
Visual Orientation and Directional Selectivity through Thalamic Synchrony

CIT
SYAa
Tb
T
Amisatacwncicn(pfpsstcdsntisittncpl
1
*T�EAmctscr
Neuroscience 142 (2006) 833–842
0d
ENTRAL SENSITIZATION IN THALAMIC NOCICEPTIVE NEURONSNDUCED BY MUSTARD OIL APPLICATION TO RAT MOLAR
OOTH PULPKrw
Ic(Stdwkcfli(tsiscap(acmn
ztsmf2atlesint1chs
. ZHANG,a1 C. Y. CHIANG,a1 Y. F. XIE,a S. J. PARK,a
. LU,a J. W. HU,a J. O. DOSTROVSKYb
ND B. J. SESSLEa,b*
Faculty of Dentistry, University of Toronto, 124 Edward Street,oronto, Ontario, Canada M5G 1G6
Department of Physiology, Faculty of Medicine, University of Toronto,oronto, Ontario, Canada M5S 1A8
bstract—We have recently demonstrated that application ofustard oil (MO), a small-fiber excitant and inflammatory
rritant, to the rat maxillary molar tooth pulp induces centralensitization that is reflected in changes in spontaneousctivity, mechanoreceptive field (RF) size, mechanical activa-ion threshold, and responses to graded mechanical stimulipplied to the neuronal RF in trigeminal brainstem subnu-leus caudalis and subnucleus oralis. The aim of this studyas to test whether central sensitization can be induced inociceptive neurons of the posterior thalamus by MO appli-ation to the pulp. Single unit neuronal activity was recordedn the ventroposterior medial nucleus (VPM) or posterior nu-lear group (PO) of the thalamus in anesthetized rats, andociceptive neurons were classified as wide dynamic rangeWDR) or nociceptive-specific (NS). MO application to theulp was studied in 47 thalamic nociceptive neurons andound to excite over 50% of the 35 VPM neurons tested and toroduce significant long-lasting (over 40 min) increases inpontaneous activity, cutaneous pinch RF size and re-ponses to graded mechanical stimuli, and a decrease inhreshold in the 29 NS neurons tested; a smaller but statisti-ally significant increase in mean spontaneous firing rate andecrease in activation threshold occurred following MO in theix WDR neurons tested. Vehicle application to the pulp didot produce any significant changes in six VPM NS neuronsested. MO application to the pulp produced pronouncedncreases in spontaneous activity, pinch RF size, and re-ponses to mechanical stimuli, and a decrease in thresholdn three of the six PO neurons. In conclusion, application ofhe inflammatory irritant MO to the tooth pulp results in cen-ral sensitization of thalamic nociceptive neurons and thiseuronal hyperexcitability likely contributes to the behavioralonsequences of peripheral inflammation manifesting asain referral, hyperalgesia and allodynia. © 2006 IBRO. Pub-
ished by Elsevier Ltd. All rights reserved.
Both authors contribute equally to this study.Correspondence to: B. J. Sessle, Faculty of Dentistry, University oforonto, 124 Edward Street, Toronto, Ontario, Canada M5G 1G6. Tel:1-416-979-4921; fax: �1-416-979-4936.-mail address: [email protected] (B. J. Sessle).bbreviations: CCI, chronic constriction injury; MinO, mineral oil; MO,ustard oil; NMDA, N-methyl-D-aspartic acid; NS, nociceptive-spe-
ific; PO, posterior nuclear group of the thalamus; RF, mechanorecep-ive field; RM ANOVA, repeated measures analysis of variance; S-R,timulus-response; V, trigeminal; Vc, subnucleus caudalis; Vo, subnu-
aleus oralis; VPL, ventroposterior lateral nucleus; VPM, ventroposte-ior medial nucleus; WDR, wide dynamic range.
306-4522/06$30.00�0.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2006.06.063
833
ey words: thalamic ventroposterior medial nucleus, poste-ior nuclear group, trigeminal, nociceptive specific neurons,ide dynamic range neurons.
n recent years great progress has been made in the study ofentral sensitization of nociceptive neurons in the spinal cordfor review, see Salter, 2004; Dubner, 2005; Woolf andalter, 2006). Much less is known about sensitization in the
rigeminal (V) brainstem nucleus but recent studies haveocumented that neurons in the subnucleus caudalis (Vc),hich is the V homologue of the spinal dorsal horn and isnown as the medullary dorsal horn, also undergo markedentral sensitization following peripheral noxious stimuli, in-ammation or nerve damage. For example, application of the
nflammatory irritant mustard oil (MO) to the tooth pulpChiang et al., 1998, 2002, 2005b) as well as other orofacialissues (Hu et al., 1992; Yu et al., 1993) induces Vc centralensitization reflected in increases in the spontaneous activ-
ty, cutaneous mechanoreceptive field (RF) size and re-ponses of Vc nociceptive neurons and decreases in theirutaneous mechanical activation threshold; these changesre dependent on brainstem glutamatergic (N-methyl-D-as-artic acid, NMDA) and purinergic P2X receptor mechanismsChiang et al., 1998, 2005b; Hu et al., 2002). In the inferiorlveolar nerve injury model comparable but more prolongedhanges also occur in Vc and in a chronic pulp inflammationodel sensitization in Vc has been reported in association withociceptive behavior (Iwata et al., 2001; Tsuboi et al., 2006).
The Vc plays a major role in mediating central sensiti-ation at the V brainstem level as evidenced by findingshat the central sensitization of nociceptive neurons inubnucleus oralis (Vo) and accompanying increased jawuscle electromyographic activity are dependent on the
unctional integrity of Vc (Cairns et al., 2001; Chiang et al.,002; Hu et al., 2002). However, limited information isvailable on central sensitization of nociceptive neurons inhe posterior thalamus. The few studies at the thalamicevel have largely concentrated on the ventroposterior lat-ral (VPL) nucleus of the ventrobasal thalamus. Severaltudies have shown that following experimentally induced
nflammation, the responses of many thalamic nociceptiveeurons are increased and their thresholds reduced andhey may display other altered properties (Guilbaud et al.,987; Dostrovsky and Guilbaud, 1988, 1990). Similarhanges in the properties on VPL nociceptive neuronsave also been observed in the sciatic nerve chronic con-triction injury (CCI) model of neuropathic pain (Guilbaud et
l., 1990, 1995; Bordi and Quartaroli, 2000). Alterations inved.

rtcccWatuuir
rnVittautstfGscTzafioHb
A
TwstPomtewwpftis(aqswwm
ac3davrgi
R
SeemtirdtBta(pcp
scwetna
trsi12nampdwss(owpdns1mbtp(eppf
S. Zhang et al. / Neuroscience 142 (2006) 833–842834
esponse properties including central sensitization of VPLhalamic neurons have also recently been reported in asso-iation with central neuropathic pain, e.g. following spinalord injury, lesions to ascending pathways, or block of gly-inergic inhibition in the spinal cord (Sherman et al., 1997;eng et al., 2000, 2003; Gerke et al., 2003). These changes
ppear not to be simply a reflection of changes occurring athe spinal level since the spinal cord injury results in anpregulation of Nav1.3 voltage-gated sodium channel sub-nits in VPL (Hains et al., 2005) and block of NMDA receptors
n VPL thalamus reduces the hyperalgesia produced by pe-ipheral inflammation (Kolhekar et al., 1997).
