
Both oxidative and nitrosative stress are associated with muscle wasting in tumour-bearing rats
7
Both oxidative and nitrosative stress are associated with muscle wasting in tumour-bearing rats Esther Barreiro a , Beatriz de la Puente a , Sı ´lvia Busquets b , Francisco J. Lo ´ pez-Soriano b, * , Joaquim Gea a,c , Josep M. Argile ´s b a Muscle and Respiratory System Research Unit, IMIM and CEXS, Universitat Pompeu Fabra, Barcelona, Spain b Departament de Bioquı ´mica i Biologia Molecular, Facultat de Biologia, Universitat de Barcelona, Diagonal 645, 08028 Barcelona, Spain c Respiratory Medicine Department, Hospital del Mar, Barcelona, Spain Received 31 January 2005; accepted 2 February 2005 Available online 21 February 2005 Edited by Barry Halliwell Abstract Reactive oxygen and nitrogen species (ROS and RNS) have been proposed as mechanisms of cancer-induced ca- chexia. In this study, we assessed using Western blot analysis the levels of total protein carbonylation (2,4-dinitrophenylhydrazine assay), both malondialdehyde- (MDA-) and 2-hydroxy-4-none- nal- (HNE-) protein adducts, Mn-superoxide dismutase (Mn- SOD), catalase, heme oxygenase-1 (HO-1) and 3-nitrotyrosine formation in gastrocnemius muscles of rats bearing the Yoshida AH-130 hepatoma. In the muscles of the tumour-bearing ani- mals, protein carbonylation as measured by total levels of car- bonyl group formation and both HNE and MDA-protein adducts, and protein tyrosine nitration were significantly greater than in control muscles. Protein levels of the antioxidant en- zymes Mn-SOD, catalase, and HO-1 were not significantly modified in the rat cachectic muscles compared to controls. The inefficiency of the antioxidant enzymes in neutralizing excessive ROS production may account for elevated markers of protein oxidation and be responsible for the development of both oxidative and nitrosative stress in cancer-induced cachexia. Ó 2005 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. Keywords: Oxidative stress; Nitrosative stress; Skeletal muscle; Cachexia 1. Introduction The development of cancer cachexia is the most common manifestation of advanced malignant disease. Indeed, ca- chexia occurs in the majority of cancer patients before death, and it is responsible for the death of 22% of cancer patients [1]. The abnormalities associated with cancer ca- chexia include anorexia, weight loss, muscle loss and atro- phy, anemia and alterations in carbohydrate, lipid, and protein metabolism [2]. The degree of cachexia is inversely correlated with the survival time of the patient and it always implies a poor prognosis [3–5]. Asthenia (lack of muscular strength), which reflects the important muscle waste that takes place in the cachectic cancer patient, is one of the most relevant characteristics of cachexia [6]. Asthenia is also characterized by a general weakness as well as physical and mental fatigue [7]. In addition, lean body mass depletion is one of the main trends of cachexia and it involves not only skeletal muscle but it also affects cardiac proteins, resulting in important alterations in heart performance. The rat ascites hepatoma Yoshida AH-130 is a suitable model system to study the mechanisms involved in the establishment of cachexia. Its growth causes rapid and pro- gressive loss of body weight and tissue waste, particularly in skeletal muscle [8]. Acceleration of tissue protein breakdown accounts for most of the waste in the AH-130 bearers [9–11]; in particular, skeletal muscle hypercatabolism is mainly due to hyperactivation of the ATP–ubiquitin-dependent proteo- lytic system [12]. High plasma levels of tumour necrosis fac- tor-a (TNF) [13] and perturbations in the hormonal homeostasis [14] may play an important role in forcing the metabolic balance towards the catabolic side. In addition to increased muscle protein degradation during cancer growth, the presence of the tumour also induces an in- creased rate of apoptosis in skeletal muscle in rats [15], which also seems to be activated by cytokines, TNF in par- ticular [16]. Oxidative stress due to greater levels of reactive oxygen species (ROS) production than those normally neutralized by intracellular antioxidant defenses has recently gained much attention for its possible involvement in cancer ca- chexia. Buck and Chojkier [17] demonstrated that muscle wasting and dedifferentiation could be prevented by inhibi- tors of nitric oxide (NO) synthesis and antioxidants. Simi- larly, Gomes-Marcondes and Tisdale [18] have shown that muscle wasting in cancer cachexia is associated with in- creased levels of malondialdehyde in gastrocnemius muscles, and that mild oxidative stress increased protein degradation in skeletal muscle by causing a greater expression of the ma- jor components of the ubiquitin–proteasome pathway. In addition, Murr et al. [19] have shown elevated neopterin (an indicator of ROS production) concentrations in the ser- um and urine of cancer patients, this elevation correlating with the disease stage, significantly predicting worse progno- sis regarding relapse and survival. Proteins are one of the major targets of oxidative stress- derived effects on tissues. Among the different modifications induced by ROS in amino acid residues, that of protein car- bonylation constitutes one of the best characterized markers of protein oxidation in several conditions and disease states, * Corresponding author. Fax: +34 934021559. E-mail address: fl[email protected] (F.J. Lo ´ pez-Soriano). 0014-5793/$30.00 Ó 2005 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2005.02.017 FEBS Letters 579 (2005) 1646–1652 FEBS 29354
