
Cancer Prev Res In vivo Nitrosomethylurea Circadian and Growth-Regulatory Genes Disrupted by...
14
2010;3:640-652. Published OnlineFirst April 27, 2010. Cancer Prev Res Ming Zhu Fang, Xun Zhang and Helmut Zarbl In vivo Nitrosomethylurea Circadian and Growth-Regulatory Genes Disrupted by Methylselenocysteine Resets the Rhythmic Expression of Updated version 10.1158/1940-6207.CAPR-09-0170 doi: Access the most recent version of this article at: Material Supplementary 0170.DC1.html http://cancerpreventionresearch.aacrjournals.org/content/suppl/2010/04/27/1940-6207.CAPR-09- Access the most recent supplemental material at: Cited Articles http://cancerpreventionresearch.aacrjournals.org/content/3/5/640.full.html#ref-list-1 This article cites by 52 articles, 16 of which you can access for free at: Citing articles http://cancerpreventionresearch.aacrjournals.org/content/3/5/640.full.html#related-urls This article has been cited by 1 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications for Cancer Research. on February 11, 2014. © 2010 American Association cancerpreventionresearch.aacrjournals.org Downloaded from Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170 for Cancer Research. on February 11, 2014. © 2010 American Association cancerpreventionresearch.aacrjournals.org Downloaded from Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Cancer Prev Res In vivo Nitrosomethylurea Circadian and Growth-Regulatory Genes Disrupted by...

2010;3:640-652. Published OnlineFirst April 27, 2010.Cancer Prev Res Ming Zhu Fang, Xun Zhang and Helmut Zarbl
In vivoNitrosomethylurea Circadian and Growth-Regulatory Genes Disrupted by Methylselenocysteine Resets the Rhythmic Expression of
Updated version
10.1158/1940-6207.CAPR-09-0170doi:
Access the most recent version of this article at:
Material
Supplementary
0170.DC1.html
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2010/04/27/1940-6207.CAPR-09-Access the most recent supplemental material at:
Cited Articles
http://cancerpreventionresearch.aacrjournals.org/content/3/5/640.full.html#ref-list-1
This article cites by 52 articles, 16 of which you can access for free at:
Citing articles
http://cancerpreventionresearch.aacrjournals.org/content/3/5/640.full.html#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
for Cancer Research. on February 11, 2014. © 2010 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
for Cancer Research. on February 11, 2014. © 2010 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170

640
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
Research Article Cancer
Prevention ResearchMethylselenocysteine Resets the Rhythmic Expression ofCircadian and Growth-Regulatory Genes Disrupted byNitrosomethylurea In vivo
Ming Zhu Fang1, Xun Zhang1, and Helmut Zarbl1,2
Abstract
Authors' AInstitute, Rand DentisInstitute oUniversityNew Jersey
Note: Supption Resea
Present adRoche, Pal
CorresponHealth ScieJersey, RoPhone: 732edu.
doi: 10.115
©2010 Am
Cancer P
Downl
Epidemiologic and animal studies indicate that disruption of circadian rhythm increases breast cancerrisk. Previously, we showed that methylselenocysteine reduced the incidence of N-nitroso-N-methylurea(NMU)–induced mammary carcinomas in Fischer 344 rats by 63%. Methylselenocysteine also increasedthe expression of Period 2 (Per2) and D-binding protein (DBP), providing evidence for a link between cir-cadian rhythm and chemoprevention. Here, we report that NMU disrupted the expression of core circadiangenes (Per1, Per2, Cry1, Cry2, and RevErbAα) and circadian-controlled genes, including melatonin receptor1α (MTNR1A), estrogen receptors (ERα and ERβ), and growth-regulatory genes (Trp53, p21, Gadd45α, andc-Myc) in mammary glands of Fischer 344 rats. By contrast, dietary methylselenocysteine (3 ppm seleni-um) given for 30 days, significantly enhanced the circadian expression of these genes (except for Cry1 andCry2). The largest effect was on the levels of the Per2, MTNR1A, and ERβmRNAs, which showed 16.5-fold,4.7-fold, and 9.5-fold increases in their rhythm-adjusted means, respectively, and 44.5-fold, 6.5-fold, and9.7-fold increases in amplitude as compared with the control diet, respectively. Methylselenocysteine alsoshifted the peak expression times of these genes to Zeitgeber time 12 (ZT12; lights off). Methylselenocys-teine also induced rhythmic expression of Trp53, p21, and Gadd45α mRNAs with peak levels at ZT12,when c-Myc expression was at its lowest level. However, methylselenocysteine had no significant effecton the circadian expression of these genes in liver. These results suggest that dietary methylselenocysteinecounteracted the disruptive effect of NMU on circadian expression of genes essential to normal mammarycell growth and differentiation. Cancer Prev Res; 3(5); 640–52. ©2010 AACR.
Introduction
In mammals, circadian rhythm is controlled by the in-teraction of positive and negative biochemical feedbackloops comprising a molecular oscillator. Heterodimersof brain and muscle arnt-like protein 1 (Bmal1, knownas Arnt1 in rat) and Clock or neuronal PAS domain pro-tein 2 (Npas2) activate transcription by binding to E-boxelements present in the promoters of circadian genes, in-
ffiliations: 1Environmental and Occupational Health Sciencesobert Wood Johnson Medical School, University of Medicinetry of New Jersey, Piscataway, New Jersey and 2Cancerf New Jersey, Robert Wood Johnson Medical School,of Medicine and Dentistry of New Jersey, New Brunswick,
lementary data for this article are available at Cancer Preven-rch Online (http://cancerprevres.aacrjournals.org/).
dress for X. Zhang: Department of Genetics and Genomics,o Alto, CA 94304.
ding Author: Helmut Zarbl, Environmental Occupationalnces Institute, University of Medicine and Dentistry of Newom 426, 170 Frelinghuysen Road, Piscataway, NJ 08854.-445-2354; Fax: 732-445-4161; E-mail: [email protected].
8/1940-6207.CAPR-09-0170
erican Association for Cancer Research.
rev Res; 3(5) May 2010
for Cancer Recancerpreventionresearch.aacrjournals.orgoaded from
cluding Period (Per), Cryptochrome (Cry), Rev-ErbAα,retinoid-related orphan nuclear receptors (ROR), andnumerous circadian-controlled genes (CCG). The Rev-ErbAα and ROR proteins, respectively, inhibit and activateBmal1 transcription (1). Posttranscriptionally modifiedPer/Cry heterodimers, in turn, repress Clock/Bmal1 tran-scriptional activity, thus limiting their own transcriptionand the expression of CCGs (2). The Clock protein wasshown to have histone acetyltransferase activity (3), sug-gesting that the activation of E-box–containing promotersis modulated at the level of the chromatin structure. Themolecular oscillator functions in most peripheral tissuesand maintains a periodicity of ∼24 hours that persistsin the absence of external cues (1, 2, 4, 5). To synchronizethe periodicity among cells in peripheral organs in vivoand adjust the phase to changing environmental signals,the intrinsic molecular oscillator is entrained by externalsignals that includes melatonin and glucocorticoids (6).In vitro studies on fibroblasts indicate that the intracellu-lar mediators of circadian gene expression include Ca2+,protein kinase C, protein kinase A, mitogen-activatedprotein kinase, and glucocorticoid receptor signalingpathways (6, 7). In mammals, the central pacemakerfor the autonomous oscillators in peripheral organs ismelatonin, a hormone whose secretion from the pineal
search. on February 11, 2014. © 2010 American Association

Methyselenocysteine Resets Circadian Gene Expression
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
gland is synchronized by light-induced signaling from thesuprachiasmatic nucleus (8, 9). In this way, the suprachias-matic nucleus coordinates the behavioral, physiologic,and biological functions of organisms and cells withlight-dark cycles, food availability, and a variety of environ-mental signals.Circadian rhythms also play a critical role in normal
cell growth, differentiation, and cellular responses to gen-otoxic stressors (2). Disruption of normal circadianrhythm by environmental exposures is associated withan increased risk of several cancer types. For example, dis-ruption of circadian rhythm by constant light exposure orpinealectomy increased the incidence of mammary ade-nocarcinoma in rodents (10, 11). Moreover, mice lackingPer2 show a significantly elevated incidence of carcino-mas after exposure to genotoxic stress (1), whereas over-expression of either Per1 or Per2 in cancer cells inhibitstheir neoplastic cell growth and increases apoptotic rates(12–14). Clinical studies also reveal an association be-tween the deregulation of Per genes and various humancancers (15–17). In addition, epidemiologic studies indi-cate that disruption of circadian rhythm by shift work(exposure to light at night) increases breast and prostatecancer risk (18), prompting the International Agency forCancer Research to classify shift work as a probable hu-man carcinogen (type 2A).Both epidemiologic and animal studies have suggested
that various forms of selenium, a dietary trace element,reduce cancer risk at multiple organ sites, includingbreast and prostate (19). Methylselenocysteine, an organ-ic form produced by plants, mediates its chemopreven-tive effects at the early stages of carcinogenesis (20–22).Our recent studies were the first to suggest an associationbetween circadian rhythm and chemoprevention. Weshowed that dietary methylselenocysteine, given for 30days after exposure to a single carcinogenic dose of N-nitroso-N-methylurea (NMU), reduced the incidence ofmammary carcinomas in Fischer 344 (F344) rats by63% at 36 weeks. Gene expression profiles of normalmammary tissue indicated that 30 days of dietary sup-plementation with methylselenocysteine significantly in-creased the levels and rhythmic expression of the corecircadian gene, Period 2 (Per2), and the circadian outputgene, D-binding protein (DBP; ref. 23). In the presentstudy, we extended these observations by demonstratingthat a single carcinogenic dose of NMU significantly dis-rupted the rhythmic expression of most core circadiangenes and CCGs during the early stages of carcinogene-sis. By contrast, chemopreventive methylselenocysteinereset and enhanced the circadian gene expression inmammary tissues of NMU-treated rats. The circadianand CCGs affected by methylselenocysteine includedcore circadian genes (Per2 and Rev-ErbAα), estrogenand melatonin receptors, and DNA damage–responsivegenes. We also found that the ablation and resetting ofcircadian expression of these genes by NMU and methyl-selenocysteine, respectively, were independent of serummelatonin levels.
