
Caterpillar D8R Dozer Engine Oil Filtration Installation ...
Upload
khangminh22Category
view
4download
0
Cardiovascular Research, 1979, 13, 547-561
REVIEW
Capillary filtration coefficient: the technique and its application to the small intestine P . D . I . R I C H A R D S O N , ' D . N . G R A N G E R ' , A N D A . E . T A Y L O R
From the Department of' Physiology, College of' Medicine, The University of' South Alabama, Mobile, Alabama
SUMMARY In this review, we discuss the physiological basis of the capillary filtration coefficient (Kf,,), and its measurement in the small intestine. First the various techniques by which Kf,, can be measured are discussed and assessed; the second part of the review considers the volumetric method of determining Kf,, in more detail, in an attempt to identify the causes of variability in published values for intestinal Kf,,. Finally, we discuss changes in intestinal Kf,, due to sym- pathetic nerve stimulation and the administration of drugs and hormones.
The ma,jority of estimates of intestinal Kf., are about 0.38 t o 0.60 (innervated) to 0.60 to 0.75 (denervated) cm"min-l.kPa-l.lOOg-l. These values are increased by most vasodilator drugs and hormones, and decreased by sympathetic nerve stimulation and by vasoconstrictor drugs.
The capillary filtration coefficient (Kf,,, CFC, Lps) has often been used as an index of the functional ex- change capacity of an organ (Landis and Gibbon, 1933; Pappenheimer and Soto-Rivera, 1948; Folkow et al., 1963). More precisely, when measured by the gravimetric technique, Kf,, is a measure of the hydraulic conductance, ie, it relates net fluid filtra- tion (or absorption) to a pressure gradient across a membrane. Kf,, is influenced by several factors; for example, the size and number of capillary pores, and the number of filtering capillaries. These factors in- variably account for the wide range of Kf,, values reported for different tissues. Values of the order of 0.1 13 (Pappenheimer and Soto-Rivera, 1948; Chen et a/., 1976), 1.50 (Drake et al., 1978; Staub, 19741, 0.22 to 1.13 (Folkow et al., 1963; Johnson and Hanson, 1966; Richardson, 1974), 2.26 (Greenway and Lautt, 1970) and 4.96 cms.rnin-l.kPa~'.100g', (Davies er a/., 1974) have been reported for skeletal muscle, lung, intestine, liver, and spleen, respectively. Several investigators have interpreted the large variation in Kf,, between tissues to be a result of comparable differences in capillary density. Al-
'Present address: Department of Physiology. The Medical College of St. Bartholomew's Hospital, Chaterhouse Square, London ECI M 684 . 'Address all correspondence to: Dr D. N. Granger, Depart- ment of Physiology, College of Medicine, University of South Alabama, Mobile, Alabama 36688, USA.
though the available histological data suggests that capillary density varies widely between tissues (eg a tenfold difference in capillary density exists between muscle and intestine: Casley-Smith et al., 1975), regional differences in capillary permeability must also be considered a potentially major determinant of the variation in Kf,, for different tissues.
As an indicator of capillary exchange capacity, the capillary filtration coefficient has been employed in studies of capillary fluid exchange (Folkow et al., 1963; Johnson and Hanson, 1966; Granger et al., 1979c) and oxygen delivery (Granger et al., 1976) in various organs, including the small intestine. In transporting organs, such as the small intestine, the importance of the capillary filtration coefficient lies not only in capillary filtration and oxygen delivery, but also in the removal of newly-absorbed fluid from the mucosal interstitium into the capillaries (Granger and Taylor, 1978).
In the first part of this review, we discuss the various techniques which have been used to measure the capillary filtration coefficient. The second part considers the problems which have given rise to variations of as much as fivefold in control K,,;s reported under very similar conditions for the feline small intestine, and the third part sum- marises the changes in intestinal capillary filtration coefficient due to physiological and pharmacological stimuli.
547
by guest on February 20, 2016
Dow
nloaded from
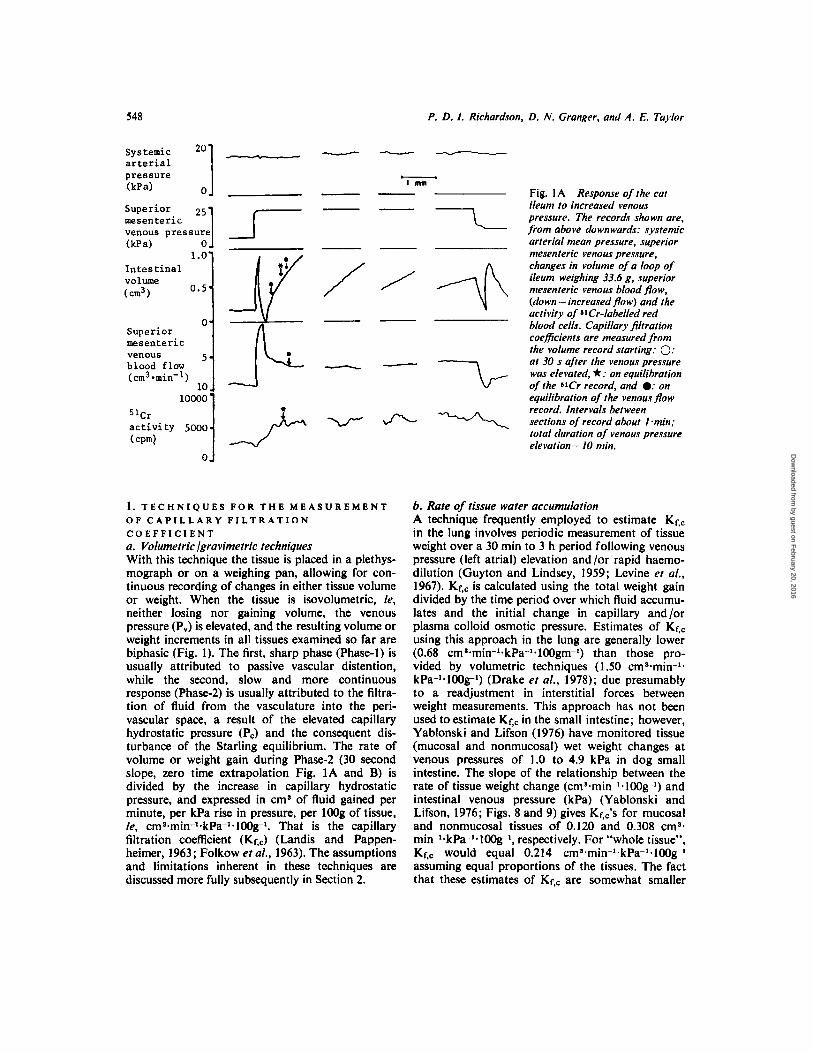
S48 P. D. I . Richardson, D. N. Granger, and A. E. Taylor
a r t e r i a l p ressure Systemic (kP4 201 0 - Super ior 25 mesenter ic venous pressuri (kP4 0
1.0 I n t e s t i n a l volume ( cm3 ) 0 . 5
0 Super ior mesenter ic venous blood flow ( 0 3 mmin-1)
5
10 10000
5 1 ~ r a c t i v i t y 5000 ( CP4
0
- I min
- - - A
1. T E C H N I Q U E S F O R T H E M E A S U R E M E N T O F C A P I L L A R Y F I L T R A T I O N C O E F F I C I E N T a . Volumetric lgravimetric techniques With this technique the tissue is placed in a plethys- mograph or on a weighing pan, allowing for con- tinuous recording of changes in either tissue volume or weight. When the tissue is isovolumetric, ie, neither losing nor gaining volume, the venous pressure (P,) is elevated, and the resulting volume or weight increments in all tissues examined so far are biphasic (Fig. 1). The first, sharp phase (Phase-1) is usually attributed to passive vascular distention, while the second, slow and more continuous response (Phase-2) is usually attributed to the filtra- tion of fluid from the vasculature into the peri- vascular space, a result of the elevated capillary hydrostatic pressure (Pc) and the consequent dis- turbance of the Starling equilibrium. The rate of volume or weight gain during Phase-2 (30 second slope, zero time extrapolation Fig. 1A and B) is divided by the increase in capillary hydrostatic pressure, and expressed in cm’ of fluid gained per minute, per kPa rise in pressure, per l00g of tissue, ie, cmS.min-l.kPa-l.100g1. That is the capillary filtration coefficient (KrJ (Landis and Pappen- heimer, 1963; Folkow et al., 1963). The assumptions and limitations inherent in these techniques are discussed more fully subsequently in Section 2.