Transcript of Both oxidative and nitrosative stress are associated with muscle wasting in tumour-bearing rats

FEBS Letters 579 (2005) 1646–1652 FEBS 29354
Both oxidative and nitrosative stress are associated with muscle wastingin tumour-bearing rats
Esther Barreiroa, Beatriz de la Puentea, Sılvia Busquetsb, Francisco J. Lopez-Sorianob,*,Joaquim Geaa,c, Josep M. Argilesb
a Muscle and Respiratory System Research Unit, IMIM and CEXS, Universitat Pompeu Fabra, Barcelona, Spainb Departament de Bioquımica i Biologia Molecular, Facultat de Biologia, Universitat de Barcelona, Diagonal 645, 08028 Barcelona, Spain
c Respiratory Medicine Department, Hospital del Mar, Barcelona, Spain
Received 31 January 2005; accepted 2 February 2005
Available online 21 February 2005
Edited by Barry Halliwell
Abstract Reactive oxygen and nitrogen species (ROS andRNS) have been proposed as mechanisms of cancer-induced ca-chexia. In this study, we assessed using Western blot analysis thelevels of total protein carbonylation (2,4-dinitrophenylhydrazineassay), both malondialdehyde- (MDA-) and 2-hydroxy-4-none-nal- (HNE-) protein adducts, Mn-superoxide dismutase (Mn-SOD), catalase, heme oxygenase-1 (HO-1) and 3-nitrotyrosineformation in gastrocnemius muscles of rats bearing the YoshidaAH-130 hepatoma. In the muscles of the tumour-bearing ani-mals, protein carbonylation as measured by total levels of car-bonyl group formation and both HNE and MDA-proteinadducts, and protein tyrosine nitration were significantly greaterthan in control muscles. Protein levels of the antioxidant en-zymes Mn-SOD, catalase, and HO-1 were not significantlymodified in the rat cachectic muscles compared to controls.The inefficiency of the antioxidant enzymes in neutralizingexcessive ROS production may account for elevated markersof protein oxidation and be responsible for the development ofboth oxidative and nitrosative stress in cancer-induced cachexia.� 2005 Federation of European Biochemical Societies. Publishedby Elsevier B.V. All rights reserved.
Keywords: Oxidative stress; Nitrosative stress; Skeletal muscle;Cachexia
1. Introduction
The development of cancer cachexia is the most common
manifestation of advanced malignant disease. Indeed, ca-
chexia occurs in the majority of cancer patients before
death, and it is responsible for the death of 22% of cancer
patients [1]. The abnormalities associated with cancer ca-
chexia include anorexia, weight loss, muscle loss and atro-
phy, anemia and alterations in carbohydrate, lipid, and
protein metabolism [2]. The degree of cachexia is inversely
correlated with the survival time of the patient and it always
implies a poor prognosis [3–5]. Asthenia (lack of muscular
strength), which reflects the important muscle waste that
takes place in the cachectic cancer patient, is one of the
most relevant characteristics of cachexia [6]. Asthenia is also
*Corresponding author. Fax: +34 934021559.E-mail address: [email protected] (F.J. Lopez-Soriano).
0014-5793/$30.00 � 2005 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2005.02.017
characterized by a general weakness as well as physical and
mental fatigue [7]. In addition, lean body mass depletion is
one of the main trends of cachexia and it involves not only
skeletal muscle but it also affects cardiac proteins, resulting
in important alterations in heart performance.
The rat ascites hepatoma Yoshida AH-130 is a suitable
model system to study the mechanisms involved in the
establishment of cachexia. Its growth causes rapid and pro-
gressive loss of body weight and tissue waste, particularly in
skeletal muscle [8]. Acceleration of tissue protein breakdown
accounts for most of the waste in the AH-130 bearers [9–11];
in particular, skeletal muscle hypercatabolism is mainly due
to hyperactivation of the ATP–ubiquitin-dependent proteo-
lytic system [12]. High plasma levels of tumour necrosis fac-
tor-a (TNF) [13] and perturbations in the hormonal
homeostasis [14] may play an important role in forcing the
metabolic balance towards the catabolic side. In addition
to increased muscle protein degradation during cancer
growth, the presence of the tumour also induces an in-
creased rate of apoptosis in skeletal muscle in rats [15],
which also seems to be activated by cytokines, TNF in par-
ticular [16].
Oxidative stress due to greater levels of reactive oxygen
species (ROS) production than those normally neutralized
by intracellular antioxidant defenses has recently gained
much attention for its possible involvement in cancer ca-
chexia. Buck and Chojkier [17] demonstrated that muscle
wasting and dedifferentiation could be prevented by inhibi-
tors of nitric oxide (NO) synthesis and antioxidants. Simi-
larly, Gomes-Marcondes and Tisdale [18] have shown that
muscle wasting in cancer cachexia is associated with in-
creased levels of malondialdehyde in gastrocnemius muscles,
and that mild oxidative stress increased protein degradation
in skeletal muscle by causing a greater expression of the ma-
jor components of the ubiquitin–proteasome pathway. In
addition, Murr et al. [19] have shown elevated neopterin
(an indicator of ROS production) concentrations in the ser-
um and urine of cancer patients, this elevation correlating
with the disease stage, significantly predicting worse progno-
sis regarding relapse and survival.
Proteins are one of the major targets of oxidative stress-
derived effects on tissues. Among the different modifications
induced by ROS in amino acid residues, that of protein car-
bonylation constitutes one of the best characterized markers
of protein oxidation in several conditions and disease states,
blished by Elsevier B.V. All rights reserved.