There have been two previous studies on the possibleole of the thalamus in mediating central sensitization ofociceptive processing arising from the orofacial region.os et al. (2000) reported changes in neuronal properties
ncluding increased spontaneous activity in the ventropos-erior medial nucleus (VPM) of the ventrobasal thalamus inhe model of CCI of the infraorbital nerve, and Kaneko etl. (2005) reported increased responses to electrical stim-lation of the pulp and decreased mechanical threshold of
he cutaneous RF after MO pulpal application. Previoustudies indicate that a major input in the rat VPM as well ashe posterior nuclear group (PO) of the thalamus arisesrom Vc (Peschanski, 1984; Gauriau and Bernard, 2004;uy et al., 2005) and we recently provided details of theomatotopic organization and response properties of no-iceptive neurons in the rat VPM (Chiang et al., 2005a).he aim of this study was to test whether central sensiti-ation can be induced in nociceptive neurons of the VPMs well as the PO by MO application to the pulp. Thendings are compared with those we have detailed previ-usly in Vc and Vo (Chiang et al., 1998; Park et al., 2001;u et al., 2002). Preliminary findings have been reportedriefly (Hu et al., 2001).
EXPERIMENTAL PROCEDURES
nimal preparation
his study was carried out in 63 Sprague–Dawley adult male ratseighing 290–420 g. The methods used for animal preparation,timulation, and neuronal recording and classification were similaro those detailed previously (Chiang et al., 1998, 2002, 2005b;ark et al., 2001; Hu et al., 2002) and so will only be brieflyutlined. The animal was anesthetized by an i.p. injection of aixture of urethane (1 g/kg) and alpha-chloralose (50 mg/kg). The
rachea and the left external jugular vein were cannulated. Toxpose the pulp of the right maxillary first molar, an occlusal cavityas prepared with a low-speed dental drill and temporarily filledith a cotton pellet soaked with saline. The animal was thenlaced in a stereotaxic apparatus, and a craniotomy was per-ormed to expose the contralateral (left) cerebral cortex overlyinghe thalamus. A supplemental dose of urethane (200–400 mg/kg,.v.) was administered 1 h after surgery and during the recordingession. The animal was immobilized with gallamine triethiodideinitial dose, 42 mg/kg; maintenance dose, 14 mg/h; i.v.) andrtificially ventilated throughout the experimental period. An ade-uate level of anesthesia was confirmed periodically by the lack ofpontaneous movements and responses to pinching the pawhen the gallamine-induced muscle paralysis was allowed toear off. In addition, pupil size and heart rate were also routinely
onitored to ensure their stability when noxious pinch stimuli were spplied. The expired % CO2 and rectal temperature were alsoontinuously monitored and maintained at physiological levels of.5–4.5% and 37–38 °C, respectively. All surgeries and proce-ures were carried out with the intent of minimizing the number ofnimals used and any suffering, and were approved by the Uni-ersity of Toronto Animal Care Committee in accordance with theegulations of the Ontario Animal Research Act (Canada) and theuidelines of the Canadian Council on Animal Care and the Amer-
can Physiological Society.
ecording and stimulation procedures
ingle neuronal activity in the contralateral thalamus was recordedxtracellularly by means of an epoxy-resin-coated tungsten micro-lectrode (15 mega-ohm; FHC, Bowdoinham, ME, USA) held by aicrodrive. As the microelectrode was advanced perpendicularly
hrough the cortex into the thalamus, low and high threshold mechan-cal stimuli were applied to the orofacial tissues to search for neuronseceiving an orofacial mechanoreceptive input. The VPM and imme-iately adjacent PO were dorsoventrally explored 2.1–3.2 mm lateralo the midline and between frontal planes P2.6 and P4.3 referred toregma (Paxinos and Watson, 1986). These neurons were assumed
o be thalamocortical neurons although not identified by antidromicctivation, since in the rat all neurons in this region project to cortexSherman and Guillery, 2001). Neuronal activity was amplified, dis-layed on oscilloscopes and also led to a window discriminatoronnected to an A/D converter (CED 1401plus, CED, UK) and aersonal computer.
A wide range of mechanical (brush, pressure and pinch)timuli was applied to the orofacial skin or intraoral mucosa tolassify each neuron as low-threshold mechanoreceptive (LTM),ide dynamic range (WDR) or nociceptive-specific (NS) (Chiangt al., 1998, 2005b; Park et al., 2001; Hu et al., 2002). Noxious
hermal heat was sometimes used to help identify nociceptiveeurons. Only WDR and NS neurons were characterized in detailnd included in the study.
The spontaneous activity, RF size, mechanical activationhreshold and responses to graded (from subthreshold to suprath-eshold) mechanical stimuli were assessed at the time intervalspecified below. After the recorded neuron’s firing became stable,ts spontaneous (baseline) activity was measured over a period of
min. As detailed in our previous reports (Chiang et al., 1998,002, 2005b; Park et al., 2001; Hu et al., 2002), the RF of eacheuron was determined through the use of a brush, blunt probend a pair of forceps. The extent of the cutaneous RF wasanually defined by brushing (in the case of a tactile RF) orinching (in the case of a pinch RF) the skin; as previouslyescribed (Chiang et al., 1994, 2005a,b). The noxious stimulationas used sparingly at the same spot so as to avoid damage to thekin or mucosa and cause neuronal sensitization. A burst re-ponse consisting of at least two spikes during each stimulus trialtouch or pinch) was accepted as the criterion for the RF boundaryf the neuron tested. A neuron was considered to have a deep RFhen the neuron responded to stimulation applied by a bluntrobe to the skin overlying muscle, bone, tendon, or temporoman-ibular joint (TMJ) and had a mechanical threshold above 5 g buto response could be evoked by the wide range of cutaneoustimuli used (Iggo, 1960; Schaible and Schmidt, 1983; Yu et al.,993). For orofacial and/or intraoral as well as body RF measure-ent, 14 designated parts throughout the orofacial region andody, including intraoral (upper and lower intraoral mucosa, andongue), perioral (upper and lower lips), face, nose, ears, fore-aws and hindpaws, and tail were tested as previously describedChiang et al., 1998, 2005b; Park et al., 2001; Hu et al., 2002). Thextent of the neuronal RF was assessed in terms of the number ofarts from which the response was evoked by light touch or firmressure applied with a dental explorer or pinch with a constantorce forceps (200 g). The activation threshold to a mechanical
timulus applied to the orofacial RF was assessed by a pair of
fimmptnWta
tt545trsoc
D
TtCmvi(om
E
T(ancsctt2cc((T0
tMpttmavmracottrc
H
RcwbdtV
m2nwatiabtpyrcrsav
AW
F(N
S. Zhang et al. / Neuroscience 142 (2006) 833–842 835
orce-monitoring forceps (with an attached strain gauge that mon-tored force levels up to 600 g/mm2). The gradual increase in the
echanical force and the responses of the tested neuron wereonitored and recorded simultaneously by the use of the Spike 2rogram (CED 1401 plus). However, the forceps did not allow uso accurately assess the mechanical activation threshold of WDReurons located in the intraoral region, therefore the threshold ofDR neurons was not systematically assessed, and only the
hresholds of a few WDR neurons having a cutaneous RF weressessed with von Frey hair filaments in this study.
