
The Serial Engagement Model 17 Years After: From TCR Triggering to Immunotherapy
Upload
independentCategory
view
0download
0
of July 12, 2016.This information is current as
Membrane RaftsAg-Independent Polarization of Plasma
Signaling during2+CD28 Sensitizes TCR Ca
RodgersJennifer N. Byrum, Jeffrey S. Van Komen and William
http://www.jimmunol.org/content/191/6/3073doi: 10.4049/jimmunol.1300485August 2013;
2013; 191:3073-3081; Prepublished online 21J Immunol
MaterialSupplementary
5.DC1.htmlhttp://www.jimmunol.org/content/suppl/2013/08/21/jimmunol.130048
Referenceshttp://www.jimmunol.org/content/191/6/3073.full#ref-list-1
, 28 of which you can access for free at: cites 55 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2013 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 12, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
CD28 Sensitizes TCR Ca2+ Signaling during Ag-IndependentPolarization of Plasma Membrane Rafts
Jennifer N. Byrum,*,†,‡ Jeffrey S. Van Komen,‡ and William Rodgers*,†,‡,x
T cells become polarized during initial interactions with an APC to form an Ag-independent synapse (AIS) composed of membrane
rafts, TCR, and TCR-proximal signaling molecules. AISs occur temporally before TCR triggering, but their role in downstream
TCR signaling is not understood. Using both human and murine model systems, we studied the signals that activate AIS formation
and the effect of these signals on TCR-dependent responses. We show that CD28 produces AISs detectable by spinning disc confocal
microscopy seconds following initial interactions between the T cell and APC. AIS formation by CD28 coincided with costimulatory
signaling, evidenced by a cholesterol-sensitive activation of the MAPK ERK that potentiated Ca2+ signaling in response to CD3
cross-linking. CD45 also enriched in AISs but to modulate Src kinase activity, because localization of CD45 at the cell interface
reduced the activation of proximal Lck. In summary, we show that signaling by CD28 during first encounters between the T cell
and APC both sensitizes TCR Ca2+ signaling by an Erk-dependent mechanism and drives formation of an AIS that modulates the
early signaling until TCR triggering occurs. Thus, early Ag-independent encounters are an important window for optimizing
T cell responses to Ag by CD28. The Journal of Immunology, 2013, 191: 3073–3081.
The activation of T cells by Ag presentation is preceded byserial interactions between motile T cells and APCs as theT cell surveys its environment for cognate peptide–MHC
(pMHC) (1–4). Recognition of pMHC triggers a cascade of in-tracellular signals that drive a global reorganization of the T cell toform an immunological synapse (IS) (5, 6). Within the context ofthese complex cellular interactions, T cells exhibit an exquisitesensitivity to Ag, where only a few cognate pMHC are necessary toproduce a robust activation (7–9). How this sensitivity is achievedis poorly understood, yet underlies features of the adaptive immuneresponse germane to vaccination, tolerance, and autoimmunity.Spatiotemporal studies of T cells during Ag presentation show
that TCR triggering measured by intracellular Ca2+ signaling ispreceded by formation of a membrane complex composed of theactin cytoskeleton, membrane rafts, TCR, and TCR proximal pro-teins (4, 10). The membrane complex occurs seconds followinginitial contact between the T cell and APC, and minutes beforemeasurable Ca2+ signaling from the TCR (10). The pre-TCR com-
plexes are both Ag- and MHC class II (MHC II) independent,because they occur in CD4+ T cells when conjugated to MHC II–deficient B cells (10), and they correlate temporally with an Ag-independent adhesion of the T cell to the APC that precedes TCRtriggering (4). Given the compositional similarities of the Ag-independent complexes and ISs, these structures have been re-ferred to as an Ag IS (AIS) (11). However, the role of the AIS inT cell activation by pMHC is not understood.In this paper, we report experiments that identified the princi-
pal signals that initiate AIS formation during initial T cell–APCinteractions, and the functional contribution of complex forma-tion to TCR signaling. Specifically, we show that AIS formation isCD28 and Lck dependent, requiring sites in each that are neces-sary for Lck interactions with CD28. Engagement of CD28 in Ag-independent conditions produced a robust activation of the MAPKERK that augmented Ca2+ release once signaling from the TCRoccurred. We also show that CD45 is targeted to AISs, and CD45downregulates proximal Lck activity in conjugates to modulateearly Ag-independent signals. Altogether, our results show thatLck-dependent CD28 costimulatory signals occur during the ear-liest stages of T cell–APC interactions, and they produce a mem-brane environment that assists in quenching these signals in theevent that Ag is not encountered.
Materials and MethodsPlasmid construction
Ten amino acids of Lck (L10)-GFP and 15 aa Src (S15)-GFP in pWay20 (12)have been described previously (13). A gene encoding CD28 followed byenhanced GFP (CD28-GFP) was cloned by first amplifying DNA encodingfull-length CD28 by RT-PCR of RNA prepared from mouse splenocytesusing iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA), using the fol-lowing primers: 59-ATGGCACTCAGGCTGCTGTT-39 (coding) and 59-GT-TGTTGTTGTTGTTGGGGCGGTACGCTGCAAAGT-39 (noncoding). TheCD28 PCR product was subcloned into the SmaI site of the GFP expressionvector pWay20 (12) upstream and in-frame with sequence encoding en-hanced GFP. For retroviral expression, the CD28-GFP gene was subclonedinto the HpaI site of the retroviral expression vector pMSCVneo (BD Bio-sciences, San Jose, CA). CD28P187,190A-GFP was generated by site directedmutagenesis using QuickChange II (Agilent Technologies, Santa Clara, CA)using the following primers: 59-AAGCCTTACCAGGCCTACGCCGCTGC-CAGAGACTTT-39 (coding) and 59-AAAGTCTCTGGCAGCGGCGTAGG-
*Department of Biochemistry and Molecular Biology, University of OklahomaHealth Sciences Center, Oklahoma City, OK 73104; †Department of Pathology, Uni-versity of Oklahoma Health Sciences Center, Oklahoma City, OK 73104; ‡OklahomaMedical Research Foundation, Oklahoma City, OK 73104; and xDepartment of Microbi-ology and Immunology, University of Oklahoma Health Sciences Center, Oklahoma City,OK 73104
Received for publication February 22, 2013. Accepted for publication July 15, 2013.
This work was supported by National Institutes of Health Grant R01 GM070001 (toW.R.), Oklahoma Center for the Advancement of Science and Technology GrantsHR08-084 and HR11-42 (to W.R.), and American Cancer Society Fellowship PF-08-178-01-LIB (to J.S.V.K.).
Address correspondence and reprint requests to Dr. William Rodgers, Department ofBiochemistry and Molecular Biology, University of Oklahoma Health Sciences Cen-ter, 940 Stanton L. Young Boulevard, BMSB 819, Oklahoma City, OK 73104. E-mailaddress: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: AIS, Ag-independent synapse; CTX, cholera toxin;DIC, differential interference contrast; FRET, fluorescence resonance energy transfer;IS, immunological synapse; L10, 10 aa Lck; MbCD, methyl-b-cyclodextrin; MHC II,MHC class II; pMHC, peptide–MHC; PP2, 4-amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo[3,4-d]pyrimidine; pTyr394, phospho-Tyr394; Rel. Fluor. Enrich., relative fluo-rescence enrichment; S15, 15 aa Src; SH3, Src homology 3; TR, Texas Red.
Copyright� 2013 by The American Association of Immunologists, Inc. 0022-1767/13/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1300485
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

CGTCGTAAGGCTT-39 (noncoding). Genes encoding the cytoplasmic do-main of CD45 with alternate membrane-targeting sequences, L10CD45-GFPor S15CD45-GFP, were constructed by first amplifying by RT-PCR DNAencoding full-length CD45, using RNA prepared from Jurkat lymphoma cellsas template. The primers for the RT-PCR were 59-ACATCAGGTACCAT-GACCATGTATTTGTGGCT-39 (CD45 coding) and 59-TGAACCTTGATT-TAAAGCTGGACTTGC-39 (CD45 noncoding). The L10- and S15- wereadded by PCR amplification of full-length CD45 using the following primers:59-ACATCATCTAGAATGGGCTGTGTCTGCAGCAAAATCTATGATCT-AC-39 (coding, L10CD45) and 59-ACATCAGTTACCATGGGGAGCAGC-AAGAGCAAGCCCAAGGACCCCAGCCA GCGCCGGAAAATCTATG-ATCTACAT-39 (coding, S15CD45) and the CD45 noncoding primer.Following amplification, the respective products were inserted upstream to andin-frame with enhanced GFP in pWay20 (Invitrogen, Carlsbad, CA) using theSmaI site of the vector. All clones were verified by DNA sequencing.
