
Archivos do Museu Nacional do Rio de Janeiro
236
ISSN 0365-4508 Nunquam aliud natura, aliud sapienta dicit Juvenal, 14, 321 In silvis academi quoerere rerum, Quamquam Socraticis madet sermonibus Ladisl. Netto, ex Hor RIO DE JANEIRO Outubro/Dezembro 2004
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Archivos do Museu Nacional do Rio de Janeiro

ISSN 0365-4508
Nunquam aliud natura, aliud sapienta dicit Juvenal, 14, 321
In silvis academi quoerere rerum, Quamquam Socraticis madet sermonibus
Ladisl. Netto, ex Hor
RIO DE JANEIRO
Outubro/Dezembro 2004

Arquivos do Museu Nacional
Universidade Federal do Rio de Janeiro
Reitor Aloísio Teixeira
Museu Nacional
Diretor Sérgio Alex K. Azevedo
Editores Pro Tempore Miguel Algel Monné Barrios
Ulisses Caramaschi
Editores de Área Alexander Wilhelm Armin Kellner
Cátia Antunes de Mello Patiu
Ciro Alexandre Ávila
Débora de Oliveira Pires
Gabriel Luiz Figueira Mejdalani
Isabel Cristina Alves Dias
João Alves de Oliveira
Marcelo Araújo de Carvalho
Maria Dulce Barcellos Gaspar de Oliveira
Marília Lopes da Costa Facó Soares
Rita Scheel Ybert
Vânia Gonçalves Lourenço Esteves
Normalização Vera de Figueiredo Barbosa
Diagramação e Arte-final Lia Ribeiro
Conselho Editorial
André Pierre Prous-Poirier Universidade Federal de Minas Gerais
David G. Reid The Natural History Museum - Reino Unido
David John Nicholas Hind Royal Botanic Gardens - Reino Unido
Fábio Lang da Silveira Universidade de São Paulo
François M. Catzeflis Institut des Sciences de VÉvolution - França
Gustavo Gabriel Politis Universidad Nacional dei Centro - Argentina
John G. Maisey Americam Museun of Natural Flistory - EUA
Jorge Carlos Delia Favera Universidade do Estado do Rio de Janeiro
J. Van Remsen Louisiana State University - EUA
Maria Antonieta da Conceição Rodrigues Universidade do Estado do Rio de Janeiro
Maria Carlota Amaral Paixão Rosa Universidade Federal do Rio de Janeiro
Maria Helena Paiva Henriques Universidade de Coimbra - Portugal
Maria Marta Cigliano Universidad Nacional La Plata - Argentina
Miguel Trefaut Rodrigues Universidade de São Paulo
Miriam Lemle Universidade Federal do Rio de Janeiro
Paulo A. D. DeBlasis Universidade de São Paulo
Philippe Taquet Museum National d'Histoire Naturelle - França
Rosana Moreira da Rocha Universidade Federal do Paraná
Suzanne K. Fish University of Arizona - EUA
W. Ronald Heyer Smithsonian Institution - EUA

ARQUIVOS
DO
MUSEU NACIONAL
VOLUME 62
NÚMERO 4
OUTUBRO/DEZEMBRO 2004
RIO DE JANEIRO
Arq. Mus. Nac. Rio de Janeiro v. 62 n.4 p.341-572 out./dez.2004

ISSN 0365-4508
Arquivos do Museu Nacional, mais antigo periódico científico do Brasil (1876), é uma publicação trimestral (março, junho, setembro e dezembro), com tiragem de 1000 exemplares, editada pelo Museu Nacional/ Universidade Federal do Rio de janeiro. Tem por finalidade publicar artigos científicos inéditos nas áreas de Antropologia, Arqueologia, Botânica, Geologia, Paleontologia e Zoologia. Está indexado nas seguintes bases de dados bibliográficos: Biological Abstracts, ISI - Thomson Scientific, Ulrich'5 International Periodicals Directory, Zoological Record, NISC Colorado e Periódica.
As normas para preparação dos manuscritos encontram- se disponíveis em cada número dos Arquivos e em htttp://acd.ufrj.br/~museuhp/publ.htm. Os artigos são avaliados por, pelo menos, dois especialistas na área envolvida e que, eventualmente, pertencem ao Conselho Editorial. O conteúdo dos artigos é de responsabilidade exclusiva do(s) respectivo(s) autor(es).
Os manuscritos deverão ser encaminhados para Museu Nacional/UFRj, Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de janeiro, RJ, Brasil.
Financiamento
CNPq Conselho Nacional de Desenvolvimento Cientifico e Tecnológico
Arquivos do Museu Nacional, the oldest Brazilian scientific publication (1876), is issued every three months (March, June, September and December). It is edited by Museu Nacional/Universidade Federal do Rio de Janeiro, with a circulation of 1000 copies. Its purpose is the edition of unpublished scientific articles in the areas of Anthropology, Archaeology, Botany, Geology, Paleontology and Zoology. It is indexed in the following bases of bibliographical data: Biological Abstracts, ISI - Thomson Scientific, Ulrich's International Periodicals Directory, Zoological Record, NISC Colorado and Periódica.
Instructions for the preparation of the manuscripts are available in each edition of the publication and at http://acd.ufrj.br/~museuhp/publ.htm. The articles are reviewed, at least, by two specialists in the area that may, eventually, belong to the Editorial Board. The authors are totally responsible for the content of the texts.
The manuscripts should be sent to Museu Nacional/ UFRJ, Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil.
© 2004 - Museu Nacional/UFRJ
Arquivos do Museu Nacional - vol.l (1876) - Rio de Janeiro: Museu Nacional.
Trimestral Até o v.59, 2001, periodicidade irregular
ISSN 0365-4508
1. Ciências Naturais - Periódicos. I. Museu Nacional (Brasil).
CDD 500.1

TmnrronnnjfnnnrTi nnniiituiíiüífflnnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004
ISSN 0365-4508
TEORIA E MÉTODOS EM ANTRACOLOGIA.
2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO 1
(Com 4 figuras)
RITA SCHEEL-YBERT 2
RESUMO: Este artigo dá continuação a uma série de textos sobre a metodologia antracológica adaptada às
regiões tropicais. São apresentadas as técnicas de trabalho de campo e de laboratório, em particular o modo
de depósito dos carvões de origem arqueológica, amostragem antracológica, concentração do material,
constituição de coleções de referência, utilização de bancos de dados antracológicos, determinação dos
carvões e dimensão dos fragmentos identificados, assim como uma discussão sobre amostras flotadas,
considerando os fragmentos que bóiam e o refugo de peneira.
Palavras-chave: antracologia, carvão, banco de dados, microscopia, flotação.
ABSTRACT: Theory and methods in anthracology. 2 - Field and laboratory techniques.
This paper presents a suite to a series of articles on the anthracological methodology in the tropics. We
discuss here the techniques of field and laboratory work, especially the way archaeological charcoal is
deposited, anthracological sampling, concentration of the material, constitution of reference collections, use
of charcoal data-bases, charcoal determination, and dimension of charcoal pieces that can be identified, as
well as a discussion on the flotation technique.
Key words: anthracology, archaeological charcoal, data-base, microscopy, flotation.
INTRODUÇÃO
A identificação de espécies a partir de material carbonizado é bastante antiga (HEER, 1866; PREJAWA, 1896; BREUIL, 1903), mas o método utilizado na época, a confecção e análise de lâminas finas, era lento e difícil, e as pesquisas não podiam almejar à reconstituição paleoambiental. A utilização da microscopia de luz refletida (WESTERN, 1963; VERNET, 1973) possibilitou a multiplicação das análises antracológicas, facilitando o estudo dos carvões e propiciando o desenvolvimento de abordagens paleoecológicas e paleoetnológicas, além de ensejar a realização de pesquisas metodológicas.
Este desenvolvimento levou os pesquisadores a aprimorar também os métodos de amostragem no campo, assim como a aprofundar uma série de questionamentos teóricos sobre a pertinência e a validade dos estudos antracológicos (cf. SCHEEL- YBERT, 2004). Ainda que a preocupação com a análise metodológica tenha se desenvolvido muito entre os antracólogos nas últimas décadas, a maior
parte dos trabalhos foi realizada em regiões mediterrâneas e temperadas da Europa (CHABAL, 1988, 1997; HEINZ, 1990; BADAL-GARCIA & HEINZ, 1991; THÉRY-PARISOT, 2001) e, em menor grau, nos Estados Unidos (SMART & HOFFMAN, 1988). Uma grande parte dos princípios estabelecidos por estes autores pode ser aplicada aos estudos de antracologia tropical, mas a definição de procedimentos metodológicos adaptados aos trópicos é muito importante, devido às especificidades de seus diversos ambientes.
A antracologia é uma disciplina relativamente nova no Brasil (SCHEEL, GASPAR & YBERT, 1996a, 1996b; SCHEEL-YBERT, 1998, 1999, 2000, 2001; SCHEEL-YBERT et al, 2003), onde a necessidade de adaptação da metodologia de coleta de campo se fez sentir desde os primeiros trabalhos. Até hoje, ainda é raro que os arqueólogos brasileiros coletem os carvões de modo sistemático, pois este material costuma ser considerado exclusivamente como uma possível fonte de datações radiocronológicas. Esta atitude, aliás, não é restrita à arqueologia brasileira, e foi observada também em Portugal
1 Submetido em 10 de fevereiro de 2004. Aceito em 22 de novembro de 2004.
Apoio financeiro: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq - PROFIX n° 540207/01-2) e da Fundação de Amparo à Pesquisa
do Estado do Rio de Janeiro (FAPERJ - E-26/152.430/02: Projeto “Soberanos da Costa”, coordenado por M.D.Gaspar)
2 Museu Nacional/UFRJ, Departamento de Antropologia. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil. E-mail: [email protected].

344 R. SCHEEL-YBERT
(FIGUEIRAL, 1994) e na Itália (FIORENTINO, 1995), por exemplo.
Este artigo tem como objetivo ressaltar a importância da coleta sistemática dos carvões e apresentar aos arqueólogos e futuros antracólogos brasileiros uma metodologia adaptada.
TÉCNICAS DE CAMPO
1. Modo de depósito dos carvões
A metodologia de amostragem antracológica deve ser adaptada em função do modo de depósito dos carvões. A distinção entre os carvões concentrados nas estruturas de combustão e os carvões dispersos nas camadas arqueológicas é fundamental, pois estes dois tipos de depósito fornecem informações distintas e, em conseqüência, devem ser coletados separadamente. Os carvões concentrados são mais visíveis durante a escavação, mas as camadas arqueológicas em geral apresentam também uma grande quantidade de carvões dispersos no sedimento (CHABAL, 1988, 1997; HEINZ, 1990; BADAL-GARCIA & HEINZ, 1991), provenientes da dispersão dos fragmentos de queima ou da limpeza periódica de fogões ou fogueiras.
Os sedimentos da maior parte dos sítios arqueológicos brasileiros contêm uma grande quantidade de fragmentos de carvão dispersos. É provável que eles tenham sido originados pela queima de lenha doméstica, mas essa hipótese ainda deve ser confirmada na maioria dos casos. No caso de sambaquis do litoral brasileiro, foi demonstrado que os carvões dispersos provinham efetivamente da lenha coletada para uso doméstico, para a qual não havia seleção (SCHEEL- YBERT, 1999,2001). A análise de fragmentos de carvão provenientes de várias coletas de madeira (lenha) sucessivas ao longo do período de ocupação, que representem uma boa amostragem da vegetação contemporânea, é uma premissa indispensável para uma boa interpretação paleoecológica baseada neste material (SCHEEL-YBERT, 2004).
Os carvões concentrados freqüentemente se originam de fogos, fogões ou fogueiras que tiveram uma curta utilização no tempo, ou representam apenas os restos da última queima neles realizada, o que implicaria numa amostra pouco significativa da vegetação como um todo. Eles podem também estar relacionados a atividades especializadas, por exemplo materiais de construção ou objetos manufaturados queimados acidentalmente, queima de cerâmica ou, em outras regiões do mundo, fornos de padaria, de metalurgia etc., que muitas vezes requerem um combustível
especial mas que podem ser reconhecidos no campo a partir de critérios arqueológicos. As estruturas contendo carvões concentrados sempre devem ser coletadas separadamente, pois seu estudo oferece importantes informações paleoetnológicas.
Em regiões temperadas, considera-se que as interpretações paleoecológicas devem ser baseadas exclusivamente nos fragmentos de carvões dispersos. Não só estes fragmentos recobrem um período de atividades mais longo, mas principalmente porque se considera que o estudo de carvões concentrados fornece resultados qualitativos e quantitativos incompletos e aberrantes do ponto de vista paleoecológico (HEINZ, 1990). Diversos autores observaram que as amostras de fogueiras (carvões concentrados) fornecem geralmente um número de taxa inferior àquele encontrado nos carvões dispersos nos sedimentos arqueológicos, e que eles são freqüentemente sub- ou super-representados (HEINZ 1990; BADAL GARCIA & HEINZ 1991; BADAL GARCIA 1992).
No entanto, CHABAL (1989a) e THIÉBAULT (1995) observaram que em certos casos os carvões de fogueiras fornecem informações bastante precisas sobre o paleoambiente vegetal; para isso, é necessário que a amostra apresente uma grande riqueza taxonômica, associada a um longo tempo de utilização. PERNAUD (1992) concorda que os carvões concentrados em certas estruturas podem fornecer uma imagem válida do ambiente vegetal, mas alerta que é somente a análise dos fragmentos dispersos que confirma, ou não, a validade das interpretações obtidas.
Na zona tropical, só existe até agora um estudo sobre esta questão (SCHEEL-YBERT, 1998). Diversas amostras de carvões concentrados de fogueiras de sambaquis foram analisadas no sudeste do Estado do Rio de Janeiro, demonstrando que eles em geral apresentam uma grande diversidade florística e permitem em todos os casos a dedução de um ambiente vegetal que é qualitativamente semelhante ao que é verificado pelo estudo dos carvões dispersos. Isso tende a mostrar que, em teoria, o estudo de carvões concentrados fornece, nos trópicos, informações paleoecológicas fiáveis, desde que se disponha de um grande número de fogueiras, contendo uma grande quantidade de fragmentos, em cada camada arqueológica. Na prática, esta condição é muito dificilmente preenchida, de modo que, em função do mínimo amostrai, as análises paleoecológicas devem ser preferencialmente baseadas no estudo dos carvões dispersos.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

TEORIA E MÉTODOS EM ANTRACOLOGIA. 2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO 345
2. Amostragem antracológica
Uma análise antracológica qualitativa e quantitativamente confiável só é possível a partir da análise de um grande número de carvões para cada estrato3, de modo que a superfície do sítio arqueológico deve ser amostrada o mais amplamente possível, para cada nível estratigráfico.
O ideal é que a amostragem antracológica seja feita concomitantemente à escavação arqueológica, a qual deve seguir o método de decapagem de camadas naturais em superfícies amplas, a fim de obter resultados que permitam boas interpretações paleoecológicas e paleoetnológicas e que levem em conta as eventuais heterogeneidades locais do sítio.
O método geralmente preconizado para a amostragem concomitante à escavação consiste na amostragem sistemática do sítio seguindo uma malha de lm2. Deve-se coletar uma amostra em cada quadrícula, em cada decapagem. A superfície a ser
amostrada deve perfazer entre V* e a totalidade de cada metro quadrado, em função da riqueza da camada em carvões (CHABAL, 1988).
Este método pode ser adaptado, seja em razão de contingências da escavação arqueológica (SCHEEL- YBERT, 1998), seja devido à abundância de carvões no sítio (SCHEEL-YBERT 85 SOLARI, no prelo).
No caso do Sambaqui Ilha da Boa Vista I (Estado do Rio de Janeiro), por exemplo, cuja escavação foi feita pelo método de decapagem de superfícies amplas, a malha de quadriculamento adotada pelos arqueólogos foi de 2 metros de lado (BARBOSA, GASPAR & BARBOSA, 1994). A amostragem antracológica foi feita em quadrículas alternadas, coletando-se, em cada decapagem, um balde de 10
litros de sedimento em seções de 30 x 30cm situadas no ângulo nordeste de cada quadrícula (Fig.l). Este material foi peneirado a seco, no campo, com malha de 4 mm e reservado para flotação e triagem posterior.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 R
Q
P
O
N
M
L
K
J
I
H
G
F
E
D
C
B
A
g ■ ■ ■ ■ ■ u u ■
g ■ ■ ■ ■ ■ u u ■
p ■ ■ ■ ■ ■ ■ u ■
■ ■ ■ 1 ■ u ■ 1 trincheira leste - oeste
. ■ . ////// • YJjOtâ ;. íSíííí
trincheira norte - sul
0 6 m
Fig.l- Esquema do quadriculamento do Sambaqui da Ilha da Boa Vista I, com localização das quadrículas amostradas e do
perfil antracológico (■) em escala exagerada. A elipse cinza escuro representa a superfície do sítio; o círculo cinza claro
representa a área de maior concentração dos vestígios de habitação (adaptado de BARBOSA, GASPAR & BARBOSA, 1994).
3 Uma discussão sobre o número mínimo de carvões da amostra antracológica e sobre a validade amostrai será apresentada em um artigo posterior desta série.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

346 R. SCHEEL-YBERT
No caso do Abrigo Santa Elina (Estado de Mato Grosso), sítio que se caracteriza pelo excelente estado de conservação dos macro-restos vegetais e pela grande abundância de carvões (SCHEEL- YBERT & SOLARI, no prelo), a malha definida durante a escavação arqueológica foi de 1 metro de lado. A amostragem antracológica foi inicialmente feita em todas as quadrículas, nas quais foi coletado sedimento em seções correspondentes a V* de cada metro quadrado, em cada decapagem. Este material foi peneirado a seco com malha de 4mm e os carvões foram recuperados por triagem manual diretamente no campo. A quantidade excessiva de carvões obtida levou a uma redução da amostragem em campanhas posteriores, que continuou sendo feita em seções de Vi de metro quadrado, mas em quadrículas alternadas. Ainda assim, uma grande quantidade de carvões foi coletada.
Na arqueologia brasileira, limitações de tempo e dinheiro freqüentemente conduziram os arqueólogos a realizar escavações em pequenas trincheiras, baseadas em decapagens artificiais e com recuperação parcial dos vestígios. Embora esta tendência esteja sendo revertida, escavações em superfícies amplas ainda são relativamente raras, de modo que a amostragem antracológica deve sempre ser feita de acordo com o modo de escavação escolhido e as condições encontradas pela equipe que estuda o sítio. Nos casos em que uma amostragem por superfícies amplas não é possível, ou em sítios nos quais as escavações tenham sido realizadas previamente, sem recuperação dos carvões, pode-se proceder à amostragem antracológica pelo método de perfis. Este método também pode ser utilizado em associação ao método de amostragem sistemática.
No caso de amostragem de solos, a coleta é feita exclusivamente ao longo de perfis, por decapagem de camadas sucessivas em níveis artificiais.
A amostragem em perfil padronizada por nossa equipe segue seções de 2m de largura por 50cm de profundidade (ou seja, lm2); a decapagem é feita em níveis artificiais de lOcm de espessura. Nos casos em que a estratificação natural da camada arqueológica estiver em discordância com os níveis artificiais, é importante sempre proceder de modo a não misturar o material proveniente de sedimentos diferentes. Deve-se também sempre coletar as amostras de carvões concentrados separadamente.
Freqüentemente é difícil manter-se constantes a horizontalidade e a espessura dos níveis artificiais
neste tipo de amostragem. Por isso, foi concebido um “guia de amostragem”, muito simples de construção e de utilização, que facilita a realização das amostragens em níveis regulares (YBERT, SCHEEL & GASPAR, 1997).
Por outro lado, deve-se sempre evitar “raspar” o sedimento com a colher durante a coleta para amostragem antracológica, a fim de não esmagar os carvões. Preferencialmente, deve-se retirar pequenos blocos de sedimento e colocá-los no balde. O mesmo deve ser observado durante a peneiragem; a utilização da pazinha para “ajudar” o sedimento a passar pela peneira destrói os carvões e impossibilita sua análise, ainda mais quando o sedimento está molhado. Deve-se evitar inclusive esmagar o sedimento contra a peneira, mesmo com as mãos, a fim de não destruir os carvões que se pretende analisar.
3. Concentração do material
De um modo geral, três métodos de recuperação dos carvões podem ser utilizados: coleta manual, peneiragem ou flotação (BADAL et al, 1989; FIGUEIRAL, 1992).
A coleta manual dos carvões só é indicada para os carvões concentrados. Ela é útil no caso da identificação de estruturas no campo, por exemplo fogueiras ou estacas queimadas, para dar informações sobre a associação de restos botânicos com outros tipos de vestígios, e também para datação. No entanto, este método não deve ser utilizado como único modo de amostragem antracológica, pois geralmente só leva em conta os fragmentos maiores, resultando inevitavelmente numa amostra distorcida.
A utilização da peneira garante uma melhor amostragem, sendo importante para revelar os carvões dispersos contidos no sedimento arqueológico. No entanto, é fundamental que todos os fragmentos de carvão retidos pela peneira sejam coletados, pois a seleção das peças maiores ou mais bem conservadas introduz um elemento de escolha subjetiva que acarretará erros de interpretação.
Em regiões tropicais, a concentração do material arqueológico deve ser feita utilizando-se peneiras de malha de 4mm (vide “Dimensão dos fragmentos
identificados”). Podem-se usar peneiras de malha inferior, mas nunca superior.
Tanto no caso de amostragem sistemática em superfícies amplas, quanto de coletas em perfil, o sedimento deve ser coletado em baldes. Deve-se sempre anotar a quantidade de baldes retirada em
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

TEORIA E MÉTODOS EM ANTRACOLOGIA. 2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO 347
cada unidade, a fim de se ter uma estimativa mais precisa do volume de sedimento amostrado. Sempre que possível, deve-se pesar o sedimento contido nos baldes. Estes dados são importantes para se comparar a abundância relativa de carvões entre as diversas camadas de um sítio ou entre sítios diferentes, o que só é possível quando se conhece o tamanho exato da amostra (superfície amostrada, volume, peso do sedimento amostrado etc.).
A peneiragem a seco, relativamente fácil, pois normalmente faz parte da cadeia operatória da escavação arqueológica, em geral é insuficiente para uma concentração eficaz da amostra antracológica. Embora o ideal seja peneirar o sedimento uma única vez, isto nem sempre é possível, devido a limitações técnicas. Por isso pode- se, por exemplo, peneirar o sedimento a seco no campo, a fim de diminuir a quantidade a ser transportada, e fazer a flotação posteriormente.
A peneiragem com água pode ser feita por imersão parcial das peneiras num tanque, piscina ou equivalente, ou lavando-se o sedimento da peneira com uma mangueira. A peneiragem com mangueira, muito útil para outras disciplinas, principalmente a zooarqueologia, costuma ser inadequada para a recuperação dos restos vegetais, que podem ser pulverizados pelo jato de água ou pelos instrumentos utilizados para revolver o material.
O maior problema envolvido na utilização de peneiras para a amostragem antracológica decorre do enorme volume de material a ser triado, em particular para os níveis ricos em conchas (no caso de sambaquis) ou em cascalho, tornando este método, em geral, lento e pouco produtivo.
A flotação é o método ideal para recuperação dos restos vegetais, pois implica em menor esforço metodológico e maior eficiência, além de ser o menos agressivo para o material. A flotação permite a recuperação de material botânico de todas as classes de tamanho preservadas no sedimento. Os fragmentos de carvão sendo de pequenas dimensões e muito leves, sua capacidade de flutuação pode ser aproveitada para separá-los do material mais pesado. Além disso, na medida em que uma parte do material flutua e a outra permanece retida pela peneira da célula de flotação, este método facilita a separação de numerosos outros restos, úteis a outras disciplinas (sementes, moluscos, ossos de peixe, cerâmica, fauna etc.).
O princípio de funcionamento de uma “célula de flotação” é bastante simples. Consiste em lavar o sedimento, depositado em uma peneira submersa
numa cuba, em uma corrente de água turbilhonante. Os carvões, liberados do sedimento, são levados à superfície da água e em direção à periferia da cuba, caindo sobre uma peneira de malha fina onde os elementos sólidos são recuperados (YBERT, SCHEEL & GASPAR, 1997; PEARSALL, 2000). Esta técnica é baseada na diferença de densidade dos resíduos orgânicos e inorgânicos. Fragmentos carbonizados de madeira, sementes ou tubérculos em geral flutuam, enquanto restos de moluscos, ossos e cerâmica, são depositados na peneira.
Existem vários modelos de células de flotação, desde o artesanal, baseado na imersão de um balde numa cuba maior contendo água, até os mais elaborados, com assistência mecânica. O modelo de uma célula de flotação desenvolvido no Brasil foi apresentado por YBERT, SCHEEL & GASPAR (1997); vários modelos utilizados nos Estados Unidos e na Europa foram apresentados por PEARSALL (2000).
Se existir uma fonte de água nas proximidades do sítio, a flotação pode ser feita diretamente no campo. Quando possível, deve-se esvaziar sistematicamente a cuba da célula de flotação após o tratamento de cada amostra, a fim de evitar contaminação entre amostras diferentes, especialmente quando existe a possibilidade de se fazer uma datação radiocarbônica posterior (vide “Análise e identificação
dos carvões”). No campo ou no laboratório, o sedimento que permanece sobre a peneira (refugo de peneira) deve ser sistematicamente triado, a fim de recuperar os fragmentos de carvão que não flutuaram (vide “Amostras flotadas: fragmentos
flutuantes e refugo de peneira’).
O material peneirado com água ou submetido a flotação deve ser seco longe de uma fonte de calor intensa para evitar sua fragmentação e deterioração da estrutura anatômica. A utilização de cones de papel jornal é muito prática para secar os fragmentos, que em seguida devem ser acondicionados em sacos plásticos, mas deve ser evitada quando se intenciona fazer a datação dos carvões. Os carvões não devem ser manipulados até sua secagem completa, a fim de evitar uma quebra acidental.
4. Amostras flotadas: fragmentos flutuantes e refugo
DE PENEIRA
Num estudo feito em diferentes frações resultantes da flotação de uma amostra do sítio de Lattara, na região mediterrânea da França, foi observado que
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

348 R.SCHEEL-YBERT
o refugo da peneira contém tantos fragmentos quanto a fração flotada (que bóia), mas que a riqueza taxonômica é muito mais elevada nesta última (CHABAL, 1989b, 1997). A autora considera esse fato como uma regra geral, e acredita que a ílotação provoque uma triagem desigual em função do peso dos fragmentos: apenas os maiores fragmentos seriam retidos na peneira (o que explicaria a menor riqueza taxonômica do refugo da peneira) e somente eles sofreriam uma refragmentação posterior (o que explicaria o mesmo número de fragmentos nas duas frações).
No estudo antracológico do Sambaqui do Forte (Cabo Frio, Estado do Rio de Janeiro), a fração flotada e o refugo da peneira de 32 amostras foram coletados e analisados separadamente (SCHEEL-YBERT, 1998). A quantidade de fragmentos em cada uma destas frações é muito variável de um nível a outro, mas os carvões flotados são em geral predominantes. No plano qualitativo, observou-se que certos taxa se encontram sempre preferencialmente no refugo da peneira (p.ex. Condalia sp) ou nos carvões flotados (p.ex. Maytenus sp, Pachystroma sp), enquanto outros podem ser, de acordo com as amostras, mais abundantes em uma fração ou em outra (p.ex. Myrtaceae tipo 1 e tipo 5). A riqueza taxonômica das duas frações é, em geral, comparável (Fig.2). Além disso, não foi notada nenhuma diferença de repartição de tamanho entre os carvões flotados e os que permaneceram na peneira, e nós consideramos que não existe nenhuma razão que possa explicar que uma refragmentação posterior afete os fragmentos de somente uma das duas frações.
A melhor forma de explicar as variações observadas são as diferenças de características físicas entre os taxa, especialmente a densidade, mas também a presença eventual de impregnações calcárias, que tendem a aumentar a densidade dos fragmentos e, logo, a impedi-los de flutuar. Nos casos em que o sedimento arqueológico está molhado, os carvões também ficam encharcados, o que aumenta sua densidade e impede a flutuação. Por outro lado, é provável que o fenômeno de vitrificação4 também esteja relacionado. De fato, os fragmentos de Condalia sp, encontrados principalmente na fração não-flotada, estavam sempre vitrificados. Durante
a ílotação de amostras de fornos de carvoeiros, os fragmentos vitrificados afundam sistematicamente, ao contrário dos não vitrificados (P.OGEREAU, com.pes., 1998). A vitrificação, que provoca a fusão das paredes celulares, diminui os espaços vazios no interior do carvão, o que resulta num aumento de densidade do carvão.
Apesar de discordar de suas hipóteses, há concordância com CHABAL (1989b, 1997) quanto à necessidade de se triar sistematicamente o refugo de peneira após a ílotação e de se analisar os fragmentos das duas frações, a fim de obter uma imagem mais representativa da composição taxonômica da amostra antracológica.
TÉCNICAS DE LABORATÓRIO
1. Análise e identificação dos carvões
A determinação taxonômica dos carvões fósseis é feita pela comparação de sua estrutura anatômica, que se conserva perfeitamente após carbonização, com aquela do lenho de espécies atuais conhecidas, seja diretamente a partir de amostras carbonizadas contidas numa coleção de referência, seja através de descrições e/ou fotografias de obras da literatura.
Existe um grande número de atlas de anatomia da madeira relativos às espécies mediterrâneas e temperadas (GREGUSS, 1959; SCHWEINGRUBER, 1978, 1990; VERNET et al, 2001 etc.), mas eles são raros nas zonas tropicais e, quando existentes, se referem mais especificamente à região amazônica (DÉTIENNE & JACQUET, 1983; DECHAMPS, 1979, 1980, 1985), ou são especializados em madeiras comerciais (MAINIERI & CHIMELO, 1989), com raras exceções (BARROS & CALLADO, 1997). Existem também livros gerais de anatomia que, embora bastante completos, não têm vocação para “atlas” (METCALFE & CHALK, 1950).
Uma importante série de trabalhos referentes às madeiras americanas foi publicada por RECORD e HESS (RECORD, 1942, 1943a, b, 1944a, b, c, d, e; HESS, 1946, 1948; RECORD & HESS, 1943, entre outros). Suas chaves de determinação são muito úteis, mas a raridade das ilustrações torna difícil a utilização destes trabalhos para a determinação antracológica. Este é o caso também das obras de
4 Fenômeno de fusão e de homogeneização das paredes celulares do carvão, que adquire um aspecto vitrificado e muito refringente. Apesar do nome, este
fenômeno não tem relação com a fusão do silício, que só ocorre em temperaturas muito altas (ca. 1300 a 1400°C). Corpos silicosos intactos são freqüentemente
encontrados no interior de células carbonizadas, e servem como critério de determinação.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

349 TEORIA E MÉTODOS EM ANTRACOLOGIA. 2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO
nível 200-2lOcm □ flotados (Ni: 61; Nsp: 23)
■ não flotados (Ni: 93; Nsp: 28)
nível 230-240cm
<
Fig.2- Proporção entre carvões flotados e não flotados em três níveis diferentes do Sambaqui do Forte (RJ), em número de fragmentos; (-») taxa mencionados no texto.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

350 R.SCHEEL-YBERT
DECHAMPS (1979, 1980, 1985). Pode-se utilizar também um grande número de artigos especializados na anatomia de espécies atuais, que é impossível citar por extenso. Para uma excelente compilação das referências bibliográficas sobre a anatomia sistemática da madeira de espécies atuais publicadas de 1900 até 1993, ver GREGORY (1994).
A análise dos fragmentos de carvão a serem identificados é feita em microscópio óptico de luz refletida com campo claro e campo escuro, a partir da quebra manual dos fragmentos segundo os três planos fundamentais da madeira: transversal, longitudinal tangencial e longitudinal radial (Fig.3). Para as identificações, uma grande importância deve ser dada aos caracteres qualitativos (disposição dos poros e do parênquima, seriação dos raios, ornamento das pontuações intervasculares, tamanho e forma das pontuações radiovasculares, presença de canais, corpos silicosos etc.), mas deve-se levar em conta também as medidas dos caracteres anatômicos. Informações sobre o estudo da anatomia da madeira podem ser encontradas na literatura especializada (BURGER 86 RICHTER, 1991; IAWA COMMITTEE, 1989; METCALFE 86 CHALK, 1950; etc.).
A utilização de técnicas de microscopia mais avançadas, como por exemplo o contraste interferencial (DIC), pode ser muito útil para uma melhor visualização dos caracteres anatômicos, em particular detalhes observados em maior aumento, como por exemplo as características das pontuações.
Observações em microscopia eletrônica de varredura (MEV) também podem ser feitas, a fim de confirmar as determinações ou para representação fotográfica das amostras. Embora BLANKENHORN, JENKINS 8& KLINE (1972) e CUTTER, CUMBIE 86 MCGINNES (1980) façam referência à boa condutividade dos carvões, que tornaria inútil qualquer preparação antes da observação em MEV, nós obtivemos melhores resultados com uma metalização prévia com platina.
Note que, como nenhum tratamento químico é efetuado, é possível obter-se, após a determinação anatômica no microscópio óptico, uma datação de 14C no mesmo fragmento (VERNET, BAZILE 8& EVIN, 1979), o que é muito interessante quando a quantidade de carvões coletada é pequena.
Plano transversal
Fig.3- Os três planos fundamentais da madeira: relação com o eixo do lenho e imagens em microscopia eletrônica de varredura (Leguminosae Caesalpinoideae: Cassia speciosa Schrad.).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

TEORIA E MÉTODOS EM ANTRACOLOGIA. 2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO 351
2. Dimensão dos fragmentos identificados
Fragmentos de carvão de apenas 0,5mm de lado podem ser identificados, especialmente no caso das gimnospermas de regiões temperadas (VERNET, BAZILE & EVIN, 1979), mas em fragmentos tão pequenos a determinação é em geral longa, difícil e imprecisa (CHABAL, 1988; SCHEEL-YBERT, 2000). Quanto menor 0 fragmento, mais difícil será sua determinação, e mais elevado será o nível taxonômico alcançado. O estudo de fragmentos muito pequenos, além de aumentar o esforço de determinação, tem como conseqüência diminuir a precisão da determinação taxonômica.
Em regiões tropicais, fragmentos cujo lado maior é inferior a 4mm são muito dificilmente identificáveis, pois eles não costumam reunir um conjunto de caracteres anatômicos suficientemente amplo para permitir sua determinação, sequer no nível de família (SCHEEL-YBERT, 2000, 2001).
A precisão de determinação possível em regiões onde a diversidade florística é menor não será provavelmente jamais alcançada em estudos nos trópicos. Contudo, uma diminuição no grau de incerteza das determinações pode ser obtida levando-se em consideração sistematicamente um grande número de caracteres anatômicos em cada amostra, o que impõe um tamanho mínimo para os fragmentos estudados e torna necessária a utilização de um máximo de dados da literatura, de uma boa coleção de referência e de um banco de dados o mais completo possível.
3. Constituição de uma coleção de referência
De modo geral, a anatomia de madeiras tropicais é muito pouco conhecida, especialmente no Brasil. Na maioria dos casos, os trabalhos existentes na literatura são insuficientes para a determinação da maioria das espécies lenhosas encontradas nas análises antracológicas. Em conseqüência, a elaboração de coleções de referência de madeiras e de carvões atuais para as diferentes associações vegetais encontradas na região de estudo é indispensável.
As amostras podem ser obtidas a partir de coletas de campo ou de doações de xilotecas. Quando se faz amostragem no campo, é indispensável coletar também material fértil (flores e frutos), a fim de produzir exsicatas que permitam a identificação taxonômica do material por especialistas.
Todas as amostras de madeira atual obtidas devem ser, na medida do possível, divididas em duas partes: uma parte deve ser conservada intacta e a
outra deve ser carbonizada visando sua inclusão na coleção de referência. As amostras carbonizadas não devem medir menos de lcm de lado.
Idealmente, a carbonização é feita em um forno a mufla, no qual as amostras são carbonizadas a 400°C durante 40 minutos. Cada amostra de madeira deve ser embrulhada em papel alumínio, com o número de referência escrito a lápis, na madeira, e com caneta permanente, no papel alumínio.
Quando não se dispõe de um forno, as amostras de madeira podem ser queimadas em uma fogueira (G.WILLCOX, com.pes., 1996), embrulhadas em papel alumínio e com as referências de cada amostra marcadas, por exemplo, em etiquetas de alumínio escritas em baixo-relevo. Embora o grafite seja resistente à queima, não se deve confiar exclusivamente na referência escrita a lápis sobre a madeira, pois pode ocorrer uma deposição de óleos e graxas que cobre a superfície da amostra.
Após carbonização, as amostras devem ser organizadas de forma a facilitar seu acesso e consulta. A melhor solução que encontramos é a utilização de organizadores com pequenas gavetas (Fig.4). Cada gaveta é etiquetada com o nome da espécie e o número de referência, mas um arquivo completo contendo todas as informações de coleta também deve ser feito.
4. Características da madeira carbonizada
A constituição de coleções de referência de madeira carbonizada é fundamental, em particular devido às eventuais modificações estruturais do lenho após combustão.
Embora não afete a organização dos tecidos do lenho, o processo de carbonização pode resultar em variações dos caracteres morfométricos (tamanho das células), as quais são devidas a uma importante reorganização estrutural da madeira submetida a fortes temperaturas. O aumento da temperatura provoca a degradação da celulose e da hemicelulose e o rearranjo do carbono numa estrutura semelhante à grafite, acompanhados da destruição da estrutura microfibrilar da madeira (MCGINNES, KANDEEL & SZOPA, 1971). Experimentos de carbonização de amostras de Quercus alba mostraram que as alterações mais significativas concernem as dimensões das amostras, com uma retração que variou de cerca de -12% a -25% e que é mais elevada no plano tangencial longitudinal (MCGINNES, KANDEEL & SZOPA, 1971).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

352 R.SCHEEL-YBERT
Fig.4- Aspecto dos organizadores contendo a coleção de referência de madeiras carbonizadas (antracoteca) do Laboratório
de Antracologia do Museu Nacional.
A análise das respostas à carbonização de seis espécies africanas pertencentes a diversas famílias demonstrou que a natureza e a magnitude das modificações são muito variáveis, e em grande parte dependentes da natureza das fibras da madeira (PRIOR & GASSON, 1993). As variações nos caracteres morfológicos e anatômicos medidos são tão mais importantes quanto a temperatura de carbonização é mais elevada. Segundo estes resultados houve, após carbonização, uma dilatação das células de parênquima axial e uma diminuição significativa da altura dos raios lenhosos em todas as espécies, enquanto que as variações do diâmetro tangencial dos poros variou entre uma diminuição de -41% até uma dilatação de +28%.
Em consequência, tanto que nosso conhecimento dos processos de carbonização e das mudanças anatômicas que intervém em sua estrutura durante sua transformação em carvão não serão completas,
deveremos utilizar os dados anatômicos quantitativos com muita prudência, especialmente quando compararmos carvões a amostras de madeira não-carbonizadas. Em todo caso, o grau de incerteza nas determinações será sempre minimizado pela utilização de uma coleção de referência de amostras carbonizadas.
5. Banco de dados antracológico
A imensa riqueza da vegetação brasileira tem por conseqüência uma maior dificuldade de identificação das espécies que no caso de regiões onde a diversidade florística é menor. Para facilitar o processo de determinação dos fragmentos antracológicos, foi elaborado um programa que permitiu a constituição de um banco de dados informatizado (SCHEEL-YBERT, SCHEEL & YBERT, 1998). Esse programa funciona em Windows, foi escrito em Microsoft Access 2.0 e
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

TEORIA E MÉTODOS EM ANTRACOLOGIA. 2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO 353
posteriormente atualizado para Access 2000 (SCHEEL-YBERT, SCHEEL & YBERT, 2002). Os caracteres anatômicos utilizados foram baseados na lista-padrão proposta pela Associação Internacional de Anatomistas da Madeira (IAWA COMMITTEE, 1989).
Este programa permite a entrada de dados anatômicos referentes a carvões atuais (ou eventualmente a madeiras não carbonizadas), aos carvões fósseis e aos dados da literatura. Ele permite a realização de pesquisas baseadas em um ou mais caracteres, eventualmente com uma margem de erro que é estabelecida durante a consulta, isto é, um número definido de características das fichas-resultado que podem ser diferentes daquelas estabelecidas na ficha- consulta; as características discordantes aparecem marcadas em vermelho. Os resultados da pesquisa são apresentados por ordem de família e de espécie, mas podem ser ordenados de outra forma.
Pode-se imprimir relatórios sob a forma de fac-símile das fichas, ou de descrições anatômicas padronizadas. Neste último caso, o resultado da pesquisa é gravado sob forma de um arquivo-texto que pode ser lido por qualquer editor de texto. Até seis imagens podem ser associadas a cada ficha anatômica, e estas também podem ser impressas. Além disso, o programa é trilingue, e todas as opções podem ser lidas em português, francês ou inglês.
Os dados anatômicos de todas as amostras da coleção de referência foram incluídos no banco de dados, assim como tipos anatômicos encontrados em amostras arqueológicas. Informações sobre a ecologia e a área de distribuição geográfica das espécies foram incluídas para a maior parte dos taxa, a partir dos dados de coleta ou de informações obtidas na literatura.
O banco de dados se mostrou extremamente útil para a determinação de fragmentos fósseis, pois é possível utilizá-lo como chave de determinação. O principal interesse deste programa, além de sua eficácia e simplicidade de uso, é o fato de ter sido especialmente concebido para a identificação antracológica. De fato, se a maior parte dos caracteres qualitativos da madeira se conservam nos carvões (com exceção dos caracteres organolépticos, como cor, cheiro, densidade etc.), os parâmetros morfométricos, especialmente o tamanho das células, a espessura das paredes, o diâmetro dos poros e das pontuações, etc., podem variar (vide “Características da madeira
carbonizada*). Por isso, a identificação taxonômica
não pode se basear exclusivamente na comparação das medidas dos caracteres anatômicos de um fragmento de carvão fóssil com aqueles de uma amostra de madeira atual não carbonizada.
CONCLUSÃO
Os estudos antracológicos fornecem, a partir das mesmas amostras, informações importantes tanto no domínio das variações paleoambientais e paleoclimáticas quanto em aspectos paleoetnológicos. Estes dois temas interpretativos não são incompatíveis e podem ser abordados a partir do mesmo material, desde que se conheça a origem de cada amostra no contexto arqueológico. A intervenção do homem pré-histórico no transporte da madeira até o sítio arqueológico não invalida as reconstituições da vegetação passada baseadas na antracologia (SCHEEL-YBERT, 1998, 2000). Por outro lado, o uso da antracologia para as reconstituições paleoambientais não deve ofuscar as informações paleoetnológicas contidas na mesma amostra (SCHEEL-YBERT, 2001).
A amostragem de carvões com objetivos paleoecológicos deve obedecer a dois requisitos básicos: eles devem provir de uma utilização doméstica para combustível, e corresponder aos resíduos de uma atividade que tenha tido uma duração temporal suficientemente longa (CHABAL, 1992). O primeiro aspecto deve-se ao caráter pouco seletivo da coleta de madeira para lenha doméstica, ao contrário do que ocorre com a utilização da madeira para finalidades específicas (material de construção, fabricação de artefatos ou combustíveis especializados). O segundo, ao fato de que existe uma correlação direta entre o tempo de duração da coleta de lenha e a superfície da área de captação de recursos, cuja vegetação será certamente mais bem representada a partir de várias coletas de lenha durante o tempo de ocupação do sítio. Por isso, as análises paleoecológicas devem ser preferencialmente baseadas nos carvões dispersos, enquanto os carvões concentrados fornecem principalmente informações paleoetnológicas.
A imensa riqueza da vegetação tropical traz alguns problemas para a determinação taxonômica das espécies em fragmentos de carvões. A precisão da determinação é sensivelmente diminuída em relação às regiões temperadas, onde a diversidade florística é menor, e onde as determinações ao nível de espécie são bastante comuns. Apesar disso, os resultados obtidos até o momento mostram que os estudos antracológicos são perfeitamente aplicáveis a regiões
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

354 R.SCHEEL-YBERT
tropicais. Reconstituições paleoambientais muito satisfatórias podem ser obtidas mesmo com identificações taxonômicas limitadas ao gênero, e a abrangência das informações paleoetnológicas que podem ser obtidas é incalculável.
O desenvolvimento da antracologia no Brasil deve se acompanhar de um aprofundamento dos estudos metodológicos e tafonômicos e de um enriquecimento das coleções de referência. Estas pesquisas poderão contribuir, posteriormente, com estudos de modelização do ecossistema que levem em conta a atividade humana.
REFERÊNCIAS BIBLIOGRÁFICAS
BADAL, E.; FIGUEIRAL, I.; HEINZ, C. & VERNET, J.L.,
1989. Charbons de bois archéologiques
méditerranéens: de la fouille à 1’interprétation. Acta
Interdisciplinaria Archaeologica, Nitra, 7:7-22.
BADAL-GARCIA, E. & HEINZ, C., 1991. Méthodes
utilisées en anthracologie pour 1’étude de sites
préhistoriques. In: WALDREN, W.H.; ENSENYAT, J.A.
& KENNARD, R.C. (Eds.) Ilnd Deya International
Conference of Prehistory. Recent developments in
Western Mediterranean prehistory: Archaeological
techniques, technology and theory. v. 1. British
Archaeological Reports International Series,
Oxford, 573:17-47.
BADAL-GARCIA, E., 1992. Lánthracologie préhistorique:
à propos de certains problèmes méthodologiques.
Bulletin de la Société Botanique de France, Paris,
Actualités botaniques, 139(2/3/4): 167-189.
BARBOSA, M.; GASPAR, M.D. & BARBOSA, D.R., 1994.
A organização espacial das estruturas habitacionais
e distribuição dos artefatos no sítio Ilha da Boa Vista
I, Cabo Frio, RJ. Revista do Museu de Arqueologia
e Etnologia, São Paulo, 4:31-38.
BARROS, C.F. & CALLADO, C.H. (Eds.), 1997.
Madeiras da Mata Atlântica: Anatomia do lenho
de espécies ocorrentes nos remanescentes
florestais do Estado do Rio de Janeiro, Brasil,
v. 1. Rio de Janeiro: Jardim Botânico do Rio de
Janeiro. 86p.
BLANKENHORN, P.R.; JENKINS, G.M. & KLINE, D.E.,
1972. Dynamic mechanical properties and
microstructure of some carbonized hardwoods. Wood
and Fiber, 4(3) :212-224.
BREUIL, H., 1903. Les fouilles dans la grotte du Mas
d’Azil (Ariège). Bulletin Archéologique, Paris, 421-
436p.
BURGER, L.M. & RICHTER, H.G. 1991. Anatomia da
madeira. São Paulo: Ed. Nobel. 154p.
CHABAL, L., 1988. Pourquoi et comment prélever les
charbons de bois pour la période antique: les
méthodes utilisées sur le site de Lattes (Hérault).
Lattara, Lattes, 1:187-222.
CHABAL, L., 1997. Forêts et sociétés en Languedoc
(Néolithique final, Antiquité tardive): L’anthracologie,
méthode et paléoécologie. Documents d’Archéologie
Française, Paris, 63:1-188.
CHABAL, L., 1989a. Étude paléoécologique à partir
des charbons de bois. In: FICHES, J.L. (Ed.)
L’oppidum d’Ambrussum et son territoire: fouilles
au quartier du Sabias (Villetelle, Hérault).
Collection des monographies du CRA, Sophia
Antipolis, 2:180-192.
CHABAL, L., 1989b. Perspectives anthracologiques sur
le site de Lattes (Hérault). Lattara, Lattes, 2:53-72.
CHABAL, L., 1992. La représentativité paléo¬
écologique des charbons de bois archéologiques
issus du bois de feu. Bulletin de la Société
Botanique de France, Paris, Actualités
botaniques, 139(2/3/4):213-236.
CUTTER, B.E.; CUMBIE, B.G. & MCGINNES, E.A., 1980.
SEM and shrinkage analyses of Southern pine wood
following pyrolysis. Wood Science and Technology,
Heidelberg, 14:115-130.
DECHAMPS, R., 1979. Étude anatomique de bois
d’Amérique du Sud. Volume I. Acanthaceae à
Lecythidaceae. Tervuren: Musée Royal de 1’Afrique
Centrale. 332p.
DECHAMPS, R., 1980. Étude anatomique de bois
d’Amérique du Sud. Volume II. Leguminosae.
Tervuren: Musée Royal de 1’Afrique Centrale. 229p.
DECHAMPS, R, 1985. Étude anatomique de bois
d’Amérique du Sud. Volume III. Linaceae à
Zygophyllaceae. Tervuren: Musée Royal de 1’Afrique
Centrale. 47lp.
DÉTIENNE, P. & JACQUET, P., 1983. Atlas
d’identification des bois de 1’Amazonie et des
régions voisines. Nogent-sur-Marne/Montpellier:
Centre Technique Forestier Tropical. 640p.
FIGUEIRAL, I., 1994. A antracologia em Portugal:
progressos e pespectivas. Actas dos Trabalhos de
Antropologia e Etnologia, Porto, 34(3/4):427-444.
FIGUEIRAL, L, 1992. Méthodes en anthracologie: étude
de sites du Bronze final et de 1’âge du Fer du nord-
ouest du Portugal. Bulletin de la Société Botanique
de France, Paris, Actualités botaniques, 139(2/
3/4): 191-204.
FIORENTINO, G., 1995. New perspectives in
anthracological analysis. Palaeoecological and
technological implications of charcoals found in the
Neolithic flintmine at La Defensola (Vieste, Apulia,
Italy). Quaternaria Nova, Roma, 5:99-128.
GREGORY, M., 1994. Bibliography of systematic wood
anatomy of Dicotyledons. IAWA Journal, Leiden,
suppl., 1:1-266.
GREGUSS, P., 1959. Holzanatomie der Europãischen
Laubhõlzer und Stráucher. Budapest: Akad.
Kiadó. 330p.
HEER, O., 1866. Pflanzen der Pfanhlbauten. Neujbl.
Naturf. Ges. Zürich auf das Jahr, Zurique,
1866:1-54.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

TEORIA E MÉTODOS EM ANTRACOLOGIA. 2 - TÉCNICAS DE CAMPO E DE LABORATÓRIO 355
HEINZ, C., 1990. Dynamique des végétations holocènes en
Méditerranée nord occidentale d’après l’anthracoanalyse
de sites préhistoriques: méthodologie et paléoécologie.
Paléobiologie Continentale, Montpellier, 16(2):1-212.
HESS, R.W., 1946. Keys to American woods. XVII. Woods
with vasicentric tracheids. XVIII. Fibers with spiral
thickenings. XIX. Special fibers in parenchyma-like
arrangement. XX. Woods with unilaterally paratracheal
parenchyma. Tropical Woods, 85:11-19.
HESS, R.W., 1948. Keys to American woods. XX. Woods
with unilaterally paratracheal parenchyma. XXL
Parenchyma in numerous concentric bands. Tropical
Woods, 94:29 52.
IAWA COMMITTEE, 1989. IAWA list of microscopic
features for hardwood identification. WHEELER, E.A.;
BAAS, P. & GASSON, P.E. (Eds.) IAWA Bulletin, N.S.,
Leiden, 10(3) :219-332.
MAINIERI, C. & CHIMELO, J.P., 1989. Fichas de
características das madeiras brasileiras. 2.ed. São
Paulo: IPT. 418p.
MCGINNES, E.A.; KANDEEL, S.A. & SZOPA, P.S., 1971.
Some structural changes observed in the
transformation of wood into charcoal. Wood and
Fiber, 3(2): 77-83.
METCALFE, C.R. & CHALK, C., 1950. Anatomy of the
dicotyledons. 2 vol. Oxford: Clarendon Press. 1500p.
PEARSALL, D.M., 2000. Paleoethnobotany: A
handbook of procedures. 2.ed. San Diego: Academic
Press. 700p.
PERNAUD, J.M., 1992. L’interprétation paléoécologique
des charbons concentrés dans des fosses dépotoirs
protohistoriques du Carroussel (Louvre, Paris).
Bulletin de la Société Botanique de France, Paris,
Actualités botaniques, 139(2/3/4):329-341.
PREJAWA, H., 1896. Die Ergebnisse der
Bohlweguntersuchungen in Grenzmoor zwischen
Oldenburd und Preussen und in Mellinghausen im
Kreise Sulingen. Mitt. Ver. Gesch. Landesskde,
Osnabrück, 21:98-178.
PRIOR, J. & GASSON, P., 1993. Anatomical changes on
charring six African hardwoods. IAWA Journal,
Leiden, 14(l):77-86.
RECORD, S.J., 1942. Keys to American woods. I. Ring-
porous woods. II. Woods with pores in ulmiform or
dendritic arrangement. Tropical Woods, 72:19-35.
RECORD, S.J., 1943a. Keys to American woods. IV.
Vessels virtually all solitary. V. Vessels with spiral
thickenings. Tropical Woods, 73:23-42.
RECORD, S.J., 1943b. Keys to American woods. VI.
Vessels with scalariform plates. VII. Vessels with very
fine pitting. IX. Woods with conspicuous rays.
Tropical Woods, 75:8 26.
RECORD, S.J., 1944a. Keys to American woods. X.
Woods with storied structure. Tropical Woods,
76:32-47.
RECORD, S.J., 1944b. Keys to American woods. XI.
Woods with resin or gum ducts. XII. Parenchyma
reticulate. Tropical Woods, 77:18-38.
RECORD, S.J., 1944c. Keys to American woods. XIII.
Woods with septate fibers. Tropical Woods, 78:35-45.
RECORD, S.J., 1944d. Keys to American woods. XIV.
Dicotyledonous woods with xylem rays virtually all
uniseriate. Tropical Woods, 79:25-34.
RECORD, S.J., 1944e. Keys to American woods. XV.
Fibers with conspicuous bordered pits. XVI. Woods
with oil (or similar) cells. Tropical Woods, 80:10-15.
RECORD, S.J. & HESS, R.W., 1943. Timbers of the
New World. New Haven: Yale University Press. 640p.
SCHEEL, R.; GASPAR, M.D. & YBERT, J.P., 1996a. A
anatomia dos carvões pré-históricos. Arqueologia
encontra respostas em restos de fogueiras e incêndios
florestais. Revista Ciência Hoje, Rio de Janeiro,
21(122):66-69.
SCHEEL, R.; GASPAR, M.D. & YBERT, J.P., 1996b.
Antracologia, uma nova fonte de informações para a
arqueologia brasileira. Revista do Museu de
Arqueologia e Etnologia, São Paulo, 6:3-9.
SCHEEL-YBERT, R., 1998. Stabilité de 1’écosystème
sur le littoral Sud-Est du Brésil à l’Holocène
Supérieur (5500-1400 ans BP). Les pêcheurs-
cueilleurs-chasseurs et le milieu végétal: apports
de 1’anthracologie. Tese de Doutorado. Montpellier:
USTL, UM II. 3 vol. 520p.
SCHEEL-YBERT, R., 1999. Paleoambiente e paleoetnologia
de populações sambaquieiras do sudeste do Estado do
Rio de Janeiro. Revista do Museu de Arqueologia e
Etnologia, São Paulo, 9:43-59.
SCHEEL-YBERT, R., 2000. Vegetation stability in the
Southeastern Brazilian Coastal area from 5500 to
1400 14C yr BP deduced from charcoal analysis.
Review of Palaeobotany and Palynology,
Amsterdam, 110:111-138.
SCHEEL-YBERT, R., 2001. Man and vegetation in the
Southeastern Brazil during the Late Holocene.
Journal of Archaeological Science, San Diego,
28(5):471-480.
SCHEEL-YBERT, R., 2004. Teoria e métodos em
antracologia. 1. Considerações teóricas e
perspectivas. Arquivos do Museu Nacional, Rio de
Janeiro, 62(1):3-14.
SCHEEL-YBERT, R. & SOLARI, M.E., no prelo. Macro-
restos vegetais do Abrigo Santa Elina: Antracologia
e Carpologia. In: VILHENA-VIALOU, A. & VIALOU,
D. (Eds.) Pré-história do Mato Grosso: uma
pesquisa brasileira-francesa pluridisciplinar. São
Paulo: EDUSP.
SCHEEL-YBERT, R.; GOUVEIA, S.E.M.; PESSENDA,
L.C.R.; ARAVENA, R.; COUTINHO, L.M. & BOULET,
R., 2003 - Holocene palaeoenvironmental evolution
in the São Paulo State (Brazil), based on anthracology
and soil 813C analysis. The Holocene, Londres, 13
(1): 73-81.
SCHEEL-YBERT, R.; SCHEEL, M. & YBERT, J.P., 1998.
Atlas Brasil: Banco de dados antracológicos e chave
computadorizada para determinação de carvões (em
Português, Inglês e Francês). Versão 1.8 [CD-ROM].
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

356 R. SCHEEL-YBERT
SCHEEL-YBERT, R.; SCHEEL, M. & YBERT, J.P., 2002.
Atlas Brasil: Banco de dados antracológicos e
chave computadorizada para determinação de
carvões. Ver.2.2 [CD-ROM].
SCHWEINGRUBER, F.H., 1978. Mikroskopische
Holzanatomie / Anatomie microscopique du bois.
Birmensdorf: Swiss Federal Institute of Forestry
Research. 226p.
SCHWEINGRUBER, F.H., 1990. Anatomy of European
woods. Bern/Stuttgart: P. Haupt publ. 800p.
SMART, T.L. & HOFFMAN, E.S., 1988. Environmental
interpretation of archaeological charcoal. In:
HASTORF, C.A. & POPPER, V.S. (Eds.) Current
Paleoethnobotany : Analytical methods and
cultural interpretation of archaeological plant
remains. Chicago/London: The University of Chicago
Press. p. 167-205.
THIÉBAULT, S., 1995. Functioning of hearths and
ancient vegetation at the Balme de Thuy (Haute-
Savoie, France). The charcoal contribution.
Quaternaria Nova, Roma, 5:129-170.
THÉRY-PARISOT, I., 2001. Économie des combustibles
au Paléolithique. Paris: CNRS. 196p.
VERNET, J.L., 1973. Étude sur 1’histoire de la végétation
du sud-est de la France au Quaternaire, d’après les
charbons de bois principalement. Paléobiologie
Continentale, Montpellier, 4(1): 1-90.
VERNET, J.L.; BAZILE, E. & EVIN, J., 1979.
Coordination des analyses anthracologiques et des
datations absolues sur charbon de bois. Bulletin
de la Société Préhistorique Française, Paris,
76(3):76-79.
VERNET, J.L.; OGEREAU, P.; FIGUEIRAL, I.;
MACHADO-YANES, C. & UZQUIANO, P., 2001. Guide
d’identification des charbons de bois
préhistoriques et récents: Sud-Ouest de l’Europe:
France, Péninsule Ibérique et íles Canaries. Paris:
CNRS Editions. 395p.
WESTERN, C., 1963. Wood and charcoal in
archaeology. In: BROTHWELL, D. & HIGGS, E.
(Eds.) Science in Archaeology. A comprehensive
survey of progress and research. Londres: Thames
and Hudson. p.150-162.
YBERT, J.P.; SCHEEL, R. & GASPAR, M.D., 1997.
Descrição de alguns instrumentos simples
utilizados para a coleta e concentração de
elementos fósseis de pequenas dimensões de
origem arqueológica ou pedológica. Revista do
Museu de Arqueologia e Etnologia, São Paulo,
7:181-189.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.343-356, out./dez.2004

TmnrronnnjfnnnrTi nnniiituiíiüífflnnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004
ISSN 0365-4508
ESTUDO POLÍNICO DE ESPÉCIES DE AESCHYNOMENEAE E
PHASEOLEAE (PAPILIONOIDEAE - LEGUMINOSAE JUSS.)
OCORRENTES NAS RESTINGAS DO ESTADO DO RIO DE JANEIRO 1
(Com 40 figuras)
FABIANA CARVALHO DE SOUZA 2>3
MARIANA ALBUQUERQUE DE SOUZA 2>3
CLÁUDIA BARBIERI FERRREIRA MENDONÇA 2>4
VANIA GONÇALVES-ESTEVES 2>5
RESUMO: Foram analisados os grãos de pólen de espécies encontradas nas restingas do Estado do Rio de
Janeiro, pertencentes às tribos: Aeschynomeneae (Benth.) Hutch. [Aeschynomene fluminensis Vell.,
Stylosanthes guianensis (Aubl.) Sw., S. viscosa (L.) Sw., Zornia latifolia Sm.] e Phaseoleae DC. [Canavalia
marítima (Aubl.) Thouras, Centrosema virginianum (L.) Benth. e Macroptilium bracteatum (Nees & Mart.) Maréchal
& Baudet.]. Pretende-se, com este trabalho, oferecer subsídios palinológicos que auxiliem à Taxonomia, à
Aeropalinologia, à Melissopalinologia, bem como, contribuir para a formação do catálogo polínico da flora
das restingas do Estado do Rio de Janeiro. Os resultados polínicos obtidos sobre as espécies das duas tribos
estudadas permitiram separá-las dentro das tribos. Os resultados obtidos mostram que as espécies da tribo
Aeschynomeneae apresentaram grãos de pólen parassincolpados ou colpados e parassincolporados; os grãos
de pólen da tribo Phaseoleae foram descritos como colporados e o tamanho, a forma dos grãos de pólen e a
ornamentação da sexina diferiram entre as espécies. Conclui-se que dentro dos gêneros existe uma certa
homogeneidade nas características polínicas, contudo, os gêneros são, morfopolinicamente, distintos. Assim,
as espécies puderam ser separadas dentro das tribos.
Palavras-chave: Palinologia, Papilionoideae-Leguminosae, Aeschynomeneae, Phaseoleae, Restinga.
ABSTRACT: Pollinic study of some species of Aeschynomeneae e Phaseoleae (Papilionoideae - Leguminosae
Juss.) occurring in “restingas” of Rio de Janeiro State.
In the present paper were analised the pollen grains of the family Leguminosae Juss. (Papilionidae) occuring in
the flora of Coastal strand vegetation (“restingas”) from Rio de Janeiro State belonging to the following tribes:
Aeschynomeneae (Benth.) Hutch. [Aeschynomene fluminensis Vell., Stylosanthes guianensis (Aubl.) Sw., S.
mscosa (L.) Sw., Zornia latifolia Sm.] and Phaseoleae DC. [Canavalia marítima (Aubl) Thouras, Centrosema
virginianum (L.) Benth. and Macroptilium bracteatum (Ness &Mart.) Marechal & Baudet.]. It was aimed to offer
palynological data to Taxonomy, Aeropalynolgy, Melissopalinolgy as well as contribute to organize a pollinic
catalog of the “restingas” of the Rio de Janeiro State. The obtained results demonstrated that: the species of
tribe Aeschynomeneae were organized by presence of parassincolpate pollen grains or colpate and
parassincolporate ones; the pollen grains of species of the tribe Phaseolae were colporated, with size, shape
and ornamentation of the sexine of pollen grains different among species. We conclude that inside the each
genus exist some homogenity in the pollinic characteristics; however, the genera are morphopollinical distinct.
Key words: Palinology, Papilionoideae-Leguminosae, Aeschynomeneae, Phaseoleae, “Restinga”
INTRODUÇÃO
Em continuidade ao estudo da família Leguminosae Juss. (Papilionoideae) ocorrentes nas restingas do Estado do Rio de Janeiro
(GONÇALVES-ESTEVES & CRESPO, 1994) apresenta-se, no momento, a palinologia de espécies pertencentes às tribos: Aeschynomeneae (Benth.) Hutch. [Aeschynomene fluminensis Vell., Stylosanthes guianensis (Aubl.) Sw., Stylosanthes
1 Submetido em 02 de julho de 2004. Aceito em 05 de novembro de 2004.
2 Museu Nacional/UFRJ, Departamento de Botânica. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil.
3 Bolsista da Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ).
4 Museu Nacional/UFRJ, Programa de Pós-Graduação em Ciências Biológicas/Botânica. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil
Bolsista da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
5 Bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)

358 F.C.SOUZA, M.A.SOUZA, C.B.F.MENDONÇA & V.GONÇALVES-ESTEVES
viscosa (L.) Sw., Zornia latifolia Sm.] e Phaseoleae DC. [Canavalia marítima (Aubl.) Thouras, Centrosema virginianum (L.) Benth. e Macroptilium
bracteatum (Nees & Mart.) Maréchal 8& Baudet.]. Espécies dessas tribos já foram objeto de estudos palinológicos desenvolvidos por VISHNU-MITTRE 85 SHARMA, 1962; BARTH, 1964; MELHEM, 1971; PIRE, 1974; MAKINO, 1978; TEWARI & NAIR, 1979; GUINET, 1981; FERGUSON & SKVARLA 1981, 1983, 1988; GONÇALVES- ESTEVES 8s CRESPO, 1994 e SILVESTRE- CAPELATO 85 MELHEM, 1997. Todos esses autores evidenciaram a variedade morfopolínica das espécies. Pretende-se, com este trabalho, oferecer subsídios palinológicos que auxiliem à taxonomia, à Aeropalinologia, à Melissopalinologia, bem como, contribuir para a formação do catálogo polínico da flora das restingas do Estado do Rio de Janeiro.
MATERIAL E MÉTODOS
O material polínico foram retiradas anteras férteis de flores em antese e/ou botões florais bem desenvolvidos. O material foi obtido de exsicatas depositadas nos herbários do Museu Nacional Rio de Janeiro (R) e da Fundação Estadual de Engenharia do Meio Ambiente Alberto Castellanos (GUA). As siglas seguem HOLMGREN, HOLMGREN 86
BAINETT (1990).
Material utilizado - BRASIL.
Aschynomeneae - Aschynomene fluminensis - RIO DE JANEIRO: Jacarepaguá, D.Sucre 6549 e outros (GUA*); ESPÍRITO SANTO: Vitória, I. Weiter 575 (R). Stylosanthes guianensis - MACAÉ: Cabiúnas, V.Esteves 609 e outros (R*). MINAS GERAIS: Alto Paraíso, H.S.Irwin s/n, 19/III/71 (RB256252); RIO DE JANEIRO: Jacarepaguá, restinga, Vaunce s/n, s/data (RB181901). S. viscosa - RIO DE JANEIRO: Saquarema, Restinga de Ipitangas, D.Araújo 9127 (GUA*); Cabo Frio, A.P.Viegas e H.P.Krug s/n, 16/X/1938 (RB112866). Zornia latifolia - RIO DE JANEIRO: Angra dos Reis, A.Castellanos 2585 (GUA*); Angra dos Reis, Ilha Grande, L.C.Giordano e outros 282 (RB); Rio de Janeiro, C.Cauvelli s/n, 17/11/1944 (RB49117).
Phaseoleae - Canavalia marítima - RIO DE JANEIRO: Maricá, Barra de Maricá, D.Araújo 4819 (GUA*); Praia Grande, C.Rizzini s/n, s/data (RB342818); Arraial do Cabo, T.Kajita e outros
s/n, 10/XII/1999 (RB268085). Centrosema
virginianum - RIO DE JANEIRO: Jacarepaguá, H.M.Chaves s/n, 22/X/1963 (GUA03693*); Restinga de Jacarepaguá, Rizzo s/n, s/data (RB 163958). Macroptilium bracteatum - RIO DE JANEIRO: Jacarepaguá, H.E.Strang s/n, 14/ VIII/1978 (GUA00169); MACAÉ: Cabiúnas, R.M.Harley 16169 (RB*).
Sempre que possível, procurou-se analisar os grãos de pólen de três espécimes de uma mesma espécie sendo um destes escolhido como padrão (indicado no material examinado por um asterisco), para as mensurações, descrições e ilustrações polínicas. Para 0 estudo sob microscópio de luz o material polínico foi preparado segundo o método acetolítico de ERDTMAN (1952) enquanto que para a obtenção das eletromicrografias em microscopia eletrônica de varredura (Zeiss DSM 960), utilizou-se material polínico não acetolisado. Os grãos de pólen foram espalhados sobre suportes metálicos previamente recobertos por fita de carbono e, em seguida, receberam uma fina camada de ouro paládio por cerca de três minutos. Do material padrão foram mensurados 25 grãos de pólen dos diâmetros polar (DP) e equatorial (DE), em vista equatorial (em Aschynomene fluminensis e em Canavalia
marítima não foram obtidas 25 medidas devido à posição preferencial de queda, em vista polar, dos grãos de pólen na lâmina). Com os resultados obtidos, foram efetuados tratamentos estatísticos calculando-se a média aritmética (x), 0 desvio-padrão da amostra (s); o desvio padrão da média (s-); 0 coeficiente de variabilidade (CV%) e 0 intervalo de confiança a 95% (IC 95%). Para as medidas dos demais caracteres, como as do diâmetro equatorial em vista polar (DEVP), do lado do apocolpo (LA), das aberturas e camadas da exina foi calculada a média aritmética a partir de 10 medidas, o mesmo ocorrendo para as medidas dos diâmetros dos grãos de pólen dos espécimes de comparação.
A terminologia adotada e as descrições polínicas seguiram os critérios de BARTH 86 MELHEM (1988), MELHEM et al. (2003) e PUNT et al.
(1999), levando-se em consideração 0 tamanho, a forma, o número de aberturas e o padrão de ornamentação da sexina. A denominação da área polar e o tamanho da abertura estão de acordo com a classificação estabelecida por FAEGRI 85
IVERSEN (1966) para 0 índice da área polar.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004

ESTUDO POLÍNICO DE ESPÉCIES DE AESCHYNOMENEAE E PHASEOLEAE, ESTADO DO RIO DE JANEIRO 359
RESULTADOS
Tribo Aeschynomeneae Aeschynomene fluminensis (Figs.1-6)
Grãos de pólen pequenos (Tabs.1-2), isopolares, subprolatos, âmbito subtriangular, sem área polar, 3-parassincolporados (Figs.l, 4), trilobados, exina microrreticulada (Figs.2-3).
Os colpos são longos, largos (Fig.5), com opérculo e membrana ornamentada, endoabertura circular (Tab.3).
A exina apresenta microrretículos conspícuos no mesocólporo (Fig.6) e menores no pólo e próximo das aberturas (Figs.4, 6); a sexina é simplescolumelada e mais espessa que a nexina.
Stylosanthes
(Figs.7-16)
Espécies estudadas - Stylosanthes guianensis -
(Figs.7-13); S. viscosa - (Figs.l4-16).
Grãos de pólen médios (Tabs.1-2), isopolares, prolatos, âmbito subcircular, sem área polar, 3- parassincolpados (Fig. 10), exina microrreticulada.
Os colpos são longos, largos (Tab.3), com opérculo e membrana ornamentada (Figs.l 1-12, 14). Em MEV, pode-se observar que a membrana apresenta dois tipos definidos de granulação: um menos conspícuo localizado no contorno do colpo e outro, mais conspícuo, sobre o colpo (Figs.10, 13, 15).
A exina apresenta microrretículos com muros estreitos e perfurações esparsas (Figs.8-9, 13, 16); a sexina é simplescolumelada e mais espessa que a nexina.
Zomia latifolia
(Figs. 17-23)
Grãos de pólen médios (Tabs.1-2), isopolares, prolatos, âmbito subcircular, área polar pequena, 3-colpados, exina microrreticulada.
Os colpos são longos, largos (Tab.3), com opérculo e membrana ornamentada (Figs.22-23). Em MEV, pode-se observar que a membrana apresenta dois tipos definidos de granulação: um menos conspícuo localizado no contorno do colpo e outro mais conspícuo, sobre o opérculo (Fig.23).
A exina possui microrretículos cujos lumens apresentam formas e tamanhos variados, sem
redução do diâmetro próximo das aberturas (Figs. 18-20); a sexina é simplescolumelada e mais espessa do que a nexina.
Tribo Phaseoleae
Canavalia marítima
(Figs.24-26)
Grãos de pólen grandes (Tab.l), isopolares, suboblatos, âmbito triangular, área polar pequena (Tab.2), 3-colporados, exina perfurada/ruguiada.
Os colpos são longos, com endoabertura lalongada e recobertos por membrana ornamentada (Fig.25). Os grãos de pólen caem na lâmina, preferencialmente, em vista polar.
A exina é perfurada no mesocólporo e rugulada na região polar (Figs.24, 26). A sexina é simplescolumelada e o teto é espesso na região do mesocólporo, tornando-se mais fino próximo da abertura (Fig.24). Os pólos apresentam uma concavidade quando observados sob MEV (Fig.25).
Centrosema virginianum
(Fig.27-34)
Grãos de pólen grandes, isopolares, prolato- esferoidais (Tab.l), âmbito triangular, área polar pequena (Tab.2), 3-colporados, exina heterobrocada.
Os colpos são longos, largos, cobertos por membrana ornamentada (Figs.30, 32, 34) e apresentam uma margem com retículos menores (Fig.31), a endoabertura é lalongada e muito ampla (Figs.33-34).
A exina apresenta muros largos e baixos, com perfurações esparsas, lumens amplos, com báculos no interior (Fig.31).
Macroptilium bracteatum
(Fig.35-40)
Grãos de pólen médios (Tab.l), isopolares, oblato- esferoidais, âmbito subcircular, área polar pequena (Tab.2), 3-colporados, exina microrreticulada.
Os colpos são longos, largos, com margem ornamentada (Figs. 38-39), endoabertura aproximadamente circular (Fig.39).
Exina com muros largos, baixos com elevações nos pontos de interseção das malhas (Fig.37), sexina mais espessa do que a nexina.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004
360 F.C.SOUZA, M.A.SOUZA, C.B.F.MENDONÇA & V.GONÇALVES-ESTEVES
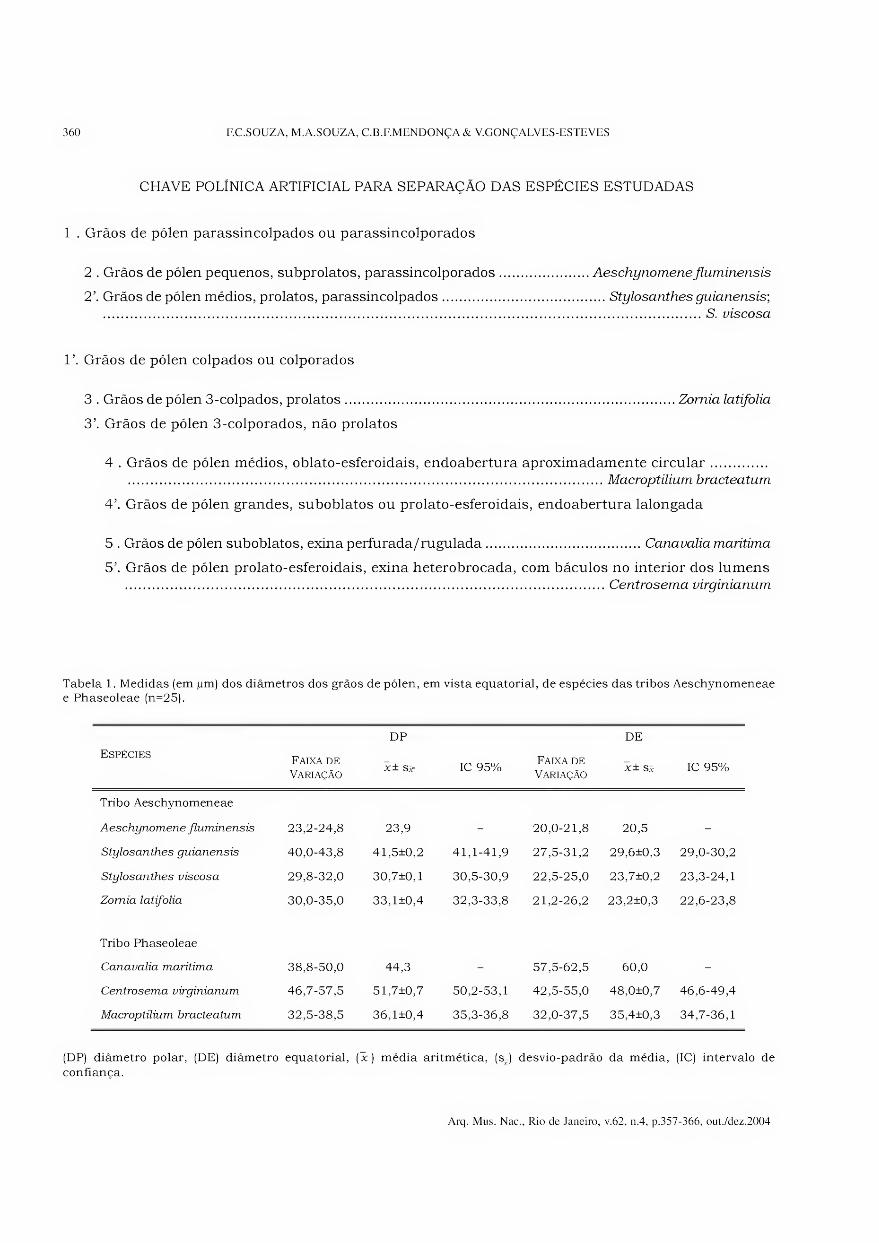
CHAVE POLÍNICA ARTIFICIAL PARA SEPARAÇÃO DAS ESPÉCIES ESTUDADAS
1 . Grãos de pólen parassincolpados ou parassincolporados
2 . Grãos de pólen pequenos, subprolatos, parassincolporados.Aeschynomene fluminensis
2\ Grãos de pólen médios, prolatos, parassincolpados.Stylosanthes guianensis;
.S. viscosa
V. Grãos de pólen colpados ou colporados
3 . Grãos de pólen 3-colpados, prolatos.Zomia latifolia
3’. Grãos de pólen 3-colporados, não prolatos
4 . Grãos de pólen médios, oblato-esferoidais, endoabertura aproximadamente circular. .Macroptüium bracteatum
4’. Grãos de pólen grandes, suboblatos ou prolato-esferoidais, endoabertura lalongada
5 . Grãos de pólen suboblatos, exina perfurada/ruguiada.Canavalia marítima
5’. Grãos de pólen prolato-esferoidais, exina heterobrocada, com báculos no interior dos lumens .Centrosema virginianum
Tabela 1. Medidas (em jum) dos diâmetros dos grãos de pólen, em vista equatorial, de espécies das tribos Aeschynomeneae e Phaseoleae (n=25).
Espécies Faixa de
Variação
DP
3c ± S3r IC 95% Faixa de
Variação
DE
X± Sx IC 95%
Tribo Aeschynomeneae
Aeschynomene fluminensis 23,2-24,8 23,9 - 20,0-21,8 20,5 -
Stylosanthes guianensis 40,0-43,8 41,5±0,2 41,1-41,9 27,5-31,2 29,6+0,3 29,0-30,2
Stylosanthes viscosa 29,8-32,0 30,7±0,1 30,5-30,9 22,5-25,0 23,7±0,2 23,3-24,1
Zomia latifolia 30,0-35,0 33,1±0,4 32,3-33,8 21,2-26,2 23,2±0,3 22,6-23,8
Tribo Phaseoleae
Canavalia marítima 38,8-50,0 44,3 - 57,5-62,5 60,0 -
Centrosema virginianum 46,7-57,5 51,7±0,7 50,2-53,1 42,5-55,0 48,0+0,7 46,6-49,4
Macroptilium bracteatum 32,5-38,5 36,1±0,4 35,3-36,8 32,0-37,5 35,4±0,3 34,7-36,1
(DP) diâmetro polar, (DE) diâmetro equatorial, (jc) média aritmética, (s-) desvio-padrão da média, (IC) intervalo de
confiança.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004
ESTUDO POLÍNICO DE ESPÉCIES DE AESCHYNOMENEAE E PHASEOLEAE, ESTADO DO RIO DE JANEIRO 361
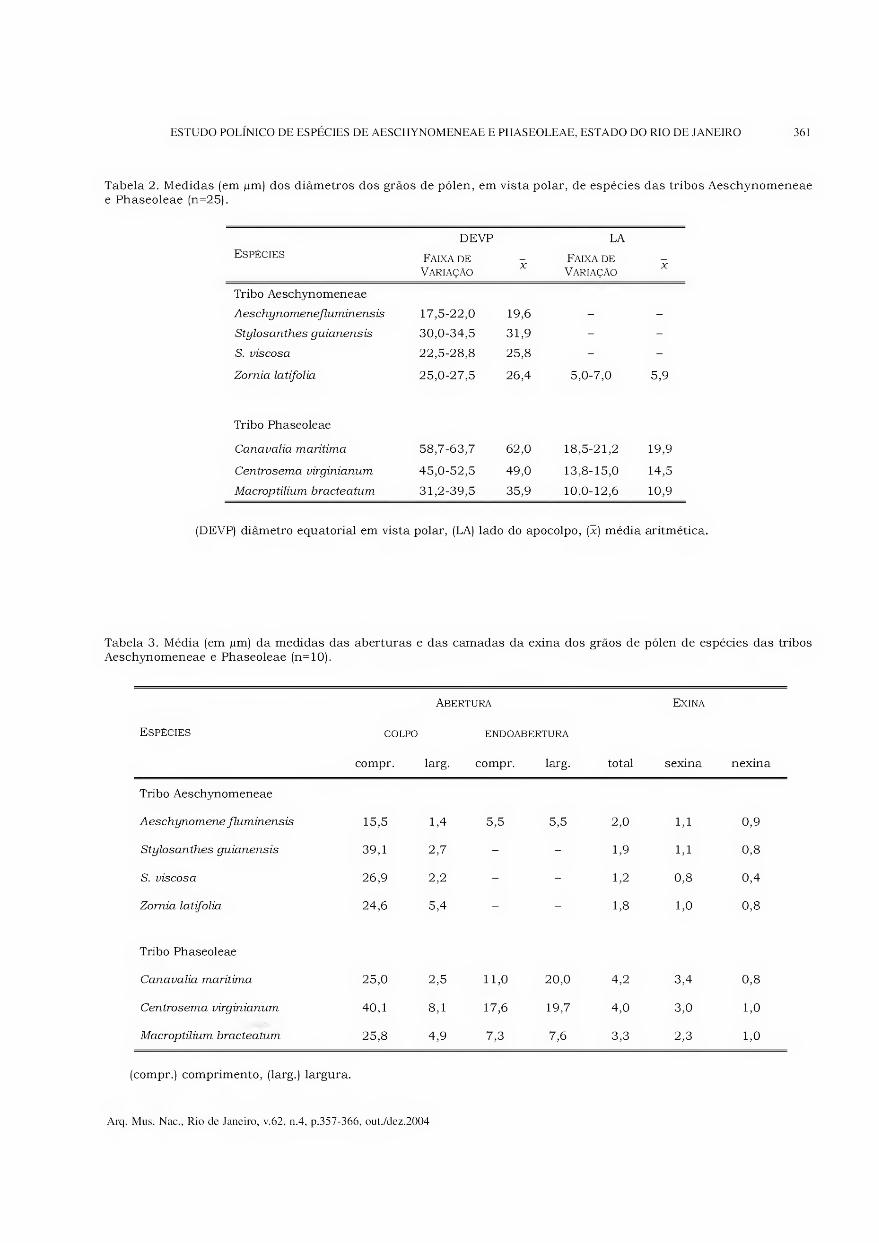
Tabela 2. Medidas (em pm) dos diâmetros dos grãos de pólen, em vista polar, de espécies das tribos Aeschynomeneae e Phaseoleae (n=25).
Espécies DEVP
Faixa de Variação
X
LA
Faixa de Variação
X
Tribo Aeschynomeneae
Aeschynomenefluminensis 17,5-22,0 19,6 - -
Stylosanthes guianensis 30,0-34,5 31,9 - -
S. viscosa 22,5-28,8 25,8 - -
Zornia latifolia 25,0-27,5 26,4 5,0-7,0 5,9
Tribo Phaseoleae
Canavalia marítima 58,7-63,7 62,0 18,5-21,2 19,9
Centrosema virginianum 45,0-52,5 49,0 13,8-15,0 14,5
Macroptilium bracteatum 31,2-39,5 35,9 10,0-12,6 10,9
(DEVP) diâmetro equatorial em vista polar, (LA) lado do apocolpo, (x) média aritmética.
Tabela 3. Média (em pm) da medidas das aberturas e das camadas da exina dos grãos de pólen de espécies das tribos Aeschynomeneae e Phaseoleae (n=10).
Espécies COLPO
compr.
Abertura
ENDOABERTURA
larg. compr. larg. total
Exina
sexina nexina
Tribo Aeschynomeneae
Aeschynomene fluminensis 15,5 1,4 5,5 5,5 2,0 1,1 0,9
Stylosanthes guianensis 39,1 2,7 - - 1,9 1,1 0,8
S. inscosa 26,9 2,2 - - 1,2 0,8 0,4
Zornia latifolia 24,6 5,4 - - 1,8 1,0 0,8
Tribo Phaseoleae
Canavalia marítima 25,0 2,5 11,0 20,0 4,2 3,4 0,8
Centrosema virginianum 40,1 8,1 17,6 19,7 4,0 3,0 1,0
Macroptilium bracteatum 25,8 4,9 7,3 7,6 3,3 2,3 1,0
(compr.) comprimento, (larg.) largura.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004
362 F.C.SOUZA, M.A.SOUZA, C.B.F.MENDONÇA & Y.GONÇALVES-ESTEVES
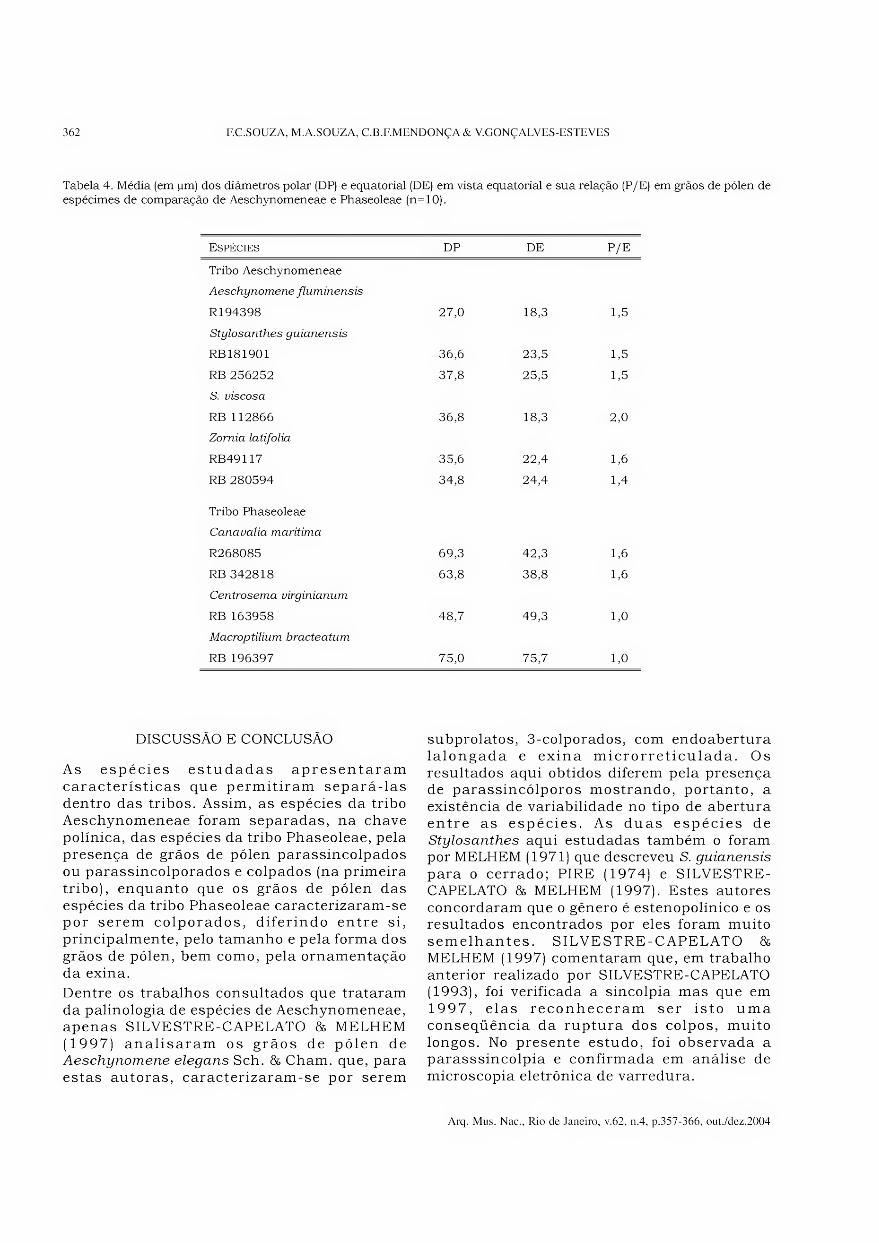
Tabela 4. Média (em jum) dos diâmetros polar (DP) e equatorial (DE) em vista equatorial e sua relação (P/E) em grãos de pólen de espécimes de comparação de Aeschynomeneae e Phaseoleae (n=10).
Espécies DP DE P/E
Tribo Aeschynomeneae
Aeschynomene fluminensis
R194398
Stylosanthes guianensis
27,0 18,3 1,5
RB181901 36,6 23,5 1,5
RB 256252 37,8 25,5 1,5
S. viscosa
RB 112866 36,8 18,3 2,0
Zornia latifolia
RB49117 35,6 22,4 1,6
RB 280594 34,8 24,4 1,4
Tribo Phaseoleae
Canavalia marítima
R268085 69,3 42,3 1,6
RB 342818 63,8 38,8 1,6
Centrosema virginianum
RB 163958 48,7 49,3 1,0
Macroptilium bracteatum
RB 196397 75,0 75,7 1,0
DISCUSSÃO E CONCLUSÃO
As espécies estudadas apresentaram características que permitiram separá-las dentro das tribos. Assim, as espécies da tribo Aeschynomeneae foram separadas, na chave polínica, das espécies da tribo Phaseoleae, pela presença de grãos de pólen parassincolpados ou parassincolporados e colpados (na primeira tribo), enquanto que os grãos de pólen das espécies da tribo Phaseoleae caracterizaram-se por serem colporados, diferindo entre si, principalmente, pelo tamanho e pela forma dos grãos de pólen, bem como, pela ornamentação da exina.
Dentre os trabalhos consultados que trataram da palinologia de espécies de Aeschynomeneae, apenas SILVESTRE-CAPELATO & MELHEM (1997) analisaram os grãos de pólen de Aeschynomene elegans Sch. & Cham. que, para estas autoras, caracterizaram-se por serem
subprolatos, 3-colporados, com endoabertura lalongada e exina microrreticulada. Os resultados aqui obtidos diferem pela presença de parassincólporos mostrando, portanto, a existência de variabilidade no tipo de abertura entre as espécies. As duas espécies de Stylosanthes aqui estudadas também o foram por MELHEM (1971) que descreveu S. guianensis
para o cerrado; PIRE (1974) e SILVESTRE- CAPELATO & MELHEM (1997). Estes autores concordaram que o gênero é estenopolínico e os resultados encontrados por eles foram muito semelhantes. SILVESTRE-CAPELATO & MELHEM (1997) comentaram que, em trabalho anterior realizado por SILVESTRE-CAPELATO (1993), foi verificada a sincolpia mas que em 1997, elas reconheceram ser isto uma conseqüência da ruptura dos colpos, muito longos. No presente estudo, foi observada a parasssincolpia e confirmada em análise de microscopia eletrônica de varredura.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004
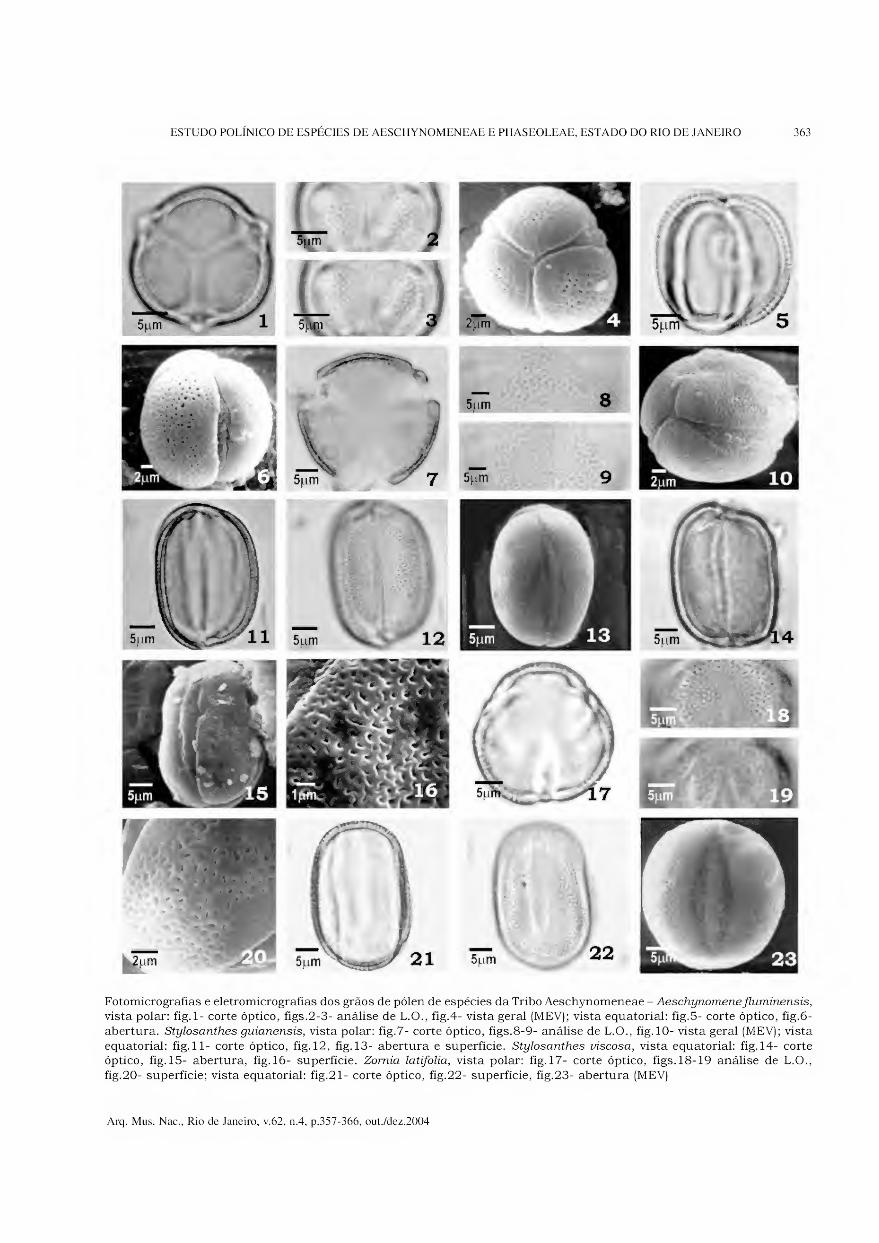
ESTUDO POLÍNICO DE ESPÉCIES DE AESCHYNOMENEAE E PHASEOLEAE, ESTADO DO RIO DE JANEIRO 363
Fotomicrografias e eletromicrografias dos grãos de pólen de espécies da Tribo Aeschynomeneae - Aeschynomene fluminensis, vista polar: fig.l- corte óptico, figs.2-3- análise de L.O., fig.4- vista geral (MEV); vista equatorial: fig.5- corte óptico, fig.6-
abertura. Stylosanthes guianensis, vista polar: fig.7- corte óptico, figs.8-9- análise de L.O., fig. 10- vista geral (MEV); vista
equatorial: fig. 11- corte óptico, fig. 12, fig. 13- abertura e superfície. Stylosanthes viscosa, vista equatorial: fig. 14- corte óptico, fig. 15- abertura, fig. 16- superfície. Zomia latifolia, vista polar: fig. 17- corte óptico, figs.18-19 análise de L.O.,
fig.20- superfície; vista equatorial: fig.21- corte óptico, fig.22- superfície, fig.23- abertura (MEV)
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004
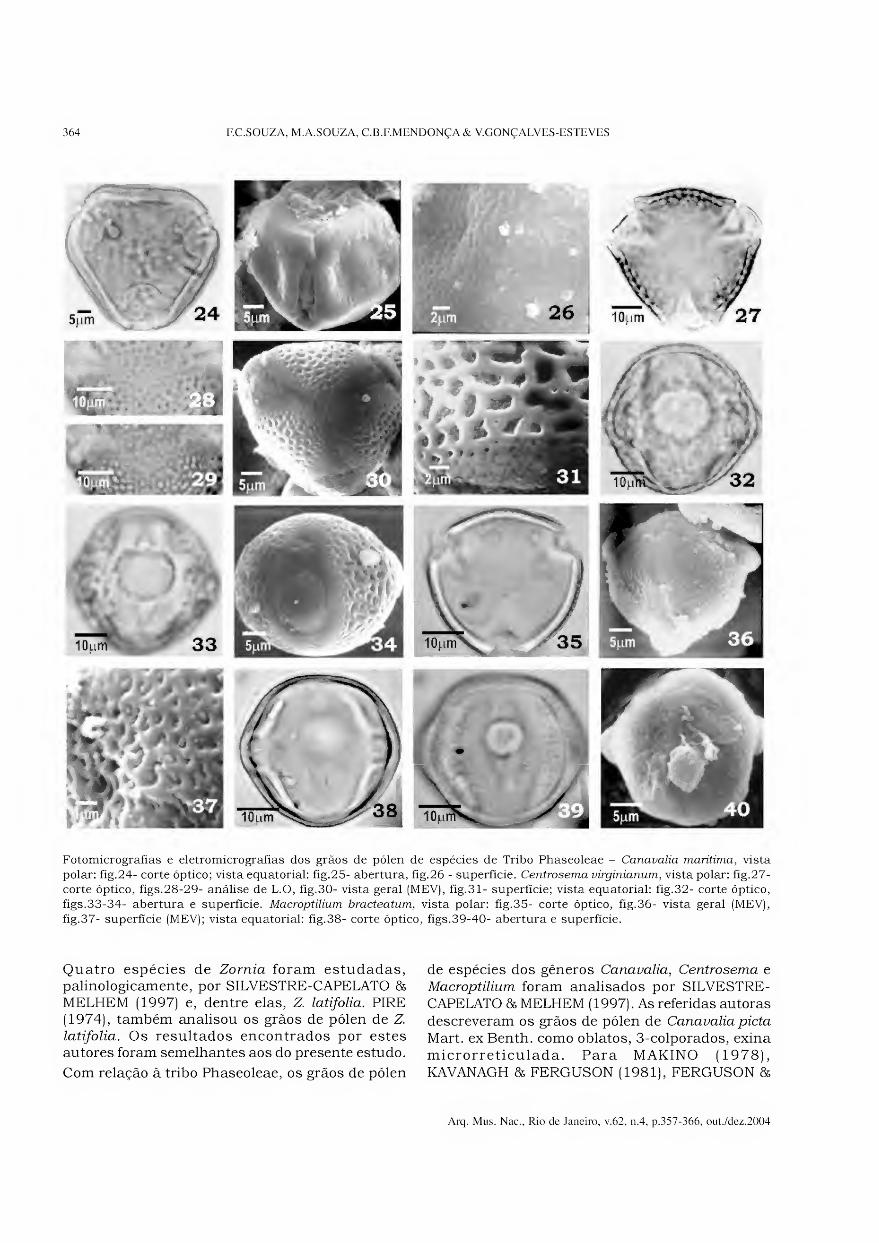
364 F.C.SOUZA, M.A.SOUZA, C.B.F.MENDONÇA & Y.GONÇALVES-ESTEVES
Fotomicrografias e eletromicrografias dos grãos de pólen de espécies de Tribo Phaseoleae - Canavalia marítima, vista
polar: fig.24- corte óptico; vista equatorial: fig.25- abertura, fig.26 - superfície. Centrosema virginianum, vista polar: fig.27- corte óptico, figs.28-29- análise de L.O, fig.30- vista geral (MEV), fig.31- superfície; vista equatorial: fig.32- corte óptico,
figs.33-34- abertura e superfície. Macroptilium bracteatum, vista polar: fig.35- corte óptico, fig.36- vista geral (MEV),
fig.37- superfície (MEV); vista equatorial: fig.38- corte óptico, figs.39-40- abertura e superfície.
Quatro espécies de Zornia foram estudadas, palinologicamente, por SILVESTRE-CAPELATO & MELHEM (1997) e, dentre elas, Z. latifolia. PIRE (1974), também analisou os grãos de pólen de Z. latifolia. Os resultados encontrados por estes autores foram semelhantes aos do presente estudo.
Com relação à tribo Phaseoleae, os grãos de pólen
de espécies dos gêneros Canavalia, Centrosema e Macroptilium foram analisados por SILVESTRE- CAPELATO & MELHEM (1997). As referidas autoras descreveram os grãos de pólen de Canavalia picta Mart. ex Benth. como oblatos, 3-colporados, exina microrreticulada. Para MAKINO (1978), KAVANAGH & FERGUSON (1981), FERGUSON &
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004

ESTUDO POLÍNICO DE ESPÉCIES DE AESCHYNOMENEAE E PHASEOLEAE, ESTADO DO RIO DE JANEIRO 365
SKVARLA (1981), os grãos de pólen eram heteropolares com exina psilada. No presente estudo, a espécie Canaualia marítima foi descrita como possuindo grãos de pólen suboblatos, isopolares, com exina perfurado-rugulada, caracterizando, assim, uma grande variação nas características polínicas nas espécies de Canavalia.
Para SILVESTRE-CAPELATO & MELHEM (1997), os grãos de pólen de Centrosema grandiflomm Benth. eram subprolatos (acetolisados) ou oblatos (não acetolisados), 3-colporados, endoabertura lolongada, operculada, exina reticulada, com muros sinuosos e lúmens menores junto das aberturas. No presente estudo, C. virginianum foi descrita como possuindo características polínicas semelhantes às citadas pelas autoras no que se refere ao tamanho do grão de pólen, quanto ao tipo de abertura e à ornamentação da sexina. A endoabertura, no entanto, foi considerada lalongada, ampla, os colpos apresentaram-se recobertos por membrana ornamentada e apresentaram uma margem com reticulação menor. A exina foi descrita como heterorreticulada, apresentando muros largos e baixos com perfurações esparsas, lúmens amplos, com báculos no interior.
Os grãos de pólen de Macroptilium erythroloma
(Benth.) Urban foram descritos por SILVESTRE- CAPELATO 86 MELHEM (1997) como prolato- esferoidais, 3-colporados, circundados por área psilada, endoabertura lolongada e exina microrreticulada. As características encontradas no presente estudo para M. bracteatum mostraram ser semelhantes àquelas de M. erythroloma diferindo, apenas, na forma dos grãos de pólen, na presença de margem ornamentada sobre os colpos, nas dimensões da endoabertura (aproximadamente circular) e na presença de muros largos, baixos com elevações nos pontos de interseção das malhas.
Embora o número de espécies estudadas de cada gênero, pertencentes as duas tribos, não tenha sido grande, foi possível concluir que os táxons de Stylosanthes apresentaram certa homogeneidade nas características polínicas. Os caracteres intergenéricos e intragenéricos, no entanto, mostraram-se, polinicamente, distintos.
AGRADECIMENTOS
Ao Laboratório de Microscopia Eletrônica da Pontifícia Universidade Católica - Rio de Janeiro (PUC-RJ), na pessoa da MSc. Maria de Fátima Lopes (in memorian), que forneceu condições de trabalho para a obtenção das eletromicrografias;
ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), à Fundação Universitária José Bonifácio e à Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), pelos auxílios concedidos.
REFERÊNCIAS BIBLIOGRÁFICAS
BARTH, O.M., 1964. Catálogo sistemático dos pólens
das plantas arbóreas do Brasil Meridional V-
Leguminosae: Papilionatae. Memórias do Instituto
Oswaldo Cruz, Rio de Janeiro, 62:95-123.
BARTH, O.M. & MELHEM, T.S., 1988. Glossário
ilustrado de palinologia, Campinas: Ed. Unicamp.
76p., 101 figs.
ERDTMAN, G., 1952. Pollen morphology and plant
taxonomy Angiosperms. Stockholm: Almqvist &
Wiksell. 539p., 261 figs.
FAEGRI, G. & IVERSEN, J., 1966. Textbook of modern
pollen analysis. 2.ed. Copenhagen: Scandinavian
University Books.
FERGUSON, I.K. & SKVARLA, J.J., 1981. The pollen
morphology of the subfamily Papilionoideae
(Leguminosae). In: POLHILL, R.M. & RAVEN, P.H.
(Eds.) Advances in legume systematics 1. London:
Kew Royal Botanic Gardens. p.859-896.
FERGUSON, I.K. & SKVARLA, J.J., 1983. The granular
interstitium in the pollen of subfamily Papilionoideae
(Leguminosae). American Journal of Botany, New
York, 70(9): 1401-1408.
FERGUSON, I.K. & SKVARLA, J.J., 1988. Pollen
morphology of the tribe Swartzieae (Subfamily
Papilionoideae: Leguminosae). 1. Introduction and
all genera excluding Aldina and Swartzia. American
Journal of Botany, New York, 75(12):1884-1897.
GONÇALVES-ESTEVES, V. & CRESPO, S.R.M., 1994.
Estudo polínico em plantas de restinga do Estado do
Rio de Janeiro - Leguminosae A.L. Juss. - Faboideae
Hutch. Boletim Museu Nacional, Nova Série,
Botânica, Rio de Janeiro (96): 1-11.
GUINET, Ph., 1981. Comparative account of pollen
characters in the Leguminosae. In: POLHILL, R.M. &
RAVEN, P.H. (Eds.) Advances in legume systematics
1. London: Kew Royal Botanic Gardens. p.789-799.
HOLMGREN, P.K.; HOLMGREN, N.H. 86 BAINETT, L.G.,
1990. Index Herbariorum. Part. 1. The Herbaria of
the world. New York: New York Botanical Garden.
KAVANAGH, T.A. 86 FERGUSON, I.K., 1981. Pollen
morphology and taxonomy of the subtribe Diocleinae
(Leguminosae: Papilionoideae: Phaseoleae). Review of
Palaeobotany and Palynology, Missouri, 32:317-367.
MAKINO, H., 1978. Palynological studies in
Leguminosae (Lotoideae) tribe Phaseoleae. Hoehnea,
São Paulo, 7:47-98.
MELHEM, T.S., 1971. Pollen grains of the “cerrado”.
Leguminosae-Lotoideae: Tribe Phaseoleae. Hoehnea,
São Paulo, 1:119-151.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004

366 F.C.SOUZA, M. A. SOUZA, C.B.F.MENDONÇA & V. GONÇALVES-ES TE VES
MELHEM, T.S.; CRUZ-BARROS, M.A.V.; CORRÊA,
A.M.S.; MAKINO-WATANABE, H.; SILVESTRE-
CAPELATO, M.S.F. & GONÇALVES-ESTEVES, V.,
2003. Variabilidade polínica em plantas de Campos
de Jordão (São Paulo, Brasil). Boletim do Instituto
de Botânica, São Paulo, 16:01-104.
PIRE, S.M.L., 1974 - Estúdio palinologico de la tribe
“Hedysareae” (Leguminosae). Bonplandia 3(12): 143-169.
PUNT, W.; BLACKMORE, S.; NILSSON, S. 85 LE THOMAS,
A., 1999. Glossary of pollen and spore terminology.
Disponível em: <http://www.biol.ruu.nl/~palaeo/
glossary/glos-int.htm>. Acesso em: 18 abr. 1999
SILVESTRE-CAPELATO, M.S.F., 1993. Palinologia das
Leguminosae da Reserva Biológica do Parque
Estadual das Fontes do Ipiranga. Campinas, 187p.
Tese (Doutorado em Biologia Vegetal) Universidade
Estadual de Campinas.
SILVESTRE-CAPELATO, M.S.F. & MELHEM, T.S.,
1997. Flora polínica da Reserva do Parque
Estadual das Fontes do Ipiranga (São Paulo,
Brasil). Família: 81 - Leguminosae. Hoehnea, São
Paulo, 24(1): 115-163.
TEWARI, R.B. 85 NAIR, P.K.K., 1979. Pollen morphology
of some Indian Papilionaceae. Journal of
Palynology, Missouri, 15(2):49-73.
VISHNU-MITTRE 85 SHARMA, B.D., 1962. Studies of
Indian pollen grains. 1. Leguminosae. Pollen et
Spores, Viena, 4(l):5-45.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.357-366, out./dez.2004

nnniTOnnnrtnnnni nnniiitiiiMiftfinnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.367-374,out./dez.2004
ISSN 0365-4508
PASSIFLORACEAE DA MATA DE ENCOSTA DO JARDIM BOTÂNICO DO
RIO DE JANEIRO E ARREDORES, RIO DE JANEIRO, RJ 1
(Com 3 figuras)
MICHAELE ALVIM MILWARD-DE-AZEVEDO 2>3 MARIA DA CONCEIÇÃO VALENTE 4
RESUMO: As Passifloraceae da mata de encosta do Jardim Botânico do Rio de Janeiro e arredores estão
representadas por cinco táxons: Passiflora amethystina Mikan, P. capsularis L., P. racemosa Brot., P.
sidaefolia M. Roemer e P. truncata Regei. Para identificação das espécies apresenta-se uma chave analítica,
seguida de descrições morfológicas, citação do material examinado, distribuição geográfica, nomes
populares, dados fenológicos e do hábitat, comentários e ilustrações.
Palavras-chave: Passifloraceae, Passiflora, taxonomia, Rio de Janeiro.
ABSTRACT: Passifloraceae in the Rio de Janeiro Botanical Garden hillside forest and surroundings, Rio de
Janeiro, RJ.
The inventory of the Passifloraceae family in the Rio de Janeiro Botanical Garden hillside forest and
surroundings was realized and five taxa was found: Passiflora amethystina Mikan, P. capsularis L., P.
racemosa Brot., P. sidaefolia M.Roemer and P. truncata Regei. A key for Identification of the species, with
their morphological descriptions, examined material, geographic distribution, popular names, phenology,
habitats, taxonomic comments and illustrations are presented.
Key words: Passifloraceae, Passiflora, taxonomy, Rio de Janeiro.
INTRODUÇÃO
A família Passifloraceae está representada por 17 gêneros (BRUMMITT, 1992; MABBERLEY, 1997) e aproximadamente 630 espécies (HOLM-NIELSEN, J0RGENSEN & LAWESSON, 1988; DEGINANI, 1999), com distribuição tropical pelas Américas e África (METCALFE & CHALK, 1965; JOLY, 1993). A maioria está subordinada ao gênero Passiflora L. (KILLIP, 1938; PURI, 1947; BARROSO et dl., 1978), que possui cerca de 400 espécies (BRIZICKY, 1961; SACCO, 1980; CERVI, 1986; ESCOBAR, 1988; DEGINANI, 1999).
A família é facilmente reconhecida por apresentar hábito escandente, folhas alternas com gavinhas axilares, pecíolo geralmente com glândulas e flores com corona e androginóforo.
A área investigada compreende 147,06ha referente à mata de encosta do Jardim Botânico do Rio de Janeiro e Horto Florestal, e 34,8ha do Parque Lage, composta por floresta ombrófila densa submontana alterada, com regiões de
afloramentos rochosos, fazendo parte da Serra Carioca e limitando-se com o Parque Nacional da Tijuca.
O estudo desta flora vascular iniciou-se com as coletas de Kuhlmann que, em 1927, apresentou um mapa fitogeográfico assinalando as espécies arbóreas do Horto Florestal (MARQUETE etal, 1994).
O presente estudo faz parte do Projeto “Vegetação das áreas do entorno do Jardim Botânico do Rio de Janeiro, Parque Lage e Horto Florestal”, desenvolvido pela equipe do Projeto “Avanços Taxonômicos e Coleções Botânicas (Programa Diversidade Taxonômica - PROTAXON)”, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Ao dar-se continuidade à divulgação, atualização e complementação da listagem de espécies, objetiva-se contribuir para a publicação de dados mais abrangentes sobre a flora local e, conseqüentemente, para o conhecimento da diversidade das espécies no Estado do Rio de Janeiro, e conseqüentemente, do Brasil.
1 Submetido em 13 de maio de 2003. Aceito em 03 de novembro de 2004.
Parte da monografia de M.A.Milward-de-Azevedo, Curso de Ciências Biológicas/Botânica, Universidade Federal do Estado do Rio de Janeiro (UNIRIO).
2 Museu Nacional/UFRJ, Programa de Pós-Graduação em Ciências Biológicas/Botânica. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil.
Bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
3 E-mail: [email protected].
4Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Rua Pacheco Leão, 915, Jardim Botânico, 22460-030, Rio de Janeiro, RJ, Brasil.

