
pH-Jump Induced Leucine Zipper Folding beyond the Diffusion Limit.
Upload
independentCategory
view
0download
0
Arabidopsis Basic Leucine-Zipper Transcription FactorsTGA9 and TGA10 Interact with Floral GlutaredoxinsROXY1 and ROXY2 and Are Redundantly Required forAnther Development1[C][W][OA]
Jhadeswar Murmu, Michael J. Bush, Catherine DeLong, Shutian Li, Mingli Xu, Madiha Khan,Caroline Malcolmson, Pierre R. Fobert, Sabine Zachgo, and Shelley R. Hepworth*
Department of Biology, Carleton University, Ottawa, Ontario, Canada K1S 5B6 (J.M., M.J.B., M.X., M.K., C.M.,S.R.H.); National Research Council Canada, Plant Biotechnology Institute, Saskatoon, Saskatchewan, CanadaS7N 0W9 (C.D., P.R.F.); and Department of Botany, University of Osnabruck, 49076 Osnabruck, Germany(S.L., S.Z.)
ROXY1 and ROXY2 are CC-type floral glutaredoxins with redundant functions in Arabidopsis (Arabidopsis thaliana) antherdevelopment. We show here that plants lacking the basic leucine-zipper transcription factors TGA9 and TGA10 have defects inmale gametogenesis that are strikingly similar to those in roxy1 roxy2mutants. In tga9 tga10mutants, adaxial and abaxial antherlobe development is differentially affected, with early steps in anther development blocked in adaxial lobes and later stepsaffected in abaxial lobes. Distinct from roxy1 roxy2, microspore development in abaxial anther lobes proceeds to a later stagewith the production of inviable pollen grains contained within nondehiscent anthers. Histological analysis shows multipledefects in the anther dehiscence program, including abnormal stability and lignification of the middle layer and defects in septumand stomium function. Compatible with these defects, TGA9 and TGA10 are expressed throughout early anther primordia butresolve to the middle and tapetum layers during meiosis of pollen mother cells. Several lines of evidence suggest that ROXYpromotion of anther development is mediated in part by TGA9 and TGA10. First, TGA9 and TGA10 expression overlaps withROXY1/2 during anther development. Second, TGA9/10 and ROXY1/2 operate downstream of SPOROCYTELESS/NOZZLE,where they positively regulate a common set of genes that contribute to tapetal development. Third, TGA9 and TGA10 directlyinteract with ROXY proteins in yeast and in plant cell nuclei. These findings suggest that activation of TGA9/10 transcriptionfactors by ROXY-mediated modification of cysteine residues promotes anther development, thus broadening our understand-ing of how redox-regulated TGA factors function in plants.
TGACG (TGA)motif-binding proteins form a distinctsubclade in the basic leucine-zipper (bZIP) super-family of Arabidopsis (Arabidopsis thaliana) transcrip-tion factors (Jakoby et al., 2002). This subclade contains10 members: TGA1 to TGA7, PERIANTHIA (PAN),and two uncharacterized members, bZIP21/TGA9(At1g08320) and bZIP65/TGA10 (At5g06839; Jakoby
et al., 2002). Past studies have identified overlappingroles for TGA transcription factors in plant diseaseresistance and stress responses (Despres et al., 2003;Zhang et al., 2003; Kesarwani et al., 2007; Fode et al.,2008; Mueller et al., 2008), indicating that these factorsoperate with a high degree of functional redundancy.To date, developmental roles for TGA factors areknown only for PAN, a key regulator of floral pattern-ing. Flowers lacking PAN typically contain an extrasepal and petal and one fewer stamen, resulting in apentamerous arrangement of floral organs (Runningand Meyerowitz, 1996; Chuang et al., 1999).
Recent studies have shed light on how posttranscrip-tional regulation of TGA transcription factors relayschanges in intracellular redox status. A pair of Cysresidues in the C terminus of TGA1 or TGA4 form anintermolecular disulfide bridge that is reduced uponexposure to salicylic acid (SA; Despres et al., 2003). Thismodification stimulates DNA binding and permits in-teraction with the BTB-ankryin protein NONEXPRES-SOR OF PATHOGENESIS-RELATED GENES1 (NPR1),a transcriptional coactivator (Despres et al., 2003; Rochonet al., 2006). PAN similarly partners with BTB-ankryinproteins, BLADE-ON-PETIOLE1 (BOP1) and BOP2, to
1 This work was supported by grants to S.R.H. from the CanadaFoundation for Innovation (grant no. 360228), the Ontario Innova-tion Trust (grant no. ER07–03–033), and the Natural Sciences andEngineering Research Council (grant no. 327195). C.D. was sup-ported by National Research Council Plant Biotechnology Institutecore funding to P.R.F.
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Shelley R. Hepworth ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
[W] The online version of this article contains Web-only data.[OA] Open Access articles can be viewed online without a sub-
scription.www.plantphysiol.org/cgi/doi/10.1104/pp.110.159111
1492 Plant Physiology�, November 2010, Vol. 154, pp. 1492–1504, www.plantphysiol.org � 2010 American Society of Plant Biologists

restrict the number of sepal-whorl organs in flowers(Hepworth et al., 2005). One of two regulatory Cysresidues in TGA1 is conserved in PAN and requiredfor activity (Li et al., 2009). Modification of thisresidue is proposed to depend on the activity ofROXY1, a CC-type glutaredoxin that interacts withPAN in plant cell nuclei (Li et al., 2009). roxy1 mutantsinitiate fewer petals than the wild type and later petalmorphogenesis can be aberrant, indicating thatROXY1 is a regulator of PAN (Xing et al., 2005; Liet al., 2009). ROXY1, together with its closest homologROXY2, is also required for anther cell specificationand microspore development (Xing and Zachgo,2008), but TGA partners in this process have yet tobe identified.The Arabidopsis anther is a bilaterally symmetrical
four-lobed structure in which pollen develops. Themitotic division of archesporial cells specified in theinterior of each lobe generates pollen mother cellssurrounded by three morphologically distinct celllayers: the tapetum, middle layer, and endothecium,underlying the epidermis (Sanders et al., 1999). Pollenmother cells divide meiotically to generate tetrads ofhaploid microspores that separate and mature intopollen grains. The tapetum contributes to microsporedevelopment by secreting enzymes necessary for mi-crospore release and by providing nutrients and struc-tural components essential for the production of viablepollen (Scott et al., 2004). Pollen grains are releasedfrom the anthers by dehiscence, a process whoseterminal step is breakage of the stomium connectingthe abaxial and adaxial anther lobes (Sanders et al.,1999; Ma, 2005). Interestingly, abaxial and adaxialanther lobe development is differentially dependenton ROXY activity. Loss of ROXY1/2 blocks sporoge-nous cell formation only in adaxial anther lobes. Inabaxial lobes, anther cell layers are specified butdevelopment of pollen mother cells and the tapetumis abnormal, leading to microspore degradation (Xingand Zachgo, 2008).We show here that mutations in TGA9 and TGA10
lead to male sterility and differential defects in abax-ial and adaxial anther lobe development similar toroxy1 roxy2 mutants. Additional defects not seen inroxy1 roxy2 mutants include abnormal stability ofthe middle layer and lack of septum and stomiumdegeneration, resulting in nondehiscent anthers. Weshow that TGA9 and TGA10 are expressed in anoverlapping fashion with ROXY1 and ROXY2 andregulate an overlapping set of genes with tapetumfunctions. In addition, ROXY proteins interact di-rectly with TGA9 and TGA10 in the nuclei of yeastand plant cells, suggesting that activation of TGA9and TGA10 transcription factors by the modificationof redox-sensitive Cys residues is needed for normalanther development. These data reveal a new rolefor TGA factors in anther patterning and dehis-cence, thus broadening our understanding of howredox-regulated TGA factors contribute to plant de-velopment.
