
UTY-specific TCR-transfer generates potential graft-versus-leukaemia effector T cells
11
UTY-specific TCR-transfer generates potential graft-versus- leukaemia effector T cells In recent years donor lymphocyte infusions (DLIs) have gained acceptance as the most successful approach to the treatment of relapsed lymphoid malignancies (Barrett, 2003; Sureda & Schmitz, 2003; Kolb et al, 2004). The key therapeutic mech- anism of the DLI is an immune response of donor T lymphocytes against leukaemic cells of a patient. This beneficial graft-versus-leukaemia (GVL) effect is accompanied by the T cell reactivity towards healthy tissues that frequently causes graft-versus-host disease (GVHD). Although GVL and GVHD are closely related, numerous observations have been made that suggest a distinct antigenic basis of these two DLI effects (Jiang et al, 1997; Michalek et al, 2003). If true, this may permit the development of strategies that augment GVL without exacerbation of GVHD. Obviously, this would signi- ficantly improve the clinical results of DLI therapy. Therefore, the identification of antigens that can induce T cell responses that specifically target leukaemic cells is a major goal of tumour immunologists. The adoptive transfer of T cells that are genetically engineered to react against such leukaemia- specific antigens is a new and promising treatment modality (Stanislawski et al, 2001). In a human leucocyte antigen (HLA)-identical setting, T cell responses are directed against minor histocompatibility anti- gens (mHags) – products of polymorphic proteins, which are different between a patient and a donor. H–Y antigens represent a separate class of mHags. These antigens are encoded by genes located on the male Y chromosome and are therefore absent in females. Recent observations have estab- lished that male recipients of female haematopoietic stem cells have the lowest risk of relapse in comparison with other patients and that this phenomenon is independent of the concomitant GVHD (Randolph et al, 2004). Presumably, this selective GVL effect is mediated by T cells recognizing certain H–Y antigens on leukaemic cells. Although no H–Y genes with a haematopoietic-restricted expression have been found (Skaletsky et al, 2003), the preferential immune recognition Roman Ivanov, 1 Samantha Hol, 1 Tineke Aarts, 1 Anton Hagenbeek, 1 Elisabeth H. Slager 2 and Saskia Ebeling 1 1 Jordan Laboratory for Haemato-Oncology, Department of Haematology, University Medical Centre Utrecht, Heidelberglaan 100, Utrecht, the Netherlands, and 2 Department of Haematology, Leiden University Medical Centre, PO Box 9600, Leiden, the Netherlands Received 26 December 2004; accepted for publication 21 February 2005 Correspondence: Saskia Ebeling, PhD, Department of Haematology, University Medical Centre Utrecht Heidelberglaan 100, 3584 CX Utrecht, the Netherlands. E-mail: [email protected] Summary Immunotherapeutic approaches that target antigens that are differentially recognized on haematopoietic and non-haematopoietic cells may specifically enhance the graft-versus-leukaemia (GVL) effect of donor lymphocyte infusion. In this study, we have characterized a new HLA-B*5201-restricted epitope of the UTY gene. Unusually, presentation of this epitope was restricted to lymphoblasts. As a result, a T cell clone specific to this epitope recognized normal and malignant male B and T lymphoblasts, while showing little reactivity towards male HLA-B*5201+ fibroblasts. Transfer of its T cell receptor (TCR) into donor T cells led to the generation of large numbers of T cells, which acquired the specificity of the original clone, its avidity and the differential pattern of reactivity towards lymphoblasts and fibroblasts. Remarkably, the specific response of TCR-transferred T cells was significantly higher than that of the original clone. This is the first demonstration of the possibility to preserve the specific pattern of a T cell response to a differentially expressed antigen after TCR-transfer and to augment the amplitude of this response concomitantly. These results indicate that it may be feasible to enhance the GVL effect of donor lymphocyte infusions in lymphoproliferative malignancies by the transfer of TCRs specific to epitopes that are differentially recognized on lymphoblasts. Keywords: H–Y antigens, T cell receptor-transfer, graft-versus-leukaemia, immunotherapy, donor lymphocyte infusion. research paper doi:10.1111/j.1365-2141.2005.05461.x ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of UTY-specific TCR-transfer generates potential graft-versus-leukaemia effector T cells

UTY-specific TCR-transfer generates potential graft-versus-leukaemia effector T cells
In recent years donor lymphocyte infusions (DLIs) have gained
acceptance as the most successful approach to the treatment of
relapsed lymphoid malignancies (Barrett, 2003; Sureda &
Schmitz, 2003; Kolb et al, 2004). The key therapeutic mech-
anism of the DLI is an immune response of donor
T lymphocytes against leukaemic cells of a patient. This
beneficial graft-versus-leukaemia (GVL) effect is accompanied
by the T cell reactivity towards healthy tissues that frequently
causes graft-versus-host disease (GVHD). Although GVL and
GVHD are closely related, numerous observations have been
made that suggest a distinct antigenic basis of these two DLI
effects (Jiang et al, 1997; Michalek et al, 2003). If true, this may
permit the development of strategies that augment GVL
without exacerbation of GVHD. Obviously, this would signi-
ficantly improve the clinical results of DLI therapy. Therefore,
the identification of antigens that can induce T cell responses
that specifically target leukaemic cells is a major goal of
tumour immunologists. The adoptive transfer of T cells that
are genetically engineered to react against such leukaemia-
specific antigens is a new and promising treatment modality
(Stanislawski et al, 2001).
In a human leucocyte antigen (HLA)-identical setting, T cell
responses are directed against minor histocompatibility anti-
gens (mHags) – products of polymorphic proteins, which are
different between a patient and a donor. H–Y antigens
represent a separate class of mHags. These antigens are
encoded by genes located on the male Y chromosome and are
therefore absent in females. Recent observations have estab-
lished that male recipients of female haematopoietic stem cells
have the lowest risk of relapse in comparison with other
patients and that this phenomenon is independent of the
concomitant GVHD (Randolph et al, 2004). Presumably, this
selective GVL effect is mediated by T cells recognizing certain
H–Y antigens on leukaemic cells. Although no H–Y genes with
a haematopoietic-restricted expression have been found
(Skaletsky et al, 2003), the preferential immune recognition
Roman Ivanov,1 Samantha Hol,1 Tineke
Aarts,1 Anton Hagenbeek,1 Elisabeth H.
Slager2 and Saskia Ebeling1
1Jordan Laboratory for Haemato-Oncology,
Department of Haematology, University Medical
Centre Utrecht, Heidelberglaan 100, Utrecht, the
Netherlands, and 2Department of Haematology,
Leiden University Medical Centre, PO Box 9600,
Leiden, the Netherlands
Received 26 December 2004; accepted for
publication 21 February 2005
Correspondence: Saskia Ebeling, PhD,
Department of Haematology, University
Medical Centre Utrecht Heidelberglaan 100,
3584 CX Utrecht, the Netherlands.
E-mail: [email protected]
Summary
Immunotherapeutic approaches that target antigens that are differentially
recognized on haematopoietic and non-haematopoietic cells may specifically
enhance the graft-versus-leukaemia (GVL) effect of donor lymphocyte
infusion. In this study, we have characterized a new HLA-B*5201-restricted
epitope of the UTY gene. Unusually, presentation of this epitope was
restricted to lymphoblasts. As a result, a T cell clone specific to this epitope
recognized normal and malignant male B and T lymphoblasts, while showing
little reactivity towards male HLA-B*5201+ fibroblasts. Transfer of its T cell
receptor (TCR) into donor T cells led to the generation of large numbers of
T cells, which acquired the specificity of the original clone, its avidity and the
differential pattern of reactivity towards lymphoblasts and fibroblasts.
