
Behavioral and Physiological Characteristics of the Antarctic Krill, Euphausia superba
16
AMER. ZOOL., 31:49-63 (1991) Behavioral and Physiological Characteristics of the Antarctic Krill, Euphausia superba 1 LANGDON B. QUETIN AND ROBIN M. ROSS Marine Science Institute, University of California at Santa Barbara, Santa Barbara, California 93106 SYNOPSIS. The antarctic krill, Euphausia superba, is considered a success in the intensely seasonal environment of the Southern Ocean because of its abundance and central role as an important food item for many of the larger carnivores in the ecosystem. The behavioral and physiological characteristics that foster this success are: (1) the ability to find concentrations of food in several types of habitat and efficiently exploit whatever food is available; (2) the close correspondence of the life cycle with seasonal cycles of food availability; and (3) a combination of physiological mechanisms that enable krill to survive the long winter period of low food availability. We evaluated the relative importance of the following four major winter-over mechanisms that have been proposed for adult krill west of the Antarctic Peninsula. The three-fold reduction in metabolic rate is the most important winter-over mechanism for these adults, although lipid utilization and shrinkage also help satisfy energy requirements in the winter. Alternate food sources did not appear to contribute significantly as a winter energy source. However, the extent, predictability and complexity of the ice cover in a region during winter may have a great influence on the relative importance of these winter-over mechanisms for different populations. Ice cover in the waters west of the Antarctic Peninsula is unpredictable and smooth surfaced when it occurs, providing the krill with little refuge from predation. In multi-year pack ice of the Weddell Sea, however, ice cover is predictable and extensive, and there is a complex undersurface that provides hiding places. In this multi-year ice, adult krill have been observed under the ice feeding, whereas west of the Antarctic Peninsula most adult krill are in the water column in the winter and are not feeding. The balance between acquiring energy and avoiding predation may be different in these two regions in the winter because of differences in predictability and complexity of the ice cover. INTRODUCTION primary production in several ways. First, The habitat of the antarctic krill, t h e s e a i c e cover limits the availability of Euphausia superba, is an environment dom- Vl S ht t o t h e phytoplankton in the water col- inated by extreme seasonal changes in day- umn > but lt also provides a habitat in which length, food availability and ice cover. In organisms can live and grow, close to what- areas where krill are most abundant, day- ever incident light is available. Despite low length ranges from constant light or near !ig ht levels, significant net primary pro- constant light in December to constant duction occurs in both the sea ice and the darkness or only a few hours of dusk in water column in late winter (Kottmeier and June. The amount of light reaching the Sullivan, 1987). Second, phytoplankton surface governs both the maximum amount production in both the water column and of energy available for photosynthesis and t h e s e a i c e increases dramatically in the the ambient temperature of the air and spring, as the melting ice creates vertical water. As temperatures decrease in the stability in the water column. Recent winter, sea ice forms—covering more than research has shown that as the ice edge 20 million km 2 of the sea surface at its max- retreats a series of blooms follows, contin- imum extent, five times the minimum in " in g fr om September until December. The spring and summer (Garrison and Siniff, increase in standing stock from these ice 1986). This annual advance and retreat of edge blooms extends to about 250 km from the ice-covered area affects total annual the ice edge (Smith et al, 1988). The pro- duction involved is substantial, and in some years can increase annual primary produc- • From the Symposium on Antarctic Marine Biology j estimates for the Southern Ocean by presented at the Annual Meeting of the American / Society of Zoologists, 27-30 December 1988, at San 60% (Smith and Nelson, 1986; Smith et al, Francisco, California. 1988). Open water phytoplankton blooms 49 at University of California, Santa Barbara on January 8, 2015 http://icb.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
14 -
download
0
Transcript of Behavioral and Physiological Characteristics of the Antarctic Krill, Euphausia superba

AMER. ZOOL., 31:49-63 (1991)
Behavioral and Physiological Characteristics of theAntarctic Krill, Euphausia superba1
LANGDON B. QUETIN AND ROBIN M. ROSS
Marine Science Institute, University of California at Santa Barbara,Santa Barbara, California 93106
SYNOPSIS. The antarctic krill, Euphausia superba, is considered a success in the intenselyseasonal environment of the Southern Ocean because of its abundance and central roleas an important food item for many of the larger carnivores in the ecosystem. Thebehavioral and physiological characteristics that foster this success are: (1) the ability tofind concentrations of food in several types of habitat and efficiently exploit whateverfood is available; (2) the close correspondence of the life cycle with seasonal cycles of foodavailability; and (3) a combination of physiological mechanisms that enable krill to survivethe long winter period of low food availability. We evaluated the relative importance ofthe following four major winter-over mechanisms that have been proposed for adult krillwest of the Antarctic Peninsula. The three-fold reduction in metabolic rate is the mostimportant winter-over mechanism for these adults, although lipid utilization and shrinkagealso help satisfy energy requirements in the winter. Alternate food sources did not appearto contribute significantly as a winter energy source. However, the extent, predictabilityand complexity of the ice cover in a region during winter may have a great influence onthe relative importance of these winter-over mechanisms for different populations. Icecover in the waters west of the Antarctic Peninsula is unpredictable and smooth surfacedwhen it occurs, providing the krill with little refuge from predation. In multi-year packice of the Weddell Sea, however, ice cover is predictable and extensive, and there is acomplex undersurface that provides hiding places. In this multi-year ice, adult krill havebeen observed under the ice feeding, whereas west of the Antarctic Peninsula most adultkrill are in the water column in the winter and are not feeding. The balance betweenacquiring energy and avoiding predation may be different in these two regions in thewinter because of differences in predictability and complexity of the ice cover.
INTRODUCTION primary production in several ways. First,The habitat of the antarctic krill, t h e s e a i c e c o v e r l i m i t s t h e availability of
Euphausia superba, is an environment dom- VlSht t o t h e phytoplankton in the water col-inated by extreme seasonal changes in day- umn> b u t lt a l s o provides a habitat in whichlength, food availability and ice cover. In organisms can live and grow, close to what-areas where krill are most abundant, day- ever incident light is available. Despite lowlength ranges from constant light or near !ight levels, significant net primary pro-constant light in December to constant duction occurs in both the sea ice and thedarkness or only a few hours of dusk in water column in late winter (Kottmeier andJune. The amount of light reaching the Sullivan, 1987). Second, phytoplanktonsurface governs both the maximum amount production in both the water column andof energy available for photosynthesis and t h e s e a i c e increases dramatically in thethe ambient temperature of the air and spring, as the melting ice creates verticalwater. As temperatures decrease in the stability in the water column. Recentwinter, sea ice forms—covering more than research has shown that as the ice edge20 million km2 of the sea surface at its max- retreats a series of blooms follows, contin-imum extent, five times the minimum in " i ng from September until December. Thespring and summer (Garrison and Siniff, increase in standing stock from these ice1986). This annual advance and retreat of edge blooms extends to about 250 km fromthe ice-covered area affects total annual the ice edge (Smith et al, 1988). The pro-
duction involved is substantial, and in someyears can increase annual primary produc-
• From the Symposium on Antarctic Marine Biology j estimates for the Southern Ocean bypresented at the Annual Meeting of the American /Society of Zoologists, 27-30 December 1988, at San 60% (Smith and Nelson, 1986; Smith et al,Francisco, California. 1988). Open water phytoplankton blooms
49
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

50 L. B. QUETIN AND R. M. Ross
peak from December through February (El-Sayed, 1985). Most investigators believethat primary production in the SouthernOcean is limited by wind-induced turbu-lence, not nutrients, and that productivityis higher in nearshore coastal regions andin regions of upwelling than in the openocean (El-Sayed, 1985; Smith et al, 1988).These pulses in primary production andtheir locations are predictable. Althoughprimary production is not zero in the win-ter (Kottmeier and Sullivan, 1987), theintense seasonality in food availabilitymeans that growth, reproduction and feed-ing in herbivores also must be seasonal, andthat organisms must be able to survive longperiods when food is scarce or non-exis-tent.
E. superba is a pelagic shrimp-like crus-tacean that grows to 50 to 60 mm in totallength and can swim as well as small fishsuch as anchovy or sardines (Hamner et al,1983). For most of its life cycle the antarc-tic krill occurs in the upper 200 m of thewater column in discrete schools or swarms,in densities up to 20,000 to 30,000 m"3.Schools vary in size from about 200 m inone horizontal dimension and severalmeters thick (Hamner, 1984) to the rare"super swarm" over 12 km long (Macaulayetal, 1984).
