
Carbon dynamics of different land use systems in NW Ethiopian
229
University of Natural Resources and Life Sciences, Vienna Institute of Forest Ecology Carbon dynamics of different land use systems in NW Ethiopian Dissertation A thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy (Dr. nat. techn.) at University of Natural Resources and Life Sciences, Vienna Submitted by MSc. Dessie Assefa Supervisor: Univ. Prof. Ph.D. Dr. Douglas L. Godbold 1 Co-supervisor: Ass. Prof. Dipl.-Biol. Dr.rer.nat. Boris Rewald 1 March 2017 Vienna, Austria 1 Institute of Forest Ecology at Department of Forest and Soil Science
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Carbon dynamics of different land use systems in NW Ethiopian

University of Natural Resources and Life Sciences, Vienna
Institute of Forest Ecology
Carbon dynamics of different land use systems in
NW Ethiopian
Dissertation
A thesis submitted in conformity with the requirements for the degree of Doctor of
Philosophy (Dr. nat. techn.) at University of Natural Resources and Life Sciences,
Vienna
Submitted by
MSc. Dessie Assefa
Supervisor: Univ. Prof. Ph.D. Dr. Douglas L. Godbold1
Co-supervisor: Ass. Prof. Dipl.-Biol. Dr.rer.nat. Boris Rewald1
March 2017
Vienna, Austria
1Institute of Forest Ecology at Department of Forest and Soil Science

I
Abstract
The amount of carbon (C) stored in the upper 100 cm soil is about three times greater than C stored in vegetation and twice as much as C present in the atmosphere. Soil organic carbon (SOC) is accumulated if inputs from leaf litterfall, root turnover, and other biomass are greater than losses by mineralization, erosion, and other processes. Potential shifts in SOC stocks and rising atmospheric CO2 concentrations are major concerns of climate change. Despite the large quantity of C stored in soil, consensus is lacking on the dynamics and the extent of land use change effects on soil C storage especially for tropical ecosystems. Several field and laboratory techniques were carried out to examine the magnitude of SOC stock changes due to land use conversion, the spatial distribution of SOC stocks, the origin, chemical composition of input materials and their decomposition rates, and substrate and temperature dependency of microbial respiration. The study was conducted in the NW Ethiopian highlands and a semi-arid savannah woodland. The land use systems studied were natural forest, eucalyptus plantation, exclosure, grazing land, and cropland. Total C and N stocks were determined along a climatic gradient. Litterfall was determined by litter traps and fine root production was estimated using sequential coring, ingrowth core and ingrowth net methods. Litter decomposition rates of leaves and fine roots were estimated using a litterbag technique. Chemical compositions of input materials were characterized by sequential extraction. Individual biomarkers were identified and quantified by gas chromatography and mass spectrometry at a molecular level. The temperature sensitivity of soil respiration was measured using a MicroResp method - glucose, lignin, and starch were added as a proxy for substrate availability and nitrogen and phosphorous were supplied as nutrient proxies to determine the factors limiting microbial respiration. Results showed that land use conversion from native forest to cropland or grazing land reduced SOC stocks by 70-90% in less than 50 years. Afforestation and exclosure led to an increase of SOC; however, the rate of increase (ca. 0.3 kg m-2 yr-1) was lower than losses (ca. 0.4 kg m-2 yr-1). The geomarker analysis (Sr/Ca and Ba/Ca ratio) and vertical distribution of carbon suggests that the major factor for SOC reduction is erosion while the C loss through mineralization ranged from 1.6-3.9 mg g soil-1 yr-1 and increased with increasing temperature. This is consistent with C availability in the soil. Soils supplemented with glucose increased heterotrophic respiration by 10-fold. The annual litterfall production in the natural forest ecosystem was about 1100 g m-2 of which leaf comprised 65%. Above-ground litter inputs in eucalyptus, cropland, and grazing land are estimated to be minor due to leaf litter raking, complete residual harvest, and overgrazing. The annual fine root production was estimated to be 700 g m-2 in natural forest and eucalyptus whereas fine root production in grazing land and cropland was about 50-60 g m-2. This illustrates that conversion of native forest to grazing land and cropland resulted in the reduction of C input into the soil by >90%. Fine roots decomposed slower than leaf litters with decomposition rate constant of 1.7 yr-1 for fine roots and 2.5 yr-1 for leaves. The variation in decomposition is due to their chemical composition. The acid-insoluble fraction in fine roots (50%) was greater than leaves (42%). Thus, fine roots contributed about 1.5 times more recalcitrant C input into the soil than leaf litters. From the biomarker analysis, the amount of suberin was 2-times that of cutin further confirming higher inputs of recalcitrant carbon from fine roots. The ratio of lignin-derived phenols of syringyls to vanillyls suggested that angiosperm plants are the predominant sources for lignin. The dominance of aliphatic lipids and lignin in SOC revealed that higher plants are a major input of SOC while microbial inputs were present as minor components (<1%). Overall, conversion of native forest to open lands in Ethiopian highland resulted in substantial loss of SOC stock due to both erosion and mineralization. The major inputs of SOC are fine roots of angiosperm plants due to their larger carbon, lignin and suberin contents and this slower decomposition. The results suggest C loss due to land use change in the Amhara region needs urgent attention. Future land use management need to raise awareness on the importance of SOC management as the basis for essential ecosystem functions and improve food security.

II
Abstrakt
Die Menge an gespeichertem Kohlenstoff (C) in den oberen 100 cm Boden ist etwa dreimal mehr als die der Vegetation und doppelt so viel wie in der Atmosphäre vorhandene C. Organischer Kohlenstoff im Boden (SOC) wird akkumuliert, wenn der C Eintrag durch Laubfall, Wurzelumsatz und andere Biomasse größer ist als die Verluste durch Mineralisierung, Erosion und andere Prozesse. Trotz der großen Wichtigkeit von Bodenkohlenstoff für globale biogeochemische Prozesse ist der Stand des Wissens über die SOC-Dynamik im Allgemeinen und die Auswirkungen von Landnutzungsänderungen im Speziellen vor allem für tropische Ökosysteme noch sehr limitiert. In dieser Studie wurden daher verschiedenste Feld- und Laboruntersuchungen durchgeführt um SOC-Bestandsveränderungen aufgrund von Landnutzungsumwandlung, die räumlichen Verteilung der SOC-Bestände, der Herkunft und chemische Zusammensetzung der Ausgangsmaterialien und ihrer Zersetzungsraten, sowie die Substrat- und Temperaturabhängigkeit der mikrobiellen Atmung zu untersuchen. Die untersuchten Landnutzungssysteme im Äthiopischen Hochland und der Savanne waren artenreiche Mischwälder, eine Eukalyptus-Plantage, eine Weideausschlußfläche, sowie Weide- und Ackerland. Gesamtbodenkohlenstoff und -stickstoff wurden entlang eines Klimagradienten bestimmt. Der Laubfall wurde mit Streusammlern, die Feinwurzelproduktion unter Verwendung von sequenzieller Bohrkernbeprobung, und Einwuchssäulen und -netzen bestimmt. Zersetzungsraten von Blättern und Feinwurzeln wurden unter Verwendung von litterbags gemessen. Die chemische Zusammensetzung der Ausgangsmaterialien wurden durch sequentielle Extraktion charakterisiert. Einzelne Biomarker wurden durch Gaschromatographie und Massenspektrometrie auf molekularer Ebene identifiziert und quantifiziert. Die Temperaturempfindlichkeit und Substratabhängigkeit der Bodenatmung wurde unter Verwendung eines MicroResp-Verfahrens gemessen. Die Ergebnisse zeigen, dass die Umwandlung von natürlichen Wäldern zu Acker- oder Weideland den Bodenkohlenstoffvorrat in weniger als 50 Jahren um 70-90% reduziert. Aufforstung und Ausschluss von Weidetieren führen zu einer Erhöhung des SOC-Vorrats. Allerdings ist die Akkumulationsrate (ca. 0,3 kg m-2 a-1) niedriger als die Verlustrate (ca. 0,4 kg m-2 a-1). Die Geomarker-Analyse (Sr:Ca- bzw. Ba:Ca-Verhältnisse) und die vertikale Verteilung von Kohlenstoff deuten darauf hin, dass Erosion der Hauptfaktor für die SOC-Abnahme war. Die heterotrophe Atmung ist durch die Kohlenstoffverfügbarkeit im Boden eingeschränkt. Die jährliche oberirdische Streuproduktion im Naturwald beträgt etwa 1100 g m-2, wobei der Blattanteil 65% betrug. Überirdische Streueinträge sind in der Eukalyptusplantage, sowie im Acker- und Weideland vernachlässigbar. Die jährliche Feinwurzelproduktion wurde in beiden Waldwirtschaftsformen auf ca. 700 g m-2 geschätzt, in Weide- und Ackerland auf etwa 50-60 g m-2. Dies zeigt, dass die Umwandlung von Wald zu Weide- bzw. Ackerland den Kohlenstoffeintrag in den Boden um >90% verringert. Die Feinwurzeln der untersuchten Bäume zersetzten sich langsamer als deren Blätter, mit einer Abbaukonstante von 1,7 a-1 für Feinwurzeln und 2,5 a-1 für Blätter. Die Zersetzungsrate ist abhängig von der chemische Zusammensetzung, insbesondere ist die säureunlösliche Fraktion in Feinwurzeln (50%) größer als in Blättern (42%). In der Schlussfolgerung tragen im untersuchten Waldsystem die Wurzeln etwa 1,5-mal mehr zum Eintrag recalcitranten Kohlenstoffs in den Boden bei als Blattstreu. Dies wird auch durch die Ergebnisse der Biomarkeranalyse gestützt. Diese zeigen, dass die Menge an Suberin (aus Wurzeln) im Boden die des Cutins (aus Blättern) um das Zweifache übersteigt. Die Biomarker zeigen zudem, dass der überwiegende Teil des Bodenkohlenstoffs von höheren Pflanzen im Allgemeinen und Angiospermen im Besonderen stammt. Insgesamt führte die Umwandlung von Wald in landwirtschaftlich genutzte Flächen im äthiopischen Hochland zu einer erheblichen Reduzierung des Bodenkohlenstoffvorrats, insbesondere durch verstärkte Erosion und Mineralisierungsraten. Der Bodenkohlenstoff stammt hauptsächlich von Feinwurzeln von Angiospermen, diese weisen größere Kohlenstoff-, Lignin- und Suberin-Gehalte auf als Blätter und zersetzen sich langsamer. Die Ergebnisse der Arbeit verdeutlichen, dass der Verlust von Bodenkohlenstoff aufgrund von Landnutzungsänderungen in der Region Amhara ein extremes Ausmaß angenommen hat und die Lösung dringend politische Aufmerksamkeit erfordert. Ein zukünftiges Landnutzungsmanagement muss vor Allem das Bewusstsein für die Wichtigkeit von Bodenkohlenstoff zur Aufrechterhaltung wesentlicher Ökosystemfunktionen stärken um u.a. die landwirtschaftliche Produktion zu stabilisieren und damit die Ernährungssicherheit in der Region zu gewährleisten.

III
Acknowledgment
I sincerely thank Prof. Douglas L. Godbold, who has always encouraged, supported my
research activities, and provided me the opportunity to become a PhD student at the Institute
of Forest Ecology, University of Natural Resources and Life Science in Austria. I appreciate
that he was always available when I needed his help and gave generous amounts of advice. I
would also like to thank Ass. Prof. Boris Rewald for his enthusiasm and fruitful discussions of
the numerous experiments and manuscripts. I am grateful to PD Hans Sandén for his support,
guidance, and useful suggestions throughout this study including his advice during numerous
field trips. I additionally thankful to Prof. Egbert Matzner and PD Gernot Bodner for their
willingness and time to serve on my comprehensive exam and defense committee.
I am thankful to Christoph Rosinger, Marcel Hirsch, and Frauke Neuman for their technical
advice and assistance with sample analysis in the laboratory. I am grateful to Dr. Karin
Wriessnig for her kind help during importing process for plant and soil samples for the last three
years as well as training me on laboratory analysis. Many thanks to Astrid Hobel and Prof. Axel
Mentler, from the Institute of Soil Science, for training me on gas chromatograph - mass
spectrometer (GC-MS), high performance liquid chromatography (HPLC), and other various
instruments in the laboratory – this thesis would not have been possible without their help!
I want to give my special thanks to Sigrid Gubo, for her dedication and help on my private
matters – making my life in Vienna very easy and enjoyable. I am also thankful to Martin
Wresowar for his quick technical support related to software whenever needed. I would like to
express my special thanks to members of the Institute of Forest Ecology and my officemates
and friends Dr. Iftekhar Ahmed and Dr. Norbu Wangdi. Many thanks should go to Dr. Mathias
Mayer, and Dr. Bradley Matthews who supported me mentally and cheered me up in the past
three years. It was a great working, talking, and laughing with members of the Institute of Forest
Ecology, which I could not mention all your names here and thank you for making my stay in
Vienna so much memorable.
I am very grateful to my wife, Nebyate Kebede, for her help during root washings. Washing
more than 5,000 root samples would not be possible without her help. Thank you for sharing
life in Vienna with me. Special thanks to Peter Kube and Christiane from Germany for their
love, encouragement, and for their frequent visits.
Finally, this work was carried out within the project (“Carbo-part”) funded by the Austrian
Ministry of Agriculture, Forestry, Environment, and Water Management. It would have not been
possible without the financial help of this project (“Carbo-part”) and the Austrian government.

IV
Table of contents
Abstract ....................................................................................................................... I
Abstrakt ...................................................................................................................... II
Acknowledgment ....................................................................................................... III
Table of contents ....................................................................................................... IV
List of figures ........................................................................................................... VIII
List of tables ............................................................................................................... X
1 General Introduction ............................................................................................. 1
1.1 Land degradation in the Amhara region: history, extent, causes, and consequences .................................................................................................................................. 1
1.2 The nature and formation of soil organic carbon ...................................................... 4
1.2.1 Soil carbon stock and temporal changes after land use change ..................................... 4
1.2.2 Mechanisms for soil organic carbon accumulation .......................................................... 6
1.2.3 Tracing the biological origin and degradation status of organic carbon in soil ................ 9
1.3 Aims and outline .....................................................................................................11
1.4 Hypotheses ............................................................................................................12
2 Soil organic carbon dynamics after land use change in Northwest Ethiopia ...... 13
2.1 Abstract ..................................................................................................................13
2.2 Introduction ............................................................................................................13
2.3 Material and methods .............................................................................................16
2.3.1 Research sites ............................................................................................................... 16
2.3.2 Soil sampling ................................................................................................................. 19
2.3.3 Bulk density determination ............................................................................................ 19
2.3.4 Total C and N stock determination ................................................................................ 20
2.3.5 Strontium, calcium and barium elemental analysis ....................................................... 21
2.3.6 Determination of soil pH ................................................................................................ 21
2.3.7 Particle size determination ............................................................................................ 21
2.3.8 Data analysis ................................................................................................................. 22
2.4 Results ...................................................................................................................23
2.4.1 Soil carbon and nitrogen stocks .................................................................................... 23
2.4.2 Vertical distributions of C and N .................................................................................... 26
2.4.3 Soil bulk density ............................................................................................................. 28
2.4.4 Strontium:calcium and barium:calcium ratios ................................................................ 29
2.4.5 Factors effecting soil carbon stocks .............................................................................. 29
2.5 Discussion ..............................................................................................................31
2.5.1 Soil organic carbon and nitrogen stocks ....................................................................... 31
2.5.2 Effect of land use change on carbon stock ................................................................... 32
2.5.3 Potentials of soil carbon gain due to afforestation and exclosure ................................. 35
2.5.4 Magnitude of soil carbon loss due to deforestation ....................................................... 36
2.6 Conclusion .............................................................................................................36

V
3 Fine root dynamics in Afromontane forest and adjacent land uses in the Ethiopian highlands .................................................................................................. 37
3.1 Abstract ..................................................................................................................37
3.2 Introduction ............................................................................................................37
3.3 Materials and Methods ...........................................................................................40
3.3.1 Site descriptions ............................................................................................................ 40
3.3.2 Root sampling ................................................................................................................ 41
3.3.3 Soil sampling ................................................................................................................. 43
3.3.4 Estimation of annual fine root production, mortality, and decomposition ...................... 43
3.3.5 Calculation of fine root turnover ..................................................................................... 47
3.3.6 Fine root vertical distribution .......................................................................................... 47
3.3.7 Carbon and nitrogen analysis ........................................................................................ 47
3.3.8 Statistical analyses ........................................................................................................ 48
3.4 Results ...................................................................................................................48
3.4.1 Fine root biomass, necromass and distribution with depth ........................................... 48
3.4.2 Seasonal variation of root stocks ................................................................................... 51
3.4.3 Fine root production, mortality, and turnover ................................................................. 52
3.4.4 Annual C and N flux into the soil ................................................................................... 57
3.5 Discussion ..............................................................................................................57
3.5.1 Effect of land use change on fine root stocks and production ....................................... 57
3.5.2 Seasonal variation of fine root mass ............................................................................. 59
3.5.3 Limitations of sampling methods ................................................................................... 60
3.5.4 Implications of fine root turnover in ecosystem carbon cycling ..................................... 63
3.6 Conclusion .............................................................................................................63
4 Fine root morphology, biochemistry and litter quality indices of fast- and slow-growing woody species in Ethiopian highland forests ............................................... 64
4.1 Abstract ..................................................................................................................64
4.2 Introduction ............................................................................................................65
4.3 Materials and methods ...........................................................................................67
4.3.1 Study site ....................................................................................................................... 67
4.3.2 Root sampling ................................................................................................................ 68
4.3.3 Root morphology ........................................................................................................... 68
4.3.4 Root biochemistry and construction costs ..................................................................... 69
4.3.5 Statistical analysis ......................................................................................................... 70
4.4 Results ...................................................................................................................71
4.4.1 Root morphology ........................................................................................................... 71
4.4.2 Root biochemistry .......................................................................................................... 73
4.4.3 Carbon cost of root production ...................................................................................... 75
4.4.4 Correlation of morphological and biochemical fine root traits ....................................... 77
4.5 Discussion ..............................................................................................................78
4.5.1 Root morphological traits and growth pattern ................................................................ 78
4.5.2 Root biochemistry and carbon cost implications for root litter quality ........................... 80

VI
4.6 Conclusion .............................................................................................................84
5 Litter production, chemistry, and turnover in a pristine forest ecosystem in the Ethiopian highland .................................................................................................... 85
5.1 Abstract ..................................................................................................................85
5.2 Introduction ............................................................................................................86
5.3 Materials and methods ...........................................................................................88
5.3.1 Description of the study site .......................................................................................... 88
5.3.2 Litterfall collection and fine root biomass determination ................................................ 89
5.3.3 Litter (leaf and root) decomposition using litterbag technique ....................................... 89
5.3.4 Chemical analysis .......................................................................................................... 90
5.3.5 Calculations of decomposition parameters, C and N input into the soil ........................ 91
5.3.6 Statistical analysis ......................................................................................................... 91
5.4 Results ...................................................................................................................92
5.4.1 Annual litterfall production ............................................................................................. 92
5.4.2 Fine root biomass, production, and turnover ................................................................. 93
5.4.3 Initial litter chemistry ...................................................................................................... 94
5.4.4 Litter decay measurement and turnover rate ................................................................ 96
5.4.5 C and N fluxes into the soil ............................................................................................ 99
5.5 Discussion ............................................................................................................ 101
5.5.1 Above-ground litter production and seasonal pattern .................................................. 101
5.5.2 Factors controlling leaf litter decomposition ................................................................ 102
5.5.3 Fine root and coarse root decomposition .................................................................... 105
5.5.4 Biochemical fluxes from litters into the soil .................................................................. 106
5.6 Conclusion ........................................................................................................... 107
6 The biological origin of soil organic carbon and response to land use change based on biomarker analysis .................................................................................. 108
6.1 Abstract ................................................................................................................ 108
6.2 Introduction .......................................................................................................... 109
6.3 Methodology......................................................................................................... 112
6.3.1 Study area ................................................................................................................... 112
6.3.2 Soil sampling, C and N analysis .................................................................................. 112
6.3.3 Sequential extraction procedures ................................................................................ 113
6.3.4 Base hydrolysis ............................................................................................................ 113
6.3.5 CuO oxidation .............................................................................................................. 114
6.3.6 Derivatization and GC/MS Analysis ............................................................................. 114
6.3.7 Origin and degradation parameters ............................................................................. 115
6.4 Results .............................................................................................................. 117
6.4.1 Soil C and N content and yields of sequential extractions .......................................... 117
6.4.2 Composition and distribution of solvent-extractable free lipids ................................... 118
6.4.3 Composition and distribution of bound lipids ............................................................... 120
6.4.4 Composition and distributions of lignin compounds .................................................... 121
6.5 Discussion ............................................................................................................ 122

VII
6.5.1 Biological origin of organic carbon in the soil .............................................................. 122
6.5.2 Degradation stage of plant organic matter .................................................................. 129
6.6 Conclusions.......................................................................................................... 132
7 Microbial soil respiration and its dependency on substrate availability and temperature in four contrasting land use systems .................................................. 133
7.1 Abstract ................................................................................................................ 133
7.2 Introduction .......................................................................................................... 134
7.3 Materials and methods ......................................................................................... 137
7.3.1 Site description ............................................................................................................ 137
7.3.2 Soil sampling ............................................................................................................... 137
7.3.3 Total C, N, and soil pH analysis .................................................................................. 138
7.3.4 Determination of moisture content and water holding capacity ................................... 138
7.3.5 Preparation of carbon and nutrient sources ................................................................ 138
7.3.6 Preparing agar and indicator solution for the MicroResp ............................................ 139
7.3.7 Soil filling and incubation ............................................................................................. 139
7.3.8 Measurement of soil respiration .................................................................................. 140
7.3.9 Quantifying CO2 efflux and biological indices .............................................................. 140
7.4 Results ................................................................................................................. 142
7.4.1 Soil chemical and physical properties in four land use systems ................................. 142
7.4.2 Basal respiration in the four land use systems ............................................................ 143
7.4.3 Microbial respiration in response to substrate availability ........................................... 144
7.4.4 Soil microbial biomass carbon ..................................................................................... 145
7.4.5 Temperature effect on CO2 efflux at different land use systems ................................. 146
7.5 Discussion ............................................................................................................ 148
7.5.1 Resource availability and carbon use efficiency of microorganisms ........................... 148
7.5.2 Biological indicators for eco-physiological stress ........................................................ 150
7.5.3 Effect of temperature on microbial decomposition ...................................................... 151
7.6 Conclusion ........................................................................................................... 153
8 Summary and conclusions ............................................................................... 154
References ............................................................................................................. 159
Appendices ............................................................................................................. 184
Appendix 1. Supplementary materials for Chapter 2 ....................................................... 184
Appendix 2. Supplementary materials for Chapter 4 ....................................................... 186
Appendix 3. Supplementary materials for Chapter 5 ....................................................... 188
Appendix 4. Supplementary materials for Chapter 6 ....................................................... 190
Appendix 5 Data not used in this thesis .......................................................................... 200

VIII
List of figures
Figure 2 1 Stocks (kg m-2, mean ±SE) of soil organic carbon (filled bars) and soil nitrogen (unfilled bars) in different land use types at a range of sites in the Amhara region of Northern Ethiopia. Shown are the pooled data of different land use systems from three highland areas (a) and two land use systems of the lowland area (b) (see legend Table 2.2). Bars without the same letters (small case letters for filled bars and capital case letters for unfilled bars) are significantly different
(mean ±SE; Scheffe, P<0.05, n (highland) = 30, and n (lowland) = 10). ................................................... 24
Figure 2 2 Vertical distribution of soil C concentrations in the highland land-use system a) and the relative cumulative SOC stock fraction across soil depth for each land use type b). Shown are the means of the pooled data of the individual sites. Different small case letters for panel (a) indicate significant differences of data points along soil depth within a land use type whereas capital case letters indicate differences between land use type for each depth (mean ±SE; Scheffe, P<0.05, n (forest) = 40, n (eucalyptus, grazing land, cropland) = 30). The panel (b) is Y = 1– ßd, where Y is the cumulative SOC fraction from the surface (proportion between 0 and 1) to soil depth (d in cm) and ß is the fitted parameter of the asymptotic nonlinear model (Gale and Grigal 1987). A Y value of 0.75 at 30 depth means that 75% of the total SOC storage is located above 30 cm soil depth. Higher ß index value indicates the relative proportion of SOC to lower
profile is greater in the specific land use, or the relative change of SOC across depth is smaller. ....... 26
Figure 2 3 Soil organic carbon stocks in relation to soil depth of different land use types in the Ambober site. Small case letters indicate a significant difference between SOC stocks at different soil depths, while capital letters indicate a significant difference between land use systems at one
depth (mean ±SE; Scheffe, P<0.05, n = 10). ................................................................................................. 28
Figure 2 4 Strontium (Sr) to calcium (Ca) and barium (Ba) to calcium ratios at different soil depths in forests (filled bars) and cropland (unfilled bars) at Katassi and Gelawdios. Within a ratio and land use bars with different letters are significantly different (mean ±SE; Scheffe, P<0.05, n = 10).
.............................................................................................................................................................................. 29
Figure 2 5 Soil carbon stocks (kg m-2) in relation to N stock (a, forest; b, cropland) and soil pH (c, forest; d, cropland). Different symbols represent a research site and values are individual samples
(n = 10)................................................................................................................................................................. 30
Figure 2 6 The relationship between soil carbon stock (kg m-2) and a) mean annual precipitation
(MAP); b) mean annual temperature. (Mean ±SE; Scheffe, P<0.05, n = 10). ........................................... 31 Figure 3 1 Cumulative root fraction distribution at Gelawdios natural forest as a function of root depth for different root categories (very fine roots vs fine roots, biomass vs necromass, and herbaceous roots vs roots from trees). Fit equation is Y = 1– ßd, where Y is the cumulative root fraction from the surface (proportion between 0 and 1) to soil depth (d in cm in the middle) and ß is the fitted parameter of the asymptotic nonlinear model (Gale and Grigal 1987). Larger β values imply deeper rooting profiles. E.g., a Y value of 0.75 at 30 cm depth means that 75% of the root biomass is located above 30 cm or, conversely, 25% of the root biomass is located below 30 cm
soil depth. ............................................................................................................................................................ 51
Figure 3 2 Seasonal variation of fine root biomass and necromass (g m-2) at Gelawdios a) forest; b) eucalyptus stand. Fine root estimates are based on coring method. Bars with different small case letters are significantly different for filled bars (biomass), and different upper case letters indicate significant differences for unfilled bars (necromass). Error bars represent mean±1SE
(p<0.05; n=10). ................................................................................................................................................... 52
Figure 3 3 Comparison of fine root production between land use types from in-growth core samples. Values for each stock were calculated based on the Decision Matrix method. Samples from grazing land and cropland was taken during the growing season only, assuming no new root production during dry time and after crop harvest. Error bars represent mean±SE (p<0.05; n=10
for forest and eucalyptus, n = 5 for grazing and cropland)........................................................................... 55

IX
Figure 3 4 Estimated fine root production (g m-2 yr-1) to a depth of 40 cm (filled bars) and 20 cm (unfilled bars) for the natural forest ecosystem with different sampling methods. In the unfilled bars, values for sequential coring and in-growth cores methods were calculated to a depth of 20 cm for uniformity with the in-growth net method because the latter was established to only 20 cm
depth. Error bars represent mean±1SE (p<0.05; n=10). .............................................................................. 56
Figure 3 5 Relationship between total root mass (biomass + necromass) with C stock a) and N stock b) for four land use systems at Gelawdios, Ethiopia. n=10 (forest and eucalyptus), and 5
(cropland and grazing land). ............................................................................................................................. 62 Figure 4 1 Non-linear regression model of relative diameter class length distribution (rDCL; cm) (n = 15) of fine roots ≤2 mm diameter of ten tropical tree and shrub species from Gelawdios forest
in the Ethiopian highlands. ................................................................................................................................ 72
Figure 4 2 Calculated amount of glucose needed to produce one gram of fine root biomass (g glucose g-1 dw) in fast- and slow-growing woody species of the Gelawdios forest, NW Ethiopia. Small case letters indicate significant differences between groups (mean±SE; Tukey, p<0.05; n = 5). .......................................................................................................................................................................... 76
Figure 4 3 Linear correlations of carbon content (C, %) in fine roots of five fast- and five slow-growing woody species of the Gelawdios forest, NW Ethiopia with a) nitrogen content (N; %); b) lignocellulose index (AIF/cell wall fraction ratio); c) acid-insoluble fractions (AIF; %); and d) root
tissue density (RTD; g cm-3) (mean±SE; Tukey, p<0.05; n = 3).................................................................. 78 Figure 5 1 Seasonal patterns of litterfall a) and monthly distributions of total litterfall, leaves, woody, reproductive organs, and miscellaneous b) (g m-2). Bars with different small case letters are significantly different. Error bars represent mean±SE (p<0.05; n=10). Each point in the monthly distributions of litterfall represents a monthly mean calculated value from ten litter collections. ........................................................................................................................................................... 93
Figure 5 2 Litter decay of four species and one mixed over one year expressed as percentage remaining of original mass in litter bags at various time intervals for a) coarse roots; b) fine roots;
c) leaves; d) coarse roots, fine roots, and leaves. ......................................................................................... 97
Figure 5 3 Linear correlations between decomposition rate (k) with a) Nitrogen (N) content; b) Carbon (C) to N ratio; c) acid insoluble fraction (AIF) to N ratio; d) extractive fractions; e) acid soluble fraction (ASF); f) AIF. Filled dots represent for fine roots and unfilled dots represent for
leaf litters. ............................................................................................................................................................ 98
Figure 5 4 Linear correlation between soil carbon stocks with a) annual fine root production; b) annual litterfall production; c) total biomass input (litterfall + fine roots). ................................................. 100 Figure 6 1 Soil C (filled bars) and N content (unfilled bars) for four land use systems from Gelawdios, Ethiopia. Values are mean±SE, n= 10. Different lower case letters indicate significant differences in soil carbon content, upper case letters significant differences in N content between land use systems. ............................................................................................................................................. 117
Figure 6 2 Carbon-normalized extract yields of solvent, base hydrolysis, and CuO oxidation
products of four land use systems from Gelawdios, Ethiopia. Values are mean±SE, n= 3. ................. 118
Figure 6 3 Lignin source parameters of the monomeric cinnamyl/vanillyl (C/V) and syringyl/vanillyl
(S/V) phenols of four land use systems. Boxes are drawn according to Goñi et al. (2000). ................. 127 Figure 7 1 Linear regression of SOC with panel a) water holding capacity (WHC), panel b) basal
respiration (BR), and panel c) microbial biomass carbon (MBc) for the four land use soils. ................ 144
Figure 7 2 Correlation of microbial biomass carbon (MBc) with basal respiration a) and soil pH b)
for the four land use soils. ............................................................................................................................... 145

X
Figure 7 3 Comparison of total C mineralized at different carbon sources with a range of recalcitrance (glucose, lignin, starch) and nutrient sources (phosphate and ammonium nitrate) in four land use systems a) forest soil; b) eucalyptus plantation soil; c) grazing land soil; and d) cropland soil. Substrates were supplemented with 25µl of each substance in solution at a concentration of 30 mg ml-1 of soil water. Control samples were amended with distilled water so that all samples were maintained at the same moisture content (ca. 60%). The average data were best fit using an Arrhenius type relationship between respiration rate and temperature (equation
8). ........................................................................................................................................................................ 147 Figure A4.1 Total ion GC–MS chromatograms (TIC) of the silylated solvent extracts of four land use systems from Gelawdios, Ethiopia. , n-Alkanols; +, n-alkanes; Δ, n-Alkanoic acids; #, carbohydrates (gl, glucose; ma, mannose; su, sucrose); MAG, Monoacylglycerides; S1-S7, steroids; T1-T5, Triterpenoids, U1, Unknown. Numbers refer to total carbon numbers in aliphatic lipid series. Detail description of each compound with its quantity, molecular formula, and
molecular weight are described in Table A4.1. ............................................................................................ 190
Figure A4.2 Total ion GC–MS chromatograms (TIC) of the methylated and silylated extracts after base hydrolysis of four land use systems from Gelawdios, Ethiopia. , n-Alkanols; +, n-alkanes; Δ, n-Alkanoic acids; iso- alkanoic acids; α- alkanoic acids; α, ω-alkanedioic acids; ω-hydroxyalkanoic acids; #, MAG, Monoacylglycerides; steroids (β-Sitpsterol); organophosphate; phenols, and U1-U5, Unknown. Numbers refer to total carbon numbers in aliphatic lipid series. Detail description of each compound with its quantity, molecular formula, and molecular weight are described in Table A4.2. ........................................................................................................................... 191
Figure A4.3 Total ion GC–MS chromatograms (TIC) of the silylated CuO oxidation products of four land use systems from Gelawdios, Ethiopia. U1-U7, Unknowns. Detail description of each compound with its quantity, molecular formula, and molecular weight are described in Table A4.3.
............................................................................................................................................................................ 192 Figure A5 1 Vertical distributions of fine root mass (g m-2) estimated from coring methods during March 2014. Each line represented one site and same letters within site are not significantly different between soil depths at p<0.005 and values are means of 20 samples ± 1SE and for
Ambober n= 10 core samples. ....................................................................................................................... 200
Figure A5 2 Fine root stock comparison between research sites in the forest ecosystem and exclosure area (Ambober) as estimated by coring method. Different small case letters are significantly different between biomass (filled bars), while upper case letters indicate significant differences between necromass (unfilled bars). (Mean ±SE; n = 20 except Ambober n = 10. Note:
* in Ambober refers to Exclosure. .................................................................................................................. 200
List of tables
Table 2 1 Site characteristics of a range of sites in the highlands or lowlands of the Amhara region
of Northwestern Ethiopia. .................................................................................................................................. 17
Table 2 2 Stocks (kg m-2, mean ±SE) of soil organic carbon (SOC) and soil nitrogen (N) in different land use types at a range of sites in the highlands or lowlands of the Amhara region of Northwestern Ethiopia. Values are for a soil depth of 50 cm. All woodlands apart from Ambober are mature forests; the site at Ambober is a developing exclosure. Different letters (abc) indicate significant difference between site for one land use and the letters (ABC) indicate significant
difference between land use types at one site (P ≤0.05, n = 10). .............................................................. 25 Table 3 1 Simplified Decision Matrix for estimating fine root production, mortality, and decomposition according to McClaugherty et al. (1982), Osawa and Aizawa (2012), and Yuan and Chen (2013). The appropriate quadrant is selected according to the direction of change in biomass (B) and necromass (N) during the interval between two sampling times. Production (P), mortality (M), and decomposition (D) for the sampling interval are calculated using the equations

XI
in the chosen quadrant. Vertical bars indicate the absolute values. Annual estimates are
calculated by summing the estimates from all sampling intervals within the year. ................................... 45
Table 3 2 Vertical distribution of fine root mass (g m-2) to a soil depth of 40 cm for native forest, eucalyptus stand, grazing land and cropland. Fine roots are categorized as tree vs herbaceous roots, biomass (live roots) vs necromass (dead roots) and very fine roots (<1 mm) vs fine roots (1-2 mm) for both native forest and eucalyptus plantations. Values for forest and eucalyptus were determined based on sequential coring. Roots from cropland and grazing land are taken from the last in-growth core harvest, assuming peak rooting time at the end of the rainy season. Roots from these land use systems are entirely herbaceous and mostly <1 mm. Values are mean±SE;
n=10 (forest and eucalyptus); n=5 (cropland and grazing land). ................................................................. 50
Table 3 3 Fine root biomass (mean, maximum), production (biomass (B), necromass (N), decomposition (D), total production (TP)), and turnover rate to a depth of 40 cm for natural forest and eucalyptus plantation. The annual productions are calculated from sequential coring data based on Decision Matrix and Maximum-Minimum methods, and the turnover rates are calculated by dividing the annual production by the mean biomass (Bmean) or by maximum biomass (Bmax).
Samples in sequential coring were collected in quarterly intervals. Values are mean±SE; n=10. ......... 54
Table 3 4 Root production estimates using in-growth cores and in-growth nets to 20 cm depth. Sampling corresponded to 1, 2, 3, 4, 6, 8, and 12-month interval times. Fine root biomass, necromass, decomposition, and total production are calculated based on the mass balance method according to Santantonio and Grace (1987), Osawa and Aizawa (2012), and Li et al. (2013). Small case letters indicate significant difference between land use types and upper case letters indicate significant difference between sampling methods. Values are mean±SE; n=10
(forest and eucalyptus), n=5 (grazing land and cropland). ........................................................................... 55
Table 3 5 Total C and N flux (g m-2 yr-1) to soils via fine roots in four land use systems at Gelawdios, Ethiopia. Element fluxes were calculated according to Xia et al. (2015) from element concentrations in the roots multiplied by annual fine root production estimated from in-growth cores. Numbers in brackets are number of samples. For elemental analysis in forests and eucalyptus, all categories were considered (i.e. three each: roots <1 mm, roots 1-2 mm, and herbaceous roots), whereas samples from grassland and cropland were all herbaceous and we
took only three samples. ................................................................................................................................... 57 Table 4 1 Morphological traits of fine roots of ten woody species. SRA, specific root area; SRL, specific root length; RTD, root tissue density. Species are grouped into fast- (FG) and slow-growing (SG) species (see Supplementary Information Table A2.2 for details). Different small case letters indicate significant trait differences between species irrespective of group, and upper case letters indicate differences between FG and SG group averages (mean±SE; Tukey, p<0.05;
nspecies = 15, ngroup = 5). ...................................................................................................................................... 72
Table 4 2 Major biochemical fractions of fine roots of ten woody species. Shown are nonpolar extractives (NPE), polar extractives (PE), extractives fraction (EF, sum of NPE and PE), acid-soluble fraction (ASF), acid insoluble fraction (AIF), and ash content. Species are grouped into fast- (FG) and slow-growing (SG) species. Different small case letters indicate significant differences between species and upper case letters indicate significance differences between FG
and SG group averages (mean±SE; Tukey, p<0.05; nspecies = 3, ngroup = 5). ............................................. 74
Table 4 3 Litter quality indices of ten woody species. Shown are carbon (C) and nitrogen (N) contents, C/N ratio, acid insoluble fraction (AIF) to N ratio, and lignocellulose index. Species are grouped into fast- (FG) and slow-growing (SG) species; see Supplementary Information Table A2.2 for details. Lignocellulose index is the ratio of AIF to cell wall fraction. Small case letters indicate significant differences between species and upper case letter indicate differences
between FG and SG group averages (mean±SE; Tukey, p<0.05; nspecies = 3, ngroups = 5). ..................... 75
Table 4 4 Estimated glucose investment for fine root biomass production of ten woody species. Species are grouped into fast- (FG) and slow-growing (SG) species; see Supplementary Information Table A2.2 for details. Small case letters indicate significant differences between species and upper case letter indicate differences between group averages (mean±SE; Tukey,
p<0.05; nspecies = 3, ngroups = 5). ......................................................................................................................... 76

XII
Table 4 5 Pearson correlation matrix of root morphological and chemical traits. Values are the Pearson (r) value of the 4 morphological and 6 chemical traits across 10 co-occurring woody species in the Gelawdios forest, NW Ethiopia (n=10). Significant correlations (p<0.05) are indicated in bold. NPE, nonpolar extractives; PE, polar extractives; EF, extractive fraction; ASF, acid-soluble fraction; AIF, acid-insoluble fraction; SRL, specific root length; SRA, specific root
area; RTD, root tissue density; AD, average root diameter. ........................................................................ 77 Table 5 1 Soil physical and chemical properties at the Gelawdios forest. Values of carbon (C),
nitrogen (N), C:N ratio, and pH are mean±SE. .............................................................................................. 88
Table 5 2 Average annual litterfall production (g m-2 yr-1) by components between July 2015 and June 2016, at Gelawdios natural forest, Ethiopia. Values with the same letter are not significantly
different (mean±SE; n = 10; p < 0.05). ............................................................................................................ 92
Table 5 3 Average fine root stock (biomass and necromass), production, and turnover rate at
Gelawdios forest, Ethiopia. Values are mean±SE (n=10; α = 0.05). .......................................................... 94
Table 5 4 Initial litter chemistry and litter quality indices of the four dominant species at Gelawdios forest, Ethiopia. AIF, acid-insoluble fraction; ASF, acid-soluble fractions; CC, carbon cost; EF, extractive fractions; NPE, non-polar extractives; and PE, polar extractives. EF is the sum of polar and non-polar extractives. Values are means±SE of three replicates. Different small case letters in the same row indicate significant differences between species (P<0.05). Upper case letters
indicate significance differences between leaves and roots of the same variable (p<0.05). .................. 95
Table 5 5 Litter decay rate coefficients and residence time for leaves, fine roots, and coarse roots of the four monospecies, two possible combinations for leaves and four species combinations for all categories. The decomposition rates (k) were estimated using a single exponential decay model as Mt = M0*e-kt according to Olson, (1963). Where Mt is the litter dry mass at time t, M0 is the initial litter mass, t is the sampling time interval, and k is the annual decay constant. Mean residence time (Rt) of litter in each treatment was estimated by the inverse of k calculated. T(0.5) is a half-life period calculated as 0.693/k, whereas the T(95) and T(99) are the time needed for 95% and 99% mass loss and calculated as 3/k and 5/k, respectively. Values in decay rate constant (k)
are mean±SE; n=3. ............................................................................................................................................ 99
Table 5 6 Mean flux of each biochemical class to soil via leaf litter, fine roots (<2 mm), and the proportion (%) of the combined flux of leaf litter and fine root flux contributed by fine roots. AIF,
acid-insoluble fraction; RCC, carbon cost; EF, extractive fractions; ASF, acid soluble fractions. ........ 100 Table 6 1 Occurrence and quantities of compounds (µg/g C) identified in the solvent extracts of
soil samples of different land use systems in Gelawdios, Ethiopia. Values are mean±SE, n= 3. ........ 119
Table 6 2 Occurrence and quantities of compounds (µg/g C) identified from base hydrolysis of
soil samples in different land use systems in Gelawdios, Ethiopia. Values are mean±SE, n= 3. ........ 120
Table 6 3 Occurrence and quantities of major compounds (µg/g C) identified in the CuO oxidation extracts of soil samples in different land use systems in Gelawdios, Ethiopia. Values are
mean±SE, n= 3 ................................................................................................................................................. 122
Table 6 4 Source and degradation parameters of major biomarker classes in different land use
systems in Gelawdios, Ethiopia. .................................................................................................................... 130 Table 7 1 Average physico-chemical characteristics of soil samples (0-10 cm depth) in four land
use systems (Mean ±SE; n=3 except texture n = 1). .................................................................................. 143
Table 7 2 Soil microbial parameters in the sampled soils at 25°C. The amount of glucose added in the substrate induced respiration (SIR) used a standard concentration designed to deliver 30 mg ml-1 of soil water at 40% of water holding capacity in the MicroResp (Campbell et al. 2003). Metabolic quotient (qCO2) was calculated as the ratio of soil basal respiration to microbial biomass carbon (MBc) (Anderson and Domsch 1978). Values denote means and standard errors of three replicates. Different letters show significant differences between land use systems
following Tukey’s HSD test (p<0.05). ............................................................................................................ 144

XIII
Table 7 3 Soil CO2 efflux and temperature sensitivity of four land use soils with different C source and nutrient addition. The amount of substrates added used a standard concentration designed to deliver 30 mg ml-1 of soil water at 40% of water holding capacity in the MicroResp (Campbell et al. 2003). Temperature sensitivity indices (Q10) were calculated as a function of soil respiration and temperature. Soil respiration = R*Q10
((T-10)/10) according to Schindlbacher et al. (2010), where
R and Q10 are fitted parameters. Values are mean±SE of triplicate values. .......................................... 146 Table A1.1 Soil texture, carbon (C) and nitrogen (N) concentrations and stocks, bulk density (BD) and soil pH at all study plots (labelled as site and land use type) and four soil depths (mean±SE;
n(texture) = 3, n(BD/pH) = 5, n(C/N) = 10). .................................................................................................. 184
Table A1.2 Concentrations (mg g-1) of calcium (Ca), strontium (Sr) and barium (Ba) at Katassi
and Gelawdios forests and croplands at four soil depth (Mean±SE; n=10). ........................................... 186 Table A2 1 Edaphic characteristics of the study site at Gelawdios forest, NW Ethiopia. Values
are Mean±SE; npH = 5, nC,N = 10, ntexture = 3. ................................................................................................ 186
Table A2 2 Scientific and local names of the ten studied woody species with family, and growth form within the Gelawdios forest, NW Ethiopia. The species are classified as either fast- or slow-growing based on previous studies and information from local experts. Species characteristics are shortly outlined. .......................................................................................................................................... 187 Table A3 1 Average annual litterfall (g dw m-2) by species/Class. Values are mean±SE (n=3). ......... 188
Table A3 2 Decay rate (% of mass loss) by species. Values are mean±SE (n=3). ............................... 189 Table A4 1 Occurrence and quantities of compounds (µg/g C) identified in the solvent extracts of soil samples at different land use systems in Gelawdios, Ethiopia. MF, molecular formula; MW,
molecular weight. Values are mean+SE; n =1-3 (see below footnotes). ................................................. 193
Table A4 2 Occurrence and quantities of compounds (µg/g C) with molecular formula (MF) and molecular weight (MW) identified from base hydrolysed of soil samples at different land use systems in Gelawdios, Ethiopia. MF, molecular formula; MW, molecular weight. Values are
mean+SE; n =1-3 (see below footnotes). ..................................................................................................... 195
Table A4 3 Occurrence and quantities of major compounds (µg/g C) identified in the CuO oxidation extracts of soil samples at different land use systems in Gelawdios, Ethiopia. MF,
molecular formula; MW, molecular weight. Values are mean+SE; n =1-3 (see below footnotes). ...... 198 Table A5 1 Proportions of fine roots (g m-2) based on diameter class, type of roots (herbaceous vs tree roots), and biomass (living) and necromass (dead) roots for different ecosystems as determined by coring method. Values are mean(±1SE); (n(highland forest) = 80, n(lowland forest) = 20, n(eucalyptus) = 30, n(exclosure) = 10). ..................................................................................... 201
Table A5 2 Fine root stock per depth (g m-2) for each research sites in the forest ecosystem and exclosure area (Ambober) as estimated by coring method. Values are mean (±SE); n = 20 except Ambober n = 10. Note: * in Ambober refers to Exclosure; na, not available ....................................... 201
Table A5 3 Carbon fractionation per depth (%) for each research site and land use systems. LC I, labile carbon one; LC II, labile carbon two (medium); RC, recalcitrant carbon. Similar fractionation for nitrogen (N). Values are mean of five samples. ........................................................ 202
Table A5 4 Total quantity of ergosterol as determined by high performance liquid chromatography
(HPLC) for Gelawdios site and Mahibere-Selassie savanna woodland. ............................................. 204
Table A5 5 Soil organic carbon row data collected at large scale based on grid coordinates. Values were determined by los of ignition. .......................................................................................... 204
Table A5 6 Soil moisture content and water holding capacity of soils during the time of carbon measurement. Values are mean±SE of the mean (n=3). .................................................................... 215

1
1 General Introduction
1.1 Land degradation in the Amhara region: history, extent, causes,
and consequences
The Amhara region is located in the northwestern part of Ethiopia between latitude 9°
to 13°45' N and longitude 36° to 40°30' E, with a total area of 170,152 km2 (Desta et al.
2000). The region is topographically divided into two main parts, namely highlands and
lowlands. The highlands are above 1,500 m above sea level (a.s.l.) and comprise the
largest part of the northern and eastern parts of the region including the highest peak
in the country (4,620 m a.s.l.). The lowlands cover mainly the western and eastern parts
of the region (31%), with altitudes from 500-1500 m a.s.l. (UNECA 1996). The highland
part comprises extensive volcanic plateaus and mountainous landscapes, generally
known for very rugged and severely broken steep slopes with angles of greater than
15%. According to UNECA (1996), about 34% of the land in the region is estimated to
feature a slope grade of over 35%.
The annual mean temperatures of the region is between 15°C and 21°C (UNECA 1996,
Ayalew et al. 2012). Relatively high temperatures occur in some valleys and in marginal
areas exhibiting arid climate and can exceed 27°C (Ayalew et al. 2012). The distribution
of rainfall largely depends upon the direction of moisture-bearing monsoon winds and
altitude. The central part of the region receives about 1600 mm of mean annual rainfall,
partially exceeding 2000 mm (Awulachew et al. 2009). The amount of rainfall is lowest
(<700 mm) in the northwest and northeast parts along the border to Sudan and Tigray
and Afar regions (Bewket and Conway 2007). About 80% of the total rain falls during
the summer months, starting in mid-June and ending early in September (UNECA
1996).
In 2007, the population of the region was estimated to be 17 million with an equal sex
ratio (CSA 2008). This represents 23% of the Ethiopian population. Of these, 87% live
in rural areas. The highlands are over-populated due to favourable climatic conditions,
whereas the lowlands are sparsely populated due to the harsh climate (Desta et al.
2000). The Amhara region, by virtue of its proximity to the ancient and medieval centres
of Ethiopian civilization, was one of the earliest settled parts of the country, with
agricultural activities dating back more than 3,000 years (UNECA 1996). From the early

2
15th to the late 19th century, the region was the centre of Ethiopia's culture and politics
(UNECA 1996).
Although accurate records on forest cover and reliable data on the rate of deforestation
are scarce (Pankhurst, 1995), an assessment by the Bureau of Agriculture indicates
that natural forest covered 0.48% of the total area, or 81,047 ha before 2007 (Bane et
al. 2007). That report also indicates that woodland and plantation forests account for
4.2% and 1.23% of land cover, respectively. The Woody Biomass Inventory and
Strategic Planning (WBISP) project (WBISP 2004) report, the only credible nation-wide
vegetation assessment in Ethiopia, shows that in the Amhara region, natural forest
covered 2.3%, woodland 4%, and shrub land 16% of the area in 2003. Since then, the
expansion of agricultural areas as a result of population growth and the need for new
farmland (Teketay et al. 2010) resulted in the permanent devastation of almost all the
highland forests. Biomass energy at the national level provides more than 99.6% of the
total domestic energy consumption: 78% from woody biomass, 8% from crop residue,
11% from animal dung and 3.1% from modern energy (WBISP 2004). About 20,000
hectares of forest are cleared annually in the Amhara region for the expansion of
farmland and fuelwood (Meseret 2016). Currently, no more forest area is available to
be cleared for cultivation in the densely populated highlands, but the practice continues
in the lowland woodlands. At present, remnants of the original vegetation are found
mainly around churches, monasteries and along very inaccessible mountainsides and
gorges. The numerous monasteries and churches have played major roles in forest
conservation (Aerts et al. 2016). To combat the deforestation trend, restoration of
vegetation cover through afforestation or establishment of exclosures (protecting it from
animal and human interference) have been widely practiced since the 1980s (Girmay
et al. 2008). In relation to afforestation, most of the planting involves exotic species,
especially eucalyptus species because of their fast growth and high economic return.
The plantation forests in the region are estimated at 44,600 ha, of which 18,000 ha are
covered by Eucalyptus species (Bekele 2011).
More than 87% of the human population is engaged in mixed farming; about 30% of
the region is devoted to agriculture (Desta et al. 2000, CSA 2008). Cropping is
predominantly rain fed and is based on inefficient methods of cultivation. Despite
inefficient utilization of the land, the region has abundant water resources, suitable for
crop production and livestock husbandry. This pertains to certain areas of the western

3
lowlands and to parts of the central plateaus. The Amhara region is one of the major
‘teff’ (Eragrostis teff) producing areas in the country. Barley, wheat, oil seeds, cotton,
maize, sorghum, and sesame are also major crops produced here in large quantities.
Nonetheless, agricultural production far from meets the food demands of the population
(UNECA 1996). Primitive farm implements and poor management of land use,
exacerbated by high soil erosion, are deemed to be the major factors for low agricultural
productivity (UNECA 1996, Desta et al. 2000, Meseret 2016). Thus, agricultural
practices have remained traditional (no modern technological intervention) and the
same plots of lands have been cultivated repeatedly until they become exhausted. Land
which was once considered unsuitable for farming by the local people is now being
brought into use.
The major environmental problem in the highlands of the Amhara region is land
degradation due to soil erosion (Meseret 2016, Molla and Sisheber 2017). The
estimated annual rate of soil loss here due to water erosion is about 119 million tons,
equivalent to 70% of the total soil loss in Ethiopia (Meseret 2016). Accordingly, most
highland farms are badly eroded, with the soil becoming too shallow and stony to
support crops. Previous studies estimate that the soil loss from arable land is 42-79 t
ha-1 yr-1 (Shiferaw and Holden 1999, Bewket and Sterk 2003, Brhane and Mekonen
2009), underlying a yield reduction of 1% to 2% per year (UNECA 1996). In 1990, land
degradation at the Ethiopian highlands reduced the production by 465,000 t of grain,
equivalent to $70 million y-1 (Girmay et al. 2008). The Amhara region started to
implement land rehabilitation measures through a comprehensive soil and water
conservation (SWC) program in the 1970s, following severe famine (Meseret 2016).
However, land degradation through soil erosion remains a serious problem (Molla and
Sisheber 2017) and is predicted to become even more severe in the future (Meseret
2016).
Livestock are integral to the farming system, supplying power for cultivation (oxen
plowing system), food, and income to households. About 35% of Ethiopia's livestock
population are located in the Amhara region (Desta et al. 2000, Meseret 2016). The
natural vegetation of the grazing land is the main source of animal feed, followed by
crop residues. The livestock density is estimated to be about 23 livestock units (LU) per
ha, whereas the carrying capacity has been estimated to be 2 LU ha-1 (Desta et al.
2000). Accordingly, overgrazing and trampling cause damage and virtually denude the

4
vegetation cover: most communal grazing lands are bare ground even during the rainy
season (Nedessa et al. 2005). The traditional practice of free grazing in the region is
thus a major cause of land degradation. Land degradation due to land-use change,
erosion and overgrazing is a negative factor in ecosystem functioning and in stabilizing
carbon (C) levels in soil. Thus, quantifying the effects of land use change and
management on C dynamics is important in terms of estimating local and global C
cycling, climate change effects, and the vulnerability of smallholder farmers. The
vulnerability of the soil organic carbon (SOC) stock is expected to be more intense and
faster in the Amhara region compared to other parts of Ethiopia due to high annual
rainfall and a steep topography facilitating losses through topsoil erosion.
1.2 The nature and formation of soil organic carbon
1.2.1 Soil carbon stock and temporal changes after land use change
Organic carbon stored in soils is the largest terrestrial carbon pool, with global
estimates ranging from 2376-2456 Pg of C in the upper 200 cm (Batjes 1996, Jobbágy
and Jackson 2000). This pool is more than four times the size of the biotic carbon pool
(550 Pg C; Eswaran et al., 1993) and three times the amount of carbon in the
atmosphere (750 Pg C; Batjes 1996, Hiederer and Köchy 2011, Fan et al. 2016).
Geographically, 32% of the total carbon pool in soil is located in the tropics; 40% of this
C is stored in forest soils (Eswaran et al. 1993). The C sink potential of global forest
ecosystems was calculated as 3.1 Pg of C per year for the period 2006-2015 (Le Quéré
et al. 2016), sequestrating about one third of anthropogenic C emissions (Fan et al.
2016). Tropical forest ecosystems account for one third of the terrestrial net primary
production (Malhi et al. 2011) and contain roughly 25% of the terrestrial biosphere C
(Becker et al. 2015). Tropical forests have more carbon stored in biomass (56%) and
less in soil (32%); boreal forests, in contrast, have on average less C stored in biomass
(20%) and more in soil (60%) (Pan et al. 2011).
The world has experienced dramatic land use changes during the past few decades,
including conversion of natural forests to agriculture, grazing land or plantations (Guo
and Gifford 2002). The sub-Saharan African region has experienced the fastest
conversion of forestland to agriculture in the past 20 years (Nkonya et al. 2013). Land
use change plays a major role in determining C storage in ecosystems (Le Quéré et al.
2016). The greatest SOC losses result from conversion of forests to open lands

5
(cropland and grazing land) (Chapter 2). Globally, average estimates showed that
conversion of forest to cropland reduces SOC by 30-35% within the first 30 years in the
top 10 cm soil (Oertel et al. 2016). A study in southern Ethiopia showed that SOC fell
by 62% in 10 years and by 75% in 53 years of cultivation after deforestation (Lemenih
et al. 2006). Carbon loss from ecosystems is mainly accomplished through erosion of
topsoil and mineralization (Yuste et al. 2007, Feng and Simpson 2008). Indeed, in the
highlands of the Amhara region, substantial amounts of topsoil are removed due to
erosion as a result of poor vegetation cover (Chapter 2; Hurni et al. 2010). A previous
study on the Ethiopian highlands showed an estimated rate of soil loss from cultivated
fields of up to 79 Mg ha-1 yr-1 (Bewket and Sterk 2003). This far exceeds the rate of soil
formation. In contrast, conversion of cropland to plantation forests increased the soil
carbon stocks by 29% worldwide (Don et al. 2011), illustrating the great potential of
recovering C stock by ecological restoration.
Another major pathway of carbon loss from terrestrial ecosystem is through respiration,
particularly soil respiration. Soil respiration is a measure of CO2 released from the soil
after decomposition of soil organic matter (SOM) by soil microbes (mineralization) and
respiration by plant roots and soil fauna (Rochette and Hutchinson 2005). Globally, soil
respiration by both plant roots and microbes releases about 90 Pg C yr−1 into the
atmosphere (Hashimoto et al. 2015). About half of the soil respiration (55%) has been
estimated to derive from metabolic activity of roots and associated mycorrhizae
(autotrophic respiration) (Saiz et al. 2007). The reminder is associated with
heterotrophic respiration from microbial communities using organic material as an
energy substrate (Ryan and Law 2005, Saiz et al. 2007). Land use conversion,
particularly from forests to open lands (e.g. cropland or grazing land), results in
substantial SOC losses that may elevate atmospheric CO2 concentrations (Batjes
1996, Lorenz et al. 2007). Historically, soils have a net loss of about 90 Pg C globally
through respiration due to cultivation (Smith 2007). Global estimates during the 1990s
showed that the annual net CO2 flux after land use change was 2.2 Pg C yr-1 (Houghton
and Goodale 2004). This value is almost entirely from the tropics, whereas outside the
tropics the average flux was a sink of 0.01 Pg C yr-1 (Houghton and Goodale 2004).
However, this CO2 flux due to land use change was reduced to 1.0 Pg C yr-1 for 2009-
2015 (Le Quéré et al. 2016). Soil carbon exchange with the atmosphere is an important
component of the global carbon cycle and this strongly influences the net carbon

6
accumulation in the atmosphere (Ryan and Law 2005). The CO2 concentration in the
atmosphere has increased from approximately 277 parts per million (ppm) in 1750, the
beginning of the industrial era, to 399 ppm in 2015 (Le Quéré et al. 2016) and is a major
concern with respect to climate change. Compared with direct anthropogenic
emissions, roughly nine times more CO2 is released from soils to the atmosphere via
soil respiration on an annual basis (Carey et al. 2016).
The level of SOC and ecosystem net exchange are determined by many factors
including temperature (Bradford et al. 2008), moisture (Meisner et al. 2015), carbon
input (Yuste et al. 2007) and nutrients such as N and P (Gnankambary et al. 2008,
Birgander et al. 2014). For example, previous studies showed that soil respiration
increases with rising temperatures (Reynolds et al. 2015, Mayer et al. 2016). However,
the magnitude of this effect remains uncertain because other studies show neutral or
even negative responses to warming, often attributed to other controlling factors (Ryan
1991, Suseela et al. 2012, Reynolds et al. 2015). For example, Carey et al. (2016)
reported that there is limited evidence of acclimation of soil respiration to experimental
warming in several major biome types. The amount of substrate availability and quality
(recalcitrance), land use management, and the below-ground community are important
to control the rate and extent of soil respiration (Chapter 7; Ryan and Law 2005,
Reichstein et al. 2005, Conant and et.al. 2011, Giesler et al. 2012).
1.2.2 Mechanisms for soil organic carbon accumulation
The organic carbon pool in the soil accumulates after the partial decomposition of
various organisms (Kögel-Knabner 2000, Kuzyakov and Domanski 2000, Angst et al.
2016). Net SOC accumulation is a result of a positive imbalance between organic
matter production and its losses through mineralization and other processes such as
topsoil erosion (Batjes and Sombroek 1997, Hairiah et al. 2001, Lorenz et al. 2007).
A major sources of SOC derive from plant litter (Chapter 3, 5; Kuzyakov and Domanski,
2000; Maeght et al., 2013). Soil fauna and microorganisms – as part of the food chain
– also provide important parent materials for SOM formation (Chapter 6; Kögel-
Knabner, 2002; Pautler et al., 2010; Wild et al., 2016). The two main inputs of plant-
derived carbon into the soil are below-ground root litter (Chapter 3) and above-ground
litterfall (Chapter 5; Jia et al., 2016). The amount of plant litter, its chemical composition
(Chapter 4), and the activity of the microbial community (Chapter 7) are essential

7
controlling factors for the formation and accumulation of soil organic matter and for
biogeochemical cycles in terrestrial ecosystems (Kögel-Knabner 2002, Solly et al.
2014). During the decomposition processes, one fraction of organic carbon is
stabilized in the soil, one part of C is incorporated into the decomposer biomass, and
another part is respired as CO2 (Wutzler and Reichstein 2008, Solly et al. 2014). The
amount and quality of litter varies considerably between regions (Macinnis-Ng and
Schwendenmann 2015), seasons (Chapter 3 and 5, Zhang et al., 2014), and with tree
species composition (Chapter 4, Becker et al., 2015). Litterfall is an important and
regular source of organic matter, making its quantification and turnover important for
understanding the productivity, temporal patterns, biogeochemical cycles, and carbon
dynamics in forest ecosystems (Vitousek 1984).
Regarding below-ground plant litter, mainly fine roots (defined as those roots with a
diameter <2 mm) are the most dynamic part of the root system (Lukac 2012). Fine
roots make up less than 5% of the total root biomass but may account for as much as
67-70% of the net primary production (NPP) of forest ecosystems and are highly
important in biogeochemical cycling (Vogt et al. 1996, Jackson et al. 1997). Even
though fine roots constitute a relatively small fraction of the total tree biomass
(Makkonen and Helmisaari 1999), the below-ground C allocation of plant litter
commonly exceeds the above-ground litterfall C input (Chapter 5; Trumbore et al.,
1995). Due to high turnover, fine roots can contribute 2-5 times more organic carbon to
the soil than the above-ground parts (Kucbel et al. 2011, Xia et al. 2015). The root-
derived C has a high potential to be stabilized long-term for soil C sequestration (Angst
et al. 2016). Nonetheless, studies on below-ground ecosystem processes are relatively
rare compared to those dealing with above-ground traits of plants. This reflects their
difficulty, the reliability of technological methods, economic limitations, and the
widespread assumption that fine roots are a rather marginal component of plants (Vogt
et al. 1998, Waisel et al. 2002, Maeght et al. 2013).
Fine root biomass is affected by climate variables such as temperature and precipitation
(Leppälammi-Kujansuu 2014), altitude, soil fertility (Finér et al. 2011), and above-
ground stand characteristics (Kucbel et al. 2011). Fine root biomass and production
generally increased with increasing mean annual temperature and precipitation, and
decreased with latitude (Yuan and Chen 2010). Interestingly, empirical evidence
indicates that the fine root biomass stock in terrestrial ecosystems is higher in

8
temperate deciduous forests (780 g m-2) than in boreal forests (600 g m-2) and tropical
evergreen or deciduous forests (570 g m-2) (Jackson et al. 1997). However, estimates
of below-ground biomass are uncertain and vary depending on methodology (Chapter
3). In the last decade, several methods have been developed to estimate fine root
production, mortality and turnover in forest ecosystems (Majdi et al. 2005, Lukac 2012,
Maeght et al. 2013). The most commonly used methods to estimate root production
and turnover are sequential coring, ingrowth cores, ingrowth nets and minirhzotron
techniques (Chapter 3; Johnson et al., 2001; Lukac and Godbold, 2001).
Litterfall (including leaves, flowers, fruits, bark and twigs) is also an important pathway
of carbon from vegetation to soil (Clark et al. 2001, Macinnis-Ng and Schwendenmann
2015). Previous studies show that total litterfall represents about 30% of forest net
primary productivity (Aragão et al. 2009). Numerous studies underline that the amount
of litterfall is heavily influenced by climatic conditions (Zhang et al. 2014, Becker et al.
2015) and vegetation diversity (Celentano et al. 2011, Moura et al. 2016). For example,
litterfall can increase with the onset of drought (Brando et al. 2008) and be higher in
diverse mixed stands compared to monocultures (Tang et al. 2010, Celentano et al.
2011).
Litter turnover rates are mainly controlled by climatic and edaphic properties such as
soil moisture and temperature (Gill et al. 1999). Tissue chemistry is another parameter
that explains a major amount of variability (Silver and Miya 2001, Solly et al. 2014).
This chemical composition may reveal further important aspects of biogeochemical
cycles (Chapter 4-7; Xia et al. 2015). Thus, the chemical compositions of roots (Chapter
4) and leaves (Chapter 5), e.g. indicated by labile and recalcitrant fractions, varies with
species and largely determines the rate of decay (Couteaux et al. 1995, Silver and Miya
2001, Sun et al. 2013) as well as the quality of C input into soil systems (Rasse et al.
2005). For example, recalcitrant tissues are characterized by relatively low
concentrations of easily degraded substrates such as lignin (Xia et al. 2015). Such
tissues are important to estimate the long-term carbon input into the soil system
(Chapter 4-6).
Beyond examining chemical composition, fine root morphological characteristics of tree
species are important with regard to plant growth strategy and carbon cycling (Comas
et al. 2002, Bardgett et al. 2014, McCormack et al. 2015, Kong et al. 2016). Root

9
morphological parameters such as root diameter (mm), specific root surface area (SRA;
cm2 g–1), specific root length (SRL; m g–1), and root tissue density (RTD; g cm–3) are
key traits in the root economics spectrum because they reflect a plant’s ecological
strategies of gaining access to soil resources (Comas et al. 2002, Weemstra et al.
2016). Some studies reported that roots of fast-growing species feature a high SRL or
low RTD (Chapter 4), both of which are associated with a resource acquisition strategy
involving short root life spans (Eissenstat 1991, Comas et al. 2002). The fine root
morphological parameters have also been interpreted to indicate the cost-benefit ratio
of roots, which determines litter quality and the root decomposition rate (Sun et al. 2013,
Collins et al. 2016). For example, root lifespan is negatively related to SRL (Silver and
Miya 2001, McCormack et al. 2012). Root traits also impact soil C cycling indirectly by
influencing the composition of the soil microbial community (Bardgett et al. 2014). In
summary, the amount of plant litter (both above- and below-ground), its biochemical
composition, and the activity of the microbial community are essential controlling factors
for the formation and accumulation of soil organic matter in terrestrial ecosystems
(Kögel-Knabner 2002). To date, information on below- or above-ground litter production
and turnover and the contribution to total ecosystem carbon budgets, particularly for
the Ethiopian highlands, remains scarce.
1.2.3 Tracing the biological origin and degradation status of organic carbon in
soil
Soil organic matter (SOM) is a chemically heterogeneous mixture of organic
compounds of plant, animal, and microbial origin exhibiting different stages of biological
oxidation (Amelung et al. 2008, Feng and Simpson 2011). Dead plant material (litter)
and animal residues are gradually decomposed until their original identity is no longer
recognizable, at which point they are considered SOM (Chapin et al. 2011). Once SOM
is incorporated into soil, however, it is difficult to infer its biological origin at the
molecular level (Amelung et al. 2008). Thus, the contribution of various biological
components to SOM accumulation remains largely unknown in general (Feng and
Simpson 2011). More specifically, the biological origin and degradation status of soil
organic carbon at the molecular level has been studied to a lesser extent in African
regions compared to temperate ecosystems. This calls for examining the molecular
composition of SOM and its degradation status to better understand the soil carbon
dynamics and biogeochemical processes here. In particular, biomarker analysis can

10
provide useful information and quantitative evidence to reconstruct the biological origin
of parent material (Kögel-Knabner 2000, Poirier et al. 2005, Otto and Simpson 2007,
Amelung et al. 2008). Biomarkers are soil organic compounds with a defined structure
carrying information of their origins, e.g. from plants, microorganisms, animals, or
anthropogenic sources in soil (Simoneit 2002, Amelung et al. 2008). The isolation and
quantification of biomarkers include: total solvent extraction of free lipids (of plant and
microbial origin), base hydrolysis to obtain bound lipids (from leaf cutin and the suberin
of roots), and copper (II) oxide (CuO) oxidation to obtain lignin-derived phenols (Otto
and Simpson 2007, Pisani et al. 2013, Spielvogel et al. 2014).
Solvent-extractable (free) lipids usually comprise less than 10% of SOM and can be
isolated using solvents that have different polarities, such as methanol,
dichloromethane, or ether (Otto et al. 2005). In contrast, ester-bound lipids are not
extractable with organic solvents, but they can be cleaved from SOM using chemolytic
methods such as base hydrolysis (Riederer et al. 1993, Nierop et al. 2005, 2006, Otto
et al. 2005). In general, soil lipids are primarily plant-derived and they range from simple
structures such as alkanes, alkanols, alkanoic acids, and steroids to complex unknown
lipids including plant waxes and biopolymers (such as suberin in the periderms of barks
and roots and cutin in leaf cuticles) (Mendez-Millan et al. 2011, Spielvogel et al. 2014).
The predominant long-chain ω-hydroxyalkanoic and α, ω-alkanedioic acids are typical
biomarkers for suberin, primarily indicating root or bark inputs into the soil (Chapter 6;
Otto et al., 2005). C16 and C18 ω-hydroxyalkanoic acids with mid-chain hydroxy or epoxy
groups are biomarkers for cutin or leaf inputs (Otto et al. 2005, Mendez-Millan et al.
2011, Spielvogel et al. 2014). Bound lipids such as suberin and cutin are more stable
than solvent-extractable lipids (Kögel-Knabner et al. 1989). Microorganisms are minor
contributors to soil lipids, such as branched short-chain (<C20) alkanoic acids,
hopanoids, and ergosterol from fungi (Kögel-Knabner 2002, Otto et al. 2005, Feng
2009).
Lignin is the second most abundant biopolymer (after cellulose and hemicellulose) in
nature and a major contributor to SOM (Kögel-Knabner 2002). Alkaline CuO oxidation
yields lignin-derived phenols classified as vanillyl, syringyl, and cinnamyl compounds
(Goñi and Hedges 1992, Pautler et al. 2013, Conti et al. 2016). The composition of
lignin monomers is commonly used to describe the major sources of plant groups
(angiosperms, gymnosperms) and tissue types (woody, non-woody) (Hedges and

11
Mann 1979a, Feng and Simpson 2011). Ratios of lignin-derived phenolic acids to their
corresponding aldehydes (Ad/Al) are useful tools for determining the stage of lignin
degradation in soils (Otto et al. 2005, Feng and Simpson 2011). Lignin is considered to
be a recalcitrant organic compound, but, polymethylene compounds (cutin, suberin) are
the most recalcitrant components of soil OM (Kögel-Knabner et al. 1989, Simpson et
al. 2005, Hamer et al. 2012). Many studies reported that enrichment of root-derived
suberin in soils suggests preferential stabilization of recalcitrant root-derived carbon
(Nierop 1998, Feng and Simpson 2007, Otto and Simpson 2007, Mendez-Millan et al.
2010). Other studies, however, found that plant roots possess on average higher
concentrations of lignin compared to leaves and stems (Abiven et al. 2005, de Leeuw
et al. 2005, Lorenz et al. 2007, Sun et al. 2013, Xia et al. 2015). This may be responsible
for a reduced mass loss rate of roots during decomposition compared to leaves
(Chapter 5).
1.3 Aims and outline
The overall objective of the thesis was to investigate the impact of land use change on
soil carbon dynamics, biochemistry, biological origin, and degradation status at the
landscape level of Ethiopian highland soils.
First, I investigated the soil carbon stocks of five land use systems that include natural
forest, eucalyptus plantations, exclosure, cropland, and grazing land. Here, the specific
objective was to determine the effect of land use conversion on SOC stocks and the
effectiveness of ecological restoration on SOC stock levels through afforestation and
natural regeneration (exclosure). Because soil carbon stock changes are the result of
differences between above- and below-ground litter inputs and losses, I subsequently
determined the above- and below-ground litter production, turnover, and changes in
response to land use conversion to estimate the amount of carbon fluxes into the soil.
The morphological traits and the intrinsic differences in the species-specific chemical
composition of ten dominant woody species were investigated to elucidate the
decomposition rate of plant materials and the quality of carbon deposition into the soil.
Subsequently, a decomposition experiment was conducted with single and mixed litters
of both fine roots and leaves of the four most dominant tree species, assuming that the
occurrence of highly degradable litter favors the breakdown of more recalcitrant litter in
the mixtures. A biomarker analysis was conducted to reconstruct the biological origin

12
of soil organic carbon based on their carbon skeleton. The extent of carbon loss by
heterotrophic respiration (CO2 efflux) was investigated following temperature change
with or without carbon and nutrient substrates.
1.4 Hypotheses
Land-use change from natural forest to cropland or grazing land results in a rapid loss
of SOC stocks, and reestablishment of perennial vegetation cover on degraded lands
restores SOC, but not to the original level.
Fine roots are more biochemically resistant to decomposition than leaf litter due to
intrinsic variations in tissue chemistry, and thus fine roots are major contributors of
recalcitrant organic carbon to soil in the Ethiopian highlands.
The efficiency of microbes in decomposing soil organic matter and the CO2 efflux to the
atmosphere is limited more by availability of C substrates and by N and P deficiencies
than by temperature.
Pre-publication of parts of this thesis
Chapter 2 – a manuscript from this section has been accepted for publication as
follows:
Assefa, D., Rewald, B., Sandén, H., Rosinger, C., Abiyu, A., Yitaferu, B., Godbold,
D.L., 2017. Deforestation and land use strongly effect soil organic carbon and
nitrogen stock in Northwest Ethiopia. Catena 153:89-99.
Chapter 4 – a manuscript from this section has been accepted with major changes for
publication in Ecosytems as:
Assefa, D., Rewald, B., Abiyu, A., Godbold, D.L., 2017. Fine root morphology,
biochemistry and litter quality indices of fast- and slow-growing woody species in the
Ethiopian highland forests.

13
2 Soil organic carbon dynamics after land use change in
Northwest Ethiopia
2.1 Abstract
Soil is the largest terrestrial organic carbon pool and can act as a source or sink for
atmospheric CO2. Although reliable soil carbon (SOC) stock measurements of major
ecosystems are essential for predicting the influence of advancing climate change,
comprehensive data on SOC stocks is still scarce for most ecosystems in subtropical
areas. In this study, SOC and N stocks of different land use systems were investigated
along a climatic gradient in Northwest Ethiopia. The land use systems ranged from dry
subtropical Afromontane forest, as the baseline, to cropland as the most degraded
system. In addition, we investigated the changes of SOC stocks after interventions to
recover vegetation cover; these were eucalyptus plantations and an exclosure to
prevent grazing. Total SOC varied between land use systems and ranged from 3.1 kg
C m-2 in croplands to 23.9 kg C m-2 in natural forest, and average N stock ranged from
0.4 kg N m-2 in croplands to 2.1 kg N m-2 in natural forest. In forests, there were a clear
vertical gradient in SOC and N stock down the soil profile, and 60% of the total SOC
and N stocks were found in the upper 10 cm soil depth. Using the Sr/Ca and Ba/Ca
ratios and the vertical distribution of the C/N ratio of the soil, the losses of SOC were
shown to be due to loss of the of the upper soil layer. Afforestation of degraded
croplands and grazing lands with eucalyptus increased SOC stocks to nearly 70% of
the natural forest levels within 30 years. Exclosure, which removed grazing pressure
and allowed regeneration of native vegetation, increased SOC in the top soil only.
Key words: land use change, eucalyptus, grazing land, exclosure, strontium
2.2 Introduction
Soil is a key factor in the global carbon cycle and can act either as a source or as a sink
to atmospheric CO2. Worldwide, soils are estimated to hold 3150 Pg of carbon (C) which
is more than four times the amount of carbon stored in terrestrial plant biomass (650
Pg C) or the atmosphere (750 Pg C) (Fan et al. 2016). The size of the pool of soil
organic carbon (SOC) is determined by the input of plant-derived carbon, the potential

14
to sequester carbon through physical and bio-chemical processes, and the loss of SOC
through heterotrophic respiration, leaching, and erosion (Jobbágy and Jackson 2000,
De Deyn et al. 2008).
Reliable SOC stock measurements of major ecosystems are essential to parameterize
models estimating net C stocks and changes in different biomes (Zimmermann et al.
2009, Powers et al. 2011). While current models are used to estimate global and
regional soil carbon pools and for predicting the influence of advancing climate change
on those pools (Jones et al. 2005), comprehensive data on SOC stocks is still scarce
for some ecosystems at scales relevant to local management as well as national carbon
inventories (Victoria et al. 2012). Because the impact of land use conversion (LUC)
varies depending on the land use type and the abiotic factors present, regional and
ecosystem level studies are important (Milne et al. 2007, Schulp et al. 2008, Schrumpf
et al. 2011). In Ethiopia, extensive soil carbon surveys are especially scarce for the
remnant natural forests of the NW Ethiopian highlands and the land use systems
established on previously naturally forested area since 1950 (Zeleke and Hurni 2001).
Natural forests and woodlands cover less than 9.5% of the Amhara region and about
60% of the total area is used as cropland and grazing land (Desta et al. 2000, Bekele
2011). Eucalyptus globules (Labill.) has been planted in the central highlands of
Ethiopia since 1895 (Pohjonen and Pukkala 1990). It is the dominant exotic species
planted in the highland areas because of its fast growth (Pohjonen and Pukkala 1990),
non-palatability to livestock, multiple use and high economic return .
Soil carbon stocks are influenced by factors such as climate, geology and weathering
history, and biotic variables such as species composition and density (Fernandez et al.
2013, Vesterdal et al. 2013). The most important human effect on the rate of changes
in SOC stock is attributed to LUC (Guo and Gifford 2002, Houghton and Goodale 2004,
Don et al. 2011), and LUC is a significant factor in global emissions of CO2 (IPCC,
2014). The contribution of LUC to anthropogenic CO2 emissions have recently been
estimated at 1.2 Pg yr-1, or about 12-15% of total anthropogenic emissions (Deng et al.
2016b). Many studies have shown that the conversion of natural vegetation to cropland
often leads to a depletion of SOC stocks (Guo and Gifford 2002, Poeplau et al. 2011,
Abu-hashim et al. 2016). For example, Del Galdo et al. (2003) reported that cropland
soils had a 48% lower SOC content in the top 10 cm compared to permanent grassland.
The reduction of original SOC stock is more intense and faster in tropical ecosystems

15
with loss of up to 60% in a few years compared to humid temperate ecosystems with
loss of 30% in 60 years of cultivation (Victoria et al. 2012). Within the complex of LUC,
two of the major environmental factors influencing SOC stocks are temperature and
moisture that affect heterotrophic soil respiration and hence SOC stocks (Wynn et al.
2006, Feng and Simpson 2008, Chen et al. 2013). In addition to affecting soil moisture
in high rainfall areas, precipitation can also have physical effects on soils such as
erosion (Guo and Gifford 2002). Guo and Gifford (2002) hypothesized that after
conversion of forest to croplands, especially in areas with high annual rainfall, topsoil
erosion causes a major loss of carbon. Mean annual precipitation in the central
highlands of Ethiopia is ca.1600 mm, but can reach up to 2000 mm within the three
main rainy months (Awulachew et al. 2009). Due to such heavy seasonal rainfall
combined with a topography of steep slopes, extensive topsoil erosion after heavy
rainfall events is common in the Ethiopian highlands (Shiferaw and Holden 1999).
To gain an insight into changes in soils, strontium (Sr) to calcium (Ca) and barium (Ba)
to calcium ratios have been widely used as markers to reconstruct environmental
history (Bullen et al. 2005, Tabouret et al. 2010) and biogeochemical properties of the
ecosystem (Capo et al. 1998, Kabata-Pendias 2010). Biologically, Sr2+ is considered to
be a non-essential element, however, it is strongly associated with and chemically very
similar to Ca (Capo et al. 1998) with a similar ionic radius and charge (Bullen et al.
2005). Strontium/Ca and Ba/Ca ratios can be a powerful tool in studies of chemical
weathering and soil genesis. These ratios often combined with measurements of
isotopic 87Sr/86Sr ratios have been used to trace the relative contributions of
atmospheric dust input and mineral weathering to soil formation (Bullen et al. 2005,
Derry and Chadwick 2007, Li et al. 2016). For example, Li et al. (2016) could show that
atmospheric dust inputs from continental Asia are an important factor in soil
development in Hainan Island of the coast of China. In study of European forest soils,
Lequy et al. (2012) showed that up to 30% of nutrient inputs to the soil resulted from
inputs of Aeolian dust.
In this study, we hypothesized that land-use change for forest to cropland results in a
rapid loss of SOC stocks, and that reestablishment of vegetation cover on degraded
lands results in a restoration of SOC. To test this hypothesis, we determined SOC
stocks of different land-use types in ecosystems typical for Eastern Africa. In addition,
we investigated the effectiveness of planting eucalyptus (afforestation) and excluding

16
grazing on degraded ecosystems (exclosure) in restoring SOC stock levels. Total C
and N stocks were estimated in five land use systems: natural forest, eucalyptus
plantation, exclosure (a degraded area excluded from animal and human interventions
for rehabilitation), cropland, and grazing land at elevations ranging from 800 to 2500 m
a.s.l. (above sea level). In addition, Sr/Ca and Ba/Ca ratios were used as putative
markers of soil loss.
2.3 Material and methods
2.3.1 Research sites
The study was conducted in the Amhara National Regional State (170,752 km2) located
in the North-central parts of Ethiopia. Elevations range from approximately 500 m a.s.l.
near to the Sudanese boarder to 4620 m in the highlands. Thirty one percent of the
Amhara region can be classified as lowlands (<1500 m) and 69% as highlands
(Awulachew et al. 2009, Ayalew et al. 2012). The mean annual temperatures varies
from 11°C in the highlands (Betrie et al. 2011) to 27°C in the lowlands (Ayalew et al.
2012). The average annual rainfall in lowland areas is around 700 mm (Bewket and
Conway 2007) whereas highlands often receive over 2000 mm of precipitation
(Awulachew et al. 2009). While Ethiopia is located in the tropics, the climate of the study
areas in the highlands are temperate with dry winters and warm summers, and are
classified as Cwb according to the Köppen-Geiger climate classification system (Peel
et al. 2007). The majority of the area has a unimodal rainfall characteristic with the main
rainfall months between June and September whereas the dry months are from
October to May with little rain in April and May. Climate data for the sites (Table 2.1)
were obtained from previous studies (Moges and Kindu 2006, Workneh and Glatzel
2008, Wassie et al. 2009, Abiyu 2012, Assaye et al. 2013).
17
Table 2 1 Site characteristics of a range of sites in the highlands or lowlands of the Amhara
region of Northwestern Ethiopia.
Site characteristics Gelawdios Katassi Tara Gedam Ambober Mahibere-Selassie
Location 11°38’25’’ N
37°48’55’’ E
11°0’05’’ N
36°44’8’’ E
12°8’47’’ N
37°44’45’’ E
12°31’15’’ N
37°31’53’’ E
12°32’20’’ N
36°35’9’’ E
Altitude (m a.s.l.) 2500 2200 2230 2230 850 Air temperature (°C)* 19a 15b 21c 20d 27e Precipitation* (mm yr-1)
1220a 2200b 1100c 1083d 965e
Soil type Cambisols Luvisols Cambisols Leptosols Fluvisols Available land use types F, EP, C, G F, EP, C F, EP, G EX, C, G, F, C Basal area g (m2 ha-1) Forest 34.1 17.7 19.8 3.4 Eucalyptus 19.0 16.7 na na Tree density (N ha-1)g Forest 6334 2293 4170 205 Eucalyptus 3031 552 na na
F = Forest; EP = Eucalyptus plantation; EX = Exclosure; G = Grazing land; C = Cropland; na,
not available. *Climate data was obtained from previous studies (dAbiyu, 2012a; bAssaye et al.,
2013; eMoges and Kindu, 2006; aWassie et al., 2009; cWorkneh and Glatzel, 2008). Inventory
data was obtained from Gebrehana (2015). Basal area was measured at 1.3 m above the
ground from a 10-15 m radius circular plot depending on forest size. Tree density includes
saplings but height >1.5 m.
The topography is typical of volcanic landscapes comprising volcanic rocks, deeply
incised by streams resulting the current ragged and undulating landforms. The ragged
topography combined with heavy rain events during the wet season (June-August)
promotes soil erosion (Bewket and Sterk 2005, Betrie et al. 2011, Shiferaw 2011). The
major soil types in the region are Alisols, Cambisols, Leptosols, Luvisols, Nitosols, and
Vertisols according to World Reference Base for soil resources (Awulachew et al. 2009,
Betrie et al. 2011, WRB 2014).
Five research sites were selected in the northwestern part of Amhara to cover major
climatic and edaphic conditions within the Amhara region (Table 2.1). The sites Katassi,
Gelawdios, Tara Gedam, Ambober, and Mahibere-Selassie have an average distance
of 120 km between each other. At each site, up to five land use types, namely natural
forest, eucalyptus plantation, exclosure (rejuvenated woodland on former grazing land),
cropland, and grazing land were identified adjacent to each other to give similar
topography, edaphic, and climate conditions for comparison of land uses at each site.
However, not all land use systems were available at each site. Geographical location,

18
climate, soil, and vegetation characteristics of each site as well as the available land
use systems are described in Table 2.1. Forests at Gelawdios, Katassi, and Tara
Gedam are dry Afromontane remnant pristine forests composed of mostly an intimate
mixture of indigenous tree species. These forests (except Katassi) are traditionally
better protected by local institutions and mostly confined to sacred groves associated
with churches and monasteries (Aerts et al. 2016). The dominant tree species for
highland areas (Gelawdios, Katassi, and Tara Gedam) are Albizia schimperiana,
Apodytes dimidiata, Calpurnia aurea, Carissa edulis, Croton macrostachyus, Ekebergia
capensis, Maytenus arbutifolia, Olea europaea, Prunus africana, and Schefflera
abyssinica. The extensive lowland semiarid forest around Mahibere-Selassie
monastery is a savannah woodland dominated by grasses with some scattered trees.
This site, which represents the typical lowland forest type of this region, is subject to
frequent man-made fires that burn off the grass cover but does not cause extensive
damage to the trees. Characteristic tree species in the lowland are Acacia polyacantha,
Balanites aegyptica, Boswellia papyrifera, Diospyros abbyssinica, Ficus sycomorus,
Pterocarpus lucens, Sterculea setigera, Oxytenanthra abyssinica, and Ziziphus
spinachrist. The Eucalyptus globules plantations at Katassi and Gelawdios were
planted on grazing land around 1985, and were thus ca. 30 years old at the time of
sampling. In Tara Gedam, Eucalyptus camaldunesis was planted on former cropland
and has been cut 3 times since planting. The date of planting is unknown but as in this
part of Ethiopia, the coppicing intervals are between 5 to 10 years, the site must be
between 20 to 40 years old and more likely 40 years old. Details of tree and basal area
densities in the four natural forests and two eucalyptus stands can be found in
Table 2.1. The exclosure at Ambober was established in 2007 on former grazing land
(Abiyu 2012). The exclosure system is commonly used to rehabilitate degraded land
by protecting land areas from further animal grazing and human interference. In
addition to preventing grazing, enrichment planting of seedlings of indigenous and
some exotic tree species was carried out. Since inception, natural revegetation has
returned to the area and it is now covered in an extensive bush woodland. The
management, utilization and protection of the site is undertaken by the local community
(Girmay et al. 2008). All studied croplands and grazing lands were converted from
natural forest within the last 50 years. Grazing lands used as common land to graze
herds of cattle, sheep, goats, and donkeys. The number of years that the cropland was
continuously cultivated after conversion of forest was obtained from local knowledge

19
(Nyssen et al. 2008). Croplands are ox-ploughed to a depth of approximately 30 cm.
Owing to the nature of climate, crops such as Eragrostis tef, Eleusine coracana,
Sorghum bicolor, Zea mays, Triticum aestivum, and Vicia faba are grown during the
wet season. Between years, the crop planted is changed.
2.3.2 Soil sampling
Soil sampling took place during two sampling campaigns, one in March 2014 (end of
the dry season), and another during the wet season in June 2015. In the natural forests,
10 sampling points were marked every 50-100 m along a transect line. At each point,
two soil samples were taken at a distances of about 2 m apart. In eucalyptus
plantations, exclosure, croplands, and grazing lands, one soil sample was taken at 10
sampling points. The distance between sampling points along transects in these land
use systems was 20-50 m according to plot size. After removing the litter layer (if
present), a soil corer (6.6 cm internal diameter) was driven to a maximum of 50 cm
depth or until the bedrock was reached. Soil samples were extracted into a Styrofoam
tray and morphologically described, using the terminology of World Reference Base for
soil resources (WRB 2014). In total, 190 soil cores were collected from all land use
systems and cores were divided into depth classes of 0-10 cm, 10-20 cm, 20-30 cm,
and 30-50 cm. For the samples from the natural forest, a composite sample for each
depth was made from the two cores taken at each sampling point. Each soil sample
was sieved using a 2 mm sieve, homogenised, and placed in a plastic bag. The
samples were then transported to Vienna for laboratory analysis.
2.3.3 Bulk density determination
Bulk soil density samples were taken at five sampling points, every second sampling
point along the transect line. The samples were taken during the onset of wet season
when the soil had no drying cracks. At the edge of each sampling plot, a profile was
dug to 60 cm. Soil samples of known volume were taken from the sides of the profile
centred at 5, 15, 25, and 40 cm depth using a stainless steel bulk density ring. Using a
trowel, the ring was removed from the horizon and the soil trimmed to the tops and
bottoms of the ring using a sharp knife. Any stones (>2 mm) were sieved out and
weighed separately. The volume of stones was quantified by displacement in a water
bath. Bulk density (soil particle <2 mm) was determined after oven dry at 105°C as a
stone free dry weights according to Don et al. (2007).

