Changing animal use at Neolithic Çatalhöyük, Turkey
27
ANCIENT NEAR EASTERN STUDIES ARCHAEOZOOLOGY OF THE NEAR EAST X Proceedings of the Tenth International Symposium on the Archaeozoology of South-Western Asia and Adjacent Areas Edited by Bea DE CUPERE, Veerle LINSEELE and Sheila HAMILTON-DYER PEETERS LEUVEN – PARIS – WALPOLE, MA. 2013 SUPPLEMENT 44
Transcript of Changing animal use at Neolithic Çatalhöyük, Turkey

ANCIENT NEAR EASTERN STUDIES
ARCHAEOZOOLOGY OF THE NEAR EAST X
Proceedings of the Tenth International Symposium on the
Archaeozoology of South-Western Asia and Adjacent Areas
Edited by
Bea DE CUPERE, Veerle LINSEELE and Sheila HAMILTON-DYER
PEETERSLEUVEN – PARIS – WALPOLE, MA.
2013
SUPPLEMENT 44

CONTENTS
Preface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ixBea DE CUPERE, Veerle LINSEELE, Sheila HAMILTON-DYER
Faunal remains from Wadi Abu Tulayha: a PPNB outpost in the steppe-desert of southern Jordan . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
Hitomi HONGO, Lubna OMAR, Hiroo NASU, Petra KRÖNNECK and Sumio FUJII
Evaluating differences in animal consumption patterns from PPNB houses at Tell Halula (Euphrates Valley, Syria) . . . . . . . . . . . . . . . . . . . . . . . . . . 27
Carlos TORNERO, Miquel MOLIST and Maria SAÑA
Changing animal use at Neolithic Çatalhöyük, Turkey . . . . . . . . . . . . . . . 45Nerissa RUSSELL, Katheryn C. TWISS, David C. ORTON and G. Arzu DEMIRERGI
Herding and settlement identity in the central Anatolian Neolithic: herding deci-sions and organisation in Çatalhöyük, elucidated through oxygen isotopes and microwear in sheep teeth . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
Elizabeth HENTON
Chalcolithic pig remains from Çamlıbel Tarlası, Central Anatolia . . . . . . . . . 101László BARTOSIEWICZ, Roz GILLIS, Linus GIRDLAND FLINK, Allowen EVIN, Thomas CUCCHI, Rus HOELZEL, Una VIDARSDOTTIR, Keith DOBNEY, Greger LARSON and Ulf-Dietrich SCHOOP
Bronze and Iron Age subsistence changes in the Upper Tigris: zooarchaeology of Operation E at Ziyaret Tepe, south-eastern Turkey . . . . . . . . . . . . . . . . 121
Tina L. GREENFIELD-JONGSMA and Haskel J. GREENFIELD
New data on the exploitation of animal resources in the Upper Tigris River area (Turkey) during the second and first millennia BC . . . . . . . . . . . . . . . . 145
Rémi BERTHON
What is inside this pit? Micro- and macrofaunal investigations at Chalcolithic and Early Bronze Age Ovçular Tepesi (Nakhchivan, Azerbaijan) . . . . . . . . . . . . 163
Zsófia E. KOVÁCS, Rémi BERTHON, Wim VAN NEER and Thomas CUCCHI
Royal tombs with horse sacrifices in Nerkin Naver, Armenia (Middle Bronze Age) 173Hakob SIMONYAN and Ninna MANASERYAN

viii CONTENTS
The mysteries of Egyptian Nile perch (Lates niloticus). The case of Tell Tweini (Syria, Middle Bronze Age–Iron Age) . . . . . . . . . . . . . . . . . . . . . . . . 209
Veerle LINSEELE, Wim VAN NEER and Joachim BRETSCHNEIDER
Zooarchaeology and social identity in Bronze Age and Iron Age Israel: a research framework . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 227
Nimrod MAROM and Guy BAR-OZ
Persian period dog burials in the Levant: new evidence from Tell el-Burak (Lebanon) and a reconsideration of the phenomenon . . . . . . . . . . . . . . . . . . . . . 243
Canan ÇAKIRLAR, Verena AMER, Jens KAMLAH and Hélène SADER
Environmental influence on animal exploitation and meat consumption during the Early Islamic period in Syria. A case study from Qasr al-Hayr al-Sharqi and al-Hadir . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265
Jacqueline STUDER, Denis GENEQUAND and Marie-Odile ROUSSET
A leopard in the Predynastic Elite Cemetery HK6 at Hierakonpolis, Egypt . . . . 283Wim VAN NEER, Bea DE CUPERE and Renée FRIEDMAN
A tale of two sites: Old Kingdom subsistence economy and the infrastructure of pyramid construction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 307
Richard W. REDDING
Animal bone assemblages from a Bronze Age palace at Tell el-Dab’a, Egypt . . . . 323Günther Karl KUNST
Tomb of the Dogs in Gebel Asyut al-gharbi (Middle Egypt, Late to Ptolemaic/Roman period): preliminary results on the canid remains . . . . . . . . . . . . . 343
Chiori KITAGAWA
Pampered puss? Cats from the Roman port of Myos Hormos at Quseir, Egypt . . . 357Sheila HAMILTON-DYER
The goats (Capra hircus L.) from Kerma (Sudan) (2050–1750 BC) – A contribu-tion to the knowledge of African goats . . . . . . . . . . . . . . . . . . . . . . . 373
Louis CHAIX
New evidence for dog butchering from prehistoric coastal sites in the Sultanate of Oman . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 403
Elena MAINI and Antonio CURCI
List of all contributors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 417

1 Marciniak and Czerniak 2007; Biehl et al. 2012. 2 Mellaart 1962, 1963, 1964, 1965, 1966, 1967, 1998; Todd 1976. 3 Hodder 1996, 1999, 2000, 2005a, 2005b, 2006a, 2006b, 2007. 4 Farid 2008.
CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY
Nerissa RUSSELL, Katheryn C. TWISS, David C. ORTON and G. Arzu DEMIRERGI
ABSTRACT
Recent work at Çatalhöyük has focused on the later levels of the Neolithic East Mound, and on the Chalcolithic West Mound. This permits us for the first time to examine animal use through virtually the whole sequence at the site. Earlier work has shown considerable stability in the balance of taxa, hunting and herding practices, and butchery through roughly the first two-thirds of the occupation on the East Mound. In the later levels of the East Mound, changes are evident. Domestic sheep/goat (mainly sheep) become even more dominant; this appears to be primarily an increase in sheep rather than a decrease in other taxa. Domestic cattle appear at this point, although wild cattle maintain their symbolic importance as seen in, for example, architectural installations. Changes in cut mark patterns, fragmentation, and body-part distributions suggest new forms of butchery, cooking, and commensality. Prelimi-nary work on West Mound materials indicates a reduction in both wild and domestic cattle and diminished symbolic salience for cattle and wild animals. Caprine herding retains its importance, with the possible introduction of a new (larger) variety of sheep.
KEYWORDS
Çatalhöyük, Neolithic, cattle, sheep, herding practices, cooking
INTRODUCTION
Çatalhöyük is a large early agricultural site in central Anatolia (Fig. 1), with the larger East Mound (Neolithic) occupied c.7300–6200 cal BC, with a population in the thousands, and the smaller West Mound occupied during the Chalcolithic period, following immedi-ately on the Neolithic occupation on the East Mound or even overlapping slightly with it.1 The better-known East Mound settlement is characterised by closely packed mudbrick houses with roof entrances, subfloor burials, wall paintings, and installations of animal parts into architectural features. James Mellaart2 first excavated the site in the 1960s. This article derives from the on-going Çatalhöyük Research Project directed by Ian Hodder3 since 1993. Here we report on the animal bones from the second major phase of excavation in the Çatalhöyük Research Project, which targeted the later levels on the East Mound: roughly VI–I in Mellaart’s levels, or South.P–T and 4040.G–I in Hodder’s levels (Fig. 2).4 We also include some very preliminary information from the West Mound excavations led by Peter Biehl and Eva Rosenstock.

46 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
5 Perkins 1969. 6 Ducos 1988. 7 Russell and Martin 2005. 8 Farid 2008.
To review briefly, Dexter Perkins studied the animal bones from Mellaart’s excavations, and argued for local cattle domestication.5 Cattle formed c.70 per cent of the assemblage, and Perkins believed that the sheep and goats, and all other fauna except dogs, were wild. Perkins never published a full report, however. Pierre Ducos later published a much more thorough analysis of a portion of the assemblage, and argued for proto-domestication of the cattle.6 Except for some mixed material from Hellenistic pits, all the animal bones from Mellaart’s excavations have since been lost. As Ducos realised, the lack of sieving created recovery bias problems.