Four graded intensities of mechanical stimuli were applied tohe tested neuron’s RF with the force-monitoring forceps (e.g. toest for pinch- or pressure-evoked responses of NS neurons to 25,0, 100, and 200 g stimuli and of WDR neurons to 5, 10, 20, and0 g stimuli; the stimulus was applied in ascending order, each fors, and successive stimuli were separated by an interval of more
han 30 s). The stimulus-response (S-R) relationship of NS neu-ons was determined by calculating the mean response (sum ofpikes during each 5 s stimulus) of all the neurons evoked by eachf the four graded stimuli. The peak firing rate response of noci-eptive neurons to the graded stimuli was also noted.
rug administration
o establish an acute chemical irritant-induced inflammation in theooth pulp, MO (0.2 �l; allyl isothiocyanate, 95%; Aldrich Chemicalo., Milwaukee, WI, USA) was applied locally to the exposedolar pulp. In our previous studies, a similar application of the
ehicle, mineral oil (MinO, 0.2 �l) to the pulp was shown to beneffective in inducing inflammation or central sensitizationChiang et al., 1998; Sunakawa et al., 1999; Park et al., 2001), sonly a limited number of experiments using MinO application wasade in this study.
xperimental paradigm
o test for neuroplastic changes indicative of central sensitizationsee above) in VPM or PO, only one neuron was tested in eachnimal. After a neuron was identified, the extent of the RF andeuronal response properties including spontaneous activity, me-hanical activation threshold and responses to graded mechanicaltimuli was first determined and served as a baseline value. Aomplete determination of the neuronal RF and response proper-ies took around 7 min (1 min for spontaneous activity, 1 min forhreshold, 2–3 min for tactile and/or pinch or pressure RF size and–3 min for responses to graded stimuli). Then, the saline-soakedotton pellet in the prepared cavity of the maxillary first molar wasarefully replaced by a segment (0.5 mm) of dental paper pointX-fine, Absorbent Point, Kerr Co., MI, USA) soaked with MO0.2 �l), and the cavity was sealed with CAVIT (ESPE, Germany).o ensure that the amount of MO solution was standardized, the.5 mm segment was cut off from the tip of the paper point and
ig. 1. A drawing of three coronal sections of the left VPM and adjacent
NS and WDR) neurons in VPM. These sections represent the rostral, middle and cS neurons and open circles and triangles WDR neurons. Numbers indicate thehen placed on a concave glass surface, and soaked with a 0.2 �lO solution delivered by a 1 �l Hamilton syringe. This freshlyrepared soaked paper point was immediately used for applica-ion to the molar pulp. Fifteen minutes after the MO application,he spontaneous activity, RF and response properties were deter-ined again. The determination of the spontaneous activity, RFnd neuronal response properties was repeated at 10 min inter-als over a 60-min period. In control experiments, after the deter-ination of the baseline spontaneous activity, RF and neuronal
esponse properties, 0.2 �l MinO was applied to the molar pulp insimilar procedure to that of the MO experiments. Then, the
avity was sealed by CAVIT. Fifteen minutes later, another seriesf complete determinations was performed in a similar manner ashat of the MO experiments. Due to technical difficulties to main-ain a stable recording over the long time period of the neuronalecording, only 50% of the nociceptive neurons could be studiedompletely for the entire 60-min period.
istological and statistical analysis
ecording sites were marked by an electrolytic lesion (anodalurrent 8.5 �A, 12 s) at the end of each experiment, and verifiedith conventional histological procedures. The recording sitesased on either the lesion location at the recording site or theiristance from a reference lesion site indicated by means of his-ological reconstruction were plotted on schematic drawings of thePM and immediately adjacent thalamic regions.
All values of neuronal response properties were expressed asean�S.E. Those of RF score were expressed as median and5th and 75th percentiles. Differences between WDR and NSeurons with respect to their neuronal properties were evaluatedith the Mann-Whitney test. Differences between baseline valuesnd values at a given time point were compared by the paired-test or signed rank test for each of the measurement time-pointsn the MO and MinO groups and evaluated by repeated measuresnalysis of variance (RM ANOVA) or ANOVA on ranks, followedy Dunnett’s test or Tukey test, respectively. Differences betweenwo groups at a corresponding time-point were treated by the ariori t-test or Mann-Whitney test. For the linear regression anal-sis of the S-R function to the graded mechanical stimuli, both theesponses and stimulus were logarithmically transformed and aonstant value of 0.1 was added before transformation of theesponses because many of these values were 0 (i.e. no re-ponse). The differences in S-R function prior to and following MOpplication were assessed by two-way RM ANOVA (SAS softwareersion 9.1). The level of significance was set at P�0.05.
RESULTS
total of 47 thalamic nociceptive neurons (40 NS, sevenDR) was recorded, of which 41 (35 NS, six WDR) neu-
regions including PO that shows the recording sites of all the nociceptive
thalamic audal portions of VPM, respectively. Filled circles and triangles representdistance from Bregma. Modified from (Paxinos and Watson, 1986).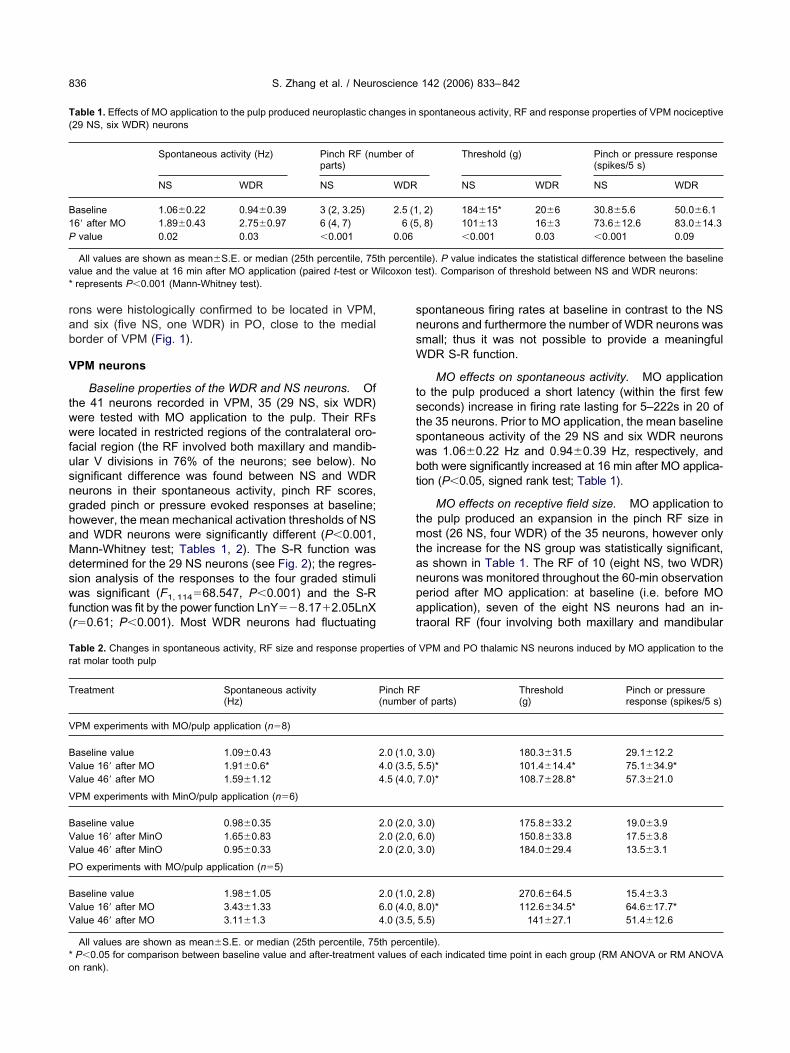
rab
V
twwfusnghaMdswf(
snsW
tstswbt
tmtanpat
T(
B1P
v*
Tr
T
V
BVV
V
BVV
P
BVV
*o
S. Zhang et al. / Neuroscience 142 (2006) 833–842836
ons were histologically confirmed to be located in VPM,nd six (five NS, one WDR) in PO, close to the medialorder of VPM (Fig. 1).