20
2.3.4 Total C and N stock determination
Before analysis, the bulk soil samples from each horizon were homogenised again, and
a subsample of approximately 3-5 g was dried at 105°C for 48h, then crushed to a fine
powder and remixed. From each subsample, a single large aliquot of about 200 mg
was taken and total carbon and nitrogen concentration were determined using a CN
elemental analyser (Truspec CNS LECO, St. Joseph, USA). The analyser was initially
calibrated using manufacturer’s standard material (Part No 502-309 purchased from
LECO), and the calibration controlled using the same standard every 30 samples.
Baseline correction was carried out every 90 samples using 5-10 empty cells. Using
the manufacturer’s standard material the instrument’s typical precision was found to be
±0.005% for carbon and ±0.001% for N.
Carbon and nitrogen stocks were determined on a weight to area basis (kg of C/N per
m2 of soil) per soil depth class and down to 50-cm. Total C stock was calculated by
summing the carbon stock of each soil horizon determined. The carbon concentration
was corrected for bulk density and soil volume, and interpolated to an area basis as
shown in the following equation:
𝑇𝐶 = ∑ 𝐶%𝑖 ∗ 𝐵𝐷𝑖 ∗ 𝑣𝑖 ∗ 0.001 𝑘𝑔𝑛𝑖 (1)
where TC is total carbon (kg C m-2); C%i is concentrations of carbon in percentage at
depth i; BDi is bulk density at depth i; and Vi is volume of soil at each horizon. Total
nitrogen (TN) was calculated in the same way.
Vertical distribution of cumulative SOC fraction (Y) from the surface to any depth (d)
was calculated according to a model developed by Gale and Grigal (1987). This model
was previously used for vertical root distributions and later adopted for vertical
distribution of SOC (Jobbágy and Jackson 2000, Deng et al. 2016b). An asymptotic
nonlinear model of the following form was used to describe the vertical SOC
distributions:
Y = 1-ßd (2)
where Y is the cumulative SOC fraction from the surface to soil depth d in centimetres
(midpoint), and ß the estimated parameter used as a measure of index of vertical SOC
distribution.

21
2.3.5 Strontium, calcium and barium elemental analysis
For analysis of strontium, calcium, and barium from the bulk soil sample of Gelawdios
and Katassi natural forest and cropland, a further 20 g subsample was taken from each
sample, dried for 3 days at 50°C, and remixed. Before analysis, residual moisture was
removed by drying at 105°C for 30 minutes. From each sample, a 500 - 600 mg aliquot
of dried soil was added into a 75 ml fusion tube with 20 ml of Aqua regia (3:1 HCl and
HNO3). To prevent the solution from foaming, 10 μl of Octanol was added.The soil
samples were shaken for 4.5 h and then digested at 125°C for 2 h. The cooled samples
were then made up to a volume of 75 ml with deionized water and filtered with Whatman
Grade 589/1 paper. The concentrations of total strontium (Sr), calcium (Ca), and barium
(Ba) were determined by inductively coupled plasma optical emission spectroscopy
(Perkin Elmer Optima 8300; ICP-OES Spectrometer, Waltham, USA) using external
calibration in line with ÖNORM L 1085. Two samples of a standard soil were used as
internal standards (ICP Multi-element Standard solution XIV, CertiPUR, purchased
from Merck KGaA, Germany).
2.3.6 Determination of soil pH
Soil pH was determined in 1:3 soil suspensions in both deionized water and 0.01 M
CaCl2 solution using a digital potentiometric pH-meter. Five samples per site and
horizon were analyzed for each land use type.
2.3.7 Particle size determination
For soil texture analysis from the 10-bulk soil sample per horizon, equal amounts of soil
of between 4-5 g were taken and thoroughly mixed. About 30 g of the composite soil
samples were taken per horizon for soil texture analysis. Soil texture was determined
by a combination of wet sieving and sedimentation methods (Kettler et al. 2001). Soil
particle were dispersed in 3% aqueous sodium hexametaphosphate (HMP, (NaPO3)n)
using a 3:1 HMP (90 mL) to soil (30 g) ratio, and shaking for 16 h (overnight). After
dispersion, the soil slurry was passed through a range of sieves of different mesh sizes
(0.5 – 0.053 mm) to separate sand particles and particulate organic matter (POM). The
collected sand particles (>0.053 mm) were dried at 105°C to constant weight, and
residual POM was then removed by ignition at 450°C for 4 h. The sand percentage was
calculated based on its fraction of the original sample mass after ignition at 450°C. The

22
solution (containing silt + clay) passing the sieve was collected in a 500 mL pre-weighed
beaker, stirred thoroughly to achieve suspension of all soil particles, and then left
undisturbed at room temperature (18-24°C) for 6h to allow silt particles to settle. After
sedimentation period, the supernatant containing the suspended clay was decanted,
and the settled silt fraction was then dried in the beaker at 105°C to constant weight.
The sand, silt, and clay percentages were calculated based on their fraction of the
original sample mass as follows.
Sand% = (Sand mass/original sample mass)*100% (3)
Silt% = (silt mass/original sample mass)*100% (4)
Clay% = 100 – (Sand% + Silt%) (5)
To assign texture classes, the USDA particle size classes of sand (2.0-0.053 mm), silt
(0.053-0.002 mm), and clay (0.002 mm) were used.
2.3.8 Data analysis
If the data was not normally distributed, it was square-root-transformed to reach
normality. Analysis of variance (ANOVA) was used to test the significance of mean
differences in total carbon and nitrogen as dependent variable and land use systems,
climate variables, and soil depths as factors. To determine differences between groups
Scheffe post hoc tests were used. Linear regression was used to evaluate the
relationships between the dependent variable (total C stock) with total N, pH, bulk
density, soil texture, mean annual precipitation (MAP) and mean annual temperature
(MAT) as independent variable. The IBM SPSS analytical software package (version
21) and SigmaPlot (version 13) were used for all statistical analyses and graphs.
Significant level determined at α = 0.05. Data are presented as mean ± standard error
(SE).

23
2.4 Results
2.4.1 Soil carbon and nitrogen stocks
Total SOC and N stocks varied considerably between land uses systems. The average
C stock (0-50 cm soil depth) was significantly higher in the forest ecosystems than other
land use systems and cropland had the lowest C stock (Fig. 2.1a). The average C
stocks per land use type can be ranked as, forest (22.0 kg C m-2) > eucalyptus
plantation (15.7 kg C m-2) > grazing land (9.3 kg C m-2) > cropland (6.8 C kg m-2),
however the difference between grazing land and cropland was not significant.
Similarly, soil N stock was also showed significant differences between the land use
systems and followed a similar trend to C stock (Fig. 2.1a). In the mature woodland
ecosystems, the lowest C and N stocks were found in the lowland semiarid forest at
Mahibere-Selassie (Table 2.2). There was no significant difference found in soil C or N
stock between Gelawdios, Tara Gedam, and Katassi. At Mahibere-Selassie, in contrast
to all the other mature forest sites, the C and N stocks of cropland were significantly
higher than that of the corresponding forest (Fig. 2.1b; P < 0.001). Total C and N stock
did not differ significantly among three land uses in Ambober (Table 2.2; Appendix 1
Table 1 (Table A1.1). Soil organic carbon stock of the eucalyptus plantation was similar
at Katassi and Tara Gedam but was significantly lower at Gelawdios compared to the
other two sites. Soil organic carbon stock of the eucalyptus plantation was on average
57% and 40% higher than the respective cropland and grazing land, but was on
average 29% lower than the SOC stock of the natural forest (Table 2.2). Similarly, soil
N stock was 47% and 29% higher in eucalyptus soil than the cropland and grazing land
soils.
The C/N ratios of soils were similar for all sites and land use types except for the
croplands. The values of the C/N ratios were between 10.9 and 13.6, and higher in
eucalyptus than the natural forests and grazing lands. The cropland at Katassi had a
very low C/N ratio (6.3) and was significantly different from all the other sites. The
highest C/N ratio was recorded at Mahibere-Selassie cropland (14.3; Table 2.2).
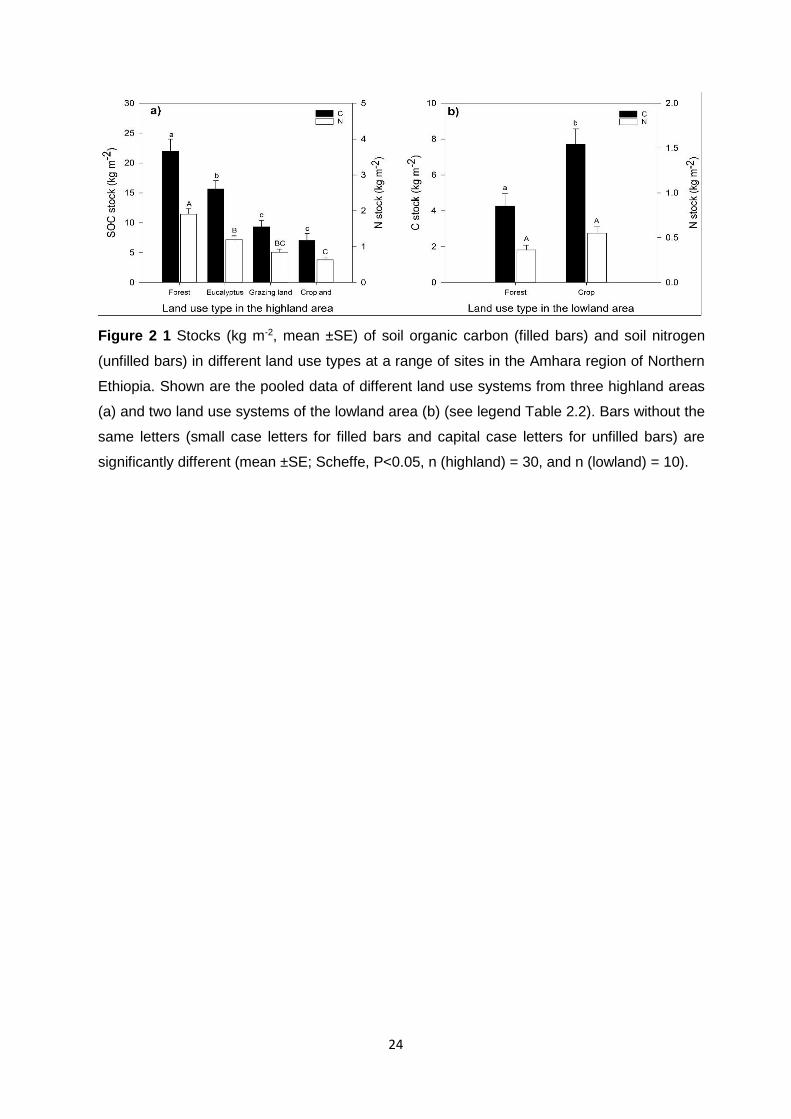
24
Figure 2 1 Stocks (kg m-2, mean ±SE) of soil organic carbon (filled bars) and soil nitrogen
(unfilled bars) in different land use types at a range of sites in the Amhara region of Northern
Ethiopia. Shown are the pooled data of different land use systems from three highland areas
(a) and two land use systems of the lowland area (b) (see legend Table 2.2). Bars without the
same letters (small case letters for filled bars and capital case letters for unfilled bars) are
significantly different (mean ±SE; Scheffe, P<0.05, n (highland) = 30, and n (lowland) = 10).
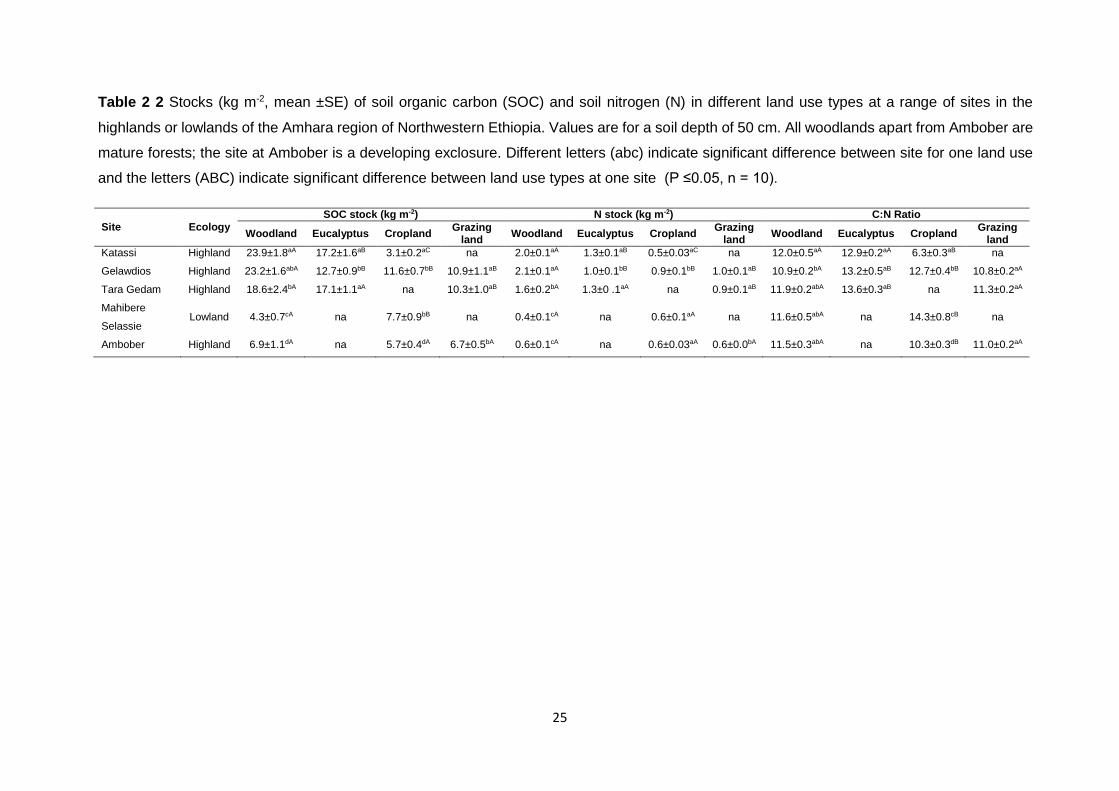
25
Table 2 2 Stocks (kg m-2, mean ±SE) of soil organic carbon (SOC) and soil nitrogen (N) in different land use types at a range of sites in the
highlands or lowlands of the Amhara region of Northwestern Ethiopia. Values are for a soil depth of 50 cm. All woodlands apart from Ambober are
mature forests; the site at Ambober is a developing exclosure. Different letters (abc) indicate significant difference between site for one land use
and the letters (ABC) indicate significant difference between land use types at one site (P ≤0.05, n = 10).
Site Ecology
SOC stock (kg m-2) N stock (kg m-2) C:N Ratio
Woodland Eucalyptus Cropland Grazing
land Woodland Eucalyptus Cropland
Grazing land
Woodland Eucalyptus Cropland Grazing
land
Katassi Highland 23.9±1.8aA 17.2±1.6aB 3.1±0.2aC na 2.0±0.1aA 1.3±0.1aB 0.5±0.03aC na 12.0±0.5aA 12.9±0.2aA 6.3±0.3aB na
Gelawdios Highland 23.2±1.6abA 12.7±0.9bB 11.6±0.7bB 10.9±1.1aB 2.1±0.1aA 1.0±0.1bB 0.9±0.1bB 1.0±0.1aB 10.9±0.2bA 13.2±0.5aB 12.7±0.4bB 10.8±0.2aA
Tara Gedam Highland 18.6±2.4bA 17.1±1.1aA na 10.3±1.0aB 1.6±0.2bA 1.3±0 .1aA na 0.9±0.1aB 11.9±0.2abA 13.6±0.3aB na 11.3±0.2aA
Mahibere
Selassie Lowland 4.3±0.7cA na 7.7±0.9bB na 0.4±0.1cA na 0.6±0.1aA na 11.6±0.5abA na 14.3±0.8cB na
Ambober Highland 6.9±1.1dA na 5.7±0.4dA 6.7±0.5bA 0.6±0.1cA na 0.6±0.03aA 0.6±0.0bA 11.5±0.3abA na 10.3±0.3dB 11.0±0.2aA
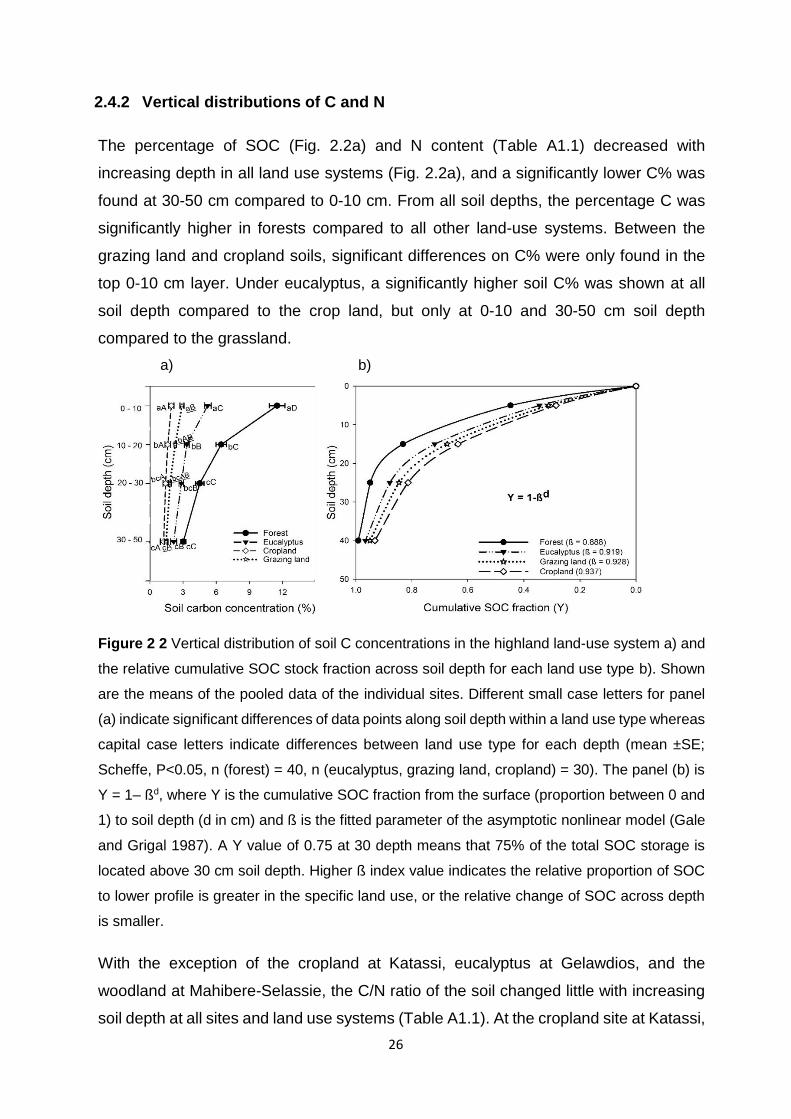
26
2.4.2 Vertical distributions of C and N
The percentage of SOC (Fig. 2.2a) and N content (Table A1.1) decreased with
increasing depth in all land use systems (Fig. 2.2a), and a significantly lower C% was
found at 30-50 cm compared to 0-10 cm. From all soil depths, the percentage C was
significantly higher in forests compared to all other land-use systems. Between the
grazing land and cropland soils, significant differences on C% were only found in the
top 0-10 cm layer. Under eucalyptus, a significantly higher soil C% was shown at all
soil depth compared to the crop land, but only at 0-10 and 30-50 cm soil depth
compared to the grassland.
a) b)
Figure 2 2 Vertical distribution of soil C concentrations in the highland land-use system a) and
the relative cumulative SOC stock fraction across soil depth for each land use type b). Shown
are the means of the pooled data of the individual sites. Different small case letters for panel
(a) indicate significant differences of data points along soil depth within a land use type whereas
capital case letters indicate differences between land use type for each depth (mean ±SE;
Scheffe, P<0.05, n (forest) = 40, n (eucalyptus, grazing land, cropland) = 30). The panel (b) is
Y = 1– ßd, where Y is the cumulative SOC fraction from the surface (proportion between 0 and
1) to soil depth (d in cm) and ß is the fitted parameter of the asymptotic nonlinear model (Gale
and Grigal 1987). A Y value of 0.75 at 30 depth means that 75% of the total SOC storage is
located above 30 cm soil depth. Higher ß index value indicates the relative proportion of SOC
to lower profile is greater in the specific land use, or the relative change of SOC across depth
is smaller.
With the exception of the cropland at Katassi, eucalyptus at Gelawdios, and the
woodland at Mahibere-Selassie, the C/N ratio of the soil changed little with increasing
soil depth at all sites and land use systems (Table A1.1). At the cropland site at Katassi,

27
the C/N ratio in the 0-10 cm layer was 8.4 and this decreased to 4.3 at 30-50 cm soil
depth. In the eucalyptus at Gelawdios, the C/N ratio of the 0-10 cm was significantly
higher than the deeper soil layers and was the highest value at any site. At the
woodland of Mahibere-Selassie, the C/N ratio of the 30-50 cm soil was 8.3 and
significantly lower than the other soil layers.
For all land use systems, more than 60% of total C stock was found in the upper 20 cm
depth (Fig 2.2b). In the estimation of relative distribution of SOC with soil depth, the
curves clearly separated for each land use, hence the largest relative proportion of SOC
in the 0-20 cm soil layer was in the natural forest and the lowest was in the cropland.
This distribution is also reflected in the calculated index values of ß, where value for
the cropland was 0.937 followed by grazing land (0.928), eucalyptus (0.919) and the
natural forest (0.888) (Fig. 2.2b).
At the Ambober site, C% in the soil was significantly higher in the 0-10 cm soil layer
between the exclosure and the cropland soils, but not at greater soil depths (Fig. 2.3).
There was no significant difference in C% in the soils between the grassland and the
other two land use types in the 0-10 cm soil layer. In all land use types, the C% in the
soil was significantly lower at 20-30 cm soil depth than at 0-10 cm soil depth.
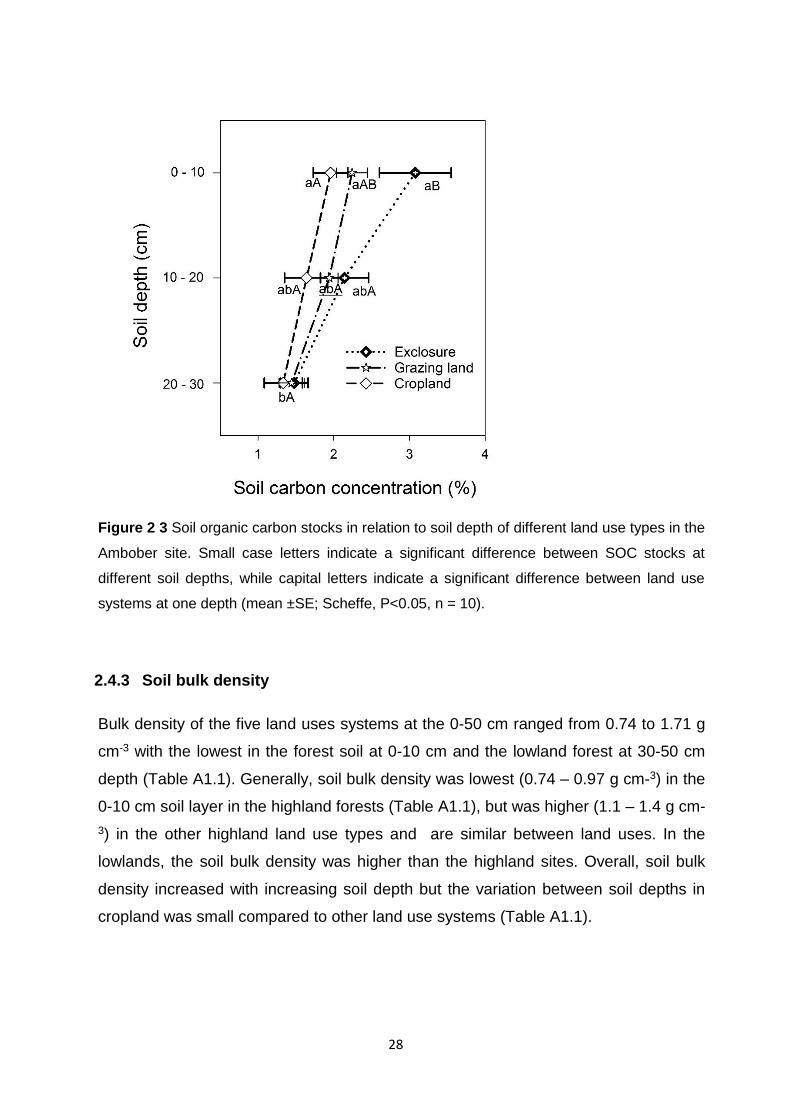
28
Figure 2 3 Soil organic carbon stocks in relation to soil depth of different land use types in the
Ambober site. Small case letters indicate a significant difference between SOC stocks at
different soil depths, while capital letters indicate a significant difference between land use
systems at one depth (mean ±SE; Scheffe, P<0.05, n = 10).
2.4.3 Soil bulk density
Bulk density of the five land uses systems at the 0-50 cm ranged from 0.74 to 1.71 g
cm-3 with the lowest in the forest soil at 0-10 cm and the lowland forest at 30-50 cm
depth (Table A1.1). Generally, soil bulk density was lowest (0.74 – 0.97 g cm-3) in the
0-10 cm soil layer in the highland forests (Table A1.1), but was higher (1.1 – 1.4 g cm-
3) in the other highland land use types and are similar between land uses. In the
lowlands, the soil bulk density was higher than the highland sites. Overall, soil bulk
density increased with increasing soil depth but the variation between soil depths in
cropland was small compared to other land use systems (Table A1.1).

29
2.4.4 Strontium:calcium and barium:calcium ratios
At both the Katassi and Gelawdios forest sites, the ratios of Sr/Ca and Ca/Ba increased
with increasing soil depth (Fig. 2.4). With the exception of Sr/Ca at Katassi, the ratios
of Sr/Ca and Ca/Ba were significantly lower in the 0-10 cm soil than in the 20-30 cm
soil. Significant differences in the ratios of both Sr/Ca and Ca/Ba were found at both
sites if the 0-10 cm and 30-50 cm soil depths are compared. In contrast, in the cropland
soil profile, the ratios of Sr/Ca and Ca/Ba did not change with soil depth at either site.
The changes in ratios of Sr/Ca and Ca/Ba shown in the forest soil profiles were primarily
due to differences in the levels of Ca between the soils layers (Table A1.2). In both
forests sites higher levels of Ca were determined in the 0-10 cm soil layer than the
deeper soil layers. In cropland soils, the levels of Ca were similar at all soil depths.
Figure 2 4 Strontium (Sr) to calcium (Ca) and barium (Ba) to calcium ratios at different soil
depths in forests (filled bars) and cropland (unfilled bars) at Katassi and Gelawdios. Within a
ratio and land use bars with different letters are significantly different (mean ±SE; Scheffe,
P<0.05, n = 10).
2.4.5 Factors effecting soil carbon stocks
Total SOC in both the forest (Fig. 2.5a) and cropland (Fig. 2.5b) showed a strong
positive correlation with N stock (R2 = 0.96 and 0.83 respectively; P<0.001). Soil pH
showed a negative correlation with SOC in the forest soil (R2 = 0.80; P<0.001; Fig. 2.5c)
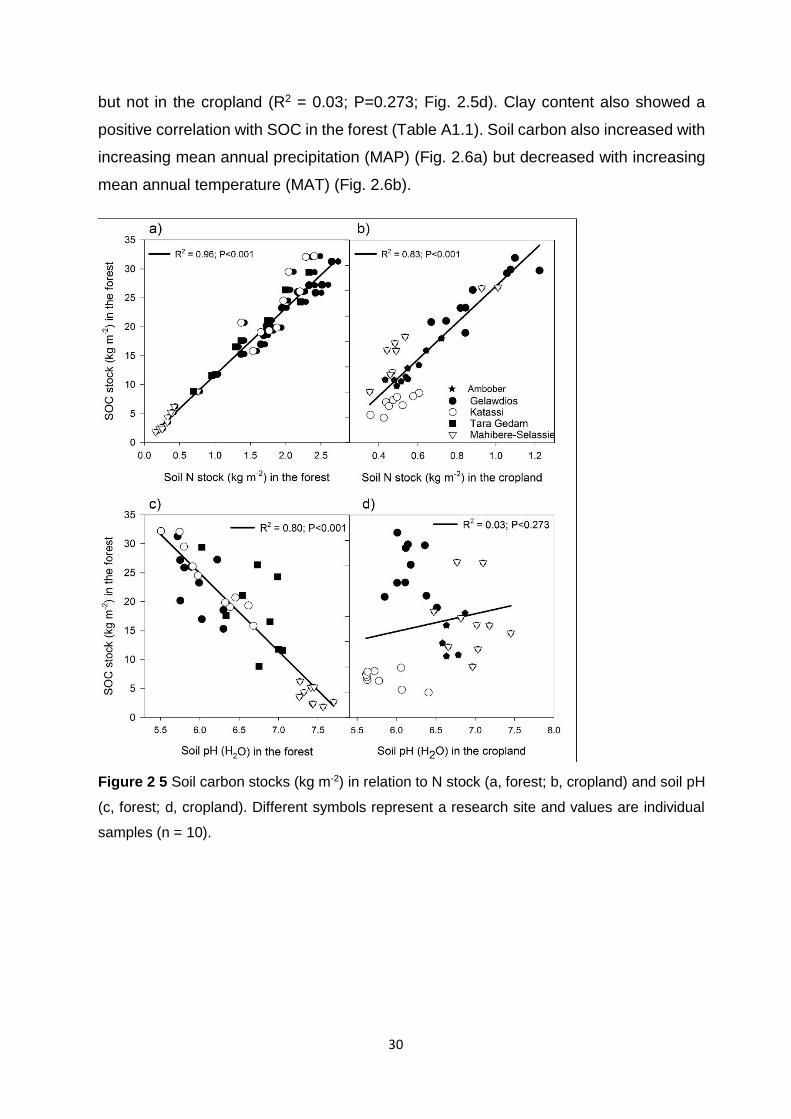
30
but not in the cropland (R2 = 0.03; P=0.273; Fig. 2.5d). Clay content also showed a
positive correlation with SOC in the forest (Table A1.1). Soil carbon also increased with
increasing mean annual precipitation (MAP) (Fig. 2.6a) but decreased with increasing
mean annual temperature (MAT) (Fig. 2.6b).
Figure 2 5 Soil carbon stocks (kg m-2) in relation to N stock (a, forest; b, cropland) and soil pH
(c, forest; d, cropland). Different symbols represent a research site and values are individual
samples (n = 10).

31
Figure 2 6 The relationship between soil carbon stock (kg m-2) and a) mean annual precipitation
(MAP); b) mean annual temperature. (Mean ±SE; Scheffe, P<0.05, n = 10).
2.5 Discussion
2.5.1 Soil organic carbon and nitrogen stocks
There were large difference in the carbon stock in soil between the different land use
systems, and differences in pattern between the highland and lowland systems. In the
highlands (Fig. 2.1a), greater SOC stocks were found in the natural forest and
eucalyptus plantation than in the grazing or croplands. In the lowlands, a greater SOC
stock was found in the cropland than in the savannah woodland (Fig. 2.1b). The overall
soil carbon stock in forests ranged from 4 kg SOC m-2 in the lowlands to 24 kg SOC m-
2 in the highlands. These values fall within the range of estimates of global tropical
means (3 - 41 kg C m-2; Batjes, 1996), and are similar to other studies in southern
Ethiopia (4 - 21 kg C m-2; Lemenih and Itanna, 2004). In the highland soils, the large
differences in SOC stock were due to higher SOC levels in the top 10 cm of soil. In the
natural forest, more than 60% of the SOC stock was in the first 10 cm of soil. Overall,
for all land use systems, more than 70-90% of total SOC stock was found in the upper
20 cm depth of soil (Fig. 2.2b, Table A1.1). This finding is consistent with other similar
studies of C distribution with soil depth (Alvarez and Lavado 1998, Tesfaye et al. 2016).
In the forests, biomass input into SOC comes from both above-ground leaf and
deadwood litter and below-ground root litter, whereas in the eucalyptus plantations, leaf

32
litter is raked and removed. In the cropland system, all above ground biomass is
removed for fodder. The grasslands are also so heavily grazed that biomass input to
soils must be primarily from root litter inputs (Kuzyakov and Domanski 2000).
Investigation of fine root biomass in the forest and eucalyptus plantations showed a
similar distribution to the SOC stock down the soil profile (Chapter 3). Jobbágy and
Jackson (2000) suggested that the vertical distributions of SOC in forests could be
explained in part by root distribution and the associated root turnover.
Soil organic carbon stock increased with increasing MAP and decreased with
increasing MAT (Fig. 2.6a and b). Other similar studies showed that soil C stores are
positively correlated with MAP and negatively correlated with MAT (Alvarez and Lavado
1998, Deng et al. 2016b). Deng et al. (2014) reported that higher temperature leads to
higher losses of soil carbon through decomposition of soil organic matter. At the global
scale, the SOC stocks showed a tendency of increasing from temperate regions to
subtropical regions (Post and Kwon 2000).
A clear relationship was found between SOC stocks and N stocks in both forest and
cropland soils (Fig. 2.5a and b). The C/N ratio in the soil was similar in all land use
types and sites (ca. 11-13) with the exception of the cropland at Katassi, which had a
C/N ratio of 6.2. The strong correlation between C and N suggests that, even in the
cropland soils, the storage of N is directly related to the levels of SOC within the soil,
and that C inputs control N stocks. The very low value of C/N ratio in the cropland soils
of Katassi at 30-50 cm soil depth (Table A1.1) corresponds to a very low level of SOC
of <1%. Decreasing C/N ratios of SOM with increasing soil depth have been shown in
many soils (Lal 1995) and have been attributed to the SOM being older and more
processed by microbes than SOM in upper soil layers (Lal 1995, Callesen et al. 2007).
The low C/N ratio in the whole soil profile of the cropland at Katassi is more
representative of subsoil layers than surface soil layers.
2.5.2 Effect of land use change on carbon stock
At the two highland sites, conversion of natural forests into croplands induced a strong
reduction of organic carbon in the soil. At Katassi and Gelawdios, the SOC stock was
reduced by 87% and 50% respectively that seems the average rate of loss of SOC is
0.42 kg m-2 yr-1 at Katassi and 0.23 kg m-2 yr-1 at Gelawdios for the 50-year period (Fig.
2.1a). However, chronosequence studies have shown that the conversion of forests to

33
cropland caused a rapid initial decrease in SOC stocks, followed by a slow decline (Wei
et al. 2014, Deng et al. 2016b). Similar results have also been reported for other sites
in Northern Ethiopia, where cultivation land had a 58% lower SOC level compared to
forest land in Northwest Tigray (Gebremariam and Kebede 2010), and 63% lower SOC
level in cropland compared to forest after a 30 years of cultivation period in the southern
highlands of Ethiopia (Solomon et al. 2002). Similarly, studies at sites in Southern
Ethiopia with a similar climate to the highland sites showed that SOC was reduced by
70% within 33 years (Lemenih et al. 2006) and 75% in a 53 year cultivation period in
the top 10 cm of soil (Lemenih et al. 2005).
Above-ground and below-ground litter inputs may be a determinant factor for the high
accumulation of C in the forest soil (Berg 2000, Smith 2007, Fernandez et al. 2013)
compared to small carbon inputs from grazing and cropland, where potential inputs are
limited and occur only during growing season. The reduction of carbon stock in the
cropland is exacerbated by complete removal of crop residue for cattle feeding. Malhi
et al. (1999) and Balesdent et al. (1998) estimated that, after forest clearance, rates of
decay of soil C were 10 times higher in cultivated soil than in forest soil, and that soil C
in some factions had become unprotected. However, our data also suggests that much
of the SOC has been lost in crop lands by direct erosion in addition to losses from
increased heterotrophic respiration after cultivation (Tosi et al. 2016). The SOC level in
cropland topsoil (0-10 cm) at Katassi (1.1 kg m-2) is less than the SOC stock of the
adjacent forest soil at the depth of 30-50 cm (3.2 kg m-2; Table A1.1). Similarly, at the
Gelawdios site, cropland carbon stock at 0-10 cm (3.1 kg m-2) was nearly comparable
to the adjacent forest carbon stock (3.7 kg m-2) at 20-30 cm depth. In addition, the
higher distribution of relative SOC stock in the lower soil profile of cropland (ß = 0.937)
than forest (ß = 0.888) indicates that the variation down the soil depth in cropland is
minor. As stated above, the C/N ratio of the cropland soil at Katassi is indicative of
subsoils rather than surface soils. This is not however the case at Gelawdios where the
cropland soil has similar C/N ratios to that of the adjacent forest.
Strontium (Sr) to calcium (Ca) and barium (Ba) to calcium ratios are widely used as
markers of environmental history (Bullen et al. 2005, Tabouret et al. 2010) and
biogeochemical properties of the ecosystem (Kabata-Pendias 2010). The Sr/Ca and
Ba/Ca ratio at 30-50 cm depth in the forest soil profile is comparable to the 0-10 cm
depth of cropland soil at both Katassi and Gelawdios (Fig. 2.3). At both sites in forests,

34
the lower Sr/Ca and Ba/Ca ratios in the upper soil profile are due to higher levels of Ca
in these soil horizons, particularly in the 0-10 cm layer. The higher levels of Ca may be
due to inputs of Ca rich leaf litter (Dauer et al. 2007) or atmospheric dust inputs (Bullen
et al. 2005, Derry and Chadwick 2007), but whatever the source of Ca, this layer is
clearly missing in the cropland soils. Taken together, both the soil profile distribution of
C and the C/N ratio as well as element ratios suggest the upper soil profile has been
lost in cropland soils. The most probable reason for the loss of the upper soil profile in
croplands is direct erosion, as if the soil C and C/N ratio had changed due to an increase
in heterotrophic respiration (Deng et al. 2014) this would have not led to a concomitant
loss of soil Ca in the cropland soils. Climate conditions in Ethiopia such as heavy rainfall
events are known to result in a high rate of soil erosion (Bewket and Sterk 2005, Betrie
et al. 2011, Shiferaw 2011). In addition, after the protective tree canopy has been
removed, the low bulk density of the upper soil layers in the natural forest may make
these soils highly sensitive to wind or water erosion (Jepsen et al. 1997).
At the highland sites, conversion of natural forest to grazing land also significantly
reduced SOC stock in the soil by 53%. Bewket and Stroosnijder (2003) showed on
other highlands sites that grazing land had 48% lower levels of SOM than natural forest.
These results are in contrast to studies of productive grasslands in tropical climates
where grassland often have similar or greater SOC storage compared to forests
(Conant et al. 2001), and even conversion of forest to grassland tends to increase C
stock in the soil (Conant et al. 2001, Guo and Gifford 2002). Guo and Gifford (2002) in
their review showed that accumulation of SOC after conversion from forest to grassland
occurred in climates between 2000-3000 mm, but not at < 2000 mm precipitation.
However, poor management of grasslands after conversion led to a decrease in SOC
even in wet tropical areas (Fearnside et al. 1998). The high levels of SOC often found
in productive grasslands are produced by high biomass inputs from roots (Kuzyakov
and Domanski 2000). At the highland sites in this study, the levels of fine root biomass
in grasslands are only 10% of those in the natural forest (Chapter 3), and thus
necromass inputs must have greatly decreased. In addition, most of the grasslands are
degraded due to overgrazing. Often the grazing land is completely denuded. Desta et
al. (2000) estimated for the grasslands that the stocking density (23 livestock unit (LU)
ha-1) is ten times the carrying capacity (2-3 LU ha-1).

35
The lowland savannah woodland at Mahibere-Selassie has a SOC stock of ca. 20% of
the highland sites. The lower SOC stocks were clearly related to the lower MAP and
higher MAT, which is likely to affect rates of SOC turnover, but also strongly affects the
vegetation structure. The basal area and stocking density of the tree of the savannah
woodland (Table 2.1) are only a fraction of that of the highland sites, and thus biomass
inputs from the trees will be low. In addition, the grass cover of the woodland is regularly
burnt, which potentially decreases SOC stocks (Knicker 2007). Furthermore, the soil of
the savannah woodland has a very low clay content and a high sand content compared
to the adjacent cropland and all the highland forest sites (Table A1.1). Exceptionally,
cropland at Mahibere-Selassie had a 60% higher SOC stock than the woodland (Fig.
2.1b). The higher SOC stock may in part be due to the higher clay content of the soil
(Ajami et al. 2016), but could also be due to the different farming systems in the
lowlands. Ajami et al. (2016) could also show a linear relationship between SOC stock
and soil clay content in loess soils in Iran after deforestation.
2.5.3 Potentials of soil carbon gain due to afforestation and exclosure
At two sites, Katassi and Tara Gedam, the soils under eucalyptus had higher SOC
stocks than either the cropland or the grazing land soil. However, at Gelawdios, the
SOC stocks were similar in the eucalyptus, grazing land, or cropland soils. Assuming
the eucalyptus plantation at Katassi and Tara Gedam are ca. 30 and 40 years old, the
rates of SOC accumulation is 0.49 kg m-2 yr-1 at Katassi and 0.17 kg m-2 yr-1 at Tara
Gedam, which is a similar to the rate of loss of SOC after forest removal. A study in
Southern Ethiopia, (Tesfaye et al. 2016) determined similar levels of SOC in 28 year
old Eucalyptus saligna plantations, and represents an annual SOC accumulation of
0.18 kg m-2 yr-1 compared to the levels of SOC determined in cropland. At Katassi and
Gelawdios, the SOC stock in the natural forest exceeded that of the eucalyptus
plantation by 29 and 55% respectively, but was similar between the two land uses at
Tara Gedam. Tesfaye et al. (2016) reported a 30% greater SOC storage in natural
forests compared to the Eucalyptus saligna plantations. These results suggest that
eucalyptus can restore SOC storage in soils even under management systems using
leaf litter raking and removal. However, the low bulk density of the surface layers of the
natural forests was not restored. The negative ecological consequences of eucalyptus
plantations also needs to be considered (Martins et al. 2013).

36
The conversion of heavily degraded grazing land to exclosure in Ambober significantly
increased SOC stock in the 0-10 cm soil layer by 42% compared to cropland after an 8
years exclosure period (Fig. 2.3). Other studies (Girmay et al. 2008, Mekuria et al. 2009,
Li et al. 2012) have also shown the positive effects of exclosure on carbon stock. For
example, Li et al., (2012) reported that on rangelands in inner Mongolia, SOC stock in
the top 10 cm layer increased from 93 to 638 g m-2 in 26 years exclosure period at the
rate of 31 g C m-2 yr-1, but these authors found no significant difference after 8 years
exclosure period (107 g m-2). The increase of SOC stock in exclosure area indicates
that an increase of vegetation growth and input of carbon can begin to restore SOC
stocks, but it takes long recovery period to obtain the approximate original SOC stock
levels.
2.5.4 Magnitude of soil carbon loss due to deforestation
Land use conversion from forest to cropland or grazing land reduced soil carbon stock
up to 88%. Using the SOC storage of current remnant forests and assuming a 40%
forest cover, the SOC stock of the Amhara region before 50 years ago was about 1.5
Gt C. Forest cover of the Amhara region is currently estimated to be about 9.5%
(Government unpublished report) giving a SOC stock of 0.3 Gt. Hence, it is clearly
important to determine if this substantial loss of 1.2 Gt has been displaced in the
landscape by erosion or lost to the atmosphere by increased SOC turnover. Our data
suggest that SOC may not only be lost to the atmosphere as CO2 (Balesdent et al.
1998) but may also be lost due to erosion. The fate of this lost SOC in the landscape is
unknown.
2.6 Conclusion
Removal of natural forests resulted in substantial loss of SOC stock, which appears to
be due to both erosion of sensitive low bulk density soil of surface layers and possibly
increased rates of SOC turnover. Afforestation with eucalyptus has the potential to
restore the levels of SOC storage partially but does not restore the physical properties
of the soil. Exclosure, and thus exclusion of the degraded land from grazing, also results
in a recovery of SOC stocks in the upper soil layer.

37
3 Fine root dynamics in Afromontane forest and adjacent
land uses in the Ethiopian highlands
3.1 Abstract
Fine roots are a major pathway of C flow from plants to soil, but information on root
dynamics in east African ecosystems is scarce. The aim of this study was to quantify
fine rootstock, production, and turnover and to examine the impact of land use change
on fine root dynamics. The study was conducted in a remnant Afromontane forest and
other land use types in NW Ethiopia using three different methods: sequential coring,
in-growth cores, and in-growth nets. Soil cores for sequential analyses were taken in
quarterly intervals, while in-growth cores and nets were harvested corresponding to 1,
2, 3, 4, 6, 8, and 12-month interval. Fine rootstocks averaged 564, 425, 56, and 46 g
m-2 in the forest, eucalyptus, grazing land, and cropland ecosystems, respectively,
based on sequential coring. The values decreased exponentially with increasing soil
depth. The root production averaged 723 and 694 g m-2 yr-1, while necromass loss due
to decomposition was 416 and 282 g m-2 yr-1, in the forest and eucalyptus, respectively,
based on sequential coring. Fine root production based on in-growth coring averaged
468, 293, 70, and 52 g m-2 yr-1 for forest, eucalyptus, grazing land, and cropland,
respectively. In general, land use conversion from forest to grazing land or cropland
reduced fine rootstock and production by 85-91%.The turnover rate of fine roots based
on mean biomass was 1.5 for forest and 2.1 for eucalyptus soil. From in-growth core
estimations, the annual C flux into the soil averaged 226, 133, 32, and 24 g m-2 yr-1 in
the forest, eucalyptus, grazing land, and cropland ecosystems.
Key words: biomass, necromass, root production, sequential coring, in-growth core, turnover
rate, carbon efflux, decision matrix
3.2 Introduction
Assessments of fine roots by simple excavation techniques were started as early as
the 18th century (Bohm 1979). Since then, in forests, most ecological studies on fine
roots, their biomass distribution and turnover have been carried out in temperate and
boreal ecosystems (Leuschner and Hertel 2003, Pinno et al. 2010, Yuan and Chen
2010, McCormack et al. 2012), while information for tropical areas in general, and

38
African ecosystems in particular, is scarce. Subsequently, worldwide and regional
ecosystem estimates of fine root production, turnover, and carbon fluxes remain
uncertain (Gill and Jackson 2000). Due to a limited understanding of below-ground
process as well as the inherent difficulties in measuring fine root turnover compared to
above-ground components of trees (Santantonio and Grace 1987, Johnson et al. 2001,
Yuan and Chen 2010), parameterising C input models is difficult for different land use
ecosystems. Although several methods have been developed for measuring fine root
production, mortality, and eventually decomposition in situ (Vogt et al. 1998), all have
drawbacks and are often confounded by artifacts (Hertel and Leuschner 2002, Lukac
2012, Andreasson et al. 2015). For example, the sequential coring method has been
widely used (Yuan and Chen 2013, Sun et al. 2015) but may miss root turnover
between sampling dates. This results in a zero estimate (Brunner et al. 2012) because
root mortality and production occur simultaneously (Hansson 2013). The sequential
coring method also relies on a calculated rather than measured estimate of fine root
production. Direct measurements of fine root production can be obtained using in-
growth cores (Lukac 2012) or in-growth nets (Lukac and Godbold 2010). One problem
with the in-growth core method is that it greatly modifies the soil physical properties and
can thus affect root growth (Makkonen and Helmisaari 1999) and either overestimate
(Neill 1992) or underestimate root production (Hertel and Leuschner 2002). In-growth
nets are often difficult to insert or remove in heavy, stony soils. Thus, to obtain more
robust estimations of fine root production and turnover rates, a combination of sampling
techniques and calculation methods should be employed (Majdi et al. 2005).
Estimates of fine root production in pan-tropical forests range from 75 to 2193 g m-2 y-
1 (Hertel and Leuschner 2002). Importantly, estimated values of fine root biomass,
production, and turnover are also influenced by environmental and seasonal factors
(Yang et al. 2004a), which may be strong factors in highly seasonal environments such
as the Ethiopian highlands. For example, fine root growth has been shown to rapidly
increase following soil rewetting (Comas et al. 2013). Moreover, soil characteristics may
also influence fine root biomass and rates of fine root turnover (Helmisaari and
Hallbäcken 1999, Godbold et al. 2003). Fine root biomass was reported to be higher
under low nutrient availability (Priess et al. 1999, Yuan and Chen 2010) or in acidified
soils (Godbold et al. 2003). Other studies, however, reported no effect of these factors

39
on fine root biomass (Ruess et al. 2006) or even lower values of fine root biomass
(Yang et al. 2004a).
Forest conversion to other land use systems is a common phenomenon in the
highlands of Ethiopia, mainly because of population growth and the need for new
agricultural lands (Nyssen et al. 2004). The conversion of natural forests often leads to
land degradation (Meshesha et al. 2014) and to losses of soil C and N stocks (Chapter
2). Much of the original forest that still remains is confined to areas around churches
and monasteries (Wassie et al. 2009, Aerts et al. 2016). Natural forests including
church forests and woodlands cover less than 9.5% of the Amhara region (Desta et al.
2000, Bekele 2011). Investigation of these church forests provides an opportunity to
determine the baseline for this landscape in terms of C storage and the C dynamics
that contribute to this storage. Most previous investigations of church forests have
focussed on above-ground aspects such as species richness (Wassie et al. 2009), and
little is known about the below-ground ecology here. To reverse the negative impacts
of deforestation and to provide a wood supply, degraded lands have often been planted
with eucalyptus. Eucalyptus globules (Labill.) plantations have been established in the
central highlands of Ethiopia since 1895 (Pohjonen and Pukkala 1990) and are
estimated to cover 172 km2 of the region (Bekele 2011). Planting of eucalyptus
increases soil C stocks after ca. 30 years (Chapter 2), but not to the levels found in
natural forests. Although the proportion of the tree biomass constituted by fine roots
(diameter <2 mm) is estimated at <5% (Vogt et al. 1996), fine roots are major
contributors to carbon input into the soil because of their rapid turnover (Vogt et al.
1998, Majdi et al. 2005, Ruess et al. 2006, Lukac 2012). Across terrestrial ecosystems,
fine root biomass is estimated to account for 14-33% of annual net primary production
(Jackson et al. 1997, Malhi et al. 2011, McCormack et al. 2015) and can contribute up
to two-fold higher organic carbon to the soil than leaf litter (Xia et al. 2015). Due to the
importance of fine root turnover in soil C storage (King et al. 2001, Leppälammi-
Kujansuu 2014), we investigated the fine root biomass and turnover on a remnant
church forest, eucalyptus plantation, grazing land, and cropland in the Ethiopian
highland landscapes of the Amhara region. The objective of this study was to assess
the fine rootstock, production, and turnover and to examine the impact of land use
change on fine root dynamics. In the tropics, where temperature permits year-round
growth, the timing of leaf production is probably a result of the availability of moisture

40
during the rainy season (Mulkey et al. 1996). Root production might also follow leaf
phenology because fine root growth depends heavily on newly fixed carbon from the
canopy (Joslin et al. 2001, Steinaker and Wilson 2008). We therefore expected that fine
root biomass and necromass would be greater during the rainy season, when trees
were physiologically active to capture nutrients, than during the dry season.
3.3 Materials and Methods
3.3.1 Site descriptions
The study was conducted at a natural remnant forest at Gelawdios and on adjacent
land use systems (eucalyptus plantation, grazing land, and cropland) situated in the
Amhara National Regional State (11°38’25’’ N 37°48’55’’ E) in northwest Ethiopia. The
altitude of the study area is 2500 m a.s.l. The monsoonal climate features a mean
annual temperature of 19°C and a mean annual precipitation of about 1200 mm, with
the main rainy season between June and September (Wassie et al. 2009). The climate
is classified as temperate with a dry winter and warm summer (Cwb) according to the
Köppen-Geiger climate classification (Peel et al. 2007). The soils of the study area are
Cambisols (WRB 2014) with weak horizon differentiation and rocks below 50 cm depth.
The forest at Gelawdios has an area of about 100 ha (Wassie et al. 2009) and is a
remnant of natural pristine forest composed mostly of a mixture of indigenous tree
species. The forest type is a dry Afromontane forest. This forest with old remnant trees
has been protected because it is a church forest. The church was built around 1500
A.D. (Wassie et al. 2009). The dominant tree species are Chionanthus mildbraedii,
Albizia schimperiana, Apodytes dimidiata, Calpurnia aurea, Croton macrostachyus,
Ekebergia capensis, Maytenus arbutifolia, and Schefflera abyssinica. The Eucalyptus
globules plantation was established on previous grazing land around 1985 and was
successively thinned. The cropland and grazing land were converted from natural forest
within the last 50 years (specific date not known). The number of years that the land
was continuously cultivated after conversion from forest was obtained from local
records and knowledge. The cropland is ploughed 2-3 times per year to ca. 30 cm depth
using an ox-drawn plough. During the sampling year (2014), the farmland was planted
with Eragrostis tef, and partly with Triticum aestivum in June and July, and was
harvested in September and October, respectively. Grazing lands are common land
used to graze herds of cattle, sheep, goats, and donkeys.

41
3.3.2 Root sampling
In the natural forest and eucalyptus plantation, ten permanent circular plots with an
area of 100 m2 each were established along a transect line with a minimum distance of
100 m between the plots. In the grazing land and cropland, plots were established along
a transect line with ca. 20 m distance between each plot. Three methods were used for
rootstock and production estimations: sequential coring, in-growth cores, and in-growth
net methods.
Sequential coring
Standing fine roots (<2 mm diameter) were determined using sequential soil coring.
The fine roots were later divided into live fine roots (biomass) and dead fine roots
(necromass). Intact soil cores were extracted with a stainless steel corer (6.6 cm
diameter) to a depth of 40 cm or until the bedrock was reached. In the eucalyptus
plantation, grazing land, and cropland, one core was taken per plot at each sampling
date from each of the ten permanent plots. Due to the greater heterogeneity in the
natural forest, two cores ca. 2 m apart were taken per plot. Soil samples were divided
into 0-10, 10-20, 20-30 and 30-40 cm soil depths. For the two cores from the natural
forest, the fine roots from each corresponding depth increment were combined to give
a single sample. Soil cores were taken in March, June, and September 2014 and in
February 2015. The timing of sampling took into account seasonal variations in rainfall
and temperature, and the planting time of the crop field. The March and February
sampling times are during the dry season, the June and September samplings times
are during the rainy season.
In-growth core and net methods
Root production was estimated from the above sequential coring as well as using in-
growth cores (Leppälammi-Kujansuu et al. 2014) and in-growth nets (Godbold et al.
2003). In the natural forest and eucalyptus plantation, five in-growth cores per plot were
established (50 cores per land use type in total) in late June 2014. One in-growth core
was positioned at the centre of the plot and the four others 2 m from the centre in N, E,
W, and S directions. Soil cores were extracted using a soil corer (6.6 cm internal
diameter*40 cm length) as described above, and the holes were lined with a 2 mm
mesh size polyethylene net and filled with the root-free soil taken from the cores. An
effort was made to restore the original soil horizons and compact the soil back to

42
approximately the original soil bulk density. In the grazing land and cropland plots, three
soil cores per plot (30 in total for each land use type) were established as described
above. One in-growth core was positioned at the centre of the plot and two others 2 m
away from the centre. One in-growth core per plot was removed from natural forest and
eucalyptus plantation in late July, August, September 2014 and February and June
2015, corresponding to 1, 2, 3, 8 and 12 months in-growth. In cropland and grazing
land, one core per plot was retrieved in July, August, and September 2014,
corresponding to 1, 2 and 3 months. Each time an in-growth core was removed, a new
in-growth core per plot (ten in total per land use) was established in a new position to
estimate root production and mortality between each sampling date. In total, the
sampling dates correspond to 1, 2, 3, 4, 6, 8, and 12-month time intervals from the
beginning of measurement. After removal, the in-growth cores were carefully divided
into 0-10, 10-20, 20-30, and 30-40 cm depths. Soil from the cores was spread on a
plastic sheet, and fine roots were handpicked from soil.
Similarly, in the natural forest and eucalyptus plantation, per plot, five in-growth nets of
10 cm width and 20 cm length with mesh size of 1 mm (Franz Eckert GmbH, D-79183
Waldkirch, Germany) were inserted into the soil about 30 cm away from the in-growth
cores; three nets per plot were inserted into the cropland and grazing land. As in an
earlier trial, it proved difficult to insert and remove the nets in the stony soil. To insert
the nets, a 10 cm wide and 5 cm thick soil column was excavated to a depth of 20 cm.
The 10 cm wide and 20 cm long nylon nets were inserted in the holes on one face and
the holes were refilled with the root-free soil and compacted to approximately the
original bulk density. In the natural forest and eucalyptus plantation, one net per plot
was removed by cutting a soil column (10 cm wide, 20 cm deep) using a long knife in
late July, August, September 2014, February 2015, and June 2015, corresponding to
1, 2, 3, 8, and 12 months in-growth. Similarly, in the cropland and grazing land, one net
per plot was removed in July, August, and September 2014, corresponding to 1, 2, and
3 months in-growth. For additional measurements between sampling dates, again, a
new net was inserted each time and an in-growth net was removed, yielding 1, 2, 3, 4,
6, 8, and 12-month time intervals. After removal, the nets were divided into 0-10 and
10-20 cm soil depths, and then roots penetrating into the nets were carefully removed
by hand.

43
For both in-growth cores and in-growth nets, the root samples were placed in separate
plastic bags and transported to Vienna (Austria) for further analysis. Samples were
stored at 4°C until processed. In Vienna, fine roots were placed on a 1 mm sieve and
soil was washed away under running tap water, and then the roots were handpicked
with forceps from the sieve. Fine roots were classified as tree roots and grass roots as
well as live and dead based on colour, breakability, wrinkled bark (texture consistency),
and other visual factors (Vogt et al. 1998, Godbold et al. 2003). Roots were further split
into two diameter classes using a digital calliper: very fine roots (<1 mm) and fine roots
(1-2 mm). All root samples were oven dried to constant weight at 70°C, weighed to the
nearest 0.01 g, and converted to g m-2.
3.3.3 Soil sampling
Soil sampling took place during March 2014 at the time of the first root sampling using
a stainless steel corer (6.6 cm diameter) as described above. Soil cores were laid onto
a Styrofoam tray and morphologically described using the terminology of the World
Reference Base for soil resources (WRB 2014). After dividing the soil corers into 0-10,
10-20, 20-30, and 30-40 cm depths, fine roots were collected for root biomass analysis
as stated above and each root-free soil sample per depth was sieved in 2 mm mesh
sieve and then homogenised. From the natural forest, two composite soil samples per
plot per depth, and from other land use systems one sample per plot per depth were
taken at ten sampling points each and placed in plastic bags in Ethiopia. The samples
were then transported to Vienna for laboratory analysis.
3.3.4 Estimation of annual fine root production, mortality, and decomposition
Using the values of standing fine root biomass and necromass from sequential coring,
two methods were used to estimate fine root production, the ‘Maximum-Minimum’ (Max-
Min) and Decision Matrix calculation techniques according to Brunner et al. (2012) and
Yang et al. (2004) . For in-growth cores and in-growth nets, fine root production (P) was
estimated using a simple ‘balancing model’ proposed by Li et al. (2013), Osawa and
Aizawa (2012), and Santantonio and Grace (1987) with some modifications.
Using the Max-Min method the annual fine root production (g m-2 yr-1) was calculated
by subtracting the lowest biomass or necromass (Bmin, Nmin) from the highest biomass

44
or necromass value (Bmax or Nmax). This method assumes a single annual pulse of fine
root production (McClaugherty et al. 1982, Brunner et al. 2012).
P (g m-2 yr-1) = Bmax- Bmin (1)
The Decision Matrix (DM) calculates the annual fine-root production by summing all
calculated productions between each pair of consecutive sampling dates throughout a
full year. The production (Pj) between two sampling dates is calculated by adding the
differences in biomass (∆B), necromass (∆N), and decomposition (D) (McClaugherty et
al. 1982, Yang et al. 2004a, Brunner et al. 2012).
Annual production (P; g m−2 yr-1) = ∑ 𝑃𝑗𝑛𝑗=1 = ∑ (∆𝐵𝑗 + ∆𝑁𝑗 + 𝐷𝑗)𝑛
𝑗=1 (2)
where the superscript j refers to a definite interval t to t + 1.
The change in standing crop of live fine roots (∆Bj) at a given time (t) equals production
(Pj) minus mortality (M)
dB/dt = dP/dt – dM/dt = ∆Bj = Bt+1 – Bt (3)
where Bt and Bt+1 represent fine root biomass at the start of the growing season and at
time t+1, respectively. All terms have the unit g m−2.
The change in standing crop of dead fine roots (∆Nj) equals mortality minus
decomposition (D).
dN/dt = dM/dt – dD/dt = ∆Nj = Nt+1 – Nt (4)
where Nt and Nt+1 represent fine root necromass at the start of the growing season and
at time t+1, respectively.
The condition under which the Decision Matrix was used as the basis for calculations
of P between sampling dates is shown in Table 3.1. In this study, all differences in
biomass and necromass from interim periods were taken into account, assuming that
the biomass and necromass pool are continuously changing (Brunner et al. 2012).
Mortality is the decrease in fine root biomass between two consecutive sampling dates.
When estimating annual root production, mortality was included to either necromass or
partly to decomposition depending on the mass change of necromass due to this
mortality. Decomposition was therefore calculated from a decrease in necromass or a
decrease in fine root biomass that was not compensated by an increase in necromass

45
(see Table 3.1 for details). To calculate the annual root production, production values
obtained from all changes in both biomass and necromass from interim periods of
consecutive sampling dates are summed from the start of sampling until the same time
point in the following year, regardless of whether the differences were statistically
significant (McClaugherty et al. 1982, Hertel and Leuschner 2002, Yuan and Chen
2013).
Table 3 1 Simplified Decision Matrix for estimating fine root production, mortality, and
decomposition according to McClaugherty et al. (1982), Osawa and Aizawa (2012), and Yuan
and Chen (2013). The appropriate quadrant is selected according to the direction of change in
biomass (B) and necromass (N) during the interval between two sampling times. Production
(P), mortality (M), and decomposition (D) for the sampling interval are calculated using the
equations in the chosen quadrant. Vertical bars indicate the absolute values. Annual estimates
are calculated by summing the estimates from all sampling intervals within the year.
Biomass
Increase Decrease
Necro
ma
ss
Increase
P = ΔB + ΔN M = ΔN D = 0
P = ΔB + ΔN or 0 M = ΔN or |-ΔB| D = |-ΔB| – ΔN or 0
Decrease P = ΔB M = ΔB D = |-ΔN|
P = 0 M = |-ΔB| D = |-ΔB| + |-ΔN|
In the in-growth cores and net methods, the last harvest at the end of the year is usually
considered as direct root production (Vogt et al. 1998, Lukac 2012). However, in a one-
year in-growth core and in in-growth net methods, biomass increases until it colonizes
the whole space and reaches an equilibrium. Nevertheless, fine root growth, death, and
decomposition also occur simultaneously between the sampling dates. Therefore, the
interim root production and decomposition between sampling dates are missed.
Moreover, production, mortality, and decomposition may vary in the growing season
due to seasonal moisture and temperature variations. To overcome this problem, the
additional in-growth cores and in-growth nets were used to estimate root production
between harvests. The underlying assumption here is that root production favoured by
soil disturbance during installation of in-growth cores and in-growth nets is
compensated by the lag time of growth of severed roots. Therefore, we used a mass
balance model from time of insertion to any given time (t) in the growing season to
estimate fine root production (P), mortality (M), and decomposition (D). The ‘mass

46
balance model’ is the same as those of Santantonio and Grace (1987), Osawa and
Aizawa (2012), and Li et al. (2013) except we considered additional growth between
harvests. The interim production of biomass (bi) between each harvest was calculated
as:
bi = bt – b0 = bt, since b0 = 0 (5)
Similarly, the interim production of necromass (ni) between each harvest was calculated
as:
ni = nt – n0 = nt, since n0 = 0 (6)
Since there are no fine roots in the in-growth cores and in-growth nets at the start of
the installation, both b0 and n0 are always zero. Then, the actual mortality for in-
growth cores and in-growth nets was calculated as:
Mt =bt-Bj where Bj = Bt+1 – Bt (7)
This means that root production between interim periods from the short-term cores and
nets should be equal to zero. If production over the short term within this time was
higher than the calculated value from the initial installation (long-term), then the
differences were assumed to represent root mortality in the long-term. This mortality
was then included to either necromass or partly to decomposition depending on the
mass change of necromass due to this mortality.
Mj = Nt+1-Nt + Dj (8)
Decomposition was therefore calculated from a decrease of necromass, or a decrease
in fine root biomass that was not compensated for by an increase in necromass, with
the following equation.
Dj = Mj + ni + Nt – Nt+1 = bi + Bt - Bt+1 + nt + Nt – Nt+1 (9)
where the small case letters bi and ni are root biomass and necromass from the interim
periods.
Annual mortality = ∑ 𝑀𝑗𝑛𝑗=1 (10)
Annual decomposition = ∑ 𝐷𝑗𝑛𝑗=1 (11)
To calculate the annual root production, all production values obtained from all changes
in both biomass, necromass, and decomposition from interim periods of consecutive

47
sampling dates were summed from the start of sampling until the same time point in
the following year (same as equation 2).
3.3.5 Calculation of fine root turnover
Root turnover rate (yr-1) was calculated as the ratio between annual root production
and average root biomass (McClaugherty et al. 1982, Aerts et al. 1992) or the highest
(maximum) biomass value (Gill and Jackson 2000). Mean residence time of roots is the
reciprocal of turnover rate.
Annual carbon inputs from fine roots into soil were calculated using fine root production
along with the C and N concentrations in the roots, assuming that there was no
translocation during root senescence. Hence, the annual fluxes of C and N into soil
(expressed in g m-2 yr-1) were estimated according to Xia et al. (2015) as:
Ia = P*C/N% (12)
Ia, annual input of C or N into the soil (g m-2 yr-1): P, annual fine root production (g m-2);
C/N, concentration of C or N in the fine roots (%).
3.3.6 Fine root vertical distribution
Vertical root distribution or cumulative root fraction (Y) from the surface to any depth
(d) was calculated according to a model developed by (Gale and Grigal 1987). An
asymptotic nonlinear model of the following form was fitted to describe the vertical root
distributions:
Y = 1-ßd (13)
where Y is the cumulative root fraction from the surface to soil depth d in centimetres
(midpoint), and ß the estimated parameter used as a measure of index of vertical root
distribution. The parameter ß can have values from 0 to 1, whereby higher values
indicate a greater proportion of roots at deeper soil (Jackson et al. 1997, Valverde-
Barrantes et al. 2007, Kucbel et al. 2011).
3.3.7 Carbon and nitrogen analysis
A very limited dry mass of fine roots and herbaceous roots was available. Accordingly,
to determine the total C and N concentrations in root materials, the original ten samples

48
were pooled by category to form nine samples each from the forest and eucalyptus,
and three samples each from grazing land and cropland. The nine samples from the
forest ecosystem were further processed to form three categories: roots <1 mm
diameter, roots from 1-2 mm diameter, and herbaceous roots. Each had three
replicates. The roots were dried at 70°C and ground to a fine powder (Fritsch
Pulverisette 5, Idar-Oberstein, Germany). Sub-samples of about 160 mg were taken
from each sample for C and N determinations using a CN elemental analyser (Truspec
CNS LECO, St. Joseph, USA). In addition, the soils from each sampling point and depth
were taken during root sampling and were dried at 105°C until constant weight. Ten
samples per land use system were analysed for total C and N concentration using 200
mg samples on the same CN elemental analyser as described above. Soil bulk density
was taken from values presented in Chapter 2.
3.3.8 Statistical analyses
Means and standard errors were calculated using the SPSS analytical software
package (version 21), and graphs were prepared using SigmaPlot (version 13). The
significance level was set at α = 0.05. Throughout the paper, error bars to the mean
are ±1SE. The relationships between different variables were analysed using one-way
analysis of variance and the Scheffe comparison method as well as simple regression
analysis. Assumptions of normality and homogeneous variance were examined by
Shapiro-Wilk’s and Levene’s test, respectively. When the assumptions of normality
were not met, the data were log10-transformed to normalize the distribution.
3.4 Results
3.4.1 Fine root biomass, necromass and distribution with depth
Fine root biomass and necromass to 40 cm soil depth for all land use systems is shown
in Table 3.2. Total weight of fine roots (biomass and necromass) in the soil layer 0–40
cm was higher in natural forest (564 g m-2) than in the eucalyptus stand (425 g m-2).
From the total root mass, living roots represented 83% in the natural forest and 79% in
eucalyptus plantation, whereas the necromass (dead roots) accounted for 17% in the
natural forest and 21% in the eucalyptus plantation (Table 3.2). The necromass,
however, was not significantly different between the forest (98±13 g m-2) and

49
eucalyptus stand (88±10 g m-2). Samples were not taken from grazing land and
cropland during the dry season because the land was entirely bare. Root biomasses
during the rainy season in these two fields were 56 and 46 g m-2, respectively, and were
very low compared to the tree ecosystems. Of this amount, living roots represent 96%
in the grazing land and 98% in the cropland. Herbaceous roots contributed ca. 30% to
the total root mass in the eucalyptus stand, whereas in the native forest the understory
herbaceous root contribution to fine root mass was negligible (1.4%). In the eucalyptus
stand, the roots of the understory vegetation were mainly from grasses. The fine roots
from grazing land and cropland were herbaceous. The biomass of smaller diameter
roots (<1 mm) accounted for ca. 50% of the total root mass both in the natural forest
and eucalyptus plantation; all roots in the grazing land and cropland were <1 mm
diameter (exception: 2.8% in the cropland were between 1-2 mm diameter) (Table 3.2).
All land use systems had relatively shallow fine root systems with >40% of the total
mass (live plus dead) situated in the upper 10 cm, decreasing to 9-11% at 30-40 cm for
vegetated ecosystems. In the cropland, all roots were found in the upper 30 cm depth.
For the pooled data of the forest ecosystem, the changes of the cumulative root fraction
by soil depth were analysed using the model of Gale and Grigal (1987). Fine root mass
declined exponentially with depth for all land use systems, with β index values ranging
from 0.789 to 0.923 (Fig. 3.1). Herbaceous roots were the shallowest roots and were
mostly concentrated in the upper 20 cm depth. When comparing diameter classes, fine
roots (1-2 mm diameter) were deeper than very fine roots (<1 mm diameter; Fig. 3.1),
as the ß-values were significantly different (P<0.001). In addition, the vertical
distribution of necromass was deeper (0.913) than the biomass (0.901; P<0.05).

50
Table 3 2 Vertical distribution of fine root mass (g m-2) to a soil depth of 40 cm for native forest,
eucalyptus stand, grazing land and cropland. Fine roots are categorized as tree vs herbaceous
roots, biomass (live roots) vs necromass (dead roots) and very fine roots (<1 mm) vs fine roots
(1-2 mm) for both native forest and eucalyptus plantations. Values for forest and eucalyptus
were determined based on sequential coring. Roots from cropland and grazing land are taken
from the last in-growth core harvest, assuming peak rooting time at the end of the rainy season.
Roots from these land use systems are entirely herbaceous and mostly <1 mm. Values are
mean±SE; n=10 (forest and eucalyptus); n=5 (cropland and grazing land).
Land use Depth Total root
mass
Live and dead roots Diameter class Plant type
Biomass Necromas
s Roots <1
mm Roots 1-2
mm Tree roots
Herbaceous roots
Forest Total 564.2±41.7 466.5±31.6 97.7±12.8 272.9±18.2 291.4±27.7 555.8±42.2 8.4±3.2
0-10 239.5±18.5 195.5±14.7 46.4±6.9 137.5±9.5 108.1±12.1 232.1±18.9 7.5±2.9
10-20 174.2±15.2 153.3±11.8 30.1±4.5 81.4±6.5 101.9±10.5 177.9±15.0 0.7±0.4
20-30 90.0±11.3 74.0±8.4 17.8±3.9 37.2±2.8 61.4±10.4 89.8±11.3 0.2±0.2
30-40 60.5±8.4 55.6±8.3 12.6±2.2 27.1±3.7 44.3±6.8 65.4±8.6 na
Eucalyptus Total 424.7±36.9 336.8±30.5 87.9±10.2 211.8±17.1 223.0±22.8 302.6±28.9 122.1±15.1
0-10 231.4±19.8 176.2±16.5 55.2±6.7 123.6±12.1 107.8±11.4 140.5±11.8 90.9±13.2
10-20 93.6±12.7 75.8±11.0 20.9±4.0 45.4±4.5 60.3±10.9 74.2±12.1 19.4±3.1
20-30 58.1±7.5 48.7±7.0 12.5±1.6 27.1±2.9 41.3±6.7 50.6±7.7 8.7±1.7
30-40 41.6±14.3 41.2±15.5 9.7±1.9 17.4±2.8 47.2±21.7 44.0±16.2 3.1±0.7
Grazing land
Total 55.5±13.3 53.2±12.7 3.4±1.6 55.5±13.3 na na 55.5±13.3
0-10 31.9±7.8 29.8±7.1 2.1±1.3 31.9±7.8 na na 31.9±7.8
10-20 13.3±5.6 13.3±5.6 na 13.3±5.6 na na 13.3±5.6
20-30 6.4±1.7 5.9±1.6 0.5±0.0 6.4±1.7 na na 6.4±1.7
30-40 4.9±2.1 4.1±1.5 0.8±1.3 4.9±2.1 na na 4.9±2.1
Cropland Total 46.1±4.4 45.3±3.9 0.8±0.6 44.8±3.4 1.3 na 46.1±4.4
0-10 31.2±3.2 30.6±6.3 0.7±0.0 31.2±3.2 na na 31.2±3.2
10-20 13.2±4.2 13.0±4.2 0.2±0.0 11.9±3.1 1.3 na 13.2±4.2
20-30 1.7±1.1 1.7±1.1 na 1.7±1.1 na na 1.7±1.1
30-40 na* na na na na na na
*na, not available
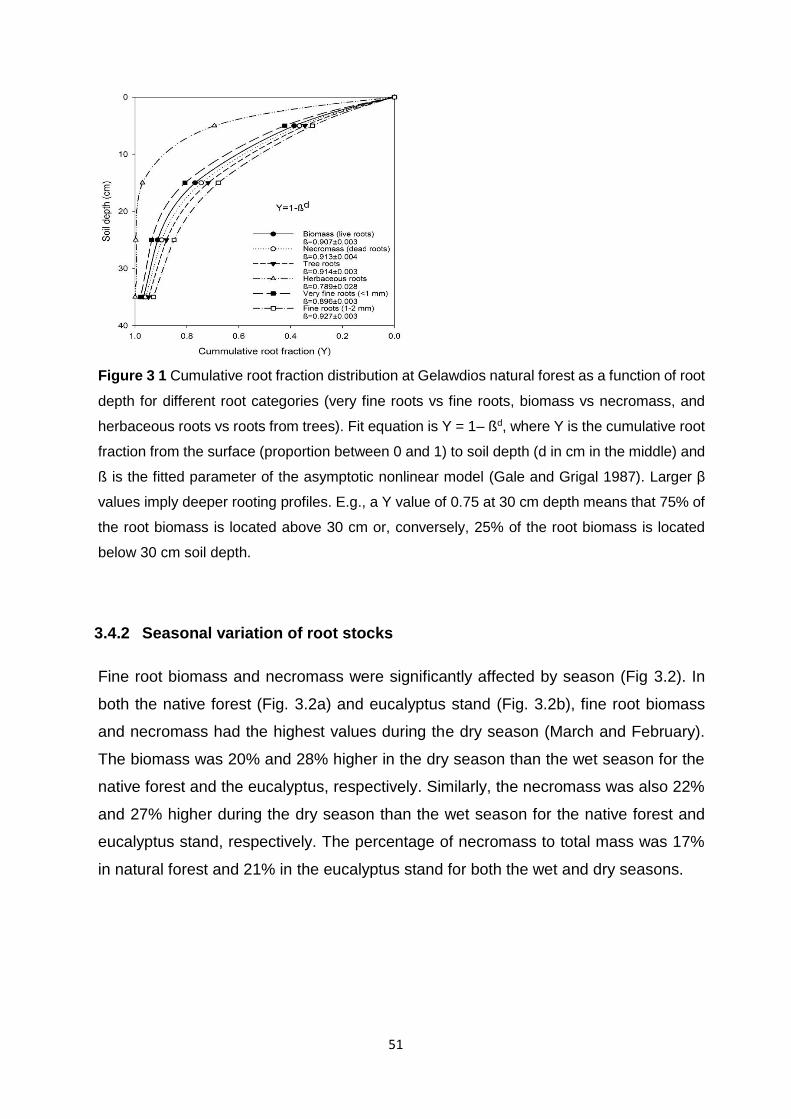
51
Figure 3 1 Cumulative root fraction distribution at Gelawdios natural forest as a function of root
depth for different root categories (very fine roots vs fine roots, biomass vs necromass, and
herbaceous roots vs roots from trees). Fit equation is Y = 1– ßd, where Y is the cumulative root
fraction from the surface (proportion between 0 and 1) to soil depth (d in cm in the middle) and
ß is the fitted parameter of the asymptotic nonlinear model (Gale and Grigal 1987). Larger β
values imply deeper rooting profiles. E.g., a Y value of 0.75 at 30 cm depth means that 75% of
the root biomass is located above 30 cm or, conversely, 25% of the root biomass is located
below 30 cm soil depth.
3.4.2 Seasonal variation of root stocks
Fine root biomass and necromass were significantly affected by season (Fig 3.2). In
both the native forest (Fig. 3.2a) and eucalyptus stand (Fig. 3.2b), fine root biomass
and necromass had the highest values during the dry season (March and February).
The biomass was 20% and 28% higher in the dry season than the wet season for the
native forest and the eucalyptus, respectively. Similarly, the necromass was also 22%
and 27% higher during the dry season than the wet season for the native forest and
eucalyptus stand, respectively. The percentage of necromass to total mass was 17%
in natural forest and 21% in the eucalyptus stand for both the wet and dry seasons.

52
Figure 3 2 Seasonal variation of fine root biomass and necromass (g m-2) at Gelawdios a)
forest; b) eucalyptus stand. Fine root estimates are based on coring method. Bars with different
small case letters are significantly different for filled bars (biomass), and different upper case
letters indicate significant differences for unfilled bars (necromass). Error bars represent
mean±1SE (p<0.05; n=10).
3.4.3 Fine root production, mortality, and turnover
The methods used for calculating annual fine root production from the sequential coring
data were the decision matrix and minimum-maximum method (Table 3.3); the mass
balance method was used for in-growth cores and in-growth net measurements (Table
3.4). Root production for all methodological approaches (sequential coring, in-growth
cores, and in-growth nets methods) clearly differed between land use types (Table 3.3
and 3.4; Fig. 3.3). For example, based on in-growth core methods, the annual root mass
produced in the forest ecosystem down to 40 cm was 468 g m-2 yr-1, which is about
37%, 87% and 90% higher than in the eucalyptus stand, grazing land, and cropland
ecosystems, respectively (Fig. 3.3). The highest annual root production was obtained
by sequential coring in the forest ecosystem (723±93 g m-2 yr-1), of which more than
58% (416±66 g m-2) was from mass loss due to decomposition (Table 3.3). Similarly,
the annual root mass production in the eucalyptus stand at the same depth was 694±95
g m-2 yr-1, necromass accounted for 14% of the total mass, and the mass loss due to
decomposition was ca. 41% (282±71 g m-2; Table 3.3). Fine root production estimated
based on the Decision Matrix was higher than the total standing mass by 22% and 39%
in the natural forest and eucalyptus, respectively. However, the annual root production
estimated by the in-growth core and in-growth net methods were ca. 43 and 62% lower

53
than the standing rootstock. The annual fine root production down to 40 cm was 60 g
m-2 yr-1 in grazing land and 52 g m-2 yr-1 in cropland based on the in-growth core method
with the mass balance calculation technique (Fig 3.3). This value was significantly lower
than in the forest and eucalyptus. Note that root growth in grazing land and cropland
was limited to the rainy season. When comparing the sampling methods, fine root
production in the forest ecosystem to 0-20 cm soil depth was highest (531±76 g m-2 yr-
1) with sequential coring, followed by the in-growth core method (329±51 g m-2 yr-1) and
the in-growth net method (218±18 g m-2 yr-1) (Fig 3.4). Except for the forest ecosystem,
the fine root production values obtained with the in-growth core and in-growth net
methods did not differ significantly in any land use system (Table 3.4). Root production
estimates using the in-growth cores and in-growth nets showed a consistent increase
over the course of the year. In the forest ecosystem, fine root production increased from
63 to 360 g m-2 from first month to the 12th month after installation of in-growth cores.
In the last four months, the average increment per month was 30 g m-2. Similarly, fine
root production as estimated with the in-growth net method increased from 42 to 191 g
m-2 with an average increment of 17 g m-2 in each of the last four months. Calculation
methods also affected the estimation of fine root production: the Decision Matrix always
showed a higher estimation than the Max-Min method (Table 3.3).
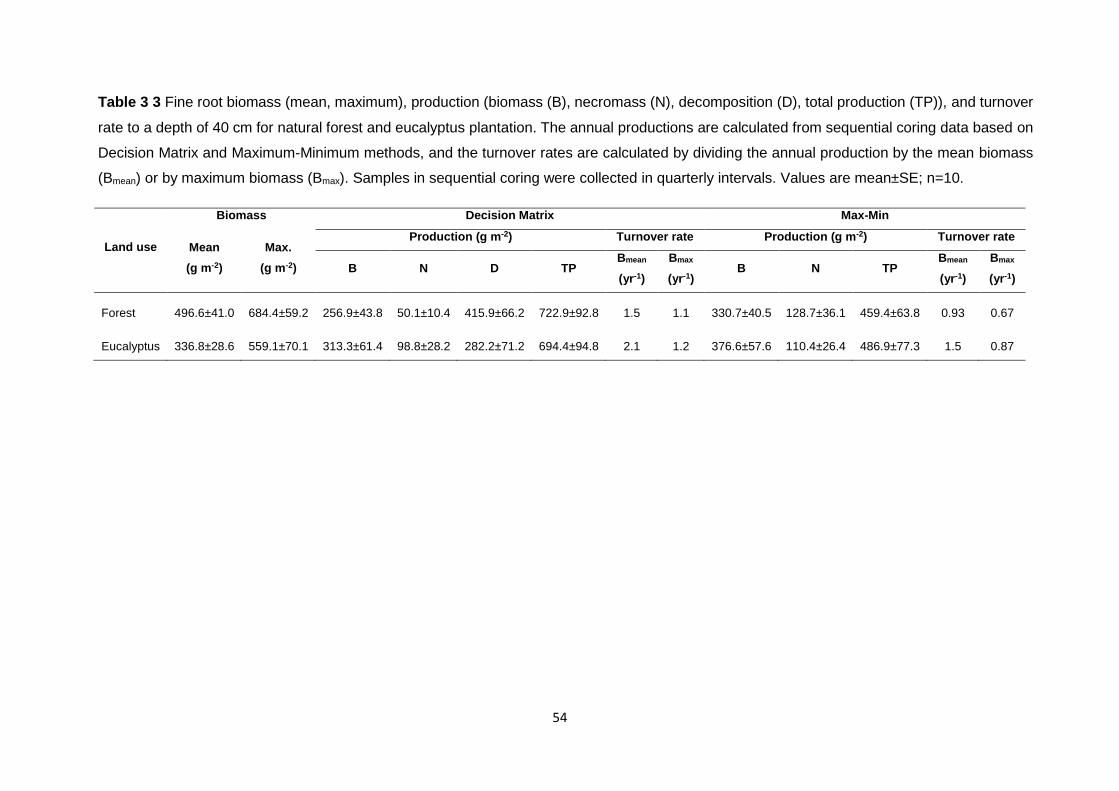
54
Table 3 3 Fine root biomass (mean, maximum), production (biomass (B), necromass (N), decomposition (D), total production (TP)), and turnover
rate to a depth of 40 cm for natural forest and eucalyptus plantation. The annual productions are calculated from sequential coring data based on
Decision Matrix and Maximum-Minimum methods, and the turnover rates are calculated by dividing the annual production by the mean biomass
(Bmean) or by maximum biomass (Bmax). Samples in sequential coring were collected in quarterly intervals. Values are mean±SE; n=10.
Land use
Biomass Decision Matrix Max-Min
Mean
(g m-2)
Max.
(g m-2)
Production (g m-2) Turnover rate Production (g m-2) Turnover rate
B N D TP Bmean
(yr-1)
Bmax
(yr-1) B N TP
Bmean
(yr-1)
Bmax
(yr-1)
Forest 496.6±41.0 684.4±59.2 256.9±43.8 50.1±10.4 415.9±66.2 722.9±92.8 1.5 1.1 330.7±40.5 128.7±36.1 459.4±63.8 0.93 0.67
Eucalyptus 336.8±28.6 559.1±70.1 313.3±61.4 98.8±28.2 282.2±71.2 694.4±94.8 2.1 1.2 376.6±57.6 110.4±26.4 486.9±77.3 1.5 0.87
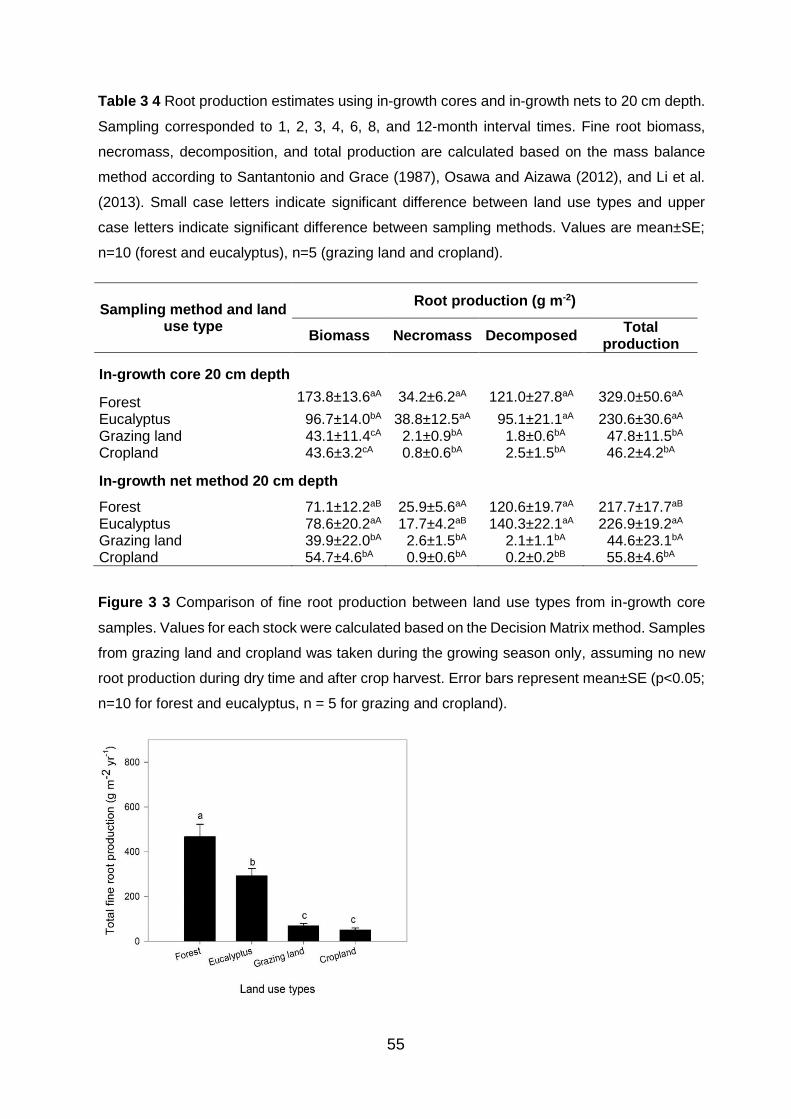
55
Table 3 4 Root production estimates using in-growth cores and in-growth nets to 20 cm depth.
Sampling corresponded to 1, 2, 3, 4, 6, 8, and 12-month interval times. Fine root biomass,
necromass, decomposition, and total production are calculated based on the mass balance
method according to Santantonio and Grace (1987), Osawa and Aizawa (2012), and Li et al.
(2013). Small case letters indicate significant difference between land use types and upper
case letters indicate significant difference between sampling methods. Values are mean±SE;
n=10 (forest and eucalyptus), n=5 (grazing land and cropland).
Sampling method and land use type
Root production (g m-2)
Biomass Necromass Decomposed Total
production
In-growth core 20 cm depth
Forest 173.8±13.6aA 34.2±6.2aA 121.0±27.8aA 329.0±50.6aA
Eucalyptus 96.7±14.0bA 38.8±12.5aA 95.1±21.1aA 230.6±30.6aA Grazing land 43.1±11.4cA 2.1±0.9bA 1.8±0.6bA 47.8±11.5bA Cropland 43.6±3.2cA 0.8±0.6bA 2.5±1.5bA 46.2±4.2bA
In-growth net method 20 cm depth
Forest 71.1±12.2aB 25.9±5.6aA 120.6±19.7aA 217.7±17.7aB Eucalyptus 78.6±20.2aA 17.7±4.2aB 140.3±22.1aA 226.9±19.2aA Grazing land 39.9±22.0bA 2.6±1.5bA 2.1±1.1bA 44.6±23.1bA Cropland 54.7±4.6bA 0.9±0.6bA 0.2±0.2bB 55.8±4.6bA
Figure 3 3 Comparison of fine root production between land use types from in-growth core
samples. Values for each stock were calculated based on the Decision Matrix method. Samples
from grazing land and cropland was taken during the growing season only, assuming no new
root production during dry time and after crop harvest. Error bars represent mean±SE (p<0.05;
n=10 for forest and eucalyptus, n = 5 for grazing and cropland).
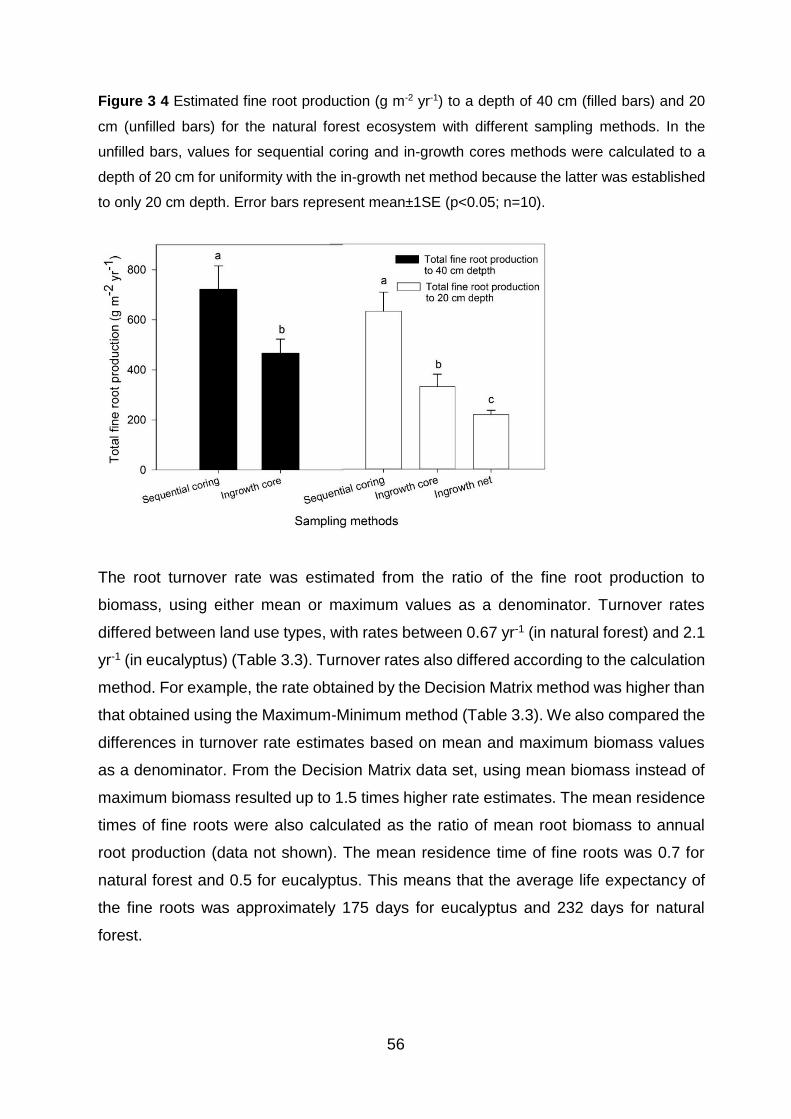
56
Figure 3 4 Estimated fine root production (g m-2 yr-1) to a depth of 40 cm (filled bars) and 20
cm (unfilled bars) for the natural forest ecosystem with different sampling methods. In the
unfilled bars, values for sequential coring and in-growth cores methods were calculated to a
depth of 20 cm for uniformity with the in-growth net method because the latter was established
to only 20 cm depth. Error bars represent mean±1SE (p<0.05; n=10).
The root turnover rate was estimated from the ratio of the fine root production to
biomass, using either mean or maximum values as a denominator. Turnover rates
differed between land use types, with rates between 0.67 yr-1 (in natural forest) and 2.1
yr-1 (in eucalyptus) (Table 3.3). Turnover rates also differed according to the calculation
method. For example, the rate obtained by the Decision Matrix method was higher than
that obtained using the Maximum-Minimum method (Table 3.3). We also compared the
differences in turnover rate estimates based on mean and maximum biomass values
as a denominator. From the Decision Matrix data set, using mean biomass instead of
maximum biomass resulted up to 1.5 times higher rate estimates. The mean residence
times of fine roots were also calculated as the ratio of mean root biomass to annual
root production (data not shown). The mean residence time of fine roots was 0.7 for
natural forest and 0.5 for eucalyptus. This means that the average life expectancy of
the fine roots was approximately 175 days for eucalyptus and 232 days for natural
forest.