www.aacrjournals.org
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
Materials and Methods
Animal maintenance and diet preparationAll protocols were reviewed by and received the approval
of the Institutional Animal Care and Use Committees. An-imal experiments were done in Association for Assessmentand Accreditation of Laboratory Animal Care International–accredited facilities at the Fred Hutchinson Cancer Re-search Center (Seattle, WA) and the University of Medicineand Dentistry of New Jersey (Piscataway, NJ). Female F344rats were from Harlan Laboratories (Indianapolis, IN).For acclimatization to a powdered ration from HarlanTekland, animals were maintained on a powdered stan-dard AIN-76A diet containing 0.1 ppm of selenium (sodi-um selenite). The methylselenocysteine-enriched diet wasmade by admixing L-Se-methylselenocysteine (SeleniumTechnologies) with the standard powdered AIN-76A dietto a final concentration of 3 ppm of selenium. Animalswere housed under controlled climate conditions and a12-hour light/12-hour dark cycle (7 a.m. on/7 p.m. off).Referring to 7 a.m. as Zeitgeber time (ZT) 0 indicates thetime of lights on; and 7 p.m. is ZT12, indicating the timeof lights off.
Animal treatment and sample collectionForty-two female F344 rats between 50 and 55 days of
age received a single i.p. injection of NMU (Ash Stevens,Inc.) at a dose of 50 mg/kg body weight. NMU was dis-solved in acidified saline at a final concentration of10 mg/mL just prior to injection. Rats were then random-ized to two groups and maintained on either the standard-ized control diet or the methylselenocysteine-enriched dietfor 30 days. An additional 21 age-matched female F344rats injected with the vehicle and maintained on the stan-dardized diet served as control group. Three rats of eachgroup were sacrificed every 4 hours for 24 hours, begin-ning at 7 a.m. (ZT0). Blood samples were collected intoheparinized-BD Vacutainers by cardiac punctures afterCO2 asphyxiation. Plasma samples were separated by se-quential centrifuge (at 1,300 rcf for 10 minutes first andthen at 2,400 rcf for 15 minutes) and stored at −80°C.All mammary glands from each side of individual rats werecarefully dissected, combined into a pool of left or rightmammary glands, frozen on dry ice, and stored at −80°C. The liver of each rat was also dissected, frozen on dryice, and stored at −80°C.
Quantitative real-time PCRTotal RNA was extracted from a small piece (∼50 mg)
of mammary tissue from the left side pool of mammaryglands and liver of each rat using Tri Reagent (Sigma) andQiagen RNeasy Mini Kit (Qiagen), and then digested withQiagen RNase-free DNase. Relative mRNA expressionlevels were determined with real-time quantitative reversetranscription-PCR (RT-PCR) using specific primers (Sup-plementary Table S1), designed with Primer ExpressSoftware V 3.0 (Applied Biosystems), and SYBR asthe reporter. For reverse transcription, 1 μg of RNA was
Cancer Prev Res; 3(5) May 2010 641
search. on February 11, 2014. © 2010 American Association

Fang et al.
642
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
reverse-transcribed to cDNA using iScript cDNA SynthesisKit (Bio-Rad Laboratories). For real-time PCR, cDNAwas amplified with the SYBR Universal PCR Master Mix(Applied Biosystems) in ABI Prism 7900 SequencingDetector according to the instructions of the manufactur-er. A no-template control was included in each assay.β-Actin was used as an endogenous control. The expres-sion level of each gene in mammary tissue of the ratsacrificed at ZT0 was used as a calibrator in each group.Three independent samples were analyzed for each timepoint. The comparative Ct method was used to analyzethe relative mRNA expression levels (24).
Western blotTotal protein was extracted from a small piece (∼200
mg) of mammary tissue from the left side pool of mam-mary glands of each animal using radioimmunoprecipita-tion assay buffer lysis buffer [50 mmol/L Tris-HCl (pH7.5), 150 mmol/L NaCl, 1 mmol/L EDTA, 0.5% sodiumdeoxycholate, 0.1% SDS] supplemented with a cocktailof protease inhibitor (1:100), phosphatase inhibitor I(1:100), and phosphatase inhibitor II (1:100; Sigma-Al-drich). Forty micrograms of a protein sample was sepa-rated on 7.5% SDS-polyacrylamide gel and transferredto nitrocellulose membrane. After blocking, the mem-brane was incubated with primary antibody, anti-ratPer2 (G-19, 1:200; Santa Cruz Biotechnology), anti-ratERβ (1:2,000; Abcam), or anti-rat melatonin receptor 1α(MTNR1A) antibody (R-18, 1:200; Santa Cruz Biotechnol-ogy) overnight at 4°C, followed by incubation with IRDye 800 CW anti-goat or anti-rabbit IgG (LI-COR). Tar-geted protein signals with fluorescence were detectedusing Odyssey Infrared Imaging System (LI-COR). Wholetissue protein from rat brain was used as a positivecontrol for Per2 and MTNR1A; rat ovary tissue proteinwas used for ERβ. The intensities of bands were deter-mined using densitometry, and β-actin was used as aninternal control for normalization.
ImmunohistochemistryA small piece (0.5 cm2) of frozen mammary tissue from
the left side pool of mammary glands of each rat was em-bedded in optimal cutting temperature compound medi-um and stored at −80°C. Cryosections (15 μm) werefixed in 4% paraformaldehyde for 20 minutes. Immuno-histochemical staining was done with anti-rat Per2(1:50), ERβ (1:200), or MTNR1A (1:50) antibody used inWestern blot, using avidin-biotin-peroxidase complex(ABC) staining systems (Santa Cruz Biotechnology; ref.25). PBS containing 1.5% normal serum instead of primaryantibody was used as a negative control in immunohisto-chemistry.
Determination of plasma melatonin levelsPlasma was separated from blood by centrifugation and
stored at −80°C. Melatonin levels were determined usingMelatonin ELISA Kit (GenWay Biotech) and SpectraMaxV5 microplate reader (Molecular Devices).
Cancer Prev Res; 3(5) May 2010
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
Sequence analyses of promoter regionsThe sequences of the promoter regions (10,000 bp 5′
upstream, exon 1, and 2,000 bp intron 1) of MTNR1A,ERβ, Per1, Per2, Cry1, Rev-ErbAα, and DBP genes wereobtained from NCBI Map Viewer. The E-box motif(CACGTG) of Bmal1 was identified within the promoterregions of these genes. The 66 bp sequences includingthe motif in the promoter regions of MTNR1A in differ-ent species were analyzed using Multiple SequenceAlignment algorithm of F. Corpet (http://bioinfo.geno-toul.fr/multalin/).
Statistical analysesIntergroup differences were evaluated using multivariate
ANOVA. Statistical significance of circadian rhythmicitywas documented by Cosinor analysis using Time Series Sin-gle Cosinor 6.3 (Expert Soft Tech; ref. 26). Period s = 24hours or s = 12 hours was considered a priori. The rhythmcharacteristics estimated by this linear least squares methodinclude mesor (rhythm-adjusted mean) and amplitude(a half of the difference between minimum and maximumof fitted cosine function). A rhythm was detected if the nullhypothesis was rejected with P < 0.05. Significance of differ-ences in protein expression levels between methylseleno-cysteine-enriched versus control diet group were analyzedwith Student's t test (mean ± SE, n = 3).
