
Machines Outperform Laypersons in Recognizing Emotions Elicited by Autobiographical Recollection
Upload
independentCategory
view
0download
0
This article was originally published in a journal published byElsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, fornon-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specificcolleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including withoutlimitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’swebsite or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Autho
r's
pers
onal
co
py
Neuropsychologia 45 (2007) 2467–2479
Bilateral damage to the mammillo-thalamic tract impairs recollection butnot familiarity in the recognition process: A single case investigation
G.A. Carlesimo a,b,∗, L. Serra b, L. Fadda a,b, A. Cherubini c, M. Bozzali c, C. Caltagirone a,b
a Neurology Clinic, Tor Vergata University, Rome, Italyb Unit of Clinical and Behavioural Neurology, IRCCS, Rome, Italy
c Neuroimaging Laboratory, Santa Lucia Foundation, IRCCS, Rome, Italy
Received 24 October 2006; received in revised form 26 March 2007; accepted 26 March 2007Available online 8 April 2007
Abstract
Focal damage confined to the hippocampus may result in recognition deficits characterized by a dissociation between impaired recollection andpreserved familiarity. Here, we report a single case of an amnesic patient with bilateral damage to the anterior part of the thalamus, who presentedwith a neuropsychological profile suggesting such a dissociation. We hypothesized that this focal damage involved the so-called Delay and Brion’scircuit, which has been theorized to subserve episodic memory processes, but at a different anatomical level than in patients with hippocampallesions. Using two independent experimental paradigms (remember/know and confidence receiver operating characteristics [ROC]) and recruitinga sex- and age-matched group of healthy controls, we demonstrated that this patient’s recognition deficits were due to a selective impairment ofrecollection with a normal familiarity process. The patient underwent an ad hoc brain MRI study, and a quantitative analysis of his MR images wasperformed. Tissue damage extended bilaterally to the mammillo-thalamic tract, with complete preservation of the medio-dorsal thalamic nuclei.Our findings support the idea that the same functional specialization hypothesized for the different sub-regions of the mesial temporal lobe mightalso extend to the thalamus. This case will be discussed in light of its implications in support of recent theories, which regard recollection andfamiliarity as independent processes associated with different neural circuits.© 2007 Elsevier Ltd. All rights reserved.
Keywords: Memory; Recognition; Recollection; Familiarity; Amnesia; Thalamus
1. Introduction
Focal lesions that involve the thalami (especially those ofvascular origin) have often been recognized as a potential causeof memory impairment in humans (e.g., Castaigne et al., 1981;Mori, Yamadori, & Mitani, 1986; von Cramon, Hebel, & Schuri,1985). Van der Werf, Jolles, Witter, & Uylings (2003) and Vander Werf, Witter, Uylings, & Jolles (2000) recently empha-sized the role of these lesions in determining a specific patternof memory impairment in a remarkable number of patients.Such a pattern is hardly distinguishable from that observedin the amnesic syndrome, which typically occurs in patientswith damage involving the mesial temporal lobe (MTL) (Squire,2004). These patients typically present with poor performance
∗ Corresponding author at: Fondazione IRCCS S. Lucia, V. Ardeatina, 306,00179 Rome, Italy. Tel.: +39 06 51501517; fax: +39 06 51501584.
E-mail address: [email protected] (G.A. Carlesimo).
on episodic memory tests when either recall or recognition tasksare employed. By contrast, they usually obtain normal scores ontests investigating short-term, semantic and procedural memory,or general intellectual functioning measures (i.e., Gentilini, DeRenzi, & Crisi, 1987; Graff-Radford, Tranel, Van Hoesen, &Brandt, 1990).
It is generally accepted that the MTL-like amnesic syndrome,which occurs in patients with thalamic lesions, is due to a dis-connection between MTL structures and nuclei of the anteriorpart of the thalami. Consistently, pathological (Castaigne etal., 1981) and neuroradiological (Gentilini et al., 1987; Graff-Radford et al., 1990; Van der Werf et al., 2003) reports suggestthat the mammillo-thalamic tract (MTT) is critically involved inthe amnesic syndrome (see Vann & Aggleton, 2004 for a review).The MTT is part of the so-called Delay and Brion’s memorycircuit (1969), which connects the hippocampal formation of theMTL to the mammillary bodies through the subiculum-fimbria-fornix complex. The MTT, in turn, connects the medial andlateral nuclei of the mammillary bodies to the anterior thalamic
0028-3932/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.doi:10.1016/j.neuropsychologia.2007.03.025

Autho
r's
pers
onal
co
py
2468 G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479
nuclei, which project to the limbic cortical areas (anterior andposterior cingulate cortex and retrosplenial cortex). Graff-Radford et al. (1990) and Parkin, Rees, Hunkin and Rose (1994)pointed out the existence of an additional MTL-thalamic con-nection, which conveys fibers from the perirhinal cortex (and theadjacent amygdala) to the medio-dorsal (MD) nucleus of the tha-lamus. The intra-thalamic portion of this fiber tract (known as theventroamygdalofugal pathway) is part of the inferior thalamicpeduncle, which terminates in the MD nucleus of the thalamus.
The fact that two distinct anatomical pathways, the first orig-inating in the hippocampus proper, the other in the perirhinalcortex in the parahippocampal gyrus, connect the MTL to thethalamus raises the question as to whether the functional spe-cialization hypothesized for the different subregions of the MTLalso extends to the thalamus. Indeed, it has been claimed thatsome structures in the MTL might be selectively involved in theunaided recall of recently stored episodic information, whileother structures might be preferentially involved in recogni-tion processes. According to the two-process model of episodicmemory (Mandler, 1980; Yonelinas, 2002), several authors haveprovided evidence that some neocortical areas in the parahip-pocampal gyrus (mainly the perirhinal cortex) are implicatedin familiarity based processes. These cortical regions receiveinputs from projections arising from virtually all of the uni-and polymodal sensory areas of the brain and contribute, atleast partially, to the episodic recognition of individual (non-associative) information. In contrast, the hippocampus (CornusAmmonis, dentate gyrus and subiculum) mainly receives cor-tical inputs from the parahippocampal gyrus and projects tomultiple cortical and subcortical brain regions. Therefore, itis more likely that the hippocampus supports the so-calledrecollection process which underlies the retrieval of associa-tive information (e.g., the temporo-spatial context of a specificepisode). Thus, since unaided recall would only be sustainedby recollection processes, while both familiarity and recollec-tion processes participate in item recognition, patients withpure hippocampal damage (sparing parahippocampal cortices)should be characterized by a more severe deficit when freerecall rather then recognition testing procedures are used. Bycontrast, patients with damage involving both the hippocam-pus and the parahippocampus should suffer from a comparabledeficit of recall and recognition. Neuropsychological evidencesconsistent with this prediction have been provided. In a meta-analysis of the available literature, Aggleton and Shaw (1996)reported that amnesic patients with damage well confined tothe hippocampus, or involving its direct subcortical projections(fornix and mammillary bodies), obtained near normal scoreson Warrington’s two-choice word- and face-recognition tests(Warrington, 1984). By contrast, a group of patients with braindamage extending beyond this so called extended hippocam-pal system performed poorly on recognition tests. There arealso more recent detailed descriptions of individuals with purehippocampal damage, who presented with severely impairedrecall but relatively normal recognition (Aggleton et al., 2005;Bastin et al., 2004; Holdstock, Mayes, Gong, Roberts, & Kapur,2005; Mayes et al., 2004). Turriziani, Fadda, Caltagirone, &Carlesimo (2004) also described a group of patients with iso-
lated damage to the hippocampus, who scored normally on arecognition test for non-associative visual information (i.e., dis-criminating between studied and unstudied faces of unknownpeople), but failed on recognition tests that required the rec-ollection of associative information (i.e., pairing a previouslystudied face with some relevant information such as its occupa-tion). Finally, recent single case studies (Aggleton et al., 2005;Bastin et al., 2004; Holdstock et al., 2002; Mayes et al., 2004)showed poor recollection but normal familiarity in patients withpure hippocampal damage. It should be noted, however, thatthe available evidence is not fully consistent with the claim ofhippocampal specialization for recollection processes. Indeed,Squire and coworkers (Wais, Wixted, Hopkins, & Squire, 2006;Wixted & Squire, 2004) failed to find a sparing of recognitionand, in particular, of the familiarity component of recognitionin amnesic patients with neuroradiological evidence of damageconfined to the hippocampal formation.
