
Behavioural consequences of two chronic psychosocial stress paradigms: Anxiety without depression
14
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of Behavioural consequences of two chronic psychosocial stress paradigms: Anxiety without depression

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Behavioural consequences of two chronicpsychosocial stress paradigms: Anxiety withoutdepression
David A. Slattery a, Nicole Uschold a, Mauro Magoni b, Julia Bar a,Maurizio Popoli b, Inga D. Neumann a, Stefan O. Reber a,*
aDepartment of Behavioural and Molecular Neurobiology, University of Regensburg, Regensburg, GermanybCenter of Neuropharmacology, Department of Pharmacological Sciences and Center of Excellence on Neurodegenerative Diseases,University of Milano, Milano, Italy
Received 12 July 2011; received in revised form 5 September 2011; accepted 6 September 2011
Psychoneuroendocrinology (2012) 37, 702—714
KEYWORDSAnxiety;Depression;Social avoidance;Social stress;CSC;SDOC
Summary Chronic stress, in particular chronic psychosocial stress, is a risk factor in theaetiology of various psychopathologies including anxiety- and depression-related disorders.Therefore, recent studies have focussed on the development of social-stress paradigms, whichare believed to be more relevant to the human situation than non-social-stress paradigms. Themajority of these paradigms have been reported to increase both anxiety- and depression-relatedbehaviour in rats or mice. However, in order to dissect the mechanisms underlying anxiety ordepression, animal models are needed, which specifically induce one, or the other, phenotype.Here, we study both short- (1 d after stressor termination) and long-term (4 d or 7 d after stressortermination) behavioural and physiological consequences of two well-validated chronic psycho-social stress models: social-defeat/overcrowding (SD/OC) and chronic subordinate colony housing(CSC). We demonstrate that SD/OC and CSC result in different physiological alterations: SD/OCmore strongly affecting body-weight development, whereas CSC more strongly affects adrenaland pituitary morphology. Both stressors were shown to flatten circadian locomotor activityimmediately after stress termination, which normalized 7 d later in SD/OC group but reversed tohyperactivity during the dark phase in the CSC group. Importantly, neither stress paradigmresulted in an increase in depression-related behaviour as assessed using the forced swim test,tail suspension test and saccharin preference test at any time-point. However, both stressparadigms lead to an anxiogenic phenotype; albeit with different temporal profiles and nottowards a novel con-specific (social anxiety). CSC exposure elevates anxiety-related behaviourimmediately after stressor termination, which lasts for at least 1 wk. In contrast, the anxiogenicphenotype only develops 1 wk after SD/OC termination. In conclusion, both models are unique for
* Corresponding author at: Chronic Stress Group, Department of Behavioural and Molecular Neuroendocrinology, University of Regensburg,Universitaetsstrasse 31, 93053 Regensburg, Germany. Tel.: +49 941 9433050; fax: +49 941 9433052.
E-mail address: [email protected] (S.O. Reber).
Available online at www.sciencedirect.com
jou rn a l home pag e : ww w. el sev ie r. com/ loca te /psyn eu en
0306-4530/$ — see front matter # 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.psyneuen.2011.09.002

Author's personal copy
Introduction
Despite substantial research efforts in the last decades, theaetiology of stress-based disorders such as major depressionand anxiety remains poorly understood. This has led to aresurgence of interest in developing more relevant animalmodels than the majority currently employed. Therefore,given the evidence purporting chronic social stress to be arisk factor for the development, not only of cardiovascularand inflammatory diseases, but also of depression and anxi-ety in vulnerable individuals (Cryan and Slattery, 2007),recent attempts have focussed on the development ofchronic social stress paradigms (Blanchard et al., 1995;Keeney and Hogg, 1999; Bartolomucci et al., 2003; Fuchsand Flugge, 2003; Stefanski et al., 2003; Engler et al., 2005;Berton et al., 2006; Reber et al., 2006, 2007; Tsankova et al.,2006; Krishnan et al., 2007; Schmidt et al., 2007). Suchparadigms are believed to be more relevant to the humansituation than non-social stress paradigms (e.g. repeatedrestraint (Cryan and Slattery, 2007)) and, thus, can betterreveal the behavioural, neuroendocrine or immunologicalconsequences of chronic stress.
Thus, the use of social defeat/subordination (De Goeijet al., 1992; Amstislavskaya and Kudryavtseva, 1997; Avgus-tinovich et al., 1997; Bartolomucci et al., 2001; Keeneyet al., 2001; Stefanski, 2001; Bartolomucci et al., 2003;Berton et al., 2006; Reber et al., 2006, 2007; Miczeket al., 2008; Savignac et al., 2010; Denmark et al., 2010)and social instability (Engler et al., 2005; Bailey et al., 2006;Schmidt et al., 2007; Kemme et al., 2008; Sterlemann et al.,2008; Engler et al., 2008; Bailey et al., 2010) paradigms in thelast decades have revealed numerous behavioural and phy-siological consequences that occur following chronic psycho-social stress exposure. The majority of these stressors havebeen reported to increase both anxiety- and depression-related behaviour (Keeney and Hogg, 1999; Blanchardet al., 2001; Berton et al., 2006; Krishnan et al., 2007;Schmidt et al., 2007). This is perhaps not surprising, giventhe high comorbidity between anxiety- and depression-related disorders, with approximately half of the patientsdiagnosed with major depressive disorder (MDD) also meetingcriteria for comorbid anxiety (Kennedy, 2008). In addition,approximately 50% of patients with generalised anxiety dis-order display symptoms of depression, while this level dropsdramatically for other anxiety disorders, such as post-trau-matic stress disorder and specific phobias (Hirschfeld, 2001;Kessler et al., 2006; Mackenzie et al., 2011). As in humans,behavioural alterations induced by the above mentionedsocial stressors persist for a long-time after the terminationof the stressor (Fuchs, 2005; Berton et al., 2006; Krishnanet al., 2007; Kennedy, 2008). Thus, such models allow us toenhance our knowledge of underlying mal-adaptationscaused by chronic stressor exposure. For example, chronicsocial defeat in mice was shown to result in an anxious-likephenotype, to induce an anhedonic state, and to reduce their
social approach, for at least one month after stress exposure.In contrast, behaviour in traditional depression tests such asthe forced swim test (FST) and tail suspension test (TST) werenot altered (Berton et al., 2006; Krishnan et al., 2007). Suchfindings demonstrate not only the comorbid phenotypeobserved after social stress exposure, but also the need inpreclinical models to assess multiple behavioural readouts.However, they also highlight a drawback of the models cur-rently used to assess stress-related psychopathologies. Inorder to really dissect the mechanisms underlying anxiety ordepression, animal models are needed, which specificallyinduce one phenotype. There are two main rationales for thisapproach: (i) despite the high comorbidity between anxietyand depression, there are a large percentage of patients whosuffer from only one or the other disease and (ii) such a model islikely to make analysis of the underlying pathophysiology ofthe disorder simpler. To date, only the social disruption modelhas been reported to cause increased anxiety in the absence ofenhanced depression-related behaviour (Kinsey et al., 2007).However, only FSTand TST, and not anhedonic-like behaviourswere assessed.
We have previously reported the physiological and immu-nological consequences of exposure to two chronic psycho-social stress paradigms: social defeat/overcrowding (SD/OC)and chronic subordinate colony housing (CSC) (Reber et al.,2006, 2007, 2008; Reber and Neumann, 2008; Veenema et al.,2008; Singewald et al., 2009; Schmidt et al., 2010; Reberet al., 2011). The SD/OC model combines exposure torepeated social defeat over 3 wk with periods of overcrowd-ing and single-housing, which adds a component of socialinstability. In contrast, the CSC model consists in housing 4experimental mice together with a dominant, and larger,resident for 3 wks. Thus, while CSC reflects a continuous formof chronic stress, SD/OC rather mimics an intermittentchronic stressor, but both cause numerous alterations instress-related physiological and immunological parameters(Reber et al., 2006, 2007, 2008; Veenema et al., 2008;Singewald et al., 2009; Schmidt et al., 2010; Reber et al.,2011). Interestingly, the increased level of pro-inflammatorycytokines observed following CSC is evident for at least 8 d(Reber et al., 2008). These results, coupled with those fromthe other chronic stress paradigms, suggest that it is likelythat SD/OC and CSC induce lasting behavioural changes. Inthis context it has repeatedly been demonstrated that CSCexposure leads to an anxious-like phenotype immediatelyafter stressor termination (Reber et al., 2007, 2008; Reberand Neumann, 2008; Veenema et al., 2008; Singewald et al.,2009). However, the effect of CSC on depression-relatedbehaviour remains unknown, and no studies have examinedthe behavioural consequences of SD/OC, at least in thecontext of a standard diet (Finger et al., 2011a,b).
Therefore, in the present study we aimed to assess theimpact of these two well-validated, clinically-relevant, psy-chosocial stress paradigms on both short- and long-termanxiety- and depression-related behaviour.
uncovering the molecular underpinnings of anxiety-related behaviour without conflicting depres-sion-based alterations.# 2011 Elsevier Ltd. All rights reserved.