368 M.A.MILWARD-DE-AZEVEDO & M.C.VALENTE
MATERIAL E MÉTODOS
Foram realizadas excursões mensais na área em
estudo, de 1991 a 1999, pelo “Projeto vegetação
das áreas do entorno do Jardim Botânico do Rio
de Janeiro, Parque Lage e Horto Florestal”,
procurando complementar as coletas realizadas
antes da implantação do mesmo.
Foi utilizado material adicional, de áreas
próximas à estudada, para completar a
descrição das espécies, quando estas se
encontravam sem flor ou fruto.
Dados sobre a distribuição geográfica,
floração e frutificação, além de outras
informações importantes, como nome vulgar
e características das espécies no hábitat
natural, foram obtidos nas etiquetas das
exsicatas dos herbários consultados (siglas de
acordo com HOLMGREN, KEUKEN &
SCHOFIELD, 1990): GUA, HB, R, RB e RFA e
acrescidos de dados descritos na bibliografia
especializada. A caracterização da vegetação
está de acordo com a classificação de
VELOSO, RANGEL FILHO & LIMA (1991).
RESULTADOS E DISCUSSÃO
Passiflora L., Sp. Pt 1: 955. 1753.
Trepadeiras herbáceas com gavinhas axilares.
Folhas alternas, pecíolos com ou sem glândulas,
lâminas membranáceas a cartáceas, simples,
lobadas a levemente lobadas para o ápice,
margem inteira a levemente serreada, 3-5
nervuras saindo da base. Estipulas foliáceas ou
linear-subuladas. Brácteas 3, ovadas a
lanceoladas, ou ausentes. Flores hermafroditas,
axilares, isoladas ou aos pares; pedicelo articulado; tubo do cálice curto-cilíndrico a
campanulado; 5 sépalas membranáceas,
oblongas a lanceoladas; 5 pétalas membranáceas,
oblongas a lanceoladas; corona filamentosa, com
1-5 séries; opérculo membranoso; límen anular,
cupuliforme ou ausente; androceu com 5 estames
livres aderidos ao ginóforo, formando o
androginóforo, anteras dorsifixas; gineceu com
3 estiletes, estigmas capitados. Frutos bagas ou
cápsulas; sementes comprimidas, numerosas,
com testa endurecida, costada, foveolada,
reticulada ou sulcada transversalmente.
Chave para identificação das espécies de Passiflora
DA MATA DE ENCOSTA DO JARDIM BOTÂNICO DO RlO DE JANEIRO E ARREDORES
1. Pecíolos sem glândulas; coronas com 1 série de filamentos; brácteas ausentes; frutos cápsulas .Passiflora capsularis
1 ’. Pecíolos com glândulas; coronas com 2-5 séries de filamentos; brácteas presentes; frutos bagas.2
2. Lâminas com ápice truncado a obtuso, glândulas entre as nervuras laterais, 3 nervuras saindo da base; estipulas linear-subuladas; testas das sementes irregularmente sulcadas. .Passiflora truncata
2\ Lâminas com ápice agudo a obtuso, glândulas nos bordos entre os lobos, 5 nervuras saindo da base; estipulas ovadas, oblíquas ou assimétricas ovadas, foliáceas; testas das sementes costadas, foveoladas ou reticuladas.3
3. Inflorescências racemosas; sépalas corniculadas no ápice, límens ausentes.Passiflora racemosa
3’. Flores solitárias; sépalas com carena no ápice ou não, límens cupuliformes.4
4. Lâminas nitidamente 3-lobadas, 5-8 glândulas nos pecíolos; sépalas com carena no ápice; coronas com 4 séries de filamentos; frutos elípticos; testas das sementes costadas.Passiflora amethystina
4\ Lâminas levemente 3-lobadas no ápice, 2 pares de glândulas nos pecíolos; sépalas sem carena no ápice; coronas com 5 séries de filamentos; frutos globosos; testas das sementes reticuladas. .Passiflora sidaefolia
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

PASSIFLORACEAE DA MATA DO JARDIM BOTÂNICO DO RIO DE JANEIRO E ARREDORES 369
1. Passiflora amethystina Mikan, Delect. Fl. etFaun.
Bras. 39. 1820. (Fig.l a-e)
Descrição - Caule cilíndrico, tricomas esparsos. Folhas com pecíolos 2,0-6,5cm compr., 5-8 glândulas curto-estipitadas, tricomas esparsos; lâminas membranáceas, 3-lobadas, 2,8-9,4cm compr. na nervura média, 2,5-7,5cm compr. na nervura lateral, 4,l-12,7cm larg., ápice de agudo a obtuso, apiculado, base subcordada a subpeltada, margem inteira a levemente serreada, glândulas nos bordos entre os lobos, 5 nervuras saindo da base, glabras em ambas as faces. Estipulas 1,5-2,7 x 0,4-l,5cm, foliáceas, ovado- assimétricas, ápice apiculado, base oblíqua. Brácteas 0,6-3,0 x 0,3-l,3cm, lanceoladas, caducas. Flores azuis, liláses, púrpuras ou alvas, axilares, solitárias; pedicelos 7,0-ll,0cm compr., glabros; sépalas 1,6-2,6 x 0,4-0,5cm, oblongo- lanceoladas, ápice obtuso, carena dorsal aristada 0,4-0,7cm compr., glabras; pétalas 1,8-2,7 x 0,3- 0,4cm, oblongas, ápice obtuso; corona com 4 séries, 2 externas ca. 2,0cm compr., filamentos liguliformes, 2 internas ca. l,2cm compr., filamentos filiformes; opérculo plicado; límen cupuliforme; androginóforo 2,0-3,0cm compr., estames 0,5-0,8cm compr., anteras 0,6-0,8 x 0,1- 0,3cm; ovário 0,4-0,6 x 0,2-0,3cm, oblongo, viloso, ferrugíneo-tomentoso, estilete 0,8-l,0cm compr. Frutos 4,0-6,2 x l,0-2,3cm, bagas, elípticos; sementes ca. 0,5 x 0,3cm, ovadas, testa costada.
Material examinado - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, encosta do Jardim Botânico do Rio de Janeiro, Lagoinha, fl., RB23561, A.Ducke e J.G.Kuhlmann s.n., 20/111/1922.
Material adicional - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, estrada da Vista Chinesa, fr, RB, J.A.Jesus 1608, 30/V/1972; fl, RB, V.G.L.Klein e M.L.Pereira 1065, 09/V/1991.
Distribuição geográfica - Bolívia, Brasil e Paraguai. No Brasil: Goiás, Distrito Federal, Bahia e nas regiões Sudeste e Sul.
Nome popular - Maracujá-azul, maracujá-de- cobra, maracujá.
Dados fenológicos - É encontrada, ocasionalmente, em flor e fruto o ano todo.
Hábitat- Floresta Ombrófila Densa e Savana. Espécie heliófila a semi-heliófila, ocorrendo principalmente em beira de estrada e em clareiras da mata.
Comentários - Passiflora amethystina é uma espécie bem distinta das outras estudadas, principalmente
por apresentar lâminas nitidamente 3-lobadas, 5-8 glândulas no pecíolo, carena no ápice das sépalas, corona com quatro séries de filamentos e semente com testa costada. SACCO (1980) caracteriza a espécie pelas folhas escurecidas quando submetidas à secagem, flores azul-purpúreas e ovário em geral ferrugíneo-viloso-tomentoso. Pertence ao subgênero Passiflora L. série Lobatae Killip.
2. Passiflora capsularis L., Sp. Pl. 1: 957. 1753. (Fig.lf-j)
Descrição - Caule anguloso, pubescente. Folhas com pecíolos 0,5-3,0cm compr., glândulas ausentes, pubescentes; lâminas membranáceas, 2- lobadas, raro 3-lobadas, l,0-8,0cm compr. na nervura média, 2,0-10,6cm compr. na nervura lateral, 2,5-8,3cm larg., ápice agudo, apiculado, base cordada, margem inteira, 3 nervuras saindo da base, pubescente em ambas as faces. Estipulas 0,4-0,7cm compr, linear-subuladas, levemente falcadas. Brácteas ausentes. Flores alvas ou cremes, axilares, solitárias ou aos pares; pedicelos l,0-5,0cm compr., pubescentes; sépalas 1,5-2,7 x 0,3-0,4cm, linear-lanceoladas, ápice agudo, sem carena, pubescentes, 3 nervuras proeminentes na face dorsal; pétalas 1,0-1,5 x 0,2cm, oblongo- lanceoladas, ápice obtuso; corona com 1 série, ca. l,2cm compr., filamentos filiformes, unidos na base por membrana delgada; opérculo plicado; límen anelar; androginóforo 0,7-l,5cm compr., estames 0,3-0,5cm compr., anteras ca. 0,4 x 0,1 cm, ovário 0,3-0,5 x 0,2-0,4cm, obovóide, levemente pubérulo, estilete 0,4-0,6cm compr. Frutos 2,3-8,5 x 0,7- 4,5cm, capsulares-loculicidas, elipsoidais ou fusiformes, violáceos; sementes ca. 0,4 x 0,2cm, elípticas, testa sulcada transversalmente.
Material examinado - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, estrada para o Horto do Instituto Brasileiro de Desenvolvimento Florestal, fl., RB340717, s.col., s.n., sem data.
Material adicional - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, Tijuca, Exclesior, fr., HB, E.Pereira 66, 21/XII/1940.
Distribuição geográfica - América Central; Colômbia, Equador, Brasil, Paraguai e Uruguai. No Brasil: Pará, Mato Grosso, Goiás, Piauí, Ceará e nas regiões Sudeste e Sul.
Nome popular - Maracujá-branco-miúdo, maracujá- branco, maracujá-mirim, maracujazinho, maracujá.
Dados fenológicos - Foi encontrada em flor nos meses de janeiro a maio, e outubro a dezembro, e em fruto em janeiro, março a maio, novembro e dezembro.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

370 M.A.MILWARD-DE-AZEVEDO & M.C.VALENTE
Hábitat - Floresta Ombrófila Densa, Vegetação de Influência Marítima, Savana e Estágio Sucessional Secundário. É uma espécie heliófila a semi- heliófila, e tem preferência por capoeiras e orla de florestas, em terrenos com boa drenagem, raramente encontrada no interior da floresta (CERVI, 1981;1992), sendo uma planta de formação secundária que sofre intensa herbivoria.
Comentários - Passiflora capsularis distingue-se das demais espécies estudadas principalmente pela ausência de glândulas no pecíolo e de brácteas,
flores com corona unisseriada e frutos capsulares. Suas folhas são geralmente bilobadas, podendo ser encontrado um curto lobo central em alguns espécimes. Pertence ao subgênero Decaloba (DC.) Rchb. seção Xeroga
(Raf.) Killip. Esta espécie pode apresentar uma segunda série de filamentos da corona de 3,0 mm compr., de coloração violeta (CERVI, 1986). Suas folhas são abortivas e suas raízes são suspeitas de prejudiciais ao gado e hemenogogas (PIO- CORRÊA, 1984).
Fig. 1- Passiflora amethystina: (a) forma da folha e aspecto geral da nervação, (b) esquema geral da flor, (c) ápice da sépala, (d) fruto, (e) semente; Passiflora capsularis: (f) forma da folha e aspecto geral da nervação, (g) esquema geral da flor, (h)
ápice da sépala, (i) fruto, (j) semente. Escalas: a, b, d, f, i = lcm; c, e, h, j = lmm; g = 2cm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

PASSIFLORACEAE DA MATA DO JARDIM BOTÂNICO DO RIO DE JANEIRO E ARREDORES 371
3. Passiflora racemosa Brot., Trans. Linn. Soc. 12: 71. 1817. (Fig.2 a-f)
Descrição - Caule cilíndrico, glabro. Folhas com pecíolos 0,8-6,0cm compr., 2 pares de glândulas sésseis, glabros; lâminas membranáceas a cartáceas, polimorfas, ovadas quando inteiras, assimétricas quando lobadas de um só lado, ou simetricamente 3-lobadas como na forma adulta com 4,0-10,5cm compr. na nervura média, 3,5-9,0cm compr. na nervura lateral, 5,5-14,6cm larg., lobos oblongos 1,1- 4,5cm larg., ápice agudo, base subpeltada ou cordada, margem inteira, glândulas nos bordos entre os lobos, 5 nervuras saindo da base, glabras em ambas as faces. Estipulas 1,2-2,0 x 0,8-l,3cm, foliáceas, ovadas, oblíquas, ápice apiculado, base obtusa. Brácteas 0,8-1,0 x 0,5-0,6cm, ovadas, ápice agudo, base obtusa, róseas a vinosas. Inflorescências axilares, racemosas. Flores alvas, rosas ou vermelhas, solitárias ou aos pares; pedicelos l,5-2,0cm compr., glabros; sépalas 3,5-4,0 x 0,8-0,9cm, oblongas, ápice corniculado, sem carena, glabras; pétalas 2,0-2,5 x 0,4-0,5cm, oblongas, ápice obtuso; corona filamentosa até a base com 3 séries, ca. 0,4cm compr.; opérculo divergindo horizontalmente, denticulado; límen ausente; androginóforo 3,5-4,8cm compr., estames 0,7-l,2cm compr., anteras 0,6-0,8 x 0,1- 0,2cm; ovário 0,6-0,7 x ca.0,2cm, oblongo, glabro, estilete 0,6-0,8cm compr. Frutos 5,0-7,0 x 2,0-3,0cm, bagas, oblongos; sementes 0,5-0,6 x 0,3-0,4cm, elípticas, testa foveolada.
Material examinado - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, matas do Jardim Botânico do Rio de Janeiro, RB, A.P.Duarte 140, 04/ VII/1946; fl., RB, Miguel e outros 4,06/111/1963; Pedra do Marinheiro, fl., GUA, M.Santos e outros 37, 24/V/ 1979; fl., fr., RB, N.M.F.Silva e outros 274, 02/VI/ 1992; fl., RB, R.Marquete e outros 1996,24/VIII/1994; fl., RB, P.Botelho e outros 60, 29/VIII/1995; RB, M.Milward e R.Marquete 32, 20/X/1999; Grotão, Pedra d’água, fl., RB, G.Martinelli 2829,01/VIII/1977.
Distribuição geográfica - Brasil: Espírito Santo e Rio de Janeiro. Introduzida na horticultura européia (KILLIP, 1938).
Nome popular - Maracujá-mirim, maracujá.
Dados fenológicos - Foi encontrada em flor nos meses de março a dezembro e em fruto em maio a junho, agosto a dezembro.
Hábitat - Floresta Ombrófila Densa e Vegetação de Influência Marítima. Espécie ombrófila a heliófila, ocorrendo em bordas de florestas e clareiras.
Comentários - Passiflora racemosa caracteriza-se
principalmente pelas inflorescências racemosas, flores com os ápices das sépalas corniculados, ausência de límen e três séries de filamentos da corona, e testa das sementes foveoladas. As lâminas são polimorfas, apresentando-se desde inteiras até 3-lobadas, alguns exemplares desta espécie são encontrados somente com lâminas inteiras, ou seja, um indivíduo jovem, geralmente, este tipo de material pode dificultar a identificação. Pertence ao subgênero Calopathanthus (Harms) Killip, que antes nomeava uma seção, devido à forma de seu opérculo. Sendo uma das mais vistosas passifloras, tem sido muito introduzida no cultivo, produzido extensamente na hibridação artificial (PIO-CORRÊA, 1984). Passiflora
racemosa é conhecida também sob o nome de P.
princeps Lodd., ambos os nomes foram propostos em 1817 (KILLIP, 1938). Comestível, medicinal e ornamental, sendo própria para pergolados de jardins residenciais (FUKS etal, 1999).
4. Passiflora sidaefolia M. Roemer, Fam. Nat. Syn.
2: 173. 1846. (Fig.2 g-1)
Descrição - Caule cilíndrico, glabro. Folhas com pecíolos 0,8-3,lcm compr., 2 pares de glândulas sésseis, glabros; lâminas membranáceas a cartáceas, levemente 3-lobadas para o ápice, 3,2-8,6cm compr. na nervura média, 2,8-6,5cm compr. na nervura lateral, 2,4-7,lcm larg., ápice agudo a obtuso, base arredondada, subtruncada ou subpeltada, margem inteira, glândulas nos bordos entre os lobos, 5 nervuras saindo da base, glabras em ambas as faces. Estipulas 1,3-3,0 x 0,5-l,8cm, foliáceas, assimétrico-ovadas, ápice apiculado a aristado, base oblíqua. Brácteas 3,5-6,0 x l,8-3,5cm, lanceolado- ovadas, imbricadas, ápice apiculado, base auriculada. Flores azuis, alvas ou alva-esverdeadas, axilares, solitárias; pedicelos ca. 2,5cm compr., glabros; sépalas 2,5-3,0 x l,0-l,2cm, oblongo- lanceoladas, ápice agudo, sem carena, glabros; pétalas 2,5-3,0 x l,0-l,3cm, oblongo-lanceoladas, ápice agudo; corona com 5 séries, 2 externas, 2,5- 3,0cm compr., filamentos ligulados, 3 internas com filamentos filiformes; opérculo membranoso, encurvado, crenado na margem; límen cupuliforme; androginóforo ca. 4,0cm compr., estames ca. l,0cm compr., anteras ca.0,6 x 0,2cm; ovário ca.0,6 x 0,3cm, ovóide, glabro, estilete ca.l,2cm compr. Frutos 2,5- 4,5 x 2,l-3,8cm, bagas, globosos; sementes 0,5-0,6 x 0,3-0,4cm, obovadas, testa reticulada.
Material examinado - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, entorno do Jardim Botânico do Rio de Janeiro, Lagoinha, fl., RB21315, D.Constantino s.n., 07/XI/1925.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

372 M.A.MILWARD-DE-AZEVEDO & M.C.VALENTE
Material adicional - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, Vista Chinesa, fr., RB, R.Marquetee outros 2171, 23/VI/1995; estrada das Paineiras, fl., fr., RB, J.P.Fontella e outros 405, 20/ 1/1970; estrada do Redentor, fl., RB, A.P.Duarte 347, 08/X/1946; fl., RB, A.P.Duarte 8743, X/1964; morro do Corcovado, fl., fr., RB, R.Marquete e outros 228, 04/X/1989; fr., RB298876, L.C.S.Silva s.n., 28/VII/1992.
Distribuição geográfica - Brasil: Minas Gerais, Rio de Janeiro e São Paulo.
Nome popular - Maracujazinho.
Dados fenológicos - Foi encontrada em flor nos meses de janeiro, e outubro a dezembro, e fruto em janeiro, junho, julho, outubro e dezembro.
Hábitat - Floresta Ombrófila Densa. Espécie heliófila, ocorrendo em orla de floresta e beira de estrada.
Comentários - As principais características diagnósticas para P. sidaefolia são as lâminas levemente 3-lobadas para o ápice, flores com dnco séries de filamentos na corona e testa da semente reticulada. De acordo com MASTERS (1872), as brácteas são um atributo de fácil reconhecimento. Pertence ao subgênero Passiflora L. série Imbricatae Killip.
Fig.2- Passiflora racemosa: (a) forma da folha jovem e aspecto geral da nervação, (b) forma da folha adulta e aspecto geral da
nervação, (c) esquema geral da flor, (d) ápice da sépala, (e) fruto, (f) semente. Passiflora sidaefolia: (g) forma da folha e aspecto geral da nervação, (h) esquema geral da flor, (i) detalhe do ramo frutífero, (j) ápice da sépala, (1) semente. Escalas: a- c, e, g-i = lcm; f, j, 1 = lmm; d = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

PASSIFLORACEAE DA MATA DO JARDIM BOTÂNICO DO RIO DE JANEIRO E ARREDORES 373
5. Passiflora tmncata Regei, Gartenflora 8: 356, est. 276. 1858. (Fig.3 a-e)
Descrição - Caule triangular, glabrescente. Folhas com pecíolos 0,5-2,8cm compr., 1 par de glândulas sésseis, glabrescente; lâminas membranáceas, levemente 3- lobadas para o ápice, 3,0-6,4cm compr. na nervura média, 3,9-7,0cm compr. na nervura lateral, 4,4-8, lcm larg., ápice truncado a obtuso, apiculado, base obtusa, margem inteira, 1-5 glândulas entre as nervuras laterais, 3 nervuras saindo da base, glabrescente em ambas as faces. Estipulas ca. 0,3cm compr, linear- subuladas. Brácteas ca. 0,1 x 0,05cm, linear- subuladas. Flores alvas, esverdeadas ou amareladas, axilares, solitárias ou aos pares; pedicelos 0,4-2,0cm compr.; sépalas 0,5-0,7 x 0,2 x 0,4cm, lanceoladas, ápice agudo a obtuso, sem carena, glabrescente; pétalas 0,3-0,4 x 0,lcm, espatulada, ápice obtuso; corona com 2 séries, a externa 0,2-0,4cm compr., filamentos falciformes, a interna ca. 0,lcm compr., filamentos filiformes; opérculo plicado, ápice curvo; límen anelar; androginóforo ca. 0,5cm compr., estames ca. 0,2cm compr., anteras ca. 0,2 x 0,lcm, ovário 0,1- 0,2 x 0,lcm, elíptico, glabrescente, estilete ca. 0,3cm compr. Frutos 1,6-3,0 x l,6-3,0cm, bagas, globosos; sementes ca. 0,5 x 0,3cm, elípticas, testa irregularmente sulcada.
Material examinado - BRASIL, RIO DE JANEIRO,
município do Rio de Janeiro, Parque Lage, bt., fr., RB, B.Kurtz e outros 166, 27/11/1991.
Material adicional - BRASIL, RIO DE JANEIRO, município do Rio de Janeiro, Vista Chinesa, bt., fl., R, A.C.Brade 10624, 01/111/1931; bt., fl., HB, RFA, E.Pereira 4264, 29/1/1959; bt., RB, V.F.Ferreira 3356, 04/V/1984; Mesa do Imperador, fl., R90277, A.C.Brade s.n., 22/1/1932; Caminho do Pico da Tijuca, bt., fl., fr., RB55798, Altamiro e outros s.n., 06/11/1946.
Distribuição geográfica - Brasil: Rio de Janeiro, São Paulo, Paraná e Santa Catarina.
Nome popular - Maracujá.
Dados fenológicos - Foi encontrada em flor nos meses de outubro a abril, e em fruto de fevereiro a abril e outubro a dezembro.
Hábitat - Floresta Ombrófila Densa. Espécie ciófila, ocorrendo principalmente no interior de mata e raramente em sua orla (SACCO, 1980).
Comentários - Passiflora tmncata distingue-se das demais espécies estudadas principalmente pelas lâminas levemente truncadas no ápice, glândulas laminares entre as nervuras laterais, flores com duas séries de filamentos na corona e testa da semente irregularmente sulcada. Pertence ao subgênero Decaloba (DC.) Rchb. seção Cieca (Medik.) DC.
Fig.3- Passiflora tmncata: (a) forma da folha e aspecto geral da nervação, (b) esquema geral da flor, (c) ápice da sépala,
(d) fruto, (e) semente. Escalas: a, b, d = lcm; c, e = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

374 M.A.MILWARD-DE-AZEVEDO & M.C.VALENTE
CONCLUSÃO
De acordo com os levantamentos feitos nos herbários citados e excursões realizadas na mata de encosta do Jardim Botânico do Rio de Janeiro e arredores, foram encontradas para a família Passifloraceae as seguintes cinco espécies Passiflom amethystina, P. capsularis, P.
racemosa, P. sidaefolia e P. tmncata. As espécies distinguem-se pelo formato das folhas, estipulas, brácteas, fruto, testa da semente e pela disposição e número de glândulas na lâmina e pecíolo, além do número e formato das séries da corona.
Os táxons estudados ocorrem, preferencialmente, em beira de estradas, orla de florestas ou em clareiras, sendo raramente encontrados no interior da mata e sofrendo intensa herbivoria. Passiflora racemosa, P.
sidaefolia e P. tmncata apresentam distribuição restrita no Brasil ocorrendo, até o momento, na região Sudeste e a última sendo também encontrada na região Sul. Passiflora amethystina ocorre em toda região Sudeste e Sul, além dos Estados da Bahia e Goiás, enquanto que P. capsularis apresenta ampla distribuição por quase todo o país.
AGRADECIMENTOS
À Dra. Claudia Petean Bove (Museu Nacional - Rio de Janeiro), pela co-orientação neste trabalho; ao Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, pelo apoio; ao Programa Institucional de Bolsas de Iniciação Científica (PIBIC/CNPq), pela bolsa concedida durante o curso de bacharelado em Ciências Biológicas (Universidade do Rio de Janeiro).
REFERÊNCIAS BIBLIOGRÁFICAS
BARROSO, G.M.; PEIXOTO, A.L.; ICHASO, C.L.F.;
GUIMARÃES, E.F. & COSTA, C.G., 1978. Sistemática de Angiospermas do Brasil. São Paulo: LTC/EDUSP. v. 1, p. 194-197, il.
BRIZICKY, G.L., 1961. The genera of Turneraceae and
Passifloraceae in the southeastem United States. Journal of the Arnold Arboretum, Cambridge, 42:204-218.
BRUMMITT, R.K., 1992. Vascular plants families and
genera. Kew: Royal Botanic Gardens. 804p. CERVI, A.C., 1981. Revision dei genero Passiflora L.
(Passifloraceae) dei Estado de Parana, Brasil
(resumo). Barcelona. 27p. Tese (Doutorado em Ciências), Universistat de Barcelona.
CERVI, A.C., 1986. Passifloraceae. In: RIZZO, S.A. (Org.) Flora do Estado de Goiás - Coleção Rizzo. Goiânia: Editora UFG, v.7, 45p.
CERVI, A.C., 1992. Passifloraceae. In: MELO, M.M.R.F. et dl. (Eds.) Flora fanerogâmica da Ilha do Cardoso
(São Paulo, Brasil). São Paulo: Instituto de Botânica,
3:11-20.
DEGINANI, N.B., 1999. Passifloraceae L. - Flora Del Valle de Lerma. Aportes Botânicos de Salta - Serie Flora,
Salta, 6:1-20.
ESCOBAR, L.K., 1988. Passifloraceae. In: PINTO, P. & LOZANO, G. (Eds.) Flora de Colombia. Bogotá:
Universidad Nacional de Colombia, v.10 (?), 138p.
FUKS, R.; GUIMARÃES, E.F.; LACANNA, M.F. & MONTEIRO, S.N., 1999. Plantas ornamentais I. In: VALENTE, M.C. & MARQUETE, N.F.S. (Eds.) Plantas
úteis das áreas do entorno do Jardim Botânico do Rio de Janeiro, Horto Florestal e Parque Lage I.
Rio de Janeiro: Ministério do Meio Ambiente, Instituto
de Pesquisas Jardim Botânico do Rio de Janeiro, p.61. (Série Estudos e Contribuições 16)
HOLMGREN, P.K.; KEUKEN, W. & SCHOFIELD, E.K., 1990.
Index herbariorum, Part 1: The Herbaria of the World. New York: New York Botanical Garden. 693p.
HOLM-NIELSEN, L.B.; J0RGENSEN, P.M. & LAWESSON,
J.E., 1988. Passifloraceae. In: HARLING, G. & ANDERSSON, L. (Eds.) Flora of Ecuador.
Copenhagen: Nordic Journal of Botany. v.31, 130p. JOLY, A.B., 1993. Botânica - Introdução à taxonomia
vegetal. São Paulo: Ed. Nacional. 777p., il.
KILLIP, E.P., 1938. The american species of Passifloraceae. Publication Field Museum of Natural History, Botanical Series, Chicago, 19(1-2): 1-613.
MABBERLEY, D.J., 1997. The Plant Book. A portable
dictionary of the vascular plants. Cambrigde:
Cambrige University Press. p.532-533. MARQUETE, N.F.S.; VALENTE, M.C.; MARQUETE, R.;
MARQUETE, O.; GUIMARÃES, E.F.; FUKS, R.; GIORDANO, L.C.; LACANNA, M.F.S.; MONTEIRO, S.N.;
MYNSSEN, C.M.; MEDEIROS, E.S. & GOMES, H.F.S., 1994. Vegetação das áreas do entorno do Jardim Botânico do Rio de Janeiro, Horto Florestal e Parque
Lage I. Rio de Janeiro: Ministério do Meio Ambiente,
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. 69p., il. (Séries Estudos e Contribuições 12)
MASTERS, M.T., 1872. Passifloraceae. In: MARTIUS, C.F.P.; EICHLER, AW. & URBAN, I. (Eds.) Flora Brasiliensis, München, Wien, Leipzig, 13(l):529-628.
METCALFE, C.R. & CHALK, L., 1965. Anatomy of the
dicotyledons I. Leaves, stem, and wood in relation to taxonomy with notes on economic uses. Oxford:
At the Clarendon Press. p.674-680. PIO-CORRÊA, M., 1984. Dicionário de plantas úteis do
Brasil e das exóticas cultivadas. Rio de Janeiro:
Imprensa Nacional, v.3, p.238-239; v.5, p.108-129. PURI, V., 1947. Studies in floral anatomy VI. Vascular
anatomy of the flower of certain species of the Passifloraceae. American Journal of Botany,
Burlington, 34:562-573.
SACCO, J.C., 1980. Passifloráceas. In: REITZ, R. (Ed.)
Flora Ilustrada Catarinense, Itajaí: Herbário Barbosa
Rodrigues. Fascículo Passifloraceae. 132p., il. VELOSO, H.P.; RANGEL FILHO, A.L.R. & LIMA, J.C.A.,
1991. Classificação da vegetação brasileira,
adaptada a um sistema universal. Rio de Janeiro:
Fundação Instituto Brasileiro de Geografia e
Estatística (IBGE), Departamento de Recursos
Naturais e Estudos Ambientais. 124p.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.367-374, out./dez.2004

wmmnraTmnnrTl nnniiitüiiiüífflnnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.375-386,out./dez.2004
ISSN 0365-4508
NEW DATA ON SOUTH AMERICAN SPECIES OF CHORDODES (NEMATOMORPHA) 1
(With 7 figures)
CRISTINA DE-VILLALOBOS 2
FERNANDA ZANCA 2
ANDREAS SCHMIDT-RHAESA 3
ABSTRACT: Taxonomic characters of both male and female horse-hair worms Chordodes brasiliensis, and
C. nobilii (Nematomorpha) and female of C. carmelitanus are re-described by scanning electron microscopy.
Sexual dimorphism was found in the cuticular pattern of C. brasiliensis, the male has three different
areolar types but the female shows five different areolar types that includes all three types of areoles as in
the males, but along the ventral and dorsal midline, two further types of areoles are present.
The female cuticle of C. carmelitanus is characterized by three types of areoles, the crown areoles always
occur in pairs on both sides along the ventral midline. Male of C. nobilii has two types of areoles, type 1 are
slightly oval or circular in shape with minute projections on the apical surface. Type 2 areoles occur individually
or in clusters and carry on their apical surface projections that can be short or longer. The female of this
species has three types of areoles. Areoles of types 1 and 2 are very similar to each other. Crowned areoles
(type 3) have quite long apical filaments and occur only along the ventral and the dorsal midline. A neotype
of nobilii is erected as the original material cannot be traced. Host data of C. brasiliensis and new distribution
records of C. brasiliensis and C. nobilii are reported.
Key words: Nematomorpha, Gordiida, Chordodes, South America, scanning electron microscopy.
RESUMO: Novos dados sobre espécies sul-americanas de Chordodes (Nematomorpha).
Neste trabalho estão sendo redescritas com microscopia de varredura, três espécies de Chordodes, C.
brasiliensis, C. carmelitanus e C. nobilii. Macho do C. brasiliensis, com cutícula provida com três tipos de
aréolas, as fêmeas apresentam dimorfismo sexual com cinco tipos de aréolas, dois tipos das quais encontram-
se na linha ventral do corpo. A fêmea de C. carmelitanus com cutícula com aréolas de três tipos, apresentam
na região ventral, longitudinalmente, uma faixa tendo aréolas coroadas pares no meio das quais saem tufos
de prologamentos muito longos. O macho de C. nobilii com dois tipos de aréolas, tipo 1 de forma oval ou
arredondadas tendo no ápice finos e curtos prolongamentos. Aréolas do tipo 2 apresentam-se isoladas, aos
pares ou em grupos tendo no ápice prologamentos curtos ou longos. As fêmeas de C. nobilii com três tipos de
aréolas. Nas aréolas do tipos 1 e 2 são semelhantes. O tipo 3 (aréolas coroadas) na linha ventral do corpo,
apresentam no ápice prolongamentos longos. Outorga-se informações sobre hospedeiros do C. brasiliensis e
novos dados da distribuição geográfica do C. brasiliensis e de C. nobilii.
Palavras-chave: Nematomorpha, Gordiida, Chordodes, América do Sul, microscópio de varredura.
INTRODUCTION
The biodiversity of horsehair worms (Nematomorpha) from Southern South America, in particular Argentina, is comparatively well known (see, among many publications, MIRALLES & DE- VILLALOBOS, 1993). However, still several species from Argentina and more from the remaining South America are insufficiently known. Scanning Electron Microscopy has become the standard method in nematomorph taxonomy, because the
fine cuticular characters can be documented reliably with this method. We investigated here representatives of the genus Chordodes from different museum collections. Chordodes icontains about 90 species and is the largest genus in Nematomorpha. The cuticular structure is usually very diverse, with several types of elevations called areoles. Characteristis for Chordodes are the so- called crowned areoles which are elevated above the surface and have an apical ring (“crown”) of shorter or longer filaments.
1 Submitted on May 26, 2004. Accepted on November 3, 2004.
2 Facultad de Ciências Naturales y Museo. Paseo dei Bosque S/N. 1900 La Plata, Argentina. E-mail: [email protected].
3 University of Bielefeld, Zoomorphology and Systematics. P.O.Box 100131, 33501 Bielefeld, Germany.

376 C.DE-VILLALOBOS, F.ZANCA & A.SCHMIDT-RHAESA
One of the best known species from South America is Chordodes brasiliensis Janda, 1894, but descriptions of this species, differ slightly between different specimens. JANDA (1894), in his original description mentions three types of areoles, an abundant ílat one, a highly elevated one with fine apical filaments and irregularly distributed solid spines. The first two types were confirmed by subsequent descriptions, whereas the spines probably represent so-called tubercle areoles in which a fingerlike projection arises from a ílat areole (MIRALLES, 1969). CAMERANO (1897a) described crowned areoles with very long filaments next to the dorsal and ventral midline in the female, a fact that was repeated in later descriptions (MIRALLES, 1976), but not confirmed by independent investigations (CARVALHO, 1946a, 1946b; CARVALHO 86 FEIO, 1950; MIRALLES 86 CAMINO, 1983; MIRALLES, 1993).
We additionally reinvestigated the holotype of Chordodes carmelitanus Carvalho 8& Feio, 1950. Scanning Electron Microscopical observations were already available from new recorded specimens (MIRALLES, 1989), but the investigation of the holotype could add new details. Finally, several specimens from the collection of the Museo de Ciências Naturales in La Plata, Argentina, correspond to description of Chordodes nobilii
Camerano, 1901. Because the holotype could not be located, we describe a neotype for this species.
MATERIAL AND METHODS
Preserved specimens from the following museums were investigated: Museo de Ciências Naturales de La Plata (MLP), Museum für Naturkunde, Berlin, Germany (MNB), Museu Nacional of Rio de Janeiro, Brazil (MNRJ), Museum Regionale di Scienze Naturali, Torino (MZUT) and Natural History Museum, London, UK (NHML). All the preserved specimens were fixed in ethanol. In the present study we investigated twenty nine specimens of Chordodes brasiliensis, one of C. carmelitanus and twenty of C. nobilii and we report new records of Chordodes brasiliensis and C. nobilii. In order to optimize the traditional observation technique, we followed the new protocol propoused by SCHMIDT- RHAESA (2002a). Body measurements were made with outstretched worms using a ruler. Diameters were measured under dissecting microscope using a caliper ruler. Fragments of worms (anterior end, mid-body and posterior end) were dehydrated in an increasing ethanol series, criticai point-dried,
mounted on bronze blocks and gold-sputter coated. Observations were performed using a JEOL SLM 1000 scanning electron microscope.
RESULTS
Chordodes brasiliensis Janda, 1894 (Figs.1-2)
1894 Chordodes brasiliensis Janda - Zool. Jahrb. Syst. Geogr. Tiere, 7:608, pis: 19-20.
Holotype (no examinated) - One male from Brazil (no further precision of locality) (JANDA, 1894).
Material examined - SEM. Midbody from: ld (Córdoba: Alta Gracia, Argentina, MLP 3575); ld
(Corrientes, Capital, Argentina, MLP 5049); l9 (Corrientes, Capital, Argentina, MLP 5052); 10d, 29 (Salta: Tala, Argentina, MZUT G2, G42); l9 (Salta Capital, Argentina, MLP 5052); ld (Rio Noro, Humboldt, Station Catherina, NHML 1921.12.13.23- 24); ld (San Luis: El Reparito, Estancia Grande, Argentina, MLP 3574); l9 (Jujuy: San Lorenzo, Argentina, MZUTG46); ld, 29 (Tucumán: LasTipas river, Argentina, MLP 3578); ld (Misiones, Parque Nacional Iguazú, Base Yawi, Argentina, MLP 4903); l9 (Misiones: Cuha Pirú stream, Argentina, MLP 5050); ld" (Rio de Janeiro, Brazil, MNRJ 5348); ld (Niterói, Brazil, MNRJ, no accession number); l9 (Japuhiba, Brazil, MNRJ no accession number); l9 (Viçosa, Brazil, MNRJ no accession number); l9 (Santa Teresa, Brazil, MNRJ no accession number). SEM from terminal end: ld (Salta: Tala, Argentina, MZUT G2), ld (Rio Noro, Humboldt, Station Catherina, NHML 1921.12.13.23-24), l9 (Corrientes, Capital, Argentina, MLP 5052). LM of midbody from 25<d (Salta: Tala, Argentina, MZUT G2.
Host - MANTODEA: Stagmatoptera hyáloptera (see CAMERANO, 1897b), Stagmatoptera sp. (see CARVALHO, 1946a), Stagmatoptera precaria (see CARVALHO, 1946b), undetermined mantid (see CARVALHO, 1946b), probably Phyllovates sp. (see SCHMIDT-RHAESA 8& EHRMANN, 2001).
d - Color of body varies between light brown to dark brown. Posterior end of males (Fig. IA) is not bilobed. The cloacal opening is oval and surrounded by slender spines (circumcloacal spines). Around the cloacal opening the cuticle is smooth with minute bristles. Anterolateral of the cloacal opening are two straight and narrow rows of bristles. They reach up to 250jum in length.
The cuticle shows three types of areoles (Fig. 1C-F). Type 1 areoles are the most abundant, they are oval
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

NEW DATA ON SOUTH AMERICAN SPECIES OF CHORDODES (NEMATOMORPHA) 377
with an irregular shape and a roughly structured surface (Fig. 1C-F). Scattered among these areoles are areoles of type 2 (tubercle areoles) with a long (15.7jum) fingerlike projection (tubercle) which is in an excentric position on the areole (Fig. 1D, E). Both areolar types are transversal to the longitudinal axis of the body. Areoles of type 3 are distinctly elevated over the other areoles (up to 21.5jum). They occur abundantly, but individualized and are scattered between the areoles of type 1 (Fig. 1C, F). Rarely they form lose clusters of up to four areoles (Fig. 1F). The apical surface of these areoles differ between specimens, is rugged (MLP 4903) (Fig. 1F) or with very fine processes (MNRJ 5348) (Fig. 1D).
9 - Color of body as in males, it varies between light brown to dark brown. The posterior end (Fig. 2A) is rounded and the cloacal opening is circular with a central terminal position. Radiating around the cloacal opening are fine grooves (Fig.2A).
The cuticle includes all three types of areoles as in the males (Fig.2B, C), but along the ventral and dorsal midline, two further types of areoles are present. Both additional types are clustered together. Two kinds of filaments, one very long and solid, the other much thinner and shorter, arise from a more or less ílat region which probably represents two areoles (Fig.2C). These central areoles are surrounded by 5-6 slightly elevated areoles which are curved towards the center (Fig.2C).
Dimensions - According to the literature (CAMERANO, 1897b; CARVALHO, 1946a; CARVALHO & FEIO, 1950; MIRALLES & CAMINO, 1983) females vary between 140 and 271mm in length and are maximally 2mm in diameter; males range from 85 to 355mm in length and from 0.7 to 2mm in diameter. Measurements of the newly reported specimens range in length from 63 to 206mm in males and from 112 to 240mm in females. The diameter varies between 0.5 to l.Omm in males and between 0.6 and 1.8mm in females
Comments - CAMERANO (1897b) designates “numerous” specimens as Chordodes brasiliensis,
in our SEM and LM reinvestigation we have analyzed all of these specimens (54cf and l9 ) and we can confirm that only 35 males belong to C. brasiliensis, and the rest of the specimens (19cf and l9 ) to Neochordodes talensis.
Our study confirmed the areolar types from previous investigations, ie., the abundant flat areoles, highly elevated areoles and tubercle areoles (see JANDA, 1894; CAMERANO, 1987a, 1987b; CARVALHO, 1946a, 1946b; CARVALHO & FEIO, 1950; MIRALLES
& CAMINO, 1983). Additionally, we could confirm the presence of further areoles in the female as noted by CAMERANO (1897a). Camerano reports paired areoles (“papille”) with long processes, but in his figures the paired character of areoles is not evident. Instead, filaments seem to originate from a broad region. This is in accordance with our fmdings, where filaments can not clearly be assigned to one areole. As paired crowned areoles with long filaments, surrounded by a circle of elevated areoles, are a common character in Chordodes (see, e.g., SCHMIDT- RHAESA, 2002a, 2002b), it can be assumed that the structures in C. brasiliensis represent modified crowned areoles. The presence of two kinds of filaments, thick and thin, however, is new and unique for C. brasiliensis. CAMERANO (1897a) figured elevated areoles surrounding the areoles with the long filaments, including a curvation towards the center, but did not mention these as a particular type of areoles. What remains questionable is the nature of the abundant elevated areoles on the cuticular surface. As these may cariy small processes, they might represent some form of crowned areoles. We think that future studies will allow a better idea about the real identity of these areoles.
Distribution - ARGENTINA - Buenos Aires: Sierra de la Ventana, Cerro Tres Picos (3cf, de Miralles & Camino, 1983), Arroyo El Loro, Rio Sauce Grande (8cf, 49 , MIRALLES, 1980). Córdoba: Alta Grada, La Granja stream (29 , MIRALLES, 1976), Alta Grada 31°40’00”S, 64°24’00”W (lcf, newrecord). Corrientes: Corrientes Capital 27°28’S, 58°49W (lcf, l9 , new record). Entre Rios: Gualeguaychú (lcf, CARVALHO, 1946a). Jujuy: San Lorenzo (29 , CAMERANO, 1897a). Misiones: Puerto Bembey (3cf, 29 , CARVALHO & FEIO, 1950), Cuha Pirú stream 27°08’00”S, 54°54’00”W (l9 , newrecord), Parque Nacional Iguazú, Base Yawi 25°36’00”S, 54°34’00”W (lcf, newrecord). Salta: Tala (35cf, 29 , CAMERANO, 1897a), Salta capital 24°51’00”S, 65°29’00”W (l9 , newrecord). San Luis: El Reparito, Estancia Grande (lcf, MIRALLES, 1976). Tucumán: LasTipasriver (lcf, 29 , MIRALLES, 1984, 1993). BRAZIL - Espírito Santo: Santa Teresa 20°06’02”S, 40°31’47”W (2cf , 39 , CARVALHO, 1946b); Rio de Janeiro: 22°55’00”S, 43°30’00”W (lcf, newrecord), Niterói22°56’00”S, 43°04’00”W (lcf, new record), Cachoeira de Macacu, Japuhiba 22°27’24”S, 42°39’24”W (l9 , new record), Minas Gerais: Viçosa 20°45’39”S, 42°52’10’W (l9 , newrecord). URUGUAY: Nueva Palmira (lcf, CARVALHO, 1946a), Tacuarembó (lcf, CARVALHO, 1946a), Mercedes (lcf, CARVALHO, 1946a), Paysandú (lcf, l9 , CARVALHO, 1946a), unspecified location (CAMERANO, 1915).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

378 C.DE-VILLALOBOS, F.ZANCA & A.SCHMIDT-RHAESA
Chordodes brasüiensis, d, SEM - fig.l- (A) posterior end (MZUT G2) unlobed; (B) posterior end (NHM 13) with a ventral
groove and two lobe like structures; (C) midbody cuticle, general view (MZUT G2); (D) midbody cuticle, detail of areoles
types 1, 2 and 3; (E) midbody cuticle, detail of areoles types 1 and 2 (MNRJ 5348); (F) midbody cuticle, lateral view showing areole type 3 (MLP 4903). Scale bars: (A-C) = lOOjum, (D-F) = lOjum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

NEW DATA ON SOUTH AMERICAN SPECIES OF CHORDODES (NEMATOMORPHA) 379
Chordodes brasiliensis, 9 , (MLP 5050), SEM - fig.2- (A) terminal end with radiating grooves (arrow) around the cloacal opening; (B) midbody cu ti cie, lateral side with type 1-3 areoles; (C) midbody cuticle, ventral midline with areolar clusters
with types 3 and 4 (crowned areoles). Scale bars: (A and C) = 100 jum, (B) = 50jum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

380 C.DE-VILLALOBOS, F.ZANCA & A.SCHMIDT-RHAESA
Chordodes camnelitanus Carvalho & Feio, 1950 (Fig.3)
1950 Chordodes carmelitanus. Carvalho & Feio. Ann. Acad. Bras. Cienc., 22:197, pis.4-6.
Holotype - l9 from Fazenda da Alegria, Carmo do Rio Claro, Minas Gerais, Brasil. (Museu Nacional of Rio de Janeiro, Brasil, no accesion number).
Material examined (holotype) - SEM of midbody. l9 (Cordoba: San Lorenzo Stream, Cura Brochero, Argentina, MLP 3579), 29 9 (Buenos Aires: Sierra de la Ventana, Argentina, MLP 3580).
Host - Unknown.
Redescription of holotype - Body length 220mm and diameter in the middle region 1.8mm. Body colour dark brown. Terminal end rounded and the cloacal opening situated terminally.
The cuticle contains three types of areoles. Areoles of type 1 and 2 are oval or rounded (Fig.3A, B), moderately elevated and with superficial or deep grooves in perpendicular direction to the body axis. The interareolar spaces between areoles type 1 and 2 are broad, structured into cord-like folds and contain scattered small protuberances. Type 1 areoles are the most abundant and among these there are scattered type 2 areoles with a short tubercle on the top that seems to originate from a depression. Crowned areoles (third type of areoles) always occur in pairs which are densely surrounded by a circle of areoles of type 1 (Fig.3B-D). They contain long slender apical filaments (up to 297pm).
Comments - Our reinvestigation in general confirms the description of CARVALHO & FEIO (1950), but differs in the interpretation. As first type of areoles, CARVALHO & FEIO (1950) described very small areoles. Observed with SEM, these areoles turn out to be irregular elevations of the cordlike structures in the interareolar space. Under lower magnifications [e.g., Fig.3D), these structures appear as areoles, but under higher magnifications, they do not possess a defined margin and are therefore not regarded as areoles by us. CARVALHO & FEIO (1950) summarized what we have called areoles type 1 and 2 as one type, but also noted the distribution of crowned areoles along the ventral midline.
Specimens from Buenos Aires and Córdoba (Argentina), reported by MIRALLES (1989) as C. carmelitanus, differ from the holotype and therefore we transferred them to the species C. nohilii (see next).
Distribution - BRAZIL - Minas Gerais: Carmo do Rio Claro (l9 , CARVALHO & FEIO, 1950).
Chordodes nobiíii Camerano, 1901 (Figs.4-6)
1901 Chordodes nobiliiCamerano. Boll. Mus. Zool. Anat. Comp. Torino, 16(410):1
Holotype - l9 from Cosquin, Cordoba, Argentina could not be located and seems to be lost.
Neotype- 1(3 (San Luis: Carpinteria, Piedra Blanca stream MLP 5290/1). Paraneotypes: 2(3 (San Luis: Carpinteria, Piedra Blanca stream MLP 5290/2).
Material examined - SEM of midbody (all reports from Argentina): Neotype 1(3 (San Luis: Carpinteria, Piedra Blanca stream MLP 5290/1), Paraneotypes 2(3 (San Luis: Carpinteria, Piedra Blanca stream MLP 5290/2), 1(3 (Buenos Aires: Sierra de la Ventana, El Loro stream, MLP 5063), 1(3 (MLP 5063), 39 (Buenos Aires: Sierra de la Ventana [as C. carmelitanus, MIRALLES, 1989], MLP 3580), 4(3
(Buenos Aires: Sierra de la Ventana, Sauce Grande stream, MLP 3582), l9 (Buenos Aires, Balcarce, MLP 5055); lcf (Cordoba, El Durazno, MLP 5291) 29 (Cordoba, MNB 4437), l9 (Cordoba: Cura Brochero, San Lorenzo stream [as C. carmelitanus,
MIRALLES, 1989], MLP 3579), 2(3 and l9 (San Luis: Villa Larca, Aguas Blancas, San Ignacio stream, MLP 5289). SEM terminal end: Neotype lcf (San Luis: Carpinteria, Piedra Blanca stream MLP 5290/1). 1(3
(Buenos Aires: Sierra de la Ventana, Sauce Grande stream, MLP 3582), lcf (Cordoba, El Durazno, MLP 5291), l9 (Buenos Aires, Balcarce, MLP 5055).
Host - Unknown.
C3 - The body color ranges from light to dark brown. The anterior end is tapered and a dark collar is always lacking. The posterior end in most of the males is not divided into lobes (Fig.4A), but in two specimens we found a clear indication of two lobes at the ventral side of the posterior end (Fig.4C). The cloacal opening is round or oval and is surrounded by slender spines undivided at the top (circumcloacal spines) (Fig.4A- C). Anterolateral of the cloacal opening are paired regions in which numerous bristles are located (bristlefields). In the anterior and posterior region of the cloacal opening are short spines.
The body cuticle is densely structured by areoles. There are two types of areoles. The most abundant type (type 1) are slightly oval or circular in shape and have a warty surface (“blackberry-like”). Irregular minute projections are distributed on the whole apical surface (Fig.5B). Type 2 areoles are slightly larger than the first type and carry on their apical surface projections which are not scattered, but clustered. These projections can be extremely
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

NEW DATA ON SOUTH AMERICAN SPECIES OF CHORDODES (NEMATOMORPHA) 381
short or slightly longer and bristlelike (Fig.5A-D). Type 2 areoles occur individually or in clusters of 3 to 10 areoles. The cuticle between the areoles is structured into cord-like folds and there are scarcely distributed tubercles emerging among type 1 areoles (Fig.5A, D).
9 - Posterior end round with a terminal cloacal opening. Radiating aroimd the cloacal opening are fine
grooves (Fig.6A). In the cuticle, the same types of areoles are present as in the male, with additional crowned areoles. Areoles of types 1 and 2 are very similar to each other (Fig.6B, C). Crowned areoles (type 3) occur only along the ventral and the dorsal midline (Fig.6D, E). They have quite long apical filaments and appear in a cluster of two areoles (Fig.6E). The surrounding areoles are from type 1 (Fig.óE).
Chordodes carmelitanus, $ holotype, SEM - fig.3- (A) cuticle of midbody with type 1 areole and tubercle areoles (type 2),
scale bar = lOOjum; (B-D) ventral midline. Areolar clusters with types 3 and 1 (crowned areoles), scale bar = lOjum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004
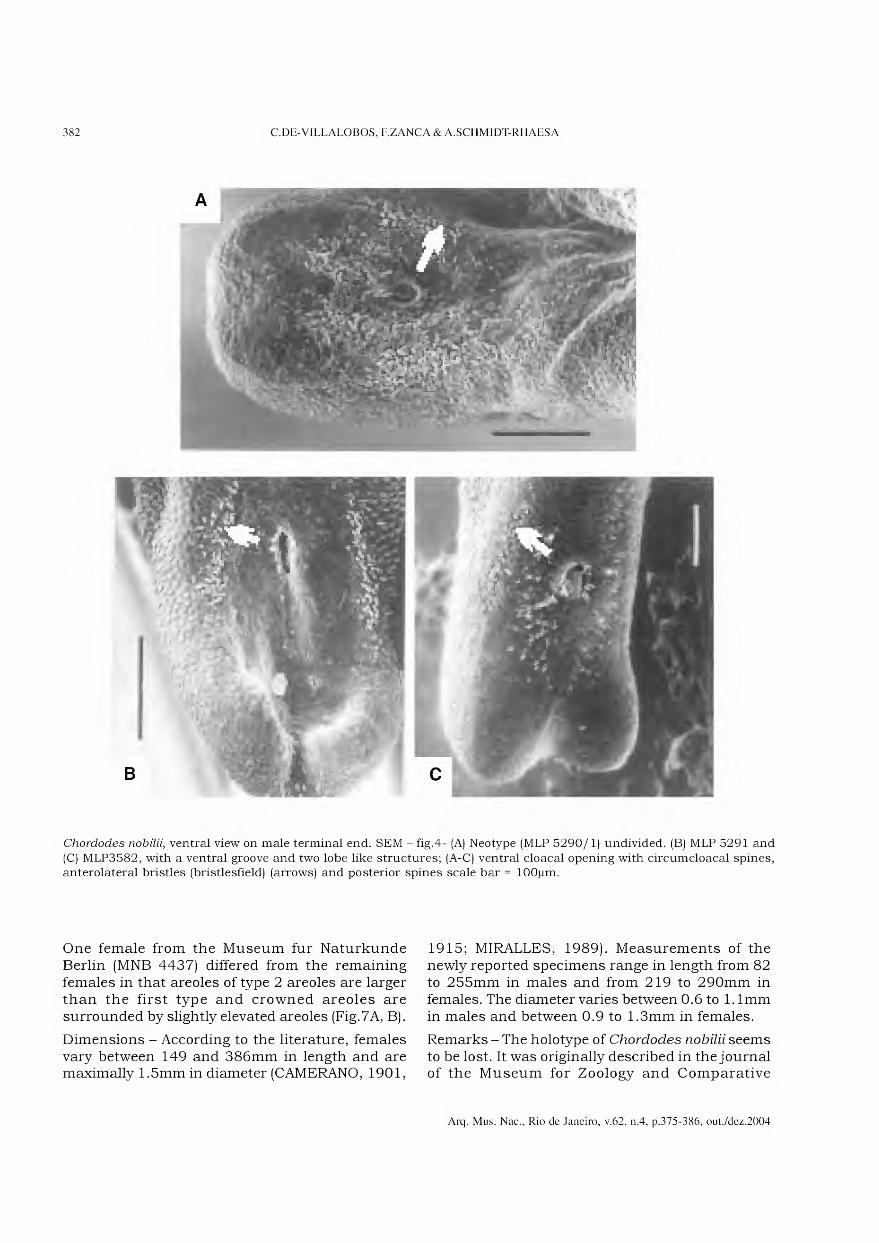
382 C.DE-VILLALOBOS, F.ZANCA & A.SCHMIDT-RHAESA
Chordodes nobilii, ventral view on male terminal end. SEM - fig.4- (A) Neotype (MLP 5290/1) undivided. (B) MLP 5291 and
(C) MLP3582, with a ventral groove and two lobe like structures; (A-C) ventral cloacal opening with circumcloacal spines, anterolateral bristles (bristlesfield) (arrows) and posterior spines scale bar = lOOjum.
One female from the Museum fur Naturkunde Berlin (MNB 4437) differed from the remaining females in that areoles of type 2 areoles are larger than the first type and crowned areoles are surrounded by slightly elevated areoles (Fig.7A, B).
Dimensions - According to the literature, females vary between 149 and 386mm in length and are maximally 1.5mm in diameter (CAMERANO, 1901,
1915; MIRALLES, 1989). Measurements of the newly reported specimens range in length from 82 to 255mm in males and from 219 to 290mm in females. The diameter varies between 0.6 to 1. lmm in males and between 0.9 to 1.3mm in females.
Remarks - The holotype of Chordodes nohilii seems to be lost. It was originally described in the journal of the Museum for Zoology and Comparative
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004
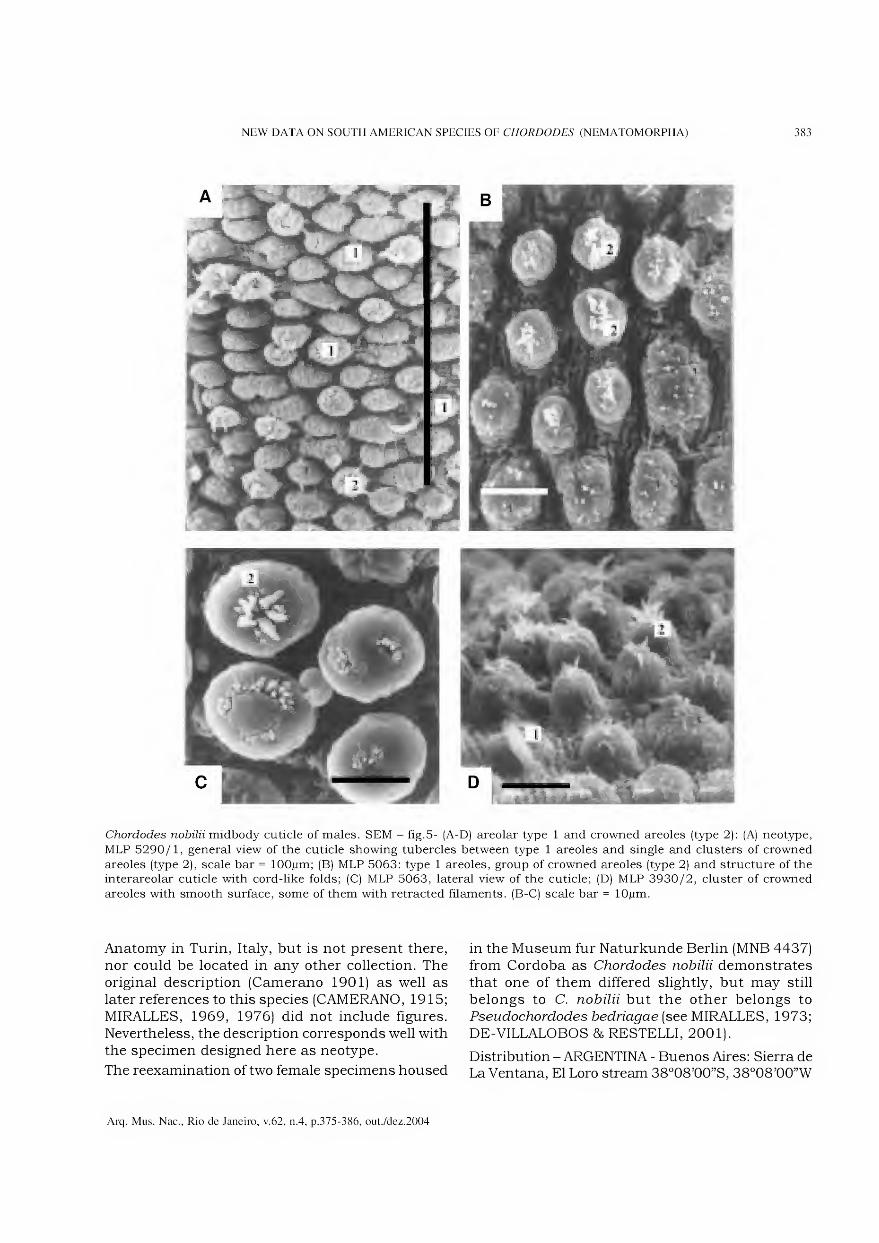
NEW DATA ON SOUTH AMERICAN SPECIES OF CHORDODES (NEMATOMORPHA) 383
Chordodes nobilii midbody cuticle of males. SEM - fig.5- (A-D) areolar type 1 and crowned areoles (type 2): (A) neotype,
MLP 5290/1, general view of the cuticle showing tubercles between type 1 areoles and single and clusters of crowned
areoles (type 2), scale bar = lOOjum; (B) MLP 5063: type 1 areoles, group of crowned areoles (type 2) and structure of the interareolar cuticle with cord-like folds; (C) MLP 5063, lateral view of the cuticle; (D) MLP 3930/2, cluster of crowned
areoles with smooth surface, some of them with retracted filaments. (B-C) scale bar = lOjum.
Anatomy in Turin, Italy, but is not present there, nor could be located in any other collection. The original description (Camerano 1901) as well as later references to this species (CAMERANO, 1915; MIRALLES, 1969, 1976) did not include figures. Nevertheless, the description corresponds well with the specimen designed here as neotype.
The reexamination of two female specimens housed
in the Museum fur Naturkunde Berlin (MNB 4437) from Cordoba as Chordodes nobilii demonstrates that one of them differed slightly, but may still belongs to C. nobilii but the other belongs to Pseudochordodes bedriagae (see MIRALLES, 1973; DE-VILLALOBOS & RESTELLI, 2001).
Distribution - ARGENTINA - Buenos Aires: Sierra de La Ventana, El Loro stream 38°08’00”S, 38°08’00”W
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

384 C.DE-VILLALOBOS, F.ZANCA & A.SCHMIDT-RHAESA
Chordodes nobilii, 9 , SEM - fig.6- (A) MLP 3580, terminal end with radiating fine grooves around the cloacal opening; C)
midbody cuticle with type 1 areoles. (B) MLP 3579, midbody cuticle with type 1 areoles, type 1 areoles and interareolar
tubercles; (C) MLP 3580, midbody cuticle with type 1 areoles, type 1 areoles with various shapes and sizes; (D) 3579,
cuticle of the ventral midline, overview of the cuticle with crowned areoles with long filaments (type 2); (E) MLP 3579, cuticle of the ventral midline, crowned areoles (type 2) in detail. Scale bars: (A, D-E) = lOOjum, (B-C) = lOjum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

NEW DATA ON SOUTH AMERICAN SPECIES OF CHORDODES (NEMATOMORPHA) 385
d* , new record), Sierra de la Ventana 39 , MIRALLES, 1989), Sierra de La Ventana, Sauce Grande stream 38°05’00”S, 62°13’00”W (4cf, new record), Balcarce 37°45’00”S, 58°18’00” (l9 , new record). Cordoba: Cordoba31°24’00”S, 64°11’00”W (l9 , new record), Cosquin (l9 , CAMERANO,1901), Cura Brochero (39 , MIRALLES, 1989), El Durazno 31°20’15”S, 64°40’09”W (lcf, new record). San Luis: Villa Larca, Aguas Blancas, San Ignacio stream (32° 38’09”S, 64°57’06”W) (2cf, l9,new record), Carpinteria, Piedra Blanca stream (1024m) (32°24’ 43”S, 64°58’47”W) (3cf, new record).
CONCLUSIONS
This study by SEM allows us to confirm the observations previously made by CARVALHO & FEIO (1950) for C. carmelitanus that in females the crown areoles with long filaments are limited to the ventral and/or dorsal and ventral groove. This particular feature in females has been mentioned for other South American species such species such as C. brasiliensis
(see JANDA, 1894, CAMERANO, 1897; CARVALHO, 1946b), C. peraccae (see DE-VILLALOBOS & ZANCA, in press), C. bdlzani (see CARVALHO & FEIO, 1950;
Chordodes nobilii, 9 (MNB 4437) - fig.7- (A) midbody cuticle, scale bar = lOjum; (B) Nematomorpha, Gordiida, Chordodes,
South America, scanning electron microscopy. Ventral groove with cluster of crowned areoles surrounded by slightly elevated areoles.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

386 C.DE-VILLALOBOS, F.ZANCA & A.SCHMIDT-RHAESA
DE-VILLALOBOS et al, 2004). C. moraisi (see CARVALHO, 1944; DE-VILLALOBOS & ZANCA, in press), C. stauiarskii (see CARVALHO & FEIO, 1950; DE-VILLALOBOS & ZANCA, in press) and in the present study for C. nobilii Although more studies are necessary on both sexes of genus Chordodes in South America, there seems to be a feature of sexual dimorphism as was considered by CARVALHO (1946b) for the species C. brasiliensis, by CAMERANO (1987) and CARVALHO AND FEIO (1950), DE-VILLALOBOS et al (2004) for C. balzani and DE-VILLALOBOS & ZANCA (in press) for C. moraisi and C.staviarskii
ACKNOWLEDGEMENTS
To Lisa Levi (Museo Regionale di Science Naturali, Torino, Italy), Cristiana Serejo (Museu Nacional - Rio de Janeiro, Brasil) and Birger Neuhaus (Museum für Naturkunde, Berlin, Germany) for the permission to investigate museum specimens. To Patrícia Sarmiento, from the Scanning Electron Microscopy Service (Museo de Ciências Naturales, La Plata, Argentina), for preparation of the material. Many thanks also to two anonymous reviewers for constructive comments on the manuscript.
LITERATURE CITED
CAMERANO, L., 1897a. Monografia dei Gordei. Memoire delle Reale Accademia delle Science di Torino,
Italy, 47:339-419. CAMERANO, L., 1897b. Vaggio dei Dott. Alfredo Borelli
nel Chaco boliviano e nella Repubblica Argentina. Bollettino dei Musei di Zoologia ed Anatomia comparata delia Reale Universita di Torino, Italy,
12(294): 1-2. CAMERANO, L., 1901. Gordii raccolti dei Dottor Filippo
Silvestri nella Repubblica Argentina en el Paraguay. Bollettino dei Musei di Zoologia ed Anatomia
Comparata delia Reale Universita di Torino, Italy,
16(410): 1-2. CAMERANO, L., 1915. Revisione dei Gordii. Memoire
delle Reale Accademia delle Science di Torino,
Italy, 66:1-66.
CARVALHO, J.C., 1946a. Gordiaceos do Museu de
Historia Natural de Montevideo. Comunicaciones Zoologicas dei Museo de Historia Natural de
Montevideo, Uruguay, 32(2): 1-7.
CARVALHO, J.C., 1946b. Uma nova espécie de Gordiáceo do Brasil e considerações sôbre Chordodes brasiliensis Janda. Boletim do Museu Nacional do
Rio de Janeiro, Brasil, 62:1-8. CARVALHO, J.C. & FEIO, J.L., 1950. Sobre alguns
Gordiaceos do Brasil e da Republica Argentina
(Nematomorpha, Gordioidea). Anais da Academia Brasileira de Ciências, Rio de Janeiro, Brasil
(2): 193-206.
DE-VILLALOBOS, C. &RESTELLI, M., 2001. Ultrastructural study of cuticle and epidermis in Pseudochordodes
bedriagae (Nematomorpha, Gordidae). Cell & Tissue Research, Germany, 305(1): 129-134.
DE-VILLALOBOS, C.; HANCOCK, E.G. & ZANCA, F., 2004.
Redescription and sexual dimorphism of Chordodes
balzaniCamerano, 1896 (Nematomorpha). Journal of Natural History, London, 38(18):2305-2313.
DE-VILLALOBOS, C. & ZANCA, F., (in press). Ultrastructural redescription of Chordodes moraisi
(Carvalho 1942) and C. staviarskii Carvalho & Feio, 1950 and reinterpretation of C. gestri Camerano,
1904 and Pseudochordodes griffinii (Camerano 1898) (Gordiida Nematomorpha). Journal of Natural History, London.
JANDA, J., 1894. Beitráge zur systematik der Gordiien.
I. Die Gordiien galiziens. II. Über das genus Chordodes. Zoologische Jahrbücher für
Systematik, Geogrphie und Biologie der Tiere, Germany, 7:595-612.
MIRALLES, D., 1969. Lista Preliminar de Gordiaceos de
Argentina y datos biológicos sobre Neochordodes uniareolatus Carvalho. Acta Zoologica Lilloana,
Argentina, 24:183-192
MIRALLES, D., 1973. Sobre la presencia de Pseudochordodes bedriagae (Camerano) en Argentina.
Neotrópica, Argentina, 19(59):101-103.
MIRALLES, D., 1976. Notas sobre Gordiaceos Argentinos. Neotrópica, Argentina, 22(68):77-80.
MIRALLES, D., 1980. Observaciones Bioecologicas sobre
Gordiaceos de Ventania. Neotrópica, Argentina, 26(76) :217-224.
MIRALLES, D., 1984. Estúdio sobre gordiaceos argentinos. Revista dei Museo de La Plata,
Argentina, 13:283-289.
MIRALLES, D., 1989. Estructura cuticular de Chordodes
carmelitanus Carvalho y Feio, 1950 nueva cita para la Argentina (Nematomorpha Chordodidae).
Neotrópica, Argentina, 35:95-99.
MIRALLES, D., 1993. Contribución al estúdio de la estructura cuticular de Gordiaceos argentinos. Neotrópica, Argentina, 39(101-102):41-44.
MIRALLES, D.B. & CAMINO, N., 1983. Sobre la colección de Gordiaceos dei Museo Argentino “Bernardino
Rivadavia”. Neotrópica, Argentina, 29(81):51-56.
MIRALLES, D. & DE-VILLALOBOS, C., 1993. Gordioidea: Distribución en la Argentina y la Región Neotropical. Fauna de Agua Dulce de la República Argentina,
Argentina, 13(2-3) :5-32. SCHMIDT-RHAESA, A., 2002a. Are the genera of
Nematomorpha monophyletic taxa? Zoologica Scripta, Germany, 31:185-200.
SCHMIDT-RHAESA, A., 2002b. Australians species of Chordodes (Nematomorpha) with a description of two
new species, remarks on the genus Chordodes and its life history. Journal of Natural History, London,
36:1569-1588. SCHMIDT-RHAESA, A. & EHRMANN, R., 2001.
Horsehair worms (Nematomorpha) as parasites of praying mantids with a discussion of their life cycle.
Zoologischer Anzeiger, Germany, 240:167-179.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.375-386, out./dez.2004

Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
ISSN 0365-4508
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF REPRESENTATIVES
OF THE SUPERFAMILIES OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE
(MOLLUSCA, CAENOGASTROPODA) 1
(With 354 figures)
LUIZ RICARDO L. SIMONE 2
ABSTRACT: As part of a larger project on the phylogeny of the Caenogastropoda superfamilies, a detailed morphological study of 19 non-marine species is provided, of the following taxa: a) Family Ampullariidae: 1) Pomacea crosseana (Hidalgo) (from Pantanal, Brazil); 2) P. curumim new species (from Xingu River, Amazon Basin, Brazil); 3) P. scalaris (Orbigny) (from Pantanal, Brazil); 4) P. canaliculata (Lamarck) (same); 5) P. lineata (Spix) (several Coastal places of Brazil); 6) P. sórdida Swainson (Rio de Janeiro, Brazil); 7) P. bridgesi (Reeve) (Pará, Brazil) (most published elsewhere); 8) Asolene megastoma (Sowerby) (Rio Grande do Sul, Brazil); 9) Felipponea neritiniformis (Dali) (Paraná, Brazil); 10) Marisaplanogyra Pilsbry (Pantanal, Brazil); b) Family Cyclophoridae: 11) Neocyclotusprominulus (Orbigny) (São Paulo, Brazil); 12) Incidostoma tupy new species (Rondônia, Brazil); 13) Aperostoma branchetiana (Moricand) (Minas Gerais, Brazil); c) Family Viviparidae: 14) Viviparus acerosus (Bourguignat) (Hungary); 15) V. contectus (Millet) (same); 16) Notopala ampullaroides (Reeve) (Australia); 17) N. essingtonensis (Frauenfeld) (same); 18) Larina cf. stangei (Adams) (same); d) Family Annulariidae: 19) Annularia sp. (Yucatan, México). Based on this study, 143 characters (192 States) were searched, polarized, and aphylogenetic analysis is performed. The single obtained cladogram is the following: ((Neocyclotus prominulus (Aperostoma blanchetiana - Incidostoma tupy)) (((Annularia sp. - Cerithioidea) ((Viviparus acerosus - V. contectus) ((Notopala ampullaroides - N. essingtonensis) Larina cf strangei))) (Marisa planogyra (Pomacea crosseana (Felipponea neritiniformis (P. scalaris (P. curumim (P. lineata ((P. canaliculata-Asolene megastoma) (P. bridgesi-P. sórdida)))))))))), length 251; ci 77; ri 91. A pool of archaeogastropod characters, the rissooideans-littorinoidean Annularia, and the Cerithioidea ground plan were used as main outgroups, however the two latter taxa are operationally included as part of the ingroup. The analysis of obtained cladogram revealed that the taxon Architaenioglossa is paraphyletic, grouping 3 branches of basal caenogastropods successively distributed on the tree as follows: Cyclophoroidea (Cyclophoridae), Ampullarioidea (Ampullariidae), and Viviparioidea (Viviparidae), since the representatives of the remainder the caenogastropods (Annularia and Cerithioidea) were terminal in the ingroup. The current concept of Ampullarioidea (Ampullariidae plus Viviparidae) was also paraphyletic.
Key words: Ampullarioidea, Viviparioidea, Cyclophoroidea, Phylogeny, Morphology
RESUMO: Morfologia Comparativa e Filogenia de representantes das superfamílias de arquitenioglossos e Annulariidae (Mollusca, Caenogastropoda).
Como parte de um projeto sobre filogenia das superfamílias de Caenogastropoda um estudo morfológico detalhado de 19 espécies não marinhas é desenvolvido dos seguintes táxons: a) Família Ampullariidae: 1) Pomacea crosseana (Hidalgo) (Pantanal, Brasil); 2) P. curumim nova espécie (Xingu River, Amazônia, Brasil); 3) P. scalaris (Orbigny) (Pantanal, Brasil); 4) P. canaliculata (Lamarck) (mesmo); 5) P. lineata (Spix) (vários locais costeiros do Brasil); 6) P. sórdida Swainson (Rio de Janeiro, Brasil); 7) P. bridgesi (Reeve) (Pará, Brasil) (maioria dos dados publicados em outro tranbalho); 8) Asolene megastoma (Sowerby) (Rio Grande do Sul, Brasil); 9) Felipponea neritiniformis (Dali) (Paraná, Brasil); 10) Marisa planogyra Pilsbry (Pantanal, Brasil); b) Família Cyclophoridae: 11) Neocyclotus prominulus (Orbigny) (São Paulo, Brasil); 12) Incidostoma tupy espécie nova (Rondônia, Brasil); 13) Aperostoma branchetiana (Moricand) (Minas Gerais, Brasil); c) Família Viviparidae: 14) Viviparus acerosus (Biurguignat) (Hungria); 15) V contectus (Millet) (mesmo); 16) Notopala ampullaroides (Reeve) (Austrália); 17) N essingtonensis (Fauenfeíd) (mesmo); 18) Larina cf. stangei (Adams) (mesmo); d) Família Annulariidae: 19) Annularia sp. (Yucatan, México). Baseando-se neste estudo, 143 caracteres (192 estados) foram levantados, polarizados, e uma análise filogenética é realizada. O cladograma único é o seguinte: ((Neocyclotus prominulus (Aperostoma blanchetiana - Incidostoma tupy)) (((Annularia sp. - Cerithioidea) ((Viviparus acerosus - V contectus) ((Notopala ampullaroides - N. essingtonensis) Larina cf strangei))) (Marisa planogyra (Pomacea crosseana (Felipponea neritiniformis (P. scalaris (P. curumim (P. lineata ((P. canaliculata - Asolene megastoma) (P. bridgesi-P. sórdida)))))))))), passos: 251; IC 77; IR 91. Um conjunto de caracteres de archaeogastrópodes, o Rissooidea Annularia, e o plano básico de Cerithioidea foram usados como principais grupos externos, entretanto, os 2 últimos táxons foram incluídos operacionalmente como parte do grupo interno. A análise do cladograma obtido revelou que o táxon Architaenioglossa é parafilético, agrupando 3 ramos de cenogastrópodes basais sucessivamente distribuídos na árvore como segue: Cyclophoroidea (Cyclophoridae), Ampullarioidea (Ampullariidae), e Viviparioidea (Viviparidae), uma vez que os representantes dos demais cenogastrópodes (Annularia e Cerithioidea) resultaram como parte terminal do grupo interno. O senso prévio de Ampullarioidea (Ampullariidae mais Viviparidae) também resultou parafilético.
Palavras-chave: Ampullarioidea, Viviparioidea, Cyclophoroidea, Filogenia, Morfologia.
1 Submitted on January 28, 2003. Accepted on November 3, 2004.
2 Museu de Zoologia da Universidade de São Paulo. Caixa Postal 42594, São Paulo, 04299-970, SP, Brasil. E-mail: [email protected].

388 L.R.L.SIMONE
INTRODUCTION
Architaenioglossa is an enigmatic taxon that encompasses non-marine mesogastropod prosobranchs, corresponding to three families: Ampullariidae (=Pilidae), Viviparidae and Cyclophoridae (s.L). HASZPRUNAR (1988) presented the more accurate sense of this taxon, regarded as the sister taxon of the branch Caenogastropoda plus Heterobranchia, in the archaeogastropod grade. PONDER AND LINDBERG (1996, 1997), in their phylogenetic analysis of the Gastropoda, advocated that the architaenioglossans were basal members of the caenogastropod branch. Despite those studies, the validity of the formal taxon Architaenioglossa is still unclear, because it is not defined by any synapomorphy, other than the non-marine habitat. It is possible that this taxon actually represents a paraphyletic or even a polyphyletic group. Besides, their relationship with other gastropods is still controversial.
The literature on some architaenioglossans is relatively rich, mainly on the ampullariids, which have species with medicai, sanitary, and gastronomic importance. However, despite of this, the taxonomy remains unclear.
The Ampullariidae are médium to large freshwater snails, with representatives in all tropical areas of the Américas, África and Asia. There are some papers describing their morphology, such as, e.g.,
SCOTT (1957) with particular attention to South American representatives. BERTHOLD (1991) made an extensive study with representatives of all genera, and included aspects on morphology, phylogeny and evolution. BERTHOLD (1991), however, did not make a formal phylogenetic analysis of the group, but this, in part, was provided by BIELER (1993). PAIN (1972) referred on the family history. DEMIAN &YOUSIF (1973) provided embryological data. CASTELLANOS & FERNÁNDEZ (1976) provide biologic and biogeographic information. COWIE (1997) addressed the nomenclatural problems with the name Ampullariidae and the genera Pomacea and Ampullaria. Revisions of South American ampullariids include studies at the Instituto Oswaldo Cruz [e.g., THIENGO, 1987, 1989, 1995).
Viviparidae are freshwater, filter-feeders found in most part of Europe, Southern Asia, north and east Australia, and the central-east region of North America. Analysis of the history and distribution of the viviparids can be found in PRASHAD (1928). As its name suggests, they are viviparous or
ovoviviparous, with females brooding the young in the pallial oviduct.
The Ampullariidae and the Viviparidae have been normally referred to the same superfamily in recent literature, sometimes as Ampullarioidea [e.g.,
BOSS, 1982; PONDER & WARÉN, 1988), other times as Viviparioidea [e.g., VAUGHT, 1989). This hypothesis is also investigated herein.
The Cyclophoridae are terrestrial, distributed worldwide, lacking gill. They are, sometimes, divided into several families [e.g., six families in VAUGHT, 1989). They are, also, the subject of several monographic papers, which include taxonomical and some anatomical data. TIELECKE (1940), for example, introduced a classification based mainly on the characters of the shell and pallial oviduct. MORRISON (1955) established a taxonomy based mainly on the male reproductive organ of the American species. Some papers also provide further modifications and further criticai analysis of unstable cyclophorid classification is profuse in the literature [e.g., SOLEM, 1956; THOMPSON, 1963, 1967).
The Annulariidae are terrestrial snails from Central and North America, with known relationships with putative basal caenogastropods such as Littorinidae. THOMPSON (1978), pointed out the unusual radula and genital system and explored the concept of the family.
As part of a larger project on the phylogeny of the Caenogastropoda, representatives of each family of all superfamilies of this taxon have been studied in a detailed morphological and comparative scenario. For each caenogastropod superfamily, the following attributes have been tested: 1) the monophyly; 2) taxonomic status; and 3) the ground plan, i.e., the attributes of their hypothetical ancestral. These three parameters can only be tested by means of a phylogenetic analysis. The Architaenioglossa is the focus of the present paper. As the group in general, and the South American members in particular, have a very confused taxonomy, this paper also provides a basis for future more detailed studies on each sub-group. Some previously published phylogenies are part of the project (SIMONE, 1999, 2000, on Terebridae; SIMONE 2001, on Cerithioidea and SIMONE in press, on Stromboidea, Cypraeoidea and Calyptraeoidea) as well as other smaller papers, serving as outgroups. The terrestrial Annulariidae, which has a representative studied herein, are undoubtedly a non-architaenioglossan
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 389
caenogastropod (see THOMPSON, 1978; PONDER & WARRÉN, 1988). Its inclusion has as main objective to test how adaptation to a non-marine environment can influence this kind of analysis.
MATERIAL AND METHODS
Although some living species of Pomacea were examined in the laboratories of the Museu de Zoologia da USP (MZSP), most of the specimens studied were fixed in 70% ethanol. The extraction of the specimens from their shells were mostly easy, in some specimens the shell was broken. The dissections were performed under a stereomicroscope following traditional techniques, with the specimens immersed in fixative. All drawings were made with aid of a camera lúcida. The hard parts, such as shells (of small specimens), jaws and radulae, were also examined in SEM, in the Laboratório de Microscopia Eletrônica of MZSP.
In the descriptions, a more detailed one is presented for the first species, P. crosseana, (it was the first species studied in the project). In the remaining species, and when possible, the description is basically comparative to P. crosseana, without any description of similar characters. A similar approach is given in the figures.
A part of the cladistic analysis was performed with the aid of the Computer programs and “Tree Gardner 2.2” (RAMOS, 1997). The last basically works as an interface of the program “Hennig86” (FARRIS, 1988), the algorithm used was mh*, bb* and ie. For the polarization of the character States the outgroup method was used.
The section of comparative morphology is organized as a phylogenetic analysis, the presentation of each character begins with abbreviated descriptive sentence followed by plesiomorphic and derived conditions(s); also included are Cl and RI (consistency and retention indices, respectively), values for the character under the most parsimonious hypotheses. Following the apomorphic state(s), a list of terminal taxa with the apomorphic condition is presented. In the next section, all character States are presented in the table 1.
The remaining Caenogastropoda studied in this project were selected as outgroups, despite sometimes they are not directly included in the data matrix. They are mainly the following: Cerithioidea (SIMONE, 2001); Littorinoidea (SIMONE, 1998); Rissooidea-Littorinoidea (SIMONE & MORACCHIOLI, 1994; SIMONE 1995a, Annularia
sp. - this study); Stromboidea (SIMONE in press c); Calyptraeoidea (SIMONE, submitted); Tonnoidea (SIMONE, 1995b); Muricoidea (SIMONE, 1995c on Thala crassa; SIMONE, 1996b on Buccinanops spp); Conoidea (SIMONE, 1999, 2000, on Terebridae). As more distant outgroups, some archaeogastropods were also analyzed (e.g., SIMONE, 1996A; 1997; LEAL 85 SIMONE, 1998; 2000). However, in the analysis of the matrix in the Computer programs, only three outgroups are included: 1) A pool of archaeogastropods, mainly based on SASAKI (1998). 2) The Annularia sp., studied herein, representing the Risooidea-Littorinoidea branch, with a species adapted to the non-aquatic environment. 3) The Cerithioidea ground plan, i.e., the node # 1 of the cerithioidean cladogram, obtained in a previous study (SIMONE, 2001). Operationally, only the archaeogastropod set of characters was utilized as the outgroup, with Annularia sp. and Cerithioidea ground plan analyzed as part of the ingroup. So that, the monophyly of the “Architaenioglossa” could be tested.
Characters were polarized separately, and the polarization is justified, if necessary, in the discussion that follows each character. In that discussion, the final cladogram obtained is considered {i.e., the allocation and optimization of some character States are discussed) although, the cladogram is only formally presented in the following section.
Three parameters considered here are known as polemic, and sometimes there are authors who do not accept th em. The FIRST is the use of the ingroup synapomorphies (those that support node 1, or the ingroup “autapomorphies” in this case). They are used herein because they are the main goal of this study, since the Architaenioglossa is still lacking a phylogenetic definition. This definition, as referred in the Introduction, is essential for the larger project on caenogastropod phylogeny. Besides, there are authors who advocate this approach [e.g., YEATES, 1992). The SECOND is to consider some multistate characters as additive (ordered). In those character States, the ordering has support from ontogeny or by comparison [i.e., each State is clearly a modification of the preceding one). For additive characters, an extra explanation is presented in the discussion of each character. Furthermore, a parallel analysis in which the character is considered unordered was performed and any change of the result is reported. The THIRD is the arbitrary choice of an optimization in the presented cladogram (Fig.353) in those few cases where more than one
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

390 L.R.L.SIMONE
optimization is equally parsimonious. It is possible to apply the heuristic options ACCTRAN (assuming convergence is more likely than reversion) or DELTRAN (vice-versa), which would produce two trees with same the topology, a duality that is avoided here. In those cases where more than one optimization is possible, they are all reported, though only one is presented in figure 353.
Abbreviations in the figures: (aa) anterior aorta, (ad) adrectal sinus, (aí) afferent gill vessel, (ag) albumen gland, (am) ampulla of anterior aorta, (an) anus, (ap) apical gland of penis shield, (ar) male pore, (as) aperture of seminal receptacle into pallial cavity, (au) auricle, (bb) bulged region of br, (bg) buccal ganglion, (bm) buccal mass, (bp) brood pouch, (br) subradular membrane, (bv) blood vessel, (ce) cerebral ganglion, (cg) capsule gland, (cm) columellar muscle, (co) collar vessel, (cp) capsules, (cv) ctenidial vein, (dc) dorsal chamber of buccal mass, (dd) duct to digestive gland, (dí) dorsal fold of buccal mass, (dg) digestive gland, (di) diaphragm-like septum of haemocoel, (dp) dorsal- basal gland of penis shield, (dq) dorsal-basal gland of penis shield aperture, (ec) esophageal crop, (ed) ejaculatory duct immerse in integument, (ef) esophageal folds, (en) endostyle, (ep) esophageal pouch, (es) esophagus, (ey) eye, (fg) food groove, (fp) female pore, (fs) foot sole, (ft) foot, (ga) gastric ventral chamber, (gb) gastric dorsal chamber, (gf) gastric fold, (gi) gill, (gm) gastric muscle, (go) gonad, (gp) pleural ganglion, (gs) gastric shield, (gt) gastric transversal septum, (ha) haemocoel, (ic) intestinal chamber, (il) intestinal loop(s) into kidney chamber, (in) intestine, (ip) intestinal loop close to pericardium, (ir) insertion of m4 in tissue on radula (to), (is) insertion of m5 in radular sac, (jw) jaw, (ka) kidney anterior chamber (kc) kidney connection between anterior and posterior chambers, (kd) kidney dorsal lobe (ki) kidney (kp) kidney posterior chamber (11) left head-foot lobe (lp) lung pneumostome, (lr) right head-foot lobe, (lu) lung, (ml to ml4) extrinsic and intrinsic odontophore muscles, (ma) mouth abductor muscle, (mb) mantle border, (mc) buccal sphincter and circular muscles of oral tube, (me) mesentery, (mj) jaws, buccal, and oral tube muscles, (mo) mouth, (ne) nephrostome, (nr) nerve ring, (nv) nerve, (oa) odontophore secondary cartilage, (oc) odontophore cartilage, (od) odontophore, (om) ommatophore, (op) operculum, (os) osphradium, (ot) oral tube, (ov) pallial oviduct, (oy) ovary, (pa) posterior aorta, (pc) pericardium, (pd) penis duct, (pe) penis, (pf) penis furrow, (pg) pedal glands anterior furrow, (ph) penis shield notch, (pi) penis sac, (pl) penis shield longitudinal furrow, (pm) penis shield middle gland, (po) solid portion of penis sac
(penis base), (pp) penis papilla, (pr) penis sac connection with penis shield, (ps) penis shield, (pt) prostate, (pu) pedal ganglion, (py) pallial cavity, (ra) radula, (rm) retractor muscle of snout, (rn) radular nucleus, (rp) reno-pericardial aperture, (rs), radular sac, (rt) rectum, (sa) salivary gland aperture, (sc) subradular cartilage, (sd) salivary duct, (se) septum between esophagus and odontophore in buccal mass, (sg) salivary gland, (sl) snout tentacle, (sn) snout, (sr) seminal receptacle, (st) stomach, (su) supra- esophageal ganglion, (sv) seminal vesicle, (sy) statocyst, (te) cephalic tentacle, (tg) integument, (to) tissue on middle region of radula preceding its exposure, (tp) distai chamber of copulatory right cephalic tentacle, (ts) testis, (ug) urinaiy gutter, (up) ureter pore, (ur) ureter, (vd) vas deferens, (ve) ventricle, (vf) vas deferens pallial furrow, (vg) vaginal duct, (vo) visceral oviduct.
Abbreviations of institutions: (AMS) Australian Museum, Sydney; (BMNH) The Natural History Museum, London; (MNHN) Muséum National d’Histoire Narurelle, Paris; (MZSP) Museu de Zoologia da Universidade de São Paulo; (ZSM) Zoologische Staatssammlung Müchen, Germany.
Some terms used in discussion merit some explanation. The word “archaeogastropod” is used in traditional sense, but it is recognized as a paraphyletic taxon. The term “basal” caenogastropods refers to those taxa which generally are in the beginning of the mesogastropods in most catalogues (e.g., Abbott, 1974; Rios, 1994), in particular the Cerithioidea, Littorinoidea and Hydrobioidea. The term “higher” caenogastropods in general refers to Tonnoidea and Neogastropoda. Where mentioned “examined species” only the ingroup species are included.
SYSTEMATICS
Family Ampullariidae Genus Pomacea Perry, 1810
Type-species Pomacea maculata Perry, 1810)
Ampullaria Lamarck, 1799:76 (Case 2996, ICZN, see Cowie, 1997). Pomacea Perry, 1810:28. Ampullarius Montfort, 1810:242.
Pomacea crosseana (Hidalgo, 1871) (Figs.l- 2, 97- 98, 115-116, 150-173)
Ampullaria crosseana Hidalgo, 1871:206-207; 1872:142-143 (pl.7, fig.l); Sowerby, 1909:348.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 391
Ampullarius crosseanus- MORRETES, 1949:66.
Type-locality: Brazil, Amazon River.
Description
Shell (Figs. 1-2). Médium to large (up to 45mm), globose. Color homogeneous pale brown, sometimes successive spiral bands pale and dark brown. Periostracum thin, velvet-like. Spire small. Suture somewhat deep. Aperture elongated antero- posteriorly.
Head-foot (Figs. 151-152, 158). Color beige, with irregular brown spots. Head broad, protruded. Cephalic tentacles stubby, slender and long, tip pointed. Ommatophore long, about XU of tentacle length, tip rounded, base at outer surface of tentacles, their base slightly overlapping. Eyes dark, at tip of ommatophores. Snout somewhat cylindrical, broad, anterior surface flattened; mouth longitudinal, in middle region of anterior surface. Two slender snout tentacles, originating laterally at anterior end of snout, about as long as cephalic tentacles, base broad gradually narrowing distally, tip somewhat pointed. Two nuchal lobes, or siphons, flap-like, projecting forward, located on both sides of head, origin at posterior region of head, between head and adjacent region of connection between head-foot and mantle. Left siphon long and broad, with secondary low fold connecting its right edge with right attachment between mantle and head-foot. Right siphon shorter and narrower than left siphon. Foot large, ample, dorso-ventrally flattened. Opercular pad large on dorsal posterior surface of foot. Anterior furrow of pedal glands narrow, borders thin, occupying almost entire anterior half of pedal margin. Columellar muscle broad, with lateral muscular expansions supporting connection of mantle border; very short (about 1/6 whorl), slightly longer at right.
Operculum (Figs.97-98, 151). Occupying entire aperture. Corneous, semi-circular, thick, flexible. Margins thin, sharp, almost circular, short expansion on inner-superior region. Nucleus almost central, slightly dislocated towards inner margin. From nucleus concentric growth lines. Inner surface smooth. Scar circular, large, close to inner margin.
Mantle organs (Figs. 153-156). Mantle border thick, generally smooth, sometimes with series of small undulations at right; color homogeneous beige. Pallial cavity occupying about half whorl. Osphradium broad, elliptical, high, located in left region just posterior and parallel to mantle border.
Osphradium base a high, thick fold (stalk), elliptical in section. Osphradium filaments on tip of this basal fold; two series of thin, slightly triangular, septum- like filaments situated alternately in each side of tall, thin, fold-like, longitudinal, central axis (Figs. 155-156). Lung large (about 1/3 of pallial cavity volume and area), outline approximately elliptical, dorso-ventrally flattened, walls thick, richly vascular, scarcely muscular. Lung vessels mostly transverse (from left to right), high, close to each other. Lung vessels connect anterior marginal vessel (from afferent gill vessel) with ctenidial vein (Fig. 155). Pneumostome slit-like, margins muscular, located in anterior-left-ventral lung region, just posterior to left siphon. Gill long and narrow, slightly broader anteriorly, surrounding right and posterior lung margins from mantle border to left posterior end of pallial cavity, close to pericardium. Gill filaments triangular, slightly low, tips slightly right- central, somewhat displaced to right; right and left margins straight (Fig. 156). Gill anterior end broadly pointed. Ctenidial vein narrow in anterior region, becomes broader short distance from its anterior end, where lung vessels connect (see below for more details). Pericardium, kidney and rectum (respectively) running along right margin of gill (Figs. 153-154), with only anterior gill region free of these structures in females; in males this region occupied by penis shield. Gonoducts lying between rectum and right margin of pallial cavity; oviduct, when mature, dislocated ventrally. Pallial gonoducts described below.
Visceral mass (Fig. 153). Globose, of about 4 whorls. Pericardium and kidney as anterior structures, bulge into pallial cavity. Stomach about 1/3 whorl posterior to pallial cavity. Digestive gland dark brown, almost black, surrounds stomach except for some ventral and dorsal areas. Gonad beige, located on digestive gland in superior and peri- columellar regions.
Circulatory and excretory systems (Figs. 154, 157). Pericardium relatively large, located just posterior to left-posterior limit of pallial cavity, partly connected to pallial floor. Afferent gill vessel with branch Crossing from right to left in anterior region of gill, running dorsal to gill and short distance beyond gill in pallial roof (Fig. 155), and then as anterior border of lung. All lung drainage runs towards ctenidial vein, edging along left gill margin and right lung margin. Ctenidial vein connects with auricle just posterior to gill posterior end. Auricle somewhat short. Ventricle massive, almost spherical, posterior to auricle. Aortas located
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