Remarkably, the specific response of TCR-transferred T cells was
significantly higher than that of the original clone. This is the first
demonstration of the possibility to preserve the specific pattern of a T cell
response to a differentially expressed antigen after TCR-transfer and to
augment the amplitude of this response concomitantly. These results indicate
that it may be feasible to enhance the GVL effect of donor lymphocyte
infusions in lymphoproliferative malignancies by the transfer of TCRs specific
to epitopes that are differentially recognized on lymphoblasts.
Keywords: H–Y antigens, T cell receptor-transfer, graft-versus-leukaemia,
immunotherapy, donor lymphocyte infusion.
research paper
doi:10.1111/j.1365-2141.2005.05461.x ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402

of leukaemic cells may be caused by overexpression or altered
processing of some H–Y antigens.
In an attempt to reveal the nature of the GVL response in
sex-mismatched settings, we stimulated female T cells with
haematopoietic cells from a male patient. A number of T cell
clones were generated that recognized Epstein–Barr virus
(EBV)-transformed B cells (EBV-LCL) of patient, but not
donor, origin, while showing no significant reactivity towards
resting haematopoietic cells and fibroblasts. Detailed analysis
of the specificity of one of these clones led to the identification
of a new cytotoxic T lymphocyte (CTL) epitope of the UTY
gene as a target of an HLA-B*5201-restricted lymphoblast-
specific CTL response. Notably, HLA-B8- and B60-restricted
epitopes of the UTY gene have been reported previously (Vogt
et al, 2000; Warren et al, 2000) and the leucoblast-specific
recognition of HLA-B8-restricted epitope has been demon-
strated (Warren et al, 2000). The results of this study indicate
that the new epitope of UTY can be considered as a leukaemia-
associated antigen because of its predominant presentation on
normal and malignant lymphoblasts. Furthermore, the feasi-
bility to generate large numbers of lymphoblast-specific CTL
through retroviral transfer of the UTY-specific T cell receptor
(TCR) was shown.
Materials and methods
Cell culture
The generation and culture of the YKII.39 T cell clone were
performed according to the previously described protocol
(Ebeling et al, 2003). Activation and expansion of B cells were
carried out using the 293-CD40-sCD40L system (Ivanov et al,
2005a). Briefly, B cells were expanded from peripheral blood
mononuclear cells (PBMC) by incubation with 1Æ2 · 104
irradiated (75 Gy) 293 cells expressing both membrane-bound
and soluble CD40L in the presence of 10 ng/ml interleukin
(IL)-10 (Peprotech, London, UK) and 200 U/ml IL-4 (Pepro-
tech). B and T cells were cultured in Roswell Park Memorial
Institute (RPMI) 1640 medium (Invitrogen, Carlsbad, CA,
USA) supplemented with 10% heat-inactivated pooled human
AB serum, 100 U/ml penicillin (Gibco, Gaithersburg, MD,
USA), 100 lg/ml streptomycin (Gibco) and 5 · 10)5 mol/l
b-mercaptoethanol (Merck, Haarlem, the Netherlands) (here-
after referred to as PSb). B cells were harvested every 3–4 d and
plated again at 6 · 104 B cells per well in a 24-well plate (Nunc,
Rochester, NY, USA), with addition of irradiated 293-CD40-
sCD40L cells, IL-4 and IL-10, as described above. Fibroblasts,
the amphotropic Phoenix packaging cell line and 293-EBNA-
B7 cell line were cultured in Dulbecco’s modified Eagles
medium (DMEM) (Invitrogen), supplemented with 10% heat-
inactivated fetal calf serum (FCS) (Integro BV, Leuvenheim,
the Netherlands) and PSb. All other cell lines were cultured in
RPMI 1640 medium (Invitrogen) with 10% FCS and PSb. Insome cases, cell lines were cultured in presence of 5 ng/ml
interferon-c (IFN-c; Peprotech) and 200 U/ml tumour
necrosis factor-a (TNF-a; R & D Systems, Minneapolis, MN,
USA) for 2 d before the test.
Cloning of HLA-B molecules and transduction of cell lines
RNA was isolated from the EBV-LCL of patient origin (EBVp)
with Trizol reagent (Invitrogen). Reverse transcription polym-
erase chain reaction (RT-PCR) was performed with HLA-B-
specific primers 5¢-GGGGTCGACATGCGGGTCAC-3¢ and
5¢-GGGGCGGCCGCTCAAGCTGTGAGAGA-3¢. The PCR
product was purified with the Qiagen Gel Extraction kit
(Qiagen, Hilden, Germany) and digested with Sal I and Not I
restriction endonucleases (New England Biolabs, Beverly, MA,
USA). The resulting fragment was subcloned in the pEGFP-N1
vector (Clontech, Palo Alto, CA, USA) and sequenced with the
BigDye Terminator v3Æ1 Cycle Sequencing Kit (Applied
Biosystems, Foster City, CA, USA). Subsequently, HLA-
B*3508 and HLA-B*5201 were cloned separately into the
retroviral pMX vector (Onishi et al, 1996) upstream of the
internal ribosomal entry sequence (IRES) and either the nerve
growth factor receptor (NGFR) or enhanced green fluorescent
protein (EGFP) marker genes.
Resulting vectors were transfected into the amphotropic
Phoenix packaging cell line with the calcium-phosphate preci-
pitationmethod (Qiagen). Viral supernatants were harvested on
the second and third days after transfection. 293-EBNA-B7 cells
were incubated for 24 h with the 1:3 diluted viral supernatant in
the presence of 6 lg/ml polybrene (Sigma-Aldrich, Steinheim,
Germany). On the third day after transduction, NGFR-positive
cells were purified by means of the MiniMACSTM separation
method (Myltenyi Biotech, Bergisch Gladbach, Germany) using
anti-NGFR antibody as described elsewhere (Weijtens et al,
2002). Fibroblasts were transduced in the same way. EGFP-
positive fibroblasts were sorted by fluorescence-activated cell
sorting (FACS). Phytohaemagglutinin (PHA) blasts and EBV-
LCLs were transduced in untreated flasks (Becton Dickinson,
Franklin Lakes, NJ, USA) coated with 12Æ5 lg/ml retronectin
(Takara, Otsu, Shiga, Japan). Cells were added at a concentra-
tion of 1 · 106 cells/ml in the culture medium, supplemented
with 300 IU/ml IL-2 (Proleukin, Chiron, Amsterdam, the
Netherlands) in case of PHA blasts. Fresh viral supernatant
was added on the second day. On the third day cells were
harvested and resuspended in freshmediumwith addition of the
corresponding cytokines. Two days after harvesting the fraction
of transduced cells was determined by flow cytometry.
Construction of mini-genes
The plasmid containing the full-length UTY sequence was used
as a template for PCR with a common sense primer
5¢-GAAGGTACCATGAAATCCTGCGCAGTG-3¢ and one of
antisense primers containing a TAA stop codon in frame with
the main open reading frame: minigene 1–334 – 5¢-TA-AGCGGCCGCTTAAGCATCCATAGGCTG-3¢; minigene 1–408 –
5¢-TAAGCGGCCGCTTATGAAAGACTCTGGCC-3¢; minigene
Generation of GVL effector T cells through TCR-transfer
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402 393

1–413 – 5¢-TAAGCGGCCGCTTACTGTACTGGATGATG-3¢;minigene 1–426 – 5¢-TAAGCGGCCGCTTACTGTAATTTCT-GTGG-3¢; minigene 1–443 – 5¢-TAAGCGGCCGCTTACTTC-TGTGCTGGATT-3¢; minigene 1–453 – 5¢-TAAGCGGCCGC-TTAAAACTGACTTTCTAA-3¢; minigene 1–468 – 5¢-TAAGC-GGCCGCTTATCGTACCTGAGCAAC-3¢; minigene 1–528 –
5¢-TAAGCGGCCGCTTAACAAGGAATACAGCC-3¢; minigene
1–596 – 5¢-TAAGCGGCCGCTTACTGACTTTTATGAAG-3¢;minigene 1–667 – 5¢-TAAGCGGCCGCTTAGCTCTCTTT-GGTAAA-3¢. These PCR products were digested with KpnI
and NotI and cloned into the pCEP4 vector. The resulting
constructs contained truncated UTY genes lacking different
parts of the carboxyl terminus of UTY.
