
Binding of the potent allergen hexahydrophthalic anhydride in the mucosa of the upper respiratory...
16
Toxicology 134 (1999) 153 – 168 Binding of the potent allergen hexahydrophthalic anhydride in the mucosa of the upper respiratory and alimentary tract following single inhalation exposures in guinea pigs and rats Christian H. Lindh a, *, Bo A.G. Jo ¨ nsson a , Gunvor Johannesson a , Xing-Dong Zhang a , Hans Welinder a , Eva B. Brittebo b a Department of Occupational and En6ironmental Medicine, Institute of Laboratory Medicine, Uni6ersity Hospital, S -221 85 Lund, Sweden b Department of Pharmaceutical Biosciences, Di6ision of Toxicology, Uppsala Uni6ersity, Box 594, S -751 24 Uppsala, Sweden Received 28 September 1998; accepted 15 February 1999 Abstract Hexahydrophthalic anhydride (HHPA; CAS No. 13149-00-3) is a highly allergenic compound commonly used in the chemical industry. Guinea pigs and rats were exposed to [ 3 H 2 ]HHPA by inhalation for 3–8 h and were killed at various intervals during 7 days. The tissue distribution of non-volatile and covalently bound radioactivity was studied by autoradiography. Tissue bound radioactivity was mainly found in the mucosa of the upper respiratory airways, whereas negligible levels were observed in the lungs. In addition, tissue bound radioactivity was present in the gastrointestinal tract and conjunctiva. Moreover, in the cortex of the kidneys in rats, but not in guinea pigs, a low level of tissue bound radioactivity was found. The radioactivity in the tissues persisted for at least 7 days after the end of exposure. Plasma proteins and soluble proteins from trachea, lung, and kidney from [ 3 H 2 ]HHPA-exposed animals were separated by gel filtration. The radioactivity in dialysed plasma was mainly found in the same fractions as albumin. The soluble proteins from trachea, lung, and kidney in both rats and guinea pigs showed a similar pattern as found in blood. The radioactivity in dialysed plasma from both guinea pigs and rats seemed to decay according to a two-compartment model. The non-extractable binding of [ 3 H 2 ]HHPA in the upper respiratory airways and Abbre6iations: CNS, central nervous system; CV, coefficient of variation; DMSO, dimethyl sulfoxide; GC – MS, gas chromatogra- phy–mass spectrometry; im, intramuscularly; iv, intravenous; HDI, hexamethylene diisocyanate; HHPA, hexahydrophthalic anhydride; HHPA – RSA, hexahydrophthalic anhydride – rat serum albumin; HHP acid, hexahydrophthalic acid; [ 3 H 2 ]HHPA, di tritium labelled hexahydrophthalic anhydride; [ 3 H 2 ]HHP acid, di tritium labelled hexahydrophthalic acid; HPLC, high performance liquid chromatography; OAA, organic acid anhydrides; PBS, phosphate buffered saline; LMW, low molecular weight; SDS – PAGE, sodium dodecylsulphate – polyacrylamide gel electrophoresis; TCA, trichloroacetic acid; TDI, toluene diisocyanate. * Corresponding author. Tel.: +46-46-173191; fax: +46-46-143702. E-mail address: [email protected] (C.H. Lindh) 0300-483X/99/$ - see front matter © 1999 Elsevier Science Ireland Ltd. All rights reserved. PII:S0300-483X(99)00032-3
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Binding of the potent allergen hexahydrophthalic anhydride in the mucosa of the upper respiratory...

Toxicology 134 (1999) 153–168
Binding of the potent allergen hexahydrophthalic anhydride inthe mucosa of the upper respiratory and alimentary tract
following single inhalation exposures in guinea pigs and rats
Christian H. Lindh a,*, Bo A.G. Jonsson a, Gunvor Johannesson a,Xing-Dong Zhang a, Hans Welinder a, Eva B. Brittebo b
a Department of Occupational and En6ironmental Medicine, Institute of Laboratory Medicine, Uni6ersity Hospital,S-221 85 Lund, Sweden
b Department of Pharmaceutical Biosciences, Di6ision of Toxicology, Uppsala Uni6ersity, Box 594, S-751 24 Uppsala, Sweden
Received 28 September 1998; accepted 15 February 1999
Abstract
Hexahydrophthalic anhydride (HHPA; CAS No. 13149-00-3) is a highly allergenic compound commonly used inthe chemical industry. Guinea pigs and rats were exposed to [3H2]HHPA by inhalation for 3–8 h and were killed atvarious intervals during 7 days. The tissue distribution of non-volatile and covalently bound radioactivity was studiedby autoradiography. Tissue bound radioactivity was mainly found in the mucosa of the upper respiratory airways,whereas negligible levels were observed in the lungs. In addition, tissue bound radioactivity was present in thegastrointestinal tract and conjunctiva. Moreover, in the cortex of the kidneys in rats, but not in guinea pigs, a lowlevel of tissue bound radioactivity was found. The radioactivity in the tissues persisted for at least 7 days after the endof exposure. Plasma proteins and soluble proteins from trachea, lung, and kidney from [3H2]HHPA-exposed animalswere separated by gel filtration. The radioactivity in dialysed plasma was mainly found in the same fractions asalbumin. The soluble proteins from trachea, lung, and kidney in both rats and guinea pigs showed a similar patternas found in blood. The radioactivity in dialysed plasma from both guinea pigs and rats seemed to decay accordingto a two-compartment model. The non-extractable binding of [3H2]HHPA in the upper respiratory airways and
Abbre6iations: CNS, central nervous system; CV, coefficient of variation; DMSO, dimethyl sulfoxide; GC–MS, gas chromatogra-phy–mass spectrometry; im, intramuscularly; iv, intravenous; HDI, hexamethylene diisocyanate; HHPA, hexahydrophthalicanhydride; HHPA–RSA, hexahydrophthalic anhydride–rat serum albumin; HHP acid, hexahydrophthalic acid; [3H2]HHPA, ditritium labelled hexahydrophthalic anhydride; [3H2]HHP acid, di tritium labelled hexahydrophthalic acid; HPLC, high performanceliquid chromatography; OAA, organic acid anhydrides; PBS, phosphate buffered saline; LMW, low molecular weight; SDS–PAGE,sodium dodecylsulphate–polyacrylamide gel electrophoresis; TCA, trichloroacetic acid; TDI, toluene diisocyanate.
* Corresponding author. Tel.: +46-46-173191; fax: +46-46-143702.E-mail address: [email protected] (C.H. Lindh)
0300-483X/99/$ - see front matter © 1999 Elsevier Science Ireland Ltd. All rights reserved.
PII: S 0300 -483X(99 )00032 -3

C.H. Lindh et al. / Toxicology 134 (1999) 153–168154
conjunctiva may be of relevance for symptoms in workers with allergy, since they mainly develop symptoms and signsfrom the nose and eyes. © 1999 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Hexahydrophthalic anhydride; Autoradiography; Inhalation; Distribution; Allergy
1. Introduction
Allergies with symptoms and signs, such asasthma and rhinitis, is a large and increasingproblem. One special group of agents inducingsuch diseases is the low molecular weight(LMW) reactive chemicals. Many of these havea vast use in the chemical industry. It is gener-ally believed that the reaction of these chemicalsto endogenous proteins is important for theirallergenic potential. Information on the distribu-tion of allergenic compounds following inhala-tion exposure is sparse, however.
The isocyanates is a group of LMW reactivechemicals that induce asthma and rhinitis, andthere are a number of reports on the distribu-tion following inhalation exposure. Kennedy etal. (1993) described in an autoradiographicstudy on guinea pigs following inhalation ofmethyl isocyanate that this chemical was mainlybound to the subepithelial region of the respira-tory tract. Toluene diisocyanate (TDI) and hex-amethylene diisocyanate (HDI) were also mainlybound to the subepithelial region of the respira-tory system of guinea pigs after inhalation expo-sure (Kennedy, 1990; Kennedy and Brown,1992) and on the apical surface in the respira-tory system (TDI; Karol et al., 1997). Moreover,an irreversible binding of isocyanates to bloodproteins, such as albumin and haemoglobin hasalso been reported (Kennedy et al., 1989; Jin etal., 1993; Schutze et al., 1995; Sepai et al.,1995a,b; Day et al., 1996).