Based on the anatomical considerations discussed above, thethalamus likely represents an extension of the functional spe-cialization already hypothesized for the MTL. According tothis theory (Aggleton & Brown, 1999), damage confined to theMTT and the anterior nuclei of the thalamus should result inmore severe and selective involvement of memory processesbased on recollection. Conversely, damage encroaching uponthe MD nucleus or its MTL afferents should produce moresevere impairment of familiarity-based recognition. Neuropsy-chological evidence in this regard is sparse and controversial.Hanley et al. (2001) described a patient with a left thalamiclesion that selectively involved the anterior thalamic nuclei withpreservation of the MD nucleus. This patient was character-ized by a striking dissociation on episodic memory tests, withsevere impairment of recall and complete preservation of recog-nition. However, when the relative contributions of recollectionand familiarity to recognition performances were investigated,both components were normal. By contrast, a patient recentlyreported by Kishiyama et al. (2005), who had a bilateral tha-lamic lesion sectioning the MTT and only partially involvingthe right MD nucleus, showed impairment of both recollection-and familiarity-based processes. Two studies also investigatedpatients with focal lesions encroaching upon the MD nuclei inwhich the MTT and anterior nuclei were unaffected (Edelstyn,Ellis, Jenkinson, & Sawyer, 2002; Zoppelt, Koch, Schwarz, &Daum, 2003). In both cases, analysis of recognition perfor-mance revealed a more severe deficit of recollection- than offamiliarity-based processes of episodic memory.
Here we report the case of an amnesic patient with bilateralthalamic damage. His neuropsychological profile (see Section2.1) documented partial sparing of recognition abilities forindividual items. This observation prompted additional inves-tigation to better characterize his peculiar recognition deficits.The patient was then enrolled in the present study together with agroup of age- and sex-matched healthy volunteers, who served ascontrols. The protocol included a neuroradiological assessmentand two different experiments (experiments 1 and 2). The aimof this study was to clarify whether or not the observed patternof memory impairment represented a dissociation between pre-served familiarity and impaired recollection, and to investigate

Autho
r's
pers
onal
co
py
G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479 2469
its relationship to the specific anatomical structures involved dueto tissue damage. The results will be discussed in terms of func-tional specialization of the thalamus as an extension of MTLspecialization.
2. Case report
G.P., a 38-year-old, right-handed man, who was employedas a lawyer in an important public company, suffered an acuteepisode of unconsciousness in February 2001. He was admittedto the hospital and regained consciousness a few hours later.He presented with severe anterograde and retrograde amne-sia. A neuropsychological examination performed several dayslater revealed a deficit of anterograde long-term memory withsubstantial preservation of his remaining cognitive functions.An MRI brain scan showed the presence of two well-definedischemic lesions involving the anterior-medial parts of the tha-lami. This patient came under our observation in July 2004,complaining of severe impairment in his day-to-day retentionof new information such as learning new routes, rememberingshort lists of items and appointments. Conversely, he seemedto have well-preserved memories of his pre-morbid life andbasic cultural knowledge. His neurological examination wasunremarkable.
2.1. Neuropsychological examination
A preliminary neuropsychological investigation was per-formed to assess the patient’s performance on tests of generalintelligence, executive functions, visual-spatial abilities, short-term memory and declarative episodic memory. An extensivebattery of tests was employed to obtain a separate evaluationof free recall and recognition abilities. For all tests adminis-tered, with the exception of recognition tests, the patient’s scoreswere first corrected for age and education and then compared tonormal ranges, according to published norms. With respect torecognition tests (for which normative data on an Italian sampleof healthy subjects were not available), the patient’s perfor-mance scores were compared directly with those obtained fromseven healthy subjects (matched for age and level of education)using Crawford and Garthwaite’s procedure (2002). Committeeapproval and written informed consent were obtained from boththe patient and the controls before the study began.
2.1.1. General intelligence, executive functions andvisual-spatial abilities
The patient’s performance on tests of general intelligence,executive functions and visual-spatial abilities are summarizedin Table 1. His WAIS full-scale IQ (110) and his performanceon Raven’s Progressive Matrices were in the medium range.However, a striking dissociation was observed between highperformance level on WAIS verbal subtests (VIQ = 133) andlow performance level on nonverbal subtests (PIQ = 76). G.P.’sscaled scores on the constructive subtests of the WAIS (BlockDesign and Object Assembly) were particularly poor. On alltests of executive functioning (Card Sorting test, Word Fluency,Trail Making test), G.P.’s scores fell in the high range of the
Table 1G.P.’s performance scores on tests of general intelligence, executive functionsand visuo-perceptual abilities
Test G.P. Maximumscore
General intelligenceWAIS-R (Wechsler, 1981)
Information 12 19Digit span 13 19Vocabulary 16 19Arithmetic 15 19Comprehension 19 19Similarities 13 19Picture completion 9 19Picture arrangement 10 19Block design 4 19Object assembly 2 19Digit symbol 9 19Verbal IQ 133Performance IQ 76Full-scale IQ 110
Raven’s colored matrices (Carlesimo,Caltagirone, & Gainotti, 1996)
26.2 (>30‰) 36
Executive functionsModified Card Sorting Test (Nocentini, Di Vincenzo, Panella,
Pasqualetti, & Caltagirone, 2002)Criteria achieved 6 (=100‰) 6Perseverative Errors 2.6 (>50‰) 0
Phonological Verbal Fluency (Carlesimoet al., 1996)
47.3 (>50‰) 17.3
Trail making test (Giovagnoli et al., 2006)A 48 s (>40‰)B 79 s (>50‰)A − B 32 s (>50‰)
Visual-spatial abilitiesVisual object and space perception battery (Warrington & James, 1991)
Object perceptionIncomplete letters 19 (>50‰) 17Silhouettes 20 (>20‰) 16Object decision 11 (<5‰) 15Progressive Silhouettes 10 (>50‰) 6
Space perception:Dot counting 10 (=100‰) 8Position discrimination 18 (=10‰) 18Number locations 10 (=100‰) 7Cube analysis 10 (=100‰) 6
Copy of Drawings (Carlesimo et al., 1996) 7.7 (<10‰) 12Copy of Drawings with Landmarks
(Carlesimo et al., 1996)65.1 (=10‰) 70
Rey’s Figure Copy (Carlesimo et al.,2002)
25.0 (<10‰) 36
For the WAIS subtests, scaled scores are reported. For the remaining tests, withthe exception of the VOSP, scores adjusted for age, education and gender arereported according to published normative data. References for the normativedata and, when available, percentile scores used are also reported.
normative population. He obtained normal scores on all subtestsof the Visual Object and Space Perception battery, except forthe Object Decision subtest, on which he performed particularlypoorly. Moreover, the patient’s performance on copying drawingtests fell in the lower part of the normal range, even though itwas not clearly pathological.

Autho
r's
pers
onal
co
py
2470 G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479
Table 2G.P.’s performance scores (adjusted for age, education and gender according topublished normative data) on tests of short-term memory and episodic recall
Test G.P. Maximumscore
Short-term memoryDigit span (Orsini et al., 1987) 7.2 (>50‰) 9Corsi span (Orsini et al., 1987) 6.2 (>50‰) 9Immediate Visual Memory (Carlesimo et
al., 1996)18.8 (>25‰) 22
Verbal recall15-word learning task (Carlesimo et al., 1996)
Immediate recall 30.4 (<10‰) 7515 min delayed recall 0 (<1‰) 15
Prose recall (Mauri et al., 1997)Immediate recall 6 (>25‰) 820 min delayed recall 2.3 (<5‰) 8
Visual-spatial recallRey’s Figure (Carlesimo et al., 2002)
Immediate reproduction 4.9 (<5‰) 3615 min delayed reproduction 1.7 (<5‰) 36
Supraspan spatial sequence learning(Spinnler & Tognoni, 1987)
4.2 (<5‰) 30.8
References for the normative data and percentile scores are also reported.
2.1.2. Short-term memoryAs expected, G.P. performed normally on short-term mem-
ory tests (Table 2). He obtained scores within the normal rangeon both digit and Corsi block span tests and on a 22-trial visualmemory task. In each trial of the latter, an abstract figure waspresented, and immediately afterward the patient was adminis-tered a recognition test and asked to identify the same figure ina four-alternative array.