G-
Fig. 1 A Response of the cat ileum to increased venous pressure. The records shown are, from above downwards: systemic arterial mean pressure, superior mesenteric venous pressure, changes in volume of a loop of ileum weighing 33.6 g, superior mesenteric venous blood pow, (down = increased flow) and the activity of 81Cr-labelled red blood cells. Capillary filtration coefficients are measured from the volume record starting: 0: at 30 s afier the venous pressure was elevated, *: on equilibration of the W r record, and 0: on equilibration of the venous flow record. Intervals between sections of record about Imin: total duration of venous pressure elevation= I0 min.
b. Rate of tissue water accumulation A technique frequently employed to estimate Kf,, in the lung involves periodic measurement of tissue weight over a 30 min to 3 h period following venous pressure (left atrial) elevation and /or rapid haemo- dilution (Guyton and Lindsey, 1959; Levine et al., 1967). Kr,c is calculated using the total weight gain divided by the time period over which fluid accumu- lates and the initial change in capillary and/or plasma colloid osmotic pressure. Estimates of Kf,c using this approach in the lung are generally lower (0.68 cma.min-l.kPa-l.lOOgm-l) than those pro- vided by volumetric techniques (1 30 cms.min-l. kPa-l.100g-l) (Drake et al., 1978); due presumably to a readjustment in interstitial forces between weight measurements. This approach has not been used to estimate Kf,c in the small intestine; however, Yablonski and Lifson (1976) have monitored tissue (mucosal and nonmucosal) wet weight changes at venous pressures of 1.0 to 4.9 kPa in dog small intestine. The slope of the relationship between the rate of tissue weight change (cma-min-l.lOOgl) and intestinal venous pressure (kPa) (Yablonski and Lifson, 1976; Figs. 8 and 9) gives Kf,;s for mucosal and nonmucosal tissues of 0.120 and 0.308 ems* rnin-l-kPa-l. 100gl, respectively. For “whole tissue”, Kf,, would equal 0.214 cms.min-l.kPa-l.lOOgl assuming equal proportions of the tissues. The fact that these estimates of Kf,c are somewhat smaller
by guest on February 20, 2016
Dow
nloaded from
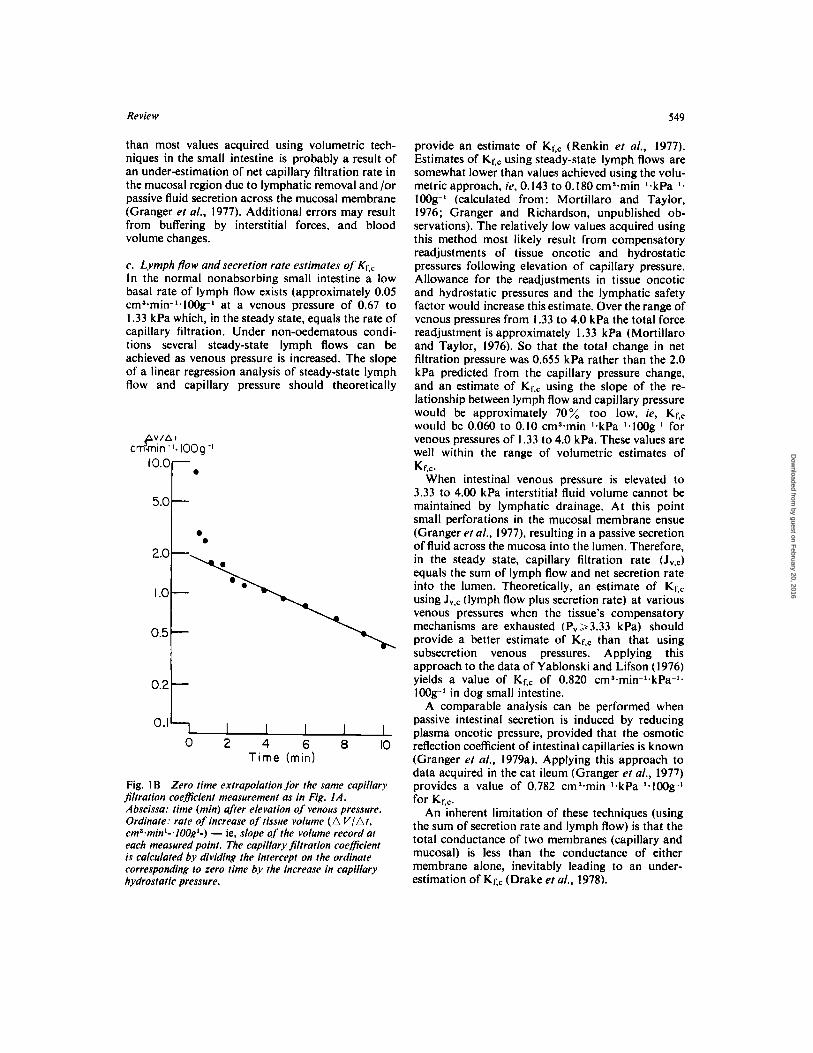
Review 549
than most values acquired using volumetric tech- niques in the small intestine is probably a result of an under-estimation of net capillary filtration rate in the mucosal region due to lymphatic removal and/or passive fluid secretion across the mucosal membrane (Granger et al., 1977). Additional errors may result from buffering by interstitial forces, and blood volume changes.
c . Lymph flow and secretion rate estimates of Kf., In the normal nonabsorbing small intestine a low basal rate of lymph flow exists (approximately 0.05 ~ m ~ * m i n - ~ ~ 1 0 0 g ~ at a venous pressure of 0.67 to 1.33 kPa which, in the steady state, equals the rate of capillary filtration. Under non-oedematous condi- tions several steady-state lymph flows can be achieved as venous pressure is increased. The slope of a linear regression analysis of steady-state lymph flow and capillary pressure should theoretically
5-0t I ..
0 2 4 6 8 1 0 T ime (min)
Fig. 1 B Zero time extrapolation for the same capillary jltration coefficient measurement as in Fig. I A, Abscissa: time (min) aJier elevation of venous pressure. Ordinate: rate of increase of tissue volume ( A VlAt , cma~minl-.lOOgl-) - ie, slope of the volume record at each measured point. The capillary filtration coefficient is calculated by dividing the intercept on the ordinate corresponding to zero time b.v the increase in capillary hydrostatic pressure.
provide an estimate of Kf,, (Renkin et al., 1977). Estimates of Kf,, using steady-state lymph flows are somewhat lower than values achieved using the volu- metric approach, ie, 0.143 to 0.180 ~ m ~ . m i n - ~ - k P a - ~ . 100g-’ (calculated from: Mortillaro and Taylor, 1976; Granger and Richardson, unpublished ob- servations). The relatively low values acquired using this method most likely result from compensatory readjustments of tissue oncotic and hydrostatic pressures following elevation of capillary pressure. Allowance for the readjustments in tissue oncotic and hydrostatic pressures and the lymphatic safety factor would increase this estimate. Over the range of venous pressures from 1.33 to 4.0 kPa the total force readjustment is approximately I .33 kPa (Mortillaro and Taylor, 1976). So that the total change in net filtration pressure was 0.655 kPa rather than the 2.0 kPa predicted from the capillary pressure change, and an estimate of Kec using the slope of the re- lationship between lymph flow and capillary pressure would be approximately 70% too low. ie, Kf,, would be 0.060 to 0.10 ~m~.min-~ .kPa-~ .100g-~ for venous pressures of 1.33 to 4.0 kPa. These values are well within the range of volumetric estimates of
When intestinal venous pressure is elevated to 3.33 to 4.00 kPa interstitial fluid volume cannot be maintained by lymphatic drainage. At this point small perforations in the mucosal membrane ensue (Granger et al., 1977), resulting in a passive secretion of fluid across the mucosa into the lumen. Therefore, in the steady state, capillary filtration rate (J,.,) equals the sum of lymph flow and net secretion rate into the lumen. Theoretically, an estimate of Kt,, using JV,, (lymph flow plus secretion rate) at various venous pressures when the tissue’s compensatory mechanisms are exhausted (P, ,3.33 kPa) should provide a better estimate of Kf,, than that using subsecretion venous pressures. Applying this approach to the data of Yablonski and Lifson (1976) yields a value of Kf,, of 0.820 ~ m ~ . m i n - ~ . k P a - ~ . I00gl in dog small intestine.
A comparable analysis can be performed when passive intestinal secretion is induced by reducing plasma oncotic pressure, provided that the osmotic reflection coefficient of intestinal capillaries is known (Granger et al., 1979a). Applying this approach to data acquired in the cat ileum (Granger et al., 1977) provides a value of 0.782 ~m~.min-~ .kPa-~ .100g-~ for Kf,,.
An inherent limitation of these techniques (using the sum of secretion rate and lymph flow) is that the total conductance of two membranes (capillary and mucosal) is less than the conductance of either membrane alone, inevitably leading to an under- estimation of KzC (Drake et al., 1978).
Kf,c.
by guest on February 20, 2016
Dow
nloaded from

550 P. D. I . Richardson, D. N . Granger, and A. E. Taylor
d. Indicator dilution methods These techniques rely upon the measurement of arteriovenous differences in the concentration of blood-borne indicators. Elevation of the venous pressure causes transcapillary filtration of protein- deficient fluid from the intravascular to perivascular compartments. Consequently, the venous blood concentration of plasma proteins and red blood cells, and the venous plasma colloid osmotic pressure ex- ceed the corresponding arterial values (Johns and Rothe, 1978). From these arteriovenous differences, and a knowledge of total blood flow, net trans- capillary fluid movement and the Kf,, may be esti- mated. Johns and Rothe (1978) calculated Kr,, values for the denervated dog intestine in the range 0.150 to 0.263 cms.min-l.kPa-l.lOOg-l. The use of multiple tracers may help to separate vascular com- pliance changes from net transcapillary fluid move- ment, and obviates the need for weight or volume measurements, which may be an advantage under some experimental conditions. The contention (Rothe et al., 1978) that gravimetric and plethysmo- graphic studies are inaccurate because of tissue leaks and instrument “drift” has not received experimental support, and as such does not constitute strong grounds for the use of the indicator dilution tech- niques. Furthermore, the indicator approach re- quires perfusion of the tissue at constant inflow, with widely varying arterial pressures if drugs are ad- ministered (3.5 to 27 kPa: Rothe et al., 1978). Moreover, the discontinuous measurements (eg of haematocrit) make the assessment of the time- course of vascular events very difficult, as may the sampling of blood from sites distant from the micro- vasculature where filtration occurs. A further difficulty is that one does not know whether the tissue is filtering under control conditions, since the technique gives no continuous information regarding volume or weight changes. I n addition, it is not possible to make any correction for the tissue’s own compensation for the alteration in forces resulting from venous pressure elevation.
e. Micropuncture techniques The hydraulic conductivity of single capillaries can be determined using the relative migration distance of red blood cells in an occluded capillary. Gore et a/ . (1976) have measured the filtration coefficient of single capillaries in rat intestinal muscle using the micro-occlusion technique. Their data suggest a re- markably high axial gradient in capillary filtration coefficient, ie, Kr,, increases by more than a factor of 10 between the arterial and venous ends of the capillary. The average Kf,, acquired by Gore et a/ . (1976) for all vessels was 0.51 pmS*pm-2.s-1*kPa-1. Using the capillary surface area estimate of Casley-
Smith et a / . (1975) for cat muscularis (4940 cm?. 100gl tissue) a value of approximately 15.0 ems. min-l*kPa-l.lOOgl would be predicted for intestinal smooth muscle. This would represent a maximal possible value since inherent in the calculation is the assumption that all capillaries in the tissue are filtering (or absorbing). The Kr,:s calculated from Yablonski and Lifson’s (1976) data (see Section Ib) suggest that the Kf,, in intestinal smooth muscle is 3 to 4 times larger than literature values for skeletal muscle. The Kf,, of the visceral smooth muscle layer is larger than one would predict from skeletal muscle values (0.1 13 ~ m ~ . m i n - ~ . k P a - ~ . 1 0 0 g ~ ) and thus may contribute more to Kr,, determination in total organs than has normally been assumed.
