
A twin-cysteine motif in the V2 region of gp120 is associated with SIV envelope trimer stabilization

Vaccine 22 (2004) 2974–2984
CCR5 targeted SIV vaccination strategy preventingor inhibiting SIV infection
W.M.J.M. Bogersa,1, L.A. Bergmeierb,1, H. Oostermeijera, P. ten Haafta,Y. Wangb, C.G. Kellyb, M. Singhc, J.L. Heeneya, T. Lehnerb,∗
a Department of Virology, Biomedical Research Primate Centre, Rijswijk, The Netherlandsb Mucosal Immunology Unit, Department of Oral Medicine and Pathology, Guy’s,
King’s and St. Thomas’ Hospital Medical and Dental Schools, London SE1 9RT, UKc Lionex Diagnostics and Therapeutics GmbH, Braunschweig, Germany
Received 19 September 2003; accepted 29 February 2004
Available online 17 June 2004
Abstract
eted theS 0 kDa heats ented SIVi nmodulateC te immunityt©
K
1
ediiataebt
T
SIVx.
za-tion
-
R5omniza-5ques-pair
iskn-ratedfromelicit
0d
Cell-surface CCR5 is a major coreceptor with CD4 glycoprotein, mediating cellular entry of CCR5 strains of HIV-1 or SIV. We targIV CCR5 coreceptor in a combined CCR5–SIV antigen immunization strategy. Rhesus macaques were immunized i.m. with the 7hock protein (HSP70) covalently linked to the CCR5 peptides, SIV gp120 and p27. Intravenous challenge with SIVmac 8980 prevnfection or decreased the viral load with the CCR5-SIV combined vaccine. CC chemokines and antibodies which block and dowCR5 were induced, as well as immune responses to the subunit SIV antigens. This novel vaccination strategy complements cogna
o SIV with innate immunity to the CCR5 coreceptor of SIV.2004 Elsevier Ltd. All rights reserved.
eywords: SIV; Vaccine; CCR5
. Introduction
CCR5 is a 7-transmembrane G-protein coupled moleculexpressed on T cells[1,2], macrophages and immature den-ritic cells[3]. CCR5 receptors play an important physiolog-
cal role in regulating the movement of mononuclear cells bynteracting with the CC chemokine ligands MIP-1�, MIP-1�nd RANTES (CCL3, 4, 5). However, the CCR5 corecep-
or has been subverted by M-tropic HIV and SIV to serves a major coreceptor to the CD4 glycoprotein receptor thatnables HIV or SIV entry[4–6]. Indeed, there are a num-er of HIV and SIV strains that use CCR5 for viral entry in
he absence of CD4[7,8]. The CCR5-dependent transmis-
∗ Corresponding author.el.: +44-207-188-3072; fax: +44-207-188-4375.
E-mail address:[email protected] (T. Lehner).1 Equal first authors.
sion suggests a novel mode of intervention in HIV orinfection by interfering with the HIV–CD4–CCR5 comple
Access of HIV to CCR5 can be inhibited by immunition in three ways: (a) by an increase in the concentraof the three CC chemokines[9–13], (b) by inducing antibodies to the extracellular domains of CCR5[14,15]and (c)by downmodulation of the cell-surface expression of CC[16]. Upregulation of CC chemokines and protection frSIV infection in macaques has been achieved by immution with SIV antigens[10,12]. Immunogenicity of CCRhas been demonstrated by xenoimmunization in maca[14,15], alloimmunization in humans[17], production of autoantibodies in persons with the homozygous 32 basedeletion of CCR5[18] and in sero-negative women at rfrom HIV infection [19]. Indeed, recently we have demostrated in non-human primates that a baculovirus-geneCCR5 construct, as well as synthetic peptides derivedthe sequences of three extracellular domains of CCR5
264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2004.02.050

W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984 2975
antibodies and T cell responses to CCR5, upregulate CCchemokines and downmodulate the cell-surface expressionof CCR5[20]. Furthermore, using 70 kDa heat shock pro-tein (HSP70) to upregulate the three CC chemokines[21],we have the means of interfering with CCR5-mediated HIVor SIV transmission.
The objectives of this investigation were to attempt toprevent SIV infection by (a) upregulating the three CCchemokines, (b) eliciting antibodies to CCR5 and (c) down-modulating the cell-surface expression of CCR5. HSP70was used as an adjuvant that induces the production of CCchemokines and peptides derived from the sequences of theextracellular domains of CCR5 were linked to HSP70 to elicitantibodies to CCR5. However, as complete absence of cell-surface expression of CCR5 cannot be accomplished, SIVgp120 and p27 subunit antigens were added in order to elicitresponses to viral antigens. The results suggest that SIV in-fection may be prevented or the viral load decreased by usinga combined strategy of innate immune CCR5 directed inhi-bition and cognate SIV directed immunity.
2. Materials and methods
2.1. Preparation of HSP70
c u-n
2
d inB asg np yl FineC n-te ver,i ithS
2
c ance)a
• er–ro–
• p–
• Pro–Thr
The N terminal first and second loop synthetic pep-tides were either covalently linked to HSP70 with glu-taraldehyde or loaded on to HSP70 in concentrations ofpeptides 10× in excess of that of HSP70. The final con-centration of each constituent in the three macaques im-munized first time by the i.m. route was as follows: HSPcovalently linked to the three peptides was 12.5�g each,and those of HSP70 loaded peptides was 25�g for the Nterminal, 35�g for the first loop and 60�g for the secondloop peptide. In the second and third i.m. immunizations,the concentrations of each peptide—HSP70 preparation wasdoubled. However, in each of the three immunizations thevaccine contained a constant amount of HSP70 covalentlylinked with glutaraldehyde to SIV gp120 (100�g) and p27(100�g).
2.4. Immunization schedule
Four groups of a total of 17 macaques were studied (Fig. 1).Group I (n = 4) was injected 100�g of HSP70 i.m. intothe arm (deltoid muscle) and 100�g into the leg (glutealmuscle) three times at 6- and 8-week intervals. Group IImacaques (n = 4) were immunized similarly, except thatthe vaccine consisted of 200�g of HSP70 linked by glu-taraldehyde to 200�g of SIV gp120 and 200�g SIV p27.G wog P70l s; Nt loop( therh ilstt l wasi es(N pro-l itedh im-m zedm
F d 17m
Mycobacterium tuberculosisHSP70 was prepared inE.oli as described earlier[22]. Four macaques were immized i.m. (3×) with 200�g of HSP70 (Group I).
.2. Preparation of SIV antigens
Recombinant SIVmac 251 gp120 was expresseaculovirus-infected cells[23] and recombinant SIV p27 wenerated in pGEX-3X as a glutathioneS-transferase fusiorotein[24]. With both preparations 100�g was covalentl
inked to HSP70 by 0.0025% glutaraldehyde (Sigmahemicals Ltd.) and 200�g was mixed with equal conce
ration of HSP70; thus, a total of 400�g of HSP70 and 200�gach of SIV gp120 and p27 were used in Group II. Howe
n Group III 100�g of the HSP70 was linked or mixed wIV gp120, p27 and the three CCR5 peptides.
