
Biancacci, Cecilia - University of the Highlands and Islands
374
UHI Thesis - pdf download summary Towards a sustainable production of Osmundea pinnatifida: insight into the cultivation and biochemical composition of the species Biancacci, Cecilia DOCTOR OF PHILOSOPHY (AWARDED BY OU/ABERDEEN) Award date: 2019 Awarding institution: University of Aberdeen Link URL to thesis in UHI Research Database General rights and useage policy Copyright,IP and moral rights for the publications made accessible in the UHI Research Database are retained by the author, users must recognise and abide by the legal requirements associated with these rights. This copy has been supplied on the understanding that it is copyright material and that no quotation from the thesis may be published without proper acknowledgement, or without prior permission from the author. Users may download and print one copy of any thesis from the UHI Research Database for the not-for-profit purpose of private study or research on the condition that: 1) The full text is not changed in any way 2) If citing, a bibliographic link is made to the metadata record on the the UHI Research Database 3) You may not further distribute the material or use it for any profit-making activity or commercial gain 4) You may freely distribute the URL identifying the publication in the UHI Research Database Take down policy If you believe that any data within this document represents a breach of copyright, confidence or data protection please contact us at [email protected] providing details; we will remove access to the work immediately and investigate your claim. Download date: 29. May. 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Biancacci, Cecilia - University of the Highlands and Islands

UHI Thesis - pdf download summary
Towards a sustainable production of Osmundea pinnatifida:
insight into the cultivation and biochemical composition of the species
Biancacci, Cecilia
DOCTOR OF PHILOSOPHY (AWARDED BY OU/ABERDEEN)
Award date:2019
Awarding institution:University of Aberdeen
Link URL to thesis in UHI Research Database
General rights and useage policyCopyright,IP and moral rights for the publications made accessible in the UHI Research Database are retainedby the author, users must recognise and abide by the legal requirements associated with these rights. This copyhas been supplied on the understanding that it is copyright material and that no quotation from the thesis may bepublished without proper acknowledgement, or without prior permission from the author.
Users may download and print one copy of any thesis from the UHI Research Database for the not-for-profitpurpose of private study or research on the condition that:
1) The full text is not changed in any way2) If citing, a bibliographic link is made to the metadata record on the the UHI Research Database3) You may not further distribute the material or use it for any profit-making activity or commercial gain4) You may freely distribute the URL identifying the publication in the UHI Research DatabaseTake down policyIf you believe that any data within this document represents a breach of copyright, confidence or data protection please contact us [email protected] providing details; we will remove access to the work immediately and investigate your claim.
Download date: 29. May. 2022

Towards a sustainable production of Osmundea pinnatifida: insight into the cultivation and biochemical composition of the species
Cecilia Biancacci
A thesis presented for the degree of Doctor of Philosophy at the University of Aberdeen
Academic Partners
The Scottish Association for Marine Science (SAMS)
University of the Highlands and Islands (UHI)
The James Hutton Institute
Under the supervision of Professor Michele S. Stanley, Professor John G. Day and Dr Gordon McDougall
2019

ii

iii
Declaration
I hereby declare that this thesis has been completed solely by Cecilia Biancacci at the Scottish
Association for Marine Science, under the supervision of Professor Michele S. Stanley, Professor
John G. Day and Dr. Gordon McDougall, during the period October 2015- September 2019. The
sources of information are specifically acknowledged. This work has not been submitted for any
previous application for a degree or personal qualification.
Cecilia Biancacci
Date 30/09/2019

iv
Abstract
Towards a sustainable production of Osmundea pinnatifida: insight into the cultivation
and biochemical composition of the species
Cecilia Biancacci
The consumption of wild harvested Osmundea pinnatifida as food in UK, Portugal and Spain has a
long tradition. However, the collection of wild material is time consuming, unsustainable and
limited by seasonality. Cultivation represents the preferred option for future exploitation of this
species. This study aimed at identifying the best rearing system for cultivation and commercial
production of O. pinnatifida. Phylogenetic analyses of populations along the West coast of Scotland
were conducted, to identify the species prior cultivation experiments, but also to explore
population variation within the frame of possible genetic pollution due to introduction of alien
species for farming purpose. The biochemical composition and variation of wild and cultivated
specimens was assessed. Giving for the first time an overview of the seasonality of the biochemical
and metabolomic profile of the species, an understanding of the correlation of biochemistry and
environmental factors, and an insight in how cultivation conditions can manipulate the
composition of the species. This information enhanced the knowledge on the metabolites present
in O. pinnatifida, with significant outcome for further commercial applications (e.g. food,
nutraceutical, pharmaceutical, cosmetic, etc.). Several cultivation methodologies were trialled,
including exploration of reproductive cycle and vegetative cultivation. Particularly, experiments in
flasks, in indoor and outdoor tank systems, PBR and on-sea out-plantation were conducted,
evaluating how cultivation parameters (e.g. light, temperature, nutrient, chemical treatments, supply
of hormones and fertilizer) affect the growth, biochemistry and morphology of the species. The
outcome was the establishment of an indoor tumbling cultivation system for the vegetative
production of O. pinnatifida tetrasporophytes. This pioneering methodology has the potential to be
scaled up and commercially applied, possibly achieving a continuous and tailored supply of the
resource that would be sustainable and independent from seasonality.

v
Acknowledgments
This work would not have been possible without the support, help, guidance and encouragement
that I have received from different people during these 4 years.
I would like to acknowledge the financial support received from IBioIC, MASTS and HIE.
Thank you to my supervisory team, Professor Michele Stanley, Professor John Day and Dr Gordon
McDougall, for their support, guidance, advices, revisions and recommendations, and for making
me a better scientist.
I am grateful for the technical assistance that I have received from Christine Beveridge, Richard
Abell, Sharon Mcneill, Debra Brennan, Alison Mair, Naomi Thomas at SAMS, and William
Allwood and Julie Sungurtas at The James Hutton Institute. Thank you to the Acadian Seaplants
for providing the Acadian Marine Plant Extract Powder (AMPEP) for the experiments presented
in Chapter 8.
To my friends, for have always been there even through miles and miles of distance.
To my family. My sister, for being my person and the best friend that I could have asked for, and
my parents, for having always supported me and for having shown me how to truly live. For their
unconditional love and for everything they have done for me.
To Gianby, for all the laughs, the long walks and talks, the support, the kindness and the love that
you have always shown. For teaching me to always look at the bright side of life and for being my
companion and friend in all the adventures.

vi
Table of Contents Chapter 1 Introduction ............................................................................................................... 1
1.1 Seaweed: a general introduction ................................................................................................... 1
1.2 Seaweed aquaculture ..................................................................................................................... 3
1.2.1 Cultivation methods ................................................................................................................ 6
1.2.2 Epiphytes in cultivated seaweeds ......................................................................................... 10
1.3 Rhodophyta .................................................................................................................................. 11
1.3.1 Introduction .......................................................................................................................... 11
1.3.2 Life cycle and reproduction ................................................................................................... 12
1.3.3 Distribution ........................................................................................................................... 13
1.3.4 Light patterns ........................................................................................................................ 14
1.3.5 Nutrients ............................................................................................................................... 16
1.3.6 Cell wall and storage of reserves in red algae ....................................................................... 17
1.3.7 Economic utilisation of red seaweeds................................................................................... 17
1.4 Scotland ........................................................................................................................................ 19
1.4.1 Scotland and aquaculture ..................................................................................................... 19
1.4.2 Scotland and seaweeds: an ancient tradition ....................................................................... 20
1.4.3 Scotland seaweed chemical industry history ........................................................................ 21
1.4.4 Scotland and commercial harvesting of wild seaweeds ....................................................... 22
1.5 Osmundea pinnatifida (Hudson) Stackhouse, 1809:79 ................................................................ 24
1.6 Project aim ................................................................................................................................... 29
1.6.1 Thesis objectives ................................................................................................................... 31
Chapter 2 Systematics and phylogeny of Osmundea pinnatifida in Scotland: investigation on
molecular biology in the context of the development of a cultivation method for the species . 33
2.1 Introduction ................................................................................................................................. 33
2.1.2 Systematics of Rhodophyceae: the order Ceramiales .......................................................... 34
2.1.3 Systematics of the Laurencia complex .................................................................................. 35
2.1.4 Taxonomy of Rhodophyceae: from morphological structures to DNA-based analyses ....... 37
2.1.5 DNA Barcoding: importance and applications ...................................................................... 38
2.2 Aims .............................................................................................................................................. 40
2.3 Materials and Methods ................................................................................................................ 41
2.3.1 Study area and sample collection ......................................................................................... 41
2.3.2 DNA extraction, PCR amplification, and sequencing ............................................................ 42
2.3.3 Multiple sequence alignments and phylogenetic analyses .................................................. 44
2.4 Results .......................................................................................................................................... 45
2.4.1 DNA extraction, PCR amplification, purification and sequencing ......................................... 45
2.4.2 Phylogenetic analyses ........................................................................................................... 47

vii
2.5 Discussion ..................................................................................................................................... 52
2.6 Conclusions .............................................................................................................................. 56
Chapter 3 Seasonal variation of biochemical composition in Osmundea pinnatifida from the West
coast of Scotland ...................................................................................................................... 58
3.1 The chemical composition of seaweeds ...................................................................................... 58
3.1.2 Red seaweeds: biochemical composition ............................................................................. 59
3.1.2.1 Red seaweeds as functional food: secondary metabolites with antimicrobial,
antioxidant and antiviral activity................................................................................................ 62
3.2 Scope ............................................................................................................................................ 63
3.2.1 Aims ........................................................................................................................................... 63
3.3 Materials and methods ................................................................................................................ 63
3.3.1 Carbohydrate analyses .......................................................................................................... 64
3.3.2 Protein analyses .................................................................................................................... 65
3.3.3 Pigments analyses ................................................................................................................. 66
3.3.4 Moisture content .................................................................................................................. 67
3.3.5 Antioxidant content .............................................................................................................. 67
3.3.6 Total fatty acid analyses ........................................................................................................ 68
3.3.7 Metal analyses ...................................................................................................................... 68
3.3.8 C:N ratio ................................................................................................................................ 69
3.3.9 Dissolved organic carbon and nitrogen (DOCN) and trace metals analyses in water samples
........................................................................................................................................................ 69
3.3.10 Statistical analyses .............................................................................................................. 70
3.4 Results .......................................................................................................................................... 71
3.4.1 Carbohydrate content ........................................................................................................... 71
3.4.2 Protein content ..................................................................................................................... 71
3.4.3 Chlorophyll-a content ........................................................................................................... 72
3.4.4 Carotenoids ........................................................................................................................... 73
3.4.5 Phycobiliproteins ................................................................................................................... 74
3.4.6 Moisture content .................................................................................................................. 74
3.4.7 Antioxidant content .............................................................................................................. 75
3.4.8 Total lipid extracted (TLE) ..................................................................................................... 76
3.4.9 Metal Analyses ...................................................................................................................... 77
3.4.10 C:N ratio .............................................................................................................................. 80
3.4.11 Dissolved organic carbon and nitrogen (DOCN) and trace metals analyses in water
samples .......................................................................................................................................... 81
3.4.12 PCA analysis of seasonal data ............................................................................................. 83
3.5 Discussion ..................................................................................................................................... 86
3.5.1 Carbohydrate content ........................................................................................................... 87

viii
3.5.2 Protein content ..................................................................................................................... 87
3.5.3 Pigment content .................................................................................................................... 88
3.5.4 Moisture content .................................................................................................................. 90
3.5.5 Antioxidant content .............................................................................................................. 90
3.5.6 Total fatty acid content ......................................................................................................... 91
3.5.7 Metals content in seaweed samples ..................................................................................... 91
3.5.8 C:N ratio in seaweed samples ............................................................................................... 93
3.5.9 Metals and DOCN analyses on water samples ...................................................................... 93
3.5.10 PCA Analysis ........................................................................................................................ 94
3.6 Conclusions .................................................................................................................................. 95
Chapter 4 Composition and seasonal variation in the metabolomic profile and flavour of
Osmundea pinnatifida ............................................................................................................... 98
4.1 Introduction ................................................................................................................................. 98
4.2 Aims ............................................................................................................................................ 101
4.3 Materials and methods .............................................................................................................. 101
4.3.1 Solvent extraction ............................................................................................................... 101
4.3.2 Oral extracts: extraction in HEPES buffer of two selected months, November and May .. 101
4.3.3 HILIC-PDA-MS analysis ........................................................................................................ 102
4.4 Results ........................................................................................................................................ 103
4.4.1 Solvent extraction ............................................................................................................... 103
4.4.2 Oral extracts: extraction in HEPES buffer of two selected months: November and May .. 118
4.5 Discussion ................................................................................................................................... 125
4.5.1 Solvent extraction ............................................................................................................... 125
4.5.1.1 ESI negative polarity ..................................................................................................... 125
4.5.1.2 ESI positive polarity ...................................................................................................... 128
4.5.1.3 Peak abundance variation: regulation trends .............................................................. 130
4.5.1.3.1 Trends for ESI negative polarity ............................................................................ 131
4.5.1.3.2 Trends for ESI positive polarity ............................................................................. 131
4.5.2 Replicating oral conditions .................................................................................................. 131
4.5.2.1 Oral extracts: extraction in HEPES buffer of two selected months, November and May
.................................................................................................................................................. 131
4.6 Conclusions ................................................................................................................................ 133
Chapter 5 Epiphytes control of O. pinnatifida cultures and spores’ production, release and
development in treated specimens ......................................................................................... 136
5.1 Introduction ............................................................................................................................... 136
5.2 Aims ............................................................................................................................................ 139
5.3 Materials and methods .............................................................................................................. 139
5.3.1 Chemical removal of epiphytes ........................................................................................... 139

ix
5.3.2 Tidal simulation as epiphytes control ................................................................................. 141
5.3.3 Epiphytes treatment of biomass grown in a tumbling tank system cultivation both indoors
and outdoors ................................................................................................................................ 143
5.3.4 In vitro spores’ production, attachment, germination and growth of juveniles of O.
pinnatifida from epiphytes treated cultures ................................................................................ 146
5.4 Results ........................................................................................................................................ 147
5.4.1 Chemical removal of epiphytes ........................................................................................... 147
5.4.1.1 Petri dish trial ............................................................................................................... 147
5.4.1.2 Flask trial ...................................................................................................................... 148
5.4.1.3 Tank trial ...................................................................................................................... 148
5.4.2 Tidal simulation as epiphytes control ................................................................................. 149
5.4.3 Impact of epiphyte treatment on tumbling cultivation system both indoor and outdoor. 152
5.4.3.1 Winter trial: indoor and outdoor cultivation ............................................................... 152
5.4.3.1.1 Biochemical composition ...................................................................................... 156
5.4.3.2 Summer trial: outdoor experiment .............................................................................. 157
5.4.3.2.1 Biochemical composition ...................................................................................... 161
5.4.4 In vitro spores’ production, attachment, germination and growth of juveniles of O.
pinnatifida from epiphytes treated cultures ................................................................................ 161
5.5 Discussion ................................................................................................................................... 166
5.6 Conclusions ................................................................................................................................ 170
Chapter 6 Effects of different media on the growth and biochemical composition of Osmundea
pinnatifida .............................................................................................................................. 172
6.1 Introduction ............................................................................................................................... 172
6.2 Aims ............................................................................................................................................ 174
6.3 Materials and methods .............................................................................................................. 174
6.3.1 Effects of three different media types on the growth and biochemical composition of
Osmundea pinnatifida adult cultures .......................................................................................... 174
6.3.2 The impact of different media types on growth and biochemical composition of Osmundea
pinnatifida juvenile plantlets ....................................................................................................... 176
6.3.3 The impact of nitrogen sources and the N:P ratio on Osmundea pinnatifida growth and
biochemical composition ............................................................................................................. 176
6.4 Results ........................................................................................................................................ 177
6.4.1 Effects of three different media types on the growth and biochemical composition of
Osmundea pinnatifida adult cultures .......................................................................................... 177
6.4.1.1 Weight and growth rate of adult cultures grown in three different media ................ 177
6.4.1.2 Effects of three different media types on the biochemical composition of Osmundea
pinnatifida adult cultures ......................................................................................................... 179
6.4.1.2.1 Antioxidant content .............................................................................................. 179
6.4.1.2.2 Carbohydrates and proteins ................................................................................. 179
6.4.1.2.3 Pigments ................................................................................................................ 180

x
6.4.2 The impact of different media types on growth and biochemical composition of Osmundea
pinnatifida juvenile plantlets ....................................................................................................... 181
6.4.2.1 Weight and growth rate of plantlets grown in either F/2 and VS media..................... 181
6.4.2.2 Biochemical composition when two different media types were employed to cultivate
Osmundea pinnatifida juvenile plantlets ................................................................................. 183
6.4.2.2.1 Antioxidant analysis .............................................................................................. 183
6.4.2.2.2 Proteins and carbohydrates analyses ................................................................... 183
6.4.2.2.3 Pigments ................................................................................................................ 184
6.4.3 The impact of nitrogen sources and N:P ratio on Osmundea pinnatifida growth and
biochemical composition ............................................................................................................. 185
6.4.3.1 Weight and growth rate of Osmundea pinnatifida adult cultures grown in different
nitrogen sources ....................................................................................................................... 185
6.4.3.2 Biochemical composition of Osmundea pinnatifida adult cultures grown in different
nitrogen sources ....................................................................................................................... 186
6.4.3.2.1 Protein and carbohydrate content ....................................................................... 186
6.4.3.2.2 Phycobiliproteins................................................................................................... 187
6.5 Discussion ................................................................................................................................... 188
6.6 Conclusions ................................................................................................................................ 192
Chapter 7 Cultivation of Osmundea pinnatifida in the Algem® Photo-bioreactor System ....... 194
7.1. Introduction .............................................................................................................................. 194
7.2 Aims ............................................................................................................................................ 196
7.3 Materials and methods .............................................................................................................. 196
7.3.1 Comparison of summer and winter conditions on the vegetative growth of O. pinnatifida
...................................................................................................................................................... 196
7.3.2 Comparison of winter conditions with a daily cycle temperature (DCT) and constant
temperature (CT) on the vegetative growth of O. pinnatifida..................................................... 197
7.3.3 Comparison of autumn and spring conditions on the vegetative growth of O. pinnatifida
...................................................................................................................................................... 198
7.4 Results ........................................................................................................................................ 199
7.4.1 Comparison of summer and winter conditions on the vegetative growth of O. pinnatifida
...................................................................................................................................................... 199
7.4.1.1 Antioxidant analyses .................................................................................................... 200
7.4.1.2 Pigment analyses.......................................................................................................... 202
7.4.2 Comparison of winter conditions with a daily cycle temperature (DCT) and a constant
temperature (CT) on the vegetative tissue growth of O. pinnatifida .......................................... 203
7.4.2.1 Pigment analyses.......................................................................................................... 204
7.4.3 Comparison of autumn and spring conditions on the vegetative growth of O. pinnatifida
...................................................................................................................................................... 206
7.4.3.1 Carbohydrates .............................................................................................................. 208
7.4.3.2 Proteins ........................................................................................................................ 209

xi
7.4.3.3 Pigments ....................................................................................................................... 210
7.4.3.4 Antioxidant analyses .................................................................................................... 211
7.5 Discussion ................................................................................................................................... 212
7.6 Conclusions ................................................................................................................................ 216
Chapter 8 Application of hormones and Acadian Marine Plant Extract Powder (AMPEP) to
cultures of O. pinnatifida and establishment of a tumbling cultivation system for the species218
8.1 Introduction ............................................................................................................................... 218
8.2 Aims ............................................................................................................................................ 220
8.3 Materials and methods .............................................................................................................. 220
8.3.1 Multi-well trials ................................................................................................................... 220
8.3.1.1 PGRs applied to cultivation of O. pinnatifida juveniles ................................................ 220
8.3.1.2 AMPEP treatments applied to cultivation of O. pinnatifida juveniles ......................... 221
8.3.2 Flask trials ............................................................................................................................ 222
8.3.2.1 Flask trials applying PGRs to adult O. pinnatifida biomass .......................................... 222
8.3.2.2 Flask trials applying AMPEP treatments to adult O. pinnatifida biomass .................... 222
8.3.3 Tank trial applying AMPEP extract to adult O. pinnatifida biomass ................................... 223
8.4 Results ........................................................................................................................................ 225
8.4.1 Multi-well trials ................................................................................................................... 225
8.4.1.1. PGRs applied to cultivation of O. pinnatifida juveniles ............................................... 225
8.4.1.2 AMPEP treatments applied to cultivation of O. pinnatifida juveniles ......................... 229
8.4.2 Flask trials ............................................................................................................................ 231
8.4.2.1. Flask trials applying PGRs to adult O. pinnatifida biomass ......................................... 231
8.4.2.1.1 Biochemical analyses of O. pinnatifida biomass cultivated applying PGRs .......... 232
8.4.2.2 Flask trials applying AMPEP treatments to adult O. pinnatifida biomass .................... 234
8.4.2.2.1 Biochemical analyses of O. pinnatifida biomass cultivated applying AMPEP
treatments............................................................................................................................ 236
8.4.3 Tank trial applying AMPEP extract to adult O. pinnatifida biomass ................................... 239
8.4.3.1 Biochemical analyses of tank trial applying the AMPEP extract to adult O. pinnatifida
biomass .................................................................................................................................... 243
8.5 Discussion ................................................................................................................................... 246
8.6 Conclusions ................................................................................................................................ 253
Chapter 9 Conclusions and future work .................................................................................. 256
9.1 Conclusions ................................................................................................................................ 256
9.2 Contribution of the work ........................................................................................................... 258
9.3 Practical implications resulting from the findings ..................................................................... 260
9.4 Limitations of the research ........................................................................................................ 261
9.5 Future work ................................................................................................................................ 261
9.6 Recommendations ..................................................................................................................... 263

xii
9.7 Concluding remarks ................................................................................................................... 264
References .............................................................................................................................. 266
Appendices ............................................................................................................................. 309
Appendix A Additional data on Chapter 3 ............................................................................ 309
A.1 Trace metals and DOCN analyses on seaweeds and seawater samples ................................... 309
A.2 Comparison between Multi-assay extraction protocol and Fournier assay for carbohydrates
determination in seasonal samples of O. pinnatifida ...................................................................... 313
A.3 PCA analyses table and abiotic factors used for the correlation analyses in Chapter 3............ 313
Appendix B Accessory data Chapter 4 ................................................................................. 319
Appendix C Additional data on cultivation experiments ...................................................... 321
C.1 Effect of temperature on biomass increase and epiphytes colonization in O. pinnatifida ....... 321
Materials and Methods ................................................................................................................ 321
Results .......................................................................................................................................... 322
C.2 Investigation on the stimulation of spores’ production applying a “heat-shock” in an indoor
tank system for O. pinnatifida ......................................................................................................... 324
Materials and methods ................................................................................................................ 324
Results .......................................................................................................................................... 324
C.3 Effect of light quality on growth rate, pigment content and reproduction in O. pinnatifida
cultures............................................................................................................................................. 326
Material and method ....................................................................................................................... 326
Results .............................................................................................................................................. 327
Discussion ......................................................................................................................................... 330
C.4 Cultivation in flasks of vegetative thalli tied onto nylon ropes ................................................. 331
Results .......................................................................................................................................... 332
Discussion ..................................................................................................................................... 333
C.5 Out-plantation of vegetative material of Osmundea pinnatifida .............................................. 334

xiii
List of figures
Figure 1.1 Women Gathering Seaweed. From: Ukiyo-e woodblock print, about 1810's, Japan, by artist
Katsukawa Shunka II ................................................................................................................................ 2
Figure 1.2 Extensive aquaculture in China, Fujian coast. Photo by George Steinmetz............................... 3 Figure 1.3 Cultivation methodologies; A) suspended long-line; B) bottom line in shallow water; C) tank
(indoor or outdoor); D) IMTA system ...................................................................................................... 9
Figure 1.4 Gracilaria sp. Cycle (modified from Lewmanomont, 1996) ..................................................... 13 Figure 1.5 Absorption spectra of photosynthetically active pigments initially studied for Porphyridium
purpureum. Spectrum of solar radiation is represented as black dotted curve. Modified from Gantt 1975
(Cole and Sheath, 2011) ........................................................................................................................... 15 Figure 1.6 Relationship between growth rate and internal nitrogen content, where “A” represents the
minimum nitrogen content for growth, the slope is the proportion between growth and internal nitrogen
content and “B” the critical value (Hanisak, 1983) .................................................................................. 16 Figure 1.7 A) Schematic view of cell wall in red algae: dark grey cylinders= cellulose microfibrillis; green
line= glucomannan; red line= sulphated glucan; grey twisted line= sulphated xylogalactans; white circle=
known link; grey circle=hypothesized link (modified from Lechat, 1998, in Fleurence and Levine, 2016).
B) Electron micrograph of P. aerugineum: p= pyrenoid; s= starch grains; g=golgi bodies; n= nucleus
(Modified from Gantt, Smithsonian Institution, in Dawes, 1998) ............................................................ 17
Figure 1.8 Scottish crofter 1880 - collecting seaweed on Skye. Photo by George Washington Wilson ..... 20 Figure 1.9 Distribution of Osmundea pinnatifida A) Worldwide (adapted from Guiry and Guiry 2018
records) and B) in UK (Hardy and Guiry, 2003) ...................................................................................... 24
Figure 1.10 Distribution of O. pinnatifida on the shore ............................................................................. 25 Figure 1.11 Samples of O. pinnatifida on the seashore from the same site in A) November, B) March and
C) June ................................................................................................................................................... 25
Figure 1.12 A) O. pinnatifida on a rocky shore (Ganavan beach), B) collected specimens ......................... 26 Figure 1.13 Transverse section of thallus at the stereo microscope; externally are visible dark pigmented
cortical cells and internally multiple layers of light pigmented medullary cells (bar=250µm) ..................... 26 Figure 1.14 Life cycle of O. pinnatifida: (a) male gametophyte (picture from Machín-Sánchez et al., 2012);
(b) female gametophyte; (e) tetrasporophyte; (f): tetraspores (bar=100µm); (g) released spore ............... 27 Figure 1.15 Ultrastructure of O. pinnatifida A) Cystocarp (1000 µm, Zeiss microscope); B)
Tetrasporocystes with visible tetraspores (scale bar=250 µm); C) Tetraspores (optical microscope); D)
Transparent trichoblast on terminal swelled tips of O. pinnatifida samples (optical microscope)
(bar=500µm) ........................................................................................................................................... 28 Figure 1.16 O. pinnatifida A) Germinated spores after release, B) Growing spore after 2 months from
release; C) developing gametophytes; D) developed gametophytes after 6 months from release .............. 28
Figure 1.17 Flow chart of the possible cultivation methodologies for Osmundea pinnatifida ...................... 30
Figure 2.1 Systematic of Rhodophyta with a focus on Osmundea genus ................................................... 34 Figure 2.2 Scheme of the triphasic Polysiphonia-type life history, from “Biology of plants”, 5th edition
(modified from Raven et al., 1992) .......................................................................................................... 35 Figure 2.3 Locations of sampling sites. In yellow Dunstaffnage, where SAMS is situated. Samples are
numbered from 1 to 6 in chronological order .......................................................................................... 41 Figure 2.4 Scheme of primers on the rbcL sequence. The primers are taken from Freshwater and Rueness
(1994) as referred in Table 2.2 ................................................................................................................. 44 Figure 2.5 A) DNA extraction Dundee sample, prepared using DNeasy Plant Mini Kit, 0.8% TBE (0.5x)
gel; B) PCR gel for Easdale sample (on the left) and Ganavan sample (on the right) with the couple of
primers FrbcL-RbcS. 5 µl of Easy Ladder I (BioLine) as reference; C) PCR gel for Easdale and South Uist
samples ................................................................................................................................................... 46 Figure 2.6 A) PCR gel for Jura sample (on the left) and South Uist samples (on the right) with the couple
of primers FrbcL-RbcS. 5 µl of Easy Ladder I (BioLine) as reference B) DNA extraction for Jura sample
with CTAB method ................................................................................................................................. 47

xiv
Figure 2.7 Purification of samples after Amicon kit procedure. L= EasyLadder I; E=Easdale;
D=Dunstaffnage; SU= South Uist; J=Jura .............................................................................................. 47 Figure 2.8 Evolutionary history inferred by using the Maximum Likelihood method based on the Jukes-
Cantor model on MEGA 7 software. Circled in green the samples from this study. The bootstrap
consensus tree inferred from 1000 replicates is taken to represent the evolutionary history of the taxa
analysed. Branches corresponding to partitions reproduced on less than 50% bootstrap replicates are
collapsed. The percentage of replicate tree in which associated taxa clustered together in the bootstrap test
(1000 replicates) are shown next to the branches. Initial tress for the heuristic search were obtained
automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated
using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with
superior log likelihood value .................................................................................................................... 49 Figure 2.9 Evolutionary history (including the Spanish entry) inferred by using the Maximum Likelihood
method based on the Jukes-Cantor model on MEGA 7 software. Circled in green the samples from this
study ....................................................................................................................................................... 49 Figure 2.10 Neighbour-Joining consensus tree obtained employing Geneious software, Clustal W
alignment, genetic distance method Jukes Cantor, and resample method Bootstrap. Circled in green the
samples here analysed .............................................................................................................................. 50 Figure 2.11 Neighbour-Joining consensus tree obtained employing Geneious software, Clustal W
alignment, genetic distance method Jukes Cantor, and resample method Bootstrap. Circled in red the
outgroup (Centroceras clavulatum, AF259490) (Geneious software) ....................................................... 50 Figure 2.12 Evolutionary history was inferred by using the Maximum Likelihood method based on the
Jukes-Cantor model. In green the samples analysed in this study; in yellow the samples for O. pinnatifida
from NCBI; in red the outgroups (Geneious software) ............................................................................ 51 Figure 3.1 Satellite image of the sampling site, indicated with a star, Dunstaffnage bay (56.4547° N,
5.4374° W) .............................................................................................................................................. 64 Figure 3.2 Variation of carbohydrates content expressed as percentage per dry weight (DW) across the
months in O. pinnatifida samples (n=3; error bar, 1SD of mean) using a Fournier (2001) modified method
................................................................................................................................................................ 71 Figure 3.3 Variation of protein content expressed as percentage per dry weight across the months
(%DW) (n=3; error bar, 1SD of mean) .................................................................................................... 72 Figure 3.4 Variation in colours of seasonal samples of O. pinnatifida from the same sampling site for A)
November, B) February, C) March, D) April, E) May, F) June, G) July, H & I) August ........................ 73 Figure 3.5 Seasonal variation of Chl-a (dark grey) and carotenoids (light grey) in O. pinnatifida samples
expressed as µg/g of dry weight, with the modified method from Torres et al. (2014) (n=3; error bar, 1SD
of mean) .................................................................................................................................................. 73 Figure 3.6 Seasonal variation of PE (dark grey) and PC content (light grey) expressed as mg/g of dry
weight in O. pinnatifida samples (n=3; error bar, 1SD of mean) ................................................................ 74 Figure 3.7 Seasonal variation of full moisture content (FMC) expressed as percentages in O. pinnatifida
samples (n=3; error bar, 1SD of mean) .................................................................................................... 75 Figure 3.8 TPC measurements expressed as µg PGE/mg (phloroglucinol equivalents) of dry weight using
the Folin-Ciocalteau method on seasonal samples (n=3; error bar, 1SD of mean) ................................... 75
Figure 3.9 FRAP assay of monthly collected samples. FRAP results are expressed in µM FeSO₄ eq/mg
dry weight (n=3; error bar, 1SD of mean) ................................................................................................ 76 Figure 3.10 Percentage of total lipid extracted expressed as percentage of dry weight from seasonal
samples of O. pinnatifida. Results are expressed as average of n=3 (Error bar, 1SD of mean).................... 77 Figure 3.11 Variation in metal content expressed as mg/kg of dry weight in seasonal samples of O.
pinnatifida (n=3) ....................................................................................................................................... 80 Figure 3.12 Nitrogen percentage of dry weight (dark grey bars), Carbon percentage of dry weight (light
grey bars) and C:N as molar ratio (dark grey line) of seasonal samples of O. pinnatifida (n=3; error bar, 1
SD of mean) ............................................................................................................................................ 80 Figure 3.13 Comparison of the trends for the DOC (light grey line) (µM) water analyses (June-August
2018) and Al values (dark grey bars) (seaweed samples, June-August 2017) for the same sampling site in
Dunstaffnage beach (n=3; error bar, 1 SD of mean) ................................................................................ 81

xv
Figure 3.14 Comparison of the trends for the TDN (light grey line) (µM) water analyses (June-August
2018) and Al values (dark grey bars) (seaweed samples, June-August 2017) for the same sampling site in
Dunstaffnage beach (n=3; error bar, 1 SD of mean) ................................................................................ 81 Figure 3.15 Correlation between salinity expressed as PSU (light grey line) and aluminium in water
samples expressed as µM (dark grey line) collected from June to August 2018 in Dunstaffnage bay. (n=3;
error bar, 1 SD of mean) ......................................................................................................................... 82 Figure 3.16 Correlation between aluminium in seawater samples (dark grey line) and precipitation data
(light grey bars) (r=0.91, p<0.05). Precipitation data for 2017-2018 are monthly averaged and obtained
from CEDA, Met Office, #918 Dunstaffnage station.............................................................................. 82 Figure 3.17 Screen plot of PCA showing the Eigenvalues and the respective component number. From
the plot trend, it is evident that the majority of data is represented by the first three components, while
from the third there is a change in the inclination of the line and the remaining components account only
for a small percentage of data .................................................................................................................. 84 Figure 3.18 The first and the second PCs were plotted since accounted for more than 75% of variance
together. ANOSIM and SIMPER analyses (PRIMER, v6) identified two groups (b and c) and one
outgroup (a, July) in relation to the similarity between the months in terms of biochemical composition . 85 Figure 3.19 BEST analyses (PRIMER, v6) identified correlation between environmental factors
(maximum temperature, precipitation, hours of light and maximum radiation) and months and importance
of the abiotic factors in influencing the seasonality of the biochemical composition of the species .......... 85 Figure 4.1 Overlay peak graph of MS profiles from August 2016 to January 2017 (Negative Mode, FSD =
7.20E7, Base peak F:FTMS-c ESI Full MS [80-1000]) ........................................................................... 104 Figure 4.2 Overlays peak graph of MS profiles from February to July 2017 (Negative mode, FSD: 9.59E7,
Base peak F:FTMS-c ESI Full MS [80--1000]) ....................................................................................... 105 Figure 4.3 Overlays peak graphs of MS profiles from August 2016 to January 2017 (Positive mode, FSD:
3.20 E8, Base Peak F:FTMS+ c ESI Full MS [80-1000]) ........................................................................ 106 Figure 4.4 Overlays peak graphs of MS profiles from February to July 2017 (Positive mode, FSD: 3.20
E8, Base Peak F:FTMS+ c ESI Full MS [80-1000]) ............................................................................... 107 Figure 4.5 Peak abundance variation over one year time period for putative components that decreased
during the summer months. Putative IDs: 1) Fumarate; 2) 1,4-Thiazane-3-carboxylic acid S-oxide, 3)
Vitamin C), 4) Chilenone A-like (possible isomer with different MS²), 5 & 6) Homocitric acid ............. 110 Figure 4.6 Peak abundance variation over one year time period for putative components that increased
during the summer months. Putative IDs: 1) Vitamin C, 2 to 5) lipid derivatives, 6) Fumarate, 7)
Chilenone B-2H, 8 to 11) Floridoside, 12) Digeneaside, 13) Chilenone A-like (possible isomer with
different MS²), 14) Trans-aconitic acid ................................................................................................... 112 Figure 4.7 Peak abundance variation over a one year time period for putative components that showed
random patterns. Putative IDs 1 & 3) Chilenone B (-2H), 2) Chilenone B, 4) Chilenone A, 5) Trans
aconitic acid .......................................................................................................................................... 113 Figure 4.8 Peak abundance variation over one year time period for putative components that showed
little variation. Putative IDs: 1) TCSO, 2) lipid derivatives ..................................................................... 114 Figure 4.9 Peak abundance variation over one year time period for putative components that showed
down regulation in summer. Putative IDs: 1 & 2) Alkaloids, 3) Amino acid derivatives, 4) valine, 5) 5-
amminopentanol ................................................................................................................................... 115 Figure 4.10 Peak abundance variation over one year time period for putative components that showed up
regulation in summer. Putative IDs: 1) Seryl tyrosine derivatives, 2) Betaine, 3) Porphyra-334, 4)
Shinorine, 5) Amminoheptanoic acid .................................................................................................... 116 Figure 4.11 Peak abundance variation over one year time period for putative components that showed
little changes. Putative IDs: 1) seryl tyrosine derivatives, 2) hydroxy Betaine derivatives, 3) Porphyra-334,
4) Palythine ........................................................................................................................................... 116 Figure 4.12 Peak abundance variation over one year time period for putative components that showed
random changes. Putative IDs: 1) Isodomoic acid, 2) Acetyl carnitine, 3) Hydroxy Betaine derivatives, 4)
Glutamine ............................................................................................................................................. 117 Figure 4.13 Peak abundance variation over one year time period for a putative component that showed
down regulation in autumn. Putative ID: 1) Palythene ........................................................................... 117

xvi
Figure 4.14 A) HEPES buffer extracted samples: in orange “May” and in pink “November”; B) solvent
extract sample ....................................................................................................................................... 119 Figure 4.15 Scanning absorbance spectrum (as absorbance units) of the HEPES extracted sample from
November ............................................................................................................................................. 119
Figure 4.16 Scanning absorbance spectrum of the HEPES extracted sample from May ........................ 120 Figure 4.17 Peak graphs of MS profiles from May oral (black), November oral (red), May (green) and
November (blue) (Positive mode, FSD = 3.90 E8, Base Peak F:FTMS+ c ESI Full MS [80-1000]). Arrow
denotes HEPES peak in oral samples .................................................................................................... 121 Figure 4.18 Peak graphs of MS profiles from May oral (black), November oral (red), May (green) and
November (blue) (Negative mode, FSD = 9.00E7, Base Peak F:FTMS+ c ESI Full MS [80-1000]). Arrow
denotes HEPES peak in oral samples .................................................................................................... 122 Figure 4.19 Main peaks identified in HILIC profiles in negative mode for Solvent and “Oral” extracted
samples (blue= November; light grey= November Oral; orange= May; black= May Oral) (n=3; error bar,
1 SD of mean) ....................................................................................................................................... 123 Figure 4.20 Abundance of main peaks identified in HILIC profiles in positive mode for Solvent and
“Oral” extracted samples (blue= November; light grey= November Oral; orange= May; black= May Oral)
(n=3; error bar, 1 SD of mean) .............................................................................................................. 124 Figure 5.1 The 4 stages of establishment in Osmundea pinnatifida spores (Axio Inverted microscope, Zeiss):
A) cell wall formation; B) polarity; C) germination; D) rhizoid growth and adhesion ............................ 138
Figure 5.2 Schematic view of the microcosm tank from the top ........................................................... 142
Figure 5.3 A) Schematic view of the system from the top; B) Schematic lateral view of the system ...... 143
Figure 5.4 Lateral view of the microcosm and reservoir tanks .............................................................. 143 Figure 5.5 Scheme of the tank tumbling system. Blue circle: air stone; grey block line: aeration pipe;
Green arrow: water inflow; grey cylinder: tambourine filter; blue arrows: circular movement of water due
to the aeration system; orange arrow: outflow system ............................................................................ 144
Figure 5.6 Indoor cultivation system from the front ............................................................................. 145
Figure 5.7 Outdoor cultivation system ................................................................................................. 146 Figure 5.8 QY measurements related to the different treatments utilised for the Petri dish trial measured
at 2 hrs, 7 days and 14 days. (control=green, 0.5% KI= blue, 25% methanol=orange, 1% NaClO= black,
0.25% Kick start=grey, 0.1% NaClO= yellow; n=3; error bar, 1 SD of mean) ....................................... 147 Figure 5.9 QY measurements related to the different treatments for the flask trial measured at 2 hrs, 7
days and 14 days. (control=green, 0.5% KI= blue, 25% methanol=orange, 0.1% NaClO= yellow; n=3;
error bar, 1 SD of mean) ....................................................................................................................... 148 Figure 5.10 QY in relation to exposure time (min) to treatment with 0.5% KI for the tank trial after 2 hrs,
1 and 2 weeks. (10 min incubation= grey, 20 min incubation=blue, 30 min incubation= orange; n=3; error
bar, 1 SD of mean) ................................................................................................................................ 149 Figure 5.11 Comparison of growth data (g, wet weight) between the two trials with different starting
biomass, 110 g (1st trial, dark grey line) and 180 g (2nd trial, light grey line). (n=3; error bar, 1 SD of mean)
.............................................................................................................................................................. 150 Figure 5.12 A) Thalli with new tips and shoots developing from the holdfast; B) Tetrasporophytes with
tetraspores; C) Tetrasporophytes after spores’ release ........................................................................... 150 Figure 5.13 Comparison of absolute growth measurements of first trial (dark grey line) and second trial
(light grey line). (n=3; error bar, 1 SD of mean) ..................................................................................... 151 Figure 5.14 Comparison of SGR measurements of of first trial (dark grey line) and second trial (light grey
line). (n=3; error bar, 1 SD of mean) ..................................................................................................... 151 Figure 5.15 Data collected with HOBO® logger for the indoor tanks from November 2017 to March
2018. Temperature is expressed in °C and Intensity in lum/ft². Temperature (dark grey bars) and Intensity
(light grey line) are plotted on the same reference axis since the values reported for the intensity in the
indoor experiment were particularly low, in set of ten ............................................................................ 152 Figure 5.16 Data collected with HOBO® logger for the outdoor tanks from November 2017 to April
2018. Temperature is expressed in °C (dark grey bars) and Intensity in lum/ft² (light grey line), unit
provided by the logger ........................................................................................................................... 152

xvii
Figure 5.17 Values relative to pH (dark grey bars) and salinity (light grey line) averaged per month for the
tanks outdoor from November 2017 to April 2018 ............................................................................... 153 Figure 5.18 Values relative to pH (dark grey bars) and salinity (light grey line) averaged per month for the
tanks Indoor from November 2017 to March 2018 ............................................................................... 153 Figure 5.19 Growth (expressed in WW) for the indoor (dark grey line) and outdoor (light grey line) tanks
trials from November 2017 to April 2018, every time corresponds to one-month interval. (n=3; error bar,
1 SD of mean) ....................................................................................................................................... 154 Figure 5.20 SGR was measured using the formula from Evans (1972) for every month and expressed as
%day⁻¹ for the Indoor trial (dark grey bars) and Outdoor one (light grey bars) (n=3; error bar, 1 SD of
mean) .................................................................................................................................................... 154 Figure 5.21 QY measured with an AquaPen every month for the cultures in the indoor (dark grey line)
and outdoor (light grey line) systems. (n=3; error bar, 1 SD of mean) .................................................... 155 Figure 5.22 Indoor biomass (A & C) and Outdoor biomass (B &D), after four months (February 2018)
into the cultivation system during the periodic weighing ........................................................................ 155 Figure 5.23 Primary axis: Chl-a (green) and carotenoids (orange) concentration expressed in µg/g for the
cultures in the indoor and outdoor tanks. Secondary axis: PE (red) and PC (blue) concentration (mg/g).
(n=3; error bar, 1 SD of mean) .............................................................................................................. 156 Figure 5.24 Protein (dark grey bars) and carbohydrate (light grey bars) content expressed in percentage
(DW) for samples undergoing Outdoor and Indoor trials. (n=3; error bar, 1 SD of mean) .................... 157 Figure 5.25 Antioxidant compounds measured with FRAP assay in samples undergoing the cultivation in
indoor (dark grey bar) and outdoor (light grey bar) tank trials. (n=3; error bar, 1 SD of mean) .............. 157 Figure 5.26 Growth (WW, g) of cultures in an outdoor tumbling system from April to September 2018.
(n=3; error bar, 1 SD of mean) .............................................................................................................. 158 Figure 5.27 SGR of culture in the outdoor tumbling system from April to September 2018. (n=3; error
bar, 1 SD of mean) ................................................................................................................................ 158 Figure 5.28 QY measurement of cultures in an outdoor tumbling system from April to September 2018.
(n=3; error bar, 1 SD of mean) .............................................................................................................. 159 Figure 5.29 Cultures during the weighing in July 2018 (A &C), wild sample in May 2018 (B) and (D) July
2018 ...................................................................................................................................................... 159 Figure 5.30 Temperature (°C) profile in the tanks in the second outdoor trial as average (blue bars),
maximum (dark grey bars) and minimum (light grey bars) ..................................................................... 160
Figure 5.31 Light profile (lum/ft²) expressed as average in the tanks in the second outdoor trial .......... 160
Figure 5.32 Salinity (light grey bars) and pH (dark grey line) measurements in the second outdoor trial 160 Figure 5.33 A) Multi-well Petri dishes with cut tips from tetrasporophytes; B) Tetraspores are visible
inside the tetrasporophytes on the extremities of the thalli of cultivated samples; empty tetrasporophytes
are visible ahead of the full tetrasporophytes; C) Haploid tetraspores (indicated by the arrows) inside the
diploid tetrasporophytes prior to release; D) Spores released prior attachment in a Petri dish and observed
under a stereo microscope ..................................................................................................................... 162 Figure 5.34 A) Live haploid spore released observed under the Axio stereomicroscope. It is possible to
see the external mucilaginous sheath; B) dead spore after release; note visible deteriorating cell wall and
loss of pigmentation .............................................................................................................................. 162 Figure 5.35 Spores under an Axio stereomicroscope (Zeiss) at different stages of development after the
release: A) Body of germinated spore; B) Rhizoid of germinated spore; C) Rounded spore without rhizoid
apparatus ............................................................................................................................................... 163 Figure 5.36 A) Developed spores after one week into the cultivation system (observed at the Axio
Inverted microscope at 10x); B & C) spores after two weeks into the cultivation system (observed at the
Axio Inverted microscope at 20x and 10x); D) Measurement of a developed spore after one month from
release (Axio Inver microscope); E) Developed juvenile after two months from the release; F) particular
of attachment disc in juvenile after two months in cultivation ............................................................... 164 Figure 5.37 A) Juveniles after two months in tissue flask, B) magnification of juveniles. Each square= 1
mm². ..................................................................................................................................................... 165

xviii
Figure 5.38 A & B) Juveniles after two months of in vitro incubation observed under a stereomicroscope
(Zeiss). C & D) Juveniles after three months of in vitro incubation observed under a stereomicroscope
(Zeiss). Visible ramification and presence of multiple branches as well as other organisms .................... 165
Figure 5.39 6x multi-well with juveniles after 5 months of cultivation .................................................. 166 Figure 6.1 Flasks (6 x 1 l) with adult cultures in the CT room under incubation: 10 °C, 12:12 photoperiod
.............................................................................................................................................................. 174
Figure 6.2 Flasks (6 x 500 ml) with juvenile cultures under incubation: 10 °C, 12 :12 photoperiod ....... 176 Figure 6.3 Different cultures under the different media treatments at the beginning and at the end of the
trial A) F/2, B) F/2 modified and, C) VS at the beginning of the experiment; D) F/2, E)F/2 modified
and, F) VS at the end of the experiment ................................................................................................ 177 Figure 6.4 Weight measurements of cultures (WW) under the three different treatments: F/2 black line,
F/2 modified dark grey line, VS light grey line. (n=3; error bar, 1 SD of mean). .................................... 178 Figure 6.5 Growth rate calculated with the equation for SGR (Evans, 1972). F/2 black line, F/2 modified
dark grey line, VS light grey line. (n=3; error bar, 1 SD of mean) ........................................................... 178 Figure 6.6 FRAP assay for the cultures under the different media. F/2 black bar, F/2 modified dark grey
bar, VS light grey bar. (n=3; error bar, 1 SD of mean) ........................................................................... 179 Figure 6.7 Carbohydrate (dark grey bars) and protein (light grey bars) content expressed as %DW for
cultures undergoing the three different treatments. (n=3; error bar, 1 SD of mean) ............................... 180 Figure 6.8 Chl-a (dark grey bars) and carotenoids (light grey bars) content of the cultures. (n=3; error bar,
1 SD of mean) ....................................................................................................................................... 180 Figure 6.9 PE (dark grey bars) and PC (light grey bars) values for cultures undergoing the three
treatments. (n=3; error bar, 1 SD of mean) ............................................................................................ 181 Figure 6.10 Juveniles cultures under the different treatments after one week A) F2; B) VS, and at the end
of the trial C) F/2 ; D) VS .................................................................................................................... 182 Figure 6.11 Weight measurements (g, WW) for cultures under two different media types: F/2 dark grey
line and VS light grey line. (n=3; error bar, 1 SD of mean) .................................................................... 182 Figure 6.12 SGR (Evans, 1972) of cultures under the different treatments. F/2 dark grey line and VS light
grey line (n=3; error bar, 1 SD of mean) ................................................................................................ 183 Figure 6.13 FRAP assay for the cultures under two media. F/2 dark grey bar and VS light grey bar. (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 183 Figure 6.14 Protein (light grey bars) and carbohydrate (dark grey bars) content expressed in % DW of the
cultures under two different media. (n=3; error bar, 1 SD of mean) ...................................................... 184 Figure 6.15 Pigments content in the cultures under the different treatments. Chlorophyll a =dark grey
bars and carotenoids= light grey bars (n=3; error bar, 1 SD of mean) ................................................... 184 Figure 6.16 PE (dark grey bars) and PC (light grey bars) content (mg/g) for the cultures under different
treatments. (n=3; error bar, 1 SD of mean) ............................................................................................ 185 Figure 6.17 Weight measurements (WW) of cultures under different nitrogen sources: F/2= blue, Urea=
orange, NH4Cl =grey and NH4NO3 =black bars (n=3; error bar, 1 SD of mean) .................................. 185 Figure 6.18 The SGR of cultures undergoing different treatment across the 7 weeks of experiment. F/2=
blue, Urea= orange, NH4Cl =grey and NH4NO3 =black bars (n=3; error bar, 1 SD of mean) ............... 186 Figure 6.19 Carbohydrate (dark grey bars) and protein (light grey bars) content expressed in percentage
(DW) for the cultures under the different treatments applied. (n=3; error bar, 1 SD of mean) ............... 187 Figure 6.20 PE (dark grey bars) and PC (light grey bars) content (mg/g) in samples undergoing different
treatments. (n=3; error bar, 1 SD of mean) ............................................................................................ 187 Figure 7.1 A) Algem®, photobioreactor used for the experiment; B) View of the open temperature
controlled chamber from the top with a 1L conical flask inside; C) Schematic of the PBR system: I=
influent, E = effluent, PP = peristaltic pump, TCC = temperature controlled chamber, MP= micro
processor, TS= Temperature sensor, H= heating system, C=cooling system, V=valve box, CH= chiller,
GMP=Gimbal mixing system. Modified from Whitton, 2006 ............................................................... 195 Figure 7.2 A) Samples under summer conditions, B) samples under winter conditions, after 2 weeks in
the PBR. (bar=1 cm) ............................................................................................................................. 199

xix
Figure 7.3 QY measurements of summer (dark grey bars) vs winter (light grey bars) cultures in the
Algem® unit (n=3; error bar, 1 SD of mean)......................................................................................... 200 Figure 7.4 SGR for cultures undergoing summer (dark grey bars) and winter (light grey bars) conditions
in the PBR (n=3; error bar, 1 SD of mean) ............................................................................................ 200 Figure 7.5 TPC analyses: comparison between field and cultivated samples in relation to summer and
winter conditions (n=3; error bar, 1 SD of mean) .................................................................................. 201 Figure 7.6 FRAP analyses: comparison between field and cultivated samples in relation to summer and
winter conditions (n=3; error bar, 1 SD of mean) .................................................................................. 201 Figure 7.7 Comparison of Chl-a (dark grey bars) and Carotenoids (light grey bars) (µg/g) contents in
biomass grown either in the Algem® for four weeks with wild harvested biomass for summer and winter
conditions, for the same months of references (n=3; error bar, 1 SD of mean) ..................................... 202 Figure 7.8 Comparison of PE (dark grey bars) and PC (light grey bars) (mg/g) contents in biomass grown
in the Algem® for four weeks with wild harvested biomass (n=3; error bar, 1 SD of mean) .................. 203 Figure 7.9 QY average measurements of cultures grown under winter conditions but two different
temperatures, daily cycle temperature (dark grey bars) and constant temperature (light grey bars) (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 204 Figure 7.10 SGR for samples under CT (light grey bars) and DCT (dark grey bars) temperature in the
Algem® PBR (n=3; error bar, 1 SD of mean) ....................................................................................... 204 Figure 7.11 Comparison of Chl-a (dark grey bars) and Carotenoids (light grey bars) (µg/g) contents in O.
pinnatifida between samples cultivated at CT, DCT and wild winter biomass for the same month of
reference (n=3; error bar, 1 SD of mean) .............................................................................................. 205 Figure 7.12 Comparison of PE (dark grey bars) and PC (light grey bars) (mg/g) contents in O. pinnatifida
between samples cultivated at CT, DCT and wild winter biomass for the same month of reference (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 205 Figure 7.13 Cultures cultivated in the Algem® PBR after 3 weeks of cultivation for A) March conditions,
B) September conditions and C) Control (bar=1cm) ............................................................................ 206 Figure 7.14 A comparison of growth, as SGR, for Control (dark grey bars) vs March (light grey bars)
conditions over a 5-weeks period (n=3; error bar, 1 SD of mean) .......................................................... 207 Figure 7.15 A comparison of growth, described as SGR, for Control (dark grey bars) vs September (light
grey bars) conditions over a 5-weeks period (n=3; error bar, 1 SD of mean) .......................................... 207 Figure 7.16 Comparison of weekly growth (SGR) between the two conditions in the Algem, September
(light grey bars) vs March (dark grey bars), over a 5-weeks period (n=3; error bar, 1 SD of mean) ......... 208 Figure 7.17 PAM measurement in reference to the QY of cultures undergoing the two different
treatments and the controls in the Algem set at Spring and Autumn conditions. Control= black, Algem
Spring= light grey and Algem Autumn =dark grey bars (n=3; error bar, 1 SD of mean) ........................ 208 Figure 7.18 Carbohydrate content expressed in percentage of samples undergoing the two different
experiments and the control (n=3; error bar, 1 SD of mean) ................................................................. 209 Figure 7.19 % DW of protein in samples undergoing the two different experiments (including controls
and wild samples for September and March) (n=3; error bar, 1 SD of mean) ........................................ 209 Figure 7.20 Comparison of Chl-a (dark grey bars) and carotenoid (light grey bars) content in samples
under Spring, Autumn and Control conditions in the PBR and wild samples for the same reference
months (n=3; error bar, 1 SD of mean) ................................................................................................. 210 Figure 7.21 Comparison of PE (grey bars) and PC (light grey bars) content in samples under Spring,
Autumn and Control conditions in the PBR and wild samples for the same reference months (n=3; error
bar, 1 SD of mean) ................................................................................................................................ 210 Figure 7.22 FRAP (antioxidant power) of cultures undergoing the two different treatments compared to
the wild samples for the same months (n=3; error bar, 1 SD of mean) .................................................. 211 Figure 7.23 TPC content (antioxidant power) of cultures undergoing the two different treatments
compared to the wild samples for the same months (n=3; error bar, 1 SD of mean) .............................. 212 Figure 8.1 Tumbling system developed for vegetative cultivation of O. pinnatifida tetrasporophytes; A)
Filters and UV system; B) Tanks under 0.5 mg/l treatment and control with 10 l media reservoirs and
dosing pumps for the nutrient supply; C) Tanks under the bathing treatment; D) Frame for supporting

xx
the tank and valve system for aeration and emptying; E) Inflow (yellow pipe) and banjo filter for the
outflow; F) cultures during the weighing ............................................................................................... 224 Figure 8.2 A) Juveniles cultures exposed to 2.5 mg BA at the beginning and B) at the end of the
experiment. Juveniles cultures exposed to IAA:BA 1:5 C) at the beginning and D) at the end of the
experiment. Juveniles cultures exposed to 2.4-D:BA:IAA 5:5:5 E) at the beginning and F) at the end of
the experiment. Each square is 1mm² .................................................................................................... 226 Figure 8.3 Results of the growth (area mm²) for the single hormone treatments applied on juveniles
cultures of O. pinnatifida (n=3; error bar, 1 SD of mean) ........................................................................ 226 Figure 8.4 Results of the growth (area, mm²) of the juveniles cultures under the single hormonal
treatment (light grey line) (2.5 mg/l BA) compared with control (dark grey line) (n=3; error bar, 1 SD of
mean) .................................................................................................................................................... 227 Figure 8.5 Results for the double hormone treatments applied on juveniles’ cultures of O. pinnatifida (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 227 Figure 8.6 Results of the growth (area, mm²) of the juveniles cultures under the double hormonal
treatment (light grey line) (IAA:BA 1:5 mg/l) compared to control (dark grey line) (n=3; error bar, 1 SD
of mean) ................................................................................................................................................ 228 Figure 8.7 Results for the triple hormone treatments applied on juveniles’ cultures of O. pinnatifida (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 228 Figure 8.8 Results of the growth (area mm²) of the juveniles’ cultures under the triple hormonal
treatment (light grey line) (2.4-D:BA:IAA 5:5:5 mg/l) compared to control (dark grey line) (n=3; error bar,
1 SD of mean) ....................................................................................................................................... 229 Figure 8.9 Measurement of the growth (area, mm²) of juveniles under the different AMPEP treatments,
measured with ImageJ®. Control = blue, 3 mg/l = light grey, 0.5 mg/l = dark grey and Bathing (20g/l) =
black bars (n=3; error bar, 1 SD of mean) ............................................................................................. 229
Figure 8.10 SGR (% day⁻¹) of juveniles undergoing different AMPEP treatments. Control= blue, 3 mg/l
= light grey, 0.5 mg/l = dark grey and Bathing (20g/l) = black bars (n=3; error bar, 1 SD of mean) ..... 230 Figure 8.11 Juveniles specimens under AMPEP treatments A) Control; B) Bathing (20 g/l); C) 0.5 mg/l;
D) 3 mg/l. Bar= 1mm .......................................................................................................................... 230
Figure 8.12 Trichoblast in juvenile specimen under AMPEP extract ..................................................... 231 Figure 8.13 Variation of weight (g, WW) across 7 weeks for cultures under different PGRs treatments.
Control: F/2 medium (blue); Single: 2.5 mg BA (orange); Double:IAA:BA (1:5) (grey); Triple: 2.4-
D:BA:IAA (5:5:5) (yellow line) (n=3; error bar, 1 SD of mean) .............................................................. 231
Figure 8.14 SGR (expressed in %day⁻¹, from Evans, 1972) of cultures under different treatments.
Control: F/2 medium (blue); Single: 2.5 mg BA (orange); Double: IAA:BA (1:5) (grey); Triple: 2.4-
D:BA:IAA (5:5:5)(yellow bars) (n=3; error bar, 1 SD of mean) .............................................................. 232 Figure 8.15 Specimens under PGRs treatments in the flask trial at the start A) Control; B) Single
treatment (2.5 mg BA); C) Double treatment (IAA:BA 1:5); D) Triple treatment (2.4D:IAA:BA 5:5:5) and
at the end of the trial E) Control; F) Single treatment; G) Double treatment; H) Triple treatment ....... 232 Figure 8.16 Carbohydrate (dark grey bars) and protein (light grey bars) content expressed in percentage in
cultures undergoing different PGRs hormonal treatments (n=3; error bar, 1 SD of mean) .................... 233 Figure 8.17 Chl-a (dark grey bars) and carotenoids (light grey bars) (µg/g) concentration in samples
undergoing different PGRs hormonal treatments (n=3; error bar, 1 SD of mean) ................................. 233 Figure 8.18 PE (dark grey bars) and PC (light grey bars) (mg/g) concentrations in cultures undergoing
different PGRs hormonal treatments (n=3; error bar, 1 SD of mean) .................................................... 234
Figure 8.19 FRAP assay (µM FeSO₄ eq/mg DW) for cultures undergoing different PGRs hormonal
treatments (n=3; error bar, 1 SD of mean) ............................................................................................ 234 Figure 8.20 SGR measurements of cultures in flask under the different AMPEP treatments. Time is
expressed in weeks (n=3; error bar, 1 SD of mean) ............................................................................... 235 Figure 8.21 Comparison of cultures weight (WW) under the three AMPEP treatments (20 g/l = orange,
0.5 mg/l = grey, 3 mg/l = yellow line) and the Control (F/2) (blue line) (n=3; error bar, 1 SD of mean)
.............................................................................................................................................................. 235

xxi
Figure 8.22 Cultures under the AMPEP treatments A) 0.5 mg/l cultures B) particular of new shoots
from the rhizoids for specimens under 0.5 mg/l; C) Control treatment and; D) Bathing (20 g/l AMPEP
treatment).............................................................................................................................................. 236 Figure 8.23 Particular of new branches (arrow) and callus (circle) formation along the thallus in cultures
under 3 mg/l AMPEP ........................................................................................................................... 236 Figure 8.24 Carbohydrate (dark grey bar) and protein (light grey bars) content expressed in percentage
(DW) of cultures under different AMPEP treatments (n=3; error bar, 1 SD of mean) ........................... 237 Figure 8.25 Chl-a (dark grey bars) and carotenoids (light grey bars) concentration (µg/g) of cultures under
different AMPEP treatments (n=3; error bar, 1 SD of mean) ................................................................ 237 Figure 8.26 PE (dark grey bars) and PC (light grey bars) concentration (mg/g) of cultures under different
AMPEP treatments (n=3; error bar, 1 SD of mean) .............................................................................. 238
Figure 8.27 FRAP analyses (µMFeSO₄eq/mg DW) of cultures under different AMPEP treatments (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 238 Figure 8.28 TPC analyses (µg/mg) of cultures under different AMPEP treatments (n=3; error bar, 1 SD
of mean) ................................................................................................................................................ 239 Figure 8.29 pH (dark grey bars) and salinity (TDS, light grey line) measurements for the tank experiment
applying AMPEP extract ....................................................................................................................... 239 Figure 8.30 Temperature (dark grey bars) and intensity measurements (average light = grey line and
maximum light = dark grey line) recorded with HOBO® loggers in the tanks under the AMPEP
treatments ............................................................................................................................................. 240 Figure 8.31 Weight (WW) of the cultures under the three treatments tested in the tank system after 15
days, 1 month, 45 days, 2 months, 75 days and 3 months (n=3; error bar, 1 SD of mean) ..................... 240 Figure 8.32 Growth rate of the cultures in the tank system applying the AMPEP extract after 15 days,
1month, 45 days, 2 months, 75 days, 3 months (n=3; error bar, 1 SD of mean) ..................................... 241 Figure 8.33 Cultures in the tank system applying the AMPEP extract after 15 days in the cultivation
system. A) 05 mg/l; B) control; C) bathing, circled callus formation; D) control cultures ..................... 241 Figure 8.34 A) cultures after one month (0.5 mg/l AMPEP); B) cultures after two months (bathing in 20
g/l AMPEP); C) new plantlets developed in cultures after 3 months (Control F/2) in the cultivation
system ................................................................................................................................................... 242 Figure 8.35 QY of the cultures under the three different treatments in the tank cultivation system at time
0, after 15 days, 1 month, 45 days, 2 months, 75 days, and 3 months (n=3; error bar, 1 SD of mean) .... 242 Figure 8.36 Content of carbohydrates in samples cultivated in tank after 1 (blue bars), 2 (grey bars) and 3
(orange bars) months of cultivation under AMPEP and control treatments (n=3; error bar, 1 SD of mean)
.............................................................................................................................................................. 243 Figure 8.37 Protein content expressed in percentage of samples after 1 (blue bars), 2 (grey bars) and 3
(orange bars) under AMPEP and control treatments (n=3; error bar, 1 SD of mean) ............................ 244 Figure 8.38 Chl-a and carotenoids content in the three sampling times under AMPEP and control
treatments (n=3; error bar, 1 SD of mean) ............................................................................................ 244 Figure 8.39 PE and PC (mg/g) content for the three sampling times under AMPEP and control
treatments (n=3; error bar, 1 SD of mean) ............................................................................................ 245 Figure 8.40 FRAP assay of samples after 1 (blue bars), 2 (grey bars) and 3 (orange bars) under AMPEP
and control treatments (n=3; error bar, 1 SD of mean) .......................................................................... 246 Figure 9.1 Flow chart that highlights the best methodology to apply for the commercial cultivation and
exploitation of the species ..................................................................................................................... 257 Figure A.1 Metal concentration (expressed in mg/kg DW) in wild (maximum value in blue and minimum
value in red) and cultivated samples (green bars) (n=3; error bar, 1SD of mean) .................................... 309 Figure A.2 Comparison between aluminium, manganese and iron concentration (mg/kg) in wild
(maximum value in blue and minimum value in red) and cultivated samples (green bars) (n=3; error bar,
1SD of mean) ........................................................................................................................................ 310 Figure A.3 Metals assessed in water samples (expressed as µM) in June (blue bars), July (orange bars) and
August (grey bars) (n=3; error bar, 1SD of mean) .................................................................................. 311

xxii
Figure A.4 Metals assessed in water samples (expressed as µg/kg) in June (blue bars), July (orange bars)
and August (grey bars) (n=3; error bar, 1SD of mean) ........................................................................... 312 Figure A.5 Correlation trends for Al (dark grey bar) and Fe (light grey bar) found in water samples (µM)
.............................................................................................................................................................. 312 Figure A.6 Comparison of carbohydrates content expressed in %(DW) with two different methods:
Multi-assay (dark grey bars) (Chen and Vaidyanathan, 2013) and Fournier (light grey bars) (2001) (n=3;
error bar, 1 SD of mean) ....................................................................................................................... 313 Figure A.7 Precipitation data (mm/m) for 2016 (dark grey line) and 2017 (light grey line) in Dunstaffnage
bay, the area of the study for the seasonal sampling (Met-office for the Dunstaffnage n°918 station) .... 314 Figure A.8 Max (dark grey line) and Min (light grey line) temperature for 2016 in Dunstaffnage bay, area
for the study for the seasonal sampling (MET Office, Dunstaffnage station, 2016-2017) ....................... 314 Figure A.9 Max (dark grey line) and Min (light grey line) temperature for 2017 in Dunstaffnage bay, area
for the study for the seasonal sampling (MET Office, Dunstaffnage station, 2016-2017) ....................... 314 Figure A.10 Radiation (W hr/m²) for 2016-2017 in Dunstaffnage bay the area for the seasonal study
(CEDA-Centre for Environmental Data Analyses) ................................................................................ 315 Figure A.11 Radiation (W hr/m²) for 2017-2018 in Dunstaffnage bay the area for the seasonal study
(CEDA-Centre for Environmental Data Analyses) ................................................................................ 315
Figure A.12 Correlation between TPC content (bars) and hours of light (line) (r=0.75) ........................ 316
Figure A.13 Correlation between FRAP (bars) and hours of light (line) (r=0.75) .................................. 316 Figure A.14 Correlation between hours of light (line) and carbohydrate content (bars) (%DW) (r=0.75)
.............................................................................................................................................................. 316
Figure A.15 Correlation between max radiation (line) and carbohydrate content (bars) (r=0.73) ........... 317 Figure A.16 Correlation between temperature (max as dark grey line and min as light grey line) and
carbohydrate content (bars) (r=0.86, max temp; r=0.82, min temp) ....................................................... 317
Figure A.17 Correlation between hours of light (bar) and Chl-a content (line) (r=-0.63) ....................... 317 Figure A.18 Correlation between temperature (max as dark grey line and min as light grey line) and Chl-a
content (bars) (r=-0.71, max temperature; r=-0.75, min temp) ............................................................... 318 Figure B.1 Negative Mode April 2017 comparison of first (black) and second (red) runs (FSD: 7.80E7,
Base Peak F: FTMS-c ESI Full MS [80-1000]) ....................................................................................... 319 Figure B.2 Positive mode April 2017 comparison of first (black) and second (red) runs (FSD: 3.45E8,
Base Peak F: FTMS+c ESI Full MS [80-1000]) ...................................................................................... 319 Figure B.3 Negative mode September 2016 comparison of first (black) and second (red) runs (FSD:
7.60E7, Base Peak F: FTMS-c ESI Full MS [80-1000]) .......................................................................... 319 Figure B.4 Positive mode September 2016 comparison of first (black) and second (red) runs (FSD:
3.50E8, Base Peak F: FTMS+c ESI Full MS [80-1000] .......................................................................... 320 Figure B.5 Negative mode comparison of August 2016 (black) and August 2017 (red) runs (FSD: 7.00E7,
Base Peak F: FTMS-c ESI Full MS [80-1000]) ....................................................................................... 320 Figure B.6 Positive mode comparison of August 2016 (black) and August 2017 (red) runs (NL: 3.40E8,
Base Peak F: FTMS+c ESI Full MS [80-1000]) ...................................................................................... 320
Figure C.1 Flasks under 4 different temperature treatments ................................................................. 321
Figure C.2 KTH medium composition ................................................................................................ 322 Figure C.3 Samples under 4 different treatments: A) 10°C after 7 and B) 28 days; C) 13°C after 7 D) and
28 days; E) 15 °C after 7 F) and 28 days; G) 20°C after 7 H) and 28 days ............................................ 323 Figure C.4 Weight growth of the samples under four treatments in relation to the time (n=3, error bar, 1
SD of mean) .......................................................................................................................................... 323 Figure C.5 Relation between weight increase and time in samples undergoing 3 different treatments:
heater (black), no nutrient (light grey) and control (dark grey bars) (n=3; error bar, 1 SD of mean) ....... 325 Figure C.6 Graph of SGR average for the treatments calculated using the formula from Evans (1972).
Heater black, no nutrient light grey and control dark grey bars (n=3; error bar, 1 SD of mean) ............. 325
Figure C.7 Flasks under culturing conditions ....................................................................................... 327

xxiii
Figure C.8 A) roped slide on the bottom of the flask for spores' collection; B) sample under red light at
the end of the experiment colonized by epiphytes; C) spores released (circled in green) outside the slide
(green light treatment) ........................................................................................................................... 327 Figure C.9 Comparison of the QY (Fv/Fm) between the different wavelengths Blue, green red and
control (grey) bars (n=3; error bar, 1 SD of mean) ................................................................................ 328 Figure C.10 SGR calculated with the formula (Evans, 1972) for Blue, green red and control (grey) bars
(n=3; error bar, 1 SD of mean) .............................................................................................................. 328 Figure C.11 Chl- a (dark grey bars) and Carotenoids content (light grey bars) expressed in µg/g for the
samples under the 4 different light treatments (n=3; error bar, 1 SD of mean) ...................................... 329 Figure C.12 PE (dark grey bars) and PC content (light grey bars) (mg/g) for the samples under different
light treatments (n=3; error bar, 1 SD of mean) ..................................................................................... 329
Figure C.13 Vegetative thalli tied to nylon ropes in A) 1 l flasks and B) 500 ml flasks .......................... 332
Figure C.14 A) 3 x 1 l flasks and B) 3 x 500 ml flasks with tied cultures ............................................... 332 Figure C.15 SGR was calculated with the formula from Evans, (1972) for 1 l cultures (dark grey bars) and
500 ml cultures (light grey bars) (n=3; error bar, 1 SD of mean) ............................................................ 332 Figure C.16 Comparison of growth (SGR) between tied (dark grey line) and untied (light grey line)
cultures (n=3; error bar, 1 SD of mean) ................................................................................................. 333 Figure C.17 A) Bags used for the trials before the out plantation; B) Deployed bags on the farm site
(winter trial) ........................................................................................................................................... 335
Figure C.18 Average temperature (°C) on the farm site relative to the winter and summer trials ........... 336
Figure C.19 Average light (Lux) on the farm site relative to the winter and summer trials..................... 336 Figure C.20 Growth (weight) of winter trial (initial weight as dark grey bars and final weight as light grey
bars) (n=3; error bar, 1 SD of mean) ..................................................................................................... 337 Figure C.21 Moisture content for deployed cultures expressed in percentage (initial FMC as dark grey
bars and final FMC as light grey bars) (n=3; error bar, 1 SD of mean) ................................................... 337
Figure C.22 Carbohydrates (% DW) of outplanted cultures (n=3; error bar, 1 SD of mean) ................ 337 Figure C.23 Chl-a (dark grey bars) and carotenoids (light grey bars) concentration of out planted cultures
(n=3; error bar, 1 SD of mean) .............................................................................................................. 338
Figure C.24 FRAP (µM FeSO₄ eq/mg DW) of out planted cultures (n=3; error bar, 1 SD of mean) .... 338
Figure C.25 Protein content (% DW) of out planted cultures (n=3; error bar, 1 SD of mean) .............. 339 Figure C.26 PE (dark grey bars) and PC (light grey bars) (mg/g) concentration in out planted cultures
(n=3; error bar, 1 SD of mean) .............................................................................................................. 339 Figure C.27 Collected bags (outside-A; inside-B) after 6 months in the out plantation site for the winter
trial ........................................................................................................................................................ 339 Figure C.28 Epibiotic organisms on collected bags including A) tunicates; B) bivalve; C) polychaetes and
D) seaweed ........................................................................................................................................... 340
Figure C.29 Alive recovered material isolated from the bags of the summer trial .................................. 340

xxiv
List of tables
Table 1.1 Table of global aquaculture production of farmed aquatic plants expressed in thousand tonnes,
live weight (modified from FAO, "The state of World Fisheries and Aquaculture”, 2018) ......................... 4 Table 1.2 Production of farmed aquatic plants in the world expressed as thousand tonnes (modified from
FAO, 2018, “The state of world fisheries and aquaculture”. nei intended as “not elsewhere included”) ..... 5 Table 2.1 Number of specific and infraspecific taxa currently recognized/total number of taxa for each
genus of the Laurencia complex (modified from Rousseau et al., 2017) ................................................... 36
Table 2.2 Primers used for the PCR reaction (Modified from: Freshwater and Rueness, 1994) ............... 43 Table 2.3 Identities residues for the samples investigated and the data from NCBI. AF259495 refers to
the entry for L. pinnatifida from France; AF281875 refers to the entry for O. pinnatifida from Ireland ....... 48
Table 2.4 Pairwise distances expressed as p-distances values obtained with MEGA 7 software............... 48
Table 2.5 Percentage of Identity inferred with Geneious® software ....................................................... 48 Table 3.1 Metals recorded in seasonal samples of O. pinnatifida. Values are expressed as mg/kg dry weight
(n=3, error, 1SD of mean) ....................................................................................................................... 79 Table 3.2 The total variance explained by PCA analyses is presented in the table below, including
eigenvalues, together with proportion and cumulative values of variance. PC1 to 10 are the principal
components identified (analyses obtained with PRIMER v6) ................................................................... 83 Table 4.1 Peaks with putative IDs and retention time for the negative ESI mode; in bold under MS² the
most abundant fragments ...................................................................................................................... 108 Table 4.2 Peaks with putative IDs and retention time for the positive ESI mode; in bold under MS² the
most abundant fragments ...................................................................................................................... 109
Table 4.3 Putatively identified components organized by regulation trends (negative ESI mode data) .. 118
Table 4.4 Putatively identified components organized by regulation trends (positive ESI mode data) ... 118
Table 5.1 Epiphytes treatments and exposure time of vegetative thalli in Petri dishes trial .................... 140
Table 5.2 Epiphytes treatments and exposure time of small volume cultures in flasks trial ................... 140
Table 5.3 Epiphytes treatments and exposure time of scale up cultures in tanks trial ............................ 141
Table 5.4 Components of the F/2 medium .......................................................................................... 145
Table 5.5 pH, salinity and temperature measurements of the first trial .................................................. 150
Table 5.6 pH, salinity and temperature measurements of the second trial ............................................. 151 Table 5.7 Biochemical composition with relative concentration of the component analysed for the
outdoor trial (n=3; error 1SD of mean) ................................................................................................. 161 Table 6.1 Media formulation trialled: F/2 recipe (from Andersen 2005), Von Stosch recipe (from
Andersen 2005); F/2 modified. The nitrogen and phosphate sources with the respective concentrations
were taken from the Von Stosch recipe. All the other elements and concentration were kept as normal as
in the original F/2 recipe. ...................................................................................................................... 175 Table 6.2 Composition of treatments applied to cultures where control refers to F/2 medium. The final
molarity of N and P was the same for all the media tested ..................................................................... 177
Table 8.1 Phytohormones found in Rhodophyta (modified from Tarakhovskaya et al., 2007) ............... 219 Table 8.2 Treatments with PGRS and relative concentrations applied on vegetative fragments of O.
pinnatifida. Control only contained F/2 medium ..................................................................................... 221
Table 8.3 AMPEP treatments applied on juveniles cultivated in 6 × multi well .................................... 222 Table 8.4 Treatments with PGRS and relative concentrations applied on cultures of O. pinnatifida. Control
only included F/2 medium .................................................................................................................... 222 Table 8.5 AMPEP treatments applied on a flask trial. Samples under Bathing treatment were left in the
solution for 1hr, then rinsed from the excess of solution and incubated in F/2 medium ........................ 223
Table 8.6 Treatments applied on a tank system .................................................................................... 223 Table A.1 Percent recovery of the certified elements for the certified reference material CRM ERM-
CD200. Cu, Zn, As, Cd and Pb are expressed in mg/kg, error as 1SD ................................................... 311 Table A.2 Data of salinity, pH, DOC and TDN for water samples collected from June to August 2018 in
Dunstaffnage bay .................................................................................................................................. 311

xxv
Table A.3 Correlation between data of metals concentration (Al, V, Mn, Fe, Zn, U) in water samples and
precipitation data obtained from CEDA for Dunstaffnage #918 station from 2017-2018 ..................... 312 Table A.4 Correlation data between metals concentration (Co, Ni, Cu, Cd, Pb) in water samples and
precipitation data obtained from CEDA for Dunstaffnage #918 station from 2017-2018 ..................... 312 Table A.5 Variables and respective principal components for PCA. Values give information about the
correlation between variables and components ...................................................................................... 313

xxvi
List of abbreviations
(18S) 18S ribosomal RNA (2,4 D) 2,4 Dichlorophenoxyacetic acid (28S) 28S ribosomal RNA (4-DG) 4-deoxygadusol (5.8S) 5.8S ribosomal RNA (A.D.) After death (AA) Arachidonic acid (Abs) Absorbance (AHL) Acylated homoserine lactone (Al) Aluminium (ALA) Llpha-linolenic (AMPEP) Ascophyllum Marine Plant Extract Powder (ANOSIM) Analyses of similarities (ANOVA) Analyses of variance (AOAC) Association of Official Analytical Chemists (AOC) Assimilable organic carbon (APC) Allophycocyanins (As) Arsenic (ATP) Adenosine triphosphate
(B₁₂) Cobalamine (BA) 6-Benzylaminopurine (Ba) Barium (BCA) Bicinchoninic acid assay (BDOC) Dissolved organic carbon (BED) Bligh and Dyer Extraction (BEST) Statistical software included in PRIMER-6 (BL) Blue light (bp) Pair base (BSA eq) Bovine Albumin Serine equivalent (C) Carbon (C:N) Carbon: nitrogen ratio (C13-NMR) (C13) Nuclear magnetic resonance (C13-NMR) Carbon-13 Nuclear Magnetic Resonance Spectroscopy (C22:6, n -3, DHA) Docosahexaenoic acid (Ca) Calcium (CB1006R) External primer (Ga Hun Boo, laboratory of Prof. Sung Min Boo) (CB44F) External primer (Ga Hun Boo, laboratory of Prof. Sung Min Boo) (CC) Corps en cerise (CCT) Collision Cell Technology (Cd) Cadmium (CEDA) Centre for environmental data (Chl-a) Chlrophyll-a (CID) Collision-induced dissociation (Co) Cobalt
(CO₂) Carbon dioxide (COB) Cytochrome b (COI) Cytochrome c-oxidase subunit 1 extended fragment (COI-5P) Cytochrome c oxidase I gene (Cox 2-3) Cytochrome oxidase subunit 2-3 intergenic spacer (Cr) Chromium (CRM) Certified Reference Material

xxvii
(Cs) Caesium (CT or CTR) Controlled Temperature room (CT) Constant temperature (CTAB) Cetyltrimethylammonium bromide (Cu) Copper (DA) Domoic Acid (DCT) Daily cycle temperature (DDA) Data dependent analyses (DDA) Data-dependent acquisition (DGDG) digalactosyldiacylglycerol) (DNA) Deoxyribonucleic acid (DOC) Dissolved organic carbon (DOCN) Dissolved organic carbon and nitrogen (DON) Dissolved organic nitrogen (DPPH) α, α-diphenyl-β-picrylhydrazyl free radical scavenging method (DW) Dry weight (EDTA) Ethylenediaminetetraacetic acid (EF2) Elongation factor 2 (EPA) Eicosapentaenoic acid (ESI) Electrospray Ionization (ETR) Electron transport rate (F) Fluorescence yield (F0) Chlorophyll fluorescence yield in dark adapted tissue (FA) Fatty acids (FAO) Food and Agriculture Organization (Fe) Iron (Fm’) maximal chlorophyll fluorescence yield (FMC) Full moisture content (Fq′/Fm′) Operating photosynthetic efficiency of photosystem II (FRAP) Ferric Reducing Antioxidant Power (FSW) filtered seawater (FT) Fourier Transform detector (FTMS) Fourier-transform mass spectrometry (FTS) Fourier Transform spectroscopy (Fv/Fm) quantum efficiency of photochemistry in PSII (FW) Fresh weight (FWHM) Full width at half maximum (GAE/g) Gallic acid equivalent/g (GC) Gas chromatography (GEq) Glucose equivalent (GG) 2-O-α-d-glucopyranosylglycerol (H1-NMR) Proton nuclear magnetic resonance (H1-NMR) Proton nuclear magnetic resonance (HCl) Hydrochloric acid (HClO4) Perchloric acid (HD) High density (HEPES) 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HF) Hydrofluoric acid (HILIC) Hydrophilic interaction chromatography (HIV) Human immunodeficiency virus (HLI) Halogenated Indole Alkaloids (HNO3) Nitric acid (HPLC) High pressure liquid chromatography (HTCO) High temperature oxidation

xxviii
(I) Iodine (IAA) Indole-3-acetic acid (ICP-MS) Quadrupole Inductively Coupled Plasma Mass Spectrometer (ICP-OES) Inductively coupled plasma optical emission spectrometry (ID) Identification (IDT) integrated DNA technologies (IMTA) Integrated Multitrophic Aquaculture (In) Indium (iP-N6) Δ2-isopentenyl)adenine (ITS1-2) Nuclear ribosomal internal transcribed spacer (JECFA) The Joint FAO/WHO Expert Committee on Food Additives (K) Potassium (kDA) Kilodaltons (KI) Potassium iodide (Km) Half-saturation constant (KTH) Kerrison, Twigg, Hughes media (L:D) Light dark photoperiod (LC-MS) Liquid chromatography–mass spectrometry (LD) Low density (LDPE) Low Density Polyethylene (LED) Light Emitting Diode (LN) Natural Logarithm (LNA) Linoleic acids (LTQ-IT) Linear Ion Trap Mass Spectrometer (MAAs) Mycosporine-like amino acids (MCL) Maximum Composite Likelihood (MEGA 7) Molecular Evolutionary Genetics Analysis (Mg) Magnesium (MGDG) Monogalactosyl-diacylglycerol (ML) Maximum likelihood (Mn) Manganese (MS) Mass spectrometry (MS) Mean Square (MUSCLE) Multiple Sequence Alignment (N) Nitrogen (N:P) Nitrogen:Phosphorous ratio
(N₂) Liquid nitrogen (NA) Sodium (NaClO) Sodium hypochlorite (NCBI) National Centre for Biotechnology Information (NDIR) Non-Dispersive infra-Red
(NH₄⁺) Ammonium (Ni) Nickel (NJ) Neighbour joining (nm) Nanometres (NO) Nitric oxide
(NO₃¯) Nitrate (P) Phosphorous (PA) Phoshatidic acid (PAM) Pulse-Amplitude-Modulation (PAR) Photosynthetically active radiation (Pb) Lead (PB) Phosphate buffer (PBR) Photo bioreactor

xxix
(PC) Phosphatidylcholine (PC) Phycocyanin (PCA) Principal Component Analyses (PCR) Polymerase Chain Reaction (PDA) Photodiode Array Detectors (PE) Phosphatidylethanolamine (PE) Phycoerythrin (PEC) Phycoerythrocyanins (PG) Phosphatidylglycerol (PGE) Phoroglucinol equivalent (PGR) Plant growth regulator (PI) Phosphatidylinositol (PL) Phospholipids (PS) Phosphatidylserine (psaA) Photosystem I P700 chlorophyll-a apoprotein A1 (PSI) Photon Systems Instruments (PSII) Photosystem II (PSU) Practical Salinity Unit (ptfe) Politetrafluoroetilene (PUFAs) Polyunsaturated fatty acids (QY) Electron quantum yield (R1) Reagent 1 (Rb) Rubidium (rbcL) Large subunit of ribulose biphosphate carboxylase (RC) Reaction centers (g) Relative centrifugal force (RD) Red light (rDNA) Ribosomal DNA (ROS) Reactive oxygen species (rpm) Revolutions per minute (rRNA) Ribosomal RNA (RT) Retention time (RT) Room temperature (RuBisCo) ribulose-1,5-bisphosphate carboxylase/oxygenase (S) Substrate (SA) Salicylic acid (SAMS) Scottish Association for Marine Science (SD) Standard deviation (SDS) sodium dodecyl sulfate (SGR) Specific growth rate (Si) Silicate (SIMPER) Similarity percentages (Sr) Strontium (SSU) rDNA sequences of the nuclear subunit (TAG) Triacylglycerol (TBE) Tris-Borate-EDTA (TCSO) 1,4-Thiazane-3-carboxylic acid S-oxide (TDN) Total dissolved nitrogen (TDS) Total dissolved salt (TFAs) Total fatty acids (TL) Total lipid (TLE) Total Lipid Extracted (TOC) Total organic a carbon (TPC) Total phenol content

xxx
(TPTZ) 2,4,6-Tris(2-pyridyl)- s-triazine (U) Uranium (UK) United Kingdom (UPW) ultra-pure water (US$ or USD) US dollar (UV) Ultraviolet (UVB) Ultraviolet B (VS or VSE) Von Stosch (W0) Initial weight (WHO) World Health Organization (Wt) Weight at the time (WW) Wet weight (yBP) Years before present (Zn) Zinc

xxxi

1
Chapter 1 Introduction
1.1 Seaweed: a general introduction
Macroalgae, also commonly called seaweeds, include macroscopic, multicellular marine red, brown
and green algae (Lee, 2008), although all seaweeds, at some stage in their life cycles, are unicellular
in the form of spores, or zygotes, and may be temporarily planktonic (Amsler and Searles, 1980).
Seaweeds are ecologically and economically important, providing essential ecosystem services and
biomass for different applications, such as food, phycocolloids, animal-feeds, soil-additives,
nutraceuticals, chemicals, cosmetics, pharmaceuticals, botanicals and as sources of natural
pigments, bioactive substances, antiviral agents etc. (Chopin, 2007).
They have been used for centuries by humans and some species continued to play large economic
and social roles (Lüning and Pang, 2003). Due to the wide variety of applications, nowadays
seaweeds are intensively cultivated, and account for almost half of the biomass of the world
mariculture production (Chopin, 2007). In 2013, 11.3 million tons of fresh algae were produced
commercially, with a market value of US$ 5.7 billion, although in 2009 the market reached 15
million tonnes of harvested and cultivated macroalgae (FAO, 2013). Farmed seaweed production
now represents 95.6% of the global seaweed supply (Chopin, 2018a). Seaweed production is not
spread uniformly across the world and while many of the properties and potential contributions of
the algae to global issues of sustainability are clear to the scientific community, the common
perception of seaweeds is still not unbiased, particularly in Western countries. Here most algae are
associated with negative concepts, also connected to environmental problems, such as green tides,
or toxic, harmful algal blooms. Even the seaweeds that cast upon the shore as “storm toss” are
considered waste and garbage. In Asian countries, such as China, Japan and Korea, seaweeds are
utilised in traditional Chinese medicines (5000 years ago), and food (4th century in Japan and 6th
century in China, Fig. 1.1) (McHugh, 2003), and are responsible for the majority of the production
(FAO, 2018) (Table 1.1).

2
Except for Iceland, Wales and Ireland, the West (including Europe and North America) has been
somehow slower to adapt to the uses of algae, compared to Asia, Polynesia, South America,
Australia and New Zeeland, and new applications and industries have developed only in the last
few decades. Globally, about 220 species of algae are consumed as food, with six genera (Laminaria,
Undaria, Porphyra, Eucheuma, Kappaphycus and Gracilaria) responsible for 94.8 % of the total seaweed
biomass production (FAO, 2018). The second main application of seaweeds is in the hydrocolloid
industry.
Figure 1.1 Women Gathering Seaweed. From: Ukiyo-e woodblock print, about 1810's, Japan, by artist Katsukawa Shunka II
The most common hydrocolloids manufactured obtained from seaweeds are alginates, agars and
carageenans (Philips and Williams, 2009; Alba and Kontogiorgos, 2017). Research has also been
carried out to evaluate macroalgae as possible alternative protein sources for farmed fish, due to
their high protein content. In the last decades, research has focused on their utilization in
bioremediation of waste waters, in particular from aquaculture systems, due to their ability to
assimilate nutrients such as phosphorous and nitrogen, and to sequester other elements, dangerous
to the ecosystem, for example heavy metals (Davis et al., 2003). Recently they have also been
investigated for their use as biofuel, due to the high biomass productivity, the possibility of utilizing
marginal, infertile land, saltwater, waste-streams as nutrient supply and combustion gas as CO₂
source (Saifullah et al., 2014).
Before being cultivated, seaweeds had been harvested by coastal communities for centuries (Fig.
1.1). Today, harvesting of natural populations of seaweed is primarily for human consumption in

3
South East Asia and for hydrocolloid (phycocolloid) production, supplying algin, agar and
carrageenan in other countries (FAO, 2014).
1.2 Seaweed aquaculture
Figure 1.2 Extensive aquaculture in China, Fujian coast. Photo by George Steinmetz
The term “aquaculture” is defined as the cultivation of marine and freshwater organisms, both
animals and plants. The seaweed industry provides a wide variety of products with an estimated
total annual value of US$ 5.5-6 billion. Food products for human consumption contribute about
US$ 5 billion of this (FAO, 2014).
In terms of price paid by weight, seaweed used for food is the most valuable, particularly in Japan
with Nori (Porphyra sp.) used for Sushi. Furthermore, in the last 15 years, world production of
farmed seaweeds has more than doubled, with expansion particularly significant in Indonesia,
related to the species Kappaphycus alvarezii and Eucheuma spp. (FAO, 2018, Table 1.2), and China
(Fig. 1.2) (e.g. with the development of high-yield strains of major cultivated species). Among Asia’s
major producers, seaweed farming production has declined only in Japan, which was overtaken by
countries such as Indonesia, Philippines and Korea (Table 1.1). However, this fall in domestic
production has been offset by imports from neighbouring countries (FAO, 2018).

4
Table 1.1 Table of global aquaculture production of farmed aquatic plants expressed in thousand tonnes, live weight (modified from FAO, "The state of World Fisheries and Aquaculture”, 2018)
Country 1990 1995 2000 2005 2010 2012 2013 2014 2015 2016
China 1,470 4,162 6,938 9,446 11,092 12,832 13,479 13,241 13,835 14,387
Indonesia 100 102 205 910 3,915 6,515 9,299 10,077 11,269 11,631
Philippines 291 579 707 1,338 1,801 1,751 1,558 1,550 1,566 1,405
Republic of Korea
411 649 374 621 901 1,022 1,131 1,087 1,197 1,351
Democratic People’s
Republic of Korea
… … … 444 444 444 444 489 489 489
Japan 565 569 528 507 432 440 418 374 400 391
Malaysia … … 16 40 208 332 269 245 261 206
Tanzania 8 39 49 77 132 157 117 140 179 119
Madagascar … … … 1 4 1 4 7 15 17
Chile … … … 16 12 4 13 13 12 15
Solomon Islands
… … … 3 7 7 12 12 12 11
Viet Nam … … … 15 18 19 14 14 12 10
Papua New Guinea
0 0 0 0 0 1 3 3 4 4
Kiribati … … … 5 5 8 2 4 4 4
India … … … 1 4 5 5 3 3 3
Others … … … 25 14 16 13 12 16 8
Total 2,845 6,100 8,817 13,450 18,895 23,475 26,780 27,270 29,275 30,050
Beyond Asia, Zanzibar (the United Republic of Tanzania) in East Africa and the Solomon Islands
in the Pacific have experienced strong growth in seaweed farming (mostly Kappaphycus alvarezii) for
export markets (FAO, 2018). In some countries, including India, Timor-Leste, the United Republic
of Tanzania, Madagascar, Fiji, Kiribati and Mozambique, seaweed farming has been recognized as
offering potential for significant production volumes (FAO, 2018). Currently, these countries each
produce from a few hundred to a few thousand tonnes annually (except Mozambique) (FAO,
2018). FAO aquaculture statistics record all farmed aquatic algae under separated species or species
groups, categorized into 13 groups according to their nature and intended uses (FAO, 2018, Table
1.2).

5
Table 1.2 Production of farmed aquatic plants in the world expressed as thousand tonnes (modified from FAO, 2018, “The state of world fisheries and aquaculture”. nei intended as “not elsewhere included”)
Species 2005 2010 2011 2012 2013 2014 2015 2016
Eucheuma spp. 987 3,481 4,616 5,853 8,430 9,034 10,190 10,519
Laminaria japonica 4,371 5,147 5,257 5,682 5,942 7,699 8,027 8,219
Gracilaria spp. 933 1,691 2,171 2,763 3,460 3,751 3,881 4,150
Undaria pinnatifida 2,440 1,537 1,755 2,139 2,079 2,359 2,297 2,070
Kappaphycus alvarezii 1,285 1,888 1,957 1,963 1,726 1,711 1,754 1,527
Porphyra spp. 703 1,072 1,027 1,123 1,139 1,142 1,159 1,353
Seaweeds nei, Algae 1,844 3,126 2,889 2,815 2,864 449 775 1.049
Porphyra tenera 584 564 609 691 722 674 686 710
Euchema denticulatum 172 259 266 288 233 241 274 214
Sargassum fusiforme 86 78 111 112 152 175 189 190
Spirulina spp. 48 97 73 80 82 86 89 89
Brown seaweeds, Phaeophyceae
30 23 28 17 16 19 30 34
Others aquatic plants 20 28 27 28 18 15 14 17
Total 13,503 18,992 20,785 23,555 26,863 27,356 29,365 30,139
The most obvious change in the species composition of world farmed aquatic algae production is
the rapid increase in the dominance of Eucheuma seaweeds (Kappaphycus alvarezii and Eucheuma spp.)
farmed in tropical and subtropical seawater and used for carageenan extraction (Table 1.2). Their
production levels exceeded those of Japanese kelp in 2010 (FAO, 2014). Species classified as
“others” and Gracilaria spp. are mostly produced in China and a large proportion of their
production is used as feed for abalone and sea cucumber culture (Table 1.2). Farmed wakame and
Porphyra spp. seaweeds are almost entirely destined for direct human consumption. In addition,
agar and carrageenan extracted from other seaweed species are also destined for human
consumption in the form of thickening agents such as those used in some beverages (FAO, 2018).
In Europe, cultivation of seaweeds is starting to develop in countries such as France, Ireland,
Norway, Scotland, Ireland, Spain and Portugal. The first commercial cultivation trials in Europe
were initiated in the early 1980s in Brittany with the seaweed Undaria pinnatifida (Wakame) (Werner
et al., 2004). To this followed spreading and diversifying of seaweed cultivation methodologies in
other countries. Seaweed aquaculture has recently been established in North America, where the
seaweed industry is comprised of small wild harvest cottage operations located along the East and
West coasts of United States and few well-established aquaculture companies in Canada. European

6
countries are now starting to focus on establishing cultivation of low-volume high-value seaweeds,
the development of new applications for algae and the identification of specific algal compounds
that can be applied as food supplements, cosmetics, biomedicine and biotechnology. Also, on
environmentally sustainable production systems, such as Integrated multi trophic aquaculture
(IMTA), within a system comprising of finfish, shellfish and/or seaweeds (Ridler et al., 2007).
Furthermore, seaweeds are becoming more widely exploited by the recent trends in lifestyles
towards products that are viewed as natural and healthy. All these aspects can lead to a prosperous
future for advancement in seaweed aquaculture (Netalgae report, 2012).
1.2.1 Cultivation methods
Seaweed production methods are often very similar between countries, but with some variations.
Some seaweeds can be cultivated vegetatively, whilst others can only be cultivated by going through
a reproductive cycle, involving alternation of generations. Some parameters are general and
fundamental for the cultivation of any seaweed and include specific requirements such as salinity
of the water, pH, nutrients, water movement, water temperature and light (FAO, 2003). During
the design of the cultivation system the type of seaweed, together with the season, and the species’
overall requirements must be taken into consideration (FAO, 2003).
Nutrients are important in determining productivity and biomass yield, but they also influence the
abundance of epiphytes. There are three main categories of nutrients: macronutrients (carbon,
nitrogen (N), phosphorous (P), micronutrients (trace elements: e.g. iron, zinc, selenium, copper,
manganese, molybdenum) and vitamins (vitamin B12, thiamine and biotin) (Lobban and Harrison,
1994). In coastal waters, the concentrations of N and P can become limiting for seaweed growth.
They will vary significantly during the year with the highest concentrations in autumn/winter and
the lowest in spring/summer (Lobban and Harrison, 1994).
Wave exposure and tidal currents are particularly important, with some species growing
predominantly in sheltered areas, while others tolerate high exposure to wave action and tidal
currents. Commonly very exposed sites are to be avoided for cultivation systems due to problems
related to the stability of the system and the maintenance of the cultivation site. Water motion is
an essential factor for algal growth and has to be considered in tank cultivation to ensure and
enhance nutrient uptake and algal productivity (Hurd, 2000). Another critical aspect for aquaculture
sites located in bays, estuaries or shallow areas, with restricted water exchanges, could be
fluctuations in salinity, which is a fundamental aspect in algae cultivation, since every species is
adapted to a specific salinity range (Lobban and Harrison, 1994). Furthermore, every species is
adapted to different temperatures and has its optimal range for the maximum growth (Lüning,

7
1990). Elevated temperature, often associated with high irradiances, can be critical for Northern
Europe seaweeds and may lead to bleaching of the thalli. For these species, selected sites should
have temperature ranges of 5-15°C for most of the year (Kerrison et al., 2015). Usually sites for
seaweed aquaculture are collocated in areas with a minimum depth of 5 meters (to guarantee the
right amount of light) and a good water exchange (FAO, 2003). Salinity should range from 33 to
35 PSU, usually resulting in optimal growth, high nutrient loading area should be preferred
(potentially also with nutrient additions, Sun et al., 2008), and density should be considered for the
growth of the cultures (Kerrison et al., 2015).
Vegetative cultivation consists of cutting small pieces from the seaweed; these are taken and placed
in an environment that will sustain their growth. This can either be on land in a tank or in bags at
sea. When they have grown to a suitable size, they are harvested, either by removing the entire
plant or by removing most of it but leaving a small piece that will grow again. When the whole
plant is removed, small pieces are again cut from it and used as seed stock for further cultivation
(Ohno and Critchey, 1993; Santelices, 1999; FAO, 2003). The seedlings that have grown better
than the other are chosen to produce new biomass. Vegetative propagation of some seaweeds,
such as Gracilaria sp. and Ascophyllum sp., has proved to be hugely successful (Ohno and Critchey,
1993; Santelices, 1999; FAO, 2003). However, cultivation involving a reproductive cycle with
alternation of generations is necessary for many seaweeds; for these, new plants cannot be grown
by taking cuttings from mature plants. This is typical for many of the brown seaweeds (e.g.
Laminaria sp.). This kind of cultivation can be divided in two main phases: 1) indoor hatchery phase
and 2) sea-based grow out phase (FAO, 2003). The first phase focuses on the microscopic life
stages of the seaweeds that are seeded, nurtured and grow-out on rafts, nets or ropes. Usually this
phase is carried out in land-based facility were all the parameters of water, temperature, light and
nutrient are carefully kept under control. In the second phase (sea-based grow-out phase) the
seaweed rafts, nets or ropes are moved to coastal waters where the seaweeds grow to maturity.
Different bottlenecks can influence this type of cultivation: the principal difficulty in this kind of
cultivation lies in the management of the transitions from spore to gametophyte to embryonic
sporophyte (FAO, 2003). Current seeding and grow-out methodology in seaweed aquaculture
includes two methods: 1) the Western method, consisting of spraying of gametophytes onto
seaweed collectors from free-living cultures; and the 2) Asian Method, with the direct application
of spore suspension to seaweed collectors (FAO, 2003).
Different cultivation methods have been widely investigated, including suspended long ropes, in
the water between wooden stakes, or tied to ropes on a floating wooden framework (a raft).
Furthermore, netting can be used instead of ropes; seaweeds can be placed on the bottom of a
pond and not fixed in any way. In more open waters, certain type of seaweed can either be forced

8
into the soft sediment on the sea bottom with a fork-like tool or held in place on a sandy bottom
by attaching it to sand-filled plastic tubes. In the off-bottom method, monofilament nylon lines, or
polypropylene ropes are stretched between wooden stakes pounded into the substrate (Fig. 1.3 B).
The floating lines method is suitable in protected areas where water current is weak, or the water
is too deep for fixed lines. Normally the floating construction or raft is used to suspend the seaweed
about 50 cm under the surface (FAO, 2003).
Generally, large-scale farming approaches use low technologies in open water situations. These
techniques were first established in Asia and have then spread rapidly. Typically, in this form of
aquaculture there are basic fixed bottom (Fig. 1.3. B) or hanging (floating) ropes (Fig. 1.3. A).
Extensive seaweed aquaculture is also applied to re-seed previously wild harvested areas in attempt
to re-establish wild populations (e.g. Chile), or it can be applied to the “ranching of seaweeds”,
relatively popular in Japan, where new, coastal constructions (such as artificial islands) are seeded
with selected seaweed (e.g. Sargassum sp.) (Werner et al., 2004). Intensive seaweed aquaculture is
relatively expensive, requires equipment and involves qualified personnel (FAO, 2003). This type
of aquaculture has been applied in South-East Asia, utilising purpose-built ponds, exclusive for
seaweed or integrated with other systems (fish, shrimps, crabs, etc.), or tanks, where the
temperature is controlled and water circulation is required (FAO, 2003). Seaweeds are grown at
high density in these systems (Fig. 1.3. C); thus salinity, oxygen, light and nutrients must be carefully
monitored. However, the cultivation of seaweed in tanks on land is more costly than nearshore
cultivation (Sanderson, 2006; Werner and Dring, 2011). Seaweeds in ponds and/or tanks can also
face contamination from other organism (algae, bacteria, animals, grazers, etc.), affecting the quality
and quantity of biomass cultured. One possible solution is the use of a bioreactor, but this is
considerably more expensive, is appropriate only for the production of high value compounds
from algae, and it is currently mainly used for the growth of microalgae (FAO, 2003; Barahona and
Rorrer 2003).
Often algae are also cultivated as feed for other aquaculture species, such as abalone (Shields and
Lupatsch, 2012) or together with other species in IMTA systems (Neori et al., 2007, 2008;
Redmond et al., 2014) (Fig. 1.3. D). Seaweed are also used as biofilter for high-nutrient waters
derived from fish/mollusc cultivation. This is especially interesting where the cultivation of
seaweeds is not necessarily feasible as the sole form of biomass to be produced (Neori et al., 2007;
Redmond et al., 2014). Furthermore, the use of seaweeds as biofilter can be applied to ecological
studies linked to marine pollution, and they can be used to reduce heavy metals, radioactivity and
chemicals contaminations (Castagna et al., 1985; Conti, 2002).