Transfection of the 293 cell line
293-EBNA-B7-B*5201 cells, plated in flat-bottom 96-well
plates (4 · 104 cells per well) 24 h in advance, were transfected
in duplicates with 1Æ2 ll Lipofectamine (Invitrogen) and
100 ng of the plasmid DNA. After 24 h, the culture medium
was substituted for 200 ll of RPMI 1640 with 10% HS and
5 · 103 T cells of the YKII.39 clone per well. After another 24 h
culture supernatant was harvested and IFN-c production was
measured by enzyme-linked immunosorbent assay (ELISA).
Peptide recognition assays
Candidate peptides were synthesized by solid phase peptide
synthesis and characterized by mass spectrometry (Pepscan
Systems, Lelystad, the Netherlands). Cells were washed twice
with RPMI 1640 medium supplemented with 2% FCS and
incubated overnight in serum-free RPMI 1640 with candidate
peptides at 10)5 mol/l concentration, if not mentioned oth-
erwise. The following day peptide-loaded cells were tested for
their ability to induce IFN-c production by YKII.39.
Chromium release assay
The chromium release assay was performed according to the
previously described protocol (Ebeling et al, 2003) with minor
modifications. Briefly, 2500 labelled target cells per well were
seeded in triplicate in 96-well round-bottomed plates (Costar,
Cambridge, MA, USA) in 200 ll of RPMI 1640 medium
supplemented with 5% FCS. Thereafter, target cells were
incubated for 4 h at 37�C with T cells at different effector to
target cell ratios. Supernatants were collected using the
Supernatant Harvesting System (Molecular Devices Corpora-
tion, Sunnyvale, CA, USA) and radioactivity was measured
with a Cobra autogamma betaplate reader (Packard, Gronin-
gen, the Netherlands).
IFN-c production assay
3 · 104 target cells were co-cultured with 3 · 104 T cells in
96-well round-bottomed plates in triplicates. After 24 h the
supernatant was harvested and IFN-c concentration was
measured by the PeliPair human IFN-c ELISA reagent set
(CLB, Amsterdam, the Netherlands), according to the manu-
facturer’s instructions.
Flow cytometric determination of cell surface markers
Flow cytometry was performed on a Calibur flow cytometer
(BD Biosciences, San Jose, CA, USA). Monoclonal antibodies
used for the flow cytometry were purchased from BD
Biosciences, except 20Æ4 anti-NGFR antibody (culture super-
natant) and goat-anti-mouse immunoglobulin (Ig) phyco-
erythrin (PE)-conjugated antibody (SBA, Birmingham, AL,
USA). Data analysis was performed using CellQuest software
(BD Biosciences).
Real time RT-PCR
RNA was isolated from cell pellets or homogenized foreskins
with Trizol reagent (Invitrogen). cDNA was prepared with the
First Strand cDNA Synthesis Kit for RT-PCR (AMV) (Roche
Diagnostics, GmbH, Mannheim, Germany) using a random
primer. Amplification reactions were performed in a 25 llfinal volume with 1X SybrGreen PCR Master Mix (Applied
Biosystems) and 300 nmol/l primers designed to specifically
amplify UTY (forward primer 5¢-TCTACAGAATGGTTCT-GATAACTGGAA-3¢, reverse primer 5¢-GGTGTCAAACA-CAACGAATAAACTTG-3¢). Amplification and detection
were performed with an ABI Prism 7700 sequence detection
system (ABI/PE, Foster City, CA, USA) under the following
conditions: 10 min at 95�C, 45 cycles of 15 s at 95�C and
1 min at 60�C. Fluorescence spectra were recorded during the
elongation phase of each PCR cycle. The Sequence Detection
Software (SDS v1.7) of the ABI-Prism 7700 was used to
generate the amplification curves for each reaction. To
differentiate specific amplicons from nonspecific products, a
DNA association curve was generated after each reaction with
the ABI Prism 7700 Sequence detection system (Applied
Biosystems). After denaturation, the temperature was
decreased from 95�C to 25�C in 1�C steps with holds for
20 s. SybrGreen fluorescence was measured during each step
and the first derivative of the fluorescence was plotted as a
function of the temperature. The cycle threshold (Ct) value
represented the refraction cycle number at which a positive
amplification reaction was measured and was set at 10 times
the standard deviation (SD) of the mean baseline emission
calculated for PCR cycles 3–15. For comparison of Ct values of
different samples correction for the amount of input material
was performed using control PCR reactions for the porpho-
bilinogen deaminase.
Cloning of the YKII.39 TCR
Usage of variable regions of TCRa (AV) and TCRb (BV)
chains in YKII.39 was analysed by RT-PCR using forward
R. Ivanov et al
394 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402

primers specific for different AV and BV gene families; an
oligonucleotide specific for either Ca or Cb was used as a
reverse primer (Arden et al, 1995). Full-length YKII.39 TCRa(AV9-2*02�AJ42�AC) was amplified with primers 5¢-TTTGGATCCGCCCACCATGAACTATTCTC-3¢ and 5¢-CCCGCGGCCGCCCTCAGCTGGACCACAGC-3¢, then cloned into Bam-
HI and NotI sites of the pMX-mTCRa-IRES-EGFP vector.
Next, EGFP marker gene in the pMX-TCRa-IRES-EGFPconstruct was substituted with the NGFR marker gene by
excising EGFP with NcoI and SalI and sequential cloning of
NGFR fragment 26–844 into NcoI and SalI sites and then NGFR
fragment 1–25 into the NcoI site. YKII.39 TCRb type could not
be found in previous experiments; therefore it was amplified
with a degenerate primer 5¢-CCCGTCGACATGGGYHSCD-
GBCTCCTMTG-3¢, where Y ¼ C + T, H ¼ A + T + C,
S ¼ C + G, D ¼ A + T + G, B ¼ T + C + G, M ¼ A + C,
and a primer 5¢-TTTGCGGCCGCTCAGAAATCCTTTCTC-3¢.The PCR product was cloned into Sal I and Not I sites of
pEGFP-N1 vector (Clontech) and sequenced with the BigDye
Terminator v3Æ1 Cycle Sequencing Kit (Applied Biosystems).
Reaction mixtures were analysed on an ABI Prism 3100 genetic
analyzer (Applied Biosystems). Sequencing of the insert
revealed BV5-5*02�BD2*01�BJ2-1�BC2 type of TCRb. ThenPCR was performed with primers 5¢-TTTCCGCGGCCAC-CATGGGCC CTGGGCTC-3¢ and 5¢-CCGTCGACCTAGCCT-CTGGAATCCTTTCTCTTGACC-3¢, the PCR product was
blunted and cloned into blunt BamHI and SalI sites of
pMX-mTCRa-IRES-EGFP vector. The sequence of all cloned
TCR genes was verified.
Results
YKII.39 recognizes an HLA-B*5201-restricted minorhistocompatibility antigen expressed on proliferatinglymphocytes
The YKII T cell line was generated through repetitive
stimulation of T cells of a female donor with chronic myeloid
leukaemia (CML) cells and, later, EBV-LCL of her HLA-
identical brother. The CD8+ T cell clone YKII.39, generated in
a limiting dilution culture of YKII, produced IFN-c in
response to stimulation with EBV-LCL of patient (EBVp)
and not donor (EBVd) origin (Fig 1A). When YKII.39
reactivity towards different subsets of the patient’s primary
leucocytes was tested, comparable reactivity was observed
towards PHA-activated T cell blasts and CD40-activated B
cells. In contrast, much lower reactivity was observed towards
CML cells, normal CD14+ cells and non-stimulated primary T
and B cells (Fig 1B). This suggested that YKII.39 is specific for
a differentially expressed mHag.
