
Astonishing Diversity of Natural Surfactants: 4. Fatty Acid Amide Glycosides, Their Analogs and...
20
ABSTRACT: FA amide glycosides are of great interest, espe- cially for the medicinal and pharmaceutical industries. These bi- ologically active natural surfactants are good prospects for future chemical preparation of compounds useful as antibiotics, anti- cancer agents, or for industry. More than 200 unusual and inter- esting natural surfactants, including their chemical structures and biological activities, are described in this review article. Paper no. L9744 in Lipids 40, 641–660 (July 2005). Fatty (carboxylic) acid amides, their analogs and derivatives are widespread in nature, although the occurrence of their glycosides is more limited (2). They are incorporated into ceramides (3), glycosphingolipids (4,5), gangliosides (6,7), N-acylated lipids (8), bacterial lipoproteins (9), and other natural complex com- pounds. The best known FA amides, such as anandamide, oleamide, N-arachidonoyldopamine, and N-acylethanolamines, as well as aminoglycosides have a high biological activity. They and primary amines attracted attention as bioregulators for the first time in 1957 when Meisinger and co-workers (10) demon- strated that N-palmitoylethanolamine is an anti-inflammatory factor contained in the lipid fraction of soybean, peanut oil, and egg yolk. More interest in FA amides with primary amines was associated with the discovery of potent neuromodulatory effects of natural N-arachidonyl-ethanolamide (anandamide) (11) and oleoylamine (oleamide) (12). Anandamide has attracted special interest because of its marked biological activities (11). Like the pharmacologically active compounds in marijuana (from Cannabis sativa), it ex- erts its effects through binding to and activation of specific cannabinoid receptors, designated CB1 and CB2, both of which are membrane-bound G-proteins. CB1 is found in the central nervous system and in some other organs, including the heart, uterus, testis, and small intestine, whereas the CB2 re- ceptor is found in the periphery of the spleen and other cells as- sociated with immunochemical functions, but not in brain (13). Recently, it has been shown that anandamide and presumably other polyunsaturated N-acylethanolamines bind to and acti- vate both the central (CB1) and peripheral (CB2) cannabinoid receptors and elicit virtually all of the known effects of cannabis. Anandamide is believed to have important anti-in- flammatory and anticarcinogenic properties, and it affects the cardiovascular system by inducing profound decreases in blood pressure and heart rate (14). cis-9,10-Octadecenamide is a primary FA amide. It was first isolated from the cerebrospinal fluid of sleep-deprived cats, and has been characterized and identified as the signaling molecule re- sponsible for causing sleep. It induces physiological sleep when injected directly into the brain of rats. Although other FA primary amides in addition to cis-9,10-octadecenamide are present natu- rally in the cerebrospinal fluid of animals, no others exhibit similar physiological activity (15). The biological effects of the other fatty acyl-ethanolamine derivatives are less clear, although they are by far the most abundant components of this lipid class. FA amide glycosides such as ceramides, gangliosides, amino- glycosides, their analogs and derivatives also show high biologi- cal activity (3–7,16–18). FA (carboxylic) amides are produced synthetically in industry in large amounts (more than 400,000 tons per annum) for use as ingredients of detergents, lubricants, inks, and many other products (19–22). However, in this review article, we are concerned with those as simple, complex of fatty (carboxylic) acid amide glycosides, which are found in nature, and some of which have profound biological functions. AMINOGLYCOSIDES AND DERIVATIVES Since Florey and co-workers (23) reported in 1940 that peni- cillin, a microbial product, is active against pathogenic mi- croorganisms, many investigations have focused on the screen- ing of natural compounds with antibiotic properties (24). A great number of antibiotics, termed aminoglycosides, have been discovered from cultures of bacteria and actinomycetes such as Bacillus, Nocardia, Micromonospora, and Strepto- myces. The aminoglycoside antibiotics show broad spectra against both Gram-positive and Gram-negative bacteria. The term ”aminoglycoside” refers to their structural aspects. These compounds are a large and diverse class of antibiotics that char- acteristically contain two or more aminosugars linked by gly- cosidic bonds to an aminocyclitol component (24,25). A few FA amide glycosides have been found. Amikacin 1 inhibits the growth of resistant bacteria producing APH(3′)-1 Copyright © 2005 by AOCS Press 641 Lipids, Vol. 40, no. 7 (2005) *Address correspondence at Department of Organic Chemistry, P.O. Box 39231, Hebrew University, Jerusalem 91391, Israel. E-mail: [email protected]; or [email protected] For previous article in this series see Reference 1. Abbreviations: fM, femtomole is 10 -15 mol; IC, inhibitory concentration; IC 50 , concentration at which growth or activity is inhibited by 50% (applies to ligand and growth inhibition); LC 50 for drugs with a cytotoxic effect, the concentration of drug at which 50% of cells die (a 50% reduction in the mea- sured protein at the end of the drug treatment as compared with that at the beginning); LD 50 , (lethal dose 50 ), the dose of a chemical that kills 50% of a sample population; MIC, minimal inhibitory concentration of antibiotic that inhibits a bacterium; MT1-MMP, membrane type 1 matrix metalloproteinase. *Address correspondence at Department of Organic Chemistry, P.O. Box 39231, Hebrew University, Jerusalem 91391, Israel. REVIEW Astonishing Diversity of Natural Surfactants: 4. Fatty Acid Amide Glycosides, Their Analogs and Derivatives Valery M. Dembitsky* Department of Organic Chemistry and School of Pharmacy, Hebrew University, Jerusalem, Israel
Transcript of Astonishing Diversity of Natural Surfactants: 4. Fatty Acid Amide Glycosides, Their Analogs and...

ABSTRACT: FA amide glycosides are of great interest, espe-cially for the medicinal and pharmaceutical industries. These bi-ologically active natural surfactants are good prospects for futurechemical preparation of compounds useful as antibiotics, anti-cancer agents, or for industry. More than 200 unusual and inter-esting natural surfactants, including their chemical structures andbiological activities, are described in this review article.
Paper no. L9744 in Lipids 40, 641–660 (July 2005).
Fatty (carboxylic) acid amides, their analogs and derivatives arewidespread in nature, although the occurrence of their glycosidesis more limited (2). They are incorporated into ceramides (3),glycosphingolipids (4,5), gangliosides (6,7), N-acylated lipids(8), bacterial lipoproteins (9), and other natural complex com-pounds. The best known FA amides, such as anandamide,oleamide, N-arachidonoyldopamine, and N-acylethanolamines,as well as aminoglycosides have a high biological activity. Theyand primary amines attracted attention as bioregulators for thefirst time in 1957 when Meisinger and co-workers (10) demon-strated that N-palmitoylethanolamine is an anti-inflammatoryfactor contained in the lipid fraction of soybean, peanut oil, andegg yolk. More interest in FA amides with primary amines wasassociated with the discovery of potent neuromodulatory effectsof natural N-arachidonyl-ethanolamide (anandamide) (11) andoleoylamine (oleamide) (12).
Anandamide has attracted special interest because of itsmarked biological activities (11). Like the pharmacologicallyactive compounds in marijuana (from Cannabis sativa), it ex-erts its effects through binding to and activation of specificcannabinoid receptors, designated CB1 and CB2, both ofwhich are membrane-bound G-proteins. CB1 is found in thecentral nervous system and in some other organs, including theheart, uterus, testis, and small intestine, whereas the CB2 re-ceptor is found in the periphery of the spleen and other cells as-
sociated with immunochemical functions, but not in brain (13).Recently, it has been shown that anandamide and presumablyother polyunsaturated N-acylethanolamines bind to and acti-vate both the central (CB1) and peripheral (CB2) cannabinoidreceptors and elicit virtually all of the known effects ofcannabis. Anandamide is believed to have important anti-in-flammatory and anticarcinogenic properties, and it affects thecardiovascular system by inducing profound decreases in bloodpressure and heart rate (14).
cis-9,10-Octadecenamide is a primary FA amide. It was firstisolated from the cerebrospinal fluid of sleep-deprived cats, andhas been characterized and identified as the signaling molecule re-sponsible for causing sleep. It induces physiological sleep wheninjected directly into the brain of rats. Although other FA primaryamides in addition to cis-9,10-octadecenamide are present natu-rally in the cerebrospinal fluid of animals, no others exhibit similarphysiological activity (15). The biological effects of the other fattyacyl-ethanolamine derivatives are less clear, although they are byfar the most abundant components of this lipid class.
FA amide glycosides such as ceramides, gangliosides, amino-glycosides, their analogs and derivatives also show high biologi-cal activity (3–7,16–18). FA (carboxylic) amides are producedsynthetically in industry in large amounts (more than 400,000tons per annum) for use as ingredients of detergents, lubricants,inks, and many other products (19–22). However, in this reviewarticle, we are concerned with those as simple, complex of fatty(carboxylic) acid amide glycosides, which are found in nature,and some of which have profound biological functions.
AMINOGLYCOSIDES AND DERIVATIVES
Since Florey and co-workers (23) reported in 1940 that peni-cillin, a microbial product, is active against pathogenic mi-croorganisms, many investigations have focused on the screen-ing of natural compounds with antibiotic properties (24). Agreat number of antibiotics, termed aminoglycosides, havebeen discovered from cultures of bacteria and actinomycetessuch as Bacillus, Nocardia, Micromonospora, and Strepto-myces. The aminoglycoside antibiotics show broad spectraagainst both Gram-positive and Gram-negative bacteria. Theterm ”aminoglycoside” refers to their structural aspects. Thesecompounds are a large and diverse class of antibiotics that char-acteristically contain two or more aminosugars linked by gly-cosidic bonds to an aminocyclitol component (24,25).
A few FA amide glycosides have been found. Amikacin 1inhibits the growth of resistant bacteria producing APH(3′)-1
Copyright © 2005 by AOCS Press 641 Lipids, Vol. 40, no. 7 (2005)
*Address correspondence at Department of Organic Chemistry, P.O. Box39231, Hebrew University, Jerusalem 91391, Israel. E-mail: [email protected]; or [email protected] previous article in this series see Reference 1.Abbreviations: fM, femtomole is 10−15 mol; IC, inhibitory concentration;IC50, concentration at which growth or activity is inhibited by 50% (appliesto ligand and growth inhibition); LC50 for drugs with a cytotoxic effect, theconcentration of drug at which 50% of cells die (a 50% reduction in the mea-sured protein at the end of the drug treatment as compared with that at thebeginning); LD50, (lethal dose50), the dose of a chemical that kills 50% of asample population; MIC, minimal inhibitory concentration of antibiotic thatinhibits a bacterium; MT1-MMP, membrane type 1 matrix metalloproteinase.*Address correspondence at Department of Organic Chemistry, P.O. Box39231, Hebrew University, Jerusalem 91391, Israel.
REVIEW
Astonishing Diversity of Natural Surfactants: 4. FattyAcid Amide Glycosides, Their Analogs and Derivatives
Valery M. Dembitsky*Department of Organic Chemistry and School of Pharmacy, Hebrew University, Jerusalem, Israel