57
3.4.4 Annual C and N flux into the soil
Total C and N fluxes into the soil were calculated from the annual root production and
concentration of C or N in the fine roots (Table 3.5). The annual C and N inputs into the
soil through fine roots in the top 40 cm as estimated from the in-growth core with the
mass balance method were highest in the natural forest (226 g C m-2 yr-1 and 7.5 g N
m-2 yr-1). The annual C flux in other land use types was highest in eucalyptus followed
by grazing land and cropland. The annual N flux followed the same trend as the C flux
(Table 3.5). The amount of fine root mass (biomass plus necromass) estimated by in-
growth cores was plotted against soil C and N stock as shown in Fig. 3.5. Fine root
mass showed the same pattern as total C and N stock (r2 = 0.76; Fig 3.5a and 0.74;
Fig 3.5b, respectively) and followed the same pattern along soil depth (data not shown).
Table 3 5 Total C and N flux (g m-2 yr-1) to soils via fine roots in four land use systems at
Gelawdios, Ethiopia. Element fluxes were calculated according to Xia et al. (2015) from element
concentrations in the roots multiplied by annual fine root production estimated from in-growth
cores. Numbers in brackets are number of samples. For elemental analysis in forests and
eucalyptus, all categories were considered (i.e. three each: roots <1 mm, roots 1-2 mm, and
herbaceous roots), whereas samples from grassland and cropland were all herbaceous and
we took only three samples.
Land use type C% in roots N% in roots Element flux (g m-2 yr-1)
C N
Forest 48.4±0.9 (n=9) 1.6±0.3 (n=9) 226.4 7.5
Eucalyptus 45.4±0.8 (n=9) 0.9±0.1 (n=9) 132.9 2.6
Grazing land 46.3±1.2 (n=3) 1.0±0.0 (n=3) 32.2 0.7
Cropland 46.3±1.2 (n=3) 1.0±0.0 (n=3) 23.9 0.5
3.5 Discussion
3.5.1 Effect of land use change on fine root stocks and production
Changes in vegetation composition because of land use conversion from native forests
to grazing land or cropland alter the overall quantity and quality of fine root stocks and
production. Hence, such conversions reduced the fine rootstock by 89 in grazing land

58
and 91% in cropland (Table 3.2). The value of fine root mass in our forest site (564 g
m-2) is comparable to that of tropical evergreen or deciduous forests (570 g m-2) and
boreal forests (600 g m-2) but lower than that of temperate deciduous forests (780 g m-
2; Jackson et al. 1997). Similarly, conversion of forest to grazing land or cropland
reduced fine root production by 85% and 89%, respectively (Fig. 3.3). In contrast, the
reverse process of these degraded lands through afforestation with exotic species
(eucalyptus) increased the fine root production by 76% using native forest as a
baseline. No similar studies are available for comparison of fine root production across
land use systems. The average production of fine roots in our study (723 g m-2 yr-1;
Table 3.3) is comparable to estimates from the mid-subtropics of China (795 g m-2 yr-1;
Yang et al. 2004), but lower than estimates from temperate forest in Germany (689-
1360 g m-2 yr-1; Hertel & Leuschner, 2002). This comparison is based on a similar
methodological approach and soil depth. Differences in fine root mass estimates
among studies may reflect different sorting processes (Ruess et al. 2006) or be linked
to above-ground stand characteristics (Finér et al. 2007, Brassard et al. 2013) or to soil
characteristics (Helmisaari and Hallbäcken 1999, Godbold et al. 2003). For example,
in relation to the sorting process, Ruess et al. (2006) reported that a thorough estimate
of fine root biomass yielded five times greater than a previous estimate (1780 g m-2 vs
221 g m-2) from the same site and using the same methodology. In the present study,
however, the strong positive correlation of fine root mass with basal area and stem
density (r2 > 0.98, p<0.05; data not shown) shows that the source of variation is more
due to vegetation characteristics.
Consistent with studies in moist tropical forests, fine root biomass and necromass
decreased exponentially with increasing depth (Valverde-Barrantes et al. 2007, Kucbel
et al. 2011). Our study found more than 70% of the fine roots in the upper 20 cm depth,
which is deeper than other estimates in the tropics of 67-78% in the uppermost layer of
0-10 cm (Kucbel et al. 2011). The vertical distribution of herbaceous roots, mainly from
grazing and cropland, showed that ca. 99% of the total root mass is located in the upper
30 cm. This underlines the superficial root systems of grasses and crops. Schenk &
Jackson (2002) also suggested that, on average, at least half of root biomass is located
in the upper 30 cm of soil for all systems globally. This reflects the fact that topsoils
provide a favourable microclimate for root development (Yang et al. 2004a). When
comparing across major biomes of the world according to Canadell et al. (1996), the

59
vertical distribution of roots in our study is more shallow (ß= 0.790 to 0.927; Fig 1) than
the average estimate for tropical evergreen or savannah forests (ß = 0.972) and
temperate conifer forests (ß = 0.976). Note that higher values of ß indicate a greater
proportion of roots in deeper soil (Jackson et al. 1997, Kucbel et al. 2011). Our study,
however, did not extend as deep as the maximum possible rooting depth, and the
estimated ß value can be highly affected by sampling depth. Canadell et al. (1996)
calculated the cumulative root fractions to a depth of 1-2 m, but our sampling depth was
a maximum of 40 cm. This does not mean that roots are limited to this depth. More
necromass at lower depth (ß = 0.913) than live roots (ß = 0.907) indicates that
necromass in the lower profile persists much longer than surface necromass.
3.5.2 Seasonal variation of fine root mass
We expected that fine root biomass and necromass would be greater during summer
(rainy season) when trees were physiologically active to capture nutrients. However,
we found peak values during the dry season and higher decomposition during the rainy
season in forest and eucalyptus ecosystems (Fig. 1). Hence, our results do not support
this hypothesis. This contrasts with many other observations reported from northern
savannah, Australia (Chen et al. 2004), California, USA (Contador et al. 2015), and
boreal forest (Yuan and Chen 2010), where the maximum values were found in the wet
season. Our result may agree with temperate forest studies, where peaks in standing
root mass were measured during winter time when trees are physiologically inactive
(Aerts et al. 1992, López et al. 2001) and when roots grew throughout the winter in the
absence of leaves (Teskey and Hinckley 1981). Steinaker and Wilson (2008) conclude
that forest leaf phenology is not a reliable index of overall vegetation phenology: they
found a significant negative correlation between leaf and root production in a Populus
tremuloides forest in North America. This is because fine root growth depends heavily
on newly fixed carbon from the canopy (Joslin et al. 2001), and photosynthates are
translocated to the roots only after the main period of growth (Priestley et al. 1976). In
other tropical forests, maximum fine root biomass was also observed during the dry
versus wet season in central Sulawesi, Indonesia (Harteveld et al. 2007), and in the
eastern Amazon, Brazil (Lima et al. 2010). Soil water availability is probably the main
factor for changes in root growth strategy (Kätterer et al. 1995, Lima et al. 2010). For
example, fine roots biomass increased at low soil moisture availability in tropical (Yavitt
and Wright 2001) and temperate forests (Hertel et al. 2013). Joslin et al. (2001) also

60
measured maximum root length in midsummer when potential evapotranspiration was
high and soil water was low. Contador et al. (2015) reported that water deficit appeared
to promote root production in deeper soil layers for mining soil water. Some species
also respond to drought by increasing root:shoot ratios (Joslin et al. 2000), and
maximum root length occurred in midsummer when potential evapotranspiration was
high and soil water was low. When water is limiting, plants should shift allocation of C
towards roots, where photosynthates can be used to increase water uptake, resulting
in greater root growth (Metcalfe et al. 2008). The other possible reason for low root
mass during wet season is that, in strongly seasonal climates, there is a strong flush of
fine roots in the spring but these roots live less than one month (Eissenstat and Yanai
1997). From our data set and the above literature, we hypothesise that higher
decomposition during the wet season and higher accumulation of necromass during
the dry season can be source of seasonal variation in fine root biomass.
3.5.3 Limitations of sampling methods
Our data suggest that fine root production estimates using sequential coring yielded
higher estimates than in-growth core and net methods. For example, root production in
the eucalyptus plantation as estimated by sequential coring was two-fold higher than
estimates from in-growth core and net methods. This agrees with a comparative study
between in-growth cores and ‘mesh’ (net) methods conducted in France (Andreasson
et al. 2015) and Germany (Godbold et al. 2003). The latter authors, for example,
reported from three years of data that fine root production is 69–89% lower for Picea
abies and 67–85% lower for understory species in the ‘meshes’ compared to the in-
growth cores. Another comparative study between sequential coring, in-growth cores
and growth chambers in northwest Germany found that annual root production is higher
in the sequential coring approach (Hertel and Leuschner 2002).
Nevertheless, all methods have limitations for root production estimation (Ruess et al.
2006). The underlying assumption is that each observed change of root mass during a
sampling interval in sequential coring is due to either production or mortality. Therefore,
estimations of root production, mortality, and decomposition with sequential coring at
steady states may result in a zero estimate (Brunner et al. 2012). From in-growth cores
and in-growth nets, many of the ingrowing roots are from damaged roots, and a lag
may occur before root production begins in the first month (Majdi et al. 2005). In

61
contrast, soil disturbance during installation may favour root production. Finally, using
root-free soils in the in-growth core method may accelerate root production in a
competition-free environment. Since root mortality and production occur simultaneously
(Hansson 2013), we may miss root turnover between sampling dates. In our study, the
time interval was 4–16 weeks. Accordingly, root growth and death between sampling
dates could not be accurately estimated. For example, in a carbon flow experiment in
a temperate forest soil, Gilbert et al. (2014) found a δ13C isotopic enrichment by fine
roots only in the rhizosphere soil at one month after labelling. To overcome the
shortcomings of sampling intervals in calculating actual root production and decay
between sampling dates (Priess et al. 1999), we placed additional in-growth cores and
nets at every retrieval time, corresponding to 1, 2, 3, 4, 6, 8, and 12-month intervals.
Our estimate of fine root production therefore increased from 268 g m-2 yr-1 to 329 g m-
2 yr-1 compared to the interval estimations involving 1, 2, 3, 8, 12-month sampling dates
but same calculation method. Still root growth and death can have occurred within
these sampling dates, and production could not be accurately estimated (Makkonen
and Helmisaari 1999).
In our study, fine root production estimates through the in-growth core and in-growth
net method showed a consistent increase over the course of the year. The root
production estimate based on in-growth cores is 22% lower than the average standing
rootstock, suggesting that only a one-year study involving the in-growth core and in-
growth net method is insufficient to reach an equilibrium stage.
Calculation methods also affect the estimation of fine root production. For example, the
Decision Matrix method showed higher values than the Max-Min method (Table 3).
Similarly, Berhongaray et al. (2013) showed that the Decision Matrix method yields
higher estimations (51 g m-2 yr-1) than the Max-Min method (41 g m-2 yr-1). The latter
method underestimates fine root production because it does not adequately account
for simultaneous processes of fine root growth, death, and decomposition occurring
continuously during the growing season.
Regarding turnover rates (death and decay), our results vary between 0.65 and 2.1 yr-
1 depending on the methodological approach and forest type. This finding provides
evidence that most of the fine root system dies and grows back at least once per year
(Lukac 2012). The inconsistency of data related to soil depth, diameter class, and

62
methodological approaches in the literature complicates comparing our turnover
estimates across studies. Nonetheless, our results are in the range of turnover rates of
European forests (0.17-3.10 yr-1) as compiled by Brunner et al. (2012). Turnover rate
estimates using mean biomass instead of maximum biomass as a denominator yielded
higher estimates (Table 3). This result is in agreement with estimates in a European
forest, with a mean difference of about 30% (Brunner et al. 2012). As in root production,
calculation methods also affect the estimation of the fine-root turnover rate. In this case,
the Decision Matrix showed a higher rate (Table 3). A comparison analysis by Brunner
et al. (2012) found that the rates calculated with the Decision Matrix were significantly
higher (approximately double, 1.14 yr-1) than those calculated with the Max-Min method
(0.57 yr-1). They suggested that the latter method is suitable for ecosystems with strong
annual fluctuations. Fine roots grow, die and decompose throughout the year. The
proportion of dead roots that have been replaced by new growth is calculated as root
production over biomass at steady state on an annual basis (McClaugherty et al. 1982).
Hence, we always underestimate root production because we miss the root growth and
death between the sampling dates. We therefore suggest that using mean biomass, as
a denominator, is more representative of average live roots than using maximum
biomass to estimate root turnover rate.
Figure 3 5 Relationship between total root mass (biomass + necromass) with C stock a) and
N stock b) for four land use systems at Gelawdios, Ethiopia. n=10 (forest and eucalyptus), and
5 (cropland and grazing land).

63
3.5.4 Implications of fine root turnover in ecosystem carbon cycling
Our fine rootstock and production estimates tended to show the same pattern of soil C
and N stocks in the soil (r2 = 0.76; P<0.001 for C and r2 = 0.74; P< 0.001 for N; Fig.
3.5). This suggests that fine roots are the major contributors of C and N input in the soil.
Similarly, Hansson et al. (2013) reported that the fine root stock distribution followed
the same pattern as the soil C and N distribution in the stands. Nonetheless, our annual
C flux estimate based on fine roots in the forest to 40 cm depth (224 g of C m-2 yr-1;
Table 3.5) was much smaller than estimates from tropical forests in Indonesia (233 g
of C m-2 yr-1 from the top 20 cm only; Hertel et al., 2008). The annual C flux from fine
roots in the eucalyptus stand, grazing land, and cropland was about 37%, 85% and
89% lower, respectively, compared to native forest. This suggests that conversion of
native forest to grazing land and cropland resulted in a ca. 85-89% C input reduction
into the soil. Previously, our carbon stock analysis (Chapter 2) showed that land use
change resulted in an up to 88% of carbon reduction at the site level. Based on the
current forest cover of the region (9.5%, unpublished government report), the total C
input to the soil through fine roots is very roughly estimated to be 3.6 mega tons
(1Mt=1012) of C yr-1. If we take the former forest cover of the country (40%, about 50
years ago; Bekele 2011) for the study area, and if there is no C translocation, then the
region lost an annual input of 11.1 Mt of C yr-1 through fine roots into soil for the last 50
years due to deforestation. This estimate is certainly conservative. The result of this
analysis suggests that forest protection against deforestation alone sequesters about
90% more carbon compared to grazing land or croplands.
3.6 Conclusion
Conversions of land use from native forest to other land use types such as grazing land
or cropland strongly reduce root production (ca. 85-89%). Root production and turnover
were also strongly affected by seasonal variations, with peak values in the dry season.
This indicates accumulation of root mass under moisture-limiting conditions. With
regard to vertical distribution, there was always a decreasing trend of root mass with
soil depth. This spatial pattern was similar to that of the C and N stock in the mineral
soil, indicating a significant contribution of roots to soil C and N pools.

64
4 Fine root morphology, biochemistry and litter quality
indices of fast- and slow-growing woody species in
Ethiopian highland forests
4.1 Abstract
Fine roots of trees are the major channels of carbon flows into the soil. Nonetheless,
the quality of carbon input into the soil via fine roots is influenced by morphological traits
and chemical composition. This study was designed to examine whether root traits
varied with species and to correlate such variations with biochemistry via proxy carbon
fractionations. Fine roots of ten tropical woody species collected from the northern
highlands of Ethiopia were analysed for root morphological indices and root chemistry
using a series of acid digestions. Fast- growing species exhibited higher specific root
length and specific root area, but lower root tissue density (RTD) than slow-growing
species. Differences in morphological parameters of fine roots in fast- and slow-growing
species reflect the ecological strategy they employ. In roots of all species, the acid-
insoluble fractions (AIF) were the highest fraction. The carbon content, AIF, and the
lignocellulose index were higher for slow-growing species and showed a strong positive
correlation with RTD. This reflects a higher carbon investment to construct fine roots of
slow growing species.
Key words: Carbon fractions, root tissue density, specific root length, acid-insoluble
fraction, carbon cost, root traits, lignin

65
4.2 Introduction
The importance of fine roots for both plant and ecosystem functioning is increasingly
recognized. At the same time, our understanding of root trait variation between plant
species and its effects on ecological processes such as biogeochemical cycling
remains limited (Xia et al. 2015, Valverde-Barrantes et al. 2016, Weemstra et al. 2016).
Despite increasing information on root traits of plants in temperate and boreal (forest)
ecosystems (Comas et al. 2002, Pinno et al. 2010), corresponding data on fine root
characteristics of tropical tree species are rare, particularly for African ecosystems.
Parameters such as specific root length (SRL) and root tissue density (RTD) of ‘fine
roots’ (< 2 mm in diameter) are key traits of a root economics spectrum. This is because
they are apparently closely linked to the carbon (C) use strategy and/or resource uptake
efficiency of trees (Comas and Eissenstat 2004, Birouste et al. 2014, Weemstra et al.
2016). From an ecological point of view, fine roots with higher RTD affect several
processes of root functioning such as soil fertility (Valverde-Barrantes et al. 2016),
respiration rate (Makita et al. 2012, Rewald et al. 2014), growth rate (Birouste et al.
2014), and longevity (McCormack et al. 2012). Similarly, other root morphology
parameters such as specific root length (SRL) have been widely used as indicators of
resource use efficiency (Comas and Eissenstat 2004, Ostonen et al. 2007).
Interestingly, SRL and RTD are not necessary correlated across phylogenetic groups
(Valverde-Barrantes et al. 2016).
The chemical composition of fine root tissue may reveal further important aspects of
carbon-use strategies of plants. For example, the fine root longevity of temperate tree
species significantly increases with decreasing C:N ratios (McCormack et al. 2012).
Beyond C:N ratios, the chemical composition of fine roots regarding labile (e.g.,
carbohydrates) and recalcitrant fractions (e.g., lignin) may further enhance our
understanding of (tree) root economic strategies (Kong et al. 2016). The pioneer work
of Kong and colleagues, however, also again emphasizes that root trait patterns are
complicated and that further studies are needed. This pertains especially to
consolidating our understanding of the relations between specific C and N fractions and
morphological parameters and to improving our trait interpretations regarding (tree) fine
root economic spectra. In particular, we need information on morphological and
chemical root traits linked to plant growth strategies (e.g. fast or slow above-ground

66
growth) (Comas et al. 2002, Comas and Eissenstat 2004) and to trait interrelationships.
Currently, evidence for correlations within and between root traits, and the wider plant
economic spectrum, is weak at best. This is probably because root system function can
be optimized using a much more diverse set of traits compared to leaves (Valverde-
Barrantes et al. 2016, Weemstra et al. 2016).
Beside their key role for plant functioning, it is increasingly recognized that fine roots
also play major roles in global biogeochemical cycles, including carbon sequestration
(Xia et al. 2015). Plant roots account for up to 48% of annual plant litter inputs (Freschet
et al. 2013) and are estimated to contribute an average of two-fold more to soil organic
C than leaf litter (Rasse et al. 2005). While root litter is thus a major source of soil
organic matter, species-specific root decomposition rates and impacts on soil organic
carbon turnover remain uncertain. The parameters that explain the largest amount of
variability in root decay are abiotic environmental factors such as temperature and
precipitation as well as root tissue chemistry (Silver and Miya 2001, Solly et al. 2014).
The chemical compositions of roots, e.g. indicated by labile and recalcitrant fractions,
varies with species and largely determines the rate of decay (Couteaux et al. 1995,
Silver and Miya 2001, Sun et al. 2013) and the quality of C input into soil systems
(Rasse et al. 2005). One approach for quantitative determination of fine root carbon
chemistry is chemical fractionation. According to Ryan et al. (1990) and Sun et al.
(2013), organic compounds of tissues can be placed into three broad fractions. 1)
Extractable labile C compounds consisting of nonpolar constituents such as fats, oils,
waxes, and polar constituents such as nonstructural carbohydrates and water-soluble
polyphenols removed using a two-stage extraction in dichloromethane and boiling
water, respectively. 2) Acid-hydrolysable structural components that are moderately
degradable C compounds consisting primarily of cellulose and hemicellulose, removed
using a two-stage digestion in 72% and 2.5% H2SO4. 3) Acid-insoluble aromatic
compounds that are highly recalcitrant C compounds conventionally referred to as
lignin (Preston et al. 2000, Xia et al. 2015) but also consisting of other highly reduced
compounds such as suberin, cutin, and tannin-protein complexes, which are the
residual of the two-stage sulfuric acid digestion minus ash mass (Sun et al. 2013). The
recalcitrant tissues are characterized by relatively low concentrations of easily
degraded substrates (Xia et al. 2015). The values obtained can be used as an estimate
of long-term carbon input into the soil system by roots.

67
This study was conducted on ten dominant woody species of a diverse, pristine
community of tropical woody species, namely a remnant church forest of the north-
central Ethiopian highland. The aim was to characterize fine root morphology,
biochemistry, and their interrelationships. The specific objectives were to determine 1)
how fine root morphology and biochemistry varies between fast- and slow-growing
woody species, and 2) if or how fine root morphological traits are correlated to root
biochemistry. We hypothesize that fast-growing species build ‘cheaper’ roots of lower
root tissue density (RTD) and that roots with a lower RTD contain less recalcitrant
carbon fractions (less lignified roots). The results are discussed in the light of root
economic strategies and the potential effects on root litter quality and quality of C input
into soil systems.
4.3 Materials and methods
4.3.1 Study site
This study was carried out at the remnant Afromontane forest of Gelawdios in the
Amhara National Regional State, north-central Ethiopia. Gelawdios (11°38’25’’N,
37°48’55’’E) is located east of Lake Tana at an altitude of 2466-2526 m above sea level.
While Ethiopia is located in the tropics, the climate of the study area is temperate with
dry winters and warm summers (Cwb) according to the Köppen-Geiger climate
classification (Peel et al. 2007). The mean annual precipitation is 1220 mm, with the
main rainy season from June to September and with low-intensity precipitation from
March to May (Wassie et al. 2009). The distribution of rainfall largely depends upon the
direction of moisture-bearing monsoon winds and altitude. The annual mean air
temperature is 19°C (Wassie et al. 2009). The soils are classified as Cambisols;
edaphic characteristics are summarized in Supplementary Information Table A1.1. The
Afromontane Gelawdios forest is a small, isolated, but pristine forest fragment (‘church
forest’) covering about 100 ha in the otherwise almost completely deforested Ethiopian
highlands (Wassie et al. 2009, Aerts et al. 2016).
Ten dominant, native woody species were studied: Allophylus abyssinicus (Hochst.)
Radlk., Apodytes dimidiata E. Mey ex. Arn., Calpurnia aurea (Ait.) Benth., Chionanthus
mildbraedii (Gilg & Schellenb.) Stearn, Combretum collinum Fresen., Dovyalis
abyssinica (A. Rich.) Warb., Ekebergia capensis (Sparm.), Maytenus arbutifolia (A.

68
Rich.) Wilczek, Podocarpus falcatus (Thunb.) Mirb., and Teclea nobilis (Del.). The
species were categorized into five fast- and five slow-growing species according to a
literature review (Fichtl and Adi 1994, Katende et al. 1995, Hedberg 2003, Bekele 2007,
Orwa et al. 2009) and information from indigenous knowledge. Species category,
characteristics, and their corresponding local names are provided in Table A2.2.
4.3.2 Root sampling
Roots were sampled in September 2014, corresponding to peak aboveground growth
and the end of the rainy season. Intact fine root branches (diameter <2 mm) were
collected from the topsoil under five randomly chosen tree individuals of each species
along a transect line; the minimum distance between sampling locations under the
same species was about 100 m. In the diverse and relatively dense forest stand, fine
root taxa for the species of interest were identified by carefully tracking coarse roots
from the tree base (Rewald et al. 2012). At each tree individual, three sample locations
were selected. At each of the locations, one intact fine root branch was carefully
extracted from a soil monolith (approx. 20 cm × 10 cm; top 20 cm of soil) with forceps;
remaining soil on roots was carefully brushed away (Wang et al. 2006). In total, fifteen
root branches per species were separately placed into sealed plastic bags, kept in a
cooling box/fridge (4-8°C), and transported to Vienna, Austria, for subsequent root
processing within 1 week after sampling. Moist paper towels were placed inside the
plastic bags to prevent desiccation.
4.3.3 Root morphology
Fine root branches were cleaned of residual soil particles with water before being
submerged and spread out individually on an A3-sized transparent tray for scanning
(Epson Expression 10000XL with transparency adapter; grey scale, 600 dpi). The PC
software WinRhizo Pro 2012b (Regent Instruments Inc., Canada) was used to
determine average root diameter (mm), total root length (cm) and root length per
diameter class (cm), total root surface area (cm2), and total root volume (cm3). Twenty
diameter classes (0-2 mm) with a class width of 0.1 mm each were set. Subsequently,
samples were dried (70°C, to constant mass) and weighed with an accuracy of ±0.1
mg. The following fine root traits were calculated: specific root area (SRA; cm2 g-1),
specific root length (SRL; m g-1), and root tissue density (RTD; g cm-3). Since root
branches had different sizes, the value of a given length per diameter class was

69
normalized using total root length, yielding a proportion of relative diameter class length
( rDCL; Zobel et al. 2007).
4.3.4 Root biochemistry and construction costs
Oven-dry root samples of each species were pooled, due to the very limited dry mass
available, and ground to powder (Fritsch Pulverisette 5, Idar-Oberstein, Germany).
Three technical replicates per species were analyzed for total C and N contents using
a CN analyser (Truspec CNS; LECO, St. Joseph, USA). Three other subsamples per
species were analysed for carbon fractions using procedures adapted from Ryan et al.
(1990), Sun et al. (2013), and Kong et al. (2016). Root carbon fractions, including non-
polar extractives (fats, oil, wax), polar extractives (carbohydrates, polyphenols), acid-
soluble structural components (cellulose, hemicellulose), acid-insoluble structural
components (mainly lignin, suberin), and ash were assessed using a series of digestion
techniques (Ryan et al. 1990).
Extractives were determined using a two-stage solvent extraction. Nonpolar fractions
(NPE) were extracted from 1 g material with 75 mL dichloromethane according to
Sluiter et al. (2005). The sample was sonicated for 30 min and the supernatant was
centrifuged at 1050 g (gravitational force) for 30 min and decanted to a dried, pre-
weighed flask. The residues were oven-dried at 60°C overnight to remove the residual
solvent (Sluiter et al. 2005), and polar fractions (PE) were extracted using hot water.
Seventy-five ml of deionized water was added in a flask containing the residue, boiled
under reflux for 3 h, and allowed to cool. After centrifugation at 1050 g for 30 min, the
supernatant was decanted into clean tubes and evaporated at 60°C until constant
weight. The residues remaining in the tubes were weighed. The two extractions
removed both polar and nonpolar extractives, considered readily decomposable,
leaving highly cross-linked cell wall components in the residue (Ryan et al. 1990, Xia
et al. 2015). The sum of polar (PE) and nonpolar (NPE) extractives is named “solvent
extractives” (EF) and the remaining residue “cell wall fraction.” Thus, the extractive
fraction is the difference between the initial weight and the weight of the cell wall fraction
plus ash (Ryan et al. 1990, Xia et al. 2015).
The cell wall fraction was subsequently divided into acid-soluble and acid-insoluble
fractions. The acid-soluble fraction (ASF), dominated by polysaccharides, was
extracted using a two-stage digestion with sulfuric acid (Ryan et al. 1990). Oven-dried

70
residues (60°C, to constant mass) were transferred to a test tube, and 3 ml of 72%
(w/w) H2SO4 was added and stirred. Test tubes were placed in a water bath (30°C, 3
h) and periodically stirred. Afterwards, the samples were transferred to 250 ml Pyrex
bottles (with Teflon-lined screw caps) by using 84 ml of distilled water, resulting in a
2.5% acid solution. The sealed bottles were autoclaved (121°C, 1 h). After cooling, the
solutions were decanted and evaporated at 60°C to constant mass. This acid-soluble
fraction (ASF) consists of hydrolyzed carbohydrates. The residue of the two-stage
sulfuric acid digestion minus the ash mass was used to determine the acid insoluble
fraction (AIF) containing structural components (lignin). The ash content of the residues
(dried at 105°C, 24 h) was determined in pre-weighed, oven-dried crucibles placed in
a muffle furnace at 575°C for 8 h (until no black residue remained). After ignition, the
crucible+ash was cooled in a desiccator and weighed. All root chemical fractions were
expressed as ash-free dry matter (DM). Litter quality indices such as C/N and AIF/N
ratios and the lignocellulose index were calculated. The lignocellulose index was
calculated as the ratio of AIF to cell wall fraction (ASF+AIF; Xia et al., 2015).
Root construction costs (CC; g glucose g dw-1) were calculated according to Vertregt
and De Vries (1987) as modified by Poorter (1994). This method is widely used on both
herbaceous and woody roots (Poorter et al. 2006, Vivin et al. 2015). CC is derived from
the C (Cdw), N (Ndw) and ash (Ashdw) contents of dry fine roots, expressed in mg g-1:
CC = (-1.041-5.077*Cdw)*(1- Ashdw)+(5.325*Ndw) (1)
4.3.5 Statistical analysis
The rDCL was plotted against diameter class according to the best fitted equation
developed by Zobel et al. (2007). The biomass and morphological data were analysed
by one-way ANOVA to determine the differences in means among species and
functional groups. Data that did not meet the assumption of normality were log or
square root transformed before analysis to reach normality. If significant differences
were found, multiple comparisons were carried out based on Tukey’s HSD test at
P<0.05. The data were also analysed using regression analysis and Pearson
correlations for examining relationships between morphological traits and biochemical
fractions. Statistical tests and analyses were performed using IBM SPSS version 21;
graphs were prepared using SigmaPlot (Version 13). All data shown are mean ±
standard error (SE).

71
4.4 Results
4.4.1 Root morphology
Morphological parameters of fine roots, i.e. average diameter, SRA, SRL, and RTD,
varied significantly among the ten examined woody species of the Gelawdios forest
(Table 4.1). Fine roots (diameter ≤ 2 mm) of the ten species had average root diameters
(AD) between 0.52-0.76 mm, with Podocarpus falcatus, Ekebergia capensis and
Teclea nobilis having significantly thicker roots. On average, slow-growing species had
significantly thicker fine roots than fast-growing species, but this finding was not
consistent on a species-level (Table 4.1). Except for the three thicker-rooted species
above, most examined species had a similar fine root diameter-class distribution, with
the majority of root length being between 0.25 and 0.40 mm in diameter (Fig. 4.1). The
specific root length (SRL) showed large variation among species, ranging from 635 cm
g-1 (Teclea nobilis) to 1695 cm g-1 (Calpurnia aurea) (Table 4.1). Specific root area
(SRA) differences were analogues to SRL: fast-growing species had on average both
significantly higher SRL and SRA values (Table 4.1). Fine roots differed also in root
tissue density (RTD), with slow-growing species unanimously featuring significantly
higher RTD than fast-growing species (Table 4.1). Average root diameter was highly
negatively correlated to SRL, and RTD was highly negatively correlated to SRA (Table
4.5).

72
Table 4 1 Morphological traits of fine roots of ten woody species. SRA, specific root area; SRL,
specific root length; RTD, root tissue density. Species are grouped into fast- (FG) and slow-
growing (SG) species (see Supplementary Information Table A2.2 for details). Different small
case letters indicate significant trait differences between species irrespective of group, and
upper case letters indicate differences between FG and SG group averages (mean±SE; Tukey,
p<0.05; nspecies = 15, ngroup = 5).
Species (Groups) Diameter (mm) SRA
(cm2 g-1) SRL
(cm g-1) RTD
(g cm-3)
Fast-growing (FG)
Apodytes dimidiata 0.55±0.02a 217±8cd 1290±78bcd 0.34±0.01a
Calpurnia aurea 0.53±0.04a 254±18d 1695±189d 0.32±0.01a
Dovyalis abyssinica 0.62±0.02ab 201±9bcd 1076±78abcd 0.33±0.01a
Maytenus arbutifolia 0.52±0.03a 244±14d 1588±172cd 0.33±0.01a
Podocarpus falcatus 0.76±0.07b 200±24bcd 1156±278abcd 0.31±0.01a
FG Average 0.59±0.05A 223±11B 1362±120B 0.33±0.01A
Slow-growing (SG)
Allophylus abyssinicus 0.55±0.01a 179±7abc 1055±66abc 0.41±0.01b
Chionanthus mildbraedii 0.59±0.04ab 165±9abc 946±83ab 0.43±0.01b
Combretum collinum 0.53±0.02a 199±8abcd 1249±96abcd 0.39±0.01b
Ekebergia capensis 0.74±0.05b 149±12ab 731±96ab 0.39±0.01b
Teclea nobilis 0.75±0.03b 142±8a 635±71a 0.39±0.01b
SG Average 0.64±0.05B 167±10A 923±111A 0.40±0.01B
Figure 4 1 Non-linear regression model of relative diameter class length distribution (rDCL;
cm) (n = 15) of fine roots ≤2 mm diameter of ten tropical tree and shrub species from Gelawdios
forest in the Ethiopian highlands.

73
4.4.2 Root biochemistry
The biochemistry of fine root tissues varied considerably both between individual
species and between slow- and fast-growing species groups (Table 4.2). For example,
the least amount of the nonpolar extractive fraction (NPE; includes fatty acids and
lipids) was present in Apodytes dimidiata fine roots (approx. 2% of dry matter), the
highest amounts in Dovyalis abyssinica and Maytenus arbutifolia (approx. 8% of dry
matter). All three species are fast growing. Thus, no significant differences of NPE were
found between species’ groups. In contrast, the amount of polar extractives (PE; incl.
sugars and phenols) was significantly greater in fast-growing species. Nonetheless,
values differed considerably between individual species (4 to 19% of dry matter), and
the difference between groups was largely driven by the high PE concentrations in the
fine roots of Calpurnia aurea and Podocarpus falcatus (both fast-growing species).
Consequently, solvent extractives (extractive fractions, EF), the sum of NPE and PE,
were significantly more abundant in fine roots of fast-growing species. The cell wall
fractions in root tissues of the ten studied species ranged from 76 to 90% of the dry
matter. In fine roots of all species except Apodytes dimidiata, the acid insoluble fraction
(AIF; referred to as lignin) were clearly the most abundant; Apodytes dimidiata had a
slightly higher acid-soluble fraction (ASF; Table 4.2). The slow-growing species
featured consistently and significantly greater AIF compared to fast-growing species;
ASF varied widely within the fine roots of both species groups. Ash contents were 1.3
to 3.1%, with significant differences between some species but no systematic
differences between fast- and slow-growing species. The acid-soluble fraction was
highly negatively correlated to EF (Table 4.5). The lignocellulose index, i.e. the
proportion of AIF among root cell wall fractions, was significantly greater in slow-
growing species; however, inter-specific differences within groups were high (Table
4.3).
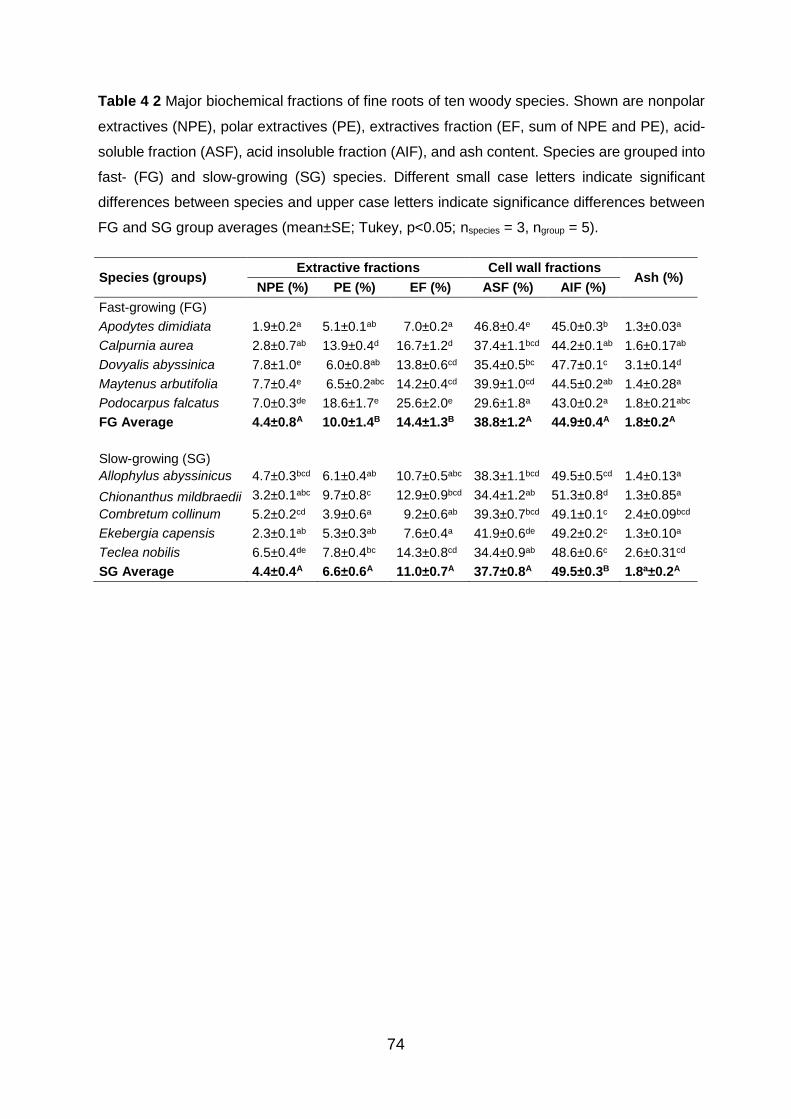
74
Table 4 2 Major biochemical fractions of fine roots of ten woody species. Shown are nonpolar
extractives (NPE), polar extractives (PE), extractives fraction (EF, sum of NPE and PE), acid-
soluble fraction (ASF), acid insoluble fraction (AIF), and ash content. Species are grouped into
fast- (FG) and slow-growing (SG) species. Different small case letters indicate significant
differences between species and upper case letters indicate significance differences between
FG and SG group averages (mean±SE; Tukey, p<0.05; nspecies = 3, ngroup = 5).
Species (groups) Extractive fractions Cell wall fractions
Ash (%) NPE (%) PE (%) EF (%) ASF (%) AIF (%)
Fast-growing (FG)
Apodytes dimidiata 1.9±0.2a 5.1±0.1ab 7.0±0.2a 46.8±0.4e 45.0±0.3b 1.3±0.03a
Calpurnia aurea 2.8±0.7ab 13.9±0.4d 16.7±1.2d 37.4±1.1bcd 44.2±0.1ab 1.6±0.17ab
Dovyalis abyssinica 7.8±1.0e 6.0±0.8ab 13.8±0.6cd 35.4±0.5bc 47.7±0.1c 3.1±0.14d
Maytenus arbutifolia 7.7±0.4e 6.5±0.2abc 14.2±0.4cd 39.9±1.0cd 44.5±0.2ab 1.4±0.28a
Podocarpus falcatus 7.0±0.3de 18.6±1.7e 25.6±2.0e 29.6±1.8a 43.0±0.2a 1.8±0.21abc
FG Average 4.4±0.8A 10.0±1.4B 14.4±1.3B 38.8±1.2A 44.9±0.4A 1.8±0.2A
Slow-growing (SG)
Allophylus abyssinicus 4.7±0.3bcd 6.1±0.4ab 10.7±0.5abc 38.3±1.1bcd 49.5±0.5cd 1.4±0.13a
Chionanthus mildbraedii 3.2±0.1abc 9.7±0.8c 12.9±0.9bcd 34.4±1.2ab 51.3±0.8d 1.3±0.85a
Combretum collinum 5.2±0.2cd 3.9±0.6a 9.2±0.6ab 39.3±0.7bcd 49.1±0.1c 2.4±0.09bcd
Ekebergia capensis 2.3±0.1ab 5.3±0.3ab 7.6±0.4a 41.9±0.6de 49.2±0.2c 1.3±0.10a
Teclea nobilis 6.5±0.4de 7.8±0.4bc 14.3±0.8cd 34.4±0.9ab 48.6±0.6c 2.6±0.31cd
SG Average 4.4±0.4A 6.6±0.6A 11.0±0.7A 37.7±0.8A 49.5±0.3B 1.8a±0.2A

75
Table 4 3 Litter quality indices of ten woody species. Shown are carbon (C) and nitrogen (N)
contents, C/N ratio, acid insoluble fraction (AIF) to N ratio, and lignocellulose index. Species
are grouped into fast- (FG) and slow-growing (SG) species; see Supplementary Information
Table A2.2 for details. Lignocellulose index is the ratio of AIF to cell wall fraction. Small case
letters indicate significant differences between species and upper case letter indicate
differences between FG and SG group averages (mean±SE; Tukey, p<0.05; nspecies = 3, ngroups
= 5).
Species (Groups) C% N% C:N Ratio AIF:N Ratio
Lignocellulose index
Fast-growing (FG) Apodytes dimidiata 44.8±0.40ab 1.38±0.01d 32.6±0.4b 32.7±0.1c 0.49±0.00a Calpurnia aurea 44.5±0.30a 1.21±0.00bc 36.8±0.3d 36.6±0.1d 0.54±0.01bc Dovyalis abyssinica 46.3±0.03bc 1.38±0.01d 33.7±0.1bc 34.7±0.1cd 0.57±0.00cde Maytenus arbutifolia 45.2±0.65ab 1.73±0.01f 26.2±0.5a 25.8±0.2a 0.53±0.01ab Podocarpus falcatus 45.3±0.40ab 1.08±0.02a 42.0±0.5f 39.9±0.6ef 0.56±0.01bcd FG Average 45.2±0.24A 1.35±0.06A 34.3±1.4A 33.9±1.3A 0.54±0.01A Slow-growing (SG) Allophylus abyssinicus 48.2±0.39d 1.26±0.01d 38.3±0.2d 39.4±0.6e 0.56±0.01bcde Chionanthus mildbraedii 49.9±0.22e 1.51±0.01b 33.0±0.3b 33.9±0.7c 0.60±0.01e Combretum collinum 47.1±0.16cd 1.36±0.01c 34.6±0.3c 36.1±0.2d 0.56±0.01bcd Ekebergia capensis 47.5±0.13cd 1.17±0.00e 40.4±0.1e 41.9±0.3f 0.54±0.00bc Teclea nobilis 47.1±0.09cd 1.74±0.01a 27.1±0.1a 28.0±0.5b 0.59±0.01de SG Average 47.9±0.29B 1.41±0.04A 34.7±1.2A 35.9±1.3A 0.57±0.01B
4.4.3 Carbon cost of root production
The calculated amount of glucose needed to produce one gram of fine root biomass
varied significantly among species (Table 4.4). For example, these carbon costs were
highest in Chionanthus mildbraedii (1.5 g glucose g-1 dw) and lowest in Calpurnia aurea
roots (1.2 g glucose g-1 dw). The calculated amount of glucose needed to synthesize
root biomass was significantly and consistently greater in slow-growing versus fast-
growing species (Fig. 4.2, Table 4.4).
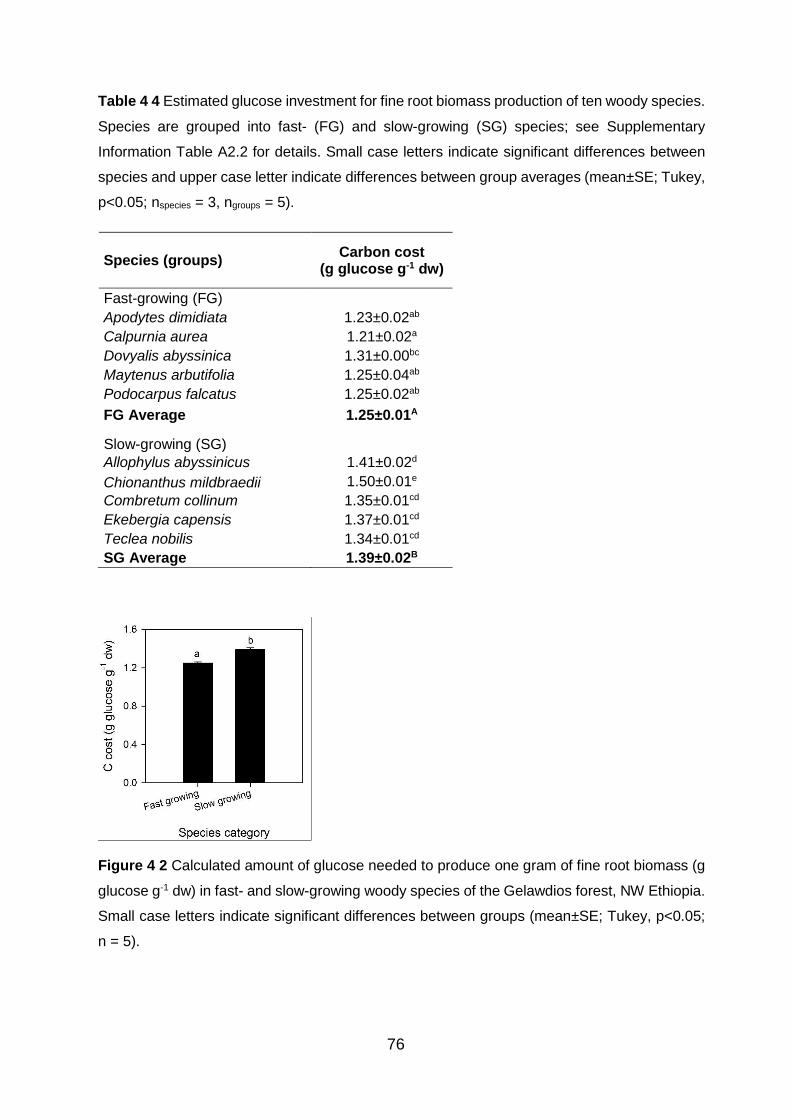
76
Table 4 4 Estimated glucose investment for fine root biomass production of ten woody species.
Species are grouped into fast- (FG) and slow-growing (SG) species; see Supplementary
Information Table A2.2 for details. Small case letters indicate significant differences between
species and upper case letter indicate differences between group averages (mean±SE; Tukey,
p<0.05; nspecies = 3, ngroups = 5).
Species (groups) Carbon cost
(g glucose g-1 dw)
Fast-growing (FG)
Apodytes dimidiata 1.23±0.02ab
Calpurnia aurea 1.21±0.02a
Dovyalis abyssinica 1.31±0.00bc
Maytenus arbutifolia 1.25±0.04ab
Podocarpus falcatus 1.25±0.02ab
FG Average 1.25±0.01A
Slow-growing (SG)
Allophylus abyssinicus 1.41±0.02d
Chionanthus mildbraedii 1.50±0.01e
Combretum collinum 1.35±0.01cd
Ekebergia capensis 1.37±0.01cd
Teclea nobilis 1.34±0.01cd
SG Average 1.39±0.02B
Figure 4 2 Calculated amount of glucose needed to produce one gram of fine root biomass (g
glucose g-1 dw) in fast- and slow-growing woody species of the Gelawdios forest, NW Ethiopia.
Small case letters indicate significant differences between groups (mean±SE; Tukey, p<0.05;
n = 5).
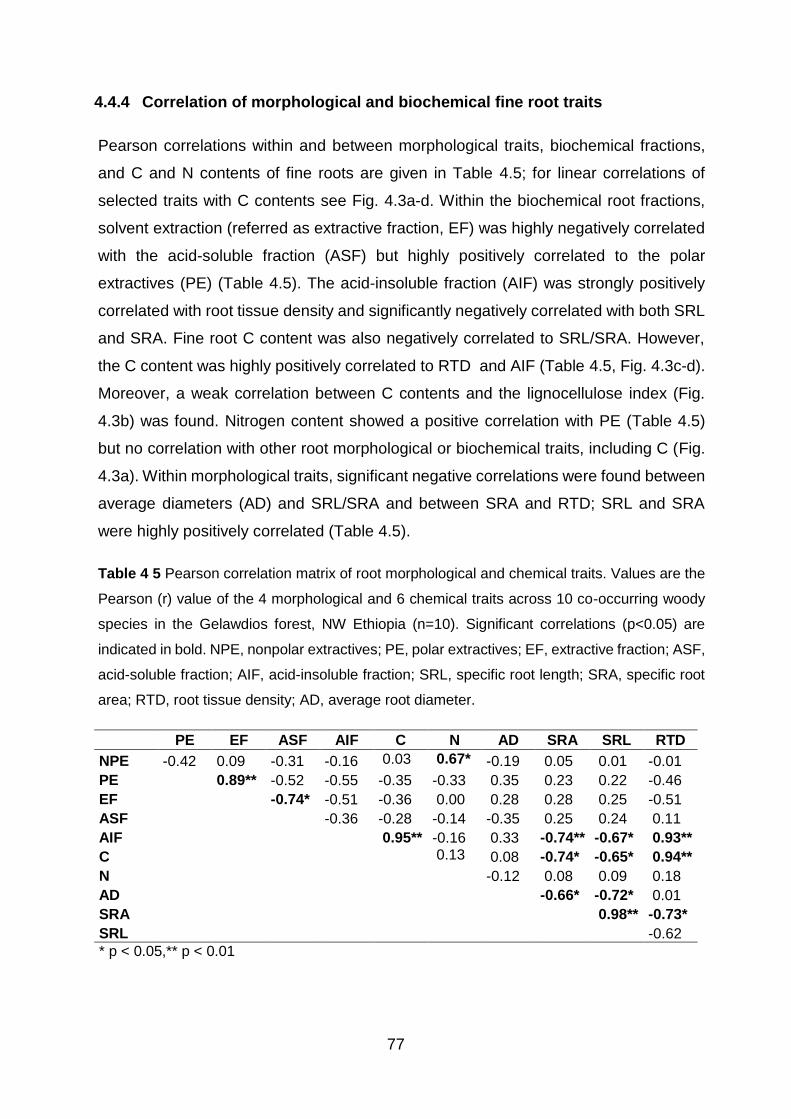
77
4.4.4 Correlation of morphological and biochemical fine root traits
Pearson correlations within and between morphological traits, biochemical fractions,
and C and N contents of fine roots are given in Table 4.5; for linear correlations of
selected traits with C contents see Fig. 4.3a-d. Within the biochemical root fractions,
solvent extraction (referred as extractive fraction, EF) was highly negatively correlated
with the acid-soluble fraction (ASF) but highly positively correlated to the polar
extractives (PE) (Table 4.5). The acid-insoluble fraction (AIF) was strongly positively
correlated with root tissue density and significantly negatively correlated with both SRL
and SRA. Fine root C content was also negatively correlated to SRL/SRA. However,
the C content was highly positively correlated to RTD and AIF (Table 4.5, Fig. 4.3c-d).
Moreover, a weak correlation between C contents and the lignocellulose index (Fig.
4.3b) was found. Nitrogen content showed a positive correlation with PE (Table 4.5)
but no correlation with other root morphological or biochemical traits, including C (Fig.
4.3a). Within morphological traits, significant negative correlations were found between
average diameters (AD) and SRL/SRA and between SRA and RTD; SRL and SRA
were highly positively correlated (Table 4.5).
Table 4 5 Pearson correlation matrix of root morphological and chemical traits. Values are the
Pearson (r) value of the 4 morphological and 6 chemical traits across 10 co-occurring woody
species in the Gelawdios forest, NW Ethiopia (n=10). Significant correlations (p<0.05) are
indicated in bold. NPE, nonpolar extractives; PE, polar extractives; EF, extractive fraction; ASF,
acid-soluble fraction; AIF, acid-insoluble fraction; SRL, specific root length; SRA, specific root
area; RTD, root tissue density; AD, average root diameter.
PE EF ASF AIF C N AD SRA SRL RTD
NPE -0.42 0.09 -0.31 -0.16 0.03 0.67* -0.19 0.05 0.01 -0.01
PE 0.89** -0.52 -0.55 -0.35 -0.33 0.35 0.23 0.22 -0.46
EF -0.74* -0.51 -0.36 0.00 0.28 0.28 0.25 -0.51
ASF -0.36 -0.28 -0.14 -0.35 0.25 0.24 0.11
AIF 0.95** -0.16 0.33 -0.74** -0.67* 0.93**
C 0.13 0.08 -0.74* -0.65* 0.94**
N -0.12 0.08 0.09 0.18
AD -0.66* -0.72* 0.01
SRA 0.98** -0.73*
SRL -0.62
* p < 0.05,** p < 0.01
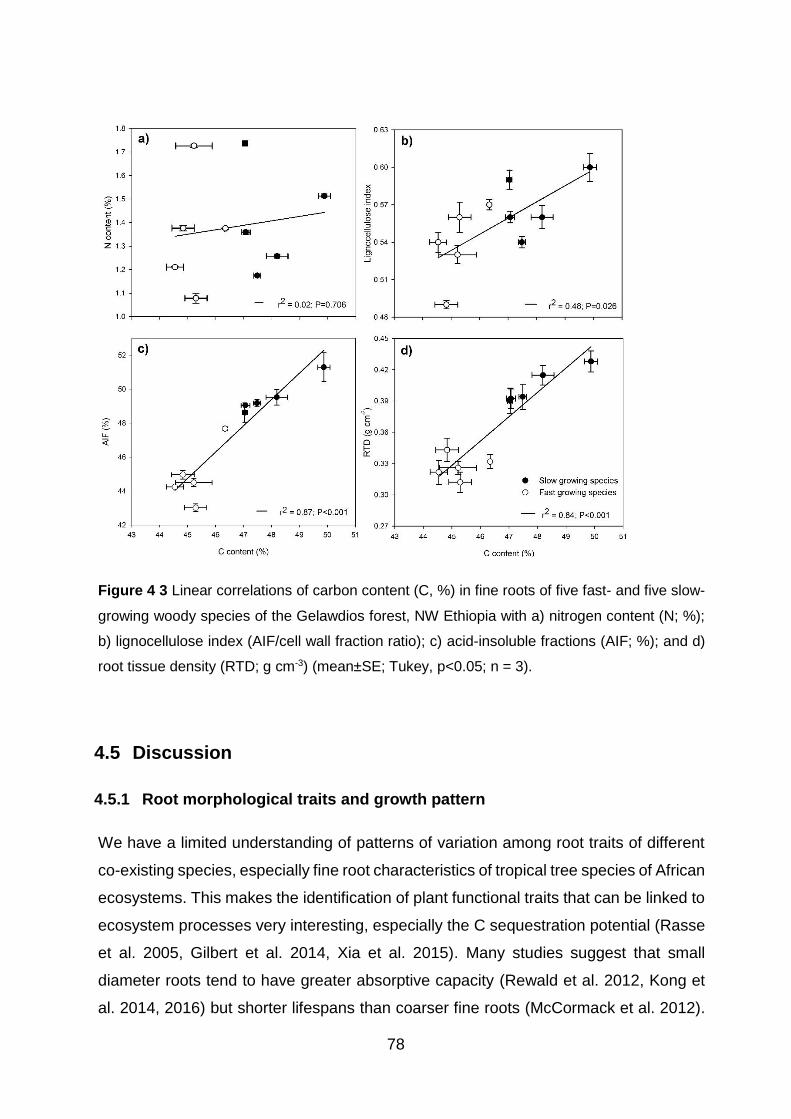
78
Figure 4 3 Linear correlations of carbon content (C, %) in fine roots of five fast- and five slow-
growing woody species of the Gelawdios forest, NW Ethiopia with a) nitrogen content (N; %);
b) lignocellulose index (AIF/cell wall fraction ratio); c) acid-insoluble fractions (AIF; %); and d)
root tissue density (RTD; g cm-3) (mean±SE; Tukey, p<0.05; n = 3).
4.5 Discussion
4.5.1 Root morphological traits and growth pattern
We have a limited understanding of patterns of variation among root traits of different
co-existing species, especially fine root characteristics of tropical tree species of African
ecosystems. This makes the identification of plant functional traits that can be linked to
ecosystem processes very interesting, especially the C sequestration potential (Rasse
et al. 2005, Gilbert et al. 2014, Xia et al. 2015). Many studies suggest that small
diameter roots tend to have greater absorptive capacity (Rewald et al. 2012, Kong et
al. 2014, 2016) but shorter lifespans than coarser fine roots (McCormack et al. 2012).

79
Such roots are also considered to play an important role in soil carbon input and nutrient
cycling. The average diameters of fine roots (0.53-0.76 mm) we recorded in Ethiopia
are in the range reported for tree species of other tropical ecosystems (0.52-1.4 mm;
Collins et al. 2016), but are considerably larger than those frequently reported for trees
in temperate forests (0.24-0.54 mm; Gu et al. 2014) and boreal forests (0.31-0.47;
Ostonen et al. 2013). For example, Pinno et al. (2010) reported – for roots of Populus
tremuloides in boreal forests – that 97% of the total root length is <1 mm diameter.
Other studies yielded similar findings: roots less than 0.5 mm in diameter accounted for
89% of the total root length in Prunus avium (Baddeley and Watson 2005) and 75% for
nine North American tree species (Pregitzer et al. 2002). In our study, about 50% of the
total fine root length was below 0.5 mm, and more than 80% of the total length were
accounted for by root segments <1 mm in diameter (Fig 4.1; Table 4.1). Species such
as Maytenus arbutifolia, Combretum collinum, and Allophylus abyssinicus are the
thinnest (90% of total root length <1 mm diameter), while Podocarpus falcatus,
Ekebergia capensis, and Teclea nobilis had the thickest values (Table 4.1). Overall, the
vast majority of fine root length was below 1 mm in diameter for all species (Fig. 4.1).
It has been shown that coarser fine roots show secondary growth, as evidenced by the
highest root tissue density (RTD), and have a lower specific root area (SRA) than finer
roots (Silver and Miya 2001, Rewald et al. 2014). In our study, fine root diameter was
negatively correlated with SRA (rpearson = -0.66; P<0.05) and SRL (rpearson = -0.72;
P<0.05; Table 4.5). Basile et al. (2007) also reported a similar negative correlation.
Comparing morphological traits with growth rates showed that average RTD is lower in
fast-growing species, whereas SRA and SRL are higher in slow-growing species. Some
of the fast-growing species such as Calpurnia aurea and Maytenus arbutifolia had much
higher SRA and SRL than the slow-growing species Ekebergia capensis and Teclea
nobilis, which exhibited the lowest SRA and SRL values (Table 4.1). Similar results
have been reported for other species, where very fine roots of fast-growing species had
much higher SRL (Pregitzer et al. 1997, Basile et al. 2007). Similarly, the RTD values
(0.31 - 0.41 g cm-3) are in the range reported for tree species of other tropical
ecosystems (0.2 - 0.6 g cm-3; Collins et al. 2016) and for temperate trees (0.32-0.83 g
cm-3; McCormack et al. 2012), but they are greater than values previously found in
other temperate forest ecosystems (Comas and Eissenstat 2004). Several authors
have suggested that a wide variety of climate and soil conditions such as temperature,

80
moisture, nutrient content, pH, and physical disturbance of the soil affect fine root
morphology (Pregitzer et al. 2002, Zobel et al. 2007, Ostonen et al. 2007). In co-existing
species of the same site, however, the morphological differences may reflect the
species’ economic spectrum and ecological strategies for resource capture under
competition (Wang et al. 2006, Collins et al. 2016, Valverde-Barrantes et al. 2016).
High SRA and SRL may facilitate faster growth and more rapid acquisition of soil
resources (Valverde-Barrantes et al. 2016). Given the apparent species-specific
differences in root morphology between co-existing species, many authors have
emphasized root morphological plasticity as an important adaptation mode to variable
growth conditions (Fransen et al. 1999, Sorgoná et al. 2007, Ostonen et al. 2013,
Gratani 2014). Based on the optimal foraging theory, Ostonen et al. (2007) identified
two main strategies of fine root adaptation to different regimes of nutrient supply: higher
C investment to increase the fine root biomass (and root length), or changing root
morphology to increased nutrient uptake efficiency through a higher specific root area.
Similar ideas have been suggested by Meinen et al. (2009) to explain differences in
root morphology between species. In relation to the litter quality index, SRA and SRL
decreased with increasing RTD (Table 4.5). The negative associations of SRA and SRL
with RTD suggest that the fast-growing species increase the total surface area of
absorbing roots for higher nutrient use efficiency and that their roots are less expensive
to construct per unit mass (Table 4.4). Similarly, Pregitzer et al. (2002) reported that
“infinitely fine” roots are the most efficient for nutrient acquisition per gram of C
expended to construct them. In contrast, slow-growing species tended to construct
more C-costly fine roots.
4.5.2 Root biochemistry and carbon cost implications for root litter quality
The fine roots of all ten species differed with respect to concentrations of C and N as
well as of chemical compounds. The C content ranged from 44% (Calpurnia aurea) to
50% (Chionanthus mildbraedii). The average C concentration for fast-growing species
was about 45% (Table 4.4). In 59 Panamanian rainforest species, C content ranged
from 42 to 52% (Martin and Thomas 2011) and 37 to 44% for 24 species in Europe
(Poorter and Bergkotte 1992). Carbon concentrations are often assumed to be about
50% of the dry mass; this value is widely used for below-ground C estimations (Gibbs
et al. 2007, Robinson 2007), but will overestimate total carbon stocks in fine roots.

81
Nitrogen contents also varied significantly in the fine roots of the 10 species. The
highest N content was determined in Teclea nobilis and Maytenus arbutifolia, which in
the former may be due to T. nobilis being a N2-fixing species (Orwa et al. 2009). The
N2-fixing status of Maytenus arbutifolia is unknown but its leaves have been shown to
contain high crude protein levels compared to 18 other species used for fodder
(Shenkute et al. 2012);16% of the crude protein in the leaves is nitrogen (Levey et al.
2000). Interestingly, however, no significance difference in N contents was found
between the fast- and slow-growing species groups (Table 4.3). This contradicts earlier
findings on seedlings (Comas et al. 2002) that N concentrations are higher in fast-
growing species. Root C:N ratios also played an important role in predicting patterns of
root decay, and these ratios are a valuable predictive tool in numerous studies of litter
decomposition at local, regional, and global scales (Silver and Miya 2001, Leppälammi-
Kujansuu et al. 2014, García-Palacios et al. 2016). The C:N ratios of roots in our study
ranged from 26 to 40, with significant differences between individual species but no
difference between growth rates. Theoretically, the optimum C:N ratio for microbial
growth, and thus decomposition, is approximately 25, but fungi and bacteria can
decompose substrates with much higher ratios (Reshi and Tyub 2007). According to
Reshi and Tyub (2007), substrates with C:N ratios of <20 decompose rapidly and NH4
is released through N-mineralization. Plant material with C:N ratios between 25-75 can
be regarded as intermediate values, and our data lie in this range. Litter with such C:N
ratio decompose quickly but N mineralization is often reduced by increased microbial
immobilization as well as protein complexation by polyphenols when the cells lyse
(Silver and Miya 2001). Roots with C:N ratios exceeding 75 are often much more
difficult to break down (Swift et al. 1979) due to greater amounts of structural woody
materials (Silver and Miya 2001).
The biochemical compositions of roots such as AIF, ASF, and EF vary with species,
and these variations determine the root litter quality and the quality of C input into soil
(Xia et al. 2015). In the present study, the AIF ranged from 43-51% (combined mean
of 47%; Table 4.2) and was consistently the highest C fraction. This value is consistent
with other studies, whose AIF averaged 49% (Hendricks et al. 2000) and 50% (Muller
et al. 1989). A recent biochemical study by Xia et al. (2015) also reported that fine roots
contained a 2.9 fold higher AIF content compared to the other fractions. Abiven et al.
(2005) also noticed large lignin-like fractions in roots of crop plants. The AIF is primarily

82
composed of highly reduced compounds such as suberin, cutin, and tannin-protein
complexes associated with lignin (Hendricks et al. 2000, Sun et al. 2013). These
compounds are thought to be highly recalcitrant and resistant to biochemical
degradation (Lorenz et al. 2007, Sun et al. 2013, Xia et al. 2015). Moreover, after
cellulose, lignin alone is one of the most abundant organic polymers in plants; the
content of lignin in wood is 20–40% of dry matter (Chen 2014). Lignin is a highly cross-
linked polymer that is resistance to chemical or biological attack; it provides mechanical
support in wood by reinforcing cell walls of xylem tissue and lignified sclerenchyma
fibres in vascular plants (Kögel-Knabner 2002, Chen 2014). Hydroxyls and many polar
groups are present in the lignin structure, resulting in strong intramolecular and
intermolecular hydrogen bonds and making the intrinsic lignin insoluble in any solvent
(Chen 2014). The presence of phenolic hydroxyl and carboxyl, however, makes the
lignin soluble in alkaline solutions (Goñi and Hedges 1992). The AIF was positively
correlated with C content (r2 = 0.87, P<0.001; Fig. 4.3c) and RTD (r2 = 0.84; P<0.001;
Fig. 4.3d) and, thus, this relationship may provide some insight into carbon investment
to fine roots. In our study, roots with a high lignin content tend to invest more carbon
per unit biomass. This can be explained by the higher glucose investment (Table 4.4)
and the higher C content within lignin compared to cellulose. For example, the
elemental composition of lignin from a red oak and yellow popular wood sample is 62
% C and 1 % N, (Jin et al. 2013). In comparison, cellulose is only 44 % C (Chen 2014).
The lignocellulose index, a ratio of AIF to the cell wall fraction (AIF+ASF), is higher in
slow-growing species, as is the total C content (Table 4.3). This suggests that both
chemical (AIF) and morphological (higher RTD) factors make the fine roots of slow-
growing species more expensive to construct in terms of C per unit mass. These
parameters (AIF, lignocellulose index, RTD) determine the decomposition dynamics
(Sun et al. 2013, Zhang and Wang 2015) and are a good indicators for root substrate
quality (Hendricks et al. 2000, Prieto et al. 2016). Input of recalcitrant materials from
the turnover of fine roots contributes to long-term CO2 sinks in soils (Xia et al. 2015).
In contrast to the AIF, the amounts of polar extractives (PE) and extractive fractions
(EF) were greater in fast-growing species. Solvent extractives are compounds of non-
structural substances, mostly low-molecular-mass compounds consisting of nonpolar
constituents such as alkaloids, fats, oils, waxes, and resins, as well as of polar
constituents such as nonstructural carbohydrates and water-soluble phenolics

83
(Pettersen 1984, Yang and Jaakkola 2011, Sun et al. 2013). Among the individual
species, Podocarpus falcatus had the highest extractive fraction (25%), Apodytes
dimidiata the lowest value (7%). Both Podocarpus falcatus and Apodytes dimidiata are
from the fast-growing species category (Table 4.2). In an investigation of 14 species
from Gelawdios forest for other chemical fractions, Podocarpus falcatus had the
greatest concentration of total phenols and condensed tannins in both roots (Tigabu
2016) and leaves (Habteyohannes 2016). Tannins are classified as hydrolyzable
tannins (esters of a sugar residues (usually D-glucose) with one or more polyphenol
carboxylic acids) and condensed tannins (polymers of flavonoids consisting mainly of
3-8 flavonoid units), and are categorized as extractives (Yang and Jaakkola 2011).
Some tannins, however, form a complex tannin-protein complex associated with the
AIF that cannot be extracted using neutral solvents (Sun et al. 2013). Moreover,
Podocarpus species are known to produce and store terpenoid resin and phenolic
resin, mainly induced by injury (Langenheim 2003). Nonetheless, the family has not
been analyzed chemically in detail and needs further investigation. Pettersen (1984)
suggested that solvent extractive materials constitute 4-10% of the dry weight of wood
of species in temperate climates and that the values may be as much as 20% of the
wood of tropical species. Preston et al. (2000), however, reported that nonpolar
extractives and water-soluble extractives together make up 40-50% of total dry litter
mass in 37 species of trees in Canadian forests. Since extractives are soluble in neutral
solvents and do not contribute to the cell wall structure (Pettersen 1984), they are
considered as labile compounds that degrade easily compared to other fractions. Sun
et al. (2013) confirmed that the extractives disappeared rapidly at the initial stages of
decomposition.
Cellulose is the most abundant organic polymer, usually accounting for 35–50% of dry
weight (Chen 2014). In most conditions, the cellulose is wrapped in hemicellulose,
which itself accounts for 20–35% dry matter (Chen 2014). In our study, the ASF that
contains mainly cellulose and hemicellulose showed no variation between functional
groups, but significant variation existed between individual species. Among the ten
studied species, Podocarpus falcatus showed the least ASF (30%), Apodytes dimidiata
the highest fraction (47%). Tigabu (2016) also found high concentrations of cellulose
and hemicellulose (41%) in fine roots of Apodytes dimidiata. These two polymers are
made of linear chains of β-1,4-linked glucose units, ranging from hundreds to ten

84
thousands of units (Kögel-Knabner 2002), and are insoluble in water, but soluble in
acidic and alkaline solutions at normal temperatures (Li et al. 2014). Due to the
chemical bonding and C content (44% in cellulose), ASF is the second fastest
decomposing fraction after the solvent extractives (Sun et al. 2013).
4.6 Conclusion
The morphological parameters of fine roots are quite variable among ten tropical
species, but reflect the general difference between fast-and slow-growing species.
Generally, fast-growing species have higher SRA and SRL but lower RTD, which are
characteristics assumed to support high rates of nutrient acquisition. Our results
provide evidence that RTD is positively correlated with AIF, C content, and C
construction cost, and is consistently higher in slow-growing species. The extractive
fractions and ASF of fine roots appeared to be unrelated to any root morphological
characteristics.

85
5 Litter production, chemistry, and turnover in a pristine
forest ecosystem in the Ethiopian highland
5.1 Abstract
Amount of plant litter and its decomposition rate determine the rate of carbon deposition
into the soil. The objective of this study is to determine the carbon input through above-
and below-ground litter production and turnover in the Ethiopian highland forests. The
litterfall and fine root production were studied for one year using litter traps and
sequential coring. Four dominant indigenous tree species were selected for chemical
analysis and the decomposition rate experiment using standard litterbag technique
from June 2015 to June 2016. Results showed that annual total litterfall production was
1090 g m-2 y-1 while fine root production was 723 g m-2 yr-1. In the decomposition
analysis, the litter mass in the litterbag declined exponentially with time for all species.
The annual decay constant (k) varied from 1.6 to 3.2 yr-1 for leaves and 1.3 to 1.8 yr-1
for fine roots indicating leaves decompose more rapidly than fine roots. Leaf litter
exhibited higher concentrations of extractive fractions (EF) and acid-soluble fractions
(ASF) that contain mainly labile carbons such as carbohydrates and cellulose than fine
roots. Fine roots contained consistently greater acid-insoluble fraction (AIF) mainly
lignin and lignin-suberin complexes than leaf litters. Thus, fine roots exhibit about 1.5
times higher recalcitrant carbon input into the soil than leaf litters. Decomposition rates
of litters were determined by EF, ASF, and AIF whereas litter quality indices such N,
C:N ratio, AIF:N ratio were poor predictors of decay in this experiment due to N is
affected by N2-fixing symbionts, N resorption, and leaching.
Keywords: litterfall production, fine roots, carbon input, litter chemistry, decomposition
rate, litterbag.

86
5.2 Introduction
Changes in the amount of soil organic carbon in the soil are the result of differences
between additions (litter inputs from above- and below-ground) and losses through
mineralization (Gaudinski et al. 2001, Conant and et.al. 2011) and erosion (Chapter 2;
Mey et al. 2015, Li et al. 2016). Above-ground fine litterfall includes leaves, flowers,
fruits, bark and twigs (Jia et al. 2016). Of the below-ground inputs, fine roots (Freschet
et al. 2013) are the major pathways of organic carbon input in to the soil through the
decomposition cycle (Berg 2000, Freschet et al. 2013, Macinnis-Ng and
Schwendenmann 2015). The quantity and quality of litter materials play an important
role in the formation and accumulation of soil organic matter and are key links in
biogeochemical cycles that connect above- and below-ground processes in terrestrial
ecosystems (Kögel-Knabner 2002, Freschet et al. 2013, Becker et al. 2015).
Previous investigations on litterfall and fine root production and their turnover have
been focused on temperate and Mediterranean climates (Guo and Sims 1999,
Celentano et al. 2011, Salete Capellesso et al. 2016). While some models have been
proposed to predict litter production (Adair et al. 2008, Cotrufo et al. 2010, Hararuk
and Luo 2014), the amount and quality of litterfall and fine root turnover varies
considerably between regions (Macinnis-Ng and Schwendenmann 2015), season
(Zhang et al. 2014), and tree species composition (Becker et al. 2015). Information on
the influence of species richness, litter composition, and the biochemical quality of
litter on carbon cycling of tropical regions remains scarce especially in the Ethiopian
highlands. Given the important role of litter inputs and decomposition to SOC storage
and ecosystem functioning, it is thus important to accurately quantify and characterize
carbon deposition through above- and below-ground litters, litter chemistry, and decay
rates more in a species rich tropical forest ecosystems (Guo and Sims 1999, Freschet
et al. 2013, Salete Capellesso et al. 2016).
The accumulation of organic detritus in the organic layer of forests are controlled by the
rate of decomposition of the plant material which is influenced by variability in litter
quality, soil moisture and temperature, and the kinds of microflora and fauna present
(Salomé et al. 2010, Yeong et al. 2016). The carbon quality of substrates may be the
predominant chemical control over decomposition (Chapin et al. 2011). Quality refers
to chemical characteristics of the litter that influence the susceptibility of litter to

87
decomposition (Karberg et al. 2008). Some studies reported that there is a 5-fold to 10-
fold range in decomposition rate of litter in a given climate, due to differences in
substrate quality (Chapin et al. 2011). Some authors reported that leaves decompose
more rapidly than woody components and deciduous leaves decompose more rapidly
than evergreen leaves and these differences in decomposition rates are a
consequence of their chemical composition (Cornelissen 1996, Chapin et al. 2011,
Prescott et al. 2011). The chemical compounds present can be categorized roughly as
labile compounds (e.g. fats, oils, waxes, nonstructural carbohydrates, and
polyphenols), moderately degradable compounds (cellulose and hemicellulose), and
highly recalcitrant compounds tissues such as lignin, suberin, cutin, and tannin-protein
complexes (Ryan et al. 1990, Sun et al. 2013, Xia et al. 2015). Litter containing high
concentrations of labile compounds (e.g. carbohydrates) tends to decompose rapidly
because these compounds can be readily metabolized by soil microorganisms or are
leached easily (Karberg et al. 2008). Labile structural compounds such as cellulose are
more stable than carbohydrates but are quickly metabolized in contrast to recalcitrant
structural compounds such as lignin because of their aromatic rings and irregular
structure (Karberg et al. 2008, Xia et al. 2015). The C:N ratio, lignin:N ratio, and
lignocellulose index have been frequently used as an index of litter quality and are good
predictors of the decomposition rate – lower values of these indices generally indicate
faster rates of decomposition (Preston et al. 2000, Girisha 2001, Moorhead et al. 2014,
Xia et al. 2015). Therefore, species difference in decomposition rate should depend on
species- and tissue-specific litter chemistry properties (Scherer-Lorenzen et al. 2007).
In this study three hypothesis are proposed to explain how initial litter quality influences
litter decomposition and carbon release. The first hypothesis suggests that leaves
decompose more rapidly than fine roots due to their chemical composition. Hence, we
expect that litter quality indices such as C:N ratio, lignin:N ratio and lignocellulose index
are lower in the leaves than fine roots. The second hypothesis suggests that species
having less acid-insoluble fractions (less lignified tissue) decompose faster than
species containing more labile compounds. The third hypothesis suggest that litter
mixtures of varying species enhance litter decomposition rates – assuming the
occurrence of highly degradable litter favors the breakdown of more recalcitrant litter in
the mixtures. Therefore, the objectives of this study were: (i) to determine the seasonal
and annual litterfall, fine root turnover, and C flux into the soil of a natural mixed forest,

88
and (ii) to investigate the in-situ decomposition rate and residence time of the detritus
material and relate to species- and tissue-specific initial litter chemistry.
5.3 Materials and methods
5.3.1 Description of the study site
This study was conducted in the remnant Afromontane forest at Gelawdios in the
Amhara National Regional State, north central part of Ethiopia. Gelawdios (11°38’25’’N,
37°48’55’’E) is located east of Lake Tana at an altitude of 2500 m above sea level.
While Ethiopia is located in the tropics, the climate of the study area is temperate with
dry winter and warm summer (Cwb) according to the Köppen-Geiger climate
classification (Peel et al. 2007). The mean annual precipitation in the area is 1200 mm
with the main rainy season from June to September and with low-intensity precipitation
from March to May (Wassie et al. 2009). The annual mean air temperature is 19°C
(Wassie et al. 2009). The soils are classified as Cambisols. The soil physical and
chemical characteristics of the site are shown in Table 4.1. The Afromontane Gelawdios
forest is a small, isolated, but pristine forest fragments (‘church forest’) covering an area
of about 100 ha in the otherwise almost completely deforested Ethiopian Highlands
(Wassie et al. 2009, Aerts et al. 2016). The dominant woody species in the forest are
Allophylus abyssinicus (Hochst.) Radlk., Apodytes dimidiata E. Mey ex. Arn., Calpurnia
aurea (Ait.) Benth., Chionanthus mildbraedii (Gilg & Schellenb.) Stearn, Combretum
collinum Fresen., Dovyalis abyssinica (A. Rich.) Warb., Ekebergia capensis (Sparm.),
Maytenus arbutifolia (A. Rich.) Wilczek, Podocarpus falcatus (Thunb.) Mirb., and
Teclea nobilis (Del.).
Table 5 1 Soil physical and chemical properties at the Gelawdios forest. Values of carbon (C),
nitrogen (N), C:N ratio, and pH are mean±SE.
Soil depth (cm)
Sand-Silt-Clay (%)
C (%) N (%) C:N ratio
pH (CaCl2)
0-10 9-39-52 11.8±0.9 1.09±0.07 10.8±0.3 5.94±0.11
10-20 5-34-61 7.2±0.7 0.70±0.07 10.3±0.2 5.55±0.10
20-30 11-46-43 4.6±0.3 0.42±0.03 11.0±0.3 5.40±0.08
30-50 9-63-28 3.5±0.3 0.30±0.02 11.7±0.4 5.35±0.07

89
5.3.2 Litterfall collection and fine root biomass determination
Litterfall was collected weekly from July 2014 to June 2015. Ten litter traps were
systematically placed in 100 m distance between each other along a transect line.
Samples were collected on a 0.5 m X 0.5 m wooden frame furnished with a 1 mm
polyamide mesh (Franz Eckert GmbH, Waldkirch, Germany); positioned 50 cm above
the ground. At the end of each month, the weekly collected litter were combined and
sorted into leaves, branches and twigs, reproductive organs (flowers, fruits, and seed),
and miscellaneous material (unidentified plant parts, mosses). The collected litter was
dried at 70°C to constant weight before weighing to the nearest 0.01g. Seasonal litter
productivity (g m-2) for the winter, spring, summer, and autumn seasons was calculated.
Winter - December, January and February are the dry months with frost in morning.
Spring - March, April and May are the autumn season in Ethiopia with occasional
showers. May is the hottest month in Ethiopia. Summer - June, July and August are the
heavy rainfall months; autumn - September, October and November are the spring
season in Ethiopia, sometimes known as the harvest season.
Methods for retrieving fine root biomass (live roots) and necromass (dead roots) stock
as well as annual root production and turnover can be found in Chapter 3. The fine
roots stock and production estimated using sequential coring with Decision Matrix
calculation technique (Chapter 3) were used for comparison of C input with leaf litter in
this chapter.
5.3.3 Litter (leaf and root) decomposition using litterbag technique
Based on dominance, four species namely, Allophylus abyssinicus, Chionanthus
mildbraedii, Combretum collinum, and Teclea nobilis were selected for chemical
analysis through sequential extraction and decomposition analysis using litterbag
technique. Since there was not enough material from the collected litter, samples of
litters for decomposition analysis and biochemical fraction analysis were collected from
fallen leaves on the forest floor. Root samples for decomposition analysis were taken
from the selected tree species by strictly tracking coarse roots from the tree base
(Rewald et al. 2012). For comparison purpose, fine roots (<2 mm diameter) and coarse
roots (2-5 mm diameter) were separately included in the litterbag during the
decomposition experiment.