.3. CCR5 peptides
The three peptides and a random unrelated 20er were pur-hased from Neosystem Laboratories (Strasbourg, Frnd the sequences of the 20ers are shown below.
N-terminal (aa 1–20): Met–Asp–Tyr–Gln–Val–Ser–SPro–ILe–Tyr–Asp–ILe–Asp–Tyr–Tyr–Thr–Ser–Glu–PCysFirst loop (aa 89–102): His–Tyr–Ala–Ala–Ala–Gln–TrAsp–Phe–Gly–Asn–Thr–Met–Cys–GlnSecond loop (aa 178–197): Cys–Ser–Ser–His–Phe–Tyr–Ser–Gln–Tyr–Gln–Phe–Trp–Lys–Asn–Phe–Gln––Leu–Lys
roup III animals (n = 4) were immunized as the other troups, except that they were given in addition to HS
inked with SIV gp120 and p27, the three CCR5 peptideerminal (aa 1–20), first loop (aa 89–102) and secondaa 178–197), one half of which was mixed and the oalf covalently linked to HSP70 using glutaraldehyde. Wh
hree macaques were immunized i.m., the fourth animammunized SC in the proximity of the iliac lymph nod3×). The macaque immunized by the SC route (Table 1,o 31) showed decreased viral load, increased T cell
iferation to HSP70 and the CCR5 peptides, and elicigher concentration of CC chemokines than in the i.m.unized animals. Group IV consisted of five unimmuniacaques.
Series A - IM Immunization
Week 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
Group I - HSP70 (n=4)
Group II - HSP70 + SIVgp120 + SIVp27 (n=4)
Group III - HSP70 + SIVgp120 + SIVp27 + 3 CCR5 peptides (n=4)
Group IV - Nil (n=5)
Autopsy
IV Challenge SIV 8980
Viral LoadImmunization
ig. 1. Immunization schedule of i.m. immunized and i.v. challengeacaques with SIV 8980.

2976 W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984
Table 1Comparative immunological analysis of the two macaques protected or with decreased virus load and two infected macaques from Group III (No. 5971, 31,4592 and 4841), at the time of challenge (week 20)
Function (A) Protected or virus load <104 (B) Infected-virus load >104
5971 31 4592 4841
1 Virus load 0 1.1× 103 1.3× 104 4.7× 106
2 Antibody titres to HSP70 3200 400 1600 4003 T cell proliferation to HSP70(SI) 6.0 11.7 2.6 1.34 Antibody titre to SIVp27 400 100 400 1005 T cell proliferation to p27 (SI) 10.7 1.4 3.6 1.26 Antibody titre to three CCR5 peptidesa 1200 2000 800 6007 T cell proliferation to three CCR5 peptides (SI) 3.3 12.7 3.9 3.08 CD8-SF at 1:5 dilution (%) 62 80 21 429 CD8 cell generated RANTES (pg/ml) 1950 2823 612 1042
10 CD8 cell generated MIP-1� (pg/ml) 654 1850 610 68411 CD8 cell generated MIP-1� (pg/ml) 3280 2089 3570 147812 CD8 cell generated three chemokines 5884 6762 4394 2720
a Combined titres (or SI) to the N terminal, first and second loop peptides.
2.5. Serum IgG and IgA antibodies
Specific serum antibodies to SIV gp120, p27, CCR5and its extracellular peptides were assayed by ELISA asdescribed previously[10,20]. Briefly plates were coatedwith a pre-determined optimal concentration of antigen(1�g/ml), CCR5 peptides (15�g/ml) and with a random20 aa peptide (R20) as a control, and they were incubatedwith doubling dilutions of test samples. Bound antibodywas detected by incubation with rabbit IgG anti-monkeyIgA (8 �g/ml) (Nordic Immunological Laboratories, Tilburg,The Netherlands) or IgG (2�g/ml; Sigma), followed byaffinity-purified goat anti-rabbit IgG-alkaline phosphataseconjugate (Sigma). The IgG and IgA antibody titres arepresented as reciprocals before and after each immuniza-tion.
2.6. T cell proliferative responses to SIV antigens andthe CCR5 peptides
T cell cultures were set up by separating mononuclearcells from defibrinated blood by Lymphoprep (Nycomed,Oslo) density gradient centrifugation before and after immu-nization from all macaques[25]. The cells were cultured for72 h without and with 1 and 10�g/ml of SIV gp120, p27,tc nd-b PMI1 dw ra dices( asc tion,f ofa ndc asei notp
2.7. Generation and assay of CD8-SF and CCchemokines
Peripheral blood mononuclear cells (PBMC) were sepa-rated and CD8+ cells were enriched by depletion of CD4+cells, B cells and monocytes by panning with CD4 (OKT4hybridoma culture supernatant) and antibodies to humanimmunoglobulin (Serotec, Oxford, UK). The enriched CD8+cells were then cultured with 10�g/ml of phytohaemag-glutinin (PHA) for 3 days. Preparation of the CD8 culturesupernatant was then carried out according to the methoddescribed previously[10,26]. PHA-stimulated CD8+ cellswere cultured at a concentration of 3× 106 cells/ml inRPMI-1640 containing 10% fetal calf serum (FCS) and10% human IL-2 preparation (TLF; Biotest, Solihull, UK).After 2 days of incubation (at 37◦C, in 5% CO2), the culturesupernatant was collected, and the cells were reconstitutedwith fresh medium; this was repeated up to three times. Ateach passage, the cellular density was adjusted to 3× 106
cells/ml. The collected supernatants were filtered througha 0.45-�m filter and stored at−70◦C for CD8-SF and theCC-chemokine assay. SIVmac acutely infected CD4+ cellswere cultured in the presence of CD8-SF at 1:2 and 1:5 dilu-tions in 96-well plates. The CD8-SF were replenished every2–3 days and on day 7 the culture supernatant was used tod theQa werea ifice ing-d inesw leaseo l.
