
Amide and urea derivatives having anti-hypercholesteremic activity ...
Upload
independentCategory
view
1download
0
Overexpression of a fatty acid amide hydrolase compromisesinnate immunity in Arabidopsis
Li Kang1, Yuh-Shuh Wang1, Srinivasa Rao Uppalapati1, Keri Wang1, Yuhong Tang1, Vatsala Vadapalli2, Barney J. Venables2,
Kent D. Chapman2, Elison B. Blancaflor1 and Kirankumar S. Mysore1,*
1Plant Biology Division, Samuel Roberts Noble Foundation, Ardmore, OK, USA, and2Center for Plant Lipid Research, Department of Biological Sciences, University of North Texas, Denton, TX, USA
Received 29 January 2008; revised 19 May 2008; accepted 5 June 2008; published online 29 July 2008.*For correspondence (fax 001 580 224 6692; e-mail [email protected]).
Summary
N-acylethanolamines are a group of lipid mediators that accumulate under a variety of neurological and
pathological conditions in mammals. N-acylethanolamine signaling is terminated by the action of diverse
hydrolases, among which fatty acid amide hydrolase (FAAH) has been well characterized. Here, we show that
transgenic Arabidopsis lines overexpressing an AtFAAH are more susceptible to the bacterial pathogens
Pseudomonas syringae pv. tomato and P. syringae pv. maculicola. AtFAAH overexpressors also were highly
susceptible to non-host pathogens P. syringae pv. syringae and P. syringae pv. tabaci. AtFAAH overexpressors
had lower amounts of jasmonic acid, abscisic acid and both free and conjugated salicylic acid (SA), compared
with the wild-type. Gene expression studies revealed that transcripts of a number of plant defense genes, as
well as genes involved in SA biosynthesis and signaling, were lower in AtFAAH overexpressors than wild-type
plants. Our data suggest that FAAH overexpression alters phytohormone accumulation and signaling which in
turn compromises innate immunity to bacterial pathogens.
Keywords: N-acylethanolamine, FAAH, fatty acid amide hydrolase, salicylic acid, non-host resistance.
Introduction
N-acylethanolamines (NAEs) comprise a class of fatty acid
amides found in trace amounts in animal tissues. One type
of NAE, namely anandamide (N-arachidonoylethanolamine)
functions in the endocannabinoid signaling pathway by
binding to a G-protein-coupled receptor, the cannabinoid
receptor 1 (CB1), in the brain. The same receptor also binds
to D9-tetrahydrocannabinol (THC), the active component of
marijuana (Cravatt et al., 2004; Devane et al., 1992).
Although this type of retrograde signaling by anandamide
has been the major function ascribed to NAEs in mammals,
NAEs are now gaining prominence not only in neuro-
transmission but also in other important physiological pro-
cesses such as immune signaling (Munro et al., 1993). One
significant breakthrough in NAE research was the cloning of
the enzymes that degrades NAEs. One of the enzymes,
called fatty acid amide hydrolase (FAAH), hydrolyzes NAE
into its corresponding free fatty acid and ethanolamine.
As more functions for NAEs in animals were being
discovered, parallel work showed that these fatty acid
amides were also endogenous constituents of plant tissues.
It appears that plants display striking similarities to mam-
malian systems in terms of their accumulation and overall
metabolism of NAEs (Blancaflor and Chapman, 2006;
Chapman, 2000, 2004). For instance, levels of NAEs in
tobacco leaves were shown to rise 10- to 50-fold when they
were challenged with fungal elicitors (Tripathy et al., 1999).
This elevation of endogenous NAEs in plants subjected to
biotic stress mirrors somewhat the accumulation of NAEs
under a variety of neurological and pathological conditions
in mammals (Berdyshev et al., 2000, 2001; Berger et al.,
2004; Schmid et al., 2002). Other biological effects of NAEs
in plants include remodeling of the cytoskeleton, alteration
of seedling growth, inhibition of phospholipase D activity,
modulation of elicitor-induced alkalinization response,
induction of defense gene expression and interaction with
ABA signaling (Austin-Brown and Chapman, 2002;
Blancaflor et al., 2003; Komis et al., 2006; Teaster et al.,
2007; Tripathy et al., 1999). Although such observations
have positioned NAEs as novel metabolites that could
govern important regulatory and signaling pathways in
336 ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd
The Plant Journal (2008) 56, 336–349 doi: 10.1111/j.1365-313X.2008.03603.x

plants (Lopez-Bucio et al., 2006), much remains to be
learned regarding their precise roles in mediating various
plant physiological processes.
The case for NAEs as potential lipid mediators in plants
has been strengthened recently by the identification of an
Arabidopsis thaliana FAAH gene (AtFAAH, At5g64440)
based upon the similarity of the amidase signature (AS)
domain and conservation of several key catalytic residues of
AtFAAH with mammalian FAAH (Shrestha et al., 2003).
When expressed as recombinant proteins in Escherichia
coli, plant FAAHs were capable of hydrolyzing NAEs in vitro
(Shrestha et al., 2003; Wang et al., 2006b). Consistent with
their in vitro NAE hydrolytic activity, seeds and seedlings of
Arabidopsis faah knockouts had elevated levels of endoge-
nous NAEs, while AtFAAH overexpressors had reduced
NAEs (Wang et al., 2006b). AtFAAH overexpressors exhib-
ited enhanced seedling growth, suggesting that the timely
depletion of NAEs might be involved in seed germination
and early seedling establishment (Wang et al., 2006b). The
availability of plants with altered FAAH expression has now
paved the way for more detailed studies of NAE metabolism
and how it affects plant development and responses to the
environment. Because of the increasing role for NAEs in
mammalian immune signaling (Buckley et al., 2006), one
logical area to probe for potential functions for NAEs in
plants would be plant immunity and defense responses,
where NAEs were first suggested to be involved (Tripathy
et al., 1999, 2003).
Here we report that overexpression of AtFAAH, a gene
involved in NAE metabolism, compromises the innate
immunity of Arabidopsis against several host and non-host
pathogens. Collectively, our data indicate that AtFAAH
intersects with plant defense pathways by altering phyto-
hormone signaling. Our results reinforce the notion that
plants, like animals, utilize fatty acid amides in mediating
immunity to pathogens and add to a growing list of
important plant processes that are modulated by NAE. This
study provides genetic evidence that altering the expression
of a gene involved in NAE metabolism can affect plant
defense responses and supports earlier pharmacological
studies implicating NAEs in plant defense signaling.
Results
Overexpressing AtFAAH compromised basal resistance in
Arabidopsis
To investigate the role of FAAH in plant disease resistance,
the Arabidopsis wild-type Columbia-0 (Col-0), a vector con-
trol line, three independent AtFAAH overexpressor lines
(OE2a-1, OE7a-1, OE11a-1) and two AtFAAH T-DNA knockout
lines (faah, salk_095108 and salk_118043), described in
Wang et al. (2006b), were used. When sprayed or infiltrated
with the virulent bacterial strains Pseudomonas syringae pv.
tomato DC3000 (P. s. tomato) and P. syringae pv. maculicola
(P. s. maculicola), all AtFAAH overexpressor lines exhibited
enhanced susceptibility manifested as larger areas of
chlorotic spots when compared with Col-0 and the vector
control line (Figure 1a and Figure S1). The faah mutant
(salk_095108) displayed a similar degree of susceptibility as
Col-0 and the vector control lines (Figure 1a). The enhanced
susceptibility of the AtFAAH overexpressors was even more
evident when considering the time in which visible symp-
toms developed. For instance, visible chlorotic lesions star-
ted to develop on leaves of Col-0, the vector control and two
knockout lines 3 days post-inoculation (dpi) with bacterial
suspension cells at 106 colony-forming units (CFU) ml)1.
However, symptoms developed on all tested AtFAAH over-
expressors as early as 2 dpi. Consistent with these symp-
toms, the bacterial population was about 10 times higher in
the leaves of AtFAAH overexpressors than in Col-0 or faah
(Figure 1b).