90
Both leaf and root decomposition was studied for a period of 1 year from June 2015 to
June 2016 using standard litter-bag techniques in a time series (Scherer-Lorenzen et
al. 2007, Karberg et al. 2008, Certini et al. 2015). The air dried litter was packed within
litter bags of polyamide mesh (Franz Eckert GmbH, Waldkirch, Germany); 1.0 mm
mesh was used on the bottom to reduce loss of small fragments and 2 mm was used
on top to allow macro-invertebrates to enter the bag (Karberg et al. 2008). The sizes of
the mesh bags were 10 cm X 10 cm for roots and 10 cm X 15 cm for leaves – adjusting
for the entered size of litter.
Four grams of either leaf or root litter materials from the four selected tree species
(Allophylus abyssinicus, Chionanthus mildbraedii, Combretum collinum, and Teclea
nobilis) were placed inside in monospecific litterbags. For leaves, a combination of two
species (2 g per species) was placed inside mixed-species litterbags. The six realized
combinations were Allophylus abyssinicus with either Chionanthus mildbraedii,
Combretum collinum, or Teclea nobilis, and Chionanthus mildbraedii with either
Combretum collinum or Teclea nobilis, and Combretum collinum with Teclea nobilis. In
four species combination (Allophylus abyssinicus, Chionanthus mildbraedii,
Combretum collinum, and Teclea nobilis), 1 g per species or 25% each were placed for
both leaves and roots. The combinations were assembled to test whether the decay of
specific litter is affected by the presence of other species, e.g. due to its palatability or
inhibitory effect on microorganisms. There were not enough materials available to
prepare all two-mixed species combinations for fine and coarse roots. Litterbags of
each treatment were prepared at five replications; retrieval took place at months of 0.5,
1, 2, 3, 6, 12 for leaves (yielding 360 samples) and months of 1, 2, 3, 6, and 12 for roots
(300 samples). Bags were placed at 45° in June 2015 at a soil depth of 3-10 cm (to
prevent their discovery and loss). At the time of retrieval, materials were carefully
brushed and cleaned to remove any attached particles that are not the original material.
The retrieved litter was carefully transferred to clean paper bags, oven-dried until
constant weight, and weighed to the nearest 0.01 g.
5.3.4 Chemical analysis
Leaf samples from forest floor of fallen leaves and root samples from each species
were transported to Vienna, for initial litter chemistry analysis. Both leaf and fine root
litter samples were dried at 70°C until constant weight and ground to powder using a

91
ball mill (Fritsch Pulverisette 5, Idar-Oberstein, Germany). Due to limited availability of
sample materials, three technical replicates per species were analyzed for total carbon
and nitrogen concentration on a CN elemental analyzer (Truspec CNS LECO, St.
Joseph, USA). Three other subsamples per species were analyzed for carbon fractions
using procedures adapted from Ryan et al. (1990), Sun et al. (2013) and Kong et al.
(2016). In brief, root carbon fractions, including non-polar extractives (fats, oil, wax),
polar extractives (carbohydrates, polyphenols), acid-soluble structural components
(cellulose, hemicellulose), acid-insoluble structural components (mainly lignin, suberin),
and ash, were assessed using a series of digestion technique (Ryan et al. 1990). Detail
procedure of chemical analysis for fine roots can be found in Chapter 4. The same
procedure was applied here for leaf litters.
5.3.5 Calculations of decomposition parameters, C and N input into the soil
Rates of litter materials decomposition were simulated by a single negative exponential
decay equation according to Olson, (1963) and Salete Capellesso et al. (2016) as:
Mt = M0*e-kt (1)
where Mt denotes litters mass at time t, M0 the initial mass and k the decomposition
rate. The rate was used to estimate the mean time needed for the litter fraction to
decompose, in days. Olson (1963) also develop a model to predict the time required
from estimated k values to reach half of the organic matter decomposition (T(0.5) =
ln(0.5)/(-k) = 0.693/k), the time required for attaining 95% mass loss (T(95) = 3/K), and
the time needed to reach 99% mass loss of the final level (T(99) = 5/k). Annual potential
C and N input (expressed in g m-2 yr-1) through plant litters were computed by
multiplying annual leaf litter or fine root mass values with its corresponding C and N
concentrations assuming that there was no translocation during leaf or root senescence
(Xia et al. 2015).
5.3.6 Statistical analysis
Statistical differences of litter production, chemical composition, and decomposition
rates between species and treatment differences were analyzed by using a one-way
ANOVA. If significant differences were found, multiple comparisons were carried out
based on Tukey’s HSD test. Assumptions of normality and homogeneous variance
were examined by Shapiro-Wilk’s and Levene’s test respectively before analysis. Data
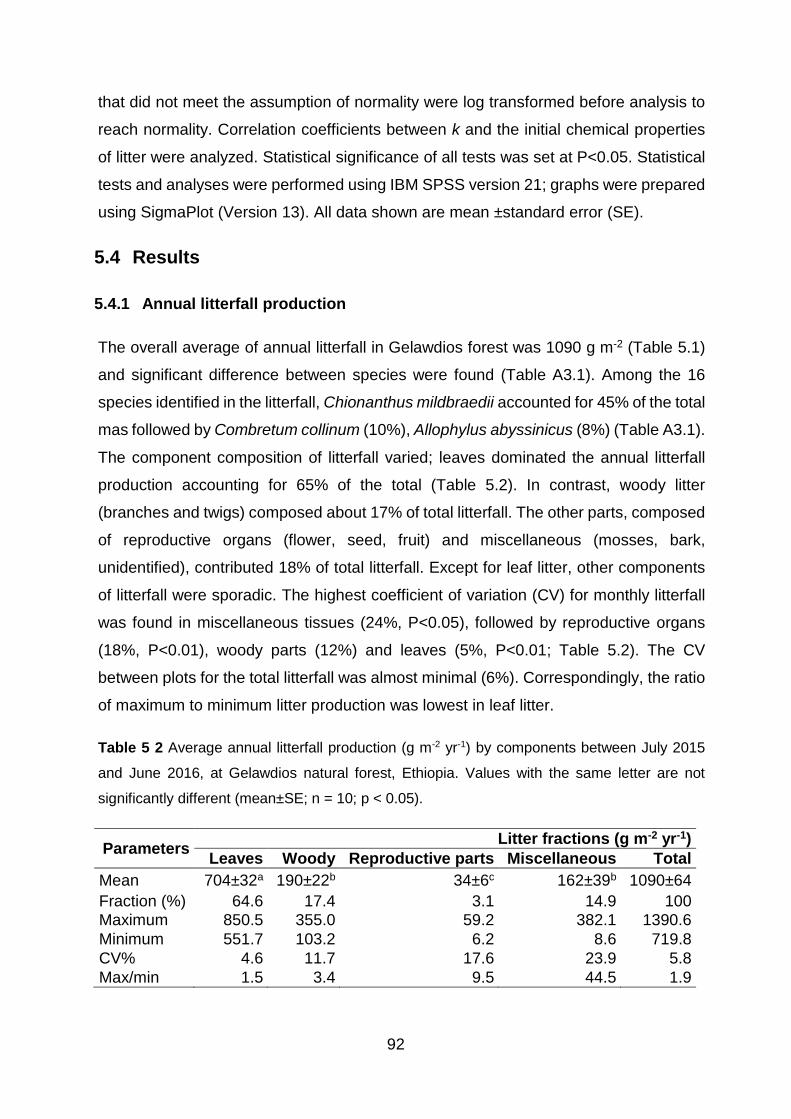
92
that did not meet the assumption of normality were log transformed before analysis to
reach normality. Correlation coefficients between k and the initial chemical properties
of litter were analyzed. Statistical significance of all tests was set at P<0.05. Statistical
tests and analyses were performed using IBM SPSS version 21; graphs were prepared
using SigmaPlot (Version 13). All data shown are mean ±standard error (SE).
5.4 Results
5.4.1 Annual litterfall production
The overall average of annual litterfall in Gelawdios forest was 1090 g m-2 (Table 5.1)
and significant difference between species were found (Table A3.1). Among the 16
species identified in the litterfall, Chionanthus mildbraedii accounted for 45% of the total
mas followed by Combretum collinum (10%), Allophylus abyssinicus (8%) (Table A3.1).
The component composition of litterfall varied; leaves dominated the annual litterfall
production accounting for 65% of the total (Table 5.2). In contrast, woody litter
(branches and twigs) composed about 17% of total litterfall. The other parts, composed
of reproductive organs (flower, seed, fruit) and miscellaneous (mosses, bark,
unidentified), contributed 18% of total litterfall. Except for leaf litter, other components
of litterfall were sporadic. The highest coefficient of variation (CV) for monthly litterfall
was found in miscellaneous tissues (24%, P<0.05), followed by reproductive organs
(18%, P<0.01), woody parts (12%) and leaves (5%, P<0.01; Table 5.2). The CV
between plots for the total litterfall was almost minimal (6%). Correspondingly, the ratio
of maximum to minimum litter production was lowest in leaf litter.
Table 5 2 Average annual litterfall production (g m-2 yr-1) by components between July 2015
and June 2016, at Gelawdios natural forest, Ethiopia. Values with the same letter are not
significantly different (mean±SE; n = 10; p < 0.05).
Parameters Litter fractions (g m-2 yr-1)
Leaves Woody Reproductive parts Miscellaneous Total
Mean 704±32a 190±22b 34±6c 162±39b 1090±64
Fraction (%) 64.6 17.4 3.1 14.9 100
Maximum 850.5 355.0 59.2 382.1 1390.6
Minimum 551.7 103.2 6.2 8.6 719.8
CV% 4.6 11.7 17.6 23.9 5.8
Max/min 1.5 3.4 9.5 44.5 1.9
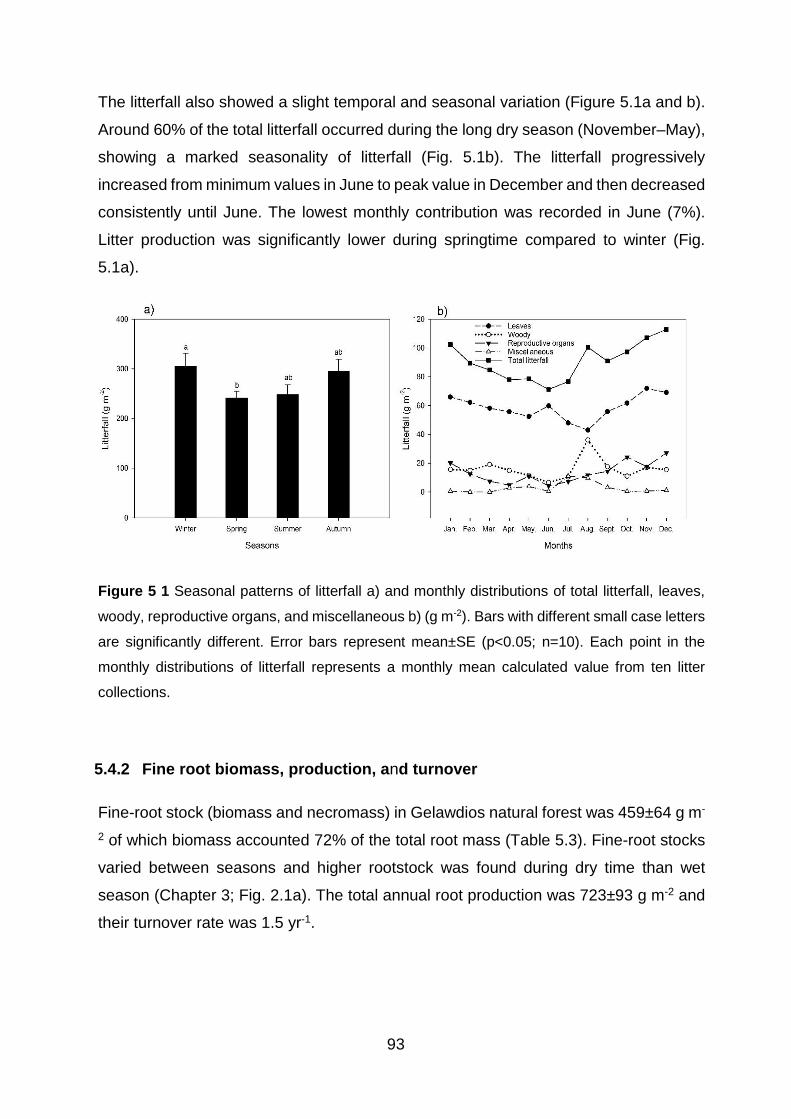
93
The litterfall also showed a slight temporal and seasonal variation (Figure 5.1a and b).
Around 60% of the total litterfall occurred during the long dry season (November–May),
showing a marked seasonality of litterfall (Fig. 5.1b). The litterfall progressively
increased from minimum values in June to peak value in December and then decreased
consistently until June. The lowest monthly contribution was recorded in June (7%).
Litter production was significantly lower during springtime compared to winter (Fig.
5.1a).
Figure 5 1 Seasonal patterns of litterfall a) and monthly distributions of total litterfall, leaves,
woody, reproductive organs, and miscellaneous b) (g m-2). Bars with different small case letters
are significantly different. Error bars represent mean±SE (p<0.05; n=10). Each point in the
monthly distributions of litterfall represents a monthly mean calculated value from ten litter
collections.
5.4.2 Fine root biomass, production, and turnover
Fine-root stock (biomass and necromass) in Gelawdios natural forest was 459±64 g m-
2 of which biomass accounted 72% of the total root mass (Table 5.3). Fine-root stocks
varied between seasons and higher rootstock was found during dry time than wet
season (Chapter 3; Fig. 2.1a). The total annual root production was 723±93 g m-2 and
their turnover rate was 1.5 yr-1.
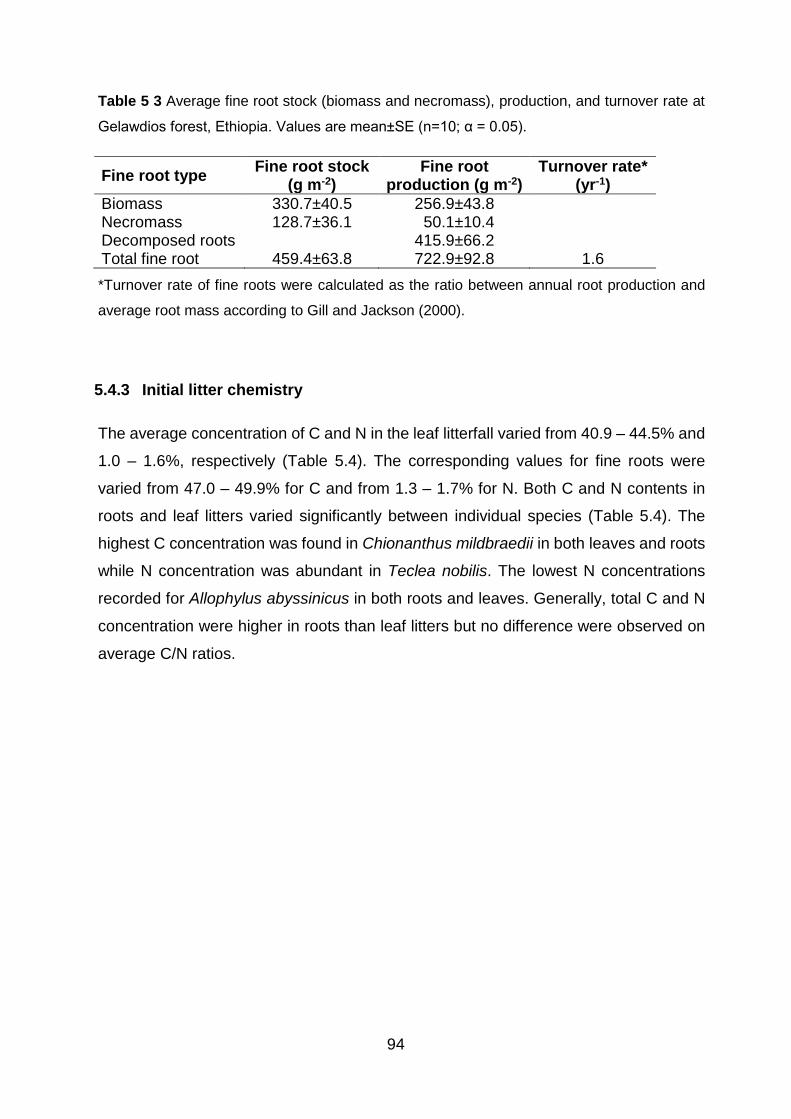
94
Table 5 3 Average fine root stock (biomass and necromass), production, and turnover rate at
Gelawdios forest, Ethiopia. Values are mean±SE (n=10; α = 0.05).
Fine root type Fine root stock
(g m-2) Fine root
production (g m-2) Turnover rate*
(yr-1)
Biomass 330.7±40.5 256.9±43.8 Necromass 128.7±36.1 50.1±10.4 Decomposed roots 415.9±66.2 Total fine root 459.4±63.8 722.9±92.8 1.6
*Turnover rate of fine roots were calculated as the ratio between annual root production and
average root mass according to Gill and Jackson (2000).
5.4.3 Initial litter chemistry
The average concentration of C and N in the leaf litterfall varied from 40.9 – 44.5% and
1.0 – 1.6%, respectively (Table 5.4). The corresponding values for fine roots were
varied from 47.0 – 49.9% for C and from 1.3 – 1.7% for N. Both C and N contents in
roots and leaf litters varied significantly between individual species (Table 5.4). The
highest C concentration was found in Chionanthus mildbraedii in both leaves and roots
while N concentration was abundant in Teclea nobilis. The lowest N concentrations
recorded for Allophylus abyssinicus in both roots and leaves. Generally, total C and N
concentration were higher in roots than leaf litters but no difference were observed on
average C/N ratios.
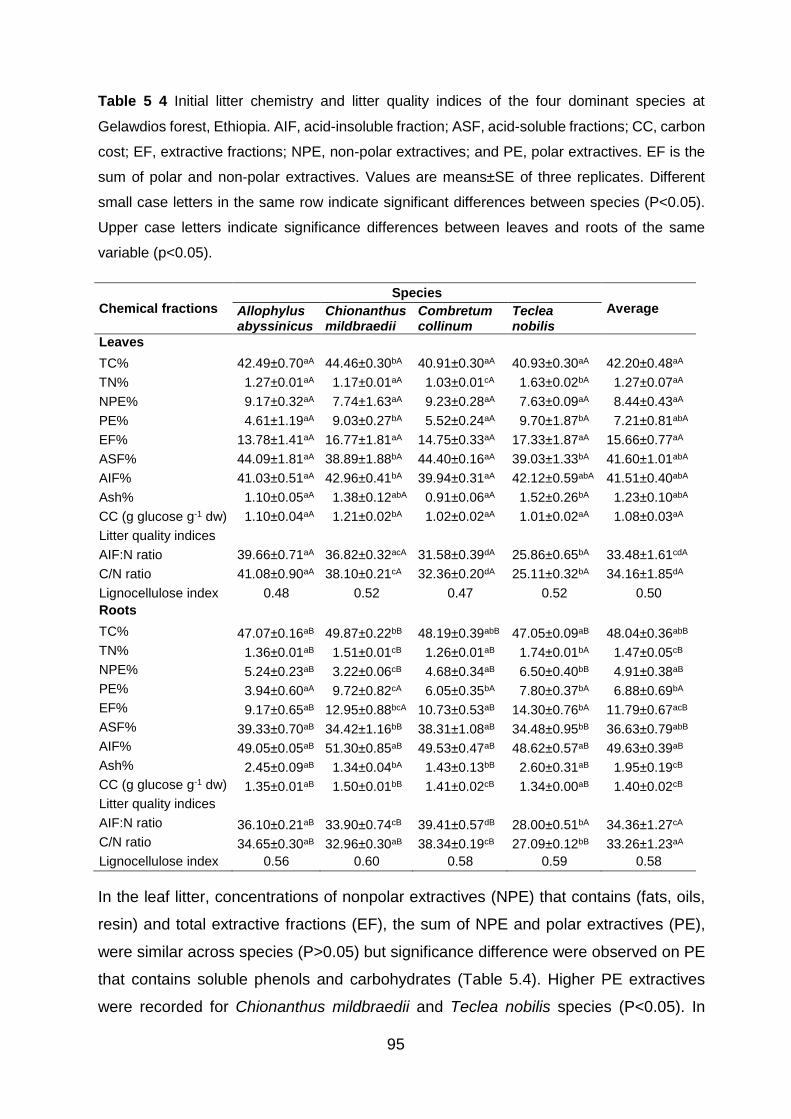
95
Table 5 4 Initial litter chemistry and litter quality indices of the four dominant species at
Gelawdios forest, Ethiopia. AIF, acid-insoluble fraction; ASF, acid-soluble fractions; CC, carbon
cost; EF, extractive fractions; NPE, non-polar extractives; and PE, polar extractives. EF is the
sum of polar and non-polar extractives. Values are means±SE of three replicates. Different
small case letters in the same row indicate significant differences between species (P<0.05).
Upper case letters indicate significance differences between leaves and roots of the same
variable (p<0.05).
Chemical fractions
Species
Average Allophylus abyssinicus
Chionanthus mildbraedii
Combretum collinum
Teclea nobilis
Leaves
TC% 42.49±0.70aA 44.46±0.30bA 40.91±0.30aA 40.93±0.30aA 42.20±0.48aA
TN% 1.27±0.01aA 1.17±0.01aA 1.03±0.01cA 1.63±0.02bA 1.27±0.07aA
NPE% 9.17±0.32aA 7.74±1.63aA 9.23±0.28aA 7.63±0.09aA 8.44±0.43aA
PE% 4.61±1.19aA 9.03±0.27bA 5.52±0.24aA 9.70±1.87bA 7.21±0.81abA
EF% 13.78±1.41aA 16.77±1.81aA 14.75±0.33aA 17.33±1.87aA 15.66±0.77aA
ASF% 44.09±1.81aA 38.89±1.88bA 44.40±0.16aA 39.03±1.33bA 41.60±1.01abA
AIF% 41.03±0.51aA 42.96±0.41bA 39.94±0.31aA 42.12±0.59abA 41.51±0.40abA
Ash% 1.10±0.05aA 1.38±0.12abA 0.91±0.06aA 1.52±0.26bA 1.23±0.10abA
CC (g glucose g-1 dw) 1.10±0.04aA 1.21±0.02bA 1.02±0.02aA 1.01±0.02aA 1.08±0.03aA
Litter quality indices
AIF:N ratio 39.66±0.71aA 36.82±0.32acA 31.58±0.39dA 25.86±0.65bA 33.48±1.61cdA
C/N ratio 41.08±0.90aA 38.10±0.21cA 32.36±0.20dA 25.11±0.32bA 34.16±1.85dA
Lignocellulose index 0.48 0.52 0.47 0.52 0.50
Roots
TC% 47.07±0.16aB 49.87±0.22bB 48.19±0.39abB 47.05±0.09aB 48.04±0.36abB
TN% 1.36±0.01aB 1.51±0.01cB 1.26±0.01aB 1.74±0.01bA 1.47±0.05cB
NPE% 5.24±0.23aB 3.22±0.06cB 4.68±0.34aB 6.50±0.40bB 4.91±0.38aB
PE% 3.94±0.60aA 9.72±0.82cA 6.05±0.35bA 7.80±0.37bA 6.88±0.69bA
EF% 9.17±0.65aB 12.95±0.88bcA 10.73±0.53aB 14.30±0.76bA 11.79±0.67acB
ASF% 39.33±0.70aB 34.42±1.16bB 38.31±1.08aB 34.48±0.95bB 36.63±0.79abB
AIF% 49.05±0.05aB 51.30±0.85aB 49.53±0.47aB 48.62±0.57aB 49.63±0.39aB
Ash% 2.45±0.09aB 1.34±0.04bA 1.43±0.13bB 2.60±0.31aB 1.95±0.19cB
CC (g glucose g-1 dw) 1.35±0.01aB 1.50±0.01bB 1.41±0.02cB 1.34±0.00aB 1.40±0.02cB
Litter quality indices
AIF:N ratio 36.10±0.21aB 33.90±0.74cB 39.41±0.57dB 28.00±0.51bA 34.36±1.27cA
C/N ratio 34.65±0.30aB 32.96±0.30aB 38.34±0.19cB 27.09±0.12bB 33.26±1.23aA
Lignocellulose index 0.56 0.60 0.58 0.59 0.58
In the leaf litter, concentrations of nonpolar extractives (NPE) that contains (fats, oils,
resin) and total extractive fractions (EF), the sum of NPE and polar extractives (PE),
were similar across species (P>0.05) but significance difference were observed on PE
that contains soluble phenols and carbohydrates (Table 5.4). Higher PE extractives
were recorded for Chionanthus mildbraedii and Teclea nobilis species (P<0.05). In

96
roots, Chionanthus mildbraedii species had the lowest NPE (P<0.05) but significantly
higher with PE than other species (P<0.05). Among all species, Combretum collinum
fine roots had the lowest PE (p<0.05). Overall, leaf litter exhibited greater
concentrations of EF and acid-soluble fractions (ASF) than fine roots (Table 5.4). By
contrast, fine roots contained consistently greater acid-insoluble fraction (AIF) than leaf
litters (P<0.001). The cell wall fractions (the sum of ASF and AIF) in root tissues ranged
from 85 to 91% of the dry matter whereas in leaves it ranged from 82 to 86%. Generally,
leaves and roots of the two deciduous species (Combretum collinum and Allophylus
abyssinicus) had higher ASF, and low AIF (lignin content) in leaves only compared to
evergreen species (Teclea nobilis and Chionanthus mildbraedii). Ash contents also
varied between species and were higher in roots than leaves except for Chionanthus
mildbraedii where no difference was found. The lignocellulose index, the proportion of
AIF to cell wall fraction, was greater in roots (58%) than leaves (50%). The AIF:N ratio
had no difference between roots and leaves. Large variation of carbon cost (the amount
of glucoses needed to produce one gram biomass) was found among plant species
and in above- versus below-ground biomass. Among the four species, carbon cost is
significantly higher in Chionanthus mildbraedii species both in leaves and in roots than
other species (P<0.05; Table 5.4). Generally, the carbon investment was significantly
higher in roots than leaves for all species (P<0.05).
5.4.4 Litter decay measurement and turnover rate
The residual litter mass (both roots and leaves) in the bags at each sampling time (in
months) declined exponentially for all species (Fig. 5.3a-d). During the first 3 months,
the weight of remaining litter mass reduced minimal but it was reduced sharply in the
3-6 months time intervals. Between 6-12 months, the weight loss due to microbial
decomposition was minimal again forming nearly a plateau. The mass loss of litter
varied between species (Table A3.2). The decay rate coefficient (k) and residence time
of decomposing materials are presented in Table 5.5. The annual mean decomposition
quotient (k) varied from 1.6±0.1 yr-1 for Chionanthus mildbraedii leaves to 3.1±0.9 yr-1
for Combretum collinum leaves. For example, at 3-months, mass loss of leaf litter was
greater for deciduous species Allophylus abyssinicus (55%) and Combretum collinum
(56%) than for evergreen species (Chionanthus mildbraedii (37%) and Teclea nobilis
(40%)). At the end of the year, more than 90% of the leaf biomass of Combretum
collinum was decomposed (Table 5.5). Generally, mass loss was average when litters
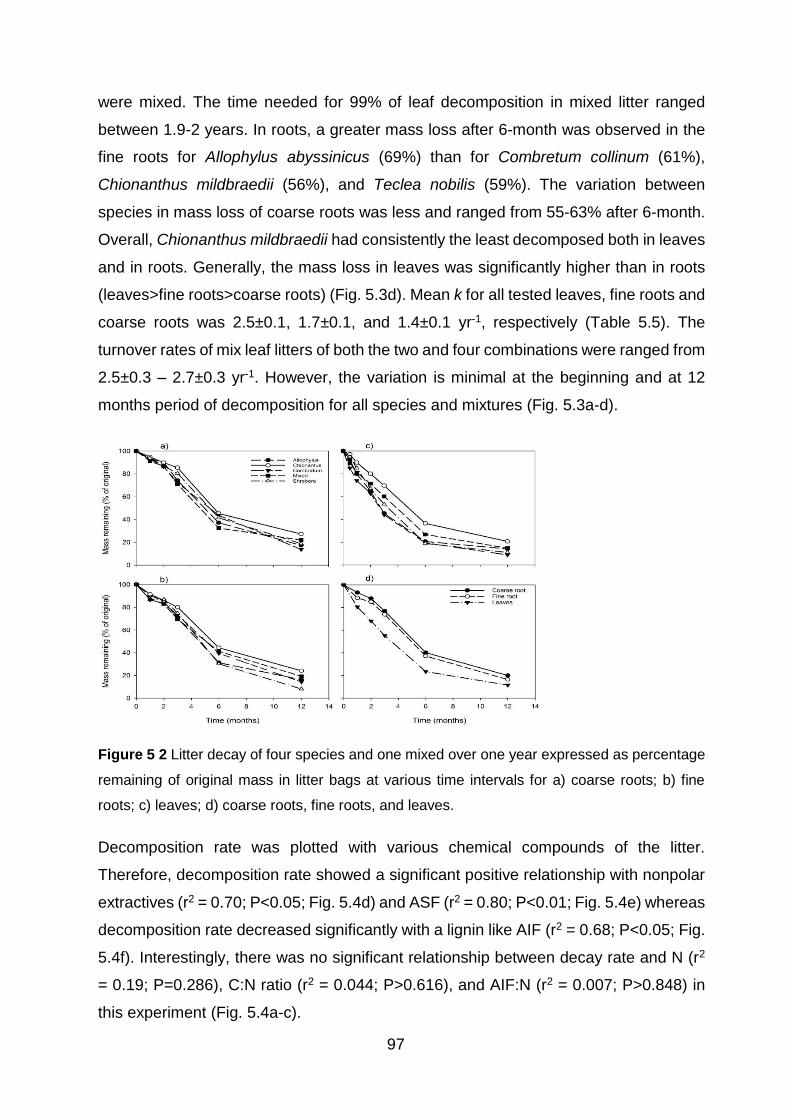
97
were mixed. The time needed for 99% of leaf decomposition in mixed litter ranged
between 1.9-2 years. In roots, a greater mass loss after 6-month was observed in the
fine roots for Allophylus abyssinicus (69%) than for Combretum collinum (61%),
Chionanthus mildbraedii (56%), and Teclea nobilis (59%). The variation between
species in mass loss of coarse roots was less and ranged from 55-63% after 6-month.
Overall, Chionanthus mildbraedii had consistently the least decomposed both in leaves
and in roots. Generally, the mass loss in leaves was significantly higher than in roots
(leaves>fine roots>coarse roots) (Fig. 5.3d). Mean k for all tested leaves, fine roots and
coarse roots was 2.5±0.1, 1.7±0.1, and 1.4±0.1 yr-1, respectively (Table 5.5). The
turnover rates of mix leaf litters of both the two and four combinations were ranged from
2.5±0.3 – 2.7±0.3 yr-1. However, the variation is minimal at the beginning and at 12
months period of decomposition for all species and mixtures (Fig. 5.3a-d).
Figure 5 2 Litter decay of four species and one mixed over one year expressed as percentage
remaining of original mass in litter bags at various time intervals for a) coarse roots; b) fine
roots; c) leaves; d) coarse roots, fine roots, and leaves.
Decomposition rate was plotted with various chemical compounds of the litter.
Therefore, decomposition rate showed a significant positive relationship with nonpolar
extractives (r2 = 0.70; P<0.05; Fig. 5.4d) and ASF (r2 = 0.80; P<0.01; Fig. 5.4e) whereas
decomposition rate decreased significantly with a lignin like AIF (r2 = 0.68; P<0.05; Fig.
5.4f). Interestingly, there was no significant relationship between decay rate and N (r2
= 0.19; P=0.286), C:N ratio (r2 = 0.044; P>0.616), and AIF:N (r2 = 0.007; P>0.848) in
this experiment (Fig. 5.4a-c).
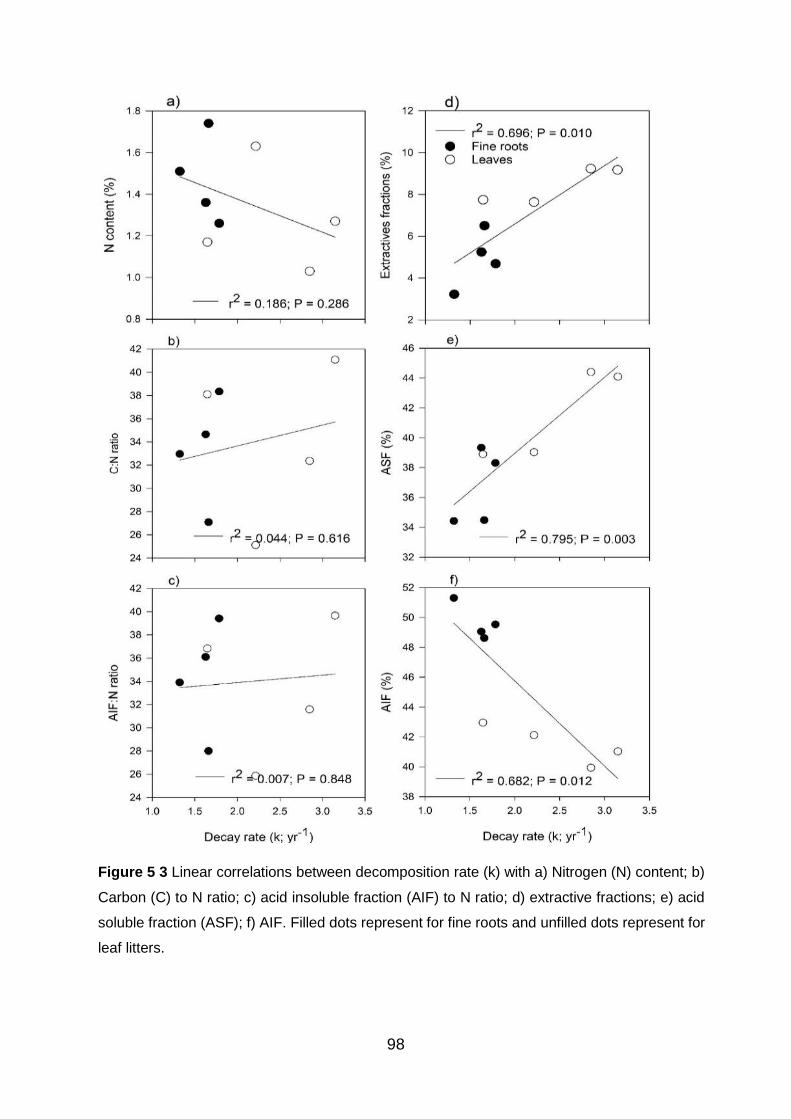
98
Figure 5 3 Linear correlations between decomposition rate (k) with a) Nitrogen (N) content; b)
Carbon (C) to N ratio; c) acid insoluble fraction (AIF) to N ratio; d) extractive fractions; e) acid
soluble fraction (ASF); f) AIF. Filled dots represent for fine roots and unfilled dots represent for
leaf litters.
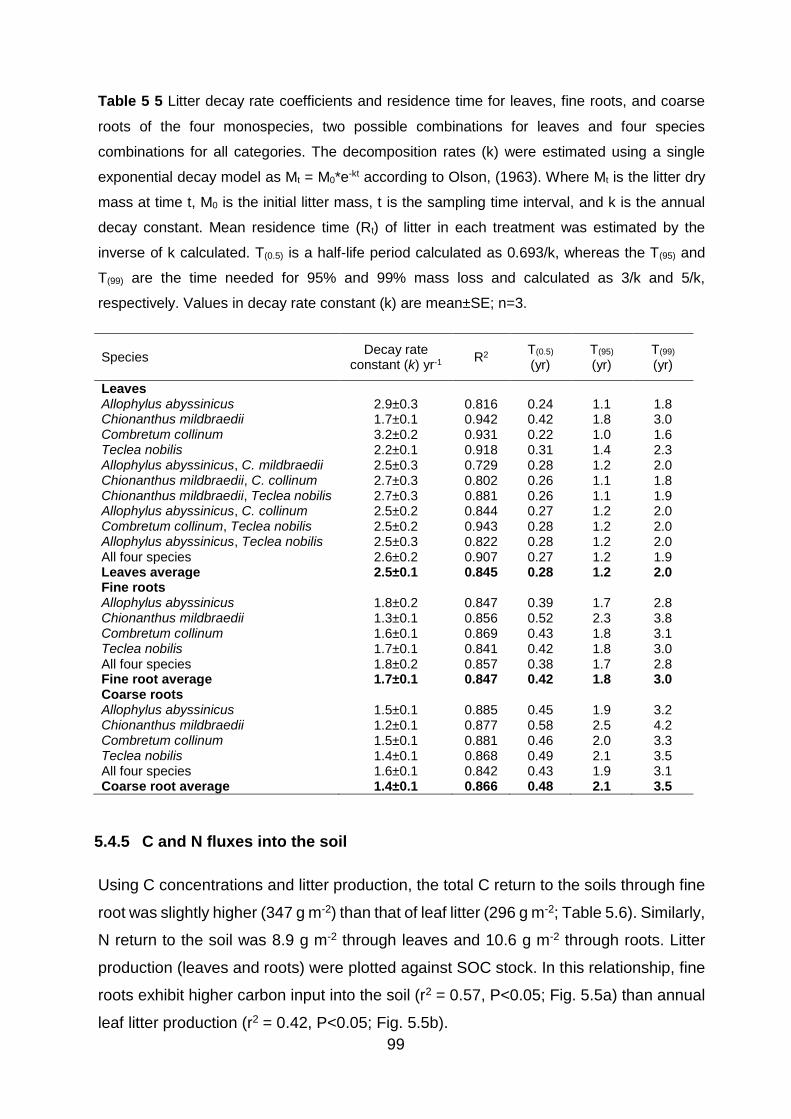
99
Table 5 5 Litter decay rate coefficients and residence time for leaves, fine roots, and coarse
roots of the four monospecies, two possible combinations for leaves and four species
combinations for all categories. The decomposition rates (k) were estimated using a single
exponential decay model as Mt = M0*e-kt according to Olson, (1963). Where Mt is the litter dry
mass at time t, M0 is the initial litter mass, t is the sampling time interval, and k is the annual
decay constant. Mean residence time (Rt) of litter in each treatment was estimated by the
inverse of k calculated. T(0.5) is a half-life period calculated as 0.693/k, whereas the T(95) and
T(99) are the time needed for 95% and 99% mass loss and calculated as 3/k and 5/k,
respectively. Values in decay rate constant (k) are mean±SE; n=3.
Species Decay rate
constant (k) yr-1 R2
T(0.5) (yr)
T(95) (yr)
T(99) (yr)
Leaves Allophylus abyssinicus 2.9±0.3 0.816 0.24 1.1 1.8 Chionanthus mildbraedii 1.7±0.1 0.942 0.42 1.8 3.0 Combretum collinum 3.2±0.2 0.931 0.22 1.0 1.6 Teclea nobilis 2.2±0.1 0.918 0.31 1.4 2.3 Allophylus abyssinicus, C. mildbraedii 2.5±0.3 0.729 0.28 1.2 2.0 Chionanthus mildbraedii, C. collinum 2.7±0.3 0.802 0.26 1.1 1.8 Chionanthus mildbraedii, Teclea nobilis 2.7±0.3 0.881 0.26 1.1 1.9 Allophylus abyssinicus, C. collinum 2.5±0.2 0.844 0.27 1.2 2.0 Combretum collinum, Teclea nobilis 2.5±0.2 0.943 0.28 1.2 2.0 Allophylus abyssinicus, Teclea nobilis 2.5±0.3 0.822 0.28 1.2 2.0 All four species 2.6±0.2 0.907 0.27 1.2 1.9 Leaves average 2.5±0.1 0.845 0.28 1.2 2.0 Fine roots Allophylus abyssinicus 1.8±0.2 0.847 0.39 1.7 2.8 Chionanthus mildbraedii 1.3±0.1 0.856 0.52 2.3 3.8 Combretum collinum 1.6±0.1 0.869 0.43 1.8 3.1 Teclea nobilis 1.7±0.1 0.841 0.42 1.8 3.0 All four species 1.8±0.2 0.857 0.38 1.7 2.8 Fine root average 1.7±0.1 0.847 0.42 1.8 3.0 Coarse roots Allophylus abyssinicus 1.5±0.1 0.885 0.45 1.9 3.2 Chionanthus mildbraedii 1.2±0.1 0.877 0.58 2.5 4.2 Combretum collinum 1.5±0.1 0.881 0.46 2.0 3.3 Teclea nobilis 1.4±0.1 0.868 0.49 2.1 3.5 All four species 1.6±0.1 0.842 0.43 1.9 3.1 Coarse root average 1.4±0.1 0.866 0.48 2.1 3.5
5.4.5 C and N fluxes into the soil
Using C concentrations and litter production, the total C return to the soils through fine
root was slightly higher (347 g m-2) than that of leaf litter (296 g m-2; Table 5.6). Similarly,
N return to the soil was 8.9 g m-2 through leaves and 10.6 g m-2 through roots. Litter
production (leaves and roots) were plotted against SOC stock. In this relationship, fine
roots exhibit higher carbon input into the soil (r2 = 0.57, P<0.05; Fig. 5.5a) than annual
leaf litter production (r2 = 0.42, P<0.05; Fig. 5.5b).

100
Table 5 6 Mean flux of each biochemical class to soil via leaf litter, fine roots (<2 mm), and the
proportion (%) of the combined flux of leaf litter and fine root flux contributed by fine roots. AIF,
acid-insoluble fraction; RCC, carbon cost; EF, extractive fractions; ASF, acid soluble fractions.
Elemental and chemical fractions
Leaves (g m-2 yr-1)
Fine roots (g m-2 yr-1)
% of roots of total litter
C 29700,4 34732,9 53,9
N 893,8 1062,8 54,3
CC 760,1 1012,2 57,1
EF 11021,5 8524,2 43,6
ASF 29278,1 26490,7 47,5
AIF 29214,7 35882,5 55,1
Figure 5 4 Linear correlation between soil carbon stocks with a) annual fine root production; b)
annual litterfall production; c) total biomass input (litterfall + fine roots).

101
5.5 Discussion
5.5.1 Above-ground litter production and seasonal pattern
The annual litter production found in this study (1090±64 g m-2 yr-1) was similar to those
recorded in previous studies in central highlands of Ethiopia (1087±219 g m2;
Lisanework and Michelsen, 1994) and in tropical China (923±129 g m-2 yr-1 to 1296±171
g m-2 yr-1; Tang et al., 2010). In general, our estimate lies in the range of estimates for
tropical forest (560-1530 g m-2 yr-1) but higher than for temperate (336-1001 g m-2 yr-1)
and boreal forests (13-576 g m-2 yr-1; Vogt et al. 1986). The proportion of leaf
components from the total above ground litterfall (65%) is similar to other tropical
studies such as 66% in central highlands of Ethiopia (Lisanework and Michelsen 1994)
and 56-61% in tropical China (Tang et al. 2010). Among 16 different species recorded
in our litterfall samples, Chionanthus mildbraedii species contributed about 45% of the
total annual litterfall (Table A3.1). Indeed, previous study in this site showed that
Chionanthus mildbraedii is a dominant species (Gebrehana 2015).
Total litterfall production was characterized by higher production during winter and
autumn (Fig. 5.1a) corresponding to dry periods of the year in Ethiopia and the rhythm
of leaf abscission (Tang et al. 2010). In the monthly distribution, litter production
increased slowly after the onset of the rainy season (June-January), and declined
towards the transition between dry and wet season (February to June) (Fig. 5.1b). The
decrease of litterfall from January to June may be due to most deciduous species (such
as Allophylus abyssinicus, and Combretum collinum) lost their entire canopy before the
warmest season. Several studies have documented similar seasonal patterns and they
found that the peak rates of litterfall usually occur during the dry season to reduce
transpiration under water stress conditions (Lisanework and Michelsen 1994,
Descheemaeker et al. 2006, Quichimbo et al. 2016, Zhou et al. 2016). Although most
of the litterfall still occurred in the dry season (Fig. 5.1a), Gelawdios forest has a weaker
seasonal variability than many temperate and cool-temperate areas (Zhang et al.
2014). The weak seasonal variability could be related to a relatively greater number of
species (16 species) than other temperate areas belonging to different tree phenology
composed of deciduous and evergreen plants that buffer the effects of season.

102
5.5.2 Factors controlling leaf litter decomposition
Leaf litter mass loss followed a simple exponential decay function (Fig. 5.3a). The
decomposition rates of leaves obtained in our study (1.7 – 3.2 year-1) are within the
range of decomposition rates previously recorded for tropical seasonal rainforests (0.5-
3.7 year-1; Gholz et al., 2000). In the first three months of decomposition, we observed
slower decomposition followed by a rapid decomposition rate between 3-6 months and
generating nearly a plateau after 6 months. This pattern was also observed by Portillo-
Estrada et al., (2016) across European forests and grasslands. Nevertheless, our
decomposition rate is slower with mass loss of 10 – 26% in the first four weeks and 20
– 37% in the first 8 weeks compared to 40 – 50% of the dry weight of litterfall
decomposed in the initial five weeks in an eastern Guatemalan forest (Ewel 1976).
Similarly, Yang et al. (2004) reported from subtropical China that leaves of Castanopsis
kawakamii, and Ormosia xylocarpa lost 91% and 88% of their initial weight in the first
150-day period, respectively. We hypothesize that the initial slower decomposition rate
in our study was generated by the combination of the following factors. First, this period
coincided with the main rainy period in Ethiopia and about 80% of the total rainfall
occurring during these three months only, which starts mid-June and ends early
September (UNECA 1996). Therefore, all airspaces are filled with water at this time
which hampers gas exchange (Karavin et al. 2016) so that microbes couldn’t get
enough air and thus reduce the decomposition rate. Some authors explained that soil
moisture is effective to a certain extent (Demessie et al. 2012), however, excess water
in the soil causes a decrease in decomposition rate because of anaerobic conditions
(Karavin et al. 2016). The second reason could be related to the invasion of
microorganisms. During the initial phase, microorganisms either need some time to
colonize the litter fully (Voříšková and Baldrian 2012) or their fungal mycelia and
microbes itself may contribute to the mass left (Portillo-Estrada et al. 2016). For
example, in a leaf litter decomposition experiment across European forests and
grasslands, Portillo-Estrada et al. (2016) noticed even litter mass increases relative to
the previous sampling during the first month of decomposition due to fungal mycelia.
The relatively sharp decline of remaining mass between 3 – 6 months may be attributed
to sufficient colonization of microbes (Voříšková and Baldrian 2012), optimal soil
temperature, moisture, and air conditions for microbial organisms (Yuste et al. 2007).
After three months of heavy rainfall, the temperature increased slowly and the

103
precipitation decreased sharply (Sisay et al. 2016) but provides sufficient soil moisture
that may explain the sharp decline in weight of the remaining litter mass during these
periods. It is known that decomposition rate is increased exponentially with increasing
temperature (Bradford 2013). According to Bradford (2013), a 10°C increase of
temperature increase decomposition rate by double. In Gelawdios, the temperature is
increased progressively from the lowest (16°C) in July and August to the maximum
24°C in February and March (Wassie et al. 2009). Thus, an 8°C increase of temperature
from the first three months to the next 6 months accelerates the decomposition rate by
1.6 times than the previous decomposition rate. After 6 months of incubation, the litter
mass loss rate decreased and generating nearly a plateau shape as observed in Fig.
5.3a-d. The slow decomposition revealed at this time can be partly explained by the
longer dry period, during which decomposition is almost impeded. In addition, the
relatively slower decay rates at later stages may be due to the decrease of the substrate
quality as a result of the removal of the labile carbon and the accumulation of
recalcitrant matter in the residual litter mass (Sun et al. 2013). The decomposition
usually begins by the most degradable fractions of the litter substrate such as soluble
carbohydrates (Sariyildiz and Anderson 2003, Hättenschwiler and Jørgensen 2010,
Moorhead et al. 2014).
Leaf decay rates varied considerably among species ranging from as little as 8% mass
remaining for Allophylus abyssinicus leaves to over 20% mass remaining for
Chionanthus mildbraedii species after one-year in the litterbag experiment (Table
A3.2). Chionanthus mildbraedii litter is consistently the least decompose species that
los only 30% of the initial weight of leaves in three months and about 21% undecayed
(Table A3.2). The slow decomposition in this species may be associated with the high
leaf toughness (Eissenstat and Yanai 1997, Pan et al. 2015, Yeong et al. 2016). For
example, Habteyohannes (2016) reported that Chionanthus mildbraedii is the toughest
leaf (10 N cm2) whereas other 13 species including Combretum collinum and Teclea
nobilis are below 6 N cm2. In addition, our chemical analysis indicated that the
Chionanthus mildbraedii litter is characterized by strongly lignified leaf tissue (Table
5.4), which could further hamper decomposition of leaf litter. Combretum collinum
decayed very rapidly with over 55% decayed in three months and over 90% decayed
within a year. Allophylus abyssinicus species also decayed faster next to Combretum
collinum. These two species (Combretum collinum and Allophylus abyssinicus) are

104
deciduous broad leaf species. Decomposition in mixed species was in general average
(Table A3.2). Similar results have been reported by previous studies that that the
presence of aspen litter increased the rate of decomposition of the birch litter in the
mixtures (Ostrofsky 2007, Gao et al. 2015). The potential mechanism for these
interactions may be resource complementarity among litter species, since higher
quality litter decomposes easily and then releases nutrient elements quickly such as N,
which could nourish microorganisms and accelerating the decomposition of lower
quality litter (Schimel and Hättenschwiler 2007, Gao et al. 2015).
Decomposition in our litterbag experiment was mainly controlled by the intrinsic
differences in recalcitrant carbon fractions such as lignin, and lignocellulose index and
C content. We found a significant positive relationship with nonpolar extractives (r2 =
0.70; P<0.05; Fig. 5.4d) and ASF (r2 = 0.80; P<0.01; Fig. 5.4e) whereas decomposition
rate decreased significantly with a lignin like AIF (r2 = 0.68; P<0.05; Fig. 5.4f). In
addition, decomposition rate was positively correlated with C content (r2 = 0.65; P<0.01)
and lignocellulose index, a ratio of AIF to cell wall fraction (AIF + ASF) (r2 = 0.77;
P<0.01) (data not shown). This shows that species having higher concentrations of
solvent and acid- soluble extractives but low structural component (lignin) are
potentially more prone to decay. This is because solvent extractives (referred as
extractive fractions, EF) contain non-structural substances of nonpolar constituents
such as alkaloids, fats, oils, waxes, resins, and polar constituents such as
carbohydrates and low-molecular-mass compounds (Pettersen 1984, Yang and
Jaakkola 2011, Sun et al. 2013). Since extractives do not contribute to the cell-wall
structure and are soluble in neutral solvents (Pettersen 1984), they are considered as
labile compounds that degrade rapidly at the initial stages of decomposition compared
to other fractions (Sun et al. 2013). ASF that contains cellulose and hemicellulose are
made of linear chains of β-1,4-linked glucose units (Kögel-Knabner 2002) and thus,
they decompose faster than AIF but slower than solvent extractives (Sun et al. 2013).
In contrast, the AIF is a highly cross-linked polymer associated with the presence of
highly reduced compounds such as suberin, cutin, and tannin-protein complexes which
have been proposed as potentially recalcitrant compounds and are resistant to
decomposition (Abiven et al. 2005, de Leeuw et al. 2005, Lorenz et al. 2007, Sun et al.
2013, Xia et al. 2015). In addition, the hydroxyls and many polar groups exist in the
lignin forms a strong intramolecular and intermolecular hydrogen bonds making the

105
intrinsic lignin resistant to decomposition (Chen 2014). Thus, our result demonstrates
that the chemical parameters (EF, ASF, AIF, and lignocellulose index) are good
indicators for substrate quality (Hendricks et al. 2000, Prieto et al. 2016), and
determines the decomposition dynamics (Sun et al. 2013, Zhang and Wang 2015).
Interestingly, Cornelissen (1996) found that leaves of deciduous species decomposed
twice as fast as those of evergreens under controlled conditions. The main reason was
that initial litter chemistry such as C:N ratios and AIF:N ratios were lower in deciduous
leaves than evergreen leaves (Prescott et al. 2004, Rahman and Tsukamoto 2013,
Pearse et al. 2014). For example, Pearse et al., (2014) found higher C:N ratio and
higher lignin concentration in evergreen oak (Quercus wislizeni) litter relative to
deciduous oak (Quercus douglasii). However, our litterbag experiment showed that N,
C:N ratio, AIF:N ratio were not different between deciduous and evergreen species for
both leaves and roots (Table 5.4). In addition, there was no significant relationship
between decay rate and N (r2 = 0.19; P=0.286), C:N ratio (r2 = 0.044; P>0.616), and
AIF:N (r2 = 0.007; P>0.848) (Fig. 5.4a-c) and these proxies are poor predictors of decay
in this experiment. The main reasons may be due to N2-fixing symbionts of species
(e.g. Teclea nobilies; Orwa et al. 2009), N resorption before leaf fall (Berg and
McClaugherty 2003, Fife et al. 2008), and N leaching and decomposition between the
time of falling and collection of the samples from the forest floor (Berg and Staaf 1981,
Berg and McClaugherty 2003).
5.5.3 Fine root and coarse root decomposition
The mass loss in fine roots (<2 mm diameter) and coarse roots (2-5 mm diameter)
followed the same trend as leaf mass loss (Fig. 5.3b-c) and the same conclusion can
be applied. Similar to leaf decomposition, Chionanthus mildbraedii species is still the
least decomposed species both in fine roots (24±5% mass remaining after 12 months)
and coarse roots (27±5%) (Table A3.2). Our data indicate that the slowly decomposing
Chionanthus mildbraedii and Teclea nobilis species resulted in greater mass loss in the
litter mixtures of fine roots but litter mixture had no effect on coarse roots.
Decomposition of the litter mixture still depends on the total quality of the litter mixture
(Gao et al. 2015). For example, Chionanthus mildbraedii fine roots had the largest root
tissue density, carbon cost, and AIF among ten other species (Chapter 4) that affects
the overall decomposition rate in the mixture. In this study, AIF and lignocellulose index

106
in Chionanthus mildbraedii have no difference with other species but root carbon cost
is significantly higher than all other species (P<0.001; Table 5.4). In general, fine roots
decomposed on average significantly slower (17±2% mass remaining after 12 months,
mean ± SD) than leaves (12±1%; p<0.001) but no significant difference with coarse
roots (20±2%) (Fig. 5.3d; Table A3.2). The decomposition rate (k) was in the order of
leaves (2.5±0.1), fine roots (1.6±0.1), and coarse roots (1.4±0.1) (Table 5.5).
5.5.4 Biochemical fluxes from litters into the soil
The total C return to the soils through fine root (347 g m-2) and leaf litter (296 g m-2;
Table 5.6) is lower than other estimates in tropical forests (436 g C m-2 yr-1 for roots or
leaves in China; Chen et al., 2005). Nevertheless, our estimate is higher than that of
boreal forest in Finland (272 g C m-2 yr-1 for fine roots and 235 g C m-2 yr-1 for above-
ground litterfall; Leppälammi-Kujansuu et al., 2014). Using N concentrations and
litterfall data, N return to the soil from leaves (8.9 g m-2) is similar to Ethiopian highland
forest (8.6 g m-2; Lisanework and Michelsen, 1994) but in the lower range of reports
from eucalyptus stand in New Zealand (8.3-13.5 g m-2; Guo and Sims, 1999). The
difference is due to either amount of litterfall or N concentration in the leaf material. Still,
N return to the soil through leaves was lower than that of roots (10.6 g m-2) in this study
(Table 5.6) and this may be due to N2-fixing symbionts in roots or N resorption or
leaching in the leaves.
ASF and AIF were the two largest biochemical fluxes to the soil (47-50%) and extractive
fractions (EF) accounted for <16% of the total litter flux in leaves and <12% in fine roots.
The finding of this study showed that fine roots exhibit higher carbon input into the soil
(r2 = 0.57, P<0.05; Fig. 5.5a) than annual leaf litter production (r2 = 0.42, P<0.05; Fig.
5.5b) and dominated the fluxes of recalcitrant carbon (an 8% difference in AIF) (Table
5.4). Out of the total carbon input, fine roots contributed about 1.5-times higher
recalcitrant carbon into the soil (Table 5.6). Xia et al., (2015) also showed that fine roots
contained 2.9-fold higher AIF and 2.3-fold more condensed tannins than leaf litter; both
are relatively difficult to decompose. By contrast, leaf litter contributed greater quantities
of labile compounds: cellulose, soluble phenolics and carbohydrates (57% in leaves vs
48% in fine roots). Thus, we hypothesized that fine roots are the major source of
recalcitrant carbon fractions entering the soil at our study site.

107
5.6 Conclusion
Litterfall in the studied areas is characterized by weak seasonality mainly attributed to
plant phenology. Fine root production is nearly equal to leaf litterfall. Nevertheless, our
results suggest that fine roots might contribute more recalcitrant SOC than leaves
because of larger amounts of AIF in the fine roots. Species with higher recalcitrant
structural compounds (AIF) are slowly decomposed whereas litter containing high
concentration of labile compounds (e.g. carbohydrates, cellulose) decompose faster.
The presence of deciduous species such as Combretum collinum and Allophylus
abyssinicus stimulate the decomposition of the less decompose Chionanthus
mildbraedii species in the mixtures litter. When it comes to species selection for
regulating global biogeochemical cycle, soil carbon deposition in the long-term, and
ecosystem function, evergreen species such as Chionanthus mildbraedii and Teclea
nobilis are preferable due to higher litter biomass production and lignin content in the
litter biomass than deciduous species.

108
6 The biological origin of soil organic carbon and
response to land use change based on biomarker
analysis
6.1 Abstract
Soil organic carbon accounts for more than four times the amount of carbon in the
atmosphere and is a fundamental part of the carbon cycle and life on Earth. This
material is primarily manufactured by plants but also by animals and microorganisms
which, as part of the food chain, return to the soil when they die and decompose. Once
decomposed and incorporated to the soil, this carbon loses its anatomical
characteristics, making it difficult to characterize and isolate its biological origin and
status of degradation. Therefore, this study aimed to reconstruct the biological origin of
soil organic carbon and to attribute it to its parent materials based on its carbon
skeleton. This effort is also a step forward in characterizing the relative stage of organic
matter degradation in different land use types. Individual biomarkers were identified
and quantified by applying chromatographic and spectrometric techniques after
sequential extraction including solvent extraction, base hydrolysis, and CuO oxidation.
The soil samples stem from four land use systems in the northern highlands of Ethiopia.
Although the identified biomarkers represented only 4% of the total organic matter, they
provided useful information on their origin, tissue type, major taxonomic group, and
stages of organic matter degradation. The overall result generally revealed a major
input of soil organic carbon derived from vascular plants; the microbial and animals
inputs were present as minor components. The relative proportion of suberin was about
2 times that of cutin in natural forest, eucalyptus, and cropland. This emphasized the
importance of below-ground biomass in the distribution of organic carbon in the soil.
The occurrences and concentrations of benzyls and lignin-derived phenols was about
4 times higher in the natural forest soil than in the other land use soils, indicating the
predominant input of recalcitrant organic carbon derived from higher plants into the
forest ecosystem. The ratio of syringyls to vanillyls (S/V) and of cinnamyl to vanillyl
(C/V) monomers – along with the lignin phenol vegetation index – indicated that non-
woody angiosperms plants are the predominant source for lignin. In all land use types,
the carbon source was mainly from C3 plants, not C4 plants. Although the extent of
organic matter degradation depends on microbial activity and biochemistry, the highest

109
suberin/cutin and C16/∑C16 ratios indicate preferential degradation of cutin.
Combining the evidence of the degradation parameters, the degradation of lignin was
enhanced in the forest soil, supported by the elevated acid/aldehyde (Ad/Al) ratios of
vanillyl and syringyl units.
6.2 Introduction
The amount of carbon (C) stored in the soil (3150 Pg C) is more than four times greater
than that in the atmosphere (750 Pg C) and is a fundamental part of the global carbon
cycle (Batjes 1996, Hiederer and Köchy 2011, Fan et al. 2016). Soil is a complex
mixture of organic compounds primarily produced by plants but also by animals and
microorganisms as part of the food chain (Amelung et al. 2008, Feng and Simpson
2011). Soil organic matter (SOM) has a mean residence time in soil ranging from days
to millennia (Amelung et al. 2008). Once organic material is incorporated into the soil,
it loses its anatomical characteristics during degradation, featuring different stages of
biological oxidation (Feng and Simpson 2007). Subsequently, morphological
identification is no longer a viable method to determine the biological origin of carbon
in the soil (Amelung et al. 2008).
Quantitative and qualitative measurements of soil organic carbon (SOC) dynamics
have been of great interest in recent years. This reflects the major significance of SOC
in the environment and its high contribution to soil productivity by improving soil fertility
and soil aggregation and based on its resistance to physical degradation (Strosser
2010, Scharlemann et al. 2014). Nonetheless, unsustainable land use management
practices, commonly by small-scale farmers, accelerate decomposition and carbon
losses through respiration and erosion (Lal 2010a, 2010b). Numerous studies have
quantified carbon dynamics, turnover, distribution, and chemical characterization in
terrestrial ecosystems based on bulk properties and elemental compositions (Schuman
et al. 2002, Jones et al. 2005, Conti et al. 2016). Limited information, however, is
available on the sources and chemical composition of input materials, except for certain
temperate ecosystems (Hedges et al. 1984, Otto et al. 2005, Otto and Simpson 2006a,
2007, Pautler et al. 2010, Li et al. 2015). The overall degradation stages of organic
matter are often unknown (Guo and Sims 1999, Girisha 2001, Otto et al. 2005). This
calls for the determination of biological origin of materials for soil carbon deposition and

110
the degradation status of carbon in various physical and chemical fractions in soils.
This is an important step forward in predicting the behavior of soil carbon and the
retention time of carbon in soils of different land use systems (Otto et al. 2005). This is
the first attempt to study the soil carbon origin of different land use systems based on
biomarker analysis at the molecular level, particularly for African ecosystems.
Structurally unique biochemicals (biomarkers) carry useful information and quantitative
evidence to reconstruct their parent organic materials as well as to determine the stage
of SOM degradation based on their carbon skeleton (Kögel-Knabner 2000, Simoneit
2002, 2005, Amelung et al. 2008, Feng and Simpson 2011, Li et al. 2015). The
composition of uncharacterized SOM is very complex, and some methodological
approaches result in signal overlap. Thus, sequential biomarker extraction with the
application of chromatographic and spectrometric techniques is one of the most
commonly used approaches to detect and quantify the concentration of specific
biomarkers at the molecular level (Poirier et al. 2005, Amelung et al. 2008, Feng and
Simpson 2011). The sequential biomarker extraction techniques include organic
solvent extraction and chemolytic methods such as base hydrolysis and CuO oxidation
because one extraction method may not provide all the required information to trace
SOM sources and processes in the soil (Kögel-Knabner 2000, Otto and Simpson 2007,
Feng 2009). Identification and quantification of individual biomarkers involves gas
chromatography and mass spectrometry (GC-MS) at the molecular level (Kögel-
Knabner 2000, Poirier et al. 2005, Otto and Simpson 2007, Amelung et al. 2008).
Extraction with organic solvents isolates unbound (free) lipids (Feng 2009) such as n-
alkanes, n-alkanols, n-alkanoic acids, steroids, hopanoids, and other terpenoids
(Kögel-Knabner 2000, Otto and Simpson 2006a, Feng and Simpson 2007, Amelung et
al. 2008). Solvent extraction thus provides a general overview of biomarkers from plant
and microbial sources (Goñi et al. 2003, Angst et al. 2016). For example, among the
compounds, even-numbered long-chain (>C20) alkanoic acids and alkanols are
common constituents of plant-derived wax lipids, whereas branched short-chain (<C20)
alkanoic acids, hopanoids, and ergosterol are indicators of microbial-derived SOM
(Feng and Simpson 2011).
As opposed to ‘free’ lipids, ester-bound soil lipids are not extractable with organic
solvents, but can be cleaved from SOM using chemolytic methods such as base

111
hydrolysis (Otto and Simpson 2007, Feng and Simpson 2011). Base hydrolysis is
mainly used to trace root or leaf origin inputs based on suberin- and cutin-derived
markers (Bull et al. 2000, Otto et al. 2005). The predominant long-chain ω-hydroxy-
alkanoic acids and α, ω-alkanedioic acids in bound soil lipids are typical biomarkers for
suberin, primarily used for tracing root or bark inputs into the soil. In turn, C16 and C18
ω-hydroxyalkanoic acids with mid-chain hydroxy or epoxy groups are biomarkers for
cutin or leaf cuticle inputs (Feng and Simpson 2011). According to Feng and Simpson
(2011), suberin- and cutin-derived compounds are recalcitrant and less prone to
microbial attack than solvent-extractable compounds.
The CuO oxidation targets ether bonds to release lignin-derived phenols (Hedges and
Ertel 1982, Otto and Simpson 2006b, Amelung et al. 2008, Feng 2009, Clemente 2012).
Lignin is made up of a polymeric network of phenols. This makes measuring and tracing
lignin directly difficult because of its high molecular weight and low extractability in soils
(Kögel-Knabner 2002, Feng and Simpson 2011). Alternatively, the lignin biopolymer,
namely lignin-derived phenols (vanillyl, syringyl, and cinnamyl) can be released from
SOM with chemolytic methods such as CuO oxidation (Hedges and Mann 1979a, Goñi
and Hedges 1992). Comparison of the abundance of acids to aldehydes (Ad/Al) in this
extract can provide information about the degradation state of lignin in a sample (Otto
and Simpson 2007). The relative abundances of the phenol structural classes can also
provide information about whether lignin is derived from angiosperm vs gymnosperm
sources or from woody vs non-woody tissues (Hedges and Mann 1979a, Goñi et al.
2000) and stages of SOM degradation (Amelung et al. 2008, Feng 2009, Clemente
2012). In addition, biomarkers can provide a general overview of root or above-ground
plant sources based on suberin- and cutin-derived compounds (Otto and Simpson
2006a). Overall, these analyses provide evidence on the origin of SOM (plants, fungi,
bacteria, animals, or anthropogenic origin) by using biomarkers as a screening tool for
bulk SOM composition. Therefore, the objectives of this study are 1) to determine and
quantify the distribution of biomarkers in different land use systems at the molecular
level; 2) to reconstruct the biological origin of soil organic carbon to their parent
materials based on their carbon skeleton; and 3) to characterize the relative stage of
organic matter degradation in the soil.

112
6.3 Methodology
6.3.1 Study area
Four land use systems (natural forest, eucalyptus plantation, cropland and grazing
land) were selected in Gelawdios, north-central Ethiopia. The study area (Gelawdios)
is geographically located at 11°38’25’’ N and 37°48’55’’ E at an elevation of 2500 m
above sea level. The topography is typical of volcanic landscapes, comprising volcanic
rocks with ragged and undulating landforms. The major soil type is classified as
Cambisols with a clay loam texture according to the World Reference Base for soil
resources (Awulachew et al. 2009, Betrie et al. 2011, WRB 2014). The mean annual
rainfall at Gelawdios is 1220 mm with a unimodal rainy season, and the average annual
temperature is 19°C (Wassie et al. 2009). Although Ethiopia is geographically located
in the tropics, the climate of the study area is temperate with a dry winter and a warm,
wet summer (Cwb) according to the Köppen-Geiger climate classification (Peel et al.
2007). The forest is a pristine Afromontane dry forest composed mostly of a mixture of
indigenous tree species; it is almost exclusively confined to sacred groves associated
with a church. The dominant tree species are Albizia schimperiana, Apodytes dimidiata,
Calpurnia aurea, Croton macrostachyus, Ekebergia capensis, Maytenus arbutifolia,
and Schefflera abyssinica. In Gelawdios forest, the number of tree species per hectare
is about 6300 (Chapter 2, Table 2.1). Due to higher tree density, undergrowth of
grasses in the forest is minor. The eucalyptus stand was established on formerly
common grazing land in 1985 and was consecutively thinned to its current density
about 3000 trees per hectare (Chapter 2, Table 2.1). Due to the large space between
trees, the undergrowth is dominated with grasses and indigenous shrub species. The
adjacent grazing land and cropland was converted from natural forest approx. 50 years
ago, although the exact date is not known. The grazing land was used as communal
grazing lands for herds of animals and it consists of highly degraded, nearly bare
ground. The cropland has been cultivated to a depth of ca. 30 cm without fallow periods;
the crop residues are harvested for animal feeding. The principal crops are ‘teff’
(Eragrostis tef), wheat (Triticum aestivum), and barley (Hodeum vulgare).
6.3.2 Soil sampling, C and N analysis
Soil samples were collected at the end of the wet season in September 2015 from all
land use types. Soil samples from the top 10 cm were collected from each land use at

113
10 sampling points marked at 50-100 m along a transect line. At each sampling point,
one sample was taken to the depth of 10 cm with a soil corer (6.6 cm diameter) after
removing the litter layer (if present). The soil samples were sieved in Ethiopia, packed
separately in a plastic bag, and transported to Vienna for laboratory analysis. Sub-
samples of soils weighing approx. 3-5 g from each sample were dried at 105°C for 48
h. From each sample, about 200 mg of soil was taken and total C and N concentrations
were determined on a CN elemental analyser (Truspec CNS LECO, St. Joseph, USA).
6.3.3 Sequential extraction procedures
Sequential chemical extractions (solvent extraction, base hydrolysis, and CuO
oxidation) were conducted on soil samples to determine total solvent extracts, bound
lipids, and lignin-derived phenols, respectively (Otto et al. 2005, Feng and Simpson
2008, Feng 2009, Clemente 2012).
Solvent extraction: Three soil samples (20 g) from each land use were first sonicated
twice for 15 min, each time with 30 ml double deionized water to remove the water-
soluble polar compounds. The water-extracted soil residues (~20 g) were then freeze-
dried and extracted with organic solvents as follows: samples were sonicated for 15
min with 50 ml of methanol, dichloromethane:methanol (1:1; v/v), and dichloromethane,
sequentially. The combined solvent extracts were passed through glass-fiber filters
(Whatman GF/A) into a round bottom flask, concentrated by rotary evaporation, and
then completely dried under nitrogen gas (N2) in 2 ml glass vials. The remaining soil
samples (non-extractable materials) were air-dried for further analysis.
6.3.4 Base hydrolysis
The air-dried soil residues from solvent extraction were then subject to base hydrolysis
to yield ester-linked lipids (Otto and Simpson, 2006a). Briefly, the residues after solvent
extraction were heated at 100°C for 3 h in Teflon-lined bombs with 20 ml of 1 M
methanolic KOH. After cooling, the extracts were acidified to pH 1 with 6 M HCl and
filtered through pre-extracted cellulose filters (Fisher P5, 5-10 µm). Again, the soil
residues were extracted twice by sonication for 15 min with 30 ml
dichloromethane:methanol (1:1 v/v) as described above. The two extracts were
combined and then filtered through glass-fiber filters (Whatman GF/A) into round
bottom flasks. Double-deionized water (50 ml) was added to each extract. Lipids were

114
recovered from the water phase by liquid–liquid extraction in a separation funnel with
50 ml diethyl ether. Anhydrous Na2SO4 was added to the combined ether phases to
remove any water. The ether extracts were concentrated by rotary evaporation,
transferred to 2 ml glass vials, and dried under N2 gas. The remaining soil samples
were air-dried for further analysis.
6.3.5 CuO oxidation
The base hydrolysis residues were air-dried and further oxidized with CuO to release
lignin-derived phenols. Soil residues (~10 g) were extracted with 1 g CuO, 100 mg
ammonium iron (II) sulfate hexahydrate [Fe(NH4)2(SO4)2·6H2O], and 15 ml of 2 M
NaOH in Teflon-lined bombs at 170°C for 2.5 h. After heating, the bombs were cooled
under running water, the liquids were decanted into Teflon centrifuge tubes (50 ml),
and the residues were washed twice each with 10 ml deionized water using a magnetic
stirrer for 10 min. The combined washings and extracts were centrifuged for 30 min at
1050 g force (Heraeus Megafuge 1.0, Hanau, Germany). The supernatant was
decanted into another Teflon centrifuge tube, acidified to pH 1 with 6 M HCl, and kept
for 1 h at room temperature in the dark to prevent reactions of cinnamic acids. After
centrifugation (at 1050 g force for 30 min), the supernatant was transferred to a
separation funnel and liquid–liquid extracted twice with 50 ml diethyl ether. The ether
extracts were concentrated by rotary evaporation, transferred to 2 ml glass vials, and
dried under N2 gas.
6.3.6 Derivatization and GC/MS Analysis
Derivatization was conducted according to Otto et al. (2005) and Feng and Simpson
(2011). In brief, the extracts were re-dissolved, and aliquots (containing ~1 mg extracts)
were derivatized for GC/MS analysis. Solvent extracts and CuO oxidation products
were each re-dissolved in 500 μl dichloromethane:methanol (1:1; v/v). Aliquots of the
extracts (100 μl) were dried in a stream of N2 and then converted to trimethylsilyl (TMS)
derivatives by 90 μl N,O-bis-(trimethylsilyl)trifluoroacetamide (BSTFA) and 10 μl
pyridine for 3 h at 70°C. After cooling, 100 μl hexane was added to dilute the extracts.
The base hydrolysis products were first methylated by reacting with 600 μl of
diazomethane in ether at 37°C for 1 h, evaporated to dryness under N2, and then
silylated with BSTFA and pyridine as described above. Oleic acid (C18:1 alkanoic acid),
tetracosane, and ergosterol were derivatized and used as external standards for

115
solvent extracts. Oleic acid methyl ester and vanillic acid were used as external
standards for base hydrolysis and CuO oxidation products, respectively. To ensure a
linear response, standards ranging from 10-1000 ppm were analyzed using the same
procedure as the biomarker extracts. GC/MS analysis was performed on an Agilent
model 6890N GC coupled to a Hewlett-Packard model 5975 quadrupole mass selective
detector.
Separation was achieved on a HP5-MS fused silica capillary column (30 m × 0.25 mm
internal diameter, 0.25 μm film thickness). The GC operating condition was as follows:
temperature held at 65°C for 2 min, increased from 65 to 300°C at a rate of 6°C min-1
with final isothermal hold at 300°C for 20 min. Helium was used as the carrier gas. The
samples were injected with a 2:1 split ratio and the injector temperature was set at
280°C. The sample (1 μl) was injected with an Agilent 7683B auto sampler. The mass
spectrometer was operated in the electron impact mode (EI) at 70 eV ionization energy
and scanned from 50 to 650 Daltons. Data were acquired and processed with the
Chemstation G1701FA software.
Individual compounds were identified by comparing the mass spectra with the National
Institute of Standards and Technology library (NIST, version 2.0), Wiley MS library data,
and standards. Concentrations of individual compounds were calculated by comparing
the peak area of the compound to that of the standard in the total ion current (TIC) and
were then normalized to the soil carbon contents (Pautler et al. 2010). The C-
normalized biomarker concentrations indicates the relative contribution of the identified
biomarkers with respect to one gram of organic carbon in the soil (Pautler et al. 2010).
All the samples were run in triplicate; values are mean±SE. If a compound was
identified in only one or two of three replicates, SE was not calculated.
6.3.7 Origin and degradation parameters
Several biomarker ratios have been proposed to distinguish between the inputs of
organic matter in the soil. The ratios of C31/(C23+C27) alkanes from solvent extraction
are commonly employed to distinguish between inputs from grasses and trees,
whereby longer chain alkanes such as C31 and C33 are derived from grass tissues (Paul
2016). The relative inputs of C3 and C4 plants can be differentiated by the carboxylic
acid ratio of C24/(n-C22 + n-C26) (Wiesenberg and Schwark 2006). A ratio >0.67
indicates C4 origin, otherwise C3 origin. This is because lipid compounds containing C24

116
alkanoic acids are from C4 plants, whereas C22 and C26 acids are from C3 plants.
Suberin and cutin are used to trace below- and above-ground plant origin, respectively.
Suberin and cutin biomarkers were summarized and calculated according to their
occurrence only in suberin or cutin, based on parameters developed by Otto & Simpson
(2006). Long-chain ω-hydroxyalkanoic acids (≥C20) and α, ω-alkanoic acids (C16-C24)
are typical biomarkers for suberin (ΣS), whereas C16 and C18 ω-hydroxyalkanoic acids
are used as cutin (ΣC) biomarker signatures. Biomarkers derived from both suberin or
cutin (ΣSvC) are ω-hydroxyalkanoic acids C16, C18, C18 di- and trihydroxy acids, and α,
ω-alkanedioic acids C16, C18.
The composition of phenolic lignin compounds obtained after CuO oxidation can be
used to calculate parameters for the origin of lignin. Major plant taxonomic groups
(gymnosperms vs angiosperms) and tissue type (woody vs non-woody tissue) can be
differentiated by the ratios of syringyl to vanillyl (S/V) and cinnamyl- to vanillyl-type
(C/V) monomers (Hedges and Ertel 1982, Goñi and Montgomery 2000). Accordingly,
the S/V ratio of angiosperm tissues has been reported as >0.6. This is because
gymnosperm wood contains only vanillyl derivatives (V: vanillin, acetovanillone, vanillic
acid), while angiosperm wood is composed of approximately equal quantities of
vanillyls and syringyls (S: syringaldehyde, acetosyringone, syringic acid). The lignin-
derived phenol (V, S, C) ratios are also used to estimate the relative contributions of
woody and non-woody angiosperms. Non-woody angiosperm tissues exhibit a V:S:C
ratio of about 1:1:1 (Hedges and Mann 1979a). To improve the detection of lignin
sources as an indicator of vegetation contribution using lignin index-phenols, another
useful parameter (lignin phenol vegetation index – LPVI) was developed by Tareq et
al. (2004). It is calculated according to the following equation.
LPVI = [{S(S+1)/(V+1)+1}*{C(C+1)/(V+1)+1}] in which S=100*S/(S+V+C);
C=100*C/(S+V+C); and V=100*V/(S+V+C).
Based on this parameter, the distinct boundaries of LPVI ranges are <1 for
gymnosperm woods, 3-27 for non-woody gymnosperm tissues, 67-415 for angiosperm
woods, and 176-2782 for non-woody angiosperms.
To assess the degradation of free lipids, bound lipids, and lignin in the soil, degradation
parameters were calculated according to Goñi and Hedges (1990), Otto & Simpson
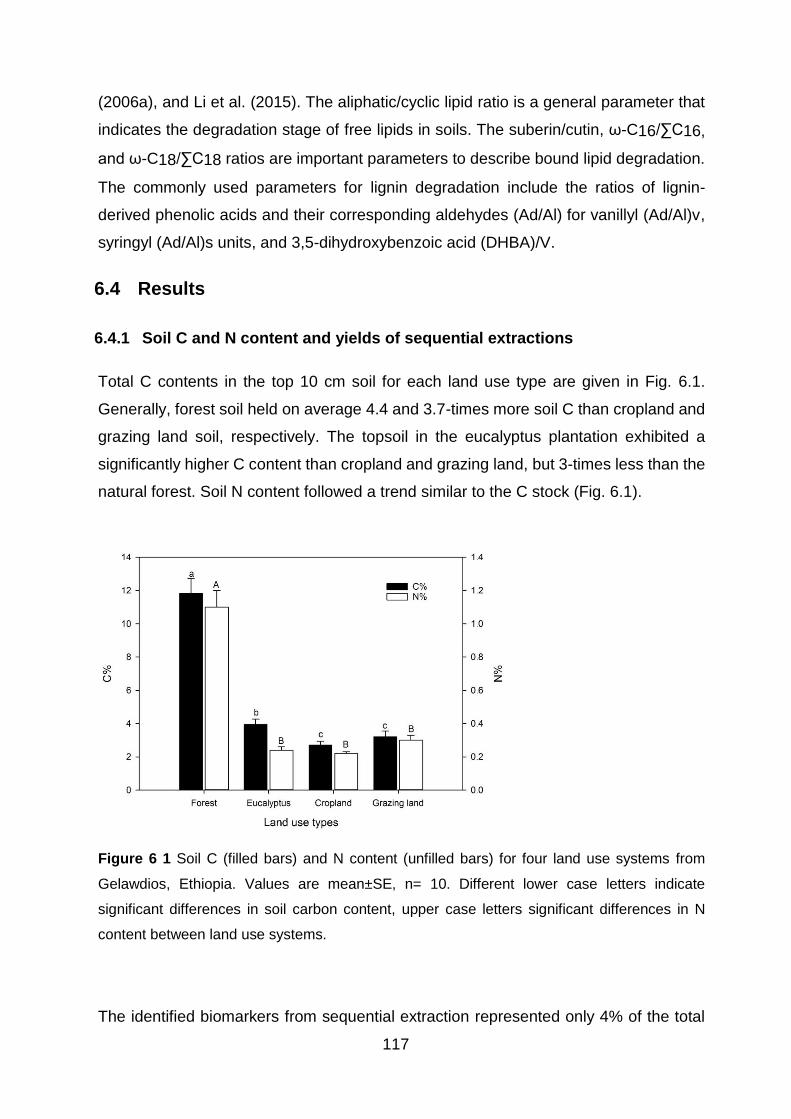
117
(2006a), and Li et al. (2015). The aliphatic/cyclic lipid ratio is a general parameter that
indicates the degradation stage of free lipids in soils. The suberin/cutin, ω-C16/∑C16,
and ω-C18/∑C18 ratios are important parameters to describe bound lipid degradation.
The commonly used parameters for lignin degradation include the ratios of lignin-
derived phenolic acids and their corresponding aldehydes (Ad/Al) for vanillyl (Ad/Al)v,
syringyl (Ad/Al)s units, and 3,5-dihydroxybenzoic acid (DHBA)/V.
6.4 Results
6.4.1 Soil C and N content and yields of sequential extractions
Total C contents in the top 10 cm soil for each land use type are given in Fig. 6.1.
Generally, forest soil held on average 4.4 and 3.7-times more soil C than cropland and
grazing land soil, respectively. The topsoil in the eucalyptus plantation exhibited a
significantly higher C content than cropland and grazing land, but 3-times less than the
natural forest. Soil N content followed a trend similar to the C stock (Fig. 6.1).
Figure 6 1 Soil C (filled bars) and N content (unfilled bars) for four land use systems from
Gelawdios, Ethiopia. Values are mean±SE, n= 10. Different lower case letters indicate
significant differences in soil carbon content, upper case letters significant differences in N
content between land use systems.
The identified biomarkers from sequential extraction represented only 4% of the total

118
organic matter in all land use systems. This means that substantial fractions of SOM
associated with mineral surfaces remain uncharacterized. Although only small fractions
could be obtained by sequential extraction of the soil, these extracts provide useful
information on their biological origin and the degradation status of organic matter based
on characteristic biomarkers. Among the sequential extraction, the highest yield was
obtained from base hydrolysis products (Fig. 6.2).
Figure 6 2 Carbon-normalized extract yields of solvent, base hydrolysis, and CuO oxidation
products of four land use systems from Gelawdios, Ethiopia. Values are mean±SE, n= 3.
6.4.2 Composition and distribution of solvent-extractable free lipids
The major chemical compounds detected in the solvent extract include a series of
aliphatic lipids (n-alkanols, n-alkanes, n-alkanoic acids), carbohydrates,
monoacylglycerides, steroids, and terpenoids (Table 6.1). The GC-MS total ion
chromatogram (TIC) of the major components in the silylated solvent extracts is shown
in Appendix 3 Figure 1 (Figure A4.1). All lists of individual compounds with their
corresponding molecular formula and weight are presented in supplementary data
Table A4.1. The C-normalized yield (mg/1 g C) obtained from solvent extracts was
highest in eucalyptus soil (8.3 mg/g C) followed by forest soil (5.7 mg/g C), cropland
(3.5 mg/g C), and grazing land (3.3 mg/g C) (Table 6.1). Aliphatic lipids accounted for
57-88% of the detected chemicals in the free lipids and were most abundant in natural
forest soil and least abundant in grazing land and cropland (Table 6.1).

119
Table 6 1 Occurrence and quantities of compounds (µg/g C) identified in the solvent extracts
of soil samples of different land use systems in Gelawdios, Ethiopia. Values are mean±SE, n=
3.
Compounda Forest Eucalyptus Cropland Grazing
land
n-Alkanols (C14-C30) 324±171 520±212 250±86 356±103
n-Alkanes (C17-C31) 222±71 391±78 139±66 58±15
n-Alkanoic acids (C9-C26) 448±184 1129±128 207±49 272±31
Iso- Alkanoic acids (C17) 24b nd nd nd
Monoacylglycerides (C19-C21) 52±26 260±19 134±46 84±2 Carbohydrates 3936±703 2429±256 1820±229 1894±192 Steroids and Terpenoids 622±267 3461±341 729±80 616±99 Unknowns 38±3 126±11 238±100 68±b Total aliphatic lipids 5007±746 4729±531 2550±430 2664±300 Identified total solvent extracts 5629±740 8190±857 3279±508 3280±394
nd, not detected
a All polar compounds were identified as their trimethylsilyl (TMS) derivatives.
b Average values detected from two samples only
The concentration of n-alkanols in the range of C14-C30 and n-alkanoic acids in the
range of C9-C26 showed a significant even-over-odd dominance, whereas n-alkanes in
the range of C17-C31 exhibited a preference of odd-numbered molecules (Table A4.1).
The combined concentrations of n-alkanols, alkanes, and alkanoic acids were in the
range of 0.6-2 mg/g C and greatest in eucalyptus soil and least in cropland soil (Table
6.1). Four carbohydrates (glucose, mannose, sucrose, and trehalose) were identified
in the solvent extracts (see details Table A4.1). Monosaccharides were detected only
in minor amounts (1% of total carbohydrates in forest, 13% in eucalyptus, 7% each in
cropland and grazing lands). Trehalose, in contrast, was found in the greatest amounts
(77-99% of total carbohydrates and 26-69% of the identified total solvent extracts). The
solvent extracts also included four monoacylglycerides, three of them being C21
compounds.
Cyclic lipids of steroids and terpenoids were detected in small amounts at
concentrations of 0.6-3.4 mg/g C; the highest concentration was detected in eucalyptus
soil (Table A4.1). The steroid cholesterol was detected in small amounts in all land use
types, whereas ergosterol was detected in minor amounts in the forest and eucalyptus
soil only. The detected phytosterols include campesterol, stigmasterol, β-sitosterol, and
sitosterone; these compounds contributed over 81-96% of total steroids in the forest,
cropland, and grazing land but only 19% in eucalyptus soil. In contrast, sesquiterpense

120
of trans-farnesol and globulol combined contributed 38% of total steroids in the
eucalyptus stand soil. Aromadendrene, γ-elemene, and ledol are also sesquiterpense,
detected in relatively higher concentration in eucalyptus soil and in minor
concentrations in cropland. The detected triterpenoids include ß- amyrin, α- amyrin,
lupeol, erythrodiol, and oleanolic acid. The latter two showed higher concentrations in
eucalyptus soil (29% of the total steroids and terpenoid extractions) but lower values in
other land use type soils (Table A4.1).
6.4.3 Composition and distribution of bound lipids
The major products identified after base hydrolysis included a series of aliphatic lipids
(n-alkanols, n-alkanoic acids, mid-chain substituted and branched acids, ω-
hydroxyalkanoic acids, α-hydroxyalkanoic acids, α,ω-alkanedioic acids, and
glycerides), with a lesser contribution from benzyls and phenols, and one steroid (Table
6.3). Compounds identified in the base hydrolysis are listed in detail in Table A4.2 and
the GC-MS chromatograms Figure A4.2. The C-normalized concentrations of total
bound lipids after base hydrolysis extraction were generally highest in the grazing land
(111.3±8.2 mg/g C), followed by cropland (97.1±4.4 mg/g C), eucalyptus soil (82.0±1.0
mg/g C), and natural forest (29.9±1.2 mg/g C) (Table 6.2). Aliphatic lipids showed a
similar pattern and represented more than 60% of the total base extracts; n-alkanoic
acids in the range of C14-C30 were detected in largest quantity (4.8-12.2 mg/g C), with
a strong even-over-odd preference (Table A4.2). Mostly even-numbered chains of α,
ω-alkanedioic acids in the range of C4-C20 were detected in minor amounts in all
samples (324-678 µg/g C). Mid-chain substituted hydroxyalkanoic acids (C16, C18, and
C19) and other α-alkanoic acids, ω- alkanoic acids, and glycerides combined were
detected in minor concentrations (1.1-2.2 mg/g C). Branched alkanoic acid (iso-C16)
was detected in low amounts (11-37 µg/g C) and was absent in grazing land soil
samples (Table 6.2). Base hydrolysis also cleaved glycerides (C19) in minor
concentrations (238-486 µg/g C) in three land use types but was absent in grazing land.
One organophosphate of C19 was also detected in all soil samples at high
concentrations (1.5-6.7 mg/g C). Benzyls and phenols ranged from 0.4-2.3 mg/g C
(Table 6.2).
Table 6 2 Occurrence and quantities of compounds (µg/g C) identified from base hydrolysis of
soil samples in different land use systems in Gelawdios, Ethiopia. Values are mean±SE, n= 3.
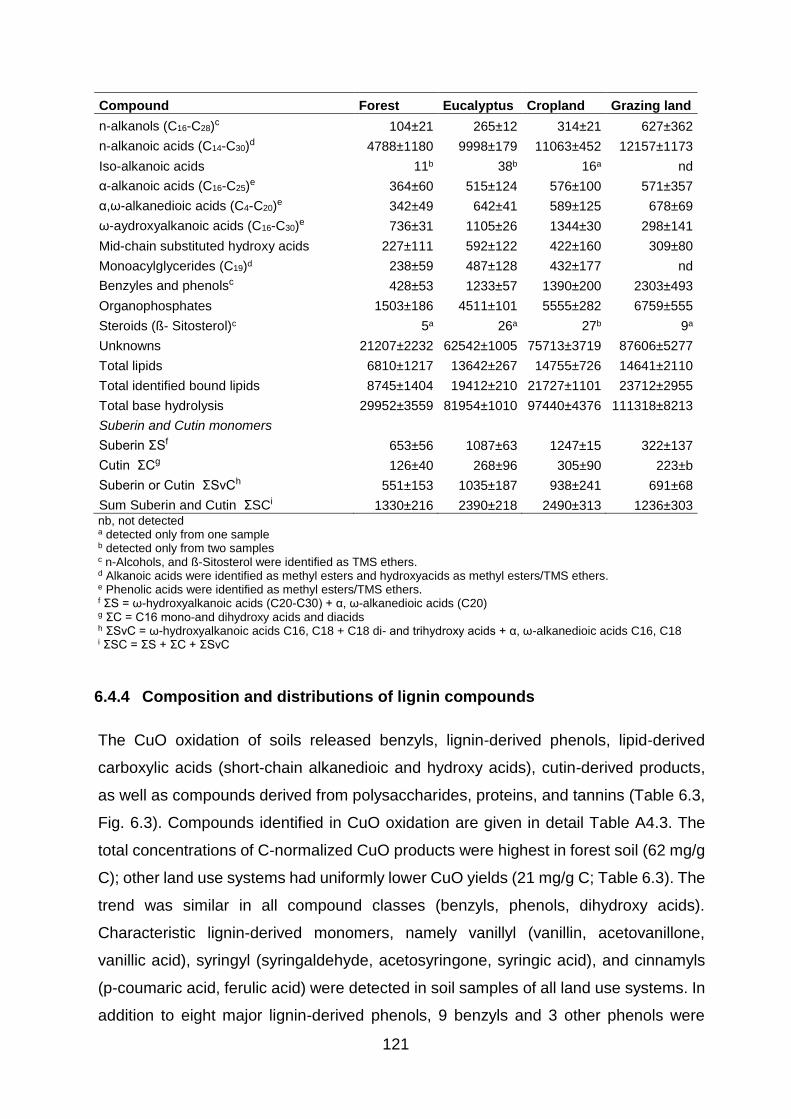
121
Compound Forest Eucalyptus Cropland Grazing land
n-alkanols (C16-C28)c 104±21 265±12 314±21 627±362
n-alkanoic acids (C14-C30)d 4788±1180 9998±179 11063±452 12157±1173
Iso-alkanoic acids 11b 38b 16a nd
α-alkanoic acids (C16-C25)e 364±60 515±124 576±100 571±357
α,ω-alkanedioic acids (C4-C20)e 342±49 642±41 589±125 678±69
ω-aydroxyalkanoic acids (C16-C30)e 736±31 1105±26 1344±30 298±141
Mid-chain substituted hydroxy acids 227±111 592±122 422±160 309±80
Monoacylglycerides (C19)d 238±59 487±128 432±177 nd
Benzyles and phenolsc 428±53 1233±57 1390±200 2303±493
Organophosphates 1503±186 4511±101 5555±282 6759±555
Steroids (ß- Sitosterol)c 5a 26a 27b 9a
Unknowns 21207±2232 62542±1005 75713±3719 87606±5277
Total lipids 6810±1217 13642±267 14755±726 14641±2110
Total identified bound lipids 8745±1404 19412±210 21727±1101 23712±2955
Total base hydrolysis 29952±3559 81954±1010 97440±4376 111318±8213
Suberin and Cutin monomers
Suberin ΣSf 653±56 1087±63 1247±15 322±137
Cutin ΣCg 126±40 268±96 305±90 223±b
Suberin or Cutin ΣSvCh 551±153 1035±187 938±241 691±68
Sum Suberin and Cutin ΣSCi 1330±216 2390±218 2490±313 1236±303 nb, not detected a detected only from one sample b detected only from two samples c n-Alcohols, and ß-Sitosterol were identified as TMS ethers. d Alkanoic acids were identified as methyl esters and hydroxyacids as methyl esters/TMS ethers. e Phenolic acids were identified as methyl esters/TMS ethers. f ΣS = ω-hydroxyalkanoic acids (C20-C30) + α, ω-alkanedioic acids (C20) g ΣC = C16 mono-and dihydroxy acids and diacids h ΣSvC = ω-hydroxyalkanoic acids C16, C18 + C18 di- and trihydroxy acids + α, ω-alkanedioic acids C16, C18 i ΣSC = ΣS + ΣC + ΣSvC
6.4.4 Composition and distributions of lignin compounds
The CuO oxidation of soils released benzyls, lignin-derived phenols, lipid-derived
carboxylic acids (short-chain alkanedioic and hydroxy acids), cutin-derived products,
as well as compounds derived from polysaccharides, proteins, and tannins (Table 6.3,
Fig. 6.3). Compounds identified in CuO oxidation are given in detail Table A4.3. The
total concentrations of C-normalized CuO products were highest in forest soil (62 mg/g
C); other land use systems had uniformly lower CuO yields (21 mg/g C; Table 6.3). The
trend was similar in all compound classes (benzyls, phenols, dihydroxy acids).
Characteristic lignin-derived monomers, namely vanillyl (vanillin, acetovanillone,
vanillic acid), syringyl (syringaldehyde, acetosyringone, syringic acid), and cinnamyls
(p-coumaric acid, ferulic acid) were detected in soil samples of all land use systems. In
addition to eight major lignin-derived phenols, 9 benzyls and 3 other phenols were
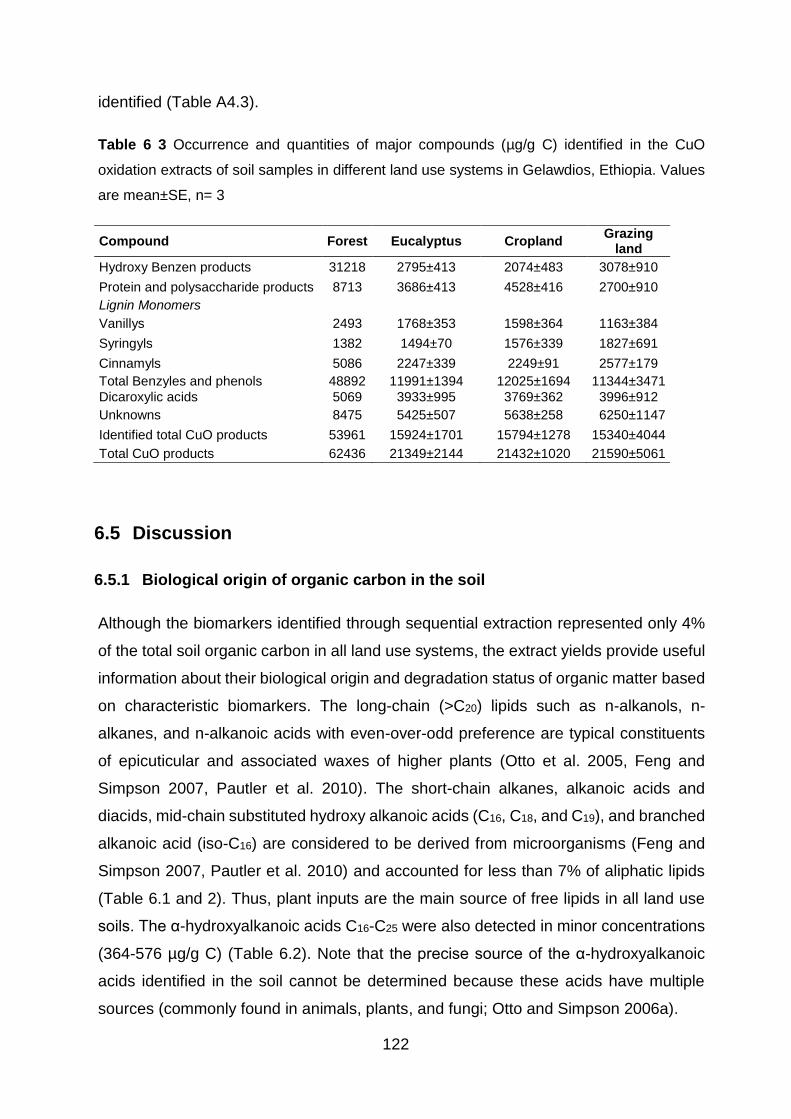
122
identified (Table A4.3).
Table 6 3 Occurrence and quantities of major compounds (µg/g C) identified in the CuO
oxidation extracts of soil samples in different land use systems in Gelawdios, Ethiopia. Values
are mean±SE, n= 3
Compound Forest Eucalyptus Cropland Grazing
land
Hydroxy Benzen products 31218 2795±413 2074±483 3078±910
Protein and polysaccharide products 8713 3686±413 4528±416 2700±910
Lignin Monomers
Vanillys 2493 1768±353 1598±364 1163±384
Syringyls 1382 1494±70 1576±339 1827±691
Cinnamyls 5086 2247±339 2249±91 2577±179
Total Benzyles and phenols 48892 11991±1394 12025±1694 11344±3471
Dicaroxylic acids 5069 3933±995 3769±362 3996±912
Unknowns 8475 5425±507 5638±258 6250±1147
Identified total CuO products 53961 15924±1701 15794±1278 15340±4044
Total CuO products 62436 21349±2144 21432±1020 21590±5061
6.5 Discussion
6.5.1 Biological origin of organic carbon in the soil
Although the biomarkers identified through sequential extraction represented only 4%
of the total soil organic carbon in all land use systems, the extract yields provide useful
information about their biological origin and degradation status of organic matter based
on characteristic biomarkers. The long-chain (>C20) lipids such as n-alkanols, n-
alkanes, and n-alkanoic acids with even-over-odd preference are typical constituents
of epicuticular and associated waxes of higher plants (Otto et al. 2005, Feng and
Simpson 2007, Pautler et al. 2010). The short-chain alkanes, alkanoic acids and
diacids, mid-chain substituted hydroxy alkanoic acids (C16, C18, and C19), and branched
alkanoic acid (iso-C16) are considered to be derived from microorganisms (Feng and
Simpson 2007, Pautler et al. 2010) and accounted for less than 7% of aliphatic lipids
(Table 6.1 and 2). Thus, plant inputs are the main source of free lipids in all land use
soils. The α-hydroxyalkanoic acids C16-C25 were also detected in minor concentrations
(364-576 µg/g C) (Table 6.2). Note that the precise source of the α-hydroxyalkanoic
acids identified in the soil cannot be determined because these acids have multiple
sources (commonly found in animals, plants, and fungi; Otto and Simpson 2006a).