9
Figure 1.3 Cultivation methodologies; A) suspended long-line; B) bottom line in shallow water; C) tank (indoor or outdoor); D) IMTA system
Another aspect to consider, when cultivating intertidal species, is their capacity to tolerate
desiccation. As the tide rises and falls, intertidal macroalgae must contend with both marine and
terrestrial conditions daily. Such alternations of submergence and emergence brings extremes in
physiological challenges (Lobban and Harrison, 1997; Helmuth and Hofmann, 2001). During high
tide, intertidal algae are underwater and generally experience reduced light levels and cool water
temperatures, while during low tides, intertidal algae may be exposed to increased light stress,
elevated air temperatures, and increased desiccation stress. When the tide returns, intertidal
organisms are rapidly submerged, rehydrated and exposed to cool water temperatures and reduced
light conditions once more. All phases of the tidal cycle affect the physiology and survival of marine
algae. Yet, few studies have tracked changes in algal performance throughout a tidal cycle. Intertidal
seaweeds resist a suite of abiotic stressors associated with submergence and emergence during the
course of a tidal cycle. Differences in the ability of macroalgae to tolerate stressors contribute to
habitat partitioning along the shore. It is well documented that tolerance to acute low tide stress is
higher for seaweeds growing higher on the shore (Bell, 1993; Hunt and Denny, 2008). Some
experiments have focused on the simulation of a tide system for macroalgae cultivation (Jones and
Dent, 1970; Fletcher and Jones, 1975; Pereira and Yarish, 2010; Mactavish and Cohen, 2014). Tidal
simulation systems constructed at a distance from coastal seawater supplies can be expensive,
elaborate, or both. Although a number of tidal simulator studies exist in the literature, there are
limitations linked to location, cost, and space associated with current designs, as well as a paucity
of information regarding relevance to field conditions (Mactavish and Cohen, 2014).

10
1.2.2 Epiphytes in cultivated seaweeds
Biotic interactions involve competition (for a resource, or space); epiphytism and allelopathy;
grazing and symbiosis (Lobban and Harrison, 1994). Epipthism is one of the most serious
problems encountered during cultivation and it can include biofouling of the crop by epiphytic
algae, or animals (Redmond et al., 2014). This may smother, overgrow, or graze the developing
crop, during the hatchery phase, or as the crop develops at sea. Grazing protozoans are important
components of natural aquatic ecosystems, and so are critical to the functioning of the marine food
web (Sherr and Sherr, 2002). In microalgal culture, contamination by protozoans and other
micrograzers can reduce algal abundance and productivity (Moreno-Garrido and Cañavate, 2001)
and so methods to screen for such contamination have been developed. Epiphytes usually
comprise undesirable nuisance algae such as diatoms, Ectocarpus sp. or Ulva sp. These algae compete
for nutrients and represent contaminants in the harvested crop. Many macrophytes are able to
deter epiphytes, either through periodic sloughing of their surface, or by production of antibiotic
chemicals (Gonzales and Goff, 1989b). Terpenoids are the most prevalent feeding deterrents in
tropical seaweeds (Lobban and Harrison, 1994), and they are one of the main compounds present
in the Laurencia complex species (Erickson, 1983; Faulkner, 1986; Bittner et al., 1985; Norte et al.,
1997; Pec et al., 2003).
Epiphytic growth on the cultured seaweed and/or on the pond’s walls can be prevented, their
growth rate slowed down, or they can sometimes be selectively killed off (Kerrison et al., 2016a,
b). Epiphyte control seems to be more easily managed in tank cultivation (Fletcher, 1995) than in
open systems, such as those typically used for large-scale seaweed cultivation (Brawley and Fei,
1987). The control over environmental conditions allowed by tank cultivation systems may be used
strategically to influence competition between maricultured seaweeds and epiphytes. The main
management tools are proper harvesting frequency and water flow (Buschmann et al., 1994),
changes in water level (Neori et al., unpublished), pulse feeding and a high stocking density
(Friedlander et al., 1991). Over-stocking and frequent slight changes in water level are particularly
effective in contrasting epiphyte growth on the pond’s walls. Since epiphytes are generally small,
they are rapidly affected by environmental stresses, a strategy that Asian mariculture operations
take advantage of by periodically raising Porphyra sp. nets out of the water and allowing the biomass
to partially dry (Neori et al., 2004). This kills much of the epiphyte biomass without significantly
harming the macrophytes. Problems with epiphyte infestation can be minimised, however, if total
crop management is implemented properly in the following manner: (1) careful selection of the
farming site with emphasis on water movement and siltation, (2) selection of quality ‘seedlings’
(free from epiphytes) to initiate cultivation, (3) use of the correct stocking density (g/m), (4) proper
culture technique, with frequent inspections of the material, and (5) good post-harvest management

11
(drying and storage) (Andersen, 2005). Many published studies have investigated macroalgal
disinfection. However, their aim has usually been the isolation of axenic laboratory cultures
(McCracken, 1989; Baweja et al., 2009; Reddy et al., 2008). The first steps are usually the selection
of source tissue with few visible epibionts, followed by physical removal of other epibiotic flora
and fauna by brushing, rubbing or using a razor blade (Baweja et al., 2009; Kerrison and Le, 2015).
This is followed by a chemical treatment to eliminate microorganisms such as bacteria, protozoa
and microalgae (Kientz et al., 2012; Baweja et al., 2009). This step should be carefully applied since
macroalgae lack a protective cuticle, and so are highly susceptible to chemical damage to their
tissues (Reddy et al., 2007; Baweja et al., 2009). Many of the methodologies for obtaining axenic
cultures utilise high concentrations and/or exposure times to harsh disinfectants, which can
severely affect the physiology of the host, leading to pigment loss or tissue degradation (Fernandes
et al., 2011). Nevertheless, as long as axenicity is obtained and the host is able to survive and
recover, these are acceptable. Disinfection during macroalgal cultivation has been reported in the
form of salinity or pH manipulation, desiccation and the use of chemicals or physical disruption
such as with a water jet (Hwang et al., 2006a, b; Xie et al., 2013).
Therefore, epithism must be more fully investigated in the context of the development of
cultivation in tank/mariculture system of new species, since it represents an obstacle to the long-
term maintenance of the cultures, growth in biomass, and quality of the harvested product.
1.3 Rhodophyta
1.3.1 Introduction
Red algae or Rhodophyta are a highly diversified group, species occur from polar, to temperate and
tropical waters of the world (Maggs et al., 2007; Robba et al., 2006). They are among the most
ancient organisms on the earth (Yoon et al., 2004, 2006; Robba et al., 2006), representing one of
the major radiations of eukaryotes (Ragan et al., 1994; Robba et al., 2006). Due to their complexity,
they can be present in many different forms and have different representative features. Their
lifecycles are intricate, and they have been valued for their unusual taste, their chemical components
and have been exploited for a long time (Cole and Sheath, 2011). The current number of known
red algal species is much greater than both the greens (ca. 1,100 species) and the brown algae (ca.
1,500 species) taken together (Graham and Wilcox, 2000), covering up to 6,000 species (Guiry and
Guiry, 2014). They can occur on the seashore, forming distinct zones, and in shallow subtidal
marine environment, where they also provide nursery for fishes. Some of the most commonly
recognized species of red algae are also the most commercially important, they can be cultivated
for food (Pereira et al., 2012), or for obtaining products and compounds that are used in a wide
range of industrial processes (Cole and Sheath, 2011).

12
1.3.2 Life cycle and reproduction
The main characteristics of this phylum are the presence of chlorophyll a, unstacked thylacoids in
plastids, plastids containing accessory pigments such as phycoerythirn, phycocyanin and
allophycocyanin, arranged in phycobilisomes, and food reserves stored as floridean starch
(Woelkerling, 1990; Graham and Wilcox, 2000; Graham et al., 2009; Harper and Saunders, 2001a,
b; Maggs et al., 2007). They also lack chloroplast endoplasmic reticulum, have pit connections
between cells of the thallus and they lack flagellated cells in the reproductive phase of their life
cycles (Woelkerling, 1990; Lee, 1999). The absence of motility has been crucial in the development
of a unique complement of reproductive structures to aid sexual reproduction and spore dispersal
(Saunders and Hommersand, 2004; Maggs et al., 2007). Male gametes are present in extracellular
mucilaginous appendages, which adjust their hydrodynamic properties, facilitating sperm
transportation. Furthermore, species-specific cell recognition proteins have been identified in male
gametes, for example, rhodobindin, which allows the attachment of the male gamete to the sessile
female gametes, called carpogonia (Delivopoulos, 2000). The female reproductive morphology
structures of carpogonium, carpogonial branch, and post fertilization development, have been
widely studied (Kylin, 1956). This was a crucial factor in red algal classification before the
introduction of DNA analyses. Red algae can have either heteromorphic or isomorphic life cycles
(Hawkes, 1990), but essentially most of them follow the same tri-phasic pattern: gametophyte,
carposporophyte and tetrasporophyte life phases (Cassano et al., 2009).
The extremely complexed life cycles of red seaweeds include the transition from unicelullarity to
complex multicellular bodies. Multicellular red algae (Rhodophyta) have indeed some of the most
complex life cycles known in living organisms, employing both sexual and asexual reproductions
(Searles, 1980; Guiry, 1987). They can have one, two or three free-living, morphological phases in
their life history and these may be morphologically similar, or different. In the cases of two or three
morphological phases, sexual and/or asexual cycles may be involved (Gabrielson and Garbary,
1986; West and Hommersand, 1981). It is well-known that reproductive processes are controlled
by one or more abiotic factors, including day length, light quality, temperature, and nutrients (Hurd
et al., 2014; García-Jiménez and Robaina, 2015). Likewise, endogenous chemical factors such as
plant growth regulators have been reported to affect reproductive events in some red seaweeds
(García-Jiménez and Robaina, 2015). Still, in the genomic era and given the high throughput
techniques at our disposal, knowledge about the endogenous molecular machinery lags far behind
that of terrestrial plants (García-Jiménez and Robaina, 2015).

13
Figure 1.4 Gracilaria sp. Cycle (modified from Lewmanomont, 1996)
In the red algae, the scheme of sexual reproduction includes the development of a specialized
female filament called the Carpogonial branch. The female gamete (carpogonium) presents a
trichogyne, which is an elongated extension responsible for receiving the male gametes. After
fertilization the zygotic nucleus develops into a diploid phase, called carposporophyte. Carpospores
develop into a second free-living phase called tetrasporophyte, which can be morphologically
similar (isomorphic alternation of generations) or different (heteromorphic alternation of
generations) from the gametophytes. Tetrasporophytic plants produce tetrasporangia by meiosis,
which release tetraspores. When released, each tetraspore will divide into four spores, giving rise
to either a male or a female haploid gametophyte (Fig. 1.4). Vegetative reproduction is also quite
common in red algae and many authors consider thallus fragmentation as the most significant kind
of vegetative reproduction in red algae due to the huge drifting biomass mats observed (Cole and
Sheath, 2011).
1.3.3 Distribution
Red algae are mainly marine, and they are geographically ubiquitous, having been reported in
tropical, temperate and cold-water regions (Cole and Sheath, 2011). They can be found at great
depths, up to 200 m, in the intertidal and in the subtidal, deeper than any other algal class, due to
the presence of their accessory pigments (Lee, 1999). However, red algae are less tolerant of low
salinity gradients compared to the Phaeophyta (Munda, 1977), below a salinity of 20‰ the number
of species decreases dramatically (Larsen and Sand-Jensen, 2006). Different species of red algae
live in the intertidal zone, influenced and controlled by the tidal levels. Algae stranded by the
retreating tides are subjected to aerial conditions until the tide returns and again different species

14
seem to be differentially susceptible to desiccation stress (Finckh, 1904), and this determines the
upper limits of several species of red algae on the shore (Hruby and Norton, 1979). Some species
of red algae thrive in relatively calm water sites, others in more turbulent localities (Conover, 1968).
Moderate water movement is beneficial, because it carries a supply of nutrients and gases to the
plants, also removing waste products. It is then not surprising that many seaweeds grow faster in
agitated, or flowing cultures than in stationary media (Whitford and Kim, 1966; Munda, 1977;
Hannach and Santelices, 1985; Parker, 1982). Muddy areas are generally inhospitable for red algae
(Norton and Mathieson, 1983) and most red algae inhabit rocky shores (Børgesen, 1908). In the
intertidal zones, red seaweeds must compete for space with ascidians, oysters, or zoanthids and
with fucoid seaweeds (Stephenson and Stephenson, 1972). The most distinctive zone is usually
found close or below the lower limit of barnacles (Stephenson and Stephenson, 1972). In cold
temperate zones, the red algal turf is often a less conspicuous feature of the shore; the most
characteristic species found in these regions are Chondrus crispus, Mastocarpus stellatus, together with
Corallina officinalis, Osmundea pinnatifida and Palmaria palmata (Lewis, 1964). Red algae are a major
source of food for a variety of herbivores, including molluscs (Carefoot, 1967; Dayton, 1971;
Kitting, 1980; Sheperd, 1973; Steneck, 1982) crustaceans (Carefoot, 1973; Nicotri, 1980) sea
urchins (Larson et al., 1980; Lawrence, 1975; Vadas, 1977; Verlaque, 1984) and fish (Horn et al.,
1982, 1985; Lundberg and Lipkin, 1979; Montgomery, 1980). Some red algae seem to encourage
the settlement and metamorphosis of the larvae of a variety of invertebrate grazers (Fretter and
Manly, 1977; Morse and Morse, 1984; Morse et al., 1979; Rumrill and Cameron, 1983; Steneck,
1982) (Cole and Sheath, 1990). The presence of an overlying algal canopy may allow the survival
of an undergrowth of red algae by providing protection from the excessive insolation or desiccation
or by inhibiting the growth of potential rivals (Kastendiek, 1982). The removal of such competitors
often enables red algae to extend the limits of their distribution (Hruby, 1976). This could also
allow the intertidal algae to colonize the lower subtidal zone (Kain, 1975). Growth rates of red
algae are influenced greatly by temperature, particularly in saturating irradiance, which is important
for the regulation of distribution of a number of red algae (Fralick and Mathieson, 1975; Yarish
and Edwards, 1982) triggering also seasonal behaviour (Breeman and ten Hoopen, 1981).
1.3.4 Light patterns
Light is the first fundamental “ingredient” for photosynthetic processes in seaweeds. The principles
of photosynthesis are similar in algae and terrestrial plants. Most of the catalytic proteins involved
in the thylakoid reactions of red algae are homologous with those in all other photosynthetic plants,
but some are analogous (Raven et al., 1990). Three kinds of pigments are directly involved in the
algal photosynthesis and they included chlorophylls, phycobiliproteins, and carotenoids (Meeks,
1974; Goodwin, 1974; Ragan, 1981; Rowan, 1989; Dring, 1990). Chlorophyll a is found in all algae

15
and plants. Seaweeds also have a wide variety of carotenoids. In red algae, the light harvesting
complex consists of phycobiliproteins, especially phycoerythrobilin. Each pigment protein
complex has a characteristic absorption spectrum that will influence the zonation of the seaweed,
making the photosynthetic apparatus of red algae unique in that only chlorophyll a occurs in these
organisms and phycobiliproteins as the major light-harvesting pigments. The phycobiliproteins
present in the red algae, greatly extend the light absorbance capacity by “filling in” where
chlorophyll absorption is low (Cole and Sheath, 2011).
Figure 1.5 Absorption spectra of photosynthetically active pigments initially studied for Porphyridium purpureum. Spectrum of solar radiation is represented as black dotted curve. Modified from Gantt 1975 (Cole and Sheath, 2011)
Phycoerythrin has a double absorption maximum at 545, 563 nm, and a shoulder at 500 nm.
Phycocyanin has a double peak at 550 nm and 617nm. Allophycocianin has a typical absorption
maximum at 650 nm (Mörschel and Rhiel, 1987; Zilinkas and Greenwald, 1986) (Fig. 1.5).
Photoinibition usually occurs at light intensities somewhat higher than light saturation (Kyle and
Ohad, 1986). This phenomenon is well documented in green plants, diatoms and dinoflagellates
(Richardson et al., 1983), but in red algae it appears to be less obvious. Usually much higher light
intensities are tolerated by red algae. However, upon exposure to desiccation conditions photo-
damage has been observed (Hodgson, 1981). A number of carotenoids have also been found in
red algae (Bjørnland et al., 1984), usually with β-carotene as the most common (Dixon, 1973).

16
1.3.5 Nutrients
Previous studies have demonstrated that C:N and N:P ratios of macroalgae are elevated in low-
level nutrient environments and that this reflects the increasing limitation of net productivity by N,
P, or both (e.g. Lapointe, 1987). C:N:P ratios of macroalgae vary as a function of taxonomic affinity
(Faganeli et al., 1986) and seasonality in nutrient availability (Chapman and Craigie, 1977; Lapointe,
1987). Among the major nutrient for algae, carbon is the most important. It seems that red algae,
along with other seaweeds, use bicarbonate as their source of carbon (Bidwell and McLachlan,
1985; Sand-Jensen and Gordon, 1984), the concentration of which in full salinity seawater rarely
limits photosynthesis (Sand-Jensen and Gordon, 1984), except in intensive cultivation (Bidwell et
al., 1985). Nitrogen is instead usually considered as the nutrient that limits the growth of an alga
(DeBoer and Ryther, 1977). This is valid also for red algae, considering that increased growth is
usually observed after the addition of nitrogen in field experiments (Hanisak, 1983) (Fig. 1.6) and
in laboratory cultures (Morgan and Simpson, 1981a; Neish et al., 1977). Seasonal variation in
nitrogen content of natural population of different red algae, e.g. Chondrus crispus (Asare and Harlin,
1983; Butler, 1936) and Porphyra tenera (Iwasaki and Matsudaira, 1954), may be linked to patterns of
growth and nitrogen availability (Cole and Sheath, 2011). Based on the ratios of nitrogen to
phosphorus concentrations (i.e. Redfield ratios), it has been suggested that phosphorus is most
likely to be the limiting nutrient, for example in coral reef environments (Wheeler and Bjornsater,
1992). Although, studies conducted on Rhodophyceae including on Gracilaria sp. (Lapointe, 1987)
have demonstrated how phosphorous can be just as an important factor, even more than nitrogen,
in winter and summer growth in temperate species.
Figure 1.6 Relationship between growth rate and internal nitrogen content, where “A” represents the minimum nitrogen content for growth, the slope is the proportion between growth and internal nitrogen content and “B” the critical value (Hanisak, 1983)

17
1.3.6 Cell wall and storage of reserves in red algae
With the exception of a few very specialized cases, the cell structure is similar in all the red algae.
Red algal thalli can be interpreted as a group of filaments held together by cell-wall interactions,
mucilage and pit-connections (pseudoparenchyma). Red algal cell walls are composed of cellulose
fibrils (rarely xylan fibrils) and a matrix of hydrocolloids. Cell wall hydrocolloid matrixes in red
algae are formed by sulfated polysaccharides classified in two main groups: agar and carrageenan.
Resulting in the high degree of complexity of cell walls in red algae (Fig. 1.7A). The most important
storage product is a polysaccharide, floridean starch (Kutzing, 1843). Grains of this material are
formed in the cytoplasm, next to the chloroplast envelope (Fig. 1.7 B) (Cole and Sheath, 2011).
Figure 1.7 A) Schematic view of cell wall in red algae: dark grey cylinders= cellulose microfibrillis; green line= glucomannan; red line= sulphated glucan; grey twisted line= sulphated xylogalactans; white circle= known link; grey circle=hypothesized link (modified from Lechat, 1998, in Fleurence and Levine, 2016). B) Electron micrograph of P. aerugineum: p= pyrenoid; s= starch grains; g=golgi bodies; n= nucleus (Modified from Gantt, Smithsonian Institution, in Dawes, 1998)
1.3.7 Economic utilisation of red seaweeds
The first recorded utilisation of seaweed as herb and food in China dates back to the 3rd century
B.C. (Read, 1936). Since then, they have been mainly used as human food and as a raw material for
the extraction of a variety of compounds with a high industrial/commercial importance to relatively
minor resources such as food and food additives in underdeveloped countries (Dixon, 1973).
However, their full agronomic and biotechnological potential has yet to be realized (Cole and
Sheath, 2011). Few studies have considered the application of red seaweed as animal feeds, since
these algae are less employed compared to the brown algae in this sector. Furthermore, the only
reported use as fertilizer is linked to the application of coralline algae as a source of lime (Dixon,
1973). At least 344 species of red algae are considered to be economic valuable (Tseng, 1981), but
to date only species of Porphyra, Gelidiella, Gloiopeltis, Euchema and Gracilaria have been cultivated
with well-established and extensive applications. As already stated, the use of algae in human food

18
has been extensive for centuries in Asian countries (China, Japan, Indo-Malayan region, Hawaii
and Philippines) (Newton, 1951; Chapman, 1970). In the West (Europe and North America) the
two most important species of red algae used as food were initially Palmaria palmata and Porphyra
spp. (usually under different colloquial name such as Dulse or Crannough), in Ireland and Scotland
(Dixon, 1973). In addition to the listed species, there are many other red algae for which there have
been used at a low level, among those Osmundea (Laurencia) pinnatifida was probably the most
common, also called Pepper Dulse and traditionally used as condiment (Dixon, 1973).
The use of red seaweeds as polyelectrolytes, pharmaceutical products, human nutrition,
antimicrobial activity, polysaccharide production, resource management, etc., was recognized and
has been investigated since the eighties (Bird and Benson, 1987). One of the main applications of
red seaweeds is still in the hydrocolloids industry. The first successful extraction of carrageenan
dates back to 1844 (Schmidt, 1844) and since then, seaweed extracts such as alginate, carrageenan
and agar have been widely used in the Western food industry, pharmaceutical and cosmetic sectors
(Nussinovitch, 1997). Agar production (US$ 132 million) is principally from two seaweeds, Gelidium
and Gracilaria, with Gracilaria been cultivated since the 1960-70s, but on a much larger scale since
the 1990s, allowing the expansion of the agar industry (FAO, 2004).
Carrageenan production (US$ 240 million) was originally dependent on wild seaweeds, especially
Chondrus crispus (Irish Moss), a small seaweed growing in cold waters, with a limited resource base.
However, since the early 1970s the industry has expanded rapidly because of the availability of
other carrageenan-containing seaweeds that have been successfully cultivated in warm-water
countries with low labour costs. Today, most of the seaweed used for carrageenan production
comes from cultivation, although there is still some demand for Irish Moss and some other wild
species from South America (FAO, 2003). The introduction of cultivation of species of Eucheuma
sp. in the Philippines during the 1970s provided the carrageenan industry with a much-enhanced
supply of raw material (FAO, 2003). Today most of the raw material comes from the two species
originally cultivated in the Philippines, but their cultivation has now spread to some other warm-
water countries, such as Indonesia and Tanzania (FAO, 2018). Limited quantities of wild Chondrus
sp. are still used; although attempts to cultivate Chondrus sp. in tanks have been successful
biologically, with valuable exploitation as food ingredient, but uneconomic as a raw material for
carrageenan (FAO, 2003; Buschmann et al., 2017). Wild species of Gigartina and Iridaea from Chile
have also been harvested and efforts have been made to find cultivation methods for these (FAO,
2003).

19
1.4 Scotland
Scotland lies on the edge of the European continental shelf, with the Atlantic Ocean to the west
and the North Sea to the east. Warm-water currents, driven along the edge of the shelf by the Gulf
Stream, keep the West coast climate warmer and wetter than that of the east coast. Waters travelling
down from the Arctic keep the north and east cooler. Scotland's coastline is almost 11,800 km
long. Its position at the edge of the continental shelf, the long coastline, its highly variable and
productive sea, and the mixing of warm and cold-water currents combine to make the waters
around Scotland a special place for marine wildlife and habitats, included seaweeds. Water depth
and exposure to wave action and tidal streams together determine which seaweeds and animals that
live on and within the seabed. From exposed vertical walls, plunging to the depths in clear waters
around Scotland's offshore islands to the sheltered kelp fringed shores of the sealochs, Scottish
seas support a wealth of rocky reef habitats (Scot. Gov., 2013).
1.4.1 Scotland and aquaculture
Nearly half of Scotland’s population lives in close proximity to the coast (within 5 km). Scotland’s
coastal economy is made up of a diverse number of sectors with connections to both terrestrial
and marine resources. For coastal communities, marine fisheries and associated industries (fish
processing) are significant, particularly in the northeast. In 2010, the value of landings by Scottish
vessels within the fisheries industry was approximately £428 million and provided direct
employment for 5,218, with Aberdeenshire employing 26% of those. Issues relating to
sustainability, such as previous over-exploitation of some stocks may have contributed to industrial
downsizing, income reduction and fewer job prospects. Coastal communities have been
experiencing a gradual trend towards part-time employment. Consequently, based on the recorded
and projected declines in fishing and fossil fuel related industries (51% drop in tonnes of products
since 2005); the future socioeconomic stability of coastal communities will depend on economic
diversification, such as the development of the aquaculture industry, predominantly on the West
coast. In 2009, the aquaculture industry was estimated to be worth £427m supporting over 1,000
full-time and 1,146 part-time jobs by farmed finfish production and 514 jobs by farmed shellfish.
More recently, provisional figures value the industry around £434m per year. Aquaculture is one
of the fastest growing food production sectors and offers employment in remote and rural
communities (Scot. Gov., 2013).
The main species cultivated in Scotland is Atlantic salmon making it the largest producer in Europe,
and in third place globally with a total production of 179,022 tonnes in 2014 (Scot. Gov., 2013).
Scottish salmon is particularly important to the country’s economy, considering that it is one of the
most significant exports, delivered to more than 50 countries. Rainbow trout, halibut and brown

20
trout are also important. Scottish shellfish production was dominated by farmed blue mussel
production at 7,683 tonnes in 2014. Pacific oyster (3,392.000 shells), native oyster (242,000 shells),
king scallop (48,000 shells) and queen scallop (18,000 shells) were also cultivated in 2014 (Scot.
Gov., 2016). A few small-scale seaweed cultivation sites have been established recently in Scotland
with the product from these seaweed farms which is likely to be used for food for human
consumption, animal feed, nutraceuticals, co-digest for anaerobic digestion plants, and
fertiliser/soil enhancer. This sector is expected to grow along with Integrated Multi-Trophic
Aquaculture. The potential for seaweed farming in Scotland is high; however, at present the
commercial seaweed industry is limited to wild harvesting by a few SMEs, or on a pilot phase
(Capuzzo and McKie, 2016).
1.4.2 Scotland and seaweeds: an ancient tradition
Eating seaweed is generally considered an unusual activity in most of Europe (Guiry and Blunden,
1991), although coastal parts of Scotland and Ireland form exceptions. Scotland had a history of
macroalgal widespread harvesting (Fig. 1.8), which was used as a raw material for the chemical
industry and has a huge potential to cultivate macroalgae.
Figure 1.8 Scottish crofter 1880 - collecting seaweed on Skye. Photo by George Washington Wilson
Seaweeds have traditionally been used in Scotland both as food and for medicinal purposes, which
has led to seaweed being investigated for their natural product content. The first reference about
the use of seaweed in Scotland goes back to 600s AD, in a questionably poem that refers to the
monks of Iona collecting Dulse (Palmaria palmata) from the rocks as food (Ross, 1996). Seaweeds
such as Dulse, especially for coastal crofters, has provided a staple part of the diet throughout the

21
Western coast, where this was eaten with oatmeal in a thick broth, or simply boiled and served with
butter as a separate dish. From the middle of the 1800s, there is some references to the use of
Dulse as medicine, against intestinal parasites. Dulse was also sold along the East coast, together
with Alaria esculenta and Laminaria saccharina. In addition, is long standing the use of species such as
Chondrus crispus, exploited in the Hebrides, primarily to make a jelly-like pudding. A number of
other species have also found their way into the diet of the people of the Highlands and Islands,
including Himanthalia elongata, which has been collected and eaten for centuries. Another main
application of seaweed was as feed for cattle (Pelvetia canaliculata and Ascophyllum nodosum). Farmers
used to employ seaweed as a fertilizer: they were abundant, nutrient-rich and alkaline, they are
particularly suited for use as a fertiliser on Scotland’s generally acidic soils. It is mainly the large,
brown species that have been used. In the past, the seaweeds were taken from winter storm-cast
and dug directly into the soil after a short period of composting. In addition, seaweed has been
used effectively in the Highlands and Islands of Scotland as a traditional medicine for many years
to treat various conditions. Several species have been traditionally used in cooking and to some
extent are still used today (Kenicer et al., 2000). Despite over 30 years of research, the majority of
species found in Scotland have yet to have their chemistry examined and are not well established
in aquaculture. Hence, further analyses and research is necessary to better develop these activities.
There are a wide variety of small, dynamic businesses currently making use of seaweed in Scotland
- mainly producing high value foodstuff and cosmetics. The largest sector using algae apart from
the alginates industry is in agriculture as a fertilizer. Seaweed use in Scotland can be seen to have a
rich history, underpinned by a boom and bust pattern. The industry has never stood still and has
always had to reinvent itself to maintain a modest success. Despite seaweed being used extensively
for their health benefits in the Highlands and Islands for many years, there is relatively little peer-
reviewed literature available on this subject. It is striking that within a region with such a rich
heritage of using natural resources more research has not been carried out.
1.4.3 Scotland seaweed chemical industry history
The Scottish seaweed industry has had three distinct historical phases:
1. The production of kelp for use in the glass and soap industries: from the 17th to the 20th century.
Extraction of soda and potash from Fucus spp., Ascophyllum spp. and Laminaria spp. provided a
ready supply of these chemicals for the British Isles. This allowed independence from the other
main centre of production in Spain. The Western Isles and Orkney archipelago were the main
centres of kelp ash production (Adams, 2016). Soda and potash were important chemicals in the
soap and glass industry and were widely used for linen bleaching. The extraction process involved

22
burning the kelp in large, often stone-lined trenches. Thanks to these applications, the Scottish
soda industry at the end of the 1700 used 400,000 tonnes of harvested seaweeds (Scot. Gov., 2013).
2. The production of kelp as a source of iodine: the Laminarians were first collected for the soda
industry they also contained, but it was not until later that they were used for the manufacture of
iodine. The discovery of iodine has represented a flourishing industry in Scotland, before its decline
after the middle of 1800’s due to the growth of the Chilean producers (Adams, 2016).
3. The use of seaweed as raw material for the alginate industry: at the end of 1800’s alginates were
discovered from Laminaria spp. (Stanford, 1893). Alginates are jelly-like carbohydrates and are used
for their water holding, gelling, emulsifying and stabilising properties. These properties turned out
to be useful in a wide range of applications, in the food industry, cosmetic, medical, paint, and
pharmaceutical industries (Pereira, 2018).
1.4.4 Scotland and commercial harvesting of wild seaweeds
Scotland has an established history of macroalgal harvesting and the potential to cultivate
macroalgae. According to the Scottish government, Scotland's wild seaweed “production” is
currently based on the harvesting and picking of wild seaweed stocks on Orkney, Shetland and the
Western Isles, with only a number of small-scale operations in these areas. This industry is
estimated to make up only a minimum part of the European market. Commercial operations are
undertaken in the Northern and Western Isles, mainly for animal feed supplements and fertilisers
(Capuzzo and McKie, 2016).
Current annual seaweed harvest is approximately 6,000 wet tonnes, with Scottish operators
harvesting a range of wild brown, red and green seaweed stocks, although the main type harvested
is Ascophyllum nodosum, with around 5,000 tonnes of wild plants harvested from intertidal waters in
the Western Isles each year (James, 2010). Smaller quantities of Fucus serratus, and kelp in Orkney
and Shetland are also harvested (Scot. Gov., 2013). “Picking” is a term given to the moderate
removal of smaller species, typically red seaweeds, which live at the top of the shore. Species such
as Mastocarpus stellatus and Chondrus crispus, the Channel Wrack (Pelvetia canaliculata), Dulse (Palmaria
palmata and Dilsea spp.), Pepper Dulse (Osmundea pinnatifida) and laverbread (Porphyra spp.) are also
picked commercially in Scotland. An unknown quantity of cast seaweed is gathered from the shore
in many of Scotland's island communities, for use as a soil conditioner or fertiliser. The Crofters
(Scotland) Act 1993 (as amended) gives crofters access to reasonable use of seaweed under
Common Grazing regulations, although it is understood that this is largely confined to the
gathering of beach-cast Laminaria spp. and other species mixed with it for spreading on machair
land in the Western Isles. As such, little information is available about the extent or size of such
gathering (Scot. Gov., 2013).

23
In Scotland, hand cutting has previously been the most common method of harvesting in the wild,
with tools such as serrated sickles or scythes used from the rocks at low tide. Mechanical harvesting
has also been used in Scottish water where feasible. Mechanized harvesting methods vary, and can
involve mowing seaweed areas with rotating blades, cutting seaweed plants with suction methods,
and dredging of areas with cutters. Modern harvesting vessels have been specifically developed for
these purposes, although harvesting methods vary. The impact of harvesting can differ according
to different considerations: the species harvested and its life cycle, the method adopted, the
proportion of harvested thallus, the level of exposure and the role in the coastal environment and
processes, the presence or absence of grazers and competitors. The method of harvesting will also
affect the regrowth of the seaweed, influencing the ecosystem related to it (Stagnol et al., 2013).
Many species, such as A. nodosum, can regenerate quickly if the frond is not cut back to the rock;
however, the removal of a seaweed in its entirety, or close to the holdfast is unlikely to permit
regrowth of a new individual (Stagnol et al., 2013).
Several studies into the impacts of harvesting techniques and regeneration of seaweeds have
observed changes in the composition of seaweed communities after harvesting (Kelly et al., 2001).
These changes are often temporary until the targeted and harvested species has undergone
sufficient regrowth. Other factors such as the frequency, magnitude and seasonality of harvesting
activities can all influence the ecological impacts of seaweed harvesting operations. The main risk
in expanding wild harvesting in Scotland is likely to be over-harvesting and the use of harvesting
practices that impair the regeneration capability of seaweed communities (i.e. removal of plants at
or close to the holdfast). These can lead to significant changes in the composition of the seaweed
communities (i.e. fewer species or changes in species proportion) and have secondary impacts on
the marine ecosystems that they inhabit (i.e. adverse impacts on biodiversity of marine fauna). By
contrast, a controlled harvesting well-managed could hasten the regeneration of some species and
seaweed communities. Frequent harvesting of the same natural seaweed communities, even if just
a proportion of the plant is harvested, may instead limit effective regrowth and contribute to long-
term loss of habitat in consequence and adversely affect future harvest yields. Other risks associated
with an uncontrolled and frequent harvesting activity are the reduction in the structural diversity
of a marine community, the spreading of non-native species, and a reduction in storm protection
with an increase of coastlines susceptibility. Currently there is no evidence that seaweed harvesting
is already affecting Scotland’s environment; however, all the problems and impacts described above
could become substantial with the future growth of this industry (Scot. Gov., 2016).
The harvest of wild macroalgae, directly or as storm cast material, in the volumes required to
produce regionally or nationally significant quantities of products is unlikely to be environmentally
acceptable or sustainable, now and in the future. It is important to find, establish and promote

24
cultivation activities in order to reduce harvesting impacts and guarantee the production of high-
value and wild-independent products.
1.5 Osmundea pinnatifida (Hudson) Stackhouse, 1809:79
Phylum Rhodophyta
Subphylum Eurhodophytina
Class Florideophyceae
Subclass Rhodymeniophycidae
Order Ceramiales
Family Rhodomelaceae
Genus Osmundea
Species Osmundea pinnatifida (Hudson) Stackhouse, 1809:79
Osmundea pinnatifida is a small red seaweed that has been reported in Europe (Britain, Bulgaria,
Faroe Islands, France, Ireland, Norway, Portugal, Spain) and Atlantic Islands (Canary Islands,
Madeira, Savage Islands); North America (Florida, North Carolina) and South America (Brazil);
Africa (Cape Verde, Ghana, Mauritania, Morocco, São Tomé and Príncipe, Senegal, Western
Sahara) Asia (Cyprus, India, Indonesia, Korea, Oman, Pakistan, Philippines, Taiwan,
Turkey,Yemen) and Australia and New Zeland (Guiry and Guiry, 2018) (Fig. 1.9 A and B).
Figure 1.9 Distribution of Osmundea pinnatifida A) Worldwide (adapted from Guiry and Guiry 2018 records) and B) in UK (Hardy and Guiry, 2003)
This species grows on open rock surfaces, confined to the eulittoral-intertidal zone of sheltered to
exposed coasts (Fig. 1.10), often covering damp slopes, either together or separately from Gelidium
pusillum. O. pinnatifida communities usually occur on shores on which a fucoid canopy is reduced in

25
extent, or even absent. Other turf-forming red seaweeds, such as Corallina officinalis, Mastocarpus
stellatus, Ceramium spp. and Callithamnion hookeri may be present, although O. pinnatifida always
dominates in this habitat, changing its morphology and colour according to the position on the
shore (Fig. 1.11).
Figure 1.10 Distribution of O. pinnatifida on the shore
Figure 1.11 Samples of O. pinnatifida on the seashore from the same site in A) November, B) March and C) June
It can be found all year round but is at its best in December through to May. It forms turf perennate
in June-September, these creeping mats of prostrate axes continue to grow outwards, increasing
the size of the turfs. Erect fronds develop from October onwards, reaching their maximum size in
February-June. According to the description by Maggs and Hommersand (1993), Osmundea (as
Laurencia) pinnatifida thalli forms extensive turfs (Fig. 1.12 A), attached by closely interwoven
stolonous holdfasts. This species presents an erect axis 2-8 cm high; the main axis is compressed,
1-2.5 mm wide and 0.6-0.8 thick. Three layers of dark pigmented cortical cells surround multiple
layers of light pigmented medullary cells (Fig. 1.13). Branches develop irregularly, alternate-
distichously to 4-5 orders, complanate (Fig. 1.12 B). In spite of being a red seaweed, O. pinnatifida
can appear of different colours: from a black or brownish purple to bleached yellow, with apices
greenish-brown in transmitted light. Fronds appear severely bleached and damaged in early
summer, due to the low tide and high temperature/light irradiance (Prathep et al., 2003). The
texture is thick and cartilaginous.

26
Figure 1.12 A) O. pinnatifida on a rocky shore (Ganavan beach), B) collected specimens
Figure 1.13 Transverse section of thallus at the stereo microscope; externally are visible dark pigmented cortical cells and internally multiple layers of light pigmented medullary cells (bar=250µm)
Male and female structures are carried by different plants (Fig. 1.14). These structures occur laterally
on the last order branchlets and consist of urn-shaped male structures and oval female structures
(approx. 1mm) without protruding pores. In tetrasporophytes, the tetrasporangia appear as tiny
dark specks (concentrated around the last order branchlets) (Fig. 1.15 B). Tetrasporangia bearing
tetraspores (Fig. 1.15 C) are borne randomly on cortical cells in the last three orders of branching,
in branchelets that are initially compressed, becoming terete. The mature tetrasporangia size varied
between 75 and 125 µm, and this has also been observed for the samples collected for this study
(pers. comm). Tetrasporangia have been recorded in October and December-June. Female plants
are rare and cystocarps have been noted only in March and June. Fertile female gametophyte
presents cystocarps (Fig. 1.15 A) laterally located, mainly in second-order branches, sessile, slightly
ovoid, with a non-protuberant ostiole. Fertile male branches show pocketshaped spermatangial pits
with apical pores, located at the bifurcations of the ultimate branchlets or laterally in series.
Spermatangial branches are of the filament-type, on the contrary of the Laurencia genus where the
spermatangial branches are of the trichoblast-type (making this one of the key factors in
recognizing and distinguishing the Osmundea genus from Laurencia) terminating in a vescicular,

27
sterile cell. It is assumed that the urn-shaped spermatangial pit with an ostiole-like upper opening
is more protective of the spermatangial trichoblasts/filaments than the open cup-shaped form.
This may have led to the evolutionary loss of the sterile branches in Osmundea clade with urn-shaped
pit structure. Transparent trichoblasts are visible on terminal swelled tips in late May in samples
from the West coast of Scotland (author pers. obs.) (Fig. 1.15 D).
Figure 1.14 Life cycle of O. pinnatifida: (a) male gametophyte (picture from Machín-Sánchez et al., 2012); (b) female gametophyte; (e) tetrasporophyte; (f): tetraspores (bar=100µm); (g) released spore

28
Figure 1.15 Ultrastructure of O. pinnatifida A) Cystocarp (1000 µm, Zeiss microscope); B) Tetrasporocystes with visible tetraspores (scale bar=250 µm); C) Tetraspores (optical microscope); D) Transparent trichoblast on terminal swelled tips of O.pinnatifida samples (optical microscope) (bar=500µm)
Once released the spores (haploid) germinate and generate gametophytes (haploids) (Fig. 1.16 A-
D) that will then merge and fertilize the cystocarp forming the carposporophyte (diploid, develops
on the haploid female thallus). This will then release diploid carpospores that generate new
terasporophytes (diploid).
Figure 1.16 O. pinnatifida A) Germinated spores after release, B) Growing spore after 2 months from release; C) developing gametophytes; D) developed gametophytes after 6 months from release
This species has a strong chemical smell and flavour, due to the peculiar chemical composition of
its biomass. The origin of its peppery flavour is unknown. However, might be hypothesized that it
derives from secondary metabolites abundant in this species (Erickson, 1983; Faulkner, 1986;

29
Bittner et al., 1985). These molecules often have a protective function and deter grazing by molluscs
and fish (Brito et al., 2002). For us, the taste, probably created by these secondary metabolites,
turns this seaweed into a new, surprising high value food ingredient. Furthermore, the peculiar
biochemical profile makes this species a valuable source of antioxidant, antiviral, antibacterial,
antifungal, anti-cancer, and anti-fouling compounds that could be exploited in nutraceutical,
cosmetic and pharmaceutical fields (Flodin et al., 1999; Iliopoulou et al., 2002a, b; Pec et al., 2003).
Currently the production is guaranteed only by wild harvesting, but this is a difficult laborious
work, especially when it is necessary to collect it in large quantities, and great care is needed when
harvesting this seaweed in order not to eradicate the species in any given area, together with other
organisms related to it. Furthermore, wild harvesting is subjected to seasonality variations. Hence,
it is fundamental to develop a sustainable cultivation method for O. pinnatifida production and
supply.
1.6 Project aim
Commercial seaweed production is “booming” with an estimated worldwide annual value of US$
6.4 billion (Cottier-Cook et al., 2016). A rising number of valuable applications for macroalgae is
driving this global seaweed demand. To meet the market needs, establishment of sustainable supply
chains for algal products are required. In Europe, products from seaweeds are breaking new
ground, especially in the gourmet food sector as low-volume high-value raw materials. O. pinnatifida
is currently only collected from the wild and marketed at a high price, dried as a peppery seasoning,
due to its unique taste. It is mainly used in food preparation and a packet (5g dry weight) retails at
approximately £12. There is no universal handbook covering how to establish new cultivation
systems for new species, such as O. pinnatifida, so novel solutions have to be developed (Fig. 1.17),
aimed at reducing the harvesting impact, improving the crop yield and understanding how to
control the cultivation cycle in order to tailor the final product according to the market needs. The
research presented in this thesis focuses on identifying the main parameters to achieve cultivation
initially in the laboratory as well as in outdoor systems. Furthermore, the biochemical composition
of the species will be investigated, taking into account seasonal variations as well as its variation
according to the cultivation parameters applied. This will give an insight into the particular
properties of this species, proving evidence that it is a good candidate for the production of
secondary metabolites that can be further exploited in the nutraceutical and pharmaceutical fields.

30
Figure 1.17 Flow chart of the possible cultivation methodologies for Osmundea pinnatifida
This industrial Ph.D. was a challenging project but represents the chance to translate research from
the academic field into a commercial reality by researchers and industry working together.

31
1.6.1 Thesis objectives
The key objectives of this research were:
To phylogenetically characterize Osmundea pinnatifida specimens’ prior cultivation
experiments as well as investigate the phylogenetic relationships between specimens from
the West coast of Scotland within the context of concerns about genetic pollution due to
the introduction of alien species for farming purposes. (Chapter 2)
To assess the seasonal variation in biochemical composition and metabolomic profile of
Osmundea pinnatifida from one selected site on the West coast of Scotland across one year
and to explore the relation between these and the abiotic factors that might be involved in
regulatory processes of the species. To then apply this knowledge to harvesting,
management, and cultivation of the resource. (Chapters 3 and 4)
To establish chemical and mechanical treatments for epiphyte management in O. pinnatifida
cultures and to explore the feasibility of cultivation of the species through a reproductive
cycle. (Chapter 5)
To investigate different cultivation methodologies and to then identify the best to apply for
commercial cultivation of O. pinnatifida. (Chapters 5, 6, 7 and 8)
To investigate different cultivation media and assess their impact on the growth and
biochemical composition of the species; to then identify the best nutrient supply for the
cultivation of O. pinnatifida. (Chapter 6)
To assess the feasibility of cultivating O. pinnatifida in the Algem® photobioreactor system
targeting the production of antioxidant compounds with interesting commercial
applications. (Chapter 7)
To assess the effects on growth, development and biochemical composition of common
auxins and cytokinins (PGRs) and the fertilizer Acadian Marine Plant Extract Powder
(AMPEP) on O. pinnatifida cultures and to develop a tumbling tank system for scale-up
commercial production of the species. (Chapter 8)

32

33
Chapter 2 Systematics and phylogeny of Osmundea pinnatifida in
Scotland: investigation on molecular biology in the context of the
development of a cultivation method for the species
2.1 Introduction
Red algae (Rhodophyta) constitute one of the major lineages of eukaryotes, originated ca. 1,500 Ma
(Yoon et al., 2004). They represent a discrete assemblage of marine and freshwater algae, which
can be distinguished from other algal lineages, on the basis of phenotypic characteristics including:
storing carbon as floridan starch; lacking in flagellae; a combination of particular photosynthetic
pigments, having chloroplasts that lack an endoplasmic reticulum, and no aggregated thylakoids
(Woelkerling, 1990; Graham and Wilcox, 2000; Harper and Saunders, 2001a, b; Maggs et al., 2007).
The Rhodophyta have traditionally been placed in a single class, the Rhodophyceae, containing two
subclasses, Bangiophycidae and Florideophycidae (Kraft, 1981; Bold and Wynne, 1985). These two
subclasses contain six and 19 orders, respectively (Woelkerling, 1990; Saunders and Hommersand,
2004). The dichotomy between subclasses Bangiophycidae and Florideophycidae is supported by
nuclear conditions (uninucleate in the Bangiophycidae vs. usually multinucleate in the
Florideophycidae), plastid number, shape and location, as well as, differing patterns of cell division,
presence of pit connection, thallus complexity, sexual reproduction, and from production of
tetrasporangia (Gabrielson and Garbary, 1986; Bold and Wynne, 1985; Ragan et al., 1994; Ragan,
1995). The taxonomic rank to which these groups have been assigned has fluctuated, as molecular
methods have been adopted (Gabrielson et al., 1985; Garbary and Gabrielson, 1990). Currently six
classes are included in the Rhodophyta phylum: Stylonematophyceae, Porphyridiophyceae,
Rhodellophyceae, Compsopogonophyceae, Bangiophyceae, and Florideophyceae (Yoon et al.,
2006). The Florideophyceae, are an assemblage of cosmopolitan red algae, with individual plants
living in marine or freshwater ecosystem, which may be currently categorized in four subclasses on
the basis of molecular and morphological data (Saunders and Hommersand, 2004). Unlike the
Bangiophyceae, the orders within the Florideophycean algae have a strong monophyletic origin
based on a single gene phylogeny and it is postulated that they evolved much later than the
Bangiophyceae (Freshwater et al., 1994; Ragan et al., 1994; Graham and Wilcox, 2000; Harper and
Saunders, 2001a, b; Maggs et al., 2007). Historically, the most widely agreed classification of red
algae was based mainly on female reproductive structures and postfertilization development (Kylin,
1923, 1925, 1932, 1956), which formed the foundation of ordinal taxonomic placement in this
highly diverse class (Maggs et al., 2007).

34
2.1.2 Systematics of Rhodophyceae: the order Ceramiales
Osmundea pinnatifida belongs to the order Ceramiales (Florideophycidae), which according to the
works of Saunders and Hommersand (2004) and Maggs et al. (2007), is the most “advanced” order
in the subclass Rhodymeniophycidae, exhibiting filamentous growth with a generally delicate form.
Shared characteristics among these algae include four-celled carpogonial branches, as well as
auxiliary cells that are lost after fertilization and borne on the supporting cell of the carpogonial
branch (Hommersand and Fredericq, 1990; Lee, 1999). Within this order, there are four families,
Ceramiaceae, Delesseriaceae, Dasyaceae, and Rhodomelaceae (Fig. 2.1).
Figure 2.1 Systematic of Rhodophyta with a focus on Osmundea genus
Ceramiales are characterized by having a triphasic Polysiphonia-type life history (Fig. 2.2), which
consists of a haploid sexual phase, the gametophyte, a diploid phase that develops directly on the
gametophytes, the carposporophyte, and a free living diploid phase, the tetrasporophyte, with
gametophytes and tetrasporophytes which are morphologically similar. Growth is uniaxial; body
plan consisting fundamentally of ecorticate axes or axes corticated by rhizoidal filaments.
Spermatangia formed singly or in clusters of 2-3, either free or separate; spermatia are released
individually, or together in a mucilaginous matrix. The female reproductive structure includes a

35
procarpic apparatus, typically consisting of a fertile periaxial cell, a sterile vegetative filament, a 3-4
celled carpogonial branch terminated by a carpogonium bearing a trichogyne. The tetrasporangia
are terminal, sessile or stipate, solitary or in clusters on determinate lateral filaments or borne on
periaxial or central cells. Tetrasporangi are spherical or subspherical, divided into four tetraspores
(Kylin, 1956; Dixon, 1973; Coomans and Hommersand, 1990; Hommersand and Fredericq, 1990;
Guiry, 1990; Maggs and Hommersand, 1993).
Figure 2.2 Scheme of the triphasic Polysiphonia-type life history, from “Biology of plants”, 5th edition (modified from Raven et al., 1992)
2.1.3 Systematics of the Laurencia complex
The genus Osmundea Stackhouse, is part of the Laurencia complex, which consists of approximately
530 species (Machín-Sánchez et al., 2018), whose ecological distribution is spread along almost all
the world’s temperate and tropical coastlines (McDermid, 1988, 1989). What makes this complex

36
distinct is that all the species included display a polysiphonous construction, with extensive
cortication and apical growth (Kylin, 1956). Over the years, the Laurencia complex has been
restructured, due to the development of molecular methods, which have assisted in species
delineation. Currently, eight related genera constitute the so-called Laurencia complex: Laurencia
sensu stricto Lamouroux; Osmundea Stackhouse; Chondrophycus (Tokida & Saito) Garbary & Harper;
Palisada (Yamada) Nam; Yuzurua (Nam) Martin-Lescanne; Laurenciella Cassano, Gil-Rodríguez,
Sentíes, Díaz-Larrea, M.C. Oliveira & M.T. Fujii; Coronaphycus Metti and, more recently, Olehopapa
Rousseau, Martin-Lescanne & Le Gall (Machín-Sánchez et al., 2018) (Table 2.1). Following these
revisions, each genus in the Laurencia complex has been morphologically and phylogenetically
delineated. With the exception of Lewis et al. (2008) and Sherwood et al. (2010), the molecular
systematics of the Laurencia complex is mainly based on the plastid marker, rbcL. This has been
widely used in the Rhodophyta to answer phylogenetic questions (Freshwater and Rueness, 1994;
Hommersand et al., 1994; Fredericq and Ramirez, 1996; Gurgel and Fredericq, 2004; Abe et al.,
2006; Martin-Lescanne et al., 2010; Cassano et al., 2012).
Table 2.1 Number of specific and infraspecific taxa currently recognized/total number of taxa for each genus of the Laurencia complex (modified from Rousseau et al., 2017)
Genus Authorship and publication Current/total
Chondrophycus (Tokida & Y.Saito) Garbary & J.T.Harper (Garbary & Harper 1998) 17 / 41
Coronaphycus (Metti et al. 2015) 2/2
Laurencia J.V.Lamour., nom. cons. (Lamouroux 1813) 172/421
Laurenciella Cassano, Gil-Rodríguez, Sentíes, Díaz-Larrea, M.C.Oliveira & M.T.Fujii
(Cassano et al. 2012)
1/1
Osmundea Stackh. (Stackhouse 1809) 21/24
Palisada K.W. Nam (2007) 24/29
Yuzurua (K.W. Nam) Martin-Lescanne (Martin-Lescanne et al. 2010) 1/1
Ohelopapa F. Rousseau, Martin-Lescanne, Payri & L. Le Gall 1/1
Laurencia complex 238/519
The genera of the Laurencia complex share typical Rhodomelacean morphology, having apical cells
sunk in apical pits at the apices of branchlets, a central cell row that is recognisable only near the
apical cell and an extensive cortex. In addition to these vegetative features, the procarp-bearing
segments, in the female reproductive structures of each genus, generally have five pericentral cells,
and the spermatangial branch pit is cup-shaped (see Figs 10-20 Nam and Choi, 2001), though it is

37
noteworthy that there are instances of pocket-shaped pits in some species of Osmundea (Martin-
Lescanne et al., 2010).
The Osmundea genus has the most distinctive combination of morphological characters within the
complex. Studies by Nam et al. (1994), Nam and Saito (1995), Garbary and Harper (1998), and
Nam (1999), has defined Osmundea as a monophyletic group, distinct from Laurencia and
Chondrophycus, with its own disjoint distribution. This definitive separation was established on the
basis of the following morphological characters:
1) The number of pericentral cells in vegetative axial filaments;
2) The type of spermatangial development;
3) The origin of tetrasporangia.
On this basis, to date, the genus Osmundea is characterised by the following combination of
characters (Nam et al., 2000):
two pericentral cells in vegetative axial filaments;
spermatangial development of filament-type;
tetrasporangia produced from random epidermal cells.
This genus includes 21 species that occur in the North American Pacific, Brazil, Atlantic Europe,
the Mediterranean Sea, India (Nam et al., 2000; Furnari et al., 2001, 2004), and also in Australia
and Northern Africa (Guiry and Guiry, 2014).
2.1.4 Taxonomy of Rhodophyceae: from morphological structures to DNA-based analyses
Interpretation of life history patterns (Guiry, 1974, 1978; Frederiq and Hommersand, 1989; Maggs
and Pueschel, 1989) and ultrastructural observations (Pueschel and Cole, 1982) have led to a
considerable revision of red algal classification (Saunders and Kraft, 1997). However, the
introduction of molecular tools capable of inferring phylogenetic relationships has had a profound
influence on red algal classification. Molecular techniques have been applied since the 1980s in red
algal taxonomy and they have enabled huge advances in the understanding of species and their
relationships. From the mid 1990’s until the present, the use of molecular DNA in algal taxonomy
and phylogenetic in general has gained popularity (Maggs et al., 2007), and for the red algae the
works of Freshwater et al. (1994), Ragan et al. (1994), Saunders and Hommersand (2004) and Yoon
et al. (2006) provided the foundation of current understanding of the taxonomic relationships of
these algae. Ragan et al. (1994) study on the divergence in nuclear small subunit (SSU) rRNA
sequences pointed out that “(…) rhodophytes are more divergent among themselves than are fungi or green algae
and green plants together.” Red algae display consistently high diversity across a variety of genes

38
including actin and the nuclear and plastid small subunit ribosomal (SSU) genes (Ragan et al., 1994;
Medlin et al., 1997). Molecular studies, revealing their genetic variation, have complemented
understanding of diversity based on their morphology and ultrastructure. The Rhodophyta have
the most gene-rich plastid genomes known, but despite their evolutionary importance, these
genomes are still poorly reported and genomic data from red algae remain scarce. To date, only
one complete nuclear genome and less than a dozen organellar genomes have been characterized
and described (Janouškovec et al., 2013).
At this time, available molecular data for the Osmundea genus is still not comprehensive, with
specimens recorded only from Europe, Brazil and California. This genus has high intraspecific
morphological variation that can lead to a misidentification of the species, as has already been
demonstrated with examples from species on the Atlantic European coasts (Magne, 1989; Maggs
and Hommersand, 1993; Nam et al., 2000) and in the Mediterranean Sea (Serio et al., 2008).
Therefore, molecular analyses represent a fundamental tool, established since the work of Nam et
al. (2000), for the resolution, especially, of the Laurencia complex.
2.1.5 DNA Barcoding: importance and applications
The DNA analyses undertaken in this project focused on DNA barcoding of O. pinnatifida. This
technique, identifying organisms based on comparisons of short, standardized DNA sequences,
(CBOL; http://barcoding.si.edu/), has been championed as a revolutionary system for the
identification and discovery of all the world’s eukaryotic organisms (Hebert et al., 2003a, b;
Saunders and Kucera, 2010). The benefits of employing this approach include the ability to develop
a database of sequences against which unknown biological samples can be compared, thus greatly
increasing the speed at which routine identification can be performed. Across all taxa, DNA
barcoding removes the reliance on morphological characters traditionally used to discriminate
species. This is of particular importance for taxonomic groups that have few phenotypic characters
on which to base identification and for which expert taxonomists are required for even routine
morphological identifications. Seaweeds are notoriously difficult to identify due to simple
morphologies, phenotypic plasticity and convergent evolution (Saunders, 2005). Molecular analyses
are therefore crucial, especially in red algae classification and identification. As described above,
their morphology can be highly variable within and between species, and conspicuous features with
which they can be readily identified are often lacking. In addition, highly convergent morphology
is commonly encountered and often conceals cryptic species that have only come to light from
molecular data. DNA barcoding has proven an effective tool for both the red and brown seaweeds
(Saunders, 2005; Robba et al., 2006; Kucera and Saunders, 2008; Saunders, 2008; McDevit and
Saunders, 2009; Saunders, 2009; Walker et al., 2009; Le Gall and Saunders, 2010) and in analysing
species diversity within the Laurencia complex (Machín-Sanchez et al., 2016).

39
There are several requirements when choosing an appropriate marker for DNA barcoding. First,
the genetic variability of the marker must be adequate for species level resolution. To this end,
species discrimination is considered successful when specimens from a single species cluster
together in a distance analysis (Hebert et al., 2003a) and the largest intraspecific divergence is less
than the smallest interspecific divergence – this difference is termed the “barcoding gap” (Meier et
al., 2008). Second, and equally important, the marker should be universally recoverable across taxa,
meaning there is high PCR and sequencing success, ideally with a near-universal set of PCR
primers. Given current technologies, it is also of benefit for the DNA barcode marker to be short
enough to be sequenced in a single read (<700bp) (Hollingsworth et al., 2009b).
The first bar code sequence to be investigated for red algae was the nuclear ribosomal internal
transcribed spacer (ITS) marker, which has been proved to be useful for species discrimination and
in detecting hybridization, in some cases for population level analyses, and for resolving
relationships amongst closely related taxa (Steane et al., 1991). The SSU, was widely applied in early
studies of red algal phylogenetic (e.g. Bird et al., 1992; Ragan et al., 1994; Saunders and Kraft, 1994),
and is still useful for exploring deeper relationships (e.g. West et al., 2008; Harper and Saunders,
2001b). Freshwater and Bailey (1998) first used the large subunit rRNA (LSU), while Harper and
Saunders (2001a, b) refined the marker by extending the portion of the gene that was amplified. Le
Gall and Saunders (2007) championed the Elongation factor 2 (EF2) as a phylogenetic marker for
red algae in a survey of Florideophyceae and Yoon et al. (2002) devised an amplification strategy
for psaA. Undoubtedly, the widest used marked in Florideophyte phylogenetic and barcoding
studies is the Ribulose-1.5-bisphosphate carboxylase large subunit (rbcL, ~1,350 bp), which was
firstly applied broadly to these algae by Freshwater et al. (1994). Alternative bar codes were
investigated by Saunders (2005) and Robba et al. (2006), proving that Cytochrome c-oxidase
subunit 1 extended fragment, ~1,232 bp (COI) can be used as a barcode marker in red algae, and
subsequent COI based studies have continued to reveal its usefulness for identifying species
(Saunders, 2008, 2009; Yang et al., 2008; Clarkston and Saunders, 2010; Le Gall and Saunders,
2010; Freshwater et al., 2010). Saunders and McDevit (2012) have reviewed the DNA barcode
marker cytochrome c oxidase subunit I (COI-5P) in detail but cautioned that primer advances were
constantly being developed. Cytochrome b (COB, ~940 bp) is a relative “newcomer” to red algal
studies and is amplified with the novel external primers CB44F and CB1006R designed by Ga Hun
Boo et al. (2013) (Saunders and Moore, 2013). Finally, Cytochrome oxidase subunit 2-3 intergenic
spacer (cox 2-3, ~350-400 bp) was first used in red algae by Zuccarello et al. (1999) and has proved
to be a useful tool at the population level (Saunders and Moore, 2013). Although most of these
markers are useful for species level identification, some are not single copy gene or they may be
long, requiring several primer series to obtain a complete sequence. Furthermore, the use of

40
different markers by different laboratories has made it difficult to make comparisons across the
red algae (Saunders, 2005).
Among all the above bar codes, the most frequently employed phylogenetic markers used are the
chloroplast encoded rbcL (1,467 base pair single copy gene that codes for the large subunit of the
ribulose 1,5-bisphosphate carboxylase/ oxygenase enzyme), and the nuclear-encoded ribosomal
cistron encoding 18S, ITS1, 5.8S, ITS2 and the 28S rDNA (e.g. Freshwater et al., 1994; Harper and
Saunders, 2001a, b). Ribosomes are organelles that have the same function (protein synthesis)
throughout the eukaryotes and prokaryotes, and their structure has been highly conserved during
evolution. The 18S and 28S rRNAs are sufficiently large (containing about 2,000 and 5,000
nucleotides, respectively) to allow valid measurement of relationships.
In this study the chloroplast encoded rbcL was chosen because it can provide good resolution of
red algal relationships at the species to genus level (e.g. Freshwater et al., 1994; Bailey and
Freshwater, 1997; de Jong et al., 1998; Muller et al., 1998). It is widely used to generate reliable
phylogenetic hypotheses in red algae at different levels of biological organization (e.g. Gelidiales:
Freshwater et al., 1994; Ceramiales/Delesseriaceae: Lin et al., 2001; Hildenbrandiales: Sherwood
and Sheath, 2003; Gracilariales: Gurgel and Fredericq, 2004; Abe et al., 2006; Martin-Lescanne et
al., 2010, Cassano et al., 2012, 2013), providing a large proportion of sequence data with a high
number of phylogenetically-informative sites, owing to its relatively higher rate of mutation in
comparison to the nuclear small subunit ribosomal marker (SSU) (Bailey and Freshwater, 1998).
The usefulness of this marker has been exemplified in the Laurencia complex where it has been
demonstrated to provide good resolution at both genus and species level (Nam et al., 2000; McIvor
et al., 2002; Abe et al., 2006, Gil-Rodriguez et al., 2009, Martin-Lescanne et al., 2010; Cassano et
al., 2012; Machín-Sanchez et al., 2016).
This chapter focuses on the identification and phylogeny of O. pinnatifida specimens obtained from
the West coast of Scotland. To assess the species identification and phylogenetic relation between
different populations and with other related species a sampling program was conducted along the
West coast of Scotland, including five sites. The delineation of the species based on the plastid-
encoded large subunit of RuBisCO (rbcL) was performed, and relationships among the samples
and other data available from NCBI for related species inferred through a rbcL phylogeny for the
different populations.
2.2 Aims
To develop an extraction protocol for DNA isolation and PCR amplification in O.
pinnatifida from the West coast of Scotland;

41
To identify phylogenetically the species prior cultivation experiments and compare the
analyses with available data on NCBI from previous studies on the species from different
European regions;
To gain a comprehension of the molecular biology of the species from the West coast of
Scotland potentially for further breeding applications;
To explore the population variation of specimens collected along the West coast of
Scotland within the frame of genetic pollution due to introduction of alien species for
farming purpose.
2.3 Materials and Methods
2.3.1 Study area and sample collection
Specimens were collected between October and January of 2015-2016 from sites on the West coast
of Scotland namely: Ganavan (2) (56.4314° N, 5.4747° W), Dunstaffnage (1) (56.4547° N, 5.4374°
W), Easdale (3) (56.2918° N, 5.6582° W), South Uist (4) (57.2642° N, 7.3312° W) and Jura (5)
(55.9042° N, 5.9414° W). In addition, samples were obtained from a site on the East coast of
Scotland in the spring of 2016 (Dundee, site number 6, 56°27'09.2"N 3°03'58.0"W) (Fig. 2.3).
Preliminary species identification was based on morphological characters, following the key
characters defined by Maggs and Hommersand (1993), without undertaking an examination of the
tissue by light microscope.
Figure 2.3 Locations of sampling sites. In yellow Dunstaffnage, where SAMS is situated. Samples are numbered from 1 to 6 in chronological order

42
Samples for molecular analyses were placed in plastic bags on collection and transported to the lab,
where they have been washed with filtered, UV sterilized and tyndalized seawater (33PSU), cleaned
manually, blotted and weighed. The sample from Dundee was also freeze dried by collaborators at
the James Hutton institute prior to dispatch to the Scottish Association for Marine Science (SAMS).
2.3.2 DNA extraction, PCR amplification, and sequencing
DNA was extracted with the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) following the
instructions of the manufactures. A cetyltrimethylammonium bromide (CTAB) DNA extraction
was also performed, on fresh and freeze-dried material. The steps of the CTAB method used are
described below, with two variations tested. In an initial trial, samples were weighted into 1, 2 and
2.5 g aliquots. In subsequent analyses, 1 g was adopted for all the further studies. Samples were
first ground in liquid nitrogen using a pestle and a mortar followed by grinding the tissue in CTAB
buffer (NaCl 1.4M; Ethylenediaminetetraacetic acid, EDTA 20 mM; Tris-HCl 100 nM; CTAB 2%
W/v). Samples were placed in 50 ml tubes with 20 ml of CTAB buffer, stored at room temperature
for 15 minutes and 100 µl of β-mercaptoethanol added to each sample. Incubated at 55 °C for 1
hour in a heated bath, allowed to cool, then Phenol: Chloroform: Isoamyl (25:24:1) was added (15
ml per sample) and the samples mixed gently 30 times by inverting the tube. They were then
centrifuged at 720 g (2,500 rpm) in a Multifuge X3FR (Thermo Fisher ®) for 10 min. The
supernatant was retained, and the pellet discarded; then 20 ml of Phenol: Chloroform: Isoamyl
(25:24:1) were added, followed by a centrifugation step at 4,200 g (6,000 rpm) for 40 min. The
supernatant was retained and 15 ml of Chloroform: Isoamyl (24:1) added to each sample. The
samples were recentrifuged at 720 g (2,500 rpm) for 10 min and the supernatant retained, 2/3 of
the volume of Isopropanol was added to each sample, mixed and stored at -20°C for 1 hour and
then centrifuged at 7,400 g (8,000 rpm) for 20 min. The isopropanol removed and the pellet washed
with 3 ml of cold ethanol (70%) and centrifuged again at 7,400 g (8,000 rpm) for 5 min. The ethanol
was then removed, and samples left to dry covered overnight or for few hours on the bench (at
room temperature). Finally, they were resuspended in 1 ml TE buffer.
The second approach employed included the addition of 522 µl of Lysis Buffer (Tris HCl 100mM;
NaCl 150 mM; EDTA 10 mM), 28 µl of Sodium dodecyl sulphate (SDS, 10%) and 10 µl of
Proteinase K (20 mg/ml) to each sample after the grinding step. This included an additional
incubation step at 56 °C for 30 minutes. Samples were then treated as described above. DNA
extractions were checked with a Nano spectrophotometer (IMPLEM Nano spectrophotometer)
using a 50 and/or 10 lid factor, in order to verify the quantity and quality of the products. Samples
were also checked on a 0.8 % Tris-borate-EDTA (TBE) agarose gel to verify whether the extraction

43
was successful or not. Subsequent PCR analyses were performed on the samples using a DNA
engine gradient Cycler (Mj Research Ptc-200, GMI).
The My-Taq ᵀᴹPlant-PCR Kit (BioLine®) was also tested on fresh material and DNA extracted
samples. Samples (0.5 mg) in triplicate of fresh tissue were weighted on an analytical balance. The
kit was used following the manufacturer’s instructions: to each sample were added 25 µl of MyTaq
Plant-PCR Mix, 2x, 1 µl of Forward and Reverse primer (FrbcL and RbcS, 20µM) and 23 µl of
water (dH₂O).
The primers used (obtained from IDT) (Table 2.2) were as detailed by Freshwater and Rueness
(1994).
Table 2.2 Primers used for the PCR reaction (Modified from: Freshwater and Rueness, 1994)
Primers Sequences
F -rbcL start: 5' -TGTGTTGTCGACATGTCT AACTCTGTAGAAG-3' (forward amplification primer)
F-577: 5' -GTATATGAAGGTCTAAAAGGTGG-3'
F-753: 5' -GGAAGATATGTATGAAAGAGC-3'
R-rbcS start: 5' -TGTGTTGCGGCCGCCCTTGTGTTAGTCTCAC-3' (reverse amplification primer)
R-1150: 5' -GCATTTGTCCGCAGTGAATACC-3'
R-753: 5' -GCTCTTTCATACATATCTTCC-3'
Initially a gradient PCR (annealing temperature from 37 °C to 55°C) was performed in order to
assess the best temperature for every couple of Primer used: 1a: F-rbcL start- 1b R753; 2a F577-
2b R1150; 3a F753- 3b R-rbcS start (Fig. 2.4). The PCR cycle was readapted from Freshwater and
Rueness (1994) and was as described below: denaturation step at 94 °C for 3 minutes, followed by
40 cycles at 94 °C for 1.5 minutes, annealing temperature at 37.5 °C for Primers F577-R1150; 47.6
°C for Primers FrbcL-R753, and 39.9 °C for Primers F753-RbcS; elongation at 72 °C for 3 minutes
and final elongation at 72 °C for 10 minutes. Subsequently, only one set of primers (1a FrbcL-3b
RbcS) was used to amplify the whole of the rbcL sequence of interested. For these primers, FrbcL-
RbcS, were employed the following parameters: 94°C for 5 minutes, 94°C for 1 minute, annealing
temperature at 42 °C for 30 seconds, 72°C for 2 minutes, repeated for 40 cycles, and a final
elongation step at 72 °C for 10 minutes. This was finally optimized for the following conditions:
94°C for 5 minutes, 94°C for 1 minute, annealing temperature at 50 °C for 30 seconds, 72°C for 2
minutes, repeated for 35 cycles, and a final elongation step at 72 °C for 10 minutes.

44
Figure 2.4 Scheme of primers on the rbcL sequence. The primers are taken from Freshwater and Rueness (1994) as referred in Table 2.2
The PCR products were analysed on a 0.8 % TBE (0.5x) agarose gel at 120 V for 22 minutes and
in order to size the PCR products, 5 µl of Easy Ladder I (Bioline®) was run alongside the PCR
products.
The PCR products were subsequently purified using an Amicon Ultra 30KDa 0.5 kit (Millipore®)
and Wizard SV Gel and PCR Clean-Up System (PROMEGA®, USA), according to the
manufacturer’s instructions. On using the Millipore kit, two extra washing steps with 450 µl of TE
(10 mM Tris-HCl, 1 mM disodium EDTA, pH 8.0) and 450 µl of dH₂O water were performed
according to the manufacture’s suggestions, in order to improve the purification of the samples.
The Wizard SV Gel and PCR Clean-Up System (PROMEGA®, USA) was used in order to cut
bands of interest from the gel, improving the result of the sequencing. A further purification step
using an Amicon Ultra 30KDa 0.5 kit (Millipore®) was also applied.
Purified PCR products were dispatched by post to the DNA Sequencing and Services Medical
Sciences Institute, School of Life Sciences, University of Dundee for sequencing.
2.3.3 Multiple sequence alignments and phylogenetic analyses
The DNA sequences obtained were first trimmed, in order to remove the region of “noise” at the
start and the end of the sequence. The DNA sequence alignments were then constructed using the
multiple alignment on Geneious software (http://www.geneious.com/) and the multiple alignment
on MEGA 7 (http://www.megasoftware.net/) using ClustalW, MUSCLE and Geneious
alignments. Phylogenetic trees have been created with Neighbour joining (NJ) method on
Geneious and Maximum likelihood (ML) method on MEGA 7, using the Jukes-Cantor model for
distance and inferring final bootstrap trees. Phylogenetic trees were performed on 2 datasets: one
including samples investigated in this study originating from Ganavan, Easdale, South Uist, Jura
and Dunstaffnage, plus two from NCBI (AF259495, O. pinnatifida France and AF281875, O.
pinnatifida Ireland), and one for all the sequence data generated plus 45 related species randomly

45
chosen from NCBI, inclusive for the Laurencia complex, following the work of Machín-Sánchez et
al. (2012). The accession numbers AF25940 (Centroceras clavulatum) and AF259491 (Ceramium
brevizonatum) were chosen as outgroups, following the work proposed by Machín-Sánchez et al.
(2012). The evolutionary history was inferred by using the Maximum Likelihood method based on
the Jukes-Cantor model. The bootstrap consensus trees were inferred on MEGA7 from 1000
replicates and taken to represent the evolutionary history of the taxa analysed. Branches
corresponding to partitions reproduced on less than 50% bootstrap replicates were collapsed. The
percentage of replicate tree in which associated taxa clustered together in the bootstrap test (1000
replicates) are shown next to the branches. Initial trees for the heuristic search were obtained
automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances
estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the
topology with superior log likelihood value. The analyses involved 50 nucleotide sequences. Codon
positions included were 1st+2nd+3d+Noncoding. There were 1,650 positions in the final dataset.
Trees were also built using Geneious, setting as genetic distance method Jukes Cantor, as tree
building method Neighbor-Joining and as resample the bootstrap method. Distances and Identities
between sequences were analysed through matrices on Geneious software both on the 50
sequences data set and on the 5 obtained sequences plus the two datasets more closely related to
them (AF259495, O. pinnatifida France and AF281875, O. pinnatifida Ireland). The intraspecific
sequence divergences within the groups were tested with the pairwise distance tool for the method
“p-distance” (identified as the proportion of nucleotides sites at which two sequences being
compared are different), Kamur and Tamura (2015) on MEGA 7, and using the matrix of distance
on Geneious.
2.4 Results
2.4.1 DNA extraction, PCR amplification, purification and sequencing
The CTAB DNA extraction and DNeasy Plant Mini Kit (Qiagen®, Hilden, Germany) were both
successful in DNA extraction on the samples tested. However, the DNeasy Plant Mini Kit (Qiagen,
Hilden, Germany) was successful only on fresh material (Fig. 2.5 C), while the CTAB method
proved to be effective on both fresh and freeze-dried material, in both the variations applied (Figs
2.5 B and 2.6). The plastid rbcL gene was successfully amplified by all the PCR cycles tested, either
using the primer pairs: FrbcL-R753, F577-R1150, and F753-RbcS, and as one single fragment using
the primer pair F-rbcL-RbcS (Freshwater and Rueness, 1994) with the parameters listed in section
2.3.2. This last primer pair (primer couple FrbCL- RbcS) was chosen for subsequent work, since
the results obtained were the same of the three separate reactions using the entire set of Primers
but the application of just one set of primers made the PCR process quicker and more efficient.
Increasing of the annealing temperature was also successful in reducing the “primer dimer” effect on

46
the reaction and in better bands definition. The reduction of the PCR cycle from 40 to 35 did not
negatively affected the PCR reaction, resulting in positively shortening the cycle’s duration, and the
results obtained from this PCR cycle were satisfactory. Despite DNA being extracted from the
Dundee sample, as confirmed by gel electrophoresis (Fig. 2.5.A), the ratios were not satisfactory
(A260/A280=1.405, A260/A230 =1.040; concentration=52.0 ng/µl-lid factor 10, read at the Nano
spectrophotometer). This suggests a problem with the starting material and the PCR reaction, with
the listed parameters, was not successful for this sample. Multiple trials were conducted but without
any success on this sample, which has then been omitted from this study. The EasyLadder I
(Bioline®) was used as reference in all the PCR runs. This ladder has five regularly spaced bands,
including sizes (bp) 100, 250, 500, 500, 1000, and 2000.
Figure 2.5 A) DNA extraction Dundee sample, prepared using DNeasy Plant Mini Kit, 0.8% TBE (0.5x) gel; B) PCR gel for Easdale sample (on the left) and Ganavan sample (on the right) with the couple of primers FrbcL-RbcS. 5 µl of Easy Ladder I (BioLine) as reference; C) PCR gel for Easdale and South Uist samples
Around 1400 bp section of the rbcL gene were amplified for all the other samples examined, except
for the Jura sample, since the sequence obtained was ~800 bp. Both purification methods were
successful for the tested samples. The Wizard SV Gel and PCR Clean-Up System (PROMEGA,
USA) was applied only if multiple bands were detectable on the gel, in order to excise the band of
interest (Fig. 2.7). The length of the PCR products obtained matched with the ̴1.400 pb reported
for the rbcL gene in O. pinnatifida (Nam et al., 2000; Martin-Lescanne et al., 2010; Cassano et al.,
2012; Machín-Sánchez et al., 2012).

47
Figure 2.6 A) PCR gel for Jura sample (on the left) and South Uist samples (on the right) with the couple of primers FrbcL-RbcS. 5 µl of Easy Ladder I (BioLine) as reference B) DNA extraction for Jura sample with CTAB method
Figure 2.7 Purification of samples after Amicon kit procedure. L= EasyLadder I; E=Easdale; D=Dunstaffnage; SU= South Uist; J=Jura
2.4.2 Phylogenetic analyses
Of the ~1,400 bp included in the rbcL sequences analysed for the 5 samples, the identities’ table
on Geneious software ®, showed the highest value for the Ganavan sample with 1,342 identical
residues, followed by South Uist, Dunstaffnage and Easdale samples respectively and the lowest
for the Jura sample with almost 580 identities for AF259495 sample (O. pinnatifida France). This
was the same for the comparison of identities between samples and the AF281875 sample (O.
pinnatifida Ireland), with the highest value reported for Ganavan (1,342 residues), followed by South
Uist, Dunstaffage and Easdale samples respectively and the lowest for Jura sample (530 identical

48
residues) (Table 2.3). The pairwise distance matrix produced by MEGA7 using the p-distance for
the 5 samples, indicates that the sample from Ganavan has a p-distance value of 0.09 and 0.05 for
AF281875 and AF259495 respectively, followed by Dunstaffnage, Easdale and South Uist samples
(Table 2.4). The sample from Jura had the largest intraspecific sequence divergences among the
samples examined, with values ranging from 0.18 (for AF281875) to 0.35 (for Easdale and
Duntaffnage) (Table 2.4). Percentage of Identities produced by Geneious are reported in Table 2.5
and showed the highest value (in relation to AF259495) among the analysed samples for Ganavan
(94.64), followed by South Uist, Jura and Dunstaffnage, and the lowest for Easdale (71.128).
Table 2.3 Identities residues for the samples investigated and the data from NCBI. AF259495
refers to the entry for L. pinnatifida from France; AF281875 refers to the entry for O. pinnatifida
from Ireland
AF259495
Ganavan AF281875
Easdale South Uist Dunstaffnage Jura
AF259495 1342 1245 990 1130 1049 577
Ganavan 1342 1202 982 1092 1030 530
AF281875 1245 1202 919 1046 994 530
Easdale 990 982 919 977 888 440
South Uist 1130 1092 1046 977 1025 521
Dunstaffnage 1049 1030 994 888 1025 480
Jura 577 530 530 440 521 480
Table 2.4 Pairwise distances expressed as p-distances values obtained with MEGA 7 software
Pairwise distance (MEGA 7) 1 2 3 4 5 6 7
1 O. pinnatifida_France_(AF259495) 0.00 0.01 0.01 0.01 0.01 0.02
2 O. pinnatifida_Ireland(AF281875) 0.00 0.01 0.01 0.01 0.01 0.01
3 South_Uist 0.15 0.09 0.01 0.01 0.01 0.02
4 Dunstaffnage 0.20 0.14 0.25 0.01 0.01 0.02
5 Easdale 0.19 0.16 0.18 0.27 0.01 0.02
6 Ganavan 0.09 0.05 0.16 0.21 0.20 0.02
7 Jura 0.25 0.18 0.31 0.35 0.35 0.29
Table 2.5 Percentage of Identity inferred with Geneious® software
Pairwise distance (Geneious) 1 2 3 4 5 6 7
1 O. pinnatifida_France_(AF259495) 94.64 100 71.128 78.283 73.003 74.838
2 Ganavan 94.64 94.945 70.192 76.236 71.934 69.372
3 O. pinnatifida_Ireland (AF281875) 100 94.945 73.452 83.433 78.693 82.555
4 Easdale 71.128 70.192 73.452 74.491 65.171 57.837
5 South Uist 78.283 76.236 83.433 74.491 69.064 66.624
6 Dunstaffnage 73.003 71.934 78.693 65.171 69.064 62.711
7 Jura 74.838 69.372 82.555 57.837 66.624 62.711

49
Bootstrap consensus trees were generated from both the two data sets of 50 sequences (Fig. 2.12)
and the 7 sequences representing the 5 samples plus two data of O. pinnatifida (AF281875 and
AF259495) that have shown the closest identity to the samples here investigated (Figs 2.8-2.11).
Figure 2.8 Evolutionary history inferred by using the Maximum Likelihood method based on the Jukes-Cantor model on MEGA 7 software. Circled in green the samples from this study. The bootstrap consensus tree inferred from 1000 replicates is taken to represent the evolutionary history of the taxa analysed. Branches corresponding to partitions reproduced on less than 50% bootstrap replicates are collapsed. The percentage of replicate tree in which associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. Initial tress for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value
Figure 2.9 Evolutionary history (including the Spanish entry) inferred by using the Maximum Likelihood method based on the Jukes-Cantor model on MEGA 7 software. Circled in green the samples from this study

50
Figure 2.10 Neighbour-Joining consensus tree obtained employing Geneious software, Clustal W alignment, genetic distance method Jukes Cantor, and resample method Bootstrap. Circled in green the samples here analysed
Figure 2.11 Neighbour-Joining consensus tree obtained employing Geneious software, Clustal W alignment, genetic distance method Jukes Cantor, and resample method Bootstrap. Circled in red the outgroup (Centroceras clavulatum, AF259490) (Geneious software)

51
Figure 2.12 Evolutionary history was inferred by using the Maximum Likelihood method based on the Jukes-Cantor model. In green the samples analysed in this study; in yellow the samples for O. pinnatifida from NCBI; in red the outgroups (Geneious software)
Both the maximum likelihood and NJ trees show the same pattern (Figs 2.9 and 2.10), with the
population from South Uist forming a sister group with the Easdale population, thus they both
form a clade with the Dunstaffnage sample. However, the Jura sample is more closely related to

52
the sequences from France and Ireland than to the other samples of the West of Scotland (Figs
2.8, 2.9, 2.10, and 2.11).
Different alignments (MUSCLE, CLUSTAL W and Geneious) were tested on the 50 sequences
data set and the alignment in Fig. 2.12 ultimately chosen. The trees obtained had some differences,
obviously reflecting the different alignment methods; however, what was important for this study
was that all the O. pinnatifida samples from the West coast of Scotland grouped together with the
other data for O. pinnatifida species obtained from NCBI (Figs 2.8, 2.9, 2.10, and 2.11). This
confirmed that the species being studied is O. pinnatifida and in addition provided an insight into
the relationship between the sampled populations. Furthermore, the values of bootstrap shown on
the nodes of the trees strengthen this observation. Therefore, even if different alignment methods
were tested, the trees here reported were considered the more suitable to represent the dataset here
analysed. They provided a clear and inclusive representation of the relationship inferred among the
Laurencia complex species (Fig. 2.12) and the populations of O. pinnatifida in Scotland (Figs 2.8, 2.9,
2.10, and 2.11).
2.5 Discussion
Methods for major seaweeds cultivation have been developed since the mid-twentieth century
(McHugh, 2003). Nowadays, the seaweed industry is booming, accounting for ~49% of the total
mariculture production (FAO, 2018). Unabated exponential growth of this industry in the last 50
years, has led to a market value of US$ 6.4 billion in 2014, providing jobs, especially in developing
and emerging economies (FAO, 2015). The farming of seaweeds for commercial purposes
undergoes different considerations, amongst which stands out species selection, based on
application as food or other production possibility (e.g. carrageenan, alginate, or biomass for
energy). Strain selection within a species might maximize properties such as growth, disease
resistance, and product quality (Roesijadi et al., 2008). Furthermore, selected strains will allow the
farmers to expand breeding seasons and enhance production (Kim et al., 2017).
The red seaweed represents a species-rich phylum with high commercial value either as food or
industrial products, such as hydrocolloids (Wattier et al., 2000). However, both the genome
characterisation (Stoebe and Kovallik, 1999) and intraspecific genetic biodiversity (Wattier et al.,
1997; Engel et al., 1999) are still poorly explored for many species (Wattier et al., 2000). Therefore,
a comprehension of the molecular biology of a species undergoing farming is fundamental to fully
understand and investigate its diversity and breeding. Additionally, the understanding of seaweed
genetics can potentially lead to engineering applications such as genome-design tools for synthetic
applications and genome-editing methods (Purnick and Weiss, 2009; Lin and Qin, 2014). Seaweed

53
genetic engineering could represent the chance to fill the gap between fundamental and applied
studies in seaweed research (Lin and Qin, 2014).
Most of the preliminary molecular biology studies on red seaweeds have focused on protocols for
DNA isolation and phylogenetic relationship among the species (Wee et al., 1992; Shioda and
Murakmi-Murofusi, 1987; Doyle and Doyle, 1987; Saunders and Bailey, 1997, 1999; Saunders and
Hommersand, 2004). DNA of high quality and quantity is a prerequisite for ensuring the efficacy
of applications, such as polymerase chain reaction (PCR). Since the earlier 2000’s, much of the
work has been focused on trying to improve the existing methods and protocols for the extraction
of genetic material, with the objective of finding a method that could be easily adapted to a range
of different species of seaweed. The application of molecular tools in studies of marine algae has
often been hindered by the difficulties of extracting suitable DNA quantity. This may be due to
polysaccharides, such as sulfated polysaccharides and carboxylic polysaccharides (Bold and Wynne,
1978; Joubert and Fleurence, 2008) in their thallus, high nuclease activity (Wee et al., 1992), and
the presence of secondary metabolites. This may act as an obstacle to DNA isolation, since DNA
often co-purifies with them, thus inhibiting downstream enzymatic reactions such as PCR
(Saunders, 2004). The release of such compounds during cell lysis leads to highly viscous
supernatants, the main source of DNA contamination.
Amongst the published DNA isolation protocols, one of the most widely used is the CTAB
extraction techniques (Doyle and Doyle, 1987). However, different methods have been tested and
applied to O. pinnatifida DNA extraction. In the literature, the most common methods reported are
namely: DNA extraction using DNeasy Plant Mini Kit (Qiagen®), both on silica dried samples
(Machín-Sànchez et al., 2012; Cassano et al., 2012) and fresh material (Nam et al., 2000; Rousseau
et al., 2017); DNeasy Plant Mini Kit (Quiagen®) plus proteinase K (Martin-Lescanne et al., 2010);
CTAB modified method on silica and ethanol treated samples (Abe et al., 2006) or DNA extraction
involving EDTA and SDS (Lewis et al., 2008). All these variants have proved to be successful in
the listed works for O. pinnatifida in obtaining good quality DNA for further analyses. In this study,
the DNeasy Plant Mini Kit (Quiagen) was compared with a modified (CTAB) extraction method
(Doyle and Doyle, 1987) and the My-Taq ᵀᴹPlant-PCR Kit (BioLine®) on both dried and fresh
material.
The Bioline My-Taq ᵀᴹPlant-PCR Kit, has been designed for direct PCR amplification of plant’s
material and has not been yet investigated on red algae. This kit includes a novel buffer system that
replaces the need for complicated extraction or purification steps (including freezing of plant
tissues with liquid nitrogen, mechanical disruption, organic extraction or column DNA
purification). It has the potential of speeding up the process, therefore it has been considered for

54
this study. However, it was not efficient in providing PCR product for O. pinnatifida samples;
therefore, it was not further investigated.
The first two methods, instead, have previously been proved to be effective for DNA extraction
in studies conducted on samples of O. pinnatifida from different geographic regions (Nam et al.,
2000; Martin-Lescanne et al., 2010; Cassano et al., 2012; Machín-Sánchez et al., 2012). Moreover,
in all of the above cited studies, the quantity and the quality of the DNA extracted was sufficient
(applying both methods) for further PCR and purification analyses. (Nam et al., 2000; Machín-
Sánchez et al., 2012; Cassano et al., 2012; Rousseau et al., 2017).
In this chapter, the type of material used for the extraction has made a difference: results were
obtained mainly from a fresh material extraction; the silica gel treatment was not considered in this
study, due to the problematics in using silica gel; with freeze-dried material proving to be the most
difficult for a successful DNA extraction. Even if DNA was extracted from a freeze-dried sample
(Fig. 2.5 A, Dundee sample), the PCR reaction failed. Furthermore, in this study DNeasy Mini
Plant kit (Qiagen®) was successful only on wet biomass from South Uist sample (Fig. 2.5 C). This
sample, compared to the others, was more squashed and brittle, since it was dispatched from the
Outer Hebrides, and this has probably helped in disrupting the tissue and extracting the DNA. The
kit was not successful on freeze dried material. To summarize, for O. pinnatifida from the Scottish
area, a DNA extraction with the CTAB modified method on fresh material appears to be the best
solution for this particular molecular investigation.
As already stated, most of the molecular approaches on O. pinnatifida bar-coding, have been based
on the plastid-encoded rbcL gene (Nam et al., 2000; McIvor et al., 2002; Abe et al., 2006; Fujii et
al., 2009, 2012; Cassano et al., 2009, 2012; Martin-Lescanne et al., 2010), utilising the Primers
proposed by Freshwater and Rueness (1994) (e.g. FrbcLstart-R753, F577-R1150 and F753-RrbcS)
(Fig. 2.4). However, other primers have been previously investigated for the rbcL region (such as
rbcLFC and rbcLRD - Nam et al., 2000) and sometimes different bar-coding investigated (such as
COI-5P – Machín-Sánchez et al., 2016). Since the majority of data available on O. pinnatifida
regarding molecular analyses and bar-coding is based on rbcL gene and the primers proposed by
Freshwater and Rueness (1994), these were chosen for this study. Furthermore, investigations
based on these components have already proved to be resolving at a genus and species level (Nam
et al., 2000; Martin-Lescanne et al., 2010; Machín-Sánchez et al., 2012).
Once obtained, the sequences were blasted on NCBI database for an initial identification of the
species. High level of identity (90-96%) and good E-values (0.00) with the records present in the
NCBI database for the species O. pinnatifida were found. Samples from a range of field sites have
then been phylogenetically analysed together with other species of the Laurencia complex in order

55
to have a more inclusive view of the phylogenetic of the populations investigated in this study
within the Laurencia complex. The Laurencia complex is a monophyletic group, as previously
reported (Garbary and Harper, 1998; Nam et al., 2000) and confirmed by other authors (Martin-
Lescanne et al., 2010; Machín-Sánchez et al., 2012, 2016), that includes eight genera (Rousseau et
al., 2017; Machín-Sánchez et al., 2018). Furthermore, the data obtained confirmed that the genus
Osmundea is distinct from Laurencia. The sequences divergences between the Laurencia complex and
for the genus Osmundea were similar to those reported by other workers (Machín-Sánchez et al.,
2012) and are comparable with intergeneric sequence divergences found in other red algal families
(Hommersand et al., 1994; Lin et al., 2001; McIvor et al., 2002; Martin-Lescanne et al., 2010;
Machín-Sánchez et al., 2012) (Tables 2.3, 2.4, and 2.5).
In this study the sample from South Uist forms a sister group with the Easdale specimen, and they
are grouped together in a clade with the samples from Ganavan, Dunstaffnage, Jura and the entries
from Asturias (North Spain) and the Canary Islands (JF781518, JF781514 and JF781516) (Fig. 2.8).
The Jura sample is closer on a phylogenetic basis to the Spanish, French and Irish samples than to
the other samples collected for this study (Figs 2.8, 2.9, 2.10, and 2.11). This was particularly
interesting, as one might anticipate that populations sampled along the West coast of Scotland
should be more related to each other than with populations from different geographic areas. It
should be noted that the Jura sequences (almost 800 bp) was shorter than those obtained from the
other samples (~1,400 bp); however, this does not compromise the results, since the divergence of
this population was confirmed by statistical analyses (Geneious® and MEGA 7 software, pairwise
distance matrices). Furthermore, sequences deposited in NCBI by other authors for Laurencia
complex species, used for the multiple alignment in this project and other studies, were of
comparable short lengths (Martin-Lescanne et al., 2010; Machín-Sànchez et al., 2012). It should be
noted that this study did not aim to be a comprehensive investigation on the population genetic of
O. pinnatifida in West Scotland.
The results from the phylogenetic methods employed, indicate that the samples collected can be
identify as O. pinnatifida species, that they are grouped in the same clade as the O. pinnatifida
sequences from GenBank (Figs 2.8, 2.9, 2.10 and 2.11), and that is it possible to recognize different
closely related subclades, on the basis of the bar-codes analyses. The samples share common
ancestors and a lower level of divergence (Tables 2.3, 2.4 and 2.5).
This study aimed to provide an overview of the phylogenetic relationships between O. pinnatifida
populations of the West coast of Scotland, in the contest of the development of a breeding system
for the target species. From a cultivation perspective, is important to understand and recognize the
molecular biology of a commercial-exploited species, in order to achieve a successful commercial
production and to prevent possible issues often associated with breeding systems (FAO, 1990).

56
The world-wide expansion in aquaculture has resulted in a significant increase in the number of
species (e.g. aquatic animals and plants), which have been displaced from their native ranges for
the purposes of aquaculture (Welcomme, 1988). These translocations can represent both a positive
improvement to cultivation systems, as well as a negative impact, determining issues such as
introduction of diseases, competition with other species, interbreeding and reduction of genetic
diversity (Rueness, 1989; Turner, 1988; FAO, 1990). Furthermore, molecular knowledge and
subsequent applications to aquaculture systems can be important for a selective breeding of aquatic
organisms (FAO, 1990) or for the application of genetic engineering tools -already investigated for
different seaweeds species (e.g. Chondrus sp., Gelidium sp., Laminaria sp., Euchema sp., etc.) in order
to improve the quality or quantity of the production (FAO, 1990) or to tailor the species for the
production of a specific compound (e.g. phycocolloids and biofuel production; Chen and Taylor,
1978; Saga et al., 1978; Zhang, 1982; Fang et al., 1983b; Patwary and van der Meer, 1992; Lin and
Qin, 2014; Radakovits et al., 2010). The Scottish government published a policy statement (2017)
regarding seaweed cultivation, to support and develop the sustainable economic growth of
aquaculture activities. The Policy 2 cited: “Only species native to the area where seaweed cultivation will take
place should be cultivated, to minimise the risk from non-native species”.
2.6 Conclusions
The best methodology for DNA extraction of O. pinnatifida from the West coast of Scotland
appears to be a CTAB modified method with fresh material. This provided good quality
and quantity of DNA for further analyses;
PCR cycle was improved from previous protocols available in order to maximise the results
and one single couple of PRIMERS was successfully selected from the sets suggested in
previous studies in order to perform the procedure in one set rather than three, as trialled
at the beginning;
According to this study, the problems regarding genetic pollution or genetic loss in
breeding systems for O. pinnatifida appear to be limited, due to the close phylogenetic
relationships between the populations present in Europe (Figs 2.8-2.12); however,
introduction of non-native species should always be avoided;
Selective breeding through strains’ selection and genetic engineering have not been
considered in this contest but should be in order to continue the presented study and
support the future development of O. pinnatifida cultivation.