(3′-aminoglycoside phosphotransferase) and AAD(2′) (2′-amino acid decarboxylase). Amikacin 1 was biologically trans-formed from kanamycin by a mutant of butirosin-producingBacillus circulans (26). Arbekacin 2 showed strong activityagainst resistant bacteria including Pseudomonas. It waslaunched into the clinical market in Japan in 1990 as achemotherapeutic agent for the treatment of infections causedby methicillin-resistant Staphylococcus aureus (27). 2″-Amino-2″-deoxyarbekacin 3 demonstrated potent activityagainst S. aureus (28). Isepamicin 4 showed activity against re-sistant bacteria with lower toxicity than gentamicin B and hasbeen useful in the clinical treatment of infection (24).
Fortimicins A 5 and B are bicyclic aminoglycoside antibi-otics that contain a fortamine moiety instead of the deoxystrep-tamine; both are produced by strain Micromonospora sp. MK-7 (29,30), and fortimicin A also is produced by M. olivoast-erospora KY 1151 (31). Fortimicin A showed excellentactivity against 352 strains of Enterobacteriaceae (a concentra-tion of 6.2 µg/mL inhibited the greatest number of strains,92.6%) and other medically significant organisms, and weakactivity against Pseudomonas aeruginosa strains (32,33). Themarine bacterium Streptomyces tenjimariensis produced theantibiotics istamycin A 6 and B 7 under select laboratory cul-ture conditions (34). Butirosin A 8 and B 9, which are amino-glycosidic antibiotics produced by a mutant of Bacillus circu-lans, showed antibacterial activity in vitro in mice (35,36).
Simple amides of uronic acids 10–12 were isolated by mildacid degradation of the lipopolysaccharides of the marine bac-terium Shewanella fidelis type strain KMM 3582T (37). Origi-nally, this bacterial strain was isolated from a sipunculan(Phascolosoma japonicum), a common inhabitant of TroitsaBay in the Gulf of Peter the Great (Sea of Japan, Russia) (38).Mycothiol 13 has been identified from the bacterial pathogenMycobacterium tuberculosis and M. bovis (39).
NUCLEOSIDE FATTY AMIDE GLYCOSIDES
Nucleosides are glycosylamines made by attaching a nucle-obase to a ribose ring. Examples of these include cytidine, uri-dine, adenoside, guanosine, thymidine, and inosine. Nucleo-sides can be acylated and/or phosphorylated by specific kinasesin the cell, producing bioactive compounds that are the molec-ular building blocks of DNA and RNA (40).
Many nucleosides are categorized as antibiotics and havenonspecific antibacterial activities (41–43). Three classes ofnucleotide antibiotics are targeted against translocase I. Thefirst class is represented by tunicamycin, which was identi-fied from the fermentation broth of Streptomyces lysosuperfi-cus nov. as having antiviral activity (44–46). Series of nucle-oside fatty amide glycosides named tunicamycins I–X,14–23, respectively, containing uracil, have been isolatedfrom S. lysosuperficus nov. (44–46). Tunicamycins showedantibacterial activities against Gram-positive bacteria, espe-cially those of the genus Bacillus [minimal inhibitory concen-tration of antibiotic that inhibits a bacterium (MIC), from 0.1to 20 µg/mL). The related compounds of tunicamycins namedstreptovirudins (series II) 24–28 (47–49), corynetoxins 29–42(50,51), and streptovirudins (series I, where uracil is replacedby dihydrouracil) 43–47 (52) have been isolated from Strep-tomyces griseoflavus subsp. thuringiensis (47–49), and fromCorynebacterium rathayi (51), respectively. Streptovirudins(series II) 24–28 have potent antibacterial activities againstthe genus Bacillus (MIC, 0.31–10 µg/mL). All corynetoxins29–42 exhibit toxicity against ryegrass (Lolium rigidum) (50).Sheep were very sensitive to corynetoxins (the lethal dosewas about 35 µg/kg bodyweight) (53).
642 REVIEW
Lipids, Vol. 40, no. 7 (2005)

FATTY AMIDE OLIGOSACCHARIDES
Among natural surfactants there are some groups of high-molec-ular-mass bioemulsifiers, including amphipathic polysaccha-rides, proteins, lipopolysaccharides, lipoproteins, and complexmixtures of these biopolymers, that are more effective at stabi-lizing oil-in-water emulsions than the low-molecular-massbioemulsifiers such as glycolipids, trehalose lipids, sophoro-
lipids, and rhamnolipids (2,54,55). Bioemulsifiers have severaladvantages over chemical surfactants, such as (i) low toxicity,(ii) biodegradability, (iii) high specificity and biocompatibility,(iv) molecular masses ranging from 500 to 1,500 Da, (v) solubil-ity and surface-active properties that are dependent on the orien-tation of residues, and (vi) occurrence in a wide variety of mi-croorganisms. These advantages should allow these compoundsto become important in industrial and environmental applica-tions. The potential commercial applications of bioemulsifiersinclude bioremediation of oil-polluted soil and water; enhancedoil recovery; replacement of chlorinated solvents used in clean-ing up oil-contaminated pipes, vessels, and machinery; use in thedetergent industry; formulations of herbicides and pesticides;and formation of stable oil-in-water emulsions for the food andcosmetic industries.
Bacterial lipopolysaccharides are the major components ofthe outer surface of Gram-negative bacteria (56). They areoften of interest in medicine for their immunomodulatory prop-erties. In small amounts they can be beneficial, but in largeramounts they may cause endotoxic shock. Although they sharea common architecture (see structures 48–53), their structuraldetails exert a strong influence on their activity. These mole-cules comprise (i) a lipid moiety, called lipid A, which is con-sidered to be the endotoxic component, (ii) a glycosidic part,consisting of a core of approximately 10 monosaccharides, and(iii) in “smooth-type” lipopolysaccharides, a third region,named O-chain, consisting of repetitive subunits of one to eightmonosaccharides responsible for much of the immunospeci-ficity of the bacterial cell.
The most common FA in lipid A have 10–16 carbon atomsalthough longer chains exist, i.e., C18 FA in Helicobacter py-lori lipid A, and C18 and C21 in Chlamydia trachomatis(57–60). Hydroxylated FA are usually found in direct acylationof the sugar except in a few cases (see below) where they ap-pear in secondary acylation. Some Bordetella strains have un-hydroxylated FA at the C-3 position (61). In Chlamydia lipidA, hydroxylated FA occur only as amide-linked substituentswhereas positions C-3 and C-3′ are both substituted by C14 FA(62,63). Another kind of FA, which has been described for afew species, is the 3-oxo FA, e.g., the 3-oxotetradecanoic acidfound in Rhodobacter capsulatus and the 27-oxooctacosanoicand heptacosane-1,27-dioic acids found in Legionella pneu-mophila (64,65). The number of acyl groups present in a mole-cule has a direct effect on its toxicity: Six acyl groups consti-tute the optimum for many isolated lipopolysaccharides. FAchain length is also critical for lipid A toxicity since C12,C12OH, C14, and C14OH FA are the moieties found in the mosttoxic lipids A (66). Unsaturated FA are rarely present in lipidA, but examples were found in Rhodopseudomonassphaeroides and other species of this genus as well as inRhodobacter capsulatus (67). Another kind of unusual FA,with a methyl branch at Cn-1, is found in some Legionella andPorphyromonas strains (68). Selected structures of lipid A iso-lated from bacteria are 48 (Escherichia coli), 49 (Yersinia ente-rocolitica, both species belonging to the Enterobacteriaceae;50 isolated from Bordetella pertussis, 51 isolated from Neisse-
REVIEW 643
Lipids, Vol. 40, no. 7 (2005)

ria meningitidis, 52 from Chlamydia trachomatis, and 53 fromRhizobium etli (56).
GUANIDINE GLYCOSIDE DERIVATIVES
A large number of biologically active molecules, includingpeptides, peptide mimetics, and guanidine-containing naturalcompounds, possess a guanidine group as an important struc-tural element (69). Guanidine-containing bioactive molecules,particularly the analogs or derivatives of natural products, arenotable targets for drug design and discovery (70–73). Becauseof the strongly basic character of the guanidine group, thisgroup is protonated under physiological conditions. As a resultof the positive charge thus imposed on the molecule, specificinteractions between ligand and receptor or enzyme and sub-strate can be formed. With the current evolution in drug dis-covery whereby combinatorial chemistry plays an increasinglyimportant role, the need for building blocks is rising as well(69).
Erylusamines A–E 54–58 are interleukin (IL)-6 receptorantagonists that were isolated from the Japanese sponge Ery-lus placenta (74,75). The cytotoxic glycolipids erylusamineTA 59, erylusine 60, and erylusidine 61, similar to ery-lusamines, have been isolated from Red Sea sponge Eryluscf. lendenfeldi (76). Pachymoside A 62 has been isolated fromcrude extract of the North Sea marine sponge Pachymatismajohnstonia (77).
644 REVIEW
Lipids, Vol. 40, no. 7 (2005)
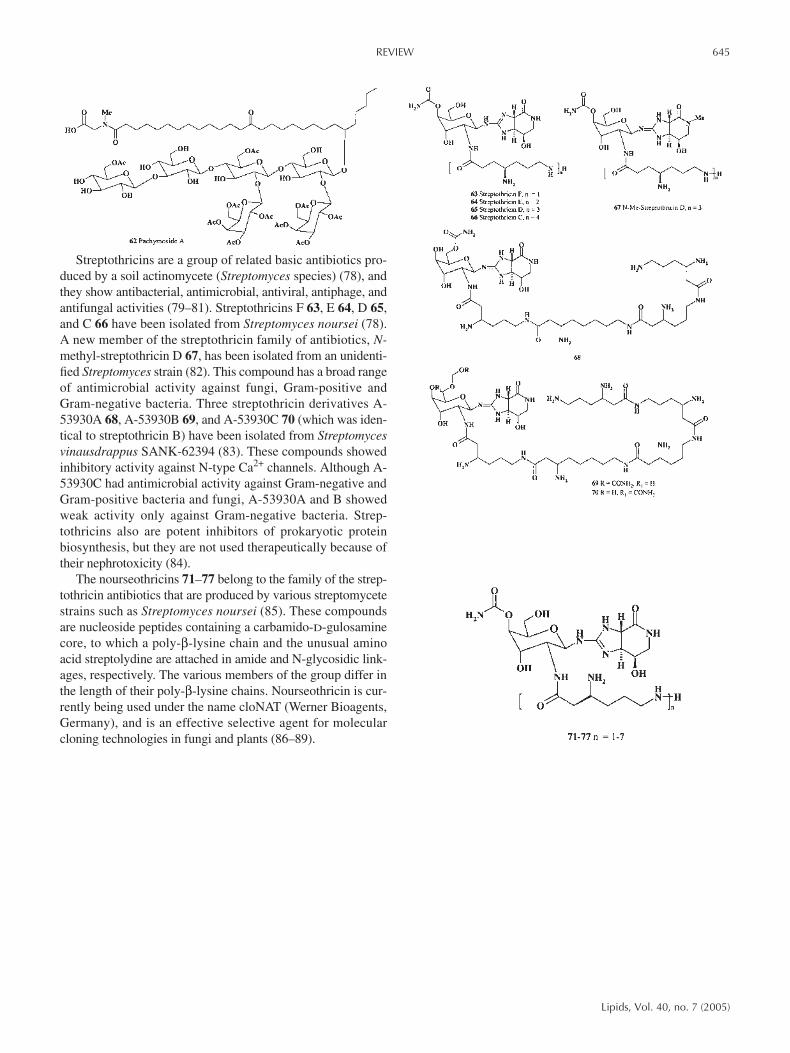
Streptothricins are a group of related basic antibiotics pro-duced by a soil actinomycete (Streptomyces species) (78), andthey show antibacterial, antimicrobial, antiviral, antiphage, andantifungal activities (79–81). Streptothricins F 63, E 64, D 65,and C 66 have been isolated from Streptomyces noursei (78).A new member of the streptothricin family of antibiotics, N-methyl-streptothricin D 67, has been isolated from an unidenti-fied Streptomyces strain (82). This compound has a broad rangeof antimicrobial activity against fungi, Gram-positive andGram-negative bacteria. Three streptothricin derivatives A-53930A 68, A-53930B 69, and A-53930C 70 (which was iden-tical to streptothricin B) have been isolated from Streptomycesvinausdrappus SANK-62394 (83). These compounds showedinhibitory activity against N-type Ca2+ channels. Although A-53930C had antimicrobial activity against Gram-negative andGram-positive bacteria and fungi, A-53930A and B showedweak activity only against Gram-negative bacteria. Strep-tothricins also are potent inhibitors of prokaryotic proteinbiosynthesis, but they are not used therapeutically because oftheir nephrotoxicity (84).
The nourseothricins 71–77 belong to the family of the strep-tothricin antibiotics that are produced by various streptomycetestrains such as Streptomyces noursei (85). These compoundsare nucleoside peptides containing a carbamido-D-gulosaminecore, to which a poly-β-lysine chain and the unusual aminoacid streptolydine are attached in amide and N-glycosidic link-ages, respectively. The various members of the group differ inthe length of their poly-β-lysine chains. Nourseothricin is cur-rently being used under the name cloNAT (Werner Bioagents,Germany), and is an effective selective agent for molecularcloning technologies in fungi and plants (86–89).
REVIEW 645
Lipids, Vol. 40, no. 7 (2005)

