
Challenges in the determination of unsubstituted food folates: impact of stabilities and conversions...
11
Challenges in the Determination of Unsubstituted Food Folates: Impact of Stabilities and Conversions on Analytical Results Hanna Sara Strandler,* ,† Johan Patring, § Margaretha Ja ̈ gerstad, § and Jelena Jastrebova § † The National Food Agency, P.O. Box 622, SE-751 26 Uppsala, Sweden § Department of Food Science, Uppsala BioCenter, Swedish University of Agricultural Sciences (SLU), P.O. Box 7051, SE-750 07 Uppsala, Sweden ABSTRACT: Tetrahydrofolate is the parent molecule of the folate coenzymes required for one carbon metabolism. Together with other unsubstituted folates such as dihydrofolate and folic acid, tetrahydrofolate represents the third pool of dietary folates following 5-methyltetrahydrofolate and formyl folates. Low intake of dietary folates and poor folate status are common problems in many countries. There is a critical need for reliable methods to determine folate in foods to accurately estimate folate intakes in populations. However, current values for folates in foods in databanks are often underestimated due to the high instability of several folate forms, especially tetrahydrofolate. The present review highlights the occurrence of unsubstituted folates in foods and their oxidation mechanisms and chemical behavior as well as interconversion reaction between tetrahydrofolate and 5,10- methylenetetrahydrofolate. The review shows also the important role of antioxidants in protecting folates during analysis and describes strategies to stabilize unsubstituted folates throughout all steps of the analytical procedure. KEYWORDS: folic acid, tetrahydrofolate, dihydrofolate, 5,10-methylenetetrahydrofolate, folate oxidation, folate stabilization ■ INTRODUCTION Folate belongs to the B vitamin group, and it is essential for cell growth, particularly during pregnancy when a low folate status (intake) increases the risk for neural tube defects (NTD) early in the pregnancy. 1 The classical human folate deficiency disease is megaloblastic anemia. 2 Other clinical aspects link a good folate status to protection against coronary heart disease, malformations, cancer, and cognitive functions, but the results from these studies are still not conclusive. 3−6 High intake of synthetic folic acid is investigated for cancer risks, but so far the results are not consistent. 6−8 Folate exerts its biological function as a transporter of C1 groups. Tetrahydrofolate accepts and donates C1 groups (methyl, formyl, formimino, and methylene) by acting as coenzyme. 9,10 The substituted tetrahydrofolates are generally more stable than tetrahydrofolate (H 4 folate), which is one of the most oxygen-sensitive forms among the native folates. If not protected from oxidation, it is only stable for up to 30 min at room temperature; the main oxidation pathway splits the molecule into two biologically inactive parts, p-aminobenzoyl- glutamic acid (pABG) and pterins. 11 Lower temperature, neutral to alkaline pH, and antioxidants stabilize H 4 folate and retard its oxidative degradation, thereby forming also other unsubstituted folates, for example, dihydrofolate (H 2 folate) and folic acid in addition to pABG and pterins. 9,12−15 The three unsubstituted folates represent the third pool of food folates following 5-methyltetrahydrofolate and formyl folates. 16 They are most common in yeast, liver, green vegetables, and cereals or products fortified with folic acid. 17 This review gives a brief background of the chemistry, biological functions, and occurrence of the unsubstituted folates in foods. Their stability and chemical behavior and factors affecting them in the food chain and during determination are highlighted. In addition, the stability and formation of the even more labile 5,10-methylenetetrahydrofolate (5,10-CH 2 - H 4 folate) is included in this review because this form easily dissociates back to H 4 folate at neutral and acidic pH and contributes to the H 4 folate pool. 18 Because of this, determi- nation could potentially lead to false results by underestimating 5,10-CH 2 -H 4 folate or overestimating H 4 folate or both. The purpose of this review is to compile current knowledge of the unsubstituted folates in foods to achieve a better understanding of their stabilities and conversions during food processing, storage, and determination. Only monoglutamate forms of the unsubstituted folates are discussed in the review as the native polyglutamate forms occurring in foods are deconjugated to monoglutamates prior to analysis. ■ BIOCHEMISTRY AND BIOLOGICAL FUNCTIONS Plants and microorganisms synthesize folates de novo through a complex metabolic route that is now fully elucidated. 19,20 In contrast, humans and other vertebrates lack a complete biosynthetic pathway and thus need dietary folates, of which plants are major sources. 9,20 Dietary folates are usually linked to a polyglutamic chain, which is hydrolyzed to monoglutamyl folate by a brush-border enzyme before absorption. During the passage through the intestinal mucosa, dietary folates are reduced and converted to 5-methyltetrahydrofolic acid (5-CH 3 - H 4 folate), which is the major form of folate present in the systemic circulation. Synthetic (oxidized) folic acid given as a supplement or through fortified foods is reduced enzymatically by folate reductase, partly during the transit through the Received: October 15, 2014 Revised: January 30, 2015 Accepted: February 2, 2015 Published: February 2, 2015 Review pubs.acs.org/JAFC © 2015 American Chemical Society 2367 DOI: 10.1021/jf504987n J. Agric. Food Chem. 2015, 63, 2367−2377
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Challenges in the determination of unsubstituted food folates: impact of stabilities and conversions...

Challenges in the Determination of Unsubstituted Food Folates:Impact of Stabilities and Conversions on Analytical ResultsHanna Sara Strandler,*,† Johan Patring,§ Margaretha Jagerstad,§ and Jelena Jastrebova§
†The National Food Agency, P.O. Box 622, SE-751 26 Uppsala, Sweden§Department of Food Science, Uppsala BioCenter, Swedish University of Agricultural Sciences (SLU), P.O. Box 7051, SE-750 07Uppsala, Sweden
ABSTRACT: Tetrahydrofolate is the parent molecule of the folate coenzymes required for one carbon metabolism. Togetherwith other unsubstituted folates such as dihydrofolate and folic acid, tetrahydrofolate represents the third pool of dietary folatesfollowing 5-methyltetrahydrofolate and formyl folates. Low intake of dietary folates and poor folate status are common problemsin many countries. There is a critical need for reliable methods to determine folate in foods to accurately estimate folate intakes inpopulations. However, current values for folates in foods in databanks are often underestimated due to the high instability ofseveral folate forms, especially tetrahydrofolate. The present review highlights the occurrence of unsubstituted folates in foodsand their oxidation mechanisms and chemical behavior as well as interconversion reaction between tetrahydrofolate and 5,10-methylenetetrahydrofolate. The review shows also the important role of antioxidants in protecting folates during analysis anddescribes strategies to stabilize unsubstituted folates throughout all steps of the analytical procedure.
KEYWORDS: folic acid, tetrahydrofolate, dihydrofolate, 5,10-methylenetetrahydrofolate, folate oxidation, folate stabilization
■ INTRODUCTION
Folate belongs to the B vitamin group, and it is essential for cellgrowth, particularly during pregnancy when a low folate status(intake) increases the risk for neural tube defects (NTD) earlyin the pregnancy.1 The classical human folate deficiency diseaseis megaloblastic anemia.2 Other clinical aspects link a goodfolate status to protection against coronary heart disease,malformations, cancer, and cognitive functions, but the resultsfrom these studies are still not conclusive.3−6 High intake ofsynthetic folic acid is investigated for cancer risks, but so far theresults are not consistent.6−8
Folate exerts its biological function as a transporter of C1groups. Tetrahydrofolate accepts and donates C1 groups(methyl, formyl, formimino, and methylene) by acting ascoenzyme.9,10 The substituted tetrahydrofolates are generallymore stable than tetrahydrofolate (H4folate), which is one ofthe most oxygen-sensitive forms among the native folates. If notprotected from oxidation, it is only stable for up to 30 min atroom temperature; the main oxidation pathway splits themolecule into two biologically inactive parts, p-aminobenzoyl-glutamic acid (pABG) and pterins.11 Lower temperature,neutral to alkaline pH, and antioxidants stabilize H4folate andretard its oxidative degradation, thereby forming also otherunsubstituted folates, for example, dihydrofolate (H2folate) andfolic acid in addition to pABG and pterins.9,12−15 The threeunsubstituted folates represent the third pool of food folatesfollowing 5-methyltetrahydrofolate and formyl folates.16 Theyare most common in yeast, liver, green vegetables, and cerealsor products fortified with folic acid.17
This review gives a brief background of the chemistry,biological functions, and occurrence of the unsubstituted folatesin foods. Their stability and chemical behavior and factorsaffecting them in the food chain and during determination arehighlighted. In addition, the stability and formation of the even
more labile 5,10-methylenetetrahydrofolate (5,10-CH2-H4folate) is included in this review because this form easilydissociates back to H4folate at neutral and acidic pH andcontributes to the H4folate pool.18 Because of this, determi-nation could potentially lead to false results by underestimating5,10-CH2-H4folate or overestimating H4folate or both. Thepurpose of this review is to compile current knowledge of theunsubstituted folates in foods to achieve a better understandingof their stabilities and conversions during food processing,storage, and determination. Only monoglutamate forms of theunsubstituted folates are discussed in the review as the nativepolyglutamate forms occurring in foods are deconjugated tomonoglutamates prior to analysis.
