

bruna cunha gondim de alencar vacinação com a proteína 2 ...
150
BRUNA CUNHA GONDIM DE ALENCAR VACINAÇÃO COM A PROTEÍNA 2 DA SUPERFÍCIE DE AMASTIGOTAS CONTRA A INFECÇÃO EXPERIMENTAL PELO Trypanosoma cruzi: EFICÁCIA DE DIFERENTES ESTRATÉGIAS VACINAIS E MECANISMOS DE IMUNIDADE Tese apresentada à Universidade Federal de São Paulo para obtenção do título de Doutor em Ciências. SÃO PAULO 2009
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of bruna cunha gondim de alencar vacinação com a proteína 2 ...

BRUNA CUNHA GONDIM DE ALENCAR
VACINAÇÃO COM A PROTEÍNA 2 DA SUPERFÍCIE DE AMASTIGOTAS
CONTRA A INFECÇÃO EXPERIMENTAL PELO Trypanosoma cruzi:
EFICÁCIA DE DIFERENTES ESTRATÉGIAS VACINAIS E
MECANISMOS DE IMUNIDADE
Tese apresentada à Universidade Federal
de São Paulo para obtenção do título de
Doutor em Ciências.
SÃO PAULO
2009

BRUNA CUNHA GONDIM DE ALENCAR
VACINAÇÃO COM A PROTEÍNA 2 DA SUPERFÍCIE DE AMASTIGOTAS
CONTRA A INFECÇÃO EXPERIMENTAL PELO Trypanosoma cruzi:
EFICÁCIA DE DIFERENTES ESTRATÉGIAS VACINAIS
E MECANISMOS DE IMUNIDADE
Orientador: Prof. Dr. Mauricio Martins Rodrigues
Tese apresentada à Universidade Federal
de São Paulo para obtenção do título de
Doutor em Ciências.
SÃO PAULO
2009

Ficha Catalográfica
de Alencar, Bruna Cunha Gondim
Vacinação com a proteína 2 da superfície de amastigotas contra a infecção experimental pelo Trypanosoma cruzi: eficácia de diferentes estratégias vacinais e mecanismos de imunidade./ Bruna Cunha Gondim de Alencar – São Paulo, 2009.
XI, 140f Tese (Doutorado) – Universidade Federal de São Paulo.
Programa de pós-graduação em Microbiologia e Imunologia. Título em inglês: Vaccination with amastigote surface protein 2
against experimental infection by Trypanosoma cruzi: efficacy of different vaccination strategies and mechanisms of immunity.
1 – Trypanosoma cruzi. 2 – Proteínas recombinantes. 3 – Adenovírus recombinante. 4 – Imunização.

UNIVERSIDADE FEDERAL DE SÃO PAULO
Departamento de Microbiologia, Imunologia e Parasitologia
(DMIP)
Centro Interdisciplinar de Terapia Gênica (Cintergen)
Programa de pós-graduação em Microbiologia e Imunologia
Chefe do Departamento: Profa. Dra. Clara Lúcia Barbieri Mestriner
Coordenador do Curso de Pós-graduação: Prof. Dr. Renato Arruda
Mortara
Auxílio financeiro: Fundação de Amparo à Pesquisa do Estado de São
Paulo (FAPESP) e Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq)

Às pessoas mais queridas
do mundo, meus pais Joyl e
Neide, meu irmão Joyl e
meu marido Daniel.

VI
AGRADECIMENTOS
Em primeiro lugar, gostaria de agradecer ao Dr. Mauricio Rodrigues,
meu orientador, por tudo que me ensinou sobre ciência, e também pela
paciência e disponibilidade nos vários anos em que estive em seu laboratório.
Agradeço também aos meus colegas de laboratório, Ronnie
Vasconcelos, Adriano Araújo, Meire Hiyane, Daniela Santoro, Fanny Tzelepis,
Carla Claser, Eduardo Silveira, Filipe Haolla, Mariana Dominguez, Laís Teixeira
e Ariane Camacho, pelo auxílio nos momentos necessários e pela amizade.
À Dra. Silvia Boscardin, pelo apoio e pelas discussões sempre repletas
de informações interessantes. Agradeço à Dra. Karina Carvalho, Fernanda
Romano e Dr. Ésper Kallás, pela constante boa vontade, pelas dicas e
protocolos de citometria, e pela convivência agradável. Ao Drs. Ricardo
Gazzinelli, Oscar Bruna-Romero, Maria Bellio, Alexandre Machado, Braulia
Caetano e Marcus Penido, pelas importantes colaborações.
Aos meus amigos, professores e funcionários da disciplina de
Parasitologia da UNIFESP, em especial ao Adílson, à Mércia e à Regiane, cuja
ajuda foi determinante em diversas ocasiões.
Às minhas amigas biomédicas, Fernanda Nakamoto, Luciana Semião,
Mariana Grings e Juliana Colucci, pela inestimável amizade, pelos conselhos e
discussões científicos ou pessoais e pela convivência de tantos anos que me é
essencial.
Às minhas queridas amigas da GM e à minha família, pelo apoio
incondicional e pela presença em todos os momentos da minha vida.

VII
SUMÁRIO
LISTA DE ABREVIATURAS ..............................................................................................................VIII
LISTA DE FIGURAS ..............................................................................................................................IX
RESUMO ................................................................................................................................................... X
INTRODUÇÃO........................................................................................................................................... 1
1.1 A DOENÇA DE CHAGAS .................................................................................................................... 2
I) Transmissão e epidemiologia ................................................................................................. 2
II) Manifestações clínicas ............................................................................................................ 3
III) Patogenia ................................................................................................................................... 4
IV) Diagnóstico ............................................................................................................................... 6
V) Tratamento ................................................................................................................................. 7
1.2 O CICLO DE VIDA DO T. CRUZI ......................................................................................................... 8
1.3 A RESPOSTA IMUNE NA INFECÇÃO EXPERIMENTAL PELO T.CRUZI................................................ 10
I) Resposta imune inata ............................................................................................................. 10
II) Resposta imune adaptativa .................................................................................................. 13
III) Alvos da resposta imune adaptativa................................................................................. 19
1.4 VACINAÇÃO CONTRA A INFECÇÃO EXPERIMENTAL PELO TRYPANOSOMA CRUZI ......................... 20
I) Proteína 2 da superfície de amastigotas (ASP-2) ............................................................ 23
1.5 DESENVOLVIMENTO DE VACINAS RECOMBINANTES VISANDO À INDUÇÃO DE UMA RESPOSTA IMUNE MEDIADA PELOS LINFÓCITOS T CD4 E CD8 ............................................................................. 27
I) Proteínas recombinantes ....................................................................................................... 27
II) DNA plasmidial ........................................................................................................................ 29
III) Vírus recombinantes ............................................................................................................. 30
IV) Imunização e reforço (Prime-boost) heterólogo............................................................ 34
OBJETIVOS ............................................................................................................................................. 40
RESULTADOS ........................................................................................................................................ 42
ARTIGOS PUBLICADOS ......................................................................................................................... 43
ARTIGO 1............................................................................................................................................... 44
CD8-T-Cell-Dependent Control of Trypanosoma cruzi Infection in a Highly Susceptible Mouse Strain after Immunization with Recombinant Proteins Based on Amastigote Surface Protein 2................................................................................................... 45
ARTIGO 2............................................................................................................................................... 55
Cross-priming of long lived protective CD8+ T cells against Trypanosoma cruzi infection: importance of a TLR9 agonist and CD4+ T cells. ............................................. 56
ARTIGO 3............................................................................................................................................... 67
Perforin and gamma interferon expression are required for CD4+ and CD8
+ T-cell-
dependent protective immunity against a human parasite, Trypanosoma cruzi, elicited by heterologous plasmid DNA prime recombinant adenovirus 5 boost vaccination. ................................................................................................................................... 68
CONSIDERAÇÕES FINAIS E CONCLUSÕES ................................................................................. 82
VACINAÇÃO CONTRA A DOENÇA DE CHAGAS...................................................................................... 83
MECANISMOS IMUNOLÓGICOS DE PROTEÇÃO CONTRA A INFECÇÃO EXPERIMENTAL PELO T. CRUZI.85
CONCLUSÕES........................................................................................................................................ 89
APÊNDICES ............................................................................................................................................ 90
APÊNDICE 1........................................................................................................................................... 91
APÊNDICE 2........................................................................................................................................... 97
APÊNDICE 3......................................................................................................................................... 110
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................................. 121
ABSTRACT............................................................................................................................................ 139

VIII
LISTA DE ABREVIATURAS
AdASP-2 Adenovírus 5 recombinante expressando a ASP-2 da cepa Y
APC Célula apresentadora de antígenos
ASP-2 Proteína 2 da superfície de amastigotas
Alum Hidróxido de alumínio
CpG ODN Oligodeoxinucleotídeo contendo dinucleotídeos CG não metilados
DAMP Padrão molecular associado a perigo
ELISA Enzyme Linked Immuno Sorbent Assay, ensaio imunoenzimático
GST Glutationa S-transferase
TLR Receptor semelhante ao Toll
hAd5 Adenovírus humano sorotipo 5
IL Interleucina
IFN Interferon
IRF Fator regulador de interferon
KLRG Receptor G semelhante à lectina de células “matadoras” (NK)
KO Knockout, camundongos apresentando deleção de gene
MHC Complexo principal de histocompatibilidade
MVA Vaccínia Ankara modificado
MyD88 Fator de diferenciação mielóide 88
NFκB Cadeia leve κ do fator nuclear intensificador de células B ativadas
NLR Receptor semelhante ao Nod
NK Célula matadora natural
PAMP Padrão molecular associado a patógenos
PCR Reação em cadeia da polimerase
PD-1 Morte programada 1
pIgSPCl.9 Plasmídio contendo o gene da ASP-2 da cepa Y de T. cruzi
PRR Receptores que reconhecem padrões
STAT Proteína transdutora de sinal e ativadora de transcrição
TCR Receptor de células T
TLR Receptores semelhantes ao Toll
TNF Fator de necrose tumoral
TRIF Adaptador contendo domínio TIR indutor de interferon β
TS Trans-sialidase

IX
LISTA DE FIGURAS
Página 9 - Figura 1: Representação esquemática do ciclo de vida do T. cruzi.
Página 22 - Tabela 1: Sumário dos diferentes antígenos e formulações usados
para a vacinação experimental contra a infecção pelo T. cruzi.
Página 25 - Figura 2: Representação esquemática da sequência deduzida de
aminoácidos da ASP-2.
Página 32 - Figura 3: Representação da estrutura do adenovírus.
Página 36 - Figura 4: Representação da geração de uma resposta imune
adaptativa específica após diferentes esquemas de imunização.

X
RESUMO
A imunização genética com um DNA plasmidial, contendo o gene que
codifica a proteína 2 da superfície de amastigotas (ASP-2), é capaz de proteger
parte dos camundongos altamente suscetíveis A/Sn contra uma infecção letal
pelo Trypanosoma cruzi. Com base neste resultado, a hipótese central desta
tese foi a de que se poderia melhorar racionalmente a eficácia da vacinação
contra a infecção experimental pelo T. cruzi, utilizando diferentes formulações
vacinais e/ou esquemas de vacinações. Para tal, empregaram-se protocolos de
imunização e reforço homólogo ou heterólogo, usando DNA plasmidial,
proteínas recombinantes, e vírus recombinantes da ASP-2 de T. cruzi.
Inicialmente, testou-se se o reforço heterólogo com a proteína ASP-2
recombinante aumentava a eficácia da vacinação com o DNA plasmidial
(pIgSPCl.9). Este protocolo não se mostrou mais eficaz, mas abriu a
possibilidade de que a imunização com a proteína ASP-2 recombinante
pudesse ser utilizada como um protocolo vacinal alternativo. Com esta
abordagem, descreveu-se que a administração de 3 doses de uma proteína
recombinante (GST-P4P7), representando os aminoácidos 261-500 da ASP-2
em fusão com a glutationa S-transferase, na presença dos adjuvantes alum e
CpG ODN 1826, era capaz de induzir altos níveis de proteção. Neste modelo
experimental, observou-se que a imunidade protetora foi de longa duração e
dependente dos linfócitos CD8 específicos para o epítopo imunodominante
TEWETGQI. Determinou-se também que o adjuvante CpG ODN 1826
melhorou a eficácia da imunidade protetora, e que linfócitos T CD4 específicos
são importantes para a indução dos linfócitos CD8 protetores.
Como o protocolo de vacinação com a proteína recombinante GST-P4P7
exigia o uso de três doses, testou-se então a possibilidade de que um
adenovírus recombinante, expressando a ASP-2 (AdASP-2), pudesse ser
utilizado num protocolo de imunização e reforço homólogo ou heterólogo
(somente duas doses). Na comparação dos protocolos de vacinação
homólogos ou heterólogos com pIgSPCl.9 e AdASP-2, observou-se que, nos
animais A/Sn, tanto a imunização homóloga quanto a heteróloga, usando o
AdASP-2, proporcionaram um alto grau de imunidade protetora contra a
infecção pelo T. cruzi. Os estudos utilizando o protocolo heterólogo (pIgSPCl.9

XI
seguido por AdASP-2), que protege 80-100% dos camundongos, mostraram
que a imunidade era de longa duração e dependente dos linfócitos T CD4 e
CD8. Esta imunidade protetora se correlacionou com a atividade citotóxica
específica in vivo, antes do desafio com o T. cruzi. Com base nestes
resultados, analisou-se a imunidade protetora após a vacinação heteróloga em
camundongos com deficiência de um importante mediador da citotoxicidade, a
perforina. Os animais deficientes (KO) da expressão de perforina vacinados se
mostraram suscetíveis à infecção experimental. Uma análise da resposta
imune celular destes camundongos mostrou que eles apresentavam uma
menor produção de IFN-γ por esplenócitos re-estimulados in vitro, na presença
do antígeno recombinante. Na análise dos linfócitos T CD8 específicos
observou-se que havia: i) uma expansão numérica normal; ii) uma redução
numérica significativa dos linfócitos multifuncionais, capazes de expressar
simultaneamente IFN-γ/CD107a e IFN-γ/TNF-α.; iii) baixa citotoxicidade in vivo;
iv) baixa expressão de CD44 e de KLRG-1.
Por fim, para definir a importância relativa do IFN-γ e da citotoxicidade
na proteção contra a infecção pelo T. cruzi após a vacinação heteróloga,
camundongos IFN-γ KO foram imunizados com pIgSPCl.9 seguido por AdASP-
2. Estes animais apresentaram um alto grau de citotoxicidade in vivo, mas
nenhuma imunidade protetora detectável.
Em resumo, ao longo deste trabalho obtiveram-se evidências que
confirmam a hipótese inicial de que, utilizando diferentes formulações vacinais
e/ou esquemas de vacinações, se poderia melhorar racionalmente a eficácia da
vacinação contra a infecção experimental pelo T. cruzi. Pela utilização de
depleções in vivo e de animais geneticamente deficientes, definiu-se a
importância dos linfócitos T CD4 e CD8, da perforina e do IFN-γ na imunidade
gerada pela vacinação com a ASP-2 de T. cruzi. Acredita-se que os dados
obtidos no decorrer desta tese serviram para: i) uma melhor compreensão dos
mecanismos imunológicos de resistência à infecção pelo T. cruzi e dos seus
antígenos alvos; ii) o desenvolvimento de vacinas recombinantes contra
infecções por protozoários intracelulares em geral e, especificamente, contra a
doença de Chagas; iii) o desenvolvimento de vacinas recombinantes capazes
de induzir uma potente resposta imune mediada por linfócitos T.

1
Introdução

2
1.1 A doença de Chagas
I) Transmissão e epidemiologia
A doença de Chagas (ou tripanossomíase americana), descrita pela
primeira vez há cem anos pelo médico brasileiro Carlos Chagas [1], é causada
pelo protozoário flagelado Trypanosoma cruzi, e existe em 21 países da
América Latina, sendo endêmica em pelo menos 15 deles [2].
A principal forma de transmissão da doença de Chagas dá-se por meio
do contato das mucosas (incluindo a gástrica) ou de lesões cutâneas com as
fezes de insetos triatomíneos da família Reduvidae, infectados pelo T. cruzi.
Além da transmissão vetorial, o homem pode contrair a doença de Chagas por
meio de transfusões de sangue, de transplantes de órgãos ou por transmissão
vertical (materno-fetal).
A doença de Chagas está frequentemente associada à pobreza e às
condições precárias de habitação, já que os insetos vetores envolvidos nos
ciclos domésticos de transmissão se alojam, preferencialmente, nas casas
construídas com madeira, barro e palha, típicas das zonas rurais menos
favorecidas. No entanto, por causa da migração da população rural para áreas
urbanas dos países latino-americanos, ocorrida nas décadas de 1970-1980, a
doença de Chagas passou a ser também uma doença urbana transmissível
pela transfusão sanguínea.
Nas últimas décadas, diversos países vêm implementando ações que
visam à redução da transmissão da doença de Chagas, pela eliminação dos
principais vetores da doença e pela sorologia pré-transfusional dos doadores
(Iniciativa do Cone Sul, lançada em 1991; Iniciativa dos países andinos,
lançada em 1996; Iniciativa dos países da América Central, lançada em 1997 e
Iniciativa da Amazônia, lançada em 2004). A iniciativa que obteve maior
sucesso até o momento foi a do Cone Sul: o Uruguai em 1997, o Chile em
1999 e o Brasil em 2006, receberam a certificação de interrupção da
transmissão pelo Triatoma infestans, o principal vetor domiciliar nessas regiões
(revisto em [2]). Além disso, hoje em dia, na maioria dos países da América
Latina, os bancos de sangue testam as amostras para detectar a doença de
Chagas, descartando as positivas [2].

3
Estas iniciativas levaram a uma diminuição da incidência da doença de
700.000 casos/ano em todo o continente em 1990, para cerca de 200.000
casos/ano em 2006 [3]. Além disso, estima-se que a prevalência da doença de
Chagas caiu de 30 milhões de pessoas em 1990 para cerca de 15 milhões em
2006, tendo o número de mortes pela doença de Chagas passado de
45.000/ano para 12.500/ano [3].
Ainda assim, acredita-se que pelo menos 28 milhões de pessoas
estejam sob risco de contrair a doença na América Latina [3]. Em algumas
regiões o controle dos vetores é difícil, uma vez que eles não vivem no
ambiente doméstico, dirigindo-se aos domicílios apenas para se alimentar. Em
outras regiões os vetores já desenvolveram resistência ao inseticida utilizado
[4]. Além disso, números crescentes de casos vêm sendo reportados na região
amazônica, frutos das infecções por alimentos contaminados ou do contato
com insetos silvestres [5]. O Trypanosoma cruzi infecta não somente o homem,
mas também outros mamíferos placentados (como roedores e cães) e
marsupiais (como gambás), os quais servem como reservatório para o
protozoário causador da doença, impossibilitando assim a erradicação desta
[5].
Por fim, nos países desenvolvidos nos quais não há transmissão natural,
a doença de Chagas vem sendo considerada um problema em potencial, pois a
imigração das populações provindas das áreas endêmicas traz o risco da
contaminação pelas transfusões sanguíneas e pelos transplantes. A partir de
2007, os bancos de sangue dos Estados Unidos passaram a fazer a sorologia
para a detecção da doença de Chagas em todas as amostras, mas tal prática
ainda não está estabelecida nos países europeus.
II) Manifestações clínicas
Após o contato com o parasita, o hospedeiro vertebrado desenvolve a
fase aguda da doença, a qual pode durar semanas ou meses e ser sintomática
ou assintomática. Os possíveis sintomas são: inflamação na região da entrada
do parasita (chagoma de inoculação), febre, dor de cabeça, mialgia,
linfadenopatia, hepato/esplenomegalia e, nos casos mais graves, miocardite e
meningo-encefalite.

4
Após o controle da parasitemia pelo sistema imune, o indivíduo passa
então à fase crônica da doença, a qual é caracterizada por uma parasitemia
subpatente. Como o parasita não é eliminado, a fase crônica da doença de
Chagas perdura por toda a vida do indivíduo. Em cerca de 66% dos casos, as
pessoas infectadas permanecem assintomáticas, apresentando a chamada
forma indeterminada da doença. Já nos outros 34% dos casos, depois de anos
ou décadas, os indivíduos desenvolvem sintomas do trato digestivo (cerca de
6%), como megacólon ou megaesôfago, do sistema nervoso periférico (cerca
de 3%), ou cardíacos (25% dos infectados) [6,7,8]. As afecções cardíacas, que
incluem problemas da condução elétrica, arritmia, cardiomiopatia e insuficiência
cardíaca, em muitos casos levam à morte.
III) Patogenia
Mesmo cem anos após a descoberta da doença de Chagas, a sua
patogenia ainda não foi completamente entendida. Questões que se referem a
como se desenvolvem as lesões e o que leva determinados pacientes a
apresentarem as diferentes formas da doença ou a permanecerem
assintomáticos, estão ainda sem uma resposta definitiva, sendo motivos de
controvérsias na comunidade científica. Duas hipóteses já foram propostas
para explicar a patogenia da doença de Chagas: (1) a hipótese da
autoimunidade e (2) a hipótese da persistência do parasita.
Por muitos anos, o fato de que o patógeno era raramente encontrado
nas lesões dos indivíduos infectados deu suporte à hipótese da autoimunidade.
Dentro desta hipótese, a autoimunidade poderia ser derivada de uma quebra
de tolerância induzida pela inflamação em resposta à infecção (ativação
bystander), da existência de antígenos do parasita com reatividade cruzada
com autoantígenos (mimetismo molecular) ou de uma ativação policlonal dos
linfócitos (revisto em [9]). Assim, a doença era vista como uma condição que se
perpetuaria mesmo após a eliminação do elemento que a deflagrou (o T. cruzi).
De fato, alguns trabalhos demonstraram a presença de anticorpos [10,11,12] e
de linfócitos T [13,14,15] que reagem contra antígenos próprios (e em alguns
casos apresentam reações cruzadas com os antígenos do parasita) em
pacientes e camundongos infectados pelo T. cruzi (revisto em [9]).

5
Nos últimos anos, contudo, a utilização de novas técnicas como a
imunohistoquímica, a hibridização in situ e principalmente a PCR, permitiu a
detecção do parasita, ou do DNA do parasita, nas lesões ou em regiões
próximas às lesões dos pacientes cronicamente infectados [16,17,18]. Além
disso, a reativação da doença de Chagas (com casos reportados de encefalite
e parasitemia detectável por PCR), nos pacientes imunossuprimidos em
decorrência da AIDS [19,20], indica que, de fato, o T. cruzi persiste, mesmo
nos pacientes que apresentam a forma indeterminada da doença. Outros
trabalhos evidenciaram a presença do T. cruzi nos pacientes cronicamente
infectados e a participação do parasita na patologia. Estes trabalhos mostraram
que o tratamento dos pacientes e dos animais cronicamente infectados com
drogas tripanosomicidas trazia uma diminuição da severidade ou uma
progressão mais lenta da doença [21,22,23]. Estes dados sugerem que,
mesmo que haja um componente autoimune, a doença de Chagas decorre da
presença do T. cruzi no tecido.
Assim, atualmente, o fato de que o T. cruzi está presente nos pacientes
cronicamente infectados é bem aceito pela maior parte da comunidade
científica. Os defensores da hipótese da autoimunidade, no entanto, acreditam
que, dada a baixa quantidade de parasitas no tecido, uma resposta imune
direcionada apenas contra o T. cruzi não seria suficiente para causar os danos
teciduais observados nos pacientes sintomáticos [9,24]. Por outro lado, alguns
especialistas acreditam que a existência de uma resposta autoimune durante a
infecção pelo T. cruzi não deve ser considerada surpreendente, uma vez que,
assim como em outras doenças crônicas, o microambiente de citocinas
inflamatórias e de dano tecidual propicia a quebra de tolerância a antígenos
sequestrados [25]. Várias objeções foram feitas à participação dos
autoanticorpos na patogênese da doença de Chagas (revisto em [26]). Em
primeiro lugar, os autoantígenos descritos como alvos dos anticorpos são, em
muitos casos, moléculas intracelulares que não estão expostas em células
intactas (como a miosina e as proteínas ribossomais). Em segundo lugar,
vários desses antígenos estão distribuídos em diferentes tecidos, tornando
difícil de explicar porque as lesões ocorreriam apenas em alguns locais. Além
disso, os autoanticorpos (principalmente a antimiosina) também foram
observados nos pacientes que sofriam de diferentes problemas cardíacos

6
como infarto do miocárdio, miocardite viral e que tinham passado por cirurgias
cardíacas [27,28]. Do mesmo modo, os autoanticorpos foram observados nos
pacientes com a forma indeterminada da doença (assintomáticos) e, em alguns
casos, também nos pacientes sadios [10,29]. Quanto à participação dos
linfócitos T autoreativos na patogênese da doença de Chagas, alguns autores
acreditam que os dados existentes são pouco extensos, difíceis de reproduzir
e, por vezes, mal-interpretados [26], não sendo suficientes para demonstrar a
importância da autoimunidade na patogênese da doença de Chagas [25,26].
Ainda que haja dúvidas em relação à principal fonte de antígenos contra
os quais a resposta imune é direcionada, a maioria dos cientistas acredita que
a doença de Chagas é o resultado de danos teciduais causados pelo próprio
T. cruzi ou por uma resposta imune inflamatória crônica alimentada (direta ou
indiretamente) por um parasitismo em baixos níveis, porém incessante [24].
Assim, em teoria, ações visando à redução desse parasitismo, como as
imunizações e os tratamentos com drogas tripanosomicidas, trariam benefícios
para os indivíduos infectados.
A persistência do parasita, nos pacientes que apresentam tanto as
formas sintomáticas quanto a forma indeterminada da doença de Chagas,
levanta uma outra dúvida ainda não esclarecida, sobre quais seriam os fatores
que influenciam o curso da infecção para as diferentes formas da doença.
Algumas hipóteses foram sugeridas, entre elas o envolvimento do backgroud
genético tanto do parasita quanto do hospedeiro. Por um lado, alguns estudos
mostraram que os parasitas encontrados nos diferentes tecidos do mesmo
hospedeiro eram geneticamente diversos, apontando para um tropismo
específico de cada clone [19,30]. Por outro lado, alguns estudos mostraram
distribuições tendenciosas de alelos relacionados à resposta imune do
hospedeiro, nos pacientes com diferentes manifestações da doença [31].
Todos esses estudos, no entanto, são ainda pouco informativos para que se
consiga entender ou intervir na patogenia da doença.
IV) Diagnóstico
O diagnóstico durante a fase aguda da doença de Chagas pode ser feito
pela observação direta do parasita no exame de sangue a fresco. Caso este

7
teste seja negativo, podem ser utilizados métodos de concentração, como
micro-hematócrito ou Strout. Na fase crônica, os métodos parasitológicos como
a hemocultura e o xenodiagnóstico apresentam baixa sensibilidade, sendo os
testes sorológicos, como a imunofluorescência indireta, a hemaglutinação
indireta ou o ELISA, os mais indicados [32].
Estas técnicas também podem ser utilizadas para avaliar a cura dos
pacientes tratados, no entanto não existem critérios 100% confiáveis. A baixa
sensibilidade dos exames parasitológicos, como o xenodiagnóstico e a
hemocultura, pode ser aumentada com várias repetições dos testes (revisto em
[33]). Já o exame sorológico convencional muitas vezes permanece positivo
por vários anos, apesar dos testes parasitológicos se mostrarem repetidamente
negativos. A técnica da lise dependente de anticorpos mediada por
complemento foi proposta como um indicador de infecção ativa, já que este tipo
de teste se mostra negativo para os pacientes considerados curados antes do
exame sorológico convencional (revisto em [34]). Mais recentemente, as
técnicas da PCR e da PCR em tempo real vêm sendo estudadas para esse fim.
Apesar de não estar claro se uma PCR positiva representa a presença de
parasitas intactos ou do DNA derivado de organismos lisados, existem
evidências de que o DNA livre do parasita só pode ser detectado por até 48h,
indicando, portanto, que a PCR é útil para a detecção da infecção ativa (revisto
em [33]).
V) Tratamento
Atualmente existem apenas duas drogas disponíveis para o tratamento
da doença de Chagas: o benzonidazol e o nifurtimox. O mecanismo de ação do
nifurtimox parece estar associado à produção de grandes quantidades de
espécies reativas do oxigênio, enquanto a ação do benzonidazol parece
envolver o estresse redutor, com modificações covalentes de macromoléculas
por intermediários nitro-redutores (revisto em [35]).
Ambas as drogas, no entanto, frequentemente causam efeitos colaterais
que levam parte dos pacientes a abandonar o tratamento. O nifurtimox induz
reações adversas em cerca de 40% dos indivíduos tratados, incluindo náuseas,
vômitos, dores abdominais, perda de peso, anorexia severa e complicações

8
neurológicas (revisto em [36]). O benzonidazol induz efeitos colaterais em uma
percentagem menor de indivíduos [36], incluindo edema, febre, rash cutâneo,
dor muscular, neuropatia periférica e neutropenia [37].
A percentagem de cura das duas drogas, nos pacientes na fase aguda
da doença, é similar (cerca de 80%), mas varia de região para região,
provavelmente devido às diferenças na suscetibilidade da cepa de T. cruzi
[33,35]. De fato, a resistência natural à droga é considerada comum [33]. Um
outro problema é que, nos pacientes na fase crônica, os dados referentes à
eficácia do tratamento indicam que mais de 80% deles não atinge cura
parasitológica [38]. Ainda assim, existem evidências de que o tratamento na
fase crônica traz benefícios para os pacientes [21,23], o que levou médicos e
cientistas de diversos países a se envolverem em um teste clínico de grandes
proporções, a fim de avaliar os benefícios do tratamento com o benzonidazol
nos pacientes na fase crônica da doença de Chagas [36].
Os dados apresentados acima indicam que, apesar das iniciativas para o
controle dos insetos vetores e a sorologia para a detecção da doença de
Chagas nos bancos de sangue haverem levado a uma redução da incidência e
da prevalência dessa enfermidade na América Latina, ainda existe um grande
número de indivíduos infectados e sob risco de infecção, para os quais o
tratamento disponível está longe de ser o ideal. Há, portanto, uma grande
necessidade de se desenvolverem novos tratamentos ou formas de prevenção,
sendo para isso indispensável investir-se na pesquisa básica e clínica que
envolva novas drogas e/ou vacinas.
1.2 O ciclo de vida do T. cruzi
O Trypanosoma cruzi é um protozoário flagelado da ordem
Kinetoplastida, a qual é caracterizada por organismos que apresentam uma
única mitocôndria denominada cinetoplasto. O T. cruzi pertence à família
Trypanosomatidae, a qual inclui outros parasitas humanos como a Leishmania
spp. e o Trypanosoma brucei. O T. cruzi é um organismo digenético (ou
heteroxeno), apresentando um hospedeiro vertebrado (mamíferos) e um
hospedeiro invertebrado (insetos da família Reduvidae).

9
Esse parasita, presente em sua forma tripomastigota metacíclica nas
fezes e na urina do inseto vetor, infecta o hospedeiro mamífero por meio das
mucosas ou por lesões cutâneas. Uma vez no hospedeiro mamífero, ele invade
células, nas quais se diferencia na forma amastigota intracelular, que é uma
forma replicativa. Após diversos ciclos de divisão binária, os amastigotas se
diferenciam novamente em tripomastigotas, que lisam a célula e caem na
corrente sanguínea, podendo assim se dirigir para outros tecidos e infectar
novas células.
O inseto vetor é infectado após se alimentar do sangue de um
hospedeiro mamífero infectado. O inseto ingere o sangue contendo formas
tripomastigotas sanguíneas do T. cruzi que, no intestino do vetor, se
diferenciam em formas epimastigotas (replicativas). Na porção posterior do
intestino, os epimastigotas sofrem o processo da metaciclogênese,
diferenciando-se em tripomastigotas metacíclicos que serão eliminados nas
fezes e na urina do inseto. A Fig. 1 oferece uma representação esquemática do
ciclo de vida do Trypanosoma cruzi.
Figura 1. Representação esquemática do ciclo de vida do T. cruzi. Adaptado de [39].

10
1.3 A resposta imune na infecção experimental pelo T. cruzi
A persistência do parasita no hospedeiro durante longos períodos é um
evento importante no ciclo de vida do T. cruzi, pois ela maximiza a
possibilidade do encontro entre o hospedeiro infectado e o inseto vetor,
permitindo assim a transmissão e a sobrevivência do parasita. Para que essa
permanência ocorra, o T. cruzi deve ser capaz de estabelecer uma situação de
equilíbrio com o sistema imune do seu hospedeiro. Por um lado, se não houver
indução da resposta imune e o parasita puder se replicar sem restrições, o
hospedeiro sucumbirá rapidamente. Por outro lado, a indução de uma resposta
imune muito potente pode levar à eliminação rápida e completa do T. cruzi. Em
ambos os casos, isto reduziria a eficiência da transmissão e os parasitas
seriam evolutivamente prejudicados e possivelmente extintos. Assim, foram
selecionados parasitas capazes de induzir uma resposta imune adequada à
sobrevivência do hospedeiro, porém incapaz de eliminar os parasitas antes da
transmissão destes para um novo hospedeiro. A elucidação dos mecanismos
imunológicos de sobrevivência do hospedeiro é de vital importância para o
desenvolvimento de intervenções que possam eventualmente desequilibrar a
relação parasita/hospedeiro em favor do segundo, e bloquear assim o ciclo
biológico do primeiro.
I) Resposta imune inata
Após a infecção, o sistema imune do hospedeiro mamífero é capaz de
responder ao T. cruzi por meio do reconhecimento de padrões moleculares
associados à patógenos (PAMPs), o qual é efetuado por receptores específicos
[40].
Nas últimas décadas foram descritos diversos receptores que
reconhecem padrões (PRRs), entre eles os receptores semelhantes ao Toll
(TLRs), localizados em membranas, e os receptores semelhantes ao NOD
(NLRs), que são citoplasmáticos. Estas moléculas estão presentes em
diferentes tipos celulares, como as células epiteliais, mas principalmente nas
células do sistema imune, como os macrófagos, as células dendríticas e os
linfócitos. Os TLRs reconhecem proteínas, lipídeos, polissacarídeos e ácidos

11
nucleicos típicos de patógenos; já os NLRs, além de reconhecerem moléculas
derivadas de patógenos (como peptídeos das paredes bacterianas, flagelina e
ácidos nucleicos), respondem também a padrões moleculares associados ao
perigo (DAMPs), como o ácido úrico [40]. A ligação dos PAMPs aos TLRs e
aos NLRs dispara vias de sinalização específicas que ativam a resposta imune
inata, induzindo atividades antimicrobianas e inflamatórias, e regulam a
resposta imune adaptativa de acordo com o tipo de patógeno a ser combatido
[41].
Já foram descritos no T. cruzi três PAMPs: as âncoras de glicosilfosfati-
dilinositol (GPI), que se ligam ao TLR2 [42]; os glicoinositolfosfolipídeos
(GIPLs), que se ligam ao TLR4 [43]; e os motivos CpG não-metilados
presentes no DNA do parasita, que seriam agonistas do TLR9 [44]. Esses
receptores parecem ser realmente importantes no reconhecimento e
consequente resposta ao T. cruzi, uma vez que camundongos deficientes do
TLR4 ou do TLR9 apresentaram níveis mais altos de parasitemia e de
mortalidade do que os camundongos normais, após a infecção por esse
parasita [43,44]. Além disso, camundongos deficientes simultaneamente do
TLR2 e do TLR9 também se mostraram mais suscetíveis à infecção pelo T.
cruzi do que animais deficientes em apenas um ou outro desses receptores
[45], evidenciando que os diversos TLRs cooperam na ativação da resposta
após o reconhecimento do parasita. Apesar de apenas três PAMPs terem sido
descritos no T. cruzi, existem evidências de que outros ligantes dos TLRs
podem estar presentes no parasita. Os camundongos deficientes da molécula
adaptadora MyD88 (comum às vias de sinalização de quase todos os TLRs
descritos, com a exceção do TLR3) se mostraram ainda mais suscetíveis do
que os camundongos duplamente deficientes do TLR2 e do TLR9,
apresentando maior mortalidade [45]. Mais do que isso, demonstrou-se que os
camundongos duplamente deficientes de MyD88 e de TRIF (uma outra
molécula adaptadora na via de sinalização de alguns TLRs, incluindo o TLR3)
eram ainda mais suscetíveis ao T. cruzi do que os MyD88 KO, apresentando
níveis mais altos de parasitemia e uma mortalidade ainda mais acelerada [46].
Por fim, dado que o sistema de reconhecimento de patógenos é altamente
redundante, já que algumas moléculas podem ser reconhecidas por vários
receptores diferentes (a flagelina, por exemplo, é reconhecida pelo TLR5, pelo

12
Ipaf e pelo Naip5, [40]), é possível que as âncoras de GPI, os GIPLs ou outras
moléculas do T. cruzi ativem outros receptores como os NLRs.
O reconhecimento do patógeno por PRRs ativa vias de sinalização que
culminam com a indução e/ou translocação de fatores de transcrição, como o
AP-1, o NFκB e os IRFs. Esses fatores de transcrição são responsáveis pela
produção de citocinas pró-inflamatórias, como a IL-12, os IFN-α/β e o TNF-α,
pelos macrófagos e pelas células dendríticas. Acredita-se que a IL-12 pode
ativar as células natural killer (NK) [47]. As células NK parecem ser importantes
nos estágios iniciais da infecção experimental pelo T. cruzi, já que a sua
depleção com anticorpos anti-asialo GM1 ou anti-NK1.1 resulta em maiores
níveis de parasitemia e de mortalidade dos camundongos C57BL/6 infectados
com as cepas CL e Y [48,49]. O mecanismo de ação das células NK pode estar
relacionado à lise de tripomastigotas livres por mecanismos independentes de
perforina [50] ou à lise de células infectadas por mecanismos independentes do
contato célula-célula e dependentes da produção de IFN-γ e óxido nítrico (NO)
[51]. Além disso, as células NK podem produzir IFN-γ nos estágios iniciais da
infecção por T. cruzi [47].
Alguns subtipos não convencionais de linfócitos T, que por apresentarem
TCRs menos diversificados são muitas vezes considerados parte da resposta
imune inata, também podem participar na resposta contra a infecção pelo T.
cruzi. Um exemplo são as células NKT restritas pelo CD1d. Duthie e
colaboradores [52] mostraram que os camundongos deficientes do CD1d, que
não possuem nenhum tipo de célula NKT, após a infecção com a cepa CL de
T. cruzi, apresentam parasitemias mais altas do que os camundongos normais,
apesar da mortalidade observada ser semelhante. Por outro lado, os mesmos
autores observaram que os camundongos Jα18-/-, deficientes no subtipo
invariante das células NKT, apresentam um fenótipo bem mais grave, com
maior morbidade e mortalidade, implicando as células iNKT na modulação da
resposta inflamatória [53]. No entanto, estes resultados podem variar quando
outras cepas de T. cruzi são utilizadas.
Um outro subtipo de linfócitos T não convencionais são os T γδ. Os
estudos deste subtipo na infecção pelo T. cruzi, contudo, são escassos e
conflitantes. Santos Lima e colaboradores [54] reportaram, utilizando

13
camundongos deficientes de Cδ e, portanto dos linfócitos T γδ, que a ausência
destas células durante a infecção seria benéfica, já que os camundongos
deficientes apresentaram uma menor mortalidade e níveis mais baixos de
inflamação nos tecidos musculares cardíaco e esquelético, após a infecção
com a cepa CL de T. cruzi. Já Nomizo e colaboradores [55] mostraram que os
camundongos tratados com anticorpos anti-Vgamma1 e, portanto, depletados
da população Vγ1+ de linfócitos T γδ, apresentavam uma menor ativação dos
linfócitos T convencionais, níveis mais altos de parasitemia e mortalidade
acelerada em relação aos camundongos normais, após a infecção com a cepa
Y de T. cruzi. As discrepâncias entre os dados podem ser causadas por
diferenças metodológicas, como o subtipo de células removido (todas as γδ vs.
somente a população Vγ1+) ou a cepa do parasita utilizada, não permitindo
assim uma conclusão definitiva.
II) Resposta imune adaptativa
A resposta imune inata é sem dúvida essencial para o controle da
infecção pelo Trypanosoma cruzi. O reconhecimento inato do parasita permite
uma resposta rápida, que retarda a multiplicação do patógeno e,
provavelmente, dispara sinais críticos para a geração da resposta adaptativa.
No entanto, na ausência de qualquer uma das principais vertentes da resposta
imune adaptativa, a resposta imune inata é incapaz de proteger os animais de
uma infecção pelo Trypanosoma cruzi. A ausência dos linfócitos B [56], T CD4
ou T CD8 [57,58] torna os camundongos extremamente suscetíveis à infecção
experimental pelo T. cruzi. Há evidências de que é a resposta imune que
mantém a parasitemia sob controle durante a fase crônica da infecção
experimental, já que o tratamento com drogas imunossupressoras (como a
ciclofosfamida), durante a fase crônica, leva a um aumento da parasitemia, a
qual se torna detectável [59].
A importância dos anticorpos na proteção contra a infecção pelo T. cruzi
foi demonstrada pela transferência passiva de anticorpos provindos do soro de
camundongos cronicamente infectados, para camundongos naive, que
apresentaram uma redução importante da parasitemia e uma mortalidade

14
significativamente retardada [60,61]. Além disso, a transferência de anticorpos
monoclonais também foi capaz de reduzir a parasitemia e atrasar a mortalidade
dos camundongos naive [62]. O mecanismo de ação desses anticorpos não foi
ainda totalmente esclarecido, mas demonstrou-se que os anticorpos protetores
eram capazes de aglutinar tripomastigotas in vitro [60], facilitar a fagocitose [63]
e atuar na citotoxicidade celular dependente de anticorpos (ADCC) [64,65].
Além disso, é possível que os anticorpos neutralizem a molécula reguladora de
complemento presente na superfície dos tripomastigotas [66], permitindo assim
o depósito da C3 e a lise do parasita pela via alternativa do complemento [34].
Outra evidência da importância das células B é que a sua ausência
aumenta a suscetibilidade de camundongos ao desafio com T. cruzi. É
importante notar, no entanto, que os camundongos desprovidos dos linfócitos B
apresentaram uma sobrevida mais longa do que aqueles deficientes dos
linfócitos T [56]. Além disso, dado que, além de produzirem anticorpos, os
linfócitos B também atuam como apresentadores de antígenos, é possível que
sua ausência seja letal na infecção pelo T. cruzi devido aos efeitos sobre a
resposta de linfócitos T. De fato, já foi demonstrado que as células B são
importantes apresentadoras de antígenos para os linfócitos T CD8 na
imunização com a proteína TS [67], e que camundongos geneticamente
deficientes dos linfócitos B apresentam alterações nos marcadores de ativação
dos seus linfócitos T [68]. Estes dados sugerem que a ausência das células B
pode prejudicar a resposta celular.
Em relação à importância dos linfócitos T CD4 e CD8 na infecção pelo
T. cruzi, diversos trabalhos já ofereceram evidências, utilizando animais
geneticamente modificados deficientes (KO) dessas subpopulações de células
[56,57,58,69]. Após a infecção do hospedeiro, algumas cepas de T. cruzi se
multiplicam no tecido e alcançam a corrente sanguínea. Quando esta
parasitemia atinge níveis críticos, uma potente resposta celular é gerada,
controlando-a e permitindo a sobrevivência dos animais.
No caso da resposta de linfócitos T CD8, a clássica curva de expansão
rápida das células específicas não se aplica durante a infecção pelo T. cruzi. Já
foi descrito que, após a infecção por este parasita, existe uma demora na
indução da resposta de linfócitos T CD8 [69,70,71] que não é comumente
observada nas infecções virais e bacterianas. Diversos grupos propuseram que

15
tal atraso seria uma estratégia que permitiria o estabelecimento de infecções
crônicas, tanto no caso do T. cruzi quanto no de outros patógenos [72,73]. A
favor desta hipótese, demonstrou-se que o adiantamento da resposta imune
por meio da imunização reduz significativamente a parasitemia, permitindo a
sobrevivência dos camundongos a um desafio letal [Vasconcelos et al.,
manuscrito em preparação].
A resposta imune dos linfócitos T CD4 também é crítica para o controle
do parasitismo agudo. No entanto, a cinética exata desta não foi documentada
por causa da dificuldade de se encontrar epítopos específicos. Enquanto no
caso dos linfócitos T CD8 foram descritos diversos epítopos que se ligam a
diferentes moléculas do MHC I de modelos experimentais [70,74,75,76], um
único epítopo para os linfócitos T CD4, proveniente de antígenos do T. cruzi, foi
documentado [77]. Mesmo assim, ainda não foi possível enumerar os linfócitos
T CD4 específicos na sua cinética de expansão.
Um subtipo de linfócitos T CD4 que foi estudado na infecção pelo
T. cruzi foi o subtipo que expressa os marcadores CD25 ou GITR. Os trabalhos
publicados apresentaram resultados conflitantes. Em um deles, o bloqueio
funcional dessas células por meio da inoculação de anti-CD25 mostrou-se
incapaz de influenciar a parasitemia e a mortalidade dos camundongos
infectados pela cepa Tulahuén de T. cruzi [78]. Outro trabalho apontou um
efeito modesto deste tratamento sobre o curso da infecção, que era
dependente da dose e da cepa de parasita utilizada [79]. Em contraste,
Mariano e colaboradores [80] observaram uma miocardite exacerbada (com um
aumento do infiltrado inflamatório e um maior número de ninhos de
amastigotas), além do aumento da parasitemia e uma mortalidade precoce dos
camundongos tratados com anti-GITR e infectados com a cepa Y de T. cruzi.
Apesar da inequívoca participação dos linfócitos T CD4 e CD8 no
controle da infecção aguda pelo T. cruzi, ainda não se sabe precisamente qual
é o mecanismo de ação destas células. Já foi extensivamente documentado
que a produção das citocinas do tipo 1 é essencial para o controle da fase
aguda da infecção e a sobrevivência dos camundongos. Um estudo foi
realizado utilizando camundongos deficientes de STAT4 (um regulador
transcricional ativado em resposta à sinalização pela IL-12 e que dirige a
diferenciação dos linfócitos Th1/Tc1) e camundongos deficientes em STAT6

16
(um regulador transcricional ativado em resposta à sinalização pela IL-4 e/ou
IL-13 e que dirige a diferenciação dos linfócitos Th2 [81]). Os camundongos
deficientes de STAT4, quando infectados pela cepa Brazil de T. cruzi,
apresentaram parasitemias extremamente altas e morreram precocemente, em
tempos similares aos camundongos deficientes de CD4 ou CD8. Já os
camundongos deficientes de STAT6 apresentaram uma parasitemia e uma
longevidade similares àquelas dos camundongos normais, porém sem
evidências da presença do T. cruzi ou de inflamação no tecido cardíaco em um
período tardio da infecção. Estes dados indicam que a diferenciação dos
linfócitos T CD4 e T CD8 em Th1 e Tc1 é essencial para que os camundongos
sobrevivam à fase aguda da infecção pelo T. cruzi. Por outro lado, a presença
dos linfócitos do tipo Th2 parece ser responsável pela persistência do parasita
na fase crônica.
Os linfócitos T CD8 Tc1, além de produzirem as citocinas do tipo1,
podem ser também altamente citotóxicos. A importância deste mecanismo foi
testada em camundongos deficientes de perforina, granzimas e Fas. Os
resultados, porém, divergiram conforme a cepa de T. cruzi. Enquanto nas
infecções com as cepas Y e Tulahuén os camundongos deficientes de
perforina apresentaram uma maior suscetibilidade [69,82,83], na infecção pela
cepa Brazil estes camundongos apresentaram parasitemia e longevidade
similares aos animais selvagens [56]. Também os camundongos deficientes de
granzima B, quando infectados pela cepa Brazil se comportaram de modo
similar aos camundongos selvagens [56]. Já os camundongos deficientes das
granzimas A e B, assim como os deficientes em Fas, quando infectados pela
cepa Tulahuén, se mostraram mais suscetíveis do que os selvagens [83]. Na
realidade, não existem evidências de que a lise de uma célula infectada pelo T.
cruzi destrua o parasita. Na infecção pelo Toxoplasma gondii, por exemplo, a
lise da célula hospedeira, mediada pela perforina, parece induzir a saída do
parasita da célula hospedeira em condições viáveis [84]. No entanto, é possível
que a lise das células infectadas pelo T. cruzi contribua para o combate ao
patógeno, por abreviar a proliferação do mesmo e expor a forma amastigota a
anticorpos, ao sistema complemento e a outros fatores circulantes, já que é
menos provável que essa forma do parasita expresse mecanismos de proteção
como a proteína reguladora de complemento (CRP) [66].

17
Entre as citocinas importantes na infecção pelo T. cruzi, pode-se citar as
do tipo 1: a IL-12, o IFN-γ e o TNF-α. Os animais deficientes da IL-12
apresentam um aumento da parasitemia e da mortalidade [85], assim como os
hamsters tratados com um bloqueador do TNF-α [86], os camundongos
deficientes do receptor de TNF-α [87,88], de IFN-γ ou do seu receptor [89].
Mais recentemente também foi descrito que os camundongos deficientes do
receptor de IFN-α/β apresentam parasitemias mais altas do que os normais,
mas não diferem no tempo da mortalidade [90].
Acredita-se que a produção dessas citocinas inflamatórias ative os
macrófagos, os quais passam a eliminar os parasitas fagocitados. O
mecanismo de ação do IFN-γ nos macrófagos parece ser, pelo menos em
parte, dependente da produção do NO. Na ausência de receptores para o IFN-
γ, o NO não é produzido por macrófagos, sugerindo que esta citocina é a
principal indutora da NO sintase 2 (NOS2). A importância do NO na resistência
à infecção pode ser dependente também da cepa de parasita, uma vez que os
animais deficientes da NOS2 são mais suscetíveis à infecção pelas cepas
Tulahuén e Y de T. cruzi [89,91], mas não à infecção pela cepa Brazil [92].
Assim, é possível que outros mecanismos independentes da NOS2, mas
também ativados pelo IFN-γ, sejam importantes na eliminação do T. cruzi.
As citocinas anti-inflamatórias parecem também participar da modulação
da resposta imune durante a infecção experimental pelo T. cruzi. Por exemplo,
camundongos KO para il-4 já foram descritos como sendo mais resistentes à
infecção pela cepa Colombiana, apresentando menor parasitemia, mortalidade
e parasitismo tissular [85], assim como os camundongos tratados com anti-IL-4
se mostraram mais resistentes à cepa Tulahuén [93]. No entanto, outro
trabalho mostrou que, apesar da diminuição no parasitismo tissular, os
camundongos il-4 KO infectados com a cepa Colombiana apresentavam uma
maior inflamação no tecido cardíaco, sugerindo que a IL-4 seria importante na
modulação da resposta Th1, impedindo o dano tissular [94]. Já no caso da IL-
10, os trabalhos mostraram que, apesar do melhor controle da parasitemia nos
camundongos il-10 KO, eles apresentaram uma maior mortalidade em
comparação aos normais, provavelmente em decorrência da secreção
excessiva do TNF-α [95,96]. Um estudo com camundongos KO cujos linfócitos

18
T não respondem ao TGF-β também demonstrou que essa citocina era
essencial para controlar a proliferação dos linfócitos T CD8 e para proteger os
camundongos do dano tissular durante a infecção experimental pelo T. cruzi
[97].
Algumas quimiocinas e seus respectivos receptores podem também
estar implicados no controle da resposta imune durante a infecção pelo
T. cruzi. A expressão do CCR5 (receptor das quimiocinas MIP1-α, MIP1-β e
RANTES) vem sendo estudada nos linfócitos T CD4 e T CD8 presentes no
tecido cardíaco de animais infectados. Marino e colaboradores [98] mostraram
que, ao contrário dos camundongos sadios, no coração daqueles infectados
pela cepa Colombiana de T. cruzi há um grande número de células CCR5+.
Além disso, eles observaram um aumento do número de células CD8+CCR5+
circulantes, nos animais infectados. Machado e colaboradores [99], trabalhando
com a cepa Y, mostraram que o T. cruzi é capaz de aumentar in vitro a
expressão do CCR5 nas células CD4+ e principalmente CD8+, enquanto Kroll-
Palhares e colaboradores [100] sugeriram que a sinalização por TNF-α/TNFR1
induz a expressão do CCR5 nos linfócitos T CD8. Alguns trabalhos [99,101]
também mostraram que os camundongos deficientes do CCR5 são mais
suscetíveis à fase aguda da infecção, apresentando maiores níveis de
parasitemia e de mortalidade que os normais.
Marino e colaboradores [98] ainda descreveram que o tratamento com o
antagonista do CCR5 Met-RANTES diminuiu, entre os dias 14 e 28 após a
infecção, o número de linfócitos T CD4, T CD8 e CCR5+ infiltrantes no coração,
além de diminuir a deposição de fibronectina e de aumentar a sobrevida dos
animais. Resultados semelhantes foram obtidos por Medeiros e colaboradores
[102], por meio do tratamento com Met-RANTES nos dias 150 a 180 após a
infecção de camundongos com a cepa Colombiana.
Em conjunto, estes dados indicam uma complexa interação entre as
diferentes células, citocinas e quimiocinas do sistema imune durante a infecção
pelo T. cruzi. Na maioria dos casos estes fatores têm papéis protetores,
levando à sobrevivência do hospedeiro, mas são fortemente regulados para
impedir uma patologia exacerbada e a eliminação completa do T. cruzi.

19
III) Alvos da resposta imune adaptativa
Os principais alvos já descritos da resposta imune humoral são as
mucinas e as trans-sialidases presentes na superfície do parasita. Os únicos
alvos de anticorpos protetores descritos até o momento são a proteína
reguladora do complemento [103] e os oligossacarídeos sialilados ligados às
glicoproteínas semelhantes às mucinas [62].
Já quanto à resposta T CD4, o único epítopo descrito está presente no
antígeno SA85.11, um membro da superfamília das trans-sialidases [77].
Apesar disso, é altamente provável que outras proteínas sejam alvos da
resposta imune mediada pelos linfócitos T CD4. De fato, há indícios da
presença de epítopos T CD4 no antígeno ASP-2, por exemplo [104].
Recentemente, foram descritos diversos alvos da resposta imune
mediada pelos linfócitos T CD8. Durante a infecção pelo T. cruzi, a resposta de
células CD8 parece se concentrar nos epítopos imunodominantes expressos
principalmente nas proteínas que fazem parte da superfamília das trans-
sialidases [70,105]. Apesar de estar claro que os linfócitos T CD8 são críticos
para a sobrevivência do hospedeiro à infecção, ainda não se sabe
precisamente porque a resposta imune destes é tão direcionada. Este
fenômeno da imunodominância pode ser decisivo não só para a sobrevivência
do hospedeiro, mas pode também ser um mecanismo do parasita para a
evasão da resposta imune, pois impede que esta seja mais ampla, o que
levaria à eliminação completa do patógeno [105,106].
Já foram descritos epítopos imunodominantes nas proteínas da
superfamília das trans-sialidases como a ASP-2, a TSA-1 e a TS [70,74,75,76].
Além destes, antígenos subdominantes foram descritos em proteínas que não
fazem parte da família das trans-sialidases, como a cruzipaína e a β-galactofu-
ranosil transferase [70] e as proteínas do flagelo PFR1 e PFR3 [107].

20
1.4 Vacinação contra a infecção experimental pelo Trypanosoma cruzi
A maioria das vacinas disponíveis para uso humano até o momento são
compostas por microorganismos mortos ou atenuados. No século passado,
este tipo de abordagem também foi utilizado para tentar proteger os
camundongos da infecção pelo T. cruzi. Os resultados, no entanto, não foram
animadores, pois, no caso das cepas atenuadas, havia grande risco de
reversão do parasita para fenótipos mais virulentos. Já as vacinas com
parasitas mortos induziam, principalmente, respostas mediadas por anticorpos,
as quais eram insuficientes para proteger os camundongos (revisto em [108]).
Assim, com as evidências descritas acima da importância dos linfócitos
T, a ideia de que uma resposta imune mediada pelos linfócitos CD4 e CD8 do
tipo I fosse capaz de refrear a multiplicação do parasita, reduzindo dessa
maneira o dano tecidual causado pela persistência do T. cruzi, floresceu entre
a comunidade científica. As atenções se voltaram, então, para as vacinas de
subunidades. Nas últimas décadas, diversos antígenos e sistemas de liberação
foram testados em infecções experimentais, tendo mostrado resultados
promissores. Entre os sistemas de liberação testados, podem ser citados: as
proteínas nativas ou recombinantes adicionadas de uma série de adjuvantes, o
DNA plasmidial e os vírus e bactérias recombinantes.
As imunizações com proteínas na presença de adjuvantes como o CpG
ODN [67,75,109], o MALP-2 (um lipopeptídeo agonista do TLR2/6, extraído das
bactérias do gênero Mycoplasma) [110], a IL-12 [111,112] ou o CFA/IFA
(adjuvante completo/incompleto de Freund) [113] mostraram-se eficazes em
controlar a parasitemia aguda, o parasitismo tecidual e a mortalidade dos
camundongos em infecções experimentais. No entanto, a imunização com
proteínas, além de induzir respostas celulares fracas (principalmente no caso
de respostas de linfócitos T CD8), depende fortemente do adjuvante ao qual
elas estão associadas, e muitos adjuvantes que são capazes de induzir fortes
respostas imunes podem ser bastante tóxicos. Assim, apesar de na última
década diversos adjuvantes novos terem sido desenvolvidos e os mecanismos
de ação de alguns deles haverem sido elucidados, hoje apenas alguns poucos
adjuvantes são aprovados para serem aplicados em seres humanos.

21
Dessa maneira, outras estratégias que não dependem tão fortemente de
adjuvantes extrínsecos vêm sendo testadas. Entre elas, a imunização com
DNA plasmidial vem obtendo bastante sucesso em vacinações contra a
infecção experimental pelo T. cruzi em camundongos. As imunizações com
genes como ts [67,114], asp-2 [115,116,117] e tcg-1, tcg-2 ou tcg-4 [118] foram
capazes de reduzir significativamente a parasitemia, o dano tecidual e a
mortalidade dos camundongos. No entanto, as vacinas de DNA se mostraram
muito menos imunogênicas nos seres humanos do que nos pequenos roedores
(revisto em [119]), trazendo a necessidade do desenvolvimento de outros
sistemas de liberação para o DNA.
Os vírus e as bactérias recombinantes também passaram a ser
utilizados para imunizar camundongos contra a infecção experimental pelo T.
cruzi. Entre os sistemas testados com sucesso, podem ser citados os
adenovírus recombinantes [120,121], os MVAs recombinantes (vírus vaccínia
modificado) [120], o vírus Sendai recombinante [122] e a Salmonella
recombinante [112,123]. Ademais, diversos estudos utilizaram estes diferentes
sistemas de liberação em protocolos de imunização e reforço heterólogos, que
serão discutidos com maior profundidade mais adiante.
Vários antígenos têm sido utilizados na vacinação contra a infecção
experimental pelo T. cruzi. Um resumo dos antígenos utilizados e as
respectivas formulações vacinais podem ser observados na Tabela 1.

22
Tabela 1
Sumário dos diferentes antígenos e formulações usados para a vacinação
experimental contra a infecção pelo T. cruzi.
Forma do antígeno
Adjuvante Antígeno/gene T. cruzi
Referência
Proteínas nativas
Alum e IL-12
PFR
[124]
Proteína nativa
CpG ODN
Cruzipaína
[109]
Proteína Rec.
Proteína Rec.
Anti-IL4/IL-12
MALP-2
Cruzipaína
Cruzipaína
[112]
[110]
Proteína Rec.
Adjuvante de Freund
TS
[113]
Proteína Rec.
CpG ODN
TS
[67]
Proteína Rec.
Alum/CpG ODN
ASP-2
[75,104]
DNA plasmidial Nenhum TSA-1 [125,126,127]
DNA plasmidial
IL-12
TSSA
[111,128] DNA plasmidial Nenhum TS [67,114,129]
DNA plasmidial
Nenhum
ASP-1
[130]
DNA plasmidial
Nenhum
ASP-2
[115,116,117,130,131]
DNA plasmidial
Nenhum
ASP-3
[132]
DNA plasmidial
Nenhum
ASP-4
[132]
DNA plasmidial
Nenhum
CRP
[133]
DNA plasmidial
Nenhum
KMP11
[134]
DNA plasmidial
Nenhum
Tc24
[126]
DNA plasmidial
IL-12/GM-CSF
TcG1
[118]
DNA plasmidial
IL-12/GM-CSF
TcG2
[118]
DNA plasmidial
IL-12/GM-CSF
TcG4
[118]
Adenovírus Nenhum TS [121]
Adenovírus
Nenhum
ASP-2
[121]
Vírus Sendai recombinante
Nenhum
ASP-2
[122]
DNA plasmidial + Rec. MVA
Nenhum ASP-2 Dutra et al., em preparação
Adenovirus rec. + MVA Rec.
Nenhum TSA [120]
Salmonella Rec. Nenhum Cruzipaína [123]

23
O mecanismo de proteção de muitas destas imunizações parece ser de
fato baseado na indução das respostas imunes do tipo Th1 e Tc1 contra os
antígenos estudados. Alguns dos trabalhos verificaram que a ausência dos
linfócitos T CD4 e/ou T CD8 reverte a proteção [67,75,111,117,121,124],
enquanto praticamente todos eles mostraram a indução de células produtoras
de citocinas tipo I, como o IFN-γ.
É importante observar, todavia, que nenhuma das vacinas contra a
infecção experimental pelo T. cruzi testadas até o momento provou ser capaz
de eliminar completamente o parasita. Contudo, uma forte resposta imune, que
controle a parasitemia e reduza a carga parasitária tecidual, pode ser suficiente
para evitar a progressão da doença para as formas crônicas sintomáticas,
mantendo os indivíduos como chagásicos assintomáticos [135]. Ainda assim, é
importante que se determine com cautela como as intervenções que modulam
a reposta imune do hospedeiro podem interferir na patogenia da doença, a qual
ainda é controversa.
Devido às características da doença, os testes clínicos de desafio
experimental nos seres humanos seriam eticamente inaceitáveis. Assim, uma
possível solução para esta questão seria a procura por correlatos imunológicos
de proteção. Desse modo, as formulações candidatas à vacina devem ser
testadas em modelos animais de infecção pelo T. cruzi, devendo os correlatos
imunológicos de proteção ser extraídos, para que se busque a indução deste
mesmo tipo de resposta imunológica nos indivíduos vacinados.
I) Proteína 2 da superfície de amastigotas (ASP-2)
Os estudos desta tese utilizaram como modelo para a vacinação
experimental o antígeno denominado Proteína 2 da Superfície de Amastigotas
(ASP-2). Este antígeno foi identificado por Pan & McMahon-Pratt [136] como
uma proteína de 83 kDa, expressa na membrana plasmática da forma
amastigota do parasita. Posteriormente, o gene que codifica essa proteína foi
clonado por Low & Tarleton [137] a partir do DNA genômico de parasitas da
cepa Brazil, tendo o sequenciamento completo mostrado que o gene
apresentava alta homologia com o grupo II da superfamília das trans-
sialidases. Com base nestes estudos iniciais, isolaram-se genes relacionados

24
com a ASP-2 a partir do cDNA de amastigotas da cepa Y de T. cruzi [115] e,
posteriormente, das cepas CL-Brener, Tulahuén, Colombiana, G e Sylvio X10/4
[131].
A sequência predita de aminoácidos da ASP-2 da cepa Y apontou a
presença de: um peptídeo sinal de transporte para o retículo endoplasmático;
dois motivos de ácido aspártico (Asp Box), típicos de sialidases bacterianas e
virais [138]; um motivo altamente conservado VTV Box, que está presente no
módulo tipo III da fibronectina e é característico dos membros da superfamília
das trans-sialidases [139]; e da região hidrofóbica C-terminal para a adição da
âncora de GPI. A expressão dessa proteína mostrou-se restrita à forma
amastigota do parasita, o que foi verificado utilizando-se tanto anticorpos
monoclonais quanto policlonais [115,131,136,137].
A figura 2 contém uma representação esquemática da sequência predita
de aminoácidos da ASP-2 de diversas cepas de T. cruzi [131]. As cepas Brazil,
Y, CL-Brener, Tulahuén, Colombiana e G expressam isoformas da ASP-2 muito
semelhantes, com identidade de aminoácidos variando entre 73,5 e 97%. Por
outro lado, as cepas G e Sylvio X10/4 apresentam isoformas da ASP-2, cujas
identidades no nível de aminoácidos, com as isoformas encontradas nas outras
cepas, são baixas, variando de 43,4 a 49%. Por outro lado, as ASP-2
observadas nas cepas G e Sylvio X10/4 apresentam uma alta identidade de
aminoácidos entre elas: 78,7% [131].