57

58
Chapter 3 Seasonal variation of biochemical composition in Osmundea
pinnatifida from the West coast of Scotland
3.1 The chemical composition of seaweeds
Several epidemiological studies have emphasized the health benefits associated with the
consumption of seaweeds (Cassolato et al., 2008; Hold and Kraan, 2011). There is renewed interest
in Europe in the formulation of health-promoting food using macroalgae as one of the ingredients,
such as, addition of Enteromorpha spp., Undaria pinnatifida, Himanthalia elongata, Porphyra umbilicalis,
and Sargassum thunbergii in snack foods, pasta, patties, bakery and meat products (Chun et al., 1999;
López-López et al., 2009). Seaweeds are good sources of protein, carbohydrates, polyunsaturated
fatty acids (PUFAs), amino acids, antioxidants, minerals, dietary fibres and vitamins (Chandini et
al., 2008; Mohamed et al., 2012). The nutritional composition of seaweeds varies within species
(Fujiwara-Arasaki et al., 1984; Nisizawa et al., 1987), geographic area, season, environmental
conditions (Kaehler and Kennish, 1996; Haroon, 2000; Chandini et al., 2008), maturity and gender
(Ito and Hori, 1989; Murakami et al., 2011). Studies on the nutraceutical properties of seaweeds
have been conducted since the 1950’s (Black, 1950b; Paine and Vadas, 1969; Munda, 1977;
Fleurence, 1999; Wong and Cheung, 2000; Rupérez and Saura-Calixto, 2001; McHugh, 2003;
Sanchez-Machado et al., 2004a; Marsham et al., 2007).
In general, seaweeds follow a similar nutrient composition pattern; total concentration of
polysaccharides ranges from 4% to 76% of dry weight (Holdt and Kraan, 2011) the highest levels
are: agar, algins carrageenans and fucoidans. These are all widely used commercially, and can exhibit
biological activities (Mak et al., 2013). The protein content is also variable, and the highest levels
are generally found in green and red seaweeds (1030% of dry weight), in comparison to brown
seaweeds (5-15% of dry weight) (Holdt and Kraan, 2011; Zhou et al., 2015). Phospholipids and
glycolipids are the major classes of lipids found in seaweeds accounting for 1-5% of cell
composition (Chojnacka et al., 2012). Phenolic compounds are another widely reported group of
compounds (Lordan et al., 2011) varying quantitatively and qualitatively for each specimen of red,
brown or green seaweeds (Machu et al., 2015). Carotenoids and phenolic are plant and seaweed
antioxidants that can rapidly neutralize free radicals and retard or decrease the extent of oxidative
deterioration (Rengasamy et al., 2015). This bioactivity has been demonstrated in red algae (Duan
et al., 2006) with the content and profile of phenolic compounds varying with the species. In red
algae for example, one of the main class of phenolic compounds are bromophenols, which have
shown antioxidant activity (Takamatsu et al., 2003). Seaweeds are also recognized as sources rich
in several elements such as Ca, Mg, Na, P and K or trace elements such as Zn, I or Mn. This
elemental richness is related to their capacity to retain inorganic compounds accounting for up to

59
36% of dry matter in some species (Lordan et al., 2011). Furthermore, seaweeds are known to have
a low calorie content, rich in minerals, vitamins, such as thiamine and riboflavin, β-carotene, trace
minerals (Kumar et al., 2008), tocopherols (which may reduce the risk of heart disease, thrombosis
and atherosclerosis, Mishra et al., 1993), steroids and dietary fibres (Lordan et al., 2011). Seaweeds
are of potential interest as added-value foods because they are claimed to contain low cholesterol
content, fight obesity, reduce blood pressure, tackle free radicals and promote healthy digestion
(Plaza et al., 2008). Furthermore, some of them have demonstrated biological properties such as
antibacterial, antiviral, antifungal, and anticoagulant activities (Alghazeer et al., 2013; Chandía and
Matsuhiro, 2008). The Chlorophyceae and the Rhodophyceae, in particular could be seen as
complementary sources of food proteins for human and animal nutrition (Fleurence, 1999; Paiva,
2017).
3.1.2 Red seaweeds: biochemical composition
Because of their substantial diversity and composition, red seaweeds (i.e. Rhodophyta) have
aroused significant interest in the food and the pharmaceutical industry as new nutritional food
sources and bioactive compounds. It is noteworthy that, among seaweeds, red algae contain high
amounts of carbohydrates, proteins and minerals (Rupérez, 2002). Specific functional properties
have been attributed to Rhodophyta proteins/peptides and polysaccharides because of their unique
composition (Cian et al., 2015).
Three main pigments are involved in light harvesting in seaweeds: chlorophylls, phycobiliproteins,
and carotenoids (Meeks, 1974; Ragan, 1981; Rowan, 1989; Dring, 1990). Chlorophyll a is found in
all algae whilst other chlorophylls, such as b and c, are not present in red algae (Lobban and
Harrison, 1997), but chlorophyll d, whose exact function is not well known, has been reported in
some red algae (Lobban and Harrison, 1997). Macroalgae, like higher plants, possess several photo-
protection mechanisms that operate together. An essential part of this mechanism relies on
carotenoids, which play a key role in preventing damage to the photosynthetic apparatus. They act
as light-harvesting pigments and play an essential role by quenching triplet chlorophyll molecules,
by scavenging singlet oxygen and by regulating the rate of thermal energy dissipation (Young,
1991). It has been indicated that the Rhodophyta exhibit a general pattern of carotenoid
composition formed by α and/or β-C and one major xanthophyll, either L or Z (Marquardt and
Hanelt, 2004). Macroalgal pigments can be affected by environmental factors and extreme
conditions, such as salinity, temperature, nutrients, and intense irradiance, which may result in a
high rate of pigment production (Boussiba et al., 1999; Zucchi and Necchi, 2001). Intertidal
macroalgae are exposed to dramatic changing irradiances and complex light patterns governed by
the daily solar cycle, the tidal rhythm and changing cloud cover (Lüning, 1990). Additionally,
nutrient concentrations, salinity, temperature, and hydration are other factors that vary during tides

60
(Benson et al., 1983). For example, when low tides occur at noon on a sunny day, very high
irradiance is frequently associated with photo-inhibition and photo-oxidative damage (Gómez et
al., 2004).
Phycobiliproteins are predominantly light-harvesting pigment-protein complexes found in
cyanobacteria, red algae, some cryptophytes and dinoflagellates (Zhao et al., 2011). They are water-
soluble pigments that form particles on the surfaces of thylakoids called phycobilisome (PBS) that
lie near the chlorophyll a reaction centre of Photosystem II (Glazer, 1989; Sun et al., 2003). They
consist of pigmented phycobilins, whose colours originate mainly from the tetrapyrrole
chromophores, bounded in various combinations to protein complexes. They include
phycoerythrobilin and phycocyanobilin. Generally, four types of phycobiliproteins are
distinguished based on their absorption maxima (Abs max): phycoerythrocyanins (PEC, Abs max
~ 575 nm), phycoerythrins (PE, Abs max ~ 495–575 nm), phycocyanins (PC, Abs max ~ 615–640
nm) and allophycocyanins (APC, Abs max ~ 650–655 nm). In commercial applications, R-
phycoerythrin is commonly used not only for applications in immunology, cell biology and flow
cytometry, as well as a dye in the cosmetics industry and in natural foods. R-phycoerythrin subunits
isolated from red algae can be used as a photosensitizer in photodynamic therapy of carcinoma
cells (Cian et al., 2012). Pigments are also fundamental to depth adaptation. Rhodophyta have the
ability to live at greater depths than other algae groups, with the potential to grow at a depth of up
to 200 m, a capability related to the presence of the accessory pigments (Lee, 2008).
Fibrillary polysaccharides are the most inert and resistant cell wall component in red algae, with
cellulose being the most important (Miller, 1997). The glycoprotein domain is not well understood,
but it consists of glycoproteins that contain “cellulose binding domains” which promote
crosslinking of polysaccharide fibres. Finally, the amorphous matrix consists of sulfated galactans,
polysaccharides, e.g. carrageenans, agarans and galactans where carrageenan and agaran structures
cooccur (DL-hybrid), that contain multiple units of the monosaccharide galactose with sulfate ester
(Jiménez-Escrig et al., 2011). These polysaccharides are also called “phycocolloids” due to their ability
to form aqueous gels (Al-Alawi et al., 2011). Carrageenans are mainly synthesized by red algae of
the Gigartinales order (Gigartina, Chondrus crispus, Eucheuma and Hypnea), although they also appear
in other Rhodophyta species (Campo et al., 2009). Their differences in chemical composition and
configuration are responsible for their interesting properties, which make them useful as gelling,
stabilizing and thickening agents in the food, pharmaceutical and cosmetics industry (Al-Alawi et
al., 2011). Agarans are mainly synthesized by red seaweeds belonging to the Pyropia, Gelidium,
Gracilaria and Pterocladia genera (Souza et al., 2012). These polysaccharides comprise the hot water-
soluble portion of the cell wall and are the main components of marine red algae (Venkatpurwar
et al., 2011).

61
The protein content of red algae, such as Porphyra, Palmaria, Gracilaria, Gelidium and Eucheuma, that
are among the major edible algae, (McLachlan et al., 1972) are at high levels (Galland-Irmouli et
al., 1999), reaching almost 47% w/w of dry matter (Sanchez-Machado et al., 2004a). This is in
contrast with green (926 g protein 100 g¯¹ dry weight) and brown algae (315 g 100 g¯¹ dry weight)
(Fleurence et al., 2012). As an example of the protein content of red algae, the species Pyropia and
Porphyra have a crude content comparable to that of soya (Norziah and Ching, 2000). The
physiological effects of dietary proteins depend on the capacity of the gastrointestinal tract to digest
protein intake. Digestibility of red algal proteins has been found to be moderate when compared
to that of animal proteins. This lower digestibility may be due to a high fibre content of red algae
in general (Cian et al., 2014; Misurcova et al., 2010). Although the structure and biological
properties of algal proteins are still relatively poorly documented, the amino acid composition
corresponding to several species of algae is known (Harnedy and FitzGerald, 2011). Amino acid
levels vary with the season and fluctuations have been linked to a number of variables, including
nutrient supply, which, in turn is related to environmental factors, such as water temperature,
available light, and salinity (Harnedy and FitzGerald, 2011).
Lipid composition of algae can also be affected by environmental factors such as light, nutrients
availability, temperature, etc. (Harwood, 1984; Thompson, 1996). Algal lipids include
phospholipids, glycolipids (glycosylglycerides) and non-polar glycerolipids (neutral lipids)
analogous to higher plants, along with betaine and some other unusual lipids that maybe
characteristic of a particular genus or species (Kumari et al., 2013). Phospholipids (PL) represent
1020% of total lipids in algae (Dembitsky and Rozentsvet, 1990; Dembitsky and Rozentsvet,
1996). The major phospholipids found in algae are phosphatidylglycerol (PG), phosphatidylcholine
(PC), phosphatidylethanolamine (PE), phosphatidylserine (PS), phosphatidylinositol (PI) and
phosphatidic acid (PA) (Kumari et al., 2013), with PC representing >60% of PL in red algae (Illijas
et al., 2009; Jones and Harwood, 1992; Kulikova and Khotimchenko, 2000). Moreover, red algae
contain small amounts of sphingolipids such as cerebrosides and ceramides detected in Chondrus
crispus, Polysiphonia lanosa, Ceratodictyon spongiosum and Halymenia sp. (Bano et al., 1990; Pettitt et al.,
1989). Marine algae also contain long chain C20 and C22 PUFAs such as arachidonic acid (AA),
eicosapentaenoic acid (EPA) and docosahexaenoic acid (C22:6, n -3, DHA). MGDG and DGDG
(monogalactosyl-diacylglycerol and digalactosyldiacylglycerol) in red algae contain AA and EPA
(Kumari et al., 2013). Triacyglycerols (TAGs) are the most prevalent neutral lipid that accumulates
in algae as a storage product and energy reservoir. Algae contain a wide variety of fatty acids and
their oxidized products (oxy-lipins), and sterols of nutritional and chemo-taxonomic importance.
In general, marine macrophytes do not exceed 24.5 % dry weight as lipids, mainly as phospholipids
and glycolipids (Holdt and Kraan, 2011). Of these, the long-chain polyunsaturated fatty acids
(PUFAs) and carotenoids are most noteworthy as functional foods (Holdt and Kraan, 2011).

62
3.1.2.1 Red seaweeds as functional food: secondary metabolites with antimicrobial, antioxidant and
antiviral activity
According to the definition given by FAO, functional foods are all those foods that are similar to
conventional food in appearance, intended to be consumed as part of a normal diet containing
biologically active compounds that offer potential for enhanced health or reduced risk of disease.
For nutraceutical and/or dietary supplements, no agreed definition is found for either and there is
still ambiguity about the regulatory requirements related to nutraceuticals (Shirwaikar and Khan,
2011). Nevertheless, a common aspect is that in all of the products the focus is on improving health
and reducing disease risk through prevention towards improvement of quality of life and well-being
contributing to an increased health and longevity (Shahidi, 2009).
A number of studies have already investigated the capacity of seaweeds to produce secondary
metabolites of antimicrobial value, including: volatile components (phenols, terpenes) (Ozdemir et
al., 2004; König et al., 1999; Awad, 2004; Glombitza et al., 1985; Xu et al., 2003, Karabay-yavasoglu
et al., 2007; Vairappan, 2003); steroids (Awad, 2000); phlorotannins (Nagayama et al., 2002) and
lipids (Ballantine et al., 1987; Freile-Pelegrin and Morales, 2004; Freile-Pelegrin and Robledo, 2013).
Edible seaweeds can contain appreciable amounts of polyphenols (Rodríguez-Bernaldo de Quirós
et al., 2010a, b), which are effective antioxidants and may have particular biological activities e.g.
inhibition of proliferation of cancer cells (Kwon et al., 2007; Yuan et al., 2005), influence in anti-
inflammatory responses (Kim et al., 2009), and influence on responses to diabetes (Lee et al., 2008,
2010a; Nwosu et al., 2011).
Halogenated compounds are produced naturally, mainly by red and brown seaweed. In many cases,
they possess biological activities of pharmacological interest, including antibacterial (Vairappan et
al., 2001a) and anti-tumoral (Fuller et al., 1992) properties. The most notable producers of
halogenated compounds in the marine environment belong to the Laurencia complex (Rhodophyta;
Faulkner, 2001). These compounds may also play multifunctional ecological roles such as acting as
a grazing deterrent (Brito et al., 2002; Flodin et al., 1999; Iliopoulou et al., 2002a, b; Suzuki et al.,
2002). Antioxidant activity has been screened in vitro, revealing that O. pinnatifida is a potential
source of biological active compounds with antioxidant properties (Campos et al., 2019; Silva et
al., 2018; Andrade et al., 2013; Barreto et al., 2012; Jiménez-Escrig, et al., 2012; Paiva et al., 2012;
Rodrigues et al., 2015a, b), although further characterizations are required. The use of natural
antioxidants in food, cosmetic, and therapeutic industry are promising alternative for synthetic
antioxidants with respect to low cost, highly compatible with dietary intake and no harmful effects
to the human body (Lobo et al., 2010). Dietary antioxidants from plants are believed to help prevent
ageing and many degenerative diseases such as cardiovascular diseases and cancers through radical
scavenging activity (Dillard and German, 2000; Wargovich, 2000; Yang et al., 2011). However, the

63
analyses of functional components, such as phenolic compounds, in edible seaweeds is still limited
(Yoshie et al., 2000).
3.2 Scope
A wide variety of seaweeds grows along the Scottish coasts and the red alga investigated here, O.
pinnatifida, is common on rocky shores, in mid to high exposed areas, particularly from October
until June. However, this particular species is perennial with pale-yellow/bleached specimens found
from July to September, after which it starts to recover (pers. obsv.). This variation is thought to
be determined by environmental parameters (e.g. light, temperature, etc.); but no data is available
regarding the link between the biochemical composition and the impact of light/temperature
exposition on the biomass. Furthermore, the few studies conducted on the biochemical
composition of O. pinnatifida in the UK or elsewhere tend to refer only to one particular sample,
without taking into consideration the impact of seasonal variation on the biochemical composition
that may occur in this species and if this variation is directly correlated to a particular abiotic factor
(Paiva et al., 2014; Marsham et al., 2007; Patarra et al., 2011; Rodrigues et al., 2015).
3.2.1 Aims
To investigate, for the first time, the seasonal variation of biochemical composition in O.
pinnatifida from the West coast of Scotland across one year;
To assess the metal content of the species in the view of legislations for human
consumption and intakes of metals with sources of food;
To develop protocols and assays for the biochemical analyses of O. pinnatifida specimens;
To identify the best harvesting season for the species in terms of biochemical composition,
for further cultivation and commercial applications (e.g. targeting interesting components
such as antioxidants);
To link the abiotic parameters (e.g. light and temperature) to the biochemical profile of the
species in order to understand the regulating processes behind the production of certain
compounds.
3.3 Materials and methods
Samples of Osmundea pinnatifida were collected monthly at low tide from Dunstaffnage bay
(56.4547° N, 5.4374° W, Fig. 3.1), a rocky shore environment, over a one-year period starting from
August 2016 until July 2017. Samples were then taken to the lab, cleaned manually from any visible
epiphytes and/or grazers, rinsed with filtered and UV sterilised seawater three times, blotted dried
and wrapped in aluminium foil. They were then plunged into N₂ until completely frozen and stored

64
at -80°C for at least 24 hours. They were then ground milled with a mortar and pestle (<2-3 mm),
the powder flushed with N₂ and then store at -80 °C whilst waiting analysis.
Figure 3.1 Satellite image of the sampling site, indicated with a star, Dunstaffnage bay (56.4547° N, 5.4374° W)
3.3.1 Carbohydrate analyses
A modified phenol-sulphuric acid method derived from that of Fournier (2001), originally
described by Dubois et al. (1956) was applied. Initially, 5 mg of lyophilised biomass were placed
into a 10 ml test-tube with a Teflon-lined screw-cap. To this, 2.5 ml of 1M H₂SO₄ was added and
the lids loosely replaced. The samples were acid-hydrolysed by placing them in an autoclave at
121°C for 15 min (TCR/40/H, Touchclave-R, LTE Scientific, UK). After cooling to RT, the
samples were vortexed (Vortex 3, IKA®) and then 2 ml of hydrolysate was transferred into an
Eppendorf tube. To reduce the likelihood of interference of particulate matter, samples were
centrifuged for 2 minutes at 15,600 g (5414 Microcentrifuge, Eppendorf®) and the supernatant
transferred into a clean Eppendorf tube. Triplicate samples (30 µl) of the hydrolysate (or glucose
stock solutions) were pipetted into a clean reaction tubes and 500 µl of 4% (w/v) phenol solution
added, dispensed using a rapid reagent dispenser. The mixture was then gently swirled and 2.5 ml
of concentrated (>96%) sulphuric acid added directly to the mixture, using a rapid reagent
dispenser. The solution was then lightly vortexed, allowed to cool to RT, and transferred into a
quartz cuvette, using a glass Pasteur pipette, and read spectrophotometrically at 490 nm (OD490).
The concentration of soluble carbohydrate present in hydrolysates was compared against a glucose
standard curve that had a linear equation (y = mx + c, where m is the slope and c is the y-intercept)
and was expressed as a glucose equivalent (GEq), using the equation:

65
𝐺𝐸𝑞 (𝑚𝑔 × 𝐿−1 ) =𝑂𝐷490 − 𝑐
𝑚
Calibration curves were generated by performing the carbohydrate quantification method on
glucose stock solutions ranging from 0-2 mg/ml. Blanks were composed of 30 µl volumes of
dH₂O. The GEq’s were then used to calculate the total soluble carbohydrate content per unit DW
biomass using the formula:
𝐶𝑎𝑟𝑏𝑜ℎ𝑦𝑑𝑟𝑎𝑡𝑒 𝐶𝑜𝑛𝑡𝑒𝑛𝑡 (%𝐷𝑊) = (𝐺𝐸𝑞 × 𝑉
𝑀) × 100
Where GEq is the outcome of the equation of the standard curve, V is the volume of 1M H₂SO₄
added and M the mg (DW) of the sample extracted. Aliquots of samples after the autoclaving step
were stored at -80 °C for possible further HPLC analyses.
3.3.2 Protein analyses
Samples (2 mg) in triplicate, of freeze-dried and ground milled material were weighted in an
analytical balance (0.0001) and put into 2 ml screw Eppendorf tubes. Phosphate Buffer (PB, pH
7.4) in volume of 24.3 µl, 1.8 ml of Reagent 1 (25% methanol in 1N NaOH) and 0.1-0.2 ml of glass
beads (Glass beads, unwashed 425-600 µm, Sigma-Aldrich) were added to the sample. Samples
were then bead-milled in a Tissue Lyser (QIAGEN) at 30 1/s for 30 minutes. They were transferred
into 2 ml capped glass vials and saponified in a block-oven at 100 °C for 30 minutes, left to cool
and vortexed before being transferred into 2 ml Eppendorf and centrifuged at 6,261 g (10,000 rpm)
for 5 minutes. 50 µl in triplicate per each sample were used for the protein assay using a Pierce™
BCA Protein Assay Kit (Thermo Fisher®) to estimate protein content in the samples and to
generate a calibration curve from a known protein standard. Samples, standards and blanks were
prepared as listed: 50 µl of sample or 50 µl of protein standard and 50 µl of R1 were placed in a 2
ml microcentrifuge tube with 1 ml of the reagent mix A+B (Reagent A: sodium carbonate, sodium
bicarbonate, bicinchoninic acid and sodium tartrate in 0.1M sodium hydroxide; Reagent B:
containing 4% cupric sulfate; 50:1 proportion). They were vortexed and put into a water-bath at
37 °C for 30 minutes. Then left to cool down and vortexed again. Samples were transferred and
read in polystyrene cuvettes at 562 nm (IMPLEM nanospectrophotometer). A calibration curve
was generated according to the manufacturer instructions for the BCA kit, using a 2 mg/ml Bovine
serum albumin standard ampule which comes with the kit. Protein (% DW) were calculated using
the formula:
𝑃𝑟𝑜𝑡𝑒𝑖𝑛 𝐶𝑜𝑛𝑡𝑒𝑛𝑡 (%𝐷𝑊) = (𝐵𝑆𝐴𝑞 × 𝑉
𝑀) × 100

66
Where BSAeq is obtained from the equation:
𝐵𝑆𝐴𝑒𝑞 =𝐴𝑏𝑠562𝑛𝑚 − 𝑐
𝑚
V is the volume of R1 added and M the mg (DW) of material extracted. In the second equation c
is the y-intercept and m the slope.
3.3.3 Pigments analyses
Pigments, including chlorophyll-a and carotenoids were determined employing a modified method
proposed originally by Torres et al. (2014). Samples (5 mg) of DW were weighted in triplicate on
an analytical balance (d=0.00001 g) and 1.5 ml of methanol (100%) were added to each sample,
centrifuged at 20,800 g at 4°C for 5 minutes and then read at A750-A666-A453-A416 (IMPLEM
nanospectrophotometer), in polystyrene cuvettes. The same samples were read for carotenoids at
A750- A480- A470-A450. The equations used were readapted by Torres et al. (2014) for red algae.
For Chl-a extracted in Methanol the formula applied was:
𝐶ℎ𝑙𝑎 (µ𝑔 × 𝑚𝑙−1) = 12.61 𝑥 𝐴𝑏𝑠666
for the carotenoids:
𝐶𝑎𝑟𝑜𝑡𝑒𝑛𝑜𝑖𝑑𝑠 (µ𝑔 × 𝑚𝑙−1) = (1000 × 𝐴𝑏𝑠470 − 1.63 × 𝐶ℎ𝑙 − 𝑎)
221
and the obtained values were converted and expressed in µg/g DW.
For the phycobiliproteins assay, phycoerythrin (PE) and phycocyanin (PC) were extracted
following the protocol proposed by Pereira et al. (2012) with modifications. For each sample 50
mg of freeze-dried ground milled material was weighted in triplicate, placed into 15 ml Falcon tubes
and 5 ml of 0.1 M PB pH 6.8 added to the tubes. These were incubated in the dark overnight at 4
°C. The samples were then centrifuged at 11,648 g (10,000 rpm) for 20 minutes. The supernatant
was collected, and absorption measurements were performed at 564, 592 and 455 nm for PE; 592,
618, and 645 nm for PC (IMPLEM nanospectrophotometer).
PC and PE concentration (mg/ml) were calculated according to the following formulae (Beer and
Eshel, 1985):
𝑃𝐸 = [(𝐴564 − 𝐴592) − (𝐴455 − 𝐴592) × 0.20] × 0.12
𝑃𝐶 = [(𝐴618 − 𝐴645) − (𝐴592 − 𝐴645) × 0.51] × 0.15
and then converted and expressed as mg/g DW.

67
3.3.4 Moisture content
Full moisture content (FMC) was estimated in duplicate after freeze drying the samples until all the
water was removed (72 hours) and was calculated by subtracting the dry sample weight (DW) from
the wet weight (WW). Calculation were done using the following formula (Zhang et al., 2010):
𝐹𝑀𝐶 = (𝑊𝑊 − 𝐷𝑊)
𝑊𝑊× 100
3.3.5 Antioxidant content
The antioxidant properties of O. pinnatifida were estimated with a Folin’s method and a Ferric
Reducing Antioxidant Power (FRAP) (Benzie and Strain, 1996) analyses. The preparation of the
samples was the same for both methods: 100 mg ± 5 mg of freeze dried and grounded samples
were weighted in triplicate. Subsequent extraction was done with 1 ml of 50% acetonitrile/UPW
(ultrapure water) containing 0.1% formic acid (w/v). Samples were vortexed thoroughly and
extracted on a blood tube rotator for 30 minutes at a constant speed of 40 rpm, vortexing the
samples every 10 minutes. They were then centrifuged at 16,100 g (13,200 rpm) at 5 °C for 10
minutes. After the extraction, the samples separate into two distinct phases. To ensure full mixing
of the two phases, they were dried in a Speed Vac rotatory evaporator, using the alcohol program
until dry. Dried samples were re-suspended in 600 µl of 20% acetonitrile/UPW vortexed and
placed in an ultrasonic bath for 10-15 minutes, chilled with iced water until fully re-suspended.
They were then centrifuged at 16,100 g (13,200 rpm) for 10 minutes at 5°C.
Total phenol assay was performed applying the Folin-Ciocalteu method. To 10 µl of sample in
duplicate, 240 µl of (UPW) and 250 µl of Folins reagent (Sigma Aldrich®) were added and the
samples were left to stand for 3 minutes. To this 500 µl 13% filtered sodium carbonate (Whatman®
filter paper, pore size 11 µm) was added and the samples were left to stand for one hour at room
temperature. They were read at A750 nm in polystyrene cuvettes (Ultrospec 3100 pro UV/Visible
spectrophotometer). A blank was prepared in parallel with 250 µl of water, 250 µl of Folins reagent
and 500 µl of 13 % filtered sodium carbonate. A standard curve for the phenol assay was prepared
by dissolving 18 mg phloroglucinol in 1.8 ml MeOH (1:10; 10 mg/ml ; 100%); 100 µl of this was
dissolved in 900 µl UPW (1 mg/ml; 10%) and 10 µl dissolved in 990 µl UPW (1%). Standards were
made from dilutions between 0.2 and 8 %. The TPC was calculated from the linear equation of the
standard curve and expressed as µg PGE/mg DW.
A further FRAP assay was performed in order to estimate more accurately the antioxidant power
of O. pinnatifida. The samples were prepared as described above. The reagents used for the reaction
were: 300 mM acetate (Na) buffer, pH 3.6 (3.1 g C₂H₃NaO₂x3H₂O in 16 ml of C₂H₄O₂/l); 10 mM

68
2,4,6-Tri(2-pyridyl)-s-triazine (TPTZ) in 40 mM HCl (0.312 g/100 ml in 0.1% HCl); 20 mM FeCl₃
x 6H₂O in UPW (0.540 g/10 ml). The working solution was made mixing 50 ml of acetate buffer
with 5 ml of TPTZ and 5 ml of ferric ionic solution. The sample was assayed undiluted at 100% as
follows: 100 µl of sample was mixed with 900 µl of the working solution, and a blank made of 100
µl of UPW and 900 µl of the working solution. A standard curve was obtained by using Iron (II)
sulfate between 100-1000 µM. The values obtained from the equation obtained by the calibration
curve were expressed in µM FeSO₄E/mg DW. Samples were read 4 minutes after the addition of
the working solution, at 593 nm (Ultrospec 3100 pro UV/Visible spectrophotometer).
3.3.6 Total fatty acid analyses
Total fatty acids were determined using a Bligh and Dyer (BED) Extraction (Bligh and Dyer, 1959).
Samples (0.3 g) were weighted in triplicate at five decimal places into glass test tubes. During the
procedure, the samples were kept on ice. One ml of 0.88 KI (w/v) in dH₂O was added to each
sample to hydrate the powder. Subsequently, 15 ml of methanol was added to each sample. Samples
were then homogenised for 20 sec using an Ultra Turrax, 7.5 ml of chloroform and 6.75 ml of
0.88% (w/v) KCI in dH₂O were added to each sample, then mixed vigorously before storage at -
20°C for 1 hour. During this time, samples were shaken for ~20 sec every 15 minutes. An additional
7.5 ml of chloroform and 6.75 ml of KCl solution were added to each sample. Samples were placed
at -20°C overnight. The following day samples were centrifuged at 1,200 rpm for 5 minutes (5810R
centrifuge, Eppendorf ®). The upper layer was carefully removed and the remaining sample filtered
into pre-weighted test tube using prewashed chloroform:methanol (2:1) Whatman® no. 1 filters.
Samples were then dried under N-EVP nitrogen evaporator until dried (~2-3 hours). Once dried
they were placed in a vacuum desiccator overnight before being weighed. Total lipid was calculated
using the equation:
𝑇𝑜𝑡𝑎𝑙 𝑙𝑖𝑝𝑖𝑑 = 𝑤𝑒𝑖𝑔ℎ𝑡 𝑜𝑓 𝑣𝑖𝑎𝑙 𝑐𝑜𝑛𝑡𝑎𝑖𝑛𝑖𝑛𝑔 𝑙𝑖𝑝𝑖𝑑 − 𝑤𝑒𝑖𝑔ℎ𝑡 𝑜𝑓 𝑒𝑚𝑝𝑡𝑦 𝑣𝑖𝑎𝑙
The percentage of total fatty acid in the samples was calculated using the equation:
%𝑇𝐿 = 𝑚𝑎𝑠𝑠 𝑜𝑓 𝑙𝑖𝑝𝑖𝑑 (𝑔)
𝑚𝑎𝑠𝑠 𝑜𝑓 𝑠𝑎𝑚𝑝𝑙𝑒 (𝑔)× 100
3.3.7 Metal analyses
The work on metal analyses on seaweed and water samples described in sections 3.3.7, 3.3.8, and
3.3.9, was conducted in conjunction with the technician Richard Abell at SAMS. Metals analyses
were performed on 0.20-0.25 g (DW) ground milled samples weighted in triplicate into acid cleaned
Teflon beakers and acid digested (acids used were high purity analytical grade; SpA grade, ROMIL
Ltd, Convent Drive, Waterbeach, Cambridge, UK). After sample weighing the organic component

69
of the seaweed was initially oxidised at room temperature with addition of 3 ml of 69% HNO3,
then heated to boiling on a Teflon coated hotplate and allowed to cool. This allowed breakdown
of the readily oxidizable component before addition of the highly reactive perchloric acid. When
cool, 1 ml of 65% HClO4 was added to the samples and the beakers were capped and heated to
~120 °C then left to reflux overnight. After the first oxidation step, the lids were removed, and
samples were evaporated to dryness with care so that they were not burnt. Once cool, 3 ml of 69%
HNO3, 1 ml of 35% HCl and 1 ml of 40% HF (3:1:1 v/v), were added with great care, and the
samples refluxed for 8 hours. They were evaporated to near dryness reducing the heat towards the
end of the evaporation, removing the samples when only a small bead of acid remained. Volume
(500 µl) of 69% HNO3 was then added and the samples dried down again. This steps effectively
removes any residual fluorides from the final sample. Finally, 2.5 ml of HNO3 was added and the
samples gently heated before dilution to 50 ml in 18.2 MΩ water. Samples were transferred to pre-
acid-cleaned LDPE bottles for storage prior to analysis. A commercially available certified
reference material (n=3) made from F. vesiculosus (CRM ERM-CD200, JRC European Commission,
Appendix A.1, Table A.1) and procedural blanks (reagents) were digested in tandem with unknown
samples to estimate the accuracy of this procedure and monitor possible contamination. Analyses
were performed with a Thermo Scientific X- Series (II) quadrupole Inductively Coupled Plasma
Mass Spectrometer (ICP-MS), equipped with Collision Cell Technology (CCT hexapole mass
analyser) and Cetac autosampler, including as reference a Standard Seaweed Trace element (ERM-
CD200, LGC). Results were expressed as mg/kg (dry weight) and error estimated as 1 standard
deviation of these triplicate measurements.
3.3.8 C:N ratio
Total organic carbon and nitrogen was also determined in triplicate per each sample (ANCAGSL
20-20 stable isotope analyzer, PDZ Europa, Sercon Ltd., Crewe, UK). Freeze-dried samples over
the range 1-1.2 mg DW biomass were combusted in pre-weighed aluminium capsules with helium
as a carrier gas. The instrument was calibrated with spirulina standards ranging from 0.1 to 3 mg.
3.3.9 Dissolved organic carbon and nitrogen (DOCN) and trace metals analyses in water samples
To investigate the increase of certain metals in the seaweed seasonal samples, such as aluminium,
especially in July, water samples were collected the following year (2017-2018) from June to August
every week at low tide from the same sampling site that was used for the seaweed collection for
the seasonal study in the previous year. Immediately after collection, salinity (PSU), temperature
and pH of water samples were measured with a Multi 340i/set (WTW) (Appendix A.1, Table A.2).
DOCN and metals were investigated in the water samples and 11 samples in total were collected
in 150 ml plastic bottles, which had been washed with 6M HCl and rinsed with UPW (18.2 MΩ)

70
prior to seawater collection. The bottle was kept in a sampling bag before and after the sampling
and rinsed with UPW (18.2 MΩ) before been brought into the analytical lab for the processing.
The work relative to DOC and DON analyses was conducted in conjunction with the technician
Sharon Mcneill at SAMS. For DOCN, 20 ml of the water samples were filtered through ashed GFF
filters (VWR 513-5242) into a 22 ml borosilicate glass clear 23x85 mm 20-400 screw neck vial
(VWR 548-0679), closed with a chromocol 20-400 mm cap (VWR CRMA20-SC) and injection 20
mm seal silicone, ptfe for 20-SC cap (VWR CRMA20-ST3S) containing 50 µl 85% phosphoric acid.
Samples were than analysed on a Shimadzu TOC-VCPH with Total Nitrogen Module by high
temperature oxidation (HTCO). DOC was analysed on a Total Organic Carbon analyzer
(Shimadzu TOC-VCPH) with Platinum catalyst (0.5% on alumina), carrier gas compressed air (150
ml/min), coupled to an NDIR (Non-Dispersive infra-Red) detector. Total Dissolved Nitrogen
(TDN) was analysed with a nitric oxide Analyser (TNM-1) using high temperature oxidation
converting total nitrogen to nitric oxide (NO) and detection by chemiluminescence. Dissolved
organic nitrogen (DON) was determined by subtraction of total inorganic nitrogen from TDN.
For metal analyses in water samples, samples were filtered through a 0.45 mm polycarbonate filter.
The filtrate was retained in separate acid cleared beakers and acidified using concentrated HCl
(Romil, Cambridge, UK) to give a final concentration of 0.024 M HCl (pH < 2). Indium (In)
internal standard was spiked into the acidified samples prior to analysis to ensure complete recovery
of analytes during analysis. Dissolved metals were analysed using a seaFAST Pico automated pre-
concentration system (Elemental Scientific Instruments Ltd, Warrington, UK). This instrument
loads a 10 ml samples onto a chelation column, removing the seawater matrix prior to elution and
collection of the remaining target metals prior to ICP-MS analysis. A certified reference material
(CRM) NASS6 was processed and analysed in the same manner as unknown samples to estimate
precision and accuracy of the analysis. All metals analysis was performed using a Thermo Scientific
X-Series (II) quadrupole ICP-MS, equipped with a collision cell and Cetac autosampler.
3.3.10 Statistical analyses
Statistical analyses were conducted applying an ANOVA single factor (p<0.05, Minitab 18) with
Tukey’s and Fisher’s post-hoc tests. To understand the relation between months and the
biochemical composition analysed, a PCA (Minitab 18 and PRIMER v6) was performed and the
first two components (considered the most representative) were plotted in an explicative biplot.
Data were standardized as required. BEST, ANOSIM and SIMPER analyses (PRIMER v6) were
performed for correlation with environmental data including temperature (MET Office from
Dunstaffnage station, 2016-2017, Appendix A.3, Figs A.8 and A.9), radiation and hours of light

71
(CEDA-Centre for Environmental Data Analyses, Appendix A.3, Figs A.10 and A.11) and
precipitation data (Met-office for the Dunstaffnage n°918 station, Appendix A.3, Fig. A.7).
3.4 Results
3.4.1 Carbohydrate content
The concentration (% DW) of carbohydrates was determined using the equation obtained from
the calibration curve of standard glucose concentrations (0.25 g/l Glucose stock). Seasonal
variation of carbohydrates in O. pinnatifida samples over a year is reported in Fig. 3.2.
Figure 3.2 Variation of carbohydrates content expressed as percentage per dry weight (DW) across the months in O. pinnatifida samples (n=3; error bar, 1SD of mean) using a Fournier (2001) modified method
There was a statistical difference (p<0.05) between September-May and November -January, also
three main groupings were seen (Sep-May-Aug-Jun-Jul; Apr-Mar-Dec-Oct-Feb; Feb-Nov-Jan)
(Tukey’s comparison, Minitab 18) (Fig. 3.2). The highest values were reported from May to
September and the lowest from October to April.
3.4.2 Protein content
The concentration of protein (%DW) was calculated using the equation obtained from the albumin
standard (2 mg/ml) calibration curve. Absorbance for the samples was corrected with the value
obtained at the same wavelength (562 nm) for the blank (n=3, SD=0.038). Seasonal variation of
protein in O. pinnatifida samples over the 12 months sampling period is reported in the Fig. 3.3.
This follows a different pattern compared to the total carbohydrates, with peaks in September and
December and lower values in the spring and summer months (June: 11.38 %). There was a
statistical difference (p<0.05) between the months, and 4 groupings were identified: Sep-Oct-Nov-
0
5
10
15
20
25
30
35
Car
bo
hyd
rate
co
nte
nt
(% D
W)
Months

72
Dec-Jan, Aug-Apr, Mar, Jul-May-Feb-Jun, which were statistically different from each other
(Tukey’s comparison test).
Figure 3.3 Variation of protein content expressed as percentage per dry weight across the months (%DW) (n=3; error bar, 1SD of mean)
3.4.3 Chlorophyll-a content
Values for chlorophyll-a content with the modified method proposed by Torres et al. (2014) are
reported as µg/g in Fig 3.5. They increased towards the spring period (especially in March) and
started to decrease towards the summer, with lower values for August, compared to the winter
month (such as November, December and January; Fig. 3.5). There was a statistical difference
(p<0.05) between March and August, whilst 4 groups were identified as being similar (Nov-Apr-
Dec-Jan; Dec-Jan-Oct; Jan-Oct-Feb-Sep; Oct-Feb-Sep-June-May-Jul-Aug) with March statistically
separated from the other months. A phenotypical difference in colour of the biomass among the
seasonal samples was also observed (Fig. 3.4).
0
5
10
15
20
25
30
35
Pro
tein
co
nte
nt
(% D
W)
Months

73
Figure 3.4 Variation in colours of seasonal samples of O. pinnatifida from the same sampling site for A) November, B) February, C) March, D) April, E) May, F) June, G) July, H & I) August
Figure 3.5 Seasonal variation of Chl-a (dark grey) and carotenoids (light grey) in O. pinnatifida samples expressed as µg/g of dry weight, with the modified method from Torres et al. (2014) (n=3; error bar, 1SD of mean)
3.4.4 Carotenoids
The modified method proposed by Torres et al. (2014), was effective for extraction of carotenoids
in O. pinnatifida samples and their values for the seasonal samples are reported as µg/g in Fig. 3.5.
The total carotenoid values follow the same pattern as that of the Chl-a measurements (Fig. 3.5).
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Ch
loro
ph
yll a
an
d c
aro
ten
oid
s co
nte
nt
(µg/
g D
W)
Months

74
There were statistical differences (p<0.05) between the months, in particular between March-
April-July. These months showed an increase in the carotenoid content compared to August-
September-October-January-February, which showed a decrease.
3.4.5 Phycobiliproteins
Phycoerythrin (PE) and phycocyanin (PC) were extracted successfully following the protocol
proposed by Pereira et al. (2011) with modification. Statistical analyses, ANOVA (one-way,
p<0.05), identified differences among the samples for PE, especially between the months of
January, December and March. In these months there was an increase in the phycobiliprotein
content, and the samples collected from July, August and May showed a decrease (Tukey’s and
Fisher’s comparison, Minitab 18). The overall trend for the PC and PE pigments was very similar,
with an increase towards December-January and in the spring months (March and April) and a
decrease in the summer months (Fig. 3.6).
Figure 3.6 Seasonal variation of PE (dark grey) and PC content (light grey) expressed as mg/g of dry weight in O. pinnatifida samples (n=3; error bar, 1SD of mean)
3.4.6 Moisture content
The percentage moisture content of O. pinnatifida varies across the months, reaching a maximum
reported value in August and a minimum value in May (Fig 3.7). There were statistical differences
between the months, especially between August, the highest peak for moisture content (88.4 %),
and May and February, which accounted for the lowest values (81.31 and 81.58 respectively)
(p<0.05).
0
0.5
1
1.5
2
2.5
3
3.5
4
PE
and
PC
co
nte
nt
(mg/
g D
W)
Months

75
Figure 3.7 Seasonal variation of full moisture content (FMC) expressed as percentages in O. pinnatifida samples (n=3; error bar, 1SD of mean)
3.4.7 Antioxidant content
Different dilutions, 1 % dilution and 10 %, for a selected sample were tried in order to estimate
the dilution factor for a more representative reading for TPC and FRAP assays. Neither gave a
strong signal, in relation to the standard curve, as such the samples were analysed undiluted. There
were statistical differences between the samples (p<0.05) especially between June-July-August and
Dec-Feb-Jan, with four groups identified within the one-year sampling period (July-August-June-
March-September; Aug-Mar-June-Sep-Oct; Jun-Mar-Sep-Oct-Nov; Mar-Sep-Oct-Nov-Apr-May-
Dec-Feb-Jan) (Fig 3.8).
Figure 3.8 TPC measurements expressed as µg PGE/mg (phloroglucinol equivalents) of dry weight using the Folin-Ciocalteau method on seasonal samples (n=3; error bar, 1SD of mean)
74
76
78
80
82
84
86
88
90
92
Mo
istu
re c
on
ten
t (%
FM
C)
Months
0
20
40
60
80
100
120
140
160
Tota
l Ph
eno
l Co
nte
nt
(µg/
mg
DW
)
Months

76
The values of the FRAP assay were calculated applying the equation obtained from the Iron (II)
sulfate standard curve (100-1000 µM) and are reported as µM FeSO₄eq/mg DW (FeSO₄Eq stands
for Iron II sulfate equivalents). There was a statistical difference between June-July and the other
months (p<0.05, Tukey’s comparison), while Fisher’s comparison identified 4 groups as statistically
similar. The main difference was reported between July and December. Furthermore, the trend
reported for FRAP analyses is similar to the one for TPC analyses, with the highest values reported
in July and the lowest in December (Fig. 3.9).
Figure 3.9 FRAP assay of monthly collected samples. FRAP results are expressed in µM FeSO₄ eq/mg dry weight (n=3; error bar, 1SD of mean)
3.4.8 Total lipid extracted (TLE)
The highest percentage for total lipid extracted was obtained for December and the lowest value
in July (Fig 3.10). Overall, the percentage of total lipid tends to remain stable across the year. There
was a statistical difference (p<0.05, Tukey’s test, Minitab 18) between the months of December
and July. Generally, the autumn-winter months had the higher concentration of lipid compare to
those of spring and summer.
0
0.5
1
1.5
2
2.5
FRA
P
(µM
FeS
O4
eq
/mg
DW
)
Months

77
Figure 3.10 Percentage of total lipid extracted expressed as percentage of dry weight from seasonal samples of O. pinnatifida. Results are expressed as average of n=3 (Error bar, 1SD of mean)
3.4.9 Metal Analyses
Table (3.1) reports the main metals values expressed in mg/kg (n=3, SD=1) in seasonal samples
of O. pinnatifida. Metal concentrations follow the same pattern across the year, with a general
increase of concentration in February, June, July, October and December, with the peak in
concentrations reached in July (Fig 3.11). There were statistical differences (p<0.05) among the
samples. July differs from September for the Al content, while March and December showed the
highest values for Cr. For Mn concentration, December, February and January differ from the
other months, accounting for the highest values. December, February and July differ from the
other months for Fe content, reporting the highest values; September and August reported the
statistically highest values for Cu concentration. December differs from the other months for the
content of Ni, showing the highest value. November, December, January and February are
statistically higher compared to May and July for Zn content. The highest content for As was
reported in February, which together with June, January and March statistically differ from the
lowest value recorded for July. No statistical differences were reported among the months for Rb.
July shows the highest value for Sr concentration that statistically differs from March. July accounts
for the highest value for Ba concentration, which differs statistically from all the months (excluded
December and June). The highest value for Pb was reported in February and the lowest in
September, with February statistically different from all the other months. Co has the highest
concentration in December, which statistically differs from the other months together with
February. Cd has the statistically highest value in December and the statistically lowest in May. Al
is the most abundant of the metals investigated and ranges from 3245.57 to 526.98. Iron is the
second in abundance and varies from 1147.57 and 316.88. Sr from 458.61 to 100.85; Mn varies
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
Tota
l lip
id e
xtra
cted
(%
DW
)
Months

78
from 261.54 to 56.16; Zn from 65.96 to 27.89; Ba from 40.76 to 6.68; As from 34.89 to 17.25; Ni
from 13.26 to 5.12; Cu from 8.07 to 1.77; Cr ranges from 4.52 to 1.25; Pb from 3.23 to 0.90; Cd
from 2.57 to 0.66; Co from 0.76 to 0.29; U from 0.44 to 0.18, and Cs from 0.15 to 0.07. Values are
expressed in mg/kg DW (Table 3.1; Fig. 3.11).
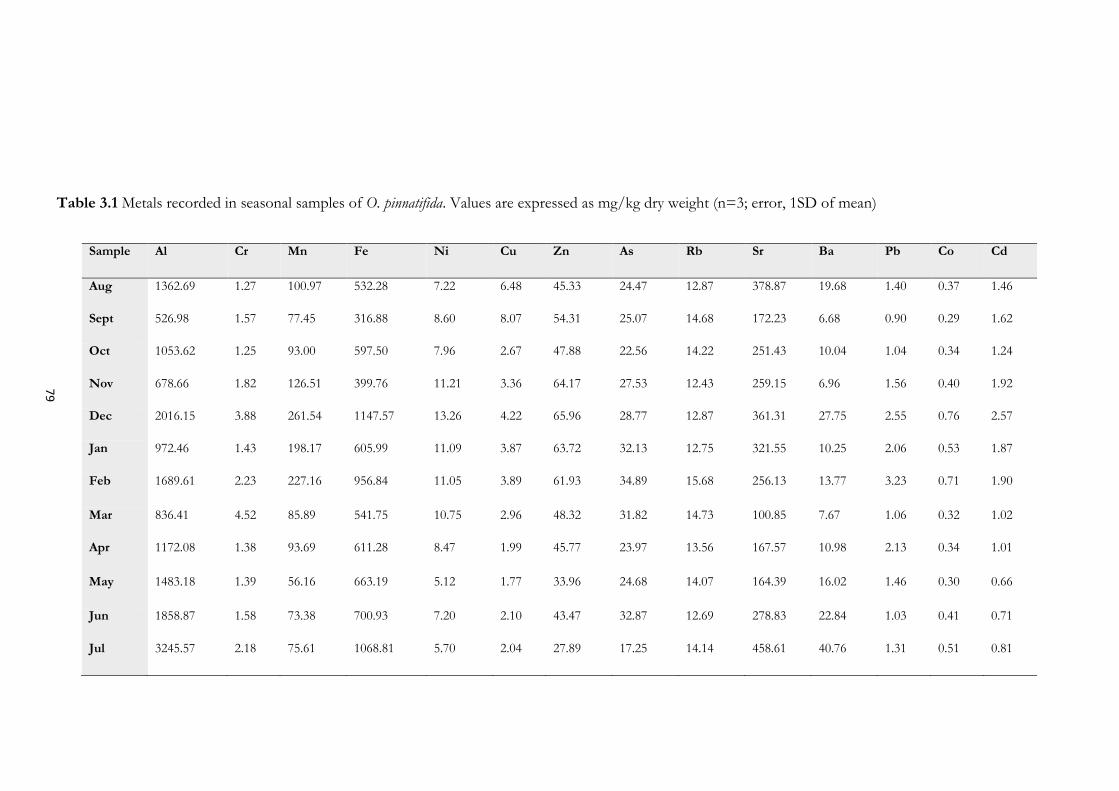
79
Table 3.1 Metals recorded in seasonal samples of O. pinnatifida. Values are expressed as mg/kg dry weight (n=3; error, 1SD of mean)
Sample Al Cr Mn Fe Ni Cu Zn As Rb Sr Ba Pb Co Cd
Aug 1362.69 1.27 100.97 532.28 7.22 6.48 45.33 24.47 12.87 378.87 19.68 1.40 0.37 1.46
Sept 526.98 1.57 77.45 316.88 8.60 8.07 54.31 25.07 14.68 172.23 6.68 0.90 0.29 1.62
Oct 1053.62 1.25 93.00 597.50 7.96 2.67 47.88 22.56 14.22 251.43 10.04 1.04 0.34 1.24
Nov 678.66 1.82 126.51 399.76 11.21 3.36 64.17 27.53 12.43 259.15 6.96 1.56 0.40 1.92
Dec 2016.15 3.88 261.54 1147.57 13.26 4.22 65.96 28.77 12.87 361.31 27.75 2.55 0.76 2.57
Jan 972.46 1.43 198.17 605.99 11.09 3.87 63.72 32.13 12.75 321.55 10.25 2.06 0.53 1.87
Feb 1689.61 2.23 227.16 956.84 11.05 3.89 61.93 34.89 15.68 256.13 13.77 3.23 0.71 1.90
Mar 836.41 4.52 85.89 541.75 10.75 2.96 48.32 31.82 14.73 100.85 7.67 1.06 0.32 1.02
Apr 1172.08 1.38 93.69 611.28 8.47 1.99 45.77 23.97 13.56 167.57 10.98 2.13 0.34 1.01
May 1483.18 1.39 56.16 663.19 5.12 1.77 33.96 24.68 14.07 164.39 16.02 1.46 0.30 0.66
Jun 1858.87 1.58 73.38 700.93 7.20 2.10 43.47 32.87 12.69 278.83 22.84 1.03 0.41 0.71
Jul 3245.57 2.18 75.61 1068.81 5.70 2.04 27.89 17.25 14.14 458.61 40.76 1.31 0.51 0.81

80
Figure 3.11 Variation in metal content expressed as mg/kg of dry weight in seasonal samples of O. pinnatifida (n=3)
3.4.10 C:N ratio
The C:N molar ratio and N and C percentages for the seasonal samples were analysed in triplicate
(Fig. 3.12). The highest value for C content is reported in February while the lowest value is in
September. The highest value for N is reported in December and the lowest in May and September.
The highest C:N ratio was recorded for July and the lowest in March.
Figure 3.12 Nitrogen percentage of dry weight (dark grey bars), Carbon percentage of dry weight (light grey bars) and C:N as molar ratio (dark grey line) of seasonal samples of O. pinnatifida (n=3; error bar, 1 SD of mean)
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
5500
Aug Sept Oct Nov Dec Jan Feb Mar Apr May Jun Jul
Met
al c
on
ten
t (m
g/kg
DW
)
Months
Al Cr Mn Fe Ni Cu Zn As Rb Sr Ba Pb Co Cd
0
2
4
6
8
10
12
14
16
18
0
5
10
15
20
25
30
35
40
45
50
C:N
mo
lar
rati
o
Nit
roge
n a
nd
Car
bo
n (
% D
W)
Months

81
3.4.11 Dissolved organic carbon and nitrogen (DOCN) and trace metals analyses in water
samples
There were no statistical differences (ANOVA, p<0.05, Tukey’s test, Minitab 18) reported for the
DOC and TDN values across the months. However, the DOC and TDN analyses follow the same
trend as that of the Al values recorded for the same months the previous year, with a peak in July
(Figs 3.13 and 3.14).
Figure 3.13 Comparison of the trends for the DOC (light grey line) (µM) water analyses (June-August 2018) and Al values (dark grey bars) (seaweed samples, June-August 2017) for the same sampling site in Dunstaffnage beach (n=3; error bar, 1 SD of mean)
Figure 3.14 Comparison of the trends for the TDN (light grey line) (µM) water analyses (June-August 2018) and Al values (dark grey bars) (seaweed samples, June-August 2017) for the same sampling site in Dunstaffnage beach (n=3; error bar, 1 SD of mean)
A positive correlation was reported between Al in seaweed samples and DOC for the seawater
(r=0.99, p<0.05) and Al in seaweed samples and TDN for the seawater (r=0.99, p<0.05).
98
99
100
101
102
103
104
105
106
0
500
1000
1500
2000
2500
3000
3500
4000
Jun July Aug
DO
C in
wat
er s
amp
les
(µM
)
Al c
on
ten
t in
sea
wee
d
(mg/
kg D
W)
Months
0
2
4
6
8
10
12
0
500
1000
1500
2000
2500
3000
3500
4000
Jun July Aug
TDN
in w
ater
sam
ple
s (µ
M)
Al c
on
ten
t in
sea
wee
d
(mg/
kg D
W)
Months

82
Statistical analyses of data on metals content of water samples showed a negative correlation
between aluminium content in water and salinity (r=-0.65, Pearson coefficient p<0.05, Fig. 3.15).
A co-variation of Al and Fe in seawater samples (r=0.99, Pearson coefficient p<0.05) was also
reported (Appendix A.2, Fig. A.5). Furthermore, a positive correlation was observed between Al
content in water samples and precipitation data (r=0.91, p<0.05) (Fig. 3.16) (data from CEDA,
Met office 2017-2018, #918 Station Dunstaffnage, Appendix A.2, Fig. A.7, Tables A.3 and A.4).
Figure 3.15 Correlation between salinity expressed as PSU (light grey line) and aluminium in water samples expressed as µM (dark grey line) collected from June to August 2018 in Dunstaffnage bay. (n=3; error bar, 1 SD of mean)
Figure 3.16 Correlation between aluminium in seawater samples (dark grey line) and precipitation data (light grey bars) (r=0.91, p<0.05). Precipitation data for 2017-2018 are monthly averaged and obtained from CEDA, Met Office, #918 Dunstaffnage station
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
25
26
27
28
29
30
31
32
33
34
1 2 3 4 5 6 7 8 9
Al i
n w
ater
sam
ple
s (µ
M)
Salin
ity
(PSU
)
Samples
0
0.2
0.4
0.6
0.8
1
1.2
0
20
40
60
80
100
120
140
160
Jun-18 Jul-18 Aug-18
Al i
n s
eaw
taer
sam
ple
s (µ
M)
Pre
cip
itat
ion
(m
m/m
)
Months

83
3.4.12 PCA analysis of seasonal data
The PCA, Eigenvalues, together with proportion and cumulative values are listed in Table 3.2 (see
Appendix A.3, Table A.5 for variables and respective principal components for PCA).
Table 3.2 The total variance explained by PCA analyses is presented in the table below, including eigenvalues, together with proportion and cumulative values of variance. PC1 to 10 are the principal components identified (analyses obtained with PRIMER v6)
PC1 PC2 PC3 PC4 PC5 PC6 PC7 PC8 PC9 PC10
Eigenvalue 5.2138 2.3024 0.8645 0.7731 0.3643 0.2624 0.1118 0.0760 0.0312 0.0005
Proportion 0.521 0.230 0.086 0.077 0.036 0.026 0.011 0.008 0.003 0.000
Cumulative 0.521 0.752 0.838 0.915 0.952 0.978 0.989 0.997 1.000 1.000
Principal component analysis was applied in this study in order to understand the correlation
between the different variables and to explain the general trend recorded from the seasonal
variation of the biochemical composition. Interpretation of the principal components is based on
finding which variables are most strongly correlated with each component. In this case, the first
principle component analyses is a measure of the concentration of PC, PE, FRAP and C:N. PE
and PC have a positive correlation while FRAP and C:N negative, so if the first ones increase the
others will decrease. The second principal component is a measure of the concentration of protein,
carotenoids and lipids. If the protein and lipid increase, the carotenoids will decrease. The third
principal component is a measure of the concentration of TPC, carbohydrates and lipids, all with
negative values of correlation. It means that if one of these value decreases the others will follow
the same trend. The fourth principal component is a measure of the concentration of TPC,
proteins, carbohydrates and carotenoids. If the proteins and TPC values increase, the carbohydrates
and carotenoids decrease. The fifth principal component is a measure of the concentration of TPC,
C:N and carotenoids. If the last two decrease than TPC values increase. However, most of the data
can be represented by the first two components that account for the 75% of variance (Table 3.5,
Fig. 3.17).
Correlation of the environmental parameters and biochemical composition was performed with
data relative to light, temperature and precipitation from MET office and CEDA. (Appendix A.3,
Figs A.7-A.11, precipitation, temperature, and radiation data from 2016 to 2017 for Dunstaffnage
bay). From this, ANOSIM (R=0.758) and SIMPER analyses (Figs 3.18 and 3.19) identified three
main groups. Group “a” only includes July. This differentiates from the other two groups in terms
of the TLE, TPC and C:N values measured. Group “b” includes October, November, December,
January, March and April, differentiates from group “c”, which includes February, May, June,
August and September, in terms of the pigments (Chl-a and carotenoids), moisture and protein

84
contents measured. Group “b” is characterized by the protein, phycobiliproteins, and TLE
contents while group “c” is commonly characterized by the concentration of carbohydrates,
antioxidant and carotenoids. BEST analyses (Fig. 3.18) demonstrated that the two abiotic factors
that express correlations within the monthly samples are the temperature and the hours of light
(No. var=2, Corr. Selections=0.528). Bivariate correlation showed that compounds such as
antioxidants and carbohydrates, are positively correlated with abiotic factors, such as light intensity
and temperature (r for antioxidant compounds and light=0.75; r for antioxidant compounds and
temperature=0.72; r for carbohydrates and light=0.75; r for carbohydrates and temperature=0.86,
see Appendix A.3, Figs A.12-A.18).
Figure 3.17 Screen plot of PCA showing the Eigenvalues and the respective component number. From the plot trend, it is evident that the majority of data is represented by the first three components, while from the third there is a change in the inclination of the line and the remaining components account only for a small percentage of data

85
Figure 3.18 The first and the second PCs were plotted since accounted for more than 75% of variance together. ANOSIM and SIMPER analyses (PRIMER, v6) identified two groups (b and c) and one outgroup (a, July) in relation to the similarity between the months in terms of biochemical composition
Figure 3.19 BEST analyses (PRIMER, v6) identified correlation between environmental factors (maximum temperature, precipitation, hours of light and maximum radiation) and months and importance of the abiotic factors in influencing the seasonality of the biochemical composition of the species

86
3.5 Discussion
According to Chojnacka et al. (2012), there are about ten thousand identified macroalgae species,
yet only about 5% thereof are used as human food, or as animal feed. In Europe, the use of edible
seaweeds for food purposes is largely historic, although this is changing and currently researchers
are studying less-known seaweed species. Not only because of their potential biological properties-
due to specific functional compounds, but mostly for their nutrient profile. They are being
considered as important alternative sources for protein and complex carbohydrates (Ibañez and
Cifuentes, 2013).
From a cultivation perspective, it is important to have a knowledge of the biochemical composition
of a target species and its nutritional profile, in order to exploit it in further applications, such as
food and/or bioactive compound. Nonetheless, investigations are fundamental to improve or
manipulate the nutritional profile in order to fit the market requirements and to exploit new species,
such as O. pinnatifida, as source of biologically active compounds with interests for further
biotechnological applications. While few cultivation trials have been undertaken for O. pinnatifida
(Silva, 2015; Gonçalves, 2018), different studies have been conducted on its nutrient levels,
bioactive properties and biochemical profile (Campos et al., 2019; Rodrigues et al., 2015, 2019;
Paiva et al., 2014; Patarra et al., 2011, 2013; Sabina and Aliya, 2009, 2011; Marsham et al., 2007;
Rizvi and Shameel, 2005). Whilst this study is the second to present the biochemical composition
of O. pinnatifida in UK, the first one was undertaken by Marsham et al. (2007). This study is the
first one to investigate the impact of seasonal variation on the biochemical/chemical composition
of O. pinnatifida, and the first to also consider the relationship this has with abiotic factors.
O. pinnatifida shows variation in its nutrient components across the months, for all the compounds
analysed. Based on previous studies conducted on O. pinnatifida, the following methods were
applied by previous authors for biochemical investigations: nitrogen content was determined by
the Kjeldahal method (Marsham et al., 2007; Denis et al., 2010; Munier et al., 2013; Rodrigues et
al., 2015); total fat content by Soxhlet extraction (AOAC, 2000; Rodrigues et al., 2015) and
Soxtherm method (Marsham et al., 2007) together with the application of GC for fatty acid (FA)
profile (Paiva et al., 2014); total sugar content was calculated by subtracting protein content and fat
content from total organic content (Rodrigues et al., 2015); soluble carbohydrates content has been
determined throughout the phenol-sulphuric acid colourimetric method (Dubois et al., 1956);
polyphenols were extracted and analysed with the Folin-Ciocalteu method (Rodrigues et al., 2015)
and run on a LC-MS system (Nwosu et al., 2011). Elements have been assessed through an acid
digestion and ICP-OES (Rodrigues et al., 2015) or with HPLC (Paiva et al., 2014). Antioxidant
activity (as 2,2-Diphenyl-1-picrylhydrazyl, DPPH*) was determined spectrophotometrically
(Andrade et al., 2013). Amino acids analyses were performed following Barbarino and Lourenco

87
(2005) method, with a modified procedure for derivatisation from Sánchez-Machado et al. (2003),
and an evaluation of amino acids profile on HPLC system (Paiva et al., 2014). However, in this
study a different approach was taken, and a new set of assays for biochemical composition were
tested and compared to those previously employed for O. pinnatifida. This was undertaken in order
to develop and achieve faster, more reliable and efficient methodologies for assessing the chemical
composition of O. pinnatifida.
3.5.1 Carbohydrate content
In Paiva et al. (2014) carbohydrates were measured employing the colorimetric method described
by Dubois et al. (1956) and the value reported (17.59 ± 0.27% of DW) is comparable to that
obtained in this study for the same month (17.09 %, January). Rodrigues et al. (2015) reported a
percentage of total sugar equivalent to 32.4 for O. pinnatifida in Portugal, which is higher compared
to those reported in this study (Fig. 3.2) for the same month (April). The second method applied
(modified from Fournier, 2001) proved to be more efficient in carbohydrates extraction from dry
biomass of O. pinnatifida, compared to the single tube multi-assay extraction (Appendix A.2, Fig.
A.6) (Slocombe et al., 2013). The carbohydrates follow almost a linear trend from January to April,
with an increase in May-June and again in August-September, followed by a decrease in October,
November and December. The highest value obtained was in September (27.78 %) and the lowest
in January (17.09 %). The level of carbohydrates is higher during the spring-summer compared to
the winter and autumn; because the seaweeds accumulate and store this type of resource to use
later in the season when the available nutrients are at lower levels (Lüning and Pang, 2003). The
trends for carbohydrates and protein content look almost inverted, with lower levels of protein
from May to August, when the carbohydrates have increased and is at its highest levels of
concentration (Figs 3.2 and 3.3).
3.5.2 Protein content
The value relative to the percentage of protein are similar to those reported by Rodrigues et al.
(2015), that estimated 21 % for O. pinnatifida sample in April. In Paiva et al. (2014), protein levels
were estimated employing a modified Kjedahl procedure (AOAC, 1990) and the application of a
conversion factor of 6.25, with a value of 20.79 %± 0.12. The same method was applied by Patarra
et al. (2011), with an average value of 20.64 (for January-February) and by Marsham et al. (2007),
that reported a value of 27.3 ± 0.1 % of protein for O. pinnatifida in the UK. These values are similar
to those obtained in this study (Fig. 3.3). Protein content of seaweeds varies not only between
species (Fleurence, 1999), but also between seasonal periods (Mishra et al., 1993). This was
confirmed in this study for O. pinnatifida seasonal samples. As already seen for other red algae
(Galland-Irmouli et al., 1999; Sanchez-Machado et al., 2003; Cian et al., 2015), O. pinnatifida has

88
high level of proteins. Higher values are reported for autumn-winter months (27.41% September
25.13 % December) while the lowest for February (12.04 %) and summer months in general (11.38
% June). The concentration of proteins follows an oscillation along the months but was more
stable compared to the variation in carbohydrate and pigment content (Figs 3.2, 3.3, 3.5 and 3.6).
Most seaweeds contain all the essential amino acids and are a rich source of the acidic residues
aspartic and glutamic acid (Fleurence et al., 2012). However, the composition of the protein was
not estimated in this study, but the data obtained from the metabolomics analyses presented in
Chapter 4, gives some hints as to the major proteins present. Similar variation in protein content,
with an increase in the winter and a decrease in the summer (especially July and August), was
reported for other species of red algae; e.g. G. turuturu (Denis et al., 2010) and P. palmata (Galland-
Irmouli et al., 1999). The same trend was also observed for the phycobiliproteins seasonal samples
of O. pinnatifida, in this study, at least for December and March-April. This is probably related to
the necessity to be able to harvest light in sufficient quantities for photosynthesis to occur during
the winter. As confirmed by other authors (Pereira et al., 2012; Kosovel and Talarico, 1979;
Campbell et al., 1999; Aguilera et al., 2002; Orduña-Rojas et al., 2002) the highest concentration of
protein and phycobiliproteins were observed in the months with the lowest light levels and the
lowest in the months with elevated sunlight levels. This association was confirmed in this study for
O. pinnatifida (Figs 3.3 and 3.6).
3.5.3 Pigment content
Initially for the pigments’ analyses in O. pinnatifida, a single tube multi-assay was considered. This
method was used for measuring the protein content and trialled for carbohydrates, but was
considered unsuitable, because it had been developed for the analyse of green algal biomass (Chen
and Vaidyanathan, 2013). Another method was tested (Torres et al., 2014), which involved
methanol extraction of the pigments from the harvested biomass. The Chl-a and carotenoids were
calculated following the equations in this paper and adapted to red algae (Torres et al., 2014). Within
this chapter, an initial trial was carried out reducing the quantity DW of biomass to be extracted
from 25 mg, as described in the paper, to 5 mg of DW. Between these two values, a range of
selected quantities of DW material were extracted and the results, in term of concentration,
obtained compared. This confirmed that the reduction in biomass extracted gave acceptable set of
results in relation to the quantity of pigments measured. The values obtained were comparable to
those estimated by Torres et al. (2014) (Fig. 3.5). Chl-a concentration changed across the months,
increasing significantly in February-March and then decreasing towards the summer (August). This
is probably related to the light intensity levels and the length of light exposure. The same pattern
was observed for the carotenoid concentration, which means that both pigments act simultaneously
in light harvesting (Chl-a) and protection (carotenoids) (Fig 3.5). Carotenoid contents in

89
macroalgae responds to high radiant energy increasing its concentration for a photo protective
action in preventing photo oxidation (Harker et al., 1999).
For the measurement of phycobiliproteins, the method proposed by Pereira et al. (2012), applied
for Gracilaria sp., was tested for seasonal samples of O. pinnatifida in this study. Only 50 mg of DW
were used for the analyses (rather than 90 mg, as proposed by the protocol). The other steps of the
protocol were kept the same (Pereira et al., 2012). PE and PC in O. pinnatifida followed the same
pattern, with an increase in December-January and March-April, and a decrease towards the
summer. It has been previously reported that high light irradiance can negatively affect
phycobiliprotein pigment concentrations in macroalgae (Aguilera et al., 2002; Zubia et al., 2014).
Another factor that plays a role in the phycobobiliproteins concentration is N limitation, since this
may cause red algae to use their phycobiliproteins as a catabolite, with the consequence of a
reduction in their light-harvesting ability (Lobban and Harrison, 1994) and a “bleaching” effect
(Penniman and Mathieson, 1987). This was confirmed in this study were the concentration of
phycobiliproteins decreases during the summer, this is also the period during which this species
undergoes N limitation, verified by the C:N data (Fig. 3.12). The concentration of PE is higher
than PC, except in August, September and May. A similar pattern, but with lower values were also
obtained for Gracilaria domingensis (Pereira et al., 2012) for PE and PC. The equations applied for
their estimations were those proposed by Beer and Eshel (1985). The results obtained were similar
to those reported for Gracilaria sp., ranging from 0.10 to 0.6 (mg/g) for PC and from 1.5 to 7
(mg/g) for PE (from Pereira et al., 2012) and to those proposed by Beer and Eshel (1985) for
different red algal species (Fig. 3.6). This is one of the most frequently cited phycobilin
quantification techniques (Beer and Eshel, 1985) that utilizes the spectral properties of crude
aqueous extracts to estimate the pigment contents of sample tissues. In particular, pigments were
investigated in this study because O. pinnatifida has a great variability in the colour (Fig. 3.4), linked
to where and when it grows. Since the colour is a fundamental parameter in the commercialization
of different products (e.g. salmon, sea urchin’s gonads, etc.), it is important to see if is it possible
to regulate the phenotype of a species in order to tailor the final product according to the market’s
needs. This might be applicable also to O. pinnatifida; however, it is still not understood if the colour
has any influence on the taste of this particular algal species. The literature reports that macroalgae
regulate their pigment contents, constitution and morphology responding to prolonged exposure
to different light levels (Machalek et al., 1996). Furthermore, when internal N reserves decrease,
seaweed start to lose their dark reddish-brown pigments, turning into a pale-yellow colour and
growth ceases (Ryther et al., 1981). This loss of colour might be correlated to the fact that pigments
are processed as alternative source of protein (Lapointe, 1981; Bird et al., 1982).

90
3.5.4 Moisture content
Moisture content reported in this study is very similar to that estimated by Paiva et al. (2014): for
the sample from Azorean area of January, they reported 90.6 %. Furthermore, Marsham et al.
(2007) reported a similar value of 86.4 ± 3.7 as well as Rodrigues et al. (2015), reporting a
percentage of 88.23 ± 0.01. In this study, the moisture content does not change much across the
months, with statistical differences reported only between August and May-February (Fig. 3.7).
3.5.5 Antioxidant content
The total phenolic content evaluated through the Folin-Ciocalteu colorimetric method has become
a standard assay in studying phenolic antioxidants. It is a convenient, simple and reproducible assay
(Huang et al., 2005). Seaweed may contain a wide variety of phenolic compounds: from simple (e.g.
phenolic acids) to more complex substances (e.g. phlorotannins) in different quantities. The lack
of an accurate and universal extraction method for phenolic analyses in seaweed is because these
compounds can be combined with other components in the biomass (such as carbohydrates and
proteins). Nevertheless, several solvents (such as methanol, ethanol, acetone and their
combinations) have been widely used for the extraction of phenolic compounds (Dai and Mumper,
2010). As already observed for other chemical compounds, the composition of phenols might also
vary, qualitatively and quantitatively, depending on the habitat, environmental parameters (light,
temperature and salinity), the season, the geographical distribution and the actual species been
investigated (Ibañezet et al., 2012; Rodríguez-Bernaldo et al., 2010a, b). Antioxidant properties
were analysed applying TPC and FRAP assays, without considering the DPPH* measurements,
already reported in previous studies for this species (Silva and Abreu, 2014; Silva et al., 2018). For
TPC, an extraction with 50% Acetonitrile/UPW containing 0.1% formic acid (w/v), followed by
Folin-Ciocalteu method was conducted in this study, while extraction with ethyl acetate plus Folin-
Ciocalteu method was applied by Rodrigues et al. (2015). They reported a TPC of 337± 22 µg/g
DW for O. pinnatifida for April, while in this study the TPC value for that month was 93.69 ± 0.04
µg/mg, with the highest value measured for August and the lowest for January (Fig 3.8). Similar
results were proposed by Campos et al. (2019) that reported a value of 172 mg Gallic acid equivalent
(GAE/100 g DW, TPC) for O. pinnatifida in March. The results obtained from TPC analyses in this
study (Fig 3.8) are higher than those reported by Silva and Abreu (2014) for O. pinnatifida cultivated
in a trial in Portugal (wild samples 87.57 and IMTA-samples 65.89 mg GAE/g). The difference
with previous authors’ work can be attributed to the different extraction methods and reference
standards applied. Furthermore, TPC and FRAP analyses in this study followed the same trend,
confirming the seasonality reported. Generally, the primary function of phenolic compounds in
land plants is as protection against ultraviolet radiation and pathogens (Manach et al., 2004).
Phenolic compounds found in algae also include the phlorotannins found in brown algae and in

91
lower amounts in some red algae (Machu et al., 2015). They are components of cell walls, but they
have also other secondary ecological functions such as radiation protection and reproduction. They
have also been investigated for their potential therapeutic properties (anticancer, antioxidative,
antibacterial, anti-allergic, anti-diabetes, anti-aging, anti-inflammatory and anti-HIV activities) (Li
et al., 2011). Polyphenolic compounds are very common constituents of human diets, there is also
increasing interest from both consumers and food manufacturers linked to these compounds
(Manach et al., 2004). It has been suggested that the phenolic compounds of this species may play
a role in its peculiar taste. A deeper understanding of their concentration and variation could be
applied to the commercial exploitation and tailoring treatments to the cultivation of this species.
But the information present here is relevant for identifying the best harvesting season for this
species and for further commercial exploitation of O. pinnatifida for biotechnological applications.
3.5.6 Total fatty acid content
With respect to the total lipid extracted, it should be noted that the method here applied extracts
all the lipids present in the sample (including pigments, such as carotenoids). The results obtained
were different from those reported by Rodrigues et al. (2015), who published a value of 0.9 for %
total fat (g/100 g dry seaweed) (analyses on gas chromatography) and Paiva et al. (2014), who
reported a lipid % of 7.53 (applying a Soxhlet extraction). However, they are similar to those
recorded by Marsham et al. (2007) that reports a percentage of 4.3 (using a Soxtherm method).
Higher values were reported for the autumn-winter months, while lower ones for the spring-
summer months, with the lowest value reported for July (Fig. 3.10). Similar results have been
obtained for other seaweeds (e.g. Undaria pinnatifida, Laminaria japonica, Fucus serratus, Egregia
menziesii, Condrocanthus canaliculatus and Ulva lobata; Gerasimenko et al., 2011; Kim et al., 1997;
Nelson et al., 2002). Composition of lipids was not analysed in this study.
3.5.7 Metals content in seaweed samples
Seaweeds play an important role in the nutrient dynamics of coastal systems and reflect changes in
water quality efficiently (Wilson, 2002). Contamination of heavy metals in the marine environment
is posing a major problem, which is directly related to anthropogenic actions (Karthick et al., 2012).
Since the species here investigated is used in food applications, it is particularly important to
investigate the metals and minerals content of the biomass. Metals were analysed for O. pinnatifida
in this study and values compared with those obtained by Rodrigues et al. (2015). The accuracy of
the digestion procedure and analyses applied in this study was confirmed by the percent recovery
of the certified elements (Appendix A.1, Table A.1). Furthermore, the step with hydrofluoric acid
was chosen following previous in-house seaweed investigations where residues (possibly silicates)
and sometimes clasts were still clearly visible after the first nitric acid/perchloric acid reflux step

92
and reproducibility of some metals was found to be inconsistent (Abell, pers comm.). These
adjustments confirmed that the methodology was high and sensitive enough to resolve any seasonal
variation within the seaweed samples.
Rodrigues et al. (2015) reported 3.7 for iron, 0.58 for zinc, 0.05 for copper and 0.12 for manganese
(as mg/10g dry weight). Higher results were obtained in this study (for the same month, April),
especially regarding the concentration of iron. Arsenic measured in this study was higher compared
to its concentration estimated in other red algae (e.g. L. papillosa: 1.48 µg/g dry weight; L. johnstonii:
5.65 µg/g dry weight, Sanchéz-Rodrigues et al., 2001) but similar to the results reported for other
species, such as G. pachidermatica (13.8 µg/g dry weight) (Sanchéz-Rodrigues et al., 2001). The other
metals estimated in Sanchéz-Rodrigues et al. (2001) are similar to those estimated in this study
(Table 3.1). Apart from the many positive aspects associated with the consumption of algae, there
are also concerns to consider, especially regarding the concentration of heavy metals. Algae are
known for their capacity of absorbing and accumulating heavy metals (e.g. arsenic, cadmium and
mercury) (Rincon et al., 2005; Brinza et al., 2007). Limits for the consumption of these
contaminants has been determined by the FAO/WHO (2008, 2011a) Expert Committee on Food
Additives (JECFA) with units expressed as µg/kg body weight per week, with the value set for 15
and 7 for arsenic and cadmium respectively. The level of arsenic (mg/kg) in this study range from
34.89 (March) to 17.25 (July), while the Cd value vary from a maximum of 2.57 (December) to 0.66
(May). These results demonstrated that arsenic and cadmium levels are not a major concern for
this species harvested in this geographical region, particularly considering the low amounts of this
seaweed consumed. It should be noted that the toxic potential of heavy metals is also dependent
on their physical state, with the arsenic being the most toxic in its inorganic form. Usually algae
convert most of the inorganic arsenic in arsenosugars and arsenophospholipids (García-Salgado et
al., 2012). Aluminium concentration in all the seasonal samples was higher than any other element
investigated, followed by iron (Table 3.1). However, the critical elements (e.g. As, Pb, Cu, Cd) were
found in moderate/low values, when compared to other seaweed species (such as Laminaria spp.,
S. latissima, A. esculenta, L. obtuse, P. palmata) (Schiener et al., 2015; Mæhre et al., 2016). However, Fe
concentration was particularly high, especially compared with other red algae (such as P. palmata,
Fe 100 mg/kg, Mæhre et al., 2014). High amounts of barium can be normally found in seaweeds
and fish; these minerals are released into aquatic ecosystems by industrial processes and can
accumulate in the organisms. The level of this metal has to be low since high concentration can
have severe and deadly effects on humans and other organisms (Apaydin et al., 2010). In this study,
the level of Ba was generally low, the exception was the month of July. Sr in its elemental form
occurs naturally in many environments and the highest level of Sr in this study was reported for
July. It has been seen that concentrations of chemical elements in seaweeds are generally low in the
warmer months, i.e. the time of the highest algae metabolic activity resulting in dilution of the

93
accumulated metals whereas in the winter with a slowing down of metabolic processes the element
content is higher (Bojanowski, 1973; Hou and Yan, 1998; Villares et al., 2002). However, this was
not completely confirmed in this study, were some metals were observed at their maximum
concentration in the spring-summer months (Fig. 3.11).
3.5.8 C:N ratio in seaweed samples
C:N values between 10 and 15 are typical for macroalgae with normal growth rates (Lobban and
Harrison, 1994). Higher values than this threshold indicate limitation of growth due to nitrogen
deficit while lower values show nitrogen accumulation (Hanisak, 1983). C:N ratio was considered
in this study because since the 1970s it has been used as an index of food quality in seaweed
(McMahon et al., 1974), a low C:N ratios (which mean high N levels) suggests relatively high levels
of protein. A positive correlation between protein and nitrogen content has previously been
reported in brown, red and green seaweeds (Kim et al., 2013; Lapointe and Ryther, 1979;
Robertson-Andersson et al., 2009). However, N and protein content may not be directly related if
the nitrogen is stored in refractory or non-proteinaceous material (McMahon et al., 1974). C:N and
C:P ratios are usually elevated in low nutrient environments, reflecting an increased limitation of
net productivity by N, P or both (Lapointe, 1987). In this study, the highest C:N ratio for O.
pinnatifida was reported for the months from May to August, the maximum values reported was for
July (15.1) and the lowest for March (8.5, Fig. 3.12). However, in general O. pinnatifida never exceeds
C:N ratio of 15, showing a general trend of normal growth rate across the whole year.
3.5.9 Metals and DOCN analyses on water samples
To better understand the high peaks reported in certain months (such as July) for certain metals,
like aluminium, water sampling followed by DOCN and metals analyses were conducted from June
2018 to August 2018 of the following year (2017-2018) and the results compared to the seasonal
seaweed specimens collected in the same area, the previous year, in 2016-2017. Interestingly,
DOCN analyses on water samples showed the same trend reported for the aluminium content in
the seaweed samples, with a peak reported in July. DOCN is an important parameter to consider
when it comes to metal analyses in water. Metals can aggregate with dissolved organic matter in the
water column, such as carbon and nitrogen, through adsorption and flocculation (Dekov et al.,
1998), and to have a complete account of the metals in the water this proportion has to be
considered. Dissolved organic carbon (DOC) present in water is primarily in the form of refractory
(i.e. poorly biodegradable) compounds with some biodegradable dissolved organic carbon (BDOC)
or assimilable organic carbon (AOC). DOC and C:N analyses conducted in this study on the water
samples suggest that there is a higher terrestrial signal of organic matter for the August sample.
C:N ratio <8 are usually from marine sources, while higher ratios (C:N >15) are likely to come

94
from terrestrial sources (Gray and Biddlestone, 1973; Premuzic et al., 1982; Ruttenberg and Goñi,
1997b). Heavy metal pollution originates from natural and anthropogenic sources; the natural
sources include rock weathering and soil erosion, and the anthropogenic sources include domestic
sewage, industrial wastewater, mining wastewater, agricultural fertilizer leachate, and coal burning
(Alvarez-Vazquez et al., 2017; Davis and Birch, 2010; Pan and Wang, 2012; Skordas et al., 2014).
Statistical analyses conducted on seawater and environmental data and evaluation of correlation
with salinity and precipitation data, suggests that the high level of Al and Fe (abundant in terrestrial
derived carbon, Krachler et al., 2010) reported first in the seaweeds samples and then in the water
samples might be related to an inland input (Appendix A.1, Tables A.3 and A.4). Dunstaffnage bay
is surrounded by granitic mountains and is located in close proximity of Loch Etive. Both of which
could supply significant amounts of aluminosilicate particles and cations to the seawater into the
area sampled (Krachler et al., 2010; Muller et al., 2012).
Interestingly, the comparison between wild and cultivated samples (obtained from tank experiment
applying F/2 medium, see Appendix A.1, Figs A.1 and A.2) in regards to certain metals, shows
some differences in particular for Fe, Co, Cu concentration (higher values are reported in the
cultivated sample compared to the wild) and As, Sr, Ba, Al, Mn and U (lower values are reported
in the cultivated sample compared to the field samples). This is probably due to the F/2 medium
used for the cultivation and the source of seawater used for its preparation which comes from
filtered, UV sterilised and tyndalysed seawater pumped from the back beach at SAMS. This can
also be particularly important from a cultivation perspective, since some of the heavy metals
showed statistically lower values in the cultivated samples compared to the wild ones. This
reduction in content of particular metals (As, Ba, Al, Mn and U, Appendix A.1, Figs A.1 and A.2)
during cultivation is interesting for the final product obtained since it can increase the number of
food and nutraceutical applications.
3.5.10 PCA Analysis
The first and the second principal components account for 75% of the variability in the data, while
the cumulative variability for the first three PC is 86 %. Variables were plotted with the first two
components together with the sampled months. Two main groups were observed in the chart and
one outgroup, July. According to the biplot (Figs 3.18 and 3.19) it seems that the summer-early
autumn months are commonly characterized by the concentration of carbohydrates, antioxidant
compounds and C:N ratio. Autumn and early-winter months are instead associated with higher
concentration of protein and lipid. Months of late winter-spring are instead grouped by the values
of pigments, including phycobiliproteins, chlorophyll-a and carotenoids. July emerges as outgroup
from these analyses. This information is useful in understanding which biochemical components
are more characteristic and specific for each month in order to comprehend how the biochemical

95
composition in O. pinnatifida varies, becoming then useful in the application of a harvesting plan
and cultivation trials.
Seaweeds are exposed to seasonal variations of abiotic factors (such as temperature, irradiance,
salinity, etc.) that can influence their metabolic responses, including photosynthesis and growth
rate, as well as levels of their principal constituents (Orduña-Rojas et al., 2002) and their
biochemical composition (Fábregas et al., 1995; Fernandéz-Reiriz et al., 1989). Moreover, the
nutritional properties of seaweeds and their seasonal oscillation are poorly known and normally are
evaluated from the chemical composition (Mabeau and Fleurence, 1993). This study considers
some of the abiotic parameters available for the sampling site in the Dunstaffnage bay, including
hours of light, radiation, temperature and rainfall data. This data was analysed together with the
biochemical components in order to verify if there was any correlation between them and the
variation of the biochemical composition of O. pinnatifida in the West coast of Scotland. The data
considered referred to the period from August 2016 to July 2017, the same time interval of the
biomass sampling.
PCA analyses helped in understanding how the biochemical profile of the species changes across
the seasons. Correlation analyses demonstrated that abiotic factors (such as temperature and
irradiance) can influence the metabolic responses and the biochemical composition of the species,
as already seen for other seaweeds (Orduña-Rojas et al., 2002; Fábregas et al., 1995; Fernandéz-
Reiriz et al., 1989). A positive correlation was found between the hours of light and the production
of antioxidant compounds as well as carbohydrates content (see Appendix A.3, Figs A.12-A.15).
Furthermore, statistical analyses provided a characterization of the months according to their
biochemical profiles, which is fundamental for timing and management of harvesting.
3.6 Conclusions
As already underlined, this study confirms that O. pinnatifida is an important source of
bioactive compounds with beneficial properties such as antioxidant, antiviral, antibacterial
and anticancer. Therefore, the information obtained from this study are fundamental to
gain a better knowledge of how the production of these compounds is regulated by the
environmental factors (such as light and temperature) and how this information can be
used and applied to a cultivation perspective in order to target the final product ensuring a
determined biochemical profile;
Protocols were successfully developed and established for the biochemical analyses of the
species or existing ones were modified and readapted;
The metal content, especially in relation to arsenic and cadmium, does not represent a
threat to human health in the vision of a commercialization of the species as food or

96
nutraceutical products; furthermore, metals’ content can be manipulated in cultivation
systems which can become particularly valuable for commercial production of the biomass
and can reduce the concentration of unwanted metals;
This study helps in identifying a harvesting plan for the species, according to the further
commercial applications of it. This is important if the final scope of the harvested biomass
of this species is, for example, the production of bioactive compounds rather than biomass
to be used in the food industry. In this case, late spring-summer months should be
preferred. On the other hand, if the main aim is instead the exploitation in the food
industry, other parameters such as carbohydrates and protein have also to be taken into
consideration. In regard to carbohydrates, the best harvesting season is summer-early
autumn, while for protein and lipids autumn-early winter. Finally, for pigments the highest
content is reported in spring. Moreover, the information obtained from this study can
become helpful to outline a legislation for collection of the species, considering not only
the abundance but also the quality of the harvested biomass.
This is the first time that the seasonal variation in biochemical composition of O. pinnatifida
across one year has been investigated and also correlated to abiotic data. This information
is paramount in understanding the biochemistry of the species and the application of this
knowledge to cultivation methods (in terms of light and temperature requirements) might
guarantee the continuos supply of a high value tailored and controlled product, exploitable
commercially;
Biochemical analyses confirmed, as already reported by previous authors, that O. pinnatifida
has a good potential for further processing or for direct food and nutraceutical applications
(Rodrigues et al., 2015; Paiva et al., 2014; Patarra et al., 2011, 2013; Sabina and Aliya, 2009,
2011; Marsham et al., 2007; Rizvi and Shameel, 2005).

97

98
Chapter 4 Composition and seasonal variation in the metabolomic
profile and flavour of Osmundea pinnatifida
4.1 Introduction
Metabolomics has now become a high-resolution biochemical phenotyping tool capable of
advancing our understanding of primary and secondary metabolism. It has made great success in
assessment of responses to environmental stress, biomarker analysis, chemotaxonomy, comparing
mutants and different growth stages, drug discovery, studying global effects of genetic
manipulation, and natural product discovery (Higashi and Saito, 2013; Kim et al., 2011; Muranaka
and Saito, 2013; Putri et al., 2013). Metabolomic analyses provide information on the regulatory
processes and metabolic responses of organisms, and it has recently helped in gaining newer
insights into the biochemical composition and functioning in seaweeds (Gaubert et al., 2019). The
continuing investigation of seaweeds for multiple biochemical, nutraceutical, pharmaceutical
applications, has greatly increased their applications in a wide array of sectors, e.g. food, feed,
fertilizer and hydrocolloids, and new, e.g. cosmetic, nutraceutical, biomaterial, and bioenergy
(Bixler and Porse, 2011; Holdt and Kraan, 2011). These developments have also led to a sudden
rise in seaweed production (FAO, 2010). Metabolomics in seaweeds is still in its infancy compared
to terrestrial plants which have been more widely investigated (Davis and Vasanthi, 2011).
However, some groups of metabolites have been studied for decades in seaweeds including
mycosporine-like amino acids, halogenated compounds (Kundel et al., 2012; La Barre et al., 2010;
Yuan et al., 2009), and lipids and their derivatives (Bouarab et al., 2004; Goulitquer et al., 2012;
Kumari et al., 2013a, b).
Metabolite concentrations in seaweed varies between and within species (Oliveira et al., 2013;
Wright et al., 2000), and can be affected by environmental factors (biotic and abiotic) (Kooke et
al., 2011; Slattery et al., 2014). Metabolite profiling in seaweeds has been carried out for decades
but has been restricted to targeted identification of one group of metabolites (Kumari et al., 2013a,
b; Kundel et al., 2012). There is a lack of information on the global metabolomics response of
seaweeds, especially in relation to changing environment conditions (Greff et al., 2017), with the
majority of present studies that focus on specific families of compounds therefore overlooking
many metabolites likely to play important ecological functions. In 2011, the first high coverage
metabolomics study on red algae was published (Nylund et al., 2011), and it described changes in
metabolites produced by Gracilaria vermiculophylla in relation to defence response. More recently an
attempt has been made to constitute a reference library for seaweed metabolites (Davis and
Vasanthi, 2011).

99
Red seaweeds, compared to green and brown, have proved to produce halogenated metabolites,
especially bromine and iodine derivatives (McConnell and Fenical, 1977; Fenical, 1981; König et
al., 1999a). The orders of Nemaliales, Gigartinales, Ceramiales, Rhodymeniales and Cryptonemiales
have been shown to be engaged in biological halogenations yielding a diverse array of organic
compounds (McConnell and Fenical, 1977). Species of the Laurencia complex (included O.
pinnatifida), have been extensively investigated and are well known source of bioactive halogenated
and non-halogenated secondary metabolites (Masuda et al., 1996; 1997a, b). More than 300
compounds have been isolated and classified in four groups: sesquiterpenoid, diterpenoid,
triterpenoid and C15 bromoethers. Laurencia species have been the most extensively studied for
the composition of these compounds (Vairappan et al., 2001a; Da Gama et al., 2002; Pereira et al.,
2003; Sudatti et al., 2006, 2011; Salgado et al., 2008). In the early 80’s, the structures and absolute
configurations of three halogenated vinyl acetylenes, which are natural products from the red alga
O. pinnatifida, were described (Gonzalez et al., 1982). These first chemical studies were concerned
with assessing the diversity of halogen-based secondary metabolite biosynthesis and structural
proposals were made with spectroscopic C13-NMR and H1-NMR (Gonzalez et al., 1982). Since
the end of the 80s, red algae, included O. pinnatifida, have been recognised as a prolific source of
unusual terpenoid and non-terpenoid C15 metabolites (Erickson, 1983; Faulkner, 1986; Bittner et
al., 1985). In 1988, for example, pinnatazane, a bridged cyclic ether sesquiterpene, was isolated
from O. pinnatifida and structurally characterized with X-ray crystallographic and spectroscopic
techniques (Atta-Ur-Rahman et al., 1988). In 1989, the products E- and Z- dihydrorhodophytin
were isolated in O. pinnatifida extracts (Norte et al., 1989). After the isolation of pinnatifidone (Bano
et al., 1988), pinnatazane (Atta-ur-Rahman et al., 1988) and laurol (Ahmad et al., 1990), pinnatifenol
was isolated in 1991 from O. pinnatifida (Ahmad and Shaiq, 1991). These compounds proved to
have interesting properties, such as the marine polyether triterpenoid dehydrothyrsiferol 298,
originally isolated from O. pinnatifida, which was shown to induce apoptosis in estrogen-dependent
and independent breast cancer cells (Norte et al., 1997; Pec et al., 2003).
Since the late eighties, O. pinnatifida has also been investigated for its antimicrobial activity
(Usmanghani et al., 1984; Bano et al., 1986). A study conducted by Usmanghani, et al. (1984) on
Arabian marine algae, underlined the significant antimicrobial activity played by O. pinnatifida
against microorganisms such as Bacillus subtilis, Bacillus megaterium and Streptococcus viridans. This has
led to new studies in the following years to better understand the chemical composition of its
constituents (Bano et al., 1987). O. pinnatifida, together with other red algae, has also been
investigated for its antileishmanial activities (Sabina and Aliya, 2011), proving that this species
influences the metabolic activity of the investigated parasite, as well as antifungal activity (Silva et
al., 2018). Recent researches focused on the bioactivity (Campos et al., 2019) and chemical,
structural and cytotoxic characterization of the species (Rodrigues et al., 2019).

100
Algae have been part of the human diet for thousands of years (e.g. 14,000 yBP in Chile; Dillehay
et al., 2008; in China, 300 A.D.; in Ireland, 600 A.D.; Newton, 1951; Tseng, 1981; Aaronson, 1986;
Turner, 2003; Gantar and Svircev, 2008; Craigie, 2010). The FAO (2014) reported that 38 % of the
23.8 million t of seaweeds in the 2012 global harvest was eaten by humans in forms recognizable
as seaweeds (e.g. kelps, nori/laver). Overall, there is a trend for increasing nutritional demand for
algal products on a global basis for application in health products and food additives. Nutritional
value is the main quality aspect for food (Picone et al., 2011). Nowadays, molecular nutrition studies
have become an important approach and alternative to the traditional food component analyses
(which focus on protein, carbohydrates, lipids, etc.), offering the possibility to detect/identify
hundreds or thousands of distinct chemical identities in food (Wishart, 2008).
Seasonal samples were collected over a 12-months period (as analysed in Chapter 3) and extracted
with 50% acetonitrile/ultrapure water (UPW) solution (as described in Chapter 3). In addition, an
extraction designed to mimic the conditions in the mouth (artificial saliva) was carried out on dried
biomass of Osmundea pinnatifida for the first time, to mimic what happens in the first phase of the
digestion in the mouth, using an HEPES buffer. This was carried out on winter (November) and
summer (May) samples in order to understand how the taste/flavour varies according to the
harvesting season. The totality of their composition was assessed using HILIC-PDA-MS analyses
in positive and negative MS modes. A wide array of techniques can be applied to obtain data for
metabolomics analyses; however, MS-based and NMR spectroscopic approaches are most popular
and particularly MS-based approaches are the most preferred techniques for metabolite profiling
(Gupta et al., 2014). This was because of their moderate to high sensitivity, high reproducibility
and superior separation potential based on electron ionisation and charge to mass ratio (Gupta et
al., 2014). The hydrophilic interaction chromatography (HILIC) technique is a relatively recent
advance in metabolomics studies, which gives better metabolite coverage than reverse phase
chromatography (Gika et al., 2013). The reference mass spectral library and software packages for
peak assignments for LC-MS data is under progress. A few software tools available publicly for
pre-processing and analysis of LC-MS data include Markerlynx, Mzmine, XCMS and MetAlign.
(Tautenhahn et al., 2012).
Osmundea pinnatifida is a delicious ingredient with a strong smell and flavour, whose origin has not
yet been investigated. However, its flavour may be linked to the secondary metabolites abundant
in this species (Erickson, 1983; Faulkner, 1986; Bittner et al., 1985). It is the taste that makes this
seaweed particularly interesting as a high value food ingredient.

101
4.2 Aims
To characterize, for the first time, the seasonal variation in the metabolomic profile of
Osmundea pinnatifida;
To explore the relationship between taste/flavour and metabolomic profile;
To understand, through the variation of metabolomic profile, the response of the seaweed
to environmental changes;
To identify seasonal patterns, due to identification of chemical classes or fragmentation
groups specific to a particular month or season;
To identify the best harvesting season for the species in terms of nutritional profile and
taste/flavour, for further cultivation and commercial applications.
4.3 Materials and methods
Samples were collected and pre-treated as described in section 3.3 Material and Method, Chapter
3.
4.3.1 Solvent extraction
Seasonal samples were extracted in triplicate using 50 % acetonitrile/UPW containing 0.1 % formic
acid as already described in Chapter 3 (section 3.3.5). This extraction procedure has been found to
give good coverage of metabolites from plants and previous work on selected seaweeds (Dailey
and Vuong, 2015; Gaubert et al., 2019).
4.3.2 Oral extracts: extraction in HEPES buffer of two selected months, November and May
Two seasonal samples, from November and May, were extracted with HEPES buffer in an attempt
to mimic an artificial saliva digestion to compare the total suite of components extracted using
solvents against those which may be present under conditions in the mouth. These two months
were chosen as representative of winter and summer conditions and because they represent the
moment at which the seaweed reaches the maximum of biomass (November) and the reproductive
season (May), before the biomass decreases and eventually bleaches out in the summer. Samples
of biomass which had been freeze-dried and ground were weighed out in triplicate (100 ± 5 mg).
Subsequent extraction was performed with 1 ml of 100 mM HEPES containing 300 mg CaCl₂ L⁻¹
at pH 6.8. Samples were vortexed for 10 seconds and extracted on a blood rotator at 40 rpm for 5
minutes at room temperature. They were then vortexed and centrifuged at 14,000 rpm for 5
minutes. The supernatant was removed, and the pellet re-extracted with 1.5 ml HEPES solution
for 5 minutes at 40 rpm on a blood rotator. Finally, after centrifugation at 5 minutes at 14,000 rpm,
the supernatants were combined and analysed in the same way as the seasonal samples, as described

102
in the following section. Furthermore, the absorbance spectra of these samples were read from 190
to 700 nm in an UltroSpec spectrometer.
4.3.3 HILIC-PDA-MS analysis
HPLC separations were performed on seasonal and HEPES extracted samples with a Dionex
U3000 UHPLC system coupled with a U3000 PDA and an LTQ-Orbitrap XL MS system
(Thermo-Fisher Ltd. Hemel Hempstead U.K). The MS and PDA systems were tuned and
calibrated according to the manufacturers recommended procedures. The HPLC was operated at
a flow rate of 300 µl/min and the column and guard column (Sequant ZicP HILIC 150 x 4.6mm,
5 µm particle size PN 1.50461.0001, Sequant ZicP HILIC Guard 20 x 2.1mm PN 1.50437.0001,
Merck Ltd.) were maintained at a temperature of 30oC. The solvent system compositions were, A:
20 mM ammonium carbonate prepared in HPLC grade water (J.T. Baker, Scientific Chemical
Supplies Ltd.), and solvent B: HPLC grade acetonitrile (J.T. Baker). Prior to injecting the sample,
the HILIC column and guard column were conditioned with 100% solvent A then 100% solvent
B for a minimum of 40 min at a flow rate of 300 µl/min. A sample injection volume of 20 µl was
employed in partial-loop mode. The gradient programme was as follows: 80-20% B 0-30 min, 20-
5% B 30-30.01 min, hold 5% B 30.01-35 min, 5-80% 35-35.01 mins, hold 80% B 35.01-45 min.
Autosampler syringe and line washes were performed with 80:20 HPLC grade water:acetonitrile
(J.T. Baker.). The HPLC column eluent was first monitored by the U3000 PDA detector where
spectra were collected in wavelength/absorbance mode from 200-600 nm with a filter bandwidth
and wavelength step of 1 nm, the filter rise time was 1 sec, the sample rate was 5 Hz. Additionally
three channel set points were employed, Channel A 280 nm, Channel B 365 nm, Channel C 520
nm, with a bandwidth of 9 nm and a sample rate of 10 Hz. The PDA detector eluate was next
transfered to a Thermo LTQ-Orbitrap XL mass spectrometry system operated under Xcalibur
software (Thermo-Fisher Ltd. UK). Mass spectra were collected with a primary full scan event (m/z
80-1000) at a mass resolution of 30,000 full width at half maximum (FWHM defined at m/z 400)
within the Fourier Transform (FT) detector, and a secondary data-dependent analysis (DDA) MS2
scan for the top3 most intense ions within the LTQ-IT. Helium was applied as a collision gas for
CID at a normalised collision energy of 45%, a trapping window width of 2 (+/- 1) m/z was
applied, an activation time of 30 ms and activation Q of 0.25 were applied, only singly charged ions
were selected for DDA, isotopic ions were also excluded. The preliminary full scan event within
the FT generated 'profile' mode spectral data, where as the LTQ-IT MS2 data were collected in
‘centroid’ mode. A scan speed of 0.1 seconds and 0.4 seconds were apllied in the LTQ-IT and FT-
MS respectively. The Automatic Gain Control was set to 1 x 105 and 1 x 106 for the LTQ-IT and
FT-MS respectively. The following settings were applied to ESI: Spray voltage -3.5 kV (ESI-) and
+4.0 kV (ESI+); Sheath gas 60; Aux gas 30; Capilary voltage -35 V (ESI-) +35 V (ESI+); Tube

103
lens voltage -100 V (ESI-) and +100 V (ESI+); Capilary temperature 280oC; ESI probe temperature
100oC.
The analyses on the 12 samples (n=3) were conducted in two different blocks; hence, two samples
were chosen from the first block of analyses and re-run with the second block of samples. The
peaks and intensity were the same for both runs, proving the robustness of the method applied
and the consistency of the analyses (see Appendix B).
Furthermore, the samples were analysed in a completely randomised order as two independent
analytical blocks respective of ESI positive and ESI negative polarities. For each analytical block,
initially three injections of a mixed sample were performed for LC-MS system conditioning. A
control blank sample was analysed at the start and end of the analytical block.
4.4 Results
4.4.1 Solvent extraction
Extracted samples were analysed and plotted with Xcalibur software (Thermo Fisher®). This
software only allows comparison of 8 plots, so each set of 12 month samples was divided into two
groups, from August to January and from February to July (Figs 4.1-4.4). Graphs of all samples
were smoothed (level 5) with the software in order to reduce the signal noise. The main peaks
detected for the negative mode are listed in the Table 4.1 and those for the positive mode in Table
4.2.

