
Biodiversity of the Genus Hypocrea/Trichoderma in Different Habitats
37
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Biodiversity of the Genus Hypocrea/Trichoderma in Different Habitats


BIOTECHNOLOGY AND BIOLOGY OF
TRICHODERMA
Vijai K. Gupta, MoniKa SchMoll, alfredo herrera-eStrella, r. S. upadhyay, irina druzhinina, Maria G. tuohy
AMSTERDAM • BOSTON • HEIDELBERG • LONDON • NEW YORK • OXFORD PARIS • SAN DIEGO • SAN FRANCISCO • SINGAPORE • SYDNEY • TOKYO

Elsevier225, Wyman Street, Waltham, MA 02451, USAThe Boulevard, Langford Lane, Kidlington, Oxford OX5 1GB, UKRadarweg 29, PO Box 211, 1000 AE Amsterdam, The Netherlands
Copyright © 2014 Elsevier B.V. All rights reserved.
No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form or by any means electronic, mechanical, photocopying, recording or otherwise without the prior written permission of the publisher.
Permissions may be sought directly from Elsevier’s Science & Technology Rights Department in Oxford, UK: phone (+44) (0) 1865 843830; fax (+44) (0) 1865 853333; email: [email protected]. Alternatively you can submit your request online by visiting the Elsevier web site at http://elsevier.com/locate/permissions, and selecting Obtaining permission to use Elsevier material.
NoticeNo responsibility is assumed by the publisher for any injury and/or damage to persons or property as a matter of products liability, negligence or otherwise, or from any use or operation of any methods, products, instructions or ideas contained in the material herein. Because of rapid advances in the medical sciences, in particular, independent verification of diagnoses and drug dosages should be made.
British Library Cataloguing in Publication DataA catalogue record for this book is available from the British Library
Library of Congress Cataloging-in-Publication DataA catalog record for this book is available from the Library of Congress
ISBN: 978-0-444-59576-8
Printed and bound in Poland14 15 16 17 18 10 9 8 7 6 5 4 3 2 1
For information on all Elsevier publications visit our web site at store.elsevier.com

v
Preface xiForeword xiiiList of Contributors xv
ABIOLOGY AND BIODIVERSITY
1 Biodiversity of the Genus Hypocrea/Trichoderma in Different Habitats
LÁSZLÓ KREDICS, LÓRÁNT HATVANI, SHAHRAM NAEIMI, PÉTER KÖRMÖCZI, LÁSZLÓ MANCZINGER, CSABA VÁGVÖLGYI,
IRINA DRUZHININA
Introduction 3Methodology of Studying Trichoderma Biodiversity 3Trichoderma Diversity in Different Habitats 5Conclusions 18Acknowledgments 18References 18
2 Ecophysiology of Trichoderma in Genomic PerspectiveLEA ATANASOVA
Trichoderma in Its Ecological Niche 25From Diversity to Genomics 27Mycotrophy of Trichoderma 28Saprotrophy of Trichoderma on Dead Wood 30Trichoderma Growth in Soil 31Rhizosphere Competence of Trichoderma 32Trichoderma versus Mycorrhizae 32Trichoderma + Bacteria = ? 33Facultative Endophytism of Trichoderma 33Animal Nourishment of Trichoderma 34Most of the Famous Trichoderma Species are Environmental
Opportunists 34Versatile Carbon Utilization Patterns Reflect Ecological
Specialization of Trichoderma spp. 35Acknowledgments 37References 37
3 DNA Barcode for Species Identification in TrichodermaLÓRÁNT HATVANI, CSABA VÁGVÖLGYI, LÁSZLÓ KREDICS,
IRINA DRUZHININA
Introduction 41The Tools 42Application of DNA Barcoding in Species-Level Identification
of Trichoderma 43Taxonomic Studies 43
Biodiversity Studies 45Identification of Industrial Trichoderma Strains 47Identification of Biocontrol Trichoderma Strains 48Identification of Trichoderma Isolates with Clinical Relevance 50Identification of Mushroom Pathogenic Trichoderma Strains 51Conclusions 51Acknowledgments 52References 52
4 Understanding the Diversity and Versatility of Trichoderma by Next-Generation Sequencing
CHRISTIN ZACHOW AND GABRIELE BERG
Introduction 57Access to Fungal and Trichoderma Diversity—Taxonomic
Profiling 58Plants Life under Control of Trichoderma—Functional Profiling 62Conclusion 63Acknowledgments 63References 63
5 Molecular Evolution of Trichoderma ChitinasesVERENA SEIDL-SEIBOTH, KATARINA IHRMARK,
IRINA DRUZHININA, MAGNUS KARLSSON
Introduction 67Phylogeny and Evolution of the GH Family 18 Gene Family in
Trichoderma 68Subgroup A Chitinases 69Subgroup B Chitinases 71Subgroup C Chitinases 74Conclusions 77Acknowledgments 77References 77
BSECRETION AND PROTEIN
PRODUCTION
6 Protein Production—Quality Control and Secretion Stress Responses in Trichoderma reeseiM. SALOHEIMO T. PAKULA N. ARO, J.J. JOENSUU
Introduction—Milestones of Trichoderma reesei 81Protein Secretome of T. reesei 82ER Quality Control and Secretion Stress Responses 84Conclusion 86References 86
Contents

CONTENTSvi
7 Heterologous Expression of Proteins in TrichodermaHELENA NEVALAINEN AND ROBYN PETERSON
Introduction 89Promoter Options 92Fusion Partners 93Extracellular Proteases 94Secretion Stress in the Frame 95Mass Production of Heterologous Protein by Fermentation 97N-glycosylation of Heterologous Proteins Produced in T. reesei 97Conclusions 98Acknowledgments 99References 99
8 Trichoderma Secretome: An OverviewSUNIL S. ADAV AND SIU KWAN SZE
Introduction 103Proteomic Analysis of Secretory Proteins 105Extraction of Extracellular Proteins for Proteomic Analysis 106Extracellular Protein Secretion by T. reesei 107Polysaccharide Degradation Machinery of T. reesei 108New Candidates in Cellulose Degradation 109Hemicellulose Hydrolyzing Enzymes 110Lignin Degradation by T. reesei 111Industrial Applications of T. reesei Cellulolytic Enzymes 111Conclusion 112References 112
9 The Secretory Pathway in the Filamentous Fungus Trichoderma
MARCO J. HERNÁNDEZ-CHÁVEZ, ROBERTO J. GONZÁLEZ-HERNÁNDEZ, JOSÉ E. TRUJILLO-ESQUIVEL, ARTURO HERNÁNDEZ-CERVANTES,
HÉCTOR M. MORA-MONTES
Introduction 115Translocation 115Cotranslational Translocation 116Post Translational Translocation 116Protein Modifications in the ER 116Vesicle Transport from ER to Golgi Complex and Trafficking
within the Golgi Cisternae 118Transport after Trafficking within the Golgi Complex 119Secreted Proteins in Trichoderma 119Concluding Remarks 120Acknowledgments 120References 120
CSECONDARY METABOLISM
10 Secondary Metabolism and Antimicrobial Metabolites of Trichoderma
ROSA HERMOSA, ROSA ELENA CARDOZA, MARÍA BELÉN RUBIO, SANTIAGO GUTIÉRREZ, ENRIQUE MONTE
Introduction 125Peptaibols 126
Diketopiperazine-Like Compounds 129Polyketides 129Pyrones 130Terpenes 131Concluding Remarks and Future Directions 133Acknowledgments 134References 134
11 Recent Advancements on the Role and Analysis of Volatile Compounds (VOCs) from Trichoderma
SHAFIQUZZAMAN SIDDIQUEE
Introduction 139Detection Techniques of VOCs 140Types of Volatiles Compounds 142Application of VOCs in Agriculture 165Conclusion 168References 168
DTOOLS
12 Molecular Tools for Strain Improvement of Trichoderma spp.
ROBERT BISCHOF AND BERNHARD SEIBOTH
Introduction 179Genetic Transformation Techniques 180Auxotrophic and Dominant Selection Markers 181Marker Recycling Strategies and Marker Free Strains 182Advanced Methods for Gene Targeting 183RNA Mediated Gene Silencing 184Promoters for Recombinant Protein Expression and Targeting 185Concluding Remarks 188References 188
13 Genetic Transformation and Engineering of Trichoderma reesei for Enhanced Enzyme Production
ANLI GENG
Introduction 193Engineering Cellulase and Hemicellulase Regulation 194Homologous and Heterologous Gene Expression and Gene
Disruption 195Protein Engineering 196Engineering Promoters 197Conclusion 198References 198
14 Applications of RNA Interference for Enhanced Cellulase Production in Trichoderma
SHAOWEN WANG AND GANG LIU
Introduction 201RNA Interference in Fungus 202Transcriptional Regulation of Cellulase Gene Expression 203

CONTENTS vii
Application of Gene Downregulation Strategy for Enhanced Cellulase Production 204
Combination of RNAi and Overexpression of the Regulating Genes 208
Conclusions and Prospects 211References 211
15 RNAi-Mediated Gene Silencing in Trichoderma: Principles and Applications
XIAOYUN SU, LINA QIN, ZHIYANG DONG
Introduction 215Molecular Mechanisms 216Advantages and Disadvantages of Using RNAi-Mediated
Gene Silencing as a Genetic Manipulation Tool in Filamentous Fungi 218
Strategies of Applying RNAi for Gene Silencing in Trichoderma and Other Filamentous Fungi 220
Conclusions 223References 224
ECELLULASES
16 Cellulase Systems in Trichoderma: An OverviewLUIS H.F. DO VALE, EDIVALDO X.F. FILHO, ROBERT N.G. MILLER,
CARLOS A.O. RICART, MARCELO V. DE SOUSA
Introduction 229Degradation of Cellulose by Cellulase Systems 230History of the Trichoderma Cellulase Research 232Structural and Functional Diversity of Trichoderma Cellulases 232Cellulase Systems and Complexes 240Acknowledgments 241References 241
17 Use of Cellulases from Trichoderma reesei in the Twenty-First Century—Part I: Current Industrial Uses and Future Applications in the Production
of Second Ethanol GenerationNICOLAS LOPES FERREIRA, ANTOINE MARGEOT,
SENTA BLANQUET, JEAN-GUY BERRIN
Overview of the Global Enzyme Market 245Industrial Cellulases 246Current Applications 249Perspectives 253Application of Trichoderma Cellulases in the Bioethanol
Industry 253References 258
18 Use of Cellulases from Trichoderma reesei in the Twenty-First Century—Part II: Optimization of
Cellulolytic Cocktails for Saccharification of Lignocellulosic Feedstocks
JEAN-GUY BERRIN, ISABELLE HERPOEL-GIMBERT, NICOLAS LOPES FERREIRA, ANTOINE MARGEOT,
SENTA HEISS-BLANQUET
Genetics of Industrial Trichoderma reesei Strains 263The T. reesei Enzyme Cocktail 264Hydrolysis of Cellulose 266Limitations in Lignocellulose Hydrolysis 267Improvement of Enzyme Cocktails by Optimization of
Enzyme Ratios 269Improvement by Supplementation of T. reesei Enzyme Cocktails 270Adapting Cellulose Cocktails to Process Conditions 275Conclusions and Perspectives 275References 275
19 Beta-Glucosidase from Trichoderma to Improve the Activity of Cellulase Cocktails
WARAWUT CHULALAKSANANUKUL
Introduction 281Cellulase Classification 282Trichoderma reesei Cellulases 282Trichoderma reesei BGLs 284BGLs from Aspergillus oryzae 284Synergism between Cellulases 286Heterologous Expression of Cellulases 286Yarrowia Lipolytica Expression Platforms 286Pichia pastoris Expression Platforms 287β-Glucosidase from Trichoderma to Improve the Activity of
Cellulase Cocktails 287Acknowledgments 288References 288
20 Regulation of Glycoside Hydrolase Expression in Trichoderma
HODA BAZAFKAN, DORIS TISCH, MONIKA SCHMOLL
Introduction 291Regulation by Environmental Parameters 292Regulatory Mechanisms 297Physiological Responses 302References 303
21 Trichoderma Proteins with Disruption Activity on Cellulosic Substrates
CHRISTIAN DERNTL, ASTRID R. MACH-AIGNER, ROBERT L. MACH
Structure and Occurrence of Cellulose in Nature 309General Aspects of Cellulose Degradation 310Cellulose Degradation by T. reesei 311Cellulolytic Enzymes in Other Trichoderma Species 314Acknowledgments 314References 314
22 Molecular Mechanism of Cellulase Production Systems in Trichoderma
KATOCH MEENU, GURPREET SINGH, R. A. VISHWAKARMA
Introduction 319Cellulase System of T. reesei 319Induction Mechanism of Cellulase Production 320Promoter Involved in Cellulase Production 320Molecular Mechanism of Cellulase Production 320

CONTENTSviii
Approaches for Refining the Cellulases Production System in T. reesei 321
References 322
23 Trichoderma in Bioenergy Research: An OverviewVIJAI K. GUPTA, ANTHONIA O’DONOVAN, MARIA G. TUOHY,
GAURI DUTT SHARMA
Introduction 325Fungal Enzyme Systems and Trichoderma Technology 326Industrial Applications of Trichoderma 327Trichoderma Enzyme Systems in Bioenergy Research 328Conclusion 332References 332
FINDUSTRIAL APPLICATIONS
24 Trichoderma Enzymes for Food IndustriesADINARAYANA KUNAMNENI, FRANCISCO J. PLOU,
ANTONIO BALLESTEROS
Introduction 339Fungus of Industrial Interest 340Trichoderma Enzymes for Industries 340Xylanases 341Cellulases 341Other Enzymes 342Food Industry 342Perspectives for Biotechnological Production of Enzymes by
Trichoderma 343References 343
25 Trichoderma: A Dual Function Fungi and Their Use in the Wine and Beer Industries
CARLOS ROBERTO FELIX, ELIANE FERREIRA NORONHA, ROBERT N. G. MILLER
Introduction 345Application in the Wine and Beer Industries 347Acknowledgments 348References 348
26 Trichoderma Enzymes for Textile IndustriesTERHI PURANEN, MARIKA ALAPURANEN, JARI VEHMAANPERÄ
Substrate 351Enzymes 352Textile Processes 353Trichoderma Enzymes in Textile Finishing Processes 355Trichoderma as a Production Host for Textile Enzymes 357Future Trends 359Acknowledgments 359References 359
27 Metabolic Diversity of TrichodermaROBERTO NASCIMENTO SILVA, ANDREI STECCA STEINDORFF,
VALDIRENE NEVES MONTEIRO
Introduction 363Global Metabolism 364Carbohydrate Metabolism and Glycoside Hydrolases 366Energy Metabolism 368Secondary Metabolism 369Metabolism and Transporters 372Acknowledgments 374References 374
28 Sequence Analysis of Industrially Important Genes from Trichoderma
AHMED M.A. EL-BONDKLY
Introduction 377Gene Sequence Analysis Fundamentals 378Genome Analysis of Trichoderma 383Industrially Genes from Trichoderma 384Sequence Analysis of Industrially Genes from
Trichoderma 384Conclusion 389References 390
29 Biosynthesis of Silver Nano-Particles by Trichoderma and Its Medical Applications
KHABAT VAHABI AND SEDIGHEH KARIMI DORCHEH
Introduction 393SNP Biosynthesis 395Mechanism 397Medical Application 399References 400
30 Role of Trichoderma Species in Bioremediation Process: Biosorption Studies
on Hexavalent ChromiumDHARA SHUKLA AND PADMA S. VANKAR
Introduction 405Hexavalent Chromium Bioremediation will Be Discussed Here
with a Case Study Representing Chromium Biosorption by Trichoderma Species 407
Conclusion 411References 412
GBIOCONTROL AND PLANT
GROWTH PROMOTION
31 Applications of Trichoderma in Plant Growth Promotion
ALISON STEWART AND ROBERT HILL
Introduction 415Trichoderma as a Plant Growth Promoter 416Consistency of Growth Promotion 418Commercialization 419

CONTENTS ix
Mechanisms of Growth Promotion 420Conclusions 425References 425
32 Molecular Mechanisms of Biocontrol in Trichoderma spp. and Their Applications in
AgricultureVIANEY OLMEDO MONFIL AND SERGIO CASAS-FLORES
Introduction 429Mycoparasitism 430Morphological Changes 430Roll of Cell Wall Degrading Enzymes 431Signal Transduction in Mycoparasitism 432ROS-Nox-Signal Transduction 433Antibiosis (Secondary Metabolites Involved in Biocontrol) 435Pyrones 436Polyketides 437Nonribosomal Peptides 437Mycotoxins Produced by Trichoderma spp. 438Synergism between Enzymes and Antibiotics 439Competition for Nutrients 439Plant Growth Promotion by Trichoderma 440Plant Root Colonization 442Induction of Systemic Resistance to Plants by Trichoderma
spp. 443Signal Transduction Pathways that Mediate Trichoderma-Plant
Communication 444Trichoderma Elicitor of Systemic Resistance in Plants 446Signal Transduction during Plant–Trichoderma Interaction in
Trichoderma 448Transgenic Plants Expressing Trichoderma Genes 448Concluding Remarks 449Acknowledgments 449References 449
33 Genome-Wide Approaches toward Understanding Mycotrophic Trichoderma Species
ALFREDO HERRERA-ESTRELLA
Introduction 455Lessons from the Genome Sequence 457Transcriptome Analyses 458The Functional Genomics View of Mycoparasitism 458High-Throughput Analysis of the Trichoderma-Plant Interaction 459Future Directions 461Concluding Remarks 462Acknowledgments 462References 462
34 Insights into Signaling Pathways of Antagonistic Trichoderma Species
SUSANNE ZEILINGER AND SABINE GRUBER
Introduction 465G Protein Signaling 465Effector Pathways of G Protein Signaling in Fungi 466Signaling Pathways and Characterized Components in
Trichoderma Species 467
Signal Transduction Components and Pathways Affecting Vegetative Growth and Conidiation 469
The Role of Signaling in Trichoderma Mycoparasitism and Biocontrol 471
Conclusions 474Acknowledgments 474References 474
35 Enhanced Resistance of Plants to Disease Using Trichoderma spp.
M.G.B. SALDAJENO, H.A. NAZNIN, M.M. ELSHARKAWY, M. SHIMIZU, M. HYAKUMACHI
Introduction 477Induced Disease Resistance in Plants 478Induced Resistance by Trichoderma spp. 481Signaling Pathways of Trichoderma-Induced Resistance 482Trichoderma spp.-Secreted Elicitors of Plant Resistance 483Engineering Plants for Disease Resistance Using Trichoderma
Genes 485Combination of Trichoderma with Other Beneficial
Microorganisms 486Other Effects of Trichoderma spp. Inoculation to the Plant 487Conclusion 487References 488
36 Enhanced Plant Immunity Using TrichodermaHEXON ANGEL CONTRERAS-CORNEJO, LOURDES MACÍAS-RODRÍGUEZ,
JESÚS SALVADOR LÓPEZ-BUCIO, JOSÉ LÓPEZ-BUCIO
Introduction 495Mechanisms of Plant Protection by Microbes 495Trichoderma-Induced Immunity 498Plant Protection Conferred by Trichoderma 500Conclusions 501Acknowledgments 501References 501
37 Genes from Trichoderma as a Source for Improving Plant Resistance to
Fungal PathogenBARBARA REITHNER AND ROBERT L. MACH
Introduction 505Trichoderma Inducing Resistance in Plants 506Transgenic Plants Expressing Trichoderma Genes Develop
Increased Resistance to Fungal Pathogens 506Trichoderma Genes Involved in Elicitation of ISR 508Conclusion 511Abbreviations 511Acknowledgments 511References 511
38 Trichoderma Species as Abiotic Stress Relievers in Plants
NAJAM W. ZAIDI, MANZOOR H. DAR, SUDHANSHU SINGH, U.S. SINGH
Introduction 515Microbes for the Management of Abiotic Stresses 516Alleviation of Abiotic Stress in Plants by Trichoderma 516