Results
Effects of NMU and methylselenocysteine on theexpression of circadian genesIn our previous study, we showed that a chemopreven-
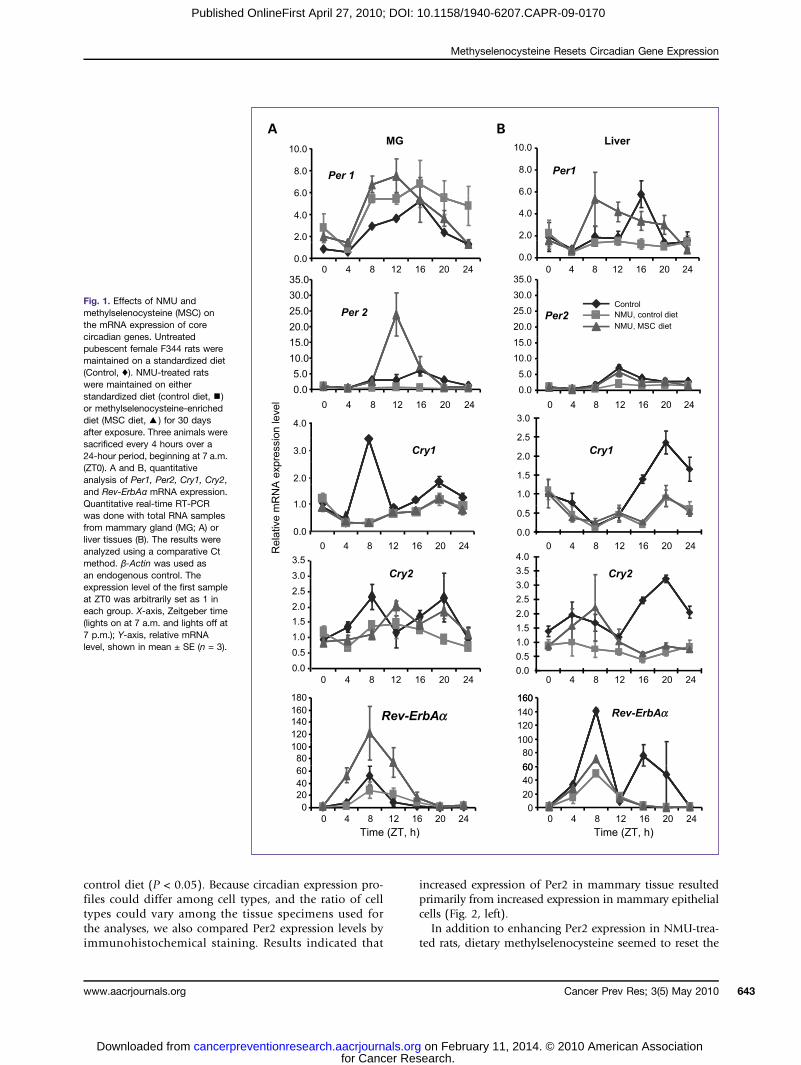
tive regimen of dietary methylselenocysteine-enricheddiet significantly increased the level and rhythm of Per2mRNA in mammary gland, but not in the liver, of NMU-treated rats. To determine if NMU was directly responsiblefor the ablation of circadian gene expression, we usedquantitative RT-PCR to generate circadian expression pro-files of circadian and CCGs in mammary gland and livertissues of rats treated with a carcinogenic dose of NMU orthe vehicle. Our results showed that NMU significantly re-duced the circadian expression of Per2, Cry1, Cry2, andRev-ErbAα genes in mammary tissues (Fig. 1A), but hadonly a modest effect in liver tissues (Fig. 1B; Table 1).By contrast, the circadian expression of Per1 was signifi-cantly reduced by NMU in liver, but not in mammarygland.The present study confirmed that in NMU-treated rats,
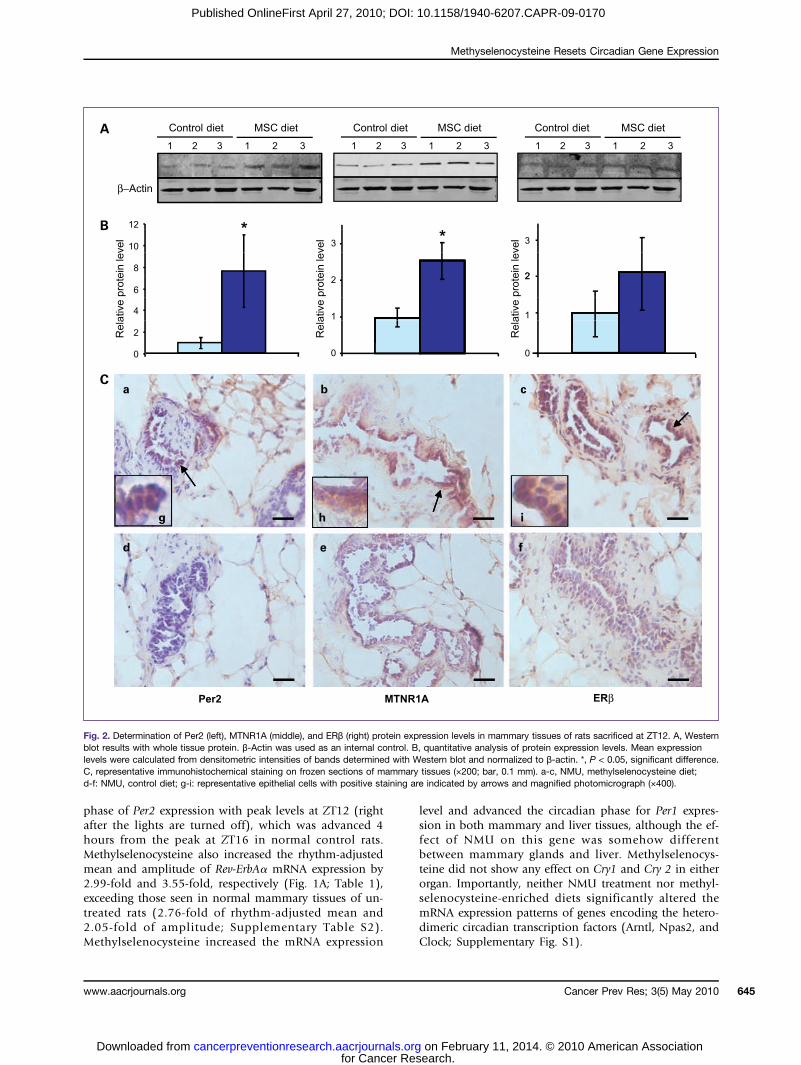
methylselenocysteine induced the rhythmic expression ofPer2, with a 16.5-fold increase of rhythm-adjusted meanand a 44.5-fold increase in amplitude of Per2 mRNA ex-pression relative to the control diet (Fig. 1A; Table 1),and a 1.5-fold increase in rhythm-adjusted mean and2.9-fold increase in amplitude as compared with the un-treated control rats (Supplementary Table S2). Westernblot analysis indicated that methylselenocysteine alsoinduced a statistically significant increase in Per2 proteinexpression in mammary tissues as compared with the
Cancer Prevention Research
search. on February 11, 2014. © 2010 American Association
Methyselenocysteine Resets Circadian Gene Expression
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
control diet (P < 0.05). Because circadian expression pro-files could differ among cell types, and the ratio of celltypes could vary among the tissue specimens used forthe analyses, we also compared Per2 expression levels byimmunohistochemical staining. Results indicated that
www.aacrjournals.org
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
increased expression of Per2 in mammary tissue resultedprimarily from increased expression in mammary epithelialcells (Fig. 2, left).In addition to enhancing Per2 expression in NMU-trea-
ted rats, dietary methylselenocysteine seemed to reset the
Fig. 1. Effects of NMU andmethylselenocysteine (MSC) onthe mRNA expression of corecircadian genes. Untreatedpubescent female F344 rats weremaintained on a standardized diet(Control, ♦). NMU-treated ratswere maintained on eitherstandardized diet (control diet,▪)or methylselenocysteine-enricheddiet (MSC diet,▴) for 30 daysafter exposure. Three animals weresacrificed every 4 hours over a24-hour period, beginning at 7 a.m.(ZT0). A and B, quantitativeanalysis of Per1, Per2, Cry1, Cry2,and Rev-ErbAα mRNA expression.Quantitative real-time RT-PCRwas done with total RNA samplesfrom mammary gland (MG; A) orliver tissues (B). The results wereanalyzed using a comparative Ctmethod. β-Actin was used asan endogenous control. Theexpression level of the first sampleat ZT0 was arbitrarily set as 1 ineach group. X-axis, Zeitgeber time(lights on at 7 a.m. and lights off at7 p.m.); Y-axis, relative mRNAlevel, shown in mean ± SE (n = 3).