Organic acid anhydrides (OAAs) are LMWreactive chemicals, which are commonly used, inthe chemical industry. Up to more than 60% ofexposed workers have been reported to developairways symptoms and/or have specific IgE andIgG antibodies against OAA-serum albuminconjugates even at very low exposures (Ven-ables, 1989; Nielsen et al., 1992; Yokota et al.,
1997). Hexahydrophthalic anhydride (HHPA) isa particularly potent allegen (Welinder et al.,1994). It is known that a large amount of theinhaled HHPA in humans is rapidly absorbedand hydrolysed to hexahydrophthalic acid (HHPacid) a compound with a low vapour pressure(Jonsson and Skerfving, 1993). However, it hasbeen described that HHPA in exposed workersalso forms adducts with plasma proteins (Jon-sson et al., 1996) and haemoglobin (Jonsson etal., 1997; Lindh and Jonsson, 1998a,b). In addi-tion, it has been found that another OAA,methyltetrahydrophthalic anhydride (MTHPA),forms lysine adducts with collagen following in-tratracheal injections of guinea pigs (Jonsson etal., 1995). However, the knowledge about thetissue distribution of OAAs and the OAA ad-ducts is still limited.
The aim of the present study was to examinethe distribution of non-volatile and non-ex-tractable radioactivity following single inhalationexposure of [3H2]HHPA in guinea pigs. The dis-tribution of the [3H2]HHPA-derived radioactivitywas confirmed by additional studies in rats. Inaddition, the binding of [3H2]HHPA/metabo-lite(s) to plasma proteins and soluble tissueproteins in guinea pigs and rats were examined.
2. Materials and methods
2.1. Synthesis of [ 3H2]HHPA
[3H2]Hexahydrophthalic acid ([3H2]HHP acid)was obtained from Amersham tritium labellingservice (Buckinghamshire, UK). The [3H2]HHPacid (portions of 100 MBq) was purified usingpreparative high-performance liquid chromatog-raphy (HPLC; model HP1050, Hewlett Packard,Palo Alto, CA) with a 25×0.46 cm Apex

C.H. Lindh et al. / Toxicology 134 (1999) 153–168 155
cyl column with 5 mm particles (Jones Chro-matography, Lakewood, CO). The mobile phasewas 0.1% trifluoroacetic acid (TFA) in water in agradient from 0–100% of 0.07% TFA in acetoni-trile in 30 min. The flow rate of the mobile phasewas 1 ml/min. The fraction containing the[3H2]HHP acid was collected, and added with 0.1mg of unlabelled HHP acid, the solution was thenevaporated. The dry residue was treated withacetyl chloride at room temperature for 18 h toform HHPA and [3H2]HHPA (Fig. 1). The acetylchloride containing the [3H2]HHPA and unla-belled HHPA was thereafter transferred into theglass vials later used in the exposure system andevaporated into dryness. The radiochemical puritywas assessed with HPLC and scintillation count-ing (LS180; Beckman Instruments, Fullerton, CA)and found to be \95%. The amount of[3H2]HHPA was less than 10% of the unlabelledHHPA as determined by mass spectrometry (MS).The specific activity was determined by gas chro-matography (GC)–MS, according to the methoddescribed by Lindh and Jonsson (1997), and scin-tillation counting.
2.2. Synthesis of HHPA-rat serum albumin(RSA) conjugate
Unlabelled HHPA (1 mg) and 0.2 GBq[3H2]HHPA were dissolved in 100 ml acetonitrile.This solution was then slowly added to a phos-phate buffered saline (PBS) solution containing
100 mg RSA. The solution was stirred for 1 h anddialysed for 10 days against PBS solution until thedialysis solution was blank. The conjugate wassterilised by filtration prior to use. The purity ofthe conjugate was checked with HPLC, using aGenesis (Jones Chromatography, Lakewood, CO)C8 column (10×0.46 cm with 4 mm particles)both directly as such, and after digestion, withPronase E (Sigma, St Louis, MO). The conjugatewas found to contain B3% free [3H2]HHP acid.
2.3. Animals
The guinea pigs used were inbred white maleDunkin–Hartley strain obtained from Sahlins(Malmo, Sweden). The rats were inbred maleBrown Norway strain obtained from Møllegaard(Ejby, Denmark). The strain and sex of the ani-mals were chosen due to their high allergenicresponsiveness (Welinder et al., 1995; Zhang etal., 1998). The animals were maintained on thefacilities with food and water provided ad libitumand on a cycle of 8 h light and 16 h dark. Theanimals were acclimated for 1 week before experi-ments. In addition, the animals used in the Bat-telle design exposure chambers were acclimatedfor increasing time periods in the chambers duringthe 5 days before the exposure. The animal anaes-thetised during the exposure was continuouslyadministered a mixture of ketamine hydrochloride(50 mg/ml; Park–Davis, Barcelona, Spain) andxylazine chloride (20 mg/ml; Bayer, Gothenburg,Sweden), prepared in a ratio of 3:2. After theexposures, blood was collected in test tubes con-taining sodium heparin by heart puncture fromthe animals. Before blood sampling the animalswere anaesthetised with 1 ml/kg im of the mixtureof ketamine hydrochloride and xylazine chloride.The animals were killed by an overdose of ke-tamine/xylazine. The experiments were approvedby the Animal Research Ethical Committee atLund University.
2.4. Inhalation exposure systems
Two different exposure equipments were usedfor the inhalation exposure study. Initially, in a
Fig. 1. Chemical structure of hexahydrophthalic anhydride(HHPA). * Indicate the position of the tritium-labelling.
C.H. Lindh et al. / Toxicology 134 (1999) 153–168156
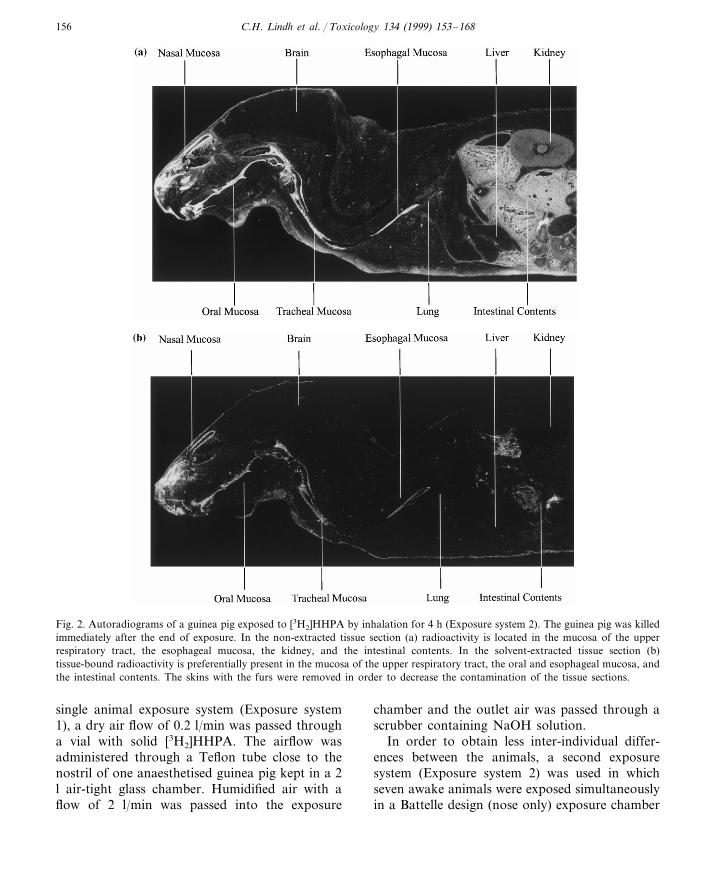
Fig. 2. Autoradiograms of a guinea pig exposed to [3H2]HHPA by inhalation for 4 h (Exposure system 2). The guinea pig was killedimmediately after the end of exposure. In the non-extracted tissue section (a) radioactivity is located in the mucosa of the upperrespiratory tract, the esophageal mucosa, the kidney, and the intestinal contents. In the solvent-extracted tissue section (b)tissue-bound radioactivity is preferentially present in the mucosa of the upper respiratory tract, the oral and esophageal mucosa, andthe intestinal contents. The skins with the furs were removed in order to decrease the contamination of the tissue sections.
single animal exposure system (Exposure system1), a dry air flow of 0.2 l/min was passed througha vial with solid [3H2]HHPA. The airflow wasadministered through a Teflon tube close to thenostril of one anaesthetised guinea pig kept in a 2l air-tight glass chamber. Humidified air with aflow of 2 l/min was passed into the exposure
chamber and the outlet air was passed through ascrubber containing NaOH solution.