CONTENTSx
Alleviation of Drought Stress in Plants by Trichoderma 517Alleviation of Salinity Stress in Plants by Trichoderma 518Alleviation of Heat Stress in Plants by Trichoderma 519Trichoderma Genes for Abiotic Stress Tolerance 520Mechanism of Abiotic Stress Tolerance Using Trichoderma 520Host Gene: Stress Tolerant Varieties 521Conclusion 522References 523
39 Advances in Formulation of Trichoderma for Biocontrol
CHRISTIAN JOSEPH R. CUMAGUN
Introduction 527Types of Formulation 528Microencapsulation 528Enhancement of Shelf Life and Application Efficiency 528Compatibility with Other Biological Systems 529
Conclusion and Future Prospects 530References 530
40 Trichoderma: A Silent Worker of Plant RhizosphereAKANKSHA SINGH, BIRINCHI K. SARMA, HARIKESH B. SINGH,
R. S. UPADHYAY
Introduction 533Diverseness Amongst Trichoderma 534Trichoderma as Inducer of Plant Defense Response 536Trichoderma as a Biofertilizer and Plant Growth Promoter 538Commercialization 538Trichoderma Genes Responsible for Playing “Big Games” 539Conclusion 540Acknowledgments 540References 540
Index 543

xi
A growing world population and the increased energy consumption caused by a higher standard of living pose a challenge on current efforts to sustain a healthy environment and counteract climate change in the future. Replacing the limited resource of fossil oil and related products with renewable, carbon dioxide-neutral resources requires a considerate strategy, as also renewable biomass is not an unlimited resource. In order to achieve a sustainable economy, the delicate balance between use of biomass/land for food production and for use in industry and as an energy resource has to be kept. Species of the genus Trichoderma can play a signifi-cant role in the strategy for a sustainable future and this book summarizes the capabilities these fungi offer.
On the one hand, the metabolic capacities of Tricho-derma are of central importance for breakdown of plant cell walls into small compounds that can be utilized by yeast not only for bioethanol production, but also as building blocks for chemical synthesis. With its potent cellulase system and its versatility for heterologous pro-teins, which facilitates complementation of this system with efficient enzymes from other organisms, Tricho-derma reesei has become one of the cornerstones for sec-ond-generation biofuel production. Several chapters of this book provide an overview of the enzyme system of Trichoderma and its optimization for efficient utilization and conversion of lignocellulosic material. Additionally, novel and established tools for enhancing cellulase pro-duction are discussed. However, besides production of second-generation biofuels from plant material, indus-trial use of Trichoderma also extends to production of silver nanoparticles and applications in beer and wine industry as well as in textile industry.
Trichoderma also serves as a versatile host for expres-sion of heterologous proteins and a broad array of tools are available for modification of the genome of this fun-gus for improvement of its production capacity. Chapters on heterologous protein production with Trichoderma, secretion and industrial strain improvement provide an
overview of the use of this fungus as a cell factory in biotechnology. The enzyme systems of Trichoderma have even been used for bioremediation, which is a further important contribution to environmental sustainability. Other products of potential relevance for industry are the secondary metabolites produced by Trichoderma spp. as well as metabolic byproducts with interesting physi-ological or chemical functions.
While T. reesei serves as a workhorse for industrial enzyme production, other species of the genus are used for plant protection in agriculture. Thereby, these fungi play an important role in establishing this important and delicate balance between food production and the use of biomass for energy production and chemical industry. Efficient and sustainable use of biomass requires protec-tion of energy plants and food crops from pathogens in order to guarantee that biomass as a limited resource can fulfill the need of both society and industry. Different species of Trichoderma act positively on plant growth and resistance of plants against disease. The chapters of this book include a thorough summary on mechanisms and application of biocontrol, the enhancing effect on plant immunity and mycoparasitism.
Considering the huge potential of Trichoderma for use in agriculture and industry, exploration of natural isolates of the genus is warranted to further increase the genomic resources to be exploited. Screening the biodiversity of different habitats and the ecophysiol-ogy of Trichoderma in a genomic perspective as well as analysis of this diversity delivers important insights into the promises the genus Trichoderma holds for the future.
In summary, this book gives a detailed overview of the field of industrial and agricultural use as well as the research with Trichoderma from industrial enzyme production to strain improvement to biocontrol and diversity.
Editors
Preface

xiii
Trichoderma exists probably since at least 100 millions of years, but it entered the scientific spotlight only in the late seventies of the last century, when the first oil shock prompted governments to look for alternatives for fossil fuel. Researchers at the US military laboratories at Natick, Massachusetts, then remembered to possess a culture of a green fungus that had been destroying all the cotton material (tents, belts, clothes) of the US soldiers during the Second World War in the South Pacific at Guadal-canal (Solomon Islands), and who was subsequently demonstrated to have exceptional cellulolytic abilities. This fungus, like any other Trichoderma isolate at that time, was then named “T. viride” because the genus was believed to consist only of a single species. It was later re-identified as “T. reesei” (in honor of one of the research-ers exploring its cellulolytic properties, Elwyn T. Reese), for a few years misnamed as T. longibrachiatum, and finally found out to be the asexual form of a very com-mon tropical ascomycete, Hypocrea jecorina. The inter-est in this organism was of outmost importance to the Trichoderma community in general, because it challenged researchers to develop a whole toolbox of molecular genetics techniques for its manipulation, finally culmi-nating in the sequencing of the genome of the original isolate QM6a and several of its cellulase-producing mutants, which comprise an invaluable aid to study this organism.
While Trichoderma is consequently known to many people only as the organism that makes cellulases, a parallel world of Trichoderma started to develop in 1932 when R. Weindling published the mycoparasitic abilities of Trichoderma “lignorum” (an illegitimate name) on plant pathogenic fungi. This biocontrol ability is due to the profound ability of Trichoderma to parasitize or even prey on other fungi, which today is known to be the innate nature of the whole genus. Weindling’s findings formed the basis for a multitude of studies on the potential use of various Trichoderma spp. for the biological control of plant pathogenic fungi, resulting in the commercializa-tion of some of them. The cellulase and the biocontrol researchers long formed two isolated communities with little information exchange, but this improved once the
need for exchange of molecular genetics research tech-niques became apparent. Most recently the genomes of several Trichoderma biocontrol species have been sequenced, and two of them (T. atroviride, T. virens) have been published.
Yet Trichoderma offers much more to science: its spe-cies are among the most frequent mitosporic fungi com-monly detected in cultivation-based surveys. They can be isolated from an innumerable diversity of natural and artificial substrata, particularly also from materi-als infested with xenobiotics, demonstrating their high opportunistic potential and adaptability to various eco-logical conditions. Consequently, it has broad impacts on mankind: one of the most stimulating recent find-ings is that some Trichoderma spp. occur or can occur as symptomless associates of plant-endophytes, thereby stimulating plant growth, delaying the onset of drought stress and preventing attacks of pathogens. Yet, there are also negative impacts of Trichoderma on mankind: in a clinical context, a pair of genetically related species (T. longibrachiatum and T. orientale) have been shown to occur as opportunistic pathogens of immunocompro-mised humans, and several Trichoderma spp. can occur as indoor molds, although their effect on human health is less severe than that of other fungal species. Finally, some species like T. aggressivum, T. pleuroticola, T. pleurotum, and T. mienum have turned their mycoparasitic abilities against commercial mushrooms like Agaricus and Pleuro-tus, thereby causing large economic losses.
All these properties make Trichoderma one of the most versatile and intriguing fungal genus, which still offers numerous aspects to be dealt with in more detail. This book has been initiated to describe the current stage of knowledge on Trichoderma from various perspectives, thereby outlining also those areas where further prog-ress is needed.
Christian P. KubicekProfessor for Biotechnology and Microbiology
Department of Chemical EngineeringVienna University of Technology
Vienna, Austria
Foreword

xv
Sunil S. Adav School of Biological Sciences, Nanyang Technological University, Singapore
Marika Alapuranen Roal Oy, Rajamäki, Finland
Miguel Alcalde Departamento de Biocatálisis, Instituto de Catálisis y Petroleoquímica, CSIC, Madrid, Spain
N. Aro VTT Technical Research Centre of Finland, Espoo, Finland
Lea Atanasova Research Area Biotechnology and Microbiology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria
Antonio Ballesteros Departamento de Biocatálisis, Instituto de Catálisis y Petroleoquímica, CSIC, Madrid, Spain
Hoda Bazafkan Health and Environment Department, Austrian Institute of Technology GmbH (AIT), Tulln, Austria
Gabriele Berg Graz University of Technology, Environmental Biotechnology, Graz, Austria
Jean-Guy Berrin Laboratoire de Biologie des Champignons Filamenteux, INRA, Polytech Marseille, Aix Marseille Université, Marseille, France
Robert Bischof Institute of Chemical Engineering, Vienna University of Technology and Austrian Centre of Industrial Biotechnology (ACIB), Vienna, Austria
Senta Blanquet IFP Energies nouvelles, Biotechnology Department, RueilMalmaison, France
Rosa Elena Cardoza Area of Microbiology, University School of Agricultural Engineers, University of León, Ponferrada, Spain
Sergio Casas-Flores División de Biología Molecular, IPICyT, San Luis Potosí, México
Warawut Chulalaksananukul Biofuels by Biocatalysts Research Unit, Chulalongkorn University, Bangkok, Thailand; Department of Botany, Chulalongkorn University, Bangkok, Thailand
Hexon Angel Contreras-Cornejo Instituto de Investigaciones QuímicoBiológicas, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México
Christian Joseph R. Cumagun College of Agriculture, University of the Philippines Los Baños, Los Baños, Laguna, Philippines
Manzoor H. Dar International Rice Research Institute, IRRI, New Delhi, India
Marcelo V. de Sousa Department of Cell Biology, University of Brasilia, Brasilia, Federal District, Brazil
Christian Derntl Research Area Gene Technology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria
Luis H.F. Do Vale Department of Cell Biology, University of Brasilia, Brasilia, Federal District, Brazil
Zhiyang Dong Institute of Microbiology, Chinese Academy of Sciences, Beijing, China
Sedigheh Karimi Dorcheh Institute for Genetic Microbiology, FriedrichSchiller University Jena, Jena, Germany
Irina Druzhinina Institute of Chemical Engineering, Vienna University of Technology, Research Area Biotechnology and Microbiology, Vienna, Austria
Ahmed M.A. El-Bondkly National Research Centre, Dokki, Giza, Egypt
M.M. Elsharkawy Laboratory of Plant Pathology, Faculty of Applied Biological Sciences, Gifu University, Gifu City, Japan
Carlos Roberto Felix Departamento de Biologia Celular, Universidade de Brasilia, Brasilia, Federal District, Brasil
Nicolas Lopes Ferreira IFP Energies nouvelles, Biotechnology Department, RueilMalmaison, France
Edivaldo X.F. Filho Department of Cell Biology, University of Brasilia, Brasilia, Federal District, Brazil
Anli Geng School of Life Sciences and Chemical Technology, Ngee Ann Polytechnic, Clementi, Singapore
Roberto J. González-Hernández Departamento de Biología, Universidad de Guanajuato, Guanajuato, México
Sabine Gruber Research Area Biotechnology and Microbiology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria
Vijai K. Gupta Molecular Glycobiotechnology Group, Department of Biochemistry, School of Natural Sciences, National University of Ireland Galway, Galway, Ireland
Santiago Gutiérrez Area of Microbiology, University School of Agricultural Engineers, University of León, Ponferrada, Spain
Lóránt Hatvani Department of Microbiology, University of Szeged, Szeged, Hungary
Senta Heiss-Blanquet IFP Energies nouvelles, Biotechnology Department, RueilMalmaison, France
Rosa Hermosa Centro HispanoLuso de Investigaciones Agrarias (CIALE), University of Salamanca, Salamanca, Spain
Arturo Hernández-Cervantes Departamento de Biología, Universidad de Guanajuato, Guanajuato, México
Marco J. Hernández-Chávez Departamento de Biología, Universidad de Guanajuato, Guanajuato, México
Isabelle Herpoel-Gimbert Laboratoire de Biologie des Champignons Filamenteux, INRA, Polytech Marseille, Aix Marseille Université, Marseille, France
List of Contributors

LIST OF CONTRIBUTORSxvi
Alfredo Herrera-Estrella Laboratorio Nacional de Genómica para la Biodiversidad, Cinvestav Sede Irapuato, Irapuato, Guanajuato, Mexico
Robert Hill BioProtection Research Centre, Lincoln University, Canterbury, New Zealand
M. Hyakumachi Laboratory of Plant Pathology, Faculty of Applied Biological Sciences, Gifu University, Gifu City, Japan
Katarina Ihrmark Uppsala BioCenter, Department of Forest Mycology and Plant Pathology, Swedish University of Agricultural Sciences, Uppsala, Sweden
J.J. Joensuu VTT Technical Research Centre of Finland, Espoo, Finland
Magnus Karlsson Uppsala BioCenter, Department of Forest Mycology and Plant Pathology, Swedish University of Agricultural Sciences, Uppsala, Sweden
Péter Körmöczi Department of Microbiology, University of Szeged, Szeged, Hungary
László Kredics Department of Microbiology, University of Szeged, Szeged, Hungary
Adinarayana Kunamneni Departamento de Biocatálisis, Instituto de Catálisis y Petroleoquímica, CSIC, Madrid, Spain
Gang Liu College of Life Science, Shenzhen University, Shenzhen, China
Jesús Salvador López-Bucio Instituto de Biotecnología, Universidad Nacional Autónoma de México, Cuernavaca, Morelos, México
José López-Bucio Instituto de Investigaciones QuímicoBiológicas, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México
Robert L. Mach Research Area Gene Technology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria
Astrid R. Mach-Aigner Research Area Gene Technology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria
Lourdes Macías-Rodríguez Instituto de Investigaciones QuímicoBiológicas, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, México
László Manczinger Department of Microbiology, University of Szeged, Szeged, Hungary
Antoine Margeot IFP Energies nouvelles, Biotechnology Department, RueilMalmaison, France
Katoch Meenu Microbial Biotechnology Division, Indian Institute of Integrative Medicine (CSIR), Jammu, Jammu and Kashmir, India
Robert N.G. Miller Department of Cell Biology, University of Brasilia, Brasilia, Federal District, Brazil
Vianey Olmedo Monfil Departamento de Biología, Universidad de Guanajuato, Guanajuato, México
Enrique Monte Centro HispanoLuso de Investigaciones Agrarias (CIALE), University of Salamanca, Salamanca, Spain
Valdirene Neves Monteiro Universidade Estadual de Goiás, Unidade Universitária de Ciências Exatas e Tecnológicas da Universidade Estadual de GoiásUnUCET, Anápolis, Goiás, Brazil
Héctor M. Mora-Montes Departamento de Biología, Universidad de Guanajuato, Guanajuato, México
Shahram Naeimi Department of Biological Control Research, Iranian Research Institute of Plant Protection, Amol, Mazandaran, Iran
H.A. Naznin Laboratory of Plant Pathology, Faculty of Applied Biological Sciences, Gifu University, Gifu City, Japan
Helena Nevalainen Department of Chemistry and Biomolecular Sciences, Macquarie University, NSW, Australia; Biomolecular Frontiers Research Centre, Macquarie University, NSW, Australia
Eliane Ferreira Noronha Departamento de Biologia Celular, Universidade de Brasilia, Brasilia, Federal District, Brasil
Anthonia O’Donovan Molecular Glycobiotechnology Group, Department of Biochemistry, School of Natural Sciences, National University of Ireland Galway, Galway, Ireland
T. Pakula VTT Technical Research Centre of Finland, Espoo, Finland
Robyn Peterson Department of Chemistry and Biomolecular Sciences, Macquarie University, NSW, Australia; Biomolecular Frontiers Research Centre, Macquarie University, NSW, Australia
Francisco J. Plou Departamento de Biocatálisis, Instituto de Catálisis y Petroleoquímica, CSIC, Madrid, Spain
Terhi Puranen Roal Oy, Rajamäki, Finland
Lina Qin Institute of Microbiology, Chinese Academy of Sciences, Beijing, China
Barbara Reithner Research Area Gene Technology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria
Carlos A.O. Ricart Department of Cell Biology, University of Brasilia, Brasilia, Federal District, Brazil
María Belén Rubio Centro HispanoLuso de Investigaciones Agrarias (CIALE), University of Salamanca, Salamanca, Spain
M.G.B. Saldajeno Laboratory of Plant Pathology, Faculty of Applied Biological Sciences, Gifu University, Gifu City, Japan
M. Saloheimo VTT Technical Research Centre of Finland, Espoo, Finland
Birinchi K. Sarma Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, Uttar Pradesh, India
Monika Schmoll Health and Environment Department, Austrian Institute of Technology GmbH (AIT), Tulln, Austria
Bernhard Seiboth Institute of Chemical Engineering, Vienna University of Technology and Austrian Centre of Industrial Biotechnology (ACIB), Vienna, Austria

LIST OF CONTRIBUTORS xvii
Verena Seidl-Seiboth Institute of Chemical Engineering, Vienna University of Technology, Research Area Biotechnology and Microbiology, Vienna, Austria
Gauri Dutt Sharma Bilaspur University, Bilaspur, Chattisgarh, India
M. Shimizu Laboratory of Plant Pathology, Faculty of Applied Biological Sciences, Gifu University, Gifu City, Japan
Dhara Shukla Facility for Ecological and Analytical Testing, Indian Institute of Technology, Kanpur, Uttar Pradesh, India
Shafiquzzaman Siddiquee Biotechnology Research Institute, University Malaysia Sabah, Kota Kinabalu, Sabah, Malaysia
Roberto Nascimento Silva Department of Biochemistry and Immunology, School of Medicine, University of São Paulo, Ribeirão Preto, São Paulo, Brazil
Akanksha Singh Department of Botany, Banaras Hindu University, Varanasi, Uttar Pradesh, India
Harikesh B. Singh Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi, Uttar Pradesh, India
Gurpreet Singh Microbial Biotechnology Division, Indian Institute of Integrative Medicine (CSIR), Jammu, Jammu and Kashmir, India
Sudhanshu Singh International Rice Research Institute, IRRI, New Delhi, India
U.S. Singh International Rice Research Institute, IRRI, New Delhi, India
Andrei Stecca Steindorff Departamento de Biologia Celular, Universidade de Brasília, Brasília, Distrito Federal, Brazil
Alison Stewart BioProtection Research Centre, Lincoln University, Canterbury, New Zealand
Xiaoyun Su Institute of Microbiology, Chinese Academy of Sciences, Beijing, China
Siu Kwan Sze School of Biological Sciences, Nanyang Technological University, Singapore
Doris Tisch Health and Environment Department, Austrian Institute of Technology GmbH (AIT), Tulln, Austria
José E. Trujillo-Esquivel Departamento de Biología, Universidad de Guanajuato, Guanajuato, México
Maria G. Tuohy Molecular Glycobiotechnology Group, Department of Biochemistry, School of Natural Sciences, National University of Ireland Galway, Galway, Ireland
R.S. Upadhyay Department of Botany, Banaras Hindu University, Varanasi, Uttar Pradesh, India
Csaba Vágvölgyi Department of Microbiology, University of Szeged, Szeged, Hungary
Khabat Vahabi Institute of General Botany and Plant Physiology, FriedrichSchiller University Jena, Jena, Germany
Padma S. Vankar Facility for Ecological and Analytical Testing, Indian Institute of Technology, Kanpur, Uttar Pradesh, India
Jari Vehmaanperä Roal Oy, Rajamäki, Finland
R.A. Vishwakarma Microbial Biotechnology Division, Indian Institute of Integrative Medicine (CSIR), Jammu, Jammu and Kashmir, India
Shaowen Wang College of Life Science, Shenzhen University, Shenzhen, China
Christin Zachow Austrian Centre of Industrial Biotechnology (ACIB GmbH), Graz, Austria; Graz University of Technology, Environmental Biotechnology, Graz, Austria
Najam W. Zaidi International Rice Research Institute, IRRI, New Delhi, India
Susanne Zeilinger Research Area Biotechnology and Microbiology, Institute of Chemical Engineering, Vienna University of Technology, Vienna, Austria

S E C T I O N A
BIOLOGY AND BIODIVERSITY

Biotechnology and Biology of Trichoderma http://dx.doi.org/10.1016/B978-0-444-59576-8.00001-1 Copyright © 2014 Elsevier B.V. All rights reserved.
3
INTRODUCTION
Members of the genus Trichoderma are cosmopolitan and prevalent components of different ecosystems in a wide range of climatic zones (Kubicek et al. 2008). The occurrence of Trichoderma species is modulated by sev-eral factors including microclimate, the availability of substrates as well as complex ecological interactions (Hoyos-Carvajal and Bissett, 2011). Survival in different geographical habitats can be related to metabolic diver-sity, high reproductive capacity and competitive capa-bilities of Trichoderma strains in nature (Cardoso Lopes et al. 2012). The aim of this chapter is to give an overview about the studies aimed at the investigation of Tricho-derma biodiversity in a wide variety of different ecologi-cal habitats.
METHODOLOGY OF STUDYING TRICHODERMA BIODIVERSITY
Methods for the Identification of Trichoderma Strains
Formerly the species-level identification of Tricho-derma/Hypocrea isolates was performed based on exclu-sively their morphological characteristics (Danielson and Davey, 1973; Summerbell, 2003; Gams and Bissett, 1998). Different media were used for culturing Tricho-derma isolates for the analysis of their morphology and culture characteristics, e.g. malt extract agar, which is appropriate for conidium production and the observa-tion of conidiophore branching, or potato dextrose agar, which proved useful for observing pigment production
C H A P T E R
1Biodiversity of the Genus Hypocrea/Trichoderma
in Different HabitatsLászló Kredics1, *, Lóránt Hatvani1, Shahram Naeimi2, Péter Körmöczi1,
László Manczinger1, Csaba Vágvölgyi1, Irina Druzhinina3
1Department of Microbiology, University of Szeged, Szeged, Hungary, 2Department of Biological Control Research, Iranian Research Institute of Plant Protection, Amol, Mazandaran, Iran, 3Institute of Chemical Engineering, Vienna University of Technology, Research Area Biotechnology and Microbiology,
Vienna, Austria*Corresponding author email: [email protected]
O U T L I N E
Introduction 3
Methodology of Studying Trichoderma Biodiversity 3Methods for the Identification of Trichoderma Strains 3Evolution of the Approach: From the Culture-Based
Method to Metagenomics 4
Trichoderma Diversity in Different Habitats 5Natural Soils, Decaying Wood and Plant Material 5Agricultural Habitats 9
Living Plants (Endophytes) 11Mushroom-Related Substrata 13Human Body 14Water-Related Environments 14Air and Settled Dust 17
Conclusions 18
Dear user
Highlight
Dear user
Highlight

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS4
A. BIOLOGY AND BIODIVERSITY
(Hoyos-Carvayal and Bissett, 2011). The preliminary identification of species based on conidiophore struc-ture, morphology as well as the size and morphology of conidia can be performed with the aid of taxonomic keys and descriptions available in the literature (Bissett, 1984, 1991a, b, c, 1992; Gams and Bissett, 1998; Chaverri and Samuels, 2003; Jaklitsch, 2009, 2011; Samuels et al., 2006b, 2012a,b). However, without professional expertise this may often lead to incorrect diagnoses due to the difficul-ties of morphology-based species identification, therefore the results of early studies must be handled with special care (Kubicek et al., 2008). In order to get around such problems and give precise species-level diagnoses, the use of biochemical and molecular methods is recommended.
Among the biochemical methods, the metabolic profiling technique of Biolog Incorporated (Hayward, California)—which provides the possibility of quanti-tative measurements of growth and the assimilation of different carbon and nitrogen sources—proved to be a useful tool to aid species identification and provide data on the ecological roles of species (Kubicek et al., 2003; Hoyos-Carvajal et al., 2009; Atanasova and Druzhinina, 2010).
A cellulose-acetate electrophoresis-based isoenzyme analysis—with the involvement of glucose-6-phosphate dehydrogenase, glucose-6-phosphate isomerase, 6-phos-phogluconate dehydrogenase, peptidases A, B and D, and phosphoglucomutase enzymes (Hebert and Beaton, 1993)—was applied by Szekeres et al. (2006) and Kredics et al. (2011a, 2012) for the identification of Trichoderma strains deriving from clinical samples and winter wheat fields, respectively.
Neuhof et al. (2007) suggested an alternative bio-chemical technique for Hypocrea⁄Trichoderma species and strains, which was developed based on intact-cell mass spectrometry for the direct detection of hydrophobins in the mycelia as well as spores of 32 Hypocrea⁄Trichoderma strains representing 29 species. The hydrophobin patterns were shown to be characteristic to species and isolates, and the method is proposed to enable the rapid and direct detection of class II hydrophobins.
Among the molecular methods, DNA-fingerprint-ing (Arisan-Atac et al., 1995), the sequence analysis of the ribosomal internal transcribed spacer (ITS) region (ITS1–5.8S rDNA–ITS2) and of segments from genes encoding for the translation elongation factor 1-alpha (tef1), endochitinase (chi18-5, formerly known as ech42), RNA polymerase II subunit (rpb2) and calmodulin (cal1) ( Kullnig-Gradinger et al., 2002; Druzhinina et al., 2008) were found to be suitable for giving precise species iden-tification of Hypocrea⁄Trichoderma strains.
The ITS-based online barcoding program TrichOKEY (www.isth.info; Druzhinina et al., 2005; Druzhinina and Kopchinskiy, 2006) provides another useful tool for the identification of Hypocrea⁄Trichoderma species. The
development and practical applications of ITS barcodes are presented and discussed in chapter 3: DNA barcode for species identification in Trichoderma. For the analy-sis of tef1, ITS and rpb2 sequences the online programme TrichoBLAST and its updated version, TrichoMARK are recommended (www.isth.info; Kopchinskiy et al., 2005). TrichoCHIT (www.isth.info), an online barcoding pro-gramme for the screening and identification of excellent chitinase producer strains of Hypocrea lixii/Trichoderma harzianum was developed by Nagy et al. (2007).
The use of species-specific primers in polymerase chain reaction can also lead to quick and precise diagno-sis. For example, Chen et al. (1999a, b) developed a PCR-based assay for the fast and specific detection of Th2 and Th4, the aggressive Trichoderma biotypes causing green mould disease of Agaricus bisporus, while the method developed by Kredics et al. (2009) allows the rapid and specific identification of Trichoderma pleurotum and Trich-oderma pleuroticola, the causal agents of green mould in the world-wide production of oyster mushroom (Pleu-rotus osteatus) even directly from the growing substrate without the need of cultivation.
Evolution of the Approach: From the Culture-Based Method to Metagenomics
Most of the studies about Trichoderma biodiversity applied the standard culture-based approach compris-ing the collection of samples, isolation of Trichoderma strains on one of the selective media described in the literature (Elad et al., 1981; Papavizas and Lumsden, 1982; Askew and Laing, 1993; Williams et al., 2003) and their maintenance in culture, which can be followed by the application of the above-mentioned species-level identification methods. The problem of this approach is that certain Trichoderma species may be easier, while others harder to isolate, therefore the diversity detected in the culture-based studies does not necessarily reflect the actual diversity of the genus in the examined habi-tat. The application of the metagenomic approach pro-vides a solution to this problem, as it is examining the habitats in situ, without the isolation and culturing of the Trichoderma strains. This approach is recently gath-ering ground in Trichoderma biodiversity studies. In the first metagenomic attempt, Hypocrea/Trichoderma- specific primers were designed for the ITS1 fragment of the rRNA gene cluster by Hagn et al. (2007). With the application of this method, the authors found only about 12 species in arable soil. Later studies demon-strated that ITS1 alone is not sufficiently diagnostic as certain species share the same allele. Based on a master alignment of ITS sequences, Meincke et al. (2010) devel-oped a novel Trichoderma-specific primer pair for diver-sity analysis, which amplifies an approximately 650 bp fragment of the ITS region suitable for identification

