
Bryostatin-1 Enhances the Maturation and Antigen-Presenting Ability of Murine and Human Dendritic...
11
2004;64:6756-6765. Cancer Res Yoonkyung Do, Venkatesh L. Hegde, Prakash S. Nagarkatti, et al. Cells Antigen-Presenting Ability of Murine and Human Dendritic Bryostatin-1 Enhances the Maturation and Updated version http://cancerres.aacrjournals.org/content/64/18/6756 Access the most recent version of this article at: Cited Articles http://cancerres.aacrjournals.org/content/64/18/6756.full.html#ref-list-1 This article cites by 45 articles, 28 of which you can access for free at: Citing articles http://cancerres.aacrjournals.org/content/64/18/6756.full.html#related-urls This article has been cited by 8 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on June 13, 2013. © 2004 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Bryostatin-1 Enhances the Maturation and Antigen-Presenting Ability of Murine and Human Dendritic...

2004;64:6756-6765. Cancer Res Yoonkyung Do, Venkatesh L. Hegde, Prakash S. Nagarkatti, et al. CellsAntigen-Presenting Ability of Murine and Human Dendritic Bryostatin-1 Enhances the Maturation and
Updated version
http://cancerres.aacrjournals.org/content/64/18/6756
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/64/18/6756.full.html#ref-list-1
This article cites by 45 articles, 28 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/64/18/6756.full.html#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 64, 6756–6765, September 15, 2004]
Bryostatin-1 Enhances the Maturation and Antigen-Presenting Ability of Murineand Human Dendritic Cells
Yoonkyung Do,1 Venkatesh L. Hegde,1 Prakash S. Nagarkatti,2 and Mitzi Nagarkatti1
Departments of 1Microbiology and Immunology and 2Pharmacology and Toxicology, Medical College of Virginia Campus, Virginia Commonwealth University,Richmond, Virginia
ABSTRACT
In this study, we investigated the effect of bryostatin-1 (Bryo-1), anantineoplastic agent, on dendritic cell (DC) maturation, activation, andfunctions. Murine bone marrow-derived DCs on culture with Bryo-1alone, Bryo-1 � calcium ionophore (CI), but not CI alone exhibitedmorphologic changes characteristic of mature DCs and expressed in-creased levels of CD40, CD80, and CD86. Moreover, Bryo-1 � CI–treatedDCs exhibited enhanced antigen-presenting ability to naive and antigen-specific T cells and alloreactive T cells. Bryo-1 � CI–mediated activationof DCs involved protein kinase C (PKC), especially PKC-�, -�, and -�, andaddition of PKC inhibitors impaired their ability to activate T cells.Bryo-1 � CI treatment of DCs did not activate mitogen-activated proteinkinase (MAPK)/extracellular signal-regulated kinase, p38 MAPK, orstress-activated protein kinase/c-Jun NH2-terminal kinase pathways. Fi-nally, treatment of DCs with Bryo-1 alone and Bryo-1 � CI, but not CIalone, induced nuclear translocation of nuclear factor �B as studied byconfocal microscopy. DCs generated from human peripheral blood mono-cytes or from human cord blood CD34� hematopoietic stem cells, whencultured with Bryo-1 � CI, also showed maturation and increased T-cellstimulatory activity. Bryo-1 � CI was more potent in inducing maturationand activation of DCs when compared with other agents such as tumornecrosis factor �, lipopolysaccharide, or phorbol 12-myristate 13-acetate � CI. Collectively, the current study shows for the first time thatBryo-1 alone or in combination with CI may promote the maturation ofDCs and therefore may be useful in development of DC-based cancerimmunotherapy.
INTRODUCTION
Bryostatin-1 (Bryo-1) is a macrocyclic lactone isolated from themarine bryozoan Bugula neritina (1). The primary interest in Bryo-1has been initiated by recognition of the potent antiproliferative effectsagainst various tumor cells (2). Such effects have been related to theability of Bryo-1 to modulate protein kinase C (PKC) activity byactivating or degrading certain isoforms of PKC. However, not all ofthe activities of Bryo-1 are PKC dependent; thus, further studies arenecessary to understand its mechanism of action (2). Some studieshave shown direct antitumor activity of Bryo-1, whereas others sug-gested that Bryo-1 modulates cellular response to other anticanceragents by acting as a sensitizer to anticancer drugs. The latter findingsare further supported by several clinical trials, in which the effect ofBryo-1 was maximal when Bryo-1 preceded other anticancer drugs(3). Bryo-1 has been shown to activate human hematopoietic progen-itor cell growth and promote T-cell activation (4, 5). Another interestin Bryo-1 recently has been stimulated by the observation that Bryo-1can facilitate the differentiation of myelogenous leukemic cells intomonocytic or dendritic-like cells (6).
In the current study, we investigated whether Bryo-1 � calciumionophore (CI) induces maturation of dendritic cells (DCs) and en-hances their antigen-presenting abilities. Unlike other well-known DCstimulators, such as lipopolysaccharide (LPS), tumor necrosis factor �(TNF-�), phorbol 12-myristate 13-acetate (PMA) � CI, and CpGoligonucleotides, Bryo-1 has the advantage that it is in clinical use asan anticancer agent, and if it can activate DCs, it can serve as a noveltool for use in DC-based immunotherapy of cancer. Our data showedthat Bryo-1 � CI treatment induced the maturation of DCs andpromoted their function to stimulate naive and Ag-specific T cells.Our data also suggested that the ability to activate DCs is PKCdependent and mediated through nuclear factor �B (NF�B) activationwithout involving mitogen-activated protein kinase (MAPK) or c-JunNH2-terminal kinase (JNK) pathways.
MATERIALS AND METHODS
Mice. Adult (6 to 8 weeks of age) female C57BL/6 (H-2b) and DBA/2(H-2d) mice were purchased from The Jackson Laboratory (Bar Harbor, ME).
Dendritic Cell Culture from Murine Bone Marrow. Murine DCs wereobtained from bone marrow cells by culturing with murine recombinantgranulocyte macrophage colony-stimulating factor (GM-CSF; 5 ng/mL;PharMingen, San Diego, CA) for 6 days (7). Briefly, bone marrow cells weredepleted of CD4� and CD8� T cells and B220� B cells by using specificmonoclonal antibodies (mAbs) and complement. The cells (1 � 106/well) wereplaced in 24-well plates supplemented with GM-CSF. The cultures were fedevery 2 days to remove nonadherent monocytes by aspirating 75% of themedium and adding fresh medium with GM-CSF. On day 6 of culture, theattached cells were dislodged by gentle pipetting and applied to 6-mL columnsof 50% FCS-RPMI 1640, to enrich the aggregated cells, which released a largenumber of phenotypically characteristic DCs after another day of culture withGM-CSF.
Dendritic Cell Culture from Human Peripheral Blood. Peripheral bloodmononuclear cells (PBMCs) were isolated from adult healthy volunteers bycentrifugation on Ficoll/hypaque (8). The nonadherent cells were collected andused for isolating T cells, as described below. The adherent cells were culturedwith GM-CSF (800 units/mL; PharMingen) and interleukin 4 (IL-4; 20 ng/ml;R&D System, Minneapolis, MN) for 1 week to generate DCs and were referredto as PBMC-DC.
