
Assessment of standing wood and fiber quality using ground and airborne laser scanning: A review
51
1 Assessment of standing wood and fiber quality using ground and airborne laser scanning: a review Martin van Leeuwen 1* , Thomas Hilker 1 , Nicholas C. Coops 1 , Gordon Frazer 2 , Michael A. Wulder 2 , Glenn J. Newnham 3 , Darius S. Culvenor 3 1-Faculty of Forest Resources Management, University of British Columbia, 2424 Main Mall, Vancouver, BC, V6T 1Z4, Canada. 2-Canadian Forest Service (Pacific Forestry Centre), Natural Resources Canada, 506 West Burnside Road, Victoria, BC, V8Z 1M5, Canada. 3-CSIRO Sustainable Ecosystems. Private Bag 10, Clayton South 3189, Australia. (*) corresponding author: Martin van Leeuwen Phone: (604) 822 6452, Fax (604) 822-9106, Email: [email protected] Pre-print of published version. Reference: Van Leeuwen, M., Hilker, T., Coops, N.C., Frazer, G., Wulder, M.A., Newnham, G.J., Culvenor, D.S. 2011. Assessment of standing wood and fiber quality using ground and airborne laser scanning: A review. Forest Ecology and Management. 261: 1467-1478. DOI: doi:10.1016/j.foreco.2011.01.032 Disclaimer: The PDF document is a copy of the final version of this manuscript that was subsequently accepted by the journal for publication. The paper has been through peer review, but it has not been subject to any additional copy-editing or journal specific formatting (so will look different from the final version of record, which may be accessed following the DOI above depending on your access situation).
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Assessment of standing wood and fiber quality using ground and airborne laser scanning: A review

1
Assessment of standing wood and fiber quality using ground and
airborne laser scanning: a review
Martin van Leeuwen1*
, Thomas Hilker1
, Nicholas C. Coops1
, Gordon Frazer2
, Michael
A. Wulder2
, Glenn J. Newnham3, Darius S. Culvenor
3
1-Faculty of Forest Resources Management, University of British Columbia, 2424 Main
Mall, Vancouver, BC, V6T 1Z4, Canada.
2-Canadian Forest Service (Pacific Forestry Centre), Natural Resources Canada, 506
West Burnside Road, Victoria, BC, V8Z 1M5, Canada.
3-CSIRO Sustainable Ecosystems. Private Bag 10, Clayton South 3189, Australia.
(*) corresponding author:
Martin van Leeuwen
Phone: (604) 822 6452, Fax (604) 822-9106, Email: [email protected]
Pre-print of published version. Reference: Van Leeuwen, M., Hilker, T., Coops, N.C., Frazer, G., Wulder, M.A., Newnham, G.J., Culvenor, D.S. 2011. Assessment of standing wood and fiber quality using ground and airborne laser scanning: A review. Forest Ecology and Management. 261: 1467-1478. DOI: doi:10.1016/j.foreco.2011.01.032 Disclaimer: The PDF document is a copy of the final version of this manuscript that was subsequently accepted by the journal for publication. The paper has been through peer review, but it has not been subject to any additional copy-editing or journal specific formatting (so will look different from the final version of record, which may be accessed following the DOI above depending on your access situation).

2
ABSTRACT
Accurate information on the wood-quality characteristics of standing timber and logs is
needed to optimize the forest production value chain and to assess the potential of
forest resources to meet other services. Physical and chemical characteristics of wood
vary with both tree and site characteristics. At the tree scale, crown development, stem
shape and taper, branch size and branch location, knot size, type and placement, and
age all influence wood properties. More broadly, at the stand level, stocking density,
moisture, nutrient availability, climate, competition, disturbance, and stand age have
also been identified as key determinants of wood quality. Such information is often
captured in polygon based forest inventory data. Other terrain-related spatial
information, such as elevation, slope and aspect, can improve assessments of site
conditions and limitations upon plant growth which impact wood quality. Light Detection
And Ranging (LiDAR) is an emerging technology, which directly measures the three-
dimensional structure of forest canopies using ground or airborne laser instruments, and
can provide highly accurate information on individual-tree and stand-level forest
structure. In this paper, we explore the potential of LiDAR and other geospatial
information sources to model and predict wood quality based on individual-tree and
stand structural metrics. We identify a number of key wood quality attributes (i.e., basic
wood density, cell perimeter, cell coarseness, tracheid length, and microfibril angle) and
demonstrate links between these properties and forest structure and site attributes.
Finally, the potential for using LiDAR in combination with other geospatial information
sources to predict wood quality in standing timber is discussed.
Key words: wood quality, fiber properties, lidar, remote sensing

3
1. INTRODUCTION
An increased focus on sustainable forestry requires forest managers to consider both
ecological and socio-economic conditions, as well as to anticipate future needs under a
changing global climate (Sayer et al., 1997). This includes recognition of the numerous
competing ecosystem services of the forest resource, such as timber production and
other non-timber forest values, such as carbon sequestration, clean water production,
regulation of various ecological processes, and biodiversity. To comply with sustainable
forest management principles, forest management has seen a requirement to move
towards harvesting trees from younger second-growth forests and plantations grown in
shorter rotations, while conserving mature first-growth forests. As a result mills now
receive and are required to process logs with a wider range of wood fiber properties. To
improve wood utilization and production efficiencies, forest managers and mill owners
need to understand how wood quality varies throughout the stem and across a forested
landscape, so that harvested logs can be matched with requirements (Zobel and Van
Buijtenen, 1989). In addition, wood density is a key input needed for accurate estimation
of forest biomass and carbon storage (Freyburger et al., 2009; Vallet et al., 2006), and
such information has potential to improve carbon accounting under schemes such as
REDD (Reducing Emissions from Deforestation and Degradation) (Herold et al., 2007).
For instance, the value of structural timber is often determined by wood properties, such
as stiffness, straightness, and stability, which are largely governed by wood density and
driven by radial (pith-to-bark) and axial (top-to-bottom) variations in cell anatomical and
chemical properties. Information on these characteristics can thus help optimize the
forest product value chain, as well as aid the decision making process around the

4
multiple forest ecosystem services, such as biodiversity conservation and carbon
storage and accumulation.
The physical and chemical characteristics of timber vary with species, tree, stand, and
site characteristics (Wilhelmsson et al., 2002; Väisänen et al., 1989; Zobel and Van
Buijtenen, 1989) and change with tree age and stand development (Downes and Drew,
2008; Kint et al., 2010). Hardwood species have a much higher wood density than
softwood species (e.g., oak species can range from 590 to 930 kg m-3
, while western
redcedar is approximately 380 kg m-3
). At the tree scale, knot size, type and placement
(Duchesne et al., 1997), crown development (Mansfield et al., 2007; Lyhykäinen et al.,
2009), branch size and branch placement (Mäkinen and Colin, 1998), stem diameter,
ring width and cambial age (Amarasekara and Denne, 2002; Wilhelmsson et al., 2002)
of the wood have all been shown to influence wood fiber properties. More broadly, at the
stand scale, stocking density, thinning (commercial and pre-commercial), moisture and
nutrient availability (including fertilization), climate, competition, and disturbance are
also important determinants of wood properties, with responses varying by location,
species and clones (Watt et al., 2006; Wilhelmsson et al. 2002; Ramirez et al., 2009).
These characteristics change over time as the result of stand development, branch
mortality, and occlusion (Kint et al., 2010; Hein et al., 2008). As a result, relationships
between wood fiber properties and individual tree and stand scale structure and its
development over time may provide an efficient way to map wood quality over broad
areas (Duchesne et al., 1997; Wilhelmsson et al., 2002; Lindström, 1997; Zobel and
Van Buijtenen, 1989).

5
One technology that can provide information on tree and stand level structure is Light
Detection And Ranging (LiDAR). LiDAR measures the three-dimensional (3-D) structure
of forests using the measured time-of-flight of laser pulses reflected from a target of
interest, while simultaneously recording position and orientation of the scanning device
(Lim et al. 2003). LiDAR instruments can be distinguished into discrete and full
waveform systems. Discrete- return systems record single- or multiple-returns from
objects that are located within the path of a single laser beam (e.g. the forest canopy,
understorey, and ground), while full- waveform instruments record the fully digitized
profile of reflected energy from objects illuminated by the laser. LiDAR data typically
represent forest structure as a 3-D point cloud, from which a range of forest inventory
attributes can be derived such as leaf area index, foliage height profiles, tree diameter,
tree or stand height, and biomass (Coops et al. 2007; Lim et al. 2003; Van Leeuwen et
al. 2010). While LiDAR has been extensively investigated and applied operationally in
the context of forest inventory (Næsset et al. 2004; Wulder et al. 2008), its potential to
derive wood quality attributes through links with tree, stand, and terrain-related indices
is a new field of research.
In this paper, we review the current state of knowledge of predicting wood quality using
tree and stand structural characteristics. First, a number of the wood fiber properties are
described and discussed in relation to their relevance to timber production and carbon
sequestration. The links between fiber properties and tree and stand level indicators are
then investigated. Finally, the potential for using both airborne and terrestrial LiDAR to

6
determine these characteristics is discussed and a suite of possible tree and stand level
structural attributes is proposed.
2. Wood anatomy and its radial and axial variation
Wood (secondary xylem) is primarily composed of cellulose, hemicelluloses, and lignin.
Of these, cellulose is the major component of wood, accounting for up to 50% of the
wood’s constituents (Raven et al. 2005). Cellulose molecules are arranged structurally
into fibrils, which in turn are organized into larger structuring elements that build the cell
wall. Lignin is the second most prevalent constituent of wood and acts as a cementing
agent between individual cells. Lignin content varies most notably between softwoods
(25-33%) and hardwoods (18-25%) and can be found throughout the cell walls (Bowyer
et al. 2007). Hemicellulose provides the critical link between the cellulose and lignin
polymers. Finally, extraneous materials can be divided into organic and inorganic
components. Organic components (e.g. oils, fats, resins, tannins, waxes, gum, and
starch) contribute to color, odor, density, resistance against decay, hygroscopy, and
flammability of wood (Twede and Selke 2005). The most abundant inorganic
constituents are calcium, potassium, nitrogen, phosphorus, and magnesium; however,
trace amounts of sodium, iron, silicon, manganese, copper, and zinc are also commonly
found (Raven et al. 2005).
Throughout the stem, these constituents are highly organized within woody cells to fulfill
a broad spectrum of functions, including mechanical support, water transport,
photosynthate translocation, and ultimately, growth. The cross-section of a stem is