5
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
by TrichOKEY and TrichoBLAST from all taxonomic clades of the genus Trichoderma The authors applied a seminested strategy for DNA amplification from soil: the first PCR- amplification was performed with a fun-gal specific forward primer and the Trichoderma-specific reverse primer, while the Trichoderma-specific forward and reverse primers were used together in the second reaction. ITS amplicons were subjected to denaturing gradient gel electrophoresis (DGGE) analysis or cloned to pGEM-T Easy vector and sequenced. The designed primer system was applied to study Trichoderma commu-nities in the rhizosphere of potatoes. However, several species are undetectable by the use of this method as the reverse primer of this system is located 30 bp upstream of the last genus-specific TrichOKEY hallmark in a still polymorphic and indel-rich area of ITS2. In a more recent study, Friedl and Druzhinina (2012) designed six reverse primers and demonstrated their high specificity and selectivity. Applied along with the forward primer ITS5 (White et al., 1990), this set of reverse primers is able to amplify the entire diagnostic region of ITS1 and 2 of all members of the genus. The strategy is that after six separate PCR amplifications from the tested soil sample—each containing the same forward and one of the reverse primers—the products are combined, puri-fied and subcloned to pGEM-T Easy vector resulting in a clone library. The sequences of the individual clones are determined and analyzed with TrichOKEY 2.0 and TrichoBLAST. Atanasova et al. (2010) applied this metage-nomic strategy to study the diversity of the Trichoderma genus in air samples.
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
Natural Soils, Decaying Wood and Plant Material
In an early study, Danielson and Davey (1973) exam-ined the Trichoderma propagules in a variety of forest soils in the southeastern U.S. and Washington State and iden-tified the isolates as Trichoderma hamatum, T. harzianum, Trichoderma koningii, Trichoderma polysporum, Trichoderma pseudokoningii and Trichoderma viride. T. koningii and T. hamatum were reported as the most widely distributed species aggregates. Trichoderma polysporum and T. viride were found to be largely restricted to cool temperate regions, T. harzianum was reported to be characteristic of warm climates, while T. hamatum and T. pseudokoningii were the dominant forms under conditions of excessive moisture. Widden and Abitbol (1980) studied the seasonal distribution of Trichoderma species in a spruce-forest soil in Canada and reported the occurrence of T. hamatum, T. harzianum, Trichoderma longibrachiatum, T. polysporum,
T. koningii, T. pseudokoningii, and T. viride. Vajna (1983) reported the isolation and morphology- as well as cul-ture characteristics-based identification of Trichoderma aureoviride, T. harzianum, T. koningii, T. longibrachiatum and T. viride from dead wood of apple twigs, oak wood and cork wood samples collected in Hungary. In the 1990s, broad studies on Trichoderma taxonomy and bio-diversity were performed by Bissett (1991a,b,c, 1992) in North America and some European regions. Trichoderma harzianum, T. polysporum and T. viride were the three taxa reported from the Hubbard Brook Experimental Forest in New Hampshire (USA), which were examined for their potential to degrade organochlorine xenobiotics (Smith, 1995). However—as already mentioned—the results of these early studies are hard to evaluate as no molecu-lar tools were available for species identification and the taxonomy of the genus Trichoderma has also changed substantially since the publication of these reports. The advent of molecular techniques resulted in a new era also in the field of Trichoderma biodiversity studies. Nev-ertheless, the results of certain recent studies should still be handled with care due to the lack of the application of molecular techniques for species identification. For instance Vasanthakumari and Shivanna (2011) reported the occurrence of Trichoderma asperellum, T. harzianum, T. koningii and T. viride from the rhizosphere and rhizo-plane of grasses of the subfamily Panicoideae in the Lak-kavalli Region of Karnataka, India, however, the isolates were identified based on morphological and cultural characteristics only.
Several studies addressed the biodiversity of the genus Hypocrea and Trichoderma in Asia. Kullnig et al. (2000) studied 76 Trichoderma strains isolated from Russia—including Siberia—and the Himalayas by ITS sequence analysis, RAPD and DNA-fingerprinting and reported the occurrence of T. asperellum, Trichoderma atroviride, Trichoderma ghanense, T. hamatum, T. harzia-num, T. koningii, Trichoderma oblongisporum, Trichoderma virens as well as some previously undetected taxa, which were later described based on morphological and physi-ological characters as well as ITS1, 2 and tef1 sequences as Trichoderma effusum, Trichoderma rossicum and Tricho-derma velutinum (Bissett et al., 2003). The T. harzianum species complex proved to be the most frequently occur-ring and genetically most diverse taxon with six dif-ferent ITS-genotypes (Kullnig et al., 2000). A follow-up study on the biodiversity of Trichoderma in Southeast Asia including Burma, Cambodia, Malaysia, Singapore, Taiwan, Thailand and Western Indonesia applied the sequence analysis of the ITS region as well as Biolog phenotype microarrays to examine 96 Trichoderma iso-lates (Kubicek et al., 2003), and revealed the occurrence of T. asperellum, T. atroviride, T. ghanense, T. hamatum, T. harzianum, Hypocrea jecorina/Trichoderma reesei, T. kon-ingii, Trichoderma spirale, T. virens and T. viride. Based on

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS6
A. BIOLOGY AND BIODIVERSITY
the results of this study, the T. harzianum complex con-tains species with high metabolic diversity and partially unique metabolic characteristics, which may explain its wide distribution over different habitats. The three new species previously detected by Kullnig et al. (2000) could also be isolated, along with additional four previously undetected taxa, which were subsequently described by Bissett et al. (2003) as Trichoderma cerinum, Trichoderma erinaceum, Trichoderma helicum and Trichoderma sinense. Zhang et al. (2005) examined the Trichoderma biodiver-sity and biogeography on 135 isolates deriving from four regions of China: provinces Hebei (North), Zheji-ang (South-East), Yunnan (West) and the Himalayan part (Tibet) and identified T. asperellum, T. atroviride, T. cerinum, Trichoderma citrinoviride, T. harzianum, T. konin-gii, T. longibrachiatum, T. sinense, T. velutinum, T. virens, T. viride, as well as two putative new species. The results of the study provided evidence for a North to South distribution of Trichoderma species in East Asia and identified Northern China as a potential center of ori-gin of a unique haplotype of T. harzianum. In a recent study, Sun et al. (2012) identified 12 taxa (T. asperellum, T. atroviride, Trichoderma brevicompactum, T. citrinoviride, T. erinaceum, T. hamatum, Trichoderma koningiopsis, H. lixii/T. harzianum, T. reesei/H. jecorina, T. spirale, Tricho-derma stromaticum, Trichoderma vermipilum and Hypocrea virens/T. virens) from Chinese forest soils by ITS barcod-ing. Tsurumi et al. (2010) explored the distribution of Trichoderma species in four countries of Asia: Indonesia, Japan, Mongolia and Vietnam through the examination of 332 isolates. Trichoderma crassum, T. hamatum, T. harzia-num and T. virens occurred in most habitats. Trichoderma atroviride, T. koningiopsis and Trichoderma stramineum were also frequent but not in cooler regions, where the occurrence of T. polysporum and Trichoderma viridescens was reported. Trichoderma brevicompactum, T. erinaceum and T. ghanense were prevalent in tropical areas. In addition to these species, potentially new taxa were also detected. Abd-Elsalam et al. (2010) isolated Tricho-derma strains from soil collected from protected areas (Rawdet Khuraim) in Saudi Arabia. Identification of the isolates by M13-microsatellite PCR and ITS barcoding revealed the presence of T. harzianum/H. lixii and the T. longibrachiatum/Hypocrea orientalis species duplet, suggesting them as pan-global taxa of Trichoderma/Hypocrea (Abd-Elsalam et al., 2010). Further species known from Asia include Hypocrea catoptron/Tricho-derma catoptron, Hypocrea cornea/T. sp., Hypocrea crassa/T. crassum, Hypocrea rugulosa, T. spirale, Hypocrea tawa/Trichoderma tawa, Hypocrea thailandica/Trichoderma thai-landicum and Hypocrea thelephoricola/Trichoderma the-lephoricola (Chaverri and Samuels, 2003), Trichoderma capillare, Trichoderma gracile, Trichoderma parareesei and Trichoderma pinnatum (Samuels et al., 2012a), Trichoderma barbatum, Trichoderma caesareum/H. sp. and Trichoderma
floccosum/H. sp. (Samuels et al., 2012b), as well as Trich-oderma arundinaceum (Degenkolb et al., 2008).
Thanks to a series of biodiversity studies, plenty of information is available about the biodiversity of the genus in Europe. Wuczkowski et al. (2003) studied the diversity of the genus Trichoderma in an original Euro-pean river-floodplain habitat, the Danube national park, which is a primeval, riparian forest area located south-east from Vienna, Austria. Besides morphological examinations, sequence analysis of the ITS region and a fragment of the tef1 gene as well as RAPD analysis were applied for the identification of the isolated Trichoderma strains. In the order of abundance, the species identified were T. harzianum, T. rossicum, T. cerinum, T. hamatum, T. atroviride, T. koningii (recognized now by TrichOKEY as T. koningiopsis) and Trichoderma sp. MA3642 from section Longibrachiatum, which was recently described by Samuels et al. (2012a) as T. capillare. Mysterud et al. (2007) examined the plant litter-associated fungi from the spring “grazing corridor” of a sheep herd in west-ern Norway and detected a wide variety of fungi includ-ing two Trichoderma isolates that the authors failed to identify by NCBI BLAST search of their ITS sequences. A search with TrichOKEY (Druzhinina et al., 2005) reveals that one of these isolates is T. hamatum while the other one belongs to the T. koningiopsis/Trichoderma ovalisporum/T. asperellum species triplet. Jaklitsch (2009, 2011) performed a wide-scale survey over 14 European countries with temperate climate to study the biodiver-sity of the Hypocrea/Trichoderma genus based on 620 specimens by examining their morphology, culture char-acteristics as well as ITS, rpb2 and tef1 sequences. Far exceeding the previous estimations about the number of Hypocrea/Trichoderma species in Europe, a total of 75 spe-cies were detected including previously described species (the holomorphs Hypocrea atroviridis/T. atroviride, Hypocrea aureoviridis/T. aureoviride, Hypocrea citrina/Trichoderma lacteum, Hypocrea epimyces/Trichoderma epimyces, Hypo-crea gelatinosa/Trichoderma gelatinosum, H. lixii/T. harzia-num, Hypocrea lutea/Trichoderma deliquescens, Hypocrea koningii/T. koningii, Hypocrea minutispora/Tricho-derma minutisporum, Hypocrea neorufa/T. sp., Hypocrea ochroleuca/T. sp., Hypocrea pachybasioides/T. polysporum, Hypocrea pilulifera/Trichoderma piluliferum, Hypocrea protopulvinata/T. sp., Hypocrea pulvinata/T. sp., Hypo-crea rufa/T.viride, Hypocrea schweinitzii/T. citrinoviride, Hypocrea sulphurea/T. sp. as well as Hypocrea species without known anamorphs: Hypocrea argillacea, Hypo-crea spinulosa, Hypocrea splendens, Hypocrea strobilina), new taxa (the holomorphs Hypocrea aeruginea/Tricho-derma aerugineum, Hypocrea albolutescens/Trichoderma albolutescens, Hypocrea atlantica/Trichoderma atlanti-cum, Hypocrea auranteffusa/Trichoderma auranteffusum, Hypocrea austriaca/Trichoderma austriacum, Hypocrea bavarica/Trichoderma bavaricum, Hypocrea calamagrostidis/

7
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
Trichoderma calamagrostidis, Hypocrea fomiticola/Trichod-erma fomiticola, Hypocrea junci/Trichoderma junci, Hypocrea luteffusa/Trichoderma luteffusum, Hypocrea luteocrystallina/ Trichoderma luteocrystallinum, Hypocrea margaretensis/ Trichoderma margaretense, Hypocrea neorufoides/Trichoderma neorufoides, Hypocrea pachypallida/Trichoderma pachypal-lidum, Hypocrea parepimyces/Trichoderma parepimyces, Hypocrea parestonica/Trichoderma parestonicum, Hypo-crea phellinicola/Trichoderma phellinicola, Hypocrea silvae- virgineae/Trichoderma silvae-virgineae, Hypocrea subeffusa/Trichoderma subeffusum and Hypocrea valdunensis/Tricho-derma valdunense; the new teleomorphs Hypocrea danica, Hypocrea rhododendri and Hypocrea sambuci; Hypocrea lon-gipilosa described as the teleomorph state of Trichoderma longipile; as well as Trichoderma alutaceum, Trichoderma dacrymycellum, Trichoderma delicatulum, Trichoderma leu-copus, Trichoderma moravicum, Trichoderma placentula, Trichoderma psychrophilum, Trichoderma subalpinum and Trichoderma tremelloides, which are the anamorphs of previously described sexually reproducing species). Among the new Hypocrea species described during the past decade, Hypocrea estonica/T. sp., Hypocrea phyllo-stachidis, Hypocrea sinuosa/Trichoderma sinuosum, Hypo-crea strictipilosa/Trichoderma strictipile, H. thelephoricola (Chaverry and Samuels, 2003), Hypocrea stilbohypoxyli (Lu and Samuels, 2003), Hypocrea parapilulifera (Lu et al., 2004), Hypocrea nybergiana (Chamberlain et al., 2004), Hypocrea voglmayrii (Jaklitsch et al., 2005), Hypocrea alcalifuscescens/T. sp. and Hypocrea parmastoi/T. sp. (Over-ton et al., 2006), Hypocrea petersenii and Hypocrea rogerso-nii (Samuels et al., 2006b), Hypocrea crystalligena (Jaklitsch et al., 2006a), Hypocrea viridescens (Jaklitsch et al., 2006b), Hypocrea alni and Hypocrea brunneoviridis (Jaklitsch et al., 2008a), Hypocrea decipiens (Jaklitsch et al., 2008b), Hypo-crea seppoi (Jaklitsch et al., 2008c) and Hypocrea rodmanii (Degenkolb et al., 2008) also occur in Europe. From the Longibrachiatum clade, Trichoderma saturnisporum is also known from Sardinia (Samuels et al., 2012a). Results of these studies demonstrated that not just a small num-ber of Trichoderma species are capable of forming a tele-omorph and suggest that the biodiversity of the genus is higher on and above the litter layer than inside the soil. Although specific associations with host fungi or trees were found, the majority of the species were sug-gested to be necrotrophic on diverse fungi on wood and bark. Since the publication of the papers of Jaklitsch (2009, 2011), the above list of European species was supplemented with additional, recently described taxa Hypocrea britdaniae, Hypocrea foliicola (Jaklitsch and Vogl-mayr, 2012), Hypocrea caerulescens, Hypocrea hispanica and Trichoderma samuelsii (Jaklitsch et al. 2012). During an ITS barcoding-based study on the species diversity of Tricho-derma in Poland, Błaszczyk et al. (2011) found that soil and decaying wood were the most diverse among the examined substrata. Species detected in forest soils were
T. atroviride, Trichoderma gamsii, T. hamatum, T. harzianum, Trichoderma tomentosum, T. viride and T. viridescens. Except for T. tomentosum, decaying wood samples also harbored these species as well as T. citrinoviride, T. koningii and T. koningiopsis. Following T. harzianum, the most abundant Trichoderma species collected from forest soil and forest wood were T. atroviride and T. viridescens, respectively. A taxon-specific metagenomic approach was applied by Friedl and Druzhinina (2012) for the assessment of the Trichoderma diversity in situ in soil samples of aspen and beech forests along the Danube floodplain. Identi-fied taxa comprised H. alni, T. asperellum, H. atroviridis/T. atroviride, T. brevicompactum, T. cerinum, T. harzianum sensu stricto, H. pachybasioides, H. pachypallida, T. pleu-roticola, Hypocrea pseudoharzianum, the species duplet H. orientalis/T. longibrachiatum, T. rossicum, H. schweinitzii, Trichoderma sp. C.P.K. 2974 and H. virens/T. virens, with the highest abundance of T. asperellum in both habitats. Two presumably new taxa, Trichoderma sp. MOTU 2B 48 from section Trichoderma and Trichoderma sp. MOTU 1A 64 from section Longibrachiatum were also detected in aspen and beech forests, respectively. The species dis-tribution proved to be uneven in the vertical profiles of the examined soils. The authors concluded that only a relatively small number of Hypocrea/Trichoderma species adapted to soil as a habitat.
Members of the genus Hypocrea/Trichoderma occurring in natural habitats of the North-American subcontinent include Hypocrea ceramica/T. sp., T. crassum, Hypocrea cremea/T. sp., Hypocrea cuneispora/T. sp., Trichoderma fertile, T. hamatum, T. longipile, T. oblongisporum, Tricho-derma pubescens, H. strictipilosa/T. strictipile, Trichoderma strigosum, Hypocrea surrotunda/Trichoderma surrotundum and T. tomentosum with conidiophore elongations and green conidia (Chaverri et al., 2003), the green asco-spored species Hypocrea ceracea/Trichoderma ceraceum, H. ceramica/Trichoderma ceramicum, Hypocrea chlorospora/Trichoderma chlorosporum, Hypocrea chromosperma/Tricho-derma chromospermum, Hypocrea cinnamomea/Trichoderma cinnamomeum, H. crassa/T. crassum, H. cremea/Tricho-derma cremeum, H. cuneispora/Trichoderma cuneisporum, H. lixii/T. harzianum, H. sinuosa/T. sinuosum, H. strictipilosa/T. strictipile and H. virens/T. virens (Chaverri and Samuels, 2003), T. ghanense, T. longibrachiatum, T. parareesei, Hypocrea pseudokoningii/T. pseudokoningii, Trichoderma saturnisporopsis, T. saturnisporum and H. schweinitzii/ T. citrinoviride from the Longibrachiatum clade (Samuels et al., 2012a), T. arundinaceum, T. brevicompactum and H. rodmanii from the T. brevicompactum clade (Degenkolb et al., 2008), H. alcalifuscescens/T. sp., Hypocrea farinosa/ T. sp. and H. sulphurea/T. sp. (Overton et al., 2006), T. bar-batum (Samuels et al., 2012b) as well as T. capillare (Samu-els et al., 2012a).
Central American species occurring in Costa Rica comprise Hypocrea candida/Trichoderma candidum,

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS8
A. BIOLOGY AND BIODIVERSITY
H. chlorospora/T. chlorosporum, Hypocrea costaricensis/T. sp., Hypocrea nigrovirens/Trichoderma nigrovirens, Hypocrea substipitata, Hypocrea tuberosa, Hypocrea virescentiflava/T. sp. (Chaverri and Samuels, 2003) and T. spirale (Chaverri et al., 2003), T. brevicompactum and Trichoderma turri-albense (Degenkolb et al., 2008), Hypocrea flaviconidia (Druzhinina et al., 2004) as well as Hypocrea eucorticioides/ T. sp. (Overton et al., 2006).
In South America, a polyphasic method based on the analysis of ITS1, 2 and tef1 sequences as well as Biolog metabolic profiling was used by Hoyos-Carvajal et al. (2009) to study the biodiversity of Trichoderma species in habitats of neotropic regions in Peru, Mexico, Gua-temala and Colombia including rainforest soils, river sand, humus and wood. A total of 182 isolates were identified from 18 species (T. asperellum, T. atroviride, T. brevicompactum, T. crassum, T. erinaceum, T. gamsii, T. hamatum, T. harzianum, H. jecorina/T. reesei, T. koningiopsis, T. longibrachiatum, T. ovalisporum, T. pubescens, T. rossicum, T. spirale, T. tomentosum, T. virens and T. viridescens) and 11 undescribed species were also discovered. The pre-dominant species were T. asperellum and T. harzianum. In a subsequent paper, Hoyos-Carvajal and Bissett (2011) reviewed the biodiversity of the genus Trichoderma in tropical American regions. The occurrence of T. asperel-lum, Trichoderma asperelloides, T. atroviride, T. brevicom-pactum, Trichoderma caribbaeum, Trichoderma caribbaeum var. aequatoriale, T. crassum, T. erinaceum, Trichoderma evansii, T. gamsii, T. hamatum, T. harzianum, H. jecorina/ T. reesei, T. koningiopsis, Trichoderma lieckfeldtiae, T. longi-brachiatum, Trichoderma neokoningii, T. ovalisporum, T. para-reesei, Trichoderma paucisporum, T. pleurotum, T. pubescens, T. rossicum, Trichoderma scalesiae, T. spirale, Trichoderma stilbohypoxyli, Trichoderma theobromicola, T. tomentosum, T. virens and T. viridescens was reported, providing a wide repertoire for the selection of biocontrol agents of crop diseases. Rivas and Pavone (2010) examined Ven-ezuelan soils and found the T. harzianum species com-plex to be the most frequently occurring taxon, followed by T. virens, T. brevicompactum, T. theobromicola, T. kon-ingiopsis, T. ovalisporum, T. asperellum, T. pleurotum and T. koningiopsis. Other species known from South America include T. strigosum and Hypocrea stromatica/Trichoderma stromaticum (Chaverri et al., 2003), Hypocrea gyrosa and H. virescentiflava/T. sp. (Chaverri and Samuels, 2003) from Brazil, Hypocrea clusiae/T. sp. from French Guyana (Chaverri and Samuels, 2003), as well as Hypocrea andi-nensis (Samuels et al., 2012a) and H. eucorticioides/T. sp. (Overton et al., 2006) from Venezuela.
Less information is available about the biodiversity of the genus in natural habitats of Africa. The distribution of Trichoderma species in soils of Embu and Taita regions in Kenya with relation to land use practices was investi-gated by Okoth et al. (2009). Species isolated from indig-enous forests of the Embu region in order of prevalence were T. harzianum, T. viride, Trichoderma aggressivum and
T. atroviride, while in the case of indigenous forests of the Taita region the detected species were T. harzianum, T. atroviride, T. koningii, T. aggressivum, T. viride and T. asperellum. The identities of the isolates were determined based on a morphological key, but unfortunately the identifications were not confirmed by molecular tech-niques, therefore results like the reported occurrence of the mushroom pathogenic species T. aggressivum in a nat-ural habitat need to be handled critically. Sadfi-Zouaoui et al. (2009) studied four different bioclimatic zones in Tunisia for Trichoderma diversity. The T. harzianum spe-cies complex proved to be the most prevalent taxon identified. Trichoderma harzianum and T. longibrachia-tum proved to be predominant in North-Tunisian forest soils. Trichoderma harzianum, T. saturnisporum and a yet unidentified Trichoderma species were detected in forest soils from central Tunisia while T. hamatum and T. har-zianum could be isolated from oasis soils in the Southern part of the country. Further species reported from Africa include Trichoderma aethiopicum, Trichoderma flagellatum and T. parareesei from Ethiopia, Trichoderma konilangbra from Uganda, H. orientalis from Zambia (Samuels et al., 2012a), H. catoptron/T. catoptron (Chaverri and Samuels, 2003), Hypocrea subcitrina (Overton et al., 2006) and T. vermipilum (Samuels et al., 2012b) from South Africa, T. arundinaceum from Namibia (Degenkolb et al., 2008), Hypocrea subsulphurea (Overton et al., 2006), as well as Trichoderma lanuginosum/H. sp., Trichoderma medusae/H. sp. from Cameroon and Trichoderma ivoriense from Ivory Coast (Samuels et al., 2012b).
Studying the biodiversity of the genus in islands as geographically separated regions may reveal important data about endemic taxa as well as inva-sive ones arriving from the nearby continents. Spe-cies reported from New Zealand include T. crassum, Hypocrea semiorbis/Trichoderma sp. and H. tawa/T. tawa (Chaverri et al., 2003), Hypocrea atrogelatinosa/T. sp., H. cremea/T. cremeum and Hypocrea macrospora (Chaverri and Samuels, 2003), Hypocrea novae-zelandiae/T. sp. and H. pseudokoningii/T. pseudokoningii (Samuels et al., 2012a), as well as H. subcitrina (Overton et al., 2006). Members of the genus occurring in Japan comprise Hypocrea aureoviridis f. macrospora and H. ceramica (Chaverri and Samuels, 2003), Hypocrea albocornea/T. sp., Hypocrea centristerilis/T. sp. and H. strictipilosa/T. strictipile (Chaverri and Samuels, 2003), H. farinosa/T. sp. and H. subsulphurea/T. sp. (Overton et al., 2006), as well as H. sulphurea/T. sp. and Hypo-crea victoriensis/T. sp. (Overton et al., 2006). The species Hypocrea melanomagna/Trichoderma melano-magnum and Hypocrea sulawesensis/T. sp. are known from Australia and Indonesia, respectively (Chaverri and Samuels, 2003). Hypocrea catoptron/T. catoptron, H. cornea/T. sp., H. rugulosa and Hypocrea straminea/ T. stramineum (Chaverri and Samuels, 2003), as well as T. parareesei, T. pinnatum/H. sp. and H. pseudokoningii/