Although krill are circumpolar in distri-bution, high concentrations of krill arefound in only a few locations (Marr, 1962;Laws, 1985). Close to the antarctic conti-nent and its associated shelf and slope, thewinds come from the east, creating thecounterclockwise East Wind Drift. Theclockwise West Wind Drift forms a ring inthe northern part of the Southern Oceanbetween the Antarctic Convergence andthe East Wind Drift. Some krill are foundin parts of the West Wind Drift, but mostare concentrated within the area coveredby the annual advance and retreat of seaice, the East Wind Drift and its associatedgyres and eddies. This distribution is sug-gestive of a close coupling between ice edgeblooms and krill populations. It is impor-tant to recognize that krill are as muchcreatures of the ice edge and the semi-enclosed bays and straits where productiv-ity is the greatest as they are of the open
ocean. The largest concentration of E.superba is found in waters to the north andeast of the Antarctic Peninsula, continuingeast into the Scotia Sea and the WeddellSea gyre (Laws, 1985).
E. superba is a "success" from several dif-ferent vantage points. First, this one spe-cies is the most important "macrozoo-planktonic" herbivore in the SouthernOcean, unlike most other oceans wheremuch of the primary production is grazedby copepods (Clarke, 1985). Although krillare capable of ingesting animal prey (Priceet al., 1988), in the summer freshly col-lected krill have dark green hepatopan-creases, suggesting that the primary foodsource at this time is phytoplankton. Thehigh biomass of krill is present year-round,and can be over 50% of" the total zooplank-ton biomass in the epipelagic layer (Hop-kins, 1985). Second, the result of this year-round dominance is that krill are theprimary food source for a large number ofspecies (Laws, 1985), and may deserve tobe called a "keystone species" for theSouthern Ocean ecosystem. In fact, in theSouthern Ocean, most vertebrates directlydepend on krill as a food source (Fig. 1).
Originally the Southern Ocean was char-acterized as a simple linear food chain: dia-toms to krill to consumers. As we learnedmore about the interactions in this ecosys-tem, however, the simple food chain con-cept had to be replaced with a complexfood web (Fig. 1). Although the base of thefood web is still phytoplankton, both dia-toms and nanoplankton must be includedas primary producers. The food webincludes herbivores both smaller and larger(salps) than krill (Lancraft et al., 1989), asubstantial link to the benthos (Smith et al.,1988), and a role for the microplankton(Hewes etal, 1985). Krill are eaten by birds,including albatrosses, petrels and threespecies of penguins (Laws, 1985), bydemersal fish (Nototheniiformes) (Kock,1985) and pelagic fish (Williams, 1985), byfour species of seals (Laws, 1985; Siniff andStone, 1985),bysquid(Nemoto^a/., 1985)and by baleen shales (Laws, 1985). Krill arefrom 33 to over 90% of the diet of theselarger marine carnivores. Annual con-sumption of krill by these predators is esti-
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from
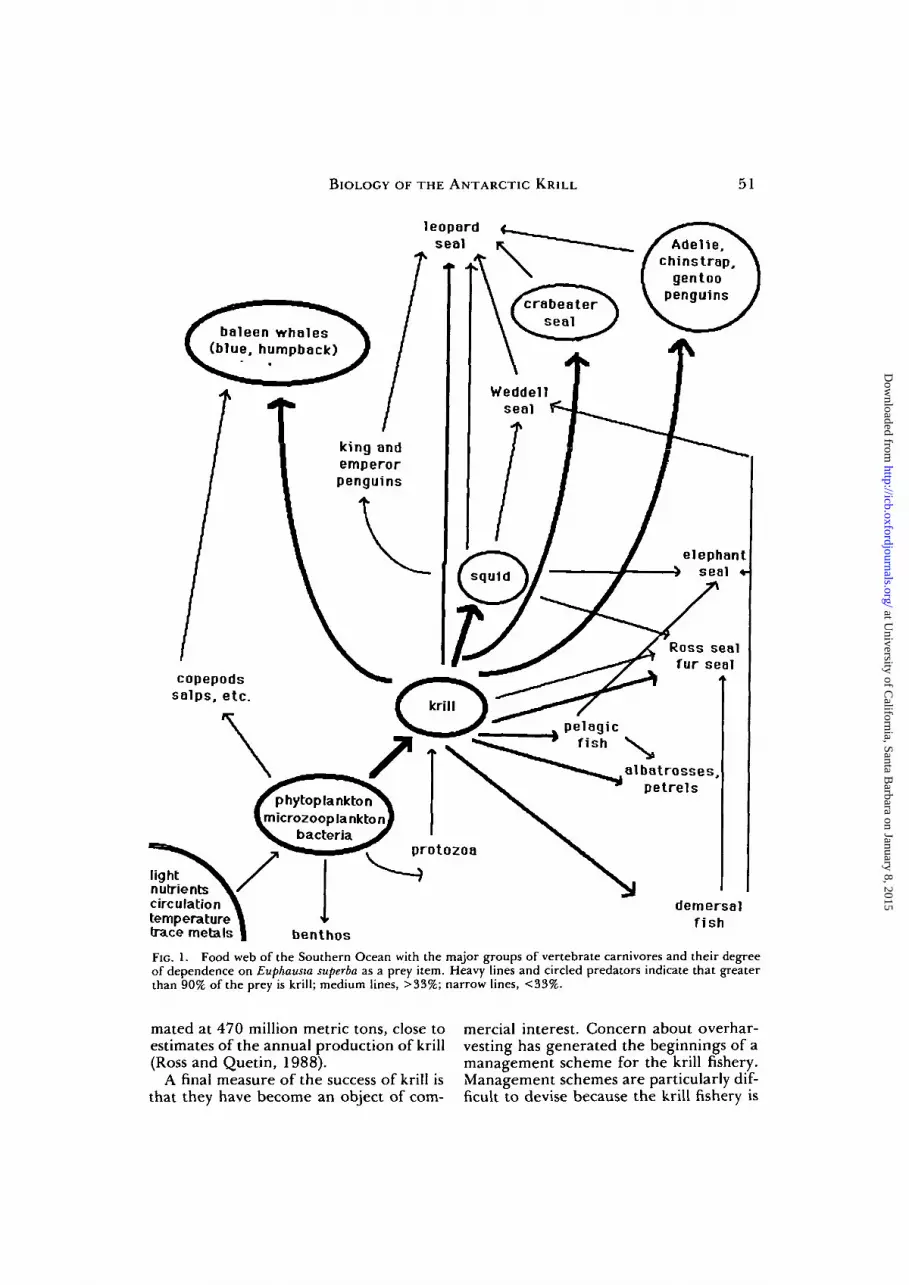
BIOLOGY OF THE ANTARCTIC KRILL 51
leopardseal
baleen whales(blue, humpback)
Weddeilseal
king andemperorpenguins
elephantseal •
Ross sealfur seal
copepodssalps, etc.
albatrossespetrels
phytoplanktonmicrozooplankton
bacteria
lightnutrientscirculationtemperaturetrace metals benthos
demersalfish
FIG. 1. Food web of the Southern Ocean with the major groups of vertebrate carnivores and their degreeof dependence on Euphausia superba as a prey item. Heavy lines and circled predators indicate that greaterthan 90% of the prey is krill; medium lines, >33%; narrow lines, <33%.
mated at 470 million metric tons, close toestimates of the annual production of krill(Ross and Quetin, 1988).
A final measure of the success of krill isthat they have become an object of com-
mercial interest. Concern about overhar-vesting has generated the beginnings of amanagement scheme for the krill fishery.Management schemes are particularly dif-ficult to devise because the krill fishery is
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

52 L. B. QUETIN AND R. M. Ross
a multi-species fishery (May et al., 1979),i.e., man is only one of many predators onkrill. Such management schemes rely onmodels of the life cycle and biology of thespecies that may or may not be correct. Forinstance, in the late 1970s scientists esti-mated from analysis of length-frequencydata that the lifespan of krill ranged fromtwo to four years (reviews: Ettershank,1984; Siegel, 1987). The combination ofhigh biomass and a short lifespan sug-gested that annual production was high,and that populations of krill could sustaina large fishery. However, a managementscheme based on this age estimate wouldhave been a poor one. In the last decade,age determinations of krill with the fluo-rescent age pigment technique and withcomputer-aided analysis of length-fre-quency data all suggest that the adult lifeof krill lasts five or six years, so the totallife span is close to eight years, over twicethe earlier estimates (Ettershank, 1984;Siegel, 1987; Berman et al, 1989). Esti-mates of total annual production of krillmust be reduced accordingly (Ross andQuetin, 1988).