TETRAMIC ACID GLYCOSIDE DERIVATIVES
A culture of a marine strain of the fungus Epicoccum purpuras-cens, isolated from the inner tissue of the jellyfish Aurelia au-rita, produced a novel tetramic acid derivative, epicoccamide78 (90). Ancorinosides A 79, B 80, C 81, and D 82, inhibitorsof membrane type 1 matrix metalloproteinase (MT1-MMP),were identified from the marine sponge Penares sollasi (91).Ancorinosides B–D inhibited MT1-MMP with IC50 values of180–500 µg/mL.
Aurantosides A 83 and B 84, cytotoxic tetramic acid amineglycosides, have been isolated from the marine spongeTheonella sp. (92). The structures of these chlorinated metabo-lites were elucidated by chemical and physical methods. Au-rantoside C 85, which is lethal to brine shrimp, was found inthe sponge Homophymia conferta (93). Other aurantosides D86, E 87 and F 88, which exhibit potent antifungal activityagainst Aspergillus fumigatus and Candida albicans, were iso-lated from the marine sponge Siliquariaspongia japonica (94).
Rubroside A 89, B 90, C 93, D 91, G 92, E 94, F 95, H 96,and also hexadecahydrorubroside D 97, aurantoside A 98, andaurantoside B 99 have also been found in Siliquariaspongiajaponica (95). The rubrosides induced numerous large intra-cellular vacuoles in 3Y1 rat fibroblasts at concentrations of0.5–1.0 µg/mL, and rubrosides A, C, D, and E were cytotoxicagainst P388 murine leukemia cells with IC50 (concentration atwhich growth or activity is inhibited by 50%) values of0.046–0.21 µg/mL. Most rubrosides show antifungal activityagainst Aspergillus fumigatus and Candida albicans.
Aurantosides G–I 100–102 have been isolated from thelithistid sponge Theonella swinhoei from Papua New Guinea(96). Compounds 100–102 represent new monochloropen-taenoyl tetramic acids with mono-, di-, and tri-N-saccharidesubstituents, respectively. Aurantosides G–I failed to show anysignificant cytotoxicity against the human colon tumor cell lineHCT-116.
646 REVIEW
Lipids, Vol. 40, no. 7 (2005)

CEREBROSIDES
Cerebrosides are a group of glycosphingolipids composed of ahexose, a long-chain aminoalcohol (sphingoid), and an amide-linked long-chain FA. Galactocerebrosides were first isolatedfrom brain tissue in the late 1800s (97). The “sphingosin” back-bone of sphingolipids was so named by J.L.W. Thudichum in1884 for its enigmatic (“Sphinx-like”) properties. These mam-malian cerebrosides include a mix of saturated and monoenoicn-alkyl and 2-hydroxy-FA, with sphingosine (4-trans-sphinge-nine, a 1,3-dihydroxy-2-aminoalcohol) as the major sphingoid(97). Over 30 years ago, glucocerebrosides were identified as aminor (<1%) lipid class in plant tissues (98). Perhaps becausethey constitute such a small portion of the total tissue lipids,plant glucocerebrosides received little attention prior to the re-cent finding (following development of the two-phase mem-brane partitioning method) that they can represent a major frac-tion of the total lipids in both the plasma membrane and tono-plast (99–102). Several reports have also implicatedglucocerebrosides as playing a role in chilling injury (101) andin acclimation to cold and water-deficit stress in different plantspecies (99,103).
Many FA amides as well as their glycosides that have beenisolated from marine microorganisms and invertebrates showhigh biological activity (104–106). Placosides A 103 and B 104have been isolated from Bahamian sponge Plakortis simplex(107). These compounds exhibit immunosuppressive proper-ties. Glycosyl ceramides 105–110 have been isolated from ma-rine sponges Agelas dispar (108), A. clathrodes (109), A.conifera (110), and A. longissima (111,112). All the isolatedcompounds were tested using the mixed leucocyte reactionassay, and only 106, 107, and 110 were shown to be im-munoactivating agents, suggesting a possible structure–activityrelationship. It was also reported that glycosyl ceramides iso-lated from sponges belonging to the genus Agelas show antitu-mor activity (108,109).
Another sponge species, Agelas mauritianus, from OkinawaIsland contained a series of glycosphingolipids termed agelas-phins: AGL-10 111 and AGL-12 112 (two major components),AGL-7A 113, AGL-7B (mixture), AGL-9A 114, AGL-9B 115,AGL-11 116, and AGL-13 117 (113). Three glycosphin-golipids, regulosides A-C 118–120, were obtained from ex-tracts of the starfish Pentaceraster regulus (114), and two re-lated compounds, temnosides A 121 and B 122, were isolatedfrom the sea urchin Temnopleurus toreumaticus; both speci-mens were collected from the Mandapam coast of India (115).
REVIEW 647
Lipids, Vol. 40, no. 7 (2005)

Six cerebrosides, acanthacerebrosides D–F 123–125 andA–C 126–128, have been isolated from the starfish Acanthasterplanci (116,117), along with two ceramide lactosides, acantha-lactosides A 131 and B 132 (118). Ophidiacerebrosides A–F133–137, the cytotoxic cerebrosides, have been isolated fromthe Mediterranean starfish Ophidiaster ophidiamus (119).Cerebrosides CE-2b 138, CE-2c 139, and CE-2d 140 werefound in the sea cucumber Cucumaria echinata (120); andcerebrosides PA-0–1 129, PA-0–5 130, PA-2–5 141, and PA-2–6 142 were isolated from Pentacta austalis (121).
Axiceramides A 143 and B 144 were major components ina marine sponge belonging to the genus Axinella (122). Cere-broside 145, which inhibits histidine decarboxylase, has beenisolated from Chondropsis sp. (123). The ethanolic extractfrom the stem bark of Dimocarpus fumatus showed in vitro cy-totoxic activity against KB cells. Fractionation of the extractgave compounds belonging to different classes, and one sphin-golipid, soyacerebroside 146, two glycosides of sitosterol, andFA were also identified (124).
A series of neurogenic cerebrosides, termitomycesphins A–F147–152, have been isolated from the edible Chinese mushroomTermitomyces albuminosus (125,126). The isolated cerebrosideswere shown to induce neuronal differentiation in rat PC12 cells.The marine fungus Microsphaeropsis olivacea, obtained from asponge Agelus sp. (Florida), contains a cerebroside 153 (127).Common plant cerebrosides 154–157, their structural diversity,biosynthesis, and functions have recently been reviewed (128).Also, a recent article reviewed the distribution, syntheses, andbiological activities of the cerebrosides (129).
648 REVIEW
Lipids, Vol. 40, no. 7 (2005)

GANGLIOSIDES
Gangliosides are a family of glycosphingolipids containing oneor more N-acetyl-neuraminic acid (sialic acid) residues in theirstructure. They are found in the plasma membrane of eukary-otic cells and confer a net negative charge on most animal cells.Localized mainly on the outer surface of cells, they are in-volved in various cellular functions including growth, differen-tiation, cell-to-cell interactions, and signal transduction (130).Gangliosides are also antigenic and participate in the pathogen-esis of certain autoimmune diseases (131,132). The expressionof gangliosides is tissue- and cell-specific and also develop-ment- and stage-specific. The species of animals is another de-terminant for ganglioside expression. Thus, it is suggested thatgangliosides may carry a specific function(s) in a certain celltype of individual animal species (133).
The biological significance of the phylogenetic patterns oflenticular gangliosides is not known. Regarding brain ganglio-sides, it has been claimed that their remarkable changes in com-position and concentration may be required for the develop-ment of brain structure and function and environmental adap-tation during evolution (134–136). Lenticular gangliosides mayperform specific function(s) in the formation and function ofeye lenses (137).
According to the National Library of Medicine (Bethesda,MD), more than 11,000 original and review articles have beenpublished on gangliosides since 1950. The structures of morethan 500 gangliosides isolated from animals, invertebrates, andmicroorganisms have been elucidated, and many novel biolog-ically active gangliosides were synthesized. This review articlewill cover the structures of only some novel gangliosides thatwere isolated from natural sources.
Two new ganglioside molecular species, named acanthagan-glioside I 158 and acanthaganglioside J 159, have been isolatedfrom the whole body of the crown-of-thorns starfish Acan-thaster planci (138). The N-acyl groups on residue are C22:0and C24:0 FA, whereas the fatty alcohol is straight (C16:0) inboth gangliosides. Two monomethylated GM3-type ganglio-side molecular species, 160 and 161, have been obtained fromthe polar lipid fraction of the chloroform/methanol extract ofthe starfish Luidia maculata (139). The ceramide moieties werecomposed of heterogeneous unsubstituted FA, 2-hydroxy FA,sphingosine, and phytosphingosine units. Compound 160, des-ignated as LMG-3, represented a new ganglioside molecularspecies, and compound 161 was a known ganglioside species.
Three ganglioside molecular species, SCG-1 162, SCG-2163, and SCG-3 164, were obtained from the lipid fraction ofthe sea cucumber Stichopus chloronotus (140). The ceramidemoieties were composed of heterogeneous long-chain base andFA units. SCG-3 is the first type of ganglioside shown to con-tain a fucopyranose in the sialosyl trisaccharide moiety. More-over, these three gangliosides exhibited neuritogenic activitytoward the rat pheochromocytoma PC12 cells in the presenceof nerve growth factor. The proportions of the neurite-bearingcells of SCG-1, SCG-2, and SCG-3 at a concentration of 3.3mg/mL were 34.1, 24.4, and 24.5%, respectively. These effectswere compared with that of the mammalian ganglioside GM1(22.1% at a concentration of 3.3 mg/mL). Gangliosides withthe same sugar moiety as that of SCG-1 have been identifiedfrom some marine invertebrates: Cucumaria japonica (141),Holothuria atra, Telenota ananas (142), Stichopus japonicus(143), Holothuria leucospilota (144), Ophiura sarsi (145),Ophiocoma echinata, Ophiomastix annulosa (146), Ophio-coma scolopendrina (147), and Anthocidaris crassispina (148).SCG-1 differs from them in the structure of the ceramide moi-ety. Gangliosides with the same sugar moiety as that of themajor component of SCG-2 also have been obtained from O.sarsi (145), O. scolopendrina (147), and Hemicentrotus pul-cherrimus (149), although the major component of SCG-2 isdifferent from them in the ceramide structure.
REVIEW 649
Lipids, Vol. 40, no. 7 (2005)

On the other hand, to the best of our knowledge, SCG-3 is anew ganglioside with a unique sialosyl trisaccharide moiety,164. The major species of gangliosides 165–167 were isolatedfrom the Japanese sea cucumber Stichopus chloronotus (140).All three species displayed neuritogenic activity against PC12cells in the presence of nerve growth factor. Structurally morecomplex ganglioside molecular species SJG-2 168–170, iso-lated from a Japanese collection of S. japonicus, also exhibitedneuritogenic activity.
MACROLACTONE FATTY AMIDE GLYCOSIDES
Vicenistatin 171 is an antitumor antibiotic produced by Strep-tomyces halstedii HC-34. Its structure is unique in that it com-prises a 20-membered lactam aglycon having an aminosugarvicenisamine. Its biological activity is also intriguing since an-titumor activity has been particularly shown againstxenographed models of certain human colon cancers. 171 and172 have been isolated as minor congeners of vicenistatin 171in the culture filtrate of the producing microorganism, andnamed as vicenistatin M 172, because the minor congener issubstituted by D-mycarose in place of vicenisamine of 171(150,151). A novel ansamitocin amide N-glycoside, N-demethyl-N-β-D-glucopyranosyl ansamitocin P-2, namedansamitocinoside P-2 173, was isolated from Actinosynnemapretiosum (152).
650 REVIEW
Lipids, Vol. 40, no. 7 (2005)