9
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
T. pseudokoningii are known from Sri Lanka (Samuels et al., 2012a). The Trichoderma communities of the island of Sardinia were studied by Migheli et al. (2009). Fifteen soil samples from different habitats including undis-turbed or extensively grazed grass steppes, forests and shrub lands were examined and the widely distributed species T. asperellum, H. atroviridis/T. atroviride, T. gamsii, T. hamatum, H. koningii/T. koningii, Hypocrea koningiopsis/ T. koningiopsis, H. lixii/T. harzianum, H. semiorbis, T. spi-rale, T. tomentosum, H. virens/T. virens, H. viridescens/ T. viridescens and T. velutinum could be identified by ITS barcoding. Only a single, potentially endemic ITS1 allele could be detected in the case of T. hama-tum, suggesting a significant reduction in the diversity of native species from the genus in Sardinia and an invasion of nonendemic species from Eurasia, Africa and the Pacific Basin. From the Longibrachiatum clade, T. saturnisporopsis is also known from Sardinia (Samuels et al., 2012a). Zachow et al. (2009) examined the fungal biodiversity of soils at different vegetation regions on Tenerife (Canary Islands). From the genus Trichoderma/Hypocrea, the species isolated and identi-fied by TrichOKEY were Trichoderma chionea, T. gamsii, T. harzianum, H. rufa/T. viride, T. spirale and T. tomen-tosum, with a clear dominance of T. harzianum. The majority of the isolates could be characterized with excellent mycoparasitic activities against the fungal plant pathogens Botrytis cinerea, Guignardia bidwellii, Rhizoctonia solani, Sclerotium rolfsii and Verticillium dahliae, suggesting the colonization of the island Tenerife by highly competitive Trichoderma species from the continent.
Agricultural Habitats
Several biotic and abiotic factors affect populations and diversity of microbial communities in agricultural ecosystems including plant species and their growth stage, total microbial competition, soil physical and chemical properties, application of pesticides or fertiliz-ers as well as the geographical region.
Trichoderma spp. can be theoretically isolated from almost all types of agricultural fields. They have sev-eral positive impacts on cultivated plants including biological control of plant diseases, inducing systemic resistance, increasing nutrient availability and uptake, promotion of plant growth, improving crop yields and degrading xenobiotic pesticides (Harman, 2006). For the reasons mentioned above, these fungi have been widely studied and commercially marketed as biofungicides, biofertilizers and soil amendments (Vinale et al. 2008).
The rhizosphere is among the common ecological niches for Trichoderma spp., which attracts them by the presence of different soil borne fungi as their prey and by rich plant root- derived nutrients (Druzhinina et al. 2011). Members of the genus were more frequently
isolated from rhizosphere and non-rhizosphere soils than from phyllospheres. Numerous Trichoderma species have been collected from different crop fields in diverse climatic zones of all continents. Members of the genus Trichoderma are among the most frequently isolated soil fungi. However, some species are ubiquitous while oth-ers are limited to specific geographical areas (Harman et al. 2004).
The majority of the research which involved the iso-lation and identification of Trichoderma strains from various agricultural and horticultural crop fields in different agro-climatic zones was undertaken in order to evaluate them for biological control potential against various phytopathogens. Therefore, only a limited num-ber of studies deal with population, abundance and diversity of the genus Trichoderma in specific crop fields or agroecosystems.
Cereal crop fields—Trichoderma spp. proved to be among the dominating fungi in cereal (rye, triticale, wheat) field soils in Poland (Pięta et al. 2000), and reported to be the most prevalent taxa among the fungal communities in winter wheat (Triticum aestivum L.) soils of Germany, where the most frequently isolated species were T. atroviride and T. viride (Hagn et al. 2003). Tricho-derma piluliferum was also isolated but surprisingly, the cosmopolitan species T. harzianum has not been obtained in this study. Season, soil type and farming management practice influenced only the distribution of T. viride iso-lates. Diversity of Trichoderma spp. was very high in soil samples of wheat fields of China (Liang et al. 2004). In another study, 11 Trichoderma species were identified by ITS-barcoding from rhizosphere soils of five winter wheat fields in the Pannonian Plain (Hungary) com-prising T. atroviride, T. brevicompactum, T. gamsii, T. har-zianum, T. koningiopsis/T. ovalisporum, the species duplet T. longibrachiatum/H. orientalis, T. pleuroticola, T. rossicum, T. spirale, T. tomentosum/T. cerinum and T. virens (Kredics et al. 2012). Trichoderma harzianum was the most abun-dant species representing various ITS haplotypes includ-ing two yet unknown ones. Other frequent species were T. virens, T. rossicum and T. atroviride, each of which grouped into two ITS-genotypes. Agricultural fields dif-fered in species composition as well as the abundance of individual Trichoderma species. Trichoderma spp. in rhizospheres of winter wheat in the Pannonian Plain were found to be common and very diverse. In contrast, Trichoderma biodiversity in agricultural soils (cultivated with various crops including wheat) of the Nile valley in Egypt was very low and contained only T. harzia-num and the anamorph of H. orientalis (Gherbawy et al. 2004). This low degree of diversity may occur due to the alkalinity of the investigated soils (pH = 7.3–8.4). Tricho-derma harzianum isolates were genetically more diverse and displayed three different ITS haplotypes and three RAPD genotypes. Furthermore, enzymatic activities and RAPD fingerprints of the isolates did not correlate with

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS10
A. BIOLOGY AND BIODIVERSITY
the habitat. Corn field soils in Egypt (Gherbawy et al. 2004) and Mexico (Sánchez-Perez, 2009) revealed only two (T. harzianum and the anamorph of H. orientalis) and three (T. harzianum, T. koningiopsis and T. virens) species, respectively, while nine Trichoderma species (T. asperel-lum, T. atroviride, T. erinaceum, T. harzianum, T. koningi-opsis, T. pleurotum, T. reesei, T. spirale and T. virens) were identified from the soils of the same crop in Venezuela (Pavone and Domenico, 2012). Trichoderma harzianum was the most prevalent species in all three cases. Based on UP-PCR and rDNA-ITS1 analysis, 42 Trichoderma iso-lates obtained from rice (Oryza sativa L.) field soils in four provinces of the Philippines belonged only to T. viride and T. harzianum, the latter comprised the majority of isolates (93%) (Cumagun et al. 2000). Similarly, T. harzianum was the most common species in rice field soils in Bangla-desh (Mostafa Kamal and Shahjahan, 1995), as well as in upland and lowland rice fields of the Philippines (Naga-mani and Mew, 1987) however, molecular identifications were not carried out in these studies. Two hundred and two Trichoderma isolates were collected from soil and phyllosphere of rice in paddy fields located at different geographical areas at the southern coast of the Caspian Sea, Iran, which belonged to six species: T. asperellum, T. atroviride, T. brevicompactum T. hamatum, T. harzianum and T. virens according to the results of ITS barcode-based identification (Naeimi et al. 2010). Like the rhizo-sphere of winter wheat in Hungary (Kredics et al. 2011a, 2012), rice paddy field habitats in Northern Iran are rich sources of potential biocontrol isolates belonging to T. atroviride, T. harzianum and T. virens, taxa that are inten-sively studied and applied in biological control of plant diseases. Trichoderma harzianum and T. virens were two most dominant species (>90%) in this region and only these two taxa were isolated from rice phyllosphere. Phylogenetic analysis revealed that T. harzianum was the most diverse species representing 14 different ITS haplo-types clustered into four groups. Trichoderma virens was the only other species from this study that showed intra-specific variation with three different genotypes in one clade. Correlation of the genotypes with sampling site or substrate (soil/phyllosphere) was not observed. In addi-tion, the results suggested that different genotypes could coexist in a single habitat (Naeimi et al. 2011).
Potato (Solanum tuberosum L.) rhizosphere—Six species: T. asperellum, T. atroviride, T. hamatum, T. harzianum, T. kon-ingii and T. tomentosum were identified by ITS barcoding and restriction fragment length polymorphism (RFLP) analysis from the rhizosphere, rhizoplane and bulk soil of potato (S. tuberosum L.) as well as onion (Allium cepa L.) in New Zealand, and similar species diversity was reported in these habitats (Bourguignon, 2008). Tricho-derma hamatum, T. harzianum and T. koningii appeared to be the most frequent species. Moreover, biodiversity analysis of Trichoderma communities in the rhizosphere
soil of different potato cultivars grown in two fields located in Southern Germany revealed that the popu-lation of Trichoderma spp. and species diversity were site-dependent, and high field heterogeneity of Tricho-derma communities was revealed by DGGE fingerprints, although differences among them were not statistically significant (Meincke et al. 2010). A study undertaken in Poland showed that Trichoderma spp. were predominant in potato field soils (Pięta et al. 2000).
Coffee (Coffea arabica) rhizosphere—Trichoderma iso-lates were recovered from the rhizosphere soils of coffee plants in forests and disturbed semiforests of Ethiopia and 134 isolates belonging to eight common species were identified by ITS-barcoding, which were the fol-lowing in order of abundance: T. harzianum and T. hama-tum (the most predominant species in both habitats), T. asperelloides, T. spirale, T. atroviride, T. koningiopsis, T. gamsii and T. longibrachiatum (Mulaw et al. 2010). Cul-tivated and uncultivated coffee regions were rich in Trichoderma populations and showed relatively high diversity, but interestingly the biodiversity indices and evenness were the same for both habitats. In addition, correlation analysis of the existence of individual Tricho-derma species to altitude and some chemical properties of sampling site soils revealed that Trichoderma spp. did not have an ecological preference. Intraspecific variation detected by phylogenetic analysis based on tef1 revealed that T. harzianum was the most diverse species. More-over, strains of T. hamatum, T. atroviride and T. spirale rep-resented new genotypes. It was concluded that the high genetic diversity of Trichoderma from coffee plantation soil and the establishment of new taxa were influenced by the variability of the host plant.
Cocoa (Theobroma cacao L.) rhizosphere—One hundred and thirty five Trichoderma isolates collected from rhizo-spheres in different locations across the Ivory Coast were identified by ITS-barcoding as T. asperellum, T. harzianum, T. virens and T. spirale (Mpika et al. 2009). The first two species were obtained from all cocoa fields and proved to be the most abundant in this habitat.
Sugar beet (Beta vulgaris L.) rhizosphere—Sixteen isolates of Trichoderma were obtained from soils of a sugar beet field in France and identified based on morphology as well as ITS and tef1 sequence analysis as T. gamsii, T. harzianum, T. tomentosum and T. velutinum (Anees et al. 2010). Trichoderma velutinum and T. gamsii were the most prevalent species.
Oilseed rape (Brassica napus L.) rhizosphere— Trichoderma spp. were among the prevalent fungi and showed high biodiversity and plant specificity in the rhizosphere and bulk soil of oilseed rape as well as strawberry (Fragaria × ananassa Duch.) in different locations of Germany (Berg et al. 2005). Diversity and abundance of Trichoderma in bulk soil was higher than in rhizosphere soil and the occurring species showed more genotypic

11
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
diversity by BOX-PCR compared to other fungal genera. Another study in France showed that Trichoderma was among the dominant fungal genera in rape rhizosphere and contributed to the mineralization of organic sulfur, which is an essential element for plant growth and pro-ductivity (Slezack-Deschaumes et al. 2012).
Common bean (Phaseolus vulgaris L.) rhizosphere— Trichoderma asperellum, T. erinaceum, T. harzianum, T. koningiopsis and T. tomentosum were identified by ITS barcoding in the rhizosphere soils of common bean fields in different areas of Brazil and high level of inter- and intraspecific diversity in terms of metabolic pro-files and assimilation of carbon sources was reported (Cardoso Lopes et al. 2012). Trichoderma asperellum and T. harzianum were the most frequent and diverse species detected.
Oil palm (Elaeis guineensis Jacq.) rhizosphere— Trichoderma harzianum, T. virens and T. koningii were the most prevalent Trichoderma species recovered from oil palm soils in Malaysia and the population of these fungi was almost the same in cultivated and unculti-vated oil palm ecosystems (Sariah et al. 2005). Popula-tions of Trichoderma spp. increased by adding empty fruit bunches to the fields, while soil pH and moisture did not affect their distribution and frequency.
Rhizosphere of other crops—Trichoderma hamatum, T. harzianum, T. koningii, T. pseudokoningii and T. viride were detected from soybean (Glycine max (L.) Merr.) soils in Poland (Pięta and Patkowska, 2003), but identities of the isolates were not confirmed by molecular techniques. Six species, (T. atroviride, T. citrinoviride, T. harzianum, T. longibrachiatum, T. koningiopsis and T. reesei) were iden-tified in Mexico from soils cultivated with Sorghum bicolor based on morphological characteristics, enzymatic activ-ity, macro- and microculture test results, rDNA restric-tion patterns (AFLP), ITS1–5.8S–ITS2 rDNA sequences and protein profiles (Larralde-Corona et al. 2008). In Japan, T. hamatum, T. harzianum, T. koningii and T. viride were identified from soils of a radish (Raphanus sativus L.) field (Mghalu et al. 2007). Eleven species of Tricho-derma were obtained from tobacco (Nicotiana tabacum L.) fields in China, among which T. harzianum, T. viride and T. hamatum were the most dominant species (Yu and Zhang, 2004). Trichoderma harzianum and T. hamatum—identified solely on the basis of classical macro- and microscopic criteria—were the most dominant species among 150 fungal species in cucumber rhizosphere soils in Switzerland (Girlanda et al. 2001). From greenhouse soils in China, T. atroviride, T. aureoviride, T. citrinoviride, T. fertile, T. harzianum, T. longibrachiatum, Trichoderma parceramosum, T. reesei, T. virens and T. viride were reported (Zhao et al. 2004). Trichoderma spp. (mostly T. harzianum based on RAPD-analysis) were isolated from rhizosphere soils of various flowers (e.g. carnation, gladiolus and lilium) and vegetables (e.g. tomato) in India
(Shanmugam et al. 2008). Trichoderma spp. were report-edly common or even the most abundant fungi obtained from various crop fields worldwide such as undisturbed cotton (Gossypium hirsutum L.) roots in USA (Baird and Carling, 1998), ginseng (Panax ginseng C.A. Meyer) rhi-zosphere in South Korea (Hyun-Sung and Lee, 1986) and arecanut palm (Areca catechu L.) rhizosphere in India (Bopaiah and Bhat, 1981).
Sadfi-Zouaoui et al. (2009) isolated Trichoderma strains from the soils of cultivated fields in North-East Tunisia and identified them as T. atroviride and T. hamatum.
In the study of Hoyos-Carvajal et al. (2009), 10 out of 29 Trichoderma species originated from agricultural related habitats in Colombia and Mexico. Trichoderma harzianum and T. asperellum were the most dominant species in this region. Distribution of the species was related to the soil and substrate type as well as to the climatic zone. Recent investigations of Trichoderma diversity in China by ITS barcoding and tef1 sequence analysis resulted in the identification of 23 species from different garden, vegeta-ble, farmland and pasture soils all over the country (Sun et al. 2012). The diversity of the genus was the highest in vegetable soils with 13 detected species (T. asperellum, T. atroviride, T. brevicompactum, T. citrinoviride, T. harzianum, T. koningiopsis, T. longibrachiatum, T. pleuroticola, T. sinense, T. stromaticum, T. velutinum, T. virens, T. viride), followed by pasture soils (eight detected species: T. asperellum, T. atroviride, T. erinaceum, T. hamatum, T. koningii, T. longibra-chiatum, T. stromaticum, T. velutinum), garden soils (eight detected species: T. asperellum, T. erinaceum, T. hamatum, T. harzianum, T. longibrachiatum, T. pleuroticola, T. tomento-sum, T. virens) and farmland soils (6 detected species: T. asperellum, T. atroviride, T. aureoviride, T. brevicompactum, T. erinaceum, T. harzianum). The distribution, proposition and frequency of the species were associated with the geographical area. Trichoderma harzianum was the most abundant and widely distributed species followed by T. asperellum and T. hamatum. According to the phyloge-netic analysis of their ITS and tef1 sequences, T. harzia-num was the most variable species in China representing 12 different ITS and 17 tef1 genotypes.
Living Plants (Endophytes)
The studies discussed above demonstrate that the occurrence of Trichoderma is general in the rhizosphere of a wide variety of soils. Certain Trichoderma strains can also colonize the plant roots and take part in sym-biotic relationships. In recent times, numerous studies were carried out to prove that some Trichoderma species can reach the inner tissues of the plants and develop an endophytic relationship.
The cocoa plant (Theobroma cacao) was in the focus of several studies, as growing this plant is very com-mon in various tropical countries of the world. Certain

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS12
A. BIOLOGY AND BIODIVERSITY
plant pathogenic fungi can cause serious crop losses. In Central- and South America, the three most common diseases of cocoa plants are black pod (Phytophtora spe-cies), witches' broom (Crinipellis perniciosa) and frosty pod rot (Moniliophtora roreri) (Bailey et al., 2006). In order to find an efficient biocontrol agent against these pathogens, the endophytic microbial community of the cocoa plant is being studied intensively. These exami-nations proved that a series of Trichoderma species may occur as endophytes of the cocoa plant, including mem-bers of the former T. koningii species aggregate such as T. ovalisporum and the new species T. caribbaeum var. aequa-toriale, T. koningiopsis (Samuels et al., 2006a) and Tricho-derma protrudens (Degenkolb et al., 2008). Trichoderma theobromicola and T. paucisporum (Samuels et al., 2006a), T. stromaticum (Samuels et al., 2012b) as well as Tricho-derma martiale (Hanada et al., 2008) were also identified as endophytic Trichoderma species of cocoa. Rubini et al. (2005) studied the diversity of endophytic fungi of the cocoa plant and successfully identified a range of fungal species, however, the prevalence of Trichoderma species was very low among the strains isolated. The endo-phytic microbiota of coffee seedlings was also reported to contain Trichoderma species including T. hamatum and T. harzianum identified by ITS sequence analysis (Posada et al., 2007).
The identification of endophytic fungi is an intensively investigated field in the case of other plants as well. Such studies may result in the description of new Trichoderma species. Chaverry et al. (2011) described Trichoderma ama-zonicum as a new species based on isolates from rubber tree (Hevea spp.), Zhang et al. (2007) described Tricho-derma taxi from Taxus mairei tree in China, while Samuels et al. (2012) described T. solani as an endophyte in tubers of Solanum hintonii in Mexico.
Six different Trichoderma species were identified by TrichOKEY as endophytic fungi of banana root (Xia et al., 2011), among which four species: T. asperellum, T. virens, T. brevicompactum and H. lixii could be found inside the roots while two species: T. atroviride and T. koningiopsis were detected only on root surface. Trichoderma asperel-lum and T. virens showed the highest frequencies in the examined samples.
Dang et al. (2010) examined the endophytic fungi of Panax notoginseng, a traditional Chinese medicinal plant. According to ITS-based identifications, a T. ovalisporum strain with antibacterial activity against Escherichia coli, Bacillus cereus, Staphylococcus aureus and Micrococcus luteus could be isolated during this study. Other well-known medicinal plants such as Salvia miltiorrhiza and Huperzia serrata were also examined. A T. atroviride strain identified by its morphology and ITS sequence analysis was isolated as an endophytic fungus, which produced tanshinone I and tanshinone IIA (Ming et al., 2012). Moreover, in an independent examination, Chen
et al. (2011) also identified T. atroviride as well as T. gam-sii strains based on their ITS sequences from Huperzia serrata.
A series of studies were aimed at the examination of the production of secondary metabolites by Trich-oderma strains. Trichoderma gamsii identified by ITS sequencing from P. notoginseng, was further examined and as a result, four new cytochalasins: trichoderones A and B and trichalasins C and D were identified by Ding et al. (2012a,b). Souza et al. (2008) studied the secondary metabolite production of a T. koningii iso-late (identified by ITS sequence analysis) derived from Strychnos cogens and described the production of koninginins A, F and E. Two new octahydronaph-thalene derivatives produced by a T. spirale strain iso-lated from Aquilaria sinensis (Li et al., 2012), as well as trichodermanin A produced by T. atroviride isolated from Cephalotaxus fortunei (Sun et al., 2011) were also reported (both producer strains were identified by ITS barcoding).
Studies on endophytic fungi of carnivorous plants also resulted in the detection of endophytic Trichoderma strains. Quilliam and Jones (2010) studied Drosera rotun-difolia plants during spring and autumn and observed seasonal distribution of endophytic fungi, however, T. viride (confirmed by ITS) could be isolated from all of the samples. Later the same authors carried out the investigation of endophytic fungi from the carnivorous plant Pinguicula vulgaris's (Quilliam and Jones, 2012). Although differences could be observed between the endophytes of the two plants, Trichoderma species were detected in both cases.
The endophytic and mycorrhizal fungi were studied also in the case of seeds and roots originated from Den-drobium nobile and Dendrobium chrysanthum belonging to the Orchidaceae family (Chen et al., 2012). The presence of Trichoderma species was proved among the 127 endo-phytic fungal isolates. Moreover, Yuan et al. (2009) inves-tigated further 288 samples from D. nobile and identified T. chlorosporum based on ITS sequence analysis among the detected species.
One of the most harmful pathogenic fungi of the potato plant is R. solani. It is capable of causing seri-ous quality and quantity damages in the potato tuber. Therefore a lot of laboratory work is aimed worldwide at finding an effective biocontrol agent against this dangerous pathogen. The biocontrol ability of endo-phytic fungi isolated from potato plants was examined in agar confrontation tests (Lahlali and Hijri, 2010). On the basis of the results of these tests it was concluded that the isolates of Epicoccum nigrum and T. atroviride (confirmed by ITS) showed the highest inhibition of R. solani. In the case of T. atroviride the mycoparasitic phenomenon was found to be determinative in the inhibition process.

