
BAMBI gene is epigenetically silenced in subset of high-grade bladder cancer
11
BAMBI gene is epigenetically silenced in subset of high-grade bladder cancer Sann Sanda Khin 1,2 , Riko Kitazawa 1 , Ne Win 3 , Than Than Aye 4 , Kiyoshi Mori 1 , Takeshi Kondo 1 and Sohei Kitazawa 1 * 1 Department of Pathology, Kobe University Graduate School of Medicine, Kobe, Japan 2 Pathology Research Division, Department of Medical Research (Lower Myanmar), Yangon, Union of Myanmar 3 National Health Laboratory, Yangon, Union of Myanmar 4 New Yangon General Hospital, Yangon, Union of Myanmar The bone morphogenetic protein (BMP) and activin membrane- bound inhibitor (BAMBI) is a transmembrane TGFRI/BMPRI- related pseudoreceptor, antagonizes transforming growth factor (TGF)-b/BMP signaling by inhibiting the formation of functional authentic receptor complexes (TGFRI/BMPRI and TGFRII/ BMPRII). On the assumption that BAMBI gene expression is epi- genetically altered during human bladder cancer progression, we screened the expression of BAMBI protein by immunohistochem- istry and the methylation status of the BAMBI promoter. In the normal or reactive urothelium, BAMBI expression was mostly overlapped with that of BMPRI, and a similar colocalization pat- tern was noted in low-grade papillary cancers. In high-grade and invasive cancers, however, mainly two reciprocal immunohisto- chemical expression patterns were observed: BAMBI-low/BMPRI- high, and BAMBI-high/BMPRI-low, indicating that BAMBI expression is controlled such that it does not interfere with the responsiveness of high-grade cancer cells to TGF-b/BMP signal- ing. Moreover, methylation of the BAMBI gene correlated signifi- cantly with negative BAMBI expression in bladder tumors. Although BAMBI overexpression significantly increased the num- ber of apoptotic cells in T24 line, knock-down small interfering RNA showed no remarkable change. Cell motility assay revealed that on treatment with either TGF-b1 or BMP2, T24 and HTB9 lines showed a marked increase in the number of migrated cells which, however, decreased significantly through the forced expression of BAMBI. Since certain subsets of aggressive tumors often promote cell motility, invasion and survival by inducing epi- thelial-to-mesenchymal transition through TGF-b/BMP in an autocrine and paracrine manner, hypermethylation of the BAMBI gene promoter that leads to BAMBI gene suppression may be one of the epigenetic events affecting the invasiveness or aggressive- ness of bladder cancers. ' 2009 UICC Key words: BAMBI; methylation; TGF-b; cell motility; bladder cancer Urinary bladder cancer is the seventh most common cancer worldwide with an annual estimated 260,000 new cases in men and 76,000 in women. 1 Of bladder tumors, 70–80% are, at the time of initial examination, detected as low-grade noninvasive or minimally invasive papillary tumors (pTa or pT1) with a high rate of recurrence, but with a low tendency for progression to advanced disease. 2,3 The remaining 20% are, despite improvements in diag- nostic methods and treatments, detected as high-grade invasive cancer with poor prognosis, 4 and seem as either superficial spread- ing flat (carcinoma in situ/pTis) or invasive nodular tumors. By conventional histopathological grading and staging of bladder can- cer, however, especially at initial biopsy obtained from the super- ficial area or urinary cytological samples, it is often difficult to foresee which tumors will recur or progress to advanced stages. Further understanding of the underlying molecular pathways and their roles in bladder carcinogenesis and progression is, therefore, needed to obtain new molecular markers for risk assessment of bladder cancer. Transforming growth factor-b (TGF-b)/bone morphogenetic protein (BMP) signaling plays an important role in a broad spec- trum of biology including cell proliferation, differentiation, migra- tion, immune response, angiogenesis and apoptosis. Activin, a member of the TGF-b superfamily, also regulates a wide variety of cellular events, including cell proliferation, differentiation and apoptosis. 5–7 The role of TGF-b/BMP in cancer development and progression is also complex and often controversial, involv- ing both aspects of tumor suppression and promotion according to tumor type and stage. The deregulation of activin/BMP signals also contributes to the development of cancer. The receptors and their ligand-binding features, as well as receptor-SMAD interac- tion, are all similar to those of TGF-b; the activation mechanism of type-I receptor, through phosphorylation in the GS (glycine- and serine-rich sequence) domain by type-II receptor, is con- served among TGF-b, activin and BMP. In general, TGF-b/BMP signaling initially plays the role as a tumor suppressor in epithe- lial cells, and then as a promoter for invasion and metastasis dur- ing the later stages of carcinoma progression. 7–11 Numerous genetic alterations in components associated with the TGF-b/ BMP signaling pathway that correlate with carcinogenesis, tumor progression and prognosis in various types of malignancies have been identified. 8,12,13 In colorectal cancers, for example, where the TGF-b/BMP signaling pathway in general negatively regu- lates tumor cell proliferation from early carcinogenesis to a later progression stage, loss-of-function mutation in TGF-b/BMP sig- naling, such as the inactivating mutation of SMAD4, has fre- quently been identified as an important early carcinogenic event. 10,14,15 On the other hand, in various types of cancers espe- cially at later cancer progression stages, TGF-b/BMPs, fre- quently produced by cancer cells themselves, promote angiogen- esis and epithelial-to-mesenchymal transition (EMT). Further- more, in a certain subset of aggressive cancers, the TGF-b/BMP signaling pathway often promotes cancer cell invasion and migration in an autocrine and paracrine manner. 9,10,13 The BMP and activin membrane-bound inhibitor (BAMBI), a transmembrane protein preserved from Xenopus to Homo sapiens, is highly homologous to TGF-b/BMP type-I receptors (TGFRI/ BMPRI), except that it lacks an intracellular serine/threonine- kinase domain. It is incorporated into complexes with TGF-b/ BMP/activin type-II receptors (TGFRII/BMPRII) and, as a pseu- doreceptor, antagonizes all TGF-b/BMP and activin signal- ing. 5,7,16–19 BAMBI is coexpressed with BMP4 in many develop- mental processes during embryogenesis in Xenopus, zebrafish and mouse. 17,20,21 Because BMP4 signaling is an essential and highly regulated process during many developmental processes, BAMBI, Additional Supporting Information may be found in the online version of this article. Grant sponsors: Ministry of Health and Welfare, Japan, Ministry of Edu- cation, Culture, Sports, Science and Technology, Japan; Grant numbers: 16590278, 19390100, 19659086, 20790283, 18590372. *Correspondence to: Division of Pathology, Kobe University Graduate School of Medicine, 7-5-1 Kusunoki-cho, Chuo-ku, Kobe 650-0017, Japan. Fax: 181-78-362-0297. E-mail: [email protected] Received 22 August 2008; Accepted after revision 16 January 2009 DOI 10.1002/ijc.24318 Published online 3 February 2009 in Wiley InterScience (www.interscience. wiley.com). Abbreviations: BAMBI, BMP and activin membrane-bound inhibitor; BMP, bone morphogenetic protein; BMPR, BMP receptor; CpG, phospho- diester-linked cytosine-guanine pair; GFP, green fluorescent protein; sFRP, secreted frizzled-related gene; MSP, Methylation-specific polymer- ase chain reaction; RT-PCR, reverse transcription-polymerase chain reac- tion; (Q-RT) PCR, quantitative real time polymerase chain reaction; TGF- b, transforming growth factor-b; TGFR, TGF-b receptor. Int. J. Cancer: 125, 328–338 (2009) ' 2009 UICC Publication of the International Union Against Cancer
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of BAMBI gene is epigenetically silenced in subset of high-grade bladder cancer

BAMBI gene is epigenetically silenced in subset of high-grade bladder cancer
Sann Sanda Khin1,2, Riko Kitazawa
1, Ne Win
3, Than Than Aye
4, Kiyoshi Mori
1, Takeshi Kondo
1and Sohei Kitazawa
1*
1Department of Pathology, Kobe University Graduate School of Medicine, Kobe, Japan2Pathology Research Division, Department of Medical Research (Lower Myanmar), Yangon, Union of Myanmar3National Health Laboratory, Yangon, Union of Myanmar4New Yangon General Hospital, Yangon, Union of Myanmar
The bone morphogenetic protein (BMP) and activin membrane-bound inhibitor (BAMBI) is a transmembrane TGFRI/BMPRI-related pseudoreceptor, antagonizes transforming growth factor(TGF)-b/BMP signaling by inhibiting the formation of functionalauthentic receptor complexes (TGFRI/BMPRI and TGFRII/BMPRII). On the assumption that BAMBI gene expression is epi-genetically altered during human bladder cancer progression, wescreened the expression of BAMBI protein by immunohistochem-istry and the methylation status of the BAMBI promoter. In thenormal or reactive urothelium, BAMBI expression was mostlyoverlapped with that of BMPRI, and a similar colocalization pat-tern was noted in low-grade papillary cancers. In high-grade andinvasive cancers, however, mainly two reciprocal immunohisto-chemical expression patterns were observed: BAMBI-low/BMPRI-high, and BAMBI-high/BMPRI-low, indicating that BAMBIexpression is controlled such that it does not interfere with theresponsiveness of high-grade cancer cells to TGF-b/BMP signal-ing. Moreover, methylation of the BAMBI gene correlated signifi-cantly with negative BAMBI expression in bladder tumors.Although BAMBI overexpression significantly increased the num-ber of apoptotic cells in T24 line, knock-down small interferingRNA showed no remarkable change. Cell motility assay revealedthat on treatment with either TGF-b1 or BMP2, T24 and HTB9lines showed a marked increase in the number of migrated cellswhich, however, decreased significantly through the forcedexpression of BAMBI. Since certain subsets of aggressive tumorsoften promote cell motility, invasion and survival by inducing epi-thelial-to-mesenchymal transition through TGF-b/BMP in anautocrine and paracrine manner, hypermethylation of the BAMBIgene promoter that leads to BAMBI gene suppression may be oneof the epigenetic events affecting the invasiveness or aggressive-ness of bladder cancers.' 2009 UICC
Key words: BAMBI; methylation; TGF-b; cell motility; bladdercancer
Urinary bladder cancer is the seventh most common cancerworldwide with an annual estimated 260,000 new cases in menand 76,000 in women.1 Of bladder tumors, 70–80% are, at thetime of initial examination, detected as low-grade noninvasive orminimally invasive papillary tumors (pTa or pT1) with a high rateof recurrence, but with a low tendency for progression to advanceddisease.2,3 The remaining 20% are, despite improvements in diag-nostic methods and treatments, detected as high-grade invasivecancer with poor prognosis,4 and seem as either superficial spread-ing flat (carcinoma in situ/pTis) or invasive nodular tumors. Byconventional histopathological grading and staging of bladder can-cer, however, especially at initial biopsy obtained from the super-ficial area or urinary cytological samples, it is often difficult toforesee which tumors will recur or progress to advanced stages.Further understanding of the underlying molecular pathways andtheir roles in bladder carcinogenesis and progression is, therefore,needed to obtain new molecular markers for risk assessment ofbladder cancer.
