ELEMENTS AND CONSTITUENTS OF A NATIONAL POLICY ON PUBLIC SAFETYAND SECURITY
Upload
independentCategory
view
1download
0
ORIGINAL ARTICLE
© Copyright 2006 by Humana Press Inc.All rights of any nature whatsoever reserved.1085-9195/(Online)1559-0283/06/46:155–164/$30.00
Cell Biochemistry and Biophysics 155 Volume 46, 2006
INTRODUCTION
The physiological responses that are impaired inmicrogravity-adapted individuals involve multiplemechanisms including hypovolemia, attenuated baro-reflex sensitivity, cardiovascular structural changes, andneurohumoral vascular tone regulation (1). Evidencefrom bed rest and postflight human studies indicatesthat inadequate vasoconstrictor responsiveness is animportant factor in orthostatic intolerance (2). Yet, thisaspect would require further investigation to be eluci-dated (3,4). Even after several decades of study, the
basic cellular and molecular mechanisms underlyingthis insufficient vasoconstrictor responsiveness are notfully understood.
Hind-limb unloading (HU) in rodents has been exten-sively used as a model to simulate cardiovascular decon-ditioning in humans. This model exhibits many of theknown cardiovascular consequences of microgravity inhumans. The HU rodents model has clearly shown that,among the multiple factors influencing arterial func-tions, the increased release of vasoactive substancesfrom the endothelium, particularly nitric oxide (NO), isa major mechanism deregulating vascular tone (5).Sangha and coworkers (6) provided functional evidencethat NO synthesis was strongly enhanced in HUrodents. It has been reported that both activity andexpression of endothelial nitric oxide synthase (eNOS)
Caveolae and Caveolae Constituents in Mechanosensing
Effect of Modeled Microgravity on Cultured Human Endothelial Cells
Enzo Spisni,1 Mattia Toni,1 Antonio Strillacci,1 Grazia Galleri,2
Spartaco Santi,3 Cristiana Griffoni,1 and Vittorio Tomasi1,*1Department of Experimental Biology, University of Bologna, Bologna, Italy; 2Department of Physiological, Biochemical and Cellular Science, University of Sassari, Sassari, Italy; and 3Institute of Organ Transplant
and Immunocytology (ITOI), Bologna Unit – CNR c/o I.O.R, Via di Barbiano 1/10, I-40138 Bologna, Italy
Abstract
Studies in modeled microgravity or during orbital space flights have clearly demonstrated that endothelialcell physiology is strongly affected by the reduction of gravity. Nevertheless, the molecular mechanisms bywhich endothelial cells may sense gravity force remain unclear. We previously hypothesized that endothelial cellcaveolae could be a mechanosensing system involved in hypergravity adaptation of human endothelial cells. Inthis study, we analyzed the effect on the physiology of human umbilical vein endothelial cell monolayers ofshort exposure to modeled microgravity (24–48 h) obtained by clinorotation. For this purpose, we evaluated thelevels of compounds, such as nitric oxide and prostacyclin, involved in vascular tone regulation and synthesizedstarting from caveolae-related enzymes. Furthermore, we examined posttranslational modifications of Caveolin(Cav)-1 induced by simulated microgravity. The results we collected clearly indicated that short microgravityexposure strongly affected endothelial nitric oxide synthase activity associated with Cav-1 (Tyr 14) phosphory-lation, without modifying the angiogenic response of human umbilical vein endothelial cells. We propose herethat one of the early molecular mechanisms responsible for gravity sensing of endothelium involves endothelialcell caveolae and Cav-1 phosphorylation.
Index Entries: Microgravity; endothelial cells; nitric oxide; Caveolin-1; caveolae.
*Author to whom all correspondence and reprint requestsshould be addressed. E-mail: [email protected]

and inducible NO synthase (iNOS) are increased in thecarotid artery and thoracic aorta of HU rodents (7).Instead, in other vascular districts, different results havebeen collected; for example, the expression of eNOS andiNOS in femoral arteries did not change during HU (7).
The endothelium is a highly heterogeneous and dis-seminated organ with a wide variety of functional propri-eties, including vascular homeostasis and vascular tonecontrol. Endothelium may adapt its functional phenotypeon the basis of various pathophysiological factors, such ashormones, growth factors, cytokines, and biomechanicalstimuli. It is well known that endothelium can respond tomechanical deformations resulting from strain associatedwith vessel stretch and from shear stress (8). Nevertheless,the mechanism by which endothelial cells recognizemechanical stimuli (mechanosensing) is not well under-stood. Several potential mechanosensing systems havebeen proposed, including cytoskeleton (9,10), ion chan-nels (11,12), junction proteins (13,14), G proteins (15), andcaveolae (16–18). Many independent studies clearly indi-cate that caveolae are involved in endothelial response toshear stress (19,20). Boyd and collaborators (19) found anincreased caveolae formation in endothelial cells exposedto chronic shear stress. They also found that caveolin-1(Cav-1), the main organizer of caveolae, dramaticallychanges its distribution after 24 h of laminar shear. Rizzoand colleagues (17) have shown an increased expressionof Cav-1 and eNOS in shear-adapted bovine aorticendothelial cells associated with an increased density ofcaveolae at the luminal plasma membrane. Other evi-dence of the involvement of caveolae in mechanotrans-duction comes from studies based on cholesteroldepletion disassembling of rafts and caveolae. Ferraroand collaborators (21) demonstrated that cholesteroldepletion strongly decreased mechanotransduction inosteoblast cultures, whereas Lungu and collaborators (22)showed that a decreased cholesterol content in caveolae,induced by Cyclosporin A, was associated with an inhibi-tion of the flow-mediated activation of eNOS. In our lab-oratory, it has been shown that the modulations of Cav-1and eNOS are among the main mechanisms involved inthe hypergravity adaptation of endothelial cells (18).Therefore, we have also proposed that endothelium maydirectly detect gravity force by using mechanisms involv-ing endothelial caveolae, because we recorded severalmodifications involving the expression and the activity ofenzymes, mainly compartmentalized into endothelial cellcaveolae, such as eNOS, Cav-1, cyclooxygenase 2, andprostacyclin synthase (PGIS) in human umbilical veinendothelial (HUVE) cells exposed to mild hypergravityconditions (3g).
