
Bacterial Community Changes during Plant Establishment at the San Pedro River Mine Tailings Site
11
1249 Mine tailings are moderately to severely impacted sites that lack normal plant cover, soil structure and development, and the associated microbial community. In arid and semiarid environments, tailings and their associated contaminants are prone to eolian dispersion and water erosion, thus becoming sources of metal contamination. One approach to minimize or eliminate these processes is to establish a permanent vegetation cover on tailings piles. Here we report a revegetation trial conducted at a moderately impacted mine tailings site in southern Arizona. A salt and drought-tolerant plant, four- wing saltbush [Atriplex canescens (Pursh) Nutt.], was chosen for the trial. A series of 3 by 3 m plots were established in quadruplicate on the test site to evaluate growth of four-wing saltbush transplants alone or with compost addition. Results show that >80% of the transplanted saltbush survived after 1.5 yr in both treatments. Enumeration of heterotrophs and community structure analysis were conducted to monitor bacterial community changes during plant establishment as an indicator of plant and soil health. e bacterial community was evaluated using denaturing gradient gel electrophoresis (DGGE) analysis of 16S rDNA PCR gene products from tailings samples taken beneath transplant canopies. Significant differences in heterotrophic counts and community composition were observed between the two treatments and unplanted controls throughout the trial, but treatment effects were not observed. e results suggest that compost is not necessary for plant establishment at this site and that plants, rather than added compost, is the primary factor enhancing bacterial heterotrophic counts and affecting community composition. Bacterial Community Changes during Plant Establishment at the San Pedro River Mine Tailings Site Karyna Rosario, Sadie L. Iverson, David A. Henderson, and Shawna Chartrand University of Arizona Casey McKeon and Edward P. Glenn Environmental Research Laboratory Raina M. Maier* University of Arizona T here is emerging concern regarding the environmental consequences of mine tailings sites, especially those in sensitive arid and semiarid riparian or wildlife refuge areas. Tailings are produced during ore processing and are characterized by little or no vegetation, poor soil structure, low nutrient content, surface crusting, and high levels of contaminants including heavy metals (Ledin and Pedersen, 1996). e impact of tailings goes beyond the immediate site because erosion and leaching processes release and spread tailings contaminants to other areas. Removal of tailings materials is costly largely because they are extensive in nature and considered to be hazardous waste. us, remediation efforts have focused on stabilization of the tailings. One approach to stabilization is phytoremediation or establish- ment of a plant cover that acts to reduce all spreading mechanisms: eolian dispersion, water erosion, and leaching. Concomitantly, a plant cover acts to stabilize soil contaminants by altering the water flux through the soil, by incorporating free contaminants into the roots and rhizosphere, and by improving ecosystem structure and function within the tailings (Cunningham et al., 1995). e chal- lenge with phytostabilization in arid and semiarid environments is that plant establishment is often limited by high levels of metals and extreme pH in addition to the other challenges that arid environ- ments pose to plants (i.e., low water availability, low organic matter, salinity, and high temperatures). In general, the development of a persistent vegetative cover depends on selecting plant species suitable to site conditions and amending tailings with organic matter that improves soil properties that favor plant growth. e improvements made to mine tailings sites through phytosta- bilization rely, in part, on activities of soil microorganisms that are essential to healthy ecosystems (Crowley et al., 1997). Bacteria play an important role in the success of phytostabilization by promoting Abbreviations: ANOVA, analysis of variance; BLM, Bureau of Land Management; C, plots containing transplants with compost; CECL, University of Arizona Chorover Enviromental Chemistry Laboratory; CFU, colony-forming unit; DGGE, denaturing gradient gel electrophoresis; HIC, University of Arizona Superfund Basic Research Program Hazard Identification Core; ICP-MS, inductively coupled plasma–mass spectrometry; KNMDS, Kruskal’s Non-Metric Multidimensional Scaling; NC, plots containing transplants with no compost, OSC, off-site control plots; SPRNCA, San Pedro River National Conservation Area; U, unplanted control plots; WQCL, University of Arizona Water Quality Center Lab. K. Rosario, S.L. Iverson, S. Chartrand, and R.M. Maier, Dep. of Soil, Water, and Environmental Science, Univ. of Arizona, Tucson, AZ 85721-0038; D.A. Henderson, Dep. of Animal Sciences, Univ. of Arizona, Tucson, AZ 85721-0038; and C. McKeon and E.P. Glenn, Environmental Research Lab, 2601 E. Airport Drive, Tucson, AZ 85706. Copyright © 2007 by the American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America. All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including pho- tocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Published in J. Environ. Qual. 36:1249–1259 (2007). doi:10.2134/jeq2006.0315 Received 7 Aug. 2006. *Corresponding author ([email protected]). © ASA, CSSA, SSSA 677 S. Segoe Rd., Madison, WI 53711 USA TECHNICAL REPORTS: BIOREMEDIATION & BIODEGRADATION Published online July 17, 2007
Transcript of Bacterial Community Changes during Plant Establishment at the San Pedro River Mine Tailings Site

1249
Mine tailings are moderately to severely impacted sites that lack normal plant cover, soil structure and development, and the associated microbial community. In arid and semiarid environments, tailings and their associated contaminants are prone to eolian dispersion and water erosion, thus becoming sources of metal contamination. One approach to minimize or eliminate these processes is to establish a permanent vegetation cover on tailings piles. Here we report a revegetation trial conducted at a moderately impacted mine tailings site in southern Arizona. A salt and drought-tolerant plant, four-wing saltbush [Atriplex canescens (Pursh) Nutt.], was chosen for the trial. A series of 3 by 3 m plots were established in quadruplicate on the test site to evaluate growth of four-wing saltbush transplants alone or with compost addition. Results show that >80% of the transplanted saltbush survived after 1.5 yr in both treatments. Enumeration of heterotrophs and community structure analysis were conducted to monitor bacterial community changes during plant establishment as an indicator of plant and soil health. Th e bacterial community was evaluated using denaturing gradient gel electrophoresis (DGGE) analysis of 16S rDNA PCR gene products from tailings samples taken beneath transplant canopies. Signifi cant diff erences in heterotrophic counts and community composition were observed between the two treatments and unplanted controls throughout the trial, but treatment eff ects were not observed. Th e results suggest that compost is not necessary for plant establishment at this site and that plants, rather than added compost, is the primary factor enhancing bacterial heterotrophic counts and aff ecting community composition.
Bacterial Community Changes during Plant Establishment at the San Pedro River Mine
Tailings Site
Karyna Rosario, Sadie L. Iverson, David A. Henderson, and Shawna Chartrand University of Arizona
Casey McKeon and Edward P. Glenn Environmental Research Laboratory
Raina M. Maier* University of Arizona
There is emerging concern regarding the environmental
consequences of mine tailings sites, especially those in
sensitive arid and semiarid riparian or wildlife refuge areas. Tailings
are produced during ore processing and are characterized by
little or no vegetation, poor soil structure, low nutrient content,
surface crusting, and high levels of contaminants including heavy
metals (Ledin and Pedersen, 1996). Th e impact of tailings goes
beyond the immediate site because erosion and leaching processes
release and spread tailings contaminants to other areas. Removal
of tailings materials is costly largely because they are extensive in
nature and considered to be hazardous waste. Th us, remediation
eff orts have focused on stabilization of the tailings.
One approach to stabilization is phytoremediation or establish-
ment of a plant cover that acts to reduce all spreading mechanisms:
eolian dispersion, water erosion, and leaching. Concomitantly, a
plant cover acts to stabilize soil contaminants by altering the water
fl ux through the soil, by incorporating free contaminants into the
roots and rhizosphere, and by improving ecosystem structure and
function within the tailings (Cunningham et al., 1995). Th e chal-
lenge with phytostabilization in arid and semiarid environments is
that plant establishment is often limited by high levels of metals and
extreme pH in addition to the other challenges that arid environ-
ments pose to plants (i.e., low water availability, low organic matter,
salinity, and high temperatures). In general, the development of a
persistent vegetative cover depends on selecting plant species suitable
to site conditions and amending tailings with organic matter that
improves soil properties that favor plant growth.
Th e improvements made to mine tailings sites through phytosta-
bilization rely, in part, on activities of soil microorganisms that are
essential to healthy ecosystems (Crowley et al., 1997). Bacteria play
an important role in the success of phytostabilization by promoting
Abbreviations: ANOVA, analysis of variance; BLM, Bureau of Land Management; C, plots
containing transplants with compost; CECL, University of Arizona Chorover Enviromental
Chemistry Laboratory; CFU, colony-forming unit; DGGE, denaturing gradient gel
electrophoresis; HIC, University of Arizona Superfund Basic Research Program Hazard
Identifi cation Core; ICP-MS, inductively coupled plasma–mass spectrometry; KNMDS,
Kruskal’s Non-Metric Multidimensional Scaling; NC, plots containing transplants with
no compost, OSC, off -site control plots; SPRNCA, San Pedro River National Conservation
Area; U, unplanted control plots; WQCL, University of Arizona Water Quality Center Lab.
K. Rosario, S.L. Iverson, S. Chartrand, and R.M. Maier, Dep. of Soil, Water, and
Environmental Science, Univ. of Arizona, Tucson, AZ 85721-0038; D.A. Henderson, Dep.
of Animal Sciences, Univ. of Arizona, Tucson, AZ 85721-0038; and C. McKeon and E.P.
Glenn, Environmental Research Lab, 2601 E. Airport Drive, Tucson, AZ 85706.
Copyright © 2007 by the American Society of Agronomy, Crop Science
Society of America, and Soil Science Society of America. All rights
reserved. No part of this periodical may be reproduced or transmitted
in any form or by any means, electronic or mechanical, including pho-
tocopying, recording, or any information storage and retrieval system,
without permission in writing from the publisher.