10
4
Figure 4.1 Overlay peak graph of MS profiles from August 2016 to January 2017 (Negative Mode, FSD = 7.20E7, Base peak F:FTMS-c ESI Full MS [80-1000])

10
5
Figure 4.2 Overlays peak graph of MS profiles from February to July 2017 (Negative mode, FSD: 9.59E7, Base peak F:FTMS-c ESI Full MS [80--1000])

10
6
Figure 4.3 Overlays peak graphs of MS profiles from August 2016 to January 2017 (Positive mode, FSD: 3.20 E8, Base Peak F:FTMS+ c ESI Full MS [80-1000])

10
7
Figure 4.4 Overlays peak graphs of MS profiles from February to July 2017 (Positive mode, FSD: 3.20 E8, Base Peak F:FTMS+ c ESI Full MS [80-1000])

10
8
Table 4.1 Peaks with putative IDs and retention time for the negative ESI mode; in bold under MS² the most abundant fragments
Peak RT m/z [M-H] MS2 Mol. Formula Putative ID
1 4.15
4.15
4.59
291
293
287
211, 203, 193, 169, 97;
275, 205, 195;
197, 189, 129, 114
C15H15O6
C15H17O6
C12H15O8
Unknown – 2H less saturated than chilenone B
Chilenone B (San-Martín et al., 1987)
Methyl furanone derivative (similar MS2 with loss of 98)
2 4.74 195 177, 109, 107, 87, 85 C10H11O4 Chilenone A (San-Martín et al., 1983)
3 5.35-8.00 112.99 95, 87, 69 C4H2O4 Fumarate as 2- ion (ChemSpider ID4573886)
4 5.35-8.00 175 115, 113 C6H7O6 Fumarate + FA/vitamin C (ChemSpider ID10189562)
5 5.35-8.00 305 287, 261, 229, 211, 201, 181,
169
C20H3402 Lipid derivatives – eicosatrienoic acid derivative
6 5.35-8.00 Various
353
351
273
None
C23H4602,
C23H4402,
C16H32O3
Lipid derivatives
methyl-docosanoic acid (Omar et al., 2018)
tricosenoic acid
hydroxy-hexadecanoic acid
7 11.37 162 118 (44), 112 (50), 100 (62), 92
(70), 75 (87)
C5H9O3NS 1, 4-Thiazane-3-carboxylic acid S-oxide (Fumio and Oka, 1963; Kuriyama et al., 1960)
8 12.3 132 71 C₅H₁₁O₃N Amino-oxy pentanoic acid
9 13.11 & 13.57 253 (299 +46) 235, 199, 187, 179, 161, 143,
131, 128, 91
C9H18O8 2-O-α-D-galactopyranosyl glycerol (Floridoside), often present as isomers floridoside
and isofloridoside – main loss is 92 amu = glycerol (Barrow et al., 1995).
10 14.61 97 53 (44) H3PO4 Phosphoric acid (ChemSpider ID979)
11 15.36 267 249, 237, 219, 161, 143, 105, 89,
87
C9H16O9 2-O-α-D-mannpyranosyl glycerol (digeneaside) (Ascencio et al., 2006; Barrow et al.,
1995)
12 15.65 132 115, 114, 88, 71 C₄H₇NO₄ Aspartic acid
13 17.89 205 187, 125 C7H9O7 Homocitric acid (ChemSpider ID26392)
14 18.64 & 19.05 173 129, 111, 85 C6H6O6 Trans-Aconitic acid (MS2 matches MassBank record) (Kurata and Amiya, 1965; Kazuya
and Takashi, 1975).
15 20.15 173 129, 85 C6H6O6 Cis- aconitic acid (different MS2 to trans form) (ChemSpider ID558863)

10
9
Table 4.2 Peaks with putative IDs and retention time for the positive ESI mode; in bold under MS² the most abundant fragments
Peak RT m/z [M-H] MS2 Mol. Formula Putative ID
1 4.40 330 312.7, 295, 277, 215, 197 C15
H23
O7N Amino acid derivative similar to isodomoic acid + 18 (H20) (Zaman et al.,
1997; Maeda et al., 1986,1987)
2 6.83 410 392, 375, 357, 324, 312 C20
H27
O8N Alkaloid (Walter and Buck, 1937)
3 8-10 383 325, 297, 285, 245, 209 C18
H26
O7N
2 Loss of 98 to palythene Possible Seryl tyrosine derivative
4 9-11 285 270, 267, 241, 226, 197 C13
H20
O5N
2 Palythene (mycosporine-like AA)
(Barceló-Villalobos et al., 2017)
5 10.67 204 144, 85, 60 C9H
17O
4N Acetyl carnitine (ChemSpider ID21243783)
6 11.10, 11.86 118 100, 59, 58 C5H
11O
2N Betaine/Valine (Blunden et al., 2010, 1986; Tyihák et al., 1994; Kumar and
Kaladharan, 2007; Černá 2011; Impellizzeri et al., 1977).
7 12.45 134 117, 116, 72, 73, 62, 63 C5H
11O
3N Hydroxy Betaine derivative
8 12.77 116 70 C5H
9O
2N Proline (Hardjani et al., 2017; Mouritsen et al.,2012, 2013; Černá 2011).
9 13.36 347 No MS² C14
H22
O8N
2 Porphyra 334 (mycosporine-like AA) (Hartman et al., 2017; Gracesa et al.,
2018)
10 13.41 146 87, 60 C7H
15O
2N Aminoheptanoic acid (ChemSpider ID198338)
11 13.93 289 274, 230, 186 C12
H20
O6N
2 Mycosporine-like AA (Asterina-330) (Carreto and Carignan, 2011)
12 14.23 333 No MS² C13
H20
O8N
2 Shinorine (mycosporine-like AA) (Cardozo et al., 2006; Carreto and Carignan
2011)
13 14.44 245 230, 227, 209, 186 C10
H16
O5N
2 Palythine (mycosporine-like AA) (Carreto 2011; Hartmann et al., 2017; Carreto
and Carignan, 2011)
14 15.22 147 130, 117 C5H
10O
3N
2 Glutamine (Cole and Sheath, 1990; Dominguez 2013)
15 20.24 104 86, 61, 60 C5H
13ON 5-aminopentanol (ChemSpider ID68156)

110
Peaks that corresponded to putatively identified components were grouped together into five
categories, depending on the trends that those compounds followed across one year of sampling.
The five main trends identified were for compounds with “Levels up in summer”, “Levels down
in summer”, “Random”, “No variation” and “Levels down in autumn”. Results are reported in the
Figs and Tables below. Graphs for each putative compound are reported according to increasing
retention time (RT).
Abundance peak graphs for the trends identified in the ESI negative mode are reported in Figs 4.5-
4.8 (y=abundance, x=months). Particularly representative of “Levels down in summer” were:
Fumarate, 1,4-Thiazane-3-carboxylic acid S-oxide, Vitamin C, Chilenone A (isomer), and
Homocitric acid (Fig. 4.5). The code at the top of the graph indicates m/z then retention time, so
for example, the code 112990_0538N represents a peak at 5.38 mins with m/z 112.990.
Figure 4.5 Peak abundance variation over one year time period for putative components that decreased during the summer months. Putative IDs: 1) Fumarate; 2) 1,4-Thiazane-3-carboxylic acid S-oxide, 3) Vitamin C), 4) Chilenone A-like (possible isomer with different MS²), 5 & 6) Homocitric acid

111
Abundance peak graphs for the putatively identified components that showed “Levels up in
summer” are reported in Fig. 4.6 (y=abundance, x=months). Particularly representative of this
class were lipid derivatives and compounds putatively linked to the osmoregulation of the species
(such as floridoside and digeneaside).

112
Figure 4.6 Peak abundance variation over one year time period for putative components that increased during the summer months. Putative IDs: 1) Vitamin C, 2 to 5) lipid derivatives, 6) Fumarate, 7) Chilenone B-2H, 8 to 11) Floridoside, 12) Digeneaside, 13) Chilenone A-like (possible isomer with different MS²), 14) Trans-aconitic acid
Abundance peak graphs for the putatively identified components that showed “Random”
variations are reported in Fig. 4.7 (y=abundance, x=months).

113
Figure 4.7 Peak abundance variation over a one year time period for putative components that showed random patterns. Putative IDs 1 & 3) Chilenone B (-2H), 2) Chilenone B, 4) Chilenone A, 5) Trans aconitic acid
Abundance peak graphs for the putatively identified components that showed “No changes” are
reported in Fig. 4.8 (y=abundance, x=months).

114
Figure 4.8 Peak abundance variation over one year time period for putative components that showed little variation. Putative IDs: 1) TCSO, 2) lipid derivatives
Abundance peak graphs for the trends identified in the ESI positive mode are reported in Figs 4.9-
4.13 (y=abundance, x=months). Particularly representative of “Levels down in summer”
compounds were valine, amino acid derivatives, and alkaloids.

115
Figure 4.9 Peak abundance variation over one year time period for putative components that showed down regulation in summer. Putative IDs: 1 & 2) Alkaloids, 3) Amino acid derivatives, 4) valine, 5) 5-amminopentanol
Abundance peak graphs for the putatively identified components that showed “Levels up in
summer” are reported in Fig. 4.10. Particularly representative were: MAAs and betaine.

116
Figure 4.10 Peak abundance variation over one year time period for putative components that showed up regulation in summer. Putative IDs: 1) Seryl tyrosine derivatives, 2) Betaine, 3) Porphyra-334, 4) Shinorine, 5) Amminoheptanoic acid
Abundance peak graphs for the putatively identified components that showed little or “No
changes” are reported in Fig. 4.11.
Figure 4.11 Peak abundance variation over one year time period for putative components that showed little changes. Putative IDs: 1) seryl tyrosine derivatives, 2) hydroxy Betaine derivatives, 3) Porphyra-334, 4) Palythine
Abundance peak graphs for the putative IDs that showed “Random changes” are reported in Fig.
4.12. Among the “Random” regulated were reported: acetyl carnitine, glutamine, hydroxyl betaine
derivatives and isodomoic acid.

117
Figure 4.12 Peak abundance variation over one year time period for putative components that showed random changes. Putative IDs: 1) Isodomoic acid, 2) Acetyl carnitine, 3) Hydroxy Betaine derivatives, 4) Glutamine
Abundance peak graph for the putatively identified component that showed “Down regulation in
autumn” is reported in Fig. 4.13.
Figure 4.13 Peak abundance variation over one year time period for a putative component that showed down regulation in autumn. Putative ID: 1) Palythene
A summary of the putative IDS with respective m/z, RT and regulation trend for the negative and
positive mode is presented in Tables 4.3 and 4.4 respectively.

118
Table 4.3 Putatively identified components organized by regulation trends (negative ESI mode data)
RT m/z [M-H] Putative ID Trend
5.38 162 1, 4-Thiazane-3-carboxylic acid S-oxide Down summer
14.94 175 Vitamin C/fumarate Down summer
15.20 195 Chilenone A-like Down summer
17.93 and 18.83 205 Homocitric acid Down summer
18.80 173 Trans aconitic acid Down summer
5.54 175 Vitamin C/fumarate Up summer
5.76, 6.40, 7.17,
7.31 and 31.72
273,305,351,353 Lipid derivatives Up summer
10.86 291 Chilenone B (-2H) Up summer
13.06, 13.10 and
13.50
253 2-O-α-D-galactopyranosyl glycerol
(Floridoside)
Up summer
15.32 267 2-O-α-D-mannpyranosyl glycerol
(digeneaside)
Up summer
18.96 195 Chilenone A-like Up summer
20.39 173 Trans aconitic acid Up summer
31.84 287 Methyl furanone derivative Up summer
4.30, 4.33, 4.41 293 Chilenone B Random
4.71 195 Chilenone A Random
18.80 173 Trans aconitic acid Random
11.91 162 TCSO No changes
33.31 305 Lipid derivatives No changes
Table 4.4 Putatively identified components organized by regulation trends (positive ESI mode data)
RT m/z [M-H] Putative ID Trend
20.60 104 5-aminopentanol Down summer
11.82 118.08 Betaine/Valine Down summer
6.33 410.18 Alkaloids Down summer
9.68 285.14 Palythene (MAA) Down autumn
11.17 118.08 Betaine/Valine Up summer
13.38 146.11 Aminoheptanoic acid Up summer
14.27 333.12 Shinorine Up summer
14.10 347.14 Porphyra 334 (MAA) Up summer
10.93 144.02 Acetyl carnitine Random
15.37 147.07 Glutamine Random
4.68 330.15 Amino acid derivatives,
similar to isodomoic
acid+18
Random
13.32 347.14 Porphyra 334 (MAA) Random
14.36 245.11 Palythine (MAA) No changes
12.53 134.08 Hydroxy Betaine derivative No changes
8.41 383.18 Seryl Tyrosine derivatives No changes
4.4.2 Oral extracts: extraction in HEPES buffer of two selected months: November and May
The HEPES extracts of the samples from the different months had clearly different colours (Fig.
4.14 A). The May samples were orange whereas the November samples were pink. Furthermore,
oral extracts differed in colour from the solvent extracts (Fig. 4.14 B).

119
Figure 4.14 A) HEPES buffer extracted samples: in orange “May” and in pink “November”; B)
solvent extract sample
Extracted samples in HEPES, as a proxy for artificial saliva, were read via scanning
spectrophotometric over 380 to 750nm and the main peaks are shown in Figs 4.15 and 4.16.
The absorbance spectra of the extracted samples showed clear differences. The orange colour of
the May samples was due to an increase in absorbance at ~460 nm. This could reflect the presence
of carotenoids, but it is doubtful how well these generally hydrophobic compounds would be
extracted in aqueous HEPES. The pink coloration obvious in the November samples, may be due
to phycobiliproteins, which are also probably present in the May samples (see peaks at around 500-
600 nm). However, carotenoids do not ionise well under ESI conditions and phycobiliproteins
have high molecular weights beyond the detection range of the MS detector at m/z 2000.
Figure 4.15 Scanning absorbance spectrum (as absorbance units) of the HEPES extracted sample from November
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
380 430 480 530 580 630 680 730 780
Ab
sorb
ance
(A
U)
nm

120
Figure 4.16 Scanning absorbance spectrum of the HEPES extracted sample from May
Extracted samples were analysed and plotted with Xcalibur software (Thermo Fisher®), and
comparison of peak graphs of MS profiles of oral and wild in positive ESI mode are shown in Fig.
4.17 and for negative ESI mode in Fig. 4.18.
The HILIC profiles of these HEPES extracted samples were compared against the solvent
extracted samples to assess which components were also more likely to be available in the mouth
and contribute to the flavour profile of the seaweed. The HEPES was carried over in the oral
samples and was the major peak in these HILIC runs. The abundance of peaks in both the solvent
and “oral” extracts in positive ESI mode are shown in Fig 4.17 and those for the negative mode in
Fig 4.18.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
375 405 435 465 495 525 555 585 615 645 675 705 735
Ab
sorb
ance
(A
U)
nm

12
1
Figure 4.17 Peak graphs of MS profiles from May oral (black), November oral (red), May (green) and November (blue) (Positive mode, FSD = 3.90 E8, Base Peak
F:FTMS+ c ESI Full MS [80-1000]). Arrow denotes HEPES peak in oral samples

12
2
Figure 4.18 Peak graphs of MS profiles from May oral (black), November oral (red), May (green) and November (blue) (Negative mode, FSD = 9.00E7, Base
Peak F:FTMS+ c ESI Full MS [80-1000]). Arrow denotes HEPES peak in oral samples1
1 Since HEPES peak was off-scale, the scale chosen was the solvent extract (9.00 E7), which allows for a more equal comparison of the data.

123
No peaks were present in the oral extracts that were not present in the total solvent extracts.
Therefore, for simplicity, only the peaks common to the two extractions (already identified in
Tables 4.1 and 4.2 - negative and positive ESI modes respectively) were examined. Those identified
in the negative ESI mode were chilenone B (m/z 291 & 293) and A (m/z 195), fumarate (m/z
112.99, 175), lipid derivatives (m/z 273, 353, 305), TCSO (m/z 162), floridoside (m/z 253),
digeneaside (m/z 267), homocitric acid (m/z 205), and trans and cis aconitic acids (m/z 173) (Fig.
4.19).
Figure 4.19 Main peaks identified in HILIC profiles in negative mode for Solvent and “Oral” extracted samples (blue= November; light grey= November Oral; orange= May; black= May Oral) (n=3; error bar, 1 SD of mean)
Those identified in the positive mode were 5-aminopenthanol (m/z 104), glutamine (m/z 130, 147),
palythine (m/z 245), shinorine (m/z 333), proline (m/z 116), aminoheptanoic acid (m/z 146),
asterina (m/z 289), betaine/valine (m/z 118), hydroxy betaine derivatives (m/z 134), acetyl carnitine
(m/z 144, 204), palythene (m/z 267, 285), porphyra 334 (m/z 347), seryl tyrosine derivative (m/z
285, 383), alkaloids (m/z 312, 410), amino acid derivatives (m/z 312, 330, 277) (Fig. 4.20).
0.00E+00
5.00E+08
1.00E+09
1.50E+09
2.00E+09
2.50E+09
3.00E+09
3.50E+09
4.00E+09
4.50E+09
m/z291
m/z293
m/z195
m/z112
m/z175
m/z273
m/z353
m/z305
m/z162
m/z162
m/z299
m/z253
m/z253
m/z299
m/z267
m/z205
m/z173
m/z205
m/z173
Ab
un
dan
ce
Compound (m/z)

124
Figure 4.20 Abundance of main peaks identified in HILIC profiles in positive mode for Solvent and “Oral” extracted samples (blue= November; light grey= November Oral; orange= May; black= May Oral) (n=3; error bar, 1 SD of mean)
In positive mode, the same compounds were present in both solvent and HEPES extracted
samples, but they varied in abundance. Some compounds did not show differences between the
two sets of samples, such as amino acids derivatives, acetyl carnitine, MAAs, alkaloids, hydroxy
betaine derivative, or proline. Others had higher levels in the solvent extracted samples, e.g.
palythene, palythine, amino acid derivatives, betaine/valine, and seryl tyrosine derivative. A few
components were higher in the HEPES extracted samples including aminoheptanoic acid at m/z
146, alkaloids at m/z 410, and the amino-acid derivative at m/z 330.
However, in negative mode, some compounds that appeared in the solvent samples were not
detected in the “Oral” samples (e.g. chilenone B -2H at m/z 291, fumarate, vitamin C, and lipid
derivatives). Solvent extracted samples had higher abundance of most compounds except for
floridoside (m/z 299), which had higher levels in the HEPES extracts of both May and November
samples. This component was identified as a formate adduct of floridoside (see Table 4.1), but the
main floridoside peak at m/z 253 was higher in the solvent extracts. This difference may be due to
isomers of this compound, one which more readily forms the formate adduct and may be more
extractable in HEPES.
There were differences in peak abundance between the May and November samples, but these
generally showed the same trends in solvent and HEPES extracted samples.
For the HEPES extracted samples in ESI positive mode, statistically higher abundances were
reported in November oral extract for amino heptanoic acid and in May oral extract for seryl
-4E+09
-2E+09
0
2E+09
4E+09
6E+09
8E+09
1E+10
1.2E+10
1.4E+10
1.6E+10
1.8E+10
2E+10
m/z
31
2
m/z
33
0
m/z
27
7
m/z
31
2
m/z
41
0
m/z
41
0
m/z
20
9
m/z
31
2
m/z
38
3
m/z
33
0
m/z
28
5
m/z
28
5
m/z
14
4
m/z
14
4
m/z
11
8
m/z
26
7
m/z
13
4
m/z
11
6
m/z
14
6
m/z
27
7
m/z
28
9
m/z
34
7
m/z
33
3
m/z
24
5
m/z
13
4
m/z
10
4
m/z
10
4
m/z
14
4
Ab
un
dan
ce
Compounds

125
tyrosine derivatives, palythene and asterina (330). For the HEPES extracted samples in ESI
negative mode, statistically higher abundances were reported in November oral extract for
chilenone B, and in May oral sample for lipid derivatives, floridoside and trans aconitic acid (Fig.
4.19).
4.5 Discussion
4.5.1 Solvent extraction
4.5.1.1 ESI negative polarity
In this study the metabolomic profile of O. pinnatifida across a one-year time period was evaluated
for the first time. Compounds were identified in both negative and positive ESI modes, as reported
in Tables 4.1 and 4.2 respectively. Interestingly, compounds that could be identified in negative
mode mainly included chilenones A & B (and their isomers and isotope), methyl furanone
derivative, lipid derivatives, fumarate, TCSO, carbohydrates (e.g. floridoside and digeneaside) and
aconitic acids (trans and cis). These compounds have been previously reported in other seaweeds
having bioactive properties, e.g. antimicrobial, antiviral, antifungal, anti-grazing and anti-
carcinogenic capabilities (San-Martín et al., 1983; Gianturco et al., 1964; de Nys et al., 1998;
Plouguerné et al., 2010; Tominaga and Oka, 1963; Kuriyama et al., 1960; Omar et al., 2018). More
specifically, Chilenone A was identified by San-Martín et al. (1983) in Laurencia chilensis and is a
dimer of 2-methyl-3(2H)-furanone, which has been reported as one of the volatile constituents of
roasted coffee (Gianturco et al., 1964). San-Martín et al. (1987) isolated Chilenone B from the same
species, which is a trimer of 2-methyl-3(2H)-furanone. Both of the above compounds are natural
sesquiterpenoids (Fraga, 1990, 2012). Methyl furanone derivatives have a role as signal molecules
and they may have anti-carcinogenic, antimicrobial, antiviral, antifungal, and anti-grazing activities
(Slaughter, 1999). In animals, they have a role as attraction signals (Slaughter, 1999), while in plants
and fruits they have been identified as aroma compounds (e.g. in strawberry) (Schieberle and
Hofmann, 1997). The most detailed information available on anti-microbial effects of furanones
in seaweed comes from studies on the Australian subtidal red alga, Delisea pulchra. This seaweed can
prevent the swarming and adhesion of a range of marine bacteria that use an intercellular signal to
change phenotype from free-living to colony through a quorum sensing mechanism. By secreting
halogenated furanones which resemble Acylated homoserine lactone (AHL) (e.g. furanone of the
3-butyl-2-one type) the seaweed prevents colonization by bacteria, interfering with their
intercellular signal (acylated homoserine lactone) (Kjelleberg et al., 1997). The production and
release of these compounds proved also to deter grazing by herbivorous animals (de Nys et al.,
1998; Dworjanyn et al., 1999).

126
Fumarate has previously shown antioxidant activities in seaweeds, and it is commonly present in
many plants and animal, being essential to tissue respiration. It is also prepared industrially and
applied as flavouring additives for beverages, baked food and baked goods (Winter, 2009). It has
been reported previously in Porphyra purpurea (UniProtKB - P80477, SDHB_PORPU).
With respect to Vitamin C, more information is required to confirm its identity in this study, such
as comparison with a standard. The Vitamin C content of algae varies markedly with season, groups
and species. Submerged species are usually richer in vitamin C irrespective of their taxonomical
grouping (Sarojini and Sarma, 1999). Maximum levels of vitamin C are reported for green and
brown algae in summer and for red algae in winter. Vitamin C is thought to affect the health of the
plants as seaweeds that show a more intense and brilliant colour are observed to have higher
content of it (Sarojini and Sarma, 1999). Furthermore, Vitamin C has a key role in the metabolic
reaction of seaweeds, being associated with the morphogenetic, vegetative and reproductive phases
(Anantharaman et al., 2011).
Lipid derivatives were only partially identified in this study with evidence for methyl-docosanoic
acid, eicosatrienoic acid, tricosenoic acid and hydroxy-hexadecanoic acid. Omar et al. (2018)
reported the presence of methyl-docosanoic acid in extracts of the red alga, Laurencia papillosa,
which had anti-microbial activity. Indeed, previous reports have revealed that FA derivatives from
seaweeds have shown strong anti-fouling activity (Bazes et al., 2009; Plouguerné et al., 2010).
Members of the Laurencia genus (Rhodophyta) are prolific producers of terpenoid and non-
terpenoid secondary metabolites that showed antibacterial and antiviral activities (Fujii et al., 2011).
Furthermore, FA derivatives also include carotenoids, that are naturally occurring fat soluble
tetraterpenoids, organic pigments that are found in phototrophic organisms and can have
remarkable biological functions and applications in human health (e.g. β-carotene, astaxanthin and
fucoxanthin) (Maeda et al., 2008; Boominathan and Mahesh, 2015).
One of the more interesting compounds identified in this study was 1,4-thiazane-3-carboxylic acid
S-oxide (TCSO), which has been previously identified in other algae, such as Undaria pinnatifida
(Tominga and Oka, 1963) and Chondria crassicaulis (Tominga and Oka, 1963; Kuriyama et al., 1960).
It is a sulfur-containing amino acid whose natural occurrence in sources other than the red alga
Chondria crassicaulis and Undaria pinnatifida had not previously been reported until now, with this
study. As a cycloallin, it has been reported as organosulfur compound in onion and garlic, reporting
several biological activities (Ichikawa et al., 2006), playing probably a role in the particular taste of
this seaweed.
Aspartate was reported in this study, which has been associated with umami taste, in addition to
glutamate, with less potency and an unknown sensory mechanism (Chandrashekar et al., 2006; Li

127
et al., 2002). Higher levels of glutamate as well as aspartate are associated with more flavourful
seaweeds (e.g. S. japonica, Mouritsen et al., 2012; Mouritsen, 2013).
Floridoside and digeneaside are common low molecular weight carbohydrates biosynthesized
during photosynthesis in marine red seaweeds (Meng et al., 1987). These compounds are thought
to have several functions, such as osmolytes and compatible solutes, with stabilizing effects on
enzymes, membranes and structural macromolecules under hypersaline concentrations (Barrow et
al., 1995). Digeneaside (2-O-α-D-mannopyranosyl glycerol) has been recognized as
chemotaxonomic marker for red algal order Ceramiales (Kirst and Bisson, 1979; Kirst, 1980; Reed,
1990). It was thought that some genera of the Ceramiales (e.g. Laurencia and Osmundea) produced
and accumulated floridoside rather than digeneaside (Barrow et al., 1995). This study suggests the
opposite to what was previously reported for this species (Kremer and Vogl, 1975; Kremer, 1978).
However, this is assuming that these two sugar glycerides ionize to a similar extent, which seems
reasonable.
Floridoside (2-O-glycerol-α-D-galactopyranoside) is a natural glycerol glycoside found in red algae
and is believed to play important roles in carbon storage, transport, assimilation and in the
regulation of osmotic balance (Kirst, 1980; Reed et al., 1980; Ekman et al., 1995). It is a compatible
solute that can be accumulated at high intracellular concentrations without interfering with the
normal functioning of the metabolism (da Costa et al., 1998; Roberts, 2005; Hagemann and Pade,
2015). Its synthesis is enhanced by high osmotic pressure conditions (Kirst and Bisson, 1979; Reed,
1985; Ekman et al., 1995). This glycoside also constitutes the major soluble pool of carbon fixed
by photosynthesis and is a precursor for cell wall polysaccharides in some species (Li et al., 2001,
2002). Apart from its in vivo role as osmolyte, floridoside has been described has having other
interesting properties. Hellio et al. (2004) reported that floridoside is able to inhibit the settlement
of cryptid larvae on the surface of underwater devices, suggesting its application as non-toxic,
natural compound for preventing biofouling, a worldwide problem estimated to cause a loss of
billions of dollars to the marine industry (Callow and Callow, 2002). Moreover, floridoside is a
potential therapeutic agent with the ability to modulate the immune response (Courtois et al., 2008;
Kim et al., 2013) and to promote bone formation (Ryu et al., 2015). The structural similarity with
2-O-α-d-glucopyranosylglycerol (GG), a compatible solute accumulated by cyanobacteria, suggests
that might also be functional in applications as moisturizing agent (Thiem et al., 1997), a non-
cariogenic, low calorie sweetener (Takenaka and Uchiyama, 2000) and a protein stabilizer
(Sawangwan et al., 2010). However, industrial applications for floridoside have not been developed
yet due to limited compound availability (Martínez-García et al., 2016).
The antioxidant capacity of algae is comprised of a variety of constituents such as carotenoids,
tocopherols, flavan-3-ols, phenolic acids, lignans, phlorotannins as well as mycosporine-like amino

128
acids (MAAs; Athukorala et al., 2003, Dunlap et al., 1998, Dunlap and Yamamoto, 1995; Yuan et
al., 2005). Natural compounds produced by seaweeds are usually referred to as secondary
metabolites since they are not involved in the basic biological functions (Cimino et al., 2001). Many
of these secondary metabolites are halogenated (McKey, 1979). Among the different classes of
marine macroalgae, red algae are the major producers of halogenated compounds (Cabrita et al.,
2010). Marine halogenated compounds range from peptides, polyketides, indoles, terpenes,
acetogenins and phenols to volatile halogenated hydrocarbons (Butler et al., 2009). Laurencia and
Osmundea (Rhodomelaceae), are considered two of the most prolific genera in this respect
(Faulkner, 2001; Wright et al., 2003). Trans-Aconitic acid and Cis- aconitic acid are both organic
acid that can become halogenated (e.g. conjugated with bromophenol) already investigated in other
algae (Kurata et al., 1998) and that have been identified in this study proving once again the
relevance of this species for further nutraceutical, pharmaceutical and cosmetic applications.
4.5.1.2 ESI positive polarity
The positive ESI mode mainly identified compounds such as amino acids (e.g. betaine, valine,
proline), amino acid derivatives (isodomoic acid, seryl tyrosine derivative), mycosporine-like amino
acids (palythene, porphyra-334, asterina-330, shironine, palythine), acetyl carnitine and alkaloids
(Table 4.2). Most of these compounds have reported osmoregulation properties and are produced
as compatible osmolytes to regulate adaptation in transient environments (Blunden, 2003;
Hartmann et al., 2017).
Domoic acid (DA) has been previously reported in other algae and originally isolated in Chondria
armata (Zaman et al., 1997). It was then investigated for its insecticidal and anthelmintic activities
(Maeda et al., 1986, 1987). Domoic acid is the most potent activator of the α-kainic receptors in
the central nervous system, which is known to act as a potent agonist to the glutamate receptor,
which conducts Na+ ion channels in the postsynaptic membrane (Debonnel et al., 1989) and can
cause poisoning. However, DA poisoning from red algae is unknown (Higa and Kuniyoshi, 2000).
Together with other FAs such as octanoic acid, nonanoic acid, decanoic acid, dodecanoic acid, and
hexadecanoic acid, the amino-heptanoic acid have been reported as precursors to seafood flavours
and as plant metabolites. But there is no information available for seaweeds (Josephson and
Lindsay, 1986) and this is the first report of their presence in red algae.
Alkaloid derivatives were identified in this study, but not specifically classified. Alkaloids in seaweed
include simple phenylamine (tyramine, hordenine) and catecholamine (dopamine) as well as sulfur-
containing bromoalkaloids, extracted from red algae (Güven et al., 2010). Pharmacologically
activities have been recognized for these compounds including antibacterial (e.g. terrestrial and
some marine bacteria), anticancer, and cytotoxic activities. Within the alkaloids, halogenated Indole

129
Alkaloids (HLI) have only been isolated in marine organisms and algae (mostly red) but not in
terrestrial plants. These alkaloids contain an indole group substituted by bromine and chlorine
atoms that are highly abundant in red algae (e.g. O. pinnatifida, Asparagopsis taxiformis, Euchema sp.,
Gracilaria sp., Chondrus Crispus) (Küpper et al., 2014; Cabrita et al., 2010; Laturnus et al., 2010;
Mithoo-Singh et al., 2017).
The amino acid composition of algae has been often studied, with red seaweeds showing lower
amounts of glutamic and aspartic acids than those recorded from other algae groups (Fleurence,
1999). Essential amino acids can constitute almost 46% of the total amino acid fraction in red algae
(e.g. P. palmata). This study reported essential amino acid such as valine (commonly reported in
seaweed, Tabarsa et al., 2012), showing the potential this seaweed has as a food additive in human
and animal nutrition. Betaine was also identified, and this compound has been studied for its
osmoregulation properties, being an osmolyte and facilitating adaptation to saline and dry
environments (Blunden, 2003). Compounds of this class have been found in many animals, plants,
algae, fungi and bacteria (Blunden and Gorden, 1986), and previously reported in red seaweeds
(e.g. Pyropia yezoensis, Mao et al., 2019).
Proline was identified in this study, which together with histidine, valine and methionine is
associated with a bitter taste (Kato et al., 1989). Proline is known to accumulate in large quantities
in terrestrial plants in response to environmental stresses (Ashraf and Foolad, 2007), and many
plants accumulate proline as a non-toxic and protective osmolyte under saline conditions (Khatkar
and Kuhad, 2000).
One of the main classes of compounds reported in this study are mycosporine-like amino acid.
MAAs are algal secondary metabolites, with absorbance maxima between 310 and 360 nm (Karsten
and Wiencke, 1999; Shick and Dunlap, 2002). More than 30 different MAAs have been identified,
with Rhodophyta accounting for the highest concentration and greatest variety among the different
algal divisions (Karsten et al., 1998; Sinha et al., 1998). MAAs have the ability to absorb UV
radiation due to their structural characteristics and to dissipate its energy without forming ROS
(Sinha et al., 1998; Tao et al., 2008). The induction of MAA synthesis is stimulated by different
light qualities, which cannot be initiated by the same photoreceptor or at least the same
chromophore (Liscum and Briggs, 1995). They have a characteristic cyclo-hexenone or -
hexenimine core, which is conjugated to the nitrogen moiety of an amino acid. This core primary
metabolism is synthesised by the shikimic acid pathway via 3-dehydroquinic acid and 4-
deoxygadusol (4-DG), a known strong antioxidant (Dunlap et al., 1997; Shick and Dunlap, 2002).
They act as UV-radiation protectors in macroalgae (e.g. P. palmata, P. tenera and Devaleraea
ramentacea), corals (e.g. Palythoa tuberculosa), marine species such as sea squirts (Lissoclinum stellatus)
and krill (Euphausia superba) and have potential in photo-protective formulations (Karsten and

130
Wiencke, 1999; Bedoux et al., 2014). They have also antioxidant properties as evidenced in a study
by Dunlap and Yamamoto (1995) for P. palmata and a role as osmoregulators has also been
recognised (Hartmann et al., 2017). The MAAs compounds reported in this study included
palythene, shironine, palythine, asterina 330, and porphyra-334, already reported in other algae
(Barceló-Villalobos et al., 2017; Hartmann et al., 2017; Carreto and Carignan, 2011). These have
potent bioactivities, for example, porphyra-334 exerts antioxidant activity and prevents cellular
damage caused by UV-induced ROS with free radical scavenging capacity (Tao et al., 2008). Several
recent publications have described the immunoregulatory and anti-inflammatory properties of
mycosporine-glycine, shinorine and porphyra-334. Particularly, porphyra-334 and shinorine have
been identified as unique natural products that can provide both a direct antioxidant defence and
stimulate enhanced cytoprotection, making these compounds commercially relevant (Suh et al.,
2014, 2017; Ryu et al., 2015).
Another interesting molecule identified with this study, which can be linked to the taste of the
seaweed and its nutritional profile is acetyl carnitine, a naturally occurring amino acid, previously
reported in brown algae (e.g. Undaria sp.). This can have applications as a diet supplement, for
thyroid control, for treatment of hypertension, and cardiovascular diseases (Jeukendrup and
Randell, 2011; Pereira, 2011). Glutamine is a central amino acid in red seaweed used as nitrogen
source for ammonium assimilation (Cole and Sheath, 1990), which has been investigated for its
relationship with the umami taste in other seaweeds (e.g. Ulva sp. and Gelidium sp., Nagahama et
al., 2009). Identification of this compound and its variation in the seasons can have considerable
relevance in understanding the properties that regulate the taste of O. pinnatifida.
4.5.1.3 Peak abundance variation: regulation trends
The metabolomics analyses conducted in this study took into consideration the variation in the
abundance of the described compounds across a full-year (Figs 4.1-4.4), in order to identify possible
trends and regulation patterns to assess the best harvesting season and link the production of
interesting secondary metabolites with seasonal changes. Secondary metabolites are often
antioxidants, whose production varies as a response of the alga to tissue desiccation or UV-
irradiation (Karsten and Wiencke, 1999). Therefore, it is not surprising that algal levels of
antioxidants (such as carotenoids and tocopherols) tend to be lower during winter and spring
months and higher in summer and autumn (Aguilera et al., 2002, Burritt et al., 2002; Yuan, 2007).
This was confirmed also in this study and by the analyses conducted on the abundance variation
across the seasons for the identified compounds (Tables 4.3 and 4.4).

131
4.5.1.3.1 Trends for ESI negative polarity
The compounds identified in negative ESI mode included osmoregulatory compounds (such as
floridoside) and antioxidant compounds (e.g. lipid derivatives, trans aconitic acid, and chilenone A
& B), which were all statistically higher in the summer months compared to the winter (Figs 4.5-
4.8). Accumulation of these compounds suggests a biological response of the seaweed to high
irradiation, day light length, desiccation, and higher temperatures that characterized the West coast
of Scotland in the summer months, especially for this intertidal species exposed to tides,
temperature, salinity, and light variation. These data confirmed what already seen with the statistical
PCA and BEST analyses in Chapter 3 (Figs 3.18 and 3.19), in terms of relations between
environmental parameters (e.g. irradiation and hours of light) and the biochemical composition of
the species. On the other hand, compounds that were down-regulated in the summer were generally
compounds that could be related to taste and palatability, such as fumarate, vitamin C, chilenone
A-like, and TCSO. This information can give pioneering and useful information on the regulation
of the taste in O. pinnatifida.
4.5.1.3.2 Trends for ESI positive polarity
In positive mode, compounds that may be related to the nutritional qualities and, hypothetically,
flavour, were statistically higher in winter and autumn months compared to spring and summer
months (e.g. valine, alkaloids, amino acid derivatives, and acetyl carnitine, Figs 4.9-4.13). On the
other hand, as already seen for the negative data, compounds that may be linked to stress resistance,
osmoregulation and UV protection were statistically higher in the summer (e.g. betaine and MAAs).
Interestingly, glutamine, identified here as m/z 147 (MS²=130), RT 15.22, and previously linked to
the umami taste in other seaweeds, showed the highest value in June, followed by March and April,
and the lowest in October. The initial hypothesis might be that since the species reaches its
maximum biomass between November and December, when it also shows the darker colour,
higher levels of glutamine would be expected in these months, rather than in June, March and
April. However, the abundance of glutamine is generally high throughout the year, although it
varies between months (Fig. 4.12), and the peculiar taste could be created by a synergetic effect of
different compounds (e.g. acetyl carnitine, TCSO, amino acid derivatives), that mainly show higher
level in late winter months, with glutamine and each other.
4.5.2 Replicating oral conditions
4.5.2.1 Oral extracts: extraction in HEPES buffer of two selected months, November and May
Under conditions that mimic those present in the mouth, the profiles of compounds changed
compared to that of solvent extracted samples. The compounds extracted in HEPES are more
likely to be similar to those released during eating and therefore responsible for the particular

132
flavour of O. pinnatifida. Except certain compounds (e.g. fumarate, vitamin C, and lipid derivatives),
the solvent and HEPES extracted samples generally contained the same main peaks listed in Tables
4.1 and 4.2 As could be expected, almost all components were reduced in abundance compared to
the solvent extraction. (Figs 4.17-4.20).
In negative mode, certain compounds were found only in solvent extracts, chilenone B -2H (m/z
291), fumarate (m/z 112, 175) and the putative lipid derivatives (m/z 273, 353). Some compounds
were higher in solvent extracts such as chilenone A (m/z 195), chilenone B (m/z 293), TCSO (m/z
162), digeneaside (m/z 267) and cis-aconitic acid (m/z 173); others had similar abundance in oral
and solvent extracts, e.g. floridoside (m/z 299) and homocitric acid (m/z 205). Certain components
were higher in oral extracts, such as lipid derivatives (m/z 305 =possibly eicosatrienoic acid in May)
and floridoside (m/z 299, 253 - in May).
In positive mode, differences in abundance of some components were reported between solvent
and oral extracts. The amino-acid derivative (m/z 312, 330), alkaloid derivative (m/z 312), seryl
tyrosine derivative (m/z 383), MAAs (m/z 267, 245), betaine/valine (m/z 118) and 5-
aminopentanol (m/z 104) were higher in the solvent extracts. The alkaloid derivative (m/z 410),
the amino acid derivative (m/z 330), MAAs (m/z 285, 289 for May), and aminoheptanoic acid (m/z
146) were higher in oral extracts. Other components including the amino acid derivative (m/z 277),
seryl tyrosine derivative (m/z 209), acetyl carnitine (m/z 144), hydroxy betaine derivatives (m/z 134,
116) and MAAs (m/z 347, 333) had similar abundances in solvent and oral extracts.
This was the first time that an artificial saliva digestion of dry biomass of O. pinnatifida was
performed to gain a better understanding of the role that certain compounds might have in
influencing and determining the taste of the species. As previously explained, some of the
compounds mentioned above, e.g. seryl tyrosine derivative, aminoheptanoic acid, floridoside, trans
and cis- aconitic acid, acetyl carnitine, TCSO, and fumarate, can have a role in influencing the taste
of O. pinnatifida as well. The fact that the same compounds can be extracted in the aqueous saliva
like HEPES buffer, strengthens the hypothesis previously discussed that these metabolites may
influence the flavour of the species. Furthermore, solvent and oral samples follow same seasonality
patterns, and if a compound is higher in May and lower in November in the solvent samples, this
is reflected also in the oral samples. This further confirms that the metabolomic profile of the
species varies with the season, therefore determining a variation in the taste and flavour of the
species according to the harvesting time.
However, further targeted analyses would need to be performed to confirm the identity and levels
of the key compounds and gain a better understanding of the variation in taste according to the
season. Also, due to time-constraints, this study only focused on major peaks in the HILIC traces
and only discussed the abundance of those components that could be at least putatively identified.

133
However, the MS data was very rich and non-targeted analysis using various data-handling software
techniques (e.g. XCMS data deconvolution and use of multivariate statistics, e.g. McDougall et al.,
2014) could compare the abundance of all peaks across the months and pick out those which
showed similar trends of expression (e.g. “Levels up in summer”), and compare them to those
identified peaks shown above. This non-targeted approach was not possible within this thesis but
could be carried out for future publications. This would provide powerful means for identifying
other, previously unknown, but possibly related compounds that could influence the flavour and
quality of O. pinnatifida.
The method here applied has strengths as well as limitations. While terpenoid, non-terpenoid C15
and halogenated metabolites were previously reported for the species (Erickson, 1983; Faulkner,
1986; Bittner et al., 1985; Atta-Ur-Rahman et al., 1988; Norte et al., 1989; Bano et al., 1988; Ahmad
et al., 1990; Ahmad and Shaiq, 1991), these compounds were not detected in this study. One reason
could be that the extraction applied is not ideal for these molecules, since they might not be
effectively extracted in 50% acetonitrile/UPW solution. Moreover, no special extraction or clean-
up procedures commonly used for pre-selection and detection of certain components were applied
in this study. Therefore, these components may be present but may be swamped out by other more
abundant metabolites. The application of the XCMS non-targeted approach (McDougall et al.,
2014) may help to overcome this and identify more minor components that alter between the
months. In addition, XCMS would assign possible formula including all elements and may help
pick out halogenated compounds previously noted in red seaweeds. However, as this study
compared monthly samples extracted in the same manner, this suggests that these previously
identified compounds do not change across the seasons and are unlikely to contribute to the
seasonal taste profile differences.
4.6 Conclusions
The seasonal variation in the metabolomic profiles of O. pinnatifida was reported for the
first time, providing valuable information regarding seasonal variation in composition as
related to flavour and confirming the importance of harvesting time for the quality of this
commercially valuable product;
The seasonal seaweed samples contained similar metabolites, but they varied in abundance
across the different months of harvest. Seasonal patterns were reported with certain
metabolites increasing over the summer months and others increasing in the autumn and
winter;

134
Particularly, components associated with stress alleviation and resistance were accumulated
during the summer months when the seaweed would be subject to increased levels of
ultraviolet light;
Components (e.g. 1,4-thiazane-3-carboxylic acid S-oxide, glutamine, acetyl carnitine, and
trans- and cis- aconitic acid) were identified that may be related to the particular flavour of
this seaweed;
Selected samples were extracted under replicating conditions in the mouth and compared
to solvent extracts, which confirmed compounds that could be related to the taste and the
seasonality of the flavour of the species, such as seryl tyrosine derivative, aminoheptanoic
acid, floridoside, trans and cis- aconitic acid, acetyl carnitine, TCSO, and fumarate;
A few components were higher in the HEPES extracted compared to the solvent samples
such as alkaloid derivative, amino acid derivative, MAAs, and aminoheptanoic acid. This
suggests that these compounds might play an important role in regulating the flavour of
the seaweed;
Differences in compounds abundance were found between November and May oral
extracts; in particular, higher concentration of alkaloids and aminoheptanoic acid were
recorded in November, suggesting a more intense flavour in this month compared to May;
however, further analyses involving a taste panel should be conducted to gain a better
understanding of flavour variations in the species;
These findings enhanced our understanding of the nutritional features and profile of O.
pinnatifida harvested at different times, of the seasonal pattern associated with the
metabolomic profile, and of the compounds that are present in the species, providing useful
information on the quality assessment of the species;
This study demonstrated that, as already anticipated by previous authors, O. pinnatifida has
a significant potential for further processing in nutraceutical, pharmaceutical and cosmetic
fields or for direct food applications.

135

136
Chapter 5 Epiphytes control of O. pinnatifida cultures and spores’
production, release and development in treated specimens
5.1 Introduction
Land-based tank system as a sustainable and reliable source of macroalgal biomass production
relies on maintaining vegetative growth with increases in yield and a good quality of the stock
material. These aspects can be heavily affected by epiphytism from other algae, or grazers (Neish
et al., 1977; Lüning and Pang, 2003; Hwang et al., 2006a), and can lead to the spread of diseases
among the cultures or degradation of the biomass (e.g. Ulva spp., Colorni, 1989; Del Campo et al.,
2002). Since the end of 1970s, epiphytic growth has been recognized as one of the greatest issues
for commercial seaweed cultivation (Ryther, 1977) as highlighted by Wheeler et al. (1981) “the control
of epiphytes and the provision of nutrients represented two major problems for macroalgae farmers”.
Seaweed are a flourishing ecosystem for a variety of different organisms. When they are transferred
to the lab for cultivation, they carry with them all the other organisms (e.g. grazers, other seaweeds,
etc.) that they are naturally associated with. Without intervention, contaminating organisms can
multiply, competing with the stock and/or causing deterioration in the quality, potentially leading
to the collapse of the culture (Borowitzka, 2007). The physical removal of contaminating organisms
by washing and hand sorting substantially reduces their impact (Baweja et al., 2009). However, this
is time-consuming and does not eliminate all of the epibionts. In addition, epibionts may be
introduced later in the cultivation cycle during seawater refreshment, by accidental cross-
contamination or by wind- carried spores/eggs in the case of outdoor tanks. Efficient methods for
the removal of such contaminants are essential. In some cases, manipulation of the culture
conditions can prevent or minimise such contamination, such as high stocking densities, low light
and nutrient levels (Lüning and Pang, 2003) or high pH and the release of alleochemicals caused
by the target cultivation species itself (Björk et al., 2004; Gross, 2010). Epiphytes can be controlled
by reducing the input of N in the cultures until the internal seaweed N reaches a critical level
(Harrison and Hurd, 2001). A chemical treatment may also be utilised to kill the contaminant. This
relies on the existence of differential susceptibility, with sensitive contaminants and a resilient
cultivation species (Guillard, 2007). Detergent, sodium hypochlorite (NaClO), reactive iodine,
formaldehyde and organic solvents have been successfully utilised for the disinfection of
macroalgal tissue to create axenic cultures (Shephard, 1970; McCracken, 1989; Baweja et al., 2009).
Other compounds such as acids, peroxides, sodium hydroxide and commercial preparations, for
example Virocid and Kick-start, are used in the aquaculture industry to disinfect equipment and
prevent or treat diseases (Togersen and Håstein, 1995; Barge, pers. comm.). These may also be
useful for decontamination in macroalgal cultures.

137
One of the more efficient way to verify that seaweeds are alive and healthy is by the monitoring of
their photosynthetic activity, using a non-invasive method such as PAM fluorometry. It has been
already applied with success to several macroalgal studies (Kolber and Falkowski, 1993; Enriquez
and Borowitzka, 2010), and is used extensively to assess stress-dependent changes in
photosynthesis of terrestrial plants, micro- and macroalgae as well as cyanobacteria (Schreiber et
al., 1986; Flameling and Kromkamp, 1998; Figueroa et al., 2006). A relative measurement of algal
photosynthetic health is obtained through measurement of the operating photosynthetic efficiency
of photosystem II (Fq′/Fm′). Higher values indicate greater photosynthetic electron flow towards
carbon fixation, while very low values indicate poor health or death (Maxwell and Johnson, 2000;
Cosgrove and Borowitzka, 2010; Kerrison et al., 2016a, b). Thus, fluorometry is a suitable method
to monitor the condition of the algal cell following a chemical treatment, which may disrupt the
delicate balance of cellular processes necessary for photosynthesis (Falkowski and Raven, 2007).
Such measurements have been used previously to assess the suitability of decontamination
treatments on Sargassum spp. (Hwang et al., 2006a, b; Kerrison et al., 2016a, b).
Chemical treatments have been successfully applied for reduction and removal of contaminants
from macroalgae (Guillard, 2007). However, this must be balanced against damaging the biomass
been cultivated (Tamiya, 1955; Richmond, 2004). A previous study by Kerrison et al. (2016a), tested
several chemical treatments on the removal of contaminates from O. pinnatifida in order to identify
the best method for decontamination of epiphytes in tank cultivation. It was not clear whether
consideration was given to the fact that O. pinnatifida is a perennial seaweed. There are three main
disadvantages associated with perennial species and their epiphytes. Firstly, epiphytes and grazers
can easily colonize its thallus and negatively affect the growth and productivity of the plant.
Secondly, once spores/gametes are released from the old thallus, this tends to be cast off from the
plant, in an attempt to remove the colonizing epiphytes. Lastly, the growth rate might be reduced
during the summer even if nutrients are sufficiently present, because the carbohydrates produced
during the photosynthesis have to be stored and used for the winter (Lüning, 1979).
Since O. pinnatifida, in its natural habitat, is exposed to tidal variations, being completely exposed
during low tide, this desiccation aspect was considered important for the cultivation process of this
species. For example, in Japan, farmers have resisted the use of chemicals on seaweed (such as
Nori) and controlled epiphytes and diseases by drying and/or freezing cultivated infected nets
(Ding and Ma, 2005). Depending on the species been cultivated, the epiphytes, especially weed
algae, can be more susceptible to desiccation compared to other species such as Pyropia sp.
(Mumford and Miura, 1988). Methods to prevent/eliminate epiphytes in tank systems are
fundamental to maximise productivity and quality.

138
Another potentially important aspect of seaweed cultivation is obtaining, settling and growth of
algal spores on ropes. Several factors can trigger spore release in algae, including desiccation,
grazing by invertebrates, abrasion and chemicals (Maggs and Callow, 2003). Spores released by red
algae are non-motile and mucilage-coated (Cole and Sheath, 2011). Once released, spores can stick
to the substratum on contact, but their initial adhesion is not always permanent, and they can
detach again (Sawada et al., 1972). Usually, tenacity of adhesion, as already reported for several red
algae, increases with time (Chamberlain, 1976; Chamberlain and Evans, 1973; Charters et al., 1973;
Suto, 1950). In general, rougher substrata enhance the settlement and survival of spores (Harlin
and Lindbergh, 1977; Ogata, 1953). These types of surfaces also protect the spores from
dislodgement by wave action/water currents/ grazer activity (Norton and Mathieson, 1983;
Brawley and Johnson, 1991) as well as desiccation, providing also evaporative cooling processes in
the intertidal region (Gonzales and Goff, 1989b). Seaweeds usually produce enormous numbers of
spores, and this is true also for red algae; however, the great majority of the spores will not survive
long enough to germinate (Maggs and Callow, 2003). Adhesion will comprise of both an initial
(temporary) and primary (permanent) attachment. This latter step is fundamental for colonizing a
new substratum (Maggs and Callow, 2003) and it is likely to depend on the residual mucilage sheath,
a common component of red algal spores (Boney, 1975; Ngan and Price, 1979) together with
adhesive substances such as glycoproteins complexes (Chamberlin and Evans, 1973, 1981-
Ceramium sp.; Pueschel, 1979-Palmaria sp.). After this, “establishment” will occur and comprises of
four stages: cell wall formation, establishment of polarity, germination and primary rhizoid growth
and adhesion (Fig. 5.1 A-D) (Fletcher and Callow, 1992).
Figure 5.1 The 4 stages of establishment in Osmundea pinnatifida spores (Axio Inverted microscope, Zeiss): A) cell wall formation; B) polarity; C) germination; D) rhizoid growth and adhesion

139
A tank system was trialled for indoor and outdoor cultivation of O. pinnatifida in order to assess the
growth of the culture in a tumbling system (in which seaweed are cultured without attachment and
left free to tumble in the tanks due to aeration provided) and the efficiency of a set of the treatments
against epiphytes in such a system. The tumbling system was specifically developed and adapted to
the species. To test the viability of spores obtained from tank cultivated O. pinnatifida
tetrasporophytes that reached this stage during the cultivation trials and after the treatment against
epiphytes, tetraspores were released and allowed to grow. Spores were taken through all the stages
detailed above and allowed to grow to juvenile then through to young plantlets.
5.2 Aims
To establish a protocol for the mechanical and chemical treatment of epiphytes in O.
pinnatifida cultures;
To develop a low-cost tidal simulation system for in tank cultivation of the species;
To apply the chemical treatment for epiphytes control to flasks and tanks culture
experiments for the species;
To assess how stressful the various treatments to be tested were to O. pinnatifida through
the use of PAM fluorometry;
To determine the parameters responsible for the spores’ production and release in cultures
of O. pinnatifida and develop a protocol for spores’ release, collection and isolation;
To determine the development of spores in vitro cultivation;
To identify the parameters responsible for the growth at a juveniles’ stage of O. pinnatifida
spores;
To evaluate the possibility of a cultivation of the species through a reproductive cycle.
5.3 Materials and methods
5.3.1 Chemical removal of epiphytes
Samples were collected in November 2016 at low tide from Seil (56.29°N -5.63°W), brought to the
laboratory into a Cool-box, washed with FSW and cleaned manually from visible epiphytes and
grazers. The treatments detailed in Table 5.1 were trialled on small fragments of O. pinnatifida thalli
(2-3 cm), over a range of exposure times.

140
Table 5.1 Epiphytes treatments and exposure time of vegetative thalli in Petri dishes trial
Treatments Time (min) Number of
Replicates
0.1 % NaClO 10 3
1 % NaClO 1 3
0.5 % KI 10 3
0.25 % Kickstart 10 3
25 % methanol 3 3
*control - 3
*untreated samples in F/2 medium
The biomass was subjected to the treatments in 6 well plates containing 20 ml of each solution.
After the allotted exposure time, the fragments were transferred into 200-300 ml of filtered, UV-
sterilised and Tyndalised seawater using sterile forceps and rinsed well in order to remove any
excess of the decontamination treatments. They were then transferred into new 30 ml Petri dishes
with 25 ml of F/2 medium and incubated at 9 °C, 12:12 (L:D) and 50 µmol/m²/sec for two weeks.
The initial PAM fluorometry measurements of the samples were taken with an AquaPen-P (PSI)
according to the manufacturer instructions, before the start of the experiment, to ensure that the
biomass was healthy. Samples were checked to verify the impact of the treatments using PAM
fluorometry, after two hours, one week and 14 days. Statistical analyses for all the trials listed in
this and subsequent sections were conducted applying an ANOVA single factor (p<0.05, Minitab
18) with Tukey’s and Fisher’s post-hoc tests.
The three treatments with the best level of recovery of the cultures were selected to treat an
increased amount of biomass, five gr of fresh material, at 250 ml. The treatments are listed in Table
5.2.
Table 5.2 Epiphytes treatments and exposure time of small volume cultures in flasks trial
Treatments Time (min) Number of Replicates
0.1 % NaClO 10 3
0.5 % KI 10 3
25 % methanol 3 3
*control - 3
*untreated samples in F/2 medium
The biomass was incubated in the treatment for the time listed on a shaking platform (Gyro-
Rocker) at 70 Rev/min. The biomass was then rinsed in 500-600 ml of filtered, UV sterilized and
Tyndalized seawater in order to remove the treatments from the tissue, placed in a new 300 ml
flasks with 250 ml of F/2 medium and incubated at 9 °C, 12:12 (L:D), 50 µmol/m²/sec for two
weeks. The initial PAM fluorometry of the samples were taken with an AquaPen-P (PSI) before

141
the start of the experiment to ensure that the biomass was healthy. Samples were checked to verify
the impact of the treatments on the biomass using PAM fluorometry, after two hours, one week
and 14 days. Tank treatments were then carried out using the parameters listed in Table 5.3.
Table 5.3 Epiphytes treatments and exposure time of scale up cultures in tanks trial
Treatments Time (min) Number of Replicates
0.5 % KI 10 1
0.5 % KI 20 1
0.5 % KI 30 1
*control - 1
*untreated samples in F/2 medium
Biomass, 100 g in triplicate of bottled-weight material, was placed in 2.5 l buckets with a lid and
with 2 l of 0.5 % KI solution (diluent FSW) for 10, 20 and 30 minutes in a shaking incubator
(75rpm/ at 10 °C). After being incubated in the treatment, the material was washed and rinsed with
four litres of filtered, UV sterilised and Tyndalised seawater in order to remove the treatment
solution. The biomass was then blotted and placed in five litres tanks with three litres of F/2
medium and incubated at 12:12, 9°C, 80 µmol/m²/sec for two weeks. The initial PAM fluorometry
of the samples was taken with an AquaPen-P (PSI) before the start of the experiment to ensure
that the samples were healthy. Samples were checked to verify the impact of the treatments on the
biomass using PAM fluorometry, after two hours, one week and 14 days.
5.3.2 Tidal simulation as epiphytes control
Samples were collected at the end of November 2016 from Seil (56.30° N, 5.65° W) (~700 g), kept
in a Cool-box during the transfer and brought to the laboratory where they were cleaned manually
from visible grazers and epiphytes. They were then washed with 0.5 % KI solution (diluent FSW)
for 30 minutes in a shaking incubator (75rpm/ at 10 °C), rinsed with filtered and sterilized seawater
and equally split into 3 x 5 l tanks (200 g per tank). The tanks were incubated at 10 °C (in a
controlled temperature room), 8:16 (L:D) photoperiod, 50-80 µmol/m²/sec in F/2, which was
changed weekly.
The biomass was then transferred into the tidal system which was personally designed and
developed for the trial. The biomass was weighted in 180 g ± 0.5 in triplicate and put into 3 x 20 l
tanks with 10 l of sterilized F/2 changed weekly (manually) and the tanks were cleaned every week
with Decon 85% and rinsed with UV-sterilized filtered seawater. QY, weight, pH, salinity and
temperature were measured periodically. Cultures were incubated at 10 °C in a controlled
temperature room, 8:16 (L:D) photoperiod, 100 µmol/m²/sec. The light went on at 9.00 am and
switched off at 5.00 pm. Light bulbs were of the Gro-Lux type (Sylvania, F8W) and were collocated

142
on top of the tanks. F/2 was added as medium. The tidal indoor system was organized as follows:
every set of tanks was in triplicate and included two tanks, one working as a microcosm and the
other as reservoir (Figs 5.2-5.4). In every tank, there was a pump (Rule Bilge pump 360 GPH)
which was regulated by a 2 Channel Digital timer. The emptying interval for each pump was 30
seconds. From 6 am to 12 pm (noon) the water was kept in the Reservoir tanks. At 12 pm the
pumps were activated and pumped the water into the Microcosm tanks, where it stayed until 18.00
pm when the pumps were activated again to pump the water back into the Reservoir tanks. The
water was kept in the Reservoir until 12.00 am (midnight) and then pumped again in the Microcosm
tanks until 6.00 am. The seaweed biomass was allowed to float freely in the tanks. A plastic fine
mesh was placed on the bottom of the tank, attached with aquarium suction cups, in order to
maintain the cultures above the level of the water remaining in the tank during the low tide (Figs
5.2 and 5.4). A plastic mesh was also placed in the microcosm tanks to separate the pumps from
the cultures in order to avoid blocking the pumping system due to biomass being sucked into them.
Aeration was provided through either air stones, or aquarium flexible pipes (Figs 5.2-5.4).
The experiment was repeated twice: the first trial went from January to April and the second from
the end of May to the end of June. Two different starting amounts of material were trialled: 180 g
for the first trial and 110 g (blotted weight) for the second, to investigate the effect of biomass
density on the growth.
Figure 5.2 Schematic view of the microcosm tank from the top

143
Figure 5.3 A) Schematic view of the system from the top; B) Schematic lateral view of the system
Figure 5.4 Lateral view of the microcosm and reservoir tanks
5.3.3 Epiphytes treatment of biomass grown in a tumbling tank system cultivation both indoors
and outdoors
Samples were collected at low tide from Seil (56.3000° N, 5.6500° W) in October 2017 and then
again in March 2018 for a second trial, for a total amount of 6 and 4 kg respectively. In both cases,
the collection and treatment of the samples were the same. Samples were kept in Cool-boxes with
wet tissues and brought to the laboratory where they were cleaned manually in order to remove all
the visible epiphytes and grazers. Secondly, samples were washed thoroughly with filtered and UV
sterilised seawater. The biomass was further cleaned with a 0.5 % KI (purchased from Sigma
Aldrich®) solution (made up in filtered and UV sterilised seawater) and left in the solution for 30
minutes with powerful aeration. It was then rinsed three times for 30 minutes in filtered and UV
sterilised seawater. The water in the tank was changed again and samples were left overnight in
filtered and UV sterilised seawater with powerful aeration and with no nutrient added for 48 hours
until the beginning of the experiment.

144
Figure 5.5 Scheme of the tank tumbling system. Blue circle: air stone; grey block line: aeration pipe; Green arrow: water inflow; grey cylinder: tambourine filter; blue arrows: circular movement of water due to the aeration system; orange arrow: outflow system
The cleaned biomass was weighted into 250 g (fresh weight) aliquots, with triplicates per treatment
(3 x indoor and 3 x outdoor) and transferred to 60 l tanks with sand filtered seawater. For both
indoor and outdoor cultivation trials, 3 x 60 l black conical tanks were used. They were placed on
top of 60 l grey buckets. Aeration was provided from the bottom through a central tube inserted
into a pipe and weighed down with a weight and air stone.
The inflow was supported by a pipe connected with taps, pouring directly into each tank. The
outflow consisted of a tambourine meshed filter (in order to avoid the loss of material) (Figs 5.5-
5.7). Water was kept in continuous circulation at a flow rate of 500 ml/min. HOBO® (onset)
loggers for register light intensity and temperature (recording interval 30 mins), were placed in one
tank inside and one outside. Tanks were cleaned twice per month with a solution of 85% Decon,
rinsed with fresh water and successively filtered seawater, and samples were weighted every month
after been spun in a spin salad to measure the fresh weight (Ross et al., 2017). QY of the cultures
was measured with an AquaPen (PSI) in order to assess the physiology of the specimens.
In the indoor system, the light was provided artificially at a photoperiod of 12:12 (L:D). Data
relative to the light intensity (lum/ft²) was plotted as was the temperature (°C). The pH and
conductivity (mS/cm) were measured every 30 minutes with a GHL ProFilux III controller. F/2
was pulse fed through an auto dosing pump (Jebao®, DP-4) releasing 20 ml three times per day of
F/2 medium at the final concentrations (M) listed in Table 5.4. The interval between pulse feeds
was set at 8 hours.
In the outdoor system pH and salinity were monitored every day manually with a Multi 340i/set
(WTW). F/2 nutrients were provided through an Easy Irrigation system (EasyGrowᵀᴹ) at the same
interval and volume as used in the indoor trial. A plastic transparent cover was used on the outside
tanks in order to avoid excessive contamination from rainwater and to keep them clean from leaves,
dust, etc. The canopy did not interfere with the light entering the tanks.

145
The experiment was repeated, but only for the outdoor system, from April to September (2018),
keeping all the parameters and methodology the same. Biochemical analyses of the samples were
conducted at the end of the experiments for all the trials, including pigments, carbohydrates,
protein, and antioxidant compounds (with the methodologies described in Chapter 3) at least on
triplicate samples per tank.
Table 5.4 Components of the F/2 medium
Figure 5.6 Indoor cultivation system from the front
F/2
Component Final concentration (M)
Phosphate stock
NaH₂PO₄.2H₂O 3.62 x 10¯⁵ Nitrate stock
NaNO₃ 8.82 x 10¯⁴ Trace elements
FeCl₃ x 6H₂O 1.17 x 10¯⁵
MnCl₂ x 4H₂O 9.10 x 10¯⁷
NA₂ EDTA 1.17 x 10¯⁵
CuSO₄ x 5H₂O 3.93 x 10¯⁸
ZnSO₄ x 7H₂O 7.65 x 10¯⁸
CoCl₂ x 6H₂O 4.20 x 10¯⁸
Na₂MoO₄ x 2H₂O 2.60 x 10¯⁸ Vitamin stock Thiamine HCl
(Vitamin B₁) 2.96 x 10¯⁷
Biotin (vitamin H) 2.05 x 10¯⁹ Cyanocobalamin
(vitamin B₁₂) 3.69 x 10¯¹0

146
Figure 5.7 Outdoor cultivation system
5.3.4 In vitro spores’ production, attachment, germination and growth of juveniles of O.
pinnatifida from epiphytes treated cultures
When tips with visible tetraspores were identified in tank cultivated material, they were cut from
the adult fronds with a sterile blade in a laminar cabinet. The material was rinsed multiple times
with FSW and placed into 12 multi well Petri dishes. The dishes were incubated at 10 °C, 12:12
photoperiod and 20-30 µmol/m²/sec with 2 ml of FSW per well until spores released occurred
(which was normally after two hours; however, the release can continue for up to 48-72 hours).
Multiple release of spores from the same tissue normally occurs. The tissue and spores released
were periodically monitored under an Axio Inverted microscope and a stereo microscope (Zeiss).
Once released, spores, either attached or not, were isolated with a Pasteur pipette in a laminar
cabinet, transferred to six multi-well Petri dishes with FSW and incubated at 12:12. 10°C, 10
µmol/m²/sec for two weeks. The FSW was changed every two days. After the first week of
incubation, F/2 medium (Guillard and Ryther, 1962; Guillard, 1975) was added to the released
spores and this was changed twice per week. After two weeks, germinated spores were transferred
into a tissue culture flasks (75 cm²) with F/2, and incubated at 12:12, 10 °C and 20 µmol/m²/sec.
Medium and the flasks were changed once per week and photographs taken with an Axio Inverted
microscope (Zeiss) to assess the growth of the cultivated juveniles. After two months of cultivation,
juveniles were transferred into 6 multi-well plates (3 juveniles per well with 4 ml of F/2) incubated
under the same conditions as described above with the medium changed every two days. Spores
were cultivated for 5 months and used for further experiments. The number of spores released was
estimated on just a few of the samples from the multi-well plates with a counting cell (Sedgewick®
rafter cell).

147
5.4 Results
5.4.1 Chemical removal of epiphytes
5.4.1.1 Petri dish trial
The 0.25% Kickstart treatment effectively killed the O. pinnatifida cultures. The biomass was pink
in colour and brittle. The QY value observed for this treatment was 0 at every time point measured,
confirming that photosynthesis was not occurring.
Biomass treated with 1% NaClO was slower to recover, when compared to the 0.1% NaClO
treatment. This concentration resulted in a greater recovery and seemed to have less impact on the
biomass. At 1% NaClO an average of 0.08 QY after 2 hours was measured (SD: 0.014), 0.18 (SD:
0.07) after 7 days and 0.15 (SD: 0, 13) after 14 days. With 0.1% NaClO a reported average QY of
0.14 (SD: 0.03) was measured after 2 hours, 0.21 (SD: 0.08) after 7 days and 0.26 (SD: 0.07) after
14 days, demonstrating that photosynthesis was occurring in both sets of treated biomass, but at a
higher rate in the 0.1% NaClO treatment.
Employing 0.5% KI seemed to be the best treatment in terms of recovery with minimal effects on
the biomass of this species. Average QY measurements of 0.26 (SD: 0.07) after 2 hours, 0.28 (SD:
0.03) after 7 days and 0.35 (SD: 0.07) after 14 days were recorded. An ANOVA single factor,
p<0.05, was applied indicating a significant difference between the treatments tested after 2 hours,
7 and 14 days.
All treatments differed significantly from the control after 7 days, while only 0.5% KI and 0.1%
NaClO did not differ from the control after 14 days (Dunnett comparison test, Minitab 18) (Fig.
5.8), demonstrating that these last two treatments were not negatively influencing the cultures.
Figure 5.8 QY measurements related to the different treatments utilised for the Petri dish trial measured at 2 hrs, 7 days and 14 days. (control=green, 0.5% KI= blue, 25% methanol=orange, 1% NaClO= black, 0.25% Kick start=grey, 0.1% NaClO= yellow; n=3; error bar, 1 SD of mean)
0
0.1
0.2
0.3
0.4
0.5
0.6
after 2 hours after 7 days after 14 days
PA
M (
QY)
Time

148
In all the chemical treatments tested, when the biomass was examined periodically under a
stereomicroscope (Zeiss), it was possible to see that grazers (e.g. isopodes) and colonizing
epiphytes (e.g. green and brown seaweeds) were dead/heavily damaged by the treatment.
5.4.1.2 Flask trial
The biomass treated with the 0.1% NaClO solution showed a slow recover of the tissue. This was
supported by the QY average measurements of 0.04 (SD: 0.02) after 2 hours, 0.07 (SD: 0.03) after
7 days and 0.14 (SD: 0.05) after 14 days. The tissue was also green in colour and brittle. Biomass
treated with a 25% methanol solution recovered slowly as supported by the QY measurements:
0.13 after 2 hours (SD: 0.12), 0.11 (SD: 0.03) after 7 days and 0.12 (SD: 0.06) after 14 days. But the
biomass morphology (e.g. texture, colour) seemed to be less affected compare to the 0.1% NaClO
treatment.
When the biomass was treated with 0.5% KI the average QY measured was 0.15 (SD: 0.03) after 2
hours, 0.23 (SD: 0.03) after 7 days and 0.26 (SD: 0.06) after 14 days. The colour and texture of the
samples seemed to have not been affected by the treatment. Grazers were killed by all the
treatments, while epiphytes were reduced visibly. An ANOVA single factor, p<0.05 was applied
demonstrating a significant difference between the treatments tested (MS Between group: 0.037,
MS within groups: 0.002), with 0.5% KI statistically different from the other treatments and this
was taken forward in the tank trial (Fig. 5.9).
Figure 5.9 QY measurements related to the different treatments for the flask trial measured at 2 hrs, 7 days and 14 days. (control=green, 0.5% KI= blue, 25% methanol=orange, 0.1% NaClO= yellow; n=3; error bar, 1 SD of mean)
5.4.1.3 Tank trial
Treatment with 0.5% KI proved to be effective in both the Petri dish and flask trials at controlling
epiphytes (and grazers), leaving the cultures overall healthy, without affecting their colour and
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
after 2 hours after 7 days after 14 days
PA
M (
QY)
Time

149
texture. In order to further optimize this treatment, the biomass was exposed to the 0.5% Kl
solution for 10, 20 and 30 minutes. The results showed that there was a statistical difference
between the 10 and 20 minutes incubation periods and the 30 minutes one after 2 hours (Tukey’s
and Fisher’s tests) (ANOVA, Single factor, p< 0.05). After 7 days there was no statistical difference
between the treatments (ANOVA single factor, p< 0.05, comparison Tukey’s and Fisher’s tests).
The 30 minutes exposure treatment statistically differed from 10 and 20 minutes treatments after
14 days (Fig. 5.10), demonstrating photosynthetically that the biomass recovered quicker from the
longer exposure time treatment. As the experiment proceeded, the cultures continued to grow and
develop tetrasporophytes, were a dark red colour and Ulva spp., Enteromorpha spp., or brown algae
were absent. Demonstrating further that the method tested was effective in removing epiphytes in
long-term tank cultures. This was supported by the PAM measurements (Fig. 5.10).
Figure 5.10 QY in relation to exposure time (min) to treatment with 0.5% KI for the tank trial after 2 hrs, 1 and 2 weeks. (10 min incubation= grey, 20 min incubation=blue, 30 min incubation= orange; n=3; error bar, 1 SD of mean)
5.4.2 Tidal simulation as epiphytes control
Measurements of weight, pH, salinity and temperature were taken every three weeks (Tables 5.5.
and 5.6). The decrease in weight was evident in all the replicates, but it must be emphasized that
the epiphytes were kept under control, and absent in both the tanks and on the cultures. The
biomass also had new shoots developing from the tips and holdfasts (Fig. 5.12 A). Spores were
released from the biomass for the entire cultivation period (Fig. 5.12 B and C). Several methods
were tried to attach the seaweed to the net or a grid (such as plastic clips, aquarium epoxy, cable
ties), however without success, except for tying cultures to nylon ropes (Appendix C.4). A tumbling
cultivation in this study was therefore applied.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
time 0 after 2 hours after 1 week after 2 weeks
PA
M (
QY)

150
Figure 5.11 Comparison of growth data (g, wet weight) between the two trials with different starting biomass, 110 g (1st trial, dark grey line) and 180 g (2nd trial, light grey line). (n=3; error bar, 1 SD of mean)
Figure 5.12 A) Thalli with new tips and shoots developing from the holdfast; B) Tetrasporophytes with tetraspores; C) Tetrasporophytes after spores’ release
A weight decrease was observed in all the tanks and in both trials (Fig. 5.11). It was noted in the
second trial that the seaweed had new shoots developing from tips and holdfast, epiphytes were
kept under control and spores were produced and released spontaneously from the biomass for
the entire length of the trial. The two trials followed the same pattern (Figs 5.11, 5.13 and 5.14),
even if the initial density and the seasons were different and did not affect the overall growth rates
measured for both (ANOVA, single way, p< 0.05). Measurements of pH, temperature, salinity and
weight were recorded every 8 days (Tables 5.5 and 5.6).
Table 5.5 pH, salinity and temperature measurements of the first trial
Parameters time 0 time 1 time 2 time 3
pH 8 7.8 8 8.04
salinity 35 30.9 32 32.4
temperature 10 13.4 12 11.5
0
20
40
60
80
100
120
140
160
180
200
0 1 2 3
wei
ght
(g W
W)
Months

151
Table 5.6 pH, salinity and temperature measurements of the second trial
Parameters time 0 time 1 time 2 time 3
pH 7.95 8.16 8.03 8.14
salinity 32.4 31.9 31.9 32.2
temperature 14 12.3 12.9 12
Figure 5.13 Comparison of absolute growth measurements of first trial (dark grey line) and second trial (light grey line). (n=3; error bar, 1 SD of mean)
Figure 5.14 Comparison of SGR measurements of of first trial (dark grey line) and second trial (light grey line). (n=3; error bar, 1 SD of mean)
The average growth from the replicates of both trials were different, but this was probably linked
to the different durations of both experiments. Higher values were recorded in the second trial
(Figs 5.11, 5.13 and 5.14). However, an ANOVA (one-way, p<0.05, Minitab 18) indicated there
was no statistical difference between the two trials. This is also clear considering the SGR average
comparison of the two trials (Fig. 5.14).
-100
-80
-60
-40
-20
0
20
0 0.5 1 1.5 2 2.5 3 3.5
Ab
solu
te g
row
th (
%d
ay⁻¹
)
Months
-0.007
-0.006
-0.005
-0.004
-0.003
-0.002
-0.001
0
0.001
0 0.5 1 1.5 2 2.5 3 3.5
SGR
(%
day
¯¹)
Months

152
5.4.3 Impact of epiphyte treatment on tumbling cultivation system both indoor and outdoor
5.4.3.1 Winter trial: indoor and outdoor cultivation
Temperature, light and pH were measured for the indoor and outdoor tanks (Figs 5.15-5.18).
While the light intensity in the indoor experiment was low and constant for all the length of the
trial (Fig. 5.15), light intensity reported for the outdoor trial showed an increase along the
experimental time, according to the seasonal variation (Fig. 5.16).
Figure 5.15 Data collected with HOBO® logger for the indoor tanks from November 2017 to March 2018. Temperature is expressed in °C and Intensity in lum/ft². Temperature (dark grey bars) and Intensity (light grey line) are plotted on the same reference axis since the values reported for the intensity in the indoor experiment were particularly low, in set of ten
Figure 5.16 Data collected with HOBO® logger for the outdoor tanks from November 2017 to April 2018. Temperature is expressed in °C (dark grey bars) and Intensity in lum/ft² (light grey line), unit provided by the logger
Values of pH and salinity for the outdoor experiment followed the same trend, increasing
simultaneously along the trial (Fig. 5.17), while pH and salinity for the indoor experiment followed
0
2
4
6
8
10
12
14
November December January February March
Tem
p (
°C)
and
Lig
ht
Inte
nsi
ty (
lum
/ft²
)
Months
0
50
100
150
200
250
300
350
400
0
2
4
6
8
10
12
November December January February March April
Ligh
t In
ten
sity
(lu
m/f
t²)
Tem
per
atu
re (
°C)
Months

153
an inverted trend, with a decrease in salinity values and an increase in pH towards the end of the
experiment (Fig. 5.18).
Figure 5.17 Values relative to pH (dark grey bars) and salinity (light grey line) averaged per month for the tanks outdoor from November 2017 to April 2018
Figure 5.18 Values relative to pH (dark grey bars) and salinity (light grey line) averaged per month for the tanks Indoor from November 2017 to March 2018
The growth of the cultures indoor started to decline right at the beginning of the experiment, and
the trial was stopped one month before the outdoor trial due to this. However, although the
outdoor tanks showed an initial decrease, they then reached a plateau and started to increase in
weight after March (time 5) (Fig. 5.19). This was also confirmed by the SGR measurements (Fig.
5.20).
27
28
29
30
31
32
33
7.7
7.8
7.9
8
8.1
8.2
8.3
8.4
November December January February March April
Salin
ity
(PSU
)
pH
Months
0
10
20
30
40
50
60
7.9
7.95
8
8.05
8.1
8.15
8.2
8.25
8.3
8.35
November December January February March
Co
nd
uct
ivit
y (m
S/cm
)
pH
Months

154
Figure 5.19 Growth (expressed in WW) for the indoor (dark grey line) and outdoor (light grey line) tanks trials from November 2017 to April 2018, every time corresponds to one-month interval. (n=3; error bar, 1 SD of mean)
Figure 5.20 SGR was measured using the formula from Evans (1972) for every month and
expressed as %day⁻¹ for the Indoor trial (dark grey bars) and Outdoor one (light grey bars) (n=3; error bar, 1 SD of mean)
QY measurements fluctuated for the outdoor tanks, reaching a peak between January and February,
while values remain stable and lower for the indoor tanks, due to the difference in the light available
(Figs 5.15 and 5.16). The cultures of the outdoor trial maintained a dark red colour and a thick
texture (Fig. 5.22 B and D), whereas for the indoor tanks the biomass changed colour, turning into
green and becoming brittle (Fig. 5.22 A and C).
0
50
100
150
200
250
300
time 0 time 1 time 2 time 3 time 4 time 5 time 6
wei
ght
(g W
W)
Month
-2
-1.5
-1
-0.5
0
0.5
Time 1 time 2 time 3 time 4 time 5 time 6
Gro
wth
rat
e (%
day
¯¹)
Month

155
Figure 5.21 QY measured with an AquaPen every month for the cultures in the indoor (dark grey line) and outdoor (light grey line) systems. (n=3; error bar, 1 SD of mean)
An ANOVA single factor (Minitab 18) recorded a statistical difference between the QY response
for the tank indoor and outdoor, with the last samples showing a better physiological response
(Fig. 5.21).
Figure 5.22 Indoor biomass (A & C) and Outdoor biomass (B &D), after four months (February
2018) into the cultivation system during the periodic weighing
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
November December January February March April
PA
M (
QY)
Months
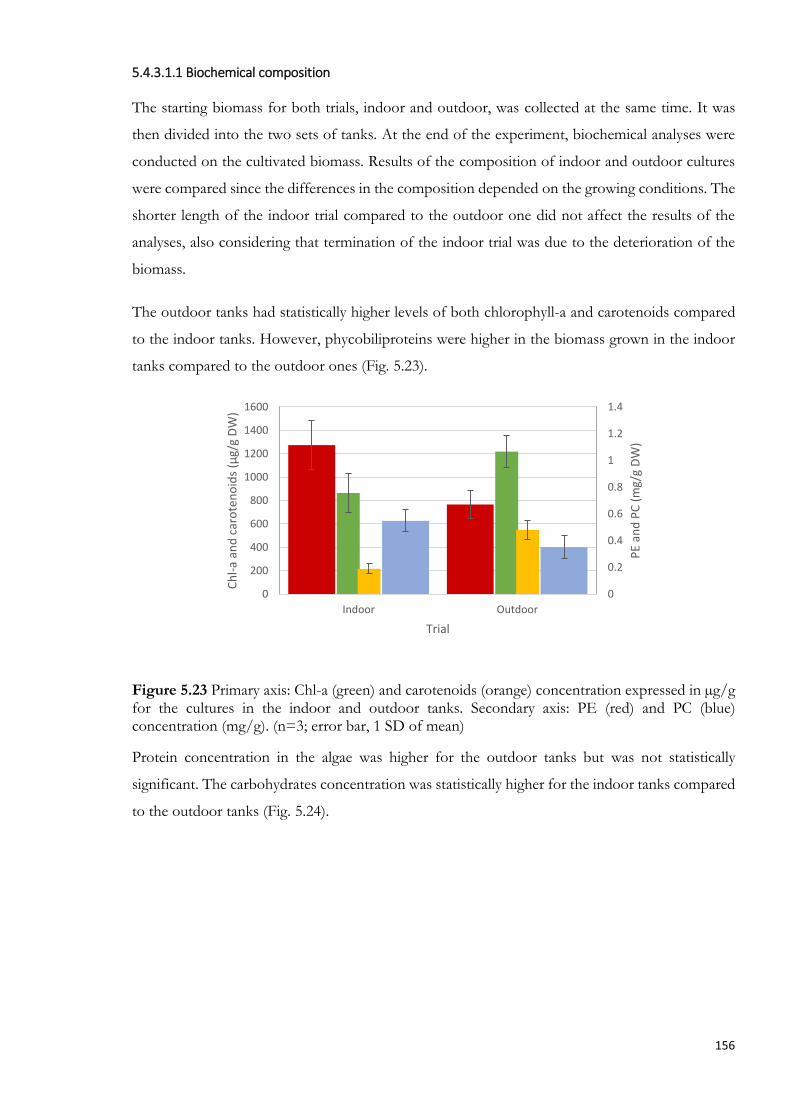
156
5.4.3.1.1 Biochemical composition
The starting biomass for both trials, indoor and outdoor, was collected at the same time. It was
then divided into the two sets of tanks. At the end of the experiment, biochemical analyses were
conducted on the cultivated biomass. Results of the composition of indoor and outdoor cultures
were compared since the differences in the composition depended on the growing conditions. The
shorter length of the indoor trial compared to the outdoor one did not affect the results of the
analyses, also considering that termination of the indoor trial was due to the deterioration of the
biomass.
The outdoor tanks had statistically higher levels of both chlorophyll-a and carotenoids compared
to the indoor tanks. However, phycobiliproteins were higher in the biomass grown in the indoor
tanks compared to the outdoor ones (Fig. 5.23).
Figure 5.23 Primary axis: Chl-a (green) and carotenoids (orange) concentration expressed in µg/g for the cultures in the indoor and outdoor tanks. Secondary axis: PE (red) and PC (blue) concentration (mg/g). (n=3; error bar, 1 SD of mean)
Protein concentration in the algae was higher for the outdoor tanks but was not statistically
significant. The carbohydrates concentration was statistically higher for the indoor tanks compared
to the outdoor tanks (Fig. 5.24).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0
200
400
600
800
1000
1200
1400
1600
Indoor Outdoor
PE
and
PC
(m
g/g
DW
)
Ch
l-a
and
car
ote
no
ids
(µg/
g D
W)
Trial

157
Figure 5.24 Protein (dark grey bars) and carbohydrate (light grey bars) content expressed in percentage (DW) for samples undergoing Outdoor and Indoor trials. (n=3; error bar, 1 SD of mean)
A statistical difference (p<0.05) was measured for the antioxidant compound concentration for the
outdoor tanks when compared to the indoor ones (Fig. 5.25).
Figure 5.25 Antioxidant compounds measured with FRAP assay in samples undergoing the cultivation in indoor (dark grey bar) and outdoor (light grey bar) tank trials. (n=3; error bar, 1 SD of mean)
5.4.3.2 Summer trial: outdoor experiment
There was an initial increase in growth when the biomass was weighted (from April to June), after
this peak the weight of the culture started to decrease and begun to stabilise again after August
(Fig. 5.26). New shoots and new tips grew on the cultured biomass which also became reproductive
from April to June. This was confirmed also by the SGR measurements (Fig. 5.27).
0
5
10
15
20
25
30
35
40
Outdoor Indoor
Pro
tein
an
d c
arb
oh
ydra
te c
on
ten
t (%
DW
)
Trial
0
0.5
1
1.5
2
2.5
3
3.5
4
Outdoor Indoor
FRA
P
(µM
FeS
O₄
eq/m
g D
W)
Trial

158
Figure 5.26 Growth (WW, g) of cultures in an outdoor tumbling system from April to September 2018. (n=3; error bar, 1 SD of mean)
Figure 5.27 SGR of culture in the outdoor tumbling system from April to September 2018. (n=3; error bar, 1 SD of mean)
QY measurements showed an initial decrease in the recorded values especially in June, followed by
a subsequent increase that reached its maximum in September (Fig. 5.28). The cultures in the
outdoor system underwent the same change in colour that can be seen in the biomass growing in
the wild (Fig. 5.29 B and D). This is directly linked to changes in day length and light intensity.
However, overall the material was healthy and thick in texture, with new growth visible (Fig. 5.29
A and C).
0
50
100
150
200
250
300
350
400
450
April May June July August September
wei
ght
(g W
W)
Month
-1.5
-1
-0.5
0
0.5
1
1.5
SGR 1 SGR 2 SGR 3 SGR 4 SGR 5
Gro
wth
rat
e (%
day
¯¹)
Month

159
Figure 5.28 QY measurement of cultures in an outdoor tumbling system from April to September 2018. (n=3; error bar, 1 SD of mean)
Figure 5.29 Cultures during the weighing in July 2018 (A &C), wild sample in May 2018 (B) and (D) July 2018
Temperature, light, pH and salinity are reported in Figs 5.30-5.32, showing an increase of
temperature especially in June and July (Fig. 5.30), a reduction of light intensity across the
experimental time (Fig. 5.31), an increase of salinity with a peak in August (Fig. 5.32), and a decrease
of pH across the months (Fig. 5.32).
0
0.1
0.2
0.3
0.4
0.5
0.6
April May June July August September
PA
M (
QY)
Month

160
Figure 5.30 Temperature (°C) profile in the tanks in the second outdoor trial as average (blue bars), maximum (dark grey bars) and minimum (light grey bars)
Figure 5.31 Light profile (lum/ft²) expressed as average in the tanks in the second outdoor trial
Figure 5.32 Salinity (light grey bars) and pH (dark grey line) measurements in the second outdoor trial
0
5
10
15
20
25
April May June July August September
Tem
per
atu
re (
°C)
Months
0
200
400
600
800
1000
1200
1400
1600
1800
2000
April May June July August September
Ligh
t In
ten
sity
(lu
m/f
t²)
Months
7.8
7.9
8
8.1
8.2
8.3
8.4
8.5
8.6
April May June July August September
31
31.2
31.4
31.6
31.8
32
32.2
32.4
32.6
pH
Months
Salin
ity
(PSU
)

161
5.4.3.2.1 Biochemical composition
Biochemical composition for the outdoor trial is presented in Table 5.7. All analyses were
conducted in triplicate per each tank on the harvested biomass at the end of the trial in September.
For the pigments, there was a statistical difference between tank number 1 and the other two, with
the former having a higher value when compared to the other two.
Table 5.7 Biochemical composition with relative concentration of the component analysed for the outdoor trial (n=3; error 1SD of mean)
Component Content (DW)
Protein 23.82 %
Carbohydrates 23.62 %
Chlorophyll a 2503 µg/g
Carotenoids 1283.6 µg/g
PE 0.68 mg/g
PC 0.25 mg/g
FRAP 1.53 µm FeSO₄/mg
5.4.4 In vitro spores’ production, attachment, germination and growth of juveniles of O.
pinnatifida from epiphytes treated cultures
The procedure for spores’ release for the successive isolation and cultivation is showed in Fig. 5.33.
Tips with visible tetraspores were cut (Fig. 5.33 A-C), and release of the spores occurred in the
multiplate (Fig. 5.33 D).
It was estimated that approximately 5,000 to 30,000 spores could be released during multiple release
events from each tip. The high variability of the released numbers and the fact that it was not
possible to obtain an average number of spores released per single plant made the survival
assessment unreliable. However, it was estimated that around 5% of spores survived and reached
the germination phase of the life cycle. The cultures obtained were not axenic.

162
Figure 5.33 A) Multi-well Petri dishes with cut tips from tetrasporophytes; B) Tetraspores are visible inside the tetrasporophytes on the extremities of the thalli of cultivated samples; empty tetrasporophytes are visible ahead of the full tetrasporophytes; C) Haploid tetraspores (indicated by the arrows) inside the diploid tetrasporophytes prior to release; D) Spores released prior attachment in a Petri dish and observed under a stereo microscope
Once released, spores that did survive (Fig. 5.34 A) either attached immediately (within 2 hours) or
remained unattached. In both cases, germination occurred within 24 hours and the attachment was
not fundamental to initiate it (pers. obsv.). Dead spores before germination were also observed
(Fig. 5.34 B).
Figure 5.34 A) Live haploid spore released observed under the Axio stereomicroscope. It is possible to see the external mucilaginous sheath; B) dead spore after release; note visible deteriorating cell wall and loss of pigmentation

163
The spores started to germinate, developing a rudimental rhizoid structure that allowed them to
adhere to the substratum. Their shape changed from rounded to oval and the division in septi of
the body became visible (indicated with A in Fig. 5.35). The basal part of the rudimental rhizoid
appendix quickly developed a swollen extremity, which was the main surface involved in anchoring
the development juvenile (indicated with B in Fig. 5.35). Moreover, even if the spores were released
at the same time, they did not develop at the same rate and while some were already attached and
germinated, others still retained their initial rounded shape or had a shorter developed appendix
(indicated with C in Fig. 5.35).
Figure 5.35 Spores under an Axio stereomicroscope (Zeiss) at different stages of development after the release: A) Body of germinated spore; B) Rhizoid of germinated spore; C) Rounded spore without rhizoid apparatus
After one week, germinated spores developed a long-shaped structure, increased in size and area,
and presented attachment disc (Fig. 5.36 A-F). Considering the different rates at which the spores
of the same age develop, it is difficult to give a general measure of the spores after defined times
in vitro cultivation.
After one month, the juveniles started to elongate and increase the size. The structure of the body
became more complex and the differentiation in cortical and medullar tissue more evident (Fig.
5.36 D). Measurements from the germinated spores were taken, in terms of the body and the
appendix, showing a clear development and enlargement of the spores (Fig. 5.36 D). After two
months of development the juveniles presented a main body, developed a complex structure, with
a cylindrical aspect and a clear distintion between the small rounded cortical cells and the medullary
one (Fig. 5.36 E). Branching was also observed, together with swolled and defined structures
anchoring the juveniles to the surface (Fig. 5.36 F).

164
Figure 5.36 A) Developed spores after one week into the cultivation system (observed at the Axio Inverted microscope at 10x); B & C) spores after two weeks into the cultivation system (observed at the Axio Inverted microscope at 20x and 10x); D) Measurement of a developed spore after one month from release (Axio Inver microscope); E) Developed juvenile after two months from the release; F) particular of attachment disc in juvenile after two months in cultivation

165
Figure 5.37 A) Juveniles after two months in tissue flask, B) magnification of juveniles. Each square= 1 mm².
Most of the juveniles that survived reached approximately 2-3mm after two months of cultivation
(Fig. 5.37 A and B). The proportion between the body and the rhizoid appendices changed, and
individuals generally had a longer cylindrical body of the former. Furthermore, the structure of the
body of the juveniles branched out, developing a structure that was charactheristic of the adult
stage of the life cycle (Fig. 5.38 A-D).
Figure 5.38 A & B) Juveniles after two months of in vitro incubation observed under a stereomicroscope (Zeiss). C & D) Juveniles after three months of in vitro incubation observed under a stereomicroscope (Zeiss). Visible ramification and presence of multiple branches as well as other organisms
After 5 months of in vitro cultivation, the length of the juveniles varied from five to ten mm (Fig.
5.39).

166
.
Figure 5.39 6x multi-well with juveniles after 5 months of cultivation
5.5 Discussion
One of the major challenges in establishing laboratory cultures of seaweed is maintaining them free
of contaminating algae (microalgae and cyanobacteria) and other epiphytes (Redmond et al., 2014).
These organisms tend to grow faster than the seaweed, compete for nutrients and may release
substances that inhibit growth, or are toxic to the alga of interest (Berland et al., 1972). Several
published studies have focused on macroalgal decontamination system; however, they usually aim
to obtain axenic cultures in the laboratory, rather than disinfect cultures used in long-term
cultivation systems (McCracken, 1989; Baweja et al., 2009; Reddy et al., 2008). The physical removal
stage (using razor blade, rubbing, or brushing the seaweeds) to remove epiphytes is followed by
chemical treatments (Baweja et al., 2009; Kientz et al., 2012). These steps can heavily affect
seaweeds that lack a protective cuticle (Reddy et al., 2008; Baweja et al., 2009). The most common
disinfectants utilised for the macroalgal culturing are: sodium hypochlorite (NaClO), reactive iodine
and organic solvents, often followed by an antibiotic mix (Baweja et al., 2009; McCracken, 1989;
Kerrison et al., 2016a, b). In these experiments the most effective treatment for epiphytes removal
on O. pinnatifida cultures was a 0.5% KI solution. This treatment significantly affected epiphytes
and grazers on the cultures, without having an impact on the seaweed. It was able to carry on
growing, showed very little loss in photosynthetic activity, as demonstrated via QY measurements,
plus it was able to develop, produce and release spores. The initial treatment prevented the
formation of epiphytes in the tank system even after months of culturing. The combination of the
chemical treatments and the tidal simulation system, proved to be an effective method in
controlling the growth of epiphytes and in preventing their proliferation in the tank cultivation
system.

167
As an intertidal species, O. pinnatifida must depend on the tides for its biological cycle (e.g.
reproduction and induction of spore release); furthermore, desiccation processes have previously
proved to be efficient in epiphytes control for other species (e.g. Pyropia sp., Ding and Ma, 2005).
Even if the tidal system did not lead to an overall increase in the biomass of the cultures (Fig. 5.11),
it was observed that while the older thalli were degenerating/becoming brittle, new shoots were
developing from the holdfast and from the tips of the branches, as the cultures were beginning to
regenerate. The cultures also produced and released spores for the entire length of the experiment.
Despite the results reported for the absolute and relative growth rates, the cultures were still alive
and growing. In this case it would probably be of more value to evaluate the quality of the culture
biomass rather than the quantity, especially in light of the slow growth rates that O. pinnatifida has
in the wild (pers. obsv.). In seaweeds in general, once the spores are released, the old thallus portion
tends to be slough off from the plant, and resources are used to support the development of new
shoots (Lüning and Pang, 2003). This is probably what happened in the O. pinnatifida tank cultures.
Further evidence to support this was provided in subsequent experiments, were O. pinnatifida
cultures required a long cultivation period to stabilise and eventually demonstrate a measureable
growth in weight.
The development of a tumbling cultivation system for this species provided a more controlled
regime where all the parameters could be monitored. The continuous tumbling of the cultures also
guaranteed a continuous exchange of nutrients, sufficient aeration as well as a uniform distribution
of light. This was the second time that an outdoor tank system for O. pinnatifida has been trialled
and the first time that an indoor system was used (Silva, 2015). The results obtained from this study
differ from those reported by Silva (2015), who conducted short outdoor tank cultivation trials in
Portugual. In the Portuguese study the SGR and productivity were positive for 5 weeks, from
November to December, and this was demonstrated further by an overall increase in the quantity
of biomass obtained. However, they excluded the summer growth experiment that had also been
carried out. Furthemore, cultures in the Portuguese study were supplied with seawater from an
IMTA system (Silva, 2015). Similar results, with fluctuations in SGR in outdoor tanks, were
reported for another Portoguese study (Gonçalves, 2018). For the experiments undertaken within
this chapter, the SGR gave negative values over the complete duration of the winter trials for the
indoor trial, while a positive value was reported at the end of the experiment for the outdoor trial;
on the other hand, positive values were reported for the first two months of the experiment for
the outdoor summer trial. Except the indoor trial, the biomass did not show visible signs of
decomposition. The indoor and outdoor cultivation systems were compared for biomass
production and quality. From this experiment, the outdoor system demonstrated that it was a
promising alternative, from a commercial point of view, and also had lower management costs
associated with it.

168
The indoor experiment was not successfully; this was considered to be due mainly to the type of
tanks used for the experiment. As already showed by previous authors (Lignell and Pedersen, 1987;
Smit et al., 1997), tank colour significantly affects the light field experienced by the algae. The tanks
used in this trial were black conical tanks and their position in the aquarium was away from the
light source (white lights) that were just hanging from the ceiling. Whilst light is fundamental for
seaweed biomass productivity and should always be sufficient to allow photosynthesis (Harrison
and Hurd, 2001). The impact of light limitation was evident from the PAM measurements of the
cultures and the pigments’ analyses. The only exception to this was for PE and PC values, which
were higher in the indoor cultures, probably because they were under stress and accumulating N
in different forms, such as pigments (Lapointe and Duke, 1984; Rico and Fernández, 1996). The
aeration and flow rates were also quite low due to limitations in the supply to the area utilised for
the experiment, whilst adequate water movement is needed to stimulate nutrients exchange and
provide carbon dioxide (Kerrison and Le, 2015). Deficit in the growth of the indoor cultures was
also reflected in accumulation of carbohydrates that are normally used by seaweeds as an energy
source (McCandless, 1981; Stiger et al., 2016).
Similar problems were also noted in the outdoor system, since in the summer water and air supplies
were limited, creating problems. In the winter trial, after four months, the cultures finally started
to stabilize and gain weight, with new shoots developing from the holdfast of the plants. Moreover,
cultures became reproductive around March-April and spontaneously released spores. Epiphytes
were kept under control during the experiment, indicating that the treatment applied previously to
the cultures was effective in their control even in the outdoor system, where the water used was
only filtered through a sand filter. The summer trial was only performed in an outdoor system,
since the conditions available indoor had already proved to be unsuitable. This time the cultures
increased in weight for the first 2-3 months, probably because of increased of light levels (Harrison
and Hurd, 2001) that occurs in Scotland in late spring months (April-May). Few issues caused a
drastic decrease of weight of the cultures, including a low water exchange rate (due to the scarce
water supply and the dependence to the tides variation), an insufficient filtration system that did
not control the inflow of contaminants (e.g. Ectocarpus sp.) and grazers (e.g. polychaetes) that
negatively affected the growth of the cultures (Redmond et al., 2014). The tanks, the filters, as well
as the pipes that supplied water to the tanks were completely infested with Ectocarpus sp., this
proliferated especially once the temperature in the tanks reached their highest levels (18-19°C).
Such high temperatures, negatively affected the cultures (as already showed in a preliminary
experiment, see Appendix C.1), considering that the biomass becomes brittle and eventually dies
off at temperature above 15 °C. Cultures started to stabilize again after August, when the light
intensity, but especially the temperatures, started to decrease. The response of the cultures was also
confirmed by the PAM measurements (Fig. 5.28).

169
The winter and summer trials showed different results in terms of biochemical composition.
Furthermore, variations in the biochemical profiles were previously observed for wild specimens
in relation to the season (Chapter 3), and these differences might be related to the particular
environmental conditions, (Kaehler and Kennish, 1996; Haroon, 2000; Chandini et al., 2008) that
have an effect on the biochemical composition of the seaweeds, especially in the summer trial.
Interestingly, the cultivation system was able to influence the morphology similarly to what
happens in the wild, with cultures showing the same variation in colour already observed in seasonal
samples.
Cultures in the tank systems investigated produced tetraspores, even after treatment against
epiphytes, showing that this chemical treatment does not negatively affect the life cycle of this
species. The reproductive cycle is considered one of the main possibilities for cultivation of a range
of species for seeded line (McHugh, 2003). It offers the advantage of having continuous genetic
recombination that makes future cultures more resistant to diseases, e.g. Ice-ice (Santelices, 1999;
FAO, 2003). But there are limitations with this methodology, such as high mortality rates, slow
growth and the costs associated with cultivation of spores until they reach the juveniles stage, step
that is usually carried out in a hatchery where normally parameters such as pH, salinity, high quality
water, light and temperature are artificially controlled with an associated cost (FAO, 2003). On the
other hand, vegetative production might guarantee a faster increase in biomass, lower production
costs, but this may also be associated with an increased susceptibility to diseases (FAO, 2003). In
the end, it will depend on the market the biomass is being produced for.
In this study, the reproductive cycle and juvenile production was performed in vitro for O.
pinnatifida for the first time from mature tetrasprophytes treated with a 0.5% KI solution cultivated
in a tank system. It was noted that tetrasporophytes could be easily obtained from the adults
cultivated in tanks even outside the natural reproductive period, which is May (Maggs and
Hommersand, 1993, Appendix C.2). Release of spores from the tips was observed once they were
cut from the adult and submerged in filtered seawater. Multiple releases from the same tip
commonly occurred. Spores were released in large numbers and quickly attached and germinated.
For the whole process to be scaled up and commercially applicable there are still several limitations
that need to be overcome. First, it will be necessary to assess the release and survival rates of the
spores, this will give an indication of how much reproductive starting material will be needed at a
commercial level. It will also be important to find an adequate substratum for the attachment of
the spores, which has already been identified as important requirement for a variety of seaweed
spores (Maggs and Callow, 2003; Hurd et al., 2014; Redmond et al., 2014). For this species, it
appears that they are able to attach to several different types of surfaces, including glass and plastic

170
Petri dishes. However, trials on different types of substrata should be conducted in order to solve
the bottleneck in survival rates, as well as the rate of the development.
5.6 Conclusions
Epiphytes and contaminants can be mitigated and controlled in O. pinnatifida cultures
applying a chemical treatment with a 0.5% KI solution and an incubation time of 30
minutes, hence this treatment should always be applied prior cultivation;
The chemical treatment coupled together with the tidal simulation system personally
developed proved to be efficient in controlling and eliminating the presence of epiphytes
on the cultures and cultivation system;
The chemical treatment does not affect the biology of the species and its capacity to
produce and release tetraspores;
Spores production and release for the species investigated is possible; however, several
limitations/problems need to be solved in order to improve the process, such as reduce
the mortality, accelerate the growth rate, control the contaminants and test different
substrata (e.g. shells) for settlement optimization;
Juveniles successfully developed from germinated spores cultivated in vitro reaching the
size of five to 10 mm in five months;
From a commercial prospective, a vegetative cultivation of this species rather than a
reproductive one seems to be more suitable at the moment for exploitability and up-scaling
of O. pinnatifida;
These preliminary tank experiments specifically developed for these trials, permitted to
design and develop a successful indoor tumbling cultivation system for the cultivation of
this species, as described in Chapter 8.

