
Apoptosis Th2 Resistance from Fas-Mediated Death-Inducing ...
Upload
independentCategory
view
2download
0
A
sFrhdrre©
K
1
Ftiuscioer
Biological control of bayoud disease in date palm: Selection ofmicroorganisms inhibiting the causal agent
and inducing defense reactions
Majida El Hassni a, Abdelbasset El Hadrami b, Fouad Daayf b, Mohamed Cherif c,Essaid Ait Barka d, Ismaıl El Hadrami a,∗
a Laboratoire de Biotechnologies, Protection et Valorisation des Ressources Vegetales ; Equipe Biotechnologies, Ecophysiologie et Valorisation desPlantes Departement de Biologie, Faculte des Sciences Semlalia, BP. 2390, 40 000 Marrakech, MOROCCO
b Department of Plant Science, 222 Agriculture Building, University of Manitoba, Winnipeg, MB, Canada R3T 2N2c Laboratoire de Phytopathologie, Institut National Agronomique de Tunisie, 43 Avenue Charles Nicolle, 1082 Cite Mahrajene, Tunis, Tunisia
d Laboratoire de Physiologie Vegetale, UFR Sciences URVVC, Universite de Reims Champagne-Ardenne, B.P. 1039, 51687 Reims Cedex 2, France
bstract
Twenty-one isolates of microorganisms, including Bacillus spp., Rhizobium spp., Ulocladium atrum, Candida guilliermondii, Pseudomonasp., Rahnella aquatilis and other bacteria not yet identified, were tested to determine their effects on the mycelial growth and the sporulation ofusarium oxysporum f.sp. albedinis (Foa), the causal agent of bayoud on date palm. The potential of these antagonists in the induction of defense
eactions in date palm seedlings was also studied. Four bacteria, B. pumilus W1, R. aquatilis W2, B. cereus X16 and n.d. S1, have exhibited aigh inhibition toward mycelial growth of Foa (70–77%), and its sporulation (80–95% of the control). Moreover, cytological alterations have been
etected in the Foa mycelium grown in the inhibition zone. Application of these antagonists into date palm seedlings has led to trigger defenseeactions with an accumulation of non-constitutive hydroxycinnamic acid derivatives, such as the sinapic derivative I2, known to play a crucialole in resistance of date palm to Foa. This reaction was more pronounced in resistant cultivar (BSTN) than in susceptible (JHL). The combinedffects of direct and indirect actions of Foa antagonists are discussed in the hope of providing a biocontrol strategy against bayoud.2006 Elsevier B.V. All rights reserved.
is (Fo
tameiota1B
eywords: Date palm; Bayoud; Biocontrol; Fusarium oxysporum f.sp. albedin
. Introduction
The vascular fusariosis commonly named bayoud, caused byusarium oxysporum f.sp. albedinis (Foa), is the most destruc-
ive fungal disease of date palm (Phoenix dactylifera L.). Thempact of this disease is most severe in North Africa partic-larly in Morocco where 2/3 of palm trees were destroyedo far (Fernandez et al., 1995). As for most vascular diseasesaused by soilborne pathogens, fighting strategies are very lim-ted or quasi-inexistent. Among these strategies, the deployment
f Foa-resistant cultivars appear to be the most suitable andconomic approach (Djerbi, 1990; Louvet, 1991) but naturalesistant genotypes are scarce with a poor quality of fruits and∗ Corresponding author. Tel.: +212 24 439997; fax: +212 24 439997.E-mail address: [email protected] (I. El Hadrami).
ctb1tGw
a); Phenols; Phytoalexins
he date palm breeding system is laborious and offered only aslong-term plan (El Hadrami et al., 2005). Alternative controleasures such as the use of Foa antagonists are, therefore, nec-
ssary and need to be explored. This strategy is based, as shownn other pathosystems involving Fusarium species, on the usef microorganisms such as bacteria, yeasts or saprophytic fungihat have either a significant potential to inhibit disease causalgents or ability to enhance defense mechanisms (M’Piga et al.,997; Larkin and Fravel, 1998; Benhamou and Nicole, 1999; Deoer et al., 1999; Cherif et al., 2002; Silva et al., 2004). In thease of fusariosis biocontrol, most of the studies have reportedhe use of non-pathogenic Fusarium isolates alone or in com-ination with bacteria such as Pseudomonas fluorescens (Sneh,
998; Duijff et al., 1999; Fuchs et al., 1997, 1999). Other biocon-rol agents belonging to the genera of Trichoderma, Penicillium,liocladium, Sporidesmium, Burkholderia, Bacillus, Serratiaere also shown to confer some levels of protection against
s2
mttdbcraLGc1ct1rtif2ittnatccteesa1ssF
aawotm
2
2
rJss
t(p
2
cpBYXgP2caDogboAim
2
sasopTioowiiuwoadrceear
oilborne pathogens including Fusarium species (Garbeva et al.,004).
Biocontrol agents achieve a better disease control via aicrobial antagonism, which implies a direct interaction with
he causal agent of disease and/or an indirect action involvinghe host (Thomashow, 1996; Benhamou et al., 2002). Theirect effect is usually a result of an antagonism between theiocontrol agent and the pathogen due to antibiosis or nutrientompetition and/or parasitism, whereas the indirect interactionesults from the enhancement of the plant defense mechanismsgainst the invading pathogen (Glick and Bashan, 1997; Vanoon et al., 1998; Larkin and Fravel, 1999; Cherif et al., 2002;etha and Vikineswary, 2002). Association of both effects
an be noticeable using some biocontrol agents (Thomashow,996; Van Loon et al., 1998; Benhamou et al., 2002). In thease of date palm, a little is known about microorganismshat can protect plants against Foa (Hatimi, 1989; Maslouhy,989) and even less with regard to the induction of defenseeactions (El Hassni et al., 2004). Recently, we have shown thathe pretreatment of date palm seedlings with a hypoaggressivesolate of F. oxysporum has provided a protection againsturther attacks by an aggressive isolate of Foa (El Hassni et al.,004). This protection was most likely achieved via an earlynduction of the plant defense reactions by the hypoaggressiveo counterpart the aggressive isolate of Foa. In this study,he hypoaggressive isolate did not exhibit any direct antago-ism/competition toward Foa; however, it was able to promotefaster accumulation of specific phenolics and induce earlier
he peroxidase activity both in the susceptible and the resistantultivars tested. The main phenolics induced were previouslyharacterized as hydroxycinnamic acid derivatives and reportedo be biomarkers of date palm resistance to bayoud (El Hadramit al., 1997; Ramos et al., 1997; Daayf et al., 2003a; El Hassnit al., 2004). Among these de novo synthesized compounds theinapic derivative I2 was reported to be the main phytoalexinccumulated in date palm in response to Foa (El Hadrami et al.,997). Trials including the studied hypoaggressive isolate aretill ongoing under confined laboratory conditions in order toet up different parameters of its use as biocontrol agent towardoa in the greenhouse and optimistically in the field.