Anxiety without depression 703

Author's personal copy
Materials and methods
Animals
Male C57BL/6 mice (Charles River, Sulzfeld, Germany) weigh-ing 19—22 g (experimental mice) or 30—35 g (dominant mice)were individually housed in standard cages for one weekbefore the experimental procedure started. All mice werekept under standard laboratory conditions (12-h light/darkcycle, lights on at 0600 h, 22 8C, 60% humidity) and had freeaccess to tap water and standard mouse diet. All experimentalprotocols were approved by the Committee on Animal Healthand Care of the local government, and performed according tointernational guidelines on the ethical use of animals.
Experimental procedure (see Fig. 1)
Experimental mice were chronically stressed by 19 d expo-sure to either the social defeat/overcrowding (SD/OC) para-digm (Reber et al., 2006) or the chronic subordinate colonyhousing (CSC) paradigm (Reber et al., 2007) or were kept asnon-stressed single housed controls (SHC). SHC, SD/OC, andCSC mice were exposed to either the saccharin preferencetest (SPT), the elevated plus-maze (EPM), or the socialpreference/avoidance test (SPAT) on day 20, to the TST orthe EPM on day 24, or the FST, EPM, or SPAT on day 28.Importantly, every behavioural test was performed in aseparate set of animals to avoid possible interactionsbetween the different tests. In another cohort of animals,24 h homecage locomotion was tracked on days �1/0, days20/21, and days 26/27, before mice were killed on day 28 toreveal changes in adrenal and pituitary weights. SD/OC andCSC mice that were tested/killed on day 24 or 28 were singlehoused after termination of respective chronic psychosocialparadigm (day 20 to day 24/28) (Fig. 1).
Chronic subordinate colony housing (CSC)paradigm
The CSC paradigm was conducted as described recently(Reber et al., 2007). Briefly, four experimental CSC mice
were housed together with a larger dominant male for 19consecutive days, in order to induce chronic psychosocialstress. Before the CSC procedure, the future dominant maleswere tested for their aggressive behaviour. Males that startedto injure their opponents by harmful bites were not used. Toavoid habituation during the chronic stressor exposure, eachlarger male was replaced by a novel larger male at days 8 and15.
Social defeat/overcrowding (SD/OC) paradigm
The SD/OC paradigm was conducted as described recently(Reber et al., 2006). Briefly, mice were exposed to unpre-dictable SD and OC during the 19-day stress period. SDconsisted in repeatedly placing mice into the home cageof a male resident for 2 h either once, or twice, a day. Inorder to avoid physical injuries, the two opponents wereseparated by a perforated partition wall immediately afterthe first attack of the resident, allowing visual, olfactory andauditory contact. The intruder was confronted with differentresidents to avoid habituation to the resident. During OC,performed on days 3, 10, 13, 14 and 18, large groups (n = 16)of experimental mice were housed together in one observa-tion cage (38 cm � 22 cm � 35 cm) for 24 h with free accessto water and food.
Body weight
To assess chronic psychosocial stress effects on body weightgain SHC, SD/OC and CSC mice were weighed on days 1, 3, 8,10, 15, 17, 19, 22, and 28.
Adrenal and pituitary weight
To assess chronic psychosocial stress effects on adrenal andpituitary weight, SHC, SD/OC and CSC mice were killed onday 28. After removal of the adrenals and the pituitary, theseorgans were pruned from fat and weighed. The left and rightadrenals were pooled for each animal. Values reportedrepresent absolute (mg) and relative to body weight (mg/g) adrenal and pituitary weights.
201-6 24 time [days]28-1 0 21 26 27
NOLDUSNOLDUSNOLDUS
SHC, SD/OC or CSCarrival
SPT TST FSTEPMSAT
EPM EPMSAT
KILLING
Figure 1 Experimental design. One week after arrival, experimental mice were chronically stressed by 19 d exposure to either thesocial defeat/overcrowding (SD/OC) paradigm or the chronic subordinate colony housing (CSC) paradigm or were kept as non-stressedsingle housed controls (SHC). SHC, SDOC, and CSC mice were exposed to either the saccharin preference test (SPT), the elevated plus-maze (EPM), or the social preference/avoidance test (SPAT) on day 20, to the tail suspension test (TST) or the EPM on day 24, or theforced swim test (FST), EPM, or SPATon day 28. Importantly, every behavioural test was performed in a separate set of animals to avoidpossible interactions between the different tests. In another set of animals 24 h homecage locomotion was tracked (using NOLDUSEthoVision system) on days �1/0, days 20/21, and days 26/27 before mice were killed on day 28 to reveal changes in adrenal andpituitary weights. SD/OC and CSC mice that were tested/killed on day 28 were single housed after termination of respective chronicpsychosocial paradigm (day 20 to day 28).
704 D.A. Slattery et al.

Author's personal copy
Home cage locomotion
To assess chronic psychosocial stress effects on locomotion,distance travelled [cm] in the home cage over 24 h wasrecorded on days �1/0, 20/21, and 26/27 in SHC, SD/OC,and CSC mice using EthoVision XT (Version 5.0.216, NoldusInformation Technology, Wageningen; the Netherlands). Dataare presented as mean + SEM of absolute distance travelledper 1 h-intervals and during last 6 h of light and first 6 h ofdark phase.
Elevated plus-maze (EPM) test
To assess chronic psychosocial stress effects on anxiety-related behaviour, SHC, SD/OC and CSC were exposedto the EPM test for 5 min (Pellow et al., 1985; Lister, 1987)on days 20, 24, of 28 as previously described (Reber andNeumann, 2008). EPM testing took place between 0800 hand 1100 h. The mouse plus-maze consists of two open(6 cm � 30 cm, 140 lx) and two closed (6 cm � 30 cm �17 cm, 25 lx) arms radiating from a central platform(6 cm � 6 cm) to form a plus-shaped figure elevated130 cm above the floor. The open arm edges were 0.3 cmin height to avoid falling. Each mouse was placed on thecentral platform facing a closed arm. The time spent on theopen and closed arms was recorded by means of a video/computer setup to allow calculation of the percentage oftime spent on the open arms of the maze. The number ofentries performed into the closed arms of the maze was takenas indicator for locomotor activity. The maze was cleanedthoroughly before each test.
Saccharin preference test (SPT)
To assess chronic psychosocial stress effects on anhedonicbehaviour, SHC, SD/OC and CSC mice were exposed to 0.05%(based on own unpublished data) saccharin solution (Sigma—Aldrich, Chemie GmbH, Steinheim, Germany) or tap waterover 24 h in a two-bottle choice tests on day 20/21 afterbeing single-housed. Both bottles were weighed before andafter the test. To exclude effects based on side preferencesbottles were switched every 3 h. Preference was calculatedbased on the following formula: [saccharin intake [g]/(sac-charin intake [g] + water intake [g])] � 100%.
Tail suspension test (TST)
To assess chronic psychosocial stress effects on depressive-like behaviour SHC, SD/OC and CSC mice were exposed to theTST (Steru et al., 1985; Swiergiel et al., 2008) on day 24.Briefly, the distal 1—2 cm of the mouse tail was taped to avertical bar, fixed on the ceiling of a plastic box (width:40 cm; depth: 40 cm; height: 80 cm), for 6 min. The durationof immobility, defined as passive hanging without anymotions, was recorded during the 6-min trial. The box wascleaned thoroughly before every test. TST testing took placebetween 0800 h and 1100 h. Predictive validity of the TSTwas confirmed by the anti-depressant imipramine in aprevious separate experiment. Imipramine (20 mg/kg; IP;Sigma—Aldrich, Chemie GmbH, Steinheim, Germany) orsaline were injected 20 min before the TST (n = 9 mice
per group). As expected (Bai et al., 2001) imipraminereduced the duration of immobility [controls: 260.06 �8.16 s (mean � S.E.M.), imipramine: 196.80 � 22.07 s,Mann—Whitney U test: P = 0.03].
Forced swim test (FST)
To assess chronic psychosocial stress effects on depressive-like behaviour, SHC, SD/OC and CSC mice were exposed to theFST (Porsolt et al., 1977; Bai et al., 2001) on day 28. Briefly,mice were placed in clear plastic cylinders (diameter: 12 cm;height: 40 cm) filled to 13 cm with 23 � 2 8C water for 6 min.The duration of immobility was recorded during the last 4 minof the 6-min trial. A mouse was regarded as immobile whenfloating motionless or making only those movements neces-sary to keep its head above the water. FS testing took placebetween 0800 h and 1100 h. Predictive validity of the FSTwasconfirmed by the anti-depressant imipramine in a previousseparate experiment. Imipramine (20 mg/kg; IP; Sigma—Aldrich, Chemie GmbH, Steinheim, Germany) or saline wereinjected 20 min before the FST (n = 9 mice per group). Asexpected (Bai et al., 2001) imipramine reduced the durationof immobility [controls: 161.00 � 5.48 s (mean � S.E.M.),imipramine: 101.89 � 17.78 s, Mann—Whitney U test:P = 0.04].