PM neurons
Baseline properties of the WDR and NS neurons. Ofhe 41 neurons recorded in VPM, 35 (29 NS, six WDR)ere tested with MO application to the pulp. Their RFsere located in restricted regions of the contralateral oro-
acial region (the RF involved both maxillary and mandib-lar V divisions in 76% of the neurons; see below). Noignificant difference was found between NS and WDReurons in their spontaneous activity, pinch RF scores,raded pinch or pressure evoked responses at baseline;owever, the mean mechanical activation thresholds of NSnd WDR neurons were significantly different (P�0.001,ann-Whitney test; Tables 1, 2). The S-R function wasetermined for the 29 NS neurons (see Fig. 2); the regres-ion analysis of the responses to the four graded stimulias significant (F1, 114�68.547, P�0.001) and the S-R
unction was fit by the power function LnY��8.17�2.05LnXr�0.61; P�0.001). Most WDR neurons had fluctuating
able 1. Effects of MO application to the pulp produced neuroplastic ch29 NS, six WDR) neurons
Spontaneous activity (Hz) Pinch RF (nuparts)
NS WDR NS
aseline 1.06�0.22 0.94�0.39 3 (2, 3.25)6= after MO 1.89�0.43 2.75�0.97 6 (4, 7)value 0.02 0.03 �0.001
All values are shown as mean�S.E. or median (25th percentile, 75alue and the value at 16 min after MO application (paired t-test or Wrepresents P�0.001 (Mann-Whitney test).
able 2. Changes in spontaneous activity, RF size and response propat molar tooth pulp
reatment Spontaneous activity(Hz)
PM experiments with MO/pulp application (n�8)
aseline value 1.09�0.43alue 16= after MO 1.91�0.6*alue 46= after MO 1.59�1.12
PM experiments with MinO/pulp application (n�6)
aseline value 0.98�0.35alue 16= after MinO 1.65�0.83alue 46= after MinO 0.95�0.33
O experiments with MO/pulp application (n�5)
aseline value 1.98�1.05alue 16= after MO 3.43�1.33alue 46= after MO 3.11�1.3
All values are shown as mean�S.E. or median (25th percentile, 75P�0.05 for comparison between baseline value and after-treatment
n rank).pontaneous firing rates at baseline in contrast to the NSeurons and furthermore the number of WDR neurons wasmall; thus it was not possible to provide a meaningfulDR S-R function.
MO effects on spontaneous activity. MO applicationo the pulp produced a short latency (within the first feweconds) increase in firing rate lasting for 5–222s in 20 ofhe 35 neurons. Prior to MO application, the mean baselinepontaneous activity of the 29 NS and six WDR neuronsas 1.06�0.22 Hz and 0.94�0.39 Hz, respectively, andoth were significantly increased at 16 min after MO applica-
ion (P�0.05, signed rank test; Table 1).
MO effects on receptive field size. MO application tohe pulp produced an expansion in the pinch RF size inost (26 NS, four WDR) of the 35 neurons, however only
he increase for the NS group was statistically significant,s shown in Table 1. The RF of 10 (eight NS, two WDR)eurons was monitored throughout the 60-min observationeriod after MO application: at baseline (i.e. before MOpplication), seven of the eight NS neurons had an in-raoral RF (four involving both maxillary and mandibular
spontaneous activity, RF and response properties of VPM nociceptive
Threshold (g) Pinch or pressure response(spikes/5 s)
NS WDR NS WDR
, 2) 184�15* 20�6 30.8�5.6 50.0�6.1, 8) 101�13 16�3 73.6�12.6 83.0�14.3
�0.001 0.03 �0.001 0.09
tile). P value indicates the statistical difference between the baselineest). Comparison of threshold between NS and WDR neurons:
VPM and PO thalamic NS neurons induced by MO application to the
of parts)Threshold(g)
Pinch or pressureresponse (spikes/5 s)
3.0) 180.3�31.5 29.1�12.25.5)* 101.4�14.4* 75.1�34.9*7.0)* 108.7�28.8* 57.3�21.0
3.0) 175.8�33.2 19.0�3.96.0) 150.8�33.8 17.5�3.83.0) 184.0�29.4 13.5�3.1
2.8) 270.6�64.5 15.4�3.38.0)* 112.6�34.5* 64.6�17.7*5.5) 141�27.1 51.4�12.6
tile).each indicated time point in each group (RM ANOVA or RM ANOVA
anges in
mber of
WDR
2.5 (16 (5
0.06
th percenilcoxon t
erties of
Pinch RF(number
2.0 (1.0,4.0 (3.5,4.5 (4.0,
2.0 (2.0,2.0 (2.0,2.0 (2.0,
2.0 (1.0,6.0 (4.0,4.0 (3.5,
th percenvalues of
dlfttetpdctrMiatai(s(
tMihesnpvt
m
hfievamtb1s(it(
Ftrtrrppd
FtarMpreH
Fvtc
S. Zhang et al. / Neuroscience 142 (2006) 833–842 837
ivisions, two the maxillary division and one the mandibu-ar division), of which three also had a convergent inputrom the perioral region, one from the nose, and one fromhe nasal bone; the remaining NS neuron had its RF limitedo the nose and septum. None of these neurons had a RFxtending beyond the orofacial region. After MO applica-ion, all eight NS neurons had an RF that expanded into theerioral region and that involved both maxillary and man-ibular divisions, and in two neurons this expansion in-luded the ear, forepaw, hindpaw and tail. Moreover, aactile RF on the eyeball, gum and upper lip skin tempo-arily appeared in four of these NS neurons. Overall, afterO application, the median RF score (eight NS neurons)
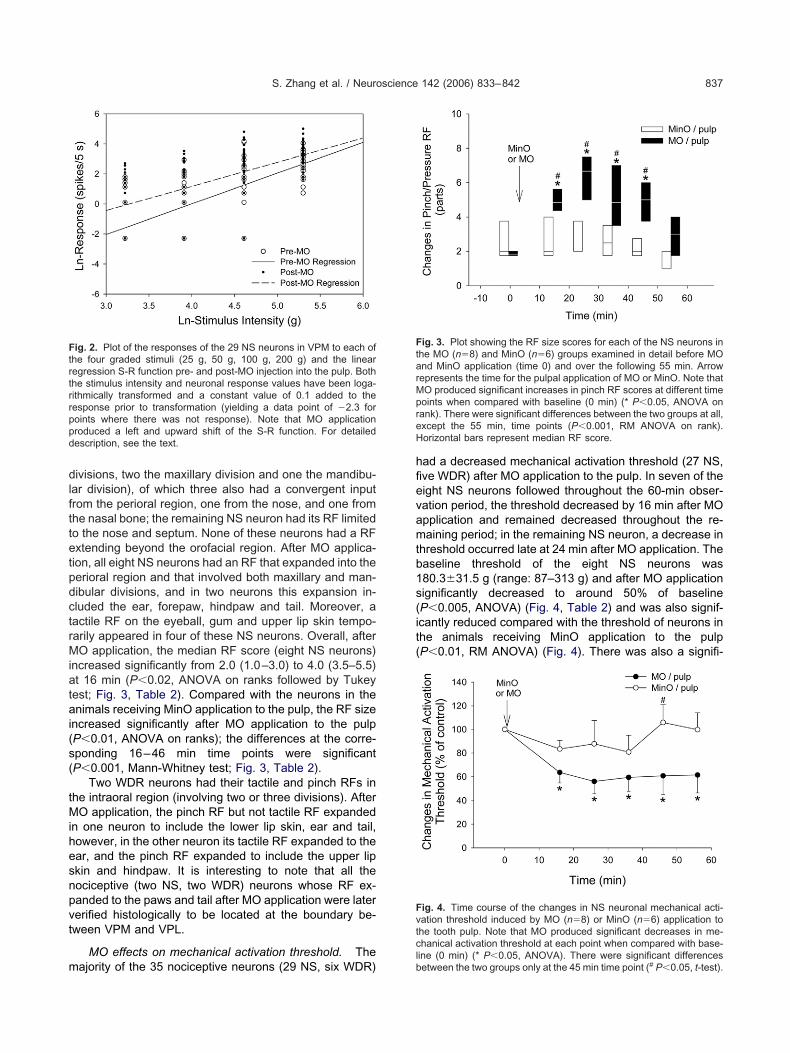
ncreased significantly from 2.0 (1.0–3.0) to 4.0 (3.5–5.5)t 16 min (P�0.02, ANOVA on ranks followed by Tukeyest; Fig. 3, Table 2). Compared with the neurons in thenimals receiving MinO application to the pulp, the RF size
ncreased significantly after MO application to the pulpP�0.01, ANOVA on ranks); the differences at the corre-ponding 16–46 min time points were significantP�0.001, Mann-Whitney test; Fig. 3, Table 2).