171

172
Chapter 6 Effects of different media on the growth and biochemical
composition of Osmundea pinnatifida
6.1 Introduction
Flask cultivation of seaweeds is an established methodology allowing for the control of culturing
parameters such as light, temperature and nutrients, in order to manage culture productivity
(Andersen, 2005). Nutrient supply is an important parameter in the management of cultivation
systems (Lignell and Pedersen, 1987), with media being usually composed of three main
components: macronutrients (generally nitrogen and phosphorous), trace elements and vitamins.
For optimal growth, macronutrients are usually required in a ratio of 16N:16Si:1P (Redfield ratio,
1934) (Parsons et al., 1984; Brzezinski, 1985), replicating the ratio found in natural seawater.
However, the N:P for seaweed can vary between 10:1 and 80:1, with an average of 30:1 (Atkinson
and Smith, 1983). Nitrogen is fundamental for the production of amino acids, nucleic acid,
enzymes, nucleoproteins and alkaloids; phosphorous is used by seaweeds in the process of
photosynthesis, respiration, cell division and enlargement, carbohydrates metabolism and has a role
in the transfer/storage of energy (Hopkins and Hüner, 2009). Typically trace metal stock solutions
consist of chloride or sulfate salts of zinc, cobalt, manganese, selenium, nickel, and iron (Andersen,
2005). Usually three vitamins, vitamin B₁₂ (cyanocobalamin), thiamine, and biotin, are added, but
very few algae need all three vitamins (Provasoli and Carlucci, 1974; Provasoli, 1966). The usual
order of vitamin requirements by algae are vitamin B₁₂ > thiamine > biotin.
In this Chapter, three different media types were investigated for the flask cultivation of vegetative
tissue of Osmundea pinnatifida. The aim was to identify the best cultivation medium for this species
in terms of growth and biomass increase, as well as biochemical composition. The commonly used
nutrient medium for indoor cultivation of red seaweeds is von Stosch enrichment medium (VSE)
(Redmond et al., 2014). Several studies have investigated the use of this medium for the cultivation
of different red seaweeds species (Guiry and Cunningham, 1984), including Gigartina sp., as well as
more recent studies; Hypnea cervicornis (Ribeiro et al., 2013), Gracilaria domingensis (J. Agardh; Mendes
et al., 2012); Gracilaria verrucosa (Mensi et al., 2011); Gracilaria sp. (Kim and Yarish, 2014); O.
pinnatifida (Silva, 2015). von Stosch enrichment medium contains nutrients such as nitrate,
phosphate, iron, manganese, EDTA, and vitamins (vitamin B₁₂, thiamine, and biotin). Compared
to other media, its application at commercial scale is limited by its high material and preparation
costs (Kim and Yarish, 2014). In order to confirm the most appropriate cultivation medium, a
comparison was carried out with one of the most widely employed algal medium (F/2) designed
for growing coastal marine algae, in particular diatoms (Guillard and Ryther, 1962; Guillard, 1975),
and a modified version of this medium, where the sources of nitrogen and phosphate were the

173
same (in concentration and type) as that used in VSE medium, while the other elements were kept
the same as F/2. The biochemical composition of the biomass produced from the cultivation in
these three media types was analysed in order to identify any impact on the biochemical profile on
this species. A comparison of media effects on growth and biochemical composition of two
different stages of cultures, adults and juveniles, was carried out again to see if the stage/age of the
cultures might affect the nutrient uptake, as already demonstrated for other seaweeds (Lobban and
Harrison, 1997; Pedersen, 1994).
In addition, the effect of different nitrogen sources on growth and biochemical composition of O.
pinnatifida was also investigated. There is, in general, a lack of available data on the effect that
nutrients, in terms of regimes, type, concentration and N/P, have on seaweed biomass and growth.
This knowledge is fundamental for the management and cultivation of seaweeds (Rueness and
Tananger, 1984). Generally, macroalgal nutrient uptake depends on several factors, such as
environmental N concentration (Harrison and Hurd, 2001) and biological factors, e.g. metabolism,
morphology, tissue type, species, age of the algae and nutritional history (Neori et al., 2004; Lobban
and Harrison, 1997; Pedersen, 1994; Rosenberg and Ramus, 1984), and typically follows a
Michaelis-Menten-type curve (Jimenez del Rio et al., 1996). The growth rate and productivity of
algae is, in part, controlled by the concentration of dissolved inorganic nitrogen (DIN) in the
aqueous medium surrounding the thallus (Chapman and Craigie, 1977; Rosenberg and Ramus,
1982; Lavery and McComb, 1991). The ability of an alga to utilise N for biomass production is
determined by the rate at which DIN can be absorbed and thereafter used in biochemical processes
(Wheeler, 1980; Koch, 1994; Sanford and Crawford, 2000). In seawater, N is available to seaweeds
in three major forms: NO₃, NH₄, and urea (Phillips and Hurd, 2004). It is most likely the key factor
in limiting growth of seaweed, especially those from temperate areas (Lobban and Harrison, 1997).
Generally, N limitation reduces growth, increases phycocolloid content, and decreases
photosynthetic activities (Chopin et al., 1990a). In many macroalgae, growth rates are reduced as
soon as the total N content of the thallus falls below the critical level of approximately 2% (Hanisak,
1983). These studies led to the concept of the “Neish effect”, reduced carrageenan content with
increasing N availability (Neish et al., 1977). However, seaweed can store nitrogen in excess, to
sustain their growth during periods of nutrient limitation/deficiency; this mechanism has important
implications for nutrient management of cultivated seaweeds (Bird et al., 1982; Lapointe, 1985).
The relationship between N, growth, biochemistry (e.g. production of carrageenan) and
photosynthesis is well documented for certain species such as Chondrus crispus (Neish et al., 1977;
Simpson et al., 1978; Chopin et al., 1995). However, there is no information available for O.
pinnatifida, the species been investigated in this thesis. This chapter details experiments carried out

174
in order to understand the impact of different media types and N sources on the growth and
biochemical composition of O. pinnatifida.
6.2 Aims
To identify the best treatment in term of medium composition that would be able to sustain
the growth and the biochemical composition of the species been investigated;
To investigate different nitrogen sources and their impact on cultivated samples in terms
of growth and biochemical composition;
To assess the influence of media composition and stage of development of the cultures
(adults and juveniles) on the biochemical profile of O. pinnatifida.
6.3 Materials and methods
6.3.1 Effects of three different media types on the growth and biochemical composition of
Osmundea pinnatifida adult cultures
The biomass for this experiment was collected in mid-November 2017 from Seil (56.30° N, 5.65°
W) at low tide. A total of six kg of fresh weight biomass was stored in a Cool-box during transport
to the laboratory, manually cleaned removing all the visible grazers and epiphytes, and rinsed
thoroughly with FSW in order to remove all the remaining debris and epiphytes. Finally, samples
were cleaned with a 0.5% solution of KI (Sigma Aldrich®) in 50 l of filtered and UV sterilised
seawater for 30 minutes. Samples were then rinsed three times for 30 minutes each time in FSW
and left to acclimate at 10 °C, 12:12 (L:D) in filtered and UV sterilised seawater for one week. The
water was changed every day.
Figure 6.1 Flasks (6 x 1 l) with adult cultures in the CT room under incubation: 10 °C, 12:12 photoperiod

175
One-litre flasks with 10 g of wet weight material (dried in a spin salad, as developed by Ross et al.
2017, before weighing) and 0.45 µm Whatman® filters connected to glass Pasteur pipettes for
aeration were used in triplicate for each treatment (Fig. 6.1). Cultures were incubated in a CT room
at 10°C, 12:12 (L:D), 80-100 µmol/m²/sec light intensity. The media treatments are detailed in
Table 6.1. Media were prepared aseptically in filtered, UV sterilized and Tyndalysed natural
seawater. All stock solutions for the different media types were autoclaved prior to use, except for
the vitamin stocks, which were filtered sterilised (0.2 µm), and the iron stock for the VS recipe
(ferrous sulfate was added aseptically to sterile distilled water); both were stored in the fridge until
required. The medium was changed twice per week in a laminar cabinet. Every week the samples
were weighted (after been dried in a spin salad) and the specific growth rate (SGR) was calculated
using the formula (Evans, 1972) where LN is natural logarithm, Wf final weight, Wi initial weight
and t time:
𝑆𝐺𝑅 =(𝐿𝑁(𝑊𝑓) − 𝐿𝑁(𝑊𝑖))𝑥100
𝑡
The experiment was run for 5 weeks in total. Statistical analyses for all the trials listed in this and
subsequent sections were conducted applying an ANOVA single factor (p<0.05, Minitab 18) with
Tukey’s, Fisher’s and Dunnett’s post-hoc tests.
Table 6.1 Media formulation trialled: F/2 recipe (from Andersen 2005), Von Stosch recipe (from Andersen 2005); F/2 modified. The nitrogen and phosphate sources with the respective concentrations were taken from the Von Stosch recipe. All the other elements and concentration were kept as normal as in the original F/2 recipe.
F/2 F/2 modified Von Stosch
Component Final concentration
(M)
Component Final concentration
(M)
Component Final concentration
(M)
Phosphate stock Phosphate stock Phosphate stock
NaH₂PO₄x2H₂O 3.62 x 10⁻⁵ Na₂ b-glycerophosphate
2.48 x 10⁻⁴ Na₂ b-glycerophosphate
2.48 x 10⁻⁴
Nitrate stock Nitrate stock Nitrate stock
NaNO₃ 8.82 x 10⁻⁴ NaNO₃ 5.00 x 10⁻³ NaNO₃ 5.00 x 10⁻³
Trace elements Trace elements Trace elements
FeCl₃x6H₂O 1.17 x 10⁻⁵ FeCl₃x6H₂O 1.17 x 10⁻⁵ FeSO₄x7H₂O 1.00 x 10⁻⁵
MnCl₂x4H₂O 9.10 x 10⁻⁷ MnCl₂x4H₂O 9.10 x 10⁻⁷ MnCl₂x4H₂O 1.00 x 10⁻⁴
NA₂ EDTA 1.17 x 10⁻⁵ NA₂ EDTA 1.17 x 10⁻⁵ Na₂EDTAx 2H₂O 1.00 x 10⁻⁴
CuSO₄x5H₂O 3.93 x 10⁻⁸ CuSO₄x5H₂O 3.93 x 10⁻⁸
ZnSO₄x7H₂O 7.65 x 10⁻⁸ ZnSO₄x7H₂O 7.65 x 10⁻⁸
CoCl₂x6H₂O 4.20 x 10⁻⁸ CoCl₂x6H₂O 4.20 x 10⁻⁸
Na₂MoO₄x2H₂O 2.60 x 10⁻⁸ Na₂MoO₄x2H₂O 2.60 x 10⁻⁸
Vitamin stock Vitamin stock Vitamin stock
Thiamine HCl
(Vitamin B₁) 2.96 x 10⁻⁷ Thiamine HCl
(Vitamin B₁) 2.96 x 10⁻⁷ Thiamine HCl
(Vitamin B₁) 5.93 x 10⁻⁶
Biotin (vitamin H) 2.05 x 10⁻⁹ Biotin (vitamin H) 2.05 x 10⁻⁹ Biotin (vitamin H) 4.09 x 10⁻⁹
Cyanocobalamin
(vitamin B₁₂) 3.69 x 10⁻¹⁰ Cyanocobalamin
(vitamin B₁₂) 3.69 x 10⁻¹⁰ Cyanocobalamin
(vitamin B₁₂) 1.48 x 10⁻⁹

176
At the end of the 5 weeks cultivating period, the biomass was harvested and protein, carbohydrates,
antioxidant compounds (FRAP assay) and pigments (including carotenoids, PE and PC and
chlorophyll-a), were analysed in triplicate for each culture with the methodologies already described
in Chapter 3.
6.3.2 The impact of different media types on growth and biochemical composition of Osmundea
pinnatifida juvenile plantlets
The previous experiment (see section 6.3.1) was repeated, but this time on juvenile plantlets of O.
pinnatifida and with only two of the media types, Von Stosch and F/2 (Table 6.1). For this
experiment, 500 ml flasks with 200 ml of medium and 2 g of material (fresh weight after spin-
drying) were used and again the experiment was performed in triplicate for each treatment. All the
other conditions and methodologies used are already described in section 6.3.1.
Figure 6.2 Flasks (6 x 500 ml) with juvenile cultures under incubation: 10 °C, 12 :12 photoperiod
The biochemical composition of the samples was determined as described in Chapter 3 and
included antioxidant compounds (FRAP assay), carbohydrates, protein, and pigments (chlorophyll-
a, carotenoids, PE and PC).
6.3.3 The impact of nitrogen sources and the N:P ratio on Osmundea pinnatifida growth and
biochemical composition
Cultures for this experiment, collected as described in section 6.3.1, were acclimated as follows,
five grams per 12 cultures were incubated for one week in filtered, UV-sterilized and tyndalyzed
seawater with a photoperiod of 12:12 (L:D), white fluorescent light (80-100 µmol/m²/sec) and at
10 °C. The experiment was conducted in triplicate in 500 ml flasks with 5 g WW of biomass and
250 ml of medium changed weekly. F/2 medium was the basic medium type (Table 6.1) with the
nitrogen source varied for the other treatments (Table 6.2):

177
Inorganic Sources: NaNO₃, NH₄Cl, NH₄NO₃
Organic source: NH₂CONH₂ (urea)
Nitrogen was added to the media at a final concentration of 824 µM (Starr and Zeikus 1993).
Phosphate was kept constant (82.4 µM, as normal concentration of F/2). With an N:P ratio of 10:1
(Table 6.2).
Table 6.2 Composition of treatments applied to cultures where control refers to F/2 medium. The final molarity of N and P was the same for all the media tested
Trace elements (ml/l)
Vitamin mix (ml/l)
Phosphate (g/l)
NaH₂PO₄x2H₂O
Nitrogen sources Weight (g/ l)
1 1 5.65 NaNO₃ (control) 75
1 1 5.65 NH₄Cl 40
1 1 5.65 NH₄NO₃ 65.9
1 1 5.65 NH₂CONH₂ 49
Blotted weight, after spin-drying was recorded every week, before the medium was changed.
Relatively daily growth rate was calculated using the formula of Evans (1972) detailed in section
6.3.1. The experiment was run for 7 weeks and the biomass was harvested for protein,
carbohydrates and phycobiliproteins analysis using the methodologies described in Chapter 3.
6.4 Results
6.4.1 Effects of three different media types on the growth and biochemical composition of
Osmundea pinnatifida adult cultures
6.4.1.1 Weight and growth rate of adult cultures grown in three different media
Cultures in this experiment demonstrated positive growth for all of the treatments investigated for
the entire length of the experiment. Samples grew new extremities, as well as developed new shoots
(Fig. 6.3); there was no change in colour, or contamination by epiphytes in the cultures undergoing
all three of the treatments.
Figure 6.3 Different cultures under the different media treatments at the beginning and at the end of the trial A) F/2, B) F/2 modified and, C) VS at the beginning of the experiment; D) F/2, E)F/2 modified and, F) VS at the end of the experiment

178
An ANOVA single factor (p<0.05) indicated a statistical difference in the first two weeks of the
experiment, with the VS treatment giving a statistically higher growth compared to the other two
treatments. However, the trend changed after this, with no statistical difference reported between
the tested treatment and a slightly higher growth reported, at the last time point, for F/2, followed
by VS and lastly F/2 modified (Fig. 6.4).
Figure 6.4 Weight measurements of cultures (WW) under the three different treatments: F/2 black line, F/2 modified dark grey line, VS light grey line. (n=3; error bar, 1 SD of mean).
The relative growth rate shows a decreasing trend, especially in the first two weeks of the
experiment (Fig. 6.5). After that, the relative growth, particularly for VS and F/2 reached a stable
state towards the end of the experiment. All three treatments were able to sustain the growth of O.
pinnatifida vegetative tissue, giving a maximum growth rate in terms of a percentage per week of 1.2
for VS, 0.86 for F/2 and 0.67 for F/2 modified.
Figure 6.5 Growth rate calculated with the equation for SGR (Evans, 1972). F/2 black line, F/2 modified dark grey line, VS light grey line. (n=3; error bar, 1 SD of mean)
9
9.5
10
10.5
11
11.5
12
Time 0 Time 1 Time 2 Time 3 Time 4 Time 5
wei
ght
(g W
W)
Week
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Time (1) Time (2) Time (3) Time (4) Time(5)
Gro
wth
rat
e (%
day
¯¹)
Week

179
6.4.1.2 Effects of three different media types on the biochemical composition of Osmundea pinnatifida
adult cultures
6.4.1.2.1 Antioxidant content
An ANOVA single factor (p<0.05) indicated a statistical difference between VS and the other
treatments, in terms of the antioxidant content (Fisher’s and Tukey’s tests), with F/2 and F/2
modified being statistically different from VS (Fig. 6.6). The highest result was reported for F/2
modified, 5.47 µM/mg FeSO₄, followed by F/2, 4.8 µM/mg FeSO₄ and last VS, 3.40 µM/mg
FeSO₄, demonstrating that the first two media statistically increase the production of antioxidant
compounds in the species.
Figure 6.6 FRAP assay for the cultures under the different media. F/2 black bar, F/2 modified dark grey bar, VS light grey bar. (n=3; error bar, 1 SD of mean)
6.4.1.2.2 Carbohydrates and proteins
An ANOVA single factor (p<0.05) and post hoc tests (Tukey’s and Fisher’s) indicated a statistical
difference in terms of carbohydrate content between VS and F/2 modified, and F/2, with the first
two showing higher values (Fig. 6.7). An ANOVA single factor (p<0.05) and post hoc tests
(Tukey’s and Fisher’s) reported no statistical difference for the protein concentration; however, VS
showed the lowest concentration compared to F/2 (highest value) and F/2 modified. Furthermore,
analyses showed that carbohydrates and protein contents followed inverted trends, while
carbohydrates content increased, the protein content decreased and vice versa (Fig. 6.7).
0
1
2
3
4
5
6
7
VS F/2 F/2 modified
FRA
P
(µM
FeS
O4
eq
/mg
DW
)
Treatment

180
Figure 6.7 Carbohydrate (dark grey bars) and protein (light grey bars) content expressed as %DW for cultures undergoing the three different treatments. (n=3; error bar, 1 SD of mean)
6.4.1.2.3 Pigments
An ANOVA single factor (p<0.05) and post hoc tests (Tukey’s and Fisher’s) indicated a statistical
difference between VS and the other two media tested, with the first one showing the lowest
concentration of Chl-a and the highest value reported for F/2 modified (Fig. 6.8). The same result
was obtained when applying the Dunnet’s test using F/2 as a control. However, no statistical
differences were observed between algae cultivated in F/2 and F/2 modified.
Figure 6.8 Chl-a (dark grey bars) and carotenoids (light grey bars) content of the cultures. (n=3; error bar, 1 SD of mean) A similar situation was recorded for the concentration of carotenoids, where an ANOVA single
factor (p<0.05, Tukey and Fisher’s tests) reported a statistical difference between VS and the other
two treatments, with the first one showing the lowest value and the highest reported for F/2
modified. No statistical difference was reported between F/2 and F/2 modified (Fig. 6.8). For the
0
5
10
15
20
25
VS F/2 F/2 modified
Car
bo
hyd
rate
s an
d p
rote
in c
on
ten
t (%
DW
)
Treatment
0
200
400
600
800
1000
1200
VS F/2 F/2 modified
Ch
loro
ph
yll a
an
d c
aro
ten
oid
s co
nte
nt
(µg/
g D
W)
Treatment

181
PE and PC content, no statistical differences were noted (ANOVA single factor p<0.05, Tukey’s
and Fisher’s tests). For PE the highest value reported was for F/2 modified, followed by VS and
finally F/2. For PC, the highest value was reported for F/2, followed by VS and finally F/2
modified (Fig. 6.9).
Figure 6.9 PE (dark grey bars) and PC (light grey bars) values for cultures undergoing the three treatments. (n=3; error bar, 1 SD of mean)
6.4.2 The impact of different media types on growth and biochemical composition of Osmundea
pinnatifida juvenile plantlets
6.4.2.1 Weight and growth rate of plantlets grown in either F/2 and VS media
From the previous experiment, F/2 and VS were confirmed as more suitable, in sustaining growth
of vegetative tissue of the species being investigated. They were used in subsequent experiments
to investigate their effects on growth and biochemical composition. For juvenile plantlets grown
in both media types (Fig. 6.10), the ANOVA single factor (p<0.05) showed no statistical difference
between the two treatments, in terms of growth (g) for the first 4 weeks, even when growth in F/2
was higher when compared to VS for the 2nd, 3rd and 4th weeks respectively. However, by week 5
there was a statistical difference between the treatments, with F/2 > VS (Fig. 6.11).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
VS F/2 F/2 modified
PE
and
PC
co
nte
nt
(mg/
g D
W)
Treatment

182
Figure 6.10 Juveniles cultures under the different treatments after one week A) F2; B) VS, and at the end of the trial C) F/2 ; D) VS
Figure 6.11 Weight measurements (g, WW) for cultures under two different media types: F/2 dark grey line and VS light grey line. (n=3; error bar, 1 SD of mean)
After an initial decrease, the SGR for F/2 treatment increased, reaching a maximum value at the
beginning of the trial (2.26 % in 7 days) and then increasing again in the 3rd week (0.37 % in 7 days)
and decreasing again after the 4th week. However, with the VS treatment there was a constant
decrease until the 4th week, then an increase in the last week. The maximum value was recorded at
the beginning of the experiment (1.57 % in 7 days), with no statistical differences reported in terms
of the growth rate (Fig. 6.12).
0
0.5
1
1.5
2
2.5
3
3.5
Time 0 Time 1 Time 2 Time 3 Time 4 Time 5
wei
ght
(g W
W)
Week

183
Figure 6.12 SGR (Evans, 1972) of cultures under the different treatments. F/2 dark grey line and VS light grey line (n=3; error bar, 1 SD of mean)
6.4.2.2 Biochemical composition when two different media types were employed to cultivate Osmundea
pinnatifida juvenile plantlets
6.4.2.2.1 Antioxidant analysis
An ANOVA single factor (p<0.05) indicated a statistical difference between F/2 and VS in terms
of antioxidant content (Fisher’s and Tukey’s tests), with F/2 > VS (Fig. 6.13).
Figure 6.13 FRAP assay for the cultures under two media. F/2 dark grey bar and VS light grey bar. (n=3; error bar, 1 SD of mean)
6.4.2.2.2 Proteins and carbohydrates analyses
An ANOVA single factor (p<0.05, Tukey’s and Fisher’s tests) demonstrated that there was no
statistical difference between the two treatments for protein content, with a slightly higher value
for VS compared to F/2 (Fig. 6.14). In the previous experiment, described in Section 6.4.1.2, no
-2
-1
0
1
2
3
4
5
SGR1 SGR2 SGR 3 SGR 4 SGR 5
Gro
wth
rat
e (%
day
¯¹)
Week
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
VS F/2
FRA
P
(µM
FeS
O4
eq
/mg
DW
)
Treatment

184
statistical difference was observed, but a higher value was reported for F/2 compared to VS. This
difference could be due to the different type, in terms of development, of algal tissue used.
Figure 6.14 Protein (light grey bars) and carbohydrate (dark grey bars) content expressed in % DW
of the cultures under two different media. (n=3; error bar, 1 SD of mean)
An ANOVA single factor (p<0.05, Tukey’s and Fisher’s Tests) indicated a statistical difference
between the treatments for the carbohydrates content, with VS > than F/2 (Fig. 6.14). This was
already estimated in the previous experiment (section 6.4.1.2). Furthermore, in this trial the
percentages reported were higher than those seen for the older thalli tissue.
6.4.2.2.3 Pigments
An ANOVA single factor (p<0.05, Tukey’s and Fisher’s Tests) indicated no statistical difference
in terms of Chl-a and carotenoids content for the cultures under F/2 and VS. However, higher
values were reported for VS (Fig. 6.15), which was different from what was seen in Section 6.4.1.2.3
It was assumed that the difference was probably due to the different typology (in term of
development) of the material investigated.
Figure 6.15 Pigments content in the cultures under the different treatments. Chlorophyll a =dark grey bars and carotenoids= light grey bars (n=3; error bar, 1 SD of mean)
0
5
10
15
20
25
30
35
VS F/2
Pro
tein
an
d c
arb
oh
ydra
tes
con
ten
t (%
DW
)
Treatment
0
100
200
300
400
500
600
700
800
VS F/2
Ch
loro
ph
yll a
an
d c
aro
ten
oid
co
nte
nt
(µg/
g D
W)
Treatment

185
An ANOVA single factor (p<0.05, Tukey’s and Fisher’s tests) demonstrated a statistical difference
between VS and F/2 in terms of PE and PC content, with VS>F/2 (Fig. 6.16). This was already
seen in the previous experiment (see section 6.4.1.2.3) and is in line with what recorded for the
protein content, where the highest concentration was reported for VS.
Figure 6.16 PE (dark grey bars) and PC (light grey bars) content (mg/g) for the cultures under different treatments. (n=3; error bar, 1 SD of mean)
6.4.3 The impact of nitrogen sources and N:P ratio on Osmundea pinnatifida growth and
biochemical composition
6.4.3.1 Weight and growth rate of Osmundea pinnatifida adult cultures grown in different nitrogen
sources
No statistical differences (single factor, p<0.05, Tukey’s, Fisher’s and Dunnett’s-F/2 as control-
tests) were reported for the different treatments in term of weight increase, with the nitrogen
sources used not having a direct effect on the weight increase of O. pinnatifida cultures (Fig. 6.17).
Figure 6.17 Weight measurements (WW) of cultures under different nitrogen sources: F/2= blue, Urea= orange, NH4Cl =grey and NH4NO3 =black bars (n=3; error bar, 1 SD of mean)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
VS F/2
PE
and
PC
co
nte
nt
(mg/
g D
W)
Treatment
0
1
2
3
4
5
6
7
Time 0 Time 1 Time 2 Time 3 Time 4 Time 5 Time 6 Time 7
wei
ght
(g W
W)
Week

186
Relatively daily growth rate measurements are reported in the Fig. 6.18, with no statistical difference
(single factor, p<0.05, Tukey’s, Fisher’s and Dunnett’s- F/2 as control- tests) reported in terms of
SGR for the different treatments, except for the 6th week were the treatment NH₄Cl was statistically
different from the control (F/2>NH₄Cl). The general pattern in all the cultures was an initial
increase in weight, followed by a slow decrease in the last 2 weeks of the experiment. However,
the NH₄Cl grown biomass decreased in weight in the 3rd week, followed by an increase in the 4th
and again a decrease in the 5th, 6th and 7th weeks. While the F/2 treatment mainly showed a positive
increase in biomass, except in the 2nd and 7th weeks (Fig. 6.18).
Figure 6.18 The SGR of cultures undergoing different treatment across the 7 weeks of experiment. F/2= blue, Urea= orange, NH4Cl =grey and NH4NO3 =black bars (n=3; error bar, 1 SD of mean)
6.4.3.2 Biochemical composition of Osmundea pinnatifida adult cultures grown in different
nitrogen sources
6.4.3.2.1 Protein and carbohydrate content
Statistical analyses (ANOVA, single factor p<0.05, Tukey’s, Fisher’s and Dunnett’s tests) did not
reveal any significant difference in terms of protein content %(DW) in the samples undergoing the
different treatments (Fig. 6.19). However, the highest percentage was reported for the only organic
source supplied (urea) and the lowest for NH₄Cl (14.97 and 13.55 %, respectively). No statistical
differences (p<0.05) were reported in terms of carbohydrates contents in cultures grown in the
different treatments (Fig. 6.19).
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
2.2
Time 1 Time 2 Time 3 Time 4 Time 5 Time 6 Time 7
Gro
wth
rat
e (%
day
⁻¹)
Week

187
Figure 6.19 Carbohydrate (dark grey bars) and protein (light grey bars) content expressed in percentage (DW) for the cultures under the different treatments applied. (n=3; error bar, 1 SD of mean)
6.4.3.2.2 Phycobiliproteins
Phycobiliproteins analyses are shown in Fig. 6.20 expressed in mg/g. ANOVA (single factor
p<0.05, Tukey’s, Fisher’s and Dunnett’s tests) revealed statistical differences for the PE
concentration between the F/2 (control) and the other treatments, with the lowest value reported
for NH₄Cl (0.12 mg/g). Statistical difference in PC content was observed between F/2 (control)
and the other treatments, with the lowest value reported for urea (0.07 mg/g).
Figure 6.20 PE (dark grey bars) and PC (light grey bars) content (mg/g) in samples undergoing different treatments. (n=3; error bar, 1 SD of mean)
0
5
10
15
20
25
30
35
40
F/2 Urea NH₄Cl NH₄NO₃
Car
bo
hyd
rate
s an
d p
rote
in c
on
ten
t (%
DW
)
Treatment
0
0.2
0.4
0.6
0.8
1
1.2
1.4
F/2 Urea NH₄Cl NH₄NO₃
PE
and
PC
co
nte
nt
(mg/
g D
W)
Treatment

188
6.5 Discussion
The impact of different media types on the cultivation of O. pinnatifida has not previously been
reported; previous cultivation studies only used VS medium, effluents from an IMTA system (Silva,
2015) or PES (Gonçalves, 2018). However, every seaweed is different in its requirements and
optimal growing cultivation, and it is a fundamental requirement to identify the best medium for
the cultivation of any selected species. Nutrients and specifically nitrogen source preference can be
determined in macroalgae by species-specific morphology, surface area, shore-line position, the
physiological capability and energetic requirement to uptake and assimilate certain nitrogen forms,
and nutritional or life history stage/age (Phillips and Hurd, 1994; Smit, 2002; Bracken and
Stachowicz, 2006). For example, young tissue has reported to have a higher uptake rates compared
to mature thalli/tissue (Wallentinus, 1984; Thomas et al., 1985; Harrison et al., 1986; Hurd and
Dring, 1990). Furthermore, besides growth rate and nutrient uptake capacity, biochemical
composition is also affected by nutrient regime, as reported by previous authors (Schiener et al.,
2015; Sode et al., 2013; Smit et al., 1997). This aspect can be particularly important for commercially
exploitable species, such as O. pinnatifida. Therefore, the effects of different media and nitrogen
sources on O. pinnatifida growth and biochemical composition were investigated in this Chapter.
In general, all the media tested were capable of sustaining the growth of O. pinnatifida cultures,
resulting in increased biomass and the control of epiphytes growth. The first two trials
demonstrated that there was a statistical difference between the treatments, and even if VS medium
has been considered more suitable for red seaweed cultivation, (Guiry and Cunningham, 1984), for
this particular species, F/2 seems to be more suitable, both for adult and juvenile plantlets. In all
cases, no epiphytes were detected for the entire length of the experiment, demonstrating that the
pre-treatment undertaken prior to cultivation was efficient in keeping the cultures clean. It also
does not affect the growth and development of the seaweeds. The cultures showed two different
patterns of growth: vertically, in elongation, with new tissue developing from the extremities, and
horizontally, with new shoots developing from the holdfast. This type of growth is also observed
in field specimens (Maggs and Hommersand, 1993). In the first trial, F/2 and VS both seemed to
perform better in terms of the growth measured than F/2 modified and as such they were the only
media types taken forward to be re-tested in the second experiment using younger specimens. A
statistical difference (p<0.05) between the two treatments, in term of growth, was confirmed with
the second trial, with F/2 performing better than VS (Fig. 6.11). In addition, adults and juveniles
seemed to have different patterns of growth in terms of weight increase as well as SGR. This
suggests that would be probably more suitable to start a culture in vitro with young material, thus
ensuring exponential growth.

189
These experiments confirmed that the morphology, age and nutritional status of material used
affect the biochemical profile of this species, as has previously been reported for other seaweeds
(Rosenberg and Ramus, 1984; Pedersen, 1994; Neori et al., 2004). In particular, the carbohydrate
concentration seems to follow the same trend for both adult and juvenile plants, with the highest
values reported for VS compared to F/2 in both cases. For the PE and PC, the situation is the
same, with the highest values reported for VS compared to F/2 in both cases. For the protein, Chl-
a and carotenoids content, the juvenile tissue contained the highest levels for all three of these
components in the VS treatment, while in the adult plants tested, the F/2 treatment resulted in the
highest values for the listed components. However, no statistical differences were observed
between F/2 and VS treatments with respect to the protein concentration in both the first and
second trial, as well as for the PE and PC content in the first trial for the different treatments tested.
Whereas, a statistical difference was recorded between the two media types for the Chl-a and
carotenoids contents only in cultivation of adults and for PE and PC content only in cultivation of
juvenile plantlets. These differences were probably due to the diverse age and therefore
typology/development of the material analysed (Lobban and Harrison, 1997; Pedersén, 1994).
Cultures grown in VS and F/2 modified followed the same trend in terms of carbohydrates, protein
and pigments concentration, differentiating from F/2. This might be due to the sources of N and
P employed in the VS and F/2 modified treatment, which differed from those used in the F/2
medium (see Table 6.1). In particular, higher concentrations of carbohydrates were reported for
VS and F/2 modified, compared to F/2, and vice versa for the protein. It has been demonstrated
experimentally that the tissue N content of macroalgal biomass increases with increasing
concentrations of N in the growth medium (Sode et al., 2013). Whilst under nitrogen limitation,
carbohydrates show the opposite trend to that of protein, phycobiliproteins and chlorophyll-a, with
an accumulation of polysaccharides (Smit et al., 1997). This has been reported previously for other
red algae (Chapman and Craigie, 1977; DeBoer et al., 1978; Bird et al., 1982; Rosenberg and Ramus,
1982a; Chopin et al., 1995; Lewis and Hanisak, 1996) and might explain the inverse trend for
carbohydrates and protein contents observed in this study.
Antioxidant analyses (FRAP) both on adults and juveniles showed the same results, with statistically
higher FRAP content of algae in F/2 modified and F/2 compared to VS in the first trial, and in
F/2 compared to VS in the second one. In the first trial, the statistically highest value was reported
for cultures cultivated with F/2 modified, followed by F/2 treatment and then VS. In the second
trial the highest value was observed for F/2 cultivated algae compared to those incubated in VS
medium. While for the juvenile plantlets this could be explained by the lower concentration of N
and P in F/2 compared to VS, as already reported for other seaweeds (Mellouk et al., 2017) and
specifically for red algae, for example Gracilaria spp. and Grateloupia filicina (Lapointe, 1985; Vergara

190
et al., 1995; Lewis and Hanisak, 1996; Athukorala et al., 2003a). For the adults this explanation does
not apply, since F/2 and F/2 modified cultivated algae both had statistically higher values for
antioxidant compounds compared to VS, but F/2 modified shares the same N and P sources and
concentration as VS.
The lower production of antioxidant compounds under VS cultivation in adults and juveniles of
O. pinnatifida was assumed to be mainly due to a shortage of microelements (such as Cu, Zn, SO₄,
etc.), since VS has a simpler trace elements’ composition compared to F/2. Uptake processes of
these metal ions are directly dependent on metabolic processes, and vary with changes in
temperature, light, and age of the seaweed (Sánchez-Rodríguez et al., 2001). As already reported
for other organism such as yeasts (Bhosale, 2004), cyanobacteria (Kobayashi et al., 1992; Tarko et
al., 2012), and plants (Bradáčová et al., 2016), trace elements play a vital role in the production of
antioxidant compounds, acting as enzymatic co-factors in antioxidative stress defence. Trace
elements are generally linked to the composition/functionality of several antioxidant compounds
identified in seaweed, phenols, sulfated polysaccharides, carotenoid pigments, sterols, catechins,
proteins, and vitamins, (Rupérez et al., 2002; Toyosaki and Iwabuchi, 2009). For example,
superoxide dismutases in eukaryotic algae have Mn or Fe, or some combination of Fe, Mn, and
Cu + Zn (Wolfe-Simon et al., 2005). Hence, the richer trace elements composition of F/2, and
consequently F/2 modified, compared to VS, might explain the difference in antioxidant content
of the adult and juvenile cultures grown in the different media types. Considering how important
the production of antioxidant compounds is in this species, particularly with respect to the
possibility of commercial exploitation in pharmaceutical and nutraceutical applications (Rodrigues
et al., 2015, 2016, 2019; Küpper et al., 2014; Silva et al., 2018), there is a clear case for using F/2,
or F/2 modified, instead of VS for the commercial cultivation of O. pinnatifida. As Kim and Yarish
(2014) have stated, economically F/2 medium has the advantage over VS of being cheaper to use,
making the media of choice convenient at a commercial scale.
It has previously been reported that red algae have a preference for NH₄ sources (DeBoer et al.,
1978; Hanisak, 1990; Haglund and Pedersén, 1988; Lobban and Harrison, 1997; Phillips and Hurd,
2004; Smit, 2002). This is probably due to the lower levels of energy required to assimilate this
nutrient (Lobban and Harrison, 1997). While NH₄ uptake often occurs by passive diffusion, nitrate
uptake typically shows saturation kinetics and is an energy-dependent process that can be described
by the Michaelis–Menten equation: 𝑉 =𝑉𝑚𝑎𝑥 (𝑆)
𝐾𝑚+𝑆 (DeBoer, 1981). Nitrogen in the form of urea,
represents an excellent source for certain species such as kelps, but it can reduce the growth of
other seaweeds (DeBoer, 1981; Navarro-Angulo and Robledo, 1999). For some species, for
example Palmaria palmata, there is a preference for N sources dependent on the N-status of the

191
fronds, absorption rate (Hanisak, 1990), with different absorption rates for different sources
directly linked to amino acid production, NH₄ (Lobban and Harrison, 1994), and on nutrient
concentrations (Martínez and Rico, 2004; Corey et al., 2013; Grote, 2016). In this study the
preference for NH₄ sources was not evident, and the growth rate of the species was not affected
either by NH₄ or NO₃. Similar experiments have been conducted on other species of red algae
(Starr and Zeikus, 1993; Navarro and Robledo, 1999) resulting in a comparable set of observations.
No significant difference was found for relative growth rate and biomass increase in this study, or
in a previous study on a different species (Navarro and Robledo, 1999). The growth rate (%day⁻¹)
ranged from 0.03 to 0.43 for F/2; from 0.001 to 0.16 for urea; from 0.05 to 0.20 for NH₄Cl and
from 0.023 to 0.8 for NH₄NO₃, with an evident decrease towards the end of the experiments for
all the treatments investigated.
The results obtained also differ from previous studies on other red seaweed in relation to the
protein content. For example, in G. cornea (Navarro and Robledo, 1999), the highest protein content
was measured when grown on either NH₄Cl or NH₄NO₃ and was lower when cultivated in either
NaNO₃ or urea. Similar results have been reported for P. palmata, with increased protein content
under NH₄ compared to NO₃ (Morgan and Simpson, 1981b; Grote, 2016). Whilst in this case the
opposite situation was true, with urea having the highest value, even if not statistically significant,
followed by NaNO₃ (F/2), NH₄NO₃ and then NH₄Cl. For the carbohydrate analyses there were
no differences between the treatments tested, showing that, according to this study, the source of
nitrogen does not influence their concentration in O. pinnatifida cultures, that might instead depend
on the P available (Hopkins and Hüner, 2009).
Phycobiliproteins represent an important mode of nitrogen storage in red algae (Lapointe, 1981;
Ryther et al., 1981), and have been demonstrated to respond the fastest to nitrogen availability in
the medium (Vergara and Niell, 1993). Furthermore, additions of N normally result in an increase
of N-containing photosynthetic pigments (Dawes, 1995; Vergara et al., 1995). Presumably because
of N limitation, red algae frequently become green during the summer (bleaching), because of
reductions in phycobiliprotein contents relative to chlorophyll-a (Penniman and Mathieson, 1987).
Both PE and PC production were affected by the different nitrogen sources and media types. For
PE, the highest production was obtained with F/2, followed by NH₄NO₃, urea and lastly NH₄Cl.
For PC, the highest content was observed in F/2 cultivated algae, followed by NH₄NO₃, NH₄Cl
and lastly urea. These results differ from those reported for another red species, G. cornea, were the
highest concentration of PE was found with NH₄Cl as the nitrogen source and PC highest with
NH₄NO₃ (Navarro and Robledo, 1999). But for both G. Cornea and O. pinnatifida in cultures
supplied with urea, the phycobiliproteins content was usually lower. This indicates a low affinity of

192
O. pinnatifida to absorb urea, which is an adaptation of certain other intertidal seaweeds linked to
their distribution (Phillips and Hurd, 2004).
6.6 Conclusions
F/2 medium is more appropriate for cultivating O. pinnatifida compared to VS. F/2 was
able to guarantee growth for the duration of the experiment, ensuring a high-quality
product. The algae were thick in texture, dark in colour, with new branches, without
contaminants and with a satisfying biochemical profile, especially in terms of antioxidant
concentration. Furthermore, F/2 is a simpler medium compared to VS and requires a lower
volume of stocks, and is therefore more economical to formulate;
When targeting the production of antioxidant compounds, F/2 modified and F/2 might
be employed instead of VS since these resulted in a statistically higher antioxidant content,
with F/2 modified resulting in the statistically highest content;
Phycobiliproteins and carbohydrates production is increased in cultivation in VS medium
while the antioxidant compounds production seems to be stimulated by F/2 and modified
F/2 media;
The growth rate of the species seems to do not be affected either by the NH₄ or NO₃
sources. On the other hand, the biochemical composition of the cultures is affected by the
treatments applied; in particular, urea treatment seemed to statistically stimulate the
production of protein and NH₄NO₃ the carbohydrates content;
The stage of development of the cultures has an impact on both the growth and the
biochemical composition of the species. Therefore, younger plantlets should be preferred
when starting a cultivation of the species investigated.

193

194
Chapter 7 Cultivation of Osmundea pinnatifida in the Algem® Photo-
bioreactor System
7.1. Introduction
Cultivation of Osmundea pinnatifida in flasks and/or tanks has yet to be established beyond the lab
scale, with only two significant studies to date having been undertaken (Silva, 2015; Gonçalves,
2018). However, establishment of a methodology for the supply and production of this red seaweed
has great potential considering that it can be exploited either as food, or as a source of biological
active compounds (such as antioxidant, antiviral, antileishmanial, antibacterial and anticancer, Bano
et al., 1986; Silva et al., 2018; Campos et al., 2019 Rodrigues et al., 2019). As already described in
Chapters 3 and 4, this species has been largely investigated for its biochemical composition since
the late eighties (Faulkner, 1986; Bittner et al., 1985; Bano et al., 1988; Norte et al., 1989a).
Secondary metabolites are produced naturally by red seaweeds, acting as grazing deterrents (Brito
et al., 2002; Flodin et al., 1999; Iliopoulou et al., 2002a, b; Suzuki et al., 2002), as well as protectants
against high irradiance and biotic stress in general (Li et al., 2011; Thomas and Kim, 2011). An
insight into how the production of these compounds is regulated in O. pinnatifida would enable the
manipulation of the cultivation conditions in order to stimulate and control their productivity. This
would be an important step forward for the commercial exploitation of this species, considering
that obtaining large amounts of biomass for large volume products is unlikely to be feasible with
such a slow-growing species. Nevertheless, low-volume high-value applications would instead
represent the perfect match for a commercial utilisation of this alga. With this scope in mind, the
Algem® unit, a lab scale photo-bioreactor (PBR) used for microalgae cultivation, has been utilised
in this study to further examine the impact of cultivation parameters on growth and biochemical
profile of the species investigated.
Cultivation systems for algae range from open to closed systems, such as photo-bioreactors, the
latter been used mainly for microalgal cultivation (Holdt et al., 2013). Photo-bioreactors (ranging
from laboratory to industrial scale) have recently attracted interest because they offer a controlled
environment, for conditions such as pH, CO₂, temperature and light, when compared to, for
example, open pond systems (Singh and Sharma, 2012). Photo-bioreactor systems (PBR) have also
been shown to increase the production of biomass as well as control contaminations in microalgae
cultivation (Ugwu et al., 2008). To date, a range of PBR types have been used in production of
various algal products (mainly from microalgae) at a laboratory scale (Ugwu et al., 2008).
Small-scale trials in the photo bioreactor Algem® were previously conducted with microalgae for
comparison of growth and production of modified strains (e.g. P. tricornutum, D’Adamo et al., 2018),

195
cultivation of Cyanobacteria (e.g. Synechocystis sp., Lea-Smith et al., 2013), Dunaliella spp. for
potential commercial production of carotenoids (Xu et al., 2016, 2018), strains of diatoms (Matthijs
et al., 2018), C. reinhardtii strains (Stoffels et al., 2017; Young and Purton, 2018), microalgae for
nutrient removal (Whitton et al., 2018), and Tetraselmis sp. (Pereira et al., 2019). However, this is the
first time that this type of bioreactor has been employed for macroalgae cultivation.
The Algem® consists of two chambers per unit (Fig. 7.1 A), with one conical flask of one liter in
each chamber, which can be linked together. The flasks are closed with hermetic lids with inlets
for filtered gas supply and the chambers can be independently regulated for temperature, light
levels and shaker speed (Fig.7.2 B).
Figure 7.1 A) Algem®, photobioreactor used for the experiment; B) View of the open temperature controlled chamber from the top with a 1L conical flask inside; C) Schematic of the PBR system: I= influent, E = effluent, PP = peristaltic pump, TCC = temperature controlled chamber, MP= micro processor, TS= Temperature sensor, H= heating system, C=cooling system, V=valve box, CH= chiller, GMP=Gimbal mixing system. Modified from Whitton, 2006
Light is provided by LEDs disposed on the bottom of the chamber, blue, red or white, with both
the intensity and light cycle fully controllable. Each chamber is gently rotated by a gimbal mixing
system. A temperature-controlled environment is created in the chamber where temperature is
regulated thanks to a heating and cooling system, and a temperature sensor in each chamber (Fig.
7.1 C). The software controlling the system can also generate geographical modelling of monthly

196
light and temperature profiles according to the coordinates chosen. The system provides data about
the pH and the growth/culture density (expressed as absorbance) of the cultures. It is a flexible
and innovative PBR designed for the cultivation of microalgae species. This chapter describes the
growth of O. pinnatifida in this system using a range of temperature and light profiles relevant to
the conditions for a particular area in Scotland.
7.2 Aims
• To investigate the possibility of cultivating a seaweed in the Algem® system;
• To investigate the capability of the system in guaranteeing a growth of the cultures and
respecting the morphology and biochemical composition of the species investigated;
• To determine the capacity of the system to replicate the environmental conditions of the
set coordinates;
• To investigate the possibility of increasing the production of antioxidant compounds in the
species examined through cultivation in the photo bioreactor system.
7.3 Materials and methods
7.3.1 Comparison of summer and winter conditions on the vegetative growth of O. pinnatifida
Biomass samples were collected in November 2017 at low tide from Seil Island (56.3000° N,
5.6500° W), kept in Cool-boxes during transport and brought to the lab where they were manually
cleaned and treated with a 0.5 % KI solution in FSW, in order to remove epiphytes and grazers, as
described in Chapter 5. They were then acclimated prior to the trials for one week at 10 °C, 12:12
light-dark cycle at 80 µmol/m²/s.
The coordinates selected for the experiment were 56.4547° N, 5.4374° W (referring to the area
used for sampling during this project, Dunstaffnage Bay). Two different conditions were simulated
in the Algem®: one representing winter conditions and the other spring-summer conditions. The
winter conditions replicated the light quality and quantity for December in this area of Scotland,
while the spring-summer condition refers to May. The temperature was set as a stable parameter
at 10 °C. In the summer condition the light (expressed in µmol/m²/s) was as described: maximum
red light was 166, average 68; blue light: 0; maximum white light 1051, average 378; total light:
maximum 1081; average 445, for a total of 17 hours of light, with levels ramping up and down
similar to that observed naturally. Aeration was provided via an aquarium air pump filtered through
Sartorius® air filters (0.2 µm). In the winter conditions the light levels (expressed in µmol/m²/s)
were as follow: maximum red light 30, average 5; blue light: 0; maximum white light: 167, average
31; total light: maximum 197; average 36, for a total of 6.50 hours of light. O. pinnatifida specimens
were blotted dried, five grams was weighed and placed into 1 l flasks with 500 ml of sterilized F/2.

197
Each condition tested was replicated in triplicate samples. During the experiment the pH, light and
growth of the cultures were monitored. QY of the cultures was assessed using an AquaPen-P (PSI),
according to manufacturer instructions, at the beginning of the experiment and weekly. The starting
QY average for the samples was 0.50 (Fv/Fm). The samples were blotted weighed weekly and the
F/2 medium changed weekly. The experiment ran for a total of four weeks. SGR was assessed
using the formula according to Evans (1972) where LN is natural logarithm, Wt weight at final
time, W0 initial weight and t time:
SGR= 𝐿𝑁(𝑊𝑡)−𝐿𝑁(𝑊0)×100
𝑡
Biochemical composition of the cultures (including pigments and antioxidant properties) was
assessed using the methods described in Chapter 3. Statistical analyses were conducted using
Minitab18, performing ANOVA (single factor, p<0.05, with post hoc tests -Tukey’s, Fisher’s and
Dunnett’s).
7.3.2 Comparison of winter conditions with a daily cycle temperature (DCT) and constant
temperature (CT) on the vegetative growth of O. pinnatifida
The experiment was repeated in a second trial using new shoots from material collected in
November 2017 from Seil, as described previously. Temperature was considered in this case and a
comparison was made between a constant temperature (CT) and a daily cycle temperature (DCT)
in order to understand if this parameter played any controlling role in the production of pigments
as well as antioxidant compounds for this species. The month selected for the coordinates and for
the variation of the temperature in relation to the DCT was December (as already considered in
the section 7.3.1). Only one winter condition was tested, with one set of triplicate flasks being
incubated at a constant temperature (10 °C) and the other set of flasks at the environmental
temperature as determined by the Algem® system according to the chosen month and hours
(varying from 0 to 10 °C), which reflected the variation of the seawater temperature, rather than
the air temperature in that specific location. The aim was to determine any significant differences
in weight increase and QY efficiency related to the temperature interval, as well as variation in the
biochemical composition. Five grams was weighed and placed into 1 l flasks with 500 ml of
sterilized F/2. The light levels employed were set to the environmental conditions for December
as described in the previous experiment. The light for both conditions was specifically: maximum
red light 30, average 5; blue light: 0; maximum white light: 167, average 31; total light: maximum
197; average 36; for a total of 6.50 hours of light (expressed in µmol/m²/s). Again, the cultures
were monitored as described in 7.3.1. The experiment ran for a total of four weeks with SGR
assessed using the formula (see section 7.3.1) according to Evans (1972).

198
At the end of the four weeks the biomass was harvested for biochemical analysis, including
chlorophyll-a, carotenoids and phycobiliproteins, using the methods described in Chapter 3.
Statistical analyses were conducted with Minitab 18, performing ANOVA (single factor, p<0.05,
with post hoc tests -Tukey’s, Fisher’s and Dunnett’s).
7.3.3 Comparison of autumn and spring conditions on the vegetative growth of O. pinnatifida
In this third trial, successive autumn and spring conditions were compared against a control.
Samples were collected and treated as already described in section 7.3.1. For the autumn
experiment, September was chosen as the month of reference whilst for the spring condition
March, on the basis of the results obtained in the seasonal study undertaken (see Chapter 3). The
coordinates set for this experiment were 56.4547° N, 5.4374° W (referring to the area used for
sampling during this project, Dunstaffnage bay). The control was set, for both runs, at 10°C and a
12:12 photoperiod with a constant light intensity set at 100 µmol/m²/sec. The light for the
September trial was maximum red light: 99; average red light 31.37; blue light: 0; maximum white
light: 545, average white light: 177; total light: maximum 644, average 208. The light for March was
maximum red light 90; average red light: 27; blue light: 0; maximum white light: 496; average white
light: 151; total light: maximum 586, average 178 (expressed in µmol/m²/s). Temperature followed
the seasonal profiles for the select area replicated by the model and mostly referred to the sea water
temperature rather than air temperature. Three replicates were set for each condition (either
autumn/spring or control). The biomass, 10 g (WW), was cultivated in 500 ml of F/2; pH and
growth (expressed in absorbance) was monitored during the lifetime of the experiment. The
medium was changed weekly, samples weighted and analysed for QY using an Aqua-Pen. The
length of the experiment was of five weeks for each set of conditions (September vs Control and
March vs Control). SGR was assessed using the formula (see section 7.3.1) according to Evans
(1972).
Biochemical composition of the cultures undergoing the cultivation trials (including pigments, and
antioxidant properties for the first two trials; pigments, antioxidant properties, carbohydrates and
protein for the last trial) was measured using the methodologies already described in Chapter 3.
Statistical analyses were conducted with Minitab 18, performing ANOVA (single factor, p<0.05,
with post hoc tests-Tukey’s, Fisher’s and Dunnett’s).

199
7.4 Results
7.4.1 Comparison of summer and winter conditions on the vegetative growth of O. pinnatifida
May and December were chosen for this trial because, from a previous seasonal study (described
in Chapter 3), the field samples related to them showed characteristic profiles clearly diversified in
term of biochemical composition.
Furthermore, even if O. pinnatifida is a perennial species, its abundance as well as quality changes
along the year, reflecting also in its biochemical composition. The two months chosen were selected
to explore the effects of two extreme conditions for this species.
There was no statistical difference observed (p<0.05) in term of increase of weight between the
two sets of conditions (Fig. 7.4). The growth levels from the Algem® system (expressed as
absorbance) varied from 0.003 to 1.82 in the summer condition, whilst for the winter conditions
this was recorded by the system to be -0.48 and 3.80. The average pH was 8.9 for the summer
simulation and 8.52 for the winter. QY was measured weekly and no statistical difference was noted
between the two conditions (Fig. 7.3).
Figure 7.2 A) Samples under summer conditions, B) samples under winter conditions, after 2 weeks in the PBR. (bar=1 cm)
A difference in the colour of the cultures depending on the conditions applied was recorded (Fig.
7.2 A and B). SGR gave a positive value for the summer conditions in the second and third week,
whilst the cultures under winter condition showed a slow decrease in their SGR (Fig. 7.4). Growth
rate (%day⁻¹) varied from -06 to 0.4 in the summer condition and from -0.1 to -0.4 in the winter
condition

200
Figure 7.3 QY measurements of summer (dark grey bars) vs winter (light grey bars) cultures in the Algem® unit (n=3; error bar, 1 SD of mean)
Figure 7.4 SGR for cultures undergoing summer (dark grey bars) and winter (light grey bars)
conditions in the PBR (n=3; error bar, 1 SD of mean)
7.4.1.1 Antioxidant analyses
The results obtained from the TPC assay were higher compared to the TPC conducted on seasonal
samples collected from Dunstaffnage beach (56.4547° N, 5.4374° W), for those months (see
Chapter 3). An average of 156.29 µg/mg (SD= 0.13) was reported for the summer condition and
124.25 µg/mg (SD=0.09) for the winter condition. However, in the wild harvested material the
highest levels for the winter were measured at 92.45 µg/mg (SD=0.05) in December and 127.47
µg/mg (SD=0.007) in July, while in May the figure was 92.73 µg/mg (SD=0.05). ANOVA (single
factor, p<0.05) showed no statistical differences between the summer and winter samples
cultivated in the Algem® unit in relation to antioxidant compounds (Tukey’s and Fisher’s
comparison tests). However, ANOVA (single factor, p<0.05) indicated statistical difference
between the winter sample from the Algem® and wild harvested winter sample, as well as for the
summer samples, with the Algem® samples being significantly higher in values compared to the
field samples (Fig. 7.5).
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
Time 0 Time 1 Time 2 Time 3 Time 4P
AM
(Q
Y)
Week
-1
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
SGR 1 SGR 2 SGR 3 SGR 4
Gro
wth
rate
(%
day
⁻¹)
Week

201
Figure 7.5 TPC analyses: comparison between field and cultivated samples in relation to summer and winter conditions (n=3; error bar, 1 SD of mean)
The statistical analyses of FRAP values showed a statistical difference between the winter sample
cultivated in the Algem® and the field samples. The same effect was observed in the summer
samples, with the Algem® samples being significantly higher in the levels measured compared to
the wild harvested samples. An average of 4.98 µM/mg (SD=0.11) was measured for the summer
condition and 3.34 µM/mg (SD=0.04) for the winter one in the Algem® units. However, in the
seasonal samples collected from the wild the highest value measured for the summer was for July
with 2.04 µM/mg (SD=0.06) and 0.84 µM/mg (SD=0.10) for February for the winter. While for
the December seasonal sample it was 0.42 µM/mg (SD=0.02) and for May 0.95 µM/mg (SD=0.01)
(Fig. 7.6). Furthermore, a statistical difference was reported also between the summer and winter
samples cultivated in the Algem®, as already reported for the wild samples.
Figure 7.6 FRAP analyses: comparison between field and cultivated samples in relation to summer and winter conditions (n=3; error bar, 1 SD of mean)
0
20
40
60
80
100
120
140
160
180
Summer Algem Summer Wild Winter Algem Winter Wild
Tota
l ph
eno
l co
nte
nt
(µg/
mg
DW
)
Samples
0
1
2
3
4
5
6
Summer Algem Summer Wild Winter Algem Winter Wild
FRA
P
(μM
FeSO
₄eq
/mg
DW
)
Samples

202
7.4.1.2 Pigment analyses
An ANOVA (single factor, p<0.05, Tukey’s and Fisher’s comparisons tests) indicated no statistical
difference between the concentration of Chl-a in the samples undergoing the summer condition
and those under winter conditions (Fig 7.7). The wild samples analysed for the seasonal variation
in biochemical composition of O. pinnatifida showed an average (n=3) of 669.6 µg/g for May and
1194.2 (n=3) for December (the months set as reference in the PBR). The values obtained for the
samples under winter conditions in the Algem® are similar to those of the wild sample collected
in December (1097.07 µg/g), while the value for the summer condition are slightly higher than
those for the natural harvested samples (938.6 µg/g). However, no statistical differences were
observed (ANOVA, single factor, p<0.05 Minitab 18) (Fig. 7.7).
Figure 7.7 Comparison of Chl-a (dark grey bars) and Carotenoids (light grey bars) (µg/g) contents in biomass grown either in the Algem® for four weeks with wild harvested biomass for summer and winter conditions, for the same months of references (n=3; error bar, 1 SD of mean)
There was no statistical difference (p<0.05) between the concentration of carotenoids in the
samples grown under the summer condition and those under winter conditions in the Algem®
system. The wild samples analysed for the seasonal variation for carotenoid composition of O.
pinnatifida showed an average (n=3) of 369.3 µg/g for May and 318.6 µg/g (n=3) for December.
No statistical differences were reported among the treatments and the wild samples (p<0.05).
Furthermore, the Algem® cultures showed the same pattern as the seasonal sample in terms of
chlorophyll-a and carotenoids concentration, with a higher concentration of carotenoids during the
summer as compared to the winter (Fig. 7.7).
There was no statistical difference (p<0.05) between the concentration of PE in the samples
incubated under the summer condition and those under winter conditions cultivated in the PBR
(Fig. 7.8). The wild samples analysed for the seasonal variation in biochemical composition of O.
pinnatifida showed an average (n=3) of 0.19 for May and 3.10 for December. A statistical difference
0
200
400
600
800
1000
1200
1400
1600
Summer Algem Summer Wild Winter Algem Winter Wild
Ch
loro
ph
yll a
an
d c
aro
ten
oid
co
nte
nt
(µg/
g D
W)
Samples

203
between the winter seasonal sample and the winter and summer Algem® samples was reported
(p<0.05).
Figure 7.8 Comparison of PE (dark grey bars) and PC (light grey bars) (mg/g) contents in biomass grown in the Algem® for four weeks with wild harvested biomass (n=3; error bar, 1 SD of mean)
There was no statistical difference (p<0.05) between the concentration of PC in the samples
incubated under the summer condition and those under winter conditions cultivated in the PBR.
The wild samples analysed for the seasonal variation in biochemical composition of O. pinnatifida
showed an average (n=3) of 0.23 for May and 0.72 for December. A statistical difference between
the winter seasonal sample and the winter and summer Algem samples was observed (p<0.05) (Fig.
7.8) with the seasonal sample having a higher value than the Algem cultivated biomass.
7.4.2 Comparison of winter conditions with a daily cycle temperature (DCT) and a constant
temperature (CT) on the vegetative tissue growth of O. pinnatifida
A minimal increase in the cultures weight or a decrease was recorded and no statistical differences
were noted after 7 days, statistical differences after 14 days, no statistical difference after 21 or after
28 days (p<0.05) (Fig. 7.10). However, new shoots were observed on the specimens and the QY
values increased during the life time of the experiment (Fig. 7.9). However, there were no statistical
differences (p<0.05) observed between the two different conditions, in relation to the temperature,
regarding the QY response (Fig. 7.9). For the second and third weeks, a positive SGR was noted
for both the conditions tested, but with no statistical differences (p<0.05) (Fig. 7.10). Growth rate
(%day⁻¹) varied from -0.02 to 1.6 for DCT conditions, and from -0.14 to 0.30 for the CT
conditions.
0
0.5
1
1.5
2
2.5
3
3.5
4
Summer Algem Summer Wild Winter Algem Winter Wild
PE
and
PC
co
nte
nt
(mg/
g D
W)
Samples

204
Figure 7.9 QY average measurements of cultures grown under winter conditions but two different temperatures, daily cycle temperature (dark grey bars) and constant temperature (light grey bars) (n=3; error bar, 1 SD of mean)
Figure 7.10 SGR for samples under CT (light grey bars) and DCT (dark grey bars) temperature in the Algem® PBR (n=3; error bar, 1 SD of mean)
7.4.2.1 Pigment analyses
For the pigment analyses of the second Algem® experiment, there was no statistical difference
(p<0.05) between the concentration of Chl-a in the samples undergoing the DCT condition and
those under CT (Fig. 7.11).
0
0.1
0.2
0.3
0.4
0.5
0.6
Time 0 Time 1 Time 2 Time 3 Time 4
PA
M (
QY)
Week
-2.5
-2
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
SGR 1 SGR 2 SGR 3 SGR 4
Gro
wth
rat
e (%
day
¯¹)
Week

205
Figure 7.11 Comparison of Chl-a (dark grey bars) and Carotenoids (light grey bars) (µg/g) contents in O. pinnatifida between samples cultivated at CT, DCT and wild winter biomass for the same month of reference (n=3; error bar, 1 SD of mean)
There was no statistical difference (p<0.05) in the concentration of carotenoids between the
samples collected from the wild in December and those undergoing both treatments (DCT and
CT) in the Algem® (Fig. 7.11).
There was a statistical difference (p<0.05) in the concentration of PE between the samples
collected from the wild in December and those grown in the Algem®, with a higher concentration
observed in samples collected from the wild (Fig. 7.12).
Figure 7.12 Comparison of PE (dark grey bars) and PC (light grey bars) (mg/g) contents in O. pinnatifida between samples cultivated at CT, DCT and wild winter biomass for the same month of reference (n=3; error bar, 1 SD of mean)
There was a statistical difference (p<0.05) in the concentration of PC between the samples
collected from the wild in December and those cultivated under both conditions in the PBR, with
a higher concentration observed in those collected from the wild (Fig. 7.12).
0
200
400
600
800
1000
1200
1400
DCT CT Wild Winter
Ch
loro
ph
yll a
an
d c
aro
ten
oid
s co
nte
nt
(µg/
g D
W)
Conditions
0
0.5
1
1.5
2
2.5
3
3.5
4
DCT CT Wild Winter
PE
and
PC
co
nte
nt
(mg/
g D
W)
Conditions

206
7.4.3 Comparison of autumn and spring conditions on the vegetative growth of O. pinnatifida
For this experiment, which compared spring with autumn conditions, a control was performed for
each condition. Samples cultivated in the PBR under the three conditions (spring (A), autumn (B)
and control (C)) are showed in Fig. 7.13. Spring and autumn conditions were chosen in order to
fulfil the study of the variation of the biochemical composition across the seasons of the species
investigated. The length of the experiment was extended for a further week in order to understand
if the trend of the growth would have changed or remained constant with the time. The average
pH was reported at 8.4 for the control, 8.25 for the spring simulation and 8.43 for the autumn one.
The maximum growth (absorbance) reported for the spring simulation was 1.32 (average), the
minimum -0.43 and the maximum 2.62; the growth reported for the autumn simulation was 1.74
(average), 0.37 (minimum) and 2.99 (minimum), while those of the control were 1.62 (average),
0.43 (minimum) and 2.56 (maximum).
Figure 7.13 Cultures cultivated in the Algem® PBR after 3 weeks of cultivation for A) March conditions, B) September conditions and C) Control (bar=1cm)
In the spring experiment, a control vs March as the reference month, indicated no statistical
difference (p<0.05) in terms of growth (g) (Fig. 7.14). Furthermore, the cultures looked healthy
and new growth was visible (Fig. 7.13 A).
The SGR was measured weekly for the control and the reference month. It was positive for the
length of the experiment, in contrast to what observed in the previous experiments, which

207
demonstrated no statistical difference between the two conditions. The exception to this was for
the second week of the trial (p<0.05).
No statistical differences were observed in terms of growth (g) between the control and the autumn
reference month, September, (p<0.05). The SGR, except for the first week, was always positive
with no statistical differences between the two conditions investigated (Fig. 7.15). Growth rate
(%day⁻¹) varied from 0.02 to 0.5 in March and from 0.05 to 0.48 in the Control of March.
Figure 7.14 A comparison of growth, as SGR, for Control (dark grey bars) vs March (light grey bars) conditions over a 5-weeks period (n=3; error bar, 1 SD of mean)
Growth rate (%day⁻¹) varied from -0.3 to 0.40 in September and from -0.4 to 0.31 in the Control
of September.
Figure 7.15 A comparison of growth, described as SGR, for Control (dark grey bars) vs September (light grey bars) conditions over a 5-weeks period (n=3; error bar, 1 SD of mean)
When compared, there was no statistical difference in the growth (g) between the reference
months, March and September, (p<0.05) (Fig. 7.16). However, for the PAM assessment there was
a statistical difference between conditions for the two months tested in the first, fourth and fifth
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
SGR 1 SGR 2 SGR 3 SGR 4 SGR 5
Gro
wth
rat
e (%
day
¯¹)
Week
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
SGR 1 SGR 2 SGR 3 SGR 4 SGR 5
Gro
wth
rat
e (%
day
¯¹)
Week

208
weeks (September > March) (Fig. 7.17). There was no statistical difference observed between the
controls from the two experiments (p<0.05).
Figure 7.16 Comparison of weekly growth (SGR) between the two conditions in the Algem, September (light grey bars) vs March (dark grey bars), over a 5-weeks period (n=3; error bar, 1 SD of mean)
Figure 7.17 PAM measurement in reference to the QY of cultures undergoing the two different treatments and the controls in the Algem set at Spring and Autumn conditions. Control= black, Algem Spring= light grey and Algem Autumn =dark grey bars (n=3; error bar, 1 SD of mean)
7.4.3.1 Carbohydrates
In these experiments, carbohydrates and proteins were also analysed, together with pigments and
antioxidant compounds. This was performed in order to have a better understanding of the impact
that the PBR has on the biochemical composition of O. pinnatifida. Results for percentage of
carbohydrates are reported in Fig. 7.18 and for the proteins in Fig. 7.19. There was no statistical
difference (p<0.05) in terms of carbohydrates content between September and March, with highest
values reported for September and slightly lower for March. This is similar to what already observed
in the seasonal samples (see Chapter 3). Furthermore, no statistical difference was noted between
-1.2
-1
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
SGR 1 SGR 2 SGR 3 SGR 4 SGR 5G
row
th r
ate
(%d
ay¯¹)
Week
0
0.1
0.2
0.3
0.4
0.5
0.6
week 0 week 1 week 2 week 3 week 4 week 5
PA
M (
QY)
Week

209
the reference months and the control (p<0.05). However, a statistical difference was observed
between the wild samples and the one cultured in the Algem® unit (p<0.05) (Fig. 7.18).
Figure 7.18 Carbohydrate content expressed in percentage of samples undergoing the two different experiments and the control (n=3; error bar, 1 SD of mean)
7.4.3.2 Proteins
No statistical difference in the protein content was observed, with the highest value recorded for
the Control run together with the March condition and the lowest for September (p<0.05). The
results reported for the protein content are similar to those observed for the same months in the
seasonal samples (see Chapter 3). No statistical difference was noted between the reference months
and the control (Fig. 7.19). However, statistical differences between the wild sample of September
and all the other samples was observed (p<0.05) (Fig. 7.19).
Figure 7.19 % DW of protein in samples undergoing the two different experiments (including controls and wild samples for September and March) (n=3; error bar, 1 SD of mean)
0
5
10
15
20
25
30
35
40
Algem Control Algem Spring Algem Autumn Wild Spring Wild Autumn
Car
bo
hyd
rate
s co
nte
nt
(% D
W)
Samples
0
5
10
15
20
25
30
35
Algem Control Algem Spring Algem Autumn Wild Spring Wild Autumn
Pro
tein
co
nte
nt
(% D
W)
Samples

210
7.4.3.3 Pigments
No statistical difference (p<0.05) was observed between March and September for Chl-a content.
However, a statistical difference was reported for the carotenoids content between the two
references months, with a highest value recorded for September, which is different to what was
observed in the seasonal field samples. A statistical difference (p<0.05) was also observed between
the control and March for the Chl-a content, with the highest value reported for the control (Fig.
7.20).
Figure 7.20 Comparison of Chl-a (dark grey bars) and carotenoid (light grey bars) content in samples under Spring, Autumn and Control conditions in the PBR and wild samples for the same reference months (n=3; error bar, 1 SD of mean)
Figure 7.21 Comparison of PE (grey bars) and PC (light grey bars) content in samples under Spring, Autumn and Control conditions in the PBR and wild samples for the same reference months (n=3; error bar, 1 SD of mean)
No statistical differences were observed among the treatments for the PE content; however, a
statistical difference was noted for the PC content between the two reference months, with the
highest value reported in September, which is different to what was observed in the seasonal field
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Algem Control Algem Spring Algem Autumn Wild Spring Wild Autumn
Ch
loro
ph
yll a
an
d c
aro
ten
oid
co
nte
nt
(µg/
g D
W)
Samples
0
0.5
1
1.5
2
2.5
Algem Control Algem Spring Algem Autumn Wild Spring Wild Autumn
PE
and
PC
co
nte
nt
(mg/
g D
W)
Samples

211
samples. A statistical difference was also reported for the PC content between the control and
March, with a highest value reported for the control (p<0.05) (Fig. 7.21).
7.4.3.4 Antioxidant analyses
The highest value for the FRAP assay was observed in September, which, together with March, are
statistically different from the control and the seasonal samples (Fig. 7.22). However, no differences
were noted between September and March samples. This is similar to what already reported for the
seasonal samples (see Chapter 3), and is a similar response observed in the TPC analyses, which
also demonstrated a statistical difference between both the controls and the reference months
(September and March).
Figure 7.22 FRAP (antioxidant power) of cultures undergoing the two different treatments compared to the wild samples for the same months (n=3; error bar, 1 SD of mean)
Furthermore, no statistical differences between the two reference months were recorded for the
TPC content (Fig.7.22). However, both the cultured samples had statistically higher antioxidant
contents compared to the wild samples for the same reference months (p<0.05) (Figs 7.22 and
7.23).
0
1
2
3
4
5
6
Algem Control Algem Spring Algem Autumn Wild Spring Wild Autumn
FRA
P
(µM
FeS
O₄e
q/m
g D
W)
Samples

212
Figure 7.23 TPC content (antioxidant power) of cultures undergoing the two different treatments compared to the wild samples for the same months (n=3; error bar, 1 SD of mean)
7.5 Discussion
The Algem® system is currently marketed as a PBR for microalgal cultivation with the possibility
to reproduce, in a controlled situation, environmental parameters of a specific area. This represents
the possibility to determine the impact that environmental factors have on both biomass
productivity and biochemical composition. Previous researches on the cultivation of microalgae in
the Algem® have mainly used the system as a controlled environment for cultivation trials, without
focusing on manipulation of biochemical profiles of the species been investigated. Nevertheless,
all the species of microalgae cultivated in this PBR have potential future commercial/
biotechnological applications as source of carotenoids, secondary metabolites, antibacterial or
nutrient removal for waste water treatments (D’Adamo et al., 2018; Lea-Smith et al., 2013; Xu et
al., 2016, 2018; Matthijs et al., 2018; Stoffels et al., 2017; Young and Purton, 2018; Whitton et al.,
2018). Similar to what has been described within this chapter, Pereira et al. (2019) investigated how
the simulation of seasonal conditions might affect the growth of Tetraselmis sp., observing statistical
differences in growth rate of the selected species for the simulated spring season compared to the
autumn in the Algem® PBR.
It is known that some seaweeds properties are dependent or can be influenced by environmental
parameters (Lüning, 1990; Lobban and Harrison, 1994), such as temperature, light regime,
photoperiod, etc., leading to the production of interesting compounds in artificial conditions.
Several studies have focused on the antioxidant properties of species belonging to the Laurencia
complex, including O. pinnatifida, investigating the capacity of the seaweed to produce secondary
metabolites of antimicrobial value. These compounds, which include volatile components, steroids,
phlorotannins, lipids and polyphenols (Vairappan et al., 2001a, b; Vairappan, 2003; Awad, 2000;
Nagayama et al., 2002; Freile-Pelegrin and Morales, 2004; Rodríguez-Bernaldo de Quirós et al.,
0
50
100
150
200
250
300
350
Algem Control Algem Spring Algem Autumn Wild Spring Wild Autumn
Tota
l ph
eno
l co
nte
nt
(µg/
mg
DW
)
Samples

213
2010a, b), are effective antioxidants and may have interesting biological activities (e.g. inhibition of
proliferation of cancer cells, Kwon et al., 2007; Yuan et al., 2005; influence in anti-inflammatory
responses, Kim et al., 2009; influence on response to diabetes, Lee et al., 2008, 2010). Since the
‘80s, red algae, including O. pinnatifida, have been investigated for the production of terpenoid
compounds, that can have pharmaceutical/nutraceutical applications (Erickson, 1983; Faulkner,
1986; Bittner et al., 1985; Norte et al., 1989a, b, 1997a, b; Bano et al., 1988; Atta-ur-Rahman et al.,
1988; Ahmad et al., 1990; Pec et al., 2003; Brito et al., 2002; Flodin et al., 1999; Iliopoulou et al.,
2002a, b; Suzuki et al., 2002). Increasing the production of antioxidant compounds by manipulating
cultivation conditions is particularly important due to the potential commercial exploitability of
these extracts for cosmetic, pharmaceutical and/or nutraceutical applications. Cultivating O.
pinnatifida in the Algem® bioreactor has demonstrated that this system has the potential to enhance
this type of antioxidant compound production in O. pinnatifida. This was demonstrated in all the
trials conducted within this chapter, especially when compared to biomass collected from the wild
for the same months with similar environmental conditions, light and temperature, as set by the
software operating the PBRs as well with the control. This suggests that the production of these
compounds in this species is influenced not only by the quantity but also by the quality of light that
the cultures receive. For all the experiments run in the Algem®, the concentration of the
antioxidant compounds was not only statistically higher but mostly more than doubled according
to TPC and five time higher for the FRAP analyses in the cultivated samples, when compared to
the wild harvested biomass and set of control conditions in the last trial. This discover is
fundamental for further commercial applications. Ideally, O. pinnatifida could be harvested even in
winter, when antioxidant compounds are lower than in summer (see Chapter 3), and the
concentration of interesting antioxidant compound could be efficiently improved in the PBR
system. Particularly, light and temperature parameters applied in the Summer and Autumn
simulations resulted in the highest antioxidant concentration. This would extremely maximise the
financial value of the species, if we consider that the content of total phenol compounds can go
from 100 mg/g of dry weight powder to 250 mg/g in only four weeks. In this study antioxidant
compound were not specifically identified and results refer to the total phenol content and the
reducing power of the species; however, as seen in Chapter 4, antioxidant compounds play an
important role in determining the flavour of the species. Therefore, increasing their concentration
in controlled cultivation conditions will make more valuable the final tailored product that could
be sold as delicacy at higher price than the wild harvested one to restaurants. Moreover, the final
product could be exploited for applications in the pharmaceutical and nutraceutical sectors.
QY results showed an increase of value during the experimental time, as the species adapted to the
incubation conditions, meaning also that the system is able to simulate the wild conditions in
relation to the selected coordinates. Moreover, the species investigated is able to respond, as it

214
would do under similar conditions in the wild. The change in colour that the cultures underwent,
especially in the summer simulation, is the same as already reported in the wild samples harvested
during the same relevant season.
Pigments analyses demonstrated that Chl-a and carotenoids concentrations do not vary statistically
between the summer and winter conditions set by the Algem® system. However, the samples
cultivated in the bioreactor unit showed the same patterns of pigment production as the wild
biomass samples, with higher concentration of Chl-a in the winter compared to the summer
simulations, and higher concentrations of carotenoid for the summer compared to the winter.
Considering that carotenoid contents in macroalgae responds to high radiant energy increasing its
concentration as they act as photo protector (Harker et al., 1999), it is perhaps not surprising that
this was also true for this study. The phycobiliproteins concentration did not appear to be
influenced by cultivation of the biomass in the Algem® system. However, similar levels were
produced when compared to the seasonal sample, with a slightly higher concentration of PE and
PC in the winter stimulated conditions when compared to the summer, as is seen in the seasonal
samples (see Chapter 3). Variation of the pigments content was also reflected in the change in
colour of the cultures grown under the different conditions (Fig. 7.2).
Previous authors have stated that temperature can strongly influence the cellular chemical
composition, uptake of nutrients, CO₂ and the growth rates of algae (Singh and Singh, 2015).
However, results obtained from the trial undertaken that compared a DCT and CT show that the
temperature seems to do not have an effect on the production of Chl-a and carotenoids. On the
other hand, PE and PC were statistically higher under CT conditions compared to DCT. This could
suggest that the production of such compounds in O. pinnatifida might be affected by the
temperature, as already seen for some cyanobacteria (Takano et al., 1995; Chaneva et al., 2007;
Grossman et al., 1995; Simeunovic et al., 2013; Maurya et al., 2014). Furthermore, the values of PE
and PC in samples cultivated in the PBR were statistically lower compared to those obtained from
the wild samples collected in December. The differences in profile showed by phycobiliproteins in
the PBR could potentially be due to the absence of blue light in the system, since usually, blue and
red light stimulated the accumulation of these pigments (Godínez-Ortega et al., 2008). Blue light
also stimulates nitrogen assimilation and the accumulation of N-compounds such as biliproteins
(Figueroa et al., 1995a, Appendix C.3). Another reason could be N limitation in the cultured
samples (Lobban and Harrison, 1994), or a high light irradiance, which can again negatively affect
phycobiliprotein pigment concentrations in macroalgae (Aguilera et al., 2002; Zubia et al., 2014).
The same situation in terms of phycobiliproteins reduction in comparison to wild samples was
confirmed in the subsequent trial. Phycobiliproteins have gained considerable importance in the
commercial sectors for pharmaceutical applications, in the food industry, immunology, cell biology,

215
flow cytometry, as fluorescent probes and cosmetics colorant (Glazer, 1994; Sekar and
Chandramohan, 2008; Sørensen et al., 2013; López-Cristoffanini et al., 2015). Understanding how
the production of these compounds is regulated in seaweeds, such as O. pinnatifida, by
environmental conditions, has the potential to facilitate the exploitation of a species for these
applications. The measured pigment content in the subsequent run, using September and March
conditions as reference months, resulted in slightly different levels when compared to the wild
biomass samples for the same months. It must also be noted that for these samples there was no
statistical difference between March and September in terms of pigments content measured (see
Chapter 3).
Protein and carbohydrate content analysed in the biomass grown in the last trial showed no
statistical difference between the samples. However, for both March and September, the total
carbohydrate content was statistically different from the values obtained for the seasonal samples,
proving a connection between the exposure to a controlled photoperiod/light intensity and the
production of these compounds. The production of carbohydrates (as already described in Chapter
3) is influenced by the light intensity and quality (Orduña-Rojas et al., 2002; Fábregas et al., 1995;
Fernandéz-Reiriz et al., 1989). This can be explained when considering the role that the
carbohydrates and their immediate derivatives have in relation to photosynthesis in algae (Raven
and Beardall, 2003). Furthermore, protein concentrations increased in the Algem® system at least
for the March conditions, having a statistical difference compared to the seasonal biomass samples
analysed for the same month. This suggests that the bioreactor system is either able to improve or
retain comparable levels of production of carbohydrates and proteins, as observed in the field.
Cultivation of biomass in the PBR has demonstrated, for this particular species, that antioxidant
activity can be controlled and even increased. As already demonstrated in Chapter 3, there is a
correlation between the quality and quantity of light and the production of antioxidant compounds.
Furthermore, as already demonstrated for other species, antioxidant compounds often include
compounds that are used by the seaweeds to protect themselves from light irradiation, photo
oxidation, UVB irradiations, etc. (such as mycosporines, bromo phenols, phycobiliproteins,
carotenoids, phenolic compounds, terpenoids) (Munir et al., 2013). It is then not surprising that
the controlled manipulation of the light parameters under specific cultivation conditions can have
an impact on the production of such compounds in this particular species. This has the potential
to be used to manipulate the culture in order to trigger or target a specific property that is then
commercially exploitable. Antioxidant compounds produced from algae have several health
benefits and they can be utilised in cosmetic, pharmaceutical and nutraceutical markets (Kim and
Yarish, 2010; Haijin et al., 2003; Veena et al., 2007; Parys et al., 2010; Yuan et al., 2005a, b; Ahn et
al., 2007; Hwang et al., 2006; Yabuta et al., 2010; Zhang et al., 2003; Munir et al., 2013). Numerous

216
species of macroalgae and microalgae are also currently referenced in the literature as sources of
bioactive compounds suitable for use as functional food ingredients (Holdt and Kraan, 2011;
Freile-Pelegrin and Robledo, 2013). The global market for antioxidants is constantly growing, with
an estimated value of about USD 63 billion in 2010, which was expected to reach at least USD 90.5
billion by 2013 (Kaur and Das, 2011; Pelegrín and Robledo, 2013).
The feasibility of using this particular PBR for small scale in vitro cultivation for this seaweed has
been demonstrated. This must be considered in terms of the slow growth rate of this species both
in the Algem® and in the wild (pers. obsv.). At a larger scale, cultivation will potentially be difficult.
Nevertheless, this methodology has the potential for a low-volume high-quality production from
an interesting alga, such as O. pinnatifida, as source of natural antioxidants. This can off-set increased
costs of cultivation in a PBR, when taken into consideration with the value of the final product.
7.6 Conclusions
Osmundea pinnatifida can be successfully cultivated in the Algem® unit with an increase of
growth and the production of healthy cultures in terms of physiology and morphology;
The morphology and colour of the samples cultivated in the unit match with that of the
wild harvested samples for the same months of reference;
The comparison of the biochemical profiles of wild and cultivated samples for the same
months demonstrated that the system is either able to maintain the same trend or even
increase the production of certain compounds;
All the trials conducted demonstrated that the system is able to result in a statistically
increased concentration of antioxidant compounds, more than doubling the content
compared to wild harvested material and making the cultivated biomass a valuable, tailored
and high-quality product;
It is possible to use this system to target and improve commercially interesting properties
giving the opportunity for commercial applications not only in the established food
market, where the high quality delicacy can be sold to restaurants instead of the wild
harvested product, but also in nutraceutical and pharmaceutical sectors, giving a promising
possibility of exploiting the increased antioxidant compounds content for low volume-
high-value applications.

217

218
Chapter 8 Application of hormones and Acadian Marine Plant Extract
Powder (AMPEP) to cultures of O. pinnatifida and establishment of a
tumbling cultivation system for the species
8.1 Introduction
Algal growth is controlled at a variety of different levels, cellular, tissue, and organismal, (Polevoi,
1982; 1986). Coupled to this, macroalgae undergo a series of morphogenic events during their life
cycles. These include germination, branching, reproduction and senescence, which need to be
under some sort of control mechanisms (Stirk et al., 2003). Our knowledge of how the algal
hormonal system impacts on seaweed metabolism and its mechanisms of action, are, at present,
scarce, but they are likely to play an important role in regulating the growth and development of
seaweeds (Bradley, 1991; Evans and Trewavas, 1991). In the 19601970s, an active search for
phytohormones in various algal taxa was performed. During this period, a wide set of compounds
with hormonal activity was discovered in Chlorophyta, Phaeophyta, and Rhodophyta
(Tarakhovskaya et al., 2007). In 1980, all known phytohormones of higher plants were found also
in algae (Niemann and Dorffling, 1980; Bajguz and Czerpak, 1996; Cooke et al., 2002; Ördög et
al., 2004; Tarakhovskaya et al., 2007). Despite their expected importance, only a very limited
number of papers has been published reporting a comprehensive analysis of phytohormone actions
in algae (Wang et al., 2014; Mikami et al., 2016). This might be due to the difficulty in establishing
a robust methodology as well as the possibility that the phytohormones detected are produced by
epiphytic microorganisms (Mori et al., 2017). For red algae, chemical analyses detected endogen
hormones, such as IAA, BA, iP-N6 -(Δ2-isopentenyl) adenine, and SA-salicylic acid, in thalli of the
Bangiophycean algae Pyropia yezoensis, Pyropia haitanensis, and Bangia fuscopurpurea (Mikami et al.,
2015).
The effect of plant growth regulators (PGRs) on cultures of Osmundea pinnatifida has never been
investigated before. However, several studies have analysed the process of plant regeneration in
other red algae such as Gigartinales (Chen and Taylor, 1978; Polne-Fuller and Gibor, 1987; Bradley
and Cheney, 1990; Huang and Fujita, 1996, 1997; Yokoya and Handro, 1996, 1997), underlying
that PGRs play a significant role in the regeneration process (Yokoya et al., 2004). An insight on
the role that hormones have on the growth and development of O. pinnatifida tissue culture is
important. In order to understand the regulatory processes and identify treatments that could
potentially increase the growth rate of this species as well as biomass production. The hormones
tested in this Chapter are amongst the most common for the control of the physiological
development of algae (Table 8.1). They include auxins and cytokinins, specifically Indole-3-acetic
acid (IAA, 3-IAA), 6-Benzylaminopurine (BA) and 2,4 Dichlorophenoxyacetic (2,4-D). The

219
presence of these auxins and cytokinins in algae is well-established (Crouch, 1990; Kiseleva et al.,
2012). Auxins were found in red algae in the 1960-1970s (Schiewer et al., 1967), and they are usually
responsible in terrestrial plants for the induction of elongation growth; differentiation of phloem
elements; apical dominance; tropisms; initiation of root formation; etc. IAA, for example, activates
the plasmalemmal H⁺-ATPase involved in the process of growth by elongation (Polevoi, 1982,
1986). On the other hand, cytokinins regulate the control of cell division; bud and leaf blade
development; senescence retardation (Tarakhovskaya et al., 2007), and are light dependent, playing
a crucial role in photosynthesis (Stirk et al., 2014a).
Table 8.1 Phytohormones found in Rhodophyta (modified from Tarakhovskaya et al., 2007)
Division Phytohormones found References
Rhodophyta IAA (genera Botryocladia, Porphyra) Provasoli and Carlucci, 1974; Sitnik et al., 2003,
Schiewer et al., 1967
Cytokinins (genus Porphyra) Stirk et al., 2003
Jasmonic acid (genus Gelidium) Arnold et al., 2001
Polyamines (genera Cyanidium, Gelidium, Grateloupia) Hamana et al., 1990; Marián et al., 2000;
Sacramento et al., 2004
Rhodomorphin (genus Griffithsia)
Waaland and Watson, 1980; Watson and
Waaland, 1983
This chapter will also investigate the efficiency of Ascophyllum Marine Plant Extract Powder
(AMPEP), extracted from Ascophyllum nodosum, a seaweed-based culture medium, in the
regeneration of O. pinnatifida. This will involve using tissue culture techniques for the production
of vegetative growth and tank cultivation. Several reports have shown the positive effect of this
seaweed extract on land-based crops for growth, yield and quality, pest and disease resistance, and
environmental stress tolerance (Featonby-Smith and van Staden, 1987; Crouch, 1990; Rayorath et
al., 2008a, b). It has been used successfully with agricultural crops and horticulture for the past 30
years (Craigie, 2011). However, the actual mechanisms of action of the AMPEP extract has not yet
been identified (Hurtado and Critchley, 2018). It is a source of naturally occurring major and minor
nutrients, carbohydrates, aminoacids, and plant growth promoting substances, which have proved
to enhance crop health, nutrition and quality. It is also known to contain the following major
macronutrients: total nitrogen (N)=0.81.5%, available phosphoric acid (P₂O₅)=12% and soluble
potash (K₂O)=1722%, these are potentially having an effect on promoting and supporting the
improvements in plant growth reported (Hurtado et al., 2009).
The efficiency of AMPEP extract in improving the daily growth rate and preventing the growth of
epiphytes was demonstrated in previous studies conducted on other red algae, K. alvarezii (Hurtado
et al., 2009; Loureiro et al., 2010; Yunque at al., 2011; Borlongan et al., 2011). Other research has
demonstrated that exogenous application of the commercial A. nodosum extract increased

220
endogenous antioxidant activity in plants, such as increased amounts of nonenzymatic antioxidant
compounds. For example, α-tocopherol, ascorbate, and β-carotene, enhanced activities of
antioxidant enzymes including glutathione reductase and superoxide dismutase, and elevated total
phenolic content (Fan et al., 2010).
Furthermore, the possibility to apply the AMPEP extract to a tank cultivation trial and compare it
with F/2 medium alone was investigated. The design and development of a tumbling indoor
cultivation system for vegetative cultivation of O. pinnatifida tetrasporophytes was undertaken. This
system was again used to compare the effects of AMPEP extract, in comparison with F/2 medium,
on the growth and biochemical composition of cultures.
Plant growth hormones and AMPEP extract were applied separately in every trial on cultures of
Osmundea pinnatifida in order to understand the effect of each compound individually on the species,
therefore not investigating the simultaneous effect of both compounds on O. pinnatifida.
8.2 Aims
To study the effects that some PGRs might have on the growth, development and
biochemical composition of O. pinnatifida (juveniles and adults);
To study the effects that AMPEP might have on the growth, development and biochemical
composition of O. pinnatifida (juveniles and adults);
To identify the best treatment for an eventual further application on a large-scale
cultivation, in order to improve the growth and the biomass production;
To apply the AMPEP extract on juveniles, adult cultures and in a cultivation system for
epiphytes mitigation;
To establish an indoor tumbling tank system for vegetative cultivation of O. pinnatifida.
8.3 Materials and methods
8.3.1 Multi-well trials
8.3.1.1 PGRs applied to cultivation of O. pinnatifida juveniles
For the PGRs and AMPEP experiments, biomass was harvested from Seil (56.30° N, 5.65° W) in
September 2018. The material was initially incubated at 10-15 °C, 12:12 photoperiod, 50-80
µmol/m²/s and enriched filtered seawater with F/2 prior to experimentation. New growing shoots
were cut from the biomass with a sterile blade under a laminar cabinet in preparation for multi-
well trials.
Every hormone treatment was carried out in triplicate on nine fragments per treatment. The
hormones used were Indole-3-acetic acid (IAA, 3-IAA), 6-Benzylaminopurine (BA) and 2,4

221
Dichlorophenoxyacetic acid (Sigma Aldrich®) in concentrations of 0.5, 2.5 and 5 mg/l as single
hormonal treatment. The hormones were combined for double and triple treatments at 1 and 5
mg/l. All solutions were made up in F/2 medium (Table 8.2). A control was run under the same
conditions, with no hormones added to the F/2 culture media. Samples were incubated at 50-80
µmol/m²/s 9-10°C, 12:12 photoperiod in 6 multi-well dishes for six weeks. The medium was
changed weekly and pictures of the plates were taken at the same time with a Nikon D5500 camera
at a standard distance from the surface, 30 cm, and a 18-55 lens without zoom. A paper grid was
place underneath the plates to act as a reference, and the images were then analysed with ImageJ®
to assess the area increase (expressed in mm²). Averages and sums were calculated, and statistical
analyses were conducted applying ANOVA (single factor, p<0.05; Fisher’s hoc test) with Minitab
18.
Table 8.2 Treatments with PGRS and relative concentrations applied on vegetative fragments of O. pinnatifida. Control only contained F/2 medium
Single Hormone
Conc (mg/l)
Double Hormones
Conc (mg/l)
Triple Hormones
Conc (mg/l)
Control
2,4-D 0.5 IAA:BA 1:1 2.4-D:BA:IAA
1:1:1 F/2 medium, no PGRS
2.5 5:1 5:1:1
5 1:5 5:5:1
IAA 0.5 5:5 1:1:5
2.5 2,4-D:IAA 1:1 1:5:1
5 5:1 1:5:5
BA 0.5 1:5 5:1:5
2.5 5:5 5:5:5
5 2.4-D:BA 1:1 1:1:1
5:1 5:1:1
1:5 5:5:1
5:5 1:1:5
1:5:1
1:5:5
5:1:5
8.3.1.2 AMPEP treatments applied to cultivation of O. pinnatifida juveniles
Cultivated 5 months old juveniles (as described in Chapter 5) were cleaned manually with a brush
prior to setting up the experiment. They were acclimated at 9 °C, 12:12 (L:D), 30 µmol/m²/sec
before the start of the trial. For the AMPEP extract, three different treatments were investigated
plus a control on juveniles cultivated in vitro in 6 multi-well plates: Bathing for one hour in 20 g/l
solution of AMPEP extract; 3 mg/l AMPEP in FSW and 0.5 mg/l AMPEP in FSW. The control
was just F/2 medium (Table 8.3). There were three multi-wells per treatment with three juveniles
per well, in total there were 54 juveniles per treatment. During the Bathing treatment, plates were
kept on a rocking plate (VWR®) at 10 °C in order to guarantee a complete mixing of the juveniles

222
with the solution. During the experiment, medium was replaced twice per week in a laminar cabinet
and the multi-well plate replaced once per week. Four ml of media were placed in each well. Growth
assessment was performed as described in section 8.3.1.1.
Table 8.3 AMPEP treatments applied on juveniles cultivated in 6 × multi well
Multi well trial N° of plantlets Treatment Replicate Duration
3 x 6 multi-well 54 3 mg/l of AMPEP 3x 30 days
3 x 6 multi-well 54 0.5 mg/l of AMPEP 3x 30 days
3 x 6 multi-well 54
Bathing in 20g/l AMPEP solution 3x 30 days
3 x 6 multi-well 54 F/2 (control) 3x 30 days
8.3.2 Flask trials
8.3.2.1 Flask trials applying PGRs to adult O. pinnatifida biomass
Flask trials utilising PGRs are detailed in Table 8.4. The control was F/2 medium only. Five grams
±0.005 of FW material were weighed and placed into 500 ml flasks with 250 ml of FSW plus
hormone or control treatment. The flasks were bubbled with air filtered through 0.45 µm Whatman
filters and incubated at 50-80 µmol/m²/s, 12:12 and 10 °C for seven weeks.
Table 8.4 Treatments with PGRS and relative concentrations applied on cultures of O. pinnatifida. Control only included F/2 medium
Treatment Hormone Concentration (mg/l)
Single Hormone BA 2.5
Double Hormones IAA:BA 1:5
Triple Hormones 2.4-D:BA:IAA 5:5:5
Control No PGRs
Pictures were taken every week using a paper grid as reference. At the end of the seven weeks the
biomass from various treatments was harvested for biochemical analyses including carbohydrates,
protein, pigments and FRAP assays. The assays were conducted according to the methodologies
described in Chapter 3. Statistical analyses were conducted applying ANOVA (single factor,
p<0.05; Fisher’s hoc test) with Minitab 18.
8.3.2.2 Flask trials applying AMPEP treatments to adult O. pinnatifida biomass
For the AMPEP flask trial, three treatments plus a control (Table 8.5) were tested on wild biomass
collected in September 2018 from Seil (56.300° N, 5.650° W), cleaned manually and treated with a
0.5 % KI solution in FSW (as described in Chapter 5). Every treatment was set up in triplicate.
During the Bathing treatment with a 20 g/l AMPEP solution, cultures were kept on a rocking plate
(VWR®) at 10 °C in order to guarantee a complete mixture of the cultures with the solution. The
biomass was weighed and 10 g (WW) were placed into 500 ml flasks with 250 ml media in triplicate.

223
The medium was changed twice per week. Aeration was provided through Whatman® filter (0.45
µm) and cultures incubated at 9°C, 12:12, 80-100 µmol/m²/s for 30 days.
For both trials (application of hormones and AMPEP extract), the biomass weight was recorded
every week after draining it with a spin salad in order to remove the excess of water from the
cultures (Ross et al., 2017), and pictures were taken. Weight (g) of the biomass was recorded and
SGR was assessed weekly applying the following formula (Evans, 1972) where LN is natural
logarithm, Wf final weight, Wi initial weight and t time:
SGR= (𝐿𝑁(𝑊𝑓)−𝐿𝑁(𝑊𝑖)×100
𝑡
Table 8.5 AMPEP treatments applied on a flask trial. Samples under Bathing treatment were left in the solution for 1hr, then rinsed from the excess of solution and incubated in F/2 medium
Flask trial Mass (FW) (g)
Treatment Replicate Duration
500 ml flask 10 3 mg/l AMPEP 3x 30 days
500 ml flask 10 0.5 mg/l AMPEP 3x 30 days
500 ml flask 10 Bathing in 20g/l solution 3x 30 days
500 ml flask 10 F/2 (control) 3x 30 days
8.3.3 Tank trial applying AMPEP extract to adult O. pinnatifida biomass
A seaweed fertilizer, AMPEP extract, was also tested in a tank experiment. Treatments and
concentrations tested are described in Table 8.6.
Table 8.6 Treatments applied on a tank system
Tank trial Mass (FW) (g) Treatment Replicate Duration
60 L tank 200 0.5 mg/l AMPEP
2x 90 days
60 L tank 200 Bathing in 20 g/l solution
2x 90 days
60 L tank 200 F/2 (control) 2x 90 days
Biomass, 3 kg in total, was collected at low tide from Seil (56.300° N, 5.650° W) at the end of
September 2018, placed in Cool boxes with wet tissues and transported to the laboratory. Here it
was manually cleaned in order to remove all visible epiphytes and grazers. The biomass was then
washed thoroughly with filtered UV sterilised seawater and cleaned further in filtered UV sterilised
seawater containing 0.5 % KI solution for 30 minutes with aeration. Finally, it was rinsed three
times, 30 minutes each time, in filtered UV sterilised seawater to remove any residual Kl (as
described in Chapter 5). The water in the tank was changed again, samples were then left overnight
in filtered UV sterilised seawater with powerful aeration and no nutrients. The seawater was
changed the next day and the biomass left in filtered UV sterilized seawater with aeration. Nutrients
were added to the tank after 48 hours. The biomass was acclimated at 10°C, 12:12 light

224
photoperiod, 100 µmol/m²/s, F/2 medium for 28 days before the growth trial. For this, the
biomass was again incubated for 48 hours without nutrients and finally placed into the cultivation
system. Approximately 200 g (±0.05) of biomass (Fig. 8.1 F) was placed into a white 60 l conical
tank with 55 l FSW final volume, there were six tanks in total and the trial lasted three months. The
tanks were supported by metal frames that permitted access to their bottoms where a valve system
for aeration and tank emptying was collocated (Fig. 8.1 D).
Figure 8.1 Tumbling system developed for vegetative cultivation of O. pinnatifida tetrasporophytes; A) Filters and UV system; B) Tanks under 0.5 mg/l treatment and control with 10 l media reservoirs and dosing pumps for the nutrient supply; C) Tanks under the bathing treatment; D) Frame for supporting the tank and valve system for aeration and emptying; E) Inflow (yellow pipe) and banjo filter for the outflow; F) cultures during the weighing
The cultivation system was located in a 10°C CT room (Fig. 8.1 B). Seawater was pumped directly
into the system having first been filtered through a sand filter and then a set of three filters: 100-
20 and 10 µm. A successive 5 µm filter was added for the final month of the experiment. The
water was also UV treated (Fig. 8.1 A). The inflow rate was 1.2 l × min⁻¹, with the seawater in the
tanks being completely replaced every hour. Lights in the CT room were Growth Lux type

225
(Sylvania®) and were kept on 24/7. The maximum radiation reported was almost 3000 lux, the
minimum 80 lux with the average being 1300 lux. HOBO® loggers were placed in two of the six
tanks to measure temperature and light. pH and salinity were measured daily with a pH and TDS
monitor (pH-2771).
Three different treatments (two replicates per treatment) were tested in the system. Two tanks were
kept as controls and were fed with F/2 medium (Guillard and Ryther, 1962). Two tanks were
treated for the entire length of the experiment with 0.5 mg/l AMPEP solution (Fig. 8.1 B). The
cultures in the remaining two tanks were bathed in a 20 g/l solution for 1 hour (in a 60 l tank at
10°C and with aeration) prior to the experiment (Fig. 8.1 C). Then the biomass was placed in the
system and fed with F/2 medium at the same rate as the control tanks. Three electric dosing pumps
(Easy irrigation System, EasyGrow ᵀᴹ) were used to feed the cultures (one for every two tanks),
releasing 180 ml per tank twice per day. The outflow of the tanks constituted of banjo filters (Fig.
8.1 E) and outflow pipes that merged with the main pipe that transported the wastewater to the
water treatment system and finally discharged. Aeration was provided from the bottom of the
system through a 2-3 cm diameter hole and this caused the cultures to tumble. A two-way valve
system allowed the tanks to be emptied directly from the bottom, aeration was switched off when
the tanks were being emptied (Fig. 8.1 D).
Filters were replaced every week and tanks were cleaned every two weeks with a 50% Decon
solution. Cultures were weighted at this time, QY measured and pictures taken. SGR was calculated
applying the formula of Evans (1972) as detailed in section 8.3.2. Every month, excess biomass
was harvested and the new plantlets, which had developed, were placed back into the tanks to
continue growing. The final biomass of the cultures was again taken to the starting biomass weight
of 200 ± 0.05 g. This was done in order to control the density of the cultures and avoid a decrease
in the growth rate due to excessive competition for light and nutrients.
8.4 Results
8.4.1 Multi-well trials
8.4.1.1. PGRs applied to cultivation of O. pinnatifida juveniles
The 2.4-D treatment, at all concentrations tested, was not particularly effective on influencing the
growth of this species. Occasionally the fragments treated with this hormone turned from red to
green in colour. Indicating that the cultures were under some sort of stress. Higher concentrations
of 2.4-D hormone again were unsuitable for this species, the exception being when it was included
in the triple treatment 2.4D:IAA:BA (5:5:5) (Fig. 8.2 E and F). This treatment performed best when
compared to the other treatments.