Published in J. Environ. Qual. 36:1249–1259 (2007).
doi:10.2134/jeq2006.0315
Received 7 Aug. 2006.
*Corresponding author ([email protected]).
© ASA, CSSA, SSSA
677 S. Segoe Rd., Madison, WI 53711 USA
TECHNICAL REPORTS: BIOREMEDIATION & BIODEGRADATION
Published online July 17, 2007

1250 Journal of Environmental Quality • Volume 36 • September-October 2007
plant growth through the mineralization of inorganic nutrients (Xu
and Johnson, 1995), production of growth regulatory substances
like phytohormones (Pishchik et al., 2002), facilitation of nutri-
ent uptake from the soil (Burd et al., 2000), improvement of soil
structure by soil aggregation (Maier et al., 2000), and reduction
of metal toxicity (Salt et al., 1999; Sandrin et al., 2000). In return,
established plants provide a nutrient-rich environment that can
stimulate bacterial activity (Glick, 2003; Kuiper et al., 2004). Th us,
monitoring changes in the microbial community can aid in assess-
ing the success of reclamation strategies.
Th e goal of this study was to use the local native shrub four-
wing saltbush to revegetate barren areas in a former silver-gold mill
site located adjacent to the San Pedro River in southern Arizona
and determine if plant establishment had a measurable infl uence
on the bacterial community. Th e site is in the San Pedro riparian
area, which was designated as a National Conservation Area in
1988 to help protect the fast disappearing desert riparian ecosystem
in the Southwest. A revegetation trial was conducted using four-
wing saltbush transplants to investigate if this species was suitable
for phytostabilization purposes. Th e specifi c objectives of the re-
vegetation trial were (i) to determine if four-wing saltbush could be
successfully established in the tailings, (ii) to evaluate if the addition
of compost amendment in the tailings was necessary to achieve
this goal, and (iii) to follow changes in the bacterial community
during the revegetation process. Bacterial community responses
beneath transplant canopies were studied by monitoring changes
in the abundance of heterotrophic bacteria (culturable counts) and
in the bacterial community structure based on DGGE community
fi ngerprints using universal bacterial primers.
Materials and Methods
Site Background and Study Area DescriptionTh e Boston mill mine tailings site (Fig. 1) is adjacent to the
San Pedro River in the San Pedro River National Conservation
Area (SPRNCA), approximately 3.2 km (2 miles) south of Fair-
bank, AZ, and 12.6 km (8 miles) southwest of Tombstone, AZ,
at an elevation of 1192 m (31°40′38.6″N, 10°10′53.4″W). It
is one of fi ve mill sites along the upper
San Pedro River, within the SPRNCA,
that were active from 1879 to 1887 for
processing silver and gold ore that was
brought from mines in Tombstone, AZ.
Th e site covers approximately 2.8 ha (7
acres) and can be divided into three areas;
an upper tier near the mill foundation,
a lower tier at a slightly lower elevation
south of the upper tier, and tailings across
the railroad tracks in the fl oodplain. Pre-
liminary data provided by the Bureau of
Land Management (BLM), which cur-
rently manages the site indicated that the
upper tier has the highest concentrations
of heavy metals. Th is area has a shallow
layer of tailings (10–20 cm) and includes
completely barren areas to the west of
the mill foundation and some areas with
patchy vegetation on the eastern side composed primarily of big
sacaton grass (Sporobolus wrighti Munro ex Scribn) and mesquite
(Prosopis spp.). Th is partial vegetation suggests that natural at-
tenuation has slowly occurred on the tailings during the past
century. For this reason the Boston mill site is considered to be
a moderately impacted mine tailings site. Nevertheless, barren
areas still pose a source of concern for eolian dispersion and water
erosion of tailings into the San Pedro River and the surrounding
region, which is a wildlife refuge and hiking area.
Experimental Design and Sampling SummaryA 24 by 30 m study area in the upper tier of the Boston Mill
site was selected for the revegetation trial (Fig. 1). One of the objec-
tives of this work was to determine whether compost amendment
was necessary for establishment of four-wing saltbush on the site.
Th erefore, two treatments were evaluated: four-wing saltbush
transplants with compost amendment (C), and unamended trans-
plants (NC). A 6 by 12 m off -site control (OSC) area (no mine
tailings) was chosen southeast of the study site. For each treat-
ment (compost, C; no compost, NC; and off -site control, OSC)
quadruplicate plots (n = 4) were established on the relevant study
area following a complete randomized design. Each plot received
fi ve transplants and plant growth was evaluated by height and
crown measurements made on each plant at time 0, 11, and 18
mo (n = 20). A soil sample was taken from each plot for chemical,
physical, and microbiological analysis before planting (n = 4). Fol-
lowing planting, root zone soil samples were taken from one plant
in each plot at 3, 4, 5, 7, 8.5, 11, 13, 16, and 18 mo for microbial
analysis (n = 4). Shoot tissue samples were collected at 18 mo to
determine plant shoot metal content. Tissue from each of the fi ve
plants in a plot was harvested and combined into a single com-
posite sample for each NC and C treatment plot (n = 4). It should
be noted that transplants did not survive in the OSC area, and so
for shoot metal content comparison four composite samples were
taken from nearby indigenous four-wing saltbush shrubs to obtain
background shoot tissue metal levels (n = 4).
For the microbial analysis a third treatment, unplanted con-
Fig. 1. Photos of the study area (A) showing an aerial view of the site and the relative locations of the transplant plot area, the control site, and their proximity to the San Pedro River. The line bisecting the site from the lower left of the photo (northwest) to the mid-right (southeast) is a railroad track. Photo (B) shows a closeup of two of the 3 by 3 m plots just after having received the transplants. The inset photo is an example of a transplant.
Rosario et al.: Bacterial Community Changes during Plant Establishment 1251
trol (U), was added to account for natural variations in the bac-
terial community. Th e U samples were initially taken from two
areas (n = 2). Th e fi rst was an area 1.7 m outside the west side
of the 24 by 30 m trial area and the second was just inside the
24 by 30 m trial area on the west side. Th irteen months follow-
ing planting, two more U sites were randomly selected on the
east side of the plot and monitored from 13 to 18 mo (n = 4).
TransplantsTh e plant chosen for this study was four-wing saltbush, which
is a perennial halophytic subshrub native to Arizona, California,
Nevada, and Utah (USDA-NRCS 2005). Th is plant is considered
drought tolerant and has been observed encroaching into other
historical mine tailings sites (USDA-SCS, 1977; Booth et al.,
1999; Arunachalam et al., 2004; Jeff erson, 2004) as well as the
Boston Mill site. Four-wing saltbush transplants were germinated
and grown in the greenhouse for 2 mo in a 2:1 sand/compost mix
in 0.375-L pots. Th e compost was a commercial composted mulch
(Sunshine mix no. 1, Sun Gro Horticulture). Th is material was also
used as the compost amendment in the fi eld trial (Tables 1 and 2).
Field StudyPlanting took place on 31 Oct. 2003. Th e study area was di-
vided into eight 7.5 by 12 m plots. A point within each plot was
selected using random X and Y coordinates and marked with a pin
fl ag to serve as a reference to later establish the 3 by 3 m treatment
areas in each plot (Fig. 1). Quadruplicate plots per treatment were
established on the study area following a complete randomized
design. To establish the treatments on the fi eld, a 3 by 3 m quadrat
was centered on a random point that was previously marked with
a pin fl ag in each of the eight 7.5 by 12 m plots. Th e four corners
of the quadrat were fl agged and one transplant was placed near the
inside of each of the quadrat corners and one in the middle. Th e
transplants were planted by digging a hole deep enough to cover
the roots, centering the transplant in the hole, and fi lling it back
with the original soil (Treatment NC) or with approximately 3.8 L
(1 gallon) of compost (Treatment C). For Treatment C, half a gal-
lon of compost was added fi rst, followed by a thin layer of soil, the
other half of the compost, and fi nally a thin layer of soil at the top.
Th e 6 by 12 m OSC area similarly received transplants in four 3
by 3 m plots that were evenly spaced in the area. After planting, all
transplants were covered with chicken wire cages for approximately
6 mo to protect against foraging animals. Over the next 8 mo the
transplants were irrigated with a total of 44 L of water per plant
using the following schedule: Day 0 and 3, 1.5 L; Day 7, 4 L; Day
14, 3 L; Day 24, 4 L; Days 40, 45, 52, 3 L; Days 83, 100, 4 L;
Days 114, 131, 181, 3 L; and on Day 218, 4 L.
Th e height (h) and crown diameters (d1 and d
2) of the
transplants were measured immediately after planting and at
11 and 18 mo to assess growth. Th e height measurement was
made from the ground to the highest point on the shrub. Th e
two crown diameter measurements were made perpendicular
to each other at the widest part of the shrub canopy. Shrub
volume was estimated by assuming an elliptical shrub shape
(V = 4/3π(h/2 × d1/2 × d
2/2) (Th omson et al., 1998).
Soil and Plant SamplingSoil samples were taken from each of the C and NC treat-
ment plots (n = 4) before planting and from the U plots
(n = 4) at the end of the trial to determine selected physical
and chemical characteristics. A 500-g surface sample (5–10
cm) was taken from each plot using a closed bucket auger and
stored in a plastic bag for analysis.
For microbiological analysis, 5-g soil samples were taken
Table 1. Selected characteristics for treatment plot tailings samples, the off -site control soil, and the compost used in this study.