RESULTS
Mutant Isolation and Characterization
TGA9 and TGA10 form a distinct subclade in thephylogenetic tree of Arabidopsis TGA bZIP familymembers (Supplemental Fig. S1), but their functionshave yet to be elucidated. Homologs of TGA9 andTGA10 in other plant species have developmentalroles. In tobacco (Nicotiana tabacum), overexpression ofNtTGA10 causes derepression of lateral branching anddefects in plant defense signaling (Schiermeyer et al.,2003). In maize (Zea mays), mutation of the TGA9homolog LIGULELESS2 causes defects in the formationof the blade-sheath boundary in leaves and delayedflowering (Walsh et al., 1998; Walsh and Freeling, 1999).To examine the functions of Arabidopsis TGA9 andTGA10, we obtained insertion lines from the SalkT-DNA collection (Fig. 1A). Lines designated tga9-1and tga9-2 contain T-DNA inserts at nucleotides 2,023and 3,325 of the coding sequence, respectively. Notranscripts were produced in these lines (Fig. 1B). Linesdesignated tga10-1 and tga10-2 contain T-DNA inserts atnucleotides 1,826 and 2,284 of the coding sequence,respectively. Partial transcripts were produced in bothlines, but the insertion in tga10-1 disrupts the bZIPdomain (Fig. 1, A and C) so as to encode a nonfunctionalprotein. No single mutants showed an obvious pheno-type. Analysis of TGA9 and TGA10 transcript by reversetranscription (RT)-PCR revealed an overlapping expres-sion pattern in flowers, suggesting that their activitiesmay overlap in floral development (Fig. 1, D and E). Toexamine this possibility, we constructed tga9 tga10 dou-ble mutants.
Male-Sterile Phenotype in tga9 tga10 Double Mutants
Strikingly, four of 72 F2 plants derived from a crossbetween tga9-1 and tga10-1 failed to set seed. Genotypingconfirmed that sterility was restricted to double mutants(Fig. 2). Examination of tga9 tga10 flowers showed nochanges in floral patterning, but anther morphologywasabnormal (Fig. 3). Stage 12 anthers in the tga9 tga10mutant were only partially filled with pollen and failedto dehisce at stage 13 (Fig. 3). Hand pollination of tga9tga10 carpels with wild-type pollen yielded a similarnumber of viable seeds (1.29% aborted seeds; n = 465) asa wild-type control cross (1.30% aborted seeds; n = 461),confirming that female fertility was normal. Unfortu-nately, expression of TGA9 and TGA10 coding sequencesunder the control of either a cauliflowermosaic virus 35Sor native promoter failed to complement the tga9 tga10mutant phenotype. However, an identical male-sterilityphenotype was seen in tga9-1 tga10-2 double mutants(data not shown), providing further evidence that theanther defect in tga9 tga10 mutants is indeed due to aloss of TGA9 and TGA10 function.
To further examine the male-sterility phenotype intga9 tga10 mutants, Alexander’s stain was used toassess pollen viability (Alexander, 1969). The anthersof late stage 12 flowers in the mutant contained pollen
TGA9 and TGA10 Are Required for Anther Development
Plant Physiol. Vol. 154, 2010 1493

that were abnormally aggregated but most stainedpurple-red, suggesting that they were potentially via-ble (Supplemental Fig. S2, A, B, D, and E). However,pollen grains that were excised from indehiscent an-thers failed to germinate on agar medium, revealingthem to be unviable (Supplemental Fig. S2, C and F;0% germination for tga9 tga10 pollen versus 70.5%germination for wild-type pollen [n = 200]). Scanningelectron micrographs of mutant pollen showed thatgrains were small and irregular, lacking the character-istic oblong shape of wild-type pollen grains. Theexine structure was also abnormal, with highly com-pressed columella, indicating defects in pollen wallmaturation (Supplemental Fig. S2, G–J).
Differential Effect of tga9 tga10 Mutation on Abaxial andAdaxial Anther Development
Anther cross-sections were used to examine thedetailed morphology of wild-type and tga9 tga10 an-
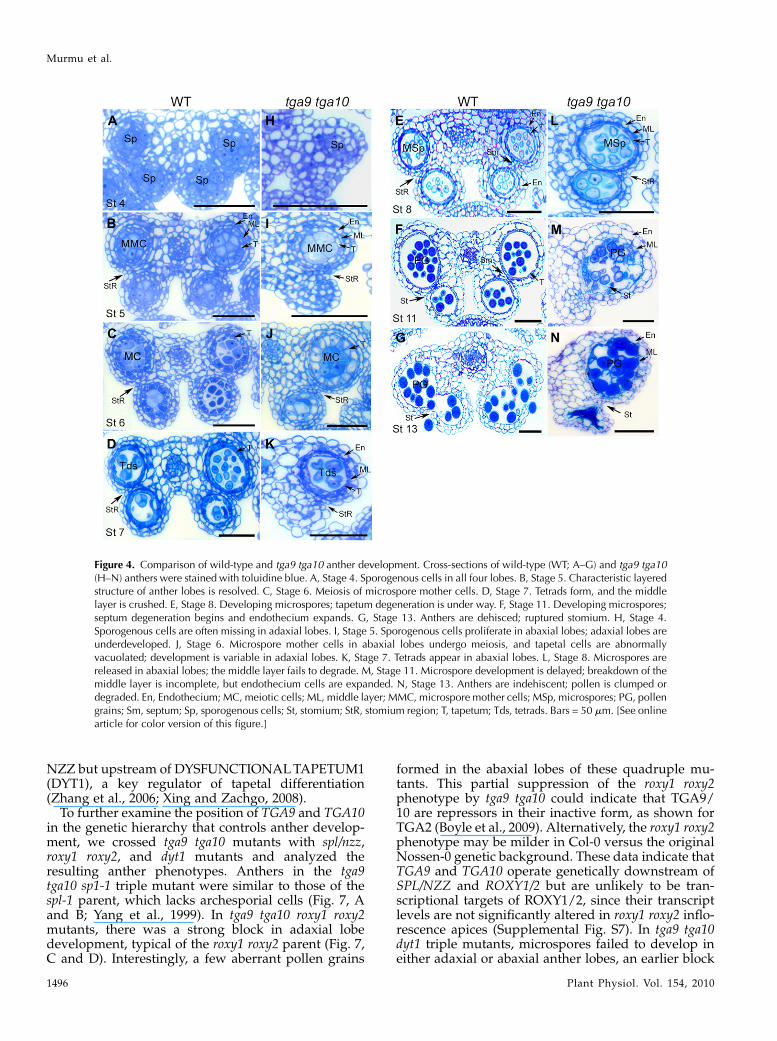
thers during development (Fig. 4) Morphological ab-normalities in the mutant were apparent from stage 4onward. In wild-type anthers, primary parietal cellsand sporogenous cells were specified in all four antherlobes and underwent cell division to form microsporemother cells surrounded by four morphologicallydistinct cell layers: the tapetum, middle layer, endo-thecium, and epidermis (Fig. 4, A and B; Sanders et al.,1999). In the double mutant, sporogenous cells formedreliably only in abaxial anther lobes (Fig. 4H). Inadaxial lobes with sporogenous cells, cell divisionswere often variable or disorganized, resulting in de-velopmental arrest. Cell division in abaxial lobesfollowed a pattern similar to the wild type, resultingin microspore mother cells surrounded by tapetum,middle, and endothecium cell layers at stage 5 (Fig. 4,B and I). During stages 6 and 7, pollen mother cells inabaxial lobes underwent meiosis to form tetrads en-capsulated by callose (Fig. 4, C, D, J, and K; Supple-mental Fig. S3). However, cells in the tapetum layer ofthe mutant at late stage 6 were expanded, with largevacuoles compared with the wild type (Fig. 4, C and J),suggesting abnormalities in tapetum function.
