
Biogeography of tropical reef fishes: the South Atlantic puzzle
11
Biogeography of tropical reef fishes: the South Atlantic puzzle J.-C. Joyeux 1 *, S. R. Floeter 2 , C. E. L. Ferreira 3 and J. L. Gasparini 11 Departamento de Ecologia e Recursos Naturais, Universidade Federal do Espı´rito Santo, Av. Fernando Ferrari S/N, Vito ´ria, ES 29060-900, Brazil, 2 Laborato ´rio de Cieˆncias Ambientais, Universidade Estadual do Norte Fluminense, Av. Alberto Lamego, 2000, Campos dos Goytacazes, RJ 28015-620, Brazil, and 3 Departamento de Oceanografia, IEAPM, Rua Kioto 253, Arraial do Cabo, RJ 28930-000, Brazil Abstract Aim To reveal underlying distributional mechanisms of tropical reef fishes in the South Atlantic. Location The tropical South Atlantic, with emphasis on the Brazilian province. Methods The disjunct distributions of thirty-five reef fish species occurring in the Brazilian Province were analysed to allow a better understanding of present biogeo- graphical patterns. To avoid potential bias because of taxonomic problems or misidentification, we only included families taxonomically well documented, with conspicuous species, and relatively easy to identify. Results The low-level differentiation between Caribbean and Brazilian species/popula- tions, as well as the presence of restricted populations of a few northern or southern species on the opposite side of the Amazon River, clearly indicate that this barrier to dispersion can be occasionally bridged. Transoceanic dispersal appears to be frequent and to occur along three routes: from the Caribbean to North East Atlantic, from northern Brazil to the Gulf of Guinea, and from Africa to southern Brazil. Intermediate stepping-stones are apparently not required along transatlantic routes because of the rarity of consecutive colonizations. However, intermediate stopovers are persistently used along short routes. In all cases, the long-term success of colonists, i.e. the establishment of stable local populations, appears to depend upon ecological factors. Main conclusions The puzzling patterns of distribution found in the South Atlantic seem primarily the outcome of allopatric speciation and of the interaction of long distance dispersal abilities and ecological processes. Successfulness of the colonization of remote sites appears to be less dependent from dispersal ability than upon persistence ability or settlement preferences. Keywords Dispersal, islands, disjunct, colonization, speciation, Amazon barrier, Brazilian prov- ince, distribution. INTRODUCTION The South Atlantic Ocean has some peculiar biogeograph- ical and ecological characteristics. The most obvious is the freshwater outflow from the Amazon River that apparently acts as a major biogeographical barrier separating the shore fish faunas of Brazil and the Caribbean (Briggs, 1995; Floeter & Gasparini, 2000). The oceanic islands of the tropical South Atlantic are widely scattered and support shelf faunas that exhibit reduced species diversity and other effects of isolation (Floeter & Gasparini, 2000). Considering that the islands vary in age, size and distance from other shores, comparative studies of their faunas can reveal *Correspondence: Departamento de Ecologia e Recursos Naturais, Universidade Federal do Espı´rito Santo, Av. Fernando Ferrari S/N, Vito ´ ria, ES 29060-900, Brazil. E-mail: [email protected] Journal of Biogeography, 28, 831–841 Ó 2001 Blackwell Science Ltd
Transcript of Biogeography of tropical reef fishes: the South Atlantic puzzle

Biogeography of tropical reef ®shes: the SouthAtlantic puzzleJ.-C. Joyeux1*, S. R. Floeter2, C. E. L. Ferreira3 and J. L. Gasparini1 1Departamento de
Ecologia e Recursos Naturais, Universidade Federal do EspõÂrito Santo, Av. Fernando Ferrari
S/N, VitoÂria, ES 29060-900, Brazil, 2LaboratoÂrio de CieÃncias Ambientais, Universidade
Estadual do Norte Fluminense, Av. Alberto Lamego, 2000, Campos dos Goytacazes, RJ
28015-620, Brazil, and 3Departamento de Oceanogra®a, IEAPM, Rua Kioto 253, Arraial do
Cabo, RJ 28930-000, Brazil
Abstract
Aim To reveal underlying distributional mechanisms of tropical reef ®shes in the SouthAtlantic.
Location The tropical South Atlantic, with emphasis on the Brazilian province.
Methods The disjunct distributions of thirty-®ve reef ®sh species occurring in theBrazilian Province were analysed to allow a better understanding of present biogeo-graphical patterns. To avoid potential bias because of taxonomic problems ormisidenti®cation, we only included families taxonomically well documented, withconspicuous species, and relatively easy to identify.
Results The low-level differentiation between Caribbean and Brazilian species/popula-tions, as well as the presence of restricted populations of a few northern or southernspecies on the opposite side of the Amazon River, clearly indicate that this barrier todispersion can be occasionally bridged. Transoceanic dispersal appears to be frequentand to occur along three routes: from the Caribbean to North East Atlantic, fromnorthern Brazil to the Gulf of Guinea, and from Africa to southern Brazil. Intermediatestepping-stones are apparently not required along transatlantic routes because of therarity of consecutive colonizations. However, intermediate stopovers are persistentlyused along short routes. In all cases, the long-term success of colonists, i.e. theestablishment of stable local populations, appears to depend upon ecological factors.
Main conclusions The puzzling patterns of distribution found in the South Atlanticseem primarily the outcome of allopatric speciation and of the interaction of longdistance dispersal abilities and ecological processes. Successfulness of the colonization ofremote sites appears to be less dependent from dispersal ability than upon persistenceability or settlement preferences.
Keywords
Dispersal, islands, disjunct, colonization, speciation, Amazon barrier, Brazilian prov-ince, distribution.
INTRODUCTION
The South Atlantic Ocean has some peculiar biogeograph-ical and ecological characteristics. The most obvious is the
freshwater out¯ow from the Amazon River that apparentlyacts as a major biogeographical barrier separating the shore®sh faunas of Brazil and the Caribbean (Briggs, 1995;Floeter & Gasparini, 2000). The oceanic islands of thetropical South Atlantic are widely scattered and supportshelf faunas that exhibit reduced species diversity and othereffects of isolation (Floeter & Gasparini, 2000). Consideringthat the islands vary in age, size and distance from othershores, comparative studies of their faunas can reveal
*Correspondence: Departamento de Ecologia e Recursos Naturais,
Universidade Federal do EspõÂrito Santo, Av. Fernando Ferrari S/N, Vito ria,
ES 29060-900, Brazil. E-mail: [email protected]
Journal of Biogeography, 28, 831±841
Ó 2001 Blackwell Science Ltd

considerable information about general patterns of evolutionand distribution (cf. Robertson, 2001). St Helena is the ofone the world's most isolated tropical islands (located from1270 km Ascension and 1870 km from Africa). Others lyingfar out from shore are Ascension, Trindade and St Paul'sRocks (Fig. 1a). Closer to the continental shores, but still farenough to demonstrate the effects of isolation, are otherisland groups located on both sides of the tropical SouthAtlantic. In addition to the oceanic islands, there aresubmerged seamounts (the Fernando de Noronha and theVito ria-Trindade chains) that remain to be explored.