Treatment plot†
pH‡ CEC§ Sand¶ Silt¶ Clay¶ TOC# TN††
cmolc kg−1 ––––––––––%–––––––––– ––g kg−1––
Off -site 8.7 8.0 62.5 26.9 10.6 4.8 0.2
NC-1 8.7 25.1 37.8 43.4 18.8 11.6 0.4
NC-2 8.4 28.7 45.3 40.9 13.8 11.0 0.7
NC-3 8.6 25.5 52.9 32.5 14.7 10.5 0.3
NC-4 9.0 24.5 74.6 18.9 6.5 10.1 0.3
C-1 8.8 22.2 47.4 40.5 12.1 8.1 0.3
C-2 8.5 30.4 26.4 47.2 26.4 21.4 0.8
C-3 9.0 32.5 61.4 30.1 8.5 8.6 0.3
C-4 8.3 28.5 45.3 38.3 16.3 11.2 0.8
U1 9.0 27.2 52.1 33.9 13.9 9.4 0.3
U2 9.0 29.1 40.8 41.6 17.7 9.7 0.3
U3 8.9 21.8 57.4 30.4 12.3 9.6 0.2
U4 9.8 25.5 41.6 39.8 18.5 17.4 0.5
Compost 8.3 ND‡‡ ND ND ND 169 3.7
† Treatment plots include no compost (NC), compost (C), and unplanted
controls (U).
‡ pH- soil to deionized water 1:2 ratio after 1 h mixing.
§ Cation exchange capacity (CEC) determined by the magnesium nitrate
method (University of Arizona Water Quality Center Lab, WQC).
¶ Texture determined by the hydrometer method (WQC).
# Total organic carbon (TOC) determined by the diff erence in total carbon
and inorganic carbon (Chorover Environmental Chemistry Lab, CECL).
†† Total nitrogen (TN) determined by high temperature combustion (CECL).
‡‡ Not determined.
Table 2. Total metal concentrations (mg kg−1) for the treatment plot tailings, the off -site control soil, and the compost used in this study.
Treatment Pb As Hg Zn Cu Fe Mn Cd Al
Off -site control† 127 (3) 7 (1) 3 (0) 989 (139) 97 (8) 10 500 (1450) 730 (69) 1 (0) 14 300 (3500)
NC‡ 11 100 ± 6100 576 ± 302 503 ± 191 5410 ± 3070 2440 ± 1370 15 600 ± 5810 44 500 ± 22 000 93 ± 50 11 500 ± 3590
C‡ 15 800 ± 958 845 ± 94 560 ± 83 8100 ± 1370 3620 ± 404 19 500 ± 4007 67 300 ± 24 000 138 ± 30 12 900 ± 4490
U‡ 14 100 ± 463 474 ± 35 178 ± 77 6570 ± 506 2290 ± 200 16 000 ± 1520 50 400 ± 7 780 86 ± 12 8000 ± 885
Compost† 21 (1) 3 (0) 2 (1) 200 (4) 28 (5) 2040 (1400) 0 0 1750 (110)
† Samples (n = 1) were analyzed in duplicates. The number in parentheses represents the deviation from the mean of the duplicates.
‡ Values are the average ± standard deviation (n = 4).

1252 Journal of Environmental Quality • Volume 36 • September-October 2007
(5–10 cm) using a sterile spatula (washed with 95% ethanol and
fl amed between samples), placed into sterile vials, and stored at
4°C within 4 h of collection for microbial analysis. Th ese samples
were collected at 0, 3, 4, 5, 7, 8.5, 11, 13, 16, and 18 mo to fol-
low bacterial community changes during plant establishment.
Th e time zero samples were taken before planting as described
above. Following planting, one plant was chosen from each treat-
ment plot (NC and C treatments, n = 4) for the microbial analy-
sis. Selected plants looked healthy, and with the exception of plot
C3, were at least 0.5 m away from any encroaching vegetation.
Soil samples were taken beneath the transplant canopy of the
chosen plant within 2 cm of the stem and approximately 5 cm
deep. For the unplanted controls, two U plots were sampled
from 3 to 11 mo (n = 2) and two more U plots were added from
13 to 18 mo (n = 4). U plots samples were taken within 5 cm of
the pin fl ag marking a random point.
Shoot tissue samples were collected 18 mo after planting
from each NC and C treatment plot to determine plant shoot
metal content. Since transplants did not survive in the off -site
control area, four composite samples were taken from nearby
indigenous four-wing saltbush shrubs to obtain background
shoot tissue metal levels.
Soil and Plant AnalysisSoil physical and chemical properties were determined by
the Water Quality Center Lab (WQCL) and by the Chorover
Enviromental Chemistry Laboratory (CECL) of the Univer-
sity of Arizona using standard soil analysis methods (Tables 1
and 2). Total metal concentrations were determined follow-
ing microwave-assisted digestion of dry soil samples (USEPA
Method 3051) and analysis by inductively coupled plasma–
mass spectrometry (ICP-MS).
For plant tissue total metal content, samples were carefully
rinsed with nanopure water to remove soil particles, dried at 65°C
for 2 d, and digested following USEPA Method 3051. Samples
were analyzed using ICP-MS by the University of Arizona Super-
fund Basic Research Program Hazard Identifi cation Core (HIC).
Heterotrophic Bacterial CountsHeterotrophic counts were performed using R2A agar
medium (Becton, Dickinson and Co., Sparks, MD) with
200 mg L−1 of cyclohexamide to prevent fungal growth. Each
sample was plated in triplicate. Briefl y, inocula were prepared
by mixing 1 g (wet wt.) of soil with 9.5 mL sterile Zwittergent
(CalBiochem, La Jolla, CA) extractant (per L H2O: 2 g NaCl,
0.0002% Zwittergent). Th e sample was vortexed for 2 min
and then serially diluted in sterile water, plated on R2A, incu-
bated at 23°C in the dark, and enumerated after 4 d.
Soil Bacterial Community DNA Extraction and Purifi cationDNA was extracted from soil bacterial communities by direct
lysis using the Fast DNA Spin Kit for Soil (Qbiogene, Carlsbad,
CA). Th e manufacturer’s protocol was followed with some modi-
fi cations. Before the extraction all sterile tubes, caps, and solu-
tions used in the protocol were exposed to UV light for 5 min in
a UVC-508 Ultraviolet Crosslinker (Ultra-Lum, Claremont, CA)
to remove any potential DNA contamination. Th e bead beating
procedure was performed by attaching the lysing matrix tubes
to a Vortex Genie II vortexer (VWR, West Chester, PA) using a
Mo Bio adaptor (Mo Bio Laboratories, Solana Beach, CA) and
vortexing the tubes at maximum speed for 10 min. Cell debris,
soil particles, and the lysing matrix beads were separated from
the DNA-containing supernatant by centrifugation at 14 000 × g
for 15 min. An alternative manufacturer’s protocol was followed
to enhance removal of soil humic materials that could inhibit
PCR amplifi cation. Samples with supernatants retaining a brown
color were assumed to contain humic material. For these samples
the Binding Matrix-DNA complex was rinsed repeatedly with
saturated 5.5 M guanidine thiocyanate (GTC, Sigma, St. Louis,
MO) until the supernatant lost its brown tint.
16S rDNA PCR for Denaturing Gradient Gel ElectrophoresisA 391 bp (including the GC clamp) portion of the 16S
rRNA gene (rDNA) encompassing the hypervariable V9 region
of the domain Bacteria was amplifi ed to target the soil bacterial
community in DNA extracts using primers 1070f and 1406r-
GC (Ferris et al., 1996). Th e PCR for DGGE was accomplished
using a modifi ed protocol from Colores et al. (2000). Briefl y,
50 μL reactions contained 1X Expand Hi Fidelity PCR Buff er
with 1.5 mM MgCl2 (Roche, Indianapolis, IN), 0.5 μM of each
primer, 0.4 g L−1 unacetylated bovine serum albumin (Sigma, St.
Louis, MO) to relieve PCR inhibition (Kreader, 1996), 200 μM
of each deoxynucleoside triphosphate, 5% dimethyl sulfoxide,
1.4 U Expand High Fidelity PCR System Enzyme (Roche, Indi-
anapolis, IN), and 0.2 to 0.28 mg L−1 soil bacterial DNA extract.
Th e reactions were exposed to 30 cycles at 95°C for 45 s, 55°C
for 45 s, and 72°C for 30 s in a GeneAmp 2400 PCR System
thermal cycler (PE Applied Biosystems, Foster City, CA).
Th e PCR products (5 μL per lane) were visualized and
evaluated using electrophoresis through a 2% GenePure LE
agarose gel (Intermountain Scientifi c Corp., Kaysville, UT)
followed by ethidium bromide staining. Products in lanes that
showed two bands, the desired product and another of slight-
ly larger size, were rejected because DGGE analysis indicated
that the extra band was associated with template overloading.
Rejected extracts were reamplifi ed using a reduced template
concentration. Most of the soil DNA extracts produced single
bands using an approximate concentration of 0.28 mg L−1,
but those requiring reamplifi cation were diluted incrementally
down to 0.2 mg L−1 until only a single band was observed.
Denaturing Gradient Gel ElectrophoresisTh e DGGE analysis was performed using a Dcode Universal
Mutation Detection System (Bio-Rad Laboratories, Hercules, CA).
Acrylamide gels (6%) were prepared with a 50 to 60% urea–for-
mamide denaturing gradient according to the manufacturer’s
protocol. Gel lanes were loaded with 25 to 35 μL of PCR product
depending on its intensity on the agarose gel. Th e gels were run at
a constant voltage of 50 V for 19 h at 60°C, stained with 3X SYBR
Green I (Molecular Probes, Eugene, OR) for 40 min, and then
imaged. To compensate for inconsistencies in electrophoresis, the

Rosario et al.: Bacterial Community Changes during Plant Establishment 1253
gels were aligned by loading an external reference ladder at regular
intervals. Th e ladder was composed of pooled PCR product from
isolated heterotrophic bacteria from tailings samples. Th e DGGE
profi les (banding patterns) were manually scored according to the
method of Konopka et al. (1999) and evaluated using Quantity
One 4.5.2 software (Bio-Rad Laboratories, Hercules, CA).