In the wild type, following the release of micro-spores at stage 8, the middle layer is crushed and thetapetum compacts as programmed cell death of theselayers takes place (Fig. 4, D and E; Sanders et al., 1999).
Figure 1. Characterization of tga9 and tga10mutant alleles, and tissueexpression of TGA9 and TGA10. A, Scale diagrams of TGA9 andTGA10 genomic sequences showing the positions of features asindicated in the key at bottom left. Black boxes indicate exons, andhorizontal arrows represent annealing positions of primers used fortranscript analysis. B, RT-PCR analysis of TGA9 transcripts in the wildtype (WT) and tga9 mutants (40 cycles). C, RT-PCR analysis of TGA10transcripts in the wild type and tga10mutants (40 cycles). Full, Productobtained using P1/P3 primer combination; Partial, product obtainedusing P1/P2 primer combination. D and E, RT-PCR analysis of TGA9and TGA10 transcripts in wild-type plant tissues (35 and 45 cycles). InB to E, GAPC control transcript used 23 cycles.
Figure 2. Sterility phenotype in tga9 tga10 double mutants. A and B,Comparison of wild-type (WT) and tga9 tga10 inflorescences. Arrowsindicate empty siliques. C, Wild-type and tga9 tga10 siliques. D and E,Wild-type and tga9 tga10 dissected siliques. Bars = 1 mm. [See onlinearticle for color version of this figure.]
Murmu et al.
1494 Plant Physiol. Vol. 154, 2010

In the mutant, the middle layer was only partlydegraded, and by stages 10 and 11, microspore devel-opment was visibly impaired (Fig. 4, L and M; Sup-plemental Fig. S4). Pollen grains in stage 13 anthersof the mutant were clumped together and irregular inappearance (Fig. 4N). Even in anthers with pollen inadaxial lobes, the septum and stomium were nonfunc-tional, blocking dehiscence (Fig. 4, K and L; Supple-mental Fig. S4). Staining of stage 11 anthers withphloroglucinol indicated that the endothecium inboth the wild type and mutant is lignified to thesame degree, suggesting that reinforcement of this celllayer, essential for dehiscence, is not disrupted (Mitsudaet al., 2005; Mizuno et al., 2007; Yang et al., 2007b).However, the residual middle layer in mutants washighly lignified, which may contribute to nondehis-cence (Supplemental Fig. S5).Similar to tga9 tga10, mutations in roxy1 roxy2 dif-
ferentially affect adaxial and abaxial lobe devel-opment. ROXY1 and ROXY2 encode two CC-typeglutaredoxins that are expressed in flowers and thatinteract with several TGA bZIP transcription factors inyeast (Li et al., 2009). Primary parietal and sporoge-nous cells fail to differentiate in roxy1 roxy2 adaxialanther lobes, whereas in abaxial anther lobes, micro-spore development terminates prematurely. Pollenmother cells enter meiosis but eventually aggregateand degrade, giving rise to empty locules (Xing andZachgo, 2008). Although the tga9 tga10 block in pollendevelopment is less severe than in roxy1 roxy2, thedifferential effect of these mutations on anther lobedevelopment suggests that ROXY-dependent promo-tion of anther development may occur in part via theregulation of TGA9 and TGA10, likely at the post-transcriptional level.
TGA9 and TGA10 Expression Overlaps with ROXY in
Developing Anthers
ROXY1 and ROXY2 are expressed in all four antherlobes beginning at stage 3 and in both meiocytes andthe tapetum at stage 6 (Li et al., 2009). To examineTGA9 expression in developing anthers, ecotypeColumbia-0 (Col-0) plants expressing a GUS reportergene fused to a 3.9-kb TGA9 promoter sequence weregenerated. Whole-mount staining of inflorescencesshowed activation of GUS expression in stage 7 flow-ers (anther stage 4). Expression peaked in stage 8flowers (anther stages 5–6) and declined in stage 9 to11 flowers (anther stages 7–11; Fig. 5, A–E). However,this same promoter driving expression of the TGA9coding sequence failed to restore fertility in tga9 tga10mutants, suggesting that additional regulatory motifspresent in the introns or 3# untranslated region areneeded for full expression. Therefore, in situ hybrid-ization was used to monitor TGA9 transcript accumu-lation in anther cross-sections. This analysis showedactivation of TGA9 in stage 2 anther primordia duringarchesporial cell specification. During stages 2 to 3 ofanther development prior to the emergence of distinctlocules, expression was in a horseshoe pattern associ-ated with the lateral and adaxial portion of primordia(Fig. 5, F and G). During stages 4 and 5, expression wasthroughout sporogenic tissue and surrounding cellslayers in adaxial and adaxial locules (Fig. 5, H and I). Atstage 6, expression localized to the tapetum andmiddlelayers, gradually fading postmeiosis with degenerationof these cell layers (Fig. 5, J–L). TGA10was expressed ina similar pattern, albeit at lower levels (SupplementalFig. S6). These data show that ROXY1/2 and TGA9/10are coexpressed during stages 3 to 6 of anther devel-opment, consistent with functional overlap.
TGA9 and TGA10 Function Genetically Downstream ofSPOROCYTELESS/NOZZLE and Promote Its Expressionin Adaxial Anther Lobes
Initiation of sporogenesis requires SPOROCYTE-LESS/NOZZLE (SPL/NZZ), a MADS box-like tran-scription factor regulated by the floral homeotic factorAGAMOUS (Ito et al., 2004). SPL/NZZ is expressed inarchesporial and sporogenous cells and later in pollenmother cells and the tapetum (Schiefthaler et al., 1999;Yang et al., 1999). In situ hybridization showed anaccumulation of SPL/NZZ transcript in the stage 2anthers of both the wild type and tga9 tga10 mutants,when archesporial cells are specified (Fig. 6, A and D;Sanders et al., 1999). SPL/NZZ transcript was uniformthroughout all four anther lobes in the wild type, butin tga9 tga10 mutants, expression was often less in-tense in one or both adaxial lobes, consistent withvariable specification of sporogenous cells in theselobes (Fig. 6, D–F; stages 2, 4–5, and 6). A similar butmore pronounced reduction of SPL/NZZ expression inadaxial anther lobes was reported for roxy1 roxy2mutants, whose activity lies downstream of SPL/
Figure 3. Flower and anther morphology in the wild type and tga9tga10mutants. Floral stages are as indicated. A, Wild-type (WT) flower.B, Wild-type anther, adaxial view. C, Wild-type dissected flower. D,tga9 tga10 flower. E, tga9 tga10 anther, adaxial view. F, tga9 tga10dissected flower; anthers are nondehiscent. Bars = 1 mm, except 100mm in B and E. [See online article for color version of this figure.]