Until recently, the tropical western South Atlantic presen-ted a conspicuous gap in our knowledge of the distributionof tropical reef ®shes (cf. Sale, 1991). In the late 1990s, anincrease in exploration and specimen collection resulted inthe publication of considerable new information about thegeographical distribution of many species (e.g. Moura et al.,1999; Floeter et al., 2001), the description of new species(e.g. Sazima et al., 1998; Rocha & Rosa, 1999; Heiser et al.,2000; Burgess, 2001; Moura et al., 2001), the ®rst compre-hensive collections for certain sites (e.g. Rosa & Moura,1997; Rocha et al., 1998; Gasparini & Floeter, 2001), andthe ®rst works on phylogeography (Bernardi et al., 2000;Bowen et al., 2001; Muss et al., 2001). Also, there is nowavailable new work on the tectonic, eustatic, climatic andoceanographic characteristics of the tropical Atlantic (e.g.Zebiak, 1993; Arz et al., 1999; Knoppers et al., 1999;Stramma & Schott, 1999). These recent advances permit usto draw a ®rst picture of some aspects of the biogeography ofthe region.
METHODS
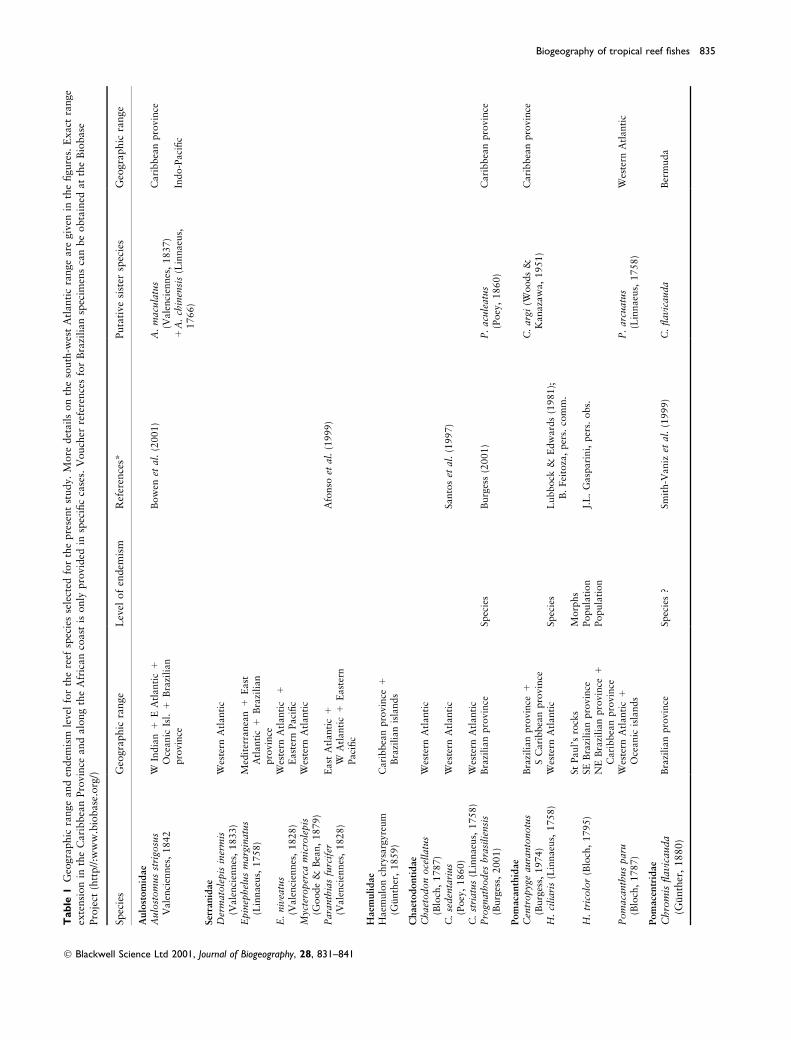
This study is based on the recorded occurrences of thirty-®ve tropical teleost species primarily associated with hardsubstrata (rocky shores, coral and coralline algae reefs),here referred to as `reef ®shes', in the South Atlanticcontinental shelf and oceanic islands (Fig. 1a). To avoidpotential bias caused by taxonomic problems or misiden-ti®cation, we only included families taxonomically welldocumented (e.g. Pomacanthidae, Labridae, Scaridae), withconspicuous (e.g. generally common, large, non-cryptic,colourful, etc.) species, and relatively easy to identify(Table 1). Species not presenting disjunct distributions (i.e. present on the coast while absent in all islands, orendemic to one lone island) were excluded because such`even' distributions are unlikely to bring much insight. Thedistribution patterns have been determined primarilythrough original studies by the authors and colleagues, aswell as compilation of data from the literature (e.g.Lubbock, 1980; Lubbock & Edwards, 1981; Edwards,1990; Afonso et al., 1999; Smith-Vaniz et al., 1999). Thespecies were considered to be resident at a particularlocation if they were suf®ciently abundant to have estab-lished self-recruiting populations or to be vagrants ifexceedingly rare and probably recruited from other sites.Although the Manuel Luis Reefs are not considered to be anoceanic island, they were included as such in this study for
their ecological and biogeographical signi®cance, as theyare the ®rst reef formation south of the Amazon plume(Rocha & Rosa, 2001a).
RESULTS AND DISCUSSION
Distribution patterns
The South Atlantic reef ®sh biogeography proved to be anintricate tapestry (Fig. 1). The thirty-®ve selected speciesdisplayed intriguing distribution patterns. The wrasseHalichoeres bathyphilus and the damsel®sh Chromis ¯avic-auda present strongly accentuated antitropical distributions(Fig. 1c). Other species such as the groupers Epinephelusniveatus and Mycteroperca microlepis and the parrot®shSparisoma atomarium show discontinuous distribution onAmerican coastal reefs (Fig. 1c). The tropical extinctiontheory (the loss of the centre of distribution) is commonlyevoked to explain this type of distribution pattern (TheÂel,1886; Briggs, 1995). Three species found in the south-westare presumed immigrants from African waters the trumpet-®sh Aulostomus strigosus, the grouper Epinephelus margin-atus and the surgeon®sh Acanthurus monroviae (Fig. 1d).Twenty-one species are indistinguishable from their Carib-bean counterparts (Fig. 1e±i). The remaining six species(Fig. 1c and i) are southern endemics the butter¯y®shPrognathodes brasiliensis, the damsel®sh Stegastes fuscus,the wrasses Bodianus insularis, Halichoeres brasiliensis andThalassoma noronhanum, and the parrot®sh Sparisomaaxillare, with close relatives in the northern or easternAtlantic [respectively, P. aculeatus, S. imbricatus, B. pul-chellus/B. rufus, H. radiatus, T. bifasciatum, and S. rubrip-inne (Table 1)].