Cells and mice
Cell lines were maintained and transfected as described previously (14, 15).Mouse strains C57BL/6 and Cd28tmMak/J (C57BL/6 background) used forpreparation of primary T cells were purchased from The Jackson Laboratory(Bar Harbor, ME). Animals were maintained under specific pathogen-freeconditions in accordance with institutional guidelines for animal welfare.
Primary T cells were harvested from 4- to 8-wk-old mice sex-matchedmice using complete media consisting of RPMI 1640 medium containing10% FCS, 0.1% 2-ME, 2% sodium pyruvate, and 1% nonessential aminoacids. Collected tissue was passed through a 70-mm nylon cell strainer,washed once with 0.14 M NH4Cl in 17 mM Tris (pH 7.2), and then washedtwice with complete media. T cells were enriched using nylon wool washedand equilibrated with Hank’s buffer containing 10% FCS. Following ad-dition to the column, cells were incubated for 1 h at 37˚C and then elutedusing Hanks buffer plus 10% FCS. Following enrichment, the T cellswere cultured overnight at a density of 3–5 3 106 cells/ml in completemedia containing 0.02 mg/ml PMA and 0.2 mg/ml ionomycin. On day 2,the medium was replaced with complete media containing 0.3 mg/ml IL-2. Cells were transduced after culturing for an additional 48–72 h.
Retroviral transductions
Phoenix-E cells (5 3 106) were seeded onto a 10-cm plate and grown inDMEM + 10% FCS + sodium pyruvate overnight. On day 2, the cells weretransfected by calcium phosphate (Invitrogen, Grand Island, NY) with 20 mgpMSCVneo containing the gene of interest. Culture supernatant was col-lected on day 4, cleared by centrifugation (1200 rpm for 5 min, Eppendorf A-4-62), and then passed through a 0.45-mm filter. Next, 6 ml viral supernatantcontaining 50 ml 8.0 mg/ml polybrene and 2 ml IL-2 was added to 5 3 106
cultured primary T cell. Following 4 h at 37˚C, the cells were sedimentedand resuspended in 10 ml complete media containing 3 ml IL-2. Viable cellswere enriched using a Ficoll pad (Mediatech, Herndon, VA) 72–96 h post-transduction.
Treatments preceding conjugation were performed at 37˚C at a celldensity of 107/ml in RPMI 1640 medium containing 50 mM HEPES (pH7.4) (RPMI-HEPES). The treatments consisted of the following: 10 mM4-amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo[3,4-d]pyrimidine (PP2;Calbiochem, La Jolla, CA) for 15 min, 10 mM methyl-b-cyclodextrin(MbCD; Sigma-Aldrich, St. Louis, MO) for 30 min, 5 mg/ml filipin(Cayman Chemical, Ann Arbor, MI) for 30 min, and 5 mm PD98059 for5 min, 50 mM b-lactose, or 50 mM sucrose (Fisher Scientific) for 20 min.Blocking of respective ligands in CH27 cells was performed by incubating106 B cells with 1.5 mg Ab at 37˚C for 10 min, followed by washing andmixing with D10 T cells. The Abs used for the blocking experiments wereas follows: clone HM48-1 (CD48), clone 16-10A1 (CD80), clone GL1(CD86), clone 3E2 (ICAM-1), and clone 3C4 (ICAM-2).
T cell conjugation
T cell–B cell conjugates were prepared as described previously (15). In brief,106 T cells were added to 2 3 106 B cells, gently mixed, then sedimented bycentrifuging at 4000 rpm (Brickmann) for 30 s. Next, the samples were in-cubated at 37˚C for 2 min, followed by gentle resuspension. In microscopyexperiments, the sample was seeded onto a poly-L-lysine–coated coverslip,then fixed with 2% paraformaldehyde, followed by washing with PBS con-taining 10mMglycine (pH 7.4) (PBS-glycine). In flow cytometry experiments,the cells were maintained and fixed in suspension. The respective treatmentswere performed, as described above, immediately before conjugation.
Cell stimulation
In immunoblotting experiments, T cell stimulation by Ab cross-linking ofCD3ε and CD28 was performed by first suspending 106 cells in 100 ml
RPMI-HEPES, followed by incubation with either 7.5 mg anti-CD28 mAb(28.2) or 7.5 mg OKT3 for 2 min at 37˚C. In stimulations by CD28 cross-linking, the samples were also incubated with secondary Ab to mouse IgGfor 2 min at 37˚C. In experiments consisting of cross-linking CD28 fol-lowed by cross-linking CD3ε, the cells were incubated with Ab to CD28for 2 min at 37˚C, followed by a 1-min incubation with secondary Ab tomouse IgG. Next, OKT3 was added, and the sample was incubated for anadditional 2 min at 37˚C.
Following the respective incubations with Abs, the samples were washedwith RPMI-HEPES, then suspended in Laemmeli sample buffer con-taining 1% 2-ME. Samples were incubated at 100˚C for 5 min, then sepa-rated by gel electrophoresis, and transferred to polyvinylidene difluoride.Total ERK1/2 and p-ERK1/2 (pThr202/pTyr204) were probed using mAbs3A7 and E10 (Cell Signaling Technologies, Danvers, MA), respectively.Immunoblots were developed using ECL (Amersham Biosciences, Rock-ford, IL), and detected using a G:Box imaging system (Syngene, Frederick,MD).
Fluorescence staining
Immunostaining for confocal microscopy was performed by incubatingfixed conjugates on coverslips with 1–5 mg/ml Ab in PBS-glycine for 45min at room temperature. Primary Abs used for labeling were as follows:mAb HI30 (Jurkats), or I3/2.3 (mouse primary T cells), each to CD45;mAb (clone 2B; BD Biosciences, San Jose, CA) to the Lck N terminus,rabbit Ab to phospho-Src family Tyr416 (Cell Signaling Technology) todetect Lck Tyr394; and rabbit polyclonal Ab to CD28 (H-93; Santa CruzBiotechnology, Santa Cruz, CA). Gammatubulin was stained using mAbclone GTU-88 (Sigma-Aldrich). Immunostaining Lck and gamma-tubulinwas preceded by permeabilizing samples with 0.1% Triton X-100 in PBS-glycine. All secondary Abs for immunostaining were from JacksonImmunoResearch Laboratories (West Grove, PA). GM1 in T cells waslabeled with cholera toxin B subunit (CTX) (Sigma-Aldrich) prior toconjugation with B cells by incubating 106 cells in 100 ml RPMI-HEPEScontaining 0.01 mg/ml biotinylated CTX for 45 min at 4˚C. The CTX waslabeled with streptavidin-conjugated Texas Red (TR) following cell con-jugation and fixation, using 9.0 mg/ml in PBS-glycine, with incubation for45 min at room temperature.
Immunostaining for flow cytometry experiments was performed usingfixed cell suspensions permeabilized with chilled (4˚C) methanol. Immu-nostaining consisted of Alexa645-labeled p-ERK1/2, incubated at a con-centration of 1 mg/ml for 1 h at room temperature. Measurement wasperformed using a FACSCalibur (BD Biosciences), and the data wereanalyzed using CellQuest. For experiments measuring T cell ERK acti-vation from conjugation to B cells, CH27 B cells were prelabeled with 4mM CellTracker Orange (CMTMR) (Life Technologies, Grand Island,NY) in RPMI-HEPES for 10 min at 37˚C, then pretreated with PD98059for 5 min at 37˚C to block interference with the p-ERK signal in T cells.This was followed by conjugation with D10 T cells expressing GFP-labeledCD28 and flow cytometry to measure Alexa645 signal in CMTMR+GFP+
events.
Calcium measurements
Cells were labeled with 2 mM Indo-1 for 30 min at 37˚C in RPMI-HEPESsupplemented with 140 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1.8 mMCaCl2, and 10 mM glucose (flux media). Next, the samples were incubatedwith 7.5 mg anti-CD28 Ab (28.2) for 45 min, followed by secondary anti-mouse Ab for 30 min, each at 4˚C. Next, the cells were suspended in fluxmedia that was maintained at 37˚C and then immediately injected intoa MoFlo (Beckman Coulter). Following establishment of a baseline, 7.5 mgOKT3 was added. Relative intracellular Ca2+ was represented by thefluorescence emission of 475 and 400 nm following excitation at 390 nm.