7
mainly composed of bark, cambium, sapwood, heartwood, and pith. The cambium is
responsible for secondary (diameter) growth and produces phloem to the outside and
secondary xylem cells to the inside (Zobel and Van Buijtenen, 1989; Raven et al. 2005).
The outer bark protects the tree against disease, damage, and dehydration, and its
thickness can vary widely with species and age. The inner bark acts as a pipeline
carrying photosynthate from leaves to the vascular cambium (secondary meristem) and
other living parts of the tree. In gymnosperms, the secondary xylem is mostly composed
of tracheids, ray cells, and resin canals. Angiosperms are more complex in their
structure, and additionally contain vessels, fibers, and longitudinal parenchyma (Zobel
and Van Buijtenen, 1989; Raven et al. 2005). Although tracheids and fibers differ in
function, both are often considered to be wood fibers (Zobel and Van Buijtenen, 1989),
and this terminology will also be followed in this paper. While most of the transport of
water, nutrients, and chemicals occurs vertically through tracheids and vessels, some
movement of resources takes place radially through the ray cells. In temperate climates,
where there are distinct growing seasons, new wood production results in annual
growth rings, most often with larger cells produced in the spring (known as earlywood)
and smaller cells produced in the summer (known as latewood or summerwood) (Zobel
and Van Buijtenen, 1989). Secondary xylem can be classified as either sapwood or
heartwood. Sapwood is responsible for the transport of water and soluble mineral
nutrients from the roots upwards. As sapwood ages, ray and parenchyma cells die and
sapwood turns into heartwood (Plomion et al. 2001). Heartwood is mainly responsible
for the stability of the tree and its high concentration of extractives provides protection
from crown rot and other forms of decay. Extractives are responsible for the darker color

8
of the heartwood when compared to the sapwood (Zobel and van Buijtenen, 1989). At
the center of the stem is the pith, a small core of soft tissue from which primary,
longitudinal growth took place. Xylem cells change markedly in their structure and
function with tree age, producing juvenile wood in the first 20 to 40 years of tree growth,
and mature wood thereafter.
Throughout the stemwood, variations in the relative amounts of cell types, chemical
composition, and cell dimensions occur between earlywood and latewood, between
heartwood and sapwood, and between species. The distribution of cell types, cell
dimensions, and chemical compositions determine wood quality attributes such as
shrink, warp, strength, stiffness and pulp and paper quality such as smoothness, and
tear strength (Cannell, 1988; Downes et al. 2002; Zobel and Van Buijtenen, 1989; Oliver
and Larson, 1996; Gartner, 2006). Genetics and environment act in concert to drive
variability in wood cell anatomical and chemical properties (Zobel and van Buijtenen,
1989; Downes and Drew, 2008; Downes et al. 2002, 2009). Genetic control over
morphological and physiological traits in plants is regulated through environmental
factors that alter plant performance and the final expression of these genetic traits (e.g.,
St. Clair, 1994a, 1994b).
3. Intrinsic and extrinsic indicators of wood quality
Assessment of wood quality can be made either through measurement of intrinsic or
extrinsic indicators. Intrinsic indicators relate to the internal, anatomical structure of
wood, and comprise supra-cellular characteristics, such as wood density, ring width, the

9
proportion of juvenile and mature wood, the percentage of sapwood and heartwood.
(Sub-) cellular characteristics, such as cell length, microfibril angle, and cell wall
thickness determine the wood’s utility for various applications. Extrinsic indicators of
wood quality relate to external characteristics of the tree, such as tree height, diameter,
stem taper, branch length, thickness, and number, and dimensions of the live crown.
Endogenous (physiology) and exogenous (ecology and environment) factors influence
wood anatomical structure and tree form. Ecological processes related to light
absorption and photosynthesis, nutrient cycling, water storage and transport,
competition (i.e., shading and crowding), and disturbance all have a profound effect on
tree growth and wood quality. Local and regional scale variations in climate and
physiography are the two primary environmental factors that control the spatiotemporal
distribution of critical resources (i.e., solar radiation, temperature, and moisture) needed
for plant growth.
To assess wood quality in standing trees, studies have explored statistical relationships
among intrinsic and extrinsic indicators, ecological and environmental site factors, and
wood quality attributes (e.g. Zobel and Van Buijtenen, 1989). Findings of these studies
form the basis of many silvicultural prescriptions designed to optimize stand productivity
(growth increment) and wood fiber characteristics and properties. However, these
studies have often been confined to relatively small experimental sites, due to budget
and time constraints; plus, comparison of results among geographical locations and
species have often been found contradictory and lacking of consistent trends (Downes
et al. 2009).

10
Recently, rapid acquisition of intrinsic wood fiber properties has become possible
through the SilviScan™ system that combines image acquisition, X-ray densitometry
and X-ray diffraction to measure wood cellular anatomy in radial and tangential planes
(Downes et al. 2002; Downes and Drew, 2008; Evans et al. 2000; Evans and Ilic, 2001).
SilviScan measures variations in wood anatomy and fiber properties from tree cores
and other wood samples, and has allowed the development of large databases of wood
anatomical properties. The wide scale collection of wood anatomical characteristics
from trees grown under a broad range of stand and site conditions facilitates research
on the statistical modeling and prediction of wood fiber properties from tree, stand, and
site measurements derived from remote sensing technology and geospatial databases.
In the following sub- section, we identify and briefly describe a number of key intrinsic
and extrinsic indicators of wood and fiber quality. Exhaustive overviews beyond the
scope of this communication can be found in Zobel and Van Buijtenen (1989) and
references therein.
3.1. Intrinsic Indicators of Wood Quality
Five intrinsic wood fiber characteristics have been widely regarded as key indicators of
wood quality in the wood science literature: wood density, cell perimeter, cell
coarseness, fiber length, and microfibril angle.
3.1.1. Basic wood density (kg/m3), defined as the ratio of dry mass to green volume
(Swenson and Enquist, 2007; Zobel and Van Buijtenen, 1989), is determined by cell

11
wall thickness, tracheid size and shape, and the ratio of latewood to earlywood. Wood
density is strongly influenced by tree age (juvenile vs. mature), the seasonal timing,
rate, and duration of secondary growth, and plays a key role in plant-level hydraulic
efficiency, protection from cavitation during drought, and mechanical support (Baas et
al. 2004; Hacke et al. 2001; Roderick and Berry, 2001; Gartner, 2006; Swenson and
Enquist, 2007). Basic wood density has been shown to vary within and among species,
geographically, and with both short- (daily) and long-term (seasonal) variations in
temperature and precipitation (Swenson and Enquist, 2007). The within-tree variability
of wood density can be observed between earlywood and latewood, where latewood is
characterized by smaller, denser wood cells (e.g. Reme and Helle, 2002), and between
juvenile and mature wood, where mature wood generally has higher density (Mansfield,
2009). Wood density is one of the most common indicators (Zobel and Van Buijtenen,
1989) of solid-wood strength and stiffness, elasticity, rupture and pulp yield (Lindström,
1996a, 1996b, 1996c; Downes and Drew, 2008).
3.1.2. Cell perimeter (μm) is calculated from the radial and tangential xylem cell
diameters (μm). Cell perimeter has a strong influence on water transport (i.e., xylem
conductivity and vulnerability to cavitation under water stress) and mechanical support
(Dean and Long, 1986; Gartner, 2006; Baas et al. 2004) and correlates with basic wood
density. Reme and Helle (2002) found that cell wall perimeters of Scandinavian grown
Scots pine and Norway spruce increased with distance from the pith, with most of the
increase occurring during the first 20 years of age, and that the perimeter gradually
decreased from earlywood to latewood. Fibers with larger perimeters and thinner walls,

12
collapse more readily and therefore make denser, stronger, and smoother paper (Wang
and Aitken, 2001; Zobel and Van Buijtenen, 1989).
3.1.3. Cell coarseness (μg/μm) is defined as the ratio of cell wall mass (μg) to cell unit
length (μm) (Gartner, 2006). Theoretical models indicate that cell coarseness is tightly
coupled to drought resilience (Hacke et al. 2001; Gartner, 2006). Cell coarseness is a
strong determinant of paper strength and stiffness (Via et al. 2004), with improved paper
qualities associated with low cell coarseness. However, a decrease in cell coarseness
correlates with lower basic wood density and improvements in paper quality attained
through lower cell coarseness comes at the expense of pulp yield (Via et al. 2004).
3.1.4. Microfibril angle (°) is defined as the angle of the cellulose microfibrils with
respect to the long axis of the individual cells (Wimmer et al., 2002), and is largely
influenced by the S2 layer of the secondary cell wall. This layer accounts for 75 - 85% of
the total cell wall thickness (Donaldson, 2008; Gierlinger and Schwanninger, 2006).
Microfibril angle (MFA) normally ranges from 5° up to 20° in outer growth rings and is
higher near the pith and in reaction wood that is formed when the tree is subjected to
mechanical stress such as wind and gravity (Donaldson, 2008; Huang et al. 2003). MFA
strongly influences the shrinkage behavior of wood (Burdon et al. 2004; Huang et al.
2003). Evans and Ilic (2001) found that MFA accounted for 86% of variation in modulus
of elasticity (a measure of the ability of wood to resist deformation under load) for
Eucalyptus Delegatensis R.T. Baker. In conifers, MFA varies both radially and vertically,
with the highest angles in juvenile wood (Donalson, 2008; Gartner, 2006). A negative

13
relationship is also found with stem height, where there is often observed variability in
the lower and upper parts of the stem, and constant MFA for the middle section
(Donaldson, 2008). Wimmer et al. (2002) found that MFA was greater in earlywood and
juvenile wood and lower in latewood and mature wood, and that longer growth periods
and higher growth rates were associated with larger MFA.
3.1.5. Fiber length (μm) is the length of the wood cell which in turn influences paper and
timber strength and stiffness. Longer fiber length, together with reduced cell
coarseness, leads to improved bonding by increasing the number of cell to cell contacts
(Via et al. 2004). Fiber length is also important attribute for timber products, since longer
fibers result in greater resistance against buckling of wood beams; however, the trait is
considered of lesser importance than wall thickness and wood density (Zobel and Van
Buijtenen, 1989). Fiber length has been found to correlate with MFA, with smaller
angles (microfibrils more aligned with cell longitudinal axis) related to longer fiber
lengths and this trend of elongating of the fibers can be found from the pith outwards to
the bark (Sheng-zuo et al. 2004).
3.2. Extrinsic tree-level indicators
Wood fiber attributes vary in response to individual tree characteristics such as age,
crown architecture, growth rate, stem lean and taper, and in response to site attributes
such as climate, topography, stand structure, and disturbance history. As a result,
variations in wood fiber properties occur within and between individual trees, species,
and regions (Zobel and van Buijtenen 1989; Burdon et al. 2004; Lindström, 1997). At