Fluvirucins A1 174, A2 175, B1 176, B2 177, and B3 178 aremacrolactam antibiotics produced by the actinomycete Microtet-raspora tyrrhenii sp. nov. (Actinomadura pusilla group) strainsQ464-31, L407-5, R-359-5, and R516-16 (153,154). All isolatedcompounds are active against the influenza A virus; B1 176 isalso active against Candida sp. (MIC, 0.91 µg/mL), and dermato-phytes (MIC > 80.6 µg/mL) (154). Fluvirucin B2 177 from theculture broth of Streptomyces inhibited PI-specific phospholipaseC in cultured A431 cell cytosol with an IC50 of 1.6 µg/mL (155).
Semisynthetically produced 3-keto-substituted macrolide an-tibiotics (ketolides) 179 and 180 were compared for concentra-tion-dependent inhibitory effects on growth rate, viable cell num-bers, and protein synthesis rates in Staphylococcus aureus cells.Inhibitory effects on 50S ribosomal subunit formation were alsoexamined, as this is a second target for these antibiotics. A con-centration range of 0.01–0.1 µg/mL was tested. Compound 179was the most effective from tested compounds with an IC50 =0.035 mg/mL. Telithromycin 180 had an IC50 of 0.08 mg/mL(156). The authors concluded that these results indicate specificstructural features of these antimicrobial agents contribute to de-fined inhibitory activities against susceptible organisms.
MISCELLANEOUS COMPOUNDS
A series of biologically active chlorinated metabolites thatare derivatives of 7-methoxytetradec-4-enoic acid are pro-duced by marine cyanobacteria belonging to the genus Lyng-bia and by some marine invertebrates. The malyngamides,which are present in extracts of L. majuscula, consist typi-cally of a FA side chain and a polar head group containingthe unusual vinyl chloride functionality. The first malyn-gamide was isolated by Moore and co-workers in 1979 fromHawaiian L. majuscula (157). The (non-chlorine-containing)FA amide glycoside malyngamide J 181 isolated from Cu-raçao collections of Lyngbia majuscula (158) was toxic toboth brine shrimp (LC50 18 µg/mL) and fish (goldfish LC5040 µg/mL) (158). Malyngamide A 182 was found in amethanolic extract of L. majuscula (157) and isolated fromthe sea hare Stylocheilus longicauda (159). The sea hares(Opisthobranchia, Anaspidea) generally consume seaweedsthat are chemically rich in secondary metabolites and con-centrate these secondary metabolites in their digestiveglands. The lipid extract of a Madagascan L. majusculayielded malyngamides Q 183 and R 184 (160). MalyngamideR 184 was modestly toxic to brine shrimp (LD50 18 µg/mL).Isomalyngamides A (isomer 182) and B 185 and the isomerof the latter, 186, were isolated from L. majuscula collectedfrom Hawaiian waters (161).
Three novel lipid nucleoside antibiotics named li-posidomycins A 187, B 188, and C 189, produced by Strepto-myces griseosporeus, strongly inhibit bacterial peptidoglycansynthesis (162). The structure of the FA component of li-posidomycin A 187 was a 3-hydroxy-7,10-hexadecadienoicacid, not previously found in nature.
REVIEW 651
Lipids, Vol. 40, no. 7 (2005)

Simocyclinones are novel natural hybrid antibiotics isolatedfrom the fermentation broth of Streptomyces antibioticus Tü6040 that combine structural elements from various groups ofknown antibiotics such as aquayamycin, fumagillin, and novo-biocin (163,164). So far, 14 different angucyclinone antibioticshave been isolated and classified in four series, A–D. The simo-cyclinones of the A-series consist solely of an angucycline ring.The components of series B are glycosylated. Compounds of theC-series have a tetraene side-chain, and those of the D-series190–193 also have a coumarin ring system. A novel family ofangucyclinone antibiotics demonstrated antibiotic activityagainst Gram-positive bacteria, as well as cytostatic effectsagainst human tumor cell lines and antibacterial and antitumoractivities (163–167). The 3-amino-4,7-dihydroxy-coumarin moi-ety is also found in the antibiotics novobiocin 194, clorobiocin195, and coumermycin A1 196, which are very potent inhibitorsof DNA gyrase (168). The aminocoumarin ring is of central im-portance for the binding of these compounds to the B subunit ofgyrase (169–171). Recent experiments have suggested that simo-cyclinone D8 193 is even more potent as a DNA gyrase inhibitorthan novobiocin (171). The aminocoumarin antibiotics novo-biocin 194, clorobiocin 195, and coumermycin A1 196 are pro-duced by different Streptomyces strains.
Ten different derivatives of glycine-containing glycolipids197–206 have been isolated from a Gram-negative, biosurfac-tant-producing, n-alkane-degrading marine bacterium Al-canivorax borkumensis (172). This Gram-negative, aerobic,rod-shaped bacterium uses a limited number of organic com-pounds, including aliphatic hydrocarbons, volatile FA, andpyruvate and its methyl ether. During cultivation on n-alkanesas sole source of carbon and energy, all strains produced bothextracellular and cell-bound surface-active glucose lipids197–206 (173). This novel class of glucolipids was producedonly by these strains, which were isolated from sediment sam-ples collected near the Isle of Borkum (North Sea). The mixedactive biosurfactants produced are characterized as glucoselipids, one of which consists of four 3-hydroxydecanoic acidslinked together by ester bonds and coupled glycosidically withthe C-1 of glucose. Other FA were found, namely, 16:0, 16:1,and 18:1, in the phospholipid fraction (173).
652 REVIEW
Lipids, Vol. 40, no. 7 (2005)

The bengamides 207–225, isolated from marine spongesbelonging to the genus Jaspidae, comprise a large family ofnatural metabolites. In 1986, a group from the University ofCalifornia–Santa Cruz reported the first examples of this class,(+) bengamide A 207 and (+) B 208, as an easily separablemixture from a small sponge collection, subsequently identi-fied as Jaspis cf. coriacea (Family Coppatiidae, Order Choris-tida) (174). These compounds display a wide and interestingrange of biological activities, including antitumor, antibiotic,and antihelmintic properties. Bengamides A 207 and B 208were more potent than bengamide P, with average IC50 valuesof 0.046, 0.011, and 2.70 fM, respectively (175). The in vitroantitumor activity against MDA-MB-435 human mammarycarcinoma was also determined for natural bengamides A, B,E, F, P, M, O, and Z and for synthetic samples of B and O. Thebest activity was observed for the natural bengamides A (IC50= 1 nM) and O (IC50 = 0.3 nM) (175).
Pathogenic strains of the soilborne fungus Periconia circi-nata produce peritoxins 227–231 with host-selective toxicityagainst susceptible genotypes of sorghum at concentrations aslow as 1.0 ng/mL (176,177). The peritoxins are low-M.W. hy-brid molecules consisting of a peptide and a chlorinatedpolyketide 228–231. Peritoxins A 228 and B 229 and peri-conins A 230 and B 231 have been isolated together with theknown metabolite circinatin 226 from culture filtrates of thefungal pathogen P. circinata.
Several unusual α-glucosidase inhibitors 232–237 containingsulfated FA have been isolated from some marine spongespecies. Penasulfate A 232, a new α-glucosidase inhibitor, wasisolated from a marine sponge Penares sp. (178); this compoundincluded a novel sulfated FA (2-piperidine-carboxylic acid).Three similar α-glucosidase inhibitors named schulzeines A 233,B 234, and C 235 were isolated from the marine sponge P.schulzei (179). Schulzeines A–C showed inhibitory activityagainst α-glucosidase, with IC50 values of 48–170 nM. Unique30- and 31-membered macrolides encompassing a prolineresidue and three sulfate groups, named penarolide sulfates A1236 and A2 237, have been isolated from a marine spongePenares sp. (180). These metabolites inhibited α-glucosidase,with IC50 values of 1.2 and 1.5 mg/mL.
REVIEW 653
Lipids, Vol. 40, no. 7 (2005)

UNUSUAL AMIDE GLYCOSIDES
Amide surfactants are widely used as functional chemicals andare indispensable for various industrial fields (181–183). Syn-thetic detergents have molecular structures and properties thatare similar to soap. Detergent molecular structures consist of along hydrocarbon chain and a water-soluble ionic group. Mostdetergents have a negative ionic group and are called anionicdetergents. Surfactants have a major impact on the performanceof modern-day cleaning systems. In addition to performance,however, there is a growing appreciation for surfactants thatare also environmentally friendly and produced from renew-able resources. Sugar-based surfactants fit these requirements,and glucosamides (183–186) are one form that is experiencingrapid growth rates. In recent years, much attention has beenpaid from the ecologial and industrial viewpoints to sugar-based surfactants such as alkyl glucoside (187–189). Some in-teresting surfactants based on amide bond(s) that have biolog-ical activities have been synthesized (183).
Some diseases are initiated by the adherence of viruses, bac-teria, or bacterial toxins to cell surface carbohydrates, a num-ber of which are components of glycosphingolipids. Studies ofthe binding of lectins indicated that many adhered weakly tomonomeric carbohydrate ligands (183,184). In addition tomore completely defining ligand specificity of the carbohy-
drate-binding pathogen, identification of multivalent carbohy-drate ligands has led to studies of their efficacy as pathogen in-hibitors. This commentary focuses on pathogens that recognizethe carbohydrate portion of glycosphingolipids. Because manyglycosphingolipid-binding pathogens have been shown to bindmultivalent saccharides, approaches for identifying and prepar-ing them as well as methods for characterizing their effective-ness as ligands were reviewed (183,184). Several multivalentsaccharide ligands were reported in the past 10 years.
Synthesis of amide glycoside derivatives of multivalent lig-ands that cross-link concanavalin A dimers of saccharides 238and 239 and a biophysical evaluation of their interaction withthe plant lectin concanavalin A have been reported (190). Tothe best of our knowledge, these structures are the first reportedof their kind. Results obtained by Dimick et al. (190) indicatethat hemagglutination assays evaluate the ability of ligands toinhibit the formation of cross-linked lattices, a process onlytangentially related to reversible ligand binding. Cluster glyco-side effects observed in agglutination assays must therefore beviewed with caution. Such effects may be relevant to the de-sign of therapeutically useful amide saccharides.
Several convergent glycodendrimer syntheses based on aro-matic cores have been reported (183). Stoddart and co-workersreported convergent synthesis of polyvalent dendritic com-pound 240 (191). Thus, carbohydrate-centered ligands are ter-minated with a total of 12 glucose residues, although the mi-croscopic environment of each is somewhat different. A relatedgroup of ligands, again derived from a convergent syntheticroute and centered on an aromatic core, present terminal glu-cose residues in nominally equivalent microenvironments.Burke and co-workers synthesized three mannosyl residuesaround a macrocyclic core 241 to produce a rigid trivalent lig-and (192).
654 REVIEW
Lipids, Vol. 40, no. 7 (2005)

A group of so-called two-component toxins are responsiblefor the pathology of the world’s most severe enterotoxic dis-eases; these toxins include the Escherichia coli heat-labiletoxin, the shiga toxin, and the cholera toxin. The toxins arestructurally and mechanistically related to the tetanus, diphthe-ria, and pertussis toxins and to the plant toxins abrin and ricin.Fan and co-workers (193) synthesized a pentavalent galacto-side ligand for the E. coli heat-labile toxin based on a macro-cyclic core 242–245. Multivalent cyclo-oligosaccharides246–250, versatile carbohydrate clusters with dual roles as mo-lecular receptors and lectin ligands, have recently been synthe-sized (194).
SUMMARY
FA amides are commonly used as foaming and wetting agents inhand dish-washing detergents, shampoos, and bar soaps, particu-
larly the diethanollauryl amide. Amphoteric or zwitterionic sur-factants have two functional group, one anionic and one cationic.In most cases it is the pH that determines which of the groupswill dominate, by favoring one or the other ionization: anionic atalkaline pH and cationic at acid pH. Near the isoelectric point,these surfactants display both charges and are truly amphoteric,often with a minimum of interfacial activity and a concomitantmaximum of water solubility. Amphoteric surfactants, particu-larly the amino acid ones, are quite biocompatible and are usedin pharmaceuticals and cosmetics. Besides their excellent sur-factant properties, i.e., wetting power, cleaning performance,foaming power, hard water tolerance, and lime soap dispersibil-ity, particular emphasis is placed on the mildness of amphotericsurfactants and on their low toxicity toward the skin and eyesand, in particular, on the improvement in dermatological and mu-cous membrane compatibility in mixtures with anionic surfac-tants in shampoos. In addition, their zwitterionic character makesthem compatible with both anionic and cationic surfactants. Am-photeric surfactants are a minor, but important ingredient of cos-metic products. Beside their activity as a surfactant, they con-tribute in combination with anionics toward the particular mild-ness and low toxicity of the products.
REFERENCES
1. Dembitsky, V.M. (2005) Astonishing Diversity of Natural Sur-factants. 3. Carotenoid Glycosides and Isoprenoid Glycolipids,Lipids 40, 535–557.
2. Dembitsky, V.M. (2004) Chemistry and Biodiversity of Biolog-ically Active Natural Glycosides, Chem. Biodivers. 1, 673–781.
3. Kester, M. (1997) Sphingolipid Metabolites and the CellularPhenotype, Trends Glycosci. Glycotechnol. 9, 447–460.
4. Yowler, B.C., and Schengrund, C.L. (2004) Glycosphin-golipids—Sweets for Botulinum Neurotoxin, Glycoconj. J. 21,287–293.
5. Morales, A., Colell, A., Mari, M., Garcia-Ruiz, C., and Fernan-dez-Checa, J.C. (2003) Glycosphingolipids and Mitochondria:Role in Apoptosis and Disease, Glycoconj. J. 20, 579–588.
6. Tettamanti, G. (2003) Ganglioside/Glycosphingolipid Turnover:New Concepts, Glycoconj. J. 20, 301–317.
7. Malykh, Y.N., Schauer, R., and Shaw, L. (2001) N-Glycolyl-neuraminic Acid in Human Tumours, Biochimie 83, 623–634.
8. Schmid, H.H.O., Schmid, P.C., and Natarajan, V. (1990) N-Acy-lated Glycerophospholipids and Their Derivatives, Prog. LipidRes. 29, 1–43.
9. Sutcliffe, I.C., and Harrington, D.J. (2004) Lipoproteins of My-cobacterium tuberculosis: An Abundant and Functionally DiverseClass of Cell Envelope Components, FEMS Microbiol. Rev. 28,645–659.
10 Kuehl, F.A., Jr., Jacob, T.A., Ganley, O.H., Ormond, R.E., andMeisinger, M.A.P. (1957) The Identification of n-(2-Hydrox-yethyl)-palmitamide as a Naturally Occurring Anti-inflamma-tory Agent, J. Am. Chem. Soc. 79, 5577–5578.
11. Devane, W.A., Hanus, L., Breuer, A., Pertwee, R.G., Steven-son, L.A., Griffin, G., Gibson, D., Mandelbaum, A., Etinger, A.,and Mechoulam, R. (1992) Isolation and Structure of a BrainConstituent That Binds to the Cannabinoid Receptor, Science258, 1946–1949.
12. Cravatt, B.F., Prospero-Garcia, O., Siuzdak, G., Gilula, N.B.,Henriksen, J., Boger, D.L., and Lerner, R.A. (1995) ChemicalCharacterization of a Family of Brain Lipids That Induce Sleep,Science 268, 1506–1509.
REVIEW 655
Lipids, Vol. 40, no. 7 (2005)

