
Population pharmacokinetics of intravenous caffeine in neonates with apnea of prematurity

Cationic Albumin–Conjugated Pegylated Nanoparticles Allow Gene
Delivery into Brain Tumors via Intravenous Administration
Wei Lu,1Qing Sun,
2Jin Wan,
3Zhenjue She,
3and Xin-Guo Jiang
1
1Department of Pharmaceutics, School of Pharmacy; 2Gene Research Center; and 3Department of Anatomy, Histology,and Embryology, Shanghai Medical School, Fudan University, Shanghai, P.R. China
Abstract
Patients with malignant gliomas have a poor prognosisbecause these tumors do not respond well to conventionaltreatments. Studies of glioma xenografts suggest that they maybe amenable to gene therapy with cytotoxic genes, such as theproapoptotic Apo2 ligand/tumor necrosis factor–relatedapoptosis-inducing ligand (Apo2L/TRAIL). Gene therapy ofgliomas ideally employs i.v. given vectors, thus excluding viralvectors as they cannot cross the brain microvascular endo-thelium or blood-brain barrier. Recently, we reported thesynthesis of cationic albumin–conjugated pegylated nano-particles (CBSA-NP) and showed their accumulation in mousebrain cells upon i.v. administration. In this study, plasmidpORF-hTRAIL (pDNA) was incorporated into CBSA-NP, andthe resulting CBSA-NP-hTRAIL was evaluated as a nonviralvector for gene therapy of gliomas. Thirty minutes aftertransfection of C6 glioma cells, CBSA-NP-hTRAIL was inter-nalized and mostly located in the cytoplasm, whereas NP-hTRAIL was entrapped in the endolysosomal compartment. At6 and 48 hours after transfection, respectively, released pDNAwas present in the nuclei and induced apoptosis. At 30minutes after i.v. administration of CBSA-NP-hTRAIL toBALB/c mice bearing i.c. C6 gliomas, CBSA-NP-hTRAIL colo-calized with glycoproteins in brain and tumor microvascula-ture and, via absorptive-mediated transcytosis, accumulatedin tumor cells. At 24 and 48 hours after i.v. administration ofCBSA-NP-hTRAIL, respectively, hTRAIL mRNA and proteinwere detected in normal brain and tumors. Furthermore,repeated i.v. injections of CBSA-NP-hTRAIL induced apoptosisin vivo and significantly delayed tumor growth. In summary,this study indicates that CBSA-NP-hTRAIL is a promisingcandidate for noninvasive gene therapy of malignant glioma.(Cancer Res 2006; 66(24): 11878-87)
Introduction
Patients with malignant glioma have a poor prognosis becausethese brain tumors respond poorly to radiation or chemotherapy,the conventional treatments of cancer (1). Instead, gene therapywith cytotoxic genes, such as the proapoptotic Apo2 ligand/tumornecrosis factor–related apoptosis-inducing ligand (Apo2L/TRAIL),may offer therapeutic promise. Most glioma cell lines express theagonist Apo2L/TRAIL receptors but no or undetectable levels of
the antagonist receptors (2). Normal cells, on the other hand, havebeen found to express antagonist Apo2L/TRAIL receptors (3, 4).Thus, Apo2L/TRAIL may allow selective killing of tumor cells only.I.t. administration of Apo2L/TRAIL has induced tumor regressionin a xenograft model with i.c. glioma implantation (5, 6).Yet, the brain capillary endothelium, which with its tight
junctions forms the brain-blood barrier (BBB) in vivo , poses amajor obstacle for delivery of exogenous genes into brain tumors(7). Viral vectors, which have been used for efficient delivery ofexogenous genes into peripheral tumors, cannot cross the BBB, andtheir in vivo delivery into brain tumors requires highly invasiveadministration routes (7–9). This makes multi-dosing difficult,while the viral vectors themselves are associated with immunoge-nicity, inflammation, oncogenic properties, and unknown long-term effects in patients (7, 10, 11).The difficulties with the viral vectors have led to the development
of nonviral vectors, such as pegylated immunoliposomes, whichallow gene delivery into the brain via i.v. administration (12–14).Targeting of the pegylated immunoliposomes to the BBB is attainedby monoclonal antibodies against either the transferrin receptor orinsulin receptor, which upon binding to their ligands triggerreceptor-mediated endocytosis (15). Upon i.v. administration ofpegylated immunoliposomes, expression of a pegylated immuno-liposome–encapsulated gene has been shown in human gliomacells implanted i.c. in mice (13).Pegylated nanoparticles, compared with liposomes, are physi-
cally and chemically more stable and allow lyophilization for long-term storage (16, 17). Cationic albumin–conjugated pegylatednanoparticles (CBSA-NP), recently developed in our laboratory,have shown brain delivery property (18, 19). A higher accumula-tion of CBSA-NP than of native albumin–conjugated pegylatednanoparticles has been found in mouse brain cells after i.v.administration of these nanoparticles loaded with a fluorescentprobe (19). Free CBSA has been reported to target delivery to theBBB in brain capillaries via absorptive-mediated transcytosis(AMT; refs. 20, 21).To determine the potential of CBSA-NP as a nonviral vector for
the systemic delivery of exogenous genes into brain, we chosehTRAIL-encoding plasmid (pORF-hTRAIL) as a model therapeuticgene for malignant glioma. The mechanisms of gene delivery byCBSA-NP-hTRAIL into normal brain and brain tumor and theintracellular fate of this nanoparticle were studied. Expression ofhTRAIL in tumors and its antitumor effect were evaluated in C6glioma tumors i.c. implanted in BALB/c mice.
Materials and Methods
Materials and animals. CBSA and copolymers of MPEG-PLA (molecular
weight = 43 kDa) and maleimide-PEG-PLA (molecular weight = 44 kDa) weresynthesized in our laboratory, as previously described (19). Plasmid pORF-
hTRAIL (v.16, 4,058 bp) was purchased from Invivogen (San Diego, CA). Fetal
bovine serum (FBS) was from Life Technologies/Invitrogen (Carlsbad, CA).
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).
Requests for reprints: Xin-Guo Jiang, Department of Pharmaceutics, School ofPharmacy, Fudan University (Fenglin Campus), P.O. Box 130, 138 Yi Xue Yuan Road,Shanghai 200032, P.R. China. Phone: 86-21-5423-7381; Fax: 86-21-5423-7381; E-mail:[email protected].
I2006 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-06-2354
Cancer Res 2006; 66: (24). December 15, 2006 11878 www.aacrjournals.org
Research Article
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

6-Coumarin was from Sigma-Aldrich (St. Louis, MO). Texas Red-transferrin,LysoTracker Blue, ethidium monoazide (EMA), Alexa Fluor 633-wheat germ
agglutinin (WGA), CellTracker CM-DiI, and PicoGreen DNA quantitative
analysis kit were from Molecular Probes (Eugene, OR). Rabbit anti-human/
mouse/rat active caspase-3 polyclonal antibody was from Chemicon(Temecula, CA). hTRAIL ELISA kit was from Biosource (Camarillo, CA).