In order to obtain less inter-individual differ-ences between the animals, a second exposuresystem (Exposure system 2) was used in whichseven awake animals were exposed simultaneouslyin a Battelle design (nose only) exposure chamber

C.H. Lindh et al. / Toxicology 134 (1999) 153–168 157
system (Cannon et al., 1983). A dry air flow of 5l/min was passed through a test tube containingthe solid [3H2]HHPA and distributed to all ani-mals. All air leaving the exposure chamber waslikewise passed through a scrubber containingNaOH solution. The exposure level in the cham-bers was determined by air sampling, using bub-blers with 5 ml 0.1 M NaOH, during theexposure. The radioactivity in the solutions wasanalysed with liquid scintillation. All exposureswere performed in a ventilated hood, and mea-sures were taken to contain the reactive radioac-tive gas in tight chambers. The radioactiveexposure of the laboratory personnel was checkedby scintillation of sampled urine. A test of theexposure system was performed by exposing sixguinea pigs for 4 h with unlabelled HHPA. Thelevels of unlabelled HHPA in air were determinedas described by Jonsson et al. (1991). The levels ofHHP acid in plasma were analysed according toLindh and Jonsson (1997).
2.5. Inhalation exposure of animals to[ 3H2]HHPA
In the first exposure system, one anaesthetisedguinea pig (360 g) were exposed for 8 h and killedimmediately after the end of exposure. The animalwas exposed to 3 mmol (N=4) HHPA/m3 with aspecific activity of 0.2 GBq/mmol HHPA. Varioustissues were collected from the animal for light-microscopic autoradiography.
In the second exposure system, one set of sevenguinea pigs (approx. 230 g) were exposed for 4 hand one set of seven rats (approx. 100 g) wereexposed for 3 h. The animals were killed after 0, 4h (two animals), 1, 2 (two animals), and 7 days.The guinea pigs were exposed to an average of 3mmol unlabelled HHPA/m3 (CV=78%; N=9)with a specific activity of 0.7 GBq/mmol HHPAand the rats were exposed to an average of 2 mmolunlabelled HHPA/m3 (CV=66%; N=12) with aspecific activity of 0.5 GBq/mmol HHPA. To min-imise stress among the rats the exposure wasterminated after 3 h. To avoid contamination ofthe tissue sections by radioactivity bound to thefur, the skin including the fur was removed fromthe animals before the animals were frozen andembedded. The embedded animals were processedfor whole-body autoradiography as described be-low. Two animals from each set were killed after4 h and 2 days, respectively, for collection of lung,kidney, palpebral conjunctiva, trachea for light-microscopic autoradiography as described below.The chosen HHPA air levels were in the samerange as the higher occupational exposures foundin Sweden.
2.6. Exposure by i6 injection
[3H2]HHPA dissolved in 85 ml (20 MBq)dimethyl sulfoxide (DMSO) was injected in thefemoral vein in the inguinal region of two anaes-thetised rats (approximately 250 g). The animals
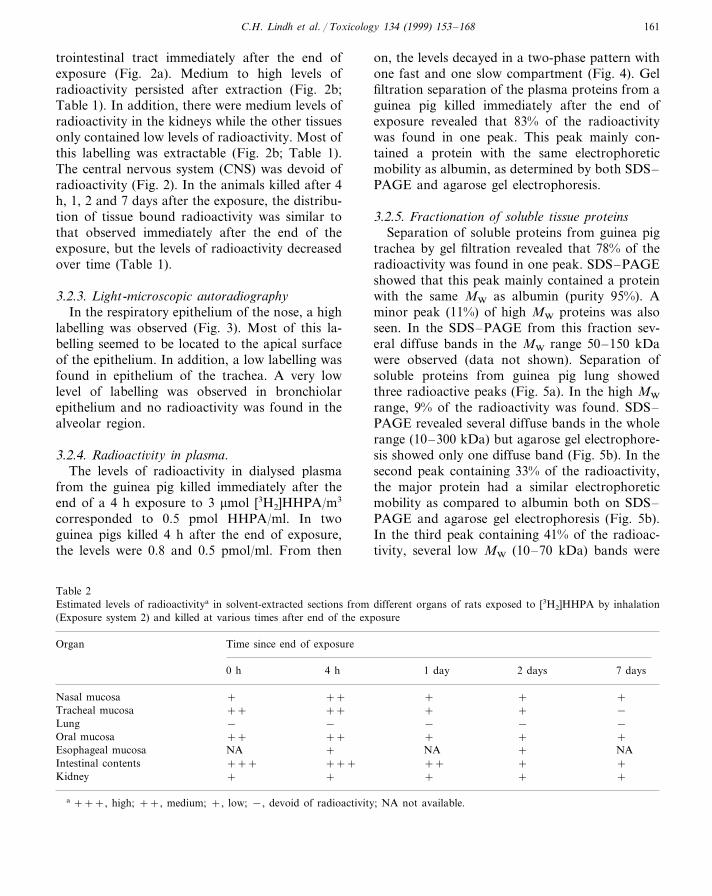
Table 1Estimated levels of radioactivitya in solvent-extracted sections from different organs of guinea pigs exposed to [3H2]HHPA byinhalation (Exposure system 2) and killed at various times after end of the exposure
Time since end of exposureOrgan
1 day 2 days0 h 7 days4 h
++ +Nasal mucosa +++ +++++ ++++Tracheal mucosa ++
− − − −Lung −+++++Oral mucosa ++
NA NAEsophageal mucosa ++ NA NA+ −Intestinal contents ++ + +
− −Kidney − −−
a +++, high; ++, medium; +, low; −, devoid of radioactivity; NA, not available.

C.H. Lindh et al. / Toxicology 134 (1999) 153–168158
Fig. 3. Light-microscopic autoradiogram (100× ) of a solvent-extracted section from the nasal passage (turbinate) of a guinea pigexposed to [3H2]HHPA by inhalation for 8 h (Exposure system 1). The animal was killed immediately after the end of exposure.There is a selective localisation of radioactivity in the nasal respiratory epithelium. Stained with hematoxylin–eosin.
were killed after 6 h and 1 day, respectively. Inaddition, [3H2]HHPA-RSA dissolved in PBS wasinjected (185 ml; 6 MBq) in the femoral vein in theinguinal region of three anaesthetised rats. Theanimals were killed after 6 h, 1 day and 2 days.The animals were then stored at −20°C until use.These animals were processed for whole-body au-toradiography as described in the next section.
