Binding of chloroquine–conjugated gold nanoparticles with bovine serum albumin

Author’s Accepted Manuscript
Biophysical and molecular docking insight into theinteraction of cytosine β-D Arabinofuranoside withHuman serum albumin
Parvez Alam, Sumit Kumar Chaturvedi, TamannaAnwar, Mohammad Khursheed Siddiqui, MohdRehan Ajmal, Gamal Badr, Mohamed H.Mahmoud, Rizwan Hasan Khan
PII: S0022-2313(15)00147-7DOI: http://dx.doi.org/10.1016/j.jlumin.2015.03.011Reference: LUMIN13252
To appear in: Journal of Luminescence
Received date: 4 December 2014Revised date: 27 February 2015Accepted date: 11 March 2015
Cite this article as: Parvez Alam, Sumit Kumar Chaturvedi, Tamanna Anwar,Mohammad Khursheed Siddiqui, Mohd Rehan Ajmal, Gamal Badr, MohamedH. Mahmoud and Rizwan Hasan Khan, Biophysical and molecular dockinginsight into the interaction of cytosine β-D Arabinofuranoside with Human seruma l b u m i n , Journal of Luminescence,http://dx.doi.org/10.1016/j.jlumin.2015.03.011
This is a PDF file of an unedited manuscript that has been accepted forpublication. As a service to our customers we are providing this early version ofthe manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting galley proof before it is published in its final citable form.Please note that during the production process errors may be discovered whichcould affect the content, and all legal disclaimers that apply to the journal pertain.
www.elsevier.com/locate/jlumin

1
Biophysical and Molecular Docking Insight into the Interaction of Cytosine β-
D Arabinofuranoside with Human Serum Albumin
Parvez Alam1, Sumit Kumar Chaturvedi
1, Tamanna Anwar
2, Mohammad Khursheed Siddiqui
1,
Mohd Rehan Ajmal1 Gamal Badr
3 and Mohamed H. Mahmoud
4 and Rizwan Hasan Khan
1*
1Interdisciplinary Biotechnology Unit, Aligarh Muslim University, Aligarh-202002, India
2Center of Bioinformatics Research and Technology, Aligarh-202002, India
3Laboratory of Immunology & Molecular Biology, Zoology Department, Faculty of Science,
Assiut University, 71516 Assiut, Egypt
4 Food Science and Nutrition Department, National Research Center, Dokki, Cairo, Egypt
*To whom correspondence should be addressed
Prof. Rizwan Hasan Khan Interdisciplinary Biotechnology Unit,
Aligarh Muslim University,
Aligarh-202002, U.P., India.
E-mail:[email protected],
Phone: 91-571-2720388
Fax: + 91-571-2721776

2
Abstract
Interaction of pharmacologically important anticancer drug Cytosine β-D Arabinofuranoside
with Human serum albumin (HSA) at physiological pH 7.4 has been studied by utilizing various
spectroscopic and molecular docking strategies. Fluorescence results revealed that Cytosine β-D
Arabinofuranoside interacts with HSA through static quenching mechanism with binding affinity
of 2.4⨉103
M-1
. The average binding distance between drug and Trp214
of HSA was found to be
2.23 nm on the basis of the theory of Förster’s energy transfer. Synchronous fluorescence data
indicated that interaction of drug with HSA changed the microenvironment around the
tryptophan residue. UV-visible spectroscopy and circular dichroism results deciphered the
complex formation and conformational alterations in the HSA respectively. Dynamic light
scattering was utilized to understand the topology of protein in absence and presence of drug.
Thermodynamic parameters obtained from Isothermal titration calorimetry (ΔH= -26.01 KJmol-1
and TΔS= -6.5 KJmol-1
) suggested the involvement of van der Waal interaction and hydrogen
bonding. Molecular docking and displacement study with site specific markers suggested that
Cytosine β-D Arabinofuranoside binds to subdomain IB of HSA which is also known as the
hemin binding site. This study will be helpful to understand the binding mechanism of Cytosine
β-D Arabinofuranoside with HSA and associated alterations.
Key words: Serum albumin, Binding, Fluorescence quenching, Molecular docking.
Abbreviations: HSA- human serum albumin, CBDA- cytosine β-D arabinofuranoside, ITC-
isothermal titration calorimetry

3
1. Introduction
Drug-protein interaction is a crucial pharmacological phenomenon that affects structure,
physiological action of carrier protein, distribution and elimination of drugs.[1; 2] Therapeutic
efficacy of drug conjointly depends upon the degree of binding affinity with plasma proteins.
Pharmacokinetic behavior of drug depends on its freely and bound form present in body .[3]
HSA serves as a transport vehicle for several endogenous compounds including fatty acids,
hemin, bilirubin, and tryptophan. Furthermore, distribution, concentration and the metabolism of
various exogenous drugs also depends upon their affinity with serum albumins.[3; 4; 5] Apart
from the roles depicted above, HSA is vital in maintaining osmolarity in plasma and intestinal
fluids and exhibits many enzymatic and antioxidant functions.[6; 7]
Human serum albumin (HSA) is most abundant, single chain, multifunctional, non-glycosylated
and negatively charged protein of molecular weight 66.5 kDa.[8] X ray crystallography revealed
that HSA is globular heart shaped protein consists of three domains viz. domain I, II and III
which are further subdivided into A and B sub domains.[9; 10] Moreover, binding and molecular
docking study depict that HSA binds to large number of small molecules due to diverse binding
sites distributed over the surface of the protein. The two best characterized binding sites for
ligands are located in hydrophobic pocket of sub domain II A and III A which corresponds to
Sudlow site I and Sudlow site II.[11; 12]
Cytosine β-D Arabinofuranoside (CBDA) is also known as Ara-C (Arabinofuranosyl Cytidine)
consists of cytosine base with arabinose sugar as shown in Fig. 1. It is used as chemotherapeutic
agent such as in treatment of acute myeloid leukemia, non-hodgkin lymphoma, acute
lymphocytic leukemia and lymphomas.[13; 14; 15] Antiviral activity of CBDA was utilized for