25
Fig. 2: Representação esquemática da sequência deduzida de aminoácidos da ASP-2 [131].
Nada se sabe ainda sobre a função biológica da ASP-2. A conservação
da sequência desta proteína em várias cepas sugere que ela tenha um papel
importante. É sabido, no entanto, que proteínas ancoradas ao GPI, ou as
moléculas de GPI livres na membrana do parasita, são importantes para os
processos de sobrevivência e de multiplicação dos amastigotas no citoplasma
da célula hospedeira [140]. No entanto, ainda não se conhecem as moléculas
que participam nestes processos. Outros estudos indicam que o motivo
VTVXNVFLYNR, comum às proteínas da superfamília das trans-sialidases,
pode ser um domínio de ligação a proteínas como a citoqueratina 18, presente
na superfície da célula [141]. A ligação do motivo VTVXNVFLYNR à
citoqueratina 18 parece ativar a cascata de sinalização ERK1/2, culminando
com uma facilitação da entrada do parasita [142]. Assim, a presença do mesmo
motivo conservado na ASP-2 e em outras proteínas da superfamília das trans-
sialidases sugere existir um papel adesivo para tais moléculas.

26
Os primeiros estudos imunológicos com a ASP-2 demonstraram que
esta proteína contém epítopos reconhecidos pelos linfócitos T CD8 que são
gerados durante a infecção de camundongos C57BL/6 [76]. Estes resultados
impeliram diversos novos estudos sobre a imunização experimental utilizando
plasmídios, proteínas e vírus recombinantes que expressam o gene da ASP-2.
Garg & Tarleton [130] demonstraram que os camundongos C57BL/6,
imunizados com o gene da asp-2 da cepa Brazil e desafiados com
tripomastigotas sanguíneos desta mesma cepa, tiveram uma redução do pico
da parasitemia e uma sobrevida de 66%. Além disso, a vacinação genética
com o gene asp-2 reduziu a inflamação e o dano tecidual no coração e no
músculo esquelético durante a fase tardia (crônica) da doença.
Em paralelo, Boscardin e colaboradores [115] imunizaram camundongos
BALB/c com um plasmídio pcDNA3 contendo o gene da ASP-2 da cepa Y,
denominado de clone 9, e a sequência de um peptídeo sinal da cadeia κ de
imunoglobulina murina (plasmídio pIgSPCl.9). Após o desafio com
tripomastigotas sanguíneos da cepa Y, os animais vacinados apresentaram
uma redução significativa do pico da parasitemia e 100% de sobrevida. Em
contraste, somente 10% dos animais do grupo controle, injetados com o
plasmídio pcDNA3, sobreviveram ao desafio letal. A imunoproteção ocorreu em
paralelo à indução de células CD4 específicas nos camundongos imunizados
com o plasmídio pIgSPCl.9 que, quando estimuladas in vitro com a proteína
recombinante His-65kDa (ASP-2 recombinante da cepa Y), secretaram IFN-γ e
NO [115].
Em um estudo subsequente, Vasconcelos et al. [117] avaliaram a
eficácia protetora da vacinação com esse antígeno após a imunização de
camundongos A/Sn (altamente suscetíveis) apenas com o plasmídio pIgSPCl.9
ou, simultaneamente, com um plasmídio contendo o gene da trans-sialidase,
expressa na forma tripomastigota de T. cruzi. Após o desafio com
tripomastigotas sanguíneos da cepa Y de T. cruzi, os animais vacinados
apresentaram uma redução significativa do pico da parasitemia. Os que foram
imunizados somente com o gene asp-2 tiveram uma sobrevida de 65% e os
imunizados com ambos os plasmídios tiveram uma sobrevida de 85%. Apesar
de não ter sido observada uma imunidade estéril, foi observada pouca ou
nenhuma patologia inflamatória no coração ou no músculo esquelético dos

27
camundongos infectados na fase tardia (crônica) da doença [117]. A imunidade
observada correlacionou-se com a presença dos linfócitos T CD4 do tipo Th1 e
dos linfócitos T CD8 Tc1. Os resultados otimistas de imunoproteção, usando
DNA plasmidial contendo o gene asp-2 de T. cruzi, estimularam os estudos que
deram origem a esta tese.
1.5 Desenvolvimento de vacinas recombinantes visando à indução de
uma resposta imune mediada pelos linfócitos T CD4 e CD8
Dada a importância dos linfócitos T CD4 e T CD8 na imunidade protetora
contra a infecção por diferentes patógenos intracelulares como Mycobacterium
tuberculosis, HIV, Plasmodium spp., Leishmania spp., T. cruzi, T. gondii, etc.,
diversos grupos vêm tentando produzir vacinas baseadas na indução de fortes
respostas imunes mediadas pelos linfócitos T CD4 e CD8. A seguir, diferentes
estratégias vacinais são analisadas quanto à sua capacidade de induzir
respostas mediadas pelos linfócitos T CD4 e T CD8.
I) Proteínas recombinantes
As proteínas nativas ou recombinantes purificadas, quando
administradas em soluções salinas são, em geral, muito pouco imunogênicas.
A geração de respostas imunes contra proteínas é, portanto, fortemente
dependente da utilização concomitante de adjuvantes. Os adjuvantes são
substâncias que, quando adicionadas a um antígeno, aumentam ou modulam a
sua capacidade de induzir uma resposta imune.
Existem diferentes classes de substâncias que mostram atividade
adjuvante, entre as quais estão os produtos bacterianos, os sais minerais, as
emulsões água-em-óleo ou óleo-em-água, as micropartículas, os ácidos
nucleicos, as saponinas e os lipossomas. No entanto, apenas algumas
substâncias adjuvantes já foram liberadas para o uso em seres humanos,
sendo que a maioria falhou devido a reações tóxicas inaceitáveis. Entre os
adjuvantes liberados para o uso humano (revisto por [143]) estão o alum (ou
hidróxido de alumínio), o MF59 (uma emulsão de óleo esqualeno em água) e o

28
AS04 (uma formulação contendo hidróxido de alumínio e monofosforil lipídio A
ou MPL).
O mecanismo de ação da maioria dos adjuvantes era desconhecido até
o final da década de 1990, a partir de quando ocorreram as descrições dos
TLRs e dos seus ligantes e, posteriormente, de outros PRRs da resposta imune
inata (como os NLRs). Estas descobertas permitiram não apenas a elucidação
do mecanismo de ação de alguns adjuvantes, mas também o desenvolvimento
de novos ligantes de PRRs e a sua utilização como adjuvantes em formulações
vacinais.
No caso do alum, sua atividade adjuvante inclui o aumento da fagocitose
pelas células dendríticas [144] e o recrutamento dos monócitos para o sítio da
injeção [145]. O mecanismo de ação desse adjuvante, largamente utilizado,
ainda é controverso apesar do grande avanço ocorrido nos últimos anos na sua
elucidação, devido à descrição dos PRRs. Alguns grupos sustentam que a
ação adjuvante do alum é independente dos TLRs, já que os camundongos
deficientes de MyD88 e de TRIF foram capazes de gerar títulos de anticorpos
semelhantes aos gerados pelos camundongos normais, após a imunização
com um antígeno adsorvido ao alum [146]. Mais recentemente, diversos
trabalhos reportaram que a indução da IL-1β pelo alum é dependente da
ativação do Nalp3 (um NLR), o qual se associa à Asc e à caspase-1 para
formar a estrutura conhecida como inflamassomo [147,148]. Ainda assim,
alguns pesquisadores acreditam que as propriedades adjuvantes do alum não
dependem do Nalp3 ou da produção da IL-1β [149], não se sabendo ao certo
se o alum ativa Nalp3 direta ou indiretamente, por exemplo pela indução de
ácido úrico (revisto em [150]).
Ao contrário do alum, cujo mecanismo de ação permanece controverso,
o adjuvante CpG ODN teve o seu mecanismo de atuação desvendado. A ação
imuno-estimulante dos dinucleotídeos CpG não-metilados, típicos do DNA
bacteriano, é dependente do TLR9, tanto nos camundongos quanto nas células
humanas [151,152]. A sinalização pelo TLR9 se dá por meio da molécula
adaptadora MyD88 [153] e culmina com a ativação de fatores de transcrição
como o NFκB e o AP-1, os quais levam à produção de diversas citocinas (como

29
o TNF-α, a IL-6 e a IL-12) e à expressão do MHC II e de moléculas co-
estimulatórias nas células apresentadoras de antígenos (revisto em [154]).
Uma outra importante ação descrita para o CpG ODN é a de estimular a
apresentação cruzada dos antígenos, o que tem grande relevância quando
este adjuvante é utilizado nas imunizações com proteínas recombinantes. A
geração da resposta imune dos linfócitos T CD8 pelas imunizações com
proteínas recombinantes é dificultada pelo fato de que os antígenos solúveis
extracelulares tendem a serem processados e apresentados, pelas células
apresentadoras de antígeno (APCs), principalmente nas moléculas de MHC II,
mas não de MHC I. A adição do CpG ODN ao antígeno, porém, parece
estimular a entrada do mesmo na via de processamento e apresentação no
MHC I, levando à chamada apresentação cruzada, tanto nas células
dendríticas quanto nos linfócitos B, o que culmina com a estimulação dos
linfócitos T CD8 [155,156,157]. Os mecanismos envolvidos na apresentação
cruzada não foram completamente elucidados, mas o CpG ODN parece agir
por meio da estabilização do mRNA do MHC I e da produção dos IFNs tipo I
[158].
II) DNA plasmidial
A vacinação com DNA plasmidial teve início na década de 1990, quando
diversos trabalhos independentes mostraram que a injeção de DNA plasmidial
desnudo era capaz de induzir uma resposta imune contra o antígeno codificado
por um transgene [159,160,161]. A injeção intramuscular do DNA plasmidial
leva à transfecção principalmente dos miócitos [162], mas, ocasionalmente,
também das células imunes residentes, como as células dendríticas e os
monócitos [163,164]. Assim, o antígeno exógeno produzido pelos miócitos
pode ser apresentado pelas APCs no MHC de classe II. Além disto, o antígeno
pode chegar à via do MHC de classe I quando produzido intracelularmente nas
APCs transfectadas diretamente, ou quando, por meio da apresentação
cruzada, é adquirido das células apoptóticas [165,166]. As vacinas de DNA,
portanto, são capazes de estimular respostas tanto dos linfócitos T CD4 quanto
dos linfócitos T CD8.

30
As vantagens desse tipo de vacinação incluem: a segurança, o baixo
custo, a fácil produção e a estabilidade. Além do que, como o DNA não gera
uma resposta imune específica contra o próprio vetor, a imunização com DNA
plasmidial induz uma resposta direcionada somente para epítopos codificados
pelo transgene. Outra vantagem é que o DNA plasmidial possui uma atividade
adjuvante inerente, devido à sua capacidade de induzir a produção dos IFNs
tipo I por vias dependentes da TANK-binding kinase 1 (TBK-1, [167]) e de
STING [168].
O grande obstáculo para as vacinas de DNA é, contudo, conseguir
reproduzir nos seres humanos as efetivas respostas imunes induzidas nos
pequenos roedores, uma vez que, em geral, as vacinas de DNA utilizadas nos
estudos clínicos não induziram altos níveis de imunidade (revisto em [169]).
Nos últimos anos, diversas abordagens vêm sendo testadas com o intuito de
aumentar a imunogenicidade das vacinas de DNA. Estas estratégias incluem
modificações no transgene para aumentar a expressão antigênica, como o uso
de sequências com códons otimizados [170,171]. Além disto, novas estratégias
de entrega do DNA vêm sendo avaliadas, incluindo a eletroporação [172,173] e
o uso de microagulhas para as imunizações transcutâneas, levando o antígeno
para as células de Langerhans (revisto em [174]). Por fim, vários tipos de
adjuvantes têm sido adicionados às vacinas de DNA, como as citocinas, as
quimiocinas e os ligantes dos TLRs [175].
III) Vírus recombinantes
Os vírus recombinantes vivos atenuados são, até agora, os vetores mais
promissores para a indução de respostas imunes mediadas pelos linfócitos T
[176]. Diversos vírus já foram modificados geneticamente para carregar genes
exógenos, entre eles os poxvírus [177,178] os adenovírus [179,180], os
flavivírus [181,182] e o influenza [183,184]. Estes vetores virais apresentam a
vantagem de infectar as células hospedeiras de maneira ativa (ao contrário do
DNA plasmidial, por exemplo, que é endocitado passivamente), sendo mais
efetivos na expressão intracelular do transgene. Assim, a proteína resultante
pode entrar na via de processamento e apresentação de antígenos para ativar
fortes respostas de linfócitos T CD8. Além disso, APCs podem fagocitar debris

31
das células infectadas expressando o transgene, ou o antígeno solúvel, de
maneira a que este também chegue à via de processamento e apresentação
para os linfócitos T CD4. Outra conveniência desses vetores virais é que eles
apresentam uma série de “sinais de perigo”, ou seja, de ligantes para
receptores como os TLRs e os NLRs, que estimulam o sistema imune inato e
potencializam as respostas imunes adaptativas.
Entre os vírus recombinantes já utilizados, o adenovírus é certamente
um dos mais potentes [185]. Os adenovírus são vírus não envelopados que
apresentam genomas de DNA dupla-fita linear de aproximadamente 38kb. Eles
já foram isolados a partir de diversas aves e mamíferos, havendo pelo menos
51 sorotipos conhecidos de adenovírus humanos, divididos em 6 subgrupos
(revisto em [186]). Os adenovírus geralmente causam afecções respiratórias,
oculares ou intestinais, cuja sintomatologia é tipicamente branda. Ainda assim,
por razões de segurança, os adenovírus que vêm sendo testados como vetores
vacinais foram modificados geneticamente, não sendo capazes de se replicar
nas células humanas normais.
O genoma dos adenovírus apresenta regiões transcritas antes da
replicação do DNA viral, denominadas de regiões early (E), e regiões
transcritas posteriormente, denominadas de late (L). As regiões E1-E4
expressam proteínas que estão, em geral, envolvidas na promoção da
replicação viral, enquanto aquelas expressas a partir das regiões L estão
relacionadas ao empacotamento e à formação do capsídeo viral (revisto em
[187]). A maioria dos vetores adenovirais apresenta deleções da E1, o que
torna o vírus incapaz de se replicar devido às funções essenciais de promoção
da replicação exercidas pela E1A. Esses vírus são cultivados em linhagens
celulares transfectadas com a região E1, que os complementam in trans e
permitem sua a replicação in vitro. Além disso, muitos vetores adenovirais
apresentam deleções também na região E3, a qual está envolvida na evasão
da resposta imune do hospedeiro. Mais recentemente, uma nova geração de
vetores adenovirais com deleção da E4 também vem sendo utilizada. A região
E4 atua na replicação viral, e a deleção de uma segunda região essencial não-
contínua com a primeira reduz a chance de ocorrerem fenômenos de
recombinação durante o crescimento in vitro e a consequente reversão para

32
um fenótipo replicativo. A deleção dessas regiões do genoma adenoviral
permite a inserção de 7-8kb de DNA exógeno (revisto em [188]).
O capsídeo dos adenovírus é uma estrutura icosaédrica formada por 240
proteínas denominadas hexons. Em cada vértice do icosaedro observa-se uma
proteína denominada penton base, ligada covalentemente a uma proteína fiber,
a qual se projeta como uma espícula (Fig. 3). A extremidade da proteína fiber
forma uma estrutura como uma maçaneta, e é chamada de knob. A fiber é a
primeira proteína do adenovírus a interagir com a célula hospedeira, mediante
a ligação da knob aos diferentes receptores das células hospedeiras. Em geral,
os adenovírus dos grupos A, C, E e F interagem com o receptor CAR
(coxsackie adenovirus receptor) [189,190]. Já os do grupo B se ligam ao CD46
e os dos grupos D e E utilizam outros receptores (revisto em [186]). Apesar
desses serem os principais receptores identificados, outros também podem ser
utilizados. O hAd5 (adenovírus sorotipo 5 humano), por exemplo, cujo principal
receptor é o CAR, é capaz de infectar células que não expressam esse
receptor [191]. Além disso, algumas regiões da penton base também interagem
com as integrinas αv [192,193], as quais atuam como co-receptores e facilitam
a internalização do vírus por vesículas recobertas por clatrina [194]. A diferença
entre os receptores utilizados por cada sorotipo de adenovírus é de grande
importância, pois certamente influencia no tropismo e, consequentemente, na
indução da resposta imune.
Fig. 3: Representação da estrutura do adenovírus. Retirado de [186].

33
Após a internalização, os adenovírus localizam-se nos endossomos,
onde a mudança do pH leva a uma desestabilização do capsídeo viral e à
liberação de algumas proteínas. Ocorre então a ruptura do endo/lisossomo
(possivelmente mediada pela proteína VI do adenovírus) e o vírus é dirigido ao
núcleo, onde seu material genético é liberado (revisto em [186]).
A infecção pelo adenovírus permite que ele seja reconhecido por PRRs
presentes no interior da célula hospedeira. Já foi descrito que os adenovírus
são capazes de induzir a produção de citocinas pró-inflamatórias e de IFNs tipo
I por vias dependentes e independentes dos TLRs, e que os mecanismos
variam bastante dependendo do tipo celular infectado. Diversos trabalhos
sugerem que os adenovírus ativam o TLR9 [195,196,197] ou algum TLR [198],
enquanto outros trabalhos também observaram a ativação de respostas imunes
na ausência da molécula adaptadora MyD88, sugerindo uma ação por outros
receptores [197,199]. Recentemente, Cheng e colaboradores [200]
descreveram a participação do receptor RIG-1 na resposta inata contra o
adenovírus. Além disto, a proteína STING parece mediar a indução do IFN-β e
da IL-6 por DNA adenoviral nos fibroblastos murinos [168]. Por fim, foi descrito
recentemente que o DNA adenoviral induz a maturação da pró-IL-1β por meio
da ativação do inflamassomo dependente do Nalp3 e da Asc [201].
Além de induzir a produção de citocinas pela resposta imune inata, os
adenovírus são grandes indutores das respostas celulares, principalmente dos
linfócitos T CD8, o que os fazem vetores atrativos para as vacinas contra os
patógenos intracelulares. A resposta de linfócitos T CD8 induzida por uma
única dose de hAd5, contra o antígeno codificado pelo transgene, é mais alta
do que a induzida por outros vetores virais como o MVA [202,203], por
exemplo, ou pelo Semliki Forest virus (SFV) [204]. As razões para tal não são
claras, mas podem estar relacionadas ao fato de que o adenovírus expressa
uma quantidade reduzida de outras proteínas (cerca de 30, [202]), além
daquela codificada pelo transgene. Esse pequeno número de proteínas,
provavelmente, dá origem a poucos peptídeos imunodominantes para competir
com aqueles derivados do transgene, o que pode permitir uma maior expansão
das células específicas para o antígeno de interesse.

34
Quanto à resposta de linfócitos T CD4 gerada pelos adenovírus, poucos
estudos foram realizados. Aparentemente, a magnitude da resposta de
linfócitos T CD4 é apenas um décimo da resposta de linfócitos T CD8 [187].
Apesar disso, essas células são importantes para o desenvolvimento da
resposta de linfócitos T CD8, pois, na ausência dos linfócitos CD4, a expansão
primária das células CD8 é mais baixa, além delas apresentarem uma menor
sobrevida e problemas na expansão secundária [205]. Também observou-se
que, nos camundongos CD4 KO, a resposta citotóxica dos linfócitos T CD8
induzida pela imunização com AdASP-2 era extremamente baixa ([105], dados
não mostrados).
Alguns fatores influenciam o tipo de resposta de linfócitos T que será
gerado, como o nível e a duração da expressão do antígeno. Após a
imunização com adenovírus, a expressão do transgene ocorre em altos níveis
por cerca de 7 dias, começando então a diminuir lentamente [206]. A maioria
dos trabalhos relata que os antígenos expressos no hAd5 ficam disponíveis por
um período de 20 a 40 dias após a imunização [206,207], apesar de haver uma
descrição da persistência de sequências do transgene por até 1 ano após a
imunização [208].
Outras questões importantes relacionadas à apresentação antigênica
permanecem sem resposta: o principal tipo celular responsável pela ativação
dos linfócitos T CD8 após a imunização com o adenovírus ainda não foi
identificado, e as contribuições relativas das formas de apresentação direta ou
cruzada não foram avaliadas. Já foi descrito que as células dendríticas são
permissivas à infecção pelo adenovírus 5 [191,209,210], sendo também
infectadas in vivo [211]. Contudo, não se sabe se elas são realmente
responsáveis pela ativação dos linfócitos CD8. É possível que a via de
inoculação vacinal influencie no tipo celular a ser infectado e,
consequentemente, também na APC que induzirá a ativação das células CD8.
IV) Imunização e reforço (Prime-boost) heterólogo
A geração de uma resposta robusta de linfócitos T geralmente requer
mais de uma dose de imunização, ou seja, uma dose inicial (prime) e uma ou
mais doses de reforço (boost). Quando se utiliza um antígeno carreado pelo

35
mesmo vetor ou sistema de liberação, tanto no prime quanto no boost, chama-
se a esse tipo de imunização de prime-boost homólogo. Essa estratégia
funciona muito bem para a indução de respostas imunes humorais, no entanto
pode não ser tão eficaz para a geração da imunidade celular, principalmente no
caso dos vetores virais. Além de induzirem respostas celulares potentes contra
o transgene, os vetores virais são também muito efetivos para induzir respostas
imunes, tanto celulares quanto humorais, contra as suas próprias proteínas.
Assim, se altos títulos de anticorpos contra o vetor viral forem gerados durante
o priming, eles podem neutralizar o vírus e impedi-lo de invadir novas células
durante o boost, prejudicando a apresentação dos antígenos. Alguns trabalhos
já sugeriram que os linfócitos T de memória, gerados na dose inicial, podem
reconhecer e eliminar rapidamente as células apresentadoras de antígenos
durante as respostas às doses de reforço, impedindo que os linfócitos T naive
sejam recrutados e expandam a resposta [212].
Para evitar este tipo de problema, um protocolo que vem sendo utilizado
com bastante sucesso para as imunizações contra infecções causadas por
patógenos intracelulares é o chamado prime-boost heterólogo, no qual as
doses de prime e de boost são compostas do mesmo antígeno, sendo este
carreado por sistemas de liberação ou por vetores diferentes (Fig. 4, retirada de
[213]).

36
Os mecanismos que levam à geração de uma potente resposta de
linfócitos T por meio da combinação de vetores ainda não foram
completamente elucidados. O aumento da magnitude da resposta pode ser
decorrente da transposição de uma regulação natural dos linfócitos T, pois a
presença do mesmo epítopo em um contexto antigênico diverso poderia
prevenir a rápida eliminação das células apresentadoras de antígenos, o que
em outras situações limitaria a expansão da resposta [212]. Além disso, a
imunização heteróloga parece subverter a hierarquia de imunodominância
estabelecida entre os epítopos, tornando a resposta mais ampla [214]. Por fim,
há evidências de que o prime-boost heterólogo melhora a funcionalidade dos
linfócitos T, induzindo um número maior de células multifuncionais [214,215].
O sucesso de um protocolo de prime-boost heterólogo depende da
ordem em que os vetores são administrados [216,217]. Alguns vetores são
efetivos tanto no priming quanto no boosting, outros apresentam maior eficácia
em primar células efetoras, enquanto um grupo diferente de vetores parece ser
melhor no boosting dessas respostas (revisto em [188]). As razões para tais
diferenças estão sendo estudadas, mas não existe ainda uma resposta
definitiva. A utilidade de cada vetor parece depender de características
Figura 4. Representação da geração de
uma resposta imune adaptativa específica
após diferentes esquemas de imunização.
Os vírus A e B expressam um antígeno
exógeno em comum.
Linha contínua: resposta contra um epítopo
exógeno expresso nos vírus A e B.
Linha pontilhada: resposta imune contra os
epítopos presentes apenas no vírus A.
Linha tracejada: resposta imune contra os
epítopos presentes apenas no vírus B.
A: resposta imune após uma única dose do
vírus A.
B: resposta imune após a imunização
homóloga com o vírus A.
C: resposta imune após o prime-boost
heterólogo com os vírus A e B.
Retirado de [213].

37
intrínsecas, como o nível e a duração da expressão do antígeno, outras
proteínas expressas pelo vetor, as células que são transfectadas/infectadas e
expressam o transgene, e os PRRs que são ativados por cada vetor.
O prime-boost heterólogo foi primeiro utilizado na década de 1990, nas
imunizações com os vírus influenza e vaccínia recombinantes expressando um
epítopo de células CD8 da proteína CS, para proteger os camundongos da
infecção experimental com o Plasmodium yoelii [216,218]. Desde então,
diferentes combinações de sistemas de liberação (incluindo imunizações com
DNA plasmidial, proteínas e vírus recombinantes) têm sido utilizadas para
induzir respostas imunes dos linfócitos T contra patógenos como o HIV
[219,220], o Plasmodium [221,222] e a Mycobacterium tuberculosis [223,224],
entre outros.
Alguns estudos mostram que uma única imunização com o vírus
vaccínia é capaz de induzir respostas de linfócitos T CD8, ainda que estas
sejam mais baixas do que aquelas induzidas por outros vírus recombinantes
[202,203]. O maior potencial da vacinação com o vírus vaccínia recombinante,
no entanto, é o boost das respostas induzidas por outros sistemas de liberação
de antígeno, protocolos que já foram testados em diversos modelos
[217,225,226,227]. Os motivos pelos quais o vírus vaccínia parece ser tão bom
em expandir as células previamente primadas, e pouco efetivo em iniciar
respostas imunes, são ainda pouco conhecidos, mas podem incluir: a
expressão antigênica em altos níveis, porém de maneira breve (por volta de
uma semana) [206]; e a expressão de mais de 200 proteínas próprias, o que
gera uma forte resposta de linfócitos T CD8 contra os inúmeros epítopos do
vetor [202,228].
Até hoje, os testes nos seres humanos empregando vaccínia ou
modified vaccinia ankara (MVA, um vaccínia atenuado) ainda não obtiveram
grande sucesso no caso da vacina contra a malária, pois o protocolo DNA/MVA
foi capaz de induzir proteção em pessoas que nunca haviam entrado em
contato com a doença [229], mas falhou em proteger os habitantes das áreas
endêmicas [230]. Mesmo um protocolo de prime-boost envolvendo dois
poxvírus, o fowlpox (FPV) e o MVA, falhou em reduzir a incidência da malária
nas crianças residentes nas áreas endêmicas [231,232].

38
Os protocolos utilizando o MVA, expressando o antígeno 85A de
Mycobacterium tuberculosis, como reforço para a vacinação com BCG têm se
mostrado seguros e capazes de induzir uma boa imunogenicidade nos seres
humanos, estimulando principalmente respostas de células T CD4 [233,234].
Ainda assim, são baixas as respostas de células T CD8 geradas por esse
protocolo de imunização nos seres humanos [234,235].
No caso da vacina contra o HIV, Harari e colaboradores [236]
recentemente descreveram a indução de respostas imunes em mais de 90%
dos voluntários imunizados com um protocolo DNA/NYVAC (NYVAC é uma
outra cepa do vírus vaccínia atenuado). Pelo menos outros 5 testes clínicos
contra o HIV envolvendo poxvírus estão em andamento: um em fase III, com
priming com o ALVAC (vírus canarypox) e boosting com a proteína env; outro
com um protocolo DNA/FPV; e os outros três com protocolos DNA/MVA, os
últimos estando na fase I ou II (revisto em [176]). Até que as informações sobre
a eficácia desses testes clínicos sejam divulgadas, a utilidade dos poxvírus nas
vacinações de seres humanos permanecerá ignorada.
O adenovírus humano sorotipo 5 é um dos mais estudados devido à sua
grande potência para induzir respostas imunes celulares. Este vetor induziu
respostas imunes de linfócitos T CD4 e T CD8 de grande magnitude (e muitas
vezes protetoras) nas imunizações homólogas ou heterólogas de modelos
animais contra diversas doenças infecciosas, como a gripe [237], a malária
[217], a hepatite C [238], a dengue [239] e a doença de Chagas [121].
Foi a busca por uma vacina contra o HIV, no entanto, que mais
impulsionou a utilização do hAd5. Diversos estudos mostraram a grande
potência deste vetor na geração de respostas imunes contra as proteínas do
HIV/SIV, principalmente de linfócitos T CD8, nos modelos com macacos rhesus
[219,240] e babuínos [241]. Outros estudos não se limitaram a testar a resposta
imune, mas também desafiaram os animais com o SHIV [242]. O modelo de
infecção pelo SHIV, entretanto, parece não ser tão fiel em predizer os
resultados obtidos nos seres humanos quanto o modelo que utiliza o SIV
(revisto em [243]). Assim, vários grupos utilizaram o protocolo DNA/hAd5
(carregando diferentes combinações de genes do SIV) para conseguir
demonstrar controle parcial da viremia e redução da perda dos linfócitos T CD4
após o desafio com o SIV [244,245,246]. Mais recentemente, Wilson e

39
colaboradores [247] também descreveram o controle da viremia e uma menor
perda dos linfócitos T CD4 em macacos imunizados com esse protocolo, após
um desafio com um SIV heterólogo.
Boa parte dos protocolos testados envolveram DNA/hAd5, mas este
vetor viral também tem sido utilizado para reforçar as imunizações com
proteínas recombinantes (em geral adicionadas de adjuvantes ligantes dos
TLRs ou sistemas de liberação), sendo capaz de induzir fortes respostas de
células T CD4 e T CD8 nos modelos animais [248,249,250].
Uma séria preocupação com a efetividade das imunizações com o hAd5
é que esta possa ser comprometida pela imunidade prévia ao vetor. De fato, foi
demonstrado que tal fenômeno ocorre em alguns modelos [251,252]. Como a
prevalência da sorologia positiva para o hAd5 é alta, tanto nos Estados Unidos
e Europa quanto na África, (respectivamente 35-40%, 60% e 90%) [253,254],
vêm sendo desenvolvidas estratégias para contornar o problema. Alguns
grupos sugeriram que a imunização com DNA como prime reduz
consideravelmente o efeito da imunidade pré-existente [219,255]. A imunização
com DNA induz uma expressão antigênica em baixos níveis, porém prolongada
[206], sem que outro antígeno seja expresso concomitantemente. Estes fatores
resultam na indução de uma resposta imune discreta, mas que parece ser
suficiente para expandir as células CD8 específicas, garantindo a
imunodominância de epítopos codificados pelo transgene durante o boost com
adenovírus.
Por outro lado, alguns grupos desenvolveram adenovírus 5
recombinantes quiméricos, contendo a proteína hexon ou partes dela,
provenientes de outros sorotipos menos prevalentes [256], já que grande parte
dos anticorpos neutralizantes é direcionada para essa proteína [257]. Além
disto, vêm se estudando sorotipos de adenovírus humanos menos prevalentes,
como o Ad35 [252,258], o Ad41 [259], o Ad26 [214,215] e o Ad48 [215]. Outra
alternativa é o uso de adenovírus de outras espécies de primatas
[180,260,261]. Apesar de tais sistemas serem promissores, reagentes e
técnicas precisam ser desenvolvidos e disponibilizados para que possam ser
utilizados em grande escala.

40
Objetivos

41
A imunização genética com DNA plasmidial, contendo o gene que
codifica a proteína 2 da superfície de amastigotas (ASP-2), é capaz de proteger
parte dos camundongos altamente suscetíveis A/Sn contra uma infecção letal
pelo T. cruzi. Com base neste resultado, o objetivo principal desta tese foi o de
melhorar racionalmente a eficácia da vacinação contra a infecção experimental
pelo T. cruzi, por meio de protocolos de imunização e reforço homólogo ou
heterólogo utilizando DNA plasmidial, proteínas recombinantes ou adenovírus
recombinante da ASP-2 de T. cruzi.
Após melhorar a eficácia da vacinação, também foi nosso objetivo
estudar a importância da participação de alguns tipos celulares (linfócitos T
CD4 ou CD8) e da expressão de determinadas moléculas (perforina e IFN-γ) na
imunidade protetora. Por último, tivemos como objetivo caracterizar funcional e
fenotipicamente os linfócitos T específicos, responsáveis pela imunoproteção
após a vacinação experimental.

42
Resultados

43
Artigos Publicados

44
Artigo 1

INFECTION AND IMMUNITY, Sept. 2005, p. 6017–6025 Vol. 73, No. 90019-9567/05/$08.00�0 doi:10.1128/IAI.73.9.6017–6025.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
CD8�-T-Cell-Dependent Control of Trypanosoma cruzi Infection in aHighly Susceptible Mouse Strain after Immunization with
Recombinant Proteins Based on Amastigote SurfaceProtein 2
Adriano F. S. Araujo,1,2 Bruna C. G. de Alencar,1,2 Jose Ronnie C. Vasconcelos,1,2 Meire I. Hiyane,1,2
Claudio R. F. Marinho,3 Marcus L. O. Penido,4 Silvia B. Boscardin,2 Daniel F. Hoft,5Ricardo T. Gazzinelli,4 and Mauricio M. Rodrigues1,2*
Centro Interdisciplinar de Terapia Genica (CINTERGEN)1 and Departmento de Microbiologia, Imunologia e Parasitologia,2
Universidade Federal de Sao Paulo-Escola Paulista de Medicina, Departamento de Imunologia, ICB, Av. Prof. LineuPrestes, 1730, Universidade de Sao Paulo,3 Sao Paulo, SP 05508-900, and Departamento de Bioquımica eImunologia, Instituto de Ciencias Biologicas, Universidade Federal de Minas Gerais e Centro de Pesquisas
Rene Rachou, FIOCRUZ, Avenida Augusto de Lima 1715, Barro Preto, 30190-002, Belo Horizonte,Minas Gerais,4 Brazil, and Division of Infectious Diseases & Immunology, Departments
of Internal Medicine & Molecular Microbiology, Saint Louis University HealthSciences Center, 3635 Vista Avenue, FDT-8N,
St. Louis, Missouri 631105
Received 4 April 2005/Returned for modification 6 May 2005/Accepted 11 May 2005
We previously described that DNA vaccination with the gene encoding amastigote surface protein 2 (ASP-2)protects approximately 65% of highly susceptible A/Sn mice against the lethal Trypanosoma cruzi infection.Here, we explored the possibility that bacterial recombinant proteins of ASP-2 could be used to improve theefficacy of vaccinations. Initially, we compared the protective efficacy of vaccination regimens using either aplasmid DNA, a recombinant protein, or both sequentially (DNA priming and protein boosting). Survival afterthe challenge was not statistically different among the three mouse groups and ranged from 53.5 to 75%. Thefact that immunization with a recombinant protein alone induced protective immunity revealed the possibilitythat this strategy could be pursued for vaccination. We investigated this possibility by using six differentrecombinant proteins representing distinct portions of ASP-2. The vaccination of mice with glutathioneS-transferase fusion proteins representing amino acids 261 to 500 or 261 to 380 of ASP-2 in the presence of theadjuvants alum and CpG oligodeoxynucleotide 1826 provided remarkable immunity, consistently protecting100% of the A/Sn mice. Immunity was completely reversed by the in vivo depletion of CD8� T cells, but notCD4� T cells, and was associated with the presence of CD8� T cells specific for an epitope located betweenamino acids 320 and 327 of ASP-2. We concluded that a relatively simple formulation consisting of arecombinant protein with a selected portion of ASP-2, alum, and CpG oligodeoxynucleotide 1826 might be usedto cross-prime strong CD8�-T-cell-dependent protective immunity against T. cruzi infection.
In spite of the significant reduction in transmission over thepast 20 years in countries such as Argentina, Brazil, Chile, andUruguay, Chagas’ disease (American trypanosomiasis) is still amajor problem for many Latin American countries. The dis-ease afflicts more than 15 million individuals, with an annualdeath toll of approximately 45,000 (reviewed in reference 18).The efficacy of conventional chemotherapy with nifurtimox orbenznidazole is low and varies widely according to the infec-tion status (acute or chronic) and the region of endemicity.Because of the low efficacy of treatment, the chronic phase ofthis infection usually persists for life. Approximately one-thirdof infected persons progress slowly to the symptomatic formsof the disease, characterized by cardiomyopathy and/ormegagastrointestinal syndromes. Asymptomatic, chronicallyinfected persons transmit the disease through blood transfu-
sions and tissue transplantation. The lack of efficient drugtreatment increases the importance of the development of newstrategies to prevent or reduce the disease.
A number of studies have been published over the past 10years demonstrating that recombinant proteins or plasmidDNA can elicit protective immunity against experimentalTrypanosoma cruzi infection (2, 3, 4, 5, 7, 8, 12, 16, 20, 21, 23,24, 28, 31, 32, 33, 34; reviewed in references 17 and 22).Vaccinated mice develop a strong cellular immune responsemediated by CD4� Th1 and CD8� Tc1, thus surviving other-wise lethal acute infection. In addition to surviving acute in-fection, chronic phase pathologies of vaccinated mice weresignificantly reduced (4, 8, 28). Although in most studies, pro-phylactic vaccinations were performed, a recent report showedthat therapeutic vaccination can also be effective in micechronically infected with certain T. cruzi strains (4). We con-sider studies on vaccination against experimental Chagas’ dis-ease important, as an efficacious vaccine may complementother strategies such as vector control and chemotherapy forprevention and treatment of the disease.
* Corresponding author. Mailing address: CINTERGEN, UNIFESP—Escola Paulista de Medicina, Rua Mirassol, 207, Sao Paulo-SP 04044-010, Brazil. Phone and fax: (55) (11) 5571-1095. E-mail: [email protected].
6017
by on October 27, 2009
iai.asm.org
Dow
nloaded from

Most experiments related to vaccination against T. cruziinfection have used BALB/c or C57BL/6 mice. Although thesemice die when challenged with the infective trypomastigotes ofcertain parasite strains, they are not as susceptible to infectionwith T. cruzi as A/Sn mice. In order to study the mechanisms ofprotective immunity required for vaccination, we have beenusing A/Sn mice. This mouse strain is highly susceptible toinfection with different strains of T. cruzi, and 100% of theanimals die after a challenge with relatively small doses of theparasite. Due to its high susceptibility, we believe that thismouse strain provides an interesting model to study the immu-nological mechanisms required to induce a high degree ofprotective immunity against infection.
Using highly susceptible A/Sn mice, we have recently de-scribed that vaccination with a plasmid expressing amastigotesurface protein 2 (ASP-2) generates specific CD4� Th1 andCD8� Tc1 immune responses. Most importantly, immuniza-tion with the asp-2 gene alone promotes the survival of ap-proximately 65% of the A/Sn mice against a lethal Trypano-soma cruzi infection (28). Protective immunity could beimproved if the animals were vaccinated simultaneously withthe genes of ASP-2 and trans-sialidase (28).
Based on the optimistic results obtained with the asp-2 gene,we attempted to further improve protective immunity in thismouse model of infection. To this end, the initial goal of thepresent study was to compare the protective efficacy of vacci-nation regimens using either plasmid DNA, recombinant pro-tein, or both sequentially (DNA priming and protein boosting).Our initial assumption was based on a number of reportsshowing that a DNA priming/protein boosting strategy couldimprove the protective efficacy in different vaccination models(14, 19, 26, 27).
During the course of these experiments, we observed thatprotective immunity was similar following immunization witheither a plasmid DNA or a recombinant protein adsorbed toalum or when both types were combined (DNA priming/pro-tein boosting). The fact that immunization with a recombinantprotein alone was partially protective was unexpected; how-ever, it revealed the possibility that this strategy could be pur-sued for experimental vaccination. Toward that goal, we usedseveral recombinant proteins representing distinct portions ofASP-2. These proteins confirmed the protective potential ofrecombinant antigens when used in a relatively simple adjuvantformulation. Also, they identified a region containing highlyprotective epitopes capable of providing protective immunityto 100% of the vaccinated A/Sn mice.
MATERIALS AND METHODS
Mice and parasites. Female 5- to 8-week-old A/Sn mice were purchased fromthe University of Sao Paulo. Parasites of the Y strain of T. cruzi were used in thisstudy (25). Bloodstream trypomastigotes were obtained from mice infected 7days earlier. The concentration of parasites was estimated and adjusted to 1,250trypomastigotes per ml. Each mouse was inoculated intraperitoneally (i.p.) with0.2 ml (250 trypomastigotes). Parasite development was monitored by countingthe number of bloodstream trypomastigotes in 5 �l of fresh blood collected fromthe tail vein (13).
Plasmid and recombinant proteins based on the sequence of the asp-2 gene.The plasmid encoding the asp-2 gene, ASP-2 DNA, was created and purified aspreviously described by Boscardin et al. (2). This plasmid contains the nucleo-tides encoding the mouse immunoglobulin � chain signal peptide in frame withthe nucleotide sequence encoding amino acids (aa) 67 to 694 of ASP-2.
The nucleotide sequences corresponding to different portions of ASP-2 (clone
9) were obtained by PCR amplification using Platinum Taq High Fidelity DNApolymerase (Invitrogen). Oligonucleotides containing the EcoRI and BamHIrestriction sites, purchased from Invitrogen, were used for amplification. Theamplified fragments were treated with the restriction enzymes EcoRI andBamHI and ligated into the pGEX-3X vector (Amershan) treated with thesesame enzymes. The recombinant plasmids were transformed into Escherichia coliDH5�. The recombinant proteins were expressed and purified as describedearlier (2). The purity of recombinant protein was determined by sodium dodecylsulfate (SDS)-polyacrylamide gel electrophoresis (PAGE). SDS molecularweight markers (Sigma) were used as protein standards.
Immunization. A/Sn mice were immunized according to protocols describedearlier (3, 29). Each mouse received 3 doses of 100 �g of plasmid DNA injectedintramuscularly at 0, 3, and 5 weeks. Immunization with recombinant His-65kDaconsisted of 3 i.p. doses of 25 �g of protein adsorbed to alum (Alhydrogel 85Superfos; Biosector, Vedbaek, Denmark) given at 0, 3, and 5 weeks. Efficientcoupling of recombinant His-65kDa to alum was obtained by mixing 2.5 �l 2%(wt/vol) Al2O3 per �g of recombinant protein for 2 h. Our DNA priming/proteinboosting regimen consisted of two doses of 100 �g of plasmid DNA injected at0 and 3 weeks. A third dose was provided 2 weeks later and consisted of an i.p.injection of 25 �g of recombinant His-65kDa adsorbed to alum.
Immunization with the recombinant proteins His-65kDa, His-25kDa, glutathi-one S-transferase (GST)-P1-P3, GST-P4-P7, GST-P4-P5, and GST-P6-P7 wasperformed as described in the legend to Fig. 2. CpG oligodeoxynucleotide(ODN) 1826 (TCCATGACGTTCCTGACGTT) was synthesized with a nucle-ase-resistant phosphorothioate backbone (Coley Pharmaceutics, Wellesley,MA).
One week after the last immunizing dose with plasmids and/or recombinantproteins, the blood was collected from the tail and the sera were analyzed for thepresence of antibodies to His-65kDa. One week later, mice were challenged withbloodstream trypomastigotes. Results are representative of two or more inde-pendent experiments.
Immunological assays. Anti-His-65kDa antibodies were detected by enzyme-linked immunosorbent assay as described earlier (2). Titers are reported as thereciprocals of the highest serum dilution producing an average optical density at492 nm greater than 0.1. The enzyme-linked immunospot assay (ELISPOTassay) was performed essentially as described earlier (2).
Hemoculture and histopathological analysis. Aliquots of 0.1 ml of blood werecollected and cultured at 28°C for 1 month in 5 ml of axenic liver infusiontryptose medium. Parasite growth was monitored microscopically every week.
Tissue specimens were collected from vaccinated and infected mice at theindicated day after challenge and fixed in paraformaldehyde for further process-ing. Paraffin-embedded tissue sections were stained with hematoxylin and eosinstain and analyzed by optical microscopy. Slides from the heart and the quadri-ceps muscle of each mouse were analyzed in a blind fashion by one of us(C.R.F.M.). Areas of inflammatory infiltrates in the myocardium, pericardium,and endocardium or in the striated muscle were analyzed and semiquantified.
In vivo depletion of CD4� and CD8� T cells. Ascites fluids containing GK1.5and 53.6.7 monoclonal antibodies were produced in BALB/c nude mice, and theconcentration of rat immunoglobulin G (IgG) was estimated as described pre-viously (15). At days 3 and 4 before challenge with trypomastigotes, mice im-munized with GST-P4-P7 were treated i.p. with a dose of 1 mg per day of rat IgG,anti-CD4, or anti-CD8. Seven days after challenge, each mouse received onemore dose of 1 mg of rat IgG, anti-CD4, or anti-CD8. The efficacy of depletionof CD4� or CD8� spleen cells before challenge was commonly higher than 98 or96%, respectively.
Peptide synthesis. The peptides KEGSKSGV (aa 94 through 101), AESLFVYI (aa 121 through 128), HEHNLFGI (aa 130 through 137), YEIVAGYI (aa140 through 147), AESWPSIV (aa 150 through 157), TEWETGQI (aa 320through 327), and PETLGHEI (aa 650 through 658) were prepared with stan-dard N�[9-fluorenylmethyloxycarbonyl] on a PSSM8 multispecific peptide syn-thesizer (Shimadzu, Kyoto, Japan) by solid-phase synthesis with a scale of 30�mol. Peptides were purified by high-pressure liquid chromatography in a Shi-madzu system. Peptides were analyzed in a C18 Vydac column (10 by 250 mm,5-�m particle diameter). The peptides were obtained in a range of 80 to 90%purity. Their identities were confirmed by using the Micro quantitative time offlight mass spectrometer equipped with an electrospray ionization source (Mi-cromass, United Kingdom).
Statistical analysis. The values of peak parasitemia of each individual mousewere log transformed before being compared by one-way analysis of variancefollowed by Tukey’s honestly significant difference (HSD) tests available at thesite http://faculty.vassar.edu/lowry/VassarStats.html. The log rank test was usedto compare the mouse survival rate after challenge with T. cruzi. Fisher’s exact
6018 ARAUJO ET AL. INFECT. IMMUN.
by on October 27, 2009
iai.asm.org
Dow
nloaded from

test was used to compare the different proportions of mice that survived infec-tion. The differences were considered significant when the P value was �0.05.
RESULTS
Initially, we compared the protective immune responses ofA/Sn mice immunized according to the group protocolspcDNA3 (I), alum (II), ASP-2 DNA (III), Alum/His-65kDa(IV), or ASP-2 DNA (V), followed by Alum/His-65kDa (DNApriming/protein boosting). After a challenge with bloodstreamtrypomastigotes of T. cruzi, we observed a significant reductionin the peak parasitemia of animals from mouse groups III, IV,and V compared to the control mice of groups I and II (P �0.05 in all cases) (Table 1). The peak parasitemias of animalsfrom mouse groups III, IV, and V were not statistically differ-ent (P � 0.05).
We also recorded the mouse mortality among the differentmouse groups after the challenge with T. cruzi. Survival at day
60 was 75, 53.31, or 73.74% in mouse groups III, IV, or V,respectively. Statistical comparisons were performed by the logrank, chi-square, or Fisher exact probability tests. In all cases,we failed to detect a significant difference among these threemouse groups.
Based on the results that immunization with a recombinantprotein adsorbed to alum provided protective immunity, weused six different recombinant proteins representing differentportions of ASP-2 for subsequent studies of vaccination. Fig-ure 1A and 1B displays a schematic representation and theSDS-PAGE analysis of the different recombinant proteins, re-spectively.
Initially, A/Sn mice were immunized with the recombinantproteins His-65kDa, GST-P1-P3, GST-P4-P7, and His-25kDa.These recombinant proteins were adsorbed to alum and in-jected i.p. in the presence of CpG ODN 1826. Control micewere immunized with the adjuvants only (alum and CpG ODN
FIG. 1. Schematic representation and SDS-PAGE analysis of the recombinant proteins based on the ASP-2 antigen. (A) Schematic represen-tation of the recombinant proteins. (B) SDS-PAGE of recombinant proteins stained with Coomassie blue. The amount of protein loaded wasapproximately 1 �g per lane.
TABLE 1. Trypomastigote-induced parasitemia and mortality in mice immunized with either ASP-2 DNA or recombinant proteinadministered individually or sequentially (DNA priming/protein boosting)
Mousegroup no. Immunogena Peak parasitemia
(log � SD)bNo. of mice alive/total
no. challengedc % Protection
I pcDNA3 6.35 � 0.08 0/16 0II Alum 6.53 � 0.05 0/8 0III ASP-2 DNA 5.42 � 0.09 12/16 75.00IV Alum/His-65kDa 5.62 � 0.32 8/15 53.31V ASP-2 DNA plus alum/His-65kDa 5.55 � 0.23 11/15 73.74
a Groups of seven or eight mice were immunized at 0, 3, and 5 weeks with pcDNA3, ASP-2 DNA, alum, or the formulation alum/His-65kDa. A fifth group of micewas injected with two doses of ASP-2 DNA at 0 and 3 weeks. A third immunizing dose was administered after 5 weeks and consisted of the formulation alum/His-65kDa.
b Two weeks after the last dose, mice were challenged i.p. with 250 bloodstream trypomastigotes. The values of peak parasitemia of mouse groups III, IV, and V werelower than those for mouse groups I and II (P � 0.05 in all cases). The peak parasitemias of groups III, IV, and V were not statistically different (P � 0.05).
c The number of mice alive was recorded 60 days after challenge. Animals from groups III, IV, and V survived longer than mice from groups I and II (P � 0.0001in all cases by the log rank test). There was no significant statistical difference among groups III, IV, and V (P � 0.05).
VOL. 73, 2005 VACCINATION AGAINST T. CRUZI INFECTION 6019
by on October 27, 2009
iai.asm.org
Dow
nloaded from

1826). To determine whether the immunizations were per-formed properly, after the third immunizing dose, the presenceof specific antibodies was determined by enzyme-linked immu-nosorbent assay using the recombinant protein His-65kDa asan antigen. The stronger antibody responses were detected inanimals immunized with recombinant protein His-65kDa orHis-25kDa. Antibody titers of mice immunized with GST-P4-P7 were higher than the titers of mice immunized withGST-P1-P3. Sera from control mice injected with adjuvant onlyhad no detectable specific antibodies to His-65kDa (data notshown).
After challenge with bloodstream trypomastigotes in miceimmunized with His-65kDa or GST-P4-P7, we observed a sig-nificant reduction in the peak parasitemia compared to ani-mals immunized with adjuvant only, GST-P1-P3, or His-25kDa(Fig. 2) (P � 0.01 in all cases). No significant difference instatistics was observed when we compared the peak para-sitemia of mice immunized with His-65kDa or GST-P4-P7(Fig. 2) (P � 0.05). Also, no difference was found when wecompared the peak parasitemias of mice immunized with GST-P1-P3 or His-25kDa or adjuvant alone (Fig. 2) (P � 0.05).
Also important was the fact that immunization with His-65kDa or GST-P4-P7 dramatically reduced mouse mortalityafter a challenge. While 100% of the control mice injected withadjuvant died before the 30th day after the challenge, 66.66%or 100% of the mice immunized with His-65kDa or GST-P4-P7, respectively, survived the challenge (Fig. 3). In contrast,survival was limited in mice immunized with recombinant pro-teins GST-P1-P3 or His-25kDa. In these cases, mouse survivalwas 6.25% or 35.74%, respectively (Fig. 3).
Statistical analysis revealed significant differences in the sur-vival of mice immunized with GST-P4-P7 compared to miceimmunized with other recombinant proteins or adjuvant only(P � 0.05 in all cases). Mice immunized with His-65kDa dis-played a survival rate significantly higher than that for othermouse groups, with the exception of the animals that receivedGST-P4-P7 (P � 0.05 in all cases). Mice immunized with His-25kDa had a survival rate higher than that for the controlanimals that received adjuvant only (P � 0.05). In contrast, thesurvival of mice injected with GST-P1-P3 or adjuvant only wasnot statistically different (P � 0.05).
The mice vaccinated with GST-P4-P7 that survived the chal-lenge were rechallenged 56 days later. Seven of eight micesurvived this second challenge, while all negative control miceimmunized with adjuvant alone died (n � 6). Parasitologicalstudies were performed on the surviving mice 77 days after thesecond challenge. We found that 57.1% of the mice (four ofseven) harbored living parasites that could be isolated fromtheir blood. These results indicated that the majority of thevaccinated mice were not able to completely eliminate theparasites.
We also evaluated the presence of parasite nests and thelevel of inflammatory response in their heart and striated mus-cle. Histological analyses failed to detect the presence of par-asite nests in the heart or striated muscle of mice vaccinatedwith GST-P4-P7. No inflammatory lesions were found in thestriated muscle of vaccinated mice. In their heart tissue, eitherno lesions or minimal lesions were found. We concluded thatmost vaccinated A/Sn mice not only survived two challenges
with T. cruzi but also controlled the inflammatory reaction inthe skeletal and heart muscles (data not shown).
The recombinant protein GST-P4-P7 expresses aa 261 to500 of ASP-2. In an attempt to further narrow the protectiveepitopes present in this recombinant protein, we immunizedmice with two other proteins representing aa 261 to 380 (GST-P4-P5) or aa 381 to 500 (GST-P6-P7) of ASP-2. After chal-lenged with trypomastigotes, mice vaccinated with GST-P4-P7or GST-P4-P5 displayed a peak parasitemia significantly lowerthan that for animals injected with GST-P6-P7 or adjuvantalone (Table 2) (P � 0.01). No statistically significant differ-ence was observed when we compared the peak parasitemia ofmice immunized with GST-P4-P7 or GST-P4-P5 (Table 2) (P� 0.05). Also, no difference was found when we compared thepeak parasitemia of mice immunized with GST-P6-P7 or ad-juvant alone (Table 2) (P � 0.05).
Statistical analysis revealed a significant difference in thesurvival of mice immunized with GST-P4-P7 or GST-P4-P5when compared to mice immunized with GST-P6-P7 or adju-vant only (P � 0.0001 in all cases). The survival of mice in-jected with GST-P6-P7 or adjuvant only was not statisticallydifferent (P � 0.05) (Table 2). These results clearly indicatedthat the protective epitopes present in GST-P4-P7 were lo-cated at the aa 261 to 380 present in the recombinant proteinGST-P4-P5.
To determine the participation of either CD4�, CD8�, orboth types of T cells in the protective immunity observed afterimmunization with the recombinant protein GST-P4-P7, wetreated vaccinated mice with anti-CD4 or anti-CD8 antibodiesprior to the challenge with T. cruzi. Treatment with monoclo-nal antibody anti-CD8 dramatically increased mouse suscepti-bility to infection compared to animals that received only ratIgG. The peak parasitemia of mice treated with anti-CD8 wassignificantly higher than that for vaccinated mice that receivedrat IgG or anti-CD4 (P � 0.05 in both cases) (Fig. 4A).
While 100% of the mice vaccinated with GST-P4-P7 andtreated with rat IgG survived the infection, 100% (10 of 10) ofanti-CD8-treated mice died (P � 0.0001). In contrast, only 4 ofthe 14 mice vaccinated with GST-P4-P7 and treated with anti-CD4 died after the challenge (P � 0.05) (Fig. 4B). We con-cluded from these experiments that, in vivo, CD8� T cells werecritical to the efficient reduction of peak parasitemia and tomouse survival after vaccination with the recombinant proteinGST-P4-P7.
Our results established that CD8� T cells provide the pro-tective immunity observed after immunization with GST-P4-P7. In order to determine whether we could detect specificCD8� T cells in the spleens of mice immunized with the dif-ferent recombinant proteins, we synthesized seven peptidesbased on the predicted amino acid sequence of the asp-2 gene.Peptides were selected by the presence of putative sites forbinding to A/Sn mouse major histocompatibility complex classI gene product H-2Kk. Peptides with putative sites for bindingto H-2Dd were not present in the predicted amino acid se-quence of the asp-2 gene (2).
The spleen cells of immunized mice were tested in the ELIS-POT assay for their ability to secrete gamma interferon(IFN-) upon in vitro stimulation in the presence of the dif-ferent peptides. During repeated experiments, we detected thepresence of peptide-specific IFN--secreting cells in the
6020 ARAUJO ET AL. INFECT. IMMUN.
by on October 27, 2009
iai.asm.org
Dow
nloaded from

spleens of mice immunized with His-65kDa or GST-P4-P7specific for the epitope represented by the synthetic peptideTEWETGQI (Table 3). In contrast, in mice immunized withGST-P1-P3 or His-25kDa, we were unable to detect the pres-ence of specific IFN--secreting cells to the peptides tested
(Table 3). The epitope TEWETGQI is located between aa 320and 327 of ASP-2. These amino acids are represented betweenGST-P4-P7 and GST-P4-P5.
The phenotype of IFN--producing cells specific for thepeptide TEWETGQI was established using a selective in vivo
FIG. 2. Trypomastigote-induced parasitemia in A/Sn mice immunized with recombinant protein His-65kDa, GST-P1-P3, GST-P4-P7, orHis-25kDa. Groups of six mice were immunized at 0, 2, and 4 weeks with His-65kDa, GST-P1-P3, GST-P4-P7, His-25kDa, or adjuvant alone. Eachmouse received i.p. 25 �g of recombinant protein adsorbed to alum in the presence of 10 �g of CpG ODN 1826. Two weeks after the last dosewas administered, mice were challenged i.p. with 250 bloodstream trypomastigotes. The parasitemia for each individual mouse is represented. Thepeak parasitemias of mice immunized with His-65kDa or GST-P4-P7 were lower than the parasitemias of animals immunized with adjuvant only,GST-P1-P3, or His-25kDa (P � 0.01 in all cases by Tukey’s HSD test). No significant difference was observed when we compared the peakparasitemias of mice immunized with His-65kDa or GST-P4-P7 (P � 0.05). Also, no difference was found when we compared the peak parasitemiasof mice immunized with GST-P1-P3 or His-25kDa or adjuvant alone (P � 0.05).
VOL. 73, 2005 VACCINATION AGAINST T. CRUZI INFECTION 6021
by on October 27, 2009
iai.asm.org
Dow
nloaded from

depletion approach. Mice immunized with GST-P4-P7 weretreated with either anti-CD4 or anti-CD8 or rat IgG. Thenumber of peptide-specific IFN--producing cells per 106
spleen cells of mice treated with rat IgG, anti-CD4, or anti-CD8 was 170.0 � 16.9, 174.6 � 6.0, or 36.0 � 8.7, respectively.Therefore, we concluded that the in vivo depletion of CD8�
cells reduced the number of peptide-specific IFN--producingcells by 79%. In contrast, depletion of CD4 cells had no impacton the number of cells specific to the epitope TEWETGQI.
To determine whether there was a correlation between theprotective immunity observed after immunization with the re-combinant proteins His-65kDa, GST-P4-P7, GST-P1-P3, orHis-25kDa and the immune response to the peptide TEWETGQI, we evaluated the number of IFN--secreting cells in thespleens of individual mice immunized with each recombinantprotein. The number of IFN--secreting cells specific to the
peptide TEWETGQI per 106 spleen cells in mice immunizedwith GST-P4-P7 or His-65kDa were 147.11 � 28.87 and 41.66� 11.86, respectively (n � 3, P � 0.01, by Tukey’s HSD test).In mice immunized with GST-P1-P3, His-25kDa, or adjuvantonly, we could not detect IFN--secreting cells specific for thepeptide TEWETGQI.
DISCUSSION
The aim of our study was to explore the possibility of usingbacterial recombinant proteins based on the ASP-2 antigen forvaccination against experimental T. cruzi infection. Our initialhypothesis was that a recombinant protein based on the ASP-2antigen could be useful as a booster injection in mice previ-ously immunized with DNA. We reasoned that this regimencould enhance the protective immune response observed withDNA vaccination alone. Contrary to this hypothesis, we foundthat the survival of mice immunized with DNA alone or with aDNA priming/protein boosting regimen was nearly identical(75 or 73.74%, respectively).
Based on the partial protection that we observed, we de-cided to pursue the question of whether recombinant proteinsof ASP-2 could be used to elicit protective immunity against aT. cruzi infection. In the subsequent experiments, we added asecond adjuvant, CpG ODN 1826, to alum. We selected thisadjuvant based on previous studies showing that CpG ODN1826 could improve Th1 immune response and protective im-munity to a T. cruzi infection induced by a purified nativeprotein (6). At this point, the precise relevance of CpG ODN1826 in our system is not clear. It is certainly not criticalbecause we can see significant protection in the absence ofCpG ODN 1826 after vaccination with His-65kDa or GST-P4-P7 (Table 1 and data not shown, respectively). At the sametime, the presence of CpG ODN 1826 did help protectiveefficacy. It is possible that the presence of CpG ODN 1826increases the activation of specific CD8� T cells. This hypoth-esis remains to be tested.
To map the protective determinants present within His-
FIG. 3. Trypomastigote-induced mortality in A/Sn mice immunizedwith recombinant proteins His-65kDa, GST-P1-P3, GST-P4-P7, orHis-25kDa. The graph shows the Kaplan-Meier curves for the survivalof 14 to 16 mice per group. Statistical analysis revealed significantdifferences in the survival of mice immunized with GST-P4-P7 com-pared to that of mice immunized with other recombinant proteins oradjuvant only (P � 0.05 in all cases by the log rank test). Mice immu-nized with His-65kDa displayed a survival rate significantly higher thanthat for other mouse groups with the exception of the animals thatreceived GST-P4-P7 (P � 0.05 in all cases). Mice immunized withHis-25kDa had a survival rate higher than that for the control animalsthat received adjuvant only (P � 0.05). In contrast, the survival of miceinjected with GST-P1-P3 or adjuvant only was not statistically different(P � 0.05).
TABLE 2. Trypomastigote-induced parasitemia and mortality inA/Sn mice immunized with recombinant protein GST-P4-P7,
GST-P4-P5, or GST-P6-P7
Immunogena Peak parasitemia(log � SD)b
No. of micealive/total no.challengedc
%Protection
Adjuvant 6.06 � 0.16 0/6 0GST-P4-P7 4.67 � 0.61 5/5 100GST-P4-P5 4.96 � 0.34 6/6 100GST-P6-P7 6.13 � 0.21 0/6 0
a Groups of five or six mice were immunized at 0, 2, and 4 weeks withGST-P4-P7, GST-P4-P5, GST-P6-P7, or adjuvant alone as described in thelegend to Fig. 2. Two weeks after the last dose was given, mice were challengedi.p. with 250 bloodstream trypomastigotes.
b The peak parasitemias in mice immunized with either GST-P4-P7 or GST-P4-P5 were lower than those for animals immunized with GST-P6-P7 or adjuvantalone (P � 0.01 in all cases by Tukey’s HSD test). The peak parasitemias of miceimmunized with either GST-P4-P7 or GST-P4-P5 were not statistically different(P � 0.05).
c Mouse survival was recorded at day 60 after infection. Animals immunizedwith GST-P4-P7 or GST-P4-P5 survived longer than mice injected with GST-P6-P7 or adjuvant alone (P � 0.0001 by the log rank test). There was nosignificant statistical difference between the survival curves of mice vaccinatedwith GST-P4-P7 or GST-P4-P5.
6022 ARAUJO ET AL. INFECT. IMMUN.
by on October 27, 2009
iai.asm.org
Dow
nloaded from