2.7. Whole-body autoradiography
The frozen animals were embedded in aqueouscarboxymethyl cellulose and frozen in a CO2/ethanol bath. Series of 20 mm sections were takensagitally on tape and were freeze-dried (Ullberg etal., 1982). To evaluate possible covalent tissue-bound radioactivity, selected tissue sections wereextracted stepwise for 1 min using 5%trichloroacetic acid (TCA), water, 50% ethanol,absolute ethanol, heptane, absolute ethanol, 50%ethanol, and water. The sections were then rinsedfor 5 min in running water and dried. The sol-vent-extracted sections and the adjacent freeze-dried non-extracted sections were then pressedagainst Hyperfilm-3H (Amersham). All films were
exposed at −20°C for 6 months. Ten or elevenautoradiograms for each animal were visuallyevaluated and the amount of radioactivity in thetissue were graded: + + + , high level of radioac-tivity, defined by white shade on the positiveprint; + + , medium level of radioactivity,defined by light grey shades; + , low level ofradioactivity, defined by dark grey shades; and− , devoid of radioactivity, defined by black.
2.8. Light-microscopic autoradiography
Various tissues from rats and guinea pigs wereprocessed for light-microscopic autoradiographyas described in Kowalski et al. (1985). The tissueswere fixed in 4% formaldehyde in PBS for 2–3days and then stored in 70% ethanol until pro-cessing. To prepare the nasal region, the headswere removed and immersed in 4% formaldehydein PBS for 2 weeks. The formaldehyde was re-moved and replaced with 30% formic acid for aperiod of 4 weeks. The nasal region was dissectedand stored in 70% ethanol. The tissues were em-bedded in Historesin (LKB, Bromma, Sweden),sectioned (4 mm) and placed on glass slides. The

C.H. Lindh et al. / Toxicology 134 (1999) 153–168 159
slides were extracted and dipped in liquid filmemulsion (NTB-2; Kodak, Rochester, NY, USA).After exposure, the sections were stained withhematoxylin and eosin and then evaluated.
2.9. Treatment of blood
The blood, collected from the animals exposedin the Battelle design exposure system, was al-lowed to cool to room temperature and was thencentrifuged at 1500×g for 10 min. The plasmawas separated from the erythrocytes and dialysedagainst five changes of PBS for 2 weeks until thedialysis solution was blank.
2.10. Fractionation of plasma proteins
The dialysed plasma was diluted (1:1) with 0.05M NH4HCO3 and then injected into a 500 ml loopconnected to an A8 KTA purifier liquid chromatog-raphy (LC) system (Pharmacia Biotech, Uppsala,Sweden) equipped with a gel filtration column(Sephadex 200, model 10/30; Pharmacia Biotech).
The mobile phase was 0.05 M NH4HCO3 with aflow of 0.5 ml/min. The proteins were detected at280 nm. Fractions were collected in 1 min inter-vals. An aliquot of 400 ml from each fraction wasadded to 10 ml of scintillation cocktail (ReadySolve HP; Beckman, Fullerton, CA, USA) priorto counting on a LS1801 Beckman scintillationequipment.
2.11. Fractionation of soluble tissue proteins
For analyses of the radioactivity bound in solu-ble proteins, about 0.5 g tissue from lung, trachea,and kidney from rats and guinea pigs exposed byinhalation were homogenised in 4 ml of PBS witha glass homogeniser. The PBS was added with theprotease inhibitors 5 mM benzamidine, 10 mMEDTA, 5 mM phenylmethansulfonyl fluoride, and5 mM N-ethylmaleimid. The homogenised solu-tions were centrifuged at 2000×g for 20 min andthe supernatants were dialysed against PBS withprotease inhibitors for 7 days until the dialysissolution was blank. The last buffer change was in0.05 M NH4HCO3. The solutions were then frac-tionated as described above for plasma proteinsexcept that these solutions were not diluted beforethe injections.
2.12. Gel electrophoresis
Sodium dodecyl sulphate–polyacrylamide gelelectrophoresis (SDS–PAGE) was performed onaliquots from the fractions obtained from the gelfiltration. The SDS–PAGE gels were from tris/glycin buffer with the acrylamide concentrationsof 12 or 4–15% and purchased from Biorad (Her-cules, CA, USA). In addition, agarose gel elec-trophoresis, as described by Jeppsson et al. (1979),was performed on aliquots from the fractions.The proteins were stained using Coomassie blue.
3. Results
3.1. Precision in inhalation of hexahydrophthalicanhydride (HHPA)
A series of six guinea pigs were exposed in the
Fig. 4. The natural logarithm of the amount of radioactivity indialysed plasma from rats and guinea pigs. Rats (n=7) wereexposed to [3H2]HHPA by inhalation for 3 h and killed atvarious times (+ ). Guinea pigs (n=7) were exposed to[3H2]HHPA by inhalation for 4 h and killed at various times(�).The decay of radioactivity seems to follow a two compart-ment model.
C.H. Lindh et al. / Toxicology 134 (1999) 153–168160
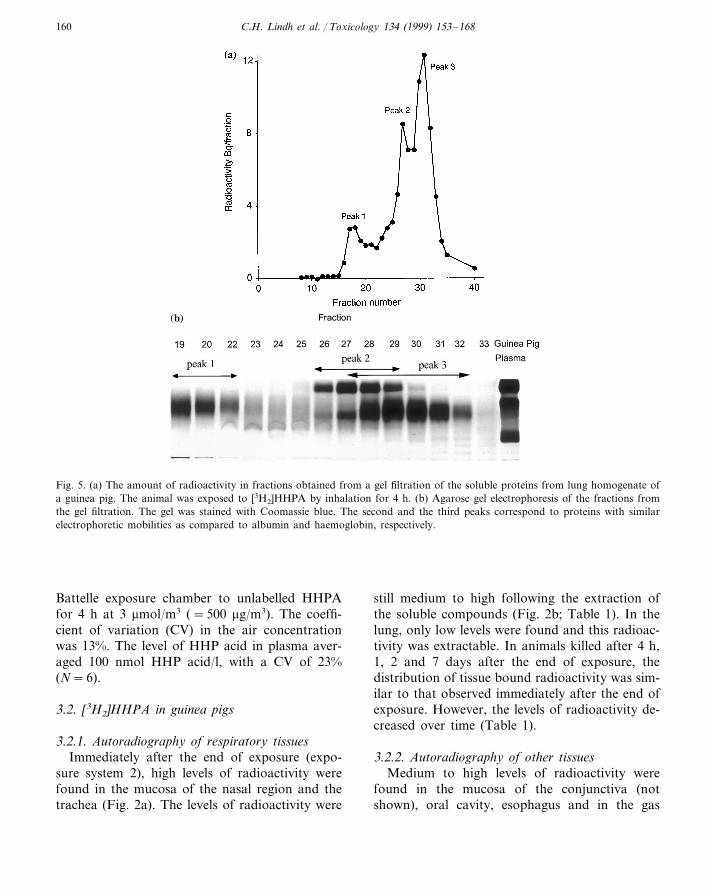
Fig. 5. (a) The amount of radioactivity in fractions obtained from a gel filtration of the soluble proteins from lung homogenate ofa guinea pig. The animal was exposed to [3H2]HHPA by inhalation for 4 h. (b) Agarose gel electrophoresis of the fractions fromthe gel filtration. The gel was stained with Coomassie blue. The second and the third peaks correspond to proteins with similarelectrophoretic mobilities as compared to albumin and haemoglobin, respectively.
Battelle exposure chamber to unlabelled HHPAfor 4 h at 3 mmol/m3 (=500 mg/m3). The coeffi-cient of variation (CV) in the air concentrationwas 13%. The level of HHP acid in plasma aver-aged 100 nmol HHP acid/l, with a CV of 23%(N=6).
3.2. [3H2]HHPA in guinea pigs
3.2.1. Autoradiography of respiratory tissuesImmediately after the end of exposure (expo-
sure system 2), high levels of radioactivity werefound in the mucosa of the nasal region and thetrachea (Fig. 2a). The levels of radioactivity were
still medium to high following the extraction ofthe soluble compounds (Fig. 2b; Table 1). In thelung, only low levels were found and this radioac-tivity was extractable. In animals killed after 4 h,1, 2 and 7 days after the end of exposure, thedistribution of tissue bound radioactivity was sim-ilar to that observed immediately after the end ofexposure. However, the levels of radioactivity de-creased over time (Table 1).