123
Since, longer chain alkanes such as C31 and C33 are derived from grass tissues (Pautler
et al. 2010; and references therein), the ratios of C31/(C23+C27) alkanes from solvent
extraction are used to distinguish between inputs from grasses and trees. The highest
value of this ratio obtained in grass land (0.27) and cropland (0.14) provides evidence
of higher inputs from grasses to soil organic carbon compared to eucalyptus soil (0.12)
and forest soil (0.10) (Table 6.4). Nonetheless, these values are very small compared
to the 0.37-0.60 reported by Marshall (2014). The lower value in our study may reflect
a lower vegetation input of cropland and grazing land in the Ethiopian highlands. All
crop residues were collected from cropland for cattle feeding. The high cattle population
– above carrying capacity in grazing land (23 livestock units ha-1 versus the carrying
capacity of 2 livestock units ha-1; Desta et al. 2000) – largely limit the input of grasses
to the soil organic carbon. Accordingly, the biological origin of the soil carbon in
cropland and grazing land could be largely from previous inputs of higher plants before
conversion of land use. Further study based on carbon dating would help determine
when this carbon was added to the soil in each land use system.
The relative inputs of C3 and C4 plants can be differentiated based on the carboxylic
acid ratio of C24/(n-C22 + n-C26) (Wiesenberg and Schwark 2006). Wiesenberg and
Schwark proposed that ratios >0.67 indicate C4 origin, otherwise C3 origin. This is
because lipid compounds containing C24 alkanoic acids are from C4 plants, while C22
and C26 acids are from C3 plants (Amelung et al. 2008). In our study, the ratios of these
compounds are all below 0.67 (Table 6.1), indicating the carbon source is mainly from
C3 plants.
The steroid cholesterol is derived from non-plant sources (Otto et al. 2005, Otto and
Simpson 2007). It contributes only minimally to organic carbon as reflected by its low
abundance in all land use types (<2% of free lipids; Table 6.1). Ergosterol, a specific
biomarker for living fungal biomass (Montgomery et al. 2000, Mille-Lindblom et al.
2004), was detected in minor amounts in forest (0.6%) and eucalyptus soil (1.1%). It
was absent in cropland and grazing land soil (Table A4.1), indicating relatively low
fungal activity (Pautler et al. 2010) or fast turnover rates of ergesterol (Wallander et al.
2013). The concentration of ergosterol in eucalyptus is about 2.6 times that of natural
forest soil (Table A4.1). This may reflect a higher association of mycorrhizal fungi in
eucalyptus versus natural forest (Burgess et al. 1993, Thomson et al. 1994, Ouaryi et
al. 2016). Since ergosterol was not detected in most soil samples, microbial inputs

124
associated with the mid-chain alkanoic acids are more likely from bacteria. Since
cropland and grazing land are open lands that receive intensive ultraviolet radiation,
photochemical degradation may be the other possible factor to decrease the ergosterol
content (Li et al. 2015). Previous studies reported that ergosterol lasts for only a few
days after cell death and decreases quickly (43% in 24 h) under photochemical
degradation (Mille-Lindblom et al. 2004, Feng and Simpson 2011).
Other cyclic lipids of phytosterols in solvent extracts include campesterol, stigmasterol,
β-sitosterol, stigmastanol, and sitosterone; they are commonly found in steroids of
higher plant waxes (Otto and Simpson 2007, Feng 2009, Pautler et al. 2010, Li et al.
2015). Sesquiterpense of aromadendrene, γ-elemene, ledol, globulol, and farnesol are
natural chemical compounds found in a variety of aromatic plants and reported as
common constituents of essential oil, mainly from eucalyptus (Song et al. 2009, Elaissi
et al. 2012, Rowshan and Tarakemeh 2013, Thanighaiarassu and Sivamani 2013). We
detected them in large quantities in eucalyptus and cropland soils (Table 6.1). Their
occurrence in the cropland may be caused by the eucalyptus trees planted adjacent to
cropland. Triterpenoids of the ß-amyrin, α-amyrin, lupeol, erythrodiol, and oleanolic acid
type are typical biomarkers for angiosperms (Otto and Simpson 2006a, Jäger et al.
2009, Pautler et al. 2010). They were found in large quantities in eucalyptus soil (Table
6.1). Particularly erythrodiol and oleanolic acid are major components in essential oils
of eucalyptus leaves (Santos et al. 1997, Jäger et al. 2009). Generally, the composition
of the aliphatic lipids, with even-over-odd predominance, revealed a major input of lipids
into SOM derived from higher plant waxes (Otto et al. 2005; references therein).
Monosaccharide carbohydrates are widely distributed among plants, animals, and
microorganism membranes and are not suitable as biomarkers specific for distinct
organisms (Otto et al. 2005, Feng and Simpson 2007, Pautler et al. 2010). By contrast,
the stress protectant trehalose is attributed to organisms such as fungi, bacteria, and
insects, but is only rarely found in plants (Otto et al. 2006, Feng et al. 2007). Therefore,
the disaccharide trehalose found in the solvent extracts is most likely derived from a
non-plant source and can be used as a microbial biomarker. Nonetheless, because the
fungal biomarker ergosterol was not detected in the cropland and grazing land soils,
occurring in small amounts in forest and eucalyptus soil only, the contribution of fungi
to SOM is unlikely.

125
Suberin is a characteristic biomacromolecule of roots and bark of vascular plants (Otto
and Simpson 2007) and is thus used to trace mainly belowground plant origin,
assuming that bark contributes little to soil organic carbon. Cutin is produced in the
epidermal cells to form a protective barrier on leaves, flowers, and fruits of vascular
plants and can be used to estimate the input of organic matter originating from shoot
biomass (Nierop 1998, Nierop et al. 2006). Due to the compositional similarity of
suberin and cutin, the differentiation of characteristic biomarkers for both sources is
difficult. Some compounds derived from plant waxes or the lingo-cellulose complex
show signal overlap (Otto and Simpson 2006a), and they were excluded here as
characteristic suberin or cutin biomarkers. Therefore, from the detected compounds,
typical suberin biomarkers (ΣS) include ω-hydroxyalkanoic acid (≥C20) and α, ω-
alkanoic acids (C16-C24). They represented 1.4% (322 µg/g C) of the identified bound
lipids in grazing land soil to 7.5% (1247 µg/g C) in forest soil (Table 6.2). Compounds
of C16 mono-and dihydroxy acids and diacids represented <1.4% (126-305 µg/g C;
Table 6.2) and are used as cutin biomarker signatures (Otto and Simpson 2006a).
Biomarkers derived from either suberin or cutin (ΣSvC) are ω-hydroxyalkanoic acids
C16, C18, C18 di- and trihydroxy acids, and α, ω-alkanedioic acids C16, C18 and
accounted for 3-6% of identified bound lipids (551-1035 µg/g C; Table 6.2). The
combined suberin and cutin represented 8-19% of aliphatic lipids. The relative
proportion of suberin was about 2 time that of cutin in forest, eucalyptus, and cropland
soils, whereas in grazing land soil, suberin and cutin concentrations were similar (Table
6.4). This shows that below-ground biomass in the forest ecosystem contributed about
twice the organic carbon than above-ground parts. In cropland, all crop residues are
harvested for either cattle fodder or fuel and, thus, the remaining part is crop roots that
may increase the contribution of suberin compared to cutin. Since forest-derived
monomers are preserved in the soil for more than 50 years, the higher suberin content
in cropland seemed to be derived from former forest vegetation (Mendez-Millan et al.
2012). The lower suberin concentration in grazing land may be because grass-derived
monomers degrade rapidly compared with forest-derived monomers of the same
compound classes in tropical soils (Hamer et al. 2012). The ω-hydroxy carboxylic acids
and α, ω-alkanedioic acids of forest origin may have been stabilized in the soils by
bonding to soil minerals. In addition, Hamer and coauthors found a more than five-times
higher concentration of x,16-diOH C16:0 compounds in forest litter than in grass
biomass, potentially explaining the greater proportion of forest-derived monomer. In

126
general, the composition of the bound soil lipids extracted by base hydrolysis such as
aliphatic lipids (6.8-14.8 mg/g C) and phenols (0.4-2.3 mg/g C; Table 6.1, 6.2) mainly
originate from suberin, cutin, and plant surface wax esters (Hedges and Ertel 1982,
Otto et al. 2005, Otto and Simpson 2006a). This reflects major inputs by vascular plants
and minor inputs by microorganisms to soil carbon.
The observed phenols (p-hydroxyacetophenone, 4-hydroxybenzoic acid, vanillyl,
acetovanillone, vanillic acid, isovanillic acid, p-coumaric acid, ferulic acid compounds)
are reported to be characteristic for lignin monomers (Otto and Simpson 2007).
Nonetheless, the phenols identified in the bound lipid fractions are likely derived from
the ester-bound aromatic constituents of suberin and the lignocellulose complex
(Kögel-Knabner 2000, Otto et al. 2005). This is because the applied base hydrolysis
cleaves esters, but not ethers (Otto et al. 2005). According to Pautler et al. (2013), the
observed phenols after alkaline hydrolysis are likely derived from suberin and/or
lignocelluloses as they are connected by ester bonds within these biopolymers (rather
than from the lignin cell wall biopolymers, which are linked by ether bonds).
CuO oxidation released mainly lignin-specific phenols from the lignin polymer. These
refer to the sum of vanillyl, syringyl, and cinnamyl phenols, which are essential
components of the cell walls of vascular plants (Table 6.3; Goñi and Hedges 1992).
These phenols accounted for more than 52% of all CuO products in all land use
systems (Table 6.3). Benzoic acid, p-hydroxybenzaldehyde, 4-hydroxybenzeneacetic
acid, pyrrol-1-carboxylic acid, and short-chain dicarboxylic acids are considered to be
the oxidation products of proteinaceous and polysaccharide material (Goñi and
Montgomery 2000, Otto et al. 2005, Tesi et al. 2014). In contrast, tannins and other
flavonoids have been suggested as the sources for 3,5-dihydroxy benzoic acid (Otto et
al. 2006). According to Otto et al. (2005) the exact source of 2,4,6-trihydroxybenzoic
acid is not known, although similar compounds such as 1,2,4- and 1,3,5-
trihydroxybenzoic acids have been reported as black carbon markers from thermal
alteration of parent structures (Kuo et al. 2008). These compounds may thus indicate
biochar residues remaining after conversion of forest to other land use types using fire
as a tool or during (illegal) charcoal production in the forest. Overall, the occurrences
and concentrations of benzyls and lignin-derived phenols are about 2 times higher in
forest soil than other land use soils. This points to a predominant input of recalcitrant
organic carbon into the forest ecosystem derived from higher plants.
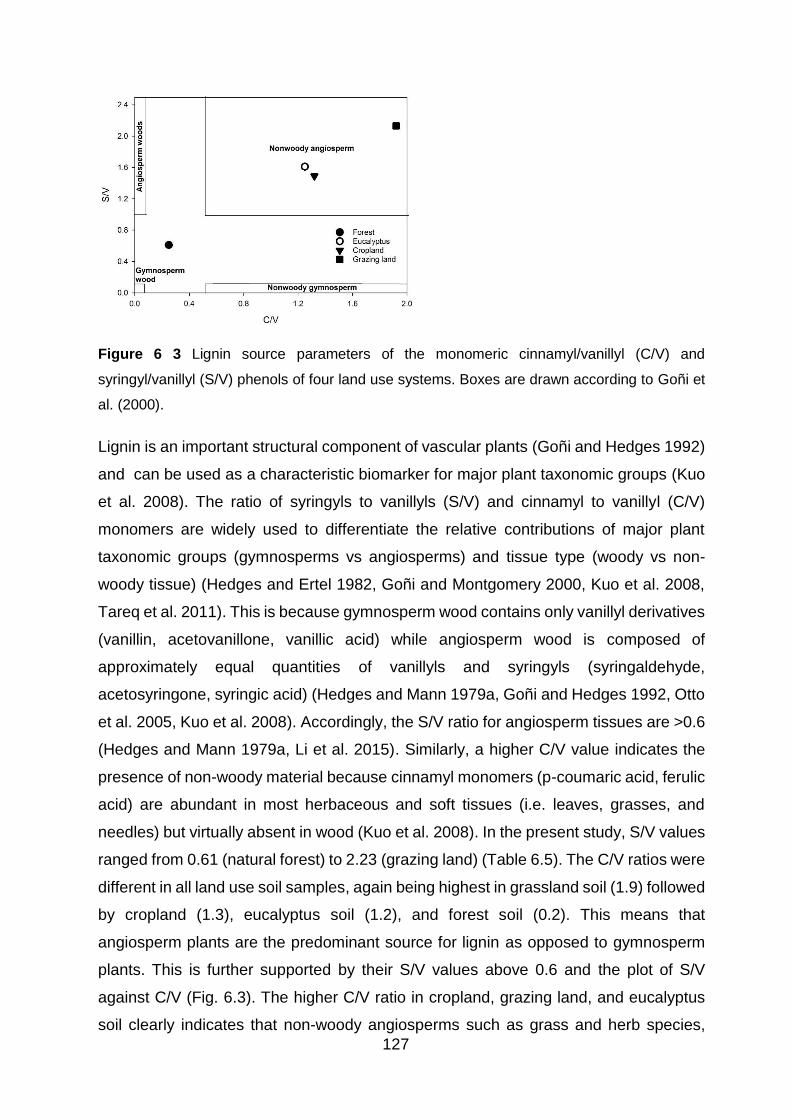
127
Figure 6 3 Lignin source parameters of the monomeric cinnamyl/vanillyl (C/V) and
syringyl/vanillyl (S/V) phenols of four land use systems. Boxes are drawn according to Goñi et
al. (2000).
Lignin is an important structural component of vascular plants (Goñi and Hedges 1992)
and can be used as a characteristic biomarker for major plant taxonomic groups (Kuo
et al. 2008). The ratio of syringyls to vanillyls (S/V) and cinnamyl to vanillyl (C/V)
monomers are widely used to differentiate the relative contributions of major plant
taxonomic groups (gymnosperms vs angiosperms) and tissue type (woody vs non-
woody tissue) (Hedges and Ertel 1982, Goñi and Montgomery 2000, Kuo et al. 2008,
Tareq et al. 2011). This is because gymnosperm wood contains only vanillyl derivatives
(vanillin, acetovanillone, vanillic acid) while angiosperm wood is composed of
approximately equal quantities of vanillyls and syringyls (syringaldehyde,
acetosyringone, syringic acid) (Hedges and Mann 1979a, Goñi and Hedges 1992, Otto
et al. 2005, Kuo et al. 2008). Accordingly, the S/V ratio for angiosperm tissues are >0.6
(Hedges and Mann 1979a, Li et al. 2015). Similarly, a higher C/V value indicates the
presence of non-woody material because cinnamyl monomers (p-coumaric acid, ferulic
acid) are abundant in most herbaceous and soft tissues (i.e. leaves, grasses, and
needles) but virtually absent in wood (Kuo et al. 2008). In the present study, S/V values
ranged from 0.61 (natural forest) to 2.23 (grazing land) (Table 6.5). The C/V ratios were
different in all land use soil samples, again being highest in grassland soil (1.9) followed
by cropland (1.3), eucalyptus soil (1.2), and forest soil (0.2). This means that
angiosperm plants are the predominant source for lignin as opposed to gymnosperm
plants. This is further supported by their S/V values above 0.6 and the plot of S/V
against C/V (Fig. 6.3). The higher C/V ratio in cropland, grazing land, and eucalyptus
soil clearly indicates that non-woody angiosperms such as grass and herb species,

128
which contain abundant cinnamyl phenols, are the predominant sources of lignin.
Although degradation might affect these ratios and can alter the original plant
composition (Tesi et al. 2014), both angiosperm woody and non-woody tissues are
equally important for SOM accumulation in the natural forest ecosystem. These lowest
S/V and C/V values in natural forest soil reflect a mixture of different angiosperm
species and to some extent gymnosperm woody and non-woody (leaves) sources. This
indicates that the forest was probably once covered by an Olive-Juniper-Podocarpus
forest as evidenced by the presence of Podocarpus falcatus species inside the natural
forest and scattered Olea africana and Juniperus procera trees in the surrounding area
and compounds of Gelawdios church. This, however, calls for further study on the
history of the forest. Gymnosperms yield higher amounts of ω-C16/∑C16 acids relative
to angiosperms (Pisani et al. 2013) and this may lower the S/V ratios, as indicated by
more abundant ω-C16 acids in the forest ecosystem (Table 6.4). Other studies also
reported lower S/V values for conifer-dominated species. For example, Li et al. (2015)
reported a low S/V value (0.23) under pine-dominated forests and 0.84 for farmland
soils. The lignin-derived phenols (V:S:C) ratios are also used to estimate the relative
contributions of woody and non-woody angiosperms; the latter exhibit a ratio of about
1:1:1 (Hedges and Mann 1979a, Kögel-Knabner 2000, Otto et al. 2006). The lignin-
derived phenols detected in our samples exhibited a V:S:C ratio of 4:2:1 (forest soil)
and 1:1:1 (eucalyptus, cropland, and grazing land soils) (Table 6.4). The relatively equal
ratio in the latter three soils further confirms that lignin-derived phenols originate mainly
from non-woody angiosperm sources. This is consistent with the S/V and C/V ratios.
The higher V:S:C ratio in the natural forest indicates that woody angiosperms contribute
significantly to soil organic carbon. However, the lignin index-phenols and their ratios
do not give a fine resolution spectrum to distinguish sources among angiosperms,
gymnosperms, and non-woody contributions. This is because of the complex mixture
of different plants as well as the different diagenetic reactivity of cinnamyl, syringyl, and
vanillyl groups (Tareq et al. 2011). To improve the detection of lignin sources, another
useful parameter (lignin phenol vegetation index – LPVI) was developed by Tareq et
al. (2004). LPVI provides a better resolution of the lignin sources based on the relative
abundances of vanillyl, syringyl, and cinnamyl-type phenols. Tareq et al. (2011)
calculated the LPVI values from lignin-phenol data of Hedges and Mann (1979). This
yielded values of 1 for gymnosperm wood, 3-27 for non-woody gymnosperm tissues,
67-415 for angiosperm wood, and 176-2782 for non-woody angiosperms (Tareq et al.

129
2004). In the present study, the LPVI values of the natural forest (92) are in the range
of angiosperm woods. In contrast, soil samples from eucalyptus (880), cropland (901),
and grazing land (2051) show characteristics for non-woody angiosperms, which is
consistent with plots of S/V vs C/V (Fig. 6.4).
6.5.2 Degradation stage of plant organic matter
Soil lipids are highly resistant to biodegradation (Kögel-Knabner 2000), but the extent
of lipid degradation depends on microbial activity and physico-chemical conditions
(Nierop et al. 2005, Otto et al. 2005). The aliphatic/cyclic lipid ratio is a general
parameter to assess the degradation of free lipids in the soil (Otto and Simpson 2006a,
Li et al. 2015). After solvent extraction, the aliphatic/cyclic lipids ratios were 22 (forest),
1.4 (eucalyptus), 3.4 (cropland), and 4.4 (grazing land). The lowest value (1.4) suggests
that cyclic lipids are better preserved in eucalyptus soil. In the natural forest soil, the
highest ratio is due to less accumulation of cyclic lipids and continuous input of aliphatic
lipids into the soil.
The suberin/cutin ratios are important parameters to describe bound lipid degradation.
Suberin is more resistant to degradation than cutin due to its contents of ω-
hydroxyalkanoic acids and phenolic units (Goñi & Hedges 1990; Otto et al. 2005; and
references therein). In this study the suberin/cutin ratios were 1.9 (forest), 1.7
(eucalyptus), 1.9 (cropland), and 1.1 (grazing land) (Table 6.4). The higher ratios in
forest, eucalyptus, and cropland soils compared to grazing land indicate preferential
degradation of cutin over suberin. Mendez-Millan et al. (2012) showed that 58-63% of
the forest-derived suberin monomers such as C18 di- and trihydroxy and epoxy acids
are preserved in the soil for more than 53 years. In contrast, cutin monomers such as
C16 mono-and dihydroxy acids, amounting to a maximum of 37-42%, are preserved
only for 20 years in pastureland. Moreover, grass-derived monomers degrade rapidly
compared with forest-derived monomers of the same compound classes due to their
chemical structure (Hamer et al. 2012). This yields the lowest ratio in grassland soil
(Table 6.4). The ω-hydroxy carboxylic acids and α, ω-alkanedioic acids of forest origin
may have been stabilized by bonding to soil minerals. The higher suberin/cutin ratio in
cropland is mainly due to the removal of above-ground biomass after crop harvest along
with selective degradation of cutin over suberin. Increased ratios of ω-C16/∑C16 and
ω-C18/∑C18 have been reported with progressing cutin degradation in marine
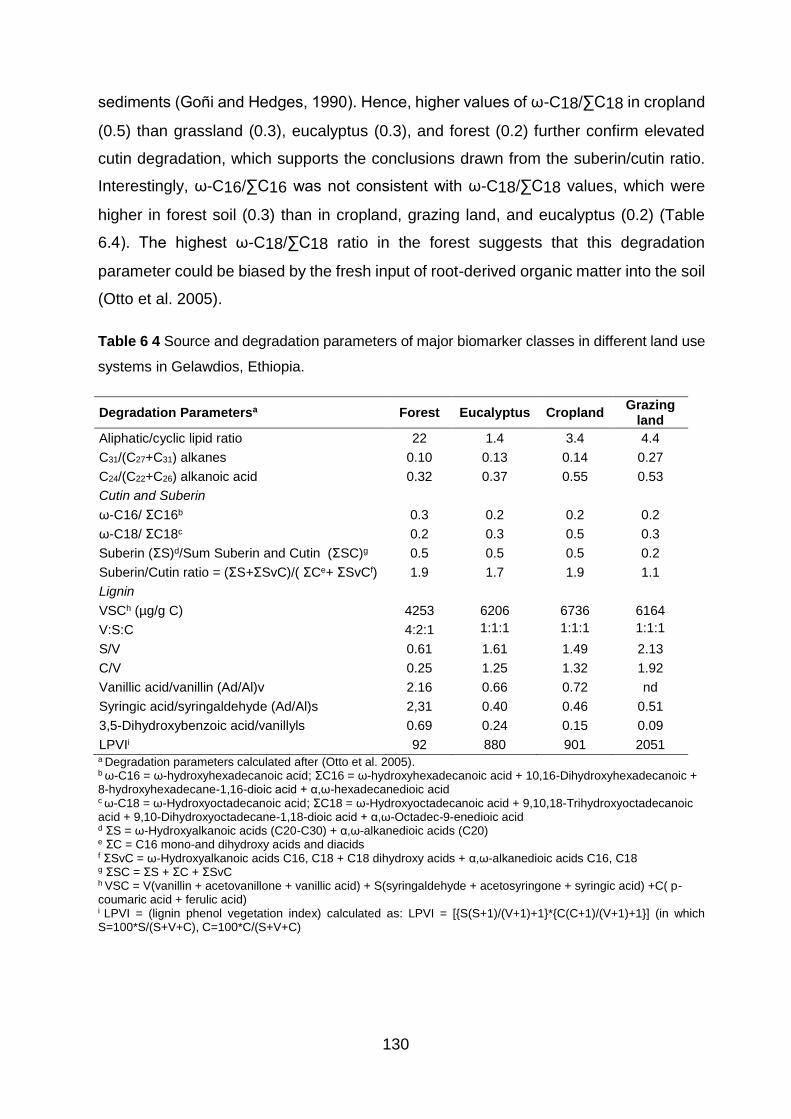
130
sediments (Goñi and Hedges, 1990). Hence, higher values of ω-C18/∑C18 in cropland
(0.5) than grassland (0.3), eucalyptus (0.3), and forest (0.2) further confirm elevated
cutin degradation, which supports the conclusions drawn from the suberin/cutin ratio.
Interestingly, ω-C16/∑C16 was not consistent with ω-C18/∑C18 values, which were
higher in forest soil (0.3) than in cropland, grazing land, and eucalyptus (0.2) (Table
6.4). The highest ω-C18/∑C18 ratio in the forest suggests that this degradation
parameter could be biased by the fresh input of root-derived organic matter into the soil
(Otto et al. 2005).
Table 6 4 Source and degradation parameters of major biomarker classes in different land use
systems in Gelawdios, Ethiopia.
Degradation Parametersa Forest Eucalyptus Cropland Grazing
land
Aliphatic/cyclic lipid ratio 22 1.4 3.4 4.4
C31/(C27+C31) alkanes 0.10 0.13 0.14 0.27
C24/(C22+C26) alkanoic acid 0.32 0.37 0.55 0.53
Cutin and Suberin
ω-C16/ ΣC16b 0.3 0.2 0.2 0.2
ω-C18/ ΣC18c 0.2 0.3 0.5 0.3
Suberin (ΣS)d/Sum Suberin and Cutin (ΣSC)g 0.5 0.5 0.5 0.2
Suberin/Cutin ratio = (ΣS+ΣSvC)/( ΣCe+ ΣSvCf) 1.9 1.7 1.9 1.1
Lignin
VSCh (µg/g C) 4253 6206 6736 6164
V:S:C 4:2:1 1:1:1 1:1:1 1:1:1
S/V 0.61 1.61 1.49 2.13
C/V 0.25 1.25 1.32 1.92
Vanillic acid/vanillin (Ad/Al)v 2.16 0.66 0.72 nd
Syringic acid/syringaldehyde (Ad/Al)s 2,31 0.40 0.46 0.51
3,5-Dihydroxybenzoic acid/vanillyls 0.69 0.24 0.15 0.09
LPVIi 92 880 901 2051 a Degradation parameters calculated after (Otto et al. 2005). b ω-C16 = ω-hydroxyhexadecanoic acid; ΣC16 = ω-hydroxyhexadecanoic acid + 10,16-Dihydroxyhexadecanoic + 8-hydroxyhexadecane-1,16-dioic acid + α,ω-hexadecanedioic acid c ω-C18 = ω-Hydroxyoctadecanoic acid; ΣC18 = ω-Hydroxyoctadecanoic acid + 9,10,18-Trihydroxyoctadecanoic acid + 9,10-Dihydroxyoctadecane-1,18-dioic acid + α,ω-Octadec-9-enedioic acid d ΣS = ω-Hydroxyalkanoic acids (C20-C30) + α,ω-alkanedioic acids (C20) e ΣC = C16 mono-and dihydroxy acids and diacids f ΣSvC = ω-Hydroxyalkanoic acids C16, C18 + C18 dihydroxy acids + α,ω-alkanedioic acids C16, C18 g ΣSC = ΣS + ΣC + ΣSvC h VSC = V(vanillin + acetovanillone + vanillic acid) + S(syringaldehyde + acetosyringone + syringic acid) +C( p-coumaric acid + ferulic acid) i LPVI = (lignin phenol vegetation index) calculated as: LPVI = [{S(S+1)/(V+1)+1}*{C(C+1)/(V+1)+1}] (in which S=100*S/(S+V+C), C=100*C/(S+V+C)

131
Lignin is among the best preserved components in the soil organic matter due to its
greater stability to biodegradation (Kuo et al. 2008). Its recalcitrance plays a significant
role in the biospheric carbon cycle (Colberg, 1988) and has been extensively used in
characterizing the degradation stage of soil organic matter (Tesi et al. 2014).
Biodegradation of lignin is mainly governed by white-rot and brown-rot fungi by
oxidative cleavage (Hedges and Mann 1979a). The yields of VSC and the ratios of
lignin-derived phenolic acids to corresponding aldehydes of vanillyl and syringyl units
(Ad/Al)v, (Ad/Al)s, and 3,5-dihydroxybenzoic acid (DHBA)/V are generally used as
indicators of the level of lignin degradation in the soil (Hedges and Mann 1979a, Goñi
and Montgomery 2000, Otto et al. 2005, Otto and Simpson 2006b). Based on total
lignin-index phenols, the yields of VSC were in the order: cropland (6.7 mg/g C), grazing
land and eucalyptus (6.2 mg/g C), and forest soil (4.2 mg/g C). Accordingly, lignin
compounds are least preserved in forest soils (Table 6.4). Since these values are
normalized to soil carbon contents, the lowest value of lignin in the natural forest can
be biased by the different inputs of organic matter into the soil. The presence of fungi
in this ecosystem, however, as indicated by the ergosterol biomarker, may contribute
to lower VSC yields in the forest but has little effect on the eucalyptus soil. The lowest
S/V (0.6) and C/V (0.2) ratios are in forest, with values in the other land use soils
exceeding 1.2 (Table 6.4). This indicates preferential degradation of syringyls and
cinnamyls monomers. The explanation is that cinnamyls link the carbohydrates and
lignin in the ligno-cellulose complex, making them more accessible to decomposition
than vanillyls (Feng & Simpson 2007, and references therein). Syringyls were also
reported to degrade faster than vanillyls in the environment, while vanillyls accumulate
more than cinnamyls and syringyls (Hedges et al. 1984, Otto et al. 2005)
Previous studies report that fresh angiosperm wood has Ad/Al ratios of 0.1-0.5 for both
syringyl and vanillyl units; higher values of 0.2-1.6 were recorded for fresh non-woody
tissues such as leaves and grasses (Goñi and Hedges 1992, Otto and Simpson 2006b,
Pisani et al. 2013). The Ad/Alv ratios for the samples analyzed in the present study were
highest in forest soil (2.2) followed by cropland and grazing land soil (0.7). In addition,
the (Ad/Al)s ratio was higher (2.3) in the forest soil than eucalyptus soil (0.4), cropland
and grazing land (0.5) (Table 6.4) suggesting the preferential degradation of
syringaldehyde over syringic acid. Finally, forest soils may be more oxidized due to the
optimal soil conditions for lignin-degrading fungi, as indicated by the presence of

132
ergosterol and the soil pH, temperature, moisture values (Pisani et al. 2013). The 3, 5-
dihydroxybenzoic acid to vanillyls ratio was in the order: forest (0.69), eucalyptus (0.24),
cropland (0.15), and grazing land (0.09) (Table 6.4). The highest value in forest soil
shows increased degradation of tannins because 3,5-dihydroxybenzoic acid after CuO
oxidation was derived from tannin oxidation (Li et al. 2015). Combining the evidence of
the degradation parameters, enhanced degradation of lignin occurred in the forest soil
as indicated by elevated acid/aldehyde (Ad/Al) ratios of vanillyl and syringyl units and
3,5-dihydroxybenzoic acid /vanillyls.
6.6 Conclusions
Overall, aliphatic lipids and lignin accounted for 46-84% of the total detected chemicals.
This demonstrated that the major input of soil organic carbon is from higher plants. The
short-chain alkanoic acids, ergosterol, and the disaccharide trehalose accounted for
less than 7% of aliphatic lipids in all land use soils, pointing to minor microbial inputs.
Ergosterol, however, a fungal biomarker, was not detected in the cropland and grazing
land soils, and occurred in only small amounts in forest and eucalyptus soil.
Accordingly, the microbial input more likely originates from bacteria. The high S/V and
C/V ratios (except for the forest ecosystem) indicate that non-woody angiosperm
tissues – mainly from C3 plants – are major contributors among the taxonomic groups.
This is because syringyl and cinnamyl monomers are abundant in most herbaceous
and soft tissues but virtually absent in wood. The lowest S/V and C/V values obtained
in forest soil reflect a mixture of different sources including woody angiosperms and
gymnosperms. A higher VSC ratio and higher LPVI values in the forest soil than in other
land use systems further provide a better resolution of the lignin sources: woody
angiosperms are equally important in contributing to soil organic carbon. The
terpenoids such as the ß-amyrin, α-amyrin, lupeol, and erythrodiol are also typical
biomarker for angiosperms. In the forest, eucalyptus, and cropland soils, root biomass
contributed C input in the soil twice compared to above-ground parts, as revealed by
their relative proportions of suberin and cutin. In addition, the highest suberin to cutin
ratio indicates that suberin is more resistant to degradation than cutin (due to contents
of ω-hydroxyalkanoic acids and high mineral bonding capacity of phenolic units in
suberin). The ratios of lignin-index phenols revealed that syringyls and cinnamyls are
degraded faster than vanillyls. In general, lignin degradation was enhanced in the forest
soil, as indicated by elevated acid/aldehyde (Ad/Al) ratios of vanillyl and syringyl units.

133
7 Microbial soil respiration and its dependency on
substrate availability and temperature in four contrasting
land use systems
7.1 Abstract
Soil organic carbon (SOC) is a fundamental part of the carbon cycle. Soil microorganisms are
the center of the biological activity regulating the breakdown and accumulation of SOC stocks
as well as the CO2 efflux to the atmosphere. Nevertheless, the extent to which substrate
availability and quality affects microbial functioning is largely unknown. This study was therefore
designed to determine the amount of CO2 effluxes from different ecosystems to the atmosphere
along a temperature gradient with or without carbon sources. Glucose, lignin, and starch were
added as proxies for substrate availability. Ammonium nitrate and calcium phosphate were also
included to determine whether microbes are limited by nutrient deficiency. The investigation is
based on the use of the MicroResp method containing a cresol red gel carrying colorimetric
indicator. This enables measuring the amount of CO2 evolved from the soil along the
temperature gradient. We found that basal respiration ranged from 0.4±0.1 to 0.9±0.1 µg CO2-
C g soil-1 h-1, which is consistent with the SOC content. Nonetheless, a strong C use efficiency
was observed at lower C content in the soil, as indicated by elevated specific basal respiration
in cropland and grazing land. Respiration rates were higher in the soils where carbon sources
(glucose, lignin, starch) were added, and microbes showed a preferential degradation of
substrates. In contrast to the conventional belief that aromatic structures are recalcitrant and
stable in soil, lignin was preferentially degraded over starch substrate. Microbial respiration in
the nutrient (N and P) addition soils was significantly lower than in soils supplied with carbon
sources, but did not differ significantly from control soil. The contribution of microbial biomass
carbon (MBc) to soil organic carbon was generally <1% of total organic carbon in the soil for all
land use types, but forest soil had twice-higher levels of MBc than other land use systems. The
ratio of basal respiration to microbial biomass carbon (metabolic quotient; qCO2) ranged from
2.1 (forest soil) to 3.9 (eucalyptus soil), reflecting higher microbial C use efficiency under
eucalyptus. The microbial communities released more CO2 per unit microbial biomass under
acidic soil conditions. Generally, microbial respiration increased with temperature in all land
use systems, but at a decreasing rate above 25°C, and the Q10 values declined at high
temperature and at lower substrate availability.
Keywords: SOC, microbial respiration, temperature sensitivity, substrate, nutrient, microbial
quotient, land use change, CO2 efflux

134
7.2 Introduction
Soil is the largest terrestrial carbon pool (3150 Pg C; Fan et al., 2016), accounting for
more than four times the amount of carbon in the atmosphere, (750 Pg C) and is a
fundamental part of carbon cycle and life on Earth (Batjes 1996, Hiederer and Köchy
2011). This pool is a result of an imbalance between organic matter production and its
decomposition, and is a source of CO2 to the atmosphere (Yuste et al. 2007). Soil
microorganisms regulate the breakdown of organic matter and determine SOC stocks
in the soil as well as the CO2 efflux to the atmosphere (Bradford 2013). These
microorganisms decompose organic matter and assimilate the produced low molecular
weight organic compounds from the soil environment. Thus, microorganisms
themselves account for 1-3% of soil organic C (Martens 1995, Wang and Sainju 2014).
Soil microbial respiration releases about 60 Pg of carbon per year to the atmosphere
as CO2 (Shao et al. 2013). The source of this respiration is microbial decomposition of
soil organic matter (Kalinina et al. 2010) to obtain energy for growth and functioning.
The size of the CO2 flux is determined by temperature (Bradford et al. 2008), moisture
(Meisner et al. 2015), carbon input (Yuste et al. 2007), and nutrients such as N and P
(Gnankambary et al. 2008, Birgander et al. 2014). Respiration rates respond more
positively to increasing temperature than do photosynthetic rates (Frank et al. 2015).
This imbalance due to elevated microbial respiration rates implies that increased
temperature will lead to an increase in the net flux from soil to the atmosphere (Bradford
2013).
The extent to which indirect vs direct temperature effects on SOC dynamics along with
substrate availability and quality in different ecosystems is largely untested (Rousk et
al. 2012). Many studies have shown that soil microbial respiration increases with
temperature (Davidson and Janssens 2006, Allison et al. 2010, Karhu et al. 2014,
Mayer et al. 2016). This information, however, is uncertain because the response of
soil microbial communities to changing temperatures has the potential to either
decrease (Bradford et al. 2008, 2010) or increase (Hartley et al. 2008) soil microbial
respiration. For example, a study conducted from the Arctic to the Amazon along the
climate gradient reported that the strongest enhancing response occurred in soil from
cold climatic regions (Karhu et al. 2014). Similarly, Hartley et al. (2008) reported that
microbial activity acclimates to long-term temperature, greatly reducing the potential for
enhanced respiration in Arctic soils. Moreover, the relationship between soil microbial

135
respiration and substrate availability (C and nutrient sources) with changing
temperature at different land use systems remains uncertain. Many studies have
limitations in that they consider the response of microbial respiration to temperature
change, substrate availability, substrate quality, and anthropogenic effects only in one
particular ecosystem. Incorporating carbon and nutrient sources in different ecosystem
soils – and incubating soils at different temperatures prior to measuring CO2 production
– overcomes this limitation and may provide more realistic outcomes that reflect actual
ecosystem responses to temperature change. This makes it important to measure
biological activity in the soil, because this reflects the capacity of soil to support soil
microorganisms, the level of microbial activity, SOM content and its decomposition.
Conversion of tropical forest to other land use systems reduces soil carbon stocks
either through loss of C input or through C mineralization by microbes (Joergensen
2010). Guillaume et al., (2016) observed up to 70% soil organic carbon (SOC) losses
in the topsoil under oil palm and rubber plantations in Indonesia compared to rainforest.
SOC losses are also associated with a decline of soil quality and functions related to
microbial activity (Guillaume et al. 2016). This calls for determining the sensitivity of
biological indicators to land-use changes as well as to substrate and temperature
fluctuations. Microbial activity depends on SOC availability (Moorhead et al. 2014).
Thus, the basal respiration per unit of SOC can be used as an indicator of C availability
(Guillaume et al. 2016). The production of enzymes requires microbial investment of
energy in the form of C and nutrients, particularly N and P (Schimel and Weintraub
2003, Hernández et al. 2010). Accordingly, their activity may increase with the
increasing availability of labile C and nutrient fertilization. Microbial activity can be
simply regulated by providing C sources and by N or P fertilization (Schlentner and
Cleve 1985, Thirukkumaran and Parkinson 2000, Hernández et al. 2010). However,
the effects of these additions depend on the enzyme of interest (produced by
microorganisms) and on the quality of the substrates (Hernández et al. 2010). Further
indices such as the metabolic quotient (basal respiration to microbial biomass ratio;
qCO2) are the most widely used and reflect the carbon-use efficiency of microbial
communities. They are also suggested to reflect the soil function of supporting microbial
growth (Anderson and Domsch 1990, Joergensen 2010, Guillaume et al. 2016).
Many methods have been developed to measure microbial biomass and microbial
respiration, including the fumigation method (Jenkinson and Powlson 1976, Vance et

136
al. 1987), the substrate-induced respiration method (Anderson and Domsch 1978), and
the adenosine triphosphate (ATP) extraction method (Tate and Jenkinson 1982). In this
study, the microbial activity and biological indices were determined using the
MicroResp method in sieved soils under laboratory incubation. MicroResp is a
microplate-based respiration system which enables analyzing basal respiration and
substrate-induced respiration simultaneously (Campbell et al. 2003, Chapman et al.
2007, Renault et al. 2013). This helps in testing a range of C sources and substrate
qualities at different temperatures. This approach measures and characterizes CO2
production by microbial communities when the microbes feed on soil organic matter
(Campbell et al. 2003). Profiling microbial respiration on various substrates and at
various temperature provides an insight into the effects of rising atmospheric CO2
concentrations and the concomitant projected climate change scenarios, which have
unknown implications for long-term soil C storage (Renault et al. 2013). We
hypothesized that the efficiency of microbes in decomposing soil organic matter, and
the amount of CO2 efflux, are limited by the availability of C substrates and by N & P
deficiencies. The chemical composition of different soil SOM pools varies, being
composed of easily degradable compounds and of compounds more resistant to
microbial attack (Hernández et al. 2010). Which chemical structures are preferentially
consumed by microbes in the field at different temperatures remains unclear. The aim
of this study was therefore to determine fluxes of carbon from the soil to the atmosphere
at different temperatures with or without carbon sources in soils of different land use
systems. We examined the effect of temperature variation with the addition of glucose,
lignin, and starch as proxies for substrate availability of contrasting recalcitrance on
CO2 efflux from soil relative to soils receiving water alone. We also examined the effect
of N and P nutrients to determine whether microbial activity is N or P limited. This was
done on soils from four contrasting land use systems: natural forest, eucalyptus
plantation, grazing land, and cropland. The amount of CO2 evolved from the soil and
its response to temperature change was measured using the MicroResp method on
sieved soils under laboratory incubation.

137
7.3 Materials and methods
7.3.1 Site description
Four land use systems (natural forest, eucalyptus plantation, cropland, grazing land)
were selected in Gelawdios, north-central Ethiopia. The study area (Gelawdios) is
located at 11°38’25’’ N and 37°48’55’’ E at an elevation of 2500 m above sea level
(a.s.l.). The topography is typical of volcanic landscapes, comprising volcanic rocks
with ragged and undulating landforms. The major soil type is classified as Cambisols
with a clay loam texture according to the World Reference Base for soil resources
(WRB 2014). The mean annual rainfall at Gelawdios is 1220 mm, with a unimodal rainy
season with the main rainfall months between June and September; the average
annual temperature is 19°C (Wassie et al. 2009). Although Ethiopia is geographically
located in the tropics, the climate of the study areas is temperate with dry winters and
warm summers (Cwb) according to the Köppen-Geiger climate classification (Peel et
al. 2007). The forest is a pristine Afromontane dry forest composed mostly of a mixture
of indigenous tree species and is almost exclusively confined to sacred groves
associated with churches. The dominant tree species are Albizia schimperiana,
Apodytes dimidiata, Calpurnia aurea, Croton macrostachyus, Ekebergia capensis,
Maytenus arbutifolia, and Schefflera abyssinica. The eucalyptus stand was established
on formerly common grazing land in 1985 and was successively thinned to its current
density about 3000 trees per hectare. The adjacent grazing land and cropland were
converted from natural forest approximately 50 years ago (exact date unknown). The
grazing land was used as communal grazing lands for herds of animals; it is highly
degraded, nearly bare ground. The cropland has been cultivated without fallow, and all
crop residues are harvested to feed cattle at home.
7.3.2 Soil sampling
Soils were collected in September 2015 from the four land use systems (natural forest,
eucalyptus plantation, grazing land, and cropland). Soils were taken in the first 10 cm
depth from three sampling points along a transect line. The distance between sampling
points was 200 m in the natural forest and 50 m in the other land use types. Soils were
sieved through a 2 mm stainless sieve in Ethiopia and transported to Vienna for further
laboratory analysis. They were stored at 4°C in zipped bags until use in the

138
experiments. All lab analyses were done at University of Natural Resources and Life
Sciences, Vienna (Austria).
7.3.3 Total C, N, and soil pH analysis
A subsample of approximately 3-5 g soil was dried at 105°C for 48 h for total carbon
and nitrogen analysis. Detailed procedures are provided in chapter 2. Total carbon and
nitrogen concentrations were determined on a CN elemental analyser (Truspec CNS
LECO, St. Joseph, USA). The analytical precision and reproducibility of the instrument
were determined based on an analysis of a calibration sample standard (Part No 502-
309 purchased from LECO). Soil pH was determined in 1:3 soil suspensions in 0.01 M
CaCl2 solution using a digital potentiometric pH-meter.
7.3.4 Determination of moisture content and water holding capacity
Soil moisture content was determined using subsamples of about 10 g soil, which were
dried at 105°C for 24 h. Another portion of soil samples (~20 g) was used to determine
water-holding capacity. The soils were soaked in water for 24 h and drained before
weighing. The soils were then dried in the oven at 105°C and re-weighed. We also
conducted preliminary runs to determine the appropriate water content in relation to
water holding capacity for maximum respiration. This prevented restricting the gaseous
exchange due to too wet or dry soil. From the available moisture content, the soils were
supplemented with deionized water to the desired water content of the soil (40, 50, 60,
70, 80, 90, and 100% of water holding capacity). Our pre-test showed that microbial
respiration was highest at 50-70% water holding capacity; below or above these values,
respiration declined. We therefore adjusted our samples to 40% water holding capacity
so that, after a carbon source in solution is added, the moisture content increased to
the required level of 50-70% water holding capacity. The soil without substrate also
received deionized water to reach 50-70% water holding capacity.
7.3.5 Preparation of carbon and nutrient sources
Three carbon sources (glucose, starch, lignin) that are ecologically relevant to soil were
selected based on the previously described studies (Campbell et al. 2003, García-
Palacios et al. 2013, Bao et al. 2015). Two additional nutrient sources (ammonium
nitrate and calcium phosphate) were included to determine whether microbes are
limited by N or P deficiencies. The concentrations of carbon and nutrient sources were

139
prepared so as to deliver 30 mg C, P or N per g of soil water (MicroResp technical
manual).
7.3.6 Preparing agar and indicator solution for the MicroResp
MicroResp detection plates were prepared according to the protocol provided by
MicroResp technical manual. In short, 3% Purified Agar (3 g per 100 ml) was prepared
in deionized water and dissolved by heating in a microwave to 120 °C until the agar
dissolves. Indicator solution was prepared in 1 L (18.25 mg cresol red, 16.77 g KCl,
0.315 g NaHCO3) and dissolved at 50°C, then filled up to end volume. Cresol is a water-
soluble indicator that changes color when it reacts with CO2. Once the temperature of
each solution has equilibrated at 60°C, the indicator solution and agar were transferred
into a beaker and mixed thoroughly with constant stirring. Then, 150 µL aliquots were
dispensed into the microplate using an 8-channel pipette. Precautions were taken
during dispensing to avoid bubbles in the wells. The plates were then stored in the dark
at room temperature in a desiccator with self-indicating soda lime on the base and a
beaker of water to keep the atmosphere CO2 free and moist.
7.3.7 Soil filling and incubation
We used sieved soil because it homogenizes the soils, removes rocks and roots, and
simplifies soil filling in the deep-well plates. The MicroResp consists of a 96-deep well
microplate to hold soil samples with a capacity of 1.2 ml, a 96-well detection plate,
MicroResp seal (silicone rubber gasket), and metal clamp to hold the three parts firmly
together (Campbell et al. 2003). Soil was placed into the deep-well plate by using a
third device made from a 300-µl well microtiter plate from which the bottom had been
removed and replaced with a Perspex sliding base. In each deep well of the microplate,
approx. 0.3 g of soil was filled from either natural forest, eucalyptus plantation, grazing
land, or cropland. Each land use type had 24 wells, six treatments with four replicates
each. After removing the filling device, the deep well plates were covered with Parafilm
for incubation, after adjusting to the desired water holding capacity for normalization.
Incubation was carried out as described by Campbell et al. (2003) and Creamer et al.
(2014) at different temperatures. In summary, microplates holding about 0.3 gm of soil
per deep well plate – at 40% of WHC, corresponding to optimal edaphic conditions for
microbial respiration – were incubated at room temperature (ca. 22°C) for 1 week in the
dark for normalizations (Bérard et al. 2016). This pre-incubation also enables recovery

140
after disturbances induced by sieving and humidity adjustment. During incubation, the
plates were covered with Parafilm to prevent water loss and soil contamination.
7.3.8 Measurement of soil respiration
We quantified microbial respiration by measuring the amounts of CO2 production during
incubation with or without substrates. After one-week normalization, four wells per land
use system were each supplemented with 25 μl of the desired substrate solutions of
glucose, starch, lignin, ammonium, phosphate, or water (control), which set them to 50-
70% water holding capacity. Three replications (plates) for each temperature scale (5,
15, 25, 35°C) were made. The plate containing the agar gel was read colorimetrically
before incubation with an absorbance microplate reader (VMAX; Molecular Devices,
Wokingham, United Kingdom) at 570 nm (t0). The plates were then sealed together with
a silicone rubber gasket with interconnecting holes between the corresponding wells.
The assembly is held together firmly with a purpose-designed clamp. In each closed
well of a 96-well microplate, moist soils with or without C, N and P substrates were
incubated for 6 h (Creamer et al. 2014) at 5, 15, 25, and 35°C in the presence of an
agar gel carrying cresol red as indicator dye. After 6 h of incubation, the plates
containing the gel were immediately re-read (t1).
7.3.9 Quantifying CO2 efflux and biological indices
The absorbance after 6 h (t1) was normalized for any differences recorded at time zero
(t0) and then converted to headspace CO2 concentration. This involved a calibration
curve obtained from a calibration dye color (detection gel) with different CO2
concentrations by using equilibrated solutions (Drage et al. 2012).
The CO2 product rate is calculated by converting the 6 h %CO2 data to µg CO2-C g soil-
1 h-1 using gas constants, constants for incubation temperature in °C (T), headspace
volume (vol) in the well (µl), fresh weight (fwt) of soil per well (g), incubation time in
hours (t), and soil sample% dry weight (dwt) (Bérard et al., 2011; Campbell et al., 2003,
MicroResp technical manual).
R (µg CO2-C) = {[(%CO2/100)*vol*(44/22.4)*(12/44)*(273/(273+T))]/soil fwt*(soil% dwt/100)}/t (1)
Soil carbon loss per annum was estimated by the following equation according to Feng
and Simpson (2008):

141
Soil carbon loss = r*365*24*(12/44)*10-6*100% (2)
where r is the measured microbial respiration rate in the units of µgCO2 g soil -1 h-1.
Valuable information such as microbial biomass, turnover time, and efficiency of carbon
use can be derived from the CO2 efflux. Active microbial biomass carbon was estimated
from glucose-induced respiration according to the method of Anderson and Domsch
(1978). Hence, microbial biomass C (MBC) can be estimated as:
MBC (µg C g-1 soil) = 40.4SIR+0.37 (3)
where SIR is the substrate induced respiration rate, mg CO2 g soil-1 h-1.
To assess the ecosystem stress and metabolic rates of soil microbial communities, we
calculated the microbial metabolic quotient (qCO2) as the amount of CO2-C produced
(basal respiration; BR) per unit of microbial biomass carbon (Bérard et al. 2011).
Metabolic quotient (qCO2) = BR/MBC (4)
The proportion of the microbial biomass carbon (MBC) to total soil organic carbon
(referred as microbial quotient; Anderson and Domsch, 1989) is determined as:
%MBc/SOC = MBC*100/SOC (5)
Temperature sensitivity (respiration rate, R at a given temperature, T) can be modelled
using a two-parametric exponential function to describe the relationships between soil
CO2 fluxes and soil temperature. We chose the Arrhenius model to describe the soil
CO2 efflux rates R as a function of soil temperature T as:
R = aeb/T (6)
where a and b are fitted parameters (Creamer et al. 2014).
Despite the complexity of soil respiration controls, many studies describe the response
of soil respiration with Q10 functions in which respiration responds exponentially to
increasing temperature (Reichstein et al. 2005, Creamer et al. 2014, Karhu et al. 2014).
The Q10 is the parameter that determines the temperature response of the soil CO2
efflux (i.e. the factor by which efflux increases for an increase in temperature (T) by

142
10°C). Hence, we calculated the Q10 values based on fitted parameter (b) values from
the whole temperature range according to Gershenson et al. (2009) as:
Q10 = e (b*10) (7)
The effects of incubation temperature during the experiment and substrate input
(treatment effect) were tested by one-way ANOVA, after checking for homogeneity of
variance using the Levene test. ANOVAs were calculated using IBM SPSS (version
21). Differences between groups were determined using Tukey’s post hoc test. Linear
regression was used to evaluate the relationships between different variables such as
SOC, water-holding capacity, soil pH, basal respiration, substrate induced respiration,
and microbial biomass carbon. SigmaPlot (version 13) was used for graphs. The
significance level was determined at α = 0.05. Data are presented as mean ±standard
error (SE).
7.4 Results
7.4.1 Soil chemical and physical properties in four land use systems
The mean soil organic carbon (SOC) was about 2.5-3.5 times higher in the forest soil
than in other land use systems (Table 7.1). Accordingly, conversion of forest to cropland
and grazing land reduced the soil carbon content by 68%-72% at a rate of ca.1.4 mg C
g soil-1 yr-1. In contrast, reafforestation of degraded grazing land with eucalyptus
plantations increased the SOC stock by 24% at the rate of 0.3 mg C g soil-1 yr-1.
Nevertheless, the rehabilitation process did not return the soil carbon content to a level
equal to the rate of soil carbon loss. The carbon content difference between cropland,
grazing land, and eucalyptus soils was not significant. The C/N ratio varied between 11
and 14, and the differences between land use systems were not significant. A similar
trend was observed for total N, being 4 times higher in the forest soil than in cropland
and eucalyptus soils, and 3 times higher than in grazing land soil. Soil pH(CaCl2) in the
top 10 cm was moderately acidic with a mean range of 4.9-5.8; values were highest in
the forest and lowest in eucalyptus soil (Table 7.1). The percent of sand content was
higher in the grazing land, whereas the clay content was very low compared to other
land use systems (Table 7.1). Soil water holding capacity (WHC) was plotted against
SOC, revealing a significant positive relationship (r2 = 0.88; P<0.001; Fig 7.1a). The
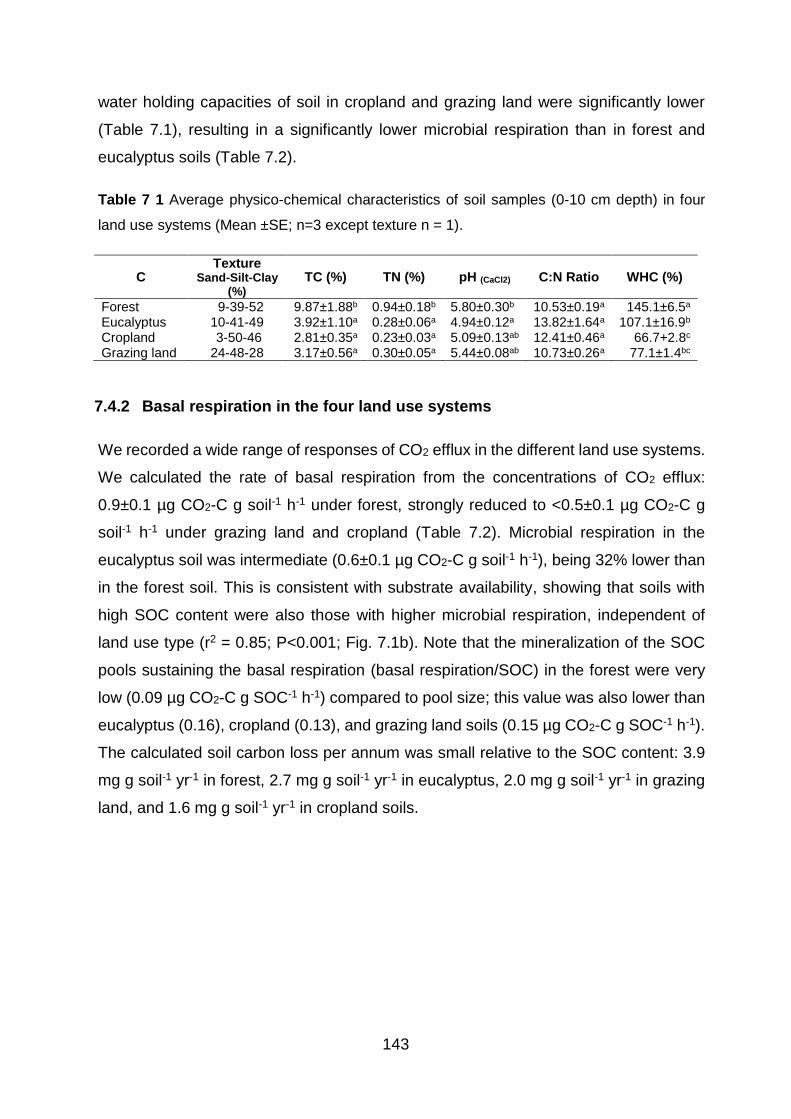
143
water holding capacities of soil in cropland and grazing land were significantly lower
(Table 7.1), resulting in a significantly lower microbial respiration than in forest and
eucalyptus soils (Table 7.2).
Table 7 1 Average physico-chemical characteristics of soil samples (0-10 cm depth) in four
land use systems (Mean ±SE; n=3 except texture n = 1).
C Texture
Sand-Silt-Clay (%)
TC (%) TN (%) pH (CaCl2) C:N Ratio WHC (%)
Forest 9-39-52 9.87±1.88b 0.94±0.18b 5.80±0.30b 10.53±0.19a 145.1±6.5a Eucalyptus 10-41-49 3.92±1.10a 0.28±0.06a 4.94±0.12a 13.82±1.64a 107.1±16.9b Cropland 3-50-46 2.81±0.35a 0.23±0.03a 5.09±0.13ab 12.41±0.46a 66.7+2.8c Grazing land 24-48-28 3.17±0.56a 0.30±0.05a 5.44±0.08ab 10.73±0.26a 77.1±1.4bc
7.4.2 Basal respiration in the four land use systems
We recorded a wide range of responses of CO2 efflux in the different land use systems.
We calculated the rate of basal respiration from the concentrations of CO2 efflux:
0.9±0.1 µg CO2-C g soil-1 h-1 under forest, strongly reduced to <0.5±0.1 µg CO2-C g
soil-1 h-1 under grazing land and cropland (Table 7.2). Microbial respiration in the
eucalyptus soil was intermediate (0.6±0.1 µg CO2-C g soil-1 h-1), being 32% lower than
in the forest soil. This is consistent with substrate availability, showing that soils with
high SOC content were also those with higher microbial respiration, independent of
land use type (r2 = 0.85; P<0.001; Fig. 7.1b). Note that the mineralization of the SOC
pools sustaining the basal respiration (basal respiration/SOC) in the forest were very
low (0.09 µg CO2-C g SOC-1 h-1) compared to pool size; this value was also lower than
eucalyptus (0.16), cropland (0.13), and grazing land soils (0.15 µg CO2-C g SOC-1 h-1).
The calculated soil carbon loss per annum was small relative to the SOC content: 3.9
mg g soil-1 yr-1 in forest, 2.7 mg g soil-1 yr-1 in eucalyptus, 2.0 mg g soil-1 yr-1 in grazing
land, and 1.6 mg g soil-1 yr-1 in cropland soils.
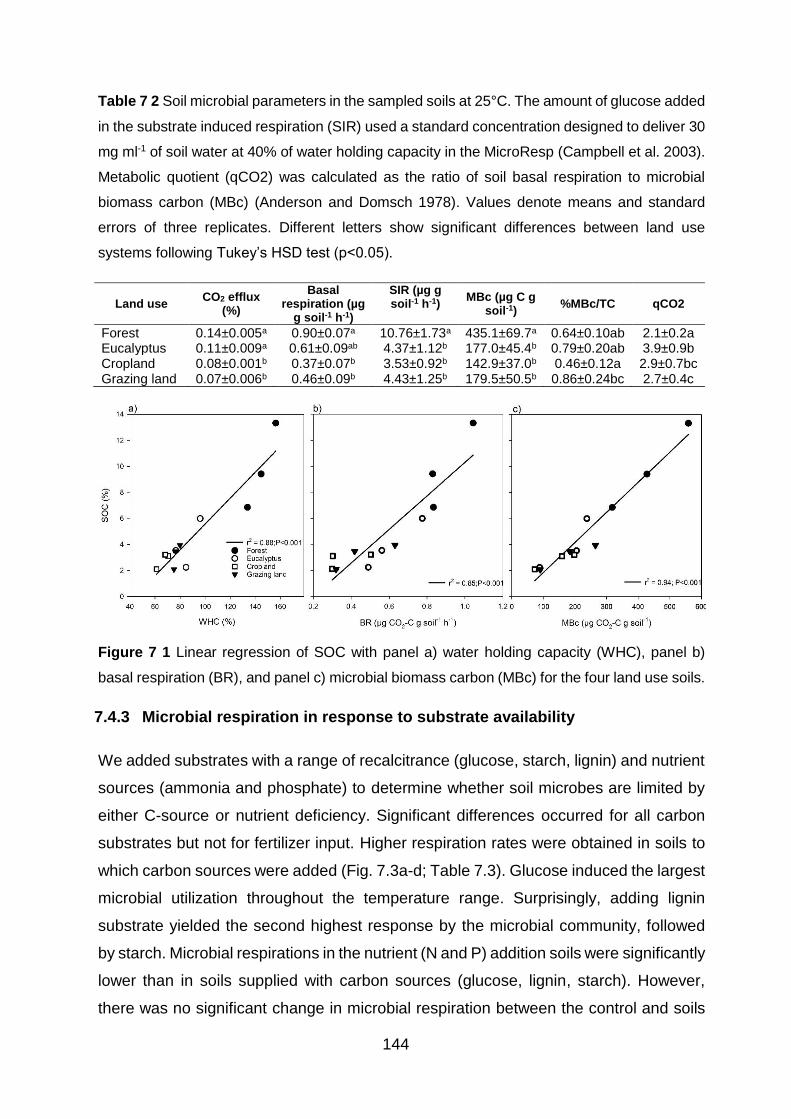
144
Table 7 2 Soil microbial parameters in the sampled soils at 25°C. The amount of glucose added
in the substrate induced respiration (SIR) used a standard concentration designed to deliver 30
mg ml-1 of soil water at 40% of water holding capacity in the MicroResp (Campbell et al. 2003).
Metabolic quotient (qCO2) was calculated as the ratio of soil basal respiration to microbial
biomass carbon (MBc) (Anderson and Domsch 1978). Values denote means and standard
errors of three replicates. Different letters show significant differences between land use
systems following Tukey’s HSD test (p<0.05).
Land use CO2 efflux
(%)
Basal respiration (µg
g soil-1 h-1)
SIR (µg g soil-1 h-1)
MBc (µg C g soil-1)
%MBc/TC qCO2
Forest 0.14±0.005a 0.90±0.07a 10.76±1.73a 435.1±69.7a 0.64±0.10ab 2.1±0.2a Eucalyptus 0.11±0.009a 0.61±0.09ab 4.37±1.12b 177.0±45.4b 0.79±0.20ab 3.9±0.9b Cropland 0.08±0.001b 0.37±0.07b 3.53±0.92b 142.9±37.0b 0.46±0.12a 2.9±0.7bc Grazing land 0.07±0.006b 0.46±0.09b 4.43±1.25b 179.5±50.5b 0.86±0.24bc 2.7±0.4c
Figure 7 1 Linear regression of SOC with panel a) water holding capacity (WHC), panel b)
basal respiration (BR), and panel c) microbial biomass carbon (MBc) for the four land use soils.
7.4.3 Microbial respiration in response to substrate availability
We added substrates with a range of recalcitrance (glucose, starch, lignin) and nutrient
sources (ammonia and phosphate) to determine whether soil microbes are limited by
either C-source or nutrient deficiency. Significant differences occurred for all carbon
substrates but not for fertilizer input. Higher respiration rates were obtained in soils to
which carbon sources were added (Fig. 7.3a-d; Table 7.3). Glucose induced the largest
microbial utilization throughout the temperature range. Surprisingly, adding lignin
substrate yielded the second highest response by the microbial community, followed
by starch. Microbial respirations in the nutrient (N and P) addition soils were significantly
lower than in soils supplied with carbon sources (glucose, lignin, starch). However,
there was no significant change in microbial respiration between the control and soils
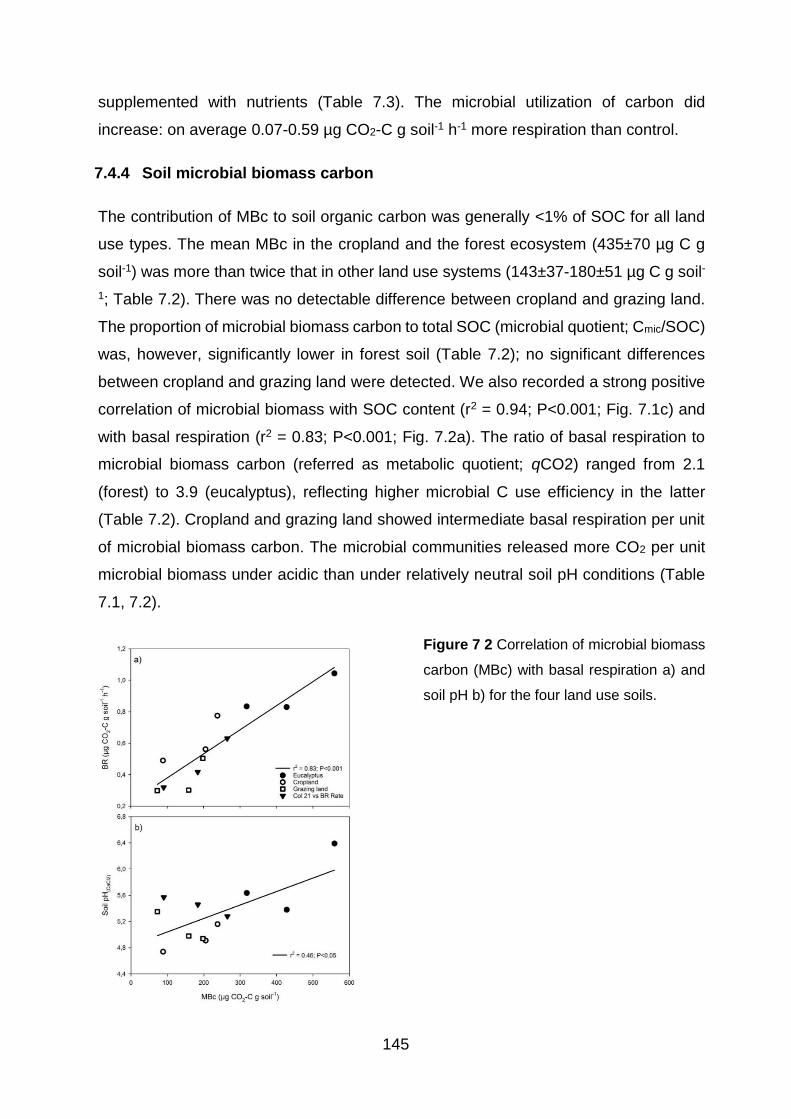
145
supplemented with nutrients (Table 7.3). The microbial utilization of carbon did
increase: on average 0.07-0.59 µg CO2-C g soil-1 h-1 more respiration than control.
7.4.4 Soil microbial biomass carbon
The contribution of MBc to soil organic carbon was generally <1% of SOC for all land
use types. The mean MBc in the cropland and the forest ecosystem (435±70 µg C g
soil-1) was more than twice that in other land use systems (143±37-180±51 µg C g soil-
1; Table 7.2). There was no detectable difference between cropland and grazing land.
The proportion of microbial biomass carbon to total SOC (microbial quotient; Cmic/SOC)
was, however, significantly lower in forest soil (Table 7.2); no significant differences
between cropland and grazing land were detected. We also recorded a strong positive
correlation of microbial biomass with SOC content (r2 = 0.94; P<0.001; Fig. 7.1c) and
with basal respiration (r2 = 0.83; P<0.001; Fig. 7.2a). The ratio of basal respiration to
microbial biomass carbon (referred as metabolic quotient; qCO2) ranged from 2.1
(forest) to 3.9 (eucalyptus), reflecting higher microbial C use efficiency in the latter
(Table 7.2). Cropland and grazing land showed intermediate basal respiration per unit
of microbial biomass carbon. The microbial communities released more CO2 per unit
microbial biomass under acidic than under relatively neutral soil pH conditions (Table
7.1, 7.2).
Figure 7 2 Correlation of microbial biomass
carbon (MBc) with basal respiration a) and
soil pH b) for the four land use soils.
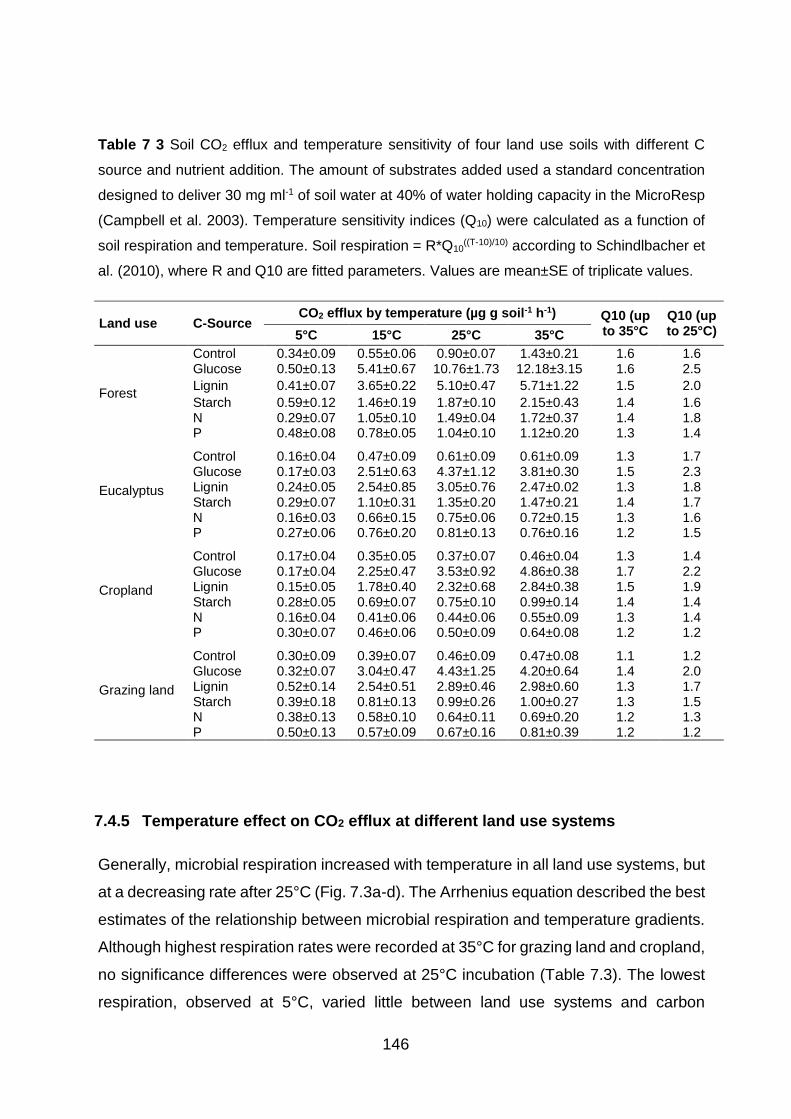
146
Table 7 3 Soil CO2 efflux and temperature sensitivity of four land use soils with different C
source and nutrient addition. The amount of substrates added used a standard concentration
designed to deliver 30 mg ml-1 of soil water at 40% of water holding capacity in the MicroResp
(Campbell et al. 2003). Temperature sensitivity indices (Q10) were calculated as a function of
soil respiration and temperature. Soil respiration = R*Q10((T-10)/10) according to Schindlbacher et
al. (2010), where R and Q10 are fitted parameters. Values are mean±SE of triplicate values.
Land use C-Source CO2 efflux by temperature (µg g soil-1 h-1) Q10 (up
to 35°C Q10 (up to 25°C) 5°C 15°C 25°C 35°C
Forest
Control 0.34±0.09 0.55±0.06 0.90±0.07 1.43±0.21 1.6 1.6 Glucose 0.50±0.13 5.41±0.67 10.76±1.73 12.18±3.15 1.6 2.5
Lignin 0.41±0.07 3.65±0.22 5.10±0.47 5.71±1.22 1.5 2.0
Starch 0.59±0.12 1.46±0.19 1.87±0.10 2.15±0.43 1.4 1.6 N 0.29±0.07 1.05±0.10 1.49±0.04 1.72±0.37 1.4 1.8 P 0.48±0.08 0.78±0.05 1.04±0.10 1.12±0.20 1.3 1.4
Eucalyptus
Control 0.16±0.04 0.47±0.09 0.61±0.09 0.61±0.09 1.3 1.7 Glucose 0.17±0.03 2.51±0.63 4.37±1.12 3.81±0.30 1.5 2.3 Lignin 0.24±0.05 2.54±0.85 3.05±0.76 2.47±0.02 1.3 1.8 Starch 0.29±0.07 1.10±0.31 1.35±0.20 1.47±0.21 1.4 1.7 N 0.16±0.03 0.66±0.15 0.75±0.06 0.72±0.15 1.3 1.6 P 0.27±0.06 0.76±0.20 0.81±0.13 0.76±0.16 1.2 1.5
Cropland
Control 0.17±0.04 0.35±0.05 0.37±0.07 0.46±0.04 1.3 1.4 Glucose 0.17±0.04 2.25±0.47 3.53±0.92 4.86±0.38 1.7 2.2 Lignin 0.15±0.05 1.78±0.40 2.32±0.68 2.84±0.38 1.5 1.9 Starch 0.28±0.05 0.69±0.07 0.75±0.10 0.99±0.14 1.4 1.4 N 0.16±0.04 0.41±0.06 0.44±0.06 0.55±0.09 1.3 1.4 P 0.30±0.07 0.46±0.06 0.50±0.09 0.64±0.08 1.2 1.2
Grazing land
Control 0.30±0.09 0.39±0.07 0.46±0.09 0.47±0.08 1.1 1.2 Glucose 0.32±0.07 3.04±0.47 4.43±1.25 4.20±0.64 1.4 2.0 Lignin 0.52±0.14 2.54±0.51 2.89±0.46 2.98±0.60 1.3 1.7 Starch 0.39±0.18 0.81±0.13 0.99±0.26 1.00±0.27 1.3 1.5 N 0.38±0.13 0.58±0.10 0.64±0.11 0.69±0.20 1.2 1.3 P 0.50±0.13 0.57±0.09 0.67±0.16 0.81±0.39 1.2 1.2
7.4.5 Temperature effect on CO2 efflux at different land use systems
Generally, microbial respiration increased with temperature in all land use systems, but
at a decreasing rate after 25°C (Fig. 7.3a-d). The Arrhenius equation described the best
estimates of the relationship between microbial respiration and temperature gradients.
Although highest respiration rates were recorded at 35°C for grazing land and cropland,
no significance differences were observed at 25°C incubation (Table 7.3). The lowest
respiration, observed at 5°C, varied little between land use systems and carbon
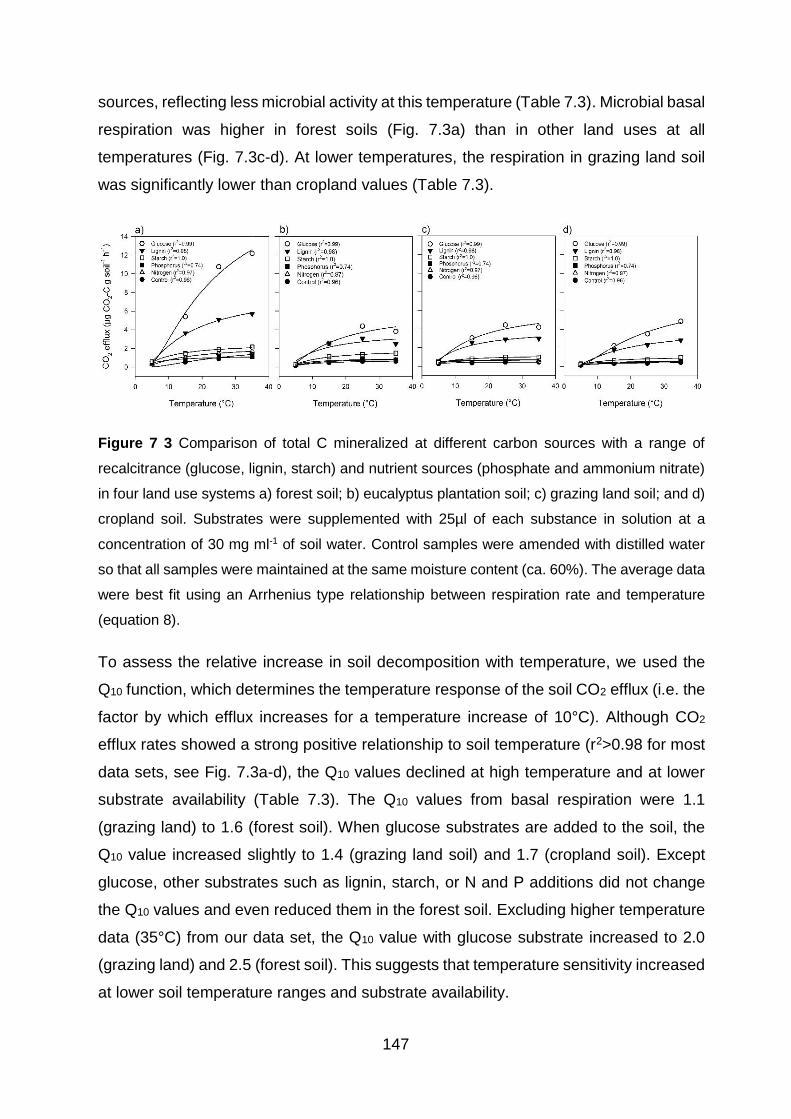
147
sources, reflecting less microbial activity at this temperature (Table 7.3). Microbial basal
respiration was higher in forest soils (Fig. 7.3a) than in other land uses at all
temperatures (Fig. 7.3c-d). At lower temperatures, the respiration in grazing land soil
was significantly lower than cropland values (Table 7.3).
Figure 7 3 Comparison of total C mineralized at different carbon sources with a range of
recalcitrance (glucose, lignin, starch) and nutrient sources (phosphate and ammonium nitrate)
in four land use systems a) forest soil; b) eucalyptus plantation soil; c) grazing land soil; and d)
cropland soil. Substrates were supplemented with 25µl of each substance in solution at a
concentration of 30 mg ml-1 of soil water. Control samples were amended with distilled water
so that all samples were maintained at the same moisture content (ca. 60%). The average data
were best fit using an Arrhenius type relationship between respiration rate and temperature
(equation 8).
To assess the relative increase in soil decomposition with temperature, we used the
Q10 function, which determines the temperature response of the soil CO2 efflux (i.e. the
factor by which efflux increases for a temperature increase of 10°C). Although CO2
efflux rates showed a strong positive relationship to soil temperature (r2>0.98 for most
data sets, see Fig. 7.3a-d), the Q10 values declined at high temperature and at lower
substrate availability (Table 7.3). The Q10 values from basal respiration were 1.1
(grazing land) to 1.6 (forest soil). When glucose substrates are added to the soil, the
Q10 value increased slightly to 1.4 (grazing land soil) and 1.7 (cropland soil). Except
glucose, other substrates such as lignin, starch, or N and P additions did not change
the Q10 values and even reduced them in the forest soil. Excluding higher temperature
data (35°C) from our data set, the Q10 value with glucose substrate increased to 2.0
(grazing land) and 2.5 (forest soil). This suggests that temperature sensitivity increased
at lower soil temperature ranges and substrate availability.

148
7.5 Discussion
7.5.1 Resource availability and carbon use efficiency of microorganisms
Soil respiration is a good indicator of overall soil biological activity and of the carbon
use efficiency by microorganisms. The mean soil respiration rates measured in this
study (0.4-0.9 µg CO2-C g soil-1 h-1) were similar to average tropical soil respiration
rates (0.42-0.83 µg CO2-C g soil-1 h-1; Joergensen, 2010). Our values, however, are
slightly higher than the mean basal respiration rate of 0.46 µg CO2-C g soil-1 h-1
measured in 62 temperate German soils as mentioned by Joergensen (2010).
We observed a clear decreasing pattern in the microbial respiration rate along a
decreasing SOC gradient (Fig. 7.1b-c). This indicates that soil C is the limiting factor
for microorganism activity. The greater soil respiration we recorded in the natural forest
suggests higher rates of soil carbon turnover than in other land use systems. Recently,
Oertel et al. (2016) reported higher soil respiration rates in forest than in other bare land
use types, and they correlated this with higher soil organic carbon contents. The
continuous input of below- and above-ground organic matter is a crucial energy source
for the microbial community in the forest ecosystem. Many studies have shown that
conversion of tropical forest to cropland or grazing land decreases the soil organic
matter content by up to 70% depending on the number of years under cultivation and
land use management (Fearnside et al. 1998, Deng et al. 2016a, Fan et al. 2016). The
lowest CO2 respiration rates recorded for cropland and grazing land soils probably
reflect their low total C content, which apparently provide insufficient food for microbes.
This may reflect the combined effect of complete removal of crop residues and poor
management practices, which remove valuable energy sources for microbial
communities. The eucalyptus soils showed an intermediate respiratory activity, but
significantly higher than that of the cropland and grazing land. This suggests that
restoring degraded sites provides greater benefits in terms of soil organic matter inputs
and microbial activity. Microbial respiration differences among land use systems were
probably also driven by soil moisture differences resulted from soil organic matter
content differences. For example, a 1% rise of SOC increased the water holding
capacity by ca. 9 times (Fig. 7.1a). Therefore, more SOC in the forest soil increased
the water holding capacity proportionally during adjustments to 40% before incubation;

149
this provides better conditions for microorganisms, potentially enhancing soil
respiration.
We supplied microorganisms with carbon sources of contrasting recalcitrance to
determine whether soil organic matter and carbon quality are an important determinant
of soil respiration. In general, soil respiration rates in the glucose-, lignin-, and starch-
supplemented soils were significantly higher than in the control soil (Table 7.3; Fig. 7.3),
further confirming that microbes are limited by carbon availability. In addition to
substrate availability, substrate quality resulted in different soil respiration rates. The
magnitude of effects on microbial respiration depends on the specific composition of
the substrate added. In general, adding labile carbon sources (glucose) stimulates
microbial respiration in all land use soils (Fig. 7.3). Surprisingly, the second highest CO2
efflux was in soils supplied with lignin substrate, indicating preferential degradation of
lignin over starch. Lignin is considered to be more recalcitrant than starch (Hernández
et al. 2010), hence the moderately labile substrate (starch) should have been depleted
faster than the more recalcitrant substrate (lignin). Moreover, starch is easily degraded
by aerobic as well as anaerobic microorganisms (Kögel-Knabner 2002), whereas white-
rot and brown-rot fungi (Goñi et al. 1993) and certain bacteria such as Streptomyces
sp. or Nocarda sp. (Brown and Chang 2014, Duboc et al. 2014) are commonly
responsible for lignin degradation. Therefore, the higher CO2 efflux in the lignin-induced
soils indicates the relative profile structure of microorganisms with lignin-specialized
fungi and bacterial dominance. Previous studies have shown that lignin decomposition
in soil can be as fast or even faster than bulk soil organic matter, and only a small
fraction of lignin (10%) seems to persist on a time scale of decades (Marschner et al.
2008, Duboc et al. 2014). Moreover, Feng et al. (2008) showed that warming can
accelerate lignin decomposition, while soil fungi, the primary decomposers of lignin in
soil, increased in abundance. Bradford (2013) and Steinweg et al. (2008) reported that
substrate limitation might shift enzyme expression toward higher affinity enzymes. That
would reduce maximum catalytic rates, favouring a slower growing microbial biomass
and a lower respiration rate. Further microbial identification and quantification would
help to elucidate microbial community structure and the taxonomy of the species.
Soil respiration in soils supplied with nutrients (N and P) was not significantly different
from control soils, suggesting that microbial activity was less affected by nutrient
deficiencies. In temperate and boreal forests, N and P has usually been found to be the

150
main limiting factor after C (Gnankambary et al. 2008, Giesler et al. 2012). On the
contrary, adding N fertilizer to soils even reduced soil respiration, and this has been
suggested to reflect microbial reallocation of C to biomass or enzyme production
(Schimel and Weintraub 2003, Allison and Vitousek 2005).
7.5.2 Biological indicators for eco-physiological stress
We calculated the microbial metabolic quotient (qCO2) – the amount of CO2 produced
per unit of microbial biomass carbon (Anderson and Domsch 1990) – to assess the
ecosystem stress and metabolic rates of soil microbial communities. This index has
been widely used as a biological indicator of ecosystem soil management practice and
disturbance (Wardle and Ghani 1995, Thirukkumaran and Parkinson 2000, Campbell
et al. 2003, Tosi et al. 2016). Disturbance supposedly increases qCO2, suggesting that
the richness of organic C from different cultures benefits respiration (Bastida et al.
2008). We found a lower metabolic quotient (qCO2) in the forest ecosystem (Table 7.3).
This points to favourable conditions for microbes and less disturbance in this
ecosystem. We also found that the eucalyptus soil had the highest qCO2, which is not
consistent with disturbance. The probable explanation is that eucalyptus species are
known to be inhibitory to soil microorganisms (Louzada et al. 1997, Martins et al. 2013).
These trees are associated with low litter decomposition and reportedly adversely affect
soil attributes (Lemenih et al. 2004). Combined, these factors may induce stress in
microbial communities, which have been shown to have high qCO2. Plant litter
decomposition studies demonstrate that a significant increase in qCO2 can eventually
occur in litter types initially resistant to decomposition (Wardle and Ghani 1995).
Differences in pH probably explain some of the variation in qCO2 because we found a
negative (but not significant) relationship between qCO2 and soil pH (r2 = 0.84; P=0.086;
data not shown). The relationship between the qCO2 and pH means that careful
management of soil pH is required to ensure a healthy and continued soil fertility (Yan
et al. 2003). Our eucalyptus plantation site has been less disturbed during the plantation
period (the last 30 years), but qCO2 was higher in the eucalyptus than at the other 3
sites, indicating that microbial stress is related more to acidity than to disturbance. This
result agrees with Wardle and Ghani (1995), who reported that qCO2 often declines
with increasing pH, which points to an effect of stress rather than disturbance levels.
Other studies by Wolters (1991) reported an increase in qCO2 after acid rain and a

151
reduction after lime treatment in beech forest soils. We can generalize that microbial
communities release more CO2 per unit microbial biomass under acidic than under
relatively neutral soil pH conditions.
7.5.3 Effect of temperature on microbial decomposition
The CO2 efflux increased with temperature but at a decreasing rate above 25°C (Fig.
7.3a-d). This relationship is best represented by the Arrhenius equation by fitting the
measured microbial CO2 efflux as a function of temperature (Lloyd and Taylor 1994).
For C substrate induced respiration, r2 values for all fitted temperature response
functions were >0.98. The Arrhenius model predicted an initial increase in soil
respiration due to the temperature sensitivity of microbial activity, but then approaching
a steady state at higher temperature (Fig. 7.3). This means that this model does not
support the scenarios in which CO2 concentration increases exponentially with
temperature. Despite the wide usage of the exponential model, it can only be valid in a
narrow temperature range because it is unrealistic to expect that biological activity
would increase without limit as a function of temperature (Tuomi et al. 2007). This
growth must cease and ultimately turn into a decrease as temperature increases above
a certain level.
The temperature sensitivity (expressed as Q10) declined at lower substrate availability
and at high temperature (Table 7.3). The metric “Q10” is a commonly used parameter
to estimate temperature sensitivity that computes the relative increase in
decomposition rate per 10°C temperature rise (Bradford 2013). The CO2 effluxes are
less sensitive to temperature change at low organic C content. For example, cropland
and grazing land soils have low Q10 values of 1.1-1.3 (Table 7.3). This may be partly
explained by thermal adaptation due to long exposure to temperature fluctuations in
these open fields. Previous studies also reported that the temperature sensitivity (Q10)
of respiration declines at lower substrate availabilities (Davidson and Janssens 2006,
Allison et al. 2010). Higher temperature sensitivity in forest and eucalyptus plantation
soils suggests the C availability and quality might affect the soil C response to warming.
Our addition of different substrates increased the Q10 value (Table 7.3), further
confirming that the temperature sensitivity of soil organic matter breakdown is also
influenced by substrate availability (Conant and et.al. 2011). The higher Q10 value after
lignin versus starch addition (Table 7.3) supports the Arrhenius kinetic theory that the

152
decomposition of more recalcitrant compounds is more sensitive to temperature
increase compared to more simple compounds (Davidson and Janssens 2006).
Similarly, Feng and Simpson (2008) reported that lignin monomers exhibited higher Q10
values than the solvent-extractable compounds, which is consistent with the above
theory. This is because less reactive and more chemically recalcitrant molecules with
greater activation energies have greater temperature sensitivity (Conant and et.al.
2011).
The temperature sensitivity (Q10) also declined at high temperature (Table 7.3).
Excluding the respiration values recorded at the 35°C incubation from our data set, we
observed a twofold variation (Q10 = 2.0-2.2). Atkin and Tjoelker (2003) reported that
adaptation of microbial respiration to warming is exhibited through a drop in Q10 values.
Empirical studies suggest that warming at high temperatures reduced microbial carbon
use efficiency from 0.31 to 0.23, also reducing the amount of assimilated C allocated
to microbial growth (Allison et al. 2010). Some studies argued that warming at high
temperatures induced a change in microbial physiology and community shifts
(Davidson and Janssens 2006, Bradford et al. 2008). For example, Bérard et al. (2011)
reported that high temperature shifts the soil microbial community from fungi towards
bacteria. One reason for the reduced CO2 efflux at higher temperature may be the
reduction of soil moisture at 35°C during incubation period: only few water droplets
were observed on the surface of the gasket. This is because warming dries the soil,
directly affecting enzyme activity and diffusion rates between microbes and their
immediate environment (Conant and et.al. 2011, Bradford 2013) as well as substrate
availability (Davidson and Janssens 2006). This returns soil respiration to a steady state
(Steinweg et al. 2008, Allison et al. 2010, Bradford et al. 2010). We did not measure
the amount of water loss during incubation and therefore cannot generalize the effect
of drought on microbial respiration. In our previous pre-test measurements, microbial
respiration was highest at 50-70% water holding capacity; below or above these values,
respiration declined.