65kDa, we used three proteins containing different domains ofASP-2. Immunization with recombinant proteins GST-P1-P3or His-25kDa representing the N- or the C-terminal domainsof ASP-2, respectively, provided limited protective immunitycompared to His-65kDa. In contrast, when we used a recom-binant protein expressing aa 261 to 500 of ASP-2 (GST-P4-P7)for vaccination, we obtained 100% protection against a lethalchallenge with T. cruzi trypomastigotes. Statistical analysis re-vealed that the protective efficacy of the recombinant proteinGST-P4-P7 was higher than that of His-65kDa. The resultsobtained with GST-P4-P7 not only confirmed the protectivepotential of recombinant proteins based on the ASP-2 of T.cruzi but also mapped highly protective epitope(s) to the cen-tral domain of this molecule. Using a shorter recombinantprotein (GST-P4-P5), we were able to narrow this highly pro-tective epitope(s) to the region between aa 261 and 380.Whether smaller recombinant proteins, or even synthetic pep-tides, can be used to vaccinate A/Sn mice remains to be eval-uated.
Protective immunity obtained after immunization with GST-P4-P7, alum, and CpG ODN 1826 was remarkable comparedto other protocols of immunization. This formulation provideda high degree of protective immunity compared to ASP-2DNA, His-65 kDa, or both in the same protocol of vaccination.Until now, more than 50 mice immunized with this formulationhave been challenged. To date, all of these immunized micehave survived T. cruzi challenge. Detailed studies on the lon-gevity of protective immune responses induced with GST-P4-P7 vaccination have yet to be performed. Nevertheless,vaccinated A/Sn mice that survived the first challenge wererechallenged 56 days later. Seven of eight mice survived infec-tion, proving that this protective immunity is highly efficient.
Although protective immunity elicited by immunization withrecombinant protein GST-P4-P7 drastically reduced the peakparasitemia and promoted mouse survival, we did not obtainsterile immunity in most animals. In spite of the fact that sterileimmunity was not achieved, we found little or no inflammationin the heart or striated muscle of mice challenged twice in thecourse of 133 days. Our observations suggest that vaccinationcontrols acute-phase parasitemia, survival, and the develop-ment of inflammatory responses in the heart and striated mus-cle. Our results reinforce previously described observationsthat immunity induced by T. cruzi ASP-2 encoding the DNAvaccine gene drastically reduce the inflammatory responses inthe heart and skeletal muscle of A/Sn mice (28).
While vaccination with GST-P4-P7 protected 100% of themice against infection, immunization with His-65kDa or ASP-2DNA never achieved this level of protective immunity. Thefact that immunization with GST-P4-P7 was more efficientthan the vaccination protocol using a larger recombinant pro-tein correlated with the number of IFN--producing cells spe-cific to the peptide TEWETGQI (Table 3). Therefore, it isplausible to propose that the shorter GST-P4-P7 recombinantprotein can induce greater numbers of protective CD8� T cellsspecific for the peptide TEWETGQI. The specific reason forthese results is currently unknown. It is possible that the pro-tective epitope TEWETGQI may be better processed by anti-gen-presenting cells after immunization with GST-P4-P7. Also,the presence of GST may provide greater T-cell help to theepitope TEWETGQI. Alternatively, we should evaluate
FIG. 4. Trypomastigote-induced parasitemia and mortality in miceimmunized with GST-P4-P7 and treated with anti-CD4 or anti-CD8antibodies. Groups of five or six mice were immunized as described inthe legend to Fig. 2 with the recombinant protein GST-P4-P7 oradjuvant alone. Mice immunized with the recombinant protein weretreated with rat IgG, anti-CD4, or anti-CD8 as described in Materialsand Methods. (A) The peak parasitemias in animals immunized withGST-P4-P7 and treated with rat IgG or anti-CD4 were significantlylower than the parasitemias of immunized animals treated with anti-CD8 (P � 0.01 in all cases by Tukey’s HSD test). The values ofparasitemia of mice immunized with GST-P4-P7 and treated withanti-CD8 were not different from those of control mice injected withadjuvant only (P � 0.05). (B) The graph shows Kaplan-Meier curvesfor the survival of 10 to 14 mice per group. Animals immunized withGST-P4-P7 and treated with rat IgG or anti-CD4 survived longer thanmice treated with anti-CD8 (P � 0.0001 by the log rank test). Therewas no significant statistical difference between the survival curves ofmice vaccinated with the recombinant protein GST-P4-P7 and treatedwith anti-CD8 or control animals injected with adjuvant alone (P �0.05).
VOL. 73, 2005 VACCINATION AGAINST T. CRUZI INFECTION 6023
by on October 27, 2009
iai.asm.org
Dow
nloaded from

whether His-65kDa contains an epitope(s) that activates reg-ulatory T cells which can inhibit the CD8�-T-cell immuneresponse to TEWETGQI.
Through depletion studies, we found that treatment withanti-CD8, but not with anti-CD4 antibodies, completely re-versed the protective immunity elicited by immunization withGST-P4-P7. The fact that depletion of CD4� T cells had alimited impact on protective immunity was not fully expected.Our previous study has shown that when treated with anti-CD4antibodies, mice vaccinated simultaneously with asp-2 and tsgenes were as susceptible to infection as anti-CD8-treatedmice (28). The results described here cannot be attributed onlyto an inefficient elimination of CD4� T cells because immu-nized mice were treated with the same antibodies and scheduleused in the previous report (28). Also, the extension of deple-tion was monitored by fluorescence-activated cell sorter anal-ysis before and after challenge. However, the most importantobservation was the fact that mice treated with anti-CD8 wereas susceptible to infection as control mice injected with adju-vant only. Therefore, it seems that in the absence CD8� Tcells, CD4� T cells were not able to provide a significantdegree of protective immunity. These results suggest a differ-ent balance in these two T-cell populations after immunizationwith DNA or the recombinant protein GST-P4-P7.
Previous studies have shown that formulations containingdistinct recombinant proteins could elicit protective immunityin BALB/c or C57BL/6 mice against a lethal challenge with T.cruzi (16, 20, 23, 32, 34). Depletions of CD4� or CD8� T cellsbefore challenge were not performed in animals vaccinatedwith cruzipain or TC52 (20, 23). In the case of vaccination withrecombinant paraflagellar rod proteins, mice genetically defi-cient in CD8 expression failed to develop protective immunityagainst T. cruzi infection (34). Our results concur with theselatter results, and both studies confirm that CD8� T cells playa critical role in protective immunity elicited by recombinantproteins, as well as DNA vaccines (12, 28; reviewed in refer-ences 17 and 22).
How are these protective CD8� T cells primed by the for-mulation containing recombinant protein, alum, and CpGODN 1826? It is plausible to speculate that this formulationuses the cross-priming route of antigen presentation (reviewedin references 1 and 9). In fact, formulations containing CpG
ODN 1826 are described as capable of inducing specific CD8�
T cells by the cross-priming route (10, 11; reviewed in refer-ence 30). Whether this route of antigen presentation is oper-ating in our case should be tested; this may provide an inter-esting model to study the development of an anti-T. cruzivaccine.
In summary, our data on the protective immune responseelicited by recombinant proteins of the ASP-2 antigen createda new model to study how recombinant proteins can be used toelicit CD8�-T-cell-mediated protective immune responses tothis intracellular pathogen. Also, we have confirmed and ex-tended the results of several other studies that have describedthe protective potential of this antigen (2, 5, 8, 28). Together,they emphasize the importance of the immune response toASP-2 for the development of immunity to T. cruzi infection.Studies on the immune response to ASP-2 may aid our under-standing of the immunopathological mechanisms leading toChagas’ disease and the development of new strategies for theprevention or treatment of the disease.
ACKNOWLEDGMENTS
This work was supported by grants from Fundacao de Amparo aPesquisa do Estado de Sao Paulo, CNPq, FAPEMIG, Brazil (EDT24.000), and The National Institutes of Health, Bethesda, Md. (RO1AI040196).
We thank Fanny Tzelepis for helping with the ELISPOT assay andGisela Sasso for the histology. A.F.S.A., M.I.H., R.T.G., and M.M.R.are recipients of fellowships from CNPq. J.R.C.V., S.B.B., andB.C.G.D.A. were recipients of fellowships from FAPESP.
REFERENCES
1. Ackerman, A. L., and P. Cresswell. 2004. Cellular mechanisms governingcross-presentation of exogenous antigens. Nat. Immunol. 5:678–684.
2. Boscardin, S. B., S. S. Kinoshita, A. E. Fujimura, and M. M. Rodrigues.2003. Immunization with cDNA expressed by amastigotes of Trypanosomacruzi elicits protective immune response against experimental infection. In-fect. Immun. 71:2744–2757.
3. Costa, F., G. Franchin, V. L. Pereira-Chioccola, M. Ribeirao, S. Schenkman,and M. M. Rodrigues. 1998. Immunization with a plasmid DNA containingthe gene of trans-sialidase reduces Trypanosoma cruzi infection in mice.Vaccine 16:768–774.
4. Dumonteil, E., J. Escobedo-Ortegon, N. Reyes-Rodriguez, A. Arjona-Torres,and M. J. Ramirez-Sierra. 2004. Immunotherapy of Trypanosoma cruzi in-fection with DNA vaccines in mice. Infect. Immun. 72:46–53.
5. Fralish, B. H., and R. L. Tarleton. 2003. Genetic immunization with LYT1or a pool of trans-sialidase genes protects mice from lethal Trypanosomacruzi infection. Vaccine 21:3070–3080.
TABLE 3. ELISPOT assay for the detection of peptide-specific IFN--secreting cells
Peptideb inculture
No. of IFN--secreting cells/106 spleen cells of mice immunized with recombinant proteinsa:
Adjuvantc His-65kDa GST-P1-P3 GST-P4-P7 His-25kDa
No peptide 3.0 � 4.2 6.0 � 0.0 5.0 � 4.2 5.0 � 1.4 5.0 � 4.2KEGSKSGV 2.0 � 0.0 0.0 � 0.0 0.0 � 0.0 5.0 � 4.2 4.0 � 2.8AESLFVYI 1.0 � 1.4 1.0 � 1.4 0.0 � 0.0 3.0 � 1.4 5.0 � 1.4HEHNLFGI 5.0 � 1.4 3.0 � 1.4 1.0 � 1.4 8.0 � 2.8 2.0 � 0.0YEIVAGYI 0.0 � 0.0 0.0 � 0.0 2.0 � 0.0 8.0 � 2.8 0.0 � 0.0AESWPSIV 0.0 � 0.0 1.0 � 1.4 0.0 � 0.0 5.0 � 1.4 1.0 � 1.4TEWETGQI 1.0 � 1.4 29.0 � 1.4d 1.0 � 1.4 113.0 � 9.8d 0.0 � 0.0PETLGHEI 3.0 � 1.4 2.0 � 2.8 0.0 � 0.0 4.0 � 2.8 4.0 � 0.0
a The number of IFN--secreting cells was determined by the ELISPOT assay.b The final concentration of each peptide in culture was 10 �g/ml.c A/Sn mice were immunized with the indicated recombinant protein as described in the legend to Fig. 2. Two weeks after the last immunization, pooled spleen cells
were collected from three mice.d The number of IFN--secreting cells in the cultures containing the peptide TEWETGQI were higher than the number in cultures containing no peptide or other
peptides (P � 0.01 by Tukey’s HSD test).
6024 ARAUJO ET AL. INFECT. IMMUN.
by on October 27, 2009
iai.asm.org
Dow
nloaded from

6. Frank, F. M., P. B. Petray, S. I. Cazorla, M. C. Munoz, R. S. Corral, andE. L. Malchiodi. 2003. Use of a purified Trypanosoma cruzi antigen and CpGoligodeoxynucleotides for immunoprotection against a lethal challenge withtrypomastigotes. Vaccine 22:77–86.
7. Fujimura, A. E., S. S. Kinoshita, V. L. Pereira-Chioccola, and M. M. Ro-drigues. 2001. DNA sequences encoding CD4� and CD8� T-cell epitopesare important for efficient protective immunity induced by DNA vaccinationwith a Trypanosoma cruzi gene. Infect. Immun. 69:5477–5486.
8. Garg, N., and R. Tarleton. 2002. Genetic immunization elicits antigen-spe-cific protective immune responses and decreases disease severity in Trypano-soma cruzi infection. Infect. Immun. 70:5547–5555.
9. Heath, W. R., G. T. Belz, G. M. Behrens, C. M. Smith, S. P. Forehan, I. A.Parish, G. M. Davey, N. S. Wilson, F. R. Carbone, and J. A. Villadangos.2004. Cross-presentation, dendritic cell subsets, and the generation of im-munity to cellular antigens. Immunol. Rev. 199:9–26.
10. Heit, A., K. M. Huster, F. Schmitz, M. Schiemann, D. H. Busch, and H.Wagner. 2004. CpG-DNA aided cross-priming by cross-presenting B cells.J. Immunol. 172:1501–1507.
11. Heit, A., T. Maurer, H. Hochrein, S. Bauer, K. M. Huster, D. H. Busch, andH. Wagner. 2003. Toll-like receptor 9 expression is not required for CpGDNA-aided cross-presentation of DNA-conjugated antigens but essential forcross-priming of CD8 T cells. J. Immunol. 170:2802–2805.
12. Katae, M., Y. Miyahira, K. Takeda, H. Matsuda, H. Yagita, K. Okumura, T.Takeuchi, T. Kamiyama, A. Ohwada, Y. Fukuchi, and T. Aoki. 2002. Coad-ministration of an interleukin-12 gene and a Trypanosoma cruzi gene im-proves vaccine efficacy. Infect. Immun. 70:4833–4840.
13. Krettli, A. U., and Z. Brener. 1976. Protective effects of specific antibodies inTrypanosoma cruzi infections. J. Immunol. 116:755–760.
14. Letvin, N. L., D. C. Montefiori, Y. Yasutomi, H. C. Perry, M. E. Davies, C.Lekutis, M. Alroy, D. C. Freed, C. I. Lord, L. K. Handt, M. A. Liu, and J. W.Shiver. 1997. Potent, protective anti-HIV immune responses generated bybimodal HIV envelope DNA plus protein vaccination. Proc. Natl. Acad. Sci.USA 94:9378–9383.
15. Li, S., M. Rodrigues, D. Rodriguez, J. R. Rodriguez, M. Esteban, P. Palese,R. S. Nussenzweig, and F. Zavala. 1993. Priming with recombinant influenzavirus followed by administration of recombinant vaccinia virus inducesCD8� T-cell-mediated protective immunity against malaria. Proc. Natl.Acad. Sci. USA 90:5214–5218.
16. Luhrs, K. A., D. L. Fouts, and J. E. Manning. 2003. Immunization withrecombinant paraflagellar rod protein induces protective immunity againstTrypanosoma cruzi infection. Vaccine 21:3058–3069.
17. Martin, D., and R. Tarleton. 2004. Generation, specificity, and function ofCD8� T cells in Trypanosoma cruzi infection. Immunol. Rev. 201:304–317.
18. Morel, C. M., and J. Lazdins. 2003. Chagas disease. Nat. Rev. Microbiol.1:14–15.
19. Nyika, A., A. F. Barbet, M. J. Burridge, and S. M. Mahan. 2002. DNAvaccination with map1 gene followed by protein boost augments protectionagainst challenge with Cowdria ruminantium, the agent of heartwater. Vac-cine 20:1215–1225.
20. Ouaissi, A., E. Guilvard, Y. Delneste, G. Caron, G. Magistrelli, N. Herbault,N. Thieblemont, and P. Jeannin. 2002. The Trypanosoma cruzi Tc52-releasedprotein induces human dendritic cell maturation, signals via Toll-like recep-tor 2, and confers protection against lethal infection. J. Immunol. 168:6366–6374.
21. Planelles, L., M. C. Thomas, C. Alonso, and M. C. Lopez. 2001. DNAimmunization with Trypanosoma cruzi HSP70 fused to the KMP11 proteinelicits a cytotoxic and humoral immune response against the antigen andleads to protection. Infect. Immun. 69:6558–6563.
22. Rodrigues, M. M., S. B. Boscardin, J. R. Vasconcelos, M. I. Hiyane, G. Salay,and I. S. Soares. 2003. Importance of CD8 T cell-mediated immune responseduring intracellular parasitic infections and its implications for the develop-ment of effective vaccines. An. Acad. Bras. Cienc. 75:443–468.
23. Schnapp, A. R., C. S. Eickhoff, D. Sizemore, R. Curtiss III, and D. F. Hoft.2002. Cruzipain induces both mucosal and systemic protection againstTrypanosoma cruzi in mice. Infect. Immun. 70:5065–5074.
24. Sepulveda, P., M. Hontebeyrie, P. Liegeard, A. Mascilli, and K. A. Norris.2000. DNA-Based immunization with Trypanosoma cruzi complement regu-latory protein elicits complement lytic antibodies and confers protectionagainst Trypanosoma cruzi infection. Infect. Immun. 68:4986–4991.
25. Silva, L. H. P., and V. Nussenzweig. 1953. Sobre uma cepa de Trypanosomacruzi altamente virulenta para o camundongo branco. Folia Clin. Biol. 20:191–203.
26. Song, M. K., S. W. Lee, Y. S. Suh, K. J. Lee, and Y. C. Sung. 2000. Enhance-ment of immunoglobulin G2a and cytotoxic T-lymphocyte responses by abooster immunization with recombinant hepatitis C virus E2 protein in E2DNA-primed mice. J. Virol. 74:2920–2925.
27. Tanghe, A., S. D’Souza, V. Rosseels, O. Denis, T. H. Ottenhoff, W. Dalemans,C. Wheeler, and K. Huygen. 2001. Improved immunogenicity and protectiveefficacy of a tuberculosis DNA vaccine encoding Ag85 by protein boosting.Infect. Immun. 69:3041–3047.
28. Vasconcelos, J. R., M. I. Hiyane, C. R. F. Marinho, C. Claser, A. M. Vieira-Machado, R. T. Gazinelli, O. Bruna-Romero, J. M. Alvarez, S. B. Boscardin,and M. M. Rodrigues. 2004. Protective immunity against Trypanosoma cruziinfection in a highly susceptible mouse strain following vaccination withgenes encoding the amastigote surface protein-2 and trans-sialidase. Hum.Gene Ther. 15:878–886.
29. Vasconcelos, J. R., S. B. Boscardin, M. I. Hiyane, S. S. Kinoshita, A. E.Fujimura, and M. M. Rodrigues. 2003. A DNA-priming protein-boostingregimen significantly improves type 1 immune response but not protectiveimmunity to Trypanosoma cruzi infection in a highly susceptible mousestrain. Immunol. Cell Biol. 81:121–129.
30. Wagner, H., A. Heit, F. Schmitz, and S. Bauer. 2004. Targeting split vaccinesto the endosome improves vaccination. Curr. Opin. Biotechnol. 15:538–542.
31. Wizel, B., N. Garg, and R. Tarleton. 1998. Vaccination with trypomastigotessurface antigen1-encoding plasmid DNA confers protection against lethalTrypanosoma cruzi infection. Infect. Immun. 66:5073–5081.
32. Wrightsman, R. A., B. D. Dawson, D. L. Fouts, and J. E. Manning. 1994.Identification of immunodominant epitopes in Trypanosoma cruzi trypomas-tigote surface antigen-1 protein that mask protective epitopes. J. Immunol.153:3148–3154.
33. Wrightsman, R. A., and J. E. Manning. 2000. Paraflagellar rod proteinsadministered with alum and IL-12 or recombinant adenovirus expressingIL-12 generates antigen-specific responses and protective immunity in miceagainst Trypanosoma cruzi. Vaccine 18:1419–1427.
34. Wrightsman, R. A., K. A. Luhrs, D. Fouts, and J. E. Manning. 2002.Paraflagellar rod protein-specific CD8� cytotoxic T lymphocytes targetTrypanosoma cruzi-infected host cells. Parasite Immunol. 24:401–412.
Editor: W. A. Petri, Jr.
VOL. 73, 2005 VACCINATION AGAINST T. CRUZI INFECTION 6025
by on October 27, 2009
iai.asm.org
Dow
nloaded from

55
Artigo 2

56
Cross-priming of long lived protective CD8+ T cells against Trypanosoma
cruzi infection: importance of a TLR9 agonist and CD4+ T cells.
A evidência de que as imunizações com a proteína recombinante
representando a porção central da ASP-2 (GST-P4P7), adicionada do alum e
do CpG ODN 1826, era capaz de proteger camundongos altamente suscetíveis
contra a infecção pelo T. cruzi, por mecanismos dependentes dos linfócitos T
CD8, estimulou o estudo de outros fatores que fossem determinantes para a
proteção induzida.
Neste trabalho, estabeleceu-se, inicialmente, que essa proteção é de
longa duração, sendo capaz de proteger os animais eficientemente por, pelo
menos, 5 meses. Passou-se então a estudar o papel do adjuvante CpG ODN
1826 e da ativação dos receptores TLR9 naquelas imunizações. Para tal,
conduziram-se vacinações com a proteína GST-P4P7 misturada ao alum, na
presença ou ausência do CpG ODN 1826. Observou-se que este adjuvante
potencializava a resposta de linfócitos T CD8 e melhorava a sua eficácia
protetora. Estudou-se, também, o papel da ativação dos receptores TLR4 por
meio do uso de camundongos portando uma mutação, a qual torna o TLR4
não-funcional. Observou-se que, em imunizações na presença do CpG ODN
1826, a ativação dos receptores TLR4 não era importante.
Para o estudo da participação dos linfócitos T CD4, foram realizados
ensaios de depleção in vivo destes linfócitos, antes das imunizações com GST-
P4P7 adicionada de alum/CpG ODN 1826, tendo também sido feitas
imunizações com uma nova proteína recombinante, contendo apenas o epítopo
da ASP-2 para linfócitos T CD8, em fusão com a GST, na presença dos
mesmos adjuvantes. Em ambos os casos, observou-se a indução de linfócitos
T CD8 secretores de IFN-γ ex vivo, os quais eram, porém, incapazes de
proteger os camundongos A/Sn contra o desafio com o T. cruzi.
Concluiu-se que o agonista do TLR9, o CpG ODN 1826, melhora a
eficiência da imunidade protetora; e que os linfócitos T CD4 são críticos para a
ativação dos linfócitos T CD8 capazes de proteger, por longos períodos, os
camundongos suscetíveis contra o desafio com Trypanosoma cruzi.

A
AToaC©
K
1
abvian(
MT
0d
Vaccine 25 (2007) 6018–6027
Cross-priming of long lived protective CD8+ T cells againstTrypanosoma cruzi infection: Importance of a
TLR9 agonist and CD4+ T cells
Bruna C.G. de Alencar a,b, Adriano F.S. Araujo a,b, Marcus L.O. Penido c,Ricardo T. Gazzinelli c, Mauricio M. Rodrigues a,b,∗
a Centro Interdisciplinar de Terapia Genica (CINTERGEN), Universidade Federal de Sao Paulo, Escola Paulista de Medicina, Brazilb Departmento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de Sao Paulo, Escola Paulista de Medicina, Brazil
c Departamento de Bioquımica e Imunologia, Instituto de Ciencias Biologicas,Universidade Federal de Minas Gerais e Centro de Pesquisas Rene Rachou, FIOCRUZ, Avenida Augusto de Lima 1715,
Barro Preto, 30190-002 Belo Horizonte, Minas Gerais, Brazil
Received 26 February 2007; received in revised form 9 April 2007; accepted 13 May 2007Available online 4 June 2007
bstract
We recently described that vaccination of mice with a gluthatione S transferase fusion protein representing amino acids 261–500 of themastigote Surface Protein-2 efficiently cross-primed protective CD8+ T cells against a lethal challenge with the human protozoan parasiterypanosoma cruzi. In this study, we initially established that this protective immunity was long lived. Subsequently, we studied the importance
+ +
f TLR9 agonist CpG ODN 1826, TLR4 and CD4 T cells for the generation of these protective CD8 T cells. We found that: (i) the TLR9gonist CpG ODN 1826 improved the efficiency of protective immunity; (ii) TLR4 is not relevant for priming of specific CD8+ T cells; (iii)D4+ T cells are critical for priming of memory/protective CD8+ T cells.2007 Elsevier Ltd. All rights reserved.lcplcwo
eywords: Vaccination; Trypanosoma cruzi; Amastigotes; CpG ODN
. Introduction
MHC class Ia-restricted CD8+ T cells are important medi-tors of adaptive immune response against infections causedy intracellular microorganisms (reviewed in Ref. [1]). Iniew of the importance of these cells for host survival follow-ng viral, bacterial or parasite challenge, it has become largelyccepted that they should be considered in the design of a
ew generation of vaccines against intracellular pathogensreviewed in Ref. [2]).∗ Corresponding author at: CINTERGEN, UNIFESP, Escola Paulista deedicina, Rua Mirassol 207, Sao Paulo 04044-010, SP, Brazil.
el.: +55 11 5571 1095; fax: +55 11 5571 1095.E-mail address: [email protected] (M.M. Rodrigues).
tpia[
tC
264-410X/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2007.05.022
Infection of humans or mice with the digenetic intracel-ular protozoan parasite Trypanosoma cruzi induces MHClass Ia-restricted CD8+ T cells specific for immunodominantarasite epitopes [3,4]. Since the seminal publication by Tar-eton et al. [5] showing that MHC class I-restricted cells wereritical for acquired immunity against experimental infectionith T. cruzi, a number of reports confirmed the importancef CD8+ T cells for host resistance following challenge withhis parasite ([3,6,7] and reviewed in Refs. [8,9]). The anti-arasitic mechanisms mediated by these cells are multiple,ncluding cytokine secretion, and possibly direct cytotoxicitygainst infected target cells ([3,10,11] and reviewed in Refs.
8,9]).Vaccination studies against experimental T. cruzi infec-ion extended these observations by providing evidence thatD8+ T cells are particularly relevant for the development of

B.C.G. de Alencar et al. / Vaccin
F2
ppmTiw
nrPwtanotmt
ilCg
2
2
mosdaiPbclCP
2s
sF[bGrTth54Ecrat
wnnptGGt9PtPAEtHpTP
2
ptco(Os(
ig. 1. Schematic representation of the Amastigote Surface Protein-2 (ASP-) of Trypanosoma cruzi and the recombinant proteins used in this study.
rotective immunity in mice immunized with recombinantlasmid DNA, proteins and viruses [12–17]. These experi-ents have given rise to the concept that a long-lived CD8+
-cell mediated immune response might be a key elementn the development of an effective vaccine against infectionith this human protozoan parasite (reviewed in Refs. [8,9]).Following this rationale, we recently described that vacci-
ation with a gluthatione S transferase (GST) fusion proteinepresenting amino acids 261–500 of the Amastigote Surfacerotein-2 (GST-P4–P7, Fig. 1), coupled to alum and admixedith the TLR 9 agonist CpG ODN 1826, provided strong pro-
ective immunity to a highly susceptible mouse strain againstlethal challenge with T. cruzi [15]. This protective immu-ity was mediated by CD8+ T cells as the in vivo depletionf these cells prior to challenge completely reversed protec-ion [15]. The epitope recognized by these CD8+ T cells was
apped to the peptide TEWETGQI (amino acids 320-327 ofhe Amastigote Surface Protein-2).
Based on these observations, this study was designed tonvestigate first whether this protective immunity was long-ived. Subsequently, we assessed whether the TLR9 agonistpG ODN 1826, TLR4 and CD4+ T cells influenced theeneration of these protective CD8+ T cells.
. Materials and methods
.1. Mice and parasites
Female 5- to 8-week-old A/Sn, C3H/HeJ and C3H/HePASice were purchased from University of Sao Paulo. Parasites
f the Y strain of T. cruzi were used in this study [18]. Blood-tream trypomastigotes were obtained from mice infected 7ays earlier. The concentration of parasites was estimatednd adjusted to 1250 ml−1. Each mouse was inoculatedntraperitoneally (i.p.) with 0.2 ml (250 trypomastigotes).arasite development was monitored by counting the num-er of bloodstream trypomastigotes in 5 �l of fresh blood
ollected from the tail vein [19]. Mouse survival was fol-owed daily. This study has been approved by the Ethicalommittee for Animal Care of the Federal University of Saoaulo.1oAd
e 25 (2007) 6018–6027 6019
.2. Plasmid and recombinant proteins based on theequence of the amastigote surface protein (asp-2) gene
Gluthatione S transferase fusion protein (GST) repre-enting amino acids 261–500 of the ASP-2 (GST-P4–P7,ig. 1) was expressed and purified as described earlier15]. The plasmid pGEX-4T-2-TEWETGQI was generatedy ligating the oligonucleotides containing the forward (5′-ATCCACCGAATGGGAAACCGGCCAGATTGG-3) and
everse (5′-AATTCCAATCTGGCCGGTTTCCCATTCGG-G-3′) sequences representing the epitope TEWETGQI and
he BamHI and EcoRI sites. These oligonucleotides wereeated at 90 ◦C for 5 min and kept at 60 ◦C for moremin. This fragment was ligated into the vetor pGEX-T-2 (GE Amershan) previously treated with BamHI andcoRI enzymes. The presence of the oligonucleotides wasonfirmed by sequencing the recombinant plasmid. Theecombinant protein denominated GST-TEWTGQI (Fig. 1)nd GST were expressed and purified according to the pro-ocols provided by the manufacturer.
The recombinant plasmid denominated pHis6-P4–P7as generated by inserting a DNA fragment representingucleotides 781–1500 of clone 9 gene (GenBank accessionumber: AY 186572) into EcoRI and SalI sites of the plasmidHis [20]. The DNA fragment was generated by PCR usinghe forward oligonucleotide (5′-CCG GAA TTC GAT GTTTG ACT GCT GG-3′) or the reverse oligonucleotide (5′-GG TCT AGA TTA CGT ATT GGA CAG GA-3′) designed
o anneal with nucleotides 781–799 or 1483–1500 of clonegene, respectively. PCR were performed using the enzymeLATINUM Pfx DNA Polymerase (Life Technologies) and
he annealing temperature used was 48 ◦C. The amplifiedCR products were cloned into the pMOSBlue vector (GE-mersham), removed by treatment with restriction enzymescoRI and SalI and ligated into the pHIS vector treated with
hese same enzymes. The recombinant protein denominatedis6-P4–P7 represented amino acids 261–500 of ASP-2. Therotein was expressed and purified as described earlier [15].he purity of recombinant proteins was determined by SDS-AGE and was represented by a single band.
.3. Immunization
Each A/Sn mouse received three i.p. doses of 25 �g ofrotein adsorbed to alum (Alhydrogel “85” Superfos Biosec-or, Vedbaek, Denmark) given at 0, 2 and 4 weeks. Efficientoupling of recombinant GST, GST-P4-P7, GST-TEWTGQIr His6-P4–P7 to alum was obtained by mixing 2.5 �l 2%w/v) Al(OH)3 per �g of recombinant protein for 2 h. CpGDN 1826 (TCCATGACGTTCCTGACGTT) was synthe-
ized with a nuclease-resistant phosphorothioate backboneColey Pharmaceutics, Wellesley, MA). When indicated,
0 �g of CpG ODN 1826 was mixed with alum containingr not the recombinant proteins before i.p. immunization of/Sn mice. Two weeks or 5 months after the last immunizingose mice were challenged with bloodstream trypomastig-
6 / Vaccin
oir
2
ddH1grttR3mog5(paU(
Aao1pwcC
TPog2a6(fiaa17pfsPaa
2
pIiord9
2
bmbwapibs
2
wAsoAuTt
3
hGplmGawtmw
020 B.C.G. de Alencar et al.
tes. Results of parasitemia are representative of two or morendependent experiments. Results of mortality are pooledesults of two or more experiments.
.4. Immunological assays
The ELISPOT assay was performed essentially asescribed earlier [20]. Briefly, the preparation of plates wasone by coating 96-well nitrocellulose plates MultiscreenA (Millipore) with 60 �l/well of sterile PBS containing0 �g/ml of the anti-mouse IFN-� MAb R4-6A2 (Pharmin-en). After overnight incubation at rt, MAb solution wasemoved by sterile aspiration and the plates were washedhree times with plain RPMI medium under sterile condi-ions. Plates were blocked by incubating wells with 100 �lPMI medium containing 10% (v/v) FBS for at least 2 h at7 ◦C. Responder cells were obtained from spleens of A/Snice. Responder cells were ressuspended to a concentration
f 106 viable cells per ml in RPMI medium (Life Technolo-ies) supplemented with 10 mM HEPES, 2 mM l-glutamine,× 10−5 M 2-mercaptoethanol, 1mM sodium pyruvate, 1%
v/v) nonessential amino acids solution, 1% (v/v) vitamin (allurchased from Life Technologies), 100 U/ml of penicillinnd streptomycin (Sigma), 10% (v/v) FBS (Hyclone, Logan,tah), and 30 U/ml of recombinant human Interleukin-2
kindly provided by Hoffman-LaRoche).Antigen presenting cells were prepared by irradiating
/Sn spleen cells for 7 min. These cells were adjusted toconcentration of 4 × 106 viable cells per ml and incubatedr not with the synthetic peptide at a final concentration of0 �M for 30 min at 37 ◦C. One hundred microliters of sus-ensions containing responders or antigen presenting cellsere pipetted to each well. The plates were incubated in static
ondition for 24 h at 37 ◦C in an atmosphere containing 5%O2.
After incubation, the bulk of cultured cells was flicked off.o remove residual cells, plates were washed 3 times withBS and 3 times with PBS-Tween. Each well received 75 �lf biotinylated anti-mouse IFN-� MAb XMG1.2 (Pharmin-en) diluted in PBS-Tween to a final concentration of�g/ml. Plates were incubated overnight at 4 ◦C. Unboundntibodies were removed by washing the plates at least
times with PBS-Tween. Peroxidase-labeled streptavidinKPL) was added at a 1:800 dilution in PBS-Tween in anal volume of 100 �l/well. Plates were incubated for 1–2 ht rt and then washed three to five times with PBS-Tweennd three times with PBS. Plates were developed by adding00 �l/well of peroxidase substrate (50 mM Tris–HCl at pH.5 containing 1 mg/ml of DAB and 1 �l/ml of 30% hydrogeneroxide solution, both from Sigma). After incubation at rt
or 15 min, the reaction was stopped by discarding the sub-trate solution and rinsing the plates under running tap water.lates were dried at rt and spots were counted with the aid ofstereomicroscope (Nikon). Results of the ELISPOT assayre representative of two or more independent experiments.
wwoim
e 25 (2007) 6018–6027
.5. In vivo depletion of CD4+ T cells
Ascites fluids containing GK1.5 and 53.6.7 mAbs wereroduced in BALB/c nude mice and the concentration of RatgG was estimated as described [21]. At days 2 and 4 beforemmunization GST-P4–P7 mice were treated i.p. with a dosef 1 mg per day of Rat IgG or anti-CD4. This treatment wasepeated before the second immunizing dose. The efficacy ofepletion of CD4+ spleen cells was commonly higher than8%.
.6. Peptide synthesis
The peptide TEWETGQI (AA 320-327) was preparedy standard N�[9-fluorenylmethyloxycarbonyl] on a PSSM8ultispecific peptide synthesizer (Shimadzu, Kyoto, Japan)
y solid-phase synthesis with a scale of 30 �mol. Peptideas purified by HPLC in a Shimadzu system. Peptides were
nalyzed in a C18 Vydac column (10 mm × 250 mm, 5 (marticle diameter). Different peptide batches were obtainedn a range of 80–90% purity. Their identities were confirmedy Q-TOF MicroTM equipped with an electrospray ionizationource (Micromass, UK).
.7. Statistical analysis
The values of peak parasitemia of each individual mouseere log transformed before being compared by one-wayNOVA followed by Tukey HSD tests available at the
ite http://faculty.vassar.edu/lowry/VassarStats.html. Valuesf ELISPOT assay were also compared by the One Waynova followed by Tukey HSD tests. The LogRank test wassed to compare the mouse survival rate after challenge with. cruzi. The differences were considered significant whenhe P value was <0.05.
. Results
In our previous study we showed that immunization ofighly susceptible A/Sn mice with the recombinant proteinST-P4–P7 coupled to alum admixed with CpG ODN 1826rovided a high degree of protective immunity against aethal challenge with T. cruzi. This protective immunity was
ediated by CD8+ T cells specific for the epitope TEWET-QI located within AA 320 and 327 of ASP-2 (Ref. [15]
nd Fig. 1). To determine whether this protective immunityas long-lived, mice vaccinated with GST-P4–P7 coupled
o alum admixed with CpG ODN 1826 were challenged 5onths after the last immunization dose. Following infectionith bloodstream trypomastigotes, a significant reductionas observed in the peak parasitemia in vaccinated mice
hen compared to animals immunized with the adjuvantsnly (Fig. 2A and B, P < 0.01). Most relevant was the fact thatmmunization with GST-P4–P7 dramatically reduced mouseortality after challenge. While 100% of the control mice
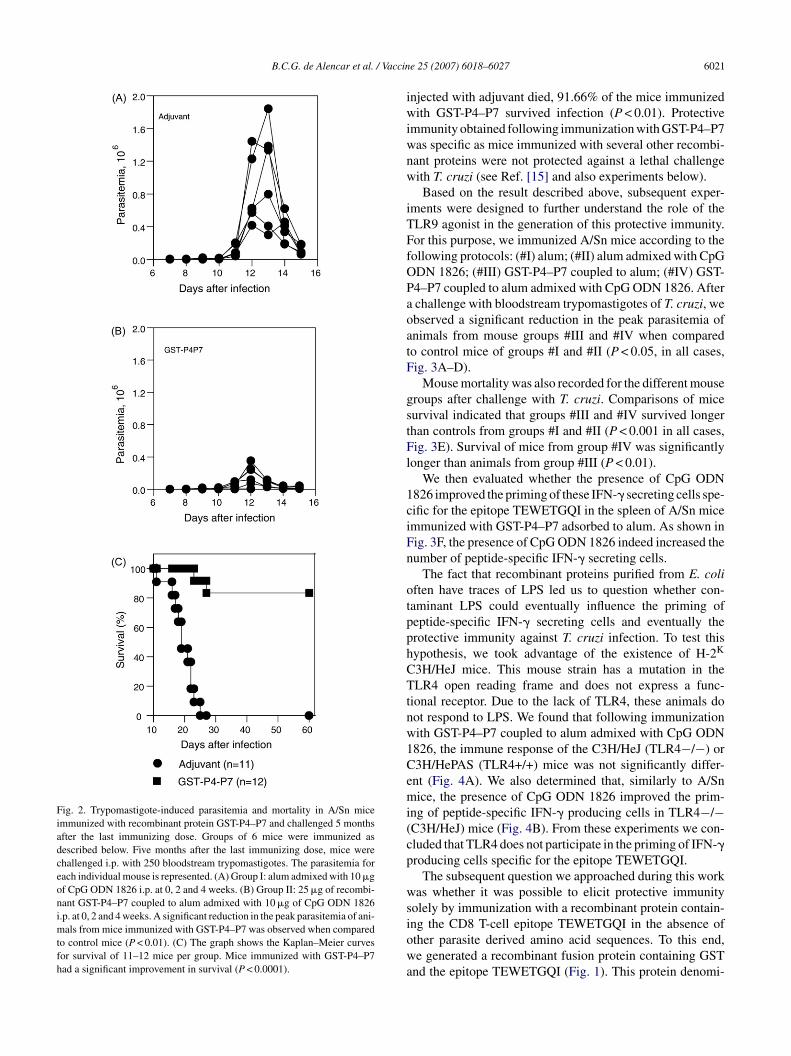
B.C.G. de Alencar et al. / Vaccin
Fig. 2. Trypomastigote-induced parasitemia and mortality in A/Sn miceimmunized with recombinant protein GST-P4–P7 and challenged 5 monthsafter the last immunizing dose. Groups of 6 mice were immunized asdescribed below. Five months after the last immunizing dose, mice werechallenged i.p. with 250 bloodstream trypomastigotes. The parasitemia foreach individual mouse is represented. (A) Group I: alum admixed with 10 �gof CpG ODN 1826 i.p. at 0, 2 and 4 weeks. (B) Group II: 25 �g of recombi-nant GST-P4–P7 coupled to alum admixed with 10 �g of CpG ODN 1826i.p. at 0, 2 and 4 weeks. A significant reduction in the peak parasitemia of ani-mals from mice immunized with GST-P4–P7 was observed when comparedto control mice (P < 0.01). (C) The graph shows the Kaplan–Meier curvesfor survival of 11–12 mice per group. Mice immunized with GST-P4–P7had a significant improvement in survival (P < 0.0001).
iwiwnw
iTFfOPaoatF
gstFl
1ciFn
otpphCTtnw1Cemi(cp
wsiowa
e 25 (2007) 6018–6027 6021
njected with adjuvant died, 91.66% of the mice immunizedith GST-P4–P7 survived infection (P < 0.01). Protective
mmunity obtained following immunization with GST-P4–P7as specific as mice immunized with several other recombi-ant proteins were not protected against a lethal challengeith T. cruzi (see Ref. [15] and also experiments below).Based on the result described above, subsequent exper-
ments were designed to further understand the role of theLR9 agonist in the generation of this protective immunity.or this purpose, we immunized A/Sn mice according to theollowing protocols: (#I) alum; (#II) alum admixed with CpGDN 1826; (#III) GST-P4–P7 coupled to alum; (#IV) GST-4–P7 coupled to alum admixed with CpG ODN 1826. Afterchallenge with bloodstream trypomastigotes of T. cruzi, webserved a significant reduction in the peak parasitemia ofnimals from mouse groups #III and #IV when comparedo control mice of groups #I and #II (P < 0.05, in all cases,ig. 3A–D).
Mouse mortality was also recorded for the different mouseroups after challenge with T. cruzi. Comparisons of miceurvival indicated that groups #III and #IV survived longerhan controls from groups #I and #II (P < 0.001 in all cases,ig. 3E). Survival of mice from group #IV was significantly
onger than animals from group #III (P < 0.01).We then evaluated whether the presence of CpG ODN
826 improved the priming of these IFN-� secreting cells spe-ific for the epitope TEWETGQI in the spleen of A/Sn micemmunized with GST-P4–P7 adsorbed to alum. As shown inig. 3F, the presence of CpG ODN 1826 indeed increased theumber of peptide-specific IFN-� secreting cells.
The fact that recombinant proteins purified from E. coliften have traces of LPS led us to question whether con-aminant LPS could eventually influence the priming ofeptide-specific IFN-� secreting cells and eventually therotective immunity against T. cruzi infection. To test thisypothesis, we took advantage of the existence of H-2K
3H/HeJ mice. This mouse strain has a mutation in theLR4 open reading frame and does not express a func-
ional receptor. Due to the lack of TLR4, these animals doot respond to LPS. We found that following immunizationith GST-P4–P7 coupled to alum admixed with CpG ODN826, the immune response of the C3H/HeJ (TLR4−/−) or3H/HePAS (TLR4+/+) mice was not significantly differ-nt (Fig. 4A). We also determined that, similarly to A/Snice, the presence of CpG ODN 1826 improved the prim-
ng of peptide-specific IFN-� producing cells in TLR4−/−C3H/HeJ) mice (Fig. 4B). From these experiments we con-luded that TLR4 does not participate in the priming of IFN-�roducing cells specific for the epitope TEWETGQI.
The subsequent question we approached during this workas whether it was possible to elicit protective immunity
olely by immunization with a recombinant protein contain-
ng the CD8 T-cell epitope TEWETGQI in the absence ofther parasite derived amino acid sequences. To this end,e generated a recombinant fusion protein containing GSTnd the epitope TEWETGQI (Fig. 1). This protein denomi-

6022 B.C.G. de Alencar et al. / Vaccine 25 (2007) 6018–6027
Fig. 3. Trypomastigote-induced parasitemia in A/Sn mice immunized withthe recombinant protein GST-P4–P7 coupled to alum in the presence orabsence of CpG ODN 1826. Groups of 6 mice were immunized i.p. at 0, 2and 4 weeks with 25 �g of GST-P4–P7 adsorbed to alum admixed or notwith 10 �g of CpG ODN 1826, or adjuvant alone. Two weeks after the lastdose, mice were challenged i.p. with 250 bloodstream trypomastigotes. Theparasitemia for each individual mouse is represented. (A) Group #I, alum; (B)group #II, alum admixed with CpG ODN 1826; (C) group #III, GST-P4–P7coupled to alum; (D) group #IV, GST-P4–P7 coupled to alum admixed withCpG ODN 1826. The peak parasitemia of animals from mouse groups #Iand #II were higher than mice of groups #III and #IV (P < 0.05, in all cases).(E) The graph shows the Kaplan–Meier curves for survival of 6–13 mice pergroup. Mice immunized with GST-P4–P7 (mouse groups #III and #IV) hada significant improvement in survival when compared to mice immunizedwith adjuvant only (P < 0.001 in all cases). Survival of animals immunizedwith GST-P4–P7 coupled to alum admixed with CpG ODN 1826 (mousegroup #IV) was significantly longer than mice from group #III (P < 0.01). (F)Groups of three mice were immunized i.p. at 0 and 2 weeks with GST-P4–P7or GST. Each mouse received i.p. 25 �g of recombinant protein adsorbedto alum admixed or not with 10 �g of CpG ODN 1826. One week after thelast dose, ELISPOT assay was performed to detect IFN-� producing cellsspecific for TEWETGQI. Results represent the mean of spot forming cells(SFC) ± S.D. of three mice per group. Asterisk denotes significantly fewerSFC (P < 0.01).
Fig. 4. Groups of three C3H/HePAS or C3H/HeJ mice were immunized i.p.at 0 and 2 weeks with GST-P4–P7 or GST. Each mouse received i.p. 25 �gof recombinant protein adsorbed to alum admixed or not with 10 �g of CpGODN 1826. One week after the last dose, ELISPOT assay was performed todmf
nCcsitBcclHd
etect IFN-� producing cells specific for TEWETGQI. Results represent theean of SFC ± S.D. of three mice per group. Asterisk denotes significantly
ewer SFC (P < 0.01).
ated GST-TEWETGQI was coupled to alum, admixed withpG ODN 1826 and used to vaccinate A/Sn mice. After ahallenge with bloodstream trypomastigotes of T. cruzi, noignificant reduction was observed in the peak parasitemian mice vaccinated with GST-TEWETGQI when comparedo control mice immunized with GST (P > 0.05, Fig. 5A and). Mortality in the two mouse groups after challenge with T.ruzi was only slightly different. Statistical comparisons indi-ated that mice immunized with GST-TEWETGQI survived
onger than mice which had received GST only (P = 0.003).owever, 100% of the GST-TEWETGQI immunized miceied following infection (Fig. 5E).
B.C.G. de Alencar et al. / Vaccin
Fig. 5. Trypomastigote-induced parasitemia and mortality in mice immu-nized with GST-TEWETGQI. Groups of 5 mice were immunized asdescribed in the legend of Fig. 2. Two weeks after the last immunizingdose, mice were challenged i.p. with 250 bloodstream trypomastigotes. Theparasitemia for each individual mouse is represented. (A) alum admixedwith CpG ODN 1826; (B) GST-TEWETGQI coupled to alum admixedwith CpG ODN 1826; (C) GST-P4–P7 coupled to alum admixed withCpG ODN 1826; (D) His6-P4–P7 coupled to alum admixed with CpGODN 1826. No significant reduction in the peak parasitemia of mice vac-cinated with GST-TEWETGQI was observed when compared to controlmice immunized with GST (P > 0.05). Peak parasitemia of mice vaccinatedwith GST-P4–P7 or His6-P4–P7 were lower than animals injected with GSTor GST-TEWETGQI (P < 0.01, in all cases). No difference was observedwhen we compared the peak parasitemia of animals vaccinated with GST-P4–P7 or His6-P4–P7 (P > 0.05). (E) The graph shows the Kaplan-Meiercurves for survival of 15 to 16 mice per group. Mice immunized with GST-TEWETGQI survived longer than mice which received GST (=0.003). Thesurvival of mice vaccinated with GST-P4–P7 or His6-P4–P7 was longer thananimals which received GST or GST-TEWETGQI (P < 0.0001, in all cases).The survival curves of mice immunized with His6-P4–P7 or GST-P4–P7were not statistically different (=0.0975). (F) Groups of three mice wereimmunized i.p. at 0 and 2 weeks with GST-P4–P7 or GST-TEWETGQI orGST. Each mouse received i.p. 25 �g of recombinant protein adsorbed toalum admixed with 10 �g of CpG ODN 1826. One week after the last dose,ELISPOT assay was performed to detect IFN-� producing cells specific forTEWETGQI. Results represent the mean of spot forming cells (SFC) ± S.D.of three mice per group.
TpHtdiGnGnpPGwind
GfbtopTtn
amcEiOfLnCssmiGpc[
sCadaasi
e 25 (2007) 6018–6027 6023
The immunogenicity of the recombinant protein GST-EWETGQI was compared to two other recombinantroteins, the conventional GST-P4–P7 and a newly producedis6-P4–P7. This last recombinant protein does not contain
he GST protein, therefore the putative CD4 epitopes areerived from the parasite antigen ASP-2 (Fig. 1). As shownn Fig. 5C and D, following challenge, mice vaccinated withST-P4–P7 or His6-P4–P7 presented a peak parasitemia sig-ificantly lower than that of animals injected with GST orST-TEWETGQI (P < 0.01, in all cases). No statistically sig-ificant difference was observed when comparing the peakarasitemia of animals vaccinated with GST-P4–P7 or His6-4–P7 (P > 0.05). Also the survival of mice vaccinated withST-P4–P7 or His6-P4–P7 was longer than that of animalshich had received GST or GST-TEWETGQI (P < 0.0001,
n all cases, Fig. 5E). The survival curves of mice immu-ized with His6-P4–P7 or GST-P4–P7 were not statisticallyifferent (=0.0975).
Subsequently, we determined whether immunization withST-TEWETGQI could elicit IFN-� secreting cells specific
or the peptide TEWETGQI. To our surprise, similar num-ers of peptide-specific IFN-� secreting cells were detected inhe spleen of A/Sn mice immunized with GST-TEWETGQIr GST-P4–P7 (Fig. 5F). These results indicated that theresence of IFN-� secreting cells specific for the epitopeEWETGQI detected by ELISPOT failed to correlate with
he limited protective immunity observed following immu-ization with the recombinant protein GST-TEWETGQI.
This unexpected finding raised the possibility that thebsence of CD4+ T cell epitopes present in the GST-P4–P7ight not be important in the expansion of IFN-� secreting
ells specific for the epitope TEWETGQI as detected by theLISPOT. Instead, these CD4+ T cells might be important
n the priming of specific memory/protective CD8+ T cells.ur hypothesis was based on recent observations obtained
rom experimental models of infections with influenza virus,isteria monocitogenes or Herpes simplex virus I and immu-ization with synthetic peptides. These studies reported thatD4+ T cells can be critically important in the generation of
pecific memory CD8+ T cells [22–26]. To evaluate this pos-ibility, we used an assay that we had recently developed toeasure the expansion of specific memory cells. This assay
s based on our published observation that immunization withST-P4–P7 coupled to alum admixed with CpG ODN 1826rimed memory peptide-specific IFN-� secreting cells thatan expand vigorously following a challenge with T. cruzi3].
As shown in Fig. 6A (white bars), immunization with aingle dose of GST-P4–P7 coupled to alum admixed withpG ODN 1826 generated an immune response that peakedt one week (185 ± 32.5 SFC per 106 spleen cells) andeclined in the subsequent 2 and 4 weeks (30.83 ± 12.3
nd 20.83 ± 20.21, respectively), as measured by ELISPOTssay. Thirteen days after challenge, the number of peptide-pecific IFN-� secreting cells in these immunized micencreased several times, to 1,150 ± 210, 1,510 ± 282.96 or
6024 B.C.G. de Alencar et al. / Vaccin
Fig. 6. Groups of three mice were immunized i.p. once with GST-P4–P7or GST. Each mouse received i.p. 25 �g of recombinant protein adsorbedto alum admixed with 10 �g of CpG ODN 1826. One, 2 or 4 weeks later,ELISPOT assay was performed to detect IFN-� producing cells specificfor TEWETGQI (open bars). Alternatively, mice were challenged with 250bloodstream trypomastigotes of T. cruzi. Thirteen days later, ELISPOT assaywas performed to detect IFN-� producing cells specific for TEWETGQI(hatched bars). Results represent the mean of SFC ± S.D. of three mice pergroup. Groups of three mice were immunized i.p. twice at 0 and 2 weeks withGST-P4–P7, GST-TEWETGQI or GST. Each mouse received i.p. 25 �g ofrecombinant protein adsorbed to alum admixed with 10 �g of CpG ODN1826. One week later, ELISPOT assay was performed to detect IFN-� pro-ducing cells specific for TEWETGQI (open bars). Alternatively, mice werechallenged with 250 bloodstream trypomastigotes of T. cruzi. Thirteen dayslater, ELISPOT assay was performed to detect IFN-� producing cells spe-cific for TEWETGQI (hatched bars). Treatment with anti-CD4 antibodieswSS
8wsmtcT
Fdn
(ocwTwltfItTtr�rsTosw(
paTAsoatIc
4
vCrti[nTt[stTb
Initially, we determined that protective immunity gen-
as performed as described in Section 2. Results represent the mean ofFC ± S.D. of three mice per group. Asterisks denote significantly fewerFC (P < 0.01).
70 ± 390.74, respectively (hatched bars). This Expansionas dependent on the immunization with GST-P4–P7, as
uch expansion of CD8+ T cells was not found in controlice immunized with GST. Based on these results, we believe
hat this assay reveals the presence of specific memory cellsapable of expanding vigorously following a challenge with. cruzi.
This assay allowed us to ask two related questions.
irstly, we tested whether these memory cells were inducedifferently following the immunization with the recombi-ant proteins GST-P4–P7 (protective) or GST-TEWETGQIeai
e 25 (2007) 6018–6027
non-protective). Secondly, we asked whether the primingf these memory cells was in fact dependent on CD4+ Tells. To approach the first question, we immunized miceith the recombinant proteins GST-P4–P7 (protective), GST-EWETGQI (non-protective) and GST (control). These miceere either challenged or not with T. cruzi. Thirteen days
ater, we estimated the number of peptide-specific T cells inhe spleen of the different mouse groups. As shown in Fig. 6B,ollowing T. cruzi infection, the number of peptide-specificFN-� producing cells in the spleen of mice immunized withhe recombinant proteins GST-P4–P7 (protective) or GST-EWETGQI (non-protective) expanded significantly more
han in mice immunized with GST (control). However, mostelevant was the fact that the number of peptide-specific IFN-producing cells in the spleen of mice immunized with the
ecombinant protein GST-P4–P7 was 3096 ± 1172, a figureeveral times higher than for mice immunized with GST-EWETGQI (1066.67 ± 676.56). It is important to pointut that mice immunized, but not challenged, presented aimilar number of peptide-specific IFN-� producing cellshether they had received GST-P4–P7 or GST-TEWETGQI
121.33 ± 20.50 and 140 ± 122.17, respectively).To determine the importance of CD4+ T cells during the
riming of memory CD8+ T cells, we treated A/Sn withnti-CD4 antibodies before immunization with GST-P4–P7.hese mice were subsequently challenged or not with T. cruzi.s shown in Fig. 6B, following infection, very few peptide-
pecific IFN-� producing cells were detected in the spleenf the vaccinated mice that had been treated with anti-CD4ntibodies (253.20 ± 192). This figure represents a reduc-ion of more than 10 times in the number of peptide-specificFN-� producing cells when compared to immunized andhallenged mice not treated with anti-CD4 antibodies.
. Discussion
A key question in the development of new or improvedaccines is how to induce protective memory T cells.ross-priming of CD8+ T cells following vaccination with
ecombinant proteins may represent an interesting strategyo achieve this goal. This type of immunization can be usedndependently or in protocols of heterologous prime-boosting27–32]. In the studies performed so far using recombi-ant proteins for cross- priming of protective CD8+ T cells,LR agonists were used as part of these vaccine formula-
ions. Among them, TLR9 agonists are of particular interest27–32]. Based on the relevance of this subject, the aim of ourtudy was to investigate certain aspects that could be impor-ant during the cross-priming of long-lived protective CD8+
cells following immunization with a recombinant proteinased on the T. cruzi antigen ASP-2.
rated by immunization with GST-P4–P7 coupled to alumdmixed with CpG ODN 1826 was long-lived, as detectedn mice vaccinated 5 months earlier (Fig. 2). This fact is

/ Vaccin
ioincp5
tt(i(trfEbvHw�crvC[ts
eappiTiCwa
Omaeantp[mplmrp
abitbmriabf
sihetAtInTGtcc(ocin
itovhooT
ipitpvlc
op
B.C.G. de Alencar et al.
mportant because it provides evidences that long-lived mem-ry CD8+ T cells against this human protozoan parasite canndeed be efficiently primed by immunization with recombi-ant proteins. The longevity of the protective immunity wasomparable to that obtained following immunization with theowerful replication-defective recombinant adenovirus type[17].Our second relevant observation was the fact that pro-
ective immunity could be elicited by vaccination withhe recombinant protein GST-P4–P7 coupled to alum onlyFig. 3). This formulation primed splenic IFN-� produc-ng cells specific for the CD8 T cell epitope TEWETGQIFig. 3F). This result was not completely expected as it con-rasts with a previous study showing that immunization withecombinant Hepatitis B antigen coupled to this adjuvantailed to prime specific CD8+ T cells as detected ex vivo byLISPOT assay [33]. The presence of contaminant LPS coulde a possible explanation for it. LPS could act as an adju-ant to prime these peptide-specific IFN-� producing cells.owever, using C3H/HeJ mice that do not respond to LPS,e were easily able to detect ex vivo peptide-specific IFN-producing cells following immunization with GST-P4–P7
oupled to alum (Fig. 4B). Based on this, we believe that theecombinant protein GST-P4–P7 itself may have some adju-ant property. As mentioned above, cross-priming of specificD8+ T cells using antigen coupled to alum is rarely seen
33], therefore the route of antigen processing and presenta-ion in vivo in our model is completely obscure at present andhould be an interesting topic to be studied in the future.
In spite of the fact that GST-P4–P7 coupled to alum couldlicit some degree of immunity, the presence of the TLR9gonist CpG ODN 1826 significantly improved the anti-arasitic effect of vaccination (Fig. 3). This improvement inrotective immunity correlated with an increase in the prim-ng of splenic IFN-� producing cells specific for the CD8-cell epitope TEWETGQI (Fig. 3E). Together, these resultsndicate that efficient cross-priming of long-lived protectiveD8+ T cells can be better achieved following vaccinationith this recombinant protein in the presence of the TLR9
gonist CpG ODN 1826.Although it has been noticed in different models that CpG
DN 1826 improves cross-priming of CD8+ T cells, the exactechanisms mediated by this adjuvant in vivo are unknown
t present. TLR-9 agonists stimulate a number of differ-nt events in dendritic cells (DC) which may be critical forntigen presentation. In vitro studies showed that a TLR9 ago-ist coupled to a soluble antigen (ovalbumin) improves theranslocation of the complex to endosomal–lysosomal com-artment of DCs, a factor that may aid cross-presentation34]. In addition, activation by TLR9 agonists leads toaturation of these cells that express increased levels of trans-
orter associated protein and MHC class I molecules. Higher
evel of transporter associated protein expression may aug-ent transport of CD8 T-cell epitopes to the endoplasmiceticulum, facilitating the formation of MHC I-peptide com-lexes presented to specific CD8+ T cells [35]. However,
i1ai
e 25 (2007) 6018–6027 6025
recent study suggests that the main mechanism mediatedy CpG ODN in vitro to improve cross-presentation is thencreased MHC I mRNA and surface protein expressionhrough a novel posttranscriptional mechanism involving sta-ilization of MHC I mRNA [36]. Whether these or otherechanisms operate in vivo to improve cross-presentation
emains to be elucidated. In addition to the mechanisms thatmprove antigen presentation by DCs, it has been shown thatTLR9 agonist can mediate cross-priming of CD8+ T cellsy B cells [37]. The participation of B cells may be an additiveactor to improve cross-priming of CD8+ T cells.
In addition to the direct consequences on the antigen pre-enting cells (DCs and/or B cells) TLR9 agonists may alsoncrease CD8+ T cell cross-priming by improving T cellelp. To investigate the role of parasite-derived CD4 T cellpitopes in the protective immunity generated by vaccina-ion with recombinant proteins based on the sequence of theSP-2 of T. cruzi, we generated a new recombinant pro-
ein containing solely the CD8 T cell epitope TEWETGQI.mmunization with GST-TEWETGQI elicited a comparableumber of splenic IFN-� producing cells specific for the CD8-cell epitope TEWETGQI as did the immunization withST-P4–P7 (Fig. 5D). In spite of that, very limited protec-
ive immunity was observed following a challenge with T.ruzi (Fig. 5B and E). In contrast, two recombinant proteinsontaining parasite-derived putative CD4+ T-cell epitopesGST-P4–P7 or His6-P4–P7) provided a significant degreef protective immunity (Fig. 5C–E). These results impli-ate parasite-derived epitopes other than TEWETGQI in thenduction of protective immunity observed following immu-ization with GST-P4–P7 or His6-P4–P7.
The lack of protective immunity observed followingmmunization with GST-TEWETGQI might be explained byhe absence of effector CD4+ T cells. However, in our previ-us study, we described that depletion of CD4+ T cells in miceaccinated with GST-P4–P7 prior to challenge with T. cruziad no impact on protective immunity. In contrast, depletionf CD8+ T cells completely reversed protection [15]. Basedn this, we consider remote the possibility that effector CD4+
cells might play a predominant role in protective immunity.The lack of protective immunity observed following
mmunization with GST-TEWETGQI correlated with a lowerriming of memory peptide-specific T cells detected follow-ng a parasite challenge (Fig. 6B). This result suggested thathe absence of CD4 T-cell epitopes could be important in theriming of memory/protective CD8+ T cells. By depleting inivo CD4+ T cells, we confirmed that this sub-population ofymphocytes was indeed critical in the priming of memoryells (Fig. 6B).
The importance of CD4+ T cells in the priming of mem-ry CD8+ T cells in our system corroborates several recentlyublished studies. Following mouse primary infection with
nfluenza virus, L. monocitogenes or Herpes simplex virus, memory CD8+ T cells failed to develop properly in thebsence of sufficient CD4+ T cell help [22–25]. Whethermmunization with other recombinant proteins and CpG
6 / Vaccin
OowoTcroivn
tfaaceboisu
A
AMo(sF
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
026 B.C.G. de Alencar et al.
DN 1826 requires appropriate CD4 help to expand mem-ry/protective CD8+ T cells is unknown at present. Also,hen these CD4+ T cells are important in providing help inur system remains to be investigated. For example, CD4+
cells specific for Herpes simplex virus 1 antigen act byonditioning antigen presenting cells early on in the immuneesponse, therefore providing an early help for proliferationf specific CD8+ T cells [25]. Differently, CD4+ T cellsnduced following L. monocitogenes infection improved sur-ival of memory specific CD8+ T cells, maintaining theirumber and function [37].
To sum up, we studied some aspects we considered impor-ant in the generation of long-lived protective CD8+ T cellsollowing immunization with a recombinant protein of ASP-2ntigen. We concluded that a TLR9 agonist and CD4+ T cellsre important in the development of these memory/protectiveells. It is possible that these studies on the immune responselicited by cross-priming following vaccination with recom-inant proteins of ASP-2 may contribute to the developmentf new strategies for the prevention or treatment of T. cruzinfection. However, due to the complex nature of this para-itic disease, we believe that it is too early to anticipate anyse of this or other strategy in humans.
cknowledgments
This work was supported by grants from Fundacao demparo a Pesquisa do Estado de Sao Paulo, CNPq, Theillennium Institute for Vaccine Development and Technol-
gy (CNPq, 420067/2005-1), and FAPEMIG (EDT 24.000)Brazil). AFSA, RTG and MMR were recipients of fellow-hips from CNPq. BCGA is a recipient of a fellowship fromAPESP.
eferences
[1] Wong P, Pamer EG. CD8 T cell responses to infectious pathogens. AnnuRev Immunol 2003;21:29–70.
[2] Robinson HL, Amara RR. T cell vaccines for microbial infections. NatMed 2005;11:S25–32.
[3] Tzelepis F, de Alencar BC, Penido ML, Gazzinelli RT, Persechini PM,Rodrigues MM. Distinct kinetics of effector CD8+ cytotoxic T cellsafter infection with Trypanosoma cruzi in naive or vaccinated mice.Infect Immun 2006;74:2477–82.
[4] Martin DL, Weatherly DB, Laucella SA, Cabinian MA, Crim MT,Sullivan S, et al. CD8+ T-Cell responses to Trypanosoma cruzi arehighly focused on strain-variant trans-sialidase epitopes. PLoS Pathog2006;2(8):e77.
[5] Tarleton RL, Koller BH, Latour A, Postan M. Susceptibility of beta 2-microglobulin-deficient mice to Trypanosoma cruzi infection. Nature1992;356:338–40.
[6] Rottenberg ME, Bakhiet M, Olsson T, Kristensson K, Mak T, Wigzell
H, et al. Differential susceptibilities of mice genomically deleted ofCD4 and CD8 to infections with Trypanosoma cruzi or Trypanosomabrucei. Infect Immun 1993;61:5129–33.[7] Tarleton RL, Sun J, Zhang L, Postan M, Glimcher L. Trypanosomacruzi infection in MHC-deficient mice: further evidence for the role of
[
[
e 25 (2007) 6018–6027
both class I- and class II-restricted T cells in immune resistance anddisease. Int Immunol 1996;8:13–22.
[8] Rodrigues MM, Boscardin SB, Vasconcelos JR, Hiyane MI, SalayG, Soares IS. Importance of CD8 T cell-mediated immune responseduring intracellular parasitic infections and its implications for thedevelopment of effective vaccines. Ann Acad Bras Cienc 2003;75:443–68.
[9] Martin D, Tarleton R. Generation, specificity, and function of CD8+ Tcells in Trypanosoma cruzi infection. Immunol Rev 2004;201:304–17.
10] Henriques-Pons A, Oliveira GM, Paiva MM, Correa AF, Batista MM,Bisaggio RC, et al. Evidence for a perforin-mediated mechanism con-trolling cardiac inflammation in Trypanosoma cruzi infection. Int J ExpPathol 2002;83:67–79.
11] Muller U, Sobek V, Balkow S, Holscher C, Mullbacher A, MuseteanuC, et al. Concerted action of perforin and granzymes is critical for theelimination of Trypanosoma cruzi from mouse tissues, but preventionof early host death is in addition dependent on the FasL/Fas pathway.Eur J Immunol 2003;33:70–8.
12] Garg N, Tarleton R. Genetic immunization elicits antigen-specificprotective immune responses and decreases disease severity in Try-panosoma cruzi infection. Infect Immun 2002;70:5547–55.
13] Katae M, Miyahira Y, Takeda K, Matsuda H, Yagita H, Okumura K,et al. Coadministration of an interleukin-12 gene and a Trypanosomacruzi gene improves vaccine efficacy. Infect Immun 2002;70:4833–40.
14] Vasconcelos JR, Hiyane MI, Marinho CRF, Claser C, Vieira-MachadoAM, Gazinelli RT, et al. Protective immunity against Trypanosomacruzi infection in a highly susceptible mouse strain following vac-cination with genes encoding the Amastigote Surface Protein-2 andtrans-sialidase. Hum Gene Ther 2004;15:878–86.
15] Araujo AF, de Alencar BC, Vasconcelos JR, Hiyane MI, Marinho CR,Penido ML, et al. CD8+-T-cell-dependent control of Trypanosoma cruziinfection in a highly susceptible mouse strain after immunization withrecombinant proteins based on amastigote surface protein 2. InfectImmun 2005;73:6017–25.
16] Miyahira Y, Takashima Y, Kobayashi S, Matsumoto Y, Takeuchi T,Ohyanagi-Hara M, et al. Immune responses against a dingle CD8+-T-Cell epitope induced by virus vector vaccination can successfullycontrol Trypanosoma cruzi infection. Infect Immun 2005;73:7356–65.
17] Machado AV, Cardoso JE, Claser C, Rodrigues MM, Gazzinelli RT,Bruna-Romero O. Long-term protective immunity induced againstTrypanosoma cruzi infection after vaccination with recombinant ade-noviruses encoding Amastigote Surface Protein-2 and trans-sialidase.Hum Gene Ther 2006;17:898–908.
18] Silva LHP, Nussenzweig V. Sobre uma cepa de Trypanosoma cruzialtamente virulenta para o camundongo branco. Folia Clin Biol1953;20:191–203.
19] Krettli AU, Brener Z. Protective effects of specific antibodies in Try-panosoma cruzi infections. J Immunol 1976;116:755–60.
20] Boscardin SB, Kinoshita SS, Fujimura AE, Rodrigues MM. Immu-nization with cDNA expressed by amastigotes of Trypanosoma cruzielicits protective immune response against experimental infection.Infect Immun 2003;71:2744–57.
21] Li S, Rodrigues M, Rodriguez D, Rodriguez JR, Esteban M, PaleseP, et al. Priming with recombinant influenza virus followed byadministration of recombinant vaccinia virus induces CD8+ T-cell-mediated protective immunity against malaria. Proc Natl Acad Sci USA1993;90:5214–8.
22] Belz GT, Wodarz D, Diaz G, Nowak MA, Doherty PC. Compromisedinfluenza virus-specific CD8+-T-cell memory in CD4+-T-cell-deficientmice. J Virol 2002;76:12388–93.
23] Sun JC, Bevan MJ. Defective CD8 T cell memory following acuteinfection without CD4 T cell help. Science 2003;300:339–42.
24] Shedlock DJ, Shen H. Requirement for CD4 T cell help in generatingfunctional CD8 T cell memory. Science 2003;300:337–9.
25] Smith CM, Wilson NS, Waithman J, Villadangos JA, Carbone FR,Heath WR, et al. Cognate CD4+ T cell licensing of dendritic cellsin CD8+ T cell immunity. Nat Immunol 2004;5:1143–8.