4
prevention of herpes virus infection [16]. It interferes with DNA synthesis and induces cancerous
cell death. CBDA is among first series of cancer drugs that altered the sugar component albeit
other drugs modify the base. It gets transformed into cytosine arabinoside triphosphate which
causes DNA damage when cells are in S phase of mitosis therefore rapidly dividing cells are
most affected. Moreover, it also inhibits DNA polymerase, RNA polymerase and Nucleotide
reductase needed for DNA synthesis[17]. Previously, our group has reported the binding
mechanism of various detergents, drugs, toxins, pollutants to serum albumins. [18; 19; 20; 21;
22; 23]
In the present study, binding of HSA with CBDA was thoroughly studied by utilizing Isothermal
Titration Calorimetry (ITC) and various spectroscopic techniques. The topology of HSA in
absence and presence of CBDA was studied by using dynamic light scattering measurements.
Moreover, displacement experiment with site specific markers and molecular docking study
have also been performed to reveal the location of CBDA binding site on HSA as well as forces
involved in the binding interaction.
2. Materials and methods
Human serum albumin (A1887), Cytosine β-D Arabinofuranoside (1768), warfarin (A2250),
hemin (H5533) and diazepam (D0406) were purchased from Sigma Aldrich, India. All of the
other reagents were of analytical grade.
2. 1 Sample preparation
HSA (100 µM) stock solution was prepared in 20 mM phosphate buffer (pH 7.4). Protein
concentration was determined spectrophotometrically using of 5.30 at 280 nm by using
Perkin-Elmer Lambda double beam UV-visible spectrophotometer.[24; 25] Drug solution (5

5
mM) was prepared in 20 mM phosphate buffer (pH 7.4) and diluted to various concentrations in
the same buffer. All experiments were carried out in 20 mM phosphate buffer (pH 7.4) at 298K.
2.2 pH determination
pH measurements were carried out on Mettler Toledo pH meter (Seven Easy S20–K) using
Expert “Pro3 in 1” type electrode. The least count of the pH meter was 0.01 pH unit.
2.3 Steady state fluorescence quenching measurement
Fluorescence measurements were performed on a Shimadzu fluorescence spectrophotometer,
model RF-5301. The fluorescence spectra were measured at 25 ± 0.1 °C with a 1 cm path length
cell. Both excitation and emission slits were set at 5 nm. Intrinsic fluorescence was measured by
exciting the protein solution at 295 nm and emission spectra were recorded in the range of 300-
450 nm. The titration of CBDA (0-115 μM) to HSA (5 μM) solution was carried out at 298 K.
The fluorescence intensity at 340 nm (tryptophan) was used to calculate the binding constant
(Kb).
2.4 Synchronous fluorescence spectroscopy
Synchronous fluorescence spectroscopy was carried out by simultaneously scanning the
excitation and emission mono-chromators. Conformational alteration around the tyrosine and
tryptophan residue of HSA can be marked when the wavelength interval (Δλ) was 15 and 60 nm
respectively. The concentration of HSA was 5 μM and CBDA varied in the range (0-115 μM).
All parameters were kept same as discussed in above section

6
2.5 Drug displacement experiment
Different site specific markers, warfarin for (site I), diazepam for (site II) and hemin for
(subdomain IB), were used for performing displacement experiments.[26; 27; 28] Titration of
CBDA was carried out with the solution having protein and site marker in the molar ratio of 1:1.
The fluorescence emission spectra were recorded in a similar way as mentioned in fluorescence
measurements and the Ksv values of drug–protein–marker were evaluated using Stern-Volmer
equation.
2.6 UV-visible absorption measurements
Absorbance measurements were carried out on Perkin-Elmer Lambda 25 double beam UV-
visible spectrophotometer attached with Peltier temperature programmer 1 (PTP-1). Cuvette of
cell length of 10 mm and scanning speed 1000 nm/min was used. The concentration of protein
was kept at 12 μM and 1:0, 1:10 and 1:20 molar ratio of HSA to CBDA were used.
2.7 Circular dichroism measurements
Far-UV CD measurements (190-250 nm) were carried out on JASCO-J 813 spcetro polarimeter
equipped with a Peltier-type temperature controller at 298 K using a quartz cell of path length of
0.1 cm. Three scans were accumulated at a scan speed of 100 nm min-1
. 1:0, 1:10 and 1:20 molar
ratio of HSA-CBDA were taken. The percent α-helical and β-sheet contents of the protein were
calculated by using online K2D3 software [29] using the CD values from 200-250 nm as
extracted from the CD spectra.

7
2.8 Dynamic light scattering (DLS) measurements
DLS measurements were carried out at 830 nm using DynaPro-TC-04 dynamic light scattering
equipment (Protein Solutions, Wyatt Technology, Santa Barbara, CA) equipped with a
temperature- controlled micro sampler. HSA (2 mg/ml) was incubated with CBDA for 12 hours.
The samples were spun at 10,000 rpm for 10 min and were filtered serially through 0.22 and 0.02
µM Whatman syringe filters directly into a 12 µl quartz cuvette. For each experiment, 20
measurements were carried out. Mean hydrodynamic radius (Rh) and polydispersity were
analyzed using Dynamics 6.10.0.10 software at optimized resolution. The Rh was estimated on
the basis of an autocorrelation analysis of scattered light intensity data based on translationa
diffusion coefficient by Stokes- Einstein relationship:
D
kTR
6h (1)
Where Rh is the hydrodynamic radius, k is Boltzmann constant, T is temperature, η is the
viscosity of water and D is diffusion coefficient.
2.9 Isothermal titration calorimetry
Isothermal titration calorimetry allows the measurement of the magnitude of the binding affinity,
and the two contributing thermodynamic terms: enthalpy (ΔH⁰) and entropy (ΔS⁰) changes.
Binding of CBDA to HSA was studied by ITC (VP-ITC (Microcal Inc., Northampton, MA) at 25
°C. Prior to the titration experiment, all solutions were degassed properly on a thermovac. The
sample cell (approximately 1.44 mL) was loaded with HSA (25 µM) and CBDA (1 mM) was
injected into the reaction cell. The titration cell was stirred continuously at 307 rpm, which
ensured rapid mixing but did not cause foaming in the protein solution. Titrations were