Study of the animal bones from the first phase of excavations (1995–1999) in the Çatal-höyük Research Project demonstrated the severity of the recovery bias. With sieving, the position of caprines and cattle are roughly reversed, with caprines forming c.70 per cent of the assemblage in the earlier levels targeted in these excavations, and cattle c.15–20 per cent. With better samples and the aid of other studies in the region in the intervening decades, we are confident that the sheep and goats are largely domestic (with the occasional wild individual) from the beginning of the occupation, and the cattle are wild through the earlier levels. All other taxa, except for dogs, appear to be wild. In the earlier levels there was little change through time, although spatial and contextual variation was evident. Notably, we documented a bias to bulls (male cattle) in feasting and special deposits.7
We have now completed analysis of material from the second phase of excavation, con-ducted from 2003 through 2008, which as previously stated targeted the East Mound’s later phases of occupation. We now have nearly a complete sequence at the site (with the latest Neolithic levels on the East Mound, from the Team Poznan [TP] area, to be reported separately). Here we report on the South and 4040 Areas (Fig. 2), including material from the earlier levels of the South Area excavated in the first phase. The South Area has a deep sounding with the longest sequence; however, it currently has a gap that may be filled with part of the 4040 sequence. Additionally, we include some very preliminary results from the on-going excavations in Trenches 5 and 7 and Building 25 (Trench 1) on the West Mound, which appear to be Early Chalcolithic I (early sixth millennium BC).
Until recently, the Çatalhöyük Research Project continued to use Mellaart’s system of building levels (0–XII, with XII being the earliest). The lower levels of the site (Mellaart’s VI–XII) matched quite well with the new excavations. However, as the focus shifted to the later levels, it became clear that Mellaart’s Levels 0–V did not match the stratigraphy very well, and a new system of stratigraphic levels was necessary (Fig. 2).8 Also, since the Çatal-höyük Research Project has excavated in several discontiguous areas of the site, the areas cannot be linked stratigraphically, so these must have independent phasing systems. In Hodder’s new phasing system, the South Area runs from South.G (early) to South.T, with virtually no material currently available from South.N–O. Large horizontal exposure was emphasised in the 4040 Area, at the expense of depth; here we have material from levels

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 47
9 Bayliss and Farid 2008, 2010.
4040.G–I. The letters of the 4040 levels do not match those of the South Area; 4040.G probably corresponds roughly to South.N–O, while 4040.H–I fall somewhere in the South.P–T sequence. A rigorous program of radiocarbon dating is currently underway,9 but the results are not yet available. Therefore the dates given in Figure 2 should be regarded as tentative and very rough approximations, especially for the later levels.
We have now recorded just over one million bones from all site areas, or about 30 per cent of the animal bone excavated. Since the bone is for the most part highly fragmented, only about 10 per cent of these specimens can be identified to taxon beyond size classes, while the rest provide mainly taphonomic information. In the first phase of excavation and analysis, we aimed to record a representative sample of context types. Based on that experi-ence, in the second phase we have targeted primary (deliberately placed or discarded where used) and secondary (discarded away from the location of use/consumption) deposits: those related to the occupation level in which they occur. Redeposited material in deposits such as fills and construction material (tertiary deposits) is less informative about human behav-iour. We were able to record the material from all indoor primary and secondary deposits from the second phase of excavation, and a substantial portion of the outdoor midden deposits. Middens accumulated in open areas between houses, created when a house was not immediately rebuilt. While some activities probably occurred in middens, they mainly contain secondary deposits dumped from activities elsewhere, particularly in or on the roofs of nearby houses. The middens contain rich, dense deposits of a variety of materials, includ-ing animal bone, deriving from a range of activities, including daily meals. We also find
Fig. 1. Map of Anatolia, showing Çatalhöyük and other sites mentioned in the text.

48 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
10 Russell and Martin 2005. 11 E.g. Russell and Martin 2005.
feasting deposits, as pockets in middens or in other locations, that are distinguished as con-centrations of large amounts of rapidly deposited, minimally processed bones, often from large animals.10
For many purposes it is necessary to lump assemblages from different levels in order to achieve a viable sample size. Based on patterning in both the faunal and other materials, we have combined them into meaningful units. For the South Area, we separate South.G, from which the only excavated material comes from an area on the edge of the early settlement; it is thus both the earliest material and contextually distinct. As noted in earlier publications,11
Fig. 2. Çatalhöyük plan, showing excavation areas with their phasing and approximate dates. Note that dating is tentative pending a radiocarbon study in progress; dates here are based on earlier work
(Cessford et al. 2005; Last and Gibson in prep).

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 49
12 Russell et al. in press. 13 Watson 1979; for more detail on all taxa identified, see Russell et al. in press.
most of the faunal variation in South.H–M appears to be contextual, due to varying balances of different context types that were excavated in each level. After the gap in South.N–O (which will be addressed in the final phase of excavations), the material from South.P–T shows some notable differences from the earlier levels, to be detailed below. Faunally, 4040.H–I are very similar to South.P–T, while 4040.G exhibits some of the changes seen in the later South levels, but in other ways more resembles the earlier levels. We make a rough distinction between the “earlier” (South.G–M) and “later” (South.P–T and 4040) levels. In the future, with firmer dating, finer distinctions may be possible. Finally, the Early Chalco-lithic I material from the West Mound, still in process of excavation, is included as it pro-vides a window into later developments at the site.
A fuller discussion of methods and results will be offered in the forthcoming report on the South and 4040 material.12 Here we highlight some of the key changes evident in the later levels of the East Mound and continuing in the Early Chalcolithic on the West Mound. We focus on the appearance of domestic cattle, changes in sheep herding practices and a sharp increase in the size of caprine flocks, and a shift to cooking more meat off the bone in the later levels.
RESULTS
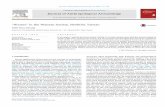
Figure 3 summarises the proportions of the major taxa through time, based on Watson’s diagnostic zones.13 South.G is contextually distinctive as the area so far excavated was on the edge of the settlement at that time, and contains dumps of heavily processed waste from daily consumption. South.I has a very small sample that is likely not representative; other-wise the assemblages from South.H–M are fairly consistent. In the later periods (South.P–T and the 4040 Area), caprines become still more abundant. Cattle appear to play a somewhat larger role in the 4040 Area than South.P–T, but this is due to a greater number of feasting and other special deposits in the 4040 Area.