The abundance and central role of krillattest to its success in the Southern Ocean.We will describe the behavioral and phys-iological characteristics that allow E. superbato thrive with a food supply that is stronglyseasonal and patchy geographically, and willdiscuss the possible role of ice in deter-mining food availability and life historystrategies. The characteristics of krill thatunderwrite its success can be grouped intothree categories: (1) krill are able to findconcentrations of food and efficientlyexploit whatever food is available, wher-ever it is available; (2) the growth andreproductive cycles of krill are keyed to theseasonal cycles of food availability and foodhabitat; and (3) a combination of physio-logical mechanisms enables krill to survivethe long winter period of low food avail-ability.
FOOD SEARCHING ANDGATHERING ABILITY
The food gathering ability depends onthe complex morphology of the feedingbasket and the variety of feeding behaviors
exhibited by E. superba. The feeding basketis formed when the six pairs of thoracicappendages and associated setae move apartduring feeding. Water and food particlesenter the food basket from the front andthe food particles become trapped on thefilter formed by the primary, secondary andtertiary setules (Hamner, 1988) (Fig. 2).The filter can trap small particles, withmean intersetal intervals for primary setaefrom 34.5 to 58.2 nm and for secondarysetae from 6.5 to 9.3 nm in adult krill(McClatchie and Boyd, 1983). The wateris forced out laterally through the filter,and the food particles are moved from thefilter to the mouth with the comb setae andtips of the filter setae (Hamner, 1988). Withthis large feeding basket and fine mesh,krill are able to capture and retain a widesize range and type of free floating parti-cles, from small phytoplankton cells tomicroplankton such as tintinnids. Krill alsohave a variety of feeding behaviors, rang-ing from compression filtration, the rhyth-mic expansion and compression of thefeeding basket, to raptorial feeding (Priceet al., 1988), to using the thoracic append-ages to scrape algae off the bottom of seaice (Hamner et al., 1983; Stretch et al.,1988). Krill can and do eat both plant andanimal matter, as is clear from the varietyof feeding behaviors observed, and thepresence in their stomachs of both tintin-nid loricae and crustacean remains. Giventhe green color of the hepatopancreases offreshly collected krill in the summer, how-ever, E. superba is primarily a herbivorewhen phytoplankton is abundant.
The necessary complements to this well-designed feeding basket and repertoire ofbehaviors are the swimming, schooling andchemosensory characteristics of krill(Hamner et al., 1983) which enable krill tosearch for concentrations of phytoplank-ton. Adult krill are strong swimmers andhave been observed moving distances ofgreater than 10 kilometers a day (Kanda etal., 1982). They may follow the retreatingice edge and associated blooms and searchout areas of high standing stock in the pro-tected bays and straits, identifying areas ofhigh phytoplankton concentration bychemical stimuli and remaining in these
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

BIOLOGY OF THE ANTARCTIC KRILL 53
•V-v:
FIG. 2. The feeding basket oiEuphausia superba, and the size of common food items in relation to the meshof the filter. P = primary filter setae, S = secondary filter setae.
areas until the food supply is depleted(Antezana and Ray, 1983). Evidence doesnot support recent arguments by Smith etal. (1988) that krill are "planktonic" orweak swimmers unable to move with aretreating ice edge.
CYCLES OF GROWTH AND REPRODUCTION
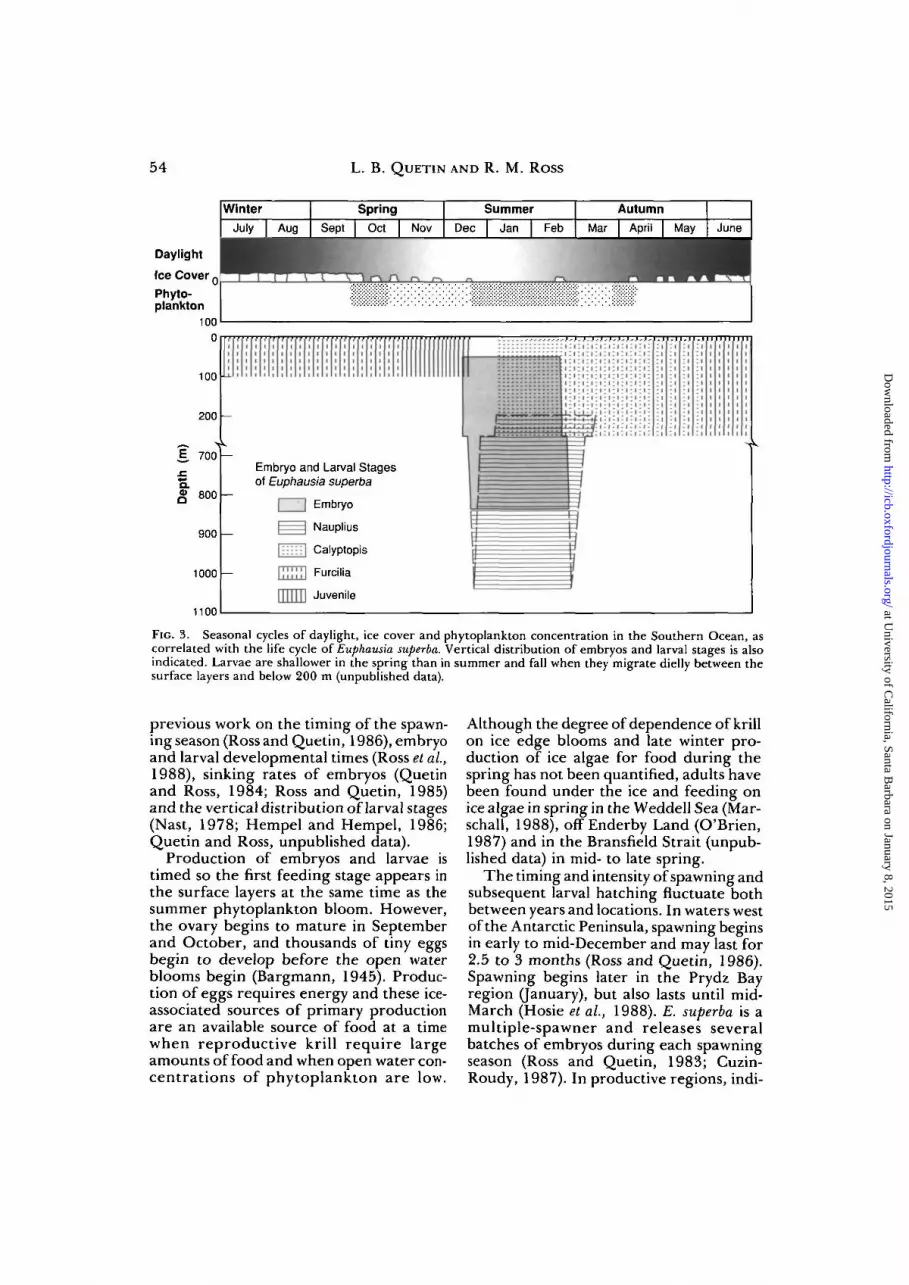
Seasonal cycles of growth and reproduc-tion in E. superba are marked and keyed toseasonal cycles of light and food in the envi-ronment (Fig. 3). Food concentrations arehigh in the water column during the tra-
ditional summer phytoplankton blooms,during the ice edge blooms in the springand occasionally in the fall either as a fallbloom of ice algae on the underside of floes(Hoshiai, 1977) or when the phytoplank-ton are concentrated in the surface layerof frazil ice (Garrison et al., 1986). Con-centrations of ice algae can also be signif-icant in the winter and are many times thosein the water column (Kottmeier and Sul-livan, 1987). The timing and vertical dis-tributions of the embryos and different lar-val stages shown in Figure 3 are based on
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from
L. B. QUETIN AND R. M. Ross
Daylight
Ice Cover
Phyto-plankton
| Oct I Nov Dec I Jan Feb Mar I April I May June
Embryo and Larval Stagesof Euphausia superba
| | Embryo
Nauplius
Calyptopis
[TUTU Furcilia
Juvenile
FIG. 3. Seasonal cycles of daylight, ice cover and phytoplankton concentration in the Southern Ocean, ascorrelated with the life cycle of Euphausia superba. Vertical distribution of embryos and larval stages is alsoindicated. Larvae are shallower in the spring than in summer and fall when they migrate dielly between thesurface layers and below 200 m (unpublished data).
previous work on the timing of the spawn-ing season (Ross and Quetin, 1986), embryoand larval developmental times (Ross et ai,1988), sinking rates of embryos (Quetinand Ross, 1984; Ross and Quetin, 1985)and the vertical distribution of larval stages(Nast, 1978; Hempel and Hempel, 1986;Quetin and Ross, unpublished data).