13
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
Mushroom-Related Substrata
The association of Trichoderma species with wild as well as cultivated mushrooms has been reported from various countries. The Trichoderma-caused green mould disease severely affects the production of both button mushroom (A. bisporus) and oyster mushroom (Pleuro-tus ostreatus), causing serious losses for growers world-wide. Overviews of the current status of A. bisporus and P. ostreatus green mould were given by Kredics et al. (2010) and Hatvani et al. (2008), respectively.
During the early appearance of mushroom green mould disease, various Trichoderma species such as T. atroviride, T. citrinoviride, T. harzianum, T. koningii and T. longibrachiatum were found to be associated with cultivated A. bisporus. However, the predominant spe-cies, causing aggressive compost colonization were identified exclusively as the T. harzianum biotypes Th2 (Seaby, 1987, 1989; Doyle, 1991) and Th4 (Castle et al. 1998) in Great Britain/Ireland and United States/Canada, respectively. The appearance of green mould due to T. harzianum biotype Th2 in other Western Euro-pean countries was subsequently reported (Hermosa et al. 1999; Mamoun et al. 2000). Based on morphologi-cal characteristics as well as the phylogenetic analyses of ITS1 and a fragment of the tef1 gene, Samuels et al. (2002) redescribed the T. harzianum biotypes Th2 and Th4, the causal agents of Agaricus green mould disease in Europe and North America, as the new species T. aggressivum f. europaeum and T. aggressivum f. aggressi-vum, respectively.
The cultivation of A. bisporus in Hungary was found to be affected mostly by T. aggressivum f. europaeum, indi-cating the spread of the Agaricus green mould epidemic from Western to Central Europe. Besides T. aggressivum, further five Trichoderma species, T. asperellum, T. atro-viride, T. ghanense, T. harzianum and T. longibrachiatum were detected in compost samples (Hatvani et al. 2007). The holotype strain of the recently described new species T. capillare (Samuels et al., 2012a) was also recovered from a Hungarian Agaricus-producing facility: it was isolated from the wall of a mushroom growing cellar (Hatvani et al., 2006).
In Poland T. aggressivum, T. atroviride, T. citrinoviride, T. harzianum, T. longibrachiatum, T. virens and T. viride were identified in association with mushrooms, with T. aggressivum being the most abundant species (60% of the isolates) (Błaszczyk et al. 2011). This finding demon-strates a change in the representation of species, as an earlier study revealed the dominance of T. harzianum in the country (Szczech et al., 2008). Green mould-affected Agaricus compost in Croatia yielded exclusively T. har-zianum, indicating a broadening spectrum of Trichoderma species being able to cause green mould disease in but-ton mushroom cultivation (Hatvani et al. 2012).
Species found in green mould-affected oyster mush-room substrate in Hungary were T. asperellum, T. atroviride, T. longibrachiatum, and the yet undescribed species Tricho-derma sp. DAOM 175924, which represented 90% of the isolates (Hatvani et al. 2007). Strains belonging to this taxon could be divided into two groups based on an A/C transversion at position 447 of the ITS2 region and corre-sponded to Trichoderma sp. K1 and K2, the Pleurotus patho-genic Trichoderma species observed in Korea (Park et al. 2004), which were subsequently described as the new species T. pleurotum and T. pleuroticola (Park et al. 2006).
The results of the comprehensive study of Komoń-Zelazowska et al. (2007) confirmed that these two species were responsible for green mould infections in Pleuro-tus farms in various countries, such as Italy, Hungary, Romania and the Netherlands. In Croatia the same spe-cies were found to cause oyster mushroom green mould, being the sole species recovered from infected Pleurotus substrate samples (Hatvani et al. 2012). Pleurotus green mould in Spain was shown to be caused exclusively by T. pleurotum (Gea, 2009). Kredics et al. (2009) devel-oped a PCR-based technique for the specific detection of T. pleurotum and T. pleuroticola, the Trichoderma patho-gens of cultivated oyster mushroom. Through the use of the newly introduced method, the presence of T. pleu-roticola was detected in high proportions in the growing substrate and on the fruiting bodies of wild Pleurotus species, which might act as potential sources of infec-tion of mushroom farms. Further Trichoderma species found in these habitats were T. atroviride, T. harzianum and T. longibrachiatum. Trichoderma pleurotum could not be detected in any of the samples tested, however, fur-ther investigations revealed that the natural substratum of oyster mushroom is a habitat of this species as well (Kredics et al. unpublished data). Recently, Kim et al. (2012) described Trichoderma mienum as a new species of the Semiorbis clade isolated from oyster mushroom and shiitake bed logs in Japan.
Trichoderma species were found to be the most fre-quent contaminants of shiitake (Lentinula edodes) cultiva-tion in Thailand. The majority of the isolates belonged to T. harzianum, but T. aureoviride, T. koningii, T. piluliferum and T. pseudokoningii were also detected in small propor-tions (Pukahuta et al. 2000). Turóczi et al. (1996) reported the isolation of T. hamatum from the fruiting bodies of Lentinula edodes. The strains showed intermediate antag-onistic properties towards phytopathogenic fungi.
Certain members of the genus Trichoderma (T. hama-tum, T. harzianum, T. koningii, T. virens and T. viride) were shown to be among the most abundant microfungi iso-lated from the surroundings of wild Termitomyces species in Thailand (Manoch et al., 2002), suggesting their poten-tial role in the stimulation of the occurrence of termite fungi. The ITS barcoding-based detection of T. hamatum, T. harzianum, T. spirale, T. virens and an unidentified

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS14
A. BIOLOGY AND BIODIVERSITY
Trichoderma sp. was reported from the nests of leafcutter ants cultivating basidiomycetous fungi from Agaricales for nutritional purposes (Rodrigues et al., 2008).
Rivera et al. (2010) reported the presence of Tricho-derma species at 16% of the moulds isolated from the ascocarps of the truffle Tuber aestivum. Wang et al. (2011) examined the microbial communities of wild Chroogom-phus rutilus, and found that T. koningiopsis strains repre-sented 28.6% of the fungal isolates.
Human Body
The role of Trichoderma species as facultative patho-gens of humans was firstly summarized by Kredics et al. (2003) and later extensively reviewed and discussed (Kredics et al., 2011b). Infections caused by Trichoderma species known from the literature include peritonitis and intra-abdominal abscess in patients undergoing continu-ous ambulatory peritoneal dialysis (CAPD), liver infec-tion, acute invasive sinusitis and disseminated infections (e.g. abdominal, lung and skin disseminations) of trans-plant recipients, brain abscess, skin infection, necrotiz-ing stomatitis and pulmonary infections of patients with hematological malignancies, fungemia by contaminated saline, rhinosinusitis, pulmonary mycetoma and fibro-sis, hypersensitivity pneumonitis, endocarditis, otitis externa, cerebrospinal fluid infection and allergic reac-tions (Kredics et al., 2011b). Trichoderma species reported in case descriptions of human infections in the literature are T. atroviride, T. citrinoviride, T. harzianum, T. koningii, T. longibrachiatum, H. orientalis, T. pseudokoningii, T. reesei, T. viride and a Hypocreaceae sp. However, it is question-able whether all of these Trichoderma species are in fact able to cause human infections, as many Trichoderma isolates recovered from clinical samples were identified based on their morphological characters only, which is frequently problematic. Although a key for the morphol-ogy-based identification of clinical Trichoderma isolates was introduced by Summerbell (2003) and the use of the morphological key of Gams and Bissett (1998) was also proposed, this may still result in incorrect identifications due to the lack of expertise. Therefore the application of biochemical and molecular techniques is suggested to confirm the species-level diagnosis of clinical Tricho-derma isolates.
As a biochemical solution, cellulose-acetate electro-phoresis-based isoenzyme analysis according to Hebert and Beaton (1993) was performed by Szekeres et al. (2006) for the identification of clinical Trichoderma iso-lates. The authors suggested this method as a cheap and efficient alternative of molecular techniques for the quick and specific identification of clinical T. lon-gibrachiatum isolates. DNA-based molecular methods applied to assess the genetic diversity of clinical Tricho-derma isolates include RFLP of the mitochondrial DNA
(Antal et al., 2006) as well as PCR-fingerprinting and ITS sequence analysis (Kuhls et al. 1999), which revealed that the reported identities of clinical Trichoderma strains were incorrect in several cases and that T. longibrachiatum was the most frequent, almost exclusive causal agent of opportunistic infections within the genus Trichoderma. However, as the species T. longibrachiatum and H. orien-talis are not distinguishable based on their ITS sequences alone, the analysis of further phylogenetic markers is needed in the cases of the clinical involvement of this species pair. For the examination of a Trichoderma strain collection of 15 clinical and 36 environmental isolates belonging to T. longibrachiatum/H. orientalis, Druzhinina et al. (2008) applied multilocus phylogenetic analysis involving the ITS region along with fragments of tef1, calmodulin (cal1) and endochitinase (chit18-5) genes. The results of this study have reinforced that H. orientalis is a sexually recombining, while T. longibrachiatum is a strictly clonal species. Hypocrea orientalis was also identi-fied as an opportunistic human pathogen, and clinical T. longibrachiatum isolates were shown not to form a par-ticular subpopulation of the species. These findings sug-gest that all strains of T. longibrachiatum and H. orientalis might be able to cause infections in humans.
Besides T. longibrachiatum and H. orientalis, the involve-ment of further four Trichoderma species was confirmed with molecular tools: T. atroviride (Ranque et al., 2008), T. citrinoviride (Kuhls et al. 1999), T. harzianum (Guarro et al., 1999; Kantarcioğlu et al., 2009) and an unknown Hypocreaceae species close to the genus Hypocrea/Tricho-derma (Druzhinina et al., 2007), which shares identical ITS and rpb2 sequences with Hypocrea peltata, a recently described sexually reproducing Hypocrea species with-out a Trichoderma anamorph (Samuels and Ismaiel, 2011).
Potential virulence factors of Trichoderma species as opportunistic pathogens of humans are suggested to include the ability to grow at elevated temperatures and neutral pH, the production of extracellular proteases and the ability to utilize amino acids as carbon and nitro-gen sources (Antal et al., 2005). Antifungal susceptibil-ity studies on clinical Trichoderma strains revealed high resistance of numerous isolates to a series of widely used antimycotics, but e.g. voriconazole can be suggested for the treatment of patients (Kredics et al., 2011b).
Water-Related Environments
Trichoderma species were shown to be abundant in different water-related environments including marine and sweet water habitats as well as water-damaged building materials.
The marine occurrence of Trichoderma species was firstly mentioned by Kohlmeyer (1974). Since that time, Trichoderma has frequently been reported in association with different marine sponge species, including Agelas

15
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
dispar collected from waters around the Island of Domi-nica (Neumann et al., 2007), Latrunculia corticata from the Red Sea at Sharm El-Sheikh, Egypt (El-Bondkly et al., 2012), Geodia corticostylifera form the South Atlantic Ocean, Brazil (Rocha et al., 2012), Mycale fibrexilis from the South China Sea nearby Yongxing Island (Zhou et al., 2011), Suberites zeteki from Rainbow Bay Marina in Pearl Harbor and Gelliodes fibrosa from Coconut Island in Kaneohe Bay on Oahu, Hawaii (Wang et al. 2008) and Psammocinia sp. from the Mediterranean Sea at Sedot-Yam, Israel (Paz et al., 2010; Gal-Hemed et al., 2011). Other sea animals from which the isolation of Trichoderma strains was reported include the gorgonian sea fan Annella sp. (Khamtong et al., 2012), the seastar Acanthaster planci (Lan et al., 2012), the blue mussel Myti-lus edulis (Ruiz et al., 2007a,b; Sallenave-Namont et al., 2000) and the cockle Cerastoderma edule (Sallenave et al., 1999; Sallenave-Namont et al., 2000), where the pres-ence of Trichoderma was shown to contribute to shellfish toxicity (Sallenave et al., 1999). Besides sea animals, a Trichoderma strain identified as T. harzianum was isolated from the green alga Chaetomorpha linum collected from the North Sea around the Island of Helgoland, Germany (Neumann, 2008).
Trichoderma species could be isolated from sedi-ments on the root of mangrove Ceriops tagal collected in the South Sea intertidal zone, China (Sun et al., 2009), as well as from marine sediments collected at different locations including the South China Sea (Burtseva et al., 2003; Song et al., 2010), the Fujian province of China (Du et al., 2009), the tideland of the Yellow Sea at Lianyun-gang, China (Sun et al., 2008), various regions of the Sea of Japan (Khudiakova et al., 2000), St. Helena Bay, South Africa (Mouton et al., 2012), marine shellfish farm-ing areas along the Western coast of France (Sallenave-Namont et al., 2000) and the Antarctic Penguin Island (Ren et al., 2009). In the Alimini Grande brackish lake in Italy, Trichoderma proved to be the dominant genus in sediment samples in a marshy area, where it is supposed to be involved in the decomposition of allochthonous plant material (mainly Phragmites australis) (De Donno et al., 2008).
Marine-derived fungi including Trichoderma species are attracting increasing interest as potential sources of metabolites (Table 1.1) with a wide range of biologi-cal activities including antibacterial activity against methicillin-resistant S. aureus (Khamtong et al., 2012), cytotoxicity against human colon carcinoma cells (Garo et al., 2003), a melanoma cell line (Sun et al., 2006), other cancer cell lines and bioactivities against HIV protease (You et al., 2010). El-Bondkly et al. (2012) used a marine Trichoderma strain for intergeneric protoplast fusion with Penicillium and Aspergillus strains to improve cellulase production. Activities of β-1,3-glucanase (Burtseva et al., 2003) and laminarinase enzymes (Burtseva et al., 2006)
as well as a tyrosinase inhibitor (Tsuchiya et al., 2008) were also studied in the case of marine-derived Tricho-derma strains. A Trichoderma strain isolated from the marine sponge G. corticostylifera was found to catalyze the bioreduction of a-chloroacetophenone (Rocha et al., 2009), the hydrolysis of benzyl glycidyl ether (Martins et al., 2011) and the asymmetric bioconversion of iodo-acetophenones to the corresponding iodophenyletha-nols (Rocha et al., 2012). The potential application of a strain (reported as T. viride) from sea water samples collected from a heavy metal-polluted area in the Medi-terranean Sea, Alexandria, Egypt was suggested for the mycoremediation of Cr (VI) from water systems (El-Kassas and El-Taher, 2009). A T. longibrachiatum strain isolated from mussels in a shellfish farming area was investigated for total lipid production, total lipid fatty acids, and phospholipid fatty acids and did not found marked differences when compared to lipid class and fatty acid profiles of terrestrial Trichoderma species (Ruiz et al., 2007a).
As the species level identification of marine Tricho-derma isolates has not been performed in the majority of these studies or it has been performed based on morpho-logical characters only, the marine occurrence of certain species reported in these articles (e.g. T. reesei, T. viride) lacks molecular evidence. In certain cases, ITS sequences were determined but their NCBI BLAST analysis resulted in an incorrect identification, e.g. Khamtong et al., 2012 identified an isolate as T. aureoviride while Song et al. (2010) another one as T. koningii, however, a TrichOKEY 2.0 analysis of the respective sequences (GenBank acces-sion numbers EU714396 and GU244589) reveals the identity of the isolates as T. harzianum and T. koningiop-sis/ovalisporum, respectively. An exact identification was provided in the study of Mohamed-Benkada et al. (2006), who identified the studied trichobrachin- producing strain as T. longibrachiatum using metabolic profiles on Biolog FF MicroPlatesTM and sequence analysis of the ITS region and the intron-rich region of the tef1 gene.
The most detailed data about Trichoderma population structure in a marine habitat were provided by the stud-ies of Paz et al. (2010) and Gal-Hemed et al. (2011). Paz et al. (2010) collected samples from the sponge Psammo-cinia sp. at a depth of 2–6 m, approximately 200 m off-shore at Sedot-Yam, Israel in the winter and summer of 2007 and isolated 400 fungal strains, among which 220 were subjected to ITS and tef1-based molecular identifi-cation. Trichoderma species with mycoparasitic potential were also recorded. Species identifications of 29 Tricho-derma strains from Psammocinia sp. were refined in the subsequent study by Gal-Hemed et al. (2011). The great-est number of isolates proved to belong to T. atroviride (9), the T. harzianum species complex (7), as well as to T. longibrachiatum (5) and the closely related H. orientalis (4). A single isolate of T. asperelloides and two putative
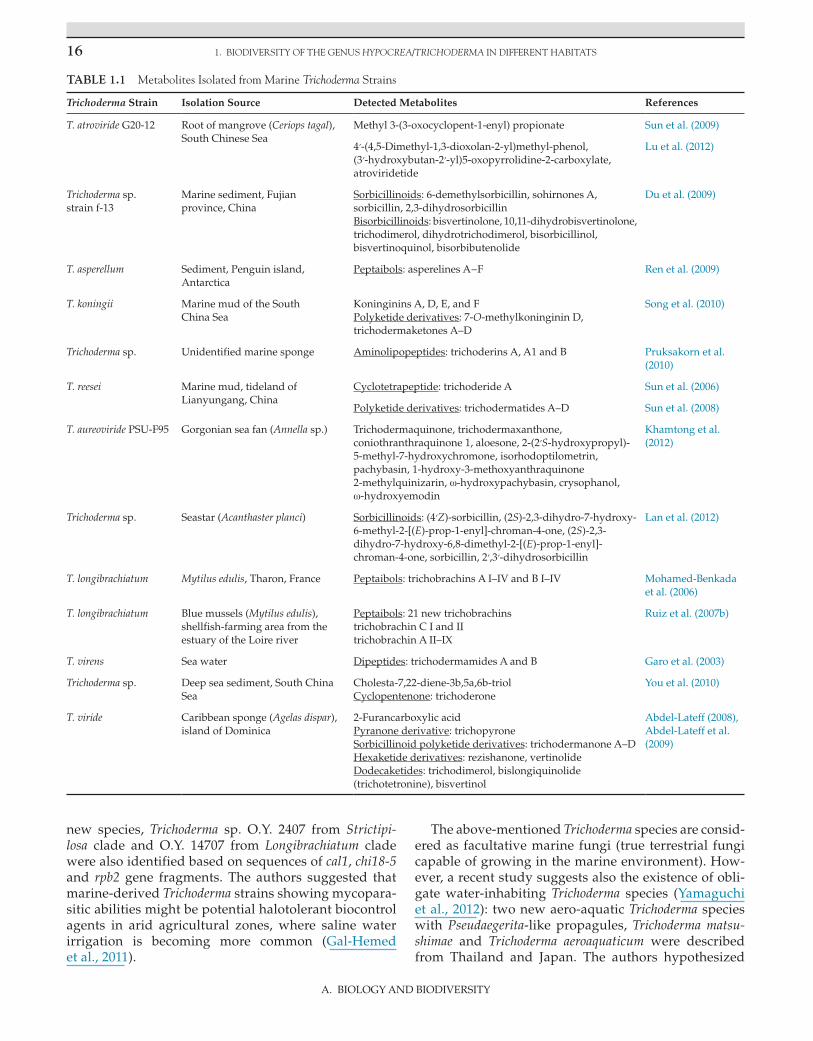
1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS16
A. BIOLOGY AND BIODIVERSITY
new species, Trichoderma sp. O.Y. 2407 from Strictipi-losa clade and O.Y. 14707 from Longibrachiatum clade were also identified based on sequences of cal1, chi18-5 and rpb2 gene fragments. The authors suggested that marine-derived Trichoderma strains showing mycopara-sitic abilities might be potential halotolerant biocontrol agents in arid agricultural zones, where saline water irrigation is becoming more common (Gal-Hemed et al., 2011).
The above-mentioned Trichoderma species are consid-ered as facultative marine fungi (true terrestrial fungi capable of growing in the marine environment). How-ever, a recent study suggests also the existence of obli-gate water-inhabiting Trichoderma species (Yamaguchi et al., 2012): two new aero-aquatic Trichoderma species with Pseudaegerita-like propagules, Trichoderma matsu-shimae and Trichoderma aeroaquaticum were described from Thailand and Japan. The authors hypothesized
TABLE 1.1 Metabolites Isolated from Marine Trichoderma Strains
Trichoderma Strain Isolation Source Detected Metabolites References
T. atroviride G20-12 Root of mangrove (Ceriops tagal), South Chinese Sea
Methyl 3-(3-oxocyclopent-1-enyl) propionate Sun et al. (2009)
4′-(4,5-Dimethyl-1,3-dioxolan-2-yl)methyl-phenol, (3′-hydroxybutan-2′-yl)5-oxopyrrolidine-2-carboxylate, atroviridetide
Lu et al. (2012)
Trichoderma sp. strain f-13
Marine sediment, Fujian province, China
Sorbicillinoids: 6-demethylsorbicillin, sohirnones A, sorbicillin, 2,3-dihydrosorbicillinBisorbicillinoids: bisvertinolone, 10,11-dihydrobisvertinolone, trichodimerol, dihydrotrichodimerol, bisorbicillinol, bisvertinoquinol, bisorbibutenolide
Du et al. (2009)
T. asperellum Sediment, Penguin island, Antarctica
Peptaibols: asperelines A−F Ren et al. (2009)
T. koningii Marine mud of the South China Sea
Koninginins A, D, E, and F Polyketide derivatives: 7-O-methylkoninginin D, trichodermaketones A–D
Song et al. (2010)
Trichoderma sp. Unidentified marine sponge Aminolipopeptides: trichoderins A, A1 and B Pruksakorn et al. (2010)
T. reesei Marine mud, tideland of Lianyungang, China
Cyclotetrapeptide: trichoderide A Sun et al. (2006)
Polyketide derivatives: trichodermatides A–D Sun et al. (2008)
T. aureoviride PSU-F95 Gorgonian sea fan (Annella sp.) Trichodermaquinone, trichodermaxanthone, coniothranthraquinone 1, aloesone, 2-(2′S-hydroxypropyl)-5-methyl-7-hydroxychromone, isorhodoptilometrin, pachybasin, 1-hydroxy-3-methoxyanthraquinone 2-methylquinizarin, ω-hydroxypachybasin, crysophanol, ω-hydroxyemodin
Khamtong et al. (2012)
Trichoderma sp. Seastar (Acanthaster planci) Sorbicillinoids: (4′Z)-sorbicillin, (2S)-2,3-dihydro-7-hydroxy-6-methyl-2-[(E)-prop-1-enyl]-chroman-4-one, (2S)-2,3-dihydro-7-hydroxy-6,8-dimethyl-2-[(E)-prop-1-enyl]-chroman-4-one, sorbicillin, 2′,3′-dihydrosorbicillin
Lan et al. (2012)
T. longibrachiatum Mytilus edulis, Tharon, France Peptaibols: trichobrachins A I–IV and B I–IV Mohamed-Benkada et al. (2006)
T. longibrachiatum Blue mussels (Mytilus edulis), shellfish-farming area from the estuary of the Loire river
Peptaibols: 21 new trichobrachins trichobrachin C I and II trichobrachin A II–IX
Ruiz et al. (2007b)
T. virens Sea water Dipeptides: trichodermamides A and B Garo et al. (2003)
Trichoderma sp. Deep sea sediment, South China Sea
Cholesta-7,22-diene-3b,5a,6b-triolCyclopentenone: trichoderone
You et al. (2010)
T. viride Caribbean sponge (Agelas dispar), island of Dominica
2-Furancarboxylic acidPyranone derivative: trichopyroneSorbicillinoid polyketide derivatives: trichodermanone A–DHexaketide derivatives: rezishanone, vertinolide Dodecaketides: trichodimerol, bislongiquinolide (trichotetronine), bisvertinol
Abdel-Lateff (2008), Abdel-Lateff et al. (2009)