Cell imaging and analysis
Confocal imaging was performed using a Zeiss (Carl Zeiss, Thornwood,NY) LSM 510 META confocal microscope equipped with a 363 oil ob-jective (NA 1.4). For imaging, GFP was excited at 488 nm, and theemission was detected between 530 and 560 nm. TR was excited at 543 nmexcitation, and wavelengths 600–660 nm were collected for imaging.
Time-lapse imaging was performed using a Zeiss Axiovert 200M mi-croscope equipped with a high-speed Yokogawa spinning disk confocaland ORCA-ERG digital cameras (Hamamatsu, Bridgewater, NJ), and anApochromat oil immersion objective (363). The samples were maintainedat 37˚C in RPMI-HEPES containing 5% FCS, using a thermo-controlledstage and heated objective. B cells were adhered to coverslips using poly-L-lysine, to which labeled T cells were added. z stacks were generated bycollecting confocal planes 0.50–0.75 mm apart. The time interval betweenstacks was 3 s.
3074 Ag-INDEPENDENT SYNAPSES IN T CELL COSTIMULATION
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

All image processing and quantitation was performed using iVision 4.0software (Bio Vision, Exton, PA). Fluorescence enrichment in cell con-jugates was quantitated by separately measuring the average fluorescenceintensity of the T cell outer membrane where it contacted the B cell, and theaverage fluorescence intensity of the remaining plasma membrane. Theratio of average intensity at the interface divided by the average intensity ofremaining outer membrane was termed relative fluorescence enrichment(Rel. Fluor. Enrich.). In time lapse experiments, Rel. Fluor. Enrich. wasmeasured for each image in the stack, and the largest value was used torepresent the amount of protein enrichment at the corresponding time point(Figs. 1, 2). For scoring AISs in conjugates, an AIS was defined as whenthe average fluorescence intensity at the cell interface was at least 2 SDsgreater than the average intensity of the remaining membrane. Rel. Fluor.Enrich. measurements and AIS scoring were performed for all T cell–B cell conjugates identified in each field of cells. Conjugates were definedas where contact of the T cell and B cell was visible in the differentialinterference contrast (DIC) image of the field. Each trial consisted of 20–30 fields, and three or more trials were averaged for each experiment.
Fluorescence resonance energy transfer (FRET) between CFP- and YFP-labeled proteins was measured and quantitated as we described previously(16, 17).
Statistical analysis
Analysis was performed using Prism 5 software (GraphPad Software, LaJolla, CA). Means and SEMs are indicated by lines and error bars, re-spectively, in the plots and were calculated as previously described (18)using data from three or more separate trials, with each trial consisting ofmeasurement of ∼30 separate conjugates. ***p , 0.001, **p , 0.01, and*p , 0.05 were determined by one-way ANOVA, using a Dunnett’s test todetermine significance relative to control sample in each experiment. Forexperiments that consisted of only two separate conditions, analysis wasby a two-tailed Student t test.
ResultsCD28 is necessary for AIS formation
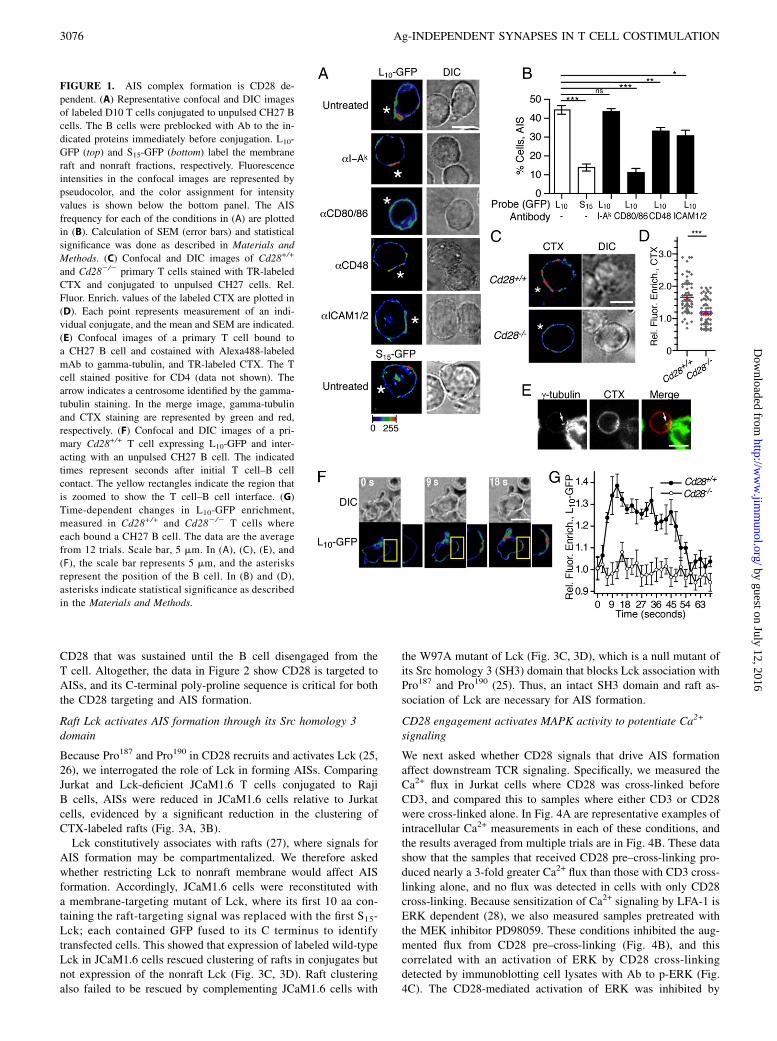
We studied AIS formation during T cell–B cell interactions bymeasuring conjugates composed of clonal D10 T cells andconalbumin-free (unpulsed) CH27 B cells. To identify T cellsurface receptors that activate AIS formation, we preblocked theB cells with Abs to CD80 and CD86, CD48, or ICAM-1, becausecross-linking CD28, CD2, or LFA-1 results in membrane capswith a lipid and protein composition similar to that of the AIS(19–22). After preblocking, conjugates were formed by gentlycosedimenting the T cells and B cells and then incubating thesamples for 2 min at 37˚C and fixation. Conjugates were imagedby confocal microscopy, and AISs were identified by enrichmentof a raft-targeted fluorescent protein that we expressed in theT cells, which consisted of the first L10-GFP (13). The L10 se-quence targets soluble proteins to the plasma membrane rafts (16),and L10-GFP enriches in AIS complexes in a cholesterol- andcytoskeleton-dependent manner (10). To determine the specificityof protein labeling in AISs, we expressed and measured a separateconstruct containing GFP anchored to the plasma membrane bythe first 15 aa Src (S15-GFP) (13). The S15-GFP neither associateswith rafts, nor enriches in AIS complexes (10, 16).In Fig. 1A are representative examples of conjugates that were
formed in each of the preblocking conditions, and plotted in Fig.1B is the fraction of conjugates that contained an AIS. Forquantitation, an AIS was defined as when the average fluorescenceintensity at the cell interface was at least 2 SDs larger than theaverage intensity of the remaining outer membrane. These datashow AIS formation was not affected by blocking MHC II but wasinhibited by blocking ligands to CD2, LFA-1, or CD28. The mostrobust inhibition occurred with blocking CD80 and CD86, whichreduced the frequency of conjugates with GFP enrichment to thatof control cells expressing S15-GFP. Similar to these results, com-paring conjugates containing either CD28-deficient (Cd282/2)or wild-type (Cd28+/+) primary T cells, each with rafts labeled by
prestaining with CTX B subunit to detect GM1, showed thatconjugates with CD28-deficient T cells exhibited significantly lessB cell–dependent clustering of CTX than conjugates with wild-type T cells (Fig. 1C, 1D).We previously demonstrated that AIS formation is rapid, be-
ginning seconds following initial T cell–B cell contact, andpeaking 20–30 s afterward (10). Similarly, capping of CTX-la-beled rafts in mouse primary T cells was evidenced in conjugatesthat were seeded and fixed immediately following mixing andsedimentation (Fig. 1E). Furthermore, all T cells containing anAIS exhibited polarization of the centrosome toward the site ofconjugation, scored by the centrosome being localized within one-third of the area of the T cell that contacted the B cell. Conversely,no AIS was observed in conjugates where the centrosome was notpolarized toward the B cell. These findings suggest AIS formationis accompanied by polarization of the microtubule-organizing centertoward the cell interface, such as has been reported in T cells un-dergoing stimulation by Ag (23).We also investigated the role of CD28 in the early dynamics of
AIS assembly by live cell imaging of transduced primary T cellsexpressing L10-GFP. The raft clustering in CD28+ cells was bothrapid and transient, peaking 9 s following initial T cell–B cellcontact, and becoming extinguished within 1 min (Fig. 1F, 1G,Supplemental Video 1). In contrast, no polarization of label wasdetected at any time point in the CD28-deficient T cells (Fig. 1E,1F, Supplemental Video 2). Using the criteria described in Fig. 1Bfor scoring AISs, the AIS frequency was 67% for wild-type T cellsthat contacted B cells during the live cell imaging, whereas noAISs were detected with the CD28-deficient T cells. Altogether,these data show that CD28 is essential for AIS formation, and thisoccurs seconds following the beginning of T cell–B cell inter-actions. The rapid and transient nature of AISs contrasts with ISsthat form in the context of Ag presentation, which require minutesto occur, and can be sustained for hours (6, 24).