■ BIOCHEMISTRY AND BIOLOGICAL FUNCTIONS
Plants and microorganisms synthesize folates de novo througha complex metabolic route that is now fully elucidated.19,20 Incontrast, humans and other vertebrates lack a completebiosynthetic pathway and thus need dietary folates, of whichplants are major sources.9,20 Dietary folates are usually linked toa polyglutamic chain, which is hydrolyzed to monoglutamylfolate by a brush-border enzyme before absorption. During thepassage through the intestinal mucosa, dietary folates arereduced and converted to 5-methyltetrahydrofolic acid (5-CH3-H4folate), which is the major form of folate present in thesystemic circulation. Synthetic (oxidized) folic acid given as asupplement or through fortified foods is reduced enzymaticallyby folate reductase, partly during the transit through the
Received: October 15, 2014Revised: January 30, 2015Accepted: February 2, 2015Published: February 2, 2015
Review
pubs.acs.org/JAFC
© 2015 American Chemical Society 2367 DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
intestinal mucosa and finally intracellularly, especially in theliver, first to H2folate and subsequently to H4folate.
9,12
Immediately upon intracellular entrance, H4folate partic-ipates in the biosynthesis of polyglutamate forms, usually up tofive to seven residues, to be retained within cells and subcellularcompartments. Polyglutamated folates are the preferredcoenzymes involved in C1 metabolism. Chain elongationincreases the anionic nature of folate coenzymes by providingα-carboxyl charges and decreases affinity for membrane carriers,thus impairing folate diffusion through hydrophobic barriers.19
The polyglutamated H4folate molecules accept C1 groupssupplied from the catabolism of certain amino acids such asglycine, serine, and histidine or from the catabolism of purines.The C1 groups are donated enzymatically to be included in thenucleotide synthesis of DNA and RNA and in the methylationof homocysteine to methionine. After donation of theirC1groups, the substituted folates regenerate to polyglutamateforms of H4folate or H2folate and are then ready to accept newC1 groups.2,9,10
■ OCCURRENCE IN FOOD
Plant foods are the major source of folate because they cansynthesize folate. This biosynthesis route has already beenexplored for biofortification of tomato fruit and rice with 25-and 50-fold increases in folate content, respectively.21,22 Othertypes of bioprocessing to enhance dietary folate aregermination, sprouting, malting, or fermentation based onyeast or lactic acid bacteria (LAB) or both. Bioprocessing iscommonly used for the manufacture of bread, yogurt,buttermilk, beer, and wine.23−26 Good dietary sources of nativefolate include liver, legumes, green leafy vegetables, citrus fruitand juice, certain berries, cereals, and egg.17 Mandatory orvoluntary fortification with folic acid is also common mostly asprophylaxis against neural tube defects because women offertile age throughout the world have difficulties meeting thedaily folate recommendations during pregnancy and lactation.27
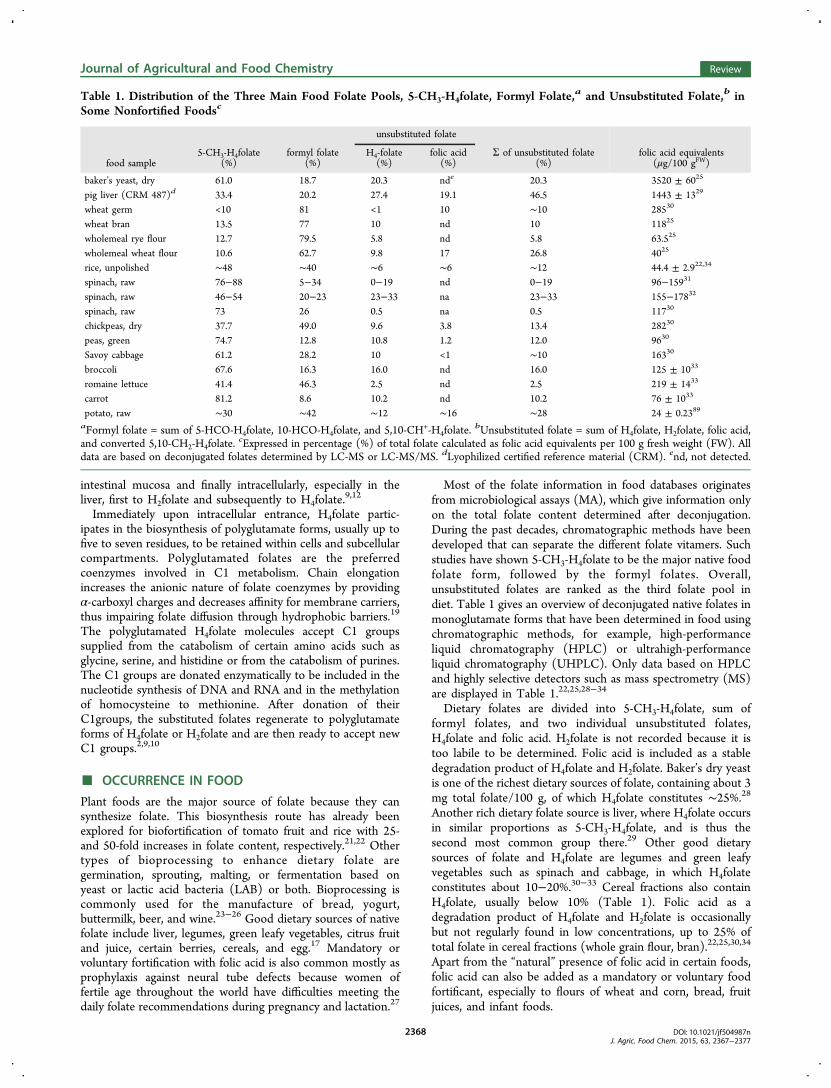
Most of the folate information in food databases originatesfrom microbiological assays (MA), which give information onlyon the total folate content determined after deconjugation.During the past decades, chromatographic methods have beendeveloped that can separate the different folate vitamers. Suchstudies have shown 5-CH3-H4folate to be the major native foodfolate form, followed by the formyl folates. Overall,unsubstituted folates are ranked as the third folate pool indiet. Table 1 gives an overview of deconjugated native folates inmonoglutamate forms that have been determined in food usingchromatographic methods, for example, high-performanceliquid chromatography (HPLC) or ultrahigh-performanceliquid chromatography (UHPLC). Only data based on HPLCand highly selective detectors such as mass spectrometry (MS)are displayed in Table 1.22,25,28−34
Dietary folates are divided into 5-CH3-H4folate, sum offormyl folates, and two individual unsubstituted folates,H4folate and folic acid. H2folate is not recorded because it istoo labile to be determined. Folic acid is included as a stabledegradation product of H4folate and H2folate. Baker’s dry yeastis one of the richest dietary sources of folate, containing about 3mg total folate/100 g, of which H4folate constitutes ∼25%.28Another rich dietary folate source is liver, where H4folate occursin similar proportions as 5-CH3-H4folate, and is thus thesecond most common group there.29 Other good dietarysources of folate and H4folate are legumes and green leafyvegetables such as spinach and cabbage, in which H4folateconstitutes about 10−20%.30−33 Cereal fractions also containH4folate, usually below 10% (Table 1). Folic acid as adegradation product of H4folate and H2folate is occasionallybut not regularly found in low concentrations, up to 25% oftotal folate in cereal fractions (whole grain flour, bran).22,25,30,34
Apart from the “natural” presence of folic acid in certain foods,folic acid can also be added as a mandatory or voluntary foodfortificant, especially to flours of wheat and corn, bread, fruitjuices, and infant foods.
Table 1. Distribution of the Three Main Food Folate Pools, 5-CH3-H4folate, Formyl Folate,a and Unsubstituted Folate,b inSome Nonfortified Foodsc
unsubstituted folate
food sample5-CH3-H4folate
(%)formyl folate
(%)H4-folate(%)
folic acid(%)
Σ of unsubstituted folate(%)
folic acid equivalents(μg/100 gFW)
baker’s yeast, dry 61.0 18.7 20.3 nde 20.3 3520 ± 6025
pig liver (CRM 487)d 33.4 20.2 27.4 19.1 46.5 1443 ± 1329
wheat germ <10 81 <1 10 ∼10 28530
wheat bran 13.5 77 10 nd 10 11825
wholemeal rye flour 12.7 79.5 5.8 nd 5.8 63.525
wholemeal wheat flour 10.6 62.7 9.8 17 26.8 4025
rice, unpolished ∼48 ∼40 ∼6 ∼6 ∼12 44.4 ± 2.922,34
spinach, raw 76−88 5−34 0−19 nd 0−19 96−15931
spinach, raw 46−54 20−23 23−33 na 23−33 155−17832
spinach, raw 73 26 0.5 na 0.5 11730
chickpeas, dry 37.7 49.0 9.6 3.8 13.4 28230
peas, green 74.7 12.8 10.8 1.2 12.0 9630
Savoy cabbage 61.2 28.2 10 <1 ∼10 16330
broccoli 67.6 16.3 16.0 nd 16.0 125 ± 1033
romaine lettuce 41.4 46.3 2.5 nd 2.5 219 ± 1433
carrot 81.2 8.6 10.2 nd 10.2 76 ± 1033
potato, raw ∼30 ∼42 ∼12 ∼16 ∼28 24 ± 0.2389
aFormyl folate = sum of 5-HCO-H4folate, 10-HCO-H4folate, and 5,10-CH+-H4folate.bUnsubstituted folate = sum of H4folate, H2folate, folic acid,
and converted 5,10-CH2-H4folate.cExpressed in percentage (%) of total folate calculated as folic acid equivalents per 100 g fresh weight (FW). All
data are based on deconjugated folates determined by LC-MS or LC-MS/MS. dLyophilized certified reference material (CRM). end, not detected.