Statistical AnalysisAll statistics were performed using the statistical software
package R (R Foundation for Statistical Computing, Vienna,
Austria). Data were all found to be normally distributed with
constant variance. Results of all F tests were considered signifi -
cant at 95% confi dence level. For the revegetation trial, data
were subjected to a two-way analysis of variance (ANOVA) for a
completely randomized design, with sample date (11 or 18 mo)
and soil treatment (compost or no compost) as independent
variables and plant volume as the dependent variable. Dead
plants were excluded from the data analysis. Th e ANOVA had
1 df due to sample date, 1 df due to soil treatment, and 64 df
due to error. Systat 11.0 software (Systat Software, San Jose, CA)
was used to conduct the analysis of variance. Plant volumes were
log transformed to equalize variances among treatments. Th e
hypotheses of unequal variance between soil treatments or sample
dates were not signifi cant (p > 0.05) for transformed data. At the
end of the trial, diff erences in shoot metal concentrations were
analyzed using one way ANOVA of a completely randomized
design. Pearson’s correlation coeffi cient (r) was used to screen
linear relationships between plant and soil metal concentrations.
For the bacterial analysis, treatment eff ects on heterotrophic cul-
turable counts as a function of time were analyzed using a nested
ANOVA model with a maximum likelihood estimator on log
transformed data. Treatment was set as a fi xed factor, while plant
and time eff ects were set as random factors. Treatment diff erences
were evaluated using pairwise t tests on weighted means calcu-
lated for each treatment level (i.e., least squares means, α = 0.05).
Th e profi les obtained from DGGE gels were evaluated in
three ways. First, ANOVA analysis of the number of bands in
each profi le was used to evaluate whether there were treatment
eff ects on species richness (the number of populations present).
Second, Dice coeffi cient analysis was used to evaluate the similar-
ities between individual profi les within each treatment plot (C1,
C2, C3, C4, NC1, NC2, NC3, NC4, U1, and U2) as a func-
tion of time. Th is analysis examined the extent of the community
structure change that occurred in each plot between consecutive
sampling times during the fi eld trial. Th e Dice coeffi cient is ex-
pressed as S = 2a/(b + c) where a represents the number of bands
present in both profi les, b refers to the number of bands present
in Profi le 1, and c represents the number of bands present in
Profi le 2. Finally, Kruskal’s Non-Metric Multidimensional Scal-
ing (KNMDS; Venables and Ripley, 2002) was used to evaluate
similarities among profi les from all treatments at each time point.
Th is nonparametric ordination method was used to visualize
and interpret changes in the bacterial community during the
revegetation trial based on binary distance where similar bacterial
communities (i.e., profi les) cluster more closely than those with
low similarity in multidimensional space. Diff erences between
bacterial communities were evaluated with a permutation test (α
= 0.05). A stress factor (sf) was also calculated to refl ect good-
ness-of-fi t of the model (sf < 0.1 was considered a good fi t). Th e
confi gurations in multidimensional space were evaluated using
three or four dimensions (D) to minimize stress. Comparisons
of the bacterial community could be performed only for samples
within the same gel. Th erefore, treatment and time eff ects were
analyzed individually due to a limited number of wells per gel.
Results
Site CharacterizationSelected physical and chemical characteristics were determined
for samples from each treatment plot: NC, C, U, and OSC (Table
1). Th e alkaline pH and the presence of organic carbon are key pa-
rameters that indicate this as a moderately stressed site. In severely
stressed sites, the pH is usually acidic (<3 to 4) and there is essen-
tially no organic matter. In this site the pH is moderately alkaline,
ranging from 8.3 to as high as 9.8 in one sample and total organic
carbon values ranged from 8.6 to 21.4 mg kg−1 with an average
of 11.6 mg kg−1. Th ese values can be compared very favorably to
the OSC, which has a pH of 8.7 and a total organic C content of
4.8 mg kg−1. Th e big diff erence between the tailings and the OSC
is the total metal concentrations, which were greater in the mine
tailings samples than the OSC sample for all metals measured ex-
cept Al (Table 2). In particular, Pb, Hg, and Cd were elevated up
to 100-fold over the OSC.
Revegetation TrialOne goal of this 18-mo revegetation fi eld trial was to evaluate
whether four-wing saltbush transplants would survive and grow
in a moderately impacted mine tailings site and if so, whether
compost addition was necessary. Results from this study show
that after 1.5 yr, more than 80% of the transplants survived in
the tailings area regardless of whether or not compost was added
during planting. A signifi cant increase in growth (p < 0.05) was
observed during the fi rst 11 mo of the study with average plant
volumes increasing from 0.00047 ± 0.000067 m3 to 0.015 ±
0.012 m3. Th ere was no signifi cant diff erence in growth between
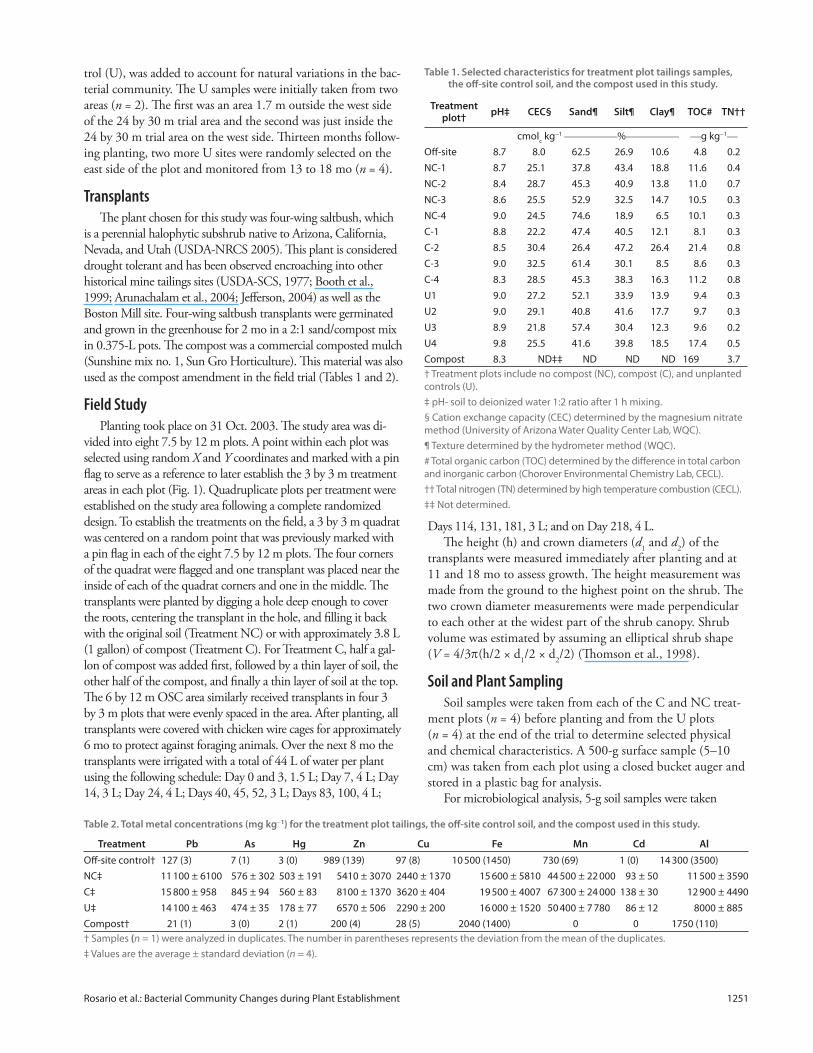
the transplants in the NC and C treatments (p > 0.05) (Fig. 2).
From 11 to 18 mo, plants did not grow further and even lost
volume to an average of 0.0080 ± 0.0067 m3, although this
volume loss was not signifi cant (p > 0.05) (Fig. 2). It should be
noted that the plants were irrigated only from 0 to 8 mo and did
not receive water for the latter part of the study. Further, during
this fi eld trial, the SPRNCA was experiencing a drought. Infor-
mation from the closest weather station, 8 miles to the northeast
in Tombstone, AZ, indicates that the area received a total of
362 mm of precipitation during the 18 mo of this study with ap-
proximately 44% of the precipitation occurring in the fi rst 8 mo.
None of the four-wing saltbush transplants survived in the
OSC plots, which was unexpected since they are indigenous
to the site. Failure to establish transplants in the OSC plots
could have been due to a fairly dense growth of indigenous
grass. Grasses, like western wheatgrass, have been found to
negatively aff ect four-wing saltbush establishment (Sabey et
1254 Journal of Environmental Quality • Volume 36 • September-October 2007
al., 1990). Th us, it is possible that naturally established grass
in this area either out-competed the transplants for nutrients
or had an inhibitory eff ect on their growth.