8
performed to ensure full occupancy of the binding sites and until the titration signal was
constant. The calorimetric data were analyzed using the MicroCal Origin 7.0 software provided
with the instrument. The enthalpy change for each injection was calculated by integrating the
area under the peaks and then subtracted with control titrations.
The other thermodynamic parameters were calculated according to the equation
∆G⁰ = -RTlnKa = ∆H⁰-T∆S⁰ (2)
where T is the absolute temperature (298 K) and R, is universal gas constant (8.3151 J mol-1
K-1
).
2.10 Molecular docking
The molecular docking study was performed using Autodock 4.2 and Autodock tools (ADT)
using Lamarckian genetic algorithm.[30] The crystal structure of HSA (PDB id: 1AO6) and three
dimensional structure of CBDA (CID: 6253) was obtained from Brookhaven Protein Data Bank
and PubChem respectively. Chain A of the protein was selected water molecules and ions were
removed and all hydrogen atoms were added. Then partial Kollman charges were assigned to
HSA. The protein was set to be rigid and there is no consideration of solvent molecules on
docking. The grid size was set to be 126, 126 and 126 along X, Y and Z axes with 0.564 Å grid
spacing. Autodock parameters used were GA population size: 150 and maximum number of
energy evolutions: 2,50,0000. 10 best solution based on docking score was retained for further
analysis, Discovery studio 3.5 were used for visualization and for the identification of residues
involved in binding. LIGPLOT program was used to analyse amino acid residues involved in the
binding between HSA and CBDA.[31]

9
3. Results and Discussion
3.1 Steady state fluorescence quenching measurement
Fluorescence spectroscopy is a valuable tool to study Protein-ligand interactions. Protein
containing aromatic amino acid residues (tyrosine, phenylalanine and tryptophan) has intrinsic
fluorescence, however among them the contribution of tryptophan is maximum.[22] In
fluorescence quenching experiment titration of drug with protein is carried out to get detailed
information of related binding parameters.[32]
The fluorescence intensity of HSA decreases with gradual addition of CBDA at 298 K as shown
in (Fig. 1A). The maximum emission wavelength of HSA was found at 340 nm and the value
decreases regularly upon increasing concentration of CBDA, indicating that interaction between
drug and HSA occurred. These results are also in accord with the previous reports.[12; 33]
The fluorescence quenching data were analyzed by Stern-Volmer equation (equation 2).[34]
F₀/F = Ksv+1 = kqτ₀ [Q] +1 (3)
where F0 and F are the steady state fluorescence intensities in the absence and presence of
quencher, respectively, KSV is the Stern-Volmer quenching constant and Q is the concentration of
quencher, kq is the bimolecular quenching constant and τ₀ is the life time of tryptophan in the
absence of quencher. Life time of tryptophan for HSA (τ₀) is 5.6 ns.[35] Stern Volmer plot is
shown in Fig. 1B. In order to establish the mechanism of quenching, kq was calculated by using
following equation
(4)

10
In static quenching, quencher physically interact with the excited molecule through chemical
bonds. However, in dynamic quenching, quencher indirectly interact with the molecule. The
bimolecular quenching constant for CBDA calculated to be 1 ⨉ 1011
M-1
s-1
which is greater than
maximum collision quenching constant 2 ⨉ 1010
M-1
s-1
.[36]
This suggests the formation of HSA-CBDA complex occurred through static quenching manner.
Value of binding constant and number of binding sites were obtained from the modified Stern-
Volmer plot as shown in Fig. 1C by using the following equation
Log (F₀/F -1) = logKb + n log [Q] (5)
where F0 and F are the steady state fluorescence intensities in the absence and presence of
quencher, respectively, Kb is binding constant, n is the number of binding sites and Q is the
concentration of quencher. Binding parameters are summarizes in Table 1, the value of binding
constant was found to be in the order of 103
which suggest that binding of CBDA binds to HSA
in static manner. [37]
3.2 Synchronous fluorescence measurements
Synchronous fluorescence spectroscopy provides information about the micro environmental
changes around tyrosine and tryptophan in HSA upon ligand binding. D-value (Δλ) between
excitation and emission wavelength was set at 15 and 60 nm for Tyr and Trp residues
respectively. [38] Change in the polarity around the micro environment of fluorophore is inferred
from the changes in maximum fluorescence emission peak (λ em max). Blue shift and red shift in
λem max signifies enhancement in hydrophobicity and polar environment around the fluorophore
(Tyr and Trp) respectively[39]. As can be inferred from the Fig. 2A, almost no red shift in
emission maximum of HSA was observed in presence of increasing concentration of CBDA
A B
C
A B C

11
when Δλ=15 nm, suggesting that the micro environment around the Tyr residues did not altered
significantly. On the other hand when Δλ=60 nm (Fig. 2B), substantial red shift (~10 nm) was
observed in emission maximum of HSA, inferring that microenvironment around the Trp residue
get significantly altered. CBDA induced conformational changes indicated microenvironment
around the Trp residue (Trp214
) get significantly perturbed and CBDA-HSA complex formation
has occured.[40] This implies that CBDA cause more conformational change near tryptophan
residue than tyrosine. Moreover the fluorescence intensity decreases regularly on addition of
CBDA in both systems which further support the occurrence of fluorescence quenching (Fig 2A
and B)
3.3 Drug displacement experiments
Majority of the drugs bind to HSA at two primary sites known as Sudlow site I and II but few
drugs and fatty acids also binds at subdomain IB of HSA. In order to determine binding site of
CBDA on HSA, competitive binding experiment with site specific markers have been carried
out. Warfarin for site I, Diazepam for site II and Hemin for the subdomain IB were used. To
trace the binding site of CBDA on HSA, KSV values were calculated by using the emitted
fluorescence intensity data in the absence and presence of markers by Stern-Volmer equation.
There was no significant change in the KSV value was observed with warfarin and diazepam
although with hemin KSV value gets markedly reduced (supplementary table 1). This difference in
KSV is sufficient enough to locate binding site on HSA as reported in earlier literature.[41] It is
evident from the displacement experiments that CBDA binds at subdomain IB on HSA.