E. Barreiro et al. / FEBS Letters 579 (2005) 1646–1652 1647
such as sepsis and chronic obstructive pulmonary disease
[20,21]. Excessive production of NO in vivo may exert
damaging effects on tissues owing to its diffusion-limited
reaction with the superoxide anion to form peroxynitrite
[22]. This powerful oxidant species and its decomposition
products are now thought to be greatly responsible for most
of the deleterious effects of excessive production of NO
[22,23]. Nitrosation and nitration reactions whereby NO is
incorporated into the target molecules are the most charac-
teristic mechanisms among the different oxidative modifica-
tions induced by peroxynitrite and other reactive nitrogen
species, RNS [23,24]. Moreover, nitration of aromatic amino
acids, lipids, and DNA bases have been shown to constitute
one of the most characteristic irreversible reactions of NO-
induced effects on tissues [22,23,25]. Specifically, nitration
of the amino acid tyrosine to form 3-nitrotyrosine is now
considered to be a biomarker for the production of RNS
in vivo [22,23].
Bearing all this in mind, we hypothesized that cancer-
induced cachexia might lead to the development of increased
levels of oxidative stress in the limb muscles of rats bearing
the ascites hepatoma Yoshida AH-130. To test our hypothesis,
firstly, we aimed at assessing whether oxidative and nitrosative
stress occur in those muscles using different indexes of protein
oxidation (protein carbonylation) and by detection of
3-nitrotyrosine immunoreactivity. Secondly, we explored mus-
cle levels of several antioxidant mechanisms that might not be
sufficiently upregulated to counterbalance the deleterious
effects of oxidative stress in those cachectic muscles.
2. Materials and methods
2.1. AnimalsMale Wistar rats (Interfauna, Barcelona, Spain), of about 6 weeks
of age, were used in the different experiments (n = 6/group). The ani-mals were maintained at 22 ± 2 �C with a regular light–dark cycle(light on from 08:00 a.m. to 08:00 p.m.) and had free access to foodand water. The diet (B.K. Universal G.J./S.L., Sant Vicenc delHorts, Barcelona, Spain) consisted of 45.5–48.5% carbohydrate(3.5% absorbable glucose, 42–45% starch), 18.5% protein and 3.1%fat (the residue was non-digestible material). The food intake wasmeasured daily. All animal manipulations were made in accordancewith the European Community guidelines for the use of laboratoryanimals.
2.2. Tumour inoculationRats were divided into two groups, namely controls and tumour
hosts. The latter received an intraperitoneal inoculum of 108 AH-130Yoshida ascites hepatoma cells obtained from exponential tumours[11]. On days 4 and 7 after tumour transplantation, the animals wereweighed and anesthetized with an i.p. injection of ketamine/xylazinemixture (3:1) (Imalgene� and Rompun�, respectively). The tumourwas harvested from the peritoneal cavity and its volume and cellularityevaluated. Tissues were rapidly excised, weighed, and frozen in liquidnitrogen.
2.3. BiochemicalsThey were all reagent grade and obtained either from Roche (Barce-
lona, Spain) or from Sigma Chemical Co. (St. Louis, MO, USA).
2.4. Total protein carbonylationTotal levels of those highly reactive carbonyl groups in the protein
side chains were detected by reaction with 2,4-dinitrophenylhydrazine(DNPH), resulting in the formation of 2,4-dinitrophenylhydrazone
(DNP) [26]. The DNP-derivatized proteins were subsequently sepa-rated by electrophoresis and further subjected to immunoblotting withselective antibodies against the DNP moieties.
2.5. Formation of aldehyde-protein adductsThe lipid peroxidation products malondialdehyde (MDA) and 4-hy-
droxy-2-nonenal (HNE) can cause further cellular damage by bindingto and modifying proteins leading to the formation of aldehyde-protein adducts. The characteristic feature of these adductions arethe introduction of carbonyl groups into the modified proteins. HNEreacts with lysine, cysteine, and histidine residues of proteins to formMichael adducts that can be stabilized and further detected using aselective antibody [27]. MDA reacts with lysine residues to form Schiffbase adducts that can also be detected in tissues using a selective anti-body [28].
2.6. Western blot analysisImmunoblotting was used to evaluate the effects of ROS and
RNS on muscle proteins and lipids according to methodologies pre-viously described [21]. Accordingly, total carbonyl groups, bothHNE- and MDA-protein adducts, Mn-superoxide dismutase (Mn-SOD), catalase, and heme oxygenase-1 (HO-1) were determined onthe one hand, and protein tyrosine nitration (3-nitrotyrosine immu-noreactivity) on the other. Frozen muscle samples (n = 6/group)were homogenized in a buffer containing HEPES 50 mM, NaCl150 mM, NaF 100 mM, Na pyrophosphate 10 mM, EDTA 5 mM,Triton-X 0.5%, leupeptin 2 lg/ml, PMSF 100 lg/ml, aprotinin2 lg/ml, and pepstatin A 10 lg/ml. Samples were then centrifugedat 1000 · g for 30 min. The pellet was discarded and the supernatantwas designated as a crude homogenate. Total muscle protein level ineach sample was spectrophotometrically determined with the Brad-ford technique using different runs of triplicates in each case and bo-vine serum albumin as the standard (Bio-Rad protein reagent, Bio-Rad Inc., Hercules, CA). The final protein concentration in eachsample was calculated from at least two almost identical Bradfordmeasurements. Equal amounts of total protein from crude musclehomogenates were always loaded (20 lg per sample/lane) onto thegels, as well as identical sample volumes/lanes. While conductingthe different Western blot analyses, the same samples were alwaysrun together and kept in the same order. Proteins were then sepa-rated by electrophoresis, transferred to polyvinylidene difluoride(PVDF) membranes, blocked with non-fat milk and incubated over-night with selective antibodies. The following antibodies were usedto detect the different antigens and