Cancer Prev Res; 3(5) May 2010 643
search. on February 11, 2014. © 2010 American Association
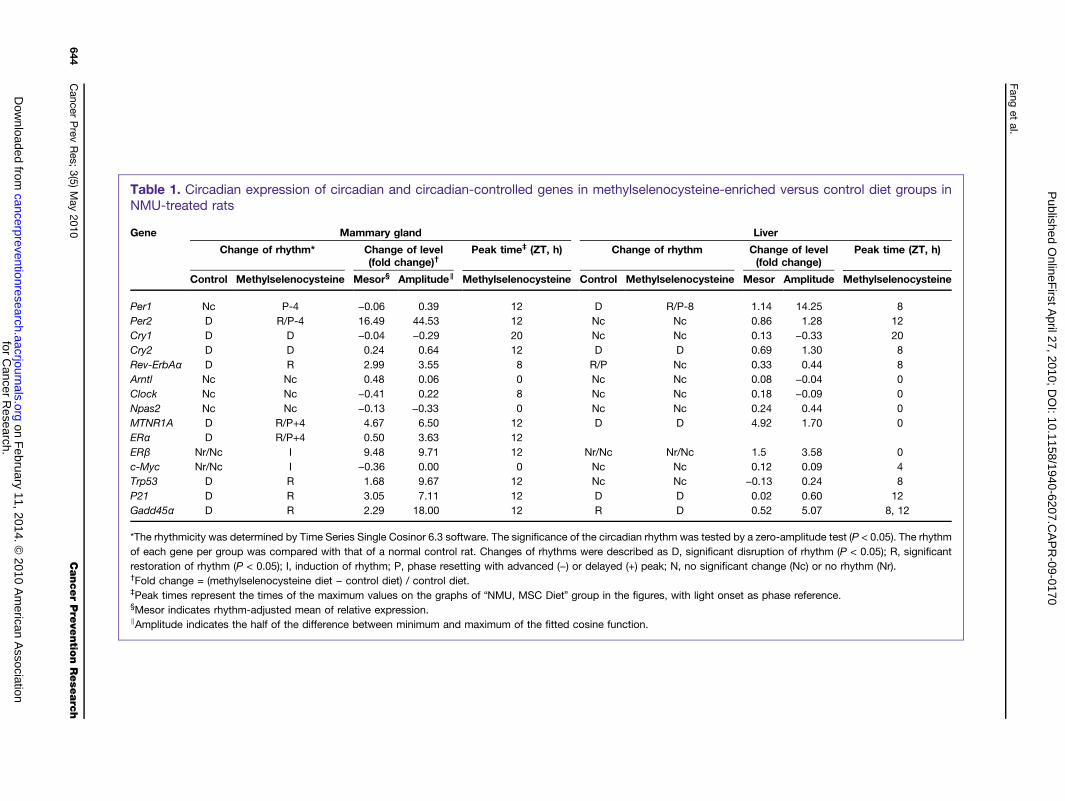
Table 1. Circadian expression of circadian and circadian-controlled genes in methylselenocysteine-enriched versus control diet groups inNMU-treated rats
Gene Mammary gland Liver
Change of rhythm* Change of level(fold change)†
Peak time‡ (ZT, h) Change of rhythm Change of level(fold change)
Peak time (ZT, h)
Control Methylselenocysteine Mesor§ Amplitude∥ Methylselenocysteine Control Methylselenocysteine Mesor Amplitude Methylselenocysteine
Per1 Nc P-4 −0.06 0.39 12 D R/P-8 1.14 14.25 8Per2 D R/P-4 16.49 44.53 12 Nc Nc 0.86 1.28 12Cry1 D D −0.04 −0.29 20 Nc Nc 0.13 −0.33 20Cry2 D D 0.24 0.64 12 D D 0.69 1.30 8Rev-ErbAα D R 2.99 3.55 8 R/P Nc 0.33 0.44 8Arntl Nc Nc 0.48 0.06 0 Nc Nc 0.08 −0.04 0Clock Nc Nc −0.41 0.22 8 Nc Nc 0.18 −0.09 0Npas2 Nc Nc −0.13 −0.33 0 Nc Nc 0.24 0.44 0MTNR1A D R/P+4 4.67 6.50 12 D D 4.92 1.70 0ERα D R/P+4 0.50 3.63 12ERβ Nr/Nc I 9.48 9.71 12 Nr/Nc Nr/Nc 1.5 3.58 0c-Myc Nr/Nc I −0.36 0.00 0 Nc Nc 0.12 0.09 4Trp53 D R 1.68 9.67 12 Nc Nc −0.13 0.24 8P21 D R 3.05 7.11 12 D D 0.02 0.60 12Gadd45α D R 2.29 18.00 12 R D 0.52 5.07 8, 12
*The rhythmicity was determined by Time Series Single Cosinor 6.3 software. The significance of the circadian rhythm was tested by a zero-amplitude test (P < 0.05). The rhythmof each gene per group was compared with that of a normal control rat. Changes of rhythms were described as D, significant disruption of rhythm (P < 0.05); R, significantrestoration of rhythm (P < 0.05); I, induction of rhythm; P, phase resetting with advanced (−) or delayed (+) peak; N, no significant change (Nc) or no rhythm (Nr).†Fold change = (methylselenocysteine diet − control diet) / control diet.‡Peak times represent the times of the maximum values on the graphs of “NMU, MSC Diet” group in the figures, with light onset as phase reference.§Mesor indicates rhythm-adjusted mean of relative expression.∥Amplitude indicates the half of the difference between minimum and maximum of the fitted cosine function.
Fang
etal.
Cancer
Prev
Res;
3(5)May
2010CancerPre
ventio
nRese
arc
h644
for Cancer R
esearch. on F
ebruary 11, 2014. © 2010 A
merican A
ssociationcancerpreventionresearch.aacrjournals.org
Dow
nloaded from P
ublished OnlineF
irst April 27, 2010; D
OI: 10.1158/1940-6207.C
AP
R-09-0170
Methyselenocysteine Resets Circadian Gene Expression
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
phase of Per2 expression with peak levels at ZT12 (rightafter the lights are turned off), which was advanced 4hours from the peak at ZT16 in normal control rats.Methylselenocysteine also increased the rhythm-adjustedmean and amplitude of Rev-ErbAα mRNA expression by2.99-fold and 3.55-fold, respectively (Fig. 1A; Table 1),exceeding those seen in normal mammary tissues of un-treated rats (2.76-fold of rhythm-adjusted mean and2.05-fold of amplitude; Supplementary Table S2).Methylselenocysteine increased the mRNA expression
www.aacrjournals.org
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
level and advanced the circadian phase for Per1 expres-sion in both mammary and liver tissues, although the ef-fect of NMU on this gene was somehow differentbetween mammary glands and liver. Methylselenocys-teine did not show any effect on Cry1 and Cry 2 in eitherorgan. Importantly, neither NMU treatment nor methyl-selenocysteine-enriched diets significantly altered themRNA expression patterns of genes encoding the hetero-dimeric circadian transcription factors (Arntl, Npas2, andClock; Supplementary Fig. S1).
Fig. 2. Determination of Per2 (left), MTNR1A (middle), and ERβ (right) protein expression levels in mammary tissues of rats sacrificed at ZT12. A, Westernblot results with whole tissue protein. β-Actin was used as an internal control. B, quantitative analysis of protein expression levels. Mean expressionlevels were calculated from densitometric intensities of bands determined with Western blot and normalized to β-actin. *, P < 0.05, significant difference.C, representative immunohistochemical staining on frozen sections of mammary tissues (×200; bar, 0.1 mm). a-c, NMU, methylselenocysteine diet;d-f: NMU, control diet; g-i: representative epithelial cells with positive staining are indicated by arrows and magnified photomicrograph (×400).
Cancer Prev Res; 3(5) May 2010 645
search. on February 11, 2014. © 2010 American Association

Fang et al.
646
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
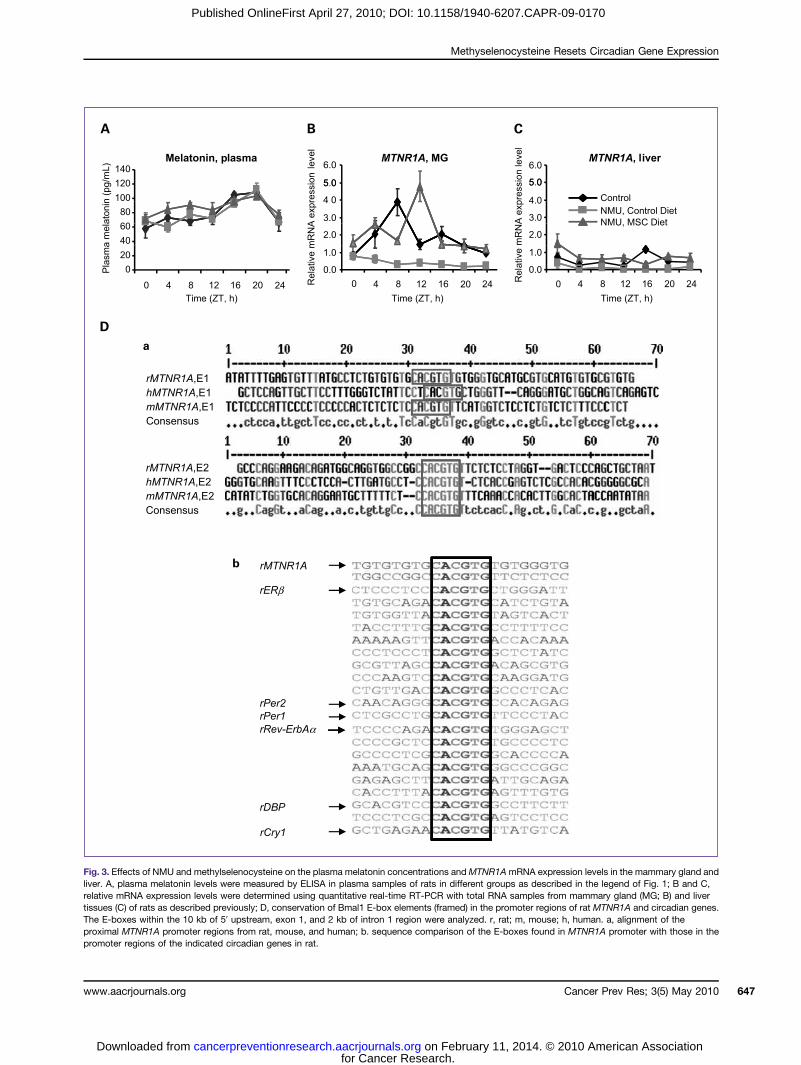
Effects of NMU and methylselenocysteine on plasmamelatonin and melatonin receptor expressionTo investigate the mechanisms by which NMU and
methylselenocysteine alter the expression levels and phasesof circadian genes, we first investigated their effects on theentrainment of circadian rhythm. We compared plasmamelatonin levels of control rats over a 24-hour period tothose of NMU-treated rats with or without a methylseleno-cysteine-enriched diet. All three groups showed normal cir-cadian patterns of plasma melatonin, with nighttime levelsattaining peak concentrations (∼111.7 pg/mL) at ZT20,and the minimum daytime levels (∼59.9 pg/mL) occurringat ZT4 (Fig. 3A). These results excluded the possibility thateither compound altered the production or stability of thehormone. Because the melatonin-mediated responses inperipheral organs are mediated through the MTNR1A (27,28), we next asked if NMU and/or methylselenocysteine af-fected the expression level ofMTNR1A in mammary tissue.The results showed that MTNR1A expression is normallyunder circadian control in rat mammary gland with peakexpression level at ZT8 (Fig. 3B). A single carcinogenic doseof NMU completely disrupted MTNR1A expression inthe mammary gland. By contrast, methylselenocysteine re-stored the levels of the MTNR1A mRNA expression to thenormal levels of untreated control rats, increasing inrhythm-adjustedmean (4.67-fold) and amplitude (6.5-fold)in mammary gland (Fig. 3B) relative to those in NMU-treated rats on control diet (Table 1; Supplementary TableS2). Dietary methylselenocysteine also shifted the phase ofpeak MTNR1A mRNA expression by 4 hours from ZT8 toZT12 (Fig. 3B). The methylselenocysteine-enriched diet alsosignificantly increased expression levels of MTNR1A proteininmammary tissues of NMU-treated rats compared with thecontrol diet (P < 0.05). Immunohistochemistry confirmedthat the differences observed byWestern blot were primarilydue to the increased MTNR1A protein expression in mam-mary epithelial cells (Fig. 2,middle) inmethylselenocysteineversus control diet group. By contrast, the rhythmicMTNR1Aexpression in normal liver was much lower, and neitherNMU nor methylselenocysteine had an appreciable effecton its expression levels in liver (Fig. 3C).Many circadian genes and CCGs are transcriptionally
regulated by the binding of Bmal1/Clock heterodimerictranscription factors to the E-box binding elements in theirpromoter regions. To examine the possibility that therhythmic expression of the melatonin receptor is underBmal1/Clock-mediated transcriptional control, we analyzedthe sequence of the promoter region of the rat MTNR1Agene and compared its conservation across species. DNAsequence comparisons identified two well-conservedE-box binding motifs (CACGTG) of Bmal1 in the promot-er regions of human (at −2926 and −2509), mouse (at−4505 and −237), and rat (at −8949 and −7047) MTNR1Agenes (Fig. 3D, a). We also found numerous copies of theE-box binding motifs of Bmal1 in the promoter regions ofERβ and various circadian genes in rat, as reported previ-ously in mouse (Fig. 3D, b; ref. 29). These findings indi-cated that the MTNR1A might be a CCG, the expression of
Cancer Prev Res; 3(5) May 2010
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
which could be entrained by exogenous signals that alterBmal1 and Clock levels or activity.