It is well known that HUVE cells are very sensitive togravitational unloading (23). In the attempt to elucidateif caveolae and caveolae constituents could also be
involved in early events after microgravity adaptation,we have grown HUVE cell monolayers for 24–48 h inmodeled microgravity obtained by using a three-dimen-sional (3D) clinostat. Our findings are consistent with thehypothesis that caveolae and Cav-1 are involved in theadaptation of endothelium to microgravity.
MATERIALS AND METHODS
Cell CultureHUVE cells were isolated from recently collected
unfrozen umbilical cords. Cells were grown in M199medium supplemented with 20% fetal calf serum, 2 mML-glutamine, 100 µg/mL endothelial cell growth supple-ments (ECGS), 5 U/mL heparin, and antibiotics (peni-cillin 100 U/mL and streptomycin 100 µg/mL), aspreviously described (18). Cells were maintained at 37ºCin a 5% CO2 incubator and used for experiments at thirdand fourth passages. Cells were subcultured using 0.05%trypsin, 0.02% ethylenediaminetetraacetic acid (EDTA)solution. Cell viability was assessed by using the trypan-blue dye exclusion assay. In this study, microgravity wasgenerated by a 3D clinostat positioned in a thermostaticroom, and particular attention was taken to exclude thepossibility of false-positive results. The effects of vibra-tion and gradient accumulation or depletion of nutrientswere almost completely eliminated by conducting con-trol experiments. Modeled microgravity conditions(named here as microgravity or 0g) were achieved byRandom Position Machine (RPM, Dutch-Space). TheRPM reproduces gravity acceleration between 10–4 and10–3. Cell cultures exposed to modeled microgravity werepositioned close to the center of the frame. This framerotates within a second rotating frame. Both frames aredriven by separate mechanisms. Rotation of each frame israndom, autonomous, and regulated by computer soft-ware. The rotation velocity of the frames was 60°/s–1.Static control cultures (named here as 1g) were posi-tioned on the basement of the RPM (24). HUVE cells weregrown on both sides of OptiCells (BioCristall Ltd.) flasks,without any coating. OptiCells were then completelyfilled (to avoid shear stress) with 10 mL of completemedium. OptiCells generate a closed cellular environ-ment with high cellular density (100 cm2/10 mL) in whichthe O2 and CO2 levels into the medium are maintainedthrough the pores of the growth membranes. As a result,oxygenation of the medium was not required. At conflu-ence, cells were exposed to modeled microgravity for24–48 h. Both exposed and control OptiCells were treatedidentically until the start of clinorotation. The temperatureand the pH of the culture medium were measured andkept constant to ascertain that the experiments were con-ducted under identical conditions. Scrupulous attentionwas made to eliminate the formation of bubbles into the
156 Spisni et al.
Cell Biochemistry and Biophysics Volume 46, 2006

OptiCells flasks during clinorotation. Immediately aftermicrogravity exposure, cells were lysed for proteinexpression or detached and used for angiogenesis assayin normal gravity. Conditioned media were collected andfrozen at –80ºC before NO and PGI2 analysis.
Antibodies and ReagentsThe primary antibodies used for immunofluorescence
studies were a monoclonal anti–Cav-1 (BD Bioscience),and a polyclonal antibody against eNOS (Santa CruzBiotechnology). The primary antibodies used for Westernblotting were a polyclonal anti–Cav-1 (BD Bioscience), apolyclonal anti-phospho-Cav-1 (Tyr 14) (Cell Signalling),a polyclonal anti-phospho-eNOS (Ser 1177) (Santa CruzBiotechnology), a polyclonal anti-PGI2 synthase (Cay-man), and polyclonal antibodies against eNOS and iNOS(Santa Cruz Biotechnology). Secondary antibodies forimmunofluorescence (FITC or Cy3 conjugated) and forWestern blotting (horseradish peroxide-conjugated) werepurchased from Sigma. Medium 199, L-glutamine, antibi-otics, heparin, bovine serum albumin (BSA), 1,4-diazabi-cyclo-(2,2,2)-octane (DABCO) were purchased fromSigma; fetal calf serum was from Life Technologies. ECLreagents were from Amersham, UK.
Western BlottingFor Western blotting analysis, confluent cells were
scraped from the OptiCells and lysed in a buffer con-taining Tris-HCl 50 mM, pH 7.5; EDTA 2 mM, NaCl 100mM, NP 40 1%, and protease inhibitors (10 mg/mLaprotinin, 10 mg/mL leupeptin, 10 mg/mL soybeantrypsin inhibitor). Samples were sonicated three timesfor 10 s on ice and centrifuged at 10,000g to discard thepellet-containing nucleus and cell debris. A smallamount of the supernatants was used for Lowry proteinassay. Sodium dodecyl sulfate (SDS)-polyacrylamidegel electrophoresis (PAGE) buffer was added to thesupernatant collected and, after boiling, 40 µg of dena-tured proteins were separated in 12% SDS-PAGE andthen transferred to nitrocellulose papers (Hybond-CExtra, Amersham). After blotting, nitrocellulose paperswere incubated with antibodies. Detection was per-formed by using the ECL procedure developed byAmersham. Quantization of the bands was performedby using densitometric image analysis software (ImageMaster, Pharmacia Biotech) and normalization wasmade against β-actin expression.
Immunoprecipitation AssayImmunoprecipitation of Cav-1 complexes was car-
ried out essentially as previously described (25). Briefly,HUVE cells exposed to micro or normal gravity werelysed in 500 µL of a buffer containing 10 mM Tris (pH7.5), 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.4%SDS, 0.5% NP40, 10% glycerol, and protease inhibitors.
Precleared supernatants were immunoprecipitatedusing anti Cav-1 polyclonal antibody (3 µg/mL) andthen incubated with protein A Sepharose. After wash-ings, immunoprecipitated samples were processed byWestern blotting to detect eNOS and PGIS.