LipofectAMINE 2000 was from Invitrogen (Carlsbad, CA). Goat anti-human
TRAIL-specific IgG and in situ apoptosis detection kit (TACS TdT kit) were
from R&D Systems (Minneapolis, MN). DNA ladder was from MBI(Fermentas International, Canada). Rabbit anti-goat IgG-Cy3 antibody,
Heparin-biotin sodium salt, streptavidin-gold (10-nm colloidal gold), and 4,6-
diamidino-2-phenylindole (DAPI) were from Sigma-Aldrich. Poly(ethylenei-
mine) (molecular weight = 1,800 Da) was from Polysciences, Inc.(Warrington, PA). Double-distilled water was purified using a Millipore
Simplicity System (Millipore, Bedford, MA). All other chemicals were analytic
reagent grades and used without further purification. The rat C6 glioma cellline (CCL-107) was obtained from the American Type Culture Collection
(Rockville, MD). The BALB/c mice (Department of Experimental Animals,
Fudan University, Shanghai, China) used in this study were treated according
to protocols approved by the ethical committee of Fudan University.Preparation of nanoparticles loaded with plasmid DNA. pORF-
hTRAIL (hereafter pDNA) was amplified in the Escherichia coli Top10,
extracted by alkaline lysis, and purified using the plasmid Giga column
isolation kit (Qiagen GmbH, Hilden, Germany). To prepare CBSA-NP-hTRAIL, pDNA and thiol-reactive pegylated nanoparticles were
subjected to a double emulsion and solvent evaporation technique
according to our previous method (19). Hereto, the internal water phaseincluded pDNA (250 Ag) in 50 AL of Tris-EDTA buffer (pH 7.5). Oil phase
was 1 mL of dichloromethane solution containing MPEG-PLA (24.0 mg)
and maleimide-PEG-PLA (2.4 mg). The sonication procedures of primary
and secondary emulsion were set as 120 W, 15 seconds (continuous) and120 W, 15 seconds (intermittent), respectively. After evaporation and
removal of unencapsulated pDNA by washing, the thiol-reactive pegylated
nanoparticles were slowly mixed with thiolated CBSA (19) at a 1:1
thiolated/maleimide ratio on a rotating plate for 8 hours. The resultingCBSA-NP-hTRAIL was purified through Sepharose CL-4B column eluted
with saline. pORF-hTRAIL–loaded MPEG-PLA nanoparticle (NP-hTRAIL)
was prepared similarly to CBSA-NP-hTRAIL, except that MPEG-PLA(26.4 mg/mL) was used only.
To allow analysis of their cellular uptake and intracellular distribution,
CBSA-NP-hTRAIL and NP-hTRAIL were labeled with the fluorescent dye
6-coumarin, as previously described (22).Furthermore, to allow for its intracellular tracking, pDNA was covalently
labeled with the fluorescent dye EMA by photolysis, as described elsewhere
(23). The preparation of 6-coumarin-labeled CBSA-NP containing EMA-
labeled pDNA (CBSA-NP-hTRAIL-EMA) was similar to that of 6-coumarin-labeled CBSA-NP-hTRAIL, except that EMA-labeled pDNA was used instead
of pDNA.
Nanoparticle characterization. Mean (volume based) diameter and
zeta potential of the nanoparticles were determined by dynamic light
scattering using a Zeta Potential/Particle Sizer Nicomp 380 ZLS (Particle
Sizing Systems, Santa Barbara, CA). For zeta potential measurements,
nanoparticle suspensions (200 Ag/mL in 0.001 mol/L HEPES buffer) were pHadjusted with 0.1 mol/L sodium hydroxide or 0.1 mol/L hydrochloric acid.
Nanoparticles were visualized with scanning (JOEL 6320FV, Philips/FEI,
Hillsboro, OR) and transmission (JOEL CM1200, Philips/FEI) electron
microscopes. Drug encapsulating and loading efficiencies of CBSA-
NP-hTRAIL and NP-hTRAIL were determined, as previously described (19).
Covalent conjugations between CBSA and nanoparticles were confirmed by
immunostaining of CBSA-NP-hTRAIL with heparin-biotin and streptavidin-
gold complexes (19).
The ability of CBSA to protonate and obtain a positive charge over a 10
to 2 pH range was determined by acid-base titration (24, 25). Briefly, 10 mg
CBSA, native bovine serum albumin (BSA), or poly(ethyleneimine) wasdissolved in 10 mL of 150 mmol/L NaCl, adjusting to pH 10 with 1 mol/L
NaOH. The pH of the solution was then monitored as aliquots of 0.1 mol/L
HCl were added.
The release of pDNA from both CBSA-NP-hTRAIL and NP-hTRAIL weredone in Tris-EDTA buffer (26). The amount of plasmid released in each time
interval was determined by the PicoGreen assay, which is very sensitive for
supercoiled and double-strained DNA, but not for single-strained DNA
and oligonucleotides (17). pDNA stability was determined by gel electro-phoresis (1% agarose containing ethidium bromide, 110 V, 90 minutes).
Cellular uptake of CBSA-NP-hTRAIL and NP-hTRAIL. CBSA-NP-
hTRAIL and NP-hTRAIL were labeled with the lipophilic fluorescent dye
6-coumarin (f0.1% loading) to allow analysis of their cellular uptake and
intracellular fate (22). C6 cells (16 � 103 per 200 AL/well) were seeded in
96-well plates in RPMI 1640, 10% FBS. After 24 hours and a 30-minute
preincubation in RPMI 1640, cells were treated with 100 Ag/mL of
6-coumarin-labeled CBSA-NP-hTRAIL or NP-hTRAIL and various inhibitors
for 30 minutes: RPMI 1640 (control), CBSA (0.1 or 1 mg/mL), polylysine
(50 or 500 Ag/mL), 0.1% w/v sodium azide, 450 mmol/L sucrose, 20 Amol/Lphenylarsinoxide, or 10 Ag/mL filipin. Cells were washed with ice-cold PBS,acid buffer [120 mmol/L NaCl, 20 mmol/L sodium barbital, 20 mmol/L
sodium acetate (pH 3)] at 4jC for 5 minutes and ice-cold PBS. Each well
received 50 AL PBS and 150 AL DMSO, and 6-coumarin fluorescence was
measured by spectrofluorometry. A standard curve was constructed by
measuring samples of 150 AL DMSO and 50 AL PBS with increasing
concentrations of the nanoparticles. Samples were tested in pentaplicate.
Nanoparticle uptake was presented as percent uptake of the control.
Intracellular tracking of nanoparticles and pDNA. C6 cells (1 � 104
per coverslip) were plated on glass coverslips coated with L-polylysine. After
24 hours and a 30-minute preincubation in RPMI 1640, cells were treated for
30 minutes with a suspension of 6-coumarin-labeled CBSA-NP-hTRAIL orNP-hTRAIL (100 Ag/mL) in RPMI 1640 with markers of endolysosomal
compartments, LysoTracker Blue (50 nmol/L), and of early and recycling
endosomes, Texas Red-transferrin (100 Ag/mL). Cells were washed, fixed,
and mounted in DakoCytomation fluorescent mounting medium. Imageswere captured with UV (LysoTracker Blue), fluorescein (6-coumarin), and
rhodamine (Texas Red-transferrin) filters using a Zeiss dual photon LSM
510 confocal microscope. Images were superimposed to determine the
intracellular nanoparticle localization.To track pDNA intracellularly, it was covalently labeled with the
fluorescent dye EMA. C6 cells plated as above and preincubated with RPMI
1640 for 30 minutes were treated with CBSA-NP-hTRAIL-EMA (100 Ag/mL)for 30 minutes. Cells were then incubated with blank RPMI 1640 and after5 hours were treated with LysoTracker Blue (50 nmol/L) for 30 minutes.
After washing, fixing, and mounting steps, slides were examined by confocal
microscopy, using a rhodamine filter for EMA-labeled DNA.In vitro gene expression and cell apoptosis determination. C6 cells
(1 � 105 per well) were cultured in six-well plates until 70% confluency was
reached. Cells were incubated with RPMI 1640 for 15 minutes and
transfected with 2 mL serum-free medium containing LipofectAMINE2000 plus 4 Ag pDNA, CBSA-NP-hTRAIL with 10 Ag encapsulated pDNA, orNP-hTRAIL with 10 Ag encapsulated pDNA (controls: medium, CBSA-NP, or
NP). After 5 hours, medium was replaced by 2 mL complete medium. Forty-
eight hours after transfection, soluble hTRAIL in the supernatant wasmeasured using a hTRAIL ELISA kit. Cells were trypsinized, stained by
propidium iodide, and analyzed by fluorescence-activated cell sorter for
apoptotic cells (FACSCalibur flow cytometry system, BD Biosciences, SanJose, CA).