2
susm nd-
hree extracellular peptides of CCR5 (10 and 40�g/ml), aontrol peptide (R20), and Concanvalin A in 96-well rouottomed plates (Costar, Cambridge, MA), containing R640 (Gibco), as described before[25]. The cells were pulseith 20�Ci/ml (25�l per well) 3H-methyl thymidine fobout 20 h. The results were expressed as stimulation inSI, ratio of counts with and without antigen), as wellounts per minute (cpm) before and after each immunizaor cultures stimulated with the optimum concentrationntigen. All cultures yielded high stimulation indices aounts with Concanavalin A, and no significant incren counts was seen with the control peptide (dataresented).
etermine the reverse transciption (RT) activity by usinguant RT kit (Amersham, UK). RANTES, MIP-1�, MIP-1�nd macrophage chemoattractant protein-1 (MCP-1)ssayed in the CD8+ cell culture supernatant using a specnzyme immunoassay (R & D Systems Europe Ltd., Abon, Oxfordshire, UK). Plasma levels of CC chemokere not assayed, as they yield variable results due to ref chemokines from platelets, which is difficult to contro
.8. Viral infection and kinetics
The challenge virus SIVmac 8980 was grown in rheonkey PBMC[27] and analyzed for CCR5 coreceptor bi

W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984 2977
ing using the “Ghost system”, as described previously[28].The MID50/ml of the stock was determined to be 2.5×104. All animals were inoculated intravenously (i.v.) with 30MID50 of the SIV 8980 stock. At regular time points after in-oculation blood sampling was performed to measure plasmaviral loads. For the detection of RNA plasma levels a highlysensitive quantitive competitive RNA-PCR (QC-RNA-PCR)was used to estimate the virus load[29]. Briefly, RNA wasextracted from 200�l EDTA plasma using guanidine isoth-iocyanate mediated lysis, followed by propan-2-ol precipita-tion. A known amount of internal standard RNA (IS-RNA)was added before the RNA extraction and was copurifiedto monitor the efficiency of the purification. The RNA wasreverse transcribed and amplified in a single reaction proto-col using RTth DNA polymerase and biotinylated primers.The IS-RNA is coamplified to monitor the amplification ef-ficiency. The amplified fragments were denatured and hy-bridized to an immobilized capture probe in a microwell plate.The amplified fragments were detected by an avidin-enzymeconjugate mediated colorimetric reaction. The amplified in-ternal standard is hybridized to a different capture probe inseparate microwells.
2.9. Evaluation of the proportion and cell-surfaceexpression of CCR5
owc lect mlp ick-il oft 2D7( con-j na bya en-L ingt 2%( plesw aliB-R ick-i n aC oft-w erea
2
es-t uner determ eacho airedS
3. Results
3.1. Serum antibodies
3.1.1. HSP70, SIV gp120 and p27All 12 macaques were immunized with HSP70 and elicited
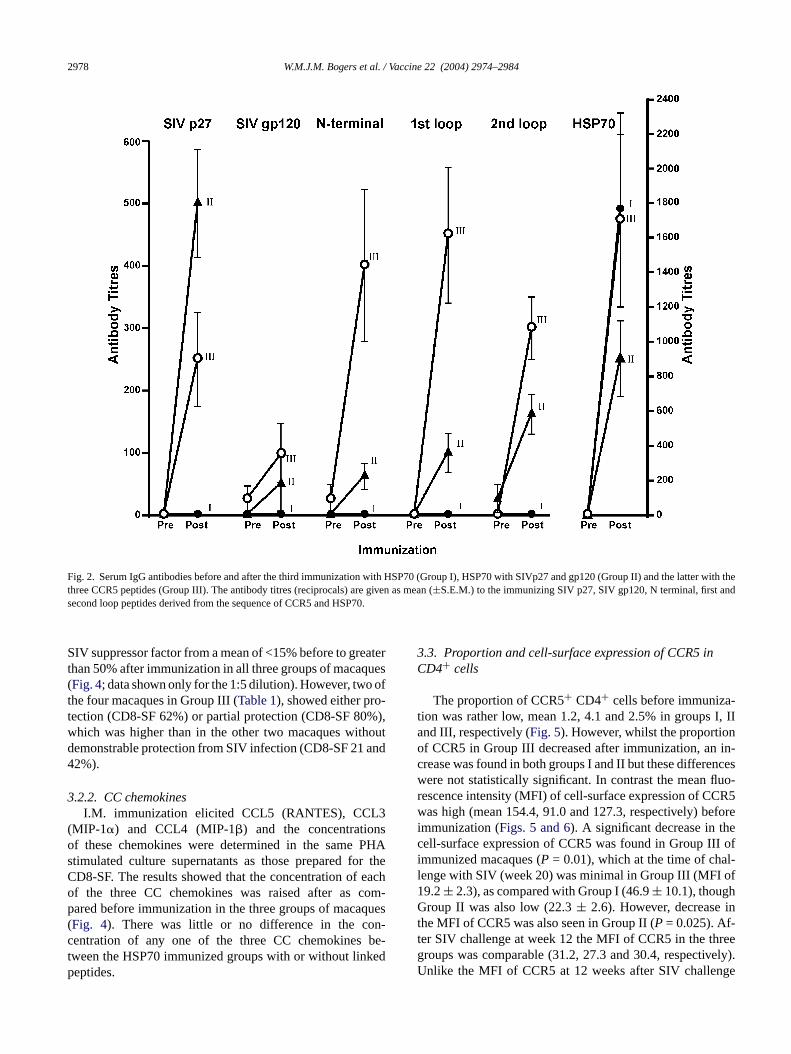
IgG antibodies to this antigen (Fig. 2), with an increase inantibody titres which was <100 before immunization. Themean (±S.E.M.) antibody titre after the third immuniza-tion, that is just before challenge in the HSP70 alone im-munized macaques (Group I) was higher (1760± 561) thanthat in Group II animals, immunized with covalently linkedSIV antigens (900± 28), but not that in Group III, withHSP70 linked SIV antigens and CCR5 peptides (1700±497); these results were not statistically significant. Antibod-ies to SIV p27 were induced only in Group II (500± 87)and Group III (250± 75) macaques and these were signifi-cantly higher than those in Group I animals (P = 0.02). Verylow antibody titres to SIV gp120 were found in Groups IIand III. Sera from Group III animals immunized with theCCR5 peptides yielded significantly raised antibody titres(P = 0.02) to the N terminal (400± 122), first loop (450± 109) and second loop (300± 50) peptides (Fig. 2). Sur-prisingly, low antibody titres to the three CCR5 peptideswere detected in Group II macaques immunized with HSP70c nti-b t pre-s
3
e-s f 5.0(sf ti-g quesi ro-l -n d byt ns.S upsI -nS sei mentw nic-i est y inG -nF
3hed
C in
Phenotypic analysis of cells was performed by flytometry with fresh whole EDTA blood. For doubolour staining, 100�l blood was incubated with 10�l ofhe appropriate monoclonal antibody, mixed in a 5.0olystyrene, round bottom tube (Falcon 2058, Becton D
nson, Lincoln Park, NY, USA), at 4◦C for 30 min, fol-owed by washing with FACS-buffer. For detectionhe CCR5 coreceptor we used monoclonal antibodyPharmingen, Woerden, The Netherlands). Appropriateugated antibody-mixtures (10�l) were incubated for 30 mit 4◦C. After staining, red blood cells were lyseddding 2 ml FACS-lysis buffer (Becton Dickinson, Etteur, NL) for 20 min at room temperature. After wash
he cells, they were fixed with 2 ml freshly preparedw/v) paraformaldehyde overnight. The sequential samere standardized by means of fluorescent beads (CITE 3 beads: FITC, PE or PerCP labelled; Becton D
nson, Eten Leur, NL). Flow cytometry was performed ooulter Epics XL-MCL cytometer using the Cell Quest sare (Becton Dickinson) and a minimum of 5000 events wnalyzed.