The micro-occlusion technique offers the unique advantage of allowing for discrimination between permeability and surface area effects on volumetric estimates of Kf,, as the measurements are made on single capillaries but this technique has not as yet been applied to mucosal capillaries of the small intestine.
J Starling forces balance Another approach used to estimate Kr,, is to measure all of the filtration forces in the Starling equilibrium equation and total organ lymph flow in the steady-state, and from these to calculate Kf,,. This approach was applied to a mesentry-intestine preparation by Hargens and Zweifach (1976) and yielded a mean value for Kf,, in the cat mesentry of 0.014 cms.min~l.kPa-l.lOOg~l, a value much lower than that reported for the cat intestine using other approaches. Erdmann et al. (1975) used a similar approach to estimate Kf,, of the lung in an un- anaesthetised sheep preparation and acquired values which are approximately one-tenth the value acquired using gravimetric techniques in isolated and intact dog lungs (Drake et al., 1978). Difficulties in the use of this technique include: (i) the assump- tion that the lymph flow ( J v , ~ ) measurements re- present total lymphatic drainage from the prepara- tion; (ii) the assumptions that tissue hydrostatic pressure = lymphatic hydrostatic pressure, = 0; and (iii) the assumption that tissue oncotic pressure = lymph oncotic pressure. Perhaps more important, as discussed by Hargens and Zweifach (1976), is the fact that some measurements were made in the mesentery (PI, P,) and some in the intestine ( K ~ , J ~ . L ) .
This approach has the advantage that it is possible to calculate Kr,, values with measurements obtained using relatively non-invasive techniques; however, there are major difficulties in making all of the necessary measurements in the intestine, a fact borne out by the calculation of a negative value for
by guest on February 20, 2016
Dow
nloaded from

Review 5 5 1
Krqc using this technique (Hargens and Zweifach, 1976) in the mesentery-intestine preparation.
Since the most widely used techniques for esti- mating Kf,c in the small intestine are the volu- metric/gravimetric methods, the next section of this Review deals with the applications and limitations of these approaches.
2. V O L U M E T R I C M E A S U R E M E N T S O F C A P I L L A R Y F I L T R A T I O N C O E F F I C I E N T The volumetric methods of determining capillary filtration coefficient suffer from two fundamental problems (Folkow and Mellander, 1970): first, there is difficulty in assessing the end-point of the passive increase in vascular volume (Phase-I of the volume response) which is due to vascular distention, as a consequence of venous pressure elevation, and second, that on elevation of the venous pressure, tissue forces continuously readjust to compensate for transcapillary filtration. The vascular (venous) dis- tention and the readjustment of tissue forces have markedly different time-courses : the former occurs within seconds and the latter within 2 to 3 min of venous pressure elevation (Granger, 1977). Ideally, therefore, the rate of increase in tissue volume used to estimate the capillary filtration coefficient should lie between the end of the vascular distention and the onset of the tissue forces readjustment. a. Blood volume shift Wallentin (1966). in a study using radioactively- labelled red blood cells, showed that the bulk of the blood volume shift on elevating the superior mesenteric venous pressure of the cat by 2.0 kPa is complete within about 30 s; this indicates that measuring the rate of volume increase from a point 30 s after the venous pressure elevation gives an acceptable basis for capillary filtration coefficient measurements. These conclusions are supported by the data of Johnson and Hanson (1966) which show a separation of the Phase-I and Phase-2 responses in dog intestine at a point well under 30 s using a semi- logarithmic plot of the change in intestinal weight against time following P, elevation from 0 to 1.33 kPa. In contrast, studies in cat ileum (Granger et al., 1979~) suggest a separation in the two phases at approximately 60 s when the rate of ileal volume change is plotted semi-logarithmically against time.
An alternative approach to the use of radio- actively-labelled red blood cells in assessing the end of the blood volume shift has been described by Granger et a / . (1979b): on elevation of the superior mesenteric venous pressure, venous outflow from the intestine falls initially to a low value and sub- sequently attains a plateau lower than the control level (see Fig. IA). The point of attainment of this plateau (“flow equilibration”) represents the end of
the blood volume shift, and measuring the rate of in- testinal volume increase from that point gives Kf., values in agreement with those determined from the end of the blood volume shift assessed using radio- actively-labelled red blood cells, or Kr,, valves obtained by the zero time extrapolation technique. Granger et al. (1979b) showed a clear positive cor- relation between the extent of venous pressure ele- vation and the time taken for Phase-I to end; how- ever, there was sufficiently wide variation between experiments to make prediction of a fixed time for the end of Phase-I liable to considerable error. The time required for Phase-I to end should theoretically be influenced by alterations in vascular tone due to drug infusions provided vascular compliance is sufficiently altered. Johns and Rothe (1978) have shown that noradrenaline decreases while papa- verine increases intestinal vascular compliance. The data of Granger et a/. (1979~) suggest that local intra-arterial infusions of adenosine nearly double the time at which there is separation of the Phase-I and Phase-2 components of ileal volume change following venous pressure elevation, presumably due to an adenosine-induced increase in vascular com- pliance. A priori, one would expect that the degree of smearing (ie, of stress-relaxation into Phase-2) would be dependent upon vascular compliance, especially of the veins and venules. Theoretically, a greater degree of “smearing” of the stress-relaxation response to P, elevation into the Phase-2 slope should occur at higher vascular compliances com- pared with lower vascular compliances. If the Phase-2 slope is measured at a fixed time after the P, elevation (eg between 30 and 60 s), this “smearing” would produce an erroneously high “Kf.,” value, and the extent of the error would be directly related to the vascular compliance.
Upon elevation of venous pressure, vascular volume increases in two distinct phases: an initial rapid change due to passive distention of the elastic vessels and a slower more prolonged increase which is considered to be the time-dependent delayed com- pliance (visco-elastic creep). The initial rapid phase is considered to represent the Phase-I tissue volume increase following P, elevation while the delayed compliance vascular phase is volumetrically in- separable from the filtration portion of Phase-2. Johns and Rothe (1978) recently demonstrated a time-dependent delayed compliance in the vascu- lature of the dog small intestine of 12.1 cm3.kg1 tissue weight for a venous pressure change of 2.7 kPa over a period from 2 to7 min after the P, eleva- tion: such a delayed compliance, of 0.090 cm3.rnin-’. kPa-l*IOOg-’ would result in an error in a volumetric determination of Kf,, of about 10% with a Kt;, of 0.752cm .rnin-’ .k Pa-’ ‘ 1 OOg’ .
by guest on February 20, 2016
Dow
nloaded from

552 P. D . I . Richardson, D. N . Granger, and A . E. Taylor
b. Readjustment of tissue forces Fluid movement across intestinal capillaries can be described by the modified Starling (1 896) equation:
J,, = Kf,, x NFP NFP = (Pc - Pt) - ~ p ( n p - nt)
where J , , NFP = net filtration pressure Pc = capillary hydrostatic pressure Pt = tissue fluid pressure q, = osmotic reflection coefficient nP = plasma oncotic pressure nt = interstitial oncotic pressure I n an isovolumetric preparation, with intact
lymphatic drainage, it can be assumed that capillary filtration is in a steady steady and equals the lym- phatic drainage (J,.L). If P, is suddenly elevated by increasing venous pressure, there is an instantaneous increase in NFP equal to the change in capillary hydrostatic pressure. As fluid enters the inter- stitium, interstitial volume increases, resulting in an increase in Pt and J,,L while nt decreases (Johnson and Richardson, 1974; Yablonski and Lifson, 1976; Mortillaro and Taylor, 1976). In addition, the move- ment of fluid from vascular to interstitial compart- ments results in an increase in np. These changes in tissue forces tend to decrease the NFP until inter- stitial volume achieves a new steady state, at which time the intestine once again becomes isovolume- tric. The timecourse of these changes is dependent on the increase in venous pressure; usually, about 10 min is required for the intestine to attain a new iso- volumetric state with venous pressure increments of between 0.66 and 2.66 kPa. The rate of volume in- crease used to calculate the Kf,, will therefore decline continuously due to the readjustment of tissue forces, as the net filtration pressure decreases during the period of venous pressure elevation. An estimate of K,,c from a rate of volume increase at a time where appreciable tissue force readjustments had taken place would therefore represent a considerable underestimate of the true K,,,. Such readjustments of tissue forces with progressive falls in NFP in all probability explain the apparent fall in Kf,, values for the dog intestine reported by Johns and Rothe (1978) to occur between 2 and 12 min after P, elevation.