Two WDR neurons had their tactile and pinch RFs inhe intraoral region (involving two or three divisions). AfterO application, the pinch RF but not tactile RF expanded
n one neuron to include the lower lip skin, ear and tail,owever, in the other neuron its tactile RF expanded to thear, and the pinch RF expanded to include the upper lipkin and hindpaw. It is interesting to note that all theociceptive (two NS, two WDR) neurons whose RF ex-anded to the paws and tail after MO application were latererified histologically to be located at the boundary be-ween VPM and VPL.
MO effects on mechanical activation threshold. The
ig. 2. Plot of the responses of the 29 NS neurons in VPM to each ofhe four graded stimuli (25 g, 50 g, 100 g, 200 g) and the linearegression S-R function pre- and post-MO injection into the pulp. Bothhe stimulus intensity and neuronal response values have been loga-ithmically transformed and a constant value of 0.1 added to theesponse prior to transformation (yielding a data point of �2.3 foroints where there was not response). Note that MO applicationroduced a left and upward shift of the S-R function. For detailedescription, see the text.
ajority of the 35 nociceptive neurons (29 NS, six WDR)lb
ad a decreased mechanical activation threshold (27 NS,ve WDR) after MO application to the pulp. In seven of theight NS neurons followed throughout the 60-min obser-ation period, the threshold decreased by 16 min after MOpplication and remained decreased throughout the re-aining period; in the remaining NS neuron, a decrease in
hreshold occurred late at 24 min after MO application. Theaseline threshold of the eight NS neurons was80.3�31.5 g (range: 87–313 g) and after MO applicationignificantly decreased to around 50% of baselineP�0.005, ANOVA) (Fig. 4, Table 2) and was also signif-cantly reduced compared with the threshold of neurons inhe animals receiving MinO application to the pulpP�0.01, RM ANOVA) (Fig. 4). There was also a signifi-
ig. 3. Plot showing the RF size scores for each of the NS neurons inhe MO (n�8) and MinO (n�6) groups examined in detail before MOnd MinO application (time 0) and over the following 55 min. Arrowepresents the time for the pulpal application of MO or MinO. Note thatO produced significant increases in pinch RF scores at different timeoints when compared with baseline (0 min) (* P�0.05, ANOVA onank). There were significant differences between the two groups at all,xcept the 55 min, time points (P�0.001, RM ANOVA on rank).orizontal bars represent median RF score.
ig. 4. Time course of the changes in NS neuronal mechanical acti-ation threshold induced by MO (n�8) or MinO (n�6) application tohe tooth pulp. Note that MO produced significant decreases in me-hanical activation threshold at each point when compared with base-
ine (0 min) (* P�0.05, ANOVA). There were significant differencesetween the two groups only at the 45 min time point (# P�0.05, t-test).
cs
ttsNorAwbAttth2iNp
rstLeaeftSudsF
M
ToN(imsgitA
P
TwsVhmttitnbdnetrRirfiTVs(Apsfta2cnps
Otrc1
Fgpc(tt
S. Zhang et al. / Neuroscience 142 (2006) 833–842838
ant decrease in the mechanical activation threshold of theix WDR neurons after MO application to the pulp (Table 1).
MO effects on responses to mechanical stimuli. Ofhe 35 nociceptive neurons tested with MO application tohe pulp, both NS and WDR neurons had increased re-ponses to graded mechanical stimuli. A total of 10 (eightS, two WDR) neurons were tested throughout the 60-minbservation period. Prior to MO application, the meanesponse of the eight NS neurons was 29.1�12.2 spikes.fter MO application, the responses of all these neuronsere significantly increased by two- to threefold of theaseline values over the 60-min period (P�0.01, RMNOVA; Fig. 5, Table 2). Compared with the neurons
ested with MinO application to the pulp, the responses ofhese neurons increased significantly after MO applicationo the pulp (P�0.002, RM ANOVA). Two WDR neuronsad baseline responses to graded pinch stimuli of 55 and9 spikes, and at 16 min after MO application, responses
ncreased to 78 and 87 spikes, respectively. Examples ofS and WDR neuronal responses to pinch stimulationrior to and after MO application are shown in Fig. 6.
After MO application, the regression analysis of theesponses of the 29 NS neurons to the four gradedtimuli was significant (F1, 114�72.304, P�0.001) andhe S-R function was fit by the power functionnY��5.26�1.61LnX (r�0.62; P�0.001). The differ-nces between the S-R functions prior to (describedbove) and after MO application were significant (for MOffect: F1, 84�10.89, P�0.005; for graded intensity ef-
ect: F3, 84�395.6, P�0.001; for MO�intensity interac-ion: F3, 84�6.91, P�0.02; two-way RM ANOVA) and the-R function was significantly shifted to the left andpward after MO application; there were also significantifferences in the responses to the 25 g, 50 g, and 100 gtimuli (P�0.05, pair-wise multiple comparison) (seeig. 2).
ig. 5. Time course of the changes in NS neuronal responses toraded mechanical stimuli induced by MO (n�8) or MinO (n�6) ap-lication to the tooth pulp. Note that MO produced significant de-reases at 16–36 time points when compared with baseline (0 min)* P�0.05, Tukey test). There were significant differences between the
pwo groups (P�0.002, RM ANOVA on rank) and at the 16 and 26 minime points (# P�0.05, t-test).
inO effects
he effects of MinO application to the pulp were examinedn six NS neurons (see examples Figs. 3–5; Table 2).one of the neurons was spontaneously active prior to
baseline) or after MinO application. All six neurons hadpsilateral orofacial RFs (five involving both maxillary and
andibular divisions, one the mandibular division) andtable mean activation thresholds and responses toraded pinch or pressure stimuli. MinO application did not
nduce any significant changes in the RF size, activationhreshold or responses to mechanical stimuli (P�0.1, RMNOVA).
O neurons
he baseline spontaneous activity of the five NS neuronsas 1.98�1.05 Hz (range: 0.02–5.82 Hz), and was notignificantly different from that of the 35 NS neurons inPM (P�0.05, Mann-Whitney test). All five NS neuronsad their pinch RF in the intraoral region (four involving theaxillary division) but unlike VPM neurons, the RF ex-
ended to include also the ears and hindpaws in three ofhe five PO neurons. MO application produced a twofoldncrease in spontaneous activity in two of the neuronsested. After MO application, the pinch RF of four POeurons expanded into the perioral region (two involvingoth maxillary and mandibular divisions, two all three Vivisions); the RF of the remaining neuron expanded to theose. In addition, the RF of three of the five neuronsxpanded to include the ears, forepaws and tail. A novelactile RF temporarily appeared in the intraoral/perioralegions and ear in three of the five NS neurons. The pinchF increased significantly from two to six (Table 2) which
s greater than the increase observed for the VPM neu-ons. The mean mechanical activation threshold of theseve NS neurons was 270.6�64.5 g (range: 79–474 g;able 2) and was not significantly different from that of thePM NS neurons (P�0.06). MO application produced aignificant decrease in mechanical activation threshold�50% of baseline values) in three NS neurons (Table 2).ll the NS neurons in PO responded to strong pressure orinch stimuli with a variable number of spikes (nine to 30pikes). The mean response of 15.4�3.3 spikes (peakrequency 5�1.1 Hz) was not significantly different fromhat of VPM NS neurons (P�0.3, Mann-Whitney test). MOpplication significantly increased the responses (range:5–125 spikes) of three NS neurons (Table 2). MO appli-ation did not produce any noticeable effects on the WDReuron. Examples of NS and WDR neuronal responses toinch stimulation prior to and after MO application arehown in Fig. 6.