/ Vaccin
[
[
[
[
[
[
[
[
[
[
[
B.C.G. de Alencar et al.
26] Kumaraguru U, Suvas S, Biswas PS, Azkur AK, Rouse BT. Concomi-tant helper response rescues otherwise low avidity CD8+ memory CTLsto become efficient effectors in vivo. J Immunol 2004;172:3719–24.
27] Heit A, Schmitz F, O’Keeffe M, Staib C, Busch DH, Wagner H,et al. Protective CD8 T cell immunity triggered by CpG-proteinconjugates competes with the efficacy of live vaccines. J Immunol2005;174:4373–80.
28] Wille-Reece U, Wu CY, Flynn BJ, Kedl RM, Seder RA. Immunizationwith HIV-1 Gag protein conjugated to a TLR7/8 agonist results in thegeneration of HIV-1 Gag-specific Th1 and CD8+ T cell responses. JImmunol 2005;174:7676–83.
29] Wille-Reece U, Flynn BJ, Lore K, Koup RA, Kedl RM, Mattapallil JJ,et al. HIV Gag protein conjugated to a Toll-like receptor 7/8 agonistimproves the magnitude and quality of Th1 and CD8+ T cell responsesin nonhuman primates. Proc Natl Acad Sci USA 2005;102:15190–4.
30] Sugauchi F, Wang RY, Qiu Q, Jin B, Alter HJ, Shih JW. Vig-orous hepatitis C virus-specific CD4+ and CD8+ T cell responsesinduced by protein immunization in the presence of Montanide ISA720
plus synthetic oligodeoxynucleotides containing immunostimulatorycytosine-guanine dinucleotide motifs. J Infect Dis 2006;193:563–72.31] Tritel M, Stoddard AM, Flynn BJ, Darrah PA, Wu CY, Wille U,et al. Prime-boost vaccination with HIV-1 Gag protein and cytosinephosphate guanosine oligodeoxynucleotide, followed by adenovirus,
[
e 25 (2007) 6018–6027 6027
induces sustained and robust humoral and cellular immune responses.J Immunol 2003;171:2538–47.
32] Wille-Reece U, Flynn BJ, Lore K, Koup RA, Miles AP, Saul A, etal. Toll-like receptor agonists influence the magnitude and quality ofmemory T cell responses after prime-boost immunization in nonhumanprimates. J Exp Med 2006;203:1249–58.
33] Hutchings CL, Gilbert SC, Hill AV, Moore AC. Novel protein andpoxvirus-based vaccine combinations for simultaneous induction ofhumoral and cell-mediated immunity. J Immunol 2005;175:599–606.
34] Heit A, Huster KM, Schmitz F, Schiemann M, Busch DH, Wagner H.CpG-DNA aided cross-priming by cross-presenting B cells. J Immunol2004;172:1501–7.
35] Cho HJ, Hayashi T, Datta SK, Takabayashi K, Van Uden JH, Horner A,et al. IFN-�� promotes priming of antigen-specific CD8+ and CD4+ Tlymphocytes by immunostimulatory DNA-based vaccines. J Immunol2002;168:4907–13.
36] Kuchtey J, Chefalo PJ, Gray RC, Ramachandra L, Harding CV.Enhancement of dendritic cell antigen cross-presentation by CpG DNA
involves type I IFN and stabilization of class I MHC mRNA. J Immunol2005;175:2244–51.37] Sun JC, Williams MA, Bevan MJ. CD4+ T cells are required for themaintenance, not programming, of memory CD8+ T cells after acuteinfection. Nat Immunol 2004;5:927–33.

67
Artigo 3

68
Perforin and gamma interferon expression are required for CD4+ and CD8+
T-cell-dependent protective immunity against a human parasite,
Trypanosoma cruzi, elicited by heterologous plasmid DNA prime
recombinant adenovirus 5 boost vaccination.
O protocolo de imunização envolvendo um DNA plasmidial ou uma
proteína recombinante baseada na ASP-2 exigia, no mínimo, três doses para
induzir um grau significativo de proteção contra o desafio pelo T. cruzi nos
camundongos A/Sn. Em vista disto, levantou-se a hipótese de que se poderia
induzir uma imunidade protetora com um protocolo mais curto (apenas duas
doses), utilizando imunizações homólogas ou heterólogas com um adenovírus
recombinante expressando a ASP-2 (AdASP-2).
Observou-se que, tanto um protocolo homólogo com AdASP-2, quanto
um protocolo heterólogo com pIgSPCl.9 seguido de AdASP-2, foram capazes
de induzir altos níveis de imunidade protetora contra a infecção pelo T. cruzi
nos camundongos A/Sn. Os estudos utilizando o protocolo heterólogo
(pIgSPCl.9/AdASP-2), que protege 80-100% dos camundongos A/Sn,
mostraram que a proteção era de longa duração e dependente de linfócitos T
CD4 e T CD8. Além disto, a imunidade protetora se correlacionou com a
presença in vivo da atividade citotóxica específica antes do desafio.
Com base nesta observação, analisou-se a imunidade protetora induzida
pelo protocolo vacinal heterólogo em camundongos deficientes da perforina,
um importante mediador da citotoxicidade. Os camundongos perforina KO
imunizados se mostraram suscetíveis à infecção pelo T. cruzi. A análise da
resposta imune celular desses animais mostrou que os seus esplenócitos
apresentavam uma menor produção de IFN-γ após o re-estímulo in vitro com a
proteína recombinante. Além disto, os seus linfócitos T CD8 apresentavam uma
expansão numérica normal, porém baixa frequência de células multifuncionais
(expressando simultaneamente IFN-γ/TNF-α ou IFN-γ/CD107a), baixa
citotoxicidade e baixa expressão de CD44 e de KLRG-1. Para se determinar a
importância relativa do IFN-γ e da citotoxicidade, camundongos IFN-γ KO foram
vacinados, tendo desenvolvido células T CD8 citotóxicas, mas não mostraram
qualquer imunidade protetora.
Assim, nos camundongos A/Sn, foi possível induzir uma imunidade
protetora contra o T. cruzi com um protocolo heterólogo envolvendo DNA
plasmidial e adenovírus recombinante (apenas duas doses), e demonstrar um
papel para a perforina e o IFN-γ nas diversas funções efetoras mediadas pelas
células T, na imunidade antiparasitária gerada por este protocolo vacinal.

INFECTION AND IMMUNITY, Oct. 2009, p. 4383–4395 Vol. 77, No. 100019-9567/09/$08.00�0 doi:10.1128/IAI.01459-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Perforin and Gamma Interferon Expression Are Required for CD4�
and CD8� T-Cell-Dependent Protective Immunity against a HumanParasite, Trypanosoma cruzi, Elicited by Heterologous
Plasmid DNA Prime-Recombinant Adenovirus 5Boost Vaccination�
Bruna C. G. de Alencar,1,2 Pedro M. Persechini,3 Filipe A. Haolla,1,2 Gabriel de Oliveira,4Jaline C. Silverio,4 Joseli Lannes-Vieira,4 Alexandre V. Machado,5 Ricardo T. Gazzinelli,5,6,7
Oscar Bruna-Romero,8 and Mauricio M. Rodrigues1,2*Centro Interdisciplinar de Terapia Genica (CINTERGEN)1 and Departmento de Microbiologia, Imunologia e Parasitologia,2
Universidade Federal de Sao Paulo-Escola Paulista de Medicina, Rua Mirassol 207, Sao Paulo, SP 04044-010, Brazil; Instituto deBiofísica Carlos Chagas Filho, Centro de Ciencias da Saude, Bloco G, Universidade Federal do Rio de Janeiro, Ilha do Fundao,
Rio de Janeiro, RJ 21941-900, Brazil3; Lab. Biologia Celular e Biologia das Interacoes, Instituto Oswaldo Cruz, FIOCRUZ,Av. Brasil no. 4365, Rio de Janeiro, RJ 21045-900, Brazil4; Departamento de Bioquímica e Imunologia, Instituto deCiencias Biologicas, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, Pampulha, Belo Horizonte,MG 31270-901, Brazil5; Centro de Pesquisas Rene Rachou, FIOCRUZ, Av. Augusto de Lima 1715, Barro Preto,Belo Horizonte, Minas Gerais 30190-002, Brazil6; Division of Infectious Disease and Immunology, Department of
Medicine, University of Massachusetts Medical School, Worcester, Massachusetts 016057; and Departamento deMicrobiologia, Instituto de Ciencias Biologicas, Universidade Federal de Minas Gerais,
Av. Antonio Carlos 6627, Pampulha, Belo Horizonte, MG 31270-901, Brazil8
Received 30 November 2008/Returned for modification 13 January 2009/Accepted 14 July 2009
A heterologous prime-boost strategy using plasmid DNA, followed by replication-defective recombinantadenovirus 5, is being proposed as a powerful way to elicit CD4� and CD8� T-cell-mediated protectiveimmunity against intracellular pathogens. We confirmed this concept and furthered existing research byproviding evidence that the heterologous prime-boost regimen using the gene encoding amastigote surfaceprotein 2 elicited CD4� and CD8� T-cell-mediated protective immunity (reduction of acute parasitemia andprolonged survival) against experimental infection with Trypanosoma cruzi. Protective immunity correlatedwith the presence of in vivo antigen-specific cytotoxic activity prior to challenge. Based on this, our second goalwas to determine the outcome of infection after heterologous prime-boost immunization of perforin-deficientmice. These mice were highly susceptible to infection. A detailed analysis of the cell-mediated immuneresponses in immunized perforin-deficient mice showed an impaired gamma interferon (IFN-�) secretion byimmune spleen cells upon restimulation in vitro with soluble recombinant antigen. In spite of a normalnumeric expansion, specific CD8� T cells presented several functional defects detected in vivo (cytotoxicity)and in vitro (simultaneous expression of CD107a/IFN-� or IFN-�/tumor necrosis factor alpha) paralleled bya decreased expression of CD44 and KLRG-1. Our final goal was to determine the importance of IFN-� in thepresence of highly cytotoxic T cells. Vaccinated IFN-�-deficient mice developed highly cytotoxic cells but failedto develop any protective immunity. Our study thus demonstrated a role for perforin and IFN-� in a numberof T-cell-mediated effector functions and in the antiparasitic immunity generated by a heterologous plasmidDNA prime-adenovirus boost vaccination strategy.
Genetic vaccination using naked DNA or recombinant vi-ruses is being pursued as alternative vaccines. This strategy canbe particularly important in the case of intracellular pathogensand neoplasic cells when the effectiveness relies heavily on thevaccination’s capacity to elicit specific immune responses me-diated by cytotoxic CD8� T cells (reviewed in references 27,48, and 49). One of the most prolific areas of genetic vaccina-tion development is the strategy known as the heterologous
prime-boost regimen. This consists in the use of two differentvectors both carrying a gene that encodes the same antigenicprotein for priming and boosting immunizations. This strategywas first proposed in the early 1990s using a combination ofrecombinant viruses (influenza and vaccinia viruses) to induceprotective immunity against malaria (34, 50). Subsequently,this approach was extended and simplified by incorporatingnaked DNA priming, followed by a booster injection of arecombinant poxviral vector (i.e., modified vaccinia Ankara);this was also used to generate sterile protective immunity inrodent malaria (53, 54). Collectively, these studies demon-strated that the heterologous prime-boost regimen provedmore effective than the repeated use of any of these vectorsindividually. In subsequent years, the heterologous plasmid
* Corresponding author. Mailing address: CINTERGEN–UNIFESP–Escola Paulista de Medicina, Rua Mirassol 207, Sao Paulo, SP 04044-010, Brazil. Phone and fax: (55) (11) 5571-1095. E-mail: [email protected].
� Published ahead of print on 3 August 2009.
4383
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

DNA prime-poxvirus vector boost regimen was adopted world-wide as a powerful mean to elicit strong effector CD8� Tc1-mediated immune responses against viral, parasitic, and neo-plastic antigens in rodents and nonhuman primates (4;reviewed in references 29, 44, and 65). Based on the preclinicalstudies, a number of clinical human trials have also been ini-tiated. However, to our knowledge, heterologous prime-boostregimens using plasmid DNA and recombinant poxviruseshave not yet provided meaningful protective immunity to hu-mans (21, 28, 42, 45).
Although there are a number of possible vector combina-tions that significantly improve cell-mediated immunity, par-ticularly of specific CD8� T cells, heterologous prime-boostvaccination using naked plasmid DNA for priming, followed bya booster injection of recombinant replication-deficient ade-novirus 5 has recently received significant attention. This strat-egy has proved successful in some relevant experimental mod-els such as simian immunodeficiency virus and malaria,providing considerable protective immunity (2, 13, 14, 26, 33,41, 62).
The fact that protective CD8� T cells could be induced inmice and nonhuman primates against both virus and parasitesmade it an interesting strategy against other microbial infec-tions. We initially attempted to build on this strategy by gen-erating protective CD4 Th1 and cytotoxic CD8� T cells againstthe infection with a human intracellular protozoan parasite,Trypanosoma cruzi. Both CD4� and CD8� T cells were de-scribed as critical for acquired immunity against experimentalinfection with T. cruzi (38, 39, 56–59). Vaccination studiesconfirmed these observations by providing evidence that CD8�
T cells are particularly important to the development of pro-tective immunity in mice immunized with recombinant plasmidDNA, proteins, or viruses (6, 19, 25, 31, 37, 43, 60). In additionto CD8� T cells, CD4� Th1 cells also play a role in immunityagainst T. cruzi after vaccination with plasmid DNA or recom-binant protein (19, 30, 60).
In our previous studies, we showed that multiple immuniza-tions of highly susceptible A/Sn mice with a gene or a recom-binant protein based on the amastigote surface protein 2(ASP-2) of T. cruzi generated protective immunity against alethal challenge with this parasite (6, 17, 19, 55, 60). In bothcases, vaccinated animals depleted of CD8� T cells prior tochallenge became highly susceptible to infection (6, 60). Pro-tective CD8� T cells were directed to the immunodominantepitope TEWETGQI located within amino acids 320 to 327 ofASP-2 (6, 19).
The present study was designed first to evaluate whether wecould reduce the number of immunizing doses required togenerate CD4� and CD8� T-cell-mediated protective immu-nity using a heterologous prime-boost regimen. A reduction inthe number of doses may greatly improve the feasibility ofhuman vaccination trials. The fact that mice immunized withrecombinant AdASP-2 showed the highest levels of in vivocytotoxicity mediated by CD8� T cells prior to challenge, andsome degree of protective immunity led us to study the role forperforin during vaccination. Subsequently, we determinedwhether the high susceptibility observed in perforin-deficientmice correlated with impaired effector T-cell immune re-sponses and the expression of relevant activation markers byspecific CD8� T cells. Our final goal was to determine the
importance of gamma interferon (IFN-�), a critical mediatorof adaptive immunity against T. cruzi infection, for protectiveimmunity in the presence of highly cytotoxic T cells.
MATERIALS AND METHODS
Mice and parasites. Female 5- to 8-week-old A/Sn, C57BL/6 wild-type (WT),C57BL/6 perforin-deficient (perforin KO), C57BL/6 CD4 deficient (CD4 KO),C57BL/6 CD8a-deficient (CD8 KO), or C57BL/6 IFN-�-deficient (IFN-� KO)mice were purchased from University of Sao Paulo, raised at the Instituto deBiofísica Carlos Chagas Filho or at the animal facilities of the Oswaldo CruzFoundation. Parasites of the Y strain of T. cruzi were used here (10). Blood-stream trypomastigotes were obtained from mice infected 7 days earlier. Theconcentration of parasites was estimated and adjusted to 750 parasites/ml. EachA/Sn mouse was inoculated intraperitoneally (i.p.) with 0.2 ml. In the case ofC57BL/6 or perforin KO mice, the concentration of parasites was estimated andadjusted to 5 � 104/ml, and 0.2 ml was injected i.p. (104 trypomastigotes/mouse).Parasite development was monitored by counting the number of bloodstreamtrypomastigotes in 5 �l of fresh blood collected from the tail vein (10). Hemocul-tures were performed with aliquots of 0.1 ml of blood collected and cultured at28°C for 1 month in 5 ml of axenic liver infusion tryptose medium. Parasitegrowth was monitored microscopically every week. For PCR, DNA was extractedfrom 0.1 ml of blood and resuspended in a final volume of 100 �l. As controls,known numbers of T. cruzi Y strain blood trypomastigotes were diluted in 0.1 mlof blood obtained from naive mice (200, 100, 50, 25, 12.5, or 0 trypomastigotes/ml). Then, 5-�l portions were used for PCRs with Platinum Taq DNA polymer-ase (Invitrogen) and the T. cruzi kDNA primers S35 (5�-AAATAATGTACGGGGGAGATGCATGA-3�) and S36 (5�-GGGTTCGATTGGGGTTGGTGT-3�).The amplification cycles consisted of an initial denaturation of 5 min at 94°C,followed by 35 cycles of 30 s at 95°C (denaturation), 30 s at 62°C (primerannealing), and 1 min at 72°C (primer extension). We observed a 330-bp band insamples containing as few as 12.5 trypomastigotes/ml, but not in parasite-freeblood DNA. Mouse survival was monitored daily. The use of animals and theexperimental procedures were approved by the Ethics Committee for AnimalCare of the Federal University of Sao Paulo.
Peptide synthesis. Peptides VNHRFTLV and TEWETGQI were prepared bystandard N�[9-fluorenylmethyloxycarbonyl] on a PSSM8 multispecific peptidesynthesizer (Shimadzu, Kyoto, Japan) by solid-phase synthesis with a scale of 30�mol. Peptide was purified by high-pressure liquid chromatography in a Shi-madzu system. Peptides were analyzed in a C18 Vydac column (10 by 250 mm,5-�m particle diameter). The peptide purity was in a range of 80 to 90% purity.Their identities were confirmed by Q-TOF Micro equipped with an electrosprayionization source (Micromass, United Kingdom). Pentamer H-2Kb/VNHRFTLVwas purchased from ProImmune, Inc. (Oxford, United Kingdom).
Recombinant plasmids and adenoviruses used for immunization. PlasmidpIgSPclone9 was obtained by inserting into the commercial vector pcDNA3(Invitrogen) the sequence encoding the signal peptide of mouse immunoglobulinkappa chain in-frame with the gene encoding T. cruzi ASP-2 (10). pAdCMV-ASP2 is an adenoviral transfer plasmid that contains a eukaryotic expressioncassette formed by the cytomegalovirus immediate-early promoter and the sim-ian virus 40 RNA polyadenylation sequences. Inside this cassette we cloned theDNA sequences encoding T. cruzi ASP-2 obtained by restriction digestion ofplasmid pIgSPclone9 (AdASP-2 [37]). Viruses and plasmids were purified asdescribed earlier, and they both lead to expression of the antigen as evaluated byin vitro transfections (10, 37). Mice were inoculated intramuscularly (i.m.) ineach tibialis anterioris muscle with 50 �g of plasmid DNA. Twenty-one dayslater, these mice received in these same spots 50 �l of viral suspension containing108 PFU of adenovirus. Immunological assays were performed 14 days after viralinoculation.
In vivo depletion of CD4� T cells were performed by treating vaccinated A/Snor C57BL/6 mice with monoclonal antibody (MAb) GK1.5. At days 6, 4, and 2before challenge with trypomastigotes, mice were injected i.p. with a dose of 0.5mg of anti-CD4 or control rat immunoglobulin G (IgG). At 7 and 14 dayspostchallenge, each mouse received one more dose of 0.5 mg of anti-CD4 or ratIgG. As determined by fluorescence-activated cell sorting (FACS) analyses, theefficacy of depletion of CD4� spleen cells before challenge was of 99.94% inanti-CD4 treated mice compared to rat IgG-treated ones. In vivo depletion ofCD8� T cells were performed by treating vaccinated A/Sn mice with MAb 53.6.7.At days 3 and 4 before challenge with trypomastigotes, mice were injected i.p.with a dose of 1 mg of anti-CD8 or control rat IgG. At 7 days after challenge,each mouse received one more dose of 1 mg of anti-CD8 or rat IgG. The efficacy
4384 DE ALENCAR ET AL. INFECT. IMMUN.
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

of depletion of CD8� spleen cells before challenge was more than 96% inanti-CD8-treated mice compared to rat IgG-treated ones.
Immunological assays. For the in vivo cytotoxicity assays, splenocytes collectedfrom naive A/Sn or C57BL/6 mice were treated with ACK buffer (0.15 M NH4Cl,10 mM KHCO3, 0.1 mM disodium EDTA [pH 7.4]) to lyse the red blood cells.These cells were divided into two populations and labeled with the fluorogenicdye carboxyfluorescein diacetate succinimidyl ester (CFSE; Molecular Probes,Eugene, OR) at a final concentration of 5 �M (CFSEhigh) or 0.5 �M (CFSElow).CFSEhigh cells were coated for 40 min at 37°C with a 1 or 2.5 �M concentrationof the H-2Kb or H-2Kk ASP-2 peptides VNHRFTLV or TEWETGQI, respec-tively. CFSElow cells remained uncoated. Then, CFSEhigh cells were washed andmixed with equal numbers of CFSElow cells before 3 � 107 to 4 � 107 total cellsper mouse were injected intravenously. Recipient animals were mice previouslyimmunized with recombinant plasmids or adenoviruses or both. Spleen cells ofrecipient mice were collected 4 h or 20 h after transfer, as indicated in the figurelegends, fixed with 1.0% paraformaldehyde, and analyzed by FACS using aFacsCanto flow cytometer (BD, Mountain View, CA). The percent specific lysiswas determined by using the formula: [1 � (% CFSEhigh immunized/% CFSElow
immunized)/(% CFSEhigh naive/% CFSElow naive)] � 100. IFN-� secretion bycultured spleen cells and enzyme-linked immunospot (ELISPOT) assay for enu-meration of IFN-� producing cells was performed essentially as described pre-viously (10, 50).
For flow cytometry analyses, we used mouse splenocytes treated with ACKbuffer. Single-cell suspensions were washed in phosphate-buffered saline (PBS),stained for 10 min at room temperature with biotinylated major histocompati-bility complex class I (MHC-I) multimer H-Kb/VNHRFTLV, and then stained30 min at 4°C with avidin-phycoerythrin (PE)- and allophycocyanin (APC)-labeled anti-CD8 antibodies (both from BD Pharmingen). For the analyses ofother cell surface markers, single cell suspension from spleens of mice weredepleted of erythrocytes, and CD8� T cells were purified by negative selectionusing a MACS separation system (Miltenyi Biotec, Bergisch Gladbach, Ger-many) according to the manufacturer’s protocols with magnetic LS columns. Asestimated by FACS analysis, �85% of the cells were CD8� T cells after enrich-ment. These cells were stained at room temperature with biotinylated MHC-Imultimer H-Kb/VNHRFTLV and then stained 20 min at room temperature withavidin-PE- and APC-labeled anti-CD8 antibodies, as well as with fluoresceinisothiocyanate (FITC)-labeled anti-CD11a, anti-CD43, anti-CD44, anti-CD62L,anti-CD127, and anti-KLRG-1 antibodies (all from BD Pharmingen). At least100,000 cells were acquired on a FacsCanto flow cytometer and then analyzedwith FlowJo (Tree Star) using a biexponential transform.
The expression of CD107a was evaluated after in vitro culture of splenocytesin the presence or absence of antigenic stimulus. Cells were washed three timesin plain RPMI and resuspended in cell culture medium consisting of RPMI 1640medium (pH 7.4) supplemented with 10 mM HEPES, 0.2% sodium bicarbonate,59 mg of penicillin/liter, 133 mg of streptomycin/liter, and 10% HyClone fetalbovine serum (HyClone, Logan, UT). The viability of the cells was evaluated byusing 0.2% trypan blue exclusion dye to discriminate between live and dead cells.The cell concentration was adjusted to 5 � 106 cells/ml in cell culture mediumcontaining also anti-CD28 (2 �g/ml), brefeldin A (10 �g/ml), monensin (5 �g/ml), and the FITC-labeled anti-CD107a (2 �g/ml, all from BD Pharmingen). Inhalf of the cultures, a final concentration of 10 �M VNHRFTLV peptide wasadded. The cells were cultivated in flat-bottom 96-well plates (Corning) in a finalvolume of 200 �l in duplicate at 37°C in a humid environment. After 12 h ofincubation, cells were stained for surface marker with anti-CD8 on ice for 20 min.At least 100,000 cells were acquired on a FacsCanto flow cytometer and thenanalyzed with FlowJo.
To detect IFN-�, tumor necrosis factor alpha (TNF-�), and interleukin-2(IL-2) by intracellular staining (ICS), in vitro cultures of splenocytes were pre-pared as described above; however, the culture medium used contained neithermonensin nor anti-CD107a. After 12 h of incubation, cells were stained for asurface marker with anti-CD8 for 20 min on ice. The cells were then washedtwice in buffer containing PBS, 0.5% bovine serum albumin, and 2 mM EDTA;fixed in 4% PBS-paraformaldehyde solution for 10 min; and permeabilized for 15min in a PBS–0.1% bovine serum albumin–0.1% saponin solution. After beingwashed twice, cells were stained for intracellular markers using APC-Cy7 anti-CD3, PE-Cy7 anti-IFN-�, Alexa 488-anti-TNF-�, and APC-anti-IL-2 for 20 minon ice. Finally, the cells were washed twice and fixed in 1% PBS-paraformalde-hyde. At least 300,000 cells were acquired on a FacsCanto flow cytometer andthen analyzed with FlowJo.
Statistical analysis. The values of parasitemia of each individual mouse werelog transformed before being compared by one-way analysis of variance, fol-lowed by Tukey HSD tests (available at http://faculty.vassar.edu/lowry/VassarStats.html). The in vivo ELISPOT cytotoxicity assay results, the numbers
of multimer-positive cells, and the percentages of cytokine- or CD107a-express-ing cells are expressed as medians (bars) and individual sample values (dots) inthe figures and were compared by using the nonparametric Mann-Whitney orKruskall-Wallis tests. The log-rank test was used to compare mouse survival rateafter challenge with T. cruzi. The differences were considered significant whenthe P value was 0.05.
RESULTS
Protective immunity of highly susceptible A/Sn mice aftervaccination with replication-defective recombinant adenovirustype 5 expressing T. cruzi ASP-2. In earlier studies we observedthat protective immunity, as measured by reduction in the peakparasitemia and delayed mouse mortality, required three dosesof plasmid DNA or recombinant protein (6, 17, 19). To deter-mine whether protective immunity could be elicited by twoimmunizing doses, we compared different protocols of vacci-nation using either homologous or heterologous prime-boostingregimens. The homologous protocols consisted of two immuniz-ing doses of either plasmid DNA (pIgSPCl.9) or a recombinantreplication-defective adenovirus type 5 (AdASP-2) containing thegene encoding ASP-2 of T. cruzi (6, 19, 60). The heterologousimmunization regimen consisted of a priming immunization withpIgSPCl.9 followed by a booster injection of the AdASP-2. Si-multaneously, a group of mice was injected with a single immu-nizing dose of AdASP-2.
Immunized mice were challenged with a lethal dose of T.cruzi bloodstream trypomastigotes after a short (14-day) or along (98-day) period after the final immunizing dose. As shownin Fig. 1B, vaccination with two doses of pIgSPCl.9 failed toreduce the magnitude of the peak parasitemia (day 11) com-pared to control mice injected with pcDNA3 followed by re-combinant Ad-gal. Mice immunized with one or two doses ofAdASP-2 displayed a peak parasitemia 3.34- or 13.61-foldlower than that of control mice (n � 6, P 0.01 in both cases).The best reduction of the peak parasitemia was observed in thegroup of mice vaccinated with the heterologous DNA prime-recombinant adenovirus boost regimen (22.88-fold reductioncompared to controls, n � 6, P 0.01).
When mouse survival was tracked after challenge (as shownon Fig. 1C), the results from the parasitemia were mainlyconfirmed. All three mouse groups immunized with AdASP-2survived longer than control animals or mice immunized withtwo doses of pIgSPCl.9. However, mouse survival differed sig-nificantly among groups of mice immunized with two doses ofAdASP-2 compared to animals injected only once (P 0.001).Protective immunity in both groups of mice boosted with therecombinant AdASP-2 was high. No significant difference wasobserved among the mouse groups primed with pIgSPCl.9 orAdASP-2 (P � 0.05). Based on these results, we selected theheterologous prime-boost vaccination regimen to further re-search the protective cells.
Protective immunity elicited by the heterologous prime-boost regimen (pIgSPCl.9/AdASP-2) was stable at least until98 days after the final immunizing dose. In ASP-2-immunizedmice challenged 14 or 98 days after the final boost vaccination,the values of parasitemia were 16.70- or 31.41-fold lower thancontrol mice, respectively (Fig. 1E, P 0.01 in both cases).However, no difference was recorded during the peak para-sitemia between the two groups of ASP-2-immunized mice(P � 0.05). Similarly, ASP-2-vaccinated mice challenged 14 or
VOL. 77, 2009 PROTECTIVE IMMUNITY AGAINST T. CRUZI INFECTION 4385
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

FIG. 1. Trypomastigote-induced parasitemia and mortality in A/Sn mice immunized with different combinations of plasmid DNA and/orreplication defective human adenovirus type 5 expressing the ASP-2 of T. cruzi. (A) A/Sn mice were immunized as described. Priming and boostingimmunizations were performed with the indicated plasmid or adenovirus i.m. at 0 and 3 weeks, respectively. Two weeks after the final immunizingdose, mice were challenged i.p. with 150 bloodstream trypomastigotes. (B) The peak parasitemia for each mouse group is represented as themean � the standard deviation (SD) (n � 6). Asterisks denote that the parasitemia of mice from groups 3, 4, and 5 was significantly lower thanthat of mice from groups 1 and 2 (P 0.01 in all cases). Mice from groups 4 and 5 had a significantly lower parasitemia than mice from group3 (P 0.05). (C) The graph shows the Kaplan-Meier curves for survival of the mice groups immunized and challenged as described above (n �9). Pooled results from two experiments are shown. Mice from group 3 survived significantly longer than animals from group 1 and 2 (P 0.0001in both cases). Mice from groups 4 and 5 survived significantly longer than the other groups (P 0.0001 in all cases). (D) A/Sn mice wereimmunized as described. Two weeks or 98 days after the final immunizing dose, mice were challenged i.p. with 150 bloodstream trypomastigotes.(E) The parasitemia for each mouse group is represented as the mean � the SD (n � 6). Asterisks indicate that ASP-2-vaccinated mice had asignificantly lower parasitemia (P 0.01) than nonimmune animals. (F) The graph shows the Kaplan-Meier curves for survival of the mouse groupsimmunized and challenged as described above (n � 6). Mice from groups vaccinated with ASP-2 survived significantly longer than nonimmunecontrol animals (P 0.001). (G) A/Sn mice were immunized as described. Before and after challenge i.p. with 150 bloodstream trypomastigotes,mice were treated as described in Materials and Methods with rat IgG or anti-CD4 MAb. (H) The parasitemia for each mouse group is representedas the mean � the SD (n � 12). Asterisks indicate that ASP-2-vaccinated mice treated with rat IgG had a significantly lower parasitemia (P 0.01)than nonimmune animals or ASP-2-vaccinated mice treated with anti-CD4 MAb. (I) The graph shows the Kaplan-Meier curves for survival ofmouse groups immunized and challenged as described above (n � 12). ASP-2-immunized mice treated with rat IgG survived significantly longerthan nonimmune animals or ASP-2-immunized mice treated with anti-CD4 MAb (P 0.0001 in both cases). ASP-2-immunized mice treated withanti-CD4 MAb survived longer than nonimmune animals. The results are pooled from two independent experiments. (J) A/Sn mice
4386 DE ALENCAR ET AL. INFECT. IMMUN.
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

98 days after infection survived longer than control animals(Fig. 1F, P 0.001 in both cases). In addition, the mortalityrates of the mice vaccinated with ASP-2 were not statisticallydifferent (P � 0.05).
To determine whether this protective immunity was depen-dent on CD4� or CD8� T cells, A/Sn mice vaccinated with theheterologous prime-boost regimen (pIgSPCl.9/AdASP-2) weretreated with anti-CD4 or CD8 MAb prior to challenge withbloodstream trypomastigotes. Treatment with anti-CD4 MAbrenders these mice more susceptible to infection. CD4-de-pleted mice presented parasitemia similar to control mice in-jected with pcDNA3/Ad-gal (Fig. 1H). Although their mor-tality rate was high (80%), they lived longer than control mice(Fig. 1I, P 0.001).
Treatment with anti-CD8 MAb renders these mice com-pletely susceptible to infection. CD8-depleted mice presentedparasitemia and survival rates similar to those of control miceinjected with pcDNA3/Ad-gal (Fig. 1K and L). In contrast,A/Sn mice vaccinated with a heterologous prime-boost regi-men (pIgSPCl.9/AdASP-2) and treated with rat IgG had sig-nificantly lower parasitemia and survived the otherwise lethalchallenge with T. cruzi.
The blood of AdASP-2-immunized animals were collectedand added to hemocultures. Parasites did not grow in any ofthe samples, indicating a very low-grade parasitemia or theabsence of parasites. PCR analysis revealed that in fact somemice had detectable parasite DNA. Comparing to knownamounts of parasite DNA equivalents, the DNA detectedranged from 12.5 to 50 per ml of blood (n � 7, t � 150 days[data not shown]).
Perforin expression is required for maximal protective im-munity following a heterologous prime-boost vaccination reg-imen in C57BL/6 mice. In order to determine the presence ofASP-2-specific cytotoxic T cells after immunization with thedifferent protocols, we performed an in vivo cytotoxicity assay(57). For that reason, we used target cells coated with thepeptide TEWETGQI, which we had previously identified as animmunodominant H-2Kk-restricted CD8 T-cell epitope (6, 19,57, 58). The assay was performed in immunized A/Sn before or13 days after challenge with bloodstream trypomastigotes. Asshown in Fig. 2, groups of mice immunized with recombinantAdASP-2 presented in vivo cytotoxicity levels of �20% before chal-lenge. In contrast, immunization with two doses of pIgSPCl.9 failedto elicit a significant degree of in vivo cytotoxicity.
After challenge with trypomastigotes, all mice groups immu-nized with the asp-2 gene (plasmid or adenovirus) showed asignificant degree of in vivo cytotoxicity compared to controlanimals injected with pcDNA3/Ad-gal or naive mice. Animalsthat received two doses of pIgSPCl.9 had slightly lower levelsof in vivo cytotoxicity, although these did not vary significantlyfrom the levels of the other groups.
These experiments were also performed in C57BL/6 micebecause we planned to use genetically modified mice in sub-sequent experiments. In this mouse strain, we used target cellscoated with the immunodominant H-2Kb-restricted epitopeVNHRFTLV (57–59). The results essentially confirmed theobservations described above except that the levels of the invivo cytotoxicity were higher for this epitope (data not shown).
The fact that mice immunized with recombinant AdASP-2showed the highest levels of in vivo cytotoxicity prior to chal-lenge and some degree of protection (Fig. 1B and C) led us topursue our second goal: whether the perforin could be criticalto immunity elicited by vaccination.
were immunized as described. Before and after challenge i.p. with 150 bloodstream trypomastigotes, mice were treated as described in Materials andMethods with rat IgG or anti-CD8 MAb. (K) The parasitemia for each mouse group is represented as the mean � the SD (n � 6). Asterisks indicatethat ASP-2-immunized mice treated with rat IgG had a significantly lower parasitemia (P 0.01) than nonimmune animals or ASP-2-vaccinated micetreated with anti-CD8 MAb. (L) The graph shows the Kaplan-Meier curves for survival of mouse groups immunized and challenged as described above(n � 8 to 9). ASP-2-immunized mice treated with rat IgG survived significantly longer (P 0.0001) than nonimmune animals or ASP-2-immunized micetreated with anti-CD8 MAb. The results are representative of two independent experiments.
FIG. 2. In vivo cytotoxic activity against target cells coated with apeptide representing the CD8 epitope in mice immunized with differ-ent protocols before or after challenge with T. cruzi. A/Sn mice wereinjected with the different immunization protocols depicted at the sideof the charter. The immunizing doses were injected i.m. at 0 and 3weeks. Two weeks after the final immunizing dose, half of the micewere injected with splenic syngeneic cells labeled with CFSE andcoated with peptide TEWETGQI. The remaining half of the animalswas challenged i.p. with 150 blood forms of T. cruzi. Thirteen dayslater, these mice were also injected with splenic syngeneic cells labeledwith CFSE and coated with the peptide TEWETGQI. The specific invivo cytotoxic activity was estimated after 20 h as described in Mate-rials and Methods. Results are expressed as medians (bars), and eachdot represents an individual mouse. Symbols denote significantlyhigher (P 0.01) specific in vivo cytotoxic activity compared to controlnaive or pcDNA-3/Ad-gal injected mice prior to infection (asterisks)or after a challenge with T. cruzi (crosses). The results are represen-tative of experiments performed twice with similar results.
VOL. 77, 2009 PROTECTIVE IMMUNITY AGAINST T. CRUZI INFECTION 4387
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

In our earlier studies, we had observed that the in vivocytotoxicity against the ASP-2-derived epitope (VNHRFTLV)was dependent on the presence of perforin since T. cruzi-infected or adenovirus-immunized perforin KO mice failed toeliminate peptide-coated cells in vivo (59; unpublished results).Based on these observations, we performed protective im-munity experiments following heterologous prime-boost(pIgSPCl.9/AdASP-2) vaccination using perforin KO mice. Be-cause these mice have a C57BL/6 genetic background, we hadto change our experimental model to C57BL/6 WT mice. Afterthe challenge with bloodstream trypomastigotes, ASP-2-vacci-nated C57BL/6 WT mice had significantly lower peak para-sitemia (20.47-fold reduction) than control mice injected withpcDNA3/Ad-gal (Fig. 3A, P 0.001). All of the C57BL/6 WTASP-2-vaccinated mice survived, whereas 46% of the controlmice died (Fig. 3E, P 0.0001). After the challenge withtrypomastigotes, the ASP-2-vaccinated perforin KO mice ani-mals still had a significantly lower peak parasitemia (4.93-foldreduction) than control mice injected with pcDNA3/Ad-gal(Fig. 3B, P 0.001). Although vaccinated perforin KO micesurvived longer than the control perforin KO mice (P 0.0001), 92.3% of the challenged mice died (Fig. 3E). C57BL/6WT ASP-2-vaccinated mice controlled better the peak para-sitemia and survived longer than vaccinated perforin KO mice(P 0.0001). These results indicated that perforin KO miceare capable of developing some degree of protective immunity,if assessed in terms of the reduction of peak parasitemia anddelayed mortality. However, it confirmed our hypothesis thatperforin expression is indeed critical for full protective immu-nity and mouse survival following heterologous prime-boostvaccination with pIgSPCl.9/AdASP-2.
To determine the participation of CD4� and CD8� T cellsin this mouse model, we performed CD4 depletion or usedCD8KO mice. We observed that animals vaccinated withpIgSPCl.9/AdASP-2 and depleted of CD4 T cells prior to in-fection still had a significantly lower peak parasitemia (10.94-fold reduction) compared to control animals injected withpcDNA3/Ad-gal (Fig. 3C). However, only 30% of ASP-2-vaccinated CD4-depleted mice survived, whereas 100% of thecontrol CD4-depleted animals died (Fig. 3F, P 0.001).
CD8 KO mice vaccinated with ASP-2 did not show anyspecific reduction in the peak parasitemia (1.32-fold reduction)compared to control mice injected with pcDNA3/Ad-gal (Fig.3D). Although ASP-2-vaccinated CD8 KO mice survivedlonger than control CD8 KO mice (P 0.05), 87.5% of themice died (Fig. 3F). Compared to the CD4-depleted vacci-nated mice, these mice were more susceptible (P � 0.001),suggesting that CD8� T cells are a slightly more importantduring the effector phase.
Characterization of the cell-mediated immune responses inimmunized C57BL/6 WT and perforin KO mice. The fact thatvaccinated perforin KO mice were highly susceptible to infec-tion prompted us to further analyze the cell-mediated immuneresponses. When restimulated in vitro with the recombinantprotein His-65 kDa (10), splenic cells from vaccinated WTmice secreted significantly higher amounts of IFN-� than theperforin KO animals (Fig. 4A). This cytokine secretion uponstimulation with the soluble antigen reflects activation ofCD4� T cells because the presence of anti-CD4 MAb in cul-
FIG. 3. Trypomastigote-induced parasitemia and mortality inC57BL/6 WT, perforin KO, CD4-depleted, or CD8 KO mice immu-nized with the heterologous prime-boost regimen. C57BL/6 WT, per-forin KO, CD4-depleted, or CD8 KO mice were immunized withpIgSPCl.9, followed by AdASP-2 both injected i.m., at 0 and 3 weeks,respectively. Control mice were injected with pcDNA3, followed byAd-gal both injected i.m., at 0 and 3. Two weeks after the lastimmunizing dose, mice were challenged i.p. with 104 bloodstreamtrypomastigotes. (A to D) The parasitemia for each mouse group isrepresented as the mean � the SD (n � 5 to 8). Asterisks denote thatparasitemia was significantly lower (P 0.01) for ASP-2-vaccinatedmice (f) than for controls nonimmune mice (F). The results arerepresentative of experiments performed twice with similar results. (Eand F) The graphs show the Kaplan-Meier curves for survival of themouse groups immunized and challenged as described above (A to D).For C57BL/6 WT and perforin KO mice, the total number of animalswas 13. In the case of CD4-depleted or CD8 KO mice, the number ofanimals was 10 or 8, respectively. Statistical analysis showed that (i)ASP-2-vaccinated C57BL/6 WT mice survived longer than all of theother mouse groups (P 0.0001 in all cases), (ii) ASP-2-vaccinatedperforin KO mice survived longer than control nonimmune perforinKO mice or CD8 KO mice (P 0.001 in both cases), (iii) ASP-2-vaccinated CD4-depleted mice survived longer than control nonim-mune CD4-depleted mice (P 0.001, and (iv) ASP-2-vaccinated CD8KO mice survived longer than control nonimmune CD8 KO mice (P 0.01).
4388 DE ALENCAR ET AL. INFECT. IMMUN.
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

ture inhibited IFN-� secretion by 80% (reference 10 anddata not shown).
We then characterized several functional and phenotypicaspects of the specific CD8� T cells. Staining of the specificCD8� T cells with the multimer H-2Kb/VNHRFTLV showedthat the frequency and total amounts of these cells in thespleen were the same for perforin KO and C57BL/6 WTmice vaccinated with the heterologous prime boost regimen(Fig. 4B).
Because specific CD8� T cells secreted IFN-� and granule-associated proteins upon ex vivo stimulation with peptides, weperformed an ELISPOT assay and stained the splenic T cellswith antibodies to CD3, CD8, CD107a, and IFN-� after in vitrostimulation with peptide. The frequency of splenic IFN-�-pro-ducing cells was 2.77 times higher in ASP-2-vaccinatedC57BL/6 WT than in perforin KO mice (Fig. 4C, P 0.001).
By FACS analysis, we found three populations of antigen-reactive CD3� CD8� T cells. These cells were positive forCD107a (a marker for T-cell degranulation and cytotoxicity[3]) or IFN-� or both. The comparison of the amount ofCD3�CD8� T cells between vaccinated C57BL/6 WT and per-forin KO mice showed a significantly higher number of cellsexpressing both markers or IFN-� only in the first group (Fig.4D, P 0.05). This difference was also observed in CD3�
CD8� T cells when we considered the total frequency ofCD107a� expressing or not IFN-� (P 0.01) but not CD107aonly (P � 0.05). Considering that the total number of CD3�
CD8� T cells stained with the multimer H-2Kb/VNHRFTLVwere similar, we concluded that the generation and/or matu-ration of antigen-specific CD3�CD8� T cells of perforin KOmice expressing both markers or IFN-�� was selectively im-paired.
Similar results were observed when we estimated the in vivocytotoxic activity against target cells coated with peptideVNHRFTLV. C57BL/6 WT mice displayed higher cytotoxicactivity (two times or more) than vaccinated perforin KO mice(Fig. 4E, P 0.01). This difference was slightly higher in short-term elimination (4 h) than in long-term elimination (20 h).
Subsequently, we used ICS to evaluate the cytokines ex-pressed by specific CD8� T cells after in vitro peptide stimu-lation. The staining of CD3� CD8� splenic cells stimulatedwith peptide in some cases was performed simultaneously withantibodies to IFN-�, TNF-�, and IL-2. These cytokines areregularly used to identify multifunctional T cells (28). Thenumber of peptide-specific T cells expressing any of thesecytokines was higher in ASP-2-immunized C57BL/6 WT micethan in perforin KO mice. Most cytokine-producing CD3�
CD8� T cells from ASP-2 immune C57BL/6 WT mice werestained for IFN-� ( 87%) or TNF-� (58%) or both markers( 45%). The frequency of CD3� CD8� T cells stained forIL-2 was only 7% and, for triple-stained cells, the frequencywas less than 5% (Fig. 5A, B, and C). The frequency ofCD3� CD8� T cells stained for either IFN-� or TNF-� wasreduced in splenic cells from ASP-2-vaccinated perforin KOmice compared to C57BL/6 WT (Fig. 5A to D, P 0.05).However, the most significant difference was observed in thefrequency of CD3� CD8� T cells simultaneously expressingIFN-� and TNF-� after peptide stimulation. ASP-2 immuneC57BL/6 WT mice had 19-fold more double-positive CD8� Tcells than perforin KO mice (P 0.005). The frequency in the
FIG. 4. Cell mediated response of C57BL/6 WT or perforin KO miceafter immunization with the heterologous prime-boost vaccination regi-men. C57BL/6 WT or perforin KO mice were immunized as described inthe legend of Fig. 3. Two weeks after the final immunizing dose, spleniccells were restimulated in vitro in the presence of medium (Med), recom-binant glutathione S-transferase (GST; 10 �g/ml) or recombinant ASP-2(His-65 kDa; 10 �g/ml [A]). When we compared all mouse groups, ASP-2-immunized WT spleens cells secreted more IFN-� upon recombinantprotein stimulation (asterisk, P 0.05 in all cases). We estimated thefrequency of specific splenic cells by staining with anti-CD8 and themultimer H-2Kb/VNHRFTLV (B) or the number of splenic IFN-� spot-forming cells (SFC) by the ex vivo ELISPOT assay (C). When we com-pared groups of mice immunized with pIgSPCl.9/AdASP-2, immunizedC57BL/6 WT mice had a higher frequency of SFC than the immunizedperforin KO mice (asterisk, P 0.001). Alternatively, these mice had theirsplenic cells cultured in the presence of anti-CD28 and FITC-labeledanti-CD107a, with or without the peptide VNHRFTLV. (D) After 12 h,cells were stained with APC-labeled anti-CD8, fixed, permeabilized, andstained with APC-Cy7-labeled anti-CD3, PE-Cy7-labeled anti-IFN-�.When we compared groups of mice immunized with pIgSPCl.9/AdASP-2,immunized C57BL/6 WT mice had a higher frequency of CD3�CD8� Tcells expressing either IFN-� or double-stained for CD107a/IFN-� thanthe immunized perforin KO mice (asterisks, P 0.01 in all cases). Finally,we estimated the in vivo cytotoxic activity by injecting immunized micewith CFSE-labeled splenic cells labeled with CFSE and coated with thepeptide VNHRFTLV. (E) After 4 or 20 h, the in vivo cytotoxic activitywas determined. When we compared groups of mice immunized withpIgSPCl.9/AdASP-2, immunized C57BL/6 WT mice displayed signifi-cantly higher in vivo cytotoxicity than perforin KO mice (asterisk, P 0.01in both cases). The results are expressed as medians (bars) and eachindividual mouse (dots) and are representative of experiments performedat least twice with similar results.
VOL. 77, 2009 PROTECTIVE IMMUNITY AGAINST T. CRUZI INFECTION 4389
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

total of responder cells was significantly higher (a fivefoldincrease) in ASP-2 immune C57BL/6 WT mice than in perforinKO mice (P 0.001). Again, considering that the total num-bers of CD3� CD8� T cells stained with the multimer H-2Kb-VNHRFTLV were similar, we concluded that the generationand/or maturation of antigen-specific CD3�CD8� T cells wasseverely impaired among ASP-2 immune perforin KO miceexpressing either IFN-� or TNF-� or both cytokines simulta-neously. Comparison of the mean fluorescence intensities ofthe different cytokines with ICS failed to detect significant
differences between the distinct CD8� T cells subpopulationsor between WT and KO mice (data not shown).
Finally, we compared the surface markers expressed on pro-tective ASP-2-specific splenic CD8� T lymphocytes elicited bythe heterologous prime-boost regimen (pIgSPCl.9/AdASP-2).For that purpose, purified CD8� T cells obtained from im-mune mice were triple stained with anti-CD8, multimerH-2Kb/VNHRFTLV, and one of several different activationmarkers. The fluorescence intensity was compared to the ex-pression of these same activation markers on naive CD8� T cells.
FIG. 5. ICS of CD8 T cells from C57BL/6 WT or perforin KO mice immunized with the heterologous prime-boost vaccination regimen.C57BL/6 WT or perforin KO mice were immunized with pcDNA3/Ad-gal or pIgSPCl.9/AdASP-2. Fourteen days after the final immunizing dose,these mice had their splenic cells cultured in the presence of anti-CD28 and brefeldin A, with or without the peptide VNHRFTLV. After 12 h,cells were stained with APC-labeled anti-CD8, fixed, permeabilized, and stained with APC-Cy7-labeled anti-CD3, PE-Cy7-labeled anti-IFN-�, andAlexa 488-labeled anti-TNF-�. (A and B) Examples of splenic CD3� CD8� cells from immunized C57BL/6 WT or perforin KO mice stained forIFN-� and TNF-�. (C) Frequency of each cell population. Asterisks indicate that C57BL/6 WT mice immunized with IgSPCl.9/AdASP-2 had ahigher frequency of CD3� CD8� T cells expressing either IFN-� or TNF-� or both cytokines than perforin KO mice immunized with pIgSPCl.9/AdASP-2 (P � 0.01 in all cases). (D) We calculate the frequency of each cell population in relation to the total amount of cells expressing anycytokine. An asterisk indicates that C57BL/6 WT mice immunized with of pIgSPCl.9/AdASP-2 had higher frequencies of CD3� CD8� T cellsexpressing both IFN-� and TNF-� than perforin KO mice immunized with pIgSPCl.9/AdASP-2 (P 0.01 in all cases). A cross indicates thatperforin KO mice immunized with IgSPCl.9/AdASP-2 had a higher frequency of CD3� CD8� T cells expressing only IFN-� than C57BL/6 WTmice immunized with pIgSPCl.9/AdASP-2. The results are presented as medians (bars) and each individual mouse (dots) and are representativeof experiments performed twice with similar results.
4390 DE ALENCAR ET AL. INFECT. IMMUN.
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