Social preference/avoidance test (SPAT)
To assess chronic psychosocial stress effects on social avoid-ance/anxiety behaviour SHC, SD/OC and CSC mice wereexposed to the SPAT (Haller and Bakos, 2002; Bertonet al., 2006) on days 20 or 28. Briefly, the experimentalmouse was placed into the SPAT box (length: 45 cm; width:27 cm; height: 27 cm; light intensity: 10—40 lx) for 30 s tohabituate to the unfamiliar environment before a smallempty wire mesh cage (length: 10 cm; width: 6.5 cm; height:5 cm) was introduced to the SPAT box for 150 s. This initialperiod gives an indication of general anxiety as it is exposureto a novel object. Afterwards, this empty cage wasexchanged for an identical cage containing an unfamiliarmale mouse for another 150 s. Total distance travelled andtime spent in the 8-cm broad contact zone (CZ) around thewire mesh cage was recorded using EthoVision XT (Version5.0.216, Noldus Information Technology, Wageningen; theNetherlands) during both 150 s trials. Social preference/avoidance was calculated based on the following formula:(time in CZ with mouse[s]/time in CZ with emptycage[s]) � 100%. A value of less than 100% was taken torepresent social avoidance, while a value above 100% wastaken to indicate social preference as used in similar studies(Berton et al., 2006). The box was cleaned thoroughly beforeevery test. The SPAT took place between 0800 h and 1100 h.
Predictive validity of the SPAT was confirmed by acute SDand the anti-depressant imipramine in a previous separateexperiment. After SD and 2 h of sensory contact with thedominant mouse, defeated mice were given imipramine(20 mg/kg; IP; Sigma—Aldrich, Chemie GmbH, Steinheim,Germany) or saline and 20 min later exposed to the SPAT(n = 8—11 mice per group). As expected the SD-inducedreduction in social preference was reversed by imipramine[SHC-saline: 130.38 � 10.67% (mean � S.E.M.), SHC-imipra-
Anxiety without depression 705

Author's personal copy
mine: 125.49 � 13.16%, SD-saline: 90.48 � 15.77% (P = 0.023vs. resp. SHC), SD-imipramine: 140.28 � 11.25% (P = 0.012vs. resp. saline), two-way ANOVA: factor SD � imipramineF1,35 = 4.60; P = 0.04].
Statistics
For statistical comparisons, the software package SPSS (ver-sion 12) was used. Data were compared using either one-wayANOVA (factor stress: SHC, SD/OC or CSC) or two-way ANOVAfor repeated measures (factor stress: SHC, SD/OC or CSC;factor time or mouse or light). Data are presented as mean +SEM. Significance was set at p � 0.05.
Results
SD/OC and CSC differentially alter body, adrenaland pituitary weight
As a first indication of a difference between the two stres-sors, we could reveal that SD/OC resulted in a reducedweight gain during stressor exposure (P < 0.001 vs. SHC;Fig. 2A; day 10: P = 0.024, day 15: P < 0.001, day 17:P < 0.001, day 19: P = 0.003; all vs. SHC; Fig. 2D) whereasCSC mice only showed a decreased body weight on day 3(P = 0.010 vs. SHC; Fig. 2D) and day 10 (P = 0.050 vs. SHC;
SHCSD/O
CCSC
body
wei
ght d
evel
opm
ent [
g]
0
1
2
3
4
5A
SHCSD/O
CCSC
0
1
2
3
4
5B
SHCSD/O
CCSC
0
1
2
3
4
5C
time [days]19171510831
body
wei
ght [
g]
022
23
24
25
26
27post stressD
***
###
******
* ##
*##
*(CSC)
*(SD/OC)***
*** **
###(CSC)
##
###(CSC)
Figure 2 Effects of chronic psychosocial stress on body weight development. To assess chronic psychosocial stress effects on bodyweight development, delta body weight gain was calculated for day 1 to day 20 (SHC: n = 94; SD/OC: n = 93; CSC: n = 94; A), for day 20to day 28 (SHC: n = 45; SD/OC: n = 45; CSC: n = 47; B), and for day 1 to day 28 (SHC: n = 45; SD/OC: n = 45; CSC: n = 47; C), during/following chronic psychosocial stressor exposure. Changes in absolute body weight from day 1 to day 20 (SHC: n = 94, white squares;SD/OC: n = 93, grey circles; CSC: n = 94, black triangles) are shown in D. Compared with SHC mice SD/OC mice showed a reduced bodyweight on day 10, 15, 17, and 19, whereas CSC mice only on day 3 and day 10. Data represent mean + SEM; *P � 0.05, **P � 0.01,***P � 0.001 vs. SHC mice; ##P � 0.01, ###P � 0.001 vs. SD/OC mice.
706 D.A. Slattery et al.

Author's personal copy
Fig. 2D) compared with SHC mice (factor stress:F (2,278) = 12.6, P < 0.001, one-way ANOVA, Fig. 2A; factorstress � time: F(12,1668) = 28.8, P < 0.001, two-way ANOVAfor repeated measures, Fig. 2D). Conversely, following stres-sor termination, both groups (SD/OC: P < 0.001; CSC:P < 0.001) gained more weight compared with SHC (factorstress: F (2,134) = 12.6, P < 0.001, one-way ANOVA, Fig. 2B),which compensated for the loss in the SD/OC group, but leadto an overall increase in the CSC (P = 0.020) compared withthe SHC group (factor stress: F (2,134) = 6.7, P = 0.002, one-way ANOVA, Fig. 2C).
Also, the absolute adrenal weight (F(2,27) = 8.0; P = 0.002,Fig. 3A) as well as absolute (F (2,25) = 7.5; P = 0.003, Fig. 3B)and relative (F (2,25) = 4.6; P = 0.020, Fig. 3D) pituitary weightwere differently affected by SD/OC and CSC. While CSCsignificantly increased absolute (P = 0.002) adrenal as wellas absolute (P = 0.008) and relative (P = 0.022) pituitaryweight on day 28, SD/OC did not.
SD/OC and CSC interfere with diurnal home-cageactivity patterns
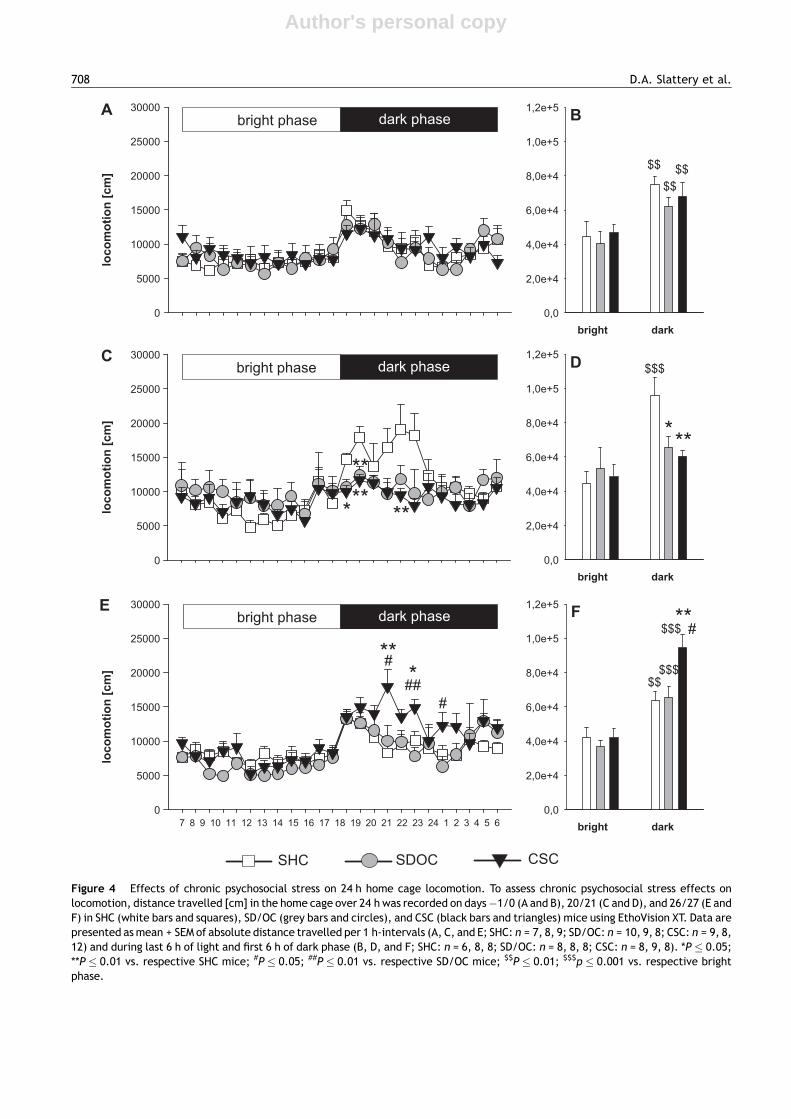
Since almost all of the anxiety and depression tests per-formed in the current study (except SPT) depend on thelocomotor activity of the mice and since exposure to stresshas been shown to disrupt circadian rhythms (Wulff et al.,2010), we assessed the effect of stress exposure on homecage locomotion before (day �1/1, Fig. 4A and B), immedi-ately after (day 20/21, Fig. 4C and D) and one week after (day26/27, Fig. 4E and F) stressor exposure. Importantly, whileSHC (P = 0.002), SD/OC (P = 0.008), and CSC (P = 0.007) miceshowed the expected increase in home cage locomotion atthe beginning of the dark phase prior to stress (factor light:F (1,19) = 31.0, P < 0.001, two-way ANOVA for repeated mea-sures, Fig. 4B; factor light: F(23,529) = 7.7, P < 0.001, two-wayANOVA for repeated measures, Fig. 4A), both stress para-digms prevented this increase as assessed on day 20/21. Thiswas indicated by a significantly decreased distance travelled