All of the estimates of tissue forces in the small intestine at various venous pressures were performed under steady state conditions, while the time course of tissue force changes subsequent to venous pressure elevation has received far less attention. Transient analysis of intestinal lymph flow and lymph oncotic pressure subsequent to venous pressure elevation suggests that 2 to 3 min are re- quired for a significant reduction in NFP due to
net fluid movement across the capillaries
lymph flow and tissue oncotic pressure (Granger et a/., 1977).
Considering the time course within which these forces readjust, the best estimate of the true Kf,, should be acquired using the rate of tissue volume change within 2 min of the increase in P,. It is im- portant to note that the time course of readjustment of tissue forces will vary depending upon the hydra- tion state of the preparation and the true value of Kf,,. Overhydrated (oedematous) preparations are characterised by minimal alterations in tissue forces subsequent to P, elevation while dehydrated pre- parations allow for greater and more rapid re- adjustments in tissue forces. Tissue force changes in normally hydrated preparations with larger capillary hydraulic conductances will occur more rapidly than in preparations or organs in which the hydraulic conductance of the capillaries is small (Granger et a/., 1978).
The zero time extrapolation (ZTE) technique was developed to obviate the influence of such tissue force readjustments on calculations of Kr,,. When the rate of tissue volume change subsequent to P, elevation is plotted semilogarithmically against time, two distinct phases of the volume change become apparent: an initial rapid component followed by a slower more prolonged component which is assumed to represent filtration (Fig. IB). By extrapolating the slow (,filtration) component, which remains linear for 8 to 10 min, to zero time, one should acquire a value which theoretically represents Kr,, before re- adjustments in tissue forces have occurred (Kedem and Katchalsky, 1958; Drake et a/., 1978). This method has been used for estimating Kf., in the isolated perfused lung (Drake et al., 1978) and more recently in the cat ileum (Granger et a/., 1979b,c).
Granger et a/. (1979b) have shown that values for Kf,, obtained using the zero time extrapolation technique in the cat ileum were not significantly different from the values obtained by measuring the rate of volume increase in response to P, elevations of 1.33 kPa or less: ( i ) at 3 0 s after the P, elevation; (ii) at the point where the accumulation of radio- actively-labelled red blood cells was complete; and (iii) where the venous blood flow record attained a plateau level during the period of P, elevation. For P, elevations > 1.33 kPa, the 3 0 s values were signi- ficantly greater than the RBC equilibration, flow equilibration, and ZTE estimates. Since the ZTE technique should eliminate the influence of tissue force readjustments on the measured Kf,, these ob- servations suggest that at the points of red cell and venous blood flow “equilibration”, tissue forces had not readjusted sufficiently to influence the measured Kf,, to a significant extent for venous pressure in- crements of, or less than 1.33 kPa.
by guest on February 20, 2016
Dow
nloaded from

Review 553
c. The nature of material accumulated during venous pressure elevation The studies in which radioactively-labelled red blood cells (RBC’s) have been used have shown that al- though RBC‘s do accumulate during the Phase-l volume response to P, elevation, any such accumu- lation during the Phase-2 response is unlikely to be significant (Wallentin, 1966; Granger et al., 1979b).
Furthermore, Johnson and Hanson (1 962) by measuring plasma protein concentrations, showed that the volume and weight gain of the Phase-2 response to P, elevation was due to the accumulation of material filtered across the exchange vessels to form a temporary oedema. Although these tech- niques have provided clear evidence that during the Phase-2 volume response to P, elevation, there is accumulation of cell-free fluid within the tissues, they are insufficiently discriminating to eliminate the possibility of a contribution to the Phase-2 volume response from continuing stress-relaxation or visco- elastic creep of the capacitance vessels. Most in- vestigators accept the possibility of a component of the Phase-2 response to P, elevation being attribut- able to continuing increases in vascular capacitance, though recent evidence (see Section 2A) suggests that this is unlikely to be a major source of error in volu- metric determinations of Kf*,. However, these data cannot be extrapolated to all tissues, and particularly not those which have sinusoidal microvascular structure, such as the liver and spleen. In the liver, for example (Greenway et al., 1969), it has been established that the underlying mechanisms associ- ated with the Phase-2 volume response to P, eleva- tion are not identical to those in intestine or skeletal muscle. Furthermore, in experiments where serial determinations of splenic venous haematocrit during P, elevation were made, it was demonstrated that the Phase-2 volume response of the isolated blood- perfused dog spleen was due to RBC sequestration with negligible plasma accumulation (B. N. Davies, P. D. I. Richardson and P. G. Withrington, un- published observations). The term “capillary filtra- tion coefficient” is therefore probably inappropriate in tissues with a sinusoidal microvasculature, such as the liver and spleen.
Since intestinal capillaries are not totally im- permeable to circulating plasma proteins, fluid filtered across the capillary wall contains measurable quantities of protein. Upon elevation of venous pressure, an enhanced transcapillary protein leakage occurs due predominantly to convection. Friedman (1976) and others (Johns and Rothe, 1978) have suggested that volumetric estimates of Kf,, signi- ficantly overestimate the true value because trans- capillary protein leakage occurs during venous pressure elevation. Using lymphatic protein flux and
lymph flow data from cat ileum (Granger et a/., 1979a, Table 2), one would estimate that, upon ele- vation of venous pressure from 1.33 to 4.0 kPa, the increased capillary protein flux accounts for <O. l % of the total volume (mass) being filtered into the interstitium. In these studies op was estimated to be 0.92 for total plasma proteins. Even when op was reduced to 0.56 by hypertonic glucose infusions (Table 3, Granger et al., 1979a), transcapillary protein leakage accounted for only 3% of the total volume change for the same increment in venous pressure. Thus, it appears that transcapillary protein leakage cannot represent a significant error in the volumetric estimate of Kf,, even when the capillaries become extremely permeable.
d. Variation of Kf,E with venouspressure The possibility that Kf,, in the intestine might be dependent upon the extent of the venous pressure increment used to determine Kf,, has been examined by Johnson and Hanson (1966) in the dog, and by Mortillaro and Taylor (1976) and Granger et al., (1979b) in the cat. In general, larger elevations of P, produced smaller values for K1;,: for example, Johnson and Hanson (1966) found Kf,, values of 2.78 and 0.83 ~m~.min-’.kPa-~.100g-~ at capillary hydrostatic pressures of 1.33 and 2.66 kPa re- spectively, while Mortillaro and Taylor (1976) found mean Kf., values of 1.872, 0.917 and 0.624 cm3* min-l.kPa-l.lOOg-l on elevating P, from zero to 1.33, 2.66, and 3.99 kPa, respectively.
This dependence of Kf,, on the magnitude of P, elevation has been ascribed to a precapillary myo- genic response to the alterations in hydrostatic pressure such that the capillary surface area is reduced by precapillary “sphincter” constriction at high venous pressures (Johnson and Hanson, 1966; Mortillaro and Taylor, 1976). Granger et al., (1979b) re-examined this problem by using varying P, elevations not from a fixed starting P,, but to a fixed final elevated P,. They found that larger changes in Pv produced smaller Kf,, values than those obtained with smaller P, elevations, but the differences were smaller than previous reports suggested : for example, elevating P, to 3.99 kPa by 1.33 kPa or less gave mean Kf., values of 1.399 (determined by the “flow equilibration method”) whereas elevations of I .33 to 2.66 kPa gave K f , i s of 0.895 and P, elevations > 2.66 kPa, 0.496cm3.rnin-’* kPa-’*100g1.
These effects may indeed be myogenic in nature: they are not likely to be due to changes in inter- stitial hydration at elevated P,, which would be ex- pected to reduce transcapillary filtration, as Granger et al. (1979b) showed a similar pattern of results with the zero time extrapolation method which should
by guest on February 20, 2016
Dow
nloaded from

554 P. D. I . Richardson, D. N. Granger, and A. E. Taylor
minimise such an effect. Similarly, this effect cannot be due to pressure-dependent changes in pre:post capillary resistance ratios as the ratio increases at increased P, (Mortillaro and Taylor, 1976) and this would be expected to cause an increase in Kr,,.
Granger et a/. (1979b) showed an additional de- pendence of Kt,, on P, changes in that when Pv was elevated by 1.33 kPa or less, the Kl;, values were greater when the final elevated pressure was 1.33 kPa (1.474) than when it was 2.66 or 3.99 kPa (0.805 and 1.399 ~ m ~ . m i n - ~ ~ k P a - ~ . l O O g ~ ~ ) , perhaps showing that the myogenic closure of precapillary sphincters to reduce the exchange vessel surface area is dependent not only upon the magnitude of venous pressure elevations, but also on the pressure range over which such elevations are made. A possible ex- planation for the reduction in Kr., as P, is elevated may lie in the blood flow distribution within the various layers of the intestinal wall at various venous pressures. Granger et al. (1979b) observed a reduc- tion in the percentage of total blood flow perfusing the mucosal-submucosal layers of the cat ileum as venous pressure was elevated, ie, 85% at Pv-O, 60% at P,=1.33, and 48% at P,=3.99 kPa. Comparable results have been observed in rat and dog intestinal preparations (R. Gore, personal com- munications; J. Friedman, personal communica- tions). Since, a priori, one would expect the capillary filtration coefficient to be higher in the mucosa be- cause of its greater capillary density (Folkow and Neal, 1971), a higher capillary filtration coefficient would be expected at lower venous pressures where the blood flow favours the mucosa. Although the mechanism responsible for the redistribution of flow remains uncertain it is conceivable that a potent myogenic response of the mucosal arterioles to P, elevation may explain these observations.