12
3.4 Fluorescence resonance energy transfer (FRET)
The efficiency of energy transfer can be used to evaluate the distance between the ligand and
fluorophore in the protein. According to Förster’s energy transfer theory the distance between
donor (Trp214
of HSA) and acceptor (CBDA), r and the efficiency of energy transfer, E can be
calculated by following equation.
(6)
where F and F○ are the fluorescence intensities of HSA in the absence and presence of CBDA,
R0 is the Förster critical distance at which the efficiency of energy transfer is 50%, R0 can be
determined by using the following equation.
(7)
where K2
is the factor related to the geometry of donor and acceptor of diploes, n is the refractive
index of the medium, is the fluorescence quantum yield of the donor in the absence of the
acceptor and J is the overlap integral of donor fluorescence emission and the acceptor
absorption, which can be calculated by using following equation.
J =∫
∫
(8)
where F (λ) is the fluorescence intensity of the donor at wavelength λ and (λ) is the molar
extinction coefficient of acceptor at wavelength λ. For HSA, K2, and n were taken as 2/3, 0.118
and 1.33, respectively.[42]

13
The FRET results are summarized in Table 2. The spectral overlap between the absorption
spectrum of acceptor (CBDA) and the fluorescence emission spectrum of donor (HSA) is shown
in Fig. 3. The binding distance (r) for the present system is 2.23 nm. Furthermore, we found that
0.5Ro<r< 1.5Ro, indicating the existence of static quenching due to complex formation between
CBDA and HSA.
3.5 UV-visible absorption spectroscopy
UV-visible spectroscopy is simple and valuable tool to study drug-protein interaction. The
absorption peak around 280 nm because of cumulative absorption of three aromatic amino acid
residues. The formation of CBDA-HSA complex is evident from UV-visible spectra
(supplementary Fig. S1). The progressive increase of absorbance along with blue shift on
increasing drug concentration governs the formation of drug-protein complex. Change in λmax
indicated the change in polarity around the tryptophan residue and hence indicates binding
between CBDA and HSA has led to the change in the conformation of protein. [43]
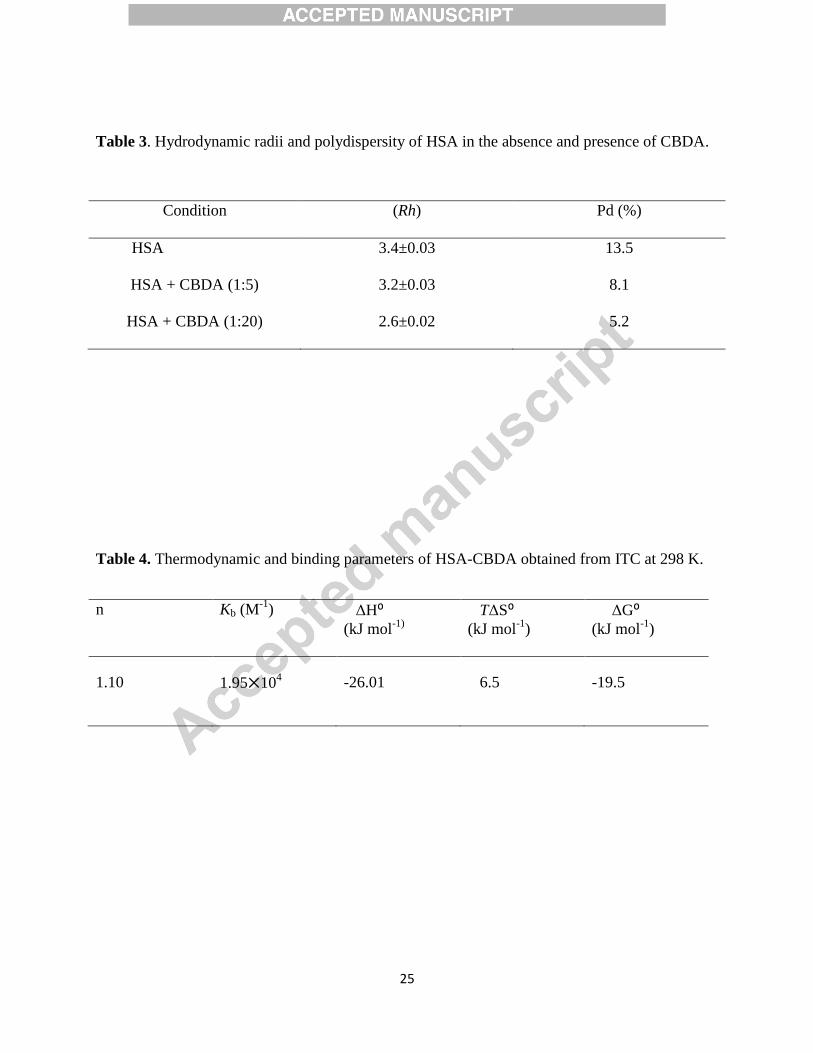
3.6 Circular dichroism analysis
CD spectroscopy is valuable tool to study secondary structure of proteins and nucleic acids.[44;
45; 46; 47; 48]
The CD experiments were carried out in order to get insight into the conformational change of
HSA induced by CBDA. Binding of ligands to globular protein modulate the intermolecular
forces liable for sustaining the secondary and tertiary structures resulting in conformational
alteration of the protein. The CD spectra of serum albumins exhibited two negative minimal
peaks in UV-region at 208 and 222 nm, characteristics of α-helical structure of the protein.[18;
49]