in CSC and SD/OC mice (factor light � stress: F(46,506) = 1.4,P = 0.047, two-way ANOVA for repeated measures, Fig. 4C)from 1900 h to 2000 h (P = 0.025 CSC vs. SHC), 2000 h to2100 h (P = 0.002 CSC vs. SHC; P = 0.007 CSC vs. SD/OC),and 2300 h to 0000 h (P = 0.008 CSC vs. SHC). Furthermore,in contrast to SHC mice (P = 0.001; factor light � stress:F(2,22) = 3.5, P = 0.047, two-way ANOVA for repeated mea-sures, Fig. 4D) CSC and SD/OC mice did not show an increasein the total distance travelled during the first 6 h of the darkphase and, thus, travelled significantly less compared withSHC mice (P = 0.007 CSC vs. SHC; P = 0.029 SD/OC vs. SHC).However, one week later, while SD/OC mice had recovered anormal diurnal activity pattern, CSC mice exhibited hyper-locomotion in the first hours of the dark phase, indicated by asignificantly increased distance travelled from 2200 h to2300 h (P = 0.011 vs. SHC, P = 0.054 vs. SD/OC; factor light -� stress: F (46,598) = 1.7, P = 0.005, two-way ANOVA forrepeated measures, Fig. 4E), 2400 h to 0100 h (P = 0.037vs. SHC, P = 0.002 vs. SD/OC), and 0200 h to 0300 h(P = 0.03 vs. SD/OC). In addition, compared with the SHC(P = 0.009) and SD/OC (P = 0.001) group, CSC mice showed asignificantly more pronounced increase in activity during thefirst 6 h of the dark phase (P = 0.011 vs. respective SHC,P = 0.016 vs. respective SD/OC; factor light � stress:F(2,21) = 4.4, P = 0.022, two-way ANOVA for repeated mea-sures, Fig. 4F). Therefore, as neither CSC nor SD/OC affectedlocomotor activity during the light phase, we decided to doall behavioural tests in the morning between 0800 h and1100 h.
SD/OC and CSC do not induce depression-relatedbehaviour
Since the most common phenotype reported followingchronic stress or exposure in rodents is an increase in depres-sion-related behaviour, we first examined depression-relatedbehaviour in three commonly used tests at different timepoints following stress exposure. Neither SD/OC nor CSC
SHCSD/O
CCSC
abso
lute
wei
ght [
mg]
0,0
3,0
3,5
4,0
4,5A adrenal
SHCSD/O
CCSC
rela
tive
wei
ght [
mg/
g]
0,000
0,100
0,125
0,150
0,175 C adrenal
SHCSD/O
CCSC
0,0
1,5
2,0
2,5
3,0B pituitary
SHCSD/O
CCSC
0,00
0,06
0,07
0,08
0,09
0,10 D pituitary
**## ****
Figure 3 Effects of chronic psychosocial stress on adrenal and pituitary weight. To assess chronic psychosocial stress effects onabsolute (A and B) and relative (C and D) adrenal (A and C) and pituitary (B and D) weight, SHC (n = 10, 9), SD/OC (n = 8, 9), and CSC(n = 12, 10) mice were killed one week following termination of chronic psychosocial stressor exposure on day 28. Afterwards thepituitary and both adrenal glands (pooled) were taken out, pruned from fat tissue, and weighed. Data represent mean + SEM;**P � 0.01 vs. SHC mice; ##P � 0.01 vs. SD/OC mice.
Anxiety without depression 707
Author's personal copy
SHC SDOC CSC
loco
mot
ion
[cm
]
0
5000
10000
15000
20000
25000
30000bright phase dark phaseA
loco
mot
ion
[cm
]
0
5000
10000
15000
20000
25000
30000bright phase dark phase
C
654321242322212019181716151413121110987
loco
mot
ion
[cm
]
0
5000
10000
15000
20000
25000
30000bright phase dark phase
E
**#
dark bright 0,0
2,0e+4
4,0e+4
6,0e+4
8,0e+4
1,0e+5
1,2e+5
dark bright 0,0
2,0e+4
4,0e+4
6,0e+4
8,0e+4
1,0e+5
1,2e+5
dark bright 0,0
2,0e+4
4,0e+4
6,0e+4
8,0e+4
1,0e+5
1,2e+5
##
B
D
F
$$
$$
#$$$
$$$
**
$$$
$$$$
*****
**
*#
***
Figure 4 Effects of chronic psychosocial stress on 24 h home cage locomotion. To assess chronic psychosocial stress effects onlocomotion, distance travelled [cm] in the home cage over 24 h was recorded on days �1/0 (A and B), 20/21 (C and D), and 26/27 (E andF) in SHC (white bars and squares), SD/OC (grey bars and circles), and CSC (black bars and triangles) mice using EthoVision XT. Data arepresented as mean + SEM of absolute distance travelled per 1 h-intervals (A, C, and E; SHC: n = 7, 8, 9; SD/OC: n = 10, 9, 8; CSC: n = 9, 8,12) and during last 6 h of light and first 6 h of dark phase (B, D, and F; SHC: n = 6, 8, 8; SD/OC: n = 8, 8, 8; CSC: n = 8, 9, 8). *P � 0.05;**P � 0.01 vs. respective SHC mice; #P � 0.05; ##P � 0.01 vs. respective SD/OC mice; $$P � 0.01; $$$p � 0.001 vs. respective brightphase.
708 D.A. Slattery et al.

Author's personal copy
altered depression-related behaviour at any time-point(Fig. 5). In more detail, neither SD/OC nor CSC mice exhib-ited altered anhedonic-like behaviour, as assessed using theSPTon day 20 (Fig. 5A). Moreover, no alteration in immobilitybehaviour in the widely-used tests of depression-relatedbehaviour, the TST (day 24) or the FST (day 28), was seen(Fig. 5B and C).
SD/OC and CSC result in a temporally-differentincrease in anxiety-related behaviour
Given the results in the depression tests, we next exam-ined whether both chronic psychosocial stressors applied inthe current study may specifically cause anxiety, without
inducing depression. In agreement with our earlier findings(Reber et al., 2007, 2008; Reber and Neumann, 2008;Veenema et al., 2008; Singewald et al., 2009), weobserved an anxious-like phenotype immediately followingCSC exposure (P = 0.001 vs. SHC; F (2,133) = 7.1, P = 0.001,Fig. 6A), which was still evident four (P = 0.026 vs. SHC;F (2,29) = 4.1, P = 0.027, Fig. 6B) and eight days later(P = 0.044 vs. SHC; F (2,29) = 5.2, P = 0.011, Fig. 6C). Con-trastingly, SD/OC exposure did not induce anxiety imme-diately following stressor termination. However, atendency was observed four days later (day 24, Fig. 6B),which reached significance on day 28 (P = 0.024 vs. SHC,Fig. 6C). An increase in anxiety-related behaviour wasreflected by a decreased percentage of time spent on
SHCSD/O
CCSC
time
imm
obile
[s]
0
60
120
180
240
300
360
SHCSD/O
CCSC
sacc
harin
pre
fere
nce
[%]
0
20
40
60
80
100
SHCSD/O
CCSC
0
60
120
180
240BA C
time
imm
obile
[s]
Figure 5 Effects of chronic psychosocial stress on depression-like behaviour. To assess chronic psychosocial stress effects ondepression-like behaviour a different set of SHC (n = 19, 18, 18), SD/OC (n = 12, 11, 18), and CSC (n = 7, 7, 18) mice was exposed to thesaccharin preference test (SPT) on day 20/21 (A), to the tail suspension test (TST) on day 24 (B), and to the forced swim test (FST) onday 28 (C). Data represent mean + SEM.
SHCSD/O
CCSC
0
10
20
30
40
50
60
*
SHCSD/O
CCSC
0
10
20
30
40
50
60
**
Exp-30/31
SHCSD/O
CCSC
% ti
me
on o
pen
arm
s
0
10
20
30
40
50
60
***
A B C
Figure 6 Effects of chronic psychosocial stress on anxiety-related behaviour. To assess chronic psychosocial stress effects on anxiety-related behaviour a different set of SHC (n = 56, 16, 13), SD/OC (n = 56, 8, 8), and CSC (n = 24, 8, 11) mice was exposed to the elevatedplus-maze (EPM) on day 20 (A), day 24 (B), and day 28 (C). Data represent mean + SEM. *P � 0.05; ***P � 0.001 vs. respective SHC mice.