Transforming growth factor-b (TGF-b)/bone morphogeneticprotein (BMP) signaling plays an important role in a broad spec-trum of biology including cell proliferation, differentiation, migra-tion, immune response, angiogenesis and apoptosis. Activin, amember of the TGF-b superfamily, also regulates a wide varietyof cellular events, including cell proliferation, differentiation and
apoptosis.5–7 The role of TGF-b/BMP in cancer developmentand progression is also complex and often controversial, involv-ing both aspects of tumor suppression and promotion accordingto tumor type and stage. The deregulation of activin/BMP signalsalso contributes to the development of cancer. The receptors andtheir ligand-binding features, as well as receptor-SMAD interac-tion, are all similar to those of TGF-b; the activation mechanismof type-I receptor, through phosphorylation in the GS (glycine-and serine-rich sequence) domain by type-II receptor, is con-served among TGF-b, activin and BMP. In general, TGF-b/BMPsignaling initially plays the role as a tumor suppressor in epithe-lial cells, and then as a promoter for invasion and metastasis dur-ing the later stages of carcinoma progression.7–11 Numerousgenetic alterations in components associated with the TGF-b/BMP signaling pathway that correlate with carcinogenesis, tumorprogression and prognosis in various types of malignancies havebeen identified.8,12,13 In colorectal cancers, for example, wherethe TGF-b/BMP signaling pathway in general negatively regu-lates tumor cell proliferation from early carcinogenesis to a laterprogression stage, loss-of-function mutation in TGF-b/BMP sig-naling, such as the inactivating mutation of SMAD4, has fre-quently been identified as an important early carcinogenicevent.10,14,15 On the other hand, in various types of cancers espe-cially at later cancer progression stages, TGF-b/BMPs, fre-quently produced by cancer cells themselves, promote angiogen-esis and epithelial-to-mesenchymal transition (EMT). Further-more, in a certain subset of aggressive cancers, the TGF-b/BMPsignaling pathway often promotes cancer cell invasion andmigration in an autocrine and paracrine manner.9,10,13
The BMP and activin membrane-bound inhibitor (BAMBI), atransmembrane protein preserved from Xenopus to Homo sapiens,is highly homologous to TGF-b/BMP type-I receptors (TGFRI/BMPRI), except that it lacks an intracellular serine/threonine-kinase domain. It is incorporated into complexes with TGF-b/BMP/activin type-II receptors (TGFRII/BMPRII) and, as a pseu-doreceptor, antagonizes all TGF-b/BMP and activin signal-ing.5,7,16–19 BAMBI is coexpressed with BMP4 in many develop-mental processes during embryogenesis in Xenopus, zebrafish andmouse.17,20,21 Because BMP4 signaling is an essential and highlyregulated process during many developmental processes, BAMBI,
Additional Supporting Information may be found in the online versionof this article.
Grant sponsors: Ministry of Health and Welfare, Japan, Ministry of Edu-cation, Culture, Sports, Science and Technology, Japan; Grant numbers:16590278, 19390100, 19659086, 20790283, 18590372.*Correspondence to: Division of Pathology, Kobe University Graduate
School of Medicine, 7-5-1 Kusunoki-cho, Chuo-ku, Kobe 650-0017, Japan.Fax: 181-78-362-0297. E-mail: [email protected]
Received 22 August 2008; Accepted after revision 16 January 2009DOI 10.1002/ijc.24318Published online 3 February 2009 in Wiley InterScience (www.interscience.
wiley.com).
Abbreviations: BAMBI, BMP and activin membrane-bound inhibitor;BMP, bone morphogenetic protein; BMPR, BMP receptor; CpG, phospho-diester-linked cytosine-guanine pair; GFP, green fluorescent protein;sFRP, secreted frizzled-related gene; MSP, Methylation-specific polymer-ase chain reaction; RT-PCR, reverse transcription-polymerase chain reac-tion; (Q-RT) PCR, quantitative real time polymerase chain reaction; TGF-b, transforming growth factor-b; TGFR, TGF-b receptor.
Int. J. Cancer: 125, 328–338 (2009)
' 2009 UICC
Publication of the International Union Against Cancer

by forming a short and direct negative feedback loop, is specu-lated to play a role in the fine-tuning of BMP4 signaling duringembryonic development.17 Little is known, however, aboutwhether or not, or how BAMBI is involved in carcinogenesis andtumor progression, except from few studies that show BAMBI neg-atively regulates TGF-b/BMP signaling in colorectal, hepatocellu-lar and gastric carcinoma.18,22,23
In the current study, on the assumption that BAMBI geneexpression is epigenetically altered during human bladder cancerprogression, we first screened the expression pattern of BAMBIprotein by immunohistochemistry and the methylation status ofthe BAMBI gene promoter with the use of clinical specimens fromJapan and Myanmar, and then examined the functional aspects ofthe BAMBI gene by forced-expression and knock-down in cancercell lines in vitro.
Material and methods
Cell lines and tissue preparation
Three human bladder cancer cell lines [T24, HTB9 (ATCC,Manassas, VA) and KU1 (Keio University, Tokyo, Japan)] andfive human colon and one breast cancer cell lines (COLO201,SW480, HT29, DLD-1, COLO320 and MCF7) (ATCC, Manassas,VA) were cultured in RPMI1640 supplemented with 10% fetal bo-vine serum, 100 U/ml penicillin and 100 U/ml streptomycin, andmaintained at 37 �C in a humidified atmosphere with 5% CO2.Formalin-fixed and paraffin-embedded tissue samples of 74 pri-mary urinary bladder tumors (41 from Japanese patients, meanage 65.4 6 4.4 years; and 33 Myanmarese patients, mean age60 6 4 years) (Table I) were used for H&E staining to determinepathological diagnosis according to the criteria of the WorldHealth Organization (2004) and the International Society of Uro-logical Pathology. This study was reviewed and approved by thelocal ethics committee at Kobe University.
Methylation analysis
DNA extraction and bisulfite modification. The bisulfite reac-tion was carried out as described.24 Briefly, 10 ll of low-meltingagarose beads, containing 0.5 lg genomic DNA derived fromeach cell line and paraffin tissue samples (n574), were formed inmineral oil and denatured with NaOH (final concentration, 0.1 M).The beads were dipped in 1 ml of freshly prepared bisulfite solu-tion containing 1 mM hydroquinone and 3.5 M NaHSO3 at pH 5.The liquid surface was sealed with mineral oil, and the samples
were incubated for 16 hr at 50 �C. The beads were then washedwith Tris-EDTA, desulfonated with 0.5 N NaOH for 30 min, neu-tralized with 1 N HCl and finally washed with Tris-EDTA. Beadfragments containing up to 0.1 lg DNA were used for polymerasechain reaction (PCR).
Methylation-specific PCR. The methylation status of p16,sFRP1, sFRP2, sFRP4, sFRP5 and BAMBI genes was analyzed bymethylation-specific polymerase chain reaction (MSP) after bisul-fite conversion of the respective DNA samples, using the sets ofprimers as previously published.25–27 Briefly, 1 ll bisulfite-modi-fied DNA was amplified in a total volume of 25 ll containing 103CoralLoad PCR buffer, 53 Q solution, 200 lM of each dNTP, 20qmol of each primer and HotStar Taq Plus DNA Polymerase(QIAGEN, Hilden, Germany). Amplification included hot-start at95 �C for 5 min, denaturing at 94 �C for 30 sec, and annealing (at56 �C for p16 and 50 �C for sFRPs and BAMBI) for 45 sec,elongation at 72 �C for 60 sec for a total of 40 cycles and a final10 min for extension. Control DNA with known methylated andunmethylated samples was used in each MSP assay to monitor thetechnical process. PCR products were analyzed by 3% agarose gelelectrophoresis and visualized under ultraviolet (UV) illumination.For BAMBI MSP, we had further confirmed the methylation statusof BAMBI by nested MSP using the new sets of primers (Fig. 1and Supplementary Table I). In nested MSP, the 30 end of the pri-mers used for the first PCR was set to the position of cytosine inthe non-CpG locus so that the bisulfite converted DNA for bothmethylated and unmethylated populations could be amplified. Inthe second PCR, the 30 end of the primers was set to the positionof cytosine in the CpG locus to selectively amplify either themethylated (M) or the unmethylated (U) DNA. The first PCRamplification included hot-start at 95 �C for 5 min, denaturing at94 �C for 30 sec, annealing at 57 �C for 45 sec and elongation at72 �C for 60 sec for a total of 30 cycles and a final 10 min forextension; the nested PCR amplification started with 95 �C for5 min, denaturing at 94 �C for 30 sec, annealing at 54 �C for45 sec, elongation at 72 �C for 60 sec for a total of 25 cycles anda final 10 min for extension. The nested PCR products were ana-lyzed by 3% agarose gel electrophoresis and visualized under UVillumination. At the same time, the first PCR product was clonedinto the TA vector (Invitrogen, Carlsbad, CA) and at least 10 inde-pendent clones were selected and sequenced using ABI PRISMTM
310 Genetic Analyzer (Perkin-Elmer, Norwalk, CT) to confirm theaccuracy of nested MSP. Methylation analysis of normal tissues orthe nontumorous surrounding urothelium was carried out for eachcase.
Immunohistochemistry and Immunocytochemistry
Formalin-fixed, paraffin-embedded tissues from the 74 urinarybladder tumor specimens were deparaffinized in xylene and rehy-drated in a series of graded ethanol. Antigens of tissue and cyto-logical slides were then retrieved by microwave treatment at 350
TABLE I – CLINICOPATHOLOGICAL DATAOF BLADDER CANCER PATIENTS
StatusBladder cancer cases
Japan, n (%) Myanmar, n (%)
No. patients 41 33Male 36 (48.6) 24 (32.4)Female 5 (6.8) 9 (12.2)
Age in years (mean) 65.4 6 4.4 60 6 4Pathologic grade
High grade 27 (36.5) 20 (27)Low grade 14 (19) 13 (17.5)
Pathologic stagepTa 17 (23) 15 (20.2)pTis – 1 (1.35)pT1 15 (20.2) 9 (12.1)pT2 9 (12.1) 8 (11)
InvasionPresence 24 (32.4) 17 (23)Absence 17 (23) 16 (21.6)
TCC 41 (55.4) 30 (40.5)Other (SCC or adenocarcinoma) – 3 (4.05)
n indicates number of cases; TCC, transitional cell carcinoma;SCC, squamous cell carcinoma. Two cases of pT1 and one case ofpT2 associate with pTis.
