
Gps Satellite Telemetry Provides New Insight into Capercaillie Tetrao urogallus Brood Movements
Upload
nationalagriculturalresearchinraCategory
view
3download
0
Characterization of a chemosensory protein (ASP3c) from honeybee(Apis mellifera L.) as a brood pheromone carrier
Loıc Briand1, Nicharat Swasdipan2, Claude Nespoulous1, Valerie Bezirard1, Florence Blon1,Jean-Claude Huet1, Paul Ebert2 and Jean-Claude Pernollet1
1Biochimie et Structure des Proteines, Unite de recherches INRA 477, Jouy-en-Josas Cedex, France; 2Department of
Biochemistry and Molecular Biology, University of Queensland, St Lucia, Australia
Chemosensory proteins (CSPs) are ubiquitous soluble smallproteins isolated from sensory organs of a wide range ofinsect species, which are believed to be involved in chemicalcommunication. We report the cloning of a honeybee CSPgene called ASP3c, as well as the structural and functionalcharacterization of the encoded protein. The protein washeterologously secreted by the yeastPichia pastoris using thenative signal peptide. ASP3c disulfide bonds were assignedafter trypsinolysis followed by chromatography and massspectrometry combined with microsequencing. The pairing(Cys(I)–Cys(II), Cys(III)–Cys(IV)) was found to be identicalto that of Schistocerca gregaria CSPs, suggesting that thispattern occurs commonly throughout the insect CSPs. CDmeasurements revealed that ASP3c mainly consists of
a-helices, like other insect CSPs. Gel filtration analysisshowed that ASP3c is monomeric at neutral pH. UsingASA, a fluorescent fatty acid anthroyloxy analogue as aprobe, ASP3c was shown to bind specifically to large fattyacids and ester derivatives, which are brood pheromonecomponents, in the micromolar range. It was unable to bindtested general odorants and other tested pheromones (sexualand nonsexual). This is the first report on a natural phero-monal ligand bound by a recombinantCSPwith ameasuredaffinity constant.
Keywords: Apis mellifera L.; brood pheromone; chemosen-sory protein; lipid-binding protein; olfaction.
In insect antennae, the first step in chemical detection is thetransport of hydrophobic signalling molecules by olfactory-binding proteins (OBPs) to receptor neurons through thesensillum lymph [1–3]. Insect OBPs are small acidic solubleproteins (13–16 kDa), highly concentrated in the sensillumlymph. They can be roughly classified as pheromone-binding proteins (PBPs) and general odorant-binding pro-teins. PBPs are supposed to be involved in sex pheromonedetection, although recent findings have brought into doubtthe currently held belief that all PBPs are specifically tunedto distinct pheromonal components [4]. In contrast, generalOBPs seem to play a more general role in olfaction bycarrying odorant molecules [5]. Although the physiologicalfunction of OBPs is not yet well understood, their essentialrole in eliciting the behavioral response and odor coding
have been demonstrated in the fruit fly [6–9] and in the fireant [10].Another class of soluble chemosensory proteins (CSPs),
which share no sequence homology with either PBPs orgeneral OBPs, has been described in insects. Such proteinshave been observed in antennae of most orders of insectssuch as Diptera [11–13], Lepidoptera [14–19], Hymenoptera[20], Coleoptera [21], Blattoidea [22], Orthoptera [23,24] andPhasmida [25–27]. Their occurrence is generally associatedwith chemosensory organs, such as legs and palpi[16,19,20,23,28,29]. They also were expressed in other sitesof the insect body, such as Drosophila melanogasterejaculatory bulb [30], Mamestra brassicae proboscis [17],labial palps of the moth Cactoblastis cactorum [14] and cellsunderlying the cuticle in Phasmatodea and Orthoptera [31].Although they have not yet been demonstrated to play anolfactory role, their tissue location and initial ligand bindingdata both support the hypothesis that CSPs are involved inchemoreception. Their natural ligands have not yet beendetermined, although binding data indicate that CSPs bindhighly hydrophobic linear molecules similar to insectpheromones and fatty acids [31,32]. CSPs do not shareany structural similarity to insect PBPs and general OBPs.They are smaller proteins (100–110 amino acid residues)containing four cysteines instead of six with conservedinterval spacing involved in two disulfide bonds [23,31].CSPs from M. brassicae and Schistocerca gregaria havebeen expressed in Escherichia coli and structurally charac-terized [31–34]. They are monomers with a high a-helicalcontent, as shown by CD and NMR spectroscopy [31,34].This was recently supported by the report of the first CSPtridimensional structure, that of the moth M. brassicae,
Correspondence to J.-C. Pernollet, Biochimie et Structure des
Proteines, Unite de recherches INRA 477, Domaine de Vilvert,
F-78352, Jouy-en-Josas Cedex, France.
Fax: 33 1 34 65 27 65, Tel.: 33 1 34 65 27 50,
E-mail: [email protected]
Abbreviations: ASA, (+/–)-12-(9-anthroyloxy)stearic acid; ASP,
antennal specific protein; BrC15-Ac, 15-bromopentadecanoic acid;
C14-Ac, myristic acid; C16-Ac, palmitic acid; C18-Ac, stearic acid;
C16-Me, methyl palmitate; C18-Me, methyl stearate; CSP, chemo-
sensory protein; OBP, odorant-binding protein; PBP, pheromone-
binding protein; RPLC, reversed phase liquid chromatography.
Enzyme: Trypsin (EC 3.4.21.4).
Note: Nucleotide sequence of ASP3c has been deposited in the
GenBank Sequence Database with accession number AF481963.
(Received 1 July 2002, accepted 30 July 2002)
Eur. J. Biochem. 269, 4586–4596 (2002) � FEBS 2002 doi:10.1046/j.1432-1033.2002.03156.x

which exhibits a novel type of a-helical fold with six helicesconnected by a–a loops [32].The honeybee (Apis mellifera L.) is able to discriminate
among a wide range of odorants [35,36]. Its OBPs, whichare evolutionary divergent from the Lepidopteran OBPs[37], were classified into three subclasses of antennal-specific proteins (ASP), namely ASP1, ASP2 and ASP3[20,38]. ASP1 has been shown to be associated with queenpheromone detection because of its higher abundance indrone, its location in sensilla placodea and ability to bind9-keto-2(E)-decenoic acid and 9-hydroxy-2(E)-decenoicacid [38,39], the most active components of the queenpheromone blend [40,41]. Based on sequence similarity,tissue-specificity and odorant binding experiments, ASP2,which does not bind any of these queen pheromonecomponents [39], was assigned to be a member of theinsect general OBP family [42]. In contrast, the ASP3subclass was classified as a CSP family due to N-terminalsequence homology [20].Recently, we purified natural ASP3c, which is com-
monly found in drones and workers and was observed asa soluble protein of 12 757.1 ± 0.3 Da. In the presentwork we report its cloning, sequencing and heterologousexpression using the yeast Pichia pastoris. Several struc-tural features of recombinant ASP3c such as its disulfidebridge pattern, secondary and quaternary structures weredetermined. We showed using a fluorescent probe bindingassay that ASP3c is able to interact with fatty acids andbrood pheromone components. This report relates the firstaffinity constant for a pheromonal ligand bound by aninsect CSP.
