
Branchial ion exchange and acid-base regulation after strenuous exercise in rainbow trout (Salmo...
16
Re,spiralion PhysioloKv (1983) 51,303- 318 303 Elsevier Biomedical Press BRANCHIAL ION EXCHANGE AND ACID-BASE REGULATION AFTER STRENUOUS EXERCISE IN RAINBOW TROUT (SALMO GAIRDNERI) GEORGE E. HOLETON +, PETER NEUMANN and NORBERT HEISLER ,4t~leilun£, Ph)'siologie, Max-Phlnck-ht.~tilut fib" experimc'ntelh" MediziH. G6tlink, en, F.R.G. Abstract. Specimens of rainbow trout (Sahno £,airdneri) were electrically stimulated to exhausting activity in a closed water recirculation system and the changes in dorsal aortic plasma pH, P(o,, Po-, 02 content, [Na+], [C1-], [K+], [Lactate ] and Ht were measured during a 24 h recovery period. Net transfer of H'. Na +, CI and ammonia between fish and environment were determined by measurement of the concentration changes in the recirculatmg water. Strcnous exercise resulted in a severe lactacidosis which was corrected by transient net transfer of H+ ions to the environmental water within 4 h, about 6 8 h before the lactate was metabolically removed. The net transfer of H ~ ions was achieved in part by branchial HCO~ CI ion exchanges. but to a larger extent by branchial exchange of H + and'or NEI2 against Na + . The excretion of ammonia, which was considerably enhanced during the first 4 h after exercise, was at least partially due to non-ionic diffusion across the gill epithelium. The observed elevation in ammonia excretion was probably the result of an exercise-induced increase in nitrogen metabolism rather than of production of ammonia for the purpose of acid-base regulation. Acid base balance Lactic acid Acidosis Metabolic acidosis Ammonia Muscular exercise Bicarbonate Nitrogen metabolism Strenuous exercise in fish generally results in production of large quantities of lactic acid as the main end-product of anaerobic glycolysis in the insufficiently perfused white musculature. The efflux of the dissociation products of lactic acid, H ~ and lactate ions from the muscle cells induces severe acid base disturbances and increases in lactate concentration of up to 30 mM in the extracellular body compartment (e.g., Secondat and Diaz, 1942; Black 1957 a,b,c: Black et al., 1959, 1962; Piiper et al., 1972: for review see Heisler, 1982a). The extracellular acid base balance is usually normalized much earlier than the lactate concentration (see Heisler, 1982a). 4cceptcd /br lmhlicalion 2<~; December 1~,;<~¢2 Deceased. 0034-5687/83:0000-0000/$03.00 :~ 1983 Elsevier Biomedical Press
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Branchial ion exchange and acid-base regulation after strenuous exercise in rainbow trout (Salmo...

Re,spiralion PhysioloKv (1983) 51,303- 318 303 Elsevier Biomedical Press
B R A N C H I A L ION E X C H A N G E A N D A C I D - B A S E R E G U L A T I O N AFTER S T R E N U O U S EXERCISE IN R A I N B O W T R O U T ( S A L M O GAIRDNERI)
G E O R G E E. H O L E T O N +, PETER N E U M A N N and N O R B E R T HEISLER
,4t~leilun£, Ph)'siologie, Max-Phlnck-ht.~tilut fib" experimc'ntelh" MediziH. G6tlink, en, F.R.G.
Abstract. Specimens of rainbow trout (Sahno £,airdneri) were electrically stimulated to exhausting
activity in a closed water recirculation system and the changes in dorsal aortic plasma pH, P(o, , Po-,
02 content, [Na+], [C1-], [K+], [Lactate ] and Ht were measured during a 24 h recovery period.
Net transfer of H ' . Na +, CI and ammonia between fish and environment were determined by
measurement of the concentration changes in the recirculatmg water.
Strcnous exercise resulted in a severe lactacidosis which was corrected by transient net transfer of
H+ ions to the environmental water within 4 h, about 6 8 h before the lactate was metabolically
removed. The net transfer of H ~ ions was achieved in part by branchial HCO~ CI ion exchanges.
but to a larger extent by branchial exchange of H + and'or NEI2 against Na + . The excretion of ammonia,
which was considerably enhanced during the first 4 h after exercise, was at least partially due to non-ionic
diffusion across the gill epithelium. The observed elevation in ammonia excretion was probably the
result of an exercise-induced increase in nitrogen metabolism rather than of production of ammonia
for the purpose of acid-base regulation.
Acid base balance Lactic acid
Acidosis Metabolic acidosis
Ammonia Muscular exercise
Bicarbonate Nitrogen metabolism
Strenuous exercise in fish generally results in production o f large quantities of lactic acid as the main end-product of anaerobic glycolysis in the insufficiently perfused white musculature. The efflux o f the dissociation products of lactic acid, H ~ and lactate ions from the muscle cells induces severe acid base disturbances and increases in lactate concentration of up to 30 mM in the extracellular body compartment (e.g., Secondat and Diaz, 1942; Black 1957 a,b,c: Black et al., 1959, 1962; Piiper et al., 1972: for review see Heisler, 1982a). The extracellular acid base balance is usually normalized much earlier than the lactate concentration (see Heisler, 1982a).
4cceptcd /br lmhlicalion 2<~; December 1~,;<~¢2
Deceased.
0034-5687/83:0000-0000/$03.00 :~ 1983 Elsevier Biomedical Press