13. Schuel, H., Burkman, L.J., Lippes, J., Crickard, K., Forester, E.,Piomelli, D., and Giuffrida, A. (2002) N-Acylethanolamines inHuman Reproductive Fluids, Chem. Phys. Lipids 121, 211–227.
14. Lambert, D.M., Vandevoorde, S., Jonsson, K.O., and Fowler,C.J. (2002) The Palmitoyl-ethanolamide Family: A New Classof Anti-inflammatory Agents? Curr. Med. Chem. 9, 663–674.
15. Fowler, C.J. (2004) Oleamide: A Member of the Endocannabi-noid Family? Br. J. Pharmacol. 141, 195–196.
16. Touw, D.J., Neef, C., Thomson, A.H., and Vinks, A.A. (2005)Cost-effectiveness of Therapeutic Drug Monitoring—A Sys-tematic Review, Ther. Drug Monit. 27, 10–17.
17. Carrasco, D.A., Vander, S.M., and Tyring, S.K. (2002) A Re-view of Antibiotics in Dermatology, J. Cutan. Med. Surg. 6,128–150.
18. Begg, E.J., Barclay, M.L., and Kirkpatrick, C.M.J. (2001) TheTherapeutic Monitoring of Antimicrobial Agents, Br. J. Clin.Pharmacol. 52 (Suppl. 1), 35S–43S.
19. Fainerman, V.B., Mobius, D., and Miller, R., eds. (2001) Sur-factants: Chemistry, Interfacial Properties, Applications, Else-vier, Amsterdam, The Netherlands, pp. 678.
20. Milton, J.R. (2004) Surfactants and Interfacial Phenomena, 3rdedn., 500 pp., John Wiley & Sons, New York.
21. Schramm, L.L., ed. (2000) Surfactants: Fundamentals and Ap-plications in the Petroleum Industry, 630 pp., Cambridge Uni-versity Press, Cambridge, United Kingdom.
22. Ash, M., and Ash, I. (1993) Handbook of Industrial Surfactants:An International Guide to More Than 16,000 Products byTradename, Application, Composition & Manufacturer, 905pp., Gower, Aldershot, Hants, England.
23. Chain, E., Florey, H.W., Gardner, A.D., Heatley, N.G., Jen-nings, M.A., Orr-Ewing, J., and Sanders, A.G. (1940) Penicillinas a Chemotherapeutic Agent, Lancet 1, 226–228.
24. Ikeda, D., and Umezawa, S. (1999) Aminoglycoside Antibi-otics, in Naturally Occurring Glycosides (Ikan, R., ed.), pp.13–42, Wiley & Sons, Chichester, England.
25. Umezawa, H., and Hooper, I.R., eds. (1982) AminoglycosideAntibiotics, Springer–Verlag, Berlin.
26. Cappelletti, L.M., and Spagnoli, R. (1983) Biological Transfor-mation of Kanamycin A to Amikacin (BBK-8), J. Antibiot.(Tokyo) 36, 328–330.
27. Yoshikawa, K., Suzuki, Y., Tanaka, M., Arihara, S., and Nigam,S.K. (1997) Three Acylated Saponins and a Related Compoundfrom Pithecellobium dulce, J. Nat. Prod. 60, 1269–1274.
28. Kondo, S., and Hotta, K. (1999) Semisynthetic AminoglycosideAntibiotics: Development and Enzymatic Modifications, J. In-fect. Chemother. 5, 1–9.
29. Okachi, R., Takasawa, S., Sato, T., Sato, S., Yamamoto, M.,Kawamoto, I., and Nara, T. (1977) Fortimicins A and B, NewAminoglycoside Antibiotics. II. Isolation, Physico-chemical andChromatographic Properties, J. Antibiot. (Tokyo) 30, 541–551.
30. Egan, R.S., Stanaszek, R.S., Cirovic, M., Mueller, S.L.,Tadanier, J., Martin, J.R., Collum, P., Goldstein, A.W., DeVault, R.L., Sinclair, A.C., et al. (1977) Fortimicins A and B,New Aminoglycoside Antibiotics. III. Structural Identification,J. Antibiot. (Tokyo) 30, 552–563.
31. Yamamoto, M., Okachi, R., Kawamoto, I., and Nara, T. (1977)Fortimicin A Production by Micromonospora olivoasterosporain a Chemically Defined Medium, J. Antibiot. (Tokyo) 30,1064–1072.
32. Girolami, R.L., and Stamm, J.M. (1977) Fortimicins A and B,New Aminoglycoside Antibiotics. IV. In vitro Study of For-timicin A Compared with Other Aminoglycosides, J. Antibiot.(Tokyo) 30, 564–570.
33. Yamashita, K., Kawabe, H., and Mitsuhashi, S. (1981) Syner-gistic Activities of Fortimicin A and β-Lactam AntibioticsAgainst Pseudomonas aeruginosa, Antimicrob. AgentsChemother. 20, 33–37.
34. Slattery, M., Rajbhandari, I., and Wesson, K. (2001) Competi-tion-Mediated Antibiotic Induction in the Marine BacteriumStreptomyces tenjimariensis, Microb. Ecol. 41, 90–96.
35. Taylor, H.D., and Schmitz, H. (1976) Antibiotics Derived froma Mutant of Bacillus circulans, J. Antibiot. (Tokyo) 29, 532–535.
36. Umezawa, S. (1974) Structures and Syntheses of Aminoglyco-side Antibiotics, Adv. Carbohydr. Chem. Biochem. 30, 111–182.
37. Kilcoyne, M., Perepelov, A., Shashkov, A.S., Nazarenko, E.L.,Ivanova, E.P., Gorshkova, N.M., Gorshkova, R.P., and Savage,A.V. (2004) Structure of an Acidic O-Specific Polysaccharidefrom Marine Bacterium Shewanella fidelis KMM 3582T Con-taining Nε—[(S)-1-Carboxyethyl]-Nα-(D-galacturonoyl)-L-Ly-sine, Carbohydr. Res. 339, 1655–1661.
38. Ivanova, E.P., Nedashkovskaya, O.I., Zhukova, N.V., Nicolau,D.V., Christen, R., and Mikhailov, V.V. (2003) Shewanellawaksmanii sp. nov., Isolated from a Sipuncula (Phascolosomajaponicum), Int. J. Syst. Evol. Microbiol. 53, 1471–1477.
39. Spies, H.S.C., and Steenkamp, D.J. (1994) Thiols of Intracellu-lar Pathogens—Identification of Ovothiol-A in Leishmaniadonovani and Structural Analysis of a Novel Thiol from My-cobacterium bovis, Eur. J. Biochem. 224, 203–213.
40. Gunaga, P., Moon, H.R., Choi, W.J., Shin, D.H., Park, J.G., andJeong, L.S. (2004) Recent Advances in 4-Thionucleosides asPotential Antiviral and Antitumor Agents, Curr. Med. Chem.11, 2585–2637.
41. Wrodnigg, T.M., and Sprenger, F.K. (2004) Bioactive Carbohy-drates and Recently Discovered Analogues as Chemotherapeu-tics, Mini Rev. Med. Chem. 4, 437–459.
42. O’Riordan, T., and Faris, M. (1999) Inhaled AntimicrobialTherapy, Respir. Care Clin. N. Am. 5, 617–631.
43. Russell, A.D. (1969) The Mechanism of Action of Some An-tibacterial Agents, Prog. Med. Chem. 6, 135–199.
44. Takatsuki, A., Arima, K., and Tamura, G. (1971) Tunicamycin,a New Antibiotic. I. Isolation and Characterization of Tuni-camycin, J. Antibiot. (Tokyo) 24, 215–223.
45. Takatsuki, A., and Tamura, G. (1971) Tunicamycin, a New An-tibiotic. II. Some Biological Properties of the Antiviral Activityof Tunicamycin, J. Antibiot. (Tokyo) 24, 224–231.
46. Takatsuki, A., and Tamura, G. (1971) Tunicamycin, a New An-tibiotic. 3. Reversal of the Antiviral Activity of Tunicamycin byAminosugars and Their Derivatives, J. Antibiot. (Tokyo) 24,232–238.
47. Thrum, H., Eckardt, K., Bradler, G., Fugner, R., Tonew, E., andTonew, M. (1975) Streptovirudins, New Antibiotics with An-tibacterial and Antiviral Activity. I. Culture Taxonomy, Fermen-tation and Production of Streptovirudin Complex, J. Antibiot.(Tokyo) 28, 514–521.
48. Tonew, E., Tonew, M., Eckardt, K., Thrum, H., and Gumpert,B. (1975) Streptovirudins—New Antibiotics with Antiviral Ac-tivity. The Antiviral Spectrum and Inhibition of Newcastle Dis-ease Virus in Cell Cultures, Acta Virol. 19, 311–317.
49. Elbein, A.D., Occolowitz, J.L., Hamill, R.L., and Eckardt, K.(1981) Streptovirudins of Series I and II: Chemical and Biologi-cal Properties, Biochemistry 20, 4210–4216.
50. Vogel, P., Petterson, D.S., Berry, P.H., Frahn, J.L., Anderton,N., Cockrum, P.A., Edgar, J.A., Jago, M.V., Lanigan, G.W.,Payne, A.L., et al. (1981) Isolation of a Group of GlycolipidToxins from Seedheads of Annual Ryegrass Lolium rigidumGaud. Infected by Corynebacterium rathayi, Aust. J. Exp. Biol.Med. Sci. 59, 455–467.
51. Frahn, J.L., Edgar, J.A., Jones, A.J., Cockrum, P.A., Anderton,N., and Culvenor, C.C.J. (1984) Structure of the Corynetoxins,Metabolites of Corynebacterium rathayi Responsible for Toxic-ity of Annual Ryegrass (Lolium rigidum) Pastures, Aust. J.Chem. 37, 165–182.
52. Eckardt, K., Thrum, H., Bradler, G., Tonew, E., and Tonew, M.(1975) Streptovirudins, New Antibiotics with Antibacterial and
656 REVIEW
Lipids, Vol. 40, no. 7 (2005)