Production of embryos and larvae istimed so the first feeding stage appears inthe surface layers at the same time as thesummer phytoplankton bloom. However,the ovary begins to mature in Septemberand October, and thousands of tiny eggsbegin to develop before the open waterblooms begin (Bargmann, 1945). Produc-tion of eggs requires energy and these ice-associated sources of primary productionare an available source of food at a timewhen reproductive krill require largeamounts of food and when open water con-centrations of phytoplankton are low.
Although the degree of dependence of krillon ice edge blooms and late winter pro-duction of ice algae for food during thespring has not been quantified, adults havebeen found under the ice and feeding onice algae in spring in the Weddell Sea (Mar-schall, 1988), off Enderby Land (O'Brien,1987) and in the Bransfield Strait (unpub-lished data) in mid- to late spring.
The timing and intensity of spawning andsubsequent larval hatching fluctuate bothbetween years and locations. In waters westof the Antarctic Peninsula, spawning beginsin early to mid-December and may last for2.5 to 3 months (Ross and Quetin, 1986).Spawning begins later in the Prydz Bayregion (January), but also lasts until mid-March (Hosie et al, 1988). E. superba is amultiple-spawner and releases severalbatches of embryos during each spawningseason (Ross and Quetin, 1983; Cuzin-Roudy, 1987). In productive regions, indi-
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

BIOLOGY OF THE ANTARCTIC KRILL 55
vidual females release a batch of about2,000 embryos about every 10 days.Because the energy required for this levelof reproduction is very high, such fecundi-ties probably only occur in specific areas ofthe Southern Ocean where food availabil-ity is high during most of the summer. Inareas of low productivity, the number ofbatches of eggs may be less or reproductionmay not occur at all (Ross and Quetin,1986). Distinct spawning grounds do exist(Marr, 1962), and they are generally locatedwhere seasonal phytoplankton blooms lastthe longest and reach the highest biomass.
After spawning, embryos sink rapidly outof the surface layers, and hatch at 850 to1,000 m after 4.5 to 6 days, depending onthe temperature structure of the water col-umn (Quetin and Ross, 1984; Ross andQuetin, 1985) (Fig. 3). The newly hatchednauplii are weak swimmers and probablycontinue to sink for a day or so, then beginto ascend to the surface (Marschall andHirche, 1984). The first three larval stages(nauplii) are found below 250 m, but theCalyptopis 1 larvae, the first stage with amouth and feeding appendages, are usu-ally found in the lighted surface layer withtheir food source—phytoplankton (Mar-schall, 1985). Krill have a long and com-plex larval sequence after the naupliarstages: three calyptopis stages and six fur-cilia stages before the metamorphosis intojuvenile at four months if food is plentiful(Ikeda, 1985), or not until nine months(mid-November) under winter conditionsof low food and temperatures (Ross et al.,1987). The availability of food once theCalyptopis 1 larvae reach the surface is crit-ical. The hatching larvae have enoughenergy stores to tolerate about 10 to 14days of starvation after metamorphosis, butfurther starvation results in eventual deatheven if food becomes available later (Rossand Quetin, 1989). Since the exact timingand length of phytoplankton blooms in theSouthern Ocean are not that predictable,not all calyptopis arriving at the surfacemay survive on the food available to them.
The ability to release many batches ofembryos during the austral summer is ofgreat survival value, giving the adults sev-eral chances to produce larvae that will
develop into the first feeding stage in anenvironment with sufficient food. Thisstrategy of producing many small embryoswithout large reserves means that the lar-vae must feed themselves soon after hatch-ing. Not only must the timing be such thatthe larvae can feed soon after reaching thesurface, but food must also be availablelong enough for the larvae to survive theearly critical stages and prepare for thelong winter.
WINTER-OVER MECHANISMS
The third set of biological characteris-tics that allow E. superba to thrive in thisintensely seasonal environment are thewinter-over mechanisms. Four majorhypotheses, shrinkage, lipid utilization,switching of food sources, and "hiberna-tion," have been proposed to explain howkrill survive the six-month winter periodof low food, low light and often extensiveice cover. To test these hypotheses and toestimate their relative importance for pop-ulations of adult E. superba west of the Ant-arctic Peninsula we compared the physio-logical condition of krill in the field atdifferent times of year (Table 1). In thispaper we will present the preliminaryresults of these experiments, but leave adetailed description of the experiments andthe complete seasonal trends for later pub-lication. Although each experiment onlyaddressed one of the hypotheses, we canuse the results of all the experiments toevaluate the relative importance of eachpotential winter-over mechanism.
Shrinkage
Krill may use their own body lipids andproteins when starved and shrink in totallength so they do not have to maintain thesame body volume (Ikeda and Dixon, 1982).In the laboratory, adult krill have survivedfor over 211 days without food, provingthat they can easily survive winter withouteating. When starved for long periods indi-viduals shrink 1 to 2% during each inter-molt period of about a month. However,Stepnik (1982), in an analysis of length-frequencies of E. superba from AdmiraltyBay, found that positive growth occurredin late winter and early spring, suggesting
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

56 L. B. QUETIN AND R. M. Ross
TABLE 1. Experiments and samples to test the four major winter-over mechanisms proposed. *
Mechanism Test Month/Year
Shrinkage
Lipid utilization
Switch food source(Carnivory)
"Hibernation"
Compare the growth rates of krill in the field inlate summer/early fall and winter
Compare lipid content at the end of the summerand in late winter
Compare rates of ingestion of phytoplanktonand fecal pellet production, summer and win-ter
Compare metabolic rates at ambient tempera-tures summer and winter
Mar/Apr 85 and Aug/Sept 85
Mar/Apr 84, 85 and Aug/Sept85
Feb/Mar 85 and Aug/Sept 85
Jan andjul 87
* All adult krill were collected from the waters west of the Antarctic Peninsula, in the Bransfield Strait andnorth of the South Shetland Islands.
that shrinkage may not occur in field pop-ulations.