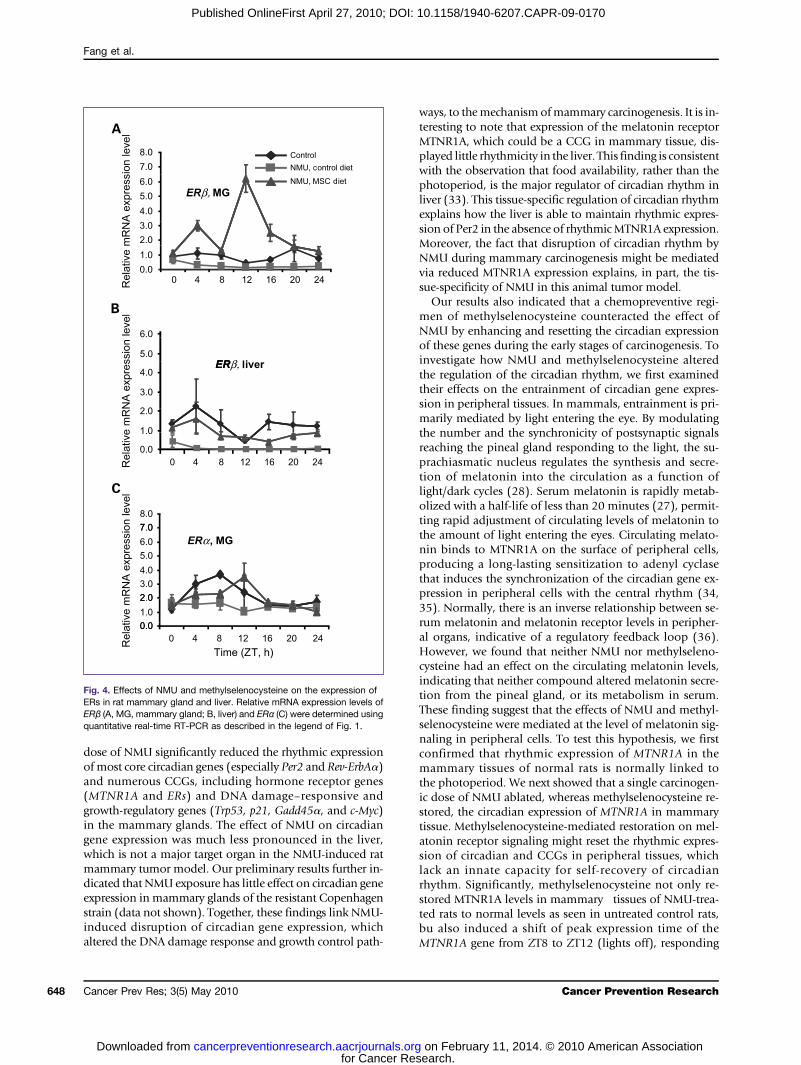
Methylselenocysteine induced circadian rhythm ofERβ and reset the phase of ERα expressionRecent studies indicated that both MTNR1A-mediated
melatonin signaling and Per2 regulate the expression levelsof estrogen receptors (ER) in vitro (29–32). We thereforeasked if NMU and methylselenocysteine altered the circa-dian expression profiles of both ERα and ERβ genes inmammary and liver tissues. We observed that the expres-sion levels of ERβ mRNA in both tissues were normallylow, with a relatively weak circadian rhythm. Nonetheless,a carcinogenic dose of NMU reduced the expression ofERβ mRNA in both tissues. More importantly, dietarymethylselenocysteine dramatically induced circadian ex-pression of ERβ mRNA in mammary gland (Fig. 4A), butnot in liver (Fig. 4B). In mammary glands of NMU-treatedrats, the methylselenocysteine-enriched diet induced a9.48-fold increase in the rhythm-adjusted mean level anda 9.71-fold increase in amplitude of ERβ mRNA expres-sion, as compared with the control diet (Table 1). Westernblot results indicated that dietary methylselenocysteine on-ly resulted in a marginal, statistically insignificant increaseof ERβ protein expression relative to the control diet. How-ever, immunohistochemical staining showed that ERβ pro-tein expression was dramatically enriched in the mammaryepithelial cells of rats on methylselenocysteine-enricheddiets as compared with rats on a control diet. ERβ proteinwas also expressed in stromal cells of mammary glands inboth groups (Fig. 2, right). Although the effect was less dra-matic, methylselenocysteine also restored NMU-disruptedcircadian expression of ERα mRNA in mammary tissuesand shifted its peak expression to ZT12 (Fig. 4C).
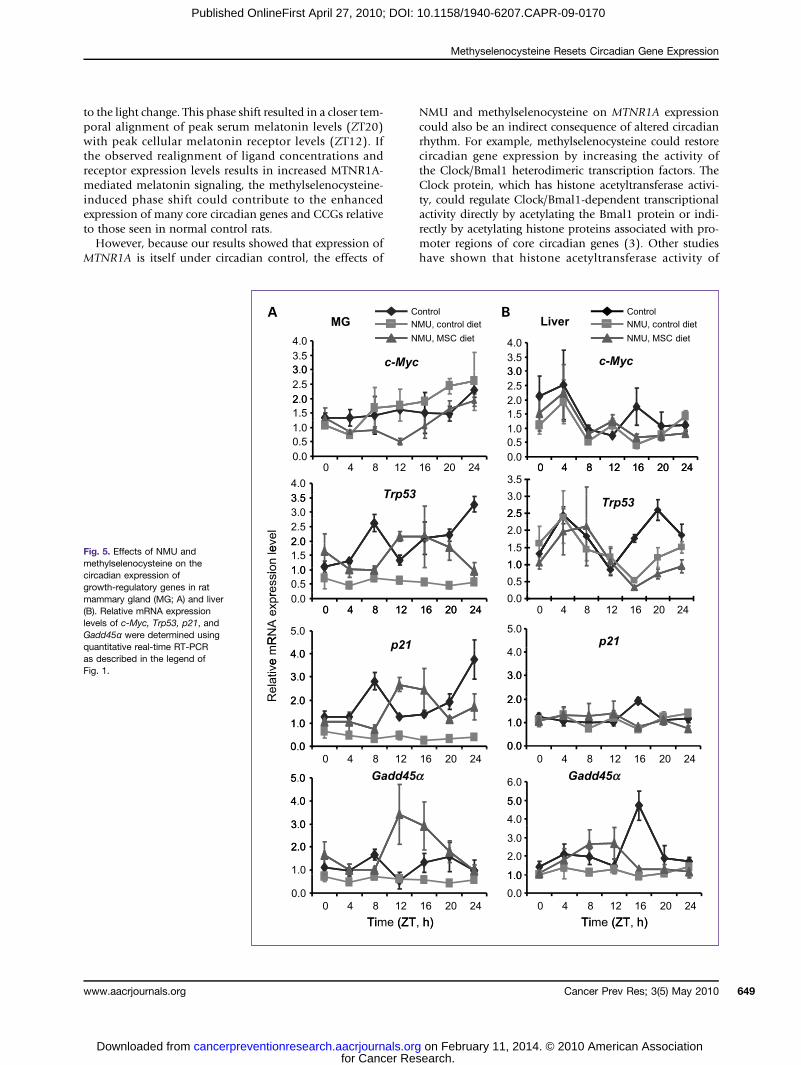
Methylselenocysteine reset the circadian expression ofgrowth-regulatory genesOur analysis further indicated that NMU significantly
reduced the circadian expression of c-Myc, Trp53, p21,and Gadd45α genes in mammary tissues. As was the casefor Per2, MTNR1A, and ERβ, dietary methylselenocysteinealso reset the phase and increased the circadian expressionof these growth-regulatory genes in the mammary gland,but not in the liver (Fig. 5; Table 1). The expression levelsof the Trp53, p21, and Gadd45α were at their peak levels atZT12, immediately after the light was turned off, whereasthe peak expression of the c-Myc proto-oncogene was at itslowest level (Fig. 5). These results indicate that NMU dis-rupted, whereas a methylselenocysteine-enriched diet re-set, the circadian expression of these genes involved incellular responses to genotoxic stress and cell cycle control.