304 ¢i. F. HOLETON ¢,: al.
In the larger spotted dogfish (Scyliorhinus slelklris) this phenomenon has been demonstrated to result from transient transepithelial net transfer of H + ions to the environmental water (Holeton and Heisler, 1983). This transfer exclusively occurred at the branchial epithelium by either excretion of H ~ ions or uptake of HCO~. The ammonia mechanism was not utilized. This could have been related to the primarily ureotelic nitrogen metabolism of the elasmobranch. But it should also
be considered that for marine animals enhancement of ionic ammonium excretion would not be an appropriate mechanism for acid-base regulation. Namely, since ionic excretion of ammonium ions is thought to be performed by carrier-mediated
ion-transfer mechanisms in exchange for Na ~ ions (e.~., Maetz, 1973; Kerstetter el a/., 1970: Evans 1977; Payan and Maetz, 1973: see also Maetz, 1974 and Evans, 1980) the elevation of NH~ excretion would be combined with an equivalent influx of Na- adding to the load of the ionoregulalory N a extrusion mechanism. In
contrast, in fl'eshwater fish extrusion of N H 4 in exchange with N a is advantageous l\)r maintaining the osmotic homeostasis of the organism.
Accordingly the present experiments about the possible role of ammonia excretion
for the acid base regulation after strenuous exercise in fish were conducted using the ammoniotelic freshwater teleost fish Salmo %airdm, ri.
Parts of the results of this study have been published in abstract form (Holeton e,t a/., 1980).
Materials and methods
Rainbow trout (Sahno ,vair~bleri), weight 935 _+ 110 g (x + SD), were obtained from commercial fish hatcheries and kept liar more than 9 weeks in a flow-through glass aquarium system with a volume of more than 150 L:fish. The water was lher- mostatted to 15 +_ 0 .2 'C and vigorously aerated so that thc PCj, of the watcr was
kept higher than 140 mm Hg and P~o lower than i).4 mm Hg. Fresh tap water which was dechlorinated in activated charcoal columns was llushcd through the aquaria at a rate of more than 200 L/(fish. day). The fish were fcd daily on coin- mercial trout food pellets until 3 5 days bet'orc experimentation.
CAT}tETER1ZATION OF THE DORSAl. AORTA
At least 36 h, in most cases more than 48 11, prior to the experiments the fish were anaesthetized in aerated urethane/water solution (20 g/L) until they lost reactivity and placed ventral side up oil an operating rack. Anaesthcsia and oxygen supply were maintained by irrigation of the gills with aerated water containing 2 3 g/L urethanc. A sharpened stainless-steel wire w.as inserted into an about 20 cnl length of PE 50 polyethylene catheter tubing, w:hich was conically pulled out at one end, such that the tip of the wire protruded ,lust beyond the tip of the catheter. The