392 L.R.L.SIMONE
posterior to ventricle; posterior aorta narrow; anterior aorta runs anteriorly on opposite side from posterior aorta. Just after separation of posterior aorta, lies anterior aorta bulb (ampulla), massive, walls muscular and thick, narrows after distance equivalent to ventricle length. Anterior aorta runs along haemocoel, with special branches in buccal mass region (see below). Kidney broad and ample (about half whorl), divided into 2 chambers. Posterior chamber most hollow, filled by intestinal loops (described below) connected to right surface by mesentery (Fig. 157). Dorsal glandular lobe of kidney dorso-ventrally flattened, cream colored, surface almost uniform, covering dorsal wall of posterior chamber (disconnected from intestine). Anterior renal chamber (“ureter”) slender and long, connected to posterior chamber by orifice at left; runs towards right attached to anterior edge of posterior kidney lobe and adjacent pallial roof, gradually narrows to pointed blind-sac. Inner surface of anterior renal chamber filled by two series of transverse septa, cream colored. Anterior series of septa larger and broader than posterior series and start between anterior and posterior renal chambers; posterior series of septa edging nephrostome. Both series of septa connected to central, longitudinal, dorsal fold. Nephrostome slit- like located in middle region between anterior and posterior renal chambers, separated from anterior renal chamber by posterior series of septa. Urinary gutter beginning ventral and at some distance from nephrostome as a tall fold, runs towards right; anteriorly attached to structures of right margin of pallial cavity, as oviduct-prostate and rectum, finishing close to right siphon.
Digestive system (Figs. 158-169) - Buccal mass (Figs. 159-161) large, occupies entire inner space of snout and about 1/3 remainder of haemocoel (Fig. 158). Buccal sphincter (mc) ventrally thick and dorsally thin. Jaw muscles (mj) thick, origin in odontophore dorsal surface, insertion around mouth inside mc. Pair of jaw plates large and thick (Figs. 150-160), outline elliptical, cutting-edge broad, composing anterior and median margins; small, broad projection in anterior region of median margin towards anterior. Pair of dorsal inner folds of buccal mass (Fig. 160) broad and tall, surface with several superficial transverse furrows. Dorsal folds cover posterior margin of jaws, running towards posterior turning medially, covering a somewhat deep dorsal chamber. Dorsal chamber with several, longitudinal, low folds. Peri-buccal and odontophoral muscles (Figs. 159, 161-167): ml)
jugal muscles, several small muscular fibers connecting buccal mass with adjacent region of haemocoel inner surface, more concentrated around mouth; mia) dorsal protractor muscle of buccal mass, somewhat thin, origin in dorsal region of mouth runs posteriorly close to median line attached to surface of buccal mass, insertion in middle levei of dorsal surface of buccal mass; mlb) two pairs of lateral protractor muscles of buccal mass, origin in lateral-dorsal region of mouth, run posteriorly edging latero-dorsal surface of buccal mass, insertion in posterior region of buccal mass, close to buccal ganglia; ma) pair of jaw abductor muscles, origin in several points of outer surface of jaw, runs immersed in mc and mj dorsally in 3 or 4 branches, after mc and mj, they unite as single pair, with insertion in adjacent region of dorsal- lateral surface of haemocoel; m2) pair of retractor muscles of buccal mass, origin in ventral-lateral surface of haemocoel adjacent to middle levei of odontophore, run dorsally, by short distance to penetrate mj fibers, insertion in inner layer of longitudinal fibers of mj in three different and somewhat equidistant, small points, located in ventral and lateral region of jaw plates; m3) superficial, thin muscle immersed in membrane covering posterior region of odontophore, with transverse fibers just anterior to radular sac; m4) large pair of ventral tensor muscles of subradular membrane, origin in inner-posterior surface of odontophore cartilages (oc), most fibers of this muscle surrounding ventral and lateral surface of cartilages, connecting with cartilage on opposite side than their origin, some fibers surrounding posterior margin of cartilages, running from posterior to anterior region edging dorsal surface of cartilages, insertion along subradular membrane (br) and in posterior-inner surface of accessory cartilages; m5) pair of dorsal tensor muscles of radula, origin wide in m4 posterior-median outer surface, surrounding m4, running towards median and anterior, gradually narrowing, insertion in middle-dorsal region of radular sac preceding its exposed (in use) area in buccal cavity; m6) horizontal muscle, with transverse fibers, connecting dorsal surface of both cartilages, insertion at some distance from dorsal edge of cartilages (Figs. 164, 166); m7) pair of very slender muscles, origin in posterior region of dorsal edge of cartilages, running posteriorly and ventrally, penetrating radular sac in region preceding its entrance in odontophore, insertion in ventral surface of radular sac like fan; m9) two pairs of
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 393
medial dorsal tensor muscles, anterior pair broad, flattened and thin, posterior pair narrower, origin in tissue (to) on middle region of radula (preceding buccal cavity), close to median line, running antero- laterally short distance to insert in posterior-dorsal edge of subradular membrane, also partly connected to adjacent outer surface of m4; mlO) pair of ventral protractor muscles of buccal mass, narrow and thin, origin in ventral inner surface of mouth, run towards posterior lying ventral surface of buccal mass, insertion in postero-ventral region of odontophore close to median line, in both sides of radular sac; mil) pair of thin and long dorsal tensor muscles of subradular membrane, origin in ventral inner surface of haemocoel in region adjacent to middle levei of odontophore, runs dorsally short distance, penetrate odontophore just anterior to radular sac, Crossing dorsal to m7, insertion along ventral surface of subradular membrane close to median line (Figs.162, 166); ml2) pair of narrow, short muscles annexed to m6 (Fig.167), origin in medial-anterior region of odontophore cartilages, just anterior to m6, running antero-laterally short distance, insertion in transition between subradular membrane and accessory cartilage just anterior to insertion of m4. Pair of odontophore cartilages large, flattened, outline elliptical. Pair of accessory cartilages thin, small, outline elliptical, in anterior-dorsal region of subradular membrane, just inside more protruded region of exposed part of radula. Subradular membrane (br) thin, strong and semi- transparent, covering inner surface of subradular cartilage, radular ribbon and adjacent areas. Subradular membrane margin mostly connected to dorsal surface of m4 pair, this pair working as tensor. Subradular cartilage forming expanded transparent projections of radula in buccal cavity, covering part of odontophore inside buccal mass. Radular sac short, little longer than odontophore. Radular nucleus broad, with pair of conspicuous blood vessels inserted in it, one terminal posterior and other ventro-anterior, this latter vessel running anteriorly along ventral surface of odontophore on median line. Radular teeth (Figs.115, 116): rachidian tooth short and broad, 7 stubby cusps, central cusps 2-3 times larger than neighboring cusps; pair of low and broad basal cusps; lateral tooth about 1.5 times broader than rachidian, curved inwards, with about 5 broad cusps, third cusp terminal and about 3 times larger than neighbor; inner marginal tooth tall, broad and flat, about same length as lateral tooth; tip bluntly
pointed, inner edge with conspicuous sub-terminal cusp, outer edge smooth or with up to 3 small, sub-terminal cusps; basal region with longitudinal concavity in outer edge where outer marginal tooth encased; outer marginal tooth tall, somewhat triangular, shorter and slender than inner marginal tooth, somewhat thick-longitudinal reinforcement along outer edge; tip sharp pointed and abruptly curved inwards, cusp small, sub-terminal on inner edge. Pair of esophageal pouches long, cylindrical, tip rounded, walls thin, located just posterior to buccal mass (Figs. 159-161). Pair of inner dorsal folds of buccal mass abruptly narrowing in levei of pouches and curving externally, penetrating into dorsal wall of pouches, at about half of pouches length, tapering gradually. Remainder of pouches inner surface smooth (Fig. 160), apparently without glands. Anterior esophagus (posterior to pouches) narrow, short, inner surface with pair of broad longitudinal folds (originating from pouches folds) and some lower, narrower secondary folds. Middle esophagus very broad, bulging, walls thin, inner surface without folds. Pair of salivary glands massive, separated from each other, located just posterior to pouches. Salivary ducts in anterior region of glands, attached to dorsal surface of buccal mass, close to median line, passing through nerve ring, penetrating buccal mass wall in about middle levei; to open lateral surface of anterior region of dorsal folds. Posterior esophagus long (about half of esophagus length), narrow, inner surface smooth. Stomach (Figs. 168-169) large, spherical, with two chambers. Ventral chamber small, ventral wall thin, about five narrow ducts to digestive gland irregularly disposed in ventral surface. Dorsal chamber larger, walls thick, muscular, inner surface folded by muscle bundles. Ventral and dorsal gastric chambers separated from each other by tall, transverse, muscular septum (part of dorsal chamber walls); esophagus and intestine connected to both cambers. Columellar- gastric muscle well-developed, thick, origin in columellar surface of mantle in levei adjacent to stomach (separated and posterior from columellar muscle), running anterio-dorsally, penetrating between esophagus insertion and intestine origin, insertion spread in gastric dorsal muscular wall, more concentrated towards posterior. Esophagus insertion and intestine origin side by side in anterior-left stomach region. Intestine wide, inner surface smooth, running anteriorly and left to renal chamber, becoming exposed inside left-posterior region of renal chamber. Intestine abruptly turning
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

394 L.R.L.SIMONE
towards posterior-right, running parallel to preceding portion, surrounding posterior and right inner surface of renal chamber. Intestine bearing chamber occupying most of outer-left inner surface of this twisted portion, where it touches pericardium; intestine having constriction, in form of transverse fold (taller right), preceding this chamber; chamber forming blind sac positioned contrary to flow, bearing 5-6 secondary smaller, blind-sacs on outer surface. Intestine with three loops, connected to renal wall by mesentery, in middle levei of right surface of renal chamber (Fig. 157). Schematic representation (in ventral view) shown in figure 154. Anterior to loops, intestine running towards anterior, exiting to pallial cavity. Anus siphoned (ie., preceded by short unattached stalk of rectum), broad, with two inner successive edges covered by narrow longitudinal folds, located close to right siphon (Figs.153, 168, 170).
Genital system - d (Figs. 170-171): Testis lies on columellar surface of visceral mass, on digestive gland, at about three whorls since visceral apex. Visceral vas deferens narrow, runs in left (inferior) edge of testis last whorl. Vas deferens becomes thicker walled in pallial cavity, running at right of and close to rectum. Posterior to anus, vas deferens Crossing over rectum, staying to left side of base of anus; where its abruptly expands as ejaculatory tube. Ejaculatory tube with loop along right wall of penis sac. Penis sac relatively large, located at left of anus; walls thin, aperture located anteriorly, turned to base of penis shield, this aperture preceded by long tubular region. From ejaculatory tube, vas deferens exits to penis sac T-shaped; anterior branch of this structure solid fold attached to dorsal wall of penis sac, preceding its aperture; Penis is posterior branch of that structure. Penis long, slender, intensely coiled, narrowing gradually; distai third very slender, about 10 times narrower than its base. Penis duct entirely closed (tubular), very coiled in penis base, gradually less coiled towards distai region. Penis duct aperture very small, in penis tip. Penis shield large (about 1 / 3 of pallial cavity volume), slightly triangular, located in superior (right) half of inner edge of mantle border, generally inside pallial cavity. Penis shield base very broad, thick, bulging due to basal gland. Basal gland aperture small, located in left-outer region of penis shield base. Penis shield gradually narrows, distai tip flat, with slightly rounded borders and central concavity. Penis shield furrow runs along central region of inner surface of shield, somewhat deep, small notch in right-basal region;
distai end of furrow Y-shaped, close to shield tip. No other penis shield gland detectable. 9 (Figs. 172- 173): Visceral structures in similar location to corresponding structures of male (Fig. 153). Ovary beige in color. Pallial structures not seen in details due to poor fixation. Relatively broad oviduct running at right of rectum for short distance and opening to large pallial oviduct. Pallial oviduct about XA of pallial cavity volume when mature; located at right and ventral to rectum, partly connected to pallial floor. Visceral oviduct inserts in right region of pallial oviduct. From this insertion broad, flat and coiled inner lumen starts, running towards anterior. Lumen immersed in capsule gland tissue. Albumen gland widely connected anteriorly to left side of posterior capsule gland, its lumen flat and broad, closed as blind-sac posteriorly. Pallial oviduct abruptly narrows in approximate anterior third levei of pallial cavity as vaginal tube. Vaginal tube long, thick walled, circular in section, at right edge of rectum. Female pore small, papilla-like, at right of, and close to anus.
Nervous system - Nerve ring very similar to those described by SCOTT (1957) and BERTHOLD (1991) for other ampullariids. Hypoathroid, located in middle levei of buccal mass. Both cerebral ganglia separated by long, thick comissure. Buccal ganglia large, located relatively close to median line in dorsal-posterior region of buccal mass, near bases of esophageal pouches. Statocysts with several small, iridescent, statoconia, located deeply in foot musculature, just ventral and slightly outside pedal ganglia.
Measurements of shells (in mm). MZSP 29522: 9 #1, 42.0 by 35.8; 9 #2, 38.6 by 31.1; d #3, 37.9 by 33.1.
Distribution - Brazil, upper Amazon and upper Parana basins.
Habitat - On Eichomia and other aquatic plants of lakes and low flow rivers.
Material examined - BRAZIL - MATO GROSSO DO SUL: Passo do Lontra, MZSP 29522, 2d, 39 , 8 young specimens, MZSP 30758, 9 (Figs. 1-2) (Simone and Lipparelli cols., V/1993).
Discussion - The shell of P. crosseana is very similar to the sympatric and more abundant P. canaliculata.
The two species are difficult to separate on shell characters alone. P. crosseana, however, is distinguishable by its elongates shell and generally uniform and paler color; although some exceptions occur. On the other hand, there are considerable
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 395
anatomical differences, as discussed in the comparison below. The simplest way for differentiate P. crosseana from P. canaliculata is by the penis shield features, which P. crosseana having flattened concave tip (Fig. 170), while P. canaliculata has longer broad tip with an apical gland (Fig.205).
Pomacea curumim new species (Figs.3-4, 57-60, 107-108, 117-118, 174-188)
Types - Holotype MZSP 30760. Paratypes - BRAZIL - PARÁ: Xingu River, Between Senador José Porfiro and Porto de Moz; 2°00’22.9-42.6”S 52°12’59-8”- 13’04.6”W, MZSP 29844, 12 specimens (sta. AMZ94-52, Zanata and others, 08/XI/1994); 2°03’40.7”-04’15.4”S 52° 14’43.5-47.4”W, MZSP 29845, 29 specimens (sta. LRP94-44, Rapp Py- Daniel and other cols., 08/XI/1994); 2°02’45.1”- 03’24.8”S 52°14’56.2”-15’03.8”W, MZSP 29846 12 specimens (sta. LRP94-45, Rapp Py-Daniel and others cols., 08/XI/1994).
Type-locality - BRAZIL - PARÁ: Xingu River, Between Senador José Porfiro and Porto de Moz; 2°04’15”S 52°14’43.5”W (Rapp Py-Danieland other cols., 08/XI/1994).
Diagnosis - Very small, globose shell, with large aperture. Lung reduced. Penis shield with single basal gland on right, an apical folded region, and notch in middle levei.
Description
Shell (Figs.3-4, 57-60). Size small (up to 12mm), globose, outline almost circular but broader near to upper suture, color pale greenish beige. Periostracum thick, smooth or sometimes with axial or spiral striae in some areas. Spire small and low, eroded, about four low, convex whorls. Last whorl large, slightly long anteriorly and somewhat conical. Aperture elliptical, slightly long antero- posteriorly. Outer lip rounded, with cut edge. Inner lip concave in superior half, without callus; convex in inferior half, with cut edge. Siphonal region bluntly angular, shortly projected anteriorly. Umbilicus deep, narrow, simple.
Head-foot (Fig. 174). Similar characters to those of preceding species but shorter antero-ventrally and broader laterally. Head very broad, tentacles lateral located. Columellar muscle very short and thick.
Operculum (Figs. 107-108, 174). Similar attributes as those of preceding species, but slightly longer antero-posteriorly. Outer surface with uniform concentric striae. Nucleus at short distance from middle levei of inner edge. Inner scar occupying
about 2/3 of inner surface, lie at short distance inner edge, and at some distance from remainder edges.
Mantle organs (Figs. 175-178, 187). Characters also similar to those of P. crosseana, distinctive or notable features following. Pallial cavity very short, less than half whorl. Penis shield occupies most of inner pallial space in males. Osphradium almost sessile, its base (stalk) in same levei of mantle (Figs. 175-177). Lung reduced, pneumostome very small pore. Gill proportionally large and straight; anterior end on mantle border or penis shield base (of males), just posterior to anterior end becomes broad, narrowing gradually. Gill filaments slightly low, curved to right, tip rounded. Ctenidial vein narrow. Between gill and right pallial structures narrow space.
Circulatory and excretory systems (Figs. 178-179). Similar attributes as those of preceding species, with following distinctive features. Heart ampulla (of anterior aorta) almost spherical and short. Kidney dorsal lobe low, separated from intestinal loops, presenting 2 regions; posterior region narrow, transverse, with several transverse furrows, edges first intestinal loop; anterior region larger (about 4/5), covered by net of vessels intensely anastomosed, large vessel covers left side of connection with anterior lobe. Kidney anterior lobe short, broad, turned forwards, inner glandular folds similar to those of P. crosseana, but with an outstanding bulged portion preceding nephrostome. Intestinal loops inside kidney chamber described below.
Digestive system - Buccal mass and odontophore characters (Figs. 180-183) similar to those of preceding species, with following differences and notes: outline shorter and circular; buccal mass in general dislocated slightly anterior to mouth; jaws with circular outline (Fig. 182); (mj) thick, short, conic, (ma) pair thinner and simple, (m4) pair with wider portion free from odontophore cartilages in their median surface, (m6) with outer surface attached to adjacent inner surface of br, (m7) pair also connected to posterior-median extremity of odontophore cartilages, (ml2) pair present. Radular teeth features similar to those of P. crosseana with following notable attributes (Figs. 117-118): rachidian tooth proportionally larger, 5-7 cusps, central cusps about 1.5 times larger than neighbor cusps, pair of basal cusps obliqúe; lateral tooth as broad as rachidian, 3-4 cusps, second cusps terminal and about double than neighbor cusps; inner and outer marginal
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

396 L.R.L.SIMONE
teeth with larger sub-terminal cusp in inner edge and no cusps in outer edge. Esophageal characters, pouches and inner folds (Fig.182) similar to those of P. crosseana, except for continuous fashion of esophageal main folds in region of pouches aperture (without notch). Esophageal pouches with narrow dorsal inner fold each, connected to adjacent dorsal surface of esophageal main folds. Salivary glands fashion also similar to those of preceding species, except for aperture located in middle region of dorsal folds of buccal mass and with form of deep slit projected towards anterior. Stomach characters (Figs. 186, 188) also similar to those of P. crosseana,
including thick muscular walls, columellar-gastric muscle and inner transverse muscular septum. Stomach differs by smaller ventral chamber (with thin walls), and fewer ducts to digestive gland. Intestine initially very broad. Region adjacent to pericardium narrow, preceded by tall fold in its dorsal surface, fold posterior portion arched, transverse, lies ventral surface, finishes towards posterior protected by posterior portion (Fig. 188). Remainder intestine runs somewhat similar to that of preceding species. In region of kidney broad and complex loop as shown in figure 179. Rectum broad and slightly short. Anus simpler, without double layer of papillate folds.
Genital system - d (Figs. 175, 187): Testis color cream, situated in upper region of digestive gland coils. Visceral and pallial vas deferens attributes similar to those of preceding species, except for broad expansion in region just at right from anus. Penis sac spherical, thin walled at left and thick muscular at right (penis base or ejaculatory tube). Penis sac amply opened in left edge towards penis shield base. Penis very shorter than that of P.
crosseana, with relative uniform width along its length. Penis tip abruptly pointed. Penis shield very large (occupies most of pallial cavity inner space) and conic. Pair of penis shield glands, basal, dorsal gland massive, with location and aperture similar to those of P. crosseana. Other gland smaller, semi- spherical, located in right side just over base. Penis shield ventral groove deep, runs about along its center; notch in right side in middle region, where penis projects. Penis shield apical region slightly broad, most folded by transverse, successive, irregular furrows (mainly in ventral and right surfaces). Penis shield ventral groove runs very narrow and shallow at left side of apical folded region. F (Figs. 184-185): Visceral structures fashion similar to those of P. crosseana. Pallial oviduct massive, also of somewhat similar features
of preceding species. Visceral oviduct ends sub- terminally in sac-like bursa copulatrix, partially separated from remainder pallial oviduct by furrow. Bursa copulatrix flattened, broad, located along left side of pallial oviduct, connected to it in middle levei. Capsule gland massive, its lumen irregularly coiled as shown in figure 185. Capsule gland lumen with 2 short coils anterior to bursa connection. Vaginal tube with about same length than capsule gland. Vaginal tube broad, walls relatively thin, inner surface with some longitudinal folds. Female pore small, at right and close to anus.
Measurements of shells (in mm) - MZSP 29845: d 1, 8.9 by 9.1; d 2, 8.8 by 7.8; 9 3, 9.2 by 8.2.
Distribution - For moment known in Xingu River (Above Amazon tributaiy), in region between Senador José Porfirio and Porto de Moz cities, Pará, Brazil.
Habitat - In clear, planar river, from 12.5 to 20m depth, 300 to 4000m distant from shore. Substrate sandy, rocky and forest.
Material examined - Types.
Etymology - The specific epithet refers to Tupy language, meaning small children [curumim). A reference to the small size in relation to normal large fashion of the other species.
Discussion - The species was initially interpreted as young samples of other species, after serial sections and exam of several specimens, the adult nature of the specimens was cleared. Complex pallial genital structures and full-developed gonad are present in most specimens, exceptionally larger than lOmm. The size and the less developed lung suggest a process of paedomorphosis. P. curumim
is certainly the smaller known species of ampulariid. On the other hand, reduction of lung has been found in ampullariids from Lake Malawi (África) (BERTHOLD, 1990) with the explanation that the animais habitat is deep (1.5 to 90m) for they reach the surface.
Pomacea scalaris (Orbigny, 1835) (Figs.5-9, 85-87, 119, 189-198)
Synonymy see CASTELLANOS & FERNANDEZ (1976: 14). Complement:
Ampullaria scalaris - ORBIGNY, 1835:31; 1843:369-371 (pl.50, figs.1-3) (see present figs.85-87); SCOTT, 1957:305-307 (pls.l, 3; pl.4, fig.5; pl.5; pl.6, fig. 10; pl.7. figs. 15-16; pis. 10, 12; pl. 13, fig.32; pl.22, fig.l); FIGUEIRAS, 1964:171; CASTELLANOS & FERNANDEZ,
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 397
1976:14-15 (pl.l, figs.l, 4, pl.2, fig.l; pl.3, fig.3); MARTÍN, 1980:47-50 (fig.4).
Ampullarius scalari - OLIVEIRA et dl., 1981:76. Pomacea scalari: - QUINTANA, 1982:111
Description
Shell (Figs.5-9, 85-87). Characteristic by a conspicuous shoulder in spire and last whorl. Color purple-brown, pale (almost white) between carina and suture. Other details in SCOTT (1957:305, pl.22, fig.l), CASTELLANOS & FERNÁNDEZ (1976:14).
Head-foot and operculum - Characters similar to those of P. crosseana. Other details in SCOTT (1957, pl.l).
Mantle organs (Figs.189-190, 193). Organization and features similar to those of P. crosseana, with remarks following. Osphradium on a stalk tall in right side and low in left side. Gill filaments narrow, pointed.
Circulatory and excretory systems (Fig. 192) - Heart attributes like those of P. crosseana. Kidney features also similar to those of P. crosseana, except for following characters. Dorsal renal lobe tall, pale brown, covered by several vessels, strongly arched (edging intestinal loops mesentery). Anterior renal chamber relatively short (less than half of posterior renal chamber length). Inner organization of septa and folds similar to those of P. crosseana, except for longitudinal fold not connected to nephrostome edge.
Digestive system (Figs.191, 194-196) - Buccal mass characters similar to those of P. crosseana,
remarkable features following. Jaw plates smaller, with circular outline (Fig. 195). Dorsal folds with smooth surface; aperture of salivary glands located between their middle and posterior thirds. Dorsal chamber (between both dorsal folds) relatively deep. Odontophore muscles: ma, 2 narrow pairs; m4 pair with smaller area attached to inner surface of cartilages; m6 narrower; mlO pair narrow; mlOa, a pair of broad muscles, origin in ventral surface of mouth, runs dorsal to mlO, insertion in middle-ventral region of odontophore. Other details of odontophore in Scott (1957, pis.3-5, 7). Radular teeth characters similar to those of P. crosseana, with following distinctions (Fig. 119): rachidian shorter, base somewhat triangular, 3-5 broad cusps, central cusps about 1.5 times larger than neighbor cusps, basal cusp narrow and obliqúe; lateral tooth little broader than rachidian, weakly curved inwards, 3-4 cusps, second cusps terminal and about 1.5 times larger than neighbor cusps; inner and outer marginal teeth slender and tall, longer than lateral tooth, inner marginal somewhat
broader than outer marginal tooth, tip narrowly rounded, sub-terminal cusp conspicuous, in inner edge [other details in Scott (1957, fig. 10)]. Esophageal pouches and inner folds with characters similar to those of P. crosseana, except for broader pouches, with larger inner fold each (Fig. 195). Stomach features also similar to those of P. crosseana, other details in Scott (1957, pl.10). Digestive gland brown in color. Intestinal loops inside renal chamber as shown in Fig. 192. Anus with single layer of papillae.
Genital system - Ó (Fig. 198): Visceral and pallial structures with characters similar to those of P.
crosseana, distinctive or notable features following. Testis beige in color. No bulged portion of vas deferens preceding anus. Penis sac with only 1 /3 of its (posterior) walls thick. Penis sac aperture long tube, only opened in anterior end, close to penis shield groove base. Penis relatively short, uniform width along its length, apex pointed. Penis shield slightly conic, tip long, narrow, curved towards right. Other details in MARTÍN (1980, fig.4). 9 (Fig.197): Visceral and pallial organs characters similar to those of P.
crosseana, with following remarkable features. Visceral oviduct connects with pallial oviduct about in middle region of its ventral side. Bursa copulatrix immerse in capsule gland, insertion of visceral oviduct in its posterior-ventral region, connection with capsule gland lumen in its anterior-ventral region. Capsule gland lumen flat, irregular, connection with bursa narrow, abruptly expands and contours bursa dorsally, after certain distance from bursa connects in middle region of broad dorsal branch of capsule gland lumen. Ventral branch of capsule gland lumen connects with vaginal tube. Vaginal tube long, narrow, walls thick. Female pore small, at right from anus.
Central nervous system - Described in SCOTT (1957, pl.12).
Measurements of shells (in mm) - MZSP 29590 (Figs 5-6):33.0 by 29.0; (Figs.7-9): 48.6 by 47.7.
Distribution - Amazon and Paraná Macrobasins.
Habitat - In freshwater courses of low flow, under aquatic plants.
Material examined - Types - BOLÍVIA: MNHN, 3 shells, syntypes. Ordinary material: BRAZIL - MATO GROSSO DO SUL: Passo do Lontra, MZSP 29590, 55 specimens (Simone and Lippareli cols., V/1993).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

398 L.R.L.SIMONE
Pomacea canaliculata (Lamarck, 1801) (Figs.11-12, 88-89, 120-121, 199-205)
Synonymy see SCOTT (1957:299); CASTELLANOS & FERNÁNDEZ (1976:13). Complement:
Ampullaria insulamm Orbigny, 1835:32; 1843:374- 375 (pl.51, figs.1-2) (see present figs.88-89); MARTÍN, 1980:47-51 (figs.2, 6).
Ampullaria canaliculata - SCOTT, 1957:299-303 (pl.22, fig.3; pl.14, figs.34-35; pl.16, fig.42; pis. 17- 21); FIGUEIRAS, 1964:170; CASTELLANOS & FERNÁNDEZ, 1976:13 (pl.l, figs.3, 6; pl.3, fig.2); MARTÍN, 1980:47-52 (figs.l, 5).
Ampullarius canaliculat - OLIVEIRA et al., 1981:78. Pomacea canaliculata - ANDREWS, 1965b:71-91
(figs.1-5, 6b, 7b); QUINTANA, 1982:109; CAZZANIGA & ESTEBENET, 1984:213-222 (fig.1-6); BERTHOLD, 1989:143, 151, 153 (figs.2, 6e); CAZZANIGA, 1990:384-387 (figs. 1-4); THIENGO et al, 1993:68-70 (figs.1-6); THIENGO, 1995:66-68 (figs.2, 8-9, 14).
Types-See MERMOD (1952:88-89, fig.149).
Description Shell (Figs.l 1-12, 88-89). Large, some specimens with about lOOmm. Color pale to dark brown, with some spiral dark bands. Spire slightly short, suture deep. Other details in SCOTT (1957: 299) THIENGO et al. (1993, fig.l).
Head-foot and operculum. Characters similar to those of P. crosseana.
Mantle organs (Figs. 199, 205). Features like those of P. crosseana, with succeeding distinctive or notable attributions. Osphradium similar to that of P. scalaris, but with taller filaments. Gill filaments narrower and pointed. Ctenidial-pulmonary vein separated into 2 parallel vessels in anterior 2/3 of its length, close with each other, pulmonary branch dorsal and broader. These 2 vessels unite with each other and run as those of preceding species. Narrow ad-rectal sinus distinguishable. Other details in ANDREWS (1965b:figs. 1-3).
Circulatory and excretory systems (Figs.202, 204) - Heart characters similar to those of P. crosseana,
ampulla of anterior aorta broad. Kidney organization also similar to that of P. crosseana,
distinguishing or remarkable features following. Dorsal renal lobe relatively small, surface most smooth, with deep notch adjacent to intestine loops; few and broad vessels present close to aperture to anterior renal chamber. Intestinal loops attributes inside renal chamber described below. Anterior renal chamber longer, with more septa, longitudinal
fold not directly attached to nephrostome wall. Urinary gutter of ordinary fashion. Other details in ANDREWS (1965b:figs. lb, 4-6)
Digestive system (Figs.200-204) - Buccal mass characters (Figs.200) comparable to those of P.
crosseana, remarkable features succeeding. Size proportionally larger. Annex cartilages thinner and smaller. Jaw plates very large and thick, with small median-anterior projection. Dorsal folds broad, smooth, with long portion covering jaws. Odontophore muscles (Fig.200): mia thinner; mlb as several thin pairs; ma as multiple, relatively narrower pairs; m3a pair of thin muscles with dorso- ventral fibers covering posterior surface of odontophore; m4 pair not so attached to inner surface of odontophore cartilages; m6 very thick; m7, mil and ml2 with insertions more anterior; mlO pair thick; mlOc absent. Esophageal characters also similar to those of preceding species, including inner folds and pouches. Differs in taller esophageal folds posterior to pouches. Radular teeth attributes similar to those of P. crosseana, distinctions following (Figs. 120-121): rachidian tooth broad and shorter, 5-7 cusps, central cusps about double than neighbor cusps, basal cusps weak; lateral tooth about as broad as rachidian, 3-4 cusps, second cusp terminal, broad, and several times larger than neighbor cusps; inner marginal tooth with about half weight and same length of lateral tooth, tip bluntly pointed, sub-terminal cusp in inner edge very small; outer marginal tooth about 3-4 times slender than inner marginal and encased in special concavity of outer edge of this tooth, tip sharp pointed, sub-terminal cusp small, in inner edge. Stomach features (Figs.201, 203) similar to those of P. crosseana, including gastric-columellar muscle, remarks following. Ventral chamber (with thin walls) very smaller, about 6 ducts to digestive gland. Gastric walls most thick muscular. Inner transverse muscular septum thick, with edges turned inwards. Intestine proximal portion relatively narrow. Digestive gland dark, almost black. Intestinal region adjacent to pericardium preceded by mosaic of several folds and chambers (Fig.201). Intestinal loops inside renal chamber complex, as shown in situ in figure 204. Anus with layer of papillae.
Genital system - d (Fig.205): Testis beige, occupies last visceral whorls up to stomach. Pallial structures similar to those if P. crosseana, with following remarks. No bulged portion of vas deferens preceding anus. Penis sac with about 2/3 thick; its posterior 1/3 with thin walls. Penis sac aperture long tube edging thick penis sac portion,
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 399
ending in penis shield groove base; this tube with aperture all along its dorsal surface. Penis very long, narrows abruptly in its middle levei, distai half very slender. Penis shield large (about half of mantle border width), almost cylindrical. Penis shield groove deep, notch in before distai third, evolved by apical gland. Apical gland only in ventral surface, with mosaic of irregular furrows and folds (Fig.205). Other details in LOPES (1956b, figs.30- 46), SCOTT (1957, fig.42), THIENGO et al (1993, figs.3-5), MARTÍN (1980, figs.l, 2, 5, 6), BERTHOLD (1989, fig.ôe). 9 : Adequate description in SCOTT (1957, figs.34-35); THIENGO et al (1993, fig.6).
Measurements of shell - MZSP 29633: 54.7 by 50.5 (Figs.l 1-12).
Distribution - Amazon and Paraná Basins.
Habitat - In freshwater curses of low energy, under aquatic plants.
Material examined - Types - ARGENTINA: Rio de La Plata, MNHN, 2 shells, syntypes of Ampullaria
insularum Orbigny. Ordinaiy material: BRAZIL - MATO GROSSO DO SUL: Passo do Lontra, MZSP 29633, 45 specimens (Simone and Lippareli cols., V/1993).
Pomacea lineata (Spix, 1827) (Figs.13-15, 64-65, 74-84,
95-96, 122, 125-126, 206-214)
Synonymy see CASTELLANOS & FERNÁNDEZ (1976). Complement:
Ampullaria fasciata-SWAINSON, 1822:21 (pl.103) [non ROISSY, 1805 nec LAMARCK, 1819); SPIX (1827:3, pl.5, fig.2) (see present figs.81-82).
Helix lineata Spix, 1827:3-4 (pl.4, fig.4; pl.5, fig.2) (see present figs.79-80).
Ampullaria fingulina Spix, 1827:2 (pl.4, fig.4) (see present fig.83-84).
Ampullariaplnysis Hupé, 1857:67 (pl. 12, fig.2) (see present fig.78).
Ampullarius lineatus - OLIVEIRA et al, 1981:77. Ampullaria testudinea Reeve, 1856 (pl.24, fig. 114)
(see present figs.74-75); SOWERBY, 1909:358; BAKER, 1913:660.
Ampullarius (Ampullarius) testudineus -
MORRETES, 1949:68. Pomacea lineata-THIENGO, 1987:564-568 (figs.l-
21); 1995:66-68 (figs.l, 4-7, 11-13, 16-17).
Description
Shell (Figs.13-15, 64-65, 74-84). Médium size (up to 45mm). Outline globose. Color of spire dark brown, almost black; last whorl greenish brown
with about 8 spiral, dark brown bands. Spire moderately tall (about half of body whorl length), slightly pointed; about 5 convex whorls. Last whorl almost spherical or weakly broader in superior region. Protoconch of 2.5 smooth whorls. Border between protoconch and teleoconch gradual and inconspicuous. Sculpture almost absent, only series of spiral lines uniformly distributed, after 5- 6 lines slightly taller line with hair. Aperture elliptical. Outer lip and siphonal region rounded. Umbilicus deep and narrow. Other details in THIENGO (1987, fig.l).
Head-foot and operculum (Figs.95-96). Features similar to those of P. crosseana. Head-foot color beige, with dark brown spots thorough exposed areas (except sole). Pedal gland furrow restrict to anterior sole edge.
Mantle organs (Figs.206, 210, 212). Similar features to those of P. crosseana, with following notable or differentiable characters. Osphradium on low stalk, relatively small, stalk slightly taller at right; osphradium base without edges covering filaments. Ctenidial and pulmonary veins separated from each other in anterior half (similar to those of P. canaliculata). Gill broader, shorter, and not so curved; its posterior end almost in middle region of pallial cavity posterior border. Gill filaments narrow, tall, tip slender and rounded, strongly curved towards right, tip blunt. Other details in THIENGO (1987, fig. 17).
Circulatory and excretory systems (Figs.208, 211)- Heart and kidney characters similar to those of P.
crosseana, remarks following. Heart proportionally smaller, ampulla about as large as ventricle. Anterior renal chamber broad and slightly short; internally 2 series of transverse septa separated by narrow longitudinal fold, all them restrict to dorsal surface. Nephrostome located in middle- posterior edge of this chamber. Posterior renal chamber broad and flat; dorsal renal lobe with circular outline, not attached to intestine, ventral surface covered by several anastomozed vessels. Connection between anterior and posterior renal chambers simple, protected at right by relatively broad vessel. Intestinal loops inside kidney chamber described below. Other details in THIENGO (1987, figs 6-8).
Digestive system (Figs.207-209, 211) - Buccal mass feature (Fig.207) similar to those described for P.
crosseana, with following distinctive or remarkable attributes. Jaw plates very large. Aperture of salivary glands located about in middle region of
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

400 L.R.L.SIMONE
dorsal folds, edged by several small folds. Dorsal folds with strong transverse furrows. Dorsal chamber between these both folds relatively deep, 2 to 4 longitudinal, somewhat tall folds running up to anterior esophagus, where faint. Odontophore muscles (Fig.207): m3 as transverse cover edging radular sac entrance in odontophore, covers also mlO insertion; mlO pair very narrow; mil pair inserted anterior, ml2 pair broad. Radular teeth attributes similar to those of P. crosseana, notable as follows (Figs.122, 125-126): rachidian broad and shorter, 5-7 cusps, central cusps 3-4 times broader than neighbor cusps; lateral tooth about as broad as rachidian, 3-4 cusps, second cusps terminal and several times larger than remainder cusps, tip bluntly pointed; inner marginal tooth with about half of rachidian width and about same lateral tooth length, basal concavity (where outer marginal encases) conspicuous and lying about 2/3 of tooth’s length, tip sharp pointed, sub-terminal cusp small and at some distance from tip; outer marginal tooth with about half of inner marginal width and about its same length, slender, weakly sigmoid, tip sharp pointed, secondary cusp at short distance from tip. Accessory pair of cartilages very thin. Branch of anterior aorta irrigating odontophore (running with mil) presents muscular fibers in its walls. Esophageal inner folds and pouches characters similar to those of P. crosseana, except for presence of 3-4 tall folds between both longitudinal folds of each pouch. Stomach features (Figs.208-209, 211) also similar to those of P. crosseana, including strong gastric-columellar muscle, remarks following. Ventral gastric chamber (with thin walls) small, about 4 ducts to digestive gland. Transverse muscular septum tall, with edge turned inwards. Intestinal origin preceded by small dorsal diverticulum. Digestive gland almost black. Intestine initially broad, its region adjacent to pericardium preceded by pair of sac-like, thick walled chambers. Ventral chamber simple, small; dorsal chamber larger, with other 3 inner sub- chambers (Figs.209, 211). Intestinal loops inside renal chamber shown in Fig.208. Anus with simple layer of papillae.
Genital system - Ó (Figs.210, 213): Visceral structures similar to those of P. crosseana, and described by Lopes (1956a) Thiengo (1987, figs.5, 9-10, 12-13). Testis pale beige. Seminal vesicle single, sac-like, small, located in posterior region of pallial vas deferens. Pallial vas deferens broad, wall thick glandular; anterior region narrower, connected to penis sac Crossing ventral to rectum.
Bulged short portion of vas deferens preceding anus present. Penis sac antero-posteriorly long; anterior half solid, posterior half thin walled, its left edge bears tubular projection united to penis shield right base; this tubular projections presents longitudinal dorsal aperture. Penis very long and coiled, base broad, gradually narrows up to very slender end long distai region. Penis duct broad and highly coiled at penis base, becomes less coiled and narrower between basal and middle penis thirds. Penis shield very large, occupying more than half of mantle edge (free portion) length, and about half of pallial cavity volume. Gill finishes on penis shield left base. Penis shield form almost conical, basal half broad, basal gland large, with normal fashion of aperture at dorsal surface. Distai half marked by abrupt reduction between it and basal half, curved towards right. Middle gland small, elliptical, located in right edge close to central furrow, in middle shield levei. Apical gland long, narrow, ventral surface transversally furrowed (more details in THIENGO, 1987). Notch between apical and middle glands, restrict to right edge. 9 (Fig.214): General organizations similar to that described for P. crosseana, and well described by Thiengo (1987: figs. 11, 15-16, 18-20). Remarks following. Ovary beige, relatively small, restricts to columellar surface of visceral whorls; ovary acina narrow, successively ramified, somewhat separated from each other. Pallial oviduct very large (more than half of pallial cavity volume when mature). Visceral oviduct very narrow, inserting in anterior end of seminal receptacle in middle levei of pallial oviduct ventral surface. Seminal receptacle large and flat, with about half of pallial oviduct width and length, located in its ventral region immerse in glandular tissue. Seminal receptacle walls iridescent muscular, outline elliptical. Narrow projection of seminal receptacle runs from its posterior end, curving abruptly towards anterior, running at left from it at about its same length; this projection wall also iridescent muscular, finishes as blind- sac. Remainder lumen of pallial oviduct bearing several loops and flattened and arched chambers obeying a shape as represented in figure 214; origin narrow, sub-terminal, in anterior end of seminal receptacle projection. Albumen gland as first loop, located along left surface of pallial oviduct. Capsule gland occupying remainder regions of pallial oviduct, without clear separation of albumen gland. Vaginal tube originating in middle region of pallial oviduct right surface, running towards anterior separated from it. Vaginal tube broad, inner lumen
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 401
also broad, its posterior region with transverse, tall, thick folds situated alternately. Its anterior region (about 2/3 of its length) with longitudinal, tall and thick folds; lateral-right fold larger, its walls curves positioned with its free edge touching its base, forming secondary inner tube. Genital pore broad and low papilla, located at short distance and slightly anterior to anus.
Measurements of shells (in mm) - MZSP 27996: #1, 35.8 by 31.2; #2, 36.6 by 30.7. MZSP 31147: d 1) 33.3 by 28.2; d 2) 34.7 by 29.6; 9 3) 32.1 by 28.5; 9 4) 32.0 by 27.1; 9 5) 32.5 by 27.5 (photo); MZSP 32197, 9 2) 44.8 by 37.5 (larger specimen).
Distribution - Central America, Amazon and Paraná Macrobasins. Coastal basins of Paraíba to Alagoas; Fernando de Noronha oceanic island.
Habitat - Similar to those of preceding species. On streams of low flow, close to margins and under aquatic plants.
Measurement of shells (in mm) - MZSP 26684: #1: 41.8 by 37.2; #2: 35.5 by 31.2.
Material examined - Types: without locality, BMNH, 1 shell, holotype of Ampullaria testudinea
Reeve. BRASIL; MNHN, 2 shells, syntypes (?) of A. lineata Spix of collection Ferussac (1837). PARAGUAY; NMHN, 2 shells, syntypes of Ampullaria physis Hupé. Remainder photos of Spix’s types are courtesy of Dr. Silvana Thiengo, Fundação Oswaldo Cruz (Figs.79-84). Ordinary material: BRAZIL - PERNAMBUCO: Fernando de Noronha Archipelago; Atalaia Beach, MZSP 31147, 23 specimens (Simone, Souza Jr. and Martins cols., 18/VII/1999). PARAÍBA: Cabedelo, MZSP 27108, ld, 29 (A.C.C. Almeida col., IV/ 1989). ALAGOAS: Ponta Verde, Sonho Verde Beach, MZSP 32197, 23 specimens (Simone col., 16/VII/ 1989). BAHIA: Ilhéus, Almada river, MZSP 26684, 2d , 29,6 shells (S.Thiengo col., 4/11/1984). SÃO PAULO: Guarapiranga Dam, near to OSEC, MZSP 27996, 11 specimens (P.Auricchio col., 21/V/1990).
Pomacea sórdida (Swainson, 1823) (Figs. 16-17, 72-73, 99-100, 123-124, 215-222)
Ampullaria sórdida Swainson, 1823 (pl.143) (no locality); PHILIPPI, 1851:38 (pl.10, fig.3); REEVE, 1856 (pl.3, fig.14); COUSIN, 1887:278; SOWERBY, 1909:357; TILLIER, 1980:24 (pl.2, fig.l).
Ampullaria intermedia Férussac, 1824 (pl.68, figs. 1- 2); 1827:135; ORBIGNY, 1835:31; 1837:371; HUPÉ, 1857:66.
Ampullaria autumnalis Reeve, 1856 (pl.4, fig. 16). Ampullaria melanicheila Reeve, 1856 (pl.5, fig.24)
(see present figs.72-73). Ampullarius (Ampullarius) sordidus - MORRETES,
1949:67; OLIVEIRA etal, 1981:76. Pomacea sórdida -LOPES, 1955:208-210 (figs. 17-
23); QUINTANA, 1982:111; THIENGO (1989:351-355, figs.1-9).
Description
Shell (Figs. 16-17, 72-73). Adequate description in Thiengo (1989, 352, fig. 1), characteristic by flatter fashion and an ampler umbilicus.
Head-foot and operculum (Figs.99-100). Similar features as those of P. crosseana.
Mantle organs (Figs.215-216, 218). Organization similar to that described for P. crosseana, with following remarks. Osphradium proportionally smaller, its basal, stalk-like portion low, lacking marginal fold covering base of filaments. Lung ampler, occupying about half of pallial roof area. Ctenidial vein separated into 2 in its posterior 1/3,
bearing pulmonary vein (although this vein apparently also receives gill blood flow). Gill weakly curved. Gill filaments tall; rod narrow, extending beyond filament; tip rounded.
Circulatory and excretory systems (Fig. 217) - Both with similar fashion than that described for P.
crosseana, with following remarkable features. Heart large, about half of kidney volume. Auricle with secondary connection to anterior-inner surface of pericardium, at right from ctenidial vein insertion. Aortas and ampulla connected to adjacent dorsal surface of pericardium. Anterior renal chamber broad and slightly short; internally 2 series of transverse septa separated by narrow longitudinal fold, all them main connected to dorsal surface and also reaching ventral surface. Nephrostome located in middle-posterior edge of this chamber. Connection between anterior and posterior renal chambers simple, protected at right by relatively broad vessel. Posterior renal chamber with tall renal dorsal lobe, this lobe covered by conspicuous, branched, anastomozed vessels. Intestine free from renal lobe.
Digestive system - Foregut characters similar to those of P. crosseana, remarks following. Dorsal chamber (between both longitudinal dorsal folds of buccal mass) with 4 longitudinal secondary folds that faint along anterior esophagus. Odontophore muscles (Figs.219, 221): mlb, generally composed by 3 narrow pairs; mld, pair parallel to mlb, but insertion more
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

402 L.R.L.SIMONE
posteriorly, in lateral-posterior region of buccal mass dorsal surface; m3 transverse, slightly thick, covering radular sac; m6 short and broad. Pair of pouches with dorsal inner fold having highly irregular surface, and lacking any longitudinal fold. Radular sac short, extending little beyond odontophore. Radular teeth of similar attributes as those of P. crosseana, notable features following (Figs.123-124): rachidian tooth with rectangular outline, 7 cusps, central cusp with about twice neighbor cusps size; lateral tooth with about 1.5 times rachidian width, 3 cusps, middle cusps terminal and several times larger than remainder cusps. Mid and hindgut features (Figs.215, 220) also similar to those of P. crosseana, distinctions following. Posterior region of esophagus smooth internally. Stomach ventral chamber very small (about 7 of dorsal chamber). Transversal septum low and restrict to ventral surface. Gastric walls slightly thinner muscular. Gastric muscle narrower. Digestive gland veiy dark, almost black. Intestine origin with smooth inner surface. Intestinal chambers in its region Crossing pericardium double (Fig.220), proximal chamber transverse and ventral, its blind- sac, narrow distai end located inside aperture of distai chamber; distai chamber longitudinal and dorsal. Remainder intestine inner surface with several transverse, low folds. Intestinal loops inside renal chamber particularly complex, obeying pattern showed in situ in figure 215.
Genital system - Ó (Figs.215, 222): Visceral and pallial structures organized similarly to those of P.
crosseana, differing by following features. Testis pale beige lying along superior and columellar surfaces of visceral whorls, fmishing half whorl posterior to pallial cavity. Pallial vas deferens thick walled (prostate) and somewhat conical; in its anterior end abruptly narrows and connects with penis sac Crossing dorsal to rectum. Penis sac almost circular, its projection connected to penis shield almost as broad as penis sac. Penis sac anterior 1/3 solid, posterior 2/3 thin walled. Penis sac projection opened longitudinally along its dorsal surface; this aperture protected posteriorly by low fold originated from penis base. Penis of moderate length and coil degree, base broader, narrows gradually up to very slender apical region. Penis duct also with moderate degree of coiling. Penis shield broad; tip blunt, thick, curved towards right. Basal penis shield gland with aperture located in its dorsal-left side and presents normally 3 closely located slits. Secondary small gland located ventrally in right edge of penis shield base. Notch of penis shield furrow located in its middle levei. Apical gland very large, composed by 5-7 successive broad
folds lying along left edge of shield. Groove, coming from penis shield longitudinal furrow, surrounds basal edge of this apical gland, runs along dorsal surface of this gland and finishes broadly opened in middle region penis shield apex. More details in LOPES (1955, figs. 19-23) and THIENGO (1989, figs.4-7). 9 : See description of Thiengo (1989, figs.8-9). Ovary, like digestive gland, almost black. Vaginal tube somewhat narrow, inner surface with some longitudinal, irregular, tall folds. Female pore a sub-terminal, ventral, transverse slit, located at right from anus.
Central nervous system - Features similar to those described for preceding ampullariids, except for lateral position of buccal ganglia.
Measurements of shells (in mm) - MZSP 32195: 9 1) 44.8 by 38.6; C? 2) 32.8 by 31.9.
Distribution - Bolivia, Brazil (RJ, SP, PR, SC), Paraguay, La Plata River (Argentina).
Habitat - Among aquatic plants of low-flow streams.
Material examined - Type: without locality, BMNH, 1 shell, holotype of Ampullaria melanocheila Reeve. Ordinary material: BRAZIL - RIO DE JANEIRO: Silva Jardim, MZSP 32195, lcf, 29 (S.Thiengo col. and leg, X/1999).
Pomacea bridgesi (Reeve, 1856) (Figs. 18-21, 67-71, 127-128)
The study on this species has been published in a separated paper: THIENGO & SIMONE, in press. However, the data on P. bridgesi have been included in the present analysis. Some shells are shown in the present paper (Figs.18-21, 67-71) including some type specimens, and radular SEM pictures (Figs. 127-128).
Genus Asolene Orbigny, 1837 Type-species - Helixplatae Maton, 1809
Asolene megastoma (Sowerby, 1825) (Figs.23-24, 90, 129, 223-229)
Synonymy see CASTELLANOS & FERNÁNDEZ (1976:18). Complement:
Ampullaria neritoides Orbigny, 1835:31; 1843:368- 369 (pl.49, figs. 1-2) (see present fig.90).
Asolene (Pomella) megastoma - SCOTT, 1957 (pl.2; pl.4, fig.6; pl.7, figs. 17-18; pls.8-9; pl.ll, fig.29; pl.13, fig.33; pl. 14, fig.36-37; pl.15; pl.23, fig.9); CASTELLANOS & FERNÁNDEZ, 1976:18 (pl. 1, fig.2; pl.4, fig.5); OLIVEIRA etal, 1981:76.
Pomella megastoma - FIGUEIRAS, 1964:173.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 403
Description Shell (Figs.23-24, 90): very large (about lOOmm). Spire short, almost plane. Aperture very ample. Umbilicus lacking. Other details in Scott (1957:314-315, pl.23, fig.9), CASTELLANOS & FERNÁNDEZ (1976:18).
Head-foot and operculum (Fig.223). Characters similar to those described for P. crosseana.
Distinctive features following. Form very broad, head relatively small. Snout somewhat an inverted cone, base narrow. Lobes (siphons) relatively short. Tentacles broader in base.
Pallial organs (Figs.224-225). Similar features to those of P. crosseana. Except for osphradium peduncled; osphradium base short and smaller (about half of osphradium area). Gill filaments narrower. Other details in SCOTT (1957, fig.20).
Circulatory and excretory systems - Heart attributes similar to those of P. crosseana, but with auricle connected to anterior wall of pericardium. Kidney closely similar to that described for P.
canaliculata. Dorsal renal lobe thick, brown, surface with irregular folds, most converging to aperture of anterior lobe; no connection with intestine. Anterior lobe long, broad, with 2 series of thick septa, pale beige, separated by longitudinal, tall fold. This longitudinal fold ventral edge free, connects only with nephrostome anterior edge. Intestinal loops inside kidney described below.
Digestive system - Buccal mass and odontophore muscles characters (Fig.226) similar to those of P.
crosseana, but with most structures thicker and proportionally larger, including jaw plates. Some distinctive and notable features follow. Odontophore muscles: mia thick; ma pair multiple; m3a pair, transverse portion anterior to radular sac covers m9; m7 pair origin located in postero- median margin of odontophore cartilages; mlO pair insertion part in m4 and part in m5, on their anterior surface (other details of odontophore in Scott, 1957, pis.7-9). Dorsal folds with very irregular-folded surface. Radular tooth similar to those of P. crosseana, with following remarks (Fig.129): rachidian broad and shorter, smooth cut- edge lacking cusps except broad, blunt central projection, basal cusps long and obliqúe; marginal tooth about as broad as rachidian, slightly sigmoid, broadly pointed tip lacking secondary cusps; inner marginal tooth 2-3 times broader than inner marginal tooth, both with very small, sub-terminal cusps in inner edge. Pair of pouches longer, inner surface with pair of opposite located longitudinal
folds, being dorsal folds broader. Esophagus characters (Fig.226) also similar to those of P.
crosseana, except for thicker muscular walls. Stomach features (Figs.227-228) also similar to those of that species, including columellar-gastric muscle and transverse muscular septum. Differs by smaller ventral chamber (with thin walls), bearing 4-5 ducts to digestive gland close with each other; and by transverse muscular septum presenting taller edge turned inwards. Intestine initially broad, inner surface smooth. Intestinal portion adjacent to pericardium preceded by 2 chambers with aperture turned towards stomach; dorsal chamber as small, simple blind sac; ventral chamber larger and more complex, located just proximal to dorsal chamber, bears 3 distai flat ramifications (Fig.227). Digestive gland dark brown, with about 3 whorls posterior to stomach. Intestinal loops inside of kidney chamber as shown in situ in figure 227, complex, wholly connected by mesentery. Rectum narrow. Anus small, with simple borders.
Genital system - d" (Fig.224): Visceral and pallial structures with characters similar to those of P.
crosseana. Distinctive or notable features following. Gonad pale cream, in columellar side of first visceral whorl. No broad region of pallial vas deferens by side of anus. Penis very slender and convolute, basal half folded by successive transverse furrows. Penis distai half narrowing gradually. Penis tip very slender. Penis sac anterior half very thick walled, posterior half very thin walled. In this posterior half dorsal fold runs from penis base to aperture of this chamber. This aperture broad, edged ventrally by thick fold with coiled distai border. Penis shield conic, very large (occupies most of pallial cavity and about half of mantle border length). Basal penis shield gland and aperture similar to those of P. crosseana, no other detectable basal glands. Penis shield ventral groove narrow, deep, running approximately along central region; secondary, smaller, obliqúe groove present at left. Penis distai region with irregular folded, massive, elliptical gland restrict to ventral side; main and secondary penis shield grooves converge in proximal margin of this distai gland and edge it in its left-proximal border. Other details in SCOTT (1957, pl. 15). 9 (Fig.229): Visceral and pallial organs similar in characters to those of P.
crosseana. Pallial oviduct very massive, greatly bulging inside pallial cavity. Visceral oviduct connects with bursa copulatrix in middle levei of left pallial oviduct side. Bursa copulatrix, just after visceral oviduct insertion, elliptical and circular in
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

404 L.R.L.SIMONE
section; bears flat, posterior, somewhat same lengthened expansion towards posterior, at some distance from visceral oviduct insertion. Capsule gland lumen flat, sinuous, complex; coils as shown in figure 229. Insertion of bursa narrow, located approximately between middle and posterior thirds of capsule gland region. Capsule gland connection with vaginal tube located approximately between middle and anterior thirds of capsule gland, towards ventral. Vaginal tube broad, walls thick, inner surface smooth, length about half of that capsule gland. Female pore small, papilla-like, located close to and at right from anus. Other details in SCOTT (1957, pl.14, figs.36-37).
Measurements of shell (Figs.23-24) - 83.0 by 87.0.
Distribution - Uruguay River (RS, Brazil; Argentina, Uruguay).
Habitat - In rivers with some degree of flow energy.
Material examined - Types - URUGUAY: Uruguay River, MNHN, 1 shell, probable holotype of Ampullaria neritoides Orbigny. Ordinary material: BRAZIL: Uruguay River; RIO GRANDE DO SUL: Itaqui, MZSP 250, 2Ó , 89 (E.Garbe col., 1915).
Genus Felipponea Dali, 1919 Type-species:
Ampullaria (Felipponea) neritiniformis Dali Felipponea neritiniformis (Dali, 1919)
(Figs. 130, 230-231)
Synonymy see CASTELLANOS & FERNÁNDEZ (1976:19). Complement:
Felipponea neritiniformis - SCOTT, 1957:317-318 (pl.6, fig. 14; pl.23, fig. 10); FIGUEIRAS, 1964:173; CASTELLANOS & FERNÁNDEZ, 1976:19 (pl.2, fig 5; pl.5, fig.15).
Asolene neritiniformis - OLIVEIRA et al, 1981:76.
N.B.: VAZ & MARTINS (in press) have provided a whole description of this species in a submitted paper. The present description is greatly reduced, restrict to some aspects only. Complementary data must be reported to that paper.
Shell. Relatively small (about 30mm). Color pale greenish beige to brown, frequently with spiral dark bands. Spire tall, eroded. Umbilicus very narrow. Other details in DALL (1919:10-11), SCOTT (1957:317-318), CASTELLANOS & FERNÁNDEZ (1976:19).
Head-foot. Similar to A. megastoma, but not so broad. Head narrower. Siphons (nuchal lobes) formed by low folds (not so tall as those of Pomacea spp).
Pallial cavity (Fig.231). Osphradium elliptical, almost sessile (stalk very short and broad).
Circulatory and excretory systems - Heart with anterior aorta ampulla similar to those of preceding species.
Digestive system - Buccal mass and odontophore muscles characters very similar to those of A.
megastoma, including m3a as fold covering radular sac, mlO pair long and narrow; differs by ma as single pair and by narrower ml pairs. Jaw plates, dorsal folds, esophageal pouches and folds similar in features as those of A. megastoma. Radular teeth features similar to those of P. crosseana, with following remarks (Fig. 130): rachidian longer, outline almost square, 5-7 small cusps, central cusp about 1.5 times larger than neighbor cusps, basal cusp weak, somewhat longitudinal; lateral tooth about as broad as rachidian, 3-4 small cusps, first cusp terminal, second cusp about 3-times larger than neighbor cusps; inner marginal tooth with about double of outer marginal tooth width, both with tip sharp pointed and sub-terminal cusps in inner edge somewhat long (other details in SCOTT, 1957, pl.23, fig. 10). Stomach characters similar to those of A. megastoma, except for thinner muscular gastric walls and by transverse muscular septum lacking long edge turned inwards. Intestinal region adjacent to pericardium preceded by pair of small, dorsal, sac-like chambers with aperture towards posterior. Intestinal loops inside renal chamber shown in figure 230.
Genital system - d : Penis shield somewhat similar to that of P. lineata, but with shorter tip lacking glands. Pair of glands in base of penis shield, being right gland smaller and more superficial. Penis shield furrow relatively deep, running almost all along its length. 9 : Pallial oviduct proportionally large, with features similar to those of A.
megastoma. Vaginal tube broad, walls glandular, origin about in half levei on capsule gland.
Material examined - BRAZIL - PARANÁ: Ivaí River, Floresta, MZSP 29713, 5 specimens, MZSP 30759, 5 specimens (A.L.Falavigna col.).
Genus Marisa Gray, 1824 Type-species: Helix comuarietis Linné, 1758
Marisa planogyra Pilsbry, 1933 (Figs.22, 25-28, 101-102, 131-132, 232-241)
Synonymy see SCOTT (1957:321). Complement: Marisa planogyra - SCOTT, 1957:321-322 (pl.23,
fig.8); CASTELLANOS & FERNÁNDEZ, 1976:21
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 405
(pl.2, fig.6; pl.5, fig.16-17); MARTÍN, 1980:47- 50 (fig.3). OLIVEIRA et al, 1981:78; MELLO, 1988:7-13 (figs.l, 3a-e); MELLO & MARINI, 1995:70-71 (fig.l).
Description
Shell (Figs.22, 25-28). Practically planispiral, with apex slightly dislocated to left. Aperture relatively small, elliptical. Other details in SCOTT (1957:321- 322), CASTELLANOS & FERNÁNDEZ (1976:21). Color usually pale beige, with dark brown spiral bands, however, some uniform colored specimens occur (Figs.22, 25-27).
Head-foot (Figs.234, 237). Characters somewhat similar to those of P. crosseana, distinctive features following. Form relatively narrower laterally and longer antero-posteriorly. Anterior furrow of pedal gland restricted to anterior foot edge. Lobes (siphons) also long; right siphon with longer longitudinal fold running along floor of pallial cavity. Columellar muscle very longer, thinner and flattened, about 1 whorl length.
Operculum (Figs.101-102, 234). Also of similar characters than that of P. crosseana, except for more rounded outline.
Pallial cavity (Figs.232-233, 235). Narrow and deep (about 3á whorl). Organs arrangement somewhat similar to that of P. crosseana, with following distinctive or notable features. Osphradium small, elliptical, on short stalk. Lung narrow and very long, almost as long as pallial cavity. Inner lung surface with transverse vessels like those of P.
crosseana. Pneumostome an obliqúe, muscular edged slit located close to anterior-left region of lung ventral surface. Gill narrow and long, edges entire lung right margin. Gill filaments short, relatively broad, right margin slightly straight, left margin convex, apex rounded. Between gill and rectum a relatively narrow space, but broader as those of preceding species.
Circulatory and excretory systems (Fig.236) - Heart relatively small and narrow. Auricle just posterior to pallial cavity posterior end. Ventricle posterior to auricle. Anterior aorta ampulla broad and long. Kidney features somewhat similar to those of P.
crosseana, with following distinctive or interesting attributes. Renal dorsal lobe slightly thin, dark brown, surface with some irregular-sized vessels; one of this vessels broader, connected to right edge of aperture to anterior renal chamber. Renal dorsal lobe free from intestine. Anterior renal chamber conic, narrow and short (about half of remainder kidney length). Anterior renal chamber with 2
same-sized dorsal series of transverse septa, separated by a tall, dorsal, longitudinal fold; this longitudinal fold not connected to nephrostome edges. Urinary gutter also similar of those of preceding species but longer; at some distance from mantle border abruptly twist and run in floor of pallial cavity (Figs.232-233).
Visceral mass (Figs.233, 236) - Length about same than that of pallial cavity, about 2.5 planispiral whorls. Stomach and adjacent digestive tubes occupy last half whorl. Gonad located in first half whorl, extending little beyond this by outer surface. Remainder visceral space occupied by dark brown digestive gland.
Digestive system - Buccal mass, odontophore muscles and esophageal characters similar to those of P. crosseana, distinctive or remarkable features following (Figs.237-238). Jaw plates thinner. Odontophore muscles (Fig.238): ma in 2 narrow pairs; m4 pair not attached to inner surface of odontophore cartilages (only covering them) and constituted by 2 thick layers (fused with each other laterally), also connects with tissue preceding exposed part of radula (to) via m9; m5 pair thicker and shorter; mlO pair narrower; ml2 pair broader. Accessory pair of cartilages softer and thinner. Radular teeth attributes similar to those of P.
crosseana, with following remarkable notes (Figs.131-132): rachidian shorter, 7-9 tall and broad cusps, central cusp larger (about double than neighbor), remainder cusps decreasing towards lateral, pair of basal cusps narrow, low and obliqúe; marginal tooth strongly curved inwards and distally, 3-4 large and tall cusps, second cusp terminal and about double sized than neighbor cusps, conspicuous longitudinal, middle reinforcement in basal half; inner and outer marginal teeth tall slender, terminally curved, similar with each other (inner slightly broader than outer marginal tooth), both several times narrower than marginal tooth, tip pointed, sub-terminal cusp with about half of tip size. Esophagus uniformly narrow (without bulged posterior portion). Esophagus inner surface with about 6 longitudinal, slightly tall folds all along its length. Stomach with similar attributes as those of P. crosseana,
including gastric-columellar muscle, distinctive features following (Figs.239-240): gastric muscular wall very thinner; transverse muscular septum absent; low longitudinal fold separates gastric chamber into esophageal and intestinal portions, inserts in posterior region of gastric-columellar muscle. Single duct to digestive gland bifurcating
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

406 L.R.L.SIMONE
at short distance from its origin, running towards opposite side. Intestine initially broad, inner surface smooth except for pair of shallow, dorsal diverticles in its limit with stomach. Intestinal portion adjacent to pericardium preceded by pair of small, sac-like chambers with apertures towards posterior; each chamber aperture edged by thick border, left chamber encased inside aperture of right chamber (Fig.239). Remainder intestine slightly narrow, running somewhat similar to that of P. crosseana. Intestinal loops inside renal chamber showed in situ in Fig.240. Rectum narrow and very long. Anus simple, small.
Genital system - Ó (Figs.232-233): Testis cream in color, located in posterior end of visceral mass first whorl. Visceral vas deferens very narrow, lying in columellar surface of visceral whorls. Pallial vas deferens also narrow and very long, lies at right from rectum. Genital organs close to mantle border, with organization somewhat similar to those preceding species. Pallial vas deferens crosses dorsal to pre- anal region of rectum without bulged portion. Penis sac most with thin walls, very short portion with thicker walls in penis base. Penis relatively short, width uniform along its length, narro ws at short distance from pointed apex. Penis sac amply opened in left region, in penis shield base. Penis shield conic, large (about half of mantle border width). Basal gland situation and aperture similar to those of preceding species. Pair of small glands in each side of basal half of penis shield, being right gland smaller and more superficial. Penis shield groove relatively shallow, runs about in central region of shield ventral surface. Penis shield groove with secondary fold in its basal-right region coming from penis sac aperture. Penis shield groove runs almost entire shield length, finishes at some distance from shield tip. Penis shield distai half narrowing gradually, conspicuous notch at right located approximately in middle shield levei. Penis shield slightly pointed, lacking glands. Other details in Mello (1988: 8-12, fig.l), Martin (1980, fig.3). 9 (Fig.241): Pallial oviduct somewhat similar to those of preceding species, but smaller and narrower. Visceral oviduct inserts sub- terminally in bursa copulatrix, inside pallial oviduct mass. Bursa copulatrix conic, long, inserts in capsule gland lumen between its middle and posterior third parts. Bursa immersed in capsule gland. Capsule gland lumen flat and as blind-sac in posterior third, remainder intensely coiled (Fig.241). Vaginal tube narrow and very long (about twice capsule gland length), runs at right from urinary gutter and rectum. Female genital pore
small, at right and close to anus. Other details in MELLO (1988:12, figs.la-e).
Measurements of shells (in mm) - MZSP 29673: CS 1, 11.7 by 32.0; cf, 12.0 by 30.0; 9 2, 11.0 by 28.8.
Distribution - Paraná Macrobasin, central South America.
Habitat - Between Eichomia and other aquatic plants.
Material examined - BRAZIL - MATO GROSSO: Poconé, MZSP 29674, 15 specimens (L.S.Rocha col., 28/111/1997). MATO GROSSO DO SUL: Passo do Lontra, MZSP 29673, 2cf, 29,1 shell (Simone and Lipparelli cols., V/1993).
Discussion - M. planogyra is very close related to M. cornuarietis (Linné, 1758), from which is very difficult separated conchologically. The anatomical differences between both species are neither well explored, except for study on their genital system (MELLO, 1988). That paper convincingly provides data for specific separation in the penis shield glands (M. planogyra has 2 glands, while M.
cornuarietis 4 glands) and length (M. cornuarietis
longer). Based on that paper, the specimens studied herein are all M. planogyra. Sexual dimorphism has been found in shell and foot color in M. cornuarietis
specimens (Demian & Ibrahim, 1972).
Superfamily Cyclophoroidea Family Cyclophoridae (=Potieridae, Amphicyclotidae)
Genus Neocyclotus Fischer & Crosse, 1886 Type-species: Cyclostoma dysoni L.Pfeiffer
Neocyclotus prominulus (Orbigny, 1840) (Figs.29-31, 91-94, 109-111, 133-134, 242-258)
Synonymy see KOBELT (1902:238). Complement: Cyclostoma prominula - ORBIGNY, 1846:362-363
(see present figs.91-94). Neocyclotus prominulus - KOBELT, 1902:238;
MORRETES, 1949:65. Incerticyclus prominulus - TORRE et al, 1942:277
(pl.39, fig.7-9). Aperostoma (?) prominula - MORRETES, 1953:47.
Description
Shell (Figs.29-31, 91-94). Size small (up to 15mm), discoid and flattened. Spire small, low. Suture deep, each whorl circular in section. Sculpture successive, uniform axial threads. Other details in KOBELT (1902:238), TORRE et al. (1942:277).
Head-foot (Figs.242, 245). Head little projected (almost inlaid). Snout stubby, most sessile, low. Mouth longitudinal, in anterior-dorsal snout surface. Pentacles shorter than snout, base at side
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 407
and posterior to snout base. Ommatophore short, located about in middle levei of outer tentacle surface. Eyes small, dark. Foot thick, relatively small. Furrow of pedal gland edging entire foot sole, deeper anteriorly. Columellar muscle very thick and short (about V3 whorl). Columellar muscle distai end somewhat bifid, with right branch flatter and taller. Males with exophalic penis located just posterior to right tentacle (described below).
Operculum (Figs. 109-111, 242). Outline circular, calcareous, thick, edge double (Fig. 111). Nucleus central, smooth; multispiral shallow furrow originated from nucleus. Outer surface opaque. Inner surface glossy, low projection central, from which scar turns around in growth (seen by previous scars). Inner scar rounded, with about V*
of opercular area.
Mantle organs (Figs.243-244, 246-247). Pallial cavity of about half whorl, relatively narrow. Mantle border thick, simple. Osphradium and gill lacking. Broad vessel (probably homologous to ctenidial vein) edges left margin of pallial cavity, anterior region with 2 branches (being collar vessel one of them), becomes broader towards posterior, inserts in left extremity of pericardium. Pericardium and kidney as left-posterior limits of pallial cavity. Between left vessel-kidney and rectum a broad smooth area without large transverse vessels (between kidney and rectum a narrower area). Rectum very broad (about Va of pallial cavity width), edging pallial cavity right border, abruptly narrows, Anus small, shortly siphoned, close to right end of mantle border. Pallial oviduct and prostate located between rectum and right border of pallial cavity, displacing it towards left (both described below).
Visceral mass (Figs.246-247). About 2.5 whorls in length. Kidney and pericardium as left-anterior limit; pallial gonoducts and rectum as right- anterior limit. Gonad located in first whorl outer surface, covering small portion of digestive gland. Stomach large and long, with about half whorl in length, half whorl posterior to pallial cavity. Stomach separates almost completely digestive gland into posterior and anterior branches, being connected with each other by thin and narrow ventral portion. Last half whorl of visceral mass filled by digestive gland (posteriorly) and intestinal loops (anteriorly). Other details below.
Circulatory and excretory systems (Figs.244, 247- 248) - Heart relatively small, located behind to left- posterior corner of pallial cavity. Narrow vessel edges anterior surface of kidney, entrance into
pericardium and inserts in auricle. Auricle and ventricle small, connected with each other by narrow and short tube, Auricle at right and slightly anterior to ventricle. Aortas bifurcate shortly after ventricle origin of common aorta, anterior aorta with about double width than posterior aorta. Broad vessel of left margin of pallial cavity runs dorsal-left to pericardium; just before this region smaller branch which edges pericardium inserts. After pericardium contoured, this vessel connects with other vessel coming from visceral mass and runs towards kidney. Kidney with 2 chambers. Anterior renal chamber totally filled by glandular tissue, somewhat triangular in section, connects with pericardium by small and narrow slit. This glandular tissue almost solid, formed by several branched, transverse, glandular septa. Posterior renal chamber simple hollow sac mostly filled by intestinal loops (described below). Nephrostome a small slit located in ventral surface of anterior renal chamber, close to posterior limit of pallial cavity. Urinary gutter starts just ventral to nephrostome, runs as shallow groove in pallial floor towards left and by left side of head-foot.
Digestive system - Buccal mass (Figs.249-251) relatively large (about 1/3 of haemocoel volume) (Fig.245). Oral tube short, walls thick, circular muscle (mouth sphincter - mc) broad. Buccal mass dorsal wall (Fig.251) thin, anterior half covered by jaws. Jaw plates very large, slightly triangular, connected with each other in median line. Surface constituted by several obliqúe striae. Jaws anterior cut-edge thin, sharp, arched. Dorsal folds tall, narrow in region just posterior to jaws, gradually become broader and newly narrow after buccal mass end. Odontophore muscles (Figs.249-250, 252-253): ml, several small and narrow muscle fibers connecting buccal mass with adjacent inner surface of haemocoel; mia, pair of narrow dorsal jugal muscles, origin in dorsal inner surface of haemocoel, in approximate middle levei of odontophore, runs towards ventral and anterior, insertion in anterior region of buccal mass dorsal surface; mlb, pair of narrow and thin jugal muscles, origin close to mia origin, runs towards ventral and posterior, insertion in each side of odontophore posterior region; mj, jaws and peribuccal muscles, relatively thick, origin in odontophore lateral-ventral surface, insertion in dorsal wall of buccal mass around jaws; ma, pair of thick and short muscles, origin in lateral-anterior inner surface of haemocoel, runs towards medial through mj fibers, insertion along outer surface of
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

408 L.R.L.SIMONE
jaws; m3d, pair of thin and narrow muscles, origin in posterior-dorsal-lateral region of buccal mass, run in surface towards ventral and anterior, insertion along ventral surface of odontophore close to median line; m4, broad pair of radular tensors, possessing some branches: m4 main body surrounding inner, ventral and lateral surfaces of cartilages, connected to cartilages only in ventral and lateral surfaces and in some sparse branches to inner surface; m4v, ventral branches, pair covering outer-lateral surface of cartilages, origin in m4 main branches posterior region, runs towards anterior, insertion along subradular membrane inner surface from median line to lateral; m4a, thinner pair covering ventrally m4 main branch, origin along their lateral surface, insertion with tissue on radular ribbon (to) in region preceding its exposure; m5, pair of dorsal tensor muscles of radula, relatively thin, origin broad on m4a, run towards medial and anterior covering m4, insertion relatively narrow in dorsal surface of radular sac preceding exposed region of radula; m6, horizontal muscle, connects dorsal surface of both cartilages along and at some distance from their medial edge; m7, pair very thin and narrow, origin in posterior end of cartilages, each runs towards medial connecting with its pair, penetrate and insert in inner surface of radular sac; mlO, pair of protractor muscles of buccal mass, origin in ventral-inner surface of mouth, runs towards posterior, insertion in ventral-anterior surface of odontophore, close to median line; mlOc, similar to mlO but running dorsal to it, insertion more anterior and dorsal to those of mlO; mil pair of narrow, thin and long muscles, origin in ventral surface of haemocoel close to median line at middle levei of odontophore, run dorsal, penetrate in odontophore in medial line, after run attached to subradular membrane, insertion along median line. Other non-muscular buccal mass structures: oc odontophore cartilages, single, large, flat pair, outline oval; ih, inner ligaments between radular sac and inner surface of m6; br, subradular membrane similar to those of ampullariids. Pair of buccal ganglia located laterally in posterior-ventral region of buccal mass (Fig.249), their comissure relatively broad. Pair of nerves penetrates in odontophore just anterior to mil penetration. Radular sac long (about 3 times longer than buccal mass length), not coiled, only twisted posteriorly. Radular nucleus thick. Radular teeth (Figs.133- 134): rachidian long and narrow, 3 stubby cusps, central cusp about double than neighbor cusps;
lateral tooth with about double of rachidian width, situated obliquely (lateral end in levei of rachidian distai roll), 3 cusps, middle cusp terminal, 3-4 times larger than neighbor cusps, base long and narrow; inner marginal tooth tall, slender, somewhat flat, 3 terminal and broad cusps, central cusp about 1.5 times larger than neighbor cusps; outer marginal tooth sigmoid, about same length than inner marginal tooth, base broad and flat, with broad projection, middle region slender and curved, distai region slightly broader, pointed, secondary cusps small, sub-terminal, on inner edge. Salivary glands somewhat triangular, separated from each other, located posterior to esophageal pouches. Salivary gland ducts long and convolute, runs on dorsal surface of anterior esophagus and buccal mass, penetrate in dorsal wall of buccal mass close to their apertures.
Anterior esophagus with pair of tall dorsal folds (continuation from those of buccal mass), which gradually faint in middle esophagus (Figs.249, 251). Pair of large, lateral pouches located just posterior to buccal mass, both connect with esophagus by narrow aperture, after aperture abruptly expand and become reniform sac. Pouches inner surface covered by several papillae. Dorsal esophageal folds cover each pouch aperture. Especial branches of anterior aorta irrigate each esophageal pouch, connected with them close to their connection with esophagus. Middle esophagus broad, inner surface with sparse, obliqúe, low folds. Posterior esophagus narrow, inner surface smooth.
Stomach (Figs.247, 254) long and broad. Esophagus inserts in stomach in middle levei of its ventral surface, but retains separation from gastric chamber by longitudinal, tall fold. Stomach mostly a simple sac, without inner especial structures, inner surface most smooth. Duct to digestive gland single, located in ventral-right region of anterior third part of stomach. Transverse groove present in anterior third of stomach, begins in aperture of duct to digestive gland, runs in ventral surface up to opposite side where faint, just in beginning of longitudinal, low fold of intestine. Digestive gland beige, with small brown spots (other details above). Intestine broad, with several loops inside posterior renal chamber as shown in Fig.247. Fecal pellets elliptical, large, fill intestine after its first loop in renal chamber. Rectum and anus described above.
Genital system - Ó : Testis (Fig.246) (described above) cream in color. Visceral vas deferens very
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 409
narrow, runs along ventral surface of stomach up
to median-ventral-posterior edge of pallial cavity.
Prostate large, located in right side of pallial cavity,
part attached to pallial floor; visceral vas deferens
inserts in middle region of prostate left side. Prostate elliptical in section, inner lumen flat
(Fig.246). Prostate aperture a small, anterior
papilla, opens towards pallial cavity floor where
groove begins. Pallial sperm groove runs short
distance up to penis base. Penis relatively small
(little longer than tentacle), located just posterior
to right tentacle (Fig.242). Penis base broad,
gradually narrows, in its middle region newly
expands and curves towards right (Figs.242, 255).
Penis tip broad, asymmetrical. Penis sperm groove
runs all along penis length, approximately in
middle of its ventral surface. Penis sperm groove
distai end on small papilla. 9 (Figs.247, 256-258):
Ovary small, occupies only columellar side of
visceral first whorl (Fig.247). Visceral oviduct very
slender, thin and long, runs along ventral surface
of stomach up to middle region of pallial cavity
posterior edge. In this region abruptly curves
towards posterior and runs obliquely to right,
augmenting very gradually. After distance
equivalent to about % whorl suddenly curves
towards posterior, running parallel to its preceding
portion. Close and anterior to its first curve inserts in seminal receptacle base. Seminal receptacle
very long, increase gradually, rounded distai tip.
Seminal receptacle aperture small slit in middle
region of pallial oviduct vaginal groove. Capsule
gland as remainder, massive region of pallial
oviduct; possesses 2 flat, thick-glandular walled
chambers. Right chamber a blind-sac connected
to left chamber posteriorly. Left chamber with
pair of tall, obliqúe folds preceding its connection
with right chamber, and several other low, narrow
folds along posterior half of its left wall. Left
chamber opens anteriorly in female pore as simple,
thick-walled slit.
Central nervous system (Figs.245, 249) - Nerve ring
located anterior to buccal mass, just posterior to
mouth. Hypoathroid type, with long comissure between both cerebral ganglia. Statocyst with
several statoliths.
Measurements of shells (in mm) - MZSP 24542:
9 1, 8.6 by 13.6; 9 2, 7.7 by 12.8; d 4, 7.4 by 12.7;
MZSP 24538, 9 : 10.4 by 15.7.
Distribution - South Rio de Janeiro and São Paulo
Atlantic Rain Forest.
Habitat - Terrestrial, under plants.
Material examined - Types - BRAZIL; MNHN, 2
shells, probable syntypes of Cyclostomaprominula
Fer. in Orbigny. Ordinary material: BRAZIL - SÃO
PAULO: São Vicente (O.Schubart col.), MZSP
24547, l9 (27/1/1960), MZSP 24545, 1 specimen (27/1/1960), Prainha, MZSP 24542, 11 specimens,
8 shells, (27/1/1958), MZSP 24539, 3 specimens
(15/11/1953), MZSP 24543, 3 specimens (27/1/
1959), MZSP 24540, 1 specimen (15/11/1953),
Paranapoã, MZSP 24548, 1 shell (1/11/1960),
Porchat Island, MZSP 24544, 3 specimens (29/1/
1959) , Itararé, MZSP 24546, 8 specimens (29/1/
1960) ; Itanhaém, Viuva Conde Farm, MZSP 24535,
1 shell (O.Schubart col., 9/IX/1941); Iporanga,
MZSP 15615, 7 specimens (Leme and Papavero
cols., 1 /XI/1983), MZSP 24538, l9 (O.Schubart
col., 30/XI/1952); Iguape, MZSP 24532, 2 shells
(Leme and Biasi cols., 1-5/XI/1968); PARANÁ:
Serra da Graciosa, MZSP 29575, 1 shell (AVC Dutra
col., X/1979).
The anatomy of Neocyclotus inca (Orbigny, 1835)
has been provided by SALGADO et al. (1989),
reporting a similar organization to N. prominulus.
These species are different mainly in the penis
fashion.
Genus Incidostoma Morrison, 1942
Type-species:
Aperostoma (Incidostoma malleatum Morrison, 1942)
Incidostoma tupy new species
(Figs.32-36, 135-136, 259-270)
Types - Holotype: MZSP 29560, l9 from type-
locality. Paratypes - BRAZIL - RONDÔNIA: Nova
Esperança, MZSP 29578, 2<d (MZ polonoroeste sta.
831888, 6-9/XII/1983), MZSP 29561, 2 specimens
(sta.831874, 6-7/XII/1983); Nova Brasília, 11°09’S
61°34W, MZSP 29561, 2 specimens (PNW 840566,
6-11 /XI / 1984); Jaru, MZSP 24553, l9 (K.Okushigue col., 10/IX/1976).
Type-locality - BRAZIL - RONDÔNIA: Santa Cruz
da Serra [MZSP 29560, Vanzolini col., (MZ poloeste
sta. 840169)].
Diagnosis - Malleated surface weakly developed. Aperture ample, separated from preceding whorl.
Outer lip free.
Description
Shell (Figs.32-36). Size about 30mm, discoid, flat,
about 4.5 whorls. Color uniform brown. Outer
surface rich of undulated and delicate growth lines;
malleated surface (i.e., several low, small
concavities sprayed by surface) almost absent, seen
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

410 L.R.L.SIMONE
in some area in outer surface preceding aperture. Walls thin. Protoconch dome-shaped, small, of 1 whorl, surface smooth and opaque, suture well- marked, transition with teleoconch unclear. Teleoconch with convex whorls, almost planispiral. Spire very low, approximate angle of 170°. Each whorl covers small area of preceding whorl. Umbilicus very ample end deep. Peristome white, somewhat thick, weakly prosocline, oval. Notch deep in superior corner, close to suture; its lips surround small orifice, anterior-left edge touches adjacent virtual suture; satellite growth lines contours notch in preceding area, gradually becoming perpendicular to suture at short distance from aperture.
Head-foot (Figs.259-260). Characters similar to those of N. prominulus, with following remarkable features. Snout slightly taller, with mouth turned ventrally. Pedal gland furrow restrict to anterior edge of foot sole. Urinary gutter at right of head deeper. Penis of males proportionally larger.
Operculum (Figs.259-260): Somewhat similar in characters to those of preceding species, except for simple (not double) outer edge, and broader last whorls of outer sculpture.
Mantle organs (Figs.261-262). Features somewhat similar to those of N. prominulus, with following distinctive or notable attributes. Lung with net of vessels more clear, with 6-7 branched vessels alternating drainage to left (ctenidial?) vessel and to ad-rectal sinus. Probable hypobranchial gland present as lobe bulging at right from kidney, part covered dorsally by rectum [normally connected to excretory features, commented below (excretory system and discussion)].
Circulatory and excretory systems (Figs. 161-162) - Heart and pericardium characters similar to those of N. prominulus, differ only in having left (ctenidial) vessel plus vessel edging kidney inserted directly to auricle. Kidney also with similar attributes as those of N. prominulus, including anterior (solid- glandular) and posterior (hollow, filled by intestinal loops) renal chambers. Differs only by more protruded nephrostome and presence of solid excretory hypobranchial gland (as described in mantle organs). Urinary gutter deeper.
Digestive system (Figs.263-264) - Buccal mass attributes similar to those of N. prominulus, with following remarks. Dorsal folds with tall middle region as stalk for aperture of salivary glands. Odontophore muscles: mlb pair thinner, closer to median line; m3 pair similar but with an additional branch connecting with each other running on
posterior odontophore surface, dorsal to radular sac; mil pair thicker. Radular teeth features similar to those of N. prominulus, with following remarkable notes (Figs. 135-136): rachidian slender, almost triangular, 3 cusps, central cusp several times larger than neighbor cusps, distai edge concave; marginal tooth with about twice rachidian length, 3 cusps, central cusp terminal and several times larger than remainder cusps, basal region long, narrow and obliqúe; inner and outer marginal teeth slightly broader. Esophageal pouches, folds and regions similar in features to those of N. prominulus, except for taller folds edging pouches aperture and more developed irrigation of blood vessel in region of these tall folds. Large blood sinus located in ventral region between esophagus and buccal mass, this sinus evolves radular sac and possesses thick branches to salivary glands, to esophageal pouches and to odontophore. Middle esophagus with 4-5 narrow, longitudinal folds. Salivary gland cluster as single mass located posterior to pouches. Stomach attributes also similar to those of N. prominulus, except for: 1) more anterior insertion of esophagus; 2) posterior region of stomach (as blind sac) longer, its inner surface covered by transverse folds (sorting area?); 3) fold continuation from esophagus taller; 4) groove running from duct to digestive gland aperture deeper. Intestinal loops, characters and digestive gland features like those described for N. prominulus.
Genital system - cf (Figs.259, 265-266): Visceral and pallial organs with characters similar to those of N. prominulus, with remarkable features following. Prostate circular in section. Sperm groove between prostate and penis base longer. Penis very larger (about same length than that of entire head- foot) . Penis base broad, gradually narrows, middle third part of uniform width. Penis distai third broad, flat, slightly triangular. Penis sperm groove runs along lateral edge of penis and also superior edge of penis tip. Penis groove ends in pointed, small papilla (Fig.266). Penis papilla stay turned inwards, bulged, semispherical structure precedes it. Immature penis simpler, with tip coiled (Figs.267-268). 9 (Figs.261, 269-270): Visceral and pallial structures with characters similar to those of N. prominulus, with following remarks. Albumen gland more differentiated, as thicker region of visceral oviduct last loop. Seminal receptacle similar located, but possessing aperture direct to pallial cavity (and not inside pallial oviduct). Seminal receptacle aperture a small papilla located in posterior edge of pallial cavity close to pallial
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 411
oviduct. Capsule gland similar to that of N.
prominulus, with same components. Genital pore
a thick edged slit.
Central nervous system (Fig.263) - Similar
characters to those of N. prominulus.
Measurements of shells (in mm) - Holotype: 17.4
by 31.3; MZSP 29561: 17.0 by 29.2; 19.3 by 30.9.
Distribution - Known in some points of Rondônia
State, Brazil, Amazon Forest.
Habitat - No data.
Material examined - Types.
Etymology - The specific epithet refers to the
ancient indigenous people living in region.
Discussion - The superior notch of outer lip clearly
allows the generic attribution of the new species,
also with the aid of generic key of TORRE et al.
(1942:187). I. tupy differs from I. malleatum
(Morrison, 1942) (without locality), I. pergrandis
(Kobelt, 1912) from Colombia and I. kobelti
(Morrison, 1942) (from Colombia) byless developed
malleated surface, ampler aperture more separated
from preceding whorl, and by more developed axial
sculpture. MORRISON (in TORRE et al, 1942)
presented the distribution of the genus for
Colombia, Ecuador and Peru, (Andes localities). The
present description expands the geographic
distribution towards east, for Brazilian territory
(Rondônia) relatively away from Andes. However,
I. malleatum, just the type species of the genus,
remains lacking a more precise locality.
The presence of the hypobranchial gland in
cyclophorids has been related to the excretion
(ANDREWS & LITTLE, 1972), but a clear gland
is found only in I. tupy (Fig.262 in middle-
superior region).
Genus Aperostoma Troschel, 1847
Type-species: A. blanchetianum Moricand
Aperostoma blanchetiana (Moricand, 1826)
(Figs.37-39, 112, 137-138, 271-275)
Synonymy see TORRE et al. (1942:244-245).
Complement:
Neocyclotus (Neocyclotus) inca - (part) KOBELT,
1902:236 (in synonymy) (non ORBIGNY, 1835).
Aperostoma (Aperostoma) blanchetianum: TORRE et
al, 1942:244-245 (pl.35, figs.7-9).
Description
Shell (Figs.37-39). Discoid and flat. Color brown
with paler spiral bands variable in localization and
number in specimens. Spire somewhat tall.
Umbilicus slightly narrow. Aperture rounded,
slightly prosocline, very weak notch in superior
corner in form perpendicular. Other details in
TORRE et al (1942:245)
Head-foot and operculum (Figs. 112). Characters
similar to those of I. tupy.
Mantle organs (Fig.271). Features similar to those
of I. tupy. Narrow fold of hypobranchial (?) gland
present edging rectum.
Circulatory and excretory systems (Fig.271) -
Characters similar to those of preceding cyclophorids, remarkable features following. Lung
vessels preceding auricle somewhat similar to those
of I. tupy, but left (ctenidial) vessel strongly curved
preceding its connection, and secondary vessel running along border between kidney and
pericardium (not running on pallial surface).
Kidney also with 2 chambers, anterior (solid-
glandular) chamber longer antero-posteriorly.
Nephrostome a slit preceded by short stalk.
Digestive system (Fig.272) - Morphological attributes in general similar to those of I. tupy, distinctive or
notable features following. Jaw plates separated
from each other, outline somewhat circular, with
small projection in median-anterior edge. Buccal
mass dorsal chamber (between both dorsal folds)
with taller longitudinal folds. Odontophore muscles:
ml, several pairs in ventral region connecting
odontophore outer surface with mouth; mlb absent.
Radular teeth with similar attributes as those of
preceding cyclophorids, remarkable features
following (Figs. 137, 138): Rachidian slender and
long, 3 cusps, central cusp about 3 times larger than
remainder cusps, distai edge concave, with an
additional central concavity; marginal tooth with 3-
4 cusps, penultimate cusp terminal and several
times larger than remainder cusps; inner and outer
marginal teeth broader, outer marginal strongly
sigmoid. Large sinus around radular sac with
branches to both pouches and salivary glands
present. Stomach with groove adjacent to duct to
digestive gland longer, surrounds transversally
entire gastric circumference with short portion
parallel to its initial region. Gastric inner surface with an additional longitudinal, low, flattened fold
located between esophagus aperture and duct to
digestive gland.
Genital system - d (Figs.274, 275): Similar
characters to those of I. tupy, with following
remarkable features. Prostate longer, broader and
curved, walls transversally folded. Sperm groove
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

412 L.R.L.SIMONE
between prostate and penis base longer. Penis large,
curve, middle third part narrower than basal and
distai regions. Penis apex broad, flat, asymmetrical,
expanded, thick. Penis sperm groove runs along
inner edge and superior edge of penis tip; finishes in penis right side. Final part of penis groove with
pair of small papillae; left papilla broader, tip
rounded, more basal located; right papilla slender,
pointed and more distai. 9 (Figs.273): Visceral and
pallial structures with characters closely similar to
those of I. tupy, including albumen gland developed
and seminal receptacle opened directly into pallial
cavity. Differs by narrower genital pore.
Measurements of shells - MZSP 16534d": 14.4 by
20.8; MZSP 16534: 14.3 by 21.5.
Distribution - Bahia and Minas Gerais States.
Habitat - No data.
Material examined - BRAZIL - MINAS GERAIS:
Buritis, Jaboticaba stream, MZSP 16534, 3cf ,
29 (sta. 642152, Vanzolini col.).
Discussion - The shell attributes of the examined specimens are closely similar to those described by
TORRE etal (1942:245) for A. blanchetiana, allowing
the specific Identification. However, there is another
very similar species, A. merrilli Morrison, 1942 (from
Bragança Railway, Amazon valley, Brazil).
Superfamily Viviparioidea
Family Viviparidae
Genus Viviparus Montfort, 1810
Type-species:
Helix fluviorum Montfort = viviparus Linné
Viviparus acerosus (Bourguignat, 1862)
(Figs.40-41, 103-104, 139-140, 276-295)
Viviparus acerosus - FECHTER & FALKNER,
1993:119 (fig.5); FALNIOWSKI etal, 1996.
Description
Shell (Figs.40-41). Globose, color greenish-brown.
Spire tall, apex slightly rounded. Suture deep, each
whorl circular in section and scarcely attached to
neighboring whorls. Body whorl harmoniously sized
with spire. Umbilicus wide. Periostracum heavy,
smooth. Sculpture lacking except growth lines and
axial undulations. Aperture circular, lips somewhat projected. Young specimens (from brood pouch)
with up to 4 whorls; protoconch smooth, of 1 whorl;
remaining whorls sculptured with several narrow,
spiral lines uniformly distributed, 2 or 3 of them markedly larger and equidistant from each other,
bearing periostracal hair.
Head-foot (Figs.276, 279-280, 291, 293). Head
slightly protruded, relatively small (about 1 /3 of
head-foot width). Snout cylindrical, anterior margin
flat. Tentacles long (about 1.5 times longer than
snout), stubby, base at side of snout base. Ommatophore short, located between basal and
middle third of outer surface of each tentacles. Pair
of siphons (lobes) similar to those of ampullariids,
but shorter. Food groove edged by thick, low
borders; running slightly to right of middle region
of pallial floor. Food groove with posterior end
fainting; anterior end in right siphon (nuchal lobe),
dividing it into two projections, anterior (dorsal)
projection with medial limit close to base of snout
(Fig.293: lr). Foot large (about 2/3 of head-foot
volume), sole simple. Opercular pad larger than
its base in dorsal foot surface, with edges projecting
beyond it. Anterior furrow of pedal glands restricted
to anterior foot, edged by thick borders. Columellar
muscle thick, length about 3/4 whorl. Head-foot
connection with mantle border very thick and richly
vascular. Male genital structures described below.
Operculum (Figs. 103-104, 276). Occupying entire
shell aperture. Horny, somewhat similar in
characters to those of ampullariids. Outline slightly
circular, with low projection in inner-superior region.
Nucleus sub-central, located closer to inner margin.
Sculptured with concentric growth lines. Inner
surface glossy; scar sub-circular, close to inner
margin, occupying about 2/3 of opercular area.
Mantle organs (Figs.277-278, 281). Pallial cavity
of about 1 whorl. Mantle border simple, thick.
Osphradium ridge-like, obliqúe, almost parallel to
mantle border, length about 1/2 of that mantle
border, very close to gill. Ctenidial vein narrow,
uniform in width throughout. Gill long and narrow,
about same length as pallial cavity; anterior end
at mantle border. Gill filaments very tall and
narrow, arched towards right; apex slightly pointed,
close to food groove. Gill filaments with short, low,
glandular portion preceding afferent gill vessel,
forming glandular ridge along left gill margin
(endostyle) (Figs.277, 281). Glandular tissue
(hypobranchial gland?) covering afferent gill vessel;
but broad space apparently lacking hypobranchial
gland between gill base and rectum. Right margin
of pallial cavity filled by oviduct in females. Ad-
rectal sinus modified in ureter, broad, running all
along pallial cavity right margin in males or edging
oviduct in females. Rectum on dorsal and left sides
of ad-rectal sinus. At right end of mantle border,
in females, presenting three openings, most
posterior and smaller is ureter pore, anus most
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 413
anterior, and female pore larger, located between anus and ureter pore.
Visceral mass (Figs.277-278). Long, with about 4 whorls, approximately smaller than pallial cavity. Kidney anterior, with stomach and adjacent intestine just posterior to it. Gonad and digestive gland occupying about 2.5 whorls posterior to stomach. Gonad brown, located in columellar surface of each whorl. Digestive gland pale green. Details of each structure below.
Circulatory and excretory systems (Figs.278, 282) - Heart relatively small, located behind posterior-left region of pallial cavity, partly posterior to anterior lobe of kidney. Auricle connection with ctenidial vein just posterior to gill; possessing portion protruding anterior to this connection. Ventricle posterior to auricle, narrow connection between both. Kidney with two chambers and ureter. Anterior kidney chamber small, triangular in section, entirely filled by glandular tissue (having only small inner hollow portion close to nephrostome). Anterior kidney chamber with four almost plane surfaces: 1) posterior surface towards pericardium, with ventral, small, middle slit as reno-pericardial pore; 2) left with pallial cavity; 3) ventral right with ureter; 4) dorsal with mantle. Nephrostome an elliptical, small pore, without muscular edges, located in middle-posterior region of ventral-anterior surface of anterior renal chamber. Posterior renal chamber simple hollow sac filled by anterior region of stomach and intestinal loops. Ureter apparently specialization of broad adrectal sinus (Figs.278, 281), runs between rectum and oviduct (in females) or right pallial cavity edge (in males). Posterior end of ureter ventral to anterior renal chamber; anterior end in muscular, papilla-like ureter pore just posterior to anus (or just posterior to female pore). No special inner glands in ureter. No urinary gutter.
Digestive system - Buccal mass just posterior to mouth, extends slightly posterior to snout inside haemocoel (Fig.280). Mouth longitudinal, ventral. Jaw plates small, thin, outline elliptical (Fig.283). Dorsal folds broad, smooth and simple. Deep dorsal chamber between both dorsal folds, inner surface smooth. Odontophore and peribuccal muscles (Figs.284-288): mc, circulatory fibers of buccal sphincter, thick and broad; ml, several pairs of small muscular fibers connecting buccal mass outer surface with adjacent regions of haemocoel, no differentiated pair; mj, pair of jaw and peribuccal muscles, origin in odontophore cartilage lateral-
anterior surface, insertion along dorsal wall of buccal mass just dorsal to jaws; m2, pair of retractor muscles of buccal mass (retractor of pharynx), origin in lateral inner surface of haemocoel posterior to buccal mass, run towards anterior, insertion in posterior-lateral surface of odontophore (Figs.284, 286), most part on membrane which covers odontophore and small part in m4; m4, pair of large ventral tensor muscle of radula and subradular membrane, origin in odontophore cartilages around mj, cover cartilages almost completely (only dorsal edge free), insertion in small lateral region of tissue on radular ribbon preceding buccal cavity (to), subradular membrane also connecting along lateral and anterior edges of m4, part of dorsal branch m4 of with weak fibers along it; m5, pair of dorsal tensor muscles of radula, originating on m4 posterior-lateral region, running medially covering m4, insertion along radular sac in region opposed to m4 insertion (each m5 does not connect with its pair); m6, horizontal muscle, similar to those of ampullariids, little shorter in length than cartilages; m7, narrow, long pair, origin in m4 dorsal branch at medial-posterior edge, run posteriorly on subradular membrane (br) inner surface fusing with each other, penetrating in radular sac, insertion along radular sac and radular nucleus (Figs.287, 288); m7a, pair of thick muscles, originating in ventral surface of haemocoel close to median line and in levei just posterior to odontophore, run dorsally and, after, anteriorly attached to radular sac ventral surface, insertion in m5 anterior edge (Figs.285, 288); m9 absent; mlO, pair of ventral protractor muscles of buccal mass, origin in ventral inner surface of mouth, run posteriorly on mc, insertion in ventral-anterior region of odontophore; mlOa, paired continuation of mlO, running close to median line and attached to membrane covering odontophore, insertion along this membrane in its medial-ventral-posterior region; mil, pair of long and relatively broad muscles, originating on haemocoel inner ventral surface in region posterior to odontophore, close to origin of m7a, running dorsally, penetrating in odontophore just anterior to radular sac, insertion along subradular membrane close to median line reducing anteriorly; ml2, small pair of muscles similar to those of ampullariids, origin in lateral- anterior inner surface of m4 and br, running towards medial and dorsal, insertion shortly on cartilages anterior-outer surface close to m6 (Fig.287). Other odontophore structures: br, subradular membrane, similar to those of
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

414 L.R.L.SIMONE
preceding species, cover inner surface of (and probably secretes) subradular cartilage, most inserted in lateral and anterior edges of m4, but also connects with radular sac (via m5 pair) and peribuccal epithelium (via mj); reinforcement of subradular membrane in its dorsal-middle region with posterior, “W”-shaped edge; oc, pair of odontophore cartilages, similar to those of preceding species, flat, outline elliptical; om, odontophore outer membrane, transparent, covering outer surfaces of odontophore, except narrow portion close to median line where is penetrated by radular sac, some muscles (m7, m7a, ml 1) and some nerves; to, tissue on radular ribbon preceding buccal cavity, length more or less equivalent to portion of radula passing through odontophore. Buccal ganglia located laterally, on each side of border of odontophore and dorsal wall of buccal mass, close and dorsal to m2 insertion. Salivary glands (Figs.280, 283, 284) clustering as single mass posterior to nerve ring (localized posterior to buccal mass) and as two narrow glandular masses through it. Salivary ducts as continuation from these two masses, run on dorsal wall of buccal mass outer surface, penetrating into wall just close to their apertures, in anterior extremities of dorsal folds, close to median line.
Radular sac about as long as buccal mass (Figs.284-286), radular sac surrounded by large blood sinus. Radular teeth (Figs. 139, 140): all teeth with similar shape, tall, as flat rod, tip rounded, curved inwards, bearing several small, sharp cusps. Rachidian shorterst tooth, with 7-9 cusps, central cusp about three times broader than neighboring cusps (but same length); lateral tooth with 7-9 cusps, central cusp terminal, about three times width of remaining cusps (but same length); inner marginal tooth similar to lateral tooth, slightly longer; outer marginal tooth similar to inner marginal tooth, differing only in uniformity of cusps (lack central differentiated cusp).
Esophagus relatively narrow and long. Anterior esophagus with pair of dorsal, broad, longitudinal folds (as continuation from dorsal folds of buccal mass), without pouches (Figs.283, 284). Middle esophagus with several (about 10) inner, narrow, low, longitudinal folds. Posterior esophagus with smooth inner surface. No detectable esophageal glands.
Stomach (Figs.278, 289-290) large, about half whorl in length, located half whorl posterior to pallial cavity; posterior half immersed in digestive gland. Esophagus insertion in posterior-left gastric extremity, as gradual enlargement, curved towards
left. In this curve very small duct to digestive gland. Stomach, after this fist curve, abruptly expands becoming almost as wide as whorl; posteriorly narrowing, without clear separation with intestine. Main duct to digestive gland located in posterior- left-ventral surface of stomach. Stomach inner surface most smooth; pair of low, narrow folds runs on its dorsal surface (as continuation from esophagus), and curve towards posterior and run along gastric dorsal wall. Another tall and complex fold, as small chamber, located in posterior extremity of stomach, at some distance from esophageal insertion and close to smaller duct to digestive gland (Fig. 189). This tall fold with outer surface possessing 4-5 secondary folds, and smooth inner surface. Narrow fold, also initiated in esophageal insertion, running further posteriorly, lying anteriorly to posterior taller fold. No clear style sac. Intestine narrowing after leaving stomach, and abruptly curving towards right and runs parallel to gastric anterior wall. This curve located adjacent to pericardium, not possessing any special inner structure (like those of ampullariids). Intestine, then, curving towards anterior in middle gastric region, and lies parallel to its preceding loop on dorsal wall of posterior renal chamber (Figs.278, 290); then exiting to pallial cavity. Rectum narrow, running along ureter outer-dorsal wall. Anus described abo ve.
Genital system -cf (Figs.291-293): Testis pale green, lies on superior and columellar surface of visceral whorls, from first whorl to pallial cavity. Visceral vas deferens very narrow, running on columella and inferior-ventral margin of testis, with successive, very narrow branches from different portions of testis. In pallial cavity, vas deferens opening to large prostate gland (Fig.292), running along pallial cavity floor for about 1 /3 of its length. Pallial vas deferens, after this distance, narrowing abruptly, becoming duct, curving towards left, penetrating into haemocoel integument. Remaining 2/3 of vas deferens lies immersed in integument, surrounded by very thick, muscular walls in several layers, forming an ejaculatory duct. Ejaculatory duct bulging into haemocoel, displacing inner structures (as esophagus) towards left; its lumen small, closer to left side. Ejaculatory tube anteriorly with broader lumen and thinner muscular walls, abruptly narrowing, penetrating through right cephalic tentacle. Vas deferens runs all along right tentacle, its inner surface relatively broad, with some inner, longitudinal, low folds. Vas deferens opens in tentacle tip as broad papilla with rounded tip. Right
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 415
tentacle possesses deep concavity located at right from genital papilla (Figs.291, 293). Papilla can retract into this concavity. 9 (Figs.294-295). Ovary greenish brown, located in same position as testis (Fig.277). Visceral oviduct very narrow, runs along columella. Pallial oviduct very large, occupying about half of pallial cavity space and also bulges posteriorly to last whorl of visceral mass, displacing kidney and intestine to left. Visceral oviduct making large loop and penetrating pallial oviduct dorsally, between posterior and middle third; connecting sub- terminally, forming small papilla inside narrow dorsal chamber. This dorsal chamber running posteriorly, continuous with ventral chamber. This dorsal chamber running to posterior end of pallial oviduct, abruptly curving ventrally and greatly increasing in size. Ventral chamber of pallial oviduct functioning as large brood pouch (although possessing some small capsules in final portion of dorsal chamber), walls thin, semi-transparent, smooth. Low and very thin fold running along dorsal- left side of entire ventral chamber, curved to right, starting close to dorsal chamber aperture, tapering posterior to genital pore. Ventral chamber (brood pouch) filled by about 20 capsules, size varying. Each capsule covered by transparent, heavy, horny membrane, filled by hyaline, yellow, jelly mass and young specimens of about 4 whorls. Vaginal tube with thick, muscular walls and some inner transverse folds, located as continuation from brood pouch. Female genital pore small, close to mantle border, with several inner folds.
Central nervous system (Figs.284) - Unlike preceding species, but similar to remainder caenogastropods, of epiathroid type, and located posterior to buccal mass. Statocysts with several statoliths. More details in Viviparus contectus.
Distribution - Europe.
Habitat - No data beyond freshwater environment. Filter feeding.
Material examined - HUNGARY - ALFÕLD: Canal Tápió, (circa 60km SE of Budapest), MZSP 29403, 1 d , 29 (J.Hemmen leg).
Viviparus contectus (Millet, 1813) (Figs. 10, 42-43, 61-62, 141-142, 296-301, 348)
Viviparus contectus - FALNIOWSKI, 1989:59 (key); FECHTER 86 FALKNER, 1993:119-119 (fig.7); ELEUTHERIADIS 86 DIMITRIADOU, 1995:41- 52; FALNIOWSKI et al, 1996; KATOH 86 RIBI, 1996:67-82.
Description
Shell (Figs. 10, 42-43, 61-62). Characters similar to those of Viviparus acerosus, differing in being broader, whorls shorter and more convex, and by paler color. Young specimens (from brood pouch) similar to those of preceding species (Figs.61-62).
Head-foot and operculum. Similar to V acerosus,
differing only in darker pigment.
Mantle organs (Fig.296). As V. acerosus.
Visceral mass. Closely similar to those of V.
acerosus, except gonad and digestive gland almost same color (pale beige) and testis shorter (restricted to last visceral whorl posterior to stomach).
Circulatory and excretory systems - Similar to Viviparus acerosus.
Digestive system (Fig.297) - Similar to V. acerosus,
except for following features. Odontophore muscles: m2 pair with small portion of insertion also connected to lateral side of anterior esophagus; m7a, mlOa pairs also present. Radular teeth similar to V acerosus (Figs. 141-142), but differing in: rachidian slightly longer, with 9-11 cusps; lateral tooth with 8-11 cusps. Stomach with same folds (including tall posterior complex fold) and ducts to digestive gland.
Genital system - Ó : Visceral and pallial structures similar to those of V. acerosus, including ejaculatory tube running in integument and modified right tentacle as penis. Differing only by shape of papilla of right tentacle tip (where vas deferens opens), being pointed and more elongated (Figs.298-299), with concavity at side of papilla deeper. 9 : Visceral and pallial organs similar to those of V acerosus, but with final region of visceral oviduct and dorsal chamber of pallial oviduct (Fig.301) shorter, and connection between both simpler. Also presenting papilla separating visceral oviduct from dorsal chamber of brood pouch. Additionally, dorsal chamber running directly towards posterior, increasing gradually. More capsules, about 30, in ventral chamber of pallial oviduct (brood pouch), chaotically organized.
Central nervous system (Fig.300, 348) - As in V.
acerosus. But commissure between both cerebral ganglia relatively long. Supra and subesophageal ganglia located far from nerve ring. Each pair of connectives uniting cerebral-pleural ganglia with pedal ganglia “V”-shaped, with dorsal portion of each connective far from each other. Cerebral ganglia separated from each other, connected by a relatively long and wide commissure, form
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