17
A. BIOLOGY AND BIODIVERSITY
TRICHODERMA DIVERSITY IN DIFFERENT HABITATS
that these fungi evolved from soil-inhabiting species of Trichoderma by adaptation to aquatic environments via the formation of bulbil-like propagules floating on water.
The abundance of Trichoderma species is also known in natural and artificial sweet water environments, e.g. in an acid mine drainage lake in Anhui Province, China (Zhang et al., 2012) or in bottled water (Ribeiro et al., 2006). In a study about the diversity and significance of mold species in Norwegian drinking water, besides Penicillium and Aspergillus, Trichoderma was also among the fungi with the highest maximum number of CFU in drinking water samples (Hageskal et al., 2006). In a sub-sequent study, a total of 123 Trichoderma strains were iso-lated from Norwegian surface-sourced drinking water (Hageskal et al., 2008). Examined samples included raw water, treated water, and water from private homes and hospital installations. Eleven known Trichoderma/Hypo-crea species and a group of unidentified Trichoderma strains representing a separate, strongly supported sub-clade within the Pachybasium A/Hamatum clade could be detected based on ITS and tef1 sequences. Trichoderma viride was the predominant species with 49% of the iden-tified strains, being present in 22% of the surface-derived water samples.
Water-damaged buildings represent a further water-related habitat from which Trichoderma species (T. atroviride, T. viride, T. hamatum as well as the clinically relevant species T. longibrachiatum, T. citrinoviride and T. harzianum) are frequently reported as the dominat-ing microfungi (Lübeck et al., 2000; Thrane et al., 2001; Ebbehøj et al. 2002). Release of spores by common indoor fungi including T. harzianum from wet wallpapered gyp-sum boards was examined by Kildesø et al. (2003). The authors concluded that these spores might be respon-sible for certain negative health effects related to build-ings contaminated with moulds. Andersen et al. (2011) studied samples from water-damaged building materi-als with visible fungal growth from private residences (houses, apartments and holiday cottages) and private businesses (shops and offices) as well as from public buildings (kindergartens, schools and offices) from all parts of Denmark and Greenland. Concrete, glass fiber, gypsum, plaster, plywood, wallpaper and wood samples were all positive for Trichoderma.
Air and Settled Dust
Air can play an important role in the dispersal of fungal spores and conidia. Based on a comprehensive review by Madsen et al. (2007), Trichoderma species could be isolated from the air of a series of different indoor and outdoor environments.
Indoor air samples positive for Trichoderma included buildings heated by wooden chips in Sweden, flats in Lithuania, homes in Germany, Poland, Taiwan, Turkey
and the USA, hospitals in Austria, Iraq, Poland, Finland and USA (from air filters) (Madsen et al., 2007). Tricho-derma was also found in air ducts of houses in Finland (Madsen et al., 2007), in air conditioners in Saudi Arabia (Madsen et al., 2007), in the air of a library and archive storage facilities in Poland (Zielińska-Jankiewicz et al., 2008), homes in New Orleans after hurricanes Katrina and Rita (Rao et al., 2007), an orchid greenhouse ( Magyar et al., 2011), a greenhouse with ornamental plants (Li and La-Mondia, 2010), child care centers in Turkey (Aydogdu and Asan, 2008) and intensive care units in Brazil, where it was reported among the most frequently occurring fil-amentous fungi (Mobin and Salmito, 2006). Trichoderma was found to be present also in the settled dust of homes in Saudi Arabia and the USA (low-traffic carpets, bed-spread/furniture surfaces), hospitals in Iraq and schools without water damage in Denmark (Madsen et al., 2007), as well as on the walls of damp homes in Croatia and in wall scrapes from basement in the USA (Madsen et al., 2007).
Outdoor isolations of Trichoderma from air samples could be realized at roofs of houses in Saudi Arabia, at rooftop of a hospital in the Netherlands, in urban areas of Kuwait including a balcony of Kuwait University, on bal-conies in Taiwan, in a coastal area in Lithuania, in Pinus nigra and Quercus forests in Turkey (Madsen et al., 2007), in the Belgrade Forest area near Istanbul ( Çolakoglu, 2003), in Trujillo, Peru (Requejo, 1975), Britain (Richards, 1956), Barcelona (Spain) (Calvo et al. 1980), Manhattan (Kansas, USA) (Kramer et al., 1959, 1964; Kramer and Pady, 1960), Sagamihara (Japan) (Takatori et al., 1994) and Israel (Barkai-Golan, 1958; Barkai-Golan and Glazer, 1962; Barkai-Golan et al., 1977).
Trichoderma was also found to be present in settled dust of roofs of houses and stationary cars in Egypt (Madsen et al., 2007).
Regarding environments related with agriculture or industry, Trichoderma could be isolated from the air of hop farms and herb processing plants in Poland, wood chip terminals in Sweden, different parts of swine farms in Finland, combine harvesters in Eng-land and storey buildings undergoing renovation in Egypt, from the air and settled dust of carpentries in Italy, from corn dust in the USA, from settled sawdust at sawmills in England and the USA, from dust blown from hay in England (Madsen et al., 2007), as well as from spots visibly contaminated with fine plastic par-ticles in a manufacturing factory for plastic caps for soft drinks (Sato, 2010).
The studies above indicate that Trichoderma conidia are commonly occurring in air samples and settled dust, however, unfortunately a species-level identi-fication has not been carried out in most of the cases. Where species names were also provided in these ear-lier studies, the occurrence of T. album, T. hamatum, T. harzianum, T. koningii, T. lignorum, T. viride, as well as

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS18
A. BIOLOGY AND BIODIVERSITY
Trichoderma inhamatum—a taxon commonly regarded a synonym of H. lixii, but also suggested as a sepa-rate species within T. harzianum sensu lato (Druzhinina et al., 2010). However, as a sequence-based molecular identification has not been carried out in these studies, they do not provide data about the real diversity of the genus and abundance of its different members in air samples. The actual Trichoderma diversity and species abundance in air samples could be revealed by metage-nomic analyses like the one of Atanasova et al. (2010), who applied the metagenomic approach to study the occurrence and diversity of members of the genus in air samples taken in the Viennese suburban area of Wie-nerwald, Austria. Sequence analysis of a total of 159 molecular operational taxonomic units (MOTUs) by TrichOKEY 2.0 revealed 15 known species with T. virens being the most abundant (52% of all MOTUs). The first European detection of tropical species (T. reesei, T. stro-maticum, T. taxi) and rare temperate species known from North-America (T. fertile, T. ceramicum) was reported in this study, supporting the possibility of long-distance spore dispersal. Other species detected were T. asperel-lum, T. citrinoviride, T. gamsii, the T. longibrachiatum/H. orientalis species duplet and T. minutisporum that all occur frequently in European ecosystems, as well as H. neorufa and Hypocrea psychrophila which are rare in Europe (Atanasova et al. 2010). Interestingly, the most frequent taxon of the genus, the T. harzianum species complex could not be detected in the examined air sam-ples, although it was abundant in soil samples from the same area. Previously it was suggested that T. harzia-num can rarely be found in the air, because its conidia may not be released easily from growth materials to the air (Madsen et al. 2007).
The prevalence of conidia in the air has a significance also from the clinical point of view: allergic diseases may be associated with airborne Trichoderma conidia ( Çolakoglu, 2003), and in the case of immunocompro-mised patients, air may also be a potential source of opportunistic Trichoderma infections like sinusitis or pneumonia. As Trichoderma species are frequently used as biocontrol agents against plant pathogenic fungi in both field agriculture and closed production systems like greenhouses, it is very important to gain informa-tion about the possible exposure of growers working in such facilities to Trichoderma conidia. Hansen et al. (2010) revealed that T. harzianum from the biocontrol product Supresivit could be detected in the air of the examined greenhouse only on the day of treatment. In a subsequent study (Hansen et al., 2012), exposure to Trichoderma could be observed for growers working in a greenhouse with senescent cucumber plants, a cabbage field and in a broccoli packing hall, however, PCR- analysis revealed that the Trichoderma isolates respon-sible for the exposure were different from the biocontrol agents applied.
CONCLUSIONS
The studies discussed above reflect that the genus Trichoderma/Hypocrea can be characterized with high adaptability to various ecological environments. How-ever, it is important to mention that the results of any study aimed at the examination of Trichoderma biodi-versity should always be evaluated in the context of the developmental stage of Trichoderma taxonomy and the species identification methods available at the time of the publication of the respective paper. Due to the constant development of the taxonomy of the genus and the description of new species, more recent exami-nations of a specific habitat may reveal higher biodi-versity of the genus and refine the results of previous studies.
Thanks to the introduction of new methods and the evolution of the approaches in biodiversity studies dur-ing the past two decades, the amount of information available about the distribution of Trichoderma species is constantly growing, therefore it can be expected that the biogeography of the genus will be understood more deeply in the near future.
AcknowledgmentsThe contribution of Csaba Vágvölgyi was realized in the frames of TÁMOP 4.2.4. A/2-11-1-2012-0001 „National Excellence Program – Elaborating and operating an inland student and researcher personal support system”. The project was subsidized by the European Union and co-financed by the European Social Fund.
ReferencesAbdel-Lateff, A., 2008. Bioproduction of sorbicillin derivatives from
marine Trichoderma sp. Z. Naturforsch. 63c, 631–635.Abdel-Lateff, A., Fisch, K., Wright, A.D., 2009. Trichopyrone and other
constituents from the marine sponge-derived fungus Trichoderma sp. Z. Naturforsch. 64c, 186–192.
Abd-Elsalam, K.A., Almohimeed, I., Moslem, M.A., Bahkali, A.H., 2010. M13-microsatellite PCR and rDNA sequence markers for identification of Trichoderma (Hypocreaceae) species in Saudi Arabian soil. Genet. Mol. Res. 9, 2016–2024.
Anees, M., Tronsmo, A., Edel-Hermann, V., Hjeljord, L.G., Héraud, C., Steinberg, C., 2010. Characterization of field isolates of Tricho-derma antagonistic against Rhizoctonia solani. Fungal Biol. 114, 691–701.
Andersen, B., Frisvad, J.C., Søndergaard, I., Rasmussen, I.S., Larsen, L.S., 2011. Associations between fungal species and water-damaged building materials. Appl. Environ. Microbiol. 77, 4180–4188.
Antal, Z., Kredics, L., Pakarinen, J., Dóczi, I., Andersson, M., Salkinoja-Salonen, M., Manczinger, L., Szekeres, A., Hatvani, L., Vágvölgyi, C., Nagy, E., 2005. Comparative study of potential virulence factors in human pathogenic and saprophytic Trichoderma longibrachiatum strains. Acta Microbiol. Immunol. Hung. 52, 341–350.
Antal, Z., Varga, J., Kredics, L., Szekeres, A., Hatvani, L., Mancz-inger, L., Vágvölgyi, C., Nagy, E., 2006. Intraspecific mitochon-drial DNA polymorphism within the emerging filamentous fungal pathogen Trichoderma longibrachiatum. J. Med. Microbiol. 55, 31–35.

REFERENCES 19
A. BIOLOGY AND BIODIVERSITY
Arisan-Atac, I., Heidenreich, E., Kubicek, C.P., 1995. Randomly ampli-fied polymorphic DNA fingerprinting identifies subgroups of Trichoderma viride and other Trichoderma sp. capable of chestnut blight biocontrol. FEMS Microbiol. Lett. 126, 249–256.
Askew, D.J., Laing, M.D., 1993. An adapted selective medium for the quantitative isolation of Trichoderma species. Plant Pathol. 42, 686–690.
Atanasova, L., Druzhinina, I.S., 2010. Review: global nutrient profil-ing by phenotype microarrays: a tool complementing genomic and proteomic studies in conidial fungi. J. Zhejiang Univ. Sci. B 11, 151–168.
Atanasova, L., Friedl, M.A., Bauer, H., Puxbaum, H., Kubicek, C.P., Druzhinina, I.S., 2010. Metagenomic study of air borne diversity of mycoparasitic fungus Trichoderma (anamorph Hypocrea). In: Abstracts of the Third Joint Conference of German Society for Hygiene and Microbiology, Hannover, Germany.
Aydogdu, H., Asan, A., 2008. Airborne fungi in child day care centers in Edirne City, Turkey. Environ. Monit. Assess. 147, 423–444.
Bailey, B.A., Bae, H., Strem, M.D., Roberts, D.P., Thomas, S.E., Crozier, J., Samuels, G.J., Choi, Ik-Young, Holmes, K.A., 2006. Fungal and plant gene expression during the colonization of cacao seedlings by endophytic isolates of four Trichoderma species. Planta 224, 1449–1464.
Baird, R., Carling, D., 1998. Survival of parasitic and saprophytic fungi on intact senescent cotton roots. J. Cotton Sci. 2, 27–34.
Barkai-Golan, R., 1958. A study of air-borne fungi in Israel. Res. Counc. Isr. 60, 247.
Barkai-Golan, R., Frank, M., Kantor, D., Karadavid, R., Tosher, D., 1977. Atmospheric fungi in the desert town of Arad and in the coastal plain of Israel. Ann. Allergy 38, 270–274.
Barkai-Golan, R., Glazer, Y., 1962. Air-borne fungi in Eilat and Tel Hashomer, Israel. J. Allergy 33, 342.
Berg, G., Zachow, C., Lottmann, J., Götz, M., Costa, R., Smalla, K., 2005. Impact of plant species and site on rhizosphere-associated fungi antagonistic to Verticillium dahliae kleb. Appl. Environ. Microbiol. 71, 4203–4213.
Bissett, J., 1984. A revision of the genus Trichoderma. I. Section Longibra-chiatum sect. nov. Can. J. Bot. 62, 924–931.
Bissett, J., 1991a. A revision of the genus Trichoderma. II. Infrageneric classification. Can. J. Bot. 69, 2357–2372.
Bissett, J., 1991b. A revision of the genus Trichoderma. III. Sect. Pachyba-sium. Can. J. Bot. 69, 2373–2417.
Bissett, J., 1991c. A revision of the genus Trichoderma. IV. Additional notes on section Longibrachiatum. Can. J. Bot. 69, 2418–2420.
Bissett, J., 1992. Trichoderma atroviride. Can. J. Bot. 70, 639–641.Bissett, J., Szakacs, G., Nolan, C.A., Druzhinina, I., Gradinger, C.,
Kubicek, C.P., 2003. New species of Trichoderma from Asia. Can. J. Bot. 81, 570–586.
Błaszczyk, L., Popiel, D., Chełkowski, J., Koczyk, G., Samuels, G.J., Sobieralski, K., Siwulski, M., 2011. Species diversity of Trichoderma in Poland. J. Appl. Genet. 52, 233–243.
Bopaiah, B.M., Bhat, N.T., 1981. Effect of continuous application of manures and fertilizers on rhizosphere microflora in arecanut palm. Plant Soil 63, 497–499.
Bourguignon, E., 2008. Ecology and Diversity of Indigenous Trichoderma Species in Vegetable Cropping Systems (Ph.D. thesis). Lincoln Uni-versity, New Zealand.
Burtseva, Y.V., Verigina, N.S., Sova, V.V., Pivkin, M.V., Zviagintseva, T.N., 2003. O-glycosylhydrolases of marine filamentous fungi: β-1,3-Glucanases of Trichoderma aureviride. Appl. Biochem. Micro-biol. 39, 475–481.
Burtseva, Y., Verigina, N., Sova, V., Pivkin, M., Zvyagintseva, T., 2006. Comparative characterization of laminarinases from the filamen-tous marine fungi Chaetomium indicum Corda and Trichoderma aure-viride Rifai. J. Appl. Phycol. 18, 375–380.
Calvo, M.A., Guarro, J., Suarez, G., Ramirez, C., 1980. Air-borne fungi in the air of Barcelona (Spain). IV. Various isolated genera. Myco-pathologia 71, 119–123.
Cardoso Lopes, F.A., Steindorff, A.S., Geraldine, A.M., Brandão, R.S., Monteiro, V.N., Júnior, M.L., Guedes Coelho, A.S., Ulhoa, C.J., Silva, R.N., 2012. Biochemical and metabolic profiles of Trichoderma strains isolated from common bean crops in the Brazilian Cerrado, and potential antagonism against Sclerotinia sclerotiorum. Fungal Biol. 116, 815–824.
Castle, A., Speranzini, D., Rghei, N., Alm, G., Rinker, D., Bissett, J., 1998. Morphological and molecular identification of Trichoderma isolates on North American mushroom farms. Appl. Environ. Microbiol. 64, 133–137.
Chamberlain, H.L., Rossman, A.Y., Stewart, E.L., Ulvinen, T., Samuels, G.J., 2004. The stipitate species of Hypocrea (Hypocreales, Hypoc-reaceae) including Podostroma. Karstenia 44, 1–24.
Chaverri, P., Castlebury, L.A., Overton, B.E., Samuels, G.J., 2003. Hypocrea/Trichoderma, species with conidiophore elongations and green conidia. Mycologia 95, 1100–1140.
Chaverri, P., Samuels, G.J., 2003. Hypocrea/Trichoderma (Ascomycota, Hypocreales, Hypocreaceae): species with green ascospores. Stud. Mycol. 48, 1–116.
Chaverri, P., Gazis, R.O., Samuels, G.J., 2011. Trichoderma amazonicum, a new endophytic species on Hevea brasiliensis and H. guianensis from the Amazon basin. Mycologia 103, 139–151.
Chen, J., Wang, Hui, Guo, Shun-Xing, 2012. Isolation and identification of endophytic and mycorrhizal fungi from seeds and roots of Den-drobium (Orchidaceae). Mycorrhiza 22, 297–307.
Chen, X.Y., Qi, Y.D., Wei, J.H., Zhang, Z., Wang, D.L., Feng, J.D., Gan, B.C., 2011. Molecular identification of endophytic fungi from medicinal plant Huperzia serrata based on rDNA ITS analysis. World J. Microbiol. Biotechnol. 27, 495–503.
Chen, X., Romaine, C.P., Ospina-Giraldo, M.D., Royse, D.J., 1999a. A polymerase chain reaction-based test for the identification of Tricho-derma harzianum biotypes 2 and 4, responsible for the worldwide green mold epidemic in cultivated Agaricus bisporus. Appl. Micro-biol. Biotechnol. 52, 246–250.
Chen, X., Romaine, C.P., Tan, Q., Schlagnhaufer, B., Ospina-Giraldo, M.D., Royse, D.J., Huff, D.R., 1999b. PCR-based genotyping of epi-demic and preepidemic Trichoderma isolates associated with green mold of Agaricus bisporus. Appl. Environ. Microbiol. 65, 2674–2678.
Çolakoglu, G., 2003. Airborne fungal spores at the Belgrad forest near the city of Istanbul (Turkey) in the year 2001 and their relation to allergic diseases. J. Basic Microbiol. 43, 376–384.
Cumagun, C.J.R., Hockenhull, J., Lübeck, M., 2000. Characterization of Trichoderma isolates from Philippine rice fields by UP-PCR and rDNA-ITS1 analysis: Identification of UP-PCR markers. J. Phyto-pathol. 148, 109–115.
Dang, L., Li, G., Yang, Z., Luo, S., Zheng, X., Zhang, K., 2010. Chemical constituents from the endophytic fungus Trichoderma ovalisporum isolated from Panax notoginseng. Ann. Microbiol. 60, 317–320.
Danielson, R.M., Davey, C.B., 1973. The abundance of Trichoderma propagules and the distribution of species in forest soils. Soil Biol. Biochem. 5, 485–494.
De Donno, A., Bagordo, F., Lugoli, F., Leopizzi, M.I., Russo, A., Napoli, C., Montagna, M.T., 2008. Spatial distribution of fungal microflora in the sediment of a brackish lake (Lake Alimini Grande, Italy) used for fish production and bathing. J. Prev. Med. Hyg. 49, 148–151.
Degenkolb, T., Dieckmann, R., Nielsen, K.F., Gräfenhan, T., Theis, C., Zafari, D., Chaverri, P., Ismaiel, A., Brückner, H., von Döhren, H., Thrane, U., Petrini, O., Samuels, G.J., 2008. The Trichoderma brevi-compactum clade: a separate lineage with new species, new pep-taibiotics, and mycotoxins. Mycol. Prog. 7, 177–219.
Ding, G., Wang, H., Li, L., Chen, A.J., Chen, L., Chen, H., Zhang, H., Liu, X., Zou, Z., 2012a. Trichoderones A and B: two pentacyclic cytochalasans from the plant endophytic fungus Trichoderma gamsii. Eur. J. Org. Chem. 2012, 2516–2519.
Ding, G., Chen, L., Chen, A., Tian, X., Chen, X., Liu, X.Z., Zhang, H., Zou, Z.M., 2012b. Trichalasins C and D from the plant endophytic fungus Trichoderma gamsii. Fitoterapia 83, 541–544.