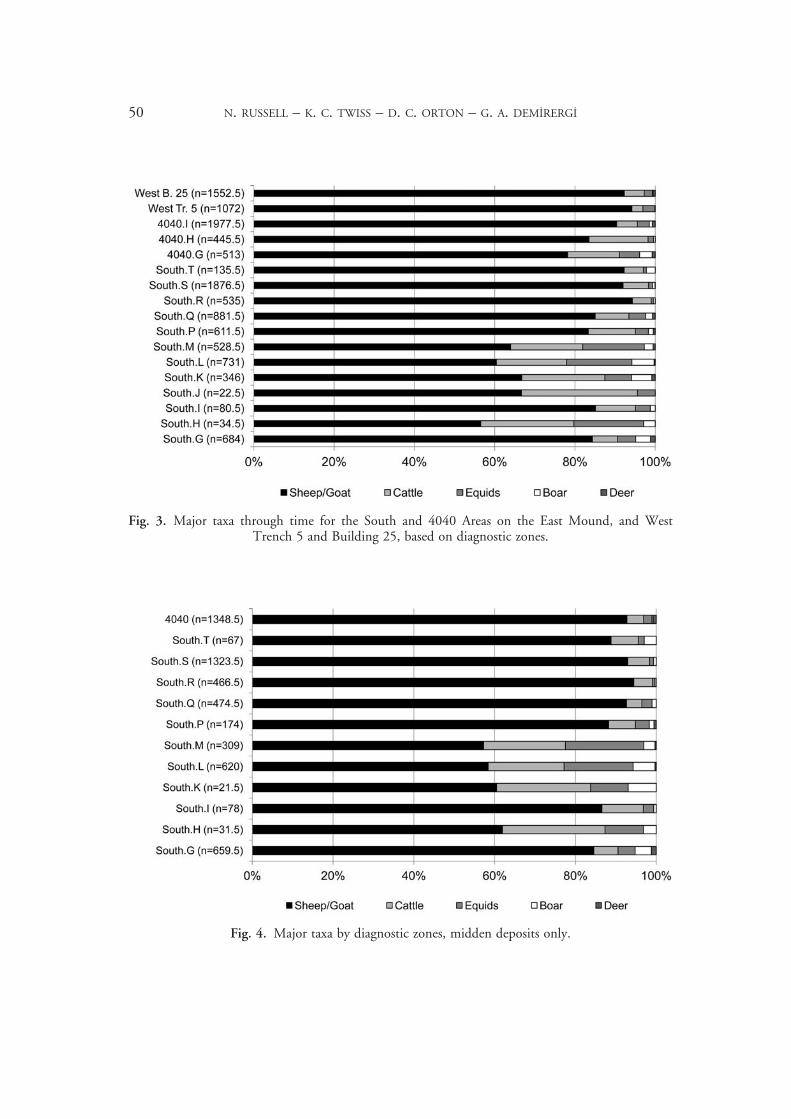
Middens, where most waste is dumped, are the best representation of overall consump-tion. Figure 4 shows the major taxa from middens through time. West Mound material does not appear because no real middens have yet been found there, and the 4040 Area phases have been lumped because the midden material is nearly all from 4040.I. Without the special deposits found in the buildings, the 4040 assemblage closely resembles those from South.P–T. To explore whether the proportional change in taxa in the later levels is the result of an increase in caprines or a decrease in cattle and other taxa, density in middens (diagnostic zones per litre) is the best measure, although not perfect. Depositional rates surely vary (indeed South.Q has middens that are remarkably dense in all materials), but discard rates should be similar enough through time that major differences are plausibly meaningful. Combining the levels into South.G (earliest site-edge deposits), South.H–M (early on-site deposits), South.P–T (later levels), and the 4040 Area, it is clear that the change in later levels is primarily a sharp increase in the numbers of caprines (Fig. 5).
50 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
Fig. 3. Major taxa through time for the South and 4040 Areas on the East Mound, and West Trench 5 and Building 25, based on diagnostic zones.
Fig. 4. Major taxa by diagnostic zones, midden deposits only.
CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 51
14 Russell and Martin 2005; Russell et al. 2005. 15 Lemppenau 1964; Greenfield 2006.
Cattle
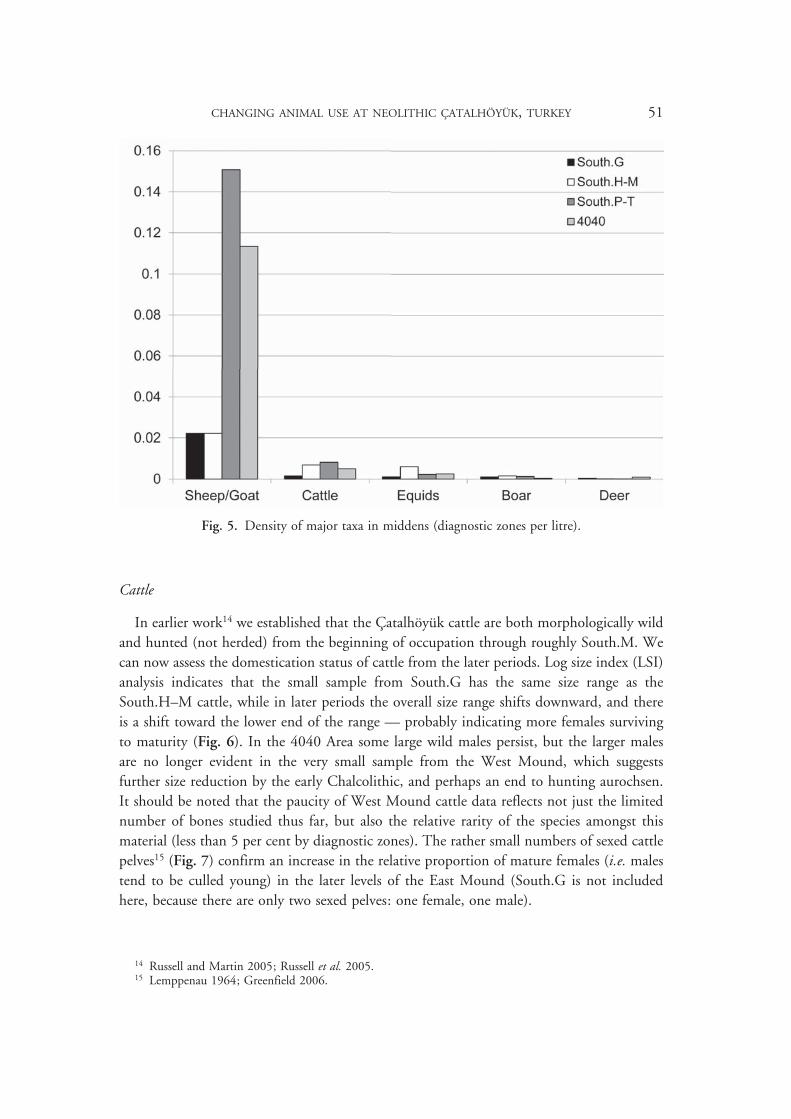
In earlier work14 we established that the Çatalhöyük cattle are both morphologically wild and hunted (not herded) from the beginning of occupation through roughly South.M. We can now assess the domestication status of cattle from the later periods. Log size index (LSI) analysis indicates that the small sample from South.G has the same size range as the South.H–M cattle, while in later periods the overall size range shifts downward, and there is a shift toward the lower end of the range — probably indicating more females surviving to maturity (Fig. 6). In the 4040 Area some large wild males persist, but the larger males are no longer evident in the very small sample from the West Mound, which suggests further size reduction by the early Chalcolithic, and perhaps an end to hunting aurochsen. It should be noted that the paucity of West Mound cattle data reflects not just the limited number of bones studied thus far, but also the relative rarity of the species amongst this material (less than 5 per cent by diagnostic zones). The rather small numbers of sexed cattle pelves15 (Fig. 7) confirm an increase in the relative proportion of mature females (i.e. males tend to be culled young) in the later levels of the East Mound (South.G is not included here, because there are only two sexed pelves: one female, one male).
Fig. 5. Density of major taxa in middens (diagnostic zones per litre).

52 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
Fig. 6. Log size index distribution of cattle measurements, using the Ullerslev cow (Degerbøl and Fredskild 1970) as the standard.
CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 53
Fig. 7. Sexed cattle pelves from the East Mound.
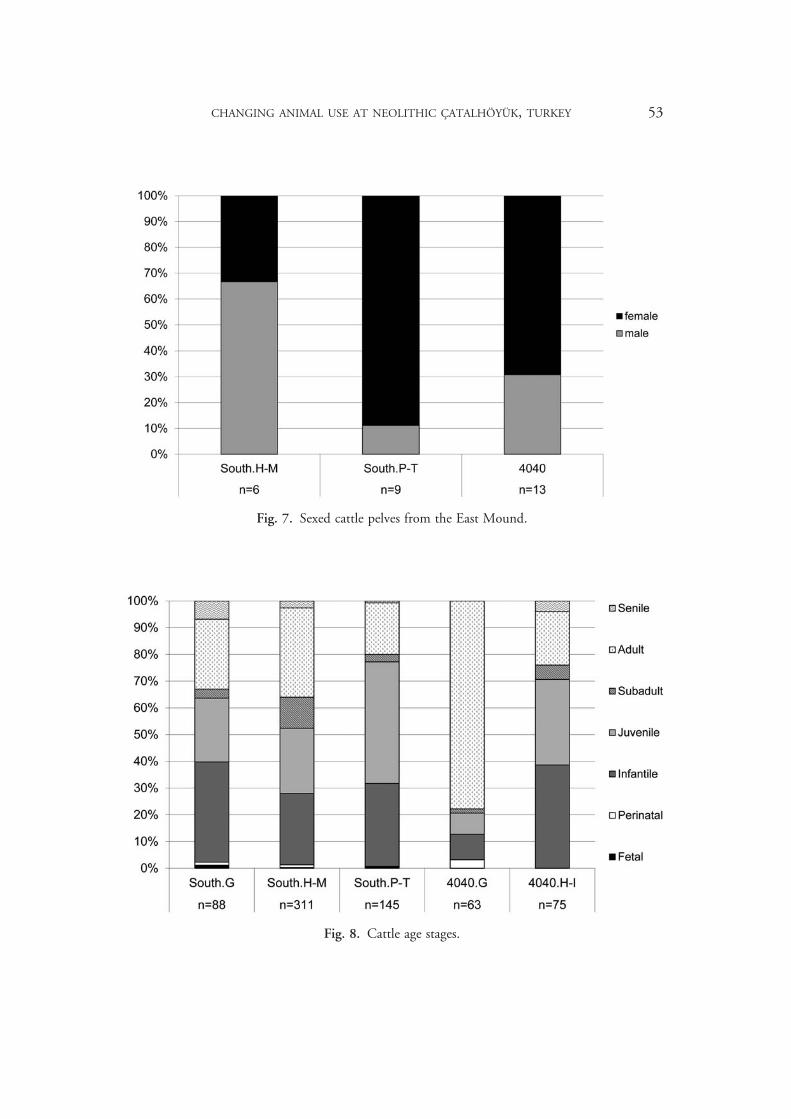
Fig. 8. Cattle age stages.