E X P E R I M E N T A L P R O C E D U R E S
Strains and materials
Escherichia coli strain DH5a was used for DNA subcloningand propagation of the recombinant plasmid. Pichiapastoris strain GS115 (his4) was used in the expressionstudy. Oligonucleotides were synthesized by MGW Biotech(France). pPIC3,5K was purchased from Invitrogen(France). Origins of chemicals are indicated in the text.
cDNA cloning of ASP3c
Antennae were collected from 7500 adult worker bees andpoly (A +) mRNA isolated using the Quick mRNAPurification Kit (Pharmacia). A cDNA library of 105
primary recombinants was generated from poly (A+)mRNA using the Capfinder (Clontech) cDNA cloningsystem and the kZAP II cloning vector (Stratagene). DNAsequencing was performed on 19 clones after in vitroexcision (Stratagene) using the services of the Australiangenome research facility.
Sequence analysis
Related protein sequences were identified using the BasicLocal Alignment Search Tool (BLAST 2.0) computed at theSwiss Institute of Bioinformatics. Sequence alignment wasperformed with CLUSTAL W using the Blosum 50 homologymatrix and per cent amino acid sequence identity wascalculated [43].
Construction of the expression vector
The cDNA encoding the precursor ASP3c with its nativesignal peptide was amplified by PCR using the followingprimers: 5¢ primer, 5¢-GAGCCCGGATCCACCATGAAGGTCTCAATAATT 3¢; 3¢ primer, 5¢-CTGACG GAATTCTTAAACATTAATGCC 3¢. These primers encoded aKozak consensus sequence as well as BamHI and EcoRIrestriction sites. The PCR-amplified fragment was clonedinto the BamHI and EcoRI sites of pPIC3,5K and theintegrity of the resulting construct was confirmed by DNAsequencing.
Transformation of Pichia pastoris and screeningfor ASP3c expression
The expression plasmid was linearized with BglII andtransferred into the Pichia pastoris yeast host by theelectroporation method as described in the manual (version3.0) of the Pichia expression Kit (Invitrogen). The selectionof multicopy integrants was achieved by using increasedlevels (0.5–2 mgÆmL)1) of G418 (Clontech, Ozyme, France).Large scale protein production was achieved as recentlydescribed [44] except that the protein was secreted for only3 days using buffered minimal MeOH medium at pH 8.0supplemented with 2% tryptone (Sigma) and 5 mM EDTA.During the induction period, MeOH was fed twice a day inorder to maintain a concentration of 0.5% v/v.
Purification of the recombinant ASP3c
ASP3c was purified by reversed phase liquid chromatogra-phy (RPLC). After removing insoluble components fromsupernatant containing recombinant proteins by filtration,the solutions were dialyzed 3 days at 4 �C, using a dialysistube with 8000 Da cut off (Servapor, Polylabo, France) andlyophilized. Purifications were performed using an Aqua-pore C8 column (Prep )10, 1.0 i.d. · 3.0 cm, Perkin Elmer,France). The lyophilized supernatant was resuspended ineluent A (25 mM ammonium acetate, pH 7.0) and thecolumn, equilibrated with the same eluent. After loading thesample, the column was washed extensively with eluent A.Elution was then achieved using a linear gradient to 33.3%eluent B (25 mM ammonium acetate, pH 7.0, 60% v/vacetonitrile in H2O) in the first 15 min, to 66.6% B in thenext 40 min and to 100% eluent B, in the last 10 min. Theflow rate was 2.5 mLÆmin)1 and the absorbance wasrecorded at 280 nm. The fractions containing purifiedproteins were pooled, dialyzed extensively against MilliQH2O and lyophilized.
Recombinant ASP3c characterization
SDS/PAGE (16%acrylamide) was performed using aMini-Protean II system (Bio-Rad, France) [45]. The molecularmass calibration kits low range and polypeptides (Bio-Rad)were used and the proteins stained with Serva blue G.ASP3c was analyzed by MALDI-TOF mass spectrometry.Two microlitres of purified ASP3c were mixed with 2 lL ofmatrix solution (saturated solution of sinapinic acid in 30%v/v acetonitrile, 0.2% v/v trifluoroacetic acid). One micro-litre of the mixture was applied to a stainless steel sampleplate and allowed to air dry. Mass calibration was made
� FEBS 2002 Brood pheromone binding by bee chemosensory protein (Eur. J. Biochem. 269) 4587

with the calibration mixture 2 (PE Biosystems) usingthioredoxin from Escherichia coli at 11 674.48 Da[M + H]+ and apomyoglobin from horse at16 952.56 Da [M + H]+. Mass spectra were obtainedusing a PE Biosystems Voyager-DE STR+ spectrometer inlinear mode. N-terminal amino acid sequence analysis ofproteins was performed by automated Edman degradationusing a Perkin-Elmer Procise 494-HT protein sequencerwith reagents and methods of the manufacturer.Oligomerization of the undenatured recombinant protein
was studied by exclusion-diffusion chromatography on a24-mL bed volume Superose 12 column (Pharmacia). Thecolumn was equilibrated in 100 mM potassium phosphate,pH 7.5, 150 mM NaCl, at 0.2 mLÆmin)1. Bovine serumalbumin (67 kDa), chicken egg ovalbumin (43 kDa),dimeric bovine b-lactoglobulin (36 kDa), bovine carbonicanhydrase (30 kDa), soybean trypsin inhibitor (21.5 kDa)and bovine ribonuclease A (13.7 kDa), purchased fromSigma, were employed as standards. A 100-lL sample ofpurified ASP3c was loaded at 0.5 mgÆmL)1 onto theSuperose column and the elution profiles were obtainedfrom on-line UV detection at 280 nm.CD spectra were recorded using a JASCO J-810
spectropolarimeter and analyzed as previously described[42]. ASP3c concentrations were determined using UVspectroscopy employing the extinction coefficient of11 200 M)1Æcm)1 at 276 nm, calculated according to Paceet al. [46]. Protein samples (� 1 mgÆmL)1 in 50 mM potas-sium phosphate buffer, pH 7.0) were placed in a 0.01-cmpath length cell. Baseline was recorded with phosphatebuffer. Secondary structure proportions were computedusing the algorithm of Deleage & Geourjon [47].
Peptide mapping and disulfide bridge assignment
In order to determine the disulfide bridge pairing, ASP3cwas digested by trypsin and the resulting peptides wereseparated by RPLC as described by Briand et al. [48]. Thefractions were manually collected. N-Terminal amino acidsequence and MALDI-TOF analysis in a reflector modewere performed as described previously.
Tryptophan quenching-based ligand binding
We tested tryptophan intrinsic fluorescence quenchingusing brominated fatty acid 15-bromopentadecanoic acid(BrC15-Ac) (Fluka, France) and palmitic acid (C16-Ac).BrC15-Ac and C16-Ac were weighed and dissolved in100% EtOH as 10 mM stock solutions. Tryptophanfluorescence was determined using an excitation wave-length of 285 nm and an emission wavelength of 326 nmwith 1 or 4 lM of ASP3c in 50 mM potassium phosphatebuffer, pH 7.5. The concentration of ASP3c was deter-mined using UV spectroscopy as previously described.Spectra were recorded with 4 lM ASP3c at 25 �C using aSFM 25 Kontron fluorometer with a 5-nm bandwidth forboth excitation and emission. For quenching experiments,successive 0.1-lL ligand aliquots were added to 1 mL of1 lM ASP3c solution using a 1-lL Hamilton syringe.Dissociation constants (Kd) were calculated from a plot offluorescence intensity vs. concentration of total ligand,obtained with a standard nonlinear regression method [49]using DELTAGRAPH 4.5 software.