14
the tree scale, age, growth rate, crown size and shape, and branchiness have been
used as indicators of wood quality, and many of these factors have been controlled
operationally through silvicultural prescriptions such as planting, thinning, pruning, and
fertilization in an attempt to improve wood quantity and quality (Jack and Long, 1996;
Oliver and Larson, 1996).
3.2.1 Age. As trees grow older, axial and radial variations in wood fiber characteristics
change markedly as a result of the maturation of woody tissue. A number of studies
have examined the relationship between basic wood density and tree social status, and
have shown that suppressed and intermediate tree classes have higher wood densities
than co-dominant and dominant tree classes within the same stand (Lindström, 1996a,
1996b, 1996c; Duchesne et al. 1997). In Lindström (1996b), variations in basic density
of Picea abies were explained using variables related to crown development, tree
height, stem taper and mean ring width, and stand variables such as stocking density
and thinning regime. At the tree level, tree height, diameter and thinning procedure were
found good explanatory variables describing the variation in basic wood density (r2 =
0.68). At the stand level, basic density was explained using solely crown related
parameters volume production, stand density and thinning procedure, yielding an
adjusted r2 of 0.99. Increases in wood density and stem straightness at older age have
been demonstrated by Suarez et al. (2010) using model simulations. Such modeling
approaches allow for estimating wood formation and quality attributes from inventory
data at fixed times and allow interpolation between, and extrapolation beyond, forest
inventories to help assess the quality of timber formed over time. This greatly aids forest

15
management and planning to optimize efficiency of timber production to meet the
required end-use properties. Similar modeling approaches are used to model stand
development and growth of individual trees. As trees develop, occlusion of older
branches by new formed branches affects photosynthetic capacity and branch mortality.
Competition among trees and mechanical stress (e.g. windiness or mechanical effects
from neighboring trees) further influences the mortality and formation of branches,
hence affect knottiness and portions of dead knots posing important implications on the
quality of formed wood (Kint et al. 2010). Hein et al. (2008) found higher portions of
sapwood for trees of younger age and closer towards the tree apex.
3.2.2. Growth rate (m3/ha/year). The rate at which an individual tree grows has an
impact on wood quality. Primary production changes daily, seasonally, and throughout
stand development due to spatiotemporal changes in solar insolation, temperature,
moisture and nutrient status, competition, disturbance, and community composition (i.e.,
mutualism) (Ryan et al. 1997; Watt et al. 2006; Lindström, 1997). Seasonal changes in
the timing, rate, and duration of secondary growth influence the characteristics and
proportions of earlywood and latewood, and the distribution of fiber properties
throughout the stem (Downes and Drew, 2008). Faster growth rates, desirable for
volume yield optimization, may be accompanied with greater earlywood portions and
hence lower overall wood densities, thus affecting the quality of the timber. Growth rates
generally decline after a certain age and with increasing tree size. This longer term
reduction in growth has implications for wood formation, production, and quality. Two
competing theories (see Mencuccini et al. 2005 for summary) have been proposed to

16
explain the apparent 'age-related growth trend’: (1) ‘cellular senescence’ theory
suggests there is a genetically controlled decline in physiological function (i.e., slowing
down of cell division, cell differentiation, and formation of secondary and tertiary cell
walls, loss of protoplasts, incomplete lignification, etc.) and (2) ‘meristem-extrinsic’
theory which suggests that declining physiological function is not genetically controlled,
but rather due to the physiological constraints imposed by tall and architecturally
complex trees (i.e., respiratory losses needed to maintain increasing stem, branch, and
root mass, as well as the physical limitations of transporting water and nutrients to the
top of a tall canopy). Effects of competition and cambial age on growth rate and fiber
properties in Picea abies were investigated by Lindström (1997) who found good
agreement between earlywood tracheid length with ring width and cambial age, while
diameter of the earlywood tracheids was correlated with cambial age, ring width and site
quality. In an earlier study, Lindström (1996b) explained variations in mean ring width
from variables as tree height and diameter, volume production and thinning regime, with
a high degree of significance (r2 = 0.95). These findings motivate research to explore if
variations in earlywood tracheid length and width may be explained using simple tree
and stand level variables.
3.2.3. Crown and Canopy Shape and Size (m). Crown shape, as determined by the
location and number of branches up the stem, as well as total branch mass, is indicative
of the number of knots present in the timber and therefore its potential use for various
wood products (Kantola et al. 2008). Dimensions of the living crown - sometimes
substituted by simpler measures as tree height, age and BDH - are widely used in

17
modeling wood formation and predicting quality attributes (Houllier et al. 1995). Crown
attributes were also found important in modeling wood density and a number of related
quality parameters in Lindström (1996a, 1996b, 1996c, 1997). Crown dimensions are
regulated by stand density and determine the photosynthetic capacity of individual trees
as well as competitive strength. Amarasekara and Denne (2002) found significant
differences in ring with between suppressed, co-dominant and dominant Corsican pines
(Pinus nigra var. maritima) with dominant trees having the widest rings. Crown depth
and variations in leaf densities within the crown determine self-occlusion and hence
mortality and productivity at different canopy strata (Kint et al. 2010). Strong
relationships between living branch area and sapwood portions have been found for
example by Hein et al. (2008) and others. Janse-Ten Klooster et al. (2007) found that
crown width was positively correlated with basic wood density over a range of species
and site conditions, and suggested that increasing wood density serves the function of
supporting the larger crowns. Wood density was also correlated with crown depth and
the number of leaders in the crown; however, this correlation was not significant for all
light conditions and soil types present in the study. The correlation between wood
density and crown architecture is generally explained by increased cambial activity and
the wider ring spacing produced by large, live crowns. Light demanding, faster growing
species also tend to have lower wood densities (Janse-Ten Klooster et al. 2007; King et
al. 2006) and this is often explained by the lower cost per unit growth (Sterck et al.
2006). Amarasekara and Denne (2002) found a higher percentage of latewood for
suppressed Corsican pine than for co-dominant and dominant individuals. Crown size,

18
competition for light, and disturbance are considered major factors explaining trends in
tree growth and stem form (Hynynen, 1995).
3.3 Extrinsic stand-level indicators
With confidence, one can state that anything affecting tree growth can also affect wood
properties, and this becomes increasingly evident for species planted outside their
natural geographic range (Zobel and Van Buijtenen, 1989). However, differences in
environmental conditions that regulate tree growth and wood formation can occur due to
topographic position, and local variations in climate (Littell et al. 2008). While detailed
examination of these influences on wood quality is difficult to undertake due to the
extensive cost of field work and subsequent wood fiber analysis, and in some cases
destructive sampling techniques, existing provenance trials and long term forestry plot
installations can provide significant insight into these factors at single plot and single
stand locations. In addition, measurements from these trials can provide unique
calibration and validation data for model development (Steward, 2006; Smithers, 1957).
Remote sensing provides an additional data source which may contain valuable
information on the spatiotemporal patterns of tree growth and wood quality over broad
geographic areas.
3.3.1. Stand / stocking density (trees/ha). Low stocking densities have been related to
shorter fiber lengths, while higher stocking densities have been found to result in lower
juvenile portions due to decreased diameter growth rates at juvenility and faster onset of
mature wood formation (Watson et al. 2003). Higher stocking densities are also related

19
to faster longitudinal (primary) growth as a result of increased competition for light.
Seeling (2001) found better drying behavior of Norway spruce grown at denser
spacings, and noted that wider spacings resulted in lower bending strength, more spiral
grain, and compression wood, and larger knot diameters. The effects of tree spacings
on growth and branch properties have also been studied for Norway spruce and
Douglas-fir by Makinen and Hein (2006) and Hein et al. (2008) respectively. Both
studies found improved branch properties at denser spacings (e.g. 1200 ha-1). In Hein
et al. (2008), decrease in branch diameters was found to be more pronounced lower in
the canopy especially maximum branch diameters which were significantly different
between 100, 200 and 1200 ha-1 stocking plots. These high stocking density plots also
showed a significantly lower portion of living branches at lower strata in the canopy and
showed lower portions of sapwood area. Sites with higher stocking densities showed a
lower cumulative branch area per unit sapwood area, compared to sites with lower
stocking densities. This may benefit the prediction of sapwood area through model
simulations, using leaf area profiles or living branch characteristics and stand density as
explanatory variables (Hein et al. 2008).
3.3.2. Nutrition and site quality. Nutrient availability has an affect on diameter growth
rate and, therefore, wood fiber structure and quality (Zobel and Van Buijtenen, 1989).
Interactions between temperature and rainfall influence seasonal moisture balance and
affect growth rate. Also, water surplus or deficit in one year may affect plant growth in
the subsequent year (Littell et al. 2008). The combined effect of these temporal and site
specific factors on growth rate have been demonstrated to alter wood quality properties

20
as basic density (Lindström 1996b; Downes and Drew 2008). In Lindström (1996b), for
example, variations in site nutrition affected growth rates and stem diameter
distributions for Norway spruce, while volume production, tree height, and stem
diameter were found significant variables and were used to describe basic density at the
tree and stand level.
3.3.3. Climate and seasonality. Continentality, physiography, and topography all
influence local to regional patterns of climate (i.e. daily, seasonal, and annual cycles of
temperature and precipitation) (Littell et al. 2008). Spring and winter temperatures,
duration of snowpack, growing-season length, and seasonal water balance
(evapotranspiration) influence ring width and earlywood and latewood proportions (Littell
et al. 2008), and the distribution of wood fiber characteristics (Downes and Drew, 2008).
Seasonal variations in precipitation have major impact on wood density and cell
dimensions. The seasonal variation in growth rate affected by climate and weather
alters fiber properties throughout the wood formed. For example, Downes et al. (2002)
explain that trees experiencing drought early in the growing season and good growth
over the summer and autumn may show a positive relationship between wood density
and ring width, arguing that seasonal distribution of growth is an important driver of
wood density and a range of fiber characteristics. In addition, Suarez et al. (2010) found
exposure to wind can be an important determinant of stem straightness which in turn
influences wood quality and yield of sawn timber.