Fluorescence Microscopy and Colocalization Analysis
To visualize the colocalization between Cav-1 andeNOS, HUVE cells, grown on OptiCells, were washedwith phosphate-buffered saline (PBS), fixed with 4%paraformaldehyde, permeabilized with PBS-Triton X-100 0.05% and then incubated with anti-Cav-1 andanti-eNOS primary antibodies diluted 1:200 in PBS-BSA 10 mg/mL. After washing, cells were incubatedwith anti-mouse FITC-conjugated and anti-rabbit CY3-coniugated secondary antibodies diluted 1:50 in PBS-BSA 10 mg/mL. Finally, cuts of OptiCells growthmembranes were mounted directly on coverslips inglycerol-PBS medium containing 50 mg/mL DABCO.The confocal imaging was performed on a Radiance2000 confocal laser scanning microscope (BioRadLaboratories), equipped with Nikon ×40 and ×63 oilimmersion objectives and with an argon/kryptonlaser. For FITC and CY3 double detection, the sampleswere simultaneously excited with the 488- and 568-nmlines of the argon/krypton laser. Optical sections wereobtained at increments of 0.1 µm in the Z-axis andwere digitized with a scanning mode format of 512 ×512 pixels and 256 gray levels. The image processingand the volume rendering were performed using theImageSpace software.
Negative controls consisted of samples not incubatedwith the primary antibody. The double-labeling immuno-fluorescence experiments were carried out avoidingcross-reactions between primary and secondary antibod-ies. In addition, different controls were performed toensure antibody specificity. The two-dimensional scatterplot diagram of each section was analyzed to evaluate thespatial colocalization of the fluorochromes. For each scat-ter plot diagram, pixels with highly colocalized fluo-rochromes (i.e., with intensity values greater than 150gray levels on a scale from 0 to 255) for both detectors,were selected to calculate the colocalization maps and cre-ate a binary image. Moreover, Pearson coefficients (27)were calculated on acquired optical sections. This coeffi-cient was calculated on 50 sections for each sample, aspreviously described (28).
PGI2 Synthase Activity DeterminationThe evaluation of PGIS enzymatic activity was assayed
by using the 6-Κeto prostaglandin F1α kit (Cayman) andrevealed by enzyme-linked immunosorbent assay. Foreach sample, the 6-keto PGF1α production was related to
Mechanosensing Role of Caveolae in Modeled Microgravity 157
Cell Biochemistry and Biophysics Volume 46, 2006
the cell number evaluated by using the acidic phos-phatase method.
NO Production AnalysisNO production was measured in conditioned media
of HUVE cells by using the Griess method for nitratequantification. The colorimetric Griess reaction (29) fornitrite was applied to measure the level of NO. Thesamples were diluted 1:2 in H2O mQ. Because the mainamount of NOx in biological fluids is found in nitrate,they were exposed to nitrate reductase (250 mU/mL)and NADPH (100 µM) for 30 min at 37°C to reducenitrate to nitrite. The nitrite-containing samples weretreated with L-glutamine dehydrogenase (670 mU/mL),2-oxoglutaric acid (4 mM), and NH4Cl (100 mM) for 10min at 37°C to consume any residual NADPH. Finally,the samples were mixed with an equal volume offreshly prepared Griess reagent (0.05% N-[1-naphthyl]ethylenediamine dihydrochloride and 0.5% sulfanil-amide in 2.5% ortho-phosphoric acid) for 10 min at37ºC. The absorbance of each colored sample was mea-sured at 540 nm. Concentrations of NO in the sampleswere determined using a calibration curve generatedwith standard NaNO2 solutions (0,1–100 µM). The sen-sitivity of the methods was 0.05 µM. Data were normal-ized against the cell number, evaluated by using theacidic phosphatase method.
3D Angiogenesis TestTo evaluate the effect of microgravity on in vitro
angiogenesis, 80,000 cells detached from OptiCell afterclinorotation were seeded on a 3D collagen gel. The gelwas prepared as previously described (25) and jelliedinto single wells of a 24-multiwell plate. Capillary for-mation was stimulated for 24 h by using human recom-binant fibroblast growth factor (bFGF) at 0.5 ng/mL.Quantitative measurement of angiogenesis wasobtained by counting the number of capillary-like struc-tures formed by HUVE cells into the gel 24 h after bFGFstimulation. For each well, six images, covering almostall the well surface, were recorded and the number ofcapillary-like ring structures was calculated.
Statistical AnalysisAll results are expressed as mean ± SEM. Differences
were analyzed by Student t-test and considered statisti-cally significant at p < 0.05 and p < 0.01.
RESULTS
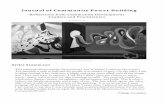
NO Synthesis is Increased in MicrogravityThe first interesting datum we collected was an
increased NO synthesis in HUVE cells grown in micro-gravity. NO increased 2.5-fold (Fig. 1A) after 24–48 h of
microgravity. To verify if an increased expression of NOsynthases was responsible for NO increase, we ana-lyzed by Western blotting the levels of iNOS and eNOSin endothelial cells monolayers exposed to normal grav-
158 Spisni et al.
Cell Biochemistry and Biophysics Volume 46, 2006
Fig. 1. (A) Nitric oxide (NO) production was evaluated inthe conditioned media, after Griess reaction, as nitrate con-centration. Each point represents the mean ± SEM of 18determinations obtained starting from the medium col-lected from six different OptiCells for each treatment (sam-ple in triplicate, n = 18). # indicates that the difference wasstatistically significant (p < 0.01). (B) Analysis of endothelialnitric oxide synthase (eNOS) and inducible nitric oxide syn-thase (iNOS) proteins expression in human umbilical veinendothelial cells exposed for 24 h to normal gravity (1g) ormicrogravity (0g) by Western blotting, starting from threedifferent OptiCells for each treatment. The images showedare representative of the results obtained in two differentexperiments. (C) Densitometric analysis of eNOS and iNOSexpression obtained by band quantification. Densitometrywas performed on six different bands for each treatment (n= 6). Both eNOS and iNOS were normalized against β-actinexpression. Band quantification results are shown settingequal to 100 the mean value obtained from the quantifica-tion and normalization of the bands obtained starting fromnormal gravity control cells.
ity or to microgravity. We found that iNOS expressionwas weak both in HUVE cells cultured in normal grav-ity and in microgravity. On the contrary, eNOS wasstrongly expressed in both the experimental conditionstested, but with no significant difference between 1gand 0g (Fig. 1B,C). This is particularly interesting con-sidering that a different mechanism—increasing eNOSmRNA expression—is involved in shear stress adapta-tion of cultured endothelial cells (30). Our results are inagreement with data collected from other researchers,showing that eNOS activity is increased by HU inrodents (31).