I.c. tumor implantation. To allow in vivo tracking of proliferation, C6
cells were labeled with the vital dye CellTracker CM-DiI, according to the
manufacturer’s protocol. C6 cells were not labeled for other in vivo studies.C6 cells (5 � 104 in 4 AL RPMI 1640) were implanted into the right striatum
(2 mm lateral to the bregma and 3 mm of depth) of BALB/c mice by using a
stereotactic fixation device with mouse adaptor (Benchmark; ref. 6).
In vivo tracking of nanoparticles. Seven days after implantation, mice
received 6-coumarin-labeled CBSA-NP-hTRAIL (control: 6-coumarin-labeled
NP-hTRAIL) via the tail vein at a dose of 60 mg/kg body weight. After
30 minutes, animals were sacrificed, and brain coronal cryostat sections
(5 Am) were prepared and viewed with a confocal microscope using
rhodamine (CellTracker CM-DiI) and fluorescein (6-coumarin) filters. WGA
specifically binds negatively charged endothelial microdomains (27).
CBSA-NP–Mediated TRAIL Gene Transfer into Glioma Cells
www.aacrjournals.org 11879 Cancer Res 2006; 66: (24). December 15, 2006
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

To detect glycoproteins, sections were stained with Alexa Fluor 633-WGA
(2.5 Ag/mL, room temperature, 1 hour) and viewed with a far-red filter. For
intracellular tracking of nanoparticles, images were taken along the z-axis
at 0.1-Am intervals, and three-dimensional images were reconstructed using
Carl Zeiss AIM software (version 3.2).CBSA-NP loaded with osmium tetroxide was prepared by adding 1 mg
osmium tetroxide to the polymer solution before emulsification. At 7 daysafter implantation, mice received CBSA-NP-osmium tetroxide via the tailvein. Thirty minutes thereafter, brain tumors were removed and preparedfor examination by a Philips CM120 transmission electron microscope, aspreviously described (28).
Gene expression in vivo . At day 7 after implantation, mice received CBSA-NP-hTRAIL, NP-hTRAIL, or saline (control) via the tail vein at a dose of 100 AgpDNA/kg body weight. Mice were sacrificed after 24 hours for in situhybridization (ISH) or after 48 hours for immunohistochemistry. For ISH,5-Am brain coronal cryostat sections were prepared after perfusing anes-thesized mice with 4% paraformaldehyde. ISH used a custom-synthesizedbiotin-labeled hTRAIL-specific oligonucleotide probe (5¶-bio-ttgccagcaggg-gctgttcatactctcttcgtc-3¶; Shenergy Biocolor BioScience and Technology Co.,Shanghai, China) and the DNADetector Chromogenic In situ Hybridizationkit (KPL, Gaithersburg, MD). After proteinase K digestion, tissue washybridized with 50% formamide hybridization cocktail (equal volumes of2 � hybridization buffer and formamide) containing biotin-labeled probe(1 ng/mL, 37jC, 18 hours) and developed with TrueBlue. No endogenousmTRAIL expression was detected upon examination by light microscopy.
Immunohistochemical detection of hTRAIL protein used goat anti-hTRAIL IgG (1:100) as primary antibody followed by Cy3-labeled secondaryantibodies (1:50). After DAPI counterstaining, slides were mounted andexamined by fluorescence microscopy (Olympus, Tokyo, Japan).
To detect tumor hTRAIL expression-time course, tumor tissues of post-implantation mice were collected at 0.5, 1, 2, 4, and 6 days after dosing. Wholeprotein extracts were prepared by direct addition of cold lysis buffer to thesamples and mechanical homogenization of tissue (29). Protein quantitationwas done using bicinchoninic acid protein assay kit (Shenergy BiocolorBioScience and Technology) and 50 Ag of protein was loaded on 10% SDS-PAGE gels. Western blot analysis of hTRAIL was according to previousdescribed method (29). The dilution of primary antibody was 1:1,000.Horseradish peroxidase–conjugated rabbit anti-IgG goat (dilution 1:2,000;Pierce Biotech, Inc., Rockford, IL) was detected by chemiluminescence usingSuper Signal West Femto Maximum Sensitivity Substrate (Pierce Biotech).
Immunohistochemistry of apoptosis markers and survival monitor-ing. At days 8, 10, and 12 after implantation, mice received CBSA-NP-hTRAIL,NP-hTRAIL, or saline (control) via the tail vein at a dose of 100 Ag pDNA/kgbody weight (1-week treatment, three doses per week). At day 14 afterimplantation, mice were sacrificed, except for 10 mice of each treatmentgroup, which were monitored for survival. The sacrificed mice were used toprepare brain coronal paraffin sections (5 Am). Slides were eitherimmunostained with anti-active caspase-3 (1:100) or subjected to terminaldeoxynucleotidyl transferase–mediated nick-end-labeling (TUNEL; TACS TdTkit) to detect nuclear DNA fragments. Slides were developed withdiaminobenzidine and counterstained with methyl green. Besides the above1-week treatment groups, an extended treatment group with the same dose ofCBSA-NP-hTRAIL at days 8, 10, 12, 15, 17, and 19 after implantation (2-weektreatment, three doses per week) was conducted for survival monitoring.
Statistical analysis. Differences between treatment groups in the in vitrocellular uptake and protein expression assays were assessed using an unpaired
Student’s t test. Survival data were presented using Kaplan-Meier plots and
were analyzed using a log-rank test. P < 0.05 was considered significant.
Results
Characterization of nanoparticles. No significant differencesin particle size were found between NP-hTRAIL (108.6 F 10.8 nm)and CBSA-NP-hTRAIL (115.7 F 13.9 nm), suggesting that CBSAconjugation did not affect particle size. No differences wereobserved either in drug encapsulating efficiency (65.0 F 6.1%versus 57.5 F 8.3%) or drug loading efficiency (0.52 F 0.05%
versus 0.46 F 0.07%). Scanning electron micrography andtransmission electron micrography (TEM) showed that CBSA-NP-hTRAIL were spherical and of equal size (Fig. 1A1 and A2).Following a two-step immunostaining process using heparin/biotin and streptavidin/gold complexes, TEM detected goldparticles on the surface of CBSA-NP-hTRAIL, indicative of cova-lent conjugations between CBSA and nanoparticle (Fig. 1A3).The average zeta potential of CBSA-NP-hTRAIL was �15.4 F 1.3mV at pH 7, but increased with decreasing pH, reaching 11.3 F 1.8mV at pH 2 (Fig. 1B1). In contrast, the zeta potential of NP-hTRAIL changed little and remained negative with decreasingpH (�18.1 F 0.9 mV at pH 7 and �11.3 F 0.9 mV at pH 2).As shown in Fig. 1B2 , poly(ethyleneimine) has a strong proton-
buffering effect at pH range from 10 to 4, which was attributed toprotonation of the primary amine groups (pH >9) and tertiary andsecondary amines (pH range, 9–4; ref. 30). However, CBSA is aweaker pH buffering system. The charge contribution of CBSAincluded primary amine group introduced by ethylenediaminefollowing CBSA synthesis (19) and carboxyl group of acidic aminoacid residues. The former caused the initial high pH protonation(pH >9), whereas the cationization of CBSA at pH 9 to 4 might beattributed to protonation of carboxyl groups. Compared withCBSA, BSA has no proton-buffering capacity at pH >7 but a weakprotonation at pH 7 to 4.Both of the particles exhibited a similar biphasic pDNA release
pattern in Tris-EDTA buffer (pH 7.4), which was characterized bya first initial rapid release (>50% of pDNA was released withinthe first day) followed by a slower and continuous release (80%was released in 10 days; Fig. 1C1). Electrophoresis result showsthe ratio between relaxed to supercoiled conformation of pDNAextracted and 1-day released from CBSA-NP-hTRAIL in compar-ison with control pDNA increased from 10:90% to 60:40%(Fig. 1C2). The relaxed/supercoiled ratio of pDNA increased to90:10% after a 10-day release. However, no DNA fragments weredetected, which has been observed before and hypothesized toreflect a protective effect of the amphiphilic PLA-PEG onshearing of pDNA (17, 26). Analysis of pDNA extracted orreleased from NP-hTRAIL shows a similar pattern as those fromCBSA-NP-hTRAIL.