As shown in Fig. 6, the phenotype of double-stained CD8�
H-2Kb/VNHRFTLV� cells from immune C57BL/6 WT animalswas CD11ahigh CD43high CD44high CD62Llow CD127low. The ac-tivation marker KLRG-1 reproducibly stained part of the cells( 60%). On the other hand, 40% of the antigen-specific CD8�
T cells did not express this marker on the surface.The pattern of expression of these surface markers on CD8�
specific T cells from ASP-2 immune perforin KO mice wasgenerally similar. Only two significant differences were repro-ducibly noted: (i) the mean intensity fluorescence of CD44 waslower on epitope-specific CD8� T cells from immune perforin
KO mice, and (ii) the frequency of specific CD8� T cells fromimmune perforin KO mice expressing the activation markerKLRG-1 was lower. The frequencies of specific CD8� T ex-pressing KLRG-1 marker in individual mice were 56.12 �11.84 and 31.67 � 14.01%, respectively, in ASP-2 immuneC57BL/6 WT mice and perforin KO mice (n � 4, P 0.05).
IFN-� KO mice immunized with a heterologous prime-boostvaccination regimen develop specific in vivo cytotoxicity butare highly susceptible to infection. Our final goal was to de-termine whether specific cytotoxicity mediated by CD8� Tcells could be elicited by the heterologous prime-boost vacci-nation regimen in the absence of IFN-�. If so, what would theimpact of the absence of this critical mediator of adaptiveimmunity be on the protective immunity of vaccinatedC57BL/6? ASP-2-vaccinated IFN-� KO mice developed highlevels of in vivo cytotoxicity (�95%, Fig. 7A). After challengewith bloodstream trypomastigotes, vaccinated IFN-� KO micehad a parasitemia and a survival rate similar to that of controlIFN-� KO mice injected with pcDNA3/Ad-gal (Fig. 7B andC, respectively). All vaccinated and control mice succumbed tochallenge before the 20th day after challenge. These resultsdemonstrated that, in the absence of IFN-�, specific cytotox-icity mediated by CD8� T cells was not sufficient to retard orreduce the parasitemia. Also, it did not delay mouse mortality,suggesting an interdependence between perforin and IFN-�during protective immunity.
DISCUSSION
As the initial aim of our study, we established an effectiveprotocol of genetic vaccination with a heterologous prime-boost regimen using plasmid DNA and recombinant replica-tion defective adenovirus both expressing the ASP-2 antigen ofthe human protozoan parasite Trypanosoma cruzi. This proto-col was an improvement over others described earlier when weused at least three doses of either plasmid DNA or recombi-nant protein to achieve significant protective immunity (6, 17,19, 60). The heterologous prime-boost vaccination was moreeffective than two doses of plasmid DNA or a single dose ofrecombinant adenovirus.
There was no significant improvement noted in the compar-ison of the heterologous prime-boost regimen with two dosesof recombinant adenovirus, since both were highly effective inour model. Nevertheless, this regimen may have a number ofadvantages for the long-term development of genetic recom-binant vaccines providing a simple solution to the problem ofwidespread immunity to the human adenoviral vector type 5(1). Priming immunization with plasmid DNA seems to besufficient for the subsequent expansion of the trans-gene spe-cific CD8� T cells in recombinant adenovirus-boosted animals(9). This expansion occurs even in animals with previous im-munity to human adenovirus 5.
Hemocultures from these vaccinated mice were all negativedenoting either very low or absent parasitemia. This result is animprovement compared to earlier studies in which we alwaysobserved parasites in hemocultures among a certain percent-age of the vaccinated A/Sn mice (6, 60). Our results from thisinitial part of the study thus confirmed the findings and appliedearlier studies of vaccination against experimental simian im-
FIG. 6. Phenotypic characterization of splenic specific CD8 T cellsinduced by immunization of C57BL/6 WT or perforin KO mice withthe heterologous prime-boost vaccination regimen. C57BL/6 WT orperforin KO mice were immunized with pIgSPCl.9/AdASP-2. Four-teen days after the final immunizing dose, these mice had their CD8�
splenic cells purified and stained with APC-labeled anti-CD8, biotin-labeled H-2Kb-VNHRFTLV, and the indicated marker-specific anti-body labeled with FITC prior to analysis by FACS. The histogramsshow the expression of the markers on CD8� H-2Kb-VNHRFTLV� Tcells (blue lines) or control naive CD8� spleen cells (red lines). Rep-resentative analyses from three or more mice studied are shown.
VOL. 77, 2009 PROTECTIVE IMMUNITY AGAINST T. CRUZI INFECTION 4391
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

munodeficiency virus and malaria infection to a human patho-gen (2, 13, 14, 26, 33, 41).
Both CD4� and CD8� T cells accounted for nonredundantmechanisms of immunity since the depletion of each one priorto challenge renders the animals susceptible to infection. Invaccinated mice without CD8� T cells, we observed a completelack of control of parasitemia in both mouse strains (A/Sn andC57BL/6). Also, no delay in mouse mortality was seen in A/Snmice, denoting a critical role for this subpopulation duringimmunity. In vaccinated C57BL/6 mice, we observed a slight
delay in mouse mortality, suggesting the presence of effectornon-CD8 immune cells. Whether they are in fact CD4� re-mains to be investigated.
In CD4� T-cell-depleted mice we still observed a significantdelay in mouse mortality compared to nonimmune controlscompatible with a CD8� T-cel-mediated immunity. However,the control of parasitemia was significantly better in C57BL/6mice. Finally, the fact that CD4� and CD8� T cells interactduring the memory immune responses makes it more difficultto evaluate precisely the effector and/or helper function ofCD4� T cells during a complex process such as protectiveimmunity in different experimental models. For that purposemore complex experimental systems will have to be developed.
In the second part of our study, we characterized the impor-tance of perforin during protective immunity elicited by DNAprime adenovirus-boost vaccination. We observed that ASP-2-vaccinated perforin KO mice developed only limited immunityto the infection that did not prevent death.
Detailed analysis of the specific immune response revealedan impaired IFN-� secretion by immune spleen cells. Althoughperforin deficiency did not impair the expansion of splenicspecific CD8� T cells, these cells had a significantly lowerfrequency of specific CD107a�/IFN-�� or IFN-��/TNF-��
cells after in vitro restimulation. Also, the in vivo cytotoxicitywas reproducibly reduced. Nevertheless, it is noteworthy thatthe in vivo cytotoxicity was present at certain levels in theperforin KO mice, indicating the presence of a perforin-inde-pendent mechanism(s) of lysis (TNF-�, FasL, etc.) yet to beidentified. Finally, the pattern of expression of certain surfacemarkers on CD8� specific T cells from immune perforin KOmice were significantly different (CD44 and KLRG-1).
Earlier studies on the characteristics of specific CD8� T cellsin perforin-deficient mice indicated a dual function for thismolecule during the homeostasis and effector phases of theimmune response. In different reports (including ours), nosignificant modification was observed in the expansion of spe-cific CD8� T cells as estimated by the multimer staining (5, 15,16). In contrast, an increase in the number of peptide-specificcytokine-expressing CD8� T cells was noted among these KOmice in certain studies (7, 64). Those authors provided evi-dence that perforin-mediated lyses of antigen-presenting cellscould account for a restriction in the expansion of specificCD8� T cells (64). Equally important, it was also noted thatduring certain chronic viral infection, perforin-mediated down-regulation of T-cell responses is critical to avoiding autoimmu-nity and immune-pathological damages (40). To date, the mostpredictable immunological impairment of the perforin KOmice has been the reduced level of cytotoxicity (in vivo or invitro) reported by distinct groups, including ours (8, 15, 32, 59).
Based on other experimental human parasitic infections, itwas not possible to predict that perforin expression would be infact critical to the protective immunity against T. cruzi infectionobserved after DNA-prime adenovirus-boost vaccination.Compared to C57BL/6 WT mice, perforin KO mice are notmore susceptible to infections with the intracellular protozoanparasites Toxoplasma gondii or Plasmodium berghei (20, 47).The fact that perforin-deficient CD8� T cells efficiently elim-inate liver stages of malaria parasites is very important (46).There is a single report stating that perforin deficiency abro-gates protective immunity against Leishmania amazonensis in-
FIG. 7. In vivo cytotoxic activity against target cells coated withpeptides representing the CD8 epitope and trypomastigote-inducedparasitemia and mortality of IFN-� KO mice immunized with heter-ologous prime-boost regimen. IFN-� KO mice were immunized asdescribed in the legend of Fig. 3. (A) Two weeks after the finalimmunizing dose, mice were injected with splenic syngeneic cells la-beled with CFSE and coated with the peptide VNHRFTLV. Thespecific in vivo cytotoxic activity was estimated after 20 h as describedin Materials and Methods. The results are expressed as medians (bars)and each individual mouse (dots, n � 3). An asterisk denotes signifi-cantly higher (P 0.01) specific in vivo cytotoxic activity of ASP-2-immunized IFN-� KO mice than control pcDNA-3/Ad-gal-injectedmice. (B) Two weeks after the final immunizing dose, mice werechallenged i.p. with 104 bloodstream trypomastigotes. The parasitemiafor each mouse group is represented as mean � the SD (n � 6).(C) The graph shows the Kaplan-Meier curves for survival of the micegroups immunized and challenged as described above (n � 6). Theseexperiments were performed twice with similar results.
4392 DE ALENCAR ET AL. INFECT. IMMUN.
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

fection elicited by vaccination with a recombinant protein (18).Therefore, we believe that the fact that perforin is critical toour system may help us to understand the role of this moleculein resistance to human parasitic infections.
The fact that perforin can be expressed in other cell types ofthe adaptive or innate immune system does not allow us toconclude that its expression on CD8� T cells is the singlerestricting factor in our system. Other types of specific lym-phocytes that may express perforin are CD4� T cells (61, 63).We observed that in our vaccination strategy CD4� T cellswere important for T. cruzi immunity after infection of A/Snand C57BL/6. CD8 KO mice were more susceptible than per-forin KO mice, indicating the presence of a non-perforin-mediated mechanisms mediated by these cells.
Other cell types may play important roles in mice immunityagainst T. cruzi infection. Natural killer (NK) cells have beendescribed as mediators of natural resistance to T. cruzi exper-imental infection. The depletion of NK cells by treatment withpolyclonal anti-asialo GM1 renders animals more susceptibleto infection (24). However, these cells are thought to act early,secreting IFN-� (11, 52). A role for cytolysis mediated byperforin has been discarded during NK direct contact-medi-ated lysis of T. cruzi or T. cruzi-infected cells (35, 36). The roleof NK cells in our system remains to be studied.
CD1d-restricted NKT cells also have been described as ca-pable of improving the resistance to T. cruzi infection (22–24).Immunization of CD1d KO mice that fail to express NKT cellsallowed us to test whether NKT cells are involved in immunityin our vaccination regimen. We found that following heterol-ogous prime-boost immunization, these mice developed pro-tective immunity similar to C57BL/6 WT mice (B. C. G. deAlencar and M. M. Rodrigues, unpublished data). Based onthis result, these cells are clearly not involved in the develop-ment of protective immunity after vaccination. Finally, �� Tcells are also not clearly associated with protective immunityduring experimental T. cruzi infection (12, 51).
Based on these observations and the fact that CD8� T cellsare critical for mouse survival after experimental vaccinationand infection, we consider it plausible that these cells representa major, but not the single, source of perforin in our system.Perforin mediates this function, allowing the full maturation ofeffector CD8� T cells. The correlation between protective im-munity and the presence of specific CD8� T cells expressingCD107a�/IFN-�� or IFN-��/TNF-�� simultaneously is im-portant to defining the type of CD8� T cells that should begenerated during vaccination protocols. Up to now, vaccinestudies aimed at determining immunity have relied heavily onthe detection of the number of CD8� T-cell using multimerstaining or IFN-� production (ELISPOT or ICS). However, aswe established, these may not be the best criteria for deter-mining the presence of immune protective CD8� T cells. Al-though we can observe reproducible differences in the numberof peptide-specific IFN-�-producing cells by ELISPOT assay,we considered that the presence of double positive for IFN-�/TNF-� or IFN-�/CD107a was more accurate to estimate theprotective immunity in our model of vaccination.
The final goal of our study was to define whether the in vivocytotoxic activity prior to challenge would ensure protectiveimmunity in the absence of IFN-�, an important mediator ofadaptive immunity during experimental T. cruzi infection. We
concluded from experiments using IFN-� KO mice that thiscytokine is critical even in the presence of high levels of in vivocytotoxicity. IFN-� KO C57BL/6 mice were more susceptiblethan CD4-depleted or CD8 KO animals, indicating that IFN-�most likely come from both sources.
A possible role for antibodies during the protective immu-nity response we observed is highly unlikely. ASP-2 is ex-pressed only by intracellular amastigotes or it is not accessibleto antibodies in the other forms of the parasite (10, 17). Also,immune sera or MAbs incubated with parasites are not able toneutralize their infectivity in vivo (M. M. Rodrigues, unpub-lished results).
In summary, we provide evidences that CD4� and CD8�
T-cell mediated immunity elicited by the DNA-prime recom-binant adenovirus-boost vaccination requires both perforinand IFN-�. The implications are that in the case of T. cruziinfection, in addition to the number of specific cells (multimerstaining) and IFN-� production (ELISPOT assay), other pa-rameters, such as the presence of double-positive IFN-�/TNF-� or IFN-�/CD107a or perforin-dependent cytotoxicity,might be crucial to determining whether immune protective Tcells are present during infection in preclinical or clinical vac-cination trials.
ACKNOWLEDGMENTS
This study was supported by grants from the Fundacao de Amparoa Pesquisa do Estado de Sao Paulo (2006/1983-4), the National Insti-tute for Vaccine Technology (INCTV-CNPq), the Millennium Insti-tute for Vaccine Development and Technology (CNPq–420067/2005-1), the Millennium Institute for Gene Therapy, and FAPEMIG (EDT24.000) (Brazil). P.M.P., A.V.M., R.T.G., O.B.-R., J.L.-V., andM.M.R. are recipients of fellowships from CNPq. B.C.G.D.A. andF.A.H. are recipients of a fellowship from FAPESP.
REFERENCES
1. Abbi, P., A. A. Lemckert, B. A. Ewald, D. M. Lynch, M. Denholtz, S. Smits,L. Holterman, I. Damen, R. Vogels, A. R. Thorner, K. L. O’Brien, A. Carville,K. G. Mansfield, J. Goudsmit, M. J. Havenga, and D. H. Barouch. 2007.Comparative seroprevalence and immunogenicity of six rare serotype recom-binant adenovirus vaccine vectors from subgroups B and D. J. Virol. 81:4654–4663.
2. Acierno, P. M., J. E. Schmitz, D. A. Gorgone, Y. Sun, S. Santra, M. S.Seaman, M. H. Newberg, J. R. Mascola, G. J. Nabel, D. Panicali, and N. L.Letvin. 2006. Preservation of functional virus-specific memory CD8� T lym-phocytes in vaccinated, simian human immunodeficiency virus-infected rhe-sus monkeys. J. Immunol. 176:5338–5345.
3. Aktas, E., U. C. Kucuksezer, S. Bilgic, G. Erten, and G. Deniz. 2009. Rela-tionship between CD107a expression and cytotoxic activity. Cell. Immunol.254:149–154.
4. Amara, R. R., F. Villinger, J. D. Altman, S. L. Lydy, S. P. O’Neil, S. L.Staprans, D. C. Montefiori, Y. Xu, J. G. Herndon, S. Wyatt, M. A. Candido,N. L. Kozyr, P. L. Earl, J. M. Smith, H. L. Ma, B. D. Grimm, M. L. Hulsey,J. Miller, H. M. McClure, J. M. McNicholl, B. Moss, and H. L. Robinson.2001. Control of a mucosal challenge and prevention of AIDS by a multi-protein DNA/MVA vaccine. Science 292:69–74.
5. Andrews, D. M., C. E. Andoniou, P. Fleming, M. J. Smyth, and M. A.Degli-Esposti. 2008. The early kinetics of cytomegalovirus-specific CD8�
T-cell responses are not affected by antigen load or the absence of perforinor gamma interferon. J. Virol. 82:4931–4937.
6. Araujo, A. F., B. C. de Alencar, J. R. Vasconcelos, M. I. Hiyane, C. R.Marinho, M. L. Penido, S. B. Boscardin, D. F. Hoft, and R. T. Gazzinelli, andM. M. Rodrigues. 2005. CD8�-T-cell-dependent control of Trypanosomacruzi infection in a highly susceptible mouse strain after immunization withrecombinant proteins based on amastigote surface protein 2. Infect. Immun.73:6017–6025.
7. Badovinac, V. P., A. R. Tvinnereim, and J. T. Harty. 2000. Regulation ofantigen-specific CD8 T cell homeostasis by perforin and interferon-�. Sci-ence 290:1354–1357.
8. Barber, D. L., E. J. Wherry, and R. Ahmed. 2003. Cutting edge: rapid in vivokilling by memory CD8 T cells. J. Immunol. 171:27–31.
9. Barouch, D. H., P. F. McKay, S. M. Sumida, S. Santra, S. S. Jackson, D. A.
VOL. 77, 2009 PROTECTIVE IMMUNITY AGAINST T. CRUZI INFECTION 4393
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

Gorgone, M. A. Lifton, B. K. Chakrabarti, L. Xu, G. J. Nabel, and N. L.Letvin. 2003. Plasmid chemokines and colony-stimulating factors enhancethe immunogenicity of DNA priming-viral vector boosting human immuno-deficiency virus type 1 vaccines. J. Virol. 77:8729–8735.
10. Boscardin, S. B., S. S. Kinoshita, A. E. Fujimura, and M. M. Rodrigues.2003. Immunization with cDNA expressed by amastigotes of Trypanosomacruzi elicits protective immune response against experimental infection. In-fect. Immun. 71:2744–2757.
11. Cardillo, F., J. C. Voltarelli, S. G. Reed, and J. S. Silva. 1996. Regulation ofTrypanosoma cruzi infection in mice by gamma interferon and interleukin 10:role of NK cells. Infect. Immun. 64:128–134.
12. Cardillo, F., A. Nomizo, and J. Mengel. 1998. The role of the thymus inmodulating �� T-cell suppressor activity during experimental Trypanosomacruzi infection. Int. Immunol. 10:107–116.
13. Casimiro, D. R., L. Chen, T. M. Fu, R. K. Evans, M. J. Caulfield, M. E.Davies, A. Tang, M. Chen, L. Huang, V. Harris, D. C. Freed, K. A. Wilson,S. Dubey, D. M. Zhu, D. Nawrocki, H. Mach, R. Troutman, L. Isopi, D.Williams, W. Hurni, Z. Xu, J. G. Smith, S. Wang, X. Liu, L. Guan, R. Long,W. Trigona, G. J. Heidecker, H. C. Perry, N. Persaud, T. J. Toner, Q. Su, X.Liang, R. Youil, M. Chastain, A. J. Bett, D. B. Volkin, E. A. Emini, and J. W.Shiver. 2003. Comparative immunogenicity in rhesus monkeys of DNA plas-mid, recombinant vaccinia virus, and replication-defective adenovirus vec-tors expressing a human immunodeficiency virus type 1 gag gene. J. Virol.77:6305–6313.
14. Casimiro, D. R., F. Wang, W. A. Schleif, X. Liang, Z. Q. Zhang, T. W. Tobery,M. E. Davies, A. B. McDermott, D. H. O’Connor, A. Fridman, A. Bagchi,L. G. Tussey, A. J. Bett, A. C. Finnefrock, T. M. Fu, A. Tang, K. A. Wilson,M. Chen, H. C. Perry, G. J. Heidecker, D. C. Freed, A. Carella, K. S. Punt,K. J. Sykes, L. Huang, V. I. Ausensi, M. Bachinsky, U. Sadasivan-Nair, D. I.Watkins, E. A. Emini, and J. W. Shiver. 2005. Attenuation of simian immu-nodeficiency virus SIVmac239 infection by prophylactic immunization withDNA and recombinant adenoviral vaccine vectors expressing Gag. J. Virol.79:15547–15555.
15. Chen, J., H. C. Hsu, A. J. Zajac, Q. Wu, P. Yang, X. Xu, S. A. McPherson,J. Li, D. T. Curiel, and J. D. Mountz. 2006. In vivo analysis of adenovirus-specific cytotoxic T lymphocyte response in mice deficient in CD28, fasligand, and perforin. Hum. Gene Ther. 17:669–682.
16. Christensen, J. E., D. Wodarz, J. P. Christensen, and A. R. Thomsen. 2004.Perforin and IFN-� do not significantly regulate the virus-specific CD8�
T-cell response in the absence of antiviral effector activity. Eur. J. Immunol.34:1389–1394.
17. Claser, C., N. M. Espíndola, G. Sasso, A. J. Vaz, S. B. Boscardin, and M. M.Rodrigues. 2007. Immunologically relevant strain polymorphism in the amas-tigote surface protein 2 of Trypanosoma cruzi. Microbes Infect. 9:1011–1019.
18. Colmenares, M., P. E. Kima, E. Samoff, L. Soong, and D. McMahon-Pratt.2003. Perforin and gamma interferon are critical CD8� T-cell-mediatedresponses in vaccine-induced immunity against Leishmania amazonensis in-fection. Infect. Immun. 71:3172–3182.
19. de Alencar, B. C., A. F. Araujo, M. L. Penido, R. T. Gazzinelli, and M. M.Rodrigues. 2007. Cross-priming of long-lived protective CD8� T cells againstTrypanosoma cruzi infection: importance of a TLR9 agonist and CD4� Tcells. Vaccine 25:6018–6027.
20. Denkers, E. Y., G. Yap, T. Scharton-Kersten, H. Charest, B. A. Butcher, P.Caspar, S. Heiny, and A. Sher. 1997. Perforin-mediated cytolysis plays alimited role in host resistance to Toxoplasma gondii. J. Immunol. 159:1903–1908.
21. Dunachie, S. J., M. Walther, J. E. Epstein, S. Keating, T. Berthoud, L.Andrews, R. F. Andersen, P. Bejon, N. Goonetilleke, I. Poulton, D. P. Web-ster, G. Butcher, K. Watkins, R. E. Sinden, G. L. Levine, T. L. Richie, J.Schneider, D. Kaslow, S. C. Gilbert, D. J. Carucci, and A. V. Hill. 2006. DNAprime-modified vaccinia virus ankara boost vaccine encoding throm-bospondin-related adhesion protein but not circumsporozoite protein par-tially protects healthy malaria-naive adults against Plasmodium falciparumsporozoite challenge. Infect. Immun. 74:5933–5942.
22. Duthie, M. S., M. Kahn, M. White, R. P. Kapur, and S. J. Kahn. 2005.Critical proinflammatory and anti-inflammatory functions of different sub-sets of CD1d-restricted natural killer T cells during Trypanosoma cruzi in-fection. Infect. Immun. 73:181–192.
23. Duthie, M. S., M. Wleklinski-Lee, S. Smith, T. Nakayama, M. Taniguchi,and S. J. Kahn. 2002. During Trypanosoma cruzi infection CD1d-restrictedNK T cells limit parasitemia and augment the antibody response to a glyco-phosphoinositol-modified surface protein. Infect. Immun. 70:36–48.
24. Duthie, M. S., and S. J. Kahn. 2006. During acute Trypanosoma cruzi infec-tion highly susceptible mice deficient in natural killer cells are protected bya single alpha-galactosylceramide treatment. Immunology 119:355–361.
25. Garg, N., and R. Tarleton. 2002. Genetic immunization elicits antigen-spe-cific protective immune responses and decreases disease severity in Trypano-soma cruzi infection. Infect. Immun. 70:5547–5555.
26. Gilbert, S. C., J. Schneider, C. M. Hannan, J. T. Hu, M. Plebanski, R.Sinden, and A. V. Hill. 2002. Enhanced CD8 T-cell immunogenicity andprotective efficacy in a mouse malaria model using a recombinant adenoviral
vaccine in heterologous prime-boost immunization regimes. Vaccine 20:1039–1045.
27. Gomez, C. E., J. L. Najera, M. Krupa, and M. Esteban. 2008. The poxvirusvectors MVA and NYVAC as gene delivery systems for vaccination againstinfectious diseases and cancer. Curr. Gene Ther. 8:97–120.
28. Goonetilleke, N., S. Moore, L. Dally, N. Winstone, I. Cebere, A. Mahmoud,S. Pinheiro, G. Gillespie, D. Brown, V. Loach, J. Roberts, A. Guimaraes-Walker, P. Hayes, K. Loughran, C. Smith, J. De Bont, C. Verlinde, D. Vooijs,S. C. Schmidt, M. Boaz, J. Gilmour, P. Fast, L. Dorrell, T. Hanke, and A. J.McMichael. 2006. Induction of multifunctional human immunodeficiencyvirus type 1 (HIV-1)-specific T cells capable of proliferation in healthysubjects by using a prime-boost regimen of DNA- and modified vacciniavirus Ankara-vectored vaccines expressing HIV-1 Gag coupled to CD8�
T-cell epitopes. J. Virol. 80:4717–4728.29. Hanke, T., N. Goonetilleke, A. J. McMichael, and L. Dorrell. 2007. Clinical
experience with plasmid DNA- and modified vaccinia virus Ankara-vectoredhuman immunodeficiency virus type 1 clade A vaccine focusing on T-cellinduction. J. Gen. Virol. 88:1–12.
30. Hoft, D. F., and C. S. Eickhoff. 2005. Type 1 immunity provides both optimalmucosal and systemic protection against a mucosally invasive, intracellularpathogen. Infect. Immun. 73:4934–4940.
31. Hoft, D. F., C. S. Eickhoff, O. K. Giddings, J. R. Vasconcelos, and M. M.Rodrigues. 2007. Trans-sialidase recombinant protein mixed with CpG mo-tif-containing oligodeoxynucleotide induces protective mucosal and systemicTrypanosoma cruzi immunity involving CD8� CTL and B cell-mediatedcross-priming. J. Immunol. 179:6889–6900.
32. Kagi, D., B. Ledermann, K. Burki, P. Seiler, B. Odermatt, K. J. Olsen, E. R.Podack, R. M. Zinkernagel, and H. Hengartner. 1994. Cytotoxicity mediatedby T cells and natural killer cells is greatly impaired in perforin-deficientmice. Nature 369:31–37.
33. Letvin, N. L., J. R. Mascola, Y. Sun, D. A. Gorgone, A. P. Buzby, L. Xu, Z. Y.Yang, B. Chakrabarti, S. S. Rao, J. E. Schmitz, D. C. Montefiori, B. R.Barker, F. L. Bookstein, and G. J. Nabel. 2006. Preserved CD4� centralmemory T cells and survival in vaccinated SIV-challenged monkeys. Science312:1530–1533.
34. Li, S., M. Rodrigues, D. Rodriguez, J. R. Rodriguez, M. Esteban, P. Palese,R. S. Nussenzweig, and F. Zavala. 1993. Priming with recombinant influenzavirus followed by administration of recombinant vaccinia virus induces CD8�
T-cell-mediated protective immunity against malaria. Proc. Natl. Acad. Sci.USA 90:5214–5218.
35. Lieke, T., S. E. Graefe, U. Klauenberg, B. Fleischer, and T. Jacobs. 2004. NKcells contribute to the control of Trypanosoma cruzi infection by killing freeparasites by perforin-independent mechanisms. Infect. Immun. 72:6817–6825.
36. Lieke, T., C. Steeg, S. E. Graefe, B. Fleischer, and T. Jacobs. 2006. Interac-tion of natural killer cells with Trypanosoma cruzi-infected fibroblasts. Clin.Exp. Immunol. 145:357–364.
37. Machado, A. V., J. E. Cardoso, C. Claser, M. M. Rodrigues, R. T. Gazzinelli,and O. Bruna-Romero. 2006. Long-term protective immunity inducedagainst Trypanosoma cruzi infection after vaccination with recombinant ad-enoviruses encoding amastigote surface protein-2 and trans-sialidase. Hum.Gene Ther. 17:898–908.
38. Martin, D. L., D. B. Weatherly, S. A. Laucella, M. A. Cabinian, M. T. Crim,S. Sullivan, M. Heiges, S. H. Craven, C. S. Rosenberg, M. H. Collins, A.Sette, M. Postan, and R. L. Tarleton. 2006. CD8� T-cell responses toTrypanosoma cruzi are highly focused on strain-variant trans-sialidaseepitopes. PLoS Pathog. 2:e77.
39. Martin, D., and R. Tarleton. 2004. Generation, specificity, and function ofCD8� T cells in Trypanosoma cruzi infection. Immunol. Rev. 201:304–317.
40. Matloubian, M., M. Suresh, A. Glass, M. Galvan, K. Chow, J. K. Whitmire,C. M. Walsh, W. R. Clark, and R. Ahmed. 1999. A role for perforin indownregulating T-cell responses during chronic viral infection. J. Virol.73:2527–2536.
41. Mattapallil, J. J., D. C. Douek, A. Buckler-White, D. Montefiori, N. L.Letvin, G. J. Nabel, and M. Roederer. 2006. Vaccination preserves CD4memory T cells during acute simian immunodeficiency virus challenge. J.Exp. Med. 203:1533–1541.
42. McConkey, S. J., W. H. Reece, V. S. Moorthy, D. Webster, S. Dunachie, G.Butcher, J. M. Vuola, T. J. Blanchard, P. Gothard, K. Watkins, C. M.Hannan, S. Everaere, K. Brown, K. E. Kester, J. Cummings, J. Williams,D. G. Heppner, A. Pathan, K. Flanagan, N. Arulanantham, M. T. Roberts,M. Roy, G. L. Smith, J. Schneider, T. Peto, R. E. Sinden, S. C. Gilbert, andA. V. Hill. 2003. Enhanced T-cell immunogenicity of plasmid DNA vaccinesboosted by recombinant modified vaccinia virus Ankara in humans. Nat.Med. 9:729–735.
43. Miyahira, Y., Y. Takashima, S. Kobayashi, Y. Matsumoto, T. Takeuchi, M.Ohyanagi-Hara, A. Yoshida, A. Ohwada, H. Akiba, H. Yagita, K. Okumura,and H. Ogawa. 2005. Immune responses against a single CD8�-T-cellepitope induced by virus vector vaccination can successfully control Trypano-soma cruzi infection. Infect. Immun. 73:7356–7365.
44. Moore, A. C., and A. V. Hill. 2004. Progress in DNA-based heterologous
4394 DE ALENCAR ET AL. INFECT. IMMUN.
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

prime-boost immunization strategies for malaria. Immunol. Rev. 199:126–143.
45. Moorthy, V. S., E. B. Imoukhuede, P. Milligan, K. Bojang, S. Keating, P.Kaye, M. Pinder, S. C. Gilbert, G. Walraven, S. M. Greenwood, and A. S.Hill. 2004. A randomised, double-blind, controlled vaccine efficacy trial ofDNA/MVA ME-TRAP against malaria infection in Gambian adults. PLoSMed. 1:e33.
46. Morrot, A., and F. Zavala. 2004. Effector and memory CD8� T cells as seenin immunity to malaria. Immunol. Rev. 201:291–303.
47. Renggli, J., M. Hahne, H. Matile, B. Betschart, J. Tschopp, and G. Corradin.1997. Elimination of Plasmodium berghei liver stages is independent of Fas(CD95/Apo-I) or perforin-mediated cytotoxicity. Parasite Immunol. 19:145–148.
48. Rice, J., C. H. Ottensmeier, and F. K. Stevenson. 2008. DNA vaccines:precision tools for activating effective immunity against cancer. Nat. Rev.Cancer 8:108–120.
49. Robinson, H. L., and R. R. Amara. 2005. T cell vaccines for microbialinfections. Nat. Med. 11:S25–S32.
50. Rodrigues, M., S. Li, K. Murata, D. Rodriguez, J. R. Rodriguez, I. Bacik,J. R. Bennink, J. W. Yewdell, A. Garcia-Sastre, R. S. Nussenzweig, M.Esteban, P. Palese, and F. Zavala. 1994. Influenza and vaccinia virusesexpressing malaria CD8� T and B cell epitopes: comparison of their immu-nogenicity and capacity to induce protective immunity. J. Immunol. 153:4636–4648.
51. Santos-Lima, E. C., and P. Minoprio. 1996. Chagas’ disease is attenuated inmice lacking gamma delta T cells. Infect. Immun. 64:215–221.
52. Sardinha, L. R., R. M. Elias, T. Mosca, K. R. Bastos, C. R. Marinho, M. R.D’Imperio Lima, and J. M. Alvarez. 2006. Contribution of NK, NK T,gamma delta T, and alpha beta T cells to the gamma interferon responserequired for liver protection against Trypanosoma cruzi. Infect. Immun. 74:2031–2042.
53. Schneider, J., S. C. Gilbert, T. J. Blanchard, T. Hanke, K. J. Robson, C. M.Hannan, M. Becker, R. Sinden, G. L. Smith, and A. V. Hill. 1998. Enhancedimmunogenicity for CD8� T-cell induction and complete protective efficacyof malaria DNA vaccination by boosting with modified vaccinia virus An-kara. Nat. Med. 4:397–402.
54. Sedegah, M., T. R. Jones, M. Kaur, R. Hedstrom, P. Hobart, J. A. Tine, andS. L. Hoffman. 1998. Boosting with recombinant vaccinia increases immu-nogenicity and protective efficacy of malaria DNA vaccine. Proc. Natl. Acad.Sci. USA 95:7648–7653.
55. Silveira, E. L., C. Claser, F. A. Haolla, L. G. Zanella, and M. M. Rodrigues.2008. Novel protective antigens expressed by Trypanosoma cruzi amastigotesprovide immunity to mice highly susceptible to Chagas’ disease. Clin. Vac-cine Immunol. 15:1292–1300.
56. Tarleton, R. L., J. Sun, L. Zhang, M. Postan, and L. Glimcher. 1996.Trypanosoma cruzi infection in MHC-deficient mice: further evidence for the
role of both class I- and class II-restricted T cells in immune resistance anddisease. Int. Immunol. 8:13–22.
57. Tzelepis, F., B. C. de Alencar, M. L. Penido, R. T. Gazzinelli, P. M. Per-sechini, and M. M. Rodrigues. 2006. Distinct kinetics of effector CD8�
cytotoxic T cells after infection with Trypanosoma cruzi in naive or vaccinatedmice. Infect. Immun. 74:2477–2482.
58. Tzelepis, F., B. C. de Alencar, M. L. Penido, C. Claser, A. V. Machado, O.Bruna-Romero, R. T. Gazzinelli, and M. M. Rodrigues. 2008. Infection withTrypanosoma cruzi restricts the repertoire of parasite-specific CD8� T cellsleading to immunodominance. J. Immunol. 180:1737–1748.
59. Tzelepis, F., P. M. Persechini, and M. M. Rodrigues. 2007. Modulation ofCD4� T-cell-dependent specific cytotoxic CD8� T cells differentiation andproliferation by the timing of increase in the pathogen load. PLoS ONE2:e393.
60. Vasconcelos, J. R., M. I. Hiyane, C. R. F. Marinho, C. Claser, A. M. Vieira-Machado, R. T. Gazinelli, O. Bruna-Romero, J. M. Alvarez, and S. B.Boscardin, and M. M. Rodrigues. 2004. Protective immunity againstTrypanosoma cruzi infection in a highly susceptible mouse strain followingvaccination with genes encoding the amastigote surface protein-2 and trans-sialidase. Hum. Gene Ther. 15:878–886.
61. Williams, N. S., and V. H. Engelhard. 1996. Identification of a population ofCD4� CTL that utilizes a perforin-rather than a Fas ligand-dependent cy-totoxic mechanism. J. Immunol. 156:153–159.
62. Wilson, N. A., J. Reed, G. S. Napoe, S. Piaskowski, A. Szymanski, J. Furlott,E. J. Gonzalez, L. J. Yant, N. J. Maness, G. E. May, T. Soma, M. R.Reynolds, E. Rakasz, R. Rudersdorf, A. B. McDermott, D. H. O’Connor,T. C. Friedrich, D. B. Allison, A. Patki, L. J. Picker, D. R. Burton, J. Lin, L.Huang, D. Patel, G. Heindecker, J. Fan, M. Citron, M. Horton, F. Wang, X.Liang, J. W. Shiver, D. R. Casimiro, and D. I. Watkins. 2006. Vaccine-induced cellular immune responses reduce plasma viral concentrations afterrepeated low-dose challenge with pathogenic simian immunodeficiency virusSIVmac239. J. Virol. 80:5875–5885.
63. Yana, F., E. Ishii, K. Kojima, A. Hasegawa, T. Azuma, S. Hirose, N. Suga, A.Mitsudome, M. Zaitsu, Y. Ishida, Y. Shirakata, K. Sayama, K. Hashimoto,and M. Yasukawa. 2003. Essential roles of perforin in antigen-specific cyto-toxicity mediated by human CD4� T lymphocytes: analysis using the com-bination of hereditary perforin-deficient effector cells and Fas-deficient tar-get cells. J. Immunol. 170:2205–2213.
64. Yang, J., S. P. Huck, S. Rebecca, S McHugh, I. F. Hermans, and F. Ronchese.2006. Perforin-dependent elimination of dendritic cells regulates the expan-sion of antigen-specific CD8 T cells in vivo. Proc. Natl. Acad. Sci. USA103:147–152.
65. Zavala, F., M. Rodrigues, D. Rodriguez, J. R. Rodriguez, R. S. Nussenzweig,and M. Esteban. 2001. A striking property of recombinant poxviruses: effi-cient inducers of in vivo expansion of primed CD8� T cells. Virology 280:155–159.
Editor: J. F. Urban, Jr.
VOL. 77, 2009 PROTECTIVE IMMUNITY AGAINST T. CRUZI INFECTION 4395
by on Septem
ber 19, 2009 iai.asm
.orgD
ownloaded from

82
Considerações finais e conclusões

83
Vacinação contra a Doença de Chagas
Durante esta tese, utilizando como modelo o antígeno ASP-2,
demonstrou-se que é possível elevar racionalmente os níveis de imunidade
protetora de longa duração, nos camundongos altamente suscetíveis à
infecção pelo T. cruzi, pelo uso de novas formulações e/ou estratégias de
vacinação. Terminou-se por demonstrar que um protocolo heterólogo
envolvendo DNA plasmidial e adenovírus recombinante, ambos expressando o
antígeno ASP-2, apresentou grande imunogenicidade e eficácia na indução de
proteção contra a infecção experimental pelo T. cruzi, sendo necessária a
administração de apenas duas doses.
Como discutido no artigo 3 por Alencar et al., 2009, este protocolo de
vacinação heteróloga pode trazer vantagens sobre a imunização homóloga,
uma vez que, de certa forma, neutraliza a imunidade ao vetor adenoviral que é
muito comum em algumas populações humanas [219,255]. É possível também
que haja diferenças nas propriedades funcionais e fenotípicas dos linfócitos T
induzidos pelas vacinações homóloga e heteróloga. Ambas as hipóteses ainda
necessitam ser testadas experimentalmente.
De qualquer forma, até o momento, as evidências são claras no sentido
de que é possível desenvolver uma forte resposta imune mediada pelos
linfócitos T do tipo Th1, reduzindo drasticamente a forma aguda e as sequelas
tardias (fase crônica) da infecção experimental pelo T. cruzi. Apesar de muitos
estudos (revistos na introdução desta tese) em modelos experimentais terem
indicado que uma vacina contra a infecção pelo T. cruzi pode ser viável,
sempre houve muito ceticismo sobre esta matéria [262].
Muitos acreditam que uma vacina profilática contra a doença de Chagas
deveria conferir proteção estéril contra o parasita. Este tipo de proteção pode,
porém, ser inalcançável, uma vez que a própria infecção seguida pelo
tratamento com o benzonidazol (e cura) não parece proteger totalmente contra
uma re-infecção [59]. Por outro lado, como mais de 60% dos indivíduos que
têm contato com o parasita são assintomáticos, talvez fosse mais realista tentar
impedir o desenvolvimento de uma sintomatologia crônica significativa.
O estudo da resposta imune dos indivíduos que tiveram contato com o
T. cruzi e são assintomáticos ou sintomáticos poderia auxiliar na compreensão
das necessidades imunológicas para se manter os indivíduos assintomáticos

84
por toda a vida. Contudo, a comparação da resposta imune celular nos
pacientes que desenvolveram ou não sintomas clínicos geraram resultados
discrepantes. Em alguns casos a alta produção de IFN-γ foi associada com a
cardiomiopatia chagásica crônica [263,264]. Nesta mesma linha, o polimorfismo
das regiões promotoras dos genes que codificam as proteínas anti-
inflamatórias BAT-1 e IL-10 indicaram que, os alelos baixos produtores, são
mais frequentes nos pacientes com a cardiomiopatia chagásica crônica
[31,265]. Em contraste, outros pesquisadores propõem que a mais alta
produção de IFN-γ se correlaciona os com sintomas clínicos mais brandos da
doença [266,267]. Também é relevante o fato de que os indivíduos curados
pelo tratamento quimioterápico durante a fase aguda da doença desenvolvem
uma forte resposta imune celular, a qual é caracterizada pela produção in vitro
de IFN-γ. Esta observação sugere que, se os linfócitos T do tipo Th1 têm um
papel patogênico, levando à exacerbação dos sintomas da miocardite
chagásica crônica, a sua ação danosa requer a presença constante de
parasitas viáveis [268]. Assim, ainda é polêmico o papel dos linfócitos Th1 no
desenvolvimento da sintomatologia crônica da doença de Chagas humana.
Mesmo no modelo de infecção de camundongos, o qual permite uma
real manipulação experimental e não se limita somente a correlações, o papel
dos linfócitos do tipo Th1/Tc1 pode ser contraditório. A infecção de animais
deficientes na expressão da STAT-6 com parasitas da cepa Brazil dá indícios
de que uma resposta imune do tipo Th1, na ausência de uma resposta Th2,
leva à eliminação quase total do T. cruzi, sem o desenvolvimento do dano
tecidual na fase tardia observado em animais normais infectados [81]. Por outro
lado, a infecção de animais deficientes na expressão da IL-4 com parasitas da
cepa Colombiana mostra que, neste caso, uma resposta imune do tipo Th1
leva a um aumento da patologia cardíaca na fase tardia da infecção. Como os
experimentos foram feitos com duas cepas diferentes e em dois modelos de
camundongos distintos, não há como se prever o que aconteceria se os
modelos fossem comparados em paralelo nas duas cepas. Portanto, ainda há
muito que se compreender sobre o desenvolvimento da imunopatologia da
miocardite crônica, seja em modelos experimentais ou nos seres humanos.

85
Acredita-se, dessa maneira, que os protocolos de vacinação
desenvolvidos nesta tese devam ser testados em modelos animais nos quais
se possa estudar melhor os efeitos sobre a fase crônica da doença. Uma
alternativa seria a utilização das cepas de T. cruzi que causam, nos
camundongos, uma cardiomiopatia com características mais semelhantes à
observada nos seres humanos, como a cepa Colombiana [269].
Alternativamente, podem-se utilizar outros modelos animais, como os hamsters
e os cães, que parecem também desenvolver cardiomiopatia [270,271].
Acredita-se, também, que novos protocolos de imunização devam ser
testados e estudados, tanto em camundongos quanto em outros modelos de
infecção, em busca de níveis de proteção mais efetivos. Neste caso, um
protocolo potencialmente interessante seria a combinação de proteína
recombinante e adenovírus, a qual não foi testada no presente trabalho. O
recente desenvolvimento de um MVA expressando a ASP-2 [Dutra et al., em
preparação] abre também a possibilidade de se testar vacinações heterólogas
com ambos os vetores virais (adenovírus e MVA). Finalmente, dada a recente
informação de que a proteção conferida pela ASP-2 é cepa-específica [272],
seria interessante que se estudasse um protocolo vacinal contendo,
simultaneamente, os antígenos ASP-2 das diversas linhagens de T. cruzi, a fim
de que se observem questões de imunodominância e de proteção contra o
desafio com as diferentes cepas do parasita.
Mecanismos imunológicos de proteção contra a infecção experimental
pelo T. cruzi.
Juntamente com a melhora na indução da imunidade protetora de longa
duração, este trabalho trouxe um melhor entendimento sobre os mecanismos
protetores induzidos por essas vacinações experimentais contra a infecção por
protozoários intracelulares, em geral, e o T. cruzi, especificamente. Em primeiro
lugar, demonstrou-se, independente da formulação utilizada, a importância dos
linfócitos T CD8 efetores na proteção induzida pela vacinação com a ASP-2.
Na vacinação homóloga com DNA plasmidial, proteína recombinante ou vírus
recombinante, a depleção dos linfócitos T CD8 antes do desafio torna os
animais completamente suscetíveis à infecção experimental [75,117,121]. O
mesmo foi observado após a vacinação heteróloga.

86
Devido à importância da resposta de linfócitos T CD8 nas infecções por
patógenos intracelulares, vêm sendo preconizadas vacinas baseadas na
indução de fortes respostas mediadas por estas células, mesmo nas infecções
em que a proteção mediada por anticorpos também possa ocorrer. Apesar do
grande interesse em se desenvolver uma imunidade protetora mediada pelos
linfócitos T CD8, ainda se tem pouco conhecimento sobre as características
dos linfócitos T que levam à proteção contra infecções em geral, e cada
infecção especificamente. Assim, a busca por possíveis correlatos de proteção
que sejam mensuráveis ex vivo é um passo muito importante para a futura
determinação da eficiência dos protocolos de vacinação. Neste sentido,
utilizou-se o esquema de imunização desenvolvido, a fim de contribuir para a
identificação das propriedades funcionais e das características fenotípicas dos
linfócitos T CD8 protetores contra a infecção pelo protozoário intracelular
(T. cruzi).
Nesta tese descreveu-se que os animais selvagens apresentaram uma
forte resposta imune mediada pelos linfócitos T CD8 multifuncionais, capazes
de produzir IFN-γ e TNF-α, expressar IFN-γ e CD107a (na superfície) e mediar
citotoxicidade in vivo. Do ponto de vista fenotípico, na superfície desses
linfócitos T CD8 específicos imunoprotetores observou-se a expressão
aumentada dos marcadores de ativação celular CD11a, CD43, CD44 e KLRG-
1. Também se observou uma expressão reduzida do CD62L. Este fenótipo
caracteriza os linfócitos CD8 do tipo TEM [273]. Mesmo após 98 dias de
imunização, os linfócitos T CD8 imunoprotetores mantiveram tais
características fenotípicas. Não se observou o desenvolvimento dos linfócitos T
CD8 com alta expressão de CD62L, característica dos linfócitos TCM. Esta
observação demonstra que os linfócitos TEM podem ser extremamente
protetores e duradouros nos animais imunizados.
Outros resultados obtidos na infecção experimental pelo T. cruzi levaram
à mesma conclusão, de que os linfócitos CD8 TEM multifuncionais, específicos
para o antígeno imunodominante (ASP-2), são responsáveis pela
imunoproteção observada na fase aguda da infecção, e são mantidos por
longos períodos [69]. Estes resultados contrastam com resultados recentes,
publicados por outros pesquisadores, que descreveram a geração de linfócitos
T CD8 específicos do tipo TCM durante a fase tardia da infecção experimental

87
de camundongos com a cepa Brazil de T. cruzi [274]. Contrastam também com
alguns resultados desses mesmos autores sugerindo que, após o tratamento
com o benzonidazol, há um aparecimento de uma população de linfócitos T
CD8 específicos do tipo TCM, e que esta sim teria propriedades imunoprotetoras
superiores às das TEM [59].
Utilizando camundongos deficientes para o gene da perforina, pôde-se
observar que as quantidades de linfócitos T CD8 específicos, determinadas
pela utilização de multímeros, podem não correlacionar com a capacidade
imunoprotetora destes. A razão é que muito linfócitos T CD8 podem não
apresentar as características multifuncionais e/ou fenotípicas importantes para
a imunoproteção. Baseando-se no que se observou nesta tese, acredita-se
que, no caso da infecção pelo T. cruzi, a busca por linfócitos específicos
multifuncionais deve nortear a análise dos diferentes protocolos de vacinação.
Contudo, estes correlatos observados devem ainda ser testados em outros
protocolos vacinais (efetivos ou não) contra a infecção experimental pelo
T. cruzi, a fim de que possam ser validados.
Assim como este trabalho, nos últimos anos, outros têm sido capazes de
trazer alguma luz sobre o que faz uma resposta CD8 ser efetiva ou não. Em
alguns modelos (como na infecção pelo HIV ou na resposta contra tumores,
revisto em [275]) não se observou uma correlação clara entre a frequência de
linfócitos T CD8 específicos e a proteção, sugerindo fortemente que tais células
são diferentes entre elas, e que a efetividade de uma resposta depende não
somente de aspectos quantitativos mas, principalmente, qualitativos.
A busca pelos aspectos qualitativos dos linfócitos T CD8 protetores
levou a análises funcionais e fenotípicas destes linfócitos. Diversas moléculas
presentes nos linfócitos T CD8 foram identificadas e utilizadas para classificá-
los. Um exemplo são as moléculas CD62L e CD127, usadas muitas vezes para
definir populações de células T CD8 efetoras (CD62Llo CD127lo), de memória
efetora (CD62Llo CD127hi) e de memória central (CD62Lhi CD127hi) [187].
Vários desses marcadores, contudo, foram definidos com base em infecções
modelo (como por LCMV ou Lysteria monocytogenes) e a sua expressão não
segue os mesmos padrões em outros cenários (revisto em [273]). Existe
também muita discussão a respeito das relações existentes entre as diferentes
populações de linfócitos T CD8 que já encontraram o antígeno (classificadas a

88
partir da expressão de marcadores). Não se sabe ao certo se elas representam
linhagens distintas totalmente diferenciadas, ou se podem se converter umas
nas outras, conforme os estímulos presentes no meio [273]. Algumas
moléculas, por exemplo, foram descritas como marcadoras de exaustão ou de
senescência celular (como o PD-1 [276] ou o KLRG-1 [277]), mas outros
trabalhos indicam que a sua expressão pode refletir um processo regulatório
contínuo e reversível [278]. Assim, a análise das características fenotípicas dos
linfócitos T CD8 ainda traz poucas respostas a respeito da sua funcionalidade e
da sua capacidade protetora.
A investigação das funções efetoras dos linfócitos T CD8 específicos
tem mostrado que a presença das células multifuncionais, que exercem mais
de uma função efetora simultaneamente (incluindo degranulação, produção de
IFN-γ, TNF-α, IL-2 ou MIP-1β), se correlaciona fortemente com a eficácia no
controle da replicação do HIV in vivo, além de tais linfócitos serem induzidos
por protocolos vacinais efetivos (revisto em [275]). Apesar dessa associação,
não se definiu ainda se a efetividade dos linfócitos T CD8 multifuncionais seria
decorrente da soma das suas funções efetoras ou se, eventualmente, a
presença simultânea dessas funções seria um marcador de células menos
diferenciadas, com um maior potencial protetor.
Nesta tese, analisou-se também a importância dos linfócitos T CD4.
Observou-se que, no protocolo de vacinação heteróloga, estes linfócitos são
críticos para a indução dos linfócitos T CD8 efetores (BCGA e MMR, dados não
publicados) e também para a imunidade protetora após o desafio experimental.
Esses linfócitos secretam IFN-γ após o re-estímulo in vitro com a ASP-2
recombinante. Tal característica se correlaciona com a imunidade protetora dos
camundongos normais, quando comparados com os animais perforina KO.
Apesar de ainda não se haver determinado formalmente se estes linfócitos T
CD4 são também do tipo multifuncionais, acredita-se que, como em outras
imunizações utilizando adenovírus recombinantes, boa parte deles apresentará
tal característica [250,279].

89
Conclusões
Ao longo deste trabalho obtiveram-se evidências que confirmam a
hipótese inicial de que, utilizando diferentes formulações vacinais e/ou
esquemas de vacinações, se poderia melhorar racionalmente a eficácia da
vacinação contra a infecção experimental pelo T. cruzi. Pela utilização de
depleções in vivo e de animais geneticamente deficientes, definiu-se a
importância dos linfócitos T CD4 e T CD8, da perforina e do IFN-γ na imunidade
gerada pela vacinação com a ASP-2 de T. cruzi.
Apesar de se estar longe de se esgotar este assunto tão complexo,
acredita-se que os dados obtidos no decorrer desta tese contribuem para: i) a
melhor compreensão dos mecanismos imunológicos de resistência à infecção
pelo T. cruzi e dos seus antígenos alvos; ii) o desenvolvimento de vacinas
recombinantes contra as infecções por protozoários intracelulares em geral e,
especificamente, contra a doença de Chagas; iii) o desenvolvimento de vacinas
recombinantes capazes de induzir potentes respostas imunes mediadas pelos
linfócitos T.

90
Apêndices

91
Apêndice 1

INFECTION AND IMMUNITY, Apr. 2006, p. 2477–2481 Vol. 74, No. 40019-9567/06/$08.00�0 doi:10.1128/IAI.74.4.2477–2481.2006Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Distinct Kinetics of Effector CD8� Cytotoxic T Cells after Infectionwith Trypanosoma cruzi in Naıve or Vaccinated Mice
Fanny Tzelepis,1 Bruna C. G. de Alencar,1 Marcus L. O. Penido,2 Ricardo T. Gazzinelli,2Pedro M. Persechini,3 and Mauricio M. Rodrigues1*
Centro Interdisciplinar de Terapia Genica (CINTERGEN) and Departmento de Microbiologia, Imunologia e Parasitologia,Universidade Federal de Sao Paulo-Escola Paulista de Medicina, Rua Mirassol, 207, Sao Paulo-SP, Brazil, 04044-0101;
Departamento de Bioquımica e Imunologia, Instituto de Ciencias Biologicas, Universidade Federal de Minas Gerais eCentro de Pesquisas Rene Rachou, FIOCRUZ, Avenida Augusto de Lima 1715, Barro Preto, 30190-002,
Belo Horizonte, Minas Gerais, Brazil2; and Instituto de Biofısica Carlos Chagas Filho,Centro de Ciencias da Saude, Bloco G, Universidade Federal do Rio de Janeiro,
Ilha do Fundao, Rio de Janeiro, 21941-900, Brazil3
Received 28 November 2005/Returned for modification 6 January 2006/Accepted 15 January 2006
The kinetics of effector CD8�-T-cell responses to specific Trypanosoma cruzi epitopes was investigated afterchallenge. Our results suggest that the delayed kinetics differs from that observed in other microbial infectionsand facilitates the establishment of the disease in naıve mice. In contrast, in vaccinated mice, the swiftCD8�-T-cell response helps host survival after challenge.
Major histocompatibility complex class Ia-restricted CD8�
T cells are critical for the survival of naıve and vaccinated miceinfected with the human protozoan parasite Trypanosoma cruzi(1, 5, 9, 11, 15, 16, 18). The antiparasitic mechanisms mediatedby these cells are multiple, including cytokine secretion andpossibly direct cytotoxicity against infected cells (6, 12, 13). Inspite of their importance for host resistance, limited informa-tion is available regarding the kinetics of effector CD8� T cellsfollowing parasite challenge. Here, we describe studies aimedat characterizing the kinetics of effector CD8�-T-cell re-sponses specific to epitopes present in the trans-sialidase (TS)or the amastigote surface protein 2 (ASP-2), prime candidatesfor vaccine development against Chagas’ disease (1, 4, 5, 18).
After infection with trypomastigotes of the Y strain, weobserved that the parasitemia of wild-type (WT) C57BL/6,CD8� knockout (KO), gamma interferon (IFN-�) KO, or per-forin KO mice was not significantly different on day 8 afterchallenge (peak parasitemia of WT mice). From days 9 to 13,WT mice quickly reduced their parasitemia, and only 9% ofthem died after challenge (Fig. 1A and B). In contrast, IFN-�KO mice were unable to control the parasitemia, dying fasterthan the other mouse groups. CD8� KO or perforin KO micewere unable to control their parasitemia at the same rate asWT animals, dying between days 15 and 21 after challenge(Fig. 1A and B). We concluded that IFN-� secreted by non-CD8� cells is important until day 13. CD8� cells and perforinare critical for survival after day 14.
Based on the observations that CD8� cells, IFN-�, and per-forin are critical for mouse survival after challenge, we fol-lowed the kinetics of specific effector CD8� T cells by using theex vivo enzyme-linked immunospot (ELISPOT) assay forIFN-� (4) and the in vivo cytotoxicity assay, which measures
the elimination of peptide-coated target cells mediated by per-forin (3). Target cells were coated with synthetic peptides rep-resenting the CD8 T-cell epitope IYNVGQVSI (TS) or VNHRFTLV (ASP-2) (Table 1). The results of the in vivo cytotoxicityassay were obtained by measuring the carboxyfluorescein diac-etate succinimidyl diester-labeled cells in the spleens of recipientmice (3). Identical results were seen when we analyzed labeledcells in lymph nodes (data not shown).
In BALB/c mice, the in vivo cytotoxicity and IFN-�-secretingcells specific to the TS peptide IYNVGQVSI were first de-tected at the peak of parasitemia (day 9). Both T-cell activitiesrose quickly until the 15th day and were kept high until the30th day after infection. During the subsequent period, theydeclined slowly but were still detectable by day 240 (Fig. 2Aand B).
A similar picture emerged when we evaluated the kinetics ofeffector cells specific to the ASP-2 peptide VNHRFTLV. InC57BL/6 mice, both T-cell activities were not detectable at thepeak of parasitemia (day 8). They rose quickly until the 16thday and were kept high until the 60th day after infection.During the subsequent period, both activities declined butwere still detectable by day 240 (Fig. 2C and D).
Treatment of T. cruzi-infected BALB/c or C57BL/6 micewith anti-CD8 monoclonal antibody resulted in a selective de-pletion of splenic CD8� T cells in vivo (�98.6%) and a com-plete reversion of the in vivo cytotoxicity response to the pep-tide IYNVGQVSI or VNHRFTLV (Fig. 3A and B, respectively).
* Corresponding author. Mailing address: CINTERGEN, UNIFESP-Escola Paulista de Medicina, Rua Mirassol, 207, Sao Paulo-SP, Brazil,04044-010. Phone and fax: (55) (11) 5571-1095. E-mail: [email protected].
TABLE 1. Peptides used in the present study
Peptide Antigen aaa MHCb
restrictionForm of the
parasite Reference
IYNVGQVSI TS 359–367 H-2Kd Trypomastigote 14VNHRFTLV ASP-2 553–560 H-2Kb Amastigote 8TEWETGQI ASP-2 320–327 H-2Kk Amastigote 1
a aa, amino acids.b MHC, major histocompatibility complex.
2477
by on August 26, 2009
iai.asm.org
Dow
nloaded from

The phenotypes of splenic IFN-�-secreting cells were deter-mined by sorting, with the aid of a FACS Vantage (BectonDickinson), CD8� cells costained with antibodies to CD62L(anti-CD8-Pe-CY5, clone 53-67, and anti-CD62L-FITC, cloneMel14; BD-Pharmingen) from C57BL/6 mice infected 80 daysearlier. ELISPOT assays performed with these purified cellsdemonstrated that splenic IFN-�-secreting cells were CD8�
CD62LLow (Fig. 4). Similar results had been described earlierfor mice infected with T. cruzi (Brazil strain) for more than 150days (10).
Finally, we compared the kinetics of effector CD8�-T-cell ex-pansion following infection of immune or naıve animals. C57BL/6 or A/Sn mice were immunized with the plasmid pIgSPclone 9(asp-2 gene) or with the recombinant protein gluthatione S-trans-ferase (GST)-P4-P7, respectively (1, 4). After challenge, micevaccinated with plasmid pIgSPclone9 (C57BL/6) or with the re-combinant protein GST-P4-P7 (A/Sn) quickly developed in vivospecific cytotoxic cells against target cells coated with the peptideVNHRFTLV or TEWETGQI, respectively (Fig. 5A and C).Control mice injected with pcDNA3 or recombinant GST pre-sented a delay in the generation of in vivo cytotoxicity.
The enumeration of specific CD8� T cells by ELISPOTshowed a similar pattern of immune response. After challenge,mice vaccinated with the plasmid pIgSPclone9 or with therecombinant protein GST-P4-P7 displayed faster expansionand significantly higher numbers of IFN-�-secreting cells (Fig.5B and D).
An interesting observation we made in naıve mice was thatthe expansion of specific splenic CD8� T cells occurred in thedays following the peak parasitemia, between days 9 and 15after challenge of BALB/c or C57BL/6 mice. The delayedkinetics of specific CD8�-T-cell expansion may be an impor-tant factor for the establishment of infection in naıve hosts.These results are in agreement with the data collected after thechallenge of naıve KO mice. Mouse survival mediated byCD8� T cells, perforin, and IFN-� occurred mainly betweendays 14 and 17 after challenge. This timing correlated closelywith the moment that specific effector CD8� T cells of C57BL/6mice reached their maximum activity (days 15 and 16).
Overall, the kinetics of T. cruzi-specific CD8� cytotoxic Tcells differs sharply from the observations made with miceinfected with lymphocytic choriomeningitis virus, Listeria mono-
FIG. 1. Trypomastigote-induced parasitemia and mortality in WT, CD8� KO, IFN-� KO, or perforin KO mice. Groups of mice were infectedintraperitoneally with 104 bloodstream trypomastigotes of the Y strain of T. cruzi. (A) Parasitemia was followed daily from days 5 to 13 afterchallenge. The results represent the mean of five or six mice � standard deviation. The asterisks denote values statistically higher than the valuesin control WT mice (P � 0.05; one-way analysis of variance). The results are representative of two independent experiments. (B) Kaplan-Meiercurves for survival of each mouse group: (i) WT, n � 32; (ii) CD8� KO, n � 8; (iii) IFN-� KO, n � 12; (iv) perforin KO, n � 16. The results werepooled from two different experiments. Statistical analysis revealed significant differences in the survival of WT mice compared to the other mousegroups (P � 0.0001 in all cases; log rank test). CD8� KO and perforin KO mice survived longer than IFN-� KO (P � 0.01 in both cases).
FIG. 2. Kinetics of peptide-specific cell-mediated immune responses in BALB/c or C57BL/6 mice after challenge with trypomastigotes of T. cruzi.BALB/c mice or C57BL/6 mice were challenged with 2.5 � 103 or 2.5 � 104 bloodstream trypomastigotes of T. cruzi, respectively. On the indicateddays, the parasitemia was monitored in these animals (closed symbols in all panels). The results represent the mean of six mice � standarddeviation (SD). (A) The in vivo cytotoxic activity of BALB/c mice against target cells coated with peptide IYNVGQVSI was determined (opensymbols). The results represent the mean of three to nine mice � SD per group. (B) IFN-�-producing spleen cells of BALB/c mice specific to thepeptide IYNVGQVSI were estimated by the ELISPOT assay (4). The results represent the mean number of spot-forming cells (SFC) per 106
splenocytes � SD (n � 4; open symbols). (C) The in vivo cytotoxic activity of C57BL/6 mice against target cells coated with peptide VNHRFTLVwas determined. The results represent the mean of three or four mice � SD per group (open symbols). (D) IFN-�-producing spleen cells ofC57BL/6 mice specific to the peptide VNHRFTLV were estimated by the ELISPOT assay. The results represent the mean number of SFC per106 splenocytes � SD (n � 4; open symbols). The results are representative of two or more independent experiments.
2478 NOTES INFECT. IMMUN.
by on August 26, 2009
iai.asm.org
Dow
nloaded from

cytogenes, and Plasmodium yoelli. In these cases, maximumCD8�-T-cell immune response was achieved between days 4and 8 following challenge and rapidly declined after that pe-riod (2, 17, 19). Studies of mice infected with -galactosidase-transgenic Toxoplasma gondii described much slower kinetics(7). The distinct kinetics can be explained by differences in thenatures of these infections (acute versus chronic).
As opposed to naıve mice, vaccinated immune animals ex-
hibited a significantly faster in vivo cytotoxic immune response.This swift immune response correlated with protective immu-nity, helping host survival after challenge.
We thank Fidel Zavala for reviewing the manuscript. We are in debtto Maria L. Juliano (UNIFESP-EPM) for providing one of the pep-tides.
This work was supported by grants from FAPESP, The MillenniumInstitute for Vaccine Development and Technology (CNPq-420067/2005-1), and FAPEMIG (EDT 24.000). R.T.G., P.M.P., and M.M.R.are recipients of fellowships from CNPq. F.T. and B.C.G.A. are recip-ients of fellowships from FAPESP.
REFERENCES
1. Araujo, A. F., B. C. de Alencar, J. R. Vasconcelos, M. I. Hiyane, C. R.Marinho, M. L. Penido, S. B. Boscardin, D. F. Hoft, R. T. Gazzinelli, andM. M. Rodrigues. 2005. CD8�-T-cell-dependent control of Trypanosomacruzi infection in a highly susceptible mouse strain after immunization with
FIG. 3. Phenotypes of the cells mediating in vivo cytotoxic activity.BALB/c (A) or C57BL/6 (B) mice were challenged 30 days before thein vivo cytotoxic assay was performed with bloodstream trypomasti-gotes of T. cruzi as described in the legend to Fig. 1. Two days earlier,infected mice were treated with Rat immunoglobulin G (RIgG) or�CD8 monoclonal antibody (18). The in vivo cytotoxic activity againsttarget cells coated with peptide IYNVGQVSI (A) or VNHRFTLV(B) was determined. The results represent the mean of three mice �standard deviation and are representative of two independent experi-ments.
FIG. 4. Phenotypes of IFN-�-producing cells. C57BL/6 mice werechallenged 80 days before the assay was performed with bloodstreamtrypomastigotes of T. cruzi as described in the legend to Fig. 1. CD8�
CD62LLow or CD8� CD62LHigh cells were 97.96% or 94.41% pure,respectively (data not shown). An ELISPOT assay was used to esti-mate the number of IFN-�-producing cells specific to the peptideVNHRFTLV in purified CD8� CD62LLow (A) or CD8� CD62LHigh
(B) cells. The results represent the mean number of spot-forming cells(SFC) per 104 splenocytes � standard deviation for triplicate cultures.The results are representative of two independent experiments.
FIG. 5. Kinetics of the CD8�-T-cell-mediated immune responsesafter challenge in mice vaccinated with the asp-2 gene or a recombi-nant protein representing ASP-2 antigen. C57BL/6 mice were immu-nized intramuscularly with three doses of 100 g plasmids pIgSPclone9(asp-2 gene; hatched bars) or pcDNA3 (control; white bars). Twoweeks after the last dose, the mice were challenged or not intraperi-toneally (i.p.) with 104 bloodstream trypomastigotes. (A) On the indi-cated days, the in vivo cytotoxic activity against target cells coated withpeptide VNHRFTLV was determined. The results represent the meanof three mice � standard deviation (SD) per group. (B) On the indi-cated days, IFN-�-producing spleen cells specific to the peptide VNHRFTLV were estimated by the ELISPOT assay. The results representthe mean number of spot-forming cells (SFC) specific to the peptideVNHRFTLV per 106 splenocytes � SD (n � 4). A/Sn mice wereimmunized with three doses of the recombinant protein GST-P4-P7(hatched bars) or GST (white bars) as described previously (1). Twoweeks after the last dose, the mice were challenged or not i.p. with 250bloodstream trypomastigotes. (C) On the indicated days, the in vivocytotoxic activity against target cells coated with peptide TEWETGQIwas determined. The results represent the mean of three mice � SDper group. (D) On the indicated days, numbers of IFN-�-producingspleen cells specific to the peptide TEWETGQI were estimated by theELISPOT assay. The results represent the mean number of SFC spe-cific to the peptide TEWETGQI per 106 splenocytes � SD (n � 4).The asterisks denote values statistically higher than the values forcontrol mice (P � 0.05; one-way analysis of variance). The results arerepresentative of two independent experiments.
2480 NOTES INFECT. IMMUN.
by on August 26, 2009
iai.asm.org
Dow
nloaded from

recombinant proteins based on amastigote surface protein 2. Infect. Immun.73:6017–6025.
2. Badovinac, V. P., A. R. Tvinnereim, and J. T. Harty. 2000. Regulation ofantigen-specific CD8� T cell homeostasis by perforin and interferon-gamma.Science 290:1354–1358.
3. Barber, D. L., E. J. Wherry, and R. Ahmed. 2003. Cutting edge: rapid in vivokilling by memory CD8 T cells. J. Immunol. 171:27–31.
4. Boscardin, S. B., S. S. Kinoshita, A. E. Fujimura, and M. M. Rodrigues.2003. Immunization with cDNA expressed by amastigotes of Trypanosomacruzi elicits protective immune response against experimental infection. In-fect. Immun. 71:2744–2757.
5. Garg, N., and R. Tarleton. 2002. Genetic immunization elicits antigen-spe-cific protective immune responses and decreases disease severity in Trypano-soma cruzi infection. Infect. Immun. 70:5547–5555.
6. Henriques-Pons, A., G. M. Oliveira, M. M. Paiva, A. F. Correa, M. M.Batista, R. C. Bisaggio, C. C. Liu, V. Cotta-De-Almeida, C. M. Coutinho,P. M. Persechini, and T. C. Araujo-Jorge. 2002. Evidence for a perforin-mediated mechanism controlling cardiac inflammation in Trypanosoma cruziinfection. Int. J. Exp. Pathol. 83:67–79.
7. Kwok, L. Y., S. Lutjen, S. Soltek, D. Soldati, D. Busch, M. Deckert, and D.Schluter. 2003. The induction and kinetics of antigen-specific CD8 T cellsare defined by the stage specificity and compartmentalization of the antigenin murine toxoplasmosis. J. Immunol. 170:1949–1957.
8. Low, H. P., M. A. Santos, B. Wizel, and R. L. Tarleton. 1998. Amastigotesurface proteins of Trypanosoma cruzi are targets for CD8� CTL. J. Immu-nol. 160:1817–1823.
9. Martin, D., and R. Tarleton. 2004. Generation, specificity, and function ofCD8� T cells in Trypanosoma cruzi infection. Immunol. Rev. 201:304–317.
10. Martin, D. L., and R. L. Tarleton. 2005. Antigen-specific T cells maintain aneffector memory phenotype during persistent Trypanosoma cruzi infection.J. Immunol. 174:1594–1601.
11. Miyahira, Y., Y. Takashima, S. Kobayashi, Y. Matsumoto, T. Takeuchi, M.Ohyanagi-Hara, A. Yoshida, A. Ohwada, H. Akiba, H. Yagita, K. Okumura,and H. Ogawa. 2005. Immune responses against a single CD8�-T-Cell
epitope induced by virus vector vaccination can successfully control Trypano-soma cruzi infection. Infect. Immun. 73:7356–7365.
12. Muller, U., V. Sobek, S. Balkow, C. Holscher, A. Mullbacher, C. Museteanu,H. Mossmann, and M. M. Simon. 2003. Concerted action of perforin andgranzymes is critical for the elimination of Trypanosoma cruzi from mousetissues, but prevention of early host death is in addition dependent on theFasL/Fas pathway. Eur. J. Immunol. 33:70–78.
13. Nickell, S. P., and D. Sharma. 2000. Trypanosoma cruzi: roles for perforin-dependent and perforin-independent immune mechanisms in acute resis-tance. Exp. Parasitol. 94:207–216.
14. Rodrigues, M. M., M. Ribeirao, V. Pereira-Chioccola, L. Renia, and F.Costa. 1999. Predominance of CD4 Th1 and CD8 Tc1 cells revealed bycharacterization of the cellular immune response generated by immunizationwith a DNA vaccine containing a Trypanosoma cruzi gene. Infect. Immun.67:3855–3863.
15. Rodrigues, M. M., S. B. Boscardin, J. R. Vasconcelos, M. I. Hiyane, G.Salay, and I. S. Soares. 2003. Importance of CD8 T cell-mediated im-mune response during intracellular parasitic infections and its implica-tions for the development of effective vaccines. Ann. Acad. Bras. Cienc.75:443–468.
16. Tarleton, R. L., B. H. Koller, A. Latour, and M. Postan. 1992. Susceptibilityof beta 2-microglobulin-deficient mice to Trypanosoma cruzi infection. Na-ture 356:338–340.
17. Tsuji, M., and F. Zavala. 2003. T cells as mediators of protective immunityagainst liver stages of Plasmodium. Trends Parasitol. 19:88–93.
18. Vasconcelos, J. R., M. I. Hiyane, C. R. F. Marinho, C. Claser, A. M. Vieira-Machado, R. T. Gazinelli, O. Bruna-Romero, J. M. Alvarez, S. B. Boscardin,and M. M. Rodrigues. 2004. Protective immunity against Trypanosoma cruziinfection in a highly susceptible mouse strain following vaccination withgenes encoding the Amastigote Surface Protein-2 and trans-sialidase. Hum.Gene Ther. 15:878–886.
19. Wherry, E. J., and R. Ahmed. 2004. Memory CD8 T-cell differentiationduring viral infection. J. Virol. 78:5535–5545.
Editor: W. A. Petri, Jr.
VOL. 74, 2006 NOTES 2481
by on August 26, 2009
iai.asm.org
Dow
nloaded from