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2368

■ CHEMICAL PROPERTIES OF UNSUBSTITUTEDFOLATES
Background information about chemical properties is essentialto understand the stability and conversions of unsubstitutedfolates during food handling and analysis. All folates comprise apteridine coupled via a carbon−nitrogen (C9−N10) bond topABG, which in turn is coupled via an amide bond through itscarboxyl moiety to the amino group of L-glutamic acid. Thepteridine moiety of folates can exist in three oxidation states:fully oxidized (folic acid) or as the reduced 7,8-dihydrofolate(H2folate) or 5,6,7,8-tetrahydrofolate (H4folate). H4folate is thecoenzymatically active form of the vitamin that accepts,transfers, and donates C1 groups, attached either at the N5
or N10 position or by bridging these positions.9,35,36 The C1groups also differ in their oxidation state, with folates existing asderivatives of formate (5-formyl-H4folate, 10-formyl-H4folate,5,10-methenyl-H4folate, and 5-formimino-H4folate), methanol(5-methyl-H4folate), or formaldehyde (5,10-methylene-H4folate).
16 In addition to these structural variables, mostnaturally occurring folates exist as γ-linked polyglutamateconjugates, with chain lengths typically in the range of five toseven glutamate residues.9 The chemical structures ofunsubstituted folates are displayed in Figure 1.
Folates are ionogenic and amphoteric molecules, which existin cationic, anionic, or zwitterionic form depending on pH. ThepH ranges, in which the various ionizable groups ofunsubstituted folates are predominantly (>50%) charged, aredisplayed in Figure 2.37−41 Ionogenic groups of particularsignificance in the pH values relevant to foods and biologicalsystems are the N5 position of H4folate (basic pKa = 4.8) andthe glutamate carboxyl groups (acidic α pKa = 3.5; γ pKa = 4.8).Folates undergo changes in ionic forms as a function of pH,which in part explains the pH dependency of folate solubilityand the stability and behavior during chromatographicseparation.9,38−41
Folates generally have minimum solubility in the mildlyacidic pH range (e.g., pH 2−4), monocationic and neutralspecies predominating, whereas solubility generally increases inproportion to the pH above this range, mainly zwitterionic andanionic species. The solubility of folates in aqueous solutions islow due to the pteridine part. An increase in temperature and
the presence of ions from buffers or salts increase thesolubility.9,35,36
UV light, air, and heating destroy the molecules by oxidationif not protected by antioxidants.41 Early works show that theeffect of pH on the stability of reduced unsubstituted folates isU-shaped with the highest stability observed between pH 1−2and 8−12, whereas the stability is lowest between pH 4 and6.9,41−43 However, even at these favorable pH intervals,H4folate and H2folate are extremely unstable with half-lives oflosses in bioactivity below 5 h.9 Folic acid, the oxidized form, ismuch more stable than reduced unsubstituted folates. Itsstability also depends on pH and it is stable under neutral andalkaline conditions, but less stable in acidic conditions (pH<5).44−48 All three unsubstituted folates are sensitive to UVlight. For more details on the stability of the unsubstitutedfolates, see Table 2.36,43,46−54
■ DEGRADATION BY AUTOXIDATIONAll unsubstituted folates undergoing autoxidation are split intheir C9−N10 bond into biologically inactive products. Figure 3gives an overview of the pathways and the major autoxidationproducts identified in the oxidation carried out in the presenceof UV light, air, or oxygen and in the absence ofantioxidants.7,15,45,55,56 The major products are pABG andvarious substituted pterins, ultimately converted into pterinsunder release of formaldehyde. Classical work has shown thatthe oxidative degradation of solutions of H4folate first producesa quinonoid isomer of H2folate as intermediate. At acidic pH,this quinonoid is oxidized to pABG and 7,8-dihydropterinunder release of formaldehyde. This dihydropterin is thenfurther oxidized to pterins. At alkaline conditions, thequinonoid-H2folate is converted to H2folate, which is furtheroxidized to pABG and 6-formyltetrapterin, followed by the
Figure 1. Structures of folic acid and the native monoglutamate folateforms.
Figure 2. Ionized groups present in unsubstituted folates at differentpH ranges and their pKa values. pH ranges where they arepredominantly charged, 50% or more, are displayed by color. pKavalues for each group are displayed where their log C curves cross thex-axis. For pKa values displayed in a table, see Patring et al.37
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2369

formation of xanthopterin and formaldehyde.15,55,56Irradiationof folic acid with UV light first produces pABG and pterine-6-carboxyaldehyde, which is converted to 6-carboxylic acid andfinally to the decarboxylated 2-amino-4-hydroxypteridine.45−48
■ MECHANISMS OF AUTOXIDATIONThe pH-dependent oxidation, affecting the rates and type ofproducts, has drawn attention to the role of ionization andprotonation of the zwitterion folate/folic acid as a mecha-nism.15,38 Akhtar and co-workers studied the effect of pH in thepH range 2−10 on photodecomposition of folic acid.46,47
According to their results folic acid exhibited best stability athighly alkaline pH 10.0, whereas its stability was lowest andsusceptibility to photolysis was highest in the acidic range (pH2−4). This clearly showed that photolysis of folic acid is a pH-dependent reaction and occurs more rapidly at low pH whenfolic acid exists predominantly as a neutral species. Morerecently, Martin et al. proposed a mechanism of photolysis offolic acid where UV radiation at 350−400 nm caused formationof pterin radicals, which were then oxidized to the knownphotoproduct 6-carboxypterin.57
According to early studies, steric hindrance is anotherphenomenon affecting the oxidative sensitivity of unsubstitutedfolates. H4folates with substituents at the N
5 position, that is, 5-CH3-H4folate and 5-HCO-H4folate, exhibit much higherstability than unsubstituted H4folate. This suggests that thestabilization effect of the N5 methyl group or the N5 formylgroup is, at least in part, due to steric hindrance in restrictingthe access of oxygen or other oxidants to the pteridine ring.The stabilizing effect of the substituent is presumably due tothe effect of the one-carbon substituent in interfering with theformation of resonance forms involved in oxidative degrada-tion.15,55
■ DEGRADATION PRODUCTS IN THE PRESENCE OFANTIOXIDANTS
In the presence of antioxidants and while protected against UVlight, the unsubstituted folates are protected against oxidation.Depending on the combination and concentration of
antioxidants, pH, time, and temperature, the oxidation ofH4folates is either fully or partly prevented. Antioxidantsdecrease the rate of oxidative degradation, making it possible todetect even early biologically active oxidation products ofH4folate, such as H2folate and folic acid. Hence, a mixture ofthese forms appears together with pABG and various pterins infoods during processing, cooking, storage, or analyticalconditions (see Figure 3).13,14,58
Thiols, for example, 2-mercaptoethanol (MCE) and 2,3-dimercaptopropanol (BAL), were the first antioxidants to beshown to retard the oxidation of H4folate.
59,60 In a classicalwork, Blakley60 found BAL to be a better antioxidant thanMCE. Later, the combination of thiols with ascorbate wasfound to be the most effective in protecting H4folate. Wilsonand Horne58 were the first group to combine the use ofascorbate and MCE as antioxidants during food folate analysis.Today, combinations of 1−2% ascorbate with MCE or BAL arecommonly used to stabilize H4folate during determination byHPLC.Although MCE is the most commonly used and inexpensive
thiol in combination with ascorbic acid/ascorbate to protectH4folate during food analysis, it is less effective and more toxicthan other thiols such as BAL, 1,4-dithiothreitol (DTT), or 2-thiobarbituric acid (TBA).22,61 Patring et al.61 compared thecombination of ascorbate (2%) and four different antioxidants(MCE, DTT, BAL, and TBA), all at 0.1%, on the stability ofH4folate solutions boiled for 1 h. BAL, DTT, and TBA were allcapable of protecting >96% of H4folate in three different buffersvarying between pH 5.1 and 7.85. DTT was the most effectiveantioxidant in acetate buffer (pH 5), DTT and BAL were themost effective in phosphate buffer (pH 6.1), and BAL and TBAwere the most effective in HEPES/CHES buffer, pH 7.85,whereas MCE was the least effective in all buffers compared tothe other thiols (degradation range 7−12%). Recently, DeBrouwer and co-workers found DTT in concentration of 0.5%to be as efficient as BAL when using phosphate buffer pH 7.2.22
Thereby, these three antioxidants (BAL, DTT, and TBA)should be considered as more efficient and environmentallyfriendly substitutes for MCE in modern analytical methods forfolate analysis, including AOAC methods.H4folate is easier to stabilize by proper combination of
antioxidants than H2folate because the initial/primary oxidationproduct of H4folate is quinonoid-H2folate, which can bereduced back to H4folate by antioxidants.