AISs require a SFK-binding polyproline sequence in the CD28cytoplasmic tail
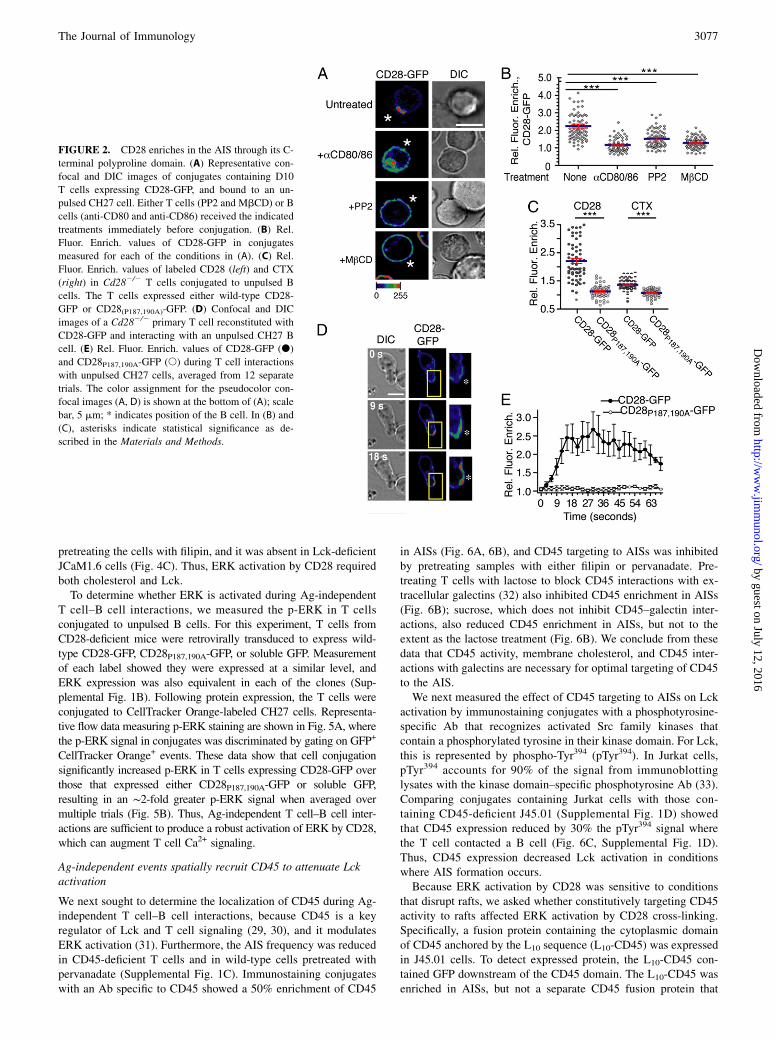
We studied CD28 localization in Ag-independent conjugates byeither immunostaining endogenous protein expressed in T cells, orby expressing a CD28 fusion protein containing GFP at its Cterminus (CD28-GFP). Both approaches showed CD28 coenrichedwith rafts in AISs, and this was inhibited by preblocking CD80 andCD86 on the B cells (Fig. 2A, 2B, Supplemental Fig. 1A). Ag-independent clustering of CD28-GFP was also blocked by pre-treating T cells with PP2 to inhibit SFK activity, and by pre-treating with MbCD to deplete membrane cholesterol (Fig. 2A,2B), each being conditions that we showed also inhibit AIS for-mation (10).We next asked whether disrupting the C-terminal polyproline
sequence in the CD28 cytoplasmic tail, containing Pro187 andPro190, affected CD28 capping and AIS formation, because thissequence recruits and activates SFKs (25), and could thereforeaccount for the PP2-sensitive nature of AIS formation. Accord-ingly, GFP-labeled CD28 containing a P187,190A double muta-tion was introduced into Cd282/2 T cells (CD28P187,190A-GFP).Measurement of fixed conjugates showed minimal clustering ofboth the mutant CD28 and CTX-labeled rafts in Ag-independentconjugates (Fig. 2C). Furthermore, in live cell imaging measure-ments of transduced T cells interacting with B cells, clustering ofGFP-labeled CD28P187,190A was not observed. In contrast, labeledwild type CD28 underwent clustering with kinetics similar toL10-GFP (Fig. 2D, 2E, Supplemental Videos 3, 4). Also in Fig. 2E,CD28-GFP enrichment was greater in both amount and durationrelative to the L10-GFP (Fig. 1F), indicating an acute activation of
The Journal of Immunology 3075
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
CD28 that was sustained until the B cell disengaged from theT cell. Altogether, the data in Figure 2 show CD28 is targeted toAISs, and its C-terminal poly-proline sequence is critical for boththe CD28 targeting and AIS formation.
Raft Lck activates AIS formation through its Src homology 3domain
Because Pro187 and Pro190 in CD28 recruits and activates Lck (25,26), we interrogated the role of Lck in forming AISs. ComparingJurkat and Lck-deficient JCaM1.6 T cells conjugated to RajiB cells, AISs were reduced in JCaM1.6 cells relative to Jurkatcells, evidenced by a significant reduction in the clustering ofCTX-labeled rafts (Fig. 3A, 3B).Lck constitutively associates with rafts (27), where signals for
AIS formation may be compartmentalized. We therefore askedwhether restricting Lck to nonraft membrane would affect AISformation. Accordingly, JCaM1.6 cells were reconstituted witha membrane-targeting mutant of Lck, where its first 10 aa con-taining the raft-targeting signal was replaced with the first S15-Lck; each contained GFP fused to its C terminus to identifytransfected cells. This showed that expression of labeled wild-typeLck in JCaM1.6 cells rescued clustering of rafts in conjugates butnot expression of the nonraft Lck (Fig. 3C, 3D). Raft clusteringalso failed to be rescued by complementing JCaM1.6 cells with
the W97A mutant of Lck (Fig. 3C, 3D), which is a null mutant ofits Src homology 3 (SH3) domain that blocks Lck association withPro187 and Pro190 (25). Thus, an intact SH3 domain and raft as-sociation of Lck are necessary for AIS formation.
CD28 engagement activates MAPK activity to potentiate Ca2+
signaling
We next asked whether CD28 signals that drive AIS formationaffect downstream TCR signaling. Specifically, we measured theCa2+ flux in Jurkat cells where CD28 was cross-linked beforeCD3, and compared this to samples where either CD3 or CD28were cross-linked alone. In Fig. 4A are representative examples ofintracellular Ca2+ measurements in each of these conditions, andthe results averaged from multiple trials are in Fig. 4B. These datashow that the samples that received CD28 pre–cross-linking pro-duced nearly a 3-fold greater Ca2+ flux than those with CD3 cross-linking alone, and no flux was detected in cells with only CD28cross-linking. Because sensitization of Ca2+ signaling by LFA-1 isERK dependent (28), we also measured samples pretreated withthe MEK inhibitor PD98059. These conditions inhibited the aug-mented flux from CD28 pre–cross-linking (Fig. 4B), and thiscorrelated with an activation of ERK by CD28 cross-linkingdetected by immunoblotting cell lysates with Ab to p-ERK (Fig.4C). The CD28-mediated activation of ERK was inhibited by
FIGURE 1. AIS complex formation is CD28 de-
pendent. (A) Representative confocal and DIC images
of labeled D10 T cells conjugated to unpulsed CH27 B
cells. The B cells were preblocked with Ab to the in-
dicated proteins immediately before conjugation. L10-
GFP (top) and S15-GFP (bottom) label the membrane
raft and nonraft fractions, respectively. Fluorescence
intensities in the confocal images are represented by
pseudocolor, and the color assignment for intensity
values is shown below the bottom panel. The AIS
frequency for each of the conditions in (A) are plotted
in (B). Calculation of SEM (error bars) and statistical
significance was done as described in Materials and
Methods. (C) Confocal and DIC images of Cd28+/+
and Cd282/2 primary T cells stained with TR-labeled
CTX and conjugated to unpulsed CH27 cells. Rel.