54 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
16 Silver 1969; Bökönyi 1972; Grant 1982; Grigson 1982. 17 Bökönyi 1970; Uerpmann 1978. 18 Twiss and Russell 2009.
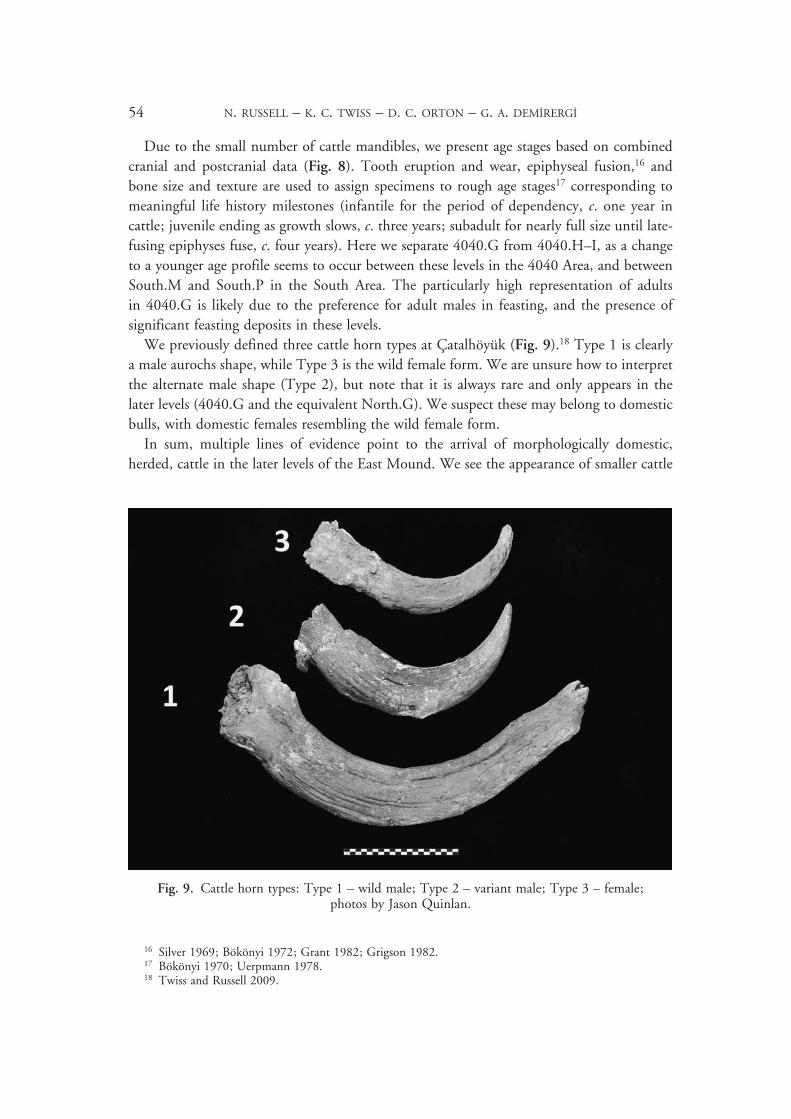
Due to the small number of cattle mandibles, we present age stages based on combined cranial and postcranial data (Fig. 8). Tooth eruption and wear, epiphyseal fusion,16 and bone size and texture are used to assign specimens to rough age stages17 corresponding to meaningful life history milestones (infantile for the period of dependency, c. one year in cattle; juvenile ending as growth slows, c. three years; subadult for nearly full size until late-fusing epiphyses fuse, c. four years). Here we separate 4040.G from 4040.H–I, as a change to a younger age profile seems to occur between these levels in the 4040 Area, and between South.M and South.P in the South Area. The particularly high representation of adults in 4040.G is likely due to the preference for adult males in feasting, and the presence of significant feasting deposits in these levels.
We previously defined three cattle horn types at Çatalhöyük (Fig. 9).18 Type 1 is clearly a male aurochs shape, while Type 3 is the wild female form. We are unsure how to interpret the alternate male shape (Type 2), but note that it is always rare and only appears in the later levels (4040.G and the equivalent North.G). We suspect these may belong to domestic bulls, with domestic females resembling the wild female form.
In sum, multiple lines of evidence point to the arrival of morphologically domestic, herded, cattle in the later levels of the East Mound. We see the appearance of smaller cattle
Fig. 9. Cattle horn types: Type 1 – wild male; Type 2 – variant male; Type 3 – female; photos by Jason Quinlan.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 55
19 De Cupere and Duru 2003; Arbuckle and Makarewicz 2009; Arbuckle 2013. 20 Payne 1987. 21 Pearson in press.
and a new male horn shape, and males in particular appear to have been culled younger. It is difficult to date the appearance of domestic cattle with precision at this point but, if we take the new horn shape as diagnostic and accept 4040.G as equivalent to South.N–O or Mellaart’s Level VI, we can infer their presence by c.6500–6400 cal BC (Fig. 2). This aligns Çatalhöyük with Erbaba in terms of the timing of the first appearance of cattle in central and south-west Anatolia, supporting Arbuckle’s argument that the adoption of domestic cattle (initially herded to the east) was resisted in central Anatolia, actually appearing earlier to the west at Bademagacı, and at Höyücek at around the same time as in central Anatolia.19 Cattle herding was practised only on a small scale, however. The adoption of domestic cat-tle coincides, intriguingly, with a sharp increase in the size of the caprine flocks, while the total number of cattle remains steady.
Sheep
Sheep always form more than 80 per cent of the Çatalhöyük caprines, with little change in the ratio through time. We will therefore focus henceforth on the sheep, sometimes elid-ing them with the caprines. The sheep are domestic throughout, in terms of size and mor-tality profile, although a few wild individuals are present, visible in the outliers on the high end of Figure 10. There is perhaps a slight size reduction in the later levels of the East Mound, and a shift to the smaller (female) end of the size range. The West Mound sheep are notably larger, suggesting the introduction of a new population.