Having such a background on the date palm × bayoud inter-ction and willing to explore antagonists as biocontrol agentsgainst Foa the present study was undertaken. The objectivesere then to select among some microorganisms from our lab-ratory collection that might be Foa antagonists in vitro, ando evaluate their effectiveness in triggering the plant defenseechanisms against bayoud.
. Materials and methods
.1. Plants
Date palm seedlings were obtained from seeds produced by
esistant (‘Bousthami Noir’, BSTN) and susceptible (‘Jihel’,HL) cultivars to bayoud. Seeds from each cultivar were firsturface sterilized before being transferred for germination intoterile peat during 2 weeks at 38 ◦C. Germinated seeds were then2
u
ransferred in pots containing a mixture of sand and peat (2:1v/v)) in a growth chamber maintained at 25 ± 2 ◦C with 16 hhotoperiod.
.2. Microorganisms
The isolates of the different microorganisms used in this workome from our laboratories’ collections and they were used inrevious biocontrol studies. These isolates belong to genera ofacillus (B. amyloliquefaciens C1; B. pumilus B2, M1, W1,1; B. subtilis B1, B3, J1; Daayf et al., 2003b) and (B. cereus16; B. thuringiensis 55T; Cherif et al., 2002); Candida (C.uilliermondii 101 and US7; Saligkarias et al., 2002a, 2002b);seudomonas sp. P1, Rahnella (R. aquatilis W2; Daayf et al.,003b); Rhizobium (Rhizobium spp. DMS and Test 2/92); Ulo-ladium atrum (U. atrum U13 and U16; Ronseaux et al., 2004);nd other non-identified bacteria (n.d. A1, n.d. A2, n.d. S1;aayf et al., 2003b). The Foa isolate ZAG was isolated from bay-ud diseased rachis of date palm harvested in a highly infestedrove in the region of Zagora, south of Morocco. This isolate haseen revealed as the most aggressive among the Foa collectionn most tested date palm cultivars (El Idrissi-Tourane, 1997).ll these microorganisms were stored within 10% glycerol ster-
le solutions at −20 ◦C and they are transferred on PDA cultureedium at 25 ± 2 ◦C before tests.
.3. In vitro tests
Antagonism between Foa and other microorganisms wastudied by co-culturing side by side on PDA plates the pathogennd other antagonists. Suspensions of each microorganism weretreaked across the center of each culture plate. After 0–48 hf incubation, two 5-mm disks of 7-day-old culture of Foa werelaced 2.5 cm apart on each side of the streaked microorganisms.he plates were then incubated at 25 ± 2 ◦C. The percentage of
nhibition was evaluated by measuring the Foa colony radiusn the side of the challenging microorganism. Colony radiusf a control grown in the absence of any other microorganismsas rated as well. Measurements were taken after 2–6 days of
ncubation and Foa mycelia samples were collected from thenterface region with other tested microorganisms, and observednder light microscope. Ten days after incubation, 5-mm disksere harvested from the periphery of Foa colonies on the sidef the tested microorganisms and suspended in an appropri-te volume of sterile distilled water. The Foa sporulation wasetermined under microscope using a hemacytometer and theesults were expressed as the number of conidia per cm2 ofolony. Two plates containing two colonies of Foa were used perxperiment for each tested Foa antagonist isolate and the wholexperiment was repeated two times. Results were expresseds the mean ± standard error (S.E.) calculated among all theeplicates.
.4. In vivo tests
Date palm seedlings (5–6-months-old) were cautiouslynearthed, cleared from soil, then the roots were carefully

wlbaowsrwidgTip
2
ib1mcWWcwandtiwpeih0as
2d
bataspnyemp
Table 1Inhibition degree of Fusarium osysporum f.sp. albedinis (Foa) mycelialgrowth ± S.E. on PDA in presence of several microorganisms after 6 days ofgrowth
Isolate Degree of inhibition (%)
Rahnella aquatilis W2 77 ± 3.65 aBacillus pumilus W1 75 ± 6.03 an.d. S1 72 ± 3.08 aBacillus cereus X16 70 ± 3.12 aBacillus pumilus Y1 44 ± 3.23 bRhizobium spp. DMS 36 ± 6.02 cn.d. A2 35 ± 2.23 cn.d. A1 30 ± 3.98 cdCandida guilliermondii US7 26 ± 0.76 dBacillus thuringiensis 55T 26 ± 3.04 dUlocladium atrum U13 26 ± 3.69 dBacillus amyloliquefaciens C1 23 ± 2.28 deUlocladium atrum U16 23 ± 3.04 deBacillus subtilis J1 23 ± 3.69 deCandida guilliermondii 101 20 ± 6.70 deBacillus subtilis B1 15 ± 5.94 ePseudomonas sp. P1 15 ± 8.94 eBacillus pumilus B2 5 ± 4.08 fBacillus pumilus M1 5 ± 4.08 fBacillus subtilis B3 5 ± 4.08 fRhizobium spp.Test 2/92 5 ± 4.08 fFoa-control 0 ± 0 f
nl
3
3
t(aiWwtamlalt
BtcrIs
ashed with sterile distilled water before use for inocu-ation with microorganisms. For the inoculation, 10 �l ofacterial suspension prepared in sterile distilled water anddjusted to 108 CFU/ml were injected into the main piv-tal root. Control plants were treated with sterile distilledater taking account the same condition as above. Forty
eedlings were used per treatment and the experiment wasepeated independently three times. After injection, seedlingsere transferred into glass tubes containing 20 ml of ster-
le distilled water. No hydroponic solutions were used; theate palm root system possesses hydrophores allowing therowth of plants only on water for more than 6 months.ubes were then covered with aluminum foil and incubated
n a growth chamber maintained at 25 ± 2 ◦C with 16 hhotoperiod.