Fluorescent fatty acid analogue-basedligand binding
Fluorophore ligand binding experiments were performedwith 1 lM ASP3c solutions in 50 mM potassium phosphatebuffer, pH 7.5. The fluorescent probe (+/–)-12-(9-anth-royloxy)stearic acid (ASA) was obtained from Sigma(France). ASA was dissolved in 10% v/v EtOH as 1 mMstock solution. Successive 0.1-lL ASA probe aliquots wereadded to 1 mL of ASP3c solution using a 1 lL Hamiltonsyringe. No cut off filter was used in the excitation beam.The excitation wavelength used for ASA was 360 nm. Oncethe binding equilibrium was reached, in approximately1 min as verified by time course experiments (not shown),the relative proportion of probe bound to ASP3c wascalculated by measuring fluorescence emission (expressed inarbitrary units). Dissociation constants (Kd) were calculatedfrom a plot of fluorescence intensity vs. concentration oftotal ligand, as described previously.
Competitive binding assay
The competitive binding assays aimed to displace fluores-cent probe with ligands were performedwith 1 lMofASP3cin 50 mM potassium phosphate buffer, pH 7.5 with 1 lMASA probe concentration. The synthetic blend correspond-ing to the major components of the queen bee mandibulargland extract was purchased from Phero Tech Inc.(Canada). It is composed of 9-keto-2(E)-decenoic acid(150 lg), 9-hydroxy-2(E)-decenoic acid (71% R-(–), 29%S-(+); 55 lg), methyl p-hydroxybenzoate (13 lg) and4-hydroxy-3-methoxyphenylethanol (1.5 lg) as defined forone queen equivalent (Qeq), the average amount ofpheromone found in the gland of mated queen [50]. Thesynthetic pheromone blend was dissolved in ethanol to afinal concentration of 10 mgÆmL)1. Other competitorligands were dissolved in 100% v/v EtOH. In order toprevent solvent competition binding [51], successive 0.1-lLfluorescent probe aliquots were added to 1 mL of ASP3csolution using a 1 lL Hamilton syringe. The EtOHconcentration in the binding-assay never exceeded 0.2%v/v leading to a maximum of relative fluorescence decay of10%. Competitor concentrations causing a fluorescencedecay to half-maximal intensity were taken as IC50 values.The apparent Kdiss values were calculated as Kdiss ¼ [IC50]/(1 + [L]/Kd) with [L] being the free fluorophore concen-tration and Kd the OBP-fluorophore complex dissociationconstant [52].
R E S U L T S
Cloning of ASP3c
In a search of putative soluble proteins involved inchemoreception, we screened a cDNA library preparedfrom honeybee antennal tissues. One clone encoded for aprotein whose N-terminal sequence matched the amino acidsequence determined on ASP3c protein purified fromhoneybee antennae [20]. Its complete cDNA sequence(Fig. 1) comprises 636 nucleotides, including an openreading frame of 393 nucleotides starting at the ATGcodon in position 40 and ending at the TAA codon atpositions 430–432. The nucleotide sequence has been
4588 L. Briand et al. (Eur. J. Biochem. 269) � FEBS 2002

deposited in the GenBank Sequence Database with acces-sion number AF481963. The open reading frame encodes a130-amino acid polypeptide. The comparison of the aminoacid sequence deduced from the cDNA sequence with thatof the N-terminal sequence of the natural ASP3c protein[20] showed that a 21-residue N-terminal signal sequence iscleaved after translation. The averagemolarmass calculatedfor the mature protein, assuming the formation of twodisulfide bridges, was 12 756.6 Da, in agreement with themeasured molar mass (12 758.3 ± 1.7 Da) of the native
protein [20]. This protein does not therefore undergo anypost-translational modification other than signal peptidecleavage and disulfide bridge formation. The calculatedisoelectric point of ASP3c was 5.9, in agreement with thosereported for other CSPs.The deduced amino acid sequence of ASP3c compared
with those of other insect CSPs and related proteins clearlyidentified ASP3c as a member of the CSP family (Fig. 2).The honeybee ASP3c protein exhibits 45% to 55% identitywith CSP-related proteins from different species, which
Fig. 1. Nucleotide and deduced amino-acid
sequences of an antennal cDNA clone fromApis
mellifera L. corresponding to ASP3c. The
nucleotides and amino acids are numbered.
The first amino acid of the ASP3c mature
sequence, indicated by a vertical arrow, is used
as a reference for amino acids numbering. The
asterisk marks the stop codon. The disulfide
bonds are indicated by a line connecting the
circled half-cystines.
Fig. 2. Sequence alignment of ASP3c with CSP isoforms and related proteins reported in other insect species.Amino-acid sequences were identified by
a BLAST search with ASP3c sequence as a query. Conserved amino acid residues are colored white with black background. Asterisks denote cysteine
residues. The conserved tryptophan residue is indicated by an arrow. Representative species are A. mellifera (line 1, EMBL accession code
AF481963), S. gregaria (line 2, EMBL accession code AF070962),M. sexta (line 3, EMBL accession code AF117599), P. americana (line 4, EMBL
accession code AF030340), L. migratoria (line 5, EMBL accession code AJ251077), D. melanogaster (line 6, EMBL accession code U05244),
H. armigera (line 7, EMBL accession code AF368375); M. brassicae (line 8, EMBL accession code AF211180), C. cactorum (line 9, EMBL
accession code U95046). Percentage identities of the predicted mature sequence proteins of 8 CSP isoforms with ASP3c are indicated.
� FEBS 2002 Brood pheromone binding by bee chemosensory protein (Eur. J. Biochem. 269) 4589

indicates a high degree of conservation between CSP-likeproteins of phylogenetically distant species. Figure 2 showsthat the four cysteines and a single conserved tryptophanare aligned for all known CSP-like sequences.
ASP3c heterologous expression
Recombinant ASP3c was secreted at high level from themethylotrophic yeast P. pastoris with its natural signalpeptide, allowing physico-chemical and functional studies.Samples of expression medium supernatants, taken atvarious time intervals, were analyzed by SDS/PAGE todetermine the optimal induction time.Only the recombinantprotein, migrating at approximately 12 kDa, was detectableby Serva blue G staining. The electrophoretic profile(Fig. 3A) reveals the protein regularly accumulating overan expression period of 3 days, while only traces of otherproteins were detected. After dialysis of culture supernatant,the recombinant protein was purified by one-step RPLC(Fig. 3B). Recombinant ASP3c eluted as a single peak at32% acetonitrile just as the natural protein did [20]. Correctprocessing of the signal sequence was verified by N-terminalanalysis of purified ASP3c, demonstrating that honeybeeinsect signal peptide was efficient for proper secretion ofheterologous ASP3c in P. pastoris. MALDI-TOF massspectrum of recombinant ASP3c (Fig. 4) showed a major
peak, together with derivatives corresponding to matrixadducts. The ASP3c mass was found to be 12 757.1 Da,which is in agreement with the theoretical and the measuredmolecular mass of the natural honeybee protein [20]. Thepurified ASP3c production reached a level of 17 mgÆL)1
over an expression period of 3 days.