Mortality profiles (Fig. 11) based on Payne’s tooth wear system20 show a slight shift in the later levels, compared to South.H–M, toward his stage C (early juvenile), and less of a peak in stage D (late juvenile); the earliest levels (South.G) resemble the later levels. This is hard to interpret in terms of herding strategies but consistent across the South and 4040 Areas; it may result from a seasonal shift in when caprines are near the settlement. There may also be fewer older adults in South.H–M, although this may be a sample size effect. The sexed pelves confirm an increase in mature females (heavier immature male cull) in the later levels, and also (with a rather small sample) in South.G (Fig. 12). Indeed, the higher male sex ratio in South.H–M is anomalous: any herded population should have more mature females than males, to preserve the breeding stock. It is therefore likely that some caprines were slaughtered and consumed away from the settlement. Possibly the sheep were grouped into larger herds in these levels (although they may have been owned by individuals or households), segregated by age and sex, with some males kept closer for slaughter. In the later periods caprines increase overall but may have been kept in smaller herds belonging to single households, or a few related households, in mixed age and sex groups. Increasing isotopic variability in the sheep from the later periods, indicating that different flocks targeted distinct parts of the landscape, supports this model.21 Isolated and perhaps inbred herds in the later periods are also indicated by a cluster of a distinctive, but
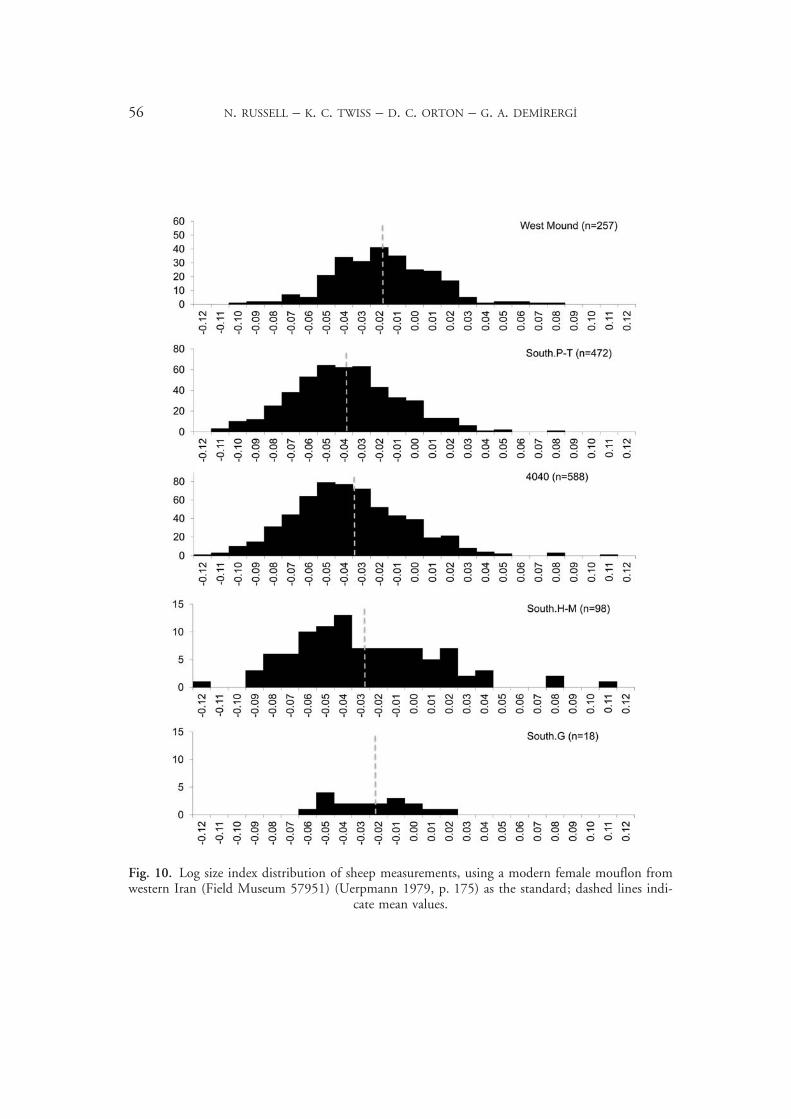
56 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
Fig. 10. Log size index distribution of sheep measurements, using a modern female mouflon from western Iran (Field Museum 57951) (Uerpmann 1979, p. 175) as the standard; dashed lines indi-
cate mean values.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 57
Fig. 11. Caprine mortality profiles based on mandibular wear (Payne stages, with approximate ages).
58 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
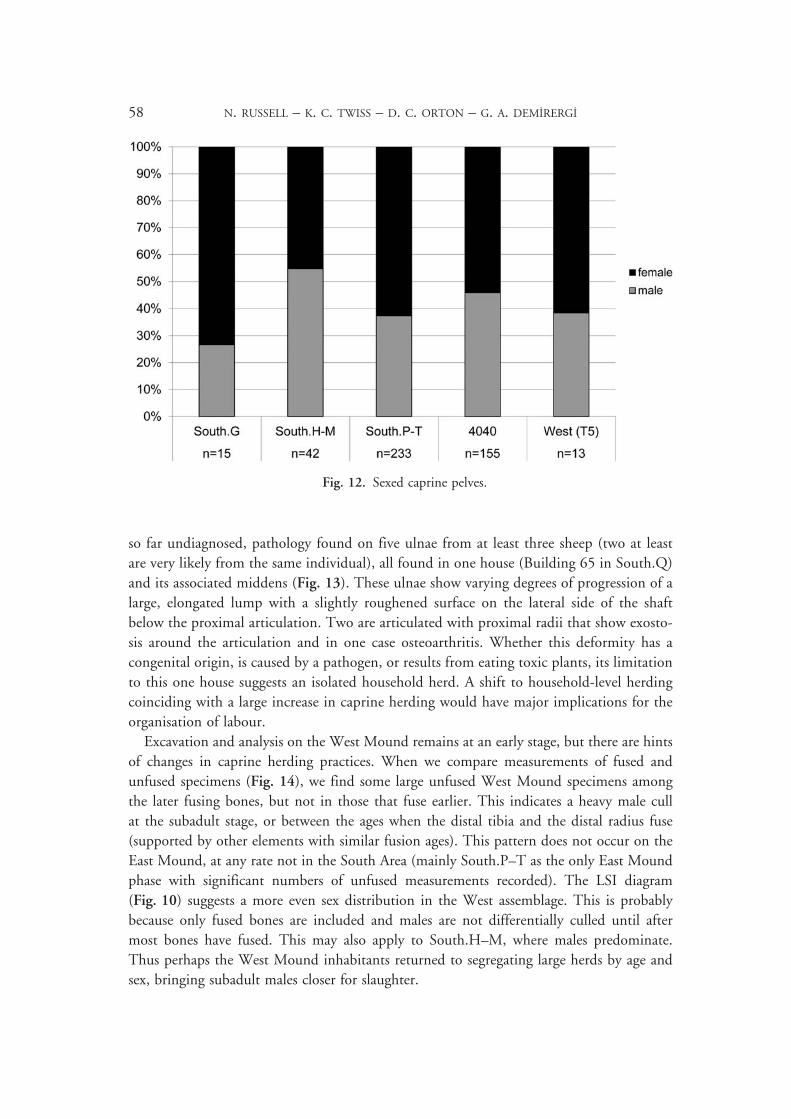
so far undiagnosed, pathology found on five ulnae from at least three sheep (two at least are very likely from the same individual), all found in one house (Building 65 in South.Q) and its associated middens (Fig. 13). These ulnae show varying degrees of progression of a large, elongated lump with a slightly roughened surface on the lateral side of the shaft below the proximal articulation. Two are articulated with proximal radii that show exosto-sis around the articulation and in one case osteoarthritis. Whether this deformity has a congenital origin, is caused by a pathogen, or results from eating toxic plants, its limitation to this one house suggests an isolated household herd. A shift to household-level herding coinciding with a large increase in caprine herding would have major implications for the organisation of labour.