At the end of the trial, shoot metal contents of transplants were
elevated from 2 to 80-fold over levels found in saltbush naturally
established in nearby areas (Table 3). Th e highest levels of metal
accumulation were for Mn (?880 mg kg−1), Pb (?310 mg kg−1),
Fe (?265 mg kg−1), Al (?230 mg kg−1), and Zn (?200 mg kg−1),
refl ecting the high total concentration of these metals in the tail-
ings (Tables 2 and 3). Th ere was a positive correlation between the
uptake of Fe and the uptake of other metals including Pb, Cu, As,
Mn, Al, and Hg (r = 0.89–0.98, Table 4). Th ere were also correla-
tions between uptake of some metals pairs including Pb and As
(r = 0.92), Pb and Hg (r = 0.93), As and Hg (r = 0.91), and As
and Mn (r = 0.99). However, there was no correlation between soil
metal concentrations and plant metal uptake for individual metals
(r = −0.22 to 0.45, data not shown)
Microbial AnalysisSoil samples were taken from beneath transplant canopies
throughout the revegetation trial to monitor changes in the bacte-
rial community. Before the trial began, heterotrophic bacterial
numbers in bulk tailings averaged 2.1 ± 0.9 × 105 CFU g−1 dry soil
(CFU, colony-forming unit). Th is is similar to other studies that
have reported heterotrophic bacteria in untreated alkaline (pH > 8)
tailings (106 CFU g−1) (Shetty et al., 1994; Noyd et al., 1995)
and in acidic (pH < 4.5) tailings (105 CFU g−1) (Moynahan et al.,
2002). Within 3 mo, a 1 to 1.5 log increase was observed in bacte-
rial numbers under transplant canopies in both the C and NC
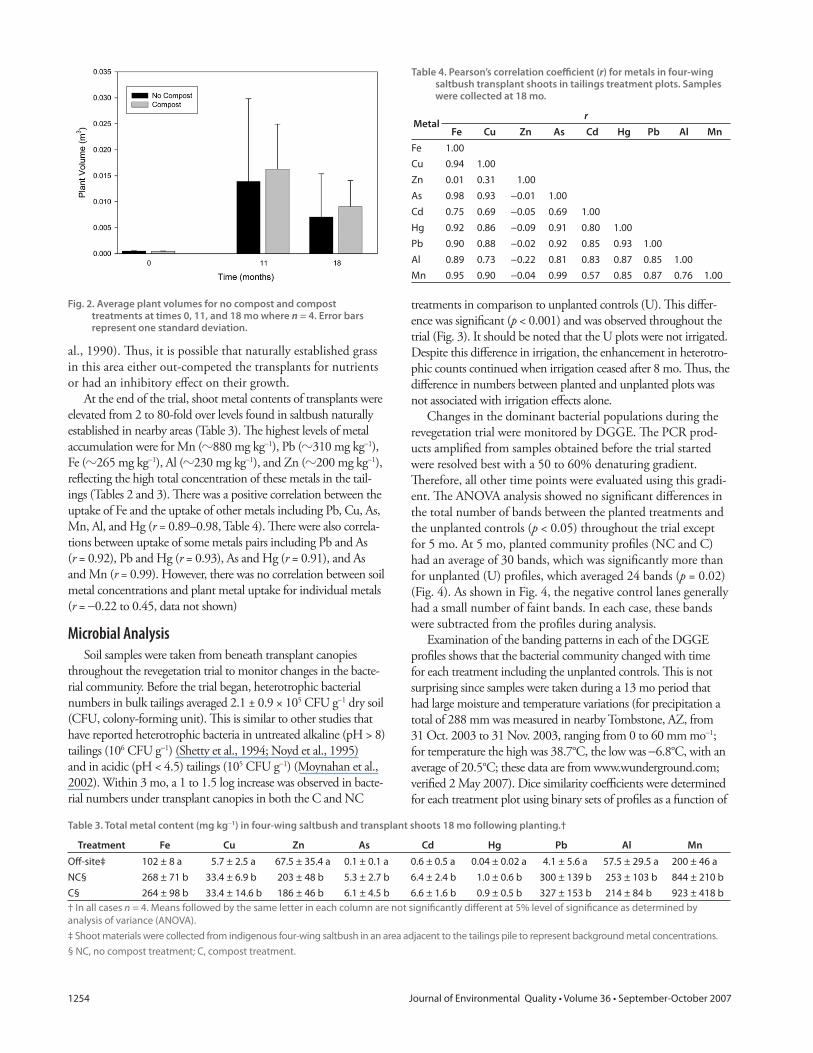
treatments in comparison to unplanted controls (U). Th is diff er-
ence was signifi cant (p < 0.001) and was observed throughout the
trial (Fig. 3). It should be noted that the U plots were not irrigated.
Despite this diff erence in irrigation, the enhancement in heterotro-
phic counts continued when irrigation ceased after 8 mo. Th us, the
diff erence in numbers between planted and unplanted plots was
not associated with irrigation eff ects alone.
Changes in the dominant bacterial populations during the
revegetation trial were monitored by DGGE. Th e PCR prod-
ucts amplifi ed from samples obtained before the trial started
were resolved best with a 50 to 60% denaturing gradient.
Th erefore, all other time points were evaluated using this gradi-
ent. Th e ANOVA analysis showed no signifi cant diff erences in
the total number of bands between the planted treatments and
the unplanted controls (p < 0.05) throughout the trial except
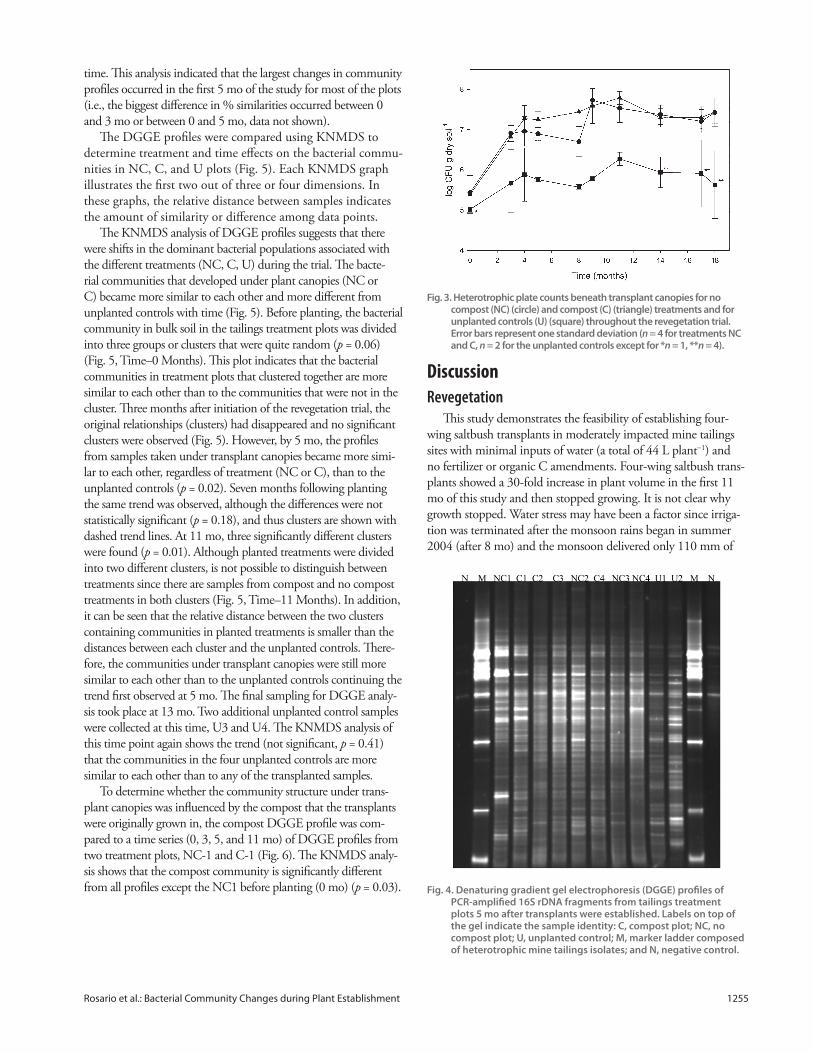
for 5 mo. At 5 mo, planted community profi les (NC and C)
had an average of 30 bands, which was signifi cantly more than
for unplanted (U) profi les, which averaged 24 bands (p = 0.02)
(Fig. 4). As shown in Fig. 4, the negative control lanes generally
had a small number of faint bands. In each case, these bands
were subtracted from the profi les during analysis.
Examination of the banding patterns in each of the DGGE
profi les shows that the bacterial community changed with time
for each treatment including the unplanted controls. Th is is not
surprising since samples were taken during a 13 mo period that
had large moisture and temperature variations (for precipitation a
total of 288 mm was measured in nearby Tombstone, AZ, from
31 Oct. 2003 to 31 Nov. 2003, ranging from 0 to 60 mm mo−1;
for temperature the high was 38.7°C, the low was −6.8°C, with an
average of 20.5°C; these data are from www.wunderground.com;
verifi ed 2 May 2007). Dice similarity coeffi cients were determined
for each treatment plot using binary sets of profi les as a function of
Fig. 2. Average plant volumes for no compost and compost treatments at times 0, 11, and 18 mo where n = 4. Error bars represent one standard deviation.
Table 3. Total metal content (mg kg−1) in four-wing saltbush and transplant shoots 18 mo following planting.†
Treatment Fe Cu Zn As Cd Hg Pb Al Mn
Off -site‡ 102 ± 8 a 5.7 ± 2.5 a 67.5 ± 35.4 a 0.1 ± 0.1 a 0.6 ± 0.5 a 0.04 ± 0.02 a 4.1 ± 5.6 a 57.5 ± 29.5 a 200 ± 46 a
NC§ 268 ± 71 b 33.4 ± 6.9 b 203 ± 48 b 5.3 ± 2.7 b 6.4 ± 2.4 b 1.0 ± 0.6 b 300 ± 139 b 253 ± 103 b 844 ± 210 b
C§ 264 ± 98 b 33.4 ± 14.6 b 186 ± 46 b 6.1 ± 4.5 b 6.6 ± 1.6 b 0.9 ± 0.5 b 327 ± 153 b 214 ± 84 b 923 ± 418 b
† In all cases n = 4. Means followed by the same letter in each column are not signifi cantly diff erent at 5% level of signifi cance as determined by
analysis of variance (ANOVA).
‡ Shoot materials were collected from indigenous four-wing saltbush in an area adjacent to the tailings pile to represent background metal concentrations.
§ NC, no compost treatment; C, compost treatment.
Table 4. Pearson’s correlation coeffi cient (r) for metals in four-wing saltbush transplant shoots in tailings treatment plots. Samples were collected at 18 mo.