2.1.3. Episodic memory: free recallThe patient’s performance on free recall tests is reported in
Table 2. His recall of verbal material was tested using a 15-wordand a prose recall test. The first test consists of five consecu-tive immediate trials, during which the examiner orally presentsa list of 15 semantically unrelated words. Immediately after-wards, the subject has to recall as many words as possible. Thistest also involves delayed recall of the same items, performed15 min after immediate recall of the last trial. In the Prose recalltest, the subject is administered a short story he has to recallimmediately after oral presentation and then 20 min later. G.P.’sscores were in the middle and lower normal range for immediaterecall of the short story and word list, respectively. However,his scores were below normal in the delayed recall of bothtests.
Recall of visual-spatial information was evaluated using theimmediate and 20 min delayed reproduction of Rey’s Figure Band the Supraspan Spatial Sequence Learning task. In the lattertask, the subject is required to reproduce an 8-block sequenceof the Corsi test immediately after presentation. If the subjectfails to reproduce the sequence correctly, the examiner repeatsthe same sequence until a learning criterion is reached (threesubsequent correct reproductions). G.P. performed poorly onboth immediate and delayed recall trials of Rey’s Figure and
did not learn the spatial sequence even after the examiner re-presented it 18 times.
2.1.4. Episodic memory: recognitionRecognition abilities were investigated using several tests
that show differences in terms of material administered (ver-bal, visual-object and visual-spatial) or paradigm employed. Inparticular, in view of the suggestion that recollection processesmake a different contribution to recognition decisions depend-ing on the kind of recognition paradigm used (i.e., larger foryes/no than for forced-choice tests) (MacMillan & Creelman,1991; but see Khoe, Kroll, Yonelinas, Dobbins, & Knight, 2000for controversial data on this issue), G.P. and the normal controlswere given yes/no, two-choice and multiple-choice recognitionmemory tests.
The 15-word recognition test investigates the yes/no recogni-tion of a word list which has been previously tested by means ofa free recall procedure. The test, which was given 15 min afterthe end of the delayed free recall test, consists of an oral presen-tation of the same list of 15 words randomly intermixed with 30new words. Subjects are requested to perform a yes/no recogni-tion test, discriminating between previously studied words andnew words.
In the two-choice Short Recognition memory test for words,taken from Warrington’s CAMDEN battery, subjects are visuallypresented with 25 words printed individually on 10 cm × 5 cmblank cards. Immediately after the study phase, 25 pairs ofwords, each consisting of a previously studied and a new word,are visually presented. Subjects are requested to decide whichof the two words was presented in the study phase. The sametesting paradigm is used in the Short Recognition memory testfor faces. In this case, black/white photos of unknown faces areused.
The multiple-choice Pictorial and Topographical Recogni-tion tests are also subtests of the CAMDEN battery. In the studyphase of both tests, 30 colored photos (of street scenes, parks andbuildings) are presented individually. During the test phase, sub-jects are presented with 30 recognition items. Each one consistsof a previously presented photo and two new ones represent-ing similar environments. The task is to identify the picturepresented in the study phase.
The patient and the normal controls were also administeredfour additional tests to assess their ability to recognize individualitems and between-item associations. In each test, the adminis-tration modalities were essentially the same, but the nature ofthe memorandum changed across tests: black and white photosof unknown faces, black and white photos of unknown build-ings, line drawings of objects and concrete words. In each test,the study phase consisted of a visual presentation of 30 pairsof items (e.g., two faces, two buildings, etc.). The patient wasinstructed to try to remember them for a successive memory task.The testing phase, which immediately followed the study phase,was articulated in two steps. The first step included a multiple-choice recognition test for individual items. The subjects wererequested to identify the item they had already seen in the studyphase, which was presented visually with three new items. Thesecond step consisted of a between-item association recognition

Autho
r's
pers
onal
co
py
G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479 2471
test. All items used as targets in step 1 of the recognition testwere presented again with the item associated with them duringthe study phase and three additional items taken from the pre-viously presented ones. The subjects were requested to selectthe item associated with the target from among other previouslystudied items. By requiring the subjects to select the associateditem from the other previously studied items, we were confidentthat their decision would not be based on a single-item familiar-ity judgment (discriminating a previously seen item from threeunseen ones) but would necessarily involve remembering thebetween-item association.
The patient’s and the healthy subjects’ performances on therecognition memory tests are reported in Table 3. G.P. scorednot dissimilar from the normal controls on the two-choice ShortRecognition memory tests for words and faces (even though aceiling effect in the performance of both G.P. and healthy con-trols could have prevented a significant difference to emerge)but he performed significantly poorer than normal controls onthe other tests. In particular, on the yes/no recognition test forthe 15-word list, he made considerably less hit rates than normalcontrols, but no false alarms. As a matter of fact, when perfor-mance on this test was scored according to the signal detectiontheory (Kamman, 1970), G.P.’s C score was in the normal con-trols’ range (indicative of normal ability to reject unstudieditems), but his memory discrimination index (d′) was signifi-cantly lower than that of the controls. In the multiple-choice
recognition tests (for both individual items and between-item associations), G.P. performed consistently worse thanNCs.
In summary, G.P. presented with a neuropsychological profiletypical of an amnesic syndrome. Indeed, impaired declarativeepisodic memory was documented by poor performance ontests of delayed free recall of verbal material and immediateand delayed recall of visual spatial information. With regardto recognition abilities, the patient’s scores on two-choice tests(for both words and faces) were normal, but his performanceson yes/no and multiple-choice tests were poor. Conversely, heshowed normal short-term memory and semantic memory abil-ities (as revealed by high scaled scores on the WAIS verbalsubtests). General intelligence, executive functions and visual-spatial perceptual abilities were also normal. G.P.’s scores werein the lower limits of the normal range or clearly pathologicalonly on tests requiring visuo-constructive abilities.
G.P.’s partial sparing of recognition abilities might be due toa normal contribution of familiarity processes to his recognitionperformances. Indeed, it has been suggested that while preservedfamiliarity may result in normal performance on two-choicerecognition tests, familiarity alone is not sufficient to supportnormal recognition in more demanding yes/no or multiple-choice recognition tests. In a two-choice recognition procedure,subjects may base their decision on a comparison of familiaritystrengths for the studied and unstudied items. Conversely, in a
Table 3G.P.’s performance scores on episodic recognition tests
Test G.P. Controls (n = 7) t p Maximum score
Yes/no recognition15-word learning task (Carlesimo et al., 1996)
Hit rates 5 13.9 (2.3) 3.6 <0.01 15False alarms 0 0.3 (0.5) 0.6 =0.30 30C 2.9 2.2 (0.8) 0.6 =0.28 2.9d′ 2.4 5.1 (0.8) 3.2 =0.01 5.8
Two-choice recognitionCAMDEN battery (Warrington, 1996)
Words 23 24.1 (0.7) 1.55 =0.10 25Faces 25 24.9 (0.4) 0.01 0.50 25
Multiple-choice recognitionCAMDEN battery (Warrington, 1996)
Pictorial 27 30.0 (0.0) 280.6 <0.001 30Topographical 17 28.9 (1.9) 6.0 <0.001 30
Single-associated item recognition (Carlesimo, Fadda, Turriziani, Tomaiuolo, & Caltagirone, 2001)Line drawings
Single 21 29.3 (1.5) 5.2 <0.001 30Associated 10 27.1 (3.1) 5.2 <0.001 30
WordsSingle 12 24.4 (2.6) 4.5 =0.002 30Associated 8 25.4 (4.8) 3.4 =0.007 30
FacesSingle 19 27.3 (1.7) 4.6 =0.002 30Associated 9 14.6 (3.8) 1.4 =0.11 30
BuildingsSingle 19 27.6 (2.6) 3.1 =0.01 30Associated 9 12.9 (6.5) 0.6 =0.30 30
For each test, mean (S.D.) scores obtained from healthy controls are reported, with t statistics according to Crawford and Garthwaite’s procedure (2002).