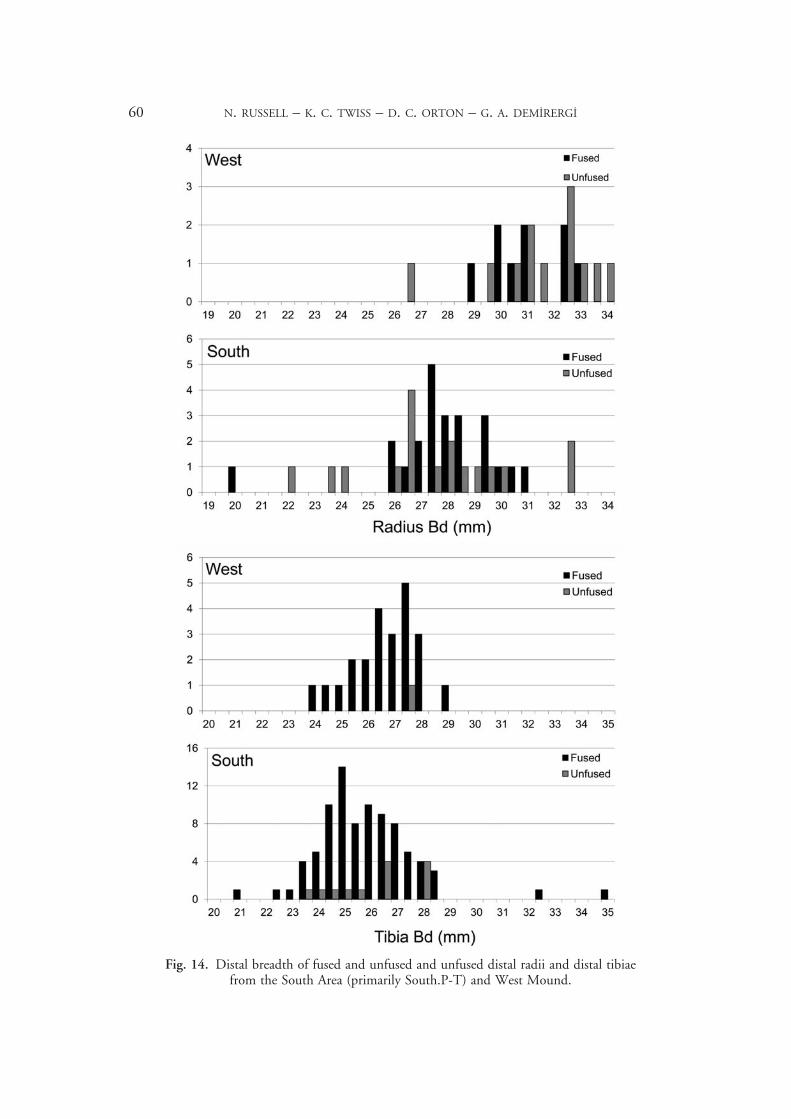
Excavation and analysis on the West Mound remains at an early stage, but there are hints of changes in caprine herding practices. When we compare measurements of fused and unfused specimens (Fig. 14), we find some large unfused West Mound specimens among the later fusing bones, but not in those that fuse earlier. This indicates a heavy male cull at the subadult stage, or between the ages when the distal tibia and the distal radius fuse (supported by other elements with similar fusion ages). This pattern does not occur on the East Mound, at any rate not in the South Area (mainly South.P–T as the only East Mound phase with significant numbers of unfused measurements recorded). The LSI diagram (Fig. 10) suggests a more even sex distribution in the West assemblage. This is probably because only fused bones are included and males are not differentially culled until after most bones have fused. This may also apply to South.H–M, where males predominate. Thus perhaps the West Mound inhabitants returned to segregating large herds by age and sex, bringing subadult males closer for slaughter.
Fig. 12. Sexed caprine pelves.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 59
Fig. 13. Sheep radii and ulnae with pathological alteration from Building 65 and associated mid-dens: a) sheep ulna 14535.F1; b) sheep ulna 14535.F11, possibly same animal as 14535.F1; c) sheep/goat ulna 15717.F438; d) sheep radius 17306.F173; e) sheep ulna 17306.F174, articulated with 17306.F173; f) sheep/goat radius 17306.F178; g) sheep/goat ulna 17306.F179, articulate with
17306.F178, and these two could possibly derive from the same animal as 17306.F173–174.
60 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
Fig. 14. Distal breadth of fused and unfused and unfused distal radii and distal tibiae from the South Area (primarily South.P-T) and West Mound.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 61
22 Martin et al. 2011. 23 Buitenhuis 1997. 24 Dewbury and Russell 2007. 25 Abe et al. 2002; Domínguez-Rodrigo and Yravedra 2009; Otárola-Castillo 2010. 26 Last 2005; Atalay and Hastorf 2006; Hodder 2011.
It is intriguing that the Çatalhöyük caprines are clearly domestic in size from the begin-ning of the occupation, since those at nearby and slightly earlier Boncuklu22 and A≥ıklı Höyük23 are wild-size, although there is evidence that they were herded at A≥ıklı. Thus the founding flocks at Çatalhöyük may have come from farther away. A second importation of exotic sheep seems to have happened in the Chalcolithic, unless the size increase is due to improved nutrition or selective breeding for larger size.
Butchery and Cooking
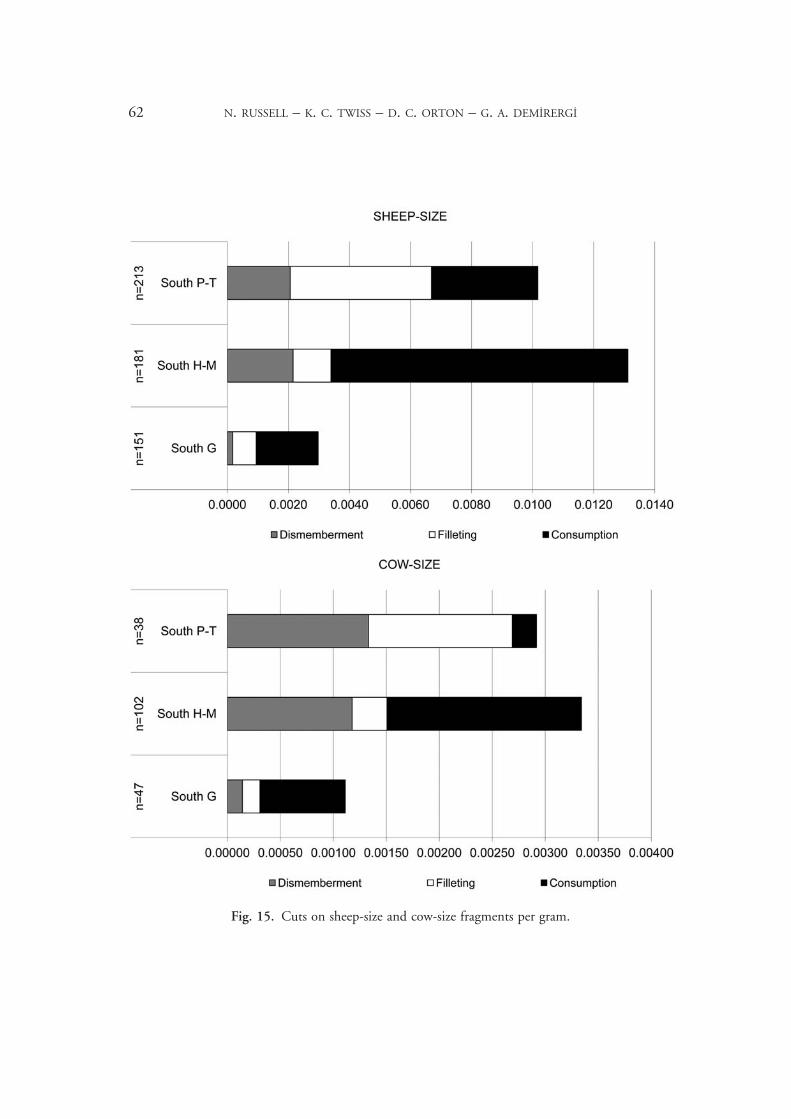
Cut marks are remarkably rare on the Çatalhöyük animal bones, probably because obsid-ian dominates the lithic assemblage and tends to leave fewer marks.24 In particular, dismem-berment cuts, which dominate most Neolithic assemblages, are very scarce. Rather, most cuts result from removing the meat from the bones either before cooking (filleting, leaving light, often diagonal, marks on shafts) or afterwards (consumption, where the softening of the bone during cooking permits deeper cuts, often with carving/sawing motions). There is a striking shift from consumption to filleting cuts between South.H–M and P–T (Fig. 15). Since many cuts occur on non-diagnostic shaft fragments, we quantify them by size class, focusing here on the sheep-size and cow-size categories, which account for most of the assemblage. Given the proportions of identified taxa, the sheep-size fragments will be over-whelmingly caprines, while the cow-size will be predominantly cattle. Quantification of cut marks is difficult when bones are fragmented.25 Since sheep-size bones tend to be more fragmented than those of large mammals at Çatalhöyük, comparing proportions of frag-ments (per cent NISP) with cut marks would be inappropriate. We have chosen to stand-ardise cut rates by quantifying in terms of cuts per gram within each size class. This means that the sheep-size rates cannot be compared with those for the cow-size (since cow-size bones weigh more), but permits better comparison across periods and cut types, regardless of differential fragmentation.
This change in treatment is seen in both size categories, suggesting that they were cooked in similar ways, with a shift from dry-heat cooking (probably baking, as there is little sign of roasting in the burning patterns) to stewing off the bone in the later levels. This uniform-ity is interesting, given that in other ways, such as fragmentation for bone grease processing, cattle tend to be treated differently, which we have interpreted as the result of their use in feasting. Cooking methods, then, were likely similar in daily meals and feasts, simply on different scales. The shift to stewing is probably related to the increasing prominence of pottery in the later levels.26
62 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
Fig. 15. Cuts on sheep-size and cow-size fragments per gram.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 63
27 Hodder, ed. in press. 28 Twiss and Russell 2009. 29 Russell and Meece 2006. 30 Mellaart 1966, 1967; Russell and Meece 2006. 31 Conolly 1999.