35 This step is the onlyone that could be reversible in the oxidation chain of H4folate,which means that once H2folate has been formed, it is furtheroxidized to folic acid or to degradation products such as pABGand pterins (see Figure 3). Several studies have reportedbetween 12 and 30% of folic acid formation from H2folateunder analytic conditions in the pH range 2.6−7.4.13,14,58,62 Asmentioned in a previous section (Occurrence in Food),H2folate is usually not analyzed in foods due to its highinstability. H2folate seems to require much higher concen-trations of antioxidants than H4folate to be protected.42,63
■ MECHANISM OF ANTIOXIDANT PROTECTIONAGAINST OXIDATION
Reducing agents such as ascorbic acids and thiols exert multipleprotective effects on folates through their action as oxygenscavenger, reducing agent, and free radical scavenger. Thedegradation of unsubstituted folates via a free radicalmechanism has been proposed by several groups.15,55,56,64
Thomas et al.48 suggested that singlet oxygen, probably
Figure 3. Pathways for interconversion between H4folate and 5,10-CH2-H4folate and degradation of unsubstituted folates exposed to airat ambient temperature;15,55,56 degradation of folic acid in the presenceof light or UV irradiation.44−46,48
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2370

produced by electronically excited states of folic acid molecule,is the oxidant agent that starts the degradation reaction. Furthersupport for free radical mechanisms is the finding that riboflavin(vitamin B2) sensitizes the photodegradation of folic acid,leading to the deactivation of the vitamin.45,47 In addition,transition metal ions such as Fe3+ or Cu2+, recognized for theirinvolvement in free radical mechanisms, have been found tocatalyze the oxidation of H4folate.
55
■ INTERCONVERSIONS BETWEEN H4FOLATE AND5,10-METHYLENE-H4FOLATE
H4folate rapidly and reversibly condensates with formaldehyde(in ∼30-fold excess) in aqueous solutions at room temperature,forming 5,10-methylenetetrahydrofolate (5,10-CH2-H4folate(see Figure 3).35,65 This is an example of interconversionreaction because it includes a reversible conversion betweentwo bioactive folate vitamers. The N5−N10-bridged compoundis found to be more stable against chemical oxidation thanH4folate, but its stability depends on an excess of form-aldehyde.11,35,65 Without excess of formaldehyde, the stabilityof 5,10-CH2-H4folate is strongly pH dependent; 5,10-CH2-H4folate appears to be completely stable at pH 9.5 or above, ineither the presence or absence of antioxidants. As soon as thepH decreases to neutral pH, 5,10-CH2-H4folate dissociatesrapidly, within 15 min, back to H4folate and formaldehyde, andheating enhances the rate of the dissociation.65,66
5,10-CH2-H4folate is one of the C1 derivatives of H4folateused for enzymatic methylation of uridinylate to thymidinylate,a key step for DNA synthesis. Hence 5,10-CH2-H4folate is anative folate form that could be present in intact living cells.Because 5,10-CH2-H4folate requires either alkaline pH (>8) oran excess of formaldehyde to remain stable, this form is notlikely to occur in foods because the pH of food is generallyneutral or slightly acidic. Hence, 5,10-CH2-H4folate mostprobably converts to H4folate or its degradation products infood samples or both.
There is only one recorded condition in food analysis whenformaldehyde can be formed in excess with potential tointerconvert H4folate into 5,10-CH2-H4folate. When Wilsonand Horne14 boiled aqueous solutions of H4folate for 10 min ina buffer (pH 7.8) containing 2% sodium ascorbate, 5,10-CH2-H4folate was formed in a yield of 25%. The authors were able toshow that heating of ascorbate led to its thermal degradationfollowed by the formation of formaldehyde, which convertedH4folate into 5,10-CH2-H4folate. This conversion did not occurwithout heating; neither did formation of formaldehyde. Morerecently, this interconversion was confirmed by de Brouwer etal. using HPLC and MS detection.62 This emphasizes the needto combine thiols and ascorbate during heat extraction, which isalso common practice today in folate determination. Theinhibitory effect of adding thiols is a formation of hemi-thioacetal with formaldehyde, thereby preventing the reactionbetween formaldehyde and H4folate.
58,66,67
■ STABILITY OF UNSUBSTITUTED FOLATE IN THEFOOD CHAIN
Because H4folate is the most vulnerable folate form, one couldexpect high losses of H4folate in the food chain, especially afterheat treatment such as thermal processing or cooking. Hence,the richest dietary sources of H4folate should occur in rawfoods with intact cell structure, where it may be bound andstabilized by proteins. A folate binding protein (FBP) is presentin milk. By adding FBP to labile folate forms in model systems,H4folate has been shown to become significantly stabilized.68
As soon as the cell integrity is compromised by freezing andthawing, mechanical stress, or thermal processing, H4folatestarts to oxidize unless protected by native antioxidants such asascorbic acid or being bound to proteins and other cellconstituents. Note that iron and copper ions leaking fromprocess equipment or process water are pro-oxidative.41
Baker’s yeast, in compressed or dried form, still containsviable cells and is therefore an example of a food ingredientwith intact cell structure. It contains about 25% of its total
Table 2. Stability of Unsubstituted Folate According to the Literature
Folic AcidAt 20 °C and 65% relative humidity, solid folic acid has a decomposition rate of 1% per year.54
Extensive degradation of solutions of folic acid at room temperature is induced by sunlight, UV, or visible light. This photolysis increases in rate at pH <5.46,47
Solutions of folic acid are stable for up to 10 h at 100 °C in the pH range 5−12, light-protected and without antioxidants, but the stability decreases at pH values <5.43
H2folateH2folate in powder is stable for up to 1 year if stored at −20 °C in oxygen-free sealed tubes.36
The extent of degradation of a stock solution of H2folate in Tris buffer, at pH 7.3, containing 0.3 M MCE and stored at 0 °C, is only 2.3% in 10 h.50
H2folate is oxidized 40 times faster at pH 4 than at pH 7. At pH 9.5, H2folate appears to be relatively stable.49
Although H2folate is relatively unstable at pH 4.0−7.0, addition of thiols and ascorbic acid appears to retard oxidation. MCE in concentration 5−10 mM is sufficient toprevent decomposition of H2folate solutions.50
H4folateH4folate is relatively stable dry in ampules at −20 °C51
The rate of degradation of an unprotected solution of H4folate increases 5-fold from 0 °C to room temperature. Increasing the temperature from 23 to 37 °C increasesthe rate of decomposition another 5-fold.51
H4folate in phosphate buffer (pH 7.5) loses half its activity in 40 min at 23 °C. MCE at a concentration of 0.01 M or bubbling with N2 gas protects H4folate for >90min.52
H4folate is stable in anaerobic conditions at room temperature.36
Aqueous solutions of H4folate stored at room temperature undergo 90% degradation in 4 h, the lability being influenced by pH and the buffer.42
Aqueous solutions of H4folate are rapidly degraded. After 5 min at room temperature, a 1 mM (6R,S)-H4folate-3HCl solution is degraded by >10% in the presence ofair. After 45 min, it is degraded by about 25%. Solutions should be made as concentrated as possible, in oxygen-free water, with lowest possible pH, and usedimmediately.51
At 100 °C, solutions of H4folate have a half-life of 2.25 min.53
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2371

folate as H4folate (Table 1). When dry baker’s yeast was storedin room temperature during 45 days, H4folate was stable. Incontrast, compressed baker’s yeast stored in a refrigerator (5°C) lost 40% of H4folate after 17 days. Drying of yeast reducedthe content of H4folate because the content of H4folate wasconsiderably higher in compressed baker’s yeast prior tostorage, but these differences reduced progressively withstorage time.28
Cereal fractions contain about 10% of its folate as H4folate.During breadmaking, yeast and fermentation could add furtherH4folate. However, several studies indicate undetectable levelsof H4folate in bread; instead they often report the occurrence offolic acid, ranging from a few percent to 50% of totalfolate.25,69,70 The breads in these studies were not made fromfolic acid fortified flours. Hence, the folic acid most probablyoriginates from oxidation of unsubstituted reduced folates aswell as from 5,10-CH2-H4folate during fermentation and ovenbaking.Green vegetables are mostly blanched and frozen before
being cooked and served. Their content of ascorbate andpossibly other antioxidants could protect the H4folate frombeing oxidized, but losses by leakage could occur. In one studyby Stea and co-workers broccoli and green peas wereinvestigated after different thermal processes.71 Blanching didnot reduce H4folate, but, in general, these vegetables retainedonly up to half of their H4folate after other thermal treatments,for example, steam cooking, microwaving, boiling, andreheating. In a recent study on the effects of industrialprocessing on folate content in green vegetables, Delchier et al.found leakage to be a major cause behind folate losses.72
Some losses of dietary folates by leakage and/or degradationseem inevitable in the food chain.71,72 Therefore, it is of utmostimportance to avoid further degradation and conversions ofunsubstituted folates during the analytical procedure asoutlined in the following section.