In calculating the capillary filtration coefficient it is generally assumed that np remains constant during the experimental manoeuvre. However, Johnson and Hanson (1966) have shown that the plasma oncotic pressure increases significantly during venous pressure elevations of less than 2.0 kPa. Theoretically, an increase in plasma oncotic pressure during P, elevation would serve to reduce the net filtration pressure, thus leading to an under- estimation of Kr,,.Since the magnitude of the increase in n,, is dependent upon the extent of the increase in venous pressure, it is conceivable that the progressive decline in Kt,, observed as P, is elevated may be due to an osmotic buffering effect at the capillary wall. On a theoretical basis, Johnson and Hanson (1966) ruled out this possibility over the venous pressure range from which the values were acquired (0 to 2.0 kPa); however, osmotic buffering may be significant for larger P, elevations (eg 2.66 to 3.99 kPa).
e. Transmission of increments in P, to the capillaries When venous pressure is elevated by a known amount, the resulting increase in capillary hydro- static pressure is smaller than the change in P,. Since Kf,,’s are expressed in cm3.min-’*kPa* IOOg-’, and the “per kPa” component of this expression properly refers to the rise in capillary hydrostatic pressure, it is important to know the proportion of an imposed rise in P, that reaches the exchange vessels. For an accurate assessment of this pro- portionate transmission, it is necessary to know the normal capillary hydrostatic pressure.
It is unacceptable to use the gravimetric technique of Pappenheimer and Soto-Rivera (1948) for deter- mination of capillary hydrostatic pressure (P,) in tissues which exhibit postcapillary /precapillary re- flexes (venous-arteriolar reflexes) (Hanson and Johnson, 1962; Folkow et al., 1963). A modification of this technique has, however, been widely em- ployed (Johnson and Hanson, 1966; Johnson and Richardson, 1974; Mortillaro and Taylor, 1976), and when Pc measurements have been made, or the percentage transmission of increments in P, to the capillaries has been estimated by a deductive tech- nique (Folkow et al., 1963). it has been assessed that about 70 to 85 % of the imposed rise in P, is trans- mitted to the exchange vessels. Most experiments in- volving Kf,, have either applied a correction factor by measuring the P, and assuming a proportionate transmission to P, of 70 to 85 % or, where the P, has not been measured (eg Greenway et al., 1969; Richardson, 1974). the Kr,, has been expressed in terms of the elevation in venous pressure. This ex- plains in part some systematic differences between experimental observations from different labora- tories.
f . Role of lymphatic drainage The importance of lymph flow in the maintenance of interstitial fluid volume has been addressed in several recent reviews (Witte and Witte, 1973; Nicoll and Taylor, 1977). It is evident from these re- ports that adequate lymphatic drainage of tissues such as the small intestine is required to maintain a physiological equilibrium of the forces governing transcapillary fluid exchange. Obstruction of in- testinal lymphatic drainage should theoretically lead to continued fluid accumulation into the interstitial spaces and ultimately result in filtration secretion. Apart from the difficulty in maintaining a true iso- volumetric state, inadequate lymphatic drainage will increase the hydration state of the preparation (oedema) and therefore alter the balance of forces across the capillaries. The ability of the tissue forces to oppose excessive capillary filtration is reduced in
by guest on February 20, 2016
Dow
nloaded from

Review 555
Table Capillary filtration coefjicients in the small intestine
References Method
Folkow et a / . (1963, 1964) I .278 Volumetric Oberg (1964)t 0.902 Volumetric Cobbold (1964) t 0.752-1.128 (n= 20) Volumetric Johnson and Hanson (1966) *$ 0.827-2.782 Volumetric Wallentin (1966) 0.752-1.128 Volumetric Haglund and Lundgren (1972) 0.391 f . 0 3 (SE) Volumetric Johnson (1973) 0.474 Volumetric Fasth and Hulten (1973) 0.376-1.128 Volumetric Biber ef a / . (1973) 0.526 *O.IS (SE), n=25) Vqlumetric Richardson ( I 974) t 0.226 fO.02 (SE, n= 44) Volumetric Richardson (1975b) 0.587 f0.06 (SE, n= 4) Volumetric Richardson (1976)t 0.451 *0.07 (SE, n= 7) Volumetric Mortillaro and Taylor (1976)t$ 0.60-4.21 Volumetric Yablonski and Lifson (1976)* 0.820 Lymph flow,
secretion rate Quillen el a/. (1978) 0.233-0.278 (n= 5) Volumetric Johns and Rothe (1978)* 0.150-0.263 Indicator dilution Granger er a/ . ( 1 9 7 9 ~ ) 0.910 f0.07 Volumetric Granger ef a/ . (1979b)$ 0.451-1.549 Volumetric
All values acquired from anaesthetised cats unless indicated to the contrary. Dog.= *. t, sympathetic innervation maintained. :, represent values obtained over a wide venous pressure range. I .O kPa= 7.52 mmHg.
overhydrated preparations. Since readjustment of tissue forces subsequent to P, elevation is less pro- nounced when lymphatic drainage is obstructed, the increase in P, will result in an increase in NFP for a longer period than when lymphatic drainage is open. This will lead to higher estimates of Kf,, than would be acquired for normally hydrated preparations with freely flowing lymphatics. In addition, it is clearly established (Granger et a/., 1977; Mortillaro and Taylor, 1976; Yablonski and Lifson, 1976) that al- though intestinal lymphatic flow at zero venous pressure is small (about 0.05 ~ m ~ ~ n i n - ~ . 1 0 0 g ~ ) , it increases with increasing P,. This means that al- though an intestinal preparation with obstructed lymphatic drainage may exhibit negligible increases in baseline volume under control conditions, eleva- tion of P, may result in a greater net retention of fluid by the tissue than would be the case if the lym- phatics were not obstructed. This would give a greater Phase-2 volume increase in response to P, elevation under obstructed-lymphatic conditions than under open-lymphatic conditions, since the fluid normally transported away from the tissue by the lymphatic system would accumulate, giving an over-estimate of Kr,,.
g. Other sources of variation in Kf,, measurements in the intestine Values for Kf,, reported from different laboratories
vary widely under control conditions (Table); in addition to the problems already discussed, varia- tions in Kr,, may arise from: (i) the nature of tissue - ie, jejunum or ileum - used in the study; (ii) the nature and route of administration of anaesthetics - the use of intraperitoneal anesthetics may affect in- testinal Kr,, and inhalation anesthetics may release suprarenalmedullary catecholamines. I n addition, (ii i) the temperature of the preparation, and blood pH, Po, and Pcoz may affect Kf,,; as may (iv) the use of plasma expanders instead of whole blood to prime the external vascular circuits (Lunde, 1967).
Furthermore, (v) plethysmographic volume- measuring techniques may affect the Kf,, if an ex- ternal hydrostatic pressure in the plethysmograph or volume-measuring system results in a “sluice” effect due to venous collapse. Such an effect would reduce the calculated Kf,, as a smaller proportion of an in- crease in P, would be transmitted to the capillary hydrostatic pressure under “sluice” as opposed to “non-sluice” conditions. In addition, a “sluice” effect can be imposed when the venous outflow pressure is below total tissue pressure. Although total tissue pressure has not been determined in the small intestine, in most tissues it ranges from 0 to 0.66 kPa (Guyton et a/., 1975). Therefore, venous outflow pressures of 0 kPa and below may impose a significant “sluice” effect and thereby lead to an underestimation of Kr,,.
by guest on February 20, 2016
Dow
nloaded from

556 P . D. 1. Richardson, D. N . Grunger, and A. E. Tavlor
3. C H A N G E S I N I N T E S T I N A L C A P I L L A R Y F I L T R A T I O N C O E F F I C I E N T In most tissues including the intestine, it is estab- lished that the Phase-2 slope is a consequence of the accumulation of cell-free fluid in the perivascular spaces. Less clearly established are the mechanisms underlying physiological and pharmacological changes in Kf,,. In general, changes in Kr,= have been attributed to alterations in the tone of the pre- capillary “sphincters”. However, with some pro- cedures such as exercise in skeletal muscle (Kjellmer, 1965; Kjellmer and Odelram, 1965) or the ad- ministration of vasoactive material such as histamine (Richardson 1975b) or bradykinin (Fasth, 1973), there is a probability that a component of the ob- served increase in Kf,, is due to increased micro- vessel permeability.
It should be emphasised that even small changes in capillary pore size can markedly influence Kf,, since flow through cylindxical channels is proportional to the fourth power of the radius (Taylor er al., 1973). The chief evidence for or against the involvement of vascular permeability changes has been to compare the increases in Kec produced by substances such as histamine with those produced by material for which there is no independent evidence of permeability changes, such as isoprenaline. If histamine produces a greater KI,, increment as compared with isopre- naline for a given increase in blood flow, it is taken as evidence that the “extra” increase in Kf,, pro- duced by histamine compared with isoprenaline must be the result of increased vascular permeability. Attractive though this approach is, it relies on the assumption that all substances produce com- parable proportionate changes in both blood flow and Kf,, unless they influence microvessel per- meability. However, Kr,, can change without any alteration in blood flow if small doses of drugs are administered (Richardson, 1974), and if the vascular smooth muscle responsible for controlling tone in precapillary resistance vessels and in precapillary “sphincters” is not continuous or homologous (Greenway et at., 1976), different proportionate changes in Kf,, and in blood flow could cocur whether the two drugs (isoprenaline, histamine) caused changes in vascular permeability or not. Similarly, different proportionate changes in blood flow and in Kf,, would be expected if there is a different distribution of pharmacological receptors (eg P-adrenoceptors; histamine receptors) in pre- capillary resistance and “sphincter” sections of the microvasculature. Clear evidence for or against in- volvement of vascular permeability changes can only be assessed using techniques such as the lymphatic protein flux method (Granger et al., 1979a).