CONCLUSIONS
We have identified three key changes in the Çatalhöyük faunal assemblage between the earlier (South.G–M) and later levels on the East Mound: a substantial increase in sheep flocks (or in sheep consumed at the site), the appearance of domestic cattle, and a change from baking meat on the bone to stewing boneless meat. There are also indications of changes in herding practices. A variety of changes in material culture also occur at this time, including increased use of pottery, cessation of clay ball use, and a shift in lithic sources.27 Some of these changes seem to be related to new and perhaps more individualised ways of constructing identity, such as the movement of motifs from large reliefs in houses onto portable pottery and stamp seals.
The advent of cattle herding led neither to an increase in cattle (whose numbers stayed about the same in the later levels of the East Mound — decreasing proportionally to caprines — and may actually have declined on the West Mound) nor to a lessening of the symbolic value of wild cattle and hunting. On the contrary, wild cattle symbolism increases when cattle herding begins, at least initially. Throughout the sequence, cattle horns sufficiently intact to age and sex are more often male and adult than are the postcranial cattle remains, indicating curation of bulls’ horns, probably especially those of wild bulls.28 This tendency is still more pronounced in special deposits. Wild bull horns are most prominent in archi-tectural installations and special deposits in 4040.G (and very likely also in South.N–O, as well as Mellaart’s Level VI,29 all roughly contemporary and dating to c.6500–6400 cal BC), although it is not clear how much of this is due to the tendency for burnt houses, which are most common in these levels, to be left more intact than other houses (Fig. 16). Small and almost surely domestic cattle are already present in 4040.G, for example in Buildings 49 and 77, both of which also contained substantial deposits of wild bulls’ horns. There is a general intensification in the special treatment of animal parts in the later levels. Whereas in the earlier levels special deposits apart from feasting remains consist primarily of cattle horns and scapulae, one or two at a time, in the later levels we find larger collections and more bodyparts (cattle mandibles, astragali, and equid first phalanges in addition to cattle horns and scapulae) and more taxa (deer, equids, and boar in addition to cattle) are included. Caprines also start to be used in feasting.
Although hunting decreases in the later levels (domestic caprines form a larger propor-tion of the assemblage, and some of the cattle are domestic), the idea of hunting is pro-moted more vigorously. The more spectacular paintings, with collections of many animals interacting with humans, are all from the later levels.30 These may or may not be hunting scenes, but they feature wild animals, perhaps exclusively. Projectile points also increase as hunting decreases.31 The increased importance of herding, and the reorganisation of labour associated with it, may have threatened the (gender) identity of some Çatalhöyük residents, who responded with an intensified performance of the idea of hunting and masculinity.

64 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
Fig. 16. Wild bull horns on pillars in Building 77 (4040.?G); photo by Jason Quinlan.
Fig. 17. Sheep and goat frontlets from Space 453, West Mound Trench 5; photo by Ingmar Franz.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 65
32 Hodder and Meskell 2011.
The intensification of special deposits of animal parts, many of them likely trophies or vouchers for hunts or feasts, suggests that feasting increased and became more competitive in the later levels of the East Mound. If these activities indeed peaked in 4040.G and con-temporary levels, this may have been a time when households or individuals used the new wealth of expanding herds to intensify competition in the old mode, based on feasting, ritual performance, and, ironically, big game hunting.32 Possibly the change in cooking methods, where meat is presented off the bone and less identifiable, made it more accepta-ble to include the meat of domestic animals in feasts. Subsequently, the dramatic feasts and trophies tapered off, perhaps as social prominence became increasingly based on the build-ing of wealth in household herds. By the time of the West Mound occupation, this wealth-based mode appears to have won out. We have not yet seen any trophies or special treat-ment of cattle body-parts on the West Mound, although some deposits may indicate feasting based on caprines, and a pair of domestic sheep and goat frontlets in Space 453 (Fig. 17) may indicate that caprine horns could now serve as vouchers for feasts.
REFERENCES
ABE, Y., MAREAN, C. W., NILSSEN, P. J., ASSEFA, Z. and STONE, E. C.2002 “The analysis of cutmarks on archaeofauna: a review and critique of quantification
procedures, and a new image-analysis GIS approach,” American Antiquity 67: 643–664.
ARBUCKLE, B. S.2013 “The late adoption of cattle and pig husbandry in Neolithic Central Turkey,” Journal
of Archaeological Science 40: 1805–1815.ARBUCKLE, B. S. and MAKAREWICZ, C. A.
2009 “The early management of cattle (Bos taurus) in Neolithic central Anatolia,” Antiquity 83: 669–686.
ATALAY, S. and HASTORF, C. A.2006 “Food, meals, and daily activities: food habitus at Neolithic Çatalhöyük,” American
Antiquity 71: 283–319.BAYLISS, A. and FARID, S.
2008 “Modelling chronology,” Çatalhöyük Research Project, Archive Report. http://www.catalhoyuk.com/downloads/Archive_Report_2008.pdf.
2010 “Modelling chronology,” Çatalhöyük Research Project, Archive Report. http://www.catalhoyuk.com/downloads/Archive_Report_2010.pdf.
BIEHL, P. F., FRANZ, I., OSTAPTCHOUK, S., ORTON, D. C., ROGASCH, J. and ROSENSTOCK, E.2012 “One community and two tells: the phenomenon of relocating tell settlements at the
turn of the 7th and 6th millennia in central Anatolia,” in Tells: Social and Environ-mental Space, edited by R. Hofmann, F.-K. Moetz, and J. Müller, pp. 53–65. Bonn: Dr. Rudolf Habelt Verlag.
BÖKÖNYI, S.1970 “A new method for the determination of the number of individuals in animal bone
material,” American Journal of Archaeology 74: 291–292.1972 “Zoological evidence for seasonal or permanent occupation of prehistoric settle-
ments,” in Man, Settlement and Urbanism, edited by P. J. Ucko, R. E. Tringham, and G. W. Dimbleby, pp. 121–126. London: Duckworth.
BUITENHUIS, H.1997 “A≥ıklı Höyük: a ‘protodomestication’ site,” Anthropozoologica 25/26: 655–662.

66 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
CESSFORD, C., BLUMBACH, P., AKOGLU, K. G., HIGHAM, T. F. G., KUNIHOLM, P. I., MANNING, S. W., NEWTON, M. W., ÖZBAKAN, M. and ÖZER, A. M.
2005 “Absolute dating at Çatalhöyük,” in Changing Materialities at Çatalhöyük: Reports from the 1995–99 Seasons, edited by I. Hodder, pp. 65–99. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archaeology at Ankara.
CONOLLY, J.1999 The Çatalhöyük Flint and Obsidian Industry: Technology and Typology in Context (Brit-
ish Archaeological Reports, International Series 787). Oxford: Archaeopress.DE CUPERE, B. and DURU, R.
2003 “Faunal remains from Neolithic Höyücek (SW-Turkey) and the presence of early domestic cattle in Anatolia,” Paléorient 29: 107–120.
DEGERBØL, M. and FREDSKILD, B.1970 The Urus (Bos primigenius Bojanus) and Neolithic Domesticated Cattle (Bos taurus
domesticus Linné) in Denmark (Det Kongelige Danske Videnskabernes Selskab Biolo-giske Skrifter, No. 17(1)). Copenhagen: Munksgaard.
DEWBURY, A. G. and RUSSELL, N.2007 “Relative frequency of butchering cutmarks produced by obsidian and flint: an exper-
imental approach,” Journal of Archaeological Science 34: 354–357.DOMÍNGUEZ-RODRIGO, M. and YRAVEDRA, J.
2009 “Why are cut mark frequencies in archaeofaunal assemblages so variable? A multi-variate analysis,” Journal of Archaeological Science 36: 884–894.
DUCOS, P.1988 Archaeozoologie Quantitative – Les Valeurs Numériques Immediates à Çatal Hüyük
(Les Cahiers du Quaternaire, No. 12). Paris: Éditions du Centre National de la Recherche Scientifique.
FARID, S.2008 “A review of the Mellaart Level system and the introduction of a new phasing system
at Çatalhöyük 2008,” Çatalhöyük Research Project, Archive Report. http://www.cat-alhoyuk.com/downloads/Archive_Report_2008.pdf.