21
4. Potential application of ground and air-based laser scanning
Recent and continuing developments in ground-based (terrestrial) and airborne laser
ranging scanners have allowed the direct measurement of 3-D forest canopy structure
at fine spatial scales. These active sensors typically employ a near-infrared laser light
source and detector, precise timing instruments, and GPS to determine the 3-D location
of reflecting targets such as stems, branches and foliage (Aschoff and Spiecker, 2004;
Wehr and Lohr, 1999; Baltsavias, 1999).
4.1 Terrestrial laser scanning
Terrestrial laser scanning (TLS) support very high laser angular shot densities, and is
capable of capturing very detailed 3-D digital models of vegetation canopies. TLS exist
as tripod mounted, or mobile scanning instruments and as point-scanning devices with
a partial or complete hemispherical field of view or frame-scanning devices. TLS
instruments are able to capture detailed structure out to a radial distance of up to 100m
(Jupp et al. 2009) and in the case of monitoring glacial movements up to several
kilometers (Maas et al. 2008b; Schwalbe et al. 2008). To date, the commercially
available TLS scanners record one, two or several discrete returns per emitted laser
pulse. Discrete-return TLS has been widely used to measure vegetation structure in a
variety of forest ecosystems. For example, discrete-return TLS has been used to
measure plot-level leaf area and gap fractions (Zheng and Moskal, 2009; Danson et al.
2007). A technique to estimate leaf area at the tree-level that takes into account non-
uniformity of the foliage distribution has been demonstrated by Huang and Pretzsch
(2010) using co-registered TLS scans. TLS has also been used to derive stem

22
diameters with root mean square errors ranging between 1.5 – 3.3 cm from field
measurements (Hopkinson et al. 2004; Maas et al. 2008a; Tansey et al. 2009) and has
been used to derive stem taper (Bienert et al. 2007), sweep and lean (Thies et al.
2004). Other areas of application include modeling radiative transfer through forest
canopies (Côté et al. 2009, 2010), the parameterization of water flow resistance models
(Antonarakis et al. 2009), and the estimation of tree volume (Hopkinson et al., 2004;
Lefsky and McHale, 2008).
In contrast to these discrete return instruments, full- waveform TLS is able to fully
digitize the returned energy of an emitted laser pulse, thereby making the instrument
more sensitive to small return intensities and allowing the modeling of secondary return
obscuration (Lovell et al. 2003; Strahler et al. 2008; Jupp and Lovell, 2007). Full-
waveform TLS has also been successfully used to estimate stand structural parameters
(Jupp et al. 2009; Strahler et al. 2008) and parametrize and validate radiative transfer
models (Ni-Meister et al. 2008). Full-waveform recording of multiple TLS wavebands,
while not commercially available, could allow future applications to go beyond the
classical analyses of vegetation structure (Lichti et al. 2008), and may provide a vital
link between plant structure and function.
The use of TLS to predict wood fiber attributes has not been broadly applied. However,
studies have applied TLS to estimate standing timber volume using estimates of bole
size and taper (Murphy, 2008; Maas et al. 2008a) and using allometric approaches
based on DBH and tree height (Hopkinson et al. 2004). Estimates of branch size,

23
arrangement, and leaf density have also been extracted from TLS (Bucksch and
Lindenberg, 2008; Bucksch and Fleck, 2009) as well as whorl heights and number of
branches per whorl (Klemmt et al. 2010). In a more physiological based approach
Samson et al. (2000) utilized a multi-wavelength LiDAR to discriminate nitrogen and
sulfur deficiencies through laser-induced fluorescence.
4.2. Airborne LiDAR (ALS)
LiDAR data used in forestry applications are commonly acquired from aircraft or
helicopter platforms. Airborne laser scanning (ALS) have been widely used for
generation of bare-earth digital elevation models (DEM) and the estimation of forest
inventory attributes, and are rapidly being adopted in operational forest management.
Typically, ALS are discrete-return scanning instruments with a footprint size of 0.1 m to 2
m (Lim et al. 2003; Wulder et al. 2008), and can achieve sub-meter accuracy of terrain
surface heights (Blair et al. 1994; Lefsky et al. 2002). The error for measuring the height
of individual trees from ALS is typically less than 1.0 m (e.g. Persson et al. 2002), and
varies with canopy height and height distribution (Hopkinson et al. 2006). When
estimating maximum or mean canopy height within plots and at full canopy closure this
error is often less than 0.5 m (Magnussen and Boudewyn, 1998; Næsset and Økland,
2002). ALS estimates of tree height are considered by some to be more accurate than
manual, field-based measurements (Næsset and Økland, 2002; Coops et al. 2007).
Full-waveform ALS capture the vertical profile of light penetrating the plant canopy, by
producing a fully digitized return intensity (energy) vector for every outgoing laser pulse.
This allows for the computation of radiation budgets within the canopy or - by

24
accounting for the effects of occlusion - vertical plant canopy profiles (Jupp et al. 2009;
Strahler et al. 2008). The range resolution of a full-waveform scanner is determined by
the sampling rate with which the sensor is able to record energy levels of the incoming
energy.
ALS has been successfully used in a variety of forestry applications for the retrieval of
crown and canopy structural attributes, such as crown width, length, height-to-first-living
branch, and biomass and biomass change over time (Chasmer et al. 2006; Riano et al.
2004; Morsdorf et al. 2004; Andersen et al. 2005; Solberg et al. 2006; Bollandsås &
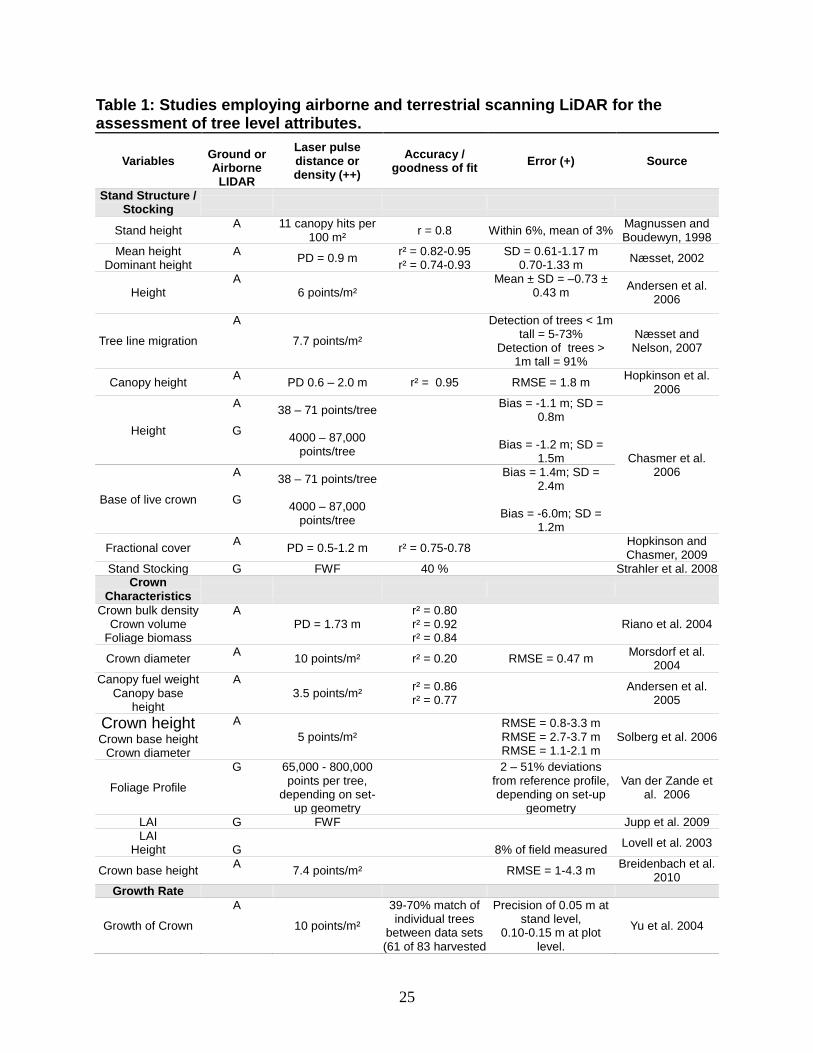
Næsset 2010). An overview of retrieved structural attributes from ALS, together with
attributes derived from TLS is listed in table 1. As these extrinsic properties have been
linked to variations in wood properties, ALS holds promise for landscape-scale mapping
of wood quality. Suarez et al. (2010) demonstrate the use of airborne LiDAR combined
with tree growth modeling to analyze the hierarchy of dominance in forest stands that is
subsequently linked to a model estimating changes in wood properties as wood density
and stem straightness. These results show strong relationships between tree
dominance and site index and wood quality parameters as wood density and stem
straightness.
25
Table 1: Studies employing airborne and terrestrial scanning LiDAR for the assessment of tree level attributes.
Variables
Ground or Airborne
LIDAR
Laser pulse distance or density (++)
Accuracy / goodness of fit
Error (+) Source
Stand Structure / Stocking
Stand height A 11 canopy hits per
100 m² r = 0.8 Within 6%, mean of 3%
Magnussen and Boudewyn, 1998
Mean height Dominant height
A PD = 0.9 m
r² = 0.82-0.95 r² = 0.74-0.93
SD = 0.61-1.17 m 0.70-1.33 m
Næsset, 2002
Height A
6 points/m² Mean ± SD = –0.73 ±
0.43 m
Andersen et al. 2006
Tree line migration
A
7.7 points/m²
Detection of trees < 1m tall = 5-73%
Detection of trees > 1m tall = 91%
Næsset and Nelson, 2007
Canopy height A
PD 0.6 – 2.0 m r² = 0.95 RMSE = 1.8 m Hopkinson et al.
2006
Height
A
G
38 – 71 points/tree
4000 – 87,000 points/tree
Bias = -1.1 m; SD = 0.8m
Bias = -1.2 m; SD =
1.5m Chasmer et al. 2006
Base of live crown
A
G
38 – 71 points/tree
4000 – 87,000 points/tree
Bias = 1.4m; SD = 2.4m
Bias = -6.0m; SD =
1.2m
Fractional cover A
PD = 0.5-1.2 m r² = 0.75-0.78 Hopkinson and Chasmer, 2009
Stand Stocking G FWF 40 % Strahler et al. 2008 Crown
Characteristics
Crown bulk density Crown volume
Foliage biomass
A PD = 1.73 m
r² = 0.80 r² = 0.92 r² = 0.84
Riano et al. 2004
Crown diameter A
10 points/m² r² = 0.20 RMSE = 0.47 m Morsdorf et al.
2004 Canopy fuel weight
Canopy base height
A 3.5 points/m²
r² = 0.86 r² = 0.77
Andersen et al.
2005
Crown height Crown base height
Crown diameter
A
5 points/m² RMSE = 0.8-3.3 m RMSE = 2.7-3.7 m RMSE = 1.1-2.1 m
Solberg et al. 2006
Foliage Profile
G 65,000 - 800,000 points per tree,
depending on set-up geometry
2 – 51% deviations from reference profile, depending on set-up
geometry
Van der Zande et al. 2006
LAI G FWF Jupp et al. 2009 LAI
Height
G
8% of field measured
Lovell et al. 2003
Crown base height A
7.4 points/m² RMSE = 1-4.3 m Breidenbach et al.
2010 Growth Rate
Growth of Crown
A
10 points/m²
39-70% match of individual trees
between data sets (61 of 83 harvested
Precision of 0.05 m at stand level,
0.10-0.15 m at plot level.
Yu et al. 2004