AtFAAH overexpressors were susceptible to a few non-host
pathogens
Results from several studies demonstrate that basal resis-
tance is often associated with non-host resistance in plants
(Bais et al., 2005; He et al., 2006). Therefore, we next exam-
ined whether the reduced basal resistance of the AtFAAH
overexpressors had any impact on non-host disease resis-
tance. Five days after infiltration with a non-host bacterial
strain, P. syringae pv. syringae (P. s. syringae), at
5 · 106 CFU ml)1 clear chlorotic lesions formed around the
leaf edges of AtFAAH overexpressors while Col-0 and faah
remained mostly symptomless (Figure 2a). Symptoms
developed on overexpressors as early as 2 days after inoc-
ulation and continued to progress while Col-0 and faah were
free of symptoms. Consistent with these observations, the
bacterial population accumulated to levels more than
50-fold higher in AtFAAH overexpressors when compared
with Col-0 and faah lines (Figure 2b). Similar results were
obtained upon inoculation with another non-host pathogen,
P. syringae pv. tabaci (P. s. tabaci; Figure 2a). The P. s. tabaci
population in AtFAAH overexpressors was almost 100-fold
higher than in Col-0 and faah lines at 3 dpi (Figure 2b).
AtFAAH overexpressors did not exhibit compromised
resistance when challenged with several other non-host
pathogens, including P. syringae pv. phaseolicola NPS3121,
P. syringae pv. glycinea, Xanthomonas campestris pv.
campestris and X. campestris pv. vesicatoria (data not
shown).
R-gene mediated resistance was not affected in AtFAAH
overexpressors
Common signaling components have been reported
between host and non-host resistance pathways (Mysore
FAAH overexpression compromises innate immunity 337
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

(a)
(b)
Figure 1. Overexpression of AtFAAH compro-
mised basal resistance in Arabidopsis.
(a) Disease symptoms on Arabidopsis wild-type
Col-0, faah knockout and AtFAAH overexpressors
(OE7a-1 and OE11a-1) were photographed
3 days post-inoculation (dpi) with Pseudomonas
syringae pv. tomato (P. s. tomato) and Pseudo-
monas syringae pv. maculicola (P. s. maculicola)
at 106 colony-forming units (CFU) ml)1. Inocu-
lated leaves are indicated by arrows.
(b) Growth of the bacteria at 3 dpi in Col-0, faah,
OE7a-1 and OE11a-1. Error bars are standard
errors from four biological replicates. Letters on
the graph donates statistically different groups
by Turkey’s group test (P > 0.005).
(a)
(b)
Figure 2. Non-host resistance was compro-
mised in AtFAAH overexpressors.
(a) Symptoms at 5 days post-inoculation (dpi)
and (b) growth (3 dpi) in Col-0, faah, OE7a-1 and
OE11a-1 after inoculation with Pseudomonas
syringae pv. syringae (P. s. syringae) and Pseu-
domonas syringae pv. tabaci (P. s. tabaci).
Leaves were infiltrated with bacterial suspen-
sions at concentration of 5 · 106 colony-forming
units (CFU) ml)1 for symptom development and
106 CFU ml)1 for growth assays. Inoculated
leaves are indicated by arrows. Letters on the
graph donates statistically different group by
Turkey’s group test (P > 0.005). Error bars are
standard errors from four biological replicates.
338 Li Kang et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

and Ryu, 2004; Peart et al., 2002). We therefore investigated
whether R-gene mediated resistance to P. s. tomato carrying
the corresponding avr gene is affected in AtFAAH overex-
pressors. The ecotype Col-0 has several R genes including
RPS2, RPM1 and RPS4. Upon inoculation with an avirulent
pathogen, P. s. tomato expressing avrRpt2 (an avirulent gene
for RPS2), the hypersensitive response (HR) on AtFAAH
overexpressors developed slightly faster than wild-type Col-0
and faah knockout. The differences were particularly clear
with a lower concentration of pathogen (Figure S2a). How-
ever, the bacterial populations were not significantly
different in any tested plant (Figure S2b). Similarly, P. s.
tomato expressing avrB (an avirulent gene for RPM1) and P.
s. tomato expressing avrRPS4 (an avirulent gene for RPS4)
caused the HR to develop faster on AtFAAH overexpressors,
but the bacterial growth was comparable to those in wild-
type plants (data not shown). These data suggest that
overexpression of AtFAAH did not compromise gene-for-
gene mediated resistance, at least for the three R-gene-
mediated pathways tested.
NAE profiles were not altered substantially in the leaves of
AtFAAH overexpressors
Previously, it was shown that several lines of Arabidopsis
AtFAAH overexpressors had a lower NAE content in seeds
and seedlings than the wild-type (Wang et al., 2006b). The
NAE profiles in leaves of 4-week-old AtFAAH overexpressors
and wild-type plants were examined to see if there was a
change in the NAE content or composition (Figure S3).
Surprisingly, the overall NAE contents in mature leaves of
AtFAAH overexpressor, OE7a-1, were similar to wild-type
leaves, particularly on a fresh weight basis (Figure S3).
Inoculation with non-host pathogen P. s. syringae or buffer
did not appear to significantly affect the overall NAE content
in the leaves of the wild-type or OE7a-1. These results
indicate that the steady-state profiles of the NAE pools in
mature leaves were not significantly altered by AtFAAH
overexpression.
AtFAAH overexpressors had low basal levels of salicylic
acid, jasmonic acid and abscisic acid
Overwhelming evidence indicates that endogenous salicylic
acid (SA) in plants is one of the key signaling molecules that
activates plant defense responses against invading patho-
gens (Durrant and Dong, 2004; Shah, 2003). We therefore
assessed whether the enhanced susceptibility of AtFAAH
overexpressors to the tested pathogens is associated with
changes in endogenous SA content. Total SA was mea-
sured in Col-0 and OE7a-1 plants before and 12 h post-
inoculation (hpi) with buffer, P. s. tomato or P. s. syringae.
Buffer-inoculated (12 hpi) or uninoculated OE7a-1 had
about two-fold less free SA and also less conjugated SA
than Col-0 (Figure 3a). Upon pathogen inoculation, the
amount of conjugated SA was still lower in OE7a-1 than Col-0
at 12 hpi whereas free SA was about two-fold higher in OE7a-
1 when compared with Col-0 (Figure 3a, lower panel).
We speculated that the initial lower SA content in AtFAAH
overexpressors might be one of the factors contributing to
their hyper-susceptibility to the tested host and non-host
pathogens. To test this hypothesis we treated OE7a-1 and
Col-0 plants with a SA analogue, benzo-(1,2,3)-thiadiazole-7-
carbothioic acid S-methyl ester (BTH), 3 days prior to
bacterial infection. It has been shown that BTH treatment
resulted in SA accumulation in plants that led to systemic
acquired resistance (SAR; Lawton et al., 1996). Our results
showed that after BTH treatment, the AtFAAH overexpres-
sor, OE7a-1, was no longer susceptible to the non-host
pathogens P. s. tabaci and P. s. syringae (Figure 3b).
Furthermore, the bacterial population in OE7a-1 was
reduced to near undetectable levels at 3 dpi and no signif-
icant differences were observed in bacterial populations
between OE7a-1 and Col-0 (data not shown). In contrast, the
virulent bacterium P. s. tomato remained 10-fold higher in
OE7a-1 than Col-0 even after BTH treatment (Figure 3c)
although the bacterial growth was slightly reduced in BTH-
treated plants. These results suggest that reduced levels of
SA may contribute to the enhanced susceptibility phenotype
of AtFAAH overexpressors.
The SA signaling pathway is reported to be antagonistic to
jasmonic acid (JA) signaling in plants (Clarke et al., 2000;
Dong, 1998). We therefore measured endogenous JA in Col-0
and OE7a-1 plants before and 24 hpi with buffer, P. s. tomato
or P. s. syringae. Buffer-inoculated (24 hpi) or uninoculated
OE7a-1 had about three-fold less JA (Figure S4a). Upon
inoculation with non-host pathogen, the amount of JA was
still lower in OE7a-1 than Col-0 at 24 hpi (Figure S4a).
Recently, it has been shown that NAE metabolism interacts
with ABA signaling in Arabidopsis seedlings (Teaster et al.,
2007). We therefore measured endogenous ABA in Col-0
and OE7a-1 plants at 24 hpi with buffer or P. s. syringae.
Buffer-inoculated (24 hpi) OE7a-1 had about 2.5-fold less
ABA (Figure S4b). Upon inoculation with non-host patho-
gen the amount of ABA was reduced in Col-0 but not in
OE7a-1 plants (Figure S4b).