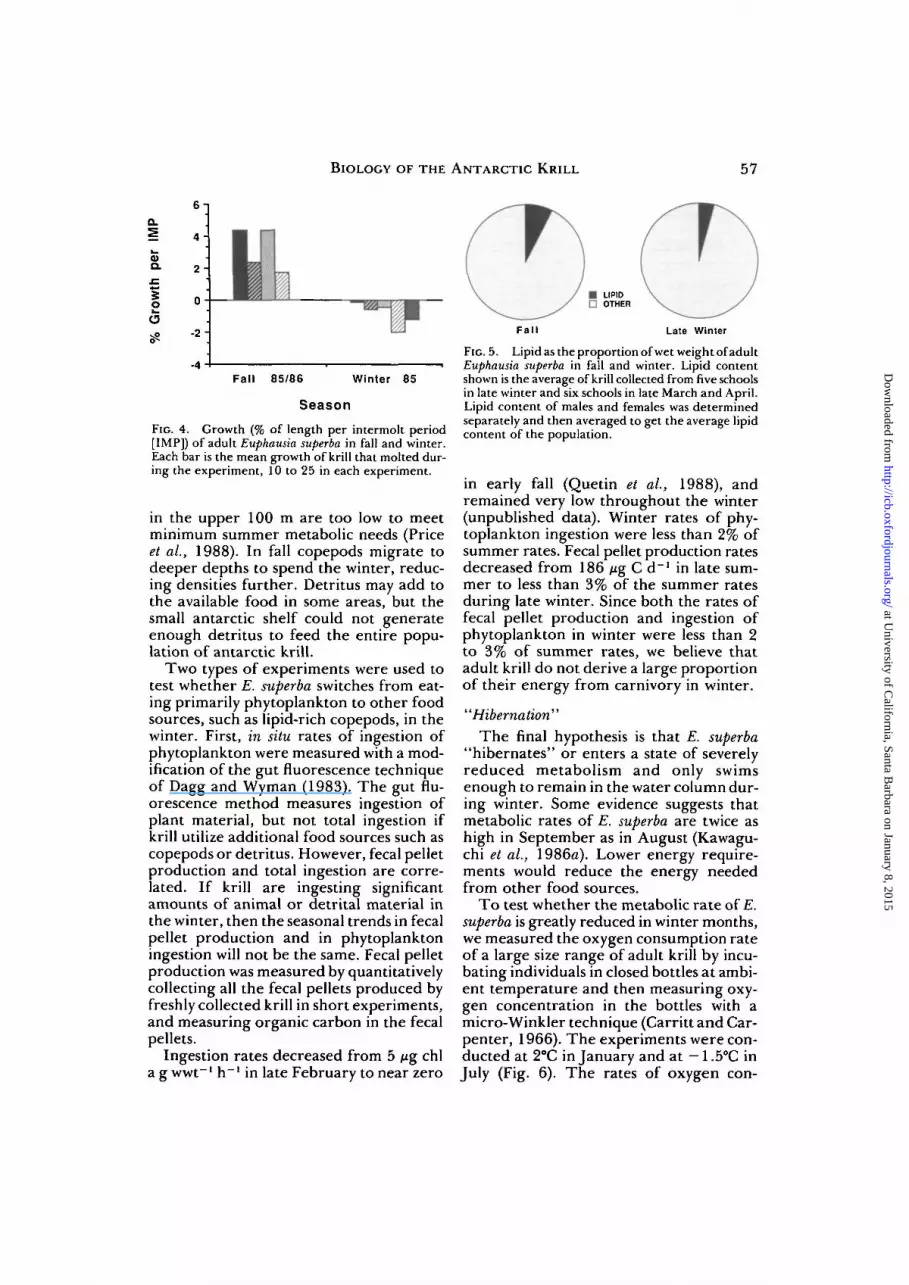
We tested the shrinkage hypothesis bymeasuring linear growth increments ofindividuals maintained in groups of 100 atambient temperatures during differenttimes of the year, eliminating the assump-tion necessary in length-frequency analysesthat the same population is sampled eachmonth. In late summer and early fall,growth was positive (1.75 to 4.40% perintermolt period) (Fig. 4), and intermoltperiods calculated for each group of krillwere 2.5 to 4.5 weeks. Growth in the win-ter was negative (-0.16 to —2.03% perintermolt period) (Fig. 4), and intermoltperiods were twice as long. Krill not onlyshrink in linear dimension during the win-ter, with a concomitant reduction in wetweight, they also lose volume, i.e., get thin-ner. The relationship between total lengthand wet weight changes with season, indi-cating that krill of any one length loseweight during fall and winter (unpublisheddata). The wet weight of a krill 42 mm longdecreases from 667 mg to 542 mg, a lossof 125 mg, during the six-month periodbetween March and September. The pro-tein and lipid in this wet weight fractionare additional sources of energy during thewinter. We now have clear evidence thatshrinkage in both total length and in pro-portional volume occurs in E. superba inthe winter west of the Antarctic Peninsula.
Lipid utilizationStored lipids may be used as an energy
source during winter. Wax esters arethought to be long-term energy reserves
that are particularly useful during winter-over periods and long-term starvation ofherbivores (Lee et al., 1970; Sargent andHenderson, 1986). So the degree of depen-dence on phytoplankton food should dic-tate whether lipid reserves for winter arelarge or small and what form they take.The lack of wax esters in antarctic krill(Clarke, 1984) and the lack of unequivocalevidence for lipid storage in some otherform suggested krill do not utilize lipids asan energy source during winter (Mauch-line and Fisher, 1969; Clarke, 1984). How-ever, previous samples of krill analyzed forlipid content did not cover the critical timesof late summer, fall and winter.
We analyzed samples of male and femalekrill in late summer-early fall and in latewinter for total lipid content, as percent ofwet weight, with a charring method (Marshand Weinstein, 1966). The average lipidcontent of the combined sexes was 7.48%in fall, dropping to 3.86% in late winter(Fig. 5). Late winter lipid content of adultswas similar to that found in immatures inDecember (Clarke, 1984) and in adults inDecember (unpublished data). Loss of halfthe total lipid during fall and winter indi-cates that lipid reserves are used during thewinter for energy.
Switch food sources
The possibility of switching to lipid-richcopepods (Boyd et al., 1984) or detritus(Kawaguchi et al., 19866) as a food sourcein winter is often mentioned. But relyingsolely on animal matter for food in the win-ter is unlikely. Even in summer, normaldensities of both copepods and protozoans
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from
BIOLOGY OF THE ANTARCTIC KRILL
oQ.
O
a
Fall 85/86 Winter 85
Season
FIG. 4. Growth (% of length per intermolt period[IMP]) of adult Euphausia superba in fall and winter.Each bar is the mean growth of krill that molted dur-ing the experiment, 10 to 25 in each experiment.
in the upper 100 m are too low to meetminimum summer metabolic needs (Priceet al, 1988). In fall copepods migrate todeeper depths to spend the winter, reduc-ing densities further. Detritus may add tothe available food in some areas, but thesmall antarctic shelf could not generateenough detritus to feed the entire popu-lation of antarctic krill.
Two types of experiments were used totest whether E. superba switches from eat-ing primarily phytoplankton to other foodsources, such as lipid-rich copepods, in thewinter. First, in situ rates of ingestion ofphytoplankton were measured with a mod-ification of the gut fluorescence techniqueof Dagg and Wyman (1983). The gut flu-orescence method measures ingestion ofplant material, but not total ingestion ifkrill utilize additional food sources such ascopepods or detritus. However, fecal pelletproduction and total ingestion are corre-lated. If krill are ingesting significantamounts of animal or detrital material inthe winter, then the seasonal trends in fecalpellet production and in phytoplanktoningestion will not be the same. Fecal pelletproduction was measured by quantitativelycollecting all the fecal pellets produced byfreshly collected krill in short experiments,and measuring organic carbon in the fecalpellets.
Ingestion rates decreased from 5 jug chla g wwt"1 h~' in late February to near zero
Late Winter
FIG. 5. Lipid as the proportion of wet weight of adultEuphausia superba in fall and winter. Lipid contentshown is the average of krill collected from five schoolsin late winter and six schools in late March and April.Lipid content of males and females was determinedseparately and then averaged to get the average lipidcontent of the population.
in early fall (Quetin et al., 1988), andremained very low throughout the winter(unpublished data). Winter rates of phy-toplankton ingestion were less than 2% ofsummer rates. Fecal pellet production ratesdecreased from 186 fig C d~' in late sum-mer to less than 3% of the summer ratesduring late winter. Since both the rates offecal pellet production and ingestion ofphytoplankton in winter were less than 2to 3% of summer rates, we believe thatadult krill do not derive a large proportionof their energy from carnivory in winter.
"Hibernation"
The final hypothesis is that E. superba"hibernates" or enters a state of severelyreduced metabolism and only swimsenough to remain in the water column dur-ing winter. Some evidence suggests thatmetabolic rates of E. superba are twice ashigh in September as in August (Kawagu-chi et al., 1986a). Lower energy require-ments would reduce the energy neededfrom other food sources.
To test whether the metabolic rate of E.superba is greatly reduced in winter months,we measured the oxygen consumption rateof a large size range of adult krill by incu-bating individuals in closed bottles at ambi-ent temperature and then measuring oxy-gen concentration in the bottles with amicro-Winkler technique (Carritt and Car-penter, 1966). The experiments were con-ducted at 2°C in January and at - 1.5°C inJuly (Fig. 6). The rates of oxygen con-
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

58 L. B. QUETIN AND R. M. Ross
10001
K - 100;
• - = ' • =
• S ©Z i 10
30 SO 60
Total Length (mm)
FIG. 6. Metabolic rate (ji\ O2 h"1) of adult Euphausiasuperba in the summer (2°C) and winter (—1.5°C). Linesrepresent the relationship between total length andoxygen consumption rate: summer, log VO2 (ml h"1)= 0.0241 TL (mm) - 2.0551, r2 = 0.70; winter, logVO, (ml h"1) = 0.0285 TL (mm) - 2.2258, r2 = 0.67.
sumption in winter were only 33% of thosein summer, a significant reduction in energyrequirements from summer to winter. Thisreduction can be partially attributed to thefact that krill are not feeding in the winter,as Ikeda and Dixon (1984) believe that theoxygen consumption rates of non-feedingkrill are only 62.5% those of feeding krill.Yet about half the decrease in energyrequirements from summer to winter isunexplained by the lack of feeding, andmay be due to other behavioral changessuch as a decrease in swimming activity orto physiological changes on a cellular levelsuch as decreases in metabolic enzymeactivity.