Anxiety without depression 709
Author's personal copy
the open arms of the EPM. Importantly, no differences inclosed arm entries reflecting locomotion were observed(data not shown; see also Fig. 4C and E).
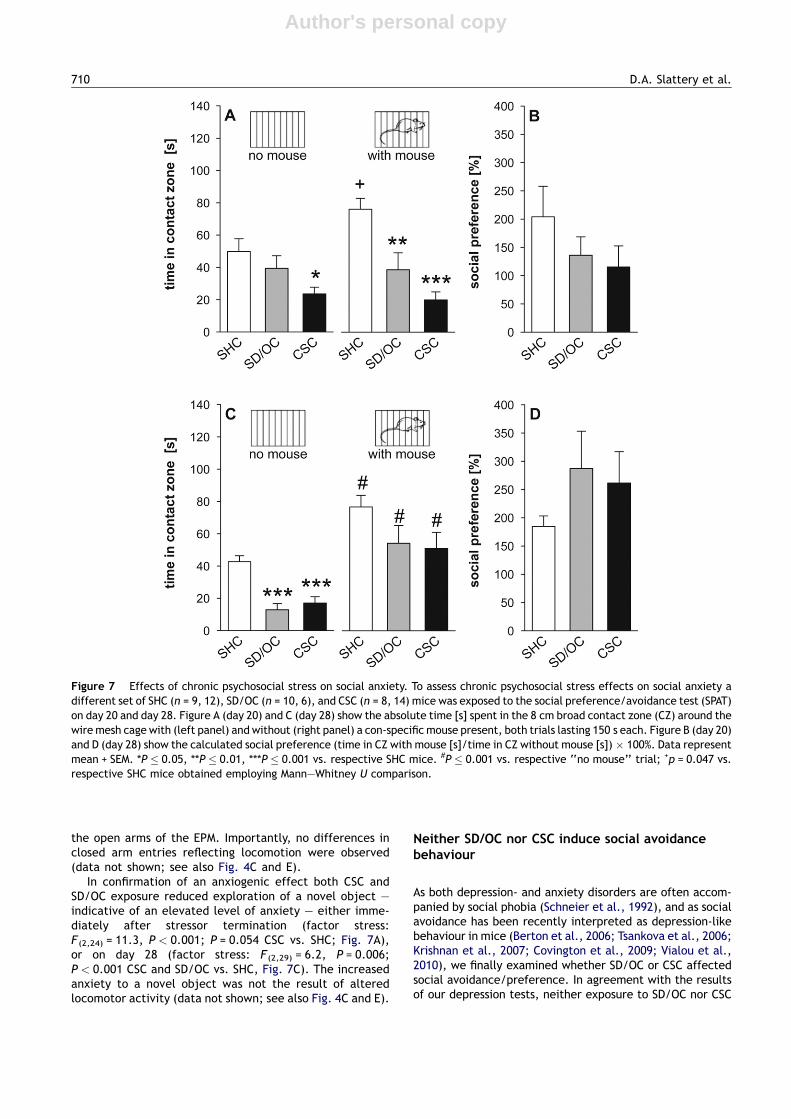
In confirmation of an anxiogenic effect both CSC andSD/OC exposure reduced exploration of a novel object —indicative of an elevated level of anxiety — either imme-diately after stressor termination (factor stress:F (2,24) = 11.3, P < 0.001; P = 0.054 CSC vs. SHC; Fig. 7A),or on day 28 (factor stress: F (2,29) = 6.2, P = 0.006;P < 0.001 CSC and SD/OC vs. SHC, Fig. 7C). The increasedanxiety to a novel object was not the result of alteredlocomotor activity (data not shown; see also Fig. 4C and E).
Neither SD/OC nor CSC induce social avoidancebehaviour
As both depression- and anxiety disorders are often accom-panied by social phobia (Schneier et al., 1992), and as socialavoidance has been recently interpreted as depression-likebehaviour in mice (Berton et al., 2006; Tsankova et al., 2006;Krishnan et al., 2007; Covington et al., 2009; Vialou et al.,2010), we finally examined whether SD/OC or CSC affectedsocial avoidance/preference. In agreement with the resultsof our depression tests, neither exposure to SD/OC nor CSC
Figure 7 Effects of chronic psychosocial stress on social anxiety. To assess chronic psychosocial stress effects on social anxiety adifferent set of SHC (n = 9, 12), SD/OC (n = 10, 6), and CSC (n = 8, 14) mice was exposed to the social preference/avoidance test (SPAT)on day 20 and day 28. Figure A (day 20) and C (day 28) show the absolute time [s] spent in the 8 cm broad contact zone (CZ) around thewire mesh cage with (left panel) and without (right panel) a con-specific mouse present, both trials lasting 150 s each. Figure B (day 20)and D (day 28) show the calculated social preference (time in CZ with mouse [s]/time in CZ without mouse [s]) � 100%. Data representmean + SEM. *P � 0.05, **P � 0.01, ***P � 0.001 vs. respective SHC mice. #P � 0.001 vs. respective ‘‘no mouse’’ trial; +p = 0.047 vs.respective SHC mice obtained employing Mann—Whitney U comparison.
710 D.A. Slattery et al.

Author's personal copy
resulted in avoidance of a novel con-specific at either timepoint assessed. Although a two-way ANOVA for repeatedmeasures failed to determine a significant over all effectof a stimulus mouse being present on day 20, separateanalyses employing Mann—Whitney U comparisons revealedthat SHC (P = 0.047) but not SD/OC and CSC mice preferredthe cage with a con-specific mouse inside to an empty one.However, this was a transient effect, as increased time spentwith the mouse compared with the empty cage (factormouse: F (1,29) = 52.6, P < 0.001, Fig. 7C) was found in SHC(P = 0.001), SD/OC (P = 0.001), and CSC (P = 0.001) mice onday 28.
Discussion
The main finding of the present study is that two well-validated psychosocial stress paradigms, SD/OC and CSC,result in increased anxiety-related but not depression-related behaviour. Moreover, the development of theseanxious-like states displays different temporal dynamics independence on the chronic stress paradigm. CSC exposureresults in increased anxiety immediately after stressor ter-mination, which lasts for at least 1 wk, while the anxious-likephenotype only develops 1 wk after SD/OC termination.Importantly, the observed behavioural alterations were nota consequence of stress-induced locomotor changes duringthe tests. Furthermore, no difference in home-cage activitywas observed during the bright-phase when the tests wereperformed. While mice subjected to both stress paradigmsdisplayed enhanced anxiety-like behaviour, they did notexhibit social avoidance of a novel con-specific; although atemporal lack of social preference was found. Moreover, bothstressors resulted in a flattened circadian rhythm immedi-ately after termination of the stressor, but this had reversedto hyperactivity in the dark-phase 1 wk later in CSC-exposedmice and had normalized in SD/OC mice. Taken together,these results demonstrate that the two models of chronicpsychosocial stress result in an anxious-like phenotype with-out comorbid depression-related behaviours in males. There-fore, both CSC and SD/OC represent attractive models todissect/differentiate the molecular underpinnings of anxi-ety- and depression-related symptoms.
Chronic stress exposure has been well-documented tolead to numerous physiological alterations includingdecreased weight gain, increased adrenal weight anddecreased thymus weight (Lucas et al., 2004; Choi et al.,2006; Savignac et al., 2010; Razzoli et al., 2011). Previously,we have reported that both CSC and SD/OC exposure lead todecreased body weight gain and increased adrenal weightduring the 19 d stress period (Reber et al., 2006, 2007).However, assessment of the body weight development duringand after stressor exposure in the current study, therebypooling all mice used in the different behavioural experi-ments (SHC: n = 94, CSC: n = 112, SD/OC: n = 93), revealedthat while there is an initial decrease in weight gain in CSCmice until day 10, this normalises over the final days ofstressor exposure (Fig. 2). Although decreased body weightgain has been shown in most studies investigating the effectsof repeated/chronic stress (Berton et al., 1998; Stefanski,1998; Keeney and Hogg, 1999; Zelena et al., 1999; Engler andStefanski, 2003), other studies have reported no alteration in
body weight development (Lucas et al., 2004; Choi et al.,2006; Savignac et al., 2010; Razzoli et al., 2011). In contrastto CSC, a reduced body weight gain over the 19 d of SD/OCexposure compared with SHC mice was observed, replicatingthe published SD/OC data (Reber et al., 2006). The weightloss observed in SD/OC animals is mainly a consequence ofthe overcrowding days. However, it should be noted thatwhile the weight recovers after stress termination, the dailySD during the paradigm is sufficient to prevent a recovery ofbody weight. Whereas a difference in weight gain wasobserved during the stress paradigms, both the CSC andthe SD/OC groups gained significantly more weight in theweek after stressor termination than unstressed controls.Similar findings have previously been reported followingsubjection to the visible burrow system (Melhorn et al.,2010) and repeated social defeat (Razzoli et al., 2011). Thisincrease in body weight after termination of chronic stressmay be an adaptive mechanism for ensuring sufficientresources in preparation for subsequent stressful eventsand, therefore, represent a general phenomenon followingprolonged stressor exposure.