416 L.R.L.SIMONE
triangular, dorso-ventrally flat; lateral edge possessing ophthalmic nerve in anterior region and tentacular nerve in posterior region. Right pleural ganglion with about half of cerebral ganglion size, located at short distance ventral from it, right cerebro-pleural connective narrow, with length equivalent to half of pleural ganglion length; form elliptical. Left pleural ganglion somewhat diffuse, with borders not so clear; left cerebro-pleural connective length equivalent to that of right one, but wider. Pedal ganglia antero-posteriorly long, located deeply inside pedal musculature, close to each other and to median line; dorsal region long and narrow, middle region wider, with a projection directed to its pair, becoming unclear if this is ganglion or commissure (both pedal ganglia separated by a shallow transversal furrow), anterior possessing a median nerve and a lateral, broad anterior projection. Cerebro-pedal connective (cl) narrow and long, connecting anterior corner of cerebral ganglia (just ventral and anterior to ophthalmic nerve) with dorsal end of pedal ganglia. Pleuro-pedal connective (pn) about twice wider that cerebro-pleural connective, boundaiy with pleural ganglia unclear, looking like their continuation. Cerebro-pedal connectives ventrally close to pleuro-pedal connectives, gradually becoming far from them towards dorsal, inserting respectively in cerebral and pleural ganglia far from each other, with a distance equivalent to that of cerebral ganglia.
Measurements of shells (in mm) - MZSP 29402: 9 1, 32.7 by 26.2; d 2, 34.5 by 25.7.
Distribution - Central Europe.
Material examined - HUNGARY - ALFÕLD: Canal Tápió (circa 60km SE of Budapest), MZSP 29402, ld, 4? (J.Hemmen col.)
Genus Notopala Cotton, 1935 Type: Paludina hanleyi Frauenfeld, 1864:339 OD
Notopala ampullaroides (Reeve, 1863) (Figs.44-46, 63, 105-106, 143-144, 302-319, 349-352)
Synonymy see SMITH (1992).
Description
Shell (Figs.44-46, 63). Somewhat similar, but shorter than those of Viviparus, of about 5.5 whorls. Color pale brown. Adult with eroded apex. Sculpture series of spiral, somewhat uniform striae, disappearing on last whorl. Aperture rounded, prosocline, peristome white. Umbilicus narrow, located at middle of inner lip. Young
specimen (found in brood pouch) (Fig.63) with up to 3.5 whorls; first whorl smooth, concave, dome- shaped; remainder of whorls almost smooth, with 3 equidistant spiral cords formed by successive small nodes, third cord located on periphery forming low carina.
Head-foot (Figs.302, 306, 315). Features similar to those of Viviparus, remarks following. Color dark grey, with some pale grey spots, sometimes coalescent. Food groove forming tall fold (Figs.302- 303), appearing abruptly in posterior end of pallial cavity; left edge taller and curved inwards; right edge low and broad. Pedal furrow wider, occupying anterior half of foot edge.
Operculum (Fig.302). Characters similar to those of Viviparus, except for nucleus being almost central, slightly towards inner edge; sculpture concentric.
Mantle organs (Figs.304, 307). Organization and components similar to Viviparus; distinctive features noted below. Mantle edge thick, smooth, color dark grey. Osphradium long and broad, located on ctenidial vein. Osphradium right edge with series of small and well-spaced projections. Ctenidial vein broad throughout. Gill filaments tall and slender, tip sharp pointed. Endostyle broad, also at right of gill.
Visceral mass (Figs.304-305). Similar to other viviparids, but slightly lower, about 2.5 whorls. Gonad occupying superior and columellar regions of each whorl. Color of both gonad and digestive gland greenish beige.
Circulatory and excretory systems (Figs.305) - Similar to Viviparus, remarks following. Heart large (about V4 of posterior renal chamber volume); auricle broad, connected to ctenidial vein posteriorly. Anterior aorta about 3-times broader than posterior aorta. Heart and posterior kidney chambers connected with each other, pericardium membrane separating them lacking (Fig.305). Posterior kidney chamber without apparent glandular tissue, greatly compressed by intestinal loop and by posterior region of pallial oviduct in females. Anterior kidney chamber solid, somewhat long and flat; posterior surface (with posterior kidney chamber) concave, triangular, with small, tight pore located in anterior-right region; its surface with pallial cavity (left) concave and broad, anterior end rounded; its surface with ureter (right) convex, with similar form of surface with pallial cavity; nephrostome located in middle region of latter part, papilla-like, with muscular, slightly tall
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 417
edges. Ureter (Figs.304-305, 307) and renal aperture as in Vivipams.
Digestive system - Most features similar to of for preceding viviparids; distinctive or remarkable characters following. Jaw plates thin, laterally located (Fig.310). Buccal mass dorsal folds tall and narrow, located close to median line. Aperture of salivary glands very small, close to median line, located in anterior end of dorsal folds. Odontophore muscles (Figs.308-309, 311-314): ma pair single, slightly thick; m2 pair with double insertion surrounding buccal ganglion; m4 insertion in tissue on radular ribbon preceding buccal cavity (to) by means of short and thin muscles; m7 pair very narrow; m7a pair well- developed, insertion with 2 branches running in opposite direction; mlO broad and thin; ml 1 pair thick and broad. Salivary glands long, slender and intensely convolute (Figs.308-309); located posterior to nerve ring; both narrowing and Crossing through nerve ring dorsally. Radular features similar to those of preceding viviparids (Figs. 143-144). Salivary ducts immersed in dorsal wall of buccal mass in posterior-lateral region (just dorsal to buccal ganglia) and run immersed in dorsal folds. Anterior esophagus with pair of inner folds continuous with dorsal folds of buccal mass; gradually these folds unite with each other and run as single, broad fold, from distance equivalent to buccal mass length, abruptly disappearing. Posterior esophagus with irregular, narrow inner folds; posterior region slender. Middle and hindgut with characters similar to those of Viviparus
(Figs.305), notable features following. Esophageal insertion in stomach sub-terminal, on its lateral- ventral surface. Stomach broad and long, somewhat conic. Stomach inner surface mostly smooth; narrow and low transverse fold running in dorsal surface, at levei of esophageal aperture; pair of narrow, low folds (origin in dorsal surface) at levei of digestive gland duct; folds running opposed to each other, surrounding at some distance duct to digestive gland; both folds approach one another in ventral surface, running parallel along intestinal surface, disappearing gradually. Duct to digestive gland single, small, located in middle region of anterior half of ventral surface of stomach. Stomach-intestinal border unclear, stomach narrowing gradually up to right region of pericardium. Digestive gland greenish beige, of 1.5 whorls posterior to stomach, and about half whorl surrounding it. Intestine, rectum and anus as in Vivipams.
Genital system - d : Similar to that of Vivipams,
remarkable features following. Testis large (Fig.317: pt), located in pallial cavity instead of visceral mass; lying along right edge of pallial cavity, bordering ureter; color beige, thick glandular walls. Pallial vas deferens originating short distance from posterior end of testis, in its ventral-left surface. Ejaculatory duct similar to those of preceding viviparids, immersed in integument (Fig.315), walls thick, with concentric muscles, bulging into haemocoel. Ejaculatory duct abruptly narrowing in base of right cephalic tentacle, walls becoming thinly muscular; it running along tentacle to its apex. Right (copulatory) cephalic tentacle base broad (Figs.315, 316), inner duct also broad; in middle levei, both tentacle and inner duct abruptly narro ws. Distai half of right tentacle strongly and irregularly curved towards left (Fig.316). 9 (Figs.318-319): Characters similar to those of Vivipams, with following distinctive or notable features. Ovary greenish beige, located in columellar and superior regions of visceral whorls, from apex to posterior end of stomach (Fig.304). Visceral oviduct very narrow, running in middle region of columellar surface of visceral mass. Pallial oviduct very large, its posterior third encroaching into renal chamber. Albumen gland solid, running in left-posterior region of posterior third of pallial oviduct, surrounding rectum. Albumen gland narrowing anteriorly, becoming duct; with visceral oviduct inserting in this region. Albumen gland narrow, with walls thick, inner surface longitudinal folded; zigzagging just within posterior edge of pallial cavity, then running towards posterior. Short distance from albumen gland posterior end, abruptly becoming broader, running anteriorly. Capsule gland posterior third of pallial oviduct, dorsal to kidney, walls thickly-glandular, inner surface longitudinally folded; special broader furrow running in ventral-left inner surface. Brood pouch anterior 2/3 of last portion of pallial oviduct, walls thin, semi-transparent inner surface smooth. About 20-30 capsules, containing specimens from 1 to 4 whorls, larger specimens located anteriorly. Anterior end of pallial oviduct with narrow duct and muscular walls; finishing in low papilla. Female pore small, sub-terminal, ventral.
Central nervous system (Figs.306, 309, 349- 352) - Located posterior to buccal mass. Cerebral ganglia flat and triangular; commissure between both cerebral ganglia (cc) with about 1 /
3 of ganglia length, located in anterior region of ganglia medial edge; a pair of small nerves located
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

418 L.R.L.SIMONE
in this commissure (Figs.350-351), running towards anterior; two pairs of broad nerves (lateral pair broader) running towards anterior, innervating dorsal-lateral surface of buccal mass (bn), these nerves as anterior vertex of ganglia; ophthalmic nerves (of) originating dorsally to these buccal mass nerves, running short distance, penetrating in ommatophores base orifice in haemocoelic surface; tentacular nerves (tn) about half-wide than ophthalmic nerves, originating in cerebral ganglia posteriorly and separated to them, running a short distance, penetrating into tentacles base. Pleural ganglia with about a third of cerebral ganglia size; both asymmetric, right ganglion (re) smaller than left (le) and possessing three nerves in anterior end, two running posteriorly (broader one as connective with visceral ganglion), and one running anteriorly; equivalent nerves inserted in left pleural ganglia in opposed side, close to connective with pedal ganglion. Connective between both cerebral and pleural ganglia (cn) very short, marked only by a single and abrupt curve in posterior vertex of cerebral ganglia. Connective between cerebral and pedal ganglia (cl) broad, separated from cerebro-pleural connectives, originated in anterior vertex, ventral to buccal mass nerves, length little shorter than cerebral ganglion length, inserting in posterior third of pedal ganglia. Pedal ganglia antero-posteriorly very long (about twice cerebral ganglia length), no clear border with pleural connective, running towards anterior penetrating in pedal musculature; pedal ganglia form similar to those of Vivipams, but shorter. Pedal commissure (uc) relatively long (about half of pedal ganglion width) and well discernible. Statocysts with about half size of pedal ganglia, located ventral and lateral to them, inside pedal musculature; internally several sand-like statoconia. Pair of visceral connectives (vc) very long (same length of haemocoel), relatively thick close to their insertion on pleural ganglia, narrowing gradually, becoming very narrow. Visceral ganglion (Fig.349: vi) relatively small, located in anterior edge of pericardium.
Measurements of shells (in mm) - AMS 202766 9 1: 26.7 by 23.2 (Figs.44-46); 9 2: 20.7 by 17.2; d 3: 18.6 by 17.5.
Material examined - AUSTRALIA: North Australia; Northern Territory, Douglas R. Crossing, 13°40.0’S 130°39.54’E, 1.5m depth, AMS 202766, 19 specimens (sta. 21437B; W.F.Ponder, C.Miller, D.L.Beechey and V.Kessner cols., 25/VI/1996).
Notopala essingtonensis (Frauenfeld, 1862) (Figs.47-48, 51, 66, 145, 320-321)
Synonymy see STODDART (1982:169); SMITH (1992).
Description
Shell (Figs.47-48, 51, 66). Characters very similar to those of N. ampullaroides, distinctions following. Outline more elongated. Color pale green. Sculpture of 3 low, spiral, equidistant, dark threads per whorl; series of low, narrow spiral striae between threads. Umbilicus very narrow. Aperture almost circular slightly prosocline. Young specimen similar to that of preceding species (Fig.66). Other details in STODDART (1982).
Head-foot and operculum. Features similar to those described by N. ampullaroides, except for darker grey color.
Mantle organs (Fig.321). Similar to those of N. ampullaroides, except for somewhat broader gill filaments. Osphradium slightly longer and narrower, bearing more small projections along right edge.
Circulatory and excretory systems - As those described for N. ampullaroides.
Digestive system (Figs. 145) - Most features closely similar to those of N. ampullaroides. Although, m6 is slightly shorter (about 2/3 of cartilage length).
Genital system - d (Fig.320): Posterior structures as described for N. ampullaroides, except for form of copulatory right cephalic tentacle, longer, narrowing gradually (slightly longer than left tentacle), with broad and low projection located in its middle- ventral surface. Copulatory tentacle inner duct broad in its base, abruptly narrowing, very slender along tentacle’s length. 9 : No available female with well-preserved visceral region was available, precluding analysis of visceral and posterior pallial structures. All other structures apparently closely similar to those described for N. ampullaroides.
Central nervous system - Similar to that of N. ampullaroides.
Measurements of shells (in mm) - AMS 153577 9 1: 27.1 by 20.8; d 2: 22.9 by 18.6; d 3: 23.6 by 18.5.
Distribution - Australia: Queensland, NT (SMITH, 1992).
Habitat - Lentic freshwater, lotic freshwater.
Material examined - AUSTRALIA - QUEENSLAND: Walkers Ck., 35 km south of Normanton, on Normanton-Karuba Rd., AMS 153577, 3cf , 39 (sta. 21169, D.F.McMichael col., 14/XII/1963).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 419
Genus Larina A. Adams, 1854 Type: L. strangei A. Adams, M)
Larina cf strangei A. Adams, 1854 (Figs.50, 52-54, 146, 322-329
Possible synonymy - Larina strangei A.Adams, 1854:41 (pl.27, fig.3); PRASHAD, 1928:179; WENZ, 1938:705 (fig.2032).
Description
Shell (Figs.50, 52-54). Similar to those of Notopala.
Distinctions following. Outline shorter and broader, somewhat discoid. Spire shorter, of 3 whorls; suture slightly deep. Color dark brown. Sculpture of irregular, spiral, low, narrow striae. Periostracum thick, smooth, except for 8 lines of periostracal hairs, each along spiral threads, clearer in young shells (Fig.50). Aperture rounded, prosocline (about 30° of longitudinal axis).
Head-foot (Fig.322). Similar to those of Notopala,
except: foot slightly larger; color pale grey; pedal gland furrow lying along anterior half of foot edge.
Operculum (Figs.323-324). Almost circular. Outer sculpture concentric, low. Nucleus almost central, slightly towards middle levei of inner edge. Inner scar occupying about V4 of inner surface area, closer to inner edge.
Mantle organs (Fig.325). Generally like those of N.
ampullaroides, remarks including following. Osphradium slightly longer, also on ctenidial vein and possessing series of small projections along its right edge. Gill filaments with broader base, narrowing abruptly; their distai 2/3 long and filiform; apex narrow and rounded. Endostyle ridge- like, running along entire gill right margin. Ad- rectal sinus conspicuous.
Circulatory and excretory systems (Fig.326) - Morphological attributes similar to those of Notopala species, with similar absence of separation between pericardium and posterior renal chamber. Anterior renal lobe broader laterally, both pores, including muscular, papilla-like pore to ureter, similar to Notopala.
Visceral mass (Fig.236) - Organization similar to preceding viviparids, but shorter, having only 1
whorl posterior to stomach. Gonad and digestive gland pale greenish beige.
Digestive system (Figs.326-327) - Foregut characters generally similar to those of Notopala.
Jaw plates somewhat thicker. Aperture of salivary gland larger, disposed longitudinally in anterior half of buccal mass dorsal folds. Odontophore muscles:
m2 pair also with double insertion surrounding buccal ganglia; m7 pair very narrow; m7a pair similar, with double branches (one running towards posterior and other anterior) in radular sac; mil pair also broad. Radular teeth shown in Fig.146. Salivary gland smaller, also convolute. Anterior esophagus slender; anterior region with pair of longitudinal folds (continuation from buccal mass dorsal folds), which gradually weaken and disappear. Middle and posterior esophagus also slender, inner surface covered by obliqúe, irregular, low, narrow folds. Middle and hindgut with similar features as those of Notopala, except for stomach having strong constriction in middle levei of right surface, internally corresponding to thin gastric shield. Pair of ducts to digestive gland, located close together in ventral-anterior surface. Digestive gland with 2 incompletely separated lobes, one posterior and other ventral to stomach, each connected to duct from stomach. Rectum with elliptical fecal pellets, lacking any apparent organization.
Genital system - d (Fig.322): Organization of posterior structures similar to those of Notopala,
remarks following. Testis also pallial, lying on right of rectum. Ejaculatory tube also muscular, immersed in integument. Right cephalic copulatory tentacle about double left tentacle length, base twisted, narrowing gradually along its length; tip somewhat rounded and slightly broader than preceding region. Vas deferens very narrow throughout right tentacle. 9 (Figs.326, 328-329): Visceral and pallial structures with similar features to those of Notopala. Albumen gland surrounding rectum behind pallial cavity. Albumen gland duct anterior and somewhat broad and thick. Visceral oviduct anterior end very narrow (Fig.328:vo), joining albumen gland duct as it exiting pallial cavity. Subsequently, albumen gland duct having long loop dorsal to ureter (about V4 of brood pouch length), lies towards posterior, running close and at right from albumen gland and ventral to rectum. Capsule gland thick glandular walled, running about % whorl in dorsal region of posterior renal chamber, twisting and lying towards anterior. Capsule gland inner surface with about 10 longitudinal, tall folds, its connection with brood pouch slightly narrow, at levei of posterior region of pallial cavity. Brood pouch as for Notopala.
Central nervous system - Nerve ring similar to Notopala. Buccal ganglia large, located laterally to posterior region of buccal mass
Measurements of shells (in mm) - AMS 338590:
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

420 L.R.L.SIMONE
9 1: 16.0 by 16.0 (photo); 9 2: 16.4 by 16.2; d 4: 8.7 by 9.1.
Distribution - Mid Coastal Queensland.
Habitat - Under rocks and wood, in small to médium rivers (Ponder, person. communic.).
Material examined - AUSTRALIA - QUEENSLAND: Dawson R. nr. Theodore, on Taroom-Theodore Rd., 24°56.49’S 150°4.18’E, 0.01-0.lm depth. AMS 338590, 2d , 139 (sta. 28911A; W.F.Ponder and D.Lindberg cols., 05/VII/1997).
Discussion - The present studied sample of Larina
is not attributed with certainty to the species of the genus, with which it is very similar, pending a review of the taxonomy of the group (PONDER, personal communication). PRASHAD (1928) suggested that Larina is the only viviparid living in an estuarine environment, the others being freshwater, and had doubts about the familial placement of the genus, since the anatomy was unknown. Despite the imprecision of the identification of the present sample must be considered, Larina appears to be clearly similar to Notopala, and may belong to the same viviparid subgroup as that genus. See additional discussion on differences in the central nervous system in the character discussion section.
Superfamily Littorinoidea Family Annulariidae
Genus Annularia Schumacher, 1817 Type-species: Turbo lincina Linné
Annularia sp. (Figs.49, 55-56, 113-114, 147-149; 330-347)
Description
Shell (Figs.49, 55-56). Of small size (up to 15mm), tall, long, apex lost. Spire of about 3 whorls convex of somewhat same size. Sculpture successive axial threads, orthocline. Aperture rounded, with projected lips. Umbilicus narrow.
Head-foot (Figs.330-332, 336). Head broad, outstanding, color beige. Snout long and broad, slightly dorso-ventrally flattened; anterior margin plane, flat, with short lateral pointed expansions. Tentacles stubby and long (about same snout length). Ommatophores small, in outer side of their bases. Tentacles tip broad, rounded, brown pigmented. Foot relatively short, borders thick. Anterior furrow of pedal glands short. Another furrow runs in pedal sole along median line, from middle-ventral region of anterior furrow to posterior third of foot sole. Opercular pad large, lateral
margins extending little beyond foot edges. Males with long penis located posterior removed and at right from head (described below). Columellar muscle thick, of 2/3 whorl, with apparently 2 regions separated by low fold. Haemocoel narrow and long, anterior half with walls thick muscular, posterior half walls thin. Pair of narrow and thin ventral retractor muscles of snout, passing through nerve ring. Some secondary thin and narrow ventral pairs of retractor muscles present, attached to haemocoel inner surface. Diaphragm-like septum in limit between head-foot haemocoel and visceral mass, thin, scarcely muscular.
Operculum (Figs. 113-114). Sub-circular, thick, calcareous, palcispiral. Pale brown basal plate connected to opercular pad. White hard fold on spiral suture, protruded outside, margins irregularly broken.
Mantle organs (Figs.333, 337). Mantle border simple, smooth slightly thick. Some specimens (about 50%) with tall septa posterior to mantle border restricting pallial cavity aperture. Pallial cavity of about 1.5 whorls. Osphradium a small, low and long fold located in anterior-left corner of cavity. Similar sized glandular fold present at left of osphradium. Gill absent. Vessel net of lung somewhat sparse, intercalated branches of adrectal and left collar main vessels (circulation described below). Rectum very broad, sometimes occupying about half of cavity volume, replete of elliptical fecal pellets. Anus siphoned, small, located close to right margin of cavity posterior removed from mantle border. Pallial gonoducts run at right and dorsal to rectum (described below). Posterior limit of pallial cavity with pericardium (left) and kidney (right).
Visceral mass (Figs.243, 333). Of about 3 whorls, 2 first whorls most filled by digestive gland. Last whorl most occupied by stomach. Gonad relatively narrow, running in columellar surface.
Circulatory and excretory systems (Figs.335, 337) - Heart relatively small, located longitudinally in left limit of pallial cavity, part connected to columellar muscle. Auricle anterior, directly connected to left collar vessel. Adrectal vessel draining to kidney. Ventricle elliptical, posterior. Kidney divided into 2 regions. Posterior region a hollow chamber mostly filled by intestinal loops, loops connected to dorsal wall by mesentery. Posterior renal region with about half whorl. Anterior kidney region with about half of posterior region size, triangular, flat; entirely filled by folded renal tissue and separated from neighbor structure
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 421
by membrane. Anterior renal region limited posteriorly by its posterior chamber, anteriorly by pallial cavity and at left by pericardium. Nephrostome a small slit in middle-dorsal region of membrane between kidney and pallial cavity. Renal lobe(s) and nephridial gland not distinguishable, probably mixed in that single folded mass. Renal tissue not connected to intestine, attached to dorsal wall of anterior kidney region and detached from ventral walls, this narrow space nephrostome connects with pallial cavity. Shallow and narrow urinary furrow starts ventral to nephrostome, in pallial cavity floor, runs edging left pallial structures, contours right insertion of mantle in head-foot and gradually faints towards ventral in short distance.
Digestive system - Mouth in center of anterior snout surface (Fig.336). Buccal mass large, half inside snout and half protruded in remainder haemocoel (Fig.336). Odontophore about 2/3of buccal mass volume. Jaw plates missing. Dorsal wall of buccal mass inner surface with pair of dorsal folds low and narrow. Aperture of salivary glands small longitudinal slits, located in median and anterior region of dorsal folds (Fig.342). Between both dorsal folds shallow and smooth surface. Buccal mass and odontophore muscles (Figs.334, 338-341): ml) several small muscular fibers of jugal muscles, particularly more concentrated around posterior region of buccal mass, no pair outstanding; mj) peribuccal muscles with some layers of circular, obliqúe and longitudinal fibers mostly originated from outer surface of cartilages; m2) absent; m3) single thin and flat muscle in posterior-dorsal region of odontophore, insertion in both lateral ends; m4) pair similar to those of preceding species but constituted by single muscular mass (and not several pairs), origin in dorsal, ventral and ventral surfaces of odontophore cartilages, surround most of these cartilages tensioning subradular membrane (br) contour mj, insertion in tissue on radula preceding its exposed area (to); m5) thick and short, origin on m4 median-posterior outer surface, runs towards dorsal, insertion in radular sac dorsal surface; m6) horizontal muscle, short, thin, restrict to anterior, subterminal region of cartilages; m7) pair narrow, thin and long, origin in m4 ventral branch, runs towards posterior attached to subradular membrane on median line, in posterior levei of odontophore connect with each other, insertion in single bundle inside radular sac region just posterior to odontophore; m9) pairs of narrow, superficial and flat muscles in dorsal-
posterior region of odontophore just anterior to m3, origin lateral in superficial membrane of odontophore, run towards posterior and medial slightly perpendicular to m4 fibers, insertion on “to” just posterior to m4 insertion; mlO) pair broad, slightly thin, origin in ventral region of mouth, run towards posterior, insertion in odontophore ventral and lateral surfaces; mil) absent; ml4) pair of superficial ventral protractor muscles of odontophore, thin and flat, origin in ventral- anterior surface of snout inner wall, run towards posterior lying odontophore ventral surface, insertion in posterior-ventral end of odontophore. Odontophore cartilages (Fig.341) long, flat and slightly arched, anterior end somewhat bifid. Septum between esophagus and odontophore tall; in this region dorsal inner folds bifurcate, outer branch very short, inner branches run along esophagus. Radular teeth (Figs.147-149): rachidian tooth tall and slightly barrow, outline triangular, cut-edge curved, cusps lacking, bluntly pointed tip, base broad and flat; lateral tooth tall, slender, outline slightly triangular, about as broad as rachidian, curved inwards, tip bluntly pointed, cusps lacking; inner marginal tooth narrow, tall, width about half of that of rachidian, uniform width along its length, base with longitudinal concavity where outer marginal tooth encases, tip broad, 4 stubby cusps in cut-edge of almost same size; outer marginal tooth broad, about twice rachidian width, base narrower than distai region, thicker reinforcement along inner edge, remainder middle and basal regions thin and flat, distai (cut) edge ample, about 50 long, slender, curved cusps which decrease gradually towards lateral. Salivary glands as 2 separated, slightly triangular masses (Fig.338) just posterior to nerve ring; their ducts somewhat broad and sinuous, run on dorsal surface of buccal mass through nerve ring, in short distance after nerve ring penetrate in dorsal wall, aperture above described. Esophagus long, broad, thin walled. Anterior esophagus with pair of low, longitudinal folds in a side (Fig.342). Posterior esophagus narrower, inner surface smooth. Stomach (Fig.343) very large and long (about 1 whorl length). Esophagus insertion in middle levei of ventral-left (inferior) surface, single duct to digestive gland at right (superior) to esophagus insertion, relatively small. Gastric shield thin, elliptical, small, located dorsal and slightly anterior to esophagus insertion. Stomach inner surface most smooth, single longitudinal fold in left (inferior) surface, from esophagus insertion (slightly dorsal to it) to
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

422 L.R.L.SIMONE
posterior gastric end. Stomach gradually narrows anteriorly, without clear separation with intestine and style sac. No style found. Digestive gland beige, covers some portions around stomach and about 1.5 whorls posterior to it. Intestine broad, with 3 loops anterior to stomach inside renal posterior chamber (Fig.337). Fecal pellets formed in first intestinal loop. Rectum and anus above described.
Genital system - d : Testis pale beige, long and very small, located in columellar surface of digestive gland first 1.5 whorls. Vas deferens very narrow, runs along ventral surface of stomach. Seminal vesicle intensely coiled (Fig.344), starts about 1 whorl posterior to pallial cavity. Seminal vesicle abruptly finishes at some distance from pallial cavity, vas deferens becomes very narrow and runs towards right and anterior, at right from rectum. Prostate long and broad (Fig.344), walls thick glandular, entirely closed (tubular). Prostate posterior end just in pallial cavity posterior limit. Prostate anterior end just posterior to anus, abruptly narrows and crosses to pallial cavity floor, runs short distance and penetrates in penis base. Penis (Figs.330, 331, 345) long, large, curved in base and apex, slightly flattened, length about same than that of pallial cavity. Muscular tissue present in anterior region of penis base connecting it with adjacent integument. Penis tip pointed. Penis duct very narrow, slightly coiled in basal half, simple in distai half, runs closer to lateral penis margin. 9 : Visceral organs Fig.343) similar located than respective structures of males, including very small and narrow gonad. Ovary color pale cream. Visceral oviduct (Figs.333, 347) with bulged portion of about % whorl followed by another portion of Vi whorl narrow and coiled, just posterior to pallial cavity. Albumen gland small, cylindrical, as thick pallial continuation of visceral oviduct. Capsule gland large, elliptical, broad, dorso-ventrally flattened, located dorsal to rectum, connected with middle levei of pallial oviduct by narrow, subterminal duct. Vaginal tube, after capsule gland duct, gradually expands, part dorsal to rectum. Aperture a longitudinal slit turned to right, posterior to anus. Inner surface of vaginal tube thick, with pair of longitudinal folds.
Nervous system (Figs.346) - Of typical epiathroid fashion, supra esophageal ganglion far from nerve ring. Anterior pedal nerve very broad. Statocysts with single and large statolyth (Figs.334, 346).
N.B. - All examined specimens possess several small pearls along visceral mass, irregular in size,
iridescent, most in larger blood sinus as renal chamber.
Measurements of shells (in mm) - MZSP 28273: 9 , 14.8 by 7.6; MZSP 28281, d 1, 10.6 by 6.1; d 2, 9.1 by 5.6.
Distribution - Full range unknown, but includes at least NE México.
Habitat - Terrestrial, on dry vegetation.
Material examined - (R.Pinto-da-Rocha and S.Casari cols.) MÉXICO - YUCATAN: Uxmal, MZSP 28266, 12 specimens, MZSP 28273, 3 specimens, MZSP 28275, 5 shells (25/IX/1996); near Blankanché Cave, MZSP 28281, 7 specimens (24/IX/1996).
Discussion - This unnamed species is very characteristic of the genus, however, a more precise identification was not possible because of the large number of species and the taxonomic confusion in the group. The generic placement of this species is based on the key to genera of HENDERSON & BARTSCH (1921:54). Although, it looks similar to Choanopoma terecostatum Thompson, 1966 (from Chiapas, México) in shell and operculum characters, but differs in its shallower suture. Despite there being relatively comprehensive revisions of the Annulariidae (e.g., BARTSCH, 1946), the anatomical knowledge of the group is still poor.
DISCUSSION OF THE CHARACTERS
Shell
1. Spire: 0= high (about 2 times aperture length); 1= low (ampullariids, cyclophorids, Larina); 2 = planispiral (Marisa) (Cl: 50; RI: 66; additive).
This character can be considered additive due to ontogeny, since Marisa presents a low spire during early development, as seen in protoconch. But when treated as non-additive nothing changes, neither in result or indices.
2. Periostracum: 0= thin; 1= thick/ (viviparids) (Cl: 100; RI: 100).
3. Suture: 0= deep; 1= shallow (plane) (F.
neritiniformis, A. megastoma) (Cl: 50; RI: 0)
4. Sculpture: 0= absent (smooth); 1= axial ridges (cyclophorids, Annularía) (Cl: 50; RI: 66).
5. Determinate growth: 0= absent; 1= present (.Annularia, cerithioideans) (Cl: 100; RI: 100).
The shell characters (1-5) are little more than 3.5% of the characters. The shell is the main structure analyzed in most literature, and is the most accessible structure that the malacologist has for
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 423
analysis. With this in mind, special attention was paid to searching for conchological characters, but most were deleted as autapomorphies. Of the 5 shell characters used, only 2 (characters 2 and 5) are non-homoplastic in this analysis.
The determinate growth (character 5), which became a non-architaenioglossan caenogastropods synapomorphy (node 7), was explored in detail by VERMEIJ 86 SIGNOR (1992). It consists of the formation of a differentiated peristome in the shell when the animal reaches the adult phase. Determinate growth is, however, also known in some Cyclophoroidea not included in this study.
Operculum
6. Outline: 0= circular; 1= with an upper-inner projection(sub-pyriform) (ampullariids, viviparids) (Cl: 50; RI: 80).
The plesiomorphic condition is that normally found in the vetigastropods, with rounded outline and multi- or paucispiral. Although somewhat circular, the ampullariids and viviparids have a modified outline, a projection in upper-inner quadrant. This is an apparent adaptation for fitting it tightly into the aperture, occluding this completely and hermetically.
7. Outer surface: 0= spiral; 1= concentric (ampullariids, viviparids, cyclophorids) (Cl: 50; RI: 50).
The members of the three architaenioglossan families in the analysis modified the outer surface of the operculum from a spiral to a concentric growth pattern. This condition was apparently reverted in the remainder basal caenogastropods, in which the paucispiral operculum is the rule.
The concentric outer sculpture is also found, by convergence, in some other caenogastropods, such as some families of Cerithioidea, in Stromboidea and in all “higher” superfamilies.
8. Type: 0= corneus; 1= calcareous (cyclophorids, Annularia) (Cl: 50;RI: 66).
The calcareous operculum is herein considered derived from the corneus one. This type resulted as a convergence between both terrestrial forms (cyclophorids and annulariids). However, calcareous opercula are known to occur in several outgroups, such as, e.g., the vetigastropod Trochoidea (Trochidae = corneus; Turbinidae, Phasianellidae = calcareous). Moreover, the ampullariids have representatives (not included here) with calcareous operculum, as in genus Pila Rõding, 1798.
Head-foot
9. Food groove: 0= absent; 1= present (viviparids) (Cl: 100; RI: 100).
The food groove running on the head-foot, in the floor of the pallial cavity, is an adaptation for filter feeding. The groove runs close to the apex of the gill filaments and conducts mucus with particles towards the mouth, by means of cilia (COOK, 1949).
Amazingly similar food grooves are found convergently in other filter-feeding caenogastropods, such as struthiolariids (Stromboidea), vermetids (Cerithioidea) and calyptraeids (Calyptraeoidea). These taxa apparently lacking any close relationship. The viviparid food groove differs from those of the other filter-feeding groups in being taller, forming almost a carina, and in having a relation with the right siphon (or nuchal lobe). This latter structure conducts the food to the mouth.
10. Right siphon: 0= absent; 1= present (ampullariids); 2= with a fold to mouth (viviparids) (Cl: 66; RI: 88; additive).
The additive optimization of this character is because of ontogeny, as very young viviparids (state 2), found inside the brood pouch, lack the fold to mouth. If this character is considered non-additive, the result is the same, but both indices change to 100.
11. Left siphon: 0= absent; 1= present (ampullariids, viviparids); 2= very long (Marisa, P. crosseana, P.
scalaris, P. curumim, P. lineata, P. bridgesi, P.
sórdida, P. canaliculata) (Cl: 40; RI: 75; additive).
The right and left siphons, also called nuchal lobes (PRASHAD, 1925; SCOTT, 1957) and pseudo- epipodia (PRASHAD, 1925), are prominent muscular flaps with the apparent function of controlling the water flow inside the pallial cavity. The left siphon is longer and works in the incurrent, and the right siphon in the excurrent flow (ANDREWS, 1965b).
In some ampullariids, the left siphon is very extensive, becoming sometimes twice as long as the shell. Its tip may be extended from the water surface, capturing air (PRASHAD, 1925:120; DEMIAN, 1965; pers.obs.). Their presence and length have also been considered by BERTHOLD (1991) and BIELER (1993, characters 37-38), and have been utilized for separating genera long ago (BINNEY, 1867:2).
Due to their similarity, the head-foot siphons of the ampullariids and viviparids were tentatively considered homologues. However, the siphons of both groups differ in some details of their bases,
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

424 L.R.L.SIMONE
with the viviparids having the base divided by a median, tall fold (clearer in the right siphon). This fold is maybe an adaptation for conducting the food to the mouth, and is lacking or inconspicuous in the ampullariids. Considering the cladogram obtained in this analysis, the siphons should have arisen in node 4 and reverted in node 7. However, another equally parsimonious optimization exists, convergence between nodes 6 and 10 (viviparids and ampullariids respectively). The first hypothesis is shown in the cladogram.
The ampullariid and viviparid siphons are clearly a modification of the head-foot, which differentiates them from the siphons of the “higher” caenogastropods. In these groups, the siphons are clearly a modification of the pallial edge (ANDREWS, 1965b:86).
The character 11 was considered additive because the State 2 is potentially a State 1 modification. The tree is the same if the character was considered non-additive, the indices, however, change to Cl: 50 and RI: 81.
12. Ommatophore in tentacles; 0= absent; 1 = present (all species) (Cl: 100; RI: 100).
The ommatophore is here defined as a secondary stalk for the eyes, positioning them away from the main axis of the cephalic tentacles. In the archaeogastropods, the ommatophore, when present, is normally located close to, but separated from the tentacle. In the architaenioglossans, as well as in the other caenogastropods that possess this structure, the ommatophore is part of the cephalic tentacle. Although, in the 2 basal branches (cyclophorids and ampullariids) the ommatophore is located on the tentacle base, while in the other caenogastropods it is located more distally.
13. Pedal furrow of pedal glands: 0= anterior margin only; 1= entire foot margin (Neocyclotusprominulus); 2= also along sole, on median line (Annularia) (Cl: 100; RI: 100; non-additive).
Pedal glands are expected to be found in all gastropods. In the caenogastropods, beyond this, possess an additional furrow lying at anterior margin of the foot, into which the anterior mucus glands open. This furrow probably is used for spreading the mucus along the foot sole. Beyond this adaptation, 2 additional modifications to the foot furrow were detected, as noted above, resulting as autapomorphies.
14. Columellar muscle: 0= 1.5 whorls; 1= 3A whorl (cyclophorids); 2= 1/3 whorl (ampullariids except Marisa) (Cl: 100; RI: 100; non-additive).
See comments after character 17.
15. Columellar muscle number: 0= 2; 1 = 1 (all species) (Cl: 100; RI: 100).
The single columellar muscle is a derived condition from the double one found in archaeogastropods. However, it is known that the State 1 is shared with a few members of the caenogastropod sister-taxon, the Heterobranchia.
16. Pair of snout tentacles: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
The snout tentacles are long projections of the lateral edges of the structure anterior surface, being almost as long as the cephalic tentacles. PRASHAD (1925) described them for Pila, calling them labial palps or anterior tentacles. These tentacles are a notable ampullariid synapomorphy, and have also been considered by BERTHOLD (1991) and BIELER (1993, character 1). Analogous structures are also found in some heterobranchs, such as the nudibranchs.
Pallial cavity
17. Pallial cavity length: 0= 1 whorl; 1= 1.5 whorls (viviparids); 2= less than Vá whorl (ampullariids except Marisa) (Cl: 100; RI: 100; non-additive).
The viviparids, because of their filter-feeding adaptations, developed a very long pallial cavity (state 1). Most ampullariids (node 12), on the other hand, presents a very short cavity (state 2), maybe a consequence of the spherical shell. This shortness is also reflected in the columellar muscle length (character 14).
18. Osphradium form and type: 0= small, circular; 1= long, ridge-like (viviparids, Annularia,
cerithioideans); 2= elliptical (ampullariids); ?= cyclophorids (Cl: 100; RI: 100; non-additive).
The osphradium commonly found in the archaeogastropods and in the heterobranchs (which have it) is in form of a small node. This state was modified in the ingroup: the cyclophorids lost it; the ampullariids developed an elliptical, bipectinate fashion (state 2); while the viviparids, as well as the remaining caenogastropods, have a long, ridge-like osphradium (state 1).
19. Osphradium leaflets: 0= absent; 1= present (ampullariids); 2= small projections on right side only (Notopala spp, Larina) (Cl: 100; RI: 100; non- additive) .
20. Osphradium: 0= sessile; 1= on a stalk (P.
crosseana, P. bridgesi, P. sórdida, P. canaliculata,
A. megastoma); 2= on ctenidial vein (Notopala spp,
Larina) (Cl: 66; RI: 85; non-additive).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 425
The osphradium is a sensory organ of olfaction, important in aquatic species. In caenogastropods, particularly, this structure is normally complex and large if compared to those of other molluscs. In terrestrial forms, the osphradium is generally reduced (e.g., annulariids), or missing (e.g., cyclophorids). The vestigial osphradium of Annularia sp., despite its small proportions, is long, which suggests the tentative attribution to the State 1 in the character 18.
The osphradium of the ampullariids has been classified as bipectinate. Bipectinate osphradia are also found in other caenogastropods, such as, e.g.,
cerithiids (Cerithioidea), strombids (Stromboidea), and “higher” caenogastropods (Calyptraeoidea, Naticoidea, Cypraeoidea, Tonnoidea, Neogastropoda), as I have observed. However, the ampullariid bipectinate osphradium is a quite different from these other caenogastropods in being located on a stalk, and in having a conspicuous longitudinal fold uniting both sides of the filaments. Those differences suggest that the bipectinate condition of the ampullariid osphradium may be due to convergence, and that this kind of modification, for increasing the surface of this sensory organ, may be a common feature among caenogastropods. Only the osphradium short length have been considered by BERTHOLD (1991) and BIELER (1993, character 2). PRASHAD (1925:95, 143) has described the osphradium of Pila
and is a quite similar to the South American species studied here.
No vestige of an osphradium was found in the examined cyclophorids. This is the reason for putting question marks on the matrix in all osphradial characters of the cyclophorids. However, it is expected that additional studies on ontogeny, and on more species, will bring new revelations.
21. GUI: 0= present; 1= absent (cyclophorids, Annularia) (Cl: 50; RI: 66).
The terrestrial, air-breading species, lost the gill. However, a vestige of its original blood supply, with an afferent vessel on the right and an efferent (homologue to the ctenidial) vessel on the left, is found in annulariids and cyclophorids. The collateral circulation between these 2 vessels is markedly increased, to form a lung.
22. Gill anterior end: 0= posterior from mantle border; 1= on mantle border (viviparids); ?= no gill (Cl: 100; RI: 100).
23. Gill outline: 0= elliptical; 1= narrow and curved (ampullariids except P. curumim) (Cl: 50; RI: 85).
24. Gill filaments: 0= triangular; 1= rounded [P.
curumim) (Cl: 100; RI: 100).
25. Gill filament tip: 0= central; 1= at right (F.
neritiniformis, P. scalaris. P. curumim, P. lineata, P.
canaliculata, A. megastoma, Viviparus spp., cerithioideans) (Cl: 25; RI: 57).
The gill of the viviparids has an additional function beyond respiration, as it is also used for filter feeding. For this task, the viviparid gill obviously presents modifications, manly an enlargement of the total length and of the filaments. The ampullariid gill, on the other hand, due to the compression of the lung (which additionally helps in the respiration), is narrow, and normally curves around the lung sac. The unusual fashion of the ampullariid gill have been considered by BERTHOLD (1991) and BIELER (1993, character 4), and was described in more detail by PRASHAD (1925:96-98) for Pila and by LUTFY & DEMIAN (1965) for Marisa comuarietis.
Once more, because of the absence of the structure in the terrestrial forms, the gill characters have been coded with a question mark on the matrix for the cyclophorids and annulariid.
26. Ctenidial vein: 0= single; 1= double (P. lineata,
P. sórdida, P. canaliculata) (Cl: 33; RI: 0).
Beyond the development of the lung (see comments after character 29), the ctenidial vein of the above Pomacea species is double in its middle and anterior parts.
27. Distance gill-rectum: 0= broad; 1= very close (Pomacea spp, A. megastoma, F. neritiniformis) (Cl: 100; RI: 100).
28. Endostyle: 0= absent; 1= present at right of gill (viviparids) (Cl: 100; RI: 100).
The name “endostyle” is used as an analogy to the structure in the Cephalochordata, and has been used in the literature on filter-feeding gastropods. The endostyle is an additional glandular ridge parallel to the gill, normally present in filter-feeding caenogastropods, such as struthiolariids (Stromboidea), turitellids-vermetids (Cerithioidea) and calyptraeids (Calyptraeoidea). In these taxa, the endostyle is different in being located along the left edge of the gill, and is apparently developed from a modification of the local mucus cells of the mantle.
As the viviparids are filter-feeding animais, they also developed a glandular ridge, here called an endostyle (Fig.304: en). However, the viviparid endostyle is unlike those of above mentioned taxa in being located along the right edge of the gill (instead of in
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

426 L.R.L.SIMONE
left side), and in being apparently developed from a modification of the gill filaments. This is suggested because of the location of the structure between the filaments and the afferent gill vessel, as well as the presence of successive septa in the endostyle, each one corresponding to a gill filament.
29. Lung sac: 0= absent; 1= present and short (ampullariids); 2= present and very long (Marisa) (Cl: 100; RI: 100; additive).
It is intuitive to deduce the evolution of the gill- less pallial cavity of air-breading snails such as the cyclophorids and annulariids (e.g., CEI, 1942). In fact the lung vessel present in both groups running on the left margin of the pallial cavity is probably homologous to the ctenidial vein, since it is located in a similar position and is connected to the auricle. However, the derivation of the net of lung vessels from the vessels and sinuses of the gill is still not clear. The pallial cavity of Cyclophoms
(TIELECKE, 1940) is similar to the cyclophorids examined herein. Some studies on the pallial blood vessel circulation [e.g., TIELECKE, 1940) shows that the circulation pattern in I. tupy and A.
blanchetiana is normal for the family, and N.
prominulus could present a derived condition. This species has the vessel homologous to the ctenidial vein running along the left edge of the lung inserting in the kidney, instead of the auricle.
The ampullariid lung differs from that in any other terrestrial gastropod, normally formed in the pallial cavity by the loss of the gill and an enlargement of the mantle vessels (see comments after character 21). The ampullariid lung is surrounded on the right by the gill and is restricted to the left region of the pallial cavity, posterior to the osphradium.
The ampullariid lung is a sac with muscular walls, its inner surface has a net of transverse brood vessels, protruding inside it. A pneumostome, with muscular edges, is the aperture of the lung sac, and is located in its anterior-left region, turned to the base of the left siphon. BERTHROLD (1991) noted that some ampullariids lack musculature surrounding the pneumostome, which may be the plesiomorphic condition, and this was considered by BIELER (1993) as character 36. The lung sac is a conspicuous synapomorphy present in all species of the family, and has also been considered by BERTHOLD (1991) and BIELER (1993, character 3).
The analysis of the morphology of the lung sac reveals that the blood inside its vessels carne from the marginal vessel located along the anterior and left edges of the organ. The origin of this vessel is
almost in the anterior end of the afferent ctenidial vessel, and afterwards, it crosses above the anterior region of the gill. This arrangement suggests that the lung sac afferent vessel may be derived from the afferent ctenidial vessel. Details of the histology of the lung are found in LUTFY & DEMIAN (1965).
The blood situated in the lung sac vessels flows to the ctenidial vein, which supplies both, the gill and the lung (ANDREWS & LITTLE, 1972: fig.ld; this study). In some species (character 26), the ctenidial vein is duplicated in its anterior and middle regions, differentiating a lung efferent vessel. However, PRASHAD (1925) has described for Pila a totally duplicated vessel, i.e., a pulmonary and ctenidial vein, wholly separated from the anterior region until they reach the auricle (see his figs. 10, 12, and fig. 1 of pl.18). This condition is assumed to be the more derived State. The additional afferent vein of Pomacea canaliculata has been advocated as a development of a horizontal division of the efferent ctenidial vein (ANDREWS, 1965b). A study on the respiratory mechanism of an ampullariid (Marisa
cornuarietis) was provided by DEMIAN (1965). BROOKS (1907) and BROOKS & MCGLONE (1908, pis. 1-7) have reported the embryological appearance of the ampullariid lung, and suggested an origin from the gill.
The long lung sac of Marisa is perhaps an adaptation to the planispiral shell, and is suggested that it has been derived from the short lung sac, present in other ampullariids. For this reason, the 3 States of character 29 are considered additive. However, nothing changes if the character is not considered additive. The lung of Pomacea has been regarded as more highly organized than that of other genera (ANDREWS, 1965b: 88), this is true in relation to Marisa, but this does not differentiate the genus from Felipponea, Pomella and Asolene; anyway, this character could be another basing the node 12.
30. Hypobranchial gland: 0= thick; 1= thin, inconspicuous (ampullariids, Notopala spp., Larina,
Annularia, cyclophorids) (Cl: 33; RI: 33).
The hypobranchial gland is normally well developed in caenogastropods, particularly in aquatic taxa. However, most ingroup species have a thin, inconspicuous hypobranchial gland, practically represented only by the mantle mucosa. ANDREWS (1965b) demonstrated that hypobranchial gland is present in Pomacea canaliculata, but only distinguishable in sections, and is unusual in covering the rectum and genital ducts. Further studies are necessary to determine, though, if the
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 427
caenogastropod hypobranchial gland is actually homologous to the hypobranchial gland of the archaeogastropods.
Cyclophorids have modified the hypobranchial gland for an accessory excretory gland (ANDREWS & LITTLE, 1972, 1982). This feature is commented in the character 44.
The basal gland, present in the base of the male penis shield of the ampullariids, has been referred to as hypobranchial gland. This nomenclature is not followed here because the gland is only present in the males. Additionally, nothing else suggests a homology of this gland with the ordinary hypobranchial gland, normally located between the gill and the rectum.
ClRCULATORY SYSTEM
31. Pericardium: 0= visceral and pallial; 1= also connected to pallial floor (ampullariids) (Cl: 100; RI: 100).
32. Ampullain anterior aorta: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
The ampullariids have the two modifications in the heart explored in the characters 31 and 32. The ampulla at the origin of the anterior aorta (Figs. 154, 179:am) is of particular interest for future physiological studies. This structure is almost as large as the ventricle, differing by its thinner walls, and by its location just after the origin of the posterior aorta. The ampulla is a long known ampullariid feature (e.g., PRASHAD, 1925: fig.10), and has also been taken into consideration by BERTHOLD (1991) and BIELER (1993, characters 5, 40).
The development of an ampulla have been advocated as an adaptation to accommodating the blood forced into the heart during the retraction of the animal into its shell (ANDREWS, 1965b).
33. Intestinal loop inside pericardium: 0= absent; 1= present (Larina, Notopalaspp.) (Cl: 100; RI: 100).
Normally there is a structural correlation between the pericardial cavity and the kidney cavity, particularly with the posterior renal chamber (when 2 chambers are present). The Australian viviparids Notopala and Larina are derived in have lost the membrane that separates both the pericardium and the posterior renal chamber. This State forms a wide cavity in which the heart and intestinal loops stay together.
Excretory system
34. Number of kidneys: 0=2; 1 = 1 (all species) (Cl: 100; RI: 100).
Most archaeogastropods possess a pair of kidneys, as well as a pair of pericardial structures and pallial organs. The caenogastropods, on the other hand, lost totally the (post-torsional) right structures, including the kidney. However, it is known that the loss of the right structures is a derived condition shared with the heterobranchs, the caenogastropods sister-taxon, and also some sparse archaeogastropod taxa.
35. Renal tissue: 0= solid glandular mass in pallial cavity roof; 1= solid glandular mass confined in a separated chamber (ampullariids); 2= a thin layer of gland and vessels (viviparids) (Cl: 100; RI: 100; non-additive).
36. Renal lobes: 0= single: 1= 2 (all species) (Cl: 100; RI: 100).
37. Intestinal loops in kidney chamber connected by a mesentery: 0= absent; 1= present (ampullariids, Annularía) (Cl: 100; RI: 100).
The clear mesentery present in some ingroup species, connecting the intestinal loops inside the posterior renal chamber to its dorsal-right surface, is an interesting feature, mainly relative to the discussion of the coelomatic condition of molluscs, as it suggests a true coelom. BERTHOLD (1991) and BIELER (1993, character 18) have considered a “repeatedly coiled” intestine as an ampullariid feature, however, but did not mention an association with the kidney. On the other hand, BOUVIER (1888, figs.2, 7) BURNE (1898) and PRASHAD (1925) have reported the coiled intestine inside the renal chamber. The ampullariid kidney function has been investigated by LITTLE (1968), but no information on the function of the intestinal loops was reported.
38. Anterior septate chamber: 0= absent; 1= present, with two equal-sized series of septa (Marisa); 2= present, with two asymmetrical series of septa (remainder ampullariids) (Cl: 100; RI: 100; additive).
The anterior chamber has been reported by BOUVIER (1888, fig.3) and PRASHAD (1925, fig.13), and was also considered by BERTHOLD (1991) and BIELER (1993, character 6). Some histological features of this kidney region are found in PRASHAD (1925:124).
The anterior chamber has been referred as “ureter” by some authors [e.g. THIENGO, 1987, fig.8). This terminology can suggest any homology with the ureter of the viviparids, however, based on investigations on the structure of viviparids (JOHANSSON, 1950), ANDREWS (1965b: 88) have
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

428 L.R.L.SIMONE
suggested that this hypothesis is not upheld. Another current name for the anterior renal chamber is “lamellar renal organ” (BURNE, 1898).
The additive optimization is based on ontogeny, since very young specimens of all ampullariids possesses State 1. However, nothing changes if the character was considered non-additive.
39. Connection between longitudinal fold of renal anterior chamber with nephrostome edge: 0= absent, 1= present (ampullariids, except Marisa) (Cl: 100; RI: 100).
40. Nephrostome: 0= in middle region of membrane between kidney and pallial cavity; 1= close to posterior end of pallial cavity (cyclophorids); 2= inside ureter (viviparids); 3= connected by an anterior chamber (ampullariids) (Cl: 100; RI: 100; non-additive).
The position of the nephrostome of the ampullariids has been shown and discussed by BURNE (1898).
41. Closed ureter: 0= absent; 1= present (viviparids) (Cl: 100; RI: 100).
42. Communication between kidney and ureter; 0= absent; 1= an open pore (viviparids); 2= a muscular papilla (Notopala spp, Larina) (Cl: 100; RI: 100; additive).
43. Urinary gutter: 0= absent; 1= a furrow along right margin of pallial cavity and head (cyclophorids); 2= a fold along rectum and pallial gonoducts (ampullariids) (Cl: 100; RI: 100; non-additive).
PRASHAD (1925), DEMIAN (1965) and LUTFY & DEMIAN (1965) have called the urinary gutter of ampullariids as “epitaenia”.
44. Collar vessel: 0= inserted in kidney; 1= inserted in auricle (A blanchetiana, I. inca) (Cl: 100; RI: 100).
The ingroup taxa are all non-marine gastropods. Adaptations to freshwater (ampullariids, viviparids) or terrestrial (cyclophorids, annulariids) environments certainly demand modifications of the kidney, because this organ must conserve electrolytes (freshwater forms) and water (terrestrial forms). These extra efforts, if compared with marine groups, certainly brings effects in the morphology. This has presumably resulted in the complexity of the ingroup kidneys, outlined in characters 35-44.
Although the members of each ingroup family have similar kidney modifications, each family is very different from each other. All taxa have two chambers, a posterior, with intestinal loops and usually connected to pericardium, and another anterior, in pallial cavity. The cyclophorids differs
in having the posterior chamber lacking visible glands, and an almost solid anterior chamber. The ampullariids have a dorsal renal lobe in the posterior chamber and the anterior chamber has two series of transverse glandular septae. The viviparids resemble the cyclophorid arrangement, but the anterior chamber is solid.
The urine transportation from the nephrostome to the exterior also involves further modifications. The ampullariids and the cyclophorids have a urinary gutter, formed by a fold running along the right edge of the pallial cavity, from the nephrostome to the region behind the head (character 43). The viviparids, on the other hand, developed a tubular ureter (character 41), which conducts the urine from the nephrostome to the region close to the mantle border; this ureter runs attached to the rectum and its pore is close to the anus.
Most of the ingroup kidney structures have obscure functions, and are an interesting area for further investigation. ANDREWS & LITTLE (1972) performed a discussion on of the excretory system of architaenioglossans, comparing their anatomy and physiology (their fig.6, p.410). The authors described a large hypobranchial gland in all examined species (no one from South America), composed by subepithelial tubular glands, secreting purines, phospholipids and mucus. Such a gland is absent, at least as a clear organ, in the examined species, except Incidostoma tupy, in which it is small and lies covered ventrally by posterior portion of rectum [Fig.262, middle-superior region, between rectum (rt)]. Further and wider discussion on the excretory strategies, in a comparative scenario, is provided by ANDREWS (e.g., 1979, 1981).
Digestive System
Buccal mass
45. Jaws: 0= pair of small plates; 1= very large (ampullariids); 2= plated fused in median line (cyclophorids); 3= absent (Annularia) (Cl: 100; RI: 100; non-additive).
The enlargement of the ampullariid jaws has been also considered by BERTHOLD (1991) and BIELER (1993, character 11).
46. Dorsal chamber of buccal mass: 0= deep; 1 = shallow (Annularia, cerithioideans) (Cl: 100; RI: 100).
47. Dorsal folds of buccal mass: 0= low, inconspicuous; 1= broad, tall (all species); 2= with transverse furrows (ampullariids) (Cl: 100; RI: 100; additive).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 429
The dorsal chamber (character 46) is located in the dorsal wall of the buccal mass, opposite the odontophore, and may be an adaptation to avoid radular auto-injury. It is particularly large in the archaeogastropods. In the caenogastropods, however, there are a pair of broad and somewhat tall folds (character 47), and the dorsal chamber is reduced. These States have been regarded as notable synapomorphies of the ingroup (e.g., Fig. 160: df, dc). Heterobranchs, on the other hand, apparently lack both the folds and chamber, with the dorsal wall of the buccal mass more muscular and with the inner surface simpler. The dorsal folds had been also reported by PRASHAD (1925), and called a “buccal gland on the roof of the buccal cavity”.
The additive optimization is based on ontogeny, since young specimens of ampullariids lack transverse furrows, acquiring them early in development. However, the result and indices are the same if the character is considered non-additive.
Odontophore
48. Mj: 0= diffuse; 1= 2 broad bands (all species) (Cl: 100; RI: 100).
49. Mia: 0= absent; 1= present (cyclophorids) (Cl: 100; RI: 100).
50. Mlb: 0= absent; 1= double [N. prominulus); 2= single mass (A. blanchetiana, I. inca) (Cl: 100; RI: 100; non-additive).
51. M2 as pair of retractor muscles of buccal mass: 0= absent; 1= present (viviparids, Annularia,
cerithioideans) (Cl: 100; RI: 100).
The m2 of the 3 above mentioned groups work as retractors of the buccal mass (retractor of “pharynx”). The pair is dissimilar to that called m2 of ampullariids, which is an abductor of the jaw plates, and is not considered homologous.
52. Ma: 0= absent; 1= present and single (cyclophorids); 2= multiple (ampullariids) (Cl: 100; RI: 100; non-additive).
PRASHAD (1925, fig.4) described 3 pairs of here denominate ‘ma’ for Pila, calling them anterior dorsolateral muscles; while BROWN & BERTHOLD (1990), describing Lanistes neritoides, called madible adductors.
53. M3: 0= absent; 1= pair with dorso-ventral fibers (N. prominulus); 2= single, with transverse (latero- lateral) fibers (P. scalaris, P. curumim, P. lineata. P.
bridgesi, P. sórdida, Annularia); 3= both (H-fashion) [Marisa, P. crosseana. F. neritiniformis, P. canaliculata,
A. megastoma) (Cl: 60; RI: 80; non-additive).
An m3 is also present in Pila, according to PRASHAD (1925, fig. 5), which he called “superficial muscles of the ventral surface of the buccal mass”.
54. M4 pair: 0= composed by several muscles; 1 = composed by 2 pairs of muscular layers surrounding odontophore cartilages (all species); 2= a single pair of large mass surrounding odontophore cartilages (viviparids; Annularia,
cerithioideans) (Cl: 100; RI: 100; additive).
The additive optimization is a try for testing if the large mass fashioned m4 is a modification (fusion) of the 2 layered m4 of 2 more basal branches. However, the result and indices do not change if the character was optimized as non-additive. This result is compatible with the suggestion, but further studies are surely necessary.
55. M4 connection with tissue on middle region of radula (to): 0= absent; 1= via m9 (ampullariids); 2= present (viviparids, Annularia, cerithioideans) (Cl: 100; RI: 100; non-additive).
Although in part of the viviparids (Larina and Notopala) a pair of m9 muscles is found (divided in some separated fibers), they have a connection between the m4 pair with the above referred tissue. Then, these species are coded in the State 2.
56. Ventral and dorsal branches of m4 pair: 0= not connected anteriorly; 1= connected anteriorly (viviparids, Annularia, cerithioideans) (Cl: 100; RI: 100).
57. Ventral tensor muscle of radula: 0= present; 1 = very reduced (mil) (all species) (Cl: 100; RI: 100).
58. Dorsal tensor muscle of radula: 0= weak; 1 = strong (m4 + m5) (all species) (Cl: 100; RI: 100).
59. M5 pair: 0= connected with odontophore cartilages; 1= on m4 (all species) (Cl: 100; RI: 100).
60. M5 pair form: 0= short and broad; 1= long and thin (all species) (Cl: 100; RI: 100).
61. M6: 0= edging most of medial surface of odontophore cartilages; 1= edging less than 2/3 of this surface (all species) (Cl: 100; RI: 100).
62. M6: 0= narrow and thin; 1= broad and thick (all species) (Cl: 100; RI: 100).
63. M7: 0= absent; 1= present (all species) (Cl: 100; RI: 100).
64. M7a pair (inserted in m5): 0= absent; 1= present [Viviparus spp) (Cl: 100; RI: 100).
65. M2 as abductor of jaw plates: 0= absent; 1 = present (ampullariids) (Cl: 100; RI: 100). See comments after character 51.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

430 L.R.L.SIMONE
66. Approximator muscles of cartilages: 0= present; 1= absent or part ofm4 (all species) (Cl: 100; RI: 100).
67. M9: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
68. M10 insertion in odontophore ventral surface: 0= posterior; 1= anterior (all species) (Cl: 100; RI: 100).
69. MIOa: 0= absent; 1= present (viviparids) (Cl: 100; RI: 100).
70. M 10c: 0= absent; 1= present (cyclophorids) (Cl: 100; RI: 100).
71. Mil: 0= broad, as ventral tensor muscle of radula; 1= narrow (all species) (Cl: 100; RI: 100).
72. M12: 0= absent; 1= present (ampullariids, Vivipams spp, cerithioideans) (Cl: 33; RI: 71).
73. M14: 0= absent; 1= present (Annularia) (Cl: 100; RI: 100).
74. Number of main cartilages: 0= more than 1 pair; 1= single pair (all species) (Cl: 100; RI: 100).
75. Pair of accessory cartilages: 0= absent; 1 = present (ampullariids) (Cl: 100; RI: 100).
The odontophore muscles and structures are an important source of comparative data (characters 48-75). The architaenioglossans present an interesting link between the archaeogastropod and other caenogastropod odontophores in several characters. An example is the pair of m4 muscles. This muscle is the largest pair from the caenogastropod odontophore, surrounding almost the entire cartilages, and apparently works as the main tensor of the subradular membrane. Analyzing the archaeogastropod odontophore, nothing similar to m4 is found. Apparently, the caenogastropod m4 is a combination of several pairs of archaeogastropod muscles, such as the several pairs that connect the cartilages to the subradular membrane (character 54), and also the pairs that connect the anterior with the posterior cartilages (character 66). The two first branches of the caenogastropods, the Cyclophoroidea and the Ampullarioidea, have the m4, composed by two layers (m4 and m4a). This State is here interpreted as an intermediate step to a single-layered and thick m4, derived from several pairs of muscles. The odontophore cartilages (character 74) are present, in general, as more than one pair in the archaeogastropods, mostly as two pairs (anterior and posterior pairs). However, all caenogastropods (including the architaenioglossans) have a single pair. This pair is homologue to the anterior pair of archaeogastropod cartilages, according to the
analysis of some muscles, such as, e.g., the horizontal muscle (m6).
A possible function of the muscles and odontophore is suggested elsewhere (SIMONE, 2001), the odontophore of the caenogastropods, including those of architaenioglossans, differs from the archaeogastropod and heterobranch odontophore in having no apparent gliding between the radula and the cartilages, provided, in those taxa, by the ventral and dorsal tensor muscles (characters 57, 58) working alternately. In the caenogastropods, the muscles apparently work pulling and stretching the subradular structures (membrane and cartilage) and the entire odontophore works grinding the food.
Most of the comparative analyses and the polarization of the odontophore characters (48-75) are based on comparisons with the archaeogastropods. Although some heterobranchs were examined, their odontophore apparently has other, still unclear, modifications, any comparative analysis is still premature.
The present discussion on odontophore features is still superficial. Further analysis will be done in the final stage of the project, when all caenogastropod superfamilies will be analyzed and each muscle will be discussed.
The ampullariids developed a secondary pair of thin cartilages (Figs. 166-167:oa), this pair of cartilages is probably a modification of the subradular membrane. These cartilages connect the ventral and dorsal branches of the pair m4. Analyzing their location, it is clear that the secondary cartilages are new acquisitions. The cyclophorids have nothing uniting both referred branches of the m4, while the remainder caenogastropods (including viviparids) both branches are United with each other (character 55). PRASHAD (1925) performed a detailed study on the odontophore of Pila globosa and have described two pairs of cartilages. What he called “superior cartilages” are apparently what is called here subradular cartilage {e.g., figs. 163, 286: sc). This structure is shown as very thick (his figs 6, 8, 9), but he referred to them as very thin in the text (PRASHAD 1925:102). Then, PRASHAD’s “superior” cartilages are not related to the accessory cartilages described herein, suggesting that Pila lacks this attribute. He called the ordinary odontophore cartilages as “lateral” cartilages.
Radula
76. Subradular cartilage: 0= narrow; 1= broad (all species) (Cl: 100; RI: 100).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 431
The subradular cartilage (sc) is most probably secreted by the subradular membrane (br), and is a transparent, chitinous pair of expansions of the radula, in buccal cavity. The subradular cartilage is a specialization of the cartilage that covers the inferior surface of the radular ribbon, which links the teeth.
The subradular cartilage is present in the archaeogastropods, but is narrow. In caenogastropods, the structure is broad, entirely covering the buccal surface of the subradular membrane that, otherwise, would be exposed directly to the buccal cavity, accessible to injuries. The heterobranchs apparently also have a broad subradular cartilage, but is sill unclear its State in the basal taxa.
77. Radular sac length: 0= long; 1= short (about samelength of buccal mass) (ampullariids) (Cl: 100; RI: 100).
The short length of the radula in ampullariids has also been considered by BERTHOLD (1991) and BIELER (1993, character 12).
78. Rachidian form: 0= narrow; 1= broad (ampullariids); 2= tall and curved (viviparids) (Cl: 100; RI: 100; non-additive).
79. Rachidian cusps: 0= few (0-3); 1= multicuspid (ampullariids, viviparids, cerithioideans) (Cl: 50; RI: 75).
80. Rachidian basal broad cusp pair: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
81. Rachidian tip: 0= broad; 1= pointed [A.
blanchetiana; I. inca; Annularia) (Cl: 50; RI: 50).
82. Lateral teeth number: 0= several (about 5); 1 = 1 (all species) (Cl: 100; RI: 100).
83. Lateral teeth tip: 0= multicuspid; 1= 1-2 cusps (A. megastoma, F. neritiniformis, cyclophorids, Annularia) (Cl: 25; RI: 40).
84. Lateral teeth shape: 0= different from rachidian; 1= tip tends to be similar shaped to rachidian (all species) (Cl: 100; RI: 100).
85. Marginal teeth number: 0= many (more than 10); 1= 2 (all species) (Cl: 100; RI: 100).
86. Marginal teeth shape: 0= similar to each other; 1= different (all species) (Cl: 100; RI: 100).
In the current literature, radular characters have generally been more important than those of the shell and operculum in comparative analyses. Several radular characters were in fact searched in present study, but they were eliminated because resulted as autapomorphies. Only the characters
77-86 are suitable, and were polarized mainly based on the analysis on the archaeogastropods. In those taxa, the typical radula is rhipidoglossate, with a central (rachidian), about 5 lateral, and many marginal teeth. Nearly all caenogastropods, however, have a taenioglossate radula (or a kind of radula derived from it), including the architaenioglossans. The taenioglossate radula has only 7 teeth per row: a central or rachidian, a pair of lateral, and two pairs of marginal teeth.
Salivary glands
87. Aperture in dorsal wall of buccal mass: 0= middle; 1= anterior (viviparids, Annularia,
cerithioideans) (Cl: 100; RI: 100).
88. Salivary glands: 0= absent; 1= present as 2 separated masses (all species); 2= a single fused mass (ampullariids, Annularia) (Cl: 66; RI: 88; additive).
The additive optimization of this character is based on ontogeny, since, as well as is known, the ontogeny of the salivary glands starts as 2 separated masses. The apparent fusion is normally due to enlargement. If the character was considered non-additive, nothing changes in the result, only the RI changes to 87.
89. Type: 0=glandular; 1= tubular (Notopala spp, Larina) (Cl: 100; RI: 100).
Salivary glands are present in other classes of the mollusks, but there are several archaeogastropod taxa lacking them (SASAKI, 1998). The polarization of salivary glands characters (87-89) is mainly based on the archaeogastropods that possess them.
The salivary glands are particularly well developed in caenogastropods. Additionally, they developed salivary ducts, a character also shared with heterobranchs.
The salivary aperture in the caenogastropods is distinct in opening generally in the anterior region of the dorsal folds of buccal mass (character 47), (character 87) in the taxa allocated after the node 5.
Esophagus
90. Position of esophagus origin: 0= in lateral- dorsal region of odontophore; 1= in its posterior region (all species) (Cl: 100; RI: 100).
The typical shape of the archaeogastropod foregut is a “V”, with the mouth at the base, and the arms are the odontophore and the esophagus respectively. This differs from the arrangement typically found in the caenogastropods, where the esophagus is more or less continuous with the
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

432 L.R.L.SIMONE
buccal mass. In this case, the esophagus is inserted in the posterior-dorsal region of the odontophore. The caenogastropod fashion is considered derived, although appears to be shared with some heterobranchs.
91. Anterior esophagus: 0= with a pair of pouches; 1= a simple tube (viviparids, Annularia,
cerithioideans) (Cl: 100; RI: 100).
92. Esophageal pouches: 0= a simple diverticulum; 1= covered with papillae (cyclophorids); 2= with a dorsal fold (ampullariids) (Cl: 100; RI: 100; non-additive).
93. Special pair of blood vessels connected to esophageal pouches: 0= absent; 1= present (ampullariids, cyclophorids) (Cl: 100; RI: 100).
The cyclophorids and the ampullariids have a pair of prominent pouches connected to the lateral surface of the anterior esophagus. Their inner surface connects with the adjacent esophageal cavity. Important branches of the anterior aorta irrigate both pouches. Something similar, but lacking the distinct blood vessels, is found in vetigastropods and neritimorphs, and other archaeogastropods (SASAKI, 1998: 150; pers.obs.). The presence of esophageal pouches is considered plesiomorphic, however, it is possible that they are not homologous structures, because they differ in several details as referred abo ve. The esophageal pouches with demarcated esophageal gland have been considered by BERTHOLD (1991) and BIELER (1993, character 16). PRASHAD (1925: 109, fig.4) described short and rounded esophageal pouches for Pila, which differ from the species studied here. PRASHAD (1925:116, fig. 11) described the arterial system in the foregut in more detail, and showed a small pair of vessels irrigating the pouches.
94. Middle esophagus: 0= narrow; 1= bulging (ampullariids except Marisa); 2= very narrow (Notopala spp, Larina) (Cl: 100; RI: 100; non- additive) .
95. Esophageal insertion in stomach: 0= in posterior region; 1= in middle region (cyclophorids, Annularia, cerithioideans) (Cl: 50; RI: 75).
The insertion of the esophagus in the posterior region of the stomach (state 0) is “U”-shaped, as happens in most archaeogastropods. The esophageal insertion in the middle region of the stomach (state 1) is apparently associated with gastric enlargement, and resulted in convergence between the cyclophorids (first branch) and the remainder of the basal caenogastropods (represented by node 7).
Stomach
96. Gastric caecum: 0= developed; 1= small, only internai (Viviparus spp); 2= absent (remainder species) (Cl: 66; RI: 50; additive).
A homologous structure to the archaeogastropod gastric caecum is not found in the ingroup species. However, the viviparids have a septum that surrounds a small chamber, and possesses a folded inner surface. This small chamber was initially interpreted as a caecum vestige (as denoted by the above States).
The gastric caecum was apparently lost in the caenogastropods and, if the small viviparid chamber is homologous to that, it is a reversion. Nevertheless, it is likely that this structure is a new viviparid acquisition. For testing this hypothesis, the character was considered additive. The result is the same if it was considered non-additive. Both indices, however, change to 100. This may be an indication that the Viviparus structure may be a new acquisition. PONDER & LINDBERG (1997) argue that gastric caecum is a vetigastropod apomorphy.
97. Stomach walls: 0= thin; 1= dorsal half muscular (Marisa, P. crosseana, P. scalaris, F. neritiniformis); 2= almost entirely muscular (remainder of ampullariids) (Cl: 100; RI: 100; additive).
98. Stomach transverse muscular septum: 0= absent; 1= present (ampullariids); 2= bending inwards (P. lineata, P. bridgesi, P. canaliculata, A.
megastoma) (Cl: 66; RI: 90; additive).
The ampullariid stomach is unusual in being highly muscular (character 97), working as a gizzard. Additionally, the stomach developed a muscular transverse septum (character 98). This septum divides the stomach of most ampullariids into 2 chambers, a larger dorsal chamber with muscular walls, and a ventral chamber with thin walls, bearing the ducts to digestive gland. This septum is also present in the genus Pila (PRASHAD, 1925).
The characters 97 and 98 are optimized as additive because of the ontogeny, since state 1 precedes 2 during development. The result is the same if both States were considered non-additive, only the RI of the character 98, changes from 90 to 85.
99. Gastric muscle: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
The gastric muscle, an ampullariid synapomorphy, is apparently reported for the first time. Its origin is in the shell columella, just posterior to that of the columellar muscle; and it runs dorsally and anteriorly, and penetrates the stomach wall just
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 433
anterior to the ventral gastric chamber. After this, the gastric muscle fibers spread like a fan along the gastric muscular walls, and divide the stomach into esophageal and intestinal branches.
The gastric muscle is small in Marisa, and broad and thick in the remainder of the studied ampullariids. Nothing is known about its occurrence in other ampullariid genera, as it is not reported in the consulted literature.
The gastric muscle probably works as a base for the musculature of the gastric wall. But, further investigations are necessary to clarify the function of this muscle, as well as the remainder of the complex structures of the ampullariid stomach, some of which are explored in characters 97-99. The ampullariid stomach is a rich source of characters for comparative analysis, but has been poorly explored. BERTHOLD (1991) and BIELER (1993) considered only a single character (17): 2 median stomach grooves.
100. Ducts to digestive gland: 0= 2; 1= 1 (cyclophorids, Annularia, Notopala spp.); 2= several (ampullariids) (Cl: 50; RI: 77; non-additive).
The plesiomorphic number of the ducts to the digestive gland, as shown by most archaeogastropods, is a pair. One of them is in the esophageal insertion, and the other in the stomach ventral surface, close to the intestinal origin. However, the number and location of the ducts are highly variable. A duct may reduce and even disappear (state 1). On the other hand, the duct may multiply, as in the case of the ampullariids (state 2). The analysis of the location of the several ducts to the digestive gland in the ampullariids suggests that they are derived from both original ducts, with both becoming close together on the ventral gastric surface. This is suggested because there are ducts turned both, anteriorly and posteriorly. Notwithstanding, it is possible that the esophageal duct could be missing, and all ducts are a result of the division of the gastric duct only. This could probably be resolved with further studies on ontogeny.
101. Style sac: 0= present; 1= absent (ampullariids, viviparids, cyclophorids) (Cl: 50; RI: 50).
Despite the absence of a gastric style sac in the archaeogastropods (the main outgroups), its presence in the caenogastropods is regarded as plesiomorphic. This polarization is based on the presence of style sacs in other classes, notably the bivalves. Thus loss is assumed in the archaeogastropod taxa, as well as the architaenioglossans. Its presence in the remaining
caenogastropods is interpreted as a reversion. On the other hand, it is possible that the caenogastropod style sac is an extraordinary convergence with those of bivalves.
Intestine
102. Intestinal loop close to kidney-pericardium region, in left part of visceral mass: 0= absent; 1 = present (ampullariids) (Cl: 100; RI: 100).
103. Dorsal chamber of intestinal region close to pericardium: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
104. Ventral chamber of intestinal region close to pericardium: 0= absent; 1= simple (ampullariids); 2= complex (P lineata, P. canaliculata, A. megastoma)
(Cl: 66; RI: 91; additive).
The ampullariids have an intestinal region near the pericardium, which contain 1 to several small, blind-sacs. Apparently the region is reported for the fist time. These chambers are disposed from longitudinally to transversally, and the aperture of each is normally turned proximally (contrary to the flow). Sometimes, the transverse chamber has its origin in the longitudinal chamber aperture. The term “complex” is used for that state with several small sub-chambers.
The formation and function of the intestinal chambers are unknown and interesting goal for further investigations.
PRASHAD (1925) have reported an intestinal “caecum” for Pila globosa, being a short blind sac in which he tried to find a crystalline style. Looking at his fig.2 (p.99) the topology of the caecum is comparable to the topology of the intestinal chambers, however, the aperture of this caecum is close to the stomach in his fig.7 of pl.17. The homology of these structures remains unclear.
105. Intestinal loops in visceral mass: 0= several; 1= a single “S” (ampullariids, viviparids, Annularia,
cerithioideans) (Cl: 100; RI: 100).
106. Intestinal loops localization: 0= immerse in digestive gland; 1= free from digestive gland (ampullariids, viviparids, Annularia, cerithioideans) (Cl: 100; RI: 100).
107. Intestinal loop in haemocoel: 0= present; 1 = absent (all species) (Cl: 100; RI: 100).
Unlike the archaeogastropods and heterobranchs, the caenogastropods tend towards a reduction of the midgut, in favor of the development of the foregut, even in herbivorous groups. Most archaeogastropods and heterobranchs have a
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

434 L.R.L.SIMONE
several-looped intestine, mostly immersed in the digestive gland, then, in visceral mass. The caenogastropods, on the other hand, including the architaenioglossans, have the midgut “Z”-shaped (an angle is the stomach and the other the pericardial or renal loop. In the cases of several looped caenogastropod intestines, the loops are inside the kidney or in the pallial cavity.
The haemocoel of the caenogastropods is separated from the visceral cavity by a thin septum. This obstructs the passage of visceral structures, what does not happen in other gastropods. In some archaeogastropod groups, such as vetigastropods and neritimorphs, there is an intestinal loop almost touching the buccal mass. Nothing similar is found in the ingroup, however, SASAKI (1998: 151) considered the condition of ampullariids comparable to archaeogastropods.
108. Intestinal region passing through kidney: 0= with single or two loops; 1= several looped (P. scálaris,
P. curumim, P. lineata, P. bridgesi, P. sórdida, P.
canaliculata, A. megastoma, Marisa) (Cl: 50; RI: 87).
The intestinal loops inside the renal chamber of the ampullariids are here apparently explored for comparative purposes for the first time. There is uniformity of the loops shape in samples of the same species, and there are conspicuous differences among species. Those differences certainly are useful for species distinction. Same ampullariids have complex renal intestinal loops that, besides their interest in systematics, are an interesting subject for further physiological analyses.
109. Fecal pellets: 0= absent; 1= present (all species) (Cl: 100; RI: 100).
Fecal pellets were also explored as phylogenetic character by previous authors (PONDER & LINDBERG, 1997; SASAKI, 1998) and are also the goal of comparative studies (MOORE, 1931). In the present study, fecal pellets resulted as a notable synapomorphy of the Caenogastropoda, differing from the archaeogastropods and heterobranchs that, characteristically, have a single continuous fecal strip.
Fecal pellets are also convergently found in some Polyplacophora and in amphibolids (Pulmonata) (PONDER & LINDBERG, 1997).
Genital System
d 110. Testis: 0= along visceral mass; 1= restrict to posterior region of visceral mass (cyclophorids) (Cl: 100; RI: 100).
111. Seminal vesicle: 0= absent; 1= present (Annularia, cerithioideans) (Cl: 100; RI: 100).
The seminal vesicle is a differentiated glandular portion of the anterior region of the visceral vas deferens, in general convolute and iridescent because of presence of spermatozoids. The vesicle is absent in the architaenioglossans, but remarkably present in remainder caenogastropods. PRASHAD (1925:127, fig.15) reported a “vesícula seminalis” for the ampullariid Pila, but of different fashion and topology, certainly a new acquisition.
112. Prostate: 0= absent; 1= present in pallial region (all species) (Cl: 100; RI: 100).
The prostate is a broad glandular portion of the caenogastropod pallial vas deferens. The gland may be opened (a furrow), as in the cerithioideans, or closed (a tube), as in architaenioglossans and annulariids. The prostate gland most probably secretes the spermatophore. The prostate of the Australian viviparids (Notopala, Larina) is difficult to be seen because of the pallial positioned testis. According to the literature (e.g., ANNANDALE & SEWELL, 1921), the prostate lies in the wall below the food groove and is actually difficult to be seen.
113. Pallial vas deferens: 0= a furrow; 1= partially closed (cyclophorids); 2= entirely closed (tubular) (ampullariids, viviparids, Annularia) (Cl: 50; RI: 60; additive).
The vas deferens is mostly absent in archaeogastropods (except Neritimorpha), and highly modified in the heterobranchs (running free into haemocoel). Those facts in part preclude the analysis of the structure in the ingroup. However, the 3 States are presently aligned in a single ordered evolutionary trend based on the ontogeny, which the 3 States appear successively during the development of the State 3-bearing groups. The result is the same if they were optimized as non- additive, both indices, however, change to 66.
114. Vas deferens aperture: 0= in posterior end of pallial cavity; 1= in dorsal end of right cephalic tentacle (viviparids); 2= in penis tip (ampullariids, Annularia); 3= in penis base (cyclophorids) (Cl: 75; RI: 85; non-additive).
The character has connection with the degree of closure of the pallial vas deferens, in such the States 0 and 1-2 are extremes (entire open and entire closed respectively), with the State 3 as intermediary.
115. Ejaculatory tube as a long muscular portion
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 435
of vas deferens immersed in integument, and protruding into pallial floor and haemocoel: 0= absent; 1= present (viviparids) (Cl: 100; RI: 100).
116. Right cephalic tentacle modified as copulatory organ, with vas deferens running along it: 0= absent; 1= present (viviparids) (Cl: 100; RI: 100).
117. Special cavity for retraction of right cephalic tentacle tip: 0= absent; 1= present (Viviparus spp) (Cl: 100; RI: 100).
The characters 115-117 and the State 2 of the character 114 explore the modification of the male pallial genital structures of viviparids, which are very different from any other gastropod. The pallial vas deferens runs in the floor of the pallial cavity, immerse in the integument, mostly surrounded by thick muscular walls disposed concentrically in many layers, protruding into the haemocoel. The right cephalic tentacle is also modified as a copulatory organ, or a penis-like structure. Although this kind of modification of the right tentacle is convergently found in several cocculiniforms and some neritimorphs, the viviparid modification is unique in having a closed duct running along its inner region. Besides, several viviparid species have special features of the copulatory tentacle, which are characteristic of the species and genera, as explored in the character 117 and others that were autapomorphic.
118. Exophalic penis (behind right tentacle): 0= absent; 1= present (cyclophorids, Annularia) (Cl: 50; RI: 66).
119. Papilla on exophalic penis tip: 0= absent; 1 = simple (N. prominulus); 2= double (A. blanchetiana,
I. inca) (Cl: 100; RI: 100; non-additive).
The cyclophorid exophalic penis (characters 118- 119) is extraordinarily similar in shape and location to the exophalic penis found in the higher caenogastropods, such as most mesogastropods and neogastropods. They are here considered homologues. But, according to the tree, the cyclophorid structure is convergent. More complex penises have been describes for other cyclophorids, having even retractile terminal papilla, e.g., in Murdochiapallidum (MORTON, 1952).
The penis of annulariids is apparently homologous to those of higher caenogastropods. However, THOMPSON (1978) referred to the penis as of pallial origin and innerved by the left pleural ganglion.
120. Penis sac: 0= absent; 1= present, with abroad
and short penis (ampullariids); 2= present, with a very long, convolute penis (P. crosseana, P.
canaliculata, A. megastoma) (Cl: 66; RI: 90; additive).
The ordered optimization is based on th ontogeny, since young specimens of taxa with State 2 penis have a State 1-like one. The result does not change if the character was considered non-additive, only the RI changes to 87.
121. Penis duct highly convolute: 0= no; 1= yes [P.
crosseana, A. megastoma, P. sórdida, P. canaliculata,
P. lineata) (Cl: 33; RI: 60).
122. Penis shield: 0= absent; 1= present (ampullariids) (Cl: 100; RI: 100).
123. Penis shield tip: 0= absent; 1= simple (ampullariids); 2= glandular (P. curumim, P. lineata,
P. bridgesi, P. sórdida, P. canaliculata, A. megastoma)
(Cl: 100; RI: 100; non-additive).
124. Penis shield middle region: 0= simple; 1= with a flap (ampullariids).
125. Penis shield middle-right gland: 0= absent; 1= present (Marisa, F. neritiniformis, P. curumim, P.
lineata, P. bridgesi, P. sórdida, P. canaliculata) (Cl: 25; RI: 57).
126. Penis shield gland in tip: 0= no gland; 1= far from middle flap (P. curumim, P. lineata), 2= close to flap (P. canaliculata, P. bridgesi, P. sórdida, A.
megastoma) (Cl: 100; RI: 100; non-additive).
Despite the fact the 3 States are considered non- additive, the position of the taxa on the tree suggests that they actually may be additive. If considered additive, nothing changes, neither in result or indices.
127. Penis shield base: 0= without gill; 1= with gill (Marisa, P. curumim, P. lineata, P. bridgesi, A.
megastoma) (Cl: 25; RI: 40).
The ampullariid modifications of the male pallial genital structures (characters 120-127) are unlike those of other gastropods. The pallial vas deferens runs along the right edge of the pallial cavity, along the side rectum, as it is found in other caenogastropods, however this is the only similarity. Close to the anus, the vas deferens crosses to a penis sac (character 120), which is half solid and half hollow. The hollow portion contains a convolute and slender penis. The penis may be shorter and broad, or very long, with a filiform apical region (character 120). The duct of the penis may or not be highly convolute (character 121). The penis is protruded protected by a special structure of the mantle border - the
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

436 L.R.L.SIMONE
penis shield (characters 122-127). The penis
shield is very large, sometimes more than half
the volume of the pallial cavity. It is retracted
inside this cavity, except during copulation. The
penis shield is much more complex than the penis itself, and has several valuable characters for
comparative analysis. Although most of the
characters are autapomorphies in the present
study, some of them are explored in the
characters (123-127). Male genital characters
have been also considered by BERTHOLD (1989,
1991) and BIELER (1993, characters 7, 48-58).
Maybe due to the invasion of freshwater
(ampullariids and viviparids) and the terrestrial
(cyclophorids) habitats, the architaenioglossans are
highly modified in their male genital system. They
possess highly specialized features not found in
any other gastropod. On the other hand, no close
correlation between these major groups is possible,
showing that most probably these structures
evolved independently.
BERTHOLD (1991) and BIELER (1993, characters
8-9) also considered two other male features as
ampullariid synapomorphies: the presence of
eupyrene and atypical sperm anchored by nurse
cells, and reduction of the spermatophore.
9
128. Pallial oviduct: 0= absent; 1= present and
opened (a glandular furrow) (cerithioideans); 2=
closed (a glandular tube) (ampullariids, viviparids,
cyclophorids, Annularia) (Cl: 66; RI: 0; additive).
The pallial oviduct is present in neritimorphs
amongst the archaeogastropods, in all
heterobranchs and all caenogastropods. Possibly
it is an indicative of a close relationship among
those three groups. Some archaeogastropods,
however, have an inconspicuous furrow running
from the female genital aperture (e.g.,
cocculiniformians), which is an analogous the
structure. In caenogastropods, the pallial oviduct
can be open (as a furrow), this condition is found
only in the cerithioideans; or closed (tubular), found
in the other groups. It is suggested that the open
condition is a step between the inconspicuous
furrow and the closed State. Though, according to
the tree, the open condition of the cerithioideans
is a reversion. If the character was optimized as
unordered, nothing changes in the result, but both
indices become 100. The indices of the whole
cladogram become the following: length: 249
(instead 252), Cl: 77 (instead 76), RI: 91 (same).
129. Brood pouch: 0= absent; 1= present as part
of oviduct (viviparids) (Cl: 100; RI: 100).
The viviparid brood pouch (or uterus), contains
capsules with young specimens from 1 to 4 whorls.
These young specimens are born already well-
formed, justifying the name of the type genus of
the family.
130. Pallial oviduct localization: 0= in pallial floor;
1= entirely at right of rectum (all species) (Cl: 100;
RI: 100).
The position of the pallial oviduct in
caenogastropods differentiates it from the pallial
oviducts of other groups that possess this structure
(the heterobranchs and neritiforms). In these
groups, the pallial oviduct runs partially or totally
in the pallial floor. Obviously, in several
caenogastropods, when the pallial oviduct is
mature, acquiring enormous proportions, a part
of it also attaches to the floor of the cavity, but it
always arises from the pallial roof.
131. Albumen gland: 0= absent; 1= loop of visceral
oviduct (cyclophorids); 2: a compact mass
(ampullariids); 3= a posterior glandular sac (Notopala
spp, Larina) (Cl: 100; RI: 100; non-additive).
The albumen glands of the different architaenio
glossan taxa are doubtless not homologous. Most
probably they are independent acquisitions from
different portions of the pallial oviduct.
132. Bursa copulatrix: 0= absent; 1= present,
connected to visceral oviduct [N. prominulus); 2= present, opening in pallial cavity (A. blanchetiana,
I. inca) (Cl: 100; RI: 100; non-additive).
The bursa copulatrix is unusual in the
cyclophorids, in which (state 2) it opens directly to
the pallial cavity. The pallial oviduct thus has two apertures: a posterior one (the bursa) and an
anterior one (the capsule gland). The plesiomorphic
condition of the bursa connected to the pallial
oviduct and not directly connected to the pallial cavity is present in some cyclophorids, such as
Cyclophorus perdix tuba (Tielecke, 1940), and
Cyclobrandia beauianus (Petit) (THOMPSON, 1967).
THOMPSON (1969) has reported a more variable condition of the bursa aperture, in a study on
Mexican and Central American cyclophorids, which
he also called a seminal receptacle.
133. Capsule gland: 0= absent; 1= present (all
species); 2= with a complex, sometimes spiral inner
duct (ampullariids) (Cl: 100; RI: 100; additive).
The additive optimization is based on ontogeny.
Young ampullariid specimens possess a very simple
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 437
capsule gland duct. However, nothing changes if
the character was considered unordered.
134. Inner blind-sac glandular chamber in capsule
gland: 0= absent; 1= present (cyclophorids) (Cl: 100;
RI: 100).
Based on the morphological disparity and different
localization, the capsule glands (character 133-
134) of the different groups are possibly not
homologous. The gland of the ampullariids is very
large, and surrounds most structures of the pallial
oviduct. Its duct is complex, sometimes spiral or
with series of blind-sac, flat chambers. The gland
of the cyclophorids is U-shaped, with a branch as
a blind sac.
135. Female pore: 0= not developed (pallial oviduct
absent or opened); 1= a slit (cyclophorids,
Annularía)-, 2= a papilla (ampullariids, viviparids)
(Cl: 50; RI: 50).
The additive optimization is based on
comparison, since State 2 appears to be a
modification of State 1. The ontogeny also
corroborates this optimization as I have observed
in young specimens. However, nothing changes
in the result or indices if the character was
considered non-additive.
136. Vaginal tube: 0= absent; 1= origin in anterior
end of pallial oviduct (ampullariids); 2= origin in
middle levei of pallial oviduct (F. neritiniformis,
P. bridgesi, P. sórdida, A. megastoma) (Cl: 50; RI:
83; additive).
The vaginal tube is long and has thick-glandular
walls. It runs parallel to the rectum and connects
the capsule gland to the female pore. This
happens in two different ways as mentioned
above. The additive condition is based on the ontogeny as I have observed, because the species,
that have State 2 in the mature stage have State
1 in immaturity. The result is the same if the
character was considered non-additive, the single
change is RI: 75.
The pallial oviduct structures have been referred
here by names, based on the nomenclature found
in the literature and on topology. However, there
is no uniformity of the designations and there is
few taxa studied on the function of each portion
(ANDREWS, 1964, 1965a; PRINCE, 1967). The
nomenclature employed here is thus provisional.
137. Brood strategy: 0= absent (eggs free); 1 =
inside pallial oviduct (viviparids); 2= a jelly mass
laid out of water (ampullariids) (Cl: 100; RI: 100;
non-additive).
The reproductive strategies of the viviparids and
ampullariids are unusual for gastropods. In the
former, the viviparous strategy raising the name
of the type genus (Viviparus) (see FALNIOWSKI
et al., 1996). In the latter, the strategy is to fix the capsules out of the water, despite the aquatic
condition.
Central Nervous System
138. Nerve ring position: 0= surrounding mouth;
1= in middle levei of buccal mass (ampullariids,
cyclophorids); 2= posterior to buccal mass
(viviparids, Annularía, cerithioideans) (Cl: 100; RI:
100; additive).
The nerve ring position in the archaeogastropods
and heterobranchs is characteristically anterior
to the buccal mass. Notwithstanding, most
caenogastropods have the nerve ring posterior
to the buccal mass. In the ampullariids and
cyclophorids, however, the position of the nerve
ring is intermediate, i.e., located in the middle
levei between the anterior and posterior ends
of the buccal mass. According to the tree, the
location of these two groups is intermediate
between the plesiomorphic condition found in
the outgroups (nerve ring anterior to buccal
mass) and the apomorphic one of the
remainder of the caenogastropods, viviparids
included. The result shows the tendency to a
posterior positioned nerve ring, present since the
basal taxa.
The additive condition of the States is based on
ontogeny. In caenogastropods the buccal mass
arises posterior to the nerve ring, and gradually
passes trough it during the development,
positioning anterior to it (BALL et al., 1997).
However, if considered non-additive, the result and
indices do not change.
139. Position of pleural ganglia: 0= close to pedal
ganglia (hypoathroid); 1= close to cerebral ganglia
(epiathroid) (viviparids, Annularía, cerithioideans)
(Cl: 100; RI: 100).
140. Connective pedal-pleural ganglia: 0= very
short; 1= long, separated from pedal-cerebral
connective (viviparids); 2= long, running close
to pedal-cerebral connective [Annularía,
cerithioideans) (Cl: 100; RI: 100; non-
additive) .
These characters (139-140) have correlations to
the hypo- and epiathroid conditions of the nerve
ring, explored by previous authors in
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

438 L.R.L.SIMONE
comparative and phylogenetic studies (e.g., HASZPRUNAR, 1988, PONDER & LINDBERG, 1997, 1998). HASZPRUNAR (1988:394-398) commented extensively about the subject and referred to the viviparid nerve ring as of dystenoid type, the same designation given by BOUVIER (1888) and ANNANDALE & SEWELL (1921:236). This type is hypoathroid on the left side and epiathroid on the right side (HASZPRUNAR, 1988:395, fig.3c).
The hypoathroid condition of the cyclophorids is a long known feature (TIELECKE, 1940, figs. 19-20). A similar condition is found in the ampullariids (BURNE, 1899; PRASHAD, 1925:fig. 17).
The nerve ring of the viviparid species studied here, show that they can be considered as epiathroid, since both pleural ganglia are located close to the cerebral ganglia (Figs.349-352). However, unlike the normal fashion of the epiathroid nerve ring, present in the other caenogastropods, the viviparid pedal connectives to the other two pairs of ganglia are separated from one another in a V-shaped fashion (Figs.300, 348-352). In the normal epiathroid condition both pair of connectives run close and parallel to each another. However, the viviparid pedal ganglia are apparently modified in a long fashion (ANNANDALE & SEWELL, 1921: figs.8- 9; FRETTER & GRAHAM, 1962, fig. 161), lacking clear boundaries. The pedal ganglia are apparently diffused along a wide length, becoming unclear where the pleuro-pedal connective and the anterior pedal nerve start. This fashion precludes the exact classification of the viviparid nerve ring.
Additionally, an interesting difference between the two groups of examined viviparids, i.e., the European Viviparus and the Australian Notopala-Larina, was found. Like the other architaenioglossans, but unlike the higher caenogastropods, the cerebral commissure is distinct and relatively long, but the Viviparus
fashion is longer than that of the Notopala, in such both cerebral ganglia are closer (Figs.348, 350:cc). The viviparid pedal ganglia are distinct in being antero-posteriorly long, but those of Notopala have a wider and more distinct pedal commissure than those of Viviparus. The connectives between the pedal ganglia with the other two pair of ganglia are shorter and broader in Notopala than in Viviparus species,
but in both cases they are separated from each other dorsally, as referred above. In both viviparid groups the pedal ganglia have a dorsal narrow and long region in such connectives to the other two pairs of ganglia insert, this region is shorter in Notopala than in Viviparus species; this dorsal region is singular in confusing with the connectives, becoming difficult to interpret where is the division between the connectives and the ganglia.
Interpreting the cladogram obtained in the present study, the epiathroid condition of the nerve ring appeared at node 5, including the viviparids (character 139). However, the connectives apparently evolved in two different ways. These ways can be seen as steps of a single evolutionary trend, as also suggested by HASZPRUNAR (1988:395). This last hypothesis is explored in character 140. This character was optimized both, as additive or non-additive, and nothing changed in the result or in the indices.
BERTHOLD (1991) and BIELER (1993) have considered other nerve ring attributes as ampullariid synapomorphies, such as presence of pleurosupraintestinal zygosis, subintestinal ganglion fused with right pleural ganglion, presence of pleural commissure, and pedal and pleural ganglia fused on both sides (BIELER, 1993, characters 19-22).
141. Statocysts: 0= with several statoconia; 1 = with single statolith (Annularia) (Cl: 100; RI: 100).
Although State 1 resulted as an Annularia
autapomorphy, it is a State found in many higher caenogastropods. Thus, the character is maintained and will be used in future comparisons.
142. Buccal ganglia: 0= close to nerve ring; 1 = separated, far from nerve ring (all species) (Cl: 100; RI: 100).
143. Buccal ganglia position: 0= close to median line; 1= lateral (viviparids, Annularia,
cerithioideans) (Cl: 100; RI: 100).
The buccal ganglia, as reported by SASAKI (1998), are not distinct in the archaeogastropods, being represented by a thickening of the buccal nerve, and located close to the nerve ring. In the caenogastropods, the pair of buccal ganglia becomes well delimited, and is located at some distance from the nerve ring, closely attached to the posterior region of buccal mass.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 439
MATRIX OF CHARACTERS
Table 1. Matrix of characters and their States, including 2 outgroups in last lines.
Character Taxon
12 3 4 5 6 7 12345 67890 12345 67890 12345 67890 12345 67890 12345 67890 12345 67890 12345 67890 12
P.crosseana 10000 11001 21021 12211 00100 01011 11012 11213 00201 02100 02311 01111 11101 11100 11
P.canaliculata 10000 11001 21021 12211 00101 11011 11012 11213 00201 02100 02311 01111 11101 11100 11
P.scalaris 10000 11001 21021 12210 00101 01011 11012 11213 00201 02100 02211 01111 11101 11100 11
P.lineata 10000 11001 21021 12210 00101 11011 11012 11213 00201 02100 02211 01111 11101 11100 11
P. curumim 10000 11001 21021 12210 00011 01011 11012 11213 00201 02100 02211 01111 11101 11100 11
Marisa 20000 11001 21001 10210 00100 00021 11012 11103 00201 01100 02311 01111 11101 11100 11
Asolene 10100 11001 11021 12211 00101 01011 11012 11213 00201 02100 02311 01111 11101 11100 11
Felipponea 10100 11001 11021 12210 00101 01011 11012 11213 00201 02100 02311 01111 11101 11100 11
P.bridgesi 10000 11001 21021 12211 00100 01011 11012 11213 00201 02100 02211 01111 11101 11100 11
P. sórdida 10000 11001 21021 12211 00100 11011 11012 11213 00201 02100 02211 01111 11101 11100 11
Neocyclotus 10010 01100 01111 00900 1???? 00001 00010 10091 00102 01111 01110 01111 11100 10101 10
Aperostoma 10010 01100 01011 00900 19999 00001 00010 10091 00112 01112 01010 01111 11100 10101 10
Incidostoma 10010 01100 01011 00900 19999 00001 00010 10091 00112 01112 01010 01111 11100 10101 10
V.contectus 01000 11012 11001 01100 01001 00100 00011 10092 11000 01100 10022 11111 11110 10110 11
V.acerosus 01000 11012 1100 1 01100 0100 1 00100 00011 10092 11000 01100 10022 11111 11110 10110 11
N.ampullaroid 01000 11012 11001 01122 01000 00101 00111 10092 12000 01100 10022 11111 11100 10110 10
N.essigntone. 01000 11012 11001 01122 01000 00101 00111 10092 12000 01100 10022 11111 11100 10110 10
Larina 11000 11012 11001 01122 01000 00101 00111 10092 12000 01100 10022 11111 11100 10110 10
Annularia 00011 00100 01201 00100 19999 00001 00010 11090 00003 11100 10222 11111 11100 10100 10
Cerithioidea 00001 00000 01001 00100 00001 00000 00010 10090 00000 11100 10022 11111 11100 10100 11
Character Taxon
7 8 9 10 11 12 13 14 345 67890 12345 67890 12345 67890 12345 67890 12345 67890 12345 67890 12345 67890 123
P.crosseana 011 11111 01011 10201 02110 21112 11111 11010 00220 00002 11110 00201 20202 12100 010
P.canaliculata 011 11111 01011 10201 02110 22212 11121 11110 00220 00002 11211 20201 20202 12100 010
P.scalaris 011 11111 01011 10201 02110 21112 11111 11110 00220 00001 01110 00201 20202 12100 010
P. lineata 011 11111 01011 10201 02110 22212 11121 11110 00220 00001 11211 11201 20202 12100 010
P. curumim 011 11111 01011 10201 02110 22112 11111 11110 00220 00001 01211 11201 20202 12100 010
Marisa 011 11111 01011 10201 02100 21012 11111 11110 00220 00001 01111 01201 20202 12100 010
Asolene 011 11111 01111 10201 02110 22212 11121 11110 00220 00002 11210 21201 20202 22100 010
Felipponea 011 11111 01111 1020 1 02110 21112 11111 11010 00220 00001 01111 0920 1 20202 22100 010
P.bridgesi 011 11111 01011 10201 02110 22212 11111 11110 00220 00001 01211 21201 20202 22100 010
P. sórdida 011 11111 01011 10201 02110 22112 11111 11110 00220 00001 11211 20201 20202 22100 010
Neocyclotus 010 10000 01111 10101 01101 20001 10000 01011 00130 00110 01000 00201 11111 00100 010
Aperostoma 010 10000 11111 10101 01101 20001 10000 01011 00130 00120 01000 00201 12111 00100 010
Incidostoma 010 10000 11111 10101 01101 20001 10000 01011 00130 00120 01000 00201 12111 00100 010
V.contectus 010 10210 01011 11101 19900 10000 10001 11010 01211 11000 01000 00211 00102 01211 011
V.acerosus 010 10210 01011 11101 19900 10000 10001 11010 01211 11000 01000 00211 00102 01211 011
N. ampullaroid 010 10210 01011 11111 19920 20001 10001 11010 01211 10000 01000 00211 30102 01211 011
N.essigntone. 010 10210 01011 11111 19920 20001 10001 11010 01211 10000 01000 00211 30102 01211 011
Larina 010 10210 01011 11111 19920 20000 10001 11010 01211 10000 01000 00211 30102 01211 011
Annularia 110 10000 11111 11201 19901 20001 00001 11010 11220 00100 01000 00201 00101 00212 111
Cerithioidea 010 10010 01011 11101 19901 20000 00001 11010 11000 00000 01000 00101 00100 00212 011
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
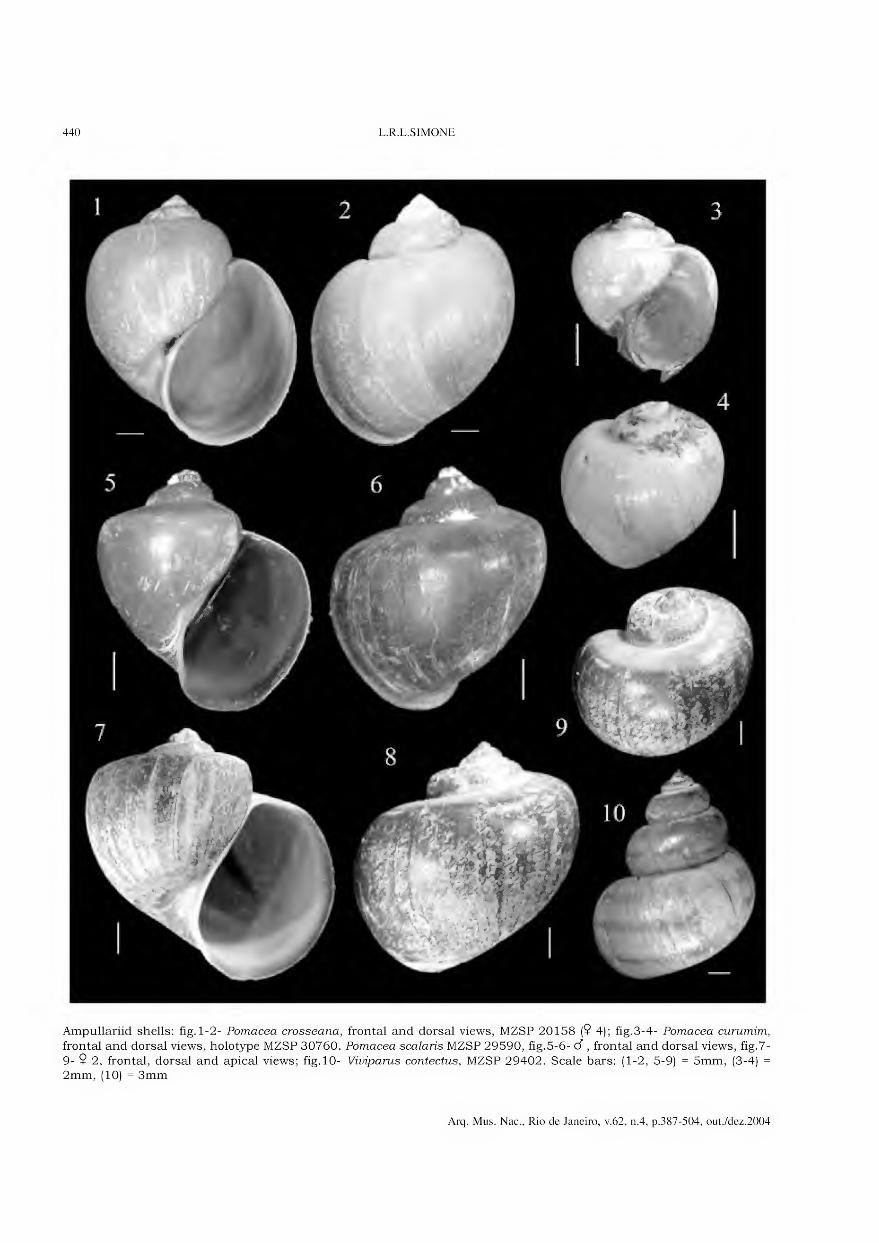
440 L.R.L.SIMONE
Ampullariid shells: fig. 1-2- Pomacea crosseana, frontal and dorsal views, MZSP 20158 (9 4); fig.3-4- Pomacea curumim,
frontal and dorsal views, holotype MZSP 30760. Pomacea scalaris MZSP 29590, fig.5-6- d , frontal and dorsal views, fig.7-
9- 9 2, frontal, dorsal and apical views; fig. 10- Viviparus contectus, MZSP 29402. Scale bars: (1-2, 5-9) = 5mm, (3-4) = 2mm, (10) = 3mm
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
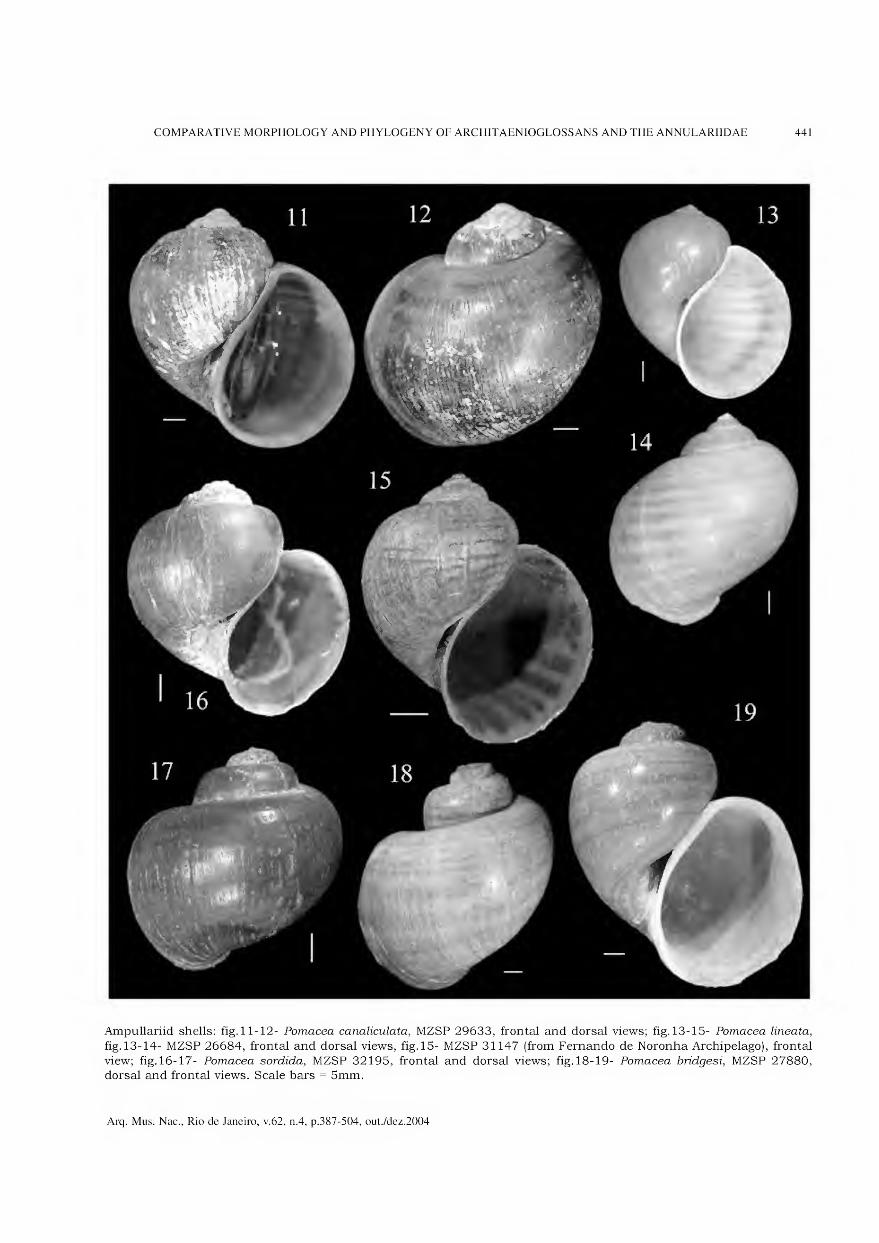
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 441
Ampullariid shells: fig.11-12- Pomacea canaliculata, MZSP 29633, frontal and dorsal views; fig.13-15- Pomacea lineata,
fig.13-14- MZSP 26684, frontal and dorsal views, fig.15- MZSP 31147 (from Fernando de Noronha Archipelago), frontal
view; fig. 16-17- Pomacea sórdida, MZSP 32195, frontal and dorsal views; fig. 18-19- Pomacea bridgesi, MZSP 27880, dorsal and frontal views. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