ION TRANSFER & ACID BASE REGULATION AFTER EXERCISE IN TROUT 305
dorsal aorta was then punctured with this assembly through the dorsal roof of the buccal cavity, the wire withdrawn and the catheter pushed into the dorsal aorta for 5 7 cm. The catheter was then shortened and connected still inside the buccal cavity to a PE 90 polyethylene catheter which was secured to and fed out of the roof of the mouth through a flange, made of a short piece of polyethylene catheter of appropriate size and put into a hole in the roof of the mouth. The flange was secured to the roof of the mouth by two atraumatic sutures. The catheter was flushed and filled with a solution of 125 250 U heparin/ml in glucose-free trout Ringer. This cannulation procedure is a modification of the method described by Soivio ez al. (1972, 1975).
Alter completion of surgery the fish were aroused by flushing the gills with aerated water until the fish made breathing attempts and became active. When they showed apparently normal ventilation, they were introduced into the experimental chamber. The chamber was flushed with fresh water for at least 36 h before the system was closed for experimentation.
EXPERIMENTAL APPARATUS
The experiments were performed in a closed water recirculation system (fig. 1) similar to that described by Heisler (1978). The water in the system was recirculated through a plexiglass box and an oxygenator with successive bubble trap. The water was thermostatted to 15 _+ 0.1 °C and was aerated at a rate of >8 L/rain with CO_~-stripped air and recirculated at a rate of about 6 L/min, For measurement of changes in the bicarbonate concentration, water from the fish system was pumped by a roller pump through three successively connected glass columns fitted with bot toms of fi'itted glass discs and therein equilibrated at constant temperature (30 + 0.05"C) with an extremely constant gas mixture of 1'~;i CO, in nitrogen, provided by a gas mixing pump (W6sthoff, Bochum, F.R.G.) selected lk)r minimal long-ternq variability (Heisler, 1978: Holeton and Heisler, 1983). Before being returned to the fish system the water thus standardized for temperature and Pc<, was fed over an electrode chain consisting of a spherical pH glass electrode and a double electrolyte bridge Ag/AgC1 reference with sleeve diaphragm, which was especially treated and selected for long-term stability (drift less than 0.001/24 h in
water equilibrated with a humidified gas of constant P~o:). This "A-bicarbonate measurement system' (Heisler, 1978) was calibrated by addition of known amounts of HCI or NaHCO~ and allowed according to the prevailing conditions (water pH ~ 7.4, water Pco~ < 0.6 mm Hg) continuous recording of changes in the water bicarbonate concentration with a resolution of 0.001 mM. [Na ~] and [C1 ] m the recirculating water were continuously monitored with Na +-sensitive glass electrodes and solid-state CI -sensitive electrodes, also selected for high long-term stability. All electrodes were connected to the recording system via high impedance precision isolation amplifiers (Model 87, Knick, Berlin, Germany) in order to prevent ground
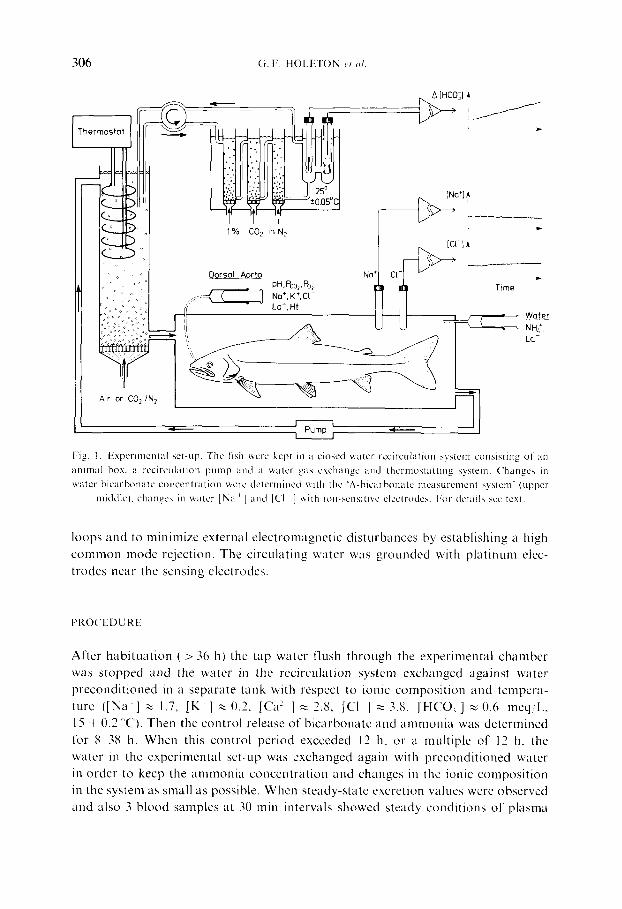
306 (;. F. HOLETON ct al.
b~ l A
T h e r m o s t a
°o ' % [ccl ~,
,, u J Dorsa l Ao r t a . . . . No+ C(-[ " ~..
. PH, Pco~, Po~ " ~ T ime
T T I o /l ' w er
Air or CO 2 /N 2
/ . i i '~ 'pu m p . i
I I
Fig. t. Experimental set-up, The fish were kept in a cMscd watcr r~:circukitkm system consisting of all
animal box, a recirculation puinp and a water gas exchange and thermostatt ing system. Changes in
water bicarbonate concentration wcrc determined with the 'A-bicarbonate illcasuremcnt system" (upper
middlcI, changcs in water [Na ~ l a n d [CI ] with ion-,,,cnsitivc elcclrodes. For de ta i l s sec tcx t .
loops and to minimize external electromagnetic disturbances by establishing a high common mode rejection. The circuhtting water was grounded with platinum elec- trodes near the sensing electrodes.
P R O C E D U R E
After habituation ( > 36 h) the tap water flush through the experimental chamber was stopped and the water in the recirculation system exchanged against water preconditioned ill a separate tank with respect to ionic composition and tempera- turc ( [ N a ' ] , ~ 1.7, [K ~] v0 .2 , [Cae ' ] ,~2 .8 , [C1 ]>~3.8, [ H C O , ] ~ 0 . 6 tneq/k, 15 + 0.2 -C). Then the control release of bicarbonate and ammonia was determined for 8 38 h. When this control period exceeded 12 h, or a multiple of 12 h, the water in the experimental set-up was exchanged again with preconditioned water in order to keep the anm~onia concentration and changes in the ionic composition in the system as small as possible. When steady-state excretion values were observed and also 3 blood samples at 3(1 rain intervals showed steady conditions of plasma

ION T R A N S F E R & A C I D BASE R E G U L A T I O N AFTER EXERCISE IN ] R O U T 3 0 7
acid base parameters, and at least 2 h after the last water exchange, the fish were forced to vigorous activity by mild electric shocks (AC, 2 4 V, 50 Hz, supplied by a transformer) for a period of 5 min.
The end of the stimulation period was defined as time zero. Blood samples were withdrawn from the dorsal aorta at time zero, 15 and 30 min, 1, 1.5, 2, 3, 4, 6, 8 and 24 h after time zero and analysed immediately for plasma pH, Pco~ and Po,, C:,o: and Ht: plasma samples were stored directly for later analysis of [Na+], [CI ] and [K+]. Plasma samples were also deproteinized, decanted and the supernatant stored for later lactate concentration analysis. Water samples were taken at the same intervals and analysed for ammonia and lactate concentration. The changes of water [HCO~], [Na +] and [C1 ] were monitored continuously throughout the experiment. Urine was collected via urethral catheters fitted to the animals in a similar way as described previously (Holeton and Heisler, 1982: Heisler, 1982b) in five hour intervals and analysed for titratable acidity, bicarbonate and ammonia concentration (Holeton and Heisler, 1983).
M E A S U R E M E N T S A N D C A L C U L A T I O N S
Heparinized blood samples were analysed for plasma pH, Pco:. and Po: using appropriate microelectrodes (Radiometer, Copenhagen) as described previously (Heisler et al. 1976a, Heisler, 1978). Oxygen content of whole blood (C~,o~) was analysed using a Lex-O,-Con Apparatus (Lexington Instr. Inc., Waltham, MA, U.S.A.). Plasma [Na ~] and [K +] were measured in plasma of non-heparinized blood by flame photometry (IL 343, Instrumentation Laboratory, Lexington, MA, U.S.A.), [CI ] by coulometric titration (Radiometer, CMT10, Copenhagen, Denmark).
Whole blood, plasma and water were deproteinized by addition of perchloric acid and the supernatant analysed for lactate concentration applying the method described by Benad6 and Heisler (1978). Changes in the water bicarbonate concen- tration were measured using the "A-bicarbonate measurement system" (see: Experi- mental apparatus, and Heisler, 1978). The ammonia concentration of the water was determined in alkalinized (pH > 12) water samples using gas-sensitive ammonia electrodes (Orion Research Inc., Cambridge, MA, U.S.A.), calibrated with alka- linized NH4CI standards. [Na ' ] and [CI ] in the water were monitored continuously using Na'-sensitive glass electrodes (lngold, Frankfurt, F.R.G.) and solid-state C1 -sensitive electrodes (Orion Research Inc., Cambridge, MA, U.S.A.) calibrated with standards of low ionic strength, similar to that of the measured water.
Plasma bicarbonate concentration was calculated by application of the Hen- derson Hasselbalch equation using pK' and ~ o: values reported for human plasma at 15 °C (Severinghaus, 1965).
The fractional water content of trout was determined by drying to constant weight at 105 ~'C to be 0.788 _+ 0.04 (.~ _+ SD).