phenomena: anti-DNP moietyantibody (Oxyblot kit, Chemicon International Inc., Temecula,CA, USA), anti-HNE antibody (Calbiochem, San Diego, CA,USA), anti-MDA antibody (Academy Bio-Medical Company, Inc.,Houston, TX, USA), anti-Mn SOD antibody (StressGen, Victoria,BC, Canada), anti-catalase antibody (Calbiochem), anti-HO-1 anti-body (Transduction Laboratories Inc., Lexington, KY), and anti-3-nitrotyrosine antibody (Cayman Chemical Inc., Ann Arbor, MI,USA). Tissue homogenates obtained from rat brain mitochondria,erythrocytes, and liver microsomes were used as positive controlsof the enzymes Mn-SOD, catalase, and HO-1, respectively. Specificproteins from all samples were detected with horseradish peroxidase(HRP)-conjugated secondary antibodies and a chemiluminescencekit. For each of the antigens, samples from the three differentgroups were always detected in the same film under identical expo-sure times. Negative control experiments in which primary antibod-ies were omitted or samples were not derivatized (carbonyl groups),and membranes were then probed only with secondary antibodieswere also conducted. Blots were scanned with an imaging densitom-eter and optical densities (OD) of specific proteins were quantifiedwith Diversity Database 2.1.1 (Bio-Rad, Philadelphia, PA, USA).Values of total protein carbonylation, both total HNE- andMDA-protein adducts, and total protein tyrosine nitration in a gi-ven sample were calculated by addition of OD of individual proteinbands in each case. Final OD obtained in each specific group of ani-mals (depicted in the corresponding histograms) corresponded to themean values of the different samples (lanes) of each of the antigensstudied. To validate equal protein loading among various lanes,PVDF membranes were stripped and re-probed with a monoclonalanti-a-tubulin antibody (Sigma) in all cases. OD in each histogramwere expressed as the ratio of the OD of the specific antigen to

Table 1Food intake, tissue weights and tumour growth in rats bearing the Yoshida AH-130 ascites hepatoma
Day 4 Day 7
Control (4) Tumour-bearing (7) Control (4) Tumour-bearing (7)
Food intake 52 ± 1 61 ± 3 123 ± 1 98 ± 1\\
Muscle weightsGastrocnemius 590 ± 16 517 ± 9\ 651 ± 10 482 ± 23\
Tibialis 195 ± 3 173 ± 4\ 235 ± 6 161 ± 5\\
Soleus 43 ± 1 41 ± 1 51 ± 2 38 ± 1\\
EDL 46 ± 1 40 ± 1\ 53 ± 2 38 ± 1\\
Heart 402 ± 9 387 ± 17 430 ± 29 330 ± 27\
Carcass weight 101 ± 2 96 ± 2 107 ± 8 89 ± 6Tumour cell content – 1886 ± 197 – 3596 ± 531
For more details see Section 2. Results are means ± S.E.M. for the number of animals indicated in parentheses. Food intake is expressed in g andrefers to the ingestion during the period of the experiment prior to sacrifice which place 4 or 7 days after tumour inoculation. Tissue weights areexpressed as mg/100 g of initial body weight. Carcass weight (body without organs or tumour) is expressed as g/100 g of initial body weight. EDL,extensor digitorum longus. Tumour cell content is expressed in millions of cells. Statistical significance of the results: \P < 0.05, \\P < 0.01 (controlversus tumour-bearing).
1648 E. Barreiro et al. / FEBS Letters 579 (2005) 1646–1652
those of a-tubulin. Besides, Red Ponceau staining of crude homog-enates on the membranes was also used to determine equal loading/transfer across lanes.
2.7. Statistical analysisData are presented as means ± S.D. Mann–Whitney U (non-para-
metric test, independent samples) test was used for assessing differencesamong the two groups (tumour-bearing animals and controls).
3. Results
3.1. Characteristics of the study animals
Table 1 indicates the main characteristics of tumour-bearing
animals. The implantation of the Yoshida AH-130 ascites hep-
atoma resulted in a decrease in muscle weights already on day
4 after tumour transplantation (12% for gastrocnemius, 11%
for tibialis, and 13% for extensor digitorum longus), and a
Fig. 1. (A) Representative examples of protein oxidation (total carbonyl grorats sacrificed on days 4 and 7 (TD4 and TD7, respectively). When samplindicated as negative control (C�). (B) OD in the histogram are expressed(optical densities, OD; arbitrary units, a.u.). Total protein carbonylation wascontrol muscles (P < 0.05).
much higher decrease on day 7 after tumour transplantation
(26% for gastrocnemius, 31% for tibialis, 25% for soleus,
28% for extensor digitorum longus).
3.2. Protein carbonylation
Total protein carbonylation. As shown in Fig. 1A, anti-DNP
antibody detected different positive protein bands with appar-
ent masses ranging from 111 to 27 kDa in the gastrocnemius of
the tumour-bearing animals. In this model, the muscles of ani-
mals sacrificed on day 7 showed higher levels of total carbonyl
content than control muscles (Fig. 1B).
HNE- and MDA-protein adducts. Fig. 2A illustrates different
HNE-protein adducts with apparent masses ranging from 70
to 25 kDa in the gastrocneumius of rats bearing the Yoshida
ascites hepatoma and their control animals. Total levels of
HNE-protein adducts were significantly greater in the muscles
of rats sacrificed on day 7 compared to control muscles (Fig.
ups) in gastrocnemius of control rats and in Yoshida tumour-bearinges were not derivatized, no protein carbonylation could be detected,as the ratio of the OD of total carbonyl groups to those of a-tubulingreater in the tumour-bearing rats sacrificed on day 7 compared with

Fig. 2. (A) Representative examples of HNE-protein adducts in the gastrocnemius of control rats and in the Yoshida tumour-bearing animalssacrificed on days 4 and 7 (TD4 and TD7, respectively). Monoclonal anti-a-tubulin antibody was used to control equal loading among various lanes.(B) OD in the histogram are expressed as the ratio of the OD of total HNE-protein adducts to those of a-tubulin (optical densities, OD; arbitraryunits, a.u.). HNE-protein adduct formation was greater in the tumour-bearing rats sacrificed on day 7 compared with control muscles (P < 0.05).