226
Among the single treatments tested, IAA seemed to initiate elongation of the vegetative tissue in
this species, whilst BA appears to regulate branching (Fig. 8.2 A and B). The combination of IAA
and BA stimulated simultaneously growth via elongation as well as branch development (Fig. 8.2
C and D).
Figure 8.2 A) Juveniles cultures exposed to 2.5 mg BA at the beginning and B) at the end of the experiment. Juveniles cultures exposed to IAA:BA 1:5 C) at the beginning and D) at the end of the experiment. Juveniles cultures exposed to 2.4-D:BA:IAA 5:5:5 E) at the beginning and F) at the end of the experiment. Each square is 1mm²
Figure 8.3 Results of the growth (area mm²) for the single hormone treatments applied on juveniles cultures of O. pinnatifida (n=3; error bar, 1 SD of mean)
0
1
2
3
4
5
6
7
8
9
Control 0.5 mg 2.4-D
2.5 mg 2.4-D
5 mg 2.4-D 0.5 mg IAA 2.5 mg IAA 5 mg IAA 0.5 mg BA 2.5 mg BA 5 mg BA
Are
a (m
m²)
Treatments
Week 1 Week 2 Week 3 Week 4 Week 5 Week 6 Week 7
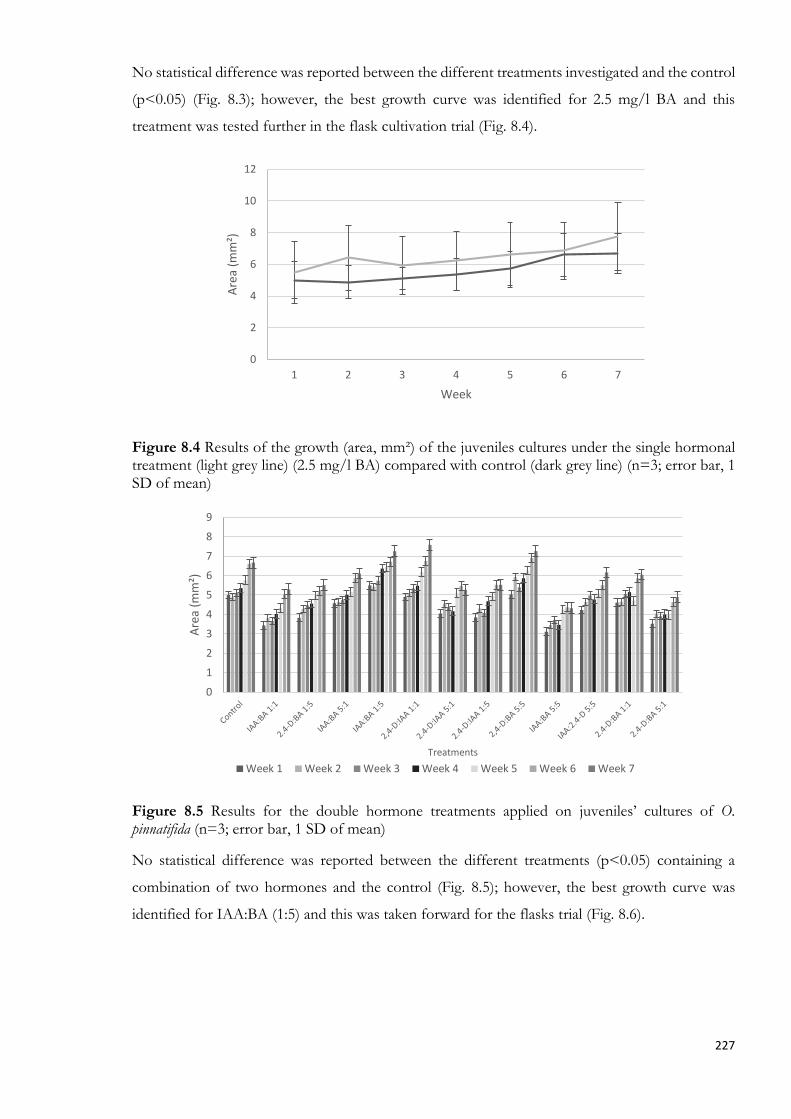
227
No statistical difference was reported between the different treatments investigated and the control
(p<0.05) (Fig. 8.3); however, the best growth curve was identified for 2.5 mg/l BA and this
treatment was tested further in the flask cultivation trial (Fig. 8.4).
Figure 8.4 Results of the growth (area, mm²) of the juveniles cultures under the single hormonal treatment (light grey line) (2.5 mg/l BA) compared with control (dark grey line) (n=3; error bar, 1 SD of mean)
Figure 8.5 Results for the double hormone treatments applied on juveniles’ cultures of O. pinnatifida (n=3; error bar, 1 SD of mean)
No statistical difference was reported between the different treatments (p<0.05) containing a
combination of two hormones and the control (Fig. 8.5); however, the best growth curve was
identified for IAA:BA (1:5) and this was taken forward for the flasks trial (Fig. 8.6).
0
2
4
6
8
10
12
1 2 3 4 5 6 7
Are
a (m
m²)
Week
0
1
2
3
4
5
6
7
8
9
Are
a (m
m²)
Treatments
Week 1 Week 2 Week 3 Week 4 Week 5 Week 6 Week 7

228
Figure 8.6 Results of the growth (area, mm²) of the juveniles cultures under the double hormonal treatment (light grey line) (IAA:BA 1:5 mg/l) compared to control (dark grey line) (n=3; error bar, 1 SD of mean)
Figure 8.7 Results for the triple hormone treatments applied on juveniles’ cultures of O. pinnatifida (n=3; error bar, 1 SD of mean)
No statistical difference was reported between the different treatments (p<0.05) containing all
three hormones and the control (Fig. 8.7); however, the best growth curve was identified for 2,4-
D:IAA:BA (5:5:5) and this was taken forward for the flasks trial (Fig. 8.8).
0
2
4
6
8
10
12
1 2 3 4 5 6 7
Are
a (m
m²)
Week
0
1
2
3
4
5
6
7
8
Control 2.4-D:BA:IAA1:1:1
2.4-D:BA:IAA5:1:1
2.4-D:BA:IAA5:5:1
2.4-D:BA:IAA1:1:5
2.4-D:BA:IAA1:5:1
2.4-D:BA:IAA1:5:5
2.4-D:BA:IAA5:1:5
2.4-D:BA:IAA5:5:5
Are
a (m
m²)
Treatments
Week 1 Week 2 Week 3 Week 4 Week 5 Week 6 Week 7

229
Figure 8.8 Results of the growth (area mm²) of the juveniles’ cultures under the triple hormonal treatment (light grey line) (2.4-D:BA:IAA 5:5:5 mg/l) compared to control (dark grey line) (n=3; error bar, 1 SD of mean)
8.4.1.2 AMPEP treatments applied to cultivation of O. pinnatifida juveniles
Results for the growth expressed as mm² and SGR relative to the application of the AMPEP
extracts to juveniles are presented respectively in Figs 8.9 and 8.10. Time is referred to weeks.
Figure 8.9 Measurement of the growth (area, mm²) of juveniles under the different AMPEP treatments, measured with ImageJ®. Control = blue, 3 mg/l = light grey, 0.5 mg/l = dark grey and Bathing (20g/l) = black bars (n=3; error bar, 1 SD of mean)
A statistical difference was reported between the 0.5 mg/l treatment and the other treatments
trialled (ANOVA, p<0.05). The lowest growth was reported for the Bathing (20 g/l) treatment
(Fig. 8.9).
0
2
4
6
8
10
12
1 2 3 4 5 6 7
Are
a (m
m²)
Week
0
0.5
1
1.5
2
2.5
3
3.5
4
Week 0 Week 1 Week 2 Week 3 Week 4
Are
a (m
m²)
Time

230
Figure 8.10 SGR (% day⁻¹) of juveniles undergoing different AMPEP treatments. Control= blue, 3 mg/l = light grey, 0.5 mg/l = dark grey and Bathing (20g/l) = black bars (n=3; error bar, 1 SD of mean)
The highest growth rate was reported for the control (week 1) and for the 0.5 mg of AMPEP
solution (for weeks 2, 3 and 4). The lowest growth rate was observed with the biomass treated with
20 g/l AMPEP solution (Fig. 8.10, week 2) (one-way ANOVA, p<0.05, Fisher’s test). After 30
days, epiphytes were found in reduced numbers in all samples treated with the AMPEP solution
(Fig. 8.11 C and D). While the untreated control showed colonization by epiphytes (Fig. 8.11 A).
This was also true for the biomass bathed in the 20 g/l solution (Fig. 8.11 B). Branching and
trichoblast formation appeared on cultures cultivated under AMPEP extract but not in the control
or in the bathed samples (Figs 8.11 C and D and 8.12).
Figure 8.11 Juveniles specimens under AMPEP treatments A) Control; B) Bathing (20 g/l); C) 0.5 mg/l; D) 3 mg/l. Bar= 1mm
-1
0
1
2
3
4
5
6
7
Week 1 Week 2 Week 3 Week 4
Gro
wth
rat
e (%
day
⁻¹)
Time

231
Figure 8.12 Trichoblast in juvenile specimen under AMPEP extract
8.4.2 Flask trials
8.4.2.1. Flask trials applying PGRs to adult O. pinnatifida biomass
Growth measurements for the hormone flask experiments are reported in Fig. 8.13. All four
treatments displayed a positive growth for the entire length of the experiment. There were no
statistical differences (p<0.05) in terms of weight increase according to the treatment applied. It is
worth noting that the phenotype of the cultures changed during the experiment, especially for the
single treatment, with some of the algal biomass turning green in colour. This phenomenon did
not appear to affect the growth of the specimens.
Figure 8.13 Variation of weight (g, WW) across 7 weeks for cultures under different PGRs treatments. Control: F/2 medium (blue); Single: 2.5 mg BA (orange); Double:IAA:BA (1:5) (grey); Triple: 2.4-D:BA:IAA (5:5:5) (yellow line) (n=3; error bar, 1 SD of mean)
A positive SGR was also seen for all the experiments and for all the treatments tested (Fig. 8.14).
The “double” treatment maintained a statistically higher SGR compare to the control in the 2nd and
3rd weeks, whilst still higher in the 4th and 6th weeks it was not statistically different. The “single”
0
1
2
3
4
5
6
7
8
9
10
Week 0 Week 1 Week 2 Week 3 Week 4 Week 5 Week 6 Week 7
wei
ght
(g W
W)
Time

232
treatment remained significantly lower than the control for the 3rd, 4th and 5th weeks, while it was
significantly higher than the control in the last week (p<0.05). Cultures had multiple branching and
tip elongation under all treatments (Fig. 8.15 E-H) and those grown under the single hormone
treatments changed colour turning green from red (Fig. 8.15 B and F).
Figure 8.14 SGR (expressed in %day⁻¹, from Evans, 1972) of cultures under different treatments. Control: F/2 medium (blue); Single: 2.5 mg BA (orange); Double: IAA:BA (1:5) (grey); Triple: 2.4-D:BA:IAA (5:5:5)(yellow bars) (n=3; error bar, 1 SD of mean)
Figure 8.15 Specimens under PGRs treatments in the flask trial at the start A) Control; B) Single treatment (2.5 mg BA); C) Double treatment (IAA:BA 1:5); D) Triple treatment (2.4D:IAA:BA 5:5:5) and at the end of the trial E) Control; F) Single treatment; G) Double treatment; H) Triple treatment
8.4.2.1.1 Biochemical analyses of O. pinnatifida biomass cultivated applying PGRs
Biochemical analyses relative to carbohydrates and protein content for the flask hormones
experiment are reported in Fig. 8.16. There was a statistical difference in terms of carbohydrates
-0.5
0
0.5
1
1.5
2
2.5
3
Week 1 Week 2 Week 3 Week 4 Week 5 Week 6 Week 7
Gro
wth
rat
e (%
day
⁻¹)
Time

233
percentage between the single hormonal treatment and the control (p<0.05) (Fig. 8.16). A statistical
difference (p<0.05, Fisher’s hoc test) was reported for the protein concentration between the triple
hormonal treatment and the control. In general, the three hormonal treatments showed higher
percentages of protein compare to the control, which accounted for the lowest rate (Fig. 8.16).
Figure 8.16 Carbohydrate (dark grey bars) and protein (light grey bars) content expressed in percentage in cultures undergoing different PGRs hormonal treatments (n=3; error bar, 1 SD of mean)
Figure 8.17 Chl-a (dark grey bars) and carotenoids (light grey bars) (µg/g) concentration in samples undergoing different PGRs hormonal treatments (n=3; error bar, 1 SD of mean)
A statistical difference (p<0.05) was reported between the double and the triple hormonal
treatments and the control for the concentration of chlorophyll-a. Only the double treatment
showed a statistical difference from the control in term of carotenoids’ concentration (Fig. 8.17).
In general, the three hormonal treatments showed higher concentrations of chlorophyll-a and
carotenoids compare to the control, which accounted for the lowest levels.
0
5
10
15
20
25
30
35
Control Single Double Triple
Car
bo
hyd
rate
s an
d p
rote
in
con
ten
t (%
DW
)
Treatment
0
200
400
600
800
1000
1200
Control Single Double Triple
Ch
loro
ph
yll a
an
d c
aro
ten
oid
s co
nte
nt
(µg/
g D
W)
Treatment

234
Figure 8.18 PE (dark grey bars) and PC (light grey bars) (mg/g) concentrations in cultures undergoing different PGRs hormonal treatments (n=3; error bar, 1 SD of mean)
Statistical differences (p<0.05) were identified between the control and the double and triple
hormonal treatment both for the PE and PC concentrations. Only the single treatment showed
lower values compare to the control. Instead, as already seen for the protein concentration, both
double and triple treatments showed higher values compare to the control (Fig. 8.18).
Figure 8.19 FRAP assay (µM FeSO₄ eq/mg DW) for cultures undergoing different PGRs hormonal treatments (n=3; error bar, 1 SD of mean)
No statistical difference was reported in terms of antioxidant content (p<0.05); however, the
three hormonal treatments showed higher values compare to the control (Fig. 8.19).
8.4.2.2 Flask trials applying AMPEP treatments to adult O. pinnatifida biomass
Growth measurements for the application of the AMPEP extract are shown in Fig. 8.20. Time is
referred as weeks.
There was no statistical difference between the treatments tested and the control in term of growth
rate or weight increase (Figs 8.20 and 8.21 respectively). In terms of weight increase, the four
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Control Single Double Triple
PE
and
PC
co
net
n (
mg/
g D
W)
Treatment
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
Control Single Double Triple
FRA
P
(µM
FeS
O₄
eq/m
g D
W)
Treatment

235
treatments performed similarly, with a slightly slower growth for the Bathing (20 g/l) treatment.
This was evident also from the SGR data (Fig. 8.21), where the Bathing treatment was the only one
to show a negative growth rate in the first week of cultivation. The control remained almost stable
for the entire experiment, while for both AMPEP treatments (0.5 and 3 mg/l), there was an
evidence of increased growth in the first week, followed then by a slight decrease and finally the
cultures settled at a constant level (Fig. 8.21).
Figure 8.20 SGR measurements of cultures in flask under the different AMPEP treatments. Time is expressed in weeks (n=3; error bar, 1 SD of mean)
The control performed better than the Bathing treatment even if this was not statistically supported
(Fig. 8.21).
Figure 8.21 Comparison of cultures weight (WW) under the three AMPEP treatments (20 g/l = orange, 0.5 mg/l = grey, 3 mg/l = yellow line) and the Control (F/2) (blue line) (n=3; error bar, 1 SD of mean)
The same trend already reported for Bathing vs control was observed for 0.5 mg/l (AMPEP) vs
control, again with no statistical differences and a reconnection of the two treatments at the last
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Control (F/2) Bathing 20 g/lAMPEP
0.5 mg/l AMPEP 3 mg/l AMPEPGro
wth
rat
e (%
day
¯¹)
TreatmentWeek 1 Week 2 Week 3 Week 4 Week 5
0
2
4
6
8
10
12
14
Week 0 Week 1 Week 2 Week 3 Week 4 Week 5
wei
ght
(g W
W)
Time

236
measurement point. Even for the 3 mg/l (AMPEP) treatment against the control, this last one
performed better, although not statistically (Fig. 8.21).
Samples cultivated under the 0.5 mg/l and 3 mg/l solutions turned green after the first week of the
experiment (Fig. 8.22 A and B). This change in colour was maintained for the length of the trial.
Cultures grown under the control treatment (Fig. 8.22 C) or those bathed in the 20 g/l solution
maintained a dark red colour (Fig. 8.22 D). Branches and ramifications even along the thallus (Fig.
8.23), that are not normally seen in O. pinnatifida grown either in cultivated biomass or in the wild,
as vegetative growth usually appears from the tips and the rhizoid, were present in cultures grown
in 0.5 mg/l and 3 mg/l AMPEP solutions.
Figure 8.22 Cultures under the AMPEP treatments A) 0.5 mg/l cultures B) particular of new shoots from the rhizoids for specimens under 0.5 mg/l; C) Control treatment and; D) Bathing (20 g/l AMPEP treatment)
Figure 8.23 Particular of new branches (arrow) and callus (circle) formation along the thallus in cultures under 3 mg/l AMPEP
8.4.2.2.1 Biochemical analyses of O. pinnatifida biomass cultivated applying AMPEP treatments
Biochemical analyses of O. pinnatifida cultures for carbohydrates and protein content grown with
the addition of AMPEP in flask cultivation system are showed in Fig. 8.24. Statistical differences
(ANOVA, p<0.05) were reported between 0.5, 3 mg/l and the other treatments for the percentage

237
of carbohydrates. The highest result was reported for 0.5 mg/l treatment and the lowest one for
the bathed samples (Fig. 8.24).
Figure 8.24 Carbohydrate (dark grey bar) and protein (light grey bars) content expressed in percentage (DW) of cultures under different AMPEP treatments (n=3; error bar, 1 SD of mean)
A statistical difference (ANOVA, p<0.05) was reported for the percentage of protein content
between the control and bathing treatment and the other two treatments, with the highest levels
reported in the control, followed by Bathing, 0.5 mg/l and finally 3 mg/l treatments (Fig. 8.24).
Statistical differences (ANOVA, p<0.05) were reported, with control and Bathing higher than the
other treatments for the chlorophyll-a and carotenoids content, with the highest levels for Chl-a
reported for the control (1264.78 µg/g), followed by Bathing (1150.45 µg/g), 3 mg/l (480.9 µg/g)
and lastly 0.5 mg/l (444.3 µg/g). For carotenoids the highest levels were reported for control (402
µg/g) followed by Bathing (385 µg/g), 3 mg/l (196 µg/g), and lastly 0.5 mg/l (188 µg/g) (Fig. 8.25).
Figure 8.25 Chl-a (dark grey bars) and carotenoids (light grey bars) concentration (µg/g) of cultures under different AMPEP treatments (n=3; error bar, 1 SD of mean)
0
5
10
15
20
25
30
35
Control (F/2) Bathing 20 g/lAMPEP
0.5 mg/l AMPEP 3 mg/l AMPEPCar
bo
hyd
rate
s an
d p
rote
in c
on
ten
t (%
DW
)
Treatment
0
200
400
600
800
1000
1200
1400
1600
Control (F/2) Bathing 20 g/lAMPEP
0.5 mg/l AMPEP 3 mg/l AMPEP
Ch
loro
ph
yll a
an
d c
aro
ten
oid
s co
nte
nt
(µg/
g D
W)
Treatment

238
The same trends were also found for the phycobiliprotein content (Fig. 8.26). The control reported
the highest levels for PE (4 mg/g) and PC (1.2 mg/g), followed by the Bathing treatment (PE 3.2
mg/g; PC 1 mg/g), 3 mg/l (PE 0.36 mg/g; PC 0.3 mg/g) and lastly 0.5 mg/l (PE 0.23mg/g; PC
0.21 mg/g). Furthermore, this difference in pigments content was reflected by the colour of the
cultures.
Figure 8.26 PE (dark grey bars) and PC (light grey bars) concentration (mg/g) of cultures under different AMPEP treatments (n=3; error bar, 1 SD of mean)
Statistical differences (ANOVA, p<0.05) were reported in terms of antioxidant compounds
between the control and the Bathing and the other two treatments, with both FRAP (Fig. 8.27)
and TPC analyses (Fig. 8.28), with highest levels reported for control, followed by Bathing, 0.5
mg/l and finally 3 mg/l treatments.
Figure 8.27 FRAP analyses (µMFeSO₄eq/mg DW) of cultures under different AMPEP treatments (n=3; error bar, 1 SD of mean)
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Control (F/2) Bathing 20 g/lAMPEP
0.5 mg/l AMPEP 3 mg/l AMPEP
PE
and
PC
co
nte
nt
(mg/
g D
W)
Treatment
0
1
2
3
4
5
6
Control (F/2) Bathing 20g/lAMPEP
0.5 mg/l AMPEP 3 mg/l AMPEP
FRA
P
(µM
FeS
O₄e
q/m
g D
W)
Treatment

239
Figure 8.28 TPC analyses (µg/mg) of cultures under different AMPEP treatments (n=3; error bar, 1 SD of mean)
8.4.3 Tank trial applying AMPEP extract to adult O. pinnatifida biomass
Data relative to pH, salinity, temperature and intensity for the tumbling cultivation system are
reported in Figs 8.29 and 8.30. While the pH increased during the experiment (even if the increase
was not statistically significant) (Fig. 8.29, dark grey bars), the salinity levels initially rose followed
by a decrease in December to then stabilize at around 28 TDS in January and 27 TDS in February
(Fig. 8.29, grey line).
Figure 8.29 pH (dark grey bars) and salinity (TDS, light grey line) measurements for the tank experiment applying AMPEP extract
The temperature slightly decreased during the experiment (Fig. 8.30, dark grey bar) even if this
decrease was not statistically significant. In general, it never surpassed the 12 °C mark and never
fell under 8 °C. The average light intensity was relevantly stable (Fig. 8.30, light grey line) and a
maximum peak was reported only once in January, again this was not statistically significant (Fig.
8.30, grey line).
0
50
100
150
200
250
300
350
Control (F/2) Bathing 20 g/lAMPEP
0.5 mg/l AMPEP 3 mg/l AMPEP
Tota
l ph
eno
l co
nte
nt
(µg/
mg
DW
)
Treatment
7.6
7.7
7.8
7.9
8
8.1
8.2
8.3
8.4
8.5
0
5
10
15
20
25
30
35
40
November December January February
pH
TDS
(mg/
l)
Months

240
Figure 8.30 Temperature (dark grey bars) and intensity measurements (average light = grey line and maximum light = dark grey line) recorded with HOBO® loggers in the tanks under the AMPEP treatments
Cultures were weighted every 15 days. Every month the biomass weight was reinstated at the same
level as the start of the experiment. The highest weight (g) for all the treatments was reported at
the end of the second month, with cultures doubling their initial weight, and the highest values
reported for the two AMPEP treatments, but with no statistical differences. A decline of the growth
rate was instead detected in the third month for all the treatment investigated, with cultures still
growing, but at a slower rate. Generally, all the treatments performed equally, with no statistical
difference reported (Fig. 8.31).
Figure 8.31 Weight (WW) of the cultures under the three treatments tested in the tank system after 15 days, 1 month, 45 days, 2 months, 75 days and 3 months (n=3; error bar, 1 SD of mean)
There was no statistical difference between the growth rate and the treatments applied (ANOVA,
p<0.05, Tukey’s test, Minitab18) (Fig. 8.32). All three treatments sustained the growth of the
0
50
100
150
200
250
300
0
2
4
6
8
10
12
14
November December January February
Ligh
t in
ten
sity
(lu
m/f
t²)
Tem
per
atu
re (
°C)
0
50
100
150
200
250
300
350
400
450
500
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
wei
ght
(g W
W)
Treatment
Time 0 15 days Month 1 Time 0 45 days Month 2 Time 0 75 days Month 3

241
cultures with positive SGR. The growth rate of the experiment was measured every 15 days. The
SGR measurements reported for all the treatments an initial decrease, followed by a subsequent
increase and a further decrease in the successive data point, for all three of the months, following
a cycling scheme. The best results were reported for all the treatments in the second month of
cultivation and the worst ones in the third month (Fig. 8.32). There was almost a doubling of the
weight of the biomass every month, with a maximum of almost 5 % day⁻¹ measured in the second
month for the Bathing treatment.
Figure 8.32 Growth rate of the cultures in the tank system applying the AMPEP extract after 15 days, 1month, 45 days, 2 months, 75 days, 3 months (n=3; error bar, 1 SD of mean)
Cultures grew vertically and horizontally (Fig. 8.33 A), developing new tips, new branches (Fig.
8.33 B) and occasionally callus formations (Fig. 8.33 C, black circle) for the entire length of the
experiment.
Figure 8.33 Cultures in the tank system applying the AMPEP extract after 15 days in the cultivation system. A) 05 mg/l; B) control; C) bathing, circled callus formation; D) control cultures
-1
0
1
2
3
4
5
6
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
Gro
wth
rat
e (%
day
⁻¹)
Treatment
15 days Month 1 45 days Month 2 75 days Month 3

242
Cultures maintained a dark purple colour and a thick texture for the length of the experiment (Figs
8.33 D and 8.34 A-C), with no difference in colour reported between the treatments.
Figure 8.34 A) cultures after one month (0.5 mg/l AMPEP); B) cultures after two months (bathing in 20 g/l AMPEP); C) new plantlets developed in cultures after 3 months (Control F/2) in the cultivation system
Pre-treatment for the prevention of epiphytes, previously described in Chapter 5, proved to be
effective for nearly the entire length of the experiment. Only one single species, Alaria sp.,
contaminated the cultures with its presence being recorded on the last 15 days of the trial.
QY of the cultures was measured every 15 days following the weighing of the cultures. Initially
there was a decrease followed by a constant increase in the PAM response of the cultures until the
second month, to then slightly decrease in the third month, this was not significant statistically (Fig.
8.35). No statistical differences were reported for the PAM measurements between the treatments
tested (p<0.05).
Figure 8.35 QY of the cultures under the three different treatments in the tank cultivation system at time 0, after 15 days, 1 month, 45 days, 2 months, 75 days, and 3 months (n=3; error bar, 1 SD of mean)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
PA
M (
QY)
Treatment
Time 0 15 days Month 1 45 days Month 2 75 days Month 3

243
8.4.3.1 Biochemical analyses of tank trial applying the AMPEP extract to adult O. pinnatifida biomass
The biochemical composition of the cultures was analysed every month, for the three treatments
and triplicate samples were taken from each. The percentage (DW) of carbohydrates for the three
treatments (after one, two and three months) is reported in Fig. 8.36. Samples after one month
reported no statistical differences. While samples for the second month reported a statistical
difference between the cultures under the control and those under the 0.5 mg/l treatments, lastly
the results for the third months showed a statistical difference between 0.5 mg/l and Bathing (20
g/l) treatments (ANOVA, p<0.05; Tukey’s post hoc test). For the three sets together, in general
after two months all the treatments showed a statistical decrease of carbohydrates concentration
(Fig. 8.36) and higher results were generally reported for the control treatment.
Figure 8.36 Content of carbohydrates in samples cultivated in tank after 1 (blue bars), 2 (grey bars) and 3 (orange bars) months of cultivation under AMPEP and control treatments (n=3; error bar, 1 SD of mean)
Protein (% DW) content is reported in Fig. 8.37. No statistical differences were reported between
the treatments, or between the three sets of samples, showing a minimum variation of protein
content in the cultures (ANOVA, p<0.05; Tukey’s post hoc test).
0
5
10
15
20
25
30
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
Car
bo
hyd
rate
co
nte
nt
(% D
W)
Treatment

244
Figure 8.37 Protein content expressed in percentage of samples after 1 (blue bars), 2 (grey bars)
and 3 (orange bars) under AMPEP and control treatments (n=3; error bar, 1 SD of mean)
Pigments contents for chlorophyll-a and carotenoids (µg/g) are reported in Fig. 8.38 and
phycobiliproteins (PE and PC, mg/g) in Fig. 8.39.
Figure 8.38 Chl-a and carotenoids content in the three sampling times under AMPEP and control treatments (n=3; error bar, 1 SD of mean)
There was no statistical difference between the treatments for the first month for Chl-a or
carotenoids content. In the second month there was a statistical difference between Bathing (20
g/l) and the control treatments for the Chl-a content, while both Bathing and 0.5 mg/l treatments
were different from the control for the carotenoids content. For the third month, there was a
statistical difference for 0.5 mg/l and the other treatments (0.5 mg/l> control> bathing). The three
months analysed together showed that the first set of samples was statistically lower than the other
two. A statistical difference was also reported between months two and three (ANOVA, p<0.05,
0
5
10
15
20
25
30
35
40
45
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
Pro
tein
co
nte
nt
(% D
W)
Treatment
0
500
1000
1500
2000
2500
3000
3500
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
Ch
loro
ph
yll a
an
d c
aro
ten
oid
sco
nte
nt
(µg/
g D
W)
Treatment
Chl-a Month 1 Chl-a Month 2 Chl-a Month 3 Car Month1 Car Month 2 Car Month 3

245
Tukey’s post hoc test). In general, a statistical increase in pigment content was reported for all the
treatments for all of the experimental period, with the highest concentrations recorded for the 0.5
mg/l treatment at the end of the experiment for both Chl-a and carotenoids.
For the first and the third sets of samples, there was no statistical difference reported between the
treatments for the phycobiliprotein content, while for the second month there was a statistical
difference between Bathing and 0.5 mg/l treatment (PE content) and between the control and 0.5
mg/l treatments (PC). (ANOVA, p<0.05, Tukey’s post hoc test). The statistical analyses between
the three sets of samples for the PE content reported a difference between the sample at month
two for the Bathing treatment and 0.5 mg/l. While for the PC content, only the samples of month
one for the Bathing treatment were statistically different from the samples of month two for the
0.5 mg/l treatment (Fig. 8.39).
Figure 8.39 PE and PC (mg/g) content for the three sampling times under AMPEP and control treatments (n=3; error bar, 1 SD of mean)
Results for antioxidant content obtained from the FRAP assay for the three sampling periods are
reported in Fig. 8.40. Statistical analyses reported no differences among the treatments for the first
set of samples, while a statistical difference between Bathing and the other two treatments
(control>0.5 mg/l> bathing) was reported for month two, and a statistical difference between 0.5
mg/l and the other two treatments (control>bathing>0.5 mg/l) reported for the third month. The
statistical analyses between the three sets, identified in particular a higher concentration of
antioxidant compound in the first month of cultivation for the cultures under Control treatment
(Fig. 8.40).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
PE
and
PC
co
nte
nt
(mg/
g D
W)
Treatment
PE Month 1 PE Month2 PE Month 3 PC Month 1 PC Month 2 PC Month 3

246
Figure 8.40 FRAP assay of samples after 1 (blue bars), 2 (grey bars) and 3 (orange bars) under AMPEP and control treatments (n=3; error bar, 1 SD of mean)
Comparing the results obtained from the biochemical analyses for the flask and the tank
experiments applying AMPEP extract, PE and PC are statistically higher in the flask experiment
for control and Bathing treatments, while the opposite is true for the 0.5 mg/l treatment. The
carbohydrates concentration was higher in the cultures cultivated in tank compared to those in the
flask experiment, as well as for the protein. The FRAP assay showed statistically higher values in
the flask experiment compared to the tank, while the opposite was seen for the Chl-a and
carotenoids concentration.
8.5 Discussion
The effects of PGRs on tissue cultures of O. pinnatifida were investigated here for the first time.
Considering the analogy with plants, differentiation in some algae might also require the
intervention of specific chemical controllers (Evans and Trewavas, 1991). There is the possibility
that the photoperiodicity and seasonality in the growth behaviour of seaweeds needs to be
controlled by substances, such as PGRs (Lüning and Tom Dieck, 1989). Endogenous PGRs
(auxins, cytokinins, abscisic acid, gibberellins, etc.) have been previously detected in a number of
algae belonging to different evolutionary lineages (Stirk et al., 2014a; Tarakhosvskaya et al., 2007;
Mikani et al., 2015). Jennings (1969) and Dawes (1971) reported stimulation of growth in algae by
auxin and kinetin. Evans (1984) discovered that gibberellins promote cell division in Ulva but acted
as growth inhibitors in other species. Cytokinins were observed to promote the branching in F.
vesiculosus and other algae (Evans and Trewavas, 1991). Besides these studies, knowledge on the
impact these compounds have is still poor, and endogenous concentration of PGRs have only
rarely been measured, making it difficult to establish an appropriate exogenous concentration for
treating algae (Evans and Trewavas, 1991). Addition of exogenous hormones to culture media in
controlled environments could enhance the establishment of seaweed cultures, with subsequent
0
1
2
3
4
5
6
Control (F/2) Bathing (20 g/l) AMPEP 0.5 mg/l AMPEP
FRA
P (
µM
FeS
O₄/
mg
DW
)
Time

247
development growth of algal biomass. Imelda et al. (1998) suggested that use of exogenous PGRs
could act synergistically with native hormones, further promoting culture growth. Positive
responses to the use of PGRs in some seaweed species have been reported by earlier researchers
(Kaczyna and Megnet, 1993; Hayashi et al., 2008; Yong et al., 2014a; Anuraj et al., 2017).
The impact of phtyoregulators, auxins and cytokinins on the growth of a limited number of red
seaweeds has been previously demonstrated (Jennings, 1971; Fries, 1974; Fries and Iwasaki, 1976).
Treatments with natural or synthetic auxins have been shown to accelerate tissue growth in cultures
and callus development and to have regulatory effects on regeneration processes (Yokoya and
Handro, 1996; García-Jiménez et al., 1998; Yokoya et al., 1999). Other researchers have
demonstrated that auxin controls thallus branching of red and brown macrophytes on the principle
of apical dominance (Yokoya and Handro, 1996; Moss, 1974). Furthermore, Aguirre-Lipperheide
et al. (1995) suggested that red seaweed species responded more to exogenously supplied PGRs
than other seaweeds. Even if numerous authors have hypothesized that the cytokinins are involved
in the regulation of algal growth (Mooney and van Staden, 1986), relatively few studies have been
carried out on the role of cytokinins as endogenous growth regulators, in particular in macroalgae,
and specifically on red seaweeds. Limitations have included complications in controlling the
mechanisms of PGR factors on seaweed growth related to the presence of marine bacteria, known
also to release plant growth promoters (Bradley, 1991), and application of seawater in growth media
and potential disturbance of active compounds contained in the media that could interfere with
the results observed (Pedersén, 1973).
Nonetheless, PGRs were investigated in this thesis to understand their influence on the growth
rate, development and biochemical composition of O. pinnatifida, at a juvenile and adult stage. It
appeared that the hormone 2,4-dichlorophenoxyacetic acid (2,4-D) is not suitable for O. pinnatifida
cultivation if used alone, while in combinations with the other PGRs tested proved to slightly
increase the growth in the triple treatment. This compound is commonly used in herbicides (Stoker
et al., 2010) and its effect, when used alone, on the vegetative tissue of O. pinnatifida was mostly
damaging. The combinations of IAA and BA hormones seemed to be involved in the elongation
of the seaweed and branching, as already seen for other seaweeds (Yokoya and Handro, 1996).
This was particularly visible in the juvenile’s growth. Single hormone treatment in flasks also
affected the colour of the cultures, turning them from red to green in colour. Analyses of the
cultures’ pigments gave further confirmation that this was in fact happening. Usually, this change
in colour is as a result of damage and/or death of the seaweed, caused by the washing out of
phycobiliproteins, which are water soluble. This leaves the remaining Chl-a as the dominant
pigment (van den Hoek et al., 1995; Fleurence and Levine, 2016). This appears to be true also for
this study; however, only the colour of the biomass changed. The overall functionality and

248
physiology of the cultures were unaffected, as shown by PAM analyses. Whilst the treatments with
the hormones did not prove to be particularly effective in increasing the growth rate of this species,
at least for the concentrations and combinations trialled in this study, they had a more visible
impact on the biochemical composition of the cultures. This indicates that they might have some
role in regulating the biochemical pathways and processes of O. pinnatifida. Proteins, pigments, Chl-
a, carotenoids and phycobiliproteins and antioxidants content all increased, showing that the supply
of hormones can affect the biochemical composition of this species, as already seen for other
seaweeds (Han et al., 2018).
The impact of AMPEP extract on the growth of O. pinnatifida was also investigated. The positive
benefits derived from Ascophyllum extracts applied for seaweed cultivation have been previously
recognized and have been reported as been biostimulant/bioeffector (van Oosten et al., 2017).
These include: effects as enhancer or inducer for the direct axis formation and growth, as abiotic
response, (Hurtado et al., 2009, 2012; Yunque et al., 2011; Tibubos et al., 2017), as mitigator to
biotic responses to damaging endophytes (Borlongan et al., 2011; Hurtado et al., 2012; Loureiro et
al., 2012), epiphytes, and epibionts (Loureiro et al., 2009; Marroig et al., 2016), and as stimulant in
in-tank cultivation (Pedra et al., 2017).
The capacity of AMPEP extract to control epiphytes colonisation was confirmed in this study,
especially for juveniles, and particularly for the 0.5 and 3 mg/l treatments. The treated biomass had
much lower levels of epiphyte contamination compared to the untreated O. pinnatifida juveniles.
Cultivation of juveniles under laboratory conditions have been observed to be particularly
susceptible to contaminants during early stages of development (pers. obsv.). The application of
this extract appeared to reduce levels of contamination of the cultures and in controlling epiphytes.
The AMPEP seems to encourage branching of the juveniles as well as the formation of trichoblasts.
These are colourless specialized hair cell types that occur on the surface of many marine and
freshwater members of the Florideophyceae (Oates and Cole, 1994; Delivopoulos, 2002). First
discovered in the Floridae (Rosenvinge, 1911), there are two kinds of trichoblasts that have evolved
within the Rhodomelaceae. One is pigmented and usually persistent, while the second is usually
colourless, deciduous, often present on the vegetative axes (Hommersand, 1963) and are eventually
shed (Garbary and Clarke, 2001). O. pinnatifida trichoblasts belong to this second type
(Hommersand, 1963). Trichoblasts are thought to be related to nutrient uptake (Oates and Cole,
1994) or with reproduction (Falkenberg, 1901; Bold and Wynne, 1985) and protection of
tetraspores (Duckett et al., 1974). Despite their significance and frequent occurrence in most of the
Rhodomelaceae, they remain one of the least investigated structures of the red algae (Delivopoulos,
2002). In this study, production of these hair-like structures seemed to be stimulated by the
AMPEP extract, and they were absent in the control. Their role in this case might be related either

249
to a form of physical protection or to an attempt to maximise nutrients absorption, which is further
supported by the change in the colour of the cultures. The colour of the juveniles was deeply
affected by the extract, changing pigmentation from red to a light green colour. This change in
colour, as already seen in Chapter 6, might be due to a N limitation, since the same effect was not
visible in cultures under F/2 medium, with consequent reductions in phycobiliprotein contents
and a greener colour of the cultivated juveniles (Penniman and Mathieson, 1987). The growth rate
for the 0.5 mg/l treatment performed statistically better than the others (p<0.05), demonstrating
that a lower concentration of this extract is more suitable for this species at this early stage of
development. Previous authors have already reported the highest efficiency of the lower
concentration of the tested extract on cultivation of other species (Hurtado et al., 2012).
The AMPEP extract proved to be an effective control of epiphytes and their proliferation
contamination also in the flask experiment. The colour of the cultures was also affected, as
previously seen in the juveniles, and this observation was confirmed by biochemical analyses. The
levels of pigments measured were statistically lower in cultures grown with the AMPEP extract
compared to the control and the bathing treatments. Trichoblasts were also visible in the adults
cultivated with the extract and developed from the apical extremities of the adults’ branches rather
than from the thallus. In this case, their role could also be in the protection of the tetraspores, as
already seen for other species (Duckett et al., 1974), beside the function of nutrient absorption
(Oates and Cole, 1994). SGR and growth showed that there was no a statistical difference among
the treatments tested in terms of increased biomass. However, it is worth considering the
application of the extract in the early stages of development of this species, when the plantlets are
more susceptible to contaminants and the growth rate is slower, to then substitute it, once they
reach a certain development stage, with a cultivation medium more suitable for adult plants (e.g.
F/2). Especially considering the effect of the extract in enhancing branching and apical elongation
(Hurtado et al., 2009, 2012; Yunque et al., 2011; Tibubos et al., 2017), which will eventually result
in an increased biomass. Biochemical analyses showed that cultures treated with the extract had
statistically higher levels of carbohydrates, probably due the P source applied (Hopkins and Huner,
2009), compared to the control and the Bathing treatments, to which corresponded a lower level
of protein. The inverse trend for carbohydrates and protein content in regard to the nutrient
applied has been reported previously (Smit et al., 1997) and discussed in Chapter 6.
The antioxidant compounds in O. pinnatifida cultures were not statistically increased by the
application of the extract, at least for the concentration trialled. Previous research has demonstrated
that this extract is able to improve the antioxidant activity of some species of red algae (Borlongan
et al., 2011; Tibubos et al., 2017). Instead, the statistically lowest values were reported for cultures

250
under the two different concentrations of the AMPEP extract tested, as already reported for certain
strains of Kappaphycus (Hurtado et al., 2012).
There have been several trials to cultivate different species of red seaweed in land-based tank
systems, such as the cultivation of Gracilaria (Buschmann et al., 1994), Gelidium (Friedlander, 2008),
Chondrus crispus (Bidwell et al., 1985), and Palmaria palmata (Corey et al., 2013; Grote 2016, 2019),
ranging from simple ponds to intensively operated tanks (Buschmann et al., 2017). One of the first
successful commercial-scale pond facilities for a red seaweed operates in the Canadian Maritimes,
to produce Chondrus crispus (Hafting et al., 2015). Commercial on-land pond cultivation has also
been developed in Israel for Gracilaria sp. (Buschmann et al., 2017). Previous attempts of cultivation
in tank of O. pinnatifida were performed in Portugal and included flask trials and small-scale outdoor
trials with some positive results (Silva, 2015; Gonçalves, 2018), as discussed in Chapter 5.
The tank experiment, comparing the AMPEP extract to F/2 medium, proved to be effective in
increasing the biomass of the cultures, which were also dark red in colour and had a thick texture.
This, for the first time, establishes a successful pioneering methodology for the on-land indoor
cultivation of this species. There are further improvements that could be made to make this system
more efficient. For example, the supply of high-quality seawater and the successive treatments used
to treat it, such as filtration, UV treatments, etc., are fundamental for O. pinnatifida cultures, as well
as a continuous flow of water through the system. In this experiment, filters (100-20-10-5 µm) were
changed every week, nonetheless, brown filamentous seaweed, similar to Ectocarpus sp. in
appearance, were found growing in the filter system. Ectocarpus sp. is a common fouling alga (Morris
and Russell, 1974), found mostly as free-floating filaments in intertidal to sublittoral zones of
temperate regions (Baweja et al., 2016). It could easily be pumped in by the water supply system.
As already seen by other authors, the cause and extent of an epiphytic outbreak can depend not
only on the quality of the cultivated strain, but also on abiotic parameters of the site, management
practise and seasonal weather fluctuations (Vairappan et al., 2006). A solution to this would be to
place the main inlet pipe further from the seashore and at a lower depth in the site. This would
reduce the likelihood of pumping in unwanted contaminates as well as overcoming variation in
flow rate due to the tide cycles. Nonetheless, the pre-filtration and UV treatment of the incoming
seawater was mostly effective, and it was only in the last 15 days of the cultivation trial that what
appeared to be Alaria sp. was discovered. This is not a common biofouling organism of other
seaweed species and is also not normally seen in tanks used during cultivation trials, being usually
cultivated on ropes (Arbona and Molla, 2006; Edwards and Watson, 2011). Spores of this species
are release between November and March (Birkett et al., 1998b), which may explain the presence
of Alaria sp. in the system, probably drawn in by the water supply system.

251
Aeration in tumbling cultivation of seaweed cultures is fundamental, as previously reported for
other species, such as Chondrus crispus (L.) Stackh (Neish and Fox, 1971; Neish and Shacklock, 1971;
Neish et al., 1977) Gracilaria, Agardhiella and other genera (DeBoer and Ryther, 1977), in order to
replicate the natural motion of the seawater and provide nutrient exchange. This should be
considered along with supplementing the cultures with CO₂, which was not investigated in this
study. This measure is normally applied to control the pH and the gas exchange of the cultures, as
well as alleviate carbon limitations. This leads to greater biological productivity with an expected
increase in the photosynthetic storage of carbon, consequently stimulating the growth of cultures
(Duarte, 1992; Duarte et al., 2005). In the absence of this, a high flow rate, such as the one adopted
here, will guarantee a continuous exchange of water, aid in gaseous exchange as well as nutrient
and inorganic carbon uptake. Seaweeds obtain the necessary nutrients for growth directly from the
medium in which they are grown in. It is important that the medium is circulated to allow this to
happen and renewed at regular intervals (FAO, 2012). Circulating the water in tanks results in the
movement of the biomass, creating a dynamic light regime that can enhance growth rates (Kübler
and Raven, 1996).
Due to the limited possibility of scaling up related to the facilities available at SAMS, scaling-up of
the production with a transfer of the cultures to bigger volumes was not tested. Instead, cultures
were brought to the original weight after one and two months (since they were doubling their
weight), in order to avoid competition for light, nutrient and an excess of biomass density in the
tanks. Seaweed with a high stocking density are usually self-shading, resulting in reduced
photosynthesis, growth rates, and consequently lower seaweed production rates (Kim et al., 2014,
Corey et al., 2014). Scaling up the cultures to larger volumes could potentially further increase the
biomass production of this seaweed.
The biochemical analyses for the tank experiment involved three sets of sampling, one been taken
every month. This was carried in order to determine if there was any variation in the biochemical
composition of the cultures not only because of the treatments been tested, but also due to the
impact the experimental period might have on growth. For all the treatments there was a general
reduction in their carbohydrate content from month one to month two, followed by an increase in
the final month with the control performing the best. This is potentially linked to the P source
supplied, which as discussed in Chapter 6 influence the carbohydrates content of the species,
compared to the other treatments tested (Hopkins and Huner, 2009). Treatments and length of the
experiment did not affect the protein content which generally remained unaltered, indicating N
was not a limiting factor. There was an increase in the pigment content, and this was supported by
the PAM data. This parameter is normally used to represent the maximum photochemical
efficiency of PSII (Baumann et al., 2009). This increase in content could be due to the quality and

252
quantity of light applied during the experiment. Light characteristics (e.g. spectral quality, quantity,
and duration) are known to have a profound effect on seaweed metabolism and development (Su
et al., 2014). Algal pigment synthesis is also regulated by various photoreceptors that absorb light
at different wavelengths (López-Figueroa and Niell, 1990, 1991). The spectrum of the Gro-Lux
lights used for this experiment includes white light, as well as high levels of blue and red radiation,
which might have influenced the pigment content.
Unlike the flask experiment, the AMPEP treatments tested in the tank system did not affect the
colour of the cultures. Instead, the cultures remained a dark red-purple colour for the entire length
of the experiment. This was evident also from the data for PE and PC concentration. The pulsing
supply of the nutrients, coupled with water motion, reduced N limitation occurring in the cultures
preventing the loss of pigments, such as phycobiliproteins. These pigments are affected by N
availability (Vergara and Niell, 1993; Dawes, 1995; Vergara et al., 1995), and as noted in the flasks,
N limitation lead to the loss of the red colouration of the cultures.
Despite what has already been observed in plants (Craigie, 2011; Khan et al., 2009) and in other
seaweeds (Yunque et al., 2011; Tibubos et al., 2017; Pedra et al., 2017), the AMPEP extract did not
improve the antioxidant content of O. pinnatifida. This was similar to the observations from the
flask experiment. The antioxidant content decreased in all of the treatments tested over the entire
length of the tank experiment. Similar results have been reported for certain colour morphotypes
of Kappaphycus, another red seaweed species. Again, with this species, there was no improvement
in the antioxidant content which decreased during the reported experimental trial (Hurtado et al.,
2012).
The differences in biochemical composition of cultures treated with AMPEP extract at the same
concentration, but cultivated in flask and tanks, could be due to several factors, not just N
limitation. Both experiments had utilised different cultivation conditions. While the temperature
was the same (9-10 °C), the light photoperiod was different (for the flask experiment: 12:12; for
the tank experiment 24/7), as well as the type of lights (white fluorescent light in the flask
experiment and Gro-lux in the tank). In addition, the cultivation systems applied were different.
Medium was static and changed twice per week in the flasks with aeration from the top. In the
tanks there was continuous water exchange and tumbling of the cultures, with aeration provided
from the bottom. Both experiments also used different biomass densities (10 g/l in the flask
experiment, 4.54 g/l in the tank experiment). Resulting differences in profiles are unsurprising since
parameters, such as light, water flow, and nutrient input, impact on the biochemical composition
of seaweeds, as already reported in other species (Dring et al., 1996; Denis et al., 2010; Chow, 2012;
Yang et al., 2015).

253
An interesting effect of the extract on cultures, either juveniles or adults, in flasks or tanks, is the
formation of accessory branches. This has been reported in previous studies for other algae such
as Ascophyllum sp. and Kappaphycus sp. (Rayorath et al., 2008a, b; Borlongan et al., 2011). The
development of new branches was detected not only from the holdfasts, which commonly follows
two patterns of growth, an apical elongation and a lateral ramification, but also from the thallus, as
observed only in the treated samples. The AMPEP extract seems also to promote the formation
of callus tissue, again as previously reported for other species (Hurtado et al., 2009; Yunque et al.,
2011). These effects might be an important consideration for long term cultivation of the
investigated species, since each of these bulges will become a new branch, contributing to increases
in weight and biomass.
Finally, AMPEP extract and the hormones were not tested in combination. Further research could
investigate the combined effects of these two very different sets of compounds. Combining both
has proved to be successful in enhancing growth and increasing the biomass in other species
(Hurtado et al., 2009, 2012; Hurtado and Critchley, 2018; Yunque et al., 2010; Tibubos et al., 2017).
It has been hypothesized that this is probably due to the interaction of the extract with compounds
such as auxin-like, gibberellin-like (Rayorath et al., 2008a, b), cytokinins, precursors of ethylene and
betaine (Mckinnon et al., 2010), which are potentially involved in enhancing plant growth responses
(Crouch, 1990).
8.6 Conclusions
The concentration and combinations of phytohormones tested did not affect the growth
of this species (for both juveniles and adults); however, they did influence developmental
growth;
It is known that phytohormones can be applied in seaweed cultivation for targeting
products such as pigments, fatty acids, and polysaccharides (Han et al., 2018). This was
confirmed in this study, since treated cultures had statistically higher levels of
phycobiliproteins, proteins, Chl-a and carotenoids;
The response of the juveniles and the adults cultivated with the AMPEP extract was
different. A possible further application of the extract could be on the juvenile stages of O.
pinnatifida in order to increase the growth rate, enhance branching, and control the
contamination of epiphytes;
AMPEP extract was able to increase pigments and carbohydrates content in adult O.
pinnatifida biomass;

254
There was no statistical difference in terms of growth and biomass increase in the tank
experiment between the three tank treatments; all were suitable for tank cultivation of this
species;
However, AMPEP extract enhanced the branching of this species, as already determined
for other algae, which will eventually lead to an increase in biomass. Its application as a pre-
treatment for long-term tank cultivation should be considered;
This was the first time that an indoor on-land system at this scale was tested for the
cultivation of this species. It proved to be successful in promoting an increase in biomass,
controlling epiphytes contamination and ensuring an ideal biochemical profile of the
cultures. There are improvements that would need to be applied; however, the proposed
system offers a commercial alternative to wild harvesting, making the production of O.
pinnatifida sustainable and independent from seasonal variation in biomass and quality.

255

256
Chapter 9 Conclusions and future work
9.1 Conclusions
Seaweed consumption and utilization as a sector continues to grow, with multiple applications for
algae and their extracts. The investigation into the cultivation of new species is expanding with
countries within Europe starting to invest in this form of aquaculture. In order to shorten the
distance between themselves and Asiatic countries, who are the leaders in the production and
consumption of seaweeds, the interest has been towards low volume but high-value species. This
will open the path for the cultivation of those species that are slow growing and “smaller” than the
more well- known kelp species. These are normally harvested in UK, but this is labour intensive,
time consuming, and care must be applied in order not to overharvest a wild population. Little is
available in term of legislation for harvesting of small species, such as O. pinnatifida, in the UK.
Advice centres on mainly cutting the seaweeds, rather than tearing them completely off the solid
substrate they are growing on. This is so part of the algae is left behind for it to regrow, and a
maximum of one third of the species can be taken from a shore in any year, again ensuring a viable
population is left behind (Angus, 2017). Investigating and establishing cultivation methodologies
to produce Pepper Dulse in Scotland, represents a sustainable and profitable alternative to wild
harvesting of this species. This seaweed is known amongst chefs as the “truffle of the sea” and it is a
valuable ingredient already sold by different companies. Furthermore, its particular biochemical
profile, rich in secondary metabolites (e.g. antioxidants) makes it a valuable species that can be
applied in nutraceutical, pharmaceutical and cosmetic fields.
If final applications such as pharmaceutical, nutraceutical, and cosmetic are to be exploited, there
is a need for fundamental knowledge on the regulation and production of compounds of potentially
interest within a chosen species. In this case O. pinnatifida.
With this in mind, the main objectives of this thesis were:
Investigate and identify the parameters that are fundamental for the cultivation of this
species in order to establish a production system both in vitro under laboratory conditions
and in tanks, indoor and outdoor;
Investigate the biochemical composition of this species and how this is regulated. How this
varies in the wild (according to different abiotic factors) and in cultivated specimens
(according to variation and manipulation of the cultivation conditions);
How cultivation conditions can be tailored to improve biomass productivity;

257
Assess the biochemistry and metabolomics profile of the species and its variation according
to the seasons. Moreover, to assess the impact that seasonality potentially has on the overall
taste profile;
Identify phylogenetically the species investigated.
The flowchart (Fig. 9.1), already represented in Chapter 1, presents the two alternative ways
for the cultivation of Osmundea pinnatifida and underlines in blue the preferred methodology.
The research conducted within this thesis identified the vegetative cultivation as the best
methodology for the commercial production and exploitation of the species and the
diagram underlines in blue boxes the steps to apply.
Figure 9.1 Flow chart that highlights the best methodology to apply for the commercial cultivation and exploitation of the species

258
9.2 Contribution of the work
Chapter 2: Several studies have previously focused on the taxonomy and phylogenetic of
O. pinnatifida, changing profoundly its classification (e.g. identifying it as member of the
genus Osmundea rather than Laurencia) (Nam, 1999; Nam et al., 2000). Most of these studies
focused on specimens from Spain and the Canary Islands. This was the first time that
different specimens from the West coast of Scotland were molecularly analysed and
compared to sequence data available for this species on the NCBI database. The best DNA
extraction method was identified, and the PCR cycle optimized. Phylogenetic analyses
confirmed what had already been reported by previous authors, that the Laurencia complex
is a monophyletic group that includes eight genera, (Garbary and Harper, 1998; Nam et al.,
2000; Martin-Lescanne et al., 2010; Machín-Sánchez et al., 2012, 2018), with the genus
Osmundea been distinct from the Laurencia genus. The sequences divergences between the
Laurencia complex and the genus Osmundea sp. were similar to those reported by other
researchers (Machín-Sánchez et al., 2012). It was comparable with intergeneric sequence
divergences found in other red algal families (Hommersand et al., 1994; Lin et al., 2001;
McIvor et al., 2002; Martine Lescanne et al., 2010; Machín-Sánchez et al., 2012). The results
obtained from the phylogenetic methods employed, indicated that the samples collected
can be identify as O. pinnatifida species, that they are grouped in the same clade as the O.
pinnatifida sequences from GenBank, and that is it possible to recognize different closely
related subclades, on the basis of the bar-codes analyses. This information can be further
applied to cultivation of the species, such as optimization of strains and DNA engineering.
Chapter 3: Even if the biochemical composition of wild specimens of O. pinnatifida from
UK has been described previously (Marsham et al., 2007), it is only for one time point in a
year. This was the first time that the biochemical composition of this species was
investigated across a whole year to elucidate the impact that seasonality has on composition.
Different extraction protocols were developed, or existing ones implemented in order to
maximise the results obtained. Analyses showed seasonal patterns and variations for all the
analysed components, which included carbohydrates, proteins, pigments (chlorophyll-a,
carotenoids, PE and PC), lipids, moisture, C:N ratio, DOCN content, antioxidants, and
metals. Statistical analyses gave for the first time an insight into the correlation between
biochemical composition and abiotic factors, highlighting important consequences for the
management of harvesting and commercial exploitation of the species.
Chapter 4: The seasonal variations in the metabolomic profile of this species across one
year has been presented for the first time. Furthermore, the flavour profile of the species
was investigated for the first time, comparing two seasonal samples to assess the effects of

259
seasonality on taste/flavour. This study confirmed that this species has great potential for
further food, nutraceutical and pharmaceutical applications, and gave an insight into
seasonality and regulation of the metabolic profile of O. pinnatifida.
Chapter 5: Epiphytes are one of the major problems in the cultivation systems of seaweed.
They were controlled and mitigated against in O. pinnatifida cultivation with the application
of a chemical treatment. This proved to be efficient, without affecting the physiology and
biology of the species, was coupled to either a tide-simulation system or applied alone. The
treatment for epiphytes mitigation and control proved to be successful in vitro, tanks
(indoor and outdoor) and on sea cultivation trials (Appendix C.5).
Spores production under laboratory conditions was feasible and multiple releases occurred.
It was possible to grow the released spores up to the juveniles’ stage in vitro cultivation.
The process was slow and needs further investigation. Treatments to control the epiphytes
contamination of this early stage were identified (Chapter 8). However, keeping in mind
the flow chart defined in Fig. 9.1, a vegetative cultivation of the species seems to be the
best option at the moment for a commercial exploitation of this seaweed.
Chapter 6: Different media types and nitrogen sources for flask cultivation of O. pinnatifida
have never been tested before. All the media trialled were able to support the growth and
the biochemical requirements of the species been investigated. N sources do not influence
the growth of the cultures but have an impact on the biochemical composition. The
medium that performed the best was F/2, which appeared to be more suitable for the
vegetative cultivation of O. pinnatifida. It was able to support the growth of the culture, to
control the epiphytes contamination, to guarantee a suitable biochemical profile and to be
more economical to be applied, compared to the other types tested. This study led to the
identification of the best nutrient requirements for cultivation in flask and in tank of O.
pinnatifida.
Chapter 7: This chapter explored the possibility to cultivate O. pinnatifida in the Algem®
photobioreactor, with pioneering and successful results. Currently this PBR is only applied
for microalgae cultivation. This study showed that the PBR tested is able to statistically
increase the concentration of antioxidants compounds in the species. Increase in growth
was successful in the system, with neither the physiology nor morphology of O. pinnatifida
negatively affected. This study provided important information for commercial
exploitation of this species, introducing a methodology for cultivation in a PBR, resulting
in an increase in biomass and a tailored biochemical profile. This may have promising
further nutraceutical and pharmaceutical applications, once the cultivated biomass is
processed and analysed.

260
Chapter 8: This was the first time that exogenous hormones and fertilizer were applied to
cultivation in flask and in tank of O. pinnatifida. While the concentration and combination
of hormones trialled in this study did not affect the growth of the cultures, they affected
the development and the biochemical composition. This study showed that the response
of the cultures to the AMPEP extract depends on the stage of development of the biomass.
AMPEP was successful in increasing the growth rate only in juvenile cultures. The extract
helped in controlling and reducing epiphytes in both adult and juvenile cultures, influencing
the biochemical composition of adults. Moreover, the extract was able to increase the
branching of the cultures, which in a long-term cultivation system, might become important
for maximising productivity of the cultures. The extract did not increase the antioxidant
content of the cultures, as reported by previous authors for other seaweeds (Tibubos et al.,
2017; Pedra et al., 2017).
The AMPEP extract was trialled in a tank experiment in comparison with F/2 medium.
For the first time an indoor tumbling tank cultivation system for the vegetative production
of O. pinnatifida tetrasporophytes was established. Cultures were cultivated for three
months, maintaining a positive growth rate for the entire length of the experiment, with
the maximum SGR recorded at almost 5%day⁻¹. PAM response positively increased in the
cultures during the trial. While there was no difference in terms of weight increases between
the treatments tested, they did modify the biochemical composition of the biomass
produced. However, despite what has already been reported by previous authors (Hurtado
et al., 2014), the extract, in the concentrations tested, was not able to increase the
antioxidant content of O. pinnatifida.
The development of this cultivation system might provide a sustainable supply of the
resource, independently from wild harvesting and seasonality. Furthermore, the system is
able to improve the biochemical profile of the species and control epiphytes
contaminations.
9.3 Practical implications resulting from the findings
As already stated by previous authors, O. pinnatifida is a promising species, not only for its explosive
taste that makes it a sought-after seasoning, but also for several nutraceutical and pharmaceutical
applications. Its value as source of antimicrobial, antiviral, antibacterial, antifungal, antifouling and
anticancer compounds has been widely recognised and investigated (Campos et al., 2019; Silva et
al., 2018; Rodrigues et al., 2019, 2016, 2015; Paiva et al., 2014; Küpper et al., 2014; Sabina and Aliya,
2011, 2009; Patarra et al., 2011; Rizvi and Shameel, 2005, 2004, 2003). Knowing how to
trigger/control the production of these compounds might have important consequences for
commercial cultivation of this species and its commercial exploitation. This project investigated

261
and established possible cultivation methodologies for the production of tailored and high-quality
cultures of O. pinnatifida, from flasks, to PBR, to tank trials. Reproductive and vegetative cultivation
were trialled, and a final vegetative indoor tumbling system developed. This could be potentially
scaled up and have further commercial applications, making the production of this species
independent from seasonality and variation in quality/quantity of biomass, tailored and sustainable.
Biochemical analyses of the species, both from the wild as well as cultivated samples, gave
important knowledge on the composition, how it varied seasonally and under cultivation
conditions. The knowledge gained can be applied for both harvesting management (understanding
how the morphology and biochemical profile change seasonally, gaining information on the growth
patterns of the species, and on when it is best to harvest the biomass for further processing) and
cultivation applications. Treatments for control and management of epiphytes in O. pinnatifida
cultures were developed and successfully identified. This aided in the establishment of cultivation
methodologies for this particular species. Metabolomics analyses highlighted unknown and rare
compounds, and their seasonal variations. Thus, providing an insight into patterns and trends of
the biochemical composition of the species according to the season. Information obtained from
these findings are fundamental in improving the knowledge available on the species biology and
ecology, as well as providing pioneering information for the on-land indoor cultivation of O.
pinnatifida, for further commercial applications.
9.4 Limitations of the research
The results obtained from this work are pioneering and meaningful. However, there are some
limitations. These are namely: the difficult in investigating and establishing cultivation
methodologies for a species that, except for its biochemical profile, has been scarcely investigated;
the slow development and growth of this species, observed also in the wild, and the complexity of
the reproductive cycle in order to achieve an efficient production through reproductive
methodologies. Other limitations related mainly to technical issues, such as a lack of experimental
areas or the adaptation of alternative arrangements, in line with what available at the institute, which
proved not ideal for the goals of the project.
9.5 Future work
Further research and development will be necessary in order to strengthen the methodologies
established in this study. This would guarantee success at the scaled-up stage for the commercial
exploitation of O. pinnatifida for food, nutraceutical, and pharmaceutical applications. Aspects,
which merit further research, have been identified on an area basis:

262
Metabolomics analyses for taste investigation: Further analyses should be conducted
again on a seasonal basis and also with cultivated material, involving a tasting panel and
application of the XCMS non-targeted approach (McDougall et al., 2014) in order to
further investigate how the flavour and taste of the species changes and which compounds
are responsible for it;
Minerals composition: Iodine was not considered in this study due to analyses logistics;
however, this element should be evaluated. Especially considering that seaweeds are one
of the best natural sources of iodine and the implication that this mineral has on the health
and diet of the human body (Yeh et al., 2014; Bocanegra et al., 2009). Bromine should be
characterised as well since O. pinnatifida is a known source of active bromine halogens
(Suzuki et al., 2005, Küpper et al., 2014).
Nutrient regime: Results obtained from Chapter 6 identified that nutrients and how they
vary, had an influence on biomass production and biochemistry. In particular, further
investigations should be undertaken in relation to the concentration of P, as well as an
analysis to determinate the absorption rate of nutrients and the impact this has during
cultivation. This would allow a more complete feeding strategy to be determined in terms
of how often media should be changed and renewed for all life stages of O. pinnatifida.
Applications of hormones and AMPEP fertilizer: Different concentrations of
hormones and AMPEP extract, compared to those investigated in this study, should be
tested. A combination of AMPEP and PGRs treatments should be undertaken in order to
further investigate the potential synergistic effects of the fertilizer and hormones on growth
and biomass productivity. Different concentrations of the AMPEP extract should be
evaluated to verify if these might result in an increase of antioxidant compounds in the
cultures as seen in other red algae (Hurtado et al., 2014).
Spores production: Studies should be conducted to further investigate methodologies for
improving spore growth and survival rates. For example, different substrata should be
evaluated as well as decontamination treatments. While smooth surfaces, such as plastic
and glass slides, should be avoided, the focus should be directed towards collectors such as
ropes, or rough substrata e.g. shells and rocks, ideally resembling a barnacle surface, which
is the common habitat of the species and probably linked to the settlement of wild spores
as well.
Indoor cultivation: The tumbling cultivation indoor system developed in this study
proved to be effective and successful for the cultivation of this species. However, in order
to guarantee a commercial application and scale-up of it, some adaptations are necessary,
as detailed in Chapter 8. The water quality should be carefully monitored, followed by an

263
efficient sterilization treatment that should include filters (down to 5µm) and UV
sterilization system in order to keep under control contaminations. Particularly,
temperature, salinity, and pH should be periodically recorded. The supplementation of the
system with CO₂ should be evaluated, considering its importance in biomass increase and
pH control. Scaling up of the system should be achieved in order to avoid nutrient
competition of the cultures and growth decrease, due to high density.
9.6 Recommendations
Although some challenges and some bottle necks still need to be solved, vegetative cultivation of
O. pinnatifida in an indoor tumbling system proved to be a promising methodology with potential
commercial applications and should be preferred to cultivation through reproductive cycle. The
following protocol highlights the steps necessary for a vegetative cultivation system of O. pinnatifida
in Scotland, taking into account all factors investigated in this thesis.
1. Collection of wild material: Wild material should be collected using scissors/knives,
between October and February, at low tides from mid-exposed sites, distant from river
inputs and human activities. Only healthy and free from contaminants cultures should be
chosen in order to reduce possibility of contamination from other organisms and
subsequently maximise the cultivation. Avoid depopulating an area and only if necessary,
remove the holdfast, otherwise leave part of the seaweed behind for it to regrow. Place the
biomass in a Cool-box with wet tissues and bring it to the laboratory for further processing.
2. Cleaning and pre-treatment of the material: Wash the harvested biomass with filtered and
sterilized seawater in order to remove debris and grazers. Carefully clean the biomass
manually from any visible epiphytes and grazers. Rinse again with filtered and sterilized
seawater. Clean the biomass with a 0.5 % KI solution in FSW for three cycles of 30 minutes
each, in a controlled temperature room at 10°C, then rinse with plenty of filtered and
sterilised seawater in order to ensure the KI solution has been removed. Leave overnight
at 10°C in FSW with aeration. Then place the biomass in acclimation tanks, at 12:12
photoperiod, 100 µmol/m²/sec growth lux light, 10 °C, powerful bubbling and with no
nutrients added for at least 48. Acclimation prior to cultivation can last up to 1 month but
nutrients (preferably F/2 medium) must be provided. If this is the case a 48-hour starvation
period prior to cultivation is recommended.
3. Tank tumbling cultivation: Preferably use white conical tanks with aeration provided from
the bottom and light source placed on top of the tank, at a distance of 40-50cm. Place the
tanks (volume can be scaled up as the production increases) in a CTR at 10°C, light
provided by Growth lux light 100µmol/m²/sec and preferably at 18:6 or 24:0 photoperiod.
Density of the cultures should be between 3 and 4 g/l. A continuous flow rate of high

264
quality filtered and sterilized seawater (FSSW) down to 5 µm is necessary. Provide strong
bubbling in order to guarantee a tumbling culture and consider input of CO₂ for pH control
and increased biomass productivity. Supply F/2 nutrient with pulse feeding at least 2-3 per
day, according to the flow rate of the system. Clean the system periodically (weekly or every
10 days). Monitor continuously pH, salinity, light intensity, and temperature. Do not allow
temperature to rise above 15°C in the tanks.
4. Harvesting and processing: Harvest older tissue periodically, leaving in the cultivation
system younger material for further growth. This would facilitate the establishment of a
semi-continuous cultivation system, ensuring continuous stock in the system and avoiding
repeated harvesting of wild populations. Finally, processing the harvested biomass for
either food or extraction of commercially valuable compounds.
9.7 Concluding remarks
Results obtained from this study are pioneering and describe a potential cultivation system for the
sustainable production of Osmundea pinnatifida. This species has a biochemical profile suitable for
human consumption as well as producing other interesting compounds (such as antioxidants) that
could be further exploited and commercially applied. Knowledge gained on the seasonality of O.
pinnatifida and variation in the biochemical profiles according to abiotic parameters, might become
useful for harvesting and cultivation management of this species. The tumbling cultivation system
developed showed promising growth rates, the potential for scaling up and commercial
development of this methodology. Establishment of a successful O. pinnatifida cultivation system
will require collaboration between industrial and research institutes, and even if might appear
challenging and it requires further development, the research presented here is promising, valuable
and commercially relevant.

265

266
References
Abe, T., Kurihara, A., Kawaguchi, S., Terada, R., and Masuda, M. (2006). Preliminary report on the molecular phylogeny of the Laurencia complex (Rhodomelaceae). Coast. Mar. Sci. 30(1): 209213.
Adams, J. (2016). An overview on seaweed uses in the UK: Past, present and future. Seaweed Resources 36: 16-21.
Aguilera, J., Bischof, K., Karsten, U., Hanelt, D., and Wiencke, C. (2002). Seasonal variation in the ecophysiological patterns in macroalgae from an Artic fjord. II. Pigment accumulation and biochemical defence systems against high light stress. Mar. Biol. 140: 1087-1095.
Aguirre-Lipperheide, M., Estrada-Rodríguez, F.J., and Evans, L.V. (1995). Facts, problems, and needs in seaweed tissue culture: an appraisal. J. Phycol. 31: 67788.
Ahmad, V.U. and Ali, M.S. (1991). Terpenoids from marine red alga Laurencia pinnatifida. Phytochemistry 30: 4172-4174.
Ahmad, V.U., Ali, M.S., and Bano, S. (1990). Marine natural products XII: a new metabolite laurol from the red alga Laurencia pinnatifida. Sci. Pharm. 58: 299-301.
Ahn, G.N., Kim, K.N., Cha, S.H., Song, C.B., Lee, J., Heo, M.S., Yeo, I.K., Lee, N.H., Jee, Y.H., Kim, J.S., Heu, M.S., and Jeon, Y.J. (2007). Antioxidant activities of phlorotannins
purified from Ecklonia cava on free radical scavenging using ESR and H₂O₂- mediated DNA damage. Eur. Food Res. Technol. 226: 71-79.
Al-Alawi, A.A., Al-Marhubi, I.M., Al-Belushi, M.S.M., and Soussi, B. (2011). Characterization of carrageenan extracted from Hypnea bryoides in Oman. Mar. Biotechnol. 13: 893899.
Alba, K. and Kontogiorgos, V. (2017). Seaweed polysaccharides (Agar, Alginate Carrageenan). 10.1016/B978-0-08-100596-5.21587-4.
Alghazeer, R., Whida, F., Abduelrhman, E., Ammoudi, F., and Azwai, S. (2013). Screening of antibacterial activity in marine green, red and brown macroalgae from the Western coast of Libya. Nat. Sci. 5: 714.
Álvarez-Vázquez, M.Á., Prego, R., Caetano, M., De Uña-Álvarez, E., Dovald, M.M., Calvo, S., and Vale, C. (2017). Contributions of trace elements to the sea by small uncontaminated rivers: Effects of a water reservoir and a wastewater treatment plant. Chemosphere 178: 173-186.
Amsler, A.C.D and Searles, R.B. (1980). Vertical distribution of seaweed spores in a water column offshore of North Carolina. J. Phycol. 16 (4): 617-619.
Anantharaman, P., Devi, G.K., Manivannan, K., and Thangavel, B. (2011). Vitamin-C content of some marine macroalgae from Gulf of Mannar marine biosphere reserve, southeast coast of India. Plant Archives. 11: 343-346.
Andersen, R.A. (2005). Phycological Methods: Algal Culturing Techniques. Academic Press, Elsevier Publ. USA, UK. pp. 578.
Andrade, P.B., Barbosa, M., Matos, R.P., Lopes, G., Vinholes, J., Mouga, T., and Valentão, P. (2013). Valuable compounds in macroalgae extracts. Food Chem. 138(2-3): 1819-1828.
Angus, S. (2017). Modern seaweed harvesting and gathering in Scotland: the legal and ecological context. Scott. Geogr. J. 133(2): 1-14.
Anuraj, A., Waman, A., Prakash, C., Dam, Roy, S., Viji, M., Baidya, M., and Chadha, N. (2017). Investigating the effects of exogenous factors on growth, photosynthetic pigments and bud induction in Gracilaria corticata var. cylindrica under in vitro conditions. IJCRAR 6(8): 3235-3246.
AOAC (1990). Official methods of analysis of the Association of Official Analytical Chemists. 15th edition. Washington, DC, Association of Official Analytical Chemists.

267
Apaydin, G., Aylikei, V., Cengiz, E., Saydan, M., Kup, N., and Tirasogu, E. (2010). Analysis of metal contents of seaweed Ulva lactuca from Istambul, Turkey by EDXRF. Turk. J. Fish. Aquat. Sc. 10: 215220.
Arbona, J.F. and Molla, M. (2006). Cultivation of brown seaweed Alaria esculenta. In: Watson, L. (ed.). Aquaculture Explained. Bord Iascaigh Mhara, Dublin. pp. 50.
Armstrong, E., Rogerson, A., and Leftley, J.W. (2000). The abundance of heterotrophic protists associated with intertidal seaweeds. Estuar. Coast Shelf. Sci. 50: 415–424.
Arnold, T.M., Targett, N.M., Tanner, C.E., Hatch, W.I., and Ferrari, K.E. (2001). Evidence for methyl jasmonate-induced phlorotannin production in Fucus vesiculosus (Phaeophyceae) J. Phycol. 37: 1026-1029.
Asare, O. and Harlin, M.M. (1983). Seasonal fluctuations in tissue nitrogen for five species of perennial macroalgae in Long Island Sound. J. Appl. Phycol. 19(2): 254-257.
Ascêncio, S.D., Orsato, A., França, R.A., Duarte, M.E., and Noseda, M.D. (2006). Complete 1H and 13C NMR assignment of digeneaside, a low-molecular-mass carbohydrate produced by red seaweeds. Carbohydr. Res. 341(5): 677-682.
Ashraf, M. and Foolad, M.R. (2007). Roles of glycine, betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 59(2): 206-216.
Athukorala, Y., Lee, K.W., Song, C.B., Ahn, C.B., Shin, T.S., Cha, Y.J., Shahidi, F., and Jeon, Y.J. (2003a). Potential antioxidant activity of marine red alga Grateloupia filicina extracts. J. Food Lipids 10: 251-265.
Atkinson, M.J and Smith, S.V. (1983). C:N:P ratios of benthic marine. Limnol. Oceanogr. 28 (3): 568-574.
Atta-ur-Rahman, V.U., Ahmad, S., Bano, S.A., Abbas, K.A., Alvi, M.S., Ali, H.S.M., Lu, and Clardy, J. (1988). Pinnatazane, a bridged cyclic ether sesquiterpene from Laurencia pinnatifida. Phytochemistry 27: 3879-3880.
Awad, N.E. (2000). Biologically active steroid from the green algae Ulva lactuca. Phytother. Res. 14: 641-643.
Awad, N.E. (2004). Bioactive brominated diterpenes from the marine red algae Jania rubens (L.) Lamx. Phytother. Res. 18: 275-279.
Bailey, J.C. and Freshwater, D.W. (1997). Molecular systematics of the Gelidiales: inferences from separate and combined analyses of plastid rbcL and nuclear SSU gene sequences. Eur. J. Phycol. 32: 343-52.
Bajguz, A. and Czerpak, R. (1996). Effect of brassinosteroids on growth and proton extrusion in the alga Chlorella vulgaris Beijerinck (Chlorophyceae). J. Plant. Growth Regul. 15: 153-156.
Ballantine, D.L., Gerwick, W.H., Velez, S.M., Alexander, E., and Guevara, P. (1987). Antibiotic activity of lipid-soluble extracts from Caribbean marine algae. Hydrobiologia 151/152: 463-469.
Bano, S., Ahmad, V.U., Perveen, S., Bano, N., Shafiuddin, and Shameel, M. (1987). Marine natural products: II. Chemical constituents of red alga Botryocladia leptopoda. Plant. Med. 53(1): 117-118.
Bano, S., Ali, M.S. and Ahmad, V.U. (1982). Marine natural products: VI. A halogenated chamigrene epoxide from the red alga Laurencia pinnatifida. Planta Med. 53: 508.
Bano, S., Ali, M.S., and Ahmad, V.U. (1988). Marine natural products: VIII. Two minors halogenated sesquiterpenoids from the red alga Laurencia pinnatifida. Sci. Pharm. (Vienna). 56: 125.
Bano, S., Bano, N., Ahmad, V.U., Shameel, M. and Amjad, S. (1986). Marine natural products: 3-formylindole from the red algae Botryocladia leptopoda. J. Nat. Prod. 49(3): 549.
Bano, S., Hayee, A., Ahmad, V.U., Shaikh, W., Usmanghani, K. and Shameel, M. (1988). Marine natural products. Part VII. Steroids from a red alga Asparagopsis sandfordiana. Polish J. Chem. 62: 905-906.

268
Bano, S., Uddin, S., and Ahmed, V.U. (1990). Marine natural products Part XV. An acetylated derivative of a new N-acylsphingosine from red alga Halymenia porphyroides. Planta Med. 56: 233-234.
Barahona, L.F. and Rorrer, G.L. (2003). Isolation of halogenated monoterpenes from bioreactor-cultured microplantlets of the macrophytic red algae Ochtodes secundiramea and Portieria hornemannii J. Nat. Prod. 66: 743-751.
Barbarino, E. and Lourenço, S.O. (2005). An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 17:447-460.
Barceló-Villalobos, M., Figueroa, F.L., Korbee, N., Álvarez-Gómez, and Abreu, M.H. (2017). Production of mycosporine-like amino acids from Gracilaria vermiculophylla (Rhodophyta) cultured through one year in an Integrated Multi-trophic Aquaculture (IMTA) System. Mar. Biotechnol. 19(3): 246-254.
Barreto, M.C., Mendonça, E., Gouveia V., Anjos, C., Medeiros, J.S., Seca, A.M.L., and Neto, A.I. (2012). Macroalgae from S. Miguel Island as a potential source of antiproliferative and antioxidant products. Arquipel. Life Mar. Sci. 29: 53-58.
Barrow, K.D., Karsten, U., King, R.J., and West, J.A. (1995). Floridoside in the genus Laurencia (Rhodomelaceae, Ceramiales)-A chemosystematic study. Phycologia. 34(4): 279-283.
Barufi, J.B., Figuerosa, F.L., and Plastino, E.M. (2015). Effects of light quality on reproduction, growth and pigment content of Gracilaria birdiae (Rhodophyta: Gracilariales) and pigment content of Gracilaria birdiae. Sci. Mar. 79(1).
Baumann, H.A., Morrison, L., and Stengel, D.B. (2009). Metal accumulation and toxicity measured by PAM-chlorophyll fluorescence in seven species of marine macroalgae. Ecotoxicol. Environ. Saf. 72(4): 1063-1075.
Baweja, P., Sahoo, D., and García-Jiménez, R.R.R. (2009). Seaweed tissue culture as applied to biotechnology: problems, achievements and prospects. Phycol. Res. 57:45-58.
Bazes, A., Silkina, A., Douzenel, P., Fay, F., Kervarec, N., Morin, D., Berge, J., and Bourgougnon, N. (2009). Investigation of the antifouling constituents from the brown alga Sargassum muticum (Yendo) fenshott. J. Appl. Phycol. 21(4): 395-403.
Bedoux, G., Hardouin, K., Marty, C., Taupin, L., Vandanjon, L. and Bourgougnon, N. (2014). Chemical characterization and photoprotective activity measurement of extracts from the red macroalga Solieria chordalis. Bot. Mar. 57: 291301.
Beer, S. and Eshel, A., (1985). Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae. Aust. J. Mar. Freshw. Res. 36: 785-92.
Bell, E.C. (1993). Photosynthetic response to temperature and desiccation of the intertidal alga Mastocarpus papillatus. Mar. Biol. 117: 337-346.
Benson, E.E., Rutter, J.C., and Cobb, A.H. (1983). Seasonal variation in frond morphology and chloroplast physiology of the intertidal alga Codium fragile (Suringar) Hariot. New Phytol. 95: 569-580.
Benzie, I.F and Strain, J.J. (1996). The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 239: 70-76.
Berland, B.R., Bonin, D.J., and Maestini, S.T. (1972). Are some bacteria toxic for marine algae? Mar. Bio. 12: 189-193.
Bhattarai, H.D., Ganti, V.S., Paudel, B., Lee, Y.K., Lee, H.K., Hong, Y.K., and Shin, H.W. (2007). Isolation of antifouling compounds from the marine bacterium, Shewanella Oneidensis SCH0402. World J. Microb. Biotechnol. 23: 243-249.
Bhosale, P. (2004). Environmental and cultural stimulants in the production of carotenoids from microorganisms. Appl. Microbiol. Biotechnol. 63: 351-361.
Bidwell, R.G.S. and Mclachlan, J. (1985). Carbon nutrition of seaweeds: Photosynthesis, photorespiration and respiration. J. exp. mar. Biol. Ecol. 86: 15-46.
Bidwell, R.G.S., McLachlan, J., and Lloyd, N.D.H. (1985). Tank cultivation of Irish Moss, Chondrus crispus Stackh. Bot. Mar. 28: 87-97.

269
Biebl, R. (1958). Temperatur und osmotische Resistenz von Meeresalgen der bretonischen Küste. Protoplasma 50: 217-242.
Bird, C.J., Rice, E.L., Murphy, C.A., and Ragan, M.A. (1992). Phylogenetic relationships in the Gracilariales (Rhodophyta) as determined by 18S rDNA sequences. Phycologia 31: 510-522.
Bird, K., Habig, C., and DeBusk, T. (1982). Nitrogen allocation and storage patterns in Gracilaria tikvahiae (Rhodophyta). J. Phycol. 18: 344-348.
Bird, K.T., and Benson, P.H. (1987). Seaweed cultivation for renewable resources. Elsevier, New York, NY. pp. 381.
Birkett, D.A., Maggs, C.A., Dring, M.J., and Boaden, P.J.S. (1998b). Infralittoral reef biotopes with kelp species: an overview of dynamic and sensitivity characteristics for conservation management of marine SACs. Natura 2000 report prepared by Scottish Association of Marine Science (SAMS) for the UK Marine SACs Project., Scottish Association for Marine Science. (UK Marine SACs Project, vol V.).
Bittner, M.L., Silva, M., Paul, V.J., and Fenical, W. (1985). A rearranged chamigrane derivative and its potential biogenetic precursor from a new species of the marine red algal genus Laurencia (Rhodomelaceae). Phytochemistry 24: 987-989.
Bixler, H.J. and Porse, H. (2011). A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 23: 321335.
Björk, M., Axelsson, L., and Beer, S. (2004). Why is Ulva intestinalis the only macroalga inhabiting isolated rockpools along the Swedish Atlantic coast? Mar. Ecol. Prog. Ser. 284: 109-116.
Bjørnland, T., Pennington, F., Haxo, F.T., and Liaaen-Jensen, S. (1984). Carotenoids of Chrysophyceae and Dinophyceae- “Coc. Min. Haltenbanken” and Gymnodinium breve (Florida red tide) 7th Int. IUPAC Symp. on Carotenoids, pp. 26
Black, W.A.P. (1950b). The seasonal variation in weight and chemical composition of the common British Laminariaceae. J. Mar. Biol. Assoc. U.K. 29: 45-72.
Bligh, E.L.G and Dyer, W.J.A. (1959). A rapid method of Total Lipid Extraction and purification. C. J. Biochem. and Physiol. 37(8): 911-917.
Blunden, G. and Gorden, S.M. (1986). Betaines and their sulphonio analogues in marine algae. In: Round, F.E. and Chapman, D.J. (eds.). Progress in Phycological Research, Vol. 4. Biopress, Bristol. pp. 39-80.
Bojanowski, R. (1973). The occurrence of major and minor chemical elements in the more common Baltic seaweed. Oceanologia 2: 81-152.
Bold, H.C. and Wynne, M.J. (1985). Introduction to the Algae. Structure and Reproduction. Englewood Cliffs. New Jersey, Prentice-Hall, xvi. pp. 720.
Boney, A.D. (1975). Mucilage sheaths of spores of red algae. J. Mar. Biol. Ass. U.K. 55: 511518.
Boo, G.H., Park, J.K. and Boo, S.M. (2013). Gelidiophycus (Rhodophyta: Gelidiales): a new genus of marine algae from East Asia. Taxon 62: 1105-1116.
Boominathan, M. and Mahesh, A. (2015). Seaweed carotenoids for cancer therapeutics. In: Kim, S.K. (ed.). Handbook of Anticancer Drugs from Marine Origin. Springer International Publishing, Switzerland.
Børgesen, F. (1908). Botany of the Faeröes. The algae-vegetation of the Faeröese coasts, with remarks on the phytogeography. Nordisk Forl. Copenhagen 3: 683-834.
Borlongan, I.A.G., Tibubos, K.R., Yunque, D.A.T., Hurtado, A.Q., and Critchley, A.T. (2011). Impact of AMPEP on the growth and occurrence of epiphytic Neosiphonia infestation on two varieties of commercially cultivated Kappaphycus alvarezii grown at different depths in the Philippines. J. Appl. Phycol. 23(3): 615-621.
Borowitzka, M.A. (2007). Culturing microalgae in outdoor ponds. In: Andersen, R.A. (ed.). Algal culturing techniques. Elsevier Academic Press, London. pp. 205218.

270
Bouarab, K., Adas, F., Gaquerel, E., Kloareg, B., Salaün, J.P., and Potin, P. (2004). The innate immunity of a marine red alga involves oxylipins from both the eicosanoid and octadecanoid pathways. Plant Physiol. 135: 1838-1848.
Boussiba, S., Bing, W., Yuan, J.P., Zarka, A., and Chen, F. (1999). Changes in pigments profile in the green alga Haeamtococcus pluvialis exposed to environmental stresses. Biotechnol. Lett. 21: 601-604.
Bracken, M.E. and Stachowicz, J.J. (2006). Seaweed diversity enhances nitrogen uptake via complementary use of nitrate and ammonium. Ecol. 87: 2397-2403.
Bradáčová, K., Weber, N.F., Moradtalab, N., Asim, M., Imran, M., Weinmann, M., and Neumann, G. (2016). Micronutrients (Zn/Mn), seaweed extracts, and plant growthpromoting bacteria as coldstress protectants in maize. Chem. Biol. Technol. Agric. 3: 19.
Bradley, P.M. (1991). Plant hormones do have a role in con-trolling growth and development of algae. J. Phycol. 27: 317-321.
Bradley, P.M. and Cheney, D.P. (1990). Some effects of plant growth regulators on tissue cultures of the marine red alga Agardhiella subulata (Gigartinales, Rhodophyta). Hydrobiologia 204/205: 353–60.
Brawley, S.H. and Adey, W.H. (1981). Micrograzers may affect macro-algal density. Nature (London). 292: 177.
Brawley, S.H. and Fei, X.G. (1987). Studies of mesoherbivory in aquaria and in an unbarricaded mariculture farm on the Chinese coast. J. Phycol. 23 (4): 614-623.
Brawley, S.H., and Johnson, L.E. (1991). Survival of fucoid embryos in the intertidal zone depends upon developmental stage and microhabitat. J. Phycol.27: 179-186.
Breeman, A.M. and ten Hoopen, A. (1981). Ecology and distribution of the subtidal red alga Acrosymphyton purpuriferum (J. Ag.) Sjøst. (Rhodophyceae, Cryptonemiales). Aquatic. Bot. 11: 143-166.
Brinza, L., Dring, M.J. and Gavrilescu, M. (2007). Marine micro- and macroalgal species as biosorbents for heavy metals. Environ. Eng. Manage J. 6: 237-251.
Brito, I., Cueto, M., Dorta, E., and Darias, J. (2002). Bromocyclocanol, a halogenated sesquiterpene with a novel carbon skeleton from the red alga Laurencia obtusa. Tetra. Lett. 43: 2551-2553.
Browne, K.L. (2001). Mariculture of the edible red alga Palmaria palmata. PhD thesis, Queen's University of Belfast, UK. pp. 186.
Brzezinski, M.A. (1985). The Si:C:N ratio of marine diatoms. Interspecific variability and the effect of some environmental variables. J. Phycol. 21: 347-357.
Burritt, D.J., Larkindale, J., and Hurd, C.L. (2002). Antioxidant metabolism in the intertidal red seaweed Stictosiphonia arbuscula following desiccation. Planta 215 (5): 829-838.
Buschmann, A.H., Carolina Camus, Infante, J., Neori, A., Israel, Á., Hernández-González, M.C., Pereda, S.V., Gomez-Pinchetti, J.L., Golberg, A., Tadmor-Shalev, N., and Critchley, A.T. (2017). Seaweed production: overview of the global state of exploitation, farming and emerging research activity. Eu. J. Phycol. 52(4): 391-406.
Buschmann, A.H., Mora, O., Gómez, P., Böttger, M., Buitano, S., Retamales, C., Vergara, P., and Gutierrez, A. (1994). Gracilaria chilensis outdoor tank cultivation in Chile: use of land-based salmon culture effluents. Aquacult. Eng. 13: 283-300.
Butler, E.C., Dong, Y., Liang, X., Kuder, T., Philp, R.P., and Krumholz (2009). Abiotic reductive dechlorination of tetrachloroethylene and trichloroethylene in anaerobic environments. Final report for the Strategic Environmental Research and Development program, University of Oklahoma.
Cabrita, M.T, Vale, C. and Rauter, A.P. (2010). Halogenated compounds from marine algae. Mar. Drugs. 8(8): 2301-2317.
Callow, M.E. and Callow, J.A. (2002). Marine biofouling: a sticky problem. Biologist 49: 15.

271
Campbell, S.J., Bité, J.S., and Burridge, T.R. (1999). Seasonal patterns in the photosynthetic capacity, tissue pigment and nutrient content of developmental stages of Undaria pinnatifida (Phaeophyta: Laminariales) in Port Phillip Bay, South- Eastern Australia. Bot. Mar. 42: 231-241.
Campo, V.L., Kawano, D.F., da Silva, D.B., and Carvalho, I. (2009). Carrageenans: Biological properties, chemical modifications and structural analysis-A review. Carbohydr. Polym. 77: 167-180.
Campos, A.M., Matos, J., Afonso, C., Gomes, R., Bandarra, N.M., and Cardoso, C. (2019). Azorean macroalgae (Petalonia binghamiae, Halopteris scoparia and Osmundea pinnatifida) bioprospection: a study of fatty acid profiles and bioactivity. Int. J. Food Sci. Technol. 54: 880-890.
Capuzzo, E. and McKie, T. (2016). Seaweed in the UK and abroadstatus, products, limitations, gaps and Cefas role. Cefas, contract report FC002I.
Cardozo, K.H.M., Carvalho, V.M., Pinto, E., and Colepicolo, P. (2006). Fragmentation of mycosporine-like amino acids by hydrogen/deuterium exchange and electrospray ionisation tandem mass spectrometry. Rapid Commun. Mass. Sp. 20: 253-258.
Cardozo, K.H.M., Vessecchi, R., Carvalho, V.M., Pinto, E., Gates, P.J., Colepicolo, P., Galembeck, S.E., and Lopes, N.P. (2008). A theoretical and mass spectrometry study of the fragmentation of mycosporine-like amino acids. Int. J. Mass. Sp. 273: 11-19.
Carreto, J. and Carignan, M. (2011). Mycosporine-like amino acids: relevant secondary metabolites. chemical and ecological aspects. Mar. Drugs 9(3): 387-446.
Cassano, V., Díaz-Larrea, J., Sentíes, A., Oliveira, M.C., Gil-Rodríguez, M.C., and Fujii, M.T. (2009). Evidence for the conspecificity of Palisada Papillosa with P. Perforata (Ceramiales, Rhodophyta) from the Western and Eastern Atlantic Ocean on the basis of morphological and molecular analyses. Phycologia 48(2): 86-100.
Cassano, V., Manchín-Sánchez, M., Neto, A. I., Oliveira, M.C., Senties, A., Díaz-Larrea, J., and Fujii, M.T. (2013). A DNA barcode approach of the Laurencia Complex (Ceramiales, Rhodophyta) in the tropical and subtropical Atlantic Ocean. Phycologia 52(4): 16.
Cassano, V., Oliveira, M.C., Gil-Rodríguez, M.C., Sentíes, A., Díaz-Larrea, J., and Fujii, M.T. (2012). Molecular support for the establishment of the new genus Laurenciella within the Laurencia complex (Ceramiales, Rhodophyta). Bot. Mar. 55(4): 349-357.
Cassolato, J.E.F., Noseda, M.D., Pujol, C.A., Pellizzari, F.M., Damonte, E.B., and Duarte, M.E.R. (2008). Chemical structure and antiviral activity of the sulfated heterorhamnan isolated from the green seaweed Gayralia oxysperma. Carbohydr. Res. 343: 3085-3095.
Castagna, A., Sinatra, F., Castagna, A.G., Stoli, S., and Zafarana, S. (1985). Trace element evaluations in marine organisms. Mar. Pollut. Bull. 16: 416-419.
Centre for Environmental Data Analysis (CEDA). Met office data relative to the years 2016-2017 and 2017-2018, #918 Station Dunstaffnage, for precipitation and radiation.
Chamberlain, A.H.L. (1976). Algal settlement and secretion of adhesive materials. In: Proceedings of the 3rd international biodegradation symposium. Sharpley, J.M. and Kaplan, A.M. (eds.). Applied Science Publ, London. pp. 417-432.
Chamberlain, A.H.L. and Evans, L.V. (1973). Aspects of spore production in the red alga Ceramium. Protoplasma 76: 139-159.
Chandía, N.P., and Matsuhiro, B. (2008). Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 42: 235-240.
Chandini, S.K., Ganesan, P., Suresh, P.V., and Bhaskar, N. (2008). Seaweeds as source of nutritionally beneficial compounds: a review. J. Food Sci. Technol. 45:1-13.
Chandrashekar, J., Hoon, M., Ryba, J.P.N., and Zuker, S.C. (2006). The receptors and cells for mammalian taste. Nature 444(7117): 288-294.
Chaneva, G., Furnadzhieva, S., Minkova, K., and Lukavsky, J. (2007). Effect of light and temperature on the cyanobacterium Arthronema africanum- a prospective phycobiliprotein-producing strain. J. Appl. Phycol. 19: 537-544.

272
Chapman, A.R.O. and Craigie, J.S. (1977). Seasonal growth in Laminaria longicruris: Relations with dissolved inorganic nutrients and internal reserves of nitrogen. Mar. Biol. 40: 197-205.
Chapman, V.J. (1970). Seaweeds and their uses. Methuen and Co. Ltd., the Camelot Press Ltd., London, UK. pp. 304.
Charnofsky, K., Towlll, L.R., and Sommerfield, M.R. (1982). Light requirements for monospore germination in Bangia atropurpurea (Rhodophyta). J. Phycol. 18: 417-422.
Charters, A.C., Neushul, M., and Coon, D. (1973). The effect of water motion on alga spore adhesion. Limnol. Oceanogr. 18: 884-896.
Chaudhury, M.K. and Daniel, S. (2006). Settlement behaviour of swimming algal spores on gradient surfaces. Biointerphases. 1(1):18-21.
Chen, L.C.M. and Taylor, A.R.A. (1978). Medullary tissue culture of the red alga Chondrus crispus. Can. J. Bot. 56: 883-886.
Chen, Y. and Vaidyanathan, S. (2012). A simple, reproducible and sensitive spectrophotometric method to estimate microalgal lipids. Anal. Chim. Acta 724: 67-72.
Chen, Y. and Vaidyanathan, S. (2013). Simultaneous assay of pigments, carbohydrates, proteins and lipids in microalgae. Anal. Chim. Acta Elsevier B.V. 776: 31-40.
Cheney, D. (1990). Genetic improvement of seaweeds through protoplasts fusion. In: Yarish, C., Penniman, C., and Van Patten, P. (eds.). Economically important marine plants of the Atlantic: their biology and cultivation. Univ. Conn Sea Grant Program, Storrs. Pp. 1525.
Choi, H.G, Kraft, G.T, Kim, H.S, Guiry, M.D, and Saunders, G.W. (2008). Phylogenetic relationships among lineages of the Ceramiaceae (Ceramiales, Rhodophyta) based on nuclear small subunit rDNA sequence data. J. Phycol. 44: 1033-1048.
Chojnacka, K., Saeid, A., Witkowska, Z., and Tuhy, L. (2012). Biologically active compounds in seaweed extracts - the prospects for the application. Open Conf. Proc. J. 3 (Suppl 1M4): 20-28.
Chopin, T. (2007). Closing remarks of the new president of the international seaweed association. Proceedings of the 19th International Seaweed Symposium, 26-31 March, Kobe, Japan. pp. 1-2.
Chopin, T. (2018a). Seaweed aquaculture–From the global, mostly Asian, picture to the opportunities and constraints of the Canadian scene. Bull. Aquacult. Assoc. Canada 1: 38.
Chopin, T., Gallant, T., and Davison, I. (1995). Phosphorus and nitrogen Nutrition in Chondrus Crispus (Rhodophyta): effects on total phosphorus and nitrogen content, carrageenan production, and photosynthetic pigments and metabolism. J. Phycol. 31(2): 283-293.
Chopin, T., Hanisak, M.D., Koehn, F.E., Mollion, J., and Moreau, S. (1990a). Studies on carrageenans and effects of seawater phosphorus concentration on carrageenan content and growth of Agardhiella subulata (C. Agardh) Kraft and Wynne (Rhodophyceae, Solieriaceae). J. Appl. Phycol. 23-16.
Chow, F. (2012). Nitrate Assimilation: the role of in vitro nitrate reductase assay as nutritional predictor. In: Applied Photosynthesis, Najafpour, M.M. (ed.). Chapter 5, InTech Publication, USA. ISBN-13: 978-953-51-0061-4. pp. 105-118.
Chun, S.S., Park, J.R., Park, J.C., Suh, J.S., and Ahn, C.B. (1999). Quality characteristics of hamburger patties added with seaweed powder. J. Korean Soc. Food Sci. Nutr. 28:140-144.
Cian, R.E., Drago, S.R., Medina, F.S., and Martínez-Augustin, O. (2015). Proteins and carbohydrates from red seaweeds: evidence for beneficial effects on gut function and microbiota. Mar. Drugs 13: 5358-5383.
Cian, R.E., Fajardo, M.A., Alaiz, M., Vioque, J., Gonzalez, R.J., and Drago, S.R. (2014). Chemical composition, nutritional and antioxidant properties of the red edible seaweed Porphyra columbina. Int. J. Food Sci. Nutr. 65: 299-305.