308 (;. F. HOLETON et al.
Results
The imposed strenuous exercise had already resulted in a maximal depression of arterial pH and bicarbonate concentra t ion by the end of the exercise period (time zero, fig. 2). The pH fell f rom about 7.8 to 7.3, and bicarbonate was reduced from
about 4.2 to 2.4 mM. In contrast , the plasma lactate concentra t ion required about 2 h to rise to an average peak concentra t ion o f about 14 mM. The pH and bicarbonate concentra t ion returned to control values within 4 h, whereas more than 10 h were required for normalizat ion o f the plasma lactate concentrat ion. Arterial Po. was slightly increased during the first hours of the recovery period (fig. 3), which resulted, after an initial reduction, probably at tr ibutable to the Bohr effect, in a parallel rise in arterial oxygen content. Arterial haematocr i t was largely increased immediately after the exercise period, recovered to control values
within 4 h and later fell below normal re-attaining pre-exercise levels after about 27 h. Plasma [K ~ ] was elevated for about 8 h, and [Na + ] followed a similar pattern
as the haematoer i t (fig. 4). [C1 ] was initially increased up to 6° , , but fell below the original level 1.5 h after exercise and did not return to the control level until 24 h after exercise.
8.0
PIle
7.5
I / { i I ~.4
bco ].51 (raM) O/
,° t [Loc,-] (raM) 1
0 L
7.0 , f , , ,
P°c02 .~ ~,.~_~_~
o ;. 8
0 2 4 t(h) 8 2/,
] i ~ . -~. [)OI'S~l] ~tOl'liC p l a s m a p H , l)~ o , - b i c a r b o n a t e a n d l a c t a t e c o n c c l l t r a l i o n t|,', d I ' t l l lCtiOll o f I]IllC
during rcco,~cry frona stlcnuotl~, muscular activity (x ± SE, n = 15). Shaded areas represent x 2 SE of lhc control values.

1ON T R A N S F E R & A C I D BASE R E G U L A T I O N A F T E R E X E R C I S E IN T R O U T 309
lO0
Pao 2 (ram Hg}
50
0 ~ - - ' ' I o 2 4 6 ; 2~
'[ {h)
Cao 2 (vot%)
0 0
4o
Ht (%)
30
20
L I ~ J J- l
2 /. 6 8 24 t lh)
i
] i 1 L _
0 2 4 6 8 24 t (hi
Fig. 3. Dorsa l aor t ic b lood oxygen par t ia l pressure (P~,o,), b lood oxygen content (Q4):) and haematoc r i t
(Ht) as a funct ion of t ime dur ing recovery from s t renuous muscu la r ac t iv i ty (x ± SE, n = 15). Shaded
areas represent X ± SE e f the cont ro l values.
The control release of bicarbonate to the environmental water at an average rate of 5.58 + 1.35 /~mol/(min.kg fish water) (fig. 5) was reversed immediately alter exercise, resulting in a net bicarbonate gain* (difference between post-exercise and control bicarbonate release) of about 1.1 mmol/kg fish water. Control values for cumulative H C O , release were re-attained about 10 h after exercise. The average control ammonia excretion rate of 4.96/~mol/(min • kg fish water) almost balanced the control bicarbonate release to a net H + excretion (AH~++,,) of -0 .62 #mol/ (min. kg fish water). This negative net H + excretion is unusual among unfed fish
* The terms "bicarbonate up take ' or "bicarbonate transfer" and s imi lar te rms arc descr ip t ively used.
a h h o u g h the mechan i sms may invob/e t ransfer of OH ions or t ransfer of HP in oppos i te d i rec t ion :
the}' are ind is t inguishab le by the me thods used.