Numerous plant defense-related genes were downregulated
in AtFAAH overexpressors
To investigate further the molecular mechanisms underly-
ing the susceptibility of AtFAAH overexpressors to non-
host pathogens, we used an Affymetrix ATH1 whole
genome microarray GeneChip to examine transcriptional
changes in AtFAAH overexpressor OE7a-1 and Col-0 at 0, 6
and 12 hpi with the non-host pathogen, P. s. syringae. We
used two approaches to interpret the microarray results.
The first approach aimed to identify the genes differentially
FAAH overexpression compromises innate immunity 339
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

expressed over the time course of inoculation by the non-
host pathogen P. s. syringae in both Col-0 and OE7a-1. The
expression profile from Col-0 or OE7a-1 at 6 and 12 hpi was
compared with those at 0 hpi of its own genotype to
determine the global changes caused by inoculation of
P. s. syringae (Figure S5a). Large numbers of genes were
differentially expressed between 0 and 6 or 12 dpi in both
Col-0 and OE7a-1 after bacterial infection. Interestingly, as
shown in Table 1, the number of genes that were differ-
entially expressed in OE7a-1 was significantly smaller than
those differentially expressed in Col-0. Many of the
defense-related genes that were induced in Col-0 in
response to P. s. syringae inoculation were not induced in
OE7a-1. These results suggest that the ability of AtFAAH
overexpressors to mount a global transcriptional response
against P. s. syringae infection is compromised.
In the second approach, gene expression at the same time
point was compared between the AtFAAH overexpressor and
the wild-type in order to identify genes intrinsically differen-
tially expressed or genes that responded in a different
(a) (b)
(c)
Figure 3. AtFAAH overexpressor had less
endogenous salicylic acid (SA) and benzo-
(1,2,3)-thiadiazole-7-carbothioic acid S-methyl
ester (BTH) treatment restored its resistance to
non-host pathogens.
(a) Salicylic acid quantification in uninoculated
Col-0 and AtFAAH overexpressor (OE7a-1) and
12 h after inoculation with buffer or Pseudomo-
nas syringae pv. tomato (P. s. tomato) or Pseu-
domonas syringae pv. syringae (P. s. syringae) at
5 · 106 colony-forming units (CFU) ml)1. Error
bars represent the standard error. The SA anal-
ysis represents an average of three independent
biological replicates.
(b) Treatment with BTH restored the resistance of
OE7a-1 to non-host pathogens Pseudomonas
syringae pv. tabaci (P. s. tabaci) and P. s. syrin-
gae. Infiltrated leaves were photographed at
5 days post-inoculation (dpi). Plants were
sprayed with buffer or BTH at 150 mM concen-
tration 72 h before infiltrating P. s. tabaci or P. s.
syringae at 5 · 106 CFU ml)1 or (c) with P. s.
tomato at 106 CFU ml)1. The graph shows
growth of P. s. tomato at 3 dpi.
Table 1 Comparison of expression profilesof Col-0 and OE7a-1 between 0 h and 6 or12 h after inoculation with the non-hostpathogen Pseudomonas syringae pv.syringae
Data set
Total no. ofgenesdifferentiallyexpressed
No. genes that wereinduced afterinoculation
No. genes that weresuppressed afterinoculation
Unique Common Unique Common
Col-0 6 h:0 h 4913 1178 1321 884 1530OE7a-1 6 h:0 h 3969 561 557Col-0 12 h:0 h 5830 1320 1615 1390 1505OE7a-1 12 h:0 h 4150 415 615
340 Li Kang et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

manner to P. s. syringae infection between OE7a-1 and Col-0.
At 0 hpi, 507 genes (223 higher and 284 lower in OE7a-1) were
intrinsically differentially expressed between OE7a-1 and
Col-0. The number of differentially expressed genes nearly
tripled at 6 hpi to 1432 differentially expressed genes (479
higher and 953 lower in OE7a-1). At 12 hpi, we observed an
even higher number of differentially expressed genes (2063)
between OE7a-1 and Col-0, with 883 higher and 1180 lower in
OE7a-1. These differentially expressed genes were classified
based on their functional annotation (Figure S6).
Using ratios of transcript levels from OE7a-1 and Col-0, a
heat map of 3126 genes differentially expressed in OE7a-1 at
any of the three time points was generated (Figure S5b). The
number of downregulated genes was slightly greater among
all the differentially expressed genes between OE7a-1 and
Col-0. Upon close examination of the microarray data, we
observed that transcripts of majority of the differentially
expressed defense-related genes (�200 genes) were less
abundant in OE7a-1 when compared with Col-0, especially at
6 and 12 hpi (Figure S5s, Table S1). Strikingly, transcripts of
around 64 genes that encode proteins with Toll and inter-
leukin-1 receptor/coiled-coil (TIR/CC) and/or nucleotide bind-
ing site (NBS) and/or leucine-rich repeat (LRR) domains
(typically present in R genes; DeYoung and Innes, 2006)
were significantly less abundant in OE7a-1 when compared
with Col-0 (Table S2).
A high degree of consistency from two independent
experiments indicates that the results generated from the
study were reproducible (Figure S5a). The correlation coef-
ficients from two independent experiments were 0.98/0.98,
0.98/0.99 and 0.98/0.99 for Col-0/OE7a-1 at 0-, 6- and 12-h
time points, respectively. To further validate the microarray
data, we selected 24 genes, along with the AtFAAH and two
reference genes, for real-time quantitative RT-PCR (qRT-
PCR) study. Despite some variations in absolute fold-differ-
ences, the qRT-PCR data were consistent with the micro-
array data (Table S3).
AtFAAH overexpressors are compromised in SA-mediated
signaling
As indicated earlier, the AtFAAH overexpressor had rela-
tively smaller amounts of total SA content than the wild-
type before and 12 h after challenging with pathogens
(Figure 3a). Consistent with these results, our microarray
data showed that transcripts of several genes involved in SA
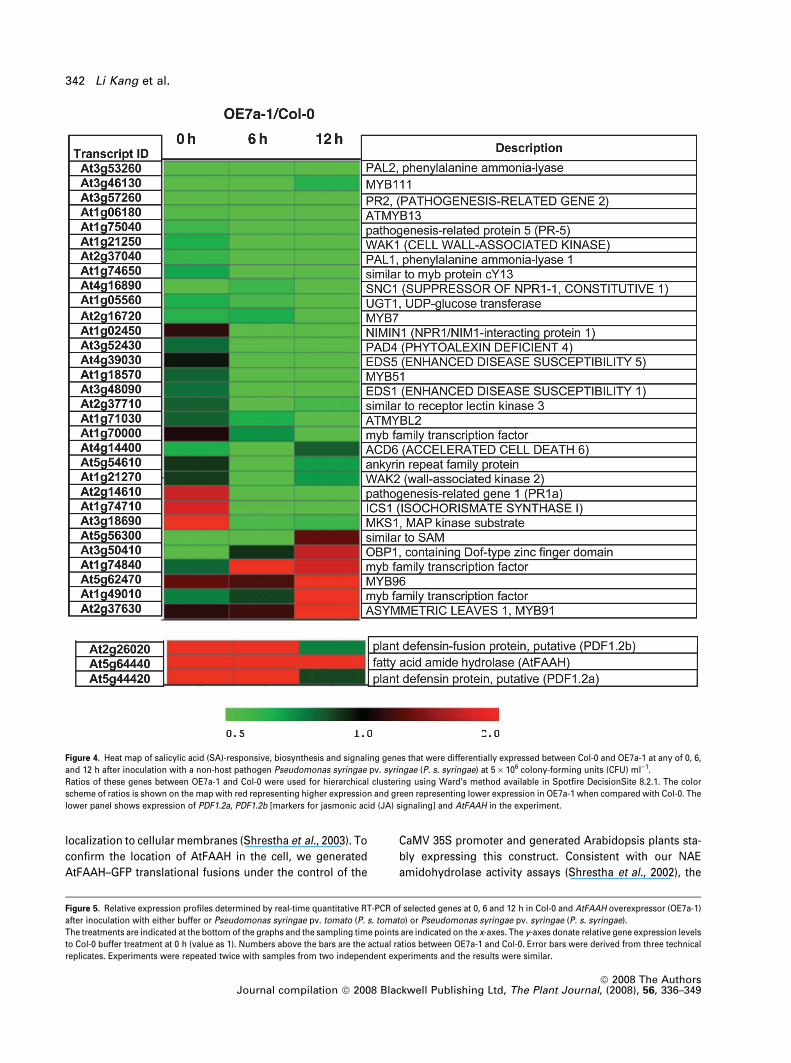
biosynthesis (ICS1, PAL1 and PAL2) were less abundant in
the AtFAAH overexpressor (OE7a-1) upon pathogen inocu-
lation when compared with Col-0 (Figure 4, Table S4). Even
though these genes were induced in both OE7a-1 and Col-0
upon bacterial inoculation, the scale of induction was much
lower in OE7a-1 when compared with Col-0. Furthermore,
transcripts of many genes involved in SA-mediated defense
signaling, such as PR1, PR2, PR5, EDS1, EDS5, PAD4, etc.,
were also less abundant in OE7a-1 when compared with Col-0
(Figure 4, Table S4).