Fluor. Enrich. values of the labeled CTX are plotted in
(D). Each point represents measurement of an indi-
vidual conjugate, and the mean and SEM are indicated.
(E) Confocal images of a primary T cell bound to
a CH27 B cell and costained with Alexa488-labeled
mAb to gamma-tubulin, and TR-labeled CTX. The T
cell stained positive for CD4 (data not shown). The
arrow indicates a centrosome identified by the gamma-
tubulin staining. In the merge image, gamma-tubulin
and CTX staining are represented by green and red,
respectively. (F) Confocal and DIC images of a pri-
mary Cd28+/+ T cell expressing L10-GFP and inter-
acting with an unpulsed CH27 B cell. The indicated
times represent seconds after initial T cell–B cell
contact. The yellow rectangles indicate the region that
is zoomed to show the T cell–B cell interface. (G)
Time-dependent changes in L10-GFP enrichment,
measured in Cd28+/+ and Cd282/2 T cells where
each bound a CH27 B cell. The data are the average
from 12 trials. Scale bar, 5 mm. In (A), (C), (E), and
(F), the scale bar represents 5 mm, and the asterisks
represent the position of the B cell. In (B) and (D),
asterisks indicate statistical significance as described
in the Materials and Methods.
3076 Ag-INDEPENDENT SYNAPSES IN T CELL COSTIMULATION
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
pretreating the cells with filipin, and it was absent in Lck-deficientJCaM1.6 cells (Fig. 4C). Thus, ERK activation by CD28 requiredboth cholesterol and Lck.To determine whether ERK is activated during Ag-independent
T cell–B cell interactions, we measured the p-ERK in T cellsconjugated to unpulsed B cells. For this experiment, T cells fromCD28-deficient mice were retrovirally transduced to express wild-type CD28-GFP, CD28P187,190A-GFP, or soluble GFP. Measurementof each label showed they were expressed at a similar level, andERK expression was also equivalent in each of the clones (Sup-plemental Fig. 1B). Following protein expression, the T cells wereconjugated to CellTracker Orange-labeled CH27 cells. Representa-tive flow data measuring p-ERK staining are shown in Fig. 5A, wherethe p-ERK signal in conjugates was discriminated by gating on GFP+
CellTracker Orange+ events. These data show that cell conjugationsignificantly increased p-ERK in T cells expressing CD28-GFP overthose that expressed either CD28P187,190A-GFP or soluble GFP,resulting in an ∼2-fold greater p-ERK signal when averaged overmultiple trials (Fig. 5B). Thus, Ag-independent T cell–B cell inter-actions are sufficient to produce a robust activation of ERK by CD28,which can augment T cell Ca2+ signaling.
Ag-independent events spatially recruit CD45 to attenuate Lckactivation
We next sought to determine the localization of CD45 during Ag-independent T cell–B cell interactions, because CD45 is a keyregulator of Lck and T cell signaling (29, 30), and it modulatesERK activation (31). Furthermore, the AIS frequency was reducedin CD45-deficient T cells and in wild-type cells pretreated withpervanadate (Supplemental Fig. 1C). Immunostaining conjugateswith an Ab specific to CD45 showed a 50% enrichment of CD45
in AISs (Fig. 6A, 6B), and CD45 targeting to AISs was inhibitedby pretreating samples with either filipin or pervanadate. Pre-treating T cells with lactose to block CD45 interactions with ex-tracellular galectins (32) also inhibited CD45 enrichment in AISs(Fig. 6B); sucrose, which does not inhibit CD45–galectin inter-actions, also reduced CD45 enrichment in AISs, but not to theextent as the lactose treatment (Fig. 6B). We conclude from thesedata that CD45 activity, membrane cholesterol, and CD45 inter-actions with galectins are necessary for optimal targeting of CD45to the AIS.We next measured the effect of CD45 targeting to AISs on Lck
activation by immunostaining conjugates with a phosphotyrosine-specific Ab that recognizes activated Src family kinases thatcontain a phosphorylated tyrosine in their kinase domain. For Lck,this is represented by phospho-Tyr394 (pTyr394). In Jurkat cells,pTyr394 accounts for 90% of the signal from immunoblottinglysates with the kinase domain–specific phosphotyrosine Ab (33).Comparing conjugates containing Jurkat cells with those con-taining CD45-deficient J45.01 (Supplemental Fig. 1D) showedthat CD45 expression reduced by 30% the pTyr394 signal wherethe T cell contacted a B cell (Fig. 6C, Supplemental Fig. 1D).Thus, CD45 expression decreased Lck activation in conditionswhere AIS formation occurs.Because ERK activation by CD28 was sensitive to conditions
that disrupt rafts, we asked whether constitutively targeting CD45activity to rafts affected ERK activation by CD28 cross-linking.Specifically, a fusion protein containing the cytoplasmic domainof CD45 anchored by the L10 sequence (L10-CD45) was expressedin J45.01 cells. To detect expressed protein, the L10-CD45 con-tained GFP downstream of the CD45 domain. The L10-CD45 wasenriched in AISs, but not a separate CD45 fusion protein that
FIGURE 2. CD28 enriches in the AIS through its C-
terminal polyproline domain. (A) Representative con-
focal and DIC images of conjugates containing D10
T cells expressing CD28-GFP, and bound to an un-
pulsed CH27 cell. Either T cells (PP2 and MbCD) or B
cells (anti-CD80 and anti-CD86) received the indicated
treatments immediately before conjugation. (B) Rel.
Fluor. Enrich. values of CD28-GFP in conjugates
measured for each of the conditions in (A). (C) Rel.
Fluor. Enrich. values of labeled CD28 (left) and CTX
(right) in Cd282/2 T cells conjugated to unpulsed B
cells. The T cells expressed either wild-type CD28-
GFP or CD28(P187,190A)-GFP. (D) Confocal and DIC
images of a Cd282/2 primary T cell reconstituted with
CD28-GFP and interacting with an unpulsed CH27 B
cell. (E) Rel. Fluor. Enrich. values of CD28-GFP (d)
and CD28P187,190A-GFP (s) during T cell interactions
with unpulsed CH27 cells, averaged from 12 separate
trials. The color assignment for the pseudocolor con-
focal images (A, D) is shown at the bottom of (A); scale
bar, 5 mm; * indicates position of the B cell. In (B) and
(C), asterisks indicate statistical significance as de-
scribed in the Materials and Methods.
The Journal of Immunology 3077
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

contained S15 in place of the L10 sequence (Fig. 6D, 6E). FRETbetween CFP-labeled L10-CD45 and L10-YFP showed a FRETsignal that was sensitive to treatment of conjugates with filipin(Fig. 6F), indicating association of L10-CD45 with nanoscopicrafts that populate macrodomains (17, 34–36). Finally, expressionof L10-CD45, but not S15-CD45, inhibited ERK phosphorylationfollowing CD28 cross-linking (Fig. 6G). Thus, although consti-tutive association of CD45 with rafts does not block AIS forma-tion (Fig. 6E), it does inhibit CD28-dependent activation of ERK.The inhibition of ERK by L10-CD45 may occur by quenchingproximal signals in raft nanodomains within the AIS. Our inter-pretation regarding Lck and ERK activation in rafts and the AIS isillustrated in Fig. 7, and discussed below.
DiscussionTCR triggering is preceded by serial interactions between the T celland APCs as the T cell surveys it environment for productiveencounters with pMHC (2–4, 37, 38). These early interactionsresult in engagement of CD28 (21, 39), and subsequent formationof an Ag-independent T cell-APC conjugate (10, 39). We showhere that the initial T cell-APC interactions also result in CD28-dependent costimulatory signals that polarize T cells and sensitizeCa2+ signaling through activation of the MAP kinase ERK (Fig. 7).Live cell imaging of T cells undergoing Ag presentation show
Ca2+ signaling by the TCR often requires minutes to be detectedfollowing initial interactions with the B cell (4, 10). Nevertheless,we showed that CD28-dependent AIS formation occurs secondsfollowing the first T cell–B cell interactions. Thus, our resultsextend those from earlier work by showing that CD28 signalingand associated costimulatory signals occur seconds after the T cellfirst contacts the APC, and temporally before TCR signaling isevidenced by intracellular Ca2+ levels. For example, Michel et al.