Autho
r's
pers
onal
co
py
2472 G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479
yes/no recognition paradigm subjects have to decide whetheror not familiarity strength exceeds an internal criterion of old-ness for each individually presented item. Depending on howstringent the established criterion is and in the absence of a nor-mal contribution of recollection to the recognition judgment,subjects may classify a high number of unstudied items as old(false alarms) and/or reject a high number of studied items asnew (missing). On the other hand, in a multiple choice recog-nition paradigm, the strategy of comparing familiarity strengthsof three or four alternatives presented simultaneously might bemuch too laborious and time consuming to permit correct recog-nition judgment. Indeed, in their meta-analysis, Aggleton andShaw (1996) considered the normal scores obtained by amnesicpatients on Warrington’s two-choice recognition tests for wordsand faces as evidence of normal contribution of familiarity torecognition.
2.2. Neuroradiological investigation
A brain MRI scan was obtained at 3.0 T (Siemens, MedicalSolutions, Erlangen, Germany). In a single session, the follow-ing pulse sequences were collected: (a) dual-echo spin echo(DE-SE) (TR = 5000 ms, TE = 20/100 ms); (b) 3D T1-weightedturbo-flash magnetization-prepared rapid-acquisition gradientecho (MPRAGE) (TR = 7.92 ms, TE = 2.4 ms, TI = 210 ms, flipangle = 15◦). For the dual-echo sequence, 52 contiguous inter-leaved axial slices were acquired with 2 mm slice thickness, witha 192 × 256 matrix over a 256 mm × 256 mm field of view, cov-ering the whole brain. The MPRAGE sequence was acquiredin a single slab, with a sagittal orientation, a 224 × 256 matrixsize over a 256 mm × 256 mm field of view, with an effectiveslice thickness of 1 mm. MRI scans were first assessed by visual
examination. With the exception of bilateral involvement of thethalamus, there were no additional abnormalities on either DE-SE or T1 weighted scans. Axial dual-echo SE and T1 weightedslices, which show the bilateral involvement of the thalamus, arepresented in Fig. 1.
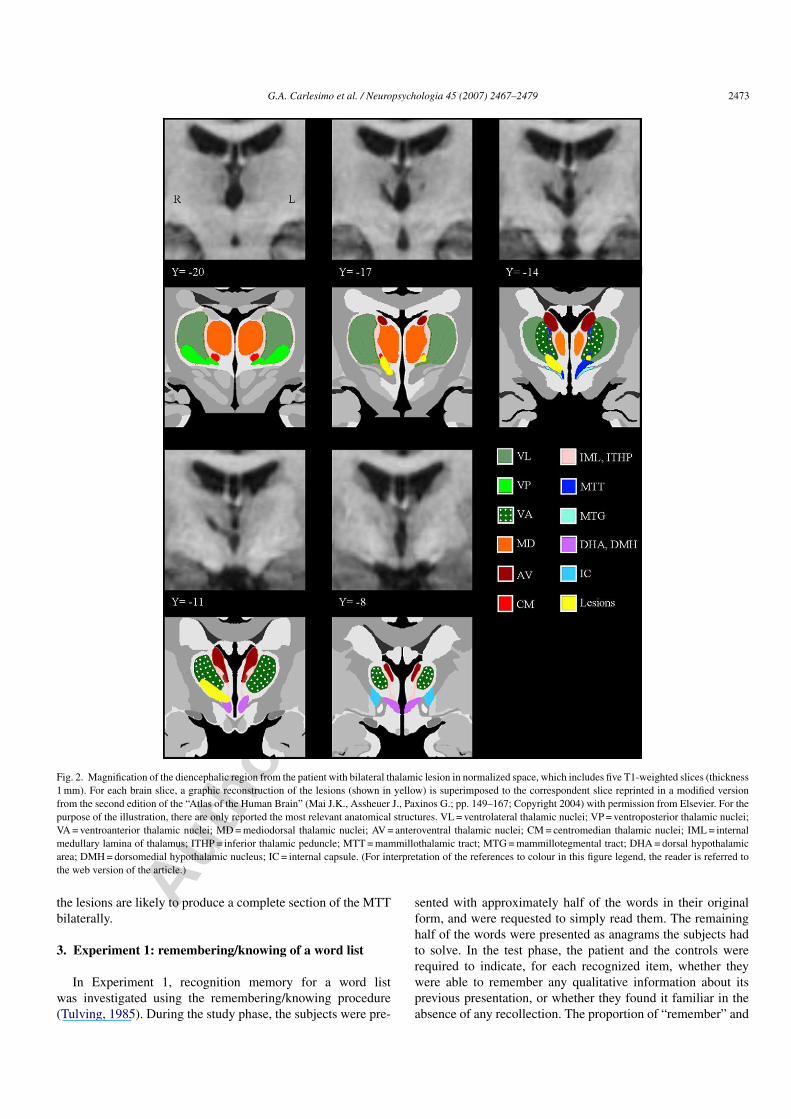
Proton density (PD) and T1 weighted images were thenco-registered and normalized to the stereotaxic space ofTalairach and Tournoux (1998) using a non linear algo-rithm (SPM2, Wellcome Dept. Cogn. Neurol., London;http://www.fil.ion.ucl.ac.uk/spm). The thalamic lesions weremanually outlined on PD and T1 weighted images using MRIcro(http://www.sph.sc.edu/comd/rorden/mricro.html), and regionsof interest (ROIs) were created for each of them. The lesion vol-ume was 316 and 217 ml on the right, and 57 and 22 ml on the leftside, considering the PD and the T1 weighted scans, respectively.As expected, estimation of the lesion load was more conserva-tive when performed on the T1 weighted than on the PD scan.Finally, an exact definition of the anatomical structures (greynuclei and fiber tracts) involved by tissue damage was madeby superimposing the ROIs obtained on the T1 weighted scanover the normalized maps derived from the Elsevier Atlas of thehuman brain (Mai, Assheuer, & Paxinos, 2004) (see Fig. 2). Theright thalamic lesion involved the MTT and, to a lesser extent,the mammillo-tegmental tract, the inferior thalamic peduncleand the internal medullary lamina. The ventral-anterior, ventral-lateral, ventral-medial and centromedian thalamic nuclei werealso damaged. The left thalamic lesion involved the MTT and theventral-anterior thalamic nucleus but it did not affect the inferiorthalamic peduncle. Interestingly, the MD nucleus was bilaterallyspared by tissue damage. Within the limitation in spatial reso-lution for such a small structure, MTT seems to be damaged ata smaller extent on the left than on the right side. Nevertheless,
Fig. 1. Four axial slices, deriving from proton density (A) and T1-weighted images (B), are presented here. They show the full extent of the patient’s tissue damage.The two ischemic lesions were selectively localized in the anterior part of the thalami, and no additional abnormality could be detected on the MRI scans. The right-sidelesion was more extensive, involving the MTT, the mammillo-tegmental tract, the inferior thalamic peduncle, the internal medullary lamina, the ventral-anterior,ventral-lateral and ventral-medial thalamic nuclei. The left side lesion involved the MTT and the ventral-anterior thalamic nucleus. The MD nucleus was bilaterallyspared (see also Fig. 2). (See text for further details. R = right; L = left.)
Autho
r's
pers
onal
co
py
G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479 2473
Fig. 2. Magnification of the diencephalic region from the patient with bilateral thalamic lesion in normalized space, which includes five T1-weighted slices (thickness1 mm). For each brain slice, a graphic reconstruction of the lesions (shown in yellow) is superimposed to the correspondent slice reprinted in a modified versionfrom the second edition of the “Atlas of the Human Brain” (Mai J.K., Assheuer J., Paxinos G.; pp. 149–167; Copyright 2004) with permission from Elsevier. For thepurpose of the illustration, there are only reported the most relevant anatomical structures. VL = ventrolateral thalamic nuclei; VP = ventroposterior thalamic nuclei;VA = ventroanterior thalamic nuclei; MD = mediodorsal thalamic nuclei; AV = anteroventral thalamic nuclei; CM = centromedian thalamic nuclei; IML = internalmedullary lamina of thalamus; ITHP = inferior thalamic peduncle; MTT = mammillothalamic tract; MTG = mammillotegmental tract; DHA = dorsal hypothalamicarea; DMH = dorsomedial hypothalamic nucleus; IC = internal capsule. (For interpretation of the references to colour in this figure legend, the reader is referred tothe web version of the article.)
the lesions are likely to produce a complete section of the MTTbilaterally.