Disulfide bridge assignment
The recombinant ASP3c protein was subjected to trypsindigestion, which was expected to cleave the polypeptidechain linked by a disulfide bridge. The tryptic peptidemixture was separated by RPLC (not shown) and analyzedby MALDI-TOF mass spectrometry and N-terminalsequencing. The calculated and experimentally determinedpeptide masses are listed in Table 1. All peptides greaterthan four residues in length have been identified by massspectrometry in the chromatogram and, in every case, themeasured mass was in perfect agreement with the calculatedvalue. The peptide C29–R35 ofmass 823.34was observed tobe linked to the peptide C36–K44 of mass 964.43 resultingin a peptide of 1784.76 (theoretical mass 1784.76), thusdemonstrating the existence of a disulfide bond betweenC29 and C36. Similarly, the peptide V46–K61 of measuredmass 1718.84 (theoretical mass 1718.86 with one disulfidebridge) appeared to have an internal disulfide bond betweenC55 and C58. All three peptides were completely sequencedby automated Edman sequencing. For the peptide C29–R35linked to peptide C36–K44, two corresponding N-terminalsequences were simultaneously observed in approximatelyequimolar amounts. For the peptide V46–K61, one conti-nuous sequence was found, despite the fact that threeputative trypsin cleavage sites exist within the peptide. Thefact that the peptide was not cleaved by trypsin supports thenotion of an internal disulfide bond that probably resultedin a compact conformation that was resistant to cleavage.
CD analysis and oligomerization
The far-UVCD spectrum of ASP3c at neutral pH (Fig. 5A)displayed a positive peak centered at 193 nm and two
Fig. 4. MALDI-TOF mass spectrometry analysis of the recombinant
ASP3c secreted byPichia pastoris. Sinapinic matrix adducts are shown.
Fig. 3. Electrophoretic analysis and purification of recombinant ASP3c.
(A) SDS/PAGE analysis of recombinant ASP3c secreted by Pichia
pastoris. Lane 1 shows standards (Low range and Polypeptide kits,
Bio-Rad, France) and lanes 2–5 are 50-lL aliquots of 0–3-days culturesupernatants. Proteins were visualized by Serva blueG250 staining. (B)
Chromatogram of ASP3c purification from the cell culture super-
natant by RPLC. Dashed line indicates the acetonitrile gradient.
4590 L. Briand et al. (Eur. J. Biochem. 269) � FEBS 2002
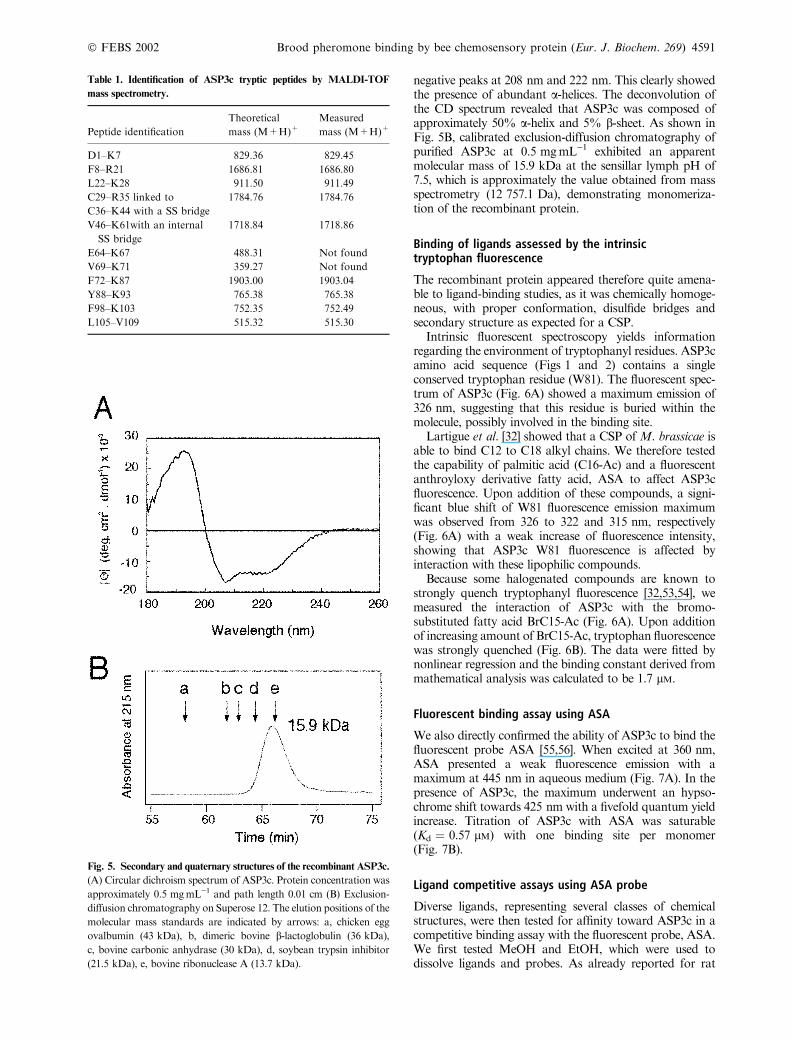
negative peaks at 208 nm and 222 nm. This clearly showedthe presence of abundant a-helices. The deconvolution ofthe CD spectrum revealed that ASP3c was composed ofapproximately 50% a-helix and 5% b-sheet. As shown inFig. 5B, calibrated exclusion-diffusion chromatography ofpurified ASP3c at 0.5 mgÆmL)1 exhibited an apparentmolecular mass of 15.9 kDa at the sensillar lymph pH of7.5, which is approximately the value obtained from massspectrometry (12 757.1 Da), demonstrating monomeriza-tion of the recombinant protein.
Binding of ligands assessed by the intrinsictryptophan fluorescence
The recombinant protein appeared therefore quite amena-ble to ligand-binding studies, as it was chemically homoge-neous, with proper conformation, disulfide bridges andsecondary structure as expected for a CSP.Intrinsic fluorescent spectroscopy yields information
regarding the environment of tryptophanyl residues. ASP3camino acid sequence (Figs 1 and 2) contains a singleconserved tryptophan residue (W81). The fluorescent spec-trum of ASP3c (Fig. 6A) showed a maximum emission of326 nm, suggesting that this residue is buried within themolecule, possibly involved in the binding site.Lartigue et al. [32] showed that a CSP ofM. brassicae is
able to bind C12 to C18 alkyl chains. We therefore testedthe capability of palmitic acid (C16-Ac) and a fluorescentanthroyloxy derivative fatty acid, ASA to affect ASP3cfluorescence. Upon addition of these compounds, a signi-ficant blue shift of W81 fluorescence emission maximumwas observed from 326 to 322 and 315 nm, respectively(Fig. 6A) with a weak increase of fluorescence intensity,showing that ASP3c W81 fluorescence is affected byinteraction with these lipophilic compounds.Because some halogenated compounds are known to
strongly quench tryptophanyl fluorescence [32,53,54], wemeasured the interaction of ASP3c with the bromo-substituted fatty acid BrC15-Ac (Fig. 6A). Upon additionof increasing amount of BrC15-Ac, tryptophan fluorescencewas strongly quenched (Fig. 6B). The data were fitted bynonlinear regression and the binding constant derived frommathematical analysis was calculated to be 1.7 lM.