(39) showed that engagement of CD28 occurs during early T cell–B cell interactions to facilitate phospholipase Cg activation andCa2+ signaling once the TCR is triggered. Our findings also un-derscore the important role of initial T cell–B cell interactions inoptimizing T cell responses to Ag by sensitizing the Ca2+ sig-
FIGURE 3. The Lck SH3 domain is required for AIS formation. (A and
C) Representative confocal and DIC images of Jurkat (Jkt) and JCaM1.6
cells stained with TR-labeled CTX and conjugated to unpulsed Raji cells.
In (C), the JCaM1.6 cells expressed Lck-GFP, S15-Lck-GFP, or LckW97A-
GFP. (B and D) The Rel. Fluor. Enrich. values of CTX measured in con-
jugates described in (A) and (B). (A and C) Scale bar, 5 mm; *, position of
the B cell. In (B) and (D) asterisks indicate statistical significance as de-
scribed in the Materials and Methods.
FIGURE 4. Pre–cross-linking CD28 activates ERK to sensitize T cell
Ca2+ signaling. (A) Intracellular Ca2+ measurements of Indo-1–labeled
Jurkat cells. The samples were stimulated by cross-linking either CD3 or
CD28, or by cross-linking CD28 prior to CD3 cross-linking. The sample
that received PD98059 was treated immediately before pre–cross-linking
CD28. (B) Ca2+ flux values averaged from separate independent trials. (C)
p-ERK (p-ERK1/2) and total ERK (ERK 1/2) analysis of whole-cell lysates
from Jurkat and JCaM1.6 cells stimulated with the indicated conditions.
The numerical values represent the signal intensity of each lane relative to
that of the unstimulated control for each blot. In (B), asterisks indicate
statistical significance as described in the Materials and Methods.
FIGURE 5. ERK activation by CD28 during Ag-independent T cell–B
cell interactions. (A) p-ERK measured in Cd282/2 T cells expressing
CD28-GFP (top panel), CD28P187,190A-GFP (middle panel), or soluble
GFP (bottom panel), and conjugated to unpulsed CH27 cells. The CH27
cells were labeled with CellTracker Orange and pretreated with PD98059
prior to conjugation. The data represent p-ERK staining of T cells (GFP+,
black line), B cells (CellTracker Orange+, gray, filled), and T cell–B cell
conjugates (GFP+CellTracker Orange+, gray line). (B) Averaged results of
the mean fluorescent intensity of p-ERK staining in GFP+CellTracker
Orange+ events. In (B) asterisks indicate statistical significance as described
in the Materials and Methods.
3078 Ag-INDEPENDENT SYNAPSES IN T CELL COSTIMULATION
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

naling. An early sensitization of TCR signaling is likely to beadvantageous in conditions where dynamic and motile T cells arerapidly sampling their environment to respond to a small andselect pool of pMHC. Such conditions exemplify the host pe-riphery.By separate complementation experiments using CD28- and
Lck-deficient T cells, we show that the C-terminal polyprolinesequence of CD28 and the SH3 domain of Lck are each critical forAIS formation. This suggests that the Ag-independent signalingis initiated by recruitment of Lck to CD28 by SH3 domain-polyproline interactions between the respective proteins, such ashas been shown to occur for Lck recruitment to the IS following Agstimulation of T cells (40). Lck undergoes activation by binding ofits SH3 domain to polyproline ligand (25), which, based on thecrystal structure of other Src kinases (41, 42), is suggested tocause displacement of an intramolecular interaction that maintainsLck in a closed, inactive conformation. Binding of the Lck SH3domain to the CD28 polyproline sequence may therefore be suf-ficient to activate Lck for the signaling that is necessary for AISformation and T cell sensitization.We first noted AISs based on an observed specific enrichment of
T cell rafts during Ag-independent interactions with B cells (10).Rafts are cholesterol-dependent lipid heterogeneities that serve asa membrane platform for cell signaling and signal regulation (17,33, 43–47). Similarly, our data show where signals for AIS for-
mation are compartmentalized to rafts. For example, AIS forma-tion was inhibited by limiting Lck expression to the non-raftmembrane fraction. Furthermore, ERK activation by CD28 cross-linking was sensitive to conditions that disrupt rafts, and by tar-geting CD45 phosphatase activity to rafts using the L10 sequence.Structurally, rafts are heterogenous in character, ranging from
submicroscopic domains that are only a few nanometers in di-ameter, to macroscopic assemblies that are microns in size.Examples of rafts macrodomains are the AIS and IS. How nano-scopic rafts are assembled into large-scale structures is not entirelyunderstood, yet evidence shows that interactions between the ac-tomyosin cytoskeleton and rafts is an important factor (48). In-terestingly, the cytoskeletal protein filamin-A binds to the C-terminal polyproline sequence of CD28, and this interaction isnecessary for IS formation in stimulated T cells (49). As a memberof a large group of proteins that simultaneously bind actin fila-ments and anchor them to the plasma membrane, filamin-A is anexcellent candidate as a factor that could promote assembly of raftmacrodomains by cytoskeleton-dependent mechanisms. WhetherCD28–filimin-A interactions function in AIS formation, and, if so,whether they compete or cooperate with those between Lck andCD28, are additional questions for study.Augmentation of TCR-dependent Ca2+ signaling by pre–cross-
linking CD28 was blocked using the MEK inhibitor PD98059,thus corroborating evidence that ERK is a critical intermediate in
FIGURE 6. CD45 localizes to the AIS to modulate
Lck signaling. (A) Confocal and DIC images of a T
cell conjugated to an unpulsed B cell and double-la-
beled with L10-GFP and TR-labeled Ab to CD45. (B)
Rel. Fluor. Enrich. values of CD45 at the cell interface
of T cell–B cell conjugates, measured in the indicated
conditions. (C) The relative signal of Lck pTyr394
where Jurkat and J45.01 cells contacted Raji B cells.
The values were calculated by dividing the average
fluorescence intensity of the pTyr394 immunostain by
the coinciding average fluorescence intensity of total
Lck immunostain. (D) Confocal and DIC images of
Jurkat cells conjugated to Raji cells and expressing
either L10- or S15-anchored CD45 cytoplasmic do-
main. The samples were imaged using the GFP label
that was on each CD45 chimera. Rel. Fluor. Erich.
values of each labeled CD45 chimera measured in T
cell-B cell conjugates are plotted in (E). (F) Average
FRET efficiencies (E%) for the indicated donor-ac-
ceptor pairs measured in the AIS of cell conjugates.
Lyn CFP/YFP is a membrane-anchored fusion protein
containing CFP and YFP separated by 3 aa. (G) p-ERK
staining measured in J45.01 cells expressing L10-
CD45, S15-CD45, or L10-GFP and stimulated by
cross-linking CD28. Measurements were gated on the
expression of the label on each protein. Scale bar,
5 mm (A, D); * indicates position of the B cell. In (B),
(C), (E), and (F) asterisks indicate statistical signifi-
cance as described in the Materials and Methods.
The Journal of Immunology 3079
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

T cell sensitization to Ag (28). Interestingly, ERK activation duringAg-independent T cell–B cell interactions can also occur subsequentto engagement of LFA-1 (28). Nevertheless, in our experimentalsystem, CD28 was the most significant in coordinating AIS forma-tion with sensitization by ERK, because AIS assembly in conjugateswas completely inhibited only by blocking CD28 interactions withCD80 and CD86, and both AIS formation and ERK activation didnot occur in T cells that lacked CD28 expression.The canonical pathway for ERK activation in T cells consists of
a TCR-dependent activation of Grb2/SOS complexes through theirrecruitment to LAT, followed by activation of Ras, then Raf, MEK,and ERK (50). The Grb/Sos recruitment occurs following phos-phorylation of LAT by ZAP-70, which is activated by Lck. Ourdata showing efficient activation of ERK by CD28 engagementsuggests a noncanonical pathway in ERK activation during Ag-independent T cell-B cell interactions. One possible mechanismis activation of ZAP-70 by CD28-associated Lck, which is pos-sible since z-chain coenriches with rafts in AISs (10). Activationof ZAP-70 in this manner may then be sufficient to cause phos-phorylation of LAT and activation of the Grb2/SOS network aspart of the MEK/ERK canonical pathway discussed above. Con-sistent with this interpretation, engagement of CD2 in Jurkat cellsresults in phosphorylation of LAT by Lck during an actin-dependent coalescence of rafts (22). Alternatively, Conche andcoworkers showed an Ag-independent activation of ERK down-stream of LFA-1 that occurs by production of cAMP and subse-quent PKA activation (28). Triggering of CD28-Lck complexformation may activate this pathway through common down-stream intermediates that are shared between LFA-1 and CD28.