.5. Analyses of phenolic compounds
Extraction of phenolic compounds was performed accord-ng to routinely-used standard protocol that has been describedy El Hadrami et al. (1997). For that, root samples harvested0, 20, and 30 days after inoculation were extracted with 80%ethanol at 4 ◦C and stored at −20 ◦C until analyzed. Phenolic
ompounds analyses were carried out using an HPLC equipmentaters 600E supplemented with a photodiode array detectoraters 990 and a Millipore software for data analysis. An effi-
ient gradient of acetonitrile-o-phosphoric acidified bidistilledater (pH 2.6) was used on an Interchrom C18 reversed phase
nalytical column (4.6 mm × 250 mm, 5 �m). Multiple chan-el wavelengths, including 280, 320 and 350 nm were useduring the elution. Phenolics were identified on the basis ofheir retention times and their characteristic spectra in compar-son with standards. When necessary, co-injection and elutionith standards were used to ensure the identity of the com-ounds. Quantities of hydroxycinnamic acid derivatives werestimated by measuring the area under the peaks and compar-ng it to chlorogenic acid used as an external standard. Rootsarvest and phenolics analyses were conducted over time at, 10, 20, and 30 days after inoculation with individual Foantagonists. HPLC analyses were performed twice from eachample.
.6. Experimental design and statistical analyses of theata
All the experiments schemes were randomized completelocks. At least two replicates per treatment were performednd each whole experiment was repeated independently two tohree times. All the collected data were submitted to ANOVAnalysis using Statistica software v. 5.5 (Statsoft, ed’99) and theignificance of differences among treatments was recorded at< 0.05. Multiple replicates of the same experiment showingo significant differences were combined within the same anal-
sis unless stated. When necessary, data were transformed tonsure the variances homogeneity. Multiple comparisons of theeans were conducted according to the Newman–Keuls test at< 0.05.brgb
.d., not determined. The values within the same column followed by the sameetter do not differ significantly (p < 0.05).
. Results
.1. In vitro tests
Based on the percentage of inhibition of the fungal growth,he tested microorganisms could be divided into three groupsTable 1). The first group represented by microorganisms suchs B. pumilus W1, B. cereus X16, R. aquatilis W2, and n.d. S1solates showing high inhibition ability toward Foa (70–77%).
ithin this group, the two isolates R. aquatilis W2 and n.d. S1ere shown to induce a strong yellowish brown pigmentation on
he Foa colonies challenged (Fig. 1). The second group exhibitedmoderate ability to inhibit Foa growth (26–44%) and includedicroorganisms such as B. pumilus Y1, B. subtilis J1, C. guil-
iermondii US7, U. atrum U13, Rhizobium spp. DMS, n.d. A1nd n.d. A2. The third group of microorganisms has shown aow inhibition of Foa mycelium growth (5–23%) and enclosedhe rest of the tested isolates.
Among all the microorganisms tested, only two isolates of. pumilus B1 and M1 have shown no significant reduction of
he Foa sporulation capacity as compared to the non-challengedontrol (Table 2). Most of the other isolates were significantlyeducing the sporulation of the pathogen by more than 50%.nterestingly, some of these isolates reducing effectively theporulation of Foa were reducing also the fungal growth rate
y more than 70% (Tables 1 and 2). There is no evidence of cor-elation between the effect of antagonists on the Foa mycelialrowth rate and the sporulation. Some isolates such as Rhizo-ium sp. Test 2/92 were completely ineffective in reducing the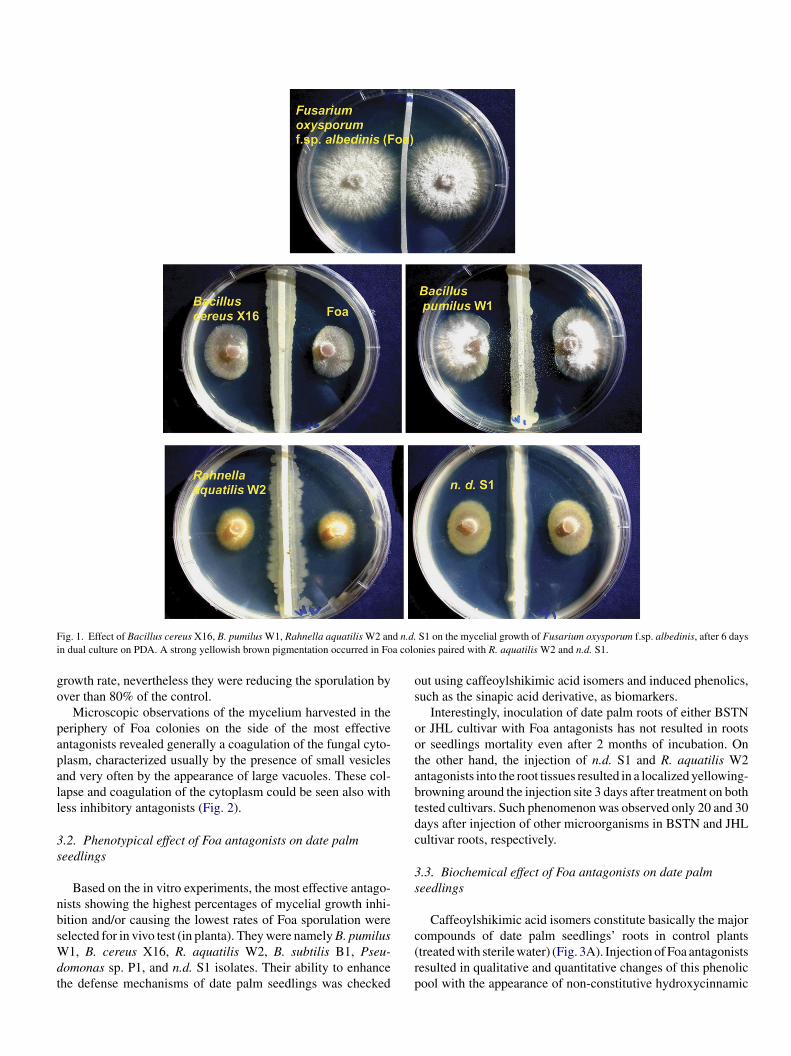
F d n.d.i a colo
go
papall
3s
nbsWdt
os
ootabtdc
3s
ig. 1. Effect of Bacillus cereus X16, B. pumilus W1, Rahnella aquatilis W2 ann dual culture on PDA. A strong yellowish brown pigmentation occurred in Fo
rowth rate, nevertheless they were reducing the sporulation byver than 80% of the control.