Exposure to both SD/OC and CSC has been shown to result inincreased adrenal weight immediately following stressor ter-mination (Reber et al., 2006, 2007), which conforms with otherforms of chronic psychosocial stress (Blanchard et al., 1993;Berton et al., 1998; Zelena et al., 1999; Schmidt et al., 2007).However, when assessed on day 28, i.e. 1 wk after stressortermination, only mice exposed to the CSC paradigm main-tained enlarged adrenals (Fig. 3). Despite the lack of a generalbody weight decrease, this clearly confirms the CSC paradigmas a reliable chronic stress model. Although not as commonlystudied as the adrenal gland, alterations in pituitary morphol-ogy have previously been reported following chronic stressexposure (Theogaraj et al., 2005; Kapitonova et al., 2010).Here, we reveal, that only CSC and not SD/OC exposure leadsto an increase in both the absolute and relative pituitaryweight determined 1 wk following termination of stressorexposure (Fig. 3). Thus, CSC results in more severe, andlong-lasting, alterations in HPA axis morphology than SD/OC; although it remains to be determined whether CSC andSD/OC lead to an alteration in pituitary morphology immedi-ately after stressor termination. Taken together, the physio-logical parameters assessed demonstrate partly differentialalterations induced by SD/OC and CSC in different temporalpatterns. Moreover, they demonstrate that reliance purely onalterations in body weight as an indicator of successful stressorexposure may lead to erroneous interpretations.
Almost all of the tests that are employed to assess altera-tions in depression- or anxiety- related behaviour rely onlocomotor activity. This can be a confounding factor if themanipulation being studied, be it pharmacological or envir-onmental, leads to alterations in general activity (Strekalovaet al., 2005; Cryan and Slattery, 2007). Therefore, prior toperforming any depression and anxiety behavioural tests, wemonitored 24 h home-cage activity prior to (day �1/0),immediately after (day 20/21) and one week (day 26/27)after stressor exposure. While no differences in basal activitywere observed, 19 d SD/OC or CSC exposure resulted in aflattened circadian-activity rhythm, mediated by a reducedactivity during the active (dark) phase (Fig. 4A—D). Whenreassessed one week later, the activity pattern of SD/OC micehad resumed that of control animals, but mice exposed to
Anxiety without depression 711

Author's personal copy
CSC exhibited a hyper-locomotion during the early hours ofthe dark phase (Fig. 4E and F). Similar flattening of thecircadian rhythm have previously been reported followingchronic stress (Mastorci et al., 2009) and may reflect aconservation of energy following stressor exposure. Impor-tantly, at all time-points assessed no differences in activitywere observed during the bright-phase. Therefore, weelected to perform all of the behavioural tests in thebright-phase to reduce the likelihood that differences inlocomotor activity could underlie any observed behaviouralchanges.
In the literature, the majority of social stress paradigmslead to both increased depression- and anxiety-related beha-viour (Keeney and Hogg, 1999; Blanchard et al., 2001; Bertonet al., 2006; Krishnan et al., 2007; Schmidt et al., 2007).However, to gain better insight into the underlying mechan-isms of these disorders, animal models that lead to either adepressed or anxious phenotype are sorely warranted. Here wecan demonstrate that both the SD/OC as well as CSC paradigmsrepresent such models. Neither stress paradigm resulted in analteration in depression-related behaviour as assessed using 3relevant tests: the SPT (day 20), the TST (day 24), and the FST(day 28; Fig. 5). The use of more than one behavioural test isimportant, as, for example, GABAB receptor knockout micewere shown to display depression-related behaviour in the FST,but not TST (Cryan and Slattery, 2010). Similarly, exposure to10 d social defeat did not alter FSTor TST behaviour, but lead toan anhedonic phenotype (Krishnan et al., 2007). We employedseparate cohorts for each test in order to prevent thatrepeated behavioural testing of the animal affects subsequentbehavioural performance (although see Chourbaji et al.,2008). Therefore, the independent and repeated demonstra-tion of unchanged depression-related behaviour strengthensthe conclusion that neither SD/OC nor CSC result in a depres-sive phenotype. However, in terms of the SD/OC paradigm thisoutcome may be dependent on the diet (Finger et al., 2011b),as animals subjected to a low, but not high, fat diet developeddepression.
We next determined whether SD/OC led to alterationsin anxiety-related behaviour, in a similar manner to thatdocumented immediately following CSC (Reber et al.,2007, 2008; Reber and Neumann, 2008; Veenema et al.,2008; Singewald et al., 2009). We could initially replicatethe increase in anxiety-related behaviour on the EPMimmediately following CSC termination and expand thisto show that anxious-like phenotype lasted for up to 1 wk(Fig. 6). However, while there was no initial effect of SD/OC exposure on anxiety-related behaviour, a gradualincrease was observable, which led to an anxious-likephenotype 1 wk after stress termination (Fig. 6). Thus,although both CSC and SD/OC result in similar anxiogeniceffects, in the absence of depressogenic effects, the phe-notypes exhibit differential temporal dynamics. To thebest of our knowledge, the only other repeated andsocial-based stress paradigm that has been reported toresult solely in an anxious-like phenotype is the socialdisruption paradigm. However, this model is only 6 d induration, and while no differences in FST or TST behaviourwere documented, it remains possible that alterations inanhedonia are present (Kinsey et al., 2007).
Recent studies assessing the effects of chronic psychoso-cial stress on behavioural outcomes have employed the SPAT
test, in which a decrease in social interest and lack of socialpreference has been reported to reflect enhanced depres-sion-related behaviour (Berton et al., 2006; Krishnan et al.,2007). However, decreased interest in the exploration of con-specifics may be more correctly interpreted as the display ofsocial avoidance or social anxiety (Kalueff et al., 2006). As weobserved the anxiety-related phenotypes on day 20 (CSC) andday 28 (CSC and SD/OC), we performed the SPATon these daysonly. Validating the test, control mice were observed tospend a longer time exploring a novel con-specific on bothtest days, indicating social preference. In contrast, miceexposed to SD/OC or CSC spent a similar time investigatingthe novel object (empty cage) and social contact (cage withthe unknown mouse), indicating a general lack of socialpreference immediately after termination of psychosocialstressor exposure. However, unlike chronic social defeat, SD/OC or CSC mice did not show active social avoidance i.e. lesstime investigating the social context than the non-socialcontext (Berton et al., 2006; Krishnan et al., 2007; Krishnanand Nestler, 2008; Covington et al., 2009). This is largely inagreement with the very recent studies by Cryan and col-leagues (Finger et al., 2011a,b). Strain differences, diet, orresident aggressiveness in the different laboratories mayunderlie the subtle differences observed. When assessed1 wk after stress or termination, despite their anxious-likephenotype, both SD/OC and CSC mice preferred to explorethe novel con-specific indicating that the deficit in socialpreference is rather transient and reverses after 1 wk ofsingle housing. In agreement with Kalueff et al. (2006),the initial lack of social preference following CSC and SD/OC is likely to reflect a temporary social deficit rather thandepressive-like behaviour, particularly as the mice did notdisplay depression-related behaviour in the other testsemployed in the current study (SPT, TST and FST). Interest-ingly, in the initial phase of the SPAT test, exposure to thenovel object can be used to determine anxiety-relatedbehaviour. Recapitulating the data from the EPM, we couldshow that CSC mice spent less time exploring the novel objecton both day 20 and day 28, while SD/OC mice only spent lesstime at the novel object on day 28, compared with controls(Fig. 7). Given that stress-related disorders are more pre-valent in females than males, it would be of interest in futurestudies to determine whether such behavioural phenotypescan be generated by chronic psychosocial stressors in femalesor are specific to males.
In conclusion, we have demonstrated that the SD/OC andCSC psychosocial stress paradigms result in increased levelsof anxiety without simultaneously increasing depression-related behaviour. This appears to be specific to the presentmodels as the majority of social defeat/subordination-basedmodels generally lead to both an anxious and depressedphenotype. Therefore, both models represent unique modelsfor uncovering the molecular underpinnings of anxiety-related behaviour without conflicting depression-basedchanges. However, differential underlying causes ofincreased anxiety in SD/OC and CSC mice are to be expected,as the physiological outcomes of SD/OC and CSC do notresemble each other, and the anxious-like phenotypes followdifferent temporal dynamics. Thus, future studies will inves-tigate the molecular and neuronal adaptations caused by theSD/OC and CSC paradigms to determine the causes of theiranxious-like phenotype.
712 D.A. Slattery et al.

Author's personal copy
Conflicts of interest
The authors declare that they have no conflicts of interest.
Funding
This study was partly funded by the DFG (RE2911/5—1).
References
Amstislavskaya, T.G., Kudryavtseva, N.N., 1997. Effect of repeatedexperience of victory and defeat in daily agonistic confrontationson brain tryptophan hydroxylase activity. FEBS Lett. 406, 106—108.