Antiviral Activity. II. Isolation, Chemical Characterization andBiological Activity of Streptovirudins A1, A2, B1, B2, C1, C2,D1, and D2, J. Antibiot. (Tokyo) 28, 274–279.
53. Jago, M.V., and Culvenor, C.C. (1987) Tunicamycin andCorynetoxin Poisoning in Sheep, Aust. Vet. J. 64, 232–235.
54. Rosenberg, E., and Ron, E.Z. (1999) High and Low MolecularMass Microbial Surfactants, Appl. Microbiol. Biotechnol. 52,154–162.
55. Lang, S., and Fischer, S. (1999) Design and Selection of Perfor-mance Surfactants in Annual Surfactants Review, (Karsa, D.R.,ed.), Vol. 2 pp. 51–103, Academic Press, Sheffield.
56. Caroff, M., and Karibian, D. (2003) Structure of BacterialLipopolysaccharides, Carbohydr. Res. 338, 2431–2447.
57. Moran, A.P., Lindner, B., and Walsh, E.J. (1997) StructuralCharacterization of the Lipid A Component of Helicobacter py-lori Rough- and Smooth-Form Lipopolysaccharides, J. Bacte-riol. 179, 6453–6463.
58. Suda, Y., Ogawa, T., Kashihara, W., Oikawa, M., Shimoyama,T., Hayashi, T., Tamura, T., and Kusumoto, S. (1997) ChemicalStructure of Lipid A from Helicobacter pylori Strain 206-1Lipopolysaccharide, J. Biochem. (Tokyo) 121, 1129–1133.
59. Qureshi, N., Kaltashov, I., Walker, K., Doroshenko, V., Cotter,R.J., Takayama, K., Sievert, T.R., Rice, P.A., Lin, J.-S.L., andGolenbok, D.T. (1997) Structure of the Monophosphoryl LipidA Moiety Obtained from the Lipopolysaccharide of Chlamydiatrachomatis, J. Biol. Chem. 272, 10594–10600.
60. Heine, H., Müller-Loennies, S., Brade, L., and Brade, H. (2003)Endotoxic Activity and Chemical Structure of Lipopolysaccha-rides from Chlamydia trachomatis Serotypes E and L-2 andChlamydophila psittaci 6BC, Eur. J. Biochem. 270, 440–450.
61. Zarrouk, H., Karibian, D., Bodie, S., Perry, M.B., Richards, J.,and Caroff, M. (1997) Structural Characterization of the LipidsA of Three Bordetella bronchiseptica Strains: Variability ofFatty Acid Substitution, J. Bacteriol. 179, 3756–3760.
62. Rund, S., Lindner, B., Brade, H., and Holst, O. (1999) StructuralAnalysis of the Lipopolysaccharide from Chlamydia trachoma-tis Serotype L2, J. Biol. Chem. 274, 16819–16824.
63. Rund, S., Lindner, B., Brade, H., and Holst, O. (2000) StructuralAnalysis of the Lipopolysaccharide from Chlamydophilapsittaci Strain 6BC, Eur. J. Biochem. 267, 5717–5726.
64. Krauss, J.H., Seydel, U., Weckesser, J., and Mayer, H. (1989)Structural Analysis of the Nontoxic Lipid-A of Rhodobactercapsulatus 37B4, Eur. J. Biochem. 180, 519–526.
65. Moll, H., Sonesson, A., Jantzen, E., Marre, R., and Zähringer,U. (1992) Identification of 27-Oxo-octacosanoic Acid and Hep-tacosane-1,27-dioic Acid in Legionella pneumophila, FEMSMicrobiol. Lett. 76, 1–6.
66. Zähringer, U., Knirel, Y.A., Lindner, B., Helbig, J.H., Sones-son, A., Marre, R., and Rietschel, E.T. (1995) The Lipopolysac-charide of Legionella pneumophila Serogroup 1 (strain Philadel-phia 1): Chemical Structure and Biological Significance, Prog.Clin. Biol. Res. 392, 113–139.
67. Mayer, H., Merkofer, T., Warth, C., and Weckesser, J. (1996)Position and Configuration of Double Bonds of Lipid A-Asso-ciated Monounsaturated Fatty Acids of Proteobacteria andRhodobacter capsulatus 37B4, J. Endotoxin Res. 3, 345–352.
68. Kumada, H., Haishima, Y., Umemoto, T., and Tanamoto, K.I.(1995) Structural Study on the Free Lipid-A Isolated fromLipopolysaccharide of Porphyromonas gingivalis, J. Bacteriol.177, 2098–2106.
69. Hannon, C.L., and Anslyn, E.V. (1993) The GuanidiniumGroup: Its Biological Role and Synthetic Analogs, in Bioor-ganic Chemistry Frontiers, (Dugas, H., ed.), Vol. 3, pp.193–221, Springer-Verlag, Berlin.
70. Berlinck, R.G.S. (1999) Natural Guanidine Derivatives, Nat.Prod. Rep. 16, 339–365.
71. Berlinck, R.G.S. (2002) Natural Guanidine Derivatives, Nat.Prod. Rep. 19, 617–649.
72. Prandota, J. (2000) Unusual Distribution, Pharmacokinetics, andMetabolism of Some Drugs and Their Implications to Pharma-cotherapy, Am. J. Ther. 7, 332–340.
73. Brown, T.D., Burris, H.A., Havlin, K.A., O’Rourke, T.J., Ro-driguez, G.I., Wall, J.G., and Weiss, G.R. (1991) New Anti-cancer Agents, Cancer Chemother. Biol. Response Modif. 12,111–146.
74. Sata, N., Asai, N., Matsunaga, S., and Fusetani, N. (1994) Ery-lusamines, IL-6 Receptor Antagonists, from the Marine Sponge,Erylus placenta, Tetrahedron 50, 1105–1110.
75. Fusetani, N., Sata, N., Asai, N., and Matsunaga, S. (1993) Isola-tion and Structure Elucidation of Erylusamine-B, a New Classof Marine Natural Products, Which Blocked an IL-6 Receptor,from the Marine Sponge Erylus placenta Thiele, TetrahedronLett. 34, 4067–4070.
76. Goobes, R., Rudi, A., and Kashman, Y. (1996) Three New Gly-colipids from a Red Sea Sponge of the Genus Erylus, Tetrahe-dron 52, 7921–7928.
77. Warabi, K., Zimmerman, W.T., Shen, J.K., Gauthier, A.,Robertson, M., Finlay, B.B., van Soest, R., and Andersen, R.J.(2004) Pachymoside A—A Novel Glycolipid Isolated from theMarine Sponge Pachymatisma johnstonia, Can. J. Chem. 82,102–112.
78. Khokhlov, A.S., and Shutova, K.I. (1972) Chemical Structureof Streptothricins, J. Antibiot. (Tokyo) 25, 501–508.
79. Riabova, I.D., Reshetov, P.D., Zhdanov, G.L., and Khokhlov,A.S. (1965) Antimicrobial Activity of Streptothricins, Antibi-otiki (Moscow) 10, 1066–1069 [in Russian].
80. Germanova, K.I., and Goncharskaia, T.I. (1969) Antiviral Prop-erties of Streptothricins A, B, C, D, F, Antibiotiki (Moscow) 14,48–51 [in Russian].
81. Germanova, K.I., and Goncharskaia, T.I. (1969) Antiphage Ac-tivity of Streptothricins A, B, C, D, F, Antibiotiki (Moscow) 14,137–140 [in Russian].
82. Kim, B.T., Lee, J.Y., Lee, Y.Y., Kim, O.Y., Chu, J.H., and Goo,Y.M. (1994) N-Methyl-streptothricin D. A New StreptothricinGroup Antibiotic from a Streptomyces spp., J. Antibiot. (Tokyo)47, 1333–1336.
83. Hisamoto, M., Inaoka, Y., Sakaida, Y., Kagazaki, T., Enokida,R., Okazaki, T., Haruyama, H., Kinoshita, T., and Matsuda, K.(1998) A-53930A and B, Novel N-Type Ca2+ Channel Block-ers, J. Antibiot. (Tokyo) 51, 607–617.
84. Hoffmann, H., Härtl, A., Bocker, H., Kahnel, H.-J., Hesse, G.,and Flemming, J. (1986) Pharmacokinetics of Nourseothricin inLaboratory Animals, Arch. Exp. Veterinarmed. 40, 699–709 [inGerman].
85. Bocker, H., and Bergter, F. (1986) Nourseothricin: Properties,Biosynthesis, Production, Arch. Exp. Veterinarmed. 40,646–657 [in German].
86. Haupt, I., Thrum, H., and Noack, D. (1986) Self-Resistance ofthe Nourseothricin Producing Strain Streptomyces noursei, J.Basic Microbiol. 26, 323–328.
87. Krügel, H., Fiedler, G., Gase, K., and Haupt, I. (1989) Strep-tothricin Resistance. in Bioactive Metabolites from Microorgan-isms (Bushell, M.E., and Gräfe, U., eds.), pp. 357–367, Elsevier,Amsterdam.
88. Krügel, H., Fiedler, G., Haupt, I., Sarfert, E., and Simon, H.(1988) Analysis of the Nourseothricin-Resistance Gene (nat) ofStreptomyces noursei, Gene 62, 209–217.
89. Krügel, H., Fiedler, G., Smith, C., and Baumberg, S. (1993) Se-quence and Transcriptional Analysis of the NourseothricinAcetyltransferase-Encoding Gene nat1 from Streptomycesnoursei, Gene 127, 127–131.
90. Wright, A.D., Osterhage, C., and Konig, G.M. (2003) Epicoc-camide, a Novel Secondary Metabolite from a Jellyfish DerivedCulture of Epicoccum purpurascens, Org. Biomolecul. Chem.1, 507–510.
91. Fujita, M., Nakao, Y., Matsunaga, S., Seiki, M., Itoh, Y., van
REVIEW 657
Lipids, Vol. 40, no. 7 (2005)