Microscopic observations of the mycelium harvested in theeriphery of Foa colonies on the side of the most effectiventagonists revealed generally a coagulation of the fungal cyto-lasm, characterized usually by the presence of small vesiclesnd very often by the appearance of large vacuoles. These col-apse and coagulation of the cytoplasm could be seen also withess inhibitory antagonists (Fig. 2).
.2. Phenotypical effect of Foa antagonists on date palmeedlings
Based on the in vitro experiments, the most effective antago-ists showing the highest percentages of mycelial growth inhi-ition and/or causing the lowest rates of Foa sporulation were
elected for in vivo test (in planta). They were namely B. pumilus1, B. cereus X16, R. aquatilis W2, B. subtilis B1, Pseu-omonas sp. P1, and n.d. S1 isolates. Their ability to enhancehe defense mechanisms of date palm seedlings was checked
c(rp
S1 on the mycelial growth of Fusarium oxysporum f.sp. albedinis, after 6 daysnies paired with R. aquatilis W2 and n.d. S1.
ut using caffeoylshikimic acid isomers and induced phenolics,uch as the sinapic acid derivative, as biomarkers.
Interestingly, inoculation of date palm roots of either BSTNr JHL cultivar with Foa antagonists has not resulted in rootsr seedlings mortality even after 2 months of incubation. Onhe other hand, the injection of n.d. S1 and R. aquatilis W2ntagonists into the root tissues resulted in a localized yellowing-rowning around the injection site 3 days after treatment on bothested cultivars. Such phenomenon was observed only 20 and 30ays after injection of other microorganisms in BSTN and JHLultivar roots, respectively.
.3. Biochemical effect of Foa antagonists on date palmeedlings
Caffeoylshikimic acid isomers constitute basically the major
ompounds of date palm seedlings’ roots in control plantstreated with sterile water) (Fig. 3A). Injection of Foa antagonistsesulted in qualitative and quantitative changes of this phenolicool with the appearance of non-constitutive hydroxycinnamic
F (Foa)i
acmstaaiwaB
maAdiiwFt
hdnfotiwti1tBdtca
ig. 2. Light microscopy observations of Fusarium oxysporum f.sp. albedinisnhibition zone in presence of Ulocladium atrum U16. Bar, 60 �m.
cid derivatives (Fig. 3B–D). Among these newly accumulatedompounds, sinapic derivative I2 was the major compound accu-ulating since the early stages of the experiment (Fig. 3B). The
ignificant accumulation of the hydroxycinnamic acid deriva-ives was accompanied by either a slight increase/maintain orreduction of caffeoylshikimic acid isomers content (Fig. 3C),
mong which some isomers tend to disappear over time depend-ng of the cultivar and the treatment (Fig. 3D). This last situationas particularly observed 30 days after the injection of n.d. S1
nd R. aquatilis W2 isolates into both cultivars and B. subtilis1 into BSTN cultivar.
The dynamic of accumulation of caffeoylshikimic acid iso-ers was different over time in response to each tested Foa
ntagonist but was also depending on the used cultivar (Fig. 4).s a high significant effect of the time after inoculation wasetected by ANOVA, mainly due to the qualitative differencesn the accumulation observed over time that were generating
nteraction between cultivars and treatments, each time pointas considered individually for statistical analysis. The effect ofoa antagonists were compared per cultivar for each consideredime point. As compared to the controls from both cultivars, the
(
at
mycelium: (A) control mycelium and (B–D) Foa mycelium sample from the
ighest increases in the accumulation of caffeoylshikimic aciderivatives were recorded, especially for R. aquatilis W2 and.d. S1, earlier after injection (10 days after inoculation) exceptor B. cereus X16, for which a high accumulation was detectablenly 30 days after inoculation of cultivar JHL. In BSTN cultivar,he highest significant accumulations of caffeoylshikimic acidsomers were recorded 10 days after the injection into the rootsith B. pumulis W1 and R. aquatilis W2 isolates, as compared
o the control (Fig. 4). The levels detected at 20 and 30 days afternjection were overall significantly lower than those observed at0 days but not significantly different from those recorded inhe healthy control except in response to n.d. S1 and B. subtilis1 isolates both at 20 and 30 days, and R. aquatilis W2 at 30ays (Fig. 4). In JHL cultivar, the situation was different withhe highest accumulations of caffeoylshikimic acid isomers, asompared to the control, observed in response to R. aquatilis W2,nd to B. cereus X16, 10 and 30 days after injection, respectively
Fig. 4).The accumulation of the sinapic acid derivative I2 dependedlso on the association Foa antagonist-date palm cultivar andhe time after injection of the antagonist into the roots (Fig. 5).