310 (; . F. H O L E ] O N c t a / .
[K*]p (raM)
oI
17° I 160
[Na']p (raM)
150
140
150 -
[ C l - ] p
(mMI
140
130
0 2 z. 6 8 24 t (h)
:ti-.k
0 2 4 6 6 ?z, t (h)
N! /
0 2 /* 6 8 2/, t (hi
l:ig. 4. Dorsa l aor t ic p lasma [K ' ] . l N a ' ] and ICI ] af ter s t r enuous muscu la r act ivi ty (x _+ SE, n = 15).
Shaded areas represent x + SE of the cont ro l values.
(see Heisler, 1982a) and may bc an indication that the fish had not yet completely achieved steady-state conditions after anaesthesia in spite of the long recovery period. Similarly long recovery periods have been found in carp (Claiborne and Heisler, 1983) and thus appear to be a common feature or freshwater fish in contrast to rather short recovery periods in marine fish. The possible minor deviation rrom control conditions would, however, not afl'ect any result of the present study significantly.
The ammonia excretion was greatly enhanced after exercise, the cumulative ammonia release returned to control levels about 24 h after exercise. The cumulative net H ' excretion as the difference between ammonia excretion and HCO~ excretion was maximal about 4 h after exercise (ca. 4.6 mmol/kg fish water) and did not return to control levels bet'orc all surplus lactate was removed from the blood

ION TRANSFER & ACID BASE REGULATION AFTER EXERCISE IN TROUT 311
5
AHCO~ w Irnmot/k'g fish ',voter)
0
0
+ aNH~ w (rnmol /kg fish water]
5
aHC03 control ~ •
/
0 10 2O 30 ~ I ~ # i ~ ~ , ,
~. a ~. control
\<TT "-.
i 0 10 2O 3O
"control ..... . - " ' " ' ~ I
AH;~w [mrnol /kg / fish water)
0 1; 2=0 30 Time (h)
Fig. 5. Changes in water bicarbonate and NH4 + lk~llowing strenuous muscular activity in rainbow trout.
Net H + excretion (AH~)~,,) is the difference bctween change of" NHJ and bicarbonate in the recircuhitmg
water (x + SE, n = 15).
(ca. 24 h). The renal contribution to this net transfer averaged 0.26 _+ 0.07 mmol/kg fish water, 2 _+ SD, equivalent to only about 6,~, of the total amount transferred.
During the first phase of recovery from the exercise-induced lactacidosis up to 1.80 mmol/kg fish water of CI was net released to the environment (fig. 6) and later recovered at the same time as the net amount of bicarbonate taken up was returned to the water after about 10 h. Similarly, sodium was net taken up from the environmental water (up to 2.8 mmol/kg fish water) and was later returned
(fig. 6). Lactate was not transferred to the environmental water at any time of the experiments_

312 ( ; F. H O L E T O N t,t ~ff
2
Net zx CI~,
0
Net zx HCO~ w
-2
( k C [ + k H C O 3 ) w 1
0 10 20
+ Net A NHa w
N e t ~ N a ~
-2 ¸
i
/i " .... ,/!~ ""'..... j i ~ ""'-....
¢
\
" " ' " . . . . . ....
............................ i
10 20
.....,.--"'"
+ + 1 (z~NHL +,~ No )w
0
(z~NH~+ ~ Na*}w I : -{&Ct-+A HCO~)w o l
FigI (~, N e t c h a n g e s ( p o s t - e x e r c i s e
. . . . . . . 0 I0 20
o ~o Time (h) 20
c o n t r o l v~.l]ues) in ~,ater ('1 , H ( ' O i , N I l 4 + and Na { f o l l o w i n g
s t r e n u o u s m u s c u l a r a c t i v i t y in r a i n b o w t r o u t ( m m o l , k g f ish w a t e r , x + SE, n = 15 ) See text .
Discussion
The ionic, osmotic and acid base equilibrium was found to be considerably disturbed alter performance of strenuous muscular activity. Haematocri t was increased im- mediately after exercise by ahnost 502i, which may partially be attributed to acidosis- induced erythrocyte swelling (Dill el al., 1937; Soivio e t a / . . 1974) and possibly also to mobilization of additional erythrocytes. Also water movements from the extra- cellular space to the intracellular compar tment of the musculature may have been involved, the water then following the osmotic gradient established by the produc- tion of lactic acid from the,osmotically less active (per molecular weight) macro-

ION T R A N S F E R & A C I D - B A S E R E G U L A T I O N AFTER EXERCISE IN T R O U T 313
+20
A [ion]
(mM)
*10
- I 0
40
Y (A [Cotions] +~ [Anions[
(mM) 20
L(3-
No+
K.
0 2 /. 6 8 24 t (h)
i i ; I ' i i J
0 2 ~. 6 8 2~ I (h)
Fig. 7. Changes in dorsal aortic plasma ion concentrations (upper panel) and the sum o1" changes o1" ionic species following strenuous muscular activity in rainbow trout.
molecule glycogen. This is suggested by the increase of the integrated activity of all the measured plasma ions by about 31 mM (+ ca. 10%) (fig, 7, lower panel) 5 30 rain after exercise. About one fourth of this increase is attributable to the rise in lactate concentration (fig. 7, upper panel). Even if this lactate would have been co-transferred with sodium across the muscle cell membranes, at least half of the ion concentration changes have to be attributed to water movements.
Alter the initial sharp increase of about 15 mM the sodium concentration dropped continuously until about 8 h after exercise, in spite of the uptake of about 3 mmol/kg fish water Na + from the environmental water during the first 6 h (fig. 6), suggesting transfer of a relatively large quantity of Na + into the intracellular body compart- ment. Plasma chloride rose initially less than [Na p ] and started to fall steeply 15 min alter exercise to attain a concentration 8 mM below the control level about 3 h alter exercise. The resulting increasing gap between [Na +] and [C1 ] was filled by lactate ions, suggesting a t ransmembrane lactate /C1 counter-ion exchange mech- anism. A Na+-Lacta te co-transfer is made unlikely by the falling plasma [Na +] and simultaneous Na + uptake. A further detailed analysis of the plasma ion concen- trations and ion fluxes between intracellular space and extracellular body compart-