Avgustinovich, D.F., Gorbach, O.V., Kudryavtseva, N.N., 1997. Com-parative analysis of anxiety-like behavior in partition and plus-maze tests after agonistic interactions in mice. Physiol. Behav.61, 37—43.
Bai, F., Li, X., Clay, M., Lindstrom, T., Skolnick, P., 2001. Intra- andinterstrain differences in models of ‘‘behavioral despair’’. Phar-macol. Biochem. Behav. 70, 187—192.
Bailey, M.T., Dowd, S.E., Parry, N.M.A., Galley, J.D., Schauer, D.B.,Lyte, M., 2010. Stressor exposure disrupts commensal microbialpopulations in the intestines and leads to increased colonizationby citrobacter rodentium. Infect. Immun. 78, 1509—1519.
Bailey, M.T., Engler, H., Sheridan, J.F., 2006. Stress induces the trans-location of cutaneous and gastrointestinal microflora to secondarylymphoid organs of C57BL/6 mice. J. Neuroimmunol. 171, 29—37.
Bartolomucci, A., Palanza, P., Costoli, T., Savani, E., Laviola, G.,Parmigiani, S., Sgoifo, A., 2003. Chronic psychosocial stresspersistently alters autonomic function and physical activity inmice. Physiol. Behav. 80, 57—67.
Bartolomucci, A., Palanza, P., Gaspani, L., Limiroli, E., Panerai, A.E.,Ceresini, G., Poli, M.D., Parmigiani, S., 2001. Social status inmice: behavioral, endocrine and immune changes are contextdependent. Physiol Behav 73, 401—410.
Berton, O., Aguerre, S., Sarrieau, A., Mormede, P., Chaouloff, F.,1998. Differential effects of social stress on central serotonergicactivity and emotional reactivity in Lewis and spontaneouslyhypertensive rats. Neuroscience 82, 147—159.
Berton, O., McClung, C.A., DiLeone, R.J., Krishnan, V., Renthal, W.,Russo, S.J., Graham, D., Tsankova, N.M., Bolanos, C.A., Rios, M.,Monteggia, L.M., Self, D.W., Nestler, E.J., 2006. Essential role ofBDNF in the mesolimbic dopamine pathway in social defeat stress.Science 311, 864—868.
Blanchard, D.C., Sakai, R.R., McEwen, B., Weiss, S.M., Blanchard,R.J., 1993. Subordination stress: behavioral, brain, and neuroen-docrine correlates. Behav. Brain Res. 58, 113—121.
Blanchard, D.C., Spencer, R.L., Weiss, S.M., Blanchard, R.J., McE-wen, B., Sakai, R.R., 1995. Visible burrow system as a model ofchronic social stress: behavioral and neuroendocrine correlates.Psychoneuroendocrinology 20, 117—134.
Blanchard, R.J., Yudko, E., Dulloog, L., Blanchard, D.C., 2001.Defense changes in stress nonresponsive subordinate males in avisible burrow system. Physiol. Behav. 72, 635—642.
Choi, D.C., Nguyen, M.M., Tamashiro, K.L., Ma, L.Y., Sakai, R.R.,Herman, J.P., 2006. Chronic social stress in the visible burrowsystem modulates stress-related gene expression in the bednucleus of the stria terminalis. Physiol. Behav. 89, 301—310.
Chourbaji, S., Brandwein, C., Vogt, M.A., Dormann, C., Gass, P.,2008. Evaluation of effects of previous exposure to an acutestressor before testing for depression-like behaviours in mice.Stress 11, 170—175.
Covington III, H.E., Maze, I., LaPlant, Q.C., Vialou, V.F., Ohnishi, Y.N.,Berton, O., Fass, D.M., Renthal, W., Rush III, A.J., Wu, E.Y.,Ghose, S., Krishnan, V., Russo, S.J., Tamminga, C., Haggarty,
S.J., Nestler, E.J., 2009. Antidepressant actions of histone dea-cetylase inhibitors. J. Neurosci. 29, 11451—11460.
Cryan, J.F., Slattery, D.A., 2007. Animal models of mood disorders:recent developments. Curr. Opin. Psychiatry. 20, 1—7.
Cryan, J.F., Slattery, D.A., 2010. GABAB receptors and depression.Current status. Adv. Pharmacol. (San Diego, Calif) 58, 427—451.
De Goeij, D.C., Dijkstra, H., Tilders, F.J., 1992. Chronic psychosocialstress enhances vasopressin, but not corticotropin-releasing fac-tor, in the external zone of the median eminence of male rats:relationship to subordinate status. Endocrinology 131, 847—853.
Denmark, A., Tien, D., Wong, K., Chung, A., Cachat, J., Goodspeed, J.,Grimes, C., Elegante, M., Suciu, C., Elkhayat, S., Bartels, B.,Jackson, A., Rosenberg, M., Chung, K.M., Badani, H., Kadri, F.,Roy, S., Tan, J., Gaikwad, S., Stewart, A., Zapolsky, I., Gilder, T.,Kalueff, A.V., 2010. The effects of chronic social defeat stress onmouse self-grooming behavior and its patterning. Behav. Brain Res.208, 553—559.
Engler, H., Bailey, M.T., Engler, A., Stiner-Jones, L.M., Quan, N.,Sheridan, J.F., 2008. Interleukin-1 receptor type 1-deficient micefail to develop social stress-associated glucocorticoid resistancein the spleen. Psychoneuroendocrinology 33, 108—117.
Engler, H., Engler, A., Bailey, M.T., Sheridan, J.F., 2005. Tissue-specific alterations in the glucocorticoid sensitivity of immunecells following repeated social defeat in mice. J. Neuroimmunol.163, 110—119.
Engler, H., Stefanski, V., 2003. Social stress and T cell maturation inmale rats: transient and persistent alterations in thymic function.Psychoneuroendocrinology 28, 951—969.
Finger, B.C., Dinan, T.G., Cryan, J.F., 2011a. The temporal impact ofchronic intermittent psychosocial stress on high-fat diet-inducedalterations in body weight. Psychoneuroendocrinology,doi:10.1016/j.psyneuen.2011.06.015, in press.
Finger, B.C., Dinan, T.G., Cryan, J.F., 2011b. High-fat diet selectivelyprotects against the effects of chronic social stress in the mouse.Neuroscience, doi:10.1016/j.neuroscience.2011.06.072, in press.
Fuchs, E., 2005. Social stress in tree shrews as an animal model ofdepression: an example of a behavioral model of a CNS disorder.CNS Spectr. 10, 182—190.
Fuchs, E., Flugge, G., 2003. Chronic social stress: effects on limbicbrain structures. Physiol. Behav. 79, 417—427.
Haller, J., Bakos, N., 2002. Stress-induced social avoidance: a newmodel of stress-induced anxiety? Physiol. Behav. 77, 327—332.
Hirschfeld, R.M., 2001. The comorbidity of major depression andanxiety disorders: recognition and management in primary care.Prim. Care Companion J. Clin. Psychiatry 3, 244—254.
Kalueff, A.V., Avgustinovich, D.F., Kudryavtseva, N.N., Murphy,D.L., 2006. BDNF in anxiety and depression. Science 312,1598—1599.
Kapitonova, M.Y., Kuznetsov, S.L., Khlebnikov, V.V., Zagrebin, V.L.,Morozova, Z., Degtyar, Y.V., 2010. Immunohistochemical charac-teristics of the hypophysis in normal conditions and chronic stress.Neurosci. Behav. Physiol. 40, 97—102.
Keeney, A.J., Hogg, S., 1999. Behavioural consequences of repeatedsocial defeat in the mouse: preliminary evaluation of a potentialanimal model of depression. Behav. Pharmacol. 10, 753—764.
Keeney, A.J., Hogg, S., Marsden, C.A., 2001. Alterations in core bodytemperature, locomotor activity, and corticosterone followingacute and repeated social defeat of male NMRI mice. Physiol.Behav. 74, 177—184.
Kemme, K., Kaiser, S., Sachser, N., 2008. Prenatal stress does not impaircoping with challenge later in life. Physiol. Behav. 93, 68—75.
Kennedy, S.H., 2008. Core symptoms of major depressive disorder:relevance to diagnosis and treatment. Dialogues Clin. Neurosci.10, 271—277.
Kessler, R.C., Coccaro, E.F., Fava, M., Jaeger, S., Jin, R., Walters, E.,2006. The prevalence and correlates of DSM-IV intermittentexplosive disorder in the National Comorbidity Survey Replica-tion. Arch. Gen. Psychiatry 63, 669—678.
Anxiety without depression 713

Author's personal copy
Kinsey, S.G., Bailey, M.T., Sheridan, J.F., Padgett, D.A., Avitsur, R.,2007. Repeated social defeat causes increased anxiety-like be-havior and alters splenocyte function in C57BL/6 and CD-1 mice.Brain Behav. Immun. 21, 458—466.