The south-western Atlantic is overall characterized byhigh endemism, about 18±20% for reef ®shes (Floeter &Gasparini, 2000). However, differences between the Carib-bean and the Brazilian provinces are at the species orsubspecies level, with no genus restricted to the south-western Atlantic. Brazilian corals, in contrast, show a levelof endemism of 33%. Twenty-three percent of thesescleractinian species are considered relict forms onlyremotely related to recent Caribbean species (Laborel,1970; LeaÄo, 1983; J. Laborel, pers. comm.). It has beenargued that the observed differentiation with respect to reef®shes is too low to be accounted for the initial establishmentof a connection of the Amazon River system to the Atlanticin the late Miocene (5±6 Ma) (cf. Hoorn et al., 1995). Wesuspect that `complete' vicariance or allopatric speciationwith stabilizing selection could not have produced suchgeneral closeness between north and south-western Atlanticichthyofaunas across the whole spectrum of ®sh species andlife histories. Thus, vicariance through parapatric speciation(separation with occasional contact) seems the most parsi-monious hypothesis to explain the low morphologicaldifferentiation in spite of the wide and old Amazon barrier(cf. Briggs, 1995; Bowen et al., 2001). The Amazon River®rst created a ®lter, not a full barrier, to dispersal withina once widespread distribution. During the Pliocene
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
832 J.-C. Joyeux et al.

Figure 1 Distribution patterns of selected south-western Atlanticreef ®shes. The letter `v' beside a number indicates that thecorresponding species is a vagrant. (a) Location of sites (excludingGough and Tristan da Cunha), (b) oceanic-scale currents,(c) antitropical distributions and an endemic of mid-Atlantic ridgeislands, (d) colonizers of the south-western Atlantic of Africanorigin. Brazilian Aulostomus is genetically similar to A. strigosus butshows intermediate morphology between A. strigosus and A.maculatus (Bowen et al., 2001), (e) colonizers of the eastern Atlanticof Caribbean or Brazilian origin 1 African population, (f) colonizersof north-eastern Brazil of Caribbean origin, (g) caribbean speciesrestricted to South Atlantic islands. 1 Although vagrant on the coastof Brazil, this species is relatively common in the Vito ria-Trindaderidge and in Trindade Island, (h) widely distributed western Atlanticspecies, and (i) south-western Atlantic endemics and species withrestricted Caribbean distribution. 1 Including subspecies S. fuscustrinidadensis. 2 Including S. lubbocki [Novelli (1998) reviewedmeristic data from all the Stegastes occurring in Brazil and found noconsistent difference between S. pictus and S. lubbocki].
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
Biogeography of tropical reef ®shes 833

(5.3±1.6 Ma) onwards, the delta experienced a rapidseaward growth caused by a major uplift and erosion ofthe Andes (Hoorn, 1994). Thus, it is possible that the ®lterinterrupted faunal interchange for the most salinity orsediment sensitive species (Muss et al., 2001), but that otherspecies breached the barrier on occasion. We hypothesize adrastic but intermittent interruption of passage during highfreshwater out¯ow periods (i.e. at the end of each glacialperiod). Another non-excluding possibility is that genetic¯ow would be completely severed during low sea-levelperiods (cf. Maslin & Mikkelsen, 1998; Arz et al., 1999;Knoppers et al., 1999). During one of the recent loweststands ()50/70 m at 12.7±11.2 ka BPBP) (Arz et al., 1999), theAmazon freshwater and material was directly exported tothe upper shelf slope (present day shelf break is at 75±80 m).This event coincided with high sedimentation rates, possibly
cutting-off passage of shallow-water forms between the twobiogeographical provinces.
It should be noted that the Brazilian province seemsisolated from the Caribbean province because of the direc-tion of prevailing currents, but that the reverse is possiblynot true (Fig. 1b). It is conceivable that species unitybetween western Atlantic, the Brazilian and the Caribbeanprovinces may be maintained in some cases through genetic¯ow from south to north. For instance, some wide-rangingBrazilian species can only be found in the southern tip of theCaribbean (Fig. 1i). The grunt Anisotremus moricandi(Ranzani), the ¯ameback angel®sh Centropyge aurantono-tus, the yellowtail damsel®sh S. pictus and the Brazilianpuffer Canthigaster sp.n., among others, distribute accord-ing to this surprising pattern (Humann, 1996; L.A. Rocha,pers. comm.). On the other hand, some Caribbean species
Figure 1 Continued.
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
834 J.-C. Joyeux et al.
Tab
le1
Geo
grap
hic
range
and
endem
ism
leve
lfo
rth
ere
efsp
ecie
sse
lect
edfo
rth
epre
sent
study.
More
det
ails
on
the
south
-wes
tA
tlan
tic
range
are
give
nin
the
®gu
res.
Exac
tra
nge
exte
nsi
on
inth
eC
arib
bea
nPro
vince
and
along
the
Afr
ican
coas
tis
only
pro
vided
insp
eci®
cca
ses.
Vouch
erre
fere
nce
sfo
rB
razi
lian
spec
imen
sca
nbe
obta
ined
atth
eB
iobas
ePro
ject
(htt
p//
:ww
w.b
iobas
e.org
/)
Spec
ies
Geo
grap
hic
range
Lev
elof
endem
ism
Ref
eren
ces*
Puta
tive
sist
ersp
ecie
sG
eogr
aphic
range
Aulo
stom
idae
Aulo
stom
us
stri
gosu
sV
alen
cien
nes
,1842
WIn
dia
n+
EA
tlan
tic+
Oce
anic
Isl.+
Bra
zili
anpro
vince
Bow
enet
al.
(2001)
A.
mac
ula
tus
(Val
enci
ennes
,1837)
+A
.ch
inen
sis
(Lin
nae
us,
1766)
Car
ibbea
npro
vince
Indo-P
aci®
c
Ser
ranid
ae
Der
mat
ole
pis
iner
mis
(Val
enci
ennes
,1833)
Wes
tern
Atl
anti
c
Epin
ephel
us
mar
ginat
us
(Lin
nae
us,
1758)
Med
iter
ranea
n+
Eas
tA
tlan
tic+
Bra
zili
anpro
vince
E.
niv
eatu
s(V
alen
cien
nes
,1828)
Wes
tern
Atl
anti
c+
Eas
tern
Pac
i®c
Myc
tero
per
cam
icro
lepis
(Goode
&B
ean,
1879)
Wes
tern
Atl
anti
c
Par
anth
ias
furc
ifer
(Val
enci
ennes
,1828)
Eas
tA
tlan
tic+
WA
tlan
tic+
Eas
tern
Pac
i®c
Afo
nso
etal
.(1
999)
Haem
uli
dae
Hae
mulo
nch
rysa
rgyr
eum
(Gu Ènth
er,
1859)
Car
ibbea
npro
vince
+B
razi
lian
isla
nds
Chaet
odonti
dae
Chae
todon
oce
llat
us
(Blo
ch,
1787)
Wes
tern
Atl
anti
c
C.
seden
tari
us
(Poey
,1860)
Wes
tern
Atl
anti
cSa
nto
set
al.