■ STABILIZATION DURING DIFFERENT STEPS OFTHE ANALYTICAL PROCEDURE
Analytical techniques used to determine folate content arebiological, such as microbiological and competitive-bindingmethods, or chemical, such as HPLC methods.9,12,18,73 Themicrobiological assay (MA) utilizes the nutritional requirementof a microorganism (Lactobacillus rhamnosus), which exhibitedhigh response to all bioactive folate forms in monoglutamic ordiglutamic forms including biologically active interconversionproducts. Hence, MA gives the sum of all bioactive forms, thatis, total folate.9 There are standard methods assessed incollaborative studies established for microbiological assays, forfortified infant formula, for cereals, and for foodstuffs.74−76 Adisadvantage of the MA is the overall time needed for analysis,almost 2 days, during which the samples are kept at 37 °C,which emphasizes the need for optimal protection byantioxidants.The competitive binding assay, using a folate-binding protein
and a radiolabeled folate to compete with the folate to bedetermined, also gives total folate content. The Biacoremethod, certified by AOAC for folic acid determinations, usesa folate binding protein isolated from bovine milk and measuresthe amount of vitamin with plasma surface resonancetechnique.77 However, the folate-binding protein does nothave the same affinity to the various folate forms.78,79
Therefore, these methods are appropriate only if it is possibleto calibrate against the same form of folate as the one present in
the sample, such as folic acid in fortified foods. They are lessuseful for food analysis with more complex matrices and severalcombinations of folate forms.However, if the focus of interest is on the individual folate
forms, chromatographic methods such as HPLC, UHPLC, orLC-MS, which separate and detect the different forms on thebasis of their chemical properties, have to be used. Figure 4gives a schematic overview of the analytical methods for folatedetermination in foods. All methods include the followingsteps: extraction, deconjugation, cleanup by filtration andcentrifugation, and detection/quantification.80 The chromato-graphic methods often include additional cleanup by affinitychromatography or solid phase extraction (SPE). It is beyondthe scope of this paper to go into all analytical details aboutchromatography-based methods for determination of foodfolates. Instead, we refer to recent reviews.12,18,30,62,73,81−83
To obtain reliable analytical results, the stabilization ofunsubstituted folates, especially H4folate, is of great importance.In Table 3 we summarize the most crucial factors in thestabilization of unsubstituted folates during analytical proce-dure. As seen from Table 3, it is possible to efficiently stabilizeunsubstituted folates by using the following protectionstrategies:
• protection from UV light, oxygen, and catalysts of freeradical reactions such as iron and copper ions throughoutthe analytical procedure;
• avoidance of repeated freeze−thaw cycles of foodsamples, food extracts, and stock solutions;
• choice of lower temperature and shorter time for eachanalytical step everywhere it does not compromise theyield of folates;
• use of combination of two antioxidants, ascorbic acid/ascorbate and a thiol (BAL, DTT, or TBA).
Sampling, Storage, and Homogenization. To obtainreliable analytical results, good laboratory practice of samplingand storage of food samples is important, by protecting themfrom UV light, air, and high temperatures. Storage shouldpreferably be carried out at −80 °C. Once the samples arehomogenized, they should be dissolved in buffers of neutral pHcontaining two antioxidants, usually 1−2% ascorbic acid/ascorbate and a thiol to protect H4folate from oxidativedegradation.58−61 Further storage after homogenization shouldbe avoided because repeated freezing and thawing results in thedegradation of H4folate.
61
Extraction and Deconjugation. Three major proceduresare used to extract folate from the food matrix: heat treatment,enzymatic extraction, or a combination of heat and enzymatictreatments. Most studies use neutral pH during extraction andother enzyme treatments. Heat treatment is usually done byboiling in a water bath (100 °C for 10 min)73,81 or heating (75°C for 1 h),80,85 when using HPLC methods, or by autoclave(120 °C/15 min), when applying MA.74−76 The enzymaticextraction is commonly performed at 37 °C for 1−4 h byadding protease and/or amylase to the sample and terminatingeach enzyme treatment by boiling (5 min).73,81 Thecombination of two antioxidants, ascorbic acid and thiol, iscrucial during sample extraction to protect the labile H4folatefrom oxidative degradation as well as to prevent interconversionof H4folate into 5,10-CH2-H4folate caused by formaldehydeformed from ascorbic acid when heated.14 Three thiols, BAL,TBA, and DTT, have been shown to be highly efficient forthese purposes, whereas the most commonly used, MCE,
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2372

appeared to be the least efficient.61 Besides efficiency, thetoxicity and environmental risks should also be consideredwhen choosing thiol to stabilize H4folate. From this point ofview, the nontoxic TBA is the best choice followed by low-toxicDTT and BAL.22,61
To limit the amount of forms for determination, mono- ordiglutamates are achieved by enzymatic deconjugation of theglutamate residues. Choice of conjugase (γ-glutamyl hydrolase)source depends on the method of determination used. ForHPLC it is necessary to use a conjugase source, usually ratplasma, human plasma, or hog kidney conjugase withmonoglutamates as the end product, whereas the bacteria inthe MA can use the diglutamates produced by chicken pancreasas well.86 The duration of deconjugation procedure usuallyvaries from 2 to 3 h to overnight incubation at 37 °C, followedby inactivation by 5 min of boiling.81 In addition to themammal-based conjugase sources hitherto used, Diaz de laGarza’s group recently evaluated two recombinant γ-glutamylhydrolases from Arabidopsis thaliana.87 These enzymes were farmore efficient in hydrolyzing folate polyglutamates in plantfood samples compared with rat plasma; in fact, thedeconjugation time could be shortened to 0.5−1 h. Moreover,the recombinants also seemed to be less sensitive towardconjugase inhibitors present in certain plant extracts.87 Therepeated heating and relatively long time of incubation duringdeconjugation (overnight incubation is most common for MA)emphasize the need to optimize the antioxidative protection ofH4folate, as mentioned above for the extraction enzymes.Moreover, blanks need to be included for all enzymes added inthe assay to subtract their endogenous folate content.Storage of food extracts after heating and deconjugation
should be avoided. Tamura and co-workers showed that storageat −70 °C for a few months caused considerable degradation ofH4folate (∼60%) in food extracts.88 Patring and co-workersalso observed losses of native H4folate in yeast extract stored at−22 °C for 4 weeks.61 With 2% sodium ascorbate and 0.1%MCE the losses amounted to 53%, whereas the replacement ofMCE with BAL reduced losses to only 19%. De Brouwer andco-workers reported stabilities of 63.7−77.7% for nativeH4folate in rice extract stored at 4 °C for 36 h, whereas thestability was >95% when stored at −20 or −80 °C with one
freeze−thaw cycle.22,31 Van Daele and co-workers used 1%ascorbic acid and 0.5% DTT for stabilization of folates instandard solutions and potato extracts. They reported stability>90% for H4folate solution when stored at 4 °C for 24 h in theautosampler or at −20 °C with three freeze−thaw cycles.89
Cleanup by SPE or Affinity Chromatography. Chro-matographic methods are more sensitive to interferingsubstances and have higher demands on sample cleanup thanMA. SPE can be used both to remove interfering substancesand to concentrate the analyte. Affinity chromatographycleanup using folate-binding protein applied on columns canbe used to increase sensitivity. However, different affinities tothe various folate forms may easily lead to considerable lossesof folate forms with low affinity.18
Folates are eluted by acidic buffers, and affinity chromatog-raphy causes the most pronounced acidification by operatingclose to pH 2 during the 10−20 min the elution takes. SPE-based cleanup procedures using strong-anion exchange (SAX)cartridges elute the folates by sodium acetate solutioncontaining sodium chloride salt (5 or 10%) and ascorbic acid(1%), which keeps the pH close to 5. However, it is stillimportant to use antioxidants in optimal amounts and UV lightprotection to prevent oxidation and degradation of reducedunsubstituted folates, especially if the samples are stored in arefrigerator or autosampler overnight until subjected intoHPLC. For instance, Kirsch and co-workers13 studied thestability of individual folate standards stored at 4 °C during 24h in buffers of different pH values (2.6, 3.4, and 7.0) containing0.1% ascorbic acid. The main interconversion products werepresented for each folate form based on UHPLC-MS/MSanalysis. H4folate showed minor interconversion (<5%) intoH2folate and folic acid, whereas H2folate showed degradation atacidic conditions and interconversion (12.7−16.7%) to folicacid.
Chromatographic Separation by HPLC and UHPLC.No antioxidants are present in the mobile phase duringchromatographic separation, which makes folates unprotectedagainst oxidation. Moreover, column temperatures might varybetween 20 and 35 °C in HPLC28,30 and 60 °C in UHPLC.34,89
The extent to which acid-catalyzed oxidations of unsubstitutedfolates occur at this stage of analysis has received little attention
Figure 4. Schematic overview of analytical methods for folate determination in foods modified from refs 12, 18, 78, and 81.