CBSA-NP-hTRAIL and NP-hTRAIL uptake. The uptake of bothCBSA-NP-hTRAIL and NP-hTRAIL by C6 cells was energydependent, as their uptake was reduced to 72.9% and 66.9%,respectively, after energy depletion by sodium azide (Fig. 2A).Phenylarsinoxide and hypertonic sucrose, inhibitors of endocytosis,also significantly decreased the intracellular uptake of CBSA-NP-hTRAIL and NP-hTRAIL. However, filipin, a specific inhibitor ofcaveolae-associated endocytosis, decreased uptake of CBSA-NP-hTRAIL but not NP-hTRAIL. Furthermore, free polycations,such as CBSA and polylysine, dose-dependently inhibited uptake ofCBSA-NP-hTRAIL, but high doses of these polycations did notinhibit uptake of NP-hTRAIL, suggesting that CBSA-NP-hTRAIL issubject to AMT.
Endolysosomal escape of nanoparticles and intracellularsorting of pDNA. To track the fate of the nanoparticlesintracellularly, C6 cells were incubated with Texas Red-transferrin(red fluorescent marker for early and recycling endosomes),LysoTracker Blue (blue fluorescent marker for secondary endo-somes and lysosomes), and the 6-coumarin-labeled nanoparticles.At 30 minutes after incubation, the superimposed image of Fig. 2B1shows that green fluorescence was both inside and outside the redand blue compartments, indicating that CBSA-NP-hTRAIL were
Cancer Research
Cancer Res 2006; 66: (24). December 15, 2006 11880 www.aacrjournals.org
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

not only inside endosomes and lysosomes but had spreadthroughout the cytoplasm. This suggested that following theiruptake CBSA-NP-hTRAIL escaped rapidly from the endolysosomalcompartments to the cytoplasm. Unlike CBSA-NP-hTRAIL, almostall green fluorescence of NP-TRAIL overlapped with blue
fluorescence, indicating NP-hTRAIL was entrapped in the second-ary endosome and lysosome rather than the early and recyclingendosomes (Fig. 2B2, superimposed image). The absence of greenfluorescence from the cytoplasm suggested a different intracellularsorting mechanism for NP-hTRAIL.
Figure 1. Characterization of CSBA-NP-hTRAIL and the pDNA (pORF-hTRAIL) encapsulated within CBSA-NP. A1, scanning electron micrograph of CBSA-NP-hTRAIL (�20K; 30 kV; bar, 200 nm). TEMs of (A2 ) CBSA-NP-hTRAIL and (A3) CBSA-NP-hTRAIL in which CBSA is labeled with 10-nm colloidal gold after atwo-step immunostaining process using heparin/biotin and streptavidin/gold complexes (1% phosphotungstic acid–negative staining; �120K; bar, 50 nm.).B1, zeta potentials of nanoparticles (200 Ag/mL) in 0.001 mol/L HEPES buffer adjusted to different pH values with either 0.1 mol/L sodium hydroxide or0.1 mol/L hydrochloric acid. Points, mean (n = 5); bars, SD. Note that CBSA-NP-hTRAIL but not NP-hTRAIL is cationic at acidic pH values. B2, acid-base titration:10 mg CBSA, BSA, or PEI1800 was dissolved in 10 mL of 150 mmol/L NaCl, adjusted to pH 10 with 1 mol/L NaOH. The solution was titrated with increasingvolume of 0.1 mol/L HCl. C1, cumulative release (% amount loaded) of pDNA from pDNA-loaded nanoparticles. Release was studied in Tris-EDTA buffer(pH 7.4) at 37jC. Points, mean (n = 4); bars, SD. C2, analysis of pDNA (pORF-hTRAIL) stability by agarose gel electrophoresis. Lane 1, DNA ladder; lane 2,original pDNA; lanes 3 and 6, pDNA extracted from CBSA-NP-hTRAIL and NP-hTRAIL, respectively; lanes 4 and 7, pDNA released from CBSA-NP-hTRAIL andNP-hTRAIL after 1 day, respectively; lanes 5 and 8, pDNA released from CBSA-NP-hTRAIL and NP-hTRAIL after 10 days, respectively. In lanes 2 to 8 , the topstrands are the relaxed conformation of pDNA, whereas the bottom strands are the supercoiled conformation.
CBSA-NP–Mediated TRAIL Gene Transfer into Glioma Cells
www.aacrjournals.org 11881 Cancer Res 2006; 66: (24). December 15, 2006
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
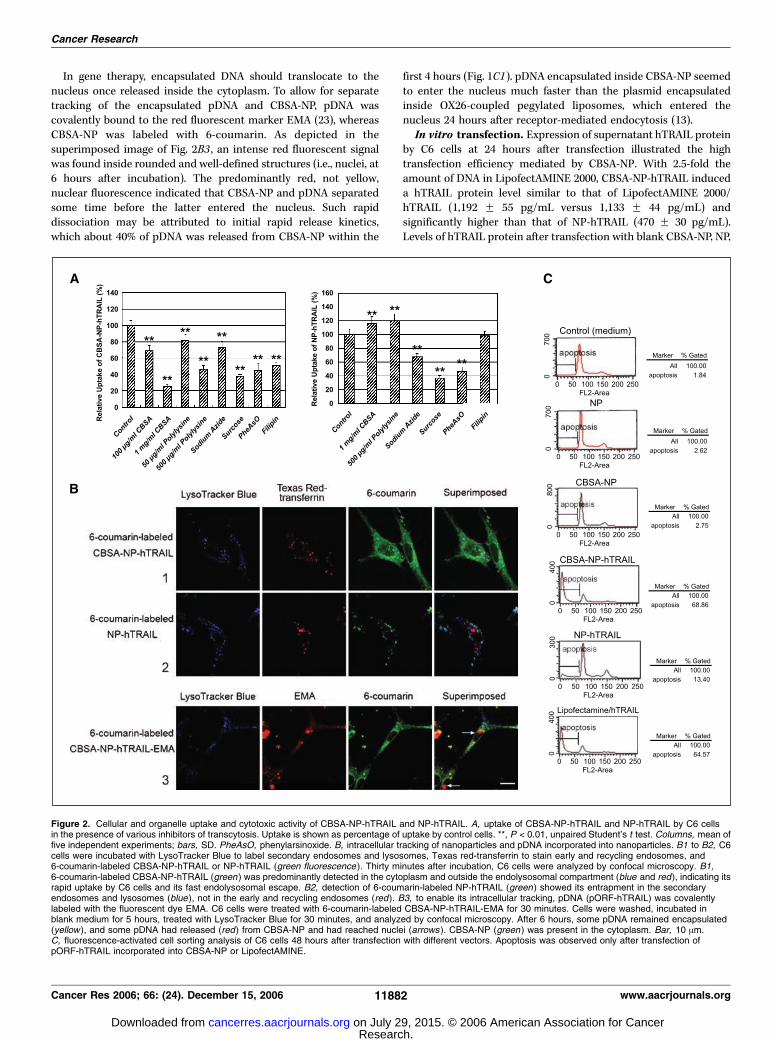
In gene therapy, encapsulated DNA should translocate to thenucleus once released inside the cytoplasm. To allow for separatetracking of the encapsulated pDNA and CBSA-NP, pDNA wascovalently bound to the red fluorescent marker EMA (23), whereasCBSA-NP was labeled with 6-coumarin. As depicted in thesuperimposed image of Fig. 2B3 , an intense red fluorescent signalwas found inside rounded and well-defined structures (i.e., nuclei, at6 hours after incubation). The predominantly red, not yellow,nuclear fluorescence indicated that CBSA-NP and pDNA separatedsome time before the latter entered the nucleus. Such rapiddissociation may be attributed to initial rapid release kinetics,which about 40% of pDNA was released from CBSA-NP within the
first 4 hours (Fig. 1C1). pDNA encapsulated inside CBSA-NP seemedto enter the nucleus much faster than the plasmid encapsulatedinside OX26-coupled pegylated liposomes, which entered thenucleus 24 hours after receptor-mediated endocytosis (13).In vitro transfection. Expression of supernatant hTRAIL protein
by C6 cells at 24 hours after transfection illustrated the hightransfection efficiency mediated by CBSA-NP. With 2.5-fold theamount of DNA in LipofectAMINE 2000, CBSA-NP-hTRAIL induceda hTRAIL protein level similar to that of LipofectAMINE 2000/hTRAIL (1,192 F 55 pg/mL versus 1,133 F 44 pg/mL) andsignificantly higher than that of NP-hTRAIL (470 F 30 pg/mL).Levels of hTRAIL protein after transfection with blank CBSA-NP, NP,