Fluorescent binding assay using ASA
We also directly confirmed the ability of ASP3c to bind thefluorescent probe ASA [55,56]. When excited at 360 nm,ASA presented a weak fluorescence emission with amaximum at 445 nm in aqueous medium (Fig. 7A). In thepresence of ASP3c, the maximum underwent an hypso-chrome shift towards 425 nm with a fivefold quantum yieldincrease. Titration of ASP3c with ASA was saturable(Kd ¼ 0.57 lM) with one binding site per monomer(Fig. 7B).
Ligand competitive assays using ASA probe
Diverse ligands, representing several classes of chemicalstructures, were then tested for affinity toward ASP3c in acompetitive binding assay with the fluorescent probe, ASA.We first tested MeOH and EtOH, which were used todissolve ligands and probes. As already reported for rat
Table 1. Identification of ASP3c tryptic peptides by MALDI-TOF
mass spectrometry.
Peptide identification
Theoretical
mass (M+H)+Measured
mass (M+H)+
D1–K7 829.36 829.45
F8–R21 1686.81 1686.80
L22–K28 911.50 911.49
C29–R35 linked to 1784.76 1784.76
C36–K44 with a SS bridge
V46–K61with an internal
SS bridge
1718.84 1718.86
E64–K67 488.31 Not found
V69–K71 359.27 Not found
F72–K87 1903.00 1903.04
Y88–K93 765.38 765.38
F98–K103 752.35 752.49
L105–V109 515.32 515.30
Fig. 5. Secondary and quaternary structures of the recombinant ASP3c.
(A) Circular dichroism spectrum of ASP3c. Protein concentration was
approximately 0.5 mgÆmL)1 and path length 0.01 cm (B) Exclusion-diffusion chromatography on Superose 12. The elution positions of the
molecular mass standards are indicated by arrows: a, chicken egg
ovalbumin (43 kDa), b, dimeric bovine b-lactoglobulin (36 kDa),c, bovine carbonic anhydrase (30 kDa), d, soybean trypsin inhibitor
(21.5 kDa), e, bovine ribonuclease A (13.7 kDa).
� FEBS 2002 Brood pheromone binding by bee chemosensory protein (Eur. J. Biochem. 269) 4591

OBP-1F [51], solvent competition effects were similar withMeOH and EtOH. We used EtOH in subsequent assaysbecause of the greater solubility of lipophilic ligands in thissolvent. Although EtOH did quench the fluorescence, thedecrease was less than 10%, when the EtOH concentrationdid not exceed 0.2%. The brominated fatty acid BrC15-Ac,previously shown to quench tryptophan fluorescence ofASP3c, was also found to efficiently compete with ASA forASP3c binding (Fig. 8). The calculated apparent dissocia-tion constant (Kdiss), deduced from the half-maximalinhibition values (IC50), was 0.65 lM. We also comparedthe influence of fatty acid chain length on ASP3c binding(Table 2). Displacement of ASA was maximal for C16-Ac.It was observed to begin with C14-Ac (Kdiss ¼ 1.64 lM),increase with C16-Ac, the best ligand for ASP3c
(Kdiss ¼ 0.51 lM) and decrease with C18-Ac(Kdiss ¼ 0.80 lM). In this series, we included two fatty acidmethyl esters (C16-Me and C18-Me), described as compo-nents of brood pheromone [57,58]. They were found tocompete with ASA (Kdiss ¼ 1.02 and 1.23 lM, respectively),but less efficiently than the corresponding nonesterified fattyacids. No binding was found to occur with floral odorantsor other components of honeybee pheromones. We assayed1,8-cineol, 2-isobutyl-3-methoxypyrazine, a-pinene andb-ionone, which are known components of floral scents[59] and 2-heptanone, geraniol, citral, 2-nonanol andisoamyl acetate, known to be honeybee nonsexual phero-mones [40]. Cuticular hydrocarbons (C22 and C30n-alcanes), involved in nestmate and kin recognition [60],and the synthetic blend corresponding to the majorcomponents of the queen bee mandibular gland extract[40] were also unable to displace ASA (not shown).
Fig. 6. Binding of fatty acid assessed by intrinsic tryptophan fluores-
cence. (A) Fluorescence emission spectra of 4 lM recombinant ASP3calone (solid squares), in presence of 10 lM palmitic acid (opensquares), in presence of 10 lM BrC15-Ac (open circles) and in presenceof 10 lM ASA (solid circles). Excitation wavelength was 285 nm andthe temperature of the cuvette was maintained at 25 �C. (B) Titrationcurve of ASP3c with BrC15-Ac; open circles show experimental data,
while the solid line is the computed binding curve; excitation wave-
length was as in (A), and ASP3c concentration was 2 lM and emissionwavelength 326 nm. Fluorescence of ASP3c alone was assigned to
100% in absence of ligand.
Fig. 7. Fluorescent binding assay using ASA. (A) Fluorescence emis-
sion spectra recorded at 25 �C of 1 lM ASA in presence of 1 lMrecombinant ASP3c (open squares); solid squares indicate the fluo-
rescence of ASA alone (1 lM) and open circles that of the proteinsolution alone (1 lM). Excitation wavelength was 360 nm (B) Titrationcurves of ASP3c with ASA; open circles show experimental data, while
solid line is the computed binding curve; excitation wavelength and
ASP3c concentration were as in (A), emission wavelength was 425 nm;
ASA probe formula is inserted.
4592 L. Briand et al. (Eur. J. Biochem. 269) � FEBS 2002

D I S C U S S I O N
In this work, we have characterized ASP3c, a Hymenop-teran soluble protein found in antennal sensilla of bothworkers and drones. As previously suggested throughN-terminal sequence [20], ASP3c is a novel member of theinsect CSP family, on the basis of deduced amino acidsequence similarity and the presence of four cysteines inconserved positions. Amino acid sequence identity amongthe CSPs from different species is high (45–55%), incontrast to insect PBPs and general OBPs, which are highlydivergent.The recombinant ASP3c, expressed using the yeastPichia
pastoris, was found to be identical to the natural honeybeeprotein according to mass spectrometry and Edmansequencing. Peptide mapping experiments assigned thedisulfide pairing (C29–C36 and C55–C58). The samecysteine pairing was exhibited by four CSP isoforms fromS. gregaria [23,31]. The high homology between CSPsindicates that disulfide bond pairing Cys(I)–Cys(II) and
Cys(III)–Cys(IV) is probably shared by all members of thisinsect protein family. Because the cysteine residues of therecombinant ASP3c formed only the predicted pair ofdisulfide bonds, it is likely that the protein was properlyfolded as corroborated by circular dichroism study. TheASP3c CD spectrum and the secondary structure propor-tions obtained by its deconvolution are similar to thoseobtained by CD or by NMR analysis with recombinantS. gregaria CSP-sg4 [31] and M. brassicae CSPMbraA6[34]. This suggests a general similar global fold for insectCSPs, whichwould be composed of six a-helices as observedin the X-ray structure of a CSP fromM. brassicae [32]. Likemany CSPs from S. gregaria [23], Carausius morosus [26]and M. brassicae [34], honeybee ASP3c was demonstratedto be a monomer by gel filtration at sensillar lymph pH.This monomeric state differs from honeybee PBPs andgeneral OBPs, which were found to be dimeric undernatural conditions [39,42].Although no natural ligand for CSPs has been identified