Altogether, further study is necessary to better understand themechanism underlying ERK activation by CD28 in Ag-freeconditions, and the hierarchy of these mechanisms in Ag-freeconditions.We show in Fig. 6 that AISs contain CD45 and, by quenching
proximal Lck, may be important for regulating the progression ofthe Ag-independent activation signals until Ag is encountered(51). Enrichment of CD45 in the AIS is interesting because itcontrasts with a documented low affinity of CD45 for raft-enrichedmembrane environments (43, 52), underscoring that proteintargeting to discrete environments is the product of multiple andunrelated parameters. In the case of CD45 targeting to AISs, weshowed that interactions with galectins contribute to some de-gree. Association of the CD45 cytoplasmic tail with the corticalcytoskeleton (53), which is enriched in AIS structures (10), maybe another factor.T cell polarization during Ag-independent T cell–B cell conjuga-
tion included relocalization of the T cell centrosome, or microtubule-organizing center, to the region of the cell underlying contact with theAPC (41, 54, 55). In T cells stimulated with Ag, reorientation of thecentrosome is thought to be critical for polarized delivery of mole-cules to the IS (55). Interestingly, while we observed Ag-independentconjugates where the centrosome was distal to the T cell–B cell in-terface, productive conjugates that exhibited an AIS was limited tothose where the centrosome was proximal or adjacent. This suggeststhat polarization of rafts and associated proteins is coupled to earlycentrosome relocalization. However, whether intracellular traffickingby centrosome-associated microtubules accounts for the accumula-tion of rafts in the AIS is not known.In summary, we show that T cell polarization to form an AIS
during initial contacts with an APC occurs as a consequence ofCD28-dependent signals that activate ERK and sensitize Ca2+
signaling. Altogether, our data move forward events associatedwith T cell costimulation, showing that they occur at the earlieststages of T cell interactions with APCs. Our findings thereforeprovide a new understanding of the spatial–temporal properties ofT cell activation by pMHC.
DisclosuresThe authors have no financial conflicts of interest.
References1. Bousso, P., and E. Robey. 2003. Dynamics of CD8+ T cell priming by dendritic
cells in intact lymph nodes. Nat. Immunol. 4: 579–585.2. Miller, M. J., A. S. Hejazi, S. H. Wei, M. D. Cahalan, and I. Parker. 2004.
T cell repertoire scanning is promoted by dynamic dendritic cell behavior andrandom T cell motility in the lymph node. Proc. Natl. Acad. Sci. USA 101:998–1003.
3. Mempel, T. R., S. E. Henrickson, and U. H. Von Andrian. 2004. T-cell priming bydendritic cells in lymph nodes occurs in three distinct phases. Nature 427: 154–159.
4. Negulescu, P. A., T. B. Krasieva, A. Khan, H. H. Kerschbaum, andM. D. Cahalan. 1996. Polarity of T cell shape, motility, and sensitivity to an-tigen. Immunity 4: 421–430.
5. Monks, C. R., B. A. Freiberg, H. Kupfer, N. Sciaky, and A. Kupfer. 1998. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature395: 82–86.
6. Grakoui, A., S. K. Bromley, C. Sumen, M. M. Davis, A. S. Shaw, P. M. Allen,and M. L. Dustin. 1999. The immunological synapse: a molecular machinecontrolling T cell activation. Science 285: 221–227.
7. Sykulev, Y., M. Joo, I. Vturina, T. J. Tsomides, and H. N. Eisen. 1996. Evidencethat a single peptide-MHC complex on a target cell can elicit a cytolytic T cellresponse. Immunity 4: 565–571.
8. Irvine, D. J., M. A. Purbhoo, M. Krogsgaard, and M. M. Davis. 2002. Directobservation of ligand recognition by T cells. Nature 419: 845–849.
9. Purbhoo, M. A., D. J. Irvine, J. B. Huppa, and M. M. Davis. 2004. T cell killingdoes not require the formation of a stable mature immunological synapse. Nat.Immunol. 5: 524–530.
10. Van Komen, J. S., S. Mishra, J. Byrum, G. R. Chichili, J. C. Yaciuk, A. D. Farris,and W. Rodgers. 2007. Early and dynamic polarization of T cell membrane raftsand constituents prior to TCR stop signals. J. Immunol. 179: 6845–6855.
FIGURE 7. Membrane remodeling and sensitization of TCR Ca2+ sig-
naling by CD28 during Ag-independent T cell–B cell interactions. Initial
and Ag-independent contacts between the T cell and APC engage CD28 to
activate Lck, leading to assembly of a raft-enriched AIS at the cell inter-
face, and ERK activation. ERK activation by Lck may be initiated by
phosphorylation of z-chain that associates with the AIS (10), followed by
ZAP-70 activation and LAT phosphorylation to recruit Grb/SOS complex
and activate Ras, Raf, and MAP kinases. The activated ERK potentiates
intracellular Ca2+ signaling once TCR binds pMHC. CD45 associated with
the AIS inactivates Lck to block ERK activation and TCR sensitization.
3080 Ag-INDEPENDENT SYNAPSES IN T CELL COSTIMULATION
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

11. Revy, P., M. Sospedra, B. Barbour, and A. Trautmann. 2001. Functional antigen-independent synapses formed between T cells and dendritic cells. Nat. Immunol.2: 925–931.
12. Lo, W., W. Rodgers, and T. Hughes. 1998. Making genes green: creating greenfluorescent protein (GFP) fusions with blunt-end PCR products. Biotechniques25: 94–96, 98.
13. Rodgers, W. 2002. Making membranes green: construction and characterizationof GFP-fusion proteins targeted to discrete plasma membrane domains. Bio-techniques 32: 1044–1046, 1048, 1050–1051.
14. Jordan, S., and W. Rodgers. 2003. T cell glycolipid-enriched membrane domainsare constitutively assembled as membrane patches that translocate to immunesynapses. J. Immunol. 171: 78–87.
15. Gordy, C., S. Mishra, and W. Rodgers. 2004. Visualization of antigen presen-tation by actin-mediated targeting of glycolipid-enriched membrane domains tothe immune synapse of B cell APCs. J. Immunol. 172: 2030–2038.
16. Chichili, G. R., and W. Rodgers. 2007. Clustering of membrane raft proteins bythe actin cytoskeleton. J. Biol. Chem. 282: 36682–36691.
17. Chichili, G. R., A. D. Westmuckett, and W. Rodgers. 2010. T cell signal regu-lation by the actin cytoskeleton. J. Biol. Chem. 285: 14737–14746.
18. Cumming, G., F. Fidler, and D. L. Vaux. 2007. Error bars in experimental bi-ology. J. Cell Biol. 177: 7–11.
19. Yang, H., and E. L. Reinherz. 2001. Dynamic recruitment of human CD2 intolipid rafts: linkage to T cell signal transduction. J. Biol. Chem. 276: 18775–18785.
20. Marwali, M. R., J. Rey-Ladino, L. Dreolini, D. Shaw, and F. Takei. 2003.Membrane cholesterol regulates LFA-1 function and lipid raft heterogeneity.Blood 102: 215–222.
21. Kovacs, B., R. V. Parry, Z. Ma, E. Fan, D. K. Shivers, B. A. Freiberg,A. K. Thomas, R. Rutherford, C. A. Rumbley, J. L. Riley, and T. H. Finkel. 2005.Ligation of CD28 by its natural ligand CD86 in the absence of TCR stimulationinduces lipid raft polarization in human CD4 T cells. J. Immunol. 175: 7848–7854.
22. Kaizuka, Y., A. D. Douglass, S. Vardhana, M. L. Dustin, and R. D. Vale. 2009.The coreceptor CD2 uses plasma membrane microdomains to transduce signalsin T cells. J. Cell Biol. 185: 521–534.