(1997)
C.
stri
atus
(Lin
nae
us,
1758)
Wes
tern
Atl
anti
cP
rogn
athodes
bra
sili
ensi
s(B
urg
ess,
2001)
Bra
zili
anpro
vince
Spec
ies
Burg
ess
(2001)
P.
acule
atus
(Poey
,1860)
Car
ibbea
npro
vince
Pom
aca
nth
idae
Cen
tropyg
eau
ranto
notu
s(B
urg
ess,
1974)
Bra
zili
anpro
vince
+S
Car
ibbea
npro
vince
C.
argi
(Woods
&K
anaz
awa,
1951)
Car
ibbea
npro
vince
H.
cili
aris
(Lin
nae
us,
1758)
Wes
tern
Atl
anti
c
StPau
l's
rock
s
Spec
ies
Morp
hs
Lubbock
&E
dw
ards
(1981);
B.
Fei
toza
,per
s.co
mm
.
H.
tric
olo
r(B
loch
,1795)
SEB
razi
lian
pro
vince
Popula
tion
J.L
.G
aspar
ini,
per
s.obs.
NE
Bra
zili
anpro
vince
+C
arib
bea
npro
vince
Popula
tion
Pom
acan
thus
par
u(B
loch
,1787)
Wes
tern
Atl
anti
c+
Oce
anic
isla
nds
P.
arcu
atus
(Lin
nae
us,
1758)
Wes
tern
Atl
anti
c
Pom
ace
ntr
idae
Chro
mis
¯av
icau
da
(Gu Ènth
er,
1880)
Bra
zili
anpro
vince
Spec
ies
?Sm
ith-V
aniz
etal
.(1
999)
C.
¯av
icau
da
Ber
muda
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
Biogeography of tropical reef ®shes 835

Tab
le1
(Conti
nued
)
Spec
ies
Geo
grap
hic
range
Lev
elof
endem
ism
Ref
eren
ces*
Puta
tive
sist
ersp
ecie
sG
eogr
aphic
range
Chro
mis
scott
i(E
mer
y,1968)
Car
ibbea
npro
vince
+B
razi
lian
pro
vince
Smit
h-V
aniz
etal
.(1
999);
Gas
par
ini
&Flo
eter
(2001);
Roch
a&
Rosa
(2001a)
Mic
rosp
athodon
chry
suru
s(C
uvi
er&
Val
enci
ennes
,1830)
Wes
tern
Atl
anti
c
Steg
aste
sfu
scus
(Cuvi
er&
Val
enci
ennes
,1830)
Bra
zili
anco
ast
Tri
ndad
eis
land
(S.
f.tr
indad
ensi
s)
Spec
ies
Subsp
ecie
sA
llen
(1991);
Gas
par
ini
etal
.(1
999)
S.im
bri
catu
s(J
enyn
s,1840)
+S.
vari
abil
is(C
aste
lnau
,1855)
Afr
ican
coas
t+
isla
nds
Bra
zili
anpro
vince
S.pic
tus
(Cas
teln
au,
1855)
Bra
zili
anpro
vince
+T
rinid
ade
&T
obag
oSp
ecie
sN
ove
lli
(1998);
L.A
.R
och
a,per
s.C
om
m.
Steg
ates
par
titu
s(P
oey
,1868)
+St
egas
tes
lubbock
iA
llen
&Sm
ith,
1992
Car
ibbea
npro
vince
Asc
ensi
on
Labri
dae
Bodia
nus
insu
lari
s(G
om
on
&L
ubbock
,1979)
Mid
-Atl
anti
cri
dge
isla
nds
Spec
ies
Gom
on
&L
ubbock
(1980)
B.
rufu
s(L
innae
us,
1758)
+B
.pulc
hel
lus
(Poey
,1860)
Wes
tern
Atl
anti
c
Wes
tern
Atl
anti
c+
SaÄo
Tom
e ÂH
alic
hoer
esbat
hyp
hil
us
(Bee
be
&T
ee-V
an,
1932)
Bra
zili
anpro
vince
Spec
ies
?Sm
ith-V
aniz
etal
.(1
999)
H.
bat
hyp
hil
us
(Bee
be
&T
ee-V
an,
1932)
Ber
muda
H.
biv
itta
tus
(Blo
ch,
1791)
Wes
tern
Atl
anti
cH
.bra
sili
ensi
s(B
loch
,1791)
Bra
zili
anpro
vince
Spec
ies
Roch
a&
Rosa
(2001b)
H.
radia
tus
(Lin
nae
us,
1758)
Car
ibbea
npro
vince+
NE
Bar
azil
ian
isla
nds
Thal
asso
ma
noro
nhan
um
(Boule
nge
r,1890)
Bra
zili
anpro
vince
Spec
ies
Roch
aet
al.
(2001)
T.
bif
asci
atum
(Blo
ch,
1791)
Car
ibbea
npro
vince
Sca
ridae
Spar
isom
aat
om
ariu
m(P
oey
,1861)
Wes
tern
Atl
anti
c
S.ax
illa
re(S
tein
dac
hner
,1878)
Bra
zili
anpro
vince
Spec
ies
Ber
nar
di
etal
.(2
000);
Moura
etal
.(2
001)
S.ru
bri
pin
ne
(Val
enci
ennes
,1839)
+Sp
aris
om
aaf
f.R
ubri
pin
ne
(Val
enci
ennes
,1839)
Car
ibbea
npro
vince
Eas
tern
Atl
anti
c
Aca
nth
uri
dae
Aca
nth
uru
sbah
ianus
(Cas
teln
au,
1855)
Bra
zili
anpro
vince
Car
ibbea
npro
vince
Popula
tion
Popula
tion
Bri
ggs
&C
aldw
ell;
(1957);
Roch
aet
al.,
per
s.co
mm
.A
.ra
ndal
li(B
rigg
s&
Cal
dw
ell,
1957)
Eas
tern
Gulf
of
Mex
ico
A.
chir
urg
us
(Blo
ch,
1787)
Wes
tan
dE
ast
Atl
anti
cA
.co
erule
us
(Blo
ch&
Schnei
der
,1801)
Wes
tern
Atl
anti
c
A.
monro
viae
(Ste
indac
hner
,1876)
Eas
tern
Atl
anti
cQ
ue Âr
oet
al.