14
Fig. 4 shows the CD spectra of HSA and HSA-CBDA. HSA exhibits two negative bands at 208
nm and 222 nm characteristic of alpha helical structure[50]. An increase in the negative CD
value without any change in the characteristic pattern of HSA was observed with gradual
increase in CBDA concentration which suggests the increment in alpha helical content in HSA
(55.80 % to 57.92% calculated from K2D3 software). From CD results it can be interpreted that
CBDA leads to stabilize the secondary structure of HSA.
3.7 Dynamic light scattering study
Dynamic light scattering was employed to determine the hydrodynamic radii of native HSA and
CBDA- HSA complex. The hydrodynamic radii of native HSA and in presence of different
molar ratios of CBDA was calculated, data represented in Fig. 5 and Table 3. The lower value of
polydispersity (5-15) indicate the presence of homogenous solution. The Rh of native HSA was
found to be 3.4 nm. [51] The Rh value of HSA starts decreasing on increasing the concentration
of CBDA. This reduction in Rh value may attributed to collapsing of protein as CBDA binds with
HSA. This response may result in a decrease in the molecular volume due to a conformational
change.[49] As CD data suggested, CBDA affects the secondary structure of HSA. This also
might be a reason that conformationally altered secondary structural components tends to
decrease hydrodynamic radius.
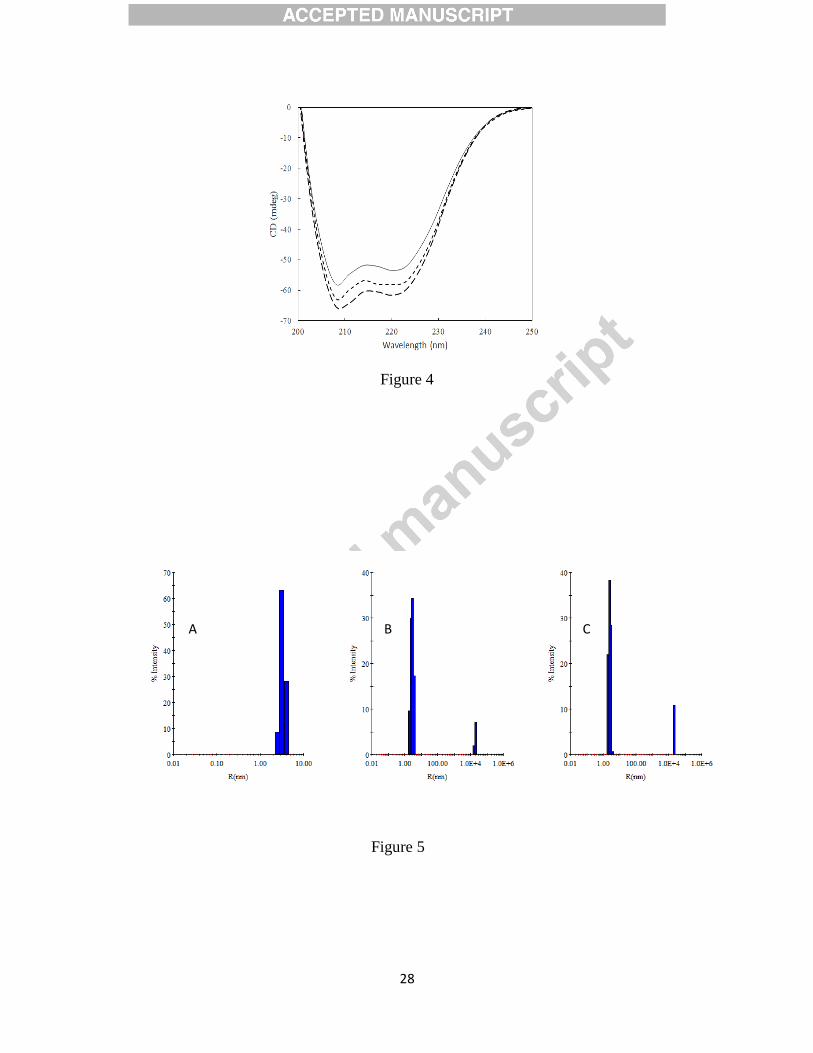
3.8 Isothermal titration calorimetry
Isothermal Titration Calorimetry was performed to determine the thermodynamic parameters for
CBDA-HSA interaction. ITC complement the fluorescence studies for the ligands which interact

15
with weak interacting forces with transport proteins as it can be carried out at higher
concentration of both protein and ligand.[52] The final data were best fitted to one binding site
model and the results are presented in Fig. 6 and Table 4. Each peak in the binding isotherms
(Fig. 6 upper panel) represents a single injection of CBDA. The negative deflection from the
baseline on addition of CBDA indicates that process is exothermic. We found that CBDA has
one binding site with affinity of order 104.
The data revealed that the binding of CBDA to HSA
shows a favorable entropical change (0<ΔS). Negative value of ΔH and positive value of ΔS
endorse the involvement of van der Waal interaction and hydrogen bonding between drug and
protein as reported earlier.[33] Moreover negative value of Gibbs’s free energy (ΔG)
demonstrates that the binding process was spontaneous. Overall the binding process was
spontaneous, enthalpy driven and entropically opposed. The differences in fluorescence and ITC
results could be ascertained to the following reason, fluorescence measures local change only
around the Trp residue whereas ITC measures global change.[53; 54]
3.9 Molecular docking study of CBDA-HSA interaction
Molecular docking has been employed to further understand the interaction of CBDA with HSA
and to confirm the binding site of CBDA on HSA. The principle regions of ligand binding on
HSA are located in hydrophobic cavities in subdomain II and III which correspond to site I and
site II respectively. Autodock 4.2 program was used to examine the binding mode of CBDA to
HSA. The docking results showed that CBDA binds within the binding pocket of subdomain IB
(Fig. 7A, B and C). It is apparent from Fig. 7 B that CBDA got insert into the hydrophobic cavity
of the protein, which indicated that the binding location was subdomain IB and this result are
also in consistent with our displacement experiments with site specific markers.