FIGURE 1 – A schema of the human BAMBI gene promoter region.CpG loci (indicated by vertical lines) are scattered in the 50-flankingregion of the BAMBI gene and form a CpG-island. TATA-box, 3 tan-dem repeats of SMAD binding elements (3XSBE) and a T-cell tran-scription factor (Tcf) are shown. Three pairs of primers 120bps for thefirst PCR and 100bps for nested PCR (horizontal gray arrows) wereused for nested methylation-specific PCR. TSS indicates transcriptionstart site.
329EPIGENETIC INACTIVATION OF BAMBI IN BLADDER CANCER

watts for 10 min in 10 mM citrate buffer (pH 6.0). The slides wereincubated in a mixture of 3% H2O2 and methanol for 10 min atroom temperature to block endogenous peroxidase activity. Aftertreatment with a blocking antibody, the primary antibodies wereadded and the slides were incubated for 60 min at room tempera-ture. Anti-human BAMBI/NMA antibody (goat IgG) raised againstNSO/nonsecreting mouse myeloma cell-derived human BAMBI(aa 1-152) (R&D Systems, Minneapolis, MN), mouse monoclonalanti-human b-catenin (IgG1, kappa) raised against recombinantC-terminal b-catenin-GST fusion protein (DAKO, Carpinteria,CA) and rabbit polyclonal antibody raised against a recombinantprotein corresponding to amino acids 466-510 mapping near thecarboxy terminus of bone morphogenetic protein receptor type-I,BMPRI(H-45), which was recommended for the detection ofBMPRIA and BMPRIB of human origin (Santa Cruz, CA) wereused as primary antibodies. Nonimmunized mouse serum wasused for preparing negative controls. The streptavidin-biotin-per-oxidase complex method was used with a MaxiTagsTM kit (Ther-moShandon, Pittsburgh, PA). Final development of the sectionswas carried out with 3,30-diaminobenzidine containing 0.03%H2O2. Immunohistochemical staining was graded as negative (nostaining) and positive (immunoreactive staining) for BAMBI andBMPRI protein expression, and as negative (no staining), membra-nous and nucleus (immunoreactive staining) for b-catenin proteinexpression.
Mutation analysis
Genomic DNA containing agarose beads (74 tissue samples)without bisulfite treatment were used for mutational analysis onexon3 of the b-catenin gene. First, nested PCR was done by usingthe following sets of primers. For the first PCR, 50-ATTTGATGGAGTTGGACATG GC-30, sense and 50-CCAGCTACTTGTTCTTGA GTGAAGG-30, antisense; for the second PCR, 50-TGGACATGGCCATGGAACCAG-30, sense and 50-TTGTTCTTGAGTGAAG GACTG-30, antisense.28 DNA (1 ll) was thenamplified in a total volume of 25 ll containing 103 CoralLoadPCR buffer, 53 Q solution, 200 lM of each dNTP, 20 qmol ofeach primer and HotStar Taq Plus DNA Polymerase. In bothPCRs, amplification started with 95 �C for 5 min, denaturing at94 �C for 30 sec, annealing at 50 �C for 45 sec, elongation at72 �C for 60 sec for a total of 35 cycles and a final 10 min forextension. PCR products were analyzed by 3% agarose gel elec-trophoresis and visualized under UV illumination. For mutationalscreening, single-strand conformation polymorphism analysis wascarried out by diluting 5 ll of the PCR product in 30 ll of loadingbuffer (0.1% bromophenol blue, 0.1% xylene cyanol and 20 mMEDTA in formamide) and then denatured at 80 �C for 5 min.Samples were run on polyacrylamide e-PAGEL electrophoresisand stained with AE-1360 Ez silver stain (ATTO, Tokyo, Japan)according to the manufacturer’s protocol. The amplified DNAshowing a positive single-strand conformation polymorphism mo-bility shift band was cloned into the TA Vector (Invitrogen, Carls-bad, CA). At least 10 independent clones were selected and therecombinant plasmids were recovered for DNA sequencing withM13F primer. The sequencing was done by using the ABI PRISMBigDye Terminator v3.1 Cycle Sequencing Kit according to themanufacturer’s instructions (Applied Biosystems, Foster City,CA). Samples were run on an ABI prism 310 Genetic Analyzer(Perkin-Elmer, Norwalk, CT).
RNA extraction, quantitative real-time PCR andreverse transcription PCR
Total RNA (1 lg), isolated from each cell line and reverse tran-scribed to produce cDNA, was then amplified and quantified bythe ABI PRISM 7300 Real Time PCR system (Applied Biosys-tems) using a set of BAMBI primers and probes (assay ID:Hs00180818_m1) and Human 18S primers and probes purchasedfrom Biosearch Tech (Applied Biosystems). The amount of
mRNA was quantified relative to that of 18S in each reaction byusing the following sets of primers:
� 18S (forward): 50d (ATGAGTCCACTTTAAATCCTTTAACGA) 30
� 18S (reverse): 50d (CTTTAATATACGCTATTGGAGCTGGAA) 30
� 18S (probe): 50FAMd (ATCCATTGGAGGGCAAGTCTGGTGC) BHQ-1 30
Small interfering RNA transfection
Cells (2 3 105/well) of each bladder cancer cell line were platedin a six-well plate in OPTI-MEM (Invitrogen, Carisbad, CA) con-taining 10% fetal bovine serum for 24 hr to yield 60–80% conflu-ency. The cell lines were then transiently transfected with BAMBIsmall interfering RNA (siRNA) or nontargeting (random) siRNA(Ambion, Austin TX) using SilencerTM siRNA Transfection Kit II(Ambion) for 48 hr. Scrambled siRNA was used as a control. Aftertransfection, total RNA was extracted for quantification of BAMBIgene expression and viable cells were counted with a hemocytom-eter and apoptotic analysis was done by using APOPercentageTM
assay (Biocolor, UK). The number of purple-red stained apoptotic cellswas counted under a light microscope at 3400 magnification.
Transient transfection studies
The plasmid constructs, pEGFP-N3-hBAMBI (kindly providedby Dr. Tetsu Akiyama) and pGL3 Basic were transfected into thecell lines using LipofectamineTM LTX Reagent (Invitrogen)according to the manufacturer’s instructions. Empty vector wasused as a control. First, the bladder cancer cell lines were culturedin 12-well plates for 24 hr to yield 60–80% confluency. The mix-ture of 1 ll (1 lg) of each plasmid DNA and 1 ll of PLUSTM rea-gent was then incubated for 5 min at room temperature. Lipofect-amine LTX (2.5 ll) was then added to diluted DNA and incubatedfor 30 min at room temperature. A DNA-lipofectamine complex(200 ll) was added to each well. After 48 hr incubation in 5%CO2 at 37 �C, apoptotic analysis was done by using Annexin V-Cy3 Apoptosis Detection Kit (BioVision, Mountain View, CA).Briefly, the cells were stained with Annexin V-Cy3 solution(1:100 dilution) for 5 min in the dark and analyzed under BZ-9000BIOREVO fluorescence microscope (KEYENCE, Osaka, Japan).The number of red apoptotic cells/HPF (high power field) underrhodamine filter and yellow to orange pEGFP-N3-hBAMBI express-ing apoptotic cells/HPF (high power field) were detected for eachcell line.
5-Aza-20-deoxycytidine (5-aza-dC) treatment
Steady-state BAMBI mRNA expression was examined inHTB9, T24 and KU1 cells with or without 5-aza-dC treatment,while changes in the methylation status of the BAMBI promoterregion were monitored by sodium bisulfite mapping. Exponen-tially growing cells were seeded at a density of 1.5 3 106 cells/60mm-diameter dish and allowed to attach overnight. The cells werethen treated with freshly prepared 1 lM 5-aza-dC (Sigma, St.Louis, MO) (day 0). After 24, 72 and 120 hr (Days 1, 3 and 5),treatment of the cells was continued by changing to a fresh me-dium containing 5-aza-dC. After 168 hr (Day 7), genomic DNAand total RNA were collected from each dish and purified bystandard methods. After confirming through sodium bisulfite map-ping that all CpG loci in the BAMBI 50-flanking region were com-pletely demethylated, RNA was used as a template for semiquanti-tative reverse transcription (RT)-PCR to evaluate BAMBI mRNAlevels.
Cell motility assay
The bladder cancer cell lines were cultured for 24 hr on 3-lmpore PET track-etched membrane inserts that formed the upperchambers of 24-well format cell culture plates (BD) to yield 80–90% confluency. Plasmid constructs pEGFP-N3-hBAMBI and
330 KHIN ET AL.
pGL3 Basic were then transfected into the cells, using Lipofect-amine LTX Reagent (Invitrogen) according to the manufacturer’sinstructions. After 48 hr transfection, the cells were treated withfreshly prepared 3 lg/ml reconstituted recombinant human TGF-b1 solution (Genzyme/Techne, USA), and incubated in 5% CO2
incubator at 37 �C. The cells were allowed to migrate for 24 hr,and the insert chambers were removed and stained with Giemsa.The membranes were air dried, detached from the inserts and fixedonto microscope slides. The migratory cells on each membraneinsert were counted under a light microscope at 3400 magnifica-tion. Empty vector was used as a control. A similar procedure wasfollowed for BMP-2-induced cell motility assay with 1 mg/ml ofrecombinant human (rh) BMP-2 (kindly provided by AstellasPharma Inc., Tokyo, Japan).
Western blotting
Five human colon and one breast cancer cell lines (COLO201,SW480, HT29, DLD-1, COLO320 and MCF7) were lysed with 1.0ml of ice-cold lysis buffer [50 mM Tris-HCl (pH 8.0), 100 mMNaCl, 0.2% Nonidet P-40, 0.2% sodium deoxycholate, 0.1% sodiumdodecyl sulfate (SDS), and 50 lM/ml aprotinin]. After sonicationfor 1 min, the lysates were centrifuged at 15,000 rpm (revolutionsper minute) for 20 min. The supernatant, equalized by protein con-centrations, was separated by 10% SDS-PAGE, transferred to thenylon membrane, Hybond-P (Amersham Biosciences, Piscataway,NJ) and immunoblotted with monoclonal anti-human b-catenin anti-
bodies (DAKO, Carpinteria, CA) against b-actin (BT-560; Biomedi-cal Technologies, Stoughton, MA). Immunocomplexes were visual-ized by the enhanced chemiluminescence Western blotting system(Amersham) according to the manufacturer’s instructions.
Statistical analysis
Statistical analysis was carried out using JMP 7.0/SAS soft-ware. Categorical variables from primary bladder tumors fromJapanese and Myanmarese cases were plotted into 2 3 2 contin-gency tables and evaluated using the Chi-square and Fisher’sExact Test. Results of apoptosis and cell motility in cell culturewere plotted as mean 6 SD and analyzed with Student’s t test. Allreported p values were two sided and considered significant whenp value was <0.05.