Figure 2. Cellular and organelle uptake and cytotoxic activity of CBSA-NP-hTRAIL and NP-hTRAIL. A, uptake of CBSA-NP-hTRAIL and NP-hTRAIL by C6 cellsin the presence of various inhibitors of transcytosis. Uptake is shown as percentage of uptake by control cells. **, P < 0.01, unpaired Student’s t test. Columns, mean offive independent experiments; bars, SD. PheAsO, phenylarsinoxide. B, intracellular tracking of nanoparticles and pDNA incorporated into nanoparticles. B1 to B2, C6cells were incubated with LysoTracker Blue to label secondary endosomes and lysosomes, Texas red-transferrin to stain early and recycling endosomes, and6-coumarin-labeled CBSA-NP-hTRAIL or NP-hTRAIL (green fluorescence ). Thirty minutes after incubation, C6 cells were analyzed by confocal microscopy. B1,6-coumarin-labeled CBSA-NP-hTRAIL (green ) was predominantly detected in the cytoplasm and outside the endolysosomal compartment (blue and red), indicating itsrapid uptake by C6 cells and its fast endolysosomal escape. B2, detection of 6-coumarin-labeled NP-hTRAIL (green ) showed its entrapment in the secondaryendosomes and lysosomes (blue ), not in the early and recycling endosomes (red). B3, to enable its intracellular tracking, pDNA (pORF-hTRAIL) was covalentlylabeled with the fluorescent dye EMA. C6 cells were treated with 6-coumarin-labeled CBSA-NP-hTRAIL-EMA for 30 minutes. Cells were washed, incubated inblank medium for 5 hours, treated with LysoTracker Blue for 30 minutes, and analyzed by confocal microscopy. After 6 hours, some pDNA remained encapsulated(yellow ), and some pDNA had released (red) from CBSA-NP and had reached nuclei (arrows ). CBSA-NP (green ) was present in the cytoplasm. Bar, 10 Am.C, fluorescence-activated cell sorting analysis of C6 cells 48 hours after transfection with different vectors. Apoptosis was observed only after transfection ofpORF-hTRAIL incorporated into CBSA-NP or LipofectAMINE.
Cancer Research
Cancer Res 2006; 66: (24). December 15, 2006 11882 www.aacrjournals.org
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

or vehicle-free culturemediumwere below the ELISA detection limit(46.8 pg/mL). As shown by fluorescence-activated cell sortinganalysis, the apoptotic effect of CBSA-NP-hTRAIL, with 2.5-foldthe amount of DNA in LipofectAMINE 2000, was similar to that ofLipofectAMINE 2000/hTRAIL and about five times higher than thatof NP-hTRAIL (Fig. 2C). Neither blank CBSA-NP nor NP causedapoptosis, showing the low toxicity of these nanoparticulate vectors.
Targeted transportation of CBSA-NP inside tumor. Todistinguish between donor and recipient cells after i.c. transplanta-tion, C6 cells were labeled with the red fluorescent, lipophilic tracerCM-DiI. CM-DiI-labeled C6 cells exhibited minimal cytotoxicity withcellular fluorescence labeling efficiency over 95% (Fig. 3A1). Sevendays after i.c. transplantation of the CM-DiI–labeled C6 cells,microscopic examination showed that they had good cell vitality andhad grown into distinct tumor tissue displaying red fluorescent cyto-lipid separation (Fig. 3A2). Thirty minutes after i.v. administration,more green fluorescence of CSBA-NP-hTRAIL than of NP-hTRAILwas detected in the tumor region, indicative of the brain tumor–specific accumulation of CBSA-NP-hTRAIL (Fig. 3A2). Three-dimensional reconstruction of brain tumor images through x-, y-,and z-axes showed colocalization of red and green fluorescence,suggesting that CBSA-NP-hTRAIL was taken in by tumor cells ratherthan remaining stagnant in the tumor interstitium (Fig. 3A3).To enter the brain via AMT, CBSA-NP-hTRAIL needs to selectively
bind to negatively charged glycoproteins in the brain capillary wall.To elucidate this interaction in vivo , a far-red fluorescent WGA wasused to detect such glycoproteins; negatively charged glycoproteinswere located in brain vasculature rather than parenchyma (Fig. 3B1to B2). Higher magnifications showed that they were expressed inboth the luminal and abluminal membranes of brain endothelialcells (Fig. 3B3 to B4). As shown in Fig. 3B3 , at 30 minutes after CBSA-NP-hTRAIL injection, the colocalization of fluorescent WGA withCBSA-NP-hTRAILwas detected not only in the luminal membrane ofendothelial cells (white arrowhead) but also in the abluminalmembrane (red arrowhead). CBSA-NP-hTRAIL was further beingtransported across the cytoplasm of endothelial cells (yellowarrowhead). This suggested that transcytosis of CBSA-NP-hTRAILacross the BBB might involve its electrostatic binding to thenegatively charged glycoproteins in the endothelial cell membrane.The transcytosis of NP-hTRAIL, however, seemed to be differentbecause NP-hTRAIL and glycoproteins in the endothelial cellmembrane did not colocalize (Fig. 3B4, arrows).In contrast to normal brain parenchyma, brain tumor expressed
higher levels of negatively charged glycoproteins, not only in tumorvasculature but also in tumor cells (Fig. 3B5 to B6). CBSA-NP-hTRAILwas found to be far more extensively distributed in tumors thanNP-hTRAIL. Moreover, white color in tumor tissue showed thestrong colocalization of CBSA-NP-hTRAIL and negatively chargedglycoproteins (Fig. 3B5). Three-dimensional reconstruction furthershowed that CSBA-NP-hTRAIL was overlapped with glycoproteins inthe tumor vessel wall, and that its transport across tumor bloodvessels involved the specific binding to anionic glycoproteins(Fig. 3B7). Virtually no colocalization between NP-hTRAIL andnegatively charged glycoproteins was detected in brain tumortissue (Fig. 3B6). Brain tumor–specific accumulation of CBSA-NP-hTRAIL was further substantiated by biodistribution data, whichshowed that increases in tissue clearance and uptake of CBSA-NP-hTRAIL were much higher in tumor than in normal brain tissues(see Supplementary Data).TEM of brain tumor vasculature proved that its pore cutoff size
was <100 nm (Fig. 3C1, arrowhead). CBSA-NP was clearly seen
adhering to membrane of endocytic vesicles of tumor endothelialcells (Fig. 3C2) and was further detected in the endolysosomalcompartment and cytoplasm of tumor cells (Fig. 3C3). Theseresults further proved that accumulation of CBSA-NP in braintumor tissue (Fig. 3A2) was accounted for by AMT.hTRAIL gene expression and tumor apoptosis. At 24 hours