YKII.39 reactivity towards EBVp was completely blocked by
antibodies against HLA Class I, HLA-B & C molecules and
CD8, but not with antibodies against HLA Class II or CD4
(Fig 1C). These results indicated that YKII.39 reacted in a
CD8-dependent manner towards an antigen presented on
either HLA-B or HLA-C. To identify the exact restriction
element of YKII.39 we transduced a random selection of 19
EBV-LCL cell lines, derived from members of two CEPH
Fig 1. YKII.39 recognizes a minor histocompatibility antigen, which is
mainly expressed on proliferating lymphocytes and presented on either
HLA-B or HLA-C. IFN-c production by YKII.39 in response to sti-
mulation with different cell types for 24 h was measured by an IFN-cELISA. Average measurements for triplicates and their standard devia-
tions are shown. (A) YKII.39 recognizes patient (EBVp) but not donor
EBV-LCL (EBVd). (B) YKII.39 reacts mainly towards activated
lymphocytesmainly. Data from independent experiments are presented.
Absolute numbers of IFN-c production differed in individual experi-
ments; therefore the IFN-c production by YKII.39 in response to sti-
mulation with different cell types is depicted in the graph as percentages
from the reactivity towards EBVp in the same experiment. (C) IFN-cproduction by YKII.39 in response to stimulation with EBVp for 24 h
was blocked by antibodies against HLA Class I, HLA-B and C molecules
and CD8, but not with antibodies against HLA Class II or CD4.
Generation of GVL effector T cells through TCR-transfer
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402 395
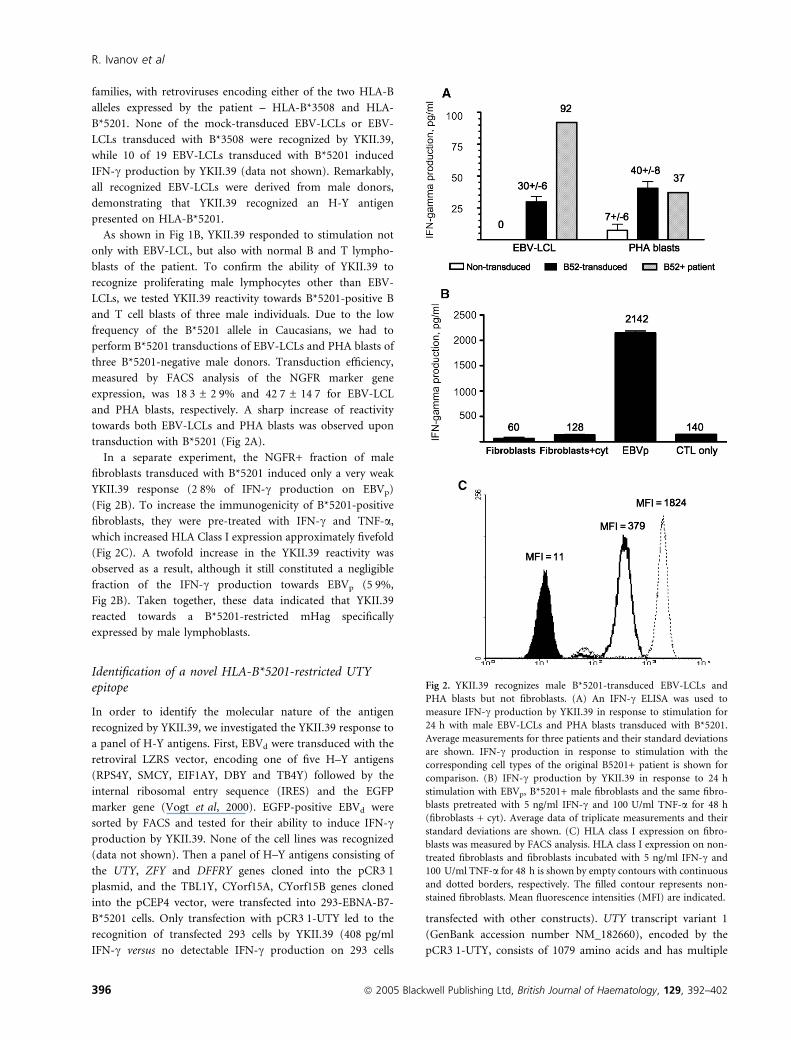
families, with retroviruses encoding either of the two HLA-B
alleles expressed by the patient – HLA-B*3508 and HLA-
B*5201. None of the mock-transduced EBV-LCLs or EBV-
LCLs transduced with B*3508 were recognized by YKII.39,
while 10 of 19 EBV-LCLs transduced with B*5201 induced
IFN-c production by YKII.39 (data not shown). Remarkably,
all recognized EBV-LCLs were derived from male donors,
demonstrating that YKII.39 recognized an H-Y antigen
presented on HLA-B*5201.
As shown in Fig 1B, YKII.39 responded to stimulation not
only with EBV-LCL, but also with normal B and T lympho-
blasts of the patient. To confirm the ability of YKII.39 to
recognize proliferating male lymphocytes other than EBV-
LCLs, we tested YKII.39 reactivity towards B*5201-positive B
and T cell blasts of three male individuals. Due to the low
frequency of the B*5201 allele in Caucasians, we had to
perform B*5201 transductions of EBV-LCLs and PHA blasts of
three B*5201-negative male donors. Transduction efficiency,
measured by FACS analysis of the NGFR marker gene
expression, was 18Æ3 ± 2Æ9% and 42Æ7 ± 14Æ7 for EBV-LCL
and PHA blasts, respectively. A sharp increase of reactivity
towards both EBV-LCLs and PHA blasts was observed upon
transduction with B*5201 (Fig 2A).
In a separate experiment, the NGFR+ fraction of male
fibroblasts transduced with B*5201 induced only a very weak
YKII.39 response (2Æ8% of IFN-c production on EBVp)
(Fig 2B). To increase the immunogenicity of B*5201-positive
fibroblasts, they were pre-treated with IFN-c and TNF-a,which increased HLA Class I expression approximately fivefold
(Fig 2C). A twofold increase in the YKII.39 reactivity was
observed as a result, although it still constituted a negligible
fraction of the IFN-c production towards EBVp (5Æ9%,
Fig 2B). Taken together, these data indicated that YKII.39
reacted towards a B*5201-restricted mHag specifically
expressed by male lymphoblasts.
Identification of a novel HLA-B*5201-restricted UTYepitope
In order to identify the molecular nature of the antigen
recognized by YKII.39, we investigated the YKII.39 response to
a panel of H-Y antigens. First, EBVd were transduced with the
retroviral LZRS vector, encoding one of five H–Y antigens
(RPS4Y, SMCY, EIF1AY, DBY and TB4Y) followed by the
internal ribosomal entry sequence (IRES) and the EGFP
marker gene (Vogt et al, 2000). EGFP-positive EBVd were
sorted by FACS and tested for their ability to induce IFN-cproduction by YKII.39. None of the cell lines was recognized
(data not shown). Then a panel of H–Y antigens consisting of
the UTY, ZFY and DFFRY genes cloned into the pCR3Æ1plasmid, and the TBL1Y, CYorf15A, CYorf15B genes cloned
into the pCEP4 vector, were transfected into 293-EBNA-B7-
B*5201 cells. Only transfection with pCR3Æ1-UTY led to the
recognition of transfected 293 cells by YKII.39 (408 pg/ml
IFN-c versus no detectable IFN-c production on 293 cells
transfected with other constructs). UTY transcript variant 1
(GenBank accession number NM_182660), encoded by the
pCR3Æ1-UTY, consists of 1079 amino acids and has multiple
Fig 2. YKII.39 recognizes male B*5201-transduced EBV-LCLs and
PHA blasts but not fibroblasts. (A) An IFN-c ELISA was used to
measure IFN-c production by YKII.39 in response to stimulation for
24 h with male EBV-LCLs and PHA blasts transduced with B*5201.