16
The CBDA-HSA complex is stabilized by hydrogen bonds between the drug and the Tyr138
,
Leu812
and Leu185
amino acid residues of the protein with bond lengths of 2.94, 2.83 and 2.92 Å,
respectively. Further, CBDA-HSA complex is also stabilized by hydrophobic interaction
between the Tyr161
, Met123
, Ile142
, and Gly189
amino acid residues of the protein and drug (Table
5). Binding energy calculated by molecular docking was -20.46 KJmol-1
which is very close to
the binding energy obtained from fluorescence and ITC experiments and further support the
spontaneity of reaction.
4. Conclusion
In view of all results it can be concluded that CBDA interacts with HSA through static
quenching mechanism and induce conformation alterations in the protein. The binding process is
spontaneous, exothermic and entropically favoured. Displacement study (with specific site
markers) and molecular docking revealed that CBDA binds at subdomain IB on HSA apart from
major drug binding site (site I and II). Hydrogen bonding and hydrophobic interactions are the
major forces in stabilizing the drug protein complex. Our study provides detailed insight to
understand the mode of interaction of other related drugs that interacts with subdomain IB.
Acknowledgements
Facilities provided by IBU, Aligarh Muslim University, Aligarh are gratefully acknowledged. PA
and SKC are gratify to Council of Scientific and Industrial Research, New Delhi for
providing fellowship in the form of JRF and SRF respectively.

17
References
[1] Y.-J. Hu, Y. Liu, T.-Q. Sun, A.-M. Bai, J.-Q. Lü, and Z.-B. Pi, Binding of anti-inflammatory drug
cromolyn sodium to bovine serum albumin. International journal of biological macromolecules
39 (2006) 280-285.
[2] S. Ranjbar, Y. Shokoohinia, S. Ghobadi, N. Bijari, S. Gholamzadeh, N. Moradi, M.R. Ashrafi-Kooshk, A.
Aghaei, and R. Khodarahmi, Studies of the interaction between isoimperatorin and human
serum albumin by multispectroscopic method: identification of possible binding site of the
compound using esterase activity of the protein. ScientificWorldJournal (2013) 305081.
[3] S. Curry, P. Brick, and N.P. Franks, Fatty acid binding to human serum albumin: new insights from
crystallographic studies. Biochim Biophys Acta 1441 (1999) 131-40.
[4] I. Petitpas, C.E. Petersen, C.E. Ha, A.A. Bhattacharya, P.A. Zunszain, J. Ghuman, N.V. Bhagavan, and S.
Curry, Structural basis of albumin-thyroxine interactions and familial dysalbuminemic
hyperthyroxinemia. Proc Natl Acad Sci U S A 100 (2003) 6440-5.
[5] N.A. Kratochwil, W. Huber, F. Muller, M. Kansy, and P.R. Gerber, Predicting plasma protein binding of
drugs: a new approach. Biochem Pharmacol 64 (2002) 1355-74.
[6] A. Salvi, P.A. Carrupt, J.M. Mayer, and B. Testa, Esterase-like activity of human serum albumin toward
prodrug esters of nicotinic acid. Drug Metab Dispos 25 (1997) 395-8.
[7] U. Kragh-Hansen, V.T. Chuang, and M. Otagiri, Practical aspects of the ligand-binding and enzymatic
properties of human serum albumin. Biol Pharm Bull 25 (2002) 695-704.
[8] T. Peters Jr, All about albumin: biochemistry, genetics, and medical applications, Academic press,
1995.
[9] D.C. Carter, and X.M. He, Structure of human serum albumin. Science 249 (1990) 302-3.
[10] D.C. Carter, and J.X. Ho, Structure of serum albumin. Adv Protein Chem 45 (1994) 153-203.

18
[11] G. Sudlow, D.J. Birkett, and D.N. Wade, The characterization of two specific drug binding sites on
human serum albumin. Mol Pharmacol 11 (1975) 824-32.
[12] N. Zaidi, M.R. Ajmal, G. Rabbani, E. Ahmad, and R.H. Khan, A comprehensive insight into binding of
hippuric acid to human serum albumin: a study to uncover its impaired elimination through
hemodialysis. PLoS One 8 (2013) e71422.
[13] W.S. Wang, C.H. Tzeng, T.J. Chiou, J.H. Liu, R.K. Hsieh, C.C. Yen, and P.M. Chen, High-dose cytarabine
and mitoxantrone as salvage therapy for refractory non-Hodgkin's lymphoma. Jpn J Clin Oncol
27 (1997) 154-7.
[14] H. Ogbomo, M. Michaelis, D. Klassert, H.W. Doerr, and J. Cinatl, Jr., Resistance to cytarabine induces
the up-regulation of NKG2D ligands and enhances natural killer cell lysis of leukemic cells.
Neoplasia 10 (2008) 1402-10.
[15] A. Pigneux, V. Perreau, E. Jourdan, N. Vey, N. Dastugue, F. Huguet, J.J. Sotto, L.R. Salmi, N. Ifrah, and
J. Reiffers, Adding lomustine to idarubicin and cytarabine for induction chemotherapy in older
patients with acute myeloid leukemia: the BGMT 95 trial results. Haematologica 92 (2007) 1327-
34.
[16] C.B. Lauter, E.J. Bailey, and A.M. Lerner, Assessment of cytosine arabinoside as an antiviral agent in
humans. Antimicrob Agents Chemother 6 (1974) 598-602.
[17] M.C. Perry, The chemotherapy source book, Lippincott Williams & Wilkins, 2008.
[18] B. Ahmad, S. Parveen, and R.H. Khan, Effect of albumin conformation on the binding of ciprofloxacin
to human serum albumin: a novel approach directly assigning binding site. Biomacromolecules 7
(2006) 1350-6.
[19] A. Varshney, M. Rehan, N. Subbarao, G. Rabbani, and R.H. Khan, Elimination of endogenous toxin,
creatinine from blood plasma depends on albumin conformation: site specific uremic toxicity &
impaired drug binding. PLoS One 6 (2011) e17230.