Results
Immunohistochemical expression of BAMBI, BMPRIand b-catenin in normal and reactive urothelium
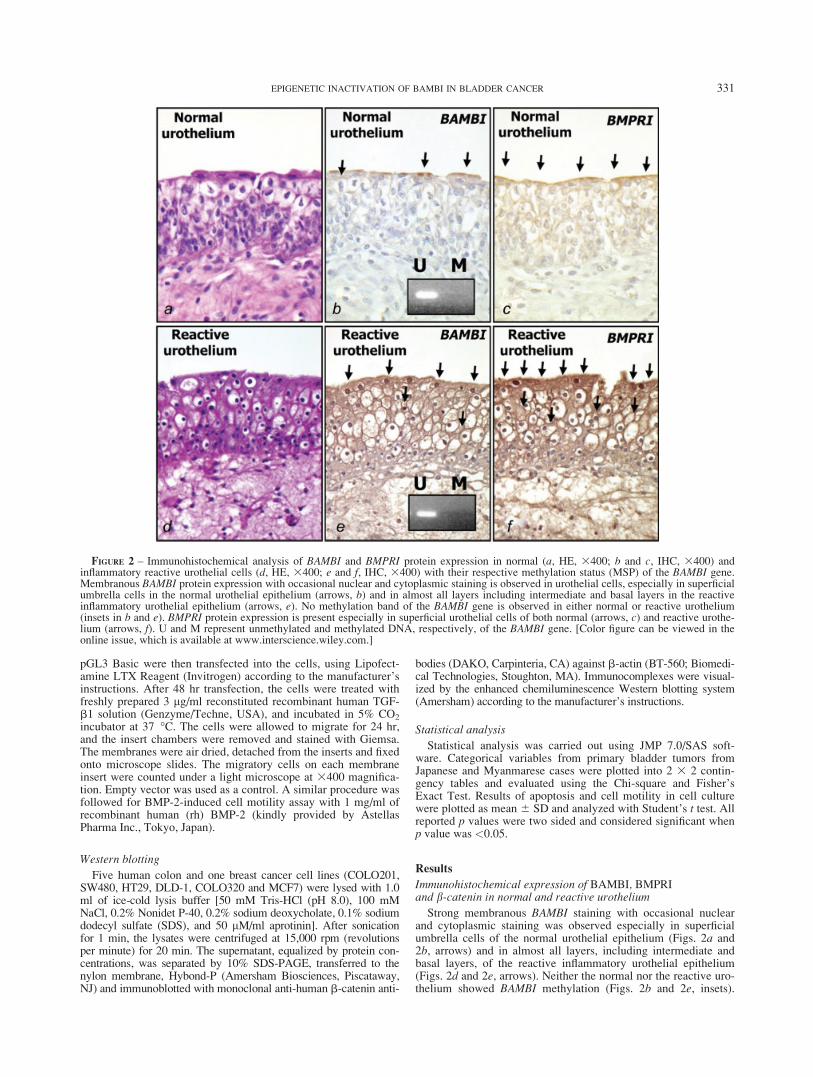
Strong membranous BAMBI staining with occasional nuclearand cytoplasmic staining was observed especially in superficialumbrella cells of the normal urothelial epithelium (Figs. 2a and2b, arrows) and in almost all layers, including intermediate andbasal layers, of the reactive inflammatory urothelial epithelium(Figs. 2d and 2e, arrows). Neither the normal nor the reactive uro-thelium showed BAMBI methylation (Figs. 2b and 2e, insets).
FIGURE 2 – Immunohistochemical analysis of BAMBI and BMPRI protein expression in normal (a, HE, 3400; b and c, IHC, 3400) andinflammatory reactive urothelial cells (d, HE, 3400; e and f, IHC, 3400) with their respective methylation status (MSP) of the BAMBI gene.Membranous BAMBI protein expression with occasional nuclear and cytoplasmic staining is observed in urothelial cells, especially in superficialumbrella cells in the normal urothelial epithelium (arrows, b) and in almost all layers including intermediate and basal layers in the reactiveinflammatory urothelial epithelium (arrows, e). No methylation band of the BAMBI gene is observed in either normal or reactive urothelium(insets in b and e). BMPRI protein expression is present especially in superficial urothelial cells of both normal (arrows, c) and reactive urothe-lium (arrows, f). U and M represent unmethylated and methylated DNA, respectively, of the BAMBI gene. [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
331EPIGENETIC INACTIVATION OF BAMBI IN BLADDER CANCER

BMPRI protein expression was observed as overlapping withBAMBI expression but seen mostly in the superficial layer (Figs.2c and 2f, arrows). Although commercially available and widelyused for the detection of BMPRIA and BMPRIB of human originby BMPRI (H-45) immunohistochemistry, we cannot exclude thepossibility that this antibody may still cross-react with othersimilar molecules. The b-catenin expression pattern was mainlylinear membranous in both the normal and reactive urothelium(data not shown). Negative controls prepared with nonimmunizedmouse serum did not show any significant staining (Supplemen-tary Fig. 4).
Immunohistochemical expression of BAMBI, BMPRIand b-catenin in primary bladder tumors
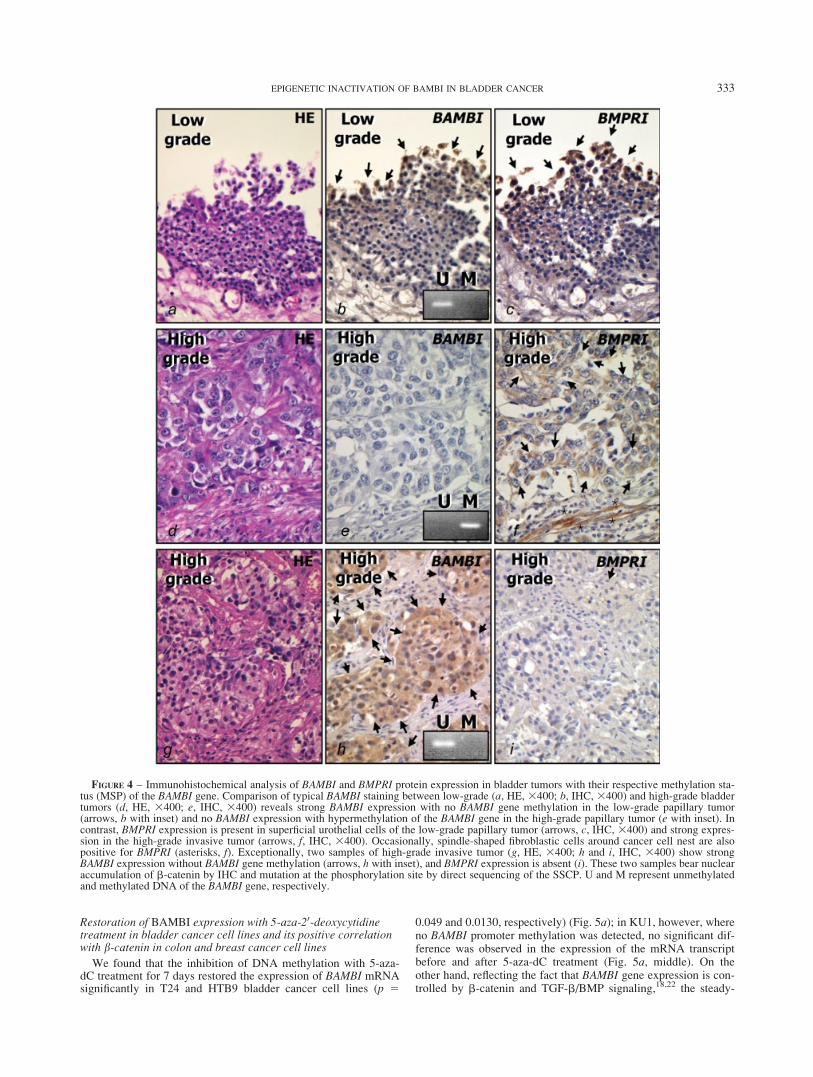
The results of immunohistochemical analyses, and epigeneticaland genetical studies are summarized in Figure 3. In 44.59% (33/74) of cases, tumor cells showed diffuse and membrano-perinu-clear as well as some nuclear localization of BAMBI expression(>25% of cells). On the other hand, BMPRI was expressed in63.5% (47/74) of cases, and a significant correlation was observedbetween strong intensity with high percentage (>75%) of BMPRIexpression and high-grade bladder tumors (p < 0.0001). A signifi-cant correlation was observed between high-grade invasive tumorsand the absence of BAMBI immunoexpression (p < 0.0001) withpositive BMPRI immunostaining (Table I and Fig. 3). The histo-pathological and immunohistochemical findings of representativecases are shown in Figure 4. Coexpression of BAMBI and BMPRIwas frequently found in low-grade tumors (Figs. 4a–4c). In high-grade invasive cancers, a BAMBI-low and BMPRI-high patternwas frequently observed (Figs. 4d–4f). The most frequent expres-sion pattern of b-catenin was the membranous 74.3% (55/74), fol-lowed by no staining 22.9% (17/74) and then nuclear staining2.7% (2/74). These two cases with strong nuclear b-catenin stain-ing were high-grade tumors with positive BAMBI and negativeBMPRI immunostaining (Figs. 4g–4i), had point mutation at thephosphorylation site in exon 3 of the b-catenin gene involving nu-cleotide change from C:G to T:A transition, and amino acid sub-stitution at codon 40 from threonine to isoleucine (T40I), asrevealed by single-strand conformational polymorphism followedby direct sequencing. Negative controls prepared with nonimmu-nized mouse serum did not show any significant staining forBAMBI, BMPRI or b-catenin (Supplementary Fig. 4).