Average measurements for three patients and their standard deviations
are shown. IFN-c production in response to stimulation with the
corresponding cell types of the original B5201+ patient is shown for
comparison. (B) IFN-c production by YKII.39 in response to 24 h
stimulation with EBVp, B*5201+ male fibroblasts and the same fibro-
blasts pretreated with 5 ng/ml IFN-c and 100 U/ml TNF-a for 48 h
(fibroblasts + cyt). Average data of triplicate measurements and their
standard deviations are shown. (C) HLA class I expression on fibro-
blasts was measured by FACS analysis. HLA class I expression on non-
treated fibroblasts and fibroblasts incubated with 5 ng/ml IFN-c and
100 U/ml TNF-a for 48 h is shown by empty contours with continuous
and dotted borders, respectively. The filled contour represents non-
stained fibroblasts. Mean fluorescence intensities (MFI) are indicated.
R. Ivanov et al
396 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402

amino acid differences with its female analogue (protein
divergence 13Æ6%) (Skaletsky et al, 2003).
YKII.39 recognizes the MQQMRHKEV peptide of UTY
To identify the epitope of UTY recognized by YKII.39, we cloned
10 truncated UTY genes, which had different parts of the
3¢ sequence missing. Only minigenes containing a sequence
coding for amino acids 454–468 induced YKII.39 activation
upon their transfection into 293-EBNA-B7-B*5201 cells
(Fig 3A). The male and female isoforms differed in this region
at three amino acid positions; moreover, two amino acids of
UTX are absent in UTY. Ten overlapping non-apeptides
spanning this region were synthesized (Fig 3B). EBVd were
loaded with these candidate peptides and tested for their ability
to induce IFN-c production by YKII.39. EBVd loaded with
peptides LMQQMRHKE, MQQMRHKEV and QQMRHKEVA
were recognized by YKII.39, while no significant reactivity was
observed towards non-treated EBVd or EBVd loaded with other
peptides (Fig 3C). Next, the response of YKII.39 to EBVd loaded
with decreasing concentrations of these three peptides was
measured. We found that peptide MQQMRHKEV was recog-
nized much more efficiently at low concentrations; a half-
maximal lysis was observed at a peptide concentration of
10)8 mol/l (Fig 3D).
Increased generation of MQQMRHKEV in normal andmalignant lymphoblasts determines their susceptibility tolysis by YKII.39
The level of UTY expression in a selection of haematopoietic
and non-haematopoietic cell types was studied to analyse
whether the differential reactivity of YKII.39 towards various
cell subsets correlates with differential levels of UTY expres-
sion. As shown in Fig 4A, relative UTY mRNA levels,
normalized for the amount of porphobilinogen deaminase
transcripts, were very high in bone marrow samples contain-
ing >80% leukaemic blasts. The UTY mRNA content was
significantly higher in leukaemic cells than in bone marrow
stromal cells and homogenized foreskins (25Æ8 ± 10Æ7 vs.
2Æ5 ± 2Æ2, P < 0Æ001). Surprisingly, non-stimulated CD3+ and
CD20+ lymphocytes isolated from healthy donors had much
higher levels of UTY mRNA expression than EBV-LCLs and
PHA blasts (35Æ2 ± 12Æ4 vs. 2Æ9 ± 2Æ6, P < 0Æ0001). In fact,
the level of UTY expression in lymphoblasts was not
significantly different from that in non-haematopoietic cells
(P ¼ 0Æ78).These findings suggest that overexpression of UTY is not the
cause of the lymphoblast-specific reactivity of YKII.39. Instead,
this differential pattern of reactivity is caused either by a more
efficient generation of the UTY epitope by lymphoblasts or by
Fig 3. YKII.39 recognizes the MQQMRHKEV peptide of the UTY gene. (A) IFN-c production by YKII.39 in response to 24 h stimulation with 293-
EBNA-B7-B*5201 cells transfected with UTY mini-genes was measured by an IFN-c ELISA. Average measurements for triplicates and their standard
deviations are shown. YKII.39 recognized only cells transfected with minigenes containing amino acids 453–468. (B) The region of UTY between
amino acids 453 and 468 contains three different amino acid residues compared with UTX (indicated with bold italics) and two amino acid deletions
(indicated by hyphens). Ten candidate non-apeptides, which were tested in the next experiment, are shown. (C) Reactivity of YKII.39 towards EBVd
loaded with candidate peptides was determined in an IFN-c ELISA. Three peptides [underlined in (B)], were found to be recognized by YKII.39. (D)
Lytic activity of YKII.39 towards EBVd loaded with increasing concentrations of peptides LMQQMRHKE, MQQMRHKEV and QQMRHKEVA was
tested in a 51Cr release assay at an effector:target ratio of 10:1.
Generation of GVL effector T cells through TCR-transfer
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402 397

an enhanced ability of these cells to trigger T cell activation due
to a higher expression of co-stimulatory and adhesion
molecules. In order to distinguish between these two possibil-
ities, we investigated YKII.39-reactivity towards B*5201-pos-
itive female fibroblasts, and resting and activated T cells loaded
with increasing concentrations of MQQMRHKEV. In the
absence of significant lytic activity towards resting T cells and
fibroblasts of male origin (Fig 4B), loading of female cells with
the antigenic peptide led to efficient lysis of all tested cell types
(Fig 4C,D). Upregulation of HLA class I expression on target
cells through treatment with IFN-c and TNF-a moderately
increased lysis, but was not essential for recognition. Together,
these data demonstrate that the lack of recognition of male
fibroblasts and resting T cells by YKII.39 is a result of highly
inefficient generation of MQQMRHKEV in these cells.
To prove susceptibility of tumour cells to lysis by YKII.39,
we investigated its reactivity towards malignant lymphoblasts.
Due to the lack of B*5201-positive samples of leukaemic cells,
we could not test the lytic activity of YKII.39 towards primary
leukaemic cells. Therefore, leukaemic cell lines were trans-
duced with the pMX-B*5201/NGFR vector and NGFR-positive
cells were immunomagnetically sorted. Specific lytic activity
was observed in EBV-LCL (92%), the Burkitt’s lymphoma cell
line Raji (54%), ALL YT cells (56%) and the cytokine-treated
myeloma cell line RPMI 8226 (49%) at Effector:Target ratios
of 30:1 (data not shown).
Retroviral transfer of the YKII.39 TCR into naı̈ve donorT cells generates vast numbers of highly effectivelymphoblast-specific CTL
Next, the immunotherapeutical potential of UTY-targeting
through TCR-transfer was evaluated. T cells of the female
donor were co-transduced with two retroviral vectors enco-
ding the a- and b-chains of the YKII.39 TCR (TCRYKII.39),
respectively (Fig 5A,B). Then TCRa/NGFR-positive T cells
were immunomagnetically purified to obtain a cell population
with 57% cells positive for both a- and b-chains of TCRYKII.39
(Fig 5C,D). Unlike mock-transduced cells, TCRYKII.39-trans-
duced donor T cells displayed profound reactivity to EBVp
(Fig 5E). The reactivity to EBVd was much lower, at a level
slightly above background. This may be caused by the presence
Fig 4. Enhanced MQQMRHKEV generation rather than UTY overexpression determines susceptibility to lysis by YKII.39. (A) Real-time RT-PCR
was performed with UTY-specific primers. Resulting Ct values were corrected for the different amount of input material by concomitant real-time
RT-PCR for porphobilinogen deaminase. The sample with the lowest corrected Ct value was considered to have one arbitrary unit of the UTYmRNA.