E. Barreiro et al. / FEBS Letters 579 (2005) 1646–1652 1649
2B). Three major MDA-protein adducts were detected in the
gastrocnemius of rats with apparent masses of 57, 52, and
35 kDa, respectively (Fig. 3A). As shown in Fig. 3B, total lev-
els of MDA-protein adducts were significantly greater in the
gastrocnemius of the tumorbearing rats sacrificed on day 7
compared to control muscles (P < 0.05).
3.3. Antioxidant enzymes
Muscle levels of the antioxidant enzymes Mn-SOD,
catalase and HO-1 in rats bearing the Yoshida ascites hepa-
toma did not show any significant modification on day 4 or
on day 7 after tumour inoculation compared to controls
(Fig. 4).
Fig. 3. (A) Representative examples of MDA-protein adducts in the gastrosacrificed on days 4 and 7 (TD4 and TD7, respectively). Monoclonal anti-a-tu(B) OD in the histogram are expressed as the ratio of the OD of total MDAunits, a.u.). MDA-protein adduct levels were greater in the tumour-bearing
3.4. Protein tyrosine nitration
Several tyrosine-nitrated protein bands were detected in the
gastrocneumius muscles of both control and cachectic Yos-
hida-bearing rats sacrificed on days 4 and 7, with apparent
masses ranging from 81 to 26 kDa (Fig. 5A). Interestingly, to-
tal muscle 3-nitrotyrosine levels were greater in the gastrocne-
mius of rats sacrificed on day 7 (Fig. 5B).
4. Discussion
The main findings of the present study are that 7 days after
the implantation of the Yoshida AH-130 ascites hepatoma in
cnemius of control rats and in the Yoshida tumour-bearing animalsbulin antibody was used to control equal loading among various lanes.-protein adducts to those of a-tubulin (optical densities, OD; arbitraryrats sacrificed on day 7 compared with control muscles (P < 0.05).
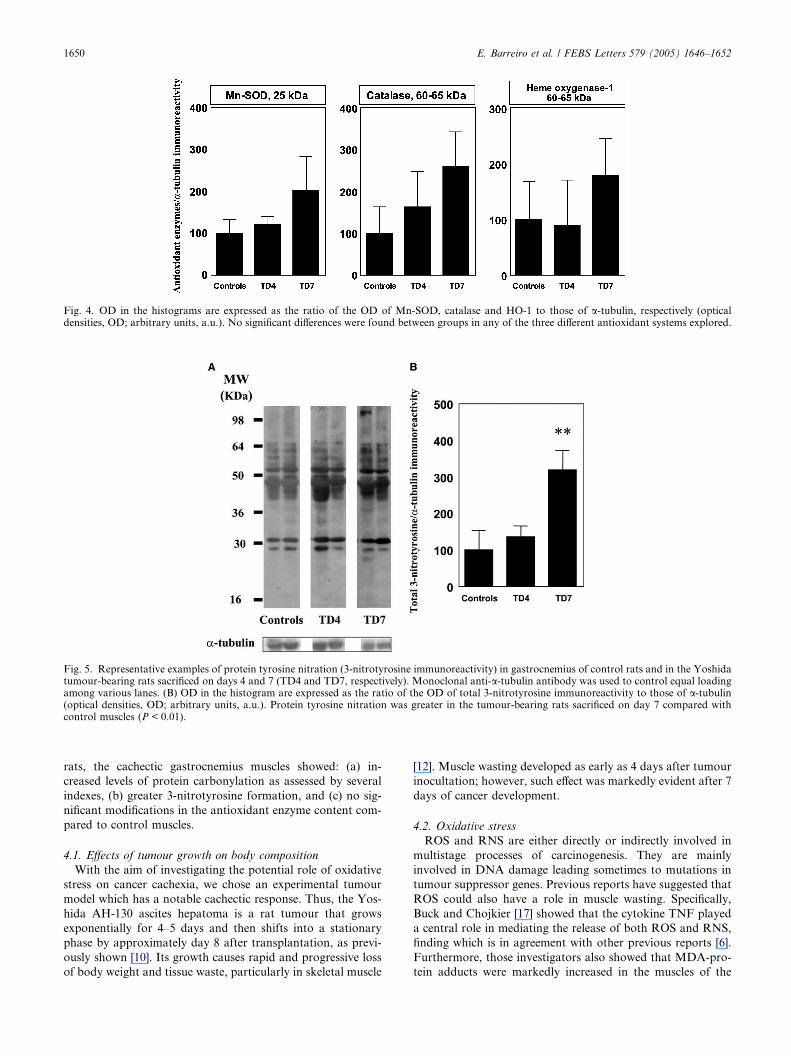
Fig. 5. Representative examples of protein tyrosine nitration (3-nitrotyrosine immunoreactivity) in gastrocnemius of control rats and in the Yoshidatumour-bearing rats sacrificed on days 4 and 7 (TD4 and TD7, respectively). Monoclonal anti-a-tubulin antibody was used to control equal loadingamong various lanes. (B) OD in the histogram are expressed as the ratio of the OD of total 3-nitrotyrosine immunoreactivity to those of a-tubulin(optical densities, OD; arbitrary units, a.u.). Protein tyrosine nitration was greater in the tumour-bearing rats sacrificed on day 7 compared withcontrol muscles (P < 0.01).