.10. Statistical analysis
The non-parametric Mann–Whitney test was used toablish if there are any significant differences in the immesponses between the three groups of macaques. Toine if the decrease in MFI between 0 and 20 weeks inf the three groups of macaques was significant, the ptudent’st-test was used.
-
ovalently linked to SIV gp120 and p27. Serum IgA aodies showed no significant changes and data are noented.
.2. T cell proliferative responses
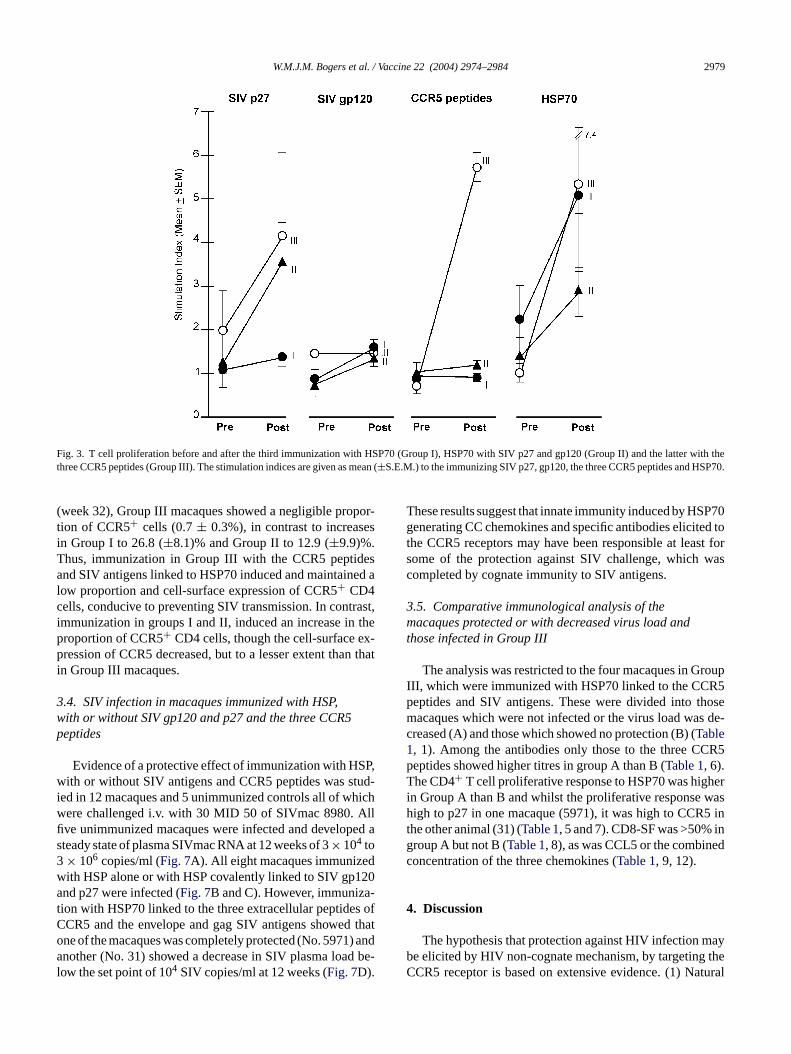
The i.m. immunization elicited T cell proliferative rponses to HSP70 with SI < 2 before and mean SI o±3.0) after immunization in Group I (Fig. 3). Similar re-ults were recorded in Group III, with SI of 1.0 (±0.2) be-ore to 5.4 (±2.0) after immunization with HSP70, SIV anens and CCR5 peptides. However, in Group II maca
mmunized with HSP70 linked to SIV antigens, the piferative responses were lower (2.9± 0.6) after immuization. This may be due to the HSP70 being altere
reatment with glutaraldehyde to link it to the SIV antigeIV p27 elicited significant responses (SI > 2) in gro
I (3.6 ± 0.9) and III macaques (4.2± 1.9) after immuization with HSP70 covalently linked to SIV p27 (Fig. 3).IV gp120 largely failed to elicit a significant respon
n groups II and III macaques, possibly because treatith glutaraldehyde may have affected the immunoge
ty of SIV gp120. Significant T cell proliferative responso the three CCR5 derived peptides were elicited onlroup III macaques (Table 1); the SI (5.7± 2.3) was sigificantly higher than in Group I or II macaques (P = 0.03;ig. 3).
.2.1. CD8-SFThe culture supernatant (at 1:2 or 1:5 dilution) of enric
D8+ T cells stimulated with PHA yielded an increase
2978 W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984
Fig. 2. Serum IgG antibodies before and after the third immunization with HSP70 (Group I), HSP70 with SIVp27 and gp120 (Group II) and the latter with thethree CCR5 peptides (Group III). The antibody titres (reciprocals) are given as mean (±S.E.M.) to the immunizing SIV p27, SIV gp120, N terminal, first andsecond loop peptides derived from the sequence of CCR5 and HSP70.
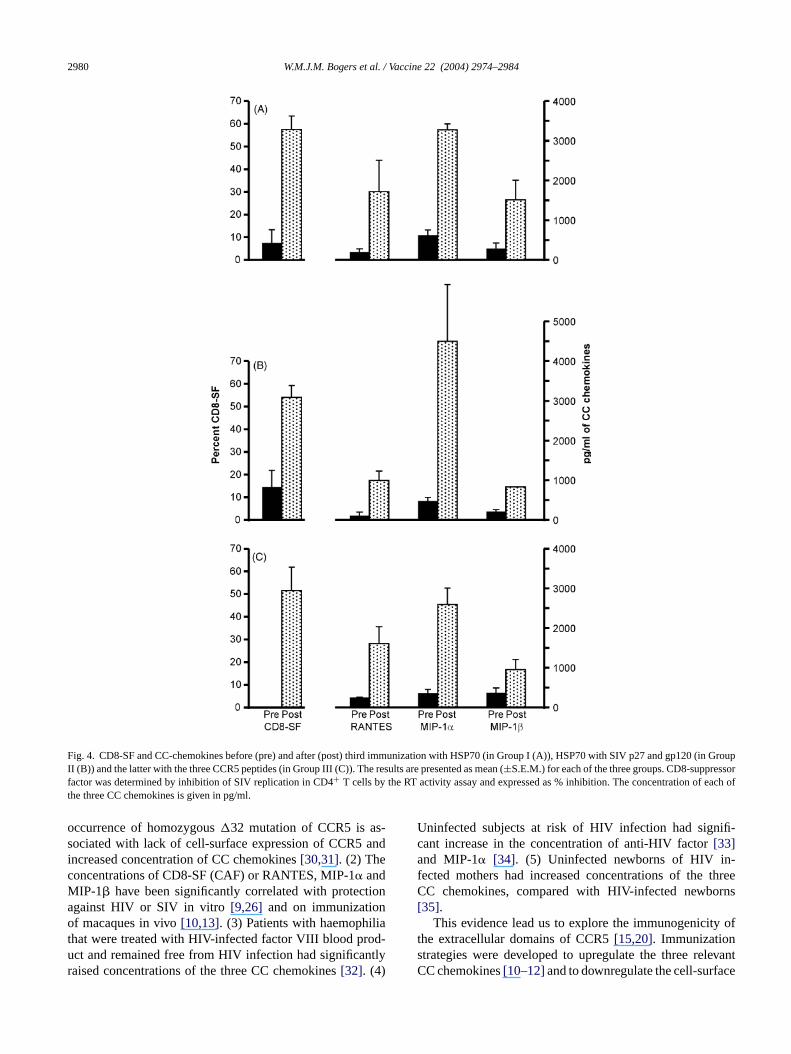
SIV suppressor factor from a mean of <15% before to greaterthan 50% after immunization in all three groups of macaques(Fig. 4; data shown only for the 1:5 dilution). However, two ofthe four macaques in Group III (Table 1), showed either pro-tection (CD8-SF 62%) or partial protection (CD8-SF 80%),which was higher than in the other two macaques withoutdemonstrable protection from SIV infection (CD8-SF 21 and42%).