Relative importance of winter-over mechanisms
We used these data to evaluate the rel-ative importance of the different hypoth-esized winter-over mechanisms for a 42-mm adult E. superba. A six-month periodof low food availability, from the last of thelate summer blooms to the beginning ofthe ice edge blooms and late winter/earlyspring production in the sea ice commu-nity, was used as the time span for all thecalculations. Summer metabolic rates wereused to predict the calories required frommid-March to mid-September. We calcu-lated the calories saved by the reductionof the metabolic rate in the winter fromthe difference between summer and winterrates. We calculated the calories in the pro-tein and lipid fraction of the wet weightloss that accompanied shrinkage (both the
ingestions h r i n k i - i
lipid
-"hibernation"
FIG. 7. Relative importance of the four major win-ter-over mechanisms proposed for Euphausia superbain the waters west of the Antarctic Peninsula. Theentire circle represents the energy requirement forthe six months of low food availability as predictedfrom the summer metabolic rates. The white segmentis the portion of the energy requirement that remainsunexplained.
decrease in total length as predicted fromaverage negative growth rates in winter andthe loss in volume) over the six-month win-ter period from the chemical composition(% protein, % lipid) of the animals, the rela-tionships between total length and wetweight in March and September, andaccepted conversion constants. Caloriesused from stored lipid reserves were esti-mated from the decrease in % lipid fromMarch to September, the wet weights inMarch and September and accepted con-version constants. The contribution ofingestion during the winter was estimatedfrom average fecal pellet production ratesin terms of carbon and accepted conver-sion constants.
We presented each savings in energy orcontribution to energy available to satisfyrequirements as a percent of the total met-abolic requirement for the six months aspredicted by summer metabolic rates (Fig.7). "Hibernation," or the lowered meta-bolic rate, was by far the most importantmechanism for successfully surviving thewinter. Over 71 % of the anticipated energyrequirements was saved by this lowering ofmetabolic rates. Direct lipid utilizationprovided almost 11 % of the energy, andutilization of the protein and lipid in thebody from shrinkage in both total lengthand in volume provided another 4% of thepredicted energy requirement. Althoughingestion was very low during winter, it
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

BIOLOGY OF THE ANTARCTIC KRILL 59
satisfied some of the energy requirements.With our preliminary evaluation of the fourhypothesized winter-over mechanisms, thesource of nearly 90% of the predictedenergy requirements for an adult krill win-tering-over west of the Antarctic Peninsulacan be identified (Fig. 7).
However, this balance may be quite dif-ferent in other regions of the SouthernOcean. For instance, in areas of lower pro-ductivity, lipid reserves may not be as high,so either additional shrinkage or weightloss may occur. The role of ice and icealgae in the availability of food during dif-ferent times of year and in the known dis-tribution patterns of E. superba has beendiscussed, but the predictability, extent andmorphology of the ice cover during thewinter may also play a role in determiningthe relative importance of the suggestedwinter-over mechanisms.
ROLE OF ICE AND ICE ALGAE
During fall, winter and spring the roleof ice and ice algae in the life history andenergetics of E. superba may be moreimportant than during austral summerbecause of the simultaneous increase in theaereal extent of sea ice and decrease inphytoplankton biomass in the water col-umn. There are few observations and evenfewer quantitative estimates of the impor-tance of sea ice and ice algae to the winter-over existence of E. superba (Table 2). Dur-ing AMERIEZ cruises in the northernWeddell Sea in fall and early spring Ainleyet al. (1988), Daly and Macaulay (1988),and Lancraft et al. (1989) found krill larvaeand juveniles associated with the ice edgeand adults more common away from theice edge in the water column or deeper inthe pack ice in the Weddell Sea. Marschall(1988) recorded distributions of adult krillunder the ice deep in the Weddell Sea withan ROV and trawl collections during springand with surface observations from the shipduring late winter in the same area. In thespring, adults were feeding on ice algae,and winter reports of krill with greenish-brown hepatopancreases from these areasimply adult krill were also feeding on icealgae during winter. Marschall (1988) alsofound adult krill more often associated with
the underside of ridged and over-rafted ice(multi-year ice) than with the smootherunderside of annual or first-year ice.
West of the Antarctic Peninsula annualice predominates and the ice habitatresembles the annual ice zone of the Wed-dell Sea (Quetin and Ross, personal obser-vation). In these waters adults are in thewater column beneath sea ice and are notclosely associated with the under surfaceof the sea ice in mid-winter (Guzman, 1983;Quetin and Ross, unpublished observa-tion). However, they do feed on ice algaein the early spring (Quetin and Ross,unpublished observation) as also reportedby O'Brien (1987) in Enderby Land. Theseobservations suggest that during winteradult krill in areas of annual or smooth iceare not directly underneath the ice and donot feed on ice algae, possibly because thereis not enough ice structure to serve as arefuge from predation and/or enough icealgae to give them an energetic advantageif they do feed. Therefore the importanceof ice algae in the winter energetics of adultkrill is small in areas of annual ice. How-ever, multi-year ice in the Weddell Sea mayhave higher algal concentrations thanannual sea ice and the inherent structuralcomplexity may enable adult krill to inhabitthe underside of the sea ice and feed onice algae without experiencing heavy pre-dation pressure.
Larvae and juveniles, on the other hand,do feed on ice algae in areas of annual orsmooth ice in both winter and spring. Guz-man (1983), Kottmeier and Sullivan (1987)and Quetin and Ross (1988) have all seenlarvae feeding on the underside of the seaice from early to late winter west of theAntarctic Peninsula as have Daly andMacaulay (1988) and Marschall (1988) inthe spring in the Weddell Sea. Most krillpredators appear to prefer eating larger oradult krill, so the relatively small size oflarvae compared to adults may give thema refuge from predation and allow them tooccupy the smooth ice zones during winterwithout high predation pressure. The dis-tribution of larvae during mid-winter sug-gests that larvae and adults do not winter-over in the same way. Possibly larvae havenot had time to accumulate sufficient lipid
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

60 L. B. QUETIN AND R. M. Ross
TABLE 2. Observations of the distribution and feeding behavior of adult and juvenile Euphausia superba in theWeddell Sea and the waters west of the Antarctic Peninsula.
Reference
Weddell SeaAinley etai, 1988
Daly and Macaulay,1988
Marschall, 1988
Area/Time
nw Weddellspr 83, fall 86
w Weddellspr 83, fall 86
e and s Weddellwin and spr 86
West of Antarctic PeninsulaGuzman, 1983
Kottmeier andSullivan, 1987
Quetin and Ross,1988, unpubl. data
Bransfield Str
Bransfield Str
Bransfield Str,Palmer Basin
Method
seabird stomachs
acoustic, nets,deck obs., div-ers
ROV
acoustic
divers
acoustic, divers
Adults
under pack ice(100 km), inopen water
in open water
under complexice, feeding
in ice free zones,not feeding
—
in water column,not feeding
Juveniles
at ice edge, underpack ice
undersurface offloes, feeding
under complex ice,feeding
under pack ice,feeding
under pack icewith ice algae
under ice, feeding
stores during a short single summer so maybe less resistant to starvation than adultkrill. Their small size, continued develop-ment during winter, and postulated lack oflipid reserves all suggest that larvae mayneed to feed in winter. Thus, larvae willhave a greater dependence on ice algae inthe winter than do adults.
Our physiological measurements of thewinter-over mechanisms of adult krill areconsistent with field observations in winter(Table 2). Adults do not feed, have loweredmetabolic rates, and negative or zerogrowth rates. Without physiological mea-surements it is impossible to evaluate theimportance of ice algae to the annual ener-getics of krill inhabiting the underside ofmulti-year ice during winter. Conclusionsabout the importance of ice algae to theenergetics of adult krill in winter in multi-year ice based on field observations to datewould be premature, especially givenquantitative physiological data for krillfound in other areas. Further studies willbe necessary to determine whether the ref-uge from predation afforded by the struc-ture of multi-year ice enables adult krill tofulfill a significant portion of their annualenergetic requirements by feeding on icealgae in winter. Recent evidence indicatesthat this is not the case in other areas.