To further confirm the microarray results and the role of
AtFAAH in SA-mediated signaling, the expression profiles of
several genes involved in SA biosynthesis or signaling, a
jasmonic acid (JA) signaling gene and a representative R
gene were examined using qRT-PCR in two separate exper-
iments with independent biological replicates for OE7a-1
and Col-0 plants treated with either buffer (control), P. s.
tomato or P. s. syringae. As expected, we observed larger
amounts of AtFAAH transcripts in OE7a-1 when compared
with Col-0 (Figure 5). It is noteworthy that the AtFAAH
expression in Col-0 was slightly induced in all treatments.
Typical marker genes for SA-mediated defense signaling,
PR1a and PR2, were highly induced in Col-0 at 12 hpi. These
genes were considerably less abundant in OE7a-1 at 6 and
12 hpi. One of the SA biosynthetic genes (ICS1) that was
tested was highly induced in Col-0 at both 6 and 12 hpi.
Consistent with the microarray data, ICS1 was not induced in
OE7a-1 except at 12 hpi with P. s. tomato. However, the
induction of ICS1 at 12 hpi in OE7a-1 was not as high as in
Col-0. The expression profiles of the WRKY transcription
factor genes (WRKY 46 and WRKY 70) and EDS5 were also
similar to that of ICS1 in all treatments in both Col-0 and
OE7a-1. The expression profiles of an RPP13-like and an
EDS-1-like genes were different from that of ICS1 and the
WRKY genes; these genes were constitutively very low in
OE7a-1 when compared with Col-0, and their expression
profiles did not change in response to mock or pathogen
inoculation in OE7a-1. On the other hand the amounts of
transcript for PDF1.2a, a marker gene for JA and ethylene
signaling, was slightly higher in OE7a-1 compared with Col-0
in most cases except at the 12-h time point in plants infected
with P. s. tomato. However, transcripts of several genes
involved in JA biosynthesis (OPR3, AOC2, AOC4, LOX1 and
LOX2) were less abundant in OE7a-1 compared with Col-0 at
one or more time points after inoculation with P. s. tomato
(Table S1). This was consistent with our observation that
OE7a-1 had less JA than Col-0 (Figure S4a).
Despite differential expression of PR genes, expression
profiles of TGA transcription factors that are involved in the
activation of PR gene expression (Zhang et al., 1999, 2003)
were not significantly different between OE7a-1 and Col-0.
However, a number of MYB and WRKY transcription factors
were expressed at much lower levels in OE7a-1 than those in
Col-0 at various sampling points (Tables S4 and S5). These
data argue for the existence of a novel mechanism through
which AtFAAH negatively affects the SA-mediated defense
signaling pathway.
AtFAAH is associated with membranes
Like mammalian FAAH, AtFAAH contains a putative
transmembrane domain near the N-terminus that predicts
FAAH overexpression compromises innate immunity 341
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349
localization to cellular membranes (Shrestha et al., 2003). To
confirm the location of AtFAAH in the cell, we generated
AtFAAH–GFP translational fusions under the control of the
CaMV 35S promoter and generated Arabidopsis plants sta-
bly expressing this construct. Consistent with our NAE
amidohydrolase activity assays (Shrestha et al., 2002), the
Figure 4. Heat map of salicylic acid (SA)-responsive, biosynthesis and signaling genes that were differentially expressed between Col-0 and OE7a-1 at any of 0, 6,
and 12 h after inoculation with a non-host pathogen Pseudomonas syringae pv. syringae (P. s. syringae) at 5 · 106 colony-forming units (CFU) ml)1.
Ratios of these genes between OE7a-1 and Col-0 were used for hierarchical clustering using Ward’s method available in Spotfire DecisionSite 8.2.1. The color
scheme of ratios is shown on the map with red representing higher expression and green representing lower expression in OE7a-1 when compared with Col-0. The
lower panel shows expression of PDF1.2a, PDF1.2b [markers for jasmonic acid (JA) signaling] and AtFAAH in the experiment.
Figure 5. Relative expression profiles determined by real-time quantitative RT-PCR of selected genes at 0, 6 and 12 h in Col-0 and AtFAAH overexpressor (OE7a-1)
after inoculation with either buffer or Pseudomonas syringae pv. tomato (P. s. tomato) or Pseudomonas syringae pv. syringae (P. s. syringae).
The treatments are indicated at the bottom of the graphs and the sampling time points are indicated on the x-axes. The y-axes donate relative gene expression levels
to Col-0 buffer treatment at 0 h (value as 1). Numbers above the bars are the actual ratios between OE7a-1 and Col-0. Error bars were derived from three technical
replicates. Experiments were repeated twice with samples from two independent experiments and the results were similar.
342 Li Kang et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

FAAH overexpression compromises innate immunity 343
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

35S::AtFAAH–GFP fusion localized primarily to the endo-
plasmic reticulum (ER) in leaf epidermal cells and cotyle-
dons. The pattern of AtFAAH–GFP localization mirrored that
of a construct fused to the HDEL motif, a recognition signal
that directs proteins to the ER (Figure 6a–c). Leaves from
plants expressing the 35::AtFAAH–GFP fusion were count-
erstained with the plasma-membrane dye FM4-64. Single
optical confocal sections at the median plane of the cells
show strong GFP labeling along the periphery of the
epidermal cells. The GFP signal coincides with regions of
FM4-64 staining indicating that in addition to the ER, AtFAAH
is localized to the plasma membrane (overlay image in
Figure 6d). Similar results were observed in four indepen-
dent transgenic lines. We then looked to see if the localiza-
tion of AtFAAH was altered after pathogen inoculation,
but the membrane localization of AtFAAH–GFP was unal-
tered 3 days after inoculation with either host (P. s. tomato)
or non-host pathogen (P. s. syringae) tested (data not
shown).
Extracts of AtFAAH–GFP fusions expressed in E. coli still
exhibited NAE hydrolase activity in vitro but activity was
about 10-fold less than AtFAAH without GFP. Furthermore,
plants expressing the 35S::AtFAAH–GFP fusion had lower
NAE hydrolase activity than plants expressing 35S::AtFAAH,
despite roughly similar levels of FAAH protein in the
transgenic plants (Sang-Chul Kim and KDC, University of
Texas, Denton, TX, unpublished data). Despite the lower
activity of the AtFAAH–GFP fusion, this construct was able to
rescue the faah line from hypersensitivity to exogenous
NAE12:0 (Wang et al., 2006b). The faah line transformed
with the 35S::AtFAAH–GFP construct exhibited enhanced
resistance to NAE levels that would typically be strongly
inhibitory to the AtFAAH knockouts (Figure 6e). However,
35S::AtFAAH–GFP expressed in the faah background resul-
ted in smaller seedlings and shorter roots compared with
35S::AtFAAH plants when grown on exogenous NAE12:0
(Figure 6e). These differences could be explained by the
lower NAE hydrolase activity of AtFAAH–GFP.
Discussion
N-acylethanolamines have diverse physiological functions
in mammals. Of particular interest here is the emerging role
of NAEs in mammalian cellular stress responses and the
immune function (Salzet et al., 2000). While there was initial
evidence that NAEs affect how plants respond to pathogens
(Chapman et al., 1998; Tripathy et al., 1999), we still lack an
appreciable understanding of the molecular mechanisms
underlying the effects of NAEs on plant immunity. To begin
to address this issue, we conducted a detailed analysis on
(a)
(d)
(e)
(b) (c) Figure 6. Localization of AtFAAH–GFP.