Methylation status of BAMBI, p16 and sFRP genes in primarybladder tumors and in cancer cell lines
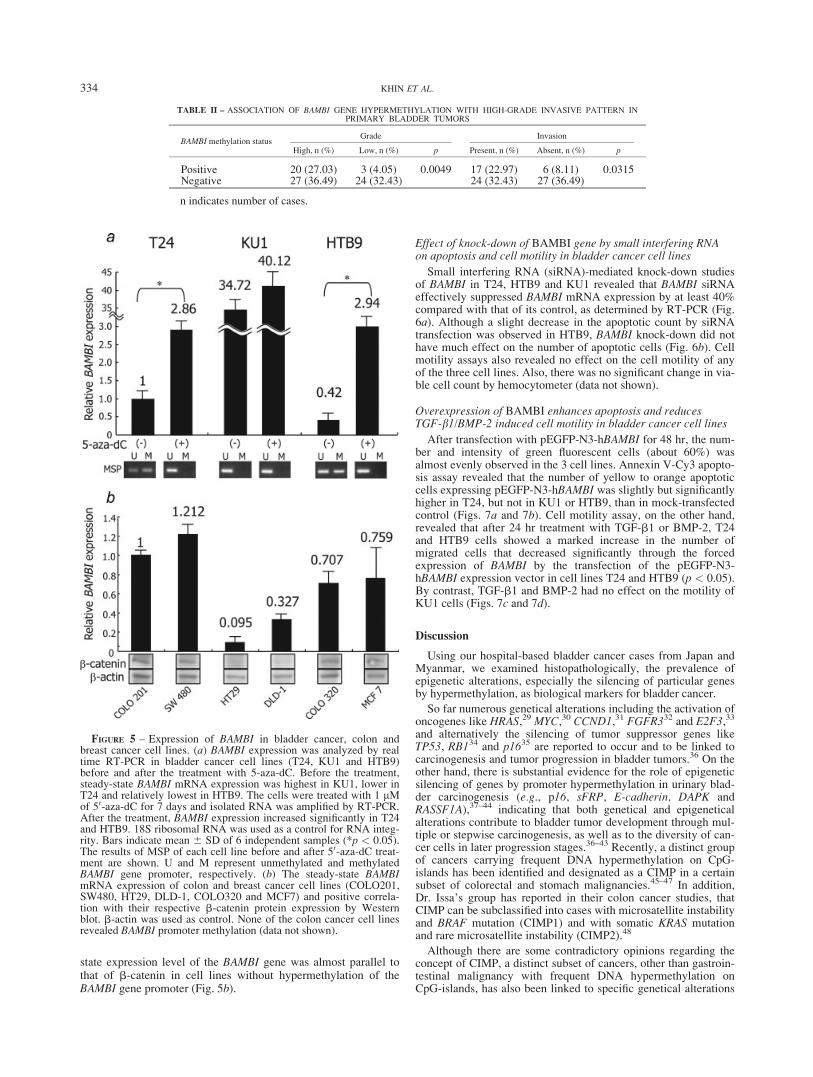
To explore whether or not the BAMBI gene is included amongCpG-island methylator phenotype (CIMP)-affected genes, theCpG-island methylator phenotype study of bladder cancer needsto be assessed. To fulfill the criteria for CIMP, we analyzed theaberrant methylation of BAMBI, p16 and sFRP genes in primarybladder tumors from Japan (n541, Fig. 3a) and Myanmar (n533,Fig. 3b), in three bladder (Fig. 3c) and five colon and one breastcancer cell lines. The prevalence of hypermethylation was 43.24%(32 of 74), 21.62% (16 of 74), 32.43% (24 of 74), 12.16% (9 of74), 50% (37 of 74) and 31.08% (23 of 74) in p16, sFRP1, sFRP2,sFRP4, sFRP5 and BAMBI, respectively. The hypermethylation ofone or more genes was significantly frequent in high-grade andinvasive tumors (p 5 0.0005 and 0.0024, respectively). Espe-cially, methylation of sFRP2 and BAMBI was significantly corre-lated with high-grade and invasive tumors (p 5 0.003 and 0.0188for sFRP2 and p 5 0.0049 and 0.0315 for BAMBI). Moreover,even in low-grade papillary tumors, the rate of methylation washigher in samples from Myanmar than in those from Japan. The fre-quent hypermethylation of the BAMBI gene was significantly corre-lated with negative BAMBI protein immunoexpression (p50.0002)and high-grade invasive primary bladder tumors (Table II). Pro-moters of these genes were also frequently hypermethylated in thebladder cancer cell lines, except that there was no methylation ofp16 in HTB9 and KU1, of sFRP5 in T24 or of BAMBI in KU1(Fig. 3c). In contrast, hypermethylation of the BAMBI promoter
region was not found in any of the five colon and one breast cancercell lines tested (data not shown). For BAMBI MSP, bisulfitesequencing data clearly reflected the MSP results: the DNA sam-ples from cases with positive MSP showed high frequency of CpGmethylation and, conversely, DNA samples from negative MSPcases revealed a low rate of or no CpG methylation.
FIGURE 3 – Summary of methylation-specific PCR (MSP) analysisof p16, sFRP1, sFRP2, sFRP4, sFRP5 and BAMBI genes; mutationalanalysis of b-catenin; BMPRI and BAMBI protein expression in the 74primary bladder carcinomas from Japan (a) and Myanmar (b), and 3bladder cancer cell lines (c). The promoter region of all genes is fre-quently methylated in a subset of high-grade primary bladder tumorsand bladder cancer cell lines; BAMBI also seems to contribute to theformation of a CpG-island methylator phenotype (CIMP). Obviously,methylation of these genes is more frequent in samples from Myanmarthan in those from Japan. The two positive cases of b-catenin muta-tion are shown with asterisks. IHC: immunohistochemistry; ICC,immunocytochemistry. [Color figure can be viewed in the onlineissue, which is available at www.interscience.wiley.com.]
332 KHIN ET AL.
Restoration of BAMBI expression with 5-aza-20-deoxycytidinetreatment in bladder cancer cell lines and its positive correlationwith b-catenin in colon and breast cancer cell lines
We found that the inhibition of DNA methylation with 5-aza-dC treatment for 7 days restored the expression of BAMBI mRNAsignificantly in T24 and HTB9 bladder cancer cell lines (p 5
0.049 and 0.0130, respectively) (Fig. 5a); in KU1, however, whereno BAMBI promoter methylation was detected, no significant dif-ference was observed in the expression of the mRNA transcriptbefore and after 5-aza-dC treatment (Fig. 5a, middle). On theother hand, reflecting the fact that BAMBI gene expression is con-trolled by b-catenin and TGF-b/BMP signaling,18,22 the steady-
FIGURE 4 – Immunohistochemical analysis of BAMBI and BMPRI protein expression in bladder tumors with their respective methylation sta-tus (MSP) of the BAMBI gene. Comparison of typical BAMBI staining between low-grade (a, HE, 3400; b, IHC, 3400) and high-grade bladdertumors (d, HE, 3400; e, IHC, 3400) reveals strong BAMBI expression with no BAMBI gene methylation in the low-grade papillary tumor(arrows, b with inset) and no BAMBI expression with hypermethylation of the BAMBI gene in the high-grade papillary tumor (e with inset). Incontrast, BMPRI expression is present in superficial urothelial cells of the low-grade papillary tumor (arrows, c, IHC, 3400) and strong expres-sion in the high-grade invasive tumor (arrows, f, IHC, 3400). Occasionally, spindle-shaped fibroblastic cells around cancer cell nest are alsopositive for BMPRI (asterisks, f). Exceptionally, two samples of high-grade invasive tumor (g, HE, 3400; h and i, IHC, 3400) show strongBAMBI expression without BAMBI gene methylation (arrows, h with inset), and BMPRI expression is absent (i). These two samples bear nuclearaccumulation of b-catenin by IHC and mutation at the phosphorylation site by direct sequencing of the SSCP. U and M represent unmethylatedand methylated DNA of the BAMBI gene, respectively.
333EPIGENETIC INACTIVATION OF BAMBI IN BLADDER CANCER
state expression level of the BAMBI gene was almost parallel tothat of b-catenin in cell lines without hypermethylation of theBAMBI gene promoter (Fig. 5b).
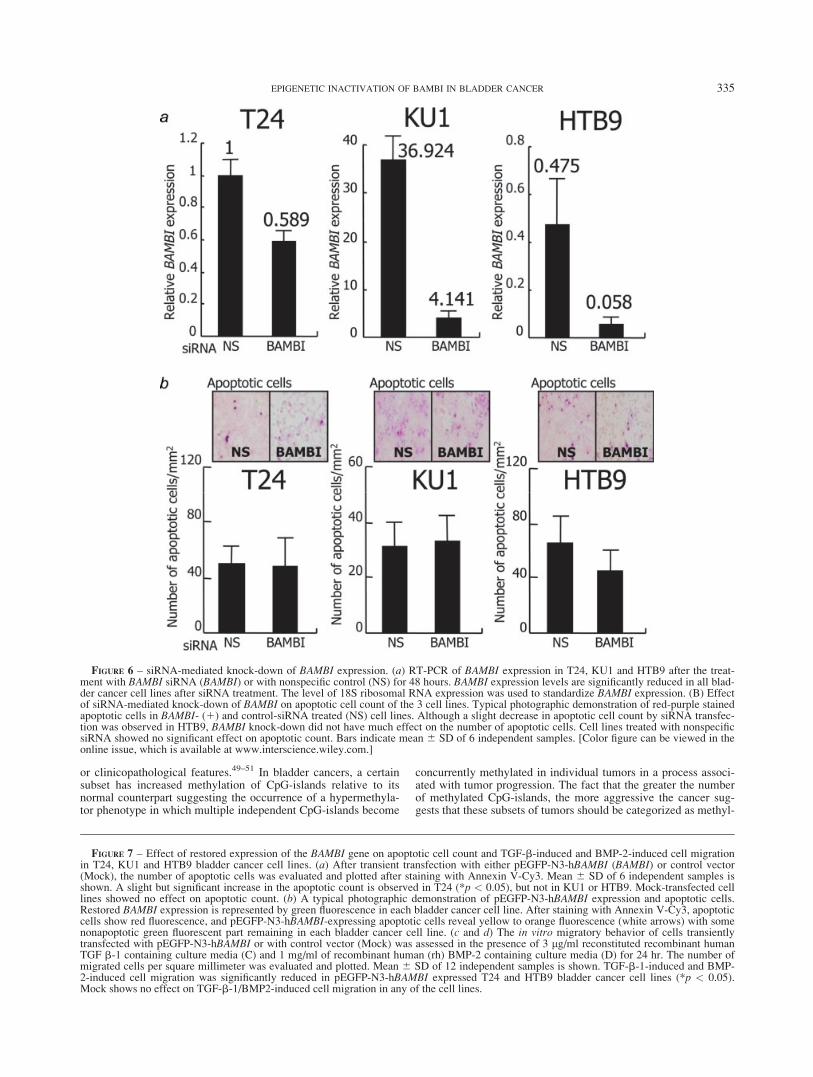
Effect of knock-down of BAMBI gene by small interfering RNAon apoptosis and cell motility in bladder cancer cell lines
Small interfering RNA (siRNA)-mediated knock-down studiesof BAMBI in T24, HTB9 and KU1 revealed that BAMBI siRNAeffectively suppressed BAMBI mRNA expression by at least 40%compared with that of its control, as determined by RT-PCR (Fig.6a). Although a slight decrease in the apoptotic count by siRNAtransfection was observed in HTB9, BAMBI knock-down did nothave much effect on the number of apoptotic cells (Fig. 6b). Cellmotility assays also revealed no effect on the cell motility of anyof the three cell lines. Also, there was no significant change in via-ble cell count by hemocytometer (data not shown).