after i.v. administration of CBSA-NP-hTRAIL, expression of hTRAILmRNA was detected throughout the cerebral cortex, periventricularregion, and tumor tissue (Fig. 4A). Moreover, hTRAIL mRNA levelsin brain tumor tissue were higher than in normal brain tissues.However, 24 hours after NP-hTRAIL administration, hTRAIL mRNAwas mainly found in ependymal cells that line the third ventricle(periventricular region). Expression of mRNA in tumor was lowercompared with those after CBSA-NP-hTRAIL administration. NohTRAIL mRNA was detected in saline control animals. At 48 hoursafter i.v. administration of CBSA-NP-hTRAIL, immunohistochem-ical analysis for the hTRAIL protein, using an antibody that did notcross-react with mTRAIL protein (31), detected specific hTRAILprotein expression in a region of the cerebral cortex inhabited byneurons and glia cells, as identified by morphology (Fig. 4B ,including inset). Focal areas of hTRAIL protein expression wereobserved in brain tumor tissues. At 48 hours after i.v. administra-tion of NP-hTRAIL, hTRAIL protein expression was undetectable inthe cerebral cortex and periventricular region, whereas low levels ofhTRAIL protein were found in brain tumor tissues. The expressionhTRAIL protein in tumor tissues is detectable 12 hours after i.v.injection of CBSA-NP-hTRAIL according to immunoblotting record(Fig. 4C1). The highest level of protein expression was found 2 daysafter dosing. The expression-time profile can remain 6 days,suggesting sustained expression effect by using nanoparticulatevectors. However, tumor hTRAIL expression following administra-tion of NP-hTRAIL was much weaker (Fig. 4C2). The absorbancelevel of hTRAIL protein 2 days after CBSA-NP-hTRAIL injection ismeasured 17-fold higher than that after NP-hTRAIL dosing.hTRAIL-mediated apoptosis was assessed by immunohistochem-
ical analysis for active caspase-3 using an antibody that detected thecleaved p17 fragment only. At 14 days after i.v. administration ofCBSA-NP-hTRAIL, p17-positive tumor cells were detected (Fig. 5A).The TUNEL assay further detected DNA fragmentation, a marker oflate apoptosis, in nuclei of tumor cells. Virtually no apoptotic cellswere detected 14 days after i.v. NP-hTRAIL treatment. The antitumoreffect of CBSA-NP-hTRAIL was also reflected in the median survivaltime of mice bearing i.c. brain tumor xenografts. After 1-weektreatment, the median survival time of CBSA-NP-hTRAIL-treatedmice (31.3 days) was significantly longer than that ofmice treatedwithNP-hTRAIL (21.5 days) or saline (20.5 days; Fig. 5B ; log-rank, P < 0.01).The extended administration of CBSA-NP-hTRAIL for 2 weeks canprolong themedian survival time to 41.5 days (P < 0.01 comparedwithCBSA-NP-hTRAIL for 1-week treatment).
Discussion
The present study shows for the first time tumor-specificnanoparticle delivery in the brain of an exogenous gene using anoninvasive, i.v. administration route. Fluorescence techniquesdeveloped in our laboratory showed that after i.v. administration,the nonviral vector CBSA-NP-hTRAIL crossed the BBB and accumu-lated in i.c. glioma xenografts via AMT. This study further showed thatthe encapsulated pDNA containing the hTRAIL genewas released andrapidly translocated to the nucleus, where hTRAILwas expressed andinduced apoptosis of tumor cells but not normal cells.
CBSA-NP–Mediated TRAIL Gene Transfer into Glioma Cells
www.aacrjournals.org 11883 Cancer Res 2006; 66: (24). December 15, 2006
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

Figure 3. In vivo uptake of CBSA-NP-hTRAIL and NP-hTRAIL into normal brain tissue and i.c. implanted C6 brain tumor. C6 cells were labeled with the vital dyeCellTracker CM-DiI and were i.c. implanted into the right striatum of BALB/c mice. Seven days after implantation, 6-coumarin-labeled CBSA-NP-hTRAIL or NP-hTRAILwas injected i.v., and after 30 minutes, the mice were sacrificed. Coronal cryostat sections of normal brain and brain tumor tissue were analyzed by confocalmicroscopy. A, i.t. accumulation of CBSA-NP-hTRAIL and NP-hTRAIL. A1, C6 cells labeled with CM-DiI (red) before i.c. implantation. A2, CBSA-NP-hTRAIL (green )accumulated more in brain tumor tissue (red) than NP-hTRAIL (green ). A3, three-dimensional reconstruction of brain tumor images showed CBSA-NP-hTRAIL (green )to be inside brain tumor cells (red). Bar, 50 Am (A1–A2 ) and 5 Am (A3 ). B, colocalization of CBSA-NP-hTRAIL and NP-hTRAIL with glycoproteins in brain vasculatureand brain tumors. Coronal cryostat sections of normal brain and brain tumor tissue were stained with Alexa Fluor 633-WGA, which specifically binds to negativelycharged residues in endothelium, and analyzed by confocal microscopy. B1 to B2, in normal brain, WGA-stained glycoproteins (purple , pseudocolor) were mainlydistributed in the endothelial membrane. B3, at higher magnification, transcytosis of CBSA-NP-hTRAIL (green ) across the BBB was shown by its binding to the luminalside (white arrowhead ), its presence inside the cytoplasm (yellow arrowhead), and its binding to the abluminal side (red arrowhead) of brain capillary endothelialcells (purple membrane ). B4, transcytosis (arrows ) of NP-hTRAIL (green ) in normal brain vasculature did not involve colocalization with glycoproteins (purple ).B5 to B7, in brain tumors, the negatively charged residues (purple ) were highly expressed in both tumor endothelial cells and tumor cells. Brain tumor accumulation of(B5) CBSA-NP-hTRAIL (green ) was higher than that of (B6 ) NP-hTRAIL (green ), with colocalization (white ) of most of CBSA-NP-hTRAIL and the anionic residues.B7, three-dimensional reconstruction of brain tumor images shows several CBSA-NP-hTRAIL (green ) specifically bound to the glycoproteins (purple ) present inthe tumor vascular wall. A, abluminal side of BBB; L, luminal side of BBB; V, blood vessel. Bar, 20 Am (B1–B7). C, absorptive-mediated transcytosis of CBSA-NPacross brain tumor microvasculature into brain tumor. Mice carrying i.c. implanted C6 brain tumors were injected i.v. with CBSA-NP loaded with osmium tetroxide atday 7 after implantation. After 30 minutes, brain tumors were removed and prepared for analysis by TEM. C1, TEM of brain tumor microvasculature revealed apore cutoff size <100 nm (arrowhead ). C2, CBSA-NP was found in endocytic vesicles of tumor endothelial cells (arrow). C3, in tumor cells, CBSA-NP was detectedinteracting with the vesicle membrane (arrow ) and free in the cytoplasm (arrowheads ). *, blood vessel; k, tumor interstitium; e, tumor endothelial cells; t, tumorcells. Bar, 200 nm (C1–C2 ) and 500 nm (C3).