3.2.2. CC chemokinesI.M. immunization elicited CCL5 (RANTES), CCL3
(MIP-1�) and CCL4 (MIP-1�) and the concentrationsof these chemokines were determined in the same PHAstimulated culture supernatants as those prepared for theCD8-SF. The results showed that the concentration of eachof the three CC chemokines was raised after as com-pared before immunization in the three groups of macaques(Fig. 4). There was little or no difference in the con-centration of any one of the three CC chemokines be-tween the HSP70 immunized groups with or without linkedpeptides.
3.3. Proportion and cell-surface expression of CCR5 inCD4+ cells
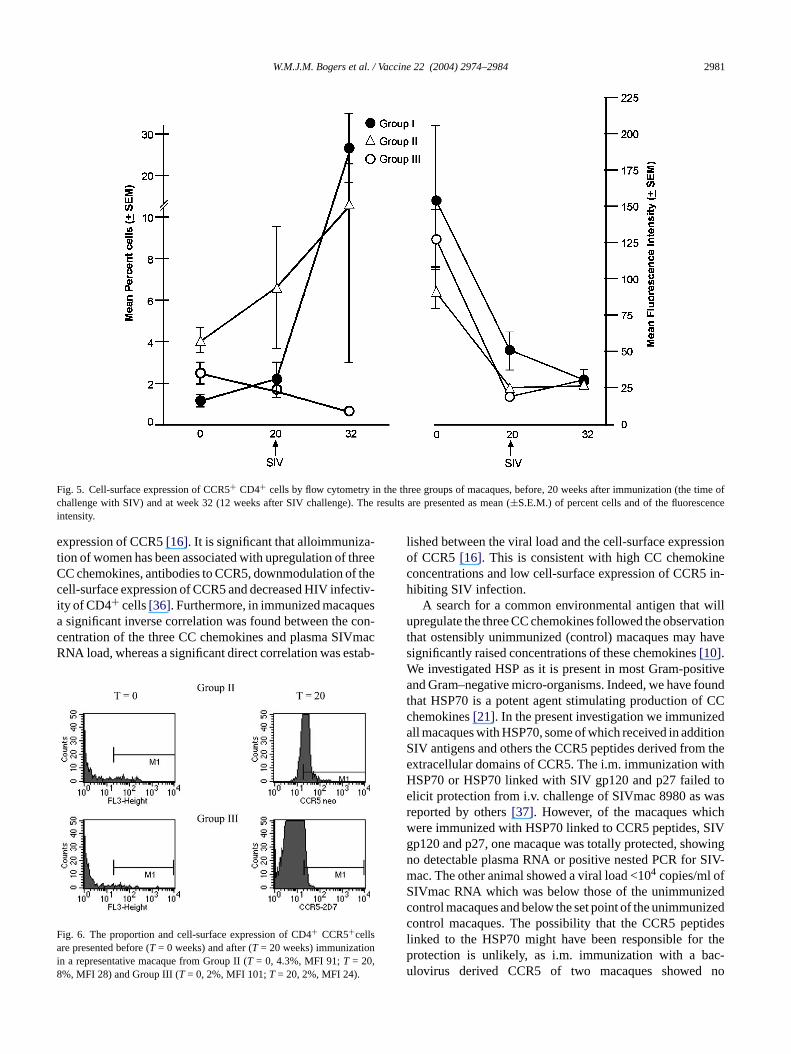
The proportion of CCR5+ CD4+ cells before immuniza-tion was rather low, mean 1.2, 4.1 and 2.5% in groups I, IIand III, respectively (Fig. 5). However, whilst the proportionof CCR5 in Group III decreased after immunization, an in-crease was found in both groups I and II but these differenceswere not statistically significant. In contrast the mean fluo-rescence intensity (MFI) of cell-surface expression of CCR5was high (mean 154.4, 91.0 and 127.3, respectively) beforeimmunization (Figs. 5 and 6). A significant decrease in thecell-surface expression of CCR5 was found in Group III ofimmunized macaques (P = 0.01), which at the time of chal-lenge with SIV (week 20) was minimal in Group III (MFI of19.2± 2.3), as compared with Group I (46.9± 10.1), thoughGroup II was also low (22.3± 2.6). However, decrease inthe MFI of CCR5 was also seen in Group II (P = 0.025). Af-ter SIV challenge at week 12 the MFI of CCR5 in the threegroups was comparable (31.2, 27.3 and 30.4, respectively).Unlike the MFI of CCR5 at 12 weeks after SIV challenge
W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984 2979
Fig. 3. T cell proliferation before and after the third immunization with HSP70 (Group I), HSP70 with SIV p27 and gp120 (Group II) and the latter with thethree CCR5 peptides (Group III). The stimulation indices are given as mean (±S.E.M.) to the immunizing SIV p27, gp120, the three CCR5 peptides and HSP70.
(week 32), Group III macaques showed a negligible propor-tion of CCR5+ cells (0.7± 0.3%), in contrast to increasesin Group I to 26.8 (±8.1)% and Group II to 12.9 (±9.9)%.Thus, immunization in Group III with the CCR5 peptidesand SIV antigens linked to HSP70 induced and maintained alow proportion and cell-surface expression of CCR5+ CD4cells, conducive to preventing SIV transmission. In contrast,immunization in groups I and II, induced an increase in theproportion of CCR5+ CD4 cells, though the cell-surface ex-pression of CCR5 decreased, but to a lesser extent than thatin Group III macaques.
3.4. SIV infection in macaques immunized with HSP,with or without SIV gp120 and p27 and the three CCR5peptides
Evidence of a protective effect of immunization with HSP,with or without SIV antigens and CCR5 peptides was stud-ied in 12 macaques and 5 unimmunized controls all of whichwere challenged i.v. with 30 MID 50 of SIVmac 8980. Allfive unimmunized macaques were infected and developed asteady state of plasma SIVmac RNA at 12 weeks of 3× 104 to3 × 106 copies/ml (Fig. 7A). All eight macaques immunizedwith HSP alone or with HSP covalently linked to SIV gp120and p27 were infected (Fig. 7B and C). However, immuniza-t s ofC thato ) anda d be-l
These results suggest that innate immunity induced by HSP70generating CC chemokines and specific antibodies elicited tothe CCR5 receptors may have been responsible at least forsome of the protection against SIV challenge, which wascompleted by cognate immunity to SIV antigens.