Confocal micrographs of epidermal cells from
Arabidopsis leaves expressing 35S::GFP (a),
35S::GFP–HDEL (b) and 35S::AtFAAH–GFP (c).
The reticulate pattern of 35S::AtFAAH–GFP is
similar to 35S::GFP–HDEL, which targets to the
endoplasmic reticulum (ER). The inset shows a
pair of guard cells. The perinuclear pattern in
AtFAAH–GFP is typical of ER localization.
(d) Single confocal optical sections of epidermal
cells from leaves of Arabidopsis plants express-
ing 35S::AtFAAH–GFP. The leaf was counter-
stained with FM4-64, which labels the plasma
membrane. The overlay image shows that the
GFP signal around the cell periphery co-localizes
with FM4-64 (yellow areas), indicating that FAAH
is also localized to the plasma membrane.
(e) The growth of faah knockouts on 30 lM
N-acylethanolamine (NAE) 12:0 is significantly
inhibited while AtFAAH overexpressors are not.
344 Li Kang et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

Arabidopsis plants with altered expression of the NAE
hydrolyzing enzyme FAAH.
Here we provided evidence that Arabidopsis plants over-
expressing FAAH have compromised immune responses to
a number of pathogens including non-host bacterial patho-
gens that typically do not cause disease in Arabidopsis.
However, resistance was not compromised against all non-
host pathogens tested, suggesting that this phenomenon
was specific for certain interactions and was not due to a
general reduction in plant fitness. In fact it should be pointed
out that AtFAAH overexpressors actually exhibited
enhanced growth phenotypes (Wang et al., 2006b). Whereas
AtFAAH overexpressors exhibited modified responses to
pathogens, faah knockouts did not show altered sensitivity
to any host or non-host pathogens tested. This observation
is consistent with our previous findings in seedlings of faah
knockouts, which did not show altered growth phenotype
other than an increased sensitivity to exogenous NAE (Wang
et al., 2006b). The lack of any obvious phenotype in the faah
knockout could be attributed to alternative competing
pathways by which NAE is catabolized in the plant. In
mammalian systems, a number of other enzymes have been
shown to participate in the degradation of NAEs, including
an acid amidase (Tsuboi et al., 2005), a second FAAH (Wei
et al., 2006), lipoxygenases (Ueda et al., 1995) and cycloox-
ygenases (Kozak et al., 2002). In this regard, bioinformatics
analyses revealed seven amidase genes in the Arabidopsis
genome that have a conserved amidase signature (AS)
domain (Kilaru et al., 2007) and may encode an alternative
FAAH. These reasons may explain why NAEs are still
degraded in the absence of AtFAAH and also explain the
lack of any visible phenotype in faah knockouts.
The NAE levels were similar between faah and Col-0
seedlings, although in seeds faah knockout plants contained
about 30% more NAE than Col-0 (Wang et al., 2006b).
Consistent with these results, faah behaved similarly to
wild-type Col-0 in response to inoculations with host and
non-host pathogens. Surprisingly, changes in NAE content
and composition in response to non-host pathogens was not
significantly different in mature leaves of FAAH overexpres-
sors compared with the wild-type (Figure S3), contrary to
what might be expected from the increased capacity for NAE
turnover in these FAAH-overexpressing plants (Wang et al.,
2006b). However, it may be that changes in NAEs are highly
localized to specific cells or subcellular compartments and
any changes may be difficult to detect relative to the bulk
steady-state levels of NAEs in leaves, given the current
methodology. This is particularly difficult given the low
levels of endogenous NAEs in mature leaves (<20 ng g)1
fresh weight for all types summed together). Infiltration and
spray application of NAEs to mature Arabidopsis plants did
not alter the resistance to pathogens, nor did it activate the
SA signaling pathway (data not shown). Moreover, FAAH
may function in ways additional to its catalytic activity to
mediate defense responses, perhaps via protein–protein
interactions. Indeed, in other work with ABA signaling in
seedlings, the action of FAAH was not entirely explained by
its effect on NAE content or composition (Teaster et al.,
2007). Overexpression of FAAH led to increased capacity for
NAE turnover, but not to significant alterations in overall
NAE content, despite a clear hypersensitivity of FAAH
overexpressors to exogenous ABA. The regulation of NAE
metabolism by FAAH, and the function of FAAH outside the
NAE pathway appear to be complex and are only beginning
to be addressed. Nonetheless it is clear that the alteration of
FAAH expression leads to marked changes in plant growth
(Wang et al., 2006b), phytohormone signaling (Teaster
et al., 2007) and the capacity for plant defense against
pathogens.
Numerous reports have put SA at center of the resistance
network against pathogens. The hyper-susceptibility of
AtFAAH overexpressors prompted us to ask whether there
was any change of SA in AtFAAH overexpressors which
would lead to compromised resistance to pathogens. Sur-
prisingly, we found that endogenous SA was at least two-
fold less in AtFAAH overexpressors when compared with
Col-0. The hyper-susceptible phenotype of AtFAAH overex-
pressors to non-host bacterial pathogens could be reversed
by treating plants with an SA analogue, BTH (Figure 3b).
This indicates that the lower SA levels in AtFAAH overex-
pressors contributed at least partially to the heightened
sensitivity to pathogens. AtFAAH overexpressors did not
compromise R-gene-mediated resistance (Figure S2), prob-
ably due to greater accumulation of SA during the resistance
response. Consistent with the reduced SA levels in AtFAAH
overexpressors was a global downregulation of SA biosyn-
thesis and SA-mediated defense signaling genes.
Recent studies have reported that lipid-based molecules
are signals for immunity in plants (Weber, 2002). Genetic
screens for Arabidopsis mutants with altered defense
responses have identified mutations in several genes puta-
tively involved in lipid metabolism (Shah, 2005). Also, work
with the Arabidopsis ssi2 (suppressor of SA-insensitivity 2)
mutant showed that deficit of 18:1 oleic acid led to constit-
utive activation of the SA-mediated resistance pathway
while JA signaling was impaired (Kachroo et al., 2001).
The enhanced susceptibility phenotype accompanied by
suppression of the SA pathway in AtFAAH overexpressors is
the reverse of the ssi2 mutant, and continues to support the
strong connection between lipid metabolism and plant
defense.
Changes in hormone levels were induced by NAE20:4 and
THC and were reported to affect spermatogenesis, fertiliza-
tion and embryo development in animals (Wang et al.,
2006a). Here, we found that overexpressing AtFAAH affected
hormone levels in plants. As mentioned above, levels of
endogenous free SA were at least two-fold lower in AtFAAH
overexpressors when compared with Col-0 (Figure 3). Sur-
FAAH overexpression compromises innate immunity 345
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

prisingly, after pathogen infection, the free SA levels
increased by more than two-fold in the AtFAAH overexpres-
sor when compared with Col-0. However, the levels of
conjugated and total SA still remained lower in the AtFAAH
overexpressor (Figure 3). Even though it has been proposed
that free SA is the mobile form of SA (Enyedi et al., 1992a,b),
it is still not clear whether conjugated SA also plays a role in
plant defense. Jasmonic acid is another phytohormone that
is known to play a role in plant defense. It has been well
documented that in plants, JA- and SA-mediated defense
responses act antagonistically (Clarke et al., 2000; Dong,
1998). Consistent with these earlier observations, we found
that transcripts of plant defensin genes PDF1.2a and PDF1.2b
(marker genes for JA signaling) were, constitutively and at
6 hpi with a non-host pathogen, more abundant in the
AtFAAH overexpressor when compared with Col-0 (Figure 5,
Table S3). However, we found that transcripts of several JA
biosynthetic genes were less abundant in the AtFAAH
overexpressor when compared with Col-0, and this was
consistent with our observation that AtFAAH overexpressors
had less JA (Figure S4). It has been observed that AtFAAH
overexpressors were hypersensitive to the phytohormone
ABA, and exogenous application of NAE12:0 induced
expression of a number of ABA-responsive genes in
Arabidopsis (Teaster et al., 2007). Recent literature suggests
a role for the ABA signaling pathway in susceptibility of
plants to disease caused by the host pathogen P. s. tomato
(Mauch-Mani and Mauch, 2005; Robert-Seilaniantz et al.,
2007; de Torres-Zabala et al., 2007). We found that the ABA
levels in AtFAAH overexpressors were about two-fold less
than in Col-0 (Figure S4B). Interestingly, ABA levels were
about two-fold less in Col-0 treated with a non-host patho-
gen when compared with buffer-treated Col-0, suggesting a
role for ABA in non-host resistance. An earlier report by
Cahill and Ward (1989) has shown that decrease in ABA
concentration occurred only in the incompatible soybean–
Phytophthora megasperma f. sp. glycinea interactions. As
reported earlier (Wang et al., 2006b), AtFAAH overexpres-
sors have enhanced vegetative growth compared with Col-0
plants, suggesting a possible involvement of growth
hormones such as auxin and cytokinin. All these data
together suggest an intricate role for AtFAAH and NAEs in
the phytohormone signaling network.