273
Cian, R.E., Lopez-Posadas, R., Drago, S.R., Sánchez de Medina, F., and Martinez-Augustin, O. (2012). Immunomodulatory properties of the protein fraction from Phorphyra columbina. J. Agric. Food Chem. 60: 8146-8154.
Cimino, G. and Ghiselin, M.T. (2001). Marine natural products chemistry as an evolutionary narrative. In: McClintock, J.B. and Baker, B.J. (eds.). Marine Chemical Ecology. CRC Press, Boca Raton, FL, USA. Pp. 115154.
Clarkston, B.E. and Saunders, G.W. (2010). A comparison of two DNA barcode markers for species discrimination in the red algal family Kallymeniaceae (Gigartinales, Florideophyceae), with a description of Euthora timburtonii sp. nov. Botany 88: 119-131.
Cole, K.M and Sheath, R.G. (2011). Biology of the Red algae (end edition). Cambridge University Press, Cambridge, New York, Port Chester, Melbourne, Sydney. pp. 517.
Cole, K.M. (1982). Rhodophycean pit plugs: an ultrastructural survey with taxonomic implications. Amer. J. Bot. 69: 703-720.
Colorni, A. (1989). Perforation disease affecting Ulva sp. cultured in Israel on the Red Sea. Dis. Aquat.Org. 7: 71-73.
Conover, J. (1968). The importance of natural diffusion gradients and transport of substances related to benthic marine plant metabolism. Bot. Mar. 11: 1-9.
Conti, M.E. (2002). SEAM (ed.). Il monitoraggio biologico della qualitá ambientale. Rome, pp. 181.
Cooke, T.J., Poli, D., Sztein, A.E., and Cohen, J.D. (2002). Evolutionary patterns in auxin action. Plant. Mol. Biol. 49: 319-338.
Coomans, R.J. and Hommersand, M.H. (1990). Vegetative growth and organization. In: Cole, K.M. and Sheath, R.G. (eds). Biology of the red algae. Univ Press, Cambridge.
Corey, P., Kim, J.K., Duston, J., and Garbary, D.J. (2014). Growth and nutrient uptake by Palmaria palmata (Palmariales, Rhodophyta) integrated with Atlantic halibut recirculating aquaculture. Algae 29: 35-45.
Corey, P., Kim, J.K., Duston, J., Garbary, D.J., and Prithiviraj, B. (2013). Bioremediation potential of Palmaria palmata and Chondrus crispus (Basin Head): effect of nitrate and ammonium ratio as nitrogen source on nutrient removal. J. Appl. Phycol. 25: 1349-1358.
Cosgrove, J. and Borowitzka, M.A. (2010). Chlorophyll fluorescence terminology: an introduction. In: Suggett, D.J., Prášil, O., and Borowitzka, M.A. (eds.). Chlorophyll a fluorescence in aquatic sciences: methods and applications. Springer, Dordrecht. pp. 117.
Cottier-Cook, E.J., Nagabhatla, N., Badis, Y., Campbell, M., Chopin, T., Day, W., Fang, J., He, P., Chad, H., Gwang Hoon, K., Yuanzi, H., Zengjie, J., Gert, K., Li, X., Feng, L., Liu, H., Liu, Y., Lu, Q., Luo, Q., and Gachon, C. (2016). Safeguarding the future of the global seaweed aquaculture industry. United Nations University. pp. 1-12.
Courtois, A., Simon-Colin Boisset, C., Berthou, C., Deslandes, E., Guézennec, J., and Bordron, A. (2008). Floridoside extracted from the red alga Mastocarpus stellatus is a potent activator of the classical complement pathway. Mar. Drugs. 6: 407-17.
Craigie, J.S. (2011). Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 23: 371-393.
Crouch, I.J. (1990). The effect of seaweed concentrate on plant growth. PhD Thesis. University of Natal, South Africa. pp. 251.
D’Adamo, S., Schiano di Visconte, G., Lowe, G., Szaub-Newton, J., Beacham, T., Landels, A., Allen, J.M., Spicer, A., and Matthijs, M. (2018). Engineering the unicellular alga Phaeodactylum tricornutum for high-value plant triterpenoid production. Plant Biotechnol. J. 1-13.
da Costa, M.S., Santos, H., and Galinski, E.A. (1998). An overview of the role and diversity of compatible solutes in Bacteria and Archaea. In: Antranikian, G. (ed.). Biotechnology of extremophiles. Springer, 1st ed. Berlin. pp. 117153.
Da Gama, B.A.P., Pereira, R.C., Carvalho, A.G.V., Coutinho, R., and Yoneshigue-Valentin, Y. (2002). The effects of seaweed secondary metabolites on biofouling. Biofouling. 18: 13-20.

274
Dai, J. and Mumper, R. J. (2010). Plant phenolics: extraction, analysis and their antioxidant and anticancer properties. Molecules, 15(10): 7313-7352.
Dailey, A. and Vuong, Q.V. (2015). Effect of extraction solvents on recovery of bioactive compounds and antioxidant properties from macadamia (Macadamia tetraphylla) skin waste. J. Food Sci. Technol. 1(1).
Davis, B. and Birch, G. (2010). Comparison of heavy metal loads in stormwater runoff from major and minor urban roads using pollutant yield rating curves. Env. Pollut. 158(8): 2541-2545.
Davis, G.D.J., and Vasanthi, A.H.R (2011). Seaweed metabolite database (SWMD): A database of natural compounds from marine algae. Bioinformation 5(8): 361-364.
Davis, T.A., Volesky B., and Mucci, A. (2003). A review of the biochemistry of heavy metal biosorption by brown algae. Water Res. 37 (18): 4311-30.
Dawes, C.J. (1971) Indole-3-acetic acid in the green algae coenocyte Caulerpa prolifera (Chlorophyceae, Siphonales). Phycologia 10: 375379.
Dawes, C.J. (1998). Marine botany. John Wiley and Sons, Inc., New York, pp. 496.
Dawes, C.J. (1995). The effect of nutrient and photon fluence on the photosynthetic responses of red and pigmented cultivars of Eucheuma denticulatum. Bot. Mar. 38: 323-327.
de Jong, Y.S.D.M., Van Der Wurff, A.W.G., Stam, W.T., and Olsen, J.L. (1998). Studies on Dasyaceae. Towards a phylogeny of the Dasyaceae (Ceramiales, Rhodophyta), based on comparative rbcL gene sequences and morphology. Eur. J. Phycol. 33: 187-201.
de Nys R., Dworjanyn S.A., and Steinberg, P.D. (1998). A new method for determining surface concentrations of marine natural products on seaweeds. Mar. Ecol. Progress Ser. 162: 79-87.
DeBoer, J.A. (1981). Nutrients. In: Lobban, C.S. and Wynne, M.J. (eds.). The Biology of Seaweeds. Blackwell Scientific, Oxford. pp. 356-391.
DeBoer, J.A. and Ryther, J.H. (1977). Potential yields from a waste-recycling algal mariculture system. In Krauss, R. (ed.). The marine plant biomass of the Pacific North-West coast. Corvallis: Oregon State Univeristy Press. pp. 231-49.
DeBoer, J.A., Guigli, H.J., Israel, T.L., and D’Elia, C.F. (1978). Nutritional studies of two red algae. I. Growth rate as a function of nitrogen source and concentration. J. Phycol. 14: 261-266.
Debonnel, G., Beauchesne, L., and de Montigny, C. (1989). Domoic acid, the alleged "mussel toxin," might produce its neurotoxic effect through kainate receptor activation: an electrophysiological study in the dorsal hippocampus. Can. J. Physiol. Pharmacol. 67(1): 29-33.
Dekov, K.M., Araújo, F., Van Grieken, R., and Subramanian, V. (1998). Chemical composition of sediments and suspended matter from the Cauvery and Brahmaputra rivers (India). Sci. Total Environ. 212: 89-105.
Del Campo, E., García-Reina, G., and Correa, J.A. (2002). Degradative disease in Ulva rigida (Chlorophyceae) associated with Acrochaete geniculate (Chlorophyceae). J. Phycol. 34: 160-166.
Delivopoulos, S. (2000). Ultrastructure of spermatiogenesis in the red alga Osmundea spectabilis var. spectabilis (Rhodomelaceae, Ceramiales, Rhodophyta). Phycologia 39(6): 517-526.
Delivopoulos, S. (2002). Ultrastructure of trichoblasts in the red alga Osmunde spectabilis var. spectabilis (Rhodomelaceae, Ceramiales). Eur. J. Phycol. 37(3): 329-338.
Dembitsky, V.M. (1996). Betaine ether linked glycerolipids–chemistry and biology. Prog. Lipid Res. 35: 1-51.
Dembitsky, V.M. and Rozentsvet, O.A (1990). Phospholipid composition of some marine red algae. Phytochemistry 29: 3149-3152.

275
Denis, C., Morançais, M., Li, M., Deniaud, E., Gaudin, P., Wielgosz-Collin, G., Barnathan, G., Jaouen, P., and Fleurence, J. (2010). Study of the chemical composition of edible red macroalgae Grateloupia turuturu from Brittany (France). Food Chem. 119(3): 913-917.
Dillard, C.J. and German, J.B. (2000). Phytochemicals: nutraceuticals and human health. J. Sci. Food Agric. 80: 1744-1756.
Ding, H.Y. and Ma, J.H. (2005). Simultaneous infection by red rot and chytrid diseases in Porphyra yezoensis Ueda. J. Appl. Phycol.17: 51-56.
Dixon, P.S. (1973). Biology of the Rhodophyta. Hafner Press, New York. pp. 285.
Doyle, J.J. and Doyle, J.L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 19: 11-15.
Dring, M.J. (1988). Photocontrol of development in algae. Ann. Rev. Plant Physiol. Plant Mol. Biol. 39: 157-174.
Dring, M.J. (1990). Light harvesting and pigment composition in marine phytoplankton and macroalgae. In: Herring, P.J., Campbell, A.K., Whitfield, M., and Maddock, L. (eds.). Light and life in the sea. Cambridge University Press, Cambridge. pp. 89-103.
Dring, M.J., Wagner, A., Boeskov, J., and Lüning, K. (1996). Sensitivity of intertidal and subtidal red algae to UVA and UVB radiation, as monitored by chlorophyll fluorescence measurements: influence of collection depth and season, and length of irradiation. Eur. J. Phycol. 31(4): 293-302.
Duan, X.J., Zhang, X.M., Li, X.M., and Wang, B.G. (2006) Evaluation of antioxidant property of extract and fractions obtained from a red alga, Polysiphonia urceolata. Food Chem. 95: 37-43.
Duarte, C.M., Middelburg, J., and Caraco, N. (2005). Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2: 1-8.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A. and Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Anal. Chem. 28(3): 350-356.
Duckett, J.G., Buchanan, J.S., Peel, M.C., and Martin, M.T. (1974). An ultrastructural study of pit connections and percurrent proliferations in the red alga, Nemalion helminthoides (Vell. in With.) Batt. New Phytol. 73: 497-507.
Dunlap, W.C. and Yamamoto, Y. (1995). Small-molecule antioxidants in marine organisms: antioxidant activity of mycosporine-glycine. Comp. Biochem. Physiol. 112(B): 105-14.
Dworjanyn, S.A., De Nys, R., and Steinberg, P.D. (1999). Localisation and surface quantification of secondary metabolites in the red alga Delisea pulchra. Mar. Biol. 133: 727-736.
Edwards, M. and Watson, L. (2011). Cultivating Laminaria digitata. In: Watson, L. (ed.). Aquaculture explained no. 26. Bord Iascaigh Mhara, Dublin. pp. 72.
Ekman, P., Yu, S., and Pedersén, M. (1995). Effects of altered salinity, darkness and algal nutrient status on floridoside and starch content, α-galactosidase activity and agar yield of cultivated Gracilaria sordida. Br. Phycol. J. 26: 123-31.
Engel, C.R., Wattier, R., Destombe, C., and Valero, M. (1999). Performance of non-motile male gametes in the sea: analysis of paternity and fertilization success in a natural population of a red seaweed, Gracilaria gracilis. Proc. R. Soc. Lond. B. Biol. Sci. 266:1879-1886.
Enriquez, S. and Borowitzka, M.A. (2010). The use of the fluorescence signal in studies of seagrasses and macroalgae. In: Suggett, D.J., Borowitzka, M.A., and Prášil, O. (eds.). Chlorophyll a fluorescence in aquatic sciences: methods and applications. Springer, Dordrecht. pp. 187-208.
Erickson, K.L. (1983). Constituents of Laurencia. In: Scheuer, P.J. (ed.). Marine Natural Products: chemical and biological perspectives. Vol. 5. Academic Press, New York, NY, USA. pp. 131257.
Evans, G.C. (1972). The quantitative analysis of plant growth. Studies in ecology. Blackwell, Oxford, 1: 1-734.
Evans, L.V. and Trewavas, A.J. (1991). Is algal development controlled by plant growth substances? J. Phycol. 27: 322-326.

276
Evans, M.L. (1984). Function of hormones at the cellular level of organisation. In: Scott, T.K., (ed.). Encyclopedia of plant Physiology, Springer-Verlag, Berlin. pp. 23-80.
Fábregas, J., Patiño, M., Arredondo-Vega, B.O., Tobar, J.L., and Otero, A. (1995). Renewal rate and nutrient concentration as tools to modify productivity and biochemical composition of cyclostat cultures of the marine microalga Dunaliella tertiolecta. Appl. Microbiol. Biotechnol. 44: 287-292.
Faganeli, J., Vukovič, A., Saleh, F.I., and Pezdič, J. (1986). C:N:P ratios and stable carbon and hydrogen isotopes in the benthic marine algae, Ulva rigida C.Ag. and Fucusvirsoides J.Ag. J. Exp. Mar. Biol. Ecol. 102 (2-3): 153-166.
Falkowski, P.G. and LaRoche, J. (1991). Acclimation to spectral irradiance in algae. J. Phycol. 27: 8-14.
Falkowski, P.G. and Raven, J.A. (2007). Aquatic photosynthesis. Princeton University Press, NJ. pp. 488.
Fan, D., Hodges, D.M., Zhang, J., Kirby, C.W., Ji, X., Locke, S.J., Critchley, A.T., and Prithiviraj, B. (2010). Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea L.) which protects Caenorhabditis elegans against oxidative and thermal stress. Food Chem. 124: 195-202.
Fang, T.C., Cui, J.J., Ou, Y.L., Dai, J.X., Wang, M.L., Liu, Q.S., and Yang, Q.M. (1983b). Breeding of the new variety “Danhai No. 1” of Laminaria japonica by using a female haploid clone of the kelp. J. Shandong Coll. Oceanol. 13(4): 63-70.
FAO, (1990). Fishery statistics: Catches and landings. Rome, Italy. pp. 502.
FAO, (2003). A guide to the seaweed industry. FAO, Fisheries technical paper, 441. Rome, Italy. pp. 118.
FAO, (2012). The State of the World Fisheries and Aquaculture. FAO Fisheries Department, Rome, Italy. pp. 230.
FAO, (2014). The state of World Fisheries and Aquaculture. Rome, Italy. pp. 243.
FAO, (2015). The State of World Fisheries and Aquaculture 2016. Rome, Italy. pp. 200.
FAO, (2018). The state of World Fisheries and Aquaculture. Rome, Italy. pp. 227.
Faulkner, D. J. (1986). Marine natural products. Nat. Prod. Rep. 3:1-33.
Faulkner, D. J. (2001). Marine natural products. Nat. Prod. Rep. 18: 1-49.
Featonby-Smith, B.C. and van Staden, J. (1987) Effects of seaweed concentrate on grain yield in barley. S. Afr. J. Bot. 53: 125-128.
Fenical, W. (1981). Natural halogenated organics. In: Duursma, E.K. and Dawson, R. (eds). Marine organic chemistry: evolution, composition, interactions, and chemistry of organic matter in seawater. Elsevier Scientific Pub, Amsterdam; New York. pp. 375-393.
Fernandes, D.R.P., Yokoya, N.S., and Yoneshigue-Valentin, Y. (2011). Protocol for seaweed decontamination to isolate unialgal cultures. Rev. Bras. Farm. 21: 313-316.
Figueroa, F.L., Aguilera, J., and Niell, F.X. (1995a). Red and blue light regulation of growth and photosynthetic metabolism in Porphyra umbilicalis (Bangiales, Rhodophyta). Eur. J. Phycol. 30: 11-18.
Figueroa, F.L., Santos, R., Conde-Álvarez, R., Mata, L., Pinchetti, J.L.G., Huovinen, P., Schuenhoff, A., and Silva, J. (2006). The use of chlorophyll fluorescence for monitoring photosynthetic condition of two tank-cultivated red macroalgae using fishpond effluents. Bot. Mar. 49: 275-282.
Flameling, I. and Kromkamp, J. (1998). Light dependence of quantum yields for PSII charge separation and oxygen evolution in eucaryotic algae. Limnol. Oceanogr. 43: 284-297.
Fletcher, A. and Jones, W.E. (1975). A simple tide simulator for maintenance of marine (intertidal) algal and lichen cultures. Br. Phycol. J. 10(3): 263-264.
Fletcher, R.L. (1995). Epiphytism and fouling in Gracilaria cultivation: An overview. J. Appl. Phycol. 7: 325-333.
Fletcher, R.L. and Callow, M.E. (1992). The settlement, attachment and establishment of marine algal spores. Br. Phycol. J. 27: 303-329.

277
Fleurence, J. (1999). Seaweed proteins: biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 10: 25-28.
Fleurence, J. and Levine, I. (eds.). (2016). Seaweed in Health and Disease Prevention. Elsevier, London. pp. 443.
Fleurence, J., Morançais, M., Dumay, J., Decottignies, P., Turpin, V., Munier, M., García-Bueno, N., and Jaouen, P. (2012). What are the prospects for using seaweed in human nutrition and for marine animals raised through aquaculture? Trends Food Sci. Technol. 27(1): 57-61.
Flodin, C., Helidoniotis, F., and Whitfield, F.B. (1999). Seasonal variation in bromophenol content and bromoperoidase activity in Ulva lactuca. Photochem. 51: 135-138.
Fournier, E. (2001). Colorimetric quantification of carbohydrates. Curr. Protoc. Food Analyt. Chem. 0(1) 1: E1.1.1-E1.1.8.
Fraga, B.M. (2012). Natural sesquiterpenoids. Nat. Prod. Rep. 29: 1334-1366.
Fralick, R.A. and Mathieson, A.C. (1975). Physiological ecology of four Polysiphonia species (Rhodophyta, Ceramiales). Mar. Biol. 29: 29-36.
Fredericq, S. and Ramírez, M.E. (1996). Systematic studies of the Phyllophoraceae (Gigartinales, Rhodophyta) based on rbcL sequence analysis. Hydrobiol. 326/327: 137-143.
Freile-Pelegrin, Y. and Morales, J.L. (2004). Antibacterial activity in marine algae from the coast of Yucatan, Mexico. Bot. Mar. 47: 140-146.
Freile-Pelegrin, Y. and Robledo, D. (2013). Bioactive Phenolic Compounds from Algae,
In: Hernández‐Ledesma, B. and Herrero, M. (eds.). Bioactive Compounds from Marine Foods: Plant and Animal Sources. Wiley-Blackwell. pp. 464.
Freshwater, D. and Rueness, J. (1994). Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence analysis. Phycologia 33(3): 187-194.
Freshwater, D.W. and Bailey, J.C. (1998). A multigene phylogeny of the Gelidiales including nuclear large subunit rRNA sequence data. J. Appl. Phycol. 10: 229-236.
Freshwater, D.W., Fredericq, S., Butler, B.S., Hommersand, M.H., and Chase, M.W. (1994). A gene phylogeny of the red algae (Rhodophyta) based on plastid rbcL. Proc. Natl. Acad. Sci. U.S.A. 91(15): 7281-7285.
Freshwater, D.W., Tudor, K., O’Shaughnessy, K., and Wysor, B. (2010). DNA barcoding in the red algal order Gelidiales: comparison of COI with rbcL and verification of the “barcoding gap.” Cryptogamie Algol. 31(4): 435-449.
Friedlander, M. (2008). Advances in cultivation of Gelidiales. J. Appl. Phycol. 20: 451-456.
Fries, L. (1974). Growth stimulation of axenic red algae by simple phenolic compounds. J. Exp. Mar. Biol. Ecol. 15: 1-9.
Fries, L. and Iwasaki, H. (1976). p-Hydroxyphenylacetic acid and other phenolic compounds as growth stimulators of the red alga Porphyra tenera. PI. Sci. Lett. 6: 299-307.
Fujii, M.T., Cassano, V., Senties, A., Diaz-Larrea, J., Machín-Sánchez, M. and Gil- Rodriguez, M.C. (2012). Comparative analysis of the corps en cerise in several species of Laurencia (Ceramiales, Rhodophyta) from the Atlantic Ocean. Braz. J. Phycol. 22(4): 795-804.
Fujii, M.T., Cassano, V., Stein, E.M., and Carvalho, L.R. (2011). Overview of the taxonomy and of the major secondary metabolites and their biological activities related to human health of the Laurencia complex (Ceramiales, Rhodophyta) from Brazil. Braz. J. Phycol. 21(2): 268-282.
Fujii, M.T., Diaz-Larrea, J., Gil-Rodriguez, M.C., Machín-Sánchez, M., Cassano, V., and Senties, A. (2009). Morphology and molecular phylogeny of the genus Osmundea (ceramiales, rhodophyta) from Canary archipelago, eastern Atlantic. Phycologia 48(4).
Fujiwara-Arasaki, T., Mino, N., and Kuroda, M. (1984). The protein value in human nutrition of edible marine algae in Japan. Hydrobiologia 116/117: 513-516.
Fuller, R.W., Cardellina, J.H., Kato, Y., Brinen, L.S., Clardy, J., Snader, K.M., and Boyd, M.R. (1992). A pentahalogenated monoterpene from the red alga Portieria hornemannii

278
produces a novel cytoxicity profile against a diverse panel of human tumour cell lines. J. Med. Chem. 35: 3007-3011.
Furnari, G., Cormaci, M., and Serio, D. (2001). The Laurencia complex (Rhodophyta, Rhodomelaceae) in the Mediterranean Sea: an overview. Cryptogamie, Algol. 22(4): 331-373.
Furnari, G., Serio, D., and Cormaci, M. (2004). Revision of Laurencia pedicularioides (Ceramiales, Rhodophyta). Taxon 53(2): 453-60.
Gabrielson, P.W. and Garbary, D.J. (1986). Systematics of red algae (Rhodophyta). CRC Critical Reviews in Plant Sciences 3: 325-366.
Gabrielson, P.W., Garbary, D.J., and Scagel, R.F. (1985). The nature of the ancestral red alga: inferences from a cladistic analysis. BioSystems 18: 335-346.
Galland-Irmouli, A.V., Fleurence, J., Lamghari, R., Lucon, M., Rouxel, C., Barbaroux, O., Bronowicki, J.P., Villaume, C., and Gueant, J.L. (1999). Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J. Nutr. Biochem. 10: 353-359.
Garbary, D.J. and Clarke, B. (2001). Apoptosis in trichoblast development in Polysiphonia harveyi (Rhodophyta). Phycol. 40: 324-329.
Garbary, D.J. and Gabrielson, P.W. (1990). Taxonomy and evolution. In: Cole, K.M. and Sheath, R.G. (eds.). Biology of the red algae. Cambridge University Press, Cambridge, UK. pp. 477-498.
Garbary, D.J. and Harper, J.T. (1998). A phylogenetic analysis of the Laurencia complex (Rhodomelaceae) of the red algae. Cryptogamie Algol. 19: 185-200.
García-Jiménez, P. and Robaina, R.R. (2015). On reproduction in red algae: further research needed at the molecular level. Front. Plant. Sci. 6: 93.
García-Jiménez, P., Rodrigo, M., and Robaina, R.R. (1998). Influence of plant growth regulators, polyamines and glycerol interaction on growth and morphogenesis of carposporelings of Grateloupia doryphora cultured in vitro. J. Appl. Phycol. 10: 95-100.
García-Salgado, S., Raber, G., Raml, R., Magnes, C., and Francesconi, K.A. (2012). Arsenosugar phospholipids and arsenic hydrocarbons in two species of brown macroalgae. Environ. Chem. 9(1): 63.
García-Salgado, S., Raber, G., Raml, R., Magnes, C., and Francesconi, K. (2012). Arsenosugar phospholipids and arsenic hydrocarbons in two species of brown macroalgae. Env. Chem. 9(1): 63.
Gaubert, J., Payri, C.E., Vieira, C., Solanki, H., and Thomas, O.P. (2019). High metabolic variation for seaweeds in response to environmental changes: a case study of the brown algae Lobophora in coral reefs. Sci. Rep. 9(1): 993.
Gerasimenko, N.I., Chaikina, E.L., Busarova, N.G., and Anisimov, M.M. (2010). Antimicrobic and hemolytic activity of low molecular metabolites of brown seaweed Laminaria cichorioides (Miyabe). Appl. Biochem. Microbiol. 46: 426-430.
Gianturco, M.A., Giammarino, A.S., Friedel, P., and Flanagan, V. (1964). The volatile constituents of coffee-IV. Tetrahedron 20(12): 2951-2961.
Gika, H.G., Theodoridis, G.A., Plumb, R.S., and Wilson, I.D. (2013). Current practice of liquid chromatography-mass spectrometry in metabolomics and metabonomics. J. Pharm. Biom. Anal. 87: 12-25.
Gil-Rodriguez, M.C., Sentìes, A., Díaz-Larrea, J., Cassano, V., and Fujii, M.T. (2009). Laurencia marilzae SP. Nov. (ceramiales, rhodophyta) from the Canary Islands, Spain, based on morphological and molecular evidence. J. Phycol. 45(1): 264-271.
Glazer, A.N. (1989). Light guides. Directional energy transfer in a photosynthetic antenna. J. Biol. Chem. 264: 14.
Glombitza, K.W., and Gerstberger, G. (1985). Phlorotannins with dibenzodioxin structural elements from the brown alga Eisenia arborea. Phyrochemurry 24: 543-551.
Godínez-Ortega, J.L., Snoeijs, P., Robledo, D., Freile-Pelegrín, Y., and Pedersén, M. (2008). Growth and pigment composition in the red alga Halymenia floresii cultured under different light qualities. J. Appl. Phycol. 20: 253-260.

279
Gómez, I., Figueroa, F.L., Ulloa, N., Morales, V., Lovengreen, C., Huovinen, P., and Hess, S. (2004). Patterns of photosynthesis in 18 species of intertidal macroalgae from southern Chile. Mar. Ecol. Prog. Ser. 270:103-116.
Gonçalves, M.S. (2018). Cultivation of Osmundea pinnatifida and Codium tomentosum, native seaweed species with commercial potential. Master Thesis, University of Porto. pp. 71.
Gonzales, M.A. and Goff, L.J. (1989b). The red algal epiphytes Microcladia coulteri and M. californica (Rhodophyceae, Ceramiaceae). II. Basiphyte specificity. J. Phycol. 25: 558-67.
González, A.G., Darias, V., and Estévez, E. (1982). Chemotherapeutic activity of polyhalogenated terpenes from Spanish algae. Planta Med. 44: 44-46.
Goulitquer, S., Potin, P., and Tonon, T. (2012). Mass spectrometry-based metabolomics to eluci- date functions in marine organisms and ecosystems. Mar. Drugs 10: 849-880.
Graham, L.E. and Wilcox, L.W. (2000) Algae. Prentice Hall Incorporated, Upper Saddle River, New Jersey.
Graham, L.E., Graham, J.M and Wilcox, L.W. (2009). Algae. Benjamin Cummings (Pearson), San Francisco, CA. pp. 616.
Gray, K.R. and Biddlestone, A.J. (1973). Composting - process parameters. The Chem. Eng. 71-76.
Greff, S., Zubia, M., Payri, C., Thomas, O.P., and Perez, T. (2017). Chemogeography of the red macroalgae Asparagopsis: metabolomics, bioactivity, and relation to invasiveness. Metabol. 13: 1-13.
Gross, E.M. (2010). Allelopathy of aquatic autotrophs. Crit. Rev. Plant. Sci. 22: 313-339.
Grossman, A., Bhaya, D., Apt, K., and Kohoe, D. (1995). Light harvesting complexes in oxygenic photosynthesis: diversity, control and evolution. Annu. Rev. Genet. 29: 231-288.
Grote, B. (2016). Bioremediation of aquaculture wastewater: evaluating the prospects of the red alga Palmaria palmata (Rhodophyta) for nitrogen uptake. J. Appl. Phycol. 28: 3075-3082.
Grote, B. (2019). Recent developments in aquaculture of Palmaria palmata (Linnaeus) (Weber and Mohr 1805): cultivation and uses. Rev. Aquac. 11: 25-41.
Grzymski, J., Johnsen, G., and Sakshug, E. (1997). The significance of intracellular self-shading on the bio-optical properties of brown, red and green macroalgae. J. Phycol. 33: 408-414.
Guaratini, T., Cardozo, K.H.M., Pinto, E., and Colepicolo, P. (2009). Comparison of diode array and electrochemical detection in the C30 reverse phase HPLC analysis of algae carotenoids. J. Brazil. Chem. Soc. 9: 1609-1616.
Guillard, R.R.L. (1975). Culture of phytoplankton for feeding marine invertebrates. In: Smith, W.L. and Chanley, M.H (eds.) Culture of marine invertebrate animals. Plenum Press, New York, USA. pp. 29-60.
Guillard, R.R.L. (2007). Purification methods for microalgae. In: Andersen, R.A. (ed.). Algal culturing techniques. Elsevier Academic Press, London. pp. 117-132.
Guillard, R.R.L. and Ryther, J.H. (1962). Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt and Detonula confervacea Cleve. Can. J. Microbiol. 8: 229-239.
Guiry, M. and Cunningham, E. (1984). Photoperiodic and temperature responses in the reproduction of the north eastern Atlantic Gigartina acicularis (Rhodophyta: Gigartinales). Phycologia 23: 357-367.
Guiry, M.D. (1974). A preliminary consideration of the taxonomic position of Palmaria palmata (Linnaeus) Stackhouse: Rhodymenia Palmata (Linnaeus) Greville. J. Mar. Biol. Assoc. U.K. 54: 509-528.
Guiry, M.D. (1978). The importance of sporangia in the classification of the Florideophycidae. In Irvine, D.E.G. and Price, J.H. (eds.). Modern approaches to the taxonomy of red and brown algae. Academic Press, London. pp. 111144.
Guiry, M.D. (1987). The evolution of the life history types in the Rhodophyta: an appraisal. Cryptogamie Algol. 8:1-12.

280
Guiry, M.D. (1990). Sporangia and spores. In: Cole, K.M. and Sheath, R.G. (eds.). Biology of the red algae. Univ Press, Cambridge. pp. 347-376.
Guiry, M.D. and Blunden, G. (1991). Seaweed resources in Europe. Uses and Potential. Wiley, Chichester. pp. 432.
Guiry, M.D. and Guiry, G.M. (2014). AlgaeBase. Worldwide electronic publication. National University of Ireland, Galway. http://www.algaebase.org
Guiry, M.D. and Guiry, G.M. (2018). AlgaeBase. Worldwide electronic publication. National University of Ireland, Galway. http://www.algaebase.org
Gupta, V., Thakur, R., Baghel, R., Reddy, C.R.K., and Jha, B. (2014). Seaweed Metabolomics: A New Facet of Functional Genomics. Adv. Bot. Res. 71:31-52.
Gurgel, C.F.D., and Fredericq, S. (2004). Systematics of the Gracilariaceae (Gracilariales, Rhodophyta): a critical assessment based on rbcL sequence analyses. J. Phycol. 40: 139-159.
Güven, K.C., Percot, A., and Sezik, E. (2010). Alkaloids in marine algae. Mar. Drugs 8: 269-284.
Häder, D.P. and Figueroa, F.L. (1997). Photo ecophysiology of macroalgae. Photochem. Photobiol. 66: 1-14.
Häder, D.P., Lebert, M., Sinha, R.P., Barbieri, E.S., and Helbling, E.W. (2002). Role of protective and repair mechanisms in the inhibition of photosynthesis in marine macroalgae. Photoch. Photobio Sci. 1: 809-814.
Hagemann, M. and Pade, N. (2015). Heterosides-compatible solutes occurring in prokaryotic and eukaryotic phototrophs. Plant Biol. 17: 927-34.
Haglund, K. and Pedersén, M. (1988). Spray cultivation of seaweeds in recirculating brackish water. Aquaculture 72: 181-189.
Haijin, M., Xiaolu, J., and Huashi, G. (2003). A k-carrageenan derived oligosaccharide prepared by enzymatic degradation containing anti-tumour activity. J. Appl. Phycol. 15: 297-303.
Hamana, K., Matsuzaki, S., Niitsu, M., Samejima, K., and Nagashima, H. (1990). Polyamines in unicellular thermoacidophilic red alga Cyanidium caldarium. Phytochemistry 29: 377-380.
Han, X., Zeng, H., Bartocci, P., Fantozzi, F., and Yan, Y. (2018). Phytohormones and effects on growth and metabolites of microalgae: a review. Fermentation 4(2): 25.
Hanelt, D., Huppertz, K., and Nultsch, W. (1993). Daily course of photosynthesis and photoinhibition in marine macroalgae investigated in the laboratory and field. Mar. Ecol. Prog. Ser. 97: 31-37.
Hanisak, M.D. (1983). The nitrogen relationships of marine macroalgae. In: Carpenter, E.J. and Capone, D.G. (eds.) Nitrogen in marine environment, Academic Press, NY. pp. 699-730.
Hannach, G. and Santelices, B. (1985). Ecological differences between the isomorphic reproductive phases of two species of Iridaea (Rhodophyta: Gigartinales). Mar. Ecol. Prog. Ser. 22: 291-303.
Hardy, F.G. and Guiry, M.D. (2003). A checklist and atlas of the seaweeds of Britain and Ireland. Published by Bri. Phycol. Soc. pp. 30.
Harker M., Berkaloff, C., Lemoine, Y., Britton, G., Young, A.J., Duval, J.C., Rmiki, N.E., and Rousseau, B. (1999). Effects of high light and desiccation on the operation of the xanthophyll cycle in two marine brown algae. Eur. J. Phycol. 34: 35-42.
Harlin, M.M., and Lindbergh, J.M. (1977). Selection of substrata by seaweeds: optimal surface relief. Mar. Bio. 40: 33-40.
Harnedy, P.A. and FitzGerald, R.J. (2011). Bioactive proteins, peptides, and amino acids from macroalgae. J. Phycol. 47: 218-32.
Haroon, A.M. (2000). The biochemical composition of Enteromorpha spp. from the Gulf of Gdańsk coast on the southern Baltic Sea. Oceanologia 42 (1): 19-28.

281
Harper, J.T. and Saunders, G.W. (2001a) Molecular systematics of the Florideophyceae (Rhodophyta) using nuclear large and small subunit rDNA sequence data. J. Phycol. 37: 1073-1082.
Harper, J.T. and Saunders, G.W. (2001b). The application of sequences of the ribosomal cistron to the systematics and classification of the Rhodophyta. Cah. Biol. Mar. 42: 25-38.
Harrison, P.J. and Hurd, C.L. (2001). Nutrient physiology of seaweeds: application of concepts to aquaculture. Cah. Biol. Mar. 42: 71-82.
Harrison, P.J., Druehl, L.D., Lloyd, K.E., and Thompson, P.A. (1986). Nitrogen uptake kinetics in three year-classes of Laminaria groenlandica (Laminariales: Phaeophyta). Mar. Biol. 93: 29-35.
Hartmann, A., Murauer A., and Ganzera M. (2017). Quantitative analysis of mycosporine-like amino acids in marine algae by capillary electrophoresis with diode-array detection. J. Pharm. Biomed. Anal. 10 (138): 153-157.
Harwood, J. (1984). Effect of the environment on the acyl lipids of algae and higher plants. In: Siegenthahler, P.A. and Eichenberger, W. (eds.) (2001). Structure, function and metabolism of plant lipids. Elsevier, Amsterdam. pp. 543550.
Hawkes, M.W. (1990). Reproductive strategies. In: Cole, K.M. and Sheath, R.G. (eds.). Biology of the Red Algae. Cambridge University Press. pp. 455-476.
Hebert, P.D.N., Cywinska, A., Ball, S.L., and DeWaard, J.R. (2003a). Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B. Biol. Sci. 270(1512): 313-321.
Hebert, P.D.N., Ratnasingham, S., and de Waard, J.R. (2003b). Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 270 (Suppl.1): S96-S99.
Hellio, C., Simon-Colin, C., Clare, A., and Deslandes, E. (2004). Isethonic acid and floridoside isolated from the red alga Grateloupia turuturu inhibit settlement of Balanus Amphitrite crypid larvae. Biofouling 20: 139-45.
Helmuth, B. and Hofmann, G.E. (2001). Microhabitats, thermal heterogeneity and patterns of physiological stress in the rocky intertidal zone. Biol. Bull. 201: 374-384.
Higa, T. and Kuniyoshi, M. (2000). Toxins associated with medicinal and edible seaweeds. J. Toxicol. Toxin Rev. 19(2): 119-137.
Hodgson, L.M. (1981). Photosynthesis of the red alga gastroclonium coulteri (rhodophyta) in response to changes in temperature, light intensity, and desiccation. J. Phycol. 17(1): 37-42.
Holdt, S.L. and Kraan, S. (2011). Bioactive compounds in seaweeds: Functional food applications and legislation. J. Appl. Phycol. 23: 543-597.
Holdt, S.L., Christensen, L., and Iversen, J.J.L. (2013). A novel closed system bubble column photobioreactor for detailed characterisation of micro- and macroalgal growth. J. Appl. Phycol. 26: 825-835.
Hollingsworth, P.M., Forrest, L.L., Spouge, J.L., Hajibabaei, M., Ratnasingham, S., Van Der Bank, M., Little, D.P. (2009b). A DNA barcode for land plants. Proc. Natl. Acad. Sci. U.S.A. 106: 12794-12797.
Hommersand, M.H. (1963). The morphology and classification of some Ceramiaceae and Rhodomelaceae. Univ. Calif. Publ. Bot. 35: 165-366.
Hommersand, M.H. and Fredericq, S. (1990). Sexual reproduction and Cystocarp development. In: Biology of the red algae. Cole, K.M and Sheath, R.G. (eds.). Cambridge University Press. pp. 305-45.
Hommersand, M.H., Fredericq, S. and Freshwater, D.W. (1994). Phylogenetic systematics and biogeography of the Gigartinaceae (Gigartinales, Rhodophyta) based on sequence analysis of rbcL. Bot. Mar. 37:193-203.
Hopkins, W.G. and Hüner, N.P.A. (2009). Introduction to plant physiology. 4th ed. John Wiley and Sons, Inc., Hoboken, NJ. pp. 523.

282
Hou, X. and Yan, X. (1998). Study on the concentration and seasonal variation of inorganic elements in 35 species of marine algae. Sci. Total Environ. 222: 141156.
Hruby, T. (1976). Observations of algal zonation resulting from competition. Estuarine and Coast Mar. Sei 4: 231-233.
Hruby, T. and Norton, T.A. (1979). Algal colonization on rocky shores in the Firth of Clyde. J. Ecol. 67: 65-77.
Huang, W. and Fujita, Y. (1997). Callus induction and thallus regeneration of the red alga Meristotheca papulosa (Rhodophyta, Gigartinales). Bot. Mar. 40: 55-61.
Huang, Z., Ito, K., Morita, I., Yokota, K., Fukushi, K., Timerbaev, A.R., Watanabe, S., and Hirokawa, T. (2005). Sensitive monitoring of iodine species in seawater using capillary electrophoresis: vertical profiles of dissolved iodine in the Pacific Ocean. J. Environ. Monit. 7: 804-808.
Hunt, L.J.H. and Denny, M.W. (2008). Desiccation protection and disruption: a trade-off for an intertidal marine alga. J. Phycol. 44(5): 1164-1170.
Hurd, C.L. (2000). Water motion, marine macroalgal physiology and production. J. Phycol. 36: 453-472.
Hurd, C.L. and Dring, M.J. (1990). Phosphate uptake by intertidal algae in relation to zonation and season. Mar. Biol. 107: 281-289.
Hurd, C.L., Harrison, P.J., Bischof, K., and Lobban, C.S. (2014). Seaweed Ecology and Physiology, 2nd ed. Cambridge: Cambridge University Press. pp. 536.
Hurtado, A. and Critchley, A. (2018). A review of multiple biostimulant and bioeffector benefits of AMPEP, an extract of the brown alga Ascophyllum nodosum, as applied to the enhanced cultivation and micropropagation of the commercially important red algal carrageenophyte Kappaphycus alvarezii and its selected cultivars. J. Appl. Phycol. 30(5): 2859-2873.
Hurtado, A.Q., Joe, M., Sanares, R. C., Fan, D., Prithiviraj, B., and Critchley, A. T. (2012). Investigation of the application of Acadian Marine Plant Extract Powder (AMPEP) to enhance the growth, phenolic content, free radical scavenging, and iron chelating activities of Kappaphycus Doty (Solieriaceae, Gigartinales, Rhodophyta). J. Appl. Phycol. 24(3): 601-611.
Hurtado, A.Q., Yunque, D.A., Tibubos, K., and Critchley, A.T. (2009). Use of Acadian marine plant extract powder from Ascophyllum nodosum in tissue culture of Kappaphycus varieties. J. Appl. Phycol. 21: 633-639.
Hwang, E.K., Ha, D.S., Baek, J.M., Wee, M.Y., and Park, C.S. (2006a). Effects of pH and salinity on the cultivated brown alga Sargassum fulvellum and associated animals. Algae 21: 317-321.
Hwang, E.K., Park, C.S., and Baek, J.M. (2006b). Artificial seed production and cultivation of the edible brown alga Sargassum fulvellum (Turner) C. Agardh: developing a new species for seaweed cultivation in Korea. J. Appl. Phycol. 18: 251-257.
Hwang, H., Chen, T., Nines, R.G., Shin, H.C., and Stoner, G.D. (2006). Photo chemoprevention of UVB-induced skin car-cinogenesis in SKH-1 mice by brown algae polyphenols. Int. J. Cancer 119: 2742-2749.
Ibañez, E. and Cifuentes, A. (2013). Benefits of using algae as natural sources of functional ingredients. J. Sci. Food. Agric. 93: 703-709.
Ibañez, E., Herrero, M., Mendiola, J.A., and Castro-Puyana, M. (2012). Extraction and characterization of bioactive compounds with health benefits from marine resources: macro and micro algae, cyanobacteria, and invertebrates. In: Hayes, M. (ed.). Marine bioactive compounds: sources, characterization and applications. Springer US, Boston, MA. pp. 5598.
Ichikawa, M., Mizuno, I., Yoshida, J., Ide, N., Ushijima, M., Kodera, Y., Hayama, M., and Ono, K. (2006). Pharmacokinetics of cycloalliin, an organosulfur compound found in garlic and onion, in rat. J. Agric. Food. Chem. 27; 54 (26): 9811-9819.

283
Iliopoulou, D., Roussis, V., Pannecouque, C., De Clercq, E., and Vagias, C. (2002a). Halogenated sesquiterpenes from the red alga Laurencia obtusa. Tetrahedron 58: 6749-6755.
Iliopoulou, D., Vagias, C., Harvala, C., and Roussis, V. (2002b). C15 Acetogenins from the red alga Laurencia obtuse. Phytochemistry 59: 111-116.
Illijas, M.I., Indy, J.R., Yasui, H., and Itabashi, Y. (2009). Lipid class and fatty acid composition of a little-known and rarely collected alga Exophyllum wentii Weber-van Bosse from Bali Island, Indonesia. J. Oleo Sci. 58: 103-110.
Imelda, J., and Chennubhotla, V.S.K. (1998). Gibberellic acid and 2,4-D as growth regulators in laboratory culture of seaweeds. Indian J. Mar. Sci. 28: 66-69.
Ito, K. and Hori, K. (1989). Seaweed: chemical composition and potential food uses. Food Rev. Int. 5(1): 101-144.
James, M.A. (2010). A review of initiatives and related R&D being undertaken in the UK and internationally regarding the use of macroalgae as a basis for biofuel production and other non-food uses relevant to Scotland. Report commissioned by the Marine Scotland, pp. 79.
Janouškovec, J., Liu, S.L., Martone, P.T., Carré, W., Leblanc, C., Collén, J., and Keeling, P.J. (2013). Evolution of red algal plastid genomes: ancient architectures, introns, horizontal gene transfer, and taxonomic utility of plastid markers. PLoS ONE 8, e59001.
Jennings, R.C. (1969). Cytokinins as endogenous plant growth regulators in the algae Ecklonia and Hypnea. Aust. J. Biol. Sci. 22: 621-627.
Jennings, R.C. (1971). Studies on the regulation of algal growth by gibberellin. Aust. J. Biol. Sci. 24: 1115-1124.
Jeukendrup, A.E. and Randell, R. (2011). Fat burners: nutrition supplements that increase fat metabolism. Obesity Rev. 12(10): 841-851.
Jimenez del Rio, M., Ramazanov, Z., and Garcia-Reina, G. (1996). Ulva rigida (Ulvales, Chlorophyta) tank culture as biofilters for dissolved inorganic nitrogen from fishpond effluents. Fifteenth International Seaweed Symposium, Springer. pp. 6166.
Jiménez-Escrig, A., Gómez- Ordóñez, E., and Rupérez, P. (2011). Seaweed as a source of novel nutraceutical: sulfated polysaccharides and peptides. Adv. Food Nutr. Res. 64 (64): 325-337.
Jiménez-Escrig, A., Gómez-Ordóñez, E., and Rupérez, P. (2012). Brown and red seaweeds as potential sources of antioxidant nutraceuticals. J. Appl. Phycol. 24(5): 1123-1132.
Jones, L.A. and Harwood, J.L. (1992). Lipid composition of the brown algae Fucus versiculosus and Ascophyllum nodosum. Phytochemistry 31: 3397-3403.
Jones, W.E. and Dent, S. (1970). Culture of marine algae using a recirculating seawater system. Helgoländer wiss. Meeresunters 20: 70-78.
Josephson, D.B. and Lindsay, R.C. (1986). Enzymic Generation of Volatile Aroma Compounds from Fresh Fish. In: Thomas, H.P and Rodney, C. (eds.). Biogeneration of aromas. American Chemical Society, Washington DC. pp. 201-219.
Joubert, Y. and Fleurence, J. (2008). Simultaneous extraction of proteins and DNA by an enzymatic treatment of the cell wall of Palmaria palmata (Rhodophyta). J. Appl. Phycol. 20: 55-61.
Kaehler, S. and Kennish, R. (1996). Summer and winter comparisons in the nutritional value of marine macroalgae from Hong Kong. Bot. Mar. 39 (1-6):11-17.
Kain, J.M. (1975) Algal recolonization of some cleared subtidal areas, The J. Ecol. 63: 739-765.
Karabay-Yavasoglu, N.U., Sukatar, A., Ozdemir, G., and Horzum, Z. (2007). Antimicrobial activity of volatile components and various extracts of the red alga Jania rubens. Phytother. Res. 21: 153-156.
Karsten, U. and Wiencke, C. (1999). Factors controlling the formation of UV-absorbing mycosporine-like amino acids in the marine red alga Palmaria palmata from Spitsbergen (Norway). J. Plant. Physiol. 155: 407-415.

284
Karsten, U., Sawall, T., and Wiencke, C. (1998). A survey of the distribution of UV-absorbing substances in tropical macroalgae. Phycol. Res. 46: 271-279.
Karthick, P., Sankar, R.S., Kaviarasan, T., and Mohanraju R. (2012). Ecological implications of trace metals in seaweeds: bio-indication potential for metal contamination in Wandoor, South Andaman Island. Egypt. J. Aquat. Res. 38: 227-231.
Kastendiek, J. (1982). Factors determining the distribution of the sea pansy, Renilla kollikeri, in a subtidal sand-bottom habitat. Oecologia 52: 340-347.
Kato, H., Rhue, M.R., and Nishimura T., (1989). Role of free amino acids and peptides in food taste. In: Teranishi, R., Buttery, R.G., and Shadidi, F. (eds.). Flavor Chemistry Trends and developments. American Chemical Society, Washington DC. pp. 158-175.
Kaur, S. and Das, M. (2011). Functional foods: an overview. Food Sci. Biotechnol. 20: 861-875.
Kelly, L., Collier, L., Costello, M.J., Diver, M., McGarvey, S., Kraan, S., Morrisey, J., and Guiry, M.D. (2001). Impact assessment of hand and mechanical harvesting of Ascophyllum nodosum on regeneration and biodiversity. Mar. Res. Ser. 19: 1-51.
Kenicer, G., Bridgewater, S., and Milliken, W. (2000). The ebb and flow of Scottish seaweed use. Bot. J. Scot. 52(2): 119-148.
Kennington, K. (2015). The plankton ecology of SEA7 (Scottish West Coast). SAHFOS Report. Plymouth, UK.
Kerrison, P.D. and Le, H.N. (2015). Environmental factors on egg liberation and germling production of Sargassum muticum. J. Appl. Phycol. 28: 481-489.
Kerrison, P.D., Le, H.N., and Hughes, A.D. (2016a). Hatchery decontamination of Sargassum muticum juveniles and adults using a combination of sodium hypochlorite and potassium iodide. J. Appl. Phycol. 28: 1169-1180.
Kerrison, P.D., Le, H.N., Twigg, G.C., Smallman, D.R., MacPhee, R., Houston, F.A.B., and Hughes, A.D. (2016b). Decontamination treatments to eliminate problem biota from macroalgal tank cultures of Osmundea pinnatifida, Palmaria palmata and Ulva lactuca. J. Appl. Phycol. 28(6): 3423-3434.
Kerrison, P.D., Stanley, M.S., Edwards, M.D., Black, K.D., and Hughes, A.D. (2015). The cultivation of European kelp for bioenergy: Site and species selection. Biomass and Bioenerg. 80: 229-242.
Khan, W., Rayirath, U.P., Subramanian, S., Jithesh, M.N., Rayorath, P., Hodges, D.M., Critchley, A.T., Craigie, J.S., Norrie, P., and Prithiviraj, B. (2009). Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 28: 386-399.
Khatkar, D. and Kuhad, M.S. (2000). Short-term salinity induced changes in two wheat cultivars at different growth stages. Biol. Plant. 43 (4): 629-632.
Kientz, B., Thabard, M., Cragg, S., Pope, J., and Hellio, C. (2012). A new method for removing microflora from macroalgal surfaces: an important step for natural product discovery. Bot. Mar. 54: 457-469.
Kim, A.R., Shin, T.S., Lee, M.S., Park, J.Y., Park, K.E., Yoon, N.Y., Kim, J.S., Choi, J.S., Jang, B.C., Byun, D.S., Park, N.K., and Kim, H.R. (2009). Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food. Chem. 57: 3483-3489.
Kim, J.H., Hudson, J.B., Huang, A.M., Bannister, K., Jin, H., Choi, T.J., Towers, G.H.N., Hong, Y.K., and DeWreede, R.E. (1997). Biological activities of seaweed extracts from British Columbia, Canada and Korea. I. Antiviral activity. Can. J. Bot. 75: 1656-1660.
Kim, J.K. and Yarish C. (2014). Development of a sustainable land based Gracilaria cultivation system. Algae 29(3): 217-225.
Kim, J.K. and Yarish, C. (2010). Antioxidant properties of tidal pool microalgae, Halochlorococcum porphyrae and Oltamannsiellopsis unicellularis from Jeju Island, Korea. Algae 25 (1): 45-56.
Kim, J.K., Yarish, C., Hwang, E.K., Park, M., and Kim, Y. (2017). Seaweed aquaculture: cultivation technologies, challenges and its ecosystem services. Algae 32 (1): 1-13.

285
Kim, T.W., Lee, K., Lee, C.K., Jeong, H.D., Suh, Y.S., Lim, W.A., Kim, K.Y., and Jeong, H.J. (2013). Interannual nutrient dynamics in Korean coastal waters. Harmful Algae 30: 15-27.
Kirst, G.O. (1980). Low mw carbohydrates and ions in Rhodophyceae: quantitative measurement of floridoside and digeneaside. Phytochemistry 19: 1107-1110.
Kirst, G.O. and Bisson, M.A. (1979). Regulation of turgor pressure in marine algae: ions and low molecular weight organic compounds. Aust. J. Plant Physiol. 6: 539-556.
Kiseleva, А.А., Tarakhovskaya, E.R., and Shyshova, M.F. (2012). Biosynthesis of phytohormones in algae. Fiziol. Rast. 59(5): 643-659.
Kjelleberg, S., Steinberg, P., Givskov, M., Gram, L., Manefield, M., and de Nys, R. (1997). Do marine natural products interfere with prokaryotic AHL regulatory systems? Aquat. Microb. Ecol. 13: 85-93.
Kobayashi, M., van de Meent, E.J., Ohoka, H., Inoue, K., Itoh, S., Amesz, J., and Watanabe, T. (1992). Pigment composition of heliobacteria and green sulphur bacteria. In: Murata, N. (ed.). Research in Photosynthesis. Kluwer Academic Publishers, Dordrecht. pp. 393-396.
Kolber, Z. and Falkowski, P.G. (1993). Use of active fluorescence to estimate phytoplankton photosynthesis in situ. Limnol. Oceanogr. 38: 1646-1665.
König, G.M., Wright, A.D., and de Nys, R. (1999a). Halogenated monoterpenes from Plocamium costatumand their biological activity. J. Nat. Prod. 62: 383-385.
Kosovel, V. and Talarico, L. (1979). Seasonal variation of phorosynthetic pigments in Gracilaria verrucosa (Huds.) Papenfuss (Florideophyceae-Gigartinales). Boll. Soc. Adriat. Sci. 63: 5-15.
Krachler, R., Krachler, R. F., von der Kammer, F., Süphandag, A., Jirsa, F., Ayromlou, S., Hofman, T., and Keppler, B.K. (2010). Relevance of peat-draining rivers for the riverine input of dissolved iron into the ocean. Sci. Total Environ. 408(11): 2402-2408.
Kraft, G.T. (1981). Rhodophyta: morphology and classification (chap. I). In: Lobban, C.S. and Wynne, M.J. (eds.). Biology of Seaweeds: Botanical Monograph, v. 17, pp. 6-51.
Krause, G.H. and Weis, E. (1991). Chlorophyll fluorescence and photosynthesis: the basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 42: 313-349.
Kremer, B.P. (1978). Patterns of photo assimilatory products in Pacific Rhodophyceae. Ca. J. Bot. 56: 1655-1659.
Kübler, J. and Raven, J. (1996). Inorganic carbon acquisition by red seaweeds grown under dynamic light regimes. Hydrobiologia. 326-327(1):401-406.
Kucera, H. and Saunders, G.W. (2008). Assigning morphological variants of Fucus (Fucales, Phaeophyceae) in Canadian waters to recognized species using DNA barcoding. Botany 86: 1065-1079.
Kulikova, I.V. and Khotimchenko, S.V. (2000). Lipids of different thallus regions of the brown alga Sargassum miyabei from the Sea of Japan. Russ. J. Mar. Biol. 26(1): 54-57.
Kumar, C.S., Raju, D., Sarada, V.L. and Rengasamy, R. (2008). Seaweed extracts control the leaf spot disease of the medicinal plant Gymnema sylvestre. Indian J. Sci. Technol. 1: 93-94.
Kumari, P., Bijo, A.J., Mantri, V.A., Reddy, C.R.K., and Jha, B. (2013b). Fatty acid profiling of tropical marine macroalgae: an analysis from chemotaxonomic and nutritional perspectives. Phytochemistry. 86: 44-56.
Kumari, P., Kumar, M., Reddy, C.R.K., and Jha, B. (2013a). Algal lipids, fatty acids and sterols, In: Dominguez, H. (ed.). Functional ingredients from algae for foods and nutraceuticals. Woodhead Publishing Ltd, Oxford, Cambridge, Philadelphia, New Delhi. pp. 87-132.
Kundel, M., Thorenz, U.R., Petersen, J.H., Huang, R.J., Bings, N.H., and Hoffmann, T. (2012). Application of mass spectrometric techniques for the trace analysis of short-lived iodine-containing volatiles emitted by seaweed. Anal. Bioanal. Chem. 402: 3345-3357.

286
Küpper, F.C., Leblanc, C., Meyer-Klaucke, W., Potin, P., and Feiters, M.C. (2014). Different speciation for bromine in brown and red algae, revealed by in vivo x-ray absorption spectroscopic studies. J. Phycol. 50 (4): 652-664.
Kurata, K., Taniguchi, K., Agatsuma, Y., and Suzuki, M. (1998). Diterpenoid feeding-deterrents from Laurencia saitoi. Phytochemistry 47: 363-369.
Kuriyama, M., Tagagi, M., and Murata, K. (1960). Ninhydrin reactive substances in marine algae. Bull. Fac. Fisheries Hokkaido Univ. 11: 58-63.
Kursar, T.A., Van Der Meer, J., and Alberte, R.S. (1983b). Light-harvesting system of the red alga Gracilaria tikvahiae. II. Phycobilissome characteristics of pigment mutants. Plant Physiol. 73: 361-369.
Kwon, H.J., Bae, S.Y., Kim, K.H., Han, C.H., Cho, S.H., Nam, S.W., Choi, Y.H., and Kim, B.W. (2007). Induction of apoptosis in HeLa cells by ethanolic extract of Corallina pilulifera. Food Chem. 104: 196-201.
Kyle, D.J. and Ohad, I. (1986). The mechanism of photoinhibition in higher plants and green algae. In: Staehelin, L.A. and Arntzen, C.J. (eds.). Encyclopedia of Plant Physiology, new series. Springer-Verlag, Berlin. pp. 468-476.
Kylin, H. (1917). Ua ber die Keimung der Florideensporen. Ark. Bot. 14: 1-25.
Kylin, H. (1923). Studien über die Entwicklungsgeschichte der Florideen. Kongl. Sven. Sk. Vetenskapsakad. Handl. 63: 1-139.
Kylin, H. (1925). The marine red algae in the vicinity of the biological station at Friday Harbor, Wash.Lunds Univ. Årrskr. N.F. Avd. 2, 21(9): 1-87.
Kylin, H. (1932). Die Florideenordnung Gigartinales. Acta Univ. Å Lund. 28(8): 1-88, Plates 1-28.
Kylin, H. (1956). Die Gattungen der Rhodophyceen. C.W.K. Gleerups, Lund, Sweden pp. xvq673.
La Barre, S., Potin, P., Leblanc, C., and Delage, L. (2010). The halogenated metabolism of brown algae (Phaeophyta), its biological importance and its environmental significance. Mar. Drugs 8: 988-1010.
Lapointe, B. (1981). The effects of light and nitrogen on growth, pigment content, and biochemical composition of Gracilaria foliifera v. agustissima (Giagartinales, Rhodophyta). J. Phycol. 17: 90-95.
Lapointe, B.E and Ryther, J.H. (1979). The effects of nitrogen and seawater flow rate on the growth and biochemical composition of Gracilaria foliifera var. angustissima in mass outdoor cultures. Bot. Mar. 22(8): 529-538.
Lapointe, B.E. (1985). Strategies for pulsed nutrient supply to Gracialria cultures in the Florida Keys: interactions between concentration and frequency of nutrient pulse. J. Exp. Mar. Biol. Ecol. 93 :211-22.
Lapointe, B.E. (1987). Phosphorus and nitrogen limited photosynthesis and growth of Gracilaria tikvahiae (Rhodophyceae) in the Florida Keys: an experimental field study. Mar. Biol. 93: 561568.
Lapointe, B.E. and Duke, C.S. (1984). Biochemical strategies for growth of Gracilaria tikvahiae (Rhodophyta) in relation to light intensity and nitrogen availability. J. Phycol. 20: 488-495.
Larsen, A. and Sand-Jensen, K. (2006). Salt tolerance and distribution of estuarine benthic macroalgae in the Kattegat- Baltic Sea area. Phycologia 45:13-23.
Laturnus, F., Svensson, T., and Wiencke, C. (2010). Release of reactive organichalogens by the brown macroalga Saccharina latissima after exposure to ultraviolet radiation. Polar Res. 29: 379-384.
Lavery, P.S. and McComb, A.J. (1991). The nutritional ecophysiology of Chaetomorpha linum and Ulva rigida in the Peel Inlet, Western Australia. Bot. Mar. 34: 251-260.
Le Gall, L. and Saunders, G.W. (2007). A nuclear phylogeny of the Florideophyceae (Rhodophyta) inferred from combined EF2, small subunit and large subunit ribosomal

287
DNA: Establishing the new red algal subclass Corallinophycidae. Mol. Phylogenetics Evol. 43: 1118-1130.
Le Gall, L. and Saunders, G.W. (2010). DNA barcoding is a powerful tool to uncover algal diversity: a case study of the Phyllophoraceae (Gigartinales, Rhodophyta) in the Canadian flora. J. Phycol. 46(2): 374-389.
Lea-Smith, D.J., Ross, N., Zori, M., Bendall, D.S., Dennis, J.S., Scott, S.A., Smith, A.G., and Howe, C.J. (2013). Thylakoid Terminal Oxidases Are Essential for the Cyanobacterium Synechocystis sp. PCC 6803 to Survive Rapidly Changing Light Intensities. Plant Physiol. 162: 485-495.
Lee, H.J., Kim, H.C., Vitek, L., and Nam, C.M. (2010a). Algae consumption and risk of type 2 diabetes: Korean National Health and Nutrition Examination Survey in 2005. J. Nutr. Sci. Vitaminol .56: 13-18.
Lee, R.E. (1999). Phycology, 3rd ed. Cambridge University Press, Cambridge, UK. pp. 614.
Lee, R.E. (2008). Phycology, 4th. Cambridge University Press. New York, NY. pp. 561.
Lee, S.H., Han, J.S., Heo, S.J., Hwang, J.Y., and Jeon, Y.J. (2010a). Protective effects of dieckol isolated form Ecklonia cava against high glucose-induced oxidative stress in human umbilical endothelial cells. Toxicol. In Vitro 24: 375-381.
Lee, S.H., Li, Y., Karadeniz, F., Kim, M.M., and Kim, S.K. (2008). α-Glucosidase and α- amylase inhibitory activities of phloroglucinol derivatives from edible marine brown alga, Ecklonia cava. J. Sci. Food. Agric. 89: 1552-1558.
Lewis, R.J. and Hanisak, D. (1996). Effects of phosphate and nitrate supply on productivity, agar content and physical properties of agar of Gracilaria strain G-16S. J. Appl. Phycol. 8(1): 41-49.
Lewis, S., Gacesa, P., Gil-Rodriguez, M.C., Valdès, F., and Frias, I. (2008). Molecular systematics of the genera Laurencia, Osmundea and Palisada (Rhodophyta) form the Canary Islands-Analysis of rDNA and RUBISCO spacer sequences. An. Jardin Bot. Madrid 65(1): 91-109.
Lewmanomont, K.S.D and Srimanobhas, V. (1996). Seagrasses of Thailand. In: Kuo, J.R.C. P., Walker, D.I., and Kirkman, H. (eds.). Seagrasses Biology: Proceedings of an international workshop, Rottnest Island, Western Australia. pp. 21-26.
Li, X., Staszewski, L., Xu, H., Durick, K., Zoller, M. and Adler, E. (2002). Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA. 99(7): 4692-4696.
Li, Y.X., Wijesekara, I., Li, Y., and Kim, S.K. (2011). Phlorotannins as bioactive agents from brown algae. Pro. Biochem. 46: 2219-2224.
Lignell, A. and Pedersén, M.F. (1987). Nutrient supply is an important operating parameter in the management of seaweed cultivation systems. Hydrobiologia 151/152: 431-441.
Lignell, A. and Pedersén, M.F. (2007). Effects of pH and inorganic carbon concentration on growth of Gracilaria secundata. Bri. Phycol. J. 24(1): 83-89.
Lin, H. and Qin, S. (2014). Tipping points in seaweed genetic engineering: scaling up opportunities in the next decade. Mar. Drugs 12: 3025-3045.
Lin, S.M., Frederiq, S., and Hommersand, M. (2001). Systematics of the Delesseriaceae (Ceramiales, Rhodophyta) based on the large subunit rDNA and rbcL sequences, including the Phycodryoideae, sub fam. Nov. J Phycol. 37: 881-899.
Liscum, E. and Briggs, W.R. (1995). Mutations in the NPH1 locus of Arabidopsis disrupt the perception of photo-tropic stimuli. Plant Cell. 7: 473-485.
Lobban, C.S. and Harrison, P.J. (1994). Seaweed Ecology and Physiology. Cambridge University Press, pp. 562.
Lobban, C.S. and Harrison, P.J. (1997). Seaweed ecology and physiology. Biologia Plantarum 40 (1): 90-90.
Lobo, V., Patil, A., Phatak, A., and Chandra, N. (2010). Free radicals, antioxidants and functional foods: impact on human health. Pharmacogn. Rev. 4: 118-126.

288
López-Cristoffanini, C., Zapata, J., Gaillard, F., Potin, P., Correa, J.A., and Contreras-Porcia, L. (2015). Identification of proteins involved in desiccation tolerance in the red seaweed Pyropia orbicularis (Rhodophyta, Bangiales). Proteomics 15: 3954-3968.
López-Figueroa, F. and Niell, F.X. (1991). Photocontrol of chlorophyll and biliprotein synthesis in seaweeds: possible photoreceptors and ecological considerations. Sci. Mar. 55: 519-527.
López-Figueroa, F. and Niell, F. X. (1990). Effects of light quality on chlorophyll and biliprotein accumulation in seaweeds. Mar. Biol. 104(2): 321-327.
López-López, I., Cofrades, S., and Jiménez-Colmenero, F. (2009). Low fat frankfurters enriched with n-3 PUFA and edible seaweed: effects of olive oil and chilled storage on physicochemical, sensory and microbial characteristics. Meat Sci. 83: 148-154.
Lordan, S., Ross, R.P., and Stanton, C. (2011). Marine bioactives and functional food ingredients: potential to reduce incidence of chronic diseases. Mar. Drugs 9: 1056-1100.
Loureiro, R.R., Reis, R.P., and Critchley, A.T. (2009). In vitro cultivation of three Kappaphycus alvarezii (Rhodphyta, Areschougiaceae) variants (green, red and brown) exposed to a commercial extract of the brown alga Ascophyllum nodosum (Fucaceae, Ochro-phyta). J. Appl. Phycol. 22: 101-104.
Lüning, K. (1979). Growth strategies of three Laminaria species (Phaeophyceae) inhabiting different depth zones in the sublittoral region of Helgoland (North Sea). Mar. Ecol. Progr. Ser. 1: 195-207.
Lüning, K. (1980). Critical levels of light and temperature regulating the gametogenesis of three Laminaria species (Phaeophyceae). J. Phycol. 16: 1-15.
Lüning, K. (1990). Seaweeds: their environment, biogeography, and ecophysiology. John Wiley and Sons. USA, NY. pp. 527.
Lüning, K. (1992). Day and night kinetics of growth rate in green, brown and red seaweeds. J. Phycol. 28: 794-803.
Lüning, K. and Diek, T. (1989). Environmental triggers in algal seasonality. Bot. Mar. 32(5): 389-397.
Lüning, K. and Dring, M.J. (1985). Action spectra and spectral quantum yield of photosynthesis in marine macroalgae with thin and thick thalli. Mar. Biol. 87:119-129.
Lüning, K. and Pang, S. (2003). Mass cultivation of seaweeds: current aspects and approaches. J. Appl. Phycol. 15: 115-119.
Mabeau, S. and Fleurence, J. (1993) Seaweed in food products: biochemical and nutritional aspects. Trends Food Sci. Technol. 4: 103-107.
Machalek, K.M., Davison, I.R., and Falkowski, P.G. (1996). Thermal and photoacclimation of photosynthesis in the brown alga, Laminaria saccharina. Plant Cell. Environ. 19: 1005-1016.
Machín-Sánchez, M., Diaz-Larrea, J., Fujii, M.T., Senties, A., Cassano, V., and Gil-Rodriguez, M.C. (2012). Morphological and molecular evidences within Osmundea (Ceramiales, Rhodophyta) from the Canary Islands, Eastern Atlantic Ocean. Afr. J. Mar. Sci. 34(1): 27-42.
Machín-Sánchez, M., Gil-Rodríguez, M.C., and Haroun, R. (2018). Phylogeography of the red algal Laurencia complex in the Macaronesia Region and nearby coastal areas: recent advances and future perspectives. Diversity 10:10.
Machín-Sánchez, M., Rousseau, F., Le Gall, L., Cassano, V., Neto, A.I., Senties, A., Fujii, M.T., and Gil-Rodriguez, M.C. (2016). Species diversity of the genus Osmundea (Ceramiales, Rhodophyta) in the Macaronesian region. J. Phycol. 52(4): 664-681.
Machu, L., Misurcova, L., Ambrozova, J.V., Orsavova, J., Mlcek, J., Sochor, J., and Jurikova, T. (2015). Phenolic content and antioxidant capacity in alga food product. Molecules 20: 1118-1133.
MacTavish, M.R. and Cohen, R.A. (2014). A simple, inexpensive, and field-relevant microcosm tidal simulator for use in marsh macrophyte studies. Appl. Plant Sci. 2(11): apps.1400058.

289
Maeda, H., Tsukui, T., Sashima, T., Hosokawa, M., and Miyashita, K. (2008). Seaweed carotenoid, fucoxanthin, as a multi-functional nutrient. Asia Pac. J. Clin. Nutr. 17(1): 196-199.
Mæhre, H.K, Edvinsen, G.K., Eilerten, K.E., and Elvevoll, E.O. (2016). Heat treatment increases the protein bioaccessibility in the red seaweed dulse (palmaria palmata), but not in the brown seaweed winged kelp (alaria esculenta). J. Appl. Phycol. 28(1): 581-590.
Mæhre, H.K., Malde, M.K., Eilertsen, K.E., and Elvevoll, E.O. (2014). Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 94: 3281-3290.
Maggs, C.A. and Pueschel, C.M. (1989). Morphology and development of Ahnfeltia plicata (Rhodophyta): proposal of Ahnfeltiales ord. nov. J. Phycol. 25: 333-351.
Maggs, C.A. and Callow, M.E. (2003). Algal Spores, Encyclopedia of life sciences. (Nature Publishing Group). pp. 1-6.
Maggs, C.A. and Hommersand, M.H. (1993). Seaweeds of the British Isles. Volume 1. Rhodophyta. Part 3A. Ceramiales. London xv: Natural History Museum Publications. pp. 1-444.
Maggs, M.A., Verbruggen, H., and De Clerck. (2007). Molecular systematics of red algae: building future structures of firm foundations. In: Brodie, J. and Lewis, J. (eds.). Unravelling the algae: the past, present and future of algal systematics. CRC Press, Boca Raton, London and New York. pp. 103-121.
Magne, F. (1989). Classification et phylogénie des Rhodophycées. Cryptogam. Algol. 10: 101-115.
Mak, W., Hamid, N., Liu, T., Lu, J., and White, W.L. (2013). Fucoidan from New Zealand Undaria pinnatifida: Monthly variations and determination of antioxidant activities. Carbohydr. Polym. 95: 606-614.
Manach, C., Scalbert, A., Morand, C., Remesy, C., and Jimenez, L. (2004) Polyphenols: food sources and bioavailability. Am. J. Clin. Nutr. 79: 727-747.
Mantri, A.V., Eswaran, K., Shanmugam, M., Ganesan, M., Veeragurunathan, V., Thiruppathi, S., Reddy, C.R.K., and Seth, A. (2017). An appraisal on commercial farming of Kappaphycs alvarezii in India: success in diversification of livelihood and prospects. J. Appl. Phycol. 29: 335-357.
Mao, Y., Chen, N., Cao, M., Chen, R., Guan, X., and Wang, D. (2019). Functional characterization and evolutionary analysis of glycine-betaine biosynthesis pathway in red seaweed Pyropia yezoensis. Mar. Drugs 17(1): 70.
Marquardt, J. and Hanelt, D. (2004). Carotenoid composition of Delesseria lancifolia and other marine red algae from polar and temperate habitats. Eur. J. Phycol. 39: 285-292.
Marroig, R.G., Loureiro, R.R., and Reis, P.R. (2016). The effect of Ascophyllum nodosum (Ochrophyta) extract powder on the epibiosis of Kappaphycus alvarezii (Rhodophyta) commercially cultivated on floating rafts. J. Appl. Phycol. 28: 2471-2477.
Marsham, S., Scott, G.W., and Tobin, M.L. (2007). Comparison of nutritive chemistry of a range of temperate seaweeds. Food Chem. 100(4): 1331-1336.
Martínez, B. and Rico, J.M. (2004). Inorganic nitrogen and phosphorous uptake kinetics in Palmaria palmata and Chondrus crispus (Rhodophyta). J. Phycol. 40 :642-650.
Martínez, B., Viejo, R.M., Rico, J.M., Rødde, R.H., Faes, V.A., Oliveros, J., and Álvarez, D. (2006). Open sea cultivation of Palmaria palmata (Rhodophyta) on the northern Spanish coast. Aquaculture 254: 376-387.
Martínez-García, M., Stuart, M.C.A., and van der Maarel, M.J.E.C. (2016). Characterization of the highly branched glycogen from the thermoacidophilic red microalgae Galdieria sulphuraria and comparison with other glycogens. Int. J. Biol. Macromolec. 89: 12-18.
Martin-Lescanne, J., Rousseau, F., De Reviers, B., Payri, C., Couloux, A., Cruaud, C., and Le Gall, L. (2010). Phylogenetic analyses of the Laurencia complex (Rhodomelaceae, Ceramiales) support recognition of five genera: Chondrophycus, Laurencia, Osmundea, Palisada and Yuzurua stat. nov. Eur. J. Phycol. 45(1): 51-61.

290
Masuda, M., Abe, T., Suzuki, T., and Suzuki, M. (1996). Morphological and chemotaxonomic studies on Laurencia composita and L. okamurae (Ceramiales, Rhodophyta). Phycologia 35(6): 550-562.
Masuda, M., Kawaguchi, S., and Phang, S.M. (1997a). Taxonomic notes on Laurencia similis and L. Papillosa (Ceramiales, Rhodophyta) from the Western pacific. Bot. Mar. 40: 229-239.
Masuda, M., Kawaguchi, S., Takahashi, Y., Matsuo, Y., and Suzuki M. (1997b). A taxonomic study of the genus Laurencia (Ceramiales, Rhodophyta) from Vietnam. I. Laurencia caduciramulosa Masuda et Kawaguchi, sp. nov. Cryptogamie, Algol. 18: 1-10.
Matthijs, M., Fabris, M., Obata, T., Foubert, I., Franco-Zorrilla, J.M., Solano, R., Fernie, A.R., Vyverman, W., and Goossens, A. (2018). The transcription factor bZIP14 regulates the TCA cycle in the diatom Phaeodactylum tricornutum. EMBO J. 36(11): 1559-1576.
Maurya, R., Ghosh, T., Paliwal, C., Shrivastav, A., Chokshi, K., Pancha, I., Ghosh, A., and Mishra, S. (2014). Biosorption of methylene blue by de-oiled algal biomass: equilibrium, kinetics and artificial neural network modelling. PLoS ONE 9: e109545.
Maxwell, K. and Johnson, G.N. (2000). Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 51: 659-668.
McConnell, O. and Fenical, W. (1977). Halogen chemistry of the red alga Asparagopsis. Phytochemistry 16(3): 367-374.
McCracken, I.R. (1989). Purifying algal cultures-a review of chemical methods. Proc. N.S. Inst. Sci. 38: 145-168.
McDermid, K.J. (1988). Laurencia from the Hawaiian Islands. In: Abbott, I.A. (ed.). Taxonomy of economic seaweeds. California Sea Grant Publication No. T-CSGCP-018, University of California, La Jolla, California. Vol.2, pp. 231-247.
McDermid, K.J. (1989). Laurencia crustiformans sp. nov. (Ceramiales, Rhodophyta) from the Hawaiian Islands. Phycologia 28: 352-359.
McDevit, D.C. and Saunders, G.W. (2009). On the utility of DNA barcoding for species differentiation among brown macroalgae (Phaeophyceae) including a novel extraction protocol. Phycol. Res. 57(2): 131-141.
McDougall, G.J., Shpiro, F., Dobson, P., Smith, P., Blake, A., and Stewart, D. (2005). Different polyphenolic components of soft fruits inhibit alpha-amylase and alpha-glucosidase. J. Agric. Food Chem. 53: 2760-2766.
McHugh, D.J. (2003). A Guide to the Seaweed Industry. FAO Fisheries Technical Paper 441, Rome, 105 p.
McIvor, L., Maggs, C.A., Provan, J., and Stanhope, M.J. (2002). rbcL sequences reveal multiple cryptic introductions of the Japanese red alga Polysiphonia harveyi. Mol. Ecol. 10: 911-919.
McKey, D. (1979). The distribution of secondary compounds within plants. In: Rosenthal, G.A. and Janzen, J.H. (eds.). Herbivores: their interactions with secondary plant metabolites. Academic Press, New York, NY. pp. 55-133.
Mckinnon, S.L., Hiltz, D., Ugarte, R., and Craft, C.A. (2010). Improved methods of analysis for betaines in Ascophyllum nodosum and its commercial seaweed extracts. J. Appl. Phycol. 22: 489-494.
McLachlan, J.L., Craigie, J.S., Chen, L.C.M, and Ogata, E. (1972). Porphyra linearis Grev. An edible species from Nova Scotia. Proc. Int. Seaweed Symp. 7: 473-476.
McMahon, R.F., Hunter, R.D., and Russell-Hunter, W.D. (1974). Variation in aufwuchs at six freshwater habitats in terms of carbon biomass and of carbon:nitrogen ratio. Hydrobiol. 45: 391-404.
Medlin, L.K., Kooistra, W.H.C.F., Potter, D., Saunders, G.W., and Andersen, R.A. (1997). Phylogenetic relationships of the ‘golden algae’ (haptophytes, heterokont chromophytes) and their plastids. Plant. Syst. and Evol. (Supplement) 11: 187-219.
Meeks, J.C. (1974). Chlorophylls. In: Stewart, W.D.P. (ed.). Algal physiology and biochemistry, Bot. Monographs. Oxford. 10:161-175.

291
Meier, R., Zhang, G., and Ali, F. (2008). The use of mean instead of smallest interspecific distances exaggerates the size of the “Barcoding Gap” and leads to misidentification. Syst. Biol. 57: 809-813.
Mellouk, Z., Benammar, I., Krouf, D., Goudjil, M., Okbi, M., and Malaisse, W. (2017). Antioxidant properties of the red alga Asparagopsis taxiformis collected on the North West Algerian coast. Exp. Ther. Med. 13(6): 3281-3290.
Mendes, L.F., Vale, L.A.S., Martins, A.P., Yokoya, N.S., Soriano, E.M., and Colepicolo, P. (2012). Influence of temperature, light and nutrients on the growth rates of the macroalga Gracilaria domingensisin. J. Appl. Phycol. 24: 1419-1426.
Meng, J., Rosell, K.G., and Srivastava, L.M. (1987). Chemical characterization of floridosides from Porphyra perforata. Carbohydr. Res. 61: 171-180.
Mensi, F., Ksouri, J., Seale, E.G., and Romdhane, M.S. (2011). Modeling laboratory culture of Gracilaria verrucosa (Hudson) Papenfuss according to nutrients concentrations and salinities levels. Bull. Inst. Natn. Scien. Tech. Mer de Salammbô, Vol. 38
Metti, Y., Millar, A. J. K., and Steinberg, P. (2015). A new molecular phylogeny of the Laurencia complex (Rhodophyta, Rhodomelaceae) and a review of key morphological characters result in a new genus, Coronaphycus, and a description of C. novus. J. Phycol. 51: 929-942.
Mikami, K., Mori, I., Matsuura, T., Ikeda, Y., Kojima, M., Sakakibara, H., and Hirayama, T. (2015). Comprehensive quantification and genome survey reveal the presence of novel phytohormone action modes in red seaweeds. J. Appl. Phycol. 28(4): 2539-2548.
Miller, I.J. (1997). The chemotaxonomic significance of the water-soluble red algal polysaccharides. Recent Res. Dev. Phytochemistry 1: 531-565.
Mishra, V.K., Temelli, F., Ooraikul, B., Shacklock, P.F., and Craigie, J.S. (1993). Lipids of the red alga, Palmaria palmata. Bot. Mar. 36: 169-174.
Misurcova, L., Kracmar, S., Klejdus, B., and Vacek, J. (2010). Nitrogen content, dietary fiber, and digestibility in algal food products. Czech J. Food Sci. 28: 27-35.
Mithoo-Singh, P.K., Keng, F.S.L., Phang, S.M., Elvidge, E.C.L., Sturges, W.T., Malin, G., and Rahman, N.A. (2017). Halocarbon emissions by selected tropical seaweeds: species-specific and compound specific responses under changing pH. PeerJ5:e2918; DOI 10.7717/peerj.2918.
Mohamed, S., Hashim, S.N., and Rahman, H.A. (2012). Seaweeds: a sustainable functional food for complementary and alternative therapy. Trends Food Sci. Technol. 23: 83-96.
Monro, K. and Poore, A.G.B. (2005). Light quantity and quality induce shade-avoiding plasticity in a marine macroalgae. J. Evolution Biol. 18: 426-435.
Mooney, P.A. and van Staden, J. (1986). Algae and cytokinins. J. Plant Physiol. 123: 1-21.
Moreno-Garrido, I. and Cañavate J.P. (2001). Assessing chemical compounds for controlling predator ciliates in outdoor mass cultures of the green algae Dunaliella salina. Aquac. Eng. 24 (2): 107-114.
Morgan, K.C. and Simpson, F.J. (1981a). Cultivation of Palmaria (Rhodymenia) palmata: effect of high concentrations of nitrate and ammonium on growth and nitrogen uptake. Aquat. Bot. 11: 167-171.
Morgan, K.C. and Simpson, F.J. (1981b). The cultivation of Palmaria palmata: effect of light intensity and temperature on growth and chemical composition. Bot. Mar. 24: 547-552.
Mori, I.C., Ikeda, Y., Matsuura, T., Hirayama, T. and Mikami, K. (2017). Phytohormones in red seaweeds: a technical review of methods for analysis and a consideration of genomic data. Bot. Mar. 60: 153-170.
Morris, O.P. and Russel, G. (1974). Interspecific differences in responses to copper by natural 6 populations of Ectocarpus. Br. Phycol. J. 9: 269-272.
Mörschel, E. and Rhiel, E. (1987). Phycobilisomes and thylakoids: the light-harvesting system of cyanobacteria and red algae. In: Harris, J.R. and Horne, R.W. (eds.). Membranous Structures. Academic Press, London. pp. 210-54.

292
Mouritsen, O.G, Williams, L., Bjerregaard, R., and Duelund, L. (2012). Seaweeds for umami flavour in the New Nordic Cuisine. Flavour 1:4.
Mouritsen, O.G. (2013). Seaweeds. Edible, available, and sustainable. The University of Chicago Press Books. pp. 304.
Muller, F.L.L. and Tankéré-Muller, S.P.C. (2012). Seasonal variations in surface water chemistry at disturbed and pristine peatland sites in the Flow Country of northern Scotland. Sci. Total Environ. 43: 351-362.
Muller, K.M., Sheath, R.G., Vis, M.L., Crease, T.J., and Cole, K.M. (1998). Biogeography and systematics of Bangia (Bangiales, Rhodophyta) based on the Rubisco spacer, rbcL gene and 18S rRNA gene sequences and morphometric analyses. I. North America. Phycologia 37: 195-207.
Mumford, T.F. Jr. and Miura, A. (1988). Porphyra as food: cultivation and economics. In: Lembi, C.A. and Waaland, J.R. (eds.). Algae and human affairs. Cambridge University Press, Cambridge. pp. 91-93.
Munda, I.M. (1977). Differences in amino acid composition of estuarine and marine fucoids. Aquat. Bot. 3: 273-280.
Munier, M., Dumay, J., Morançais, M., Jaouen, P., and Fleurence, J. (2013). Variation in the biochemical composition of the edible seaweed Grateloupia turuturu Yamada harvested from two sampling sites on the Brittany coast (France): the influence of storage method on the extraction of the seaweed pigment r- phycoerythrin. J. Chem. Article ID 568548. pp. 8.
Munir, N., Sharif, N., Naz, S., and Mazoor, F. (2013). Algae: a potent antioxidant source. SJMR 1(3): 22-31.
Murakami, K., Yamaguchi, Y., Noda, K., Fujii, T., Shinohara, N., Ushirokawa, T., Sugawa-Katayama, Y., and Katayama, M. (2011). Seasonal variation in the chemical composition of a marine brown alga, Sargassum horneri (Turner) C. Agardh. J. Food Compos. Anal. 24: 231-236.
Nagahama, T., Fujimoto, K., Takami, S., Kinugawa, A., and Narusuye, K. (2009). Effective amino acid composition of seaweeds inducing food preference behaviours in Aplysia kurodai. Neurosci. Res. 64: 243-250.
Nagayama, K., Iwamura, Y., Shibata, T., Hirayama, I., and Nakamura, T. (2002). Bactericidal activity of phlorotannins from the brown alga Ecklonia kurome. J. Antimicrob. Chemother. 50: 889-893.
Nam, K.W. (1999). Morphology of Chondrophycus undulata and C. parvipapillata and its implications for the taxonomy of the Laurencia (Ceramiales, Rhodophyta) complex. Eur. J. Phycol. 34: 455-468.
Nam, K.W. and Choi, H.G. (2001). Morphology of Laurencia clavata and L. elata (Ceramiales, Rhodophyta) in relation to generic circumscription in the Laurencia complex. Eur. J. Phycol 36: 285-294.
Nam, K.W. and Saito, Y. (1995). Vegetative and reproductive anatomy of some Laurencia (Ceramiales, Rhodophyta) species with a description of L. maris-rubri sp. nov. from the Red Sea. Phycologia 34(2): 157-165.
Nam, K.W., Maggs, C.A., and Garbary, D.J. (1994). Resurrection of the genus Osmundea with an emendation of the generic delineation of Laurencia (Ceramiales, Rhodophyta). Phycologia 33(5): 384-395.
Nam, K.W., Maggs, C.A., McIvor, L. and Stanhope, M.J. (2000b). Taxonomy and Phylogeny of Osmundea (Rhodomelaceae, Rhodophyta) in Atlantic Europe. Phycologia 36: 759-772.
Nam, K.W., Maggs, C.A., Mcivor, L., Stanhope, M.J., and Lamouroux, H. (2000). Taxonomy and Phylogeny of Osmundea (Rhodomelaceae, Rhodophyta). J. Phycol. 36: 759-772.
Navarro-Angulo, L. and Robledo, D. (1999). Effects of nitrogen source, N:P ratio and N-pulse concentration and frequency on the growth of Gracilaria cornea (Gracilariales, Rhodophyta) in culture. Hydrobiol. 398-399: 315-320.

293
Neish, A.C., Shacklock, P.F., Fox, C.H. and Simpson, F.J. (1977). The cultivation of Chondrus crispus. Factors affecting growth under greenhouse conditions. Can. J. Bot. 55: 2263-2271.
Nelson, M.M., Phleger, C.F., and Nichols, P.D. (2002). Seasonal lipid composition in macroalgae of the North eastern Pacific Ocean. Bot. Mar. 45: 58-65.
Neori, A. (2008). Essential role of seaweed cultivation in integrated multi-trophic aquaculture farms for global expansion of mariculture: an analysis. J. Appl. Phycol. 20: 567-570.
Neori, A., Chopin, T., Troell, M., Buschmann, A.H., Kraemer, G.P., Halling, C., Shpigel, M. and Yarish, C. (2004). Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration in modern mariculture. Aquac. 231: 361-391.
Neori, A., Krom, M.D, and Rijn, J.V. (2007). Biogeochemical processes in intensive zero-effluent marine fish culture with recirculating aerobic and anaerobic bio filters. J. Exp. Mar. Biol. Ecol. 349: 235-247.
Netalgae (www.netalgae.eu) (2012). Seaweed industry in Europe, Report.
Ngan, Y. and Price, I R. (1979). Systematic significance of spore size in the Florideophyceae (Rhodophyta). Br. Phycol. J. 14: 285-303.
Niemann, D.I. and Dorffling, K. (1980). Growth inhibitors and growth promoters in Enteromorpha compressa (Chlorophyta). J. Phycol. 16: 383-389.
Nisizawa, K., Noda, H., Kikuchi, R., and Watanabe, T. (1987). The main seaweed foods in Japan. Hydrobiologia 151152(1): 5-29.
Norte, M., Fernández, J.J, and Runao, J.Z. (1989b). Three new bromo ethers from the red alga Laurencia obtuse. Tetrahedron 45: 5987-5994.
Norte, M., Fernández, J.J., and Souto, M. (1997b). New polyehter squalene derivatives from Laurencia. Tetrahedron 53: 4649-4654.
Norte, M., Fernández, J.J., Cataldo, F., and González, G. (1989a). E-dihydrorhodophytin, a C15 acetogenin from the red alga Laurencia pinnatifida. Phytochemistry 28: 647-649.
Norte, M., Fernández, J.J., Souto, M.L., Gavín, J.A., and García-Grávalos, M.D. (1997a). Thyrsenols A and B, two unusual polyether squalene derivatives. Tetrahedron 53(9): 3173-3178.
Norton, T.A. and Mathieson, A.C. (1983). The biology of unattached seaweeds. Prog. Phycol. Res. 2: 333-386.
Norziah, M.H. and Ching, C.Y. (2000). Nutritional composition of edible seaweed Gracilaria raggi-an edible species of nori from Nova Scotia. Food Chem. 68: 69-76.
Nussinovitch, A. (1997). Hydrocolloid applications: gum technology in the food and other industries. Blackie Academic and Professional, New York, NY. pp. 354.
Nwosu, F., Morris, J., Lund, V.A., Stewart, D., Ross, H.A., and Mcdougall, G.J. (2011). Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 126(3): 1006-1012.
Nylund, G.M., Weinberger, F., Rempt, M., and Pohnert, G. (2011). Metabolomic assessment of induced and activated chemical defence in the invasive red alga Gracilaria vermiculophylla. PLoS ONE 6: e29359.
Oates, B.R. and Cole, K. (1994). Comparative studies on hair cells of two agarophyte red algae, Gelidium vagum (Gelidiales, Rhodophyta) and Gracilaria pacifica (Gracilariales, Rhodophyta). Phycologia 33: 420-433.
Ogata, E. (1953). Some experiments on the settling of spores of red algae Bull. Soc. Plant Ecol. 3 (3): 128-134.
Ohno, M. and Critchley, A. T. (1993). Seaweed cultivation and marine ranching. Kanagawa International Fisheries Training Center, Japan International Cooperation Agency (JICA). Kanagawa, Japan, pp. 151.
Oliveira, A.S., Sudatti, D.B., Fujii, M.T., Rodrigues, S.V., and Pereira, R.C. (2013). Inter- and intrapopulation variation in the defensive chemistry of the red seaweed Laurencia dendroidea (Ceramiales, Rhodophyta). Phycologia: 52: 130-136.

294
Omar, H., Al-Judaibiand, A., and El-Gendy, A. (2018). Antimicrobial, antioxidant, anticancer activity and phytochemical analysis of the red alga, Laurencia papillosa. Int. J. pharmacol. 14 (4): 572-583.
Ördög, V., Stirk, W.A., van Staden, J., Novak, O., and Strnad, M. (2004). Endogenous cytokinins in three genera of microalgae from the Chlorophyta. J. Phycol. 40: 88-95.
Orduña-Rojas, J., Robledo, D., and Dawes, C.J. (2002). Studies on the tropical agarophyte Gracilaria cornea J. Agardh (Rhodophyta, Gracilariales) from Yucatán, Mexico. I. Seasonal physiological and biochemical responses. Bot. Mar. 45: 453-458.
Ozdemir, G., Karabay, N.U., Dalay, M.C., and Pazarbasi, B. (2004). Antibacterial activity of volatile component and various extracts of Spirulina platensis. Phytother. Res. 18: 754-757.
Paine, R.T. and Vadas, R.L. (1969). Calorific values of benthic marine algae and their postulated relation to invertebrate food preference. Mar. Biol. 4: 79-86.
Paiva, L., Lima, E., Neto, A.I., Marcone, M. and Baptista, J. (2017). Nutritional and functional bioactivity value of selected Azorean macroalgae: Ulva compressa, Ulva rigida, Gelidium microdon, and Pterocladiella capillacea. J. Food Sci. 82(7): 1757-1764.
Paiva, L., Lima, E., Patarra, R.F., Neto, A.I., and Baptista, J. (2014). Edible Azorean macroalgae as source of rich nutrients with impact on human health. Food Chem. 164: 128-135.
Paiva, L.S., Patarra, R.F., Neto, A.I., Lima, E.M.C., and Baptista, J.A.B. (2012). Antioxidant activity of macroalgae from the Azores. Arquipelago. Life Mar. Sci. 29: 1-6.
Pan, K. and Wang, W.X. (2012). Trace metal contamination in estuarine and coastal environments in China. Sci. Total Environ. 421: 3-16.
Papenfuss, G.F. (1947). Generic names of algae proposed for conservation. I. Madrono 9(1): 8-17.
Parker, H.S. (1982). Effects of simulated current on the growth rate and nitrogen metabolism of Gracilaria tikvahiae (Rhodophyta). Mar. Biol. 69: 137-145.
Parsons, T.R., Maita, Y. and Lalli, C.M. (1984). A manual of chemical and biological methods for seawater analysis. Pergamon Press, Oxford. pp. 173.
Parys, S., Kehraus, S., Krick, A., Glombitza, K.W., Carmeli, S., Klimo, K., Gerhäuser, C. and König, G.M. (2010). In vitro chemopreventive potential of fucophlorethols from the brown alga Fucus vesiculosus L. by antioxidant activity and inhibition of selected cytochrome P450 enzymes. Phytochemistry 71: 221-229.
Patarra, R.F., Leite, J., Pereira, R., Baptista, J., and Neto, A.I. (2013). Fatty acid composition of selected macrophytes. Nat. Prod. Res. 27(7): 665-669.
Patarra, R.F., Paiva, L., Neto, A.I., Lima, E., and Baptista, J. (2011). Nutritional value of selected macroalgae. J. Appl. Phycol. 23: 205-208.
Patwary, M.U. and van der Meer, J.P. (1992). Genetics and breeding of cultivated seaweeds. Korean J. Phycol. 7: 281-318.
Pec, M.K., Aguirre, A., Moser-Their, K., Fernandez, J.J., Souto, M.L., Dota, J., Diaz-Gonzalez F., and Villar J. (2003). Induction of apoptosis in estrogen dependent and independent breast cancer cells by the marine terpenoid dehydrothyrsiferol. Biochem. Pharmacol. 65: 1451-1461.
Pedersén, M.F. (1973). Identification of a cytokinin, 6-(3 methyl-2-butenylamino) purine, in seawater and the effect of cytokinins on brown algae. Physiol. Plant 28: 101-105.
Pedersén, M.F. (1994). Transient ammonium uptake in the macroalga Ulva-lactuca (Chlorophyta)-nature, regulation, and the consequences for choice of measuring technique. J. Phycol. 30: 980-986.
Pedra, A.G.L.M., Ramlov, F., Maraschin, M., and Hayashi, L. (2017). Cultivation of the red seaweed Kappaphycus alvarezii with effluents from shrimp cultivation and brown seaweed extract: effects on growth and secondary metabolism. Aquacult. 479: 297-303.

295
Penniman, C.A. and Mathieson, A.C. (1987). Variation in chemical composition of Gracilaria tikuahiae McLachlan (Gigartinales, Rhodophyta) in the Great Bay Estuary, New Hampshire. Eot. Mar. 30: 525-34.
Pereira, D.C. Trigueiro, T.G., Colepicolo, P., and Marinho-Soriano, E. (2012). Seasonal changes in the pigment composition of natural population of Gracilaria domingensis (Gracilariales, Rhodophyta). Rev. Bras. Farmacogn. 22(4): 874-880.
Pereira, H., Páramo, J., Silva, J., Marques, A., Barros, A., and Maurício, D. (2018). Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: from an agar plate to 100-m3 industrial photobioreactors. Sci. Rep. 8(1): 5112.
Pereira, H., Silva, J., Santos, T., Gangadhar, K.N., Raposo, A., Nunes, C., Coimbra, M.A., Gouvela, L., Barreira, L., and Varela, J. (2019). Nutritional potential and toxicological evaluation of Tetraselmis sp. CTP4 microalgal biomass produced in industrial photobioreactors. Molecules 24(17): 3192
Pereira, L. (2011). A review of the nutrient composition of selected edible seaweeds. In: Pomin, V.H (ed.). Seaweed: ecology, nutrient composition and medicinal uses. Nova Scotia Publishers, Inc. pp. 15-47.
Pereira, L. (2018). Biological and therapeutic properties of the seaweed polysaccharides. Int. Biol. Rev. 2(2):1-50.
Pereira, R. and Yarish, C. (2008). Mass production of marine macroalgae. In: Jørgensen, S.E. and Fath, B.D. (eds.). Encyclopedia of ecology, vol 3, Ecological engineering. Elsevier, Oxford. Pp. 2236-2247.
Pereira, R. and Yarish, C. (2010). The role of Porphyrain sustainable culture systems: physiology andapplications. In: Israel, A. and Einav, R. (eds.). Seaweeds and their role in a globally changingenvironment. Springer, Heidelberg. pp. 339-354.
Pereira, R., Yarish, C., and Critchley, A. (2012). Seaweed aquaculture for human foods, land based. In: Costa-Pierce, B.A. (ed.). Ocean farming and sustainable aquaculture science and technology. Encyclopedia of sustainability science and technology. Springer, New York. pp. 9109-9128.
Pettit, T.R., Johnes, A.L., and Harwood, J.L. (1989). Lipid of the red algae, Chondrus and Polysiphonia lanosa. Phytochemistry 28: 399-405.
Philips, G.O. and Williams, P.A. (eds.) (2009). Handbook of Hydrocolloids. Woodhead Publishing. pp. 948.
Phillips, J.C. and Hurd, C.L. (2004). Kinetics of nitrate, ammonium, and urea uptake by four intertidal seaweeds from New Zealand. J. Phycol. 40: 534-545.
Picone, G., Engelsen, S.B., Savorani, F., Testi, S., Badiani, A., and Capozzi, F. (2011). Metabolomics as a powerful tool for molecular quality assessment of the fish Sparus aurata. Nutrients 3: 212-227.
Plaza, M., Cifuentes, A., and Ibáñez, E. (2008). In the search of new functional food ingredients from algae. Trends Food Sci. Tech. 19: 31-39.
Plouguerné, E., Georgantea, P., Ioannou, E., Vagias, C., Roussis, V., Hellio, C., Kraffe, E., and Stiger-Pouvreau, V. (2010). Anti-microfouling activity of lipidic metabolites from the invasive brown alga Sargassum muticum (Yendo) Fensholt. Mar. Biotechnol. 12(1): 52-61.
Polevoi, V.V. (1982). Fitogormony (Phytohormones), Leningrad: Leningr. Gos. Univ.
Polevoi, V.V. (1986). Rol’ auksina v sistemakh regulyatsii u ras- tenii (The Role of Auxin in Regulatory Systems of Plants), Leningrad: Nauka.
Polne-Fuller, M. and Gibor, A. (1987). Calluses and callus-like growth in seaweeds: induction and culture. Hydrobiologia 151/152: 131-138.
Prathep, A., Marrs, R.H., and Norton, T.A. (2003). Spatial and temporal variations in sediment accumulation in an algal turf and their impact on associated fauna. Mar. Biol. 142 (2): 381-390.
Premuzic, E.T., Benkovitz, C.M., Gaffney, J.S., Walsh, J.J. (1982). The nature and distribution of organic matter in the surface sediments of world oceans and seas. Org. Geochem. 4: 63-77.