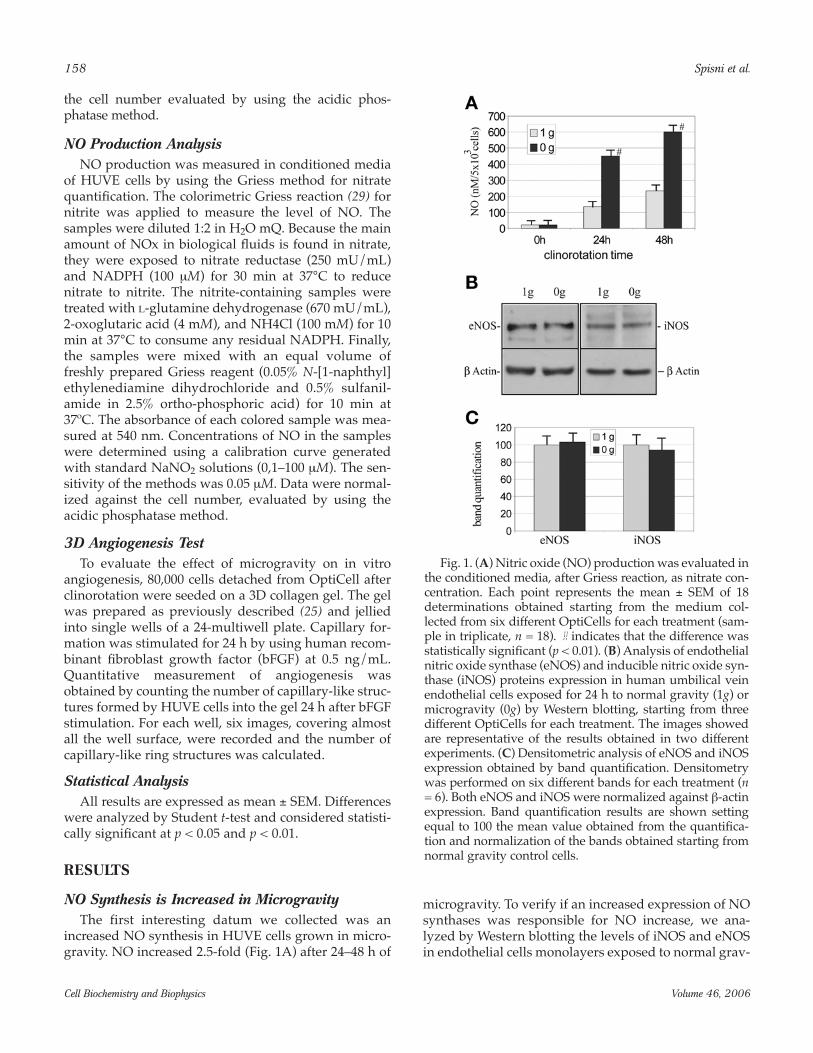
Regulation of eNOS ActivityA well-described molecular mechanism regulating
eNOS activity in endothelium is its localization intocaveolae and its protein–protein interaction with Cav-1(32). In vivo and in vitro studies have clearly shown thatCav-1 is the major negative regulatory protein for eNOSand that Cav-1 expression is inversely correlated to NOproduction (33). Moreover, these findings are supportedby genetic data showing that Cav-1 knockout mice haveincreased endothelium-dependent relaxations and NOlevels in blood (34,35). Thus we analyzed if theincreased NO production was linked to a decreasedCav-1 expression in HUVE cell monolayers cultured inmicrogravity for 24 h (Fig. 2A). Results confirmed thatthe increased NO synthesis did not correlate withdecreased Cav-1 expression. The regulation of eNOS byphosphorylation is poorly understood. Among the dif-ferent known phosphorylation sites on eNOS, Ser-1179in bovine (Ser-1177 in humans) has been characterizedmost extensively (36). Ser-1177 phosphorylation is stim-ulatory for both basal and agonist-mediated NO releaseand seems to be the major site for Akt phosphorylation(36). We found that in HUVE cells, Ser-1177 phosphory-lation is not affected after 24 h of microgravity treatment(Fig. 2A,B). Cav-1 was first identified as a substrate forthe v-Src tyrosine kinase, which phosphorylates Cav-1on Tyr 14. Moreover, it has been hypothesized that Cav-1 phosphorylation at Tyr14 may override or downregu-late the activity of the caveolin-scaffolding domain (37),the region of Cav-1 involved in eNOS binding and inhi-bition. Interestingly, after 24 h, microgravity augmentedCav-1 Tyr14 phosphorylation in HUVE cells (Fig. 2A).Our densitometric analysis results indicate that Cav-1phosphorylation increased 1.8-fold after 24 h of clinoro-tation (Fig. 2B).
Other caveolar enzymes, such as PGIS, are involvedin the regulation of vascular tone and are also bound toCav-1. To verify if microgravity may affect the release ofother vasoactive substances, we measured PGI2 pro-duction in conditioned media of HUVE cell monolayersafter clinorotation. Results (Fig. 2C) indicated that only
NO synthesis was affected after 24 h of microgravityexposure. To verify the hypothesis that Cav-1 Tyr 14phosphorylation could inactivate the CSD activity andin particular the coupling between Cav-1 and eNOS, we
Mechanosensing Role of Caveolae in Modeled Microgravity 159
Cell Biochemistry and Biophysics Volume 46, 2006
Fig. 2. (A) Western blotting analysis of Cav-1, Ser 1177phospho eNOS, Tyr 14 phospho Cav-1 expressions inhuman umbilical vein endothelial cells grown for 24 hunder normal (1g) or microgravity (0g) conditions. Imagesare representative of the results obtained from two inde-pendent experiments, consisting of three differentOptiCells for each treatment (n = 6). (B) Densitometricanalysis of the Western blotting results obtained. p-Cav-1was normalized against Cav-1, whereas p-eNOS was nor-malized against β-actin expression. Densitometry was per-formed on six different bands for each treatment (n = 6).Band quantification results are shown setting equal to 100the mean value obtained from the quantification and nor-malization of the bands obtained from normal gravity con-trol cells. (C) Prostacyclin (PGI2) production was evaluatedin the conditioned media as 6-keto PGF1α concentration, byusing enzyme-linked immunosorbent assay. Normalizationof the values shown was made relating the 6-keto PGF1αconcentration to the total number of cells in each Opticell,evaluated by using the acidic phosphatase method. Eachpoint represents the mean ± SEM of 18 determinationsobtained starting from the supernatants collected from sixdifferent OptiCells dosed in triplicate. *Indicates that differ-ence was statistically significant (p < 0.05).