so far, several roles have been proposed for insect CSPsbased on their tissue localization. For instance, p10 protein,expressed during leg regeneration in the cockroach Peri-planeta americana, has been proposed to be involved in limbregeneration [29,30]. Because of their localization in anten-nae, tarsi and labrum, it has been hypothesized that the classof CSPs could be involved in CO2 detection or taste [14].However, binding of neither radioactively labeled bicar-bonate nor glucose with CSPs of S. gregaria has beenobserved [23]. Recently, using a fluorescent-binding assay,CSP-sg4 from S. gregaria was observed to bind odorantswith a low affinity, whereas carboxylic acids and linearalcohols of 12, 14 and 18 carbons, as well as ethyl esters ofthe fatty acids, failed to displace the fluorescent probe [61].However, the structural analogy of CSPs with varioustransport proteins of lipidic compounds [34] suggested alipid carrier function possibly involving pheromones orother lipids, such as cuticular compounds. Diverse tritiatedpheromonal analogues and fatty acids were observed tobind the CSP of the Lepidopteran M. brassicae [15,17,18].Moreover, fluorescence quenching and modeling studiesshowed that theM. brassicaeCSPMbraA6 was able to bindbrominated alkyl alcohols or fatty acids [32].The hypothesis of lipid association is well supported by
our data. Amino acid sequence alignment revealed that thebee ASP3c contains a single conserved tryptophan residue(W81). Because tryptophanyl residues are frequentlyinvolved in ligand binding, the binding of ligands can bemonitored by a significant decrease in the intrinsic proteinfluorescence due to energy transfer from excited tryptophanresidues. Palmitic acid and the fluorescent probe ASA wereshown to weakly affect W81 fluorescence. In contrast,among halogenated compounds, which are known tostrongly quench tryptophanyl residues [53,54], a bromo-substituted fatty acid, BrC15-Ac was shown to efficientlyquench W81, as observed with M. brassicae CSPMbraA6[32]. We observed also that BrC15-Ac was able to displaceASA, suggesting that brominated fatty acid and ASA bothassociated with W81 in the same ligand binding site.The ligand binding activity of ASP3c was further
investigated using displacement of ASA, a fatty acid probewith an anthroyloxy fluorophore. Anthroyloxy derivativesemission maxima are only weakly affected by solventpolarity. Instead, they are sensitive to rotational steric
Fig. 8. Competitive binding assays of ASA with several ligands. EtOH
(r), C14-Ac (n), BrC15Ac (m), C16Ac (s), C18-Ac (h), C16-Me (d)
andC18-Me (j); fluorescence of ASA-ASP3c complexwas assigned to
100% in absence of competitor; experimental conditions were as
described in Fig. 6.
Table 2. Affinity of ligands for ASP3c measured with ASA as fluores-
cent competitive probe. d, maximal percentage of displacement reached
at high ligand concentration; IC50, ligand concentration provoking a
decay of fluorescence of half-maximal intensity; Kdiss, apparent dis-
sociation constant obtained byKdiss ¼ [IC50]/(1 + [L]/Kd) with [L] forthe free probe concentration and Kd the measured dissociation con-
stant of ASP3c-ASA complex.
Ligand d IC50 (lM) Kdiss (lM)
C14-Ac 27 4.5 1.64
BrC15-Ac 48 1.8 0.65
C16-Ac 58 1.4 0.51
C18-Ac 30 2.2 0.80
C16-Me 45 2.8 1.02
C18-Me 26 3.4 1.23
� FEBS 2002 Brood pheromone binding by bee chemosensory protein (Eur. J. Biochem. 269) 4593

hindrance at the level of the anthroyloxy moiety [62]. Whenbound to ASP3c, the maximal emission wavelength of ASAsignificantly decreased, revealing rotational constraints anda narrow binding site. Odorants, sexual/nonsexual phero-mones, fatty acids and fatty acid methyl ester derivatives(components of brood pheromone) were tested for theirability to displace ASA. As already observed with CSPM-braA6 [32], we demonstrated that ASP3c was indeed able tobind diverse fatty acids with dissociation constants in themicromolar range with a chemical specificity for aliphaticchains of 16–18 carbons. Methyl ester derivatives were alsoobserved to bind ASP3c, opposite to the S. gregaria CSP,which does not bind lipids either [61]. Moreover, the affinityconstant of BrC15-Ac deduced from ASA competition wasclose to that obtained from tryptophan fluorescencequenching. The only slightly lower affinity of the esterifiedfatty acid, compared to unsubstituted ones, suggests that thecarbonyl group of fatty acids is not essential for bindingASP3c. Themicromolar affinity of the tested fatty acids andfatty acid methyl esters for ASP3c is similar to thenanomolar to micromolar binding affinities observed forplant and vertebrate lipid binding proteins [63,64]. More-over, these apparent dissociation constants are very close tothose reported for the binding of pheromones and odorantsonto insect PBPs and general OBPs [4,42,65], suggesting aphysiological role of ASP3c in the transport of fatty acidsand their derivatives. Interestingly, ASP3c efficiently bindsfatty acid ester components of the brood pheromone,although it was found to be unable to bind nonfatty acidgeneral odorants and other tested pheromones (sexual andnonsexual). This pheromone is known to be involved in theregulation of behavioral sequence including feeding ofthe larvae, capping of the cells and thermoregulation of thebrood area in the colony.The recombinant protein ASP3c is produced in sufficient
quantity to provide enough material for the crystallizationtrials, which are currently under way. Moreover, we expectto locate the binding sites using site-directed mutagenesisaiming to clearly define the relationships between thestructure and the function of this honeybee CSP.
R E F E R E N C E S
1. Pelosi, P. (1996) Perireceptor events in olfaction. J. Neurobiol. 30,
3–19.
2. Steinbrecht, R.A. (1998) Odorant-binding proteins: expression
and function. Ann. N. Y. Acad. Sci. 855, 323–332.
3. Krieger, J. & Breer, H. (1999) Olfactory reception in invertebrates.
Science 28, 720–723.
4. Campanacci, V., Krieger, J., Bette, S., Sturgis, J.N., Lartigue, A.,
Cambillau, C., Breer, H. & Tegoni, M. (2001) Revisiting the
specificity of Mamestra brassicae and Antheraea polyphemus
pheromone-binding proteins with a fluorescence binding assay.
J. Biol. Chem. 276, 20078–20084.
5. Pelosi, P. &Maida, R. (1995) Odorant-binding proteins in insects.
Comp. Biochem. Physiol. 111B, 503–514.
6. Kim, M.S., Repp, A. & Smith, D.P. (1998) LUSH odorant-
binding protein mediates chemosensory responses to alcohols in
Drosophila melanogaster. Genetics 150, 711–721.
7. Kim,M.S. & Smith, D.P. (2001) The invertebrate odorant-binding
protein LUSH is required for normal olfactory behavior in Dro-
sophila. Chem. Senses 26, 195–199.
8. Wang, Y., Wright, N.J., Guo, H., Xie, Z., Svoboda, K., Malinow,
R., Smith, D.P. & Zhong, Y. (2001) Genetic manipulation of the
odor-evoked distributed neural activity in the Drosophila mush-
room body. Neuron 29, 267–276.
9. Carlson, J.R. (2001) Viewing odors in the mushroom body of the
fly. Trends Neurosci. 24, 497–498.
10. Krieger, M.J. & Ross, K.G. (2002) Identification of a major gene
regulating complex social behavior. Science 295, 328–332.
11. McKenna, M.P., Hekmat-Scafe, D.S., Gaines, P. & Carlson, J.R.
(1994) Putative Drosophila pheromone-binding proteins
expressed in a subregion of the olfactory system. J. Biol. Chem.