Metalr
Fe Cu Zn As Cd Hg Pb Al Mn
Fe 1.00
Cu 0.94 1.00
Zn 0.01 0.31 1.00
As 0.98 0.93 −0.01 1.00
Cd 0.75 0.69 −0.05 0.69 1.00
Hg 0.92 0.86 −0.09 0.91 0.80 1.00
Pb 0.90 0.88 −0.02 0.92 0.85 0.93 1.00
Al 0.89 0.73 −0.22 0.81 0.83 0.87 0.85 1.00
Mn 0.95 0.90 −0.04 0.99 0.57 0.85 0.87 0.76 1.00
Rosario et al.: Bacterial Community Changes during Plant Establishment 1255
time. Th is analysis indicated that the largest changes in community
profi les occurred in the fi rst 5 mo of the study for most of the plots
(i.e., the biggest diff erence in % similarities occurred between 0
and 3 mo or between 0 and 5 mo, data not shown).
Th e DGGE profi les were compared using KNMDS to
determine treatment and time eff ects on the bacterial commu-
nities in NC, C, and U plots (Fig. 5). Each KNMDS graph
illustrates the fi rst two out of three or four dimensions. In
these graphs, the relative distance between samples indicates
the amount of similarity or diff erence among data points.
Th e KNMDS analysis of DGGE profi les suggests that there
were shifts in the dominant bacterial populations associated with
the diff erent treatments (NC, C, U) during the trial. Th e bacte-
rial communities that developed under plant canopies (NC or
C) became more similar to each other and more diff erent from
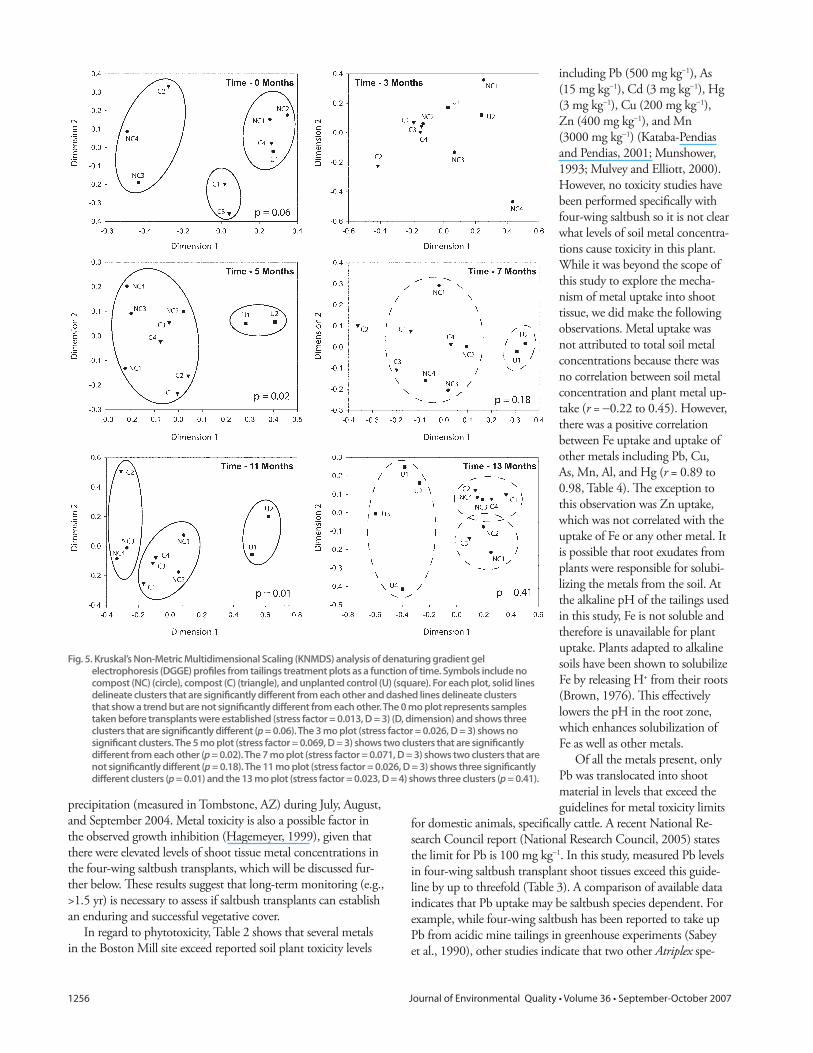
unplanted controls with time (Fig. 5). Before planting, the bacterial
community in bulk soil in the tailings treatment plots was divided
into three groups or clusters that were quite random (p = 0.06)
(Fig. 5, Time–0 Months). Th is plot indicates that the bacterial
communities in treatment plots that clustered together are more
similar to each other than to the communities that were not in the
cluster. Th ree months after initiation of the revegetation trial, the
original relationships (clusters) had disappeared and no signifi cant
clusters were observed (Fig. 5). However, by 5 mo, the profi les
from samples taken under transplant canopies became more simi-
lar to each other, regardless of treatment (NC or C), than to the
unplanted controls (p = 0.02). Seven months following planting
the same trend was observed, although the diff erences were not
statistically signifi cant (p = 0.18), and thus clusters are shown with
dashed trend lines. At 11 mo, three signifi cantly diff erent clusters
were found (p = 0.01). Although planted treatments were divided
into two diff erent clusters, is not possible to distinguish between
treatments since there are samples from compost and no compost
treatments in both clusters (Fig. 5, Time–11 Months). In addition,
it can be seen that the relative distance between the two clusters
containing communities in planted treatments is smaller than the
distances between each cluster and the unplanted controls. Th ere-
fore, the communities under transplant canopies were still more
similar to each other than to the unplanted controls continuing the
trend fi rst observed at 5 mo. Th e fi nal sampling for DGGE analy-
sis took place at 13 mo. Two additional unplanted control samples
were collected at this time, U3 and U4. Th e KNMDS analysis of
this time point again shows the trend (not signifi cant, p = 0.41)
that the communities in the four unplanted controls are more
similar to each other than to any of the transplanted samples.
To determine whether the community structure under trans-
plant canopies was infl uenced by the compost that the transplants
were originally grown in, the compost DGGE profi le was com-
pared to a time series (0, 3, 5, and 11 mo) of DGGE profi les from
two treatment plots, NC-1 and C-1 (Fig. 6). Th e KNMDS analy-
sis shows that the compost community is signifi cantly diff erent
from all profi les except the NC1 before planting (0 mo) (p = 0.03).
Discussion
RevegetationTh is study demonstrates the feasibility of establishing four-
wing saltbush transplants in moderately impacted mine tailings
sites with minimal inputs of water (a total of 44 L plant−1) and
no fertilizer or organic C amendments. Four-wing saltbush trans-
plants showed a 30-fold increase in plant volume in the fi rst 11
mo of this study and then stopped growing. It is not clear why
growth stopped. Water stress may have been a factor since irriga-
tion was terminated after the monsoon rains began in summer
2004 (after 8 mo) and the monsoon delivered only 110 mm of
Fig. 3. Heterotrophic plate counts beneath transplant canopies for no compost (NC) (circle) and compost (C) (triangle) treatments and for unplanted controls (U) (square) throughout the revegetation trial. Error bars represent one standard deviation (n = 4 for treatments NC and C, n = 2 for the unplanted controls except for *n = 1, **n = 4).
Fig. 4. Denaturing gradient gel electrophoresis (DGGE) profi les of PCR-amplifi ed 16S rDNA fragments from tailings treatment plots 5 mo after transplants were established. Labels on top of the gel indicate the sample identity: C, compost plot; NC, no compost plot; U, unplanted control; M, marker ladder composed of heterotrophic mine tailings isolates; and N, negative control.
1256 Journal of Environmental Quality • Volume 36 • September-October 2007
precipitation (measured in Tombstone, AZ) during July, August,
and September 2004. Metal toxicity is also a possible factor in
the observed growth inhibition (Hagemeyer, 1999), given that
there were elevated levels of shoot tissue metal concentrations in
the four-wing saltbush transplants, which will be discussed fur-
ther below. Th ese results suggest that long-term monitoring (e.g.,
>1.5 yr) is necessary to assess if saltbush transplants can establish
an enduring and successful vegetative cover.
In regard to phytotoxicity, Table 2 shows that several metals
in the Boston Mill site exceed reported soil plant toxicity levels
including Pb (500 mg kg−1), As
(15 mg kg−1), Cd (3 mg kg−1), Hg
(3 mg kg−1), Cu (200 mg kg−1),
Zn (400 mg kg−1), and Mn
(3000 mg kg−1) (Kataba-Pendias
and Pendias, 2001; Munshower,
1993; Mulvey and Elliott, 2000).
However, no toxicity studies have
been performed specifi cally with
four-wing saltbush so it is not clear
what levels of soil metal concentra-
tions cause toxicity in this plant.
While it was beyond the scope of
this study to explore the mecha-
nism of metal uptake into shoot
tissue, we did make the following
observations. Metal uptake was
not attributed to total soil metal
concentrations because there was
no correlation between soil metal
concentration and plant metal up-
take (r = −0.22 to 0.45). However,
there was a positive correlation
between Fe uptake and uptake of
other metals including Pb, Cu,
As, Mn, Al, and Hg (r = 0.89 to
0.98, Table 4). Th e exception to
this observation was Zn uptake,
which was not correlated with the
uptake of Fe or any other metal. It
is possible that root exudates from
plants were responsible for solubi-
lizing the metals from the soil. At
the alkaline pH of the tailings used
in this study, Fe is not soluble and
therefore is unavailable for plant
uptake. Plants adapted to alkaline
soils have been shown to solubilize
Fe by releasing H+ from their roots
(Brown, 1976). Th is eff ectively
lowers the pH in the root zone,
which enhances solubilization of
Fe as well as other metals.