442 L.R.L.SIMONE
Ampullariid shells: fig.20-21- Pomacea bridgesi, MZSP 32196, frontal and dorsal views; fig.22- Marisa planogyra, MZSP 29674, specimens with uniform color, right view; fig.23-24- Asolene megastoma, MZSP 250, frontal and dorsal views;
fig.25-28- Marisa planogyra, MZSP 29674, fig.25-27- specimen with uniform color (same than figure 22), left, frontal and obliqúe views; fig.28- specimen with bands in color, dorsal view. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
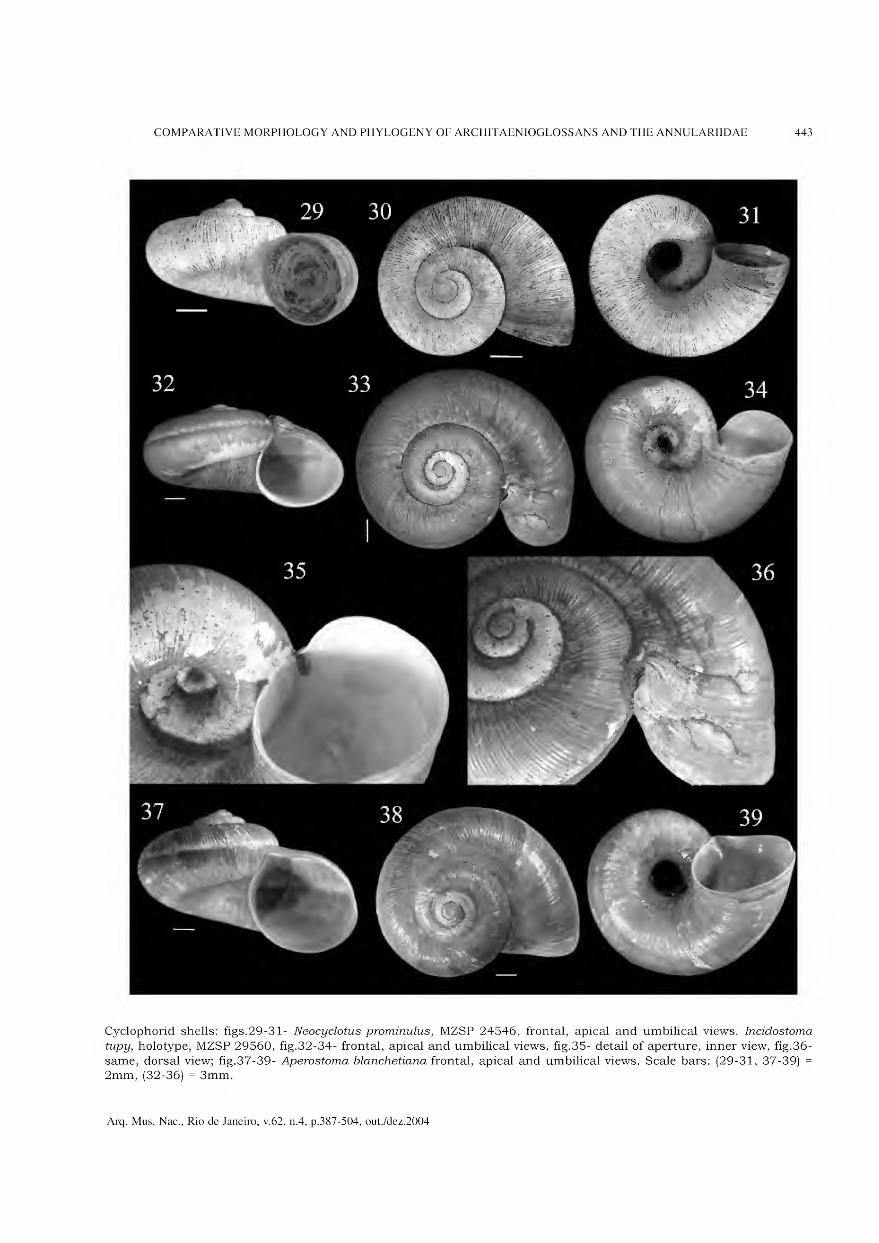
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 443
Cyclophorid shells: figs.29-31- Neocyclotus prominulus, MZSP 24546, frontal, apical and umbilical views. Incidostoma
tupy, holotype, MZSP 29560, fig.32-34- frontal, apical and umbilical views, fig.35- detail of aperture, inner view, fig.36-
same, dorsal view; fig.37-39- Aperostoma blanchetiana frontal, apical and umbilical views. Scale bars: (29-31, 37-39) = 2mm, (32-36) = 3mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
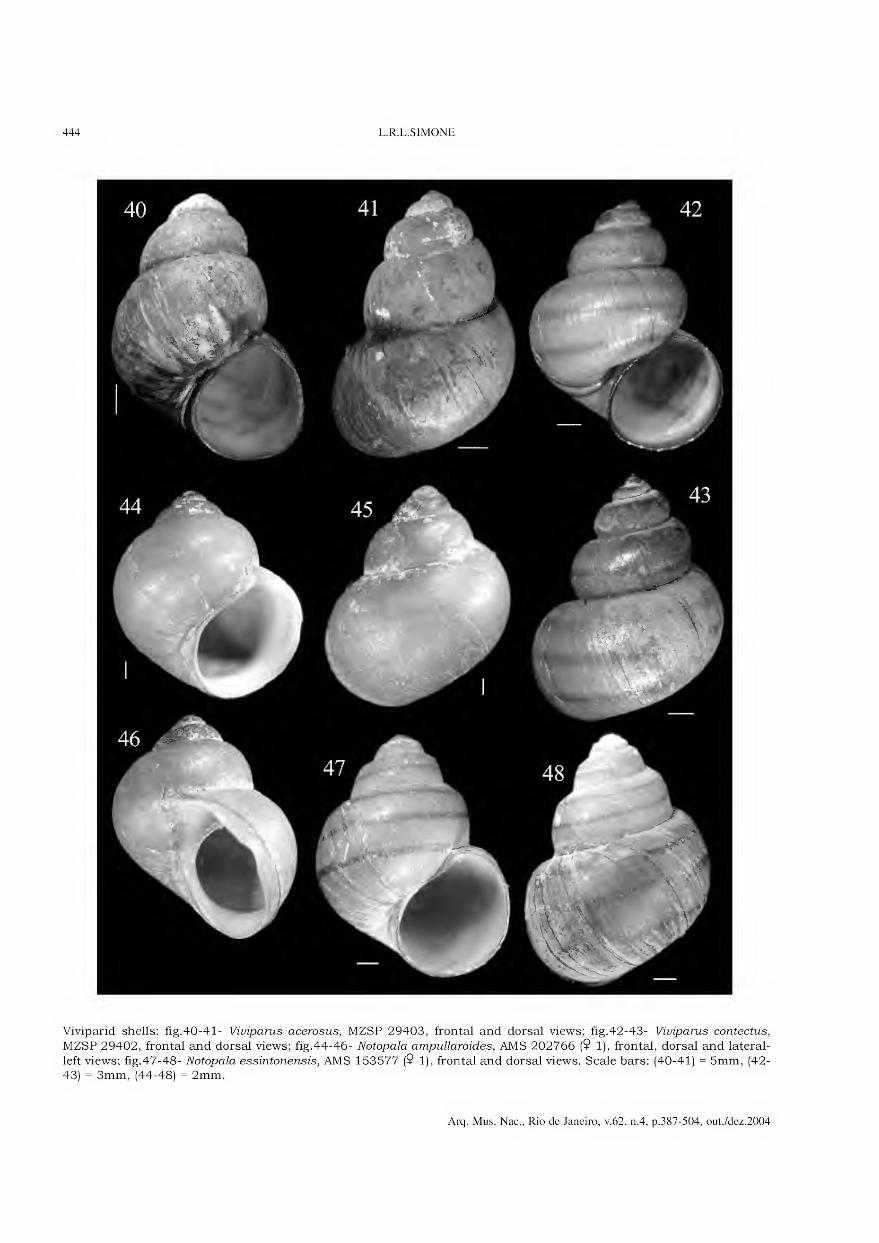
444 L.R.L.SIMONE
Viviparid shells: fig.40-41- Viviparus acerosus, MZSP 29403, frontal and dorsal views; fig.42-43- Viviparus contectus,
MZSP 29402, frontal and dorsal views; fig.44-46- Notopala ampullaroides, AMS 202766 (9 1), frontal, dorsal and lateral-
left views; fig.47-48- Notopala essintonensis, AMS 153577 (9 1), frontal and dorsal views. Scale bars: (40-41) = 5mm, (42- 43) = 3mm, (44-48) = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 445
Shells: fig.49- Annularia sp, MZSP 28266, SEM, frontal view; fig.50- Larina cf strangei, AMS 338590, SEM, yoimg specimen
extracted from brood pouch, dorsal view; fig.51- Notopala essingtonensis, AMS 153577, lateral-left view; fig.52-54- Larina
cf strangei, AMS 338590, frontal, dorsal and lateral-left views; fig.55-56- Annularia sp., MZSP 28266, frontal and dorsal views. Scale bars: (49) = lmm, (50-56) = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

446 L.R.L.SIMONE
Shells in SEM: Pomacea curumim, paratypes, fig.57-58- specimens with intact periostracum, frontal and dorsal views; fig. 59- 60- specimen with most periostracum removed, frontal and dorsal views; fig.61-62- Viviparus contectus, young specimen
removed from brood pouch, dorsal and apical views; fig.63- Notopala ampullaroides, young specimen removed from brood pouch, dorsal view; fig.64-65-. Pomacea lineata, MZSP 31147 (from Fernando de Noronha Archipelago); fig.64- young specimen, frontal view; fig.65- same, detail of apex, apical view; fig.66- Notopala essingtonensis, young specimen removed from brood
pouch, dorsal view. Scale bars: (57-60) = 0.6mm, (61-62) = 0.5mm, (63, 65-66) = 0.4mm, (64) = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
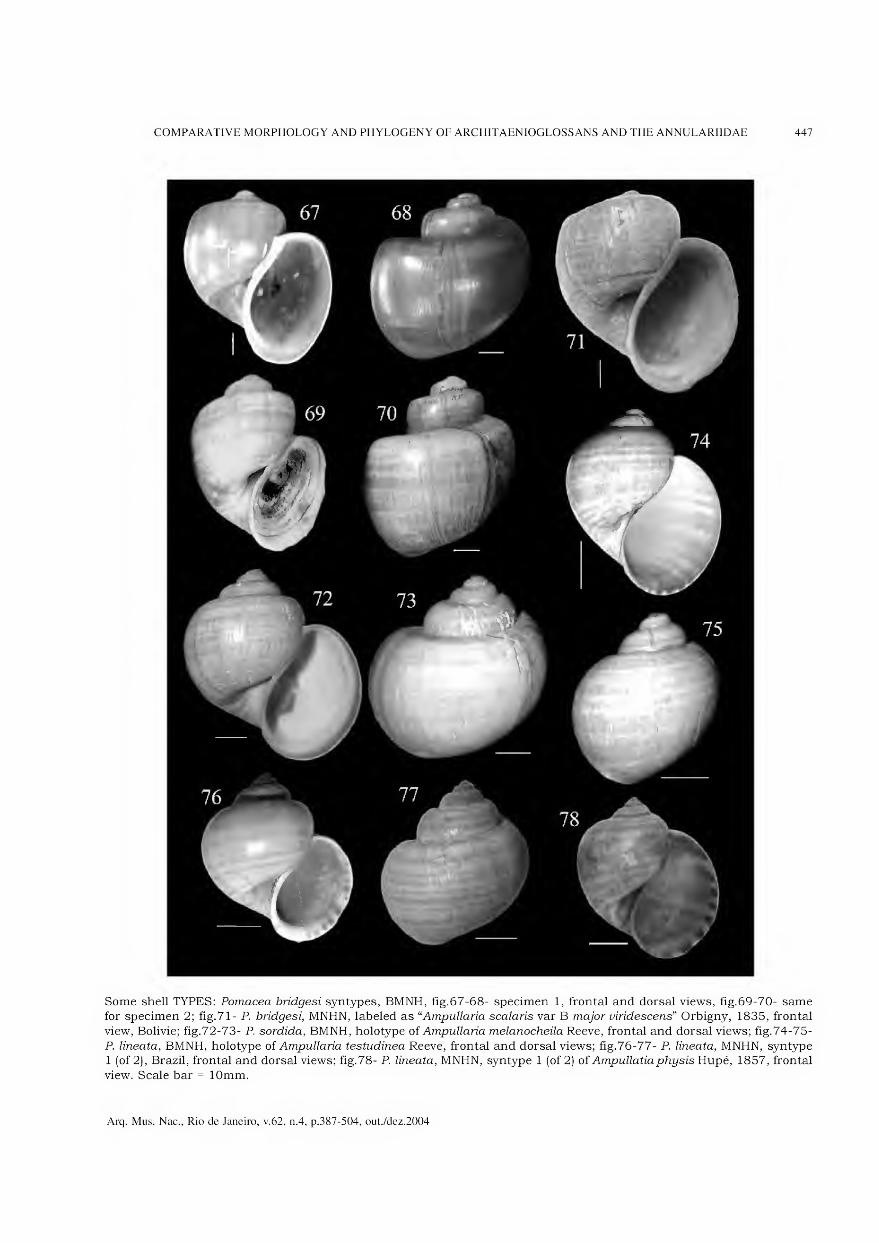
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 447
Some shell TYPES: Pomacea bridgesi syntypes, BMNH, fig.67-68- specimen 1, frontal and dorsal views, fig.69-70- same for specimen 2; fig.71- P. bridgesi, MNHN, labeled as “Ampullaria scalaris var B major viridescens” Orbigny, 1835, frontal
view, Bolivie; fig.72-73- P. sórdida, BMNH, holotype of Ampullaria melanocheila Reeve, frontal and dorsal views; fig.74-75- P. lineata, BMNH, holotype of Ampullaria testudinea Reeve, frontal and dorsal views; fig.76-77- P. lineata, MNHN, syntype 1 (of 2), Brazil, frontal and dorsal views; fig.78- P. lineata, MNHN, syntype 1 (of 2) of Ampullatiaphysis Hupé, 1857, frontal view. Scale bar = lOmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

448 L.R.L.SIMONE
Pomacea lineata TYPES: fig.79-80- MNHN, syntype 2 (of 2, see figs.76-77), Brazil, frontal and dorsal views; fig.81-82- ZSM, Spix’s types of Pomus lineatus fasciatus, Bahia; fig.83-84- ZSM, Spix’s types of Pomus fingulinus, Bahia. Figs.81-84
courtesy of Silvana Thiengo (Fundação Oswaldo Cruz, Rio de Janeiro). Scale bars = 10mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 449
Some shell TYPES: fig.85-87- Pomacea scalaris, 3 syntypes, MNHN, Bolívia; fig.88-89- P. canaliculata, MNHN, Rio de la Plata, syntypes of Pila insularum Orbigny, fig.88- frontal view of syntype 1, fig.89- dorsal view of syntype 2; fig.90- Asolene
megastoma, MNHN, probable type of Ampullaria neritoides Orbigny, Rio Uruguay, frontal view. Neocyclotus prominulus,
MNHN, 2 probable syntypes of Cyclostoma prominula Fer. in Orbigny, Brazil, coll. Deshayes, fig.91-92- specimen 1, umbilical and frontal views, fig.93-94- same for specimen 2. Scale bars: (85-90) = lOmm, (91-94) = 3mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

450 L.R.L.SIMONE
Opercula: fig.95-96- Pomacea lineata, outer and inner views; fig.97-98- Pomacea crosseana, outer and inner views; fig.99- 100- Pomacea sórdida, outer and inner views; fig. 101-102- Marisaplanogyra, outer and inner views; fig. 103-104- Viviparus
acerosus, outer and inner views; fig. 105-106- Notopala ampullaroides, outer and inner views. Scale bars: (95-100, 103- 104) = 3mm, (101-102, 105-106) = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
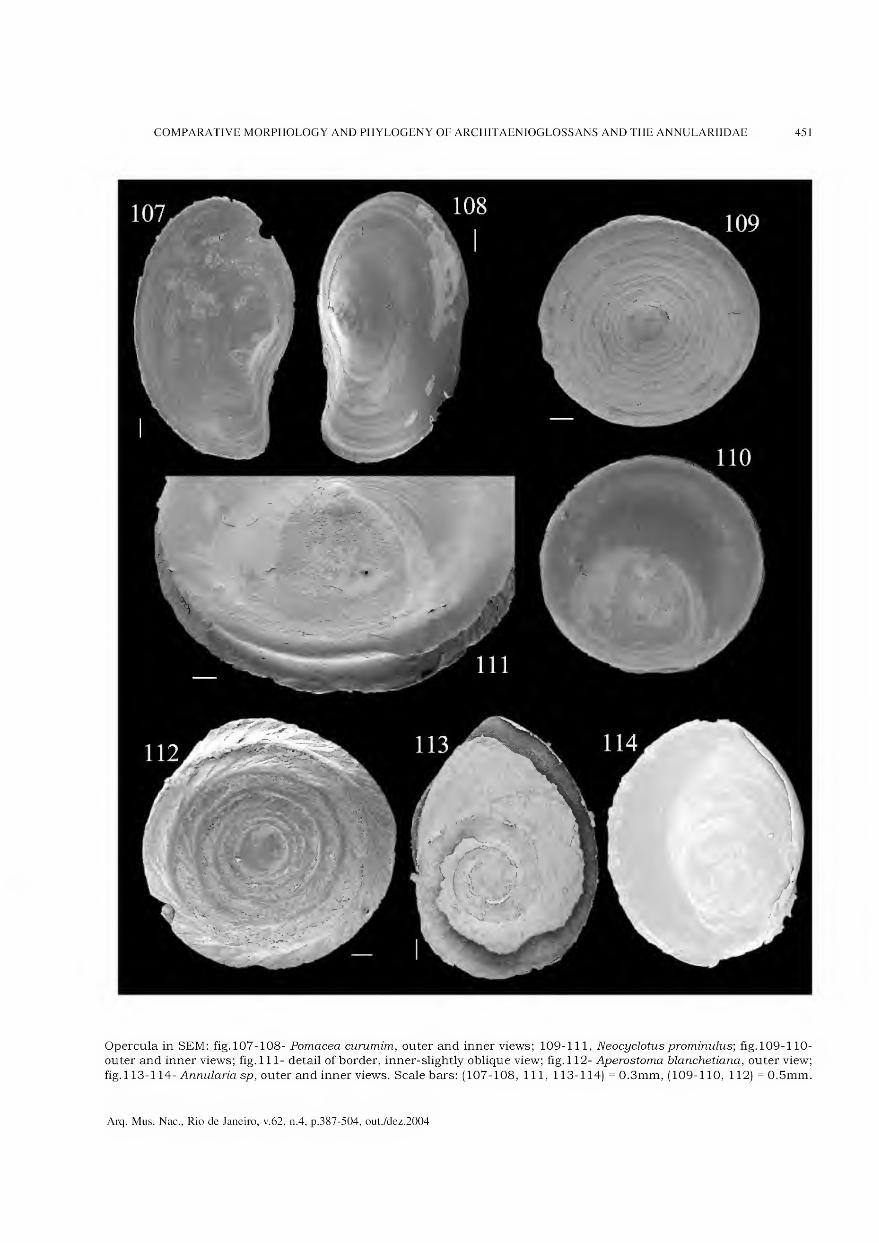
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 451
Opercula in SEM: fig. 107-108- Pomacea curumim, outer and inner views; 109-111, Neocyclotusprominulus; fig. 109-110- outer and inner views; fig. 111- detail of border, inner-slightly obliqúe view; fig. 112- Aperostoma blanchetiana, outer view;
fig. 113-114- Annularia sp, outer and inner views. Scale bars: (107-108, 111, 113-114) = 0.3mm, (109-110, 112)= 0.5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

452 L.R.L.SIMONE
Pomacea radulae in SEM: fig.115-116- Pomacea crosseana; fig.117-118- P. curumim; fig.119- P. scalaris; fig.120-121- P canaliculata; fig. 122- P. lineata; fig. 123- P. sórdida. Scale bars: (115-116, 119-123) = 200jum, (117-118) = 60jum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 453
Ampullariid radulae in SEM: fig. 124- Pomacea sórdida; fig. 125-126- P. lineata (from Fernando de Noronha Archipelago); fig. 127-128- P. bridgesi; fig. 129- Asolene megastoma; fig. 130- Felipponea neritiniformis; fig. 131-132- Marisa planogyra.
Scale bars: (124-128, 130) = 200|um, (129) = 400jum, (131-132) = lOOjum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
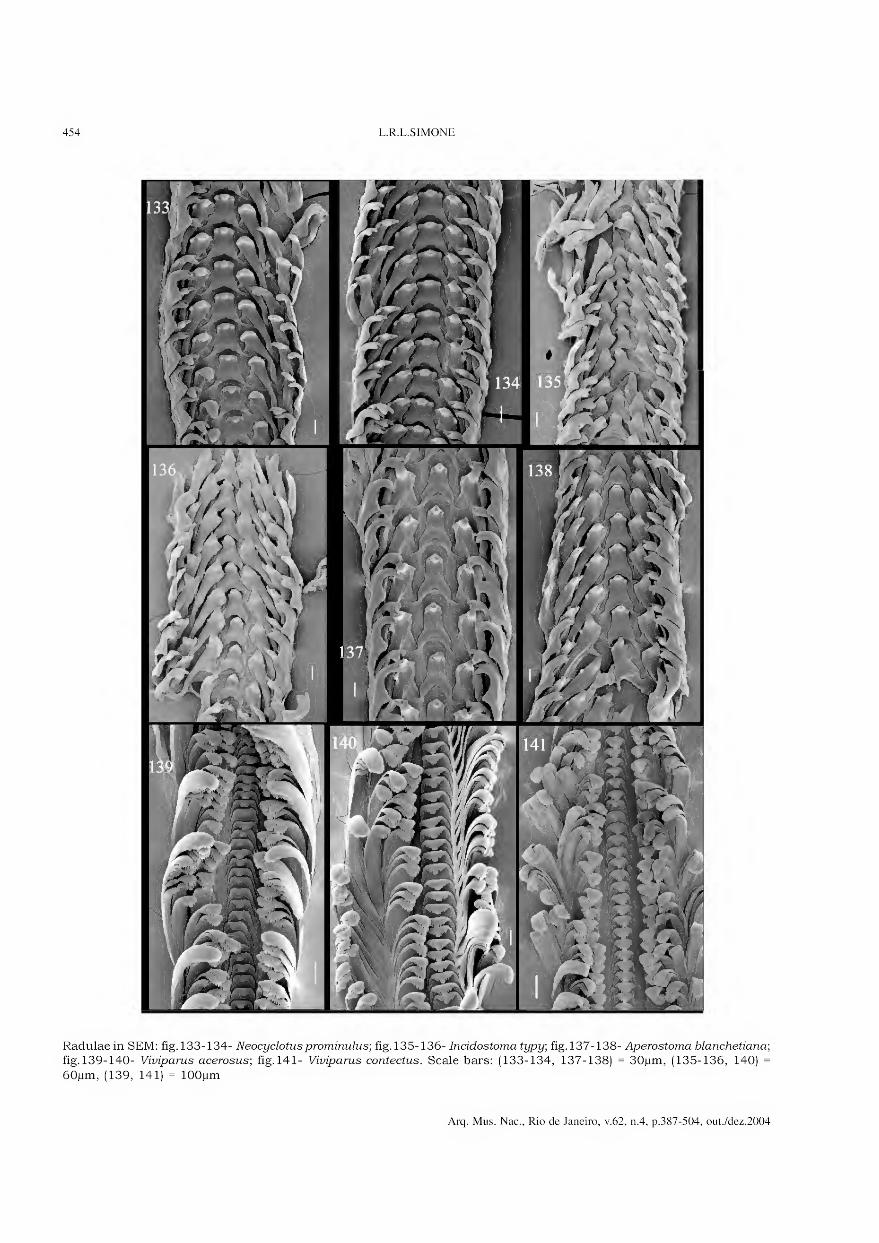
454 L.R.L.SIMONE
Radulae in SEM: fig. 133-134- Neocyclotusprominulus; fig. 135-136- Incidostoma typy; fig. 137-138- Aperostoma blanchetiana; fig. 139-140- Viviparus acerosus; fig. 141- Viviparus contectus. Scale bars: (133-134, 137-138) = 30jum, (135-136, 140) =
60jum, (139, 141) = lOOjum
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
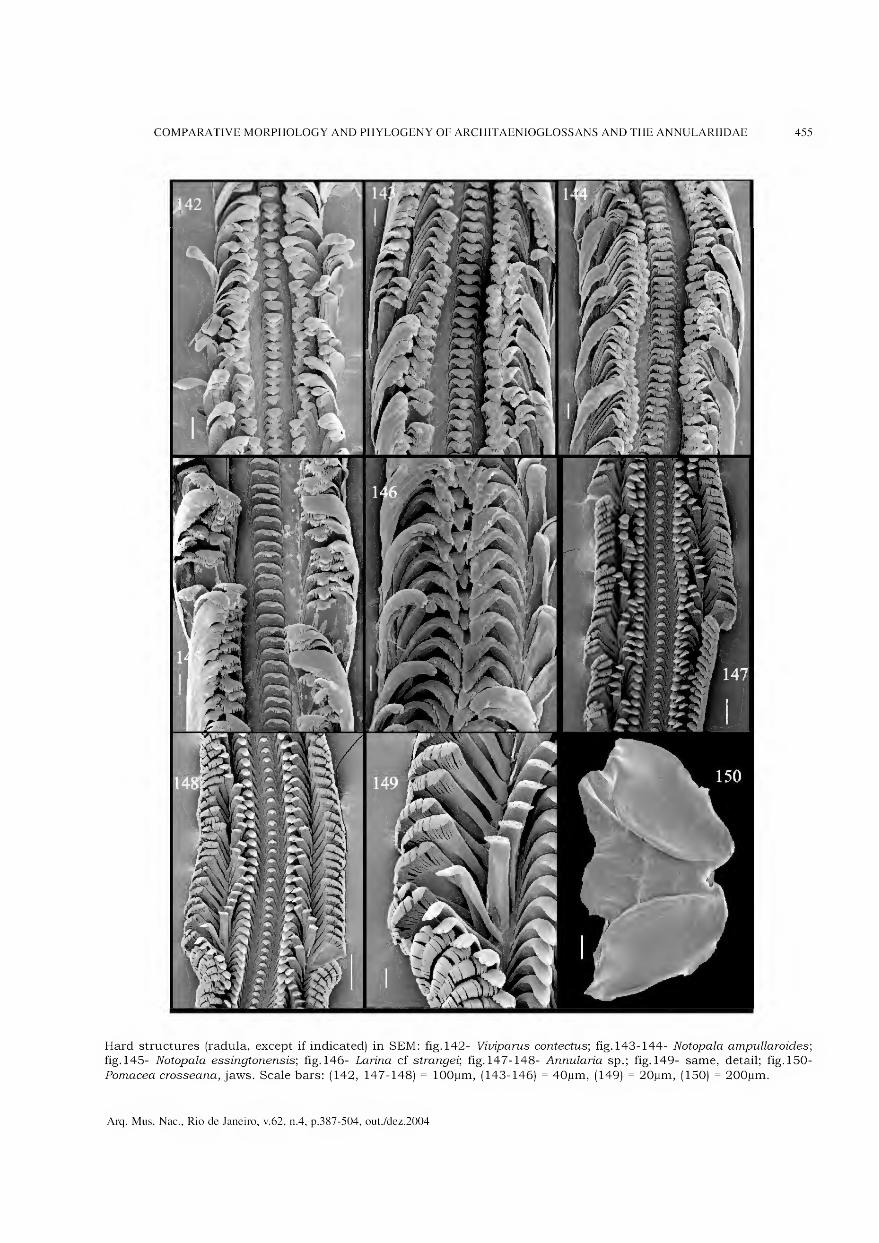
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 455
Hard structures (radula, except if indicated) in SEM: fig. 142- Viiriparus contectus; fig. 143-144- Notopala ampullaroides; fig.145- Notopala essingtonensis; fig. 146- Larina cf strangei; fig. 147-148- Annularia sp.; fig. 149- same, detail; fig. 150-
Pomacea crosseana, jaws. Scale bars: (142, 147-148) = lOOjum, (143-146) = 40jum, (149) = 20jum, (150) = 200jum.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

456 L.R.L.SIMONE
Pomacea crosseana anatomy: fig.151- head-foot, 9 , frontal view; fig.152- same, lateral-right view; fig.153- pallial cavity roof in inner-ventral view, and visceral mass; fig. 154- anterior region of visceral mass and adjacent portion of pallial
cavity, ventral view, ventral walls of pericardium and kidney removed, an schematic representation of in situ intestinal
loops inside kidney shown in a separated detail; fig. 155- detail of pallial cavity in region of lung, ventral wall of lung most removed and deflected posteriorly (to left in fig.). Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
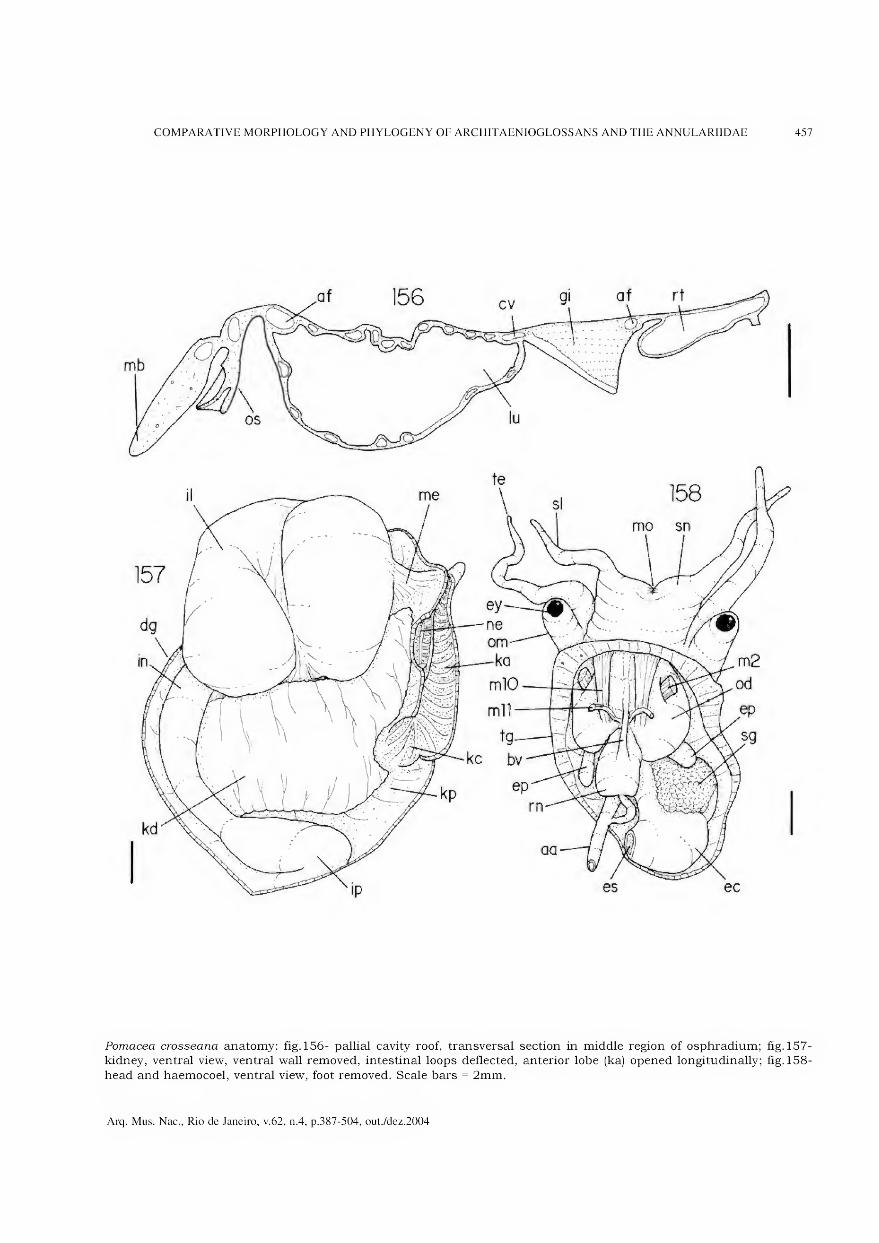
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 457
Pomacea crosseana anatomy: fig.156- pallial cavity roof, transversal section in middle region of osphradium; fig. 157- kidney, ventral view, ventral wall removed, intestinal loops deflected, anterior lobe (ka) opened longitudinally; fig. 158-
head and haemocoel, ventral view, foot removed. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

458 L.R.L.SIMONE
mo
mIO m2
Pomacea crosseana anatomy: fig. 159- foregut, buccal mass in lateral-left view, remainder structures in ventral view;
fig. 160- same, ventral view, odontophore removed, esophagus and left esophageal pouch (right in fig.) opened longitudinally;
fig. 161- buccal mass, dorsal view; fig. 162- same, ventral view; fig. 163- same, dorsal view, dorsal wall and esophagus removed. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
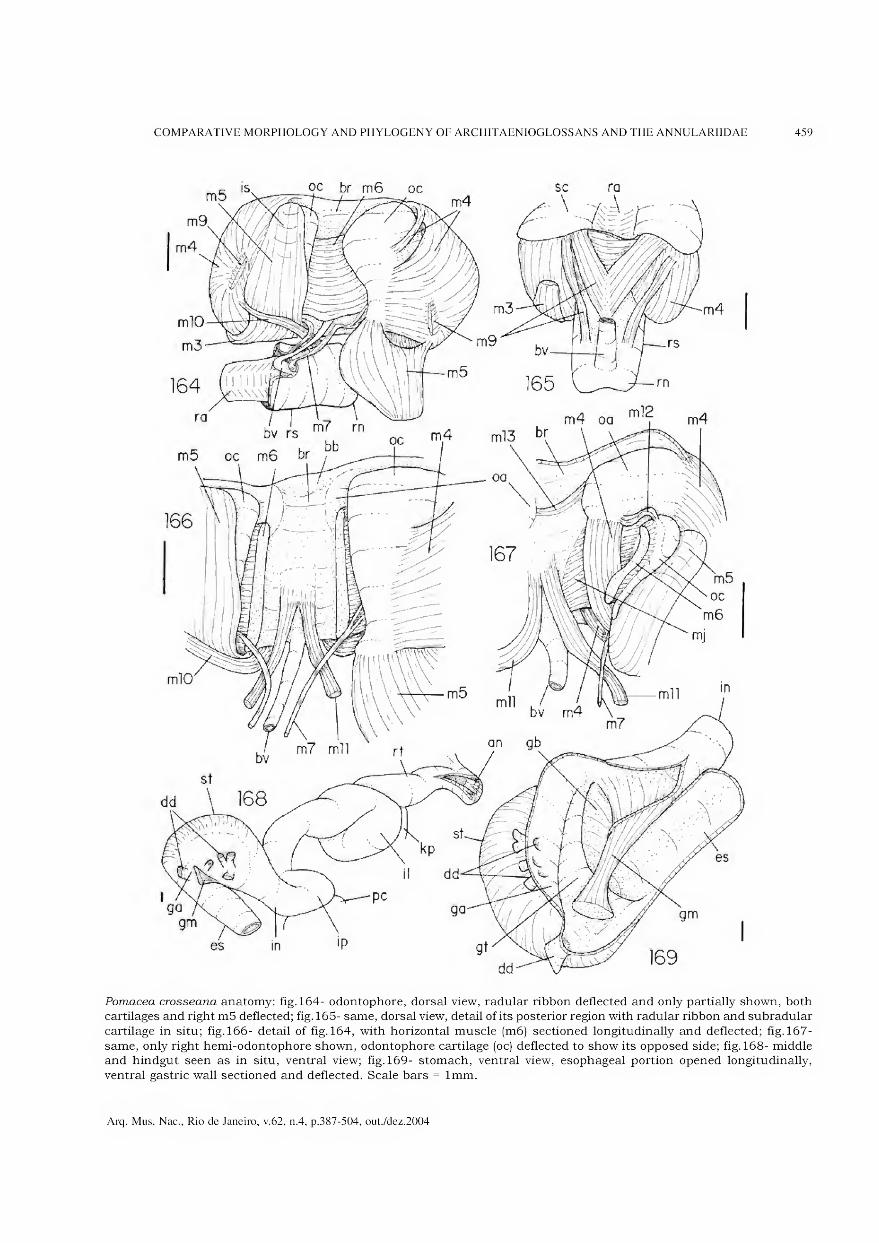
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 459
Pomacea crosseana anatomy: fig.164- odontophore, dorsal view, radular ribbon deflected and only partially shown, both
cartilages and right m5 deflected; fig. 165- same, dorsal view, detail of its posterior region with radular ribbon and subradular
cartilage in situ; fig. 166- detail of fig.164, with horizontal muscle (m6) sectioned longitudinally and deflected; fig. 167-
same, only right hemi-odontophore shown, odontophore cartilage (oc) deflected to show its opposed side; fig. 168- middle and hindgut seen as in situ, ventral view; fig. 169- stomach, ventral view, esophageal portion opened longitudinally,
ventral gastric wall sectioned and deflected. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

460 L.R.L.SIMONE
Pomacea crosseana anatomy: fig. 170- detail of pallial cavity in region of penis shield; fig. 171- same, with penis shield deflected
externally, penis sac opened, penis stretched; fig. 172-173- pallial oviduct, ventral and dorsal views. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 461
Pomacea curumim anatomy: fig. 174- head-foot, 9 , frontal view; fig. 175- pallial cavity roof, d, inner-ventral view, penis shield deflected outside; fig. 176- pallial cavity roof, transversal section in middle region of osphradium; fig. 177- detail of
osphradium and adjacent region, ventral view; fig. 178- kidney, ventral view, ventral wall removed, intestinal loops deflected,
anterior lobe (ka) opened longitudinally; fig. 179- anterior region of visceral mass and adjacent region of pallial cavity, ventral view, ventral walls of kidney and pericardium removed, special attention to fashion of renal intestinal loops;
fig. 180- foregut, lateral-right view. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

462 L.R.L.SIMONE
Pomacea curumim anatomy: fig. 181- buccal mass, lateral-right view; fig. 182- foregut, ventral view, odontophore removed, esophagus opened longitudinally right dorsal fold (left in fig.) sectioned in the levei of esophageal pouch; fig. 183-
odontophore, dorsal view, radular ribbon extracted and deflected, both cartilages deflected, left cartilage in slightly apical
view, horizontal muscle (m6) sectioned longitudinally and part deflected, left m5 also deflected; fig. 184- pallial oviduct and adjacent structures of pallial roof, ventral view; fig. 185- same, seen as a transparent structure; fig. 186- midgut,
ventral view as in situ; fig. 187- right-anterior region of pallial cavity roof, male, penis shield deflected, penis sac opened
and penis stretched; fig. 188- midgut most opened longitudinally, ventral view, ventral gastric wall deflected, its transversal septum (gt) partially sectioned. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
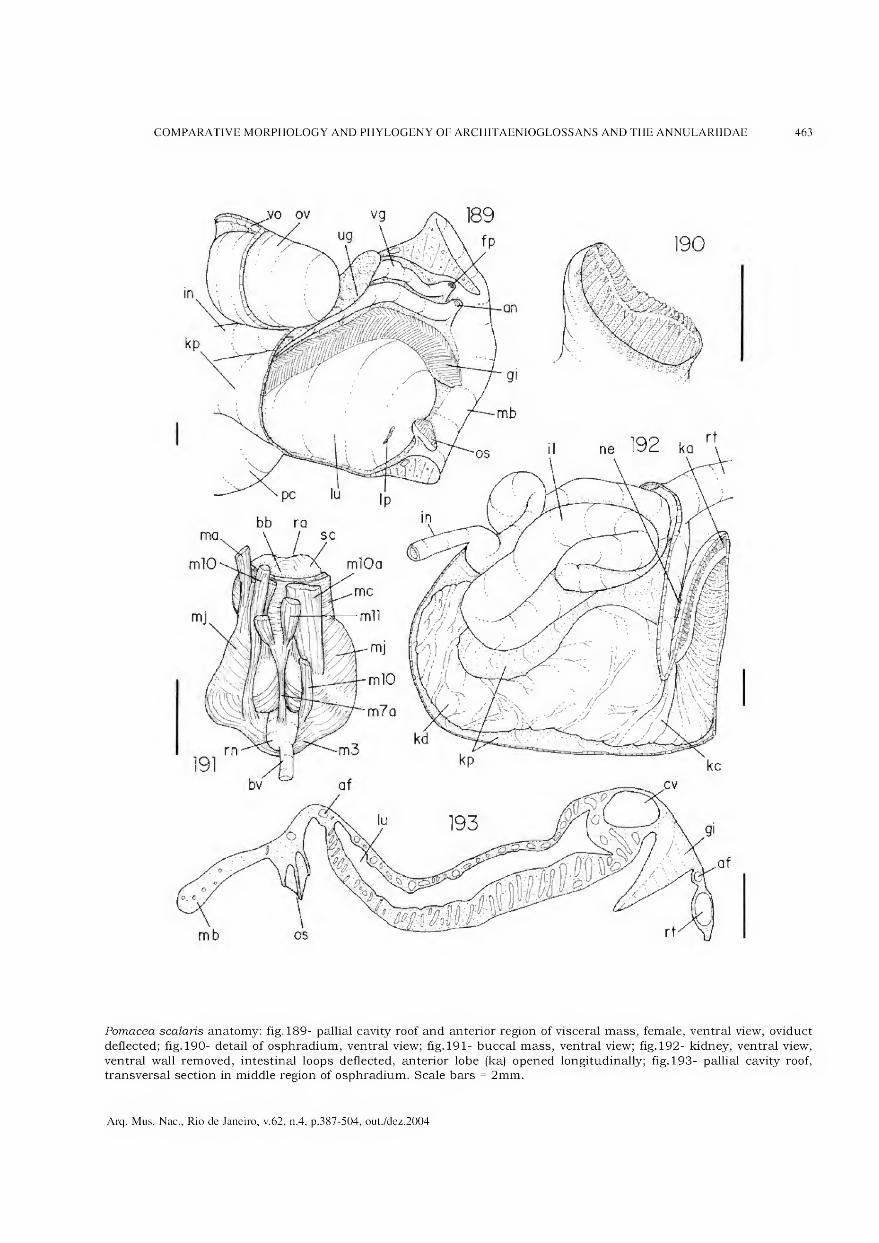
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 463
Pomacea scalaris anatomy: fig. 189- pallial cavity roof and anterior region of visceral mass, female, ventral view, oviduct
deflected; fig. 190- detail of osphradium, ventral view; fig. 191- buccal mass, ventral view; fig. 192- kidney, ventral view,
ventral wall removed, intestinal loops deflected, anterior lobe (ka) opened longitudinally; fig. 193- pallial cavity roof, transversal section in middle region of osphradium. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

464 L.R.L.SIMONE
Pomacea scalaris anatomy: fig. 194- foregut, lateral-right view; fig. 195- same, ventral view, odontophore removed, esophagus
and left esophageal pouch (right in fig.) opened longitudinally; fig. 196- odontophore, dorsal view, radular ribbon removed,
deflected and only partially shown, both cartilages and right m5 deflected; fig. 197- pallial oviduct, ventral view, seen as a transparent structure. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 465
Fig.198- Pomacea scalaris anterior region of male reproductive system, penis shield deflected outside, penis sac opened, penis stretched. Scale bar = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
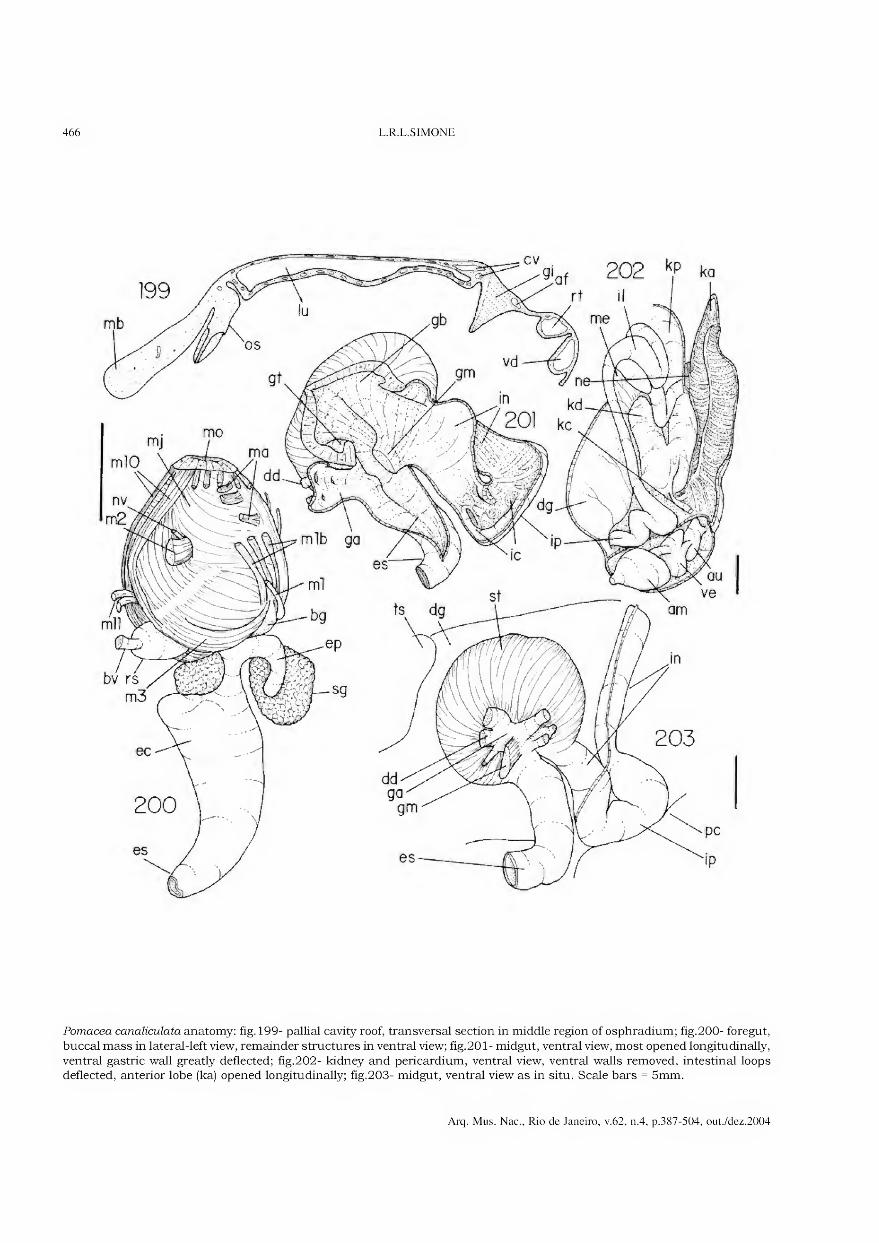
466 L.R.L.SIMONE
Pomacea canaliculata anatomy: fig. 199- pallial cavity roof, transversal section in middle region of osphradium; fig.200- foregut,
buccal mass in lateral-left view, remainder structures in ventral view; fig.201- midgut, ventral view, most opened longitudinally,
ventral gastric wall greatly deflected; fig.202- kidney and pericardium, ventral view, ventral walls removed, intestinal loops deflected, anterior lobe (ka) opened longitudinally; fig.203- midgut, ventral view as in situ. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
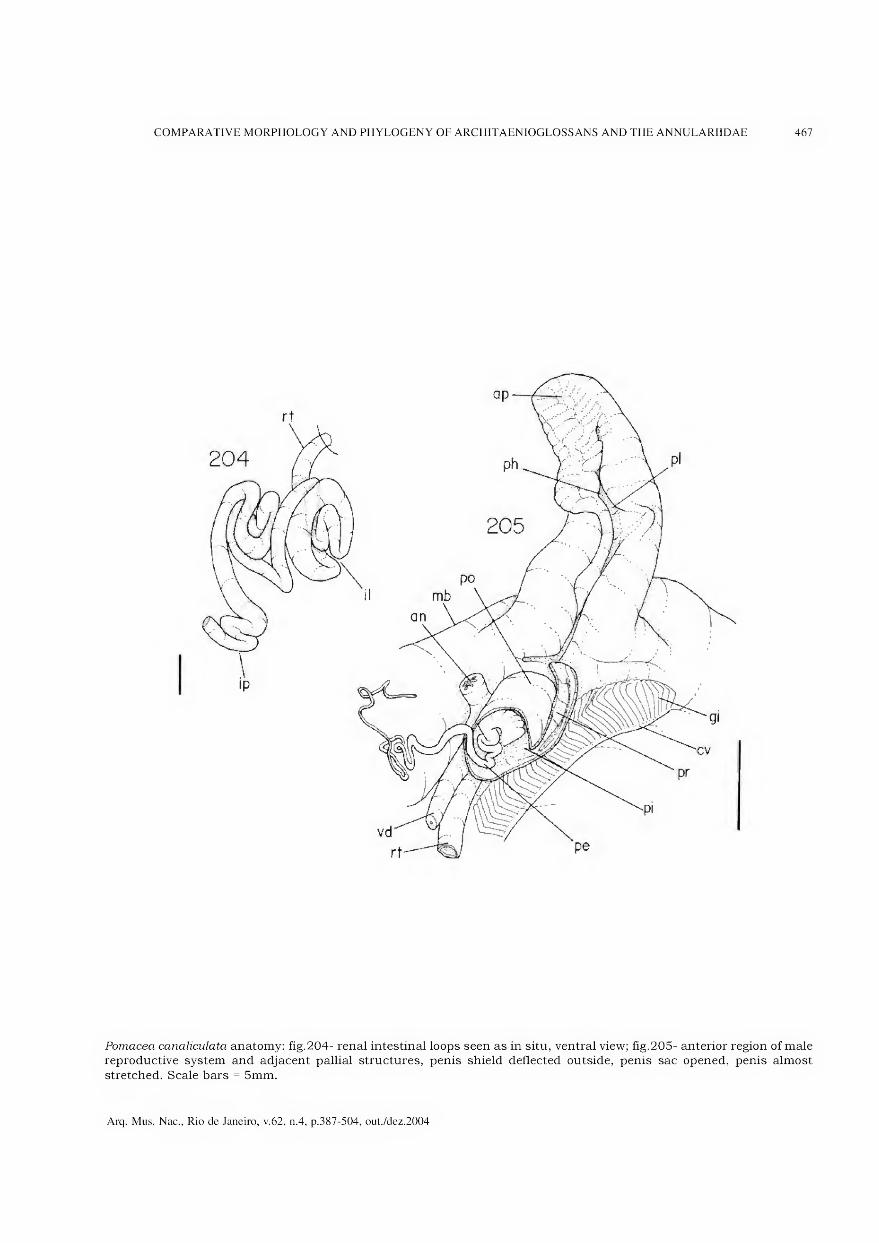
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 467
Pomacea canaliculata anatomy: fig.204- renal intestinal loops seen as in situ, ventral view; fig.205- anterior region of male reproductive system and adjacent pallial structures, penis shield deflected outside, penis sac opened, penis almost
stretched. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

468 L.R.L.SIMONE
Pomacea lineata anatomy, mainland samples: fig.206- pallial cavity roof, transversal section in middle region of osphradium; fig.207- foregut, ventral-slightly left view; fig.208- middle and hindgut, ventral view as in situ; fig.209- midgut most
opened longitudinally, ventral view. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
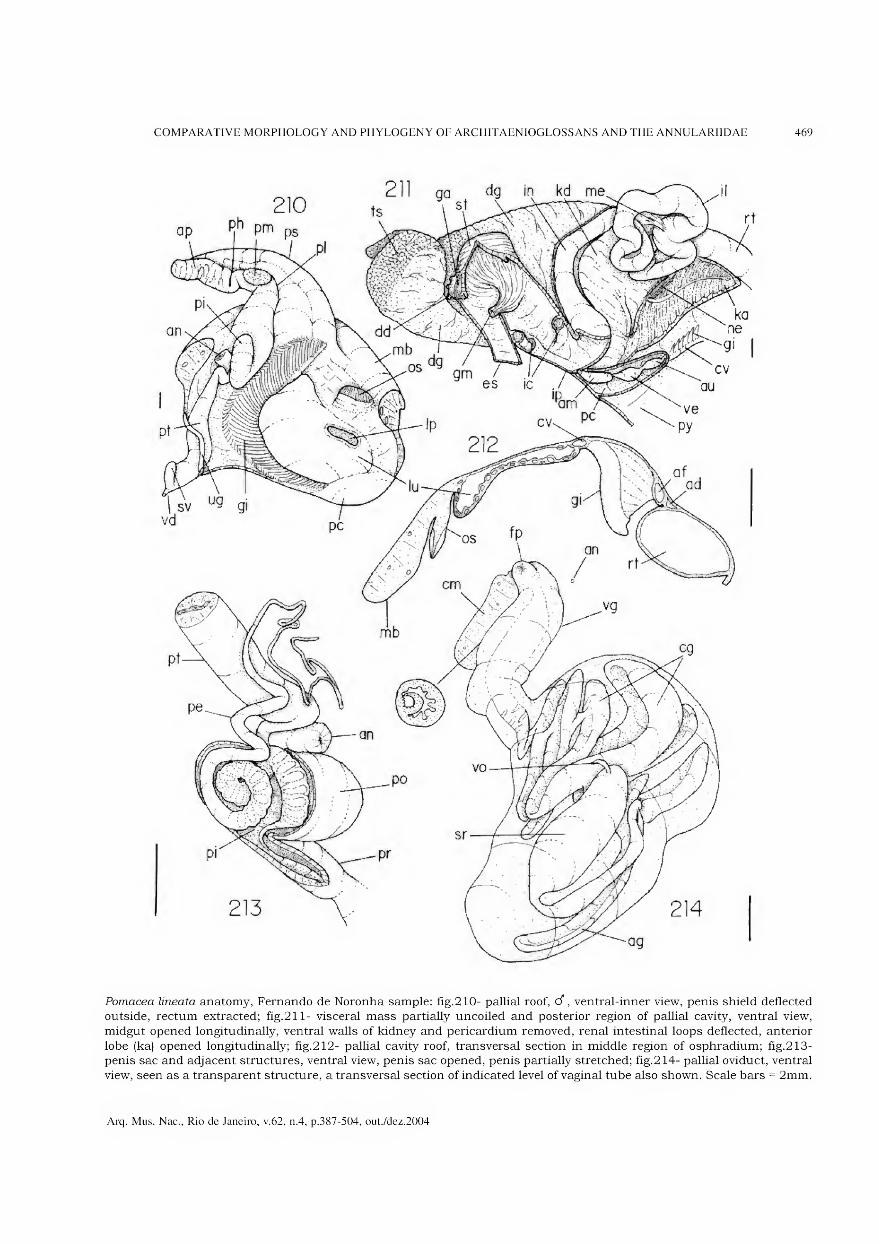
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 469
Pomacea lineata anatomy, Fernando de Noronha sample: fig.210- pallial roof, d", ventral-inner view, penis shield deflected outside, rectum extracted; fig.211- visceral mass partially uncoiled and posterior region of pallial cavity, ventral view,
midgut opened longitudinally, ventral walls of kidney and pericardium removed, renal intestinal loops deflected, anterior
lobe (ka) opened longitudinally; fig.212- pallial cavity roof, transversal section in middle region of osphradium; fig.213- penis sac and adjacent structures, ventral view, penis sac opened, penis partially stretched; fig.214- pallial oviduct, ventral
view, seen as a transparent structure, a transversal section of indicated levei of vaginal tube also shown. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
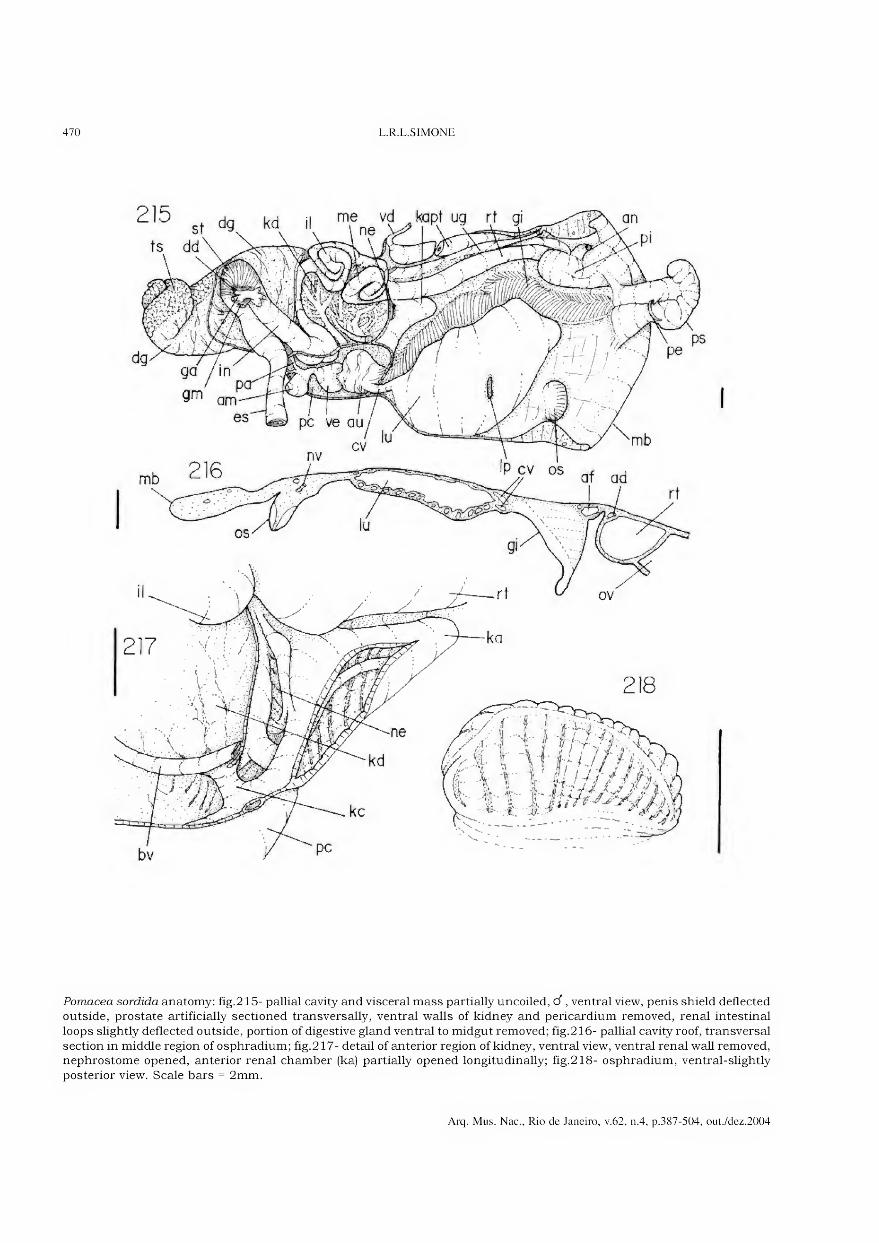
470 L.R.L.SIMONE
■ ° st kd,
ts dd\
me vd topt ug rt gj
Pomacea sórdida anatomy: fig.215- pallial cavity and visceral mass partially uncoiled, d, ventral view, penis shield deflected outside, prostate artificially sectioned transversally, ventral walls of kidney and pericardium removed, renal intestinal
loops slightly deflected outside, portion of digestive gland ventral to midgut removed; fig.216- pallial cavity roof, transversal
section in middle region of osphradium; fig.217- detail of anterior region of kidney, ventral view, ventral renal wall removed, nephrostome opened, anterior renal chamber (ka) partially opened longitudinally; fig.218- osphradium, ventral-slightly posterior view. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
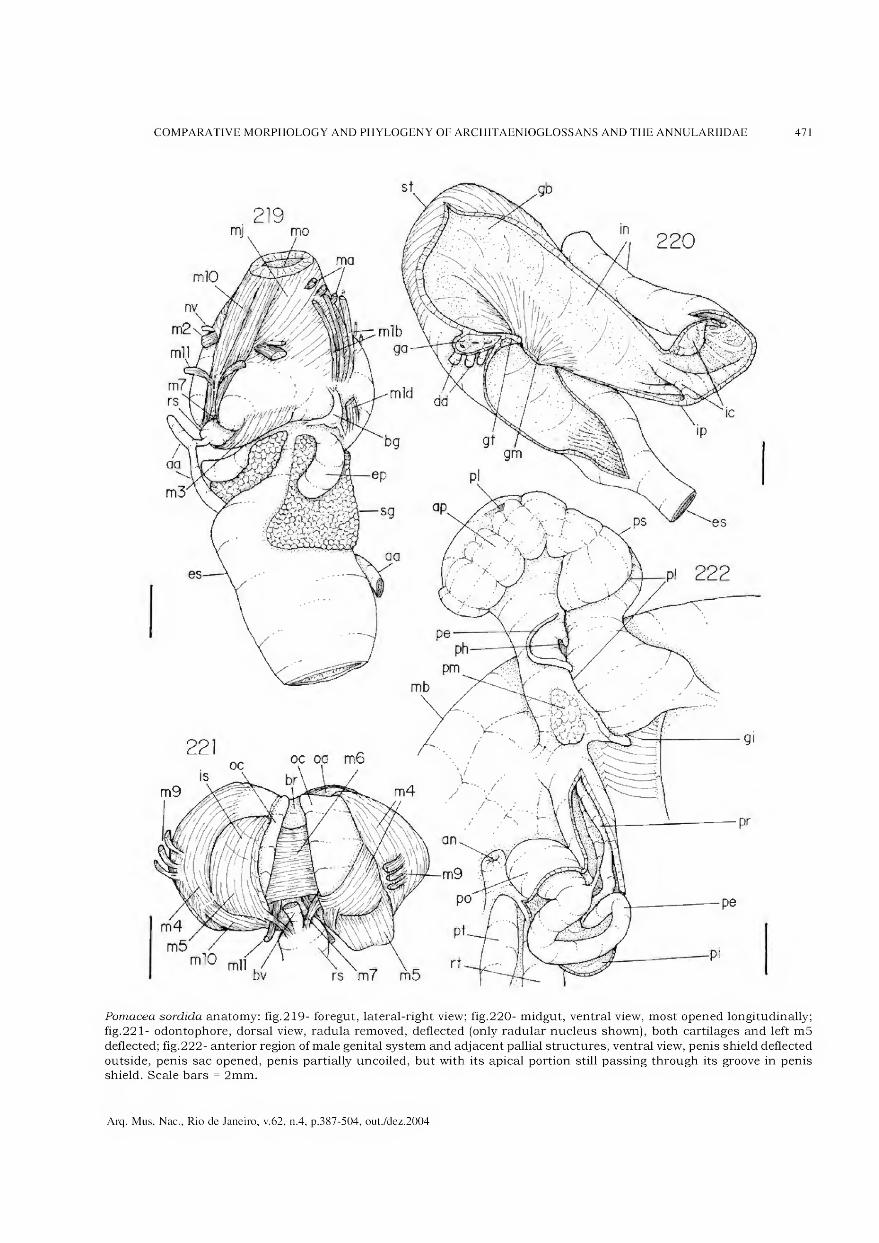
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 471
Pomacea sórdida anatomy: fig.219- foregut, lateral-right view; fig.220- midgut, ventral view, most opened longitudinally; fig.221- odontophore, dorsal view, radula removed, deflected (only radular nucleus shown), both cartilages and left m5
deflected; fig.222- anterior region of male genital system and adjacent pallial structures, ventral view, penis shield deflected
outside, penis sac opened, penis partially uncoiled, but with its apical portion still passing through its groove in penis shield. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

Asolene megastoma anatomy: fig.223- head-foot, d, frontal view; fig.224- pallial cavity, d, ventral-inner view, penis shield deflected outside, penis sac opened, penis partially stretched, prostate posterior region sectioned; fig.225- pallial
cavity roof, transversal section in middle region of osphradium; fig.226- foregut, lateral-left view. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 473
227
fp mb an
Asolene megastoma anatomy: fig.227- midgut, ventral view as in situ, chambers of intestine (ic) seen as if intestine was a
transparent structure; fig.228- stomach and adjacent portions of stomach and intestine, ventral view, entirely opened
longitudinally; fig.229- pallial oviduct and adjacent region of pallial cavity, ventral view, oviduct seen as a transparent structure. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

474 L.R.L.SIMONE
Felipponea neritiniformis anatomy: fig.230- midgut, ventral view as in situ, chambers of intestine (ic) seen as if intestine was a transparent structure; fig.231- osphradium, ventral view. Scale bars = lmm. Marisa planogyra anatomy: fig.232- anterior
portion of pallial cavity, d, ventral-inner view, penis shield deflected outside, penis sac opened, penis partially stretched;
fig.233- pallial cavity and visceral mass partially uncoiled, male, ventral view, penis shield deflected outside; fig.234- head- foot, d , frontal view; fig.235- pallial cavity roof, transversal section in middle region of lung. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 475
Marisaplanogyra anatomy: fig.236- kidney and adjacent structures, ventral view, ventral walls of kidney and pericardium removed, renal intestinal loops deflected outside and only partially shown, anterior renal chamber opened longitudinally;
fig.237- head and haemocoel, ventral view, foot removed; fig.238- foregut, lateral-left view; fig.239- midgut, ventral view,
most opened longitudinally, ventral gastric wall deflected; fig.240- midgut, ventral view as in situ; fig.241- pallial oviduct posterior region, not fully mature, ventral view, seen as a transparent structure. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
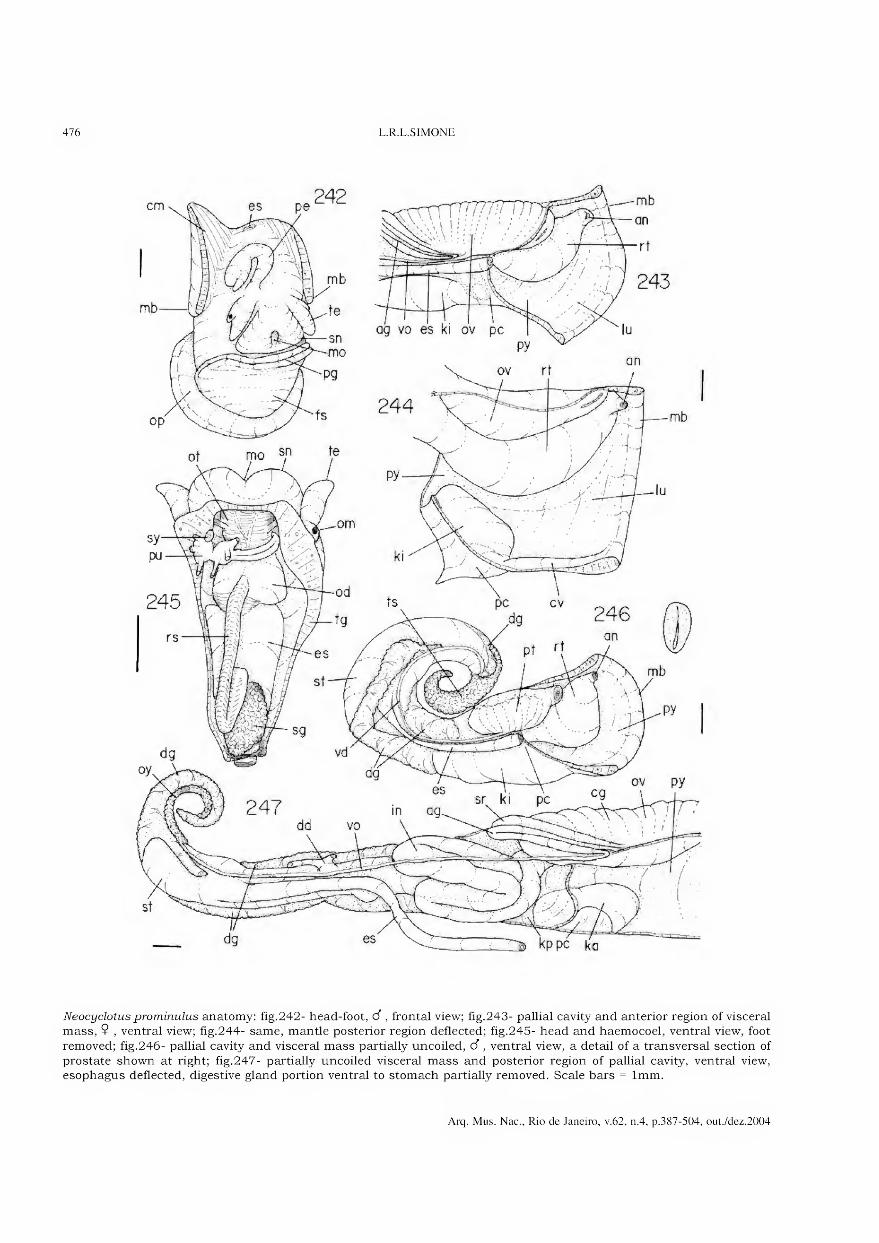
476 L.R.L.SIMONE
Neocyclotusprominulus anatomy: fig.242- head-foot, d, frontal view; fig.243- pallial cavity and anterior region of visceral mass, $ , ventral view; fig.244- same, mantle posterior region deflected; fig.245- head and haemocoel, ventral view, foot
removed; fig.246- pallial cavity and visceral mass partially uncoiled, d , ventral view, a detail of a transversal section of
prostate shown at right; fig.247- partially uncoiled visceral mass and posterior region of pallial cavity, ventral view, esophagus deflected, digestive gland portion ventral to stomach partially removed. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 477
Neocyclotusprominulus anatomy: fig.248- kidney and adjacent structures, ventral view, ventral wall of pericardium removed; fig.249- foregut, lateral-right view; fig.250- buccal mass and anterior region of esophagus, ventral view; fig.251- foregut,
ventral view, odontophore removed, esophagus and right esophageal pouch (left in fig.) opened longitudinally; fig.252-
odontophore, dorsal view, radula removed, deflected and only partially shown, both cartilages and left m5 deflected; fig.253- same, m6 sectioned longitudinally, only middle and right structures shown, m4 partially separated from cartilage. Scale bars = 0.5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
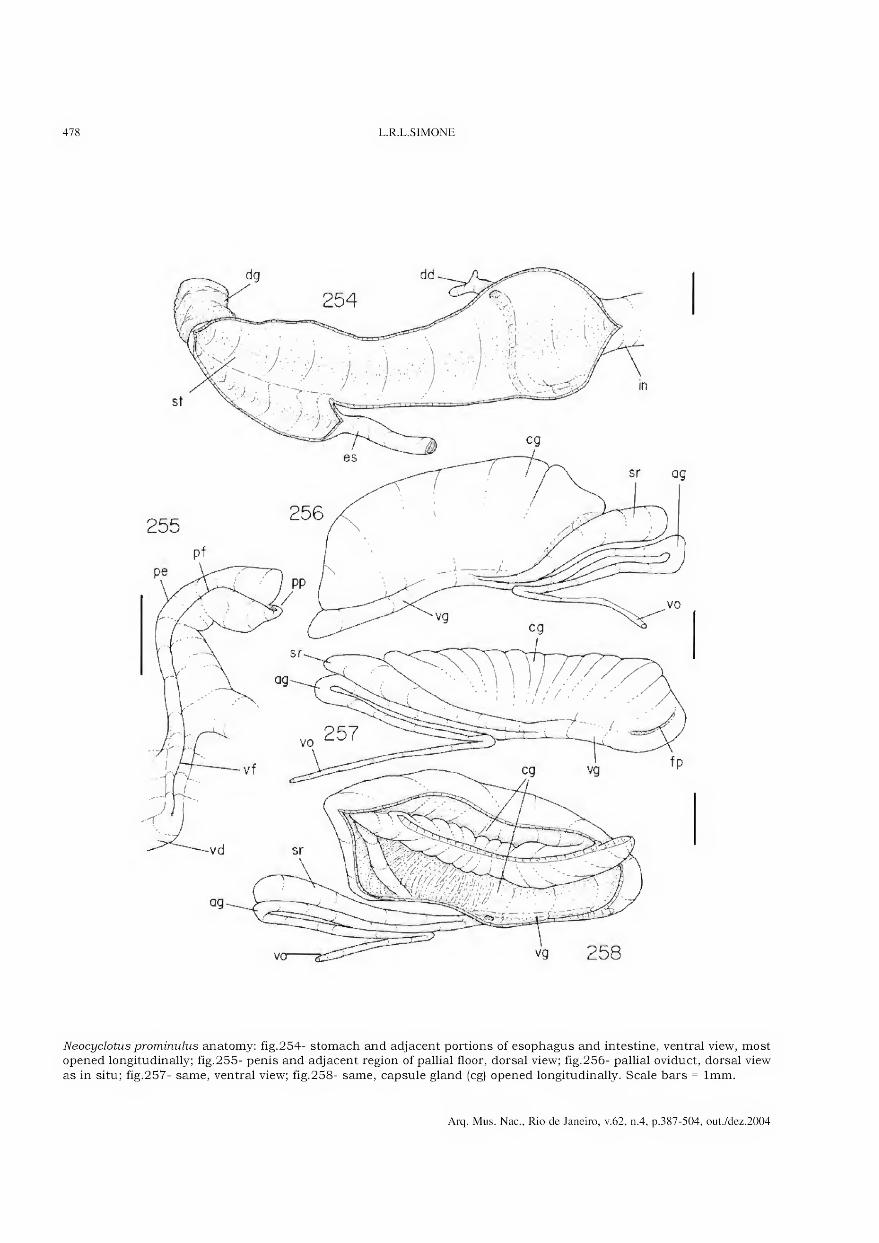
478 L.R.L.SIMONE
Neocyclotus prominulus anatomy: fig.254- stomach and adjacent portions of esophagus and intestine, ventral view, most opened longitudinally; fig.255- penis and adjacent region of pallial floor, dorsal view; fig.256- pallial oviduct, dorsal view
as in situ; fig.257- same, ventral view; fig.258- same, capsule gland (cg) opened longitudinally. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 479
Incidostoma tupy anatomy: fig.259- head-foot, d, frontal view; fig.260- head-foot, 9 , frontal-slightly left view; fig.261- pallial cavity (lung), 9 , ventral-inner view; fig.262- region between visceral mass and pallial cavity, ventral view, ventral
wall of pericardium and portion of rectum ventral to kidney removed. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
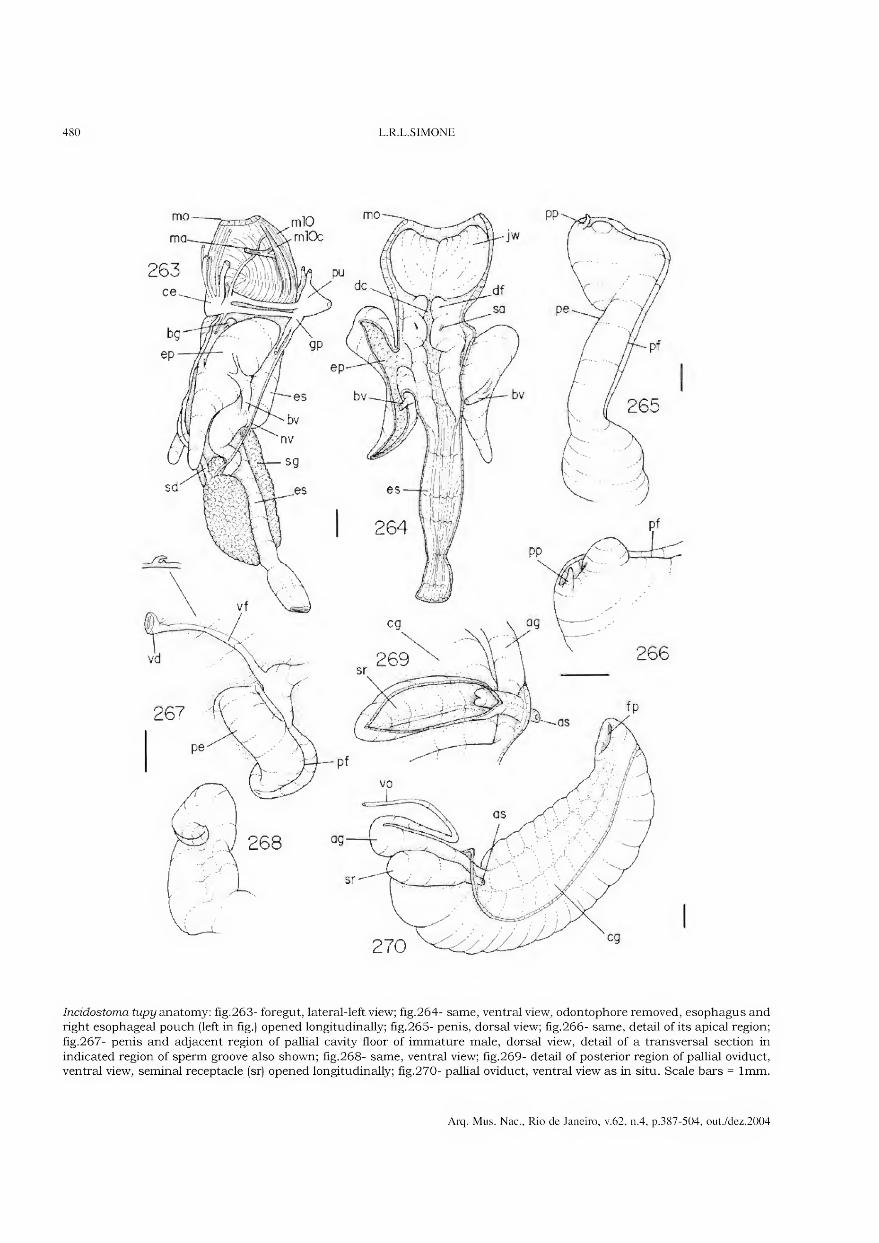
480 L.R.L.SIMONE
Incidostoma tupy anatomy: fig.263- foregut, lateral-left view; fig.264- same, ventral view, odontophore removed, esophagus and right esophageal pouch (left in fig.) opened longitudinally; fig.265- penis, dorsal view; fig.266- same, detail of its apical region;
fig.267- penis and adjacent region of pallial cavity floor of immature male, dorsal view, detail of a transversal section in
indicated region of sperm groove also shown; fig.268- same, ventral view; fig.269- detail of posterior region of pallial oviduct, ventral view, seminal receptacle (sr) opened longitudinally; fig.270- pallial oviduct, ventral view as in situ. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 481
Aperostoma blanchetiana anatomy: fig.271- kidney and adjacent structures, ventral view, ventral wall of pericardium removed; fig.272- foregut, lateral-left view; fig.273- pallial oviduct, ventral view, posterior structures shown as in situ;
fig.274- penis and adjacent region of pallial floor, dorsal view; fig.275- same, detail of its apical region. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
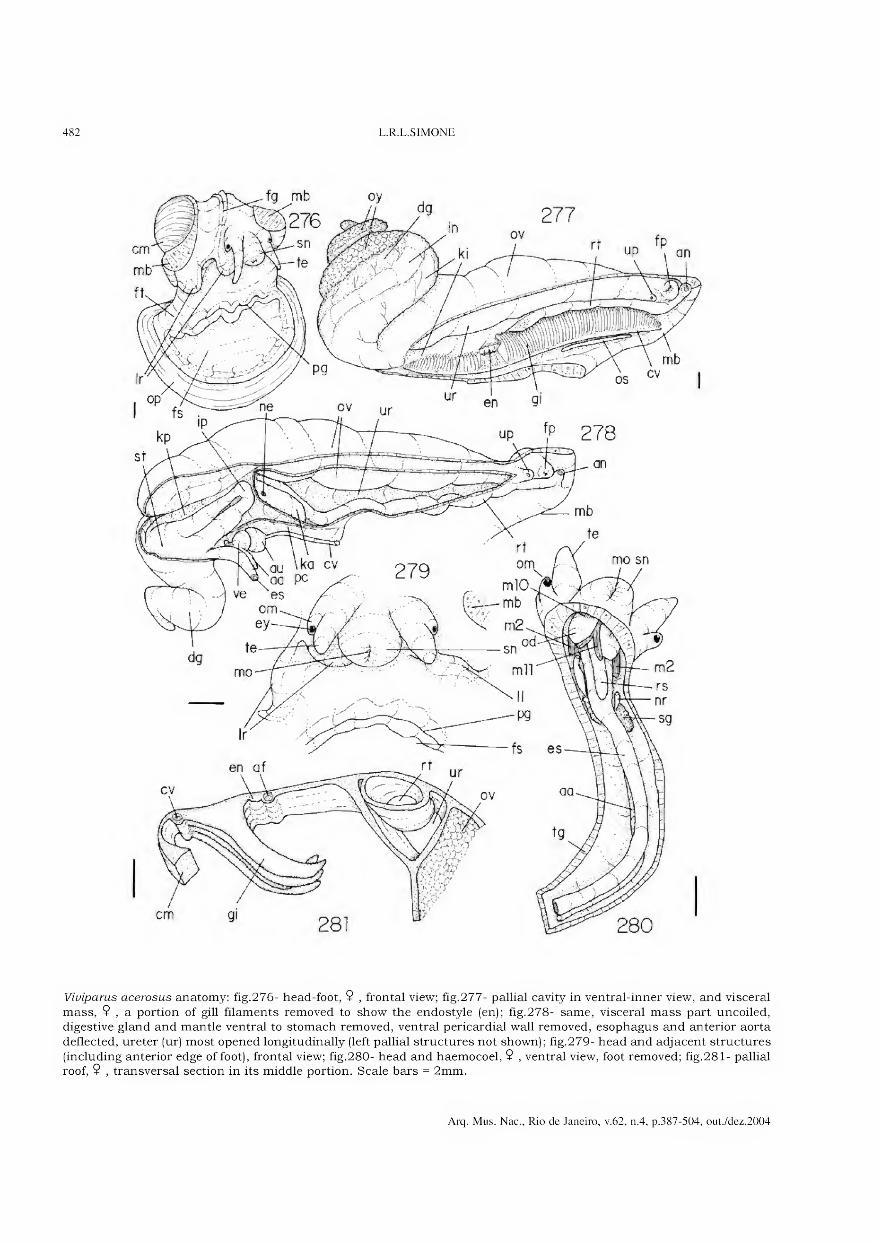
482 L.R.L.SIMONE
Viviparus acerosus anatomy: fig.276- head-foot, 9 , frontal view; fig.277- pallial cavity in ventral-inner view, and visceral
mass, 9 , a portion of gill filaments removed to show the endostyle (en); fig.278- same, visceral mass part uncoiled, digestive gland and mantle ventral to stomach removed, ventral pericardial wall removed, esophagus and anterior aorta
deflected, ureter (ur) most opened longitudinally (left pallial structures not shown); fig.279- head and adjacent structures
(including anterior edge of foot), frontal view; fig.280- head and haemocoel, 9 , ventral view, foot removed; fig.281- pallial roof, 9 , transversal section in its middle portion. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 483
Viviparus acerosus anatomy: fig.282- kidney and adjacent structures, ventral view, ventral wall of pericardium removed, ureter opened longitudinally; fig.283- foregut, ventral view, odontophore removed, esophagus partially opened longitudinally;
fig.284- foregut, lateral-left view; fig.285- odontophore, ventral view; fig.286- same, dorsal view; fig.287- same, dorsal
view, radular ribbon extracted, both cartilages deflected, m6 sectioned longitudinally, lateral structures weakly shown. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

484 L.R.L.SIMONE
Viviparus acerosus anatomy: fig.288- odontophore, dorsal view, radular ribbon removed and deflected downwards, both cartilages and left m5 deflected; fig.289- stomach opened longitudinally, ventral view, distai region not shown; fig.290-
midgut, ventral view, seen as in situ; fig.291- head and haemocoel, d, ventral view, foot removed, some transversal
sections along vas deferens artificially done, right tentacle (left in fig.) opened longitudinally; fig.292- middle region of male genital system, dorsal view, seen as in situ; fig.293- head, d , lateral-right view. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
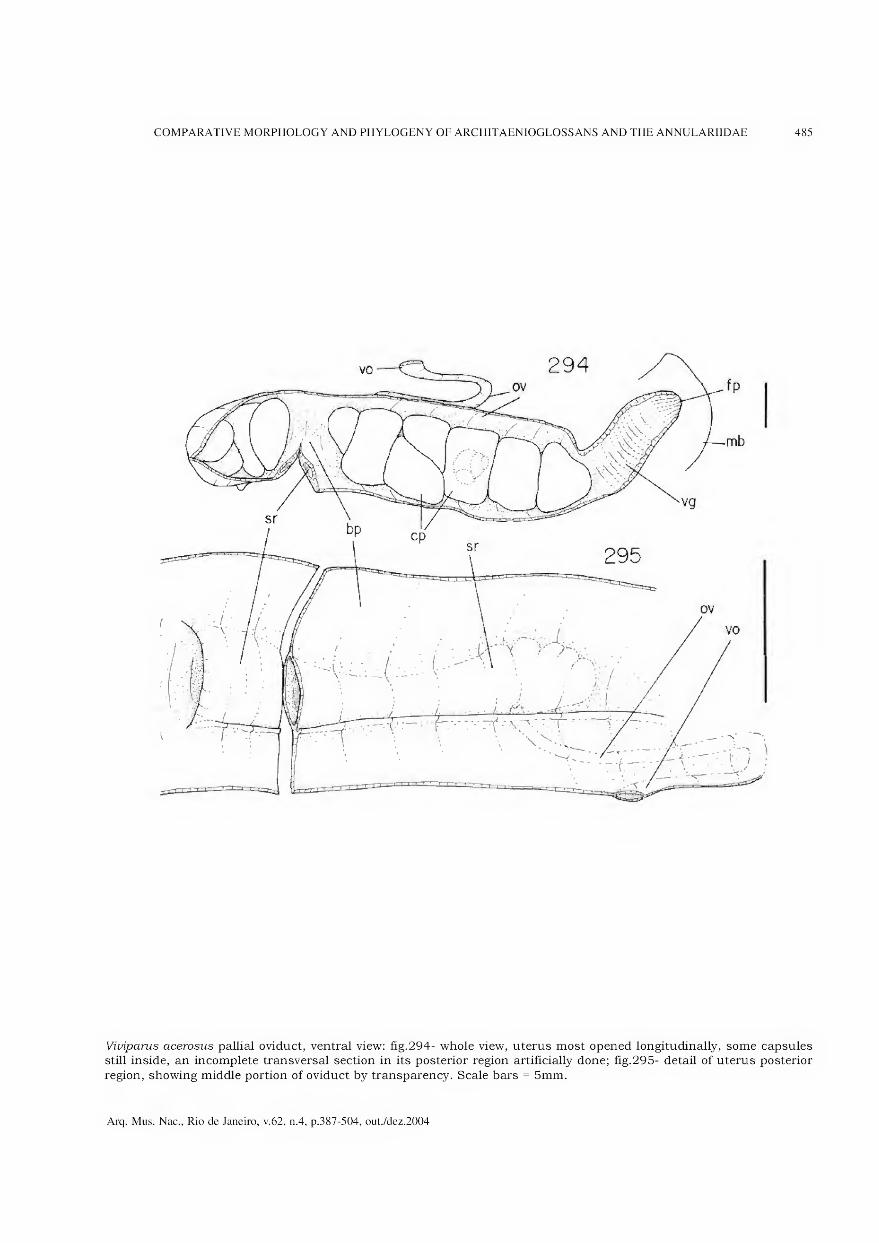
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 485
Viviparus acerosus pallial oviduct, ventral view: fig.294- whole view, uterus most opened longitudinally, some capsules still inside, an incomplete transversal section in its posterior region artificially done; fig.295- detail of uterus posterior
region, showing middle portion of oviduct by transparency. Scale bars = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
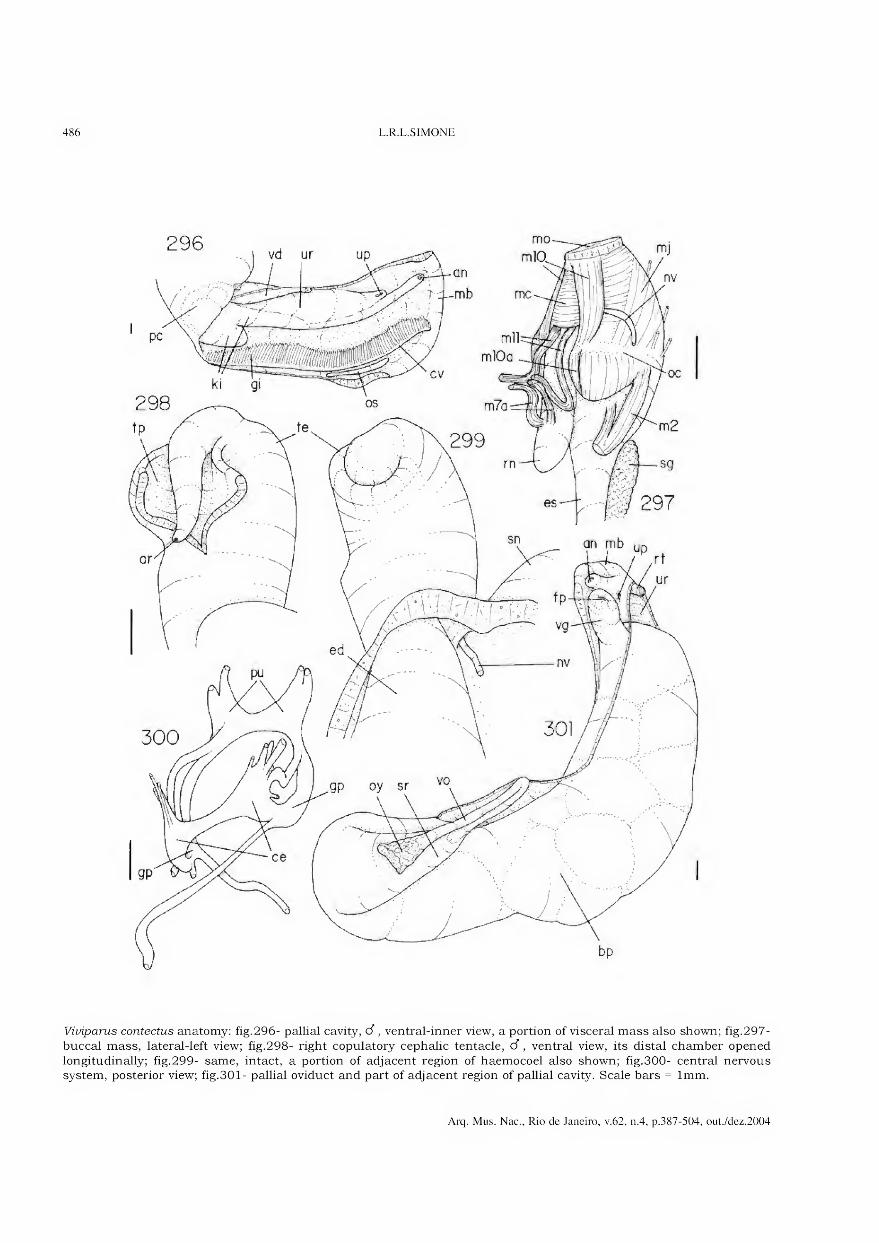
486 L.R.L.SIMONE
296 mo
Viviparus contectus anatomy: fig.296- pallial cavity, d, ventral-inner view, a portion of visceral mass also shown; fig.297-
buccal mass, lateral-left view; fig.298- right copulatory cephalic tentacle, d, ventral view, its distai chamber opened
longitudinally; fig.299- same, intact, a portion of adjacent region of haemocoel also shown; fig.300- central nervous system, posterior view; fig.301- pallial oviduct and part of adjacent region of pallial cavity. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
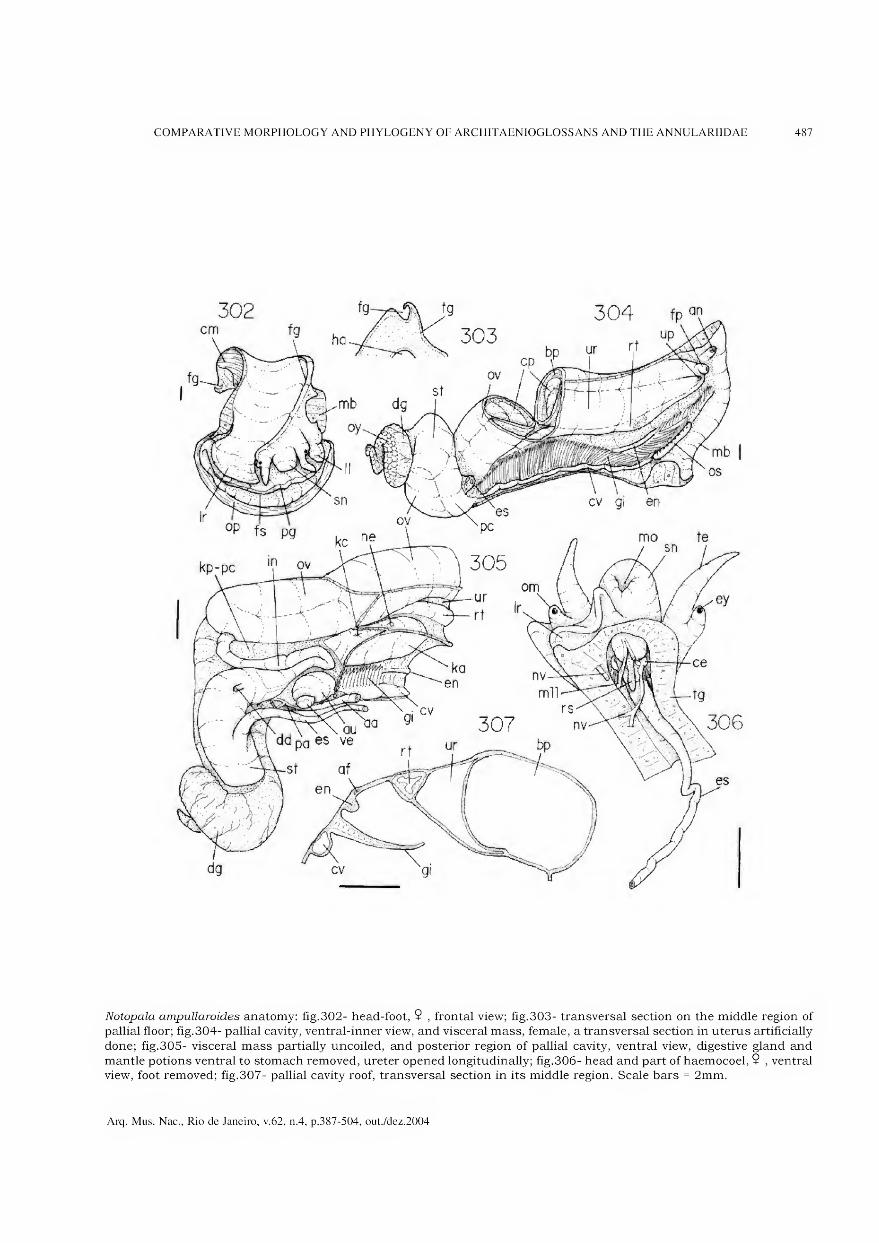
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 487
Notopala ampullaroides anatomy: fig.302- head-foot, $ , frontal view; fig.303- transversal section on the middle region of pallial floor; fig.304- pallial cavity, ventral-inner view, and visceral mass, female, a transversal section in uterus artificially
done; fig.305- visceral mass partially uncoiled, and posterior region of pallial cavity, ventral view, digestive gland and
mantle potions ventral to stomach removed, ureter opened longitudinally; fig.306- head and part of haemocoel, 9 , ventral view, foot removed; fig.307- pallial cavity roof, transversal section in its middle region. Scale bars = 2mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
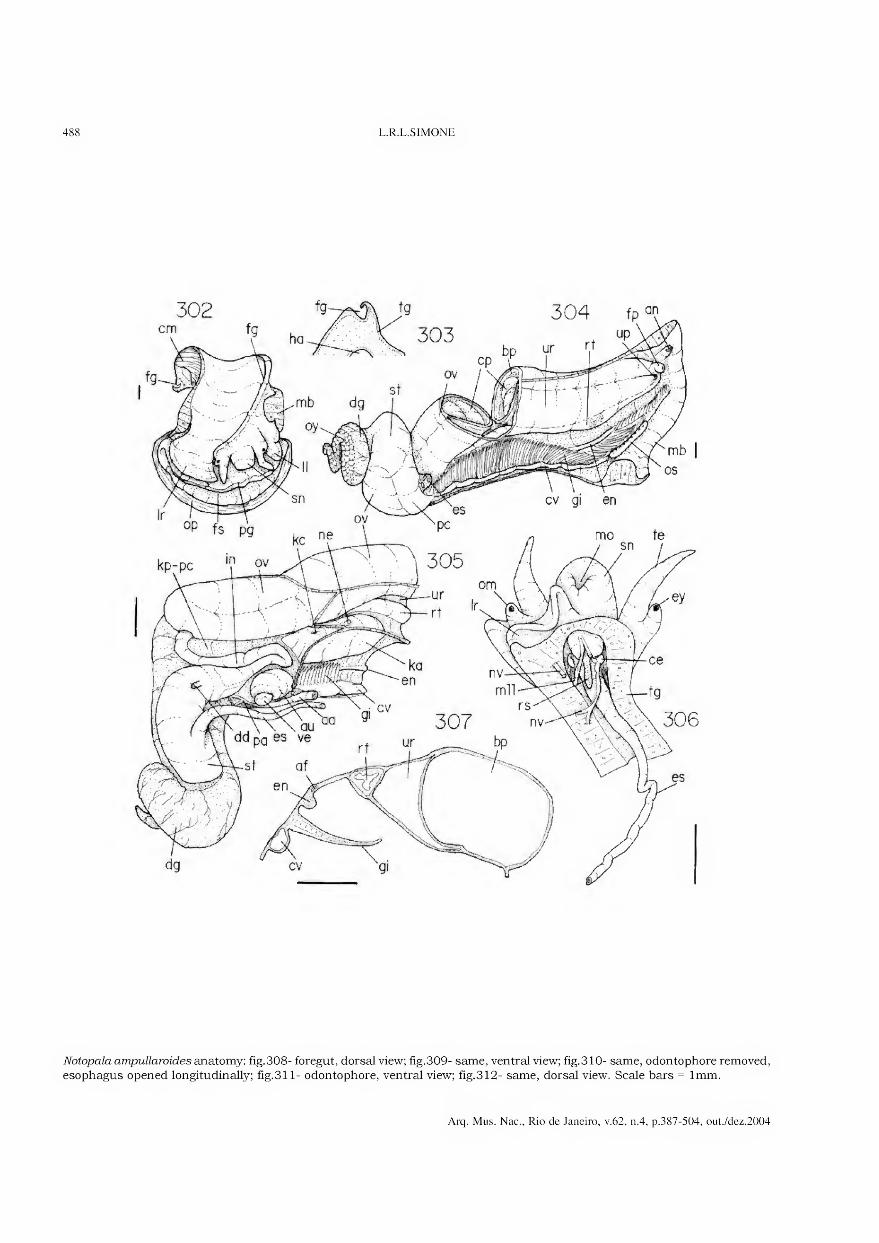
488 L.R.L.SIMONE
Notopala ampullaroides anatomy: fig.308- foregut, dorsal view; fig.309- same, ventral view; fig.310- same, odontophore removed, esophagus opened longitudinally; fig.311- odontophore, ventral view; fig.312- same, dorsal view. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 489
br mil m4
ed-
JJ
t ag
ui
m (
'^9
Notopala ampullaroides anatomy: fig.313- odontophore, dorsal view, radular ribbon removed and deflected, both cartilages and left m5 deflected; fig.314- same, m6 sectioned longitudinally and both cartilages still more deflected; fig.315- head
and part of haemocoel, d, ventral view, foot removed, some transversal sections along vas deferens done; fig.316- right
copulatory cephalic tentacle, d , ventral view, some adjacent head structures also shown; fig.317- pallial portion of male genital system, and some adjacent structures, ventral view; fig.318- pallial oviduct, ventral view as in situ; fig.319- same, transversal section in its posterior region. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