a. Nerve stimulation The influence of periarterial sympathetic nerve stimulation (Folkow et al., 1964) and of reflex acti- vation of the sympathetic nerves to the intestinal vasculature (&erg, 1964) are well established. In- creases in sympathetic discharge result in decreases in Kf,,, presumably the result of precapillary “sphincter” constriction resulting in a decrease in the functional exchange vessel area with consequent reduction in Kc,,. Less clear is the normal physio- logical role of the sympathetic innervation: com- parisons between different experimental series (Table) reveal that Kf,, is generally lower in pre- parations with an intact sympathetic innervation than in denervated preparations, though the dis- tinction is not clear-cut and in many instances where the preparations were denervated, they were also effectively adrenalectomised (Folkow et al., 1963 et seq.). A potentially important consideration may relate to the level of anaesthesia in the various preparations, and the surgical trauma involved. Richardson (1975b) observed that acute periarterial nerve section resulted in an increase in Kr,,, but the Kf,, was lower in isolated blood-perfused prepara- tions than in comparable preparations in which the superior mesenteric artery and periarterial nerves were intact (Richardson, 1976). It seems likely that under experimental conditions, there is a degree of sympathetic tone maintaining the Kec at lower levels than is the case in denervated preparations. but that the picture may be obscured by extensive handling, and more traumatic surgery to the in- testine. In pharmacological investigations, the possi- bility of alterations in systemic arterial pressure evoking baroreceptor-modulated changes in regional vasoconstrictor tone, including that of the pre- capillary “sphincters” is important, since baro- receptor reflex changes in Kr,, (oberg, 1964) will generally tend to counteract the direct drug-induced changes in Kf,, resulting in smaller experimentally- determined changes in Kf., than would be expected in the absence of the baroreceptor reflex effects (Richardson, 1974).
b. Shock The gastrointestinal circulation is particularly com- promised during haemorrhagic hypotension. Hag- lund and Lundgren (1972, 1973) have extensively analysed the effects of haemorrhagic hypotension on intestinal Kf,, and have shown that it increases by 70 % during the hypotensive period, due presumably to relaxation of precapillary sphincters by local tissue metabolites. Superimposed vasoconstrictor fibre stimulation is only seldom able to decrease Kf,, below the pre-shock value. Although hypo- tension is believed to cause mucosal oedema and
by guest on February 20, 2016
Dow
nloaded from

Re view 557
passive fluid filtration into the lumen by increasing capillary hydrostatic pressure and an increased capillary surface area (Kf,,) (Cook et at., 1971), a concomitant increase in capillary permeability may also occur.
c. a and B stimulation and blockade Intra-arterial noradrenaline (Folkow et al., 1963 ; Granger et at., 1979b: about 15 n g m r 3 blood) and intravenous phenylephrine ( I pg-kg'min-', Rich- ardson, 1974) and noradrenaline (0.2pg.kgl.min-' Richardson, 1973) reduce the Kf,, and superior mesenteric venous blood flow at constant or in- creased systemic blood pressure (BP), indicating constriction of the precapillary resistance vessels and precapillary "sphincters". These observations are in accordance with a reduction in neRb extraction, in- dicating a reduced capillary surface area in the dog intestine on infusion of noradrenaline (Shepherd et at., 1973). The vasoconstrictor responses to phenyle- phrine are blocked by an a-adrenoceptor blocking drug, phentolamine (1-2 mgekg', i.v.), which itself causes an increase in Kf,, in the sympathetically- innervated preparation (Richardson, 1974). The in- crease in Kf,, due to phentolamine may be the result of a reduction in vasoconstrictor tone, or to in- herent vascular smooth-muscle relaxant properties of phentolamine. Intra-arterial (Folkow et at., 1963, 0.1 pgmin-') and intravenous isoprenaline increase the Kec. an effect which on i.v. infusion is dose- dependent over the range 0.1 to 1.0 pg.kg-'*min-' (Richardson, 1974). The lower doses of isoprenaline (0.1 to 0.2 pg.kg'*min-', i.v.1 increase the Kf,, by 30 to 60%, but do not alter blood flow or BP; higher doses show evidence of precapillary resis- tance vessel as well as precapillary sphincter vaso- dilatation. These effects of isoprenaline are blocked by pretreatment with propranolol (100 pg'kg-l i.v.) but not by the selective B,-adrenoceptor antagonist practolol.
d. Autacoids Histamine Low doses (0.1 to 1.0 pg-kg-l-min-l) of histamine infused intravenously cause reductions in Kf,, with variable changes in BP and blood flow; higher doses (up to 40 pg-kg-lmin-l i.v.) or smaller doses after histaminase inhibition with amino- guanidine (10 mgakg', i.v.) cause increases in Kr,,. These increases in Kf,, could be due either to pre- capillary sphincter dilatation, or to increases in microvessel permeability, or both. The reductions in Kf,, on i.v. infusion of low doses of histamine are reversed by a-adrenoceptor blockade with phento- lamine (2 mgakg', i.v.) but not by sympathetic de- nervation or adrenalectomy (Richardson, 1973, 1974, 1975b). Although these effects suggest that histamine might cause a fall in Kr,, via the release of
catecholamines from the intestine, more recent evi- dence indicates that histamine can cause contraction of the isolated portal vein of the rabbit and that this effect can be blocked by phentolamine (Cook and Macleod, 1978). All effects of histamine on in- testinal Kf,, are blocked by the H , receptor anta- gonist mepyramine ( 1 to 2 mg-kg', i.v.), but not by burimamide (5 mg.kgl i.v.), suggesting that the vascular actions are mediated primarily via H, and not via H, receptors, as is the case in other vascular responses to histamine (Black et at., 1975; Richard- son and Withrington, 1977). 5-hydroxytryptamine 5 HT is released pathologically (pancreatic and gastrointestinal tumors) from the gastrointestinal tract (Burks and Long, 1966; Owman et at., 1973) so changes in Kf,, induced by 5HT are potentially important under these condi- tions. Intravenous 5HT (5-50 pg*kg lmin-l) causes a fall in Kf,, and variable effects on blood flow and BP (Richardson, 1974). These effects of 5HT are blocked by methysergide (250 pg'kg-', i.v.) (Richard- son, 1974). The responses are different when 5HT is injected or infused into the superior mesenteric artery: only increases in blood flow and in Kf,, being observed, though 5HT was observed to con- tract isolated mesenteric arteries and veins (Biber, et at., 1973a,b). The responses of the intestinal vasculature to 5HT therefore appear to depend on the dose and route of administration. Bradykinin Intra-arterial bradykinin (1 -20 n g a r S of arterial blood) causes an increase in blood flow through the small and large intestine; in addition, it increases the Kf,, an effect which is apparent at doses too low to produce significant changes in blood flow or in the motility of the gastrointestinal tract. This has led to the suggestion that bradykinin causes an increase in Kf,, primarily by increasing microvascular permeability (Fasth, 1973 ; Fasth and Hulten, 1973a,b). Bradykinin is released in patho- logical conditions involving the gastrointestinal tract such as the carcinoid and dumping syndromes, and septic shock (Seki et at., 1972). Large doses (up to10 pgekglmin-l i.v.) of bradykinin infused intra- venously also increases the Kf,, (Richardson, 1973), the high doses being necessitated by the rapid de- activation of bradykinin in the blood and pulmonary vasculature (Vane, 1969; Douglas, 1975).
In general, the autocoids increase the Kf,, and reduce the regional vascular resistance. The potential importance of these responses lies in the pathological release of the autacoids, either separately or together in a group. Concomitant release of these substances would result in increases in Kf,, (mediated in all probability by changes in vascular permeability in addition to precapillary sphincter dilatation) and increases in total blood flow through the intestine.
by guest on February 20, 2016
Dow
nloaded from

558 P. D. I . Ricliard.son, D. N. Granger, and A. E. Taylor
e. Vasoconstrictor peptides Angiotensin Intravenous infusions of angiotensin (25 to 160 ng.kg-'*min-') cause dose-dependent re- ductions in Kf,, indicating precapillary sphincter constriction and, in general, reductions in blood flow at increased BP, indicating precapillary re- sistance vessel constriction. These effects are not attenuated by a-adrenoceptor blockade (Richard- son, 1974).
Vasopressin Vasopressin infused into the superior mesenteric artery causes dose-dependent reductions in Kf,, and increases in regional vascular resistance (Quillen et al., 1977), effects qualitatively similar to those produced by intravenous infusions (Richard- son, 1973). Quillen et al. (1977) have shown that vasopressin causes an increase in the ratio of pre- capillary: postcapillary resistance, resulting in net water reabsorption from the intestine into the vas- culature; this effect might be related to the physio- logical release of ADH consequent upon increases in plasma osmolality. The effect would be enhanced by a reduction in the inflow resistance of the intrahepatic portal venous vascular bed (Richardson and Withrington, 1978) which would reduce outflow resistance from the intestine. The observed vascular effects of vasopressin in the intestine there- fore have a potential physiological role in fluid balance.
f. Vasodilator drugs Intravenous infusions of vascular smooth muscle re- laxant drugs such as aminophylline (200 to 400 pg. kg-'min-', Richardson, 1974), sodium nitroprus- side (10 pg*kg-'min-l, i.v.) and sodium nitrite (0.5 rng.kg'.min-', i.v.) (Richardson, 1973) all cause in- creases in intestinal Kf,, with reductions in intestinal vascular resistance (increases in blood flow at re- duced BP), indicating precapillary resistance vessel and, since there is no independent evidence that these vasodilator drugs increase vascular permea- bility, precapillary sphincter dilation.
g. Gastrointestinal and pancreatic hormones Secretin Secretin administered intra-arterially ( I to 25 mU.cm-s blood) or intravenously (100 to 500 m u . kg-lmin-l) (Biber et al., 1974; Richardson, 1976) causes increases in Kec and intestinal blood flow. At low doses of secretin, corresponding approximately to high physiological levels of the hormone, there are substantial increases in Kf,, with little change in regional vascular resistance (Richardson, 1976). This raises the possibility that during digestion when the hormone is circulating in elevated levels, the Kt;, of the intestine is increased.