Dendritic Cell Culture from Human Cord Blood. In some experiments,DCs were generated from CD34� hematopoetic stem cells (HSCs) isolatedfrom human cord blood provided by St. Louis Cord Blood Bank (SSMCardinal Glennon Children’s Hospital/St. Louis University School of Medi-cine, St. Louis, MO). Such DCs were referred to as CD34�HSC-DC. Briefly,mononuclear cells were enriched by centrifugation on Ficoll/hypaque. Adher-ent cells were depleted, and CD34� cells were isolated using MACS techniqueaccording to the manufacturer’s recommendations (Miltenyi Biotech, Auburn,CA). The cells then were cultured for 2 weeks with 10% RPMI 1640 supple-mented with 5% autologous plasma and cytokines such as GM-CSF (800units/mL; PharMingen) and TNF-� (50 units/mL; PharMingen) as describedpreviously (9).
Treatment of DC with Bryostatin-1 and Other Activating Agents. DCsgenerated as described previously were cultured for an additional 24 hours inmedium with vehicle alone (control DCs), Bryo-1 (10 nmol/L; Biomol, Plym-outh Meeting, PA) � CI (A23187, 0.5 �g/mL; Sigma, St. Louis, MO; Bryo-1 � CI DC), Bryo-1 alone (Bryo-1 DC), or CI alone (CI DC). For comparison,we also used DCs cultured with LPS (10 �g/mL; Sigma), TNF-� (50 to 500
Received 12/22/03; revised 7/1/04; accepted 7/15/04.Grant support: NIH grants R01AI053703, R01ES09098, R01 AI058300, R21
DA014885, R01DA016545, and R01HL058641.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Requests for reprints: Mitzi Nagarkatti, Department of Microbiology and Immunol-ogy, Medical College of Virginia Campus, Virginia Commonwealth University, Rich-mond, VA 23298. Phone: 804-828-1555; Fax: 804-828-0686; E-mail: [email protected].
©2004 American Association for Cancer Research.
6756
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

units/mL; PharMingen), or PMA (10 ng/mL; Sigma) � CI (0.5 �g/mL; Sigma)for 24 hours and were referred to as LPS DC, TNF-� DC, and PMA � CI DC,respectively. It should be noted that in our preliminary experiments, weincluded various concentrations of Bryo-1 and PMA, and based on the results,we selected the optimal concentration of these agents. In particular, it wasobserved that DCs treated with a high concentration of PMA for 24 hoursresulted in a lower yield of viable cells when compared with treatment withBryo-1.
Cell Surface Antigen Detection with Monoclonal Antibodies Using FlowCytometry. Phenotypic analysis of DCs was carried out by double-stainingwith phycoerythrin (PE)-conjugated and FITC-conjugated mAbs as describedelsewhere following incubation with Fc-block (PharMingen; ref. 10). Thefollowing are the mAbs used against mouse DCs: PE-anti-CD80, PE-anti-CD86, PE-anti-CD40, and FITC-anti-CD11c (PharMingen). For PBMC-DCsand CD34�HSC-DCs, the following mAbs were used: PE-anti-CD86, PE-anti-CD80, FITC-anti-HLA (PharMingen), and PE-anti-CD83 (Immunotech; Beck-man Coulter, Fullerton, CA). The cells were analyzed by flow cytometry(model 752, EPICS V; Coulter Electronics, Miami, FL). The data weredepicted as expression indices, which are the products of percentage ofmarker-positive cells multiplied by the mean fluorescence intensity (% posi-tive � MFI). For comparison, the value obtained for cells exposed to TNF-�or LPS was normalized to 1.
FITC-Dextran Uptake. Murine bone marrow-derived Bryo-1 DCs wereharvested, washed, and incubated with FITC-conjugated dextran (1 mg/mL;Sigma) for 30 minutes at either 37°C or 4°C. The cells were harvested, washed,and analyzed by flow cytometry (model 752, EPICS V; Coulter Electronics).
Allogeneic Mixed Lymphocyte Reaction. Murine control DCs, Bryo-1DCs, CI DCs, Bryo-1 � CI DCs, TNF-� DCs, or PMA � CI DCs wereirradiated at 2000 rad and mixed with responder murine splenic T cells toobtain various T cell:DC ratios in 96-well plates. Briefly, DCs were activatedwith medium (control), Bryo-1, CI, Bryo-1 � CI, TNF-�, or PMA � CI for 24hours as described previously, and the next day, the cells were thoroughlywashed with prewarmed medium, irradiated, and then mixed with allogeneic Tcells. After 3 days of culture, the cells were pulsed with [3H]thymidine duringthe last 8 hours. DNA synthesis was determined by [3H]thymidine incorpora-tion using a liquid scintillation counter.
To study allogeneic mixed lymphocyte reaction (allo-MLR) using humanDCs, CD34�HSC-DCs activated with medium (control), Bryo-1 � CI, LPS, orPMA � CI for 24 hours as described previously were washed and irradiatedbefore mixing with allogeneic T cells (2 � 105/well) in 96-well tissue cultureplates. The allogeneic T cells were isolated from cord blood by using theMACS technique according to the manufacturer’s recommendations (MiltenyiBiotech), the details of which are described below. After 5 days of incubation,the cultures were pulsed with [3H]thymidine during the last 8 hours. DNAsynthesis was determined by [3H]thymidine incorporation using a liquid scin-tillation counter.
Purification of T Cells. Murine T cells used in allo-MLR were purified bypassing the spleen cells of DBA/2 mice over nylon wool columns, followed bydepletion of B cells and macrophages using specific antibodies � complement,as described previously (11). Thus, the cells purified were �95% CD3� asdetermined by flow cytometric analysis. Human PBMC-derived and cordblood-derived T cells were isolated using magnetic beads and columns pro-vided by Miltenyi Biotec. The T cells were �95 to 98% pure as determined bythe expression of CD3 using a flow cytometer.
Antigen-Pulsing and Ag Presentation. In antigen-presentation experi-ments, 18 hours before treatment of DCs with Bryo-1 � CI, the cells werecultured with conalbumin (50 �g/mL). The DCs were harvested, washed,irradiated, and used as Ag-pulsed DCs. In experiments aimed at testing theability of DCs to present tumor-specific antigens, whole LSA tumor cell lysatewas prepared by repeated freezing/thawing cycles and was added directly tothe murine DC cultures 18 hours before treatment with Bryo-1 or otheractivating agents. Next, the DCs were washed, irradiated, and used as lym-phosarcoma (LSA)-pulsed DCs.
Antigen-Specific T-Cell Proliferation. To study conalbumin-specific T-cell proliferation, Ag-specific T cells were generated by injecting C57BL/6mice with 100 �g of conalbumin mixed with complete Freund’s adjuvant(Sigma) in both rear footpads to activate T cells in popliteal lymph nodes(LNs) as described previously (12). Eight to 10 days later, the draining LNswere collected and used as a source of conalbumin-specific T cells. To
determine the proliferative responsiveness of murine Ag-specific T cellsfollowing contact with DCs, the DCs (Ag pulsed or unpulsed; 1 � 105/well)were treated with medium alone or Bryo-1 � CI, irradiated, and then mixedwith Ag-specific T cells (3 � 105/well). After 72 hours, the cultures werepulsed with [3H]thymidine during the last 8 hours. DNA synthesis wasdetermined by [3H]thymidine incorporation using a liquid scintillationcounter.
To study human T-cell responsiveness to tetanus toxoid (TT), PBMC-DCswere cultured with TT (10 Lf/mL; Aventis, Bridgewater, NJ) overnight. Thenext day, the DCs were harvested, washed, and incubated with medium,Bryo-1 � CI, PMA � CI, or TNF-�, irradiated, and used as TT-pulsed DCs.Various numbers of irradiated PBMC-DCs were mixed in 96-well plates withautologous T cells (1 � 105/well) isolated from peripheral blood of samedonor. After 5 days, the cultures were pulsed with [3H]thymidine during thelast 8 hours. DNA synthesis was determined by [3H]thymidine incorporationusing a liquid scintillation counter.