Apart from genes involved in phytohormone synthesis
and signaling, transcript profiling revealed that transcripts of
several other plant defense-related genes were found to be
less abundant in the AtFAAH overexpressor when compared
with Col-0. Plants encode a large number of R proteins that
contain a region of LRRs, a putative nucleotide binding site,
an N-terminal leucine zipper and coiled-coil or Toll and
interleukin-1 receptor domains (Martin et al., 2003). Tran-
scripts of a large number of putative and well-characterized
R genes or their homologs were less abundant in the
AtFAAH overexpressor compared with Col-0 (Table S2,
Figure 5). Further investigation is needed to understand
the biological implications of downregulation of these R
genes in AtFAAH overexpressors.
Similar to its mammalian counterpart, AtFAAH–GFP
fusion proteins were associated with the cell membrane
(Figure 6c,d). The membrane localization of FAAH is
required to facilitate channeling of fatty acid amide (FAA)
through membrane, and is considered to be important for its
function (McKinney and Cravatt, 2005). In the future, we will
attempt to express AtFAAH under the control of the native
promoter since overexpression using the 35S promoter
could lead to mislocalization of the protein. Nonetheless, our
observation that AtFAAH–GFP is localized to the ER is
consistent with localization studies of mammalian FAAH
(Oddi et al., 2005). The conservation of the function and
subcellular localization of FAAH in plants and animals
suggested the possibility of a similar signal transduction
system. Identification of the NAE receptors will lead to
further understanding of the role of AtFAAH and NAEs in
plant defense.
In conclusion, we have presented data showing compro-
mised immune responses of plants overexpressing AtFAAH.
The hypersensitivity of AtFAAH overexpressors to a number
of host and non-host pathogens can be explained in part by
alterations in phytohormone synthesis and signaling.
Although there are obviously a number of other molecular
targets through which NAE and FAAH exert their effects, our
study presents an important entry point for probing further
into the mechanisms of functioning of NAE in plant immu-
nity and adds to a growing list of plant processes that are
clearly influenced by this understudied group of lipids.
Experimental procedures
Plant material and bacterial pathogens
Arabidopsis thaliana wild-type Col-0 and transgenic 35S::AtFAAHlines (OE2a-1, OE7a-1 and OE11a-1) were grown in growthchambers at 20�C under short-day condition (8 h light). Four- tofive-week-old plants were used for inoculation in all experiments.Bacterial pathogens were grown in King’s B (KB) medium contain-ing appropriate antibiotics at 30�C.
Pathogen inoculation and RNA isolation
For bacterial inoculation, bacteria were grown in KB medium withappropriate antibiotics for 16 h and were pelleted by centrifugationat 3000 g. Bacterial pellets were washed three times and suspendedin sterile water. Leaves were inoculated with a needle-less syringefor symptom development and growth assay. Leaf disks frominoculated leaves were ground in water and serial dilutions of eachsample were plated on a KB agar plate for quantification of patho-gens in plant tissues. Leaves were inoculated by spray inoculationfor some experiments. To reduce variation in infection, vacuuminfiltration was used for experiments involving microarray analysisand quantification of SA/JA/ABA. A final concentration of 0.01% of
346 Li Kang et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

Silwet L-77 (OSI, Danbury, CT) was added to facilitate infection inspraying and vacuum infiltration experiments. Leaf tissues col-lected from at least 10 individual plants at designated times werepooled for RNA extraction. Total RNA was extracted and purifiedusing the RNeasy MinElute Kit (Qiagen, http://www1.qiagen.com/),according to the manufacturer’s instructions.
Quantification of NAE
An average of 4–5 g of mature leaf material was collected from eachof the control (0 h) and infiltrated plants. The treated plants werevacuum infiltrated with buffer control or P. s. syringae at5 · 106 CFU ml)1. To reduce variations, only fully expanded matureleaves were harvested for analysis. The NAE extraction, enrichmentand identification/quantification by gas chromatography/massspectrometry (GC/MS) were performed as described (Venableset al., 2005) with a few modifications (see Appendix S1).
Quantification of SA, JA and ABA
Free SA was extracted from Arabidopsis leaves (�300 mg) and wasanalyzed as described (Schmelz et al., 2003; Appendix S1). The SAand JA were separated on a RTx-5 column (30 m · 0.25 mm ·0.25 mm, Restek, http://www.restek.com/) using conditions des-cribed earlier (Uppalapati et al., 2005). Free SA was quantified fromthe methylene chloride:1 propanol layer. The remaining (top) waterphase was used for the extraction of conjugated SA. To quantifyconjugated SA, the water phase was acid hydrolyzed (37% con-centrated HCl) for 1 h at 95�C to release free SA. The sample wascooled to room temperature, and after adding an internal standardwas re-extracted and quantified for free SA as described (Schmelzet al., 2003). The SA and JA quantification experiments wererepeated at least three times with independent samples using threetechnical replicates during each experiment. The ABA was extractedusing methods similar to those described by Walker-Simmonset al., 2000; Appendix S1).
Microarray analyses
The Affymetrix GeneChip� Arabidopsis ATH1Genome Array(Affymetrix, http://www.affymetrix.com/) was used in the expres-sion analysis. For microarray experiments, total RNA was extractedfrom leaf tissues from two independent biological replicates. Foreach replicate, inoculated leaves from eight plants were pooled forRNA isolation, and 10 lg of total RNA was used for microarraylabeling reaction. Labeling, hybridization and scanning wereperformed according to the manufacturer’s instructions (Affyme-trix). For every chip, the signal level for each gene was obtainedfrom 11 probe sets and data were collected and normalized againstbackground using MAS5 implemented in GCOS1.4 (Affymetrix).Data normalization between chips was conducted using robustregression (Dozmorov and Centola, 2003). To select genes differ-entially expressed between treatment conditions, an expressionratio of 2 and above with a cut-off P-value (Student’s t-test) of 0.05were used as the selection criteria. The ‘presence/absence’ calldetermined by the MAS5 algorithm for each gene signal was alsoused as a criterion for selection, and only genes whose signals weretruly present in the samples were selected.
For clustering analyses, ratios of selected genes were used.Hierarchical clusters were generated for genes using Ward’s method(Ward, 1963) available in Spotfire DecisionSite 8.2.1 (Spotfire Inc.,http://spotfire.tibco.com/). Gene ontology analysis for selected
genes was obtained using web-based tools at TAIR for functioncategory (http://arabidopsis.org/tools/bulk/go/index.jsp).
Real-time quantitative RT-PCR analysis
The RT reaction was performed with 2 lg of total RNA and 0.5 lg ofpoly (dT)16 primer at 37�C using Omniscript RT Kit (Qiagen) andRNase inhibitor (Promega, http://www.promega.com/) in a finalvolume of 20 ll. Gene-specific primers for PCR were designed usingthe PRIMEREXPRESS software (Applied Biosystems, http://www3.appliedbiosystems.com/) and synthesized by Integrated DNATechnologies (http://eu.idtdna.com/). Primer sequences are listed inTable S5.
Quantitative RT-PCR was performed with an ABI PRISM 7900(Applied Biosystems) using PowerSYBR Green master mix (AppliedBiosystems). Relative quantification of PCR products was per-formed via a standard curve method using Arabidopsis ElongationFactor 1a as an endogenous control. The relative expression levelsof designated calibrator (Col-0, buffer treated at 0 h) were analyzedaccording to the manufacturer’s instructions (user’s bulletin no. 2;Applied Biosystems).