Cholecystokinin-Pancreozymin ( CCK-PZ) Intra-arterial (Biber et at., 1974) and intravenous (Richardson: unpublished observations) adminis- trations of CCK-PZ produce effects similar to those seen with secretin - ie of a decrease in intestinal vascular resistance and an increase in Kf,c. Glucagon Glucagon is well-established to cause in- creases in intestinal blood flow in the dog (Kock et al., 1970a,b), and cat (Fasth and Hulten, 1971), though its effects on intestinal Kf., have only re- ceived slight attention. Intravenous glucagon (250 ng. kg'min-') causes a reduction in intestinal Kf,, an effect reversed by a-adrenoceptor blockade by phentolamine (Richardson, 1975a). Gastrin The effect of gastrin on intestinal Kf,, does not appear to have been established, though the synthetic pentapeptide analogue, pentagastrin, causes reductions in Kf,, without significant changes in blood flow or BP at a dose of 100 ng.kg-'min-l i.v. (Richardson, 1975a).
Release of the gastrointestinal hormones during digestion would therefore be expected to elicit in- creases in Kf,, which may be accompanied by small increases in intestinal blood flow. The increased capillary perfusion indicated by the increase in Kf,, might be expected to increase the absorptive activity of the intestine in digestion, and a tentative con- clusion from the levels of the hormone infused is that these observations are physiologically significant.
h. Purines Adenosine (10 p g ~ c m - ~ La.) has been shown to decrease intestinal vascular resistance, decrease Kec (Granger et al., 1978), and cause redistribution of blood flow within the intestine away from func- tional exchange vessels. Capillary hydrostatic pressure is also increased indicating a possible myo- genic reduction in Kf,, and mucosal perfusion. Adenosine does not alter lymphatic protein flux, which suggests that vascular permeability remains unchanged.
Conclusion
The more recent estimates of Kr,c in the small in- testine have been about 0.60 to 0.75 cm3.min-'. kPa-'.100g1 in preparations in which the sym- pathetic nerves have been divided, and about 0.38 to 0.60 cms.min-'.kPa-'.lOOg- I with the sym- pathetic innervation intact. Despite considerable variations from these ranges which have been re- ported, the Kdc remains an important variable in the Starling equilibrium, and consequently in capillary exchange. The precise choice of technique to measure capillary filtration coefficient in the intestine will depend upon the requirements of individual
by guest on February 20, 2016
Dow
nloaded from

Review 559
studies. However, volumetric and gravimetric tech- niques seem likely to continue to predominate: in these, variability can be reduced ( i ) by using small venous pressure elevations ( Q 1.33 kPa), (i i) by using a control venous pressure approximating normal portal venous pressure, (i i i) by assessing the end of the passive blood volume shift in every deter- mination and (iv) by ensuring an uninterrupted lymphatic drainage.
Much important information remains lacking re- garding intestinal KI:, - in particular whether it alters in response to “physiological” stimuli such as low frequency nerve stimulation and concentrations of vasoactive hormones established to occur under physiological conditions. Further research is re- quired to establish the factors leading to physio- logical, pathological and therapeutic changes in capillary filtration coefficient.
Parts of the studies described in this review were supported by grants from National Heart, Lung and Blood Institute 15680 and from the Medical Re- search Council (U.K.). Dr Richardson thanks Dr Charles Baugh, Dean of Basic Medical Sciences for extending an invitation to Dr Richardson to work at the University of South Alabama, and The Well- come Trust for a grant towards his travelling ex- penses. The authors are grateful for the clerical assistance of Ms Sue Van Devender and Cardyn Brewer.
References
Biber, B., Fara, J.. and Lundgren. 0. (1973a). Intestinal vascular responses to 5-hydroxytryptamine. Acta Physio- logica Scandinavica, 81, 526-534.
Biber, B., Fara, J., and Lundgren, 0. (1973b). Vascular re- actions in the small intestine during vasodilation. Acta Physiologica Scandinavica, 89, 449-456.
Biber, B., Fara, J., and Lundgren, 0. (1974). A pharma- cological study of intestinal vasodilator mechanisms in the cat. Acta Physiologica Scandinavica, 90, 673-683.
Black, J . W., Owen, D. A. A., and Parsons, M. E. (1975). An analysis of the depressor responses to histamine in the cat and dog: involvement of both H, and H , receptors. British Journal of Pharmaco1og.v. 54, 3 19-324.
Burks, T. F.. and Long, J. P. (1966). 5-hydroxytryptamine release into dog intestinal vasculature. American Journal
Casely-Smith, J. R., O’Donoghue, P. J.. and Crocker. K. W. J., (1975). The quantitative relationships between fenestrae in jejunal capillaries and connective tissue channels: proof of “tunnel capillaries”. Microvuscular Research, 9, 78-100.
Chen, H. I . . Granger, H J., and Taylor, A. E. (1976). In - teraction of capillary. interstitial and lymphatic forces in the canine hindpaw. Circulation Research, 39, 245-254.
Cook, B. H., Wilson, E. R., and Taylor, A. E. (1971). Intestinal fluid loss in hemorrhagic shock. American Journal of Physiolugy, 221, 1494-1498.
Cook. D. A., and Macleod, K. M. (1978). Response of rabbit portal vein to histamine. Briti.th Journal of Pharmacology, 62,165- 170.
of Physiolog~, 211, 619-625.
Davies, B. N., Richardson, P. D. I., and Withrington, P. G. (1974). The effects of post-ganglionic sympathetic nerve stimulation on the “capillary filtration coefficient” of the isolated, blood-perfused dog spleen. Journal of Physiology, 241,46-47P.
Douglas, W. W. (1975). Autacoids (Section V). In The Pharmacological basis oJ Therapeutics. Edited by L. S. Goodman, and A. Gilman, Macmillan: New York.
Drake, R., Gaar, K. A., and Taylor, A. E. (1978). Estimation of the filtration coefficient of pulmonary exchange vessels. American Journal of Physiology, 234, H266-H274.
Erdmann, A. J., Vaughn. T. R., Brigham, K. L., Woolverton, W. C., and Staub, N. C. (1975). Effect of increased vascular pressure on lung fluid balance in unanesthetized sheep. Circulation Research, 37, 27 1-284.
Fasth, S. (1973). The effect of bradykinin on gastrointestinal motility and circulation. MD Thesis, University of Goteborg.
Fasth, S., and Hulten, L. (1971). The effect of glucagon on intestinal motility and blood flow. Acta Physiologica Scandinavica, 83, 169-1 73.
Fasth, S., and Hulten, L. (1973a). The effect of bradykinin on intestinal motility and blood flow. Acta Chirurgica Scandin- avica 139,699-705.
Fasth, S., and Hulten, L. (1973b). The effect of bradykinin on the consecutive vascular sections of the small and large intestine. Acta Chirurgica Scandinavica, 139, 707-715.
Folkow. B., Lewis, D. H., Lundgren, 0.. Mellander, S., and Wallentin, 1. (1964). The effect of sympathetic vaso- constrictor fibres on the distribution of capillary blood in the intestine. Acta Physiologica Scandinavica, 61, 458-466.
Folkow, B., Lundgren, 0.. and Wallentin, 1. (1963). Studies on the relationship between flow resistance, capillary filtration coefficient and regional blood volume in the in- testine of the cat. Acta Physiologica Scandinavica, 51, 27C283.
Folkow, B., and Mellander, S. (1970). Measurements of capillary filtration coefficient and its use in studies of the control of capillary exchange. In Capillary Permeability. Alfred Benzon Symposium I I : Munksgaard, Copenhagen.
Folkow, B., and Neil, E. (1971). Gastrointestinal and liver circulation (Chapter 26). In Circulation. Oxford Uni- versity Press: London.
Friedman, J. .I. (1976). Transcapillary protein leakage and fluid movement: effect of venous pressure. Microvascular Research, 12, 275-290.
Gore, R. W., Schoknecht, W., and Bohlen, H. G. (1976). Filtration coefficients of single capillaries in rat intestinal muscle. In Microcirculation, Vol. I . pp. 331-332. Edited by J . Grayson and W. Zingg, Plenum: New York.
Granger, D. N. (1977). Interstitial fluid volume regulation in the feline small intestine: an analysis of the interactions between transmucosal flows and lymph flow. Ph.D. Thesis, University Medical Center, Jackson, Mississippi.
Granger, D. N., Brace, R. A,, Parker, R. E., and Taylor, A. E. (1979a). Analysis of the permeability characteristics of intestinal capillaries. Circulation Research, 44, 335-343,
Granger, D. N., Gabel, J. C., Drake, R. E., and Taylor, A. E. (1978). Physiologic basis for the clinical use of albumin solutions Surgery, Gynaccology and Obstetrics, 146, 97- 104.
Granger, D. N., Mortillaro, N. A., and Taylor, A. E. (1977). Interactions of intestinal lymph flow and secretion. American Journal of Physiology. 232, E 13-E 18.
Granger, D. N., Richardson, P. D. I . , and Taylor, A. E. (1979b). Volumetric assessment of the capillary filtration coefficient in the cat small intestine. European Journal
Granger. D. N., and Taylor, A. E. (1978). Effects of solute- coupled transport on lymph flow and oncotic pressures in
Physiolog,v, 381, 25-33.
by guest on February 20, 2016
Dow
nloaded from

560
the cat ileum. American Journal of Physiologv, 235,
Granger, D. N., Valleau, J. D., Parker, R. E., Lane, R. S.. and Taylor, A. E. (1979~). Effects of adenosine on in- testinal vascular hemodynamics, oxygen delivery, and capillary fluid exchange. American JCJiimal of Physiology, 235, H707-H719, 1978.