153
7.6 Conclusion
Our study demonstrates that soil respiration increases with temperature, but that the
rate of this increase drops above 25°C. This indicates that expectations of increasing
biological activity without limit as a function of temperature are erroneous. Low C
contents in soil after land use conversion from forest to cropland and grazing land
restrict soil respiration rates and make the microbial communities in these soils strongly
C limited. Greater soil respiration in the natural forest is evidence for high rates of
biological activity and soil carbon turnover. The MBc was twice as high in the forest soil
as in the other land use systems, which reflects the higher energy supply for
microorganisms. In general, soil respiration rates were significantly higher in substrate-
supplemented soils than the control soil, further confirming that microbes are limited by
C sources. Substrate quality also yielded different soil respiration rates, pointing to
preferential degradation of substrates by microbes. Nonetheless, the response of
microbes to nutrient addition was not significant, meaning that they are not limited by
nutrient deficiency. The carbon use efficiency of microbes is higher under acidic
conditions.

154
8 Summary and conclusions
The study was conducted in the Amhara Regional State located in the northcentral
parts of Ethiopia. Five research sites (Katassi, Gelawdios, Tara Gedam, Ambober, and
Mahibere-Selassie) were selected at an average distance of 120 km between each
other. Elevations range from approximately 800 m a.s.l. at Mahibere-Selassie to 2500
m at Gelawdios. At each site, up to five-land use types (natural forest, eucalyptus
plantation, exclosure (rejuvenated woodland on former grazing land), cropland, and
grazing land) were identified adjacent to each other to give similar topography, edaphic,
and climate conditions for comparison of land uses at each site. However, not all land
use systems were available at each site. The highland forests are Afromontane
remnant pristine forests mostly confined to sacred groves associated with churches.
The lowland semiarid forest around Mahibere-Selassie monastery is a savannah
woodland dominated by grasses with some scattered trees. The eucalyptus plantations
were established around 1985 with Eucalyptus globules at Katassi and Gelawdios on
grazing land and Eucalyptus camaldunesis at Tara Gedam on former cropland. The
exclosure was established at Ambober in 2007 on former grazing land. All studied
croplands and grazing lands were converted from natural forest within the last 50 years.
This thesis has addressed several major aspects of the biogeochemical cycle based
on field and laboratory investigations of different land use systems in NW Ethiopia. The
methodologies and results from experimental studies have been presented and
discussed individually in chapters two to seven. This final chapter serves to summarize,
integrate the data presented, and relate the findings to a wide range of research
questions raised during the inception of this research.
Land use plays a major role in determining the C storage of soils. In Ethiopian highland
ecosystems, greater SOC stocks were found in the natural forest than in any other land
use systems. Following land use conversion from native forest to cropland or grazing
land, SOC stocks were reduced by 88% at site level in <50 years. In contrast,
converting degraded cropland or grazing land into perennial vegetation, e.g.
afforestation with eucalyptus, is found to increase SOC stocks by increasing C input
from plant litter. Surprisingly, the effects of exclosures (natural vegetation restoration)
to increase SOC stocks was not significant over 8 years establishment in subsoils (10-

155
30 cm depth), questioning the suitability of the natural regeneration for quick recovery
of SOC stocks.
Major factors underlying the reduction of carbon stock are erosion and enhanced
mineralization. In the studied areas, the level of SOC stock in topsoil (0-10 cm) of
cropland or grazing land was less than the SOC stock of the adjacent forest soil at the
depth of 30-50 cm. The geochemical markers such as Sr/Ca and Ba/Ca ratio at 30-50
cm depth in the forest soil profiles are comparable to the 0-10 cm depth of cropland
soil. Both factors suggest that the upper soil profile has been lost by erosion in cropland
and grazing land. Concerning mineralization, the possible maximum annual carbon loss
rate through heterotrophic respiration was estimated to be 3.9 mg g soil-1 yr-1 in forests,
2.7 mg g soil-1 yr-1 in eucalyptus stand, 2.0 mg g soil-1 yr-1 in grazing lands, and 1.6 mg
g soil-1 yr-1 in croplands. This is consistent with substrate availability showing that soils
with high SOC content have higher microbial respiration independent of land use type.
However, this estimation was based on the optimum soil moisture content and
temperature throughout the year, while this optimum condition may occur only for few
days a year. The loss of SOC by heterotrophic respiration is smaller than the effect of
erosion (6.5 mg g soil-1 yr-1 on average) and can be compensated by litter input in to
the soil.
Nevertheless, land use conversions from native forest to grazing land or cropland alter
the overall quantity and quality of litter inputs. The annual above litterfall production in
the forest ecosystem was about 1100 g dry weight (dw) m-2 of which leaves comprised
65%. The above-ground C input, mainly through leaves, was estimated about 300 g m-
2. Above-ground litter inputs in eucalyptus, cropland, and grazing land are estimated to
be minor due to leaf litter raking, complete residual harvest, and overgrazing,
respectively. Similarly, the average fine root production was estimated to be about 700
g dw m-2 in natural forest and eucalyptus stands whereas fine root production in grazing
land and cropland was about 50-60 g dw m-2. Therefore, the amount of annual C input
into the soil by fine roots alone was about 340 g m-2 in natural forest and eucalyptus
stands and ca. 30 g m-2 in grazing land and croplands. Overall, the results illustrate that
conversion of native forest to grazing land and cropland resulted in the reduction of C
input into the soil by >90%. Compared to the above leaf litterfall, fine roots contributed
about 15% more C into the soil, emphasizing their importance for SOC accumulation.

156
The litter chemistry analysis revealed that acid soluble fraction (ASF; i.e. mainly
cellulose) and acid insoluble fraction (AIF; i.e. mainly lignin) were the two compound
groups contributing most to C input into the soil. ASF and AIF were equally present in
leaf extracts (ca. 42% each) whereas ASF in roots accounted for 37% and AIF
accounted for 50% of total extracts. Solvent extractives (i.e. extractive fractions mainly
carbohydrates) accounted for <16% of the total fluxes in leaves and <12% in fine roots
and the rest were ash. The AIF from roots contains a highly cross-linked polymer with
highly reduced compounds of suberin and tannin-protein complexes, which are
resistant to degradation. By contrast, leaf litter contributed considerably more labile C
compounds such as cellulose, soluble phenolics and carbohydrates to the soil than fine
roots. Thus, fine root estimated to contribute about 1.5 times more recalcitrant C input
into the soil than leaf litter. Suberin and cutin can be used as markers to estimate the
input of organic matter originating from root biomass and shoot specific contribution
into the soil. The biomarker analysis revealed that the amount of suberin was 2-times
that of cutin – further confirming the greater input of recalcitrant carbon by fine roots.
Based on the biomarker analysis of soils, aliphatic lipids and lignin accounted about 46-
84% of the total detected compounds and revealed a major input of soil organic carbon
derived from higher plants. The short-chain alkanoic acids, ergosterol, and a
disaccharide trehalose accounted only for less than 7% of aliphatic lipids in all soils
indicating the microbial inputs were present as minor components. Cholesterol is
derived from non-plant source such as soil fauna, fungi, and algae and has minor
contribution to soil organic carbon as reflected by its low abundance in all land use
types. From the CuO oxidation yields, the ratio of syringyls to vanillyls (S/V) and
cinnamyls to vanillyls (C/V) monomers are widely used to differentiate the relative
contributions of major plant taxonomic groups (gymnosperms vs angiosperms), tissue
type (woody vs non-woody tissue), and diagenetic state of lignin. Accordingly, S/V
values varied in the range of 0.61 to 2.23 suggesting that angiosperm plants are the
predominant sources for lignin compared to gymnosperm plants because S/V values
above 0.6 are considered as angiosperm origin. Similarly, a higher value of C/V ratio is
indicative of the presence of non-woody material because cinnamyl monomers (p-
coumaric acid, ferulic acid) are abundant in most herbaceous and soft tissues (i.e.
leaves, grasses) but virtually absent from wood. The lignin-derived phenols exhibited a
V:S:C ratio of 4:2:1 (forest soil); 1:1:1 (eucalypt, cropland, and grazing land soils).

157
Relatively equal V:S:C: ratio in eucalyptus, cropland, and grazing land soils further
confirming that lignin-derived phenols originate mainly from non-woody angiosperm
sources, which is consistent with S/V and C/V ratios. The higher V:S:C ratio in the
natural forest indicates that woody angiosperms have significant contributions to SOC.
The ratios of lignin-derived phenolic acids and 3, 5-dihydroxybenzoic acid (DHBA)/V
are generally used as indicators of the level of lignin and tannin degradation in the soil,
respectively. The lowest ratio of S/V (0.6) and C/V (0.2) was in forest, while the other
land use soils show a ratio greater than 1.2, indicating enhanced degradation of lignin
in the forest soil, and preferential degradation of syringyls and cinnamyls from the lignin-
derived phenols. This is because cinnamyls linking carbohydrates and lignin in the
ligno-cellulose complex and are therefore more accessible to decomposition than
vanillyls. The 3, 5-dihydroxybenzoic acid to vanillyls was in the order of forest soil
(0.69), eucalyptus (0.24), cropland soil (0.15), and grazing land soil (0.09). The highest
value in forest soil shows increased degradation of tannins as 3, 5-dihydroxybenzoic
acid in the CuO oxidation was derived from tannin oxidation.
A litterbag experiment revealed that leaf litter decomposed faster than fine root litter
with a decomposition rate constant (k) of 2.5 yr-1 for leaves and 1.7 yr-1 for fine roots.
Fine root morphological traits such as specific root surface area (SRA), specific root
length (SRL), and root tissue density (RTD; g cm–3) are important with regard to plant
growth strategy, roots life span, and root decomposition rate. This study demonstrates
that roots of fast-growing species have low RTD. The calculated amount of glucose
needed to produce a gram of tissue biomass ranged between 1.2 and 1.5 g glucose g-
1 dw and was significantly and consistently greater in slow-growing species compared
to fast-growing species. Fine root C contents were negatively correlated to specific root
length or area (SRL/SRA), but positively correlated to RTD and AIF and were largely
consistent with a range of species. This difference between growth patterns determines
the decomposition dynamics and C deposition in the soil and suggests that species
having lower RTD and low structural components (low lignin) decompose more rapidly
Due to species-specific morphological and litter chemistry differences, decomposition
of litter materials in the litterbag experiment varied largely by species. Among the
studied species, the deciduous species (Allophylus abyssinicus, and Combretum
collinum) decomposed faster than evergreen species (Chionanthus mildbraedii and
Teclea nobilis). Furthermore, the presence of litter from deciduous species in litter

158
mixtures, i.e. the normal situation in mixed forests, increased the rate of decomposition
of other slower decomposing litter types.
Overall, these findings are fundamental to improving our understanding of
biogeochemical cycles under different land use systems and its impact on the global C
balance. This thesis may support decision-making on land use and management of soil
carbon by raising awareness and upgrading existing knowledge on the SOC dynamics
and importance of SOC management as the basis for essential ecosystem functions.
Decisions on soil carbon management requires a better understanding of the spatial
variation in SOC stocks and their C source as well as the potential to use management
approaches to reduce losses and/or increase stocks. In the Amhara region, there is
both a need and an opportunity to influence land use policy and management to
improve soil C quantity and quality. This can be evidenced by mass mobilization of
farmers by the government for soil and water conservation campaign work in the whole
regions since decades. However, this campaign focused only on physical and biological
conservation activities and ignoring soil C management. Therefore, soil conservation
efforts need to emphasize and promote land use practices that focus on the possible
greatest increase or at least stabilization in SOC through afforestation, exclosure or
reduction of losses due to erosion, and above all sustainable management approaches.
Promoting land uses that maintain biomass C input is more beneficial to overall carbon
stocks that improves soil quality resulting in greater productivity and ultimately
increased food security.

159
References
Abiven, S., S. Recous, V. Reyes, and R. Oliver. 2005. Mineralisation of C and N from root, stem and leaf residues in soil and role of their biochemical quality. Biology and Fertility of Soils 42:119–128.
Abiyu, A. 2012. The role of seed dispersal, exclosures, nurse shrubs and trees around churches and farms for restoration of ecosystem diversity and productivity in the Ethiopian highlands. University of Natural Resources and Life Science, Vienna, Austria.
Abu-hashim, M., M. Elsayed, and A.-E. Belal. 2016. Effect of land-use changes and site variables on surface soil organic carbon pool at Mediterranean Region. Journal of African Earth Sciences 114:78–84.
Adair, E., W. Parton, S. del Grosso, W. Silver, M. Harmon, S. Hall, I. Burke, and S. Hart. 2008. Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates. Global change biology 14:2636–2660.
Aerts, R., C. Bakker, and H. De Caluwe. 1992. Root turnover as determinant of the cycling of C, N, and P in a dry heathland ecosystem. Biogeochemistry 15:175–190.
Aerts, R., K. Van Overtveld, E. November, A. Wassie, A. Abiyu, S. Demissew, D. D. Daye, K. Giday, M. Haile, and S. TewoldeBerhan. 2016. Conservation of the Ethiopian church forests: Threats, opportunities and implications for their management. Science of the Total Environment 551:404–414.
Ajami, M., A. Heidari, F. Khormali, M. Gorji, and S. Ayoubi. 2016. Environmental factors controlling soil organic carbon storage in loess soils of a subhumid region, northern Iran. Geoderma 281:1–10.
Allison, S. D., and P. M. Vitousek. 2005. Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biology and Biochemistry 37:937–944.
Allison, S. D., M. D. Wallenstein, and M. A. Bradford. 2010. Soil-carbon response to warming dependent on microbial physiology. Nature Geoscience 3:336–340.
Alvarez, R., and R. S. R. Lavado. 1998. Climate, organic matter and clay content relationships in the Pampa and Chaco soils, Argentina. Geoderma 83:127–141.
Amelung, W., S. Brodowski, A. Sandhage-Hofmann, and R. Bol. 2008. Combining biomarker with stable isotope analyses for assessing the transformation and turnover of soil organic matter. Advances in agronomy 100:155–250.
Anderson, J. P. E., and K. H. Domsch. 1978. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biology and Biochemistry 10:215–221.
Anderson, T.-H., and K. H. Domsch. 1989. Ratios of microbial biomass carbon to total organic carbon in arable soils. Soil Biology and Biochemistry 21:471–479.
Anderson, T.-H., and K. H. Domsch. 1990. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry 22:251–255.
Andreasson, F., M. Gonzalez, L. Augusto, and M. R. Bakker. 2015. Comparison of ingrowth cores and ingrowth meshes in root studies: 3 years of data on Pinus pinaster and its understory. Trees:1–16.
Angst, G., S. John, C. W. Mueller, I. Kögel-Knabner, and J. Rethemeyer. 2016. Tracing the sources and spatial distribution of organic carbon in subsoils using a multi-biomarker approach. Nature Publishing Group.
Aragão, L. E. O. C., Y. Malhi, D. B. Metcalfe, J. E. Silva-Espejo, E. Jiménez, D. Navarrete, S. Almeida, A. C. L. Costa, N. Salinas, O. L. Phillips, L. O. Anderson, E. Alvarez, T. R. Baker, P.

160
H. Goncalvez, J. Huamán-Ovalle, M. Mamani-Solórzano, P. Meir, A. Monteagudo, S. Patiño, M. C. Peñuela, A. Prieto, C. A. Quesada, A. Rozas-Dávila, A. Rudas, J. A. Silva Jr., and R. Vásquez. 2009. Above- and below-ground net primary productivity across ten Amazonian forests on contrasting soils. Biogeosciences 6:2759–2778.
Assaye, Y., Y. G. Selassie, and B. Ayele. 2013. A socio-economic contribution of high land bamboo (Yushania Alpina) for household livelihood in Banja District, Northwestern Ethiopia. Journal of Agriculture and Biodiversity Research 2:151–159.
Atkin, O. K., and M. G. Tjoelker. 2003. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends in Plant Science 8:343–351.
Awulachew, S. B., M. McCartney, T. S. Steenhuis, and A. A. Ahmed. 2009. A review of hydrology, sediment and water resource use in the Blue Nile Basin. International Water Management Institute (IWMI), Colombo, Sri Lanka.
Ayalew, D., K. Tesfaye, G. Mamo, B. Yitaferu, and W. Bayu. 2012. Outlook of future climate in northwestern Ethiopia. Agricultural Sciences 3:608–624.
Baddeley, J. A., and C. A. Watson. 2005. Influences of root diameter, tree age, soil depth and season on fine root survivorship in Prunus avium. Plant and Soil 276:15–22.
Balesdent, J., E. Besnard, D. Arrouays, and C. Chenu. 1998. The dynamics of carbon in particle-size fractions of soil in a forest-cultivation sequence. Plant and Soil 201:49–57.
Bane, J., S. Nune, A. Mekonnen, and R. Bluffstone. 2007. Policies to increase forest cover in Ethiopia Contents. Page 239Proceedings of a Policy Workshop organized. Addis Ababa, Ethiopia, Ethiopia.
Bao, X., J. Yu, W. Liang, C. Lu, J. Zhu, and Q. Li. 2015. The interactive effects of elevated ozone and wheat cultivars on soil microbial community composition and metabolic diversity. Applied Soil Ecology 87:11–18.
Bardgett, R. D., L. Mommer, and F. T. De Vries. 2014. Going underground: root traits as drivers of ecosystem processes. Trends in Ecology & Evolution 29:692–699.
Basile, B., D. R. Bryla, M. L. Salsman, J. Marsal, C. Cirillo, R. S. Johnson, and T. M. DeJong. 2007. Growth patterns and morphology of fine roots of size-controlling and invigorating peach rootstocks. Tree physiology 27:231–241.
Bastida, F., A. Zsolnay, T. Hernández, and C. García. 2008. Past, present and future of soil quality indices: A biological perspective. Geoderma 147:159–171.
Batjes, N. H. 1996. Total carbon and nitrogen in the soils of the world. European journal of soil science 47:151–163.
Batjes, N. H., and W. G. Sombroek. 1997. Possibilities for carbon sequestration in tropical and subtropical soils. Global change biology 3:161–173.
Becker, J., H. Pabst, J. Mnyonga, and Y. Kuzyakov. 2015. Annual litterfall dynamics and nutrient deposition depending on elevation and land use at Mt. Kilimanjaro. Biogeosciences 12:5635–5646.
Bekele, M. 2011. Forest plantations and woodlots in Ethiopia. First edition. Regional Soil Conservation Unit, Swedish International Development Authority, Nairobi, Kenya.
Bekele, T. A. 2007. Useful trees and shrubs of Ethiopia: identification, propagation and management for 17 agroclimatic zones. Page (B. Tengnäs, E. Kelbesa, S. Demissew, and P. Maundu, Eds.). RELMA in ICRAF Project, World Agroforestry Centre, Eastern Africa Region, Nairobi, Kenya.
Bérard, A., T. Bouchet, G. Sévenier, A. L. Pablo, and R. Gros. 2011. Resilience of soil microbial communities impacted by severe drought and high temperature in the context of Mediterranean heat waves. European Journal of Soil Biology 47:333–342.
Bérard, A., L. Capowiez, S. Mombo, E. Schreck, C. Dumat, F. Deola, and Y. Capowiez. 2016. Soil

161
microbial respiration and PICT responses to an industrial and historic lead pollution: a field study. Environmental Science and Pollution Research 23:4271–4281.
Berg, B. 2000. Litter decomposition and organic matter turnover in northern forest soils. Forest Ecology and Management 133:13–22.
Berg, B., and C. McClaugherty. 2003. Plant litter : decomposition, humus formation, carbon sequestration. Springer Berlin Heidelberg, Berlin, Heidelberg.
Berg, B., and H. Staaf. 1981. Leaching, accumulation and release of nitrogen in decomposing forest litter. Ecological Bulletins 33:163–178.
Berhongaray, G., I. A. Janssens, J. S. King, and R. Ceulemans. 2013. Fine root biomass and turnover of two fast-growing poplar genotypes in a short-rotation coppice culture. Plant and Soil 373:269–283.
Betrie, G. D., Y. A. Mohamed, A. van Griensven, and R. Srinivasan. 2011. Sediment management modelling in the Blue Nile Basin using SWAT model. Hydrology and Earth System Sciences 15:807–818.
Bewket, W., and D. Conway. 2007. A note on the temporal and spatial variability of rainfall in the
drought‐prone Amhara region of Ethiopia. International Journal of Climatology 27:1467–1477.
Bewket, W., and G. Sterk. 2003. Assessment of soil erosion in cultivated fields using a survey methodology for rills in the Chemoga watershed, Ethiopia. Agriculture, Ecosystems & Environment 97:81–93.
Bewket, W., and G. Sterk. 2005. Dynamics in land cover and its effect on stream flow in the Chemoga watershed, Blue Nile basin, Ethiopia. Hydrological Processes 19:445–458.
Bewket, W., and L. Stroosnijder. 2003. Effects of agroecological land use succession on soil properties in Chemoga watershed, Blue Nile basin, Ethiopia. Geoderma 111:85–98.
Birgander, J., J. Rousk, and P. A. Olsson. 2014. Comparison of fertility and seasonal effects on grassland microbial communities. Soil Biology and Biochemistry 76:80–89.
Birouste, M., E. Zamora-Ledezma, C. Bossard, I. M. Pérez-Ramos, and C. Roumet. 2014. Measurement of fine root tissue density: a comparison of three methods reveals the potential of root dry matter content. Plant and Soil 374:299–313.
Bohm, W. 1979. Methods of studying root systems. Page (D. Vj. D. Billings, A. F. Golley, W. O. L. Lange, and O. R. J. S. Olson, Eds.) Springer-Verlag Berlin Heidelberg. New York.
Bradford, M. A. 2013. Thermal adaptation of decomposer communities in warming soils. Frontiers in microbiology 4.
Bradford, M. A., C. A. Davies, S. D. Frey, T. R. Maddox, J. M. Melillo, J. E. Mohan, J. F. Reynolds, K. K. Treseder, and M. D. Wallenstein. 2008. Thermal adaptation of soil microbial respiration to elevated temperature. Ecology letters 11:1316–1327.
Bradford, M. A., B. W. Watts, and C. A. Davies. 2010. Thermal adaptation of heterotrophic soil respiration in laboratory microcosms. Global change biology 16:1576–1588.
Brando, P. M., D. C. Nepstad, E. A. Davidson, S. E. Trumbore, D. Ray, and P. Camargo. 2008. Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: results of a throughfall reduction experiment. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 363:1839–48.
Brassard, B. W., H. Y. H. Chen, X. Cavard, J. Laganière, P. B. Reich, Y. Bergeron, D. Paré, Z. Yuan, J. Laganiere, P. B. Reich, Y. Bergeron, D. Pare, and Z. Yuan. 2013. Tree species diversity increases fine root productivity through increased soil volume filling. Journal of Ecology 101:210–219.
Brhane, G., and K. Mekonen. 2009. Estimating Soil Loss Using Universal Soil Loss Equation (USLE) for Soil Conservation planning at Medego Watershed, Northern Ethiopia. Marsland Press

162
Journal of American ScienceJournal of American Science 5:58–69.
Brown, M. E., and M. C. Chang. 2014. Exploring bacterial lignin degradation. Current Opinion in Chemical Biology 19:1–7.
Brunner, I., M. R. Bakker, R. G. Björk, Y. Hirano, M. Lukac, X. Aranda, I. Børja, T. D. Eldhuset, H. S. Helmisaari, C. Jourdan, B. Konôpka, B. C. López, C. Miguel Pérez, H. Persson, and I. Ostonen. 2012. Fine-root turnover rates of European forests revisited: an analysis of data from sequential coring and ingrowth cores. Plant and Soil 362:357–372.
Bull, I. D., C. J. Nott, P. F. van Bergen, P. R. Poulton, and R. P. Evershed. 2000. Organic geochemical studies of soils from the Rothamsted classical experiments—VI. The occurrence and source of organic acids in an experimental grassland soil. Soil Biology and Biochemistry 32:1367–1376.
Bullen, T. D., S. W. Bailey, B. Thomas D., B. Scott W., T. D. Bullen, and S. W. Bailey. 2005. Identifying calcium sources at an acid deposition-impacted spruce forest: a strontium isotope, alkaline earth element multi-tracer approach. Biogeochemistry 74:63–99.
Burgess, T. I., N. Malajczuk, and T. S. Grove. 1993. The ability of 16 ectomycorrhizal fungi to increase growth and phosphorus uptake of Eucalyptus globulus Labill. and E. diversicolor F. Muell. Plant and Soil 153:155–164.
Callesen, I., K. Raulund-Rasmussen, and J. Westman. 2007. Nitrogen pools and c:n ratios in well-drained nordic forest soils related to climate and soil texture. Boreal environment research 12:681–692.
Campbell, C. D., S. J. Chapman, C. M. Cameron, M. S. Davidson, and J. M. Potts. 2003. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Applied and environmental microbiology 69:3593–3599.
Canadell, J., R. B. Jackson, J. B. Ehleringer, H. A. Mooney, O. E. Sala, and E.-D. Schulze. 1996. Maximum rooting depth of vegetation types at the global scale. Oecologia 108:583–595.
Capo, R. C., B. W. Stewart, and O. A. Chadwick. 1998. Strontium isotopes as tracers of ecosystem processes: theory and methods. Geoderma 82:197–225.
Carey, J. C., J. Tang, P. H. Templer, K. D. Kroeger, T. W. Crowther, A. J. Burton, J. S. Dukes, B. Emmett, S. D. Frey, M. A. Heskel, L. Jiang, M. B. Machmuller, J. Mohan, A. M. Panetta, P. B. Reich, S. Reinsch, X. Wang, S. D. Allison, C. Bamminger, S. Bridgham, S. L. Collins, G. de Dato, W. C. Eddy, B. J. Enquist, M. Estiarte, J. Harte, A. Henderson, B. R. Johnson, K. S. Larsen, Y. Luo, S. Marhan, J. M. Melillo, J. Peñuelas, L. Pfeifer-Meister, C. Poll, E. Rastetter, A. B. Reinmann, L. L. Reynolds, I. K. Schmidt, G. R. Shaver, A. L. Strong, V. Suseela, and A. Tietema. 2016. Temperature response of soil respiration largely unaltered with experimental warming. Proceedings of the National Academy of Sciences of the United States of America 113:13797–13802.
Celentano, D., R. A. Zahawi, B. Finegan, R. Ostertag, R. J. Cole, and K. D. Holl. 2011. Litterfall Dynamics Under Different Tropical Forest Restoration Strategies in Costa Rica. Biotropica 43:279–287.
Certini, G., L. S. Vestgarden, C. Forte, and L. Tau Strand. 2015. Litter decomposition rate and soil organic matter quality in a patchwork heathland of southern Norway 1:207–216.
Chapin, F. S., P. A. (Pamela A. . Matson, P. M. Vitousek, M. C. Chapin, F. S. Chapin III, P. A. (Pamela A. . Matson, and H. A. Mooney. 2011. Principles of Terrestrial Ecosystem Ecology. Springer, New York Dordrecht Heidelberg London.
Chapman, S. J., C. D. Campbell, and R. R. E. Artz. 2007. Assessing CLPPs Using MicroRespTM A Comparison with Biolog and multi-SIR*. J Soils Sediments 7:406–410.
Chen, G.-S., Y.-S. Yang, J.-S. Xie, J.-F. Guo, R. Gao, and W. Qian. 2005. Conversion of a natural

163
broad-leafed evergreen forest into pure plantation forests in a subtropical area: Effects on carbon storage. Annals of Forest Science 62:659–668.
Chen, H. 2014. Chemical Composition and Structure of Natural Lignocellulose. Pages 25–71Biotechnology of Lignocellulose. Springer Netherlands, Dordrecht.
Chen, S., Y. Huang, J. Zou, and Y. Shi. 2013. Mean residence time of global topsoil organic carbon depends on temperature, precipitation and soil nitrogen. Global and Planetary Change 100:99–108.
Chen, X., D. Eamus, and L. B. Hutley. 2004. Seasonal patterns of fine-root productivity and turnover in a tropical savanna of northern Australia. Journal of Tropical Ecology 20:221–224.
Clark, D. A., S. Brown, D. W. Kicklighter, J. Q. Chambers, J. R. Thomlinson, and J. Ni. 2001. Measuring net primary production in forests: concepts and field methods. Ecological Applications 11:356–370.
Clemente, J. S. 2012. Soil Organic Matter Composition Impacts its Degradability and Association with Soil Minerals. University of Toronto.
Collins, C. G., S. J. Wright, and N. Wurzburger. 2016. Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees. Oecologia 180:1037–1047.
Comas, L., T. Bouma, and D. Eissenstat. 2002. Linking root traits to potential growth rate in six temperate tree species. Oecologia 132:34–43.
Comas, L. H., S. R. Becker, V. M. V Cruz, P. F. Byrne, and D. A. Dierig. 2013. Root traits contributing to plant productivity under drought. Frontiers in plant science 4:442.
Comas, L. H., and D. M. Eissenstat. 2004. Linking fine root traits to maximum potential growth rate among 11 mature temperate tree species. Functional Ecology 18:388–397.
Conant, R. T., K. Paustian, and E. T. Elliott. 2001. Grassland management and conversion into grassland: effects on soil carbon. Ecological Applications 11:343–355.
Conant, T. R., and et.al. 2011. Temperature and soil organic matter decomposition rates – synthesis of current knowledge and a way forward. Global change biology 17:3392–3404.
Contador, M. L., L. H. Comas, S. G. Metcalf, W. L. Stewart, I. Porris Gomez, C. Negron, and B. D. Lampinen. 2015. Root growth dynamics linked to above-ground growth in walnut (Juglans regia). Annals of botany 116:49–60.
Conti, G., E. Kowaljow, F. Baptist, C. Rumpel, A. Cuchietti, N. P. Harguindeguy, and S. Díaz. 2016. Altered soil carbon dynamics under different land-use regimes in subtropical seasonally-dry forests of central Argentina. Plant and Soil:1–13.
Cornelissen, J. H. C. 1996. An Experimental Comparison of Leaf Decomposition Rates in a Wide Range of Temperate Plant Species and Types. The Journal of Ecology 84:573.
Cotrufo, M. F., J. Ngao, F. Marzaioli, and D. Piermatteo. 2010. Inter-comparison of methods for quantifying above-ground leaf litter decomposition rates. Plant and Soil 334:365–376.
Couteaux, M.-M., P. Bottner, and B. Berg. 1995. Litter decomposition, climate and liter quality. Trends in Ecology & Evolution 10:63–66.
Creamer, R. E., R. P. O. Schulte, D. Stone, A. Gal, P. H. Krogh, G. Lo Papa, P. J. Murray, G. Pérès, B. Foerster, M. Rutgers, J. P. Sousa, A. Winding, G. Lo Papa, P. J. Murray, G. Pérès, B. Foerster, M. Rutgers, G. Lo Papa, P. J. Murray, G. Pérès, B. Foerster, M. Rutgers, J. P. Sousa, A. Winding, G. Lo Papa, P. J. Murray, G. Pérès, B. Foerster, and M. Rutgers. 2014. Measuring basal soil respiration across Europe: Do incubation temperature and incubation period matter? Ecological Indicators 36:409–418.
CSA. 2008. Summary and Statistical Report of the 2007 Population and Housing Census. Addis Ababa, Eethiopia, Eethiopia.

164
Dauer, M. J., J. Chorover, A. O. Chadwick, J. Oleksyn, G. M. Tjoelker, E. S. Hobbie, B. P. Reich, and M. D. Eissenstat. 2007. Controls over leaf and litter calcium concentrations among temperate trees. Biogeochemistry:175–187.
Davidson, E. A., and I. A. Janssens. 2006. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440:165–173.
Demessie, A., B. R. Singh, R. Lal, and L. T. Strand. 2012. Leaf litter fall and litter decomposition under Eucalyptus and coniferous plantations in Gambo District, southern Ethiopia. Acta Agriculturae Scandinavica, Section B - Soil & Plant Science 62:467–473.
Deng, L., G. Liu, and Z. Shangguan. 2014. Land-use conversion and changing soil carbon stocks in China’s ‘Grain-for-Green’ Program: a synthesis. Global change biology 20:3544–3556.
Deng, L., G. Wang, G. Liu, and Z. Shangguan. 2016a. Effects of age and land-use changes on soil carbon and nitrogen sequestrations following cropland abandonment on the Loess Plateau, China. Ecological Engineering 90:105–112.
Deng, L., G. Zhu, Z. Tang, and Z. Shangguan. 2016b. Global patterns of the effects of land-use changes on soil carbon stocks. Global Ecology and Conservation 5:127–138.
Derry, L. A., and O. A. Chadwick. 2007. Contributions from Earth’s Atmosphere to Soil. Elements 3.
Descheemaeker, K., B. Muys, J. Nyssen, J. Poesen, D. Raes, M. Haile, and J. Deckers. 2006. Litter production and organic matter accumulation in exclosures of the Tigray highlands, Ethiopia. Forest Ecology and Management 233:21–35.
Desta, L., M. Kassie, S. Beninc, and J. Penderd. 2000. Land degradation in the highlands of Amhara region and strategies for sustainable land management. Page in J. P. and S. K. E. M.A. Jabbar, editor. Policies for sustainable land management in the highlands of Ethiopia. International Livestock Research Institute, Addis Ababa, Eethiopia.
De Deyn, G. B., J. H. C. Cornelissen, and R. D. Bardgett. 2008. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol Lett 11:516–531.
Don, A., J. Schumacher, and A. Freibauer. 2011. Impact of tropical land‐use change on soil organic carbon stocks–a meta‐analysis. Global change biology 17:1658–1670.
Don, A., J. Schumacher, M. Scherer-Lorenzen, T. Scholten, and E.-D. Schulze. 2007. Spatial and vertical variation of soil carbon at two grassland sites—implications for measuring soil carbon stocks. Geoderma 141:272–282.
Drage, S., D. Engelmeier, G. Bachmann, A. Sessitsch, B. Mitter, and F. Hadacek. 2012. Combining microdilution with MicroRespTM: Microbial substrate utilization, antimicrobial susceptibility and respiration. Journal of Microbiological Methods 88:399–412.
Duboc, O., M.-F. Dignac, I. Djukic, F. Zehetner, M. H. Gerzabek, and C. Rumpel. 2014. Lignin decomposition along an Alpine elevation gradient in relation to physicochemical and soil microbial parameters. Global Change Biology 20:2272–2285.
Eissenstat, D. M. 1991. On the relationship between specific root length and the rate of root proliferation: a field study using citrus rootstocks. New Phytologist 118:63–68.
Eissenstat, D. M., and R. D. Yanai. 1997. The Ecology of Root Lifespan. Advances in Ecological Research 27:1–60.
Elaissi, A., Z. Rouis, N. A. Ben Salem, S. Mabrouk, Y. ben Salem, K. B. H. Salah, M. Aouni, F. Farhat, R. Chemli, F. Harzallah-Skhiri, and M. L. Khouja. 2012. Chemical composition of 8 eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complementary and Alternative Medicine 12:81.
Eswaran, H., E. Van Den Berg, and P. Reich. 1993. Organic Carbon in Soils of the World. Soil Science Society of America Journal 57:192.
Ewel, J. J. 1976. Litter Fall and Leaf Decomposition in a Tropical Forest Succession in Eastern

165
Guatemala. The Journal of Ecology 64:293.
Fan, S., F. Guan, X. Xu, D. I. Forrester, W. Ma, and X. Tang. 2016. Ecosystem Carbon Stock Loss after Land Use Change in Subtropical Forests in China. Forests 7.
Fearnside, P. M., R. I. Barbosa, and R. Imbrozio Barbosa. 1998. Soil carbon changes from conversion of forest to pasture in Brazilian Amazonia. Forest Ecology and Management 108:147–166.
Feng, X. 2009. The Molecular Composition of Soil Organic Matter (SOM) and Potential Responses to Global Warming and Elevated CO2. University of Toronto.
Feng, X., L. L. Nielsen, and M. J. Simpson. 2007. Responses of soil organic matter and microorganisms to freeze–thaw cycles. Soil Biology and Biochemistry 39:2027–2037.
Feng, X., E. J. Simpson, K. P. Wilson, D. D. Williams, and M. J. Simpson. 2008. Increased cuticular carbon sequestration and lignin oxidation in response to soil warming. Nature Geoscience 1:836–839.
Feng, X., and M. J. Simpson. 2007. The distribution and degradation of biomarkers in Alberta grassland soil profiles. Organic Geochemistry 38:1558–1570.
Feng, X., and M. J. Simpson. 2008. Temperature responses of individual soil organic matter components. Journal of Geophysical Research: Biogeosciences (2005–2012) 113.
Feng, X., and M. J. Simpson. 2011. Molecular-level methods for monitoring soil organic matter responses to global climate change. Journal of Environmental Monitoring 13:1246–1254.
Fernandez, D. P., J. C. Neff, C. Huang, G. P. Asner, and N. N. Barger. 2013. Twentieth century carbon stock changes related to Piñon-Juniper expansion into a black sagebrush community. Carbon balance and management 8:13.
Fichtl, R., and A. Adi. 1994. Honeybee flora of Ethiopia. Margraf Verlag, Weikersheim, Germany.
Fife, D. N., E. K. S. Nambiar, and E. Saur. 2008. Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment. Tree physiology 28:187–96.
Finér, L., H.-S. Helmisaari, K. Lõhmus, H. Majdi, I. Brunner, I. Børja, T. Eldhuset, D. Godbold, T. Grebenc, and B. Konôpka. 2007. Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosystems 141:394–405.
Finér, L., M. Ohashi, K. Noguchi, and Y. Hirano. 2011. Factors causing variation in fine root biomass in forest ecosystems. Forest Ecology and Management 261:265–277.
Frank, D., M. Reichstein, M. Bahn, K. Thonicke, D. Frank, M. D. Mahecha, P. Smith, M. Velde, S. Vicca, and F. Babst. 2015. Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Global change biology 21:2861–2880.
Fransen, B., J. Blijjenberg, and H. De Kroon. 1999. Root morphological and physiological plasticity of perennial grass species and the exploitation of spatial and temporal heterogeneous nutrient patches. Plant and Soil 211:179–189.
Freschet, G. T., W. K. Cornwell, D. A. Wardle, T. G. Elumeeva, W. Liu, B. G. Jackson, V. G. Onipchenko, N. A. Soudzilovskaia, J. Tao, and J. H. C. Cornelissen. 2013. Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide. Journal of Ecology 101:943–952.
Del Galdo, I., J. Six, A. Peressotti, and M. Francesca Cotrufo. 2003. Assessing the impact of land‐use change on soil C sequestration in agricultural soils by means of organic matter fractionation and stable C isotopes. Global change biology 9:1204–1213.
Gale, M. R., and D. F. Grigal. 1987. Vertical root distributions of northern tree species in relation to successional status. Canadian Journal of Forest Research 17:829–834.

166
Gao, J., F. Kang, T. Li, X. Song, W. Zhao, X. Yu, and H. Han. 2015. Assessing the Effect of Leaf Litter Diversity on the Decomposition and Associated Diversity of Fungal Assemblages. Forests 6:2371–2386.
García-Palacios, P., R. Milla, M. Álvaro-Sánchez, N. Martín-Robles, and M. Maestro. 2013. Application of a high-throughput laboratory method to assess litter decomposition rates in multiple-species experiments. Page Soil Biology and Biochemistry.
García-Palacios, P., I. Prieto, J.-M. Ourcival, and S. Hättenschwiler. 2016. Disentangling the Litter Quality and Soil Microbial Contribution to Leaf and Fine Root Litter Decomposition Responses to Reduced Rainfall. Ecosystems 19:490–503.
Gaudinski, J., S. Trumbore, E. Davidson, A. Cook, D. Markewitz, and D. Richter. 2001. The age of fine-root carbon in three forests of the eastern United States measured by radiocarbon. Oecologia 129:420–429.
Gebrehana, K. 2015. Biodiversity and Growth Rates of Natural Forests in the Amhara Region of Ethiopia. University of Natural Resources and Life Sciences (BOKU), Austria, Vienna.
Gebremariam, M., and F. Kebede. 2010. Land use change effect on soil carbon stock, above-ground biomass, aggregate stability and soil Crust: A case from Tahtay Adyabo, North Western Tigray, Northern Ethiopia. Journal of the drylands 2:220–225.
Gershenson, A., N. E. Bader, and W. Cheng. 2009. Effects of substrate availability on the temperature sensitivity of soil organic matter decomposition. Global Change Biology 15:176–183.
Gholz, H. L., D. A. Wedin, S. M. Smitherman, M. E. Harmon, and W. J. Parton. 2000. Long-term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition. Global Change Biology 6:751–765.
Gibbs, H. K., S. Brown, J. O. Niles, and J. A. Foley. 2007. Monitoring and estimating tropical forest carbon stocks: making REDD a reality. Environmental Research Letters 2:45023.
Giesler, R., C. Esberg, A. Lagerström, and B. J. Graae. 2012. Phosphorus availability and microbial respiration across different tundra vegetation types. Biogeochemistry 108:429–445.
Gilbert, K. J., T. J. Fahey, J. C. Maerz, R. E. Sherman, P. Bohlen, J. J. Dombroskie, P. M. Groffman, and J. B. Yavitt. 2014. Exploring carbon flow through the root channel in a temperate forest soil food web. Soil Biology and Biochemistry 76:45–52.
Gill, R. A., and R. B. Jackson. 2000. Global patterns of root turnover for terrestrial ecosystems. New Phytologist 147:13–31.
Gill, R., I. C. Burke, D. G. Milchunas, and W. K. Lauenroth. 1999. Relationship Between Root Biomass and Soil Organic Matter Pools in the Shortgrass Steppe of Eastern Colorado. Ecosystems 2:226–236.
Girisha, G. K. 2001. Effects of litter and woody debris quality on decomposition and nutrient release in exotic forests in New Zealand. Lincoln University.
Girmay, G., B. R. Singh, H. Mitiku, T. Borresen, and R. Lal. 2008. Carbon stocks in Ethiopian soils in relation to land use and soil management. Land Degradation & Development 19:351–367.
Gnankambary, Z., U. Ilstedt, G. Nyberg, V. Hien, and A. Malmer. 2008. Nitrogen and phosphorus limitation of soil microbial respiration in two tropical agroforestry parklands in the south-Sudanese zone of Burkina Faso: the effects of tree canopy and fertilization. Soil Biology and Biochemistry 40:350–359.
Godbold, D. L., H.-W. Fritz, G. Jentschke, H. Meesenburg, and P. Rademacher. 2003. Root turnover and root necromass accumulation of Norway spruce (Picea abies) are affected by soil acidity. Tree physiology 23:915–921.
Goñi, M. A., and J. I. Hedges. 1990. The diagenetic behavior of cutin acids in buried conifer needles

167
and sediments from a coastal marine environment. Geochimica et Cosmochimica Acta 54:3083–3093.
Goñi, M. A., and J. I. Hedges. 1992. Lignin dimers: Structures, distribution, and potential geochemical applications. Geochimica et Cosmochimica Acta 56:4025–4043.
Goñi, M. A., and S. Montgomery. 2000. Alkaline CuO oxidation with a microwave digestion system: Lignin analyses of geochemical samples. Analytical Chemistry 72:3116–3121.
Goñi, M. A., B. Nelson, R. A. Blanchette, and J. I. Hedges. 1993. Fungal degradation of wood lignins: Geochemical perspectives from CuO-derived phenolic dimers and monomers. Geochimica et Cosmochimica Acta 57:3985–4002.
Goñi, M. A., M. J. Teixeira, and D. W. Perkey. 2003. Sources and distribution of organic matter in a river-dominated estuary (Winyah Bay, SC, USA). Estuarine, Coastal and Shelf Science 57:1023–1048.
Goñi, M. A., M. B. Yunker, R. W. Macdonald, and T. I. Eglinton. 2000. Distribution and sources of organic biomarkers in arctic sediments from the Mackenzie River and Beaufort Shelf. Marine Chemistry 71:23–51.
Gratani, L. 2014. Plant Phenotypic Plasticity in Response to Environmental Factors. Advances in Botany 2104:17.
Gu, J., Y. Xu, X. Dong, H. Wang, and Z. Wang. 2014. Root diameter variations explained by anatomy and phylogeny of 50 tropical and temperate tree species. Tree Physiology 34:415–425.
Guillaume, T., D. Maranguit, K. Murtilaksono, and Y. Kuzyakov. 2016. Sensitivity and resistance of soil fertility indicators to land-use changes: New concept and examples from conversion of Indonesian rainforest to plantations. Ecological Indicators 67:49–57.
Guo, L. B., and R. M. Gifford. 2002. Soil carbon stocks and land use change: a meta analysis. Global change biology 8:345–360.
Guo, L. B., and R. E. H. Sims. 1999. Litter production and nutrient return in New Zealand eucalypt short-rotation forests: implications for land management. Agriculture, Ecosystems & Environment 73:93–100.
Habteyohannes, L. 2016. Leaf Traits of Church Forest Trees in North West Ethiopia. University of Natural Resources and Life Sciences.
Hairiah, K., S. M. Sitompul, M. van Noordwijk, and C. Palm. 2001. Carbon stocks of tropical land use systems as part of the global C balance. Effects of forest conversion and options for clean development activities. International Centre for research in Agroforestry, ICRAF.
Hamer, U., C. Rumpel, and M.-F. Dignac. 2012. Cutin and suberin biomarkers as tracers for the turnover of shoot and root derived organic matter along a chronosequence of Ecuadorian pasture soils. European Journal of Soil Science 63:808–819.
Hansson, K., H.-S. Helmisaari, S. P. Sah, and H. Lange. 2013. Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW Sweden. Forest Ecology and Management 309:58–65.
Hararuk, O., and Y. Luo. 2014. Improvement of global litter turnover rate predictions using a Bayesian MCMC approach. Ecosphere 5:art163.
Harteveld, M., D. Hertel, M. Wiens, and C. Leuschner. 2007. Spatial and temporal variability fo fine root abundance and growth in tropical moist forests and agroforestry systems (Sulawesi, Indonesia). ECOTROPICA 13:111–120.
Hartley, I. P., D. W. Hopkins, M. H. Garnett, M. Sommerkorn, and P. A. Wookey. 2008. Soil microbial respiration in arctic soil does not acclimate to temperature. Ecology letters 11:1092–1100.
Hashimoto, S., N. Carvalhais, A. Ito, M. Migliavacca, K. Nishina, and M. Reichstein. 2015. Global spatiotemporal distribution of soil respiration Global spatiotemporal distribution of soil

168
respiration modeled using a global database Global spatiotemporal distribution of soil respiration Global spatiotemporal distribution of soil respiration. BGD Biogeosciences Discuss 12:4331–4364.
Hättenschwiler, S., and H. B. Jørgensen. 2010. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. Journal of Ecology 98:754–763.
Hedberg, I. 2003. Flora of Ethiopia and Eritrea Pt. 1. Apiaceae to Dipsacaceae.
Hedges, J. I., and J. R. Ertel. 1982. Characterization of lignin by gas capillary chromatography of cupric oxide oxidation products. Analytical Chemistry 54:174–178.
Hedges, J. I., and D. C. Mann. 1979a. The characterization of plant tissues by their lignin oxidation products. Geochimica et Cosmochimica Acta 43:1803–1807.
Hedges, J. I., and D. C. Mann. 1979b. The lignin geochemistry of marine sediments from the southern Washington coast. Geochimica et Cosmochimica Acta 43:1809–1818.
Hedges, J. I., H. J. Turin2, and J. R. Ertel. 1984. Sources and distributions of sedimentary organic matter in the Columbia River drainage basin, Washington and Oregon’. Ltmnol. Oceanogr 29:35–46.
Helmisaari, H.-S., and L. Hallbäcken. 1999. Fine-root biomass and necromass in limed and fertilized Norway spruce (Picea abies (L.) Karst.) stands. Forest Ecology and Management 119:99–110.
Hendricks, J. J., J. D. Aber, K. J. Nadelhoffer, and R. D. Hallett. 2000. Nitrogen Controls on Fine Root Substrate Quality in Temperate Forest Ecosystems. Ecosystems 3:57–69.
Hernández, D. L., S. E. Hobbie, T. Richard, C. D. L. Hernández, and S. E. Hobbie. 2010. The effects of substrate composition, quantity, and diversity on microbial activity. Plant Soil 335:397–411.
Hertel, D., M. A. Harteveld, and C. Leuschner. 2008. Conversion of a tropical forest into agroforest alters the fine root-related carbon flux to the soil. Soil Biology and Biochemistry 41:481–490.
Hertel, D., and C. Leuschner. 2002. A comparison of four different fine root production estimates with ecosystem carbon balance data in a Fagus–Quercus mixed forest. Plant and Soil 239:237–251.
Hertel, D., T. Strecker, H. Müller-Haubold, and C. Leuschner. 2013. Fine root biomass and dynamics in beech forests across a precipitation gradient - is optimal resource partitioning theory applicable to water-limited mature trees? Journal of Ecology 101:1183–1200.
Hiederer, R., and M. Köchy. 2011. Global Soil Organic Carbon Estimates and the Harmonized World Soil Database. EUR 25225 EN. Publications Office of the European Union. Citeseer, Joint Research Center. Ispra, Italy.
Houghton, R. A., and C. L. Goodale. 2004. Effects of land-use change on the carbon balance of terrestrial ecosystems. Ecosystems and land use change 153:85–98.
Hurni, H., S. Abate, A. Bantider, B. Debele, E. Ludi, B. Portner, B. Yitaferu, and G. Zeleke. 2010. Land Degradation and Sustainable Land Management in the Highlands of Ethiopia.
Jackson, R., H. A. Mooney, and E.-D. Schulze. 1997. A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences 94:7362–7366.
Jäger, S., H. Trojan, T. Kopp, M. N. Laszczyk, and A. Scheffler. 2009. Pentacyclic Triterpene Distribution in Various Plants – Rich Sources for a New Group of Multi-Potent Plant Extracts. Molecules 14:2016–2031.
Jenkinson, D. S., and D. S. Powlson. 1976. The effects of biocidal treatments on metabolism in soil—V: A method for measuring soil biomass. Soil Biology and Biochemistry 8:209–213.
Jepsen, R., J. Roberts, and W. Lick. 1997. Effects of bulk density on sediment erosion rates. Water, Air, & Soil Pollution 99:21–31.
Jia, B., G. Zhou, and Z. Xu. 2016. Forest litterfall and its composition: a new data set of observational

169
data from China. Ecology 97:1365–1365.
Jin, W., K. Singh, and J. Zondlo. 2013. Pyrolysis Kinetics of Physical Components of Wood and Wood-Polymers Using Isoconversion Method. Agriculture 3:12–32.
Jobbágy, E. G., and R. B. Jackson. 2000. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological applications 10:423–436.
Joergensen, R. G. 2010. Organic Matter and Micro-Organisms in Tropical Soils. Pages 17–44. Springer Berlin Heidelberg.
Johnson, M. G., D. T. Tingey, D. L. Phillips, and M. J. Storm. 2001. Advancing fine root research with minirhizotrons. Environmental and Experimental Botany 45:263–289.
Jones, C., C. McConnell, K. Coleman, P. Cox, P. Falloon, D. Jenkinson, and D. Powlson. 2005. Global climate change and soil carbon stocks; predictions from two contrasting models for the turnover of organic carbon in soil. Global change biology 11:154–166.
Joslin, J. D., M. H. Wolfe, and P. J. Hanson. 2000. Effects of altered water regimes on forest root systems. New Phytologist 147:117–129.
Joslin, J. D., M. H. Wolfe, and P. J. Hanson. 2001. Factors controlling the timing of root elongation intensity in a mature upland oak stand. Plant and Soil 228:201–212.
Kabata-Pendias, A. 2010. Trace elements in soils and plants. Fourth. CRC press, USA.
Kalinina, O., S. V Goryachkin, N. A. Karavaeva, D. I. Lyuri, and L. Giani. 2010. Dynamics of carbon pools in post-agrogenic sandy soils of southern taiga of Russia. Carbon Balance Manag 5:1.
Karavin, N., E. Yalman, Z. Kizir, A. Kocamaz, and B. Kartal. 2016. Variation in leaf litter decomposition rate according to salinity and water regime in Juglans regia L. Int. J. Adv. Res. Biol. Sci. International Journal of Advanced Research in Biological Sciences 3:158–162.
Karberg, N. J., N. A. Scott, C. P. Giardina, N. J. Karberg, N. A. Scott, C. P. Giardina, and C. M. Hoover. 2008. Methods for Estimating Litter Decomposition. Pages 103–111Field measurements for forest carbon monitoring.
Karhu, K., M. D. Auffret, J. A. J. Dungait, D. W. Hopkins, J. I. Prosser, B. K. Singh, J.-A. Subke, P. A. Wookey, G. I. Ågren, and M.-T. Sebastià. 2014. Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 513:81–84.
Katende, A. B., A. Birnie, and B. Tengnas. 1995. Useful trees and shrubs for Uganda : identification, propagation, and management for agricultural and pastoral communities. Regional Soil Conservation Unit, Kampala.
Kätterer, T., A. Fabião, M. Madeira, C. Ribeiro, and E. Steen. 1995. Fine-root dynamics, soil moisture and soil carbon content in a Eucalyptus globulus plantation under different irrigation and fertilisation regimes. Forest Ecology and Management 74:1–12.
Kettler, T. A., J. W. Doran, and T. L. Gilbert. 2001. Simplified method for soil particle-size determination to accompany soil-quality analyses. Soil Science Society of America Journal 65:849–852.
King, J., K. Pregitzer, D. Zak, J. Sober, J. Isebrands, R. Dickson, G. Hendrey, and D. Karnosky. 2001. Fine-root biomass and fluxes of soil carbon in young stands of paper birch and trembling aspen as affected by elevated atmospheric CO2 and tropospheric O3. Oecologia 128:237–250.
Knicker, H. 2007. How does fire affect the nature and stability of soil organic nitrogen and carbon? A review. Biogeochemistry 85:91–118.
Kögel-Knabner, I. 2000. Analytical approaches for characterizing soil organic matter. Organic Geochemistry 31:609–625.
Kögel-Knabner, I. 2002. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biology and Biochemistry 34:139–162.

170
Kögel-Knabner, I., F. Ziegler, M. Riederer, and W. Zech. 1989. Distribution and decomposition pattern of cutin and suberin in forest soils. Zeitschrift für Pflanzenernährung und Bodenkunde 152:409–413.
Kong, D. L., J. J. Wang, P. Kardol, H. F. Wu, H. Zeng, X. B. Deng, and Y. Deng. 2016. Economic strategies of plant absorptive roots vary with root diameter. Biogeosciences 13:415–424.
Kong, D., C. Ma, Q. Zhang, L. Li, X. Chen, H. Zeng, and D. Guo. 2014. Leading dimensions in absorptive root trait variation across 96 subtropical forest species. New Phytologist 203:863–872.
Kucbel, S., P. Jaloviar, and J. Špišák. 2011. Quantity, vertical distribution and morphology of fine roots in Norway spruce stands with different stem density. Plant Root 5:46–55.
Kuo, L.-J., P. Louchouarn, and B. E. Herbert. 2008. Fate of CuO-derived lignin oxidation products during plant combustion: application to the evaluation of char input to soil organic matter. Organic Geochemistry 39:1522–1536.
Kuzyakov, Y., and G. Domanski. 2000. Carbon input by plants into the soil. Review. Journal of Plant Nutrition and Soil Science 163:421–431.
Lal, R. 1995. Soils and global change. Lewis Publishers.
Lal, R. 2010a. Managing soils for a warming earth in a food-insecure and energy-starved world. Journal of Plant Nutrition and Soil Science 173:4–15.
Lal, R. 2010b. Managing Soils and Ecosystems for Mitigating Anthropogenic Carbon Emissions and Advancing Global Food Security. BioScience 60:708–721.
Langenheim, J. H. 2003. Plant resins : chemistry, evolution, ecology, and ethnobotany. Timber Press, Portland, Cambridge.
de Leeuw, J. W., G. J. M. Versteegh, and P. F. van Bergen. 2005. Biomacromolecules of algae and plants and their fossil analogues. Pages 209–233Plants and Climate Change. Springer Netherlands, Dordrecht.
Lemenih, M., and F. Itanna. 2004. Soil carbon stocks and turnovers in various vegetation types and arable lands along an elevation gradient in southern Ethiopia. Geoderma 123:177–188.
Lemenih, M., E. Karltun, and M. Olsson. 2005. Soil organic matter dynamics after deforestation along a farm field chronosequence in southern highlands of Ethiopia. Agriculture, Ecosystems & Environment 109:9–19.
Lemenih, M., B. Lemma, and D. Teketay. 2006. Changes in soil carbon and total nitrogen following reforestation of previously cultivated land in the highlands of Ethiopia. SINET: Ethiopian Journal of Science 28:99–108.
Lemenih, M., M. Olsson, and E. Karltun. 2004. Comparison of soil attributes under Cupressus lusitanica and Eucalyptus saligna established on abandoned farmlands with continuously cropped farmlands and natural forest in Ethiopia. Forest Ecology and Management 195:57–67.
Leppälammi-Kujansuu, J. 2014. Norway spruce fine root dynamics and carbon input into soil in relation to environmental factors. University of Helsinki.
Leppälammi-Kujansuu, J., L. Aro, M. Salemaa, K. Hansson, D. B. Kleja, and H.-S. Helmisaari. 2014. Fine root longevity and carbon input into soil from below- and aboveground litter in climatically contrasting forests. Forest Ecology and Management 326:79–90.
Lequy, É., S. Conil, and M.-P. Turpault. 2012. Impacts of Aeolian dust deposition on European forest sustainability: A review. Forest Ecology and Management 267:240–252.
Leuschner, C., and D. Hertel. 2003. Fine root biomass of temperate forests in relation to soil acidity and fertility, climate, age and species. Pages 405–438Progress in botany. Springer.
Levey, D. J., H. A. Bissell, and S. F. O’Keefe. 2000. Conversion of Nitrogen to Protein and Amino

171
Acids in Wild Fruits. Journal of Chemical Ecology 26:1749–1763.
Li, F., B. Pan, D. Zhang, X. Yang, H. Li, S. Liao, A. Ghaffar, H. Peng, and B. Xing. 2015. Organic matter source and degradation as revealed by molecular biomarkers in agricultural soils of Yuanyang terrace. Scientific Reports 5:11074.
Li, S., L. Bashline, L. Lei, and Y. Gu. 2014. Cellulose synthesis and its regulation. The Arabidopsis book 12:e0169.
Li, X., J. Zhu, H. Lange, and S. Han. 2013. A modified ingrowth core method for measuring fine root production, mortality and decomposition in forests. Tree physiology 33:18–25.
Li, Y., X. Zhao, Y. Chen, Y. Luo, and S. Wang. 2012. Effects of grazing exclusion on carbon sequestration and the associated vegetation and soil characteristics at a semi-arid desertified sandy site in Inner Mongolia, northern China. Canadian Journal of Soil Science 92:807–819.
Li, Z., X. Nie, X. Chang, L. Liu, L. Sun, and J. Poesen. 2016. Characteristics of Soil and Organic Carbon Loss Induced by Water Erosion on the Loess Plateau in China. PLOS ONE 11:e0154591.
Lima, T. T. S., I. S. Miranda, and S. S. Vasconcelos. 2010. Effects of water and nutrient availability on fine root growth in eastern Amazonian forest regrowth, Brazil. New Phytologist 187:622–630.
Lisanework, N., and A. Michelsen. 1994. Litterfall and nutrient release by decomposition in three plantations compared with a natural forest in the Ethiopian highland. Forest ecology and Management 65:149–164.
Lloyd, J., and J. A. Taylor. 1994. On the temperature dependence of soil respiration. Functional Ecology 8:315–323.
López, B., S. Sabaté, and C. A. Gracia. 2001. Annual and seasonal changes in fine root biomass of a Quercus ilex L. forest. Plant and Soil 230:125–134.
Lorenz, K., R. Lal, C. M. Preston, and K. G. J. Nierop. 2007. Strengthening the soil organic carbon pool by increasing contributions from recalcitrant aliphatic bio(macro)molecules. Geoderma 142:1–10.
Louzada, J. N. C., J. Schoereder, and P. De Marco. 1997. Litter decomposition in semideciduous forest and Eucalyptus spp. crop in Brazil: a comparison. Forest Ecology and Management 94:31–36.
Lukac, M. 2012. Fine root turnover. Pages 363–373Measuring Roots. Springer.
Lukac, M., and D. L. Godbold. 2001. A modification of the ingrowth-core method to determine root production in fast growing tree species. Journal of Plant Nutrition and Soil Science 164:613–614.
Lukac, M., and D. L. Godbold. 2010. Fine root biomass and turnover in southern taiga estimated by root inclusion nets. Plant and Soil 331:505–513.
Macinnis-Ng, C., and L. Schwendenmann. 2015. Litterfall, carbon and nitrogen cycling in a southern hemisphere conifer forest dominated by kauri (Agathis australis) during drought. Plant Ecology 216:247–262.
Maeght, J.-L., B. Rewald, and A. Pierret. 2013. How to study deep roots—and why it matters. Frontiers in plant science 4.
Majdi, H., K. Pregitzer, A.-S. Moren, J.-E. Nylund, and G. I. Ågren. 2005. Measuring fine root turnover in forest ecosystems. Plant and Soil 276:1–8.
Makita, N., Y. Kosugi, M. Dannoura, S. Takanashi, K. Niiyama, A. R. Kassim, and A. R. Nik. 2012. Patterns of root respiration rates and morphological traits in 13 tree species in a tropical forest. Tree Physiology 32:303–312.

172
Makkonen, K., and H.-S. Helmisaari. 1999. Assessing fine-root biomass and production in a Scots pine stand–comparison of soil core and root ingrowth core methods. Plant and Soil 210:43–50.
Malhi, Y., D. D. Baldocchi, and P. G. Jarvis. 1999. The carbon balance of tropical, temperate and boreal forests. Plant, cell & environment 22:715–740.
Malhi, Y., C. Doughty, and D. Galbraith. 2011. The allocation of ecosystem net primary productivity in tropical forests. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 366:3225–3245.
Marschner, B., S. Brodowski, A. Dreves, G. Gleixner, A. Gude, P. M. Grootes, U. Hamer, A. Heim, G. Jandl, R. Ji, K. Kaiser, K. Kalbitz, C. Kramer, P. Leinweber, J. Rethemeyer, A. Schäffer, M. W. I. Schmidt, L. Schwark, and G. L. B. Wiesenberg. 2008. How relevant is recalcitrance for the stabilization of organic matter in soils? Journal of Plant Nutrition and Soil Science 171:91–110.
Marshall, M. H. M. 2014. Characterization of Natural Organic Matter in Bentonite Clays and Clay-rich Sediments. University of Toronto.
Martens, R. 1995. Current methods for measuring microbial biomass C in soil: potentials and limitations. Biology and Fertility of Soils 19:87–99.
Martin, A. R., and S. C. Thomas. 2011. A reassessment of carbon content in tropical trees. PLoS One 6:e23533.
Martins, C., T. Natal-da-Luz, J. P. Sousa, M. J. Gonçalves, L. Salgueiro, and C. Canhoto. 2013. Effects of essential oils from Eucalyptus globulus leaves on soil organisms involved in leaf degradation. PloS one 8:e61233.
Mayer, M., H. Sandén, B. Rewald, D. L. Godbold, and K. Katzensteiner. 2016. Increase in heterotrophic soil respiration by temperature drives decline in soil organic carbon stocks after forest windthrow in a mountainous ecosystem. Functional Ecology.
McClaugherty, C. A., J. D. Aber, and J. M. Melillo. 1982. The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology:1481–1490.
McCormack, M. L., T. S. Adams, E. A. H. Smithwick, and D. M. Eissenstat. 2012. Predicting fine root lifespan from plant functional traits in temperate trees. New Phytologist 195:823–831.
McCormack, M. L., I. A. Dickie, D. M. Eissenstat, T. J. Fahey, C. W. Fernandez, D. Guo, H. H.-S. Helmisaari, E. A. Hobbie, C. M. Iversen, R. B. Jackson, J. Leppälammi-Kujansuu, R. J. Norby, R. P. Phillips, K. S. Pregitzer, S. G. Pritchard, B. Rewald, and M. Zadworny. 2015. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytologist 207:505–518.
Meinen, C., D. Hertel, and C. Leuschner. 2009. Biomass and morphology of fine roots in temperate broad-leaved forests differing in tree species diversity: is there evidence of below-ground overyielding? Oecologia 161:99–111.
Meisner, A., J. Rousk, and E. Bååth. 2015. Prolonged drought changes the bacterial growth response to rewetting. Soil Biology and Biochemistry 88:314–322.
Mekuria, W., E. Veldkamp, and M. Haile. 2009. Carbon stock changes with relation to land use conversion in the lowlands of Tigray, Ethiopia. Page Proceedings of Conference on International Research on Food Security, Natural Resource Management and Rural Development; Proceedings of the 36th Meeting of the Italian Society of Agronomy. Tropentag, University of Hamburg.
Mendez-Millan, M., M.-F. Dignac, C. Rumpel, and S. Derenne. 2011. Can cutin and suberin biomarkers be used to trace shoot and root-derived organic matter? A molecular and isotopic approach. Biogeochemistry 106:23–38.
Mendez-Millan, M., M.-F. Dignac, C. Rumpel, D. P. Rasse, G. Bardoux, and S. Derenne. 2012. Contribution of maize root derived C to soil organic carbon throughout an agricultural soil profile

173
assessed by compound specific 13 C analysis. Organic Geochemistry 42:1502–1511.
Mendez-Millan, M., M.-F. Dignac, C. Rumpel, D. P. Rasse, and S. Derenne. 2010. Molecular dynamics of shoot vs. root biomarkers in an agricultural soil estimated by natural abundance 13 C labelling. Soil Biology and Biochemistry 42:169–177.
Meseret, D. 2016. Land Degradation in Amhara Region of Ethiopia: Review on Extent, Impacts and Rehabilitation Practices. Journal of Environment and Earth Science 6:120–130.
Meshesha, D. T., A. Tsunekawa, M. Tsubo, S. A. Ali, and N. Haregeweyn. 2014. Land-use change and its socio-environmental impact in Eastern Ethiopia’s highland. Regional Environmental Change 14:757–768.
Metcalfe, D. B., P. Meir, L. E. O. C. Aragão, A. C. L. da Costa, A. P. Braga, P. H. L. Gonçalves, J. de Athaydes Silva Junior, S. S. de Almeida, L. A. Dawson, Y. Malhi, and M. Williams. 2008. The effects of water availability on root growth and morphology in an Amazon rainforest. Plant and Soil 311:189–199.
Mey, D., J. Sartohadi, D. Mardiatno, and M. A. Marfai. 2015. Prediction of soil organic carbon loss due to erosion in Girindulu Watersheds. Journal of Degraded and Mining Lands Management 2:327–334.
Mille-Lindblom, C., E. von Wachenfeldt, and L. J. Tranvik. 2004. Ergosterol as a measure of living fungal biomass: persistence in environmental samples after fungal death. Journal of Microbiological Methods 59:253–262.
Milne, E., R. Al Adamat, N. H. Batjes, M. Bernoux, T. Bhattacharyya, C. E. P. C. Cerri, C. E. P. C. Cerri, K. Coleman, M. Easter, and P. Falloon. 2007. National and sub-national assessments of soil organic carbon stocks and changes: The GEFSOC modelling system. Agriculture, Ecosystems & Environment 122:3–12.
Moges, Y., and M. Kindu. 2006. Effects of fencing and ground cultivation on natural regeneration of Boswellia papyrifera in Metema Wereda, Ethiopia. Journal of the drylands 1:45–51.
Molla, T., and B. Sisheber. 2017. Estimating soil erosion risk and evaluating erosion control measures for soil conservation planning at Koga watershed in the highlands of Ethiopia. Solid Earth 8:13–25.
Montgomery, H. J., C. M. Monreal, J. C. Young, and K. A. Seifert. 2000. Determinination of soil fungal biomass from soil ergosterol analyses. Soil Biology and Biochemistry 32:1207–1217.
Moorhead, D., G. Lashermes, S. Recous, and I. Bertrand. 2014. Interacting Microbe and Litter Quality Controls on Litter Decomposition: A Modeling Analysis. PLoS ONE 9:e108769.
Moura, P. M., T. D. Althoff, R. A. Oliveira, J. S. Souto, P. C. Souto, R. S. C. Menezes, and E. V. S. B. Sampaio. 2016. Carbon and nutrient fluxes through litterfall at four succession stages of Caatinga dry forest in Northeastern Brazil. Nutrient Cycling in Agroecosystems 105:25–38.
Mulkey, S. S., K. Kitajima, and S. J. Wright. 1996. Plant physiological ecology of tropical forest canopies. Trends in Ecology & Evolution 11:408–412.
Muller, R. N., P. J. Kalisz, and J. O. Luken. 1989. Fine root production of astringent phenolics. Oecologia 79:563–565.
Nedessa, B., J. Ali, and I. Nyborg. 2005. Exploring Ecological and Socio-Economic Issues for the Improvement of Area Enclosure Management A Case Study from Ethiopia.
Neill, C. 1992. Comparison of Soil Coring and Ingrowth Methods for Measuring Belowground Production. Ecology 73:1918–1921.
Nierop, K. G. J. 1998. Origin of aliphatic compounds in a forest soil. Organic Geochemistry 29:1009–1016.
Nierop, K. G. J., B. Jansen, J. A. Hageman, and J. M. Verstraten. 2006. The Complementarity of Extractable and Ester-Bound Lipids in a Soil Profile Under Pine. Plant and Soil 286:269–285.

174
Nierop, K. G. J., D. F. W. Naafs, and P. F. van Bergen. 2005. Origin, occurrence and fate of extractable lipids in Dutch coastal dune soils along a pH gradient. Organic Geochemistry 36:555–566.
Nkonya, E., J. Koo, E. Kato, and Z. Guo. 2013. Trends and patterns of land use change and international aid in sub-Saharan Africa.
Nyssen, J., J. Poesen, J. Moeyersons, J. Deckers, M. Haile, and A. Lang. 2004. Human impact on the environment in the Ethiopian and Eritrean highlands—a state of the art. Earth-Science Reviews 64:273–320.
Nyssen, J., H. Temesgen, M. Lemenih, A. Zenebe, N. Haregeweyn, and M. Haile. 2008. Spatial and temporal variation of soil organic carbon stocks in a lake retreat area of the Ethiopian Rift Valley. Geoderma 146:261–268.
Oertel, C., J. Matschullat, K. Zurba, F. Zimmermann, and S. Erasmi. 2016. Greenhouse gas emissions from soils—A review. Chemie der Erde - Geochemistry 76:327–352.
Olson, J. S. 1963. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 44:322–331.
Orwa, M. A., J. Kindt, R, and R. S. C. Anthony. 2009. Agroforestry Database: a tree reference and selection guide version 4.0. World Agroforestry Centre, Kenya.
Osawa, A., and R. Aizawa. 2012. A new approach to estimate fine root production, mortality, and decomposition using litter bag experiments and soil core techniques. Plant and Soil 355:167–181.
Ostonen, I., Ü. Püttsepp, C. Biel, O. Alberton, M. R. Bakker, K. Lõhmus, H. Majdi, D. Metcalfe, A. F. M. Olsthoorn, A. Pronk, E. Vanguelova, M. Weih, and I. Brunner. 2007. Specific root length as an indicator of environmental change. Plant Biosystems 141:426–442.
Ostonen, I., K. Rosenvald, H.-S. Helmisaari, D. Godbold, K. Parts, V. Uri, K. Lõhmus, B. Rewald, and C. Nabais. 2013. Morphological plasticity of ectomycorrhizal short roots in Betula sp and Picea abies forests across climate and forest succession gradients: its role in changing environments. Frontiers in microbiology 4:1–10.
Ostrofsky, M. L. 2007. A comment on the use of exponential decay models to test nonadditive processing hypotheses in multispecies mixtures of litter. Journal of the North American Benthological Society 26:23–27.
Otto, A., R. Gondokusumo, and M. J. Simpson. 2006. Characterization and quantification of biomarkers from biomass burning at a recent wildfire site in Northern Alberta, Canada. Applied Geochemistry 21:166–183.
Otto, A., C. Shunthirasingham, and M. J. Simpson. 2005. A comparison of plant and microbial biomarkers in grassland soils from the Prairie Ecozone of Canada. Organic Geochemistry 36:425–448.
Otto, A., and M. J. Simpson. 2006a. Sources and composition of hydrolysable aliphatic lipids and phenols in soils from western Canada. Organic Geochemistry 37:385–407.
Otto, A., and M. J. Simpson. 2006b. Evaluation of CuO oxidation parameters for determining the source and stage of lignin degradation in soil. Biogeochemistry 80:121–142.
Otto, A., and M. J. Simpson. 2007. Analysis of soil organic matter biomarkers by sequential chemical degradation and gas chromatography–mass spectrometry. Journal of separation science 30:272–282.
Ouaryi, A., A. Boularbah, H. Sanguin, M. Hafidi, E. Baudoin, L. Ouahmane, C. Le Roux, A. Galiana, Y. Prin, and R. Duponnois. 2016. High potential of symbiotic interactions between native mycorrhizal fungi and the exotic tree Eucalyptus camaldulensis for phytostabilization of metal-contaminated arid soils. International Journal of Phytoremediation 18:41–47.

175
Pan, X., Y.-B. Song, C. Jiang, G.-F. Liu, X.-H. Ye, X.-F. Xie, Y.-K. Hu, W.-W. Zhao, L. Cui, J. H. C. Cornelissen, M. Dong, and A. Prinzing. 2015. Evolutionary Position and Leaf Toughness Control Chemical Transformation of Litter, and Drought Reinforces This Control: Evidence from a Common Garden Experiment across 48 Species. PLOS ONE 10:e0143140.
Pan, Y., R. A. Birdsey, J. Fang, R. Houghton, P. E. Kauppi, W. A. Kurz, O. L. Phillips, A. Shvidenko, S. L. Lewis, J. G. Canadell, P. Ciais, R. B. Jackson, S. W. Pacala, A. D. McGuire, S. Piao, A. Rautiainen, S. Sitch, and D. Hayes. 2011. A Large and Persistent Carbon Sink in the World’s Forests. Science 333:988–993.
Paul, E. A. 2016. The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization.
Pautler, B. G., J. Austin, A. Otto, K. Stewart, S. F. Lamoureux, and M. J. Simpson. 2010. Biomarker assessment of organic matter sources and degradation in Canadian High Arctic littoral sediments. Biogeochemistry 100:75–87.
Pautler, B. G., P. T. Sanborn, A. J. Simpson, and M. J. Simpson. 2013. Molecular characterization of organic matter in Canadian Arctic paleosols for paleoecological applications. Organic Geochemistry 63:122–138.
Pearse, I. S., R. C. Cobb, and R. Karban. 2014. The phenology-substrate-match hypothesis explains decomposition rates of evergreen and deciduous oak leaves. Journal of Ecology 102:28–35.
Peel, M. C., B. L. Finlayson, and T. A. McMahon. 2007. Updated world map of the Köppen-Geiger climate classification. Hydrology and Earth System Sciences Discussions Discussions 4:439–473.
Pettersen, R. C. 1984. The Chemical Compostition of Wood. Pages 57–126The Chemistry of Solid Wood.
Pinno, B. D., S. D. Wilson, D. F. Steinaker, K. C. J. Van Rees, and S. A. Mcdonald. 2010. Fine root dynamics of trembling aspen in boreal forest and aspen parkland in central Canada. Annals of Forest Science 67:1–6.
Pisani, O., K. M. Hills, D. Courtier-Murias, A. J. Simpson, N. J. Mellor, E. A. Paul, S. J. Morris, and M. J. Simpson. 2013. Molecular level analysis of long term vegetative shifts and relationships to soil organic matter composition. Organic Geochemistry 62:7–16.
Poeplau, C., A. Don, L. Vvesterdal, J. Leifield, B. Van Wesemael, J. Schumacher, A. Gensior, C. Poeplau, L. Vesterdal, J. Leifeld, B. Van Wesemael, J. Schumacher, and A. Gensior. 2011. Temporal dynamics of soil organic carbon after land-use change in the temperate zone - carbon response functions as a model approach. Global Change Biology 17:2415–2427.
Pohjonen, V., and T. Pukkala. 1990. Eucalyptus globulus in Ethiopian forestry. Forest Ecology and Management 36:19–31.
Poirier, N., S. P. Sohi, J. L. Gaunt, N. Mahieu, E. W. Randall, D. S. Powlson, and R. P. Evershed. 2005. The chemical composition of measurable soil organic matter pools. Organic Geochemistry 36:1174–1189.
Poorter, H. 1994. Construction costs and payback time of biomass: a whole plant perspective. Pages 111–127in J. Roy and E. Garnier, editors.A whole plant perspective on carbon-nitrogen interactions.
Poorter, H., and M. Bergkotte. 1992. Chemical composition of 24 wild species differing in relative growth rate. Plant, cell & environment 15:221–229.
Poorter, H., S. Pepin, T. Rijkers, Y. De Jong, J. R. Evans, and C. Körner. 2006. Construction costs, chemical composition and payback time of high-and low-irradiance leaves. Journal of Experimental Botany 57:355–371.
Portillo-Estrada, M., M. Pihlatie, J. F. J. Korhonen, J. Levula, A. K. F. Frumau, A. Ibrom, J. J. Lembrechts, L. Morillas, L. Horváth, S. K. Jones, and Ü. Niinemets. 2016. Climatic controls on

176
leaf litter decomposition across European forests and grasslands revealed by reciprocal litter transplantation experiments. Biogeosciences 13:1621–1633.
Post, W. M., and K. C. Kwon. 2000. Soil carbon sequestration and land‐use change: processes and potential. Global change biology 6:317–327.
Powers, J. S., M. D. Corre, T. E. Twine, and E. Veldkamp. 2011. Geographic bias of field observations of soil carbon stocks with tropical land-use changes precludes spatial extrapolation. Proceedings of the National Academy of Sciences 108:6318–6322.
Pregitzer, K. S., J. L. DeForest, A. J. Burton, M. F. Allen, R. W. Ruess, and R. L. Hendrick. 2002. Fine root architecture of nine North American trees. Ecological Monographs 72:293–309.
Pregitzer, K. S., M. E. Kubiske, C. K. Yu, and R. L. Hendrick. 1997. Relationships among root branch order, carbon, and nitrogen in four temperate species. Oecologia 111:302–308.
Prescott, C. E., L. Vesterdal, C. M. Preston, and S. W. Simard. 2004. Influence of initial chemistry on decomposition of foliar litter in contrasting forest types in British Columbia. Canadian Journal of Forest Research 34:1714–1729.
Prescott, C. E., L. M. Zabek, C. L. Staley, and R. Kabzems. 2011. Decomposition of broadleaf and needle litter in forests of British Columbia : influences of litter type , forest type , and litter mixtures. Canadian Journal of Forest Research 30:1742–1750.
Preston, C. M., J. A. Trofymow, the, and the C. I. D. E. Working Group. 2000. Variability in litter quality and its relationship to litter decay in Canadian forests. Canadian Journal of Botany 78:1269–1287.
Priess, J., C. Then, and H. Fölster. 1999. Litter and fine-root production in three types of tropical premontane rain forest in SE Venezuela. Plant Ecology 143:171–187.
Priestley, C. A., P. B. Catlin, and E. A. Olsson. 1976. The Distribution of 14C-Labelled Assimilates in Young Apple Trees as Influenced by Doses of Supplementary Nitrogen. Annals of Botany 40:1163–1170.
Prieto, I., A. Stokes, and C. Roumet. 2016. Root functional parameters predict fine root decomposability at the community level. Journal of Ecology 104:725–733.
Le Quéré, C., R. M. Andrew, J. G. Canadell, S. Sitch, J. I. Korsbakken, G. P. Peters, A. C. Manning, T. A. Boden, P. P. Tans, R. A. Houghton, R. F. Keeling, S. Alin, O. D. Andrews, P. Anthoni, L. Barbero, L. Bopp, F. Chevallier, L. P. Chini, P. Ciais, K. Currie, C. Delire, S. C. Doney, P. Friedlingstein, T. Gkritzalis, I. Harris, J. Hauck, V. Haverd, M. Hoppema, K. Klein Goldewijk, A. K. Jain, E. Kato, A. Körtzinger, P. Landschützer, N. Lefèvre, A. Lenton, S. Lienert, D. Lombardozzi, J. R. Melton, N. Metzl, F. Millero, P. M. S. Monteiro, D. R. Munro, J. E. M. S. Nabel, S. Nakaoka, K. O&apos;Brien, A. Olsen, A. M. Omar, T. Ono, D. Pierrot, B. Poulter, C. Rödenbeck, J. Salisbury, U. Schuster, J. Schwinger, R. Séférian, I. Skjelvan, B. D. Stocker, A. J. Sutton, T. Takahashi, H. Tian, B. Tilbrook, I. T. van der Laan-Luijkx, G. R. van der Werf, N. Viovy, A. P. Walker, A. J. Wiltshire, and S. Zaehle. 2016. Global Carbon Budget 2016. Earth System Science Data 8:605–649.
Quichimbo, P., D. Veintimilla, Y. Carrión, and L. Jiménez. 2016. Litterfall production under pine plantations in the southern Andes region of Ecuador (Producción de hojarasca bajo plantaciones de pino en los Andes del Sur del Ecuador). Enfoque UTE 33:14–2514.
Rahman, M. M., and J. Tsukamoto. 2013. Leaf traits, litter decomposability and forest floor dynamics in an evergreen- and a deciduous-broadleaved forest in warm temperate Japan. Forestry 86:441–451.
Rasse, D. P., C. Rumpel, and M.-F. Dignac. 2005. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant and Soil 269:341–356.
Reichstein, M., J.-A. Subke, A. C. Angeli, and J. D. Tenhunen. 2005. Does the temperature sensitivity of decomposition of soil organic matter depend upon water content, soil horizon, or

177
incubation time? Global Change Biology 11:1754–1767.
Renault, P., M. M Ben-Sassi, and A. Berard. 2013. Improving the MicroRespTM substrate-induced respiration method by a more complete description of CO2 behavior in closed incubation wells. Geoderma 207–208.
Reshi, Z., and S. Tyub. 2007. Detritus and decomposition in ecosystems. New India Pub. Agency, New Delhi, India.
Rewald, B., C. Meinen, M. Trockenbrodt, J. E. Ephrath, and S. Rachmilevitch. 2012. Root taxa identification in plant mixtures–current techniques and future challenges. Plant and Soil 359:165–182.
Rewald, B., A. Rechenmacher, and D. L. Godbold. 2014. It’s complicated: intraroot system variability of respiration and morphological traits in four deciduous tree species. Plant physiology 166:736–745.
Reynolds, L. L., B. R. Johnson, L. Pfeifer-Meister, and S. D. Bridgham. 2015. Soil respiration response to climate change in Pacific Northwest prairies is mediated by a regional Mediterranean climate gradient. Global Change Biology 21:487–500.
Riederer, M., K. Matzke, F. Ziegler, and I. Kögel-Knabner. 1993. Occurence, distribution and fate of the lipid plant biopolymers cutin and suberin in teperate forest soils. Org. Geochem. 20:1063–1076.
Robinson, D. 2007. Implications of a large global root biomass for carbon sink estimates and for soil carbon dynamics. Proceedings of the Royal Society of London B: Biological Sciences 274:2753–2759.
Rochette, P., and G. L. Hutchinson. 2005. Measurement of Soil Respiration in situ: Chamber Techniques. USDA-ARS / UNL Faculty:247–286.
Rousk, J., S. D. Frey, and E. Bååth. 2012. Temperature adaptation of bacterial communities in experimentally warmed forest soils. Global Change Biology 18:3252–3258.
Rowshan, V., and A. Tarakemeh. 2013. Chemical composition of Scaligeria meifolia Boiss. essential oil from Iran. Herla Rolonica 59.
Ruess, R. W., R. L. Hendrick, J. C. Vogel, and B. Sveinbjornsson. 2006. The role of fine roots in the functioning of Alaskan boreal forests. Page in M. W. O. F. Stuart Chapin Keith van Cleve, Leslie A. Viereck, David L. Verbyla, editor. Alaska’s Changing Boreal Forest. Oxford University Press: New York, New York.
Ryan, M. G. 1991. A simple method for estimating gross carbon budgets for vegetation in forest ecosystems. Tree physiology 9:255–266.
Ryan, M. G., and B. E. Law. 2005. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 73:3–27.
Ryan, M. G., J. M. Melillo, and A. Ricca. 1990. A comparison of methods for determining proximate carbon fractions of forest litter. Canadian Journal of Forest Research 20:166–171.
Saiz, G., K. Black, B. Reidy, S. Lopez, and E. P. Farrell. 2007. Assessment of soil CO 2 efflux and its components using a process-based model in a young temperate forest site. Geoderma 139:79–89.
Salete Capellesso, E., K. L. Scrovonski, E. M. Zanin, L. U. Hepp, C. Bayer, and T. L. Sausen. 2016. Effects of forest structure on litter production, soil chemical composition and litter–soil interactions. Acta Botanica Brasilica:1–7.
Salomé, C., N. Nunan, V. Pouteau, T. Z. Lerch, and C. Chenu. 2010. Carbon dynamics in topsoil and in subsoil may be controlled by different regulatory mechanisms. Global change biology 16:416–426.
Santantonio, D., and J. C. Grace. 1987. Estimating fine-root production and turnover from biomass

178
and decomposition data: a compartment–flow model. Canadian Journal of Forest Research 17:900–908.
Santos, G. G., J. C. N. Alves, J. M. L. Rodilla, A. P. Duarte, A. M. Lithgow, and J. G. Urones. 1997. Terpenoids and other constituents of Eucalyptus globulus. Phytochemistry 44:1309–1312.
Sariyildiz, T., and J. M. Anderson. 2003. Interactions between litter quality, decomposition and soil fertility: a laboratory study. Soil Biology and Biochemistry 35:391–399.
Scharlemann, J. P., E. V. Tanner, R. Hiederer, and V. Kapos. 2014. Global soil carbon: understanding and managing the largest terrestrial carbon pool. Carbon Management 5:81–91.
Schenk, H. J., and R. B. Jackson. 2002. The Global Biogeography of Roots. Ecological Monographs 72:311–328.
Scherer-Lorenzen, M., J. L. Bonilla, C. Potvin, M. Scherer-Lorenzen, Á. J. L. Bonilla, and C. Potvin. 2007. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 116:2108–2124.
Schimel, J. P., and S. Hättenschwiler. 2007. Nitrogen transfer between decomposing leaves of different N status. Soil Biology and Biochemistry 39:1428–1436.
Schimel, J. P., and M. N. Weintraub. 2003. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: a theoretical model. Soil Biology and Biochemistry 35:549–563.
Schindlbacher, A., C. De Gonzalo, E. Díaz‐pinés, P. Gorría, B. Matthews, R. Inclán, S. Zechmeister‐boltenstern, A. Rubio, and R. Jandl. 2010. Temperature sensitivity of forest soil organic matter decomposition along two elevation gradients. Temperature J. Geophys. Res 115:10.
Schlentner, R. E., and K. Van Cleve. 1985. Relationships between CO2 evolution from soil, substrate temperature, and substrate moisture in four mature forest types in interior Alaska. Canadian Journal of Forest Research 15:97–106.
Schrumpf, M., E. D. Schulze, K. Kaiser, and J. Schumacher. 2011. How accurately can soil organic carbon stocks and stock changes be quantified by soil inventories? Biogeosciences Discussions 8:1193–1212.
Schulp, C. J. E., G.-J. Nabuurs, P. H. Verburg, and R. W. de Waal. 2008. Effect of tree species on carbon stocks in forest floor and mineral soil and implications for soil carbon inventories. Forest Ecology and Management 256:482–490.
Schuman, G. E., H. H. Janzen, and J. E. Herrick. 2002. Soil carbon dynamics and potential carbon sequestration by rangelands. Environmental pollution 116:391–396.
Shao, P., X. Zeng, D. J. P. Moore, and X. Zeng. 2013. Soil microbial respiration from observations and Earth System Models. Environmental Research Letters 8:34034.
Shenkute, B., A. Hassen, T. Assafa, N. Amen, and A. Ebro. 2012. Identification and nutritive value of potential fodder trees and shrubs in the mid rift valley of Ethiopia. Journal of Animal & Plant Sciences 22:1126–1132.
Shiferaw, A. 2011. Estimating soil loss rates for soil conservation planning in the Borena Woreda of South Wollo Highlands, Ethiopia. Journal of Sustainable Development in Africa 13:87–106.
Shiferaw, B., and S. Holden. 1999. Soil erosion and smallholders’ conservation decisions in the highlands of Ethiopia. World development 27:739–752.
Silver, W. L., and R. K. Miya. 2001. Global patterns in root decomposition: comparisons of climate and litter quality effects. Oecologia 129:407–419.
Simoneit, B. R. T. 2002. Molecular Indicators (Biomarkers) of Past Life. The Anatomical Record 268:186–195.