(1990);
Gola
ni
&So
nin
(1996);
Moura
(2000);
C.E
.L.
Fer
reir
a,per
s.obs.
*G
ener
alre
fere
nce
suse
dfo
rth
eW
este
rnA
tlan
tic
are
Bo Èhlk
e&
Chap
lin
(1968),
Hum
ann
(1996)
and
Ran
dal
l(1
996),
and
for
the
Eas
tern
Atl
anti
car
e:Q
ue Âr
oet
al.
(1990),
Deb
eliu
s(1
997)
and
Edw
ards
etal
.(2
001).
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
836 J.-C. Joyeux et al.

are restricted to a few sites south of the Amazon (e.g. thedamsel®sh Chromis scotti and the wrasse Halichoeresradiatus), generally on isolated locations (Fig. 1g). Suchdistributions are a clear indication that the isolation betweenBrazil and the Caribbean is occasionally bridged ± one wayor the other. Periods of isolation can be interrupted bysporadic gene ¯ow through larval transport by currents,active swimming abilities (Stobutski & Bellwood, 1997),rafting (Mora, 2001) or even through the colonization ofdeep-water sponge habitats under the Amazon River plume(Collette & Rutzler, 1977; Uyeno et al., 1983; Rocha et al.,2000).
Long-distance dispersal
Larvae of a number of species can successfully travelenormous distances before reaching suitable settling groundand colonizing new areas. Five species have thus apparentlymade the eastward transatlantic crossing. Two of them(Chaetodon sedentarius at Azores and Madeira, and theangel®sh Pomacanthus paru in the Gulf of Guinea) wereonly found vagrants in the East Atlantic, presumablyindicating recent arrival (Fig. 1e). The other three (Creole-®sh Paranthias furcifer, B. pulchellus and Sparisomaaff. rubripinne) established restricted populations (Afonsoet al., 1999; Bernardi et al., 2000) (Fig. 1e). Two specieswith now resident western populations most probablycrossed the other way (cf. Bowen et al., 2001) (Fig. 1d).However, as shown by the relative commonness of vagrantsfrom distant source populations, the establishment offounding populations appears much rarer than recruitment.In addition to the two recorded stray species in the EastAtlantic, the African acanthurid Acanthurus monroviae wasrecently found as a vagrant in south-east Brazil (Moura,2000; C.E.L.F. pers. obs.) and in the eastern Mediterranean(Golani & Sonin, 1996) (Fig. 1d). Other examples includethe thirteen species that were recorded only as juveniles inBermuda (including Chromis scotti; Fig. 1g) (Smith-Vanizet al., 1999). This pattern (comparative number of speciescrossings, limited distribution in colonized areas) ®ts withmodern and fossil records for shell-bearing mollusks(Vermeij & Rosenberg, 1993). Invaders of the East Atlanticcould have crossed the ocean from northern Brazil to theGulf of Guinea via the Equatorial Under Current (Stramma& Schott, 1999; Muss et al., 2001) (Fig. 1b and e), possiblyfavoured by ENSO-type or ENSO-induced events (Venegaset al., 1996; Stramma & Schott, 1999). Alternatively, otherspecies [e.g. C. sedentarius and Adudefduf taurus (MuÈ ller &Troschel)] (Debelius, 1997; Edwards et al., 2001) appear tohave crossed from the Caribbean to the Macaronesianislands off northern African via the Gulf Stream. In spite ofthe westward ¯owing South Equatorial Current, invasionsfrom Africa to the west are rarer, as evidenced by thelimited present distribution of the few putative colonizers ofSouth America (Fig. 1d). Stepping stones do not appearnecessary for transatlantic passage in either direction. Theinsular hog®sh B. insularis is endemic and the lone memberof its genus on the mid-Atlantic Ridge islands (Fig. 1c),
although its sister species B. pulchellus crossed the tropicalAtlantic (Fig. 1e). It is unlikely that two or more consecu-tive and successful colonizations can occur. The presence ofvarious species at oceanic islands is thus to be seen asre¯ecting the species' ability to disperse and colonize, not asa pre-requisite for further transport.
There has been an increasing attention given to theevolutionary signi®cance of dispersal and retention of earlylife-history stage (e.g. Jones et al., 1999; Swearer et al.,1999). Overall, reef ®sh seem to have evolved and success-fully dealt with the dispersion/retention duality. As evi-denced by established populations at geologically recentvolcanic islands such as Ascension (1.5 Myr) and bytransoceanic vagrants, species may be dispersed far away,thus colonizing new areas and avoiding extinction. How-ever, mass recruitment is essentially local (Jones et al., 1999;Swearer et al., 1999). Recruitment in isolated environmentsnot only allows the persistence of endemic populations(Robertson, 2001), but also may permit allopatric speci-ation. A notable example is the possibly on-going differen-tiation of Holacanthus ciliaris (Fig. 2a±b) and Chromismultilineata (Guichenot) through possible founder effectsand inbreeding at St Paul's (cf. Edwards & Lubbock, 1983).It would be of interest to know if H. ciliaris colour morphsperceive themselves as different `species', as they apparentlyoccupy different microhabitats (B. Feitoza and Osmar LuizJr, pers. comm.).
The type of spawning mode and the larval duration of theplanktonic phase are often assumed to be major constraintson the dispersal abilities of marine species (see review inSale, 1991, for a counter-example see Victor & Wellington,2000). Among the Brazilian endemic reef ®shes, 74% aresmall benthic demersal spawners, which evidently could notreadily transgress the Amazon barrier (Floeter & Gasparini,2000). A duality is readily apparent on oceanic islandswhere benthic spawners are more frequently endemic(through lack of genetic input), and pelagic spawnersproportionately more speciose (through higher dispersal),than at continental shelves (Floeter & Gasparini, 2000).Recent expeditions to St Paul's Rocks (B. Feitoza et al.,unpubl. data) noted the rarefaction of the semi-albinoticsH. ciliaris, and the comparatively higher abundance of thelocal blue morph (Fig. 2a) (cf. also Lubbock & Edwards,1981). That such island populations can undergo large¯uctuations within a few years appears to illustrate thatreversal to ancestral population composition is unlikelywithout integrating new immigrants. Thus, concurrentlywith the high-observed frequency of vagrants, we infer thatlong-distance dispersal processes are more common thangenerally predicted. For both pelagic and benthic spawners,however, short route dispersal would be eased by thepresence of stepping stones, and the south-western Atlanticis no exception to the rule. Trindade Island presents higherreef ®sh diversity than would be expected from its distancefrom the coast because of the seamounts of the Vito ria-Trindade chain (Gasparini & Floeter, 2001). Further north,St Paul's Rocks' ichthyofauna could be considered as animpoverished caricature of Fernando de Noronha (Fig. 1h),
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
Biogeography of tropical reef ®shes 837

thus showing the dependency of the former's fauna to thelatter's colonization.