3.5. Comparative immunological analysis of themacaques protected or with decreased virus load andthose infected in Group III
The analysis was restricted to the four macaques in GroupIII, which were immunized with HSP70 linked to the CCR5peptides and SIV antigens. These were divided into thosemacaques which were not infected or the virus load was de-creased (A) and those which showed no protection (B) (Table1, 1). Among the antibodies only those to the three CCR5peptides showed higher titres in group A than B (Table 1, 6).The CD4+ T cell proliferative response to HSP70 was higherin Group A than B and whilst the proliferative response washigh to p27 in one macaque (5971), it was high to CCR5 inthe other animal (31) (Table 1, 5 and 7). CD8-SF was >50% ingroup A but not B (Table 1, 8), as was CCL5 or the combinedconcentration of the three chemokines (Table 1, 9, 12).
4
ayb theC tural
ion with HSP70 linked to the three extracellular peptideCR5 and the envelope and gag SIV antigens showedne of the macaques was completely protected (No. 5971nother (No. 31) showed a decrease in SIV plasma loa
ow the set point of 104 SIV copies/ml at 12 weeks (Fig. 7D).
. Discussion
The hypothesis that protection against HIV infection me elicited by HIV non-cognate mechanism, by targetingCR5 receptor is based on extensive evidence. (1) Na
2980 W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984
Fig. 4. CD8-SF and CC-chemokines before (pre) and after (post) third immunization with HSP70 (in Group I (A)), HSP70 with SIV p27 and gp120 (in GroupII (B)) and the latter with the three CCR5 peptides (in Group III (C)). The results are presented as mean (±S.E.M.) for each of the three groups. CD8-suppressorfactor was determined by inhibition of SIV replication in CD4+ T cells by the RT activity assay and expressed as % inhibition. The concentration of each ofthe three CC chemokines is given in pg/ml.
occurrence of homozygous�32 mutation of CCR5 is as-sociated with lack of cell-surface expression of CCR5 andincreased concentration of CC chemokines[30,31]. (2) Theconcentrations of CD8-SF (CAF) or RANTES, MIP-1� andMIP-1� have been significantly correlated with protectionagainst HIV or SIV in vitro[9,26] and on immunizationof macaques in vivo[10,13]. (3) Patients with haemophiliathat were treated with HIV-infected factor VIII blood prod-uct and remained free from HIV infection had significantlyraised concentrations of the three CC chemokines[32]. (4)
Uninfected subjects at risk of HIV infection had signifi-cant increase in the concentration of anti-HIV factor[33]and MIP-1� [34]. (5) Uninfected newborns of HIV in-fected mothers had increased concentrations of the threeCC chemokines, compared with HIV-infected newborns[35].
This evidence lead us to explore the immunogenicity ofthe extracellular domains of CCR5[15,20]. Immunizationstrategies were developed to upregulate the three relevantCC chemokines[10–12]and to downregulate the cell-surface
W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984 2981
Fig. 5. Cell-surface expression of CCR5+ CD4+ cells by flow cytometry in the three groups of macaques, before, 20 weeks after immunization (the time ofchallenge with SIV) and at week 32 (12 weeks after SIV challenge). The results are presented as mean (±S.E.M.) of percent cells and of the fluorescenceintensity.
expression of CCR5[16]. It is significant that alloimmuniza-tion of women has been associated with upregulation of threeCC chemokines, antibodies to CCR5, downmodulation of thecell-surface expression of CCR5 and decreased HIV infectiv-ity of CD4+ cells[36]. Furthermore, in immunized macaquesa significant inverse correlation was found between the con-centration of the three CC chemokines and plasma SIVmacRNA load, whereas a significant direct correlation was estab-
Fig. 6. The proportion and cell-surface expression of CD4+ CCR5+cellsa ni8
lished between the viral load and the cell-surface expressionof CCR5[16]. This is consistent with high CC chemokineconcentrations and low cell-surface expression of CCR5 in-hibiting SIV infection.
A search for a common environmental antigen that willupregulate the three CC chemokines followed the observationthat ostensibly unimmunized (control) macaques may havesignificantly raised concentrations of these chemokines[10].We investigated HSP as it is present in most Gram-positiveand Gram–negative micro-organisms. Indeed, we have foundthat HSP70 is a potent agent stimulating production of CCchemokines[21]. In the present investigation we immunizedall macaques with HSP70, some of which received in additionSIV antigens and others the CCR5 peptides derived from theextracellular domains of CCR5. The i.m. immunization withHSP70 or HSP70 linked with SIV gp120 and p27 failed toelicit protection from i.v. challenge of SIVmac 8980 as wasreported by others[37]. However, of the macaques whichwere immunized with HSP70 linked to CCR5 peptides, SIVgp120 and p27, one macaque was totally protected, showingno detectable plasma RNA or positive nested PCR for SIV-mac. The other animal showed a viral load <104 copies/ml ofSIVmac RNA which was below those of the unimmunizedcontrol macaques and below the set point of the unimmunizedcontrol macaques. The possibility that the CCR5 peptidesl thep ac-u no
re presented before (T = 0 weeks) and after (T = 20 weeks) immunization a representative macaque from Group II (T = 0, 4.3%, MFI 91;T = 20,%, MFI 28) and Group III (T = 0, 2%, MFI 101;T = 20, 2%, MFI 24).
inked to the HSP70 might have been responsible forrotection is unlikely, as i.m. immunization with a blovirus derived CCR5 of two macaques showed

2982 W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984
Unimmunized
Immunized
Pla
sma
Vir
alL
oad
(RN
AE
q/m
l)
Immunized10
10
10
10
10
10
10
8
7
6
4
3
2
1
5
10
Immunized
2 4 6 8 10 120
10
10
10
10
10
10
10
8
7
6
4
3
2
1
5
10
10
10
10
10
10
10
10
8
7
6
4
3
2
1
5
10
2 4 6 8 10 120
10
10
10
10
10
10
10
8
7
6
4
3
2
1
5
10
Weeks Post-Challenge
2 4 6 8 10 120Weeks Post-Challenge
2 4 6 8 10 120
(A)
(B)
(C)
(D)
Fig. 7. Plasma virus load of SIVmac 8980 was determined at 0, 2, 4, 6, 8, 10 and 12 weeks in 17 macaques by RNA-PCR, after i.v. inoculation with 30 MID 50of the SIV 8980 stock. The results are presented as RNA Eq/ml in the 5 non-immunized (Group A) and immunized macaques with HSP70 (Group B), HSP70with SIV gp120 and p27 (Group C) and the latter with the three CCR5 peptides (Group D). The protected macaque and one with decreased virus load are bothin Group D. The set point of 104 SIV copies/ml is shown by the horizontal line.
protection when challenged i.v. with SIVmac 8980 (unpub-lished), as was also reported in immunization with DNA ofCCR5[38].