Generation of LSA Tumor-Specific T Cells. LSA tumor-specific T cellswere generated as described previously (13). Briefly, LSA tumor cells(1 � 106) were injected into syngeneic C57BL/6 mice, and 5 days later, themice were treated with 1,3-bis(chloroethyl)-1-nitrosourea (50 mg/kg bodyweight). Three days later, T cells were purified from the spleens and used asLSA-specific T cells.
Cytotoxicity against LSA Tumor Cells. To test whether treatment ofBryo-1 � CI enhances the ability of DCs to generate cytolytic T cells againstLSA tumor cells, LSA-specific T cells were obtained as described previouslyand were incubated for 5 days with either control DC or Bryo-1 � CI DCpulsed with LSA tumor cell lysate as described previously. Next, the cytolyticT cells were tested in a 4-hour 51Cr release assay against LSA tumor cells asdescribed previously (13).
Western Blot Analysis. DCs were lysed by repeated freeze/thawing, andprotein concentration in the supernatant was determined using the BCAprotein assay kit (Pierce, Rockford, IL). Total protein (15 to 40 �g) wasseparated by SDS-PAGE and transferred to a membrane. Blots wereblocked with 5% (w/v) nonfat dry milk in Tris-buffered saline/Tween [0.5mol/L NaCl, 20 mmol/L Tris-HCl, and 0.1% (v/v) Tween 20 (pH 7.6)] andprobed with the primary antibody. Next, blots were washed and incubatedwith horseradish peroxidase-conjugated secondary antibody and visualizedwith enhanced chemiluminescence reagent (Amersham Biosciences, Piscat-away, NJ).
Use of Inhibitors PKC, p44/p42 MAPK, or p38 MAPK. Murine DCswere preincubated with various concentrations of pan-PKC inhibitor bisin-dolylmaleimide I (Calbiochem, San Diego, CA), PKC-� inhibitor Rottlerin(Calbiochem), PD98059 (p44/p42 MAPK inhibitor), or SB203580 (p38MAPK inhibitor) for 1 hour at 37°C and then treated either with medium(control DC) or Bryo-1 � CI for 24 hours as described previously.
Preparation of Nuclear Extract and Electrophoretic Mobility ShiftAssay. DCs were incubated for 2 hours with LPS, TNF-�, or Bryo-1 � CI,and nuclear extracts were prepared using commercial nuclear extract kit(Active Motif, Carlsbad, CA). The reactions were started by adding 10 �gof nuclear protein extract to a reaction mixture containing 3 �g ofpoly(dI�dC) (Pharmacia), 4 �L of 5� binding buffer [50 mmol/L HEPES(pH 7.9), 50 mmol/L KCl, 12.5 mmol/L MgCl2, 0.5 mmol/L EDTA, 50%glycerol v/v, 5 mmol/L ATP, and 5 mmol/L DTT[, and [�-32P]ATP-labeleddouble-stranded DNA (dsDNA) probe (25 fmol; ref. 14). All of the DNAbinding reactions were conducted for 20 minutes at room temperature in afinal volume of 20 �L except for supershift, in which the reaction wasconducted on ice for 30 minutes. Cold competitor oligonucleotides wereadded to the reaction mixture before the radiolabeled probe. For supershiftassay, protein extracts were incubated with polyclonal anti-p50 (Santa CruzBiotechnology, Santa Cruz, CA) for 30 minutes on ice, followed by theaddition of the radiolabeled probe for another 30 minutes on ice. The entiresample then was loaded on a 5% nondenaturing polyacrylamide gel inTris-borate-EDTA buffer. After electrophoresis, gels were dried, and sep-arated protein-DNA complexes were visualized by autoradiography usingKodak X-Omat AR film (Rochester, NY). The following dsDNA oligo-nucleotides were used in electrophoretic mobility shift assay analysis aslabeled or competitor probes: NF�B sense, 5�-AGT TGA GGG GAC TTTCCC AGG-3�; NF�B antisense, 5�-GCT CCT GGG AAA GTC CCC
6757
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

TCA-3�; NF�B mutant sense, 5�-AGT TGA GGC GAC TTT CCC AGG-3�;and NF�B mutant antisense, 5�-GCT CCT GGG AAA GTC GCC TCA-3�.
Detection of Nuclear Translocation of NF�B by Confocal Microscopy.DCs stimulated for 2 hours with medium alone, LPS, Bryo-1, Bryo-1 � CI,or CI alone were washed and resuspended at 1 � 106 cells/mL, and 100 �Lof this cell suspension were layered onto polylysine-coated slides. Cellswere allowed to adhere for 30 minutes at 37°C. Next, cells were fixed in3.7% buffered paraformaldehyde at room temperature for 15 minutes andpermeabilized in 0.2% Triton X-100 in PBS for 5 minutes. The Fc receptorswere blocked as before. The primary antibodies, goat anti-NF�Bp65 (C-19)or rabbit anti-NF�Bp50 (C-20; Santa Cruz Biotechnology), were added atappropriate concentrations for 1 hour at room temperature. Cells subse-quently were washed twice in PBS and stained with the appropriatefluorescent-labeled antibodies, Cy3 antigoat IgG, or Cy2 antirabbit IgG(Jackson ImmunoResearch, West Grove, PA) at room temperature for 1hour. Cells were washed twice with PBS, and nuclei were stained withHoechst 33258 (Molecular Probes, Eugene, OR) for 1 minute before slideswere mounted in ProLong Antifade mounting medium (Molecular Probes)and examined using a Zeiss LSM510 META laser scanning confocalmicroscope (Carl Zeiss, Oberkochen, Germany). Images were acquiredusing thin (�0.8 �m) optical sections through X-Y axis. Images werecollected simultaneously and sequentially to rule out bleed-through fromone fluorescence channel to another.
Statistical Analysis. The statistical comparisons between different studygroups were carried out using Student’s t test, and differences of P � 0.05were considered to be significant. Each experiment was repeated at leastthree times.
RESULTS
Effect of Bryo-1 � CI Treatment on DC Morphology andExpression Levels of Costimulatory Molecules. DCs were culturedfrom mouse bone marrow cells with GM-CSF for 6 days. Next, theywere treated either with Bryo-1 � CI (Bryo-1 � CI DCs) or mediumalone (control DCs) for 24 hours and observed for morphology. Wegenerally observed larger proportions of cells with dendrites in Bryo-1 � CI DCs when compared with control DCs (Fig. 1A and B). TheDCs also were treated with PMA � CI (PMA � CI DCs) or TNF-�(TNF-� DCs; Fig. 1C and D), which served as positive controls.These cultures also showed enhanced numbers of cells with DCmorphology when compared with the control culture.
Next, the expression of costimulatory molecules was examined bydouble-staining the DCs with FITC-CD11c and PE–anti-costimula-tory molecules. The CD11c� population was gated, and the expres-sion level of costimulatory molecules was compared between differ-ent treatment groups (Fig. 1E). The data showed that there was asignificant up-regulation of CD40, CD80, and CD86 molecule ex-pression on Bryo-1 � CI DCs when compared with control DCs. Itshould be noted that the increased levels of costimulatory moleculeexpression on Bryo-1 � CI DCs were comparable with those seen onLPS DCs except for Bryo-1 � CI DCs, which expressed higher levelsof CD86 and MHC class II molecules than LPS DCs (data not shown).