97
Apêndice 2

Infection with Trypanosoma cruzi Restricts the Repertoire ofParasite-Specific CD8� T Cells Leading to Immunodominance1
Fanny Tzelepis,*† Bruna C. G. de Alencar,*† Marcus L. O. Penido,‡ Carla Claser,*†
Alexandre V. Machado,‡¶ Oscar Bruna-Romero,§ Ricardo T. Gazzinelli,‡¶
and Mauricio M. Rodrigues2*†
Interference or competition between CD8� T cells restricted by distinct MHC-I molecules can be a powerful means to establishan immunodominant response. However, its importance during infections is still questionable. In this study, we describe thatfollowing infection of mice with the human pathogen Trypanosoma cruzi, an immunodominant CD8� T cell immune response isdeveloped directed to an H-2Kb-restricted epitope expressed by members of the trans-sialidase family of surface proteins. To determinewhether this immunodominance was exerted over other non-H-2Kb-restricted epitopes, we measured during infection of heterozygotemice, immune responses to three distinct epitopes, all expressed by members of the trans-sialidase family, recognized by H-2Kb-, H-2Kk-,or H-2Kd-restricted CD8� T cells. Infected heterozygote or homozygote mice displayed comparably strong immune responses to theH-2Kb-restricted immunodominant epitope. In contrast, H-2Kk- or H-2Kd-restricted immune responses were significantly impaired inheterozygote infected mice when compared with homozygote ones. This interference was not dependent on the dose of parasite or thetiming of infection. Also, it was not seen in heterozygote mice immunized with recombinant adenoviruses expressing T. cruzi Ags. Finally,we observed that the immunodominance was circumvented by concomitant infection with two T. cruzi strains containing distinctimmunodominant epitopes, suggesting that the operating mechanism most likely involves competition of T cells for limiting APCs. Thistype of interference never described during infection with a human parasite may represent a sophisticated strategy to restrict primingof CD8� T cells of distinct specificities, avoiding complete pathogen elimination by host effector cells, and thus favoring hostparasitism. The Journal of Immunology, 2008, 180: 1737–1748.
M ajor histocompatibility complex class Ia-restrictedCD8� T cells are important mediators of the adaptiveimmune response against infections caused by intra-
cellular microorganisms. Pathogens have a number of potentialamino acid sequences that can bind to MHC-I molecules and pro-vide targets for specific CD8� T cells. In contrast, hosts have avast number of TCR clonally distributed on these CD8� T cells torecognize a variety of MHC class I (MHC-I)3-peptide complexes.Despite this large possible combination, pathogen-specific CD8�
T cells seem to recognize preferentially a small number ofepitopes. This preference, known as immunodominance, may oc-cur due to different mechanisms including the formation of stableMHC-I peptide complexes on the surface of APC, higher amountsor higher affinity specific T cell precursors or competition of Tcells for APC. The full biological implications of immunodomi-nance during effector and memory immune responses are unknownand are being thoroughly studied during infection with viruses orbacteria (reviewed in Refs. 1–6).
Infection of humans or mice with the digenetic intracellular pro-tozoan parasite Trypanosoma cruzi also induces MHC class Ia-restricted CD8� T cells specific for parasite epitopes (7, 8). This Tcell subpopulation is critical for host survival following infectioneven when small challenge doses of parasites are used to initiateinfection (Refs. 9–11 and reviewed in Ref. 12). Despite the factthat CD8� T cell mediated immune response is critical for hostsurvival during acute infection, T. cruzi manages to survive withinthe host and establishes a lifelong chronic infection. Parasite per-sistence is considered a key element in the development of symp-toms that occur many years or decades after initial infection inapproximately one-third of these chronically infected individuals(13–15). Thus, understanding the specificity, magnitude, and lon-gevity of CD8� T cell immune responses may greatly help us toexplain the complex immunopathology caused by T. cruzi and topropose new means for intervention for Chagas’ disease, whichaffects �20 million people in Latin America.
Studies on the immunodominance of CD8� T cell immune re-sponses during T. cruzi infection were thought not to be simple toaccomplish due to the large parasite genome which contains�12,000 genes (16). Nevertheless, the description of epitopes tar-gets of powerful CD8� T cell immune responses elicited duringT. cruzi infection in distinct inbred mouse strains allowed us to
*Centro Interdisciplinar de Terapia Genica and †Departamento de Microbiologia,Imunologia e Parasitologia, Escola Paulista de Medicina, Universidade Federal deSao Paulo, Sao Paulo, Brazil; ‡Departamento de Bioquımica e Imunologia and §De-partamento de Microbiologia, Instituto de Ciencias Biologicas, Universidade Federalde Minas Gerais, Belo Horizonte, Brazil; and ¶Centro de Pesquisas Rene Rachou,Fundacao Oswaldo Cruz, Belo Horizonte, Brazil
Received for publication May 14, 2007. Accepted for publication November25, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from Fundacao de Amparo a Pesquisa do Estadode Sao Paulo (FAPESP), and The Millennium Institute for Vaccine Development andTechnology (Conselho Nacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)-420067/2005-1), Rede Mineira de Estrutura e Funcao de Biomoleculas (Fun-dacao de Amparo a Pesquisa do Estado de Minas Gerais-24000). F.T., B.C.G.d.A.,and C.C. are recipients of fellowships from FAPESP. A.V.M., O.B.-R., R.T.G., andM.M.R. are recipients of fellowships from CNPq.2 Address correspondence and reprint requests to Dr. Mauricio M. Rodrigues, CentroInterdisciplinar de Terapia Genica, Escola Paulista de Medicina, Universidade Fed-eral de Sao Paulo, Rua Mirassol, 207, Sao Paulo-SP, Brazil, 04044-010. E-mail ad-dress: [email protected] Abbreviations used in this paper: MHC-I, MHC class I; TS, trans-sialidase; SFC,spot forming cell; ASP-2, amastigote surface protein 2.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

initiate studies on the immunodominance phenomenon (7, 8).Surprisingly, these studies suggest that CD8 response is highlyconfined to epitopes expressed by different members of a largefamily of T. cruzi surface Ags named trans-sialidases (TS).Thus, studies performed in our laboratory and elsewhere, showthat C57BL/6, BALB/c, and more recently B10.A mice infectedwith T. cruzi develop a strong and long-lasting CD8� T cellsspecific for, respectively, H-2Kb-, H-2Kd-, and H-2Kk-re-stricted epitopes expressed by members of the TS family ofsurface Ags.
The purpose of the present study was to determine the mechanismsthat established the immunodominance during naturally acquired im-munity following experimental infection with T. cruzi. Toward thatgoal, we compared the magnitude of CD8� T cell immune responsesto a number of different epitopes from members of the TS family,which are recognized by H-2Kb-restricted T cells. We found that theepitope VNHRFTLV was immunodominant following infection ofC57BL/6 mice with parasites of two different strains. Subsequently,we investigated whether this immunodominance could be exerted onthe immune response to CD8 T cell epitopes restricted by differentMHC-I molecules. We accomplished that by comparing specific im-mune responses to three distinct H-2Kb-, H-2Kd-, or H-2Kk-restrictedepitopes following T. cruzi infection of homozygote (C57BL/6,BALB/c, and B10.A) or heterozygote (F1) mice. Our results providestrong evidence that homozygote (C57BL/6) and heterozygote micedevelop similar immune responses to immunodominant H-2Kb-re-stricted epitope VNHRFTLV. However, immune responses toH-2Kd- or H-2Kk-restricted epitopes were severely diminished in het-erozygote mice. This interference/competition was restricted to T.cruzi infection because following immunization with recombinant ad-enovirus, heterozygote mice responded to these same T. cruziepitopes, as well as or better than homozygote strains. Finally, thepossible mechanism mediating this strong immunodominance wasevaluated by simultaneous infection with two T. cruzi strains contain-ing distinct immunodominant epitopes. These mice responded equallywell to the “dominant” and “subdominant” epitopes suggesting thatthe operating mechanism most likely involves competition of T cellsfor limiting APCs. We believe that this competition between T cellsof different specificities is a sophisticated strategy that T. cruzi devel-oped to restrict CD8� T cell responses and escape complete elimi-nation by host effector cells, and thus favoring host parasitism.
Materials and MethodsMice and parasites
Female 8- to 10-wk-old wild-type C57BL/6, BALB/c, B10.A, F1 C57BL/6 � BALB/c, F1 C57BL/6 � B10.A, F1 B10.A � BALB/c mice wereobtained from Federal University of Sao Paulo. Experimental procedureswere approved by the Committee of Ethics of Federal University of SaoPaulo.
Parasites of Y, G, or CL-Brener strains of T. cruzi were used in thisstudy (17, 18). Bloodstream trypomastigotes of Y strain were obtainedfrom plasma of A/Sn mice infected 7 days earlier. Tissue culture trypo-mastigotes of the G or CL-Brener strain were used to infect mice. Theconcentration of parasites was adjusted and each mouse was inoculated i.p.with 0.2 ml containing the indicated amount of trypomastigotes. C57BL/6,B10.A, F1 C57BL/6 � BALB/c, F1 C57BL/6 � B10.A, F1 B10.A �BALB/c were challenged, in most experiments, with 104 bloodstream try-pomastigotes i.p. BALB/c mice were challenged with only 2.5 � 103 par-asites because they are more susceptible to infection and succumb to chal-lenge doses higher than 5 � 103 parasites.
Peptide synthesis
Peptides VNHRFTLV, TEWETGQI, TsKb-18 (ANYDFTLV), TsKb-20(ANYKFTLV) were prepared by standard N�[9-fluorenylmethyloxycar-bonyl] on a PSSM8 multispecific peptide synthesizer (Shimadzu) by solid-phase synthesis with a scale of 30 �M. Peptide was purified by HPLC ina Shimadzu system. Peptides were analyzed in a C18 Vydac column (10 �
250 mm, 5-�m particle diameter). Different peptide batches were obtainedin a range of 80–90% purity. Their identities were confirmed by Q-TOFMicro equipped with an electrospray ionization source (Micromass). Pep-tide IYNVGQVSI was purchased from Neosystem. As estimated by HPLCanalysis, peptide IYNVGQVSI was �90% pure.
Immunological assays
For the in vivo cytotoxicity assays, splenocytes of the different mousestrains were divided into two populations and labeled with the fluorogenicdye CFSE (Molecular Probes) at a final concentration of 5 �M (CFSEhigh)or 0.5 �M (CFSElow). CFSEhigh cells were pulsed for 40 min at 37°C with1–2.5 �M of H-2Kb ASP-2 peptide (VNHRFTLV), or H-2Kb TsKb-18peptide (ANYDFTLV), H-2Kb TsKb-20 peptide (ANYKFTLV), or H-2Kd
TS peptide (IYNVGQVSI) peptide or H-2Kk ASP-2 peptide (TEWETGQI). CFSElow cells remained unpulsed. Subsequently, CFSEhigh cellswere washed and mixed with equal numbers of CFSElow cells before in-jecting i.v. 15–20 � 106 total cells per mouse. Recipient animals were micethat had been infected or not with T. cruzi. Spleen cells of recipient micewere collected 20 h after transfer, fixed with 3.7% paraformaldehyde andanalyzed by FACS, using a FACSCalibur Cytometer (BD Biosciences).Percentage of specific lysis was determined using the formula: 1 �((% CFSEhigh infected/% CFSElow infected)/(%CFSEhigh naive/% CFSElow
naive)) � 100%. In experiments using adenovirus, we estimated the per-centage of CFSEhigh or CFSElow cells in immunized, not infected mice.ELISPOT assay for enumeration of IFN-�-producing cells was performedessentially as described earlier (19).
MHC-I tetramer IYNVGQVSI/Kd or pentamer VNHRFTLV/Kb were syn-thesized at the Tetramer Core Facility (Emory University, Atlanta, GA) orwere purchased from ProImmune, respectively. Mouse splenocytes were an-alyzed. Single-cell suspensions of splenocytes were washed in PBS, stained for15 min at 37 °C with tetramers, then stained 30 min at 4°C with labeledanti-CD8 Abs as well as anti-CD4, -CD11b, and -B220. At least 1,000,000cells were acquired on a FACSCalibur flow cytometer (BD Pharmingen) thenanalyzed with FlowJo (Tree Star) using a biexponential transform.
Recombinant adenoviruses and immunization
pAdCMV-TS is an adenoviral transfer plasmid that contains an eukaryoticexpression cassette formed by the CMV immediate-early promoter and theSV40 RNA polyadenylation sequences. Inside this cassette, we cloned theDNA sequences encoding T. cruzi TS protein signal peptide and catalyticdomain obtained by restriction enzyme digestion of plasmid p154/13 (ad-eno-TS). Equivalently, pAdCMV-amastigote surface protein-2 (ASP2) en-codes ASP2 sequences obtained by restriction digestion of plasmid pIgSPclone 9 (adeno-ASP-2). Viruses were generated and purified as describedearlier (20).
Mice were inoculated i.m. in each tibialis anterioris muscle with 50 �lof viral suspension containing 5 � 107 PFU of each adenovirus. Immu-nological assays were performed 15 days after viral inoculation.
Statistical analysis
Values were compared by one-way ANOVA followed by Tukey honestlysignificant difference tests available at http://faculty.vassar.edu/lowry/VassarStats.html. Differences were considered significant when the p valuewas �0.05.
ResultsImmunodominance of the CD8� T cell immune response to theH-2Kb-restricted epitope VNHRFTLV in C57BL/6 mice infectedwith T. cruzi of Y or G strains
Three epitopes recognized by H-2Kb-restricted CD8� T cells weredescribed as targets of strong and long-lasting immune responsesfollowing infection of C57BL/6 mice with T. cruzi (Table I, Refs.
Table I. Peptides used in the study
Peptide/Epitope Sequence H-2K Restriction Ref.
VNHRFTLV VNHRFTLV b 7TsKb-18 ANYDFTLV b 8TsKb-20 ANYKFTLV b 8IYNVGQVSI IYNVGQVSI d 7TEWETGQI TEWETGQI k 7
1738 COMPETITIVE T CELL PRIMING DURING PROTOZOAN INFECTION
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

7 and 8). These epitopes are expressed by members of the TSfamily of surface proteins/Ags. The characterization of the im-mune response to the epitope VNHRFTLV was performed by our
group following infection of C57BL/6 mice with parasites of the Ystrain of T. cruzi (7). The immune responses to the other twoepitopes (TsKb-18 and TsKb-20) were described in C57BL/6 mice
FIGURE 1. Specific CD8� T cell-mediated immune responses in C57BL/6 mice challenged with T. cruzi. C57BL/6 mice were challenged or not i.p. with 104
bloodstream trypomastigotes of the Y strain (A–C), or 105 trypomastigote culture forms of the G or CL-Brener strains (D and E) of T. cruzi. A, At the indicateddays, the in vivo cytotoxic activity against target cells coated with peptides VNHRFTLV, TsKb-18, or TsKb-20 were determined as described in Materials andMethods. Results represent the mean of four mice � SD per group. �, In vivo cytotoxicity against target cells coated with peptide VNHRFTLV was significantlyhigher than TsKb-18- or TsKb-20-coated cells (p � 0.01). B, At the indicated days, IFN-�-producing spleen cells specific for peptides VNHRFTLV, TsKb-18,or TsKb-20 were estimated ex vivo by ELISPOT assay. Results represent the mean number of SFC per 106 splenocytes � SD (n � 4) following in vitro stimulationwith the indicated peptide or medium only. �, At day 15, the numbers of SFC specific for peptide VNHRFTLV or TsKb-20 were significantly higher than thenumber of TsKb-18-specific SFC (p � 0.01). �, At days 30 and 60, the number of SFC specific for peptide VNHRFTLV was significantly higher than the numberof TsKb-18- or TsKb-20-specific SFC (p � 0.01). C, Ex vivo ELISPOT assay was performed using in vitro the indicated concentrations of peptides VNHRFTLVor TsKb-20 to stimulate spleen cells from mice infected 15 days earlier. The data was normalized against the maximal values obtained with each peptide in eachindividual mouse. This value was considered 100%. Results of the titration curve of each individual mouse are shown. D, The numbers of IFN-�-producing spleencells specific for peptides VNHRFTLV, TsKb-18, or TsKb-20 were estimated ex vivo by ELISPOT assay in noninfected mice or animals infected with parasitesof the G or CL-Brener strain 15 days earlier. Results represent the mean number of SFC per 106 splenocytes � SD (n � 4) following in vitro stimulation withthe indicated peptide or medium only. �, The number of SFC specific for peptide VNHRFTLV was significantly higher than the number of TsKb-18- orTsKb-20-specific SFC (p � 0.01). †, The number of SFC specific for peptides TsKb-18 or TsKb-20 were significantly higher than the number of specific SFCfor peptide VNHRFTLV (p � 0.01). E, The in vivo cytotoxic activity against target cells coated with peptides VNHRFTLV, TsKb-18, or TsKb-20 was determinedin mice challenged with trypomastigotes of the G or CL-Brener strains. Results represent the mean of four mice � SD per group. �, In vivo cytotoxicity against target cellscoated with peptide VNHRFTLV was significantly higher than TsKb-18- or TsKb-20-coated cells (p � 0.01). †, In vivo cytotoxicity against target cells coated with peptideTsKb-18 was significantly higher than VNHRFTLV or TsKb-20-coated cells (p � 0.01). Results are representative of two or more independent experiments.
1739The Journal of Immunology
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

infected with parasites of the Brazil, CL, and Y strain (8). TsKb-18(ANYDFTLV) and TsKb-20 (ANYKFTLV) epitopes display asingle amino acid substitution (D3K) but are recognized byCD8� T cells of distinct specificities (Table I and Ref. 8).
To evaluate the immunodominance pattern among these threeepitopes after infection of C57BL/6 mice with T. cruzi, we esti-mated in vivo peptide-specific cytotoxicity and ex vivo the number
of peptide-specific IFN-�-producing cells following infection withparasites of the Y strain. In this mouse model, specific immuneresponses were first detected 9 days following infection. The max-imum in vivo cytotoxicity occurred from days 15 to 90 followinginfection (7). Fig. 1A shows the kinetics of the in vivo cytotoxicityspecific for target cells coated with peptide VNHRFTLV. Morethan 96% specific lyses were detected at day 15, 30, or 60 after
FIGURE 2. Comparison of specific CD8� T cell-mediated immune responses in homozygote or heterozygote mice challenged with T. cruzi. Mice ofthe indicated strains were challenged i.p. with 104 bloodstream trypomastigotes of Y strain of T. cruzi, except for BALB/c mice which received 2500trypomastigotes. Fifteen days later, specific immune responses were estimated in vivo by cytotoxicity (A, C, and E) or ex vivo by ELISPOT (B, D, andF). A and B, �, The in vivo cytotoxicity or the number of SFC from heterozygote F1 C57BL/6 � BALB/c mice specific for peptide IYNVGQVSI weresignificantly lower than the values of homozygous BALB/c mice (p � 0.01 in both cases). C and D, �, The in vivo cytotoxicity or the number of SFC fromheterozygote F1 C57BL/6 � B10.A mice specific for peptide TEWETGQI were significantly lower than the values of homozygous B10.A mice (p � 0.01in both cases). E and F, The in vivo cytotoxicity or the number of SFC from heterozygote F1 B10.A � BALB/c mice specific for peptides TEWETGQIor IYNVGQVSI was significantly lower than the values of homozygous B10.A or BALB/c mice, respectively (p � 0.01 in both cases). Results arerepresentative of two or more independent experiments.
1740 COMPETITIVE T CELL PRIMING DURING PROTOZOAN INFECTION
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

challenge. These values were significantly higher than cytotoxicitylevels specific for target cells coated with peptides TsKb-18 orTsKb-20. In these cases, the maximum specific cytotoxicities were52.0 � 2.3 or 63.8 � 4.0, respectively. The numbers of splenicIFN-�-producing cells specific for peptides VNHRFTLV orTsKb-20 detected ex vivo by ELISPOT assay were quite similar atday 15 postinfection. However, at days 30 and 60 postinfection,the number of IFN-�-producing cells specific for peptide VNHRFTLV was more than three times higher than cells specific forTsKb-20 (Fig. 1B). Despite the fact that TsKb-18 peptide has asingle amino acid substitution when compared with TsKb-20, veryfew IFN-�-producing cells specific for this epitope were detectedex vivo at day 15, 30, or 60 postinfection (Fig. 1B). Analysis of theimmune T cell affinities for the peptides VNHRFTLV or TsKb-20failed to provide any significant degree of difference (Fig. 1C).
We also estimated the number of IFN-�-producing cells de-tected ex vivo following infection with a second strain of T. cruzi(G strain). We found that the number of IFN-�-producing cellsspecific for peptide VNHRFTLV was significantly higher than thenumber of cells specific for peptides TsKb-18 or TsKb-20 (Fig.1D). In mice infected with the G strain, the values of the in vivocytotoxicity specific for target cells coated with peptide VNHR
FTLV were also significantly higher than cytotoxicity levels spe-cific for target cells coated with peptide TsKb-18 or TsKb-20 ( p �0.001, Fig. 1E).
Similar experiments performed with C57BL/6 mice infected with athird T. cruzi strain (CL-Brener) provided a distinct result. In theseanimals, we found that the number of IFN-�-producing cells specificfor peptide VNHRFTLV was significantly lower than the number ofcells specific for peptides TsKb-18 or TsKb-20 (Fig. 1D). In miceinfected with the CL-Brener strain, the values of the in vivo cytotox-icity specific for target cells coated with peptide VNHRFTLV werealso significantly lower than cytotoxicity levels specific for target cellscoated with peptide TsKb-18 or TsKb-20 (Fig. 1E).
In addition, a number of other peptides from T. cruzi previouslydescribed as capable of binding to H-2Kb were also tested ex vivo byELISPOT using splenic cells from C57BL/6 infected 15 or 30 daysearlier with parasites of the Y strain (21–23). None of them stimulatedIFN-�-producing cells (data not shown). Based on these observations,it seems that the CD8� T cell immune response to VNHRFTLV isimmunodominant during infection of C57BL/6 mice with parasites ofthe Y or G strain of T. cruzi. In contrast, during infection of C57BL/6mice with parasites of the CL-Brener strain, CD8� T cell immuneresponse to TsKb-18 and TsKb-20 are immunodominants.
FIGURE 3. Specific CD8� T cell-mediated immune responses in heterozygote F1 C57BL/6 � BALB/c mice challenged with different doses of T. cruzi.Groups of heterozygote F1 C57BL/6 � BALB/c mice were challenged i.p. with 104 or 105 bloodstream trypomastigotes of the Y strain of T. cruzi. Thein vivo cytotoxic activities against target cells coated with peptides VNHRFTLV or IYNVGQVSI were estimated 15 (A) or 30 days (B) following infection.Results represent the mean of four mice � SD per group. �, The in vivo cytotoxicity against target cells coated with peptide IYNVGQVSI was significantlylower than against cells coated with peptide VNHRFTLV (p � 0.01). No statistically significant difference was detected when we compared the immuneresponse in mice which received 104 or 105 parasites. C, IFN-�-producing spleen cells specific for peptides VNHRFTLV or IYNVGQVSI were estimatedex vivo by ELISPOT 15 days following infection. Results represent the mean number of SFC per 106 splenocytes � SD (n � 4) following in vitrostimulation with the indicated peptide or medium only. �, The number of SFC specific for peptide IYNVGQVSI was significantly lower than VNHRFTLV-specific SFC (p � 0.05 in both doses). No statistically significant difference was detected in the number of SFC specific for peptide IYNVGQVSI of micewhich received 104 or 105 parasites (p � 0.05). D, F1 C57BL/6 � BALB/c mice were challenged or not (day 0) i.p. with 104 bloodstream trypomastigotesof T. cruzi. At the indicated days, IFN-�-producing spleen cells specific for peptides VNHRFTLV or IYNVGQVSI were estimated ex vivo by ELISPOT.Results represent the mean number of peptide-specific SFC per 106 splenocytes � SD (n � 4). �, The number of SFC specific for peptide VNHRFTLVwas significantly higher than IYNVGQVSI-specific SFC (p � 0.01).
1741The Journal of Immunology
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

T. cruzi-infected heterozygote mice displays significantlyreduced CD8� T cell immune responses to certain parasitesepitopes
This immunodominance could be explained by several nonmutu-ally exclusive reasons. The first possibility is that during T. cruziinfection the number of MHC-I-VNHRFTLV complexes formedon the surface of APC is higher than the other two epitopes. Al-ternatively, this complex could be exposed for a longer period. Athird possibility is that C57BL/6 mice have a greater number of, orhigher affinity, precursor T cells specific for MHC I-VNHRFTLVcomplexes. Also, it is possible that a competition occurs betweenspecific T cells for APC. This last possibility could be caused, orincreased, by the first three and may provide a much strongermechanism of immunodominance. In this case, the immunodom-inant response would “interfere” with priming of immune re-sponses specific for epitopes restricted by distinct MHC-I mole-cules (24).
To investigate some of these possibilities, we took advantage ofthe fact that we had previously identified two other epitopes rec-ognized by H-2Kk- (TEWETGQI) or H-2Kd (IYNVGQVSI)-re-stricted CD8� T cells from T. cruzi infected mice (Table I and Ref.7). The presence of strong immune responses to epitopes restrictedby distinct MHC-I alleles allowed us to investigate a possible in-terference of one CD8� T cell-mediated immune response on theothers, or simultaneously on one another. To evaluate that possi-bility, we infected heterozygote and homozygote mice and com-
pared the immune responses to three distinct epitopes. Fifteen daysafter challenge, homozygote mice (C57BL/6 or BALB/c) dis-played �90% in vivo cytotoxicity specific for target cells coatedwith peptide VNHRFTLV (C57BL/6) or IYNVGQVSI (BALB/c).Heterozygote mice (F1 C57BL/6 � BALB/c) also presented an invivo cytotoxicity against target cells coated with peptide VNHRFTLV above 95%. In contrast, only 12.92 � 5.91% of the targetcells coated with peptide IYNVGQVSI were eliminated in vivo(Fig. 2A). Analysis of the number of splenic peptide-specific IFN-�-producing cells revealed a similar picture. Although homozygotemice (C57BL/6 or BALB/c) had a significant number of T cellsspecific for the respective MHC-I-restricted epitope, heterozygotemice responded well only to H-2Kb-restricted epitope VNHRFTLV (Fig. 2A). In F1 C57BL/6 � BALB/c mice, the number ofT cells specific for H-2Kd-restricted epitope IYNVGQVSI wasonly 18.51% of the number detected in BALB/c mice.
Similar experiments were performed by comparing CD8� T cellimmune responses of homozygote mice (C57BL/6 or B10.A) withheterozygote F1 C57BL/6 � B10.A. Homozygote mice presentedan immune response capable of eliminating in vivo �90% of pep-tide-coated target cells. Heterozygote animals also displayed an in
FIGURE 4. Staining of T. cruzi-specific CD8� T cells in infected ho-mozygote and heterozygote mice. Spleen cells were collected from ho-mozygote or heterozygote mice infected or not 35 days earlier with T.cruzi. The numbers of total splenic CD8� costained with either VNHRFTLV-Kb pentamer (A) or IYNVGQVSI-Kd tetramer (B) are shown. Dataare representative of three mice � SD. These experiments were performedtwice with similar results.
FIGURE 5. Recognition of epitopes VNHRFTLV and IYNVGQVSI byspecific T cells as measured by the ELISPOT assays. C57BL/6 (n � 4) orBALB/c (n � 4) mice were challenged i.p. with 104 or 2.5 � 103 blood-stream trypomastigotes of the Y strain of T. cruzi, respectively. Fifteendays after challenge, ex vivo ELISPOT assay was performed using in vitrothe indicated peptide concentrations. Spleen cells from C57BL/6 orBALB/c mice were restimulated with peptides VNHRFTLV or IYNVGQVSI, respectively. The data were as normalized against the maximalvalues obtained with each peptide in each individual mouse. This value wasconsidered 100%. Results of the titration curve of each individual mouse isshown.
1742 COMPETITIVE T CELL PRIMING DURING PROTOZOAN INFECTION
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

vivo cytotoxicity against target cells coated with peptide VNHRFTLV of 86.39 � 6.90%. In contrast, in these animals only 29.5 �10.6% of target cells coated with peptide TEWETGQI were elim-inated in vivo (Fig. 2C).
Analysis of the number of splenic peptide-specific IFN-�-pro-ducing cells revealed that homozygote mice had a significant num-ber of T cells specific for the respective MHC-I-restricted epitope.Heterozygote mice responded to the H-2Kb-restricted epitopeVNHRFTLV similarly to homozygote animals (Fig. 2D). In con-trast, in F1 C57BL/6 � B10.A mice, the number of TEWETGQI-specific cells was 8.59% of the number detected in homozygoteB10.A mice.
The last group of heterozygote mice we studied (F1 B10.A �BALB/c) presented an interesting picture. When measured in vivoby cytotoxicity, the elimination of target cells coated with peptideTEWETGQI or IYNVGQVSI was lower in heterozygote micethan in homozygote ones, suggesting that immune responses ofboth specificities interfered with one another (Fig. 2F). Analysis ofthe number of splenic peptide-specific IFN-�-producing cells re-vealed that homozygote mice had a significantly higher number of
T cells specific for the respective MHC-I-restricted epitope. Het-erozygote mice had �40% inhibition of the immune response tothe H-2Kk-restricted epitope TEWETGQI when compared withhomozygote animals. However, a drastic reduction of 92.75% wasseen in the immune response to H-2Kd-restricted epitope IYNVGQVSI (Fig. 2F).
The impaired immune responses observed against these parasiteepitopes in heterozygote mice could be dependent on the dose ofT. cruzi used for challenge. To test this hypothesis, we challengedeach mouse with 105 parasites, a dose 10 times higher than used inthe previous experiments. An increase in the dose of parasite chal-lenge did not improve the limited immune response observed in F1
C57BL/6 � BALB/c mice to the epitope IYNVGQVSI. Both, thein vivo cytotoxicity or the number of IFN-�-producing cells weresimilar in mice challenged with 104 or 105 parasites (Fig. 3, A–C).In contrast, the number of IFN-�-producing cells specific for theimmunodominant epitope VNHRFTLV increased in mice chal-lenged with 105 parasites (Fig. 3C). Also, the reduced immuneresponses observed against the epitope IYNVGQVSI in F1
C57BL/6 � BALB/c mice was not due to a delay in the priming
FIGURE 6. MHC class I expression in splenicAPC cells from homozygote or heterozygote mice.Splenic cells were stained with anti-H-2Kd-FITC oranti-H-2Kb-FITC and anti-B220-PE or anti-CD11c-PE. The frequency of each cell population is indi-cated in the corners. The mean fluorescence (MF) ofthe MHC I molecules (H-2Kb or H-2Kd) of the dou-ble-positive cells are also shown. Infected F1
C57BL/6 � BALB/c mice were infected 30 daysbefore with 104 blood stream trypomastigotes. Re-sults of each mouse group (infected and nonin-fected) were reproduced at least twice with identicalresults.
1743The Journal of Immunology
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

and/or expansion of specific CD8� T cells. The kinetics of IFN-�-producing T cells specific for VNHRFTLV showed a vigorousexpansion during the first 30 days following infection while cellsspecific for IYNVGQVSI expanded very little in that period (Fig.3C). Specific immune response measured by in vivo cytotoxicity tothe epitope IYNVGQVSI or VNHRFTLV was performed in miceinfected 60 days earlier. Although the immune response to VNHRFTLV was still high (91.1 � 0.5%, n � 3) at that point in time, theimmune response to the epitope IYNVGQVSI was still negative(0.0 � 0.0, n � 3, data not shown).
Finally, we used the staining of specific T cells to visualizewhether in fact there was a significantly lower number of CD8� Tcells specific for the epitope IYNVGQVSI in F1 C57BL/6 �BALB/c mice. As shown in Fig. 4A, the total number of CD8�
spleen cells from infected F1 C57BL/6 � BALB/c mice stainedwith the pentamer VNHRFTLV/Kb was only 18.35% lower thanthe number of cells in C57BL/6. When similar analysis was per-formed to estimate the number CD8� spleen cells stained with thetetramer IYNVGQVSI/Kd, we found that infected F1 C57BL/6 �BALB/c mice had 91.5% less cells than BALB/c mice (Fig. 4B).
Analysis of the immune T cell affinities for the peptides VNHRFTLV or IYNVGQVSI were performed using immune T cellsfrom C57BL/6 or BALB/c mice, respectively. This analysis pro-vided evidence that the affinity of T cells specific for the H-2Kd-restricted epitope IYNVGQVSI was slightly higher than the affin-ity of the T cells specific for the H-2Kb-restricted epitopeVNHRFTLV, a fact that cannot explain the immunodominance ofthe later (Fig. 5).
Analysis of the expression of MHC-I molecules H-2Kb andH-2Kd were performed using spleen cells of C57BL/6, BALB/c,and F1 C57BL/6 � BALB/c mice (Fig. 6). The expression of eitherH-2Kd or H-2Kb molecules on the surface of F1 C57BL/6 �BALB/c mice was lower than the homozygote animals. Neverthe-less, in normal F1 C57BL/6 � BALB/c mice, the expression ofH-2Kd molecules (nonresponder allele) in B220� or CD11c� orother cell types were higher than the expression of H-2Kb mole-cules (responder allele). The same picture was observed when weanalyzed spleen cells from infected F1 C57BL/6 � BALB/c.Therefore the absence of immune response in the F1 C57BL/6 �BALB/c to the H-2Kd-restricted epitope IYNVGQVSI cannot beattributed to a significantly lower expression of H-2Kd moleculeson APCs. Other experiments provided below also contribute to thisinterpretation.
Immune responses of heterozygote mice to T. cruzi epitopesfollowing immunization with recombinant adenovirus are notimpaired
A possible explanation for the impaired immune response to theH-2Kd-restricted epitope IYNVGQVSI in heterozygote mice could bethe development of a severely skewed T cell repertoire in the presenceof MHC-I H-2Kb or H-2Kk molecules. Also, the number of H-2Kd
molecules on the surface of cells from heterozygote mice could beresponsible for the reduced response to the epitope IYNVGQVSI. Totest these hypotheses in vivo, we immunized homozygote (BALB/c)and heterozygote (F1 C57BL/6 � BALB/c or F1 B10.A � BALB/c)mice with a recombinant adenovirus expressing the TS protein, theAg which contains the epitope IYNVGQVSI (20). Following immu-nization with recombinant adenovirus, the peak CD8� T cell immuneresponse is at 15 days (25). The immune responses of heterozygotemice measured by the in vivo cytotoxicity against target cells coatedwith the peptide IYNVGQVSI was higher than homozygote BALB/c(Fig. 7A). Therefore, after immunization with a recombinant virus, theheterozygote mice immune response to the peptide IYNVGQVSI wasnot impaired.
The limited immune response to epitope TEWETGQI in het-erozygote (F1 C57BL/6 � B10.A) could be explained by a reducedAg presentation in vivo to H-2Kk-restricted T cells specific for theepitope TEWETGQI caused by some intrinsic property of theH-2Kb-restricted epitope VNHRFTLV. Alternatively, T cellsspecific for VNHRFTLV could inhibit the activation of T cellsspecific for TEWETGQI by a direct interaction between CD8� Tcells. To approach these questions, we used a second recombinantadenovirus expressing the ASP-2 of T. cruzi. This Ag containsboth of these epitopes (20). Homozygote (C57BL/6 or B10.A) orheterozygote (F1 C57BL/6 � B10.A) mice immunized with therecombinant adenovirus adeno-ASP-2 developed immune responsespecific for H-2Kb-restricted epitope VNHRFTLV significantlyhigher than the response to H-2Kk-restricted epitope TEWETGQI.Nevertheless, the immune response to the epitope TEWETGQI wasidentical in homozygote or heterozygote mice (Fig. 7B). These resultsindicated that although the immune response to the epitope VNHRFTLV seems to be stronger following immunization with adeno-ASP-2 virus, it did not interfere with the smaller response to theepitope TEWETGQI.
FIGURE 7. Immune responses of homozygote and heterozygote miceto T. cruzi epitopes following immunization with recombinant adenovirus.The indicated mouse strains were immunized i.m. 15 days earlier withrecombinant adenovirus expressing a control protein (Adeno-�gal), T.cruzi trans-sialidase (adeno-TS) (A), or amastigote Surface Protein-2 (Ad-eno-ASP-2) (B). Specific immune responses to the indicated peptides wereestimated by the in vivo cytotoxicity assay. Results represent the mean offour mice � SD per group. �, The in vivo cytotoxicity of heterozygote F1
C57BL/6 � BALB/c or F1 B10.A � BALB/c mice against target cellscoated with peptide IYNVGQVSI were significantly higher than homozy-gote BALB/c mice (p � 0.01 in both cases). Results are representative ofthree independent experiments.
1744 COMPETITIVE T CELL PRIMING DURING PROTOZOAN INFECTION
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

Comparison of the CD4 T cell dependence on the immune re-sponse following infection with T. cruzi or immunization with re-combinant adeno-ASP-2 was performed using CD4 knockoutmice. We observed that these mice had a severe reduction on thein vivo cytotoxic immune response to the epitope VNHRFTLVfollowing infection or immunization proving that both cytotoxicimmune responses were dependent on CD4� T cell activation(data not shown).
Immune responses of C57BL/6 mice to T. cruzi epitopesfollowing infection with two parasite strains containing distinctimmunodominant epitopes
Precisely how this immunodominant response to VNHRFTLVepitope interferes with T cells of other specificities during T. cruziinfection was not clear. It was possible that T cells specific for theepitope VNHRFTLV prevented the presentation of the otherepitopes when both were present in the same APCs. In that case,we predicted that strong responses to both epitopes (dominant andsubdominant) would be induced when the epitopes were expressedin different APCs. To test this hypothesis, we infected C57BL/6mice with two different parasite strains expressing distinct immu-
nodominant epitopes. As described above, infection with the Ystrain of T. cruzi induced a strong response to the epitope VNHRFTLV, but not to TsKb-18, as measured by in vivo cytotoxicity orELISPOT assay (Fig. 8). In contrast, infection with parasites of theCL-Brener strain elicited strong immune responses to the epitopeTsKb-18, but not to VNHRFTLV (Fig. 8). As we predicted, miceinfected with both parasite strains at the same time displayedstrong immune response to both epitopes (Fig. 8). This result sug-gests that separation of the “dominant” and “subdominant”epitopes can circumvent the immunodominance.
DiscussionInterference or competition between CD8� T cells restricted bydistinct MHC-I molecules have been described following immu-nization with different Ags (24, 26–28). It is a powerful meansused by CD8� T cells specific for a particular MHC-I-peptidecomplex to restrict expansion of T cells of all other specificitiesestablishing an immunodominant response. Whether this type ofinterference/competition between CD8� T cells restricted for dif-ferent MHC-I molecules could in fact be critical during infectionsis still a matter of debate (29–31). Here, we demonstrate thatCD8� T cell immune response specific for H-2Kb-restrictedepitope VNHRFTLV is immunodominant during infection ofC57BL/6 mice with T. cruzi of Y and G strains. In addition, wehave demonstrated that CD8� T cells specific for the immuno-dominant epitope VNHRFTLV greatly interferes with the gener-ation of T cells restricted for other MHC-I molecules. Thus, fol-lowing infection of heterozygote mice with T. cruzi, a significantreduction in the priming of CD8� T cells restricted for H-2Kd orH-2Kk molecules was observed providing evidence that interfer-ence/competition is an important factor down-modulating immuneresponse during experimental infection with this human pathogen.In the case of infection of F1 B10.A � BALB/c mice, a mutualinterference takes place reducing both specific responses to theH-2Kd- or H-2Kk-restricted T cells.
Precisely how this immunodominant response to VNHRFTLVepitope interferes with T cells of other specificities is not clear. Theinterference observed following infection is not related to theepitopes themselves or caused by a skewed T cell repertoire inheterozygote mice as F1 C57BL/6 � BALB/c mice immunizedwith recombinant adenovirus developed immune responses higherthan the homozygote BALB/c to the H-2Kd-restricted epitopeIYNVGQVSI.
A few nonmutually excluding hypotheses can be proposed. In-terference could be due to the presence of CD4�CD25� regulatoryT cells. Thus, Haeryfar et al. (32) found that CD4� CD25� reg-ulatory T cells selectively suppressed responses to the most im-munodominant CD8� T cell epitopes in three distinct systems.However, in a recent study, Kotner and Tarleton (33) describedthat depletion of these cells had little impact on the immune re-sponse of CD8� T cells specific for T. cruzi immunodominant orsubdominant epitopes. Alternatively, it could be possible thatCD8� T cells specific for VNHRFTLV inhibit expansion/prolif-eration of T cells of other specificities by acting directly on re-sponder T cells. This possibility also seems unlikely because afterimmunization with adeno-ASP-2, heterozygote and homozygotemice presented a strong response to VNHRFTLV and also gener-ated immune responses of a similar degree to TEWETGQI epitope.Also, during concomitant infection with two parasites strains, theimmune response to VNHRFTLV did not interfere with the ex-pansion of TsKb-18 specific cells.
The explanation that we favor is that CD8� T cells specific forVNHRFTLV compete with T cells of other specificities in limitingAPC resources during priming with T. cruzi (23, 26, 27). This
FIGURE 8. Specific CD8� T cell-mediated immune responses inC57BL/6 mice challenged with one or two strains T. cruzi. C57BL/6 micewere challenged or not i.p. with 104 bloodstream trypomastigotes of the Ystrain and/or 105 trypomastigote culture forms of the CL-Brener strain of T.cruzi. A, Fifteen days after infection, the in vivo cytotoxic activity againsttarget cells coated with peptides VNHRFTLV or TsKb-18 were determined.Results represent the mean of four mice � SD per group. B, Fifteen days afterinfection, IFN-�-producing spleen cells specific for peptides VNHRFTLV orTsKb-18 were estimated ex vivo by ELISPOT assay. Results represent themean number of SFC per 106 splenocytes � SD (n � 4) following in vitrostimulation with the indicated peptide or medium only. �, Immune responsespecific for the peptide VNHRFTLV was significantly higher than to peptideTsKb-18 (p � 0.01). †, The immune response specific for the peptide TsKb-18was significantly higher than to peptide VNHRFTLV (p � 0.01). Results arerepresentative of two independent experiments.
1745The Journal of Immunology
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

hypothesis is supported by the experiment where we infectedC57BL/6 mice with two parasite strains containing distinct immu-nodominant epitopes. In this scenario, distinct parasite strains aremost likely processed by different APCs, allowing strong immuneresponse to be developed against both epitopes.
To generate an immunodominant response, H-2Kb-VNHRFTLV complexes should be formed in a larger number, or first,leading to a rapid expansion of specific T cells. These T cellswould use the available APC resources precluding or reducingactivation of T cells of other specificities. The possibility that moreH-2Kb-VNHRFTLV complexes are formed has no experimentalsupport. In fact, the observation that homozygote mice developpowerful immune responses to H-2Kd- IYNVGQVSI or H-2Kk-TEWETGQI complexes suggests that the number of complexesformed on the surface of APCs from heterozygote mice should besufficient to promote a strong T cell response. Alternatively, thenumber of T cell precursors and/or their affinities could be factorsthat accelerate the immune response causing an immunodomi-nance to the epitope VNHRFTLV. In experiments designed toevaluate whether CD8� T cells from infected BALB/c had alower affinity for H-2Kd-IYNVGQVSI complexes than cellsfrom infected C57BL/6 mice for H-2Kb-VNHRFLTV, we havefound that this is not the case (Fig. 5). Therefore, it seems thatthe TCR affinity cannot explain the discrepant activation inheterozygote mice.
After contact with APCs, the expansion of T cells with higheraffinity or more precursors for the MHC-I-peptide complex wouldovercome T cells specific for other parasite peptides. However,following immunization with recombinant adenovirus, we werenot able to detect any interference in heterozygote mice when com-pared with homozygote ones, indicating that the affinity/more pre-cursor hypotheses are not a likely explanation for the studied com-petition phenomenon.
Hence, in our opinion, the most plausible explanation is that thisimmunodominance is regulated by the timing and duration ofH-2Kb-VNHRFTLV display on the surface of APCs. Earlier stud-ies have shown that prolonged Ag exposure influences the gener-ation of immune responses improving priming of T cells specificfor immunodominant epitopes (34–36). Indeed, VNHRFTLVepitope is present in proteins expressed by trypomastigotes (infec-tive forms) and amastigotes (intracellular stages) of the Y strain ofT. cruzi (37, 38). This fact may accelerate and improve immuneresponse by providing an advantage for T cells specific for H-2Kb-VNHRFTLV complexes on the surface of APC. In contrast, theepitopes IYNVGQVSI or TEWETGQI are expressed only by try-pomastigotes (infective forms) or amastigotes (intracellularstages), respectively (38, 39). This last hypothesis would explainthe differences observed following infection or immunization ofheterozygote mice with adenovirus-expressing parasite Ag. Impor-tantly, evidence exists that immunodominance may be dictated bythe infective strain of T. cruzi. Thus, while expressing the VNHRFTLV epitope (21), the Brazil strain of T. cruzi induced an im-munodominant response specific for TsKb-20 and TsKb-18epitopes (8). This strain-dependent switch has no immediate ex-planation, but it indicates the importance of parasites in controllingthe immunodominant CD8� T cell response. In the case of theCL-Brener strain, we found during sequence analysis of the cDNAof ASP-2 that the deduced AA sequence was VNYDFTIV (aminoacid substitutions are underlined). A synthetic peptide representingthis epitope was not recognized by CD8� T cells specific for theepitope VNHRFTLV (data not shown).
Although previous studies provided compelling evidence ofcompetition between CD8� T cells for APCs expressing simulta-neously two distinct MHC-I molecules, the substrate they compete
for is not yet known. This competition occurs early in the activa-tion process of T cells (24), and involves membrane proteins re-quired for full T cell activation and/or soluble factor(s) such ascytokines (1, 24, 40). Despite its potential as a mechanism of im-munodominance, during mouse infection with lymphocytic cho-riomeningitis virus, this interference/competition was not apparent(29, 30). Also, ablation of immunodominant epitopes from Listeriamonocytogenes did not cause an increase in the immune responsesto subdominant epitopes during infection, suggesting that therewas no apparent interference/competition among CD8� T cells(31). Very recent reports suggest the opposite, that there is inter-ference between CD8� T cells of different MHC-I specificitiesduring viral infection with HIV, SIV. Severe acute respiratory syn-drome and vaccinia (41–44). Thus, we believe that our observationreinforces the idea that a competitive mechanism for CD8� T cellimmunodominance occurs during infectious diseases with differentpathogens.
Although we do not yet know precisely the mechanism used togenerate this strong non-MHC specific immunodominance, wewould like to suggest that it has an important biological implica-tion for a chronic parasitic disease. In the case of self-resolvingcommon acute viral or bacterial infections, immunodominance fo-cuses the response on few determinants. It was suggested that thisstrategy successfully maximizes the power of effector immune re-sponses and, at the same time, minimizes the risk of autoimmunity(6). In the case of T. cruzi infection (Chagas’ disease), immu-nodominance may have a similar function and may guarantee hostsurvival for a long period of time. However, on the negative side,the immunodominance described here seems to hamper T. cruzi-infected mice (homozygotes or heterozygotes) in their develop-ment of stronger, and/or broader, specific immune responses. Inaddition, an immune response relying mainly on few immunodom-inant CD8� T cell epitopes may favor the escape of immune re-sponse associated with parasite mutation or any particular anti-genic variation process. An example supporting this hypothesiswas described in a chronic viral infection with hepatitis C virus. Astrong immunodominant human CD8� T cells response was de-veloped causing the selection of an escape mutant (45). Regard-less, we favor the hypothesis that either restriction or avoidance ofmaximal host effector functions is a critical step to prevent parasiteelimination, allowing parasite persistence and leading to a chroniclifelong infection.
One important immunological aspect to be considered in therational design of a vaccine for Chagas’ disease is the pathogenesisand the intense myocarditis composed mainly of CD8� T cellselicited during T. cruzi infection. Thus, following vaccination withplasmid DNA or recombinant proteins, we generated memoryCD8� T cells that, after a challenge with T. cruzi, displayed a veryfast anamnestic immune response (7). These cells were specific forimmunodominant epitopes VNHRFTLV or TEWETGQI (7). Sucha strong immune response did not cause any discernible immuno-pathology. It in fact led to a reduced parasite development andprevented the establishment of chronic phase tissue pathologies(46, 47). Also, some of these vaccinated mice completely clearedthe parasite and established sterile immunity (46, 47). These find-ings are also supported by our vaccination studies when we ob-tained reduced parasitism and limited disease development by in-creasing the immune responses to these same immunodominantepitopes (20, 46, 47). Thus, the development of a restricted T cellresponse confined to a limited number of epitopes may be impor-tant to avoid elicitation of potentially self-reactive or pathogenic Tcell clones.
In summary, our results provide evidence of a competitive ex-pansion of CD8� T cells restricted by different MHC-I molecules.
1746 COMPETITIVE T CELL PRIMING DURING PROTOZOAN INFECTION
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

T. cruzi infection may be an interesting model to evaluate themolecular basis for CD8� T cells competition and the biologicalrole of immunodominance during infection with intracellular pro-tozoan parasites, which are major causes of life-threateningchronic infection in humans. Among the protozoan parasites,CD8� T cells have also been shown to greatly influence immunityagainst Plasmodium, Toxoplasma gondii, and Leishmania sp. (48–53). Whether this competitive priming and expansion of T cellsrestricted by different MHC-I molecules also occurs during infec-tion with these parasites also needs to be addressed.
AcknowledgmentsWe thanks Dr. Dan Hoft (St. Louis University, St. Louis, MO) for provid-ing the H-2Kd-IYNVGQVSI tetramer and Karina Inacio Carvalho (CentroInterdisciplinar de Terapia Genic (CINTERGEN), Sao Paulo, Brazil) forhelping with the FACS softwares. FACS analyses were possible due to theuse of equipment provided by BD Biosciences (Sao Paulo, Brazil). We arein debt with Dr. Victor Nussenzweig (New York University, New York, NY),Dr. Moriya Tsuji (Rockefeller University, New York, NY), Dr. George dosReis (Federal University of Rio de Janeiro, Rio de Janeiro, Brazil), Dr. MilenaSoares (Centro de Pesquisas Goncalo Muniz/Fundacao Oswaldo Cruz, BeloHorizonte, Brazil), and Dr. Esper G. Kallas (CINTERGEN-Universidade Fed-eral de Sao Paulo, Sao Paulo, Brazil) for helpful discussion and reviewing themanuscript.
DisclosuresThe authors have no financial conflict of interest.
References1. Sette, A., and R. Sundaran. 2006. The phenomenon of immunodominance: spec-
ulations on the nature of immunodominance. In Immunodominance: The Choiceof the Immune System. J. A. Frelinger, ed. Wiley-VCH Verlag, Weinheim, pp.57–71.
2. Gaddis, D. E., M. J. Fuller, and A. J. Zajac. 2006. CD8 T-cell immunodominance,repertoire, and memory. In Immunodominance: The Choice of the Immune Sys-tem. J. A. Frelinger, ed. Wiley-VCH Verlag, Weinheim, pp. 109–145.
3. Porter, B. B., and J. T. Harty. 2006. Listeria monocytogenes infection and theCD8� T-cell hierarchy. In Immunodominance: The Choice of the Immune System.J. A. Frelinger, ed. Wiley-VCH Verlag, Weinheim, pp. 149–162.
4. Kedl, R. M., J. W. Kappler, and P. Marrack. 2003. Epitope dominance, compe-tition and T cell affinity maturation. Curr. Opin. Immunol. 15: 120–127.
5. Yewdell, J. W., and M. Del Val. 2004. Immunodominance in TCD8�: meetingreview responses to viruses: cell biology, cellular immunology, and mathematicalmodels. Immunity 21: 149–153.
6. Yewdell, J. W. 2006. Confronting complexity: real-world immunodominance inantiviral CD8� T cell responses. Immunity 25: 533–543.
7. Tzelepis, F., B. C. de Alencar, M. L. Penido, R. T. Gazzinelli, P. M. Persechini,and M. M. Rodrigues. 2006. Distinct kinetics of effector CD8� cytotoxic T cellsafter infection with Trypanosoma cruzi in naive or vaccinated mice. Infect. Im-mun. 74: 2477–2482.
8. Martin, D. L., D. B. Weatherly, S. A. Laucella, M. A. Cabinian, M. T. Crim,S. Sullivan, M. Heiges, S. H. Craven, C. S. Rosenberg, M. H. Collins, et al. 2006.CD8� T-cell responses to Trypanosoma cruzi are highly focused on strain-varianttrans-sialidase epitopes. PLoS Pathog. 2: e77.
9. Tarleton, R. L., B. H. Koller, A. Latour, and M. Postan. 1992. Susceptibility of�2-microglobulin-deficient mice to Trypanosoma cruzi infection. Nature 356:338–340.
10. Tarleton, R. L., J. Sun, L. Zhang, M. Postan, and L. Glimcher. 1996. Trypano-soma cruzi infection in MHC-deficient mice: further evidence for the role of bothclass I- and class II-restricted T cells in immune resistance and disease. Int.Immunol. 8: 13–22.
11. Rottenberg, M. E., M. Bakhiet, T. Olsson, K. Kristensson, T. Mak, H. Wigzell,and A. Orn. 1993. Differential susceptibilities of mice genomically deleted ofCD4 and CD8 to infections with Trypanosoma cruzi or Trypanosoma brucei.Infect. Immun. 61: 5129–5133.
12. Martin, D., and R. Tarleton. 2004. Generation, specificity, and function of CD8�
T cells in Trypanosoma cruzi infection. Immunol. Rev. 201: 304–317.13. Tarleton, R. L. 2003. Chagas disease: a role for autoimmunity? Trends Parasitol.
19: 447–451.14. Kierszenbaum, F. 2005. Where do we stand on the autoimmunity hypothesis of
Chagas disease? Trends Parasitol. 21: 513–516.15. Albareda, M. C., S. A. Laucella, M. G. Alvarez, A. H. Armenti, G. Bertochi,
R. L. Tarleton, and M. Postan. 2006. Trypanosoma cruzi modulates the profile ofmemory CD8� T cells in chronic Chagas’ disease patients. Int. Immunol. 18:465–471.
16. El Sayed, N. M., P. J. Myler, D. C. Bartholomeu, D. Nilsson, G. Aggarwal,A. N. Tran, E. Ghedin, E. A. Worthey, A. L. Delcher, G. Blandin, et al. 2005. Thegenome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Sci-ence 309: 409–415.
17. Silva, L. H. P., and V. Nussenzweig. 1953. Sobre uma cepa de Trypanosomacruzi altamente virulenta para o camundongo branco. Folia Clin. Biol. 20:191–203.
18. Yoshida, N. 1983. Surface antigens of metacyclic trypomastigotes of Trypano-soma cruzi. Infect. Immun. 40: 836–839.
19. Miyahira, Y., K. Murata, D. Rodriguez, J. R. Rodriguez, M. Esteban,M. M. Rodrigues, and F. Zavala. 1995. Quantification of antigen specific CD8�
T cells using ELISPOT assay. J. Immunol. Methods 181: 145–154.20. Machado, A. V., J. E. Cardoso, C. Claser, M. M. Rodrigues, R. T. Gazzinelli, and
O. Bruna-Romero. 2006. Long-term protective immunity induced againstTrypanosoma cruzi infection after vaccination with recombinant adenovirusesencoding amastigote surface protein-2 and trans-sialidase. Hum. Gene Ther. 17:898–908.
21. Low, H. P., M. A. Santos, B. Wizel, and R. L. Tarleton. 1998. Amastigote surfaceproteins of Trypanosoma cruzi are targets for CD8� CTL. J. Immunol. 160:1817–1823.
22. Fralish, B. H., and R. L. Tarleton. 2003. Genetic immunization with LYT1 or apool of trans-sialidase genes protects mice from lethal Trypanosoma cruzi in-fection. Vaccine 21: 3070–3080.
23. Wrightsman, R. A., K. A. Luhrs, D. Fouts, and J. E. Manning. 2002. Paraflagellarrod protein-specific CD8� cytotoxic T lymphocytes target Trypanosoma cruzi-infected host cells. Parasite Immunol. 24: 401–412.
24. Willis, R. A., J. W. Kappler, and P. C. Marrack. 2006. CD8 T cell competitionfor dendritic cells in vivo is an early event in activation. Proc. Natl. Acad. Sci.USA 103: 12063–12068.
25. Yang, T. C., J. Millar, T. Groves, N. Grinshtein, R. Parsons, S. Takenaka,Y. Wan, and J. L. Bramson. 2006. The CD8� T cell population elicited byrecombinant adenovirus displays a novel partially exhausted phenotype associ-ated with prolonged antigen presentation that nonetheless provides long-termimmunity. J. Immunol. 176: 200–210.
26. Grufman, P., E. Z. Wolpert, J. K. Sandberg, and K. Karre. 1999. T cell compe-tition for the antigen-presenting cell as a model for immunodominance in thecytotoxic T lymphocyte response against minor histocompatibility antigens. Eur.J. Immunol. 29: 2197–2204.
27. Kedl, R. M., W. A. Rees, D. A. Hildeman, B. Schaefer, T. Mitchell, J. Kappler,and P. Marrack. 2000. T cells compete for access to antigen-bearing antigen-presenting cells. J. Exp. Med. 192: 1105–1113.
28. Kedl, R. M., B. C. Schaefer, J. W. Kappler, and P. Marrack. 2002. T cells down-modulate peptide-MHC complexes on APCs in vivo. Nat. Immunol. 3: 27–32.
29. Butz, E. A., and M. J. Bevan. 1998. Massive expansion of antigen-specific CD8�
T cells during an acute virus infection. Immunity 8: 167–175.30. Probst, H. C., T. Dumrese, and M. F. van den Broek. 2002. Cutting edge: com-
petition for APC by CTLs of different specificities is not functionally importantduring induction of antiviral responses. J. Immunol. 168: 5387–5391.
31. Vijh, S., I. M. Pilip, and E. G. Pamer. 1999. Noncompetitive expansion of cy-totoxic T lymphocytes specific for different antigens during bacterial infection.Infect. Immun. 67: 1303–1309.
32. Haeryfar, S. M., R. J. DiPaolo, D. C. Tscharke, J. R. Bennink, and J. W. Yewdell.2005. Regulatory T cells suppress CD8� T cell responses induced by directpriming and cross-priming and moderate immunodominance disparities. J. Im-munol. 174: 3344–3351.
33. Kotner, J., and R. Tarleton. 2007. Endogenous CD4�CD25� regulatory T cellshave limited role in control of Trypanosoma cruzi infection in mice. Infect. Im-mun. 75: 861–869.
34. Skoberne, M., R. Holtappels, H. Hof, and G. Geginat. 2001. Dynamic antigenpresentation patterns of Listeria monocytogenes-derived CD8 T cell epitopes invivo. J. Immunol. 167: 2209–2218.
35. Badovinac, V. P., B. B. Porter, and J. T. Harty. 2002. Programmed contraction ofCD8� T cells after infection. Nat. Immunol. 3: 619–626.
36. Stock, A. T., S. N. Mueller, A. L. van Lint, W. R. Heath, and F. R. Carbone. 2004.Cutting edge: prolonged antigen presentation after herpes simplex virus-1 skininfection. J. Immunol. 173: 2241–2244.
37. Giordano, R., D. L. Fouts, D. Tewari, W. Colli, J. E. Manning, and M. J. Alves.1999. Cloning of a surface membrane glycoprotein specific for the infective formof Trypanosoma cruzi having adhesive properties to laminin. J. Biol. Chem. 274:3461–3468.
38. Boscardin, S. B., S. S. Kinoshita, A. E. Fujimura, and M. M. Rodrigues. 2003.Immunization with cDNA expressed by amastigotes of Trypanosoma cruzi elicitsprotective immune response against experimental infection. Infect. Immun. 71:2744–2757.
39. Abuin, G., L. H. Freitas-Junior, W. Colli, M. J. Alves, and S. Schenkman. 1999.Expression of trans-sialidase and 85-kDa glycoprotein genes in Trypanosomacruzi is differentially regulated at the post-transcriptional level by labile proteinfactors. J. Biol. Chem. 274: 3461–3468.
40. Melchionda, F., T. J. Fry, M. J. Milliron, M. A. McKirdy, Y. Tagaya, andC. L. Mackall. 2005. Adjuvant IL-7 or IL-15 overcomes immunodominance andimproves survival of the CD8� memory cell pool. J. Clin. Invest. 115:1177–1185.
41. Altfeld, M., E. T. Kalife, Y. Qi, H. Streeck, M. Lichterfeld, M. N. Johnston,N. Burgett, M. E. Swartz, A. Yang, G. Alter, et al. 2006. HLA alleles associatedwith delayed progression to AIDS contribute strongly to the initial CD8� T cellresponse against HIV-1. PLoS Med. 3: e403.
42. Newberg, M. H., K. J. McEvers, D. A. Gorgone, M. A. Lifton, S. H. Baumeister,R. S. Veazey, J. E. Schmitz, and N. L. Letvin. 2006. Immunodomination in theevolution of dominant epitope-specific CD8� T lymphocyte responses in simianimmunodeficiency virus-infected rhesus monkeys. J. Immunol. 176: 319–328.
1747The Journal of Immunology
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