The SIV non-cognate mechanism of protection againstSIV infection common to the totally or partially protectedmacaque (Table 1) is an increase in the concentration of CD8-SF (62 and 80%), compared with those found in the infectedmacaques (21 and 42%). This is an in vitro biological assay ofinhibition of SIV infection generated in culture supernatantsof stimulated CD8-enriched cells[9,26]. The same super-natant yielded increased concentrations of RANTES (1950and 2823 pg/ml) in the fully or partially protected, comparedwith the infected macaques (612 and 878 pg/ml). The non-cognate immunity might be complemented by raised anti-bodies to the N terminal, first and second loop of CCR5 inpreventing SIV or HIV transmission by blocking and internal-ization of CCR5 receptors[39,40]. Indeed, we have demon-strated that CCR5 antibodies are complementary to the CCchemokines in inhibiting CCR5-mediated SIV or HIV in-fection[20]. Thus, HIV transmission can be suppressed withlow concentrations of CC chemokines that are enhanced withCCR5 antibodies, or conversely with low titres of CCR5 an-tibodies that can be enhanced by CC chemokines[20].
Cognate SIV-specific T cell proliferation to SIVp27 wasfound predominantly in the fully protected macaque, sug-
gesting that CD4+ T cell helper function is required. HSP70-specific T cell proliferation was found in both the fully andpartially protected macaques, in contrast to the infected an-imals and this is consistent with HSP70 stimulating produc-tion of CCL5[21] which might be boosted by SIV (or HIV)infection as the virion expresses HSP70[41]. It is of con-siderable interest to note that HSP70 has been used here invivo as a dual functioning agent, acting as an immunologicaladjuvant and generating CC chemokines[21]. The resultsof this investigation suggests that SIV non-cognate immu-nity which blocks and downmodulates cell-surface expres-sion of CCR5, complemented with SIV specific immunitymay prevent SIV infection or decrease the viral load. Thisis consistent with vaginal mucosal or paramucosal immu-nization with HSP70 linked to CCR5 peptides, SIV gp120and p27, showing significant clearance of the virus to unde-tectable levels when challenged vaginally with SHIV 89.6P[42].
Acknowledgements
This work was supported by grants from the EuropeanCommunity Biomed Grant (No. BMH4 CT97-2345) and theGuy’s and St. Thomas’ Charitable Foundation.

W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984 2983
References
[1] Ostrowski MA, Justement SJ, Catanzaro A, Hallahan CA, EhlerLA, Mizell SB, et al. Expression of chemokine receptors CXCR4and CCR5 in HIV-1-infected and uninfected individuals. J Immunol1998;161:3195–201.
[2] Sallusto F, Lenig D, Mackay CR, Lanzavecchia A. Flexible programsof chemokine receptor expression on human polarized T helper 1 and2 lymphocytes. J Exp Med 1998;187:875–83.
[3] Sallusto F, Schaerli P, Loetscher P, Schaniel C, Lenig D, MackayCR, et al. Rapid and corordinated switch in chemokine receptor ex-pression during dendritic cell maturation. Eur J Immunol 1998;28:2760–9.
[4] Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M, et al.Identification of a major co-receptor for primary isolates of HIV-1.Nature 1996;381:661–6.
[5] Dragic T, Litwin V, Allaway GP, Martin SR, Huang Y, NagashimaKA, et al. HIV-1 entry into CD4+ cells is mediated by the chemokinereceptor CC-CKR-5. Nature 1996;381:667–73.
[6] Chen Z, Zhou P, Ho DD, Landau NR, Marx PA. Genetically divergentstrains of simian immunodeficiency virus use CCR5 as a coreceptorfor entry. J Virol 1997;71:2705–14.
[7] Endreas MJ, Clapham PR, Marsh M, Ahuja M, Turner JD, McK-night A, et al. CD4-independent infection by HIV-2 is mediated byfusin/CXCR4. Cell 1996;87:745–56.
[8] Hoffman TL, LaBranche CC, Zhang W, Gabriella C, Robinson J,Chaiken I, et al. Stable exposure of the coreceptor-binding site in aCD4-independent HIV-1 envelope protein. Proc Natl Acad Sci USA1999;96:6359–64.
[9] Cocchi F, DeVico AL, Garzino-Demo A, Arya SK, Gallo RC, Lusso-e
L,ph
ne in
A,-im-
USA
iuliti-ated
ssonenu-Gen
W,ed
inNIH;
etptorues.
t al.co-non-
r T.andIV-1
[18] Ditzel HJ, Rosenkilde MM, Garred P, Wang M, Koefoed K, PedersenC, et al. The CCR5 receptor acts as an alloantigen in CCR5�32 ho-mozygous individuals: Identification of chemokine- and HIV-1 block-ing human antibodies. Proc Natl Acad Sci USA 1998;95:5241–345.
[19] Lopalco L, Barassi C, Pastori C, Longhi R, Burastero SE, TambussiG, et al. CCR5-reactive antibodies in seronegative partners of HIV-seropositive individuals downmodulate surface CCR5 in vivo andneutralize the infectivity of R5 strains of HIV-1 in vitro. J Immunol2000;164:3426–33.
[20] Lehner T, Doyle C, Wang Y, Babahmady K, Tao L, Bergmeier L, et al.Immunogenicity of the extracellular domains of the CCR5 chemokinereceptor and the effects on SIV or HIV infectivity in vitro. J Immunol2001;166:7446–55.
[21] Lehner T, Mitchell E, Bergmeier L, Singh M, Spallek R, CranageM, et al. Heat shock proteins generate�-chemokines which func-tion as innate adjuvants enhancing adaptive immunity. Eur J Immunol2000;30:594–603.
[22] Mehlert A, Young DB. Biochemical and antigenic characterization ofthe Mycobacterium tuberculosis71 kD antigen, a member of the 70kD heat-shock protein family. Mol Microbiol 1989;3:125–30.
[23] Doyle CB, Bhattacharyya U, Kent KA, Stott EJ, Jones IM. Regionsrequired for CD4 binding in the external glycoprotein gp120 of simianimmunodeficiency virus. J Immunol 1995;69:1256–60.
[24] Mills HR, Berry N, Burns NR, Jones IM. Simple and efficient pro-duction of the core antigen of HIV-1, HIV-2 and SIV using pGEXexpression vectors inE. coli. AIDS 1992;6:429–37.
[25] Lehner T, Bergmeier LA, Tao L, Panagiotidi C, Klavinskis LS, Hus-sain L, et al. Targeted lymph node immunization with simian im-munodeficiency virus p27 antigen to elicit genital, rectal and uri-nary immune responses in nonhuman primates. J Immunol 1994;153:
cance
a W,het-etro-
rnirusinde-
L. AvirusVirol
-sionnce
R,esis-ell
t al.tion.
ky J,withad
lat-HIV-Proc
R,V-
P. Identification of RANTES, MIP-1� and MIP-1� as the major HIV- suppressive factors produced by CD8+ T cells. Scienc1995;270:1811–5.