3. Experiment 1: remembering/knowing of a word list
In Experiment 1, recognition memory for a word listwas investigated using the remembering/knowing procedure(Tulving, 1985). During the study phase, the subjects were pre-
sented with approximately half of the words in their originalform, and were requested to simply read them. The remaininghalf of the words were presented as anagrams the subjects hadto solve. In the test phase, the patient and the controls wererequired to indicate, for each recognized item, whether theywere able to remember any qualitative information about itsprevious presentation, or whether they found it familiar in theabsence of any recollection. The proportion of “remember” and

Autho
r's
pers
onal
co
py
2474 G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479
“know” responses was used to estimate the proportion of itemsaccepted on the basis of recollection and familiarity (Yonelinas,Kroll, Dobbins, Lazzara, & Knight, 1998). In agreement with agreat deal of previous literature on the generation effect (e.g.,Slamecka & Graf, 1978; Verfaellie & Treadwell, 1993), weexpected that recognition accuracy and, in particular, recollec-tion scores would be higher for words that had been anagrammedthan for words that had been simply read.
3.1. Subjects and procedure
During the study phase, the patient and the seven healthycontrols (all males, mean age 37.4 ± 3.4; education 16.4 ± 1.3)were presented with 50 five-letter words (frequency: 27.2 permillion; range: 1–83; Bortolini, Tagliavini, & Zampilli, 1971),half in their original form and half as anagrams of words. Ana-grams were generated by keeping the second and the fourth letter(underlined) of each word in its original position and randomlyreversing two of the remaining three letters (e.g., ertno = treno),thus obtaining anagrams with only one possible solution. Thesubjects were requested to simply read the words or solve theanagrams. Any time a subject read a word or correctly solved ananagram, the subsequent trial was presented. Any time an ana-gram was incorrectly solved, subjects were encouraged to tryagain for 1 min. When the time was up, they were provided withthe correct solution. In the test phase, the subjects were admin-istered a list of items that included all the words previouslypresented intermixed with 25 new words, and were required torespond “remember”, “know”, or “new” to each item. They weretold to respond “remember” if they could recollect the specificepisode during which the word had been previously presented.They were told to respond “know” if they were sure the item hadbeen presented during the study phase, but could not recollectthe specific episode during which it had been previously pre-sented. The subjects were told to respond “new” if they thoughtthe item had not been presented during the study phase.
3.2. Results
The proportion of “remember” and “know” responses madeby the patient and the controls are reported in Table 4.
The controls produced a significantly higher percentage of“remember” (t = 5.22; p = 0.002) and a similar rate of “know”responses (t = 0.11; p > 0.10) when words that had to be ana-grammed were compared to those that had to be simply read.By contrast, the patient showed only a mild increase of “remem-ber” and a remarkable increase of “know” responses whenhe compared the words to be read with those to be ana-
grammed. In a direct comparison between the patient and thecontrols (using Crawford and Garthwaite’s procedure (2002)),there was no significant difference for words that had beensimply read (t = 0.47 and 0.70 for “remember” and “know”responses, respectively; p > 0.10 in both cases). Conversely, thepatient showed a statistically significance trend toward pro-ducing less “remember”(t = 1.80; p = 0.06) and more “know”(t = 1.54, p = 0.09) responses for anagrammed words. Finally, inno case did G.P. produce a false alarm.
Since the subjects were instructed to respond “remember”when an item was recollected, the probability of a “remem-ber” response was used as an estimate of recollection (i.e.,R = ‘remember’). Since true recollection of an unstudied itemwas impossible, recollection was estimated using a thresholdmodel that assumes new items never exceed a memory thresh-old, so that new items may be accepted as old only on thebasis of a random guess. Thus, true recollection is measuredby subtracting the proportion of false “remember” responsesfrom the proportion of true “remember” responses and thendividing by the opportunity to observe a true “remember”response, R = (Rold − Rnew)/(1 − Rnew). Because the subjectswere instructed to respond “know” when an item was familiar inthe absence of recollection [i.e., ‘know’ = F (1 – R)], familiaritywas estimated as the probability of a “know” response given theitem was not recollected [i.e., F = “know”/(1 – R)]. Yonelinaset al. (1998) incorporated response bias into the estimates byusing a model in which familiarity is assumed to reflect a signal-detection process. In such a model, false alarms arise becausethe familiarity of some of the new items exceeds the partici-pants’ response criterion. Thus, familiarity is measured as thedifference between the average familiarity of old and new items(i.e., d′). In the present case, however, due to the complete lackof false alarms in G.P.’s performance, applying signal detectionmodels to the results is problematic, because d′ approaches infin-ity as false alarms approach 0. In this case, a more appropriateway of estimating familiarity is probably to look at the estimatesof familiarity for old and new items and then estimate overallfamiliarity as the difference between these values. Accordingly,F was calculated as (Kold/1 − Rold) − (Knew/1 − Rnew).
Estimates of recollection and familiarity in the patient andNCs are presented in Fig. 3. Consistently with the pattern of“remember” and “know” responses as a function of the studycondition, the normal subjects’ recollection scores were higherfor anagrammed than for read words (t = 5.37; p = 0.002), with acomparable familiarity score in the two cases (t = 0.69; p > 0.10).Again, the patient’s performance on words that had been readduring the study phase did not differ from that of the normal con-trols. For both “remember” and “know” responses, the difference
Table 4Experiment 1: proportion of ‘remember’ and ‘know’ responses produced by G.P. and healthy controls for words that had been simply read or anagrammed duringthe study phase, and for new words
Read items Anagram items New items
G.P. Controls G.P. Controls G.P. Controls
Remember response 0.07 0.22 (0.30) 0.16 0.64 (0.25) 0.00 0.05 (0.07)Know response 0.15 0.24 (0.12) 0.41 0.13 (0.17) 0.00 0.10 (0.10)

Autho
r's
pers
onal
co
py
G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479 2475
Fig. 3. Experiment 1: Estimates of recollection (left panel) and familiarity (right panel) derived from the recognition test for a word list using the remembering/knowprocedure.
between the patient’s score and the controls’ average score didnot approach statistical significance (t = 0.39 and 0.42, respec-tively; p > 0.10 in both cases). On the contrary, when wordswhose anagram had been solved during the study phase wereconsidered, the contribution of recollection to the recognitionperformance was significantly lower in the patient than in thecontrols (t = 1.87, p = 0.05). Finally, the patient’s contribution offamiliarity to recognition performance was even higher than thatof NCs. However, the statistical comparison did not approachsignificance (t = 1.20, p > 0.10).
3.3. Discussion
Consistently with a large body of experimental literature onnormal subjects (for a review, see Yonelinas, 2002), our healthycontrols produced higher recollection scores for words whoseanagram had to be solved than for words that simply had to beread during the study phase. Conversely, the same experimentalmanipulation did not significantly affect familiarity scores. In thepsychological literature, the advantage, in terms of recollection,for anagrammed compared to simply read words is commonlyreferred to as the so-called generation effect (e.g., Slamecka& Graf, 1978; Verfaellie & Treadwell, 1993). Such an effecthas been generally interpreted as a result of more distinctiveencoding operations. Solving an anagram enhances the numberof potential cues for retrieval more than simply reading a word.This is thought to be more beneficial for an associative kindof memory process (such as recollection) than for a memoryoperation, like the feeling of familiarity, which strictly refersto the individual item to be remembered (Gardiner, 1988; seeYonelinas, 2002).
G.P. showed a different pattern of performance when com-pared to normal controls. Indeed, even though his memory scoreswere consistently higher for words that had been solved thanfor words that had been read, the gain was much larger forthe “know” (+0.26) than for the “remember” (+0.09) responses.As a consequence, in the anagram condition he produced less“remember” and more “know” responses and he obtained a sig-nificantly lower recollection, but a higher familiarity, score thanhealthy controls. These results are consistent with our predic-tion. G.P.’s recognition performance was indeed characterized
by a deficient contribution of recollection processes. The factthat such a deficit emerged only for words that had been ana-grammed during the study phase can be explained by the factthat simply reading the words probably engaged very superfi-cial stimulus encoding, thus giving rise to very poor recollectionscores also in the group of healthy subjects.
The other finding of the present experiment, namely com-parable familiarity in G.P. and healthy controls for the wordsthat had been read or anagrammed, is quite consistent with thehypothesis of a sparing of familiarity processes in the recognitionperformance of this patient.