Fig. 4. OD in the histograms are expressed as the ratio of the OD of Mn-SOD, catalase and HO-1 to those of a-tubulin, respectively (opticaldensities, OD; arbitrary units, a.u.). No significant differences were found between groups in any of the three different antioxidant systems explored.
1650 E. Barreiro et al. / FEBS Letters 579 (2005) 1646–1652
rats, the cachectic gastrocnemius muscles showed: (a) in-
creased levels of protein carbonylation as assessed by several
indexes, (b) greater 3-nitrotyrosine formation, and (c) no sig-
nificant modifications in the antioxidant enzyme content com-
pared to control muscles.
4.1. Effects of tumour growth on body composition
With the aim of investigating the potential role of oxidative
stress on cancer cachexia, we chose an experimental tumour
model which has a notable cachectic response. Thus, the Yos-
hida AH-130 ascites hepatoma is a rat tumour that grows
exponentially for 4–5 days and then shifts into a stationary
phase by approximately day 8 after transplantation, as previ-
ously shown [10]. Its growth causes rapid and progressive loss
of body weight and tissue waste, particularly in skeletal muscle
[12]. Muscle wasting developed as early as 4 days after tumour
inocultation; however, such effect was markedly evident after 7
days of cancer development.
4.2. Oxidative stress
ROS and RNS are either directly or indirectly involved in
multistage processes of carcinogenesis. They are mainly
involved in DNA damage leading sometimes to mutations in
tumour suppressor genes. Previous reports have suggested that
ROS could also have a role in muscle wasting. Specifically,
Buck and Chojkier [17] showed that the cytokine TNF played
a central role in mediating the release of both ROS and RNS,
finding which is in agreement with other previous reports [6].
Furthermore, those investigators also showed that MDA-pro-
tein adducts were markedly increased in the muscles of the

E. Barreiro et al. / FEBS Letters 579 (2005) 1646–1652 1651
tumour-induced cachectic animals [17]. However, the adminis-
tration of the antioxidant DD-a-tocopherol clearly abrogated
such an increase, and prevented the muscle wasting of those
animals [17]. Other authors have also reported an increase in
MDA in the blood of advanced gastric cancer patients [29]
as well as high levels of this type of aldehyde in skeletal muscle
during cancer cachexia [18].
Given the fact that the DNPH assay indiscriminately detects
total protein carbonyl groups formed by diverse species such
as hydroxyl and alkylperoxyl radicals, MDA, and HNE [30],
and that lipid peroxidation is a common phenomenon of oxi-
dative stress [31,32], we decided to specifically detect whether
both MDA- and HNE-protein adducts were increased in the
cachectic muscles of our tumour-bearing rats. Indeed, both
MDA- and HNE-protein adducts were found to be greater
in the gastrocnemius of these animals compared with controls,
suggesting that peroxidation of polyunsaturated membrane
lipids is likely to be a continuing event during the muscle wast-
ing process.
Total mucle protein carbonylation levels, as measured by
the DNPH assay, were also increased in the cachectic gas-
trocnemius muscles. Based on this, we can speculate that,
in our experimental model of cancer cachexia, active oxida-
tive cellular damage must occur (probably via TNF) during
the process of muscle wasting. Actually, soon after TNF
exposure, ROS can alter contractile regulation reducing mus-
cle force, while late effects of that cytokine on tissues lead to
ROS-mediated upregulation of the ubiquitin/proteasome
pathway (through NF-jb activation) accelerating protein
degradation and muscle mass loss [33]. Indeed, it is thought
that the importance of oxidative stress as a contributing fac-
tor to cancer-induced cachexia is based on the increased sus-
ceptibility of oxidized proteins to undergo proteolytic
degradation [34,35].
Another important aspect considered in the present study
was the assessment of whether reduced food intake in the
cachectic animals might have any effect on the levels of muscle
protein carbonylation. Since hypophagia is present in Yoshida
AH-130 tumour bearers on day 7, experiments using pair-fed
rats were used, and no significant differences in those levels
were detected with respect to control muscles (results not
shown). Eventually, it is also worth considering that minimum
levels of ROS were present in control muscles as shown by
those oxidized protein bands of similar sizes to the ones ob-
served in cachectic muscles (Figs. 1–3). This is actually consis-
tent with the fact that relatively low levels of ROS are required
for normal force production [36].
Muscle content of the three different antioxidant systems
analyzed in the current study were not significantly modified
in the advanced cachexia state with respect to control mus-
cles, albeit protein levels of both Mn-SOD and catalase
tended to increase in the cachectic gastrocnemius. Therefore,
we can conclude from our data that these three different
antioxidant mechanisms do not seem to have major protec-
tive effects against oxidative stress in this model of cancer-
induced cachexia. One could speculate from these findings
that cancer conditions might alter either the content or the
activity of those enzymes, or even modify their regulatory
mechanisms responsible for clearly upregulating their levels.
For instance, MacMillan-Crow et al. [37–39] have already
shown in different studies that peroxynitrite leads to nitra-
tion and inactivation of Mn-SOD in vivo under several con-
ditions including human pancreatic ductal adenocarcinoma.
In our study, the absence of a significant increase in the con-
tent of these antioxidant mechanisms along with the ex-
pected lack of increased Mn-SOD activity would certainly
lead to greater levels of muscle oxidative stress in the
cachectic animals.