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2373

over the years. Strandler and co-workers found a concentration-dependent stability of H4folate under these conditions, pHaround 3 and a temperature of 35 °C.90 At a concentration of 5μg/mL the losses of H4folate were 20% after 30 min, whereas ata lower concentration of 0.5 μg/mL H4folate was degradedcompletely after 10 min.90 Usually the concentrations ofH4folate in injected standard and food extracts are in the rangeof 1−50 ng/mL, that is, much lower than those studied byStrandler and co-workers (for references, see Table 1). Thisindicates a considerable risk for losses of H4folate during thepassage of the chromatographic column, which can lead todeviations from linearity in the lower region of calibration curveand false-negative results for H4folate. Therefore, specialconsideration should be given to the on-column stability ofH4folate in the development and optimization of new HPLC orUHPLC methods to determine folates in view of theconcentration-dependent stability of H4folate. Shorter retentiontime and lower column temperature are preferable to improvethe on-column stability of H4folate.HPLC or UHPLC with Different Detectors. When using
HPLC, the choice of calibrants becomes more crucial becauseonly the forms that are used as calibrants can be identified inthe sample. Due to interconversion and stability problems,there are limitations to which folate forms can be found in foodextracts, and this has been reviewed in previous sections of thisreview. Determination can be made by using UV, fluorescence,MS, or electrochemical detection.12,73,81,82 The reduced folateshave native fluorescence of different intensities and wavelengthmaxima.91 Folic acid does not fluorescence and is betterdetermined with UV detection.81 This means that acombination of UV and fluorescence detection at different
excitation/emission wavelengths is necessary to quantify allfolate forms. When using both detectors connected in sequence(UV detector followed by fluorescence detector), specialattention should be paid to the stability of H4folate duringthe passage of UV detector flow cell because of the high risk fordegradation of H4folate due to UV light and increasedtemperature in the flow cell. The stability check can beperformed by comparing the fluorescence response for H4folatewith and without UV detector connected before thefluorescence detector. If the fluorescence response for H4folateis lower without the UV detector, this indicates that there arelosses of H4folate in the UV detector flow cell. In such a case itis preferable to use split and connect detectors in parallel.Higher specificity is achieved by using mass spectrometric
detectors. Analytes are ionized, giving a specific mass to chargepattern, and folates can be determined in both positive andnegative modes.28 Quantification is usually performed withexternal calibration in HPLC methods when using UV andfluorescence detection. In methods with mass spectrometricdetection the use of stable isotopes as internal standards, suchas deuterium-labeled (2H4)
31or 13C-labeled (13C5) isotopes offolates,22,89 can considerably improve the relative recovery, butit cannot affect the absolute recovery.92 Although the relativerecovery of H4folate is about 100%, the absolute recovery may bemuch lower, about 37−70%,22,30,33,34,89 which may lead tonegative false results for H4folate if concentrations in foodsamples are close to the limit of quantification. Moreover, theuse of internal standards or external standards will not correctfor oxidations or conversions into other bioactive folate formsduring analysis, as recently discussed by Ringling and Rychlik,30
for instance, when native 5,10-CH2-H4folate dissociates to
Table 3. Crucial Factors in the Stabilization of Unsubstituted Folates during All Steps of the Analytical Procedurea
factor analytical steps protection strategies
presence ofoxygen
all degass with nitrogen or argon
UV light all protect from UV light, e.g., by using yellow filter on lamps or yellow lamps
freeze−thawcycle
storage of food samples avoid multiple freeze−thaw cycles, no more than onestorage of food extracts prior to analysis avoid freeze−thaw step, use storage at 4 °C instead
presence ofimpurities
all use reagents of high purity, avoid presence of pro-oxidative catalysts, e.g., copper and iron ionscheck the purity of folate standards and correct concentrations of stock solutions for purity84
temperature storage of food samples prior samplepretreatment
as low a temperature as possible, −80 °C22,34,88
storage of stock solutions −80 °Cextraction by heat treatment as low a temperature as possible, e.g., 75 °C instead of 100 °C61,85
storage of food extracts prior to analysis 4−6 °Cseparation on HPLC column avoid using elevated column temperatures if possible61,90
detection by UV followed by fluorescence use UV-DAD detector with efficient cooling system
time storage of samples and extracts as short as possibleextraction by heat treatment and/or enzymes as short as possibledeconjugation as short as possible, avoid overnight incubationseparation on the HPLC column shorter retention times are preferable
oxidation preparation of stock and working solutions,extraction and elution buffer.
use a combination of two antioxidants, ascorbic acid/ascorbate and TBA, for MA or ascorbicacid/ascorbate and BAL or DTT for HPLC methods
all steps of sample pretreatment make fresh working solutions on daily basisaOnly H4folate and folic acid are considered, because there are no studies presenting data on H2folate or the conversion product 5,10-CH2-H4folatein foods due obviously to their lack of stability as already discussed in the sections above.
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2374

H4folate and H4folate is oxidized to H2folate and folic acid.Hence, conversions and oxidations may cause some over-estimations of H4folate and folic acid because the changes arenot exactly the same in the food matrix as in the standards.It is important to consider that the bioactive unsubstituted
folates analyzed in foods, H4folate and folic acid, reflect the sumof oxidative and interconversion changes of H4folate, H2folate,and 5,10-CH2H4folate originally present in the intact cells ofvegetable, animal, or microbiological origin such as yeast andLAB. In addition, folic acid in foods either depends on theoxidation of H4folate or H2folate or is a consequence ofsupplementation/fortification or both. Analysts monitoringfolic acid content in fortified foods need to consider thatparts of the analyzed folic acid could be due to oxidation ofunsubstituted reduced folates or conversion from 5,10-CH2-H4folate to H4folate and its further oxidation. Also worthmentioning is that the relatively stable folic acid in fortifiedfoods might be underestimated due to photodecomposition ifnot protected from UV light during sample pretreatment.When it comes to UV light sensitivity, both oxidized andreduced folates are sensitive to degradation.
■ AUTHOR INFORMATIONCorresponding Author*(H.S.S.) Phone: +4618175757. Fax: +4618105848. E-mail:[email protected].
NotesThe authors declare no competing financial interest.
■ ABBREVIATIONS USED5-CH3-H4folate 5-methyltetrahydrofolate5-HCO-H4folate 5-formyltetrahydrofolate10-HCO-H4folate 10-formyltetrahydrofolate10-HCO-H2folate 10-formyldihydrofolate10-HCO-folic acid 10-formylfolic acid5,10-CH+-H4folate 5,10-methenyltetrahydrofolate5,10-CH2-H4folate 5,10-methylenetetrahydrofolateAOAC Association of Analytical ChemistsBAL 2,3-dimercapto-1-propanolC1 one-carbonCHES 2-(N-cyclohexylamino)ethanesulfonic acidCH2O formaldehydeDAD diode array detectorDTT 1,4-dithiothreitolECD electrochemical detectionFBP folate binding proteinFW fresh weightH2folate dihydrofolateH4folate tetrahydrofolateHEPES N-(2-hydroxyethyl)-piperazine-N-2-etha-
nesulfonic acidIS internal standardLAB lactic acid bacteriaMA microbiological assayMCE 2-mercaptoethanolNTD neural tube defectpABG p-aminobenzoylglutamic acidSAX strong anion exchangeSPE solid phase extractionTBA 2-thiobarbituric acidUHPLC ultrahigh-performance liquid chromatogra-
phy
■ REFERENCES(1) MRC. Vitamin Study Research Group. Prevention of neural tubedefects: results of the Medical Research Council Vitamin Study. Lancet1991, 338, 131−137.(2) Lucock, M. Folic acid: nutritional biochemistry, molecularbiology, and role in disease processes. Mol. Gen. Metab. 2000, 71,121−138.(3) Clarke, R.; Halsey, J.; Bennett, D.; Lewington, S. Homocysteineand vascular disease: review of published results of the homocysteine-lowering trials. J. Inherit. Metab. Dis. 2011, 34, 83−91.(4) De-Regil, L.; Fernandez-Gaxiola, A.; Dowswell, T.; Pena-Rosas, J.Effects and safety of periconceptional folate supplementation forpreventing birth defects. Cochrane Database Systematic Rev. 2010, 10,CD007950.(5) WCRF/AICR (World Cancer Research Fund/American Institutefor Cancer Research Publisher). Food, nutrition, physical activity andthe prevention of cancer: a global perspective; Washington DC, USA;www.dietandcancerreport.org (accessed March 2014).(6) Durga, J.; van Boxtel, M. P. J.; Schouten, E. G.; Kok, F. J.; Jolles,J.; Katan, M. B.; Verhoef, P. Effect of 3-year folic acid supplementationon cognitive function in older adults in the FACIT trial: a randomised,double blind, controlled trial. Lancet 2007, 369, 208−216.(7) Qin, X.; Cui, Y.; Shen, L.; Sun, N.; Zhang, Y.; Li, J.; Xu, X.; Wang,B.; Xu, X.; Huo, Y.; Wang, X. Folic acid supplementation and cancerrisk: a meta-analysis of randomized controlled trials. Int. J. Cancer2013, 133, 1033−1041.(8) Wien, T. N.; Pike, E.; Wisløff, T.; Staff, A.; Smeland, S.; Klemp,M. Cancer risk with folic acid supplements: a systematic review andmeta-analysis. BMJ Open 2012, 2e000653, DOI: 10.1136/bmjopen-2011-005.(9) Gregory, J. F. Chemical and nutritional aspects of folate research:analytical procedures, methods of folate synthesis, stability, andbioavailability of dietary folates. In Advances in Food and NutritionResearch; Kinsella, J. E., Ed.; Academic Press: New York, 1989; pp 1−101.(10) Nelson, D. L.; Cox, M. M. Chapter 22.2. Biosynthesis of aminoacids. In Lehninger Principles of Biochemistry, 5th ed.; Freeman: NewYork, 2008; pp 882−897.(11) Hawkes, J. G.; Villota, R. Folates in foods: reactivity, stabilityduring processing, and nutritional implications. Crit. Rev. Food Sci.Nutr. 1989, 28, 439−538.(12) Ball, G. F. M. Folate. In Vitamins in Foods: Analysis,Bioavailability, and Stability; Ball, G., Ed.; CRC Press: Boca Raton,FL, USA, 2006; pp 231−273.(13) Kirsch, S. H.; Knapp, J.-P.; Herrmann, W.; Obeid, R.Quantification of key folate forms in serum using stable-isotopedilution ultra performance liquid chromatography-tandem massspectrometry. J. Chromatogr., B 2010, 878, 68−75.(14) Wilson, S. D.; Horne, D. W. Evaluation of ascorbic acid inprotecting labile folic acid derivatives. Proc. Natl. Acad. Sci. U.S.A.1983, 80, 6500−6504.(15) Reed, L. S.; Archer, M. C. Oxidation of tetrahydrofolic acid byair. J. Agric. Food Chem. 1980, 28, 801−805.(16) Eto, I.; Krumdieck, C. L. Determination of three different poolsof reduced one-carbon-substituted folates: I. A study of thefundamental chemical reactions. Anal. Biochem. 1980, 109, 167−184.(17) USDA National Nutrient Database for Standard Reference,release 25; Nutrient Data Laboratory Home Page, 2012; http://www.ars.usda.gov/ba/bhnrc/ndl.(18) Quinlivan, E. P.; Hanson, A. D.; Gregory, J. F. The analysis offolate and its metabolic precursors in biological samples. Anal. Biochem.2006, 348, 163−184.(19) Ravanel, S.; Douce, R.; Rebeille, F. Metabolism of folates inplants. In Advances in Botanical Research; Rebeille, F., Douce, R., Eds.;Academic Press: London, UK, 2011; Vol. 59, pp 67−106.(20) Hanson, A. D.; Gregory, J. F. Folate biosynthesis, turnover, andtransport in plants. Annu. Rev. Plant Biol. 2011, 62, 105−125.