DISCUSSION
ur previous studies have shown that MO application tohe tooth pulp can induce NMDA and P2X-dependent neu-oplastic changes reflecting central sensitization in noci-eptive neurons of the V brainstem complex (Chiang et al.,998, 2005b; Park et al., 2001; Hu et al., 2002). The
resent study has provided new data showing that pulpal
aVfcntn
sV(a
sarpoBinmpp
Frbs ons.
S. Zhang et al. / Neuroscience 142 (2006) 833–842 839
pplication of MO also induces central sensitization inPM and PO nociceptive neuron that is reflected in pro-
ound changes in the neuronal properties, including in-rease in spontaneous activity, enlargement of the cuta-eous RF, decrease in cutaneous mechanical activationhreshold, and enhancement of responses to cutaneousoxious mechanical stimuli.
Previous studies have demonstrated that electricaltimulation of the tooth pulp activates some neurons inPM and that these neurons also have cutaneous RFs
Woda et al., 1975; Shigenaga and Inoki, 1976; Kaneko et
ig. 6. Examples of changes in responses to noxious pinch stimulusegions, at baseline and 13–25 min after MO application to the tooth puaseline activity, following MO application. For all post-stimulus histogtimulus marker: 200 g for both NS neurons, 80 g for both WDR neur
l., 2005). However, a possible confound with these earlier i
tudies is that electrical stimulation of the tooth pulp canlso spread to excite sensory afferents innervating sur-ounding tissue (Sessle, 1979; Rajaona et al., 1986). Ap-lication of MO to the pulp as used in the present studynly activates A delta and C-fiber tooth pulp afferents (seeyers and Narhi, 1999; Sunakawa et al., 1999). Interest-
ngly, over half (20/35) of the VPM and one of six POociceptive neurons tested in the present study were im-ediately activated by MO application to the molar toothulp, suggesting that they received excitatory afferent in-uts from the tooth pulp. The neuroplastic changes reflect-
the center of RF of NS and WDR neurons in thalamic VPM and POhe pinch-evoked responses were increased, compared with precedingrtical bar�20 Hz; x axis�30 s; bin width�1 s. Upper traces indicate
applied tolp. Note trams, ve
ng central sensitization observed in the VPM and PO NS

nitasi2aa
gacgo
ce1tfsoaacicint
hewaaRwtmisbmsDtCctctat2
Vz
PVelespTatvctl1rbpdsFcvamaKrtdtopmtfarfisosccm
sosaufnbVboC
S. Zhang et al. / Neuroscience 142 (2006) 833–842840
eurons following MO application to the pulp, however,nvolved sensory inputs arising from non-pulp tissues inhe intraoral cavity and the facial skin and thus are indic-tive of generalized alterations in processing of orofacialensory inputs. This sensitization of widespread afferentnputs, which is also observed in Vc (Chiang et al., 1998,005b; Hu et al., 2002), is thought to contribute to second-ry hyperalgesia (Klede et al., 2003; Dubner, 2005; Woolfnd Salter, 2006).
The response properties of the NS neurons in PO wereenerally similar to those of the VPM NS neurons. MOpplication resulted in significant increase in RF size, de-rease in activation threshold and increase in responses toraded pinch stimuli (Table 2) similar to the sensitizationbserved in the VPM NS neurons.
Our findings in VPM are consistent with findings ofentral sensitization in VPL nociceptive neurons (Guilbaudt al., 1987, 1990, 1995; Dostrovsky and Guilbaud, 1988,990; Bordi and Quartaroli, 2000), and also supported byhose of the recent study by Kaneko et al. (2005) whoound increased VPM neuronal responses to electricaltimulation of the pulp and decreased mechanical thresh-ld to stimulation of the cutaneous RF after MO pulpalpplication. Alterations in VPM neuronal responses werelso observed in a rat model of orofacial pain produced byonstriction of the infraorbital nerve (Vos et al., 2000),ncluding increased responses to noxious stimuli and in-reased spontaneous activity. The findings of these stud-es collectively indicate that central sensitization of VPMociceptive neurons occurs in neuropathic and inflamma-ory pain models.
A major difficulty in interpreting all of the studies thatave reported sensitization at the thalamic level is to whatxtent the observed changes are simply a reflection of theell-documented sensitization that occurs at the spinalnd V brainstem levels. The changes we observed in VPMre generally similar to the changes (i.e. expansion of theFs, decreased thresholds and increased responses) thate have documented in Vc neurons following MO applica-
ion to the tooth pulp (Chiang et al., 1998, 2005b) and thusay be largely due to sensitization of the Vc neurons. This
s supported by our other recent findings that VPM centralensitization can be markedly attenuated by synapticlockade of Vc (Park et al., 2006) and by reports that theain nociceptive projection to VPM arises in Vc (Peschan-
ki, 1984; Gauriau and Bernard, 2004; Guy et al., 2005;ostrovsky and Craig, 2006; Messlinger et al., 2006), al-
hough a contribution relayed via Vo is also likely (Dehazeron et al., 2004; Guy et al., 2005). This viewpoint isonsistent with our previous findings that MO-induced cen-ral sensitization in both Vc and Vo are similar, and that Voentral sensitization and accompanying jaw muscle elec-romyographic activity can be considerably attenuated bypplication to Vc of NMDA receptor or P2X receptor an-agonists or the synaptic blocker CoCl2 (Cairns et al.,001; Chiang et al., 2002; Hu et al., 2001, 2002).
It is unlikely that all of the changes we observed inPM and PO were entirely a reflection of central sensiti-
ation in Vc since the neuroplastic changes in VPM and wO were greater and lasted longer than those occurring inc (Chiang et al., 1998, 2005b; Hu et al., 2002). Forxample, the RFs of some of the NS neurons which were
ocated in PO or at the boundary between VPL and VPMxpanded to include the paws and tail, and the RFs ofome VPM neurons which included an intraoral RF ex-anded to include the whole intra- and extra-oral regions.hese findings suggest that additional sensory integrationnd modulation may occur in sensory thalamus, andhereby contribute to central sensitization and to the de-elopment of hyperalgesia and spread and referral ofraniofacial pain, including the pain associated with pulpi-is. This view is consistent with findings that trigeminotha-amic neurons release glutamate in VPM (Eaton and Salt,990) and that NMDA receptors are present on thalamicelay neurons (Salt, 1986); thus, thalamic neurons couldecome sensitized by NMDA-dependent mechanisms,erhaps through processes similar to those that have beenemonstrated in Vc (Chiang et al., 1997, 1998, 2003) andpinal dorsal horn (Dubner, 2005; Woolf and Salter, 2006).urther support for thalamic NMDA receptor involvementomes from findings that the nociceptive responses of ratentrobasal thalamus neurons can be reduced by locallypplied NMDA and selective metabotropic glutamateGlu1 and mGlu5 antagonists (Eaton and Salt, 1990; Saltnd Turner, 1998; Salt and Binns, 2000). Furthermore,olhekar et al. (1997) reported that injection of the NMDA
eceptor antagonist D-APV into VPL attenuated the acutehermal hyperalgesia and mechanical hyperalgesia pro-uced by intraplantar injection of carrageenan, suggestinghe involvement of thalamic NMDA receptors in the devel-pment and maintenance of hyperalgesia associated witheripheral inflammation. Nonetheless, other mechanismsay also occur in thalamus that could contribute to sensi-
ization of VPM neurons. For example, Hains et al. (2005)ound that following spinal cord contusion injury there was
an upregulation of Nav1.3 sodium channels within neu-ons in VPL, which would be expected to lead to increasedring rates and would explain the associated increasedpontaneous firing rates, hyperresponsiveness to innocu-us and noxious stimuli and the increased receptive fieldsizes that they observed. It is unlikely, however, that suchhanges could occur rapidly enough to account for thehanges demonstrated in our study which occur withininutes after MO injection to the tooth pulp.