Significant seasonal, geographical andinterannual variations occur in the ice cover
of the Southern Ocean (Zwally^ al., 1983),and have been used to predict variationsin primary production associated withreceding marginal ice zones (Smith et al.,1988). These interannual variations in icecover and spring and summer marginal icezone productivity may have importanteffects on the population dynamics, ratesof reproduction, and recruitment of E.superba. Ice algae is probably an importantfood source for krill in spring before thesummer phytoplankton blooms develop. Inthe spring larvae continue to develop andgrow and the ovaries and testes of the adultsstart to mature in preparation for thereproductive season, thus increasing ener-getic requirements. West of the AntarcticPeninsula, where krill are concentrated andwhere most of our research occurs, inter-annual variability in ice extent can be sig-nificant even when interannual variabilityin ice extent in the Southern Ocean as awhole is low (Smith et al., 1988). The win-ter of 1985 was particularly mild west ofthe Antarctic Peninsula, and even in Augustand September the Bransfield Strait wasnot frozen and the R/VPolar Duke did notencounter pack ice until well south ofAnvers Island (Fig. 8a). The winter of 1987was cold, with the pack ice extending over150 km north of the South Shetland Islandsas early as June (Fig. 8b). Interannual vari-ability in the extent of the annual sea ice
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

BIOLOGY OF THE ANTARCTIC KRILL 61
FIG. 8. The extent of ice cover in the waters west of the Antarctic Peninsula and the northwestern WeddellSea during two winters in the 1980s: (a) August and September 1985; (b) June andjuly 1987.
and associated blooms may be an impor-tant factor in the ability of larvae to developand in the distribution and reproductivesuccess of adults, as seen in the Arctic whereinterannual differences in egg productionrates of copepods appear to be a functionof warm and cold years and the extent ofthe sea ice (Smith and Vidal, 1986).
CONCLUSIONS
The antarctic krill, E. superba, is verysuccessful in this environment of extremes,successful because it is able to cope with afood supply that is both patchy and sea-sonal. The success of this species has itsroots in a combination of biological andphysiological factors, not the least of whichis a suite of winter-over mechanisms thatallow it to survive a long period of very lowfood availability.
ACKNOWLEDGMENTS
Discussions with many colleagues whowork in the Antarctic have been useful aswe explored the differences betweenregions of the Southern Ocean. We alsothank the captains and crews of the researchvessels R/V Hero (USAP 82 to 84) andR/VPolar Duke (USAP 85 to 89) who havebeen of invaluable help in our search for
antarctic krill. E. Kirsch, M. O. Amsler anda vast number of volunteers helped con-duct experiments, take samples and ana-lyze data from numerous cruises. Ourresearch has been supported by theNational Science Foundation, Division ofPolar Programs, Grant No. DPP82-18356to Quetin and Ross, and Grant No. DPP85-18872 to Ross and Quetin.
REFERENCES
Ainley, D. G., W. R. Fraser, and K. L. Daly. 1988.Effects of pack ice on the composition of micro-nektonic communities in the Weddell Sea. In D.Sahrhage (ed.), Antarctic ocean and resources vari-ability, pp. 140-146. Springer-Verlag, Berlin,Heidelberg.
Antezana, R. and K. Ray. 1983. Aggregation ofEuphausia superba as an adaptive group strategyto the Antarctic ecosystem. In S. B. Schnack (ed.),On the biology of krill Euphausia superba, pp. 199-215. Alfred-Wegener-Institute for PolarResearch, Bremerhaven, FRG.
Bargmann, H. E. 1945. The development and life-history of adolescent and adult krill, Euphausiasuperba. Discovery Rep. 23:103-176.
Berman, M. S., A. L. McVey, and G. Ettershank.1989. Age determination of antarctic krill usingfluorescence and image analysis of size. Polar Biol.9:267-271.
Boyd, C. M., M. Heyraud, and C. N. Boyd. 1984.Feeding of the antarctic krill Euphausia superba.J. Crust. Biol. 4(Spec. No. 1):123-141.
Carritt, D. E. andJ.H. Carpenter. 1966. Comparison
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

62 L. B. QUETIN AND R. M. Ross
and evaluation of currently employed modifica-tions of the Winkler methods for determiningdissolved oxygen in sea water: A NASCO report.J. Mar. Res. 24:286-317.
Clarke, A. 1984. Lipid content and composition ofantarctic krill, Euphausia superba Dana. J. Crust.Biol. 4(Spec. No. l):285-294.
Clarke, A. 1985. Food webs and interactions: Anoverview of the Antarctic ecosystem. In W. N.Bonner and D. W. H. Walton (eds.), Antarctica,pp. 329-350. Pergamon Press, Oxford, NewYork, Toronto, Sydney, Paris, Frankfurt.
Cuzin-Roudy,J. 1987. Gonad history of the antarctickrill Euphausia superba Dana during its breedingseason. Polar Biol. 7:237-244.
Dagg, M. J. and K. D. Wyman. 1983. Natural inges-tion rates of the copepods Neocalanus plumchrusand N. cnstatus calculated from gut contents. Mar.Ecol. Prog. Ser. 13:37-46.
Daly, K. L. and M. C. Macaulay. 1988. Abundanceand distribution of krill in the ice edge zone ofthe Weddell Sea, austral spring 1983. Deep-SeaRes. 35:21-41.
El-Sayed, S. Z. 1985. Plankton of the Antarctic seas.In W. N. Bonner and D. W. H. Walton (eds.),Antarctica, pp. 135-153. Pergamon Press, Oxford,New York, Toronto, Sydney, Paris, Frankfurt.
Ettershank, G. 1984. A new approach to the assess-ment of longevity in the antarctic krill Euphausiasuperba. J. Crust. Biol. 4(Spec. No. l):295-305.
Garrison, D. L., C. W. Sullivan, and S. F. Ackley.1986. Sea ice microbial communities in Antarc-tica. BioScience 36:243-250.
Garrison, D. L. and D. B. Siniff. 1986. An antarcticperspective. BioScience 36:238-242.
Guzman, O. 1983. Distribution and abundance ofantarctic krill (Euphausia superba) in the Brans-field Strait. In S. B. Schnack (ed.), On the biologyof krill Euphausia superba, pp. 169-190. Alfred-Wegener-Institute for Polar Research, Bremer-haven, FRG.
Hamner, W. M. 1984. Aspects of schooling inEuphausia superba. J. Crust. Biol. 4(Spec. No. 1):67-74.
Hamner, W. M. 1988. Biomechanics of filter feedingin the antarctic krill Euphausia superba: Reviewof past work and new observations. J. Crust. Biol.8:149-163.
Hamner, W. M., P. P. Hamner, S. W. Strand, and R.W. Gilmer. 1983. Behavior of antarctic krill,Euphausia superba: Chemoreception, feeding,schooling, and molting. Science 220:433-435.
Hempel, I. and G. Hempel. 1986. Field observationson the developmental ascent of larval Euphausiasuperba (Crustacea). Polar Biol. 6:121-126.
Hewes, C. D., O. Holm-Hansen, and E. Sakshaug.1985. Alternate carbon pathways at lower tro-phic levels in the Antarctic food web. In W. R.Siegfried, P. R. Condy, and R. M. Laws (eds.),Antarctic nutrient cycles and food webs, pp. 277—283.Springer-Verlag, Berlin, Heidelberg, New York,Tokyo.
Hopkins, T. L. 1985. Food web of an antarctic mid-water ecosystem. Mar. Biol. 89:197-212.
Hoshiai, T. 1977. Seasonal change of ice commu-
nities in the sea ice near Syowa Station, Antarc-tica. In M. J. Dunbar (ed.), Polar oceans, pp. 307-317. Arctic Inst. North America, Calgary, Can-ada.
Hosie, G. W., T. Ikeda, and M. Stolp. 1988. Distri-bution, abundance and population structure ofthe antarctic krill (Euphausia superba Dana) in thePrydz Bay region, Antarctica. Polar Biol. 8:213-224.
Ikeda, T. 1985. Life history of antarctic krill Euphau-sia superba: A new look from an experimentalapproach. Bull. Mar. Sci. 37:599-608.
Ikeda, T. and P. Dixon. 1982. Body shrinkage as apossible over-wintering mechanism of the antarc-tic krill, Euphausia superba Dana. J. Exp. Mar.Biol. Ecol. 62:143-151.
Ikeda, T. and P. Dixon. 1984. The influence of feed-ing on the metabolic activity of antarctic krill(Euphausia superba Dana). Polar Biol. 3:1-9.
Kanda, K., K. Takagi, and Y. Seki. 1982. Movementof the larger swarms of antarctic krill Euphausiasuperba population off Enderby Land during1976-1977 season. J. Tokyo Univ. Fish. 68:25-42.
Kawaguchi, K., S. Ishikawa, and O. Matsuda. 1986a.The overwintering strategy of antarctic krill(Euphausia superba Dana) under the coastal fastice off the Ongul Islands in Lutzow-Holm Bay,Antarctica. Mem. Natl. Inst. Polar Res. (Spec.Issue) 44:67-85.