Discussion
Epidemiologic and animal studies indicate that disrup‐tion of the circadian rhythm is associated with an increasedrisk of breast cancer (18). In the present study, we showedthat exposure of pubescent female F344 rats to a carcinogenic
Cancer Prevention Research
search. on February 11, 2014. © 2010 American Association
Methyselenocysteine Resets Circadian Gene Expression
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
Fig. 3. Effects of NMU and methylselenocysteine on the plasma melatonin concentrations andMTNR1AmRNA expression levels in the mammary gland andliver. A, plasma melatonin levels were measured by ELISA in plasma samples of rats in different groups as described in the legend of Fig. 1; B and C,relative mRNA expression levels were determined using quantitative real-time RT-PCR with total RNA samples from mammary gland (MG; B) and livertissues (C) of rats as described previously; D, conservation of Bmal1 E-box elements (framed) in the promoter regions of rat MTNR1A and circadian genes.The E-boxes within the 10 kb of 5′ upstream, exon 1, and 2 kb of intron 1 region were analyzed. r, rat; m, mouse; h, human. a, alignment of theproximal MTNR1A promoter regions from rat, mouse, and human; b. sequence comparison of the E-boxes found in MTNR1A promoter with those in thepromoter regions of the indicated circadian genes in rat.
Cancer Prev Res; 3(5) May 2010www.aacrjournals.org 647
for Cancer Research. on February 11, 2014. © 2010 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Fang et al.
648
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
dose of NMU significantly reduced the rhythmic expressionof most core circadian genes (especially Per2 and Rev-ErbAα)and numerous CCGs, including hormone receptor genes(MTNR1A and ERs) and DNA damage–responsive andgrowth-regulatory genes (Trp53, p21, Gadd45α, and c-Myc)in the mammary glands. The effect of NMU on circadiangene expression was much less pronounced in the liver,which is not a major target organ in the NMU-induced ratmammary tumor model. Our preliminary results further in-dicated that NMU exposure has little effect on circadian geneexpression in mammary glands of the resistant Copenhagenstrain (data not shown). Together, these findings link NMU-induced disruption of circadian gene expression, whichaltered the DNA damage response and growth control path-
Cancer Prev Res; 3(5) May 2010
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
ways, to themechanismofmammary carcinogenesis. It is in-teresting to note that expression of the melatonin receptorMTNR1A, which could be a CCG in mammary tissue, dis-played little rhythmicity in the liver. This finding is consistentwith the observation that food availability, rather than thephotoperiod, is the major regulator of circadian rhythm inliver (33). This tissue-specific regulation of circadian rhythmexplains how the liver is able to maintain rhythmic expres-sion of Per2 in the absence of rhythmicMTNR1A expression.Moreover, the fact that disruption of circadian rhythm byNMU during mammary carcinogenesis might be mediatedvia reduced MTNR1A expression explains, in part, the tis-sue-specificity of NMU in this animal tumor model.Our results also indicated that a chemopreventive regi-
men of methylselenocysteine counteracted the effect ofNMU by enhancing and resetting the circadian expressionof these genes during the early stages of carcinogenesis. Toinvestigate how NMU and methylselenocysteine alteredthe regulation of the circadian rhythm, we first examinedtheir effects on the entrainment of circadian gene expres-sion in peripheral tissues. In mammals, entrainment is pri-marily mediated by light entering the eye. By modulatingthe number and the synchronicity of postsynaptic signalsreaching the pineal gland responding to the light, the su-prachiasmatic nucleus regulates the synthesis and secre-tion of melatonin into the circulation as a function oflight/dark cycles (28). Serum melatonin is rapidly metab-olized with a half-life of less than 20 minutes (27), permit-ting rapid adjustment of circulating levels of melatonin tothe amount of light entering the eyes. Circulating melato-nin binds to MTNR1A on the surface of peripheral cells,producing a long-lasting sensitization to adenyl cyclasethat induces the synchronization of the circadian gene ex-pression in peripheral cells with the central rhythm (34,35). Normally, there is an inverse relationship between se-rum melatonin and melatonin receptor levels in peripher-al organs, indicative of a regulatory feedback loop (36).However, we found that neither NMU nor methylseleno-cysteine had an effect on the circulating melatonin levels,indicating that neither compound altered melatonin secre-tion from the pineal gland, or its metabolism in serum.These finding suggest that the effects of NMU and methyl-selenocysteine were mediated at the level of melatonin sig-naling in peripheral cells. To test this hypothesis, we firstconfirmed that rhythmic expression of MTNR1A in themammary tissues of normal rats is normally linked tothe photoperiod. We next showed that a single carcinogen-ic dose of NMU ablated, whereas methylselenocysteine re-stored, the circadian expression of MTNR1A in mammarytissue. Methylselenocysteine-mediated restoration on mel-atonin receptor signaling might reset the rhythmic expres-sion of circadian and CCGs in peripheral tissues, whichlack an innate capacity for self-recovery of circadianrhythm. Significantly, methylselenocysteine not only re-stored MTNR1A levels in mammary tissues of NMU-trea-ted rats to normal levels as seen in untreated control rats,bu also induced a shift of peak expression time of theMTNR1A gene from ZT8 to ZT12 (lights off), responding
Fig. 4. Effects of NMU and methylselenocysteine on the expression ofERs in rat mammary gland and liver. Relative mRNA expression levels ofERβ (A, MG, mammary gland; B, liver) and ERα (C) were determined usingquantitative real-time RT-PCR as described in the legend of Fig. 1.
Cancer Prevention Research
search. on February 11, 2014. © 2010 American Association
Methyselenocysteine Resets Circadian Gene Expression
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
to the light change. This phase shift resulted in a closer tem-poral alignment of peak serum melatonin levels (ZT20)with peak cellular melatonin receptor levels (ZT12). Ifthe observed realignment of ligand concentrations andreceptor expression levels results in increased MTNR1A-mediated melatonin signaling, the methylselenocysteine-induced phase shift could contribute to the enhancedexpression of many core circadian genes and CCGs relativeto those seen in normal control rats.However, because our results showed that expression of
MTNR1A is itself under circadian control, the effects of
www.aacrjournals.org
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
NMU and methylselenocysteine on MTNR1A expressioncould also be an indirect consequence of altered circadianrhythm. For example, methylselenocysteine could restorecircadian gene expression by increasing the activity ofthe Clock/Bmal1 heterodimeric transcription factors. TheClock protein, which has histone acetyltransferase activi-ty, could regulate Clock/Bmal1-dependent transcriptionalactivity directly by acetylating the Bmal1 protein or indi-rectly by acetylating histone proteins associated with pro-moter regions of core circadian genes (3). Other studieshave shown that histone acetyltransferase activity of
Fig. 5. Effects of NMU andmethylselenocysteine on thecircadian expression ofgrowth-regulatory genes in ratmammary gland (MG; A) and liver(B). Relative mRNA expressionlevels of c-Myc, Trp53, p21, andGadd45α were determined usingquantitative real-time RT-PCRas described in the legend ofFig. 1.
Cancer Prev Res; 3(5) May 2010 649
search. on February 11, 2014. © 2010 American Association

Fang et al.