TGA9 and TGA10 Are Required for Anther Development
Plant Physiol. Vol. 154, 2010 1495
NZZ but upstream of DYSFUNCTIONALTAPETUM1(DYT1), a key regulator of tapetal differentiation(Zhang et al., 2006; Xing and Zachgo, 2008).
To further examine the position of TGA9 and TGA10in the genetic hierarchy that controls anther develop-ment, we crossed tga9 tga10 mutants with spl/nzz,roxy1 roxy2, and dyt1 mutants and analyzed theresulting anther phenotypes. Anthers in the tga9tga10 sp1-1 triple mutant were similar to those of thespl-1 parent, which lacks archesporial cells (Fig. 7, Aand B; Yang et al., 1999). In tga9 tga10 roxy1 roxy2mutants, there was a strong block in adaxial lobedevelopment, typical of the roxy1 roxy2 parent (Fig. 7,C and D). Interestingly, a few aberrant pollen grains
formed in the abaxial lobes of these quadruple mu-tants. This partial suppression of the roxy1 roxy2phenotype by tga9 tga10 could indicate that TGA9/10 are repressors in their inactive form, as shown forTGA2 (Boyle et al., 2009). Alternatively, the roxy1 roxy2phenotype may be milder in Col-0 versus the originalNossen-0 genetic background. These data indicate thatTGA9 and TGA10 operate genetically downstream ofSPL/NZZ and ROXY1/2 but are unlikely to be tran-scriptional targets of ROXY1/2, since their transcriptlevels are not significantly altered in roxy1 roxy2 inflo-rescence apices (Supplemental Fig. S7). In tga9 tga10dyt1 triple mutants, microspores failed to develop ineither adaxial or abaxial anther lobes, an earlier block
Figure 4. Comparison of wild-type and tga9 tga10 anther development. Cross-sections of wild-type (WT; A–G) and tga9 tga10(H–N) anthers were stained with toluidine blue. A, Stage 4. Sporogenous cells in all four lobes. B, Stage 5. Characteristic layeredstructure of anther lobes is resolved. C, Stage 6. Meiosis of microspore mother cells. D, Stage 7. Tetrads form, and the middlelayer is crushed. E, Stage 8. Developing microspores; tapetum degeneration is under way. F, Stage 11. Developing microspores;septum degeneration begins and endothecium expands. G, Stage 13. Anthers are dehisced; ruptured stomium. H, Stage 4.Sporogenous cells are often missing in adaxial lobes. I, Stage 5. Sporogenous cells proliferate in abaxial lobes; adaxial lobes areunderdeveloped. J, Stage 6. Microspore mother cells in abaxial lobes undergo meiosis, and tapetal cells are abnormallyvacuolated; development is variable in adaxial lobes. K, Stage 7. Tetrads appear in abaxial lobes. L, Stage 8. Microspores arereleased in abaxial lobes; the middle layer fails to degrade. M, Stage 11. Microspore development is delayed; breakdown of themiddle layer is incomplete, but endothecium cells are expanded. N, Stage 13. Anthers are indehiscent; pollen is clumped ordegraded. En, Endothecium; MC, meiotic cells; ML, middle layer; MMC, microspore mother cells; MSp, microspores; PG, pollengrains; Sm, septum; Sp, sporogenous cells; St, stomium; StR, stomium region; T, tapetum; Tds, tetrads. Bars = 50 mm. [See onlinearticle for color version of this figure.]
Murmu et al.
1496 Plant Physiol. Vol. 154, 2010

than for either parent (Fig. 7, E and F). Collectively,these data indicate that TGA9 and TGA10 functiondownstream of SPL/NZZ and ROXY1/2 and up-stream or in parallel with DYT1 in the genetic hierar-chy that controls anther development.
TGA9/10 and ROXY1/2 Regulate an Overlapping Set ofGenes with Functions in Early and MiddleTapetal Development
Both ROXY1/2 and TGA9/10 are coexpressed in thetapetum during stages 4 to 7 of anther development,
and their genetic interactions are consistent with par-allel activities downstream of SPL/NZZ in antherdevelopment. Microarray analysis has identified a setof genes functioning primarily in tapetal developmentand pollen wall formation whose expression is se-verely down-regulated in roxy1 roxy2 inflorescenceapices (Xing and Zachgo, 2008). To examine the pos-sibility that some of these genes might also be targetsof TGA9/10 activity, we used RT-PCR to monitor thetranscripts of representative genes acting during theearly, middle, and late stages of tapetal developmentin wild-type and tga9 tga10 inflorescence apices (Fig. 7,
Figure 5. Expression of TGA9 in wild-type inflo-rescences and flowers. Expression was monitoredusing a TGA9::GUS reporter gene (A–E) or by insitu hybridization (F–L). A to E, GUS activity isfirst detected in stage 7 flowers, peaks at stage 8,and declines during stages 9 to 10. E, Stage 10flower cross section; expression persists in thelocule wall (arrowheads). TGA9 in situ probehybridized to anther cross-sections. F, Stage 2 to 3anthers. Expression laterally and along the adaxialface of primordia (arrows) is shown. G, Stage 2.Anther primordia. H, Stage 4. Expression in allcell layers. I and J, Stages 5 to 6. Expression isrestricted to the tapetum and middle layer. K andL, Stages 8 to 11. Expression declines. Bars = 100mm except in A (0.5 mm) and F to L (25 mm). LW,locule wall; ML, middle layer; T, tapetum. [Seeonline article for color version of this figure.]
Figure 6. In situ analysis of SPL/NZZ expressionin wild-type (WT) and tga9 tga10 anthers. Antherstages are as indicated. A to C, The wild type. SPLis uniformly expressed in adaxial versus abaxialanther lobes. D to F, The tga9 tga10 mutant. SPLexpression is reduced in adaxial anther lobes(asterisks). Sp, Sporogenous cells. Bars = 25 mm.[See online article for color version of this figure.]
TGA9 and TGA10 Are Required for Anther Development
Plant Physiol. Vol. 154, 2010 1497

G and H). The expression of genes at the top of thehierarchy required for early archesporial cell forma-tion and specification of anther cell layers, representedby SPL/NZZ and SQUAMOSA PROMOTER-BINDINGPROTEIN-LIKE8 (SPL8; Unte et al., 2003), were notmarkedly altered in tga9 tga10 inflorescence apices.
In contrast, a marked reduction was observed in thetranscripts of DYT1 and MALE-STERILITY1 (MS1),tapetal genes operating before and during the meiosisof pollen mother cells (Wilson et al., 2001; Zhang et al.,2006; Ito et al., 2007; Yang et al., 2007a). Similar to tga9tga10 mutants, ms1 mutants show an abnormally vac-uolated tapetum and defects in microspore develop-ment, resulting in pollen with an altered exine pattern(Ito et al., 2007; Yang et al., 2007a). No obvious changeswere observed in the expression of the MYB transcrip-tion factors encoded by MYB35/DEFECTIVE IN TA-PETAL DEVELOPMENT AND FUNCTION1 (TDF1),orMYB33 andMYB65, of which double mutants showtapetum hypertrophy leading to premeiotic abortionof pollen development (Millar and Gubler, 2005; Zhuet al., 2008). Tapetal genes with post-meiotic functionsin pollen coat formation, represented by MS2 andFACELESS POLLEN (FLP), were also expressed atwild-type levels in tga9 tga10 mutant apices (Aartset al., 1997; Ariizumi et al., 2003). Quantitative RT-PCRin a separate experiment using roxy1 roxy2 mutantapices as a control confirmed these trends, showing asignificant reduction in DYT1 and MS1 transcript in
tga9 tga10mutants (Supplemental Fig. S8; Supplemen-tal Materials and Methods S1).