3 1 4 el. F. H O L E T O N e t a / .
ments is not possible because of the lack of precise knowledge of the magnitude of water movements and the changes in compartment volumes in the fish.
The el'flux kinetics of H- and lactate ions from the muscle cells as the site of anaerobic lactic acid production were faster than in most other fish species studied,
which may be attributable to a higher rate of muscle perfusion in the more active salmonids. Peak lactate concentration values were attained about 2 h after exercise, whereas in non-salmonid fishes peak values were not observed before 4 8 h after exercise (e.g. Secondat and Diaz, 1942: Piiper et al. 1972; Holeton and Heisler,
1983: for review see: Heisler, 1982a). The efflux of H + ions was extremely IEst: the amount of H ~ buffered in the extracellular space attained a maximum already at the end of the 5 rain exercise period. These restllts confirm the general observation that the time constant for the elimination of surplus H ~ ions fi'om vertebrate muscle
cells is at least 8 10 times larger than tklr lactate ions (e4,'. Black et a/. 1959; Piiper el a/., 1972; Holeton and Heisler, 1978, 1982:Benad6 and Heisler, 1978).
In spite of the fast efflux kinetics of H + ions, the amount of H ~ buffered m the extracellular space after exercise is relatively small. Up to about 3.5 mmol/kg fish water of lactate (Cp • V<.) was present ill the extracellular space, whereas only about 0.6 mmol/kg fish water of H ' ions* was sufficient to lower the pH or bicarbonate concentration in the small volume of poorly buffered extracellular space so lar that a new transient equilibrium for the distribution of H" ions between intra- and
extracellular body compartments was attained; thus the driving l\/rce for further H- transfer was removed ('equilibrium limitation', Holeton and Hcisler, 1983). This amount is small in comparison with the quantity of 4.6 mmol/kg fish water of H + ions that were temporarily transferred to the environmental water. Transfer of this amount allowed normalization of the acid-base status of the fish by 3 4 h alter exercise, when the plasma lactate concentration was still considerably elewited. The final quantitative elimination of the originally produced lactic acid by further
aerobic processing took 6 8h longer. Net transfer of H ~ ions to the water utilized at least two different mechanisms.
After exercise the control release of bicarbonate was terminated and later restarted at an initially lower rate resulting in a net gain of about 1.1 mmol/kg fish water. This is only about 15<'~> of the net bicarbonate gain from the environment found in So'liorhinus after strenuous exercise (Holeton and Heisler, 1983). The ureotelic dogfish, however, did not utilize the ammonia mechanism at all, whereas in the ammoniotelic Salmo ,gairdneri the net excretion of ammonia increased by about 3.5 mmol/kg fish water (fig. 5, difference between post-exercise and control ammonia release curve).
The net H ~ ion excretion can readily be calculated from the changes in bicar- bonate and ammonium ion concentrations of the environmental water. The mech- anisms involved in the excretion, however, cannot be deduced from these para-
• Es t imate based on the red t ic t ion o f b i c a r b o n a l e in p l a sma , an ex t race l lu la r space o f 2 5 " . o f the b o d y
w a t e r a n d a n o n - b i c a r b o n a t e buf fe r value o f 10 m e q / p H • k b l o o d (for detai ls see H o l e t o n a n d Heisler ,
1983).

ION T R A N S F E R & ACID BASE R E G U L A T I O N AFTER EXERCISE IN TROUT 315
meters. Some additional information can be gained from the changes of Na + and C1- in the water as the claimed counter-ions for H + or NH4 + and HCO/ transfer
across the gill epithelium. During the first hours of the recovery period CI was actually net excreted into
the environmental water suggesting that bicarbonate was taken up in exchange (fig. 6). The concentration of sodium in the water was net reduced, which in turn could indicate either H + or NH4 + excretion. The quantities, however, were not
H + equivalent to the net changes in HCO, and N 4 in the environmental water. Up to 0.75 mmol/kg fish water more CI (ca. + 70°<;) was net released than the water bicarbonate was redticed, and up to 1.2 mmol/kg fish water more NH4 + (ca. + 45 , )
was net released than Na + taken tip (fig. 6, panels 3 and 6). On the background of I : I ion exchange ratios to be claimed for conservation of electroneutrality for the HCO3/C1 and NH4+/Na + ion exchange mechanisms, the observed discrepancies can only be explained on the basis of a NH2 CI co-transfer mechanism, or, more likely, as the result of non-ionic diffusion similar to the mechanism in the mam-
malian kidney (Pitts, 1964). Then ionization of NH, to NH4 + in the environmental water would have produced equimolar quantities of HCO~, elevating the water bicarbonate concentration and leading to the observed mismatch between net changes of CI and H C O , in the water. But also the fraction of the water NH4' equivalent to the net change in water Na + must not necessarily have been transferred
by ionic exchange with sodium. The obtained results could as well be explained on the basis of H+/Na * counter-ion exchange. Then the H + ions transferred would have balanced the amount of bicarbonate produced as a result of non-ionic diffusion
of NH~, Evidence that non-ionic diffusional transfer of NH~ is possible in physiological
conditions has also been provided by Maetz (1973). Based on the correlation between the partial pressure difference and net ammonia flux reported for goldfish, the rate of ammonia elimination by non-ionic diffusion in trout can be estimated to be at least 5 /~mol/min • kg*, a rate sufficient to cover at least the steady-state excretion. If during recovery from exercise the plasma ammonia concentration
would be elevated, which is likely according to the largely increased production rate, then even a considerable fraction of the post-exercise elevated ammonia excretion observed in the present study may be the result of passive non-ionic diffusion along the partial pressure gradient, in spite of the fact that the proportion of physically dissolved ammonia was reduced according to the lowered pH under these conditions in plasma.
It must be questioned whether the elevation of ammonia production and excretion
after exercise is advantageous for the fish. If NH4 and H + are competing for the same carrier sites for the exchange against sodium, as has been suggested previously (e.g. Maetz, 1973, 1974; Evans, 1980), then the direct elimination of H ~ ions would
• Calculated by usc of the Henderson Hasse lba lchequa t ion (pK 9.6) and the diffusion law assuming
a total plasma ammonia concentration of 0.3 raM, and plasma and water pH values as well as water
ammonia concentrations determined in thc prescnt study.