269, 16340–16347.
12. Pikielny, C.W., Hasan, G., Rouyer, F. & Rosbash, M. (1994)
Members of a family of Drosophila putative odorant-binding
proteins are expressed in different subsets of olfactory hairs.
Neuron 12, 35–49.
13. Ozaki,M.,Morisaki,K., Idei,W.,Ozaki,K.&Tokunaga,F. (1995)
A putative lipophilic stimulant carrier protein commonly found in
the taste and olfactory systems. A unique member of the phero-
mone-binding protein superfamily. Eur. J. Biochem. 230, 298–308.
14. Maleszka, R. & Stange, G. (1997) Molecular cloning, by a novel
approach, of a cDNA encoding a putative olfactory protein in the
labial palps of the moth Cactoblastis cactorum. Gene 202, 39–43.
15. Bohbot, J., Sobrio, F., Lucas, P. &Nagnan-LeMeillour, P. (1998)
Functional characterization of a new class of odorant-binding
proteins in the mothMamestra brassicae. Biochem. Biophys. Res.
Commun. 253, 489–494.
16. Picimbon, J.-F., Dietrich, K., Angeli, S., Scaloni, A., Krieger, J.,
Breer, H. & Pelosi, P. (2000) Purification and molecular cloning of
chemosensory proteins from Bombyx mori. Arch. Insect Biochem.
Physiol. 44, 120–129.
17. Nagnan-Le Meillour, P., Cain, A.H., Jacquin-Joly, E., Francois,
M.C., Ramachandran, S., Maida, R. & Steinbrecht, R.A. (2000)
Chemosensory proteins from the proboscis ofMamestra brassicae.
Chem. Senses 25, 541–553.
18. Jacquin-Joly, E., Vogt, R.G., Francois, M.C. & Nagnan-Le
Meillour, P. (2001) Functional and expression pattern analysis of
chemosensory proteins expressed in antennae and pheromonal
gland ofMamestra brassicae. Chem. Senses 26, 833–844.
19. Picimbon, J.F., Dietrich, K., Krieger, J. & Breer, H. (2001)
Identity and expression pattern of chemosensory proteins in
Heliothis virescens (Lepidoptera, Noctuidae). Insect Biochem. Mol.
Biol. 31, 1173–1181.
20. Danty, E., Arnold, G., Huet, J.-C., Huet, D., Masson, C. &
Pernollet, J.-C. (1998) Separation, characterization and sexual
heterogeneity of multiple putative odorant-binding proteins in the
honeybee Apis mellifera L. (Hymenoptera: Apidea). Chem. Senses
23, 83–91.
21. Wojtasek, H., Hansson, B.S. & Leal, W.S. (1998) Attracted or
repelled? a matter of two neurons, one pheromone binding pro-
tein, and a chiral center. Biochem. Biophys. Res. Commun. 250,
217–222.
22. Picimbon, J.-F. & Leal, W.S. (1999) Olfactory soluble proteins of
cockroaches. Insect Biochem. Molec. Biol. 29, 973–978.
23. Angeli, S., Ceron, F., Scaloni, A., Monti, M., Monteforti, G.,
Minnocci, A., Petacchi, R. & Pelosi, P. (1999) Purification,
structural characterization, cloning and immunocytochemical
localization of chemoreception proteins from Schistocerca
gregaria. Eur. J. Biochem. 262, 745–754.
24. Picimbon, J.-F., Dietrich, K., Breer, H. & Krieger, J. (2000)
Chemosensory proteins of Locusta migratoria (Orthoptera: Acri-
didae). Insect Biochem. Molec. Biol. 30, 233–241.
25. Tuccini, A., Maida, R., Rovero, P., Mazza, M. & Pelosi, P. (1996)
Putative odorant-binding protein in antennae and legs of Car-
ausius morosus (Insecta, Phasmatodea). Insect Biochem. Molec.
Biol. 1, 19–24.
26. Mameli, M., Tuccini, A., Mazza, M., Petacchi, R. & Pelosi, P.
(1996) Soluble proteins in chemosensory organs of phasmids.
Insect Biochem. Mol. Biol. 26, 875–882.
4594 L. Briand et al. (Eur. J. Biochem. 269) � FEBS 2002

27. Marchese, S., Angeli, S., Andolfo, A., Scaloni, A., Brandazza, A.,
Mazza,M., Picimbon, J.-F., Leal, W.S. & Pelosi, P. (2000) Soluble
proteins from chemosensory organs of Eurycantha calcarata (In-
sects, Phasmatodea). Insect Biochem. Mol. Biol. 30, 1091–1098.
28. Nomura, A., Kawasaki, K., Kubo, T. & Natori, S. (1992) Puri-
fication and localization of p10, a novel protein that increases in
nymphal regenerating legs of Periplaneta americana (American
cockroach). Int. J. Dev. Biol. 36, 391–398.
29. Kitabayashi, A.N., Arai, T., Kubo, T. & Natori, S. (1998)
Molecular cloning of cDNA for p10, a novel protein that increases
in the regenerating legs of Periplaneta americana (American
cockroach). Insect Biochem. Mol. Biol. 28, 785–790.
30. Dyanov, H.M. &Dzitoeva, S.G. (1995)Method for attachment of
microscopic preparations on glass for in situ hybridization, PRINS
and in situ PCR studies. Biotechniques 18, 822–824.
31. Picone, D., Crescenzi, O., Angeli, S., Marchese, S., Brandazza, A.,
Ferrara, L., Pelosi, P. & Scaloni, A. (2001) Bacterial expression
and conformational analysis of a chemosensory protein from
Schistocerca gregaria. Eur. J. Biochem. 268, 4794–4801.
32. Lartigue, A., Campanacci, V., Roussel, A., Larsson, A.M., Jones,
T.A., Tegoni, M. & Cambillau, C. (2002) X-Ray structure and
ligand binding study of a moth chemosensory protein. J. Biol.
Chem. 277, 32094–32098.
33. Campanacci, V., Spinelli, S., Lartigue, A., Lewandowsky, C.,
Brown, K., Tegoni, M. & Cambillau, C. (2001) Recombinant
chemosensory protein (CSP2) from the mothMamestra brassicae:
crystallization and preliminary crystallographic study. Acta Cryst.
D57, 137–139.
34. Campanacci, V., Mosbah, A., Bornet, O., Wechselberger, R.,
Jacquin-Joly, E., Cambillau, C., Darbon, H. & Tegoni, M. (2001)
Chemosensory protein from the moth Mamestra brassicae.
Expression and secondary structure from 1H and 15NNMR.Eur.
J. Biochem. 268, 4731–4739.
35. Hildebrand, J.G. & Shepherd, G.M. (1997) Mechanisms of
olfactory discrimination: converging evidence for common prin-
ciples across phyla. Annu. Rev. Neurosci. 20, 595–631.
36. Laska, M., Galizia, C.G., Giurfa, M. & Menzel, R. (1999)
Olfactory discrimination ability and odor structure-activity
relationships in honeybees. Chem. Senses 24, 429–438.
37. Vogt, R.G., Callahan, F.E., Rogers, M.E. & Dickens, J.C. (1999)
Odorant binding protein diversity and distribution among the
insect orders, as indicated by LAP, an OBP-related protein of the
true bug Lygus lineolaris (Hemiptera, Heteroptera). Chem. Senses
24, 481–495.