Localization of AtFAAH–GFP
Arabidopsis plants (ecotype Col-0) and faah knockouts were trans-formed with 35S::AtFAAH-GFP construct (see Appendix S1) usingthe floral dip method (Clough and Bent, 1998). Green fluorescentprotein from the epidermal cells of leaves from 4-week-old plantswas imaged with a Leica TCS SP2 AOBS laser confocal scanningmicroscope (Leica Microsystems, http://www.leica-microsys-tems.com/) using the 488-nm line of an Ar laser and emission wasdetected at 520 nm. Leaves were counterstained with membranedye FM4-64 and imaged using the 543 nm line of a He/Ne laser.
Acknowledgements
We thank Drs Jian-Min Zhou and Xiaoyan Tang for various bacterialstrains, Dr Carol Bender for the GC column and internal standardsfor SA quantification and Drs Jyoti Shah and Marilyn Roossinck forcritical review of the manuscript. This work was supported by theSamuel Roberts Noble Foundation and in part by the Department ofEnergy (DOE) Biosciences (DE-FG02-05ER15647).
Supporting Information
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Spray inoculation symptoms.Figure S2. RPS2-AvrRpt2-mediated resistance is not affected inAtFAAH overexpressors.Figure S3. Quantification of N-acylethanolamine in AtFAAH over-expressor and Col-0.Figure S4. Quantification of jasmonic acid and ABA in AtFAAHoverexpressor and Col-0.Figure S5. Heat maps of differentially expressed genes.Figure S6. Functional category of differentially expressed genes.Table S1. Differentially expressed defense related genes.Table S2. Toll and interleukin-1 receptor/coiled-coil (TIR/CC)–nucle-otide binding site (NBS) – leucine-rich repeat (LRR) type of R genesdifferentially expressed.Table S3. Comparison of microarray and real-time qRT-PCR data.Table S4. Expression values of list of salicylic acid-related genes.
FAAH overexpression compromises innate immunity 347
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

Table S5. Primer sets used for real-time quantitative RT-PCR.Appendix S1. Supplementary methods.Please note: Wiley-Blackwell Publishing are not responsible for thecontent or functionality of any supporting materials supplied by theauthors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
References
Austin-Brown, S.L. and Chapman, K.D. (2002) Inhibition of phos-pholipase D alpha by N-acylethanolamines. Plant Physiol. 129,1892–1898.
Bais, H.P., Prithiviraj, B., Jha, A.K., Ausubel, F.M. and Vivanco, J.M.
(2005) Mediation of pathogen resistance by exudation of antimi-crobials from roots. Nature, 434, 217–221.
Berdyshev, E.V., Schmid, P.C., Dong, Z. and Schmid, H.H. (2000)Stress-induced generation of N-acylethanolamines in mouseepidermal JB6 P+ cells. Biochem. J. 346, 369–374.
Berdyshev, E.V., Schmid, P.C., Krebsbach, R.J., Kuwae, T., Huang,
C., Ma, W.Y., Dong, Z. and Schmid, H.H. (2001) Role ofN-acylethanolamines in cell signaling. World Rev. Nutr. Diet. 88,207–214.
Berger, C., Schmid, P.C., Schabitz, W.R., Wolf, M., Schwab, S. and
Schmid, H.H. (2004) Massive accumulation of N-acylethanolam-ines after stroke. Cell signalling in acute cerebral ischemia?J. Neurochem. 88, 1159–1167.
Blancaflor, E.B. and Chapman, K.D. (2006) Similarities betweenendocannabinoid signaling in animal systems and N-acyletha-nolamine metabolism in plants. In Communication in Plants –Neuronal Aspects of Plant Life (Baluska, F., Mancuso, S. andVolkmann, D., eds). Heidelberg, Germany: Springer Verlag,pp. 205–219
Blancaflor, E.B., Hou, G. and Chapman, K.D. (2003) Elevated levels ofN-lauroylethanolamine, an endogenous constituent of desiccatedseeds, disrupt normal root development in Arabidopsis thalianaseedlings. Planta, 217, 206–217.
Buckley, N.E., Burbridge, D., Buranapramest, M., Ferguson, T. and
Paau, R.Y. (2006) Experimental methods to study the role of theperipheral cannabinoid receptor in immune function. MethodsMol. Med. 123, 19–40.
Cahill, D.M. and Ward, E.W.B. (1989) Rapid localized changes inabscisic acid concentrations in soybean in interactions withPhytophthora megasperma f. sp. glycinea or after treatment withelicitors. Physiol. Mol. Plant Pathol. 35, 4583–4593.
Chapman, K.D. (2000) Emerging physiological roles for N-acyl-phosphatidylethanolamine metabolism in plants: signaltransduction and membrane protection. Chem. Phys. Lipids, 108,221–229.
Chapman, K.D. (2004) The occurrence, metabolism and prospectivefunctions of N-acylethanolamines in plants. Prog. Lipid Res. 43,302–327.
Chapman, K.D., Tripathy, S., Venables, B. and Desouza, A.D. (1998)N-Acylethanolamines: formation and molecular composition of anew class of plant lipids. Plant Physiol. 116, 1163–1168.
Clarke, J.D., Volko, S.M., Ledford, H., Ausubel, F.M. and Dong, X.
(2000) Roles of salicylic acid, jasmonic acid, and ethylene incpr-induced resistance in Arabidopsis. Plant Cell, 12, 2175–2190.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16, 735–743.
Cravatt, B.F., Saghatelian, A., Hawkins, E.G., Clement, A.B., Bracey,
M.H. and Lichtman, A.H. (2004) Functional disassociation of thecentral and peripheral fatty acid amide signaling systems. Proc.Natl Acad. Sci. USA, 101, 10821–10826.
Devane, W.A., Hanus, L., Breuer, A., Pertwee, R.G., Stevenson, L.A.,
Griffin, G., Gibson, D., Mandelbaum, A., Etinger, A. and
Mechoulam, R. (1992) Isolation and structure of a brain constituentthat binds to the cannabinoid receptor. Science, 258, 1946–1949.
DeYoung, B.J. and Innes, R.W. (2006) Plant NBS-LRR proteins inpathogen sensing and host defense. Nat. Immunol. 7, 1243–1249.
Dong, X. (1998) SA, JA, ethylene, and disease resistance in plants.Curr. Opin. Plant Biol. 1, 316–323.
Dozmorov, I. and Centola, M. (2003) An associative analysis of geneexpression array data. Bioinformatics, 19, 204–211.
Durrant, W.E. and Dong, X. (2004) Systemic acquired resistance.Annu. Rev. Phytopathol. 42, 185–209.
Enyedi, A.J., Yalpani, N., Silverman, P. and Raskin, I. (1992a)Localization, conjugation, and function of salicylic acid in tobaccoduring the hypersensitive reaction to tobacco mosaic virus. Proc.Natl Acad. Sci. USA, 89, 2480–2484.
Enyedi, A.J., Yalpani, N., Silverman, P. and Raskin, I. (1992b) Signalmolecules in systemic plant resistance to pathogens and pests.Cell, 70, 879–886.
He, P., Shan, L., Lin, N.C., Martin, G.B., Kemmerling, B.,
Nurnberger, T. and Sheen, J. (2006) Specific bacterial suppres-sors of MAMP signaling upstream of MAPKKK in Arabidopsisinnate immunity. Cell, 125, 563–575.
Kachroo, P., Shanklin, J., Shah, J., Whittle, E.J. and Klessig, D.F.
(2001) A fatty acid desaturase modulates the activation of defensesignaling pathways in plants. Proc. Natl Acad. Sci. USA, 98, 9448–9453.
Kilaru, A., Blancaflor, E.B., Venables, B.J., Tripathy, S., Mysore, K.S.
and Chapman, K.D. (2007) The N-acylethanolamine-mediatedregulatory pathway in plants. Chem. Biodivers. 4, 1933–1955.
Komis, G., Quader, H., Galatis, B. and Apostolakos, P. (2006)Macrotubule-dependent protoplast volume regulation in plas-molyzed root-tip cells of Triticum turgidum: involvement ofphospholipase D. New Phytol. 171, 737–750.
Kozak, K.R., Crews, B.C., Morrow, J.D., Wang, L.H., Ma, Y.H., Wein-
ander, R., Jakobsson, P.J. and Marnett, L.J. (2002) Metabolism ofthe endocannabinoids, 2-arachidonylglycerol and anandamide,into prostaglandin, thromboxane, and prostacyclin glycerol estersand ethanolamides. J. Biol. Chem. 277, 44877–44885.