4. Experiment 2: receiver operating characteristic for aword and a face list
In Experiment 2, recognition memory for words and pre-viously unknown faces was examined using the confidencereceiver operating characteristic (ROC) procedure (Yonelinaset al., 1998). The control subjects and the patient underwenta study phase with lists of words and faces. Then, they wereadministered a recognition memory test. The task was not onlyto discriminate between studied and unstudied items, but also torate the confidence of any recognition response. Responses wereplotted on ROCs and a nonlinear regression method was used toderive parameter estimates of recollection and familiarity.
4.1. Subjects and procedure
In Experiment 2, the patient and 15 healthy subjects (allmales, mean age 35.1 ± 4.8; education 15.6 ± 1.5) were sub-mitted to two recognition memory tasks involving words andunknown faces, respectively. The former task employed 160five-letter words (frequency: 27.1 per million; range 2–331;Bortolini et al., 1971); the second task, 160 colored photos ofhuman faces (80 males, 80 females). In the study phase of theword recognition test, the participants were required to read 80words individually presented on a Personal Computer screenand to classify each word as pleasant or unpleasant. During thetest phase, immediately following the study phase, the subjectswere visually presented with a list containing all the studieditems intermixed with 80 new words. For each presented item,
Autho
r's
pers
onal
co
py
2476 G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479
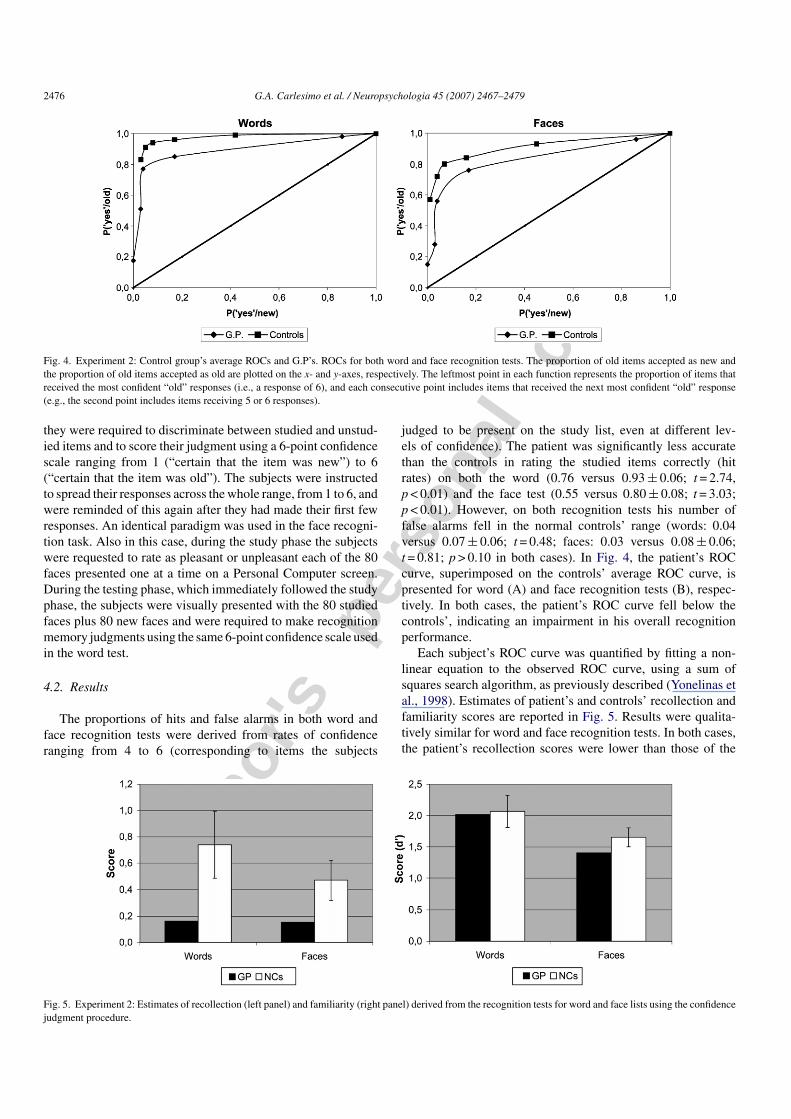
Fig. 4. Experiment 2: Control group’s average ROCs and G.P’s. ROCs for both word and face recognition tests. The proportion of old items accepted as new andthe proportion of old items accepted as old are plotted on the x- and y-axes, respectively. The leftmost point in each function represents the proportion of items thatreceived the most confident “old” responses (i.e., a response of 6), and each consecutive point includes items that received the next most confident “old” response(e.g., the second point includes items receiving 5 or 6 responses).
they were required to discriminate between studied and unstud-ied items and to score their judgment using a 6-point confidencescale ranging from 1 (“certain that the item was new”) to 6(“certain that the item was old”). The subjects were instructedto spread their responses across the whole range, from 1 to 6, andwere reminded of this again after they had made their first fewresponses. An identical paradigm was used in the face recogni-tion task. Also in this case, during the study phase the subjectswere requested to rate as pleasant or unpleasant each of the 80faces presented one at a time on a Personal Computer screen.During the testing phase, which immediately followed the studyphase, the subjects were visually presented with the 80 studiedfaces plus 80 new faces and were required to make recognitionmemory judgments using the same 6-point confidence scale usedin the word test.
4.2. Results
The proportions of hits and false alarms in both word andface recognition tests were derived from rates of confidenceranging from 4 to 6 (corresponding to items the subjects
judged to be present on the study list, even at different lev-els of confidence). The patient was significantly less accuratethan the controls in rating the studied items correctly (hitrates) on both the word (0.76 versus 0.93 ± 0.06; t = 2.74,p < 0.01) and the face test (0.55 versus 0.80 ± 0.08; t = 3.03;p < 0.01). However, on both recognition tests his number offalse alarms fell in the normal controls’ range (words: 0.04versus 0.07 ± 0.06; t = 0.48; faces: 0.03 versus 0.08 ± 0.06;t = 0.81; p > 0.10 in both cases). In Fig. 4, the patient’s ROCcurve, superimposed on the controls’ average ROC curve, ispresented for word (A) and face recognition tests (B), respec-tively. In both cases, the patient’s ROC curve fell below thecontrols’, indicating an impairment in his overall recognitionperformance.
Each subject’s ROC curve was quantified by fitting a non-linear equation to the observed ROC curve, using a sum ofsquares search algorithm, as previously described (Yonelinas etal., 1998). Estimates of patient’s and controls’ recollection andfamiliarity scores are reported in Fig. 5. Results were qualita-tively similar for word and face recognition tests. In both cases,the patient’s recollection scores were lower than those of the
Fig. 5. Experiment 2: Estimates of recollection (left panel) and familiarity (right panel) derived from the recognition tests for word and face lists using the confidencejudgment procedure.

Autho
r's
pers
onal
co
py
G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479 2477
controls (words: t = 2.67, p < 0.01; faces: t = 1.72; p = 0.05). Bycontrast, the contribution of familiarity to G.P.’s recognition per-formance did not differ from that of the controls (words: t = 0.48;faces: t = 0.39; p > 0.10 in both cases).
4.3. Discussion
The results of experiment 2 are consistent with those obtainedin experiment 1 and clearly indicate the influence of defectiverecollection on G.P.’s recognition memory performance. Indeed,for both word and face recognition the confidence level of hisrecognition decisions suggests that he had poor access to mem-ory of the study event in which the item had been previouslypresented. In agreement with experiment 1, the results of experi-ment 2 confirm that the contribution of familiarity to the patient’srecognition performance was similar to that observed in thenormal controls.
5. General discussion
Here, we report the case of a patient who developed anamnesic syndrome as a consequence of bilateral ischemic dam-age selectively involving the anterior part of the thalami. Hisneuropsychological profile was indistinguishable from that usu-ally observed in patients with MTL damage. Indeed, G.P.exhibited deficient anterograde episodic memory, which wasparticularly evident in free recall paradigms and following adelay interval between stimulus presentation and memory test,in the context of substantially preserved general intelligence,executive functions, linguistic abilities and short-term memory.Brain MRI demonstrated that the ischemic infarcts had lesionedthe MTT bilaterally, substantially sparing the MD nucleus. Thisis consistent with a large body of evidence based on both autop-tical series (Castaigne et al., 1981) and neuroradiological data(Gentilini et al., 1987; Graff-Radford et al., 1990; Van der Werf etal., 2003) documenting that an amnesic syndrome clinically andneuropsychologically indistinguishable from that observed inMTL amnesic patients is generally observed in thalamic patientsfollowing damage to the MTT.