316 (;. F. HOLETON et al.
be preferable, since it would not require breakdown of valuable amino acids for the acid base regulation. If, as suggested by the present results, a significant fraction is eliminated by non-ionic diffusion from the fish, this fraction would not contribute to the acid base regulation at all. Since the majority of previous studies of the acid base regulation in fish during acidic stress situations could not demonstrate any contribution of the ammonia mechanism (Cross et al.. 1969; Heisler el d[.,
1976b; Heisler, 1978; Heisler and Neumann, 1977: Heisler et al., 1981; Holeton and Heisler, 1983; Cameron and Kormanik, 1982a,b; Heisler 1982b), the elevation of ammonia excretion after exercise in the present study may very well be correlated to effects of strenuous exercise other than lactacidosis. Similar as in carp (Cyprim~s
cal7)io) and in Tilapia mossamhica (Driedzic and Hochachka, 1976: Kutty, 1972), the ammonia production of muscle fibres during anaerobic contractions may be enhanced as a result of the increased metabolism. Spillover of the ammonia into the blood (Driedzic and Hochachka, 1976) and elimination from the organism would then be the result of increased amino acid metabolism and not primarily related to acid base regulation.
In conclusion, severe post-exercise lactacidosis in trout is corrected to normal by transepithelial net H ~ ion transfer to the environmental water already 6 -8 h before the surplus lactic acid is removed from the organism by further aerobic metabolism. Part of this transfer is performed by branchial HCO, /CI ion exchange, the remainder by NH4' /Na ~ and/or H ~/Na + ion exchanges. A considerable fraction of the largely increased ammonia production is eliminated by non-ionic diffusion. The elevation of ammonia excretion during the first 4 h after exercise is attributed to exercise-induced production rather than to production lk)r the purpose of acid base regulation.
References
Benadd, A..I.S. and N. Heisler (1978). Comparison of the cfflux rates of hydrogen and lactate ions
fronl isolated muscles in vilro. Rc.vfir. Physiol. 32:369 380.
Black, E.C. (1957a). Alterations in tile blood level of lactic acid in certain salmonid fishes following
muscular activity. 1. Kamloops trout. ,S'almo t,,airthu,ri. ,1. f:Ls'h, Rc.~. Bd. ('all. 14: 117 134.
Bhtck, E. C (1957b). Alterations in the blood levcl of lactic acid in ccrtain salmonid fishes Following
muscular activity. II. Lake Irout. Salvelimls pmmavcu,~h. J. Fish. R~'s. Bd. ('aJz. 14:645 649.
Black, E.C. (1957c). Alterations in the blood level of lactic acid in certain salmonid ['ishcs I'ollo~ing
nluscular activity. [I1. Sockeye salmon, Om'orhvncluls ,erl,-a..I. Fish. Res. Bd. Can. 14:807 814.
Black, E. ('.. W.G. ('hiu, F. D. Forbes and A. Hanslip (1959). Changes in pH, carbonate and lactate
of the blood of yearling Kamloops trout, S(limo ,c, air~hwrL during and l'ollowing severe nluscular activity. ,]. FLail. Re,~. Bd. ('an. 16:391 4(12.
Black, E . ( . , A.R. Connor. K.C. Lull and W. (7. ( 'hiu (19621. Changes in glycogen, pyruvatc and lactate in rainbow trout :,S'ahm* ,vairdncriJ during and follo~ving muscuktr activity. J. l"ish. Rc~.
Bd. Call. 19:409 436.
('allleron, J. N, and G. A. Kormanik (1982a). lntra- and cxtracclltt[al- acid base statt>, as at function of
tcmpcraturc in the fi'esh~,,atcr channel catfish, lcl¢l/llrll,v plm~hm~. J. E\p, Biol. 99:127 142.
Camcron, J .N. and G.A. Kormanik (Ic)S2bL The acid base responses of gill,,, and kidne?s to infused acid and base loads in the channel catl'ish. Ic/a/urle~/~u*~clatu,~..I. Evp. Biol. 9t): 143 160.