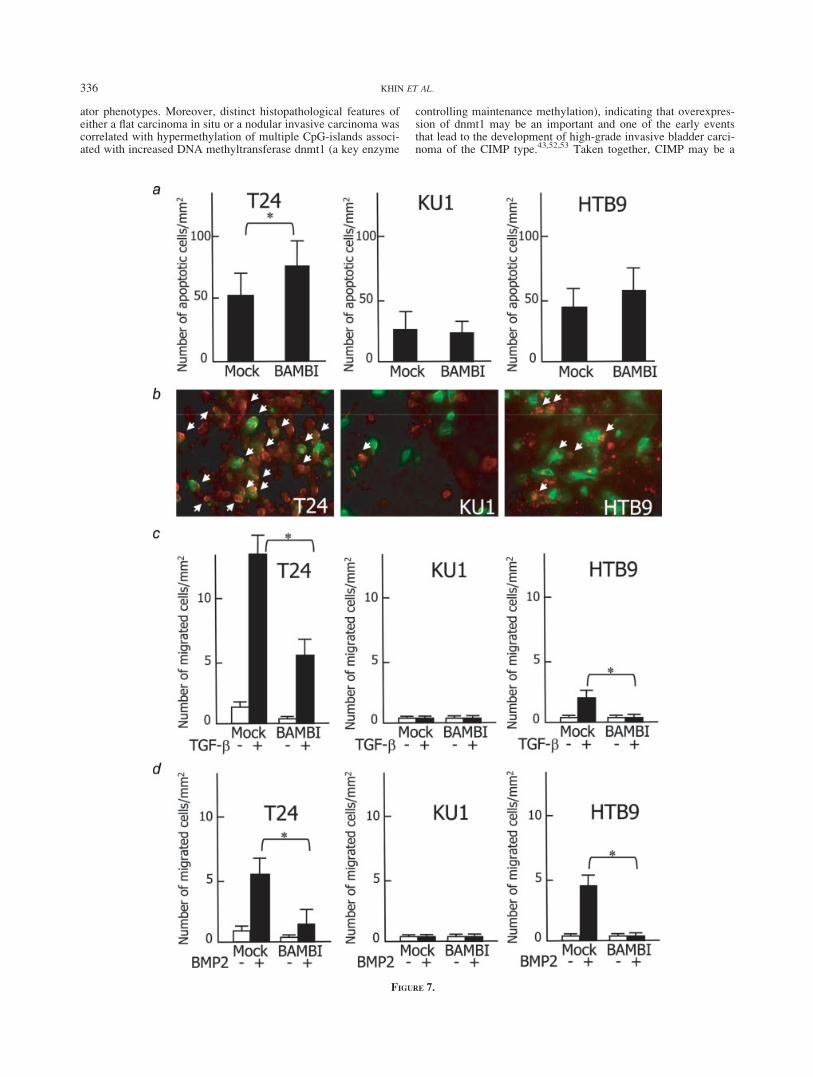
Overexpression of BAMBI enhances apoptosis and reducesTGF-b1/BMP-2 induced cell motility in bladder cancer cell lines
After transfection with pEGFP-N3-hBAMBI for 48 hr, the num-ber and intensity of green fluorescent cells (about 60%) wasalmost evenly observed in the 3 cell lines. Annexin V-Cy3 apopto-sis assay revealed that the number of yellow to orange apoptoticcells expressing pEGFP-N3-hBAMBI was slightly but significantlyhigher in T24, but not in KU1 or HTB9, than in mock-transfectedcontrol (Figs. 7a and 7b). Cell motility assay, on the other hand,revealed that after 24 hr treatment with TGF-b1 or BMP-2, T24and HTB9 cells showed a marked increase in the number ofmigrated cells that decreased significantly through the forcedexpression of BAMBI by the transfection of the pEGFP-N3-hBAMBI expression vector in cell lines T24 and HTB9 (p < 0.05).By contrast, TGF-b1 and BMP-2 had no effect on the motility ofKU1 cells (Figs. 7c and 7d).
Discussion
Using our hospital-based bladder cancer cases from Japan andMyanmar, we examined histopathologically, the prevalence ofepigenetic alterations, especially the silencing of particular genesby hypermethylation, as biological markers for bladder cancer.
So far numerous genetical alterations including the activation ofoncogenes like HRAS,29 MYC,30 CCND1,31 FGFR332 and E2F3,33
and alternatively the silencing of tumor suppressor genes likeTP53, RB134 and p1635 are reported to occur and to be linked tocarcinogenesis and tumor progression in bladder tumors.36 On theother hand, there is substantial evidence for the role of epigeneticsilencing of genes by promoter hypermethylation in urinary blad-der carcinogenesis (e.g., p16, sFRP, E-cadherin, DAPK andRASSF1A),37–44 indicating that both genetical and epigeneticalalterations contribute to bladder tumor development through mul-tiple or stepwise carcinogenesis, as well as to the diversity of can-cer cells in later progression stages.36–43 Recently, a distinct groupof cancers carrying frequent DNA hypermethylation on CpG-islands has been identified and designated as a CIMP in a certainsubset of colorectal and stomach malignancies.45–47 In addition,Dr. Issa’s group has reported in their colon cancer studies, thatCIMP can be subclassified into cases with microsatellite instabilityand BRAF mutation (CIMP1) and with somatic KRAS mutationand rare microsatellite instability (CIMP2).48
Although there are some contradictory opinions regarding theconcept of CIMP, a distinct subset of cancers, other than gastroin-testinal malignancy with frequent DNA hypermethylation onCpG-islands, has also been linked to specific genetical alterations
TABLE II – ASSOCIATION OF BAMBI GENE HYPERMETHYLATION WITH HIGH-GRADE INVASIVE PATTERN INPRIMARY BLADDER TUMORS
BAMBI methylation statusGrade Invasion
High, n (%) Low, n (%) p Present, n (%) Absent, n (%) p
Positive 20 (27.03) 3 (4.05) 0.0049 17 (22.97) 6 (8.11) 0.0315Negative 27 (36.49) 24 (32.43) 24 (32.43) 27 (36.49)
n indicates number of cases.
FIGURE 5 – Expression of BAMBI in bladder cancer, colon andbreast cancer cell lines. (a) BAMBI expression was analyzed by realtime RT-PCR in bladder cancer cell lines (T24, KU1 and HTB9)before and after the treatment with 5-aza-dC. Before the treatment,steady-state BAMBI mRNA expression was highest in KU1, lower inT24 and relatively lowest in HTB9. The cells were treated with 1 lMof 50-aza-dC for 7 days and isolated RNA was amplified by RT-PCR.After the treatment, BAMBI expression increased significantly in T24and HTB9. 18S ribosomal RNA was used as a control for RNA integ-rity. Bars indicate mean 6 SD of 6 independent samples (*p < 0.05).The results of MSP of each cell line before and after 50-aza-dC treat-ment are shown. U and M represent unmethylated and methylatedBAMBI gene promoter, respectively. (b) The steady-state BAMBImRNA expression of colon and breast cancer cell lines (COLO201,SW480, HT29, DLD-1, COLO320 and MCF7) and positive correla-tion with their respective b-catenin protein expression by Westernblot. b-actin was used as control. None of the colon cancer cell linesrevealed BAMBI promoter methylation (data not shown).
334 KHIN ET AL.
or clinicopathological features.49–51 In bladder cancers, a certainsubset has increased methylation of CpG-islands relative to itsnormal counterpart suggesting the occurrence of a hypermethyla-tor phenotype in which multiple independent CpG-islands become
concurrently methylated in individual tumors in a process associ-ated with tumor progression. The fact that the greater the numberof methylated CpG-islands, the more aggressive the cancer sug-gests that these subsets of tumors should be categorized as methyl-
FIGURE 6 – siRNA-mediated knock-down of BAMBI expression. (a) RT-PCR of BAMBI expression in T24, KU1 and HTB9 after the treat-ment with BAMBI siRNA (BAMBI) or with nonspecific control (NS) for 48 hours. BAMBI expression levels are significantly reduced in all blad-der cancer cell lines after siRNA treatment. The level of 18S ribosomal RNA expression was used to standardize BAMBI expression. (B) Effectof siRNA-mediated knock-down of BAMBI on apoptotic cell count of the 3 cell lines. Typical photographic demonstration of red-purple stainedapoptotic cells in BAMBI- (1) and control-siRNA treated (NS) cell lines. Although a slight decrease in apoptotic cell count by siRNA transfec-tion was observed in HTB9, BAMBI knock-down did not have much effect on the number of apoptotic cells. Cell lines treated with nonspecificsiRNA showed no significant effect on apoptotic count. Bars indicate mean 6 SD of 6 independent samples. [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
FIGURE 7 – Effect of restored expression of the BAMBI gene on apoptotic cell count and TGF-b-induced and BMP-2-induced cell migrationin T24, KU1 and HTB9 bladder cancer cell lines. (a) After transient transfection with either pEGFP-N3-hBAMBI (BAMBI) or control vector(Mock), the number of apoptotic cells was evaluated and plotted after staining with Annexin V-Cy3. Mean 6 SD of 6 independent samples isshown. A slight but significant increase in the apoptotic count is observed in T24 (*p < 0.05), but not in KU1 or HTB9. Mock-transfected celllines showed no effect on apoptotic count. (b) A typical photographic demonstration of pEGFP-N3-hBAMBI expression and apoptotic cells.Restored BAMBI expression is represented by green fluorescence in each bladder cancer cell line. After staining with Annexin V-Cy3, apoptoticcells show red fluorescence, and pEGFP-N3-hBAMBI-expressing apoptotic cells reveal yellow to orange fluorescence (white arrows) with somenonapoptotic green fluorescent part remaining in each bladder cancer cell line. (c and d) The in vitro migratory behavior of cells transientlytransfected with pEGFP-N3-hBAMBI or with control vector (Mock) was assessed in the presence of 3 lg/ml reconstituted recombinant humanTGF b-1 containing culture media (C) and 1 mg/ml of recombinant human (rh) BMP-2 containing culture media (D) for 24 hr. The number ofmigrated cells per square millimeter was evaluated and plotted. Mean 6 SD of 12 independent samples is shown. TGF-b-1-induced and BMP-2-induced cell migration was significantly reduced in pEGFP-N3-hBAMBI expressed T24 and HTB9 bladder cancer cell lines (*p < 0.05).Mock shows no effect on TGF-b-1/BMP2-induced cell migration in any of the cell lines.