26
trees detected) LAI Increment A 3.1-9.8 points/m² r² = 0.87-0.93 Solberg et al. 2006
Age / Volume / Biomass
Forest successional stage
A 0.26 points/m²
Class accuracies 73-100%
Falkowski et al.
2009
Canopy gap dynamics
A 0.03 & 0.19 points/m²
Delineation accuracy for gaps
> 5m² = 96%
Vepakomma et al. 2008
Timber volume A
10 points/m² RMSE = 16-25% Bias = 8.2-24.3%
Maltamo et al. 2004
Biomass Volume
A
PD = 0.7 m r² = 0.32 -0.82 r² = 0.39-0.83
RMSE = 29-44 Mg/ha RMSE = 48-53 m
3/ha
Popescu et al. 2004
Aboveground biomass
A
PD = 3-5 m r² = 0.83 RMSE = 26.0 t/ha Hyde et al. 2007
Biomass change A
3.4-4.7 points/m² R²=0.87 RMSE=4.4 Bollandsås & Næsset 2010
DBH Biomass
A
2.6 points/m² r² = 0.87 r² = 0.88
RMSE = 18% RMSE = 47%
Popescu 2007
Volume
A PD = 0.9 m r² = 0.8-0.93 18.3-31.9 m
3 ha
-1 Næsset 2002
Basal Area
G FWF r² = 0.86-0.97 Strahler et al. 2008
DBH G SD = 2.3 cm Tansey et al. 2009
Volume
G r² = 0.75–0.90 Lefsky et al. 1999
(+) RMSE = Root Mean Square Area; SD = Standard Deviation (++) PD = Point Distance; FWF = Full Waveform
4.3 Complementarily and integration of ground-based and airborne LiDAR systems
ALS and TLS systems have their own inherent strength and weakness, and should be
considered complementary technologies. While ALS can estimate structural
parameters, such as stand height, gap probability and canopy volume, a number of
limitations exist for characterizing vegetation structure lower in the canopy (Hopkinson
et al. 2004; Lovell et al. 2003). First, the ability of ALS to accurately characterize forest
vegetation canopy height profiles depends on the point density of laser returns (Lovell et
al. 2003). Consequently, the vegetation understorey is often under-represented by
airborne LiDAR due to the top-down perspective and resultant bias towards the upper
part of the canopy. The infrequency of laser returns from mid and understorey layers is
particularly apparent in forests with extremely dense overstory canopies. Second, ALS

27
is often restricted to near-nadir view angles (±20°), which are less suitable for the
estimation of canopy clumping and leaf angle distribution (Chen et al. 2003; Ni-Meister
et al. 2008) both of which have important ecological and economical implications
(Zheng and Moskal, 2009). Finally, ALS is limited in its ability to characterize the woody
component of vegetation, because the vertical projection of the stem contains very
limited information on bole size (diameter) and shape.
One of the key limitations of TLS is the restricted range within which tree and stand
structural characteristics can be obtained. Depending on stand density, the typical range
of TLS acquisitions is less than 100 m from the sensor position and due to decreasing
point densities with distance, processing TLS data may be limited to range distances
not exceeding 10 – 30 m (Liang et al. 2009; Maas et al. 2008a). Also, since static TLS
features a radial field of view, hit densities decline markedly with distance from the
sensor due to near field obscuration. This bias has to be taken in account when trying to
derive stand-level estimates from TLS data (Hilker et al. 2010). Whereas mobile TLS
improves sampling compared to static TLS (Rutzinger et al., 2010), the issue remains
that TLS-returns are typically biased towards lower parts of the canopy, and as a result
the upper crown structure and tree heights are often difficult to assess (Hilker et al.
2010; Chasmer et al. 2006). In contrast to ALS, most TLS are able to describe
vegetation structure from a broad range of view angles and perspectives (Côté et al.
2009, 2010; Jupp et al. 2009; Strahler et al. 2008), which allows for the analysis of
canopy clumping (Chen, 1996) and the effects of variations in the project function of
different canopy elements.

28
Recent work of Chasmer et al. (2006), Lindberg et al. (2010) and Hilker et al. (2010)
show that integration of TLS and ALS data to a common coordinate frame enhances the
detail of structural information obtained and overcomes challenges mentioned above
with respect to either technique. Such co-registration successfully enables the study of
tree-level structure by combining the complementary values of ALS and TLS in one data
set and thus facilitates studies on tree allometry using LIDAR remote sensing or
enhances leaf area estimates at various canopy strata (Huang and Pretzsch, 2010).
Another important advantage gained through the integration of TLS and ALS data is that
TLS allows calibration and validation of ALS data in an accurate and rapid manner (e.g.
Lindberg et al. 2010).
4.4. Modeling of wood fiber properties from LiDAR data
Tree-level parameters with demonstrated explanatory power for wood physical
properties include DBH, tree social status, light competition, tree height, and growth,
crown dimensions, and branch free bole sections as well as number of branches,
branch angle and diameters (see also Table 2). These parameters have been the focus
of various modeling approaches to estimate wood quality attributes ranging from gross
defects such as knots and spiral grain to cellular level properties. Using height-over-age
growth curves, several branchiness parameters – such as number and size and
insertion angle – can be derived from simple forest inventory parameters, while canopy
characteristics are widely used to model growth and between tree interactions (Houllier
et al. 1995). An important advancement is yet to be made through the application of

29
LiDAR remote sensing derived stand and tree-level attributes to parametrize wood
quality and tree growth models. Recent contributions include those of Suarez et al.
(2010) who demonstrate the use of the Canadian TASS model in combination with the
Timber Quality for Conifers (ConTQ) model to derive estimates of timber properties from
LiDAR data. In addition, a variety of recent studies have focused on quantifying
branchiness, crown depth, and base height, and whorl heights from LiDAR data
(Breidenbach et al. 2010; Klemmt et al. 2010; Lindberg et al. 2010).

30
Table 2: Use of LiDAR for measurement of extrinsic indicators of wood quality and stand and site conditions.
Measurement Source of information (+) Related extrinsic attributes
Related wood anatomical and wood quality characteristics
Branchiness, branch diameter
TLS Knots Heterogeneity among all fiber properties and wood density, impacts on strength, sapwood portions
Crown dimensions
TLS/ALS Social status, vigor, growth rate, branchiness
Branch free bole section, wood density
Stem diameter & growth
TLS Stem volume, stem biomass, aboveground biomass
Ring width, wood density, sapwood vs. heartwood portions, mature vs. juvenile portions, fiber length, cell coarseness, cell perimeter
Tree height & growth
ALS/TLS Stem volume, stem biomass, aboveground biomass, site quality
Ring width, wood density, sapwood vs. heartwood portions, mature vs. juvenile portions
Vertical distribution of foliage (foliage profile)
ALS/TLS Biomass, crown length, and branchiness
Branch free bole section, heterogeneity among fiber properties, uniformity of timber
Gap fraction ALS/TLS Stocking density, growth rate, availability of sunlight with canopy depth (branchiness), biomass
Heterogeneity among all fiber properties and wood density, impacts on strength, ring width
Stocking density ALS/TLS Stocking density Ring width, wood density, juvenile vs. mature portions
Terrain: slope & aspect
ALS Local climate, growth rate, site quality, hydrology
Ring width, formation of reaction wood, variations in wood density
Precipitation & temperature
Weather data/ GIS
Seasonal distribution of growth Ring width, and proportions of earlywood and latewood, wood density
Site nutrient availability
Soil sampling/ GIS
Growth rate, site quality Ring width
(+) TLS = Terrestrial Laser Scanning; ALS = Airborne Laser Scanning; GIS = Geo-Information Systems
5. Summary and outlook for future research
This paper reviews some of the key indicators of wood quality, discusses the main
endogenous and exogenous controls on plant growth and wood anatomy, and explores
the potential of TLS and ALS remote sensing technologies to capture a suite of forest
and terrain-related metrics needed to predict local and regional variations in wood
quality (Table 2). TLS allows tree-level observations of individual stems and crown
attributes at a high level of detail. For instance, TLS observations have been used to
measure the diameter at breast height and taper of individual tree boles (Strahler et al.
2008; Thies et al. 2004), plus canopy and crown properties (Jupp et al. 2009; Hilker et

31
al. 2010). These types of measurements currently allow for the accurate estimation of
stem volume, above ground biomass, and many crown attributes such as branching
structure, branch diameter, and crown volume and shape. We expect that the
integration of LiDAR measurements with other ancillary data sources related to local
and regional variations in climate and species composition may provide a strong
foundation for predictive modeling of plant growth rates and wood properties at tree,
stand, and regional scales.
ALS provides opportunities for much broader spatial characterizations of wood quantity
and quality at the stand and landscape level. TLS, on the other hand, provides
unparalleled spatial descriptions of forest canopy structure at local scales, and may
ultimately be used to calibrate and validate the lower density point clouds, structural
metrics, and wood quality predictions obtained from ALS. Among the most important
quantities measured by LiDAR is the vertical distribution of foliage within the canopy.
The vertical distribution and density (cover) of foliage within a stand are strong
indicators of stand age, height, density, successional status, stand developmental
stage, site productivity, competition (shading and crowding), and disturbance history, all
of which are expected to influence tree- and stand-level growth responses and wood
properties. Cumulative foliage area on a per-tree basis has been found to correlate with
sapwood cross-sectional area (Waring et al., 1982). Another parameter of interest that
can be routinely obtained from ALS is the canopy gap fraction. Several studies (e.g.
Danson et al. 2007; Ni-Meister et al. 2008) have demonstrated the negative correlation
between stocking density and LiDAR measurements of canopy gap fraction. Stocking

32
density determines the potential growing space available to each individual tree, and is
known to influence wood density, juvenile wood portions, and cell perimeter and
coarseness. Finally, bare-earth or digital elevation models constructed from LiDAR
ground returns allow for the extraction of topographical indices (e.g., slope, aspect,
elevation), which capture local and regional variations in critical plant resources, such
as solar radiation, moisture, and temperature. Resource availability is critical to plant
growth and the development of wood anatomical structure.
In addition to single-date acquisitions, multi-temporal LiDAR datasets may prove useful
for the assessment of vegetation dynamics (i.e., growth, mortality, and disturbance), and
can therefore be a valuable addition to traditional canopy characterizations. For
instance, daily, seasonal, and annual cycles of secondary growth influence radial and
axial patterns of ring width, proportions of earlywood and latewood, wood density, fiber
size and shape, and many other wood anatomical characteristics (Downes and Drew,
2008). Using multi-temporal LiDAR acquisitions, information on the deposition of fiber
throughout the tree's life spans can be derived and may be used as input in wood
quality models and can be used to compute, for example, sapwood portions, branch
size or the fraction of living branches interpolated between the individual LiDAR
acquisitions. This yields important information to forecast the wood quality formed
through growth, or to estimate the effects of sylvicultural interventions on wood
formation (Hein et al. 2008; Kint et al. 2010). To these means, formulation of
standardized acquisition methodologies as well as protocols are needed that enable
comparison of data from scanners from different manufacturers and model updates. In