296
Provasoli, L. (1966). Culture and collections of algae. In: Watanabe, A. and Hattori, A. (eds.). Media and prospects for the cultivation. Jap. Soc. Plant Physiol., Tokyo. pp. 63-75.
Provasoli, L. and Carlucci, A.F. (1974). Vitamins and growth regulators. In: Stewart, W.D.P. (ed.). Algal physiology and biochemistry. Berkeley, Los Angeles. pp. 741-787.
Pueschel, C.M. (1979). Ultrastructure of tetrasporogenesis in Palmaria palmata (Rhodophyta). J. Phycol. 15(4): 409-424.
Pueschel, C.M. and Cole, K.M. (1982). Rhodophycean pit plugs an ultrastructural survey with taxonomic implications. Amer. J. Bot. 69: 703-720.
Purnick, P.E.M. and Weiss, R. (2009). The second wave of synthetic biology: From modules to systems. Nat. Rev. Mol. Cell Biol. 10: 410-422.
Radakovits, R., Jinkerson, R.E., Darzins, A., and Posewitz, M.S. (2010). Genetic engineering of algae for enhanced biofuel production. Eukaryot. Cell 9: 486-501.
Ragan, M.A. (1981). Chemical constituents of seaweeds. In: Lobban, C.S. and Wynne, M. J (eds.). The biology of Seaweeds. Blackwell Scientific, Oxford. pp. 589-626.
Ragan, M.A. and Gutell, R.R. (1995). Are red algae plants? Bot. J. Linean. Soc. 118: 81-105.
Ragan, M.A. Bird C.J., Riceo, E.L., Gutell, R.R., Murphy, C.A., and Singh, R.K. (1994). A molecular phylogeny of the marine red algae (Rhodophyta) based on the nuclear small-subunit ribosomal-RNA gene. Proc. Natl. Acad. Sci. USA 91: 7276-7280.
Ragan, M.A., Bird, C.J., Rice, E.L., Gutell, R.R., Murphy, C.A., and Singh, R.K. (1994). A molecular phylogeny of the marine red algae (Rhodophyta) based on the nuclear small subunit rRNA gene. Proc. Natl. Acad. Sci. USA. 91(15): 7276-80.
Raven, J.A. and Beardall, J. (2003). Carbohydrate Metabolism and Respiration in Algae. In: Larkum, A.W.D., Douglas, S.E., and Raven, J.A. (eds.). Photosynthesis in Algae. Advances in photosynthesis and respiration. Springer, Dordrecht. vol 14.
Raven, J.A., Johnston, A.M., and Macfarlene, J.J. (1990). Carbon metabolism. In: Cole, K.M., and Sheath, R.G. (eds.). Biology of red algae. Cambridge University Press, Cambridge. pp. 171-202.
Raven, P.H., Evert, R.F., and Eichhorn, S.E. (1992). Biology of plants, 5th Edition. Worth Publishers, New York. pp. 791.
Rayorath, P., Khan, W., Palanisamy, R., MacKinnon, S.L., Stefanova, R., Hankins, S.D., Critchley, A.T., and Prithiviraj, B. (2008b). Extracts of the brown seaweed Ascophyllum nodosum induce gibberellic acid (GA3)-independent amylase activity in barley. J. Plant Growth Regul. 27: 370-379.
Rayorath, P., Mundaya, J.N., Farid, A., Wajahatullah, K., Ravishankar, P., Hankins, S.D., Critchley, A.T., and Balakrishnan, P. (2008a). Rapid bioassays to evaluate the plant growth promoting activity of Ascophyllum nodosum (L.) Le Jol. Using a model plant, Arabidopsis thaliana (L) Heynh. J. Appl. Phycol. 20: 423-429.
Read, B.E. (1936). Chinese medicinal plants from the Pen Ts’ao Kang Mu A.D. 1596, 3rd ed. Peking Nat. Hist. Bull. pp. 300.
Reddy, C.R.K., Jha, B., Fujita, Y., and Ohno, M. (2008). Seaweed micropropagation techniques and their potentials: an overview. J. Appl. Phycol. 20: 609-617.
Redmond, S., Kim, J.K., Yarish, C., Pietrak, M., and Bricknell, I. (2014). Culture of Sargassum in Korea: techniques and potential for culture in the U.S. Marine Sea Grant College program, Orono, Maine. pp. 13.
Reed, R.H. (1990). Solute accumulation and osmotic adjustement. In: Cole, K.M and Sheath, R.G. (eds.). Biology of the red algae. Cambridge University Press, Cambridge. pp. 147-170.
Rengasamy, K.R.R., Kulkarni, M.G., Stirk, W.A., and Van Staden, J. (2015). Eckol- a new plant growth stimulant from the brown seaweed Ecklonia maxima J. Appl. Phycol. 27: 581-587.
Ribeiro, A.L.N.L., Tesima, K.E., Souza, J.M.C., and Yokoya, N.S. (2013). Effects of nitrogen and phosphorus availabilities on growth, pigment and protein contents in Hypnea cervicornis J. Agardh (Gigartinales, Rhodophyta). J. Appl. Phycol. 25: 1151-1157.

297
Richardson, K., Beardall, J., and Raven, J.A. (1983). Adaptation of unicellular algae to irradiance: An analysis of strategies. New Phytol. 93(2): 157-191.
Richmond, A. (2004). Biological principles of mass cultivation. In: Richmond, A. (ed.). Handbook of microalgal culture. Biotechnology and applied phycology. Blackwell Science Ltd, Cornwall. pp. 125-177.
Rico, J.M. and Fernándes, C. (1996). Seasonal nitrogen metabolism in an intertidal population of Gelidium latifolium (Gelidiaceae, Rhodophyta). Eur. J. Phycol. 31: 149-155.
Ridler, N., Wowchuk, M., Robinson, B., Barrington, K., Chopin, T., Robinson, S., Page, F., Reid, G., Szemerda, M., Sewuster, J., and Boyne-Travis, S. (2007). Integrated multi-trophic aquaculture (IMTA): a potential strategic choice for farmers. Aquacul. Econ. Manage. 11(1): 99-110.
Rincon, J., Gonzalez, F., Ballester, A., Blazquez, M.L. and Munoz, J.A. (2005). Biosorption of heavy metals by chemically activated alga Fucus vesiculosus. J. Chem. Technol. Biotechnol. 80: 1403-1407.
Rizvi, M.A. and Shameel, M. (2005). Pharmaceutical biology of seaweeds from the Karachi coast of Pakistan. Pharmaceut. Biol. 43(2): 97-107.
Robba, L., Russell, S., and Barker, G.L. (2006). Assessing the Use of the Mitochondrial Cox 1 Marker for use in DNA barcoding of red algae (Rhodophyta). Am. J. Bot. 93(8): 1101-1108.
Roberts, M.F. (2005). Organic compatible solutes of halotolerant and halophilic micro- organisms. Saline Syst. 1: 1-30.
Robertson-Andersson, D., Wilson, D., Bolton, J., Anderson, R., and Maneveldt, G. (2009). Rapid assessment of tissue nitrogen in cultivated Gracilaria gracilis (Rhodophyta) and Ulva lactuca (Chlorophyta). Afr. J. Aquat. Sci. 34: 169-172.
Rodrigues, D., Costa-Pinto, A.R., Sousa, S., Vasconcelos, M.W., Pintado, M.M., Pereira, L., Rocha-Santos, T.A.P., da Costa, J.P., Silva, A.M.S., Duarte, A.C., Gomes, A.M.P., and Freitas, A.C. (2019). Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization. Mar. Drugs 17 (209): 1-15.
Rodrigues, D., Freitas, A.C., Pereira, L., Rocha-Santos, T.A., Vasconcelos, M.W., Roriz, M., Rodríguez-Alcalá, L.M., Gomes, A.M., and Duarte, A.C. (2015a). Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 183: 197-207.
Rodrigues, D., Sousa, S., Silva, A., Amorim, M., Pereira, L., Rocha-Santos, T.A.P., and Freitas, A.C. (2015b). Impact of enzyme- and ultrasound-assisted extraction methods on biological properties of red, brown, and green seaweeds from the central West coast of Portugal. J. Agric. Food Chem. 63(12): 3177-3188.
Rodrigues, D., Walton, G., Sousa, S., RochaSantos, T.A.P., Duarte, A.C., Freitas, A.C., and Gomes, A.M.P. (2016). In vitro fermentation and prebiotic potential of selected extracts from seaweeds and mushrooms. Lwt Food Sci. Technol. 73: 131-139.
Rodríguez-Bernaldo de Quirós A., Frecha, F.S., Vidal, P.A., and López, H.J. (2010b). Antioxidants compounds in edible brown seaweeds. Eur. Food Res. Technol. 231(3): 495-498.
Rodríguez-Bernaldo de Quirós, A., Lage-Yusty, M.A., and López-Hernández, J. (2010a). Determination of phenolic compounds in macroalgae for human consumption. Food Chem. 121(2): 634-638.
Roesijadi, G., Copping, A.E., and Huesemann, M.H. (2008). Techno-economic feasibility analysis of offshore seaweed farming for bioenergy and biobased products. Independent research and development report, IR Number: PNWD-3931.
Rosell, K.G. and Srivastava L.M. (1987). Fatty acids as antimicrobial substances in brown algae. Hydrobiologia 151: 471-475.
Rosenberg, G. and Ramus, J. (1984). Uptake of inorganic nitrogen and seaweed surface-area-volume ratios. Aquat. Bot. 19: 65-72.

298
Ross, M.E., Stanley, M.S., Day, J.G., and Semião, A.J.C. (2017). A comparison of methods for the non-destructive fresh weight determination of filamentous algae for growth rate analysis and dry weight estimation, J. Appl. Phycol. 1-12.
Ross, W.A. (1996). A little book of Celtic verse. Belfast: The Appletree Press.
Rousseau, F., Gey, D., Kurihara, A., Maggs, C.A., Martin-Lescanne, J., Payri, C., de Reviers B., Sherwood, A.R. and Le Gall, L. (2017). Molecular phylogenies support taxonomic revision of three species of Laurencia (Rhodomelaceae, Rhodophyta), with the description of a new genus. Eur. J. Taxon. 269: 1-19.
Rowan, K.S. (1989). Photosynthetic pigments of algae, Cambridge University Press, UK. pp. 334.
Rüdiger, W. and López-Figueroa, F. (1992). Photoreceptors in algae. Ann. Rev. Photochem. Photobiol. 55: 949-954.
Rueness, J. (1989). Sargassum muticum and other introduced Japanese macroalgae: biological pollution of European coasts. Mar. Poll. Bull. 20:173-176.
Rueness, J. and Tananger, T. (1984). Growth in culture of four red algae from Norway with potential for mariculture. Hydrobiologia 116-117(1): 303-307.
Rupérez, P. (2002). Mineral content of edible marine seaweeds. Food Chem. 79: 23-26.
Rupérez, P., Ahrazem, O., and Leal, J.A. (2002). Potential antioxidant capacity of sulfated polysaccharides from edible marine brown seaweed Fucus vesiculosus. J. Agric. Food Chem. 50: 840-845.
Rupérez, P., and Saura-Calixto, F. (2001). Dietary fibre and physicochemical properties of edible Spanish seaweeds. Eur. Food Res. Technol. 212: 349-354.
Ruttenberg, K.C. and Goñi, M. (1997b). Phosphorus distribution, C:N:P ratios, and δ¹³Coc in arctic, temperate, and tropical coastal sediments: tools for characterizing bulk sedimentary organic matter Article. Mar. Geol. 139(1).
Ryther, J.H. (1977). Preliminary results with a pilot-plan waste recycling-marine aquaculture system. In: D'Itri, F.M. (ed.). Wastewater Renovation and Reuse. Maral Dekker Inc, New York. pp. 89-132.
Ryther, J.H., Corwin, N., DeBusk, T.A., and Williams, L.D (1981). Nitrogen uptake and storage by the red alga Gracilaria tikvahiae (McLachlan, 1979). Aquaculture 26: 107-115.
Ryu, B.M., Li, Y.X., Kang, K.H., Kim, S.K., and Kim, D.G. (2015). Floridoside from Laurencia undulata promotes osteogenic differentiation in murine bone marrow mesenchymal cells. J. Funct. Foods. 19: 505-11.
Ryu, J., Kwon, M.J., and Nam, T.J. (2015). Nrf2 and NF-kB signalling pathways contribute toporphyra-334-mediated inhibition of UVA-induced inflammation in skin fibroblasts. Mar. Drugs 13: 4721-4732.
Sabina, H. and Aliya, R. (2009). Seaweed as a new source of flavone, scutellarein 4 '-methyl ether. Pak. J. Bot. 41(4): 1927-1930.
Sabina, H., and Aliya, R. (2011). Bioactive assessment of selected marine red algae against leishmania major and chemical constituents of Osmundea Pinnatifida. Pak. J. Bot. 43(6): 3053-3056.
Sacramento, A.T., García-Jiménez, P., Alcázar, R., Tiburcio, A.F., and Robaina, R.R. (2004). Influence of Polyamines on the Sporulation of Grateloupia (Halymeniaceae, Rhodophyta). J. Phycol. 40: 887-894.
Saga, N., Uchida, T., and Sakai, Y. (1978). Clone Laminaria from a single isolated cell Bull. Jap. Soc. Scient. Fish. 44: 87.
Saifullah, A.Z.A., Karim, Md. A., and Ahmad-Yazid, A. (2014). Microalgae: An Alternative Source of Renewable Energy. AJER 3(3): 330-338.
Salgado, L.T., Viana, N.B., Andrade, L.R., Leal, R.N., Da Gama, B.A.P., Attias M., Pereira, R.C., and Amado Filho, G.M. (2008). Intra-cellular storage, transport and exocytosis of halogenated compounds in marine red alga Laurencia obtusa. J. Struct. Biol. 162: 345-355.
San Martin, A., Rovirosa, J., Muñoz, O., Chen, M.H.M., Guneratne, R.D., and Clardy, J. (1983). The isolation and structure determination of chilenone A, a putative dimer of 2-

299
methyl-3(2H)-furanone from the marine alga Laurencia chilensis. Tetrahedron Lett. 24 (38): 4063-4066.
Sánchez-Machado, D.I., López-Cervantes, J., López-Hernández J., and Paseiro-Losada, P. (2004a). Fatty acids, total lipid, protein and ash contents of processed edible seaweeds. Food Chem. 85 (3): 439-444.
Sánchez-Machado, D.I., López-Cervantes, J., López-Hernández, J., Paseiro-Losada, P., and Simal-Lozano, J. (2003). High-performance liquid chromatographic analysis of amino acids in edible seaweeds after derivatization with phenyl isothiocyanate. Chromatographia 58: 159-163.
Sánchez-Rodríguez, I., Huerta-Díaz, M.A., Choumiline, E., Holguíın-Quiñones, O., and Zertuche-González, J.A. (2001). Elemental concentrations in different species of seaweeds from Loreto Bay, Baja California Sur, Mexico: implications for the geochemical control of metals in algal tissue. Env. Pollut. 114: 146-160.
Sanderson, J.C. (2006). Reducing the environmental impacts of sea-cage fish farming through cultivation of seaweed. PhD Thesis. Open University, Scottish Association for Marine Sciences, Oban. pp. 339.
Sand-Jensen, K. and Gordon, D.M. (1984). Differential ability of marine and freshwater
macrophytes to utilize HCO₃ and CO₂. Mar. Biol. 80: 247253.
Sanford, L.P. and Crawford, S.M. (2000). Mass transfer versus kinetic control across solid-water boundaries. Limnol. Oceanogr. 45: 1180-1186.
Santelices, B. (1999). A conceptual framework for marine agronomy. Hydrobiol. 398/399: 15-23.
Sarojini, Y. and Sarma, N.S. (1999). Vitamin C content of some macroalgae of Visakhapatnam, east coast of India. Ind. J. Mar. Sci. 28: 408-412.
Saunders, G.W. (2004). Gel purification of red algal genomic DNA: an inexpensive and rapid method for isolation of polymerase chain reaction-friendly DNA. J. Phycol. 29(2): 251-254.
Saunders, G.W. (2005). Applying DNA barcoding to red macroalgae: a preliminary appraisal holds promise for future applications. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 360 (1462): 1879-1888.
Saunders, G.W. (2008). A DNA barcode examination of the red algal family Dumontiaceae in Canadian waters reveals substantial cryptic species diversity. I. The foliose Dilsea–Neodilsea complex and Weeksia. Botany 86(7): 773-789.
Saunders, G.W. (2009). Routine DNA barcoding of Canadian Gracilariales (Rhodophyta) reveals the invasive species Gracilaria vermiculophylla in British Columbia. Mol. Ecol. Resour. 9: 140-150.
Saunders, G.W. and Bailey, J.C., (1999). Molecular systematic analyses indicate that the enigmatic Apophlaea is a member of the Hildenbrandiales (Rhodophyta, Florideophycidae) J. Phycol. 35: 171-175.
Saunders, G.W. and Hommersand, M.H. (2004). Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 91: 1494-1507.
Saunders, G.W. and Kraft, G.T. (1997). A molecular perspective on red algal evolution: focus on the Florideophycidae. Plant Syst. Evol. 11 (Supplement): 115-138.
Saunders, G.W. and Kucera, H. (2010). An evaluation of rbcL, tufA, UPA, LSU and ITS as DNA barcode markers for the marine green macroalgae. Cryptogamie Algol. 31(4): 487-528.
Saunders, G.W. and Mcdevit, D.C. (2012). Methods for DNA barcoding photosynthetic protists emphasizing the macroalgae and diatoms. Methods Mol. Biol. 858: 207-222.
Saunders, G.W. and Moore, T. (2013). Refinements for the amplification and sequencing of red algal DNA barcode and RedToL phylogenetic markers: a summary of current primers, profiles and strategies. Algae 28(1): 31-43.
Sawada T., Koga S., and Uchiyama, S. (1972). Some observations on carpospores adherence in Polysiphonia japonica Harvey. Sci. Bull. Fac. Agr. Kyushu Univ. 26: 223-226.

300
Sawangwan, T., Goedl, C., and Nidetzky, B. (2010). Glycosylglycerol and glucosylglycerate as enzyme stabilizers. Biotechnol. J. 5: 187-191.
Schieberle, P. and Hofmann, T. (1997). Evaluation of the character impact odorants in fresh strawberry juice by quantitative measurements and sensory studies on model mixtures. J. Agr. Food Chem.45: 227-232.
Schiener, P., Black, K.D., Stanley, M.S., and Green, D.H. (2015). The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta, J. Appl. Phycol. 27: 363-373.
Schiewer, V. (1967). Auxinvorkommen und Auxinstoffwechsel bei Mehrzelligen Ostseealgen: 1. Zum Vorkommen von Indol-3-Essigsäure. Planta 74: 313-323.
Schmidt, C. (1844). Ueber Pflanzenschleim und Bassorin. Justus Liebigs Annalen der Chemie 51(1): 29-62.
Schreiber, U., Schliwa, U., and Bilger, W. (1986). Continuous recording of photo-chemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 10: 51-62.
Serio, D., Petrocelli, A., Cormaci, M., Cecere, E., and Furnari, G. (2008). First record of Osmundea oederi (Gunnerus) G. Furnari comb. nov. (Rhodomelaceae, Rhodophyta) from the Mediterranean Sea. Cryptogamie, Algol. 29: 119-127.
Shahidi, F. (2009). Nutraceuticals and functional foods: Whole versus processed foods. Trends Food Sci. Technol. 20: 376-387.
Shephard, D.C. (1970). Axenic culture of Acetabularia in a synthetic medium. In: Prescott, D.M. (ed.). Methods in cell physiology. Academic Press, New York. Vol 4., pp. 46-69.
Sherr, E.B. and Sherr, B.F. (2002). Significance of predation by protists in aquatic microbial foodwebs. Antonie Van Leeuwenhoek 81: 293-308.
Sherwood, A.A. and Sheath, R.G. (2003). Systematics of the Hilden-brandiales (Rhodophyta): gene sequence and morphometric analyses of global collections. J. Phycol. 39: 409-422.
Sherwood, A.R., Sauvage, T., Kurihara, A., Conklin, K.Y., and Presting, G.G. (2010). A comparative analysis of COI, LSU and UPA marker data for the Hawaiian florideophyte Rhodophyta: implications for DNA barcoding of red algae Cryptogamie, Algol. 31(4): 451.
Shick, J.M. and Dunlap, W.C. (2002). Mycosporine-like amino acids and related Gadusols: biosynthesis, accumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 64: 223-262.
Shields, R. and Lupatsch, I. (2012). Algae for aquaculture and animal feeds. In: Posten, C. and Walter, C. (eds.). Microalgal biotechnology: integration and economy. Walter de Gruyter, Berlin, Germany. pp. 79-96.
Shioda, M. and Murakmi-Murofusi, K. (1987). Selective inhibition of DNA polymerase by polysaccharide purified from slime of Physarum polycephalum. Biochem. Biophys. Re. Commun. 146: 5279-5280.
Shirwaikar, A.P.V. and Khan, S. (2011). The changing face of nutraceuticals-An overview. Int. J. Pharm. Life Sci. 2: 925-932.
Silva, P. and Abreu, M.H. (2014). Cultivation trials in land based IMTA conditions of macroalgae with pharmacological potential: Osmundea pinnatifida, Codium tomentosum and Bifurcaria bifurcata. Aquaculture Europe 2014. pp. 1-3.
Silva, P., Fernandes, C., Barros, L., Ferreira, I.C.F.R., Pereira, L., and Gonçalves, T. (2018). The antifungal activity of extracts of Osmundea pinnatifida, an edible seaweed, indicates its usage as a safe environmental fungicide or as a food additive preventing post-harvest fungal food contamination. Food Func. 9 (12): 6188-6196.
Silva, P.J.F. (2015). Screening of biotechnological potential of Osmundea pinnatifida: cultivation trials and biological activities. Master Thesis. University of Coimbra. pp. 92.
Simeunovic, J., Beslin, K., Svircev, Z., Kovac, D., and Babic, O. (2013). Impact of nitrogen and drought on phycobiliprotein content in terrestrial cyanobacterial strains. J. Appl. Phycol. 25(2): 597-607.

301
Simpson, F.J., Shacklock, P.F., Robson, D., and Neish, A.C. (1978). Factors affecting cultivation of Chondrus crispus (Florideophyceae). Proc. Seaweed Spp. 9: 509-513.
Singh, R.N. and Sharma, S. (2012). Development of suitable photobioreactor for algae productionA review. Renew Sust. Energ. Rev. 16: 2347-53.
Singh, S.P. and Singh, P. (2015). Effect of temperature and light on the growth of algae species: a review. Renew. Sust. Energ. Rev. 50: 431-444.
Sinha, R.P, Klisch, M., Grniger, A., and Häder, D.P. (1998). Ultraviolet absorbing/screening substances in cyanobacteria, phytoplankton and macroalgae, J. Photochem. Photobiol. B: Biol. 47: 83-94.
Sitnik, K.M, Musatenko, L.I., Vasyuk, V.A., Vedenicheva, N.P., Generalova, V.M., Martin, G.G., and Nesterova, A.N. (2003). Gormonal’nii kompleks roslin i gribiv (Hormonal complex in plants and fungi), Kiiv: Akademperiodika.
Skordas, K., Kelepertzis, E., Kosmidis, D., Panagiota, P., and Vafidis, D. (2014). Assessment of nutrients and heavy metals in the surface sediments of the artificially lake water reservoir Karla, Thessaly, Greece. Environ. Earth Sci. 73(8).
Slattery, M. and Lesser, M.P. (2014). Allelopathy in the tropical alga Lobophora variegate (phaeophyceae): mechanistic basis for a phase shift on mesophotic coral reef? J. Phycol. 505: 493-505.
Slaughter, C.J. (1999). The naturally occurring furanones: formation and function from pheromone to food. Biol. Rev. Camb. Philos. Soc. 74(3): 259-76.
Slocombe, S.P., Ross, M., Thomas, N., McNeill, S., and Stanley, M.S. (2013). A rapid and general method for measurement of protein in micro-algal biomass. Bioresour. Technol. 129: 51-57.
Smit, A.J. (2002). Nitrogen uptake by Gracilaria gracilis (Rhodophyta): adaptations to a temporally variable nitrogen environment, Bot. Mar. 45: 196-209.
Smit, A.J., Robertson, B.L. and du Preez, D.R. (1997). Influence of ammonium-N pulse concentrations and frequency, tank condition and nitrogen starvation on growth rate and biochemical composition of Gracilaria gracilis. J. Appl. Phycol. 8: 473-481.
Sode, S., Bruhn, A., Balsby, T.J., Larsen, M.M., Gotfredsen, A. and Rasmussen, M.B. (2013). Bioremediation of reject water from anaerobically digested wastewater sludge with macroalgae (Ulva lactuca, Chlorophyta). Bioresour. Technol. 146: 426-435.
Sørensen, L., Hantke, A., and Eriksen, N.T. (2013). Purification of the photosynthetic pigment C-phycocyanin from heterotrophic Galdieria sulphuraria. J. Sci. Food Agric. 93(12): 2933-2938
Souza, B.W.S., Cerqueira, M.A, Bourbon, A.I., Pinheiro, A.C., Martins, J.T, Teixeira, J.A., Coimbra, M.A., and Vicente, A.A. (2012). Chemical characterization and antioxidant activity of sulfated polysaccharide from the red seaweed Gracilaria birdiae. Food Hydrocoll. 27(2): 287-292.
Stagnol, D., Renaud, M. and Davoult, D. (2013). Effects of commercial harvesting of intertidal macroalgae on ecosystem biodiversity and functioning. Estuar. Coast Shelf Sci. 130: 99-110.
Starr, R.C. and Zeikus, J.A. (1993). UTEX–the culture collection of algae at the University of Texas at Austin. The list of cultures. J. Phycol. 29: 1-106.
Steane, D.A, McClure, B.A., Clarke, A.E, and Kraft, G.T (1991). Amplification of the polymorphic 5.8S rRNA gene from selected Australian gigartinalean species (Rhodophyta) by polymerase chain reaction. J. Phycol. 27: 758-62.
Stegenga, H., Bolton, J.J., and Anderson, R.J. (1997). Seaweeds of the South African West Coast. Creda Press, Cape Town. pp. 655.
Stiger, V., Bourgougnon, N., and Deslandes, E. (2016). Carbohydrates from seaweeds. In: Fleurence, J. and Levine, I. (eds.). Seaweed in health and disease prevention. Academic Press, USA. pp. 223-274.
Stirk, W.A., Bálint, P., Tarkowská D., Novák, O., Maróti, G., Ljung, K., Turečková, V., Strnad, M., Ördög, V., and van Staden, J. (2014a). Effect of light on growth and

302
endogenous hormones in Chlorella minutissima (Trebouxiophyceae). Plant Physiol. Biochem. 79: 66-76.
Stirk, W.A., Novák, O., Strnad, M. and Van Staden, J. (2003). Cytokinins in macroalgae. Plant Growth Regul. 41: 13-24.
Stoebe, B. and Kovallik, K.V. (1999). Gene-cluster analysis in chloroplast genomics. TIG 15: 344-347.
Stoffels, L., Taunt, H.N., Charalambous, B., and Purton, S. (2017). Synthesis of bacteriophage lytic proteins against Streptococcus pneumoniae in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 15(9): 1130-1140.
Stoker, T.E. and Kavlock, R.J. (2010). Pesticides as endocrine-disrupting chemicals. In: Robert, K.(ed.). Hayes Handbook of Pesticide Toxicology (3rd Edition). Academic press, Orlando, FL. pp. 551-569.
Su, N., Wu, Q., Shen, Z., Xia, K., and Cui, J. (2014). Effects of light quality on the chloroplastic ultrastructure and photosynthetic characteristics of cucumber seedlings. Plant Growth Regul. 73(3): 227-235.
Sudatti, D.B., Fujii, M.T., Rodrigues, S.V., Turra, A., and Pereira, R.C (2011). Effect of abiotic factors on growth and chemical defense content in Laurencia dendroidea J. Agardh (Ceramiales, Rhodophyta) cultivated clones. Mar. Biol. 158: 1439-1446.
Sudatti, D.B., Rodrigues, S.V., and Pereira, R.C. (2006). Quantitative GC-ECD analysis of halogenated metabolites: determination of elatol on surface and within-thallus of Laurencia obtusa. J. Chem. Ecol. 32: 835-843.
Suh, S.S., Hwang, J., Park, M., Seo, H.H., Kim, H.S., Lee, J.H., Moh, S.H., and Lee, T.K. (2014). Anti-inflammation activities of mycosporine-like amino acids (MAAs) in response to UV radiation suggest potential anti-skin aging activity. Mar. Drugs 12(10): 5174-5187.
Suh, S.S., Oh, S.K., Lee, S.G., Kim, I.C, and Kim, S. (2017). Porphyra-334, a mycosporine-like amino acid, attenuates UV-induced apoptosis in HaCaT cells. Acta Pharm. 27; 67(2):257-264.
Sun, L., Wang, S.M., Chen, L.X., and Gong, X.Q. (2003). Promising Fluorescent Probes from Phycobiliproteins. IEEE J. Select. Quantum Electron 9: 177-188.
Sun, S., Wang, F., Li, C. L., Qin, S., Zhou, M. J., Ding, L.P., Pang, S., Duan, D., Wang, G., Yin, B., Yu, R., Jiang, P., Liu, Z., Zhang, G.T., Fei, X., and Zhou, M. (2008). Emerging challenges: massive green algal blooms in the yellow sea. Nat. Proc. hdl:10101/npre.2008.2266.1.
Suto, S. (1950). Studies on shedding, swimming and fixing of the spores of seaweeds. Bull. Jpn. Soc. Sci. Fish. 16: 1-9.
Suzuki, M., Kawamoto, T., Vairappan, C.S., Ishii, T., Abe, T., and Masuda, M. (2005). Halogenated metabolites from Japanese Laurencia spp. Phytochemistry 66(23): 2787-2793.
Suzuki, M., Yamada, H., and Kurata, K. (2002). Dictyterpenoids A and B, two novel diterpenoids with feeding-deterrent activity from the brown alga Dictyota okamurae. J. Nat. Prod. 65: 121-125.
Tabarsa, M., Rezaei, M., Ramezanpour, Z., and Waaland, J.R. (2012). Chemical compositions of the marine algae Gracilaria Salicornia (Rhodophyta) and Ulva lactuca (Chlorophyta) as potential food source. J. Sci. Food Agric. 92: 2500-2506.
Takamatsu, S., Hodges, T.W., Rajbhandari, I., Gerwick, W.H., Hamann, M.T., and Nagle, D.G. (2003). Marine natural products as novel antioxidant prototypes. J. Nat. Prod. 66: 605-608.
Takano, R., Hayashi, K., Hara, S., and Hirase, S. (1995). Funoran from the red seaweed, Gloiopeltis complanata: polysaccharides with sulfated agarose structure and its precursor structure. Carbohydr. Polym. 27: 305-311.
Takenaka, F. and Uchiyama, H. (2000). Synthesis of α-glucosylglycerol by α-glucosidase and some of its characteristics. Biosci Biotech Biochem. 64: 1321-1326.

303
Tamiya, H. (1955). Growing Chlorella for food and feed. In: Proc. World Symposium on Applied Solar Energy. Phoenix, Arizona, Stanford Research, Menlo Park, California. pp. 231.
Tao, C., Sugawara, T., Maeda, S., Wang, X. and Hirata, T. (2008). Antioxidative activities of a mycosporine-like amino acid, porphyra-334. Fish. Sci. 74: 1166-1172.
Tarakhovskaya, E.R., Maslov, Y.I., and Shishova, M.F. (2007). Phytohormonesin algae. Russ. J. Plant Physiol. 54(2): 163-170.
Tarko, T., Duda-Chodak, A., and Kobus, M. (2012). Influence of growth medium composition on synthesis of bioactive compounds and antioxidant properties of selected strains of Arthrospira cyanobacteria. Czech J. Food Sci. 30: 258-26.
Tautenhahn, R., Patti, G.J., Rinehart, D., and Siuzdak, G. (2012). XCMS Online: a web-based platform to process untargeted metabolimc data. Anal. Chem. 84(11): 5035-5039.
The Scottish Government (2013). Environmental Assessment (Scotland) Act 2005. Seaweed Policy Statement Consultation Document. Environmental report.
The Scottish Government (2016). Scotland’s aquaculture. Topic Sheet No. 106 V2.
Thiem, J., Scheel, O., and Schneider, G. (1997). Cosmetic preparations with an effective amount of glycosylglycerides as skin moisturizers. EP 0 770378, Germany.
Thomas, N.V. and Kim, S.K. (2011). Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharm. 32: 325-335.
Thomas, T.E., Harrison, P.J., and Taylor, E.B. (1985). Nitrogen uptake and growth of the germlings and mature thalli of Fucus distichus. Mar. Biol. 84: 267-274.
Thompson Jr.G.A. (1996). Lipids and membrane function in green algae. Biochim. Biophys. Acta 1302: 17-45.
Tibubos, K.R., Hurtado, A.Q, and Critchley, A.T. (2017). Direct formation of axes in new plantlets of Kappaphycus alvarezii (Doty) Doty, as influenced by the use of AMPEP K+, spindle inhibitors, and plant growth hormones. J. Appl. Phycol. 29(5): 2345-2349.
Togersen, Y. and Håstein, T. (1995). Disinfection in aquaculture. Scientific and technical review of the Office International des Epizooties 14: 419-434.
Tominaga, F. and Oka, K. (1963). On the isolation and identification of 1,4-Thiazane-3-Carboxylic Acid S-Oxide from the Brown Alga Undaria pinnatifida. J. Biochem. 54(3):222-224.
Torres, P.B., Chow, F., Furlan, C.M., Mandelli, F., Mercadante, A., and Dos Santos, D.Y.A.C. (2014). Standardization of a protocol to extract and analyze chlorophyll a and carotenoids in Gracilaria tenuistipitata Var. Liui. Zhang and Xia (Rhodophyta). Braz. J. Oceanogr. 62(1): 57-63.
Toyosaki, T. and Iwabuchi, M. (2009). New antioxidant protein in seaweed (Porphyra yezoenis Ueda.). Int. J. Food Sci. Nutr. 60: 46-56.
Tsekos, I., Niell, F.X., Aguilera, J., Figueroa, F.L., and Delivopoulos S.G. (2002). Ultrastructure of the thallus stage of Porphyra leucosticta (Rhodophyta) grown in red, blue and green light. Phycol. Res. 50: 251-264.
Tseng, C.K. (1981). Marine Phycoculture in China. Proc. Int. Seaweed Symp. 10: 123-152.
Turner, G.E. (ed.) (1988). Codes of practice and manual of procedures for consideration of introductions and transfers of marine and freshwater organisms. FAO, EIFAC/CECPI Occas. Pap. 23: 44.
Ugwu, C.U., Aoyagi, H., and Uchiyama, H. (2008). Photobioreactors for mass cultivation of algae. Bioresour. Technol. 99: 4021-4028.
Usmanghani, K., Shameel, M., Khan, K., and Mahmood, Z.A. (1984). Antibacterial and antifungal activities of marine algae from Karachi seashore of Pakistan. Fitotrap. 55: 73-77.
Vairappan, C.S (2003). Potent antibacterial activity of halogenated metabolites from Malaysian red algae, Laurencia majuscule (Rhodomelaceae, Ceramiales). Biomolecular. Eng. 20: 255-259.

304
Vairappan, C.S, Suzuki, M., Motomura, T., and Ichimura, T. (2001b). Pathogenic bacteria associated with lesions and thallus bleaching symptoms in the Japanese kelp Laminaria religiosa Miyabe (Laminariales, Phaeophyceae). Hydrobiologia 445: 183-191.
Vairappan, C.S. (2006). Seasonal occurrences of epiphytic algae on the commercially cultivated red alga Kappaphycus alvarezii (Solieriaceae, Gigartinales, Rhodophyta). J. Appl. Phycol. 18: 611-617.
Vairappan, C.S., Suzuki, M., Abe, T., and Masuda, M. (2001a). Antibacterial halogenated metabolites from the Malaysian Laurencia species. Phytochemistry 58: 291-297.
van den Hoek, C., Mann, D.G., and Jahns, H.M. (1995). Algae. An Introduction to Phycology. Cambridge University Press, Cambridge. pp. 623.
van Oosten, M.J., Pepe, O., De Pascale, S., Silletti, S., and Maggio, A. (2017). The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 4: 5.
Veena, C.K., Josephine, A., Preetha, S.P., and Varalakshmi, P. (2007). Beneficial role of sulfated polysaccharides from edible seaweed Fucus vesicuolosus in experimental hyperoxaluria. Food Chem. 100: 1552-1559.
Venkatpurwar, V., Shiras A., and Pokharkar V. (2011). Porphyran capped gold nanoparticles as a novel carrier for delivery of anticancer drug: in vitro cytotoxicity study. Int. J. Pharm. 409: 314-320.
Vergara, J.J. and Niell, F.X. (1993). Effects of nitrate availability and irradiance on internal nitrogen constituents in Corallina elongata (Rhodophyta). J. Phycol. 29: 285-293.
Vergara, J.J., Bird, K.T., and Niell, F.X. (1995). Nitrogen assimilation following NH⁴+ pulses in the red alga Gracilariopsis lemaneiformis: effect on C metabolism. Mar. Ecol. Progr. Ser. 122: 253-263.
Villares, R., Puente, X., and Carballeira, A. (2002). Seasonal variation and background levels of heavy metals in two green seaweeds. Environ. Pollut. 119: 79-90.
Waaland, S.D. and Watson, B.A. (1980). Isolation of a cell-fusion hormone from Griffithsia pacifica Kylin, a red alga. Planta 149: 493-497.
Walker, R.H., Brodie, J., Russell, S., Irvine, L.M., and Orfanidis, S. (2009). Biodiversity of coralline algae in the Northeast Atlantic including Corallina caespitose sp. nov. (Corallinoideae, Rhodophyta). J. Phycol. 45(1): 287-297.
Wallentinus, I. (1984). Comparisons of nutrient uptake rates for Baltic macroalgae with different thallus morphologies. Mar. Biol. 80: 215-225.
Wang, C., Lei, A., Zhou, K., Hu, Z., Hao, W., and Yang, J. (2014). Growth and nitrogen uptake characteristics reveal outbreak mechanism of the opportunistic macroalga Gracilaria tenuistipitata. PLoS ONE 9(10): e108980.
Wargovich, M.J. (2000). Anticancer properties of fruits and vegetables. HortScience 35 (4): 573-575.
Watson, B.A. and Waaland, S.D. (1983). Partial purification and characterization of a glycoprotein cell fusion hormone from Griffithsia pacifica, a red alga. Plant Physiol. 71: 327-332.
Wattier, R., Dallas, J.F., Destombe, C., Saumitou-Laprade, P., and Valero, M. (1997). Single-locus microsatellites in Gracilariales (Rhodophyta): high level of genetic variability within Gracilaria gracilis and conservation in related species. J. Phycol. 33: 868-880.
Wattier, R., Prodöhl, P., and Maggs, C. (2000). DNA isolation protocol for red seaweed (Rhodophyta). Plant Mol. Biol. Rep. 18(3): 275-281.
Welcomme, R.L. (ed.) (1988). International introductions of inland aquatic species (Vol. 294). Food and agriculture organization of the united nations (FAO). Rome, Italy. pp. 318.
Werner, A. and Dring, M.J. (2011). Cultivating Palmaria palmata. ‘Aquaculture Explained’ Series No. 27 Bord Iascaigh Mhara (BIM), Dublin.
Werner, A., Clarke, D., and Kraan, S. (2004). Strategic review of the feasibility of seaweed aquaculture in Ireland. Report, edited by Irish seaweed Centre, Martin Ryana Institute, National Univeristy of Irlenad (Galway).

305
West, J.A. and Hommersand, M.H. (1981). Rhodophyta: life histories. In: Lobban, C.S. and Wynne, M.J. (eds.). The biology of seaweeds. Blackwell Scientific, Oxford, UK. pp. 133-193.
West, J.A., Scott, J.L., West, K.A., Karsten, U., Clayden, S.L., and Saunders, G.W. (2008). Rhodachlya madagascarensis gen. et sp. nov.: a distinct acrochaetioid represents a new order and family (Rhodachlyales ord. nov., Rhodachlyaceae fam. nov.) of the Florideophyceae (Rhodophyta). Phycologia 47: 203-212.
Wheeler, P.A. and Bjornsater, B.R. (1992). Seasonal fluctuations in tissue nitrogen, phosphorus, and N:P for five macroalgal species common to the Pacific North-West Coast. J. Phycol. 28: 1-6.
Wheeler, W.N., Neushul, M., and Harger, B.W.W. (1981). Development of a coastal marine farm and its associated problems. In: Levring, T. (ed.). Proceedings of the tenth International Seaweed Symposium, Walter de Gruyter, Berlin. pp. 631-636.
Whitford, L.A. and Kim, C.S. (1966). The effect of light and water movement on some species of marine algae. Revue Algol. 3: 251-254.
Whitton, R. (2016). Algae Reactors for Wastewater Treatment. EngD Thesis, pp. 197.
Whitton, R., Santinelli, M., Pidou, M., Ometto, F., Henderson, R., Roddick, F., Jarvis, P., Villa, R., and Jefferson, B. (2018). Tertiary nutrient removal from wastewater by immobilised microalgae: impact of wastewater nutrient characteristics and hydraulic
retention time (HRT). H₂Open J. 1(1): 12-25.
WHO, (2008). Population nutrient intake goals for preventing diet-related chronic diseases. Recommendations for preventing dental diseases. In: World Health Organization. Diet, nutrition and the prevention of chronic diseases. Report of a joint WHO/FAO expert consulta. WHO Technical Report Series), 916. pp. 160.
WHO, (2011a). Evaluation of certain contaminants in food: seventy- second report of the Joint FAO/WHO Expert Committee on Food Additives. WHO, Geneva. Technical Report Series No. 959. pp. 115.
Wilson, S. (2002). Nutritional value of detritus and algae in blenny territories on the Great Barrier Reef. J. Exp. Mar. Biol. Ecol. 271: 155-169.
Winter, R. (2009). A consumer’s dictionary of food additives, 7th edition. Three River Press, New York (USA), pp. 595.
Wishart, D.S. (2008). Metabolomics: application to food science and nutrition research. Trends Food Sci. Tech. 19: 482-493.
Woelkerling, W.J. (1990). An introduction. In: Cole, K.M. and Sheath, R.G. (eds.). Biology of the Red Algae. Cambridge Univ. Press, New York. pp. 1-6.
Wolfe-Simon, F., Grzebyk, D., Schofield, O., and Falkowski, P. (2005). The role and evolution of superoxide dismutases in Algae. J. Phycol. 41(3): 453-465.
Wong, K.H. and Cheung, P.C.K. (2000). Nutritional evaluation of some subtropical red and green seaweeds. Part I. Proximate composition, amino acid profiles and some physicochemical properties. Food Chem. 71(4): 475-482.
Wright, J.T., De Nys, R., and Steinberg, P.D. (2000). Geographic variation in halogenated furanones from the red alga Delisea pulchra and associated herbivores and epiphytes. Mar. Ecol. Prog. Ser. 207: 227-241.
Xie, E.Y., Liu, D.C., Jia, C., Chen, X., and Li, Y.B. (2013). Artificial seed production and cultivation of the edible brown alga Sargassum naozhouense Tseng et Lu. J. Appl. Phycol. 25: 513-522.
Xu, N., Fan, X., Yan, X., Li, X., Niu, R., and Tseng, C.K. (2003). Antibacterial bromophenols from the marine red algae Rhodomela confervoides. Phytochemistry 62: 1221-1226.
Xu, Y., Ibrahim, I.M., and Harve, P.J. (2016). The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina(chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 106: 305-315.

306
Xu, Y., Ibrahim, I.M., Wosu, I.C., Ben-Amotz, A., and Harvey, P.J. (2018). Potential of New Isolates of Dunaliella Salina for Natural β-Carotene Production. Biology 7(1): 1-14.
Yabuta, Y., Fujimura, H., Kwak, C.S., Enomoto, T., and Watanabe, F. (2010). Antioxidant activity of the phycoeryth-robilin compound formed from a dried Korean purple laver (Porphyra sp.) during in vitro digestion. Food Sci. Technol. Res. 16: 347-351.
Yang, E.C., Kim, M.S., Paul, J.L., Geraldino, P.J.L., Sahoo, D., Shin, J.M, and Boo, S.M. (2008). Mitochondrial cox1 and plastid rbcL genes of Gracilaria vermiculophylla (Gracilariaceae, Rhodophyta). J. Appl. Phycol. 20: 161-168.
Yang, H., Xu, Z., Fan, M., Gupta, R., Slimane, R.H., Bland, A.E., and Wright, I. (2008). Progress in carbon dioxide separation and capture: A review. J. Env. Sci. 20: 14-27.
Yang, Y., Chai, Z., Wang, Q., Chen, W., He, Z., and Jiang, S. (2015). Cultivation of seaweed Gracilaria in Chinese coastal waters and its contribution to environmental improvements. Algal Res. 9: 236-244.
Yang, Y., Chan, S.W., Hu, M., Walden, R., and Tomlinson, B. (2011). Effects of Some Common Food Constituents on Cardiovascular Disease. ISRN Cardiology, 397136.
Yarish, C. and Edwards, P. (1982). A field and cultural investigation of the horizontal and seasonal distribution of estuarine red algae of New Jersey. Phycologia 21 :112-124.
Yeh, T.S., Hung, N.H., and Lin, T.C. (2014). Analysis of iodine content in seaweed by GC-ECD and estimation of iodine intake. JFDA 22(2): 189-196.
Yokoya, N.S. and Handro, W. (1996). Effects of auxins and cytokinins on tissue culture of Grateloupia dichotoma (Gigartinales, Rhodophyta). Hydrobiologia 326/327: 393-400.
Yokoya, N.S. and Handro, W. (1997). Thallus regeneration and growth induced by plant growth regulators and light intensity Grateloupia dichotoma (Rhodophyta). In: Kitamura, T. (ed.). Proceedings of I.T.I.T., AIST/MITI and NEDO, Takamatsu, Japan. pp. 83-86.
Yokoya, N.S., West, J.A., and Luchi, A.E. (2004). Effects of plant growth regulators on callus formation, growth and regeneration in axenic culture of Gracilaria tenuistipitata and Gracilaria perplexa (Gracilariales, Rhodophyta). Phycol. Res. 52: 244-254.
Yong, W.T.L., Ting, S.H., Yong, Y.S., Thien, V.Y., Wong, S.H., Chin, W.L., Rodrigues, K.F., and Anton, A. (2014a). Optimization of culture conditions for the direct regeneration of Kappaphycus alvarezii (Rhodophyta, Solieriaceae). J. Appl. Phycol. 26: 1597-1606.
Yoon, H.S., Hackett, J.D., and Bhattacharya, D. (2002b). A single origin of the peridinin- and fucoxanthin-containing plastids in dinoflagellates through tertiary endosymbiosis. Proc. Natl. Acad. Sci. U.S.A. 99:11724-11729.
Yoon, H.S., Hackett, J.D., Ciniglia, C., Pinto, G., and Bhattacharya, D. (2004). A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 21: 809-818.
Yoon, H.S., Müller, K.M., Sheath, R.G., Ott, F.D., and Bhattacharya, D. (2006) Defining the major lineages of red algae (Rhodophyta). J. Phycol. 42: 482-492.
Yoshie, Y., Wang, W., Petillo, D., and Suzuki, T. (2000). Distribution of catechins in Japanese seaweeds. Fisheries Sci. 66: 998-1000.
Young, A. (1991). Control of the light-harvesting function of chloroplast membranes by aggregation of the LHCII chlorophyll-protein complex. FEBS Lett. 292: 1-4.
Young, D.N., Howard, B.M., and Fenical, W. (1980). Subcellular localization of brominated secondary metabolites in the red alga Laurencia snyderae. J. Phycol. 16: 182-185.
Young, R., and Purton, S. (2018). CITRIC: cold-inducible translational read-through in the chloroplast of Chlamydomonas reinhardtii using a novel temperature-sensitive transfer RNA. Microb. Cell Fact 17: 1-12.
Yuan, Y.V. (2007). Antioxidants from edible seaweeds. ACS Symp. Ser. 956: 268-301
Yuan, Y.V., Bone, D.E., and Carrington, M.F. (2005a). Antioxidant activity of dulse (Palmaria palmata) extract evaluated in vitro. Food Chem. 91: 485-494.
Yuan, Y.V., Carrington, M.F., and Walsh, NA. (2005b). Extracts from dulse (Palmaria palmata) are effective antioxidants and inhibitors of cell proliferation in vitro. Food Chem. Toxicol. 43: 1073-1081.

307
Yuan, Y.V., Westcott, N.D., Hu, C., and Kitts, D.D. (2009). Mycosporine- like amino acid composition of the edible red alga Palmaria palmata (dulse) harvested from the West and East coasts of Grand Manan Island, New Brunswick. Food Chem. 112: 321-328.
Yunque, D.A., Tibubos, K.R., Hurtado, A.Q., and Critchley, A.T. (2011). Optimization of culture conditions for tissue culture production of young plantlets of carrageenophyte Kappaphycus. J. Appl. Phycol. 23: 433-438.
Zaman, L., Arakawa, O., Shimosu, A., Onoue, Y., Nishi, S., Shida, Y., and Noguchi, T. (1997). Two new isomers of domoic acid from a red alga, Chondria armata. Toxicon 35: 205-212.
Zertuche González, J.A., García Lepe, G., Pacheco Ruiz, I., Gendrop, V., and Guzmán, J.M. (2001). Open water Chondrus crispus Stackhouse cultivation. J. Appl. Phycol. 13: 249-253.
Zhang, D. (1982). Study on protoplasts preparation, culture and fusion of somatic cells from two species of green algae Ulva linza and Monostroma angicava Kjellm. J. Shandong College of Oceanol. 13: 57-65.
Zhang, Q., Li, N., Zhou, G., Lu, X., Xu, Z., and Li, Z. (2003). In vivo antioxidant activity of polysaccharide fraction from Porphyra haitanesis (Rhodophyta) in aging mice. Pharmacol. Res. 48:151-155.
Zhao, K.H., Porra, R.J., and Scheer, H. (2011). Phycobiliproteins, In: Roy, S., Llewellyn, C.A., Egeland, E.S., and Johnson, G. (eds.). Phytoplankton pigments: characterization, chemotaxonomy and applications in oceanography. Cambridge Univ. Press, Cambridge. pp. 375-411.
Zhou, A.Y., Robertson, J., Hamid, N., Ma, Q., and Lu, J. (2015). Changes in total nitrogen and amino acid composition of New Zealand Undaria pinnatifida with growth, location and plant parts. Food Chem. 186: 319-325.
Zubia, M., Yolanda, F.P., and Robledo, D. (2014). Photosynthesis, pigment composition and antioxidant defences in the red alga Gracilariopsis tenuifrons (Gracilariales, Rhodophyta) under environmental stress. J. Appl. Phycol. 26(5): 2001-2010.
Zuccarello, G.C., Burger, G., West, J.A., and King, R.J. (1999). A mitochondrial marker for red algal interspecific relationships. Mol. Ecol. 8: 1443-1447.
Zuccarello, G.C., Sandercock, B., and West, J.A. (2002). Diversity within red algal species: variation in world-wide samples of Spyridia filamentosa (Ceramiaceae) and Murrayella periclados (Rhodomelaceae) using DNA markers and breeding studies. Eu. J. Phycol. 37: 403-17.
Zucchi, M.R. and Necchi, O. (2001). Effects of temperature, irradiance and photoperiod on growth and pigment content in some freshwater red algae in culture. Phycol. Res. 49: 103-114.

308

309
Appendices
Appendix A Additional data on Chapter 3
A.1 Trace metals and DOCN analyses on seaweeds and seawater samples
a) Comparison of metal analyses of cultivated and wild samples
Metals and minerals analyses were conducted (as already described in Chapter 3) on cultivated
samples kept in a tank system with F/2 medium. Results showed that the concentration of certain
metals that could represent a threat to human consumption, was significantly lowered by the
cultivation conditions. This was particularly true for Al, As, Ba and Cd, while other metals, such as
Fe, are incremented with the cultivation (Fig. A.1). This is important because, even if there is no a
concern in terms of metals content for the consumption of the species (as already underlined in
Chapter 3), knowing that it is possible to control and particularly reduce/increase the concentration
of certain metals in a cultivation system can turn particularly useful for a commercial exploitation
of the species as food and nutraceutical product.
The results of the cultivated samples were compared with the highest and lowest value obtained
from the seasonal samples. Results are showed in Figs A.1 and A.2.
Figure A.1 Metal concentration (expressed in mg/kg DW) in wild (maximum value in blue and minimum value in red) and cultivated samples (green bars) (n=3; error bar, 1SD of mean)
For every month, was taken the highest and lowest value regarding the metals analysed and
compared with the average obtained from n=3 cultivated samples (indicated in green in Figs A.1
and A.2).
-20.00
0.00
20.00
40.00
60.00
80.00
100.00
Co Ni Cu Zn As Cd Cs Ba Pb U Li B Be
Met
als
con
cen
trat
ion
(mg/
kg D
W)
Metals

310
Figure A.2 Comparison between aluminium, manganese and iron concentration (mg/kg) in wild (maximum value in blue and minimum value in red) and cultivated samples (green bars) (n=3; error bar, 1SD of mean)
ANOVA single factor (p< 0.05) (Minitab 18, Tukey’s test comparison) was performed on the data
showing statistical differences between the highest values and the lowest values obtained for
cultivated samples for Al, Mn, Fe, Co, Ni, Zn, As, Cd, Cs, Ba and Pb (Figs A.1 and A.2). The
highest level reported for As in wild samples was statistically different from the value reported for
the cultivated samples, which was lower than the lowest value reported in the wild. Ba and Zn were
statistically lowered in the cultivated samples (Fig. A.1). Al content statistically lowered in cultivated
samples, in comparison with the highest value reported (Fig. A.2). While Fe content was increased
in the cultivation system (Fig. A.2). No statistical difference was reported for Li, Be, and B. For
Cu, the cultivated sample statistically differed from the lowest seasonal value (Fig. A.1). While for
the Uranium concentration, all the three values statically differ between each other (Fig. A.1).
b) Reference material for metals’ analyses in seaweed seasonal samples
The percent recovery of the certified elements expressed as:
%𝑟𝑒𝑐𝑜𝑣𝑒𝑟𝑦 =𝑎𝑛𝑎𝑙𝑦𝑡𝑒 𝑐𝑜𝑛𝑐𝑒𝑛𝑡𝑟𝑎𝑡𝑖𝑜𝑛 𝑚𝑒𝑎𝑠𝑢𝑟𝑒𝑑
𝑐𝑒𝑟𝑡𝑖𝑓𝑖𝑒𝑑 𝑎𝑛𝑎𝑙𝑦𝑡𝑒 𝑐𝑜𝑛𝑐𝑒𝑛𝑡𝑟𝑎𝑡𝑖𝑜𝑛× 100
is shown in Table A.1. For Cu, Zn, As and Cd the deviation is <10% from the certified value and
<18% for Pb at the lowest sub mg/kg concentration. The low concentration of Pb in the sample
may account for the poorer recovery. These results suggest that the accuracy of the digestion and
analysis was high and sensitive enough to resolve any seasonal variation within the seaweed
samples.
0.00300.00600.00900.00
1200.001500.001800.002100.002400.002700.003000.003300.003600.003900.004200.004500.00
Al Mn Fe
Met
als
con
cen
trat
ion
(m
g/kg
)
Metals

311
Table A.1 Percent recovery of the certified elements for the certified reference material CRM ERM-CD200. Cu, Zn, As, Cd and Pb are expressed in mg/kg, error as 1SD
c) DOC and TDN and Metals analyses on water samples
The Table (A.2) reports the data (expressed in µM) for the salinity, pH, DOC and TDN recorded
in seawater samples and discussed in Chapter 3, collected from June to August 2018 in
Dunstaffnage bay. Concentration of metals in water samples are reported in Figs A.3 and A.4. Fig.
A.5 represents the correlation between Al and Fe content in water samples. Correlations between
the various metals are reported in Tables A.3 and A.4.
Table A.2 Data of salinity, pH, DOC and TDN for water samples collected from June to August 2018 in Dunstaffnage bay
Date sal (PSU)
pH Dissolved organic carbon (DOC) (µM)
Total dissolved nitrogen (TDN) (µM)
01/06/2018 31.25 8.53 104.13 8.45
13/06/2018 31.6 8.6 102.19 9.77
18/06/2018 31.5 8.5 98.27 7.52
28/06/2018 30.7 8.43 104.72 8.86
03/07/2018 29.9 8.38 117.24 13.96
12/07/2018 29.9 8.33 108.67 8.08
17/07/2018 31.1 8.36 102.80 9.90
26/07/2018 33 8.51 87.99 8.41
01/08/2018 32 8.44 90.63 7.48
10/08/2018 28.9 8.47 120.98 8.69
15/08/2018 30 8.4 92.29 8.27
Figure A.3 Metals assessed in water samples (expressed as µM) in June (blue bars), July (orange bars) and August (grey bars) (n=3; error bar, 1SD of mean)
0
2
4
6
8
10
12
14
Al V Mn Fe Zn U
Met
als
con
cen
trat
ion
(µ
M)
Metals
Sample Cu error Zn error As error Cd error Pb error
CRM ERM-CD200 1.8 0.3 24.2 3.1 60.2 16.4 0.9 0.2 0.4 0.1
Certified Values 1.7 0.2 25.3 1.7 55.0 4.0 1.0 0.1 0.5 0.1
Recovery (%) 104.0 95.6 109.4 90.0 82.3

312
Figure A.4 Metals assessed in water samples (expressed as µg/kg) in June (blue bars), July (orange bars) and August (grey bars) (n=3; error bar, 1SD of mean)
Figure A.5 Correlation trends for Al (dark grey bar) and Fe (light grey bar) found in water samples (µM)
Table A.3 Correlation between data of metals concentration (Al, V, Mn, Fe, Zn, U) in water samples and precipitation data obtained from CEDA for Dunstaffnage #918 station from 2017-2018
Al V Mn Fe Zn U Precipitation
Al 1
V -0.86664 1
Mn -0.99824 0.894724 1
Fe 0.990063 -0.78786 -0.97997 1
Zn 0.917468 -0.99359 -0.93946 0.852411 1
U 0.968354 -0.71468 -0.95183 0.993828 0.789147 1
precipitation 0.865985 -1 -0.89414 0.78706 0.993446 0.713771 1
Table A.4 Correlation data between metals concentration (Co, Ni, Cu, Cd, Pb) in water samples and precipitation data obtained from CEDA for Dunstaffnage #918 station from 2017-2018
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
Co Ni Cu Cd PbM
etal
s co
nce
ntr
atio
n (
µM
)
Metals
0
0.5
1
1.5
2
2.5
June July August
Al a
nd
Fe
(µM
)
Months
Co Ni Cu Cd Pb precipitation
Co 1
Ni -0.60098 1
Cu 0.876508 -0.142 1
Cd 0.967868 -0.78265 0.727295 1
Pb 0.805435 -0.01033 0.991281 0.630519 1
precipitation -0.92302 0.247198 -0.99425 -0.79661 -0.97147 1

313
A.2 Comparison between Multi-assay extraction protocol and Fournier assay for carbohydrates
determination in seasonal samples of O. pinnatifida
A single tube method for a multi-assay biochemical analyses, already applied on microalgae
(Slocombe et al., 2013; Chen and Vaidyanathan, 2012, 2013), was tested and compared with the
Fournier method (2001) for carbohydrates extraction. Content of carbohydrates extracted with the
two methods is shown in Fig. A.6.
Figure A.6 Comparison of carbohydrates content expressed in %(DW) with two different methods: Multi-assay (dark grey bars) (Chen and Vaidyanathan, 2013) and Fournier (light grey bars) (2001) (n=3; error bar, 1 SD of mean)
The single tube assay was not efficient in extracting carbohydrates from freeze dried samples of O.
pinnatifida, resulting in a statistically lower content compare to the values obtained applying the
method proposed by Fournier (2001) which was applied for carbohydrates analyses in this thesis.
A.3 PCA analyses table and abiotic factors used for the correlation analyses in Chapter 3
Table A.5 Variables and respective principal components for PCA. Values give information about the correlation between variables and components
Variable PC1 PC2 PC3 PC4 PC5
Carbohydrates -0.267 0.292 -0.455 -0.422 0.268
Proteins 0.225 0.439 -0.241 0.459 -0.386
Chl-a 0.349 -0.299 -0.321 -0.018 0.149
Carotenoids 0.126 -0.510 -0.376 -0.409 -0.400
PE 0.367 -0.220 0.119 0.248 -0.075
PC 0.369 -0.225 -0.197 0.244 0.328
Lipid 0.274 0.403 -0.447 -0.097 -0.231
TPC -0.320 -0.127 -0.450 0.467 0.429
FRAP -0.375 -0.265 -0.138 0.165 -0.270
CN -0.389 -0.157 -0.125 0.262 -0.418
0
5
10
15
20
25
30
35
Car
bo
hyd
rate
s co
nte
nt
(% D
W)

314
Environmental data included in the PCA analyses described in Chapter 3 are shown in Figs A7-
A11 and include precipitation data for 2016-2017 (Fig. A.7), temperature (Fig. A.8, 2016; Fig. A.9,
2017), and radiation (Fig. A.10, 2016-2017; Fig. A.11, 2017-2018).
Figure A.7 Precipitation data (mm/m) for 2016 (dark grey line) and 2017 (light grey line) in Dunstaffnage bay, the area of the study for the seasonal sampling (Met-office for the Dunstaffnage n°918 station)
Figure A.8 Max (dark grey line) and Min (light grey line) temperature for 2016 in Dunstaffnage bay, area for the study for the seasonal sampling (MET Office, Dunstaffnage station, 2016-2017)
Figure A.9 Max (dark grey line) and Min (light grey line) temperature for 2017 in Dunstaffnage bay, area for the study for the seasonal sampling (MET Office, Dunstaffnage station, 2016-2017)
00.05
0.10.15
0.20.25
0.30.35
0.40.45
0.5
pre
cip
itat
ion
(m
m/m
)
Months
02468
101214161820
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Tem
per
atu
re (
°C)
Months
02468
101214161820
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Tem
per
atu
re (
°C)
Months

315
Figure A.10 Radiation (W hr/m²) for 2016-2017 in Dunstaffnage bay the area for the seasonal study (CEDA-Centre for Environmental Data Analyses)
Figure A.11 Radiation (W hr/m²) for 2017-2018 in Dunstaffnage bay the area for the seasonal study (CEDA-Centre for Environmental Data Analyses)
Bivariate correlation was conducted between antioxidants, carbohydrates, proteins, pigments and
the abiotic factors, and the most representative results are reported in Figs A.12-A.18.
0
500
1000
1500
2000
2500
00
:00
01
:00
02
:00
03
:00
04
:00
05
:00
06
:00
07
:00
08
:00
09
:00
10
:00
11
:00
12
:00
13
:00
14
:00
15
:00
16
:00
17
:00
18
:00
19
:00
20
:00
21
:00
22
:00
23
:00
Rad
iati
on
(W
hr/
m²)
HoursJanuary February March April May June
July August September October November December
0
200
400
600
800
1000
1200
1400
1600
1800
2000
00
:00
,
01
:00
,
02
:00
,
03
:00
,
04
:00
,
05
:00
,
06
:00
,
07
:00
,
08
:00
,
09
:00
,
10
:00
,
11
:00
,
12
:00
,
13
:00
,
14
:00
,
15
:00
,
16
:00
,
17
:00
,
18
:00
,
19
:00
,
20
:00
,
21
:00
,
22
:00
,
23
:00
,
Rad
iati
on
(W
hr/
m²)
Hours
January February March April May June
July August September October November December

316
Figure A.12 Correlation between TPC content (bars) and hours of light (line) (r=0.75)
Figure A.13 Correlation between FRAP (bars) and hours of light (line) (r=0.75)
Figure A.14 Correlation between hours of light (line) and carbohydrate content (bars) (%DW) (r=0.75)
0
20
40
60
80
100
120
140
160
0
5
10
15
20
25
TPC
(µ
g/m
g D
W)
hrs
of
ligh
t
Months
0
5
10
15
20
25
0
0.5
1
1.5
2
2.5
Hrs
of
ligh
t
FRA
P
(µM
FeSO
₄/m
g D
W)
Months
0
5
10
15
20
25
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
0
5
10
15
20
25
30
35
Car
bo
hyd
rate
s co
nte
nt
(% D
W)
Months
Hrs
of
Ligh
t

317
Figure A.15 Correlation between max radiation (line) and carbohydrate content (bars) (r=0.73)
Figure A.16 Correlation between temperature (max as dark grey line and min as light grey line) and carbohydrate content (bars) (r=0.86, max temp; r=0.82, min temp)
Figure A.17 Correlation between hours of light (bar) and Chl-a content (line) (r=-0.63)
0
5
10
15
20
25
30
35
0
500
1000
1500
2000
2500
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Car
bo
hyd
rate
s co
nte
nt
(%D
W)
Rad
iati
on
(W
hr/
m²)
Months
0
5
10
15
20
25
30
35
0
5
10
15
20
25
Aug Sep Oct Nov Dec Jan Feb Mar Apr May Jun Jul
Tem
per
atu
re (
°C)
Car
bo
hyd
rate
s co
nte
nt
(% D
W)
Months
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
5
10
15
20
25
Ch
l-a
con
ten
t (µ
g/g
DW
)
Hrs
of
ligh
t
Months

318
Figure A.18 Correlation between temperature (max as dark grey line and min as light grey line) and Chl-a content (bars) (r=-0.71, max temperature; r=-0.75, min temp)
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
5
10
15
20
25
Ch
l-a
(µg/
g D
W)
Tem
per
atu
re (
°C)
Months

319
Appendix B Accessory data Chapter 4
The analyses on the 12 samples (in three replicates) were conducted in two different blocks, two
samples were chosen from the first run and re-run the second time together with the remaining
block of samples. Results for the comparison of the same month analysed in two different runs for
the positive and negative modes are reported in Figs B.1-B.4. The peaks were the same for both
runs, proving the robustness of the method applied and the consistency of the analyses.
Figure B.1 Negative Mode April 2017 comparison of first (black) and second (red) runs (FSD: 7.80E7, Base Peak F: FTMS-c ESI Full MS [80-1000])
Figure B.2 Positive mode April 2017 comparison of first (black) and second (red) runs (FSD: 3.45E8, Base Peak F: FTMS+c ESI Full MS [80-1000])
Figure B.3 Negative mode September 2016 comparison of first (black) and second (red) runs (FSD: 7.60E7, Base Peak F: FTMS-c ESI Full MS [80-1000])
RT: 2.19 - 23.07 SM: 5B
4 6 8 10 12 14 16 18 20 22
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relat
ive Ab
unda
nce
NL:7.80E7
Base Peak F: FTMS - c ESI Full ms [80.00-1000.00] MS 008_a1_neg
NL:7.80E7
Base Peak F: FTMS - p ESI Full ms [80.00-1000.00] MS ap1
RT: 2.19 - 23.07 SM: 5B
4 6 8 10 12 14 16 18 20 22
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relat
ive Ab
unda
nce
NL:3.45E8
Base Peak F: FTMS + c ESI Full ms [80.00-1000.00] MS 008_a1_pos
NL:3.45E8
Base Peak F: FTMS + p ESI Full ms [80.00-1000.00] MS ap1
RT: 2.19 - 23.07 SM: 5B
4 6 8 10 12 14 16 18 20 22
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relat
ive Ab
unda
nce
NL:7.60E7
Base Peak F: FTMS - c ESI Full ms [80.00-1000.00] MS 003_s1_neg
NL:7.60E7
Base Peak F: FTMS - p ESI Full ms [80.00-1000.00] MS sep1

320
Figure B.4 Positive mode September 2016 comparison of first (black) and second (red) runs (FSD: 3.50E8, Base Peak F: FTMS+c ESI Full MS [80-1000]
The set of analysed samples included a sampling period from August 2016 to July 2017. To verify
if there was a difference between the same months in different years, the sample from August 2016
was compared with the sample from August 2017 and results for the positive and negative modes
are reported in Figs B.5 and B.6. The same peaks were reported in both samples.
Figure B.5 Negative mode comparison of August 2016 (black) and August 2017 (red) runs (FSD: 7.00E7, Base Peak F: FTMS-c ESI Full MS [80-1000])
Figure B.6 Positive mode comparison of August 2016 (black) and August 2017 (red) runs (NL: 3.40E8, Base Peak F: FTMS+c ESI Full MS [80-1000])
RT: 2.19 - 23.07 SM: 5B
4 6 8 10 12 14 16 18 20 22
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relat
ive Ab
unda
nce
NL:3.50E8
Base Peak F: FTMS + c ESI Full ms [80.00-1000.00] MS 003_s1_pos
NL:3.50E8
Base Peak F: FTMS + p ESI Full ms [80.00-1000.00] MS sep1
RT: 2.19 - 23.07 SM: 5B
4 6 8 10 12 14 16 18 20 22
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relat
ive Ab
unda
nce
NL:7.00E7
Base Peak F: FTMS - c ESI Full ms [80.00-1000.00] MS 004_AG1_Neg
NL:7.00E7
Base Peak F: FTMS - p ESI Full ms [80.00-1000.00] MS aug1
RT: 2.19 - 23.07 SM: 5B
4 6 8 10 12 14 16 18 20 22
Time (min)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relat
ive Ab
unda
nce
NL:3.40E8
Base Peak F: FTMS + c ESI Full ms [80.00-1000.00] MS 004_ag1_pos
NL:3.40E8
Base Peak F: FTMS + p ESI Full ms [80.00-1000.00] MS aug1

321
Appendix C Additional data on cultivation experiments
C.1 Effect of temperature on biomass increase and epiphytes colonization in O. pinnatifida
The aim of this preliminary trial was to assess the best temperature for an increase in biomass and
resistance to epiphytes in O. pinnatifida cultures.
Materials and Methods
Samples were collected from Ganavan Beach (56°26'21.0"N, 5°28'15.8"W) and Dunstaffnage
beach (56°27'07.2"N, 5°26'44.6"W) on rocky mid-sheltered shores, after morphological
identification following the key characters given by Maggs and Hommersand (1993). Samples were
transported to the lab, manually cleaned to remove visible epiphytes and grazers, thereafter, washed
with seawater. The seawater for the experiment was collected from the Algal Culture room, SAMS
facility, filtered (100 µm-25 µm-0.45 µm) and sterilized with UV and then tyndalized at 75 °C for
20 min for three days consecutively. After that, KTH medium (Fig. C.2) was added in a laminar
flow cabinet. Flasks were replaced with sterilized ones during the medium change. Air was filtered
through 0.45 µm (Whatman® filter) into 500 ml flasks and 0.30 g of WW specimens placed in each
flask with KTH medium. The flasks were divided in four different conditions in a controlled
temperature rooms, for a total of 3 flasks for each condition: 13 °C, 10 °C, 15 °C and 20 °C (Fig.
C.1). The photoperiod was 16:8 (L:D) of white light with an intensity of 50-100 µmol/m²/sec. The
length of the experiment was of 4 weeks. The medium was changed weekly, and the weighing of
the samples conducted at this point.
Figure C.1 Flasks under 4 different temperature treatments

322
Figure C.2 KTH medium composition
Results
At the end of the experiment, the 10 °C temperature seemed the most suitable for epiphytes control
and growth increase (Fig. C.3 A and B). Under the 15 °C condition, different green filamentous
epiphytes (e.g. Enteromorpha sp.) were present in the cultures and started to affect the tissue texture
(Fig. C.3 E and F). The 20 °C treatment is unsuitable for a long-term maintenance of the cultures;
after 4 weeks, it was impossible to detect the weight of the specimens since the tissue was almost
completely disrupted, brittle and colonized by green epiphytes (Fig. C.3 G and H). The best
treatment seemed to be the 10 °C, and this temperature was chosen for further experiments;
however, no statistical differences (p<0.05, Minitab 18) were recorded in terms of weight increase
among the treatments investigated, except for the 20 °C treatment (Fig. C.4).

323
Figure C.3 Samples under 4 different treatments: A) 10°C after 7 and B) 28 days; C) 13°C after 7 D) and 28 days; E) 15 °C after 7 F) and 28 days; G) 20°C after 7 H) and 28 days
Figure C.4 Weight growth of the samples under four treatments in relation to the time (n=3, error bar, 1 SD of mean)
Discussion
Four different temperatures were tested in order to assess the adaptation and resistance of the
species. The results showed that O. pinnatifida specimens from Scotland are susceptible to
temperature above 15°C. Cultures at 20°C were negatively affected, decreasing their weight and
being easily colonized by epiphytes. At the end of the experiment, the material under this condition
was heavily damaged and measurements of the weight were impossible. Epiphytes colonization
was higher also in specimens under 15°C, compared to those under 10 and 13 °C. The best
treatment, in term of morphology (Fig. C.3) and weight increase (Fig. C.4) appears to be the 10 °C
condition. Even if the species has been recorded worldwide, it must undergo phenotypical
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
weight (0) weight (1) weight (2) weight(3) weight(4)
Wei
ght
(g W
W)
Week
10-13 °C 10° C 15 °C 20 °C

324
adaptations, according to the geographical distribution and temperature where it grows.
Furthermore, must be considered that different strains can be adapted to different conditions, as
already seen for other species (e.g. Palmaria palmata, Manríquez-Hernàndez et al., 2016). From this
study emerged that tolerance to increase of temperature above 13 °C is very low in O. pinnatifida
specimens from the West coast of Scotland.
C.2 Investigation on the stimulation of spores’ production applying a “heat-shock” in an
indoor tank system for O. pinnatifida
Three different treatments were tested in order to investigate potential regulation factors for spore
production in O. pinnatifida cultures in an indoor tank system.
Materials and methods
The Algal Culture Room used for the trials is a controlled temperature room with an average
temperature of 13 °C, monitored with a digital thermometer during the period of one month. The
material for the experiment was collected manually from Easdale Island, North-West Scotland
(56°17'25.5"N 5°39'30.2"W) on a rocky shore during low tide in February 2016. A total amount of
almost 2 kg (FW) was collected. The experimental set-up consisted of 9 Plexiglas, rectangular tanks
of 5 l. They were pre-washed with a 50% DECON solution in water and rinsed to remove any
residual DECON. Before the start of the experiment, the biomass was manually cleaned from
visible epiphytes and grazers and then washed with 0.5ml/l of Sodium Hypochlorite 5% for 2
minutes and then rinsed with FSW for 10 minutes. The biomass was then acclimated for 15 days
at 13°C in FSW and vigorous aeration before the start of the experiment.
The seawater for the experiment was filtered (100 µm-25 µm-0.45 µm) and sterilized with UV and
then tyndalized at 75 ° C for 75 min for 3 days consecutively. Biomass (50 g, FW) was placed in
each tank. Three different treatments in triplicate were tested: “control” (nutrient added. KTH),
“no nutrient added”, and “heat shock” in nutrient added. For this treatment, an aquarium heater
was set for each tank at 20 °C for 3 days, then the heaters were turned off and removed from the
tanks, leaving the temperature to cool down again. The photoperiod chosen for the experiment
was 16:8 (L:D) with white light at an intensity of 80-100 µmol/m²/sec. The aeration system
consisted of 2 tubes for each tank, sunk to the bottom thanks to aquarium stones. The trial run for
4 weeks. Weekly change of the Medium (KTH) and weighing of cultures was performed. Samples
were examined under a SteREO Discovery V8 stereomicroscope (Zeiss) periodically to assess
formation of tetrasporocystes and spores release.
Results
The best growth rate was reported for the “Control” treatment. Cultures under the “no nutrient”
treatment showed a thick texture and mature tetrasporocystes. However, the colour changed

325
turning into green. The treatment using the “heat shock” was not effective in terms of induction
of the sporulation. Cultures under this treatment showed the lowest growth rate (Fig. C.5).
Moreover, the heat shock itself resulted in an increase in epiphytes from Enteromorpha sp..
Figure C.5 Relation between weight increase and time in samples undergoing 3 different treatments: heater (black), no nutrient (light grey) and control (dark grey bars) (n=3; error bar, 1 SD of mean)
Figure C.6 Graph of SGR average for the treatments calculated using the formula from Evans (1972). Heater black, no nutrient light grey and control dark grey bars (n=3; error bar, 1 SD of mean)
No differences in tetrasporocystes formation and sporulation were detected among the treatments
tested, neither a triggering factor (except the cultivation itself) identified. Tetrasporocystes
appeared in cultures under all the treatments after 7 days into the cultivation system. Spores were
naturally released from biomass under the three treatments after two weeks into the cultivation
system. After three weeks most of the tetrasporocystes of biomass under the three treatments were
empty, or with few spores inside. Since the first week of experiment, new plantlets were growing
in all the tanks especially from the holdfast of the plants. Epiphytes were visible especially in the
samples treated with the “heat-shock”.
0
10
20
30
40
50
60
0 1 2 3 4
Wei
ght
(g W
W)
Week
-2
-1.5
-1
-0.5
0
0.5
1
1 2 3 4
Gro
wth
rat
e (%
day
¯¹)
Time (week)
Heater No nutrient Control

326
Conclusions
A “heat-shock” is not effective in stimulating the formation of tetrasporocystes in O.
pinnatifida, which seems to occur once the biomass is put into cultivation systems, even
outside the normal reproductive season (which is May);
Sodium hypochlorite at the tested concentration (5%) and exposure time (2 minutes) is not
efficient in controlling epiphytes in O. pinnatifida cultures;
Absence of nutrient does not statistically reduce the growth rate of the species investigated,
can reduce the contamination of epiphytes, but results in a green colour of the cultures,
probably due to the degradation of pigments (e.g. phycobiliproteins) under N limitation
(Penniman and Mathieson, 1987; Rico and Fernandez, 1996; Harrison and Hurd, 2001). It
may be applied as pre-treatment or alternated to nutrient supply in cultivation systems for
epiphytes management.
C.3 Effect of light quality on growth rate, pigment content and reproduction in O. pinnatifida
cultures
Material and method
Four different wavelengths –red, green, blue and white fluorescent- were tested during this trial.
The light intensity at the source was 150 µmol/m²/sec for the blue and white light, 200
µmol/m²/sec for the red and the green. The flasks for each treatment were placed at the same light
intensity (80 µmol/m²/sec). A tray was placed under the flasks undergoing the same treatment to
avoid the influence of the other lights placed below or above it (Fig. C.7). Flasks (500 ml) containing
five grams of blotted material and 500 ml of autoclaved F/2 were used in triplicate for each
treatment. Aeration was provided throughout air filters (0.45 µm) and sterilized glass pipettes. QY
was assessed using an AquaPen-P (PSI) at the beginning of the experiment and weekly. The starting
QY average for the samples was 0.52 (Fv/Fm) (n=3) (Fig. C.9). The photoperiod was 12:12 (L:D)
and the trial was carried on in a controlled temperature room at an average temperature of 10°C.
The experiment run for 8 weeks. Weight, QY measurements and spore production/release were
assessed weekly (Fig. C.8). Pigment analyses were performed upon completion of the experimental
period. Pigments (including Chl-a, carotenoids, Pe and PC) were analysed as already described in
Chapter 3.

327
Figure C.7 Flasks under culturing conditions
Results
Spores appeared after one week in cultures under all the four treatments, with no difference in
timing. Spores were released spontaneously after one week from appearance in the
tetrasporocystes, without difference between the treatments (Fig. C.8 A-C).
Figure C.8 A) roped slide on the bottom of the flask for spores' collection; B) sample under red light at the end of the experiment colonized by epiphytes; C) spores released (circled in green) outside the slide (green light treatment)
A B C

328
Statistical difference (p<0.05) were reported between the treatments in term of PAM response
between green and red light in the second week, and between blue and white and red and green in
the fourth week, with the first two showing the highest results and the last two the lowest ones.
No differences were reported for the other weeks of experiment (Fig. C.9).
Figure C.9 Comparison of the QY (Fv/Fm) between the different wavelengths Blue, green red and control (grey) bars (n=3; error bar, 1 SD of mean)
The statistical analyses performed for the SGR (Fig. C.10), according to the various lights, shows
that there is a statistical difference between the red and white light treatment (p <0.05, Fisher’s
Test, Minitab 18).
Figure C.10 SGR calculated with the formula (Evans, 1972) for Blue, green red and control (grey) bars (n=3; error bar, 1 SD of mean)
0
0.1
0.2
0.3
0.4
0.5
0.6
time 0 time 1 time 2 time 3 time 4 time 5 time 6 time 7
PA
M (
QY)
Week
0
0.2
0.4
0.6
0.8
1
1.2
1.4
SGR (1) SGR (2) SGR(3) SGR(4) SGR(5) SGR(6)
Gro
wth
rat
e (%
day
¯¹)
Week

329
Figure C.11 Chl- a (dark grey bars) and Carotenoids content (light grey bars) expressed in µg/g for the samples under the 4 different light treatments (n=3; error bar, 1 SD of mean)
For the Chl-α analyses, the Tukey’s and Fisher’s tests showed significant differences in
concentration between the samples under the blue and white (control) lights, red and white and
green and white (Fig. C.11). The highest value (p<0.05) was reported for the samples undergoing
the treatment with the green light (1730.5 µg/g). For the carotenoids, the Tukey’s and Fisher’s tests
(p<0.05) showed significant differences in content between the samples under the blue and white
(control) light, red and white and green and white. The highest value was reported for the samples
undergoing the treatment with the red light (627.6 µg/g) (Fig. C.11).
Figure C.12 PE (dark grey bars) and PC content (light grey bars) (mg/g) for the samples under different light treatments (n=3; error bar, 1 SD of mean)
Statistical analyses showed no differences (p<0.05) in the PC concentration for the samples
undergoing the different treatments. The highest value was reported for the control (white
fluorescent light). Statistical analyses showed no differences in the PE concentration for the
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Blue Red Green Control
Ch
l-a
and
car
ote
no
id c
on
ten
t (µ
g/g
DW
)
Light treatment
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Blue Red Green Control
PE
and
PC
co
nte
nt
(mg/
g D
W)
Light Treatment

330
samples undergoing the different treatments with the highest value reported for the green light
treatment (Fig. C.12).
Discussion
Light is the primary energy source for algae. These organisms can adapt to the light (photo
acclimation) and avoid the excess of it (photo protection) (Falkowski and La Roche, 1991). As
other intertidal macroalgae (Lüning, 1990; Häder et al., 2002), O. pinnatifida is exposed to changing
irradiances and complex light patterns (Gómez et al., 2004). Protection against the harmful effects
of super excitation due to saturating light intensity is a fundamental survival mechanism of
seaweeds in the intertidal zone (Häder et al., 2002; Cardozo et al., 2006, 2008; Guaratini et al.,
2009). While during low photon flux conditions the algae have to harvest maximum light.
Alterations in red algae pigment content may be related to the size and number of chromophores,
as well as the number, structure or size of phycobilisomes, which are modified to optimize light
absorption (Kursar et al., 1983b; López-Figueroa and Niell, 1990, 1991). Furthermore, the synthesis
of chlorophyll and biliprotein can be regulated by phytochrome (red/far-red light) and blue light
photoreceptors (López-Figueroa and Niell, 1991). In this experiment, Chl-α and carotenoids
production was stimulated by the light quality (green, red and blue), compared to the control state
(Fig. C.12). Particularly, the Chl-a production seems to be stimulated by the green light while the
carotenoids production increases under the red light. Barufi et al. (2015) reported for Gracilaria
birdiae an increase in Chl-α production related to green and blue lights, which is similar (for the
green light) to what was estimated in this experiment (Fig. C.12). An increase in phycobiliprotein
and chlorophyll-a (Chl a) content was observed in Chondrus crispus cultivated under blue, red, and
green light in comparison with white light controls (Franklin et al., 2001). The accessory pigment
synthesis on the other hand, was stimulated only for PE production by Green light (Fig. C.13),
while the PC production was not affected by the different treatments. This is in contrast with a
study conducted on G. birdiae (Barufi et al., 2015), that showed an increase in accessory pigments
due to the different treatments, especially under the blue light. The same observation was shown
in Porphyra leucosticta (Tsekos et al., 2002). Usually, blue and red light stimulated the accumulation
of phycobiliproteins (Godínez-Ortega et al., 2008). Blue light also stimulated nitrogen assimilation
and the accumulation of such N-compounds as biliproteins (Figueroa et al., 1995a). In this
experiment, the red-light treatment showed the highest increase in weight, followed by the green
light, the blue and, as last, the control (white fluorescent) (Fig. C.11). However, must be underlined
that especially the samples undergoing the cultivation in Red light were heavily affected by
epiphytes (Fig. C.9 B). Nevertheless, the statistical difference in weight increase remained even
after the manual cleaning of the samples and the re-weighing of them. PAM response was not
deeply affected by the wavelength applied, showing in general a stable response of the cultures, and

331
only a slight decrease in the last two weeks of the experiment, which was not statistical relevant.
Light can also control metabolic, growth and reproductive processes (Rüdiger and Figueroa, 1992;
Dring, 1988; Lüning, 1992; Monro and Poore, 2005) such as spores’ germination, as already seen
in other red algae (e.g. Bangia atropurpurea, Charnofsky et al., 1982) and gamete formation in brown
algae, such as Laminaria (Lüning, 1980). Moreover, according to the action spectra for
photosynthesis in red algae, photosynthetic activity is lower under blue light than under red, owing
to the low efficiency of electron transport in photosystem II under blue light because most
chlorophyll (about 85%) in red algae is located in photosystem I. Therefore, the absence of blue
spectra could stimulate the investment of light energy for reproduction instead of photosynthesis
and stimulate the development of tetrasporophytes of some red algae, such as G. birdiae (Lüning
and Dring, 1985; Grzymski et al., 1997). In this study, however, tetrasporangium differentiation
occurred simultaneously in all the cultures under the four different treatments one week after the
beginning of the experiment. Neither spermatangial conceptacles nor cystocarps were observed.
The light quality seems to not influence reproduction and spore release in O. pinnatifida. Depending
which one is the final target of the cultivation of the species, these results are important in
understanding how the quality and quantity of pigments of O. pinnatifida can be effectively
manipulated with the manipulation of the wavelength.
C.4 Cultivation in flasks of vegetative thalli tied onto nylon ropes
Materials and methods
Material was harvested from Easdale, Seil (56°17'25.5"N 5°39'30.2"W) in November 2017, cleaned
and treated as described in Chapter 5, and placed into cultivation after acclimation in tank at 10 °C,
12:12 (L:D), with filtered and UV sterilized seawater. The experiment was done in triplicate. Two
different volumes were used, 1 l and 500 ml flasks, with 1 l and 500 ml of F/2 respectively.
Vegetative thalli were manually attached onto cleaned and dried (50% Decon solution in UPW)
nylon ropes hanging into the flasks. Two different starting biomasses were chosen, 5 g for 500 ml
volume (Figs C.13 A and C.14 B) and 10 g for the 1 l volume (Figs C.13 B and C.14 A). Medium
was changed twice per week and cultures blotted weighed every week, after been drained in a spin-
salad; cultures were spun until no more water was released from the material. Weight of the rope
was subtracted from the total wet weight of the culture. Cultures were incubated at 10 °C, 12:12
(L:D) photoperiod, 80 µmol/m²/sec. Aeration was provided through 0.45 µm (Whatman® filter).

332
Figure C.13 Vegetative thalli tied to nylon ropes in A) 1 l flasks and B) 500 ml flasks
Figure C.14 A) 3 x 1 l flasks and B) 3 x 500 ml flasks with tied cultures
Results
Cultures, in both 1 l and 500 ml flasks, grew for all the length of the experiment. They did not
detach from the rope and the holdfast of some of the plant started to attach to the nylon rope
toward the end of the experiment (pers. obsv.). No macroscopic epiphytes were detected on the
cultures. Regarding the SGR, the highest rate was reported by the 500 ml flasks (Fig. C.15),
especially after the second week from the start of the experiment. However, no statistical difference
(ANOVA, single factor, p<0.05, Minitab 18) was reported for the SGR between the 500 ml and 1
l flasks (Fig. C.15). This indicates that the density (at least for the values here tested) does not have
an influence on the growth rate of the species.
Figure C.15 SGR was calculated with the formula from Evans, (1972) for 1 l cultures (dark grey bars) and 500 ml cultures (light grey bars) (n=3; error bar, 1 SD of mean)
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
1 2 3 4
Gro
wth
rat
e (%
day
¯¹)
Week

333
Furthermore, the SGR of free-floating cultures from a previous experiment was compared with
those of tied cultures, with the same volume, media and incubation condition. No statistical
difference (p<0.05) was reported among the two treatments, with slightly higher value reported for
the free-floating cultures (Fig. C.16).
Figure C.16 Comparison of growth (SGR) between tied (dark grey line) and untied (light grey line) cultures (n=3; error bar, 1 SD of mean)
Discussion
Line rope farming is an established technique for seaweed cultivation that was pioneered in China
in the 1950s, for the cultivation of brown seaweeds. The methodology has been than broaden and
applied to other genera such as Gracilaria sp.. Ropes can be made of nylon or polypropylene and
the seaweeds can be either insert in the rope, untwisting it, or they can be tied with strings (FAO,
2012). This system is applied for the vegetative propagation of plants as well for the growth of
sporophytes on sea (FAO, 2012). It was here experimented in vitro for the vegetative cultivation
of O. pinnatifida. From this study emerged that O. pinnatifida can grow in both tumbling cultures and
attached to a substratum, following a different trend compare to free-floating cultures in the growth
curve. Since this species normally grows on the seashore, attached to rock and barnacles’ surfaces,
its attachment was considered important in this study from a cultivation prospective, and it is not
surprising that it might have influence on the growth rate and pattern of the cultures. Towards the
end of this trial, some plants started to attach to the rope with their holdfast, and new branches
were evident since from the first week of culturing. New tissue developed both from tips and from
the holdfast. The quality of the cultures was not negatively affected by the methodology, showing
no epiphytes growing on the surface neither in the flasks. However, this methodology is time
consuming and laborious, and might be difficult to apply on a large scale. Every attempt conducted
in this project aimed at attaching the material onto a tough surface failed; however, this should be
further investigated as potentially important for the growth and increase of biomass of the species.
Indeed, the growth increase reported with this study ranged from 1.90% day⁻¹ to 3.92 for 1 l flasks,
-1
-0.5
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
0 1 2 3
Gro
wth
rat
e (
%d
ay¯¹
)
Week

334
and from 1.66 % day⁻¹ to 2.95 for 500 ml flasks, which is among the highest reported for this
species in this project.
C.5 Out-plantation of vegetative material of Osmundea pinnatifida
Scope
Cultivation of seaweed in open water is an established technique for several species (FAO, 2012).
Here was trialled the possibility to cultivate O. pinnatifida in mesh bags on a seaweed farming site,
testing different depths and densities. This has been already trialled for other red algae (e.g.
Palmaria) with some positive results (Browne, 2001; Martinez et al., 2006). One of the main
problems of this methodology is the contamination that derives by biofouling organisms, such as
other algae and invertebrates, that normally colonise an object that is immersed in the sea for a
certain length of time (Armstrong et al., 2000). The activity of biofouling organisms can affect the
growth and survival rate of the cultures (Sanderson, 2006). They can also interfere with the
physiology of the seaweed (e.g. reducing photosynthetic rate, Sanderson, 2006). This section
investigated the possibility of growing O. pinnatifida inside mesh bags in the sea, trialling a low-cost
system with the potential of creating biomass quickly, with minimum effort.
Materials and methods
Samples were collected from Easdale, Seil (56°17'25.5"N 5°39'30.2"W) in the mid of November
2017. They were kept in a Cool-box and brought to the lab where they were left into a 60 l tank
with FSW, at 10 °C and 12:12 (L:D) photoperiod. They were successively cleaned with a 0.5 % KI
as described in Chapter 5. 12 x 20 cm tubular plastic nets, 5-6 cm diameter and 3-4 mm mesh size
were prepared with the material (Fig. C.17A). Two depths were chosen for the experiment: 30 cm
and 1 m from the surface. Two densities were chosen for the experiment: 15 g/20 cm and 60 g/20
cm.
The colour of the meshes was not chosen in accordance to the treatment. Of the six bags at 15g/20,
three bags were deployed at 30 cm from the surface and 3 at 1m from the surface. Of the six bags
at 60 g/20 cm (Fig. C.17 A), three bags were deployed at 30 cm in depth and 3 at 1 m. Bags were
disposed randomly relatively to the density at the target depth, along a 3 m rope in length at
intervals of ~50 cm. They were cable tied to a horizontal rope of 0.5 cm of thickness.
The site chosen for the experiment is the largest experimental seaweed farm in the UK and is used
for upscaling studies as it approaches a size that could be commercially viable. The location is
56.49N, 5.47W in the Lynn of Lorne off the island of Lismore. It is composed of a single 100 x
100 m grid system for growing of seaweed. The depth range is 15-25 m and the site is very exposed

335
to south-westerly winds from the Atlantic with a mean current speed of 0.1 m/s at 5 m below the
surface.
Two out plantation trials were tested with the described conditions, one from November 2017 to
April 2018 (Fig. C.17 B) and one from April 2018 to September 2018.
Figure C.17 A) Bags used for the trials before the out plantation; B) Deployed bags on the farm site (winter trial)
Temperature, light intensity, pH and salinity are monitored in the site of deployment. Here are
reported only the data relative to the temperature and light (Figs C.18 and C.19).
Biochemical analyses were carried on specimens including moisture, pigments, carbohydrates,
protein and antioxidant compounds following the methodologies already described in Chapter 3.
Analyses were done only for samples from the first trial because, unfortunately, the material
collected from the second trial was not analyseable.
Results
Average temperature and light (lux) for the farm site relative to the two trials are reported in Figs
C.19 and C.20. Evident is the oscillation of the recorded parameters according to the season, with
a decrease of temperature in the winter (6-8 °C, Fig. C.18) and an increase in the light intensity
from May to July (Fig. C.19).

336
Figure C.18 Average temperature (°C) on the farm site relative to the winter and summer trials
Figure C.19 Average light (Lux) on the farm site relative to the winter and summer trials
a) Winter trial
Deployed bags, 12/12, were collected after the end of the trial and they were intact. Both the low-
density bags (LD) (at either 30cm and 1 m of depth) showed an increase in weight (0.16 % day⁻¹),
whilst those with a higher density (HD) (at either 30cm and 1 m of depth) a slight decrease (-0.06
%day⁻¹) (Fig. C.20). Epiphytes were present attached outside of the meshes, but they did not
contaminate the cultures in the bags (Fig. C.27 A and B). The ones at low density showed a better
texture and colour, due to the fact that were less packed and more equally distributed in the bag
compared to the HD ones (Fig. C.20). Moisture content was estimated as already described in
Chapter 3. No differences in term of moisture content were found between the two densities as
well as the two depths tested (Fig. C.21).
0
2
4
6
8
10
12
14
16
18.8.16 26.11.16 6.3.17 14.6.17 22.9.17 31.12.17 10.4.18 19.7.18 27.10.18 4.2.19
Tem
per
atu
re (
°C)
Date
0
2000
4000
6000
8000
10000
12000
18.8.16 26.11.16 6.3.17 14.6.17 22.9.17 31.12.17 10.4.18 19.7.18 27.10.18 4.2.19
Ligh
t In
ten
sity
(Lu
x)
Date

337
Figure C.20 Growth (weight) of winter trial (initial weight as dark grey bars and final weight as light grey bars) (n=3; error bar, 1 SD of mean)
Figure C.21 Moisture content for deployed cultures expressed in percentage (initial FMC as dark grey bars and final FMC as light grey bars) (n=3; error bar, 1 SD of mean)
Biochemical composition
For carbohydrates content, only the culture at 30 cm and high density were statistically different
from the other (p<0.05) (Fig. C.22) while for chlorophyll-a and carotenoids no statistic differences
were reported (Fig. C.23) as well as for FRAP content (Fig. C.24).
Figure C.22 Carbohydrates (% DW) of outplanted cultures (n=3; error bar, 1 SD of mean)
0
10
20
30
40
50
60
70
80
30 cm depth LD 30 cm depth HD 1m LD 1m HD
wei
ght
(g W
W)
Treatment
82
83
84
85
86
87
88
89
90
91
30 cm depth LD 30 cm depth HD 1m LD 1m HD
Mo
istu
re c
on
ten
t (%
FM
C)
Treatment
0
5
10
15
20
25
30
35
40
30 cm Lowdensity
30 cm Highdensity
1 m low density 1m high density
Car
bo
hyd
rate
co
nte
nt
(% D
W)
Treatment

338
Figure C.23 Chl-a (dark grey bars) and carotenoids (light grey bars) concentration of out planted cultures (n=3; error bar, 1 SD of mean)
Figure C.24 FRAP (µM FeSO₄ eq/mg DW) of out planted cultures (n=3; error bar, 1 SD of mean)
For protein, a Fisher’s test (p<0.05) reported a statistical difference between cultures at 1 m and
low density and the other (Fig. C.25), whilst for PE content the cultures at 30 cm and low density
were statistically different from the other (p<0.05) (Fig. C.26). No statistical differences were
reported instead for PC (Fig. C.26).
0
200
400
600
800
1000
1200
30 cm low density 30 cm high density 1 m low density 1 m high density
Ch
l-a
and
car
ote
no
id c
on
ten
t (µ
g/g
DW
)
Treatment
0
0.5
1
1.5
2
2.5
3
3.5
30 cm Lowdensity
30 cm highdensity
1 m low density 1 m high density
FRA
P
(µM
FeS
O₄
eq/m
g D
W)
Treatment

339
Figure C.25 Protein content (% DW) of out planted cultures (n=3; error bar, 1 SD of mean)
Figure C.26 PE (dark grey bars) and PC (light grey bars) (mg/g) concentration in out planted cultures (n=3; error bar, 1 SD of mean)
Figure C.27 Collected bags (outside-A; inside-B) after 6 months in the out plantation site for the winter trial
b) Summer trial
Of the deployed bags, 11/12, were recovered. Unfortunately, the bags were completely covered
inside and outside of several epibiontic organisms including bryozoans, snails, bivalves (e.g.
0
5
10
15
20
25
30
30 cm low density 30 cm highdensity
1 m low density 1 m high density
Pro
tein
co
nte
nt
(% D
W)
Treatment
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
30 cm low density 30 cm highdensity
1 m low density 1 m high density
Ph
yco
bili
pro
tein
co
nte
nt
(mg/
g D
W)
Treatment

340
mussels, and scallops), worms (e.g. polychaetes), crustacean (isopodes, amphipodes, etc.), other
seaweed (Ulva spp, Palmaria spp, etc.) and tunicates (Fig. C.28 A-D). Few new plantlets were
however visible and free from epiphytes (Fig. C.29), but the majority of the cultures went
grazed/lost. Therefore, no biochemical analyses neither growth estimation were performed on
these samples.
Figure C.28 Epibiotic organisms on collected bags including A) tunicates; B) bivalve; C) polychaetes and D) seaweed
Figure C.29 Alive recovered material isolated from the bags of the summer trial
Discussion
This was the first time that an out plantation in mesh bags of Osmundea pinnatifida in the open sea
was trialled. Normally, red seaweed used for the production of carrageenans are cultivated on lines
off-bottom or long-lines as well as tube netting, these methodologies apply to species such as
Euchema spp, Gracilaria spp and Kappaphycus spp (FAO, 2012). These species have usually a high
growth rate and are grown vegetatively (FAO, 2012). They are tied to the lines and harvested up
to 8 times per year (e.g. E. cottonii, FAO 2012), proving that these methodologies are successful for
mariculture of these red algae species. The system applied in this study did not include a tie-tie

341
technique of the single plants to the rope. This because Osmundea pinnatifida individuals are small
and thin, difficult to tie to a rope, in order to be sure that the material will remain in place, (also
considering the exposure of the site selected for the experimentation). Therefore, wild specimens,
collected and cleaned as described in Chapter 5, were placed into plastic meshed and hung vertically
from one of the ropes of the seaweed farm grid in place. The size of the meshes was big enough
to guarantee an exchange of nutrient and water, as well as oxygen and light, but to preserve the
cultures from passing through it. This was done following the type of tube netting normally used
for Gracilaria edulis, a technique that permits to this species to grow throughout the net and being
harvested (Mantri et al., 2017). The same pattern of growth was also seen for O. pinnatifida in this
trial, however with a slower growth compared to Gracilaria species (Mantri et al., 2017).
Furthermore, this methodology was applied with success also to other species of red algae, such as
P. palmata (Martínez et al., 2006; Browne, 2001) and Chondrus crispus (Zertuche-González et al.,
2001). While the cultures at LD were evenly distributed in the bag, those at HD resulted more
pressed, and especially the individuals in the middle of the bag were affected by this and showed a
greener colour, probably due to a limitation of light/nutrient diffusion. This is probably also one
of the reasons why an increase in weight was recorded for the LD cultures but not for the HD
ones, with indeed a slight decrease of biomass probably due to the density. Regarding the epiphyte’s
coverage, must be acknowledge that there were no epiphytes directly on the cultures tissue in both
the trials conducted. This proves that the treatment applied prior to the out plantation is effective
in keeping epiphytes under control even in an environment such as the open sea, where
contaminations can be multiple and severe. However, the cultures from the summer trial suffered
of the activity of different epibiotic and grazers, which means that the treatment was not efficient
in controlling the effect of those animals on the cultures. The site chosen for this trial, Port
A’Bhuiltin, is an established seaweed farm used for Saccharina latissima and Alaria esculenta cultivation
and biofouling species were already present and recorded on these species as well. High levels of
biofouling in the summer months normally occur because irradiance, day length and temperatures
are high, determining an increase in phytoplankton density, which provides a food source for
invertebrates (Kennington, 2015). Nonetheless, the few individuals recovered from the summer
trial still alive in the bags were clearly new young juveniles, of a dark colour and thick texture, sign
of a partial recover and regrowth of the cultures (Fig. C.29).
The depth and density at which the cultures were kept do not seem to have influence on the
biochemical composition of the species. It could be said that the optimal seasonal for out plantation
of O. pinnatifida in this investigated site goes from November to late April. After that, the
environmental conditions of the site together with the high impact of the biofouling and grazers
make the cultivation of this species impossible, due to the loss of biomass and the contaminations
determined by all the epibiotic organisms already described. Must be also underlined that even in

342
the wild the highest biomass and best conditions of the species are achieved between October and
April-May. Furthermore, it could be that the length here chosen for the trial was too long and that
shorter cultivation periods are necessary. However, this would unlikely match with the slow growth
rate of the species that, even in nature, requires long time for growing (pers. obsv.). Further studies
are also necessary to determine the appropriate stocking density, which could affect differently the
growth rate of the species and its increase in biomass. As expected, the growth rate tents to decrease
with increasing number of fronds per bags, probably due to a reduction of nutrient supply, light
irradiance (self-shading phenomenon) and reduced turbulence (Browne, 2001; Martínez et al.,
2006- P. palmata; Zertuche-González et al., 2001- Chondrus crispus). Furthermore, the bags applied
for these trials appear to be suitable for the purpose, they resisted throughout the winter and
summer, without torn out or breaking. Results from these trials suggests that cultivation on sea is
feasible for O. pinnatifida in the West coast of Scotland, however, should be limited to a certain
period and further research is necessary.




