38. Danty, E., Michard-Vanhee, C., Huet, J.-C., Genecque, E.,
Pernollet, J.-C. &Masson, C. (1997) Biochemical characterization,
molecular cloning and localization of a putative odorant-binding
protein in the honey beeApis melliferaL. (Hymenoptera: Apidea).
FEBS Lett. 414, 595–598.
39. Danty, E., Briand, L., Michard-Vanhee, C., Perez, V., Arnold, G.,
Gaudemer, O., Huet, D., Huet, J.-C., Ouali, C., Masson, C. &
Pernollet, J.-C. (1999) Cloning and expression of a queen phero-
mone-binding protein in the honeybee: an olfactory-specific,
developmentally regulated protein. J. Neuroscience 19, 7468–7475.
40. Free, J.B. (1987) Pheromones of Social Bees. Chapman & Hall,
London.
41. Brockmann, A., Bruckner, D. & Crewe, R.M. (1998) The EAG
response spectra of workers and drones to queen honeybee man-
dibular gland components: the evolution of a social signal.
Naturwissenschaften 85, 283–285.
42. Briand, L., Nespoulous, C., Huet, J.-C., Takahashi, M. &
Pernollet, J.-C. (2001) Ligand binding and physico-chemical
properties of ASP2, a recombinant odorant-binding protein from
honeybee (Apis mellifera L.). Eur. J. Biochem. 268, 752–760.
43. Thompson, J.D., Higgins, D.G. &Gibson, T.J. (1994) CLUSTAL
W: improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–
4680.
44. Briand, L., Perez, V., Huet, J.-C., Danty, E., Masson, C. &
Pernollet, J.-C. (1999) Optimization of the production of a
honeybee odorant-binding protein by Pichia pastoris. Prot. Expr.
Purif. 15, 362–369.
45. Sallantin, M., Huet, J.-C., Demarteau, C. & Pernollet, J.-C. (1990)
Reassessment of commercially available molecular weight stan-
dards for peptide sodium dodecyl sulfate-polyacrylamide
gel electrophoresis using electroblotting and microsequencing.
Electrophoresis 11, 34–36.
46. Pace, C.N., Vajdos, F., Fee, L., Grimsley, G. & Gray, T. (1995)
How to measure and predict the molar absorption coefficient of a
protein. Protein Sci. 4, 2411–2423.
47. Deleage, G. & Geourjon, C. (1993) An interactive graphic pro-
gram for calculating the secondary structure content of proteins
from circular dichroism spectrum. Comput. Applic. Biosci. 9, 197–
199.
48. Briand, L., Nespoulous, C., Huet, J.-C. & Pernollet, J.-C. (2001)
Disulfide pairing and secondary structure of ASP1, an olfactory-
binding protein from honeybee (Apis mellifera L). J. Peptide Res.
58, 540–545.
49. Norris, A.W. & Li, E. (1998) Retinoid protocols. In Methods in
Molecular Biology, (Redfern, C.P.F., eds), Vol. 89, pp. 123–139.
Humana Press Inc, Totowa, NJ, USA
50. Slessor, K.N., Kalinski, L.-A., King, G.G.S., Borden, J.H. &
Winston, M.L. (1988) Semiochemicals basis of the retinue
response to queen honey bees. Nature 332, 354–356.
51. Briand, L., Nespoulous, C., Perez, V., Remy, J.-J., Huet, J.-C. &
Pernollet, J.-C. (2000) Ligand-binding properties and structural
characterization of a novel rat odorant-binding protein variant.
Eur. J. Biochem. 267, 3079–3089.
52. Ramoni, R., Vincent, F., Grolli, S., Conti, V., Malosse, C., Boyer,
F.D., Nagnan-Le Meillour, P., Spinelli, S., Cambillau, C. &
Tegoni, M. (2001) The insect attractant 1-octen-3-ol is the natural
ligand of bovine odorant-binding protein. J. Biol. Chem. 276,
7150–7155.
53. Jain, M.K. & Maliwal, B.P. (1985) The environment of trypto-
phan in pig pancreatic phospholipase A2 bound to bilayers.
Biochim. Biophys. Acta 814, 135–140.
54. de Foresta, B., Champeil, P. & Le Maire, M. (1990) Different
classes of tryptophan residues involved in the conformational
changes characteristic of the sarcoplasmic reticulum Ca2(+)-
ATPase cycle. Eur. J. Biochem. 194, 383–388.
55. Mei, B., Kennedy, M.W., Beauchamp, J., Komuniecki, P.R. &
Komuniecki, R. (1997) Secretion of a novel, developmentally
regulated fatty acid-binding protein into the perivitelline fluid of
the parasitic nematode, Ascaris suum. J. Biol. Chem. 272, 9933–
9941.
56. Lechner, M., Wojnar, P. & Redl, B. (2001) Human tear lipocalin
acts as an oxidative-stress-induced scavenger of potentially
harmful lipid peroxidation products in a cell culture system.
Biochem. J. 356, 129–135.
57. Le Conte, Y., Arnold, G., Trouiller, J. & Masson, C. (1990)
Identification of a brood pheromone in honeybees.
Naturwissenschaften 77, 334–336.
58. Mohammedi, A., Paris, A., Crauser, D. & Le Conte, Y. (1998)
Effect of alphatic esters on ovary development of queenless bees
(Apis mellifera L.). Naturwissenschaften 83, 455–458.
59. Knudsen, J.T., Tollsten, L. &Bergstrom, L.G. (1993) Floral scents
– a checklist of volatile compounds isolated by head-space tech-
niques. Phytochemistry 33, 253–280.
60. Salvy, M., Martin, C., Bagneres, A.G., Provost, E., Roux, M., Le
Conte, Y. & Clement, J.-L. (2001) Modifications of the cuticular
hydrocarbon profile of Apis mellifera worker bees in the presence
of the ectoparasitic mite Varroa jacobsoni in brood cells.
Parasitology 122, 145–159.
� FEBS 2002 Brood pheromone binding by bee chemosensory protein (Eur. J. Biochem. 269) 4595

61. Ban, L., Zhang, L., Yan, Y. & Pelosi, P. (2002) Binding properties
of a locust’s chemosensory protein. Biochem. Biophys. Res. Com-
mun. 293, 50–54.
62. Matayoshi, E.D. & Kleinfeld, A.M. (1981) Emission wavelength-
dependent decay of the 9-anthroyloxy-fatty acid membrane
probes. Biophys. J. 352, 212–235.
63. Douliez, J.P., Michon, T. & Marion, D. (2000) Steady-state
tyrosine fluorescence to study the lipid-binding properties of a
wheat non-specific lipid-transfer protein (nsLTP1). Biochim.
Biophys. Acta 1467, 65–72.
64. Curry, S., Brick, P. & Franks, N.P. (1999) Fatty acid binding to
human serum albumin: new insights from crystallographic studies.
Biochim. Biophys. Acta 1441, 131–140.
65. Kowcun, A., Honson, N. & Plettner, E. (2001) Olfaction in the
gypsy moth, Lymantria dispar: effect of pH, ionic strength, and
reductants on pheromone transport by pheromone-binding pro-
teins. J. Biol. Chem. 276, 44770–44776.
4596 L. Briand et al. (Eur. J. Biochem. 269) � FEBS 2002
Copyright © 2022 FDOKUMEN




