What is difficult to account for in the results of G.P.’sgeneral neuropsychological examination is his asymmetricalperformance pattern on the WAIS. Indeed, his scores wereconsiderably lower on nonverbal than on the verbal subtests.Specifically, his performance was very poor on the visual-constructive subtests, consistent with his low scores on copyingdrawings. These additional findings could be explained in termsof deficits in planning abilities, which are usually related tofrontal lobe dysfunction. However, there was no detectable MRIabnormality affecting the frontal lobes, and the patient had highscores on a number of cognitive tests exploring frontal lobefunctions. In turn, the reduction in visuo-contructive abilitiescould be due to a deficit in high-level integration of visual-spatial perception and praxic abilities. This hypothesis seemsmuch more likely, and is supported by neuroimaging evidenceof more extensive damage in the right hemisphere, which isspecialized for high-level spatial integration (Vogel, Bowers, &Vogel, 2003).
What aroused our interest in this patient was the possibil-ity of extending the dissociation between impaired recollectionbut preserved familiarity components of recognition memory,which has already been observed in amnesic patients with purehippocampal damage, to a patient with an amnesic syndromeof diencephalic origin. This expectation was based on two linesof evidence. The first one was the partial sparing of recognitionmemory in this patient. In fact, a detailed analysis of recognitionmemory abilities documented that G.P.’s performance was notdifferent from that of matched healthy controls on two-choicerecognition tests for both verbal and visual stimuli, while hescored poorly on a yes/no test for a word list and on a vari-ety of multiple-choice recognition tests for verbal and visualitems. As previously noted, it has been suggested that recol-lection processes could play a less critical role in two-choicethan in yes/no and multiple-choice recognition paradigms, thusaccounting for selective sparing of performance in two-choicerecognition paradigms in a patient with hypothetical sparing offamiliarity but impaired recollection abilities. The second lineof evidence suggesting the possibility of extending a neuropsy-chological pattern previously described in patients with purehippocampal damage to a thalamic patient is that, as discussedabove, this patient’s ischemic lesions damaged the MTT bilat-erally, which, according to the anatomical model put forwardby Aggleton and Brown (1999), is part of the “recollection cir-cuit” connecting the hippocampus proper to the anterior thalamicnuclei. Instead, the MD nucleus, which participates in the “famil-iarity circuit” since it is connected to the perirhinal cortex viathe ventroamygdalofugal pathway, was substantially spared bythe parenchymal damage. The ventroamygdalofugal pathwaywas surely preserved on the left side, where the inferior thala-mic peduncle was intact. We presume that the same structurewas also preserved on the right side. Indeed, the right inferiorthalamic peduncle was clearly involved by the lesion only in itsposterior part. Conversely, it was relatively preserved in its ante-rior portion, where the ventroamygdalofugal pathway is betterrecognizable. Nevertheless, due to spatial resolution limitationsof our imaging data, we cannot exclude that the ventroamyg-dalofugal pathway might be, at least in a minimal part, involvedby the lesion on the right side. G.P.’s results in two independentexperimental paradigms, both intended to estimate the contribu-tion of recollection and familiarity to recognition performance,were substantially consistent with the hypothesis of the study.Indeed, on both the remembering/know (Tulving, 1985) and theconfidence receiver operating characteristics (Yonelinas et al.,1998) procedures, our patient demonstrated poor contributionof recollection to his own recognition performance. By contrast,the contribution of familiarity to performance was not differentfrom that of the normal controls on both the ROCs and R/Kparadigm.
There are some published reports on assessment of the dif-ferential impairment of recollection and familiarity in patientswith thalamic lesions (Edelstyn et al., 2002; Hanley et al., 2001;Kishiyama et al., 2005; Zoppelt et al., 2003). However, thesmall number of subjects studied and the heterogeneity of theirlesion localization make a comparison across studies particu-larly difficult. Further, the approaches used to characterize tissue

Autho
r's
pers
onal
co
py
2478 G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479
damage are also heterogeneous and often based on simple visualinspection. For example, Edelstyn et al. reached different con-clusions regarding the thalamic nuclei involved in a focal lesionin their patient QX by using two different neuroradiologicalassessments (Edelstyn et al., 2002; Edelstyn, Hunter, & Ellis,2006). Among previously reported thalamic patients, in whichthe contribution of recollection and familiarity to the recognitionperformance was investigated in detail, patient R.G.’s ischemicdamage (Kishiyama et al., 2005) was closest in location to thatof G.P. In fact, also in R.G. ischemic lesions encroached onthe MTT bilaterally. Compared to our patient, however, R.G.displayed a less clear dissociation between familiarity and rec-ollection. Indeed, he had substantial impairment of recollectionand a smaller, but consistent, deficit in familiarity. It should benoted, however, that according to the authors’ description, R.G.’sthalamic damage, it also involved the right MD nucleus, thuspossibly accounting for his mild familiarity impairment.
In conclusion, the present study shows that discrete damageconfined to some strategic structures outside the MTL can resultin an amnesic syndrome with peculiar characteristics. Our resultssupport the idea that recollection and familiarity are not onlybased on different MTL sub-regions, but involve a complex net-work of gray matter structures interconnected by specific whitematter tracts (Aggleton & Brown, 1999; Yonelinas, Otten, Shaw,& Rugg, 2005). In any case, further studies on larger populationsof patients are needed to produce more homogeneous results andto better clarify the specific role of the different brain structuresinvolved in episodic memory.
Acknowledgments
We would like to thank the Elsevier Academic press forthe permission to reproduce images from the Elsevier Atlas ofthe human brain (Mai J.K., Assheuer J., Paxinos G.; Copyright2004).
References
Aggleton, J. P., & Brown, M. W. (1999). Episodic memory, amnesia, and thehippocampal-anterior thalamic axis. Behavioural and Brain Sciences, 22,425–444.
Aggleton, J. P., & Shaw, C. (1996). Amnesia and recognition memory: a re-analysis of psychometric data. Neuropsychologia, 34, 51–62.
Aggleton, J. P., Vann, S. D., Denby, C., Dix, S., Mayes, A. R., Roberts, N., etal. (2005). Sparing of the familiarity component of recognition memory ina patient with hippocampal pathology. Neuropsychologia, 43, 1810–1823.
Bastin, C., Linden, M., Charnallet, A., Denby, C., Montaldi, D., Roberts, N., et al.(2004). Dissociation between recall and recognition memory performancein an amnesic patient with hippocampal damage following carbon monoxidepoisoning. Neurocase, 10, 330–344.
Bortolini, V., Tagliavini, C., & Zampilli, A. (1971). Lessico di frequenze dellalingua italiana contemporanea [Frequency of words in the contemporaryItalian language]. Milan, Italy: Garzanti.
Carlesimo, G. A., Buccione, I., Fadda, L., Graceffa, A., Mauri, M., Lo Russo,S., et al. (2002). Standardizzazione di due test di memoria per uso clinico:Breve racconto e Figura di Rey. Nuova Rivista di Neurologia, 12, 3–13.
Carlesimo, G. A., Caltagirone, C., & Gainotti, G. (1996). The Mental Deteriora-tion Battery: normative data, diagnostic reliability and qualitative analysesof cognitive impairment. European Neurology, 36, 378–384.
Carlesimo, G. A., Fadda, L., Turriziani, P., Tomaiuolo, F., & Caltagirone, C.(2001). Selective sparing of face learning in a global amnesic patient. Journalof Neurology Neurosurgery and Psychiatry, 71, 340–346.
Castaigne, P., Lhermitte, F., Buge, A., Escourolle, R., Hauw, J. J., & Lyon-Caen, O. (1981). Paramedian thalamic and midbrain infarct: clinical andneuropathological study. Annals of Neurology, 10, 127–148.
Crawford, J. R., & Garthwaite, P. H. (2002). Investigation of the single case inneuropsychology: confidence limits on the abnormality of test scores andtest score differences. Neuropsychologia, 40, 1196–1208.
Delay, J., & Brion, S. (1969). Le Syndrome de Korsakoff. Oaris: Masson.Edelstyn, N. M., Ellis, S. J., Jenkinson, P., & Sawyer, A. (2002). Contribution of
the left dorsomedial thalamus to recognition memory: a neuropsychologicalcase study. Neurocase, 8, 442–452.