335EPIGENETIC INACTIVATION OF BAMBI IN BLADDER CANCER
ator phenotypes. Moreover, distinct histopathological features ofeither a flat carcinoma in situ or a nodular invasive carcinoma wascorrelated with hypermethylation of multiple CpG-islands associ-ated with increased DNA methyltransferase dnmt1 (a key enzyme
controlling maintenance methylation), indicating that overexpres-sion of dnmt1 may be an important and one of the early eventsthat lead to the development of high-grade invasive bladder carci-noma of the CIMP type.43,52,53 Taken together, CIMP may be a
FIGURE 7.
336 KHIN ET AL.

secondary event resulting from preceding genetical alterations thatultimately cause increased activity of dnmt1 or of other enzymeswith de novo DNA methyltransferase.49,50 It is still unclear, how-ever, whether such secondarily augmented methyltransferase ac-tivity, maintenance or de novo, triggers hypermethylation of mul-tiple CpG-islands in a promoter-specific or nonspecific manner.
Consistent with previous reports,43,52,53 we also observed that acertain group of high-grade bladder cancers with invasive growthpatterns from Japan and Myanmar, and bladder cancer cell linescarried frequent hypermethylation of multiple gene promoters(Figs. 3a–3c). Although genetic alteration of b-catenin is rare inbladder cancer,54 epigenetic inactivation of sFRPs, secreted antag-onists of Wnt signaling, is responsible for the invasive phenotypeof bladder cancer.43 Since hypermethylation of the BAMBI genewas frequently associated with hypermethylation of p16 tumorsuppressor gene (one of the most frequently silenced genes inCIMP), and Wnt-related genes sFRP1, sFRP2, sFRP4 and sFRP5,and thus BAMBI can be regarded as a member of CIMP-affectedgenes in high-grade bladder cancer. Previous studies have reportedthat Wnt/b-catenin signaling activates the transcription of BAMBI,and that BAMBI expression is aberrantly elevated in most colorec-tal carcinomas reflecting the activation status of b-catenin.18,22
We also confirmed that BAMBI gene promoter hypermethylationwas not observed and positive correlation of its expression with b-catenin in either the 9 CIMP-high clinical samples or the 5colorectal and 1 breast cancer cell lines tested (data not shown).Moreover, inactivation or blockage of the TGF-b/BMP signalingpathway in colon epithelium is one of the important initial steps incarcinogenesis, and TGF-b/BMP signaling serves mostly as an in-hibitory factor for growth and survival throughout the progressionof colorectal cancer. Continuous BAMBI expression due to acti-vated b-catenin (through the direct binding of TCF/LEF transcrip-tion factor to BAMBI promoter region) or TGF-b/BMP signalingitself (through the evolutionary conserved BMP-responsive ele-ments in BAMBI promoter, by the direct binding of SMAD3 andSMAD4 to its promoter region) favors the growth and survival ofcolorectal cancer cells by acting as a negative feedback systemagainst TGF-b/BMP signaling.18,19,22,55 It follows, therefore, thatcolorectal cancer cells expressing the BAMBI gene with hypome-thylation of the BAMBI gene promoter have a better chance of sur-viving in the tumor microenvironment. To the contrary, the effectof the TGF-b/BMP signaling pathway at later progression stagesin some aggressive tumors, such as in malignant melanoma,56
ovarian57 and bladder cancers,58,59 often promotes cancer cellinvasion and migration by inducing EMT in an autocrine and para-crine manner. We speculate, therefore, that the choice of hyperme-thylated genes in CIMP is more a flexible than a rigid phenom-enon resulting from a clonal selection of the specific tumor celltype according to the tumor microenvironment.
In the normal or reactive urothelium, on the other hand, immu-nohistochemical localization of BAMBI expression was mostlyoverlapped with that of BMPRI (Fig. 2), supporting a previous
report17 that BAMBI may be physiologically important as a directfine-tuner of TGF-b/BMP by counteracting and bufferizing its ac-tivity, and thereby, keeping its signal transduction within a limitedrange. Among cases of low-grade papillary noninvasive urothelialcancer, because the immunohistochemical colocalization patternof BAMBI and BMPRI was similar to that of the normal and reac-tive urothelium (Figs. 4a–4c), such physiological bufferizingaction of BAMBI may remain functionally intact in such low-gradecancers. In high-grade and invasive cancers, however, 2 reciprocalimmunohistochemical expression patterns were mainly observedbetween BAMBI and BMPRI (Figs. 4d–4i): BAMBI-low andBMPRI-high, and BAMBI-high and BMPRI-low, indicating thatBAMBI expression is controlled such that it does not interfere withcellular demand for TGF-b/BMP signaling defined by the authen-tic level of TGF-b/BMP receptors. Taken together with the factthat the role of TGF-b/BMP signaling in cancer is both tumor sup-pressive and progressive according to the difference of intercellu-lar signal transduction systems followed by the activation ofauthentic receptors, we speculate that when cancer cells need tobe irresponsive to tumor-suppressive TGF-b/BMP signaling,BAMBI expression is kept high while authentic receptors for TGF-b/BMP are kept low to accomplish full blocking. Conversely,when TGF-b/BMP signaling is favorable for cancer cell survivalor motility, cells with suppressed BAMBI expression by promoterhypermethylation have a better chance to survive. To support thishypothesis, in the BAMBI high and BMPRI low KU1 cell, whereepidermal growth factor (EGF) stimulates anchorage-independentgrowth through the amplified and overexpressed its cognate recep-tor (EGFR),60,61 TGF-b stimulation showed almost no effect oncell motility (Fig. 7c). On the other hand, in BAMBI low andBMPRI high bladder cancer cell lines T24 and HTB9, TGF-btreatment enhanced cell motility, which was significantly sup-pressed by forced BAMBI expression (Fig. 7c). Together with casestudies from Japan and Myanmar, these results of bladder cancercell lines indicate that among high-grade bladder cancers at least 2types can be differentiated in response to TGF-b/BMP signaling:one with irresponsive to TGF-b/BMP and the other in whichTGF-b/BMP signaling positively promotes the aggressive charac-ter of the tumor cells. This differentiation may be useful for futuretherapeutic strategy based on epigenetics.
In conclusion, since certain subsets of aggressive tumors oftenpromote cell motility, invasion and survival (e.g., EMT) by TGF-b/BMP signaling, BAMBI gene suppression by promoter hyper-methylation with negative BAMBI immunoexpression may be oneof the important epigenetic events affecting the invasiveness oraggressiveness of bladder cancers.
Acknowledgements
The authors thank Mr. Shuichi Matsuda, Ms. Noriko Sakamotoand Ms. Miki Zenigami for their excellent technical assistance.
References
1. Lopez-Beltran A, Sauter G, Gasser T, Hartmann A, Schmitz-DragerBJ, Helpap B, Ayala AG, Tamboli P, Knowles MA, Sidransky D,Cordon-Cardo C, Jones PA, et al. Infiltrating urothelial carcinoma. In:Eble JN, Sauter G, Epstein JI, Sesterhenn IA, eds. World Health Orga-nization classification of tumours: pathology and genetics of tumoursof the urinary system and male genital organsed. Lyon: IARC Press,2004. 93–109.
2. Frigerio S, Padberg BC, Strebel RT, Lenggenhager DM, MessthalerA, Abdou M-T, Moch H, Zimmermann DR. Improved detection ofbladder carcinoma cells in voided urine by standardized microsatelliteanalysis. Int J Cancer 2007;121:329–38.
3. Olsson AY, Feber A, Edwards S, Te Poele R, Giddings I, Merson S,Cooper CS. Role of E2F3 expression in modulating cellular prolifera-tion rate in human bladder and prostate cancer cells. Oncogene2007;26:1028–37.
4. Stoehr R, Wissmann C, Suzuki H, Knuechel R, Krieg RC, KlopockiE, Dahl E, Wild P, Blaszyk H, Sauter G, Simon R, Schmitt R, et al.Deletions of chromosome 8p and loss of sFRP1 expression are pro-
gression markers of papillary bladder cancer. Lab Invest 2004;84:465–78.
5. Massague J. How cells read TGF-beta signals. Nat Rev Mol Cell Biol2000;1:169–78.
6. Siegel PM, Massague J. Cytostatic and apoptotic actions of TGF-betain homeostasis and cancer. Nat Rev Cancer 2003;3:807–21.
7. Chen YG, Lui HM, Lin SL, Lee JM, Ying SY. Regulation of cell pro-liferation: apoptosis and carcinogenesis by activin. Exp Biol Med2002;227:75–87.
8. Bierie B, Moses HL. Tumour microenvironment: TGFbeta: the mo-lecular Jekyll and Hyde of cancer. Nat Rev Cancer 2006;6:506–20.
9. Leivonen S-K, Kahari V-M. Transforming growth factor-b signalingin cancer invasion and metastasis. Int J Cancer 2007;121:2119–24.
10. Massague J. TGFbeta in cancer. Cell 2008;134:215–30.11. Deli A, Kreidl E, Santifaller S, Trotter B, Seir K, Berger W, Schulte-
Hermann R, Rodgarkia-Dara C, Grusch M. Activins and activinantagonists in hepatocellular carcinoma. World J Gastroenterol 2008;14:1699–709.
337EPIGENETIC INACTIVATION OF BAMBI IN BLADDER CANCER

12. Akhurst RJ, Balmain A. Genetic events and the role of TGF beta inepithelial tumour progression. J Pathol 1999;187:82–90.
13. Bierie B, Moses HL. TGF-beta and cancer. Cytokine Growth FactorRev 2006;17:29–40.
14. Derynck R, Akhurst RJ, Balmain A. TGF-beta signaling in tumor sup-pression and cancer progression. Nat Genet 2001;29:117–29.
15. Rojas A, Meherem S, Kim Y-H, Washington MK, Willis JE, Marko-witz SD, Grady WM. The aberrant methylation of TSP1 suppressesTGF-b1 activation in colorectal cancer. Int J Cancer 2008;123:14–21.
16. Balemans W, Van Hul W. Extracellular regulation of BMP signalingin vertebrates: a cocktail of modulators. Dev Biol 2002;250:231–50.
17. Onichtchouk D, Chen YG, Dosch R, Gawantka V, Delius H, Mas-sague J, Niehrs C. Silencing of TGF-beta signalling by the pseudore-ceptor BAMBI. Nature 1999;401:480–5.
18. Sekiya T, Oda T, Matsuura K, Akiyama T. Transcriptional regulationof the TGF-beta pseudoreceptor BAMBI by TGF-beta signaling. Bio-chem Biophys Res Commun 2004;320:680–4.
19. Karaulanov E, Knochel W, Niehrs C. Transcriptional regulation ofBMP4 synexpression in transgenic Xenopus. EMBO J 2004;23:844–56.
20. Grotewold L, Plum M, Dildrop R, Peters T, Ruther U. Bambi is coex-pressed with Bmp-4 during mouse embryogenesis. Mech Dev 2001;100:327–30.