The fold difference between this sample and other samples was calculated; the UTY mRNA content expressed in arbitrary units is shown. Three
different samples of each category were tested. Average results for three samples and their standard deviations are shown. (B) YKII.39 exerted lytic
activity in a 51Cr release assay on male B*5201-positive PHA blasts (solid squares) but neither on resting cells (solid triangles) of the same patient nor
on B*5201-transduced male fibroblasts (solid circles). IFN-c and TNF-a treatment of target cells slightly increased lysis of fibroblasts (open circles)
and did not influence lysis of resting T cells (data not shown). (C) Lysis of MQQMRHKEV-loaded B*5201-positive female T cells by YKII.39 was
tested in a 51Cr release assay at an effector:target ratio 10:1. PHA blasts are indicated with solid squares, resting T cells – with solid circles, cytokine-
treated resting T cells – with open circles. (D) YKII.39 lytic activity on EGFP-positive fraction of B*5201-tranduced female fibroblasts loaded with
MQQMRHKEV was measured in a 51Cr release assay at effector:target ratio 10:1. Solid circles represent fibroblasts not treated with cytokines, open
circles – cytokine-treated fibroblasts. Mock-transduced female fibroblasts (solid triangles) were tested as a negative control.
R. Ivanov et al
398 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402

of EBV-specific T cells in the polyclonal population of
TCRYKII.39-transduced cells. The EBVp-specific reactivity was
completely confined to the CD8+ fraction, confirming the
necessity of co-receptor engagement for T cell activation via
TCRYKII.39, which was found in blocking experiments.
Remarkably, the amount of IFN-c produced by TCRYKII.39-
transduced T cells was significantly higher than that of YKII.39
(5455 ± 1286 pg/ml vs. 721 ± 134 pg/ml, P ¼ 0Æ003).In a separate experiment, we tested the reactivity of
TCRYKII.39-transduced donor cells in response to allogeneic
fibroblasts transduced with B*5201. YKII.39 had demonstrated
very low reactivity towards these fibroblasts, as shown in
Fig 2B. TCRYKII.39-transduced T cells produced some IFN-c in
response to stimulation with B*5201-positive fibroblasts,
although it was not significantly different from the reactivity
towards B*5201-negative fibroblasts (Fig 5F). Therefore, the
observed response was mediated by alloreactive T cells in the
absence of a significant B*5201-restricted UTY-specific
response. The differential pattern of recognition mediated
through TCRYKII.39 is hence preserved in TCRYKII.39-trans-
duced cells. This finding suggests that YKII.39 and TCRYKII.39-
transduced T cells require a similar amount of peptide
Fig 5. Donor T cells transduced with YKII.39 TCR preserve the specificity, the sensitivity and the differential pattern of recognition of the original
clone. Donor T cells were co-transduced with vectors pMX-TCRaYKII.39/NGFR and pMX-TCRbYKII.39. Staining with antibodies against TCRVb5Æ3and NGFR was performed on mock-transduced (A), transduced (B), transduced and immunomagnetically purified (C) donor T cells. (D) The
TCRbYKII.39 expression on TCR-transduced cells was measured by FACS analysis after staining with PE-conjugated BV5Æ3-specific antibody. Empty
contours represent staining of the original YKII.39 clone. For the Mean Fluorescence Intensity (MFI) estimation only BV5Æ3-positive TCR-transducedcells were gated (marker M1) to enable valid comparison with the original clone. The MFI of YKII.39 was 57Æ0 and the MFI of TCRYKII.39-transduced
cells was 40Æ2. (E) CD4+ and CD8+ fractions of TCR-transduced donor T cells were obtained by immunomagnetic depletion of CD8+ and CD4+ cells,
respectively. Reactivity of YKII.39 and different fractions of TCRYKII.39-transduced T cells to EBVp and EBVd was tested in an IFN-c ELISA. Average
results for triplicates and their standard deviations are shown in the figure. (F) Reactivity of CD8+ TCRYKII.39-transduced T cells towards non-
transduced versus B*5201-transduced fibroblasts, either pretreated with 5 ng/ml IFN-c and 200 U/ml TNF-a or not, was tested in an IFN-c ELISA.
IFN-c production in response to stimulation with EBVp and EBVd is shown for comparison. (G) Reactivity of YKII.39 and TCRYKII.39-transduced
T cells to EBVd loaded with increasing amounts of MQQMRHKEV was compared in a 51Cr release assay at the effector:target ratio of 10:1.
Generation of GVL effector T cells through TCR-transfer
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402 399

presented on the target cell surface for activation. Indeed, we
found that both the original T cell clone and T cells transduced
with TCRYKII.39 reacted to EBVd loaded with the
MQQMRHKEV peptide at the minimal concentration of
10)9 mol/l (Fig 5G).
Discussion
H–Y antigens were initially identified as targets of immune
responses leading to the rejection of a male graft in female
patients (Scott et al, 1997). The increased severity of GVHD in
female to male SCT is associated with responses to H–Y
antigens (Goulmy et al, 1996). Recently, it was shown that
anti-H–Y responses also contribute to the GVL effect, which is
independent from the parallel GVHD reactivity (Randolph
et al, 2004). This important observation indicates that certain
H–Y antigens should be able to elicit responses that target
leukaemic cells in the absence of significant collateral damage.
However, all H–Y antigens characterized so far as targets of
T cell responses are ubiquitously expressed (Wang et al, 1995;
Pierce et al, 1999; Vogt et al, 2000, 2002). Moreover, mapping
of the Y chromosome did not reveal any antigens with a
haematopoietic-restricted expression (Skaletsky et al, 2003).
This indicates that either overexpression, post-transcriptional
regulation or altered processing of some H–Y antigens leads to
a differential pattern of recognition of their epitopes on
leukaemic cells, resulting in GVL reactions.
We have recently found evidence to support this notion. The
male isoform of the ribosomal protein S4 (RPS4Y) induces an
HLA-B*5201-restricted CTL response that specifically targets
proliferating B and T cells (Ivanov et al, 2005b). Despite the
constantly high levels of RPS4 mRNA in different cell types, a
significantly larger amount of RPS4 protein was found in
malignant and normal lymphoblasts. This observation pro-
vides a proof of principle that an immune response to a
ubiquitously expressed antigen may result in a differential
recognition of leukaemic cells. A second H–Y antigen to
corroborate this notion is UTY. Warren et al (2000) demon-
strated a significant cell-type specificity of presentation of UTY
by cultured human cells. In that study an increased level of
UTY expression and/or processing in haematopoietic cells was
thought to play a role in the preferential recognition of UTY
on blast cells.
In the present study, a new HLA-B*5201-restricted epitope
of UTY was identified. UTY encodes a tetratricopeptide repeat
protein of unknown function. Tetratricopeptide repeats were
reported to facilitate protein–protein interactions and the
assembly into high-order complexes (Blatch & Lassle, 1999).
As a result of the alternative splicing, three transcripts are
generated of the UTY gene, resulting in UTY proteins with
different carboxyl termini. The sequence encoding the
MQQMRHKEV peptide recognized by YKII.39 can be found
in all three isoforms. This peptide, as well as its female
analogue, has a Valine residue in the ninth position that can
serve as an anchor residue for binding to B*5201, according to
published data (Rammensee et al, 1995). However, compared
with its female analogue, it has three amino acid substitutions
and two amino acid deletions, making the existence of a female
counterpart peptide and its recognition by YKII.39 rather
improbable.