Therefore, we concluded that ROXY1/2 and TGA9/10 function at similar levels in the genetic hierarchythat controls anther development and are likely tocoregulate a set of genes expressed in the tapetumbefore and during meiosis of pollen mother cells,represented by DYT1 and MS1. This result is compat-ible with the overlapping expression patterns ofROXY1/2 and TGA9/10 in the tapetum during stages4 to 7 of anther development.
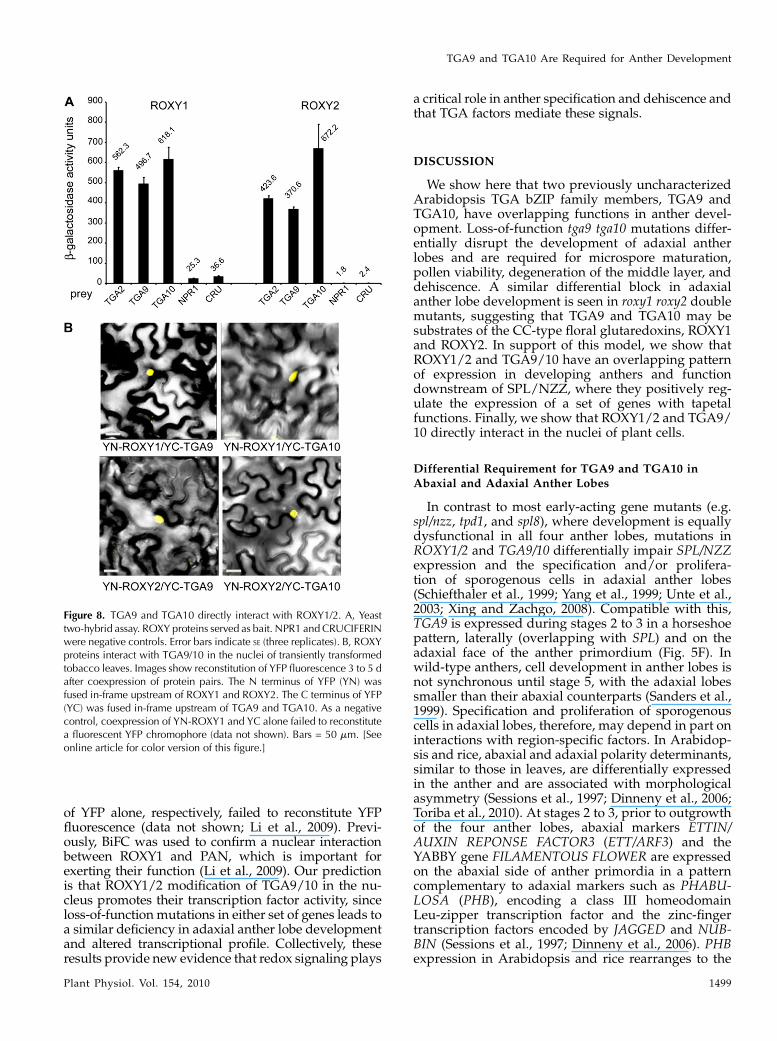
TGA9/10 and ROXY1/2 Interact in Yeast and in Planta
Yeast two-hybrid and bimolecular fluorescence com-plementation (BiFC) assays indicate that ROXY1 inter-acts with several TGA transcription factors in yeast andin planta, including PAN, TGA2, TGA3, and TGA7 (Liet al., 2009). We first used a two-hybrid assay to testfor a direct interaction between ROXY1/2 and TGA9/10, obtaining a positive result (Fig. 8A). As a furthertest, we used BiFC to examine the interaction of theseproteins in planta. This assay showed reconstitution ofyellow fluorescent protein (YFP) fluorescence in thenuclei of transformed tobacco leaf cells, indicating thatROXYproteins directly interact with TGA9 and TGA10in plant cells (Fig. 8B). Conversely, ROXY1/2 (fused tothe N terminus of YFP) or TGA9/10 (fused to the Cterminus of YFP) coexpressed with the C or N terminus
Figure 7. Double mutant analysis and RT-PCR analysis of anther transcripts in the wild type and tga9 tga10 mutants. A to F,Cross-sections of stage 11 anthers stained with toluidine blue for the indicated genotypes. Bars = 50 mm. G, RT-PCR analysis ofanther transcripts (35 cycles) in wild-type (WT) and mutant inflorescence apices. Numerical values for transcript down-regulation in roxy1 roxy2mutants (Xing and Zachgo, 2008) are shown. H, Genetic framework for control of anther development(modified from Wilson and Zhang, 2009) showing overlap for tapetal genes down-regulated in roxy1 roxy2 and tga9 tga10mutants. Shaded ovals, Down-regulated in roxy1 roxy2; dashed perimeter, down-regulated in tga9 tga10; shaded ovals withdashed perimeter; down-regulated in both. Genes not discussed in the text are as follows: TAPETAL DETERMINANT1 (TPD1;Yang et al., 2003), EXCESS MICROSPOROCYTES1/EXTRA SPOROGENOUS CELLS (EMS1/EXS; Canales et al., 2002; Zhao et al.,2002),MYB103/MYB80 (Higginson et al., 2003; Zhang et al., 2007),MYB99 (Ito et al., 2007), ABORTEDMICROSPORES (AMS;Sorensen et al., 2003), DEFECTIVE IN EXINE FORMATION1 (DEX1; Paxson-Sowders et al., 2001), NO EXINE FORMATION1(NEF1; Ariizumi et al., 2004), and CALLOSE SYNTHASE5 (CalS5; Dong et al., 2005). [See online article for color version of thisfigure.]
Murmu et al.
1498 Plant Physiol. Vol. 154, 2010
of YFP alone, respectively, failed to reconstitute YFPfluorescence (data not shown; Li et al., 2009). Previ-ously, BiFC was used to confirm a nuclear interactionbetween ROXY1 and PAN, which is important forexerting their function (Li et al., 2009). Our predictionis that ROXY1/2 modification of TGA9/10 in the nu-cleus promotes their transcription factor activity, sinceloss-of-functionmutations in either set of genes leads toa similar deficiency in adaxial anther lobe developmentand altered transcriptional profile. Collectively, theseresults provide new evidence that redox signaling plays
a critical role in anther specification and dehiscence andthat TGA factors mediate these signals.
DISCUSSION
We show here that two previously uncharacterizedArabidopsis TGA bZIP family members, TGA9 andTGA10, have overlapping functions in anther devel-opment. Loss-of-function tga9 tga10 mutations differ-entially disrupt the development of adaxial antherlobes and are required for microspore maturation,pollen viability, degeneration of the middle layer, anddehiscence. A similar differential block in adaxialanther lobe development is seen in roxy1 roxy2 doublemutants, suggesting that TGA9 and TGA10 may besubstrates of the CC-type floral glutaredoxins, ROXY1and ROXY2. In support of this model, we show thatROXY1/2 and TGA9/10 have an overlapping patternof expression in developing anthers and functiondownstream of SPL/NZZ, where they positively reg-ulate the expression of a set of genes with tapetalfunctions. Finally, we show that ROXY1/2 and TGA9/10 directly interact in the nuclei of plant cells.