F L) tre1 ffeoylh
Sacawso
aBb
ig. 3. HPLC chromatograms of soluble phenolics in date palm roots (cv. JH0, 20 and 30 days after injection of the Foa antagonist, respectively. 1–3, caydroxycinnamic acid derivatives.
uch accumulation was much more important 20 and 30 daysfter the injection. In cultivar BSTN, high accumulation of I2ompound was observed with most of the Foa antagonists tested
s compared to the control. The lowest accumulation levelsere observed in response to B. cereus X16 and Pseudomonasp. P1. In cultivar JHL, among all the Foa antagonists tested,nly R. aquatilis W2 and n.d. S1 isolates induced a significant
Wis(
ated with n.d. S1 isolate: (A) healthy control and (B–D) phenolic profiles atshikimic acid isomers; I2, sinapic derivative; 4, ferulic derivative; 5–6, other
ccumulation of I2 compound comparatively to the control.y comparing the accumulation of sinapic acid derivative I2 inoth tested cultivars, only Foa antagonist isolates R. aquatilis
2 and n.d. S1 were able to induce a similar pattern. However,solate B. subtilis B1 seemed to be highly efficient in inducinguch derivative at a high level in the bayoud resistant cultivarBSTN) than in the susceptible (JHL).

Table 2Effect of microorganisms on the sporulation of Fusarium oxysporum f.sp. albe-dinis (Foa) after 10 days of incubation on PDA
Isolate Number of conidia ((×105) per cm2)
Foa-control 55.11 ± 2.51 aBacillus pumilus M1 51.73 ± 8.38 aBacillus pumilus B2 50.81 ± 5.98 an.d. A1 27.43 ± 5.42 bBacillus subtilis B3 25.98 ± 7.38 bUlocladium atrum U13 19.56 ± 11.24 bcn.d. A2 16.84 ± 7.38 bcdBacillus cereus X16 10.57 ± 5.55 cdPseudomonas sp. P1 7.48 ± 2.68 cdBacillus pumilus W1 7.48 ± 3.38 cdRhizobium spp. DMS 6.74 ± 2.88 cdUlocladium atrum U16 5.07 ± 1.03 cdCandida guilliermondii US7 4.60 ± 2.18 cdBacillus amyloliquefaciens C1 4.57 ± 1.02 cdBacillus subtilis J1 4.53 ± 1.44 cdBacillus pumilus Y1 4.52 ± 0.89 cdRhizobium spp.Test 2/92 4.25 ± 2.88 cdBacillus thuringiensis 55T 3.9 ± 0.71 cdBacillus subtilis B1 2.38 ± 1.3 dn.d. S1 2.28 ± 1.08 dCandida guilliermondii 101 2.26 ± 0.93 dRahnella aquatilis W2 2.23 ± 1.93 d
nl
4
oattimosoodMoFindosavn
ogFo
brgentpctoszFiatwwcdpcatsapeoGacRbaoa
mfdpsmaaagpamwp
.d., not determined. The values within the same column followed by the sameetter do not differ significantly (p < 0.05).
. Discussion
Suppression of Fusarium wilt has been reported on a varietyf host plants (Alabouvette et al., 1996; Sneh, 1998; Fuchs etl., 1997, 1999). Examination of soils in many cases has shownhat even if the density of inoculum was high and the condi-ions for disease settlement were favorable, disease severity andncidence has remained low on susceptible plants. Varieties oficroorganisms, such as non-pathogenic Fusarium spp. and flu-
rescent Pseudomonas, have been shown to be involved in thisoil suppressive ability of Fusariosis (Sneh, 1998). In the casef date palm, several studies have reported the non-receptivityf certain soils (i.e. region of Marrakech city in Morocco) to theevelopment of bayoud (Sedra, 1985; Louvet, 1991; Sedra andaslouhy, 1995). This non-receptivity has been related, later
n, to the phenomena of competition between the causal agentoa and other non-pathogenic Fusarium species predominant
n these soils, and to the antagonism of several bacteria, acti-omycete and fungi (Maslouhy, 1989). The present study iseveloped within this context to test the antagonistic effectsf several microorganisms, such as Bacillus spp., Rhizobiumpp., U. atrum, C. guilliermondii, Pseudomonas sp., R. aquatilis,gainst Foa, and to detect whether they are direct antagonists initro or they act via a stimulation of date palm defense mecha-isms.
The in vitro tests have shown that simultaneous co-culture
f these microorganisms and Foa does not stop completely itsrowth. However, their culture two days prior the transfer ofoa was accompanied by the formation of a more or less zonef inhibition depending of Foa antagonist used. This direct inhi-aaBt
ition effect of Foa might be the result of antifungal compoundseleased by the antagonists into the culture media. Such sug-estions have also been reported in other studies (Ait Barkat al., 2002). In some cases of interaction with Foa antago-ists, the sporulation of Foa colonies was highly reduced par-icularly in the border where the inhibition is stirring. Suchhenomenon could be related, directly, to the effect of toxicompounds released by Foa antagonist, and/or, indirectly, tohe non-sufficient or abnormal growth of Foa under the antag-nists’ effects, that cannot let it sporulate. The decline of Foaporulation was also observed even if there was no inhibitionone by some Foa antagonists. Similar results were reported forusarium roseum var. sambucinum, the causal agent of dry rot
n potato tubers, challenged by B. thuringiensis 55T (Cherif etl., 2002). In this case, despite the absence of an inhibition zone,he destructive effect of the pathogen by B. thuringiensis 55Tas high, resulting in serious damage of the hyphae associatedith series of degradation events. This suggests that fungitoxic
ompounds and/or hydrolytic enzymes produced by antagonistsiffuse for a short distance into the agar that disorganize theathogen hyphae before any physical contact. Such disordersan lead also to a higher reduction of the pathogen sporulations observed in the present study. Microscopic observations ofhe Foa mycelium, harvested on the side of the antagonist, havehown a coagulation of cytoplasm characterized by the appear-nce of small vesicles and vacuoles, reminding other cases ofathogens submitted to the activity of antagonists (Benhamout al., 1999; Ait Barka et al., 2002), or treated by fungicidesr chitosan (Robertson and Fuller, 1990; Benhamou, 1992; Elhaouth et al., 1992; Ait Barka et al., 2004). Other changes such
s the appearance of a yellowish brown pigmentation of the Foaolonies were observed in presence of some antagonists such as. aquatilis W2 and n.d. S1. These colony color changes mighte due to the effect of diffusible metabolites produced by Foantagonists and/or to the activity of anatagonists’ phenoloxidaser peroxidase activities as shown in other systems (Badalyan etl., 2002).