To further clarify the contribution of Bryo-1 and CI on DC matu-
Fig. 1. The morphology and expression of surface markers on murine bonemarrow-derived DCs following Bryo-1 � CI treatment. DCs were cultured withBryo-1 � CI for 24 hours as described in Materials and Methods. A–D, The cellswere observed under a phase contrast microscope. The top and bottom panels showmagnification of 10� and 40�, respectively. E, The DCs (1 � 106) were harvested,washed, and double-stained with FITC-conjugated CD11c and PE-conjugated anti-CD40, -CD80, or -CD86 mAbs. The CD11c� population was gated, and the expres-sion of each costimulatory molecule was indicated. The mean fluorescence intensity(MFI, number in parenthesis) was indicated in each histogram.
6758
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

ration, we also treated DCs with either Bryo-1 alone, CI alone, or thecombination of Bryo-1 � CI (Fig. 1E). The data showed that additionof Bryo-1 alone to DCs induced similar up-regulation of costimula-tory molecules as treatment of DCs with Bryo-1 � CI. CI alone alsofailed to induce significant phenotypic changes in DCs. We corrobo-rated the maturation of DCs by Bryo-1 � CI using FITC-dextranuptake (Fig. 2). The data showed Bryo-1 � CI DCs had decreasedability to uptake FITC-dextran when compared with the control DCs,which indicated that treatment of DCs with Bryo-1 � CI induced theirmaturation.
Effect of Bryo-1 Treatment on Ability of DC to Stimulate theProliferative Responsiveness of Allogeneic and Ag-Specific TCells. To test the effect of Bryo-1 � CI treatment on functionalproperties of DCs, we performed allo-MLR (Fig. 3A and B). To this
end, responder T cells obtained from DBA/2 mice (H-2d) were stim-ulated with DCs from C57BL/6 mice (H-2b). The data showed thatBryo-1 � CI DCs stimulated the proliferative responsiveness ofallogeneic T cells stronger than control DCs (Fig. 3A). Furthermore,Bryo-1 DCs were able to stimulate allogeneic T cells similar toBryo-1 � CI DCs, although to a lesser extent at high stimulator:effector ratios. It also was observed that CI DCs were similar to thecontrol DCs in that they were not effective in further stimulatingallogeneic T cells. Thus, in functional studies such as allo-MLR, DCsstimulated with Bryo-1 � CI induced better T-cell responsivenessthan Bryo-1 alone, whereas DCs treated with CI alone failed to induceenhanced T-cell activation (Fig. 3A). Thus, in all of the experiments,we used the combination of Bryo-1 � CI to activate DCs. Moreover,the proliferative response of T cells was stronger when Bryo-1 � CIDCs were used for stimulation when compared with the use of TNF-�DCs or PMA � CI DCs (Fig. 3B).
When the DCs were pulsed with conalbumin antigen followed byBryo-1 � CI treatment and then mixed with conalbumin-specific Tcells, Bryo-1 � CI DCs induced stronger Ag-specific T-cell prolifer-ation than control DCs (Fig. 3C and D). Furthermore, when naive Tcells were used instead of sensitized Ag-specific T cells, Bryo-1 � CIDCs were able to induce significant proliferative responsiveness ofnaive T cells to conalbumin stronger than control DCs (Fig. 3E).
We also investigated the proliferative responsiveness of tumor-specific T cells following stimulation with DCs pulsed by tumor celllysate. The data shown in Fig. 3F indicate that Bryo-1 � CI DCscould induce strong proliferative responsiveness of LSA tumor-specific T cells, whereas control DCs, TNF-� DCs, or PMA � CIDCs were not as effective. It should be noted that LSA tumor-specificT cells did not respond to DCs pulsed with other syngeneic tumorlysates, such as EL-4, as also shown previously (13, 15).
Fig. 2. Endocytosis by murine bone marrow-derived DCs. The DCs (2 � 105) wereharvested and incubated with FITC-conjugated dextran (1 mg/mL) for 30 minutes at either37°C or 4°C. The filled histogram represent cells incubated at 4°C, and the emptyhistograms represent similar cells incubated at 37°C. The percentage of cells showingpositive fluorescence is indicated in each histogram. The number in parenthesis indicatesthe increase in mean fluorescence intensity (MFI).
Fig. 3. The effect of Bryo-1 on murine bonemarrow-derived DC-induced proliferative respon-siveness of allogenic T cells and Ag-specific Tcells. A and B, Various numbers of DCs ofC57BL/6 mice treated with various activators orvehicle (control) were irradiated and mixed withallogeneic T cells from DBA/2 mice at 37°C for 72hours and then pulsed with [3H]thymidine duringthe last 8 hours. C and D, show Ag-specific re-sponses in which the DCs from C57BL/6 micewere pulsed with conalbumin (50 �g/mL) and ei-ther treated with vehicle alone (control DC) or withBryo-1 � CI (Bryo-1 � CI DCs). The DCs(1 � 105/well) were irradiated and mixed withconalbumin-specific T cells (3 � 105/well). TheDCs that were not pulsed with conalbumin alsowere included as control (unpulsed DCs). The cul-tures were incubated at 37°C for 72 hours andpulsed with [3H]thymidine during the last 8 hours.E, Naive T cells (8 � 105/well) from C57BL/6wild-type mice were mixed with conalbumin-pulsed DCs (1 � 105/well). F, The DCs werepulsed with whole LSA tumor cell lysate, treatedwith different DC activators, and then mixed withLSA tumor-specific T cells. The cultures were in-cubated at 37°C for 72 hours and pulsed with[3H]thymidine during the last 8 hours. Vertical barsrepresent mean � SEM of triplicate cultures (A–F).
6759
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
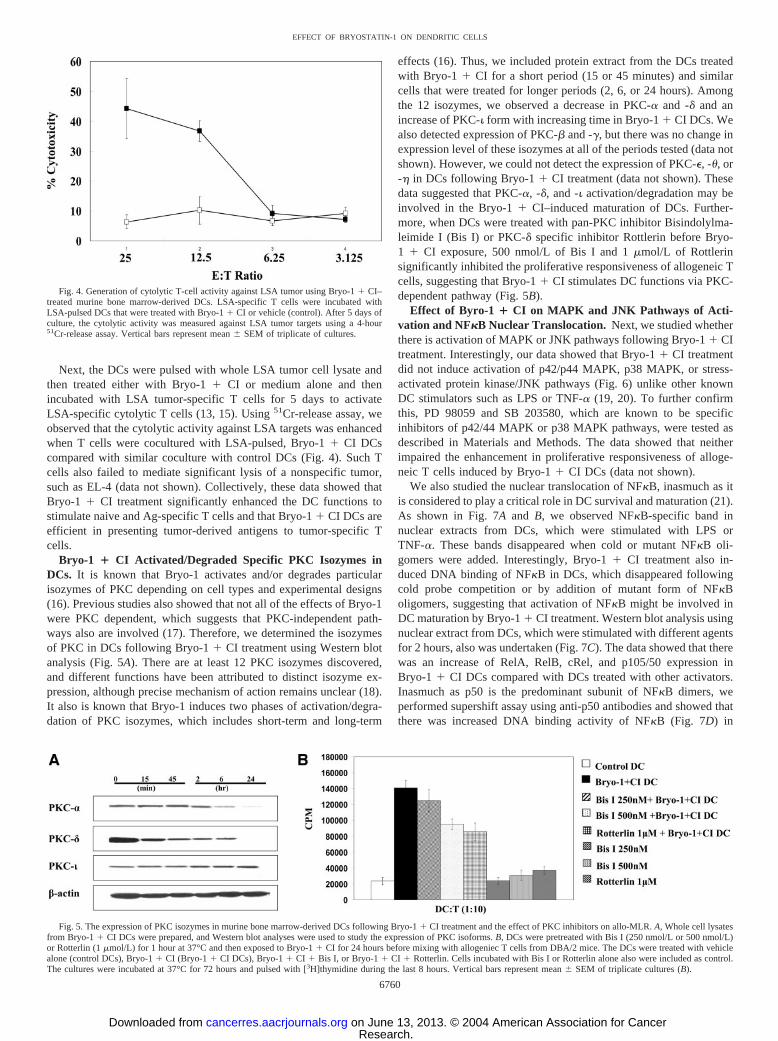
Next, the DCs were pulsed with whole LSA tumor cell lysate andthen treated either with Bryo-1 � CI or medium alone and thenincubated with LSA tumor-specific T cells for 5 days to activateLSA-specific cytolytic T cells (13, 15). Using 51Cr-release assay, weobserved that the cytolytic activity against LSA targets was enhancedwhen T cells were cocultured with LSA-pulsed, Bryo-1 � CI DCscompared with similar coculture with control DCs (Fig. 4). Such Tcells also failed to mediate significant lysis of a nonspecific tumor,such as EL-4 (data not shown). Collectively, these data showed thatBryo-1 � CI treatment significantly enhanced the DC functions tostimulate naive and Ag-specific T cells and that Bryo-1 � CI DCs areefficient in presenting tumor-derived antigens to tumor-specific Tcells.