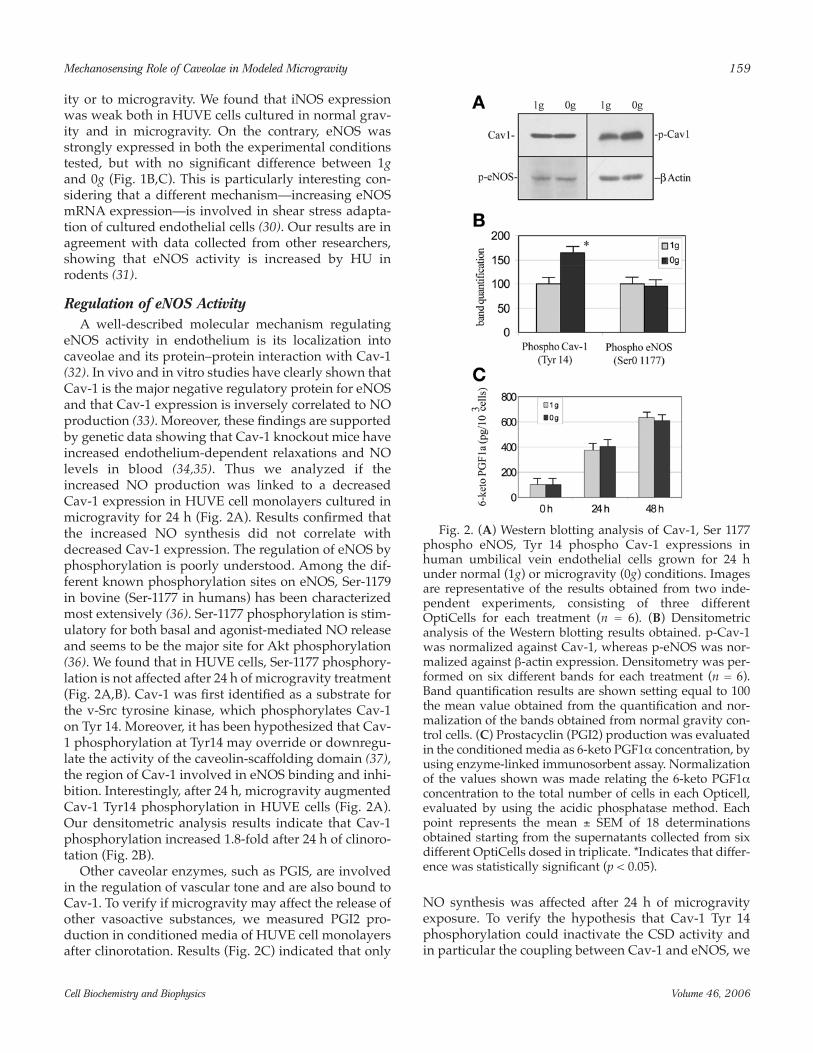
performed confocal microscopy analysis of Cav-1/eNOS colocalization. Figure 3 shows that 24 h aftermicrogravity exposure the colocalization betweeneNOS and Cav-1 at the plasma membrane level wasclearly diminished (compare panels D and H with pan-els L and P, respectively). Analysis on 50 different sec-tions indicates that Pearson’s correlation coefficientdecreases from 0.78 to 0.59 in modeled microgravityconditions. The results of the colocalization analysis
were confirmed also by the immunoprecipitation exper-iments (Fig. 4A,B), showing that the amount of eNOSrecovered after Cav-1 immunoprecipitation wasreduced in HUVE cells after 24 h of microgravity expo-sure. The selectivity of this uncoupling phenomenonhas been revealed by the examination of PGIS/Cav-1coimmunoprecipitation, because it is known that alsoPGIS is an enzyme that bind and coimmunoprecipitatewith Cav-1 (Fig. 4A).
160 Spisni et al.
Cell Biochemistry and Biophysics Volume 46, 2006
Fig. 3. Confocal microscopy localization of Cav-1 (FITC, represented in green) and endothelial nitric oxide synthase(eNOS) (Cy3, represented in red) in human umbilical vein endothelial cells exposed for 24 h to normal gravity (1g, A–H)or to microgravity (0g, I–P). Color images result from integrating different optical sections. (A–D) and (I–L) were obtainedby using the ×40 oil immersion objectives, whereas (E–H) and (M–P) were obtained by using the ×63 oil immersion objec-tives. Whereas in (A–D) and (I–L) a wide number of cells are shown, (E–H) and (M–P) show single-cell images.Superposition of the green (Cav-1) and red (eNOS) signals is shown (C,G,K,O); yellow indicates that eNOS and Cav-1 colo-calize. Scatter plot colocalization maps (D,H,L,P) were obtained as described under Materials and Methods and representthe colocalization levels in a binary black and white image (bar = 5 µm).

Microgravity and AngiogenesisBecause NO has been described as an angiogenic fac-
tor, the angiogenic response of HUVE cells was exam-ined, after 24 h of microgravity, by using an in vitro 3Dangiogenesis test based on capillary-like structures for-mation into a collagen gel (Fig. 5A,B). We decided tostimulate the angiogenic response by using bFGF toexclude the effects of the reciprocal regulation betweenNO and vascular endothelial growth factor duringangiogenesis (38). Results of the count of the number ofcapillary-like ring structures formed in each well areshown in Fig. 5C, indicating that the formation of capil-
lary-like structures, after 24 h of stimulation by bFGF,was not affected by microgravity exposure.