Soest, R.W.M., and Fusetani, N. (2001) Ancorinosides B–D, In-hibitors of Membrane Type 1 Matrix Metalloproteinase (MT1-MMP), from the Marine Sponge Penares sollasi Thiele, Tetra-hedron 57, 1229–1234.
92. Matsunaga, S., Fusetani, N., Kato, Y., and Hiroda, H. (1991)Bioactive Marine Metabolites. 38. Aurantoside-A and Auranto-side-B—Cytotoxic Tetramic Acid Glycosides from the MarineSponge Theonella sp., J. Am. Chem. Soc. 113, 9690–9692.
93. Wolf, D., Schmitz, F.J., Qiu, F., and Kelly-Borges, M. (1999)Aurantoside C, a New Tetramic Acid Glycoside from theSponge Homophymia conferta, J. Nat. Prod. 62, 170–172.
94. Sata, N.U., Matsunaga, S., Fusetani, N., and van Soest, R.W.M.(1999) Aurantosides D, E, and F: New Antifungal TetramicAcid Glycosides from the Marine Sponge Siliquariaspongiajaponica, J. Nat. Prod. 62, 969–971.
95. Sata, N.U., Wada, S., Matsunaga, S., Watabe, S., van Soest,R.W.M., and Fusetani, N. (1999) Bioactive Marine Metabolites.Part 87. Rubrosides A–H, New Bioactive Tetramic Acid Glyco-sides from the Marine Sponge Siliquariaspongia japonica, J.Org. Chem. 64, 2331–2339.
96. Ratnayake, A.S., Davis, R.A., Harper, M.K., Veltri, C.A., And-jelic, C.D., Barrows, L.R., and Ireland, C.M. (2005) Auranto-sides G, H, and I: Three New Tetramic Acid Glycosides from aPapua New Guinea Theonella swinhoei, J. Nat. Prod. 68,104–107.
97. Hakomori, S. (1983) in Handbook of Lipid Research, Vol. 3,Sphingolipid Biochemistry (Kanfer, J.N., and Hakomori, S.,eds.), pp. 1–37, Plenum Press, New York.
98. Sastry, P.S., and Kates, M. (1964) Lipid Components of Leaves.V. Galactolipids, Cerebrosides, and Lecithin of Runner BeanLeaves, Biochemistry 3, 1271–1280.
99. Lynch, D.V., and Steponkus, P.L. (1987) Plasma MembraneLipid Alterations Associated with Cold-Acclimation of WinterRye Seedlings (Secale cereale L-cv. puma), Plant Physiol. 83,761–767.
100. Rochester, C.P., Kjellbom, P., Andersson, B., and Larsson, C.(1987) Lipid Composition of Plasma Membranes Isolated fromLight-grown Barley (Hordeum vulgare) Leaves—Identificationof Cerebroside as a Major Component, Arch. Biochem. Biophys.255, 385–391.
101. Yoshida, S., Washio, K., Kenrick, J., and Orr, G. (1988) Ther-motropic Properties of Lipids Extracted from Plasma Membraneand Tonoplast Isolated from Chilling-sensitive Mung Bean(Vigna radiata [L] Wilczek), Plant Cell Physiol. 29,1411–1416.
102. Lynch, D.V. (1993) Sphingolipids, in Lipid Metabolism inPlants (Moore, T.S., Jr., ed.), pp. 285–308, CRC Press, BocaRaton, FL.
103. Norberg, P., and Liljenberg, C. (1991) Lipids of Plasma Mem-branes Prepared from Oat Root Cells—Effects of Induced WaterDeficit Tolerance, Plant Physiol. 96, 1136–1141.
104. Burja, A.M., Banaigs, B., Abou-Mansour, E., Burgess, J.G., andWright, P.C. (2001) Marine Cyanobacteria—A Prolific Sourceof Natural Products, Tetrahedron 57, 9347–9377.
105. Kobayashi, J., and Ishibashi, M. (1999) Marine Natural Prod-ucts and Marine Chemical Ecology, in Comprehensive NaturalProducts Chemistry (Mori, K., ed.), Vol. 8, pp. 415–649, Else-vier, Amsterdam.
106. Dembitsky, V.M., and Srebnik, M. (2002) Natural HalogenatedFatty Acids: Their Analogues and Derivatives, Prog. Lipid Res.41, 315–367.
107. Costantino, V., Fattorusso, E., Mangoni, A., Di Rosa, M., andIanaro, A. (1997) Glycolipids from Sponges. 6. Plakoside A andB, Two Unique Prenylated Glycosphingolipids with Immuno-suppressive Activity from the Marine Sponge Plakortis simplex,J. Am. Chem. Soc. 119, 12465–12470.
108. Costantino, V., Fattorusso, E., Mangoni, A., Di Rosa, M., Ia-
naro, A., and Maffia, P. (1996) Glycolipids from Sponges. IV.Immunomodulating Glycosyl Ceramides from the MarineSponge Agelas dispar, Tetrahedron 52, 1573–1578.
109. Costantino, V., Fattorusso, E., and Mangoni, A. (1995) Glyco-lipids from Sponges. 3. Glycosyl Ceramides from the MarineSponge Agelas conifera, Liebigs Ann. Chem. 8, 2133–2136.
110. Costantino, V., Fattorusso, E., and Mangoni, A. (1995) Glyco-lipids from Sponges. 1. Glycosyl Ceramide Composition of theMarine Sponge Agelas clathrodes, Liebigs Ann. Chem. 8,1471–1475.
111. Cafieri, F., Fattorusso, E., Mahajnah, Y., and Mangoni, A.(1994) Longiside, a Novel Digalactosylceramide from theCaribbean Sponge Agelas longissima, Liebigs Ann. Chem. 12,1187–1189.
112. Cafieri, F., Fattorusso, E., Mangoni, A., and Taglialatela-Scafati,O. (1995) Glycolipids from Sponges. 2. Glycosyl CeramideComposition of the Marine Sponge Agelas longissima, LiebigsAnn. Chem. 8, 1477–1481.
113. Natori, T., Morita, M., Akimoto, K., and Koezuka, Y. (1994)Agelasphins, Novel Antitumor and Immunostimulatory Cere-brosides from the Marine Sponge Agelas mauritianus, Tetrahe-dron 50, 2771–2784.
114. Venkannababu, U., Bhandari, S.P.S., and Garg, H.S. (1997)Regulosides A–C: Glycosphingolipids from the Starfish Pen-taceraster regulus, Liebigs Ann. Chem. 6, 1245–1247.
115. Babu, U.V., Bhandari, S.P.S., and Garg, H.S. (1997) Temno-sides A and B, Two New Glycosphingolipids from the SeaUrchin Temnopleurus toreumaticus of the Indian Coast, J. Nat.Prod. 60, 732–734.
116. Kawano, Y., Higuchi, R., Isobe, R., and Komori, T. (1988) Bio-logically Active Glycosides from Asteroidea. 13. Glycosphin-golipids from the Starfish Acanthaster planci. 2. Isolation andStructure of 6 New Cerebrosides, Liebigs Ann. Chem. 1, 19–24.
117. Sugiyama, S., Honda, M., and Komori, T. (1988) BiologicallyActive Glycosides from Asteroidea. 15. Asymmetric Synthesisof Phytosphingosine and Phytosphingosine Anhydro Base—As-signment of the Absolute Stereochemistry, Liebigs Ann. Chem.7, 619–625.
118. Kawano, Y., Higuchi, R., Isobe, R., and Komori, T. (1988) Bio-logically Active Glycosides from Asteroidea. 17. Glycosphin-golipids from the Starfish Acanthaster planci. 3. Isolation andStructure of 2 New Ceramide Lactosides, Liebigs Ann. Chem.12, 1181–1183.
119. Jin, W., Rinehart, K.L., and Jares-Erijman, E.A. (1994) Ophidi-acerebrosides—Cytotoxic Glycosphingolipids Containing aNovel Sphingosine from a Sea Star, J. Org. Chem. 59, 144–147.
120. Higuchi, R., Inagaki, K., Togawa, K., Miyamoto, T., and Ko-mori, T. (1994) Constituents of Holothuroideae. 4. Isolation andStructure of 3 New Cerebrosides, Ce-2b, Ce-2c and Ce-2d, fromthe Sea Cucumber Cucumaria echinata, Liebigs Ann. Chem. 1,79–84.
121. Higuchi, R., Inagaki, K., Togawa, K., Miyamoto, T., and Ko-mori, T. (1994) Constituents of Holothuroideae. 5. Isolation andStructure of Cerebrosides from the Sea Cucumber Pentacta aus-tralis, Liebigs Ann. Chem. 7, 653–658.
122. Costantino, V., Fattorusso, E., Mangoni, A., Aknin, M., andGaydou, E.M. (1994) Axiceramide-A and Axiceramide-B, 2Novel Tri-α-glycosylceramides from the Marine Sponge Ax-inella sp., Liebigs Ann. Chem. 12, 1181–1185.
123. Endo, M., Nakagawa, M., Hamamoto, Y., and Ishihama, M.(1986) Pharmacologically Active Substances from Southern Pa-cific Marine Invertebrates, Pure Appl. Chem. 58, 387–394.
124. Voutquenne, L., Lavaud, C., Massiot, G., Sevenet, T., and Hadi,H.A. (1999) Cytotoxic Polyisoprenes and Glycosides of Long-chain Fatty Alcohols from Dimocarpus fumatus, Phytochem-istry 50, 63–69.
125. Qi, J., Ojika, M., and Sakagami, Y. (2000) Termitomycesphins
658 REVIEW
Lipids, Vol. 40, no. 7 (2005)

A–D, Novel Neuritogenic Cerebrosides from the Edible Chi-nese Mushroom Termitomyces albuminosus, Tetrahedron 56,5835–5841.
126. Qi, J., Ojika, M., and Sakagami, Y. (2001) Neuritogenic Cere-brosides from an Edible Chinese Mushroom. Part 2: Structuresof Two Additional Termitomycesphins and Activity Enhance-ment of an Inactive Cerebroside by Hydroxylation, Bioorg.Med. Chem. 9, 2171–2177.
127. Keusgen, M., Yu, C.M., Curtis, J.M., Brewer, D., and Ayer,S.W. (1996) A Cerebroside from the Marine Fungus Mi-crosphaeropsis olivacea (Bonord.) Höhn, Biochem. Syst. Ecol.24, 465–468.
128. Sperling, P., and E. Heinz, E. (2003) Plant Sphingolipids: Struc-tural Diversity, Biosynthesis, First Genes and Functions,Biochim. Biophys. Acta 1632, 1–15.
129. Tan, R.X., and Chen, J.H. (2003) The Cerebrosides, Nat. Prod.Rep. 20, 509–534.
130. Colombaioni, L., and Garcia-Gil, M. (2004) SphingolipidMetabolites in Neural Signalling and Function, Brain Res. BrainRes. Rev. 46, 328–355.
131. Paparounas, K. (2004) Anti-GQ1b Ganglioside Antibody in Pe-ripheral Nervous System Disorders: Pathophysiologic Role andClinical Relevance, Arch. Neurol. 61, 1013–1016.
132. Gleeson, A. (1994) Glycoconjugates in Autoimmunity,Biochim. Biophys. Acta 1197, 237–255.
133. Diatlovitskaia, E.V., and Kandyba, A.G. (2004) BioeffectorSphingolipids as Stimulators of Cell Growth and Survival,Bioorg. Khim. 30, 227–233 [in Russian].
134. Avrova, N.F. (1985) Evolutionary Approach to the Analysis ofStructure and Function of Gangliosides, Neurochem. Res. 10,1547–1554.
135. Irwin, L.N. (1984) Phylogeny and Ontogeny of Vertebrate BrainGangliosides, Adv. Exp. Med. Biol. 174, 319–329.
136. Kappel, T., Hilbig, R., and Rahmann, H. (1993) Variability inBrain Ganglioside Content and Composition of EndothermicMammals, Heterothermic Hibernators and Ectothermic Fishes,Neurochem. Int. 22, 555–566.
137. Panzetta, P., and Allende, M.L. (2000) Ganglioside ExpressionDuring Differentiation of Chick Retinal Cells in vitro, Neu-rochem. Res. 25, 163–169.
138. Miyamoto, T., Yamamoto, A., Wakabayashi, M., Nagaregawa,Y., Inagaki, M., Higuchi, R., Iha, M., and Teruya, K. (2000) Bi-ologically Active Glycosides from Asteroidea, Two New Gan-gliosides, Acanthagangliosides I and J from the Starfish Acan-thaster planci, Eur. J. Org. Chem., 2295–22301.
139. Kawatake, S., Inagaki, M., Isobe, R., Miyamoto, T., andHiguchi, R. (2002) Isolation and Structure of MonomethylatedGM3-Type Ganglioside Molecular Species from the StarfishLuidia maculate, Chem. Pharm. Bull. (Tokyo) 50, 1386–1389.
140. Yamada, K., Hamada, A., Kisa, F., Miyamoto, T., and Higuchi,R. (2003) Constituents of Holothuroidea, 13. Structure of Neu-ritogenic Active Ganglioside Molecular Species from the SeaCucumber Stichopus chloronotus, Chem. Pharm. Bull. (Tokyo)51, 46–52.
141. Chekareva, N.V., Smirnova, G.P., and Kochetkov, N.K. (1991)Gangliosides of the Holothuria Cucumaria japonica Semper,Bioorg. Khim. 17, 398–402.
142. Smirnova, G.P. (1996) Gangliosides from Holothuria atra andTelenota ananas, Bioorg. Khim. 22, 134–139.
143. Kaneko, M., Kisa, F., Yamada, K., Miyamoto, T., and Higuchi,R. (2001) Constituents of Holothuroidea, 8. Structure of Neuri-togenic Active Ganglioside from the Sea Cucumber Stichopusjaponicus, Eur. J. Org. Chem. 3171–3174.
144. Yamada, K., Matsubara, R., Kaneko, M., Miyamoto, T., andHiguchi, R. (2001) Constituents of Holothuroidea. 10. Isolationand Structure of a Biologically Active Ganglioside MolecularSpecies from the Sea Cucumber Holothuria leucospilota, Chem.Pharm. Bull. (Tokyo) 49, 447–452.
145. Smirnova, G.P., Chekareva, N.V., and Kochetkov, N.K. (1986)Gangliosides of Ophiura, Ophiura sarsi, Bioorg. Khim. 12,507–513.
146. Smirnova, G.P., Chekareva, N.V., and Kochetkov, N.K. (1991)Gangliosides from 2 Species of Ophiura, Ophiocoma echinataand Ophiomastix annulosa Clark, Bioorg. Khim. 17, 387–397.
147. Inagaki, M., Shibai, M., Isobe, R., and Higuchi, R. (2001) Con-stituents of Ophiuroidea. 1. Isolation and Structure of ThreeGanglioside Molecular Species from the Brittle Star Ophiocomascolopendrina, Chem. Pharm. Bull. (Tokyo) 49, 1521–1525.
148. Kubo, H., Irie, A., Inagaki, F., and Hoshi, M. (1990) Ganglio-sides from the Eggs of the Sea Urchin, Anthocidaris crassispina,J. Biochem. (Tokyo) 108, 185–192.
149. Ijuin, T., Kitajima, K., Song, Y., Kitazume, S., Inoue, S., Hal-sam, S.M., Morris, H.R., Dell, A., and Inoue, Y. (1996) Isola-tion and Identification of Novel Sulfated and NonsulfatedOligosialyl Glycosphingolipids from Sea Urchin Sperm, Glyco-conj. J. 13, 401–413.
150. Shindo, K., Kamishohara, M., Odagawa, A., Matsuoka, M., andKawai, H. (1993) Vicenistatin, a Novel 20-Membered Macro-cyclic Lactam Antitumor Antibiotic, J. Antibiot. (Tokyo) 46,1076–1081.
151. Matsushima, Y., Nakayama, T., Fujita, M., Bhandari, R.,Eguchi, T., Shindo, K., and Kakinuma, K. (2001) Isolation andStructure Elucidation of Vicenistatin M, and Importance of theVicenisamine Aminosugar for Exerting Cytotoxicity ofVicenistatin, J. Antibiot. (Tokyo) 54, 211–219.
152. Lu, C., Bai, L., and Shen, Y. (2004) A Novel Amide N-Glyco-side of Ansamitocins from Actinosynnema pretiosum, J. An-tibiot. (Tokyo) 57, 348–350.
153. Naruse, N., Konishi, M., Oki, T., Inouye, Y., and Kakisawa, H.(1991) Fluvirucins A1, A2, B1, B2, B3, B4 and B5, New An-tibiotics Active Against Influenza A Virus. III. The Stereochem-istry and Absolute Configuration of Fluvirucin A1, J. Antibiot.(Tokyo) 44, 756–761.
154. Tomita, K., Oda, N., Hoshino, Y., Ohkusa, N., and Chikazawa,H. (1991) Fluvirucins A1, A2, B1, B2, B3, B4 and B5, New An-tibiotics Active Against Influenza A Virus. IV. Taxonomy onthe Producing Organisms, J. Antibiot. (Tokyo) 44, 940–948.
155. Ui, H., Imoto, M., and Umezawa, K. (1995) Inhibition of Phos-phatidylinositol Specific Phospholipase C Activity by Flu-virucin B2, J. Antibiot. (Tokyo) 48, 387–390.
156. Champney, W.S., and Tober, C.L. (2001) Structure–ActivityRelationships for Six Ketolide Antibiotics, Curr. Microbiol. 42,203–210.
157. Cardellina, J.H., Marner, F.-J., and Moore, R.E. (1979) Malyn-gamide A, a Novel Chlorinated Metabolite of the MarineCyanophyte Lyngbya majuscula, J. Am. Chem. Soc. 101,240–242.
158. Wu, M., Milligan, K.E., and Gerwick, W.H. (1997) Three NewMalyngamides from the Marine Cyanobacterium Lyngbya ma-juscula, Tetrahedron 53, 15983–15990.
159. Gallimore, W.A., and Scheuer, P.J. (2000) Malyngamides O andP from the Sea Hare Stylocheilus longicauda, J. Nat. Prod. 63,1422–1424.
160. Milligan, K.E., Marquez, B., Williamson, R.T., Davies-Cole-man, M., and Gerwick, W.H. (2000) Two New Malyngamidesfrom a Madagascan Lyngbya majuscula, J. Nat. Prod. 63,965–968.
161. Kan, Y., Sakamoto, B., Fujita, T., and Nagai, H. (2000) NewMalyngamides from the Hawaiian Cyanobacterium Lyngbyamajuscula, J. Nat. Prod. 63, 1599–1602.
162. Ubukata, M., Kimura, K., Isono, K., Nelson, C.C., Gregson,J.M., and McCloskey, J.A. (1992) Structure Elucidation of Li-posidomycins, a Class of Complex Lipid Nucleoside Antibi-otics, J. Org. Chem. 57, 6392–6403.
163. Schimana, J., Fiedler, H.P., Groth, I., Sussmuth, R., Beil, W.,Walker, M., and Zeeck, A. (2000) Simocyclinones, Novel Cy-
REVIEW 659
Lipids, Vol. 40, no. 7 (2005)