Bryo-1 � CI Activated/Degraded Specific PKC Isozymes inDCs. It is known that Bryo-1 activates and/or degrades particularisozymes of PKC depending on cell types and experimental designs(16). Previous studies also showed that not all of the effects of Bryo-1were PKC dependent, which suggests that PKC-independent path-ways also are involved (17). Therefore, we determined the isozymesof PKC in DCs following Bryo-1 � CI treatment using Western blotanalysis (Fig. 5A). There are at least 12 PKC isozymes discovered,and different functions have been attributed to distinct isozyme ex-pression, although precise mechanism of action remains unclear (18).It also is known that Bryo-1 induces two phases of activation/degra-dation of PKC isozymes, which includes short-term and long-term
effects (16). Thus, we included protein extract from the DCs treatedwith Bryo-1 � CI for a short period (15 or 45 minutes) and similarcells that were treated for longer periods (2, 6, or 24 hours). Amongthe 12 isozymes, we observed a decrease in PKC-� and -� and anincrease of PKC-� form with increasing time in Bryo-1 � CI DCs. Wealso detected expression of PKC-� and -�, but there was no change inexpression level of these isozymes at all of the periods tested (data notshown). However, we could not detect the expression of PKC-�, -, or- in DCs following Bryo-1 � CI treatment (data not shown). Thesedata suggested that PKC-�, -�, and -� activation/degradation may beinvolved in the Bryo-1 � CI–induced maturation of DCs. Further-more, when DCs were treated with pan-PKC inhibitor Bisindolylma-leimide I (Bis I) or PKC-� specific inhibitor Rottlerin before Bryo-1 � CI exposure, 500 nmol/L of Bis I and 1 �mol/L of Rottlerinsignificantly inhibited the proliferative responsiveness of allogeneic Tcells, suggesting that Bryo-1 � CI stimulates DC functions via PKC-dependent pathway (Fig. 5B).
Effect of Byro-1 � CI on MAPK and JNK Pathways of Acti-vation and NF�B Nuclear Translocation. Next, we studied whetherthere is activation of MAPK or JNK pathways following Bryo-1 � CItreatment. Interestingly, our data showed that Bryo-1 � CI treatmentdid not induce activation of p42/p44 MAPK, p38 MAPK, or stress-activated protein kinase/JNK pathways (Fig. 6) unlike other knownDC stimulators such as LPS or TNF-� (19, 20). To further confirmthis, PD 98059 and SB 203580, which are known to be specificinhibitors of p42/44 MAPK or p38 MAPK pathways, were tested asdescribed in Materials and Methods. The data showed that neitherimpaired the enhancement in proliferative responsiveness of alloge-neic T cells induced by Bryo-1 � CI DCs (data not shown).
We also studied the nuclear translocation of NF�B, inasmuch as itis considered to play a critical role in DC survival and maturation (21).As shown in Fig. 7A and B, we observed NF�B-specific band innuclear extracts from DCs, which were stimulated with LPS orTNF-�. These bands disappeared when cold or mutant NF�B oli-gomers were added. Interestingly, Bryo-1 � CI treatment also in-duced DNA binding of NF�B in DCs, which disappeared followingcold probe competition or by addition of mutant form of NF�Boligomers, suggesting that activation of NF�B might be involved inDC maturation by Bryo-1 � CI treatment. Western blot analysis usingnuclear extract from DCs, which were stimulated with different agentsfor 2 hours, also was undertaken (Fig. 7C). The data showed that therewas an increase of RelA, RelB, cRel, and p105/50 expression inBryo-1 � CI DCs compared with DCs treated with other activators.Inasmuch as p50 is the predominant subunit of NF�B dimers, weperformed supershift assay using anti-p50 antibodies and showed thatthere was increased DNA binding activity of NF�B (Fig. 7D) in
Fig. 4. Generation of cytolytic T-cell activity against LSA tumor using Bryo-1 � CI–treated murine bone marrow-derived DCs. LSA-specific T cells were incubated withLSA-pulsed DCs that were treated with Bryo-1 � CI or vehicle (control). After 5 days ofculture, the cytolytic activity was measured against LSA tumor targets using a 4-hour51Cr-release assay. Vertical bars represent mean � SEM of triplicate of cultures.
Fig. 5. The expression of PKC isozymes in murine bone marrow-derived DCs following Bryo-1 � CI treatment and the effect of PKC inhibitors on allo-MLR. A, Whole cell lysatesfrom Bryo-1 � CI DCs were prepared, and Western blot analyses were used to study the expression of PKC isoforms. B, DCs were pretreated with Bis I (250 nmol/L or 500 nmol/L)or Rotterlin (1 �mol/L) for 1 hour at 37°C and then exposed to Bryo-1 � CI for 24 hours before mixing with allogeniec T cells from DBA/2 mice. The DCs were treated with vehiclealone (control DCs), Bryo-1 � CI (Bryo-1 � CI DCs), Bryo-1 � CI � Bis I, or Bryo-1 � CI � Rotterlin. Cells incubated with Bis I or Rotterlin alone also were included as control.The cultures were incubated at 37°C for 72 hours and pulsed with [3H]thymidine during the last 8 hours. Vertical bars represent mean � SEM of triplicate cultures (B).
6760
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

Bryo-1 � CI DCs. Studies also were performed to confirm nucleartranslocation of p50 and RelA subunits of NF�B in vehicle, LPS,Bryo-1, CI, and Bryo-1 � CI–treated DCs by staining with thespecific antibodies and visualization by confocal microscopy (Fig.7E). In untreated DCs, RelA and p50 were localized in the cytosolicregion of the cells, whereas following treatment with LPS, Bryo-1, orBryo-1 � CI, there was nuclear translocation and colocalization ofboth the subunits as depicted by the yellow staining in the last columnfollowing merging of the images. Furthermore, CI treatment alone didnot lead to nuclear staining for p50 and RelA. It was interesting tonote that following treatment with Bryo-1 alone, p50 and RelA werefound in the nucleus and cytosol, whereas following treatment withBryo-1 � CI or LPS almost all of the p50 and RelA translocated to thenucleus.