Lawton, K.A., Friedrich, L., Hunt, M., Weymann, K., Delaney, T.,
Kessmann, H., Staub, T. and Ryals, J. (1996) Benzothiadiazoleinduces disease resistance in Arabidopsis by activation of thesystemic acquired resistance signal transduction pathway. PlantJ. 10, 71–82.
Lopez-Bucio, J., Acevedo-Hernandez, G., Ramirez-Chavez, E.,
Molina-Torres, J. and Herrera-Estrella, L. (2006) Novel signals forplant development. Curr. Opin. Plant Biol. 9, 523–529.
Martin, G.B., Bogdanove, A.J. and Sessa, G. (2003) Understandingthe functions of plant disease resistance proteins. Annu. Rev.Plant Biol. 54, 23–61.
Mauch-Mani, B. and Mauch, F. (2005) The role of abscisicacid in plant-pathogen interactions. Curr. Opin. Plant Biol. 8, 409–414.
McKinney, M.K. and Cravatt, B.F. (2005) Structure and func-tion of fatty acid amide hydrolase. Annu. Rev. Biochem. 74, 411–432.
Mellersh, D.G. and Heath, M.C. (2003) An investigation into theinvolvement of defense signaling pathways in components of thenonhost resistance of Arabidopsis thaliana to rust fungi alsoreveals a model system for studying rust fungal compatibility.Mol. Plant Microbe Interact. 16, 398–404.
Munro, S., Thomas, K.L. and Abu-Shaar, M. (1993) Molecular char-acterization of a peripheral receptor for cannabinoids. Nature,365, 61–65.
348 Li Kang et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349

Mysore, K.S. and Ryu, C.M. (2004) Nonhost resistance: how muchdo we know? Trends Plant Sci. 9, 97–104.
Oddi, S., Bari, M., Battista, N., Barsacchi, D., Cozanni, I. and
Maccarrone, M. (2005) Confocal microscopy and biochemicalanalysis reveal spatial and functional separation betweenanandamide uptake and hydrolysis in human keratinocytes. Cell.Mol. Life Sci. 62, 386–395.
Peart, J.R., Lu, R., Sadanandom, A. et al. (2002) Ubiquitin ligase-associated protein SGT1 is required for host and nonhostdisease resistance in plants. Proc. Natl Acad. Sci. USA, 99,10865–10869.
Robert-Seilaniantz, A., Navarro, L., Bari, R. and Jones, J.D. (2007)Pathological hormone imbalances. Curr. Opin. Plant Biol. 10, 372–379.
Salzet, M., Breton, C., Bisogno, T. and Di Marzo, V. (2000) Com-parative biology of the endocannabinoid system possible role inthe immune response. Eur. J. Biochem. 267, 4917–4927.
Schmelz, E.A., Engelberth, J., Alborn, H.T., O’Donnell, P.,
Sammons, M., Toshima, H. and Tumlinson, J.H. III (2003)Simultaneous analysis of phytohormones, phytotoxins, andvolatile organic compounds in plants. Proc. Natl Acad. Sci. USA,100, 10552–10557.
Schmid, P.C., Wold, L.E., Krebsbach, R.J., Berdyshev, E.V. and
Schmid, H.H. (2002) Anandamide and other N-acylethanolaminesin human tumors. Lipids, 37, 907–912.
Shah, J. (2003) The salicylic acid loop in plant defense. Curr. Opin.Plant Biol. 6, 365–371.
Shah, J. (2005) Lipids, lipases, and lipid-modifying enzymes in plantdisease resistance. Annu. Rev. Phytopathol. 43, 229–260.
Shrestha, R., Noordermeer, M.A., van der Stelt, M., Veldink, G.A.
and Chapman, K.D. (2002) N-acylethanolamines are metabolizedby lipoxygenase and amidohydrolase in competing pathwaysduring cottonseed imbibition. Plant Physiol. 130, 391–401.
Shrestha, R., Dixon, R.A. and Chapman, K.D. (2003) Molecularidentification of a functional homologue of the mammalian fattyacid amide hydrolase in Arabidopsis thaliana. J. Biol. Chem. 278,34990–34997.
Teaster, N.D., Motes, C.M., Tang, Y. et al. (2007) N-Acylethanol-amine metabolism interacts with abscisic acid (ABA) signaling inArabidopsis thaliana. Plant Cell, 19, 2454–2469.
de Torres-Zabala, M., Truman, W., Bennett, M.H., Lafforgue, G.,
Mansfield, J.W., Rodriguez Egea, P., Bogre, L. and Grant, M.
(2007) Pseudomonas syringae pv. tomato hijacks the Arabidopsisabscisic acid signaling pathway to cause disease. EMBO J. 26,1434–1443.
Tripathy, S., Venables, B.J. and Chapman, K.D. (1999) N-Acyletha-nolamines in signal transduction of elicitor perception. Attenua-tion of alkalinization response and activation of defense geneexpression. Plant Physiol. 121, 1299–1308.
Tripathy, S., Kleppinger-Sparace, K., Dixon, R.A. and Chapman, K.D.
(2003) N-acylethanolamine signaling in tobacco is mediated by amembrane-associated, high-affinity binding protein. Plant Phys-iol. 131, 1781–1791.
Tsuboi, K., Sun, Y.X., Okamoto, Y., Araki, N., Tonai, T. and Ueda, N.
(2005) Molecular characterization of N-acylethanolamine-hydro-lyzing acid amidase, a novel member of the choloylglycinehydrolase family with structural and functional similarity to acidceramidase. J. Biol. Chem. 280, 11082–11092.
Ueda, N., Yamamoto, K., Yamamoto, S. et al. (1995) Lipoxygenase-catalyzed oxygenation of arachidonylethanolamide, a cannabi-noid receptor agonist. Biochem. Biophys. Acta, 1254, 127–134.
Uppalapati, S.R., Patricia, A., Weng, H.P., Palmer, D.A., Mitchell,
R.E., Jones, W. and Bender, C.L. (2005) The phytotoxin coronatineand methyl jasmonate impact multiple phytohormone pathwaysin tomato. Plant J. 42, 201–217.
Venables, B.J., Waggoner, C.A. and Chapman, K.D. (2005) N-Acy-lethanolamines in selected legumes. Phytochemistry, 66, 1913–1918.
Walker-Simmons, M.K., Rose, P.A., Hogge, L.R. and Abrams, S.R.
(2000) Abscisic acid (ABA) immunoassay and gas chromatogra-phy/mass spectrometry verification. In Methods in MolecularBiology, Vol. 141: Plant Hormone Protocols (Tucker, G.A. andRoberts, J.A., eds). Totowa, NJ: Humana Press Inc., pp. 33–47
Wang, H., Dey, S.K. and Maccarrone, M. (2006a) Jekyll and hyde:two faces of cannabinoid signaling in male and female fertility.Endocr. Rev. 27, 427–448.
Wang, Y.S., Shrestha, R., Kilaru, A., Wiant, W., Venables, B.J.,
Chapman, K.D. and Blancaflor, E.B. (2006b) Manipulation ofArabidopsis fatty acid amide hydrolase expression modifies plantgrowth and sensitivity to N-acylethanolamines. Proc. Natl Acad.Sci. USA, 103, 12197–12202.
Ward, J.H. (1963) Hierarchical grouping to optimize an objectivefunction. J. Am. Stat. Assoc. 58, 236–244.
Weber, H. (2002) Fatty acid-derived signals in plants. Trends PlantSci. 7, 217–224.
Wei, B.Q., Mikkelsen, T.S., McKinney, M.K., Lander, E.S. and Crav-
att, B.F. (2006) A second fatty acid amide hydrolase with variabledistribution among placental mammals. J. Biol. Chem. 281,36569–36578.
Zhang, Y., Fan, W., Kinkema, M., Li, X. and Dong, X. (1999) Inter-action of NPR1 with basic leucine zipper protein transcriptionfactors that bind sequences required for salicylic acid induction ofthe PR-1 gene. Proc. Natl Acad. Sci. USA, 96, 6523–6528.
Zhang, Y., Tessaro, M.J., Lassner, M. and Li, X. (2003) Knockoutanalysis of Arabidopsis transcription factors TGA2, TGA5, andTGA6 reveals their redundant and essential roles in systemicacquired resistance. Plant Cell, 15, 2647–2653.
FAAH overexpression compromises innate immunity 349
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 56, 336–349
Copyright © 2022 FDOKUMEN




