[10] Lehner T, Wang Y, Cranage M, Bergmeier LA, Mitchell E, Taoet al. Protective mucosal immunity elicited by targeted iliac lymnode immunization with a subunit SIV envelope and core vaccimacaques. Nat Med 1996;2:767–75.
[11] Wang Y, Tao L, Mitchell E, Bogers WMJM, Doyle C, Bravery Cet al. Generation of CD8 suppressor factor and� chemokines, induced by xenogeniec immunization, in the prevention of simianmunodeficiency virus infection in macaques. Proc Natl Acad Sci1998;95:5223–8.
[12] Heeney JL, van Gils M, Teeuwsen VJP, Bogers WMJM, De GMorghen C, Radaelli A, et al.�-Chemokines and neutralizing anbody titres correlate with heterologous protection of HIV-1 vaccinmacaques. Proc Natl Acad Sci USA 1998;95:10803–8.
[13] Ahmed RKS, Nilsson C, Wang Y, Lehner T, Biberfeld G, ThorstenR. �-Chemokine production in macaques vaccinated with live attated virus correlates with protection against SIVsm challenge. JVirol 1999;80:1569–74.
[14] Lehner T, Wang Y, Bravery CA, Doycle C, Mitchell E, Bogerset al. Antibodies to CCR5 receptors and�-chemokines, generatby xenogeneic immunization, in the prevention of SIV infectionmacaques. In: Abstract of the NCVGD meeting. Bethesda:May 1997.
[15] Lehner T, Wang Y, Doyle C, Tao L, Bergmeier LA, Mitchell E,al. Induction of inhibitory antibodies to the CCR5 chemokine receand their complementary role in preventing SIV infection in macaqEur J Immunol 1999;29:2427–35.
[16] Lehner T, Wang Y, Cranage M, Tao L, Mitchell E, Bravery C, eUp-regulation of�-chemokines and down-modulation of CCR5receptors inhibit simian immunodeficiency virus transmission inhuman primates. Immunology 2000;99:569–77.
[17] Wang Y, Underwood J, Vaughan R, Harmer A, Doyle C, LehneAllo-immunization elicits CCR5 antibodies, SDF-1 chemokinesCD8 – suppressor factors that inhibit transmission of R5 and X4 Hin women. Clin Exp Immunol 2002;129:493–501.
1858–68.[26] Walker CM, Moody DJ, Stites DP, Levy JA. CD8+ lymphocytes
control HIV infectionin vitro by suppressing virus replication. Scien1996;234:1563.
[27] Heeney JL, Haldenman L, ten Haaft P, Dubbes R, KoornstrTeeuwsen V, et al. Vaccine protection and reduced virus load fromerologous macaque-propagated SIV challenge. AIDS Res Hum Rvirus 1999;10:5117–21.
[28] Trkola A, Ketas T, Kewalramani VN, Endorf F, Binley JM, KatingeH, et al. Neutralization sensitivity of human immunodeficiency vtype 1 primary isolates to antibodies and CD4-based reagents ispendent of coreceptor usage. J Virol 1998;72:1876–85.
[29] ten Haaft P, Verstrepen B, Uberla K, Rosenwerth B, Heeney Jpathogenic threshold of virus load defined in immunodeficiencyon simian-human immunodeficiency virus infected macaques. J1998;72:10281–5.
[30] Dean M, Carrington M, Winkler C, Huttley GA, Smith MW, Allikmets R, et al. Genetic restriction of HIV-1 infection and progresto AIDS by a deletion allele of the CKR5 structural gene. Scie1996;273:1856–62.
[31] Lui R, Paxton WA, Choe S, Ceradini D, Martin SR, Horuket al. Homozygous defect in HIV-1 coreceptor accounts for rtance of some multiply-exposed individuals to HIV-1 infection. C1996;86:367–77.
[32] Zagury D, Lachgar A, Chams V, Fall LS, Bernard J, Zagury J-F, eC-C chemokines, pivotal in protection against HIV types 1 infecProc Natl Acad Sci USA 1998;95:3857–61.
[33] Stranford A, Skurnick J, Kouria D, Osmond D, Chang S-Y, Sninset al. Lack of infection in HIV-exposed individuals is associateda strong CD8+ cell noncytotoxic anti-HIV response. Proc Natl AcSci USA 1999;96:1030–5.
[34] Garzino-Demo A, Moss RB, Margolick JB, Cleghorn F, Sill A, Btner WA, et al. Spontaneous and antigen-induced production ofinhibotory�-chemokines are associated with AIDS-free status.Natl Acad Sci USA 1999;96:11986–91.
[35] Wasik T, Bratosiewicz J, Wierzbicki A, Whiteman V, RutsteinStarr S, et al. Protective role of�-chemokines associated with HI

2984 W.M.J.M. Bogers et al. / Vaccine 22 (2004) 2974–2984
specific Th response against perinatal HIV transmission. J Immunol1999;162:4355–64.
[36] Wang Y, Tao L, Mitchell E, Bravery C, Berlingieri P, Armstreong P, etal. Allo-immunization elicits CD8+ T cell-derived chemokines, HIVsuppressor factors and resistance to HIV infection in women. Nat Med1999;5:1004–9.
[37] Warren JT, Levinson MA. AIDS preclinical vaccine development: bi-ennial survey of HIV, SIV and SHIV challenge studies in vaccinatednonhuman primates. J Med Primatol 1999;28:249–73.
[38] Zuber B, Hinkula J, Vodros D, Lundholm P, Nilsson C, Morner A,et al. Induction of immune responses and break of tolerance by DNAagainst the HIV-1 coreceptor CCR5 but not protection from SIVsmchallenge. Virology 2000;278:400–11.
[39] Mack M, Lucklow B, Nelson PJ, Cihak J, Simmons G, Clapham PR,et al. Aminooxypentane-RANTES induced CCR5 internalization butinhibits recylcing: a novel inhibitory mechanism of HIV infectivity. JExp Med 1998;187:1215–24.
[40] Signoret N, Pelchen-Matthews A, Mack M, Proudfoot AE, Marsh M.Endocytosis and recycling of the HIV coreceptor CCR5. J Cell Biol2000;151:1281–93.
[41] Gurer C, Cimarelli A, Luban J. Specific incorporation of heat shockprotein 70 family members into primate lentiviral virions. J Virol2002;76:4666–70.
[42] Bogers WMJM, Bergmeier LA, Ma J, Ooestermeijer H, Wang Y, KellyCG. A novel HIV-CCR5 receptor vaccine strategy in the control ofmucosal SIV/HIV infection. AIDS 2004;18:25–36.
Copyright © 2022 FDOKUMEN




