Effect of Bryo-1 � CI Treatment on Activation of HumanPBMC-DCs. To study the effect of Bryo-1 on human PBMC-DCs,we carried out several experiments using DCs isolated and culturedfrom human PBMCs. First, when PBMC-DCs treated with differentactivators were observed for morphology, we found similar morpho-logic changes as seen in mouse bone marrow-derived DCs (Fig. 8A).In particular, Bryo-1 � CI treatment induced prominent dendriticprocesses in PBMC-DCs compared with control DCs or DCs treatedwith PMA � CI or TNF-�.
It was noted that PBMC-DCs treated with Bryo-1 � CI showedsignificant up-regulation of HLA-DP/DQ/DR, CD80, CD83, andCD86 compared with the controls (Fig. 8B). For the most part,treatment of PBMC-DCs with Bryo-1 � CI also had a more pro-nounced effect on the activation of DCs compared with the DCstreated with other well-known DC stimulators, such as PMA � CI orTNF-�.
To test the effect of Bryo-1 � CI on the ability of PBMC-DCs tostimulate the proliferative responsiveness of Ag-specific T cells, theDCs were pulsed with TT antigen, treated with Bryo-1 � CI orvarious activators, and then mixed with autologous peripheral blood Tcells (Fig. 8C). The data showed that Bryo-1 � CI DCs induced T-cellproliferation stronger than other groups of DCs.
Effect of Bryo-1 � CI Treatment on the Activation of CordBlood-Derived CD34�HSC-DCs. Previous studies showed thatfunctional DCs could be generated from CD34�HSC and therebysuggested promising implications of CD34�HSC-DCs in future clin-ical applications (22). In line with these efforts, we also generatedDCs from cord blood-derived CD34�HSC and studied the effect of
Bryo-1 � CI on maturation and functional enhancement. We observedan increase in the expression of CD80, CD83, CD86, and HLA onCD34�HSC-DCs treated with Bryo-1 � CI compared with controlDCs (Fig. 9A). Moreover, Bryo-1 � CI HSC-DCs expressed CD83and CD86 stronger than PMA � CI DCs or LPS DCs, suggestingthat the mode of action of Bryo-1 � CI might be different com-pared with other known stimulators of DCs. When Bryo-1 � CIDCs were tested in allo-MLR, dramatic enhancement in T-cellproliferation was observed compared with other treatment groups(Fig. 9B).
DISCUSSION
Bryostatins have generated a wide interest because of their antin-eoplastic activity combined with low toxicity (2). Moreover, Bryo-1has been shown to induce cell differentiation and increase the sensi-tivity of tumor cells to cytotoxicity caused by other drugs (3, 23). Inthe current study, we tested whether Bryo-1 � CI can induce thematuration of DCs and promote their antigen-priming activities to Tcells. Our data showed that Bryo-1 � CI could activate the DCs asshown by: (1) up-regulation of costimulatory molecules; (2) decreasein the ability of DCs to uptake FITC-dextran; (3) enhancement ofproliferative responsiveness of allogeneic T cells and Ag-specific Tcells; and (4) enhancement of cytolytic activity against LSA tumorcells. We also showed that the maturation of DCs by Bryo-1 � CI wasPKC dependent, probably via PKC-�, -�, and -�. However, it did notinduce any MAPK or JNK pathways unlike other known DC stimu-lators. NF�B translocation also was detected in DCs followed byBryo-1 � CI treatment. Collectively, this study suggests a novel usefor Bryo-1 � CI in the maturation and functional activation of DCs.
Bryo-1 currently is tested in a number of Phase I and II studiessponsored by National Cancer Institute as a single agent or in com-bination with other chemotherapeutic agents to manage a wide rangeof cancers (24). Preliminary studies from Phase II trials at MemorialSloan-Kettering Cancer Center published in 2001 suggested thatBryo-1 treatment enhanced antitumor activity in combination withTaxol® against esophageal cancer (25). Other studies also supported apromising clinical application of Bryo-1 for the management of var-ious human cancers (26, 27). Although most of these clinical studiesdid not provide the exact mechanism by which Bryo-1 enhanced theantitumor activity, it has been suggested that Bryo-1 acts as a sensi-tizer to other anticancer drugs. Some studies reported a dose-depen-
Fig. 6. The involvement of MAPK or stress-activated protein kinase/JNK pathways in murinebone marrow-derived DCs following Bryo-1 � CItreatment. At indicated time points, whole cell ly-sates of Bryo-1 � CI DCs were analyzed by West-ern blot analysis for MAPK or stress-activated pro-tein kinase/JNK pathways.
6761
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

dent increase in TNF-� and IL-6 concentrations in plasma following1-hour intravenous infusion of Bryo-1, whereas others could notdetect any increase in such cytokines in plasma (26, 27). In anotherPhase I study, Bryo-1 was shown to increase IL-2–induced lympho-kine-activated killer cell activity and IL-2–induced lymphocyte pro-
liferation (28). However, nothing is known about the effect of Bryo-1on DCs.
Previous studies showed that various stimuli could initiate thematuration process of DCs in vitro. These include the proinflamma-tory cytokines such as TNF-� and bacterial products such as LPS (29,
Fig. 7. Nuclear translocation of NF�B and itssubunits in murine bone marrow-derived DCs fol-lowing different modes of stimulation. The nuclearprotein from DCs treated with LPS, TNF-�, andBryo-1 � CI was extracted and incubated with32P-labeled NF�B, mutant, cold oligonucleotides,or antibody (p50). The specific band of NF�B wasindicated with filled arrowhead (A, B, and D). Theempty arrowhead depicts supershift of NF�B usinganti-p50 antibody (D). The nuclear extracts wereprepared from DCs activated by various agents for2 hours, and Western blot analysis was used toanalyze distinct subunits of NF�B (C). E, showsconfocal microscopic analysis of vehicle, LPS,Bryo-1, Bryo-1 � CI, or CI-treated DCs by No-marski differential interference contrast (first col-umn) and following staining with anti-RelA anti-body and Cy3 antigoat antibody (second column),anti-p50 antibody and Cy2 antirabbit antibody(third column), nuclear staining with Hoechst(fourth column), and following merging of the im-ages (fifth column). The yellow coloration follow-ing merge depicts colocalization of RelA and p50in the last column.
6762
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

30). Ligation of CD40/CD40L and the viral double-stranded RNAalso has been shown to stimulate maturation of human DCs (31, 32).Furthermore, it was shown that when ex vivo-generated human DCswere pulsed with tumor-specific peptides or whole tumor cell lysates,
they induced antitumor responses in cancer patients, suggesting thepromising use of DCs in vaccination and immunotherapy of cancer(33). Unfortunately, despite such possibilities, there are still majorlimitations in the application of DCs in clinical settings. It is com-
Fig. 8. Effect of Bryo-1 � CI on the activation ofhuman PBMC-DCs. A, The morphology of humanPBMC-derived DCs is depicted as seen under a phasecontrast microscope. B, The DCs treated with differ-ent activators were harvested, washed, and stainedwith various FITC- and PE-conjugated mAbs. Thedata are depicted as expression indices, which are theproducts of percentage of marker-positive cells mul-tiplied by the mean fluorescence intensity (MFI) asdescribed in Materials and Methods. For the sake ofcomparison, the value obtained for cells exposed toTNF-� has been normalized to 1. Vertical bars rep-resent mean � SEM of two or three experiments.C, PBMC-DCs were pulsed with TT (10 Lf/mL) andeither treated with medium alone (control DC) orwith various activators. Various numbers of irradi-ated PBMC-DCs then were mixed with PBMC-derived T cells as described in Materials and Meth-ods. The cultures were incubated at 37°C for 5 daysand pulsed with [3H]thymidine during the last 8hours. Vertical bars represent mean � SEM of trip-licate cultures.