Long-term success of colonists
Some of the observed features cannot easily be linked todispersal abilities or possible routes. Of special interest is theabsence of the family Acanthuridae (surgeon®shes) fromSt Paul's Rocks, probably the only tropical island in theworld missing this conspicuous reef associated family(Floeter et al., 2001). Similarly, the wide-ranging butter¯y-®sh Chaetodon striatus and the hog®sh B. pulchellus werenot recorded at Atol das Rocas while both are present inneighbouring Fernando de Noronha. These local absencescould be related to the small size of the islands, and thushabitat availability together with possible post-settlement
dif®culties. The genus Scarus (family Scaridae), which occursat oceanic sites in the Indo-Paci®c and North Atlantic(Smith-Vaniz et al., 1999), is absent from the South Atlanticoceanic islands. On the other hand, all four scarid speciesthat do occur on South Atlantic oceanic islands belong to thegenus Sparisoma. This pattern could re¯ect the broaderfeeding habits of this genus (Bernardi et al., 2000). TheFrench angel®sh P. paru is common in the Caribbean, on theBrazilian coast, Atol das Rocas, Fernando de Noronha andSt Paul's Rocks. Surprisingly, on Trindade Island this speciesis apparently replaced by H. tricolor. The island is the onlyone in the South Atlantic that harbors a resident H. tricolorpopulation (Gasparini & Floeter, 2001). These pelagic-spawning species certainly do not have larval dispersalrestrictions, because the acanthurids, C. striatus, B. pulchel-lus and P. paru have successfully colonized very isolated sites
Figure 2 Holacanthus ciliaris colourmorphs from St Paul's Rocks: (a) blue morph,and (b) semi-albinotic morph. In situ photo-graphs by Osmar Luiz Jr.
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
838 J.-C. Joyeux et al.

such as Ascension, Bermuda, and Africa. Thus, specialecological requirements for proper larval or post-settlementdevelopment, including habitat and predation rates, pre-sumably determine the long-term success of colonists.
It has been argued that competition could hinder orprevent colonization of high diversity locations. EasternPaci®c ®sh and gastropod species have apparently beenunable to colonize the Indo-western Paci®c, while theopposite is frequent (Vermeij & Rosenberg, 1993; Briggs,1995). As is known from shell-bearing mollusks (cf. Vermeij& Rosenberg, 1993), western Atlantic ®shes that havecolonized Africa generally have extremely limited distribu-tions in the eastern Atlantic (Briggs, 1995) (Fig. 1d and e),most often occurring on the offshore islands of Cape Verdeand SaÄo Tome archipelago, where competition is probablylower.
Extinctions are usually believed to be rare events occurringonly on a long time-span. Even in the smallest and mostisolated islands, endemic ®shes have survived through evolu-tionary time under very restricted areas in high-risk situations(Robertson, 2001). However, local extinction processes ofapparently established populations are probably more fre-quent than generally expected. Historical disappearance orsevere decline of population of reef ®sh species caused bynatural (e.g. El NinÄo) or man-made disturbance (e.g. reefdegradation, intensive aquarium collection, over®shing) isnow documented (Smith-Vaniz et al., 1999; Hawkins et al.,2000), and has been related to the restricted geographicalrange of the species (Hawkins et al., 2000). The survival ofpotentially valuable morphs or endemic species on south-western oceanic islands is under serious threat as theaquarium trade is moving in (J.L. Gasparini & S.R. Floeter,pers. obs.). Present-day distribution patterns may or may notprovide indications of the local long-term history of species.An interesting feature found in the western Atlantic is that ofspecies present in the entire Caribbean Basin but restricted tooffshore localities in the south-western Atlantic (Fig. 1i).Assuming the latter are relict populations from a oncewidespread distribution, the pattern does not ®t with theobservation that generally the northern component of adisjunct antitropical or antiequatorial population is the ®rstto disappear (Briggs, 1995). However, one can argue thatinferences from observations in the Paci®c are dif®cult totranspose into the Atlantic because the West Indies areconsiderably more extensive and diverse than the Brazilianprovince and thus more ef®cient at providing tropical habitatsafter range reduction, especially during glacial periods.
CONCLUSIONS
The puzzling patterns of distribution found in the SouthAtlantic seem primarily the outcome of allopatric speciationand of the interaction of long distance dispersal abilities andecological processes. One emerging idea is that persistenceability or settlement preferences (see OÈ hman et al., 1998) aremore important than dispersal ability. Given enough timemany species can reach extremely isolated reefs. Thedocumented arrival of vagrants clearly indicates this capa-
bility, even within the short period of recent scienti®cinvestigations. However, not all vagrants can complete theirlife history there. No stepping stones are required alongtransoceanic routes because of the rarity of consecutivecolonizations. However, intermediate stopovers are persist-ently used along short routes. Finally, some patterns cannotbe given a satisfactory explanation. For example, why thespot®n hog®sh B. pulchellus is present at the SaÄo TomeÂarchipelago when its closest relative (Sazima & Gasparini,1999), and always sympatric, the Spanish hog®sh B. rufus isnot? Just chance?
The puzzle will certainly remain unclear for a long time. Agreat effort will be required to resolve central issues such asthe importance of speciation, long distance transport,colonization success, etc. that shaped the biogeography ofSouth Atlantic reef ®shes.
ACKNOWLEDGMENTS
This study is dedicated to John C. Briggs for his lifetimecontribution to the science of biogeography and for inspiringmany generations of researchers. The work greatly bene®tedfrom discussions or comments on earlier drafts from thefollowing colleagues: John Brigg, Alasdair Edwards, Benja-min Victor, Brian Bowen, Julian Caley, Phil Munday, andHoward Choat. Unpublished information was kindly provi-ded by Bertran Feitoza (Universidade Federal da ParaõÂba,JoaÄo Pessoa, PB, Brazil), Osmar Luiz Jr, Luiz Rocha(University of Florida, Department of Fisheries and AquaticSciences, Gainesville, FL, USA), and Jacques Laborel (Che-min des grands Bassins, La Ciotat, France).
REFERENCES
Afonso, P., Porteiro, F.M., Santos, R.S., Barreiros, J.P., Worms,J. & Wirtz, P. (1999) Coastal marine ®shes of SaÄo TomeÂIsland (Gulf of Guinea). ArquipeÂlago, 17A, 65±92.
Allen, G.R. (1991) Damsel®shes of the world, 271 p. Mergus,Melle, Germany.
Arz, H.W., PaÈtzold, J. & Wefer, G. (1999) Climatic changesduring the last deglaciation recorded in sediment cores fromthe north-eastern Brazilian Continental Margin. Geo-MarineLetters, 19, 209±218.