43. Rutigliano, J. A., T. J. Ruckwardt, J. E. Martin, and B. S. Graham. 2007. Relativedominance of epitope-specific CD8� T cell responses in an F1 hybrid mousemodel of respiratory syncytial virus infection. Virology 362: 314–319.
44. Kastenmuller, W., G. Gasteiger, J. H. Gronau, R. Baier, R. Ljapoci, D. H. Busch,and I. Drexler. 2007. Cross-competition of CD8� T cells shapes the immu-nodominance hierarchy during boost vaccination. J. Exp. Med. 204: 2187–2198.
45. Tester, I., S. Smyk-Pearson, P. Wang, A. Wertheimer, E. Yao, D. M. Lewinsohn,J. E. Tavis, and H. R. Rosen. 2005. Immune evasion versus recovery after acutehepatitis C virus infection from a shared source. J. Exp. Med. 201: 1725–1731.
46. Vasconcelos, J. R., M. I. Hiyane, C. R. F. Marinho, C. Claser, A. M.Vieira-Machado, R. T. Gazinelli, O. Bruna-Romero, J. M. Alvarez,S. B. Boscardin, and M. M. Rodrigues. 2004. Protective immunity againstTrypanosoma cruzi infection in a highly susceptible mouse strain following vac-cination with genes encoding the amastigote surface protein-2 and trans-siali-dase. Hum. Gene Ther. 15: 878–886.
47. Araujo, A. F., B. C. de Alencar, J. R. Vasconcelos, M. I. Hiyane, C. R. Marinho,M. L. Penido, S. B. Boscardin, D. F. Hoft, R. T. Gazzinelli, and M. M. Rodrigues.2005. CD8�-T-cell-dependent control of Trypanosoma cruzi infection in a highlysusceptible mouse strain after immunization with recombinant proteins based onamastigote surface protein 2. Infect. Immun. 73: 6017–6025.
48. Rodrigues, M. M., S. B. Boscardin, J. R. Vasconcelos, M. I. Hiyane, G. Salay,and I. S. Soares. 2003. Importance of CD8 T cell-mediated immune responseduring intracellular parasitic infections and its implications for the developmentof effective vaccines. An. Acad. Bras. Cienc. 75: 443–468.
49. Tsuji, M., and F. Zavala. 2003. T cells as mediators of protective immunityagainst liver stages of Plasmodium. Trends Parasitol. 19: 88–93.
50. Morrot, A., and F. Zavala. 2004. Effector and memory CD8� T cells as seen inimmunity to malaria. Immunol. Rev. 201: 291–303.
51. Kwok, L. Y., S. Lutjen, S. Soltek, D. Soldati, D. Busch, M. Deckert, andD. Schluter. 2003. The induction and kinetics of antigen-specific CD8 T cells aredefined by the stage specificity and compartmentalization of the antigen in murinetoxoplasmosis. J. Immunol. 170: 1949–1957.
52. Belkaid, Y., E. Von Stebut, S. Mendez, R. Lira, E. Caler, S. Bertholet,M. C. Udey, and D. Sacks. 2002. CD8� T cells are required for primary immu-nity in C57BL/6 mice following low-dose, intradermal challenge with Leishma-nia major. J. Immunol. 168: 3992–4000.
53. Bertholet, S., R. Goldszmid, A. Morrot, A. Debrabant, F. Afrin, C. Collazo-Custodio,M. Houde, M. Desjardins, A. Sher, and D. Sacks. 2006. Leishmania antigens arepresented to CD8� T cells by a transporter associated with antigen processing-inde-pendent pathway in vitro and in vivo. J. Immunol. 177: 3525–3533.
1748 COMPETITIVE T CELL PRIMING DURING PROTOZOAN INFECTION
on Novem
ber 4, 2009 w
ww
.jimm
unol.orgD
ownloaded from

110
Apêndice 3

ST
FJJMa
b
c
d
e
f
g
Bh
i
j
a
ARRAA
KTDAC
1
ptc
Rf
0d
Vaccine 27 (2009) 5644–5653
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
train-specific protective immunity following vaccination against experimentalrypanosoma cruzi infection
ilipe A. Haolla a,b, Carla Claser a,b, Bruna C.G. de Alencar a,b, Fanny Tzelepis a,b,osé Ronnie de Vasconcelos a,b, Gabriel de Oliveira c, Jaline C. Silvério d, Alexandre V. Machado e,oseli Lannes-Vieira d, Oscar Bruna-Romero f, Ricardo T. Gazzinelli e,g,h, Ricardo Ribeiro dos Santos i,j,
ilena B.P. Soares i,j, Mauricio M. Rodrigues a,b,∗
Centro Interdisciplinar de Terapia Gênica (CINTERGEN), Universidade Federal de São Paulo-Escola Paulista de Medicina, Rua Mirassol, 207, São Paulo, SP 04044-010, BrazilDepartmento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de São Paulo-Escola Paulista de Medicina, Rua Mirassol, 207, São Paulo, SP 04044-010, BrazilLab. Biologia Celular, Instituto Oswaldo Cruz, FIOCRUZ, Av. Brasil n0 4365, Rio de Janeiro, RJ 21045-900, BrazilLab. de Biologia das Interacões, Instituto Oswaldo Cruz, FIOCRUZ, Av. Brasil n0 4365, Rio de Janeiro, RJ 21045-900, BrazilCentro de Pesquisas René Rachou, FIOCRUZ, Avenida Augusto de Lima 1715, Barro Preto, Belo Horizonte, MG 30190-002, BrazilDepartamento de Microbiologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, Pampulha, Belo Horizonte, MG 31270-901, BrazilDepartamento de Bioquímica e Imunologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, Pampulha,elo Horizonte, MG 31270-901, BrazilDivision of Infectious Disease and Immunology, Department of Medicine, University of Massachusetts Medical School, Worcester, MA 01655, USACentro de Pesquisas Goncalo Moniz, FIOCRUZ, Rua Waldemar Falcão, 121, Salvador, BA 40295-001, BrazilHospital São Rafael, Av. São Rafael, 2152, São Marcos, Salvador, BA 41253-190, Brazil
r t i c l e i n f o
rticle history:eceived 3 June 2009eceived in revised form 28 June 2009ccepted 8 July 2009vailable online 25 July 2009
eywords:rypanosoma cruziNA vaccinedenovirus vaccine
a b s t r a c t
Immunisation with Amastigote Surface Protein 2 (asp-2) and trans-sialidase (ts) genes induces protectiveimmunity in highly susceptible A/Sn mice, against infection with parasites of the Y strain of Trypanosomacruzi. Based on immunological and biological strain variations in T. cruzi parasites, our goal was to val-idate our vaccination results using different parasite strains. Due to the importance of the CD8+ T cellsin protective immunity, we initially determined which strains expressed the immunodominant H-2Kk-restricted epitope TEWETGQI. We tested eight strains, four of which elicited immune responses to thisepitope (Y, G, Colombian and Colombia). We selected the Colombian and Colombia strains for our studies.A/Sn mice were immunised with different regimens using both T. cruzi genes (asp-2 and ts) simultane-ously and subsequently challenged with blood trypomastigotes. Immune responses before the challenge
D8 were confirmed by the presence of specific antibodies and peptide-specific T cells. Genetic vaccinationdid not confer protective immunity against acute infection with a lethal dose of the Colombian strain. Incontrast, we observed a drastic reduction in parasitemia and a significant increase in survival, followingchallenge with an otherwise lethal dose of the Colombia strain. In many surviving animals with late-stagechronic infection, we observed alterations in the heart’s electrical conductivity, compared to naive mice.In summary, we concluded that immunity against T. cruzi antigens, similar to viruses and bacteria, may
e a n
be strain-specific and hav. Introduction
Genetic vaccination is being pursued as an efficient, long-term,rotective form of immunisation against Trypanosoma cruzi infec-ion (reviewed in Refs. [1,2]). This strategy is particularly interesting,onsidering the importance of the CD4+ Th1 and CD8+ cytotoxic T
∗ Corresponding author at: CINTERGEN, UNIFESP-Escola Paulista de Medicina,ua Mirassol, 207, São Paulo, SP 04044-010, Brazil. Tel.: +55 11 5084 8807;
ax: +55 11 5084 8807.E-mail address: [email protected] (M.M. Rodrigues).
264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2009.07.013
egative impact on vaccine development.© 2009 Elsevier Ltd. All rights reserved.
cells during naturally acquired or vaccine-induced immunity to thishuman intra-cellular, protozoan parasite (reviewed in Refs. [3–5]).
Recently, we studied the immunisation of highly susceptibleA/Sn mice with a heterologous prime-boost immunisation regi-men, using plasmid DNA followed by a recombinant adenovirusthat expressed the Amastigote Surface Protein-2 (ASP-2) of T. cruzi.This immunisation protocol generated protective immunity against
a lethal challenge with parasites of the Y strain [6]. Vaccinated ani-mals depleted of CD4+ or CD8+ T cells prior to parasitic challengewere highly susceptible to infection, indicating a critical role ofthese T cells during protective immunity. Protective CD8+ T cellswere specific to the immunodominant epitope TEWETGQI, located
cine 2
avpp
pcmcsps
btdg[fDsgi2eowwvtsis
2
2
2tCdttebmtmtvhosAhuT<
2
f
F.A. Haolla et al. / Vac
t amino acids (AA) 320 and 327 of ASP-2. In this mouse model,accination with T. cruzi genes/antigens not only diminished acutehase parasitemia and mortality, but also reduced chronic phaseathology.
The results obtained with ASP-2 vaccination suggested that thisrotein could be part of a subunit prophylactic or therapeutic vac-ine against Chagas disease. Nevertheless, the fact that T. cruzi, likeost microorganisms, displays strain polymorphism necessitates
ritical interpretation of our results. The hypothesis of strain-pecific immunity is very plausible and could be applied to mostathogenic microorganisms (reviewed Ref. [7]). Therefore, a similarcenario for T. cruzi infection could be possible.
Although plausible, to the best of our knowledge, there haseen no proof suggesting that acquired or vaccine-induced, pro-ective immunity against T. cruzi is strain-specific. Recent studiesescribing the epitopes recognised by protective CD8+ T cells sug-est that the immunodominant epitopes are, in fact, strain-specific8–10]. Our studies on genetic vaccination with asp-2 genes clonedrom different parasite strains also confirmed this hypothesis [11].ue to the possible immunological variations found in parasites
trains isolated from different regions of the Americas, the firstoal of this study was to determine which strains presented themmunodominant, H-2Kk-restricted CD8+ T cell epitope of ASP-
(TEWETGQI). We then selected two strains that expressed thispitope. These strains corresponded to divergent T. cruzi lineage Ir II. Using these strains, we evaluated whether our earlier resultsere valid in susceptible A/Sn mice. These mice were vaccinatedith two genes [asp-2 and trans-sialidase (ts)] that were pre-
iously described as being protective against a challenge withhe Y strain of T. cruzi. Essentially, we confirmed our hypothe-is that both naturally acquired and vaccine-induced protectivemmunity during experimental T. cruzi infection can be strain-pecific.
. Materials and methods
.1. Mice and parasites
Female 5–8-week-old A/Sn (H-2a), B10.A (H-2a) C57BL/6 (H-b) were purchased from the Federal University of São Paulo orhe University of São Paulo. Parasites of the Sylvio X10/4, Dm28c,L-Brener, Tulahuen, or G strains of T. cruzi trypomastigotes wereerived from infected LLC-MK2, as described [11]. Parasites fromhe Colombia (COL, Ref. [12]) and Colombian [13] strains hadheir DNA extracted and rRNA genes sequenced, as describedarlier [14]. Bloodstream trypomastigotes of the Y, COL and Colom-ian strains were obtained from experimentally infected A/Snice. The challenge was performed intraperitoneally (i.p.) with
he indicated number of trypomastigotes and parasite develop-ent was monitored by counting the number of bloodstream
rypomastigotes in 5 �l of fresh blood collected from the tailein [15]. The use of animals and the experimental proceduresave been approved by the Ethics Committee for Animal Caref the Federal University of São Paulo. The values of peak para-itemia of each individual mouse were log compared by One-Waynova followed by Tukey HSD tests available at the websitettp://faculty.vassar.edu/lowry/VassarStats.html. LogRank test wassed to compare mouse survival rate after challenge with T. cruzi.he differences were considered significant when the P value was0.05.
.2. Peptides
Peptides TEWETGQI and TsKb-18 (ANYDFTLV) were purchasedrom Genscript (Piscataway, NJ). Peptide purity was in a range of
7 (2009) 5644–5653 5645
80–90% purity. Their identities were confirmed by Q-TOF MicroTM
equipped with an electrospray ionization source (Micromass, UK).
2.3. Recombinant plasmids and adenoviruses used forimmunisation
Plasmids pIgSPclone9 and p154/13 expressing ASP-2 and TS ofT. cruzi, respectively, were generated as described earlier and bothlead to expression of the recombinant antigen [15–17]. pAdCMV-asp-2 and pAdCMV-ts are adenoviral transfer plasmids that containan eukaryotic expression cassette formed by the cytomegalovirusimmediate-early promoter and the SV40 RNA polyadenylationsequences. Inside this cassette we cloned the DNA sequencesencoding T. cruzi ASP-2 or TS (AdASP-2 and Ad-TS, Ref. [17]). Virusesand plasmids were purified as described earlier [13–15]. Mice wereinoculated intra-muscularly (i.m.) in each tibialis anterioris musclewith 100 �g of plasmid DNA (pIgSPclone9 and p154/13). Twenty-one days later, these mice received in these same spots a seconddose of 100 �g of plasmid DNA or 50 �l of viral suspension contain-ing a total of 4 × 108 plaque forming units of adenovirus (AdASP-2and Ad-TS). Immunological assays and challenge were performed14 days after viral inoculation. Some groups of mice received twoi.m. doses of 4 × 108 plaque forming units of adenovirus (AdASP-2and Ad-TS). In this case the doses were administered 8 weeks apart.
2.4. Immunological assays
For the in vivo cytotoxicity assays, splenocytes collected fromnaive B10.A, C57BL/6 or A/Sn mice were divided into two popu-lations and labeled with the fluorogenic dye carboxyfluoresceindiacetate succinimidyl diester (CFSE, Molecular Probes, Eugene,Oregon, USA) at a final concentration of 5 �M (CFSEhigh) or 0.5 �M(CFSElow). CFSEhigh cells were coated for 40 min at 37 ◦C with2.5 �M of peptide TEWETGQI or 1.0 �M of peptide TsKb-18. CFSElowcells remained uncoated. Subsequently, CFSEhigh cells were washedand mixed with equal numbers of CFSElow cells before injectingintravenously (i.v.) (30–40) × 106 total cells per mouse. Recipientanimals were mice previously immunised with recombinant plas-mids or adenoviruses or both. Spleen cells of recipient mice werecollected 20 h after transfer as indicated on the legend of the figures,fixed with 1.0% paraformaldehyde and analyzed by fluorescence-activated cell sorting (FACS), using a FacsCanto flow cytometer (BD,Mountain View, CA). Percentage of specific lysis was determinedusing the formula:
1 − %CFSEhigh immunised/%CFSElow immunised%CFSEhigh naive/%CFSElow naive
× 100%
ELISPOT assay for enumeration of peptide-specific IFN-� producingcells was performed essentially as described earlier [15]. Antibodiesto recombinant ASP-2 (His-65 kDa) or TS were detected by ELISA. Tothis aim, ELISA plates were coated with 250 ng/well of each recom-binant protein and the assay was performed as previously described[15,16].
2.5. Electrocardiogram (ECG) and histology
Mice were i.p. tranquilized with diazepan (20 mg/kg) and trans-ducers were carefully placed under the skin in accordance withchosen preferential derivation (DII). Traces were recorded usinga digital system (Power Lab 2/20) connected to a bio-amplifier
in 2 mV for 1 s (PanLab Instruments). Filters were standardizedbetween 0.1 and 100 Hz and traces were analyzed using the Scopesoftware for Windows V3.6.10 (PanLab Instruments). We measuredheart rate (beats per minute—bpm), duration of the PR, QRS, QTintervals and P wave in ms (millisecond) on 222 dpi. The rela-
5 cine 2
tarckrbtRaSF
2
llvacf
3
3np
rmuamttb
ceeCeXtTfiei
shlaGmoii
itC
646 F.A. Haolla et al. / Vac
ionship between the QT interval and RR interval was individuallyssessed. To obtain physiologically relevant values for the heartate-corrected QT interval (QTc) throught Bazzet’s formula. Chroni-ally infected vaccinated A/Sn mice and age-matched controls wereilled under anesthesia at 120 days post-infection. Hearts wereemoved and fixed in 4% buffered formalin. Sections were analyzedy light microscopy after paraffin-embedding and standard hema-oxylin and eosin staining or Picrosirus-Hematoxilin (Picrrosiriused Staining—EasyPath® Erviegas). The percentage of the coloredreas (red) was determined with the aid of the software imageJ.elected tissue sections were photographed with the aid of NikonE2 microscope couple to a Zeiss camera.
.6. Statistical analysis
The values of peak parasitemia of each individual mouse wereog transformed before being compared by One-Way Anova fol-owed by Tukey HSD tests available at the site http://faculty.assar.edu/lowry/VassarStats.html. Values of ELISPOT assay werelso compared by One Way Anova. The LogRank test was used toompare mouse survival rate after challenge with T. cruzi. The dif-erences were considered significant when the P value was <0.05.
. Results
.1. Specificity of the CD8+ T cell immune response duringaturally acquired immunity in mice infected with differentarasite strains
Our initial studies on the specificity of the CD8+ T cell immuneesponse were performed in infected B10.A mice. We selected this
ouse strain because the A/Sn mice that we initially planned tose in our vaccination studies were highly susceptible to infectionnd died before developing a strong immune response [6]. Bothouse strains have the H-2a haplotype, a recombinant H-2 haplo-
ype that is homozygous to the H-2Kk and H-2Dd alleles. Therefore,he MHC1a-restricted CD8+ T cells of H-2a mice recognise epitopesound to either H-2Kk or H-2Dd molecules.
To uncover the specificity of the naturally acquired CD8+ Tell immune responses, we infected B10.A mice with parasites ofight different strains. The strains used were isolated from differ-nt endemic regions of South America (Brazil, Argentina, Chile, andolombia) and belonged to both of the divergent phylogenetic lin-ages of T. cruzi. The G, Colombian, DM28c, Tulahuén, and Sylvio/10-4 strains belong to T. cruzi lineage I; Strains Y and COL belong
o T. cruzi lineage II, and CL-Brener is a hybrid of both lineages [14].wenty days later, both infected mice and naive controls were usedor in vivo cytotoxic or ex vivo ELISPOT assays to detect specificmmune responses to the H-2Kk-restricted epitope TEWETGQI. Thispitope is immunodominant, and expressed by several proteinsncluding the ASP-2 of T. cruzi [8,10,18,19].
The timing for these experiments was based on earlier studieshowing that the CD8+ T cell immune response to T. cruzi epitopesas delayed kinetics; being detected at ∼14 days after the chal-
enge [7,8,10]. Mice infected with parasites of the Y, G, Colombian,nd COL strains displayed strong responses to the peptide TEWET-QI, as measured by both assays (Fig. 1A and B, respectively). Theagnitude of the immune responses of mice infected with each
f these two parasite strains was comparable. In contrast, micenfected with parasites of the other four strains failed to develop
mmune responses to this epitope (P > 0.05 in all cases).To confirm that the differences observed during themmune response to TEWETGQI were not due to poor infec-ivity/immunogenicity by certain T. cruzi strains, we infected57BL/6 mice. The genetic background of this mouse strain is
7 (2009) 5644–5653
identical to B10.A mice, but they have a different H-2 haplotype(H-2b). Infection with the parasites of the CL-Brener, Sylvio X/10-4or DM28-c strains, which failed to induce immune responses tothe TEWETGQI epitope in B10.A mice, elicited strong responses tothe H-2Kb-restricted peptide TsKb-18, as estimated by the in vivocytotoxic assay or the ELISPOT assay (Fig. 1C and D). Their immuneresponses were also comparable in magnitude. C57BL/6 micechallenged with other parasite strains also displayed statisticallyhigher immune responses than naive mice (Tulahuén, Y or G),though they were significantly lower than the ones describedabove.
A single strain, Tulahuén, elicited poor immune responses to thetwo peptides in both mouse strains. The inability of these mice togenerate an immune response to this strain was not due to a lackof infectivity/immunogenicity. BALB/c mice (H-2d) infected withTulahuén trypomastigotes responded strongly to a third epitope,IYNVGQVSI, as previously described. This epitope is immunodom-inant and expressed on the TS antigen of T. cruzi (data not shown[20,21]).
Together, these results support our hypothesis and previousobservations that the specificity of the acquired immune responsesto T. cruzi that are mediated by mouse CD8+ T cells vary accordingto the parasite strain used for infection.
3.2. Protective immunity in highly susceptible A/Sn micefollowing genetic vaccination with asp-2 and ts
Based on the results above, we restricted our study to three ofthe eight parasite strains that were distinct from the Y strain andresponded to the TEWETGQI epitope. The G strain, isolated froman opossum, was described as poorly infective, leading to low lev-els of parasitemia [22]. Therefore, we selected the Colombian andCOL strains. To confirm the phylogenetic origins of these strains,we sequenced the rRNA genes obtained from the blood of infectedmice. As shown in Fig. 2, the results confirmed that Colombian andCOL parasites belong to divergent T. cruzi phylogenetic lineages Iand II, respectively.
To determine whether protective immunity against these twostrains could be elicited by genetic immunisation, we comparedtwo different vaccination protocols, using either homologous orheterologous prime-boosting regimens. The homologous proto-col consisted of two immunising doses of plasmid DNA (pIgSPCl.9and p154/13) or recombinant replication-defective adenovirus type5 (AdASP-2 and Ad-TS). The heterologous immunisation regimenconsisted of a priming immunisation with plasmids pIgSPCl.9 andp154/13, followed by a booster injection of the AdASP-2 and Ad-TS.In parallel, control mice were injected with the homologous proto-col consisting of two immunising doses of plasmid DNA (pcDNA3)or recombinant adenovirus (Ad�-gal). The control for the heterolo-gous immunisation regimen consisted of a priming immunisationwith pcDNA3, followed by the administration the Ad�-gal.
These vaccination protocols were selected based on our previ-ous finding that they could elicit protective immunity after a lethalchallenge with parasites of the Y strain of T. cruzi [6]. We simultane-ously vaccinated mice with asp-2 and ts. This decision was based onour earlier observation that using both genes improved protectiveimmunity in A/Sn mice [24].
To monitor the immunisation efficiency, we determined theantibody titres to recombinant ASP-2 or TS (Fig. 3). Groups of micethat were vaccinated with T. cruzi genes presented high levelsof specific antibodies, as detected by ELISA. The antibody lev-
els were significantly higher in groups of mice immunised withrecombinant adenovirus (2 doses) or heterologous prime-boost(DNA–adenovirus), in comparison to animals that received immu-nisations with plasmids only. To determine whether CD8 immuneresponses were elicited, we selected only the group of mice that
F.A. Haolla et al. / Vaccine 27 (2009) 5644–5653 5647
Fig. 1. Specific CD8+ T cell mediated immune responses in B10.A or C57BL/6 mice challenged with different strains of T. cruzi.B10.A (Panels A and B) or C57BL/6 (Panels C and D) mice were challenged or not i.p. with bloodstream trypomastigotes (104 per mouse) of the Y, Colombian or COL strains ortrypomastigote culture forms (105 per mouse) of Sylvio X/10 Cl.4, Tulahuén, CL-Brener, DM28c or G strains.After 20 days, the in vivo cytotoxic activity against target cells coated with peptides TEWETGQI (Panel A) or TsKb-18 (Panel B) were determined as described in Section 2.Results represent the mean of four mice ± SD per group. Asterisks denote that in vivo cytotoxicity of infected mice against target cells coated with peptide TEWETGQI orTsKb-18 was significantly higher than control naive mice (P < 0.01).After 20 days, IFN-� producing spleen cells specific for peptides TEWETGQI (Panel B) or TsKb-18 (Panel D) were estimated ex vivo by ELISPOT assay. Results represent them 6 in vitrt infect(R
rma(
rpw
FoPgg
ean number of spot forming cells (SFC) per 10 splenocytes ± SD (n = 4) followinghe numbers of SFC specific for peptide TEWETGQI or TsKb-18 of spleen cells fromP < 0.01).esults are representative of two or more independent experiments.
eceived the DNA–adenovirus regimen. As shown in Fig. 4, theseice presented high levels of in vivo-specific cytotoxicity (panel A)
nd peptide-specific IFN-� producing cells, as detected by ELISPOT
Panel B).Immunised mice in this experiment were injected with theecombinant adenovirus (2 doses) or heterologous prime-boostrotocol. T. cruzi gene-injected and control mice were challengedith a lethal dose of T. cruzi bloodstream trypomastigotes of the
ig. 2. Sequence of the divergent domain D7 of the large subunit of T. cruzi rRNA 24Sbtained from distinct parasite strains.arasites from the COL and Colombian strains had their DNA extracted and rRNAenes sequenced as described earlier [14]. Their sequences were compared to rRNAenes sequences from Genbank AF301912 and AF239981.
o stimulation with the indicated peptide or medium only. The asterisks denote thated mice were significantly higher than the number of SFC of control naive animals
Colombian strain. As shown in Fig. 5A, parasitemia in all fourgroups progressed steadily. No statistically significant differencewas observed among the groups. When mouse survival was fol-lowed after the challenge (as shown in Fig. 5B), the group of miceimmunised with the heterologous prime-boost regimen survivedsignificantly longer than the other three. Nevertheless, all miceeventually died, and we concluded that no protective immunitywas elicited following genetic vaccination against parasites of theColombian strain.
In subsequent experiments, immunised mice were injected withthe homologous protocol consisting of two immunising doses ofplasmid DNA (pIgSPCl.9 and p154/13) or recombinant replication-defective type 5 adenovirus (AdASP-2 and Ad-TS). A third groupof animals received the heterologous immunisation regimen, con-sisting of a priming immunisation with plasmids pIgSPCl.9 andp154/13, followed by a booster injection with AdASP-2 and Ad-TS.
Vaccinated and control mice were challenged with a lethal doseof bloodstream trypomastigotes of the COL strain (Fig. 6). All threegroups of mice immunised with T. cruzi genes (groups 4, 5 and 6)displayed peak parasitemias significantly lower than the controls(groups 1, 2 and 3; Fig. 6, Panel A; P < 0.01 in all cases). When mouse

5648 F.A. Haolla et al. / Vaccine 27 (2009) 5644–5653
Fig. 3. Specific antibody response in A/Sn mice genetically vaccinated with ASP-2and TS.Antibody titers to recombinant TS (Panel A) or ASP-2 (Panel B) were determinedin groups of mice (n = 6) administered with T. cruzi genes, as described in Section2i(w
sfitc
anf
Fig. 4. Specific CD8+ T cell mediated immune responses in A/Sn mice geneticallyvaccinated with ASP-2 and TS.A/Sn mice were immunised with the heterologous prime-boost (DNA–adenovirus)protocol, as described in Section 2. After 14 days, the in vivo cytotoxic activityagainst target cells coated with peptides TEWETGQI (Panel A) were determined asalso described in Section 2. Results represent the mean of 4 mice ± SD per group.Asterisks denote that in vivo cytotoxicity of ASP2/TS immunised mice against targetcells coated with peptide TEWETGQI were significantly higher than in control miceinjected with pcDNA and Ad�-gal (P < 0.01).After 14 days, IFN-� producing spleen cells specific for peptide TEWETGQI (PanelB) were estimated ex vivo by ELISPOT assay. Results represent the mean numberof spot forming cells (SFC) per 106 splenocytes ± SD (n = 4) following in vitro stim-ulation with the indicated peptide or medium only. The asterisks denote that the
. Antibody titers as detected by ELISA were significantly higher in groups of micemmunised with recombinant adenovirus (2 doses) or heterologous prime-boostDNA–adenovirus) when compared to animals that received two immunisationsith plasmids (P < 0.01 in both cases).
urvival after the challenge was followed (Fig. 6, Panel B), the resultsrom the parasitemia were largely confirmed. Only 1 of the 18 micemmunised with T. cruzi genes succumbed to the acute lethal infec-ion. In contrast, all 18 control mice died by the 30th day after the
hallenge.Immunised mice that survived the acute infection were evalu-ted for cardiac disturbances at 120 days post-infection. Althougho differences were found when the statistical analyses were per-
ormed (Table 1), qualitative analysis based on cardiac conduction
numbers of SFC specific for peptide TEWETGQI of spleen cells from ASP-2/TS immu-nised mice were significantly higher than the number specific SFC of control miceinjected with pcDNA and Ad�-gal (P < 0.01).Results are representative of two independent experiments.
disturbances were detected on the individual electrocardiograms(ECGs; Fig. 7A and B and Table 2). ECG traces observed from the
plasmid DNA-vaccinated animals suggested the presence of atrialelectrical conduction disturbances due to overload (branch block-age), first or second degree atrioventricular blockage (AVB1 or 2),and sinusal bradycardia. Three of the six mice vaccinated twice withrecombinant AdASP-2/Ad-TS (group 5) did not suffer significant car-
F.A. Haolla et al. / Vaccine 27 (2009) 5644–5653 5649
Table 1ECG parameters measured.
Mouse group Immunol. status n Heart rate (n of beats/min) P wave duration (ms) PR segment (ms) QRS segment (ms) QTc
Naive 8 521.6 ± 73.3 16.5 ± 1.9 38.5 ± 7.2 14.2 ± 2.6 31.4 ± 10.44 pIgSPCl.9 (2x) 4 463.7 ± 84.4 24.8 ± 11.5 43.9 ± 15.0 15.0 ± 12.5 37.1 ± 6.55 41.76 18.5
V
do6
ttdctescnf
4
tt2iwrrwssPVwstrtMsdst
TS
C
SVEBAADIS
t
AdASP-2 (2x) 6 454.8 ± 41.7pIgSPCl.9/AdASP-2 5 413.3 ± 122.2
alues represent the means ± SD.
iac alterations at this stage of the disease (Fig. 7C and Table 2). Nonef the mice vaccinated twice with the heterologous protocol (group) developed AVB2 (Table 2).
Histological analyses of the heart tissue following hema-oxilin/eosin staining did provide evidence of a link betweenhe observed electrical disturbances and cellular infiltrates. Toetermine whether the accumulation of fibrosis or tissue repairorrelated with the electrical disturbances detected, we stainedhe interstitial collagen with picrosirius red. The stained area wasstimated and the results are presented in Fig. 8A. There was aignificant increase in the stained areas in all groups (4, 5 and 6),ompared to naive age-matched controls. In contrast, mice immu-ised with two doses of recombinant adenovirus displayed slightly
ewer stained areas (P < 0.01).
. Discussion
In this study, we tested whether immunity against T. cruzi infec-ion in mice could be specific to the parasite strain. We foundhat a single inbred mouse strain (B10.A) responded to the H-Kk-restricted CD8+ T cell epitope TEWETGQI, after experimental
nfection with four of the eight parasite strains tested. Mice infectedith the other four parasite strains failed to develop significant
esponses to this epitope. The heterogeneity among the immuneesponses of CD8+ T cells from B10.A mice to the TEWETGQI epitopeas also reproduced with other immunodominant epitopes. As
hown in Fig. 1C and D, C57BL/6 mice infected with only three of theeven parasite strains responded strongly to the TsKb-18 epitope.revious studies have shown that the immunodominant epitopeNHRFTLV is strongly recognised during infection of C57BL/6 miceith the Y or G strains of T. cruzi, but not with the CL-Brenner
train [10]. In accordance with these observations, we found thathe immunodominant H-2Kd-restricted epitope YNVGQVSI wasecognised by T cells of BALB/c mice, after infection with three ofhe seven strains tested (Y, Colombian and Tulahuen; CC, FT and
R unpublished results). The reason for the heterogeneity in thepecificity CD8+ T cell immune responses following infection withifferent parasite strains is still unknown. It may be related to AAequence polymorphism, levels of expression of the different epi-opes, immunodominance, or immune evasion mechanisms.
able 2ummary of the ECG records.
ardiac alterations Naive (n = 8) Group 4 (n
RAb 0 0RAb 0 0XTb 0 0RADb 0 0VB-1b 0 0VB-2b 0 3EFLEXb 0 0
NFRAb 0 0TSDb 0 0
a ECG record of A/Sn mouse genetically vaccinated as described in the legend of Fig. 6 ab Abbreviations: (i) SAR: sinus arrhythmia; (ii) VAR: ventricular arrhythmia; (iii) EXT: v
ricular block; (vi) AVB2: 2◦ degree atrioventricular block; (vii) DEFLEX: Q wave deflection
± 20.2 52.5 ± 5.0 12.5 ± 2.6 52.5 ± 17.1± 3.3 42.9 ± 5.8 14.5 ± 3.4 58.3 ± 29.7
We then selected two of the T. cruzi strains that expressed theTEWETGQI epitope (Colombian and COL) to test whether protec-tive immunity, elicited by the genetic vaccination with the antigensASP-2 and TS, could confer protective immunity in highly suscep-tible A/Sn mice against a lethal challenge with these parasites.Our present results demonstrated that the resulting protectiveimmunity elicited by vaccination with these antigens/genes wasstrain-specific. Vaccinated mice succumbed to a challenge withparasites of the Colombian strain. In contrast, the vast majority ofvaccinated animals survived a challenge with parasites of the COLstrain. Like earlier studies, these results demonstrated that immuni-sation with ASP-2 via distinct delivery systems can elicit protectiveimmunity in C57BL/6, BALB/c and A/Sn mice, after a challenge withthe Y or Brazil strain of T. cruzi [6,11,15,17–19,23,24]. In the case ofTS, successful vaccination studies have been performed in BALB/cor C57BL/6, but not in A/Sn mice, using the Y or Tulahuén strains[16,17,19,21,24–26].
Two possible, non-excluding explanations can account for thevariation observed. First, it is possible that these discrepanciescould be related to the antigenic polymorphism in the parasites ofthe Colombian strain, e.g., the absence of epitope(s) critical for theprotective immunity. Comparison of the predicted AA sequencesfor the ASP-2 of Colombian and Y strains revealed high similarity(96.8%, Ref. [11]). In the case of the TS of T. cruzi, the transcriptsof parasites of the Colombian strain have not yet been isolatedand sequenced. However, similar to the ASP-2, the genes encod-ing the catalytic region of TS used in our study (AA 1–678) didnot show high sequence variability among the five other strains.The identities among the predicted AA sequences of TS (Y strain)and other strains were 96, 84, 92, and 97%, in the CL (XP 807431),Sylvio X-10/4 (P23253), FALLS (CAA40511) and Tulahuén “/isolate=”Tul2 (AAA99443) strains, respectively. Nevertheless, these smallpolymorphic regions may still be critical for inducing protectiveimmunity.
Based on the fact that both strains express the important targetepitope TEWETGQI, we should still be able to detect some degree
of protective immunity mediated by CD8+ T cells, following a chal-lenge with the Colombian strain. The results obtained after inducingan immune response to infection with both strains also eliminatethe possibility that they differ in their capacity to generate similarantigen available for processing and presentation.= 4)a Group 5 (n = 6)a Group 6 (n = 5)a
0 00 00 00 20 23 00 00 00 0
nd challenged with 250 blood forms of T. cruzi 120 days earlier.entricular extrasystolis; (iv) BRAD: sinus bradicardya; (v) AVB1: 1◦ degree atrioven-
(viii) STSD: ST segment depression.

5650 F.A. Haolla et al. / Vaccine 27 (2009) 5644–5653
Fig. 5. Parasitemia and mortality in A/Sn (H-2a) mice immunised with differentcombination of plasmid DNA and/or rec. adenovirus expressing the ASP-2 and TS ofT. cruzi and challenged with trypomastigotes of the Colombian strain.A/Sn mice were immunised as described in Section 2.Two weeks after the final immunising dose, mice were challenged i.p. with 250bloodstream trypomastigotes of the Colombian strain.Panel A: the parasitemia for each mouse group is represented as a mean ± SD (n = 6 or10). The parasitemias of the different mouse groups were not statistically different.Panel B: the graph shows the Kaplan–Meier curves for the survival of the mice groupsimmunised and challenged as described above (n = 6 or 10). Mice from group 4 (�)survived significantly longer than animals from group 1 (�), 2 (�,) or 3 (�) (P < 0.01in all cases, LogRank test).Pooled results from two experiments are shown.
Fig. 6. Parasitemia and mortality in A/Sn (H-2a) mice immunised with differentcombinations of plasmid DNA and/or rec. adenovirus expressing the ASP-2 and TSof T. cruzi and challenged with trypomastigotes of the COL strain.A/Sn mice were immunised as described in Section 2.Two weeks after the final immunising dose, mice were challenged i.p. with 250bloodstream trypomastigotes of the COL strain.Panel A: the parasitemia for each mouse group is represented as mean ± SD (n = 6).The parasitemias of mice groups 4, 5 and 6 were statistically lower than mice fromgroups 1, 2 and 3 (P < 0.01 in all cases).
Panel B: the graph shows the Kaplan–Meier curves for the survival of the mice groupsimmunised and challenged as described above (n = 6). Mice from groups 4, 5 and 6survived significantly longer than animals from groups 1, 2 and 3 (P < 0.01 in all cases,LogRank test). Representative results of two independent experiments.Antigenic polymorphism, as the cause of strain-specific immu-nity, has been described for many viral and bacterial pathogens,leading to selection of antigenic variants following the use ofvaccines [27,28]. In the case of protozoan parasites, it is notclear whether strain-specific immunity plays a major role dur-

F.A. Haolla et al. / Vaccine 27 (2009) 5644–5653 5651
Fig. 7. Examples of ECG from mice which survived the lethal challenge with trypo-mastigotes of the COL strain of T. cruzi.Vaccinated and challenged A/Sn mice are from groups 4, 5 and 6 of Fig. 6.Panels A and B: ECG record of a vaccinated A/Sn mouse of group 4 (A) or group 6(B) and challenged with 250 blood forms of T. cruzi 120 days earlier. The electricdisturbances indicated by the arrows are AVB 2.Panel C: ECG record of a vaccinated A/Sn mouse of group 5. No alterations wererP2a
itianh
sbtasarfotmodrthCtcaaotcati
dwf
Fig. 8. Red picrosirius staining of heart sections from vaccinated A/Sn mice chal-lenged 120 days earlier.Vaccinated and challenged A/Sn mice are from groups 4, 5 and 6 of Fig. 6.Heart tissues sections were stained with red picrosirius/hematoxillin. Sections wereobserved using microscopy (400×) and the fibrosis areas were estimated using theNIH ImageJ software.Panel A: the fibrosis area of tissues obtained from mice of groups 4, 5 and 6 werehigher than the naive mice (P < 0.01). However, the fibrosis area of heart sectionsobtained from mice of 5 were slightly lower than for groups 4 and 6 (P < 0.01).Panel B: example of heart section stained with red picrosirius from vaccinated andchallenged mice.
ecorded.anel C: ECG record of a vaccinated A/Sn mouse from group 5 and challenged with50 blood forms of T. cruzi 120 days earlier. Fifty percent of these mice did not presentny detectable electric disturbance.
ng infection or vaccination, contributing to parasitic evasion ofhe immune response. Nevertheless, there are examples suggest-ng that this may be the case during natural infections [29,30]. Inddition, during experimental vaccination, strain-specific immu-ity, attributable to the primary sequence of the vaccine antigens,as been observed [31–33].
The second possible explanation for the observed strain-pecificity is that beyond antigenic polymorphism, other forms ofiological variability can account for the observed differences. Dis-inct parasite strains may have different tissue tropisms during thecute phase, multiplying in different target organs. Although manytudies have attempted to describe the fate of the T. cruzi parasitesfter experimental infection using histological analyses, only oneecent study has tracked the parasites of the Brazil strain, via trans-ection with a reporter gene. This study found that after i.p. infectionf A/J mice, luciferase-expressing T. cruzi multiply predominantly inhe large intestine [34]. Parasite growth was uncontrolled in those
ice, and they all died. No information of this nature is available onther T. cruzi strains. Therefore, it is possible that parasites of theistinct strains used in this study (Y, COL and Colombian) vary, withegard to tissue tropism. This fact could explain the differences inhe protective immunity observed. In a finding that supports thisypothesis, Leavey and Tarleton (2003, Ref. [35]) described howD8+ T cells could not be properly activated after penetration intohe mouse muscle. In these animals, this unresponsive phenotypeontrasted with the spleen CD8+ T, which could still be readilyctivated after muscle penetration. Therefore, in our experiment,lthough effector-specific CD8+ T cells were present in the spleenf mice infected with parasites of each strain, it is possible that inhe case of the animals infected with the Colombian strain, theseells were not functional at the sites of parasite multiplication, suchs within the myocardial muscle. In fact, experimental mouse infec-ion with parasites of the Colombian strain is well known to cause
ntense myocardial infection [36–42].In the chronic disease phase (120 dpi), the cardiac electrical con-uction system of surviving A/Sn mice infected with the COL strainas evaluated. We observed ECG alterations in most animals. This
act contrasted with our previous experiments in vaccinated A/Sn
Panel C: example of heart section stained with red picrosirius from naive mice.

5 cine 2
mmvgcatitTndp[
mirtcscfmc
noddsecbmld
mwsacbebm
cpiih
A
àM(TfRfF
[
[
[
[
[
[
[
[
[
[
[
[
652 F.A. Haolla et al. / Vac
ice infected with parasites of the Y strain [6]. Although theseice survived the lethal infection with the COL strain of T. cruzi, the
accinated A/Sn mice exhibited an increased PR interval (ms), sug-estive of atrioventricular or branch blockage, somewhat similar tohronic Chagasic cardiomyopathy. In accordance with Punukollu etl. [43], the most common symptoms in the chronic phase of infec-ions are cardiac alterations such as ventricular increase, cardiacnsufficiency and myocarditis. We verified electrical disturbances inhe PR and QTc intervals by ECG, possibly caused by the infection.his process may have resulted in destruction of the conductingervous tissues or in local inflammation that led to electrolyticisequilibrium and altered muscle fibre contraction, induced byro-inflammatory cytokines such as IFN-�, TNF-� and nitric oxide44–46].
Few studies have quantified the cardiac area affected witharkers that stain the collagen and fibrin produced during the
nflammation and tissue repair processes [47–51]. Using picrosiriused staining, we found that all groups showed greater deposi-ion of collagen and fibrin, as well as destroyed cardiomyocytes,orroborating our ECG results. Most of the vaccinated A/Sn micetill developed late-stage symptoms, which is compatible with thehronic Chagasic cardiomyopathy. Thus, our work suggested that,or highly susceptible A/Sn animals, an immune response of higher
agnitude or breadth would be required to protect against thehronic stage symptoms.
The fact that immunity can be strain-specific will impose aumber of limitations that should be considered during the devel-pment of prophylactic or therapeutic vaccines against Chagasisease. Members of the TS super-family of antigens, like the oneescribed here, may not be the best candidates for a universal,imple recombinant vaccine such as, for example, hepatitis B. Nev-rtheless, they still may be part of a more complex formulation thatan be immunogenic against most strains of T. cruzi. Recent recom-inant vaccines, such as the human papillomavirus vaccine, are aixture of antigens from four distinct virus types [52]. Neverthe-
ess, they are not expected to have universal coverage against allifferent types of the human papillomavirus.
Our results also do not exclude that other conserved antigensay show better performance, individually or in combinationith ours. To date, there are a number of antigens that are being
tudied for their potential use in vaccines against T. cruzi infections,s was recently reviewed [53]. Some of these antigens are quiteonserved; others may have a limited number of alleles that coulde used simultaneously. Nevertheless, their relative abilities tolicit protective immunity against T. cruzi infection will have toe assessed against multiple parasite strains, in different mouseodels.
Finally, we should mention that A/Sn mice are highly sus-eptible to infection. Most mouse strains, as well as the humanopulation, have a higher innate and/or acquired resistance to
nfection. It is possible that they may develop better or broadermmune responses to recombinant vaccines than was describedere.
cknowledgments
This work was supported by grants from Fundacão de AmparoPesquisa do Estado de São Paulo (2006/1983-4), CNPq, Theillennium Institute for Vaccine Development and Technology
CNPq - 420067/2005-1), The Millennium Institute for Gene
herapy, FAPEMIG (EDT 24.000) (Brazil) and The National Instituteor Vaccine Development and Technology (INCTV-CNPq). AVM,TG, OBR, JLV, RRS, MBPS and MMR were recipients of fellowshipsrom CNPq. CC, BCGA, FT and JRC are recipients of fellowships fromAPESP.
[
7 (2009) 5644–5653
References
[1] Garg N, Bhatia V. Current status and future prospects for a vaccine against Amer-ican trypanosomiasis. Expert Rev Vaccines 2005;4(December (6)):867–80.
[2] Dumonteil E. DNA vaccines against protozoan parasites: advances and chal-lenges. J Biomed Biotechnol 2007;2007(6):90520.
[3] Tarleton RL. Immune system recognition of Trypanosoma cruzi. Curr OpinImmunol 2007;19(August (4)):430–4.
[4] Martin D, Tarleton R. Generation, specificity, and function of CD8+ T cells inTrypanosoma cruzi infection. Immunol Rev 2004;201(October):304–17.
[5] Rodrigues MM, Alencar BC, Claser C, Tzelepis F. Immunodominance: a newhypothesis to explain parasite escape and host/parasite equilibrium leadingto the chronic phase of Chagas’ disease? Braz J Med Biol Res 2009;42(March(3)):220–3.
[6] de Alencar BC, Persechini PM, Haolla FA, Oliveira G, Silvério JC, Lannes-VieiraJ, et al. Perforin and Interferon-� expression are required for CD4+ and CD8+
T cell-dependent protective immunity against a human parasite (Trypanosomacruzi) elicited by a heterologous plasmid DNA prime-recombinant adenovirus5 boost vaccination; Infect Immun 2009 (in press).
[7] Grebe KM, Yewdell JW, Bennink JR. Heterosubtypic immunity to influenza Avirus: where do we stand? Microbes Infect 2008;10(July (9)):1024–9.
[8] Tzelepis F, de Alencar BC, Penido ML, Gazzinelli RT, Persechini PM, RodriguesMM. Distinct kinetics of effector CD8+ cytotoxic T cells after infection withTrypanosoma cruzi in naive or vaccinated mice. Infect Immun 2006;74(April(4)):2477–81.
[9] Martin DL, Weatherly DB, Laucella SA, Cabinian MA, Crim MT, Sullivan S, et al.CD8+ T-Cell responses to Trypanosoma cruzi are highly focused on strain-varianttrans-sialidase epitopes. PLoS Pathog 2006;2(August (8)):e77.
10] Tzelepis F, de Alencar BC, Penido ML, Claser C, Machado AV, Bruna-Romero O, etal. Infection with Trypanosoma cruzi restricts the repertoire of parasite-specificCD8+ T cells leading to immunodominance. J Immunol 2008;180(February(3)):1737–48.
[11] Claser C, Espíndola NM, Sasso G, Vaz AJ, Boscardin SB, Rodrigues MM. Immuno-logically relevant strain polymorphism in the amastigote surface protein 2 ofTrypanosoma cruzi. Microbes Infect 2007;9(July (8)):1011–9.
12] Dost CK, Albuquerque S, Hemleben V, Engels W, Prado Jr JC. Molecular geneticcharacterization of different Trypanosoma cruzi strains and comparison of theirdevelopment in Mus musculus and Calomys callosus. Parasitol Res 2002;88(July(7)):609–16.
13] Frederici EE, Albelmann WB, Neva FA. Chronic and progressive, 1964. Myocardi-tis and myositis in C3H mice infected with Trypanosoma cruzi. Am J Trop MedHyg 1964;13(March):272–80.
14] Briones MR, Souto RP, Stolf BS, Zingales B. The evolution of two Trypanosomacruzi subgroups inferred from rRNA genes can be correlated with the inter-change of American mammalian faunas in the Cenozoic and has implications topathogenicity and host specificity. Mol Biochem Parasitol 1999;104(November(2)):219–32.
15] Boscardin SB, Kinoshita SS, Fujimura AE, Rodrigues MM. Immunizationwith cDNA expressed by amastigotes of Trypanosoma cruzi elicits protectiveimmune response against experimental infection. Infect Immun 2003;71(May(5)):2744–57.
16] Costa F, Franchin G, Pereira-Chioccola VL, Ribeirão M, Schenkman S, RodriguesMM. Immunization with a plasmid DNA containing the gene of trans-sialidasereduces Trypanosoma cruzi infection in mice. Vaccine 1998;16(May (8)):768–74.
[17] Machado AV, Cardoso JE, Claser C, Rodrigues MM, Gazzinelli RT, Bruna-RomeroO. Long-term protective immunity induced against Trypanosoma cruzi infec-tion after vaccination with recombinant adenoviruses encoding amastigotesurface protein-2 and trans-sialidase. Hum Gene Ther 2006;17(September(9)):898–908.
18] Araújo AF, de Alencar BC, Vasconcelos JR, Hiyane MI, Marinho CR, PenidoML, et al. CD8+-T-cell-dependent control of Trypanosoma cruzi infection in ahighly susceptible mouse strain after immunization with recombinant pro-teins based on amastigote surface protein 2. Infect Immun 2005;73(September(9)):6017–25.
19] de Alencar BC, Araújo AF, Penido ML, Gazzinelli RT, Rodrigues MM.Cross-priming of long lived protective CD8+ T cells against Trypanosomacruzi infection: importance of a TLR9 agonist and CD4+ T cells. Vaccine2007;25(August (32)):6018–27.
20] Giddings OK, Eickhoff CS, Smith TJ, Bryant LA, Hoft DF. Anatomical route ofinvasion and protective mucosal immunity in Trypanosoma cruzi conjunctivalinfection. Infect Immun 2006;74(October (10)):5549–60.
21] Hoft DF, Eickhoff CS, Giddings OK, Vasconcelos JR, Rodrigues MM.Trans-sialidase recombinant protein mixed with CpG motif-containingoligodeoxynucleotide induces protective mucosal and systemic Trypanosomacruzi immunity involving CD8+ CTL and B cell-mediated cross-priming. JImmunol 2007;179(November (10)):6889–900.
22] Yoshida N. Surface antigens of metacyclic trypomastigotes of Trypanosoma cruzi.Infect Immun 1983;40(May (2)):836–9.
23] Garg N, Tarleton RL. Genetic immunization elicits antigen-specific protectiveimmune responses and decreases disease severity in Trypanosoma cruzi infec-
tion. Infect Immun 2002;70(October (10)):5547–55.24] Vasconcelos JR, Hiyane MI, Marinho CR, Claser C, Machado AM, Gazzinelli RT, etal. Protective immunity against Trypanosoma cruzi infection in a highly suscep-tible mouse strain after vaccination with genes encoding the amastigote surfaceprotein-2 and trans-sialidase. Hum Gene Ther 2004;15(September (9)):878–86.

cine 2
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
F.A. Haolla et al. / Vac
25] Fujimura AE, Kinoshita SS, Pereira-Chioccola VL, Rodrigues MM. DNA sequencesencoding CD4+ and CD8+ T-cell epitopes are important for efficient protectiveimmunity induced by DNA vaccination with a Trypanosoma cruzi gene. InfectImmun 2001;69(September (9)):5477–86.
26] Vasconcelos JR, Boscardin SB, Hiyane MI, Kinoshita SS, Fujimura AE, RodriguesMM. A DNA-priming protein-boosting regimen significantly improves type 1immune response but not protective immunity to Trypanosoma cruzi infec-tion in a highly susceptible mouse strain. Immunol Cell Biol 2003;81(April(2)):121–9.
27] Boni MF. Vaccination and antigenic drift in influenza. Vaccine 2008;26(July(Suppl. 3)):C8–14. Review.
28] Dagan R. Impact of pneumococcal conjugate vaccine on infections causedby antibiotic-resistant Streptococcus pneumoniae. Clin Microbiol Infect2009;15(April (Suppl. 3)):16–20.
29] Pattaradilokrat S, Cheesman SJ, Carter R. Linkage group selection: towards iden-tifying genes controlling strain specific protective immunity in malaria. PLoSONE 2007;2(September (9)):e857.
30] Wijayalath WA, Cheesman S, Rajakaruna J, Handunnetti SM, Carter R, Pathi-rana PP. Evidence for strain-specific protective immunity against blood-stageparasites of Plasmodium cynomolgi in toque monkey. Parasite Immunol2008;30(November–December (11–12)):630–6.
31] Crewther PE, Matthew ML, Flegg RH, Anders RF. Protective immune responsesto apical membrane antigen 1 of Plasmodium chabaudi involve recognition ofstrain-specific epitopes. Infect Immun 1996;64(August (8)):3310–7.
32] Flück C, Smith T, Beck HP, Irion A, Betuela I, Alpers MP, et al. Strain-specific humoral response to a polymorphic malaria vaccine. Infect Immun2004;72(November (11)):6300–5.
33] Lyon JA, Angov E, Fay MP, Sullivan JS, Girourd AS, Robinson SJ, et al. Protec-tion induced by Plasmodium falciparum MSP1(42) is strain-specific, antigenand adjuvant dependent, and correlates with antibody responses. PLoS ONE2008;3(July (7)):e2830.
34] Hyland KV, Asfaw SH, Olson CL, Daniels MD, Engman DM. Biolumines-cent imaging of Trypanosoma cruzi infection. Int J Parasitol 2008;38(October(12)):1391–400.
35] Leavey JK, Tarleton RL. Cutting edge: dysfunctional CD8+ T cells reside innonlymphoid tissues during chronic Trypanosoma cruzi infection. J Immunol2003;170(March (5)):2264–8.
36] dos Santos RR, Rossi MA, Laus JL, Silva JS, Savino W, Mengel J. Anti-CD4 abro-gates rejection and reestablishes long-term tolerance to syngeneic newbornhearts grafted in mice chronically infected with Trypanosoma cruzi. J Exp Med1992;175(January (1)):29–39.
37] Soares MB, Silva-Mota KN, Lima RS, Bellintani MC, Pontes-de-Carvalho L,Ribeiro-dos-Santos R. Modulation of chagasic cardiomyopathy by interleukin-
4: dissociation between inflammation and tissue parasitism. Am J Pathol2001;159(August (2)):703–9.38] Michailowsky V, Silva NM, Rocha CD, Vieira LQ, Lannes-Vieira J, Gazzinelli RT.Pivotal role of interleukin-12 and interferon-gamma axis in controlling tissueparasitism and inflammation in the heart and central nervous system duringTrypanosoma cruzi infection. Am J Pathol 2001;159(November (5)):1723–33.
[
[
7 (2009) 5644–5653 5653
39] Camandaroba E, Thé TS, Pessina DH, Andrade SG. Trypanosoma cruzi: clones iso-lated from the Colombian strain, reproduce the parental strain characteristics,with ubiquitous histotropism. Int J Exp Pathol 2006;87(June (3)):209–17.
40] Rocha NN, Garcia S, Giménez LE, Hernández CC, Senra JF, Lima RS, et al. Charac-terization of cardiopulmonary function and cardiac muscarinic and adrenergicreceptor density adaptation in C57BL/6 mice with chronic Trypanosoma cruziinfection. Parasitology 2006;133(December (Pt 6)):729–37.
[41] Medeiros GA, Silvério JC, Marino AP, Roffê E, Vieira V, Kroll-Palhares K, et al.Treatment of chronically Trypanosoma cruzi-infected mice with a CCR1/CCR5antagonist (Met-RANTES) results in amelioration of cardiac tissue damage.Microbes Infect 2009;11(February (2)):264–73.
42] Sales Jr PA, Golgher D, Oliveira RV, Vieira V, Arantes RM, Lannes-Vieira J, etal. The regulatory CD4+CD25+ T cells have a limited role on pathogenesis ofinfection with Trypanosoma cruzi. Microbes Infect 2008;10(May (6)):680–8.
43] Punukollu G, Gowda RM, Khan IA, Navarro VS, Vasavada BC. Clinical aspects ofthe Chagas’ heart disease. Int J Cardiol 2007;115(February (3)):279–83.
44] Kroll-Palhares K, Silvério JC, Silva AA, Michailowsky V, Marino AP, Silva NM, etal. TNF/TNFR1 signaling up-regulates CCR5 expression by CD8+ T lymphocytesand promotes heart tissue damage during Trypanosoma cruzi infection: ben-eficial effects of TNF-alpha blockade. Mem Inst Oswaldo Cruz 2008;103(June(4)):375–85.
45] Chandra M, Tanowitz HB, Petkova SB, Huang H, Weiss LM, Wittner M, et al.Significance of inducible nitric oxide synthase in acute myocarditis caused byTrypanosoma cruzi (Tulahuen strain). Int J Parasitol 2002;32(June (7)):897–905.
46] Cardioprotective effects of verapamil on myocardial structure and functionin a murine model of chronic Trypanosoma cruzi infection (Brazil Strain): anechocardiographic study.
47] Chandra M, Shirani J, Shtutin V, Weiss LM, Factor SM, Petkova SB, et al. Int JParasitol 2002;32(February (2)):207–15.
48] Caldas IS, Talvani A, Caldas S, Carneiro CM, de Lana M, da Matta Guedes PM, etal. Benznidazole therapy during acute phase of Chagas disease reduces parasiteload but does not prevent chronic cardiac lesions. Parasitol Res 2008;103(July(2)):413–21.
49] Sanchez JA, Milei J, Yu ZX, Storino R, Wenthold Jr R, Ferrans VJ. Immunohisto-chemical localization of laminin in the hearts of patients with chronic chagasiccardiomyopathy: relationship to thickening of basement membranes. Am HeartJ 1993;126(December (6)):1392–401.
50] Abdalla GK, Faria GE, Silva KT, Castro EC, Reis MA, Michelin MA. Trypanosomacruzi: the role of PGE2 in immune response during the acute phase of experi-mental infection. Exp Parasitol 2008;118(April (4)):514–21.
[51] Garzoni LR, Adesse D, Soares MJ, Rossi MI, Borojevic R, de Meirelles Mde N.Fibrosis and hypertrophy induced by Trypanosoma cruzi in a three-dimensionalcardiomyocyte-culture system. J Infect Dis 2008;197(March (6)):906–15.
52] Munoz N, Manalastas Jr R, Pitisuttithum P, Tresukosol D, Monsonego J, AultK, et al. Safety, immunogenicity, and efficacy of quadrivalent human papillo-mavirus (types 6,11, 16, 18) recombinant vaccine in women aged 24–45 years:a randomised double-blind trial. Lancet 2009;373(June (9679)):1949–57.
53] Cazorla SI, Frank FM, Malchiodi EL. Vaccination approaches against Try-panosoma cruzi infection. Expert Rev Vaccines 2009;July (8):921–35.

121
Referências Bibliográficas

122
1. Chagas C (1909) Nova tripanozomiase humana. Estudos sobre a morfolojia e o ciclo evolutivo de Schizotrypanum cruzi n. gen., n. sp., ajente etiolójico de nova entidade morbida do homem. Mem Inst Oswaldo Cruz 1: 159-218.
2. Moncayo A, Silveira AC (2009) Current epidemiological trends for Chagas disease in Latin America and future challenges in epidemiology, surveillance and health policy. Mem Inst Oswaldo Cruz 104 Suppl 1: 17-30.
3. WHO/TDR (2007) World Health Organization - Grupo de trabajos científicos sobre la enfermidad de Chagas. TDR/GTC/09, Geneve.
4. Coura JR, Dias JC (2009) Epidemiology, control and surveillance of Chagas disease: 100 years after its discovery. Mem Inst Oswaldo Cruz 104 Suppl 1: 31-40.
5. Dias JC (2009) Elimination of Chagas disease transmission: perspectives. Mem Inst Oswaldo Cruz 104 Suppl 1: 41-45.
6. Coura JR, Anunziato N, Willcox HP (1983) [Chagas' disease morbidity. I - Study of cases originating in various states of Brazil, observed in Rio de Janeiro]. Mem Inst Oswaldo Cruz 78: 363-372.
7. Coura JR, de Abreu LL, Pereira JB, Willcox HP (1985) [Morbidity in Chagas' disease. IV. Longitudinal study of 10 years in Pains and Iguatama, Minas Gerais, Brazil]. Mem Inst Oswaldo Cruz 80: 73-80.
8. Pereira JB, Willcox HP, Coura JR (1985) [Morbidity in Chagas' disease. III. Longitudinal study of 6 years, in Virgem da Lapa, MG, Brazil]. Mem Inst Oswaldo Cruz 80: 63-71.
9. Marin-Neto JA, Cunha-Neto E, Maciel BC, Simoes MV (2007) Pathogenesis of chronic Chagas heart disease. Circulation 115: 1109-1123.
10. Cunha-Neto E, Duranti M, Gruber A, Zingales B, De Messias I, et al. (1995) Autoimmunity in Chagas disease cardiopathy: biological relevance of a cardiac myosin-specific epitope crossreactive to an immunodominant Trypanosoma cruzi antigen. Proc Natl Acad Sci U S A 92: 3541-3545.
11. Kaplan D, Ferrari I, Bergami PL, Mahler E, Levitus G, et al. (1997) Antibodies to ribosomal P proteins of Trypanosoma cruzi in Chagas disease possess functional autoreactivity with heart tissue and differ from anti-P autoantibodies in lupus. Proc Natl Acad Sci U S A 94: 10301-10306.
12. Giordanengo L, Maldonado C, Rivarola HW, Iosa D, Girones N, et al. (2000) Induction of antibodies reactive to cardiac myosin and development of heart alterations in cruzipain-immunized mice and their offspring. Eur J Immunol 30: 3181-3189.
13. Cunha-Neto E, Coelho V, Guilherme L, Fiorelli A, Stolf N, et al. (1996) Autoimmunity in Chagas' disease. Identification of cardiac myosin-B13 Trypanosoma cruzi protein crossreactive T cell clones in heart lesions of a chronic Chagas' cardiomyopathy patient. J Clin Invest 98: 1709-1712.
14. Leon JS, Godsel LM, Wang K, Engman DM (2001) Cardiac myosin autoimmunity in acute Chagas' heart disease. Infect Immun 69: 5643-5649.
15. Iwai LK, Juliano MA, Juliano L, Kalil J, Cunha-Neto E (2005) T-cell molecular mimicry in Chagas disease: identification and partial structural analysis of multiple cross-reactive epitopes between Trypanosoma cruzi B13 and cardiac myosin heavy chain. J Autoimmun 24: 111-117.
16. Monteon-Padilla V, Hernandez-Becerril N, Ballinas-Verdugo MA, Aranda-Fraustro A, Reyes PA (2001) Persistence of Trypanosoma cruzi in chronic chagasic cardiopathy patients. Arch Med Res 32: 39-43.
17. Elias FE, Vigliano CA, Laguens RP, Levin MJ, Berek C (2003) Analysis of the presence of Trypanosoma cruzi in the heart tissue of three patients with chronic Chagas' heart disease. Am J Trop Med Hyg 68: 242-247.

123
18. Benvenuti LA, Roggerio A, Sambiase NV, Fiorelli A, Higuchi Mde L (2005) Polymerase chain reaction in endomyocardial biopsies for monitoring reactivation of Chagas' disease in heart transplantation: a case report and review of the literature. Cardiovasc Pathol 14: 265-268.
19. Burgos JM, Begher SB, Freitas JM, Bisio M, Duffy T, et al. (2005) Molecular diagnosis and typing of Trypanosoma cruzi populations and lineages in cerebral Chagas disease in a patient with AIDS. Am J Trop Med Hyg 73: 1016-1018.
20. Cordova E, Boschi A, Ambrosioni J, Cudos C, Corti M (2008) Reactivation of Chagas disease with central nervous system involvement in HIV-infected patients in Argentina, 1992-2007. Int J Infect Dis 12: 587-592.
21. Apt W, Arribada A, Zulantay I, Sanchez G, Vargas SL, et al. (2003) Itraconazole or allopurinol in the treatment of chronic American trypanosomiasis: the regression and prevention of electrocardiographic abnormalities during 9 years of follow-up. Ann Trop Med Parasitol 97: 23-29.
22. Garcia S, Ramos CO, Senra JF, Vilas-Boas F, Rodrigues MM, et al. (2005) Treatment with benznidazole during the chronic phase of experimental Chagas' disease decreases cardiac alterations. Antimicrob Agents Chemother 49: 1521-1528.
23. Viotti R, Vigliano C, Lococo B, Bertocchi G, Petti M, et al. (2006) Long-term cardiac outcomes of treating chronic Chagas disease with benznidazole versus no treatment: a nonrandomized trial. Ann Intern Med 144: 724-734.
24. Dutra WO, Menezes CA, Villani FN, da Costa GC, da Silveira AB, et al. (2009) Cellular and genetic mechanisms involved in the generation of protective and pathogenic immune responses in human Chagas disease. Mem Inst Oswaldo Cruz 104 Suppl 1: 208-218.
25. Tarleton RL (2003) Chagas disease: a role for autoimmunity? Trends Parasitol 19: 447-451. 26. Kierszenbaum F (2005) Where do we stand on the autoimmunity hypothesis of Chagas
disease? Trends Parasitol 21: 513-516. 27. De Scheerder IK, De Buyzere M, Delanghe J, Maas A, Clement DL, et al. (1991) Humoral
immune response against contractile proteins (actin and myosin) during cardiovascular disease. Eur Heart J 12 Suppl D: 88-94.
28. Nomura Y, Yoshinaga M, Haraguchi T, Oku S, Noda T, et al. (1994) Relationship between the degree of injury at operation and the change in antimyosin antibody titer in the postpericardiotomy syndrome. Pediatr Cardiol 15: 116-120.
29. Girones N, Rodriguez CI, Carrasco-Marin E, Hernaez RF, de Rego JL, et al. (2001) Dominant T- and B-cell epitopes in an autoantigen linked to Chagas' disease. J Clin Invest 107: 985-993.
30. Manoel-Caetano Fda S, Carareto CM, Borim AA, Miyazaki K, Silva AE (2008) kDNA gene signatures of Trypanosoma cruzi in blood and oesophageal mucosa from chronic chagasic patients. Trans R Soc Trop Med Hyg 102: 1102-1107.
31. Costa GC, da Costa Rocha MO, Moreira PR, Menezes CA, Silva MR, et al. (2009) Functional IL-10 gene polymorphism is associated with Chagas disease cardiomyopathy. J Infect Dis 199: 451-454.
32. Gomes YM, Lorena VM, Luquetti AO (2009) Diagnosis of Chagas disease: what has been achieved? What remains to be done with regard to diagnosis and follow up studies? Mem Inst Oswaldo Cruz 104 Suppl 1: 115-121.
33. Britto CC (2009) Usefulness of PCR-based assays to assess drug efficacy in Chagas disease chemotherapy: value and limitations. Mem Inst Oswaldo Cruz 104 Suppl 1: 122-135.
34. Krettli AU (2009) The utility of anti-trypomastigote lytic antibodies for determining cure of Trypanosoma cruzi infections in treated patients: an overview and perspectives. Mem Inst Oswaldo Cruz 104 Suppl 1: 142-151.