Granger, H. J., Goodman, A. T., and Granger, D. N. (1976). Role of resistance and exchange vessels in local micro- vascular control of skeletal muscle oxygenation in the dog. Circulation Research, 38, 379-385.
Greenway, C. V., and Lautt, W. W. (1970). Effects of hepatic venous pressure on transsinusoidal fluid transfer in the liver of the unanesthetized cat. Circularion Research,
Greenway, C. V., and Lautt, W. W. (1972). Effects of adre- naline, isoprenaline and histamine on transsinusoidal fluid filtration in the cat liver. Brirish Journal of Pharniu- cology, 44, 185-191.
Greenway, C. V., Scott, G. D., and Zink, J. (1976). Sites of autoregulatory escape of blood flow in the mesenteric vascular bed. Journal of Physiology, 259, 1-12.
Greenway, C. V., Stark, R. D., and Lautt, W. W. (1969). Capacitance responses and fluid exchange in the cat liver during stimulation of the hepatic nerves. Circularion Research, 25,277-284.
Guyton, A. C., and Lindsey. A. W. (1959). Effect of elevated left atrial pressure and decreased plasma protein concen- tration on the development of pulmonary edema. Circic- Iarion Research, 7, 649-657.
Guyton, A. C., Taylor, A. E., and Granger, H. J. (1975). Circulatory Physiology 11: Dynamics and control of the body fluids, Chapter 4. Saunders: Philadelphia, Pa.
Hagland, U., and Lundgren, 0. (1972). Reactions within consecutive vascular sections of the small intestine of the cat during prolonged hypotension. Acra Pl~ysitilogicrr Scandinavica, 84, 151-163.
Hagland. U., and Lundgren, 0. (1973). The effects of vaso- constrictor fibre stimulation on consecutive vascular sections of cat small intestine during hemorragic hypo- tension. Acra Physiologica Scandinavica, 88, 95-1 08.
Hanson, K. M., and Johnson, P. C. (1962). Evidence for local arteriovenous reflex in intestine. Juurnal of Applied Physiology, 17,509-51 3.
Hargens, A. R., and Zweifach, B. W. (1976). Transport be- tween blood and peripheral lymph in intestine. Micro- vascular Research, 11,89- 101.
Johns, B. L., and Rothe, C. F. (1978). Delayed vascular compliance and fluid exchange in the canine intestine. American Journal of Physiology, 234, H66CH669.
Johnson, L. (1973). Capillary filtration in isolated, homo- logously perfused small intestine. Acra Chirrrrgica Scan- dinavica, 139, 72-78.
Johnson. P. C., and Hanson, K. M. (1962). Effect of arterial pressure on arterial and venous resistance of intestine. Journal of Applied Physiology, 17, 503-508.
Johnson, P. C., and Hanson, K. M. (1966). Capillary filtra- tion in the small intestine of the dog. Circularion Research, 19,166-773.
Johnson, P. C., and Richardson, D. R. (1974). The influence of venous pressure on filtration forces in the intestine. Micro vascular Research, 7, 296-306.
Kedem, 0.. and Katchalsky, A. (1958). Thermodynamic analysis of the permeability of biological membranes to non-electrolytes. Biochimica 1’1 Biopli,vsicn Acra, 27,
Kjellmer, 1. (1965). On the competition hetween metabolic vasodilation and neurogenic vasoconstriction in skeletal niuscle. Acla Pliysiulogica Scandinavica, 63, 450-459.
E429-E436.
26, 697-703.
229-246.
Kjellmer, I . , and Odekdm, H. (1965). The effect of some physiological vasodilators on the vascular bed of skeletal muscle. Acta Physiologica Scandinavica, 63, 94- 102.
Kock, N. G., Tibblin, S., and Schenk, W. G. (1970a). Hemodynamic responses to glucagon : an experimental study of central, visceral and peripheral effects. Annals of Surgery, 171, 373-379.
Kock, N. G., Tibblin, S., and Schenk, W. G. (1970b). Mesenteric blood flow response to glucagon administration during temporary exclusion of the liver. Archives IJJ
Surgery, 100,28&283. Landis, E. M., and Gibbon, J. H. (1933). The effects of
temperature and of tissue pressure on the movement of fluid through the human capillary wall. Journal of Clinical Invesligafion, 12, 105-138.
Levine, 0. R., Mellins, R. B., Senior, R. M., and Fishman, A. P. (1967). The application of Starling’s Law of capillary exchange to the lungs. Journal of Clinical Invesrigariun. 46, 934944.
Lunde, P. K. M. (1967). The influence of perfusate composi- tion on edema development in isolated perfused rabbit lungs. Universitetsforlaget : Oslo. Norwegian Research for Science and the Humanities.
Mortillaro, N. A., and Taylor, A. E. (1976). Interaction of capillary and tissue forces in the cat small intestine. Circularion Research, 39, 348-358.
Nicoll, P. A., and Taylor, A. E. (1977. Lymph formation and flows. Annual Review of Physiology, 39, 73-95.
bberg, B. (1964). Effects of cardiovascular reflexes on net capillary fluid transfer. Aria Physiologica Scanclanavia, Supplement 229,5-98.
bberg, B.. and Rosell, S. (1967). Sympathetic control of consecutive vascular sections in canine subcutaneous adipose tissue. Acra Physiologica Scandinavica, 71, 47-56.
Owman, C., Hakanson, R., and Sundler, F. (1973). Occur- rence and function of amines in endocrine cells producing polypetide hormones. Federarion Proceedings, 32. 1785- 1791.
Pdppenheimer, J. R., and Soto-Rivera, A. (1948). Effective osmotic pressure of the plasma proteins and other quan- tities associated with the capillary circulation in the hindlimbs of cats and dogs. American Journal oJ Ph.ssi- olugy, 152,471491.
Quillen, E. W., Granger, D. N., and Taylor, A. E. (1977). The effects of arginine vasopressin on capillary filtration in the cat ileum. Casrroenrerology, 73, 1290-1295.
Renkin, E. M., Joyner, W. L., Sloop, C. H., and Watson, P. D. (1977). lnfluence of venous pressure on plasma-lymph transport in the dog’s paw; convective and dissipative mechanisms. Microvascular Research, 14, 191-204.
Richardson, P. D. I. (1973). Pharmacological responses of the vasculature of the mammalian small intestine with particular regard to the responses of the microcirculation. Ph.D. Thesis, University of London.
Richardson, P. D. I. (1974). Drug-induced changes i n capillary filtration coefficient and blood flow in the in- nervated small intestine of the anaesthetized cat. Brirish Journal of Pharmacology, 52,48 1498 .
Richardson, P. D. I. (1975a). The effects of glucagon and pentagastrin on capillary filtration coefficient in the in- nervated jejunum of the anaesthetised cat. Bririsli Journul of Pliarmacology, 54, 225P.
Richardson, P. D. 1. (1975b). An investigation of the niech- anisms responsible for a reduction in capillary filtration coefficient in the innervated cat jejunum on intravenous infusion of histamine. Journal of Pharniacy and Pharma- co1og.r. 27. 868 -870.
Richardson, P. D. I . (1976). The actions of natural secretin on the small intestinal vasculature of the anaesthetized cat. Bririsli Journal of P/iarniac~oliig.r, 58, 127- 136.
by guest on February 20, 2016
Dow
nloaded from

561
Richardson, P. D. I., and Withrington, P. G. (1977). A comparison of the effects of bradykinin, 5-hydroxy- tryptamine and histamine on the hepatic arterial and portal venous vascular beds of the dog: histamine H, and H, receptor populations. British Journal of Pharmacolog),, 60.123-133.
Richardson, P. D. I . , and Withrington, P. G. (1978). The effects of intra-arterial and intraportal injections of vaso- pressin on the simultaneously-perfused hepatic arterial and portal venous vascular beds of the dog. Circulation Research, 43, 496-502.
Rothe, C . F., Johns, B. L., and Bennett, T. D. (1978). Vascular capacitance of dog intestine using mean transit time of indicator. American Journal uf P/r,rsiolugy, 234,
Seki, T.. Nakajima, T., and Erdos. E. G. (1972). Colon kallikrein. its relation to the plasma enzyme. Biucheniical Pharmacology. 21, 1227-1235.
Shepherd, A. P.. Mailman, P. D., Burks, T. F.. and Granger, H. J. (1973). Effects of norepinephrine and sympathetic
H7-H 13.
stimulation on extraction of oxygen and ""Rb in perfused canine bowel. Circulation Research, 33, 166-1 74.
Starling, E. H. (1896). On the absorption of fluid from the connective tissue spaces. Journalof Physiology. 19,312-326.
Staub, N. C. (1974). Pulmonary edema. Physiological Rlvirws, 54, 678-8 I I.
Taylor. A. E., Gibson, W. H., Granger, H. J.. and Guyton. A. C. (1973). The interaction between intracapillary and tissue forces in the overall regulation of interstitial fluid volume. Lympho/ogy, 6, 192-208.
Wallentin, 1. (1966). Importance of tissue pressure for the fluid equilibrium between the vascular and interstitial compartments in the small intestine. Acta Physiologica Scanclanavia, 68, 304-3 15.
Witte. M. H.. and Witte, C . L. (1973). Lymph formation, lymph absorption. The formula of edema. Lymphology,
Yablonski, M. E., and Lifson, N. (1976). Mechanism of pro- duction of intestinal secretion by elevated venous pressure. Joiirnal of Clinical Investigation. 51, 904-91 5 .
6, 101-109.
by guest on February 20, 2016
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