179
Simoneit, B. R. T. 2005. A review of current applications of mass spectrometry for biomarker/molecular tracer elucidations. Mass Spectrometry Reviews 24:719–765.
Simpson, M. J., B. Chefetz, A. P. Deshmukh, P. G. Hatcher, and P. G. Hatcher. 2005. Comparison of polycyclic aromatic hydrocarbon distributions and sedimentary organic matter characteristics in contaminated, coastal sediments from Pensacola Bay, Florida. Marine Chemistry 59:139–163.
Sisay, K., C. Thurnher, and H. Hasenauer. 2016. Daily climate data for the Amhara region in Northwestern Ethiopia. International Journal of Climatology.
Sluiter, A., R. Ruiz, C. Scarlata, J. Sluiter, and D. Templeton. 2005. Determination of extractives in biomass. Page Laboratory Analytical Procedure (LAP). National Renewable Energy Laboratory, 1617 Cole Boulevard, Golden, Colorado.
Smith, P. 2007. Land use change and soil organic carbon dynamics. Nutrient Cycling in Agroecosystems 81:169–178.
Solly, E. F., I. Schöning, S. Boch, E. Kandeler, S. Marhan, B. Michalzik, J. Müller, J. Zscheischler, S. E. Trumbore, and M. Schrumpf. 2014. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant and Soil 382:203–218.
Solomon, D., F. Fritzsche, M. Tekalign, J. Lehmann, and W. Zech. 2002. Soil organic matter composition in the subhumid Ethiopian highlands as influenced by deforestation and agricultural management. Soil Science Society of America Journal 66:68–82.
Song, A., Y. Wang, and Y. Liu. 2009. Study on the chemical constituents of the essential oil of the leaves of Eucalyptus globulus Labill from China. Asian Journal of Traditional Medicines 4.
Sorgoná, A., M. R. Abenavoli, P. G. Gringeri, and G. Cacco. 2007. Comparing Morphological Plasticity of Root Orders in Slow-and Fast-growing Citrus Rootstocks Supplied with Different Nitrate Levels. Annals of botany 100:1287–1296.
Spielvogel, S., J. Prietzel, J. Leide, M. Riedel, J. Zemke, and I. Kögel-Knabner. 2014. Distribution of cutin and suberin biomarkers under forest trees with different root systems. Plant and Soil 381:95–110.
Steinaker, D. F., and S. D. Wilson. 2008. Phenology of fine roots and leaves in forest and grassland. Journal of Ecology 96:1222–1229.
Steinweg, J. M., A. F. Plante, R. T. Conant, E. A. Paul, and D. L. Tanaka. 2008. Patterns of substrate utilization during long-term incubations at different temperatures. Soil Biology and Biochemistry 40:2722–2728.
Strosser, E. 2010. Methods for determination of labile soil organic matter: an overview. Journal of Agrobiology 27:49–60.
Sun, T., L. Dong, Z. Mao, and Y. Li. 2015. Fine root dynamics of trees and understorey vegetation in a chronosequence of Betula platyphylla stands. Forest Ecology and Management 346.
Sun, T., Z. Mao, and Y. Han. 2013. Slow decomposition of very fine roots and some factors controlling the process: a 4-year experiment in four temperate tree species. Plant and Soil 372:445–458.
Suseela, V., R. T. Conant, M. D. Wallenstein, and J. S. Dukes. 2012. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Global Change Biology 18:336–348.
Swift, M. J. (Michael J., O. W. Heal, and J. M. (Jonathan M. Anderson. 1979. Decomposition in terrestrial ecosystems. University of California Press, Berkeley and Los Angeles.
Tabouret, H., G. Bareille, F. Claverie, C. Pécheyran, P. Prouzet, and O. F. X. Donard. 2010. Simultaneous use of strontium: calcium and barium: calcium ratios in otoliths as markers of habitat: Application to the European eel (Anguilla anguilla) in the Adour basin, South West

180
France. Marine Environmental Research 70:35–45.
Tang, J.-W., M. Cao, J.-H. Zhang, and M.-H. Li. 2010. Litterfall production, decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: a 10-year study. Plant and Soil 335:271–288.
Tareq, S. M., H. Kitagawa, and K. Ohta. 2011. Lignin biomarker and isotopic records of paleovegetation and climate changes from Lake Erhai, southwest China, since 18.5kaBP. Quaternary International 229:47–56.
Tareq, S. M., N. Tanaka, and K. Ohta. 2004. Biomarker signature in tropical wetland: lignin phenol vegetation index (LPVI) and its implications for reconstructing the paleoenvironment. Science of The Total Environment 324:91–103.
Tate, K. R., and D. S. Jenkinson. 1982. Adenosine triphosphate measurement in soil: An improved method. Soil Biology and Biochemistry 14:331–335.
Teketay, D., M. Lemenih, T. Bekele, Y. Yemshaw, S. Feleke, W. Tadesse, Y. Moges, T. Hunde, and D. Nigussie. 2010. Forest resources and challenges of sustainable forest management and conservation in Ethiopia. Page in T. T. Frans Bongers, editor. Degraded forests in Eastern Africa: management and restoration. Earthscan, London, Newyork.
Tesfaye, M. A., F. Bravo, R. Ruiz-Peinado, V. Pando, and A. Bravo-Oviedo. 2016. Impact of changes in land use, species and elevation on soil organic carbon and total nitrogen in Ethiopian Central Highlands. Geoderma 261:70–79.
Tesi, T., I. Semiletov, G. Hugelius, O. Dudarev, P. Kuhry, and Ö. Gustafsson. 2014. Composition and fate of terrigenous organic matter along the Arctic land–ocean continuum in East Siberia: Insights from biomarkers and carbon isotopes. Geochimica et Cosmochimica Acta 133:235–256.
Teskey, R. O., and T. M. Hinckley. 1981. Influence of temperature and water potential on root growth of white oak. Physiologia Plantarum 52:363–369.
Thanighaiarassu, R. R., and P. Sivamani. 2013. Analysis of Chemical Composition of Four Essential Oils by GC-MS and its Antibacterial Activity against Human Pathogenic Bacteria. International Journal of Pharmaceutical & Biological Archives 4:195–200.
Thirukkumaran, C. M., and D. Parkinson. 2000. Microbial respiration, biomass, metabolic quotient and litter decomposition in a lodgepole pine forest floor amended with nitrogen and phosphorous fertilizers. Soil Biology and Biochemistry 32:59–66.
Thomson, B. D., T. S. Grove, N. Malajczuk, and G. E. S. J. Hardy. 1994. The effectiveness of ectomycorrhizal fungi in increasing the growth of Eucalyptus globulus Labill. in relation to root colonization and hyphal development in soil. New Phytologist 126:517–524.
Tigabu, Y. 2016. Investigation of fine root trait differences among woody species and between functional types in church forest of Gelawdios, Amhara region, Ethiopia. University of Natural Resources and Life Sciences.
Tosi, M., O. S. Correa, M. A. Soria, J. A. Vogrig, O. Sydorenko, and M. S. Montecchia. 2016. Land-use change affects the functionality of soil microbial communities: A chronosequence approach in the Argentinian Yungas. Applied Soil Ecology 108:118–127.
Trumbore, S. E., E. A. Davidson, P. Barbosa de Camargo, D. C. Nepstad, and L. A. Martinelli. 1995. Belowground cycling of carbon in forests and pastures of Eastern Amazonia. Global biogeochemical cycles 9:515–528.
Tuomi, M., P. Vanhala, K. Karhu, H. Fritze, and J. Liski. 2007. Heterotrophic soil respiration—Comparison of different models describing its temperature dependence. Ecological modelling:182–190.
UNECA. 1996. Sustainable agriculture and environmental rehabilitation programme (SAERP): household level socio-economic survey of the Amhara region. Addis Ababa, Eethiopia,

181
Eethiopia.
Valverde-Barrantes, O. J., C. B. Blackwood, K. R. Kramer-Walter, P. J. Bellingham, T. R. Millar, R. D. Smissen, S. J. Richardson, and D. C. Laughlin. 2016. Root traits are multidimensional: specific root length is independent from root tissue density and the plant economic spectrum: Commentary on Kramer-Walter et al. (2016). Journal of Ecology 104:1311–1313.
Valverde-Barrantes, O. J., J. W. Raich, and A. E. Russell. 2007. Fine-root mass, growth and nitrogen content for six tropical tree species. Plant and Soil:357–370.
Vance, E. D., P. C. Brookes, and D. S. Jenkinson. 1987. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry 19:703–707.
Vertregt, N., and F. W. T. P. De Vries. 1987. A rapid method for determining the efficiency of biosynthesis of plant biomass. Journal of Theoretical Biology 128:109–119.
Vesterdal, L., N. Clarke, B. D. Sigurdsson, and P. Gundersen. 2013. Do tree species influence soil carbon stocks in temperate and boreal forests? Forest Ecology and Management 309:4–18.
Victoria, R., S. Banwart, H. Black, J. Ingram, H. Joosten, E. Milne, E. Noellemeyer, and Y. Baskin. 2012. The benefits of soil carbon. United Nations Environment Programme, Nairobi, Kenya.
Vitousek, P. M. 1984. Litterfall, Nutrient Cycling, and Nutrient Limitation in Tropical Forests. Ecology 65:285–298.
Vivin, P., M. Castelan-Estrada, and J.-P. Gaudillere. 2015. Seasonal changes in chemical composition and construction costs of grapevine tissues. VITIS-Journal of Grapevine Research 42:5.
Vogt, K. A., C. C. Grier, and D. J. Vogt. 1986. Production, Turnover, and Nutrient Dynamics of Above- and Belowground Detritus of World Forests. Pages 303–377in A. Macfadyen and C. D. Ford, editors.Advances in Ecological Research. Volume 15. Academic Press, New York.
Vogt, K. A., D. J. Vogt, and J. Bloomfield. 1998. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. plant soil 200:71–89.
Vogt, K. A., D. J. Vogt, P. A. Palmiotto, P. Boon, J. O’Hara, and H. Asbjornsen. 1996. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. Plant and Soil 187:159–219.
Voříšková, J., and P. Baldrian. 2012. Fungal community on decomposing leaf litter undergoes rapid successional changes. The ISME Journal 7:477–486.
Waisel, Y., A. Eshel, T. Beeckman, and U. Kafkafi. 2002. Plant roots: the hidden half. CRC Press.
Wallander, H., A. Ekblad, D. L. Godbold, D. Johnson, A. Bahr, P. Baldrian, R. G. Björk, B. Kieliszewska-Rokicka, R. Kjøller, and H. Kraigher. 2013. Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in forests soils–A review. Soil Biology and Biochemistry 57:1034–1047.
Wang, J., and U. M. Sainju. 2014. Soil Carbon and Nitrogen Fractions and Crop Yields Affected by Residue Placement and Crop Types.
Wang, Z., D. Guo, X. Wang, J. Gu, and L. Mei. 2006. Fine root architecture, morphology, and biomass of different branch orders of two Chinese temperate tree species. Plant and Soil 288:155–171.
Wardle, D. A., and A. Ghani. 1995. A critique of the microbial metabolic quotient (qCO2) as a bioindicator of disturbance and ecosystem development. Soil Biology and Biochemistry 27:1601–1610.
Wassie, A., F. J. Sterck, D. Teketay, and F. Bongers. 2009. Tree regeneration in church forests of Ethiopia: effects of microsites and management. Biotropica 41:110–119.
WBISP. 2004. Woody Biomass Inventory and Strategic Planning Terminal report.

182
Weemstra, M., L. Mommer, E. J. W. Visser, J. van Ruijven, T. W. Kuyper, G. M. J. Mohren, and F. J. Sterck. 2016. Towards a multidimensional root trait framework: a tree root review. New Phytologist 211:1159–1169.
Wei, X., M. Shao, W. Gale, and L. Li. 2014. Global pattern of soil carbon losses due to the conversion of forests to agricultural land. Scientific reports 4:4062.
Wiesenberg, G. L. B., and L. Schwark. 2006. Carboxylic acid distribution patterns of temperate C3 and C4 crops. Organic Geochemistry 37:1973–1982.
Wild, B., N. Gentsch, P. Čapek, K. Diáková, R. J. E. Alves, J. Bárta, A. Gittel, G. Hugelius, A. Knoltsch, P. Kuhry, N. Lashchinskiy, R. Mikutta, J. Palmtag, C. Schleper, J. Schnecker, O. Shibistova, M. Takriti, V. L. Torsvik, T. Urich, M. Watzka, H. Šantrůčková, G. Guggenberger, and A. Richter. 2016. Plant-derived compounds stimulate the decomposition of organic matter in arctic permafrost soils. Nature Publishing Group.
Wolters, V. 1991. Biological processes in two beech forest soils treated with simulated acid rain—a laboratory experiment with Isotoma tigrina (insecta, collembola). Soil Biology and Biochemistry 23:381–390.
Workneh, S., and G. Glatzel. 2008. Land use changes and soil organic carbon and soil nitrogen in the Northwestern Highlands of Ethiopia. University of Natural Resources and Applied Life Sciences, Vienna, Vienna.
WRB. 2014. World Reference Base for Soil Resources 2014. Page International soil classification system for naming soils and creating legends for soil maps. Food and Agriculture Organization of the United Nations, Rome, Italy.
Wutzler, T., and M. Reichstein. 2008. Colimitation of decomposition by substrate and decomposers &ndash; a comparison of model formulations. Biogeosciences 5:749–759.
Wynn, J. G., M. I. Bird, L. Vellen, E. Grand‐Clement, J. Carter, S. L. Berry, E. Grand-Clement, J. Carter, S. L. Berry, E. Grand‐Clement, J. Carter, S. L. Berry, E. Grand-Clement, J. Carter, and
S. L. Berry. 2006. Continental‐scale measurement of the soil organic carbon pool with climatic, edaphic, and biotic controls. Global biogeochemical cycles 20:1–12.
Xia, M., A. F. Talhelm, and K. S. Pregitzer. 2015. Fine roots are the dominant source of recalcitrant plant litter in sugar maple‐dominated northern hardwood forests. New Phytologist 208:715–726.
Yan, T., L. Yang, and C. D. Campbell. 2003. Microbial biomass and metabolic quotient of soils under different land use in the Three Gorges Reservoir area. Geoderma 115:129–138.
Yang, G., and P. Jaakkola. 2011. Wood chemistry and isolation of extractives from wood. Literature study for BIOTULI project-Saimaa University of Applied Sciences. Saimaa University of Applied Sciences.
Yang, Y.-S., G.-S. Chen, P. Lin, J.-S. Xie, and J.-F. Guo. 2004a. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in subtropical China. Annals of Forest Science 61:617–627.
Yang, Y. S., J. F. Guo, G. S. Chen, J. S. Xie, L. P. Cai, and P. Lin. 2004b. Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a natural forest in subtropical China. Annals of Forest Science 61:465–476.
Yavitt, J. B., and S. J. Wright. 2001. Drought and Irrigation Effects on Fine Root Dynamics in a Tropical Moist Forest, Panama. Biotropica 33:421–434.
Yeong, K. L., G. Reynolds, and J. K. Hill. 2016. Leaf litter decomposition rates in degraded and fragmented tropical rain forests of Borneo. Biotropica 48:443–452.
Yuan, Z. Y., and H. Y. H. Chen. 2010. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: literature review and meta-analyses. Critical Reviews in Plant Sciences 29:204–221.

183
Yuan, Z. Y., and H. Y. H. Chen. 2013. Simplifying the decision matrix for estimating fine root production by the sequential soil coring approach. Acta Oecologica 48:54–61.
Yuste, J. C., D. D. Baldocchi, A. Gershenson, A. Goldstein, L. Misson, and S. Wong. 2007. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Global change biology 13:2018–2035.
Zeleke, G., and H. Hurni. 2001. Implications of land use and land cover dynamics for mountain resource degradation in the Northwestern Ethiopian highlands. Mountain Research and Development 21:184–191.
Zhang, H., W. Yuan, W. Dong, and S. Liu. 2014. Seasonal patterns of litterfall in forest ecosystem worldwide. Ecological Complexity 20:240–247.
Zhang, X., and W. Wang. 2015. The decomposition of fine and coarse roots: their global patterns and controlling factors. Scientific Reports 5:9940.
Zhou, J., X. Lang, B. Du, H. Zhang, H. Liu, Y. Zhang, and L. Shang. 2016. Litterfall and nutrient return in moist evergreen broad-leaved primary forest and mixed subtropical secondary deciduous broad-leaved forest in China. European Journal of Forest Research 135:77–86.
Zimmermann, M., P. Meir, M. R. Silman, A. Fedders, A. Gibbon, Y. Malhi, D. H. Urrego, M. B. Bush, K. J. Feeley, K. C. Garcia, G. C. Dargie, W. R. Farfan, B. P. Goetz, W. T. Johnson, K. M. Kline, A. T. Modi, N. M. Q. Rurau, B. T. Staudt, and F. Zamora. 2009. No Differences in Soil Carbon Stocks Across the Tree Line in the Peruvian Andes. Ecosystems 13:62–74.
Zobel, R. W., T. B. Kinraide, and V. C. Baligar. 2007. Fine root diameters can change in response to changes in nutrient concentrations. Plant and Soil 297:243–254.

184
Appendices
Appendix 1. Supplementary materials for Chapter 2
Table A1.1 Soil texture, carbon (C) and nitrogen (N) concentrations and stocks, bulk density
(BD) and soil pH at all study plots (labelled as site and land use type) and four soil depths
(mean±SE; n(texture) = 3, n(BD/pH) = 5, n(C/N) = 10).
Soil depth (cm)
Sand-Silt-Clay (%)
C (%) N (%) BD
(g cm-3) pH
(H2O) pH
(CaCl2) C stock (kg m-2)
N stock (kg m-2)
C:N Ratio
Katassi Forest
0-10 10-30-60 13.33±1.3 0.98±0.06 0.76±0.08 6.11±0.11 5.72±0.11 9.97±0.99 0.74±0.05 13.36±0.77
10-20 16-44-40 7.48±0.77 0.64±0.05 0.85±0.07 6.15±0.13 5.68±0.14 6.17±0.63 0.52±0.04 11.63±0.42
20-30 24-48-28 5.93±0.78 0.54±0.06 0.92±0.06 6.15±0.14 5.69±0.15 4.86±0.64 0.44±0.05 10.84±0.67
30-50 9-58-33 3.16±0.29 0.30±0.03 1.03±0.06 6.16±0.23 5.53±0.26 4.84±0.44 0.45±0.04 10.64±0.23
Katassi Eucalyptus
0-10 5-69-26 5.20±0.27 0.38±0.02 1.08±0.04 5.40±0.04 4.49±0.06 5.32±0.28 0.39±0.02 13.64±0.25
10-20 3-62-35 3.87±0.36 0.30±0.03 1.13±0.05 5.34±0.09 4.39±0.04 3.73±0.35 0.29±0.03 13.04±0.26
20-30 6-59-35 3.55±0.38 0.28±0.03 1.19±0.06 5.27±0.06 4.31±0.03 3.43±0.37 0.27±0.03 12.59±0.21
30-50 8-58-34 3.06±0.51 0.25±0.04 1.32±0.06 5.33±0.06 4.34±0.02 4.69±0.94 0.38±0.07 12.02±0.35
Katassi Cropland
0-10 4-30-67 1.12±0.07 0.13±0.01 1.06±0.08 5.88±0.10 5.06±0.10 1.12±0.07 0.13±0.01 8.42±0.31
10-20 10-32-58 0.75±0.05 0.12±0.01 1.09±0.03 5.82±0.09 4.82±0.09 0.78±0.06 0.12±0.01 6.36±0.35
20-30 4-28-68 0.61±0.06 0.10±0.01 1.12±0.02 5.75±0.10 4.81±0.11 0.59±0.06 0.10±0.01 6.21±0.46
30-50 4-28-68 0.43±0.08 0.09±0.01 1.20±0.04 5.81±0.13 4.84±0.14 0.71±0.13 0.16±0.01 4.34±0.63
Gelawdios Forest
0-10 9-39-52 11.83±0.9 1.09±0.07 0.74±0.01 6.26±0.08 5.94±0.11 8.41±0.67 0.77±0.05 10.81±0.26
10-20 4-34-61 7.17±0.74 0.70±0.07 0.89±0.03 5.96±0.08 5.55±0.10 5.72±0.59 0.55±0.06 10.34±0.15
20-30 11-46-43 4.63±0.33 0.42±0.03 1.02±0.05 5.84±0.07 5.40±0.08 3.74±0.27 0.34±0.03 11.03±0.26
30-50 9-63-28 3.49±0.29 0.30±0.02 1.18±0.11 5.84±0.08 5.35±0.07 5.30±0.44 0.45±0.03 11.70±0.42
Gelawdios Eucalyptus
0-10 10-41-49 3.95±0.31 0.24±0.02 1.24±0.03 6.12±0.15 5.08±0.13 4.84±0.38 0.30±0.02 16.33±0.78
10-20 13-50-37 2.22±0.14 0.18±0.01 1.30±0.02 5.94±0.10 4.88±0.10 2.84±0.17 0.24±0.02 12.26±0.46
20-30 9-47-44 1.76±0.18 0.15±0.02 1.34±0.01 5.73±0.17 4.76±0.09 2.27±0.24 0.20±0.02 11.72±0.38
30-50 10-43-47 1.60±0.12 0.15±0.01 1.36±0.03 5.88±0.17 4.83±0.13 3.92±0.29 0.36±0.03 10.90±0.19
Gelawdios Cropland
0-10 3-50-46 2.71±0.12 0.22±0.01 1.23±0.03 5.99±0.05 5.13±0.06 3.07±0.14 0.25±0.01 12.69±0.56
10-20 3-55-42 2.58±0.18 0.21±0.02 1.26±0.05 6.13±0.08 5.31±0.09 2.70±0.19 0.22±0.02 12.36±0.50
20-30 2-52-46 2.35±0.11 0.18±0.01 1.24±0.02 6.26±0.06 5.38±0.08 2.21±0.10 0.17±0.01 13.19±0.44
30-50 6-59-35 2.12±0.16 0.17±0.01 1.33±0.02 6.30±0.09 5.43±0.10 3.67±0.27 0.29±0.02 12.67±0.50
Gelawdios Grazing land
0-10 24-48-28 3.21±0.34 0.30±0.03 1.22±0.06 5.95±0.12 5.37±0.07 3.65±0.38 0.34±0.04 10.78±0.08
10-20 32-45-24 2.47±0.21 0.23±0.02 1.20±0.04 6.16±0.11 5.44±0.07 2.68±0.23 0.25±0.02 10.71±0.18
20-30 25-34-41 2.03±0.21 0.19±0.02 1.28±0.04 6.39±0.08 5.56±0.04 1.95±0.20 0.18±0.02 10.77±0.25
30-50 39-47-14 1.52±0.24 0.14±0.02 1.29±0.04 6.53±0.06 5.65±0.03 2.66±0.42 0.24±0.04 10.96±0.33
Tara Gedam Forest
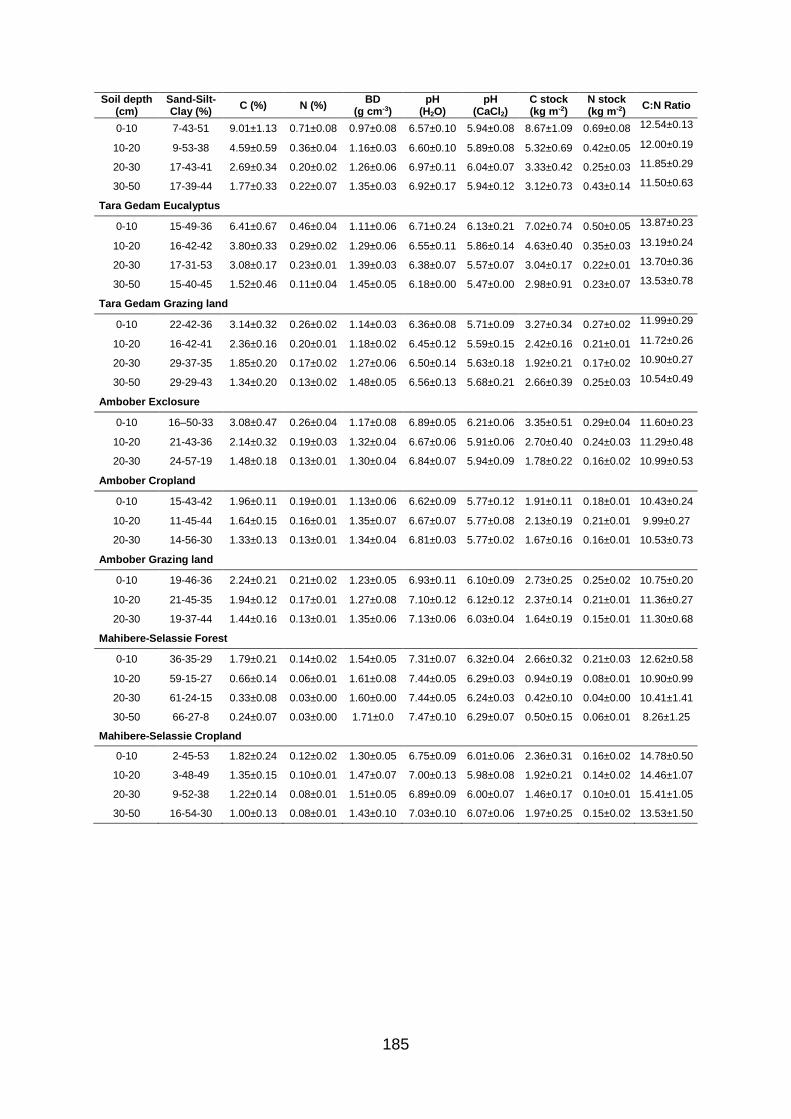
185
Soil depth (cm)
Sand-Silt-Clay (%)
C (%) N (%) BD
(g cm-3) pH
(H2O) pH
(CaCl2) C stock (kg m-2)
N stock (kg m-2)
C:N Ratio
0-10 7-43-51 9.01±1.13 0.71±0.08 0.97±0.08 6.57±0.10 5.94±0.08 8.67±1.09 0.69±0.08 12.54±0.13
10-20 9-53-38 4.59±0.59 0.36±0.04 1.16±0.03 6.60±0.10 5.89±0.08 5.32±0.69 0.42±0.05 12.00±0.19
20-30 17-43-41 2.69±0.34 0.20±0.02 1.26±0.06 6.97±0.11 6.04±0.07 3.33±0.42 0.25±0.03 11.85±0.29
30-50 17-39-44 1.77±0.33 0.22±0.07 1.35±0.03 6.92±0.17 5.94±0.12 3.12±0.73 0.43±0.14 11.50±0.63
Tara Gedam Eucalyptus
0-10 15-49-36 6.41±0.67 0.46±0.04 1.11±0.06 6.71±0.24 6.13±0.21 7.02±0.74 0.50±0.05 13.87±0.23
10-20 16-42-42 3.80±0.33 0.29±0.02 1.29±0.06 6.55±0.11 5.86±0.14 4.63±0.40 0.35±0.03 13.19±0.24
20-30 17-31-53 3.08±0.17 0.23±0.01 1.39±0.03 6.38±0.07 5.57±0.07 3.04±0.17 0.22±0.01 13.70±0.36
30-50 15-40-45 1.52±0.46 0.11±0.04 1.45±0.05 6.18±0.00 5.47±0.00 2.98±0.91 0.23±0.07 13.53±0.78
Tara Gedam Grazing land
0-10 22-42-36 3.14±0.32 0.26±0.02 1.14±0.03 6.36±0.08 5.71±0.09 3.27±0.34 0.27±0.02 11.99±0.29
10-20 16-42-41 2.36±0.16 0.20±0.01 1.18±0.02 6.45±0.12 5.59±0.15 2.42±0.16 0.21±0.01 11.72±0.26
20-30 29-37-35 1.85±0.20 0.17±0.02 1.27±0.06 6.50±0.14 5.63±0.18 1.92±0.21 0.17±0.02 10.90±0.27
30-50 29-29-43 1.34±0.20 0.13±0.02 1.48±0.05 6.56±0.13 5.68±0.21 2.66±0.39 0.25±0.03 10.54±0.49
Ambober Exclosure
0-10 16–50-33 3.08±0.47 0.26±0.04 1.17±0.08 6.89±0.05 6.21±0.06 3.35±0.51 0.29±0.04 11.60±0.23
10-20 21-43-36 2.14±0.32 0.19±0.03 1.32±0.04 6.67±0.06 5.91±0.06 2.70±0.40 0.24±0.03 11.29±0.48
20-30 24-57-19 1.48±0.18 0.13±0.01 1.30±0.04 6.84±0.07 5.94±0.09 1.78±0.22 0.16±0.02 10.99±0.53
Ambober Cropland
0-10 15-43-42 1.96±0.11 0.19±0.01 1.13±0.06 6.62±0.09 5.77±0.12 1.91±0.11 0.18±0.01 10.43±0.24
10-20 11-45-44 1.64±0.15 0.16±0.01 1.35±0.07 6.67±0.07 5.77±0.08 2.13±0.19 0.21±0.01 9.99±0.27
20-30 14-56-30 1.33±0.13 0.13±0.01 1.34±0.04 6.81±0.03 5.77±0.02 1.67±0.16 0.16±0.01 10.53±0.73
Ambober Grazing land
0-10 19-46-36 2.24±0.21 0.21±0.02 1.23±0.05 6.93±0.11 6.10±0.09 2.73±0.25 0.25±0.02 10.75±0.20
10-20 21-45-35 1.94±0.12 0.17±0.01 1.27±0.08 7.10±0.12 6.12±0.12 2.37±0.14 0.21±0.01 11.36±0.27
20-30 19-37-44 1.44±0.16 0.13±0.01 1.35±0.06 7.13±0.06 6.03±0.04 1.64±0.19 0.15±0.01 11.30±0.68
Mahibere-Selassie Forest
0-10 36-35-29 1.79±0.21 0.14±0.02 1.54±0.05 7.31±0.07 6.32±0.04 2.66±0.32 0.21±0.03 12.62±0.58
10-20 59-15-27 0.66±0.14 0.06±0.01 1.61±0.08 7.44±0.05 6.29±0.03 0.94±0.19 0.08±0.01 10.90±0.99
20-30 61-24-15 0.33±0.08 0.03±0.00 1.60±0.00 7.44±0.05 6.24±0.03 0.42±0.10 0.04±0.00 10.41±1.41
30-50 66-27-8 0.24±0.07 0.03±0.00 1.71±0.0 7.47±0.10 6.29±0.07 0.50±0.15 0.06±0.01 8.26±1.25
Mahibere-Selassie Cropland
0-10 2-45-53 1.82±0.24 0.12±0.02 1.30±0.05 6.75±0.09 6.01±0.06 2.36±0.31 0.16±0.02 14.78±0.50
10-20 3-48-49 1.35±0.15 0.10±0.01 1.47±0.07 7.00±0.13 5.98±0.08 1.92±0.21 0.14±0.02 14.46±1.07
20-30 9-52-38 1.22±0.14 0.08±0.01 1.51±0.05 6.89±0.09 6.00±0.07 1.46±0.17 0.10±0.01 15.41±1.05
30-50 16-54-30 1.00±0.13 0.08±0.01 1.43±0.10 7.03±0.10 6.07±0.06 1.97±0.25 0.15±0.02 13.53±1.50
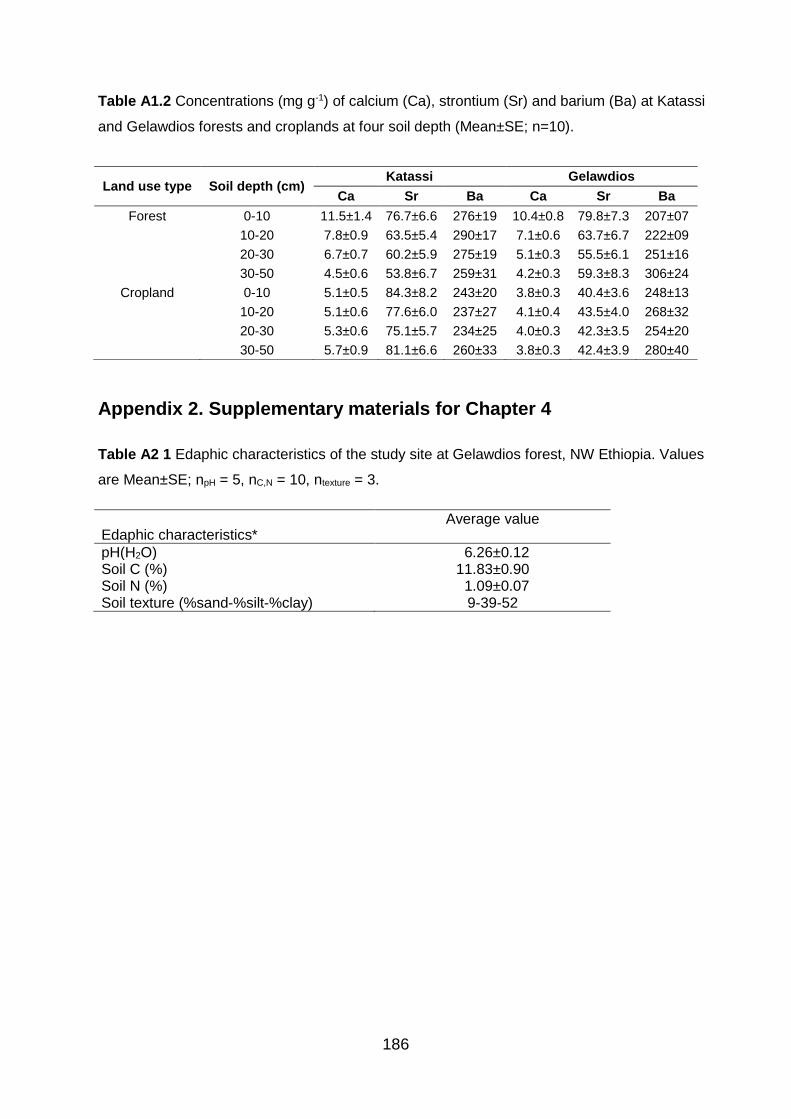
186
Table A1.2 Concentrations (mg g-1) of calcium (Ca), strontium (Sr) and barium (Ba) at Katassi
and Gelawdios forests and croplands at four soil depth (Mean±SE; n=10).
Appendix 2. Supplementary materials for Chapter 4
Table A2 1 Edaphic characteristics of the study site at Gelawdios forest, NW Ethiopia. Values
are Mean±SE; npH = 5, nC,N = 10, ntexture = 3.
Edaphic characteristics* Average value
pH(H2O) 6.26±0.12 Soil C (%) 11.83±0.90 Soil N (%) 1.09±0.07 Soil texture (%sand-%silt-%clay) 9-39-52
Land use type Soil depth (cm) Katassi Gelawdios
Ca Sr Ba Ca Sr Ba
Forest 0-10 11.5±1.4 76.7±6.6 276±19 10.4±0.8 79.8±7.3 207±07 10-20 7.8±0.9 63.5±5.4 290±17 7.1±0.6 63.7±6.7 222±09 20-30 6.7±0.7 60.2±5.9 275±19 5.1±0.3 55.5±6.1 251±16 30-50 4.5±0.6 53.8±6.7 259±31 4.2±0.3 59.3±8.3 306±24
Cropland 0-10 5.1±0.5 84.3±8.2 243±20 3.8±0.3 40.4±3.6 248±13 10-20 5.1±0.6 77.6±6.0 237±27 4.1±0.4 43.5±4.0 268±32 20-30 5.3±0.6 75.1±5.7 234±25 4.0±0.3 42.3±3.5 254±20 30-50 5.7±0.9 81.1±6.6 260±33 3.8±0.3 42.4±3.9 280±40
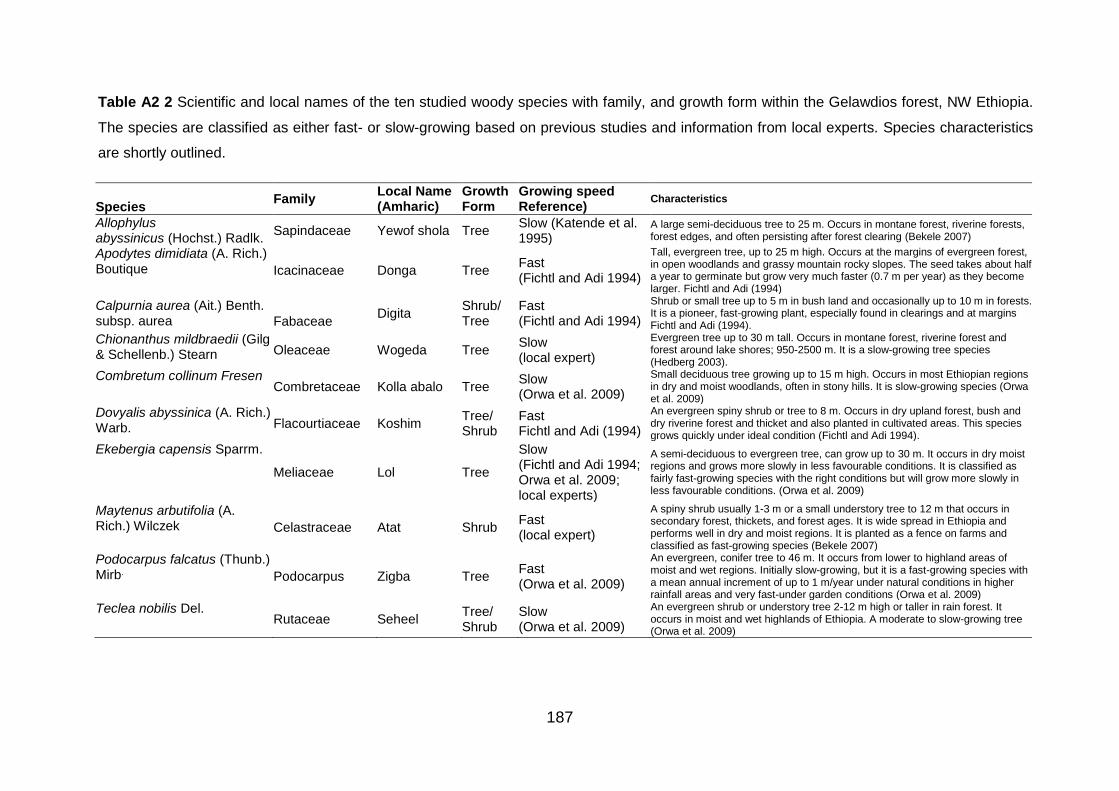
187
Table A2 2 Scientific and local names of the ten studied woody species with family, and growth form within the Gelawdios forest, NW Ethiopia.
The species are classified as either fast- or slow-growing based on previous studies and information from local experts. Species characteristics
are shortly outlined.
Species Family
Local Name (Amharic)
Growth Form
Growing speed Reference)
Characteristics
Allophylus abyssinicus (Hochst.) Radlk.
Sapindaceae Yewof shola Tree Slow (Katende et al. 1995)
A large semi-deciduous tree to 25 m. Occurs in montane forest, riverine forests, forest edges, and often persisting after forest clearing (Bekele 2007)
Apodytes dimidiata (A. Rich.) Boutique Icacinaceae Donga Tree
Fast (Fichtl and Adi 1994)
Tall, evergreen tree, up to 25 m high. Occurs at the margins of evergreen forest, in open woodlands and grassy mountain rocky slopes. The seed takes about half a year to germinate but grow very much faster (0.7 m per year) as they become larger. Fichtl and Adi (1994)
Calpurnia aurea (Ait.) Benth. subsp. aurea Fabaceae
Digita Shrub/ Tree
Fast (Fichtl and Adi 1994)
Shrub or small tree up to 5 m in bush land and occasionally up to 10 m in forests. It is a pioneer, fast-growing plant, especially found in clearings and at margins Fichtl and Adi (1994).
Chionanthus mildbraedii (Gilg & Schellenb.) Stearn Oleaceae Wogeda Tree
Slow (local expert)
Evergreen tree up to 30 m tall. Occurs in montane forest, riverine forest and forest around lake shores; 950-2500 m. It is a slow-growing tree species (Hedberg 2003).
Combretum collinum Fresen Combretaceae Kolla abalo Tree
Slow (Orwa et al. 2009)
Small deciduous tree growing up to 15 m high. Occurs in most Ethiopian regions in dry and moist woodlands, often in stony hills. It is slow-growing species (Orwa et al. 2009)
Dovyalis abyssinica (A. Rich.) Warb. Flacourtiaceae Koshim
Tree/ Shrub
Fast Fichtl and Adi (1994)
An evergreen spiny shrub or tree to 8 m. Occurs in dry upland forest, bush and dry riverine forest and thicket and also planted in cultivated areas. This species grows quickly under ideal condition (Fichtl and Adi 1994).
Ekebergia capensis Sparrm.
Meliaceae Lol Tree
Slow (Fichtl and Adi 1994; Orwa et al. 2009; local experts)
A semi-deciduous to evergreen tree, can grow up to 30 m. It occurs in dry moist regions and grows more slowly in less favourable conditions. It is classified as fairly fast-growing species with the right conditions but will grow more slowly in less favourable conditions. (Orwa et al. 2009)
Maytenus arbutifolia (A. Rich.) Wilczek Celastraceae Atat Shrub
Fast (local expert)
A spiny shrub usually 1-3 m or a small understory tree to 12 m that occurs in secondary forest, thickets, and forest ages. It is wide spread in Ethiopia and performs well in dry and moist regions. It is planted as a fence on farms and classified as fast-growing species (Bekele 2007)
Podocarpus falcatus (Thunb.) Mirb. Podocarpus Zigba Tree
Fast (Orwa et al. 2009)
An evergreen, conifer tree to 46 m. It occurs from lower to highland areas of moist and wet regions. Initially slow-growing, but it is a fast-growing species with a mean annual increment of up to 1 m/year under natural conditions in higher rainfall areas and very fast-under garden conditions (Orwa et al. 2009)
Teclea nobilis Del. Rutaceae Seheel
Tree/ Shrub
Slow (Orwa et al. 2009)
An evergreen shrub or understory tree 2-12 m high or taller in rain forest. It occurs in moist and wet highlands of Ethiopia. A moderate to slow-growing tree (Orwa et al. 2009)
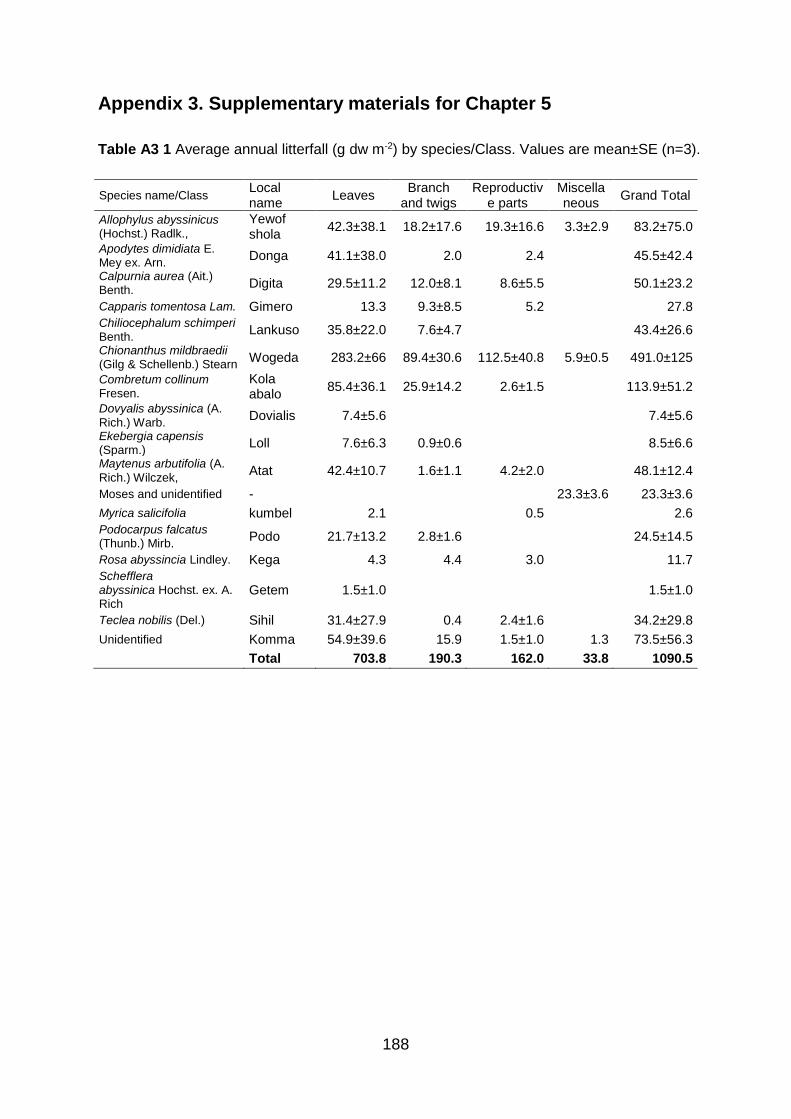
188
Appendix 3. Supplementary materials for Chapter 5
Table A3 1 Average annual litterfall (g dw m-2) by species/Class. Values are mean±SE (n=3).
Species name/Class Local name
Leaves Branch
and twigs Reproductiv
e parts Miscellaneous
Grand Total
Allophylus abyssinicus (Hochst.) Radlk.,
Yewof shola
42.3±38.1 18.2±17.6 19.3±16.6 3.3±2.9 83.2±75.0
Apodytes dimidiata E. Mey ex. Arn.
Donga 41.1±38.0 2.0 2.4 45.5±42.4
Calpurnia aurea (Ait.) Benth.
Digita 29.5±11.2 12.0±8.1 8.6±5.5 50.1±23.2
Capparis tomentosa Lam. Gimero 13.3 9.3±8.5 5.2 27.8
Chiliocephalum schimperi Benth.
Lankuso 35.8±22.0 7.6±4.7 43.4±26.6
Chionanthus mildbraedii (Gilg & Schellenb.) Stearn
Wogeda 283.2±66 89.4±30.6 112.5±40.8 5.9±0.5 491.0±125
Combretum collinum Fresen.
Kola abalo
85.4±36.1 25.9±14.2 2.6±1.5 113.9±51.2
Dovyalis abyssinica (A.
Rich.) Warb. Dovialis 7.4±5.6 7.4±5.6
Ekebergia capensis (Sparm.)
Loll 7.6±6.3 0.9±0.6 8.5±6.6
Maytenus arbutifolia (A.
Rich.) Wilczek, Atat 42.4±10.7 1.6±1.1 4.2±2.0 48.1±12.4
Moses and unidentified - 23.3±3.6 23.3±3.6
Myrica salicifolia kumbel 2.1 0.5 2.6
Podocarpus falcatus (Thunb.) Mirb.
Podo 21.7±13.2 2.8±1.6 24.5±14.5
Rosa abyssincia Lindley. Kega 4.3 4.4 3.0 11.7
Schefflera abyssinica Hochst. ex. A. Rich
Getem 1.5±1.0 1.5±1.0
Teclea nobilis (Del.) Sihil 31.4±27.9 0.4 2.4±1.6 34.2±29.8
Unidentified Komma 54.9±39.6 15.9 1.5±1.0 1.3 73.5±56.3
Total 703.8 190.3 162.0 33.8 1090.5
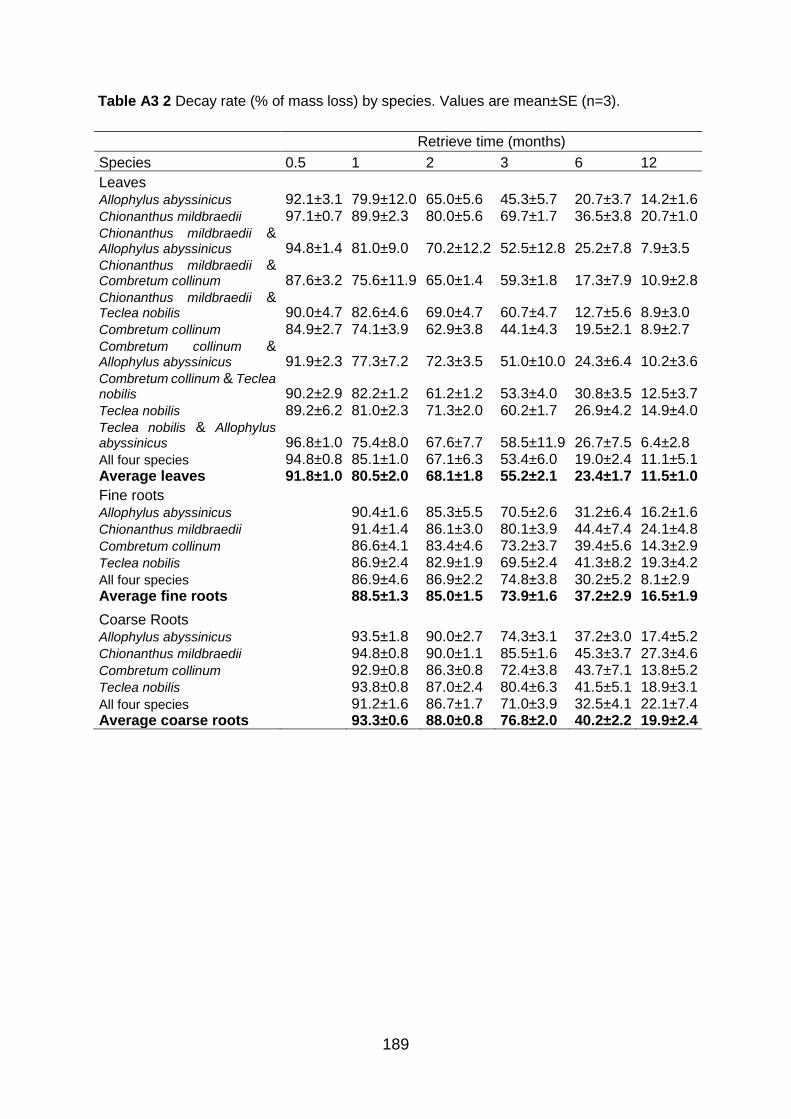
189
Table A3 2 Decay rate (% of mass loss) by species. Values are mean±SE (n=3).
Retrieve time (months)
Species 0.5 1 2 3 6 12
Leaves Allophylus abyssinicus 92.1±3.1 79.9±12.0 65.0±5.6 45.3±5.7 20.7±3.7 14.2±1.6 Chionanthus mildbraedii 97.1±0.7 89.9±2.3 80.0±5.6 69.7±1.7 36.5±3.8 20.7±1.0 Chionanthus mildbraedii & Allophylus abyssinicus 94.8±1.4 81.0±9.0 70.2±12.2 52.5±12.8 25.2±7.8 7.9±3.5 Chionanthus mildbraedii & Combretum collinum 87.6±3.2 75.6±11.9 65.0±1.4 59.3±1.8 17.3±7.9 10.9±2.8 Chionanthus mildbraedii & Teclea nobilis 90.0±4.7 82.6±4.6 69.0±4.7 60.7±4.7 12.7±5.6 8.9±3.0 Combretum collinum 84.9±2.7 74.1±3.9 62.9±3.8 44.1±4.3 19.5±2.1 8.9±2.7 Combretum collinum & Allophylus abyssinicus 91.9±2.3 77.3±7.2 72.3±3.5 51.0±10.0 24.3±6.4 10.2±3.6 Combretum collinum & Teclea nobilis 90.2±2.9 82.2±1.2 61.2±1.2 53.3±4.0 30.8±3.5 12.5±3.7 Teclea nobilis 89.2±6.2 81.0±2.3 71.3±2.0 60.2±1.7 26.9±4.2 14.9±4.0 Teclea nobilis & Allophylus abyssinicus 96.8±1.0 75.4±8.0 67.6±7.7 58.5±11.9 26.7±7.5 6.4±2.8 All four species 94.8±0.8 85.1±1.0 67.1±6.3 53.4±6.0 19.0±2.4 11.1±5.1 Average leaves 91.8±1.0 80.5±2.0 68.1±1.8 55.2±2.1 23.4±1.7 11.5±1.0
Fine roots Allophylus abyssinicus 90.4±1.6 85.3±5.5 70.5±2.6 31.2±6.4 16.2±1.6 Chionanthus mildbraedii 91.4±1.4 86.1±3.0 80.1±3.9 44.4±7.4 24.1±4.8 Combretum collinum 86.6±4.1 83.4±4.6 73.2±3.7 39.4±5.6 14.3±2.9 Teclea nobilis 86.9±2.4 82.9±1.9 69.5±2.4 41.3±8.2 19.3±4.2 All four species 86.9±4.6 86.9±2.2 74.8±3.8 30.2±5.2 8.1±2.9 Average fine roots 88.5±1.3 85.0±1.5 73.9±1.6 37.2±2.9 16.5±1.9
Coarse Roots Allophylus abyssinicus 93.5±1.8 90.0±2.7 74.3±3.1 37.2±3.0 17.4±5.2 Chionanthus mildbraedii 94.8±0.8 90.0±1.1 85.5±1.6 45.3±3.7 27.3±4.6 Combretum collinum 92.9±0.8 86.3±0.8 72.4±3.8 43.7±7.1 13.8±5.2 Teclea nobilis 93.8±0.8 87.0±2.4 80.4±6.3 41.5±5.1 18.9±3.1 All four species 91.2±1.6 86.7±1.7 71.0±3.9 32.5±4.1 22.1±7.4 Average coarse roots 93.3±0.6 88.0±0.8 76.8±2.0 40.2±2.2 19.9±2.4
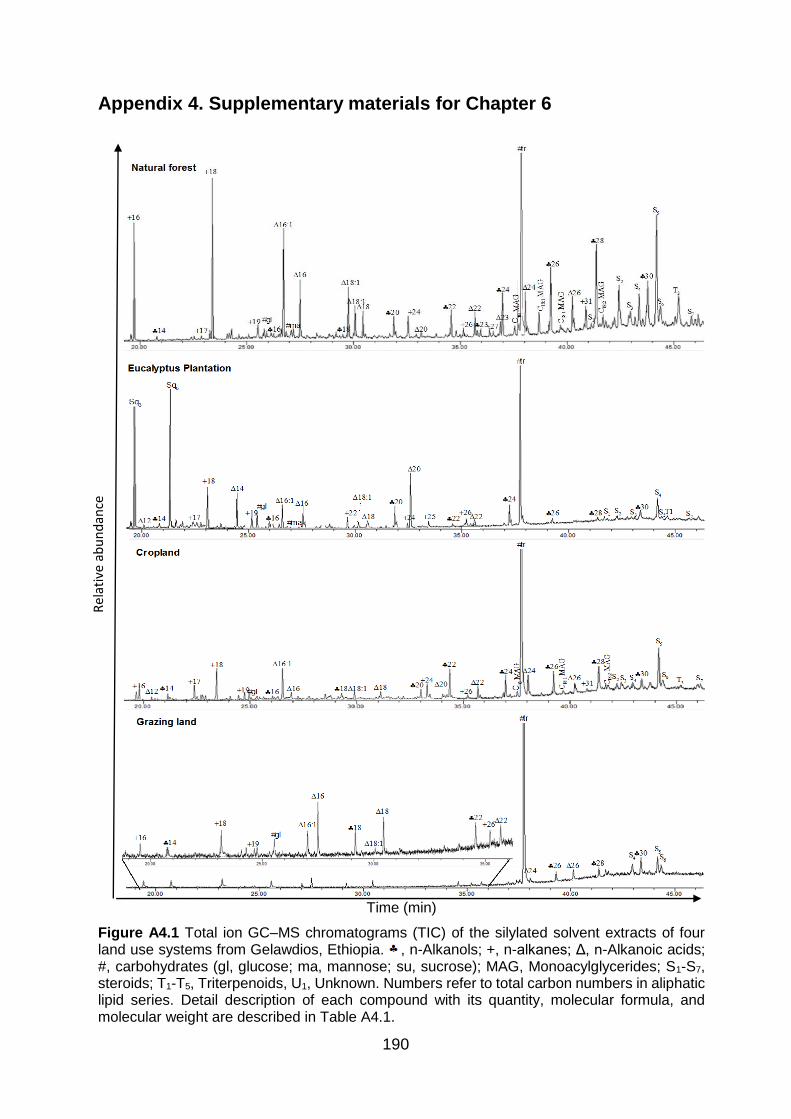
190
Appendix 4. Supplementary materials for Chapter 6
Time (min)
Figure A4.1 Total ion GC–MS chromatograms (TIC) of the silylated solvent extracts of four land use systems from Gelawdios, Ethiopia. , n-Alkanols; +, n-alkanes; Δ, n-Alkanoic acids; #, carbohydrates (gl, glucose; ma, mannose; su, sucrose); MAG, Monoacylglycerides; S1-S7, steroids; T1-T5, Triterpenoids, U1, Unknown. Numbers refer to total carbon numbers in aliphatic lipid series. Detail description of each compound with its quantity, molecular formula, and molecular weight are described in Table A4.1.
Rel
ativ
e ab
un
dan
ce
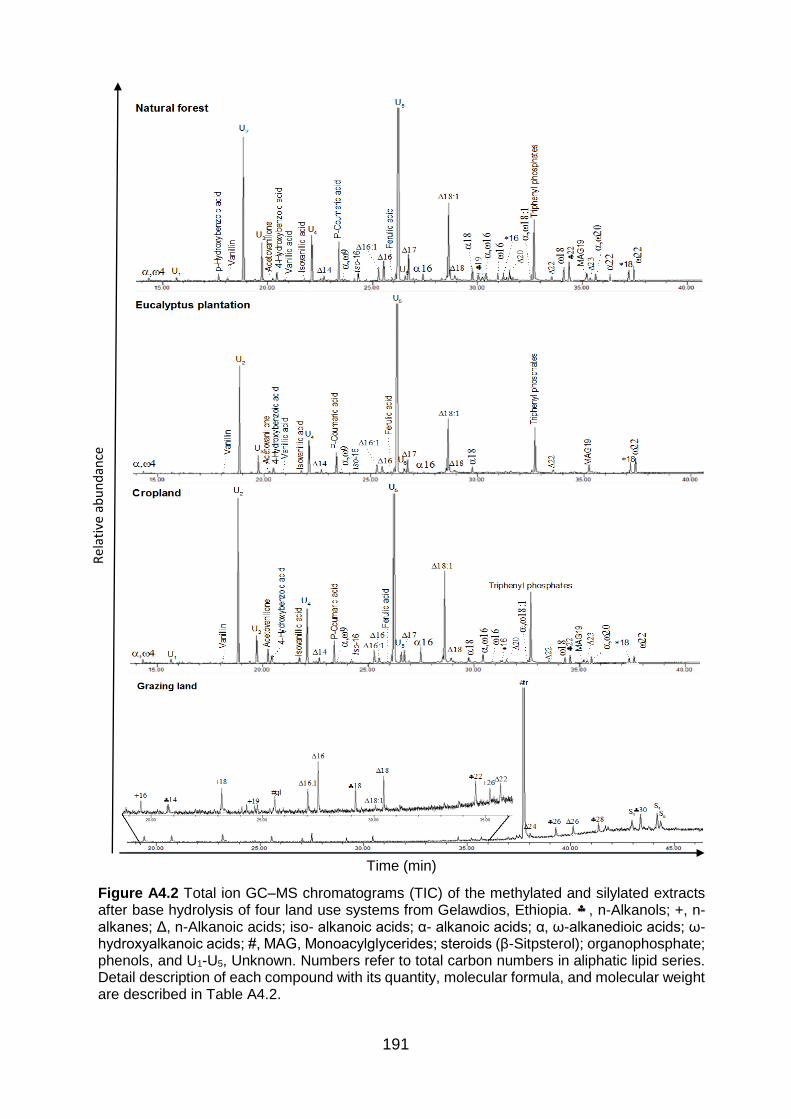
191
Time (min)
Figure A4.2 Total ion GC–MS chromatograms (TIC) of the methylated and silylated extracts after base hydrolysis of four land use systems from Gelawdios, Ethiopia. , n-Alkanols; +, n-alkanes; Δ, n-Alkanoic acids; iso- alkanoic acids; α- alkanoic acids; α, ω-alkanedioic acids; ω-hydroxyalkanoic acids; #, MAG, Monoacylglycerides; steroids (β-Sitpsterol); organophosphate; phenols, and U1-U5, Unknown. Numbers refer to total carbon numbers in aliphatic lipid series. Detail description of each compound with its quantity, molecular formula, and molecular weight are described in Table A4.2.
Rel
ativ
e ab
un
dan
ce
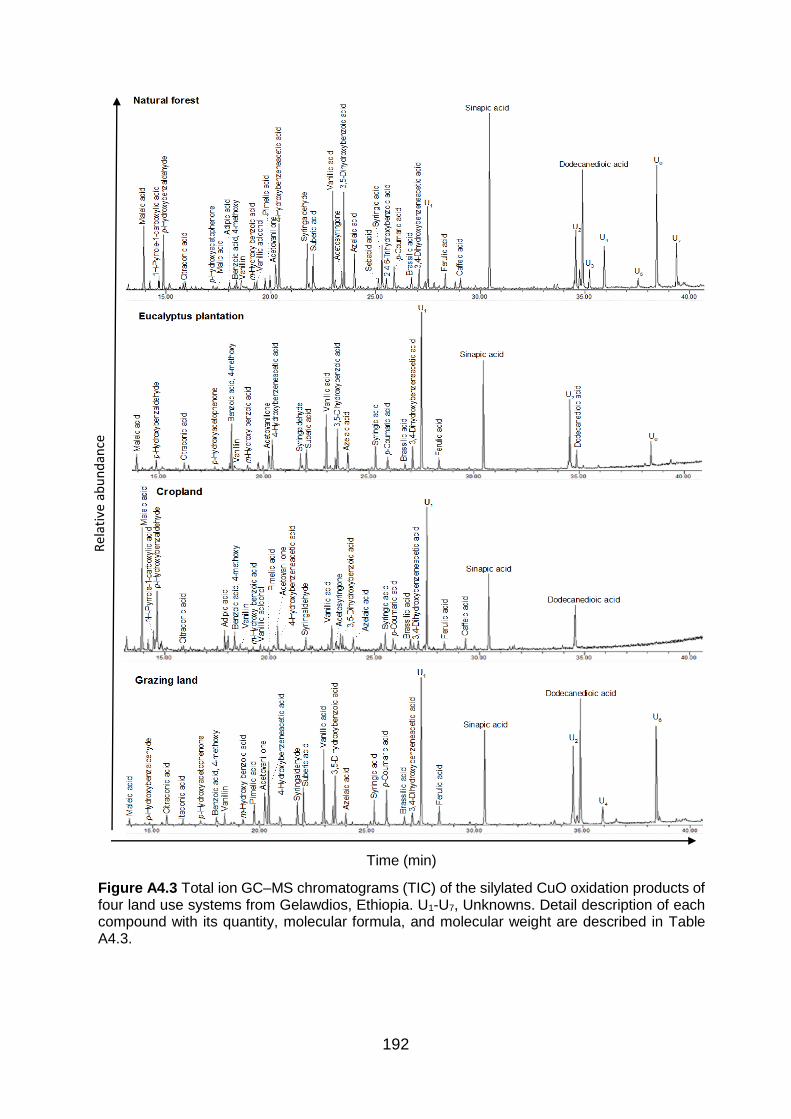
192
Time (min)
Figure A4.3 Total ion GC–MS chromatograms (TIC) of the silylated CuO oxidation products of four land use systems from Gelawdios, Ethiopia. U1-U7, Unknowns. Detail description of each compound with its quantity, molecular formula, and molecular weight are described in Table A4.3.
Rel
ativ
e ab
un
dan
ce
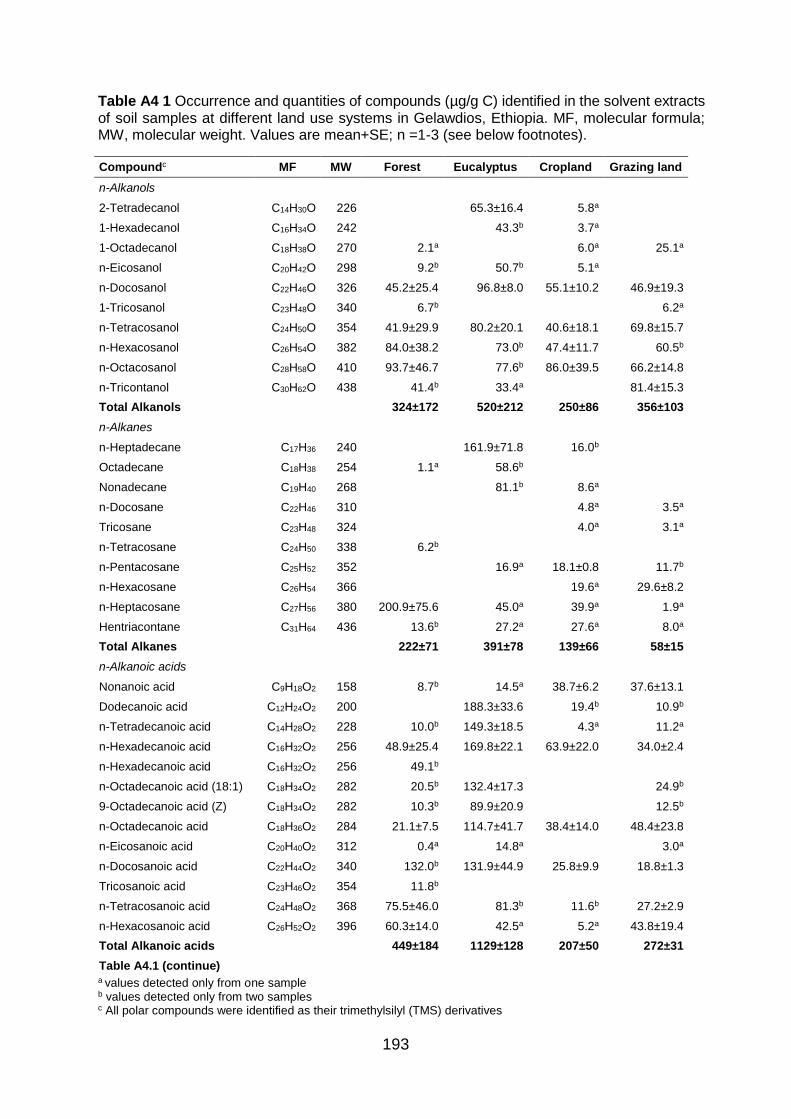
193
Table A4 1 Occurrence and quantities of compounds (µg/g C) identified in the solvent extracts of soil samples at different land use systems in Gelawdios, Ethiopia. MF, molecular formula; MW, molecular weight. Values are mean+SE; n =1-3 (see below footnotes).
Compoundc MF MW Forest Eucalyptus Cropland Grazing land
n-Alkanols
2-Tetradecanol C14H30O 226 65.3±16.4 5.8a
1-Hexadecanol C16H34O 242 43.3b 3.7a
1-Octadecanol C18H38O 270 2.1a 6.0a 25.1a
n-Eicosanol C20H42O 298 9.2b 50.7b 5.1a
n-Docosanol C22H46O 326 45.2±25.4 96.8±8.0 55.1±10.2 46.9±19.3
1-Tricosanol C23H48O 340 6.7b 6.2a
n-Tetracosanol C24H50O 354 41.9±29.9 80.2±20.1 40.6±18.1 69.8±15.7
n-Hexacosanol C26H54O 382 84.0±38.2 73.0b 47.4±11.7 60.5b
n-Octacosanol C28H58O 410 93.7±46.7 77.6b 86.0±39.5 66.2±14.8
n-Tricontanol C30H62O 438 41.4b 33.4a 81.4±15.3
Total Alkanols 324±172 520±212 250±86 356±103
n-Alkanes
n-Heptadecane C17H36 240 161.9±71.8 16.0b
Octadecane C18H38 254 1.1a 58.6b
Nonadecane C19H40 268 81.1b 8.6a
n-Docosane C22H46 310 4.8a 3.5a
Tricosane C23H48 324 4.0a 3.1a
n-Tetracosane C24H50 338 6.2b
n-Pentacosane C25H52 352 16.9a 18.1±0.8 11.7b
n-Hexacosane C26H54 366 19.6a 29.6±8.2
n-Heptacosane C27H56 380 200.9±75.6 45.0a 39.9a 1.9a
Hentriacontane C31H64 436 13.6b 27.2a 27.6a 8.0a
Total Alkanes 222±71 391±78 139±66 58±15
n-Alkanoic acids
Nonanoic acid C9H18O2 158 8.7b 14.5a 38.7±6.2 37.6±13.1
Dodecanoic acid C12H24O2 200 188.3±33.6 19.4b 10.9b
n-Tetradecanoic acid C14H28O2 228 10.0b 149.3±18.5 4.3a 11.2a
n-Hexadecanoic acid C16H32O2 256 48.9±25.4 169.8±22.1 63.9±22.0 34.0±2.4
n-Hexadecanoic acid C16H32O2 256 49.1b
n-Octadecanoic acid (18:1) C18H34O2 282 20.5b 132.4±17.3 24.9b
9-Octadecanoic acid (Z) C18H34O2 282 10.3b 89.9±20.9 12.5b
n-Octadecanoic acid C18H36O2 284 21.1±7.5 114.7±41.7 38.4±14.0 48.4±23.8
n-Eicosanoic acid C20H40O2 312 0.4a 14.8a 3.0a
n-Docosanoic acid C22H44O2 340 132.0b 131.9±44.9 25.8±9.9 18.8±1.3
Tricosanoic acid C23H46O2 354 11.8b
n-Tetracosanoic acid C24H48O2 368 75.5±46.0 81.3b 11.6b 27.2±2.9
n-Hexacosanoic acid C26H52O2 396 60.3±14.0 42.5a 5.2a 43.8±19.4
Total Alkanoic acids 449±184 1129±128 207±50 272±31
Table A4.1 (continue) a values detected only from one sample b values detected only from two samples c All polar compounds were identified as their trimethylsilyl (TMS) derivatives