Kawaguchi, K., O. Matsuda, S. Ishikawa, and Y. Naito.1986A. A light trap to collect krill and othermicronektonic and planktonic animals under theAntarctic coastal fast ice. Polar Biol. 6:37-42.
Kock, K.-H. 1985. Krill consumption by Antarcticnotothenioid fish. In W. R. Siegfried, P. R. Condy,and R. M. Laws (eds.), Antarctic nutrient cycles andfood webs, pp. 437-444. Springer-Verlag, Berlin,Heidelberg, New York, Tokyo.
Kottmeier, S. T. and C. W. Sullivan. 1987. Late win-ter primary production and bacterial productionin sea ice and seawater west of the Antarctic Pen-insula. Mar. Ecol. Prog. Ser. 36:287-298.
Lancraft, T. M., J. J. Torres, and T. L. Hopkins.1989. Micronekton and macrozooplankton in theopen waters near antarctic ice edge zones(AMER1EZ 1983 and 1986). Polar Biol. 9:225-233.
Laws, R. M. 1985. The ecology of the SouthernOcean. Am. Sci. 73:26-40.
Lee, R. F., J. C. Nevenzel, and G.-A. Paffenhofer.1970. Wax esters in marine copepods. Science167:1510-1511.
Macaulay, M. C, T. S. English, and O. A. Mathisen.1984. Acoustic characterization of swarms ofantarctic krill (Euphausia superba) from ElephantIsland and Bransfield Strait. J. Crust. Biol. 4(Spec.No. 1): 16-44.
Marr, J. W. S. 1962. The natural history of theantarctic krill (Euphausia superba Dana). Discov-ery Rep. 32:33-464.
Marschall, H.-P. 1985. Structural and functionalanalyses of the feeding appendages of krill larvae.In W. R. Siegfried, P. R. Condy, and R. M. Laws(eds.), Antarctic nutrient cycles and food webs, pp.
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

BIOLOGY OF THE ANTARCTIC KRILL 63
346-354. Springer-Verlag, Berlin, Heidelberg,New York, Tokyo.
Marschall, H.-P. 1988. The overwintering strategyof antarctic krill under the pack-ice of the Wed-dell Sea. Polar Biol. 9:129-135.
Marschall, H.-P. and H.-J. Hirche. 1984. Develop-ment of eggs and nauplii of Euphausia superba.Polar Biol. 2:245-250.
Marsh,J. B. and D. B. Weinstein. 1966. Simplechar-ring method for determination of lipids. J. LipidRes. 7:574-576.
Mauchline, J. and L. R. Fisher. 1969. The biologyof euphausiids. Adv. Mar. Biol. 7:1-454.
May, R. M., J. R. Beddington, C. W. Clark, S. J. Holt,andR. M. Laws. 1979. Management of multispe-cies fisheries. Science 205:267-277.
McClatchie, S. and C. M. Boyd. 1983. Morphologicalstudy of sieve efficiencies and mandibular sur-faces in the antarctic krill, Euphausia superba. Can.J. Fish. Aquat. Sci. 40:955-967.
Nast, F. 1978. The vertical distribution of larval andadult krill (Euphausia superba Dana) on a timestation south of Elephant Island, South Shet-lands. Meeresforsch. 27:103-118.
Nemoto, T., M. Okiyama, and M. Takahashi. 1985.Aspects of the roles of squid in food chains ofmarine Antarctic ecosystems. In W. R. Siegfried,P. R. Condy, and R. M. Laws (eds.), Antarctic nutri-ent cycles and food webs, pp. 415-420. Springer-Verlag, Berlin, Heidelberg, New York, Tokyo.
O'Brien, D. P. 1987. Direct observations of thebehavior of Euphausia superba and Euphausia crys-tallorophias (Crustacea: Euphausiacea) under packice during the Antarctic spring of 1985. J. Crust.Biol. 7:437-448.
Price, H. J., K. R. Boyd, and C. M. Boyd. 1988.Omnivorous feeding behavior of the antarctickrill Euphausia superba. Mar. Biol. 97:67-77.
Quetin, L. B. and R. M. Ross. 1984. Depth distri-bution of developing Euphausia superba embryos,predicted from sinking rates. Mar. Biol. 79:47-53.
Quetin, L. B. and R. M. Ross. 1988. Summary ofWINCRUISE II to the Antarctic Peninsula dur-ing June and July 1987. Ant. J., U.S. 23:149-151.
Quetin, L. B., R. M. Ross, and M. O. Amsler. 1988.Field ingestion rates of Euphausia superba. EOS68:1785.
Ross, R. M. and L. B. Quetin. 1983. Spawning fre-quency and fecundity of the antarctic krillEuphausia superba. Mar. Biol. 77:201-205.
Ross, R. M. and L. B. Quetin. 1985. The effect ofpressure on the sinking rates of the embryos ofthe antarctic krill Euphausia superba. Deep-SeaRes. 32:799-807.
Ross, R. M. and L. B. Quetin. 1986. How productiveare antarctic krill? BioScience 36:264-269.
Ross, R. M. and L. B. Quetin. 1988. Euphausia superba:
A critical review of estimates of annual produc-tion. Comp. Biochem. Physio. 90B:499-505.
Ross, R. M. and L. B. Quetin. 1989. Energetic costto develop to the first feeding stage of Euphausiasuperba Dana, and the effect of delays in foodavailability.J. Exp. Mar. Biol. Ecol. 133:103-127.
Ross, R. M., L. B. Quetin, M. O. Amsler, and M. C.Elias. 1987. Larval and adult antarctic krill,Euphausia superba, winter-over at Palmer Station.Ant.J., U.S. 22:205-206.
Ross, R. M., L. B. Quetin, and E. Kirsch. 1988. Effectof temperature on developmental times and sur-vival of early larval stages of Euphausia superbaDana. J. Exp. Mar. Biol. Ecol. 121:55-71.
Sargent, J. R. and R.J. Henderson. 1986. Lipids. InE. D. S. Corner and O. Hara (eds.), The biologicalchemistry of marine copepods, pp. 59-108. Claren-don Press, Oxford.
Siegel, V. 1987. Age and growth of antarcticEuphausiaceae (Crustacea) under natural condi-tions. Mar. Biol. 96:483-495.
Siniff, D. B. and S. Stone. 1985. The role of theleopard seal in the tropho-dynamics of the Ant-arctic marine ecosystem. In W. R. Siegfried, P.R. Condy, andR. M. Laws (eds.), Antarctic nutrientcycles and food webs, pp. 555-560. Springer-Ver-lag, Berlin, Heidelberg, New York, Tokyo.
Smith, S. J. and J. Vidal. 1986. Variations in thedistribution, abundance, and development ofcopepods in the southeastern Bering Sea in 1980and 1981. Cont. Shelf Res. 5:215-239.
Smith, W. O., Jr., N. K. Keene, and J. C. Comiso.1988. Interannual variability in estimated pri-mary productivity of the antarctic marginal icezone. In D. Sahrhage (ed.), Antarctic ocean andresources variability, pp. 131-139. Springer-Ver-lag, Berlin, Heidelberg.
Smith, W. O.,Jr. and D. M. Nelson. 1986. Impor-tance of ice edge phytoplankton production inthe Southern Ocean. BioScience 36:251-257.
Stepnik, R. 1982. All-year populational studies ofEuphausiaceae (Crustacea) in the Admiralty Bay(King George Island, South Shetland Islands,Antarctic). Pol. Polar Res. 3:49-68.
Stretch, J. J., P. P. Hamner, W. M. Hamner, W. C.Michel, J. Cook, and C. W.Sullivan. 1988. For-aging behavior of antarctic krill Euphausia superbaon sea ice microalgae. Mar. Ecol. Prog. Ser. 44:131-139.
Williams, R. 1985. Trophic relationships betweenpelagic fish and euphausiids in Antarctic waters.In W. R. Siegfried, P. R. Condy, and R. M. Laws(eds.), Antarctic nutrient cycles and food webs, pp.452-459. Springer-Verlag, Berlin, Heidelberg,New York, Tokyo.
Zwally, H. J., C. L. Parkinson, andj. C. Comiso. 1983.Variability of antarctic sea ice and changes incarbon dioxide. Science 220:1005-1012.
at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from

at University of C
alifornia, Santa Barbara on January 8, 2015
http://icb.oxfordjournals.org/D
ownloaded from
