4.3. Reactive nitrogen species
Buck and Chojkier [17] showed in their study that treatment
of cachectic mice with nitro-LL-arginine, a NO synthase inhibi-
tor, prevented weight loss as well as the deleterious effects of
TNF on muscle. On the grounds that excessive NO production
could be one contributing mechanism to cancer-induced ca-
chexia, we explored whether 3-nitrotyrosine formation was in-
creased in cachectic muscles. In effect, protein tyrosine
nitration was markedly greater in the muscles of tumour-bear-
ing rats with advanced cachexia (day 7). These findings led us
to the conclusion that muscle wasting, at least under experi-
mental cancer conditions, is strongly associated with nitrosa-
tive stress. This is specially important since nitration of
tyrosine residues has been shown to trigger an increased degra-
dation by the proteasome of the modified proteins [40]. Even-
tually, we also conclude that cachexia per se was associated
with the development of nitrosative stress in the cancer-in-
duced cachectic muscles since hypophagia (pair-fed animals)
did not show any significant increase in protein tyrosine nitra-
tion (data not shown).
4.4. Concluding remarks
In conclusion, in this study we provide first evidence of the
existence of a clear association between experimental cancer-
induced cachexia and the development of both oxidative and
nitrosative stress. The levels of three major antioxidant mech-
anisms were also explored, showing no significant increase
within the cachectic muscle fibers. This indicates that altera-
tions in those antioxidant systems might contribute to worsen
the levels of both oxidative stress and muscle wasting. This
hypothesis requires further investigation, since some antioxi-
dant proteins can be pharmacologically induced, suggesting a
potential novel therapeutic target for cancer-induced cachexia.
Acknowledgments: The authors are thankful to Mr. Francesc Sanchezfor his technical assistance. This work was supported by grants fromthe Instituto de Salud Carlos III, Ministerio de Sanidad y Consumo(03/0100), from the Direccion General de Investigacion Cientıfica yTecnica (BFI 2002-02186) from the Spanish Ministry of Educationand Science, from the Generalitat de Catalunya (SGR/00108), andfrom Red Respira (RTIC C03/11, Fondo de Investigacion Sanitaria,Instituto de Salud Carlos III).
References
[1] Warren, S. (1932) The immediate causes of death in cancer. Am.J. Med. Sci. 184, 610–615.
[2] Argiles, J.M., Alvarez, B. and Lopez-Soriano, F.J. (1997) Themetabolic basis of cancer cachexia. Med. Res. Rev. 17, 477–498.
[3] DeWys, W. (1985) Management of cancer cachexia. Semin.Oncol. 12, 452–460.
[4] Harvey, K.B., Bothe, A. and Blackburn, G.L. (1979) Nutritionalassessment and patient outcome during oncological therapy.Cancer 43, 2065–2069.
[5] Nixon, D.W., Heymsfield, S.B., Kutner, M.M., Ansley, J. andLawson, D.H. (1980) Protein-calorie undernutrition in hospital-ized cancer patients. Am. J. Med. 68, 491–497.

1652 E. Barreiro et al. / FEBS Letters 579 (2005) 1646–1652
[6] Argiles, J.M., Garcıa-Martınez, C., Llovera, M. and Lopez-Soriano, F.J. (1992) The role of cytokines in muscle wasting: itsrelation with cancer cachexia. Med. Res. Rev. 12, 637–652.
[7] Adams, R. and Victor, M. (1981) Asthenia in: Principles ofNeurology (Adams, R. and Victor, M., Eds.), pp. 341–345,McGraw-Hill, New York.
[8] Llovera, M., Garcıa-Martınez, C., Agell, N., Lopez-Soriano, F.J.and Argiles, J.M. (1995) Muscle wasting associated with cancercachexia is linked to an important activation of the ATP-dependent ubiquitin-mediated proteolysis. Int. J. Cancer 61,138–141.
[9] Baccino, F.M., Tessitore, L., Bonelli, G. and Isidoro, C. (1986)Protein turnover states of tumor cells and host tissues in anexperimental model. Biomed. Biochim. Acta 45, 1585–1590.
[10] Tessitore, L., Bonelli, G. and Baccino, F.M. (1987) Earlydevelopment of protein metabolic perturbations in the liver andskeletal muscle of tumor-bearing rats. Biochem. J. 241, 153–159.
[11] Tessitore, L., Costelli, P., Bonetti, G. and Baccino, F.M. (1993)Cancer cachexia, malnutrition, and tissue protein turnover inexperimental animals. Arch. Biochem. Biophys. 306, 52–58.
[12] Llovera, M., Garcıa-Martınez, C., Agell, N., Marzabal, M.,Lopez-Soriano, F.J. and Argiles, J.M. (1994) Ubiquitin geneexpression is increased in skeletal muscle of tumor-bearing rats.FEBS Lett. 338, 311–318.
[13] Costelli, P., Llovera, M., Lopez-Soriano, J., Carbo, N., Tessitore,L., Lopez-Soriano, F.J., Baccino, F.M. and Argiles, J.M. (1995)Lack of effect of eicosapentaenoic acid in preventing cancercachexia and inhibiting tumor growth. Cancer Lett. 97, 25–32.
[14] Tessitore, L., Costelli, P. and Baccino, F.M. (1993) Humoralmediation for cancer cachexia in tumor-bearing rats. Br. J. Cancer67, 15–23.
[15] Van Royen, M., Carbo, N., Busquets, S., Alvarez, B., Quinn,L.S., Lopez-Soriano, F.J. and Argiles, J.M. (2000) DNA frag-mentation occurs in skeletal muscle during tumor growth: a linkwith cancer cachexia. Biochem. Biophys. Res. Commun. 270,533–537.
[16] Carbo, N., Busquets, S., Van Royen, M., Alvarez, B., Lopez-Soriano, F.J. and Argiles, J.M. (2002) TNF-alpha is involved inactivating DNA fragmentation in skeletal muscle. Br. J. Cancer86, 1012–1016.
[17] Buck, M. and Chojkier, M. (1996) Muscle wasting and dediffer-entiation induced by oxidative stress in a murine model ofcachexia is prevented by inhibitors of nitric oxide synthesis andantioxidants. EMBO J. 15, 1753–1765.