23. Angus, K. L., and G. M. Griffiths. 2013. Cell polarisation and the immunologicalsynapse. Curr. Opin. Cell Biol. 25: 85–91.
24. Lee, K. H., A. D. Holdorf, M. L. Dustin, A. C. Chan, P. M. Allen, andA. S. Shaw. 2002. T cell receptor signaling precedes immunological synapseformation. Science 295: 1539–1542.
25. Holdorf, A. D., J. M. Green, S. D. Levin, M. F. Denny, D. B. Straus, V. Link,P. S. Changelian, P. M. Allen, and A. S. Shaw. 1999. Proline residues in CD28and the Src homology (SH)3 domain of Lck are required for T cell costimula-tion. J. Exp. Med. 190: 375–384.
26. Rudd, C. E., and H. Schneider. 2003. Unifying concepts in CD28, ICOS andCTLA4 co-receptor signalling. Nat. Rev. Immunol. 3: 544–556.
27. Rodgers, W., B. Crise, and J. K. Rose. 1994. Signals determining protein tyrosinekinase and glycosyl-phosphatidylinositol-anchored protein targeting to a glyco-lipid-enriched membrane fraction. Mol. Cell. Biol. 14: 5384–5391.
28. Conche, C., G. Boulla, A. Trautmann, and C. Randriamampita. 2009. T celladhesion primes antigen receptor-induced calcium responses through a tran-sient rise in adenosine 39,59-cyclic monophosphate. Immunity 30: 33–43.
29. Koretzky, G. A., J. Picus, T. Schultz, and A. Weiss. 1991. Tyrosine phosphataseCD45 is required for T-cell antigen receptor and CD2-mediated activation ofa protein tyrosine kinase and interleukin 2 production. Proc. Natl. Acad. Sci.USA 88: 2037–2041.
30. Zikherman, J., C. Jenne, S. Watson, K. Doan, W. Raschke, C. C. Goodnow, andA. Weiss. 2010. CD45-Csk phosphatase-kinase titration uncouples basal andinducible T cell receptor signaling during thymic development. Immunity 32:342–354.
31. Zhang, M., M. Moran, J. Round, T. A. Low, V. P. Patel, T. Tomassian,J. D. Hernandez, and M. C. Miceli. 2005. CD45 signals outside of lipid raftsto promote ERK activation, synaptic raft clustering, and IL-2 production. J.Immunol. 174: 1479–1490.
32. Chen, I. J., H. L. Chen, and M. Demetriou. 2007. Lateral compartmentaliza-tion of T cell receptor versus CD45 by galectin-N-glycan binding andmicrofilaments coordinate basal and activation signaling. J. Biol. Chem. 282:35361–35372.
33. Chichili, G. R., R. C. Cail, and W. Rodgers. 2012. Cytoskeletal modulation oflipid interactions regulates Lck kinase activity. J. Biol. Chem. 287: 24186–24194.
34. Lillemeier, B. F., J. R. Pfeiffer, Z. Surviladze, B. S. Wilson, and M. M. Davis.2006. Plasma membrane-associated proteins are clustered into islands attachedto the cytoskeleton. Proc. Natl. Acad. Sci. USA 103: 18992–18997.
35. Lillemeier, B. F., M. A. Mortelmaier, M. B. Forstner, J. B. Huppa, J. T. Groves,and M. M. Davis. 2010. TCR and Lat are expressed on separate protein islands onT cell membranes and concatenate during activation. Nat. Immunol. 11: 90–96.
36. Wilson, B. S., S. L. Steinberg, K. Liederman, J. R. Pfeiffer, Z. Surviladze,J. Zhang, L. E. Samelson, L. H. Yang, P. G. Kotula, and J. M. Oliver. 2004.Markers for detergent-resistant lipid rafts occupy distinct and dynamic domainsin native membranes. Mol. Biol. Cell 15: 2580–2592.
37. Bousso, P., and E. Robey. 2003. Dynamics of CD8+ T cell priming by dendriticcells in intact lymph nodes. Nat. Immunol. 4: 579‑585.
38. Schneider, H., J. Downey, A. Smith, B. H. Zinselmeyer, C. Rush, J. M. Brewer,B. Wei, N. Hogg, P. Garside, and C. E. Rudd. 2006. Reversal of the TCR stopsignal by CTLA-4. Science 313: 1972–1975.
39. Michel, F., G. Attal-Bonnefoy, G. Mangino, S. Mise-Omata, and O. Acuto. 2001.CD28 as a molecular amplifier extending TCR ligation and signaling capa-bilities. Immunity 15: 935–945.
40. Tavano, R., G. Gri, B. Molon, B. Marinari, C. E. Rudd, L. Tuosto, and A. Viola.2004. CD28 and lipid rafts coordinate recruitment of Lck to the immunologicalsynapse of human T lymphocytes. J. Immunol. 173: 5392–5397.
41. Ueda, H., M. K. Morphew, J. R. McIntosh, and M. M. Davis. 2011. CD4+ T-cellsynapses involve multiple distinct stages. Proc. Natl. Acad. Sci. USA 108:17099–17104.
42. Xu, W., S. C. Harrison, and M. J. Eck. 1997. Three-dimensional structure of thetyrosine kinase c-Src. Nature 385: 595–602.
43. Xavier, R., T. Brennan, Q. Li, C. McCormack, and B. Seed. 1998. Membranecompartmentation is required for efficient T cell activation. Immunity 8: 723–732.
44. Bi, K., Y. Tanaka, N. Coudronniere, K. Sugie, S. Hong, M. J. van Stipdonk, andA. Altman. 2001. Antigen-induced translocation of PKC-u to membrane rafts isrequired for T cell activation. Nat. Immunol. 2: 556–563.
45. Kabouridis, P. S., A. I. Magee, and S. C. Ley. 1997. S-acylation of LCK proteintyrosine kinase is essential for its signalling function in T lymphocytes. EMBO J.16: 4983–4998.
46. Zhang, W., R. P. Trible, and L. E. Samelson. 1998. LAT palmitoylation: itsessential role in membrane microdomain targeting and tyrosine phosphorylationduring T cell activation. Immunity 9: 239–246.
47. Filipp, D., J. Zhang, B. L. Leung, A. Shaw, S. D. Levin, A. Veillette, andM. Julius. 2003. Regulation of Fyn through translocation of activated Lck intolipid rafts. J. Exp. Med. 197: 1221–1227.
48. Chichili, G. R., and W. Rodgers. 2009. Cytoskeleton-membrane interactions inmembrane raft structure. Cell. Mol. Life Sci. 66: 2319–2328.
49. Tavano, R., R. L. Contento, S. J. Baranda, M. Soligo, L. Tuosto, S. Manes, andA. Viola. 2006. CD28 interaction with filamin-A controls lipid raft accumulationat the T-cell immunological synapse. Nat. Cell Biol. 8: 1270–1276.
50. Smith-Garvin, J. E., G. A. Koretzky, and M. S. Jordan. 2009. T cell activation.Annu. Rev. Immunol. 27: 591–619.
51. Freiberg, B. A., H. Kupfer, W. Maslanik, J. Delli, J. Kappler, D. M. Zaller, andA. Kupfer. 2002. Staging and resetting T cell activation in SMACs. Nat.Immunol. 3: 911–917.
52. Rodgers, W., and J. K. Rose. 1996. Exclusion of CD45 inhibits activity ofp56lckassociated with glycolipid-enriched membrane domains. J. Cell Biol. 135:1515–1523.
53. Cairo, C. W., R. Das, A. Albohy, Q. J. Baca, D. Pradhan, J. S. Morrow,D. Coombs, and D. E. Golan. 2010. Dynamic regulation of CD45 lateral mo-bility by the spectrin-ankyrin cytoskeleton of T cells. J. Biol. Chem. 285: 11392–11401.
54. Kupfer, A., G. Dennert, and S. J. Singer. 1985. The reorientation of the Golgiapparatus and the microtubule-organizing center in the cytotoxic effector cell isa prerequisite in the lysis of bound target cells. J. Mol. Cell. Immunol. 2: 37–49.
55. Stinchcombe, J. C., E. Majorovits, G. Bossi, S. Fuller, and G. M. Griffiths. 2006.Centrosome polarization delivers secretory granules to the immunological syn-apse. Nature 443: 462–465.
The Journal of Immunology 3081
by guest on July 12, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