DISCUSSION
It is well known that changes in gravity force maydeeply influence eukaryotic cell behavior. One of theaims of this study was to verify which molecular mech-anisms are involved in the endothelial cells early adap-tation to the decreased gravity. We have recently shownthat caveolae and caveolar enzymes are involved in theendothelial cell adaptation to hypergravity conditions(18), and we have hypothesized that caveolae may havea gravisensing role, at least in these cells. The strongincrease in NO synthesis we observed, in the experi-mental conditions we adopted, is a piece of data gener-ally confirmed by literature in most in vitro and in vivomodels for microgravity simulation. Nevertheless, dif-ferent mechanisms may regulate eNOS activity and NOsynthesis in endothelial cells. Among them, the modu-lation of eNOS expression, the posttranslational regula-tion of the enzyme and its association with regulativeproteins are the well-described regulatory mechanisms(39,40). Among them, the Ser 1177 phosphorylation of
Mechanosensing Role of Caveolae in Modeled Microgravity 161
Cell Biochemistry and Biophysics Volume 46, 2006
Fig. 4. (A) Immunoprecipitation of Cav-1 complexes inhuman umbilical vein endothelial cells exposed to normalgravity (1g) or microgravity (0g) for 24 or 48 h. Cell lysateswere immunoprecipitated (IP) with polyclonal antibody toCav-1, and the resulting immunocomplexes were ana-lyzed for the presence of endothelial nitric oxide synthase(eNOS, upper lanes) or prostacyclin synthase (PGIS)(lower lanes) by Western blotting. Image in (A) is repre-sentative of two different sets of experiments, consisting ofthree different OptiCells for each treatment. (B)Densitometric analysis of the eNOS and PGIS bandsobtained by Western blotting after Cav-1 immunoprecipi-tation. Quantification was performed on six differentbands for each treatment (n = 6). Numerical data wereobtained setting equal to 100 the mean value obtainedfrom the quantification of the bands obtained after theimmunoprecipitation of normal gravity control cellslysates. *Indicates that the difference was statistically sig-nificant (p < 0.05).
Fig. 5. Effect of microgravity on in vitro angiogenesis,evaluated by using three-dimensional collagen gel angio-genesis test. After 24 h of normal gravity or microgravityconditioning, HUVE cells were seeded on three-dimen-sional collagen gel and stimulated with 0.5 ng/mL bFGFfor 24 h (A,B). After 24 h, human umbilical vein endothe-lial (HUVE) cells started the formation of capillary-likering structures (indicated by arrows) into the gel.Quantitative analysis of the angiogenic response of HUVEcells (C) was made counting the number of capillary-likering structures, formed in each well. For each conditiontested, 12 wells were analyzed (n = 12). Data are expressedas the mean ± SEM (bar = 60 µM).

eNOS and the binding of Cav-1 scaffolding domain tothe reductase domain of eNOS are believed to be themajor posttranslational mechanisms modulating theenzyme activity (38). Our results showed that, althoughSer 1177 phosphorylation was not affected, eNOSinhibitory binding to Cav-1 was significantly reducedby microgravity. The Tyr 14 Cav-1 phosphorylation,probably induced by Src kinase, seems to be the molec-ular key of Cav-1/eNOS complex dissociation. It hasbeen hypothesized that Tyr 14-phosphorylated Cav-1may function like a growth factor receptor that recruitsSH2 domain-containing proteins to the plasma mem-brane (41). Moreover, Colonna and Podestà (42) haverecently observed that Cav-1 Tyr 14 phosphorylation isassociated with a cytoskeleton rearrangement in Y1adrenal cells. A marked cytoskeletal reorganization hasbeen also observed in HUVEC, by Carlsson and collab-orators (23) in response to modeled microgravityobtained by using the rotating wall vessel bioreactor.Yet, after 24 h of clinorotation, we could not detect anycytoskeleton modification (data not shown).
Even if the increased NO synthesis seems to be acommon feature of microgravity adaptation, the molec-ular mechanisms involved in this upregulation could bedifferent in different cell types. Indeed, although mod-eled microgravity caused an increased expression ofeNOS mRNA in microvascular endothelial cells (43), inHUVE cells, the increased NO synthesis mainly actedon posttranslational mechanisms. Our data clearlyshowed that the colocalization between Cav-1 andeNOS is significantly decreased after 24 h of clinorota-tion. Immunoprecipitation assay confirms that theuncoupling of Cav-1 and eNOS may be the cause of theincreased eNOS activity we measured in microgravity.Thus it is very likely that Cav-1 phosphorylation on Tyr14 may be the molecular event causing eNOS dissocia-tion. Interestingly, we observed that the bindingbetween Cav-1 and PGIS (also a caveolar enzymebound to Cav-1 scaffolding domain) is not affected bymicrogravity-induced Cav-1 phosphorylation. This isnot surprising because we have previously shown thatthe scaffolding domain of Cav-1 has structural featuresin agreement with the hypothesis of a multiple func-tional domain (44). Moreover, whereas for PGIS thebinding to Cav-1 seems to be mainly linked to its sub-cellular compartmentalization and does not change itsactivity (25), for eNOS the binding to Cav-1 is a funda-mental regulative step leading to its inactivation.
Our hypothesis that caveolae could behave asendothelial cell mechanosensors and gravisensors issubstantiated by the early Cav-1 Tyr-14 phosphorylationwe have observed during microgravity adaptation. Thishypothesis is also confirmed by findings showing thatcholesterol depletion inhibited eNOS activation induced
by shear stress (22) and reduced mechanotransductionpathways induced by hydrostatic pressure (21).
NO is a vasoactive compound considered to be aproangiogenic factor. The evaluation of the angiogenicresponse of endothelial cells grown in microgravity is ofgreat interest for space medicine and bioengineering,especially for the generation of vascularized tissues byusing bioreactors (45). Moreover, the effect of micro-gravity on the angiogenic response is still debated. Forexample, although Carlsson and collaborators (23)found an increased proliferation in HUVE cells exposedto microgravity, Morbidelli and coauthors (46) observedthat microgravity strongly decreased proliferation inporcine aortic endothelial cells. Our results clearly indi-cate that microgravity, obtained by clinorotation, doesnot modify the angiogenic competence of HUVE cells. Itis likely that the increased angiogenesis stimulationresulting from the increased NO synthesis was counter-balanced by the modification of other signaling path-ways induced by microgravity.