Differential Requirement for TGA9 and TGA10 inAbaxial and Adaxial Anther Lobes
In contrast to most early-acting gene mutants (e.g.spl/nzz, tpd1, and spl8), where development is equallydysfunctional in all four anther lobes, mutations inROXY1/2 and TGA9/10 differentially impair SPL/NZZexpression and the specification and/or prolifera-tion of sporogenous cells in adaxial anther lobes(Schiefthaler et al., 1999; Yang et al., 1999; Unte et al.,2003; Xing and Zachgo, 2008). Compatible with this,TGA9 is expressed during stages 2 to 3 in a horseshoepattern, laterally (overlapping with SPL) and on theadaxial face of the anther primordium (Fig. 5F). Inwild-type anthers, cell development in anther lobes isnot synchronous until stage 5, with the adaxial lobessmaller than their abaxial counterparts (Sanders et al.,1999). Specification and proliferation of sporogenouscells in adaxial lobes, therefore, may depend in part oninteractions with region-specific factors. In Arabidop-sis and rice, abaxial and adaxial polarity determinants,similar to those in leaves, are differentially expressedin the anther and are associated with morphologicalasymmetry (Sessions et al., 1997; Dinneny et al., 2006;Toriba et al., 2010). At stages 2 to 3, prior to outgrowthof the four anther lobes, abaxial markers ETTIN/AUXIN REPONSE FACTOR3 (ETT/ARF3) and theYABBY gene FILAMENTOUS FLOWER are expressedon the abaxial side of anther primordia in a patterncomplementary to adaxial markers such as PHABU-LOSA (PHB), encoding a class III homeodomainLeu-zipper transcription factor and the zinc-fingertranscription factors encoded by JAGGED and NUB-BIN (Sessions et al., 1997; Dinneny et al., 2006). PHBexpression in Arabidopsis and rice rearranges to the
Figure 8. TGA9 and TGA10 directly interact with ROXY1/2. A, Yeasttwo-hybrid assay. ROXY proteins served as bait. NPR1 and CRUCIFERINwere negative controls. Error bars indicate SE (three replicates). B, ROXYproteins interact with TGA9/10 in the nuclei of transiently transformedtobacco leaves. Images show reconstitution of YFP fluorescence 3 to 5 dafter coexpression of protein pairs. The N terminus of YFP (YN) wasfused in-frame upstream of ROXY1 and ROXY2. The C terminus of YFP(YC) was fused in-frame upstream of TGA9 and TGA10. As a negativecontrol, coexpression of YN-ROXY1 and YC alone failed to reconstitutea fluorescent YFP chromophore (data not shown). Bars = 50 mm. [Seeonline article for color version of this figure.]
TGA9 and TGA10 Are Required for Anther Development
Plant Physiol. Vol. 154, 2010 1499

notch region at stage 4, presaging the site of stomiumdevelopment (Dinneny et al., 2006; Toriba et al., 2010).Mutations in ETT, whose activity is auxin responsive,and the auxin transport mutants pin formed1 and pinoidalso perturb anther structure biased toward the loss ofadaxial features (Bennett et al., 1995; Sessions et al.,1997), indicating that adaxial-abaxial patterning reliesin part on a gradient of auxin, as in leaves (Pekkeret al., 2005; Chitwood et al., 2007). The existence ofregion-specific factors in the anther, therefore, maylessen the requirement for ROXY-TGA9/10 activity inabaxial anther lobes.
TGA9 and TGA10 Contribute to Tapetal Development
The tapetum is essential for normal microspore andpollen development, as it provides nutrients and se-cretes enzymes and structural components for thepollen coat. Indeed, mutations in tapetally expressedgenes often cause defects in microspore developmentleading to male sterility (Scott et al., 2004; Ma, 2005).Consistent with this, transcript analysis in tga9 tga10mutants identified a significant reduction inDTY1 andMS1, representative genes operating in the tapetumbefore and during meiosis (Wilson et al., 2001; Zhanget al., 2006; Ito et al., 2007). During this time, TGA9/10are strongly expressed in the middle layer and tape-tum (Fig. 5, stages 5–6). Unexpectedly, tapetal geneswith postmeiotic roles in pollen coat developmentrepresented by MS2 and FLP1 (Aarts et al., 1997;Ariizumi et al., 2003) were not markedly affected inthis assay. MS2 is down-regulated 164.6-fold in roxy1roxy2 mutants, and we expected to see a similar resultfor tga9 tga10 but did not. Microspores degeneratecompletely in roxy1 roxy2 but continue to develop intga9 tga10 mutants, which may account for this differ-ence. Tapetal programmed cell death is initiated atabout the time that microspores are released fromtetrads (Sanders et al., 1999), during which time TGA9and TGA10 expression in the tapetum declines.
TGA9 and TGA10 Are Required for Anther Dehiscence
Dehiscence is a multistep program that begins withdegeneration of the middle layer (stages 7–8) and thetapetum (stages 8–11) and is accompanied by theexpansion and reinforcement of the endothecium(stage 11). Degradation of the septum between anthercompartments and stomium differentiation are com-plete by the end of stage 12. At stage 13, breakage ofthe stomium releases the mature pollen (Sanders et al.,1999, 2005). Our results show that multiple steps of thedehiscence program require TGA9/10. First, dissolu-tion of the middle layer is incomplete in the doublemutant. Second, thickening and lignification of theendothecium, essential for dehiscence (Steiner-Langeet al., 2003; Mitsuda et al., 2005; Yang et al., 2007b), issimilar to the wild type at stage 11, but the middlelayer remnant becomes ectopically lignified, whichmay interfere with dehiscence (Supplemental Fig. S5).
Septums in the mutant are disorganized and fail todegenerate, even when immature pollen grains formin adaxial lobes (Supplemental Fig. S4), and finally,stomium-like cells form at the notch between loculesbut are nonfunctional (Supplemental Fig. S4).
Relatively few nondehiscent mutants have been de-scribed to date (Ma, 2005; Wilson and Zhang, 2009).Mutations that impair jasmonic acid (JA) biosynthe-sis or signal transduction tend to delay rather thanblock anther dehiscence (Xie et al., 1998; Sanders et al.,2000; Ishiguro et al., 2001; Mandaokar and Browse,2009). Jasmonate treatment fails to rescue dehiscencein tga9 tga10 mutants (data not shown), suggestingthat TGA9/10 function independently or downstreamof a jasmonate signal. Auxin similarly regulates thetiming of dehiscence, by at least two mechanisms:ARF6 and ARF8 promote dehiscence via JA biosyn-thesis, and premature lignification of the endotheciumoccurs in auxin receptor mutants (Nagpal et al., 2005;Cecchetti et al., 2008; Tabata et al., 2010). Mutation ofthe tapetally expressed RECEPTOR-LIKE PROTEINKINASE2 (RPK2) gene blocks dehiscence, but its modeof action is unknown (Mizuno et al., 2007). In rpk2mutants, the middle layer fails to differentiate from thetapetum, which grows hypertrophically during mei-osis and does not fully degenerate. Reinforcement ofthe endothecium and degeneration of the septum andstomium are also blocked in this mutant, suggestingimportant functions for the middle layer in dehis-cence.