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2375

(21) Díaz de la Garza, R. I.; Gregory, J. F., III; Hanson, A. D. Folatebiofortification of tomato fruit. Proc. Natl. Acad. Sci. U.S.A. 2007, 104,4218−4222.(22) De Brouwer, V.; Storozhenko, S.; Van de Steene, J. C.; Wille, S.M. R.; Stove, C. P.; van der Straeten, D.; Lambert, W. E. Optimisationand validation of a liquid chromatography-tandem mass spectrometrymethod for folates in rice. J. Chromatogr., A 2008, 1215, 125−132.(23) Jagerstad, M.; Piironen, V.; Walker, C.; Ros, G.; Carnovale, E.;Holasova, M.; Nau, H. Increasing natural food folates throughbioprocessing and biotechnology. Trends Food Sci.Technol. 2005, 16,298−306.(24) Koehler, P.; Hartmann, G.; Wieser, H.; Rychlik, M. Changes offolates, dietary fiber, and proteins in wheat as affected by germination.J. Agric. Food Chem. 2007, 55, 4678−4683.(25) Patring, J.; Wandel, M.; Jagerstad, M.; Frølich, W. Folatecontent of Norwegian and Swedish flours and bread analysed by use ofliquid chromatography-mass spectrometry. J. Food Compos. Anal. 2009,22, 649−656.(26) Kariluoto, S.; Edelmann, M.; Herranen, M.; Lampi, A.-M.;Shmelev, A.; Salovaara, H.; Korhola, M.; Piironen, V. Production offolate by bacteria isolated from oat bran. Int. J. Food Microbiol. 2010,143, 41−47.(27) FFI (Food Fortification Initiative); http://www.ffinetwork.org(accessed January 2015).(28) Patring, J. D. M.; Jastrebova, J. A. Application of liquidchromatography-electrospray ionisation mass spectrometry for deter-mination of dietary folates: effects of buffer nature and mobile phasecomposition on sensitivity and selectivity. J. Chromatogr., A 2007,1143, 72−82.(29) Vishnumohan, S.; Arcot, J.; Pickford, R. Naturally-occurringfolates in foods: method development and analysis using liquidchromatography−tandem mass spectrometry (LC−MS/MS). FoodChem. 2011, 125, 736−742.(30) Ringling, C.; Rychlik, M. Analysis of seven folates in food byLC−MS/MS to improve accuracy of total folate data. Eur. FoodRes.Technol. 2013, 236, 17−28.(31) Freisleben, A.; Schieberle, P.; Rychlik, M. Specific and sensitivequantification of folate vitamers in foods by stable isotope dilutionassays using high-performance liquid chromatography-tandem massspectrometry. Anal. Bioanal. Chem. 2003, 376, 149−156.(32) Zhang, G. F.; Storozhenko, S.; Van der Straeten, D.; Lambert,W. E. Investigation of the extraction behavior of the mainmonoglutamate folates from spinach by liquid chromatography−electrospray ionization tandem mass spectrometry. J. Chromatogr., A2005, 1078, 59−66.(33) Wang, C.; Riedl, K. M.; Schwartz, S. J. Fate of folates duringvegetable juice processing deglutamylation and interconversion.Food Res. Int. 2013, 53, 440−448.(34) De Brouwer, V.; Storozhenko, S.; Stove, C. P.; Van Daele, J.;Van der Straeten, D.; Lambert, W. E. Ultra-performance liquidchromatography-tandem mass spectrometry (UPLC-MS/MS) for thesensitive determination of folates in rice. J. Chromatogr., B 2010, 878,509−513.(35) Blakley, R. L. The Biochemistry of folic acid and relatedpteridines. In Frontiers of Biology; North Holland Publishing:Amsterdam, The Netherlands, 1969; pp 1−99.(36) Temple, C., Jr.; Montgomery, J. A. Chemical and physicalproperties of folic acid and reduced derivatives. In Folates and Pterins;Blakley, R. L., Benkovic, S. J., Eds.; Wiley: New York, 1984; Vol. 1, pp62−104.(37) Patring, J.; Lanina, S. A.; Jastrebova, J. A. Applicability of alkyl-bonded ultra-pure silica stationary phases for gradient reversed-phaseHPLC of folates with conventional and volatile buffers under highlyaqeous conditions. J. Sep. Sci. 2006, 29, 889−904.(38) Kallen, R. G.; Jencks, W. P. The dissociation constants oftetrahydrofolic acid. J. Biol. Chem. 1966, 241, 5845−5850.(39) Kallen, R. Tetrahydrofolic acid and formaldehyde. In Methods inEnzymology; Wright, D. M. A. L., Ed.; Academic Press: New York,1971; Vol. 66, pp 705−716.