33
addition, better insight into technical considerations behind scanning technology is of
critical importance though often unavailable due to proprietary knowledge involved
(Næsset, 2009).
Investigated trends between tree- and stand-level mensurational data and wood
anatomical structure, together with the proven results of LiDAR remote sensing to
retrieve a broad range of forest inventory parameters, motivate research that seeks to
explore the potential of LIDAR to gain an understanding of the spatial distribution of
wood fiber quantity and quality. This knowledge can be used to support decision
making, and to evaluate competing forest functions and management scenarios. The
link between wood density and other wood characteristics and properties holds potential
to address issues of accounting and accountability of harvested timber resources, and
can be used for monitoring carbon offsets attained through forest growth. Where sites
are managed for timber production, such information can be further used to facilitate in-
stand sorting of timber according to its fiber properties to improve processing efficiency
and to better meet end-use requirements.
However, management of several competing forest functions requires quantification of
their societal- and intrinsic values. Therefore, in parallel to research on the quantification
and spatial mapping of wood quality attributes, research on the measurement of other
forest values is needed together with forest management policies that balance socio-
economic aspects of forest use with ecological constraints in order to assure sustained
resource utilization, conservation of biodiversity, and sustained ecosystem functioning.

34
This includes considering costs of maximizing current yield and growth placed on future
generations.
This review was intended to help guide future research on the application of LiDAR
remote sensing to derive forest structure metrics that relate to wood quality and
quantity. We anticipate that LIDAR derived structure may be linked to supra-cellular
characteristics and include juvenile and mature wood portions, ring width, and wood
density. Since many wood anatomical characteristics, including wood density, correlate
well with tree size, growth rate, stand structure, and site conditions, we expect that
LIDAR-derived measurements of canopy height, density, and topography will furnish
strong predictor variables for modeling the spatial distribution of wood quality at tree,
stand, and landscape scales.

35
Acknowledgements
Components of this research were partly funded by a NSERC Accelerator grant to Prof.
Dr. Nicholas C. Coops and by the Government of Canada through the Lodgepole Pine
Partnership Project funded by the Canadian Forest Service and the Canadian Wood
Fibre Centre. The authors would like to acknowledge Prof. Dr. Shawn D. Mansfield and
two anonymous reviewers for their helpful comments.

36
7. References Amarasekara, H., Denne, M.P., 2002. Effects of crown size on wood characteristics of
Corsican pine in relation to definitions of juvenile wood, crown formed wood and core
wood. For. 75, 51-61.
Andersen, H.-E., McGaughey, R.J., Reutebuch, S.E., 2005. Estimating forest canopy
fuel parameters using LIDAR data. Remote Sens. Env. 94, 441-449.
Andersen, H.-E., Reutebuch, S.E., McGaughey, R.J., 2006. A rigorous assessment of
tree height measurements obtained using airborne LIDAR and conventional field
methods. Can. J. Remote Sens. 32, 355-366.
Antonarakis, A.S., Richards, K.S., Brasington, J., Bithell, M., 2009. Leafless roughness
of complex tree morphology using terrestrial lidar. Water Resour. Res. 45, W10401.
Aschoff, T., Spiecker, H., 2004. Algorithms for the automatic detection of trees in laser
scanner data. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 36 part 8/W2, 66-
70.
Baas, P., Ewers, F.W., Davis, S.D., Wheeler, E.A., 2004. Evolution of xylem physiology,
in: Hemsley, A.R., Poole, I., The evolution of plant physiology: from whole plants to
ecosystems. Society of London. Palaeobotany Specialist Group, pp. 512.
Baltsavias, E.P., 1999. Airborne laser scanning: existing systems and firms and other
resources. ISPRS J. Photogramm. Remote Sens. 54, 164-198.
Bienert A., Scheller, S., Keane, E., Mohan, F., Nugent, C., 2007. Tree detection and
diameter estimations by analysis of forest terrestrial laser scanner point clouds. Int.
Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 36/3-W52, 50-55.

37
Bienert, A., Scheller, S., Keane, E., Mullooly, G., Mohan, F., 2006. Application of
terrestrial laser scanners for the determination of forest inventory parameters. Int. Arch.
Photogramm. Remote Sens. Spat. Inf. Sci. 36 part 5.
Blair, J., Coyle, D., Bufton, J., Harding, D., 1994. Optimization of an airborne laser
altimeter for remote sensing of vegetation and tree canopies. Proceedings of
IGARSS’94, vol. 2, 939-941.
Bollandsås, O. M., Næsset, E., 2010. Using delta values of multi-temporal first-return
small footprint airborne laser scanner data to predict change of tree biomass in
mountain spruce forests. Silvilaser 2010, September 14-17, Freiburg, Germany.
Bowyer, J.L., Haygreen, J.G., Shmulsky, R., 2007. Forest products and wood science:
an introduction (5th edition). Blackwell, Ames, Iowa, pp 558.
Breidenbach, J., Næsset, E., Lien, V., Gobakken, T., Solberg, S., 2010. Towards an
inventory of quality attributes of individual trees using airborne laser scanning and
multispectral data. Silvilaser 2010, September 14-17, Freiburg, Germany.
Bucksch, A., Lindenbergh, R., 2008. CAMPINO - A skeletonization method for point
cloud processing. ISPRS J. Photogramm. Remote Sens. 63, 115-127.
Bucksch, A., Fleck, S., 2009. Automated detection of branch dimensions in woody
skeletons of leafless fruit tree canopies. SilviLaser 2009, October 14 – 16, Austin,
Texas.
Burdon, R.D., Kibblewhite, R.P., Walker, J.C.F., Megraw, R.A., Evans, R. & Cown, D.J.,
2004. Juvenile versus mature wood: a new concept, orthogonal to corewood versus
outerwood, with special reference to Pinus radiata and P. taeda. For. Sci. 50, 399-415.

38
Cannell M.G.R., Sheppard, L.J., Milne, R., 1988. Light use efficiency and woody
biomass production of poplar and willow. For. 61, 125-136.
Chasmer, L., Hopkinson, C., Treitz, P., 2006. Investigating laser pulse penetration
through a conifer canopy by integrating airborne and terrestrial lidar. Can. J. Remote
Sens. 32, 116-125.
Chen, M.J., 1996. Optically-based methods for measuring seasonal variation of leaf
area index in boreal conifer stands. Ag. For. Met. 80, 135-163.
Chen, J.M., Liu, J., Leblanc, S.G., Lacaze, R., Roujean, J.L., 2003. Multi-angular optical
remote sensing for assessing vegetation structure and carbon absorption. Remote
Sens. Env. 84, 516-525.
Côté, J., Widlowski, J., Fournier, R.A., Verstraete, M.M., 2009. The structural and
radiative consistency of three-dimensional tree reconstructions from terrestrial lidar.
Remote Sens. Env. 113, 1067 – 1081.
Côté, J., Fournier, R.A., Luther, J.E., Van Lier, O.R., 2010. Forest structure
characterization of balsam fir (Abies balsamea (L.) Mill.) stands with terrestrial LiDAR
and fine-scale architectural modelling. Silvilaser 2010, September 14-17, Freiburg,
Germany.
Coops, N.C., Hilker, T., Wulder, M.A., St-Onge, B., Newnham, G., Siggins, A., Trofymow,
J.A., 2007. Estimating canopy structure of Douglas-fir forest stands from discrete-
return LiDAR. Trees 21, 295-310.
Danson, F.M., Hetherington, D., Morsdorf, F., Koetz, B., Allgöwer, B., 2007. Forest
canopy gap fraction from terrestrial laser scanning. IEEE Geosci. Remote Sens. Lett.
4, 157-160.

39
Dean, T.J., Long, J.N., 1986. Validity of constant-stress and elastic-instability principles
of stem formation in Pinus contorta and Trifolium pretense. Ann. Bot. 58, 833-840.
Donaldson, L., 2008. Microfibril angle: measurement, variation and relationships - a
review. IAWA J. 29, 345-386.
Downes, G.M., Drew, D.M., 2008. Climate and growth influences on wood formation
and utilization. South. For. 70, 155-167.
Downes, G.M., Drew, D., Battaglia, M., Schulze, D., 2009. Measuring and modelling
stem growth and wood formation: an overview. Dendrochronologia 27, 147-157.
Downes, G.M., Wimmer, R., Evans, R., 2002. Understanding wood formation: gains to
commercial forestry through tree-ring research. Dendrochronologia 20, 37-51.
Duchesne, I., Wilhelmsson, L., Spangberg, K., 1997. Effects of in-forest sorting of
Norway spruce (Picea abies) and Scots pine (Pinus sylvestris) on wood and fibre
properties. Can. J. For. Res. 27, 790-795.
Evans, R., Ilic, J., 2001. Rapid prediction of wood stiffness from microfibril angle and
density. For. Prod. J. 51, 53-57.
Evans, R., Stringer, S., Kibblewhite, R.P., 2000. Variation of microfibril angle, density
and fibre orientation in twenty-nine Eucalyptus nitens trees. Appita J. 53, 450–457.
Falkowski, M.J., Evans, J.S., Martinuzzi, S., Gessler, P.E., Hudak, A.T., 2009.
Characterizing forest succession with lidar data: an evaluation for the Inland
Northwest, USA. Remote Sens. Env. 113, 946-956.
Freyburger, C., Longuetaud, F., Mothe, F., Constant, T., Leban, J.-M., 2009. Measuring
wood density by means of X-ray computer tomography. Ann. For. Sci. 66, art. nr. 804.