Degeneration of specific cell types during dehis-cence relies on programmed cell death (Wu andCheung, 2000; Sanders et al., 2005), which in senes-cence and plant defense are dependent on reactiveoxygen species as signaling intermediates (Apel andHirt, 2004; Gechev et al., 2006). Hence, it is unsurpris-ing that the dehiscence program relies on TGA9/10,whose activity is likely to be redox regulated. Jasmo-nates, produced by plants in response to biotic andabiotic stresses, are known to promote the productionof reactive oxygen species such as hydrogen peroxide(H2O2), which can be stress indicators but also impor-tant development signals (Buchanan and Balmer, 2005;
Figure 9. Model for redox control of TGA9/10 activity by ROXY1/2. Intheir inactive form, one or more oxidized C-terminal Cys residues blockTGA9/10 transcription factor activity (a dithiol bridge is shown) by anunknown mechanism. In response to redox changes triggered by adevelopmental signal, ROXY1 and ROXY2 reduce TGA9 and TGA10,converting them into a transcriptionally active form.
Murmu et al.
1500 Plant Physiol. Vol. 154, 2010

Gapper and Dolan, 2006; Huffaker and Ryan, 2007;Wasternack, 2007). Indeed, the promoter elements towhich TGA factors bind respond to various signals,including auxin, JA, and H2O2 (Xiang et al., 1996, 1997;Chen and Singh, 1999). Microarray studies show thatTGA factors contribute in large part to the plant’stranscriptional response to stress, as mimicked bytreatment with oxylipins such as 12-oxo-phytodienoicacid and JA (Mueller et al., 2008). In plant defense, theSA-inducible CC-type glutaredoxin ROXY19/GRX480interacts with TGA factors and plays a role in SA/JAcross talk by repressing JA-responsive PDF1.2 genetranscription (Ndamukong et al., 2007). Thus, TGA9/10 may respond to redox changes induced by thelocalized production of auxin, JA, or H2O2. Indeed,significant changes in the redox status of protein thiolsare seen by two-dimensional gel electrophoresis fol-lowing treatment of Arabidopsis tissues with JA(Alvarez et al., 2009). These reversible modifications,often in the form of disulfide bridges, can function asredox switches to control protein activity, as shown forTGA1 and TGA4 (Despres et al., 2003).
Posttranscriptional Control of TGA9 and TGA10 Activity
by Glutaredoxin
In plants, there are now several examples of disul-fide bridge formation or protein S-glutathionylationof Cys residues as mechanisms for regulating tran-scription factor activity (Dietz, 2008; Paulsen andCarroll, 2010). Our results support a model wherebymodification of TGA9/10 by ROXY1/2 in responseto a developmental signal such as JA biosynthesispromotes transcription factor activity (Fig. 9). Site-directed mutagenesis shows that Cys residues in theconserved C terminus of TGA1 (Cys-260 and Cys-266)and PAN (Cys-340) are crucial for function (Despreset al., 2003; Li et al., 2009). Transcriptional activationby TGA1 requires an interaction with NPR1, a BTB-ankryin transcriptional coactivator (Rochon et al.,2006). This interaction relies on the reduction of Cys-266 and Cys-260 to disrupt an intramolecular disulfidebridge, modifications that are induced in SA-treatedplants by an unknown glutaredoxin (Despres et al.,2003). In contrast, PAN interacts constitutively withBTB-ankryin cofactors BOP1 and BOP2 in yeast and inthe nuclei of Arabidopsis leaf mesophyll protoplasts(Hepworth et al., 2005; Xu et al., 2010). A single Cysresidue in PAN (Cys-340, equivalent to Cys-266 inTGA1) is crucial for function, making it unclear if PANis regulated by an intermolecular disulfide bridge orby S-glutathionylation. Genetic analysis shows thatthe pan phenotype (extra petals) is epistatic to the roxy1roxy2 phenotype (fewer petals), suggesting thatROXY1 negatively regulates PAN activity (Li et al.,2009). In support of a monothiol mechanism for PANregulation, complementation studies show that onlyone of three Cys residues (Cys-49) in the active site ofROXY1 is crucial for its in vivo activity (Xing et al.,2005). Alignments show that TGA10 alone has an
equivalent to Cys-340/Cys-260 in PAN/TGA1, respec-tively, and that both TGA9 and TGA10 have a uniqueC-terminal Cys residue (Cys-429 and Cys-435, respec-tively; Supplemental Fig. S9). Our inability at presentto complement the tga9 tga10 mutant phenotype withTGA9 or TGA10 cDNAs prevents us from testing ifthese residues are important for activity.
Functional Redundancy among TGA Family Members
Biochemical studies show that bZIP factors bind toDNA as dimers (Lamb and McKnight, 1991), support-ing the notion that TGA9 and TGA10 may functionredundantly as homodimers or heterodimers. More-over, the tga9 tga10 phenotype is less severe than that ofroxy1 roxy2 in that pollen formation is not completelyblocked (Xing and Zachgo, 2008). A likely possibility isthat other TGAs expressed in the anther work inconjunction with TGA9 and TGA10 as downstreameffectors of ROXY1 and ROXY2. Candidates includeTGA2, TGA6, and PAN, which interact with ROXY1with differing affinities in yeast or in plants (Xiang et al.,1997; Li et al., 2009). The best candidate for redundancyis TGA2, but tight linkage to TGA10 hampers thegenetic testing of this model. Given the broad expres-sion pattern of TGA9, PAN, and TGA2 in tissues otherthan the flower (Fig. 1, D and E; Chuang et al., 1999; Liet al., 2009), it is likely that genetic redundancy con-tinues to mask additional roles for this redox-regulatedfamily of transcription factors in plant development.
MATERIALS AND METHODS
Plant Materials and Growth Conditions
Plants were grown in growth chambers on agar plates and/or in soil at
21�C in 24 h of light (100 mmol m22 s21). The wild type was the Col-0 ecotype
of Arabidopsis (Arabidopsis thaliana) unless otherwise stated. T-DNA insertion
lines tga9-1 (SALK_057609), tga9-2 (SALK_091349), tga10-1 (SALK_124227),
and tga10-2 (SALK_039672) were obtained from the Arabidopsis Biological
Resource Center. The double mutant tga9-1 tga10-1 was characterized in this
study. The position of T-DNA inserts was determined by sequencing, and
homozygous mutants were identified by PCR genotyping (http://signal.salk.
edu). Mutants roxy1 roxy2, spl-1, and dyt1were described previously (Xing and
Zachgo, 2008). All mutant combinations were constructed by crossing and
confirmed by genotyping. Floral and anther development stages were deter-
mined according to Smyth et al. (1990) and Sanders et al. (1999), respectively.
Supplemental Tables S1 and S2 list the sequences of most primers used in this
study. All other primer sequences are available by request.
Construction of TGA9::GUS Reporter Lines
A TGA9::GUS translational fusion gene was created using the strategy
described by Hepworth et al. (2002). A sequence representing the putative
promoter of TGA9 (nucleotides –3,895 to +5) was amplified by PCR using
Arabidopsis bacterial artificial chromosome T27G7 DNA as the template.
Wild-type plants were transformed with pTGA9::GUS by floral dipping
(Clough and Bent, 1998) using the Agrobacterium tumefaciens strain C58C1
pGV101 pMP90 (Koncz and Schell, 1986). Basta-resistant transformants were
selected on soil using the herbicide Finale (AgrEvo).