Of all the metals present, only
Pb was translocated into shoot
material in levels that exceed the
guidelines for metal toxicity limits
for domestic animals, specifi cally cattle. A recent National Re-
search Council report (National Research Council, 2005) states
the limit for Pb is 100 mg kg−1. In this study, measured Pb levels
in four-wing saltbush transplant shoot tissues exceed this guide-
line by up to threefold (Table 3). A comparison of available data
indicates that Pb uptake may be saltbush species dependent. For
example, while four-wing saltbush has been reported to take up
Pb from acidic mine tailings in greenhouse experiments (Sabey
et al., 1990), other studies indicate that two other Atriplex spe-
Fig. 5. Kruskal’s Non-Metric Multidimensional Scaling (KNMDS) analysis of denaturing gradient gel electrophoresis (DGGE) profi les from tailings treatment plots as a function of time. Symbols include no compost (NC) (circle), compost (C) (triangle), and unplanted control (U) (square). For each plot, solid lines delineate clusters that are signifi cantly diff erent from each other and dashed lines delineate clusters that show a trend but are not signifi cantly diff erent from each other. The 0 mo plot represents samples taken before transplants were established (stress factor = 0.013, D = 3) (D, dimension) and shows three clusters that are signifi cantly diff erent (p = 0.06). The 3 mo plot (stress factor = 0.026, D = 3) shows no signifi cant clusters. The 5 mo plot (stress factor = 0.069, D = 3) shows two clusters that are signifi cantly diff erent from each other (p = 0.02). The 7 mo plot (stress factor = 0.071, D = 3) shows two clusters that are not signifi cantly diff erent (p = 0.18). The 11 mo plot (stress factor = 0.026, D = 3) shows three signifi cantly diff erent clusters (p = 0.01) and the 13 mo plot (stress factor = 0.023, D = 4) shows three clusters (p = 0.41).

Rosario et al.: Bacterial Community Changes during Plant Establishment 1257
cies, A. numularia, and A. lentiformis, did not shoot accumulate
Pb from alkaline and acidic tailings, respectively (Williams et al.,
1994; Jordan et al., 2002; Mendez et al., 2007). Th e observed
shoot accumulation of Pb in this study is undesirable as wildlife
may be negatively impacted. Th us, other native species that
naturally establish in the tailings area, such as big sacaton grass or
other Atriplex species, should be investigated.
Microbial Community AnalysisOne way in which the introduction of plants into mine
tailings sites improves ecosystem function is by enhancing mi-
crobial activity, which in turn enhances soil productivity and
supports plant establishment and succession. Th us, we hypoth-
esized that following planting the presence of root exudates
and compost C inputs would lead to an increase in culturable
heterotrophic bacterial numbers in planted treatments. Th is
hypothesis was confi rmed with a 1 to 1.5 log increase in het-
erotrophic numbers (CFUs) in planted treatment plots. Th is
increase occurred whether or not compost was added, sug-
gesting that the plants, rather than added compost, enhanced
the heterotrophic bacterial community beneath transplant.
canopies. Th is is further supported by the data showing that the
compost community DGGE profi le was quite diff erent from
the NC and C community profi les (Fig. 6). Note that both of
these communities had exposure to the compost. For the NC
treatments the plants were grown in compost before transplant-
ing and for the C treatments the plants were both grown in the
compost and were amended with compost during transplanting
We also hypothesized that plant establishment would lead
to an increase in species richness in the Boston Mill site. Th is
hypothesis was not confi rmed. An ANOVA analysis compar-
ing the number of bands among DGGE profi les suggests
that there was no signifi cant increase (p > 0.05) in richness
between planted areas and the unplanted controls (except at
5 mo, p = 0.02). While DGGE profi les do not provide total
richness indices, as one band may represent more than one
species (Jackson et al., 2000), these profi les can serve as a
screening tool to observe gross diff erences in species richness
between multiple samples. Interestingly, some studies have
suggested that root exudates can serve as selective compounds
decreasing richness indices rather than increasing them
(Marilley et al., 1998; Kozdrój and van Elsas, 2000).
As just discussed, an increase in heterotrophic bacterial numbers
is not equivalent to an increase in species richness, nor is it equiva-
lent to an increase in community functional diversity (e.g., nutrient
cycling, response to environmental stress). In fact it has been sug-
gested that heterotrophic bacterial numbers recover more readily
than functional diversity during reclamation (Moynahan et al.,
2002). Th is may be due to a slower response in the bacterial ge-
netic pool. Although species richness is not equivalent to functional
diversity, there is some indication that there may be a critical level
of species richness below which function is impaired (Griffi ths et
al., 1997). Th is is an important issue because low functional diver-
sity might compromise the ability of the bacterial community to
normally degrade organic matter, and thus, aff ect nutrient cycling.
One impact would be an accumulation of organic matter in the
site. Th is question can only be resolved by long-term monitoring
of the site to investigate if there is organic matter accumulation.
Spatial heterogeneity and seasonal changes often makes it dif-
fi cult to fi nd signifi cant diff erences among treatments in fi eld stud-
ies. In this study, although KNMDS results were not signifi cant
for every time point, a consistent trend was observed regarding
bacterial community structure from 5 mo on. In general, DGGE
profi les from planted treatments were more similar to each other
than to the unplanted controls. In contrast, there was no consistent
diff erence in profi les taken beneath transplant canopies in compost
and no compost treatments (similar to the plant growth results).
Th us, KNMDS analysis suggests that the establishment of plants,
in either the absence or presence of compost amendment, caused
a similar change in the bacterial community structure beneath
transplant canopies. Several other groups studying bacterial com-
munity structure have also observed community changes during
plant growth (e.g., Marschner et al., 2002; Kang and Mills, 2004;
Lerner et al., 2006). It would be intriguing to determine whether
the community structure changes observed in this study could be
used to develop biomarkers to predict the stage and success of phy-
tostabilization in arid and semiarid mine tailings sites.
Temporal analysis of unplanted control DGGE profi les in-
dicates that shifts occurred in the bulk soil bacterial community
(e.g., the Dice similarity coeffi cient for time 0 and time 13 mo
was 62%). Th e magnitude of this shift is actually quite similar to
that found for the NC (58.5%) and the C (52.6%) treatments
suggesting that there are natural seasonal changes in the bacte-
rial community structure in both the control and treated plots.
Such temporal shifts in soil communities have also been observed
previously in biological soil crusts (Nagy et al., 2005) and in an
upland grassland soil in the UK (Griffi ths et al., 2003). In the
latter study, profi les obtained from 5 to 10 cm and from 10 to 15
cm changed more than profi les obtained from 0 to 5 cm. In the
present study, samples were obtained from approximately 5 cm.
Th e results of this study along with similar fi ndings from
other groups show that natural temporal variations in the bac-
Fig. 6. Kruskal’s Non-Metric Multidimensional Scaling (KNMDS) analysis of the denaturing gradient gel electrophoresis (DGGE) profi le from the compost used to germinate and grow the transplants (compost only) and the profi les from one of the no compost plots (NC1) and one of the compost plots (C1) at times 0, 3, 5, and 11 mo. The plot (stress factor = 0.021, D = 4) (D, dimension) shows two signifi cantly diff erent clusters (p = 0.025).

1258 Journal of Environmental Quality • Volume 36 • September-October 2007
terial community can aff ect the interpretation of results. For
example, there was a single time point, 5 mo, where we observed
signifi cant diff erences in the number of bands among the DGGE
profi les of unplanted and planted areas. If this was the only time
point analyzed, this fi nding would have suggested that there were
increases in bacterial diversity associated with planted areas but
not unplanted controls. Th erefore, to assess reclamation success
through microbial community changes it is important to moni-
tor changes at various times throughout the year.
ConclusionsPrevious work has shown that reclamation strategies such as
liming, seeding, and compost addition can increase heterotro-
phic numbers and change community structure in both acidic
and alkaline mine tailings (Noyd et al., 1995; Mummey et al.,
2002; Moynahan et al., 2002; Kelly et al., 2003). Here we have
shown that the establishment of four-wing saltbush transplants
in the Boston Mill mine tailings site caused similar changes in
the bacterial community as detected by heterotrophic counts
and DGGE profi le analysis. In this moderately impacted mine
tailings site, compost amendment was not essential to either
establishment of plants or to the development of the bacterial
community. Study results also suggest that increased heterotro-
phic bacterial abundance and shifts in the dominant bacterial
populations did not translate into an increased bacterial species
richness. Long-term monitoring needs to be performed to inves-
tigate whether the observed changes are sustained over time and
to determine whether the observed shifts in community structure
refl ect potential increases in functional diversity. In summary,
the practical implication of this work is that reclamation eff orts
in alkaline mine tailings should take into consideration that the
addition of compost may not provide any benefi t for plant es-
tablishment. Th is should be a cost-reducing advantage for imple-
menting phytostabilization in moderately impacted mining sites.
AcknowledgmentsTh is research was supported by Grant 2 P42 ES04940-11
from the National Institute of Environmental Health Sciences
Superfund Basic Research Program, NIH. Our thanks to Karl Ford
and William L. Auby of the Bureau of Land Management for their
support and cooperation in giving us access to the Boston Mill
Site. We also wish to thank Michael Kopplin of the University of
Arizona Superfund Basic Research Program Hazard Identifi cation
Core for performing all ICP-MS total metal analyses.
ReferencesArunachalam, S.K., C. Hinz, and G. Aylmore. 2004. Soil physical properties
aff ecting root growth in rehabilitated gold mine tailings. p. 1–7. In SuperSoil 2004: 3rd Australian New Zealand Soils Conf., Univ. of Sydney, Australia.