Like the previously reported HLA-B8-restricted UTY-speci-
fic CTL, YKII.39 displayed a differential pattern of recognition
(Warren et al, 2000). Only B and T lymphoblasts were
recognized, while resting haematopoietic cells and fibroblasts
did not induce activation of YKII.39. Quantitative analysis of
UTY expression showed that, although the level of UTYmRNA
in lymphoblasts was higher than in skin or bone marrow
stromal cells, it was lower than in non-recognized resting
haematopoietic cells. Thus YKII.39-reactivity is not correlated
to UTY expression levels. Furthermore, elevated expression of
co-stimulatory and adhesion molecules on lymphoblasts is not
the cause of their specific recognition either, since loading of
fibroblasts and resting T cells with the antigenic peptide
induced their lysis by YKII.39 at levels similar to, or higher
than, those of PHA blasts. Thus, provided that sufficient
amounts of the MQQMRHKEV peptide are presented, resting
cells are equally susceptible to YKII.39 as blast cells. These data
suggest that inefficient generation of MQQMRHKEV in
resting cells is the cause of their YKII.39-resistance. Immuno-
proteasome involvement in the generation of this epitope does
not explain lymphoblast recognition, because neither fibro-
blasts treated with IFN-c, nor CML cells differentiated into
dendritic cell-like cells, were recognized. However, other
factors involved in antigen-processing may be responsible for
this phenomenon, such as the lymphoblast-restricted activity
of aminopeptidases that participate in peptide-trimming
downstream of the proteasome (Rock et al, 2004). Alternat-
ively, considering the similar recognition pattern of another
UTY epitope (Warren et al, 2000), post-transcriptional regu-
lation of expression of the UTY protein or its turnover may be
responsible for the accumulation of the UTY protein in
lymphoblasts.
Retroviral transfer of the TCR of the original clone into large
numbers of donor T cells provides an attractive solution to this
problem (Schumacher, 2002). However, tissue-specific recog-
nition of ubiquitously expressed antigens depends on a subtle
balance between the avidity of antigen-specific T cells and the
level of the antigen presented by target cells. Primary T cells of
the HLA-identical donor transduced with TCRYKII.39 consist of
T lymphocytes with different functional properties. More
importantly, they have variable levels of expression of the
exogenous TCR, which is shown to have an impact on T cell
reactivity (Heemskerk et al, 2004). Nevertheless, we demon-
strated the preservation of the avidity and the recognition
pattern of YKII.39 in donor T cells transduced with TCRYKII.39.
Both TCRYKII.39-transduced T cells and YKII.39 demonstrated
the same efficient recognition of lymphoblasts in the absence
of significant recognition of fibroblasts, which represent
principal substrates of GVL and GVHD reactions, respectively.
Although the preservation of the fine peptide specificity after
R. Ivanov et al
400 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402

TCR-transfer was reported previously (Schaft et al, 2003;
Rubinstein et al, 2003), this report represents the first
description of the preserved differential pattern of recognition
of a ubiquitously expressed tumour-associated antigen after
transfer of a human TCR into peripheral blood lymphocytes.
In other studies, either antigens with a very restricted pattern
of expression were targeted (Clay et al, 1999; Morgan et al,
2003; Heemskerk et al, 2004) or a murine TCR was used (Liu
et al, 2000; Stanislawski et al, 2001).
Differentially recognized H–Y antigens represent a promis-
ing target for immunotherapy. Unlike many mHags that are
present in a small proportion of the patient population, H–Y
antigens are shared by all male patients. A universally attractive
H–Y epitope has to be presented on an HLA allele that is
frequent in the population. HLA-B*5201 is present in 34% of
Japanese and 27% of Chinese individuals (Imanishi et al,
1992), although it is relatively rare (2–6%) in Caucasians. For
the latter patients, differentially processed UTY epitopes
presented on more frequent HLA alleles need to be identified
to make the clinical application of UTY-targeting immuno-
therapies more feasible. The significant divergence of UTY and
its female counterpart UTX ensures an abundance of potential
T cell epitopes in UTY.
In conclusion, we have identified a new epitope of the UTY
antigen that is able to elicit a male lymphoblast-specific CTL
response because of its superior generation in proliferating B
and T cells. Transfer of the anti-UTY TCR into donor T cells
resulted in generation of large numbers of lymphoblast-specific
effector cells. To our knowledge, this is the first demonstration
that a differential pattern of recognition of a ubiquitously
expressed antigen is preserved in a heterogeneous population
of TCR-transduced cells. Our findings support the feasibility of
immunogenetherapy of lymphoproliferative diseases by gen-
etically engineered T cells redirected towards the patient’s
lymphoblasts. Use of T cells transduced with the UTY-specific
TCR may be especially beneficial in case of DLI for male
patients transplanted previously with haematopoietic stem
cells of a female sibling.
Acknowledgements
The authors are grateful to Dr Pierre G. Coulie and Dr Benoit
van den Eynde (Institute of Cellular Pathology, Universite de
Louvain, Brussels, Belgium) for the 293-EBNA-B7 cell line;
Dr Ton N.M. Schumacher (Netherlands Cancer Institute,
Amsterdam, the Netherlands) for the pMX vector; Dr Garry
Nolan (Stanford University, Palo Alto, CA, USA) for the
amphotrophic packaging cell line Phoenix; M. Otten and
F. Melman for excellent technical assistance.
References
Arden, B., Clark, S.P., Kabelitz, D. & Mak, T.W. (1995) Human
T-cell receptor variable gene segment families. Immunogenetics,
42, 455–500.
Barrett, J. (2003) Allogeneic stem cell transplantation for chronic
myeloid leukemia. Seminars in Hematology, 40, 59–71.
Blatch, G.L. & Lassle, M. (1999) The tetratricopeptide repeat: a
structural motif mediating protein–protein interactions. Bioessays,
21, 932–939.
Clay, T.M., Custer, M.C., Sachs, J., Hwu, P., Rosenberg, S.A. &
Nishimura, M.I. (1999) Efficient transfer of a tumor antigen-reactive
TCR to human peripheral blood lymphocytes confers anti-tumor
reactivity. Journal of Immunology, 163, 507–513.
Ebeling, S.B., Ivanov, R., Hol, S., Aarts, T.I., Hagenbeek, A., Verdonck,
L.F. & Petersen, E.J. (2003) HLA-DRB1*16-restricted recognition of
myeloid cells, including CD34+ CML progenitor cells. British Journal
of Haematology, 121, 721–729.
Goulmy, E., Schipper, R., Pool, J., Blokland, E., Falkenburg, J.H.,
Vossen, J., Gratwohl, A., Vogelsang, G.B., van Houwelingen, H.C. &
van Rood, J.J. (1996) Mismatches of minor histocompatibility
antigens between HLA-identical donors and recipients and the de-
velopment of graft-versus-host disease after bone marrow trans-
plantation. New England Journal of Medicine, 334, 281–285.
Heemskerk, M.H., Hoogeboom, M., Hagedoorn, R., Kester, M.G.,
Willemze, R. & Falkenburg, J.H. (2004) Reprogramming of virus-
specific T cells into leukemia-reactive T cells using T cell receptor
gene transfer. Journal of Experimental Medicine, 199, 885–894.
Imanishi, T., Akaza, T., Kimura, A., Tokunaga, K. & Gojobori, T.
(1992) Allele and Haplotype Frequencies for HLA and Complement
Loci in Various Ethnic Groups. Oxford University Press, Oxford, pp.
1065–1220.
Ivanov, R., Aarts, T., Hagenbeek, A., Hol, S. & Ebeling, S. (2005a)
B-cell expansion in the presence of the novel 293-CD40L-sCD40L
cell line allows the generation of large numbers of efficient xeno-
antigen-free APC. Cytotherapy, 7, 62–73.
Ivanov, R., Aarts, T., Hol, S., Doornenbal, A., Hagenbeek, A., Petersen,
E. & Ebeling, S. (2005b) Identification of a 40S ribosomal protein
S4-derived H-Y epitope able to elicit a lymphoblast-specific cytotoxic
T lymphocyte response. Clinical Cancer Research, 11, 1694–1703.
Jiang, Y.Z., Mavroudis, D.A., Dermime, S., Molldrem, J., Hensel, N.F.