194
Table A4.1 continued
Compoundc MF MW Forest Eucalyptus Cropland Grazing land
Iso- Alkanoic acids
Iso-Heptadecanoic acid C17H34O2 270 24.5b
Monoacylglycerides
(±)-2,3-Dihydroxypropyl hexadecanoate (C16)
C19H38O4 330 9.2b 19.1a 37.12b
Monostearin (C18:1) C21H42O4 358 16.1b 134.4±26.1 97.0±19.14 64.9±8.9
Linolein, 1-mono (C18:1) C21H38O4 354 20.6±9.5 31.9a 9.3a
2-Monolinolenin (C18:2) C21H36O4 352 6.4a 74.3±16.0 9.7a
Total Monoacylglycerides 52±26 260±19 134±46 84±2
Carbohydrates
D-Glucose C6H12O6 180 16.2b 90.1±32.9 37.8±16.6 46.7±14.7
Mannose C6H12O6 180 16.9±7.0 211.9±78.4 78.4±25.3 46.0±14.0
Sucrose C12H22O11 342 7.7b 19.6a 19.5b 33.0±11.3
Trehalose C12H22O11 342 3895±712.5 2107.4±217.1 1684.1±191 1768.2±157.8
Total Carbohydrates 3936±703 2429±256 18120±229 1894±192
Steroids and Terpenoids
Aromadendrene C15H24 204 0.5a 124.9b 13.8a
γ-Elemene C15H24 204 183.0±81.1 13.5a
Ledol C15H26O 222 157.2±32.9 9.3a
(-)-Globulol C15H26O 222 72.0±7.5 17.1b 11.8a
trans-Farnesol C15H26O 222 752.7±64.9 48.2a
Cholesterol C27H46O 386 11.4b 98.8±33.0 62.2b 23.6b
Ergostrol C28H44O 396 36.8b 94.7a
Campesterol C28H48O 400 32.9b 117.3±28.7 87.9b 104.7±0.3
Stigmasterol C29H48O 412 56.0b 243.1±30.4 110.2±17.4 138.9±18.3
ß-Sitosterol C29H50O 414 232.2±93.8 243.2±6.0 237.9±46.1 142.9±25.7
Stigmastanol C29H52O 416 119.4±34.2
ß- Amyrin C30H50O 426 81.2b 161.8±1.8 98.9±16.3
α- Amyrin C30H50O 426 14.7b 66.1±17.9
Lupeol C30H50O 426 9.7a 87.3b 13.3a 28.0a
Sitosterone C29H48O 412 13.3a 73.6b 17.0a
Erthrodiol C30H50O2 442 47.4b 343.6±100.3
Oleanolic acid C30H48O3 456 85.4±52.5 642.1±132.5 46.9b
Total Steroids and Terpenoids 622±267 3461±341 729±80 616±616
Unknownd 37.6±3.4 126.2±10.8 238.1±99.6 67.7b
Total aliphatic lipids 5007±746 4729±531 2550±430 2664±300
Total solvent extracts 5629±740 8190±857 3279±508 3280±394 a values detected only from one sample b values detected only from two samples c All polar compounds were identified as their trimethylsilyl (TMS) derivatives d Identified as Glycocholic acid (C26H43NO6)
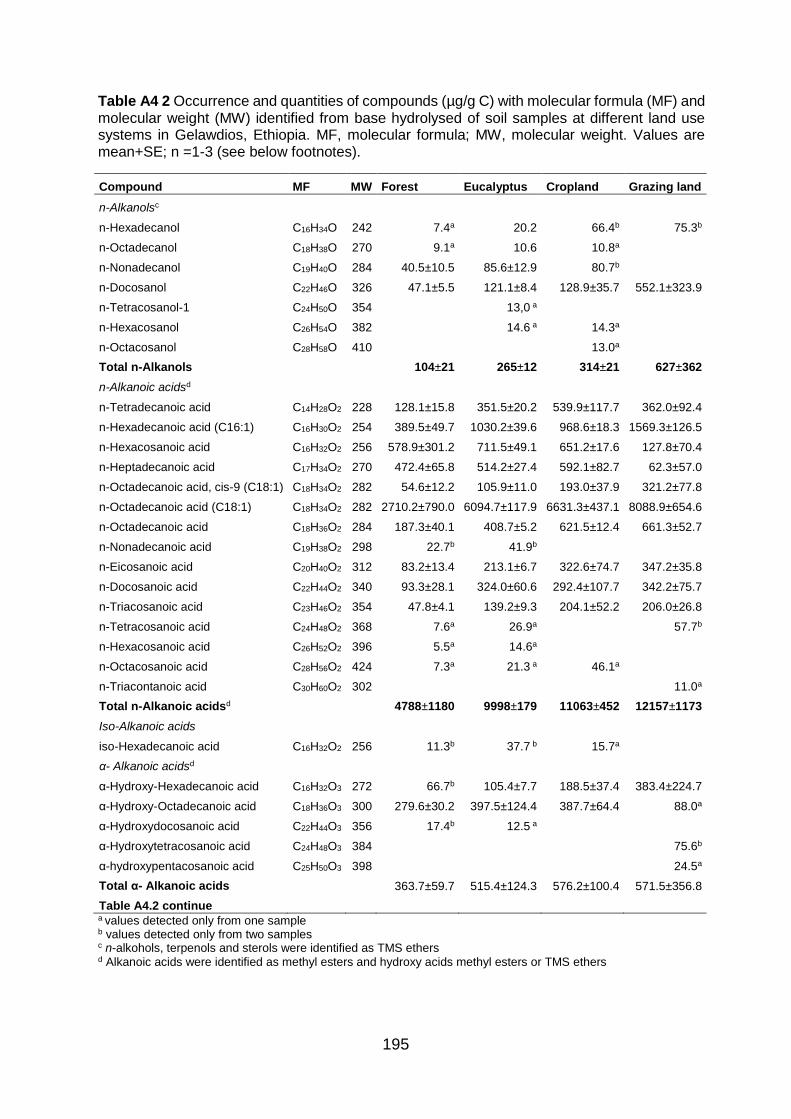
195
Table A4 2 Occurrence and quantities of compounds (µg/g C) with molecular formula (MF) and molecular weight (MW) identified from base hydrolysed of soil samples at different land use systems in Gelawdios, Ethiopia. MF, molecular formula; MW, molecular weight. Values are mean+SE; n =1-3 (see below footnotes).
Compound MF MW Forest Eucalyptus Cropland Grazing land
n-Alkanolsc
n-Hexadecanol C16H34O 242 7.4a 20.2 66.4b 75.3b
n-Octadecanol C18H38O 270 9.1a 10.6 10.8a
n-Nonadecanol C19H40O 284 40.5±10.5 85.6±12.9 80.7b
n-Docosanol C22H46O 326 47.1±5.5 121.1±8.4 128.9±35.7 552.1±323.9
n-Tetracosanol-1 C24H50O 354 13,0 a
n-Hexacosanol C26H54O 382 14.6 a 14.3a
n-Octacosanol C28H58O 410 13.0a
Total n-Alkanols 104±21 265±12 314±21 627±362
n-Alkanoic acidsd
n-Tetradecanoic acid C14H28O2 228 128.1±15.8 351.5±20.2 539.9±117.7 362.0±92.4
n-Hexadecanoic acid (C16:1) C16H30O2 254 389.5±49.7 1030.2±39.6 968.6±18.3 1569.3±126.5
n-Hexacosanoic acid C16H32O2 256 578.9±301.2 711.5±49.1 651.2±17.6 127.8±70.4
n-Heptadecanoic acid C17H34O2 270 472.4±65.8 514.2±27.4 592.1±82.7 62.3±57.0
n-Octadecanoic acid, cis-9 (C18:1) C18H34O2 282 54.6±12.2 105.9±11.0 193.0±37.9 321.2±77.8
n-Octadecanoic acid (C18:1) C18H34O2 282 2710.2±790.0 6094.7±117.9 6631.3±437.1 8088.9±654.6
n-Octadecanoic acid C18H36O2 284 187.3±40.1 408.7±5.2 621.5±12.4 661.3±52.7
n-Nonadecanoic acid C19H38O2 298 22.7b 41.9b
n-Eicosanoic acid C20H40O2 312 83.2±13.4 213.1±6.7 322.6±74.7 347.2±35.8
n-Docosanoic acid C22H44O2 340 93.3±28.1 324.0±60.6 292.4±107.7 342.2±75.7
n-Triacosanoic acid C23H46O2 354 47.8±4.1 139.2±9.3 204.1±52.2 206.0±26.8
n-Tetracosanoic acid C24H48O2 368 7.6a 26.9a 57.7b
n-Hexacosanoic acid C26H52O2 396 5.5a 14.6a
n-Octacosanoic acid C28H56O2 424 7.3a 21.3 a 46.1a
n-Triacontanoic acid C30H60O2 302 11.0a
Total n-Alkanoic acidsd 4788±1180 9998±179 11063±452 12157±1173
Iso-Alkanoic acids
iso-Hexadecanoic acid C16H32O2 256 11.3b 37.7 b 15.7a
α- Alkanoic acidsd
α-Hydroxy-Hexadecanoic acid C16H32O3 272 66.7b 105.4±7.7 188.5±37.4 383.4±224.7
α-Hydroxy-Octadecanoic acid C18H36O3 300 279.6±30.2 397.5±124.4 387.7±64.4 88.0a
α-Hydroxydocosanoic acid C22H44O3 356 17.4b 12.5 a
α-Hydroxytetracosanoic acid C24H48O3 384 75.6b
α-hydroxypentacosanoic acid C25H50O3 398 24.5a
Total α- Alkanoic acids 363.7±59.7 515.4±124.3 576.2±100.4 571.5±356.8
Table A4.2 continue a values detected only from one sample b values detected only from two samples c n-alkohols, terpenols and sterols were identified as TMS ethers d Alkanoic acids were identified as methyl esters and hydroxy acids methyl esters or TMS ethers
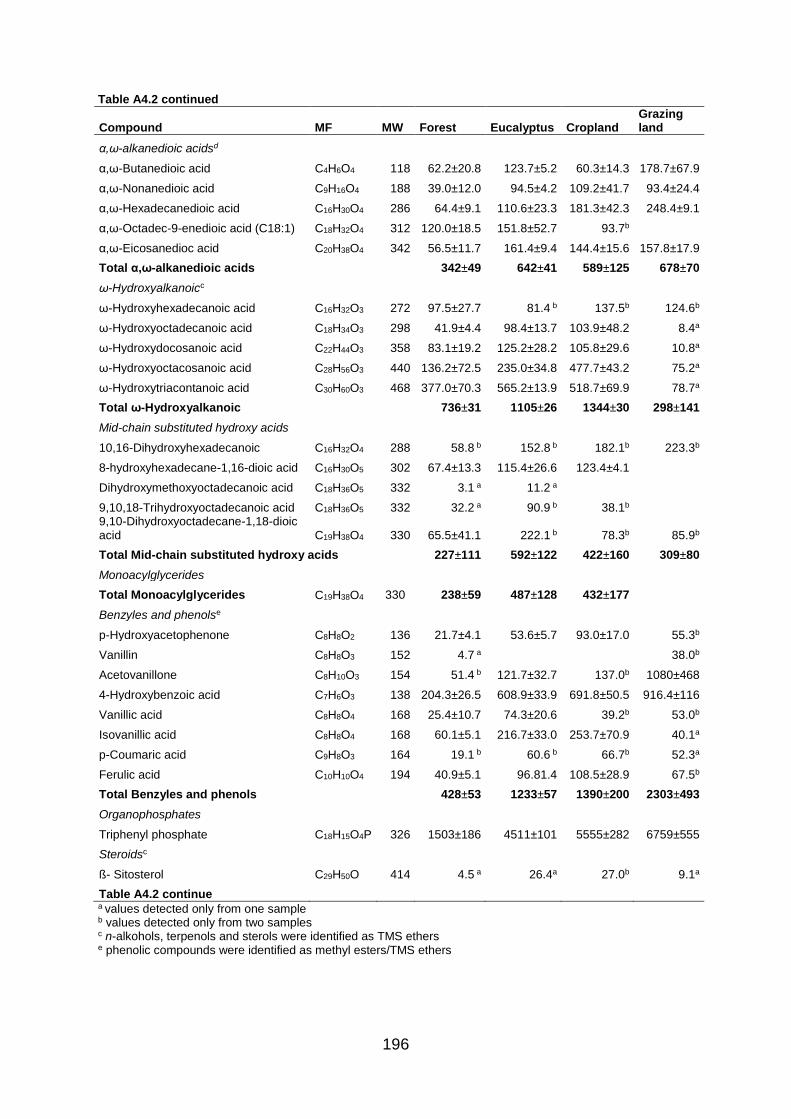
196
Table A4.2 continued
Compound MF MW Forest Eucalyptus Cropland Grazing land
α,ω-alkanedioic acidsd
α,ω-Butanedioic acid C4H6O4 118 62.2±20.8 123.7±5.2 60.3±14.3 178.7±67.9
α,ω-Nonanedioic acid C9H16O4 188 39.0±12.0 94.5±4.2 109.2±41.7 93.4±24.4
α,ω-Hexadecanedioic acid C16H30O4 286 64.4±9.1 110.6±23.3 181.3±42.3 248.4±9.1
α,ω-Octadec-9-enedioic acid (C18:1) C18H32O4 312 120.0±18.5 151.8±52.7 93.7b
α,ω-Eicosanedioc acid C20H38O4 342 56.5±11.7 161.4±9.4 144.4±15.6 157.8±17.9
Total α,ω-alkanedioic acids 342±49 642±41 589±125 678±70
ω-Hydroxyalkanoicc
ω-Hydroxyhexadecanoic acid C16H32O3 272 97.5±27.7 81.4 b 137.5b 124.6b
ω-Hydroxyoctadecanoic acid C18H34O3 298 41.9±4.4 98.4±13.7 103.9±48.2 8.4a
ω-Hydroxydocosanoic acid C22H44O3 358 83.1±19.2 125.2±28.2 105.8±29.6 10.8a
ω-Hydroxyoctacosanoic acid C28H56O3 440 136.2±72.5 235.0±34.8 477.7±43.2 75.2a
ω-Hydroxytriacontanoic acid C30H60O3 468 377.0±70.3 565.2±13.9 518.7±69.9 78.7a
Total ω-Hydroxyalkanoic 736±31 1105±26 1344±30 298±141
Mid-chain substituted hydroxy acids
10,16-Dihydroxyhexadecanoic C16H32O4 288 58.8 b 152.8 b 182.1b 223.3b
8-hydroxyhexadecane-1,16-dioic acid C16H30O5 302 67.4±13.3 115.4±26.6 123.4±4.1
Dihydroxymethoxyoctadecanoic acid C18H36O5 332 3.1 a 11.2 a
9,10,18-Trihydroxyoctadecanoic acid C18H36O5 332 32.2 a 90.9 b 38.1b 9,10-Dihydroxyoctadecane-1,18-dioic acid C19H38O4 330 65.5±41.1 222.1 b 78.3b 85.9b
Total Mid-chain substituted hydroxy acids 227±111 592±122 422±160 309±80
Monoacylglycerides
Total Monoacylglycerides C19H38O4 330 238±59 487±128 432±177
Benzyles and phenolse
p-Hydroxyacetophenone C8H8O2 136 21.7±4.1 53.6±5.7 93.0±17.0 55.3b
Vanillin C8H8O3 152 4.7 a 38.0b
Acetovanillone C8H10O3 154 51.4 b 121.7±32.7 137.0b 1080±468
4-Hydroxybenzoic acid C7H6O3 138 204.3±26.5 608.9±33.9 691.8±50.5 916.4±116
Vanillic acid C8H8O4 168 25.4±10.7 74.3±20.6 39.2b 53.0b
Isovanillic acid C8H8O4 168 60.1±5.1 216.7±33.0 253.7±70.9 40.1a
p-Coumaric acid C9H8O3 164 19.1 b 60.6 b 66.7b 52.3a
Ferulic acid C10H10O4 194 40.9±5.1 96.81.4 108.5±28.9 67.5b
Total Benzyles and phenols 428±53 1233±57 1390±200 2303±493
Organophosphates
Triphenyl phosphate C18H15O4P 326 1503±186 4511±101 5555±282 6759±555
Steroidsc
ß- Sitosterol C29H50O 414 4.5 a 26.4a 27.0b 9.1a
Table A4.2 continue a values detected only from one sample b values detected only from two samples c n-alkohols, terpenols and sterols were identified as TMS ethers e phenolic compounds were identified as methyl esters/TMS ethers
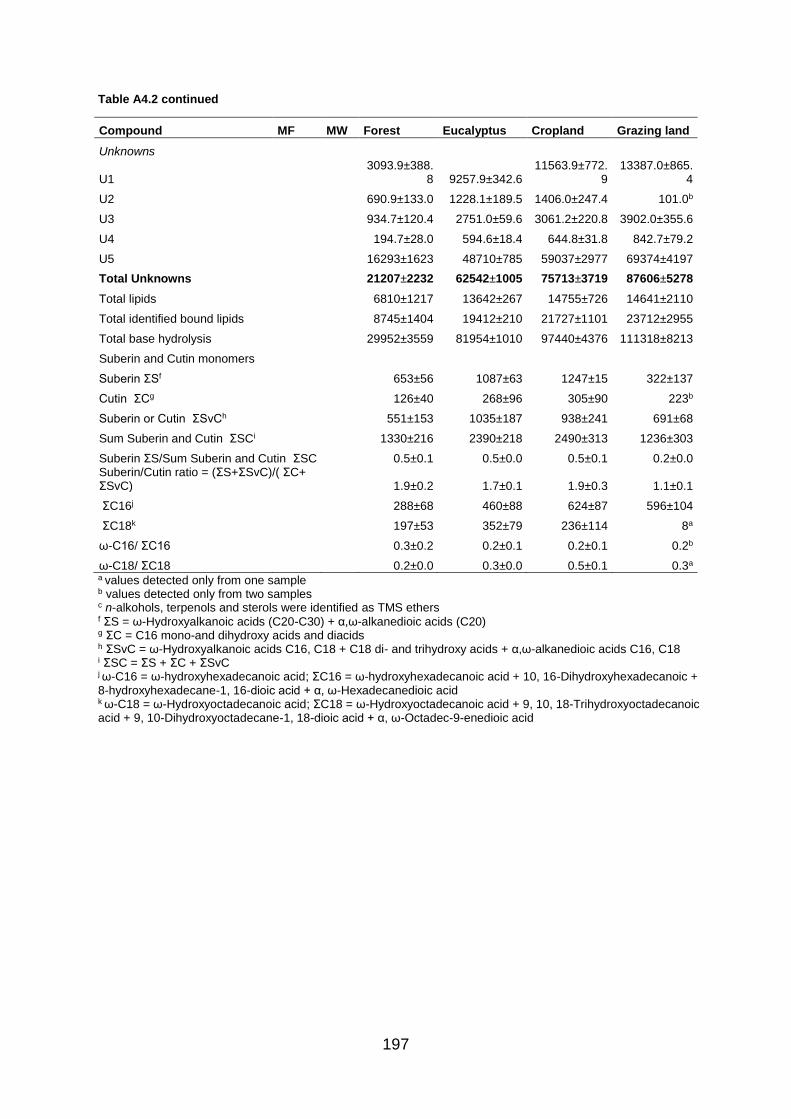
197
Table A4.2 continued
Compound MF MW Forest Eucalyptus Cropland Grazing land
Unknowns
U1 3093.9±388.
8 9257.9±342.6 11563.9±772.
9 13387.0±865.
4
U2 690.9±133.0 1228.1±189.5 1406.0±247.4 101.0b
U3 934.7±120.4 2751.0±59.6 3061.2±220.8 3902.0±355.6
U4 194.7±28.0 594.6±18.4 644.8±31.8 842.7±79.2
U5 16293±1623 48710±785 59037±2977 69374±4197
Total Unknowns 21207±2232 62542±1005 75713±3719 87606±5278
Total lipids 6810±1217 13642±267 14755±726 14641±2110
Total identified bound lipids 8745±1404 19412±210 21727±1101 23712±2955
Total base hydrolysis 29952±3559 81954±1010 97440±4376 111318±8213
Suberin and Cutin monomers
Suberin ΣSf 653±56 1087±63 1247±15 322±137
Cutin ΣCg 126±40 268±96 305±90 223b
Suberin or Cutin ΣSvCh 551±153 1035±187 938±241 691±68
Sum Suberin and Cutin ΣSCi 1330±216 2390±218 2490±313 1236±303
Suberin ΣS/Sum Suberin and Cutin ΣSC 0.5±0.1 0.5±0.0 0.5±0.1 0.2±0.0 Suberin/Cutin ratio = (ΣS+ΣSvC)/( ΣC+ ΣSvC) 1.9±0.2 1.7±0.1 1.9±0.3 1.1±0.1
ΣC16j 288±68 460±88 624±87 596±104
ΣC18k 197±53 352±79 236±114 8a
ω-C16/ ΣC16 0.3±0.2 0.2±0.1 0.2±0.1 0.2b
ω-C18/ ΣC18 0.2±0.0 0.3±0.0 0.5±0.1 0.3a a values detected only from one sample b values detected only from two samples c n-alkohols, terpenols and sterols were identified as TMS ethers f ΣS = ω-Hydroxyalkanoic acids (C20-C30) + α,ω-alkanedioic acids (C20) g ΣC = C16 mono-and dihydroxy acids and diacids h ΣSvC = ω-Hydroxyalkanoic acids C16, C18 + C18 di- and trihydroxy acids + α,ω-alkanedioic acids C16, C18 i ΣSC = ΣS + ΣC + ΣSvC j ω-C16 = ω-hydroxyhexadecanoic acid; ΣC16 = ω-hydroxyhexadecanoic acid + 10, 16-Dihydroxyhexadecanoic + 8-hydroxyhexadecane-1, 16-dioic acid + α, ω-Hexadecanedioic acid k ω-C18 = ω-Hydroxyoctadecanoic acid; ΣC18 = ω-Hydroxyoctadecanoic acid + 9, 10, 18-Trihydroxyoctadecanoic acid + 9, 10-Dihydroxyoctadecane-1, 18-dioic acid + α, ω-Octadec-9-enedioic acid
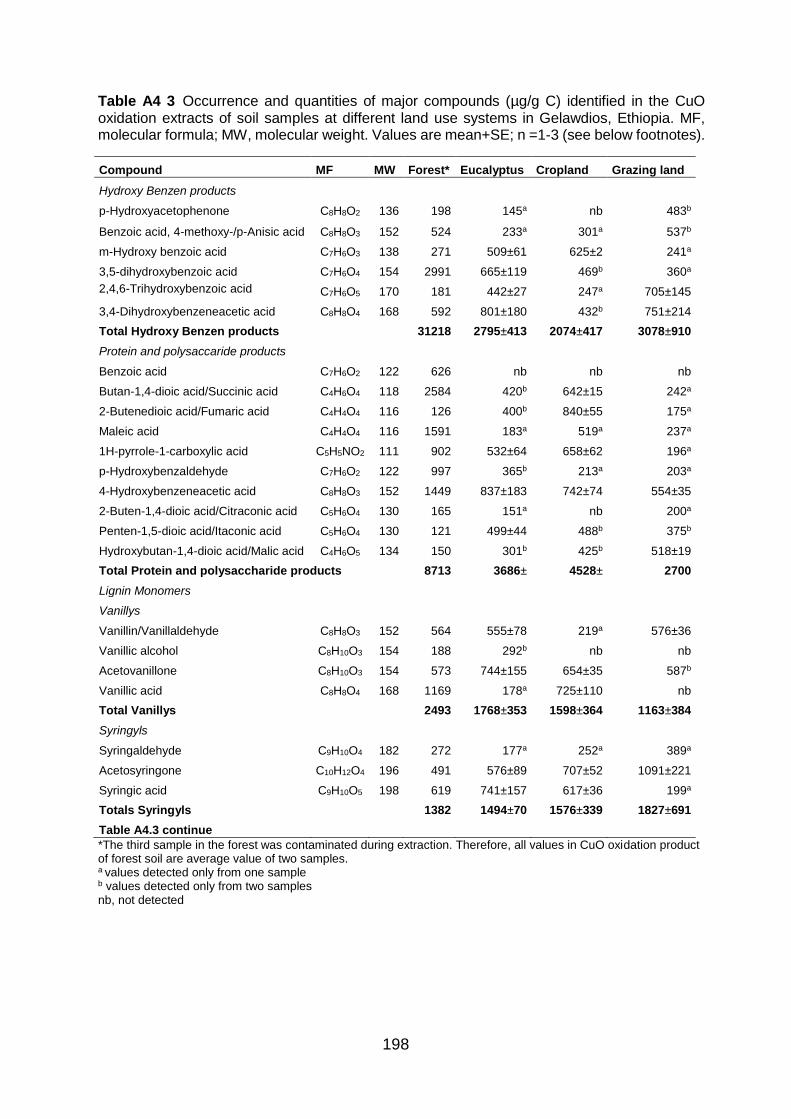
198
Table A4 3 Occurrence and quantities of major compounds (µg/g C) identified in the CuO oxidation extracts of soil samples at different land use systems in Gelawdios, Ethiopia. MF, molecular formula; MW, molecular weight. Values are mean+SE; n =1-3 (see below footnotes).
Compound MF MW Forest* Eucalyptus Cropland Grazing land
Hydroxy Benzen products
p-Hydroxyacetophenone C8H8O2 136 198 145a nb 483b
Benzoic acid, 4-methoxy-/p-Anisic acid C8H8O3 152 524 233a 301a 537b
m-Hydroxy benzoic acid C7H6O3 138 271 509±61 625±2 241a
3,5-dihydroxybenzoic acid C7H6O4 154 2991 665±119 469b 360a
2,4,6-Trihydroxybenzoic acid C7H6O5 170 181 442±27 247a 705±145
3,4-Dihydroxybenzeneacetic acid C8H8O4 168 592 801±180 432b 751±214
Total Hydroxy Benzen products 31218 2795±413 2074±417 3078±910
Protein and polysaccaride products
Benzoic acid C7H6O2 122 626 nb nb nb
Butan-1,4-dioic acid/Succinic acid C4H6O4 118 2584 420b 642±15 242a
2-Butenedioic acid/Fumaric acid C4H4O4 116 126 400b 840±55 175a
Maleic acid C4H4O4 116 1591 183a 519a 237a
1H-pyrrole-1-carboxylic acid C5H5NO2 111 902 532±64 658±62 196a
p-Hydroxybenzaldehyde C7H6O2 122 997 365b 213a 203a
4-Hydroxybenzeneacetic acid C8H8O3 152 1449 837±183 742±74 554±35
2-Buten-1,4-dioic acid/Citraconic acid C5H6O4 130 165 151a nb 200a
Penten-1,5-dioic acid/Itaconic acid C5H6O4 130 121 499±44 488b 375b
Hydroxybutan-1,4-dioic acid/Malic acid C4H6O5 134 150 301b 425b 518±19
Total Protein and polysaccharide products 8713 3686± 4528± 2700
Lignin Monomers
Vanillys
Vanillin/Vanillaldehyde C8H8O3 152 564 555±78 219a 576±36
Vanillic alcohol C8H10O3 154 188 292b nb nb
Acetovanillone C8H10O3 154 573 744±155 654±35 587b
Vanillic acid C8H8O4 168 1169 178a 725±110 nb
Total Vanillys 2493 1768±353 1598±364 1163±384
Syringyls
Syringaldehyde C9H10O4 182 272 177a 252a 389a
Acetosyringone C10H12O4 196 491 576±89 707±52 1091±221
Syringic acid C9H10O5 198 619 741±157 617±36 199a
Totals Syringyls 1382 1494±70 1576±339 1827±691
Table A4.3 continue
*The third sample in the forest was contaminated during extraction. Therefore, all values in CuO oxidation product of forest soil are average value of two samples. a values detected only from one sample b values detected only from two samples nb, not detected
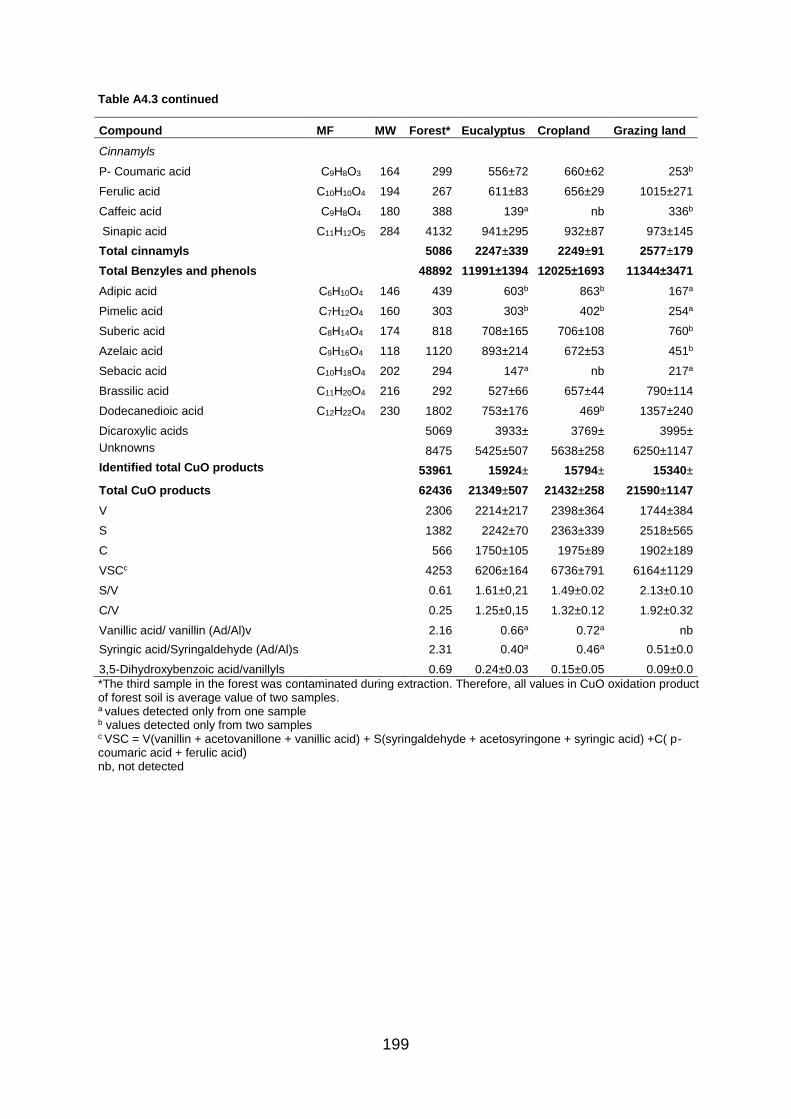
199
Table A4.3 continued
Compound MF MW Forest* Eucalyptus Cropland Grazing land
Cinnamyls
P- Coumaric acid C9H8O3 164 299 556±72 660±62 253b
Ferulic acid C10H10O4 194 267 611±83 656±29 1015±271
Caffeic acid C9H8O4 180 388 139a nb 336b
Sinapic acid C11H12O5 284 4132 941±295 932±87 973±145
Total cinnamyls 5086 2247±339 2249±91 2577±179
Total Benzyles and phenols 48892 11991±1394 12025±1693 11344±3471
Adipic acid C6H10O4 146 439 603b 863b 167a
Pimelic acid C7H12O4 160 303 303b 402b 254a
Suberic acid C8H14O4 174 818 708±165 706±108 760b
Azelaic acid C9H16O4 118 1120 893±214 672±53 451b
Sebacic acid C10H18O4 202 294 147a nb 217a
Brassilic acid C11H20O4 216 292 527±66 657±44 790±114
Dodecanedioic acid C12H22O4 230 1802 753±176 469b 1357±240
Dicaroxylic acids 5069 3933± 3769± 3995±
Unknowns 8475 5425±507 5638±258 6250±1147
Identified total CuO products 53961 15924± 15794± 15340±
Total CuO products 62436 21349±507 21432±258 21590±1147
V 2306 2214±217 2398±364 1744±384
S 1382 2242±70 2363±339 2518±565
C 566 1750±105 1975±89 1902±189
VSCc 4253 6206±164 6736±791 6164±1129
S/V 0.61 1.61±0,21 1.49±0.02 2.13±0.10
C/V 0.25 1.25±0,15 1.32±0.12 1.92±0.32
Vanillic acid/ vanillin (Ad/Al)v 2.16 0.66a 0.72a nb
Syringic acid/Syringaldehyde (Ad/Al)s 2.31 0.40a 0.46a 0.51±0.0
3,5-Dihydroxybenzoic acid/vanillyls 0.69 0.24±0.03 0.15±0.05 0.09±0.0
*The third sample in the forest was contaminated during extraction. Therefore, all values in CuO oxidation product of forest soil is average value of two samples. a values detected only from one sample b values detected only from two samples c VSC = V(vanillin + acetovanillone + vanillic acid) + S(syringaldehyde + acetosyringone + syringic acid) +C( p-coumaric acid + ferulic acid) nb, not detected
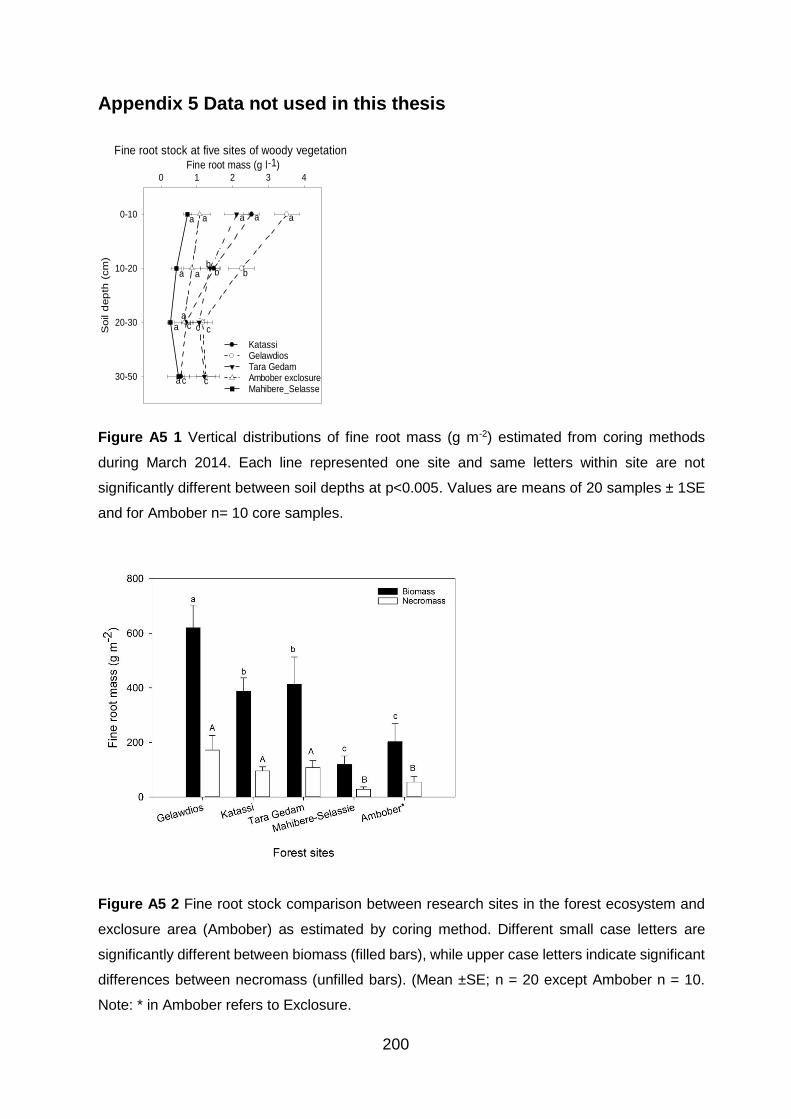
200
Appendix 5 Data not used in this thesis
Figure A5 1 Vertical distributions of fine root mass (g m-2) estimated from coring methods
during March 2014. Each line represented one site and same letters within site are not
significantly different between soil depths at p<0.005. Values are means of 20 samples ± 1SE
and for Ambober n= 10 core samples.
Figure A5 2 Fine root stock comparison between research sites in the forest ecosystem and
exclosure area (Ambober) as estimated by coring method. Different small case letters are
significantly different between biomass (filled bars), while upper case letters indicate significant
differences between necromass (unfilled bars). (Mean ±SE; n = 20 except Ambober n = 10.
Note: * in Ambober refers to Exclosure.
Fine root stock at five sites of woody vegetation Fine root mass (g l-1)
0 1 2 3 4
Soil d
ep
th (
cm
)
0-10
10-20
20-30
30-50
Katassi Gelawdios Tara Gedam Ambober exclosure Mahibere_Selasse
a
b
c
c
a
c
c
b
a
b
c
a
a
a
a
a
a
a
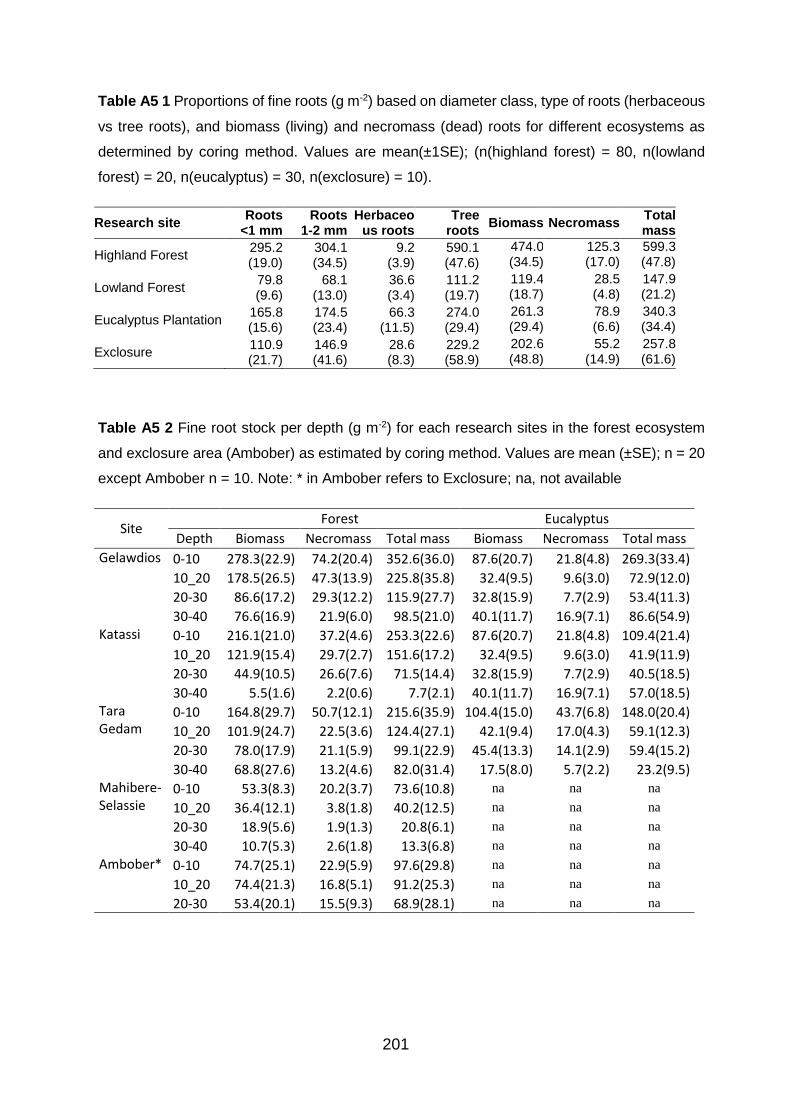
201
Table A5 1 Proportions of fine roots (g m-2) based on diameter class, type of roots (herbaceous
vs tree roots), and biomass (living) and necromass (dead) roots for different ecosystems as
determined by coring method. Values are mean(±1SE); (n(highland forest) = 80, n(lowland
forest) = 20, n(eucalyptus) = 30, n(exclosure) = 10).
Research site Roots
<1 mm Roots
1-2 mm Herbaceo
us roots Tree
roots Biomass Necromass
Total mass
Highland Forest 295.2 (19.0)
304.1 (34.5)
9.2 (3.9)
590.1 (47.6)
474.0 (34.5)
125.3 (17.0)
599.3 (47.8)
Lowland Forest 79.8 (9.6)
68.1 (13.0)
36.6 (3.4)
111.2 (19.7)
119.4 (18.7)
28.5 (4.8)
147.9 (21.2)
Eucalyptus Plantation 165.8 (15.6)
174.5 (23.4)
66.3 (11.5)
274.0 (29.4)
261.3 (29.4)
78.9 (6.6)
340.3 (34.4)
Exclosure 110.9 (21.7)
146.9 (41.6)
28.6 (8.3)
229.2 (58.9)
202.6 (48.8)
55.2 (14.9)
257.8 (61.6)
Table A5 2 Fine root stock per depth (g m-2) for each research sites in the forest ecosystem
and exclosure area (Ambober) as estimated by coring method. Values are mean (±SE); n = 20
except Ambober n = 10. Note: * in Ambober refers to Exclosure; na, not available
Site Forest Eucalyptus
Depth Biomass Necromass Total mass Biomass Necromass Total mass
Gelawdios 0-10 278.3(22.9) 74.2(20.4) 352.6(36.0) 87.6(20.7) 21.8(4.8) 269.3(33.4)
10_20 178.5(26.5) 47.3(13.9) 225.8(35.8) 32.4(9.5) 9.6(3.0) 72.9(12.0)
20-30 86.6(17.2) 29.3(12.2) 115.9(27.7) 32.8(15.9) 7.7(2.9) 53.4(11.3)
30-40 76.6(16.9) 21.9(6.0) 98.5(21.0) 40.1(11.7) 16.9(7.1) 86.6(54.9) Katassi 0-10 216.1(21.0) 37.2(4.6) 253.3(22.6) 87.6(20.7) 21.8(4.8) 109.4(21.4)
10_20 121.9(15.4) 29.7(2.7) 151.6(17.2) 32.4(9.5) 9.6(3.0) 41.9(11.9)
20-30 44.9(10.5) 26.6(7.6) 71.5(14.4) 32.8(15.9) 7.7(2.9) 40.5(18.5)
30-40 5.5(1.6) 2.2(0.6) 7.7(2.1) 40.1(11.7) 16.9(7.1) 57.0(18.5) Tara Gedam
0-10 164.8(29.7) 50.7(12.1) 215.6(35.9) 104.4(15.0) 43.7(6.8) 148.0(20.4)
10_20 101.9(24.7) 22.5(3.6) 124.4(27.1) 42.1(9.4) 17.0(4.3) 59.1(12.3)
20-30 78.0(17.9) 21.1(5.9) 99.1(22.9) 45.4(13.3) 14.1(2.9) 59.4(15.2)
30-40 68.8(27.6) 13.2(4.6) 82.0(31.4) 17.5(8.0) 5.7(2.2) 23.2(9.5) Mahibere-Selassie
0-10 53.3(8.3) 20.2(3.7) 73.6(10.8) na na na
10_20 36.4(12.1) 3.8(1.8) 40.2(12.5) na na na
20-30 18.9(5.6) 1.9(1.3) 20.8(6.1) na na na
30-40 10.7(5.3) 2.6(1.8) 13.3(6.8) na na na
Ambober* 0-10 74.7(25.1) 22.9(5.9) 97.6(29.8) na na na
10_20 74.4(21.3) 16.8(5.1) 91.2(25.3) na na na
20-30 53.4(20.1) 15.5(9.3) 68.9(28.1) na na na
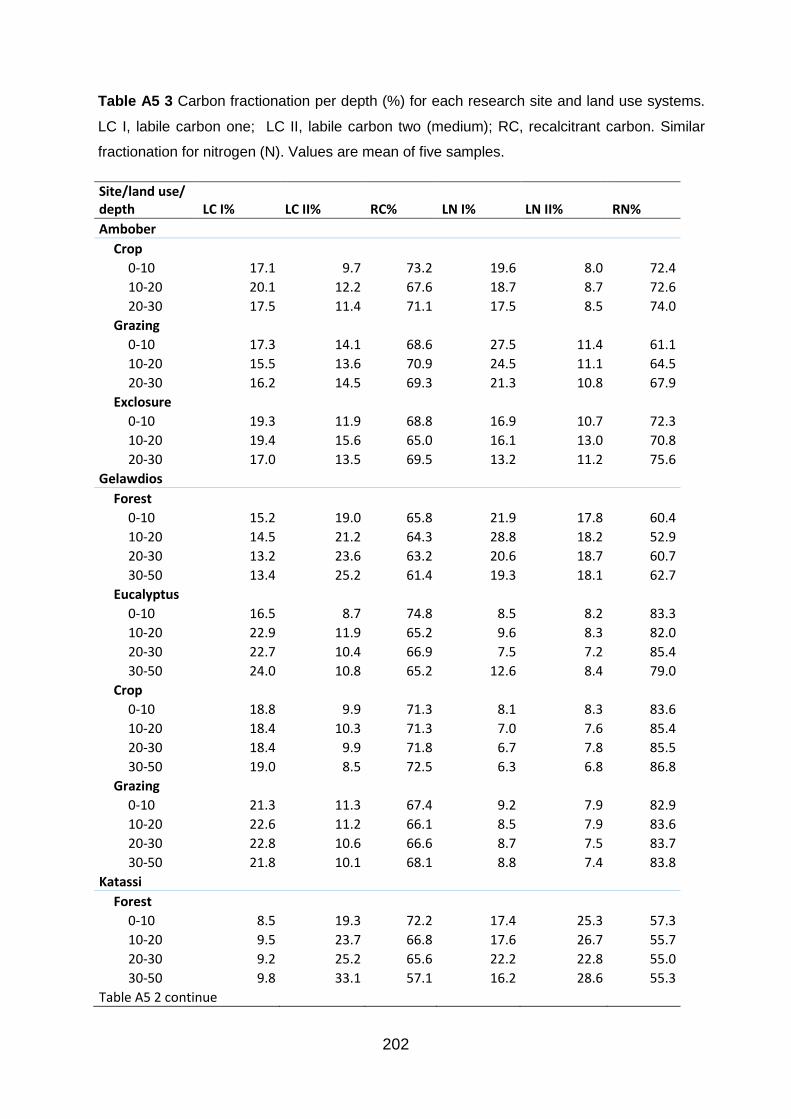
202
Table A5 3 Carbon fractionation per depth (%) for each research site and land use systems.
LC I, labile carbon one; LC II, labile carbon two (medium); RC, recalcitrant carbon. Similar
fractionation for nitrogen (N). Values are mean of five samples.
Site/land use/ depth LC I% LC II% RC% LN I% LN II% RN%
Ambober
Crop
0-10 17.1 9.7 73.2 19.6 8.0 72.4
10-20 20.1 12.2 67.6 18.7 8.7 72.6
20-30 17.5 11.4 71.1 17.5 8.5 74.0
Grazing
0-10 17.3 14.1 68.6 27.5 11.4 61.1
10-20 15.5 13.6 70.9 24.5 11.1 64.5
20-30 16.2 14.5 69.3 21.3 10.8 67.9
Exclosure
0-10 19.3 11.9 68.8 16.9 10.7 72.3
10-20 19.4 15.6 65.0 16.1 13.0 70.8
20-30 17.0 13.5 69.5 13.2 11.2 75.6
Gelawdios
Forest
0-10 15.2 19.0 65.8 21.9 17.8 60.4
10-20 14.5 21.2 64.3 28.8 18.2 52.9
20-30 13.2 23.6 63.2 20.6 18.7 60.7
30-50 13.4 25.2 61.4 19.3 18.1 62.7
Eucalyptus
0-10 16.5 8.7 74.8 8.5 8.2 83.3
10-20 22.9 11.9 65.2 9.6 8.3 82.0
20-30 22.7 10.4 66.9 7.5 7.2 85.4
30-50 24.0 10.8 65.2 12.6 8.4 79.0
Crop
0-10 18.8 9.9 71.3 8.1 8.3 83.6
10-20 18.4 10.3 71.3 7.0 7.6 85.4
20-30 18.4 9.9 71.8 6.7 7.8 85.5
30-50 19.0 8.5 72.5 6.3 6.8 86.8
Grazing
0-10 21.3 11.3 67.4 9.2 7.9 82.9
10-20 22.6 11.2 66.1 8.5 7.9 83.6
20-30 22.8 10.6 66.6 8.7 7.5 83.7
30-50 21.8 10.1 68.1 8.8 7.4 83.8
Katassi
Forest
0-10 8.5 19.3 72.2 17.4 25.3 57.3
10-20 9.5 23.7 66.8 17.6 26.7 55.7
20-30 9.2 25.2 65.6 22.2 22.8 55.0
30-50 9.8 33.1 57.1 16.2 28.6 55.3
Table A5 2 continue
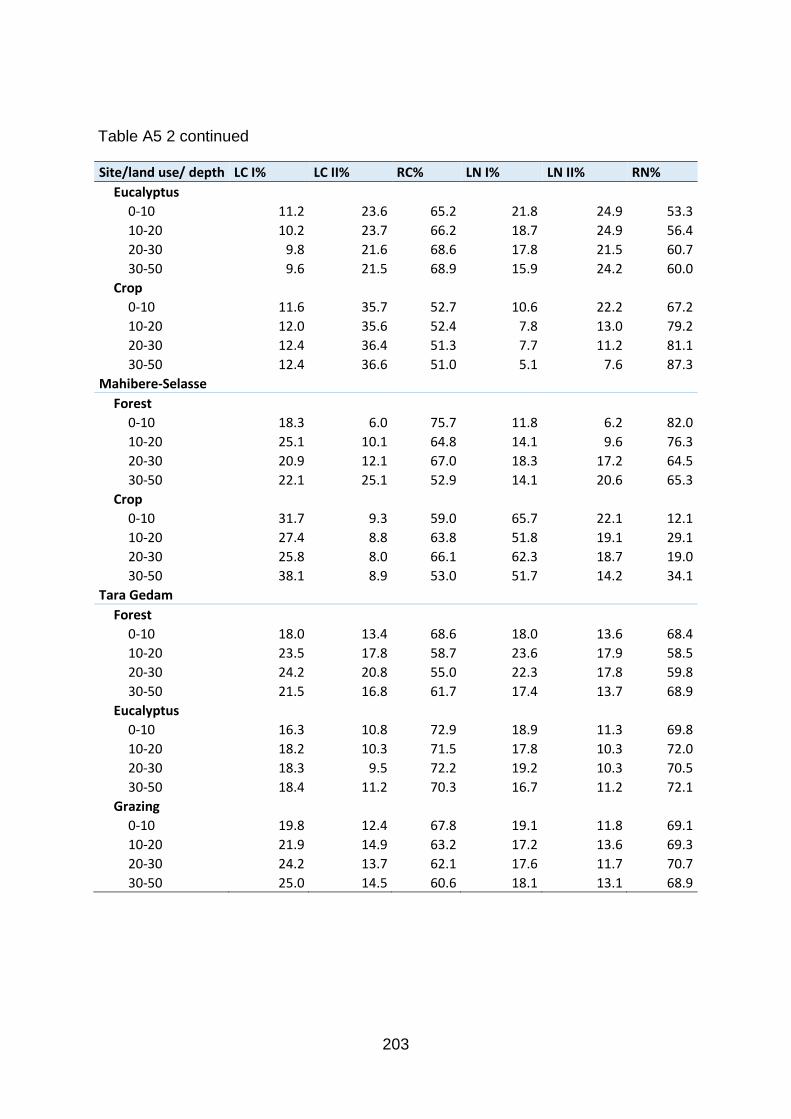
203
Table A5 2 continued
Site/land use/ depth LC I% LC II% RC% LN I% LN II% RN%
Eucalyptus
0-10 11.2 23.6 65.2 21.8 24.9 53.3
10-20 10.2 23.7 66.2 18.7 24.9 56.4
20-30 9.8 21.6 68.6 17.8 21.5 60.7
30-50 9.6 21.5 68.9 15.9 24.2 60.0
Crop
0-10 11.6 35.7 52.7 10.6 22.2 67.2
10-20 12.0 35.6 52.4 7.8 13.0 79.2
20-30 12.4 36.4 51.3 7.7 11.2 81.1
30-50 12.4 36.6 51.0 5.1 7.6 87.3
Mahibere-Selasse
Forest
0-10 18.3 6.0 75.7 11.8 6.2 82.0
10-20 25.1 10.1 64.8 14.1 9.6 76.3
20-30 20.9 12.1 67.0 18.3 17.2 64.5
30-50 22.1 25.1 52.9 14.1 20.6 65.3
Crop
0-10 31.7 9.3 59.0 65.7 22.1 12.1
10-20 27.4 8.8 63.8 51.8 19.1 29.1
20-30 25.8 8.0 66.1 62.3 18.7 19.0
30-50 38.1 8.9 53.0 51.7 14.2 34.1
Tara Gedam
Forest
0-10 18.0 13.4 68.6 18.0 13.6 68.4
10-20 23.5 17.8 58.7 23.6 17.9 58.5
20-30 24.2 20.8 55.0 22.3 17.8 59.8
30-50 21.5 16.8 61.7 17.4 13.7 68.9
Eucalyptus
0-10 16.3 10.8 72.9 18.9 11.3 69.8
10-20 18.2 10.3 71.5 17.8 10.3 72.0
20-30 18.3 9.5 72.2 19.2 10.3 70.5
30-50 18.4 11.2 70.3 16.7 11.2 72.1
Grazing
0-10 19.8 12.4 67.8 19.1 11.8 69.1
10-20 21.9 14.9 63.2 17.2 13.6 69.3
20-30 24.2 13.7 62.1 17.6 11.7 70.7
30-50 25.0 14.5 60.6 18.1 13.1 68.9
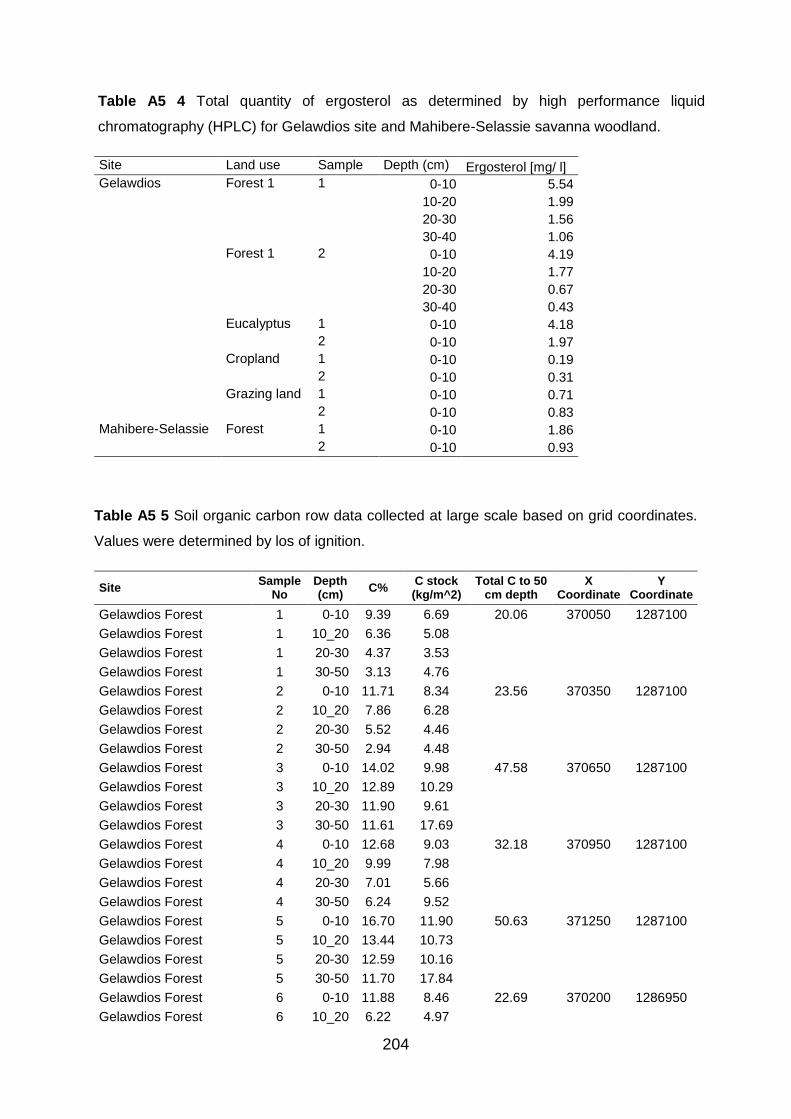
204
Table A5 4 Total quantity of ergosterol as determined by high performance liquid
chromatography (HPLC) for Gelawdios site and Mahibere-Selassie savanna woodland.
Site Land use Sample Depth (cm) Ergosterol [mg/ l]
Gelawdios Forest 1 1 0-10 5.54
10-20 1.99
20-30 1.56
30-40 1.06
Forest 1 2 0-10 4.19
10-20 1.77
20-30 0.67
30-40 0.43
Eucalyptus 1 0-10 4.18
2 0-10 1.97
Cropland 1 0-10 0.19
2 0-10 0.31
Grazing land 1 0-10 0.71
2 0-10 0.83
Mahibere-Selassie Forest 1 0-10 1.86
2 0-10 0.93
Table A5 5 Soil organic carbon row data collected at large scale based on grid coordinates.
Values were determined by los of ignition.
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Gelawdios Forest 1 0-10 9.39 6.69 20.06 370050 1287100
Gelawdios Forest 1 10_20 6.36 5.08
Gelawdios Forest 1 20-30 4.37 3.53
Gelawdios Forest 1 30-50 3.13 4.76
Gelawdios Forest 2 0-10 11.71 8.34 23.56 370350 1287100
Gelawdios Forest 2 10_20 7.86 6.28
Gelawdios Forest 2 20-30 5.52 4.46
Gelawdios Forest 2 30-50 2.94 4.48
Gelawdios Forest 3 0-10 14.02 9.98 47.58 370650 1287100
Gelawdios Forest 3 10_20 12.89 10.29
Gelawdios Forest 3 20-30 11.90 9.61
Gelawdios Forest 3 30-50 11.61 17.69
Gelawdios Forest 4 0-10 12.68 9.03 32.18 370950 1287100
Gelawdios Forest 4 10_20 9.99 7.98
Gelawdios Forest 4 20-30 7.01 5.66
Gelawdios Forest 4 30-50 6.24 9.52
Gelawdios Forest 5 0-10 16.70 11.90 50.63 371250 1287100
Gelawdios Forest 5 10_20 13.44 10.73
Gelawdios Forest 5 20-30 12.59 10.16
Gelawdios Forest 5 30-50 11.70 17.84
Gelawdios Forest 6 0-10 11.88 8.46 22.69 370200 1286950
Gelawdios Forest 6 10_20 6.22 4.97
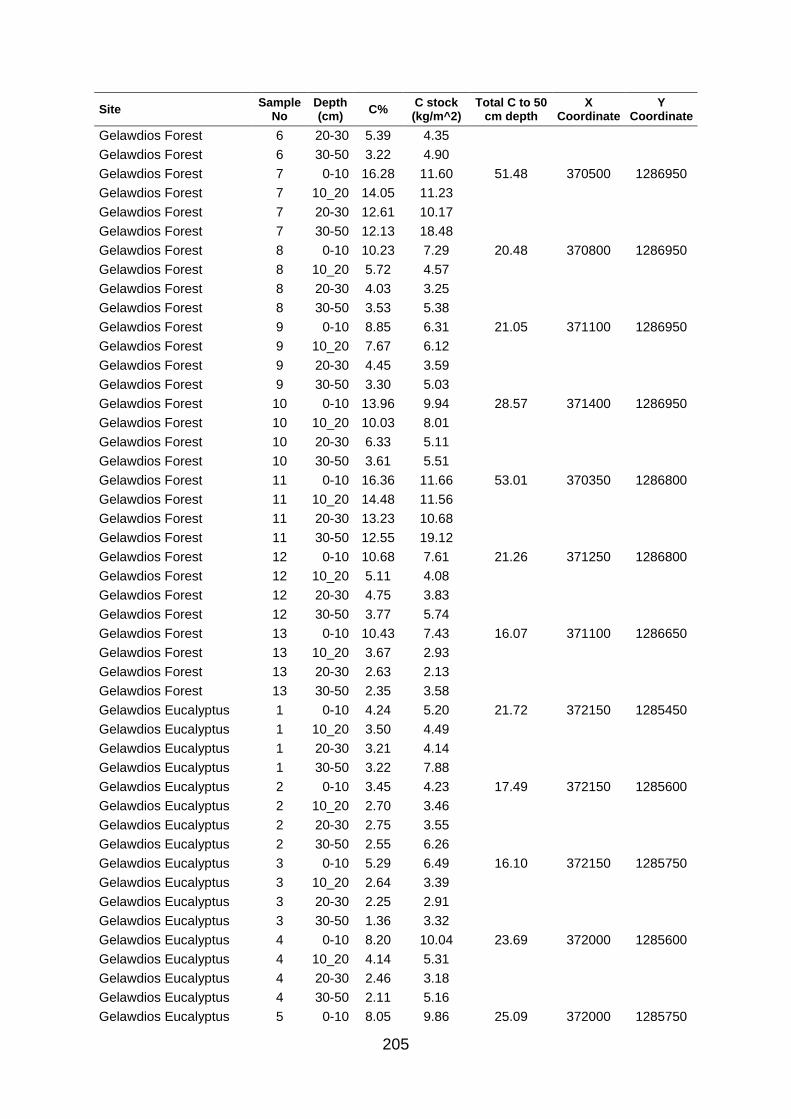
205
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Gelawdios Forest 6 20-30 5.39 4.35
Gelawdios Forest 6 30-50 3.22 4.90
Gelawdios Forest 7 0-10 16.28 11.60 51.48 370500 1286950
Gelawdios Forest 7 10_20 14.05 11.23
Gelawdios Forest 7 20-30 12.61 10.17
Gelawdios Forest 7 30-50 12.13 18.48
Gelawdios Forest 8 0-10 10.23 7.29 20.48 370800 1286950
Gelawdios Forest 8 10_20 5.72 4.57
Gelawdios Forest 8 20-30 4.03 3.25
Gelawdios Forest 8 30-50 3.53 5.38
Gelawdios Forest 9 0-10 8.85 6.31 21.05 371100 1286950
Gelawdios Forest 9 10_20 7.67 6.12
Gelawdios Forest 9 20-30 4.45 3.59
Gelawdios Forest 9 30-50 3.30 5.03
Gelawdios Forest 10 0-10 13.96 9.94 28.57 371400 1286950
Gelawdios Forest 10 10_20 10.03 8.01
Gelawdios Forest 10 20-30 6.33 5.11
Gelawdios Forest 10 30-50 3.61 5.51
Gelawdios Forest 11 0-10 16.36 11.66 53.01 370350 1286800
Gelawdios Forest 11 10_20 14.48 11.56
Gelawdios Forest 11 20-30 13.23 10.68
Gelawdios Forest 11 30-50 12.55 19.12
Gelawdios Forest 12 0-10 10.68 7.61 21.26 371250 1286800
Gelawdios Forest 12 10_20 5.11 4.08
Gelawdios Forest 12 20-30 4.75 3.83
Gelawdios Forest 12 30-50 3.77 5.74
Gelawdios Forest 13 0-10 10.43 7.43 16.07 371100 1286650
Gelawdios Forest 13 10_20 3.67 2.93
Gelawdios Forest 13 20-30 2.63 2.13
Gelawdios Forest 13 30-50 2.35 3.58
Gelawdios Eucalyptus 1 0-10 4.24 5.20 21.72 372150 1285450
Gelawdios Eucalyptus 1 10_20 3.50 4.49
Gelawdios Eucalyptus 1 20-30 3.21 4.14
Gelawdios Eucalyptus 1 30-50 3.22 7.88
Gelawdios Eucalyptus 2 0-10 3.45 4.23 17.49 372150 1285600
Gelawdios Eucalyptus 2 10_20 2.70 3.46
Gelawdios Eucalyptus 2 20-30 2.75 3.55
Gelawdios Eucalyptus 2 30-50 2.55 6.26
Gelawdios Eucalyptus 3 0-10 5.29 6.49 16.10 372150 1285750
Gelawdios Eucalyptus 3 10_20 2.64 3.39
Gelawdios Eucalyptus 3 20-30 2.25 2.91
Gelawdios Eucalyptus 3 30-50 1.36 3.32
Gelawdios Eucalyptus 4 0-10 8.20 10.04 23.69 372000 1285600
Gelawdios Eucalyptus 4 10_20 4.14 5.31
Gelawdios Eucalyptus 4 20-30 2.46 3.18
Gelawdios Eucalyptus 4 30-50 2.11 5.16
Gelawdios Eucalyptus 5 0-10 8.05 9.86 25.09 372000 1285750
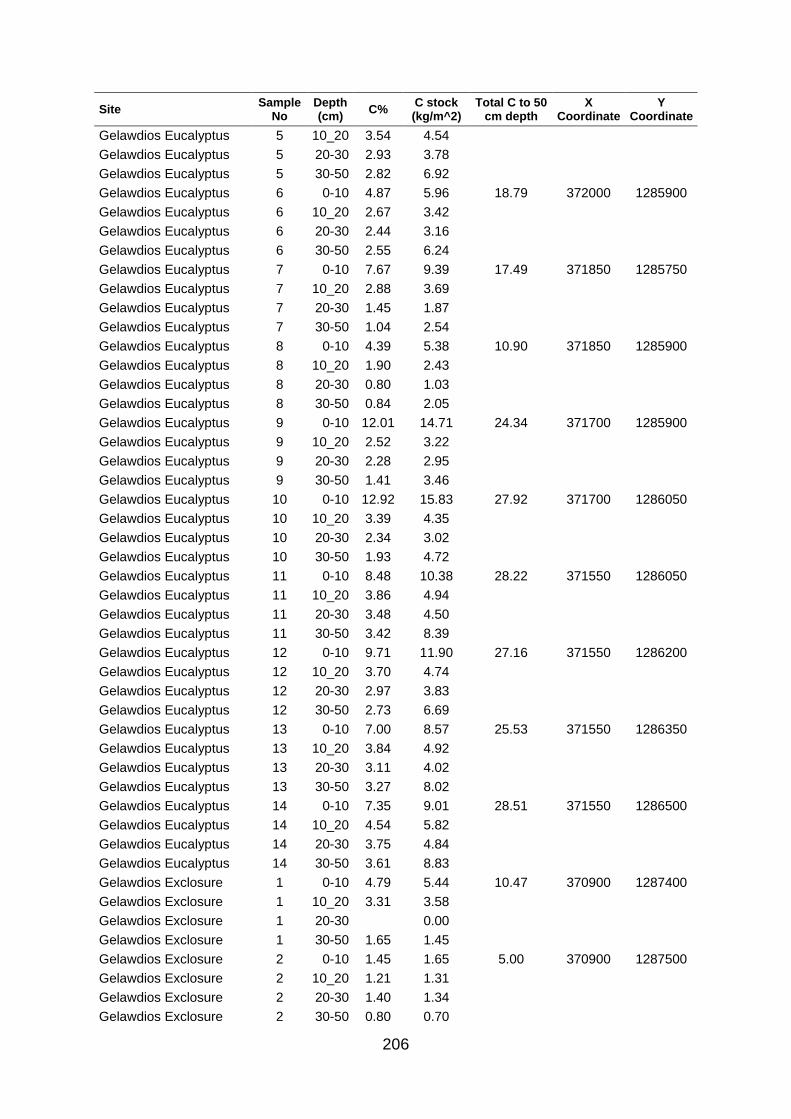
206
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Gelawdios Eucalyptus 5 10_20 3.54 4.54
Gelawdios Eucalyptus 5 20-30 2.93 3.78
Gelawdios Eucalyptus 5 30-50 2.82 6.92
Gelawdios Eucalyptus 6 0-10 4.87 5.96 18.79 372000 1285900
Gelawdios Eucalyptus 6 10_20 2.67 3.42
Gelawdios Eucalyptus 6 20-30 2.44 3.16
Gelawdios Eucalyptus 6 30-50 2.55 6.24
Gelawdios Eucalyptus 7 0-10 7.67 9.39 17.49 371850 1285750
Gelawdios Eucalyptus 7 10_20 2.88 3.69
Gelawdios Eucalyptus 7 20-30 1.45 1.87
Gelawdios Eucalyptus 7 30-50 1.04 2.54
Gelawdios Eucalyptus 8 0-10 4.39 5.38 10.90 371850 1285900
Gelawdios Eucalyptus 8 10_20 1.90 2.43
Gelawdios Eucalyptus 8 20-30 0.80 1.03
Gelawdios Eucalyptus 8 30-50 0.84 2.05
Gelawdios Eucalyptus 9 0-10 12.01 14.71 24.34 371700 1285900
Gelawdios Eucalyptus 9 10_20 2.52 3.22
Gelawdios Eucalyptus 9 20-30 2.28 2.95
Gelawdios Eucalyptus 9 30-50 1.41 3.46
Gelawdios Eucalyptus 10 0-10 12.92 15.83 27.92 371700 1286050
Gelawdios Eucalyptus 10 10_20 3.39 4.35
Gelawdios Eucalyptus 10 20-30 2.34 3.02
Gelawdios Eucalyptus 10 30-50 1.93 4.72
Gelawdios Eucalyptus 11 0-10 8.48 10.38 28.22 371550 1286050
Gelawdios Eucalyptus 11 10_20 3.86 4.94
Gelawdios Eucalyptus 11 20-30 3.48 4.50
Gelawdios Eucalyptus 11 30-50 3.42 8.39
Gelawdios Eucalyptus 12 0-10 9.71 11.90 27.16 371550 1286200
Gelawdios Eucalyptus 12 10_20 3.70 4.74
Gelawdios Eucalyptus 12 20-30 2.97 3.83
Gelawdios Eucalyptus 12 30-50 2.73 6.69
Gelawdios Eucalyptus 13 0-10 7.00 8.57 25.53 371550 1286350
Gelawdios Eucalyptus 13 10_20 3.84 4.92
Gelawdios Eucalyptus 13 20-30 3.11 4.02
Gelawdios Eucalyptus 13 30-50 3.27 8.02
Gelawdios Eucalyptus 14 0-10 7.35 9.01 28.51 371550 1286500
Gelawdios Eucalyptus 14 10_20 4.54 5.82
Gelawdios Eucalyptus 14 20-30 3.75 4.84
Gelawdios Eucalyptus 14 30-50 3.61 8.83
Gelawdios Exclosure 1 0-10 4.79 5.44 10.47 370900 1287400
Gelawdios Exclosure 1 10_20 3.31 3.58
Gelawdios Exclosure 1 20-30 0.00
Gelawdios Exclosure 1 30-50 1.65 1.45
Gelawdios Exclosure 2 0-10 1.45 1.65 5.00 370900 1287500
Gelawdios Exclosure 2 10_20 1.21 1.31
Gelawdios Exclosure 2 20-30 1.40 1.34
Gelawdios Exclosure 2 30-50 0.80 0.70
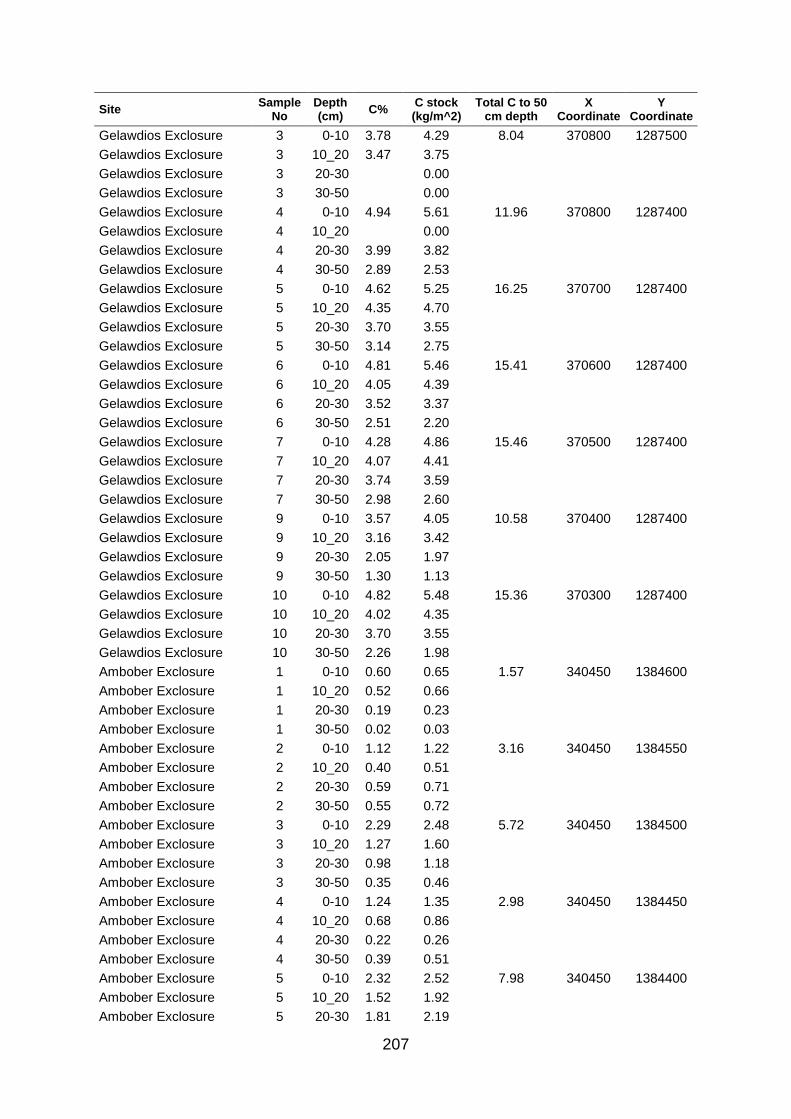
207
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Gelawdios Exclosure 3 0-10 3.78 4.29 8.04 370800 1287500
Gelawdios Exclosure 3 10_20 3.47 3.75
Gelawdios Exclosure 3 20-30 0.00
Gelawdios Exclosure 3 30-50 0.00
Gelawdios Exclosure 4 0-10 4.94 5.61 11.96 370800 1287400
Gelawdios Exclosure 4 10_20 0.00
Gelawdios Exclosure 4 20-30 3.99 3.82
Gelawdios Exclosure 4 30-50 2.89 2.53
Gelawdios Exclosure 5 0-10 4.62 5.25 16.25 370700 1287400
Gelawdios Exclosure 5 10_20 4.35 4.70
Gelawdios Exclosure 5 20-30 3.70 3.55
Gelawdios Exclosure 5 30-50 3.14 2.75
Gelawdios Exclosure 6 0-10 4.81 5.46 15.41 370600 1287400
Gelawdios Exclosure 6 10_20 4.05 4.39
Gelawdios Exclosure 6 20-30 3.52 3.37
Gelawdios Exclosure 6 30-50 2.51 2.20
Gelawdios Exclosure 7 0-10 4.28 4.86 15.46 370500 1287400
Gelawdios Exclosure 7 10_20 4.07 4.41
Gelawdios Exclosure 7 20-30 3.74 3.59
Gelawdios Exclosure 7 30-50 2.98 2.60
Gelawdios Exclosure 9 0-10 3.57 4.05 10.58 370400 1287400
Gelawdios Exclosure 9 10_20 3.16 3.42
Gelawdios Exclosure 9 20-30 2.05 1.97
Gelawdios Exclosure 9 30-50 1.30 1.13
Gelawdios Exclosure 10 0-10 4.82 5.48 15.36 370300 1287400
Gelawdios Exclosure 10 10_20 4.02 4.35
Gelawdios Exclosure 10 20-30 3.70 3.55
Gelawdios Exclosure 10 30-50 2.26 1.98
Ambober Exclosure 1 0-10 0.60 0.65 1.57 340450 1384600
Ambober Exclosure 1 10_20 0.52 0.66
Ambober Exclosure 1 20-30 0.19 0.23
Ambober Exclosure 1 30-50 0.02 0.03
Ambober Exclosure 2 0-10 1.12 1.22 3.16 340450 1384550
Ambober Exclosure 2 10_20 0.40 0.51
Ambober Exclosure 2 20-30 0.59 0.71
Ambober Exclosure 2 30-50 0.55 0.72
Ambober Exclosure 3 0-10 2.29 2.48 5.72 340450 1384500
Ambober Exclosure 3 10_20 1.27 1.60
Ambober Exclosure 3 20-30 0.98 1.18
Ambober Exclosure 3 30-50 0.35 0.46
Ambober Exclosure 4 0-10 1.24 1.35 2.98 340450 1384450
Ambober Exclosure 4 10_20 0.68 0.86
Ambober Exclosure 4 20-30 0.22 0.26
Ambober Exclosure 4 30-50 0.39 0.51
Ambober Exclosure 5 0-10 2.32 2.52 7.98 340450 1384400
Ambober Exclosure 5 10_20 1.52 1.92
Ambober Exclosure 5 20-30 1.81 2.19
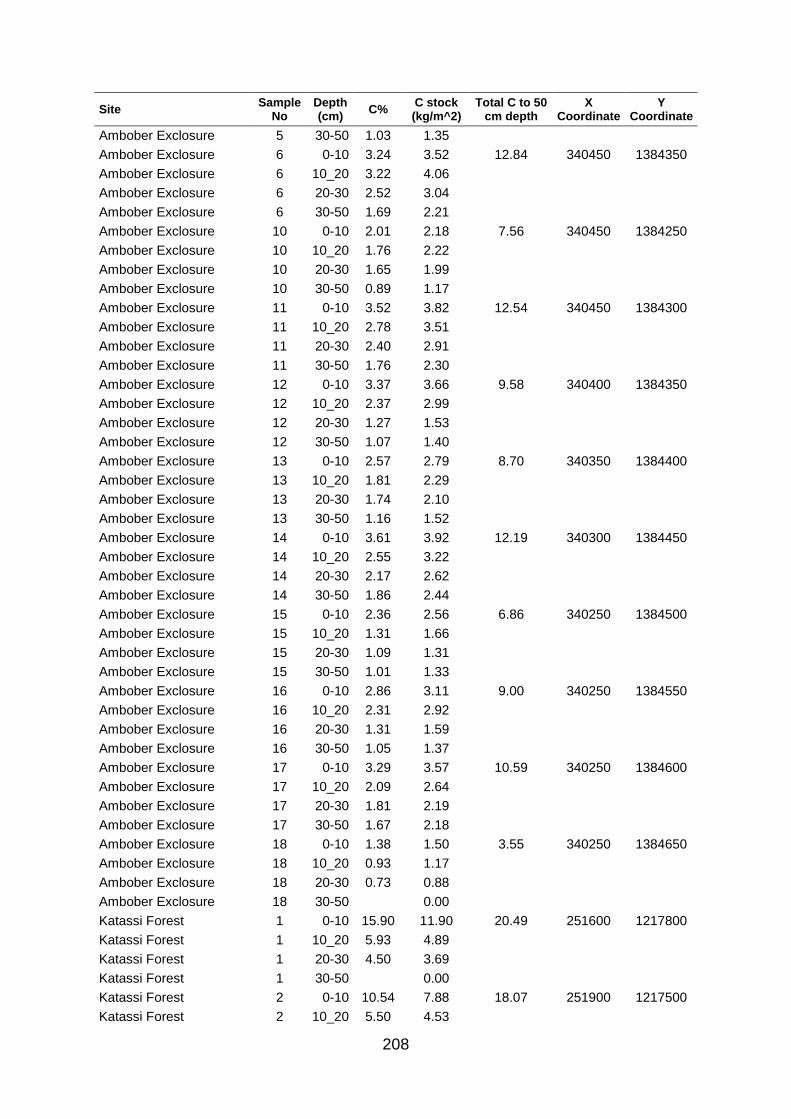
208
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Ambober Exclosure 5 30-50 1.03 1.35
Ambober Exclosure 6 0-10 3.24 3.52 12.84 340450 1384350
Ambober Exclosure 6 10_20 3.22 4.06
Ambober Exclosure 6 20-30 2.52 3.04
Ambober Exclosure 6 30-50 1.69 2.21
Ambober Exclosure 10 0-10 2.01 2.18 7.56 340450 1384250
Ambober Exclosure 10 10_20 1.76 2.22
Ambober Exclosure 10 20-30 1.65 1.99
Ambober Exclosure 10 30-50 0.89 1.17
Ambober Exclosure 11 0-10 3.52 3.82 12.54 340450 1384300
Ambober Exclosure 11 10_20 2.78 3.51
Ambober Exclosure 11 20-30 2.40 2.91
Ambober Exclosure 11 30-50 1.76 2.30
Ambober Exclosure 12 0-10 3.37 3.66 9.58 340400 1384350
Ambober Exclosure 12 10_20 2.37 2.99
Ambober Exclosure 12 20-30 1.27 1.53
Ambober Exclosure 12 30-50 1.07 1.40
Ambober Exclosure 13 0-10 2.57 2.79 8.70 340350 1384400
Ambober Exclosure 13 10_20 1.81 2.29
Ambober Exclosure 13 20-30 1.74 2.10
Ambober Exclosure 13 30-50 1.16 1.52
Ambober Exclosure 14 0-10 3.61 3.92 12.19 340300 1384450
Ambober Exclosure 14 10_20 2.55 3.22
Ambober Exclosure 14 20-30 2.17 2.62
Ambober Exclosure 14 30-50 1.86 2.44
Ambober Exclosure 15 0-10 2.36 2.56 6.86 340250 1384500
Ambober Exclosure 15 10_20 1.31 1.66
Ambober Exclosure 15 20-30 1.09 1.31
Ambober Exclosure 15 30-50 1.01 1.33
Ambober Exclosure 16 0-10 2.86 3.11 9.00 340250 1384550
Ambober Exclosure 16 10_20 2.31 2.92
Ambober Exclosure 16 20-30 1.31 1.59
Ambober Exclosure 16 30-50 1.05 1.37
Ambober Exclosure 17 0-10 3.29 3.57 10.59 340250 1384600
Ambober Exclosure 17 10_20 2.09 2.64
Ambober Exclosure 17 20-30 1.81 2.19
Ambober Exclosure 17 30-50 1.67 2.18
Ambober Exclosure 18 0-10 1.38 1.50 3.55 340250 1384650
Ambober Exclosure 18 10_20 0.93 1.17
Ambober Exclosure 18 20-30 0.73 0.88
Ambober Exclosure 18 30-50 0.00
Katassi Forest 1 0-10 15.90 11.90 20.49 251600 1217800
Katassi Forest 1 10_20 5.93 4.89
Katassi Forest 1 20-30 4.50 3.69
Katassi Forest 1 30-50 0.00
Katassi Forest 2 0-10 10.54 7.88 18.07 251900 1217500
Katassi Forest 2 10_20 5.50 4.53

209
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Katassi Forest 2 20-30 3.91 3.21
Katassi Forest 2 30-50 3.20 2.45
Katassi Forest 3 0-10 6.75 5.05 13.17 251900 1216900
Katassi Forest 3 10_20 4.43 3.66
Katassi Forest 3 20-30 3.14 2.58
Katassi Forest 3 30-50 2.46 1.88
Katassi Forest 4 0-10 11.32 8.47 29.48 251900 1218100
Katassi Forest 4 10_20 10.34 8.53
Katassi Forest 4 20-30 8.56 7.02
Katassi Forest 4 30-50 7.14 5.46
Katassi Forest 5 0-10 14.70 11.00 23.94 252200 1217200
Katassi Forest 5 10_20 7.70 6.35
Katassi Forest 5 20-30 4.52 3.71
Katassi Forest 5 30-50 3.77 2.88
Katassi Forest 6 0-10 9.07 6.79 19.04 252500 1218100
Katassi Forest 6 10_20 5.90 4.87
Katassi Forest 6 20-30 5.06 4.15
Katassi Forest 6 30-50 4.24 3.24
Katassi Forest 7 0-10 11.65 8.72 19.17 252500 1217500
Katassi Forest 7 10_20 5.29 4.37
Katassi Forest 7 20-30 4.20 3.44
Katassi Forest 7 30-50 3.45 2.64
Katassi Forest 8 0-10 10.69 8.00 18.84 252500 1216900
Katassi Forest 8 10_20 6.16 5.08
Katassi Forest 8 20-30 4.03 3.30
Katassi Forest 8 30-50 3.21 2.45
Katassi Forest 9 0-10 10.76 8.05 19.90 252800 1217200
Katassi Forest 9 10_20 6.64 5.48
Katassi Forest 9 20-30 4.57 3.75
Katassi Forest 9 30-50 3.42 2.61
Katassi Forest 10 0-10 13.45 10.07 23.22 253100 1217500
Katassi Forest 10 10_20 7.27 6.00
Katassi Forest 10 20-30 4.79 3.93
Katassi Forest 10 30-50 4.23 3.23
Katassi Forest 11 0-10 10.86 8.12 22.73 252800 1217800
Katassi Forest 11 10_20 7.41 6.12
Katassi Forest 11 20-30 5.59 4.59
Katassi Forest 11 30-50 5.11 3.90
Katassi Forest 12 0-10 12.59 9.42 22.91 253100 1218100
Katassi Forest 12 10_20 6.92 5.71
Katassi Forest 12 20-30 5.31 4.36
Katassi Forest 12 30-50 4.48 3.42
Katassi Forest 13 0-10 16.65 12.46 25.28 253400 1217800
Katassi Forest 13 10_20 6.99 5.77
Katassi Forest 13 20-30 4.88 4.00
Katassi Forest 13 30-50 4.00 3.06
Katassi Forest 14 0-10 10.61 7.94 22.05 253400 1217200
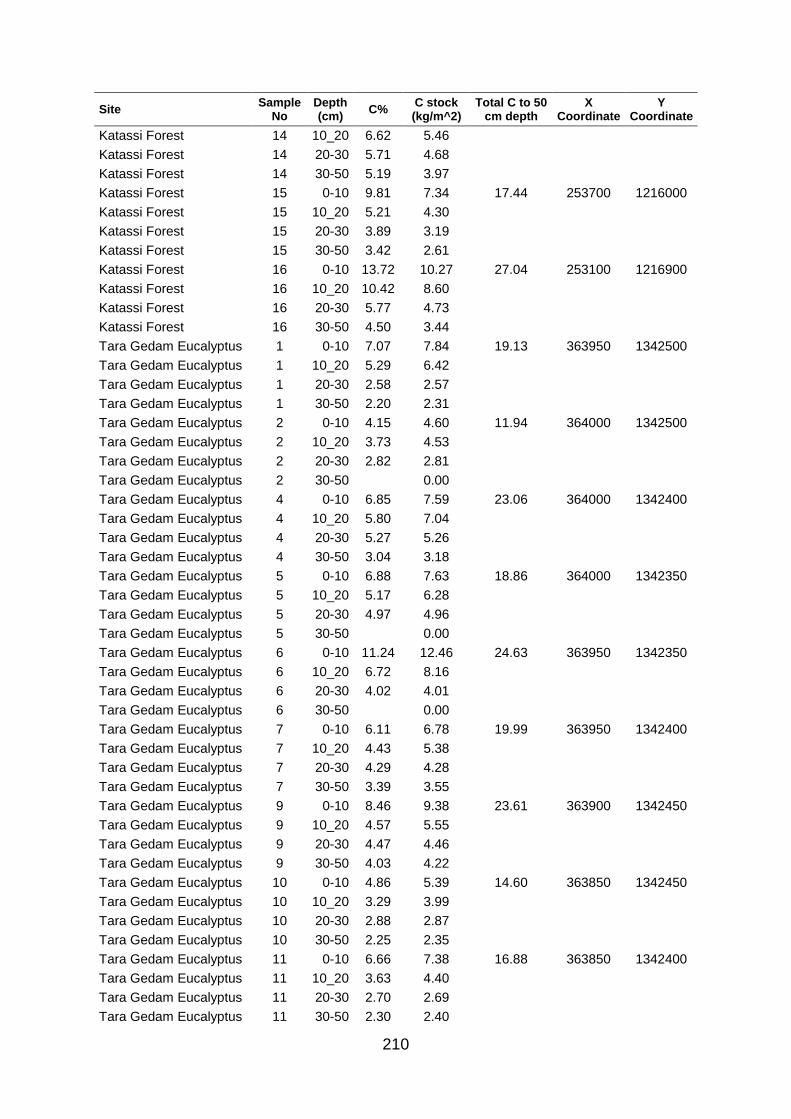
210
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Katassi Forest 14 10_20 6.62 5.46
Katassi Forest 14 20-30 5.71 4.68
Katassi Forest 14 30-50 5.19 3.97
Katassi Forest 15 0-10 9.81 7.34 17.44 253700 1216000
Katassi Forest 15 10_20 5.21 4.30
Katassi Forest 15 20-30 3.89 3.19
Katassi Forest 15 30-50 3.42 2.61
Katassi Forest 16 0-10 13.72 10.27 27.04 253100 1216900
Katassi Forest 16 10_20 10.42 8.60
Katassi Forest 16 20-30 5.77 4.73
Katassi Forest 16 30-50 4.50 3.44
Tara Gedam Eucalyptus 1 0-10 7.07 7.84 19.13 363950 1342500
Tara Gedam Eucalyptus 1 10_20 5.29 6.42
Tara Gedam Eucalyptus 1 20-30 2.58 2.57
Tara Gedam Eucalyptus 1 30-50 2.20 2.31
Tara Gedam Eucalyptus 2 0-10 4.15 4.60 11.94 364000 1342500
Tara Gedam Eucalyptus 2 10_20 3.73 4.53
Tara Gedam Eucalyptus 2 20-30 2.82 2.81
Tara Gedam Eucalyptus 2 30-50 0.00
Tara Gedam Eucalyptus 4 0-10 6.85 7.59 23.06 364000 1342400
Tara Gedam Eucalyptus 4 10_20 5.80 7.04
Tara Gedam Eucalyptus 4 20-30 5.27 5.26
Tara Gedam Eucalyptus 4 30-50 3.04 3.18
Tara Gedam Eucalyptus 5 0-10 6.88 7.63 18.86 364000 1342350
Tara Gedam Eucalyptus 5 10_20 5.17 6.28
Tara Gedam Eucalyptus 5 20-30 4.97 4.96
Tara Gedam Eucalyptus 5 30-50 0.00
Tara Gedam Eucalyptus 6 0-10 11.24 12.46 24.63 363950 1342350
Tara Gedam Eucalyptus 6 10_20 6.72 8.16
Tara Gedam Eucalyptus 6 20-30 4.02 4.01
Tara Gedam Eucalyptus 6 30-50 0.00
Tara Gedam Eucalyptus 7 0-10 6.11 6.78 19.99 363950 1342400
Tara Gedam Eucalyptus 7 10_20 4.43 5.38
Tara Gedam Eucalyptus 7 20-30 4.29 4.28
Tara Gedam Eucalyptus 7 30-50 3.39 3.55
Tara Gedam Eucalyptus 9 0-10 8.46 9.38 23.61 363900 1342450
Tara Gedam Eucalyptus 9 10_20 4.57 5.55
Tara Gedam Eucalyptus 9 20-30 4.47 4.46
Tara Gedam Eucalyptus 9 30-50 4.03 4.22
Tara Gedam Eucalyptus 10 0-10 4.86 5.39 14.60 363850 1342450
Tara Gedam Eucalyptus 10 10_20 3.29 3.99
Tara Gedam Eucalyptus 10 20-30 2.88 2.87
Tara Gedam Eucalyptus 10 30-50 2.25 2.35
Tara Gedam Eucalyptus 11 0-10 6.66 7.38 16.88 363850 1342400
Tara Gedam Eucalyptus 11 10_20 3.63 4.40
Tara Gedam Eucalyptus 11 20-30 2.70 2.69
Tara Gedam Eucalyptus 11 30-50 2.30 2.40
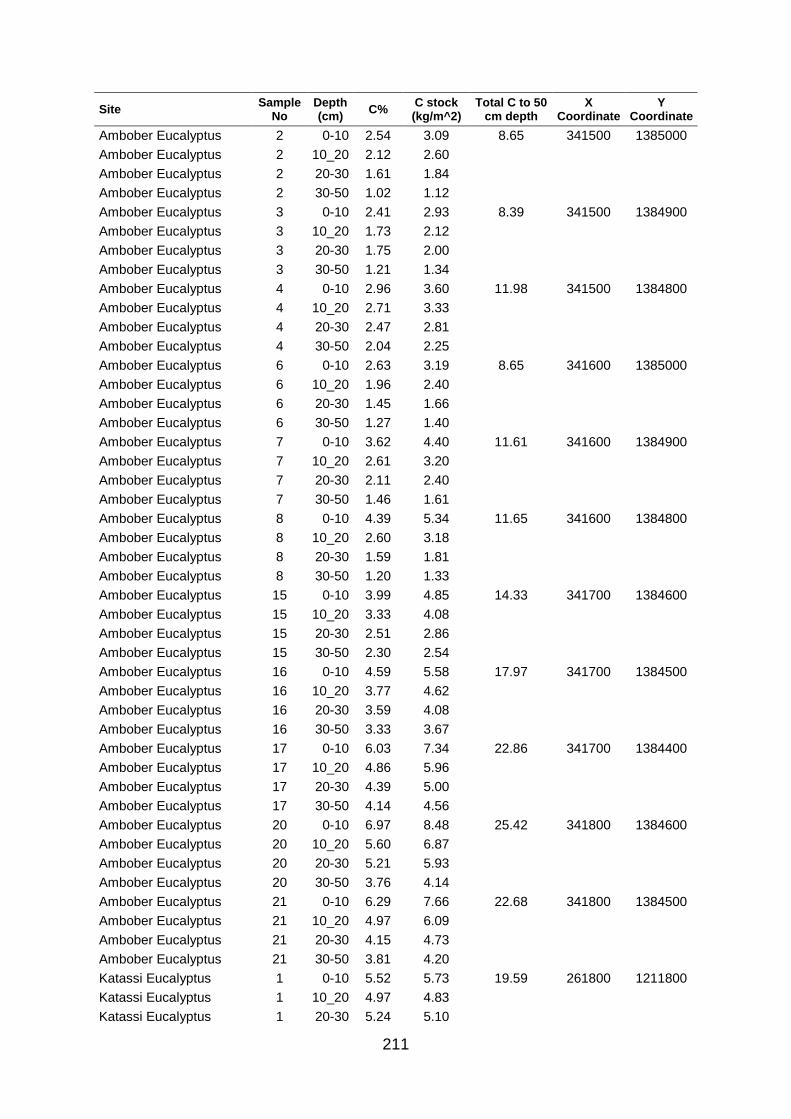
211
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Ambober Eucalyptus 2 0-10 2.54 3.09 8.65 341500 1385000
Ambober Eucalyptus 2 10_20 2.12 2.60
Ambober Eucalyptus 2 20-30 1.61 1.84
Ambober Eucalyptus 2 30-50 1.02 1.12
Ambober Eucalyptus 3 0-10 2.41 2.93 8.39 341500 1384900
Ambober Eucalyptus 3 10_20 1.73 2.12
Ambober Eucalyptus 3 20-30 1.75 2.00
Ambober Eucalyptus 3 30-50 1.21 1.34
Ambober Eucalyptus 4 0-10 2.96 3.60 11.98 341500 1384800
Ambober Eucalyptus 4 10_20 2.71 3.33
Ambober Eucalyptus 4 20-30 2.47 2.81
Ambober Eucalyptus 4 30-50 2.04 2.25
Ambober Eucalyptus 6 0-10 2.63 3.19 8.65 341600 1385000
Ambober Eucalyptus 6 10_20 1.96 2.40
Ambober Eucalyptus 6 20-30 1.45 1.66
Ambober Eucalyptus 6 30-50 1.27 1.40
Ambober Eucalyptus 7 0-10 3.62 4.40 11.61 341600 1384900
Ambober Eucalyptus 7 10_20 2.61 3.20
Ambober Eucalyptus 7 20-30 2.11 2.40
Ambober Eucalyptus 7 30-50 1.46 1.61
Ambober Eucalyptus 8 0-10 4.39 5.34 11.65 341600 1384800
Ambober Eucalyptus 8 10_20 2.60 3.18
Ambober Eucalyptus 8 20-30 1.59 1.81
Ambober Eucalyptus 8 30-50 1.20 1.33
Ambober Eucalyptus 15 0-10 3.99 4.85 14.33 341700 1384600
Ambober Eucalyptus 15 10_20 3.33 4.08
Ambober Eucalyptus 15 20-30 2.51 2.86
Ambober Eucalyptus 15 30-50 2.30 2.54
Ambober Eucalyptus 16 0-10 4.59 5.58 17.97 341700 1384500
Ambober Eucalyptus 16 10_20 3.77 4.62
Ambober Eucalyptus 16 20-30 3.59 4.08
Ambober Eucalyptus 16 30-50 3.33 3.67
Ambober Eucalyptus 17 0-10 6.03 7.34 22.86 341700 1384400
Ambober Eucalyptus 17 10_20 4.86 5.96
Ambober Eucalyptus 17 20-30 4.39 5.00
Ambober Eucalyptus 17 30-50 4.14 4.56
Ambober Eucalyptus 20 0-10 6.97 8.48 25.42 341800 1384600
Ambober Eucalyptus 20 10_20 5.60 6.87
Ambober Eucalyptus 20 20-30 5.21 5.93
Ambober Eucalyptus 20 30-50 3.76 4.14
Ambober Eucalyptus 21 0-10 6.29 7.66 22.68 341800 1384500
Ambober Eucalyptus 21 10_20 4.97 6.09
Ambober Eucalyptus 21 20-30 4.15 4.73
Ambober Eucalyptus 21 30-50 3.81 4.20
Katassi Eucalyptus 1 0-10 5.52 5.73 19.59 261800 1211800
Katassi Eucalyptus 1 10_20 4.97 4.83
Katassi Eucalyptus 1 20-30 5.24 5.10
212
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Katassi Eucalyptus 1 30-50 4.54 3.92
Katassi Eucalyptus 2 0-10 4.92 5.11 12.42 262200 1211800
Katassi Eucalyptus 2 10_20 3.35 3.26
Katassi Eucalyptus 2 20-30 2.74 2.67
Katassi Eucalyptus 2 30-50 1.60 1.38
Katassi Eucalyptus 3 0-10 3.11 3.23 10.84 262000 1211600
Katassi Eucalyptus 3 10_20 2.94 2.86
Katassi Eucalyptus 3 20-30 2.98 2.90
Katassi Eucalyptus 3 30-50 2.15 1.85
Katassi Eucalyptus 4 0-10 6.30 6.54 20.35 262400 1211600
Katassi Eucalyptus 4 10_20 5.47 5.32
Katassi Eucalyptus 4 20-30 4.87 4.75
Katassi Eucalyptus 4 30-50 4.34 3.74
Katassi Eucalyptus 5 0-10 6.58 6.83 16.08 261800 1211400
Katassi Eucalyptus 5 10_20 4.09 3.98
Katassi Eucalyptus 5 20-30 3.24 3.16
Katassi Eucalyptus 5 30-50 2.45 2.11
Katassi Eucalyptus 6 0-10 6.42 6.67 17.75 262200 1211400
Katassi Eucalyptus 6 10_20 4.92 4.78
Katassi Eucalyptus 6 20-30 4.18 4.07
Katassi Eucalyptus 6 30-50 2.59 2.24
Katassi Eucalyptus 7 0-10 9.95 10.33 31.11 262600 1211400
Katassi Eucalyptus 7 10_20 8.37 8.14
Katassi Eucalyptus 7 20-30 7.75 7.55
Katassi Eucalyptus 7 30-50 5.90 5.09
Katassi Eucalyptus 8 0-10 5.46 5.67 11.50 262000 1211200
Katassi Eucalyptus 8 10_20 2.91 2.83
Katassi Eucalyptus 8 20-30 1.94 1.89
Katassi Eucalyptus 8 30-50 1.28 1.11
Katassi Eucalyptus 9 0-10 8.32 8.64 26.42 262400 1211200
Katassi Eucalyptus 9 10_20 6.77 6.58
Katassi Eucalyptus 9 20-30 6.37 6.20
Katassi Eucalyptus 9 30-50 5.79 4.99
Katassi Eucalyptus 10 0-10 9.01 9.35 32.88 262600 1211200
Katassi Eucalyptus 10 10_20 8.64 8.40
Katassi Eucalyptus 10 20-30 8.40 8.18
Katassi Eucalyptus 10 30-50 8.05 6.94
Katassi Eucalyptus 11 0-10 5.76 5.98 14.64 262200 1211000
Katassi Eucalyptus 11 10_20 4.14 4.03
Katassi Eucalyptus 11 20-30 3.20 3.12
Katassi Eucalyptus 11 30-50 1.76 1.52
Katassi Eucalyptus 12 0-10 8.47 8.79 26.66 262600 1211000
Katassi Eucalyptus 12 10_20 6.15 5.98
Katassi Eucalyptus 12 20-30 6.50 6.34
Katassi Eucalyptus 12 30-50 6.44 5.55
Tara Gedam Forest 9 0-10 7.89 7.70 16.86 362600 1343800
Tara Gedam Forest 9 10_20 3.14 3.69
Tara Gedam Forest 9 20-30 2.90 3.64

213
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Tara Gedam Forest 9 30-50 1.86 1.83
Tara Gedam Forest 12 0-10 7.74 7.56 17.91 362900 1344100
Tara Gedam Forest 12 10_20 3.78 4.44
Tara Gedam Forest 12 20-30 3.03 3.81
Tara Gedam Forest 12 30-50 2.14 2.10
Tara Gedam Forest 16 0-10 8.91 8.70 16.34 363200 1343800
Tara Gedam Forest 16 10_20 3.03 3.55
Tara Gedam Forest 16 20-30 2.01 2.52
Tara Gedam Forest 16 30-50 1.59 1.56
Tara Gedam Forest 17 0-10 5.00 4.88 13.71 363200 1343500
Tara Gedam Forest 17 10_20 3.25 3.82
Tara Gedam Forest 17 20-30 2.22 2.79
Tara Gedam Forest 17 30-50 2.26 2.22
Tara Gedam Forest 18 0-10 10.17 9.94 18.95 363200 1343200
Tara Gedam Forest 18 10_20 3.25 3.82
Tara Gedam Forest 18 20-30 2.32 2.92
Tara Gedam Forest 18 30-50 2.32 2.28
Tara Gedam Forest 24 0-10 10.90 10.65 19.69 363800 1343200
Tara Gedam Forest 24 10_20 7.71 9.05
Tara Gedam Forest 24 20-30 0.00
Tara Gedam Forest 24 30-50 0.00
Tara Gedam Forest 25 0-10 9.66 9.44 22.39 363800 1342900
Tara Gedam Forest 25 10_20 5.01 5.88
Tara Gedam Forest 25 20-30 3.52 4.43
Tara Gedam Forest 25 30-50 2.69 2.64
Tara Gedam Forest 27 0-10 6.93 6.77 14.99 364100 1343500
Tara Gedam Forest 27 10_20 4.20 4.93
Tara Gedam Forest 27 20-30 2.61 3.29
Tara Gedam Forest 27 30-50 0.00
Tara Gedam Forest 29 0-10 14.10 13.78 45.00 364100 1342900
Tara Gedam Forest 29 10_20 12.33 14.47
Tara Gedam Forest 29 20-30 11.49 14.44
Tara Gedam Forest 29 30-50 2.36 2.31
Tara Gedam Forest 32 0-10 13.53 13.22 47.99 364400 1343200
Tara Gedam Forest 32 10_20 11.07 13.00
Tara Gedam Forest 32 20-30 10.05 12.63
Tara Gedam Forest 32 30-50 9.31 9.14
Tara Gedam Forest 36 0-10 12.45 12.16 46.23 364700 1343500
Tara Gedam Forest 36 10_20 10.51 12.33
Tara Gedam Forest 36 20-30 9.64 12.12
Tara Gedam Forest 36 30-50 9.80 9.62
Mahibere-Selasse Forest 1 0-10 1.96 2.93 3.58 239000 1381000
Mahibere-Selasse Forest 1 10_20 0.30 0.44
Mahibere-Selasse Forest 1 20-30 0.13 0.17
Mahibere-Selasse Forest 1 30-50 0.04 0.04
Mahibere-Selasse Forest 2 0-10 2.50 3.74 13.37 236000 1381000
Mahibere-Selasse Forest 2 10_20 2.68 3.89
Mahibere-Selasse Forest 2 20-30 2.53 3.21
Mahibere-Selasse Forest 2 30-50 2.45 2.53
Mahibere-Selasse Forest 3 0-10 0.85 1.27 3.31 233000 1381000
Mahibere-Selasse Forest 3 10_20 0.83 1.21
Mahibere-Selasse Forest 3 20-30 0.48 0.61
Mahibere-Selasse Forest 3 30-50 0.22 0.23

214
Site Sample
No Depth (cm)
C% C stock (kg/m^2)
Total C to 50 cm depth
X Coordinate
Y Coordinate
Mahibere-Selasse Forest 4 0-10 2.61 3.90 9.92 233000 1378000
Mahibere-Selasse Forest 4 10_20 1.80 2.61
Mahibere-Selasse Forest 4 20-30 1.71 2.16
Mahibere-Selasse Forest 4 30-50 1.21 1.25
Mahibere-Selasse Forest 5 0-10 2.31 3.45 12.16 230000 1381000
Mahibere-Selasse Forest 5 10_20 2.74 3.97
Mahibere-Selasse Forest 5 20-30 2.44 3.09
Mahibere-Selasse Forest 5 30-50 1.60 1.65
Mahibere-Selasse Forest 6 0-10 1.35 2.02 2.65 227000 1381000
Mahibere-Selasse Forest 6 10_20 0.35 0.51
Mahibere-Selasse Forest 6 20-30 0.05 0.07
Mahibere-Selasse Forest 6 30-50 0.05 0.05
Mahibere-Selasse Forest 7 0-10 1.11 1.67 4.99 224000 1381000
Mahibere-Selasse Forest 7 10_20 0.98 1.42
Mahibere-Selasse Forest 7 20-30 0.79 1.00
Mahibere-Selasse Forest 7 30-50 0.88 0.91
Mahibere-Selasse Forest 8 0-10 1.90 2.85 5.87 224000 1384000
Mahibere-Selasse Forest 8 10_20 1.27 1.85
Mahibere-Selasse Forest 8 20-30 0.55 0.69
Mahibere-Selasse Forest 8 30-50 0.46 0.48
Mahibere-Selasse Forest 9 0-10 2.22 3.32 9.75 227000 1384000
Mahibere-Selasse Forest 9 10_20 1.79 2.60
Mahibere-Selasse Forest 9 20-30 1.75 2.22
Mahibere-Selasse Forest 9 30-50 1.56 1.61
Mahibere-Selasse Forest 10 0-10 0.24 0.35 1.39 230000 1384000
Mahibere-Selasse Forest 10 10_20 0.20 0.29
Mahibere-Selasse Forest 10 20-30 0.24 0.30
Mahibere-Selasse Forest 10 30-50 0.44 0.45
Mahibere-Selasse Forest 11 0-10 0.49 0.74 2.78 233000 1384000
Mahibere-Selasse Forest 11 10_20 0.52 0.76
Mahibere-Selasse Forest 11 20-30 0.60 0.76
Mahibere-Selasse Forest 11 30-50 0.51 0.52
Mahibere-Selasse Forest 12 0-10 1.05 1.57 5.37 236000 1384000
Mahibere-Selasse Forest 12 10_20 0.73 1.06
Mahibere-Selasse Forest 12 20-30 1.17 1.48
Mahibere-Selasse Forest 12 30-50 1.21 1.25
Mahibere-Selasse Forest 13 0-10 0.97 1.45 5.22 236000 1387000
Mahibere-Selasse Forest 13 10_20 1.01 1.47
Mahibere-Selasse Forest 13 20-30 0.95 1.20
Mahibere-Selasse Forest 13 30-50 1.07 1.10
Mahibere-Selasse Forest 14 0-10 2.01 3.01 9.02 233000 1387000
Mahibere-Selasse Forest 14 10_20 2.14 3.10
Mahibere-Selasse Forest 14 20-30 1.22 1.55
Mahibere-Selasse Forest 14 30-50 1.32 1.37
Mahibere-Selasse Forest 15 0-10 0.49 0.73 1.70 230000 1387000
Mahibere-Selasse Forest 15 10_20 0.32 0.46
Mahibere-Selasse Forest 15 20-30 0.25 0.32
Mahibere-Selasse Forest 15 30-50 0.18 0.19
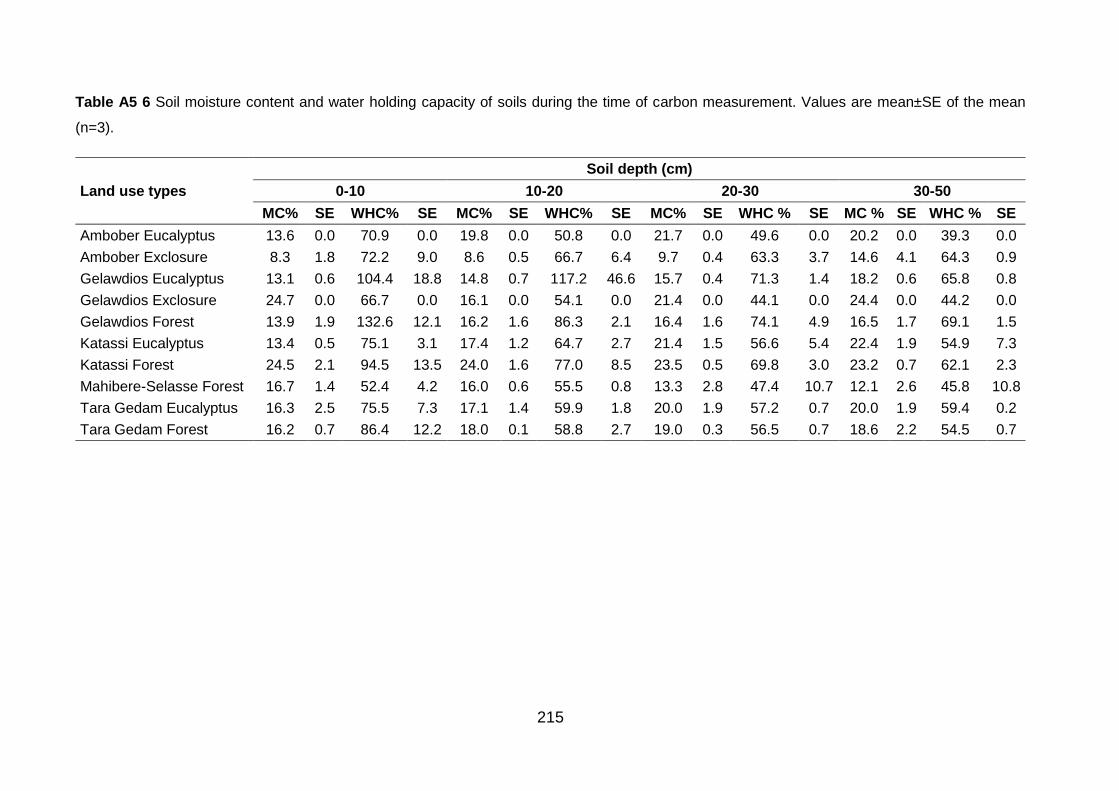
215
Table A5 6 Soil moisture content and water holding capacity of soils during the time of carbon measurement. Values are mean±SE of the mean
(n=3).
Land use types
Soil depth (cm)
0-10 10-20 20-30 30-50
MC% SE WHC% SE MC% SE WHC% SE MC% SE WHC % SE MC % SE WHC % SE
Ambober Eucalyptus 13.6 0.0 70.9 0.0 19.8 0.0 50.8 0.0 21.7 0.0 49.6 0.0 20.2 0.0 39.3 0.0
Ambober Exclosure 8.3 1.8 72.2 9.0 8.6 0.5 66.7 6.4 9.7 0.4 63.3 3.7 14.6 4.1 64.3 0.9
Gelawdios Eucalyptus 13.1 0.6 104.4 18.8 14.8 0.7 117.2 46.6 15.7 0.4 71.3 1.4 18.2 0.6 65.8 0.8
Gelawdios Exclosure 24.7 0.0 66.7 0.0 16.1 0.0 54.1 0.0 21.4 0.0 44.1 0.0 24.4 0.0 44.2 0.0
Gelawdios Forest 13.9 1.9 132.6 12.1 16.2 1.6 86.3 2.1 16.4 1.6 74.1 4.9 16.5 1.7 69.1 1.5
Katassi Eucalyptus 13.4 0.5 75.1 3.1 17.4 1.2 64.7 2.7 21.4 1.5 56.6 5.4 22.4 1.9 54.9 7.3
Katassi Forest 24.5 2.1 94.5 13.5 24.0 1.6 77.0 8.5 23.5 0.5 69.8 3.0 23.2 0.7 62.1 2.3
Mahibere-Selasse Forest 16.7 1.4 52.4 4.2 16.0 0.6 55.5 0.8 13.3 2.8 47.4 10.7 12.1 2.6 45.8 10.8
Tara Gedam Eucalyptus 16.3 2.5 75.5 7.3 17.1 1.4 59.9 1.8 20.0 1.9 57.2 0.7 20.0 1.9 59.4 0.2
Tara Gedam Forest 16.2 0.7 86.4 12.2 18.0 0.1 58.8 2.7 19.0 0.3 56.5 0.7 18.6 2.2 54.5 0.7




