3.2.2. Autoradiography of other tissuesMedium to high levels of radioactivity were
found in the mucosa of the conjunctiva (notshown), oral cavity, esophagus and in the gas
C.H. Lindh et al. / Toxicology 134 (1999) 153–168 161
trointestinal tract immediately after the end ofexposure (Fig. 2a). Medium to high levels ofradioactivity persisted after extraction (Fig. 2b;Table 1). In addition, there were medium levels ofradioactivity in the kidneys while the other tissuesonly contained low levels of radioactivity. Most ofthis labelling was extractable (Fig. 2b; Table 1).The central nervous system (CNS) was devoid ofradioactivity (Fig. 2). In the animals killed after 4h, 1, 2 and 7 days after the exposure, the distribu-tion of tissue bound radioactivity was similar tothat observed immediately after the end of theexposure, but the levels of radioactivity decreasedover time (Table 1).
3.2.3. Light-microscopic autoradiographyIn the respiratory epithelium of the nose, a high
labelling was observed (Fig. 3). Most of this la-belling seemed to be located to the apical surfaceof the epithelium. In addition, a low labelling wasfound in epithelium of the trachea. A very lowlevel of labelling was observed in bronchiolarepithelium and no radioactivity was found in thealveolar region.
3.2.4. Radioacti6ity in plasma.The levels of radioactivity in dialysed plasma
from the guinea pig killed immediately after theend of a 4 h exposure to 3 mmol [3H2]HHPA/m3
corresponded to 0.5 pmol HHPA/ml. In twoguinea pigs killed 4 h after the end of exposure,the levels were 0.8 and 0.5 pmol/ml. From then
on, the levels decayed in a two-phase pattern withone fast and one slow compartment (Fig. 4). Gelfiltration separation of the plasma proteins from aguinea pig killed immediately after the end ofexposure revealed that 83% of the radioactivitywas found in one peak. This peak mainly con-tained a protein with the same electrophoreticmobility as albumin, as determined by both SDS–PAGE and agarose gel electrophoresis.
3.2.5. Fractionation of soluble tissue proteinsSeparation of soluble proteins from guinea pig
trachea by gel filtration revealed that 78% of theradioactivity was found in one peak. SDS–PAGEshowed that this peak mainly contained a proteinwith the same MW as albumin (purity 95%). Aminor peak (11%) of high MW proteins was alsoseen. In the SDS–PAGE from this fraction sev-eral diffuse bands in the MW range 50–150 kDawere observed (data not shown). Separation ofsoluble proteins from guinea pig lung showedthree radioactive peaks (Fig. 5a). In the high MW
range, 9% of the radioactivity was found. SDS–PAGE revealed several diffuse bands in the wholerange (10–300 kDa) but agarose gel electrophore-sis showed only one diffuse band (Fig. 5b). In thesecond peak containing 33% of the radioactivity,the major protein had a similar electrophoreticmobility as compared to albumin both on SDS–PAGE and agarose gel electrophoresis (Fig. 5b).In the third peak containing 41% of the radioac-tivity, several low MW (10–70 kDa) bands were
Table 2Estimated levels of radioactivitya in solvent-extracted sections from different organs of rats exposed to [3H2]HHPA by inhalation(Exposure system 2) and killed at various times after end of the exposure
Time since end of exposureOrgan
1 day 2 days0 h 7 days4 h
+ +Nasal mucosa + ++++ −+++Tracheal mucosa ++
− − −Lung −−+++++Oral mucosa ++
+ NAEsophageal mucosa NA + NA+ +Intestinal contents +++ +++ ++
+ +Kidney + ++
a +++, high; ++, medium; +, low; −, devoid of radioactivity; NA not available.
C.H. Lindh et al. / Toxicology 134 (1999) 153–168162
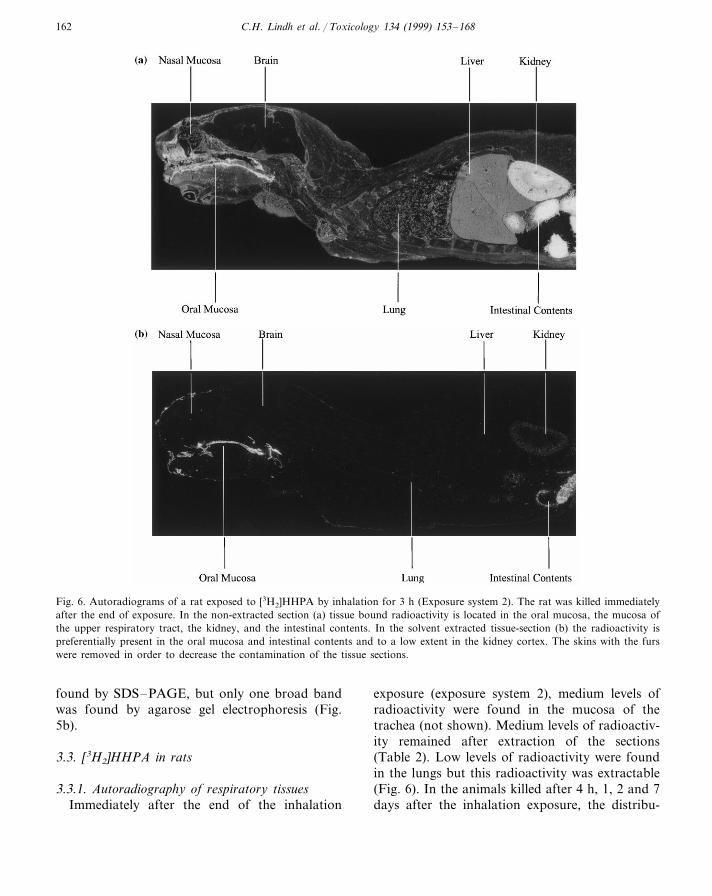
Fig. 6. Autoradiograms of a rat exposed to [3H2]HHPA by inhalation for 3 h (Exposure system 2). The rat was killed immediatelyafter the end of exposure. In the non-extracted section (a) tissue bound radioactivity is located in the oral mucosa, the mucosa ofthe upper respiratory tract, the kidney, and the intestinal contents. In the solvent extracted tissue-section (b) the radioactivity ispreferentially present in the oral mucosa and intestinal contents and to a low extent in the kidney cortex. The skins with the furswere removed in order to decrease the contamination of the tissue sections.
found by SDS–PAGE, but only one broad bandwas found by agarose gel electrophoresis (Fig.5b).
3.3. [3H2]HHPA in rats
3.3.1. Autoradiography of respiratory tissuesImmediately after the end of the inhalation
exposure (exposure system 2), medium levels ofradioactivity were found in the mucosa of thetrachea (not shown). Medium levels of radioactiv-ity remained after extraction of the sections(Table 2). Low levels of radioactivity were foundin the lungs but this radioactivity was extractable(Fig. 6). In the animals killed after 4 h, 1, 2 and 7days after the inhalation exposure, the distribu-
C.H. Lindh et al. / Toxicology 134 (1999) 153–168 163
tion of tissue bound radioactivity was similar tothat observed immediately after the end of theexposure, but the levels of radioactivity decreasedover time (Table 2). In most animals, low levelswere found in the mucosa of the nasal region.However, in the animal killed after 4 h afterexposure a medium labelling was found at thissite. No selective localisation of tissue bound ra-dioactivity was found in the respiratory tissuesfollowing iv injection of [3H2]HHPA in thefemoral vein (not shown).