21. Tsang M, Kim R, de Caestecker MP, Kudoh T, Roberts AB, DawidIB. Zebrafish nma is involved in TGFbeta family signaling. Genesis2000;28:47–57.
22. Sekiya T, Adachi S, Kohu K, Yamada T, Higuchi O, Furukawa Y,Nakamura Y, Nakamura T, Tashiro K, Kuhara S, Ohwada S, AkiyamaT. Identification of BMP and activin membrane-bound inhibitor(BAMBI), an inhibitor of transforming growth factor-beta signaling,as a target of the beta-catenin pathway in colorectal tumor cells.J Biol Chem 2004;279:6840–6.
23. Sasaki T, Sasahira T, Shimura H, Ikeda S, Kuniyasu H. Effect of Nmaon growth inhibition by TGF-beta in human gastric carcinoma celllines. Oncol Rep 2004;11:1219–23.
24. Kitazawa S, Kitazawa R, Maeda S. Identification of methylated cyto-sine from archival formalin-fixed paraffin-embedded specimens. LabInvest 2000;80:275–6.
25. Kitazawa S, Kitazawa R, Obayashi C, Yamamoto T. Desmoid tumorwith ossification in chest wall: possible involvement of BAMBI pro-moter hypermethylation in metaplastic bone formation. J Bone MinerRes 2005;20:1472–7.
26. Norrie MW, Hawkins NJ, Todd AV, Meagher AP, O’Connor TW,Ward RL. Inactivation of p16INK4a by CpG hypermethylation is nota frequent event in colorectal cancer. J Surg Oncol 2003;84:143–50.
27. Zou H, Molina JR, Harrington JJ, Osborn NK, Klatt KK, Romero Y,Burgart LJ, Ahlquist DA. Aberrant methylation of secreted frizzled-related protein genes in esophageal adenocarcinoma and Barrett’sesophagus. Int J Cancer 2005;116:584–91.
28. Doglioni C, Piccinin S, Demontis S, Cangi MG, Pecciarini L, Chiar-elli C, Armellin M, Vukosavljevic T, Boiocchi M, Maestro R. Altera-tions of beta-catenin pathway in non-melanoma skin tumors: loss ofalpha-ABC nuclear reactivity correlates with the presence of beta-cat-enin gene mutation. Am J Pathol 2003;163:2277–87.
29. Zhang ZT, Pak J, Huang HY, Shapiro E, Sun TT, Pellicer A, Wu XR.Role of Ha-ras activation in superficial papillary pathway of urothelialtumor formation. Oncogene 2001;20:1973–80.
30. Christoph F, Schmidt B, Schmitz-Drager BJ, Schulz WA. Over-expression and amplification of the c-myc gene in human urothelialcarcinoma. Int J Cancer 1999;84:169–73.
31. Dinney CP, McConkey DJ, Millikan RE, Wu X, Bar-Eli M, Adam L,Kamat AM, Siefker-Radtke AO, Tuziak T, Sabichi AL, GrossmanHB, Benedict WF, et al. Focus on bladder cancer. Cancer Cell2004;6:111–6.
32. Sibley K, Cuthbert-Heavens D, Knowles MA. Loss of heterozygosityat 4p16.3 and mutation of FGFR3 in transitional cell carcinoma.Oncogene 2001;20:686–91.
33. Feber A, Clark J, Goodwin G, Dodson AR, Smith PH, Fletcher A,Edwards S, Flohr P, Falconer A, Roe T, Kovacs G, Dennis N, et al.Amplification and overexpression of E2F3 in human bladder cancer.Oncogene 2004;23:1627–30.
34. Chatterjee SJ, Datar R, Youssefzadeh D, George B, Goebell PJ, SteinJP, Young L, Shi SR, Gee C, Groshen S, Skinner DG, Cote RJ.Combined effects of p53, p21, and pRb expression in the progressionof bladder transitional cell carcinoma. J Clin Oncol 2004;22:1007–13.
35. Chapman EJ, Harnden P, Chambers P, Johnston C, Knowles MA.Comprehensive analysis of CDKN2A status in microdissected urothe-lial cell carcinoma reveals potential haploinsufficiency, a high fre-quency of homozygous co-deletion and associations with clinical phe-notype. Clin Cancer Res 2005;11:5740–7.
36. Schulz WA. Understanding urothelial carcinoma through cancer path-ways. Int J Cancer 2006;119:1513–8.
37. Chan MW, Chan LW, Tang NL, Tong JH, Lo KW, Lee TL, CheungHY, Wong WS, Chan PS, Lai FM, To KF. Hypermethylation of mul-
tiple genes in tumor tissues and voided urine in urinary bladder cancerpatients. Clin Cancer Res 2002;8:464–70.
38. Cote RJ, Laird PW, Datar RH. Promoter hypermethylation: a newtherapeutic target emerges in urothelial cancer. J Clin Oncol2005;23:2879–81.
39. Dhawan D, Hamdy FC, Rehman I, Patterson J, Cross SS, Feeley KM,Stephenson Y, Meuth M, Catto JW. Evidence for the early onset ofaberrant promoter methylation in urothelial carcinoma. J Pathol2006;209:336–43.
40. Friedrich MG, Weisenberger DJ, Cheng JC, Chandrasoma S, Sieg-mund KD, Gonzalgo ML, Toma MI, Huland H, Yoo C, Tsai YC,Nichols PW, Bochner BH, et al. Detection of methylated apoptosis-associated genes in urine sediments of bladder cancer patients. ClinCancer Res 2004;10:7457–65.
41. Cairns P. Gene methylation and early detection of genitourinary can-cer: the road ahead. Nat Rev Cancer 2007;7:531–43.
42. Catto JW, Azzouzi AR, Rehman I, Feeley KM, Cross SS, Amira N,Fromont G, Sibony M, Cussenot O, Meuth M, Hamdy FC. Promoterhypermethylation is associated with tumor location, stage, and subse-quent progression in transitional cell carcinoma. J Clin Oncol2005;23:2903–10.
43. Marsit CJ, Karagas MR, Andrew A, Liu M, Danaee H, Schned AR,Nelson HH, Kelsey KT. Epigenetic inactivation of SFRP genes andTP53 alteration act jointly as markers of invasive bladder cancer.Cancer Res 2005;65:7081–5.
44. Yates DR, Rehman I, Abbod MF, Meuth M, Cross SS, Linkens DA,Hamdy FC, Catto JW. Promoter hypermethylation identifies progres-sion risk in bladder cancer. Clin Cancer Res 2007;13:2046–53.
45. Etoh T, Kanai Y, Ushijima S, Nakagawa T, Nakanishi Y, Sasako M,Kitano S, Hirohashi S. Increased DNA methyltransferase 1 (DNMT1)protein expression correlates significantly with poorer tumor differen-tiation and frequent DNA hypermethylation of multiple CpG islandsin gastric cancers. Am J Pathol 2004;164:689–99.
46. Kanai Y, Ushijima S, Kondo Y, Nakanishi Y, Hirohashi S. DNAmethyltransferase expression and DNA methylation of CPG islandsand peri-centromeric satellite regions in human colorectal and stom-ach cancers. Int J Cancer 2001;91:205–12.
47. Toyota M, Ahuja N, Ohe-Toyota M, Herman JG, Baylin SB, Issa JP.CpG island methylator phenotype in colorectal cancer. Proc NatlAcad Sci USA 1999;96:8681–6.
48. Shen L, Toyota M, Kondo Y, Lin E, Zhang L, Guo Y, Hernandez NS,Chen X, Ahmed S, Konishi K, Hamilton SR, Issa JP. Integratedgenetic and epigenetic analysis identifies three different subclasses ofcolon cancer. Proc Natl Acad Sci USA 2007;104:18654–9.
49. Issa JP. CpG island methylator phenotype in cancer. Nat Rev Cancer2004;4:988–93.
50. Ting AH, McGarvey KM, Baylin SB. The cancer epigenome-compo-nents and functional correlates. Genes Dev 2006;20:3215–31.
51. Jones PA, Baylin SB. The fundamental role of epigenetic events incancer. Nat Rev Genet 2002;3:415–28.
52. Nakagawa T, Kanai Y, Ushijima S, Kitamura T, Kakizoe T, HirohashiS. DNA hypermethylation on multiple CpG islands associated withincreased DNA methyltransferase DNMT1 protein expression duringmultistage urothelial carcinogenesis. J Urol 2005;173:1767–71.
53. Salem C, Liang G, Tsai YC, Coulter J, Knowles MA, Feng A-C, GroshenS, Nichols PW, Jones PA. Progressive Increases in de Novo Methylationof CpG Islands in Bladder Cancer. Cancer Res 2000;60:2473–6.
54. Stoehr R, Krieg RC, Knuechel R, Hofstaedter F, Pilarsky C, Zaak D,Schmitt R, Hartmann A. No evidence for involvement of beta-cateninand APC in urothelial carcinomas. Int J Oncol 2002;20:905–11.
55. Togo N, Ohwada S, Sakurai S, Toya H, Sakamoto I, Yamada T,Nakano T, Muroya K, Takeyoshi I, Nakajima T, Sekiya T, YamazumiY, et al. Prognostic significance of BMP and activin membrane-boundinhibitor in colorectal cancer. World J Gastroenterol 2008;14:4880–8.
56. Rothhammer T, Poser I, Soncin F, Bataille F, Moser M, Bosserhoff AK.Bone morphogenic proteins are overexpressed in malignant melanomaand promote cell invasion and migration. Cancer Res 2005;65:448–56.
57. Theriault BL, Shepherd TG, Mujoomdar ML, Nachtigal MW. BMP4induces EMT and Rho GTPase activation in human ovarian cancercells. Carcinogenesis 2007;28:1153–62.
58. Izadifar V, de Boer WI, Muscatelli-Groux B, Maille P, van der KwastTH, Chopin DK. Expression of transforming growth factor beta1 andits receptors in normal human urothelium and human transitional cellcarcinomas. Hum Pathol 1999;30:372–7.
59. Hung T-T, Wang H, Kingsley EA, Risbridger GP, Russell PJ. Molec-ular profiling of bladder cancer: Involvement of the TGF-b pathwayin bladder cancer progression. Cancer Lett 2008;265:27–38.
60. Bryan RT, Hussain SA, James ND, Jankowski JA, Wallace DM. Mo-lecular pathways in bladder cancer, part 1. BJU Int 2005;95:485–90.
61. Ishikawa J, Maeda S, Sugiyama T, Nishimura R, Mizoguchi A, Kami-dono S. EGF stimulates anchorage-independent growth of a humanbladder carcinoma cell line (KU1) with an amplified and over-expressed EGF receptor gene. Int J Cancer 1989;44:1000–4.
338 KHIN ET AL.




