GRANT, A.1982 “The use of tooth wear as a guide to the age of domestic ungulates,” in Ageing and
Sexing Animal Bones from Archaeological Sites (British Archaeological Reports, British Series 109), edited by B. Wilson, C. Grigson, and S. Payne, pp. 91–108. Oxford: Archaeopress.
GREENFIELD, H. J.2006 “Sexing fragmentary ungulate acetabulae,” in Recent Advances in Ageing and Sexing
Animal Bones, edited by D. Ruscillo, pp. 68–86. Oxford: Oxbow.GRIGSON, C.
1982 “Sex and age determination of some bones and teeth of domestic cattle: a review of the literature,” in Ageing and Sexing Animal Bones from Archaeological Sites (British Archaeological Reports, British Series 109), edited by B. Wilson, C. Grigson, and S. Payne, pp. 7–23. Oxford: Archaeopress.
HODDER, I., ed.1996 On the Surface: Çatalhöyük 1993–95. Cambridge: McDonald Institute for Archaeo-
logical Research; London: British Institute of Archaeology at Ankara.HODDER, I.
1999 “Renewed work at Çatalhöyük”, in Neolithic in Turkey: The Cradle of Civilization — New Discoveries, edited by N. Ba≥gelen and M. Özdogan, pp. 157–164. Istanbul: Arkeoloji ve Sanat Yayınları.
HODDER, I., ed.2000 Towards Reflexive Method in Archaeology: The Example at Çatalhöyük. Cambridge:
McDonald Institute for Archaeological Research; London: British Institute of Archae-ology at Ankara.
2005a Changing Materialities at Çatalhöyük: Reports from the 1995–99 Seasons. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archae-ology at Ankara.

CHANGING ANIMAL USE AT NEOLITHIC ÇATALHÖYÜK, TURKEY 67
2005b Inhabiting Çatalhöyük: Reports from the 1995–99 Seasons. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archaeology at Ankara.
2006a Çatalhöyük Perspectives: Themes from the 1995–99 Seasons. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archaeology at Ankara.
HODDER, I.2006b The Leopard’s Tale: Revealing the Mysteries of Çatalhöyük. London: Thames and Hud-
son.HODDER, I., ed.
2007 Excavating Çatalhöyük: South, North and KOPAL Area Reports from the 1995–99 Seasons. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archaeology at Ankara.
HODDER, I.2011 “Human-thing entanglement: towards an integrated archaeological perspective,”
Journal of the Royal Anthropological Institute 17: 154–177.HODDER, I., ED.
in press Substantive Technologies at Çatalhöyük: Reports from the 2000–2008 Seasons. Los Angeles: Cotsen Institute of Archaeology, University of California, Los Angeles.
HODDER, I. and MESKELL, L.2011 “A curious and sometimes a trifle macabre artistry,” Current Anthropology 52: 235–263.
LAST, JONATHAN
2005 “Pottery from the East Mound,” in Changing Materialities at Çatalhöyük: Reports from the 1995–99 Seasons (McDonald Institute Monographs), edited by I. Hodder, pp. 101–138. Cambridge: McDonald Institute for Archaeological Research; Lon-don: British Institute of Archaeology at Ankara.
LAST, J. and GIBSON, C., ED.in prep. The Sixth Millennium BC at Çatalhöyük – Excavations on the West Mound 1998–
2003. Los Angeles: Cotsen Institute of Archaeology, University of California, Los Angeles.
LEMPPENAU, U.1964 Geschlechts- und Gattungsunterschiede am Becken Mitteleuropäischer Wiederkäuer.
Unpublished PhD diss. Universität München.MARCINIAK, A. and CZERNIAK, L.
2007 “Social transformations in the Late Neolithic and the Early Chalcolithic periods in central Anatolia,” Anatolian Studies 57: 115–130.
MARTIN, L., MIDDLETON, C. and EDWARDS, Y. H.2011 “Early Neolithic hunter-foragers in Central Anatolia: preliminary results from Bon-
cuklu Höyük in the Konya Basin,” Paper presented at the Archaeozoology of South-west Asia meetings, Brussels.
MELLAART, J.1962 “Excavations at Çatal Hüyük, 1961: first preliminary report,” Anatolian Studies 12:
41–65.1963 “Excavations at Çatal Hüyük, 1962: second preliminary report,” Anatolian Studies
13: 43–103.1964 “Excavations at Çatal Hüyük, 1963: third preliminary report,” Anatolian Studies 14:
39–119.1965 “Çatal Hüyük West,” Anatolian Studies 15: 135–156.1966 “Excavations at Çatal Hüyük, 1965: fourth preliminary report,” Anatolian Studies
16: 165–191.1967 Çatal Hüyük: A Neolithic Town in Anatolia. London: Thames and Hudson.1998 “Çatal Hüyük: the 1960s seasons,” in Ancient Anatolia: Fifty Years’ Work by the
British Institute of Archaeology at Ankara, edited by R. J. Matthews, pp. 35–41. London: British Institute of Archaeology at Ankara.
OTÁROLA-CASTILLO, E.2010 “Differences between NISP and MNE in cutmark analysis of highly fragmented
faunal assemblages,” Journal of Archaeological Science 37: 1–12.

68 N. RUSSELL – K. C. TWISS – D. C. ORTON – G. A. DEMIRERGI
PAYNE, S.1987 “Reference codes for wear states in the mandibular cheek teeth of sheep and goats,”
Journal of Archaeological Science 14: 609–614.PEARSON, J. A.
in press “Human and animal diets as evidenced by stable carbon and nitrogen isotope analy-sis,” in Humans and Landscapes of Çatalhöyük: Reports from the 2000–2008 Seasons, edited by I. Hodder. Los Angeles: Cotsen Institute of Archaeology, University of California, Los Angeles.
PERKINS, D., JR.1969 “Fauna of Çatal Hüyük: evidence for early cattle domestication in Anatolia,” Science
164(3876): 177–179.RUSSELL, N. and MARTIN, L.
2005 “The Çatalhöyük mammal remains,” in Inhabiting Çatalhöyük: Reports from the 1995–99 Seasons (McDonald Institute Monographs), edited by I. Hodder, pp. 33–98. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archaeology at Ankara.
RUSSELL, N., MARTIN, L. and BUITENHUIS, H.2005 “Cattle domestication at Çatalhöyük revisited,” Current Anthropology 46: S101–S108.
RUSSELL, N. and MEECE, S.2006 “Animal representations and animal remains at Çatalhöyük,” in Çatalhöyük Perspec-
tives: Reports from the 1995–99 Seasons (McDonald Institute Monographs), edited by I. Hodder, pp. 209–230. Cambridge: McDonald Institute for Archaeological Research; London: British Institute of Archaeology at Ankara.
RUSSELL, N., TWISS, K. C., ORTON, D. C. and DEMIRERGI, G. A.in press “More on the Çatalhöyük mammal remains,” in Humans and Landscapes of Çatal-
höyük: Reports from the 2000–2008 Seasons, edited by I. Hodder. Los Angeles: Cot-sen Institute of Archaeology, University of California, Los Angeles.
SILVER, I. A.1969 “The ageing of domestic animals,” in Science in Archaeology: A Survey of Progress and
Research, edited by D. R. Brothwell and E. S. Higgs, pp. 283–302. London: Thames and Hudson.
TODD, I. A.1976 Çatal Hüyük in Perspective. Menlo Park: Cummings.
TWISS, K. C. and RUSSELL, N.2009 “Taking the bull by the horns: ideology, masculinity, and cattle horns at Çatal-
höyük (Turkey),” Paléorient 35: 19–32.UERPMANN, H.-P.
1978 “The “KNOCOD” system for processing data on animal bones from archaeological sites,” in Approaches to Faunal Analysis in the Middle East (Peabody Museum Bulle-tins), edited by R. H. Meadow and M. A. Zeder, pp. 149–167. Cambridge: Pea-body Museum, Harvard University.
1979 Probleme der Neolithisierung des Mittelmeerraums (Beihefte zum Tübinger Atlas des vorderen Oriente, Reihe B, Nr. 28). Wiesbaden: Dr. Ludwig Reichert Verlag.
WATSON, J. P. N.1979 “The estimation of the relative frequencies of mammalian species: Khirokitia 1972,”
Journal of Archaeological Science 6: 127–137.
Nerissa RUSSELL Katheryn C. TWISS
Cornell University, USA SUNY Stony Brook, USA
David C. ORTON G. Arzu DEMIRERGI
Cambridge University, UK SUNY Stony Brook, USA