490 L.R.L.SIMONE
Anatomy of some Australian viviparids - Notopála essingtonensis: fig.320- right copulatory cephalic tentacle, d , ventral
view, some adjacent head structures also shown; fig.321- pallial cavity roof, d, transversal section in its middle region;
Larina cf strangev. fig.322- head-foot, d, frontal view; fig.323- operculum, outer view; fig.324- same, inner view; fig.325- pallial cavity roof, 9 , transversal section in its middle region. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 491
Larina cf strangei anatomy: fig.326- visceral mass partially uncoiled and posterior region of pallial cavity, ventral view,
structures ventral to stomach removed or deflected, albumen gland transversal sectioned, ureter opened longitudinally;
fig.327- foregut, lateral-left view; fig.328- posterior 2/3 portion of pallial oviduct, ventral view as in situ; fig.329- same, transversal section in its posterior region. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
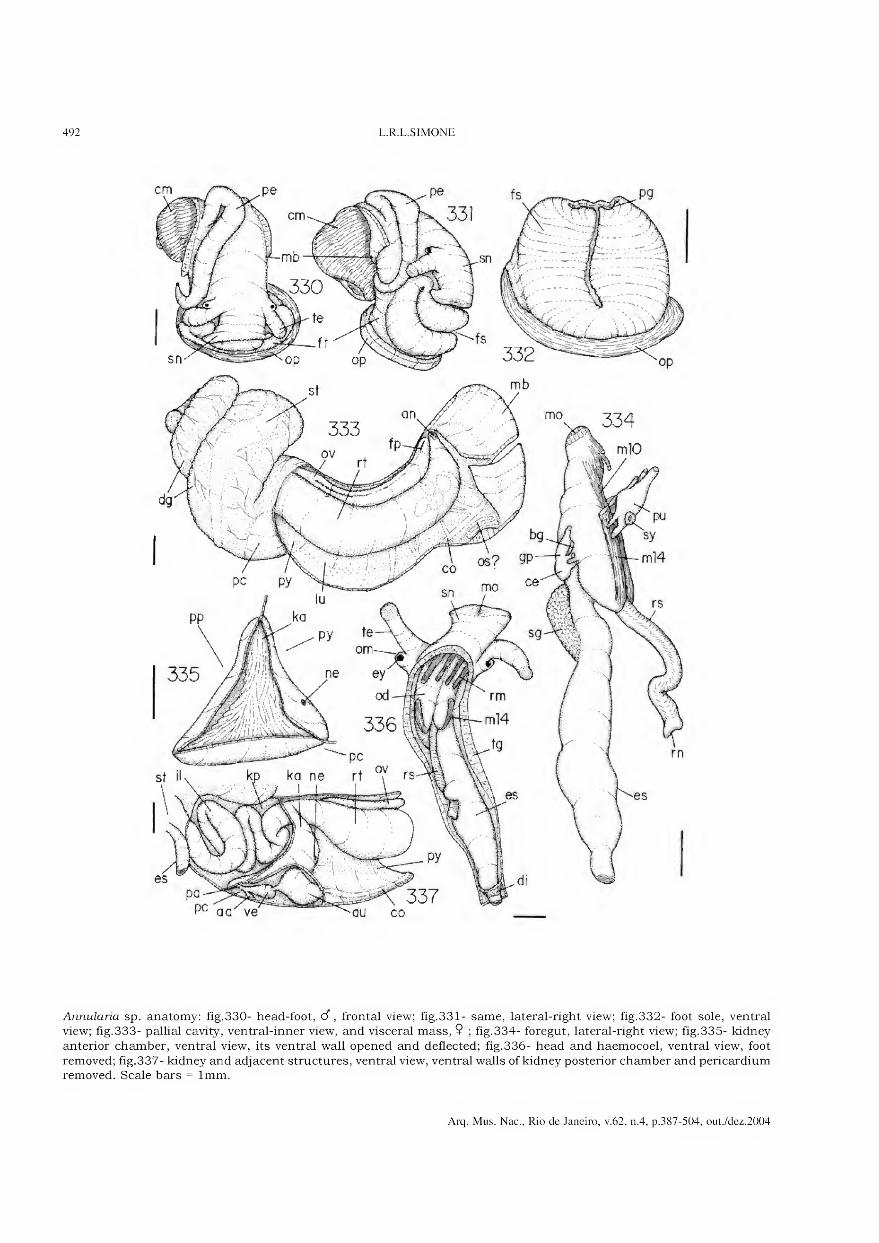
492 L.R.L.SIMONE
Annularia sp. anatomy: fig.330- head-foot, d", frontal view; fig.331- same, lateral-right view; fig.332- foot sole, ventral view; fig.333- pallial cavity, ventral-inner view, and visceral mass, 9 ; fig.334- foregut, lateral-right view; fig.335- kidney
anterior chamber, ventral view, its ventral wall opened and deflected; fig.336- head and haemocoel, ventral view, foot
removed; fig.337- kidney and adjacent structures, ventral view, ventral walls of kidney posterior chamber and pericardium removed. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
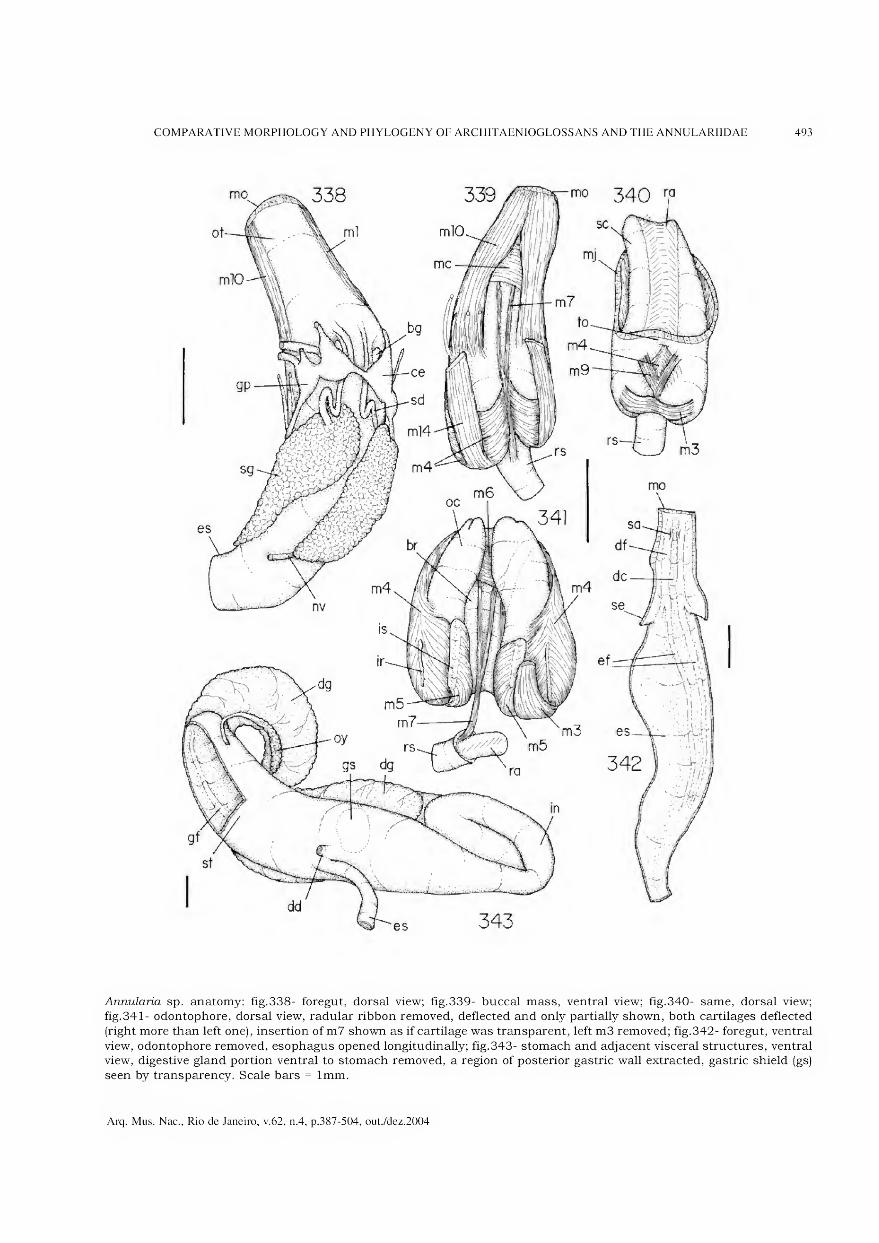
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 493
Annularia sp. anatomy: fig.338- foregut, dorsal view; fig.339- buccal mass, ventral view; fig.340- same, dorsal view; fig.341- odontophore, dorsal view, radular ribbon removed, deflected and only partially shown, both cartilages deflected
(right more than left one), insertion of m7 shown as if cartilage was transparent, left m3 removed; fig.342- foregut, ventral
view, odontophore removed, esophagus opened longitudinally; fig.343- stomach and adjacent visceral structures, ventral view, digestive gland portion ventral to stomach removed, a region of posterior gastric wall extracted, gastric shield (gs)
seen by transparency. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
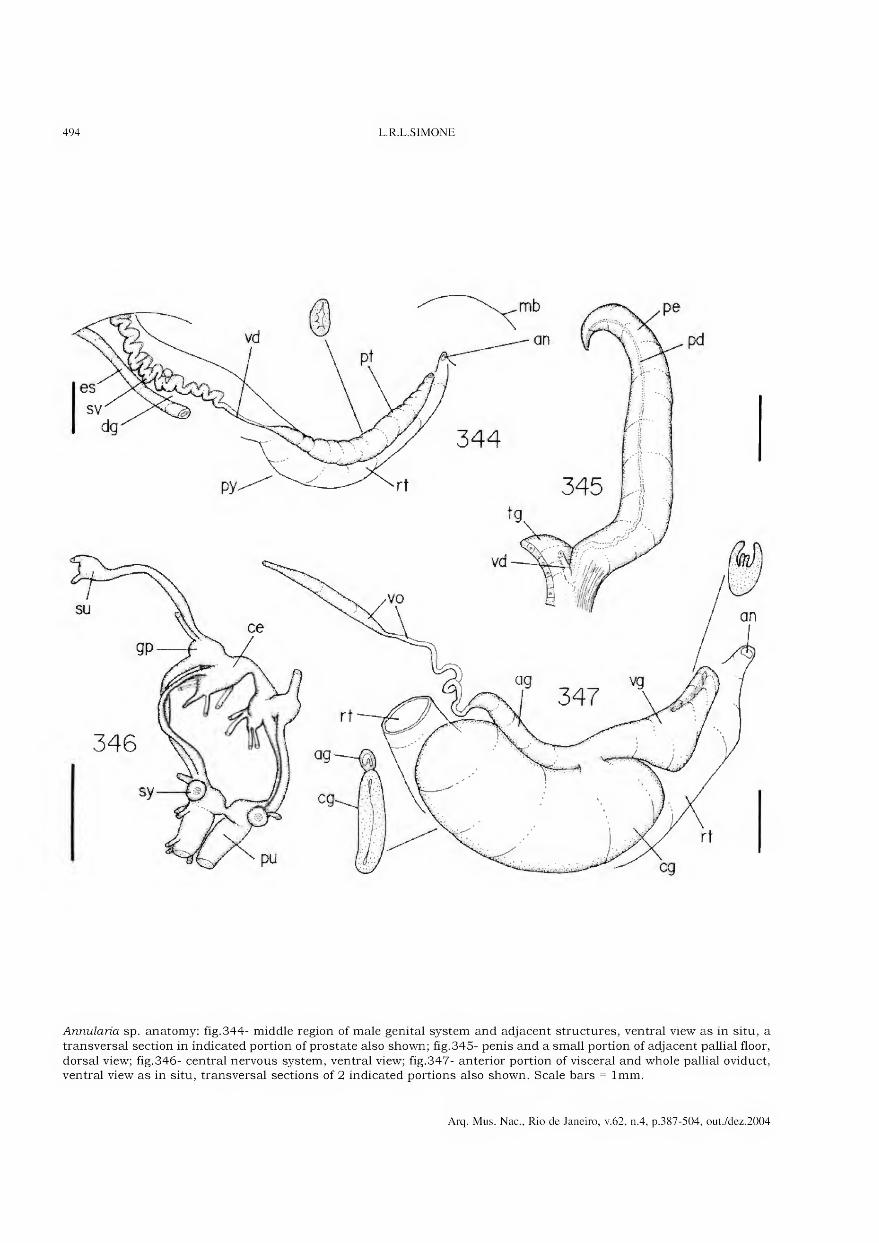
494 L.R.L.SIMONE
Annularia sp. anatomy: fig.344- middle region of male genital system and adjacent structures, ventral view as in situ, a
transversal section in indicated portion of prostate also shown; fig.345- penis and a small portion of adjacent pallial floor,
dorsal view; fig.346- central nervous system, ventral view; fig.347- anterior portion of visceral and whole pallial oviduct, ventral view as in situ, transversal sections of 2 indicated portions also shown. Scale bars = lmm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
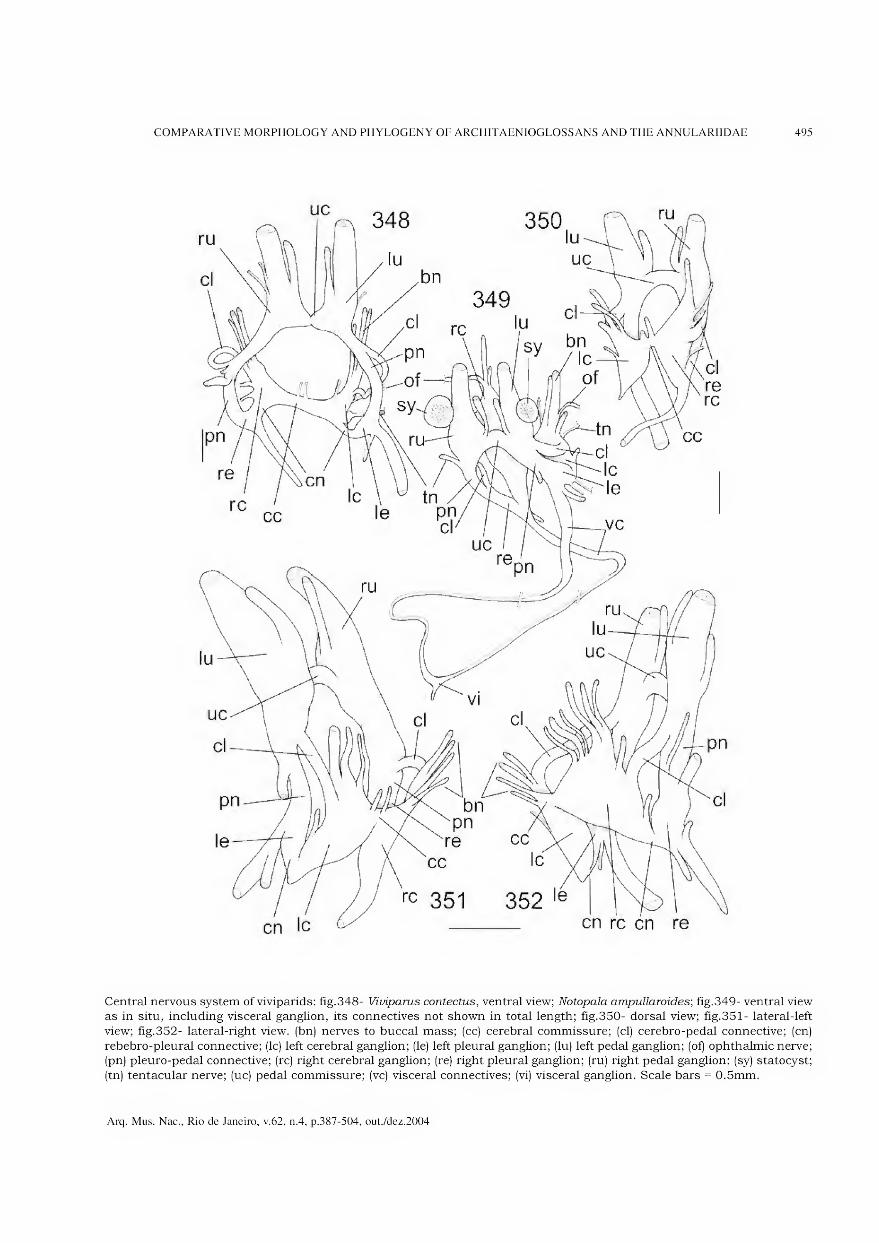
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 495
Central nervous system of viviparids: fig.348- Viviparus contectus, ventral view; Notopala ampullaroides; fig.349- ventral view as in situ, including visceral ganglion, its connectives not shown in total length; fig.350- dorsal view; fig.351- lateral-left
view; fig.352- lateral-right view. (bn) nerves to buccal mass; (cc) cerebral commissure; (cl) cerebro-pedal connective; (cn)
rebebro-pleural connective; (lc) left cerebral ganglion; (le) left pleural ganglion; (lu) left pedal ganglion; (of) ophthalmic nerve; (pn) pleuro-pedal connective; (rc) right cerebral ganglion; (re) right pleural ganglion; (ru) right pedal ganglion; (sy) statocyst;
(tn) tentacular nerve; (uc) pedal commissure; (vc) visceral connectives; (vi) visceral ganglion. Scale bars = 0.5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

496 L.R.L.SIMONE
1 7 12 15 18 30 34 3647 48 54
57 58 59 60 61 62 63 66 68 71 74
11111111111 76 82 84 85 86 88 90 93 96 101 107
111111112 1 1 109 113 128 130 133 135 138 142
11111
4 8 1421 40 43 45 49 52 70 83 92 95 100 110 114 118 119 131 132 134 |i i i 43 50 81 119 132
ffrrf 5 6 7 1011 46 4 g 13 2137 45 53 7273 79 81 83 88 100118 141
—we### • • mAnnularia
Neocyclotus prominulus
'Aperostoma blanchetiana
Incidostoma tupy
6 1011 55 72 79
105 106 113 114
1 25 5 1 54 5 5 5 6 87 91 112 138 1 39 140 1 43
□ •■■■■■■■ ■ ■ ■ ■ 011221111 2 1 1 1
10 0 0 0 1 IT [“Ti 95 101 111 140
• □ ■ ■ 10 12
2 9 10 17 22 28 35 40 41 42
fülfffflf 6978 IR 115 lÍ6lÍ9lÍ5lh ■ ■■■■■•■ 12 11112 1
30113 114 128 135
30 64 96 117
vwr
sp
'Cerithioidea
c:
11 16 18 19 23 29 31 32 35 37 38 40 43 45 52
53 65 67 75 77 78 80 88 92 97 99 100 102 103 ■ ■■■■■■•■■■■ ■ ■ 311111122112 1 1
104 120 122 123 124 125 131 133 135 136 137 ■ ■■■■■■■•■ 1 1 1 1 1 1 2 2 2 1.
1 29 108 127
fftr 20120121 125
»ID i 2 i n
Iviparus contectus
'iviparus acerosus
'Notopala ampullaroides
'Notopala essingtoensis
'Larina cf. strangei
Marisa planogyra
'Pomacea crosseana
'Felipponea neritiniformis
| = exclusive synapomorphy 0 = ingroup convergence | | = reversion
°23 24
97 123
M 0 1 26 98 104
126 127
yy 121
ff f 98 121
25 r+^^Pomacea bridgesi
Pomacea sórdida
Pomacea testudinea
Pomacea canaliculata
'Asolene megastoma
Fig.353- cladogram with synapomorphies of each node shown. Uppermost number indicates the character, the lower the State. Note that two outgroups (Annularia sp and Cerithioidea ground plan) resulted inside the ingroup. Length: 252; CI= 76; RI= 91.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004
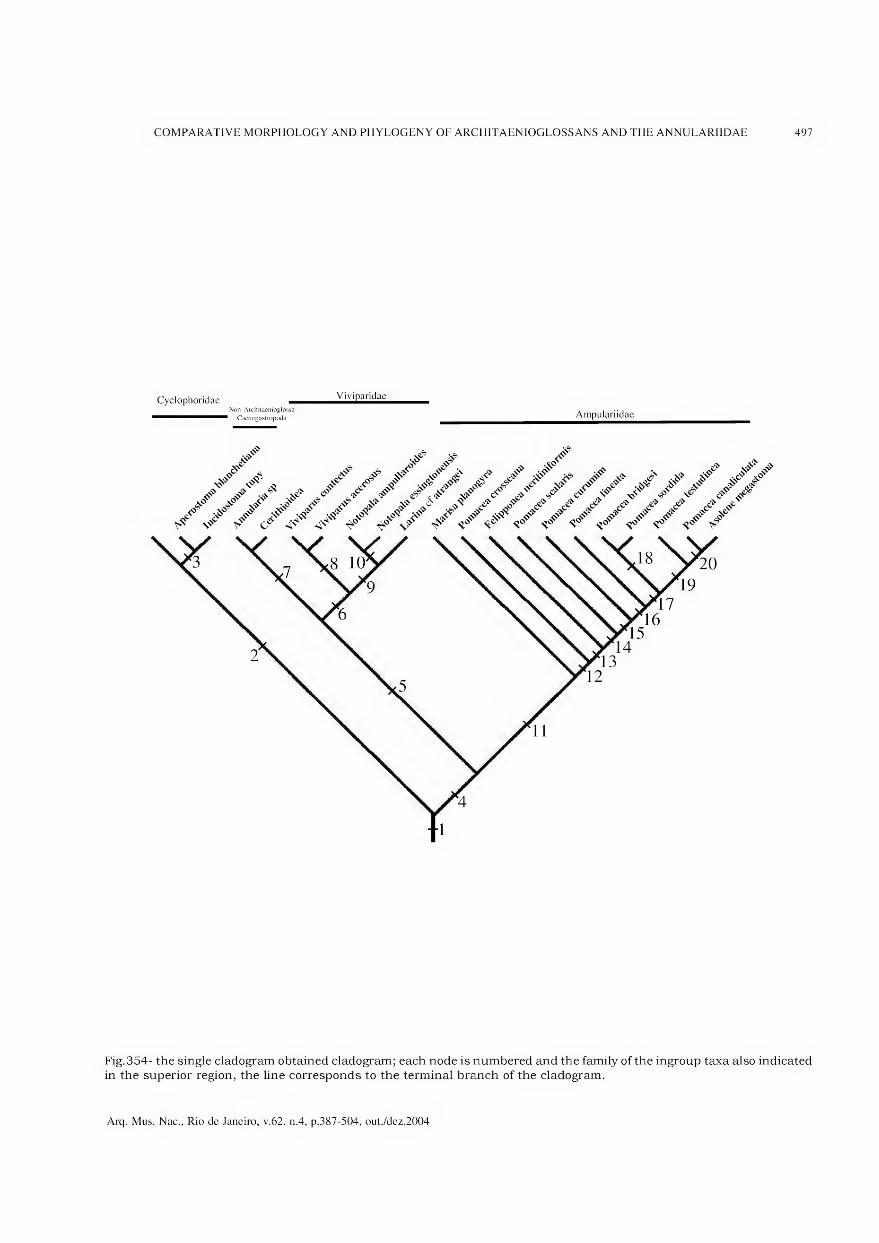
COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 497
Cyclophoridae _ViviParidae Non-Architaenioglossa
Caenogastropoda Ampulariidae
Fig.354- the single cladogram obtained cladogram; each node is numbered and the family of the ingroup taxa also indicated in the superior region, the line corresponds to the terminal branch of the cladogram.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

498 L.R.L.SIMONE
DISCUSSION OF THE CLADOGRAM
As referred in the Material and Methods section, each character was polarized one by one, analyzing each case separately, and deciding which outgroups are selected for separating its States, for polarization, and, in the cases of multistate characters, for deciding the optimization is additive or not. Those parameters can be easily changed in the Computer programs, and where changes were made, any modification in the results or indices was reported. Taking into consideration that whether all outgroups are analyzed effectively as outgroups, the monophyly of the ingroup is not tested, since it will absolutely result reunited in a branch. For testing the monophyly of any taxon, it is important to insert in the analysis some outgroups belonging to close taxa, which will be analyzed operationally as part of the ingroup. A more distant taxon (in present study a set of archaeogastropod taxa) was used as the effective outgroup. If the Architaenioglossa is actually monophyletic, the caenogastropod taxa should appear outside of the architaenioglossans clade.
This procedure was applied in the present study. The Annularia sp., as well as the Cerithioideaground plan, appeared inside the architaenioglossans taxa. Both these taxa are undoubtedly considered caenogastropods. The present result suggests that the taxon “Architaenioglossa” is non-monophyletic. The two non-architaenioglossans taxa formed a single terminal branch, and the architaenioglossans formed three successive branches, indicating its paraphyly.
Most nodes and branches (Figs.353-354) are analyzed in the following text. The more important synapomorphies that support them are also reported, but are discussed in the previous section.
The node number 1 (Fig.354) is supported by 41 synapomorphies (Fig.353). From those, the outstanding are: ommatophore located on tentacles (character 12); osphradium elongated and more complex (18); decrease of the hypobranchial gland (30); duplication of the lobe of the single kidney (34, 36); appearance of a pair of inner dorsal folds in buccal mass (47); jaw muscles separated in a pair of bundles (48); formation of the m4 pair of odontophore muscles from several pairs (54); reduction of ventral tensor muscle of the radula (57); enlargement of dorsal tensor muscle of the radula (58); m5 pair of odontophore muscles becoming longer and inserted on m4 (instead of on the cartilage) (59, 60); horizontal muscle becoming shorter and broader (61,62); arising the pair m7 (63); loss of approximator muscles of
cartilages (or incorporation to m4) (66); loss of the posterior pair of odontophore cartilages (74); enlargement of subradular cartilage (76); reduction of the radular teeth to 7 per row (82, 85); esophagus origin in posterior region of buccal mass (90); loss of gastric caecum (96); absence of visceral structures in haemocoel (107); fecal pellets (109); pallial vas deferens at least partially closed (113); large and complex pallial oviduct located at right from rectum (128, 130); capsule gland (133); developed female pore (135); nerve ring positioned more posterior from mouth (138); and buccal ganglia positioned far removed from the nerve ring (142). Being the node 1 followed by the 3 architaenioglossan branches plus the remainder caenogastropods (represented by Annularia and Cerithioidea ground plan), it is a quite possible that it represents the order Caenogastropoda as a whole. This affirmation is considered still premature, since another study concerned to the phylogeny of the entire order is in progress. The present study is part of it. For a secure affirmation on the caenogastropod synapomorphies and monophyly depends on a deeper analysis of remainder superfamilies of the group and of the sister-group taxon, the Heterobranchia. Those studies are still being performed.
HASZPRUNAR (1988) regarded Architaenioglossa as non-caenogastropod prosobranchs (archaeo gastropods). He considered it as the sister taxon of the Caenogastropoda plus Heterobranchia branch. PONDER 86 LINDBERG (1996, 1997), on the other hand, considered the architaenioglossans as basal caenogastropods. However, as also pointed out by HASZPRUNAR (1988), those authors suggested that there is a possibility that the architaenioglossans are paraphyletic (1997:225) as corroborated with the present result. Harasewych et al. (1997) performed a molecular analysis (18S rDNA and cytochrome c oxidase I), included the viviparid Cipangopaludinajaponica (von Martens, 1860), with this architaenioglossan in the first branch of the Caenogastropoda.
The node 2, supported by 21 synapomorphies, represents the family Cyclophoridae, the single one reunited in the superfamily Cyclophoroidea. From the synapomorphies, those more remarkable are: calcareous operculum (character 8); loss of pallial structures for becoming a lung (21); posterior located nephrostome (40); urinary gutter (43); fusion of jaw plates (45); odontophore muscle pair mia and mlOc (49, 70); esophageal pouches covered internally by papillae (92); the exophalic penis (114, 118, 119); albumen gland as a loop
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 499
(131); and special conformation of pallial oviduct (132, 134). Despite the very well supported node, the cyclophorids are a very diverse and widespread group. They have some endemic supra-specific taxa that were not analyzed in present study. It is possible that the family synapomorphies would be diluted when a larger number of representatives will be analyzed. However, a set of synapomorphies of the taxon is here presented by the first time, and is considered important for defining the taxon and for analyzing its relationship with the remainder gastropods.
The node 3 separates Aperostoma and Incidostoma from Neocyclotus. It is supported by 5 synapomorphies, being the more important the different organization of the lung vessels (44), the modification of the odontophore pair of muscles mia (50); the duplication of the penis papilla (119); and the bursa copulatrix opening directly to the pallial cavity instead of opening to the pallial oviduct (132).
The node 4 encompasses the remainder ingroup except the cyclophoroideans. It is supported by 10 synapomorphies, being the more remarkable the following: the head-foot siphons (10, 11, which reverted in the node 7); the connection of the m4 with tissue on the radula (55); the odontophore muscle pair ml2 (72); the closure of the pallial vas deferens (113, 114). The arrangement of the taxa on the cladogram shows that the cyclophorids are the more basal taxon of the caenogastropods, and that the remainder taxa represent another branch, uniting the ampullariids and remainder caenogastropods.
For operational reasons, we will analyze the node 11 to 18, and after them we return to numerical order.
The node 11 represents the Ampullariidae, the single family of the superfamily Ampullarioidea. The node is supported by 40 synapomorphies, being the more remarkable: snout tentacles (character 16); osphradium with bipectinated and pedunculated fashion (18, 19); lung sac (29); ampulla in anterior aorta (32); intestinal loops inside kidney (37); anterior septate renal chamber (38); multiple jaw ditator muscles (ma) (52); odontophore muscle pairs m7b and m9 (65, 67); pair of accessory odontophore cartilages (75); muscular stomach walls (97); gastric muscle (99); multiplication of ducts to digestive gland (100); special arrangement of intestinal chambers in its portion close to pericardium (103, 104); penis and penis shield structures (120-127); enormous capsule gland surrounding most of remainder pallial oviduct structures (131, 133); special brooding strategy (137). The ampullariids
have been the goal of some important papers on their anatomy (e.g., Scott, 1957), and even including phylogenetic approach (BERTHOLD, 1989, 1991; BIELER, 1993). However, several of here presented synapomorphies are apparently introduced by the fist time. They are the cases, for example, of the gastric muscle (that connect the stomach with shell columella), the intestinal chambers close to pericardium, and the renal intestinal loops. BIELER (1993), based on the data searched by BERTHOLD (1991), selected 70 ampullariid characters, and obtained 3 equally parsimonious trees. The author showed 3 cladograms (that paper figs. 1-3), the first based on the BERTHOLD’s preferences, and the other 2 based on the data matrix. All cladograms are so different from each other and from that obtained herein that it is impracticable to compare. Bieler searched 23 synapomorphies for the family, most of them taken into consideration in the present study and presented in preceding discussion.
The remaining nodes, 12-20, refer to the internai organization of the family Ampullariidae. The first branch is Marisa, separated from the remaining branches by 8 synapomorphies. The lasting taxa are, in a broad sense, representatives of the genus Pomacea. However, species belonging to the genera Asolene and Felipponea, considered by some authors as subgenera of Pomacea, where mixed. The scattered organization of the ampullariids in the cladogram is considered provisional, since some organs and structures have been under-analyzed [e.g., the kidney, pallial oviduct). This happened because of the insufficient or poorly-preserved material available. For this reason no taxonomical change is proposed. However, on the basis of this result, it appears that a reanalysis of ampullariid taxonomy may necessary.
The node 5 is supported by 13 synapomorphies, including: the appearance of the pair of retractor muscle of buccal mass (m2) (character 51); the simplification and enlargement of the odontophore muscle pair m4 (54-56); the loss of the esophageal pouches (91); the prostate gland located in pallial cavity (112); the nerve ring positioned posterior to buccal mass (138); the approach of the pleural ganglia to the cerebral ganglia (epiathroid organization) (139, 140); and the lateral position of buccal ganglia in buccal mass (143). This branch represents the caenogastropods except the Cyclophoroidea and the Ampullarioidea, encompassing the Viviparioidea and the Sorbeoconcha. PONDER & LINDBERG (1997:225) introduced this taxon, comprising the Cerithioidea
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

500 L.R.L.SIMONE
and the rest of the caenogastropods. The current literature usually considers the Viviparidae as Ampullarioidea [e.g., BOSS, 1982;VAUGHT, 1989). The viviparids actually share some characters with the ampullariids, such as the head-foot left and right siphons, the type of operculum, and the freshwater environment. Those characters were here considered homologous between the representatives of both groups. However, despite this, the two groups are paraphyletic. This resultindicates that the viviparids should be included in a monotypic superfamily Viviparioidea.
The node 6 is supported by 18 synapomorphies, and represents the viviparids. The more important synapomorphies are: adaptations to filter feeding (characters 9, 10, 17, 22, 28); ureter (40, 41); male pallial ducts running by integument and opening in the right cephalic tentacle (114-117); brood pouch in pallial oviduct (129, 137). An internai organization of the viviparids was obtained, represented by the nodes 8 and 9, separating respectively the Viviparus sample from the Australian sample. As this organization is considered provisional, then no deeper analysis or taxonomic implications is here proposed. The Australian viviparids studied here belong to the subfamily Bellamyinae (ANNANDALE & SEWELL, 1921; ROHRBACH, 1937; VAIL, 1977; SMITH, 2000), characterized mainly by the testis positioned in the pallial cavity, by side of the rectum (Fig.317:pt); this character is not explored here because the studied material is not well-preserved enough for observing this character in all samples. The Australian sample of Notopála and Larina are separated from each other (node 10) by a single synapomorphy (single gastric duct to digestive gland, character 100, State 1, convergent with other branches).
The node 7 encompasses supposedly outgroups, representing the “true” caenogastropods, or the Sorbeoconcha, in this study. The node is supported by 10 synapomorphies, being half of them reversions. The more remarkable are: the determinate shell growth (character 5); the reversion of the left and right head-foot siphons (10-11); the reduction of the dorsal chamber of buccal mass (46); the gastric style sac (101); the male seminal vesicle (111); and the approach of the pairs of connectives pedal-cerebral to pedal- pleural ganglia in the nerve ring (140).
As reported above, a more un-depth study on the caenogastropod phylogeny is still been performed, which this study is part. Apparently, the architaenioglossans can be regarded as true Caenogastropoda, being its basal branches.
However, further analysis on caenogastropod phylogeny is still in progress, and depends on the study of remainder superfamilies, as well as of more outgroups. On the other hand, the present studied sample is considered sufficient for analyzing the monophyly and phylogenetic relationship of the contemplated supra-generic taxa, just the main goals of this study.
CONCLUSIONS
1. The taxon Architaenioglossa is not monophyletic, but so paraphyletic, then it should not be used as a formal taxon.
2. The architaenioglossans can be regarded as the 3 first branches of the order Caenogastropoda, respectively: Cyclophoroidea (Cyclophoridae); Ampullarioidea (Ampullariidae) and Viviparioidea (Viviparidae).
3. The combination among the architaenioglossans and remaining caenogastropods are supported by 41 morphological synapomorphies.
4. The Cyclophoroidea, Ampullarioidea and Viviparioidea are supported respectively by 21, 40 and 18 synapomorphies.
5. The Viviparioidea are a separated taxon from the Ampullarioidea.
6. The three architaenioglossan branches presented internai, well-resolved organizations; however, they are considered provisional and no systematic change was proposed.
7. The analysis of all organs and systems are useful for phylogenetic analysis.
ACKNOWLEDGMENTS
To the researchers who provided specimens for this study; Jen Hemmen (Germany), for the gift of the Viviparus samples; Dr. Silvana Thiengo (FIOCRUZ), for the samples of Pomacea sórdida
and P. bridgesi and for permission of using some photos of Spix’s type specimens, she obtained in the Zoologische Staatssammlung Müchen, Germany; Jorge Vaz (São Paulo), for the samples of Felipponea neritiniformis; the team of ictiologists of MZSP, for the P. curumim samples; Dr. Ricardo Pinto-da-Rocha and Dr. Sonia Casari, for the samples of Annularia sp. For loaned material I thank Dr. Winton Ponder (AMS, Australia), for the samples of Notopala and Larina, and also for complete revision of the paper. For Thomas Lippareli, in
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 501
that time in Campo Grande, Mato Grosso do Sul, for providing me a field trip to Pantanal, where we collected several of the lots here studied. I also thank Dr. Cláudio Mantovani Martins (São Paulo), for providing histological examination in 2 specimens of Pomacea curumim and in helping the adult nature of the sample; Dr. Philippe Bouchet (MNHN, Paris), and Dr. John Taylor and John Todd (BMNH), for the permission of examination and photographing type specimens; Dr. José L.M. Leme, for the guidance. This research was mostly supported by a governmental grand of Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP), proc. #96/6756-2, 00/ 11074-5, 00/11357-7.
LITERATURE CITE D
ADAMS, A., 1854. Descriptions of a new genus and of
several new species of gastropodous Mollusca, from
the Cumingian collection. Proceedings of the
Zoological Society of London 1853:41-42.
ANDREWS, E.B., 1964. The functional anatomy and
histology of the reproductive system of some pilid
gastropod molluscs. Proceedings of the
Malacological Society of London 36:121-140.
ANDREWS, E.B., 1965a. The functional anatomy of the
gut of the prosobranch gastropod Pomacea
canaliculata and some other pilids. Proceedings of
the Zoological Society of London 145:19-36.
ANDREWS, E.B., 1965b. The functional anatomy of the
mantle cavity, kidney and blood system of some pilid
gastropods (Prosobranchia). Journal of Zoology,
146(l):70-94.
ANDREWS, E.B., 1979. Fine structure in relation to
function in the excretory system of two species of
Viviparus. Journal of Molluscan Studies,
45(2): 186-206.
ANDREWS, E.B., 1981. Osmoregulation and excretion
in prosobranch gastropods; part 2: structure in
relation to function. Journal of Molluscan Studies,
47(3):248-289.
ANDREWS, E.B., 1982. Osmoregulation and excretion
in prosobranch gastropods part 2: structure in
relation to function. Journal of Molluscan Studies,
47(3):248-289.
ANDREWS, E.B. & LITTLE, C., 1972. Structure and
function in the excretory systems of some terrestrial
prosobranch snails (Cyclophoridae). Journal of
Zoology, 168(3) :395-422.
ANDREWS, E.B. & LITTLE, C., 1982. Renal structure
and function in relation to habitat in some
cyclophorid land snails from Papua New Guinea.
Journal of Molluscan Studies 48(2):124-143.
ANNANDALE, N. & SEWELL, R, 1921. The banded pond
snail of índia (Viviparus bengalensis). Records of the
Indian Museum 22:215-292.
BALL, A.D.; ANDREWS, E.B. & TAYLOR, J.D., 1997. The
ontogeny of the pleurembolic proboscis in Nucella
lapillus (Gastropoda: Muricidae). Journal of
Molluscan Studies 63:87-99.
BARTSCH, P., 1946. The operculate land mollusks of
the family Annulariidae of the Island of Hispaniola
and the Bahama Archipelago. Bulletin of the United
States National Museum 192:1-264, 38 pis.
BAKER, M.D.F., 1913. The land and fresh-water
mollusks of the Stanford Expedition to Brazil.
Proceedings of the Academy of Natural Sciences
of Philadelphia 65:618-672.
BERTHOLD, T., 1989. Comparative conchology and
functional morphology of the copulatory organ of the
Ampullariidae (Gastropoda, Monotocardia) and their
bearing upon phylogeny and palaeontology.
Abhandlungen des Naturwissenschaftlichen
Vereins in Hanburg (NF) 28:141-164.
BERTHOLD, T., 1990. Phylogenetic relationship, adaptations
and biogeographic origin of the Ampullariidae (Mollusca,
Gastropoda) endemic to Lake Malawi, África.
Verhandlungen des Naturwissenschaftlichen Vereins
in Hamburg (NF) 31/32:47-84.
BERTHOLD, T., 1991. Vergleichende Anatomie, Phylogenie
und Historische Biogeographie der Ampullariidae.
Abhandlungen des Naturwissenschaftlichen Vereins
in Hanburg (NF) 29:1-256.
BERTONI, A.W., 1926. Sobre moluscos dei Paraguay.
Revista de la Sociedad Cientifica dei Paraguay
2(l):71-73.
BIELER, R., 1993. Ampulariid phylogeny - book review
and cladistic re-analysis. Veliger, 36(3) :291-299.
BINNEY, W.G., 1867. Land and fresh-water shells of
North America; part 3. Smithsonian Miscellaneous
Collections, 144(3): 1-120.
BOSS, K.J., 1982. Mollusca. In: PARKER, S.P. [Ed.]
Synopsis and classification of living animais. New
York: McGraw-Hill Book Company. v.2, p.947-1166.
BOUVIER, E.L., 1888. Étude sur 1’organisation des
ampullaires. Memoires de la Societe Philomath
Paris 100:63-85, pl.9.
BROOKS, W.K., 1907. The origin of the lung in
Ampullaria. Nautilus: 11-12.
BROOKS, W.K. & MCGLONE, B., 1908. The origin of
the lung of Ampullaria. Papers from the Marine
Biological Laboratory at Tortugas. Carnegie Institute
Publication 102:97-104, pis. 1-7.
BROWN, D.S. & BERTHOLD, T., 1990. Lanistes neritoides
sp.n. (Gastropoda) from West Central África:
description, comparative anatomy and phylogeny.
Verhandlungen des Naturwissenschaftlichen
Vereins in Hamburg (NF) 31/32:119-152.
BURNE, R.H., 1898. A reno-pericardiac pore in
Ampullaria urceus Müll. Proceedings of the
Malacological Society of London 3:49-52.
BURNE, R.H., 1899. Note on the nervous sytem of
Ampullaria sp. Proceedings of the Malacological
Society of London 3:317-319.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

502 L.R.L.SIMONE
CASTELLANOS, Z.J.A. & FERNÁNDEZ, D., 1976.
Mollusca Gasteropoda Ampullariidae. In: RINGUELET,
R. A. Fauna de agua dulce de la República Argentina
15(1): 1-33.
CAZZANIGA, N.J., 1990. Sexual dimorphism in Pomacea
canaliculata (Gastropoda: Ampullariidae). Veliger
33(4):384-388.
CAZZANIGA, N.J. & ESTEBENET, A.L., 1984. Revision
y notas sobre los hábitos alimentarios de los
Ampullariidae (Gastropoda). Historia Natural
4(22):213-224.
CEI, G., 1942. Considerazioni filogenetiche sopra alcune
particolari strutture degli organi respiratori in
Cyclostoma elegans Müll. Bolletino di Zoologia 8(5-
6):175-181.
COOK, P.M., 1949. A ciliary feeding mechanism in
Viviparus viviparus (L.). Proceedings ot the
Malacological Society of London 27:265-271.
COTTON, B.C., 1935. Recent Australian viviparidae and
a fóssil species. Records of the South Australian
Museum 5:339-344.
COUSIN, A., 1887. Faune malacologique de la pépublique
de 1’Equateur. Bulletin de la Société Zoologique
de France 12:187-287., pls.3-4.
COWIE, R.H., 1997. Pila Rõding, 1798 and Pomacea Perry,
1810 (Mollusca, Gastropoda): proposed placement on
the Official List, and Ampullariidae Gray, 1824:
proposed confirmation as the nomenclaturally valid
synonym of Pilidae Preston, 1915. Bulletin of
Zoological Nomenclature 54(2):83-88.
DALL, W.H., 1919. A new form of Ampullaria. Nautilus
33:10-11.
DEMIAN, E.S., 1965. The respiratory system and the
mechanism of respiration in Marisa comuarietis (L.).
Arkiv for Zoologi 17(8):539-560.
DEMIAN, E.S. & IBRAHIM, A.M., 1972. Sexual
dimorphism and sex ratio in the snail Marisa
comuarietis (L.). Bulletin of the Zoological Society
of Egypt 24:52-63.
DEMIAN, E.S. & YOUSIF, F., 1973. Embryonic
development and organogenesis in the snail Marisa
comuarietis (Mesogastropoda: Ampullariidae).
Malacologia 12(1): 151-174.
DOELLO-JURADO, M., 1924. Algunos moluscos
fluviátilos interesantes de Bolivia. Physis 7:238-240.
ELEUTHERIADIS, N. & DIMITRIADOU, M.L., 1995. The
life cycle, population dynamics, growth and
secondary production of the snail Viviparus contectus
(Millet) (Gastropoda: Prosobranchia) in the marshes
of the River Strymonas, Macedonia, northern Greece.
Malacologia 37(l):41-52.
FALNIOWSKI, A., 1989. Prosobranchs (Prosobranchia,
Gastropoda, Mollusca) of Poland. 1. Neritidae,
Viviparidae, Valvatidae, Bithyniidae, Rissoidae,
Aciculidae. Zeszyty Naukowe Uniwersytetu
Jagiellonskiego Prace Zoologiczne 35:1-168.
FALNIOWSKI, A.; MAZAN, K. & SZAROWSKA, M., 1996.
Embryonic shells of Viviparus - what they may tell
us about taxonomy and phylogeny? (Gastropoda:
Architaenioglossa: Viviparidae). Malakologische
Abhandlungen 18(l):35-42.
FARRIS, J.S., 1988. Hennig 86, version 1.5. Distributed
by the author (Computer program). Port Jefferson
Station, N.Y.
FECHTER, R. & FALKNER, G., 1993. Moluscos:
Moluscos europeos marinos y de interior. In:
STEINBACH, G. [Ed.] Blume Naturaleza.
Barcelona, 287p.
FÉRUSSAC, A.E.J.P.J.F.A., 1824 - Zoologie. In:
FREYCINET, L.C.D. Voyage au tour du monde...
exécuté sur FUranie et la Physicienne, pendant...
1817-1820... par M.L. de Freycinet, 1824-1842.
FÉRUSSAC, A.E.J.P.J.F.A., 1822-1827. Dict. Class
d’Histoire Naturelle:135.
FIGUEIRAS, A., 1964 - La malacofauna dulceacuicola
dei Uruguay; ensayo de catálogo sistemático y
simonímico. Comunicaciones de la Sociedad
Malacologica dei Uruguay 1(7): 161-202.
FRAUENFELD, G.R., 1862. Zusammenstellung der
Arten der Gattung Vivipara Lmk. in der Kaiserlichen
Sammlung. Verh. Zool.Bot.Ges.Wein 12:1162-1170.
FRETTER, V. & GRAHAM, A., 1962. British prosobranch
molluscs, their functional anatomy and ecology.
Ray Society. London, i-xvi, 755p.
HARASEWYCH, M.G.; ADAMKEWICZ, S.L.; BLAKE, J.A.;
SAUDEK, D.; SPRIGGS, T. & BULT, C.J., 1997.
Neogastropod phylogeny: a molecular perspective.
Journal of Molluscan Studies 63:327-351.
HASZPRUNAR, G., 1988. On the origin and evolution
of major gastropod groups, with special reference
to the Streptoneura. Journal of Molluscan Studies
54:367-441.
HENDERSON, J.B. & BARTSCH, P., 1921. A
classification of the American operculate land
mollusks of the family Annulariidae. Proceedings
of the United States National Museum
58(2327):49-82.
HIDALGO, J.G., 1871. Description d’un Ampullaria
nouveau, provenant du fleuve Amazones. Journal
de Conchyliologie 19:206-207.
HIDALGO, J.G., 1872. Descriptions d’especes nouvelles.
Journal de Conchyliologie 20:142-144, pl.7.
HUPÉ, M.H., 1857. Mollusques (3). In: CASTELNAU,
F. Animaux nouveaux ou rares récueillis pedant
1’expedition dans les parties centrales de 1’Amerique
du Sud, de Rio de Janeiro à Lima au Peru. Paris,
7(3): 1-96.
JOHANSSON, J. 1950. On the embryology of Viviparus
and its significance for the phylogeny of the
Gastropoda. Arkiv for Zoologi 1:173-177.
KATOH, M. & RIBI, G., 1996. Genetic evidence for natural
hybridization and apparent introgression between
freshwater snail species (Viviparus ater and V contectus).
Journal of Evolutionary Biology 9(l):67-82
KOBELT, W., 1902. Cyclophoridae. Das Tierreich 16:i-
xxxix, 1-662, 1 map.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

COMPARATIVE MORPHOLOGY AND PHYLOGENY OF ARCHITAENIOGLOSSANS AND THE ANNULARIIDAE 503
LAMARCK, J.B.P.A.M., 1799. Prodrome d’une nouvelle
classification des coquilles, comprenant une
rédaction appropriée des caractères génériques, et
1’établissement d’un grand nombre de genres
nouveaux. Mémoires de la Societé d’Histoire
Naturelle de Paris 1:63-91.
LAMARCK, J.B.P.A.M., 1819. Histoire naturelle des
animaux sans vertèbres, Paris, 6(l):83-88.
LITTLE, C., 1968. Aestivation and ionic regulation in
two species of Pomacea (Gastropoda, Prosobranchia).
Journal of Experimental Biology 48:569-585.
LEAL, J.H. & SIMONE, L.R.L., 1998. Propilidium
curumim, a new species of Lepetidae (Gastropoda,
Patellogastropoda) from off Southern and
southeastern Brazil. Bulletin of Marine Science
(Miami) 63(1):157-165.
LEAL, J.H. & SIMONE, L.R.L., 2000. Copulabyssia riosi, a
new deep-sea limpet (Gastropoda: Pseudococculinidae)
from the continental slope off Brazil with comments
on the systematics of the genus. Nautilus (Sanibel)
114(2):59-68.
LOPES, H.S., 1955. Sôbre duas espécies do gênero Pomacea
Perry, com um estudo da genitália em ambos os sexos
(Mesogastropoda, Architaenioglossa, Mollusca). Revista
Brasileira de Biologia 15(2):203-210.
LOPES, H.S., 1956a. Sôbre Pomacea lineata (Spix, 1827)
(Mesogastropoda, Architaenioglossa, Mollusca).
Revista Brasileira de Biologia 16(3):375-380.
LOPES, H.S., 1956b. Sôbre Pomacea canaliculata
(Lamarck, 1822) (Mesogastropoda, Architaenioglossa,
Mollusca). Revista Brasileira de Biologia 16(4):535-
542.
LUTFY, R.G. & DEMIAN, E.S., 1965. The histology of
the respiratory organs of Marisa cornuarietis (L.).
Arkiv for Zoologi 18(5):51 71
MARTIN, S.M., 1980. Contribución al studio histologico
dei complejo peniano de algunas ampularias
argentinas. Neotropica 26(75) :47-52.
MELLO, M.D.A., 1988. Morfologia do aparelho genital
de Marisa planogyra Pilsbry, 1933 e M. cornuarietis
(Linné, 1767 (Gastropodae - Ampullariidae). Revista
Nordestina de Biologia 6(1):7-13.
MELLO, M.D.A. & MARINI, A.C., 1995 - Gênero Marisa
(Gray, 1824), p.70-79. In: BARBOSA, F.S. (Org.? Ed.?)
Tópicos em Malacologia Médica. Editora Fiocruz.
Rio de Janeiro, 314p.
MERMOD, G., 1952. Les types de la collection Lamarck
au Museum de Geneve III. Revue Suisse de Zoologie
59(2):23-97.
MONTFORT, P.D., 1810. Conchyliologie systématique
et classification méthodique des coquilles. Paris,
2 vols., 676p.
MOORE, H.B., 1931. The systematic value of a study of
molluscan faeces. Journal of the Malacological
Society of London, 19:281-290.
MORRETES, F.L., 1949. Ensaio de catálogo dos
moluscos do Brasil. Arquivos do Museu Paranaense
7:1-216.
MORRETES, F.L., 1953. Adenda e corregenda ao ensaio
de catálogo dos moluscos do Brasil. Arquivos do
Museu Paranaense 10:37-76.
MORRISON, J.P.E., 1955. Notes on American
cycloporid land snails, with two new names, eight
new species, three new genera, and the family
Amphicyclotidae separated on animal characters.
Journal of the Washington Academy of Sciences
45(5): 149-162.
MORTON, J.E., 1952. A preliminary study of the land
operculate Murdochia pallidum (Cyclophoridae,
Mesogastropoda). Transactions of the Royal
Society of New Zealand 80(l):69-79.
OLIVEIRA, M.P.; REZENDE, G.J.R& CASTRO, G.A., 1981.
Catálogo dos moluscos da Universidade Federal de
Juiz de Fora. Juiz de Fora: MEC-UFJF. 520p.
ORBIGNY, A., 1835. Synopsis terrestrium et fluviatilum
molluscorum, in suo per American Meridionalem
itinerere. Magasin de Zoologie 5(6): 1-44.
ORBIGNY, A. 1834-1847. Voyage dans 1’Amérique
Méridionale, 5(3): mollusqués. Paris, 758p, 85 pis.
PAIN, T., 1972. The Ampullariidae, an historical survey.
Journal of Conchology 27(7):453-462.
PERRY, G., 1810-1811. Arcana; or the Museum of
Natural History. Stratford, London, 84 unnumbered
pis. + associated text.
PHILIPPI, R.A., 1851. Die gattung Ampullaria. INMartini
& Chemnitz. Systematisches Conchylien
Cabinet: 1-74, 24 pis.
PONDER, W.F. & LINDBERG, D.R., 1996. Gastropod
phylogeny - challenges for the 90s. In: TAYLOR,
J. [Ed.] Origin and evolutionary radiation of
the Mollusca. Oxford University Press. London,
p.135-154.
PONDER, W.F. & LINDBERG, D.R., 1997. Towards a
phylogeny of gastropod molluscs: an analysis using
morphological characters. Zoological Journal of the
Linnean Society 119:83-265.
PONDER, W.F. & WARÉN, A., 1988. Classification of
the Caenogastropoda and Heterostropha - A list of
the family group and the higher category names.
In: PONDER, W.F. [Ed.] Prosobranch Phylogeny.
Malacological Review supp. 4:288-326.
PRASHAD, B., 1925. Anatomy of the common Indian
apple-snail, Pila globosa. Memoirs of the Indian
Museum 8(3):91-152, pls.16-18.
PRASHAD, B., 1928. Recent and fóssil Viviparidae.
A study in distribution, evolution and palaeo
geography. Memoirs of the Indian Museum
8(4):153-252, pl.19.
PRINCE, G.V., 1967. Über Lebensweise, Fortpflanzung
und Genitalorgane des terrestrischen Prosobranchiers
Cochlostoma septemspirale. Archiv für
Mollusckenkunde 96(1-2): 1-18
QUINTANA, M.G., 1982. Catálogo preliminar de la
malacofauna dei Paraguay. Revista dei Museo
Argentino de Ciências Naturales “Bernardino
Rivadavia” (Zoologia) 11(3):61-158.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

504 L.R.L.SIMONE
RAMOS, T.C., 1997. Tree Gardner, version 2.2.
Distributed by the author (Computer program). São
Paulo.
REEVE, L.A., 1856. Monograph of the genus Ampullaria.
Conchologia Iconica 10:28 pis.
ROHRBACH, F., 1937. Oekogische und morphologische
untersuchungen an Viviparus (Bellamya) capillatus
Fauenfeld und V. (B.) unicolor Oliver, unter
Berücksichtigung anderer tropischer Formen und im
Hinblick auf phyletische Beziehungen. Archiv für
Mollusckenkunde 69:177-218.
ROISSY, A.F.P.M., 1805. In: BUFFON, G.L.L. (Ed.)
Histoire naturelle des mollusques. Paris, vols. 5-6.
SALGADO, N.C.; COELHO, A.C.S. & BARROS, H.M.,
1989. Prosobrânquios terrestres do Brasil.
Cyclophoridae: Neocyclotus (N.)inca (D’Orbigny, 1835)
(Mollusca, Gastropoda, Mesogastropoda). Boletim do
Museu Nacional, Nova Série, Zoologia, Rio de
Janeiro (335): 1-15.
SASAKI, T., 1998. Comparative anatomy and phylogeny
of the recent Archaeogastropoda (Mollusca:
Gastropoda). Bulletin of the University Museum,
University of Tokyo 38:1-223.
SCOTT, M.I.H., 1957. Estúdio morfologico y taxonomico
de los Ampullariidos de la Republica Argentina.
Revista dei Museo Argentino de Ciências Naturales
“Bernardino Rivadavia”, Ciências Zoologicas
3(5):231-333, 23 pis.
SIMONE, L.R.L., 1997. Morphology of the Western
Atlantic Haliotidae (Gastropoda, Vetigastropoda) with
description of a new species from Brazil. Malacologia,
39(l-2):59-75.
SMITH, B.J., 1992. ‘Non-marine Mollusca’. In:
HOUSTON, W.W.K. [Ed.] Zoological Catalogue of
Australia. Canberra, AGPS vol. 8, xii + 405p.
SMITH, D.G., 2000. Notes on the taxonomy of
introduced Bellamya (Gastropoda: Viviparidae)
species in northeastern North America. Nautilus
114:31-37.
SOLEM, A., 1956. The helicoid cyclophorid mollusks of
México. Proceedings of the Academy of Natural
Sciences of Philadelphia 108:41-59, pls.5-6.
SOWERBY, G.B., 1909. Notes on the family
Ampullariidae with list of species, varieties, and
synonyms, also descriptions of four new species.
Proceedings of the Malacological Society of
London 8:345-362.
SPIX, J.A., 1827. Testacea Fluviatilia. Dr. F. Paula
Schrank & C.F.P. Martius [Ed.]. Lipsiae, 36p, 28 pis.
STODDART, J.A., 1982. Western Australian viviparids
(Prosobranchia: Mollusca). Journal of the
Malacological Society of Australia 5(3-4): 167-173.
SWAINSON, W., 1822-1823. Zoological Illustrations.
James Moyes. London, vol.3: 181 pis.
THIENGO, S.C., 1987. Observation on the morphology
of Pomacea lineata (Spix, 1827) (Mollusca,
Ampullariidae). Memórias do Instituto Oswaldo
Cruz 82(4):563-570.
THIENGO, S.C., 1989. On Pomacea sórdida (Swainson,
1823) (Prosobranchia, Ampullariidae). Memórias do
Instituto Oswaldo Cruz 84(3) :351-355.
THIENGO, S.C., 1995. Família Pilidae Connoly, 1927,
p.50-69. In: BARBOSA, F.S. (Ed.?Org.?) Tópicos em
Malacologia Médica. Rio de Janeiro: Editora
Fiocruz, 314p.
THIENGO, S.C.; BORDA C.E. & BARROS-ARAÚJO, J.L.,
1993. On Pomacea canaliculata (Lamarck, 1822)
(Mollusca; Pilidae: Ampullariidae). Memórias do
Instituto Oswaldo Cruz 88(1):67-71.
THIENGO, S.C. & SIMONE, L.R.L., in press. Anatomy and
systematics of Pomacea bridgesi (Caenogastropoda,
Ampullariidae), from North Brazil. Memórias do
Instituto Oswaldo Cruz.
THOMPSON, F.G., 1963. Systematic notes on the land
snails of the genus Tomocyclus (Cyclophoridae).
Breviora 181:1-11.
THOMPSON, F.G., 1966. A new pomatiasid from
Chiapas, México. Nautilus 80(l):24-28.
THOMPSON, F.G., 1967. A new cyclophorid land snail
from the West Indies (Prosobranchia), and the
discussion of a new subfamily. Proceedings of the
Biological Society of Washington 80:13-18.
THOMPSON, F.G., 1969. Some Mexican and Central
American land snails of the family Cyclophoridae.
Zoologica, New York Zoological Society: 35-77, pis. 1-7.
THOMPSON, F.G., 1978. A new genus of operculate land
snails from Hispaniola with comments on the status
of family Annulariidae. Nautilus 92(l):41-54.
TIEKECKE, H., 1940. Anatomie, phylogenie und
tiergeographie der Cyclophoriden. Archiv für
Naturgeschichte, Zeitschrift für Systematische
Zoologie , N.F., 9(3):317-371
TILLIER, S., 1980. Gastéropodes terrestres et fluviatiles
de Guyane Française. Mémoires du Muséum National
D’Histoire Naturelle, série A, Zoologie 118:1-189.
TORRE, C.; BARTSCH, P. & MARRISON, J.P.E., 1942.
The cyclophorid operculate land mollusks of America.
Bulletin of the United States National Museum
181:1-306, 42 pis.
VAIL, V.A., 1977. Comparative reproductive anatomy of
3 viviparid gastropods. Malacologia 16:519-540.
VAUGHT, K,C., 1989. A classification of the living
Mollusca. ABBOTT, R.T. & BOSS, K.J. [Eds.]
American Malacologists. Melbourne, 189p.
VAZ, J.F. & MARTINS, C.M., in press. Anatomia interna
e distribuição de Felipponea neritiniformis Dali
(Ampullariidae, Gastropoda, Mollusca).
VERMEIJ, G.J. & SIGNOR, P.W., 1992. The geographic,
taxonimic and temporal distribution of determinate
growth in marine gastropods. Biological Journal of
the Linnean Society 47:233-247.
WENZ, W., 1938. Gastropoda. In: SCHINDEWOLF, O.H.
(Ed.?Org.?) Handbuch der Paláozoologie. Verlag von
Gebrüder Borntraeger. Berlin, i-xii , 1639p.
YEATES, D., 1992. Why remove autapomorphies?
Cladistics 8:387-389.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.387-504, out./dez.2004

Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004
ISSN 0365-4508
A NEW SCINAX WAGLER, 1830 OF THE S. ROSTRATUS GROUP
FROM CENTRAL BRAZIL (AMPHIBIA, ANURA, HYLIDAE) 1
(With 8 figures)
LEONCIO P. LIMA2 3
ROGÉRIO P. BASTOS 2 3
ARIOVALDO A. GIARETTA 4
ABSTRACT: A new species of the genus Scinax belonging to the S. rostratus species group is described from
the State of Goiás, Brazil. Scinax constrictus sp.nov. is characterized by small snout-vent length (males
ranging 20.8-29.4-mm; females 28.2-35.6mm), protuberant nostrils, vocal sac pigmented, superior eyelid, calcar, and jaw tubercles present, dermal constriction on the shoulders in adults, two separate tubercles
above the cloaca in females, and a discrete anal flap in males; an open land, non-forest species. A description
of the advertisement call is provided.
Key words: Amphibia, Anura, Hylidae, Scinax constrictus sp.nov., taxonomy.
RESUMO: Nova espécie de Scinax Wagler, 1830 do grupo de S. rostratus do Brasil Central (Amphibia, Anura, Hylidae).
Uma nova espécie do gênero Scinax pertencente ao grupo de S. rostratus é descrita do Estado de Goiás,
Brasil. Scinax constrictus sp.nov. caracteriza-se por apresentar pequeno comprimento rostro-anal (machos
20,8-29,4mm; fêmeas 28,2-35,6mm), narinas protuberantes, saco vocal pigmentado, presença de tubérculos na pálpebra superior, calcâneo e na mandíbula, constricção dérmica sobre os ombros dos adultos, dois
tubérculos separados acima da cloaca nas fêmeas e um discreto flap anal nos machos; espécie de área
aberta. O canto de anúncio é descrito.
Palavras-chave: Amphibia, Anura, Hylidae, Scinax constrictus sp.nov., taxonomia.
INTRODUCTION
Scinax Wagler, 1830 is one of the most diversified genus among the neotropical hylid frogs, including 85 species (CARAMASCHI, 2004; FROST, 2002). POMBAL & GORDO (1991) recognized the priority of Scinax in relation to Ololygon Fitzinger, 1843, and DUELLMAN & WIENS (1992) diagnosed the genus based on externai morphology, osteology, and larval/reproductive features, defining several species groups. Among them, the S. rostratus group is defined by characteristics as the presence of an acuminate snout, pointed tubercle on heel, and head-down calling position (DUELLMAN, 1972; FAIVOVICH, 2002). Currently, nine species are recognized in this group (FROST, 2002): S. boulengeri(Cope, 1887), S. garbei (Miranda-Ribeiro, 1926), S. joZi/z Lescure & Martin, 2001, S. kennedyi
(Pyburn, 1973), S. nebulosus (Spix, 1824), S. pedromedinae (Henle, 1991), S. proboscideus
(Brongersma, 1933), S. rostratus (Peters, 1863), and S. sugillatus (Duellman, 1973). Herein, we describe a new species of the S. rostratus group from the Brazilian Cerrado biome.
MATERIAL AND METHODS
Specimens used in the description or examined for comparisons are deposited in MNRJ (Museu Nacional, Rio de Janeiro, Brazil), ZUEC (Museu de História Natural, Universidade Estadual de Campinas, Brazil), and ZUFG (Universidade Federal de Goiás, Brazil).
The following measurements were taken to the nearest 0.0lmm with calipers: SVL (snout-vent length), HW (head width), HL (head length), THL (thigh length), TBL (tibia length), and FL (foot length). Sixothers variables, ED (eye diameter), IOD (interorbital distance), IND (internarial distance),
1 Submitted on August 25, 2003. Accepted on October 6, 2004.
2 Universidade Federal de Goiás, Instituto de Ciências Biológicas, Departamento de Biologia Geral. Caixa Postal 131, Goiânia, 74001-970, GO, Brasil.
E-mail: [email protected].
3 Bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
4 Universidade Federal de Uberlândia, Instituto de Biociências. 28400-902, Uberlândia, MG, Brasil.

506 L.P.LEMA, R.P.BASTOS & A.A.GIARETTA
END (eye-nostril distance), NSD (nostril-snout distance), and TD (tympanum diameter), were measured using an ocular micrometer in a Zeiss stereomicroscope. All measurements are in millimeters and followed DUELLMAN (2001) and CEI (1980). Webbing formula notation was described following SAVAGE & HEYER (1967), as modified by MYERS & DUELLMAN (1982).
Advertisement calls of six males were recorded with a DAT Sony TCD-D100 or DAT Tascam recorder and microphone ECM-MS 907 or Sennheiser ME66, respectively. The vocalizations were edited at a sampling frequency of 12kHz and 16 bits resolution with a PC-Pentium Computer, and analyzed with Avisoft-Sonagraph Light and Cool Edit 96 softwares. The sonogram was produced with 256 points, overlap of 87.5%, and in flat top.
RESULTS
Scinax constrictus sp.nov. (Figs.1-8)
Holotype - BRAZIL - GOIÁS: Municipality of Palmeiras (16°48’S; 49°55’W), Sítio dos Pinheiros, MNRJ 31205, adultcf, L.P.Lima col., 26/11/2001.
Paratopotypes - MNRJ 31206-31225, 19 adult cf and one adult 9 , L.P.Lima col., 26/11/2000; ZUFG 883-886, four adult cf, L.P.Lima col., 12/11/2000;
ZUFG 942-943, two adult cf , L.P.Lima col., 24/III/2002; ZUFG 958, adult d , L.P.Lima col., 13/11/2000; ZUFG 961-962,two adultcf, L.P.Lima col., 04/XI/2000; ZUFG 979, adult cf, L.P.Lima col., 30/X/2000; ZUFG 980-981, two adult cf, L.P.Lima col., 13/X/2000; ZUFG 992-995, four adult cf, L.P.Lima col., 24/III/2001; ZUFG 1008- 1012, 1014-1022, 13 adult cf and one adult 9 , collected with the holotype; ZUFG 1665 (Fig. 1), adult cf, L.P.Lima, R.C.D.Pinheiro e R.P.Bastos cols., 07/11/2003.
Referred specimens - BRAZIL - GOIÁS: Municipality ofGuapó: ZUFG 947-948, two adultcf, R.F.Juliano col., 24/III/2000. Municipality of Itaberaí: ZUFG 1300-1303, four adult cf, R.P.Bastos, L.P.Lima e D.Brandão cols., 12/1/2001. Municipality of Jussara: ZUFG 1192, adultcf, R.P.Bastos, L.P.Lima e D.Brandão cols., 13/1/2001. Municipality of Mossâmedes: ZUFG 821-822, 845, two adult cf and one adult 9 , Fazenda Água Fria, R.P.Bastos e R.F.Juliano cols., 23/1/2000. Municipality of Pirenópolis: ZUEC 10273-10276, 10277-10285, 11 adult d and two adult 9 , A.J.Cardoso, A.S.Rand e A.A.Giaretta cols., 04/XII/1992. Municipality of Pontalina: MNRJ 31226-31227, adultcf and adult 9 , Fazenda Lagoa Grande, L.D.Guimarães, C.F.B.Haddad, J.P.Pombal Jr. and R.P.Bastos cols., 16/XII/1998; ZUFG 288-289, two adult cf , L.D.Guimarães e R.P.Bastos cols., 17/111/1997;
Fig. 1- Scinax constrictus sp.nov., paratopotype (ZUFG 1665), adult male in life.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004

A NEW SCINAX WAGLER, 1830 OF THE S. ROSTRATUS GROUP FROM CENTRAL BRAZIL 507
ZUFG 515- 522, eight adult Ó, L.D.Guimarães e R.P.Bastos cols., 26/X/1998. Municipality of Porangatu: ZUFG 1402, adult cf, Leôncio P.Lima e Leopoldo P.Lima cols., 21/XII/2000. Municipality of Uirapuru: ZUFG 702, adult d, M.B.Alcantara col., 01/IV/1999.
Diagnosis - A small species (Figs.2-3) of the Scinax rostmtusgroup; SVLof males 20.8-28.4mm, females 28.2-35.6mm; protuberant nostril; vocal sac pigmented; upper eyelid, calcar, and jaw tubercles present; nostrils not projected from head contour in dorsal view; a dorsolateral dermal constriction on the shoulders in adults; two separate tubercles above the cloaca in females and a discrete anal flap in males; an open area inhabitant species.
Comparisons with others species - Scinax constrictus
sp.nov. most resembles S. nebulosus (characters in parentheses) in general aspect, but it can be distinguished generally by the smaller SVL [x
=26. lmm, range=24-30mm in males; x=38.0mm, range= 35-40mm in females; LUTZ, 1968, DUELLMAN, 1972); presence of discrete anal flap in males (no anal flap; LUTZ, 1968); presence of one row of small tubercles on the edge of lower jaw (absent or indistinct row; LUTZ, 1968, DUELLMAN, 1972); advertisement call with higher or lower dominant frequency (x=2867.3Hz, range=2685.6-3129.8Hz according to DE LA RIVA, MÁRQUEZ & BOSCH, 1994; x=4500Hz, range=4200-4800Hz according to
HÕDL, 1977), generally lower call duration (x =
277ms, range=190-385ms according to HÕDL, 1977, or x=240.6ms, range=201.1-305ms according to DE LA RIVA, MÁRQUEZ & BOSCH, 1994), and higher note number (4-7 notes according to DUELLMAN, 1972, or one note by call according to DE LA RIVA, MÁRQUEZ & BOSCH, 1994).
Scinax constrictus sp.nov. differs from S. boulengeri
(characters in parentheses) by the smaller SVL (x=41.1mm, range=35.5-48.7mm in males; SVL maximum = 53mm in females; LEÓN, 1969; DUELLMAN, 1972); presence of discrete anal flap in males (no anal flap; DUELLMAN, 1972), posterior surface of legs brown or pale green (greenish yellow or orange; DUELLMAN, 1972); advertisement call with lower dominant frequency [x =2840Hz, range=2520-3182Hz; LEÓN, 1969), lower call duration (x=350ms, range=240-470ms; LEÓN, 1969), and higher note number (one note; LEÓN, 1969).
From S. garbei (characters in parentheses), S. constrictus sp.nov. differs by the smaller SVL (range=30-42mm in males, range=38-48mm in females; RODRIGUEZ & DUELLMAN, 1994); posterior surface of legs brown or pale green (yellow or orange; DUELLMAN, 1972); advertisement call with higher note number (one note; DUELLMAN, 1970) and lower duration ( x=690ms, range=230- 1860ms; DUELLMAN, 1970).
Scinax constrictus sp.nov., holotype (MNRJ 31205): fig.2- dorsal view, fig.3- ventral view.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004

508 L.P.LIMA, R.P.BASTOS & A.A.GIARETTA
Scinax constrictus sp.nov. can be distinguished from S. jolyi (characters in parentheses; LESCURE & MARTY, 2001) by the smaller SVL [x=37.7mm,
range=36.5-39.1 in males; x =41,85mm, range=40.0- 43.7mm in females); presence of one row of small tubercles on the edge of lower jaw (no row); advertisement call with only pulsioned structure (advertisement call with harmonic and pulsioned structure).
Scinax constrictus sp.nov. differs from S. kennedyi
(characters in parentheses; PYBURN, 1973) by the smaller SVL (x=33.0mm, range=31.5-35.3 in males; SVL of one female=37.3mm); presence of one row of small tubercles on the edge of lower jaw (no row); posterior surface of legs brown or pale green (orange); advertisement call with lower duration (x = 1770ms, range=690-2100ms) and higher dominant frequency (x =1200Hz, range= 1050-1600Hz).
From S.pedromedinae(characters in parentheses), S. constrictus sp.nov. differs by, generally, the smaller SVL (x=26.4mm, range=22.2-28.9mm in males; x=29.4mm, range=27.5-31.5mm in females; DUELLMAN & WIENS, 1993); jaw tubercles less evident (line of tubercles evident; DUELLMAN & WIENS, 1993); open land species (primary rainforest species; HENLE, 1991); advertisement call with higher duration (90-100ms; DUELLMAN & WIENS, 1993).
Scinax constrictus sp.nov. differs from S. proboscideus (characters in parentheses; DUELLMAN, 2001) by the smaller SVL (x =37.4mm, range=33.0-39.8mm in males; SVL=37.0mm in one female); absence of a fleshy proboscis (presence); concealed parts of the legs brown or pale green (yellow); advertisement call with higher note number (x =1.3, range=l-3) and higher dominant frequency (x=2012Hz).
Scinax constrictus sp.nov. can be distinguished from S. rostratus (characters in parentheses; DUELLMAN, 2001) by the smaller SVL (x=45.7mm in males; x=48.0mm in females); presence of a line of tubercles on lower jaw (no line); presence of tubercle on hell (no tubercle); concealed parts of the legs brown or pale green (orange or dark yellow); advertisement call with higher dominant frequency (x=920Hz) and lower duration (x =690, range=230-860ms).
Scinax constrictus sp.nov. differs from S. sugillatus
(characters in parentheses; DUELLMAN, 1973) by the smaller SVL (x=39.9mm, range=38.7-42.0mm in males; SVL of one female=45.5mm in); jaw tubercles less evident (line more evident); higher
note number (1-2 notes); advertisement call with higher dominant frequency (x =2023Hz, range=1017-2904Hz), and lower duration (x=390, range=280-600ms).
Description of the holotype - Head 12.5% longer than wide; snout elongate, pointed in dorsal view (Fig.4) and acute in lateral view (Fig.5); nostrils not projected from head contour in dorsal view, projected in lateral view; canthus rostralis distinct; tympanum evident, circular; supratympanic fold evident with enlarged tubercles; lower eyelid bordered by a tinny white stripe; tongue slightly cordiform, free behind; vomerine teeth in two small groups between choanae; single subgular vocal sac; five tubercles along the edge of the lower jaw; upper arm strong, with 2-4 tubercles on outer side; fingers size I<II~IV<III (Fig.6); fingers I and II free, II and III, and III and IV linked at base by web; nuptial pads present but poorly developed; outer metacarpal tubercle cordiform, inner metacarpal tubercle oval; subarticular tubercles rounded; finger disks wider than long; toes size I<II<V~III<IV (Fig.7); foot webbing formula I 2 2/3-2 II 1-2 III 1- 2 IV 2-1 V; inner metatarsal tubercle rounded, the outer with half size of the inner; subarticular tubercles rounded; supernumerary tubercles discrete on hands and feet; outer margin of tarsus and foot with a ridge of tubercles; calcar tubercle prominent; a discrete anal flap formed by two enlarged tubercles; a dermal groove from the cloaca to venter of thighs; dorsum and head smooth with many enlarged tubercles; loreal and shoulder regions more tuberculate; body constricted at the levei of shoulders; dorsal surfaces of arms and legs with scattered tubercles; venter granular; ventral surfaces of legs and arms smooth.
Color in life - Males and females showed dorsum color ranging from beige or pale green to brown or dark brown, with black irregular spots. Granules of dorsum brown, yellow or orange. Tympanum bronze. A triangular spot with apex directed posteriorly, bordered by a yellow line, between the eyes. Upper surfaces of arms and legs brown with irregular stripes dark brown. Concealed parts of the legs brown or pale green. Belly white to light brown. Throat whitish with brown flecks. íris pale to dull bronze with black median vertical streak.
Color in preservative (70% alcohol) - Color similar to those in life, however more dark. The colors yellow or orange of the dorsal granules, yellow of tympanum, and yellow of line around the spot between the eyes become opaque and brown.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004

A NEW SCINAX WAGLER, 1830 OF THE S. ROSTRATUS GROUP FROM CENTRAL BRAZIL 509
Measurements of holotype - SVL 24.2; HL 8.2; HW 7.2; ED 2.5; TD 1.3; IOD 2.5; END 2.5; NSD 1.0; IND 1.7; THL 12.2; TBL 13.9; FL 10.5.
Variation - About 82% of specimens (males and females) have the triangular spot between eyes. Skin on dorsum varied from slightly tuberculate to tuberculate. The dorsum may be brown or dark brown. Most of the specimens (87%) showed limbs with three dark brown transverse bars. Nuptial pads present in 95% of the adult males. Mean, standard deviation, and range of measurements of 82 males and six females are presented in table 1.
Advertisement call - The call showed pulsioned structure (Fig.8). Average call duration 213.4±24.0ms (range=180.2-242.2ms; n=30 calls of five males); 9.3±0.9 notes (range=6.0-10.0 notes; n=30 calls), each with average duration of 17.08±2.01ms (range=14.5-20.4ms; n=150 notes); notes given at intervals of 6.9±0.7ms (range=6.0- 8.0ms; n=30 calls). Each note is constituted, in average, by 7.0±2.0 pulses (range=4.0-9.0 pulses; n=150 notes) with average duration of 2.4±0.4ms (range=2.0-4.0ms; n=150 pulses). Repetition call rate was 18.0±3.5 calls/min (range=14.0-23.0 calls/min, n=5 males); average dominant frequency
5
Scinax constrictus sp.nov., holotype (MNRJ 31205): fig.4- dorsal view of head, fig.5- lateral view of head, fig.6- ventral view of hand, fig.7- ventral view of foot. Scale bar = 5mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004
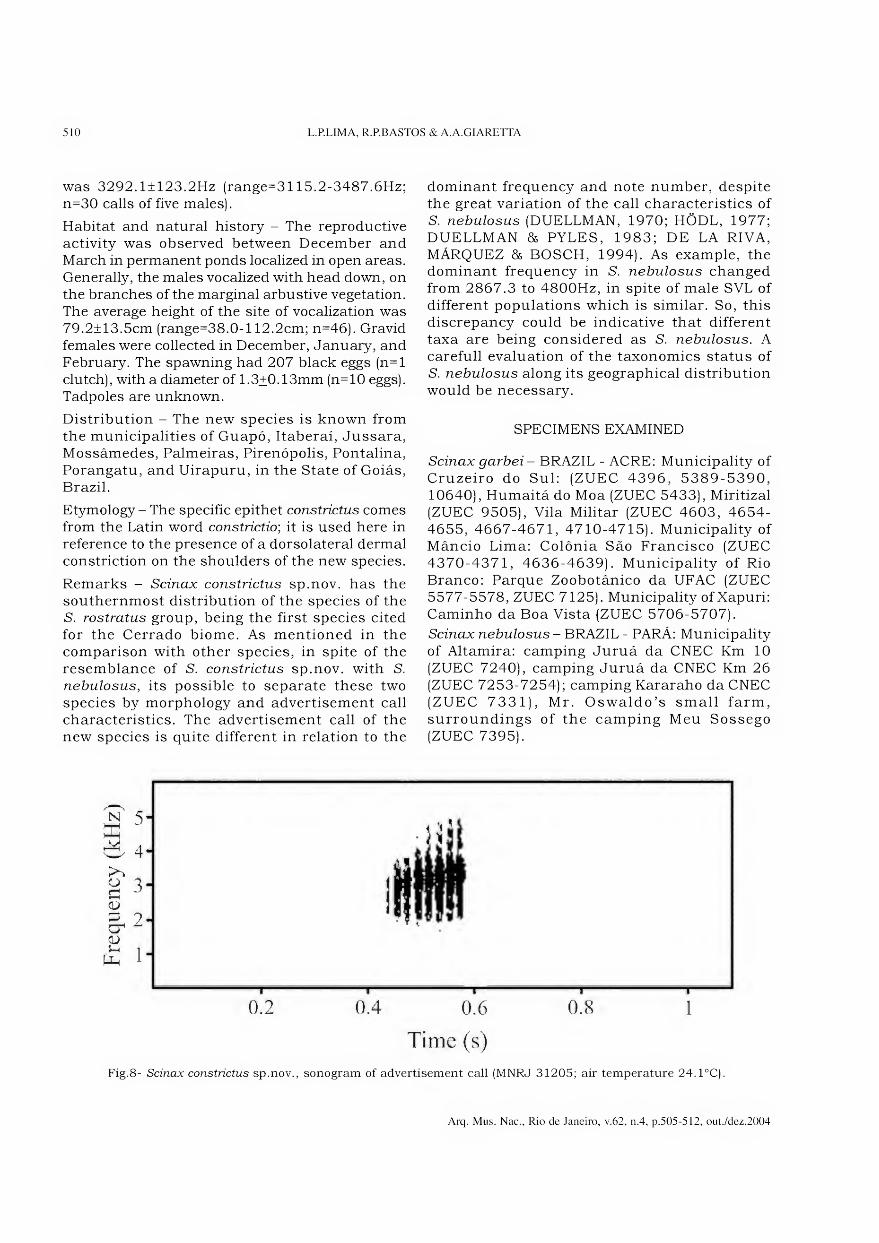
510 L.RLIMA, R.P.BASTOS & A.A.GIARETTA
was 3292. l±123.2Hz (range=3115.2-3487.6Hz; n=30 calls of five males).
Habitat and natural history - The reproductive activity was observed between December and March in permanent ponds localized in open areas. Generally, the males vocalized with head down, on the branches of the marginal arbustive vegetation. The average height of the site of vocalization was 79.2±13.5cm (range=38.0-l 12.2cm; n=46). Gravid females were collected in December, January, and February. The spawning had 207 black eggs (n=l clutch), with a diameter of 1.3+0.13mm (n=10 eggs). Tadpoles are unknown.
Distribution - The new species is known from the municipalities of Guapó, Itaberaí, Jussara, Mossâmedes, Palmeiras, Pirenópolis, Pontalina, Porangatu, and Uirapuru, in the State of Goiás, Brazil.
Etymology - The specific epithet constrictus comes from the Latin word constrictio; it is used here in reference to the presence of a dorsolateral dermal constriction on the shoulders of the new species.
Remarks - Scinax constrictus sp.nov. has the southernmost distribution of the species of the S. rostratus group, being the first species cited for the Cerrado biome. As mentioned in the comparison with other species, in spite of the resemblance of S. constrictus sp.nov. with S. nebulosus, its possible to separate these two species by morphology and advertisement call characteristics. The advertisement call of the new species is quite different in relation to the
dominant frequency and note number, despite the great variation of the call characteristics of S. nebulosus (DUELLMAN, 1970; HÕDL, 1977; DUELLMAN & PYLES, 1983; DE LA RIVA, MÁRQUEZ & BOSCH, 1994). As example, the dominant frequency in S. nebulosus changed from 2867.3 to 4800Hz, in spite of male SVL of different populations which is similar. So, this discrepancy could be indicative that different taxa are being considered as S. nebulosus. A carefull evaluation of the taxonomics status of S. nebulosus along its geographical distribution would be necessary.
SPECIMENS EXAMINED
Scinax garbei - BRAZIL - ACRE: Municipality of Cruzeiro do Sul: (ZUEC 4396, 5389-5390, 10640), Humaitá do Moa (ZUEC 5433), Miritizal (ZUEC 9505), Vila Militar (ZUEC 4603, 4654- 4655, 4667-4671, 4710-4715). Municipality of Mâncio Lima: Colônia São Francisco (ZUEC 4370-4371, 4636-4639). Municipality of Rio Branco: Parque Zoobotânico da UFAC (ZUEC 5577-5578, ZUEC 7125). Municipality of Xapuri: Caminho da Boa Vista (ZUEC 5706-5707).
Scinax nebulosus - BRAZIL - PARÁ: Municipality of Altamira: camping Juruá da CNEC Km 10 (ZUEC 7240), camping Juruá da CNEC Km 26 (ZUEC 7253-7254); camping Kararaho da CNEC (ZUEC 7331), Mr. Oswaldo’s small farm, surroundings of the camping Meu Sossego (ZUEC 7395).
0.2 0.4 0.6 0.8 1
Time (s) Fig.8- Scinax constrictus sp.nov., sonogram of advertisement call (MNRJ 31205; air temperature 24.1°C).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004
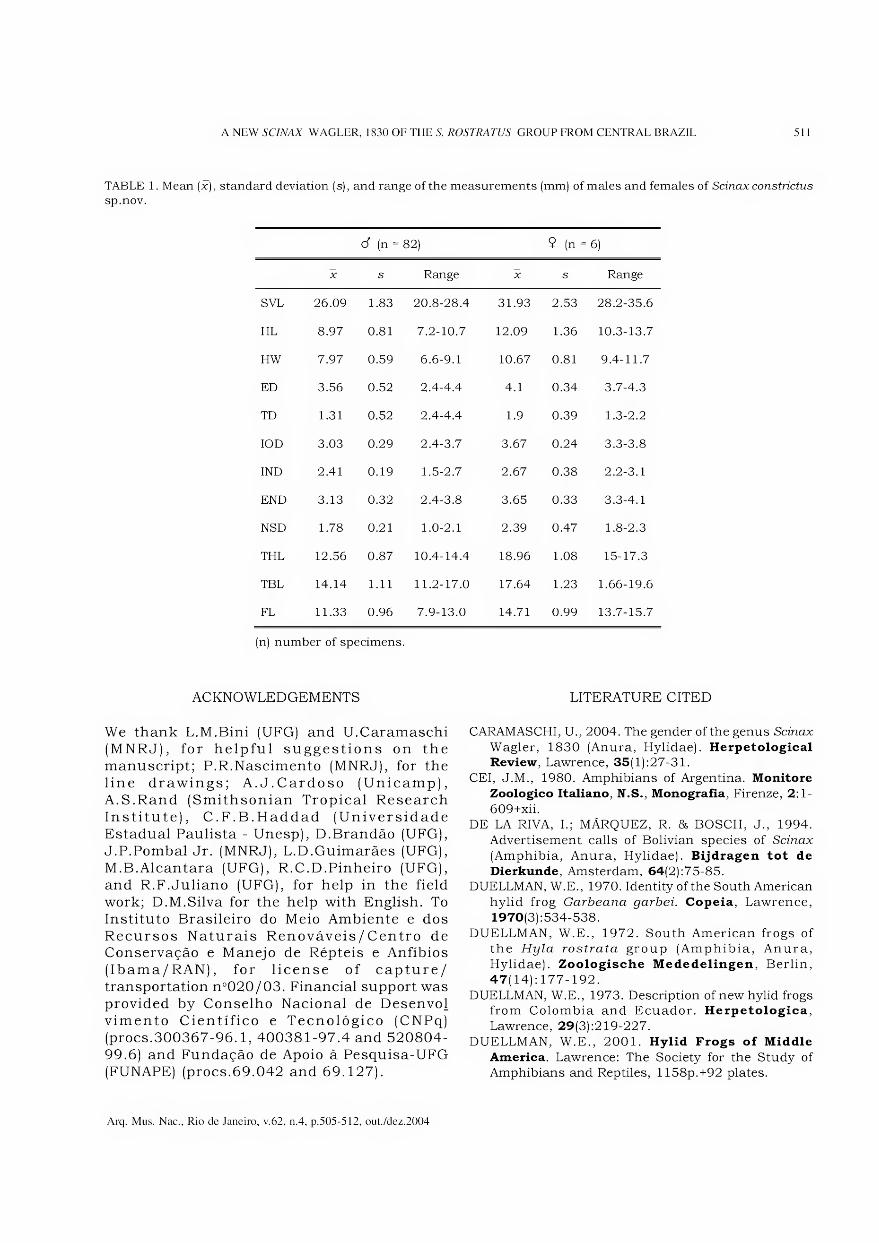
A NEW SCINAX WAGLER, 1830 OF THE S. ROSTRATUS GROUP FROM CENTRAL BRAZIL 511
TABLE 1. Mean (x), standard deviation (s), and range of the measurements (mm) of males and females of Scinax constrictus
sp.nov.
cf (n = 82) $ (n = 6)
X s Range X s Range
SVL 26.09 1.83 20.8-28.4 31.93 2.53 28.2-35.6
HL 8.97 0.81 7.2-10.7 12.09 1.36 10.3-13.7
HW 7.97 0.59 6.6-9.1 10.67 0.81 9.4-11.7
ED 3.56 0.52 2.4-4.4 4.1 0.34 3.7-4.3
TD 1.31 0.52 2.4-4.4 1.9 0.39 1.3-2.2
IO D 3.03 0.29 2.4-3.7 3.67 0.24 3.3-3.8
IND 2.41 0.19 1.5-2.7 2.67 0.38 2.2-3.1
END 3.13 0.32 2.4-3.8 3.65 0.33 3.3-4.1
NSD 1.78 0.21 1.0-2.1 2.39 0.47 1.8-2.3
THL 12.56 0.87 10.4-14.4 18.96 1.08 15-17.3
TBL 14.14 1.11 11.2-17.0 17.64 1.23 1.66-19.6
FL 11.33 0.96 7.9-13.0 14.71 0.99 13.7-15.7
(n) number of specimens.
ACKNOWLEDGEMENTS
We thank L.M.Bini (UFG) and U.Caramaschi (MNRJ), for helpful suggestions on the manuscript; P.R.Nascimento (MNRJ), for the line drawings; A.J.Cardoso (Unicamp), A.S.Rand (Smithsonian Tropical Research Institute), C.F.B.Haddad (Universidade Estadual Paulista - Unesp), D.Brandão (UFG), J.P.Pombal Jr. (MNRJ), L.D.Guimarães (UFG), M.B.Alcantara (UFG), R.C.D.Pinheiro (UFG), and R.F.Juliano (UFG), for help in the field work; D.M.Silva for the help with English. To Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis/Centro de Conservação e Manejo de Répteis e Anfíbios (Ibama/RAN), for license of capture/ transportation n°020/03. Financial support was provided by Conselho Nacional de Desenvol vimento Científico e Tecnológico (CNPq) (procs.300367-96.1, 400381-97.4 and 520804- 99.6) and Fundação de Apoio à Pesquisa-UFG (FUNAPE) (procs.69.042 and 69.127).
LITERATURE CITED
CARAMASCHI, U., 2004. The gender of the genus Scinax
Wagler, 1830 (Anura, Hylidae). Herpetological
Review, Lawrence, 35(1):27-31.
CEI, J.M., 1980. Amphibians of Argentina. Monitore
Zoologico Italiano, N.S., Monografia, Firenze, 2:1-
609+xii.
DE LA RIVA, I.; MÁRQUEZ, R. & BOSCH, J., 1994.
Advertisement calls of Bolivian species of Scinax
(Amphibia, Anura, Hylidae). Bijdragen tot de
Dierkunde, Amsterdam, 64(2):75-85.
DUELLMAN, W.E., 1970. Identity of the South American
hylid frog Garbeana garbei. Copeia, Lawrence,
1970(3) :534-538.
DUELLMAN, W.E., 1972. South American frogs of
the Hyla rostrata group (Amphibia, Anura,
Hylidae). Zoologiscbe Mededelingen, Berlin,
47(14): 177-192.
DUELLMAN, W.E., 1973. Description of new hylid frogs
from Colombia and Ecuador. Herpetologica,
Lawrence, 29(3) :219-227.
DUELLMAN, W.E., 2001. Hylid Frogs of Middle
America. Lawrence: The Society for the Study of
Amphibians and Reptiles, 1158p.+92 plates.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004

512 L.RLIMA, R.P.BASTOS & A.A.GIARETTA
DUELLMAN, W.E. & PYLES, R.A., 1983. Acoustic
resource partitioning in anuran communities.
Copeia, Lawrence, 1983(3):639-649.
DUELLMAN, W.E. & WIENS, J.J., 1992. The status of
the hylid frog genus Ololygon and the recognition of
Scinax Wagler, 1830. Occasional Papers of the
Museum of Natural History, University of Kansas,
Lawrence, 151:1-23.
DUELLMAN, W.E. 85 WIENS, J.J., 1993. Hylid frogs of
the genus Scinax Wagler, 1830, in Amazonas Ecuador
and Peru. Occasional Papers of the Museum of
Natural History, University of Kansas, Lawrence,
153:1-57.
FAIVOVICH, J., 2002. A cladistic analysis of Scinax
(Anura: Hylidae). Cladistics, Ann Arbor,
18(2002) :367-393.
FROST, D.R., 2002. Amphibian Species of the World:
anonline reference. V.2.21 (15 July 2002). Eletronic
database available at http://research.amnh.org/
herpetology/amphibia/index.html. Acessed on: 26
February 2003.
HENLE, K., 1991. Ololygon pedromedinae sp.nov., ein
neuer Knickzehenlaubfrosch (Hylidae) aus Peru.
Salamandra, Bonn, 27(l):76-82.
HÕDL, W., 1977. Call differences and calling sites
segregation in anuran species from Central
Amazonian floating meadows. Oecologia, Berlin,
28:351-363.
LEÓN, J.R., 1969. The systematics of the frogs of the
Hyla rubra group in Middle America. University of
Kansas Publications, Museum of Natural History,
Lawrence, 18(6):505-545.
LESCURE, J. & MARTY, C., 2001. Atlas des amphibiens
de Guyane. Patrimoines Naturels, Paris, 45:1-388.
LUTZ, B., 1968. New Brazilian forms of Hyla. The Pearce
Sellards Series, Texas Memories Museum, Austin,
10:1-18.
MYERS, C.W. & DUELLMAN, W.E., 1982. A new species of
Hyla from Cerro Colorado, and other tree frog records
and geographical notes from western Panama. American
Museum Novitates, New York , 2752:1-25.
POMBAL JR., J.P. & GORDO, M., 1991. Duas novas
espécies de Hyla da Floresta Atlântica no Estado de
São Paulo (Amphibia, Anura). Memórias do Instituto
Butantan, São Paulo, 53(1):135-144.
PYBURN, W.F., 1973. A new hylid frog from the llanos
of Colombia. Journal of Herpetology, Houston,
7(2):297-301.
RODRIGUEZ, L.O. & DUELLMAN, W.E., 1994. Guide to
the frogs of the Iquitos region, Amazonian Peru.
University of Kansas, Museum of Natural History,
Special Publication, Austin, 22:1-80, 12 pis.
SAVAGE, J.M. & HEYER, W.R., 1967. Variation and
distribution of the tree-frog genus Phyllomedusa in
Costa Rica, Central America. Beitraege zur
Neotropischen Fauna, Stuttgart, 5:11-131.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.505-512, out./dez.2004

nrnmnfsminml nnrtmiwnfffinnni ^1
Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.513-518, out./dez.2004
ISSN 0365-4508
REGISTRO DA ATIVIDADE DE ORGANISMOS
EM UM CAULE DO PERMIANO DO RIO GRANDE DO SUL, BRASIL 1
(Com 6 figuras)
ROBSON TADEU BOLZON 2’3 INÊS AZEVEDO 2-4
LUCIANO G AND IN MACHADO 5
RESUMO: Neste trabalho são analisadas perfurações e galerias identificadas em um fragmento de lenho
proveniente da Formação Serra Alta, Permiano Superior da Bacia do Paraná. No exemplar ocorrem áreas
com uma preservação anatômica excelente refletindo em células bem preservadas. A destruição do tecido
lenhoso produzida por insetos foi acompanhada pela degradação dos tecidos por ação de fungos e bactérias.
Nas regiões onde a atividade de organismos se manifesta, a sílica precipitou a partir das bordas formando
estruturas com anéis concêntricos indicando que estas estavam vazias antes da mineralização, o que sugere
um processo diagenético posterior.
Palavras-chave: Icnofósseis, caules fósseis, Permiano, Bacia do Paraná.
ABSTRACT: Activity of Organisms in a Stem of the Rio Grande do Sul State, Brazil.
Borings and galleries were analised in one fóssil wood fragment from Serra Alta Formation, Upper Permian,
Paraná Basin. Some regions were well preserved allowing for the observation of well preserved wall cells. The
wood destruction by insects was accompained by the degradation of wood cell by fungi and bactéria. In
these regions the silica precipited from the borders to the center forming concentric rings that were empty
before mineralization, suggesting a subsequent diagenetic process.
Key words: Ichnofossils, fóssil wood, Permian, Paraná Basin.
INTRODUÇÃO
O registro geológico demonstra que, a partir do Devoniano, ficaram mais evidentes as interações entre as plantas vasculares e os animais (LABANDEIRA, 1997, 1998). Em vegetais fósseis, as atividades biológicas incluem diferentes processos, que resultam em galerias, perfurações, degradação por ação de fungos e bactérias e coprólitos. Perfurações resultantes da atividade de insetos são encontradas em folhas, pecíolos e caules fósseis. Em folhas fósseis, manifestam-se principalmente como fitofagia e galerias. É comum, além disso, a presença de coprólitos no interior de caules fósseis.
No Brasil, poucos são os registros de atividades biológicas em vegetais fósseis. GUERRA- SOMMER (1995) registrou a ocorrência de fitofagia em folhas de Glossopteridales do
Eopermiano da Bacia do Paraná, no sul do Brasil, atribuindo aos Ortopterídeos (Protorthoptera). ADAMI-RODRIGUES et al.
(2003) analisaram folhas fósseis de Glossopteris
Brongniart, 1828 e Cordaites Unger, 1850, das formações Rio Bonito e Irati/Serra Alta, do Estado do Rio Grande do Sul, atacadas por insetos herbívoros.
O presente estudo teve como objetivo identificar os tipos de atividades biológicas encontradas em um fragmento de lenho mineralizado do Permiano Inferior da Bacia do Paraná, relacionando-as ao possível agente causador e a parâmetros de natureza tafonõmica.
MATERIAL E MÉTODOS
O material corresponde a um fragmento de lenho com xilema secundário (Gymnospermae),
1 Submetido em 10 de março de 2004. Aceito em 21 de outubro de 2004.
2 Universidade Federal do Paraná, Departamento de Geologia, Centro Politécnico, Setor de Ciências da Terra. Caixa Postal 19.001, Curitiba, 81531-970, PR, Brasil.
3 E-mail: [email protected].
4 E-mail: [email protected].
5 Bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).