The use of biological agents to enhance the plant defenseechanisms against pathogens represents an ecologically
riendly alternative to pesticides repeatedly used to control plantiseases. This strategy has more significance against soilborneathogens such as bayoud on date palm, for which fightingtrategies are very restricted or quasi-unavailable. The deploy-ent of such strategies should have a great impact taking into
ccount the perennial aspect of the crop, the socio-economicalnd ecological issues to use chemicals within date palm grovesnd the long-term solution that represent the breeding pro-ram. The success of a biological control agent in turning-onlant defense mechanisms against pathogens depends of theirbility to establish metabolically active populations that couldediate host protection and/or compete directly or indirectlyith the pathogens for nutrient resources. Against soilborneathogens, several studies have reported the use of endophytic
nd plant growth-promoting bacteria, including Bacillus spp.nd Pseudomonas spp. (Thomashow, 1996; M’Piga et al., 1997;enhamou and Nicole, 1999; Jeun et al., 2004). However, lit-le is known about the potential of R. aquatilis as a biocontrol

F ultivap �) 10d
apalaAitirc
hsinstt
ig. 4. Time course of caffeoylshikimic acids in date palm roots of resistant cumilus W1, B. subtilis B1, R. aquatilis W2, n.d. S1 and Pseudomonas sp. P1. (iffer significantly (p < 0.05). Bars represent the standard errors to the means.
gent (Laux et al., 2002). These bacteria, especially the endo-hytic, have been reported to be responsible for an increase,t early stages of the interaction, of mRNA encoding pheny-alanine ammonia-lyase (PAL) and chalcone synthase, whichre key enzymes in the phenylpropanoid pathway (Zdor andnderson, 1992). The findings of the present study showing an
ncrease in the accumulation of hydroxycinnamic acid deriva-
ives after injection of Foa antagonists into date palm roots aren accordance with theses results. In fact, treatment of date palmoots with Foa antagonists, B. pumilus W1, B. subtilis B1, B.ereus X16, R. aquatilis W2, Pseudomonas sp. P1 and n.d. S1HIor
r (BSTN) and susceptible (JHL) after treatment with Bacillus cereus X16, B.days, ( ) 20 days, (�) 30 days. The histograms having the same letter do not
as resulted in the changes in the accumulation of caffeoyl-hikimic acid isomers, the remarkably constitutive compoundsn date palm roots. It has led also to the accumulation of severalon-constitutive hydroxycinnamic acid derivatives such as theinapic acid derivative I2. Such hydroxycinnamic acid deriva-ives are known to play a major role in date palm resistanceo bayoud and to exhibit a fungitoxic effect toward Foa (El
adrami et al., 1997; Ramos et al., 1997; Daayf et al., 2003a).ndeed, these studies have shown a decrease in the accumulationf the constitutive caffeoylshikimic acid isomers in date palmoot along with an increase in the accumulation of newly induced

F r (BSW ays, (s
so1cFctHTeh
tRsd
oo
ig. 5. Time course of sinapic derivative I2 in date palm roots of resistant cultiva1, B. subtilis B1, R. aquatilis W2, n.d. S1 and Pseudomonas sp. P1. (�) 10 d
ignificantly (p < 0.05). Bars represent the standard errors to the means.
inapic acid derivatives such as I2 in response to infection with F.xysporum f.sp. albedinis (El Hadrami et al., 1997; Ramos et al.,997). The isolation and purification of these newly synthesizedompounds (I2) has led to study their biological activity towardoa (Ramos et al., 1997). A highly significant inhibition of Foaonidia germination accompanied by a reduction of the germubes growth were detected in presence of these compounds (El
adrami et al., 1997; Ramos et al., 1997; Daayf et al., 2003a).heir de novo synthesis on site in response to infection andffectiveness at lower doses in vitro beside their phenolic natureas led to suggest them as phytoalexins in date palm althoughasao
TN) and susceptible (JHL) after treatment with Bacillus cereus X16, B. pumilus) 20 days, (�) 30 days. The histograms having the same letter do not differ
heir late accumulation into the roots (El Hadrami et al., 1997;amos et al., 1997; Daayf et al., 2003a). Much more studies are
till needed to determine their exact chemical structure and roleuring bayoud infection as well as their biological effect in vivo.
In conclusion, this study has investigated the selection of a setf microorganisms that can act as antagonists against Foa basedn their ability to inhibit more or less strongly its mycelial growth
nd/or sporulation. Among the in vitro selected Foa antagonists,ome isolates of Bacillus spp., Pseudomonas spp., and Rahnellaquatilis were having also the ability to induce defense reactionsf date palm without causing any seedlings mortality. Further
sotw
A
PM
R
A
A
A
B
B
B
B
B
C
D
D
D
D
D
E
E
E
E
E
F
F
F
G
G
G
H
J
L
L
L
L
M
M
R
tudies are conducted in order to highlight the in planta effectf these antagonists on the endophytic development of Foa, ando set up the ecological conditions required for their settlementithin the date palm rhizosphere.
cknowledgments
Financial support for this work was provided by PRAD 02-02,rotars II No. P51/14, AI (France-Maroc) 01/23 and AI (Tunisie-aroc) 24/02.