Bernardi, G., Robertson, D.R., Clifton, K.E. & Azzurro, E.(2000) Molecular systematics and evolutionary ecology of theAtlantic parrot®sh genus Sparisoma. Molecular Phylogeneticsand Evolution, 15, 292±300.
BoÈ hlke, J.E. & Chaplin, C.C.G. (1968) Fishes of the Bahamasand adjacent tropical waters. Livingston Publishers,Wynnwood.
Bowen, B.W., Bass, A.L., Garcia-Rodriguez, A., Rocha, L.A. &Robertson, D.R. (2001) Phylogeography of the trumpet®sh,Genus Aulostomus: ring species complex on a planetary scale.Evolution, 55, 1029±1039.
Briggs, J.C. (1995) Global biogeography. Developments inpaleontology and Stratigraphy, Vol. 14. Elsevier, Amsterdam.
Briggs, J.C. & Caldwell, D.K. (1957) Acanthurus randalli, anew surgeon®sh from the Gulf of Mexico. Bulletin of theFlorida State Museum, Biological Sciences, 2, 43±51.
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
Biogeography of tropical reef ®shes 839

Burgess, W.E. (2001) Prognathodes brasiliensis, a new species ofbutter¯y®sh (Teleostei: Chaetodontidae) from southernBrazil. Tropical Fish Hobbyist, 6, 56±63.
Collette, B.B. & Rutzler, K. (1977) Reef ®shes over spongebottoms off the mouth of the Amazon River. Proceedingsof the Third International Coral Reef Symposium, 1,305±310.
Debelius, H. (1997) Mediterranean and Atlantic ®sh guide.IKAN-Unterwasserarchiv, Frankfurt.
Edwards, A.J. (1990) Fish and ®sheries of Saint Helena Island.Centre for Tropical Coastal Management Studies, Universityof Newcastle upon Tyne, England.
Edwards, A.J. & Lubbock, R. (1983) Marine zoogeography ofSt. Paul's Rocks. Journal of Biogeography, 10, 65±72.
Edwards, A.J., Gill, A.C. & Abohweyere, P.O. (2001) Irvine'smarine ®shes of tropical West Africa, pp. 150. (in press)
Floeter, S.R. & Gasparini, J.L. (2000) The south-westernAtlantic reef ®sh fauna: composition and zoogeographicpatterns. Journal of Fish Biology, 56, 1099±1114.
Floeter, S.R., GuimaraÄes, R.Z.P., Rocha, L.A., Ferreira, C.E.L.,Rangel, C.A. & Gasparini, J.L. (2001) Geographic variationin reef-®sh assemblages along the Brazilian coast. GlobalEcology and Biogeography 10, 423±433.
Gasparini, J.L. & Floeter, S.R. (2001) The shore ®shes ofTrindade Island, western South Atlantic. Journal of NaturalHistory (in press).
Gasparini, J.L., Moura, R.L. & Sazina, I. (1999) Stegastestrindadensis n. sp. (Pisces: Pomacentridae), a new damsel®shfrom Trindade Island, off Brazil. Boletim do Museu deBiologia Mello LeitaÄo (Nova SeÂrie), 10, 3±11.
Golani, D. & Sonin, O. (1996) The occurrence of the tropicalwest African ®shes Acanthurus monroviae (Acanthuridae)and Arius parkii (Aridae) in the Levant. Journal of Ichthy-ology and Aquatic Biology, 2, 1±3.
Gomon, M.F. & Lubbock, R. (1980) A new hog®sh of the genusBodianus (Teleostei, Labridae) from islands of the mid-Atlantic ridge. Northeast Gulf Science, 3 [for 1979], 104±111.
Hawkins, J.P., Roberts, C.M. & Clark, V. (2000) The threat-ened status of restricted-range coral reef ®sh species. AnimalConservation, 3, 81±88.
Heiser, J.B., Moura, R.L. & Robertson, D.R. (2000) Two newspecies of creole wrasse (Labridae: Clepticus) from oppositesides of the Atlantic. Journal of Ichthyology and AquaticBiology, 4, 67±76.
Hoorn, C. (1994) A reconstruction of the paleo-Amazon Riversystem (middle-late Miocene, NW Amazonia). Paleogeogra-phy, Paleoclimatology and Palaeoecology, 112, 187±238.
Hoorn, C., Guerrero, J., Sarmiento, G.A. & Lorente, M.A. (1995)Andean tectonics as a cause for changing drainage patternsin Miocene northern South America. Geology, 23, 237±240.
Humann, P. (1996) Reef ®sh identi®cation: Florida, Caribbean,Bahamas, 2nd edn, pp. 424. New World Publications,Jacksonville.
Jones, G.P., Milicich, M.J., Emslie, M.J. & Nunow, C. (1999)Self-recruitment in a coral reef ®sh population. Nature, 402,802±804.
Knoppers, B., Ekau, W. & Figueiredo, A.G. (1999) The coastand shelf of east and northeast Brazil and material transport.Geo-Marine Letters, 19, 171±178.
Laborel, J. (1970) Les peuplements des madreÂporaires des coà testropicales Do BreÂsi. Annals of the University of Abidjan,Series E, 2, 1±260.
LeaÄo, Z.M.A.N. (1983) Abrolhos ± o refu gio pleistoceÃnico deuma fauna terciaÂria de corais. CieÃncias da Terra, 8, 22±24.
Lubbock, R. (1980) The shore ®shes of Ascension Island.Journal of Fish Biology, 17, 283±303.
Lubbock, R. & Edwards, A. (1981) The ®shes of Saint Paul'srocks. Journal of Fish Biology, 18, 135±157.
Maslin, M. & Mikkelsen, N. (1998) Timing of the lateQuaternary Amazon Fan Complex mass-transport deposits.Geological evolution of ocean basins: results from the oceandrilling program, Vol. 131 (eds A. Cramp, C.J. Macleod, S.V.Lee and E.J.W. Jones), pp. 129±150. Special Publications,Geological Society, London.
Mora, C. (2001) Dispersal of reef ®shes by rafting. ReefEncounter, 29, 16±17.
Moura, R.L. (2000) Non-indigenous reef ®shes in the south-western Atlantic. Abstracts of the Ninth International CoralReef Symposium, 1, 288.
Moura, R.L., Gasparini, J.L. & Sazima, I. (1999) New recordsand range extensions of reef ®shes in the western southAtlantic, with comments on reef ®sh distribution along theBrazilian coast. Revista Brasileira de Zoologia, 16, 513±530.
Moura, R.L., Figueiredo, J.L. & Sazima, I. (2001) A newparrot®sh (Scaridae) from Brazil, and revalidation of Spari-soma frondosum (Agassiz, 1831), S. amplum (Ranzani,1842), S. Axillare (Steindachner, 1878) and Scarus trispinosusValenciennes, 1840. Bulletin of Marine Science (in press).