How cells may couple gravity force changes to sig-naling mechanisms is still an open question. Our resultsdemonstrate that both Cav-1 Tyr 14 phosphorylationand eNOS activation are caveolar mechanisms involvedin early microgravity adaptation of HUVE cells. Webelieve that caveolae can behave as gravity sensors, atleast in endothelial cells. Our studies open up the possi-bility that caveolae and Cav-1 could be deeply involvedin the impairment of vascular functions observed in themicrogravity adaptation of mammals. Yet, we can notexclude that other signaling pathways, dependent orindependent from Cav-1, also play a role in micrograv-ity impairment of vascular tone. Our conclusion is that,in the conditions we tested, Cav-1 and caveolae areresponsible for the early activation of the signallingmechanisms involved in endothelial cells adaptation tothe decreased gravity force.
ACKNOWLEDGMENTS
We thank Mr. Gavino Campus and Prof. Proto Pippiafor technical support in the use of three-dimensional cli-nostat. Supported by DCMC project (2006) fromAgenzia Spaziale Italiana (Roma) to VT.
REFERENCES
1. Watenpaugh, D. E., and Hargens, A. R. The cardiovascularsystem in microgravity. Handbook of Physiology.Environmental Physiology. American Physiology Society,Bethesda, MD, 1996.
2. Blomqvist, C. G., Buckey, J. C., Gaffney, F. A., Lane, L. D.,Levine, B. D., and Watenpaugh, D. E. (1994) Mechanismsof post-flight orthostatic intolerance. J. Gravit. Physiol. 1,122–124.
162 Spisni et al.
Cell Biochemistry and Biophysics Volume 46, 2006

3. Bonnin, P., Ben Driss, A., Benessiano, J., Maillet, A., Pavy leTraon, A., and Levy, B. I. (2001) Enhanced flow-dependentvasodilatation after bed rest, a possible mechanism fororthostatic intolerance in humans. Eur. J. Appl. Physiol. 85,420–426.
4. Buckey, J. C. Jr., Lane, L. D., Levine, B. D., et al. (1996)Orthostatic intolerance after spaceflight. J. Appl. Physiol. 81,7–18.
5. Vaziri, N. D., Ding, Y., Sangha, D. S., and Purdy, R. E. (2000)Upregulation of NOS by simulated microgravity, potentialcause of orthostatic intolerance. J. Appl. Physiol. 89,338–344.
6. Sangha, D. S., Vaziri, N. D., Ding, Y., and Purdy, R. E. (2000)Vascular hyporesponsiveness in simulated microgravity:role of nitric oxide-dependent mechanisms. J. Appl. Physiol.88, 507–517.
7. Ma, J., Kahwaji, C. I., Ni, Z., Vaziri, N. D., and Purdy, R. E.(2003) Effects of simulated microgravity on arterial nitricoxide synthase and nitrate and nitrite content. J. Appl.Physiol. 94, 83–92.
8. Barakat, A., and Lieu, D. (2003) Differential responsivenessof vascular endothelial cells to different types of fluidmechanical shear stress. Cell. Biochem. Biophys. 38, 323–343.
9. Chen, K. D., Li, Y. S., Kim, M., et al. (1999) Mechano-transduction in response to shear stress. Roles of receptortyrosine kinases, integrins, and Shc. J. Biol. Chem. 274,18393–18400.
10. Wang, N., Butler, J. P., and Ingber, D. E. (1993)Mechanotransduction across the cell surface and throughthe cytoskeleton. Science 260, 1124–1127.
11. Drummond, H. A., Gebremedhin, D., and Harder, D. R.(2004) Degenerin/epithelial Na+ channel proteins.Components of a vascular mechanosensor. Hypertension44, 643–648.
12. Tavernarakis, N., and Driscoll, M. (2001) Mech-anotransduction in Caenorhabditis elegans: the role ofDEG/ENaC ion channels. Cell. Biochem. Biophys. 35, 1–18.
13. Shay-Salit, A., Shushy, M., Wolfovitz, E., et al. (2002) VEGFreceptor 2 and the adherens junction as a mechanical trans-ducer in vascular endothelial cells. Proc. Natl. Acad. Sci.USA 99, 9462–9467.
14. Shyy, J. Y., and Chien, S. (2002) Role of integrins inendothelial mechanosensing of shear stress. Circ. Res. 91,769–775.
15. Gudi, S., Huvar, I., White, C. R., et al. (2003) Rapid activa-tion of Ras by fluid flow is mediated by Galpha(q) andGbetagamma subunits of heterotrimeric G proteins inhuman endothelial cells. Arterioscler. Thromb. Vasc. Biol. 23,994–1000.
16. Park, H., Go, Y. M., Darji, R., et al. (2000) Caveolin-1 regu-lates shear stress-dependent activation of extracellular sig-nal-regulated kinase. Am. J. Physiol. Heart Circ. Physiol. 278,1285–1293.
17. Rizzo, V., Sung, A., Oh, P., and Schnitzer, J. E. (1998) Rapidmechanotransduction in situ at the luminal cell surface ofvascular endothelium and its caveolae. J. Biol. Chem. 273,26323–26329.
18. Spisni, E., Bianco, M. C., Griffoni, C., et al. (2003)Mechanosensing role of caveolae and caveolar con-
stituents in human endothelial cells. J. Cell. Physiol. 197,198–204.
19. Boyd, N. L., Park, H., Yi, H., et al. (2003) Chronic shearinduces caveolae formation and alters ERK and Aktresponses in endothelial cells. Am. J. Physiol. Heart. Circ.Physiol. 285, 1113–1122.
20. Rizzo, V., Morton, C., DePaola, N., Schnitzer, J. E., andDavies, P. F. (2003) Recruitment of endothelial caveolaeinto mechanotransduction pathways by flow conditioningin vitro. Am. J. Physiol. Heart. Circ. Physiol. 285, 1720–1729.
21. Ferraro, J. T., Daneshmand, M., Bizios, R., and Rizzo, V.(2004) Depletion of plasma membrane cholesterol damp-ens hydrostatic pressure and shear stress-inducedmechanotransduction pathways in osteoblast cultures.Am. J. Physiol. Cell. Physiol. 286, 831–839.
22. Lungu, A. O., Jin, Z G., Yamawaki, H., Tanimoto, T., Wong,C., and Berk, B. C. (2004) Cyclosporin A inhibits flow-mediated activation of endothelial nitric oxide synthasevia altering cholesterol content in caveolae. J. Biol. Chem.279, 48794–48800.