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS20
A. BIOLOGY AND BIODIVERSITY
Doyle, O., 1991. Trichoderma green mould update. Irish Mushroom Rev. 3, 13–17.
Druzhinina, I.S., Chaverri, P., Fallah, P., Kubicek, C.P., Samuels, G.J., 2004. Hypocrea flaviconidia, a new species from Costa Rica with yel-low conidia. Stud. Mycol. 50, 401–407.
Druzhinina, I.S., Kopchinskiy, A.G., 2006. TrichOKEY. In: Meyer, W., Pearce, C. Medimond (Eds.), A DNA Oligonucleotide BarCode Program for the Identification of Multiple Sequences of Hypocrea and Trichoderma, International Proceedings of the 8th International Mycological Congress. Cairns, Australia, vol. 2. Bologna, Italy.
Druzhinina, I.S., Kopchinskiy, A.G., Komoń, M., Bissett, J., Szakacs, G., Kubicek, C.P., 2005. An oligonucleotide barcode for species identification in Trichoderma and Hypocrea. Fungal Genet. Biol. 42, 813–828.
Druzhinina, I.S., Komoń-Zelazowska, M., Kredics, L., Hatvani, L., Antal, Z., Belayneh, T., Kubicek, C.P., 2008. Alternative reproduc-tive strategies of Hypocrea orientalis and genetically close but clonal Trichoderma longibrachiatum, both capable to cause invasive mycoses of humans. Microbiol. - SGM 154, 3447–3459.
Druzhinina, I.S., Kubicek, C.P., Komoń-Zelazowska, M., Mulaw, T.B., Bissett, J., 2010. The Trichoderma harzianum demon: complex specia-tion history resulting in coexistence of hypothetical biological spe-cies, recent agamospecies and numerous relict lineages. BMC Evol. Biol. 10, 94.
Druzhinina, I.S., LaFe, K., Willinger, B., Komoń-Zelazowska, M., Ammirati, J., Kubicek, C.P., Rogers, J.D., 2007. An unknown Hypocreaceae species isolated from human lung tissue of a patient with non-fatal pulmonary fibrosis. Clin. Microbiol. Newslett. 29, 180–184.
Druzhinina, I.S., Seidl-Seiboth, V., Herrera-Estrella, A., Horwitz, B.A., Kenerley, C.M., Monte, E., Mukherjee, P.K., Zeilinger, S., Grigoriev, I.V., Kubicek, C.P., 2011. Trichoderma: the genomics of opportunistic success. Nat. Rev. Microbiol. 9, 749–759.
Du, L., Zhu, T., Li, L.Y., Cai, S., Zhao, B., Gu, Q., 2009. Cytotoxic sor-bicillinoids and bisorbicillinoids from a marine-derived fungus Trichoderma sp. Chem. Pharm. Bull. 57, 220–223.
Ebbehøj, N.E., Hansen, M.Ø., Sigsgaard, T., Larsen, L., 2002. Building-related symptoms and molds: a two-step intervention study. Indoor Air 12, 273–277.
Elad, Y., Chet, I., Henis, Y., 1981. A selective medium for improving quantitative isolation of Trichoderma spp. from soil. Phytoparasitica 9, 59–67.
El-Bondkly, A.M.A., Mervat, M., El-Gendy, A., 2012. Cellulase produc-tion from agricultural residues by recombinant fusant strain of a fungal endophyte of the marine sponge Latrunculia corticata for pro-duction of ethanol. Anton. Leeuw. Int. J. G. 101, 331–346.
El-Kassas, H.Y., El-Taher, E.M., 2009. Optimization of batch process parameters by response surface methodology for mycoremediation of chrome-VI by a chromium resistant strain of marine Trichoderma viride. Am.-Eurasian J. Agric. Environ. Sci. 5, 676–681.
Friedl, M.A., Druzhinina, I.S., 2012. Taxon-specific metagenomics of Trichoderma reveals a narrow community of opportunistic species that regulate each other’s development. Microbiology 158, 69–83.
Gal-Hemed, I., Atanasova, L., Komoň-Zelazowska, M., Druzhinina, I.S., Viterbo, A., Yarden, O., 2011. Marine isolates of Trichoderma spp. as potential halotolerant agents of biological control for arid-zone agriculture. Appl. Environ. Microbiol. 77, 5100–5109.
Gams, W., Bissett, J., 1998. Morphology and identification of Tricho-derma. In: Kubicek, C.P., Harman, G.E. (Eds.), Trichoderma and Glio-cladium, Basic Biology, Taxonomy and Genetics, vol. 1. Taylor & Francis, London, UK, pp. 3–34.
Garo, E., Starks, C.M., Jensen, P.R., Fenical, W., Lobkovsky, E., Clardy, J., 2003. Trichodermamides A and B, cytotoxic modified dipeptides from the marine-derived fungus Trichoderma virens. J. Nat. Prod. 66, 423–426.
Gea, F.J., 2009. First report of Trichoderma pleurotum on oyster mush-room crops in Spain. J. Plant Pathol. 91, 504.
Gherbawy, Y., Druzhinina, I., Shaban, G.M., Wuczkowsky, M., Yaser, M., El-Naghy, M.A., Prillinger, H., Kubicek, C.P., 2004. Trichoderma populations from alkaline agricultural soil in the Nile valley, Egypt, consist of only two species. Mycol. Prog. 3, 211–218.
Girlanda, M., Perotto, S., Moenne-Loccoz, Y., Bergero, R., Lazzari, A., Defago, G., Bonfante, P., Luppi, A.M., 2001. Impact of biocontrol Pseudomonas fluorescens CHA0 and a genetically modified deriva-tive on the diversity of culturable fungi in the cucumber rhizo-sphere. Appl. Environ. Microbiol. 67, 1851–1864.
Guarro, J., Antolin-Ayala, M.I., Gene, J., Gutierrez-Calzada, J., Nieves-Diez, C., Ortoneda, M., 1999. Fatal case of Trichoderma harzianum infection in a renal transplant recipient. J. Clin. Microbiol. 37, 3751–3755.
Hageskal, G., Knutsen, A.K., Gaustad, P., de Hoog, G.S., Skaari, I., 2006. Diversity and significance of mold species in Norwegian drinking water. Appl. Environ. Microbiol. 72, 7586–7593.
Hageskal, G., Vrålstad, T., Knutsen, A.K., Skaar, I., 2008. Exploring the species diversity of Trichoderma in Norwegian drinking water sys-tems by DNA barcoding. Mol. Ecol. Resour. 8, 1178–1188.
Hagn, A., Pritsch, K., Schloter, M., Munch, J.C., 2003. Fungal diversity in agricultural soil under different farming management systems, with special reference to biocontrol strains of Trichoderma spp. Biol. Fertil. Soils 38, 236–244.
Hagn, A., Wallisch, S., Radl, V., Charles Munch, J., Schloter, M., 2007. A new cultivation independent approach to detect and monitor common Trichoderma species in soils. J. Microbiol. Methods 69, 86–92.
Hanada, R.E., de Jorge Souza, T., Pomella, A.W., Hebbar, K.P., Pereira, J.O., Ismaiel, A., Samuels, G.J., 2008. Trichoderma martiale sp. nov., a new endophyte from sapwood of Theobroma cacao with a potential for biological control. Mycol. Res. 112, 1335–1343.
Hansen, V.M., Meyling, N.V., Winding, A., Eilenberg, J., Madsen, A.M., 2012. Factors affecting vegetable growers’ exposure to fungal bio-aerosols and airborne dust. Ann. Occup. Hyg. 56, 170–181.
Hansen, V.M., Winding, A., Madsen, A.M., 2010. Exposure to bioaero-sols during the growth season of tomatoes in an organic greenhouse using Supresivit (Trichoderma harzianum) and Mycostop (Streptomy-ces griseoviridis). Appl. Environ. Microbiol. 76, 5874–5881.
Harman, G.E., 2006. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 96, 190–194.
Harman, G.E., Howell, C.R., Viterbo, A., Chet, I., Lorito, M., 2004. Trich-oderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56.
Hatvani, L., Antal, Z., Manczinger, L., Druzhinina, I.S., Kubicek, C.P., Szekeres, A., Vágvölgyi, C., Nagy, E., Kredics, L., 2006. Monitoring the occurrence of Trichoderma species during compost production and cultivation of Agaricus bisporus in Hungary. Acta Microbiol. Immunol. Hung. 53, 272.
Hatvani, L., Antal, Z., Manczinger, L., Szekeres, A., Druzhinina, I.S., Kubicek, C.P., Nagy, A., Nagy, E., Vágvölgyi, C., Kredics, L., 2007. Green mould diseases of Agaricus and Pleurotus are caused by related but phylogenetically different Trichoderma species. Phytopa-thology 97, 532–537.
Hatvani, L., Kocsubé, S., Manczinger, L., Antal, Z., Szekeres, A., Druzhinina, I.S., Komon-Zelazowska, M., Kubicek, C.P., Nagy, A., Vágvölgyi, C., Kredics, L., 2008. The green mould disease global threat to the cultivation of oyster mushroom (Pleurotus ostreatus): a review. In: Gruening, M. (Ed.), Science and Cultivation of Edible and Medicinal Fungi: Mushroom Science, Proceedings of the 17th Congress of the International Society for Mushroom Science, vol. 27. , pp. 485–495 Cape Town, South Africa, 20–24 May 2008.
Hatvani, L., Sabolić, P., Kocsubé, S., Kredics, L., Czifra, D., Vágvölgyi, C., Kaliterna, J., Ivić, D., Đermić, E., Kosalec, I., 2012. The first report on mushroom green mould disease in Croatia. Arh. Hig. Rada Toksikol 63, 481–487.

REFERENCES 21
A. BIOLOGY AND BIODIVERSITY
Hebert, P.D.N., Beaton, M.J., 1993. Methodologies for Allozyme Analy-sis Using Cellulose Acetate Electrophoresis. a Practical Handbook. Helena Laboratories, Beaumont, TX.
Hermosa, M.R., Grondona, I., Monte, E., 1999. Isolation of Trichoderma harzianum Th2 from commercial mushroom compost in Spain. Plant Dis. 83, 591.
Hoyos-Carvajal, L., Bissett, J., 2011. Biodiversity of Trichoderma in neo-tropics. In: Grillo, O., Venora, G. (Eds.), The Dynamical Processes of Biodiversity – Case Studies of Evolution and Spatial Distribution, Intech, pp. 303–320.
Hoyos-Carvajal, L., Orduz, S., Bissett, J., 2009. Genetic and metabolic biodiversity of Trichoderma from Colombia and adjacent neotropic regions. Fungal Genet. Biol. 46, 615–631.
Hyun-Sung, S., Lee, M., 1986. Environmental factors and the distribution of soil microorganisms in ginseng field. Kor. J. Microbiol. 24, 184–193.
Jaklitsch, W.M., 2009. European species of Hypocrea part I. The green-spored species. Stud. Mycol. 63, 1–91.
Jaklitsch, W.M., 2011. European species of Hypocrea part II: species with hyaline ascospores. Fungal Diversity 48, 1–250.
Jaklitsch, W.M., Komon, M., Kubicek, C.P., Druzhinina, I.S., 2005. Hypo-crea voglmayrii sp. nov. from the Austrian alps represents a new phy-logenetic clade in Hypocrea/Trichoderma. Mycologia 97, 1365–1378.
Jaklitsch, W.M., Komon, M., Kubicek, C.P., Druzhinina, I.S., 2006a. Hypocrea crystalligena sp. nov., a common European species with a white-spored Trichoderma anamorph. Mycologia 98, 499–513.
Jaklitsch, W.M., Samuels, G.J., Dodd, S.L., Lu, B.-S., Druzhinina, I.S., 2006b. Hypocrea rufa/Trichoderma viride: a reassessment, and descrip-tion of five closely related species with and without warted conidia. Stud. Mycol. 56, 135–177.
Jaklitsch, W.M., Kubicek, C.P., Druzhinina, I.S., 2008a. Three European species of Hypocrea with reddish brown stromata and green asco-spores. Mycologia 100, 796–815.
Jaklitsch, W.M., Põldmaa, K., Samuels, G.J., 2008b. Reconsideration of Protocrea (Hypocreales, Hypocreaceae). Mycologia 100, 962–984.
Jaklitsch, W.M., Gruber, S., Voglmayr, H., 2008c. Hypocrea seppoi, a new stipitate species from Finland. Karstenia 48, 1–11.
Jaklitsch, W.M., Stadler, M., Voglmayr, H., 2012. Blue pigment in Hypo-crea caerulescens sp. nov. and two additional new species in sect. Trichoderma. Mycologia 104, 925–941.
Jaklitsch, W.M., Voglmayr, H., 2012. Hypocrea britdaniae and H. foliicola: two remarkable new European species. Mycologia 105, 1213–1221.
Kantarcioğlu, A.S., Celkan, T., Yücel, A., Mikami, Y., Kurugoglu, S., Mitani, H., Altas, K., 2009. Fatal Trichoderma harzianum infection in a leukemic pediatric patient. Med. Mycol. 47, 207–215.
Khamthong, N., Rukachaisirikul, V., Tadpetch, K., Kaewpet, M., Phong-paichit, S., Preedanon, S., Sakayaroj, J., 2012. Tetrahydroanthraqui-none and xanthone derivatives from the marine-derived fungus Trichoderma aureoviride PSU-F95. Arch. Pharm. Res. 35, 461–468.
Khudiakova, Y.V., Pivkin, M.V., Kuznetsova, T.A., Svetashev, V.I., 2000. Fungi in sediments of the Sea of Japan and their biologically active metabolites. Microbiology 69, 608–611.
Kildesø, J., Würtz, H., Nielsen, K.F., Kruse, P., Wilkins, K., Thrane, U., Gravesen, S., Nielsen, P.A., Schneider, T., 2003. Determination of fungal spore release from wet building materials. Indoor Air 13, 148–155.
Kim, C.S., Shirouzu, T., Nakagiri, A., Sotome, K., Nagasawa, E., Maekawa, N., 2012. Trichoderma mienum sp. nov., isolated from mushroom farms in Japan. Anton. Leeuw. Int. J. G. 102, 629–641.
Kohlmeyer, J., 1974. On the Definition and Taxonomy of Higher Marine Fungi. vol. 5. Veröffentl. Inst. Meeresforsch, Bremerhaven S263–S286.
Komoń-Zelazowska, M., Bissett, J., Zafari, D., Hatvani, L., Manczinger, L., Woo, S., Lorito, M., Kredics, L., Kubicek, C.P., Druzhinina, I.S., 2007. Genetically closely related but phenotypically divergent Trichoderma species cause world-wide green mould disease in oys-ter mushroom farms. Appl. Environ. Microbiol. 73, 7415–7426.
Kopchinskiy, A.G., Komoń, M., Kubicek, C.P., Druzhinina, I.S., 2005. TrichoBLAST: a multiloci database of phylogenetic markers for Trichoderma and Hypocrea powered by sequence diagnosis and simi-larity search tools. Mycol. Res. 109, 658–660.
Kramer, C.L., Pady, S.M., Rogerson, C.T., Ouye, C.G., 1959. Kansas aeromycology II. Materials, methods, and general results. Trans. Kans. Acad. Sci. 62, 184–199.
Kramer, C.L., Pady, S.M., 1960. Kansas aeromycology. XI. Fungi Imper-fecti. Trans. Kans. Acad. Sci. 63, 229–238.
Kramer, C.L., Pady, S.M., Wiley, B., 1964. Kansas aeromycology. XIV. Diurnal studies 1961–1962. Trans. Kans. Acad. Sci. 67, 442–459.
Kredics, L., Láday, M., Körmöczi, P., Manczinger, L., Rákhely, G., Vágvölgyi, Cs., Szekeres, A., 2011a. Trichoderma communities of the winter wheat rhizosphere. Agr. Vidékfejl. Szeml. 6, S413–S418.
Kredics, L., Antal, Z., Dóczi, I., Manczinger, L., Kevei, F., Nagy, E., 2003. Clinical importance of the genus Trichoderma. Acta Microbiol. Immunol. Hung. 50, 105–117.
Kredics, L., Hatvani, L., Manczinger, L., Vágvölgyi, C., Antal, Z., 2011b. Chapter 62. Trichoderma. In: Liu, D. (Ed.), Molecular Detection of Human Fungal Pathogens, Taylor & Francis Group, London, pp. 509–526.
Kredics, L., García Jimenez, L., Naeimi, S., Czifra, D., Urbán, P., Manczinger, L., Vágvölgyi, C., Hatvani, L., 2010. A challenge to mushroom growers: the green mould disease of cultivated cham-pignons. In: Méndez-Vilas, A. (Ed.), Microbiology Book Series„ Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology”, FORMATEX, Bada-joz, pp. 295–305.
Kredics, L., Kocsubé, S., Nagy, L., Komoń-Zelazowska, M., Manczinger, L., Sajben, E., Nagy, A., Vágvölgyi, C., Kubicek, C.P., Druzhinina, I.S., Hatvani, L., 2009. Molecular identification of Trichoderma spe-cies associated with Pleurotus ostreatus and natural substrates of the oyster mushroom. FEMS Microbiol. Lett. 300, 58–67.
Kredics L., Láday, M., Körmöczi, P., Manczinger, L., Rákhely, G., Vágvölgyi, C., Szekeres, A., 2012. Genetic and biochemical diversity among Trichoderma isolates in soil samples from winter wheat fields of the Pannonian Plain. Acta Biol. Szeg. 56, 141–149.
Kubicek, C., Bissett, J., Druzhinina, I., Kulling-Grandiger, C., Szakacs, G., 2003. Genetic and metabolic diversity of Trichoderma: a case study on South East Asian isolates. Fungal Genet. Biol. 38, 310–319.
Kubicek, C.P., Komon-Zelazowska, M., Druzhinina, I.S., 2008. Fungal genus Hypocrea/Trichoderma: from barcodes to biodiversity. J. Zheji-ang Univ. Sci. B 9, 753–763.
Kuhls, K., Lieckfeldt, E., Börner, T., Guého, E., 1999. Molecular reiden-tification of human pathogenic Trichoderma isolates as Trichoderma longibrachiatum and Trichoderma citrinoviride. Med. Mycol. 37, 25–33.
Kullnig, C., Szakacs, G., Kubicek, C.P., 2000. Molecular identification of Trichoderma species from Russia, Siberia and the Himalaya. Mycol. Res. 104, 1117–1125.
Kullnig-Gradinger, C.M., Szakacs, G., Kubicek, C.P., 2002. Phylog-eny and evolution of the fungal genus Trichoderma: a multigene approach. Mycol. Res. 106, 757–767.
Lahlali, R., Hijri, M., 2010. Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 311, 152–159.
Lan, W.J., Zhao, Y., Xie, Z.L., Liang, L.Z., Shao, W.Y., Zhu, L.P., Yang, D.P., Zhu, X.F., Li, H.J., 2012. Novel sorbicillin analogues from the marine fungus Trichoderma sp. associated with the seastar Acan-thaster planci. Nat. Prod. Commun. 7, 1337–1340.
Larralde-Corona, C.P., Santiago-Mena, M.R., Sifuentes-Rincón, A.M., Rodríguez-Luna, I.C., Rodríguez-Pérez, M.A., Shirai, K., Narváez-Zapata, J.A., 2008. Biocontrol potential and polyphasic character-ization of novel native Trichoderma strains against Macrophomina phaseolina isolated from sorghum and common bean. Appl. Micro-biol. Biotechnol. 80, 167–177.

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS22
A. BIOLOGY AND BIODIVERSITY
Liang, C., Li, B., Lu, G., 2004. Gliocladium and Trichoderma in agricul-tural soil. J. Zhejiang Univ. Agric. Life Sci. 30, 415.
Li, D.-L., Chen, Y.-C., Tao, M.-H., Li, H.-H., Zhang, W.-M., 2012. Two new octahydronaphthalene derivatives from Trichoderma spirale, an endophytic fungus derived from Aquilaria sinensis. Helv. Chim. Acta 95, 805–809.
Li, D.-W., LaMondia, J., 2010. Airborne fungi associated with ornamen-tal plant propagation in greenhouses. Aerobiologia 26, 2615–2628.
Lu, X., Tian, L., Chen, G., Xu, Y., Wang, H.-F., Li, Z.-Q., Pei, Y.-H., 2012. Three new compounds from the marine-derived fungus Tricho-derma atroviride G20-12. J. Asian Nat. Prod. Res. 14, 647–651.
Lu, B.S., Samuels, G.J., 2003. Hypocrea stilbohypoxyli and its Trichoderma koningii-like anamorph: a new species from Puerto Rico on Stilbohy-poxylon moelleri. Sydowia 55, 255–266.
Lu, B., Druzhinina, I.S., Fallah, P., Chaverri, P., Gradinger, C., Kubicek, C.P., Samuels, G.J., 2004. Hypocrea/Trichoderma species with pachy-basium-like conidiophores: teleomorphs for T. minutisporum and T. polysporum and their newly discovered relatives. Mycologia 96, 310–342.
Lübeck, M., Poulsen, S.K., Lübeck, P.S., Jensen, D.F., Thrane, U., 2000. Identification of Trichoderma strains from building materials by ITS1 ribotyping, UP-PCR fingerprinting and UP-PCR cross hybridiza-tion. FEMS Microbiol. Lett. 185, 129–134.
Madsen, A.M., Hansen, V.M., Meyling, N.V., Eilenberg, J., 2007. Human exposure to airborne fungi from genera used as biocontrol agents in plant production. Ann. Agric. Environ. Med. 14, 5–24.
Magyar, D., Eszéki, E.R., Oros, G., Szécsi, Á., Kredics, L., Hatavni, L., Körmöczi, P., 2011. The air spora of an orchid greenhouse. Aerobio-logia 27, 121–134.
Mamoun, M.L., Iapicco, R., Savoie, J.-M., Olivier, J.M., 2000. Green mould disease in France: Trichoderma harzianum Th2 and other species caus-ing damages on mushroom farms. Mushroom Sci. 15, 625–632.
Manoch, L., Dethoup, T., Kokaew, J., 2002. Diversity of microfungi from soil and debris at termite mounds in Thailand. Book of Abstracts, the 7th International Mycological Congress, p. 165.
Martins, M.P., Mouad, A.M., Boschini, L., Regali Seleghim, M.H., Durães Sette, L., Meleiro Porto, A.L., 2011. Marine fungi Aspergillus sydowii and Trichoderma sp. catalyze the hydrolysis of benzyl glyc-idyl ether. Mar. Biotechnol. 13, 314–320.
Meincke, R., Weinert, N., Radl, V., Schloter, M., Smalla, K., Berg, G., 2010. Development of a molecular approach to describe the compo-sition of Trichoderma communities. J. Microbiol. Methods 80, 63–69.
Mghalu, M.J., Tsuji, T., Kubo, N., Kubota, M., Hyakumachi, M., 2007. Selective accumulation of Trichoderma species in soils suppressive to radish damping-off disease after repeated inoculations with Rhizoctonia solani, binucleate Rhizoctonia and Sclerotium rolfsii. J. Gen. Plant Pathol. 73, 250–259.
Migheli, Q., Balmas, V., Komoń-Zelazowska, M., Scherm, B., Fiori, S., Kopchinskiy, A., Kubicek, C.P., Druzhinina, I., 2009. Soils of a Medi-terranean hot spot of biodiversity and endemism (Sardinia, Tyrrhe-nian Islands) are inhabited by pan-European, invasive species of Hypocrea/Trichoderma. Environ. Microbiol. 11, 35–46.
Ming, Q., Han, T., Li, W., Zhang, Q., Zhang, H., Zheng, C., Huang, F., Rahman, K., Qin, L., 2012. Tanshinone IIA and tanshinone I produc-tion by Trichoderma atroviride D16, an endophytic fungus in Salvia miltiorrhiza. Phytomedicine 19, 330–333.
Mobin, M., Salmito, M.A., 2006. Fungus microbiota in air conditioners in intensive care units in Teresina, Piauí. Rev. Soc. Bras. Med. Trop. 39, 556–559, (Article in Portuguese).
Mohamed-Benkada, M., Montagu, M., Biard, J.-F., Mondeguer, F., Verite, P., Dalgalarrondo, M., Bissett, J., Pouchus, Y.F., 2006. New short peptaibols from a marine Trichoderma strain. Rapid Commun. Mass Spectrom. 20, 1176–1180.
Mostafa Kamal, M.D., Shahjahan, A.K.M., 1995. Trichoderma in rice field soils and their effect on Rhizoctonia solani. Bangladesh J. Bot. 24, 75–79.
Mouton, M., Postma, F., Wilsenach, J., Botha, A., 2012. Diversity and characterization of culturable fungi from marine sediment collected from St. Helena Bay, South Africa. Microb. Ecol. 64, 311–319.
Mpika, J., Kébé, I.B., Issali, A.E., N’Guessan, F.K., Druzhinina, I.S., Komon-Zélazowska, M., Kubicek, C.P., Aké, S., 2009. Antagonist potential of Trichoderma indigenous isolates for biological control of Phytophthora palmivora the causative agent of black pod disease on cocoa (Theo-broma cacao L.) in Côte d’Ivoire. Afr. J. Biotechnol. 8, 5280–5293.
Mulaw, B.T., Kubicek, C.P., Druzhinina, I.S., 2010. The rhizosphere of Coffea Arabica in its native highland forests of Ethiopia provides a niche for a distinguished diversity of Trichoderma. Diversity 2, 527–549.
Mysterud, I., Høiland, K., Koller, G., Stensrud, Ø., 2007. Molecular characterization and evaluation of plant litter-associated fungi from the spring ‘grazing corridor’ of a sheep herd vulnerable to alveld disease. Mycopathologia 164, 201–215.
Naeimi, S., Khodaparast, S.A., Javan-Nikkhah, M., Vágvölgyi., Cs., Kredics, L., 2011. Species patterns and phylogenetic relationships of Trichoderma strains in rice fields of southern Caspian Sea, Iran. Cereal Res. Commun. 39, 560–568.
Naeimi, S., Okhovvat, S.M., Javan-Nikkhah, M., Kredics, L., Khosravi, V., 2010. Biological control of Rhizoctonia solani AG1-1A, the causal agent of rice sheath blight with Trichoderma strains. Phytopathol. Mediterr. 49, 287–300.
Nagamani, A., Mew, T.W., 1987. Trichoderma in Philippine rice field soils. Int. Rice Res. Newslett. 12, 25.
Nagy, V., Seidl, V., Szakacs, G., Komoń-Zelazowska, M., Kubicek, C.P., Druzhinina, I.S., 2007. Application of DNA bar codes for screen-ing of industrially important fungi: the haplotype of Trichoderma harzianum sensu stricto indicates superior chitinase formation. Appl. Environ. Microbiol. 73, 7048–7058.
Neuhof, T., Dieckmann, R., Druzhinina, I.S., Kubicek, C.P., Nakari-Setala, T., Penttilä, M., von Döhren, H., 2007. Direct identification of hydrophobins and their processing in Trichoderma using intact-cell MALDI-TOF MS. FEBS J. 274, 841–852.
Neumann, K., Abdel-Lateff, A., Wright, A.D., Kehraus, S., Krick, A., König, G.M., 2007. Novel sorbicillin derivatives with an unprec-edented carbon skeleton from the sponge-derived fungus Tricho-derma species. Eur. J. Org. Chem. 2007, 2268–2275.
Neumann, K., 2008. Marine-derived Fungi: A Source for Structurally New and Bioactive Secondary Metabolites (Dissertation), Rhein-ischen Friedrich-Wilhelms-Universität Bonn.
Okoth, S.A., Okoth, P., Wachira, P.M., Roimen, H., 2009. Spatial distri-bution of Trichoderma spp. in Embu and Taita regions, Kenya. Trop. Subtrop. Agroecosyst. 11, 291–301.
Overton, B.E., Stewart, E.L., Geiser, D.M., 2006. Taxonomy and phylo-genetic relationships of nine species of Hypocrea with anamorphs assignable to Trichoderma section Hypocreanum. Stud. Mycol. 56, 39–65.
Park, M.S., Bae, K.S., Yu, S.H., 2004. Molecular and morphological analysis of Trichoderma isolates associated with green mold epi-demic of oyster mushroom in Korea. J. Huazhong Agric. Univ. 23, 157–164.
Park, M.S., Bae, K.S., Yu, S.H., 2006. Two new species of Trichoderma associated with green mold of oyster mushroom cultivation in Korea. Mycobiology 34, 111–113.
Papavizas, G.C., Lumsden, R.D., 1982. Improved medium for isolation of Trichoderma spp. from soil. Plant Dis. 66, 1019–1020.
Pavone, M., Domenico, F., 2012. Biocontrol de Rhizoctonia solani Kühn por Trichoderma spp (Ph.D. thesis). Universidad Central de Venezuela.
Paz, Z., Komoň-Zelazowska, M., Druzhinina, I.S., Aveskamp, M.M., Shnaiderman, A., Aluma, Y., Carmeli, S., Ilan, M., Yarden, O., 2010. Diversity and potential antifungal properties of fungi associated with a Mediterranean sponge. Fungal Diversity 42, 17–26.