Muss, A., Robertson, D.R., Stepien, C.A., Wirtz, P. & Bowen,B.W. (2001) Phylogeography of Ophioblennius: the role ofocean currents and geography in reef ®sh evolution. Evolu-tion, 55, 561±572.
Novelli, R. (1998) Revis~ao TaxonoÃmica das espeÂcies de Steg-astes Jenyns, 1842 (Pomacentridae, Teleostei) ocorrentes noBrasil. PhD Thesis, Universidade Estadual do Norte Flumin-ense Campos, RJ, p. 129.
OÈ hman, M.C., Munday, P.L., Jones, G.P. & Caley, M.J. (1998)Settlement strategies and distribution patterns of coral-reef®shes. Journal of Experimental Marine Biology and Ecology,225, 219±238.
QueÂro, J.C., Hureau, J.C., Karrer, C., Post, A. & Saldanha, L.(eds) (1990) Check-list of the ®shes of the eastern tropicalAtlantic (CLOFETA), 1492 p. JNICT, Lisbon; SEI, Paris;UNESCO, Paris (3 volumes).
Randall, J.E. (1996) Caribbean reef ®shes, 3rd edn, p. 358.T.H.F., Neptune City, NJ.
Robertson, D.R. (2001) Population maintenance among trop-ical reef ®shes: inferences from small-island endemics.Proceedings of the National Academy of Science, 98,5667±5670.
Rocha, L.A. & Rosa, I.L. (1999) New species of Haemulon(Teleostei: Haemulidae) from the NE Brazilian Coast. Copeia,1999, 447±452.
Rocha, L.A. & Rosa, I.L. (2001a) Baseline assessment of reef®sh assemblages of Parcel Manuel Luiz Marine State Park,MaranhaÄo, north-east Brazil. Journal of Fish Biology, 58,985±998.
Rocha, L.A. & Rosa, R.S. (2001b) Halichoeres brasiliensis(Bloch, 1791), a valid wrasse species (Teleostei: Labridae)
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
840 J.-C. Joyeux et al.

from Brazil, with notes on the Caribbean species Halichoeresradiatus (Linnaeus, 1758). Journal of Ichthyology andAquatic Biology, 4(4), 161±166.
Rocha, L.A., Rosa, I.L. & Rosa, R.S. (1998) Peixes recifais dacosta da ParaõÂba, Brasil. Revista Brasileira de Zoologia, 15,553±566.
Rocha, L.A., Rosa, I.L. & B.M. Feitoza (2000) Sponge-dwelling®shes of northeastern Brazil. Environmental Biology ofFishes, 59, 453±458.
Rocha, L.A., GuimaraÄes, R.Z.P. & Gasparini, J.L. (2001)Redescription of the Brazilian Wrasse Thalassoma noronha-num (Boulenger, 1890) (Teleostei: Labridae). Journal ofIchthyology and Aquatic Biology, 4, 105±108.
Rosa, R.S. & Moura, R.L. (1997) Visual assessment of reef ®shcommunity structure in the Atol das Rocas BiologicalReserve, off NE Brazil. Proceedings of the Eighth Interna-tional Coral Reef Symposium, 1, 983±986.
Sale, P.F. (ed.) (1991) The ecology of ®shes on coral reefs.Academic Press, San Diego.
Santos, R.S., Porteiro, F.A. & Barreiros, J.P. (1997) Marine®shes of the Azores. Annotated checklist and bibliography.ArquipeÂlago. Suppl. 1, xxv + 242 pp.
Sazima, I. & Gasparini, J.L. (1999) A presumed hybrid of thehog®shes Bodianus pulchellus and Bodianus rufus in thewestern South Atlantic. Journal of Ichthyology and AquaticBiology, 3, 89±92.
Sazima, I., Gasparini, J.L. & Moura, R.L. (1998) Grammabrasiliensis, a new basslet from the western South Atlantic(Perciformes: Grammatidae). Journal of Ichthyology andAquatic Biology, 3, 39±43.
Smith-Vaniz, W., Collette, B.B. & Luckhurst, B.E. (1999) Fishesof Bermuda: history, zoogeography, annotated checklist, andidenti®cation keys. ASIH, Special Publication Number 4,Kansas.
Stobutski, I.C. & Bellwood, D.R. (1997) Sustained swimmingabilities of the late pelagic stages of coral reef ®shes. MarineEcology, Progress Series, 149, 35±41.
Stramma, L. & Schott, F. (1999) The mean ¯ow ®eld of thetropical Atlantic Ocean. Deep-Sea Research II 46, 279±303.
Swearer, S.E., Caselle, J.E., Lea, D.W. & Warner, R.R. (1999)Larval retention and recruitment in an island population ofcoral-reef ®sh. Nature, 402, 799±802.
TheÂel, H. (1886) The voyage of H. M. S. Challenger. Zoology,14, 1±290.
Uyeno, T., Matsuura, K. & Fujii, E. (1983) Fishes Trawled OffSurinam and French Guiana. Japan Marine Fishery ResourceResearch Center, Tokyo.
Venegas, S.A., Mysak, L.A. & Straub, D.N. (1996) Evidence forinterannual and interdecadal climate variability in the SouthAtlantic. Geophysical Research Letters, 23, 2673±2676.
Vermeij, G.J. & Rosenberg, G. (1993) Giving and receiving ±the tropical Atlantic as donor and recipient for invadingspecies. American Malacological Bulletin, 10, 181±194.
Victor, B.C. & Wellington, G.M. (2000) Endemism and thepelagic larval duration of reef ®shes in the eastern Paci®cOcean. Marine Ecology, Progress Series, 205, 241±248.
Zebiak, S.E. (1993) Air±sea interaction in the EquatorialAtlantic region. Journal of Climate, 6, 1567±1586.
BIOSKETCHES
Dr Jean-Christophe Joyeux is Professor of Ecology at theUniversidade Federal do EspõÂrito Santo and is currentlyleading various projects on larval ®sh ecology.
Sergio R. Floeter is a PhD candidate at the UniversidadeEstadual do Norte Fluminense. He is currently leading anmulti-institutional team to build a reference data base forthe tropical Atlantic reef ®shes.
Dr Carlos Eduardo L. Ferreira works as marine resear-cher at IEAPM Institute (Brazilian Navy), and has beendealing with various aspects of the ecology of Brazilianreefs and ®shes for more than a decade.
JoaÄo Luiz Gasparini is af®liated to the UniversidadeFederal do EspõÂrito Santo, and has worked for the last15 years on the taxonomy, systematics and naturalhistory of Brazilian reef ®shes.
Ó Blackwell Science Ltd 2001, Journal of Biogeography, 28, 831±841
Biogeography of tropical reef ®shes 841




