eferences
it Barka, E., Eullaffroy, P., Clement, C., Vernet, G., 2004. Chitosan improvesdevelopment, and protects Vitis vinifera L. against Botrytis cinerea. PlantCell Rep. 22, 608–614.
it Barka, E., Gognies, S., Nowak, J., Audran, J.C., Belarbi, A., 2002. Inhibitoryeffect of endophyte bacteria on Botrytis cinerea and its influence to promotethe grapevine growth. Biol. Control 24, 135–142.
labouvette, C., Lemanceau, P., Steinberg, C., 1996. Biological control ofFusarium wilts: opportunities for developing a commercial product. In: Hall,R. (Ed.), Principles and Practice of Managing Soilborne Plant Pathogens.American Phytopathological Press, St. Paul Minn, pp. 192–212.
adalyan, S.M., Innocenti, G., Garibyan, N.G., 2002. Antagonistic activity ofxylotrophic mushrooms against pathogens of cereals in dual culture. Phy-topathol. Mediterr. 41, 200–225.
enhamou, N., 1992. Ultrastructural and cytochemical aspects of chitosan onFusarium oxysporum f.sp. radicis-lycopersici, agent of tomato crown androot rot. Phytopathology 82, 1185–1193.
enhamou, N., Garand, C., Goulet, A., 2002. Ability of nonpathogenic Fusar-ium oxysporum strain F047 to induce resistance against Pythium ultimuminfection in cucumber. Appl. Environ. Microbiol. 68, 4044–4060.
enhamou, N., Nicole, M., 1999. Cell biology of plant immunization againstmicrobial infection: the potential of induced resistance in controlling plantdiseases. Plant Physiol. Biochem. 37, 703–719.
enhamou, N., Rey, P., Picard, K., Tirilly, Y., 1999. Ultrastructural and cyto-chemical aspects of the interaction between the mycoparasite Pythiumoligandrum and soilborne plant pathogens. Phytopathology 89, 506–517.
herif, M., Sadfi, N., Benhamou, N., Boudabbous, A., Boubaker, A., Hajlaoui,M.R., Tirilly, Y., 2002. Ultrastructure and cytochemistry of in vitro interac-tions of the antagonistic bacteria Bacillus cereus X16 and B. thuringiensis55T with Fusarium roseum var. sambucinum. J. Plant Pathol. 84, 83–93.
aayf, F., El Bellaj, M., El Hassni, M., J’aiti, F., El Hadrami, I., 2003a. Elicitationof soluble phenolics in date palm (Phoenix dactylifera L.) callus by Fusariumoxysporum f. sp. albedinis culture medium. Environ. Exp. Bot. 49, 41–47.
aayf, F., Adam, L., Fernando, W.G.D., 2003b. Comparative screening of bac-teria for biological control of potato late blight (strain US-8), using in-vitro,detached-leaves, and whole plant testing systems. Can. J. Plant Pathol. 25,276–284.
e Boer, M., Van der Sluis, I., Van Loon, L.C., Bakker, P.A.H.M., 1999. Com-bining fluorescent Pseudomonas spp. strains to enhance suppression ofFusarium wilt of radish. Eur. J. Plant Pathol. 105, 201–210.
jerbi, M., 1990. Methodes de diagnostic du Bayoud du palmier dattier. Bull.OEPP/EPPO Bull. 20, 607–613.
uijff, B.J., Recorbet, G., Bakker, P.A.H.M., Loper, J.E., Lemanceau, P., 1999.Microbial antagonism at the root level is involved in the suppression ofFusarium wilt by the combination of nonpathogenic Fusarium oxysporumF047 and Pseudomonas putidia WCS358. Phytopathology 89, 1073–1079.
l Ghaouth, A., Arul, J., Asselin, A., Benhamou, N., 1992. Anti-fungual
activity of chitosan on post-harvest pathogens: induction of morphologi-cal and cytological alterations in Rhizopus stolonifer. Mycol. Res. 96, 769–779.l Hadrami, A., El Idrissi-Tourane, A., El Hassni, M., Daayf, F., El Hadrami,I., 2005. Toxin-based in vitro selection and its potential application to date
R
palm for resistance to the bayoud Fusarium wilt—a review. C. R. Biol. 328,732–744.
l Hadrami, I., Ramos, T., El Bellaj, M., El Idrissi-Tourane, A., Macheix, J.J.,1997. A sinapic derivative as induced defense compound of date palm againstFusarium oxysporum f.sp. albedinis, the agent causing Bayoud disease. J.Phytopathol. 145, 329–333.
l Hassni, M., J’aiti, F., Dihazi, A., Ait Barka, E., Daayf, F., El Hadrami, I., 2004.Enhancement of defense responses against Bayoud disease by treatment ofdate palm seedlings with an hypoaggressive Fusarium oxysporum isolate. J.Phytopathol. 152, 182–189.
l Idrissi-Tourane, A., 1997. Importance des polyamines libres et conjugueeset des inhibiteurs de leur biosynthese dans les mecanismes de defense dupalmier dattier (Phoenix dactylifera L.) vis-a-vis de Fusarium oxysporumf.sp. albedinis, agent causal du Bayoud. These de troisieme cycle, UniversiteCadi Ayyad, Marrakech, p. 123.
ernandez, M., Lourd, M., Ouinten, M., Tantaoui, A., Geiger, J.P., 1995. Le Bay-oud du palmier dattier, une maladie qui menace la phoeniciculture. Phytoma469, 36–39.
uchs, J.G., Moenne-Loccoz, Y., Defago, G., 1997. Nonpathogenic Fusariumoxysporum strain F047 induces resistance to Fusarium wilt in tomato. PlantDis. 81, 492–496.
uchs, J.G., Moenne-Loccoz, Y., Defago, G., 1999. Ability of nonpathogenicFusarium oxysporum F047 to protect tomato against Fusarium wilt. Biol.Control 14, 105–110.