REFERENCES 23
A. BIOLOGY AND BIODIVERSITY
Pięta, D., Pastucha, A., Patkowska, E., 2000. Bacteria and fungi commu-nities occurring in the soil after the cultivation of cereal and potato. Electron. J. Pol. Agric. Univ. Serv. Hortic. 3 (2).
Pięta, D., Patkowska, E., 2003. The role of antagonistic fungi and bac-teria limiting the occurrence of some phytopathogens inhabiting the soybean soil environment. Electron. J. Pol. Agric. Univ. Serv. Hortic. 6 (2)http://www.ejpau.media.pl/volume6/issue2/horticulture/art-04.pdf.
Posada, F., Aime, M.C., Peterson, S.W., Rehner, S.A., Vega, F.E., 2007. Inoculation of coffee plants with the fungal entomopathogen Beau-veria bassiana (Ascomycota: Hypocreales). Mycol. Res. 111, 748–757.
Pruksakorn, P., Arai, M., Kotoku, N., Vilchčze, C., Baughn, A.D., Moodley, P., Jacobs Jr., W.R., Kobayashi, M., 2010. Trichoderins, novel aminolipopeptides from a marine sponge-derived Tricho-derma sp., are active against dormant mycobacteria. Bioorg. Med. Chem. Lett. 20, 3658–3663.
Pukahuta, C., Limtong, S., Suwanarit, P., Nutalaya, S., 2000. Species diversity of Trichoderma contaminating shiitake production houses in Thailand. Kasetsart J. Nat. Sci. 34, 478–485.
Quilliam, R.S., Jones, D.L., 2010. Fungal root endophytes of the car-nivorous plant Drosera rotundifolia. Mycorrhiza 20, 341–348.
Quilliam, R.S., Jones, D.L., 2012. Evidence for host-specificity of cultur-able fungal root endophytes from the carnivorous plant Pinguicula vulgaris (Common Butterwort). Mycol. Prog. 11, 583–585.
Ranque, S., Garcia-Hermoso, D., Michel-Nguyen, A., Dumon, H., 2008. Isolation of Trichoderma atroviride from a liver transplant. J. Mycol. Med. 18, 234–236.
Rao, C.Y., Riggs, M.A., Chew, G.L., Muilenberg, M.L., Thorne, P.S., Van Sickle, D., Dunn, K.H., Brown, C., 2007. Characterization of air-borne molds, endotoxins, and glucans in homes in New Orleans after hurricanes Katrina and Rita. Appl. Environ. Microbiol. 73, 1630–1634.
Ren, J., Xue, C., Tian, L., Xu, M., Chen, J., Deng, Z., Proksch, P., Lin, W., 2009. Asperelines A−F, Peptaibols from the marine-derived fungus Trichoderma asperellum. J. Nat. Prod. 72, 1036–1044.
Requejo, H.V., 1975. Microflora atmosférica de la ciudad de Trujillo (Peru). III. Géneros aislados durante el argo 1971. Mycopathologia 36, 15–20.
Ribeiro, A., Machado, A.P., Kozakiewicz, Z., Ryan, M., Luke, B., Buddie, A.G., Venancio, A., Lima, N., Kelly, J., 2006. Fungi in bottled water: a case study of a production plant. Rev. Iberoam. Micol. 23, 139–144.
Richards, M., 1956. A census of mould spores in the air over Britain in 1952. Trans. Br. Mycol. Soc. 39, 431–441.
Rivas, M., Pavone, D., 2010. Diversidad de Trichoderma spp. en planta-ciones de Theobroma cacao l. del estado Carabobo, Venezuela, y su capacidad biocontroladora sobre Crinipellis pernicosa (Stahel) Singer. Interciencia 35, 777–783.
Rivera, C.S., Blanco, D., Oria, R., Venturini, M.E., 2010. Diversity of culturable microorganisms and occurrence of Listeria monocytogenes and Salmonella spp. in Tuber aestivum and Tuber melanosporum asco-carps. Food Microbiol. 27, 286–293.
Rocha, L.C., Ferreira, H.V., Pimenta, E.F., Berlinck, R.G., Seleghim, M.H., Javaroti, D.C., Sette, L.D., Bonugli, R.C., Porto, A.L., 2009. Bioreduction of alpha-chloroacetophenone by whole cells of marine fungi. Biotechnol. Lett. 31, 1559–1563.
Rocha, L.C., Luiz, R.F., Rosset, I.G., Raminelli, C., Seleghim, M.H.R., Sette, L.D., Porto, A.L.M., 2012. Bioconversion of iodoacetophe-nones by marine fungi. Mar. Biotechnol. 14, 396–401.
Rodrigues, A., Bacci Jr, M., Mueller, U.G., Ortiz, A., Pagnocca, F.C., 2008. Microfungal “weeds” in the leafcutter ant symbiosis. Microb. Ecol. 56, 604–614.
Rubini, M.R., Silva-Ribeiro, R.T., Pomella, A.W., Maki, C.S., Araújo, W.L., Dos Santos, D.R., Azevedo, J.L., 2005. Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological con-trol of Crinipellis perniciosa, causal agent of Witches' Broom Disease. Int. J. Biol. Sci. 1, 24.
Ruiz, N., Dubois, N., Wielgosz-Collin, G., Robiou du Pont, T., Bergé, J.-P., Pouchus, Y.F., Barnathan, G., 2007a. Lipid content and fatty acid composition of a marine-derived Trichoderma longibrachiatum strain cultured by agar surface and submerged fermentations. Process. Biochem. 42, 676–680.
Ruiz, N., Wielgosz-Collin, G., Poirier, L., Grovel, O., Petit, K.E., Mohamed-Benkada, M., Robiou du Pont, T., Bissett, J., Vérité, P., Barnathan, G., Pouchus, Y.F., 2007b. New trichobrachins, 11-residue peptaibols from a marine strain of Trichoderma longibrachiatum. Pep-tides 28, 1351–1358.
Sadfi-Zouaoui, N., Hannachi, I., Rouaissi, M., Hajlaoui, M., Rubio, M., Monte, E., Boudabous, A., Hermosa, M., 2009. Biodiversity of Trich-oderma strains in Tunisia. Can. J. Microbiol. 55, 154–162.
Sallenave, C., Pouchus, Y.F., Bardouil, M., Lassus, P., Roquebert, M.F., Verbist, J.F., 1999. Bioaccumulation of mycotoxins by shellfish: contamination of mussels by metabolites of a Trichoderma koningii strain isolated in the marine environment. Toxicon 37, 77–83.
Sallenave-Namont, C., Pouchus, Y.F., Robiou du Pont, T., Lassus, P., Verbist, J.F., 2000. Toxigenic saprophytic fungi in marine shellfish farming areas. Mycopathologia 149, 21–25.
Samuels, G.J., Dodd, S.L., Gams, W., Castlebury, L.A., Petrini, O., 2002. Trichoderma species associated with the green mold epi-demic of commercially grown Agaricus bisporus. Mycologia 94, 146–170.
Samuels, G.J., Ismaiel, A., 2011. Hypocrea peltata: a mycological Dr Jekyll and Mr Hyde? Mycologia 103, 616–630.
Samuels, G.J., Suarez, C., Solis, K., Holmes, K.A., Thomas, S.E., Ismaiel, A., Evans, H.C., 2006a. Trichoderma theobromicola and T. paucisporum: two new species isolated from cacao in South America. Mycol. Res. 110, 381–392.
Samuels, G.J., Dodd, S.L., Lu, B.-S., Petrini, O., Schroers, H.-J., Druzhinina, I.S., 2006b. The Trichoderma koningii aggregate species. Stud. Mycol. 56, 67–133.
Samuels, G.J., Ismaiel, A., Mulaw, T.B., Szakacs, G., Druzhinina, I.S., Kubicek, C.P., Jaklitsch, W.M., 2012a. The Longibrachiatum clade of Trichoderma: a revision with new species. Fungal Diversity 55, 77–108.
Samuels, G.J., Ismaiel, A., de Souza, J., Chaverri, P., 2012b. Trichoderma stromaticum and its overseas relatives. Mycol. Prog. 11, 215–254.
Sánchez-Perez, M.I., 2009. Aislamiento y caracterización molecular y agronómica de Trichoderma spp. Nativos del norte de Tamaulipas (Doctoral dissertation). Tesis de Maestría en Ciencias en Biotec-nología Genómica. Centro de Biotecnología Genómica. Instituto Politécnico Nacional, pp. 187
Sariah, M., Choo, C.W., Zakaria, H., Norihan, M.S., 2005. Quantifica-tion and characterisation of Trichoderma spp. from different ecosys-tems. Mycopathologia 159, 113–117.
Sato, J., 2010. Distribution of filamentous fungi in a manufacturing fac-tory for plastic caps for soft drinks. Biocontrol Sci. 15, 87–90.
Seaby, D.A., 1987. Infection of mushroom compost by Trichoderma species. Mushroom J. 179, 355–361.
Seaby, D.A., 1989. Further observations on Trichoderma. Mushroom J. 197, 147–151.
Shanmugam, V., Sharma, V., Ananthapadmanaban, 2008. Genetic related-ness of Trichoderma isolates antagonistic against Fusarium oxysporum f. sp. dianthi inflicting carnation wilt. Folia Microbiol. 53, 130–138.
Slezack-Deschaumes, S., Piutti, S., Vong, P.C., Benizri, E., 2012. Dynamics of cultivable arylsulfatase-producing bacterial and fungal communi-ties along the phenology of field-grown rape. Eur. J. Soil Biol. 48, 66–72.
Smith, W.H., 1995. Forest occurrence of Trichoderma species: emphasis on potential organochlorine (xenobiotic) degradation. Ecotoxicol. Environ. Saf. 32, 179–183.
Song, F., Dai, H., Tong, Y., Ren, B., Chen, C., Sun, N., Liu, X., Bian, J., Liu, M., Gao, H., Liu, H., Chen, X., Zhang, L., 2010. Trichoderma ketones A–D and 7-O-Methylkoninginin D from the marine fungus Trichoderma koningii. J. Nat. Prod. 73, 806–810.

1. BIODIVERSITY OF THE GENUS HYPOCREA/TRICHODERMA IN DIFFERENT HABITATS24
A. BIOLOGY AND BIODIVERSITY
Souza, A.D., Rodrigues-Filho, E., Souza, A.Q., Pereira, J.O., Calgarotto, A.K., Maso, V., Marangoni, S., Da Silva, S.L., 2008. Koninginins, phospholipase A2 inhibitors from endophytic fungus Trichoderma koningii. Toxicon 51, 240–250.
Summerbell, R.C., 2003. Ascomycetes: Aspergillus, Fusarium, Sporothrix, Piedra, and relatives. In: Howard, D.H. (Ed.), Pathogenic Fungi in Humans and Animals, Marcel Dekker Inc., New York, pp. 437–449.
Sun, S., Tian, L., Wang, Y., Wu, H., Lu, X., Pei, Y., 2009. A novel natural product from the fermentation liquid of marine fungus Trichoderma atroviride G20-12. Asian J. Tradit. Med. 4, 123–127.
Sun, Y., Tian, L., Huang, J., Ma, H.-Y., Zheng, Z., Lv, A.-L., Yasukawa, K., Pei, Y.-H., 2008. Trichodermatides A–D, novel polyketides from the marine-derived fungus Trichoderma reesei. Org. Lett. 10, 393–396.
Sun, Y., Tian, L., Huang, Y.F., Sha, Y., Pei, Y.H., 2006. A new cyclotetrapep-tide from marine fungus Trichoderma reesei. Pharmazie 61, 809–810.
Sun, R., Liu, Z., Fu, K., Fan, L., Chen, J., 2012. Trichoderma biodiversity in China. J. Appl. Genet. 53, 343–354.
Sun, P.X., Zheng, C.J., Li, W.C., Jin, G.L., Huang, F., Qin, L.P., 2011. Trichodermanin A, a novel diterpenoid from endophytic fungus culture. J. Nat. Med. 65, 381–384.
Szczech, M., Staniaszek, M., Habdas, H., Uliński, Z., Szymański, J., 2008. Trichoderma spp. - the cause of green mold on polish mush-room farms. Veg. Crops Res. Bull. 69, 105–114.
Szekeres, A., Láday, M., Kredics, L., Varga, J., Antal, Z., Hatvani, L., Manczinger, L., Vágvölgyi, C., Nagy, E., 2006. Rapid identification of clinical Trichoderma longibrachiatum isolates by cellulose-acetate electrophoresis mediated isoenzyme analysis. Clin. Microbiol. Infect. 12, 369–375.
Takatori, M., Shida, T., Akiyama, K., Takatori, K., 1994. Airborne fungi during the last ten years in Sagamihara. Aerugi 43, 1–8, (Article in Japanese).
Thrane, U., Poulsen, S.B., Nirenberg, H.I., Lieckfeldt, E., 2001. Identifi-cation of Trichoderma strains by image analysis of HPLC chromato-grams. FEMS Microbiol. Lett. 203, 249–255.
Tsuchiya, T., Yamada, K., Minoura, K., Miyamoto, K., Usami, Y., Kobayashi, T., Hamada-Sato, N., Imada, C., Tsujibo, H., 2008. Puri-fication and determination of the chemical structure of the tyrosi-nase inhibitor produced by Trichoderma viride strain H1-7 from a marine environment. Biol. Pharm. Bull. 31, 1618–1620.
Tsurumi, Y., Inaba, S., Susuki, S., Kamijo, S., Widyastuti, Y., Hop, D., Balijinova, T., Sukarno, N., Nakagiri, A., Susuki, K., Ando, K., 2010. Distribution of Trichoderma species in four countries of Asia. 9th Inter-national Mycological Congress, Edinburgh, Scotland, August 2010.
Turóczi, G., Fekete, C., Kerényi, Z., Nagy, R., Pomázi, A., Hornok, L., 1996. Biological and molecular characterization of potential biocon-trol strains of Trichoderma. J. Basic Microbiol. 36, 63–72.
Vajna, L., 1983. Trichoderma species in Hungary. Acta Phytopathol. Acad. Sci. Hung. 18, 291–301.
Vasanthakumari, M.M., Shivanna, M.B., 2011. Fungal assemblages in the rhizosphere and rhizoplane of grasses of the subfamily Panicoi-deae in the Lakkavalli region of Karnataka, India. Microbes Envi-ron. 26, 228–236.
Vinale, F., Sivasithamparam, K., Ghisalberti, E.L., Marra, R., Woo, S.L., Lorito, M., 2008. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 40, 1–10.
Wang, G., Li, Q., Zhu, P., 2008. Phylogenetic diversity of culturable fungi associated with the Hawaiian sponges Suberites zeteki and Gelliodes fibrosa. Anton. Leeuw. Int. J. G. 93, 163–174.
Wang, P., Liu, Y., Yin, Y., Jin, H., Wang, S., Xu, F., Zhao, S., Geng, X., 2011. Diversity of microorganisms isolated from the soil sample surround Chroogomphus rutilus in the Beijing Region. Int. J. Biol. Sci. 7, 209–220.
White, T.J., Bruns, T., Lee, S., Taylor, J.W., 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenet-ics. In: Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J. (Eds.), PCR Protocols: A Guide to Methods and Applications, Academic Press, New York, pp. 315–322.
Widden, P., Abitbol, J.-J., 1980. Seasonality of Trichoderma species in a spruce-forest soil. Mycologia 72, 775–784.
Williams, J., Clarkson, J.M., Mills, P.R., Cooper, R.M., 2003. A selec-tive medium for quantitative reisolation of Trichoderma harzianum from Agaricus bisporus compost. Appl. Environ. Microbiol. 69, 4190–4191.
Wuczkowski, M., Druzhinina, I., Gherbawy, Y., Klug, B., Prillinger, H., Kubicek, C.P., 2003. Species pattern and genetic diversity of Tricho-derma in a mid-European, primeval floodplain-forest. Microbiol. Res. 158, 125–133.
Xia, X., Lie, T.K., Qian, X., Zheng, Z., Huang, Y., Shen, Y., 2011. Spe-cies diversity, distribution, and genetic structure of endophytic and epiphytic Trichoderma associated with banana roots. Microb. Ecol. 61, 619–625.
Yamaguchi, K., Tsurumi, Y., Suzuki, R., Chuaseeharonnachai, C., Sri-Indrasutdhi, V., Boonyuen, N., Okane, I., Suzuki, K., Nakagiri, A., 2012. Trichoderma matsushimae and T. aeroaquaticum: two aero-aquatic species with Pseudaegerita-like propagules. Mycologia 104, 1109–1120.
You, J., Dai, H., Chen, Z., Liu, G., He, Z., Song, F., Yang, X., Fu, H., Zhang, L., Chen, X., 2010. Trichoderone, a novel cytotoxic cyclo-pentenone and cholesta-7,22-diene-3b, 5a, 6b-triol, with new activi-ties from the marine-derived fungus Trichoderma sp. Ind. Microbiol. Biotechnol. 37, 245–252.
Yu, Z., Zhang, K., 2004. Population dynamics of Trichoderma species in the rhizosphere of tobacco and four species from China. J. Zhejiang Univ. Agric. Life Sci. 30, 468.
Yuan, Z.L., Chen, Y.C., Yang, Y., 2009. Diverse non-mycorrhizal fungal endophytes inhabiting an epiphytic, medicinal orchid (Dendrobium nobile): estimation and characterization. World J. Microbiol. Bio-technol. 25, 295–303.
Zachow, C., Berg, C., Müller, H., Meincke, R., Komon-Zelazowska, M., Druzhinina, I.S., Kubicek, C.P., Berg, G., 2009. Fungal diversity in the rhizosphere of endemic plant species of Tenerife (Canary Islands): relationship to vegetation zones and environmental fac-tors. ISME J. 3, 79–92.
Zhang, C., Druzhinina, I., Kubicek, C.P., Xu, T., 2005. Trichoderma bio-diversity in China: evidence for a North to South distribution of species in East Asia. FEMS Microbiol. Lett. 251, 251–257.
Zhang, L., Hao, C., Wang, L., Li, S., Feng, C., 2012. Characteristics of the eukaryotic community structure in acid mine drainage lake in Anhui Province, China. Wei Sheng Wu Xue Bao 52, 875–884, (Arti-cle in Chinese).
Zhang, C.L., Liu, S.P., Lin, F.C., Kubicek, C.P., Druzhinina, I.S., 2007. Trichoderma taxi sp. nov., an endophytic fungus from Chinese yew Taxus mairei. FEMS Microbiol. Lett. 270, 90–96.
Zhao, Z., Sun, X., Yang, R., Yang, H., Lu, G., 2004. Diversity of Trich-oderma in greenhouse soil. J. Zhejiang Univ. Agric. Life Sci. 30, 467.
Zhou, K., Zhang, X., Zhang, F., Li, Z., 2011. Phylogenetically diverse cultivable fungal community and polyketide synthase (PKS), non-ribosomal peptide synthase (NRPS) genes associated with the South China sea sponges. Microb. Ecol. 62, 644–654.
Zielińska-Jankiewicz, K., Kozajda, A., Piotrowska, M., Szadkowska-Stańczyk, I., 2008. Microbiological contamination with moulds in work environment in libraries and archive storage facilities. Ann. Agric. Environ. Med. 15, 71–78.




