ION T R A N S F E R & ACID BASE REGULATION AFTER EXERCISE IN TROUT 317
Claiborne, J.B. and N. Hcisler (1983). Acid-base regulation and ion transfers in the carp (Cvpriml,s earpio) during and after exposure to environmental hypercapnia. J. Exp. Biol. (in press).
Cross, C. E.. B. S. Packer, J. M. Linta, H. V. Murdaugh, Jr., and E. D. Robin (1969). H + buffering and excretion in response to acute hypercapnia in the dogfish Squalus acanthias. Am..1. Physiol. 216:
440 452. Dill. D. B., H.T. Edwards and W. V. Consolazio (1937). Blood as a physicochemical system. XI. Man
at rest. J. Biol. Chem, 118:635 648. Driedzic, W. R. and P. Hochachka (1976). Control of energy metabolism in fish white muscle. Am. J.
Phy.~'io/. 230:579 582. Evans. D.H. (1977). Further evidence for N a + , ' N H f exchange in marine teleost fish. J. E.vp. Biol.
70:213 220. Evans. D .H. (1980). Kinetic studies of ion transport by fish gill epithelium. Am. J. Physiol. 238:
R224 R230. Heislcr. N., H. Weitz and A . M . Weitz (1976a). Extracellular and intracellular pH with changes of
temperature in the dogfish Scrliorhinu.s stellaris. Re.v~ir. Ftlr.siol. 26:249 263.
Heisler, N., H. Weitz and A. M. Wcitz (1976b). Hypercapnia and resultant bicarbonate transfer processes in an elasmobranch fish (Scyliorhbu¢s .slellori.s). Bull. Eur, Ptlvsiopathol. Respir. 12:77 85.
Heisler, N. and P. Neumann (1977). lnlluence of sea-water pH upon bicarbonate uptake induced by
hypercapnia in an elasmobranch fish (Scyliorhhms ste/l~ri,~), p/liigers Arch. 368, Suppl. : R I9. Heislcr, N. (1978). Bicarbonate exchanges between body compar tments after changes of temperature
in the larger spotted dogfish (Scyliorhitm~ .stellaris). Re,spir. P/o'siol. 33:145 160. Heisler, N,, G .F . Holeton, D.P. Toews (1981). Regulation of gill ventilation and acid-base status ill
hyperoxia-induced hypercapnia in the larger spotted dogfish (Scy/iorhimls stel/arA). Physiologi.st 24 : 308.
Heisler. N. (1982a). Transepithelial ion transfer processes as mechanisms l`or fish acid base regulation
in hypcrcapnia and lactacidosis. Call. J. Zoo[. 6(1:1108 1122. Heisler, N. (1982b). lntracellular and extracellular acid base regulation in the tropical fresh-water teleost
fish 5[|'tlhl'tlllclllL~ tlltlrlHOl'tltlLV in response to the transition from water breathing to air breathing. J. £vp. Biol. 9 9 : 9 28.
Holeton, G. F. and N. Heisler (1978). Acid base regulation by bicarbonate exchange in the gills after
exhausting exercise in the larger spotted dogfish S~T/iorhinus szelhlris. Physiologi.~t 21: 56. Holeton, G. F., P. Neumann and N. Heisler (1980). Mechanisms of transient acid base regulation after
s trenuous activity in rainbow trout. Pttr.~ioh~gist 23: 932. Holeton. G .F . and N. Heisler (1983). Contribution of net ion transfer mechanisms to the acid base
regulation alter exhausting activity in the larger spotted dogfish (Scvliorhmus stelhtris). J. kLvp. Biol. (in press).
Kcrstetter, T. H., L. B. Kirschner and D.D. Rafuse (1970). On the mechanism of sodium ion transport by the irrigated gills of rainbow trout (Sahm~ gair~hleri). J. Gen. Physiol. 56:342 359.
Kutty, M. N. (1972). Respiratory quotient and ammonia excretion in Tilapia mossamhica. Marim' Biol. 16:126 133.
Maetz. J. (1973). Na ~ / Nl q~ , N a + / H + exchanges and NH~ movement across the gills of ('ara,ssiu~ auratus. J. E.\-p. Biol. 58:255 275.
Maetz, J. (1974). Aspects of adaptation to hypo-osmotic and hyper-osmotic environments. In: Bio- chemical and Biophysical Perspectives in Marine Biology. edited by D. C. Malins and J. R. Sargent.
New York, Academic Press, pp. 1-167. Payan, P. and J. Maetz (1973). Branchial sodium transport mechanism in Scvliorhinus catlicttla:
evidence ['or Na +,NH4 ~ and Na ~,'H ~ exchanges - and ['or a role of carbonic anhydrase. J. Exp. Biol. 58:487 502.
Piiper, J., M. Meyer and F. Drces (1972). Hydrogen ion balance in the elasmobranch Scyliorhimts stclhtris after exhausting activity. Re.v~ir. Physiol. 16:290 303.
Pitts. R, F. {It)64). Renal production and excretion of ammonia. ,4m. J. Mcd. 36:720 742.

318 ( ; F. H O t E T O N e: ed.
Secondat. M. and D. Diaz (1942). Recherches sur la lactacidemic chez le poisson d 'cau doucc. C.R. Ac~ld. Sci. Pelris 215:71 73.
Scveringhaus. J. (19(~5). Blood gas concentrations. 111 : Handbook of Physiology, Section 3 : Respiration,
Vol. 11, edited by W.O. Fenn and tl. Rahn. Washington, American Physiological Society. pp. 1475 1487.
Soivio, A., K. Westman and K. Nyhohn (1972). hnproved method of dorsal aorta catheterization: haemalological effects followed l\~r three weeks in rainbow trout :.S'alltto .e~tir~h~eH). kTtzJ~isl~ l:i,sh.
Re,~'. 1: 1l 21. Soivio, A., K. Westman and K. Nyhohn {1974}. Changes in haemalocrit values in blood samples
treated with and without oxygen: a comparative study with four salmonid species..I, l:'i~h. Biol.
6:763 76~). Soivio, A., K. Nyholm and K. Westman (1975). A technique for repcatcd sampling of the blood of
individual resting fish. J. E.vp. Bi~l. 63:207 217.




