The few VPM WDR neurons tested in this study did nothow as marked sensitization following pulpal applicationf MO as the NS neurons. Although this difference isimilar to what we observed in Vc where WDR neuronslso undergo less marked sensitization than NS neuronsnder the same conditions (Chiang et al., 1998), the dif-erence may however reflect the lower number of WDReurons that were tested with pulpal application of MO inoth regions, and the fact that most WDR neurons in bothPM and Vc have fluctuating spontaneous firing rates ataseline which hinder accurate determination of thresh-lds and responses prior to and after MO application (seehiang et al., 1998, 2005a). These factors may explain
hy in the present study not all of the neuronal properties
oticuoc
ftcNintt2eut
Ilsocples
AattS
B
B
C
C
C
C
C
C
C
C
D
D
D
D
D
E
G
G
G
G
G
G
H
H
H
H
S. Zhang et al. / Neuroscience 142 (2006) 833–842 841
f VPM WDR neurons were found to be significantly al-ered following MO application (see Table 1). The lowerncidence of WDR neurons compared with NS neuronsould be related to the anesthetic regimen used, but isnlikely since a similar anesthetic state has been used inur Vc studies (Chiang et al., 1997, 1998, 2003) where aonsiderable proportion of the neurons were WDR.
To our knowledge this is the first report of the S-Runction of NS neurons in the rat VPM. Over the rangeested, the responses increased progressively with in-reasing mechanical stimulus intensity indicating that VPMS neurons can encode the intensity changes of noxious
nputs, and thus may be involved in the sensory-discrimi-ative aspects of pain. Interestingly, during central sensi-
ization induced by pulpal application of MO, the S-R func-ion was significantly shifted to the left and upwards (Fig.), indicating that the pinch/pressure threshold was low-red as shown in Fig. 4; these changes that could contrib-te to the occurrence of allodynia and hyperalgesia underhis condition.
CONCLUSION
n summary, this study has clearly demonstrated that fol-owing application of the inflammatory irritant MO to aingle molar tooth pulp, the cutaneous RFs and responsesf nociceptive neurons in both VPM and PO are signifi-antly increased. These changes in the RF and responseroperties of thalamocortical neurons to noxious stimuli
ikely contribute to the behavioral consequences of periph-ral inflammation manifesting as pain referral, hyperalge-ia and allodynia.
cknowledgments—The authors thank Dr. J.-C. Lee for his valu-ble statistical assistance and K. MacLeod and S. Carter for theirechnical assistance. This study was supported by National Insti-ute of Dental and Craniofacial Research grant DE-04786 to B. J.essle.
REFERENCES
ordi F, Quartaroli M (2000) Modulation of nociceptive transmission byNMDA/glycine site receptor in the ventroposterolateral nucleus ofthe thalamus. Pain 84:213–224.
yers MR, Narhi MV (1999) Dental injury models: experimental toolsfor understanding neuroinflammatory interactions and polymodalnociceptor functions. Crit Rev Oral Biol Med 10:4–39.
airns BE, Sessle BJ, Hu JW (2001) Temporomandibular-evoked jawmuscle reflex: role of brain stem NMDA and non-NMDA receptors.Neuroreport 12:1875–1878.
hiang CY, Hu B, Hu JW, Dostrovsky JO, Sessle BJ (2002) Centralsensitization of nociceptive neurons in trigeminal subnucleus oralisdepends on integrity of subnucleus caudalis. J Neurophysiol 88:256–264.
hiang CY, Hu B, Park SJ, Zhang S, Kwan CL, Hu JW, Dostrovsky JO,Sessle BJ (2003) Purinergic and NMDA-receptor mechanisms un-derlying tooth pulp sitmulation-induced central sensitization in tri-geminal nociceptive neurons. In: Proceedings of the 10th WorldCongress on Pain (Dostrovsky JO, Carr DB, Koltzenberg M, eds),pp 345–354. Seattle: IASP Press.
hiang CY, Hu JW, Sessle BJ (1994) Parabrachial area and nucleus
raphe magnus-induced modulation of nociceptive and nonnociceptivetrigeminal subnucleus caudalis neurons activated by cutaneous ordeep inputs. J Neurophysiol 71:2430–2445.
hiang CY, Hu JW, Sessle BJ (1997) NMDA receptor involvement inneuroplastic changes induced by neonatal capsaicin treatment intrigeminal nociceptive neurons. J Neurophysiol 78:2799–2803.
hiang CY, Park SJ, Kwan CL, Hu JW, Sessle BJ (1998) NMDAreceptor mechanisms contribute to neuroplasticity induced in cau-dalis nociceptive neurons by tooth pulp stimulation. J Neurophysiol80:2621–2631.
hiang CY, Zhang S, Park SJ, Hu JW, Dostrovsky JO, Sessle BJ(2005a) Mechanoreceptive field and response properties of noci-ceptive neurons in ventral posteromedial thalamic nucleus of therat. Thalamus Relat Syst 3:41–51.
hiang CY, Zhang S, Xie YF, Hu JW, Dostrovsky JO, Salter MW,Sessle BJ (2005b) Endogenous ATP involvement in mustard-oil-induced central sensitization in trigeminal subnucleus caudalis(medullary dorsal horn). J Neurophysiol 94:1751–1760.
e Chazeron I, Raboisson P, Dallel R (2004) Organization of dience-phalic projections from the spinal trigeminal nucleus oralis: ananterograde tracing study in the rat. Neuroscience 127:921–928.
ostrovsky JO, Craig AD (2006) Ascending projection systems. In:Textbook of pain (McMahon SB, Koltzenburg M, eds), pp 187–203.Edinburgh: Elsevier Churchill Livingstone.
ostrovsky JO, Guilbaud G (1988) Noxious stimuli excite neurons innucleus submedius of the normal and arthritic rat. Brain Res460:269–280.
ostrovsky JO, Guilbaud G (1990) Nociceptive responses in medialthalamus of the normal and arthritic rat. Pain 40:93–104.
ubner R (2005) Plasticity in central nociceptive pathways. In: Thepaths of pain (Merskey H, Loeser JD, Dubner R, eds), pp 101–115.Seattle: IASP Press.
aton SA, Salt TE (1990) Thalamic NMDA receptors and nociceptivesensory synaptic transmission. Neurosci Lett 110:297–302.
auriau C, Bernard JF (2004) A comparative reappraisal of projec-tions from the superficial laminae of the dorsal horn in the rat: theforebrain. J Comp Neurol 468:24–56.
erke MB, Duggan AW, Xu L, Siddall PJ (2003) Thalamic neuronalactivity in rats with mechanical allodynia following contusive spinalcord injury. Neuroscience 117:715–722.
uilbaud G, Benoist JM, Gautron M (1995) Contribution of the sciaticand saphenous nerve to the ventrobasal thalamic neuronal re-sponses to pinch in rats with a chronic sciatic nerve constriction: astudy using anesthetic blocks and nerve section. Neurosci Lett187:197–200.
uilbaud G, Benoist JM, Jazat F, Gautron M (1990) Neuronal respon-siveness in the ventrobasal thalamic complex of rats with an ex-perimental peripheral mononeuropathy. J Neurophysiol 64:1537–1554.