3.3.2. Autoradiography of other tissuesMedium to high levels of radioactivity was
found in the conjunctiva, oral cavity, gastrointes-tinal tract, liver and kidneys immediately after theend of exposure (Figs. 6 and 7). Some of theradioactivity was non-extractable (Fig. 6b; Table2). In the other tissues, there were only low levelsof radioactivity and most of this radioactivity wasextractable (Fig. 6). The CNS was devoid ofradioactivity (Fig. 6). In the animals killed 4 h, 1,2 and 7 days after exposure the distribution of
tissue bound radioactivity was similar to thatobserved immediately after the end of the expo-sure. However, the levels of radioactivity decayedover time (Table 2). Following iv injection of[3H2]HHPA, high levels of non-extractable ra-dioactivity was only found at the injection site. Ivinjection of the [3H2]HHPA-RSA conjugate re-sulted in a low general labelling of the tissues withno selective localisation of radioactivity. The CNSwas devoid of radioactivity (not shown).
3.3.3. Light-microscopic autoradiographyIn the trachea, a weak but distinct labelling of
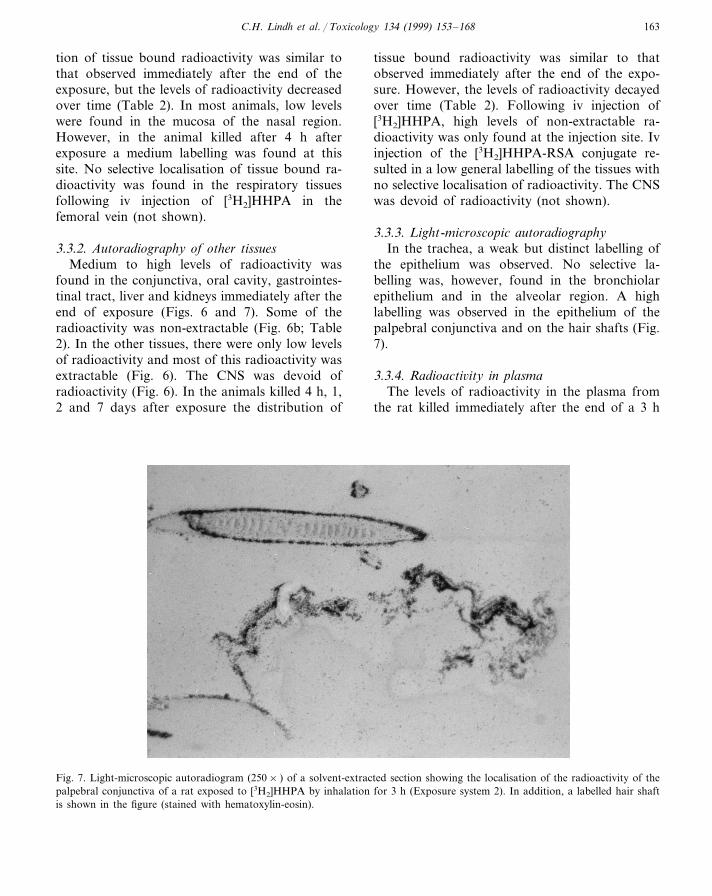
the epithelium was observed. No selective la-belling was, however, found in the bronchiolarepithelium and in the alveolar region. A highlabelling was observed in the epithelium of thepalpebral conjunctiva and on the hair shafts (Fig.7).
3.3.4. Radioacti6ity in plasmaThe levels of radioactivity in the plasma from
the rat killed immediately after the end of a 3 h
Fig. 7. Light-microscopic autoradiogram (250× ) of a solvent-extracted section showing the localisation of the radioactivity of thepalpebral conjunctiva of a rat exposed to [3H2]HHPA by inhalation for 3 h (Exposure system 2). In addition, a labelled hair shaftis shown in the figure (stained with hematoxylin-eosin).
C.H. Lindh et al. / Toxicology 134 (1999) 153–168164
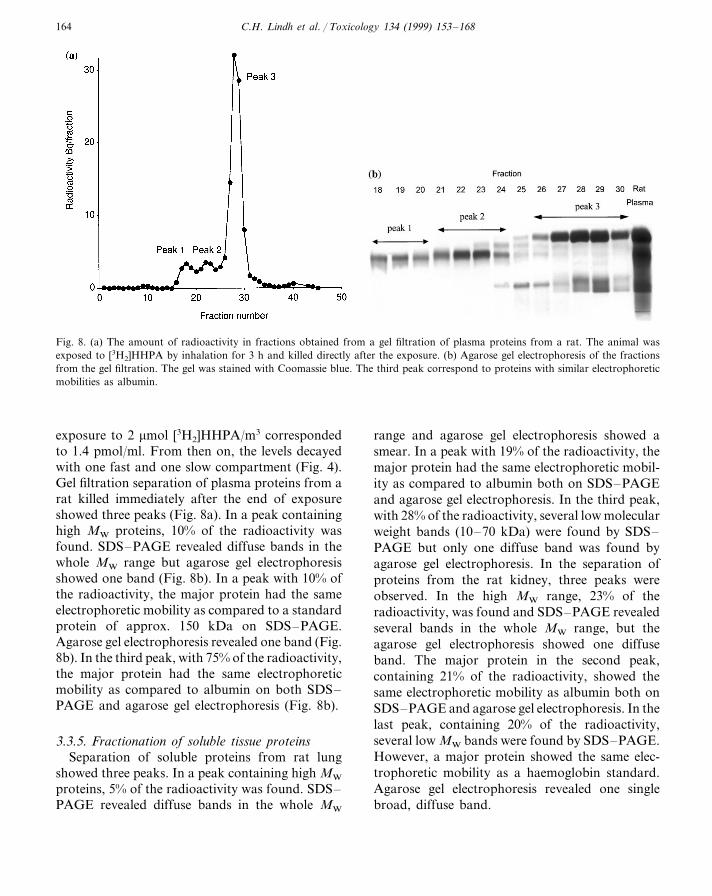
Fig. 8. (a) The amount of radioactivity in fractions obtained from a gel filtration of plasma proteins from a rat. The animal wasexposed to [3H2]HHPA by inhalation for 3 h and killed directly after the exposure. (b) Agarose gel electrophoresis of the fractionsfrom the gel filtration. The gel was stained with Coomassie blue. The third peak correspond to proteins with similar electrophoreticmobilities as albumin.
exposure to 2 mmol [3H2]HHPA/m3 correspondedto 1.4 pmol/ml. From then on, the levels decayedwith one fast and one slow compartment (Fig. 4).Gel filtration separation of plasma proteins from arat killed immediately after the end of exposureshowed three peaks (Fig. 8a). In a peak containinghigh MW proteins, 10% of the radioactivity wasfound. SDS–PAGE revealed diffuse bands in thewhole MW range but agarose gel electrophoresisshowed one band (Fig. 8b). In a peak with 10% ofthe radioactivity, the major protein had the sameelectrophoretic mobility as compared to a standardprotein of approx. 150 kDa on SDS–PAGE.Agarose gel electrophoresis revealed one band (Fig.8b). In the third peak, with 75% of the radioactivity,the major protein had the same electrophoreticmobility as compared to albumin on both SDS–PAGE and agarose gel electrophoresis (Fig. 8b).
3.3.5. Fractionation of soluble tissue proteinsSeparation of soluble proteins from rat lung
showed three peaks. In a peak containing high MW
proteins, 5% of the radioactivity was found. SDS–PAGE revealed diffuse bands in the whole MW
range and agarose gel electrophoresis showed asmear. In a peak with 19% of the radioactivity, themajor protein had the same electrophoretic mobil-ity as compared to albumin both on SDS–PAGEand agarose gel electrophoresis. In the third peak,with 28% of the radioactivity, several low molecularweight bands (10–70 kDa) were found by SDS–PAGE but only one diffuse band was found byagarose gel electrophoresis. In the separation ofproteins from the rat kidney, three peaks wereobserved. In the high MW range, 23% of theradioactivity, was found and SDS–PAGE revealedseveral bands in the whole MW range, but theagarose gel electrophoresis showed one diffuseband. The major protein in the second peak,containing 21% of the radioactivity, showed thesame electrophoretic mobility as albumin both onSDS–PAGE and agarose gel electrophoresis. In thelast peak, containing 20% of the radioactivity,several low MW bands were found by SDS–PAGE.However, a major protein showed the same elec-trophoretic mobility as a haemoglobin standard.Agarose gel electrophoresis revealed one singlebroad, diffuse band.