19
[20] E. Ahmad, G. Rabbani, N. Zaidi, B. Ahmad, and R.H. Khan, Pollutant-induced modulation in
conformation and beta-lactamase activity of human serum albumin. PLoS One 7 (2012) e38372.
[21] A.B. Khan, J.M. Khan, M.S. Ali, R.H. Khan, and D. Kabir-ud, Spectroscopic approach of the interaction
study of amphiphilic drugs with the serum albumins. Colloids Surf B Biointerfaces 87 (2011) 447-
53.
[22] A. Varshney, Y. Ansari, N. Zaidi, E. Ahmad, G. Badr, P. Alam, and R.H. Khan, Analysis of binding
interaction between antibacterial ciprofloxacin and human serum albumin by spectroscopic
techniques. Cell Biochem Biophys 70 (2014) 93-101.
[23] N. Zaidi, E. Ahmad, M. Rehan, G. Rabbani, M.R. Ajmal, Y. Zaidi, N. Subbarao, and R.H. Khan,
Biophysical insight into furosemide binding to human serum albumin: a study to unveil its
impaired albumin binding in uremia. J Phys Chem B 117 (2103) 2595-604.
[24] K. Wallevik, Reversible denaturation of human serum albumin by pH, temperature, and guanidine
hydrochloride followed by optical rotation. J Biol Chem 248 (1973) 2650-5.
[25] O.H. Lowry, N.J. Rosebrough, A.L. Farr, and R.J. Randall, Protein measurement with the Folin phenol
reagent. J Biol Chem 193 (1951) 265-75.
[26] J. Ghuman, P.A. Zunszain, I. Petitpas, A.A. Bhattacharya, M. Otagiri, and S. Curry, Structural basis of
the drug-binding specificity of human serum albumin. Journal of molecular biology 353 (2005)
38-52.
[27] K. Yamasaki, T. Maruyama, U. Kragh-Hansen, and M. Otagiri, Characterization of site I on human
serum albumin: concept about the structure of a drug binding site. Biochim Biophys Acta 1295
(1996) 147-57.
[28] F. Zsila, Circular dichroism spectroscopic detection of ligand binding induced subdomain IB specific
structural adjustment of human serum albumin. J Phys Chem B 117 (2013) 10798-806.

20
[29] C. Louis-Jeune, M.A. Andrade-Navarro, and C. Perez-Iratxeta, Prediction of protein secondary
structure from circular dichroism using theoretically derived spectra. Proteins: Structure,
Function, and Bioinformatics 80 (2011) 374-381.
[30] D.S. Goodsell, G.M. Morris, and A.J. Olson, Automated docking of flexible ligands: applications of
AutoDock. J Mol Recognit 9 (1996) 1-5.
[31] A.C. Wallace, R.A. Laskowski, and J.M. Thornton, LIGPLOT: a program to generate schematic
diagrams of protein-ligand interactions. Protein Eng 8 (1995) 127-34.
[32] P. Bourassa, S. Dubeau, G.M. Maharvi, A.H. Fauq, T.J. Thomas, and H.A. Tajmir-Riahi, Binding of
antitumor tamoxifen and its metabolites 4-hydroxytamoxifen and endoxifen to human serum
albumin. Biochimie 93 (2011) 1089-101.
[33] T. Chatterjee, A. Pal, S. Dey, B.K. Chatterjee, and P. Chakrabarti, Interaction of virstatin with human
serum albumin: spectroscopic analysis and molecular modeling. PLoS One 7 (2012) e37468.
[34] J.R. Lackowicz, Principles of fluorescence spectroscopy. Plenum Press,(New York, 1983) Chapter 5
(1983) 111-150.
[35] D. Pushparaju Yeggoni, A. Rachamallu, M. Kallubai, and R. Subramanyam, Cytotoxicity and
Comparative Binding Mechanism of Piperine with Human Serum Albumin and α-1-Acid
Glycoprotein. Journal of Biomolecular Structure and Dynamics (2014) 1-53.
[36] M.-F. Zhang, Z.-Q. Xu, Y.-S. Ge, F.-L. Jiang, and Y. Liu, Binding of fullerol to human serum albumin:
spectroscopic and electrochemical approach. Journal of Photochemistry and Photobiology B:
Biology 108 (2012) 34-43.
[37] J. Min, X. Meng-Xia, Z. Dong, L. Yuan, L. Xiao-Yu, and C. Xing, Spectroscopic studies on the
interaction of cinnamic acid and its hydroxyl derivatives with human serum albumin. Journal of
Molecular Structure 692 (2004) 71-80.

21
[38] C. Sun, J. Yang, X. Wu, X. Huang, F. Wang, and S. Liu, Unfolding and refolding of bovine serum
albumin induced by cetylpyridinium bromide. Biophys J 88 (2005) 3518-24.
[39] E. Rahnama, M. Mahmoodian-Moghaddam, S. Khorsand-Ahmadi, M.R. Saberi, and J. Chamani,
Binding site identification of metformin to human serum albumin and glycated human serum
albumin by spectroscopic and molecular modeling techniques: a comparison study. J Biomol
Struct Dyn (2014).
[40] S.R. Feroz, S.B. Mohamad, N. Bujang, S.N. Malek, and S. Tayyab, Multispectroscopic and molecular
modeling approach to investigate the interaction of flavokawain B with human serum albumin. J
Agric Food Chem 60 (2012) 5899-908.
[41] Z. Chi, and R. Liu, Phenotypic characterization of the binding of tetracycline to human serum
albumin. Biomacromolecules 12 (2010) 203-209.
[42] L. Trnková, I. Boušová, V. Staňková, and J. Dršata, Study on the interaction of catechins with
human serum albumin using spectroscopic and electrophoretic techniques. Journal of Molecular
Structure 985 (2011) 243-250.
[43] M.-G. Wen, X.-B. Zhang, J.-N. Tian, S.-H. Ni, H.-D. Bian, Y.-L. Huang, and H. Liang, Binding interaction
of xanthoxylin with bovine serum albumin. Journal of solution chemistry 38 (2009) 391-401.
[44] M. Shakir, M.S. Khan, S.I. Al-Resayes, U. Baig, P. Alam, R.H. Khan, and M. Alam, In vitro DNA binding,
molecular docking and antimicrobial studies on a newly synthesized poly (o-
toluidine)–titanium dioxide nanocomposite. RSC Advances 4 (2014) 39174-39183.
[45] P. Kumar, B. Baidya, S.K. Chaturvedi, R.H. Khan, D. Manna, and B. Mondal, DNA binding and
nuclease activity of copper (II) complexes of tridentate ligands. Inorganica Chimica Acta 376
(2011) 264-270.
[46] C.-Z. Lin, M. Hu, A.-Z. Wu, and C.-C. Zhu, Investigation on the differences of four flavonoids with
similar structure binding to Human serum albumin. Journal of Pharmaceutical Analysis (2014).