uilbaud G, Neil A, Benoist JM, Kayser V, Gautron M (1987) Thresh-olds and encoding of neuronal responses to mechanical stimuli inthe ventro-basal thalamus during carrageenin-induced hyperalge-sic inflammation in the rat. Exp Brain Res 68:311–318.
uy N, Chalus M, Dallel R, Voisin DL (2005) Both oral and caudalparts of the spinal trigeminal nucleus project to the somatosensorythalamus in the rat. Eur J Neurosci 21:741–754.
ains BC, Saab CY, Waxman SG (2005) Changes in electrophysio-logical properties and sodium channel Nav1.3 expression in tha-lamic neurons after spinal cord injury. Brain 128:2359–2371.
u B, Chiang CY, Dostrovsky JO, Sessle BJ, Hu JW (2001) Centralsensitization in ventrobasal thalamic nociceptive neurons inducedby mustard oil application to rat tooth pulp. Soc Neurosci Abstr27:734.
u B, Chiang CY, Hu JW, Dostrovsky JO, Sessle BJ (2002) P2Xreceptors in trigeminal subnucleus caudalis modulate central sen-sitization in trigeminal subnucleus oralis. J Neurophysiol 88:1614–1624.
u JW, Sessle BJ, Raboisson P, Dallel R, Woda A (1992) Stimulation
of craniofacial muscle afferents induces prolonged facilitatory ef-
I
I
K
K
K
M
P
P
P
P
R
S
S
S
S
S
S
S
S
S
S
T
V
W
W
W
W
Y
S. Zhang et al. / Neuroscience 142 (2006) 833–842842
fects in trigeminal nociceptive brain-stem neurones. Pain 48:53–60.
ggo A (1960) Non-myelinated afferent fibres from mammalian skeletalmuscle. J Physiol 155:52P–53P.
wata K, Imai T, Tsuboi Y, Tashiro A, Ogawa A, Morimoto T, MasudaY, Tachibana Y, Hu JW (2001) Alteration of medullary dorsal hornneuronal activity following inferior alveolar nerve transection inrats. J Neurophysiol 86:2868–2877.
aneko M, Sunakawa M, Matsui Y, Suda H (2005) Responsiveness oftooth pulp-driven neurons in thalamic ventral posteromedial andmediodorsal nuclei following experimental pulpitis and naloxoneadministration in rats. J Oral Biosci 47:135–148.
lede M, Handwerker HO, Schmelz M (2003) Central origin of sec-ondary mechanical hyperalgesia. J Neurophysiol 90:353–359.
olhekar R, Murphy S, Gebhart GF (1997) Thalamic NMDA receptorsmodulate inflammation-produced hyperalgesia in the rat. Pain 71:31–40.
esslinger K, Dostrovsky JO, Strassman AM (2006) Anatomy andphysiology of head pain. In: The headaches (Olesen J, GoadsbyPJ, Ramadan NM, Tfelt-Hansen P, Welch KMA, eds), pp 95–109.Philadelphia: Lippincott Williams & Wilkins.
ark SJ, Chiang CY, Hu JW, Sessle BJ (2001) Neuroplasticity inducedby tooth pulp stimulation in trigeminal subnucleus oralis involvesNMDA receptor mechanisms. J Neurophysiol 85:1836–1846.
ark SJ, Zhang S, Chiang CY, Hu JW, Dostrovsky JO, Sessle BJ(2006) Central sensitization induced in thalamic nociceptiveneurons by tooth pulp stimulation is dependent on the functionalintegrity of trigeminal brainstem subnucleus caudalis but notsubnucleus oralis. Brain Res [Epub ahead of print] doi:10.1016/j.brainres.2006.06.115.
axinos G, Watson C (1986) The rat brain in stereotaxic coordinates.Sydney: Academic Press.
eschanski M (1984) Trigeminal afferents to the diencephalon in therat. Neuroscience 12:465–487.
ajaona J, Dallel R, Woda A (1986) Is electrical stimulation of the ratincisor an appropriate experimental nociceptive stimulus? ExpNeurol 93:291–299.
alt TE (1986) Mediation of thalamic sensory input by both NMDAreceptors and non-NMDA receptors. Nature 322:263–265.
alt TE, Binns KE (2000) Contributions of mGlu1 and mGlu5 receptorsto interactions with N-methyl-D-aspartate receptor-mediated re-sponses and nociceptive sensory responses of rat thalamic neu-rons. Neuroscience 100:375–380.
alt TE, Turner JP (1998) Modulation of sensory inhibition in theventrobasal thalamus via activation of group II metabotropic glu-tamate receptors by 2R,4R-aminopyrrolidine-2,4-dicarboxylate.
Exp Brain Res 121:181–185.alter MW (2004) Cellular neuroplasticity mechanism mediating painpersistence. J Orofac Pain 18:318–324.
chaible HG, Schmidt RF (1983) Responses of fine medial articularnerve afferents to passive movements of knee joints. J Neuro-physiol 49:1118–1126.
essle BJ (1979) Is the tooth pulp a ‘pure’ source of noxious input? In:Advances in pain research and therapy, Vol. 3 (Bonica JJ, Liebe-skind JC, Albe-Fessard DG, eds), pp 245–260. New York: Raven.
herman SE, Luo L, Dostrovsky JO (1997) Altered receptive fields andsensory modalities of rat VPL thalamic neurons during spinalstrychnine-induced allodynia. J Neurophysiol 78:2296–2308.
herman SM, Guillery RW (2001) Exploring the thalamus. New York:Academic Press.
higenaga Y, Inoki R (1976) Effect of morphine on single unit re-sponses in ventrobasal complex (VB) and posterior nuclear group(PO) following tooth pulp stimulation. Brain Res 103:152–156.
unakawa M, Chiang CY, Sessle BJ, Hu JW (1999) Jaw electromyo-graphic activity induced by the application of algesic chemicals tothe rat tooth pulp. Pain 80:493–501.
suboi Y, Sessle BJ, Hu JW (2006) Pulp exposure induces centralsensitization and nociceptive behavior in rats. J Dent Res SpecialIssue B, IADR Abstracts 2585.
os BP, Benoist JM, Gautron M, Guilbaud G (2000) Changes inneuronal activities in the two ventral posterior medial thalamicnuclei in an experimental model of trigeminal pain in the rat byconstriction of one infraorbital nerve. Somatosens Mot Res 17:109–122.
eng HR, Lee JI, Lenz FA, Schwartz A, Vierck C, Rowland L, Dough-erty PM (2000) Functional plasticity in primate somatosensorythalamus following chronic lesion of the ventral lateral spinal cord.Neuroscience 101:393–401.
eng HR, Lenz FA, Vierck C, Dougherty PM (2003) Physiologicalchanges in primate somatosensory thalamus induced by deaffer-entation are dependent on the spinal funiculi that are sectioned andtime following injury. Neuroscience 116:1149–1160.
oda A, Azerad J, Guilbaud G, Besson JM (1975) A microphysiologicstudy of thalamic projection of the cat’s dental pulp (author’stransl). Brain Res 89:193–213.
oolf CJ, Salter MW (2006) Plasticity and pain: role of the dorsal horn.In: Textbook of pain (McMahon SB, Koltzenburg M, eds), pp 91–105. Edinburgh: Elsevier Churchill Livingstone.
u XM, Sessle BJ, Hu JW (1993) Differential effects of cutaneous anddeep application of inflammatory irritant on mechanoreceptive fieldproperties of trigeminal brain stem nociceptive neurons. J Neuro-
physiol 70:1704–1707.(Accepted 20 June 2006)(Available online 24 August 2006)
Copyright © 2022 FDOKUMEN




