Krishnan, V., Han, M.-H., Graham, D.L., Berton, O., Renthal, W.,Russo, S.J., LaPlant, Q., Graham, A., Lutter, M., Lagace, D.C.,Ghose, S., Reister, R., Tannous, P., Green, T.A., Neve, R.L.,Chakravarty, S., Kumar, A., Eisch, A.J., Self, D.W., Lee, F.S.,Tamminga, C.A., Cooper, D.C., Gershenfeld, H.K., Nestler, E.J.,2007. Molecular adaptations underlying susceptibility and resis-tance to social defeat in brain reward regions. Cell 131, 391—404.
Krishnan, V., Nestler, E.J., 2008. The molecular neurobiology ofdepression. Nature 455, 894—902.
Lister, R.G., 1987. The use of a plus-maze to measure anxiety in themouse. Psychopharmacology (Berl.) 92, 180—185.
Lucas, L.R., Celen, Z., Tamashiro, K.L., Blanchard, R.J., Blanchard,D.C., Markham, C., Sakai, R.R., McEwen, B.S., 2004. Repeatedexposure to social stress has long-term effects on indirect mar-kers of dopaminergic activity in brain regions associated withmotivated behavior. Neuroscience 124, 449—457.
Mackenzie, C.S., Reynolds, K., Chou, K.L., Pagura, J., Sareen, J., 2011.Prevalence and correlates of generalized anxiety disorder in a na-tional sample of older adults. Am. J. Geriatr. Psychiatry 19, 305—315.
Mastorci, F., Vicentini, M., Viltart, O., Manghi, M., Graiani, G.,Quaini, F., Meerlo, P., Nalivaiko, E., Maccari, S., Sgoifo, A.,2009. Long-term effects of prenatal stress: changes in adultcardiovascular regulation and sensitivity to stress. Neurosci.Biobehav. Rev. 33, 191—203.
Melhorn, S.J., Krause, E.G., Scott, K.A., Mooney, M.R., Johnson,J.D., Woods, S.C., Sakai, R.R., 2010. Meal patterns and hypotha-lamic NPY expression during chronic social stress and recovery.Am. J. Physiol. 299, R813—R822.
Miczek, K.A., Yap, J.J., Covington 3rd., H.E., 2008. Social stress,therapeutics and drug abuse: preclinical models of escalated anddepressed intake. Pharmacol. Ther. 120, 102—128.
Pellow, S., Chopin, P., File, S.E., Briley, M., 1985. Validation of open:closed arm entries in an elevated plus-maze as a measure ofanxiety in the rat. J. Neurosci. Methods 14, 149—167.
Porsolt, R.D., Bertin, A., Jalfre, M., 1977. Behavioral despair in mice:a primary screening test for antidepressants. Arch. Int. Pharma-codyn. Ther. 229, 327—336.
Razzoli, M., Carboni, L., Andreoli, M., Ballottari, A., Arban, R., 2011.Different susceptibility to social defeat stress of BalbC andC57BL6/J mice. Behav. Brain Res. 216, 100—108.
Reber, S.O., Birkeneder, L., Veenema, A.H., Obermeier, F., Falk, W.,Straub, R.H., Neumann, I.D., 2007. Adrenal insufficiency andcolonic inflammation after a novel chronic psycho-social stressparadigm in mice: implications and mechanisms. Endocrinology148, 670—682.
Reber, S.O., Neumann, I.O., 2008. Chronic subordinate colony (CSC)housing enhances state anxiety and reduces hypothalamic argi-nine vasopressin, but not oxytocin, synthesis. Ann. N. Y. Acad. Sci.1148, 184—195.
Reber, S.O., Obermeier, F., Straub, R.H., Falk, W., Neumann, I.D.,2006. Chronic intermittent psychosocial stress (social defeat/over-crowding) in mice increases the severity of an acute DSS-inducedcolitis and impairs regeneration. Endocrinology 147, 4968—4976.
Reber, S.O., Obermeier, F., Straub, R.H., Veenema, A.H., Neumann,I.D., 2008. Aggravation of DSS-induced colitis after chronic sub-ordinate colony (CSC) housing is partially mediated by adrenalmechanisms. Int. J. Biol. Stress 11, 225—234.
Reber, S.O., Peters, S., Slattery, D.A., Hofmann, C., Scholmerich, J.,Neumann, I.D., Obermeier, F., 2011. Mucosal immunosuppressionand epithelial barrier defects are key events in murine psychoso-cial stress-induced colitis. Brain Behav. Immun. 25, 1153—1161.
Savignac, H.M., Hyland, N.P., Dinan, T.G., Cryan, J.F., 2010. Theeffects of repeated social interaction stress on behavioural and
physiological parameters in a stress-sensitive mouse strain.Behav. Brain Res. 216, 576—584.
Schmidt, D., Reber, S.O., Botteron, C., Barth, T., Peterlik, D.,Uschold, N., Mannel, D.N., Lechner, A., 2010. Chronic psychoso-cial stress promotes systemic immune activation and the devel-opment of inflammatory Th cell responses. Brain Behav. Immun.24, 1097—1104.
Schmidt, M.V., Sterlemann, V., Ganea, K., Liebl, C., Alam, S., Harbich,D., Greetfeld, M., Uhr, M., Holsboer, F., Muller, M.B., 2007. Persis-tent neuroendocrine and behavioral effects of a novel, etiologicallyrelevant mouse paradigm for chronic social stress during adoles-cence. Psychoneuroendocrinology 32, 417—429.
Schneier, F.R., Johnson, J., Hornig, C.D., Liebowitz, M.R., Weissman,M.M., 1992. Social phobia. Comorbidity and morbidity in anepidemiologic sample. Arch. Gen. Psychiatry 49, 282—288.
Singewald, G.M., Nguyen, N.K., Neumann, I.D., Singewald, N.,Reber, S.O., 2009. Effect of chronic psychosocial stress-inducedby subordinate colony (CSC) housing on brain neuronal activitypatterns in mice. Stress 12, 58—69.
Stefanski, V., 1998. Social stress in loser rats: opposite immunologicaleffects in submissive and subdominant males. Physiol. Behav. 63,605—613.
Stefanski, V., 2001. Social stress in laboratory rats: behavior, immunefunction, and tumor metastasis. Physiol. Behav. 73, 385—391.
Stefanski, V., Peschel, A., Reber, S., 2003. Social stress affectsmigration of blood Tcells into lymphoid organs. J. Neuroimmunol.138, 17—24.
Sterlemann, V., Ganea, K., Liebl, C., Harbich, D., Alam, S., Holsboer,F., Muller, M.B., Schmidt, M.V., 2008. Long-term behavioral andneuroendocrine alterations following chronic social stress inmice: Implications for stress-related disorders. Horm. Behav.53, 386—394.
Steru, L., Chermat, R., Thierry, B., Simon, P., 1985. The tail suspen-sion test: a new method for screening antidepressants in mice.Psychopharmacology (Berl.) 85, 367—370.
Strekalova, T., Spanagel, R., Dolgov, O., Bartsch, D., 2005. Stress-induced hyperlocomotion as a confounding factor in anxiety anddepression models in mice. Behav. Pharmacol. 16, 171—180.
Swiergiel, A.H., Leskov, I.L., Dunn, A.J., 2008. Effects of chronic andacute stressors and CRF on depression-like behavior in mice.Behav. Brain Res. 186, 32—40.
Theogaraj, E., John, C.D., Christian, H.C., Morris, J.F., Smith, S.F.,Buckingham, J.C., 2005. Perinatal glucocorticoid treatment pro-duces molecular, functional, and morphological changes in the ante-rior pituitary gland of the adult male rat. Endocrinology 146, 4804—4813.
Tsankova, N.M., Berton, O., Renthal, W., Kumar, A., Neve, R.L.,Nestler, E.J., 2006. Sustained hippocampal chromatin regulationin a mouse model of depression and antidepressant action. Nat.Neurosci. 9, 519—525.
Veenema, A.H., Reber, S.O., Selch, S., Obermeier, F., Neumann, I.D.,2008. Early life stress enhances the vulnerability to chronicpsychosocial stress and experimental colitis in adult mice. Endo-crinology 144, 2727—2736.
Vialou, V., Robison, A.J., LaPlant, Q.C., Covington, H.E., Dietz, D.M.,Ohnishi, Y.N., Mouzon, E., Rush, A.J., Watts, E.L., Wallace, D.L.,Iniguez, S.D., Ohnishi, Y.H., Steiner, M.A., Warren, B.L., Krishnan,V., Bolanos, C.A., Neve, R.L., Ghose, S., Berton, O., Tamminga,C.A., Nestler, E.J., 2010. [Delta]FosB in brain reward circuitsmediates resilience to stress and antidepressant responses. Nat.Neurosci. 13, 745—752.
Wulff, K., Gatti, S., Wettstein, J.G., Foster, R.G., 2010. Sleep andcircadian rhythm disruption in psychiatric and neurodegenerativedisease. Nat. Rev. 11, 589—599.
Zelena, D., Haller, J., Halasz, J., Makara, G.B., 1999. Social stress ofvariable intensity: physiological and behavioral consequences.Brain Res. Bull. 48, 297—302.
714 D.A. Slattery et al.




