40
Gartner, B.L., 2006. Predictions of wood structural patterns in trees by using ecological
models of plant water relations, in: Characterization of the cellulosic cell Wall. Stokke,
D.D., Groom, L. H. (Eds), Blackwell Publishing, Ames, Iowa, pp. 274.
Gierlinger, N., Schwanninger, M., 2006. Chemical imaging of poplar wood cell walls by
confocal Raman microscopy. Plant Phys. 140, 1246-1254.
Hacke, U.G., Sperry, J.S., Pockman, W.T., Davis, S.D., McCulloh, K.A., 2001. Trends in
wood density and structure are linked to prevention of xylem implosion by negative
pressure. Oecologia 126, 457-461.
Hein, S., Weiskittel, A. R., Kohnle, U., 2008. Effect of wide spacing on tree growth,
branch and sapwood properties of young Douglas-fir [Pseudotsuga menziesii (Mirb.)
Franco] in south-western Germany. Eur. J. Forest Res. 127, 481-493.
Herold, M., Johns, T., 2007. Linking requirements with capabilities for deforestation
monitoring in the context of the UNFCCC-REDD process. Env. Res. Lett. 2, art. nr.
045025.
Hilker, T., Van Leeuwen, M., Coops, N., Wulder, M., Newnham, G., Jupp, D., Culvenor,
D., 2010. Comparing canopy metrics derived from terrestrial and airborne laser
scanning in a Douglas-fir dominated forest stand. Trees – Struct. Funct. 24, 819-832.
Hopkinson, C., Chasmer, L., 2009. Testing LiDAR models of fractional cover across
multiple forest ecozones. Remote Sens. Env. 113, 275-288.
Hopkinson, C., Chasmer, L., Lim, K., Trietz, P., Creed, I., 2006. Towards a universal lidar
canopy height indicator. Can. J. Remote Sens. 32, 139-152.
Hopkinson C., Chasmer, L., Young-Pow, C., Treitz, P., 2004. Assessing forest metrics
with a ground-based scanning LiDAR. Can. J. Remote Sens. 34, 573-583.

41
Houllier, F., Leban, J.-M., Colin, F., 1995. Linking growth modelling to timber quality
assessment for Norway spruce. For. Ecol. Manag. 74, 91-102.
Huang C.-L., Lindström, H., Nakada, R., Ralston, J., 2003. Cell wall structure and wood
properties determined by acoustics - A selective review. Holz als Roh - und Werkstoff
61, 321-335.
Huang, P., Pretzsch, H., 2010. Using terrestrial laser scanner for estimating leaf areas of
individual trees in a conifer forest. Trees 24, 609-619.
Hyde, P., Nelson, R., Kimes, D., Levine, E., 2007. Exploring LiDAR-RaDAR synergy--
predicting aboveground biomass in a southwestern ponderosa pine forest using
LiDAR, SAR and InSAR. Remote Sens. Env. 106, 28-38.
Hynynen, J., 1995. Predicting the growth response to thinning for Scots pine stands
using individual-tree growth models. Silva Fennica 29, 225-246.
Jack, S.B., Long, J.N., 1996. Linkages between silviculture and ecology: an analysis of
density management diagrams. For. Ecol. Manag. 86, 205-220.
Janse-Ten Klooster, S., Thomas, E.J.P., Sterck, F.J., 2007. Explaining interspecific
differences in sapling growth and shade tolerance in temperate forests. J. Ecol. 95,
1250-1260.
Jupp, D.L.B., Culvenor, D.S., Lovell, J.L., Newnham, G.J., Strahler, A.H., & Woodcock,
C.E., 2009. Estimating forest LAI profiles and structural parameters using a ground-
based laser called ‘Echidna’. Tree Phys. 29, 171-181.
Jupp, D.L.B., Lovell, J.L., 2007. Airborne and ground-based LiDAR systems for forest
measurement: background and principles. CSIRO Marine and Atmospheric Research
Paper 017.

42
Kantola, A., Härkönen, S., Mäkinen, H., Mäkelä, A., 2008. Predicting timber properties
from tree measurements at felling: evaluation of RetroSTEM model and TreeViz
software for Norway spruce. For. Ecol. Manag. 255, 3524-3533.
King D.A., Davies, S.J., Tan, S., Noor, N.S.M., 2006. The role of wood density and stem
support costs in the growth and mortality of tropical trees. J. Ecol. 94, 1365-2745.
Kint, V., Hein, S., Campioli, M., Muys, B., 2010. Modelling self-pruning and branch
attributes for young Quercus robur L. and Fagus sylvatica L. trees. Forest Ecol.
Manag. 260, 2023-2034.
Klemmt, H.-J., Seifert, T., Seifert, S., Kunneke, A., Wessels, B., Pretzsch, H., 2010.
Assessment of branchiness in a Pinus pinaster plantation by terrestrial laser scanner
data as a link between exterior and interior wood properties. Silvilaser 2010,
September 14-17, Freiburg, Germany.
Lefsky, M.A., Cohen, W.B., Acker, S.A., Parker, G.G., Spies, T.A., Harding, D., 1999.
Lidar remote sensing of the canopy structure and biophysical properties of Douglas-fir
western hemlock forests. Remote Sens. Env. 70, 339-361.
Lefsky M.A., Cohen W.B., Parker G.G., Harding D.J., 2002. LiDAR remote sensing for
ecosystem studies. Biosci. 52, 19–30.
Lefsky, M., McHale, M., 2008. Volume estimates of trees with complex architecture from
terrestrial laser scanning. J. Appl. Remote Sens. 2, art. nr. 023521.
Liang, X., Litkey, P., Hyyppä, J., Kaartinen, H., Vastaranta, M., Holopainen, M., 2009.
Automatic stem location mapping using TLS for plot-wise forest inventory. Silvilaser
2009, October 14-16, Texas, USA.

43
Lichti, D., Pfeifer, N., Maas, H.G., 2008. ISPRS Journal of Photogrammetry and Remote
Sensing theme issue "Terrestrial Laser Scanning". ISPSR J. Photogramm. Remote
Sens. 63, 1-3.
Lim, K., Treitz, P., Wulder, M., St-Onge, B., Flood, M., 2003. LiDAR remote sensing of
forest structure. Prog. Phys. Geogr. 27, 88-106.
Lindberg, E., Holmgren, J., Olofsson, K., Olsson, H., 2010. Estimation of stem attributes
using a combination of terrestrial and airborne laser scanning. Silvilaser 2010,
September 14-17, Freiburg, Germany.
Lindström, H., 1996a. Basic density in Norway spruce. Part I. A literature review. Wood
and Fiber Sci. 28, 15-27.
Lindström, H., 1996b. Basic density of Norway spruce. Part II. Predicted by stem taper,
mean growth ring width, and factors related to crown development. Wood and Fiber
Sci. 28, 240-251.
Lindström, H., 1996c. Basic density in Norway spruce. Part III. Development from pith
outwards. Wood and Fiber Sci. 28, 391-405.
Lindström, H., 1997. Fiber length, tracheid diameter, and latewood percentage in
norway spruce: development from pith outwards. Wood and Fiber Sci. 29, 21-34.
Littell, J.S., Peterson, D.L., Tjoelker, M., 2008. Douglas-fir growth in mountain
ecosystems: water limits tree growth from stand to region. Ecol. Monogr. 78, 349-368.
Lovell, J.L., Jupp, D.L.B., Culvenor, D.S., Coops, N.C., 2003. Using airborne and
ground-based ranging lidar to measure canopy structure in Australian forests. Can. J.
Remote Sens. 29, 607-622.

44
Lyhykäinen, H.T., Mäkinen, H., Mäkelä, A., Pastila, S., Heikkilä, A., Usenius, A., 2009.
Predicting lumber grade and by-product yields for Scots pine trees. For. Ecol. Manag.
258, 146-158.
Maas, H.-G., Bienert, A., Scheller, S., Keane, E., 2008a. Automatic forest inventory
parameter determination from terrestrial laser scanner data. Int. J. Remote Sens. 29,
1579-1593.
Maas, H.-G., Schwalbe, E., Dietrich, R., Baessler, M., Ewert, H., 2008b. Determination
of spatio-temporal velocity fields on glaciers in West-Greenland by terrestrial image
sequence analysis. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 37/B8, 1419-
1424.
Magnussen, S., Boudewyn, P., 1998. Derivations of stand heights from airborne laser
scanner data with canopy-based quantile estimators. Can. J. For. Res. 28, 1016-1031.
Mäkinen, H., Colin, F., 1998. Predicting branch angle and branch diameter of Scots pine
from usual tree measurements and stand structural information. Can. J. For. Res. 28,
1686-1696.
Mäkinen, H., Hein, S., 2006. Effect of wide spacing on increment and branch properties
of young Norway spruce. Eur. J. For. Res. 125, 239-248.
Maltamo, M., Eerikäinen, K., Pitkänen, J., Hyyppä, J., Vehmas, M., 2004. Estimation of
timber volume and stem density based on scanning laser altimetry and expected tree
size distribution functions. Remote Sens. Env. 90, 319-330.
Mansfield, S.D., Parish, R., Di Lucca, C.M., Goudie, J., Kang, K.-Y., Ott, P., 2009.
Revisiting the transition between juvenile and mature wood: a comparison of fibre

45
length, microfibril angle and relative wood density in lodgepole pine. Holzforschung 63,
449-456.
Mansfield, S.D., Parish, R., Goudie, J.W., Kang, K. Ott, P., 2007. The effects of crown
ratio on the transition from juvenile to mature wood production in lodgepole pine in
western Canada. Can. J. For. Res. 37, 1450-1459.
Mencuccini, M., Martínez-Vilalta, J., Vanderklein, D., Hamid, H.A., Korakaki, E., Lee, S.,
Michiels, B., 2005. Size-mediated ageing reduces vigour in trees. Ecol. Lett. 8, 1183-
1190.
Morsdorf, F., Meier, E., Koetz, B., Klaus, I., Dobbertin, M., Allgoewer, B., 2004. LIDAR-
based geometric reconstruction of boreal type forest stands at single tree level for
forest and wildland fire management. Remote Sens. Env. 92, 353-362.
Murphy, G., 2008. Determining stand value and log product yields using terrestrial lidar
and optimal bucking: a case study. J. For. 106, 317-324.
Næsset, E., 2002. Predicting forest stand characteristics with airborne scanning laser
using a practical two-stage procedure and field data. Remote Sens. Env. 90, 88-99.
Næsset, E., 2009. Effects of different sensors, flying altitudes, and pulse repetition
frequencies on forest canopy metrics and biophysical stand properties derived from
small-footprint airborne laser data. Remote Sens. Env. 113, 148-159.
Næsset, E., Gobakken, T., Holmgren, J., Hyyppa, H., Hyyppa, J., Maltamo, M., Nilsson,
M., Olsson, H., Persson, A., Soderman, U., 2004. Laser scanning of forest resources:
the Nordic experience. Scand. J. For. Res. 19, 482-499.
Næsset, E., Nelson, R., 2007. Using airborne laser scanning to monitor tree migration in
the boreal-alpine transition zone, Remote Sens. Env. 110, 357-369.