Brown, J.C. 1976. Screening plants for iron effi ciency. In M.J. Wright (ed.) Proc. Workshop on Plant Adaptation to Mineral Stress in Problem Soils. Cornell Univ. Agric. Exp. Stn., Ithaca, NY.
Booth, D.T., J.K. Gores, G.E. Schuman, and R.A. Olson. 1999. Shrub densities on pre-1985 reclaimed mine lands in Wyoming. Restor. Ecol. 7:24–32.
Burd, G.I., D.G. Dixon, and B.R. Glick. 2000. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 46:237–245.
Colores, G.M., R.E. Macur, D.M. Ward, and W.P. Inskeep. 2000. Molecular analysis of surfactant-driven microbial population shifts in hydrocarbon-
contaminated soil. Appl. Environ. Microbiol. 66:2959–2964.
Crowley, D.E., S. Alvey, and E.S. Gilbert. 1997. Rhizosphere ecology of xenobiotic-degrading microorganisms. In E.L. Kruger et al. (ed.) Phytoremediation of soil and water contaminants. ACS Symp. Ser. 664. Am. Chemical Soc., Washington, DC.
Cunningham, S.D., R.W. Berti, and J.W. Huang. 1995. Phytoremediation of contaminated soils. Trends Biotechnol. 13:393–397.
Ferris, M.J., G. Muyzer, and D.M. Ward. 1996. Denaturing gradient gel electrophoresis profi les of 16S rRNA-defi ned populations inhabiting a hot spring mat community. Appl. Environ. Microbiol. 62:340–346.
Glick, B.R. 2003. Phytoremediation: A synergistic use of plant and bacteria to clean up the environment. Biotechnol. Adv. 21:383–393.
Griffi ths, B.S., K. Ritz, and R.E. Wheatley. 1997. Relationship between functional diversity and genetic diversity in complex microbial communities. p. 1–9. In H. Insam and A. Rangger (ed.) Microbial communities: Functional versus structural approaches. Springer, New York.
Griffi ths, R.I., A.S. Whiteley, A.G. O’Donnell, and M.J. Bailey. 2003. Infl uence of depth and sampling time on bacterial community structure in an upland grassland soil. FEMS Microbiol. Ecol. 43:35–43.
Hagemeyer, J. 1999. Ecophysiology of plant growth under heavy metal stress. p. 157–180. In M.N.V. Prasad and J. Hagemeyer (ed.) Heavy metal stress in plants: From molecules to ecosystems. Springer, New York.
Jackson, C.R., E.E. Roden, and P.F. Churchill. 2000. Denaturing gradient gel electrophoresis can fail to separate 16S rDNA fragments with multiple base diff erences. Mol. Biol. Today 1:49–51.
Jeff erson, L.V. 2004. Implications of plant density on the resulting community structure of mine site land. Restor. Ecol. 12:429–438.
Jordan, F.L., M. Robin-Abbott, R.M. Maier, and E.P. Glenn. 2002. A comparison of chelator-facilitated metal uptake by an halophyte and a glycophyte. Environ. Toxicol. Chem. 21:2698–2704.
Kang, S., and A.L. Mills. 2004. Soil bacterial community structure changes following disturbance of the overlying plant community. Soil Sci. 169:55–65.
Kataba-Pendias, A., and H. Pendias. 2001. Trace elements in soils and plants. CRC Press, Boca Raton, FL.
Kelly, J.J., M.M. Häggblom, and R.L. Tate. 2003. Eff ects of heavy metal contamination and remediation on soil microbial communities in the vicinity of a zinc smelter as indicated by analysis of microbial phospholipid fatty acid profi les. Biol. Fertil. Soils 38:65–71.
Konopka, A., T. Bercot, and C. Nakatsu. 1999. Bacterioplankton community diversity in a series of thermally stratifi ed lakes. Microb. Ecol. 38:126–135.
Kozdrój, J., and J.D. van Elsas. 2000. Response of the bacterial community to root exudates in soil polluted with heavy metals assessed by molecular and cultural approaches. Soil Biol. Biochem. 32:1405–1417.
Kreader, C. 1996. Relief of amplifi cation inhibition in PCR with bovine serum albumin or T4 gene 32 protein. Appl. Environ. Microbiol. 62:1102–1106.
Kuiper, I., E.L. Lagendijk, G.V. Bloemberg, and B.J. Lugtenberg. 2004. Rhizoremediation: A benefi cial plant–microbe interaction. Mol. Plant Microbe Interact. 17:6–15.
Ledin, M., and K. Pedersen. 1996. Th e environmental impact of mine wastes: Roles of microorganisms and their signifi cance in treatment of mine wastes. Earth Sci. Rev. 41:67–108.
Lerner, A., Y. Herschkovitz, E. Baudoin, S. Nazaret, Y. Moenne-Loccoz, Y. Okon, and E. Jurkevitch. 2006. Eff ect of Azospirillum brasilense inoculation on rhizobacterial communities analyzed by denaturing gradient gel electrophoresis and automated ribosomal intergenic spacer analysis. Soil Biol. Biochem. 38:1212–1218.
Maier, R.M., I.L. Pepper, and C.P. Gerba. 2000. p. 61–89. Environmental microbiology. Academic Press, San Diego, CA.
Marilley, L., G. Vogt, M. Blanc, and M. Aragno. 1998. Bacterial diversity in the bulk soil and rhizosphere fractions of Lolium perenne and Trifolium repens as revealed by PCR restriction analysis of 16S rDNA. Plant Soil 198:219–224.
Marschner, P., C. Neumann, A. Kania, L. Weiskopf, and R. Lieberei. 2002. Spatial and temporal dynamics of the microbial community structure in the rhizosphere of cluster roots of white lupin (Luminus albus L.). Plant Soil 246:167–174.
Mendez, M.O., E.P. Glenn, and R.M. Maier. 2007. Phytostabilization potential of quailbush for mine tailings: Growth, metal accumulation and microbial community changes. J. Environ. Qual. 36:245–253.
Moynahan, O.S., C.A. Zabinski, and J.E. Gannon. 2002. Microbial community structure and carbon utilization diversity in a mine tailings revegetation study. Restor. Ecol. 10:77–87.

Rosario et al.: Bacterial Community Changes during Plant Establishment 1259
Mulvey, P.J., and G.L. Elliott. 2000. Toxicities in soils. p. 252–257. In P.E.V. Charman and B. W. Murphy (ed.) Soils: Th eir properties and management. Oxford Univ. Press, South Melbourne, Australia.
Mummey, D.L., P.D. Stahl, and J.S. Buyer. 2002. Microbial biomarkers as an indicator of ecosystem recovery following surface mine reclamation. Appl. Soil Ecol. 21:251–259.
Munshower, F.F. 1993. Practical handbook of disturbed land revegetation. Lewis Publ., Boca Raton, FL.
Nagy, M.L., A. Perez, and F. Garcia-Pichel. 2005. Th e prokaryotic diversity of biological soil crusts in the Sonoran Desert (Organ Pipe Cactus National Monument, AZ). FEMS Microbiol. Ecol. 54:233–245.
National Research Council. 2005. Mineral tolerance of animals. National Academies Press, Washington, DC.
Noyd, R.K., F.L. Pfl eger, M.R. Norland, and M.J. Sadowosky. 1995. Native prairie grasses and microbial community responses to reclamation of tactonite iron ore tailing. Can. J. Bot. 73:1645–1654.
Pishchik, V.N., N.I. Vorobyev, I.I. Chernyaeva, S.V. Timifeeva, A.P. Kozhemyakov, Y.V. Alexeev, and S.M. Lukin. 2002. Experimental and mathematical simulation of plant growth promoting rhizobacteria and plant interaction under cadmium stress. Plant Soil 243:173–186.
Sabey, B.R., R.L. Pendleton, and B.L. Webb. 1990. Eff ect of municipal sewage sludge application on growth of two reclamation shrub species in copper mine spoils. J. Environ. Qual. 19:580–586.
Salt, D.E., N. Benhamou, M. Leszczyniecka, I. Raskin, and I. Chet. 1999. A
possible role for rhizobacteria in water treatment by plant roots. Int. J. Phytorem. 1:67–69.
Sandrin, T.R., A.M. Chech, and R.M. Maier. 2000. A rhamnolipid biosurfactant reduces cadmium toxicity during naphthalene biodegradation. Appl. Environ. Microbiol. 66:4585–4588.
Shetty, K.G., M.K. Banks, B.A. Hetrick, and A.P. Schwab. 1994. Biological characterization of a southeast Kansas mining site. Water Air Soil Pollut. 78:160–177.
Th omson, E.F., S.N. Mirza, and J. Afzal. 1998. Technical note: Predicting the components of aerial biomass of four-wing saltbush from shrub height and volume. J. Range Manage. 51:323–325.
USDA-NRCS. 2005. Th e PLANTS database. Version 3.5. Available at http://plants.usda.gov (verifi ed 3 May 2007). Data compiled from various sources by Mark W. Skinner. National Plant Data Center, Baton Rouge, LA.
USDA-SCS. 1977. Notice of release of ‘Corto’ Australian saltbush for soil stabilization and cover. USDA-SCS Plant Science Div., Washington, DC, and Arizona Agric. Exp. Stn., Tucson, AZ.
Venables, W.N., and B.D. Ripley. 2002. Modern applied statistics with S. 4th ed. p. 385–389. Springer Statistics and Computing Series, New York.
Williams, T.P., J.M. Bubb, and J.N. Lester. 1994. Th e occurrence and distribution of trace metals in halophytes. Chemosphere 28:1189–1199.
Xu, J.G., and R.L. Johnson. 1995. Root growth, microbial activity, and phosphatase activity in oil-contaminated, remediated and uncontaminated soils planted to barley and fi eld pea. Plant Soil 173:3–10.




