22
[47] C. Hebia, L. Bekale, P. Chanphai, J. Agbebavi, and H.A. Tajmir-Riahi, Trypsin inhibitor complexes with
human and bovine serum albumins: TEM and spectroscopic analysis. Journal of Photochemistry
and Photobiology B: Biology 130 (2014) 254-259.
[48] J.M. Khan, A. Qadeer, E. Ahmad, R. Ashraf, B. Bhushan, S.K. Chaturvedi, G. Rabbani, and R.H. Khan,
Monomeric banana lectin at acidic pH overrules conformational stability of its native dimeric
form. PloS one 8 (2014) e62428.
[49] S.K. Chaturvedi, E. Ahmad, J.M. Khan, P. Alam, M. Ishtikhar, and R.H. Khan, Elucidating the
interaction of limonene with bovine serum albumin: a multi-technique approach. Molecular
BioSystems (2014).
[50] P. Alam, G. Rabbani, G. Badr, B.M. Badr, and R.H. Khan, The Surfactant-Induced Conformational and
Activity Alterations in Rhizopus niveus Lipase. Cell Biochemistry and Biophysics (2014) 1-8.
[51] J.M. Khan, S.K. Chaturvedi, and R.H. Khan, Elucidating the mode of action of urea on mammalian
serum albumins and protective effect of sodium dodecyl sulfate. Biochemical and biophysical
research communications 441 (2013) 681-688.
[52] P.D. Ross, and S. Subramanian, Thermodynamics of protein association reactions: forces
contributing to stability. Biochemistry 20 (1981) 3096-102.
[53] H. Watanabe, S. Tanase, K. Nakajou, T. Maruyama, U. Kragh-Hansen, and M. Otagiri, Role of Arg-410
and Tyr-411 in human serum albumin for ligand binding and esterase-like activity. Biochem. J
349 (2000) 813-819.
[54] A. Das, and G.S. Kumar, Binding studies of aristololactam-[small beta]-d-glucoside and daunomycin
to human serum albumin. RSC Advances 4 (2014) 33082-33090.

23
Legend to Figures
Figure 1: (A) CBDA-induced fluorescence quenching of HSA. The concentration of HSA was 5
μM and the concentration of CBDA was varied from 0-115 μM. The intrinsic fluorescence of the
protein was measured in 20 mM sodium phosphate buffer, pH 7.4 at 298 K upon excitation at
295 nm. (B) Stern-Volmer plot for HSA-CBDA interactions (C) Modified Stern-Volmer plot for
the HSA-CBDA interactions at 298 K.
Figure 2: Synchronous fluorescence spectra (A) Δλ 15 (B) Δλ 60, HSA (5 μM) in presence of
increasing concentration of CBDA (0-115 μM).
Figure 3: The spectral overlap of the fluorescence emission spectrum of HSA (curve a) with the
absorption spectrum of CBDA (curve b), CBDA: HSA = 1:1.
Figure 4: Far-UV CD spectra of HSA alone (─) or in complex and in presence of 1:5 (- -) and
1:20 (__
__
) molar ratio of HSA to CBDA.
Figure 5: Hydrodynamic radii pattern of HSA in the absence of CBDA (A) and presence of HSA
+ CBDA (1:5) (C) HSA + CBDA (1:20).
Figure 6: Isothermal titration calorimetry of CBDA-HSA interaction. In the top panel, the heat
released per unit time (μcal/sec) was plotted vs time (min) where each peak corresponds to the
injection of an aliquot of ligand. In the bottom panel, the heat of reaction per injection
(kcal/mole).
Figure 7: (A) Best conformation of CBDA docked to HSA (B) Cartoon representation of
residues involved in binding of CBDA to HSA (C) Ligplot representations of the interactions
between HSA and CBDA.

24
Table 1. Stern-Volmer quenching constants and binding parameters for HSA-CBDA interactions
at 298 K.
Temp
(K)
Ksv
(M-1
)
kq
(M-1
s-1
)
Kb
(M-1
)
n
ΔG⁰
(kJ mol-1
)
R2
298 1.9⨉103 1.9⨉10
11 2.4⨉10
3 1.02 -19.18 0.99
Table 2. (FRET) parameters obtained from HSA-CBDA binding.
J (M-1
cm-1
nm4) R0 (nm) r (nm) EFRET
6.35⨉10-16
1.54 2.23 0.10
25
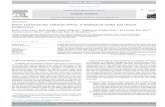
Table 3. Hydrodynamic radii and polydispersity of HSA in the absence and presence of CBDA.
Condition (Rh) Pd (%)
HSA 3.4±0.03 13.5
HSA + CBDA (1:5) 3.2±0.03 8.1
HSA + CBDA (1:20) 2.6±0.02 5.2
Table 4. Thermodynamic and binding parameters of HSA-CBDA obtained from ITC at 298 K.
n
Kb (M-1
)
ΔH⁰
(kJ mol-1)
TΔS⁰
(kJ mol-1
)
ΔG⁰
(kJ mol-1
)
1.10 1.95⨉104 -26.01 6.5 -19.5

26
Table 5. Molecular docking parameters of HSA-CBDA complex.
.
Figure 1
Binding Site Amino Acids Forces Involved ΔG (kJ mol-1
)
Site IB Tyr138
H-Bonding -20.46
Leu185
H-Bonding
Leu182
H-Bonding
Tyr161
Hydrophobic
Met123
Hydrophobic
Ile142
Hydrophobic
Gly189
Hydrophobic
A B
C

27
Figure 2
Figure 3
a
b
B A
28
Figure 4
Figure 5
A
B C
29
Figure 6

30
Figure 7
Highlights
Comprehensive insight into the interaction of CBDA with HSA.
The interaction process is spontaneous and exothermic.
The main governing forces for stabilising HSA- CBDA complex are van der Waal
interaction and hydrogen bonding.
CBDA binds at subdomain IB on HSA.
A
C
A
B
Copyright © 2022 FDOKUMEN