Cancer Research
Cancer Res 2006; 66: (24). December 15, 2006 11884 www.aacrjournals.org
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

Unlike poly(ethyleneimine) with strong proton bufferingcapacity, CBSA has weaker. This indicated endolysosomal escapeof CBSA-NP cannot be explained as the ‘‘proton sponge effect,’’which caused swelling and rupture of lysosomes (32). Acid-basetitration curve of CBSA indicated initial protonation of theprimary amine groups followed by cationization of the carboxylgroups when increasing HCl volume. Due to the decreasing pH
from physiologic to lysosomal environment, further cationiza-tion of the carboxyl groups by the excess protons from theenvironment being transferred to CBSA resulted in increase inpositive charges of CBSA, even conversion of zeta potential ofCBSA-NP-hTRAIL from negative to positive. The positive-chargedCBSA-NP-hTRAIL could enhance its interaction with the negativelycharged endolysosomal membrane, which probably destabilized
Figure 4. In vivo expression of the hTRAIL gene innormal brain and i.c. implanted C6 brain tumor. C6cells were i.c. implanted into the right striatum ofBALB/c mice. At day 7 after implantation, micereceived CBSA-NP-hTRAIL, NP-hTRAIL, or saline(control) i.v. A, at 24 hours after the i.v.administration, half of the mice were anesthesizedand perfused with 4% paraformaldehyde. Coronalcryostat sections were prepared of normal brain andbrain tumor tissue. Using a biotin-labeledhTRAIL -specific oligonucleotide probe and thechromogen True Blue as developer, expression ofhTRAIL mRNA was detected in cerebral cortex,periventricular region, and brain tumor tissue after i.v.administration of CBSA-NP-hTRAIL. Expression ofhTRAIL mRNA after i.v. administration of NP-hTRAILwas lower and was observed in the periventricularregion and brain tumor tissue only. B, at 48 hoursafter the i.v. administration, the remaining mice weresacrificed, and coronal cryostat sections wereprepared of normal brain and brain tumor tissue.Expression of the hTRAIL protein was detected bybinding of goat anti-human TRAIL-specific IgGfollowed by anti-goat IgG-Cy3 antibody (red ). Cellnuclei were counterstained with DAPI. Following i.v.administration of CBSA-NP-hTRAIL, the hTRAILprotein was detected in normal brain and in focalareas in brain tumor. After i.v. injection of NP-hTRAIL, low levels of hTRAIL were found in braintumor only. Bar, 50 Am (A and B ). C, Westernblotting analysis. Expression of hTRAIL protein intumor at 0 (control), 0.5, 1, 2, 4, and 6 days followingi.v. injection of CBSA-NP-hTRAIL (C1 ) andNP-hTRAIL (C2). Immunoblotting of h-actin wasused as internal control and confirmation of equalloading of the samples.
CBSA-NP–Mediated TRAIL Gene Transfer into Glioma Cells
www.aacrjournals.org 11885 Cancer Res 2006; 66: (24). December 15, 2006
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

the membrane at the point of contact, followed by extrusion intocytoplasm (33). This hypothesis was indirectly proven by thedemonstration that at 30 minutes after i.v. administration, NP-hTRAIL, which has a negative zeta potential at both physiologicand acidic pH, was mostly detected in secondary endosomes andlysosomes, not in cytoplasm.
Several lines of evidence support the notion that CBSA-NP-hTRAILpasses through the BBB and tumor vascular endothelial cells by AMT.First, the electrostatic binding between CBSA-NP and the anionicmicro-domains of brain capillaries could be proven by the positiveimmuno-gold staining results in vitro (Fig. 1A3), as heparan sulfatesare themajor components of negatively charged glycoproteins located
Figure 5. Effects of CBSA-NP-hTRAIL andNP-hTRAIL on apoptosis and growth of i.c.implanted C6 brain tumors. C6 cells were i.c.implanted into the right striatum of BALB/c mice.The mice received 1-week treatment of CBSA-NP-hTRAIL, NP-hTRAIL, or saline (control; i.e.,dosing at days 8, 10, and 12 after implantationfollowing i.v. injection). At day 14 after implantation,mice were sacrificed for immunohistochemistry,except for 10 mice of each treatment group, whichwere monitored for survival. A, coronal paraffinsections of brain tumors were immunostained forthe presence of active caspase-3 or DNAfragments (TUNEL). Slides were developed withdiaminobenzidine and counterstained with methylgreen. CBSA-NP-hTRAIL but not NP-hTRAILinduced apoptosis of brain tumor cells in vivo . Bar,50 Am. B, Kaplan-Meier survival curve of miceharboring i.c. C6 gliomas. Besides the above1-week treatment groups, an extended treatmentgroup with the same dose of CBSA-NP-hTRAIL atdays 8, 10, 12, 15, 17, and 19 after implantation(2-week treatment) was conducted for survivalmonitoring. Mice received 1-week treatmentof CBSA-NP-hTRAIL, designated as CBSA-NP-hTRAIL (1w), survived significantly longerthan mice that received i.v. administrations ofNP-hTRAIL or saline (P < 0.01, log-rank analysis).Mice treated with CBSA-NP-hTRAIL for 2 weeks,designated as CBSA-NP-hTRAIL (2w), survivedsignificantly longer than CBSA-NP-hTRAIL (1w)treated group (P < 0.01, log-rank analysis).
Cancer Research
Cancer Res 2006; 66: (24). December 15, 2006 11886 www.aacrjournals.org
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

in those microvessels (19, 27). Second, our findings of CSBA-NP-hTRAIL uptake by C6 glioma cells showed that this uptake was energydependent and was specifically inhibited by free polycations, such asCBSA and polylysine. Third, we showed AMTof CBSA-NP-hTRAIL bythe BBB in normal brain tissue. By using far-red fluorescent WGA,which specifically binds to glycoprotein microdomains in the braincapillary wall (27), CBSA-NP-hTRAIL and glycoproteins were found tocolocalize on the luminal and abluminal membranes of the brainendothelium. CBSA-NP-hTRAIL was also detected in the endothelialcytoplasm. Fourth, we observed AMT of CBSA-NP-hTRAIL byendothelial cells of the brain tumor vasculature. Glycoproteins werefound to be highly expressed in brain tumor endothelial cells and tocolocalize with CBSA-NP-hTRAIL. Furthermore, TEM showed CBSA-NP to be interacting with the luminal surface of the membrane of abrain tumor endothelial cell. This is in line with a previous report thatthe preferential uptake of cationic molecules by angiogenic tumorepithelium occurred mostly through vesicular organelles (34). Finally,we showed that AMT of CBSA-NP-hTRAIL resulted in its uptake bybrain tumor cells. In brain tumors, green fluorescent CBSA-NP-hTRAILwas detected inside red fluorescent tumor cells, but not in theinterstitium. TEM showedCBSA-NP in an endocytic vesicle and free inthe cytoplasm of tumor cells. CBSA-NP-hTRAIL further colocalizedwith glycoproteins, highly expressed by brain tumor cells. Increasedtumor uptake of CBSA-NP-hTRAIL was evidenced by analysis ofbiodistribution (see Supplementary Data).TRAIL selectively induces apoptosis of glioma cells via its
cognate DR4 and DR5 receptors, whereas normal cells are
protected by virtue of expression of the DcR1 and DcR2antagonistic receptors (reviewed in ref. 5). In this study, TRAILdelivered via systemic administration of CBSA-NP-hTRAIL had amoderate effect on median survival of tumor-bearing mice(41 versus 22 days for control animals) compared with the increasein median survival reported for TRAIL delivered via retroviralvector (>100 versus 36 days for control animals; ref. 6). Still,retroviral vectors require i.t. delivery via craniotomy or intracarotidarterial infusion, and the associated safety risks preclude retroviralclinical use of vectors (35). The nontoxic nature of nanoparticles,on the other hand, allows repeated i.v. administrations of CBSA-NP-hTRAIL and, thus, along with the known bystander effect ofTRAIL (36), improves antitumor action.In conclusion, this study showed the feasibility of systemic
administration of CBSA-NP-hTRAIL as a nonviral vector for genetherapy of glioma. The current findings encourage further studiesinto the application of nonviral vectors for noninvasive genetherapy of malignant glioma.
Acknowledgments
Received 6/28/2006; revised 9/26/2006; accepted 10/5/2006.Grant support: National Natural Science Foundation of China grant 30472095
(X. Jiang).The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. Olivia Cholewa (Molecular Probes Detection Technologies in Eugene,OR) for valuable technical assistance with fluorescent tracking and Dr. Anita Frijhofffor writing and editing contributions.