514 R.T.BOLZON, I.AZFVF.nO & L.G.MACHADO
coletado em um afloramento da rodovia São Gabriel - Batovi (km 18), Formação Serra Alta (ALVES, 1999; MERLOTTI, 2000), do Permiano da Bacia do Paraná, Estado do Rio Grande do Sul. O afloramento é constituído por siltitos oliva a verde-oliva ou cinzentos, com laminação plano- paralela irregular, argilitos maciços, raros arenitos intercalados e concreções silicosas freqüentemente acompanhadas por veios de sílica (GUERRA, 1976; GUERRA-SOMMER, 1978; MERLOTTI, 2000). De acordo com SCHNEIDER et al. (1974), esta seqüência está incluída litoestratigraficamente na Formação Serra Alta, de idade Neopermiano, Kazaniano (DAEMON & QUADROS, 1970). A deposição dos sedimentos provavelmente ocorreu abaixo do nível de ação de ondas, em ambiente marinho de águas calmas (SCHNEIDER et al, 1974).
O lenho composto por medula e xilema secundário possui 90mm de comprimento, com diâmetro maior de 80mm e diâmetro menor de 50mm, sendo a diferença no diâmetro ocasionada provavelmente pela perda de tecido (Fig. 1). O exemplar foi cortado nos três planos anatômicos (transversal, longitudinal e radial) e as secções analisadas em microscópio estereoscópico e ótico. O material está depositado na Coleção de Paleobotânica da Universidade Federal do Rio Grande do Sul (UFRGS, PB965).
Características do Lenho
Os cortes transversais indicam um lenho eustélico com medula cêntrica, xilema primário endarco e lenho secundário picnoxílico. A medula pequena (4mm) está mal preservada e rompida. Na periferia da medula observa-se a provável presença de canais secretores. As células do parênquima possuem parede fina. O xilema secundário é homogêneo e constituído de traqueóides e raios; as pontoações intertraqueoidais são unisseriadas e, às vezes, bisseriadas opostas.
O tecido radial está mal preservado e mostra-se homogêneo, com os raios unisseriados e muito baixos com poucas células de altura (2-4-9) e as pontoações do campo de cruzamento pequenas e em número de 4 ou mais. Os anéis de crescimento, em número de 10, são marcados pela redução gradual do diâmetro dos lumens das células axiais do lenho inicial para o tardio, que é estreito (1 a 7 células de largura). A identificação taxonômica mais específica ficou prejudicada, devido a não
visualização de características diagnósticas do exemplar, em especial, da medula. O lenho eustélico e a presença de canais secretores periféricos na medula sugerem a inclusão no Grupo Solenoide (MUSSA, 1986).
Características das Atividades Biológicas
O xilema secundário do exemplar foi destruído pela atividade orgânica em várias regiões. As perfurações aparecem normalmente preenchidas por sílica e são facilmente visíveis pela coloração esbranquiçada em plano transversal (Fig.l) e longitudinal (Fig.2). No plano transversal aparecem em número e forma variável, desde circulares a ovais, até alongadas e irregulares. As alongadas sugerem a presença de duas ou mais perfurações sobrepostas. No plano anatômico transversal, as regiões com perfurações e consequentemente com o tecido lenhoso destruído ocorrem em número superior a 200. Em todo o xilema secundário, o número médio de perfurações corresponde a 12 por cm2, com mínimo de 8 e máximo de 18 por cm2. O diâmetro das perfurações varia de 0,85 a 2,3mm. Fraturas radiais e tangenciais são encontradas no tecido lenhoso originadas, possivelmente antes da silicificação. Estes padrões de fraturas podem ser comumente observados em situações post-mortem
nos caules atuais.
A observação microscópica dos espaços vazios remanescentes após as perfurações no xilema secundário evidencia que essas estão preenchidas principalmente por bandas de sílica (Figs.3-4). A precipitação ocorreu da periferia para o centro, determinando a formação de anéis concêntricos. Os limites destas áreas possuem células com parede parcialmente preservada (Fig.5).
Nas secções longitudinais, as perfurações no xilema secundário são tubiformes, com diâmetro constante e extremidades arredondadas, e uma altura média de 6,15mm. Normalmente as perfurações mostram fundo cego, mas algumas possuem intercomunicações diagonais ao eixo anatômico longitudinal formando galerias (Fig.2). Na periferia de algumas galerias foram observadas estruturas com morfologia análoga a hifas de fungos, similar as já descritas para lenhos fósseis (BUURMAN, 1972). Também são observados traqueóides com paredes incompletas (Fig. 6) e concentrações de material amarelo escuro a marrom, correspondendo possivelmente a fragmentos de tecido lenhoso.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.513-518, out./dez.2004
REGISTRO DA ATIVIDADE DE ORGANISMOS EM UM CAULE DO PERMIANO DO RIO GRANDE DO SUL, BRASIL 515
Fig. 1- Secção transversal do lenho; fig.2- secção longitudinal mostrando as intercomunicações entre as galerias; fig.3-
secção transversal em microscopia ótica de galeria preenchida por sílica bandeada; fig.4- detalhe da figura anterior. Células junto à borda da galeria com paredes incompletas; fig.5- detalhe da galeria com células do lenho tardio e células
de raio bem preservadas; fig.6- secção longitudinal com células traqueóides bem preservadas adjacentes à galeria.
Concentração de substância marrom escura junto à borda, correspondendo provavelmente a fragmentos de tecido lenhoso. Escalas: 1-2 = mm; 3 = 500pm; 4-6 = 200pm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.513-518, out./dez.2004

516 R.T.BOLZON, I.AZFVF.nO & L.G.MACHADO
DISCUSSÃO
A atividade biológica identificada no lenho estudado inclui perfurações, galerias e outras manifestações, realizadas por insetos, fungos e provavelmente bactérias, embora seja difícil precisar, entre estes, os grupos responsáveis.
Poucos dados sobre atividades biológicas em lenhos fósseis são conhecidos no Brasil, até o momento. As galerias e as perfurações observadas podem ser classificadas como icnitos de alimentação (Fodinichnia) do tipo Xylichnia, que reúnem perfurações de insetos produzidas em madeiras. FERNANDES et al (2002) fizeram referência ao material em estudo e classificaram as perfurações como estruturas de bioerosão.
No final do Paleozoico, os artrópodes dos grupos Diplopoda, Arthropleura, Collembola e Coleoptera foram considerados como possíveis habitantes das madeiras em decomposição e os Orthoptera (Blattodea), Diplopoda, Arthropleura, Collembola, Acarida e Coleóptera, seriam xilófagos (SCOTT 8& TAYLOR, 1983; TAYLOR & TAYLOR, 1993).
LABANDEIRA (1998) apresentou alguns critérios para identificar no registro fóssil, a herbivoria, ou seja, as atividades ocorridas durante a vida do vegetal e diferenciar das ocorridas após a morte do vegetal, detritivoria. Considerou como mais importante no reconhecimento da herbivoria, à detecção da resposta do vegetal, incluindo a reação dos tecidos que implicam na multiplicação e no aumento anormal das células. De acordo com os critérios indicados pelo autor, o material estudado apresenta evidências de detritivoria.
Segundo STUBBLEFIELD & TAYLOR (1986), as galerias originadas por insetos em tecidos lenhosos, em geral, variam pouco em tamanho, não são vazias e formam uma rede anastomosada de espaços sugerindo escavações.
CICHAN 85 TAYLOR (1982) registraram evidências de atividade biológica, na forma de pelotas fecais de coleópteros e túneis conectando galerias verticais em lenhos mineralizados de Premnoxylon
Pierce & Hall 1953, do Carbonífero dos Estados Unidos. ZHOU & ZHANG (1989) também apontam coprólitos deste mesmo grupo de insetos, em lenhos de Protocupressinoxylon Eckhold, 1922, do Jurássico da China. Os ácaros perfuradores de madeira seriam muitas vezes responsáveis por túneis, preservados em madeiras permineralizadas ou petrificadas, contendo muitas vezes, coprólitos em seu interior (DEREK 8s TAYLOR, 2002). O registro fóssil indica que este grupo de ácaros ocorre
desde o Neocarbonífero (ASH, 2000; DEREK & TAYLOR, 2002). ASH (2000) descreveu um caule de Filicales do Triássico do Arizona contendo cavidades irregulares de várias formas e tamanhos, além de coprólitos pequenos (40-50pm de diâmetro e 80 a lOOpm de comprimento) preenchidos por minerais, atribuídos a ácaros Oribatida.
A ocorrência de fósseis de insetos das ordens Mecopteoidea (Família Permochoristidae); Neuropteris (Família Permithonidae); Coleoptera (Família Permocupididae) e Homoptera (Famílias Pereboridae, Prosbolecicadidae, Prosbolidae e Fulguringuidae) é registrada para o Permiano do Rio Grande do Sul (Formação Irati/Serra Alta, localidade de Mina do Leão, km 90 BR290), conforme PINTO (2000).
As comparações entre caules atuais atacados por larvas de coleópteros e 0 lenho fóssil analisado mostram semelhanças no padrão de destruição, principalmente com relação ao diâmetro das perfurações circulares e as intercomunicações entre elas. Essas semelhanças e os dados do registro fóssil permitem sugerir que as perfurações e as galerias foram realizadas por larvas de coleópteros, provavelmente após a morte do vegetal. Além disso, em madeiras atuais, as galerias ocasionadas pela atividade de invertebrados (larvas de insetos e moluscos) são rapidamente colonizadas por fungos e bactérias que intensificam a destruição do lenho.
Segundo KIM 8s SINGH (2000), em ambientes úmidos, as bactérias são mais tolerantes às condições anóxicas ou com pouco oxigênio. O padrão de decomposição da madeira por bactérias é diferente do padrão ocasionado por fungos. As bactérias atacam principalmente os elementos não lignificados da madeira e podem ser reconhecidas três diferentes formas de decomposição das paredes celulares: túneis tortuosos, erosão produzindo valas e cavidades que ocorrem na direção perpendicular ao eixo longitudinal da célula. Para os autores, a degradação por fungos é 0 resultado da ação de alguns ascomicetos, deuteromicetos e basidiomicetos. Os ascomicetos e deuteromicetos causam a podridão branda, mais comum em solos e, às vezes, associada com a degradação por bactérias. Já sob condições de oxigênio reduzido, a madeira pode ser atacada e degradada por fungos basidiomicetos e bactérias, causadores da podridão branca e da podridão parda. Na podridão branca ocorre a degradação e a corrosão dos componentes das paredes celulares. No segundo tipo, a celulose é rapidamente despolimerizada e, embora a parede fique muito porosa, a forma original da célula é mantida.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.513-518, out./dez.2004

REGISTRO DA ATIVIDADE DE ORGANISMOS EM UM CAULE DO PERMIANO DO RIO GRANDE DO SUL, BRASIL 517
STUBBLEFIELD & TAYLOR (1986), ao estudarem amostras de caules de Araucarioxylon Kraus, 1870 e Vertebraria (Royle) Schopf, 1965 do Permiano da Antártica, sugeriram seu ataque por fungos. As regiões atacadas do lenho ficaram mais claras em comparação com as células adjacentes, refletindo uma redução da quantidade de matéria orgânica na parede de algumas células. Os autores observaram que nas margens das escavações, as células estavam parcialmente degradadas e as paredes das células do tecido lenhoso de regiões próximas foram modificadas.
A colonização por fungos e provavelmente bactérias ocorreu no lenho fóssil estudado, pois algumas regiões com tecido lenhoso destruído indicam um padrão semelhante ao encontrado em lenhos com áreas de podridão. As hifas de fungos foram observadas na periferia das galerias e teriam penetrado no tecido lenhoso após a sua formação. A decomposição por fungos e a degradação por bactérias dos constituintes das paredes celulares do lenho alteraram a morfologia original de muitas perfurações e as dimensões de algumas galerias.
Os processos relacionados com a diagênese de um lenho resultam na preservação ou destruição do tecido lenhoso. As células ao redor das perfurações no caule estudado estão preservadas e indicam que não ocorreram mudanças morfológicas significativas durante os processos diagenéticos. As fraturas radiais, tangenciais e oblíquas sugerem o ressecamento do tecido lenhoso após a formação das galerias. Estas fraturas estão preenchidas por minerais similares aos que ocorrem nas galerias.
A preservação do tecido lenhoso, muitas vezes, ocorre da superfície da parede, onde a sílica precipita, para o interior da célula (LEO & BARGHOORN, 1976). Assim, em grandes espaços (SCURFIELD & SEGNIT, 1984) ou em estruturas celulares grandes, como um elemento de vaso (BOLZON & OLIVEIRA, 2000), os minerais precipitam em bandas.
No material estudado, a atividade biológica ocorreu antes do sepultamento final. A análise da mineralogia permitiu inferir que as perfurações e as galerias estavam vazias no início do processo de silicificação do lenho, pois durante a fossildiagênese, a precipitação da sílica, que iniciou provavelmente após o sepultamento, ocorreu da borda para o centro formando anéis concêntricos.
CONCLUSÃO
Um grande número de indícios de atividade de perfuração foi identificado em regiões do tecido lenhoso de um fragmento de caule proveniente de níveis do Permiano, provavelmente produzidas por insetos. No início do processo de silicificação estas perfurações e galerias estavam vazias, e posteriormente, com a precipitação da sílica, formaram-se estruturas com bandas concêntricas. Associada à ação dos insetos, ocorreu à degradação por fungos e bactérias.
AGRADECIMENTOS
Ao Departamento de Paleontologia e Estratigrafia da Universidade Federal do Rio Grande do Sul, pelo empréstimo do material, e ao Departamento de Geologia da Universidade Federal do Paraná.
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMI-RODRIGUES, K.; IANUZZI, R.; ALVES DE
SOUZA, P. & PINTO, I.D., 2003. Quantitative analisys
of herbivory on Gondwana floras of the Neopalaeozoic
of Rio Grande do Sul State, Brazil. In: CONGRESSO
BRASILEIRO DE PALEONTOLOGIA, 18., Brasília.
Resumos..., Brasília: SBP, p.33.
ALVES, L.S.R., 1999. Anéis de crescimento como
indicadores paleoclimáticos no Neopermiano da
Bacia do Paraná (Fm. Irati e Serra Alta). Porto Alegre.
27 lp. Tese (Doutorado em Geociências), Instituto de
Geociências, Universidade Federal do Rio Grande do Sul.
ASH, S., 2000. Evidence of oribatid mite herbivory in
the stem of a Late Triassic tree fern from Arizona.
Journal of Paleontology, 74(6):1065-1071.
BOLZON, R.T. & OLIVEIRA, E.V., 2000. Associação de
lenhos fósseis da região de Uruguaiana, Estado do
Rio Grande do Sul, Brasil: considerações sobre
geologia e tafonomia. Revista da Universidade de
Guarulhos, 5(n.especial): 160-163.
BUURMAN, P., 1972. Mineralization of fóssil wood.
Scripta Geologica, 12:1-43.
CICHAN, M.A. & TAYLOR, T.N., 1982. Wood-boring in
Premnoxylon: plant-animal interactions in the
Carboniferous. Palaeogeography, Palaeoclimatology,
Palaeoecology, 39:123-127.
DAEMON, R.F. & QUADROS, L.P., 1970. Bioestratigrafia
do Neoproterozóico da Bacia do Paraná. In:
CONGRESSO BRASILEIRO DE GEOLOGIA, 24.,
Brasília, Anais..., Brasília: SBG, p.359-412.
DEREK, K.W. & TAYLOR, E.L., 2002. Evidence of wood-
boring mites from the Paleozoic and Mesozoic of
Antarctica. Disponível em: <http://www.2002.
botanyconference.org/section7 / abstracts /
18.shtml>. Acesso em: 23 out. 2003.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.513-518, out./dez.2004

518 R.T.BOLZON, I.AZEVEDO & L.G.MACHADO
FERNANDES, A.C.S.; BORGHI, L.; CARVALHO, I.S. &
ABREU, C.J., 2002. Guia dos Icnofósseis de
Invertebrados do Brasil. Rio de Janeiro: Interciência.
p. 189-190.
GUERRA, M., 1976. Vertebraria (Roylé) Schopf 1965 Un
genre présent dans la Formacion Irati du Rio Grande do
Sul (Brésil). In: CONGRÈS NATIONAL DES SOCIÉTÉS
SAVANTES, 101., Lille, Actes..., Lille: Fase. I, p.51-68.
GUERRA-SOMMER, M., 1978. Ocorrência do gênero
nórdico de conífera no Gandwana sul brasileiro.
Pesquisas, 10:65-76.
GUERRA-SOMMER, M., 1995. Fitofagia em
glossopterideas na paleoflora da Mina do Faxinai
(Formação Rio Bonito, Artinskiano, Bacia do Paraná).
Pesquisas, 22(1-2): 58-63.
LABANDEIRA, C.C.,1997. Insect mouthparts:
ascertaineing the paleobiology of insect feeding
strategies. Annual Review of Ecology and
Systematics, 28:153-193.
LABANDEIRA, C.C., 1998. Early history of arthropod and
vascular plant associations. Annual Review of Earth
and Planetary Sciences, 26:329-377.
MERLOTTI, S., 2000. Os lenhos fósseis das formações
Irati e Serra Alta (Permiano Superior, Bacia do
Paraná) do Rio Grande do Sul, Brasil - registro de
dois novos espécimes. Revista de Geociências de
Guarulhos, 5(n.especial):37-39.
MUSSA, D., 1986. As formas Gondwânicas do Grupo
Solenoide e sua distribuição estratigráfica. Anais da
Academia Brasileira de Ciências, 58(l):61-88.
KIM, Y.S. & SING, A.P., 2000. Micromorphological
characteristics of wood biodegradation in wet
environments: A Review. LAWA Journal, 21(2): 135-155.
LEO, R.F. & BARGHOORN, E.S., 1976. Silicification of
wood. Botanical Museum Leaflets, 25:1-47.
PINTO, I.D., 2000. Insetos fósseis. In: HOLZ, M. &
DEROSS, L. F. (Eds.) Paleontologia do Rio Grande
do Sul. Porto Alegre: EDUFRGS. p. 131-147.
SCOTT, A.C. & TAYLOR, T.N., 1983. Plant/animal
interactions during the Upper Carboniferous.
Botanical Review, 49(3):259-307.
SCHNEIDER, R.L.; MÜLHLMANN, H.; TOMMASI, E.;
MEDEIROS, R.A.; DAEMON, R.F. & NOGUEIRA,
A.A., 1974. Revisão estratigráfica da Bacia do
Paraná. In: CONGRESSO BRASILEIRO DE
GEOLOGIA, 25., Porto Alegre. Anais..., Porto Alegre:
SBG, v. 1, p.41-65.
SCURFIELD, G. & SEGNIT, E.R., 1984. Petrifaction of
wood by silica minerais. Sedimentary Geology,
39:149-167.
STUBBLEFIELD, S.P. & TAYLOR, T.N., 1986. Wood decay
in silicified gymnosperms from Antarctica. Botanical
Gazette, 147(1):116-125.
TAYLOR, T.N. & TAYLOR, E.L., 1993. The Biology
and Evolution of Fóssil Plants. New York: Pratice
Hall. 939p.
ZHOU, Z. & ZHANG, B., 1989. A sideritic
Protocupressinoxylon with insect borings and frass
from the Middle Jurassic, Henan, China. Review of
Palaeobotany and Palynology, 59:133-143.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.513-518, out./dez.2004

wmmnraTmnnrTl nnniiitüiiiüífflnnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004
ISSN 0365-4508
DUAS NOVAS ICNOESPÉCIES DE BIFUNGITES DESIO, 1940
NA FORMAÇÃO PIMENTEIRA,
DEVONIANO DA BACIA DO PARNAÍBA, BRASIL 1
(Com 5 figuras)
SONIA AGOSTINHO 2 MARIA SOMÁLIA SALES VIANA 2
ANTONIO CARLOS SEQUEIRA FERNANDES 3
RESUMO: Este trabalho descreve duas novas icnoespécies, Bifungites munizi isp.nov. e Bifungitespiauiensis isp.nov., para os arenitos da Formação Pimenteira (Devoniano, Bacia do Parnaíba), no Estado do Piauí, Brasil. Bifungites munizi isp.nov. caracteriza-se por apresentar, em um mesmo exemplar, dois corpos terminais
de morfologias diferentes, um maior, reniforme ou em forma de âncora e outro menor de formato globóide, circular, ambos interligados por um eixo central retilíneo de seção semi-cilíndrica. Bifungites piauiensis
isp.nov. caracteriza-se pela presença de um eixo basal retilíneo, subcilíndrico, prolongando-se em extremidades
em forma de setas cujas bases apresentam projeções para trás com ângulos de 10° a 35°. As duas novas icnoespécies apresentam características morfológicas e dimensões que as diferenciam das demais formas
conhecidas na literatura, acrescentando dois novos icnotaxa às duas icnoespécies de Bifungites já formalizadas
para o Devoniano no Brasil.
Palavras-chave: Icnologia, Formação Pimenteira, Devoniano, Bacia do Parnaíba, Bifungites.
ABSTRACT: Two new ichnospecies of Bifungites Desio, 1940 from the Pimenteira Formation, Devonian of Parnaíba Basin, Brazil.
This paper describes two new ichnospecies, Bifungites munizi isp.nov. and Bifungites piauiensis isp.nov.,
that occur in sandstones of the Pimenteira Formation (Devonian, Parnaíba Basin), in the Piauí State, Brazil.
Bifungites munizi isp.nov. is morphologically characterized by different terminations, one of which is reniform and the other is dumbbell-like. Bifungites piauiensis isp.nov. shows a horizontal, straight and subcircular
shaft, with double arrow-shaped terminations which bases have projections with angles of 10° to 35°. The
morphological characteristics and dimensions of the two new ichnospecies allow their differenciation from other ichnospecies of Bifungites, resulting in two new ichnotaxa of this ichnogenus in the Brazilian Devonian.
Key words: Ichnology, Pimenteira Formation, Devonian, Parnaíba Basin, Bifungites.
INTRODUÇÃO
A Bacia do Parnaíba localiza-se principalmente na região Nordeste, abrangendo uma área com cerca de óOO.OOOkm2 e ocupando parte dos estados do Piauí, Maranhão, Tocantins, Pará, Ceará, Bahia e Goiás. Também conhecida como Bacia do Maranhão é, sobretudo, uma bacia paleozoica, embora depósitos mesozoicos e cenozoicos pouco espessos cubram grandes
áreas de sua extensão. A Bacia do Parnaíba desenvolveu-se diretamente sobre um substrato composto principalmente por rochas metamórficas oriundas de processos tectonomagmáticos não mais antigos que o Mesoproterozóico, sobre os quais se superpõem grabens (GÓES & FEIJÓ, 1994) preenchidos no Neoproterozóico (Formação Riachão) e no Cambriano-Ordoviciano (Formação Mirador). Segundo GÓES & FEIJÓ (1994), a coluna
1 Submetido em 01 de agosto de 2003. Aceito em 27 de setembro de 2004.
Apoio: Instituto Virtual de Paleontologia-RJ/Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (IVP-RJ/FAPERJ) e PRH-
26/Agência Nacional de Petróleo (ANP)/Financiadora de Estudos e Projetos (FINEP).
2 Universidade Federal de Pernambuco, Centro de Tecnologia e Geociências, Departamento de Geologia. Rua Acadêmico Hélio Ramos s/n, Cidade Univer¬
sitária, 50670-000, Recife, PE, Brasil. E-mails: [email protected] e [email protected].
3 Museu Nacional/UFRJ, Departamento de Geologia e Paleontologia. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil.
Universidade do Estado do Rio de Janeiro, Faculdade de Geologia, Departamento de Estratigrafia e Paleontologia. Rua São Francisco Xavier, 524, Maracanã,
20559-900, Rio de Janeiro, RJ, Brasil. E-mail: [email protected].
Bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).

520 S.AGOSTINHO, M.S.S.VIANA & A.C.S.FERNANDES
sedimentar foi subdividida em cinco seqüências depositadas do Siluriano ao Cretáceo, com uma sedimentação predominantemente siliciclástica, ocorrendo subordinadamente calcário, anidrita e sílex, além de diabásio e basalto, representativos de eventos magmáticos do Neotriássico ao Eocretáceo. As cinco seqüências deposicionais são: (i) Seqüência Siluriana (Grupo Serra Grande), (ii) Seqüência Devoniana (Grupo Canindé), (iii) Seqüência Carbonífero- triássica (Grupo Balsas), (iv) Seqüência Jurássica (Grupo Mearim) e (v) Seqüência Cretácea (formações Grajaú, Codó, Corda e Itapecuru). Na Seqüência Devoniana, o Grupo Canindé é constituído pelas formações Itaim, Pimenteira, Cabeças e Poti, sendo representativo da maior ingressão marinha na Bacia do Parnaíba, com o término da sedimentação atribuído ao soerguimento provocado pelos reflexos de uma Orogenia Eo-herciniana. Os icnofósseis estudados procedem de um afloramento da Formação Pimenteira, a qual é composta por espessas camadas de folhelhos cinza-escuros a pretos, depositadas em um paleoambiente nerítico dominado por tempestades as quais depositaram camadas de arenito muito fino, durante o Givetiano- Frasniano (■q.v. DELL A FÁVERA, 1990).
A Formação Pimenteira tem revelado um conteúdo icnofossilífero bastante significativo, auxiliando em seus estudos estratigráficos e de interpretação paleoambiental. Em seus estratos, até o presente, foram identificados os icnogêneros Arenicolites, Asteriacites,
Asterosoma, Bifungites, Chondrites, Cruziana,
Cylindrichnus, Diplichnites, Diplocraterion,
Helminthopsis, Lophochtenium, Isopodichnus,
Macaronichnus, Merostomichnites, Neoskolithos,
Nereites, Palaeohelminthoida, Palaeophycus,
Phycosiphon, Planolites, Rhizocorallium,
Rosselia, Rusophycus, Scolicia, Skolithos,
Spirophyton, Subphyllocorda, Teichichnus e Zoophycos, como resultado dos estudos de AGOSTINHO (2001), AGOSTINHO & VALENÇA (1999), AGOSTINHO, CORRÊA & FERNANDES (2003), AGOSTINHO, VIANA & FERNANDES (2003), AGOSTINHO et dl. (2001), ASSIS & FERNANDES (1980), BRITO (1977), CAMPANHA (1974), CAMPELO & VIANA (2003), FERNANDES, AGOSTINHO & VIANA (2003), FERREIRA & FERNANDES (1983), FONSECA & MELO (1987), KEGEL (1961, 1966), MUNIZ (1988), NASCIMENTO et al. (2003), RODRIGUES
(2001), SILVA (1999), YOUNG & BORGHI (2003) e YOUNG, BORGHI & FERNANDES (2002), em grande parte sintetizados por FERNANDES et al.
(2002). Destes autores, somente ASSIS & FERNANDES (1980), FERNANDES, AGOSTINHO & VIANA (2003) e SILVA (1999) procederam a estudos mais detalhados das ocorrências do icnogênero Bifungites.
Os icnofósseis descritos neste trabalho são procedentes de duas localidades situadas no Estado do Piauí (Figs.1-3) e encontram-se depositados na coleção de macrofósseis do Departamento de Geologia do Centro de Tecnologia e Geociências (DG-CTG) da Universidade Federal de Pernambuco (UFPE).
SISTEMÁTICA
Icnogênero Bifungites Desio, 1940
Icnoespécie-tipo - Bifungites fezzanensis Desio, 1940 por monotipia.
Diagnosis - Escavações com tubos verticais em forma de n invertido, com as estruturas da base geralmente preservadas em epirrelevo convexo (positivo) em forma de halteres e com dois corpos terminais, globulares a triangulares (em seta), interligados por um eixo central (HÁNTZSCHEL, 1975).
Observações: Bifungites foi originalmente reconhecido por DESIO (1940) como um novo tipo de icnofóssil em forma de halteres presente em rochas da Líbia, sendo em seguida interpretado como moldes de aberturas de tubos em forma de U de Rhizocorallium (DUBOIS & LESSERTISSEUR, 1964) ou de Corophioides (OSGOOD, 1970) e, posteriormente, registrado em associação a tubos em U sem spreiten (GUTSCHICK, SUTTNER 86 SWITEK, 1962; RODRIGUEZ 86 GUTSCHICK, 1970). Sua caracterização como tubos duplos verticais em forma de n
invertido, sem spreiten, terminando na base em câmaras ou expansões saculiformes ligadas entre si por um tubo horizontal, foi defendida por GUTSCHICK 86 LAMBORN (1975), classificando etologicamente Bifungites como icnito de habitação (Domichnia), gerado por organismos sedentários de corpo mole, suspensívoros, como anelídeos ou artrópodes. Posteriormente, FILLION 86 PICKERILL (1984), ao analisarem exemplares do icnogênero Arthraria
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

DUAS NOVAS ICNOESPÉCIES DE BIFUNGITES, FORMAÇÃO PIMENTEIRA, DEVONIANO DA BACIA DO PARNAÍBA, BRASIL 521
procedentes de Newfoundland, Canadá, teceram comentários a respeito da distinção entre este último icnogênero, Diplocraterion e Bifungites, face às suas semelhanças; ressaltaram ainda o fato de autores como OSGOOD (1970) e SEILACHER (1955) terem interpretado, respectivamente, Bifungites como a base de um tubo em U dotado
de spreiten ou uma escavação do mesmo tipo truncada, e de outros autores, como os já citados anteriormente, como escavações em forma de n
invertido, sem spreiten. De acordo com FILLION & PICKERILL (1984), considerando esta última interpretação e o fato da presença ou ausência de spreiten ser uma característica significativa na
44° A 42° 42° 6 41°
Fig. 1- (A) localização do afloramento de São João Vermelho, no Município de São Raimundo Nonato e (B) dos afloramentos situados nas margens do rio Sambito, Município de Pimenteiras, Estado do Piauí.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

522 S.AGOSTINHO, M.S.S.VIANA & A.C.S.FERNANDES
Fig.2- (A) vista geral do afloramento em São João Vermelho, com destaque em primeiro plano para o nível arenítico
superior onde se encontram os exemplares de Bifungites munizi isp.nov.; (B) vista geral das exposições de arenitos da Formação Pimenteira nas margens do rio Sâmbito, próximo à localidade de Oiti.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

DUAS NOVAS ICNOESPÉCIES DE BIFUNGITES, FORMAÇÃO PIMENTEIRA, DEVONIANO DA BACIA DO PARNAÍBA, BRASIL 523
Li tolo eia
r-i síitito (si
Icnoeeneros
W* Planolites/ Pataeophycus
Estruturas Sedimentares
Acamamento ondulado
m Arenito (a) i Skolithos (?)
U Arenieoltíes O Estratificação cruzada
Hummoeky
Cimento 4—^ Bifungites // Laminação plano paralela Carbonático
Laphoctenium Superfície ondulada
V Fóssil Corporal (Tentaculita)
^ Phycosiphon ©
Acamamento lenticular
Aeamamento fiaser
Concreções ferruginasa
Fig.3- (A) seção estratigráfica do afloramento de São João Vermelho e (B) do afloramento no rio Sambito, próximo à localidade de Oiti.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

524 S.AGOSTINHO, M.S.S.VIANA & A.C.S.FERNANDES
classificação icnogenérica, Bifungites não deve ser colocado como um sinônimo júnior de Diplocraterion. Caso se comprovasse a presença de tubos verticais no material-tipo de Bifungites
(ressalta-se que o material-tipo se perdeu durante a Segunda Guerra Mundial), este seria considerado mais bem como um sinônimo júnior de Arthraria.
Caso os exemplares previamente atribuídos a Bifungites possuam tubos verticais sem spreiten
entre si, estes deveriam ser renomeados como um icnogênero novo e distinto. Até que tal material se torne disponível para análise, FILLION 8& PICKERILL (1984) consideram as escavações em U ou em 71 invertido, sem spreiten entre os tubos, como Bifungites. Este icnogênero ocorre na icnofácies Cruziana e distribui-se do Cambriano Inferior ao Carbonífero (TURNER 85 BENTON, 1983). No Brasil, Bifungites foi registrado por NETTO (2000) para a Formação Santa Bárbara (pré- cambriana, da Bacia de Camaquã), mas cuja identificação foi equivocada, segundo comunicação pessoal de Renata Guimarães Netto (UNISINOS); para as formações Inajá (devoniana, da Bacia de Jatobá; MUNIZ, 1976, 1979), Ponta Grossa (devoniana; FERNANDES, 1996, 2001; FERNANDES 8s MELO, 1985), Rio Bonito/Palermo e Rio do Sul (permianas; BALISTIERI 8s NETTO, 2001; BALISTIERI, NETTO 86 WEINSCHUTZ, 2001), que posteriormente revelou-se tratar de exemplares de Diplocraterion, de acordo com comunicação pessoal de Renata Guimarães Netto, da Bacia do Paraná; e nas formações Pimenteira (AGOSTINHO 8s VALENÇA, 1999; AGOSTINHO, CORRÊA 86
FERNANDES, 2003; AGOSTINHO, VIANA 86
FERNANDES, 2003; ASSIS 8& FERNANDES, 1980; CAMPELO 86 VIANA, 2003; FERNANDES, AGOSTINHO 86 VIANA, 2003; FERREIRA 86
FERNANDES, 1983; SILVA, 1999) eLongá (MUNIZ, 1982), ambas devonianas, da Bacia do Parnaíba. Entre os espécimens estudados, entretanto, foram reconhecidas formalmente somente duas icnoespécies: B. cruciformes Muniz, 1982 e B.
paranaensis Fernandes 85 Melo, 1985, ficando os demais registros referenciados como Bifungites isp.
Bifungites munizi isp.nov. (Fig.4a-d)
Holotypus - DG-CTG-UFPE 5.689.
Paratypi- DG-CTG-UFPE 5.690 e 5.691.
Locus typicus- Os três exemplares foram coletados em afloramento localizado nos arredores do povoado de São João Vermelho (coordenadas
08°34’57”S e 42°26’55,2”W), no Município de São Raimundo Nonato, na área compreendida pelo Parque Nacional da Serra da Capivara (Fig.la). O acesso à cidade de São Raimundo Nonato pode ser feito pelas rodovias BR-020 (que liga São Raimundo Nonato a Coronel José Dias e São João do Piauí) e BR-324 (também PI-140, que liga São Raimundo Nonato a Canto do Buriti); a localidade de São João Vermelho situa-se a 26km de Coronel José Dias, percorrendo-se 8 km a partir desta cidade pela BR-020 até a guarita 020, também conhecida como guarita do desfiladeiro e, a partir dela, em estrada de terra por 18km até o afloramento próximo ao povoado. O afloramento (Fig.2a) encontra-se representado por uma seqüência de arenitos silticos laminados e arenitos finos intercalados, apresentando marcas de onda de crista reta e laminação sub-horizontal (Fig.3a). Nos arenitos, além dos exemplares de Bifungites
munizi isp.nov., são observadas outras formas de Bifungites, além de prováveis espécimens de Phycosiphon incertum von Fischer-Ooster, 1858 e amostras com exemplares que podem ser atribuídos aos icnogêneros Arenicolites, Lockeia e Palaeophycus / Planolites.
Stratum typicum- Arenitos finos a silticos finamente laminados da Formação Pimenteira.
Derivatio nomini - Em homenagem a Geraldo da Costa Barros Muniz, professor da Universidade Federal de Pernambuco que se dedicou ao estudo dos icnofósseis das bacias do Nordeste brasileiro.
Diagnosis - Um corpo terminal maior em forma de âncora em uma das extremidades e outro menor de contorno circular na extremidade oposta, interligados por um eixo central retilíneo de seção semi-cilíndrica.
Diagnosis - Burrows preserved as convex epireliefs
(epichnia) with a horizontal, straight, and subcircular
shaft between two different terminations, the bigger one being reniform-like and the other, smaller in size,
is dumbbell-like.
Descrição - Exemplares completos preservados em hiporrelevo convexo, representados pelos corpos terminais e pelo eixo central. Os corpos terminais apresentam diferenças morfológicas e de dimensões entre si, sendo 0 maior em forma de âncora e, 0
menor, de contorno circular. O eixo central é retilíneo, subcilíndrico, sem variação de largura ao longo de seu comprimento. As dimensões em milímetros como comprimento total (C), comprimento do eixo central (c), diâmetro do eixo (d), distância entre as extremidades do corpo
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

DUAS NOVAS ICNOESPÉCIES DE BIFUNGITES, FORMAÇÃO PIMENTEIRA, DEVONIANO DA BACIA DO PARNAÍBA, BRASIL 525
terminal em forma de âncora (Dl), distância entre as extremidades do corpo terminal de contorno circular (D2), comprimento do corpo terminal em forma de âncora (cctl) e de contorno circular (cct2), e as relações C/c, Dl/de D2/d, do holótipo e dos parátipos encontram-se relacionadas na tabela 1. Não foram observadas marcas de inserções dos tubos verticais nos corpos terminais.
Discussão - Os exemplares de Bifungites munizi
isp.nov. apresentam morfologia e dimensões acentuadamente diferentes dos demais exemplares de Bifungites presentes no mesmo afloramento, bem como das icnoespécies de Bifungites já descritas em formações brasileiras. No primeiro caso, os exemplares associados apresentam corpos terminais triangulares ou em forma de seta, ou com contorno arredondado (Fig.4a), identificados como Bifungites
isp. O exemplar de Bifungites isp., descrito por ASSIS & FERNANDES (1980) da Formação Pimenteira, do Estado de Goiás, é uma forma de dimensões menores, apresentando C/c = 1,55 e D/d = 1,75 e possuindo terminações cuja morfologia, em virtude da sua má preservação, varia de globular a triangular. Bifungites
munizi isp.nov. difere também de B. piauiensis
isp.nov. por esta última icnoespécie apresentar corpos terminais bem definidos em forma de seta, os quais possuem ainda algumas variações em sua forma e também com índices em grande parte com dimensões menores. Por outro lado, um confronto descritivo entre B. zmmzzz isp.nov. e B.
paranaensis permite também diferenciá-los pelo fato de B. paranaensis possuir corpos terminais bem definidos em forma de seta, os quais apresentam uma base plana, embora suas relações C/c e D/d aproximem-se de B. munizi
isp.nov. Bifungites cruciformis, outra icnoespécie devoniana, mas descrita para a Formação Longá, difere por sua vez de B. munizi isp.nov. por apresentar terminações tipicamente em forma de
cruz em ambas extremidades, que possuem eixo e braços subcilíndricos e de contorno subarredondado. É preciso ressaltar que a icnoespécie ora proposta possui variação de forma nos corpos terminais em um mesmo exemplar (um em forma de âncora ou reniforme e outro subesférico), variação que não é observada em nenhuma das icnoespécies citadas acima. Exemplares portadores de terminações reniformes também são conhecidos na literatura estrangeira. OSGOOD (1970) atribuiu os exemplares do Ordoviciano Superior de Cincinnati (Estados Unidos) à icnoespécie Corophioides biclavata (Miller, 1875) e MÉLOU & PLUSQUELLEC (1975) descreveram espécimens do Ordoviciano Superior da França como ?Bifungites isp. Os exemplares descritos por OSGOOD (1970), entretanto, possuem as terminações semelhantes, mais aproximadas à forma de halteres, sendo assim definidas pelo autor e o eixo central acentuadamente mais estreito ou de menor diâmetro que os exemplares de B. munizi isp.nov.; por outro lado, os exemplares descritos por MÉLOU & PLUSQUELLEC (1975), apesar de apresentarem corpos terminais morfologicamente diferentes entre si, possuem as terminações reniformes com contornos menos circulares com projeções laterais mais acentuadas, diferindo significativamente da forma apresentada pelas terminações reniformes de B. munizi isp.nov. SEILACHER (1983) atribuiu ambas as ocorrências à icnoespécie Bifungites biclavatus
(Miller, 1875), assinalando que as formas com terminações reniformes seriam características do Ordoviciano Superior, enquanto que as dotadas de terminações em forma de seta, identificadas como B.
fezzanensis Desio, 1940, seriam características do Devoniano, estendendo-se ao Carbonífero. A ocorrência de B. munizi isp.nov. em São João Vermelho (PI), portanto, estende a distribuição estratigráfica das formas dotadas de terminações reniformes ao Devoniano Médio.
Tabela 1. Dimensões (mm) dos exemplares de Bifungites munizi isp.nov.
Exemplares C c d Dl D2 cctl cct2 C/c Dl/d D2/d
Holótipo DG-CTG-UFPE 5.689 35,85 18,92 6,95 19,77 15,23 7,97 9,55 1,89 2,84 2,19
Parátipos DG-CTG-UFPE 5.690 34,74 18,65 5,35 18,87 12,78 7,18 7,82 1,86 3,52 2,38
DG-CTG-UFPE 5.691 30,16 15,80 5,89 18,67 14,10 8,54 6,72 1,90 2,83 2,39
(C) comprimento total; (c) comprimento do eixo; (d) diâmetro do eixo; (Dl) distância entre as extremidades do corpo
terminal em forma de âncora; (D2) distância entre as extremidades do corpo terminal em forma de halteres; (cctl)
comprimento do corpo terminal em forma de âncora; (cct2) comprimento do corpo terminal em forma de halteres. índices de relações: C/c, Dl/d e D2/d.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

526 S.AGOSTINHO, M.S.S.VIANA & A.C.S.FERNANDES
Fig.4- Bifungites munizi isp.nov.: (A) aspecto geral da amostra com os três exemplares estudados; [B) holótipo, exemplar DG-CTG-UFPE 5.689; (C) parátipo, exemplar DG-CTG-UFPE 5.690; (D) parátipo, exemplar DG-CTG-UFPE 5.691.
Bifunqites piauiensis isp.nov. (Fig.5a-c)
Holotypus - DG-CTG-UFPE 5.654.
Paratypi - DG-CTG-UFPE 5.647, 5.664, 5.665 e 5.652.
Locus typicus - Os exemplares foram coletados nos afloramentos situados na localidade de Oiti, Município de Pimenteiras, com as coordenadas de 06°16’S e 41°35W (Fig. lb). Nessa localidade, um meandro do rio Sambito forma uma cachoeira com cerca de 200m de extensão em rochas da Formação Pimenteira (Fig.2b), as quais estão representadas por arenitos finos a sílticos que se intercalam (Fig.3b) e exibem um rico conteúdo icnofossilífero. No afloramento foram também observados exemplares de Asteriacites stelliforme (Miller & Dyer, 1878), Diplichnites isp., ?Granularia
isp., Lophoctenium isp., Palaeophycus tubularis
Hall, 1847, Planolites beverleyensis (Bilings, 1862) e Scolicia isp., além de tentaculitídeos (AGOSTINHO & VIANA, 2001; CAMPELO & VIANA, 2003; SILVA, 1999).
Stratum typicum - Arenitos finos a sílticos, finamente laminados, da Formação Pimenteira.
Designatio nominis - Em virtude de ser procedente do Estado do Piauí.
Diagnosis - Escavações preservadas em epirrelevo convexo (epichnia) com eixo central retilíneo, aparentemente subcilíndrico, com corpos terminais em forma de setas, cujas bases apresentam projeções para trás com ângulos de 10° a 35°.
Diagnosis - Burrows preserved as convex epiréliefs
(epichnia) with a horizontal, straight, and
subcircular shaft, with double arrow-shaped
terminations, which bases have projections with
angles of 10o to 35°.
Descrição - Normalmente preservados como epirrelevos convexos [epichnia), dos cinco espécimes completos mensurados (Tab.2), quatro deles apresentaram comprimento compreendido entre 60mm e 73mm, e o restante medindo 40mm. Os corpos terminais apresentam-se freqüentemente em forma de seta. Ainda nos mesmos cinco
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

DUAS NOVAS ICNOESPÉCIES DE BIFUNGITES, FORMAÇÃO PIMENTEIRA, DEVONIANO DA BACIA DO PARNAÍBA, BRASIL 527
Fig.5- Bifungites piauiensis isp.nov.: (A) holótipo (exemplar superior) e parátipo (exemplar inferior), amostra DG-CTG- UFPE 5.654; (B), parátipo, amostra DG-CTG-UFPE 5.647; (C) parátipo, amostra DG-CTG-UFPE 5.664.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004
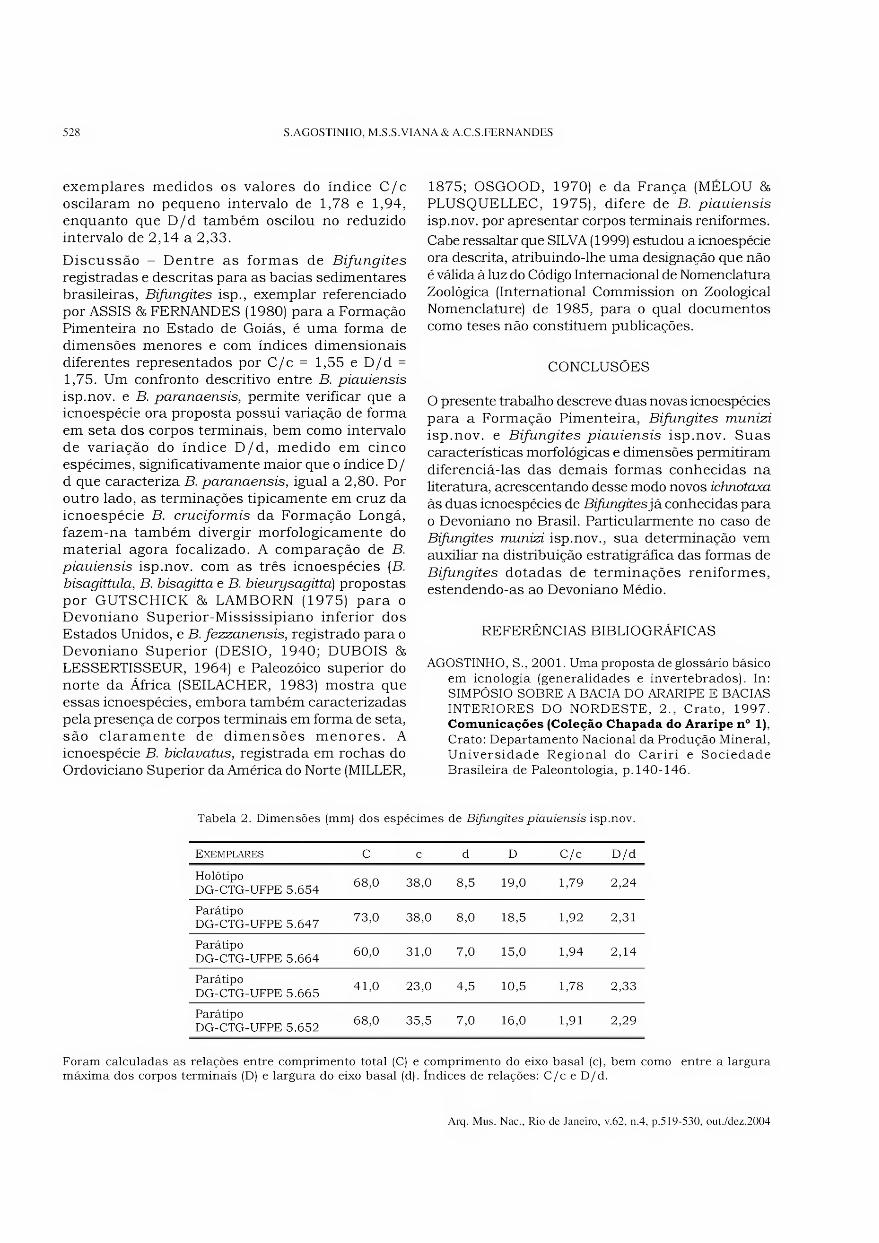
528 S.AGOSTINHO, M.S.S.VIANA & A.C.S.FERNANDES
exemplares medidos os valores do índice C/c oscilaram no pequeno intervalo de 1,78 e 1,94, enquanto que D/d também oscilou no reduzido intervalo de 2,14 a 2,33.
Discussão - Dentre as formas de Bifungites
registradas e descritas para as bacias sedimentares brasileiras, Bifungites isp., exemplar referenciado por ASSIS & FERNANDES (1980) para a Formação Pimenteira no Estado de Goiás, é uma forma de dimensões menores e com índices dimensionais diferentes representados por C/c = 1,55 e D/d = 1,75. Um confronto descritivo entre B. piauiensis
isp.nov. e B. paranaensis, permite verificar que a icnoespécie ora proposta possui variação de forma em seta dos corpos terminais, bem como intervalo de variação do índice D/d, medido em cinco espécimes, significativamente maior que o índice D / d que caracteriza B. paranaensis, igual a 2,80. Por outro lado, as terminações tipicamente em cruz da icnoespécie B. cruciformis da Formação Longá, fazem-na também divergir morfologicamente do material agora focalizado. A comparação de B.
piauiensis isp.nov. com as três icnoespécies (B.
bisagittula, B. bisagitta e B. bieurysagitta) propostas por GUTSCHICK & LAMBORN (1975) para o Devoniano Superior-Mississipiano inferior dos Estados Unidos, e B. fezzanensis, registrado para o Devoniano Superior (DESIO, 1940; DUBOIS & LESSERTISSEUR, 1964) e Paleozoico superior do norte da África (SEILACHER, 1983) mostra que essas icnoespécies, embora também caracterizadas pela presença de corpos terminais em forma de seta, são claramente de dimensões menores. A icnoespécie B. biclavatus, registrada em rochas do Ordoviciano Superior da América do Norte (MILLER,
1875; OSGOOD, 1970) e da França (MELOU & PLUSQUELLEC, 1975), difere de B. piauiensis
isp.nov. por apresentar corpos terminais reniformes.
Cabe ressaltar que SILVA (1999) estudou a icnoespécie ora descrita, atribuindo-lhe uma designação que não é válida à luz do Código Internacional de Nomenclatura Zoológica (International Commission on Zoological Nomenclature) de 1985, para o qual documentos como teses não constituem publicações.
CONCLUSÕES
O presente trabalho descreve duas novas icnoespécies para a Formação Pimenteira, Bifungites munizi
isp.nov. e Bifungites piauiensis isp.nov. Suas características morfológicas e dimensões permitiram diferenciá-las das demais formas conhecidas na literatura, acrescentando desse modo novos ichnotaxa
às duas icnoespécies de Bifungitesjá conhecidas para o Devoniano no Brasil. Particularmente no caso de Bifungites munizi isp.nov., sua determinação vem auxiliar na distribuição estratigráfica das formas de Bifungites dotadas de terminações reniformes, estendendo-as ao Devoniano Médio.
REFERÊNCIAS BIBLIOGRÁFICAS
AGOSTINHO, S., 2001. Uma proposta de glossário básico
em icnologia (generalidades e invertebrados). In:
SIMPÓSIO SOBRE A BACIA DO ARARIPE E BACIAS
INTERIORES DO NORDESTE, 2., Crato, 1997.
Comunicações (Coleção Chapada do Araripe n° 1),
Crato: Departamento Nacional da Produção Mineral,
Universidade Regional do Cariri e Sociedade
Brasileira de Paleontologia, p. 140-146.
Tabela 2. Dimensões (mm) dos espécimes de Bifungites piauiensis isp.nov.
Exemplares C c d D C/c D/d
Holótipo DG-CTG-UFPE 5.654
68,0 38,0 8,5 19,0 1,79 2,24
Parátipo DG-CTG-UFPE 5.647
73,0 38,0 8,0 18,5 1,92 2,31
Parátipo DG-CTG-UFPE 5.664
60,0 31,0 7,0 15,0 1,94 2,14
Parátipo DG-CTG-UFPE 5.665
41,0 23,0 4,5 10,5 1,78 2,33
Parátipo DG-CTG-UFPE 5.652
68,0 35,5 7,0 16,0 1,91 2,29
Foram calculadas as relações entre comprimento total (C) e comprimento do eixo basal (c), bem como entre a largura máxima dos corpos terminais (D) e largura do eixo basal (d). índices de relações: C/c e D/d.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

DUAS NOVAS ICNOESPÉCIES DE BIFUNGITES, FORMAÇÃO PIMENTEIRA, DEVONIANO DA BACIA DO PARNAÍBA, BRASIL 529
AGOSTINHO, S.; CAMPELO, F.M.C.A.; MELO, K.J.V.; VIANA, M.S.S. & FERNANDES, A.C.S., 2001. Perfil
estratigráfico da Formação Pimenteira (Devoniano da
Bacia do Parnaíba) no leito do rio Sambito (Pimenteiras-PI). In: CONGRESSO BRASILEIRO DE
P&D EM PETRÓLEO E GÁS, 1., Natal. Resumos,
Natal: Universidade Federal do Rio Grande do Norte e Sociedade Brasileira de Química, p.56.
AGOSTINHO, S.; CORRÊA, L.M.S.A. & FERNANDES,
A.C.S., 2003. Os icnofósseis da Formação Pimenteira (Devoniano da Bacia do Parnaíba) no Município de
Miranorte, Estado do Tocantins. In: CONGRESSO
BRASILEIRO DE PALEONTOLOGIA, 18., Brasília. Boletim de Resumos, Brasília: Sociedade Brasileira
de Paleontologia, p.34.
AGOSTINHO, S. & VALENÇA, L.M.M., 1999. Icnofósseis da região de Pimenteiras, PI, Formação Pimenteira,
Devoniano da Bacia do Parnaíba. In: CONGRESSO
BRASILEIRO DE PALEONTOLOGIA, 16., Crato. Boletim de Resumos, Crato: Sociedade Brasileira de
Paleontologia e Universidade Regional do Cariri, p. 11.
AGOSTINHO, S. & VIANA, M.S.S., 2001. Ocorrência de Tentaculita na Formação Pimenteira, Devoniano da Bacia do Parnaíba. Revista Brasileira de
Paleontologia, Rio de Janeiro (2):96. AGOSTINHO, S.; VIANA, M.S.S. & FERNANDES, A.C.S.,
2003. Um novo registro do icnogênero Bifungites na
Formação Pimenteira (Devoniano), Bacia do Parnaíba, Brasil. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 18., Brasília. Boletim de
Resumos, Brasília: Sociedade Brasileira de Paleontologia, p.35.
ASSIS, J.F.P. & FERNANDES, A.C.S., 1980. A ocorrência
de Bifungites Desio, 1940 na Formação Pimenteiras, Devoniano da Bacia do Maranhão. Anais da Academia Brasileira de Ciências, Rio de Janeiro,
52(2):335-338. BALISTIERI, P.R.M.N. & NETTO, R.G., 2001.
Reconstrução paleoambiental da sucessão
sedimentar Mafra/Rio do Sul (Permo-Carbonífero, Bacia do Paraná, SC) com base nas assembléias de traços fósseis. Revista Brasileira de Paleontologia,
Rio de Janeiro (2): 143-144. BALISTIERI, P.R.M.N.; NETTO, R.G. & WEINSCHUTZ,
L.C., 2001. Paleoicnologia da porção superior do
Grupo Itararé (base da Formação Rio do Sul, Permo- Carbonífero, Bacia do Paraná), na região de Mafra, SC. Revista Brasileira de Paleontologia, Rio de
Janeiro (2):88-89. BRITO, I.M., 1977. Ocorrência de bióglifos no Devoniano
Inferior do Município de Tocantínia, Goiais. Anais da Academia Brasileira de Ciências, Rio de Janeiro, 49(3):461-464.
CAMPANHA, V.A., 1974. Caracterização ichnofaciológica do
Membro Picos, Formação Pimenteiras, na região de São Miguel do Tapuio. Piauí. Anais da Academia Brasileira
de Ciências, Rio de Janeiro, 46(3/4) :593-600.
CAMPELO, F.M.A.C. & VIANA, M.S.S., 2003. A palaeoenvironmental interpretation of the Pimenteira Formation, Parnaíba Basin, Northeast Brazil, based
on ichnofossils. In: LATINAMERICAN CONGRESS OF
SEDIMENTOLOGY, 3., Belém. Abstracts, Belém: Museu Paraense Emílio Goeldi e Universidade Federal
do Pará, p. 172-173. DELLA FÁVERA, J.C., 1990. Tempestitos da Bacia do
Parnaíba. Porto Alegre. 242p. Tese (Doutorado em
Geociências), Programa de Pós-Graduação em
Geociências, Instituto de Geociências/Universidade Federal do Rio Grande do Sul.
DESIO, A., 1940. Vestigia problemática paleozoiche delle Líbia. Annali dei Museo Libico di Storia Naturale,
Trípoli, 2:47-92.
DUBOIS, P. & LESSERTISSEUR, J., 1964. Note sur
Bifungites, trace problematique du Devonien du Sahara. Bulletin de la Société Géologique de
France, Paris, 6(7):626-631.
FERNANDES, A.C.S., 1996. Os icnofósseis do Ordoviciano, Siluriano e Devoniano da Bacia do
Paraná. Rio de Janeiro. 183p. Tese (Doutorado em
Ciências - Geologia), Programa de Pós-Graduação em Geologia, Instituto de Geociências/Universidade
Federal do Rio de Janeiro.
FERNANDES, A.C.S., 2001. A paleoicnofauna brasileira de artrópodes: estado atual de seu conhecimento.
Acta Geologica Leopoldensia, São Leopoldo, 24(52/
53):359-372. FERNANDES, A.C.S. & MELO, J.H.G., 1985.
Ocorrência do icnogênero Bifungites na Formação
Ponta Grossa, Devoniano do Estado do Paraná. Anais da Academia Brasileira de Ciências, Rio
de Janeiro, 57(2):203-207.
FERNANDES, A.C.S.; AGOSTINHO, S. & VIANA, M.S.S., 2003. O icnogênero Bifungites e sua nova ocorrência
na Formação Pimenteira (Devoniano), Bacia do
Parnaíba, Brasil. In: CONGRESSO NACIONAL DE GEOLOGIA, 6., Monte da Caparica, Portugal. Ciências da Terra (Universidade Nova de Lisboa),
Lisboa, n. especial 5, p.A72-A75. (CD-ROM). FERNANDES, A.C.S.; BORGHI, L.; CARVALHO, I.S. &
ABREU, C.J., 2002. Guia dos icnofósseis de invertebrados do Brasil. Rio de Janeiro: Editora
Interciência, 260p.
FERREIRA, C.S. & FERNANDES, A.C.S., 1983.
Notícias sobre alguns icnofósseis da Formação Pimenteira, Devoniano no Estado de Goiás. Anais da Academia Brasileira de Ciências, Rio de
Janeiro, 55(1):140. FILLION, D & PICKERILL, R.K., 1984. On Arthraria
antiquata Billings, 1872 and its relationship to Diplocraterion Torell, 1870 and Bifungites Desio, 1940. Journal of Paleontology, Tulsa,
58(3) :683-696.
FONSECA, V.M.M. & MELLO, J.H.G., 1987. Ocorrência de Tropidoleptos carinatus (Conrad) (Brachiopoda,
Orthida) na Formação Pimenteira, e sua importância
paleobiogeográfica. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 10., Rio de Janeiro. Anais,
Rio de Janeiro, Sociedade Brasileira de Paleontologia, 2:505-537.
GÓES, A.M.O. & FEIJÓ, F.J., 1994. Bacia do Parnaíba. Boletim de Geociências da Petrobrás, Rio de
Janeiro, 8(l):57-68.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

530 S.AGOSTINHO, M.S.S.VIANA & A.C.S.FERNANDES
GUTSCHICK, R.C. & LAMBORN, R., 1975. Bifungitestrace fossils from Devonian-Mississipian rocks of
Pennsylvania and Montana, U.S.A. Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam,
18:193-212. GUTSCHICK, R.C.; SUTTNER, L.J. & SWITEK, M.J.,
1962. Biostratigraphy of transitional Devonian- Mississipian Sappington Formation of Southwest
Montana. In: ANNUAL FIELD CONFERENCE, 13.
Billings Geological Society Guidebook, p.79-89.
HÀNTZSCHEL, W., 1975. Trace Fossils and Problemática. In: TEICHERT, C. (Ed.) Treatise on
Invertebrate Paleontology. Boulder: Geological Society of America & University of Kansas. Part W,
Miscellanea, Supplement 1, 269p.
KEGEL, W., 1961. Rasto de trilobita (?Homalonotus) do Devoniano Inferior do Piauí. Anais da Academia Brasileira de Ciências, Rio de Janeiro,
33(2):163-167. KEGEL, W., 1966. Rastos do Devoniano da Bacia do
Parnaíba. Boletim Departamento Nacional da
Produção Mineral. Divisão de Geologia e
Mineralogia, Rio de Janeiro (233): 1-32. INTERNATIONAL COMMISSION ON ZOOLOGICAL
NOMENCLATURE, 1985. International Code of Zoological Nomenclature. 3.ed. London:
International Trust for Zoological Nomenclature, 338p.
MÉLOU, M. & PLUSQUELLEC, Y., 1975. Sur Bifungites? (Problemática) du “Grès de Kermeur”, Ordovicien de
la presqu’ ile de Crozon (Finistère, France). Neues Jahrbuch für Geologie und Palãontologie, Monatshefte, Stuttgart, p.456-479.
MILLER, S.A., 1875. Some new species of fossils from
the Cincinnati Group and remarks upon some described forms. Cincinnati Quarterly Journal of
Science, Cincinnatti, 2(4):349-355. MUNIZ, G.C.B., 1976. Macrofósseis devonianos da
Formação Inajá no Estado de Pernambuco. Recife.
165p. Tese (Livre-Docência), Universidade Federal de
Pernambuco. MUNIZ, G.C.B., 1979. Novos ichnofósseis devonianos
da Formação Inajá, no Estado de Pernambuco. Anais da Academia Brasileira de Ciências, Rio de Janeiro,
51(1):121-132. MUNIZ, G.C.B., 1982. Ichnofósseis Devonianos da
Formação Longá, no Estado do Piauí. In: CONGRESSO BRASILEIRO DE GEOLOGIA, 32., Salvador. Anais, Salvador: Sociedade Brasileira de
Geologia, 4:1305-1316. MUNIZ, G.C.B., 1988. Merostomichnites piauiensis
ichnosp. nov. do Devoniano do estado do Piauí
(Membro Picos, Formação Pimenteira). Estudos Pesquisas, Universidade Federal de Pernambuco,
Recife (9):49-53.
NASCIMENTO, M.C.; MOURA, C.R.; CAMPELO, F.M.A.C.; AGOSTINHO, S. & VIANA, M.S.S., 2003.
Paleoenvironmental significance of trilobite traces in
the Parnaíba Basin, NE of Brazil. In: LATINAMERICAN CONGRESS OF SEDIMENTOLOGY, 3., Belém.
Abstracts, Belém: Museu Paraense Emílio Goeldi e
Universidade Federal do Pará, p. 176-177. NETTO, R.G., 2000. Paleoicnologia do Rio Grande do
Sul. In: HOLZ, M. & DE ROS, L.F. (Eds.)
Paleontologia do Rio Grande do Sul. Porto Alegre: Centro de Investigação do Gonduana e Universidade
Federal do Rio Grande do Sul. p.25-43.
OSGOOD, R.G., 1970. Trace fossils of the Cincinnati area. Paleontographica Americana, Ithaca,
6:281-444.
RODRIGUES, A.S., 2001. Revisão dos icnofósseis descritos por Wilhelm Kegel para as formações
Cabeças e Longá (Devoniano Médio e Superior da
Bacia do Parnaíba). In: JORNADA DE INICIAÇÃO CIENTÍFICA DA UNIVERSIDADE FEDERAL DO
RIO DE JANEIRO, 23., Rio de Janeiro. Resumos,
Rio de Janeiro: Universidade Federal do Rio de Janeiro, p.124.
RODRIGUEZ, J. & GUTSCHICK, R.C., 1970. Late
Devonian-Early Mississipian ichnofossils from western Montana and northern Utah. In: CRIMES,
T.P. & HARPER, J.C. (Eds.) Trace Fossils. Liverpool:
Seel House Press, p.407-438. (Geological Journal Special Issue 3)
SEILACHER, A., 1955. Spuren und Fazies in
Unterkambrium. In: SCHINDEWOLF, O.H. &
SEILACHER, A., Beitráge zur Kenntnis des Kambriums in der Salt Range (Pakistan). Abhandlungen.
Mathematisch-naturwissenschaftliche Klasse. Akademie der Wissenschaften und der Literatur,
Mainz, 10:11-143.
SEILACHER, A., 1983. Upper Paleozoic trace fossils from the Gilf Kebir-Abu Ras area in southwestern Egypt. Journal of African Earth Sciences,
Johannesburg, l(l):21-34. SILVA, S.M.O.A., 1999. Icnofósseis da Formação
Pimenteira, Devoniano da Bacia do Parnaíba,
Município de Pimenteiras, Piauí. Recife. 46p.
Dissertação (Mestrado em Geociências), Programa de
Pós-Graduação em Geociências, Centro de
Tecnologia/Universidade Federal de Pernambuco. TURNER, B.R. & BENTON, M.J., 1983. Paleozoic trace
fossils from the Kufra Basin, Libya. Journal of
Paleontology, Tulsa, 57(3):447-460. YOUNG, C.G.K. & BORGHI, L., 2003. An event of forced
regression in the Pimenteira Formation (Devonian,
Parnaíba Basin, Brazil). In: LATINAMERICAN CONGRESS OF SEDIMENTOLOGY, 3., Belém.
Abstracts, Belém: Museu Paraense Emílio Goeldi e
Universidade Federal do Pará, p. 168-170. YOUNG, C.G.K.; BORGHI, L. & FERNANDES, A.C.S.,
2002. Icnofósseis na Formação Pimenteira
(Devoniano, Bacia do Parnaíba) em testemunhos de sondagem. Paleontologia em Destaque, São
Leopoldo (40):21.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.519-530, out./dez.2004

TmnfTOnrominnrTi nnniiitiiiMifttinnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004
ISSN 0365-4508
A NEW ACTINOPTERYGIAN FISH
FROM THE LATE PERMIAN OF THE PARANÁ BASIN, SOUTHERN BRAZIL 1
(With 5 figures)
FRANCISCO JOSÉ DE FIGUEIREDO 2
BARTIRA C.M.C. CARVALHO 3
ABSTRACT: Angatubichthys mendesi gen. et sp. nov., an early actinopterygian fish from the Upper Permian
of Paraná Basin (Corumbataí Formation), is described from four complete and articulated specimens preserved
in a reddish siltstone obtained in an outcrop in Angatuba municipality, State of São Paulo. It is a small fish
distinguished from the other palaeoniscoids by a combination of features: short and deep head; wrinkled
median postrostral; nasal and dermosphenotic not in contact; suboperculum larger than operculum;
conspicuous radiating ridges of ganoin on cleithrum and supracleithrum; longitudinal bands of ganoin on
dentalosplenial; maxillary canal present; ethmoidal commissure on rostropremaxilloantorbital; reduced
number of branchiostegal rays; rows of deep smooth rhomboidal scales on flanks; and caudal fin heterocercal,
deeply forked with almost equal-sized lobes. Angatubichthys mendesi gen. et sp.nov. appears to be more
advanced than the so-called basal actinopterygians (Cheirolepis, Mimia, Moythomasia, Polypterus) due to
various anatomical features (e.g., absence of pineal plate, presence of dermopterotic, deep scales on the
flanks, presence of suborbitals, reduction of the number of branchiostegal rays, and reduced number of
pectoral fin rays) but it wants synapomorphies of platysomid and neopterygian fishes.
Key words: Angatubichthys mendesi gen. et sp.nov., actinopterygian fish, Upper Permian, Paraná Basin.
RESUMO: Um novo peixe actinopterígio do Neopermiano da Bacia do Paraná, Sul do Brasil
Angatubichthys mendesi gen. et sp.nov., um primitivo peixe actinopterígio do Neopermiano da Bacia do Paraná
(Formação Corumbataí), é descrito com base em quatro espécimes completos e articulados preservados em
siltito avermelhado que aflora no município de Angatuba, Estado de São Paulo. Trata-se de um peixe de
pequeno porte distinto de outros palaeoniscóides por uma combinação de caracteres: cabeça curta e alta; pós-
rostral mediano de superfície enrugada; nasal e dermoesfenótico sem contato; subopérculo maior que o opérculo;
conspícuas estrias radiantes de ganoína sobre o cleitro e supracleitro; faixas longitudinais de ganoína sobre o
dentalo-esplenial; presença de comissura etmoideana e canal maxilar; número reduzido de raios de
branquiostégios; fileiras de escamas rômbicas e lisas nos flancos; nadadeira caudal heterocerca profundamente
bifurcada, com lobos de tamanho aproximadamente igual. Angatubichthys mendesi gen. et sp.nov. parece ser
mais avançado que actinopterígios basais (Cheirolepis, Mimia, Moythomasia, Polypterus) devido a presença de
vários aspectos anatômicos [e.g., ausência de forame pineal, presença de dermopterótico, escamas altas nos
flancos, presença de suborbitais, redução do número de raios de branquiostégios e número reduzido de raios
da nadadeira peitoral), mas carece de sinapomorfias para platisomídeos e neopterígios.
Palavras-chave: Angatubichthys mendesi_gen. et sp.nov., peixe actinopterígio, Neopermiano, Bacia do Paraná.
INTRODUCTION
The Brazilian Paleozoic paleoichthyofauna is relatively diversified but the record of well- preserved specimens is extremely meager (RICHTER, 1991; DIAS, 1995). Concerning the actinopterygian fishes, most of the specimens are represented by scales, bony fragments, and isolated teeth [e.g., RICHTER, 1985; MEZZALIRA, 1989).
Few studies were based on almost complete and articulated fishes (DUNKLE & SCHAEFFER, 1956; BELTAN, 1978; MALABARBA, 1988; RICHTER, 1991; SANTOS, 1991; DIAS, 1995; FIGUEIREDO, DIAS 86 COSTA-CARVALHO, 1998; VEGA, DIAS 86 RAGONHA, 1997; VEGA-DIAS, DIAS 8& RICHTER, 2000; and RICHTER, 2002) and most of these fóssil material were obtained from the Brazilian portion of the Paraná Basin.
1 Submitted on April 4, 2003. Accepted on October 26, 2004.
2 Universidade do Estado do Rio de Janeiro, Instituto de Biologia, Departamento de Biologia Animal e Vegetal. Rua São Francisco Xavier, 524, Maracanã,
20559-900, Rio de Janeiro, RJ, Brasil. E-mail: [email protected].
Museu Nacional/UFRJ, Programa de Pós-Graduação em Ciências Biológicas/Zoologia. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil.
3 Geosoft. Edifício Avenida Central, Avenida Rio Branco, 156, Centro, 22043-900, Rio de Janeiro, RJ, Brasil.

532 F.J.FIGUEIREDO & B.C.M.C.CARVALHO
SANTOS (1991) reported the occurrence of actinopterygian fishes for the Permian of the State of São Paulo. The material consists of a poorly preserved platysomid fish from the Tatuí Formation (see FIGUEIREDO, DIAS & COSTA-CARVALHO, 1998), and nearly complete palaeoniscoid fishes from the Corumbataí Formation. PETRI (2001) pointed out that the latter material is remarkable due to the quality of preservation and taxonomic novelty. Unfortunately, the late paleontologist Rubens da Silva Santos (1918-1996) passed away without formally describing it.
In this paper, we describe the ray-finned fish from the Corumbataí Formation originally reported by Silva Santos (SANTOS, 1991), comparing it with other South American taxa and briefly discussing its affinities.
GEOLOGICAL SETTING
The Paraná Basin is an extensive rock unit placed at the Central-Southeastern South America, covering part of the political territories of Brazil, Argentina, Uruguay, and Paraguay and encompassing an area of about l,200,000km2 (MILANI et al, 1998). Sedimentation took place during the Paleozoic and Mesozoic, and the stratigraphic record ranges from the Late Ordovician to the Late Cretaceous (LAVINA, 1991).
The following stratigraphic groups are recognised in the Paraná Basin: Paraná (Devonian); Tubarão (Upper Carboniferous-Middle Permian); Passa Dois (Upper Permian); São Bento (Triassic-Jurassic); and Bauru (Jurassic-Cretaceous) (ALMEIDA & MELO, 1981; PETROBRAS, 1998).
SCHNEIDER et al (1974) argued that the Passa Dois Group is composed in the Southern Paraná Basin by the Irati, Serra Alta, Teresina, and Rio do Rasto formations, and in the States of São Paulo, Goiás, and Mato Grosso, by the Irati and Corumbataí formations.
In São Paulo, the Corumbataí Formation corresponds to the Serra Alta, Teresina, and Serrinha (in part) formations. It consists of mudstone, siltstone, and shale of dark grey colour in the basal portion and reddish in the top (SCHNEIDER et al, 1974; ALMEIDA & MELO, 1981; MILANI, FRANÇA & SCHNEIDER, 1994).
The predominant climatic conditions during sediment deposition of the Corumbataí Formation were dry, with low rainfall, which gradually produced scattered water pools (RAGONHA,
1989a). The paleoenvironment was probably transitional from neritic to lacustrine (MILANI, FRANÇA & SCHNEIDER, 1994).
The fóssil record of the Corumbataí Formation is rich in bryophytes, pteridophytes, gymnosperms, sponges, mollusks, crustaceans, reptiles, and fish remains (MENDES, 1952; FÚLFARO, 1970; MEZZALIRA, 1989). Regarding the latter, it is common to find stratigraphic leveis with a considerable amount of disarticulated bones, dipnoan dental plates, petalodontid teeth, palaeoniscoid scales, xenacanthid and hybodontid remains, besides ctenacanthid spines and coelacanth teeth (RAGONHA, 1989b; TOLEDO, BRITO & BERTINI, 1997). At the moment, the palaeoniscoid fish Tholonotus braziliensis Dunkle & Schaeffer, 1956 is the only relatively complete and articulated vertebrate known from that formation (DUNKLE & SCHAEFFER, 1956).
MATERIAL AND METHODS
The fóssil fish material herein studied consists of four specimens preserved in a reddish siltstone. All specimens are housed at the Paleozoological Collection of the Departamento de Biologia Animal e Vegetal of the Universidade do Estado do Rio de Janeiro and are referred in this paper by the abbreviation Pz.DBAV.UERJ followed by the catalogue number.
The specimens were mechanically prepared with sharp Steel needles and Consolidated with PARALOID 67 dissolved in ethyl acetate. Casts of the specimens were made using CAF 73 (a silicone rubber). The specimens and casts were coated with a smoke of ammonium chloride to enhance skeletal details for photographs. Observations were made using a binocular microscope Olympus SZ 40. Line drawings were made from magnified photographic prints.
The anatomical terminology mainly follows GARDINER & SCHAEFFER (1989). The scale series formula follows WESTOLL (1944): D/P, A, Cl, C2, where, D is the number of scale series anterior to the origin of the dorsal fin; P, number of scales series anterior to the pelvic fin; A, number of scale series anterior to the anal fin; Cl, number of scale series anterior to origin of the caudal peduncle and C2, the number of scale series anterior to the end of the epichordal lobe of the caudal fin.
As far as possible, problems concerning homologies are discussed along the text. The
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 533
proposed diagnosis is based on the unique combination of primitive and derived characters that, although not rigorously in accordance with cladistic principies, this allows the identification of individual taxa when autapomophies are wanting (ARRATIA, 1997).
Anatomical Abbreviations - r and 1 are used before anatomical terms to indicate right and left side respectively. (ANG) angular; (APR) anterior postrostral; (AUP) pars autopalatina of palatoquadrate; (ba.s.) basal shield; (BBR) basibranchial; (BR.R) branchiostegal rays; (CH) ceratohyal; (CL) cleithrum; (CLA) clavicle; (DH) dermohyal; (DPAL) dermopalatine; (DPT) dermopterotic; (DSPH) dermosphenotic; (DSPL) dentalosplenial; (e.ps.a.) canal for efferent pseudobranchial artery; (ENPT) endopterygoid; (f.e.a.) fenestra exonarina anterior; (ep. f.) epaxial fulcra; (ethm.c.) ethmoidean commissure; (f.e.p.) fenestra exonarina posterior; (FR) frontal; (fr.f.) fringing fulcra; (HBR1) first hypobranchial; (HP) hypohyal; (iorb.s.c.) infraorbital sensory canal; (JU) jugal; (l.l.s.c.) lateral line sensory canal; (LA) lachrymal; (LEXS) lateral extrascapular; (LG) lateral guiar; (md.s.c.) mandibular sensory canal; (MEXS) medial extrascapular; (MG) median guiar; (MPR) median postrostral; (MX) maxilla; (mx.c.) maxillary canal; (NA) nasal; (OP) operculum; (p.pl.) posterior pit-line of parietal bone; (PA) parietal; (PAS) parasphenoid; (PO) postorbital; (POP) preoperculum; (pop.s.c.) preopercular sensory canal; (PSCL) presupracleithrum; (PT) post-temporal; (QJ) quadratojugal; (R+PMX+ANT) rostropremaxilloantorbital; (S) sympletic; (SCL) supracleithrum; (scl.b.) sclerotic bone; (SO) suborbital; (SOP) suboperculum; (sorb.s.c.) supraorbital sensory canal; (st.c.) supratemporal commissure; (t) teeth; (t.c.) temporal sensory canal; (t.pl.) transverse pit-line of parietal bone; (v.pl.) vertical pit-line of quadratojugal.
SYSTEMATIC PALEONTOLOGY
Osteichthyes Huxley, 1880 Actinopterygii Woodward, 1891
Actinopteri Cope, 1871 Family indeterminate
Angatubichthys gen.nov.
Diagnosis - Fusiform palaeoniscoid fish of small size recognised by the following combination of features: head length contained about five times
in the maximum body length; short and obtuse snout; operculum smaller than suboperculum; broad and spatulate median postrostral, ornamented with delicate rugae on the anterior border; ethmoidean commissure included into the rostropremaxilloantorbital; maxillary canal present; nasal separate from dermosphenotic; presence of dermopterotic; circumorbital ring including postorbital, jugal, lachrymal and two suborbitals; laniariform teeth on the oral border of jaws; six branchiostegal rays; subequal medial and lateral extrascapular bones; suspensorium vertically inclined; post- temporal and cleithrum ornamented with conspicuous ridges; presupracleithrum present; dentalosplenial ornamented with fine longitudinal ridges; endopterygoid separated; pectoral fin twice as long as the pelvic fin; rows of smooth rhombic scales along the lateral side of the body, anteriormost ones three times as high as long; caudal fin heterocercal with almost equal-sized lobes; fin-rays closely articulated and fulcra are present along the leading edges of all fins.
Type-species - Angatubichthys mendesi sp.nov.
Etymology - A combination of the name of the type-locality (Angatuba) and the Greek word, ichthys, fish.
Angatubichthys mendesi sp.nov. (Figs.l, 2a, 2b, 3-5)
Diagnosis - As for the genus (by monotypy).
Holotype - Pz.DBAV.UERJ 059, a complete specimen, 66mm maximum length (ML); Westoll Formula (WF): 26/11, 20, 39, 53.
Etymology - The specific epithet is in honor of the late Brazilian paleontologist Josué Camargo Mendes, from the Instituto de Geociências of the Universidade de São Paulo, Brazil, who collected the material.
Additional material - Pz.DBAV.UERJ 60, complete specimen, 93mm ML [WF: 27/11, 20, 41, 57]; Pz.DBAV.UERJ 61 a (part) and b (counterpart), incomplete specimen, 57,9mm estimated ML [WF: 27/12, 21, 38, 5]; Pz.DBAV.UERJ 58, complete specimen 37mm ML.
Horizon and type-locality - Upper third of the Corumbataí Formation; Upper Permian (Kazanian). The outcrop is located between kilometres 200 and 210 of the Raposo Tavares Highway (SP-270), about 6km south of the municipality of Angatuba, São Paulo State.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

534 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
DESCRI PTION
Angatubichthys mendesi (Fig.l) reaches about 90mm of maximum length. The body is elegantly fusiform, lateraly compressed and elongate, appearing to be subtrigonal in transverse section. The head is slightly longer than deep and the orbit is situated well forward. The cranium is brachycephalous and the dorsal profile is almost straight. The snout is obtuse, not projecting beyond the mouth opening. The dorsal fin is situated nearer to the caudal fin than to the head and its origin is approximately opposite to origin of the anal fin. The pelvic fin originates at about midway point between the pectoral and pelvic fin bases. The lateral line runs along the middle line of the body. There are 38 to 41 rows of rhombic and macromeric scales ahead of the hinge-line.
Most of the dermocranial bones have smooth surfaces, but show conspicuous growth lines apparently corresponding to concentric accretions of ganoin on the borders. Only rostropremaxilloantorbital, median postrostral, dentalosplenial, and the bones of the pectoral girdle are ornamented conspicually. The sensory canais are totally included in dermal bones and exit on the surface through minute pores.
The snout region is composed of the following bones: median postrostral, anterior postrostral, rostropremaxilloantorbital, and nasal. The median postrostral (Figs.2b, 3-4; MPR) is a large spatulate bone forming most of the snout anterior to the orbit. This bone is long, about one third of head length. It lacks teeth and tubercles, but is ornamented with rugae on the anterior portion. In addition, there
are marked growth lines near the borders. The obtuse posterior border meets the frontais through an obliqúe and straight suture.
This bone meets the nasais laterally and the rostropremaxilloantorbital anteroventrally. There is in its anterior third a lateral notch corresponding to the anterior limit of the fenestra exonarina anterior (Figs.2b, 3; f.e.a.).
In Pz.DBAV.UERJ 059 there is a small anamestic and reniform bone, slightly displaced from its anatomical position and lying anteriorly and below the median postrostral above the rostropremaxilloantorbital. It was interpreted as an anterior postrostral (Fig.2b; APR). Probably this bone lies on a shallow depression between the anterior margins of the median postrostral and nasal. Its presence is not a regular feature in the specimens observed. This bone was observed by DUNKLE & SCHAEFFER (1956) in Tholonotus braziliensis and apparently corresponds to the lat-rostral of Cheirolepis
canadensis Whiteaves, 1881 (see ARRATIA & CLOUTIER, 1996).
Presence of supranumerary bones in the rostral region is considered to be a primitive condition for actinopterygian fishes since it also occurs among sarcopterygian fishes and in the primitive Cheirolepis Agassiz, 1835. Therefore, the condition found in Angatubichthys (i.e., reduced number of bones in rostral region) is derived.
The rostropremaxilloantorbital (Figs.2b, 3; R+PMX+ANT) is a small bone that forms the anterodorsal limit of the mouth opening. It is a complex and squarish bone, articulated dorsally with the median postrostral and nasal, and
Fig.l- Angatubichthys mendesi gen. et sp.nov. Holotype (Pz.DBAV.UERJ 059). Scale bar = lcm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004
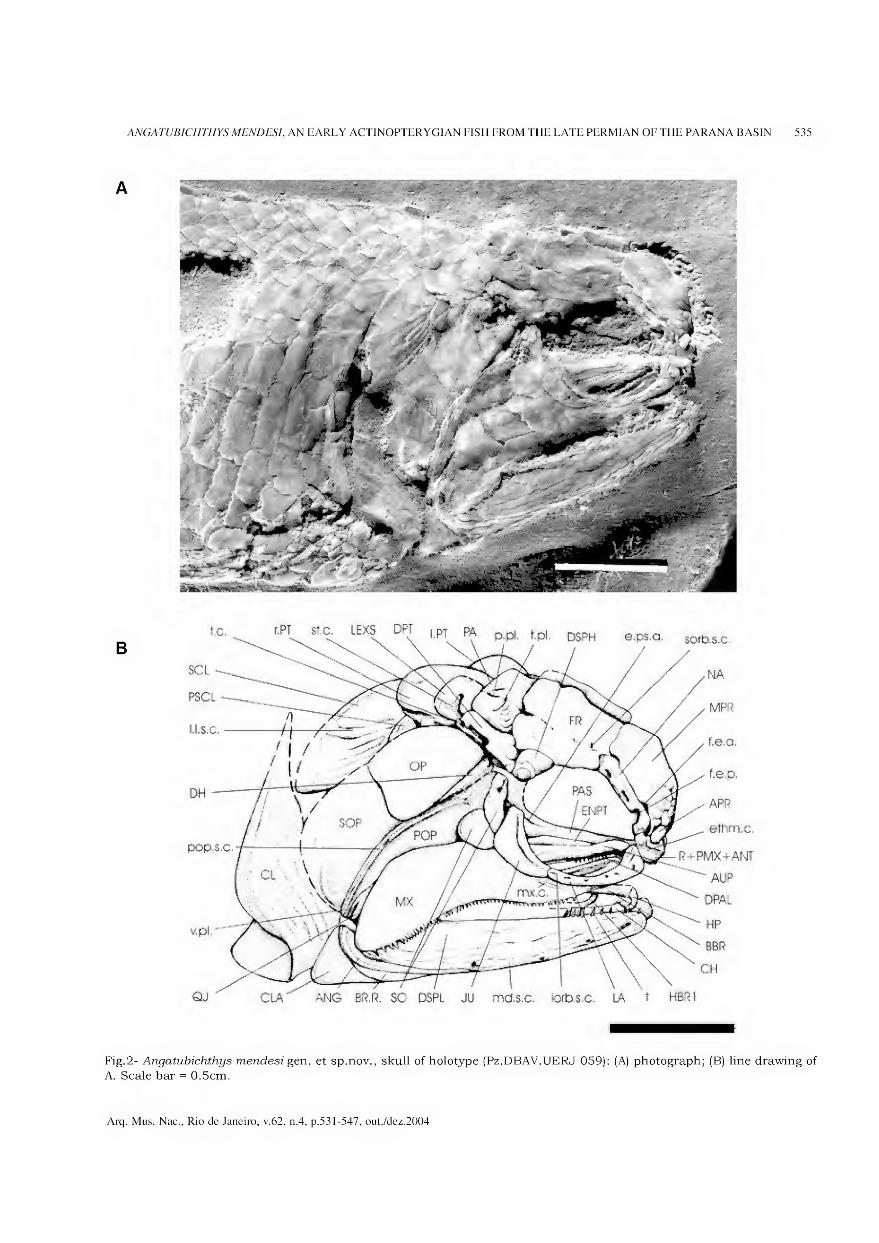
ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 535
B
Fig.2- Angatubichthys mendesi gen. et sp.nov., skull of holotype (Pz.DBAV.UERJ 059): (A) photograph; (B) line drawing of
A. Scale bar = 0.5cm.
t.C.
SCL
P5CL
lls.c.
pOp-S-C.
e.ps.o. soib.s.c.
NA
MPR
f.e.a
f.e.p.
APR
©íhm.c.
R+PMX+ANT
AUP
OPAL
HP
BGft
CH
HBRT
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004
536 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
posteriorly with the lachrymal above and maxilla below. It possesses a well developed neurodermal component marked by a tripartite pattern of sensory canais. The supraorbital and infraorbital sensory canais join together into the middle portion of the bone producing a well developed ethmoidean commissure (Figs.2b, 3; ethm.c.). The shape and arrangement of the bone correspond to that of Rhadinichthys fusiformis Lund & Poplin, 1997 (see LUND 85 POPLIN, 1997; POPLIN & LUND, 1997). In the holotype, there are in the oral margin of the rostropremaxilloantorbital three round teeth-like structures, but we are unable to establish hitherto if they are true teeth or tubercles.
The nasal (Figs.2b, 3-4; NA) is a paired bone, straight and elongate, with length equal to that of the frontal. It meets the whole lateral margin of the median postrostral forming the anterodorsal limit of the orbit. It is inclined in relation to the basicranian axis meeting the frontal posteriorly through a limbate suture. The anterior third is capitate and in this region there is a notch corresponding to the posterior border of the fenestra exonarina anterior. In the orbital margin, another notch occurs, deeper than the anterior, for the fenestra exonarina posterior
(Figs.2b, 3; f.e.p.). The supraorbital sensory canal runs along the bone in the midline exiting on the surface through few minute pores. In Cheirolepis the nasal is divided into two bony elements. This condition is considered primitive to basal actinopterygians (POPLIN & LUND, 1995) whereas the condition of Angatubichthys is derived.
The frontal (Figs.2b, 3-4; FR) and postrostral are the largest bones of the skull roof. The frontal is subrectangular, with the longitudinal margins approximately parallel, and length equivalent to the orbital diameter. It is also wide, reaching the greatest width, near the dermosphenotic bone. Its width is about 2/3 of its length. The posterior margin of the frontal is slightly wider than the anterior. Posterodorsally, it meets the dermosphenotic, posteriorly with a small portion of the dermopterotic, and medially with its antimere. The bone slightly overlapps the anterior margin of the parietal bone. A rounded medial projection in the middle third of the bone shows that this bone was slightly asymmetrical in some individuais.
The supraorbital sensory canal (Figs.2b, 3-4; sorb.s.c.) runs the frontal totally bone enclosed, exiting on surface through at least three large pores.
PT
PSCL
pop.s.c.
íorb.s,a
LG MG md.s.c CLA BR.R.
DSPH
sorb.s,c.
MPR
f.è.p.
NA
f.e.o.
R+PMX+ANT
LA
Fig.3- Angatubichthys mendesi gen. et sp.nov., skull of Pz.DBAV.UERJ 60. Scale bar = 0.5cm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 537
Fig.4- Angatubichthys mendesi gen. et sp.nov., skull of Pz.DBAV.UERJ 58. Scale bar = 0.25cm.
The frontal possesses a well-developed cleft, followed by a deep depression, in its posterior margin, corresponding to the projection of the supraorbital sensory canal into the parietal. This bone forms most of the dorsal margin of the orbit, separating the nasal from the dermosphenotic, as in Polypterus Lacepède, 1803, Calamoichthys Smith, 1865, and Cosmoptychius Traquair, 1877 and many other actinopterygians. Supraorbital bones are lacking.
The parietal bone (Figs.2b, 3, and 4; PA) is a paired and subquadrangular bone, about 1.5 times wider than long and 2.5 times shorter than the frontal. The lateral border is twice as long as the medial. Its surface is smooth, except near the anteroventral border, where there are conspicuous cuneiform striae. In the central portion of the bone there is a pair of pit-lines: an arciform transverse (Figs.2b, 3; t.pl.) and, a posterior one, running longitudinally in relation to the main body axis (Fig.2b; p.pl.), almost at right angle with the first. An anterior pit-line is lacking.
The parietal meets the dermopterotic laterally. Posteromedially, it joins the medial extrascapular and posterolaterally, the lateral extrascapular.
In Cheirolepis and in most of the sarcopterygian fishes, the parietal is a long bone, about the same length as the frontal. This condition is considered primitive to ray-finned fishes (POPLIN & LUND,
1995; ARRATIA& CLOUTIER, 1996). Angatubichthys
and most of advanced actinopterygian fishes present derived condition.
The dermosphenotic (Figs.2b, 3, and 4; DSPH) is a small triangular bone that forms the posterodorsal limit of the orbit. The loop of the temporal sensory canal (otic branch) runs along the bone towards the postorbital. In most of the primitive actinopterygians there is an ypsiloid dermosphenotic. In Angatubichthys, the long orbital branch and the posterior process are lacking.
Medially, the dermosphenotic bone meets the frontal and posteriorly, the dermopterotic. In Pz.DBAV.UERJ 061 there is a gap between the dermosphenotic and the jugal, which is occupied by a postorbital bone.
The dermopterotic (Figs.2b, 3-4; DPT) is a subrectangular bone approximately three times as longer than wide. It forms the main part of the lateral border of the skull roof. The longitudinal margins are approximately parallel, except for the anterior third where the bone is wider and shows a crenulate border to the dermosphenotic and the frontal bones. A small space anteroventrally placed above the dermohyal is interpreted as a spiracular region. The temporal sensory canal pierces the bone along its longitudinal axis close to the lateral border.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

538 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
Direct evidence from embryology of certain extant actinopterygians (e.g., PoZi/pferus Lacepède, 1803; Polyodon Lacepède, 1797; Acipenser Linnaeus, 1758; and Arma Linnaeus, 1766) shows that fusion of intertemporal with supratemporal occurs during early ontogeny (PEHRSON, 1922, 1940, 1944, 1947; JOLLIE, 1980, 1984; GRANDE & BEMIS, 1998) and this condition is considered derived for basal actinopterygians (GARDINER & SCHAEFFER, 1989). Based on that, a compound dermopterotic bone occurs in Angatubichthys.
There is a pair of subcircular extrascapulars, median and lateral, lying on the posterior limit of the skull roof. They are of same size, laminate and ílat. The lateral extrascapular (Figs.2b, 3-4; LEXS) bears a tripartite outline due to the passage of sensory canais. The main sensory canal of the lateral line (Figs.2b, 3-5; l.l.s.c.) pierces this element producing a supratemporal commissure (Fig.2b; st.c.). It pierces the bone transversely Crossing over to the medial extrascapular. Anteromedially, the lateral extrascapular meets the curved posterolateral border of the parietal bone and anterolaterally the dermopterotic. The lateral extrascapular also covers the anterolateral margin of the post-temporal bone.
Anteriorly, the medial extrascapular (Fig.4; MEXS) meets the parietal bone and laterally, the lateral extrascapular. Posteriorly, it overlapps the medial portion of the post-temporal bone.
The parasphenoid (Figs.2b, 4; PAS) is the only basicranial bone preserved. It is partially preserved in lateral view and can be seen within the orbit of the Pz.DBAV.UERJ 59 forming the floor of the anterior braincase. Laterally, it has a wing-shaped process that gradually tapers forwards. In the base of the ascending process there is a longitudinal groove for the pseudobranchial efferent artery (Fig.2b; e.ps.a.) as described by GARDINER (1973; fig.5, p. 115; 1984) in Mimia toombsi Gardiner & Bartram, 1977 of the Gogo Formation, Australia.
The anteriormost portion of parasphenoid is laterally covered by the pars autopalatina of the palatoquadrate. The rest contacts the medial lamina of the endopterygoid bone.
There are fragments of an interorbital bony septum extending towards the parasphenoid in the posterodorsal corner of the orbit in Pz.DBAV.UERJ 059 and 060.
The anteriormost element of the circumorbital series, the lachrymal (Figs.2b, 3 LA), is a long and slightly arched bone occupying the
anteroventral corner of the orbit. Anteriorly, it is dilated, while slender rear.
The infraorbital sensory canal (Figs.2b, 3; iorb.s.c.) runs longitudinally through the median portion of the lachrymal. It is recognised on the surface of the bone by a delicate tubular inflated area and by the presence of at least three exiting pores.
Anteriorly, the lachrymal meets the rostropremaxilloantorbital and posteriorly the jugal. The ventral border of lachrymal lies on a depression on the dorsal margin of the maxilla, hence the dorsal margin of the maxilla is partially covered.
The second element of the circumorbital series, the jugal (Figs.2b, 4; JU), is a crescent-shaped bone occupying of about1 /4 of the posteroventral corner of the orbit. Its membranodermal component is large and laminate. The infraorbital sensory canal pierces the bone close to the orbital rim.
The jugal meets the postorbital dorsally and posteriorly, the dorsal and ventral suborbitals.
The third element, the postorbital (Fig.4; PO) is a small, roughly trapezoidal bone only preserved in Pz.DBAV.UERJ 61. This bone forms with jugal, the posterior limit of the orbit. The infraorbital sensory canal pierces it dorsoventrally. The postorbital also occurs in sarcopterygians (SCHULTZE, 1993) and some basal actinopterygians [e.g., Pteronisculus
magnus Nielsen, 1942; see NIELSEN, 1942). The bone preserved in Angatubichthys is homochiral to that of Pteronisculus White, 1937.
The suborbitals (Figs.2b, 3-4; SO) are two anamestic dermal plates lacking ornamentation. The dorsal suborbital is a guttate and laminate bone. Anteriorly, it abuts the postorbital; ventrally, the ventral suborbital, and posteriorly, the preoperculum. The ventral suborbital is an ovoid bone, with a convex posterior border and anterior border almost straight. Anteriorly, it meets the jugal, ventrally the maxilla and posteriorly the preoperculum. Two suborbital bones occur in the most primitive actinopterygians [e.g., Mimia
Gardiner & Bartram, 1977; Moythomasia Gross, 1950), showing what it seems to be the primitive condition (GARDINER & SCHAEFFER, 1989).
There are four partially preserved sclerotic bones in the orbital region (Fig.3; scl.b.) of examined specimens. As in other paleoniscoid fishes, they are flimsy and arched structures.
The dermohyal (Figs.2b, 4; DH) is a triangular and elongate dermal bone, lying obliquely between preoperculum and operculum. It covers the lateral
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 539
face of hyomandibula near to the spiracular region. Apparently, the primitive condition for basal actinopterygians is to bear two dermohyals (or two antopercular bones) (GARDINER & SCHAEFFER, 1989). In this case, Angatubichthys presents derived condition in relation to Cheirolepis and Moythomasia.
The maxilla (Figs.2b, 3-4; MX) is a large bone, laminate and triangular. As in other paleoniscoid fishes it has a shallow and elongate anterior suborbital portion and a high and well-developed postorbital portion. Posteriorly, it is very inclined, covering part of the posterior end of the lower jaw. The oral margin contains laniariform teeth in the outer row and apparently delicate conical teeth in an inner row.
There is in Pz.DBAV.UERJ 059 a very reduced longitudinal sensory canal on the maxilla lying beneath the lachrymal bone. It is interpreted herein as a maxillary canal (Fig.2b; mx.c.) because of its resemblance with the feature occurring in the semionotid Lepidotes elvensis Blainville, 1818 (THIES, 1989).
The preoperculum (Figs.2b, 3-4; POP) is a malleiform bone consisting of a dorsal and well- developed subhorizontal arm and a ventral and slender vertical arm. The morphological pattern is similar to that of primitive actinopterygians such as Mimia and Moythomasia (see GARDINER, 1984).
The dorsal arm is projected towards the orbit. Anterodorsally, it meets the dorsal suborbital and below the ventral suborbital. At the levei of the posterodorsal angle of the maxilla, the preoperculum becomes narrow and vertical. This vertical arm ends before the posteroventral angle of maxilla. The preopercular sensory canal (Figs.2b, 3-4; pop.s.c.) runs close to the posterior margin of the bone. The subhorizontal arm of the preoperculum meets the dermohyal posteriorly, and, in part, the operculum. The posterior margin of the vertical arm articulates throughout its extension with the suboperculum.
The operculum (Figs.2b, 3-4; OP) is a sub- rectangular bone. The anterior border is straight and contacts the dermohyal. The posterodorsal margin is convex and the ventral margin shows a sigmoidal contact with the suboperculum. It partially covers the anterior margin of presupracleithrum, supracleithrum, and part of the lateral border of the post-temporal.
The suboperculum (Figs.2b, 3-4; SOP) is roughly trapezoidal and slightly larger than the operculum.
The posteroventral margin is more developed, covering most of the vast smooth inner lamina of cleithrum and meeting the posteriormost branchiostegal ray ventrally.
There are six branchiostegal rays (Figs.2b, 3-4; BRR) lying in series below angular and dentalosplenial. Ahead of the anteriormost branchiostegal ray there is a pair of guiar plates. The lateral guiar (Fig.3; LG) is triangular and slender in lateral view. An imprint in front of the lateral guiar is interpreted as the median guiar (Fig.3; MG).
The dentalosplenial (Figs.2b, 3-4 DSPL) is the largest bone of lower jaw. It is an elongate and shallow bone preserved with the ypsilateral angular, forming with it the outer surface of the lower jaw. Oral margin is almost straight and the ventral border slightly convex. Its greatest depth corresponds to about */ of its length. Its anteriormost end is slightly rounded.
There are laniariform teeth (Figs.2b, 4; t) along the oral border of the dentalosplenial. In Pz.DBAV.UERJ 058A, interpreted as ayoung fish, apparently there is a double series of teeth. The inner series appears to consist of very small conical teeth, while more acuminate teeth form the outer.
The dentalosplenial is densely ornamented on the outer surface with bands apparently corresponding to the sinuous accretion of ganoin. The mandibular sensory canal (Figs.2b, 3-4; md.s.c.) runs across the bone close to the ventral margin. From that, spaced and minute pores exit on surface.
The angular (Figs.2b, 3-4; ANG) is a fistuliform dermal bone that forms the posteroventral margin of the lower jaw. It typically carries in its midline the posterior section of the mandibular sensory canal.
The mandibular suspensorium is roughly triangular, large, and somewhat inclined. It is approximately as long as the maxilla. Considering shape and arrangement of preoperculum, the articular region between hyomandibula and otic endocranium is almost vertical. In Pz.DBAV.UERJ 059, the quadrate portion of palatoquadrate was partially preserved behind suboperculum. This portion appears to possess a large and convex articular area to lower jaw as in the acrolepid Namaichthys schroederi Gardiner, 1962. The contact zone with the lower jaw lies just behind the orbit. The design of the bones and their relationship with the parasphenoid follow the
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

540 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
pattern in Australosomus kochi Nielsen, 1949 (see NIELSEN, 1949).
The pars autopalatina (Fig.2b; AUP) of palatoquadrate is slender and narrow. Its anteriormost end is capitate. Most of the medial border abuts against the endopterygoid (Fig.2b; ENPT) that appears through the orbital space, as a narrow and elongate bone.
A dentigerous plate with pointed teeth meeting the ventral face of the pars autopalatina is interpreted as a dermopalatine (Fig.3b; DPAL). It is similar to that described by GARDINER (1962) in Namaichthys schroederi.
In Pz.DBAV.UERJ 060 the articular area of the palatoquadrate is poorly preserved, but among bone remains there is a nodular bone between ventral extremity of the vertical branch of preoperculum and the dorsal end of angular tentatively interpreted as a sympletic (S?, Fig.3). The presence of this bone among early actinopterygians is questioned by certain authors (GARDINER & SCHAEFFER, 1989) that claim this is a synapomorphy of Neopterygii. However, other [e.g., OLSEN, 1984; VÉRAN, 1988) have been surveying anatomical evidences that indicate its early occurrence among primitive actinopterygians.
A small bone identified as quadratojugal (Figs.2b, 3-4; QJ) lies between the end of the vertical arm of the preoperculum and the posteroventral border of the maxilla. It is a triangular bone with a sharp dorsal extremity and a large base bearing conspicuous vertical pit-line (Figs.2b, 3; v.pl.). A vertical pit-line on quadratojugal was considered a relevant feature by BRITO (1997) to recognize that bone in various lineages of actinopterans.
The following bones of the hyobranchial apparatus are preserved: dermohyal (seen above), ceratohyal, hypohyal, basibranchiai, and hypobranchiai.
The hypohyal (Fig.2b; HP) and the ceratohyal (= neocerato-hyal of VÉRAN, 1988; Fig.2b, CH) are partially preserved in lateral view in Pz.DBAV.UERJ 059. The first is a small triangular bone, slightly arched, with the round extremities synchondrally joined with the basibran chiai plate. The second is seemingly a slender and long bone, but only its proximal capitate end can be seen.
Conceming the branchial apparatus, a basibranchial plate (Fig.2b; BBR) and the first hypobranchial (Fig.2b; HBR1) are preserved, in lateral view, in the gape of Pz.DBAV.UERJ 059. Both bones are somewhat elongate, the basibranchial being a wide and depressed plate and the hypobranchial slightly arched in the proximal portion.
The post-temporal (Figs.2b, 3-4; PT) is an ovoid and laminate dermal bone. The main sensory canal of the lateral line runs along a reduced portion of bone close to ventral margin. Anteroventrally, it is joined with the presupracleithrum and dorsoventrally with supracleithrum.
The presupracleithrum (Figs.2b, 3-4; PSCL) is preserved in all specimens. It is a small triangular bone lying on the anterodorsal corner of supracleithrum. Typically, it belongs to the pectoral girdle (see BJERRING, 1985) and not to the spiracular series as argued by certain authors {e.g.,
LEHMAN, 1966). It occurs homoplastically in certain taxa of primitive actinops such as Pteronisculus White, 1937, Paramblypterus
Sauvage, 1888, Watsonulus Piveteau, 1934, Boreosomus Stensiõ, 1925 [passim LEHMAN, 1966).
The supracleithrum (Figs.2b, 3; SCL) is spatulate, well ossified and deep (1.8 times the height of the frontal and 1.2 times the height of the operculum). Ventrally, it is articulated with the dorsal process of the cleithrum. The bone is partially covered by operculum and suboperculum and is densely ornamented with radial ridges on the anterior margin.
The main sensory canal of the lateral line crosses the supracleithrum obliquely, running from the anterodorsal region to the median point of the convex posterior margin of the bone.
The cleithrum (Figs.2b, 3-4; CL) is the largest dermal bone of the pectoral girdle. It is slightly sigmoid and typically bears a well-developed spine-shaped dorsal process. The portion below the spine-shaped process corresponding to the externai lamina of the cleithrum is arched and ornamented with radial ridges. This rounded prominence covers the radiais and the proximal portion of the lepidotrichia of the pectoral fin. It projects medially into a concave margin that is covered by the suboperculum. Its posterior margin is sinuous, convex below the spine- shaped process and concave ventrally. This last portion projects backwards in an outer lamina. The anteriormost part of cleithrum contacts the clavicle through an obliqúe suture.
The clavicle (Figs.2b, 3-4; CLA) is triangular and projected forwards; it is partially hidden by the posterior corner of lower jaw and branchiostegal rays.
In primitive actinopterygians the clavicles are in close contact along the ventral midline. A narrow interclavicle is distinguished behind gulars (LUND, POPLIN 86 McCARTHY, 1995). In Angatubichthys
we were not able to determine the presence of an interclavicle, probably because it is hidden by the
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 541
clavicle in all specimens examined.
The pectoral fin is directly associated with the cleithrum. It is relatively long, and the largest ray reaches the eighth scale row counting from the posterior margin of the cleithrum. It is approximately twice as long as the pelvic fin. It carries at least ten segmented and distally ramified rays. A reduced number of fin-rays (<25) in the pectoral fin is considered derived condition for basal actinopterygians (TAVERNE, 1997). Small and sharp fringing fulcra (Fig.4; fr.f.) are present along the leading edge of the fin.
The pelvic girdle is not visible. The pelvic fin is very small and triangular. Its base is relatively short; the length of the anterior margin corresponds to the half the length of anterior margin of anal fin. We counted at least 12 segmented rays. The pelvic fin is preceded by a modified subcircular scute.
The dorsal fin is triangular and moderate in size. The anterior margin is equivalent to the anterior border of the anal fin. The base of dorsal fin is approximately equal to the length of the anterior margin of pelvic fin. We count 21 segmented and distally ramified rays. The largest one bears about 11 articulate segments. Fringing fulcra occur on anterior border of fin preceded by at least three basal sagitate scutes. It was not possible to observe
any endoskeletal supports for the dorsal fin because it is hidden by rows of scales.
The anal and dorsal fins are placed at the posterior half of the body; the base of the anal fin is 1.5-2 times longer than the anterior margin of the pelvic fin. It carries at least 31 segmented and distally ramified rays. The anterior margin is covered with small fringing fulcra and the base is preceded by a small basal sagital scute. Support elements were not observed.
The caudal fin (Fig.5) bears at least 50 distally ramified and segmented rays. In its dorsal margin there are 13-16 epaxial fulcra (Fig.5; ep.f.). They are long rod-like structures that progressively decrease in size toward the tail end. In the caudal base there are three basal sagitate scutes (Fig.5; ba.s.).
In the base of ventral hypochordal lobe there are two basal scutes and a series of small fringing fulcra (= “anterior lepidotrichia” of PEARSON & WESTOLL, 1979; fr. f., Fig.5).
The body is entirely covered with ganoid scales of smooth surface. In epaxial area scales are rhombic and equilaterals (4 longitudinal rows starting from back), the same for hypoaxial area (3 longitudinal rows up from abdômen). At the midline of body scales are higher than longer decreasing gradually in size backwards.
Fig.5- Angatubichthys mendesi gen. et sp.nov., caudal skeleton of holotype (Pz.DBAV.UERJ 59). Scale bar = lcm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