arbeva, P., Van Veen, J.A., Van Elsas, J.D., 2004. Microbial diversity insoil: selection of microbial populations by plant and soil type and impli-cations for disease suppressiveness. Ann. Rev. Phytopathol. 42, 243–270.
etha, K., Vikineswary, S., 2002. Antagonistic effects of Streptomyces vio-laceusniger strain G10 on Fusarium oxysporum f. sp. cubense race 4: indirectevidence for the role of antibiosis in the antagonistic process. J. Ind. Micro-biol. Biotechnol. 28, 303–310.
lick, B.R., Bashan, Y., 1997. Genetic manipulation of plant growth promot-ing bacteria to enhance biocontrol of phytopathogens. Biotechnol. Adv. 15,353–378.
atimi, A., 1989. Etude de la receptivite des sols de deux palmeraies marocainesau Bayoud. These de troisieme cycle, Universite Cadi Ayyad, Marrakech, p.58.
eun, Y.C., Park, K.S., Kim, C.H., Fowler, W.D., Kloepper, J.W., 2004. Cyto-logical observations of cucumber plants during induced resistance elicitedby rhizobacteria. Biol. Control 29, 34–42.
arkin, P.R., Fravel, D.R., 1999. Mechanisms of action and dose responserelationships governing biological control of Fusarium wilt of tomato bynonpathogenic Fusarium spp. Phytopathology 89, 1152–1161.
arkin, R.P., Fravel, D.R., 1998. Efficacy of various fungal and bacterial bio-control organisms for control of Fusarium wilt of tomato. Plant Dis. 82,1022–1028.
aux, P., Baysal, O., Zeller, W., 2002. Biological control of fire blight byusing Rahnella aquatilis Ra39 and Pseudomonas spec. R1. Acta Hort. 590,225–230.
ouvet, J., 1991. Que devons-nous faire pour lutter contre le Bayoud? Physiolo-gie des arbres et Arbustes en zones arides et semi arides: Groupes d’Etudesd’Arbre, Paris-France, pp. 337–346.
’Piga, P., Belanger, R.R., Paulitz, T.C., Benhamou, N., 1997. Increased resis-tance to Fusarium oxysporum f.sp. radicis-lycopersici in tomato plantstreated with the endophytic bacterium Pseudomonas fluorescens strain63–28. Physiol. Mol. Plant Pathol. 50, 301–320.
aslouhy, A., 1989. Contribution a l’etude in vitro et in situ des antagonistesde Fusarium oxysporum f.sp. albedinis, agent causal du Bayoud. These detroisieme cycle, Universite Cadi Ayyad, Marrakech, p. 98.
amos, T., El Bellaj, M., El Idrissi-Tourane, A., Daayf, F., El Hadrami,I., 1997. Les phenolamides des Rachis de palmes, composants de lareaction de defense du palmier dattier vis-a-vis de Fusarium oxys-
porum f.sp. albedinis, agent du Bayoud. J. Phytopathol. 145, 487–493.obertson, R.W., Fuller, M.S., 1990. Effects of the demethylase inhibitor, cypro-conazole, on hyphal tip cells of Sclerotium rolfsii. II. An electron microscopestudy. Exp. Mycol. 14, 124–135.

R
S
S
S
S
S
S
T
onseaux, S., Clement, C., Ait Barka, E., 2004. Biological control of Botrytiscinerea on Vitis vinifera L. plants by the use of fungal antagonist Ulo-cladium atrum. In: Disease, Biocontrol, Organizing, Scientific Committees(Ed.), Proceedings of International Workshop on Development of Biocon-trol Agents of Diseases for Commercial Applications in Food ProductionSystems, Sevilla-Spain, March, pp. 24–27.
aligkarias, I.D., Gravanis, F.T., Epton, H.A.S., 2002a. Biological control ofBotrytis cinerea on tomato plants by the use of epiphytic yeasts Candidaguilliermondii strains 101 and US7 and Candida oleophila strain I-182: I.In vivo studies. Biol. Control 25, 143–150.
aligkarias, I.D., Gravanis, F.T., Epton, H.A.S., 2002b. Biological control ofBotrytis cinerea on tomato plants by the use of epiphytic yeasts Candidaguilliermondii strains 101 and US7 and Candida oleophila strain I-182: II.
A study on mode of action. Biol. Control 25, 151–161.edra, My H., 1985. Potentiel infectieux et receptivite de quelques sols depalmeraies a la fusariose vasculaire du palmier dattier (bayoud) causee parFusarium oxysporum f.sp. albedinis (Kill. et Maire) Malencon. These detroisieme cycle. Agronomie, I.A.V. Hassan II, Rabat, p. 88.
V
Z
edra, My H., Maslouhy, My A., 1995. La fusariose vasculaire du palmierdattier (Bayoud). II. Action inhibitrice des filtrats de culture de six microor-ganismes antagonistes isoles des sols de la palmeraie de Marrakech sur ledeveloppement in vitro de Fusarium oxysporum f. sp. albedinis. Al Awamia90, pp. 1–8.
ilva, H.S.A., Romeiro, R.S., Macagnan, D., Vieira, B.A.H., Pereira, M.C.B.,Mounteer, A., 2004. Rhizobacterial induction of systemic resistance intomato plants: non-specific protection and increase in enzyme activities.Biol. Control 29, 288–295.
neh, B., 1998. Use of non-pathogenic or hypovirulent fungal strains to pro-tect plants against closely related fungal pathogens. Biotechnol. Adv. 16,1–32.
homashow, S.L., 1996. Biological control of plant root pathogens. Curr. Opin.
Biotechnol. 7, 343–347.an Loon, L.C., Bakker, P.A.H.M., Pieterse, C.M.J., 1998. Systemic resistanceinduced by rhizosphere bacteria. Ann. Rev. Phytopathol. 36, 453–483.
dor, R.E., Anderson, A.J., 1992. Influence of root colonizing bacteria on thedefense responses of bean. Plant Soil 140, 99–107.
Copyright © 2022 FDOKUMEN




