& Barrett, A.J. (1997) Preferential usage of T cell receptor (TCR) V
beta by allogeneic T cells recognizing myeloid leukemia cells:
implications for separating graft-versus-leukemia effect from graft-
versus-host disease. Bone Marrow Transplantation, 19, 899–903.
Kolb, H.J., Schmid, C., Barrett, A.J. & Schendel, D.J. (2004) Graft-
versus-leukemia reactions in allogeneic chimeras. Blood, 103, 767–
776.
Liu, X., Peralta, E.A., Ellenhorn, J.D. & Diamond, D.J. (2000) Tar-
geting of human p53-overexpressing tumor cells by an HLA A*0201-
restricted murine T-cell receptor expressed in Jurkat T cells. Cancer
Research, 60, 693–701.
Michalek, J., Collins, R.H., Durrani, H.P., Vaclavkova, P., Ruff, L.E.,
Douek, D.C. & Vitetta, E.S. (2003) Definitive separation of graft-
versus-leukemia- and graft-versus-host-specific CD4+ T cells by
virtue of their receptor beta loci sequences. Proceedings of the
National Academy of Sciences of the U.S.A, 100, 1180–1184.
Morgan, R.A., Dudley, M.E., Yu, Y.Y., Zheng, Z., Robbins, P.F., The-
oret, M.R., Wunderlich, J.R., Hughes, M.S., Restifo, N.P. & Rosen-
berg, S.A. (2003) High efficiency TCR gene transfer into primary
human lymphocytes affords avid recognition of melanoma tumor
antigen glycoprotein 100 and does not alter the recognition of
autologous melanoma antigens. Journal of Immunology, 171, 3287–
3295.
Generation of GVL effector T cells through TCR-transfer
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402 401

Onishi, M., Kinoshita, S., Morikawa, Y., Shibuya, A., Phillips, J.,
Lanier, L.L., Gorman, D.M., Nolan, G.P., Miyajima, A. & Kitamura,
T. (1996) Applications of retrovirus-mediated expression cloning.
Experimental Hematology, 24, 324–329.
Pierce, R.A., Field, E.D., den Haan, J.M., Caldwell, J.A., White, F.M.,
Marto, J.A., Wang, W., Frost, L.M., Blokland, E., Reinhardus, C.,
Shabanowitz, J., Hunt, D.F., Goulmy, E. & Engelhard, V.H. (1999)
Cutting edge: the HLA-A*0101-restricted H-Y minor histocompat-
ibility antigen originates from DFFRY and contains a cysteinylated
cysteine residue as identified by a novel mass spectrometric tech-
nique. Journal of Immunology, 163, 6360–6364.
Rammensee, H.G., Friede, T. & Stevanoviic, S. (1995) MHC ligands
and peptide motifs: first listing. Immunogenetics, 41, 178–228.
Randolph, S.S., Gooley, T.A., Warren, E.H., Appelbaum, F.R. & Rid-
dell, S.R. (2004) Female donors contribute to a selective graft-ver-
sus-leukemia effect in male recipients of HLA-matched, related
hematopoietic stem cell transplants. Blood, 103, 347–352.
Rock, K.L., York, I.A. & Goldberg, A.L. (2004) Post-proteasomal
antigen processing for major histocompatibility complex class I
presentation. Nature Immunology, 5, 670–677.
Rubinstein, M.P., Kadima, A.N., Salem, M.L., Nguyen, C.L., Gilland-
ers, W.E., Nishimura, M.I. & Cole, D.J. (2003) Transfer of TCR
genes into mature T cells is accompanied by the maintenance of
parental T cell avidity. Journal of Immunology, 170, 1209–1217.
Schaft, N., Willemsen, R.A., de Vries, J., Lankiewicz, B., Essers, B.W.,
Gratama, J.W., Figdor, C.G., Bolhuis, R.L., Debets, R. & Adema, G.J.
(2003) Peptide fine specificity of anti-glycoprotein 100 CTL is pre-
served following transfer of engineered TCR alpha beta genes into
primary human T lymphocytes. Journal of Immunology, 170, 2186–
2194.
Schumacher, T.N. (2002) T-cell-receptor gene therapy. Nature Reviews
Immunology, 2, 512–519.
Scott, D.M., Ehrmann, I.E., Ellis, P.S., Chandler, P.R. & Simpson, E.
(1997) Why do some females reject males? The molecular basis for
male-specific graft rejection. Journal of Molecular Medicine, 75, 103–
114.
Skaletsky, H., Kuroda-Kawaguchi, T., Minx, P.J., Cordum, H.S., Hil-
lier, L., Brown, L.G., Repping, S., Pyntikova, T., Ali, J., Bieri, T.,
Chinwalla, A., Delehaunty, A., Delehaunty, K., Du, H., Fewell, G.,
Fulton, L., Fulton, R., Graves, T., Hou, S.F., Latrielle, P., Leonard, S.,
Mardis, E., Maupin, R., McPherson, J., Miner, T., Nash, W., Ngu-
yen, C., Ozersky, P., Pepin, K., Rock, S., Rohlfing, T., Scott, K.,
Schultz, B., Strong, C., Tin-Wollam, A., Yang, S.P., Waterston, R.H.,
Wilson, R.K., Rozen, S. & Page, D.C. (2003) The male-specific
region of the human Y chromosome is a mosaic of discrete sequence
classes. Nature, 423, 825–837.
Stanislawski, T., Voss, R.H., Lotz, C., Sadovnikova, E., Willemsen,
R.A., Kuball, J., Ruppert, T., Bolhuis, R.L., Melief, C.J., Huber, C.,
Stauss, H.J. & Theobald, M. (2001) Circumventing tolerance to a
human MDM2-derived tumor antigen by TCR gene transfer. Nature
Immunology, 2, 962–970.
Sureda, A. & Schmitz, N. (2003) Allogeneic stem cell transplantation
after reduced-intensity conditioning in lymphoid malignancies.
Annals of Hematology, 82, 1–13.
Vogt, M.H., Goulmy, E., Kloosterboer, F.M., Blokland, E., de Paus,
R.A., Willemze, R. & Falkenburg, J.H. (2000) UTY gene codes for an
HLA-B60-restricted human male-specific minor histocompatibility
antigen involved in stem cell graft rejection: characterization of the
critical polymorphic amino acid residues for T-cell recognition.
Blood, 96, 3126–3132.
Vogt, M.H., van den Muijsenberg, J.W., Goulmy, E., Spierings, E.,
Kluck, P., Kester, M.G., van Soest, R.A., Drijfhout, J.W., Willemze,
R. & Falkenburg, J.H. (2002) The DBY gene codes for an HLA-
DQ5-restricted human male-specific minor histocompatibility
antigen involved in graft-versus-host disease. Blood, 99, 3027–
3032.
Wang, W., Meadows, L.R., den Haan, J.M., Sherman, N.E., Chen, Y.,
Blokland, E., Shabanowitz, J., Agulnik, A.I., Hendrickson, R.C. &
Bishop, C.E. (1995) Human H-Y: a male-specific histocompat-
ibility antigen derived from the SMCY protein. Science, 269, 1588–
1590.
Warren, E.H., Gavin, M.A., Simpson, E., Chandler, P., Page, D.C.,
Disteche, C., Stankey, K.A., Greenberg, P.D. & Riddell, S.R. (2000)
The human UTY gene encodes a novel HLA-B8-restricted H-Y
antigen. Journal of Immunology, 164, 2807–2814.
Weijtens, M., van Spronsen, A., Hagenbeek, A., Braakman, E. &
Martens, A. (2002) Reduced graft-versus-host disease-inducing
capacity of T cells after activation, culturing, and magnetic cell
sorting selection in an allogeneic bone marrow transplantation
model in rats. Human Gene Therapy, 13, 187–198.
R. Ivanov et al
402 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 129, 392–402




