6763
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

monly observed that the DCs pulsed with an antigen and injected invivo are not effective, undergo apoptosis, or sometimes show oppositeeffects. This is because of, first, the environmental factors that alreadyexisted in the immune-deficient host, and, second, the nature of DCs,which can act either as an immune tolerant or immune adjuvant(34–36). Other known DC stimulators, which make them an immuneadjuvant, have been reported and shown efficient in vivo, but unfor-tunately, not all of them can be used in human study (30, 37). CD40Lhas been considered a promising agent, although no study regardingthis has been reported yet in humans (31). On the basis of the currentstudy, we propose that Bryo-1 may serve as a novel and strong DCstimulator with the potential of being used in human clinical trials.This study showed that Bryo-1 � CI treatment induced DC matura-tion and functional enhancement better than other known DC stimu-lators, such as LPS, TNF-�, and PMA � CI. Inasmuch as Bryo-1 hasalready been used in human studies, the current findings provideattractive alternatives for the use of Bryo-1 in DC-based vaccine orimmunotherapy.
Bryo-1 is one of the first drugs to target a signaling pathway,although the relationship between its various activities and PKCmodulation is not uniformly established. Bryo-1 has been shown to actas PKC stimulator or PKC inhibitor depending on dose, time oftreatment, or experimental conditions (38, 39). In the current study,
we found that Bryo-1–mediated effect on DCs depended on PKCpathway. PKC is composed of at least 12 isozymes that can be furthercategorized into subfamilies according to their structures and cofactorregulation (18). The types of PKC isozyme that Bryo-1 activates alsoare diverse, depending on cell types or treatments. A previous studyshowed that Bryo-1 substantially regulated PKC-�, -�, and -� in NIHmouse fibroblast cells, whereas PMA did not distinguish betweenthese isozymes (40). Other studies showed that Bryo-1 down-regu-lates PKC-� in epithelial cells and PKC-� and -� in human T cells(41, 16). Bryo-1 also induces a unique biphasic response in PKC-�,depending on concentrations leading to its degradation in NIH mousefibroblast cells, B16F10 melanocytes, and HeLa cells (40, 42, 43).PKC-� recently was discovered, and its exact function is unknown(44). Further studies are required to reveal the relationship betweenspecific PKC isozymes and functions of DCs.
Because NF�B plays a critical role in DC maturation and survival,observation of the translocation of NF�B following Bryo-1 � CItreatment is not surprising. Moreover, nuclear extract from Bryo-1 � CI DCs showed enhanced expression of NF�B subunits com-pared with DCs treated with other activating agents, suggesting dif-ferential regulation of NF�B by Bryo-1 � CI in DCs, especially atearly time points. In the current study, we also noted that Bryo-1 � CIdid not induce MAPK or JNK pathways. This is in contrast to
Fig. 9. Bryo-1 � CI treatment up-regulates theexpression of surface markers on humanCD34�HSC-DCs and enhances their function tostimulate allogeneic T cells. Human CD34�HSC-DCs from cord blood were cultured and treated asdescribed in Materials and Methods. A, The DCstreated with different activators were harvested,washed, and stained with various FITC- and PE-conjugated mAbs. The data were plotted as expres-sion indices as described in Fig. 8. Vertical barsrepresent mean � SEM of two or three experiments.B, CD34�HSC-DCs were treated with vehicle alone(control DC) or with various activators. Variousnumbers of irradiated CD34�HSC-DCs then weremixed with T cells isolated from allogeneic cordblood (2 � 105/well) as described in Materials andMethods. The cultures were incubated at 37°C for 5days and pulsed with [3H]thymidine during the last 8hours. Vertical bars represent mean � SEM of trip-licate cultures.
6764
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

previous studies showing the involvement of MAP/extracellular sig-nal-regulated kinase/MAPK in the differentiation of monocytes orhuman lymphoid leukemia cells by Bryo-1 (45, 46). Other activatorsof DCs, such as TNF-� or LPS, also have been shown to activateMAPK or JNK pathways (19, 20). These studies suggest that Bryo-1 � CI may use a distinct signaling pathway to activate DCs com-pared with the widely tested agents such as LPS or TNF-�. It shouldbe noted that LPS has been shown to act primarily through Toll-likereceptor 4 (47). Thus, further studies are necessary to address whetherBryo-1 � CI acts through Toll-like receptors or through independentmechanisms.
In conclusion, this study shows for the first time that Bryo-1 � CIcan promote maturation of murine and human DCs and enhance theirantigen-presenting properties to activate T cells. Thus, Bryo-1 mayexhibit unique properties in activity as an anticancer agent and im-munostimulant, which is highly beneficial for the management ofcancer.
REFERENCES
1. Pettit GR. The bryostatins. Fortschr Chem Org Naturst 1991;57:153–95.2. Hale KJ, Hummersone MG, Manaviazar S, Frigerio M. The chemistry and biology of
the bryostatin antitumour macrolides. Nat Prod Rep 2002;19:413–53.3. Clamp A, Jayson GC. The clinical development of the bryostatins. Anticancer Drugs
2002;13:673–83.4. May WS, Sharkis SJ, Esa AH, et al. Antineoplastic bryostatins are multipotential
stimulators of human hematopoietic progenitor cells. Proc Natl Acad Sci USA1987;84:8483–7.
5. Hess AD, Silanskis MK, Esa AH, Pettit GR, May WS. Activation of human Tlymphocytes by bryostatin. J Immunol 1988;141:3263–9.
6. Roddie PH, Horton Y, Turner ML. Primary acute myeloid leukaemia blasts resistantto cytokine-induced differentiation to dendritic-like leukaemia cells can be forced todifferentiate by the addition of bryostatin-1. Leukemia 2002;16:84–93.
7. Inaba K, Inaba M, Romani N, et al. Generation of large numbers of dendritic cellsfrom mouse bone marrow cultures supplemented with granulocyte/macrophage col-ony-stimulating factor. J Exp Med 1992;176:1693–702.
8. Romani N, Reider D, Heuer M, et al. Generation of mature dendritic cells from humanblood. An improved method with special regard to clinical applicability. J ImmunolMethods 1996;137–51.
9. Rosenzwajg M, Canque B, Gluckman JC. Human dendritic cell differentiation path-way from CD34� hematopoietic precursor cells. Blood 1996;87:535–44.
10. Seth A, Pyle RH, Nagarkatti M, Nagarkatti PS. Expression of the J11d marker onperipheral T lymphocytes of MRL-lpr/lpr mice. J Immunol 1988;141:1120–5.
11. Steinman RM, Gutchinov B, Witmer MD, Nussenzweig MC. Dendritic cells are theprincipal stimulators of the primary mixed leukocyte reaction in mice. J Exp Med1983;157:613–27.
12. McKallip RJ, Do Y, Fisher MT, Robertson JL, Nagarkatti PS, Nagarkatti M. Role ofCD44 in activation-induced cell death: CD44-deficient mice exhibit enhanced T cellresponse to conventional and superantigens. Int Immunol 2002;14:1015–26.
13. Nagarkatti M, Kaplan AM. The role of suppressor T cells in BCNU-mediatedrejection of a syngeneic tumor. J Immunol 1985;135:1510–7.
14. Lahiri DK, Ge Y. Electrophoretic mobility shift assay for the detection of specificDNA-protein complex in nuclear extracts from the cultured cells and frozen autopsyhuman brain tissue. Brain Res Brain Res Protoc 2000;5:257–65.