46
Naesset, E., Økland, T., 2002. Estimating tree height and tree crown properties using
airborne scanning laser in a boreal nature reserve. Remote Sens. Env. 79, 105-115.
Ni-Meister, W., Strahler, A.H., Woodcock, C.E., Schaaf, C.B., Jupp, D.L.B., Yao, T.,
Zhao, F., Yang, X.Y., 2008. Modeling the hemispherical scanning, below-canopy lidar
and vegetation structure characteristics with a geometric-optical and radiative-transfer
model. Can. J. Remote Sens. 34, S385-S397.
Oliver, C.D., Larson, B.C., 1996. Forest stand dynamics. Wiley, New York, pp. 544.
Persson, A., Holmgren, J., Söderman, U., 2002. Detecting and measuring individual
trees using an airborne laser scanner. Photogramm. Eng. Remote Sens. 68, 925-932.
Plomion, C., Leprovost, G., Stokes, A., 2001. Wood formation in trees. Plant Phys. 127,
1513-1523.
Popescu, S.C., 2007. Estimating biomass of individual pine trees using airborne lidar.
Biomass and Bioenergy 31, 646-655.
Popescu, S.C., Wynne, R.H., Scrivani, J.A., 2004. Fusion of small-footprint lidar and
multispectral data to estimate plot- level volume and biomass in deciduous and pine
forests in Virginia, USA . For Sci. 50, 551-565.
Ramirez, M., Rodriguez, J., Peredo, M., Valenzuela, S., Mendonca, R., 2009. Wood
anatomy and biometric parameters variation of Eucalyptus globulus clones. Wood Sci.
and Technol. 43, 131-141.
Raven, P.H., Evert, R.F., Eichhorn, S.E., 2005. Biology of Plants (seventh edition).
Freeman and Company, New York. pp. 686.
Reme, P.A., Helle, T., 2002. Assessment of transverse dimensions of wood tracheids
using SEM and image analysis. Eur. J. Wood and Wood Prod. 60, 277-282.

47
Riano, D., Chuvieco, E., Condes, S., Gonzalez-Matesanz, J., Ustin, S.L., 2004.
Generation of crown bulk density for Pinus sylvestris L. from lidar. Remote Sens. Env.
92, 345-352.
Roderick, M.L., Berry, S.L., 2001. Linking wood density with tree growth and
environment: a theoretical analysis based on the motion of water. New Phytol. 149,
473-485.
Rutzinger, M., Pratihast, A.K., Oude Elberink, S., Vosselman, G., 2010. Detection and
modelling of 3D trees from mobile laser scanning data. Int. Arch. Photogramm. Remote
Sens. Spat. Inf. Sci. 38 part 5, 520-525.
Ryan, M.G., Lavigne, M.B., Gower, S.T., 1997. Annual carbon cost of autotrophic
respiration in boreal forest ecosystems in relation to species and climate. J. Geophys.
Res. 102, 28,871-28,883.
Sayer, J.A., Vanclay, J.K., Byron, N., 1997. Technologies for sustainable forest
management: challenges for the 21st century. In: Sayer, J.A. (Ed.) The Earthscan
Reader in Forestry and Development, Chapter 13. Earthscan, London, pp. 264-279.
Schwalbe, E., Maas, H.-G., Dietrich, R., Ewert, H., 2008. Glacier velocity determination
from multi temporal terrestrial long range scanner point clouds. Int. Arch. Photogramm.
Remote Sens. Spat. Inf. Sci 37/B5, 457-462.
Seeling, U., 2001. Transformation of plantation forests -- expected wood properties of
Norway spruce (Picea abies (L.) Karst.) within the period of stand stabilization. For.
Ecol. Manag. 151, 195-210.
Sheng-zuo, F., Wen-zhong, Y., Xiang-xiang, F., 2004. Variation in microfibril angle and
its correlation to wood properties in poplars. J. For. Res. 15, 261-267.

48
Samson, G., Tremblay, N., Dudelzak, A.E., Babichenko, S.M., Dextraze, L., Wollring, J.,
2000. Nutrient stress of corn plants: Early detection and discrimination using a
compact multiwavelength fluorescent lidar. Proceedings of EARSeL-SIG Workshop
LIDAR, June 16 – 17, Dresden, Germany.
Smithers L.A., 1957. Thinning in lodgepole pine stands in Alberta. In: Tech. Note 52,
Canadian Department of Northern Affairs and National Resources, pp. 26.
Solberg, S., Næsset, E., Bollandsås, O.M., 2006. Single tree segmentation using
airborne laser scanner data in a structurally heterogeneous spruce forest.
Photogramm. Eng. Remote Sens. 72, 1369-1378.
St. Clair, J.B., 1994a. Genetic variation in tree structure and its relation to size in
Douglas-fir. 1. Biomass partitioning, foliage efficiency, stem form, and wood density.
Can. J. For. Res. 24, 1226-1235.
St. Clair, J.B., 1994b. Genetic variation in tree structure and its relation to size in
Douglas-fir. 2. Crown form, branch characters, and foliage characters. Can. J. For.
Res. 24, 1236-1247.
Sterck, F.J., Poorter, L., Schieving, F., 2006. Leaf traits determine the growth-survival
trade-off across rain forest tree species. Am. Nat. 167, 758-765.
Stewart, J.D., Jones, T.N., Noble, R.C., 2006. Long-term Lodgepole pine silviculture
trials in Alberta: history and current status. Canadian Forest Service, Natural
Resources Canada.
Strahler, A.H., Jupp, D.L.B., Woodcock, C.E., Schaaf, C.B., Yao, T., Zhao, F., Yang, X.,
Lovell, J., Culvenor,D., Newnham, G., Ni-Miester, W., Boykin-Morris, W., 2008.

49
Retrieval of forest structural parameters using a ground-based lidar instrument
‘Echidna®’. Can. J. Remote Sens. 34, S426-S440.
Suarez, J.C., Gardiner, B.A., Di Luca, M., Goudie, J., Polsson, K., 2010. Consequences
of stand structure on timber quality. Silvilaser 2010, September 14-17, Freiburg,
Germany.
Swenson, N.G., Enquist, B.J., 2007. Ecological and evolutionary determinants of a key
plant functional trait: wood density and its community-wide variation across latitude
and elevation. Am. J. Bot. 94, 451-459.
Tansey, K., Selmes, N., Anstee, A., Tate, N.J., Denniss, A., 2009. Estimating tree and
stand variables in a Corsican Pine woodland from terrestrial laser scanner data. Int. J.
Remote Sens. 30, 5195–5209.
Thies, M., Pfeifer, N., Winterhalder, D., Gorte, B.G.H., 2004. Three-dimensional
reconstruction of stems for assessment of taper, sweep, and lean based on laser
scanning of standing trees. Scandinavian J. For. Res. 19, 571-581.
Twede, D., Selke, S.E.M., 2005. Cartons, crates and corrugated board: handbook of
paper and wood packaging technology. DEStech Publications, Lancaster. pp. 528.
Väisänen, H., Kellomäki, S., Oker-Blom, P., Valtonen, E., 1989. Structural development
of Pinus sylvestris stands with varying initial density: A preliminary model for quality of
sawn timber as affected by silvicultural measures. Scand. J. For. Res. 4, 233-238.
Vallet, P., Dhôte, J., Moguédec, G.L., Ravart, M., Pignard, G., 2006. Development of
total aboveground volume equations for seven important forest tree species in France.
For. Ecol. Manag. 229, 98-110.

50
Van der Zande, D., Hoet, W., Jonckheere, I., Van Aardt, J., Coppin, P., 2006. Influence
of measurement set-up of ground-based LiDAR for derivation of tree structure. Ag. For.
Met. 141, 147-160.
Van Leeuwen, M., Nieuwenhuis, M., 2010. Retrieval of forest structural parameters
using LiDAR remote sensing. Eur. J. For. Res. 129, 749-770.
Vepakomma, U., St-Onge, B., Kneeshaw, D., 2008. Spatially explicit characterization of
boreal forest gap dynamics using multi-temporal lidar data. Remote Sens. Env. 112,
2326-2340.
Via, B.K., Stine, M., Shupe, T.F., Chi-Leung, S., Groom, L., 2004. Genetic improvement
of fiber length and coarseness based on paper product performance and material
variability – a review. IAWA J. 25, 401-414.
Wang, T., Aitken, S.N., 2001. Variation in xylem anatomy of selected populations of
lodgepole pine. Can. J. For. Res. 31, 2049-2057.
Waring, R.H., Schroeder, P.E., Oren, R., 1982. Application of the pipe model theory to
predict canopy leaf area. Can. J. For. Res. 12, 556–560.
Watson, P., Garner, C., Robertson, R., Reath, S., Gee, W., Hunt, K., 2003. The effects
of initial tree spacing on the fibre properties of plantation-grown coastal western
hemlock. Can. J. For. Res. 33, 2460-2468.
Watt, M.S., Moore, J.R., Façon, J., Downes, G.M., Clinton, P.M., Coker, G., Davis, M.R.,
Simcock, R., Parfitt, R.L., Dando, J., Mason, E.G., Bown, H.E., 2006. Modelling the
influence of stand structural, edaphic and climatic influences on juvenile Pinus radiata
dynamic modulus of elasticity. For. Ecol. Manag. 229, 136-144.

51
Wehr, A., Lohr, U., 1999. Airborne laser scanning – and introduction and overview.
ISPRS J. Photogramm. Remote Sens. 54, 68-82.
Wilhelmsson, L., Arlinger, J., Spangberg, K., Lundqvist, S.-O., Grahn, T., Hedenberg,
O., Olsson, L., 2002. Models for predicting wood properties in stems of Picea abies
and Pinus sylvestris in Sweden. Scand. J. For. Res. 17, 330-350.
Wimmer, R., Downes, G.M., Evans, R., 2002. Temporal variation of microfibril angle in
Eucalyptus nitens grown in different irrigation regimes. Tree Physiology 22, 449-457.
Wulder, M.A., Bater, C.W., Coops, N.C., Hilker, T., White, J.C., 2008. The role of LiDAR
in sustainable forest management. For. Chron. 84, 807-826.
Yu, X.W., Hyyppa, J., Kaartinen, H., & Maltamo, M., 2004. Automatic detection of
harvested trees and determination of forest growth using airborne laser scanning.
Remote Sens. Env. 90, 451-462.
Zheng G., Moskal, L.M., 2009. Retrieving Leaf Area Index (LAI) using remote sensing:
Theories, methods and sensors. Sens. 9, 2719-2745.
Zobel, B.J., Van Buijtenen, J.P., 1989. Wood variation - its causes and control. Springer
Verlag: Berlin, pp 363.




