C.H. Lindh et al. / Toxicology 134 (1999) 153–168 165
4. Discussion
HHPA, a highly allergenic compound, isknown to form adducts with blood proteins inhumans (Jonsson et al., 1996, 1997; Lindh andJonsson 1998a,b). This highly reactive compoundis also known to be rapidly hydrolysed into HHPacid, which is excreted in the urine (Jonsson andSkerfving, 1993). The present results showed thata single inhalation exposure to [3H2]HHPA gener-ated a rapid and selective localisation of radioac-tivity in the mucosa of the upper respiratory andalimentary tract and in the conjunctiva of guineapigs and rats. The [3H2]HHPA-derived radioactiv-ity at these sites could only partially be extractedby organic solvents and water which suggests that[3H2]HHPA and/or HHPA-metabolite(s) was co-valently bound to tissue macromolecules. Cova-lent binding of chemicals at these sites is afrequent finding both after inhalation and sys-temic administration of radio-labelled chemicalsthat are metabolically activated to reactive prod-ucts (Bond et al., 1988; Brittebo, 1993). Iv injec-tion of [3H2]HHPA in rats did not, however,result in a binding of radioactivity at these sitesbut instead close to the injection site. Thus, thepreferential binding of HHPA in the upper res-piratory and alimentary mucosa and conjunctivais most likely related to a high local exposure of[3H2]HHPA.
In the lung of guinea pigs and rats a lowhomogeneously distributed radioactivity was ob-served following exposure to [3H2]HHPA. More-over, most of this radioactivity was extractable.The low level of radioactivity in the lung may bedue to scrubbing of HHPA in the upper respira-tory tract, leading to low levels of HHPA in thelower parts of the conducting airways and in thealveolar air. In contrast, following exposure toradio-labelled TDI and HDI, non-extractable ra-dioactivity was observed in the terminal bronchi-oles by autoradiography (Kennedy, 1990). Duringthe extraction of tissue sections of HHPA-exposedanimals, both HHP acid and adducts of solubleproteins may be removed. Gel filtration of lungsof HHPA-exposed animals indicates that there areseveral adducted proteins in lung tissue, but it isnot clear whether these originate from a forma-
tion in the lung, or if they are transported to thelung via the circulation. The major HHPA ad-ducts corresponded to serum albumin andhaemoglobin, which may indicate that the pul-monary adducts mainly originate from the blood.On the other hand, since plasma exudation is acommon mechanism in the lungs, formation ofplasma protein adducts in the airway lumenshould be considered in further studies.
None of the previous distribution studies onallergenic chemicals have reported a selective la-belling in the conjunctiva or the upper alimentarytract following inhalation exposure. A somewhatincreased level of radioactivity has, however, beenobserved in the esophagus and stomach of ratsexposed to [14C]TDI (Kennedy et al., 1994). In themucosa of the oral cavity and esophagus of[3H2]HHPA-exposed rats and guinea pigs, the ra-dioactivity could only be partially extracted byorganic solvents and was thus in part bound totissue. In addition, a high level of radioactivitywas also present in the contents of the stomachand intestines, and parts of this radioactivity re-mained after extraction. The origin of the gas-trointestinal radioactivity is unclear. Inpreliminary autoradiographic studies of[3H2]HHPA-exposed guinea pigs, high levels ofradioactivity were observed in the furs. The la-belling of the fur may possibly be related to highlevels of cysteine. Thus, the gastrointestinal ra-dioactivity may be due to ingestion of radioactiv-ity present on the fur by the animals during theirsurvival interval. In vitro synthesised conjugatesbetween HHPA and N-acetyl cysteine are un-stable and HHPA may slowly be transferred fromcysteine to lysine (B.A.G. Jonsson, personalcommunication).
A low but distinct localisation of [3H2]HHPA-derived radioactivity was also observed in thecortex of the rat kidney after solvent extraction,whereas no similar localisation was observed inthe guinea pig. Since HHPA is reactive, it isunlikely that HHPA is distributed as such to therat kidneys. However, unstable conjugates be-tween HHPA and cysteine residues can be formedand then transported to the kidney (see above).This alone will, however, not explain the differ-ence between rats and guinea pigs. Such a species

C.H. Lindh et al. / Toxicology 134 (1999) 153–168166
difference may be explained by different concen-trations of the cysteine-containing substance or aselective binding to a specific rat protein. In themale rat kidney cortex, there is for instance aselective synthesis of a2m-globulin protein,whereas male guinea-pigs do not produce thisprotein.
Gel filtration of plasma from guinea pigs andrats indicated that HHPA may be predominantlybound to serum albumin. This would be an inter-esting finding since conjugates between acid anhy-drides and serum albumin are commonly used inassays detecting specific antibodies against theseanhydrides (Welinder and Nielsen, 1991). How-ever, the identity of the HHPA-albumin adducthas yet to be confirmed. The amount of radioac-tivity in the plasma from both rats and guineapigs seemed to decay according to a two compart-ment model. The half-life of the slower phase inthe rats was estimated to some days which may bein agreement with the 3 days half-life of serumalbumin in rats (Schreiber et al., 1971). The half-life of the adducts in the slower phase in guineapigs seemed to be longer than found in rats butthere are, based on the current available informa-tion, no data on the half-life of serum albumin inguinea pigs. However, it must be emphasised thatthe decay of the slow compartments were basedon only one data point each. The fast phase mayoriginate from clearance of adducts from otherproteins or chemically modified albumin. In fact,the latter was observed in the experiment using ivinjection of the radioactive conjugate betweenHHPA and serum albumin in rats. In this experi-ment, a fast phase was observed although purealbumin was used in the synthesis of the conju-gate. The levels of adducts found in plasma fromrats and guinea pigs following the exposure toHHPA was an order of magnitude lower thanthose found in workers exposed to similar airlevels (Jonsson et al., 1996). The relatively lowlevel of adducts in the guinea pig plasma seemedreasonable since the animals were only exposedfor a short period, whereas the workers wererepeatedly exposed. On the other hand, the levelsof free HHP acid in guinea pig plasma werecomparable to those of exposed volunteers (Jon-sson and Skerfving, 1993).
It might be argued against the use of only asingle animal for each time period after exposurein the clearance experiments. However, the Bat-telle exposure chamber seemed to provide a uni-form exposure, as determined by both airsampling and biological monitoring using theHHP acid levels in plasma. The animals wereacclimated during 5 days in the exposure chamberbefore the experiments. Low stress levels probablycontributed to the low individual differences.
In conclusion, a selective binding of[3H2]HHPA-derived radioactivity, possiblyHHPA-protein adducts, was observed in the mu-cosa of the upper respiratory and alimentary tractand conjunctiva of rats and guinea pigs followinga single inhalation exposure to [3H2]HHPA. Theselective tissue localisation of potential HHPAadducts at these sites may have relevance to thesymptoms of the allergic workers, since theymainly develop symptoms and signs from the noseand conjunctiva and less often from the lowerrespiratory airways. However, it remains to beestablished which HHPA-protein conjugate that isthe main allergen in the process of the develop-ment of allergy.
Acknowledgements
The authors gratefully acknowledge the supportby Dr M. Dahlback, Astra Draco AB, Lund, inkindly providing us with the exposure equipment.We also gratefully acknowledge the help from DrD.L. Liu with the iv injections. Excellent technicalassistance was given by Ms A. Bostrom and MsM. Mattsson. This work was supported by grantsfrom the Swedish Medical Research Council, theSwedish Council for Work Life Research, theVardal Foundation, the European Union (Biomed1), and the Medical Faculty at Lund University.
References
Bond, J.A., Harkema, J.R., Russell, V.I., 1988. Regionaldistribution of xenobiotic metabolizing enzymes in therespiratory airways of dogs. Drug Metab. Disp. 16, 116–124.