650
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
Clock protein can be counteracted by Sirt1, a NAD+-dependent histone deacetylase, thereby linking cellularenergy balance to Clock/Bmal1-regulated transcriptionof circadian genes and CCGs (37). Moreover, the inhibi-tion of Sirt1-associated histone deacetylase activity withsplitomicin or nicotinamide increased histone 3 acetyla-tion levels, and further increased the circadian oscillationof circadian gene expression (37). Based on these find-ings, we hypothesize that NMU and methylselenocysteineregulate the circadian expression of circadian genes andCCGs at the epigenetic level during carcinogenesis andchemoprevention. This hypothesis is supported by severallines of evidence. Inorganic selenium, which has antioxi-dant activity, alters NADP/NADPH and NAD/NADH-coupled redox cycling (21). Methylselenocysteine couldtherefore alter the redox state of these cofactors to inhibitthe histone deacetylase activity of Sirt1 (37, 38), furtherenhancing the rhythmic expression of circadian genes.Consistent with this possibility, our previous studies sug-gested that NMU might mediate part of its carcinogeniceffects through epigenetic mechanisms that alter DNAconformation (39, 40). In addition, the majority of thecircadian genes deregulated in human cancers harbor in-
Cancer Prev Res; 3(5) May 2010
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
creased DNA methylation and/or deacetylated histones intheir promoter regions (15–17). Recent studies alsoshowed that selenite can inhibit DNA hypermethylationand histone deacetylation in prostate cancer cells in vitro(41). More importantly, our preliminary in vitro experimentsalso showed that methylselenocysteine increases histoneacetylation in mammary epithelial cells (data not shown).The present study also provides insight into the me-
chanisms by which resetting of circadian gene expressionby methylselenocysteine can contribute to chemopreven-tion. Previous studies showed that dietary methylseleno-cysteine supplements did not prevent the initiation ofcarcinogenesis, but suppressed the outgrowth of premalig-nant lesions (21). Other studies showed that ovariectomyafter carcinogen exposure prevents mammary carcinogene-sis in pubescent female rats, implicating estrogens in thepromotion of initiated cells (42). Significantly, the core cir-cadian protein Per2 binds to and increases the degradationof ERα protein and also indirectly regulates the transcriptionof ERβ (29, 31). These findings suggest that disruption ofcircadian gene expression could enhance, whereas restora-tion of circadian rhythm could inhibit, estrogen-mediatedpromotion of mammary carcinogenesis. ERα and ERβ have
Fig. 6. Hypothetical model for the interaction of NMU and methylselenocysteine with the circadian control network during the early stages ofcarcinogenesis. The proposed model combines the changes in gene expression observed in the present study with known regulating networks(blue arrows), comprising and modulating the activity of the molecular oscillator in peripheral mammalian cells (5). The rhythmic expression of the melatoninreceptor (MTNR1A), itself a CCG that plays a crucial role in the entrainment and synchronization of peripheral circadian rhythm with a central pacemaker,was abolished by NMU and reset by methylselenocysteine without altering the expression of Bmal1 and Clock. These data are consistent with thepossibility that NMU and methylselenocysteine regulate rhythmic expressions of circadian and CCGs by modifying the activity of Clock/Bmal1. AlteredBmal1/Clock activity would also modulate the expression of ERβ and other growth-regulatory genes. Hypothetically positive or negative inputs from NMUand methylselenocysteine are indicated by dotted red or green lines, respectively. E, estrogen; ERE, estrogen-response element; M, melatonin;CK, casein kinase; PKA, protein kinase A; CREB, cAMP-response element binding protein; HAT, histone acetyltransferase; HDAC, histone deacetylase.
Cancer Prevention Research
search. on February 11, 2014. © 2010 American Association

Methyselenocysteine Resets Circadian Gene Expression
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
opposing effects on estrogen-mediated cell growth of mam-mary epithelial cells, whereas overexpression of Per2 inhib-ited breast cancer cell growth (43, 44). Consistent with thesein vitro study results, we found that methylselenocysteinedramatically enhanced circadian expression of Per2 andERβ at mRNA and protein levels in mammary epithelialcells in vivo. Although we observed a less dramatic effect ofmethylselenocysteine on ERα, the large increase in ERβ le-vels would elevate the ERβ/ERα ratio in mammary cells. Ithas been shown that when coexpressed in breast cancercells, ERβ antagonizes the expression of ERα-mediatedexpression of growth-regulatory genes, including c-Mycand cyclin D1. Our in vivo studies also found that methyl-selenocysteine-enriched diets inhibited the expression ofc-Myc in mammary tissues in vivo, with the lowest levelof c-Myc expression occurring at ZT12, whereas Per2 andERβ expression levels were at their highest levels. Takentogether, these findings suggest that enhanced circadianexpression of ERβ protein might suppress the promotionphase of mammary carcinogenesis by inhibiting estrogen-induced cell growth.Another mechanism by which methylselenocysteine-
mediated effects on circadian rhythm could contribute tothe suppression of mammary carcinogenesis is through al-tered responses to genotoxic stressors (45, 46). Disruptingnormal circadian rhythm though shift work, exposure tochemical carcinogens, or genetic manipulation wouldtherefore be expected to negatively affect the ability of cellsto respond to and repair DNA damage. Consistent withsuch a mechanism, our data showed that disruption of cir-cadian rhythm by NMU inhibited the rhythmic expressionof DNA damage–responsive and growth-regulatory genes(Tpr53, p21, Gadd45α, and c-Myc). By contrast, a dietaryregimen of methylselenocysteine reset and enhanced therhythmic expression of core circadian genes, Per2 andRev-ErbAα, and CCGs, including these DNA damage–re-sponsive genes. Interestingly, the peak expression levelsof the Trp53, p21, and Gadd45α genes corresponded tem-porally with the lowest level of c-Myc gene expression, sug-gesting that enhanced circadian gene expression bymethylselenocysteine also plays a role in coordinating cel-lular growth with DNA damage responses.Together, our findings suggest that disruption and re-
setting of circadian rhythm by NMU and chemopreven-tive methylselenocysteine, respectively, are linked to themechanisms of mammary tumor promotion (Fig. 6).Disruption of circadian rhythm by shift work, exposureto light at night, and blindness has also been associatedwith increased rates of breast, prostate, and colon can-cers (18). Our findings thus have potential implicationsfor mitigating the increased risk of breast and prostate can-cers associatedwith shift work (47). Numerous studies have
www.aacrjournals.org
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
suggested that normal dietary levels of methylselenocys-teine could reduce the risk of several cancers without excesstissue accumulation or toxicity (19). Methylselenocysteineis a naturally occurring amino acid synthesized in plantsgrown in selenium-containing soil. The highest levels arefound in Brazil nuts (up to 200 μg each), garlic, wild leeks,onions, and broccoli. Lower levels are found in somemeatsand seafood. Our chemoprevention studies in rats indicat-ed that dietary methylselenocysteine at 3 ppm of seleniumfor up to 8 months yielded no apparent toxicity (23). Thislevel of dietary supplementation with methylselenocys-teine corresponds to a daily dose of ∼200 to 300 μg in hu-man, which is well below the recommended maximal dailyintake level of selenium (400 μg/d) established by the Insti-tute of Medicine (48). Significant toxicity in humans is typ-ically seen only after prolonged intake exceeding 800 μg/d(49). The average dietary intake of selenium in Americansand Europeans is ∼100 μg daily, with many populationsaround the world routinely ingesting 400 to 600 μg of sele-nium daily without apparent adverse effects (50). Multipleclinical trials with 200 and 400 μg/d of inorganic seleniumformonths to years have shown no toxicity (51, 52). Anoth-er study reported no obvious toxicity with doses from1,600 to 3,200 μg/d given for 12 months (53). We there-fore expect that a dietary supplement of ∼200 μg ofmethylselenocysteine per day, a level currently foundin many multivitamin preparations, would be effectivein resetting circadian rhythm in humans. This predictionis currently being tested in shift workers in preparationfor a prospective intervention trial to evaluate the effica-cy of methylselenocysteine in mitigating the increasedrisk of cancer associated with the disruption of circadianrhythm by shift work.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Dr. Michael Gallo for helpful discussion and Jennifer Williamsfor critical reading of the manuscript.
Grant Support
NIH grants U19ES011387, P30ES005022, and P30ES007033; fundsfrom the Environmental and Occupation Health Sciences Institute, a jointinstitute of the University of Medicine and Dentistry of New Jersey andRutgers, the State University of New Jersey.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
Received 08/16/2009; revised 02/02/2010; accepted 02/15/2010;published OnlineFirst 04/27/2010.
References
1. Fu L, Lee CC. The circadian clock: pacemaker and tumour suppres-sor. Nat Rev Cancer 2003;3:350–61.2. Chen-Goodspeed M, Lee CC. Tumor suppression and circadian
function. J Biol Rhythms 2007;22:291–8.
3. Doi M, Hirayama J, Sassone-Corsi P. Circadian regulator CLOCK is ahistone acetyltransferase. Cell 2006;125:497–508.
4. Ko CH, Takahashi JS. Molecular components of the mammaliancircadian clock. Hum Mol Genet 2006;15:271–7.
Cancer Prev Res; 3(5) May 2010 651
search. on February 11, 2014. © 2010 American Association

Fang et al.
652
Published OnlineFirst April 27, 2010; DOI: 10.1158/1940-6207.CAPR-09-0170
5. Mackey SR. Biological rhythms workshop IA: Molecular Basis ofRhythms Generation. In: Stillman BSD, Grodzicker T, editors. ColdSpring Harbor Symposia on Quantitative Biology. Cold SpringHarbor: Cold Spring Harbor Laboratory Press; 2007, p. 7–19.
6. Balsalobre A. Clock genes in mammalian peripheral tissues. CellTissue Res 2002;309:193–9.
7. Shim HS, Kim H, Lee J, et al. Rapid activation of CLOCK byCa2+-dependent protein kinase C mediates resetting of themammalian circadian clock. EMBO Rep 2007;8:366–71.
8. De Leersnyder H, Bresson JL, de Blois MC, et al. β1-Adrenergic an-tagonists and melatonin reset the clock and restore sleep in a circadi-an disorder, Smith-Magenis syndrome. J Med Genet 2003;40:74–8.