23. Carlsson, S. I., Bertilaccio, M. T., Ballabio, E., and Maier, J.A. (2003) Endothelial stress by gravitational unloading:effects on cell growth and cytoskeletal organization.Biochim. Biophys. Acta 1642, 173–179.
24. Schwarzenberg, M., Pippia, P., Meloni, M. A., Cossu, G.,Cogoli-Greuter, M., and Cogoli, A. (1999) Signal transduc-tion in T lymphocytes—a comparison of the data fromspace, the free fall machine and the random positioningmachine. Adv. Space Res. 24, 793–800.
25. Spisni, E., Griffoni, C., Santi, S., et al. (2001) Co-localizationprostacyclin (PGI2) synthase-caveolin-1 in endothelialcells and new roles for PGI2 in angiogenesis. Exp. Cell. Res.266, 31–43.
26. Tabas, I., Zha, X., Beatini, N., Myers, J. N., and Maxfield, F.R. (1994) The actin cytoskeleton is important for the stim-ulation of cholesterol esterification by atherogenic lipopro-teins in macrophages. J. Biol. Chem. 269, 22547–22556.
27. Manders, E. M. M., Verbeek, F. J., and Aten, J. A. (1993)Measurement of co-localization of objects in dual colorconfocal images. J. Microsc. 169, 375–382.
28. Riccio, M., Dembic, M., Cinti, C., and Santi, S.Multifluorescence labeling and co-localization analyses.In Methods in Molecular Biology: Cell Cycle Control andDysregulation Protocols, Vol. 85 (Giordano, A., Romano,G., eds.). Humana Press Inc., New York, 2004, pp.171–177.
29. Hevel, J. M., and Marletta, M. A. (1994) Nitric-oxide syn-thase assays. Meth. Enzymol. 233, 250–258.
30. Ranjan, V., Xiao, Z., and Diamond, S. L. (1995) ConstitutiveNOS expression in cultured endothelial cells is elevated byfluid shear stress. Am. J. Physiol. 269, 550–555.
31. Ma, J., Kahwaji, C. I., Ni, Z., Vaziri, N. D., and Purdy R. E.(2003) Effects of simulated microgravity on arterial nitricoxide synthase and nitrate and nitrite content. J. Appl.Physiol. 94, 83–92.
32. Sbaa, E., Frerart, F., and Feron, O. (2005) The double regu-lation of endothelial nitric oxide synthase by caveolae andcaveolin: a paradox solved through the study of angiogen-esis. Trends Cardiovasc. Med. 15, 157–162.
Mechanosensing Role of Caveolae in Modeled Microgravity 163
Cell Biochemistry and Biophysics Volume 46, 2006

33. Bernatchez, P. N., Bauer, P. M., Yu, J., Prendergast, J. S., He,P., and Sessa, W. C. (2005) Dissecting the molecular controlof endothelial NO synthase by caveolin-1 using cell-per-meable peptides. Proc. Natl. Acad. Sci. USA 102, 761–766.
34. Drab, M., Verkade, P., Elger, M., et al. (2001) Loss of caveo-lae, vascular dysfunction, and pulmonary defects in cave-olin-1 gene-disrupted mice. Science 293, 2449–2452.
35. Rafani, B., Engelman, J. A., Wang, X. B., et al. (2001)Caveolin-1 null mice are viable but show evidence ofhyperproliferative and vascular abnormalities. J. Biol.Chem. 276, 38121–38138.
36. Bauer, P. M., Fulton, D., Boo Y. C., et al. (2003)Compensatory phosphorylation and protein-protein inter-actions revealed by loss of function and gain of functionmutants of multiple serine phosphorylation sites inendothelial nitric-oxide synthase. J. Biol. Chem. 278,14841–14849.
37. Williams, T. M., Medina, F., Badano, I., et al. (2004) Caveolin-1 gene disruption promotes mammary tumorigenesis anddramatically enhances lung metastasis in vivo. Role of Cav-1 in cell invasiveness and matrix metalloproteinase (MMP-2/9) secretion. J. Biol. Chem. 279, 51630–51646.
38. Tai S. C., Robb G. B., and Marsden, P. A. (2004) Endothelialnitric oxide synthase: a new paradigm for gene regulationin the injured blood vessel. Arterioscler. Thromb. Vasc. Biol.24, 405–412.
39. Fleming, I., and Busse, R. (2003) Molecular mechanismsinvolved in the regulation of the endothelial nitric oxide
synthase. Am. J. Physiol. Regul. Integr. Comp. Physiol. 284,1–12.
40. Fulton, D., Fontana, J., Sowa, G., et al. (2002) Localizationof endothelial nitric-oxide synthase phosphorylated onserine 1179 and nitric oxide in Golgi and plasma mem-brane defines the existence of two pools of active enzyme.J. Biol. Chem. 277, 4277–4284.
41. Minshall, R. D., Sessa, W. C., Stan, R. V., Anderson, R. G.,and Malik, A. B. (2003) Caveolin regulation of endothelialfunction. Am. J. Physiol. Lung Cell. Mol. Physiol. 285,L1179–1183.
42. Colonna, C., and Podesta, E. J. (2005) ACTH-induced cave-olin-1 tyrosine phosphorylation is related to podosomeassembly in Y1 adrenal cells. Exp. Cell. Res. 304, 432–442.
43. Cotrupi, S., Ranzani, D., and Maier, J. A. (2005) Impact ofmodeled microgravity on microvascular endothelial cells.Biochim. Biophys. Acta 1746, 163–168.
44. Spisni, E., Tomasi, V., Cestaio, A., and Tosatto, S. C. (2005)Structural insights into the function of human caveolin 1.Biochem. Biophys. Res. Commun. 338, 1383–1390.
45. Unsworth, B. R., and Lelkes, P. I. (1998) Growing tissues inmicrogravity. Nat. Med. 4, 901–907.
46. Morbidelli, L., Monici, M., Marziliano, N., et al. (2005)Simulated hypogravity impairs the angiogenic response ofendothelium by up-regulating apoptotic signals. Biochem.Biophys. Res. Commun. 334, 491–499.
164 Spisni et al.
Cell Biochemistry and Biophysics Volume 46, 2006
Copyright © 2022 FDOKUMEN