[18] Gomes-Marcondes, M.C. and Tisdale, M.J. (2002) Induction ofprotein catabolism and the ubiquitin–proteasome pathway bymild oxidative stress. Cancer Lett. 180, 69–74.
[19] Murr, C., Fuith, L.C., Widner, B., Wirleitner, B., Baier-Bitterlich,G. and Fuchs, D. (1999) Increased neopterin concentrations inpatients with cancer: indicator of oxidative stress. Anticancer Res.19, 1721–1728.
[20] Barreiro, E., Comtois, A.S., Mohammed, S., Lands, L. andHussain, S.N.A. (2002) Role of heme oxygenases in sepsis-induced diaphragmatic contractile dysfunction and oxidativestress. Am. J. Physiol. Lung Cell. Mol. Physiol. 283, L476–L484.
[21] Barreiro, E., Gea, J., Corominas, J.M. and Hussain, S.N.A.(2003) Nitric oxide synthases and protein oxidation in thequadriceps femoris muscles of COPD patients. Am. J. Respir.Cell. Mol. Biol. 29, 771–778.
[22] Beckman, J. and Koppenol, W. (1996) Nitric oxide, superoxide,and peroxynitrite: the good, the bad, and the ugly. Am. J. Physiol.271, C1424–C1437.
[23] van der Vliet, A., Eiserich, J.P., Shigenaga, M.K. and Cross, C.E.(1999) Reactive nitrogen species and tyrosine nitration in therespiratory tract. Am. J. Respir. Crit. Care Med. 160, 1–9.
[24] Gow, A.J., Farkouh, C.R., Munson, D.A., Posencheg, M.A. andIschiropoulos, H. (2004) Biological significance of nitric-oxidemediated protein modifications. Am. J. Physiol. 287, L262–L268.
[25] Barreiro, E., Comtois, A.S., Gea, J., Laubach, V.E. and Hussain,S.N.A. (2002) Protein tyrosine nitration in the ventilatorymuscles. Am. J. Respir. Cell. Mol. Biol. 26, 438–446.
[26] Levine, R.L., Williams, J.A., Stadtman, E.R. and Shacter, E.(1994) Carbonyl assays for determination of oxidatively modifiedproteins. Methods Enzymol. 233, 346–357.
[27] Friguet, B., Stadtman, E.R. and Szweda, L.I. (1994) Modificationof glucose-6-phosphate dehydrogenase by 4-hydroxy-2-nonenal.J. Biol. Chem. 269, 21639–21643.
[28] Requena, J.R., Fu, X., Ahmed, M.U., Jenkins, A.J., Lyons, T.J.and Thorpe, S.R. (1996) Lipoxidation products as biomarkers ofoxidative damage to proteins during lipid peroxidation reactions.Nephrol. Dial. Transplant. 11, 48–53.
[29] Chevari, S., Andial, T., Benke, K. and Shtrenger, I.A. (1992) Freeradical reactions and cancer. Vopr. Med. Khim. 38, 4–5.
[30] Burchman, P. and Kuhan, Y.T. (1997) Diminished susceptibilityto proteolysis after protein modification by the lipid peroxidationproduct malondialdehyde: inhibitory role for crosslinked andnoncrosslinked adducted proteins. Arch. Biochem. Biphys. 340,331–337.
[31] Esterbauer, H., Schaur, R.J. and Zollner, H. (1991) Chemistryand biochemistry of 4 hydroxynonenal, malondialdehyde andrelated aldehydes. Free Radic. Biol. Med. 11, 81–128.
[32] Gutteridge, J.M.C. (1995) Lipid peroxidation and antioxidants asbiomarkers of tissue damage. Clin. Chem. 41, 1819–1828.
[33] Reid, M.B. and Li, Y.P. (2001) Cytokines and oxidative signalingin skeletal muscle. Acta Physiol. Scand. 171, 225–232.
[34] Cervera, J. and Levine, R.L. (1988) Modulation of the hydro-phobicity of glutamine synthetase by mixed-function oxidation.FASEB J. 2, 2591–2595.
[35] Davies, K.J. (1987) Protein damage and degradation by oxygenradicals. I. General aspects. J. Biol. Chem. 262, 9895–9901.
[36] Reid, M.B., Haack, K.E., Franchek, K.M., Valberg, P.A.,Kobzik, L. and West, M.S. (1992) Reactive oxygen in skeletalmuscle: I. Intracellular oxidant kinetics and fatigue in vitro. J.Appl. Physiol. 73, 1797–1804.
[37] MacMillan-Crow, L.A., Crow, J.P. and Thompson, J.A. (1998)Peroxynitrite-mediated inactivation of manganese superoxidedismutase involves nitration and oxidation of critical tyrosineresidues. Biochemistry 37, 1613–1622.
[38] MacMillan-Crow, L.A. and Thompson, J.A. (1999) Tyrosinemodifications and inactivation of active site manganese superox-ide dismutase mutant (Y34F) by peroxynitrite. Arch. Biochem.Biophys. 366, 82–88.
[39] MacMillan-Crow, L.A. and Cruthirds, D.L. (2001) Invitedreview: manganese superoxide dismutase in disease. Free Radic.Res. 34, 325–336.
[40] Souza, J., Choi, I., Chen, Q., Weisse, M., Daikhin, E., Yudkoff,M., Obin, M., Ara, J., Horwitz, J. and Ischiropoulos, H. (2000)Proteolytic degradation of tyrosine nitrated proteins. Arch.Biochem. Biophys. 380, 360–366.




