tostatic Angucyclinone Antibiotics Produced by Streptomycesantibioticus Tü 6040 I. Taxonomy, Fermentation, Isolation andBiological Activities, J. Antibiot. (Tokyo) 53, 779–787.
164. Schimana, J., Walker, M., Zeeck, A., and Fiedler, H.P. (2001)Simocyclinones: Diversity of Metabolites Is Dependent on Fer-mentation Conditions, J. Ind. Microbiol. Biotechnol. 27,144–148.
165. Eustaquio, A.S., Luft, T., Wang, Z.X., Gust, B., Chater, K.F.,Li, S.M., and Heide, L. (2003) Novobiocin Biosynthesis: Inacti-vation of the Putative Regulatory Gene NovE and HeterologousExpression of Genes Involved in Aminocoumarin Ring Forma-tion, Arch. Microbiol. 180, 25–32.
166. Galm, U., Schimana, J., Fiedler, H.P., Schmidt, J., Li, S.M., andHeide, L. (2002) Cloning and Analysis of the SimocyclinoneBiosynthetic Gene Cluster of Streptomyces antibioticus TO6040, Arch. Microbiol. 178, 102–114.
167. Holzenkampfer, M., and Zeeck, A. (2002) Biosynthesis ofSimocyclinone D8 in an 18O2-Rich Atmosphere, J. Antibiot.(Tokyo) 55, 341–342.
168. Li, S.M., and Heide, L. (2005) New Aminocoumarin Antibioticsfrom Genetically Engineered Streptomyces Strains, Curr. Med.Chem. 12, 419–427.
169. Lewis, R.J., Singh, O.M., Smith, C.V., Skarzynski, T., Maxwell,A., Wonacott, A.J., and Wigley, D.B. (1996) The Nature of In-hibition of DNA Gyrase by the Coumarins and the Cyclothi-alidines Revealed by X-Ray Crystallography, EMBO J. 15,1412–1420.
170. Tsai, F.T., Singh, O.M., Skarzynski, T., Wonacott, A.J., Wes-ton, S., Tucker, A., Pauptit, R.A., Breeze, A.L., Poyser, J.P.,O’Brien, R., et al. (1997) The High-Resolution Crystal Struc-ture of a 24-kDa Gyrase B Fragment from E. coli Complexedwith One of the Most Potent Coumarin Inhibitors, Clorobiocin,Proteins 28, 41–52.
171. Flatman, R.H., Howells, A.J., Heide, L., Fiedler, H.P., andMaxwell, A. (2005) Simocyclinone D8, an Inhibitor of DNAGyrase with a Novel Mode of Action, Antimicrob. AgentsChemother. 49, 1093–1100.
172. Abraham, W.R., Meyer, H., and Yakimov, M. (1998) NovelGlycine Containing Glucolipids from the Alkane Using Bac-terium Alcanivorax borkumensis, Biochim. Biophys. Acta 1393,57–62.
173. Yakimov, M.M., Golyshin, P.N., Lang, S., Moore, E.R., Abra-ham, W.R., Lunsdorf, H., and Timmis, K.N. (1998) Alcanivo-rax borkumensis gen. nov., sp. nov., a New, Hydrocarbon-De-grading and Surfactant-Producing Marine Bacterium, Int. J.Syst. Bacteriol. 48, 339–348.
174. Quinoa, E., Adamczeski, M., Crews, P., and Bakus, G.J. (1986)Bengamides, Heterocyclic Anthelmintics from a Jaspidae Ma-rine Sponge, J. Org. Chem. 51, 4494–4497.
175. Thale, Z., Kinder, F.R., Bair, K.W., Bontempo, J., Czuchta,A.M., Versace, R.W., Phillips, P.E., Sanders, M.L., Wattanasin,S., and Crews, P. (2001) Bengamides Revisited: New Structuresand Antitumor Studies, J. Org. Chem. 66, 1733–1741.
176. Macko, V., Stimmel, M.B., Wolpert, T.J., Dunkle, L.D., Ack-lin, W., Bänteli, R., Jaun, B., and Arigoni, D. (1992) Structureof the Host-specific Toxins Produced by the Fungal PathogenPericonia circinata, Proc. Natl. Acad. Sci. USA 89, 9574–9578.
177. Churchill, A.C., Dunkle, L.D., Silbert, W., Kennedy, K.J., andMacko, V. (2001) Differential Synthesis of Peritoxins and Pre-cursors by Pathogenic Strains of the Fungus Periconia circinata,Appl. Environ. Microbiol. 67, 5721–5728.
178. Nakao, Y., Maki, T., Matsunaga, S., van Soest, R.W.M., and
Nobuhiro, F. (2004) Penasulfate A, a New α-Glucosidase In-hibitor from a Marine Sponge Penares sp., J. Nat. Prod. 67,1346–1350.
179. Takada, K., Uehara, T., Nakao, Y., Matsunaga, S., van Soest,R.W.M., and Fusetani, N. (2004) Schulzeines A–C, New α-Glu-cosidase Inhibitors from the Marine Sponge Penares schulzei,J. Am. Chem. Soc. 126, 187–193.
180. Nakao, Y., Maki, T., Matsunaga, S., van Soest, R.W.M., andFusetani, N. (2000) Penarolide Sulfates A1 and A2, New α-Glu-cosidase Inhibitors from a Marine Sponge Penares sp., Tetrahe-dron 56, 8977–8987.
181. Infante, M.R., Pérez, L., Pinazo, A., Clapés, P., Morán, M.C.,Angelet, M., García, M.T., and Vinardell, M.P. (2004) AminoAcid-Based Surfactants, C.R. Chimie 7, 583–592.
182. Fernández-Pérez, M., and Otero, C. (2001) Enzymatic Synthe-sis of Amide Surfactants from Ethanolamine, Enzyme Microb.Technol. 28, 527–536.
183. Lundquist, J.J., and Toone, E.J. (2002) The Cluster GlycosideEffect, Chem. Rev. 102, 558–578.
184. Schengrund, C.-L. (2003) “Multivalent” Saccharides: Develop-ment of New Approaches for Inhibiting the Effects of Glyco-sphingolipid-Binding Pathogens, Biochem. Pharmacol. 65,699–707.
185. Han, F., and Zhang, G. (2004) New Family of Gemini Surfac-tants with Glucosamide-Based Trisiloxane, Colloid. Surf. A:Physicochem. Engineer. Aspects 237, 79–85.
186. Turner, M.W. (2003) The Role of Mannose-Binding Lectin inHealth and Disease, Mol. Immunol. 40, 423–429.
187. Abert, M., Mora, N., and Lacombe, J.M. (2002) Synthesis andSurface-Active Properties of a New Class of Surfactants De-rived from D-Gluconic Acid, Carbohydr. Res. 337, 997–1006.
188. Larpent, C., Laplace, A., and Zemb, T. (2004) MacrocyclicSugar-Based Surfactants: Block Molecules Combining Self-Ag-gregation and Complexation Properties, Angew. Chem. Int. Ed.Engl. 43, 3163–3167.
189. Kumpulainen, A.J., Persson, C.M., and Eriksson, J.C. (2004)Headgroup and Hydrocarbon Tail Effects on the Surface Ten-sion of Sugar-Based Surfactant Solutions, Langmuir 20,10935–10942.
190. Dimick, S.M., Powell, S.C., McMahon, S.A., Moothoo, D.N.,Naismith, J.H., and Toone, E.J. (1999) On the Meaning ofAffinity: Cluster Glycoside Effects and Concanavalin A, J. Am.Chem. Soc. 121, 10286–10296.
191. Colonna, B., Harding, V.D., Nepogodiev, S.A., Raymo, F.M.,Spencer, N., and Stoddart, J.F. (1998) Synthetic CarbohydrateDendrimers—Part 7—Synthesis of Oligosaccharide Den-drimers, Chem. Eur. J. 4, 1244–1254.
192. Burke, S.D., Zhao, Q., Schuster, M.C., and Kiessling, L.L.(2000) Synergistic Formation of Soluble Lectin Clusters by aTemplated Multivalent Saccharide Ligand, J. Am. Chem. Soc.122, 4518–4519.
193. Fan, E., Zhang, Z., Minke, W.E., Hou, Z., Verlinde, C.L.M.J.,and Hol, W.G.J. (2000) High-Affinity Pentavalent Ligands ofEscherichia coli Heat-Labile Enterotoxin by Modular Structure-Based Design, J. Am. Chem. Soc. 122, 2663–2664.
194. Mellet, C.O., Defaye, J., and Garcia-Fernandez, J.M. (2002)Multivalent Cyclooligosaccharides: Versatile CarbohydrateClusters with Dual Role as Molecular Receptors and Lectin Lig-ands, Chem. Eur. J. 8, 1982–1990.
[Received March 24, 2005; accepted June 25, 2005]
660 REVIEW
Lipids, Vol. 40, no. 7 (2005)




