Edelstyn, N. M., Hunter, B., & Ellis, S. J. (2006). Bilateral dorsolateral thalamiclesions disrupts conscious recollection. Neuropsychologia, 44, 931–938.
Gardiner, J. M. (1988). Functional aspects of recollective experience. Memoryand Cognition, 16, 309–313.
Gentilini, M., De Renzi, E., & Crisi, G. (1987). Bilateral paramedian thalamicartery infarcts: report of eight cases. Journal of Neurology, Neurosurgeryand Psychiatry, 50, 900–909.
Giovagnoli, A. R., Del Pesce, M., Mascheroni, S., Simoncelli, M., Laiacona, M.,& Capitani, E. (2006). Trail making test: normative values from 287 normaladult controls. Italian Journal of Neurological Sciences, 17, 305–309.
Graff-Radford, N. R., Tranel, D., Van Hoesen, G. W., & Brandt, J. P. (1990).Diencephalic amnesia. Brain, 113, 1–25.
Hanley, J. R., Davies, A. D., Downes, J. J., Roberts, J. N., Gong, Q. Y., &Mayes, A. R. (2001). Remembering and knowing in a patient with preservedrecognition and impaired recall. Neuropsychologia, 39, 1003–1010.
Holdstock, J. S., Mayes, A. R., Gong, Q. Y., Roberts, N., & Kapur, N. (2005).Item recognition is less impaired than recall and associative recognitionin a patient with selective hippocampal damage. Hippocampus, 15, 203–215.
Holdstock, J. S., Mayes, A. R., Roberts, N., Cezayirli, E., Isaac, C. L., O’Reilly,R. C., et al. (2002). Under what conditions is recognition spared relative torecall after selective hippocampal damage in humans? Hippocampus, 12,341–351.
Kamman, R. (1970). Workbook for General Psychology. In Signal DetectionTheory and Recognition Memory: Part I (Basic Calculation). Practice-Hall.
Khoe, W., Kroll, N. E., Yonelinas, A. P., Dobbins, I. G., & Knight, R. T. (2000).The contribution of recollection and familiarity to yes-no and forced-choicerecognition tests in healthy subjects and amnesics. Neuropsychologia, 38,1333–1341.
Kishiyama, M. M., Yonelinas, A. P., Kroll, N. E., Lazzara, M. M., Nolan, E. C.,Jones, E. G., et al. (2005). Bilateral thalamic lesions affect recollection- andfamiliarity-based recognition memory judgments. Cortex, 41, 778–788.
MacMillan, N. A., & Creelman, C. D. (1991). Detection theory: A user’s guide.New York, NY: Cambridge University Press.
Mai, J. K., Assheuer, J., & Paxinos, G. (2004). Atlas of the human brain. SanDiego: Academic Press.
Mandler, G. (1980). Recognizing: the judgement of previous occurrence. Psy-chological Review, 87, 252–271.
Mauri, M., Carlesimo, G. A., Graceffa, A. M. S., Loasses, A., Lo Russo, S.,Sinofriani, E., et al. (1997). Standardizzazione di due nuovi test di memoria:apprendimento di liste di parole correlate e non correlate semanticamente.Archivio di Psicologia, Neurologia e Psichiatria, 58, 621–645.
Mayes, A. R., Holdstock, J. S., Isaac, C. L., Montaldi, D., Grigor, J., Gummer, A.,et al. (2004). Associative recognition in a patient with selective hippocam-pal lesions and relatively normal item recognition. Hippocampus, 14, 763–784.
Mori, E., Yamadori, A., & Mitani, Y. (1986). Left thalamic infarction and dis-turbance of verbal memory: a clinicoanatomical study with a new method ofcomputed tomographic stereotaxic lesion localization. Annals of Neurology,20, 671–676.
Nocentini, U., Di Vincenzo, S., Panella, M., Pasqualetti, P., & Caltagirone, C.(2002). La valutazione delle funzioni esecutive nella pratica neuropsicolog-ica: dal Modified Card sorting Test al Modified Card Sorting Test-RomaVersion. Dati di standardizzazione. Nuova Rivista di Neurologia, 12,14–24.

Autho
r's
pers
onal
co
py
G.A. Carlesimo et al. / Neuropsychologia 45 (2007) 2467–2479 2479
Orsini, A., Grossi, D., Capitani, E., Laiacona, M., Papagno, C., & Vallar, G.(1987). Verbal and spatial immediate memory span. Normative data from1355 adults and 1112 children. The Italian Journal of Neurological Sciences,8, 539–548.
Parkin, A. J., Rees, J. E., Hunkin, N. M., & Rose, P. E. (1994). Impairmentof memory following discrete thalamic infarction. Neuropsychologia, 32,39–51.
Slamecka, N. J., & Graf, P. (1978). The generation effect: delineation of aphenomenon. Journal of Experimental Psychology: Human Learning andMemory, 4, 592–604.
Spinnler, H., & Tognoni, G. (1987). Standardizzazione e taratura italiana di testsneuropsicologici. Italian Journal of Neurological Science, 8, 35–38.
Squire, L. R. (2004). Memory systems of the brain: a brief history and currentperspective. Neurobiology of Learning and Memory, 82, 171–177.
Talairach, J., & Tournoux, P. (1998). Co-planar stereotaxic atlas of the humanbrain. New York: Thieme Medical Publishers.
Tulving, E. (1985). Memory and consciousness. Canadian Psychologist, 26,1–12.
Turriziani, P., Fadda, L., Caltagirone, C., & Carlesimo, G. A. (2004). Recog-nition memory for single items and for associations in amnesic patients.Neuropsychologia, 42, 426–433.
Van der Werf, Y. D., Jolles, J., Witter, M. P., & Uylings, H. B. (2003). Contri-butions of thalamic nuclei to declarative memory functioning. Cortex, 39,1047–1062.
Van der Werf, Y. D., Witter, M. P., Uylings, H. B., & Jolles, J. (2000). Neu-ropsychology of infarctions in the thalamus: a review. Neuropsychologia,38, 613–627.
Vann, S. D., & Aggleton, J. (2004). The mammillary bodies: two memorysystems in one? Nature Reviews, 5, 35–44.
Verfaellie, M., & Treadwell, J. R. (1993). Status of recognition memory inamnesia. Neuropsychology, 7, 5–13.
Vogel, J. J., Bowers, C. A., & Vogel, D. S. (2003). Cerebral lateralization ofspatial abilities: A meta-analysis. Brain and Cognition, 52, 197–204.
von Cramon, D. Y., Hebel, N., & Schuri, U. (1985). A contribution to theanatomical basis of thalamic amnesia. Brain, 108, 993–1008.
Wais, P. E., Wixted, J. T., Hopkins, R. O., & Squire, L. R. (2006). The hip-pocampus supports both the recollection and the familiarity components ofrecognition memory. Neuron, 49, 459–466.
Warrington, E. K. (1984). The recognition memory tests. Windsor, UK: NFER-Nelson.
Warrington, E. K. (1996). The Camden memory tests manual. Hove, East Sussex:Psychology Press.
Warrington, E. K., & James, M. (1991). The visual object and space perceptionbattery. Bury St. Edmunds: Thames Valley Test Company.
Wechsler, D. (1981). WAIS-R, Wechsler adult intelligence scale revised. NewYork: The Psychological Corporation.
Wixted, J. T., & Squire, L. R. (2004). Recall and recognition are equally impairedin patients with selective hippocampal damage. Cognitive and AffectiveBehavioural Neuroscience, 4, 58–66.
Yonelinas, A. P. (2002). The nature of recollection and familiarity: a review of30 years of research. Journal of Memory and Language, 46, 441–517.
Yonelinas, A. P., Kroll, N. E., Dobbins, I., Lazzara, M., & Knight, R. T.(1998). Recollection and familiarity deficits in amnesia: Convergence ofremember-know, process dissociation, and receiver operating characteristicdata. Neuropsychology, 12, 323–339.
Yonelinas, A. P., Otten, L. J., Shaw, K. N., & Rugg, M. D. (2005). Separating thebrain regions involved in recollection and familiarity in recognition memory.Journal of Neurosciences, 16, 3002–3008.
Zoppelt, D., Koch, B., Schwarz, M., & Daum, I. (2003). Involvement of themediodorsal thalamic nucleus in mediating recollection and familiarity.Neuropsychologia, 41, 1160–1170.
Copyright © 2022 FDOKUMEN




