15. Nagarkatti M, Toney DM, Nagarkatti PS. Immunomodulation by various nitrosoureasand its effect on the survival of the murine host bearing a syngeneic tumor. CancerRes 1989;49:6587–92.
16. Isakov N, Galron D, Mustelin T, Pettit GR, Altman A. Inhibition of phorbol ester-induced T cell proliferation by bryostatin is associated with rapid degradation ofprotein kinase C. J Immunol 1993;150:1195–204.
17. Curiel RE, Garcia CS, Farooq L, Aguero MF, Espinoza-Delgado I. Bryostatin-1 andIL-2 synergize to induce IFN-� expression in human peripheral blood T cells:implications for cancer immunotherapy. J Immunol 2001;167:4828–37.
18. Stabnel S. Protein kinase C—an enzyme and its relatives. Semin Cancer Biol1994;5:277–84.
19. Ardeshna KM, Pizzey AR, Devereux S, Khwaja A. The PI3 kinase, p38 SAP kinase,and NF-�B signal transduction pathways are involved in the survival and maturationof lipopolysaccharide-stimulated human monocyte-derived dendritic cells. Blood2000;96:1039–46.
20. Yanagawa Y, Iijima N, Iwabuchi K, Onoe K. Activation of extracellular signal-related kinase by TNF-� controls the maturation and function of murine dendriticcells. J Leukoc Biol 2002;71:125–32.
21. Ouaaz F, Arron J, Zheng Y, Choi Y, Beg AA. Dendritic cell development andsurvival require distinct NF-�B subunits. Immunity 2002;16:257–70.
22. Caux C, Massacrier C, Vanbervliet B, et al. CD34� hematopoietic progenitors fromhuman cord blood differentiate along two independent dendritic cell pathways inresponse to granulocyte-macrophage colony-stimulating factor plus tumor necrosisfactor �: II. Functional analysis. Blood 1997;90:1458–70.
23. Lilly M, Tompkins C, Brown C, Pettit G, Kraft A. Differentiation and growthmodulation of chronic myelogenous leukemia cells by bryostatin. Cancer Res 1990;50:5520–5.
24. Philip PA, Zonder JA. Pharmacology and clinical experience with bryostatin 1: anovel anticancer drug. Expert Opin Investig Drugs 1999;8:2189–99.
25. GPC Biotech receives Orphan Medicinal Product Designation. Expert Rev AnticancerTher 2002;2:246.
26. Philip PA, Rea D, Thavasu P, et al. Phase I study of bryostatin 1: assessment ofinterleukin 6 and tumor necrosis factor � induction in vivo. The Cancer ResearchCampaign Phase I Committee. J Natl Cancer Inst 1993;85:1812–8.
27. Jayson GC, Crowther D, Prendiville J, et al. A phase I trial of bryostatin 1 in patientswith advanced malignancy using a 24 hour intravenous infusion. Br J Cancer1995;72:461–8.
28. Scheid C, Prendiville J, Jayson G, et al. Immunomodulation in patients receivingintravenous Bryostatin 1 in a phase I clinical study: comparison with effects ofBryostatin 1 on lymphocyte function in vitro. Cancer Immunol Immunother 1994;39:223–30.
29. Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen by culturedhuman dendritic cells is maintained by granulocyte/macrophage colony-stimulatingfactor plus interleukin 4 and downregulated by tumor necrosis factor �. J Exp Med1994;179:1109–18.
30. Lyakh LA, Koski GK, Telford W, Gress RE, Cohen PA, Rice NR. Bacterial lipopo-lysaccharide, TNF-�, and calcium ionophore under serum-free conditions promoterapid dendritic cell-like differentiation in CD14� monocytes through distinct path-ways that activate NK-� B. J Immunol 2000;165:3647–55.
31. Caux C, Massacrier C, Vanbervliet B, et al. Activation of human dendritic cellsthrough CD40 cross-linking. J Exp Med 1994;180:1263–72.
32. Cella M, Salio M, Sakakibara Y, Langen H, Julkunen I, Lanzavecchia A. Maturation,activation, and protection of dendritic cells induced by double-stranded RNA. J ExpMed 1999;189:821–9.
33. Nestle FO, Alijagic S, Gilliet M, et al. Vaccination of melanoma patients withpeptide- or tumor lysate-pulsed dendritic cells. Nat Med 1998;4:328–32.
34. Vicari AP, Caux C, Trinchieri G. Tumour escape from immune surveillance throughdendritic cell inactivation. Semin Cancer Biol 2002;12:33–42.
35. Kao JY, Gong Y, Chen CM, Zheng QD, Chen JJ. Tumor-derived TGF-a� reduces theefficacy of dendritic cell/tumor fusion vaccine. J Immunol 2003;170:3806–11.
36. Bachleitner-Hofmann T, Stift A, Friedl J, et al. Stimulation of autologous antitumorT-cell responses against medullary thyroid carcinoma using tumor lysate-pulseddendritic cells. J Clin Endocrinol Metab 2002;87:1098–104.
37. Hartmann G, Weiner GJ, Krieg AM. CpG DNA: a potent signal for growth, activa-tion, and maturation of human dendritic cells. Proc Natl Acad Sci USA 1999;96:9305–10.
38. Lorenzo PS, Bogi K, Hughes KM, et al. Differential roles of the tandem C1 domainsof protein kinase C � in the biphasic down-regulation induced by bryostatin 1. CancerRes 1999;59:6137–44.
39. Hennings H, Blumberg PM, Pettit GR, Herald CL, Shores R, Yuspa SH. Bryostatin1, an activator of protein kinase C, inhibits tumor promotion by phorbol esters inSENCAR mouse skin. Carcinogenesis 1987;8:1343–6.
40. Szallasi Z, Smith CB, Pettit GR, Blumberg PM. Differential regulation of proteinkinase C isozymes by bryostatin 1 and phorbol 12-myristate 13-acetate in NIH 3T3fibroblasts. J Biol Chem 1994;269:2118–24.
41. Lee HW, Smith L, Pettit GR, Bingham Smith J. Dephosphorylation of activatedprotein kinase C contributes to downregulation by bryostatin. Am J Physiol. 1996;271:C304–11.
42. Szallasi Z, Du L, Levine R, et al. The bryostatins inhibit growth of B16/F10melanoma cells in vitro through a protein kinase C-independent mechanism: disso-ciation of activities using 26-epi-bryostatin 1. Cancer Res 1996;56:2105–11.
43. Basu A, Akkaraju GR. Regulation of caspase activation and cis-diamminedichloro-platinum(II)-induced cell death by protein kinase C. Biochemistry 1999;38:4245–51.
44. Lu Y, Jamieson L, Brasier AR, Fields AP. NF-�B/RelA transactivation is required foratypical protein kinase C e-mediated cell survival. Oncogene 2001;20:4777–92.
45. Lin H, Chen C, Li X, Chen BD. Activation of the MEK/MAPK pathway is involvedin bryostatin1-induced monocytic differenciation and up-regulation of X-linked in-hibitor of apoptosis protein. Exp Cell Res 2002;272:192–8.
46. Wall NR, Mohammad RM, Al-Katib AM. Mitogen-activated protein kinase is re-quired for bryostatin 1-induced differentiation of the human acute lymphoblasticleukemia cell line Reh. Cell Growth Differ 2001;12:641–7.
47. Beutler B. Tlr4: central component of the sole mammalian LPS sensor. Curr OpinImmunol 2000;12:20–6.
6765
EFFECT OF BRYOSTATIN-1 ON DENDRITIC CELLS
Research. on June 13, 2013. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from




