542 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
The row of scales immediately above and two immediately below the lateral line are similar in size. Behind the pelvic fin the scales that compose these three rows gradually become equilateral toward hinge line, where they are small and elongated.
The lateral line scales are crossed at midline by the sensory canal which is totally enclosed. Their anterior and posterior borders show a moderately developed notch. There are about 38-41 middle flank scales, at least 30 of which are from lateral line. Anteriormost they are rhombic and deep and more equilateral near caudal peduncle.
Posterior to the hinge line there is a gradual change of direction and shape of scales in relation to the main longitudinal body axis (Fig.5). They become inclined and lozenge-shaped.
There are about 24 modified sagital dorsal fin scales in the back, from the occiput to the origin of the dorsal fin.
DISCUSSION AND CONCLUSIONS
Angatubichthys mendesi and other South American palaeoniscoid fishes
Comparisons between Anagatubichthys mendesi
and palaeoniscoid fishes previously described from Brazil on the basis of either nearly complete or well- preserved specimens, indicate that Tholonotus
braziliensis (?Acrolepididae) shares an amount of general similarities. However, it can be distinguished from Angatubichthys by the presence of the following features: an arched dorsal profile (semionotid-like); large frontais with narrow anterior portion and excluded from the orbit; small postrostral; dermopterotic obliquely truncated in its anterior portion, but with a posterolateral expansion; dermosphenotic covering the orbit and contacting the nasal; operculum larger than suboperculum; weak or absent dentition; pectoral and anal fins with 17 rays; dorsal fin placed in the middle of the body and anterior to anal fin; and scales ornamented with wrinkles.
Tholonosteon santacatarinae Beltan, 1978, is a large palaeoniscoid from the Upper Carboniferous of the State of Santa Catarina, about which few anatomical structures relevant to systematics are known, mainly due to the fact that skull is lacking. But this species can be easily distinguished from Angatubichthys mendesi by suitable features such as: an arched dorsal profile; 36 soft rays in pectoral fin; dorsal fin with base longer than deep, rising in
the middle of the body and composed of 25 rays; caudal fin deeply forked and bearing 68 rays, besides the presence of 20 dorsal scutes and scales along the body ornamented with delicate longitudinal rugae.
Additionally to describing Tholonosteon santacatarinae,
BELTAN (1978) reported the occurrence of various species of palaeoniscoid fishes from the Upper Carboniferous of Uruguay based on partially complete specimens. Among those taxa, Gondwanichthys
maximus Beltan, 1978 (Acrolepididae) and Carbinolepis
uruguayensis Beltan, 1978 (Acrolepididae) promptly differ from Angatubichthys for their supranumerary extra-scapular and grooved scales; Mesonichthys
antipodus Beltan, 1978 (Acrolepididae), for a fenestra exonarina anterior higher on snout and operculum larger than suboperculum; Elonichthys macropercularis
Beltan, 1978 (Elonichthyidae), for the presence of ornamented scales, oval rostral and large number of branchiostegal rays (15); Rhadinichthys rionigher
Beltan, 1978 (Rhadinichthyidae), for a dermosphenotic contacting the nasal and scales with parallel grooves; Itararichthys microphthalmus Beltan, 1978 (Pygopteridae), for a reduced orbit, a single sclerotic bone, and supranumerary extrascapulars; Daphnaechelus formosus Beltan, 1978 (Amblypte ridae), for skull higher than long, small or lacking rostral, operculum larger than suboperculum, and triangular and small parietal.
Furthermore, BELTAN (1989) erected two new monotypical palaeoniscoid families: Monesedei phidae and Irajapintoseidae. The first includes Monesedeiphus Beltan, 1989, an aberrant palaeoniscoid recognised mainly for the presence of 15 extrascapulars and 12 branchiostegal rays, five suborbitals, and skull roof bones ornamented with tubercles and rugae of ganoin. The family Irajapintoseidae includes Irajapintoseidon Beltan, 1989, that possesses a robust postrostral with V- shaped posterior suture, nasal in contact with the dermosphenotic, operculum twice as large as suboperculum, 12 extrascapulars, and scales ornamented. Therefore, both differ considerably from Angatubichthys mendesi in various diagnostic anatomical features.
Santosichthys mafrensis Malabarba, 1988 (?Elonichthyidae) is known mainly from a skull cast. It differs from Angatubichthys mendesi mainly by the presence of a prominent snout; narrow frontais ornamented with rugae; long dermopterotic forming a digitate projection towards the frontal; well-developed trapezoidal dermosphenotic forming the posterodorsal limit of the orbit; narrow and
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 543
slender maxilla; presence of supraorbital bone; high number of branchiostegal rays (16); and rhomboid scales ornamented with wrinkles and indented posterior border.
RICHTER (1991) pointed out the occurrence of eight new palaeonisciform fishes mostly from Santa Catarina State, not yet formally described. Examining restorations made by her, we are unable to find any remarkable similarities with Angatubichthys mendesi The pattern of the dermal bones skull, particularly the non-contacting condition between nasal and dermosphenotic is enough to separate these fishes from Angatubichthys.
COX & HUTCHINSON (1991) pointed out the presence of an early actinopterygian fish to the Late Permian of Pedra do Fogo Formation: Brazilichthys
macrognathus Cox & Hutchinson, 1991. It is represented by a poorly preserved cranium which can be distinguished from that of Angatubichthys
by the presence of skull bones ornamented with ridges and tubercles, large maxilla with convex orbital portion, two rows of large teeth on maxilla and mandible, the inner row bearing stout and spaced fang-like teeth; unusual premaxilla, and nasal contacting dermosphenotic bone.
VEGA, DIAS & RAGONHA (1997) registered the occurrence of palaeonisciform fishes for the Rio do Rasto Formation, Permo-Triassic of the Paraná Basin, district of Santo Antônio da Platina, State of Paraná. The material consists of seven incomplete fishes preserved on the same bedding plane. In all specimens most of the skull bones are lacking, constraining an accurate Identification. Three different morphotypes were described suggesting new taxa (VEGA-DIAS, DIAS & RICHTER, 2000). A high number of scale rows separate these taxa from Angatubichthys.
BORDAS (1944) reported the occurrence of the following species from the Triassic of Argentina: Pseudobeaconia bracaccinii Bordas, 1944 and Pseudobeaconia elegans Bordas, 1944 (Redfieldiidae), Cleithrolepis cuyana Bordas, 1944 (Cleithrolepididae), and Mendocinichthys
(=Mendocinia) brevis (Bordas, 1944) (Perleidiidae). All these taxa share a deep body and arched dorsal profile as in various so-called “subholostean” fishes, therefore differing from Angatubichthys mendesi.
More recently, RICHTER (2002) described a complete palaeoniscoid fish from Late Permian of the State of Santa Catarina (Rio do Rasto Formation): Rubius
pascoalensis Richter, 2002. The fish possesses an anvil-shaped dermosphenotic meeting the nasal and the intertemporal does not reach the nasal. In addition, the operculum is larger than the suboperculum. These features are enough to separate Rubius Richter, 2002 from Angatubichthys.
Other palaeoniscoid fishes were reported from the Paleozoic of the Paraná Basin. Some remains were tentatively identified as belonging to Acrolepis
Agassiz, 1833 and Elonichthys Giebel, 1848 (BELTAN, 1978; RICHTER, 1985). However, these are based on fragmentary material consisting of isolated bones, scales, and teeth, limiting adequate Identification and comparisons. But, Angatubichtys mendesi, when compared with well known oversea species of Acrolepididae and Elonichthyidae such as Acrolepis sedgwicki Agassiz, 1834 and Elonichthys robisoni Agassiz, 1843, shares only plesiomorphous similarities. Otherwise, the families Acrolepididae and Elonichthyidae, presumably represented in the Brazilian Paleozoic strata, are hitherto paraphyletic assemblages (GARDINER & SCHAEFFER, 1989). Their scales have been reported to get three times higher than long as in the flanks of Angatubichthys mendesi. It is probable that many scales assigned to taxa of these families belong to representatives of several non related groups.
The early actinopterygian scale structure is, in general, conservative (retaining many plesiomorphies). An attempt by ALDINGER (1937) to define groups based on certain patterns of canaliculi distribution and ornamentation have been recently questioned (GARDINER & SCHAEFFER, 1989, SCHULTZE, 1996; but see RICHTER & SMITH, 1995 for a new approach to assess scale features of actinopterygians). We assume that the current knowledge of the morphological diversity of scales is still in its infancy and will not allow a correct assessment of isolated scales of most of taxa.
Taking all of the current information about anatomy and systematics of South American palaeoniscoids there is little doubt that they are poorly known. Probably most of taxa form paraphyletic assemblages waiting for instant review. In addition, there is large amount of missing data to generate noise in a premature cladistic analysis. For similar reasons, Angatubichthys mendesi does not fit any species diagnosis within palaeoniscoid families described for the Paleozoic of África too (BROUGH, 1931; GARDINER, 1962, 1969; JUBB & GARDINER, 1975).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

544 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
Comments on the phylogenetic relationships of Angatubichthys
The most influent approach concerning phylogenetic classification of lower actinopterygian fishes is that by GARDINER 85 SCHAEFFER (1989). In order to tentatively place Angatubichthys
in this system, the characters used by them are discussed independently, but not integrated into a cladistic analysis because of the large amount of missing data.
In this context, based on the available morphological data, Angatubichtys mendesi seems to occupy a more derived position in relation to various actinopterygians below the Chondrostei- Neopterygii divergence. For instance, the presence of dermopterotic bone (synapomorphy for the node H of GARDINER & SCHAEFFER, 1989, Fig.10) places it in a derived position in relation to assemblages such as “Pteronisculus group”, “Boreosomus group”, and “ Watsonichthys group”. A reduced number of branchiostegal rays and a dermosphenotic contacting less than 1/ of the dermopterotic (synapomorphies for the node I of GARDINER & SCHAEFFER, 1989, Fig.10) place it in a polytomy with a clade formed by “Redfieldius
group” + “Hoplolepis group” and another, formed by “Mesopoma group” + (“Aeduella group” + (“Platysomus group” + (“Bobasatrania group” + “Dorypterus group”))). Angatubichthys mendesi
lacks the skeletal apomophies listed for the first clade (node J of GARDINER & SCHAEFFER, 1989, Fig. 12), or for the second (node K of GARDINER & SCHAEFFER, 1989, Fig. 12). It seems to be closer to the group where Canobius is included (“Mesopoma group”).
The “Mesopoma group “ of GARDINER & SCHAEFFER (1989) that includes Mesopoma
Traquair, 1890, Canobius Traquair, 1881 and Styracopterus Aldinger, 1937 is defined by the presence of an almost vertical suspensorium, reduced preoperculum, reduced posterodorsal processes of maxilla, suboperculum subequal or larger than the operculum, and T-shaped dermosphenotic (in Mesopoma and Styracopterus).
No formal taxonomy was given for this group. Despite of general resemblances shared, only the presence of a suboperculum larger than operculum suggests putative relationship of Angatubichthys
with taxa of this clade.
Unfortunately, due to the preservation of the specimens examined, the correspondence between the number of dermal rays and endoskeletal
supports of the fins, very important to clarify the systematic position of a fish as a stem actinopteran or a neopterygian (PATTERSON, 1982; CO ATES, 1999) was inaccessible to direct observation.
More recently, COATES (1999) tested the hypotheses of phylogenetic relationships for lower actinopteygians of GARDINER & SCHAEFFER (1989) including new anatomical data and additional taxa. The trees obtained are similar in various aspects to those of GARDINER & SCHAEFFER (1989). Most of the morphological features used in his analysis were observed in well- preserved specimens showing details of endocranium and brain. Such exceptional preservation is found in relatively few fóssil taxa. Unfortunately, the available morphological data of Angatubichthys permits only comparisons of dermal elements of the skull, scales and fins.
In sum, the phylogenetic relationships of various taxa of lower actionopterygians have not yet been resolved. Considering an array of current phylogenetic approaches (KAZANTZEVA, 1971; KAZANTSEVA-SELEZNEVA, 1981; LONG, 1988; GARDINER & SCHAEFFER, 1989; LUND, POPLIN & McCARTHY, 1995; TAVERNE, 1997; COATES, 1998; 1999), Angatubichthys mendesi appears to be more advanced in relation to so-called basal actinopterygians (e.g., Cheirolepis Agassiz, 1835; Mimia Gardiner & Bartram, 1977; Moythomasia
Gross, 1950; Osorioichthys Casier, 1952; Howqualepis Long, 1988; Polypterus Lacepède, 1803). It is suggested by several derived skeletal features [e.g., absence of plate pineal, presence of dermopterotic and suborbitals, high scales on the flanks, reduced number of branchiostegals and pectoral fin rays). Nevertheless, Angatubichthys
mendesi wants synapomorphies of more advanced actinopterygian fishes of the platysomid and neopterygian clades.
It is hoped that additional informative specimens will allow the phylogenetic position of Angatubichthys and other lower actinopterygians from South America to be refined in the future.
ACKNOWLEDGEMENTS
Thanks to Valería Gallo, Paulo M. Brito, Hugo R. Secioso Santos (UERJ), Gloria Arratia (Museum für Naturkunde der Humboldt Universitát, Berlin), Michael Coates (Chicago University), Martha Richter (British Museum of Natural History, London) and Alex Kellner (Museu Nacional - Rio de Janeiro), for profitable suggestions and criticai
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 545
reading of early drafts of this paper. Denis Serrette (Muséum National d’Histoire Naturelle, Paris) kindly contributed with the photographs.
LITERATURE CITE D
ALDINGER, H., 1937. Permische Ganoidfische aus Ostgrõnland. Meddelelsen om Gronland, Copenhagen, 102(3): 1-392.
ALMEIDA, F.F.M. & MELO, M.S., 1981. A Bacia do Paraná e o vulcanismo mesozoico. In: SÃO PAULO, SECRETARIA DA INDÚSTRIA E COMÉRCIO, CIÊNCIA E TECNOLOGIA, Mapa Geológico do Estado de São Paulo. Escala 1:500 000. São Paulo, v.l, p.46-81.
ARRATIA, G., 1997. Basal Teleosts and Teleostean phylogeny. Paleo Ichthyologica, München, 7:5-168.
ARRATIA, G. & CLOUTIER, R., 1996. Reassessment of the morphology of Cheirolepis canadensis
(Actinopterygii). In: SCHULTZE, H.P. & CLOUTIER, R. (Eds.) Devonian Fishes and Plants of Miguasha, Quebec, Canada. München: Verlag Dr. Friedrich Pfeil, p. 165-197.
BELTAN, L., 1978. Découverte d’une ichthyofaune dans le Carbonifére supérieur d’Uruguay. Rapports avec les faunes ichthyologiques contemporaines des autres régions du Gondwana. Annales de la Societé Géologique du Nord, Lille, 97:351-355.
BELTAN, L., 1989. New Permian Actinoterygian Families from Uruguay. Acta Musei Reginahradecensis S.A., Scientiae Naturales, Praha, 22(1989):79-86.
BJERRING, H.C., 1985. The question of a presupracleithrum in brachiopterygian fishes. Acta Zoologica, Stockholm, 66:171-174.
BORDAS, A.F., 1944. Peces Triásicos de La Quebrada de Santa Clara (Mendozay San Juan). Revista de la Sociedad Argentina de Ciências Naturales, La Plata, 19:453-460.
BRITO, P.M., 1997. Révision des Aspidorhynchidae (Pisces, Actinopterygii) du Mésozoique: ostéologie, relations phylogénétiques, données environnementales et biogéografiques. Geodiversitas, Paris, 19(4) :681-772.
BROUGH, J., 1931. On fóssil fish from the Karoo System, and some general considerations on the bony fishes of the Triassic period. Proceedings of the Zoological Society of London, London, 1931:235-296.
COATES, M.I., 1998. Actinopterygians from the Namurian of Bearsden, Scotland, with comments on early actinopterygian neurocrania. Zoological Journal of the Linnean Society, London, 122:27-59.
COATES, M.I., 1999. Endocranial preservation of a Carboniferous actinopterygian from Lancashire, UK, and the interrelationships of primitive actinopterygians. Philosophical Transactions of the Royal Society of London, London, Series B, 354:435-467.
COX, C.B. & HUTCHINSON, P., 1991. Fishes and amphibians from the Late Permian Pedra do Fogo
Formation of Northern Brazil. Palaeontology, London, 4(3) :561-573.
DIAS, E.V., 1995. Nova espécie de peixe (Actinopterygii, Platysomidae) do Permiano- Triássico da Bacia do Paraná, Brasil. Dissertação de Mestrado, Universidade Federal do Rio Grande do Sul, Instituto de Geociências, Porto Alegre. 88p.
DUNKLE, D.H. & SCHAEFFER, B., 1956. Preliminary description of a palaeoniscoid fish from the Late Palaeozoic of Brazil. Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, 9(l):5-22.
FIGUEIREDO, F.J.; DIAS, E.V. & COSTA-CARVALHO, B.C.M., 1998. New genus and species of Platysomid fish from the Permian of Paraná Basin, Brazil. Journal of Vertebrate Paleontology, Illinois, 18(3/ supplement):41A.
FÚLFARO, V.J., 1970. Contribuição ao conhecimento da Região de Angatuba. DNPM/DGM, Rio de Janeiro, Boletim 253, 83p.
GARDINER, B.G., 1962. Namaichthys schroederi Gürich and other Palaezoic fishes from South África. Palaeontology, London, 5(1):9-21.
GARDINER, B.G., 1969. New palaeoniscoid fish fron the Witteberg series of South África. Zoological Journal of the Linnean Society, London, 48:423-452.
GARDINER, B.G., 1973. Interrelationships of teleostomes. In: GREENWOOD, P.H.; MILES, R.S. & PATTERSON, C. (Eds.) Interrelationships of Fishes. London: Academic Press, p. 105-135.
GARDINER, B.G., 1984. The relationships of the palaeoniscid fishes, a review based on new specimens of Mimia and Moythomasia from the Upper Devonian of Western Australia. Bulletin of the British Museum of Natural History (Geology), London, 37:173-428.
GARDINER, B.G. & SCHAEFFER, B., 1989. Interrelationships of lower actinopterygian fishes. Zoological Journal of the Linnean Society, London, 97:135-187.
GRANDE, L., & BEMIS, W.E., 1998. A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy. An empirical search for interconnected patterns of natural history. Society of Vertebrate Paleontology Memoir 4, supplement to Journal of Paleontology, London, 18(l):l-690.
JOLLIE, M., 1980. Development of head and pectoral girdle skeleton and scales in Acipenser. Copeia, Washington, 1980:226-249.
JOLLIE, M., 1984. Development of the head and pectoral skeleton of Polypterus with a note on scales (Pisces: Actinopterygii). Journal of Zoology, London, 204:469-507.
JUBB, R.A. & GARDINER, B.G., 1975. A preliminary catalogue of identificable fóssil fish material from South África. Annals of the South African Museum, Pretória, 67(11):381-440.
KAZANTSEVA, A.A., 1971. On the taxonomy of the Palaeonisciformes. Trudy Paleontologicheskogo Instituta Akademia Nauk, SSSR, Moskva, 130:160-167.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

546 FJ.FIGUEIREDO & B.C.M.C.CARVALHO
KAZANTSEVA-SELEZNEVA, A.A., 1981. A phylogeny of lower actinopterygians. Voprosy Ikhtiologii Moskva,
21(4):579-594.
LAVINA, E.L., 1991. Geologia Sedimentar e paleogeografia do Neopermiano e Eotriássico
(Intervalo Kazaniano-Scythiano) da Bacia do
Paraná. Ph.D. Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre, 2v., 457p.
LEHMAN, J.P., 1966. Actinopterygii. In: PIVETEAU, J.P.
(Ed.) Traité de Paléontologie. Paris: Masson et Cie.
Éditeus, 4, 3. p. 1-242.
LONG, J.A., 1988. New palaeoniscid fishes from the Late
Devonian and Early Carboniferous of Victoria. Memoirs of the Association of the Australasian
Palaeontologists, Sydney, 7:1-64.
LUND, R.; POPLIN, C. & McCARTHY, K., 1995. Preliminary
analysis of the interrelationships of some Palaeozoic Actinopterygii. In: LEVRIÈVRE, H.; WENZ, S.; BLIECK,
A. & CLOUTIER, R. (Coords.) Premiers vertébrés et vertébrés inférieurs. Paris, 4-9 septembre, 1995,
Geobios, Mémoire spécial, 19:215-220. LUND, R. & POPLIN, C., 1997. The Rhadinichthyids
(Paleoniscoid Actinopterygians) from the Bear Gulch
limestone of Montana (USA, Lower Carboniferous). Journal of Vertebrate Paleontology, Illinois, 17(3):466-486.
MALABARBA, M.C.L., 1988. A new genus and species of stem group actinopteran fish from the Lower Permian
of Santa Catarina State, Brazil. Zoological Journal of the Linnean Society, London, 94:287-299.
MENDES, J.C., 1952. A Formação Corumbataí na região do rio Corumbataí - Estratigrafia e descrição dos lamelibrânquios. Boletim da Faculdade de Filosofia,
Ciências e Letras da Universidade de São Paulo,
São Paulo, 145(8): 1-119. MEZZALIRA, S., 1989. Os Fósseis do Estado de São
Paulo. 2.ed., Secretaria do Meio Ambiente, Instituto Geológico, São Paulo. 136p.
MILANI, E.J.; FRANÇA, A.B. & SCHNEIDER, R.L., 1994.
Bacia do Paraná. Boletim de Geociências da
Petrobrás, Rio de Janeiro, 8(l):69-82. MILANI, E.J.; FACCINI, U.F.; SCHRERER, C.M.;
ARAÚJO, L.M. & CUPERTINO, J. A., 1998. Sequences and Stratigraphic Hierarchy of the Paraná Basin (Ordovician to Cretaceous), Southern Brazil. Boletim
IG - USP, Série Científica, 29:125-173. NIELSEN, E., 1942. Studies on Triassic fishes from East
Greenland. I. Glaucolepis and Boreosomus.
Palaeozoologica Groenlandica, Copenhagen, 1:1-403. NIELSEN, E., 1949. Studies on Triassic fishes from East
Greenland. II. Australosomus and Birgeria.
Meddelelser Groenlandica, Copenhagen, 1(1): 1-309.
OLSEN, P.E., 1984. The skull and pectoral girdle of the parasemionotid fish Watsonulus eugnatoides from
the Early Triassic Sakamena Group of Madagascar, with comments on the relationships of holostean fishes. Journal of Vertebrate Paleontology,
Kansas, 4:481-499. PATTERSON, C., 1982. Morphology and Interrelationships
of Primitive Actinopterygians Fishes. American Zoologist, Washington, 22:241-259.
PEARSON, D.M. & WESTOLL, T.S., 1979. The Devonian actinopterygian Cheirolepis Agassiz. Transactions of
Royal Society of Edinburgh, 70:337-399.
PEHRSON, T., 1922. Some points in the cranial development of teleostomian fishes. Acta Zoologica,
Stockholm, 3:1-63.
PEHRSON, T., 1940. The development of dermal bones in the skull of Amia calva. Acta Zoologica,
Stockholm, 21:1-50.
PEHRSON, T., 1944. Some observations on the
development and morphology of the dermal bones in the skull of Acipenser and Polyodon Acta Zoologica,
Stockholm, 25:27-48. PEHRSON, T., 1947. Some new interpretations of the
skull in Polypterus. Acta Zoologica, Stockholm,
28:399-455. PETRI, S., 2001. As pesquisas paleontológicas no Brasil.
Revista Brasileira de Paleontologia, Rio de Janeiro,
1:9-136. PETROBRÁS, 1998. Geology of the Southeastern
Paraná Basin - Field Guide & Virtual Field Trip
CD-ROM, 1998 AAPG International Conference & Exhibition, Rio de Janeiro, 08-11/novembro, 47p.
POPLIN, C. & LUND, R., 1995. Fates on the rostral, portrostral and premaxillary in the early history of actinopterygians. In: LEVRIÈVRE, H.; WENZ, S.;
BLIECK, A. & CLOUTIER, R. (Eds.) Premiers
vertébrés et vertébrés inférieurs. Paris, 4-9
septembre, 1995, Geobios, Mémoire Spécial,
19:225-230. POPLIN, C. & LUND, R., 1997. Evolution of the
premaxillary in the primitive fóssil actinopterygians.
Geodiversitas, Paris, 19(3):557-565.
RAGONHA, E.W., 1989a. Placas dentárias de Dipnoi do Grupo Passa Dois (P-Tr) da Bacia do Paraná.
Apreciações ambientais, climáticas cronológicas e estratigráficas. Anais do XI Congresso Brasileiro de Paleontologia, Curitiba, PR, 1:195-206.
RAGONHA, E.W., 1989b. Ictiodorulite de base bulbosa.
Evidências de possível processo articulatório. Anais do XI Congresso Brasileiro de Paleontologia,
Curitiba, 1:177-181.
RICHTER, M., 1985. Situação da pesquisa paleoictiológica no Paleozoico brasileiro. Coletânea de Trabalhos Paleontológicos, Sér.Geol. 27,
Brasília, Sec. Paleontologia e Estratigrafia, 2:105-110. RICHTER, M., 1991. A new marine ichthyofauna from
the Permian of the Parana Basin of Southern
Brasil. King’s College London, University of London, PhD Thesis, 173p.
RICHTER, M., 2002. A ray-finned fish (Osteichthyes)
from the Late Permian of the State of Santa Catarina (Paraná Basin), Southern Brazil. Revista Brasileira
de Paleontologia, Rio de Janeiro, 3:56-61. RICHTER, M. & SMITH, M.M. 1995. A microstructural
study of the ganoine tissue of selected lower vertebrates. Zoological Journal of the Linnean
Society, London, 114:173-212. SANTOS, R.S., 1991. Paleonisciformes do Estado de São
Paulo. Resumos do XII Congresso Brasileiro de
Paleontologia, São Paulo, SP, p.ll.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004

ANGATUBICHTHYS MENDESI, AN EARLY ACTINOPTERYGIAN FISH FROM THE LATE PERMIAN OF THE PARANA BASIN 547
SCHNEIDER, R.L.; MUHLMAN, H.; TOMMASI, E.; MEDEIROS, R.A.; DAEMON, R.H.F. & NOGUEIRA,
A.A., 1974. Revisão estratigráfica da Bacia do Paraná. Anais do XXVIII Congresso Brasileiro de Geologia,
SBG, Porto Alegre, RS, p.41-65.
SCHULTZE, H.P., 1993. Patterns of Diversity in the skull of jawed fishes. In: HANKEN, J. & HALL, B.K. (Eds.) The Skull: Patterns of Structural and Systematic
Diversity, v.2. Chicago and London: University of
Chicago Press, p. 189-254. SCHULTZE, H.P., 1996. The scales of mesozoic
actinopterygians. In: ARRATIA, G. & VIOHL, G. (Eds.) Mesozoic Fishes - Systematics and Palaeoecology. Procedings International Meeting, Eischstátt, 1993, München, Verlag Dr. Friedrich Pfeil, p.83-93.
TAVERNE, L., 1997. Osorioichthys marginis,
“Paléonisciforme” du Famennien de Belgique et la
phylogénie des Actinoptérygiens dévoniens (Pisces). Bulletin de LPInstitut Royal des Sciences Naturelles de Belgique, Sciences de la Terre,
Bruxelles, 67:57-78. THIES, D., 1989. Sinneslinien bei dem Knochenfisch
Lepidotes elvensis (Blainville, 1818) (Actinopterygii,
Semionotiformes) aus dem Oberlias (Unter-Toarcium) von Grimmen in der DDR. Neues Jahrbuch für Geologie und Palaontologie, Monatshefte,
1989(11):692-704.
TOLEDO, C.E.V.; BRITO, P.M.M. & BERTINI, R.J., 1997. Chronological meaning about the presence of
petalodonts (Holocephali incertae sedis) in the
Corumbataí Formation of São Paulo State (Brazil). Boletim de Resumos do XV Congresso Brasileiro
de Paleontologia, São Pedro, SP, p.80. VEGA, C.S.; DIAS, E.V. & RAGONHA, E.W., 1997.
Ocorrência de novos Actinopterygii da Formação
Rio do Rasto, Permo-Triássico da Bacia do Paraná, Brasil. Boletim de Resumos do XV Congresso Brasileiro de Paleontologia, São
Pedro, SP, p.87.
VEGA-DIAS, C.; DIAS, E.V. & RICHTER, M., 2000. Actinopterygian remains from the Rio do Rasto Formation, Upper Permian of the Paraná Basin,
Brazil. Acta Geologica Leopoldensia, Porto Alegre,
23(51):21-31.
VÉRAN, M., 1988. Les éléments accessoires de 1’arc
hyoidien des Poissons Téléostomes (Acanthodiens et Osteichthyens) fossiles et actuales. Mémoires du Muséum National d’Histoire Naturelle, Sciences
de la Terre, Paris, 54:1-98. WESTOLL, T.S., 1944. The Haplolepididae, a new
family of Late Carboniferous bony fishes - A study in taxonomy and evolution. Bulletin of the American Museum of Natural History, New York,
83:1-121.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.531-547, out./dez.2004


nnniTOnnnrtnnnni nnniiitiiiMiftfinnni Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004
ISSN 0365-4508
ON A NEW MANIRAPTORAN DINOSAUR (THEROPODA) FROM THE UPPER
CRETACEOUS OF NEUQUÉN, PATAGÔNIA, ARGENTINA 1
(With 41 figures)
JORGE O. CALVO 2
JUAN D. PORFIRI 2
ALEXANDER W. A. KELLNER 3
ABSTRACT: The “Futalognko” site, situated at the margins of the Los Barreales lake about 90 km northwest
of Neuquén city (Patagônia), revealed several new dinosaur specimens, all from the Portezuelo Formation (Turonian-Early Coniacian). Among those are the remains of new theropod dinosaur referable to a new
species of the maniraptoran Unenlagia, Unenlagia paynemüi n. sp. The material is represented by a pubis,
humerus, ilium, and a dorsal vertebra, plus a pedal phalanx and a claw that tentatively referred to this new
species. It shares with Unenlagia comahuensis a posteriorly oriented distai end of the pubis that shows a convex anterior margin and differs by possessing more gracile bones, angle between the anterior rim of the
deltopectoral crest and the humerus shaft more closed, the presence of a small anterior process on the
distai end of pubis, and the distai end of the postacetabular blade of ilium broader and rounded, with a shallower brevis fossa.
Key words: Dinosauria, Maniraptora, Portezuelo Formation, Patagônia, Argentina
RESUMO: Sobre um novo dinossauro Maniraptora (Theropoda) do Cretáceo Superior de Neuquén, Patagônia,
Argentina.
Novos materiais de dinossauros, todos procedentes da Formação Portezuelo (Turoniano-Coniaciano Inferior),
foram encontrados nos últimos anos na localidade denominada de “Futalognko”, situada às margens do lago Barreales a cerca de 90 km noroeste da cidade de Neuquén (Patagônia). Entre estes se destaca uma
nova espécie do maniraptora Unenlagia, denominado aqui de Unenlagia paynemili n. sp. O material é
representado pelo úmero, púbis, ílio e uma vértebra dorsal, além de uma falange e uma ungueal, atribuídos por tentativa a essa nova espécie. A principal característica compartilhada com Unenlagia comahuensis é a
margem anterior convexa do púbis, com a parte distai orientada posteriormente. As principais diferenças
observadas na nova espécie são os ossos mais gráceis, o ângulo entre a crista deltopeitoral e eixo do úmero mais fechado, a presença de uma projeção anterior no púbis, e a porção pós-acetabular do ílio mais larga e
arredondada, com a brevis fossa mais rasa.
Palavras-chave: Dinosauria, Maniraptora, Formação Portezuelo, Patagônia, Argentina
INTRODUCTION
In 2000, the team of the National University of Comahue has discovered an important fóssil deposit about 90km northwest of the Neuquén city (Patagônia), Argentina (CALVO etal, 2002b). Called the Futalognko site, this quarry is placed at Costa Dinosaurio Locality, northern coast of the Barreales lake. Due to the richness of fossils a new Paleontological Center of the National University of Comahue was built there. Today, this complex is named Barreales Lake Paleontological Center
(CePaLB) and consists the first permanent educational paleontological site in South America fully dedicated to field research in vertebrate paleontology.
The fossiliferous beds of the Futalognko site are from the Turonian- Early Coniacian Portezuelo Formation of the Rio Neuquén Group (LEANZA & HUGO, 2001) and yielded several important vertebrate fossils. Among those are titanosaurid sauropods (CALVO et al, 2001, CALVO & GRILL, 2003), new material from the theropod Megaraptor
namunhuaiquii Novas, 1998 (CALVO etal., 2002a;
1 Submitted on April 16, 2004. Accepted on June 4, 2004.
2 Universidad Nacional dei Comahue, Centro Palentológico Lago Barreales, Costa Dinosaurio, Ruta Prov. 51, km 65. Neuquén, Patagônia, Argentina.
E-mail: [email protected].
3 Museu Nacional/UFRJ, Departamento de Geologia e Paleontologia. Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil. E-mail: [email protected] .br
Fellow of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). Associated Researcher - American Museum of Natural History, New York.

550 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
PORFIRI86 CALVO, 2003), theropod teeth (POBLETE 8& CALVO, 2003; VERALLI 8& CALVO, 2003) ornithopods (PORFIRI 85 CALVO, 2002), turtles, osteichthyan fishes (GALLO, CALVO 85 KELLNER, 2003), crocodylomorphs, pterosaurs (KELLNER et al, 2004), and plants (PRÁMPARO etal, 2003).
During the field season of 2002, another interesting material was recovered from this site. It consists of isolated remains, including the left humerus and left pubis that share several features with the maniraptoran Unenlagia comahuensis Novas 85
Puerta, 1997 (CALVO, PORFIRI 85 KELLNER, 2003). The latter was found at the Sierra dei Portezuelo, about 50km south of the Futalognko site (NOVAS 8& PUERTA, 1997). In 2003, one dorsal vertebra and the posterior end of one right ilium, also sharing several features with Unenlagia
comahuensis, were recovered from that site. The purpose of this paper is to report this material, which is referred to a new species of Unenlagia (U.
paynemilin.sp.). An isolated phalanx and ungueal, also recovered from the site, are tentatively referred to the new taxon, which is described here.
GEOLOGICAL SETTING
The Neuquén Basin of western Argentina has deposits ranging from the Upper Triassic to the Tertiary. These deposits include a sequence of marine and continental strata, including the Neuquén Group, which is composed of sediments formed in a continental, fluvial environment that range from Albian to Campanian age (ULIANA 85
DELLAPE, 1981; LEGARRETA 85 GULISANO, 1989; CALVO, 1991; CALVO 85 SALGADO, 1995).
The material of the new species, Unenlagia
paynemili n.sp., comes from continental deposits from the top of the Portezuelo Formation, Rio Neuquén Subgroup, Neuquén Group (CAZAU 85
ULIANA, 1973; LEANZA,1999). The Rio Neuquen Subgroup is composed of two formations, at the base the Portezuelo and at the top the Plottier formations. The Portezuelo Formation is characterized by yellowish sandstones, red and green claystones with a poor participation of conglomerates, which have been interpreted as Turonian - Early Coniacian (LEANZA 8& HUGO, 2001). The depositional environment is interpreted as a meandering river system that runs over a flattened area by an exuberant vegetation in humid climate conditions (SÁNCHEZ etal, 2003). Fossils were recovered from a fine conglomerate and are mostly unarticulated.
SYSTEMATIC PALEONTOLOGY
Theropoda Marsh, 1881 Coelurosauria Gauthier, 1986 Maniraptora Gauthier, 1986
Unenlagia Novas 85 Puerta, 1997
Type-species - Unenlagia comahuensis Novas 85
Puerta, 1997
Unenlagia paynemili n.sp.
Etymology - In honor to Maximino Paynemil, the chief of the Paynemil Indian community that lives in the region of Loma de La Lata, where the Futalognko site is situated.
Holotype - Left humerus and left pubis, regarded as belonging to the same individual, housed at the Museo de Geologia y Paleontologia de la Universidad Nacional dei Comahue under the number MUCPv-349.
Referred specimens - A dorsal vertebra (MUCPv-416) and the posterior end of a right ilium (MUCPv-409). Tentatively, a pedal phalanx (MUCPv-415) and a pedal claw (MUCPv-343) are also referred to this species.
Locality and horizon - Futalognko site at the Centro Paleontológico Lago Barreales situated about 90km northwest of the Neuquén city, Neuquén province, Patagônia, Argentina; Portezuelo Formation (Turonian-Early Coniacian), Rio Neuquén Group.
Diagnosis - Species of Unenlagia distinguishable from Unenlagia comahuensis by possessing more gracile bones, the angle between the anterior rim of the deltopectoral crest and the humerus shaft about 116° (opposed to 140° of Unenlagia
comahuensis), a small anterior process on the distai end of pubis, and the distai end of the postacetabular blade of ilium broader and rounded, and a shallower brevis fossa.
DESCRIPTIONS AND COMPARISONS
Since 2002 a quarry map of the Futalognko site showing the relation of all collected fóssil remains is being made, including the elements attributed to Unenlagia paynemili n.sp., which were found isolated. The pubis and the humerus were lying about two and half meters from each other (close to the remains of the giant sauropod) and are regarded as belonging to the same individual due to their proximity in the quarry and similarity in size when compared to the material of the larger Unenlagia
comahuensis. Furthermore, both bones have a
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 551
distinct brownish color, different from the other sauropod and theropod elements (e.g., Megaraptor
namunhuaiquü) collected at the site. Although the size of the ilium (and the bone color) is also compatible with the humerus and pubis, it was found more distant (ca. 23 meters from the pubis) and is regarded as belonging to a different individual. The last element attributed to Unenlagia paynemili
n.sp. is an incomplete dorsal vertebrae that belongs to a proportionally larger individual (also larger than the type material of Unenlagia comahuensis) and was found in 2003 by surface collecting (probably washed out from the quarry during the flooding of the Barreales lake). Two more bones are described and are tentatively referred to Unenlagia paynemili
n.sp.: a pedal phalanx and an ungual, also found isolated (respectively 5 and 12.5 meters from the pubis). Except for the humerus and pubis, all remaining specimens are numbered separately.
The humerus (MUCPv-349) belongs to the left side and except for some parts of the proximal and distai articulations is complete with a total length of 217mm (Figs. 1-10). It is a long and slender pneumatic bone, with the thickness of the córtex around 2mm. The proximal part is flattened anteroposteriorly. The deltopectoral crest is expanded and restricted to the proximal third of the humerus, a condition common to other maniraptorans. It is thin and inclined about 116 degrees relative to the shaft. The preserved part of the internai tuberosity is anteroposteriorly compressed and longitudinally elongated as in Unenlagia comahuensis, Deinonychus antirrhopus Ostrom 1969, Archaeopteryx lithographica Meyer 1860, and some other maniraptorans (OSTROM, 1969a, b; WELLNHOFER, 1974; 1993; NOVAS & PUERTA, 1997). The shaft is essentially straight and long differing from the bowed condition present in some maniraptorans like Deinonychus antirrhopus
(OSTROM, 1969b). It is slightly compressed anteroposteriorly with an oval cross section over most its length (middle part = 17.5 x 14mm) and gets more triangular closer to the distai articulation. A blunt longitudinal ridge (length 20mm) is observed above the radial condyle (partially preserved) at the dorsal margin close to the posterior surface (Figs. 1-4, 7, 8).
Overall, the humerus of Unenlagia paynemili n.sp. is similar to the one of Unenlagia comahuensis (Figs.9- 10). Both have the proximal part, that includes the deltopectoral crest, strongly deflected relative to the shaft. The deltopectoral crest in those taxa is quadrangular in anterior view, similar to oviraptorids (BARSBOLD, MARYANSKA & OSMÓLSKA, 1990) and most dromeosaurids (OSTROM, 1969a, b), but differs
from the distinct and long deltopectoral crest of alvarezsaurids (e.g., NOVAS, 1996) and the less pronounced condition reported in Archaeopteryx
lithographica (WELLNHOFER, 1974; 1993) and in the Ornithomimosauria (BARSBOLD & OSMÓLSKA, 1990). The humerus of Unenlagia paynemili n.sp. differs from Unenlagia comahuensis mainly by being about 20% smaller and a more gracile element. Furthermore, the angle between the ventral margin of the deltopectoral crest in the new species is 116° while in Unenlagia comahuensis it is wider, about 140° (Figs.9-10). The latter also shows a developed blunt ridge on the dorsal margin posterior to the deltopectoral crest that is not observed in the new species (Figs.9-10).
Unenlagia paynemili n.sp., left humerus (MUCPv-349) in dorsal view - fig. 1- picture; fig.2- drawing. (dpc) deito
pectoral crest, (itub) internai tuberosity, (rad) radial condyle,
(rid) ridge. Scale bars = 50mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004
552 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
Unenlagia paynemili n.sp., left humerus (MUCPv-349) - fig.3- picture, posterior view; fig.4- drawing, posterior view;
fig.5- picture, ventral view; fig.6- drawing, ventral view. (dpc) deito pectoral crest, (itub) internai tuberosity, (rid) ridge. Scale bars = 50mm.
The left pubis (MUCPv-349) of Unenlagia paynemili
n.sp. is almost complete (length: 267mm) while the right one is represented only by a thin layer of bone at the tip of the distai half, forming the pubic boot (Figs.11-19). Overall, the pubis is a slender and comparatively thin bone, with an anteroposteriorly expanded proximal end that is slightly displaced medially (anterior view). In proximal view, the iliac articulation is slightly concave, long and narrow. The shaft has a convex anterior surface, with the distai part directed posteroventrally. In anteroposterior view, the lateral margin is sigmoidal, with the proximal half concave and the distai half convex. The distai end is expanded anteroposteriorly, with both pubes fusing in the midline forming a pubic boot, and showing a small anterior projection (Figs. 11-16). The pubic apron is deflected posteriorly and extends for about 135mm, reaching half the public
shaft (Figs. 13-18). Apparently this flange is too narrow to meet at the midline with the opposite element. Except for the proximal region, most of the pubic flange extends medially from the anterior margin of the shaft.
Although no complete ilium is available for Unenlagia paynemili n.sp., the comparisons with other maniraptorans, particularly Unenlagia
comahuensis indicate that the pubes is positioned vertically. The pubic apron of the new form is extended for about half of the public shaft like in most maniraptorans but differing from the smaller pubic apron diagnostic of the Avialae (NORELL, CLARK 86 MAKOVICKY, 2001). Contrary to some other maniraptorans (e.g., Velociraptor Osborn, 1924), the opposite flange forming the pubic apron apparently does not meet in the midline, a condition also reported in Unenlagia comahuensis (NOVAS 86 PUERTA, 1997).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 553
Unenlagia paynemili n.sp., left humerus (MUCPv-349) - fig.7-
picture; fig.8- drawing. (dpc) deito pectoral crest; (itub) internai tuberosity; (rad) radial condyle; (rid) ridge. Scale bars = 50mm.
The comparisons of the pubes of Unenlagia paynemili
n.sp. with other maniraptorans show that it is more similar to Unenlagia comahuensis, both sharing at least one feature: the posteriorly oriented distai end of the pubis that shows a convex anterior margin (Figs.20-21). Although NOVAS & PUERTA (1997) reconstructed the pubis of Unenlagia comahuensis
in a straight fashion in lateral view, a closer examination of the holotype indicates that the pubis (particularly the left one) had the distai part directed posteriorly as in Unenlagia paynemili n.sp., this character is considered a synapomorphy of Unenlagia. This feature differs from the straight pubes found in most other theropods and from the anteriorly curved distai pubic end of the oviraptorids (BARSBOLD, MARYANSKA & OSMÓLSKA, 1990).
The main difference between the pubes of U paynemili
n.sp. and U. comahuensis is the presence of an anterior component in the pubic boot in the new
species absent in the latter. More basal theropods such as Camotaurus Bonaparte, 1985 (BONAPARTE, NOVAS & CORIA, 1990), Pycnonemosaums Kellner & Campos, 2002 (KELLNER & CAMPOS, 2002), Sinraptor
Currie & Zhao, 1993 (CURRIE & ZHAO, 1993) and many coelurosaurs like the Oviraptorosauria (BARSBOLD, MARYANSKA & OSMÓLSKA, 1990) have the anterior portion of the pubes more developed. Other maniraptorans like Velociraptor also show a small anterior extension of the pubic boot (NORELL & MAKOVICKY, 1999) but differ from Unenlagia
paynemili n.sp. by being dorsoventrally expanded.
In posterior view, the shaft is bowed at the third distai end as in Unenlagia comahuensis, differing from other maniraptorans where the lateral margin is bowed more proximally like Velociraptor
mongolensis Osborn, 1924 (NORELL & MAKOVICKY, 1999); or straight like Achillobator giganticus Perle, Norell 85 Clark, 1999 (PERLE, NORELL 85 CLARK, 1999j; Tyrannosaurus rex Osborn, 1905 (OSBORN, 1905; BROCHU, 2003j; Aviminus portentosus
Kurzanov, 1981; Coelurosauria indet. (MARTILL et
al, 2000). This character is just present in Unenlagia
comahuensis and Unenlagia paynemili n.sp. and it might be another synapomorphy uniting those taxa.
Only the posterior portion of the right ilium (MUCPv-409) was found (length of preserved portion: 103mm), including the posterior part of the acetabulum (Figs.22-31). In dorsal view, the dorsal margin of the preserved part is sigmoid, with the posterior end displaced laterally (Figs.24-25). The postacetabular blade forms a thin lamina of bone that shows a strong lateral concavity corresponding to a medial convexity. Overall it is short and acuminate, showing a marked inflection on the dorsal margin posterior to the acetabulum. The lateral surface is smooth. Close to the dorsal margin before the beginning of the inflected border of the postacetabular portion a rugose area is present; two distinct knob-like structures are present, possibly for muscle attachment. Some weakly grooves and ridges, likely for muscle scars, are also present at the most posterior end (on the lateral and medial surfaces). Ventrally, the brevis fossa is narrow and shallow. Medially a marked ridge forms the medial shelf of the brevis fossa that is directed posterodorsally running divergent from the ventral margin of the postacetabular blade. A rounded depression is present above the posterior part of the acetabulum. The preserved surface of the acetabulum forms a rugose surface that broadens posteriorly, ending in a thick posterior edge (“lip”) that is part of the ischiadic peduncle.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

554 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
Comparison of the proximal part of the left humerus of Unenlagia comahuensis (MCF PVPH 78 - left) and Unenlagia
paynemili n.sp. (MUCPv-349 - right) - fig.9- picture, fig. 10- drawing. (dpc) deito pectoral crest, (itub) internai tuberosity, (rid) ridge. Scale bars = 50mm.
Comparing the preserved part of the ilium with other coelurosaurs, Unenlagia paynemili n.sp. shows the postacetabular part of the ilium acuminate, a feature considered synapomorphic of the Maniraptora (NORELL, CLARK & MAKOVICKY, 2001) that contrasts to the more square outline observed in other theropods such as Sinraptor
(CURRIE & ZHAO, 1993). The brevis fossa is shelf- like, a common feature within Maniraptora. Among coelurosaurs, Unenlagia paynemili n.sp. shares one feature, at the ilium, with Unenlagia
comahuensis - the marked inflection of the dorsal margin of the postacetabular blade that is here considered a diagnostic feature of the genus Unenlagia. The main differences between those species are the more rounded (and less) acuminate posterior edge and the shallower brevis fossa of Unenlagia paynemili n.sp. that also has a narrower and more open posterior portion of the acetabulum. Furthermore Unenlagia comahuensis shows a well developed ridge dividing the brevis fossa that is absent in Unenlagia paynemili n.sp.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004
ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 555
Unenlagiapaynemilin.sp., left pubis (MUCPv-349) - fig. 11- picture, lateral view; fig. 12- drawing, lateral view; figl3- picture, anterior view; fig. 14- drawing, anterior view. (pro) bone projection, (puap) pubic apron, (pub) pubic boot. Scale bars = 50mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

Unenlagia paynemili n.sp., left pubis (MUCPv-349) - fig.15- picture, medial view; fig.16- drawing, medial view; fig.17- picture, posterior view; fig.18- drawing, posterior view. (pro) bone projection, (puap) pubic apron; (pub) pubic boot.
Scale bars = 50mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004
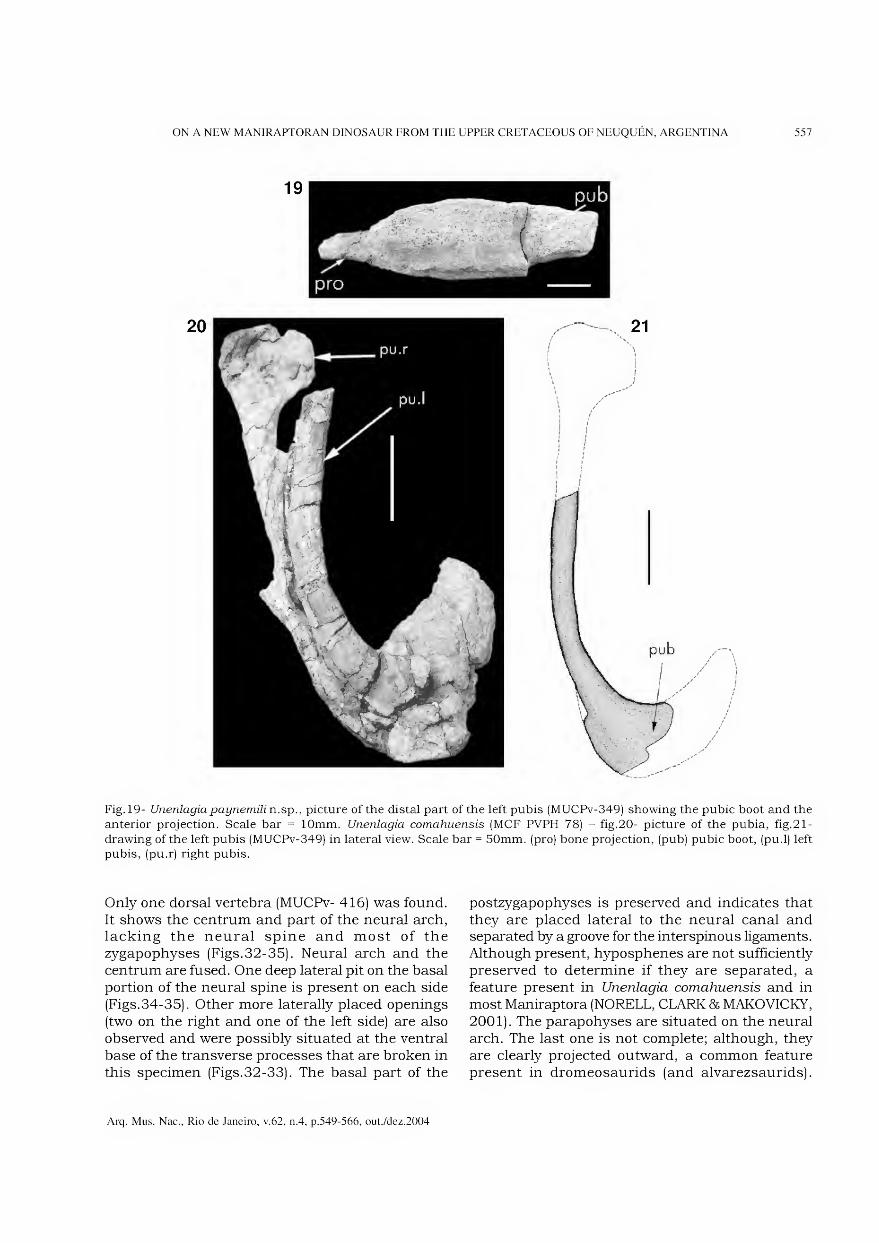
ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 557
Fig. 19- Unenlagiapaynemili n.sp., picture of the distai part of the left pubis (MUCPv-349) showing the pubic boot and the anterior projection. Scale bar = 10mm. Unenlagia comahuensis (MCF PVPH 78) - fig.20- picture of the pubia, fig.21-
drawing of the left pubis (MUCPv-349) in lateral view. Scale bar = 50mm. (pro) bone projection, (pub) pubic boot, (pu.l) left pubis, (pu.r) right pubis.
Only one dorsal vertebra (MUCPv- 416) was found. It shows the centrum and part of the neural arch, lacking the neural spine and most of the zygapophyses (Figs.32-35). Neural arch and the centrum are fused. One deep lateral pit on the basal portion of the neural spine is present on each side (Figs.34-35). Other more laterally placed openings (two on the right and one of the left side) are also observed and were possibly situated at the ventral base of the transverse processes that are broken in this specimen (Figs.32-33). The basal part of the
postzygapophyses is preserved and indicates that they are placed lateral to the neural canal and separated by a groove for the interspinous ligaments. Although present, hyposphenes are not sufficiently preserved to determine if they are separated, a feature present in Unenlagia comahuensis and in most Maniraptora (NORELL, CLARK & MAKOVICKY, 2001). The parapohyses are situated on the neural arch. The last one is not complete; although, they are clearly projected outward, a common feature present in dromeosaurids (and alvarezsaurids).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

558 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
28
Unenlagia paynemili n.sp., right ilium (MUCPv-409) - fig.22- picture, lateral view; fig.23- drawing, lateral view; fig.24-
picture, dorsal view; fig.25- drawing, dorsal view; fig.26- picture, medial view; fig.27- drawing, medial view; fig.28- picture,
ventral view; fig.29- drawing, ventral view. (bfos) brevis fossa, (isp) ischiadic peduncle; (knb) knob-like protuberance; (mus) muscle scars, (rid) ridge. Scale bars = 30mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 559
Picture comparing the posterior part of the right ilium of Unenlagia comahuensis (MCF PVPH 78 - top) and Unenlagia paynemili n.sp. (MUCPv-409 - bottom) - fig.30- lateral view, fig.31- medial view. (rid) ridge. Scale bars = 30mm.
The centrum is amphiplatyan, with the anterior articulation surface rounded and the posterior surface slightly wider than higher. Close to the contact with the neural arch, one pleurocoel followed by a small depression is present on each side. A shallow groove followed by a small foramen is present of the ventral side of the centrum.
The most interesting feature of the MUCPv- 416 is the presence of a lateral pit close to the base of the neural spine, which has been reported in the posterior dorsais of Unenlagia comahuensis (NOVAS & PUERTA, 1997). Other dorsais of the holotype of this species, however, do not show this feature (pers. obs.)
A complete pedal phalanx (MUCPv-415) was also found in the same quarry and is tentatively referred to Unenlagia paynemili n.sp. (Figs.36-37). The specimen is complete likely represents the first phalanx of the right digit I. The distai condyles are asymmetric and are separated by a marked groove. The lateral condyle shows a developed sub-circular fossa (possible for ligaments) while in the medial condyle this fossa is less developed. Proximally the articulation with the metatarsal is formed by two asymmetric surfaces separated by a blunt ridge, with the lateral one larger than the medial. Dorsally a bony ridge is positioned on the medial side. It
extends about one third of the total length of the phalanx, and it was likely the attachment surface for a muscle or ligament. An ungueal phalanx (MUCPv-343) is also tentatively referred to Unenlagia paynemili n.sp. (Figs.38-39). Itis strongly curved and laterally compressed, 65mm long on the dorsal border, and about 29mm high at the proximal end. On each side there is a groove that runs on the middle part of the claw, starting proximally close to the basal portion, reaching the dorsal margin close to the tip. Two additional parallel sulcus are present on one side. The upper sulcus runs from the proximal region, near the articular facets up to the middle groove. The lower sulcus starts at the base of the middle groove and reaches the ventral region of the claw anterior to the flexor tubercle. The ventral rim is lateroventrally rounded. It presents a flexor tubercle well developed placed below the articular facets, the latter being asymmetrical.
The lateral compression and the curvature of the Unenlagia paynemili n.sp. claw resemble that present in digit I of Rahonavis Forster, Sampson, Chiappe, & Krause, 1998. Although no foot is known from Unenlagia
comahuensis, it should be noted that FORSTER et cã.
(1998) suggest that Unenlagia comahuensis should have had a similar claw to Rahonavis.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004
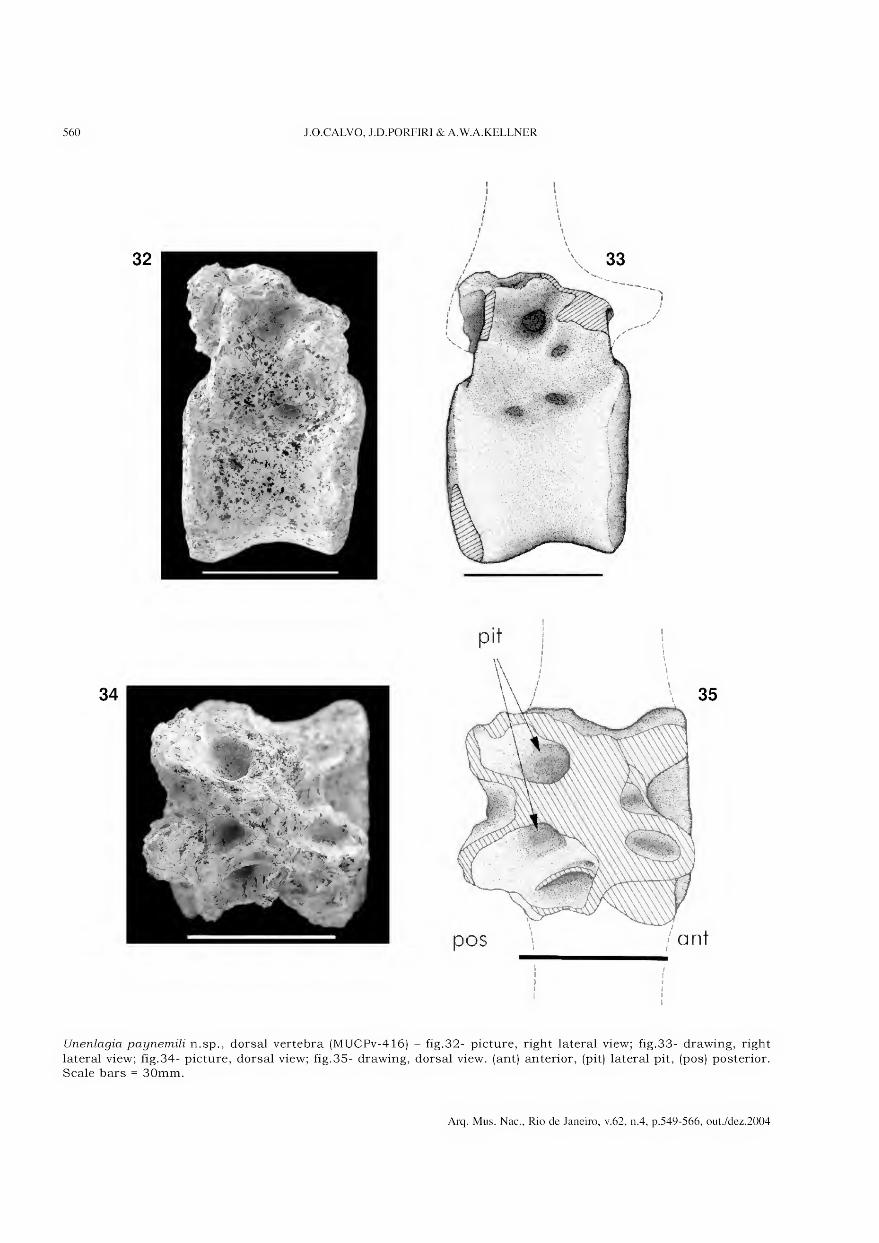
560 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
í I i J
[ i
Unenlagia paynemili n.sp., dorsal vertebra (MUCPv-416) - fig.32- picture, right lateral view; fig.33- drawing, right
lateral view; fig.34- picture, dorsal view; fig.35- drawing, dorsal view. (ant) anterior, (pit) lateral pit, (pos) posterior. Scale bars = 30mm.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004
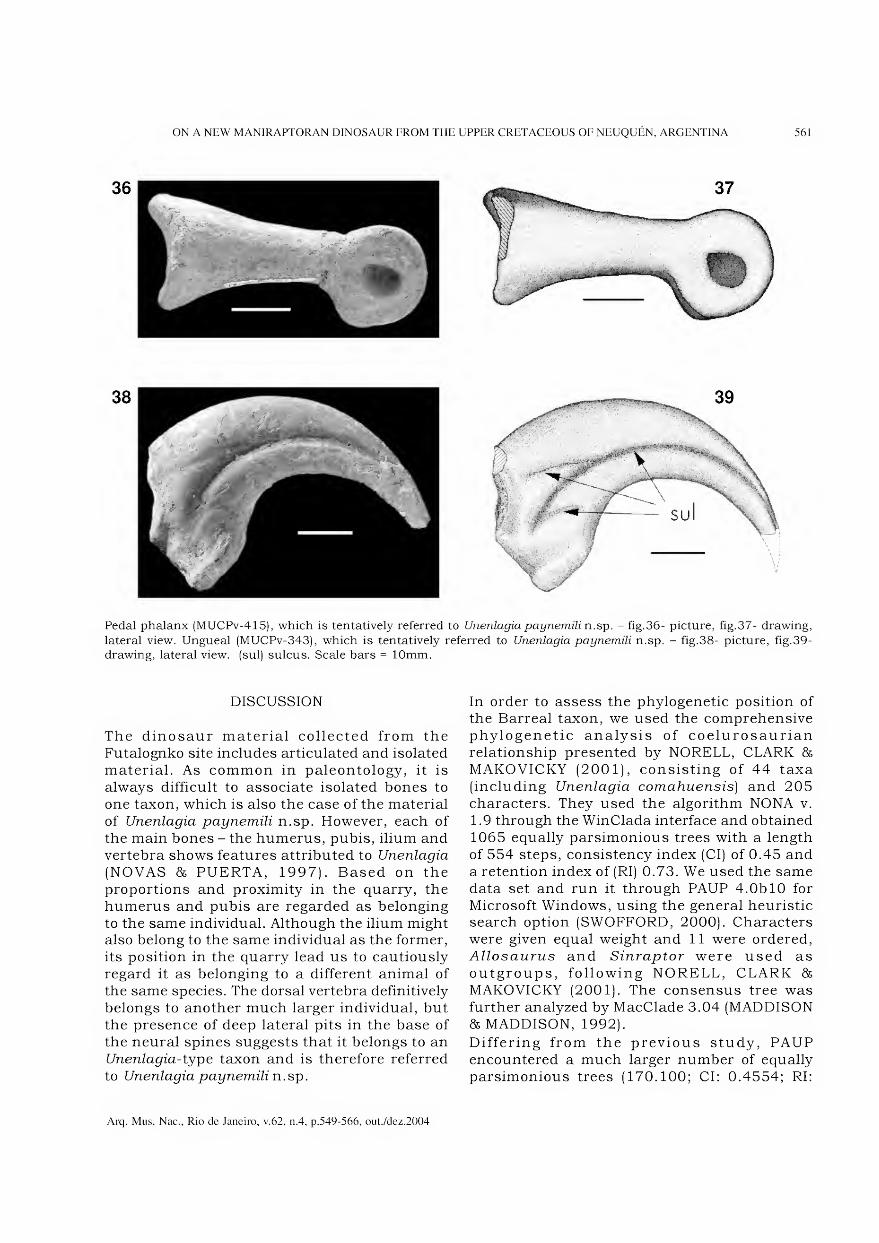
ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 561
Pedal phalanx (MUCPv-415), which is tentatively referred to Unenlagia paynemili n.sp. - fig.36- picture, fig.37- drawing,
lateral view. Ungueal (MUCPv-343), which is tentatively referred to Unenlagia paynemili n.sp. - fig.38- picture, fig.39- drawing, lateral view. (sul) sulcus. Scale bars = 10mm.
DISCUSSION
The dinosaur material collected from the Futalognko site includes articulated and isolated material. As common in paleontology, it is always difficult to associate isolated bones to one taxon, which is also the case of the material of Unenlagia paynemili n.sp. However, each of the main bones - the humerus, pubis, ilium and vertebra shows features attributed to Unenlagia
(NOVAS 86 PUERTA, 1997). Based on the proportions and proximity in the quarry, the humerus and pubis are regarded as belonging to the same individual. Although the ilium might also belong to the same individual as the former, its position in the quarry lead us to cautiously regard it as belonging to a different animal of the same species. The dorsal vertebra definitively belongs to another much larger individual, but the presence of deep lateral pits in the base of the neural spines suggests that it belongs to an Unenlagia-type taxon and is therefore referred to Unenlagia paynemili n.sp.
In order to assess the phylogenetic position of the Barreai taxon, we used the comprehensive phylogenetic analysis of coelurosaurian relationship presented by NORELL, CLARK 86
MAKOVICKY (2001), consisting of 44 taxa (including Unenlagia comahuensis) and 205 characters. They used the algorithm NONA v. 1.9 through the WinClada interface and obtained 1065 equally parsimonious trees with a length of 554 steps, consistency index (Cl) of 0.45 and a retention index of (RI) 0.73. We used the same data set and run it through PAUP 4.0b 10 for Microsoft Windows, using the general heuristic search option (SWOFFORD, 2000). Characters were given equal weight and 11 were ordered, Allosaurus and Sinraptor were used as outgroups, following NORELL, CLARK 85
MAKOVICKY (2001). The consensus tree was further analyzed by MacClade 3.04 (MADDISON 86 MADDISON, 1992). Differing from the previous study, PAUP encountered a much larger number of equally parsimonious trees (170.100; Cl: 0.4554; RI:
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

562 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
0.7150) with a shorter length of 549 steps. The consensus tree also differs from the one published by NORELL, CLARK & MAKOVICKY (2001), by recovering a monophyletic Dromaeosauridae (including Unenlagia comahuensis) in sister group relationship with the Avialae (compare figsures 40 and 41). Within Dromeosauridae, PAUP recovered a trichotomy formed by Unenlagia, Sinornithosaurus and all other dromaeosaurids (which in the previous study formed a polytomy). Furthermore, PAUP resolved the relationship between Troodontidae, with Sinornithoides followed stepwise by Byronosaurus, Troodon and Saurornithoides.
The next step was to include the information available for Unenlagia paynemili n.sp. using the same data matrix and run it through PAUP. Only 10 characters could be scored for the new species (141/ 162/ 180: State 0; 105-106/ 160/ 176-178: State 1; 179: State 2), all remaining are unknown (1-104/ 107-140/ 142-159/ 161/ 163-175/ 181- 205: ?). We also changed character 179, concerning the pubic shaft, which according to NORELL, CLARK 86 MAKOVICKY (2001) could vary from being straight (state 0) or curved anteriorly (state 1). The direct observation of the holotype of Unenlagia
comahuensis, as well as the Barreai specimen, leads us to include a third state: distai end of pubis curved posteriorly (state 2). This modified data set was run through PAUP and obtained the same number of equally parsimonious trees which were expectedly one step longer (550 steps; Cl: 0.4564; RI: 0.7158). The topology of the tree did not change, and differed only by showing a monophyletic grouping formed by Unenlagia
comahuensis and the Barreai specimen (Fig.41). Based on the comparisons and phylogenetic analysis, we opted to consider the Barreai specimen congeneric with Unenlagia comahuensis,
proposing the new species Unenlagia paynemili
n.sp. Although it is quite possible that a generic differentiation might be applied in the future for the Barreai taxon, we prefer to wait until more complete and articulated material comes to light.
According to the hypothesis of coelurosaurian relationships published by NORELL, CLARK 8& MAKOVICKY (2001), Unenlagia paynemili n.sp. presents two maniraptoran synapomorphies: the acuminate postacetabular ala of the ilium and vertical placed pubis (proximal part). The new Barreai taxon also shows a synapomorphy that unites all maniraptoran except Omitholestes: the brevis fossa shelf-like. It further has a comparatively
small anterior projection of the pubic boot also absent or slightly projected in Avialae and Dromeosauridae opposed to the well anterior projection known in most other theropods (e.g.,
Allosaurus, Deinonychus antirrhopus). Unenlagia
paynemili n.sp. does not have at least one synapomorphy of the Avialae: reduced pubic apron. Furthermore it shares one synapomorphic feature with Unenlagia comahuensis: the posteriorly oriented distai end of the pubis.
Among the diagnostic features of Unenlagia
comahuensis, NOVAS 86 PUERTA (1997) point out the presence of lateral pits at the base of the neural spines of the posterior dorsais and anterior sacrals and the inflected margin of postacetabular iliac blade. Those features are present in the referred specimens of Unenlagia paynemili n.sp. and might be considered two additional synapomorphies uniting those taxa. It should be noted, however, that the two additional dorsal vertebrae of the holotype of Unenlagia comahuensis lack lateral pits.
The main differences between Unenlagia
comahuensis and the new Barreai taxon is the different angle of the distai margin of the deltopectoral crest, the more rounded posterior edge of the ilium and the projection on the anterior part of the pubis. Furthermore the postacetabular portion of the ilium of Unenlagia paynemili n.sp. has a shallower brevis shelf with a smooth surface lacking the ridge present in Unenlagia comahuensis.
CONCLUSIONS
The new Barreales lake species shares one synapomorphy with Unenlagia comahuensis: the particular posterior inclination of the distai end of the pubis. One more character is shared by those taxa and is regarded as a potential synapomorphy of Unenlagia: the inflected dorsal margin of the postacetabular iliac blade. Although the dorsal vertebra of the Barreai species also shows the lateral pits at the base of the neural spine observed in Unenlagia comahuensis (NOVAS 86 PUERTA, 1997), this feature is absent in the additional two dorsal vertebrae of the type material and has to be seen with caution. Although the Barreales lake material possibly represents a new genus, we regard classify this species within the genus Unenlagia until more material allowing a better characterization is found. The several observed anatomical differences, however, lead us to propose a new species, Unenlagia paynemili n.sp.
The phylogenetic position of Unenlagia is disputed
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 563
with some authors regarding this taxon in sister group relationships with the Avialae (NOVAS & PUERTA, 1997) member of the Avialae (FORSTER et al, 1998) or a dromeosaurid not closely related to Avialae (NORELL & MAKOVICKY, 1999; NORELL, CLARK & MAKOVICKY, 2001). Unfortunately the incomplete nature of the Unenlagia paynemili n.sp.
does not resolve this dispute. Since we used the data basis of NORELL, CLARK & MAKOVICKY (2001), we end up as having both species of Unenlagia as members of the Dromeosauridae (Fig.41). Although
the analysis of the coelurosaurian phylogenetic hypothesis published by NORELL, CLARK & MAKOVICKY (2001) is not the purpose of the paper, the fact that PAUP produced different results than NONA, particularly recovering shorter and more resolved parsimonious trees is somewhat intriguing and should be object of further studies. In any case, the discovery of this new species of Unenlagia indicates a higher diversity of those enigmatic Unenlagia-type maniraptorans.
F
F
Altosaurus
Sinraptor
Ornitholestes
Microvônator
Caudipteryx
Ingenia
Oviraptor mongoliensis
Oviraptor philoceratops
Conchoraptor
Oviraptoríd IGM 100/42
Chirostenotes
Avimimus
Segnosaurus
Erlikosaurus
Alxasaurus
Byronosaurus
Sinornithoides
Troodon
Saurornithoides mongoliensis
Saurornitboides junior
Dromaeosaurus
Deinonychus
Vefociraptor
Unenlagia comahuensis
Adasaurus
Utahraptor
Sauromitholestes
Achillobator
IGM 100/1015
Sinomíthosaurus
Rahonavis
Archaeoptaryx
Confuciusornis
Alvarezsaurus calvoi
Patagonykus
Monanykus
Shuvuuia
Garudimimus
Peiçoanimimus
Harpymimus
Struthiomimus
Gallimimus
Tyrannosaurus
Albertosaunis
Fig.40- Strict consensus cladogram published by NORELL, CLARK & MAKOVICKY (2001).
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

564 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
Allosaurus
Sinraptor
Omitholestes
Microvenator
Caudipteryx
Ingeniã
Oviraptor mongoltensis
Oviraptor philoceratops
Conchoraptor
Oviraptoríd IGM 100/42
Chirostenotes
Avimímus
— Segfrtosat/ms
Erlikosaurus
Alxasaurus
Sinornithoides
Byronosaurus
Troodon
Sauromithoides mongoliensis
Saurornithoides junior
-Sinornithosaurus
- Dromaeosaurus
- Deinonychus
-- Velociraptor
— Adasaurus
— Utah raptor
- Saurornithotestes
-Achillobator
- IGM 100/1015
Unenlagia comahuensis
Unenlagia paynemili ÇRahonavis
Archaeopteryx
Con fuciusornis
-Aívarezsaurus calvoi
I— Patagünykus
Hj— Mononykus
Shuvuuia
- Garudimimus
- Pelecanimimus
—— Harpymimus
_r Struthlomimus
*— Gallimimus
Tyrannosaurus
Albertosaurus £
Fig.41- Strict consensus cladogram based on the data set of NORELL, CLARK & MAKOVICKY (2001) run through PAUP
4.0b 10, including the Unenlagia paynemili n.sp. See text for details.
ACKNOWLEDGMENTS
For access to the holotype of Unenlagia comahuensis, we thank Prof. Rodolfo Coria and Geol. Alberto Garrido (Carmen Funes Museum, Plaza Huincul, Neuquén). Our special thanks to the following members of Lake Barreales Paleontological Center (CePaLB) crew that helped during the excavation and preparation of the specimen: Federico Poblete, Elias Gómez, Horacio Saez, Magdalena Perini, and Cláudio Veralli. Matías A.Garcia (Neuquén) is
thanked for the drawings that illustrate this paper. This research was funded as folio ws: Dino Project (supported by Duke Energy Argentina through Duke Foundation with United Way International, and the collaboration of Daniel Andino Trailers, Pan American Energy, Topsy S.A., Unifon S.A., Skanska S.A., Petrobras Energy), T-021 Project (supported by the Universidad Nacional dei Comahue, Chevron- Texaco), and Agencia Nacional de Promoción Científica y Tecnológica (PICT 07-08277), all to J.O. Cal vo. A.W.A.Kellner thanks the Fundação de
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

ON A NEW MANIRAPTORAN DINOSAUR FROM THE UPPER CRETACEOUS OF NEUQUÉN, ARGENTINA 565
Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ - project E-26/152.442/2002) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for funding the trips to Neuquén, Argentina (2002-2004).
LITERATURE CITE D
BARSBOLD, R. & OSMÓLSKA, H., 1990. Ornitho mimosauria. In: WEISHAMPEL, D.B.; DODSON, P. &
OSMÓLSKA, H. (Eds.) The Dinosauria, Berkeley, p.225-244.
BARSBOLD, R.; MARYANSKA, T. & OSMÓLSKA, H., 1990.
Oviraptorosauria. In: WEISHAMPEL, D.B.; DODSON, P. & OSMÓLSKA, H. (Eds.) The Dinosauria, Berkeley,
p.249- 258. BONAPARTE, J.F.; NOVAS, F.E. & CORIA, R.A., 1990.
Camotaums sastrei Bonaparte, the horned, lightly build
carnosaur from the middle Cretáceos of Patagônia. Contributions Science, Natural History Museum of Los Angeles County, Los Angeles, 416:1-42.
BROCHU, C.A., 2003. Osteology of Tyrannosaurus rex: Insights from a Nearly Complete Skeleton and High- Resolution Computed Tomographic Analysis of the Skull. Journal of Vertebrate Paleontology, Illinois,
22(4): 1-138. CALVO, J.O., 1991. Huellas Fósiles de dinosaurios en la
Formación Rio Limay (Albiano-Cenomaniano) Picún Leufú. Província dei Neuquén. Argentina. (Ornithischia-
Saurischia: Saurópoda-Terópoda) Ameghiniana,
Buenos Aires, 28(3-4):241-258. CALVO, J.O.; PORFIRI, J. & KELLNER, A.W.A., 2003. A
close relative of Unenlagia comahuensis (Theropoda,
Maniraptora) from the Upper Cretaceous of Neuquén, Patagônia, Argentina. In: Boletim de Resumos, 18.
Congresso Brasileiro de Paleontologia, Brasília:
Universidade de Brasília, p.82-83. CALVO, J.O.; PORFIRI, J.; VERALLI, C. & NOVAS, F.
2002a. Megaraptor namunhuaiquii (Novas, 1998), a
new light about its phylogenetic relationships. In: VARGAS, A.; YÁNEZ, J.V. & RUBI LAR, D.R. (Eds.),
Sociedad Paleontológica de Chile. Primer Congreso Latinoamericano de Paleontologia de Vertebrados, Santiago do Chile, p.20.
CALVO, J.O.; PORFIRI, J.; VERALLI, C. & POBLETE, F.,
2001. One of the largest titanosaurid sauropods ever
found, Upper Cretaceous, Neuquén, Patagônia, Argentina. 61 Annual Meeting Society of Vertebrate
Paleontology, Illinois, 21(3):37A.
CALVO, J.O.; PORFIRI, J.; VERALLI, C.; POBLETE, F. &
KELLNER, A.W.A., 2002b. Futalognko Paleontological
Site, one of the most amazing Continental Cretaceous
Environments of Patagônia, Argentina. In: VARGAS, A.; YÁNEZ & RUBILAR, D.R. (Eds.), Sociedad Paleontológica de Chile. Primer Congreso Latinoamericano de
Paleontologia de Vertebrados, Santiago do Chile, p. 19.
CALVO, J.O. & SALGADO, L., 1995. Rebbachisaurus tessonei sp.nov. a new sauropod of the Albian-
Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia, Lisboa, 11:13-33.
CAZAU, L.B. & ULIANA, M.A., 1973. El Cretácico superior continental de la Cuenca Neuquina. V Congreso Geológico Argentino, Buenos Aires, Actas
3:131-163. CURRIE, P. & ZHAO, X., 1993. A new carnosaur
(Dinosauria, Theropoda) from the Jurassic of
Xinjiang, People’s Republic of China. Canadian Journal of Barth Sciences, Otawa, 30(10-
11):2037-2081. FORSTER, C.A.; SAMPSON, S.D.; CHIAPPE, L.M. &
KRAUSE, D.W., 1998. The theropod Ancestry of
Birds: New evidence from the Late Cretaceous of Madagascar. Science, Washington. 279:1915- 1919.
GALLO, V.; CALVO, J.O. & KELLNER, A.W.A., 2003.
First occurrence of a teleostean fish in the Portezuelo Formation (Upper Cretaceous),
Neuquén Group, Patagônia - Argentina.
Universidade do Estado do Rio de Janeiro, III Simpósio Brasileiro de Paleontologia de
Vertebrados, Rio de Janeiro, p.29.
KELLNER, A.W.A. & CAMPOS, D.A., 2002. On a Theropod Dinosaur (Abelisauria) from the Continental Cretaceous of Brazil. Arquivos do Museu Nacional,
Rio de Janeiro, 60(3): 163-170. KELLNER, A.W.A.; CALVO, J.O.; SAYÃO, J.M. &
PORFIRI, J., 2004. First Pterosaur from the
Portezuelo Formation, Neuquén Group, Patagônia, Argentina. IV Simpósio Brasileiro de Paleontologia
de Vertebrados. Rio Claro, p.29-30.
LEANZA, H.A., 1999. The Jurassic and Cretaceous Terrestrial Beds from Southern Neuquén Basin,
Argentina. Miscelânea, Tucumán, 4:1-30.
LEANZA, H.A. & HUGO, C.A., 2001. Cretaceous red beds from Southern Neuquén Basin (Argentina): age,
distribution and stratigraphic discontinuities. In:
Leanza, H.A. (Eds.) Asociación Paleontológica Argentina. VII International Symposium on
Mesozoic Terrestrial Bcosystems, Buenos Aires,
7:117-122. LEGARRETA, L. & GULISANO, C., 1989. Análisis
estratigráfico secuencial de la Cuenca Neuquina
(Triásico superior-Terciario inferior). In: CHEBLI, G.
& SPALLETTI, L. (Eds.) Cuencas Sedimentarias Argentinas. Serie Correlación Geológica.
Universidad Nacional de Tucumán, Tucumán,
6:221-243. MADDISON, W.P. & MADDISON, N.D.R., 1992.
MacClade, 3,04. Sinauer Associates, Inc. Sunderland,
Massachusetts.
MARTILL, D.; FREY, E.; SUES, H.D. & CRUIKSHANK,
A., 2000. Skeletal remains of a small theropod
dinosaur with associated soft structures from the Lower Cretaceous Santana Formation of northeastern
Brazil. Canadian Journal of Barth Sciences,
Canada, 37:891-900. NORELL, M.A. & MAKOVICKY, P.J., 1999. Important
Features of the Dromaeosaurid Skeleton II: Information from Newly Collected Specimens of
Velociraptor mongoliensis. American Museum
Novitates, New York, 3282:1-45.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

566 J.O.CALVO, J.D.PORFIRI & A.W.A.KELLNER
NORELL, M.A.; CLARK, J.M. & MAKOVICKY, P.J., 2001. Phylogenetic relationships among coelurosaurian theropods. New Perspectives on the Origin and Barly
Evolution of Birds: Proccedings of the International Symposium in Honor of John H. Ostrom. Peabody
Museum of Natural History, New Haven, 49-67.
NOVAS, F.E., 1996. Alvarezsauridae, Cretaceous basal birds from Patagônia and Mongolia. Proceedings of the Gondwanan Dinosaur Symposium. Memoirs
Queensland Museum, Brisbane, 39(3):675-702. NOVAS, F., 1998. Megaraptor namunhuaiquii, Gen. et
sp.nov., a large-clawed, late Cretaceous theropod from Patagônia. Journal of Vertebrate Paleontology, Illinois, 18(l):4-9.
NOVAS, F.E. 85 PUERTA, P., 1997. New evidence
concerning avian origins from the late Cretaceous of Patagônia. Nature, London, 387:390-392.
OSBORN, H.F., 1905. Tyrannosaurus and other
Cretaceous carnivorous dinosaur (second communication). Bulletin of the American Museum
of Natural History, New York, 21:259-265.
OSTROM, J.H., 1969a. A new theropod dinosaur from
the Lower Cretaceous of Montana. Postilla. Peabody Museum Natural History, New Haven, 128:1-17.
OSTROM, J.H., 1969b. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower
Cretaceous of Montana. Peabody Museum Natural History, New Haven, Bulletin 30:1-165.
PERLE, A.; NORELL, M. 85 CLARK, J., 1999. A new
maniraptoran Theropod-Achillobator giganteus
(Dromaeosauridae) from the Upper Cretaceous of Burkland, Mongolia. National University of
Mongolia, Mongolia, 1-104.
POBLETE, F. 85 CALVO, J., 2003. Upper Turonian
Dromaeosaurid teeth from Futalognko quarry,
Barreales Lake, Neuquén, Patagônia, Argentina. XIX Jornadas Argentinas de Paleontologia de Vertebrados, Buenos Aires p.24.
PORFIRI, J. 85 CALVO, J.O., 2002. A new record of an
ornithopod dinosaur from the Upper Cretaceous of Neuquén, Patagônia, Argentina. In: VARGAS, A.;
YÁNEZ, J.V. 85 RUBILAR, D.R. (Eds.), Sociedad Paleontológica de Chile. Primer Congreso Latinoamericano de Paleontologia de Vertebrados.
Santiago do Chile, p.45. PORFIRI, J.D. 85 CALVO, J.O., 2003. “Megaraptores” en
Lago Barreales, Neuquén, Patagônia. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina y Simposio de Tafonomía y
Paleoecología. Santa Rosa, La Pampa. p.17. PRÁMPARO, M.B.; PASSALIA, M.G.; HEREDIA, S.; 85
CALVO, J.O., 2003. Hallazgo de una macroflora en el
cretácico superior dei Grupo Neuquén, Lago Barreales, Neuquén. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina y Simposio
de Tafonomía y Paleoecología. Santa Rosa, La
Pampa. p.18. SÁNCHEZ, M.; CARDOZO, J.; HEREDIA, S. 85 CALVO,
J. 2003. Sedimentología de la Formación Portezuelo
(Grupo Neuquén), Cretácico Superior, en lago Los Barreales, Provinda dei Neuquén. 2das. Jornadas Regionales en Ciências de la Tierra. San Juan.
Actas, p.32. SWOFFORD, D.L., 2000. PAUP: Phylogenetic Analysis
Using Parsimony. Version 4,0B10. Sinauer
Associates. Inc. Sunderland, Massachusetts. ULIANA, M.A. 85 DELLAPE, D., 1981. Estratigrafia y
evolución paleoambiental de la Sucesión
Maastrichtiano-Eoterciaria dei engolfamiento Neuquino (Patagônia Septentrional). VIII Congreso Geológico Argentino, San Luis. Actas 3:673-711.
VERALLI, C. 85 CALVO, J.O., 2003. New findings of carcharodontosaurid teeth (Upper Turonian) on
Futalognko quarry, North Barreales Lake, Neuquén, Argentina. XIX Jornadas Argentinas de Paleontologia de Vertebrados, Buenos Aires, p.31.
WELLNHOFER, P., 1974. Das Funfte Skelettexmplar von
Archaeopteryx. Paleontographica, Stuttgart, 147(A): 169-216.
WELLNHOFER, P., 1993. Das siebte Exemplar von
Archaeopteryx aus den Solnhofener Schichten.
Archaeopteryx, Eichstátt, 11:1-48.
Arq. Mus. Nac., Rio de Janeiro, v.62, n.4, p.549-566, out./dez.2004

Arquivos do Museu Nacional, Rio de Janeiro, v.62, n.4, out./dez.2004
ISSN 0365-4508
SUMÁRIO / CONTENTS
Artigos originais / Original articles
Antropologia/ Anthropology
Teoria e métodos em antracologia; 2 - Técnicas de campo e de laboratório.
Theory and methods in anthracology. 2 - Field and laboratory techniques.
R.SCHEEL-YBERT. 343
Botânica / Botany
Estudo polínico de espécies de Aeschynomeneae e Phaseoleae (Papilionoideae - Leguminosae Juss.) ocorrentes nas restingas do
Estado do Rio de Janeiro.
Pollinic study of some species of Aeschynomeneae e Phaseoleae (Papilionoideae - Leguminosae Juss.) occurring in "restingas"
of Rio de Janeiro State.
F.C.SOUZA, M.A.SOUZA, C.B.F.MENDONÇA SC V.GONÇALVES-ESTEVES. 357
Passifloraceae da Mata de Encosta do Jardim Botânico do Rio de Janeiro e arredores, Rio de Janeiro, RJ.
Passifloraceae in the Rio de Janeiro Botanical Garden hillside forest and surroundings, Rio de Janeiro, RJ.
M.A.MILWARD-DE-AZEVEDO SC M.C.VALENTE. 367
Zoologia / Zoology
Novos dados sobre espécies sul-americanas de Chordodes (Nematomorpha).
New data on South American species of Chordodes (Nematomorpha).
C.DE-VILLALOBOS, F.ZANCASC A.SCHMIDT-RHAESA. 375
Morfologia Comparativa e Filogenia de representantes das superfamílias de arquitenioglossos e Annulariidae (Mollusca,
Caenogastropoda).
Comparative Morphology and Phylogeny of representatives of the superfamilies of architaenioglossans and the Annulariidae
(Mollusca, Caenogastropoda).
L.R.L. SIMONE. 387
Nova espécie de Scinax Wagler, 1830 do grupo de 5. rostratus do Brasil Central (Amphibia, Anura, Hylidae).
A new Scinax Wagler, 1830 of the 5. rostratus group from Central Brazil (Amphibia, Anura, Hylidae).
L.P.LIMA, R.P.BASTOS SC A.A.GIARETTA. 505
Paleontologia / Paleontology
Registro da atividade de organismos em um caule do Permiano do Rio Grande do Sul, Brasil.
Activity of Organisms in a Stem of the Rio Grande do Sul State, Brazil
R. B.BOLZON, I.AZEVEDO SC L.G.MACHADO.
Duas novas icnoespécies de Bifungites Desio, 1940 na Formação Pimenteira, Devoniano da Bacia do Parnaíba, Brasil.
Two new ichnospecies of Bifungites Desio, 1940 from the Pimenteira Formation, Devonian of Parnaíba Basin, Brazil
S. AGOSTINHO, M.S.S.VIANA SC A.C.S.FERNANDES.
Um novo peixe actinopterígio do Neopermiano da Bacia do Paraná, Sul do Brasil.
A new actinopterygian fish from the Late Permian of the Paraná Basin, Southern Brazil
F.J.FIGUEIREDO SC B.C.M.C.CARVALHO.
Sobre um novo dinossauro Maniraptora (Theropoda) do Cretáceo Superior de Neuquén, Patagônia, Argentina.
On a new Maniraptoran dinosaur (Theropoda) from the Upper Cretaceous of Neuquén, Patagônia, Argentina.
J.O.CALVO, J.D.PORFIRI SC A.W.A.KELLNER.
513
519
531
549

CONSULTORES CIENTÍFICOS
COLABORADORES DAS PUBLICAÇÕES EDITADAS PELO MUSEU NACIONAL EM 2004
Adalécio Kovaleski
Empresa Brasileira de Pesquisa Agropecuária
Fernando D’Incao
Fundação Universidade Federal do Rio Grande - RS
Adelita Aparecida Sartori Paoli
Universidade Estadual Paulista
Francisco de Assis Ribeiro Santos
Universidade Estadual de Feira de Santana - BA
Adriana Assis Arantes Universidade Federal de Uberlândia
Francisco Langeani Neto
Universidade Estadual Paulista
Adrian Martin Pohlit
Instituto Nacional de Pesquisas da Amazônia
Genise Vieira Somner
Universidade Federal Rural do Rio de Janeiro
Aírton Rodrigues Pinto Júnior
Pontifícia Universidade Católica do Paraná Geraldo Wilson Fernandes Universidade Federal de Minas Gerais
Alan Lane de Melo
Universidade Federal de Minas Gerais
Gloria Arratia
Museum für Naturkunde der Humboldt Universitát - Alemanha
Alejo Mesa Larramhebere
Universidade Estadual Paulista
Gustavo A. Schimidt Melo
Museu de Zoologia - USP
Andréa Luizi Ponzo Pereira
Universidade Federal de Juiz de Fora
Helena Piccoli Romanowski
Universidade Federal do Rio Grande do Sul
Angela Mascelani Museu Casa do Pontal - RJ
Ismar S. Carvalho
Universidade Federal do Rio de Janeiro
Angelo Barbosa Monteiro Machado Universidade Federal de Minas Gerais
Ivone Rezende Diniz Universidade de Brasília
Anthony Érico da Gama Guimarães
Fundação Instituto Oswaldo Cruz - RJ João Graciano Mendonça Filho Universidade Federal do Rio de Janeiro
Ariovaldo A. Giaretta
Universidade Federal de Uberlândia - MG
João Ubiratan Moreira dos Santos
Universidade Federal Rural da Amazônia
Ben Hanelt
Louisiana State University -EUA
JOAQUÍN ARROYO-CABRALES
Instituto Nacional de Antropologia e Historia - México
Carlos Eduardo de Viveiros Grelle
Universidade Federal do Rio de Janeiro
John Janovy
University of Nebraska - EUA
Carlos Frederico Duarte da Rocha Universidade do Estado do Rio de Janeiro
Jorge Luiz Nessimian
Universidade Federal do Rio de Janeiro
Carlos S. Carbonell Universidad de La Republica - Uruguai
José Albertino Rafael
Instituto Nacional de Pesquisas da Amazônia
Célio Fernando Baptista Haddad
Universidade Estadual Paulista
José Fernando Andrade Baumgratz
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
César Leandro Schultz
Universidade Federal do Rio Grande do Sul
José H. Leal
The Bailey-Matthews Shell Museum - EUA
Christopher Fielitz
Emory & Henry College - EUA José Jurberg
Fundação Instituto Oswaldo Cruz - RJ
Cibele Schwanke
Universidade do Estado do Rio de Janeiro
José W. Thomé
Pontifícia Universidade Católica - RS
Cristiana Barreto Center for Brazilian Studies - Reino Unido
JULIAN FaIVOVICH
American Museum of Natural History - EUA
Cristiane Pujol Universidade de Brasília
Julio Antonio Lombardi Universidade Federal de Minas Gerais
Dilma Solange Napp
Universidade Federal do Paraná Júlio César Garavello Universidade Federal de São Carlos - SP
Elizabeth Travassos Lins
Universidade Federal do Estado do Rio de Janeiro
Kenneth Anthony
James Cook University - Australia
Eurípedes Barsanulfo Menezes
Universidade Federal Rural do Rio de Janeiro
Lin Chau Ming
Universidade Estadual Paulista

Luciana de Carvalho
Univer Cidade - RJ
Luciane Marinoni
Universidade Federal do Paraná
Luís Amilton Foerster
Universidade Federal do Paraná
Luiz Flamarion B. Oliveira
Universidade Federal do Rio de Janeiro
Marcela Laura Monné Freire
Museu de Zoologia - USP
Marcelo F. Napoli
Universidade Federal da Bahia
Marcos Tavares
Museu de Zoologia - USP
Marcus Nadruz
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Margareth Ferreira de Sales
Universidade Federal Rural de Pernambuco
Margot Guerra Sommer
Universidade Federal do Rio Grande do Sul
Maria Cecília Arango Jaramillo
Universidad Católica de Oriente
Maria Cláudia Malabarba
Pontifícia Universidade Católica - RS
Maria das Graças Veloso Marinho
Universidade Federal de Campina Grande - PB
Maria de Nazaré do Carmo Bastos
Museu Paraense Emílio Goeldi - PA
Mario Vivo
Museu de Zoologia - USP
Marisa Afonso
Museu de Arqueologia e Etnologia - USP
Marise Sardenberg Salgado de Carvalho
Companhia de Pesquisa de Recursos Minerais - RJ
Martha Richter
Universidade Federal do Rio de Janeiro
Mary Elizabeth C. Bernardes de Oliveira
São Paulo - Brasil
Mirna Martins Casagrande
Universidade Federal do Paraná
Naércio Aquino Menezes
Universidade de São Paulo
Nicolas C. Czaplewski
Oklahoma Museum of Natural Histoiy - EUA
Nelson Papavero
Museu de Zoologia - USP
Nilda Marquete Ferreira da Silva
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Paulo Henrique Franco Lucinda
Universidade Federal do Tocantins
Paulo Knauss de Mendonça
Centro de Estudos Gerais - RJ
Paul W. Sammarco
Louisiana Universities Marine Consortium - EUA
P. B. Mordan
The Natural History Museum - Inglaterra
Regina Helena P. Andreata
Universidade Santa Úrsula - RJ
Renato Contin Marinoni
Universidade Federal do Paraná
Renato Neves Feio
Universidade Federal de Viçosa - MG
Ricardo Coutinho
Instituto de Estudos do Mar Almirante Paulo Moreira - RJ
Ricardo Macedo Corrêa e Castro
Universidade de São Paulo
Ricardo S. Rosa
Universidade Estadual do Oeste do Paraná
Ronaldo Fernandes
Museu Nacional - UFRJ
Rosaly Ale Rocha
Instituto Nacional de Pesquisas da Amazônia
Rosana Conrado Lopes
Universidade Federal do Rio de Janeiro
Ross MacPhee
American Museum of Natural History - EUA
Sheila Merlotti
Universidade Federal de Santa Catarina
Silvia Terezinha Sfoggia Miotto
Universidade Federal do Rio Grande do Sul
Tânia Andrade Lima
Museu Nacional - UFRJ
Tânia Lindner Dutra
Universidade do Vale do Rio dos Sinos - RS
Therezinha SantAnna Melhem
Instituto de Botânica - SP
Timothy Peter Moulton
Universidade do Estado do Rio de Janeiro
Toby Vincent Barrett
Instituto Nacional de Pesquisas da Amazônia
Valéria Gallo
Universidade do Estado do Rio de Janeiro
Valéria Veloso
Universidade Federal do Estado do Rio de Janeiro
Zailiang Ji
University of Victoria - EUA

Associação Amigos do Museu Nacional
SAMN
www.sam n.kit.net [email protected] 2568-8262 - ramal: 203 2568-1347 - ramal: 26
A Associação Amigos do Museu Nacional (SAMN) é uma
instituição sem fins lucrativos. Fundada em 13 de janeiro
de 1937 por professores do Museu Nacional, em 23 de
dezembro de 1966 foi reconhecida como de utilidade
pública pela Lei Estadual n°1200.
Os objetivos da SAMN ligam-se ao enriquecimento da cul¬
tura nacional - incentivar o ensino das Ciências Naturais e
Antropológicas através de visitas, cursos, concursos, etc.;
enriquecer as coleções científicas do Museu Nacional; pro¬
mover expedições científicas e atividades técnico-científi¬
cas no país; incentivar o respeito ao índio e a suas mani¬
festações culturais, à natureza e à preservação de suas ri¬
quezas e, finalmente, promover atividades esportivas, re¬
creativas e sociais.
A atual Diretoria da SAMN está trabalhando no sentido
de revitalizar a Sociedade, objetivando, inclusive, maior
integração com as escolas que visitam o Museu Nacional.
Com vistas a promover essa integração, está sendo lançada
uma campanha de adesões, de caráter permanente. A
filiação à SAMN poderá ser feita por fax (ficha de inscri¬
ção abaixo) ou on line (ficha de inscrição na homepage).
The Friends od the Brazilian National Museum (SAMN)
is a non-profit institution. It was founded on January 13,
1937 and on December 23, 1966 it was recognized as an
institution of public utility by State Law # 1200.
The objectives of SAMN are linked to the enrichment
of the national culture - motivation of the teaching of
Natural and Anthropological Sciences through visits,
courses, contests etc.; enrichment of the scientific
collections of Museu Nacional; promotion of scientific
expeditions and technical-scientific activities in the
country; motivation of the respect to the Native and
their culture, to Nature and the preservation of its
wealth and, finally, promotion of sports, recreational
and social activities.
The current Directory of SAMN works hard for the
revitalization of the Society, also aiming at the
integration with the many schools that visit the Museu
Nacional.
The affiliation to SAMN can be made by fax (application
form below) or online (application form on the
homepage)
Endereço / Address
Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brasil
r
NOME/NAME: .
ENDEREÇO/ADDRESS: .
TEL/PHONE #: .
E-MAIL:.
NASCIMENTO/BIRTH:././
ID / ÓRGÃO EXP.:.
ATIVIDADES/ACTIVITIES:.
CEP/CODE:.
CEL/MOBILE: .
HOMEPAGE:.
CPF/CNPJ / CGC:.
1
G J

INSTRUCTIONS TO AUTHORS
The authors are totally responsible for the content of the texts
1 - The MUSEU NACIONAL/UFRJ publishes reports of original scientific research in the field
of Natural and Anthropological Sciences in the following publications: Arquivos do Museu
Nacional (ISSN 0365-4508); Publicações Avulsas do Museu Nacional (ISSN 0100-6304);
Relatório Anual do Museu Nacional (ISSN 0557-0689); Boletim do Museu Nacional, Nova
Série - Antropologia (ISSN 0080-3189), Botânica (ISSN 0080-3197), Geologia (ISSN 0080-
3200) and Zoologia (ISSN 0080-312X); Série Livros (ISBN 85-7427). All are indexed in the
following data base: Biological Abstracts, ISI - Thomson Scientific, Ulrich’s International
Periodicals Directory, Zoological Record, NISC Colorado, Periódica and C.A.B. International.
2- The manuscript must be submitted in three high-quality copies, printed on one side of
A4 paper, double-spaced, font Times New Roman (12), together with an electronic ver-
sion (Word for Windows) not formatted. The manuscripts can be submitted in Portu-
guese or English (for other languages consult the Editorial Board). All foreign expres-
sions have to be italicized.
3- The text has to be preceded by the Identification of the author (name, institutional affili-
ation, and complete address).
4- An abstract followed by the title and key works in both languages have to be provided in
English and Portuguese.
5- All illustrations, designated in the text as figure (Fig. 1, Fig.2, etc.), must have scale bars. Titles
or captions must be typed apart. Illustrations should be digitalized.
6- All citations must indicate author and year of publication, with the author(s’) last name(s)
in upper case followed by the publications date of the cited document, separated by a
comma and in brackets; i.e.: (PEREIRA, 1996).
7- The references must be presented at the end of the text in alphabetical order, as follows
(adapted from the ABTN regulations):
Books/ chapters in books:
LIMA, D.A., 1982 - Present-day forest refuges in Northeastern Brazil. In: PRANCE, G.T.
(Ed.) Biological diversification in the tropics. New York: Columbia University Press,
p.245-251.
Periodicals:
MORA, O.A.; SIMÕES, M.J. & SASSO, W.S., 1987 - Aspectos ultra-estruturais dos
fibroblastos durante a regressão da cauda dos girinos. Revista Brasileira de Biologia,
Rio de Janeiro, 47(4):615-618, figs.1-2.
Papers published in scientific meetings:
VENTURA, P.E.C., 1985 - Avifauna de Morro Azul do Tinguá, Miguel Pereira, Rio de
Janeiro. In: CONGRESSO BRASILEIRO DE ZOOLOGIA, 12., Campinas. Resumos...,
Campinas: Universidade Estadual de Campinas, p.273.
Documents obtained at the internet:
POMERANCE, R., 1999 - Coral mortality, and global climate change. Available:
<http://www.gov/topical/global/envinon/99031002.htm>. Capturedon 18abr. 1999.
8- A total of 50 (fifty) copies, per article, are offered free of charge for the author(s).
9- All correspondence and manuscripts must be sent to the following address:
Comissão de Publicações
Museu Nacional/UFRJ
Quinta da Boa Vista, São Cristóvão
20940-040 - Rio de Janeiro, RJ, Brasil
Tels.: (0xx21) 2568 1347 ramal 22
E-mail: <[email protected]>

INSTRUÇÕES PARA AUTORES
O conteúdo dos artigos é de inteira responsabilidade do(s) autor(es)
1- 0 MUSEU NACIONAL/UFRJ edita, nas áreas das Ciências Naturais e Antropológicas:
Arquivos do Museu Nacional (ISSN 0365-4508); Publicações Avulsas do Museu Nacional
(ISSN 0100-6304); Relatório Anual do Museu Nacional (ISSN 0557-0689); Boletim do Mu¬
seu Nacional, Nova Série - Antropologia (ISSN 0080-3189), Botânica (ISSN 0080-3197),
Geologia (ISSN 0080-3200) e Zoologia (ISSN 0080-312X); Série Livros (ISBN 85-7427).
Indexadas nas bases de dados: Biological Abstracts, ISI - Thomson Scientific, UlriclTs
International Periodicals Directory, Zoological Record, NISC Colorado, Periódica e C.A.B.
International.
2- Os originais devem ser apresentados em três vias, em papel A4, espaço duplo, em uma só
face do papel, bem como em disquete no programa Word for Windows, em fonte Times
New Roman (corpo 12), sem qualquer tipo de formatação. Os manuscritos podem ser
encaminhados em português ou inglês (outro idioma ficará a critério da Comissão Edito¬
rial). Os termos estrangeiros no texto deverão ser grafados em itálico.
3- Os textos deverão ser precedidos de identificação do autor (nome e instituição de vínculo
com endereço completo).
4- Deverão constar Resumo e Abstract, juntamente com título e palavras-chave em portugu¬
ês e inglês.
5- As ilustrações, designadas no texto como figura (Fig.l, Fig.2, etc.), deverão conter escalas
com as unidades abreviadas (legendas à parte). Digitalizadas e salvas individualmente
(inclusive quando apresentadas em pranchas).
6- As citações no texto devem ser indicadas pelo sistema autor-data que compreende o
sobrenome do(s) autor(es), em caixa alta, seguido do ano de publicação do documento,
separado por vírgula e entre parêntese. Ex.: (PEREIRA, 1996).
7- As referências bibliográficas (adaptadas das normas da ABNT) deverão ser apresentadas
no final do texto, em ordem alfabética única dos autores.
Livro:
LIMA, D.A., 1982 - Present-day forest refuges in Northeastern Brazil. In: PRANCE, G.T.
(Ed.) Biological diversification in the tropics. New York: Columbia University Press,
p.245-251.
Periódico:
MORA, O.A., SIMÕES, M.J. & SASSO, W.S., 1987 - Aspectos ultra-estruturais dos
fibroblastos durante a regressão da cauda dos girinos. Revista Brasileira de Biolo¬
gia, Rio de Janeiro, 47(4):615-618, figs.1-2.
Trabalhos apresentados em encontros científicos:
VENTURA, P.E.C., 1985 - Avifauna de Morro Azul do Tinguá, Miguel Pereira, Rio de
Janeiro. In: CONGRESSO BRASILEIRO DE ZOOLOGIA, 12., Campinas. Resumos...,
Campinas: Universidade Estadual de Campinas, p.273.
Documentos disponíveis na internet:
POMERANCE, R., 1999 - Coral mortality, and global climate change. Disponível em:
<http://www.gov/topical/global/envinon/99031002.htm>. Acesso em: 18 abr. 1999.
8- Serão fornecidos ao(s) autor(es) 50 (cinqüenta) exemplares por artigo.
9- A correspondência editorial e os artigos deverão ser enviados para:
Comissão de Publicações Museu Nacional/UFRJ
Quinta da Boa Vista, São Cristóvão
20940-040 - Rio de Janeiro, RJ, Brasil
Tels.: (0xx21) 2568 1347 - ramal 22 E-mail: <[email protected]>


MUSEU NACIONAL Universidade Federal do Rio de Janeiro
Quinta da Boa Vista, São Cristóvão 20940-040 - Rio de Janeiro, RJ, Brasil
Impresso na Gráfica da UFRJ




