9. Liu C, Weaver DR, Jin X, et al. Molecular dissection of two distinctactions of melatonin on the suprachiasmatic circadian clock. Neuron1997;19:91–102.
10. Hamilton T. Influence of environmental light and melatonin uponmammary tumour induction. Br J Surg 1969;56:764–6.
11. Aubert C, Janiaud P, Lecalvez J. Effect of pinealectomy andmelatoninon mammary tumor growth in Sprague-Dawley rats under differentconditions of lighting. J Neural Transm 1980;47:121–30.
12. Gery S, Gombart AF, Yi WS, Koeffler C, Hofmann WK, Koeffler HP.Transcription profiling of C/EBP targets identifies Per2 as a geneimplicated in myeloid leukemia. Blood 2005;106:2827–36.
13. Hua H, Wang Y, Wan C, et al. Circadian gene mPer2 overexpressioninduces cancer cell apoptosis. Cancer Sci 2006;97:589–96.
14. Canaple L, Kakizawa T, Laudet V. The days and nights of cancercells. Cancer Res 2003;63:7545–52.
15. Chen ST, Choo KB, Hou MF, Yeh KT, Kuo SJ, Chang JG. Deregu-lated expression of the PER1, PER2 and PER3 genes in breastcancers. Carcinogenesis 2005;26:1241–6.
16. Yeh KT, Yang MY, Liu TC, et al. Abnormal expression of period 1(PER1) in endometrial carcinoma. J Pathol 2005;206:111–20.
17. Yang MY, Chang JG, Lin PM, et al. Downregulation of circadianclock genes in chronic myeloid leukemia: alternative methylation pat-tern of hPER3. Cancer Sci 2006;97:1298–307.
18. Stevens RG. Circadian disruption and breast cancer: from melatoninto clock genes. Epidemiology 2005;16:254–8.
19. Clark LC, Combs GF, Jr., Turnbull BW, et al. Nutritional Prevention ofCancer Study Group. Effects of selenium supplementation for cancerprevention in patients with carcinoma of the skin. A randomizedcontrolled trial. JAMA 1996;276:1957–63.
20. El-Bayoumy K, Sinha R. Mechanisms of mammary cancer chemopre-vention by organoseleniumcompounds.Mutat Res 2004;551:181–97.
21. Jackson MI, Combs GF, Jr. Selenium and anticarcinogenesis: under-lying mechanisms. Curr Opin Clin Nutr Metab Care 2008;11:718–26.
22. Medina D, Thompson H, Ganther H, Ip C. Se-methylselenocysteine:a new compound for chemoprevention of breast cancer. Nutr Cancer2001;40:12–7.
23. Zhang X, Zarbl H. Chemopreventive doses of methylselenocysteinealter circadian rhythm in rat mammary tissue. Cancer Prev Res 2008;1:119–27.
24. Fang MZ, Chen D, Sun Y, Jin Z, Christman JK, Yang CS. Reversal ofhypermethylation and reactivation of p16INK4a, RARβ, and MGMTgenes by genistein and other isoflavones from soy. Clin CancerRes 2005;11:7033–41.
25. Fang MZ, Jin Z, Wang Y, et al. Promoter hypermethylation andinactivation of O(6)-methylguanine-DNA methyltransferase inesophageal squamous cell carcinomas and its reactivation in celllines. Int J Oncol 2005;26:615–22.
26. Iurisci I, Filipski E, Reinhardt J, et al. Improved tumor control throughcircadian clock induction by Seliciclib, a cyclin-dependent kinaseinhibitor. Cancer Res 2006;66:10720–8.
27. Pandi-Perumal SR, Trakht I, Srinivasan V, et al. Physiological effectsof melatonin: role of melatonin receptors and signal transductionpathways. Prog Neurobiol 2008;85:335–53.
28. Stehle JH, von Gall C, Korf HW. Melatonin: a clock-output, aclock-input. J Neuroendocrinol 2003;15:383–9.
29. Cai W, Rambaud J, Teboul M, et al. Expression levels of estrogenreceptor β are modulated by components of the molecular clock.Mol Cell Biol 2008;28:784–93.
30. Cos S, Gonzalez A, Martinez-Campa C, Mediavilla MD, Alonso-
Cancer Prev Res; 3(5) May 2010
for Cancer Recancerpreventionresearch.aacrjournals.orgDownloaded from
Gonzalez C, Sanchez-Barcelo EJ. Estrogen-signaling pathway: a linkbetween breast cancer and melatonin oncostatic actions. CancerDetect Prev 2006;30:118–28.
31. Gery S, Virk RK, Chumakov K, Yu A, Koeffler HP. The clock genePer2 links the circadian system to the estrogen receptor. Oncogene2007;26:7916–20.
32. Horard B, Rayet B, Triqueneaux G, Laudet V, Delaunay F, Vanacker JM.Expression of the orphan nuclear receptor ERRα is under circadian reg-ulation in estrogen-responsive tissues. J Mol Endocrinol 2004;33:87–97.
33. Stokkan KA, Yamazaki S, Tei H, Sakaki Y, Menaker M. Entrainment ofthe circadian clock in the liver by feeding. Science 2001;291:490–3.
34. Jilg A, Moek J, Weaver DR, Korf HW, Stehle JH, von Gall C. Rhythmsin clock proteins in the mouse pars tuberalis depend on MT1 mela-tonin receptor signalling. Eur J Neurosci 2005;22:2845–54.
35. von Gall C, Weaver DR, Moek J, Jilg A, Stehle JH, Korf HW. Melatoninplays a crucial role in the regulation of rhythmic clock gene expressionin the mouse pars tuberalis. Ann N Y Acad Sci 2005;1040:508–11.
36. Witt-Enderby PA, Radio NM, Doctor JS, Davis VL. Therapeutic treat-ments potentially mediated by melatonin receptors: potential clinicaluses in the prevention of osteoporosis, cancer and as an adjuvanttherapy. J Pineal Res 2006;41:297–305.
37. Nakahata Y, Kaluzova M, Grimaldi B, et al. The NAD+-dependentdeacetylase SIRT1 modulates CLOCK-mediated chromatin remodel-ing and circadian control. Cell 2008;134:329–40.
38. Rutter J, Reick M, Wu LC, McKnight SL. Regulation of clock andNPAS2 DNA binding by the redox state of NAD cofactors. Science2001;293:510–4.
39. Jin Z, Houle B, Mikheev AM, Cha RS, Zarbl H. Alterations in H-ras1promoter conformation during N-nitroso-N-methylurea-inducedmammary carcinogenesis and pregnancy. Cancer Res 1996;56:4927–35.
40. Zarbl H. Toxicogenomic analyses of genetic susceptibility to mam-mary gland carcinogenesis in rodents: implications for human breastcancer. Breast Dis 2007;28:87–105.
41. Xiang N, Zhao R, Song G, Zhong W. Selenite reactivates silencedgenes by modifying DNA methylation and histones in prostatecancer cells. Carcinogenesis 2008;29:2175–81.
42. Tsukamoto R, Mikami T, Miki K, et al. N-methyl-N-nitrosourea-induced mammary carcinogenesis is promoted by short-termtreatment with estrogen and progesterone mimicking pregnancy inaged female Lewis rats. Oncol Rep 2007;18:337–42.
43. Williams C, Edvardsson K, Lewandowski SA, StromA, Gustafsson JA.A genome-wide study of the repressive effects of estrogen receptor βon estrogen receptor α signaling in breast cancer cells. Oncogene2008;27:1019–32.
44. Matthews J, Gustafsson JA. Estrogen signaling: a subtle balancebetween ERα and ERβ. Mol Interv 2003;3:281–92.
45. Oklejewicz M, Destici E, Tamanini F, Hut RA, Janssens R, van derHorst GT. Phase resetting of the mammalian circadian clock byDNA damage. Curr Biol 2008;18:286–91.
46. Kondratov RV, Antoch MP. Circadian proteins in the regulation of cellcycle andgenotoxic stress responses. TrendsCell Biol 2007;17:311–7.
47. Hansen J. Increased breast cancer risk among women who workpredominantly at night. Epidemiology 2001;12:74–7.
48. Monsen ER. Dietary reference intakes for the antioxidant nutrients:vitamin C, vitamin E, selenium, and carotenoids. J Am Diet Assoc2000;100:637–40.
49. Yang G, Zhou R. Further observations on the human maximum safedietary selenium intake in a seleniferous area of China. J Trace ElemElectrolytes Health Dis 1994;8:159–65.
50. Combs GF, Jr. Impact of selenium and cancer-prevention findings onthe nutrition-health paradigm. Nutr Cancer 2001;40:6–11.
51. Orzano AJ, Strickland PO, Tallia AF, et al. Improving outcomes forhigh-risk diabetics using information systems. J Am Board FamMed 2007;20:245–51.
52. Tsavachidou D, McDonnell TJ, Wen S, et al. Selenium and vitamin E:cell type- and intervention-specific tissue effects in prostate cancer.J Natl Cancer Inst 2009;101:306–20.
53. Orzano AJ, Scott J, Hudson SV, et al. Strategies for conductingcomplex clinical trials in diverse community practices. Med Care2007;45:1221–6.
Cancer Prevention Research
search. on February 11, 2014. © 2010 American Association




