124
35. Urbina JA (2009) Ergosterol biosynthesis and drug development for Chagas disease. Mem Inst Oswaldo Cruz 104 Suppl 1: 311-318.
36. Marin-Neto JA, Rassi A, Jr., Avezum A, Jr., Mattos AC, Rassi A (2009) The BENEFIT trial: testing the hypothesis that trypanocidal therapy is beneficial for patients with chronic Chagas heart disease. Mem Inst Oswaldo Cruz 104 Suppl 1: 319-324.
37. McKerrow JH, Doyle PS, Engel JC, Podust LM, Robertson SA, et al. (2009) Two approaches to discovering and developing new drugs for Chagas disease. Mem Inst Oswaldo Cruz 104 Suppl 1: 263-269.
38. Cancado JR (2002) Long term evaluation of etiological treatment of chagas disease with benznidazole. Rev Inst Med Trop Sao Paulo 44: 29-37.
39. Macedo AM, Oliveira RP, Pena SD (2002) Chagas disease: role of parasite genetic variation in pathogenesis. Expert Rev Mol Med 4: 1-16.
40. Ishii KJ, Akira S (2007) Toll or toll-free adjuvant path toward the optimal vaccine development. J Clin Immunol 27: 363-371.
41. Rasmussen SB, Reinert LS, Paludan SR (2009) Innate recognition of intracellular pathogens: detection and activation of the first line of defense. Apmis 117: 323-337.
42. Campos MA, Almeida IC, Takeuchi O, Akira S, Valente EP, et al. (2001) Activation of Toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. J Immunol 167: 416-423.
43. Oliveira AC, Peixoto JR, de Arruda LB, Campos MA, Gazzinelli RT, et al. (2004) Expression of functional TLR4 confers proinflammatory responsiveness to Trypanosoma cruzi glycoinositolphospholipids and higher resistance to infection with T. cruzi. J Immunol 173: 5688-5696.
44. Bartholomeu DC, Ropert C, Melo MB, Parroche P, Junqueira CF, et al. (2008) Recruitment and endo-lysosomal activation of TLR9 in dendritic cells infected with Trypanosoma cruzi. J Immunol 181: 1333-1344.
45. Bafica A, Santiago HC, Goldszmid R, Ropert C, Gazzinelli RT, et al. (2006) Cutting edge: TLR9 and TLR2 signaling together account for MyD88-dependent control of parasitemia in Trypanosoma cruzi infection. J Immunol 177: 3515-3519.
46. Koga R, Hamano S, Kuwata H, Atarashi K, Ogawa M, et al. (2006) TLR-dependent induction of IFN-beta mediates host defense against Trypanosoma cruzi. J Immunol 177: 7059-7066.
47. Sardinha LR, Elias RM, Mosca T, Bastos KR, Marinho CR, et al. (2006) Contribution of NK, NK T, gamma delta T, and alpha beta T cells to the gamma interferon response required for liver protection against Trypanosoma cruzi. Infect Immun 74: 2031-2042.
48. Duthie MS, Kahn SJ (2006) During acute Trypanosoma cruzi infection highly susceptible mice deficient in natural killer cells are protected by a single alpha-galactosylceramide treatment. Immunology 119: 355-361.
49. Cardillo F, Voltarelli JC, Reed SG, Silva JS (1996) Regulation of Trypanosoma cruzi infection in mice by gamma interferon and interleukin 10: role of NK cells. Infect Immun 64: 128-134.
50. Lieke T, Graefe SE, Klauenberg U, Fleischer B, Jacobs T (2004) NK cells contribute to the control of Trypanosoma cruzi infection by killing free parasites by perforin-independent mechanisms. Infect Immun 72: 6817-6825.
51. Lieke T, Steeg C, Graefe SE, Fleischer B, Jacobs T (2006) Interaction of natural killer cells with Trypanosoma cruzi-infected fibroblasts. Clin Exp Immunol 145: 357-364.
52. Duthie MS, Wleklinski-Lee M, Smith S, Nakayama T, Taniguchi M, et al. (2002) During Trypanosoma cruzi infection CD1d-restricted NK T cells limit parasitemia and augment the antibody response to a glycophosphoinositol-modified surface protein. Infect Immun 70: 36-48.

125
53. Duthie MS, Kahn M, White M, Kapur RP, Kahn SJ (2005) Critical proinflammatory and anti-inflammatory functions of different subsets of CD1d-restricted natural killer T cells during Trypanosoma cruzi infection. Infect Immun 73: 181-192.
54. Santos Lima EC, Minoprio P (1996) Chagas' disease is attenuated in mice lacking gamma delta T cells. Infect Immun 64: 215-221.
55. Nomizo A, Cardillo F, Postol E, de Carvalho LP, Mengel J (2006) V gamma 1 gammadelta T cells regulate type-1/type-2 immune responses and participate in the resistance to infection and development of heart inflammation in Trypanosoma cruzi-infected BALB/c mice. Microbes Infect 8: 880-888.
56. Kumar S, Tarleton RL (1998) The relative contribution of antibody production and CD8+ T cell function to immune control of Trypanosoma cruzi. Parasite Immunol 20: 207-216.
57. Rottenberg ME, Bakhiet M, Olsson T, Kristensson K, Mak T, et al. (1993) Differential susceptibilities of mice genomically deleted of CD4 and CD8 to infections with Trypanosoma cruzi or Trypanosoma brucei. Infect Immun 61: 5129-5133.
58. Tarleton RL, Grusby MJ, Postan M, Glimcher LH (1996) Trypanosoma cruzi infection in MHC-deficient mice: further evidence for the role of both class I- and class II-restricted T cells in immune resistance and disease. Int Immunol 8: 13-22.
59. Bustamante JM, Bixby LM, Tarleton RL (2008) Drug-induced cure drives conversion to a stable and protective CD8+ T central memory response in chronic Chagas disease. Nat Med 14: 542-550.
60. Krettli AU, Brener Z (1976) Protective effects of specific antibodies in Trypanosoma cruzi infections. J Immunol 116: 755-760.
61. Kierszenbaum F (1980) Protection of congenitally athymic mice against Trypanosoma cruzi infection by passive antibody transfer. J Parasitol 66: 673-675.
62. Franchin G, Pereira-Chioccola VL, Schenkman S, Rodrigues MM (1997) Passive transfer of a monoclonal antibody specific for a sialic acid-dependent epitope on the surface of Trypanosoma cruzi trypomastigotes reduces infection in mice. Infect Immun 65: 2548-2554.
63. Lages-Silva E, Ramirez LE, Krettli AU, Brener Z (1987) Effect of protective and non-protective antibodies in the phagocytosis rate of Trypanosoma cruzi blood forms by mouse peritoneal macrophages. Parasite Immunol 9: 21-30.
64. Okabe K, Kipnis TL, Calich VL, Dias da Silva W (1980) Cell-mediated cytotoxicity to Trypanosoma cruzi. I. Antibody-dependent cell mediated cytotoxicity to trypomastigote bloodstream forms. Clin Immunol Immunopathol 16: 344-353.
65. Lima-Martins MV, Sanchez GA, Krettli AU, Brener Z (1985) Antibody-dependent cell cytotoxicity against Trypanosoma cruzi is only mediated by protective antibodies. Parasite Immunol 7: 367-376.
66. Norris KA (1998) Stable transfection of Trypanosoma cruzi epimastigotes with the trypomastigote-specific complement regulatory protein cDNA confers complement resistance. Infect Immun 66: 2460-2465.
67. Hoft DF, Eickhoff CS, Giddings OK, Vasconcelos JR, Rodrigues MM (2007) Trans-sialidase recombinant protein mixed with CpG motif-containing oligodeoxynucleotide induces protective mucosal and systemic trypanosoma cruzi immunity involving CD8+ CTL and B cell-mediated cross-priming. J Immunol 179: 6889-6900.
68. Cardillo F, Postol E, Nihei J, Aroeira LS, Nomizo A, et al. (2007) B cells modulate T cells so as to favour T helper type 1 and CD8+ T-cell responses in the acute phase of Trypanosoma cruzi infection. Immunology 122: 584-595.
69. Tzelepis F, de Alencar BC, Penido ML, Gazzinelli RT, Persechini PM, et al. (2006) Distinct kinetics of effector CD8+ cytotoxic T cells after infection with Trypanosoma cruzi in naive or vaccinated mice. Infect Immun 74: 2477-2481.

126
70. Martin DL, Weatherly DB, Laucella SA, Cabinian MA, Crim MT, et al. (2006) CD8+ T-Cell responses to Trypanosoma cruzi are highly focused on strain-variant trans-sialidase epitopes. PLoS Pathog 2: e77.
71. Tzelepis F, Persechini PM, Rodrigues MM (2007) Modulation of CD4+ T cell-dependent specific cytotoxic CD8+ T cells differentiation and proliferation by the timing of increase in the pathogen load. PLoS One 2: e393.
72. Davenport MP, Belz GT, Ribeiro RM (2009) The race between infection and immunity: how do pathogens set the pace? Trends Immunol 30: 61-66.
73. Padilla AM, Simpson LJ, Tarleton RL (2009) Insufficient TLR activation contributes to the slow development of CD8+ T cell responses in Trypanosoma cruzi infection. J Immunol 183: 1245-1252.
74. Rodrigues MM, Ribeirao M, Pereira-Chioccola V, Renia L, Costa F (1999) Predominance of CD4 Th1 and CD8 Tc1 cells revealed by characterization of the cellular immune response generated by immunization with a DNA vaccine containing a Trypanosoma cruzi gene. Infect Immun 67: 3855-3863.
75. Araujo AF, de Alencar BC, Vasconcelos JR, Hiyane MI, Marinho CR, et al. (2005) CD8+-T-cell-dependent control of Trypanosoma cruzi infection in a highly susceptible mouse strain after immunization with recombinant proteins based on amastigote surface protein 2. Infect Immun 73: 6017-6025.
76. Low HP, Santos MA, Wizel B, Tarleton RL (1998) Amastigote surface proteins of Trypanosoma cruzi are targets for CD8+ CTL. J Immunol 160: 1817-1823.
77. Millar AE, Kahn SJ (2000) The SA85-1.1 protein of the Trypanosoma cruzi trans-sialidase superfamily is a dominant T-cell antigen. Infect Immun 68: 3574-3580.
78. Kotner J, Tarleton R (2007) Endogenous CD4(+) CD25(+) regulatory T cells have a limited role in the control of Trypanosoma cruzi infection in mice. Infect Immun 75: 861-869.
79. Sales PA, Jr., Golgher D, Oliveira RV, Vieira V, Arantes RM, et al. (2008) The regulatory CD4+CD25+ T cells have a limited role on pathogenesis of infection with Trypanosoma cruzi. Microbes Infect 10: 680-688.
80. Mariano FS, Gutierrez FR, Pavanelli WR, Milanezi CM, Cavassani KA, et al. (2008) The involvement of CD4+CD25+ T cells in the acute phase of Trypanosoma cruzi infection. Microbes Infect 10: 825-833.
81. Tarleton RL, Grusby MJ, Zhang L (2000) Increased susceptibility of Stat4-deficient and enhanced resistance in Stat6-deficient mice to infection with Trypanosoma cruzi. J Immunol 165: 1520-1525.
82. Nickell SP, Sharma D (2000) Trypanosoma cruzi: roles for perforin-dependent and perforin-independent immune mechanisms in acute resistance. Exp Parasitol 94: 207-216.
83. Muller U, Sobek V, Balkow S, Holscher C, Mullbacher A, et al. (2003) Concerted action of perforin and granzymes is critical for the elimination of Trypanosoma cruzi from mouse tissues, but prevention of early host death is in addition dependent on the FasL/Fas pathway. Eur J Immunol 33: 70-78.
84. Persson EK, Agnarson AM, Lambert H, Hitziger N, Yagita H, et al. (2007) Death receptor ligation or exposure to perforin trigger rapid egress of the intracellular parasite Toxoplasma gondii. J Immunol 179: 8357-8365.
85. Michailowsky V, Silva NM, Rocha CD, Vieira LQ, Lannes-Vieira J, et al. (2001) Pivotal role of interleukin-12 and interferon-gamma axis in controlling tissue parasitism and inflammation in the heart and central nervous system during Trypanosoma cruzi infection. Am J Pathol 159: 1723-1733.
86. Bilate AM, Salemi VM, Ramires FJ, de Brito T, Russo M, et al. (2007) TNF blockade aggravates experimental chronic Chagas disease cardiomyopathy. Microbes Infect 9: 1104-1113.

127
87. Castanos-Velez E, Maerlan S, Osorio LM, Aberg F, Biberfeld P, et al. (1998) Trypanosoma cruzi infection in tumor necrosis factor receptor p55-deficient mice. Infect Immun 66: 2960-2968.
88. Aliberti JC, Souto JT, Marino AP, Lannes-Vieira J, Teixeira MM, et al. (2001) Modulation of chemokine production and inflammatory responses in interferon-gamma- and tumor necrosis factor-R1-deficient mice during Trypanosoma cruzi infection. Am J Pathol 158: 1433-1440.
89. Holscher C, Kohler G, Muller U, Mossmann H, Schaub GA, et al. (1998) Defective nitric oxide effector functions lead to extreme susceptibility of Trypanosoma cruzi-infected mice deficient in gamma interferon receptor or inducible nitric oxide synthase. Infect Immun 66: 1208-1215.
90. Costa VM, Torres KC, Mendonca RZ, Gresser I, Gollob KJ, et al. (2006) Type I IFNs stimulate nitric oxide production and resistance to Trypanosoma cruzi infection. J Immunol 177: 3193-3200.
91. Rodrigues MM, Ribeirao M, Boscardin SB (2000) CD4 Th1 but not Th2 clones efficiently activate macrophages to eliminate Trypanosoma cruzi through a nitric oxide dependent mechanism. Immunol Lett 73: 43-50.
92. Cummings KL, Tarleton RL (2004) Inducible nitric oxide synthase is not essential for control of Trypanosoma cruzi infection in mice. Infect Immun 72: 4081-4089.
93. Hiyama K, Hamano S, Nakamura T, Nomoto K, Tada I (2001) IL-4 reduces resistance of mice to Trypanosoma cruzi infection. Parasitol Res 87: 269-274.
94. Soares MB, Silva-Mota KN, Lima RS, Bellintani MC, Pontes-de-Carvalho L, et al. (2001) Modulation of chagasic cardiomyopathy by interleukin-4: dissociation between inflammation and tissue parasitism. Am J Pathol 159: 703-709.
95. Holscher C, Mohrs M, Dai WJ, Kohler G, Ryffel B, et al. (2000) Tumor necrosis factor alpha-mediated toxic shock in Trypanosoma cruzi-infected interleukin 10-deficient mice. Infect Immun 68: 4075-4083.
96. Hunter CA, Ellis-Neyes LA, Slifer T, Kanaly S, Grunig G, et al. (1997) IL-10 is required to prevent immune hyperactivity during infection with Trypanosoma cruzi. J Immunol 158: 3311-3316.
97. Martin DL, Postan M, Lucas P, Gress R, Tarleton RL (2007) TGF-beta regulates pathology but not tissue CD8+ T cell dysfunction during experimental Trypanosoma cruzi infection. Eur J Immunol 37: 2764-2771.
98. Marino AP, da Silva A, dos Santos P, Pinto LM, Gazzinelli RT, et al. (2004) Regulated on activation, normal T cell expressed and secreted (RANTES) antagonist (Met-RANTES) controls the early phase of Trypanosoma cruzi-elicited myocarditis. Circulation 110: 1443-1449.
99. Machado FS, Koyama NS, Carregaro V, Ferreira BR, Milanezi CM, et al. (2005) CCR5 plays a critical role in the development of myocarditis and host protection in mice infected with Trypanosoma cruzi. J Infect Dis 191: 627-636.
100. Kroll-Palhares K, Silverio JC, Silva AA, Michailowsky V, Marino AP, et al. (2008) TNF/TNFR1 signaling up-regulates CCR5 expression by CD8+ T lymphocytes and promotes heart tissue damage during Trypanosoma cruzi infection: beneficial effects of TNF-alpha blockade. Mem Inst Oswaldo Cruz 103: 375-385.
101. Hardison JL, Wrightsman RA, Carpenter PM, Kuziel WA, Lane TE, et al. (2006) The CC chemokine receptor 5 is important in control of parasite replication and acute cardiac inflammation following infection with Trypanosoma cruzi. Infect Immun 74: 135-143.
102. Medeiros GA, Silverio JC, Marino AP, Roffe E, Vieira V, et al. (2009) Treatment of chronically Trypanosoma cruzi-infected mice with a CCR1/CCR5 antagonist (Met-RANTES) results in amelioration of cardiac tissue damage. Microbes Infect 11: 264-273.

128
103. Krautz GM, Kissinger JC, Krettli AU (2000) The targets of the lytic antibody response against Trypanosoma cruzi. Parasitol Today 16: 31-34.
104. de Alencar BC, Araujo AF, Penido ML, Gazzinelli RT, Rodrigues MM (2007) Cross-priming of long lived protective CD8+ T cells against Trypanosoma cruzi infection: importance of a TLR9 agonist and CD4+ T cells. Vaccine 25: 6018-6027.
105. Tzelepis F, de Alencar BC, Penido ML, Claser C, Machado AV, et al. (2008) Infection with Trypanosoma cruzi restricts the repertoire of parasite-specific CD8+ T cells leading to immunodominance. J Immunol 180: 1737-1748.
106. Rodrigues MM, Alencar BC, Claser C, Tzelepis F (2009) Immunodominance: a new hypothesis to explain parasite escape and host/parasite equilibrium leading to the chronic phase of Chagas' disease? Braz J Med Biol Res 42: 220-223.
107. Wrightsman RA, Luhrs KA, Fouts D, Manning JE (2002) Paraflagellar rod protein-specific CD8+ cytotoxic T lymphocytes target Trypanosoma cruzi-infected host cells. Parasite Immunol 24: 401-412.
108. Garg N, Bhatia V (2005) Current status and future prospects for a vaccine against American trypanosomiasis. Expert Rev Vaccines 4: 867-880.
109. Frank FM, Cazorla SI, Sartori MJ, Corral RS (2008) Elicitation of specific, Th1-biased immune response precludes skeletal muscle damage in cruzipain-vaccinated mice. Exp Mol Pathol 84: 64-70.
110. Cazorla SI, Frank FM, Becker PD, Corral RS, Guzman CA, et al. (2008) Prime-boost immunization with cruzipain co-administered with MALP-2 triggers a protective immune response able to decrease parasite burden and tissue injury in an experimental Trypanosoma cruzi infection model. Vaccine 26: 1999-2009.
111. Katae M, Miyahira Y, Takeda K, Matsuda H, Yagita H, et al. (2002) Coadministration of an interleukin-12 gene and a Trypanosoma cruzi gene improves vaccine efficacy. Infect Immun 70: 4833-4840.
112. Schnapp AR, Eickhoff CS, Sizemore D, Curtiss R, 3rd, Hoft DF (2002) Cruzipain induces both mucosal and systemic protection against Trypanosoma cruzi in mice. Infect Immun 70: 5065-5074.
113. Fontanella GH, De Vusser K, Laroy W, Daurelio L, Nocito AL, et al. (2008) Immunization with an engineered mutant trans-sialidase highly protects mice from experimental Trypanosoma cruzi infection: a vaccine candidate. Vaccine 26: 2322-2334.
114. Fujimura AE, Kinoshita SS, Pereira-Chioccola VL, Rodrigues MM (2001) DNA sequences encoding CD4+ and CD8+ T-cell epitopes are important for efficient protective immunity induced by DNA vaccination with a Trypanosoma cruzi gene. Infect Immun 69: 5477-5486.
115. Boscardin SB, Kinoshita SS, Fujimura AE, Rodrigues MM (2003) Immunization with cDNA expressed by amastigotes of Trypanosoma cruzi elicits protective immune response against experimental infection. Infect Immun 71: 2744-2757.
116. Fralish BH, Tarleton RL (2003) Genetic immunization with LYT1 or a pool of trans-sialidase genes protects mice from lethal Trypanosoma cruzi infection. Vaccine 21: 3070-3080.
117. Vasconcelos JR, Hiyane MI, Marinho CR, Claser C, Machado AM, et al. (2004) Protective immunity against trypanosoma cruzi infection in a highly susceptible mouse strain after vaccination with genes encoding the amastigote surface protein-2 and trans-sialidase. Hum Gene Ther 15: 878-886.
118. Bhatia V, Garg NJ (2008) Previously unrecognized vaccine candidates control Trypanosoma cruzi infection and immunopathology in mice. Clin Vaccine Immunol 15: 1158-1164.
119. Liu MA, Ulmer JB (2005) Human clinical trials of plasmid DNA vaccines. Adv Genet 55: 25-40.

129
120. Miyahira Y, Takashima Y, Kobayashi S, Matsumoto Y, Takeuchi T, et al. (2005) Immune responses against a single CD8+-T-cell epitope induced by virus vector vaccination can successfully control Trypanosoma cruzi infection. Infect Immun 73: 7356-7365.
121. Machado AV, Cardoso JE, Claser C, Rodrigues MM, Gazzinelli RT, et al. (2006) Long-term protective immunity induced against Trypanosoma cruzi infection after vaccination with recombinant adenoviruses encoding amastigote surface protein-2 and trans-sialidase. Hum Gene Ther 17: 898-908.
122. Duan X, Yonemitsu Y, Chou B, Yoshida K, Tanaka S, et al. (2009) Efficient protective immunity against Trypanosoma cruzi infection after nasal vaccination with recombinant Sendai virus vector expressing amastigote surface protein-2. Vaccine 27: 6154-6159.
123. Cazorla SI, Becker PD, Frank FM, Ebensen T, Sartori MJ, et al. (2008) Oral vaccination with Salmonella enterica as a cruzipain-DNA delivery system confers protective immunity against Trypanosoma cruzi. Infect Immun 76: 324-333.
124. Wrightsman RA, Manning JE (2000) Paraflagellar rod proteins administered with alum and IL-12 or recombinant adenovirus expressing IL-12 generates antigen-specific responses and protective immunity in mice against Trypanosoma cruzi. Vaccine 18: 1419-1427.
125. Wizel B, Garg N, Tarleton RL (1998) Vaccination with trypomastigote surface antigen 1-encoding plasmid DNA confers protection against lethal Trypanosoma cruzi infection. Infect Immun 66: 5073-5081.
126. Dumonteil E, Escobedo-Ortegon J, Reyes-Rodriguez N, Arjona-Torres A, Ramirez-Sierra MJ (2004) Immunotherapy of Trypanosoma cruzi infection with DNA vaccines in mice. Infect Immun 72: 46-53.
127. Zapata-Estrella H, Hummel-Newell C, Sanchez-Burgos G, Escobedo-Ortegon J, Ramirez-Sierra MJ, et al. (2006) Control of Trypanosoma cruzi infection and changes in T-cell populations induced by a therapeutic DNA vaccine in mice. Immunol Lett 103: 186-191.
128. Miyahira Y, Akiba H, Katae M, Kubota K, Kobayashi S, et al. (2003) Cutting edge: a potent adjuvant effect of ligand to receptor activator of NF-kappa B gene for inducing antigen-specific CD8+ T cell response by DNA and viral vector vaccination. J Immunol 171: 6344-6348.
129. Costa F, Franchin G, Pereira-Chioccola VL, Ribeirao M, Schenkman S, et al. (1998) Immunization with a plasmid DNA containing the gene of trans-sialidase reduces Trypanosoma cruzi infection in mice. Vaccine 16: 768-774.
130. Garg N, Tarleton RL (2002) Genetic immunization elicits antigen-specific protective immune responses and decreases disease severity in Trypanosoma cruzi infection. Infect Immun 70: 5547-5555.
131. Claser C, Espindola NM, Sasso G, Vaz AJ, Boscardin SB, et al. (2007) Immunologically relevant strain polymorphism in the Amastigote Surface Protein 2 of Trypanosoma cruzi. Microbes Infect 9: 1011-1019.
132. Silveira EL, Claser C, Haolla FA, Zanella LG, Rodrigues MM (2008) Novel protective antigens expressed by Trypanosoma cruzi amastigotes provide immunity to mice highly susceptible to Chagas' disease. Clin Vaccine Immunol 15: 1292-1300.
133. Sepulveda P, Hontebeyrie M, Liegeard P, Mascilli A, Norris KA (2000) DNA-Based immunization with Trypanosoma cruzi complement regulatory protein elicits complement lytic antibodies and confers protection against Trypanosoma cruzi infection. Infect Immun 68: 4986-4991.
134. Planelles L, Thomas MC, Alonso C, Lopez MC (2001) DNA immunization with Trypanosoma cruzi HSP70 fused to the KMP11 protein elicits a cytotoxic and humoral immune response against the antigen and leads to protection. Infect Immun 69: 6558-6563.
135. Cazorla SI, Frank FM, Malchiodi EL (2009) Vaccination approaches against Trypanosoma cruzi infection. Expert Rev Vaccines 8: 921-935.

130
136. Pan AA, McMahon-Pratt D (1989) Amastigote and epimastigote stage-specific components of Trypanosoma cruzi characterized by using monoclonal antibodies. Purification and molecular characterization of an 83-kilodalton amastigote protein. J Immunol 143: 1001-1008.
137. Low HP, Tarleton RL (1997) Molecular cloning of the gene encoding the 83 kDa amastigote surface protein and its identification as a member of the Trypanosoma cruzi sialidase superfamily. Mol Biochem Parasitol 88: 137-149.
138. Roggentin P, Rothe B, Kaper JB, Galen J, Lawrisuk L, et al. (1989) Conserved sequences in bacterial and viral sialidases. Glycoconj J 6: 349-353.
139. Frasch AC (2000) Functional diversity in the trans-sialidase and mucin families in Trypanosoma cruzi. Parasitol Today 16: 282-286.
140. Garg N, Postan M, Mensa-Wilmot K, Tarleton RL (1997) Glycosylphosphatidylinositols are required for the development of Trypanosoma cruzi amastigotes. Infect Immun 65: 4055-4060.
141. Magdesian MH, Giordano R, Ulrich H, Juliano MA, Juliano L, et al. (2001) Infection by Trypanosoma cruzi. Identification of a parasite ligand and its host cell receptor. J Biol Chem 276: 19382-19389.
142. Magdesian MH, Tonelli RR, Fessel MR, Silveira MS, Schumacher RI, et al. (2007) A conserved domain of the gp85/trans-sialidase family activates host cell extracellular signal-regulated kinase and facilitates Trypanosoma cruzi infection. Exp Cell Res 313: 210-218.
143. Reed SG, Bertholet S, Coler RN, Friede M (2009) New horizons in adjuvants for vaccine development. Trends Immunol 30: 23-32.
144. Morefield GL, Sokolovska A, Jiang D, HogenEsch H, Robinson JP, et al. (2005) Role of aluminum-containing adjuvants in antigen internalization by dendritic cells in vitro. Vaccine 23: 1588-1595.
145. Kool M, Soullie T, van Nimwegen M, Willart MA, Muskens F, et al. (2008) Alum adjuvant boosts adaptive immunity by inducing uric acid and activating inflammatory dendritic cells. J Exp Med 205: 869-882.
146. Gavin AL, Hoebe K, Duong B, Ota T, Martin C, et al. (2006) Adjuvant-enhanced antibody responses in the absence of toll-like receptor signaling. Science 314: 1936-1938.
147. Eisenbarth SC, Colegio OR, O'Connor W, Sutterwala FS, Flavell RA (2008) Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature 453: 1122-1126.
148. Li H, Willingham SB, Ting JP, Re F (2008) Cutting edge: inflammasome activation by alum and alum's adjuvant effect are mediated by NLRP3. J Immunol 181: 17-21.
149. Franchi L, Nunez G (2008) The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1beta secretion but dispensable for adjuvant activity. Eur J Immunol 38: 2085-2089.
150. Marrack P, McKee AS, Munks MW (2009) Towards an understanding of the adjuvant action of aluminium. Nat Rev Immunol 9: 287-293.
151. Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, et al. (2000) A Toll-like receptor recognizes bacterial DNA. Nature 408: 740-745.
152. Takeshita F, Leifer CA, Gursel I, Ishii KJ, Takeshita S, et al. (2001) Cutting edge: Role of Toll-like receptor 9 in CpG DNA-induced activation of human cells. J Immunol 167: 3555-3558.
153. Hacker H, Vabulas RM, Takeuchi O, Hoshino K, Akira S, et al. (2000) Immune cell activation by bacterial CpG-DNA through myeloid differentiation marker 88 and tumor necrosis factor receptor-associated factor (TRAF)6. J Exp Med 192: 595-600.

131
154. Kumagai Y, Takeuchi O, Akira S (2008) TLR9 as a key receptor for the recognition of DNA. Adv Drug Deliv Rev 60: 795-804.
155. Maurer T, Heit A, Hochrein H, Ampenberger F, O'Keeffe M, et al. (2002) CpG-DNA aided cross-presentation of soluble antigens by dendritic cells. Eur J Immunol 32: 2356-2364.
156. Heit A, Huster KM, Schmitz F, Schiemann M, Busch DH, et al. (2004) CpG-DNA aided cross-priming by cross-presenting B cells. J Immunol 172: 1501-1507.
157. Di Pucchio T, Chatterjee B, Smed-Sorensen A, Clayton S, Palazzo A, et al. (2008) Direct proteasome-independent cross-presentation of viral antigen by plasmacytoid dendritic cells on major histocompatibility complex class I. Nat Immunol 9: 551-557.
158. Kuchtey J, Chefalo PJ, Gray RC, Ramachandra L, Harding CV (2005) Enhancement of dendritic cell antigen cross-presentation by CpG DNA involves type I IFN and stabilization of class I MHC mRNA. J Immunol 175: 2244-2251.
159. Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, et al. (1993) Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 259: 1745-1749.
160. Fynan EF, Webster RG, Fuller DH, Haynes JR, Santoro JC, et al. (1993) DNA vaccines: protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proc Natl Acad Sci U S A 90: 11478-11482.
161. Wang B, Ugen KE, Srikantan V, Agadjanyan MG, Dang K, et al. (1993) Gene inoculation generates immune responses against human immunodeficiency virus type 1. Proc Natl Acad Sci U S A 90: 4156-4160.
162. Danko I, Williams P, Herweijer H, Zhang G, Latendresse JS, et al. (1997) High expression of naked plasmid DNA in muscles of young rodents. Hum Mol Genet 6: 1435-1443.
163. Dupuis M, Denis-Mize K, Woo C, Goldbeck C, Selby MJ, et al. (2000) Distribution of DNA vaccines determines their immunogenicity after intramuscular injection in mice. J Immunol 165: 2850-2858.
164. Chattergoon MA, Kim JJ, Yang JS, Robinson TM, Lee DJ, et al. (2000) Targeted antigen delivery to antigen-presenting cells including dendritic cells by engineered Fas-mediated apoptosis. Nat Biotechnol 18: 974-979.
165. Rubartelli A, Poggi A, Zocchi MR (1997) The selective engulfment of apoptotic bodies by dendritic cells is mediated by the alpha(v)beta3 integrin and requires intracellular and extracellular calcium. Eur J Immunol 27: 1893-1900.
166. Albert ML, Sauter B, Bhardwaj N (1998) Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature 392: 86-89.
167. Ishii KJ, Kawagoe T, Koyama S, Matsui K, Kumar H, et al. (2008) TANK-binding kinase-1 delineates innate and adaptive immune responses to DNA vaccines. Nature 451: 725-729.
168. Ishikawa H, Ma Z, Barber GN (2009) STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 461: 788-792.
169. Kutzler MA, Weiner DB (2008) DNA vaccines: ready for prime time? Nat Rev Genet 9: 776-788.
170. Ramakrishna L, Anand KK, Mohankumar KM, Ranga U (2004) Codon optimization of the tat antigen of human immunodeficiency virus type 1 generates strong immune responses in mice following genetic immunization. J Virol 78: 9174-9189.
171. Yan J, Yoon H, Kumar S, Ramanathan MP, Corbitt N, et al. (2007) Enhanced cellular immune responses elicited by an engineered HIV-1 subtype B consensus-based envelope DNA vaccine. Mol Ther 15: 411-421.
172. Capone S, Zampaglione I, Vitelli A, Pezzanera M, Kierstead L, et al. (2006) Modulation of the immune response induced by gene electrotransfer of a hepatitis C virus DNA vaccine in nonhuman primates. J Immunol 177: 7462-7471.

132
173. Luckay A, Sidhu MK, Kjeken R, Megati S, Chong SY, et al. (2007) Effect of plasmid DNA vaccine design and in vivo electroporation on the resulting vaccine-specific immune responses in rhesus macaques. J Virol 81: 5257-5269.
174. Vandervoort J, Ludwig A (2008) Microneedles for transdermal drug delivery: a minireview. Front Biosci 13: 1711-1715.
175. Abdulhaqq SA, Weiner DB (2008) DNA vaccines: developing new strategies to enhance immune responses. Immunol Res 42: 219-232.
176. Radosevic K, Rodriguez A, Lemckert A, Goudsmit J (2009) Heterologous prime-boost vaccinations for poverty-related diseases: advantages and future prospects. Expert Rev Vaccines 8: 577-592.
177. Ourmanov I, Kuwata T, Goeken R, Goldstein S, Iyengar R, et al. (2009) Improved survival in rhesus macaques immunized with modified vaccinia virus Ankara recombinants expressing simian immunodeficiency virus envelope correlates with reduction in memory CD4+ T-cell loss and higher titers of neutralizing antibody. J Virol 83: 5388-5400.
178. Mooij P, Balla-Jhagjhoorsingh SS, Beenhakker N, van Haaften P, Baak I, et al. (2009) Comparison of human and rhesus macaque T-cell responses elicited by boosting with NYVAC encoding human immunodeficiency virus type 1 clade C immunogens. J Virol 83: 5881-5889.
179. Barouch DH, Liu J, Lynch DM, O'Brien KL, La Porte A, et al. (2009) Protective efficacy of a single immunization of a chimeric adenovirus vector-based vaccine against simian immunodeficiency virus challenge in rhesus monkeys. J Virol 83: 9584-9590.
180. Gao G, Wang Q, Dai Z, Calcedo R, Sun X, et al. (2008) Adenovirus-based vaccines generate cytotoxic T lymphocytes to epitopes of NS1 from dengue virus that are present in all major serotypes. Hum Gene Ther 19: 927-936.
181. Monath TP, Liu J, Kanesa-Thasan N, Myers GA, Nichols R, et al. (2006) A live, attenuated recombinant West Nile virus vaccine. Proc Natl Acad Sci U S A 103: 6694-6699.
182. Tao D, Barba-Spaeth G, Rai U, Nussenzweig V, Rice CM, et al. (2005) Yellow fever 17D as a vaccine vector for microbial CTL epitopes: protection in a rodent malaria model. J Exp Med 201: 201-209.
183. de Goede AL, Boers PH, Dekker LJ, Osterhaus AD, Gruters RA, et al. (2009) Characterization of recombinant influenza A virus as a vector for HIV-1 p17Gag. Vaccine 27: 5735-5739.
184. He Q, Martinez-Sobrido L, Eko FO, Palese P, Garcia-Sastre A, et al. (2007) Live-attenuated influenza viruses as delivery vectors for Chlamydia vaccines. Immunology 122: 28-37.
185. Buchbinder SP, Mehrotra DV, Duerr A, Fitzgerald DW, Mogg R, et al. (2008) Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 372: 1881-1893.
186. Russell WC (2009) Adenoviruses: update on structure and function. J Gen Virol 90: 1-20. 187. Yang TC, Millar JB, Grinshtein N, Bassett J, Finn J, et al. (2007) T-cell immunity generated
by recombinant adenovirus vaccines. Expert Rev Vaccines 6: 347-356. 188. Limbach KJ, Richie TL (2009) Viral vectors in malaria vaccine development. Parasite
Immunol 31: 501-519. 189. Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, et al. (1997)
Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 275: 1320-1323.
190. Roelvink PW, Lizonova A, Lee JG, Li Y, Bergelson JM, et al. (1998) The coxsackievirus-adenovirus receptor protein can function as a cellular attachment protein for adenovirus serotypes from subgroups A, C, D, E, and F. J Virol 72: 7909-7915.
191. Adams WC, Bond E, Havenga MJ, Holterman L, Goudsmit J, et al. (2009) Adenovirus serotype 5 infects human dendritic cells via a coxsackievirus-adenovirus receptor-

133
independent receptor pathway mediated by lactoferrin and DC-SIGN. J Gen Virol 90: 1600-1610.
192. Wickham TJ, Mathias P, Cheresh DA, Nemerow GR (1993) Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell 73: 309-319.
193. Mathias P, Wickham T, Moore M, Nemerow G (1994) Multiple adenovirus serotypes use alpha v integrins for infection. J Virol 68: 6811-6814.
194. Patterson S, Russell WC (1983) Ultrastructural and immunofluorescence studies of early events in adenovirus-HeLa cell interactions. J Gen Virol 64: 1091-1099.
195. Cerullo V, Seiler MP, Mane V, Brunetti-Pierri N, Clarke C, et al. (2007) Toll-like receptor 9 triggers an innate immune response to helper-dependent adenoviral vectors. Mol Ther 15: 378-385.
196. Iacobelli-Martinez M, Nemerow GR (2007) Preferential activation of Toll-like receptor nine by CD46-utilizing adenoviruses. J Virol 81: 1305-1312.
197. Zhu J, Huang X, Yang Y (2007) Innate immune response to adenoviral vectors is mediated by both Toll-like receptor-dependent and -independent pathways. J Virol 81: 3170-3180.
198. Hartman ZC, Kiang A, Everett RS, Serra D, Yang XY, et al. (2007) Adenovirus infection triggers a rapid, MyD88-regulated transcriptome response critical to acute-phase and adaptive immune responses in vivo. J Virol 81: 1796-1812.
199. Basner-Tschakarjan E, Gaffal E, O'Keeffe M, Tormo D, Limmer A, et al. (2006) Adenovirus efficiently transduces plasmacytoid dendritic cells resulting in TLR9-dependent maturation and IFN-alpha production. J Gene Med 8: 1300-1306.
200. Cheng G, Zhong J, Chung J, Chisari FV (2007) Double-stranded DNA and double-stranded RNA induce a common antiviral signaling pathway in human cells. Proc Natl Acad Sci U S A 104: 9035-9040.
201. Muruve DA, Petrilli V, Zaiss AK, White LR, Clark SA, et al. (2008) The inflammasome recognizes cytosolic microbial and host DNA and triggers an innate immune response. Nature 452: 103-107.
202. Millar J, Dissanayake D, Yang TC, Grinshtein N, Evelegh C, et al. (2007) The magnitude of the CD8+ T cell response produced by recombinant virus vectors is a function of both the antigen and the vector. Cell Immunol 250: 55-67.
203. de Andrade BP, Gazzinelli RT, Del Val M, Bruna-Romero O (2007) Protective immunization against murine cytomegalovirus infection using adenoviruses and poxviruses expressing hepatitis B virus chimeras. Int Microbiol 10: 261-269.
204. Naslund TI, Uyttenhove C, Nordstrom EK, Colau D, Warnier G, et al. (2007) Comparative prime-boost vaccinations using Semliki Forest virus, adenovirus, and ALVAC vectors demonstrate differences in the generation of a protective central memory CTL response against the P815 tumor. J Immunol 178: 6761-6769.
205. Yang TC, Millar J, Groves T, Zhou W, Grinshtein N, et al. (2007) On the role of CD4+ T cells in the CD8+ T-cell response elicited by recombinant adenovirus vaccines. Mol Ther 15: 997-1006.
206. Geiben-Lynn R, Greenland JR, Frimpong-Boateng K, Letvin NL (2008) Kinetics of recombinant adenovirus type 5, vaccinia virus, modified vaccinia ankara virus, and DNA antigen expression in vivo and the induction of memory T-lymphocyte responses. Clin Vaccine Immunol 15: 691-696.
207. Hovav AH, Panas MW, Rahman S, Sircar P, Gillard G, et al. (2007) Duration of antigen expression in vivo following DNA immunization modifies the magnitude, contraction, and secondary responses of CD8+ T lymphocytes. J Immunol 179: 6725-6733.

134
208. Tatsis N, Fitzgerald JC, Reyes-Sandoval A, Harris-McCoy KC, Hensley SE, et al. (2007) Adenoviral vectors persist in vivo and maintain activated CD8+ T cells: implications for their use as vaccines. Blood 110: 1916-1923.
209. Mercier S, Gahery-Segard H, Monteil M, Lengagne R, Guillet JG, et al. (2002) Distinct roles of adenovirus vector-transduced dendritic cells, myoblasts, and endothelial cells in mediating an immune response against a transgene product. J Virol 76: 2899-2911.
210. Lore K, Adams WC, Havenga MJ, Precopio ML, Holterman L, et al. (2007) Myeloid and plasmacytoid dendritic cells are susceptible to recombinant adenovirus vectors and stimulate polyfunctional memory T cell responses. J Immunol 179: 1721-1729.
211. Zhang Y, Chirmule N, Gao GP, Qian R, Croyle M, et al. (2001) Acute cytokine response to systemic adenoviral vectors in mice is mediated by dendritic cells and macrophages. Mol Ther 3: 697-707.
212. Cockburn IA, Chakravarty S, Overstreet MG, Garcia-Sastre A, Zavala F (2008) Memory CD8+ T cell responses expand when antigen presentation overcomes T cell self-regulation. J Immunol 180: 64-71.
213. Rocha CD, Caetano BC, Machado AV, Bruna-Romero O (2004) Recombinant viruses as tools to induce protective cellular immunity against infectious diseases. Int Microbiol 7: 83-94.
214. Liu J, O'Brien KL, Lynch DM, Simmons NL, La Porte A, et al. (2009) Immune control of an SIV challenge by a T-cell-based vaccine in rhesus monkeys. Nature 457: 87-91.
215. Liu J, Ewald BA, Lynch DM, Denholtz M, Abbink P, et al. (2008) Magnitude and phenotype of cellular immune responses elicited by recombinant adenovirus vectors and heterologous prime-boost regimens in rhesus monkeys. J Virol 82: 4844-4852.
216. Li S, Rodrigues M, Rodriguez D, Rodriguez JR, Esteban M, et al. (1993) Priming with recombinant influenza virus followed by administration of recombinant vaccinia virus induces CD8+ T-cell-mediated protective immunity against malaria. Proc Natl Acad Sci U S A 90: 5214-5218.
217. Gilbert SC, Schneider J, Hannan CM, Hu JT, Plebanski M, et al. (2002) Enhanced CD8 T cell immunogenicity and protective efficacy in a mouse malaria model using a recombinant adenoviral vaccine in heterologous prime-boost immunisation regimes. Vaccine 20: 1039-1045.
218. Rodrigues M, Li S, Murata K, Rodriguez D, Rodriguez JR, et al. (1994) Influenza and vaccinia viruses expressing malaria CD8+ T and B cell epitopes. Comparison of their immunogenicity and capacity to induce protective immunity. J Immunol 153: 4636-4648.
219. Casimiro DR, Chen L, Fu TM, Evans RK, Caulfield MJ, et al. (2003) Comparative immunogenicity in rhesus monkeys of DNA plasmid, recombinant vaccinia virus, and replication-defective adenovirus vectors expressing a human immunodeficiency virus type 1 gag gene. J Virol 77: 6305-6313.
220. Goonetilleke N, Moore S, Dally L, Winstone N, Cebere I, et al. (2006) Induction of multifunctional human immunodeficiency virus type 1 (HIV-1)-specific T cells capable of proliferation in healthy subjects by using a prime-boost regimen of DNA- and modified vaccinia virus Ankara-vectored vaccines expressing HIV-1 Gag coupled to CD8+ T-cell epitopes. J Virol 80: 4717-4728.
221. Webster DP, Dunachie S, Vuola JM, Berthoud T, Keating S, et al. (2005) Enhanced T cell-mediated protection against malaria in human challenges by using the recombinant poxviruses FP9 and modified vaccinia virus Ankara. Proc Natl Acad Sci U S A 102: 4836-4841.
222. Rodriguez A, Goudsmit J, Companjen A, Mintardjo R, Gillissen G, et al. (2008) Impact of recombinant adenovirus serotype 35 priming versus boosting of a Plasmodium

135
falciparum protein: characterization of T- and B-cell responses to liver-stage antigen 1. Infect Immun 76: 1709-1718.
223. McShane H, Pathan AA, Sander CR, Keating SM, Gilbert SC, et al. (2004) Recombinant modified vaccinia virus Ankara expressing antigen 85A boosts BCG-primed and naturally acquired antimycobacterial immunity in humans. Nat Med 10: 1240-1244.
224. Vordermeier HM, Villarreal-Ramos B, Cockle PJ, McAulay M, Rhodes SG, et al. (2009) Viral booster vaccines improve Mycobacterium bovis BCG-induced protection against bovine tuberculosis. Infect Immun 77: 3364-3373.
225. Meseda CA, Elkins KL, Merchlinsky MJ, Weir JP (2002) Prime-boost immunization with DNA and modified vaccinia virus ankara vectors expressing herpes simplex virus-2 glycoprotein D elicits greater specific antibody and cytokine responses than DNA vaccine alone. J Infect Dis 186: 1065-1073.
226. Nigam P, Earl PL, Americo JL, Sharma S, Wyatt LS, et al. (2007) DNA/MVA HIV-1/AIDS vaccine elicits long-lived vaccinia virus-specific immunity and confers protection against a lethal monkeypox challenge. Virology 366: 73-83.
227. Forbes EK, Sander C, Ronan EO, McShane H, Hill AV, et al. (2008) Multifunctional, high-level cytokine-producing Th1 cells in the lung, but not spleen, correlate with protection against Mycobacterium tuberculosis aerosol challenge in mice. J Immunol 181: 4955-4964.
228. Harrington LE, Most Rv R, Whitton JL, Ahmed R (2002) Recombinant vaccinia virus-induced T-cell immunity: quantitation of the response to the virus vector and the foreign epitope. J Virol 76: 3329-3337.
229. McConkey SJ, Reece WH, Moorthy VS, Webster D, Dunachie S, et al. (2003) Enhanced T-cell immunogenicity of plasmid DNA vaccines boosted by recombinant modified vaccinia virus Ankara in humans. Nat Med 9: 729-735.
230. Moorthy VS, Imoukhuede EB, Milligan P, Bojang K, Keating S, et al. (2004) A randomised, double-blind, controlled vaccine efficacy trial of DNA/MVA ME-TRAP against malaria infection in Gambian adults. PLoS Med 1: e33.
231. Bejon P, Mwacharo J, Kai O, Mwangi T, Milligan P, et al. (2006) A phase 2b randomised trial of the candidate malaria vaccines FP9 ME-TRAP and MVA ME-TRAP among children in Kenya. PLoS Clin Trials 1: e29.
232. Bejon P, Ogada E, Mwangi T, Milligan P, Lang T, et al. (2007) Extended follow-up following a phase 2b randomized trial of the candidate malaria vaccines FP9 ME-TRAP and MVA ME-TRAP among children in Kenya. PLoS One 2: e707.
233. Beveridge NE, Price DA, Casazza JP, Pathan AA, Sander CR, et al. (2007) Immunisation with BCG and recombinant MVA85A induces long-lasting, polyfunctional Mycobacterium tuberculosis-specific CD4+ memory T lymphocyte populations. Eur J Immunol 37: 3089-3100.
234. Hawkridge T, Scriba TJ, Gelderbloem S, Smit E, Tameris M, et al. (2008) Safety and immunogenicity of a new tuberculosis vaccine, MVA85A, in healthy adults in South Africa. J Infect Dis 198: 544-552.
235. Brookes RH, Hill PC, Owiafe PK, Ibanga HB, Jeffries DJ, et al. (2008) Safety and immunogenicity of the candidate tuberculosis vaccine MVA85A in West Africa. PLoS One 3: e2921.
236. Harari A, Bart PA, Stohr W, Tapia G, Garcia M, et al. (2008) An HIV-1 clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunctional, and long-lasting T cell responses. J Exp Med 205: 63-77.
237. Gao W, Soloff AC, Lu X, Montecalvo A, Nguyen DC, et al. (2006) Protection of mice and poultry from lethal H5N1 avian influenza virus through adenovirus-based immunization. J Virol 80: 1959-1964.

136
238. Matsui M, Moriya O, Akatsuka T (2003) Enhanced induction of hepatitis C virus-specific cytotoxic T lymphocytes and protective efficacy in mice by DNA vaccination followed by adenovirus boosting in combination with the interleukin-12 expression plasmid. Vaccine 21: 1629-1639.
239. Khanam S, Pilankatta R, Khanna N, Swaminathan S (2009) An adenovirus type 5 (AdV5) vector encoding an envelope domain III-based tetravalent antigen elicits immune responses against all four dengue viruses in the presence of prior AdV5 immunity. Vaccine 27: 6011-6021.
240. Santra S, Seaman MS, Xu L, Barouch DH, Lord CI, et al. (2005) Replication-defective adenovirus serotype 5 vectors elicit durable cellular and humoral immune responses in nonhuman primates. J Virol 79: 6516-6522.
241. Casimiro DR, Tang A, Chen L, Fu TM, Evans RK, et al. (2003) Vaccine-induced immunity in baboons by using DNA and replication-incompetent adenovirus type 5 vectors expressing a human immunodeficiency virus type 1 gag gene. J Virol 77: 7663-7668.
242. Shiver JW, Fu TM, Chen L, Casimiro DR, Davies ME, et al. (2002) Replication-incompetent adenoviral vaccine vector elicits effective anti-immunodeficiency-virus immunity. Nature 415: 331-335.
243. Baroncelli S, Negri DR, Michelini Z, Cara A (2008) Macaca mulatta, fascicularis and nemestrina in AIDS vaccine development. Expert Rev Vaccines 7: 1419-1434.
244. Letvin NL, Mascola JR, Sun Y, Gorgone DA, Buzby AP, et al. (2006) Preserved CD4+ central memory T cells and survival in vaccinated SIV-challenged monkeys. Science 312: 1530-1533.
245. Mattapallil JJ, Douek DC, Buckler-White A, Montefiori D, Letvin NL, et al. (2006) Vaccination preserves CD4 memory T cells during acute simian immunodeficiency virus challenge. J Exp Med 203: 1533-1541.
246. Wilson NA, Reed J, Napoe GS, Piaskowski S, Szymanski A, et al. (2006) Vaccine-induced cellular immune responses reduce plasma viral concentrations after repeated low-dose challenge with pathogenic simian immunodeficiency virus SIVmac239. J Virol 80: 5875-5885.
247. Wilson NA, Keele BF, Reed JS, Piaskowski SM, MacNair CE, et al. (2009) Vaccine-induced cellular responses control simian immunodeficiency virus replication after heterologous challenge. J Virol 83: 6508-6521.
248. Tritel M, Stoddard AM, Flynn BJ, Darrah PA, Wu CY, et al. (2003) Prime-boost vaccination with HIV-1 Gag protein and cytosine phosphate guanosine oligodeoxynucleotide, followed by adenovirus, induces sustained and robust humoral and cellular immune responses. J Immunol 171: 2538-2547.
249. Wille-Reece U, Flynn BJ, Lore K, Koup RA, Miles AP, et al. (2006) Toll-like receptor agonists influence the magnitude and quality of memory T cell responses after prime-boost immunization in nonhuman primates. J Exp Med 203: 1249-1258.
250. Elvang T, Christensen JP, Billeskov R, Thi Kim Thanh Hoang T, Holst P, et al. (2009) CD4 and CD8 T cell responses to the M. tuberculosis Ag85B-TB10.4 promoted by adjuvanted subunit, adenovector or heterologous prime boost vaccination. PLoS One 4: e5139.
251. Barouch DH, Pau MG, Custers JH, Koudstaal W, Kostense S, et al. (2004) Immunogenicity of recombinant adenovirus serotype 35 vaccine in the presence of pre-existing anti-Ad5 immunity. J Immunol 172: 6290-6297.
252. Ophorst OJ, Radosevic K, Havenga MJ, Pau MG, Holterman L, et al. (2006) Immunogenicity and protection of a recombinant human adenovirus serotype 35-based malaria vaccine against Plasmodium yoelii in mice. Infect Immun 74: 313-320.

137
253. Nwanegbo E, Vardas E, Gao W, Whittle H, Sun H, et al. (2004) Prevalence of neutralizing antibodies to adenoviral serotypes 5 and 35 in the adult populations of The Gambia, South Africa, and the United States. Clin Diagn Lab Immunol 11: 351-357.
254. Kostense S, Koudstaal W, Sprangers M, Weverling GJ, Penders G, et al. (2004) Adenovirus types 5 and 35 seroprevalence in AIDS risk groups supports type 35 as a vaccine vector. AIDS 18: 1213-1216.
255. Yang ZY, Wyatt LS, Kong WP, Moodie Z, Moss B, et al. (2003) Overcoming immunity to a viral vaccine by DNA priming before vector boosting. J Virol 77: 799-803.
256. Roberts DM, Nanda A, Havenga MJ, Abbink P, Lynch DM, et al. (2006) Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature 441: 239-243.
257. Sumida SM, Truitt DM, Lemckert AA, Vogels R, Custers JH, et al. (2005) Neutralizing antibodies to adenovirus serotype 5 vaccine vectors are directed primarily against the adenovirus hexon protein. J Immunol 174: 7179-7185.
258. Radosevic K, Wieland CW, Rodriguez A, Weverling GJ, Mintardjo R, et al. (2007) Protective immune responses to a recombinant adenovirus type 35 tuberculosis vaccine in two mouse strains: CD4 and CD8 T-cell epitope mapping and role of gamma interferon. Infect Immun 75: 4105-4115.
259. Ko SY, Cheng C, Kong WP, Wang L, Kanekiyo M, et al. (2009) Enhanced induction of intestinal cellular immunity by oral priming with enteric adenovirus 41 vectors. J Virol 83: 748-756.
260. Reyes-Sandoval A, Fitzgerald JC, Grant R, Roy S, Xiang ZQ, et al. (2004) Human immunodeficiency virus type 1-specific immune responses in primates upon sequential immunization with adenoviral vaccine carriers of human and simian serotypes. J Virol 78: 7392-7399.
261. Tatsis N, Blejer A, Lasaro MO, Hensley SE, Cun A, et al. (2007) A CD46-binding chimpanzee adenovirus vector as a vaccine carrier. Mol Ther 15: 608-617.
262. Camargo EP (2009) Perspectives of vaccination in Chagas disease revisited. Mem Inst Oswaldo Cruz 104 Suppl 1: 275-280.
263. Gomes JA, Bahia-Oliveira LM, Rocha MO, Martins-Filho OA, Gazzinelli G, et al. (2003) Evidence that development of severe cardiomyopathy in human Chagas' disease is due to a Th1-specific immune response. Infect Immun 71: 1185-1193.
264. Gomes JA, Bahia-Oliveira LM, Rocha MO, Busek SC, Teixeira MM, et al. (2005) Type 1 chemokine receptor expression in Chagas' disease correlates with morbidity in cardiac patients. Infect Immun 73: 7960-7966.
265. Ramasawmy R, Cunha-Neto E, Fae KC, Muller NG, Cavalcanti VL, et al. (2006) BAT1, a putative anti-inflammatory gene, is associated with chronic Chagas cardiomyopathy. J Infect Dis 193: 1394-1399.
266. Laucella SA, Postan M, Martin D, Hubby Fralish B, Albareda MC, et al. (2004) Frequency of interferon- gamma -producing T cells specific for Trypanosoma cruzi inversely correlates with disease severity in chronic human Chagas disease. J Infect Dis 189: 909-918.
267. Alvarez MG, Postan M, Weatherly DB, Albareda MC, Sidney J, et al. (2008) HLA Class I-T Cell Epitopes from trans-Sialidase Proteins Reveal Functionally Distinct Subsets of CD8 T Cells in Chronic Chagas Disease. PLoS Negl Trop Dis 2: e288.
268. Bahia-Oliveira LM, Gomes JA, Cancado JR, Ferrari TC, Lemos EM, et al. (2000) Immunological and clinical evaluation of chagasic patients subjected to chemotherapy during the acute phase of Trypanosoma cruzi infection 14-30 years ago. J Infect Dis 182: 634-638.
269. Rocha NN, Garcia S, Gimenez LE, Hernandez CC, Senra JF, et al. (2006) Characterization of cardiopulmonary function and cardiac muscarinic and adrenergic receptor density

138
adaptation in C57BL/6 mice with chronic Trypanosoma cruzi infection. Parasitology 133: 729-737.
270. Bilate AM, Salemi VM, Ramires FJ, de Brito T, Silva AM, et al. (2003) The Syrian hamster as a model for the dilated cardiomyopathy of Chagas' disease: a quantitative echocardiographical and histopathological analysis. Microbes Infect 5: 1116-1124.
271. Guedes PM, Veloso VM, Caliari MV, Carneiro CM, Souza SM, et al. (2007) Trypanosoma cruzi high infectivity in vitro is related to cardiac lesions during long-term infection in Beagle dogs. Mem Inst Oswaldo Cruz 102: 141-147.
272. Haolla FA, Claser C, de Alencar BC, Tzelepis F, de Vasconcelos JR, et al. (2009) Strain-specific protective immunity following vaccination against experimental Trypanosoma cruzi infection. Vaccine 27: 5644-5653.
273. Wiesel M, Walton S, Richter K, Oxenius A (2009) Virus-specific CD8 T cells: activation, differentiation and memory formation. Apmis 117: 356-381.
274. Bixby LM, Tarleton RL (2008) Stable CD8+ T cell memory during persistent Trypanosoma cruzi infection. J Immunol 181: 2644-2650.
275. Appay V, Douek DC, Price DA (2008) CD8+ T cell efficacy in vaccination and disease. Nat Med 14: 623-628.
276. Day CL, Kaufmann DE, Kiepiela P, Brown JA, Moodley ES, et al. (2006) PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 443: 350-354.
277. Voehringer D, Koschella M, Pircher H (2002) Lack of proliferative capacity of human effector and memory T cells expressing killer cell lectinlike receptor G1 (KLRG1). Blood 100: 3698-3702.
278. Trautmann L, Janbazian L, Chomont N, Said EA, Gimmig S, et al. (2006) Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat Med 12: 1198-1202.
279. Darrah PA, Patel DT, De Luca PM, Lindsay RW, Davey DF, et al. (2007) Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nat Med 13: 843-850.

139
ABSTRACT
Genetic immunization with plasmidial DNA containing the gene encoding
the amastigote surface protein 2 (ASP-2) protects part of the highly susceptible
A/Sn mice against a lethal challenge with Trypanosoma cruzi. Based on these
results, in this thesis, we tested the hypothesis that the efficacy of the
vaccination against T. cruzi experimental infection could be rationally improved
by the use of different vaccine formulations and/or vaccinations strategies. For
that purpose, we employed homologous or heterologous priming and boosting
strategies, with plasmidial DNA, recombinant proteins, and adenovirus
expressing T. cruzi ASP-2. Initially, we tested whether boosting with a
recombinant protein of ASP-2 could improve the efficacy of the priming
vaccination with plasmidial DNA (pIgSPCl.9). This protocol was not more
efficient, but opened the possibility that the immunization with the recombinant
protein could be used as an alternative vaccine strategy. Using this approach,
we described that the administration of 3 doses of a recombinant protein
representing the amino acids 261-500 of ASP-2 in fusion o the gluthatione S-
transferase (GST-P4P7), in the presence of the adjuvants alum and CpG ODN
1826, was capable of eliciting high levels of protective immunity. In this
experimental model, protective immunity was: i) long-lasting; and ii) dependent
on the effector CD8 T lymphocytes specific for the immunodominant epitope
TEWETGQI. We also established that the adjuvant CpG ODN 1826 improved
the efficacy of protective immunity, and that CD4 T lymphocytes were important
for the induction of protective CD8 T cells.
Because the vaccination strategy employing the recombinant protein
GST-P4P7 required 3 doses, we tested the possibility that a recombinant
adenovirus expressing ASP-2 (AdASP-2) could be used in a homologous or
heterologous priming and boosting protocol with two doses only. By comparing
homologous or heterologous priming and boosting with pIgSPCl.9 and AdASP-
2, we found that homologous or heterologous immunizations with AdASP-2 led
to a high degree of protective immunity against T. cruzi infection, in highly
susceptible A/Sn mice. Follow up studies using the heterologous priming and
boosting strategy (pIgSPCl.9 followed by AdASP-2), which protects 80-100% of
the mice, showed that protective immunity was long-lived and dependent on the

140
activation of CD4 or CD8 T cells. Protective immunity correlated with antigen-
specific in vivo cytotoxic activity prior to the challenge with T. cruzi. Based on
this observation, we evaluated the protective immunity following the
heterologous vaccination in mice genetically deficient for an important mediator
of cytotoxicity, perforin. Vaccinated perforin knock-out (KO) mice were
extremely susceptible to the experimental infection. The analyses of the
immune response of these mice showed that their splenocytes produced less
IFN-γ when re-stimulated in vitro with recombinant antigen. Analysis of specific
CD8 T lymphocytes showed: i) a normal numeric expansion; ii) a significantly
lower frequency of polyfunctional cells expressing simultaneously IFN-γ/CD107a
or IFN-γ/TNF-α.; iii) lower in vivo cytotoxicity; iv) lower expression of CD44 and
KLRG-1.
Finally, to define the relative importance of IFN-γ and cytotoxic T
lymphocytes in the protection against T. cruzi infection following heterologous
vaccination, IFN-γ KO mice were immunized with pIgSPCl.9 followed by
AdASP-2. These mice developed a high level of in vivo cytotoxicity, but no
detectable protective immunity.
In summary, during this work we gathered evidences that confirmed our
initial hypothesis that we could rationally improve the efficacy of vaccination
against T. cruzi experimental infection using different vaccine formulations
and/or vaccination strategies. Using in vivo depletions and KO mice, we defined
the importance of CD4 cells, CD8 cells, perforin and IFN-γ during immunity
generated by vaccination with ASP-2 of T. cruzi. We believe that our findings
will provide: i) a better understanding of the immunologic mechanisms and the
target antigens during the resistance to T. cruzi infection; ii) to aid the
development of recombinant vaccines against intra-cellular parasitic protozoan
infections in general and Chagas’ disease, specifically; iii) to aid the
development of recombinant vaccines capable of eliciting potent immune
response mediated by T lymphocytes.




