References1. Rich JN, Bigner DD. Development of novel targetedtherapies in the treatment of malignant glioma. Nat RevDrug Discov 2004;3:430–46.
2. Rieger J, Naumann U, Glaser T, et al. APO2 ligand: anovel lethal weapon against malignant glioma? FEBSLett 1998;427:124–8.
3. KeaneMM, Ettenberg SA, NauMM, et al. Chemotherapyaugments TRAIL-induced apoptosis in breast cell lines.Cancer Res 1999;59:734–41.
4. Pan G, Ni J, Wei YF, et al. An antagonist decoy receptorand a death domain-containing receptor for TRAIL.Science 1997;277:815–8.
5. Hawkins CJ. TRAIL and malignant glioma. VitamHorm 2004;67:427–52.
6. Roth W, Isenmann S, Naumann U, et al. LocoregionalApo2L/TRAIL eradicates intracranial human malignantglioma xenografts in athymic mice in the absence ofneurotoxicity. Biochem Biophys Res Commun 1999;265:479–83.
7. Pardridge WM. The blood-brain barrier: bottleneck inbrain drug development. NeuroRx 2005;2:3–14.
8. Culver KW, Ram Z, Wallbridge S, et al. In vivo genetransfer with retroviral vector-producer cells for treat-ment of experimental brain tumors. Science 1992;256:1550–2.
9. Nilaver G, Muldoon LL, Kroll RA, et al. Delivery ofherpesvirus and adenovirus to nude rat intracerebraltumors after osmotic blood-brain barrier disruption.Proc Natl Acad Sci U S A 1995;92:9829–33.
10. Dewey RA, Morrissey G, Cowsill CM, et al. Chronicbrain inflammation and persistent herpes simplex virus1 thymidine kinase expression in survivors of syngeneicglioma treated by adenovirus-mediated gene therapy:implications for clinical trials. Nat Med 1999;5:1256–63.
11. McMenamin MM, Byrnes AP, Charlton HM, et al. Agamma34.5 mutant of herpes simplex 1 causes severeinflammation in the brain. Neuroscience 1998;83:1225–37.
12. Shi N, Pardridge WM. Noninvasive gene targeting tothe brain. Proc Natl Acad Sci U S A 2000;97:7567–72.
13. Zhang Y, Jeong Lee H, Boado RJ, et al. Receptor-
mediated delivery of an antisense gene to human braincancer cells. J Gene Med 2002;4:183–94.
14. Zhang Y, Schlachetzki F, Pardridge WM. Global non-viral gene transfer to the primate brain followingintravenous administration. Mol Ther 2003;7:11–8.
15. Shi N, Boado RJ, Pardridge WM. Receptor-mediatedgene targeting to tissues in vivo following intravenousadministration of pegylated immunoliposomes. PharmRes 2001;18:1091–5.
16. Gref R, Minamitake Y, Perracchia MT, et al. Biode-gradable long-circulating polymeric nanoparticles. Sci-ence 1994;263:1600–3.
17. Perez C, Sanchez A, Putnam D, et al. Poly(lactic acid)-poly(ethylene glycol) nanoparticles as new carriers for thedelivery of plasmid DNA. J Control Release 2001;75:211–4.
18. Lu W, Tan YZ, Hu KL, et al. Cationic albuminconjugated pegylated nanoparticle with its transcytosisability and little toxicity against blood-brain barrier. Int JPharm 2005;295:247–60.
19. Lu W, Zhang Y, Tan YZ, et al. Cationic albumin-conjugated pegylated nanoparticles as novel drug carrierfor brain delivery. J Control Release 2005;107:428–48.
20. Kumagai AK, Eisenberg JB, Pardridge WM. Absorp-tive-mediated endocytosis of cationized albumin and ah-endorphin-cationized albumin chimeric peptide byisolated brain capillaries. Model system of blood-brainbarrier transport. J Biol Chem 1987;262:15214–9.
21. Thole M, Nobmanna S, Huwyler J, et al. Uptake ofcationzied albumin coupled liposomes by culturedporcine brain microvessel endothelial cells and intactbrain capillaries. J Drug Target 2002;10:337–44.
22. Davda J, Labhasetwar V. Characterization of nano-particle uptake by endothelial cells. Int J Pharm 2002;233:51–9.
23. Roy I, Ohulchanskyy TY, Bharali DJ, et al. Opticaltracking of organically modified silica nanoparticles asDNA carriers: a nonviral, nanomedicine approach forgene delivery. Proc Natl Acad Sci U S A 2005;102:279–84.
24. Benns JM, Choi JS, Mahato RI, et al. pH-sensitivecationic polymer gene delivery vehicle: N -Ac-poly(L-histidine)-graft-poly(L-lysine) comb shaped polymer.Bioconjug Chem 2000;11:637–45.
25. Tang MX, Szoka FC. The influence of polymerstructure on the interactions of cationic polymers withDNA and morphology of the resulting complexes. GeneTher 1997;4:823–32.
26. Cohen H, Levy RJ, Gao J, et al. Sustained delivery andexpression of DNA encapsulated in polymeric nano-particles. Gene Ther 2000;7:1896–905.
27. Vorbrodt AW. Ultracytochemical characterization ofanionic sites in the wall of brain capillaries. J Neurocytol1989;18:359–68.
28. Panyam J, Sahoo SK, Prabha S, et al. Fluorescenceand electron microscopy probes for cellular and tissueuptake of poly(D,L-lactide-co-glycolide) nanoparticles.Int J Pharm 2003;262:1–11.
29. Grataroli R, Vindrieux D, Selva J, et al. Characteriza-tion of tumour necrosis factor-a-related apoptosis-inducing ligand and its receptors in the adult humantestis. Mol Hum Reprod 2004;10:123–8.
30. Lee M, Rentz J, Han SO, et al. Water-solublelipopolymer as an efficient carrier for gene delivery tomyocardium. Gene Ther 2003;10:585–93.
31. Wiley SR, Schooley K, Smolak PJ, et al. Identificationand characterization of a new member of the TNFfamily that induces apoptosis. Immunity 1995;3:673–82.
32. Boussif O, Lezoualc’h F, Zanta MA, et al. A versatilevector for gene and oligonucleotide transfer into cells inculture and in vivo : polyethylenimine. Proc Natl AcadSci U S A 1995;92:7297–301.
33. Panyam J, Zhou WZ, Prabha S, et al. Rapid endo-lysosomal escape of poly(DL-lactide-co-glycolide) nano-particles: implications for drug and gene delivery.FASEB J 2002;16:1217–26.
34. Thurston G, McLean JW, Rizen M, et al. Cationicliposomes target angiogenic endothelial cells in tumorsand chronic inflammation in mice. J Clin Invest 1998;101:1401–13.
35. El-Aneed A. An overview of current delivery systemsin cancer gene therapy. J Control Release 2004;94:1–14.
36. Kagawa S, He C, Gu J, et al. Antitumor activity andbystander effects of the tumor necrosis factor-relatedapoptosis-inducing ligand (TRAIL) gene. Cancer Res2001;61:3330–8.
CBSA-NP–Mediated TRAIL Gene Transfer into Glioma Cells
www.aacrjournals.org 11887 Cancer Res 2006; 66: (24). December 15, 2006
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

2006;66:11878-11887. Cancer Res Wei Lu, Qing Sun, Jin Wan, et al. AdministrationGene Delivery into Brain Tumors via Intravenous
Conjugated Pegylated Nanoparticles Allow−Cationic Albumin
Updated version
http://cancerres.aacrjournals.org/content/66/24/11878
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2006/12/15/66.24.11878.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/66/24/11878.full.html#ref-list-1
This article cites 36 articles, 13 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/66/24/11878.full.html#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on July 29, 2015. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN



