(40) Poe, M. Acidic dissociation constants of folic acid, dihydrofolicacid, and methotrexate. J. Biol. Chem. 1977, 25, 3724−3728.(41) Gregory, J. F. Vitamins. In Food Chemistry; Damodaran, S.,Parkin, K. L., Fennema, O. R., Eds.; Dekker: New York, 2008; pp498−503.(42) O’Broin, J.; Temperley, I.; Brown, J.; Scott, J. Nutritionalstability of various naturally occurring monoglutamate derivatives offolic acid. Am. J. Clin. Nutr. 1975, 28, 438−444.(43) Paine-Wilson, B.; Chen, T. S. Thermal destruction of folacin:effect of pH and buffer ions. J. Food Sci. 1979, 44, 717−722.(44) Mnkeni, A. P.; Beveridge, T. Thermal destruction ofpteroylglutamic acid in buffer and model food systems. J. Food Sci.1982, 47, 2038−2041.(45) Lowry, O. H.; Bessey, O. A.; Crawford, E. J. Photolytic andenzymatic transformations of pteroylglutamic acid. J. Biol. Chem. 1949,180, 389−398.(46) Akhtar, M. J.; Khan, M. A.; Ahmad, I. Photodegradation of folicacid in aqueous solution. J. Pharm. Biomed. Anal. 1999, 19, 269−275.(47) Akhtar, J. M.; Khan, A. M.; Ahmad, I. Effect of riboflavin on thephotolysis of folic acid in aqueous solution. J. Pharm. Biomed. Anal.2000, 23, 1039−1044.(48) Thomas, A. H.; Suarez, G.; Cabrerizo, F. M.; Martino, R.;Capparelli, A. L. Study of the photolysis of folic acid and 6-formylpterin in acid aqueous solutions. J. Photochem. Photobiol. A:Chem. 2000, 135, 147−154.(49) Poe, M. Dihydrofolate reductase from a methotrexate-resistantstrain of Escherichia coli: dihydrofolate monooxygenase activity.Biochem. Biophys. Res. Commun. 1973, 54, 1008−1014.(50) Hillcoat, B. L.; Nixon, P. F.; Blakley, R. L. Effect of substratedecomposition on the spectrophotometric assay of dihydrofolatereductase. Anal. Biochem. 1967, 21, 178−189.(51) Schircks laboratories data sheets on folic acid (16.203)dihydrofolic acid (16.206) and tetrahydrofolic acid (16.207).(52) Scrimgeour, K. G. Methods for reduction, stabilization, andanalyses of folates. In Methods in Enzymology; Donald, B., McCormick,L. D. W., Eds.; Academic Press: New York, 1980; Vol. 66, pp 517−523.(53) Chen, T. S.; Cooper, R. G. Thermal destruction of folacin: effectof ascorbic acid, oxygen and temperature. J. Food Sci. 1979, 44, 713−716.(54) Tripet, F. Y.; Kesselring, U. W. Stability of solid folic acid inrelation to temperature and humidity. Pharm. Acta Helv. 1975, 50,318−322.(55) Blair, J. A.; Pearson, A. J. Kinetics and mechanism of theautoxidation of the 2-amino-4-hydroxy-5,6,7,8-tetrahydropteridines. J.Chem. Soc., Perkin Trans. 2 1974, 1, 80−88.(56) Blair, J. A.; Farrar, G. Chapter 6. Oxidation of tetrahydrofolatesand tetrahydrobiopterin by molecular oxygen. In AtmosphericOxidation and Antioxidants; Scott, G., Ed.; Elsevier: Amsterdam, TheNetherlands, 1993; pp 171−182.(57) Martin, C. B.; Walker, D.; Soniat, M. Density functional theorystudy of possible mechanisms of folic acid photodecomposition. J.Photochem. Photobiol. A: Chem. 2009, 208, 1−6.(58) Wilson, S. D.; Horne, D. W. High-performance liquidchromatographic determination of the distribution of naturallyoccurring folic acid derivatives in rat liver. Anal. Biochem. 1984, 142,529−535.(59) Osborn, M. J.; Huennekens, F. M. Enzymatic reduction ofdihydrofolic acid. J. Biol. Chem. 1958, 233, 969−974.(60) Blakley, R. L. Spectrophotometric studies on the combination offormaldehyde with tetrahydropteroylglutamic acid and other hydro-pteridines. Biochem. J. 1960, 74, 71−82.(61) Patring, J. D. M.; Johansson, M.; Yazynina, E.; Jastrebova, J.Evaluation of impact of different antioxidants on stability of dietaryfolates during food sample preparation and storage of extracts prior toanalysis. Anal. Chim. Acta 2005, 553, 36−42.(62) De Brouwer, V.; Zhang, G. F.; Storozhenko, S.; Straeten, D. V.;Lambert, W. E. pH stability of individual folates during critical sample
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2376

preparation steps in prevision of the analysis of plant folates.Phytochem. Anal. 2007, 18, 496−508.(63) Zakrezewski, S. F. Evidence for the chemical interactionbetween 2-mercaptoethanol and tetrahydrofolate. J. Biol. Chem. 1966,241, 2957−2961.(64) Pearson, A. J. Kinetics and mechanisms of the autooxidation oftetrahydropterins. Chem. Ind. 1974, 233−239.(65) Osborn, M. J.; Talbert, P. T.; Huennekens, F. M. The structureof “active formaldehyde” (N5,N10-methylene tetrahydrofolic acid). J.Am. Chem. Soc. 1960, 82, 4921−4927.(66) Blakley, R. L. The reaction of tetrahydropteroylglutamic acidand related hydropteridines with formaldehyde. Biochem. J. 1959, 72,707−710.(67) Kallen, R. G.; Jencks, W. P. The mechanism of the condensationof formaldehyde with tetrahydrofolic acid. J. Biol. Chem. 1966, 241,5851−5863.(68) Jones, M. L.; Nixon, P. F. Tetrahydrofolates are greatlystabilized by binding to bovine milk folate-binding protein. J. Nutr.2002, 132, 2690−2694.(69) Pfeiffer, C. M.; Rogers, L. M.; Gregory, J. F. Determination offolate in cereal-grain food products using trienzyme extraction andcombined affinity and reversed-phase liquid chromatography. J. Agric.Food Chem. 1997, 45, 407−413.(70) Konings, E. J.; Roomans, H. H.; Dorant, E.; Goldbohm, R. A.;Saris, W. H.; van den Brandt, P. A. Folate intake of the Dutchpopulation according to newly established liquid chromatography datafor foods. Am. J. Clin. Nutr. 2001, 73, 765−776.(71) Stea, T. H.; Johansson, M.; Jagerstad, M.; Frølich, W. Retentionof folates in cooked, stored and reheated peas, broccoli and potatoesfor use in modern large-scale service systems. Food Chem. 2007, 101,1095−1107.(72) Delchier, N.; Ringling, C.; Le Grandois, J.; Aoude-Werner, D.;Galland, R.; George, S.; Rychlik, M.; Renard, C. Effects of industrialprocessing on folate content in green vegetables. Food Chem. 2013,139, 815−824.(73) Nollet, L. M. L.; Toldra, F. Food Analysis by HPLC, 3rd ed.;CRC Press: Boca Raton, FL, USA, 2012.(74) Official Methods of Analysis of AOAC INTERNATIONAL (2003)17th Ed., AOAC INTERNATIONAL: Gaithersburg, MD, USA,Official Method 992.05.(75) Official Methods of Analysis of AOAC INTERNATIONAL (2005)18th Ed., AOAC INTERNATIONAL: Gaithersburg, MD, USA.Official Method 2004.05.(76) CEN, EN 14131:2003 Foodstuffs. Determination of folate bymicrobiological assay, 2003.(77) Bostrom Caselunghe, M.; Lindeberg, J. Biosensor-baseddetermination of folic acid in fortified food. Food Chem. 2000, 70,523−532.(78) Stralsjo, L.; Arkbage, K.; Witthoft, C.; Jagerstad, M. Evaluationof a radioprotein-binding assay (RPBA) for folate analysis in berriesand milk. Food Chem. 2002, 79, 525−534.(79) Nygren-Babol, L.; Jagerstad, M. Folate-binding protein in milk:a review of biochemistry, physiology, and analytical methods. Crit. Rev.Food Sci. Nutr. 2012, 52, 410−425.(80) Strandler, H. S. Determination of Folate for Food CompositionData, Swedish University of Agricultural Sciences, Uppsala, Sweden,2012.(81) Eitenmiller, R. R.; Ye, L.; Landen, W. O. Chapter 10. Folate andfolic acid. In Vitamin Analysis for the Health and Food Sciences, 2nd ed.;CRC Press: Boca Raton, FL, USA, 2008; pp 443−505.(82) Bagley, P. J.; Selhub, J. Analysis of folate form distribution byaffinity followed by reversed-phase chromatography and electro-chemical detection. Clin. Chem. 2000, 46, 404−411.(83) Ja gerstad, M.; Jastrebova, J. Occurrence, stability, anddetermination of formyl folates in foods. J. Agric. Food Chem. 2013,61, 9758−9768.(84) van den Berg, H.; Finglas, P. M.; Bates, C. FLAIRintercomparisons on serum and red cell folate. Int. J. Vitam. Nutr.Res. 1994, 64, 288−293.
(85) Yazynina, E.; Johansson, M.; Jagerstad, M.; Jastrebova, J. Lowfolate content in gluten-free cereal products and their mainingredients. Food Chem. 2008, 111, 236−242.(86) Tamura, T.; Shin, Y. S.; Williams, M. A.; Stokstad, E. L. R.Lactobacillus casei response to pteroylpolyglutamates. Anal. Biochem.1972, 49, 517−521.(87) Ramos-Parra, P. A.; Urrea-Lopez, R.; Diaz de la Garza, R. I.Folate analysis in complex food matrices: use of a recombinantArabidopsis γ-glutamyl hydrolase for folate deglutamylation. Food Res.Int. 2013, 54, 177−185.(88) Tamura, T.; Mizuno, Y.; Johnston, K. E.; Jacob, R. A. Foodfolate assay with protease, α-amylase, and folate conjugase treatments.J. Agric. Food Chem. 1997, 45, 135−139.(89) van Daele, J.; Blancquaert, D.; Kiekens, F.; van Der Straeten, D.;Lambert, W. E.; Stove, C. P. Folate profiling in potato (Solanumtuberosum) tubers by ultrahigh-performance liquid chromatography−tandem mass spectrometry. J. Agric. Food Chem. 2014, 62, 3092−3100.(90) Strandler, H. S.; Axelsson, M.; Jastrebova, J. Food folate analysisby HPLC; effects of on-column degradation and interconversion offolates. Presented at the 1st International Vitamin Conference,Copenhagen, Denmark, 2010.(91) Gounelle, J. C.; Ladjimi, H.; Prognon, P. A rapid ans specificextraction procedure for folate determination in rat liver and analysisby high-performance liquid chromatography with fluorometricdetection. Anal. Biochem. 1989, 176, 406−411.(92) Wieling, J. LC-MS-MS experiences with internal standards.Chromatographia 2002, 55, S107−S113.
■ NOTE ADDED AFTER ASAP PUBLICATIONThis article published February 26, 2015 with errors in theTOC graphic, Figure 2, and Figure 3. The corrected filesreposted February 27, 2015.
Journal of Agricultural and Food Chemistry Review
DOI: 10.1021/jf504987nJ. Agric. Food Chem. 2015, 63, 2367−2377
2377




