C.H. Lindh et al. / Toxicology 134 (1999) 153–168 167
Brittebo, E.B., 1993. Metabolism of xenobiotics in the nasalolfactory mucosa: Implications for local toxicity. Phar-macol. Toxicol. 72, 50–52.
Cannon, W.C., Blanton, E.F., McDonald, K.E., 1983. Theflow past chamber: an improved nose-only exposure sys-tem for rodents. Am. Ind. Hyg. Assoc. J. 44, 923–928.
Day, B.W., Jin, R., Karol, M.H., 1996. In vivo and in vitroreactions of toluene diisocyanate isomers with guinea pighemoglobin. Chem. Res. Toxicol. 9, 568–573.
Jeppsson, J.O., Laurell, C.B., Franzen, B., 1979. Agarose gelelectrophoresis. Clin. Chem. 25, 629–638.
Jin, R., Day, B.W., Karol, M.H., 1993. Toluene diisocyanateprotein adducts in the brochoalveolar lavage of guineapigs exposed to vapors of the chemical. Chem. Res. Tox-icol. 6, 906–912.
Jonsson, B.A.G., Skerfving, S., 1993. Toxicokinetics and bi-ological monitoring in experimental exposure of humansto gaseous hexahydrophthalic anhydride. Scand. J. WorkEnviron. Health 19, 183–190.
Jonsson, B.A.G, Welinder, H., Skarping, G., 1991. Determi-nation of hexahydrophthalic anhydride in air using gaschromatography. J. Chromatogr. 558, 247–256.
Jonsson, B.A.G., Wishnok, J.S., Skipper, P.L., Stillwell,W.G., Tannenbaum, S.R., 1995. Lysine adducts betweenmethyltetrahydrophthalic anhydride and collagen inguinea pig lung. Toxicol. Appl. Pharmacol. 135, 156–162.
Jonsson, B.A.G., Lindh, C.H., Zhang, X.D., Welinder, H.,1996. Protein adducts of highly sensitising organic acidanhydrides (OAA) in man. Allergy Suppl. 31, 51–62.
Jonsson, B.A.G., Lindh, C.H., Welinder, H., 1997.Hemoglobin adducts and specific immunoglobulin G inhumans as biomarkers of exposure to hexahydrophthalicanhydride. Biomarkers 2, 239–246.
Karol, M.H., Jin, R., Lantz, R.C., 1997. Immunohistochemi-cal detection of toluene diisocyanate (TDI) adducts ofpulmonary tissues of guinea pigs following inhalation ex-posure. Inhal. Toxicol. 9, 63–83.
Kennedy, A.L., Brown, W.E., 1992. Isocyanates and lungdisease: experimental approaches to molecular mecha-nisms. Occup. Med. 7, 301–329.
Kennedy, A.L., Stock, M.F., Alarie, Y., Brown, W.E., 1989.Uptake and distribution of 14C during and following in-halation exposure to radioactive toluene diisocyanate.Toxicol. Appl. Pharmacol. 100, 280–292.
Kennedy, A.L., Singh, G., Alarie, Y., Brown, W.E., 1993.Autoradiographic analyses of guinea pig airway tissuesfollowing inhalation exposure to 14C-labelled methyl iso-cyanate. Fundam. Appl. Toxicol. 20, 57–67.
Kennedy, A.L., Wilson, T.R., Stock, M.F., Alarie, Y.,Brown, W.E., 1994. Distribution and reactivity of in-haled 14C-labelled toluene diisocyanate (TDI) in rats.Arch. Toxicol. 68, 434–443.
Kennedy, A.L., 1990. The in vivo reactivity of inhaled iso-cyanates: Comparative analysis of cellular and macro-
molecular targents. Ph.D. Thesis. Carnegie Mellon Uni-versity.
Kowalski, B., Brittebo, E.B., Brandt, I., 1985. Epithelialbinding of 1,2-dibromethane (EDB) in respiratory andupper alimentary tracts. Cancer Res. 45, 2616–2645.
Lindh, C.H., Jonsson, B.A.G., 1997. Determination of hex-ahydrophthalic acid and methylhexahydrophthalic acid inplasma after derivatisation with pentafluorobenzyl bro-mide using gas chromatography and mass spectrometricdetection. J. Chromatogr. 691, 331–339.
Lindh, C.H., Jonsson, B.A.G., 1998a. Quantification methodof human hemoglobin adducts from hexahydrophthalicanhydride and methylhexahydrophthalic anhydride. J.Chromatogr. 710, 81–90.
Lindh, C.H., Jonsson, B.A.G., 1998b. Human hemoglobinadducts following exposure to hexahydrophthalic anhy-dride and methylhexahydrophthalic anhydride. Toxicol.Appl. Pharmacol. 153, 152–160.
Nielsen, J., Welinder, H., Horstmann, V., Skerfving, S.,1992. Allergy to methyltetrahydrophthalic anhydride inepoxy resin workers. Br. J. Ind. Med. 49, 769–775.
Schreiber, G., Urban, J., Zahringer, J., Reutter, W., Frosch,U., 1971. The secretion of serum protein and the synthe-sis of albumin and total protein in regenerating rat liver.J. Biol. Chem. 246, 4531–4538.
Schutze, D., Sepai, O., Lewalter, J., Miksche, L., Henschler,D., Sabbioni, G., 1995. Biomonitoring of workers ex-posed to 4,4%-methylenedianiline or 4,4%-methyl-enediphenyl diisocyanate. Carcinogenesis 16, 573–582.
Sepai, O., Henschler, D., Sabbioni, G., 1995a. Albumin ad-ducts, hemoglobin adducts and urinary metabolites inworkers exposed to 4,4%-methylenediphenyl diisocyanate.Carcinogenesis 16, 2583–2587.
Sepai, O., Schutze, D., Heinrich, U., Hoymann, H.G., Hen-schler, D., Sabbioni, G., 1995b. Hemoglobin adducts andurine metabolites of 4,4%-methylenedianiline after 4,4%-methylenediphenyl diisocyanate exposure to rats. Chem.-Biol. Interact. 97, 185–198.
Ullberg, S., Larsson, B., Tjalve, H., 1982. Autoradiography.In: Glenn, H.J. (Ed.), Biological Applications of Radio-tracers. CRC Press, Boca Raton, FL, pp. 56–108.
Venables, K.M., 1989. Low molecular weight chemicals, hy-persensitivity, and direct toxicity: the acid anhydrides.Br. J. Ind. Med. 46, 222–232.
Welinder, H., Nielsen, J., 1991. Immunologic tests of specificantibodies to organic acid anhydrides. Allergy 46, 601–609.
Welinder, H.E., Jonsson, B.A.G., Nielsen, J.E., Ottosson,H.E., Gustavsson, C.A., 1994. Exposure–response rela-tionships in the formation of specific antibodies to hex-ahydrophthalic anhydride in exposed workers. Scand. J.Work Environ. Health 20, 459–465.
Welinder, H., Zhang, X.D., Gustavsson, C., Bjork, B., Sker-fving, S., 1995. Structure–activity relationships of or-ganic acid anhydrides as antigens in an animal model.Toxicology 103, 127–136.

C.H. Lindh et al. / Toxicology 134 (1999) 153–168168
Yokota, K., Johyama, Y., Yamaguchi, K., Fujiki, Y.,Takeshita, T., Morimoto, K., 1997. Specific antibodiesagainst methyltetrahydrophthalic anhydride and risk fac-tors for sensitization in occupationally exposed subjects.Scand. J. Work Environ. Health 23, 214–220.
Zhang, X.D., Welinder, H., Jonsson, B.A.G., Skerfving,S., 1998. Antibody responses in rats after im-munization with organic acid anhydrides. A model ofpredictive testing. Scand. J. Work Environ. Health 24,220–227.
..




