Pollen Germination and Viability Assays
For the in vitro assay of pollen germination, pollen grains from the anthers
of late stage 12 flowers were excised into water and transferred to pollen germi-
TGA9 and TGA10 Are Required for Anther Development
Plant Physiol. Vol. 154, 2010 1501

nation medium presolidified onto a microscope slide (Fan et al., 2001). Slides
were incubated at 22�C in a high-humidity chamber (Johnson-Brousseau and
McCormick, 2004), and pollen tubes were observed after 24 h by light
microscopy. The germination of more than 300 pollen grains was monitored
for each genotype. Pollen viability was also assessed with Alexander’s stain
(Alexander, 1969).
Histological Analysis of Developing Anthers
Floral buds were fixed overnight at 4�C in 2% (v/v) glutaraldehyde as
described (Ogawa et al., 2009). Tissues were briefly rinsed in 25 mM NaPO4
buffer (pH 6.8) and dehydrated with ethanol prior to embedding in LRWhite
resin (London Resin Company). Sections (1 mm) were heat fixed onto glass
slides and stained with 0.05% (w/v) toluidine blue prior to imaging. To detect
callose, dissected anthers were treated with 0.05% (w/v) aniline blue in 0.067
M phosphate buffer (pH 8.5) and imaged under UV illumination. To detect
lignin, paraffin-embedded anther sections were dewaxed and dehydrated
prior to successive incubations in 2% phloroglucinol (in 95% ethanol) and 6 N
HCl for color development.
GUS Staining and in Situ Hydridization
Staining, sectioning, and embedding of GUS-stained tissues was per-
formed essentially as described by Sieburth and Meyerowitz (1997). Sections
(10 mm) were affixed to glass slides and dewaxed with tert-butanol prior to
imaging. In situ hybridization and antisense probe synthesis was performed
as described previously (Hepworth et al., 2005).
RT-PCR
Total RNA was isolated from inflorescence apices using the Trizol method
(Invitrogen). Total cDNA was synthesized with SuperScript III reverse tran-
scriptase (Invitrogen) using 1mg of RNA as the template. PCRwas subsequently
performed using 2 mL of diluted cDNA as the template and Taq as the
polymerase (Invitrogen). Primers bZIP21-F2 and bZIP21-R1, and bZIP21-F2 and
bZIP21-R2, were used for amplification of TGA9 transcripts designated full and
partial, respectively. Primers bZIP65cDNA-F1 and bZIP65cDNA-R2, and
bZIP65cDNA-F1 and bZip65R1-3 were used for amplification TGA10 transcripts
designated full and partial, respectively. GAPC encoding glyceraldehyde
3-phosphate dehydrogenase served as a control transcript (Hepworth et al.,
2005).
Yeast Two-Hybrid Assay
We used the GAL4-based yeast two-hybrid system described by Kohalmi
et al. (1998). To generate bait and prey constructs, the full coding sequences of
TGA9, TGA10, ROXY1, and ROXY2 were amplified by PCR from cloned cDNA
template. PCR productswere subcloned into pCR-BluntII-TOPO (Invitrogen) and
sequenced to ensure fidelity. Recognition sites for the restriction enzymes SalI and
NotI were incorporated into the 5# ends of primers to facilitate the subsequent
directional cloning of ROXY1/2 and TGA9/10 cDNA inserts into the corre-
sponding sites of bait (pBI-880) and prey (pBI-881) plasmids, respectively.
Construction of TGA2 in pBI-880, yeast transformation, and b-galactosidase
assays were described previously (Despres et al., 2000).
BiFC Assay
BiFC assays were performed as described by Li et al. (2009). pE-SPYNE and
pE-SPYCE (Walter et al., 2004) were used to generate expression vectors using
a Gateway cloning strategy (Invitrogen). The N terminus of YFP was cloned
in-frame upstream of ROXY1 and ROXY2 cDNAs. The C terminus of YFP was
fused in-frame to the N termini of TGA9 and TGA10 cDNAs. Fusion genes
were expressed under the control of a cauliflower mosaic virus 35S promoter.
Infiltrated Nicotiana benthamiana leaves were examined for reconstitution of
YFP fluorescence 3 to 5 d after inoculation using a LSM 510 Meta confocal
microscope (Carl Zeiss).
Scanning Electron Microscopy
Samples were prepared for scanning electron microscopy as described
previously (Hepworth et al., 2005) and imaged using a VegaII XMU VPSEM
apparatus (Tescan).
Sequence data have been deposited in GenBank under accession numbers
HQ132743 (TGA9 cDNA) and HQ132742 (TGA10 cDNA).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Phylogenetic tree for Arabidopsis TGA tran-
scription factors.
Supplemental Figure S2. Analysis of pollen in wild-type and tga9 tga10
mutant plants.
Supplemental Figure S3. Callose deposition in wild-type and tga9 tga10
mutant plants.
Supplemental Figure S4. Comparison of wild-type and tga9 tga10 late-
stage anther lobe development.
Supplemental Figure S5. Endothecium lignification of wild-type and tga9
tga10 anthers.
Supplemental Figure S6. In situ analysis of TGA10 expression in devel-
oping anthers.
Supplemental Figure S7. RT-PCR analysis of TGA9 and TGA10 transcript
in wild-type (Nossen-0) and roxy1 roxy2 apices.
Supplemental Figure S8. Quantitative RT-PCR analysis of tapetum tran-
scripts in wild-type, tga9 tga10, and roxy1 roxy2 apices.
Supplemental Figure S9.Alignment of selected TGA transcription factors.
Supplemental Table S1. List of primers.
Supplemental Table S2. List of primers for RT-PCR transcript analysis of
anther development genes.
Supplemental Materials and Methods S1.
ACKNOWLEDGMENTS
We thank Laurian S. Robert and Ann-Fook Yang at the Eastern Cereal and
Oilseed Research Centre, Agriculture and Agri-Food Canada, Ottawa, for use
of their ultramicrotome facility.
Received May 14, 2010; accepted August 27, 2010; published August 30, 2010.
LITERATURE CITED
Aarts MGM, Hodge R, Kalantidis K, Florack D, Wilson ZA, Mulligan BJ,
Stiekema WJ, Scott R, Pereira A (1997) The Arabidopsis MALE
STERILITY 2 protein shares similarity with reductases in elongation/
condensation complexes. Plant J 12: 615–623
Alexander MP (1969) Differential staining of aborted and nonaborted
pollen. Stain Technol 44: 117–122
Alvarez S, ZhuM, Chen S (2009) Proteomics of Arabidopsis redox proteins
in response to methyl jasmonate. J Proteomics 73: 30–40
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative
stress, and signal transduction. Annu Rev Plant Biol 55: 373–399
Ariizumi T, Hatakeyama K, Hinata K, Inatsugi R, Nishida I, Sato S, Kato
T, Tabata S, Toriyama K (2004) Disruption of the novel plant protein
NEF1 affects lipid accumulation in the plastids of the tapetum and exine
formation of pollen, resulting in male sterility in Arabidopsis thaliana.
Plant J 39: 170–181
Ariizumi T, Hatakeyama K, Hinata K, Sato S, Kato T, Tabata S, Toriyama
K (2003) A novel male-sterile mutant of Arabidopsis thaliana, faceless
pollen-1, produces pollen with a smooth surface and an acetolysis-
sensitive exine. Plant Mol Biol 53: 107–116
Bennett SRM, Alvarez J, Bossinger G, Smyth DR (1995) Morphogenesis in
pinoid mutants of Arabidopsis thaliana. Plant J 8: 505–520
Boyle P, Le Su E, Rochon A, Shearer HL, Murmu J, Chu JY, Fobert PR,
Despres C (2009) The BTB/POZ domain of the Arabidopsis disease
resistance protein NPR1 interacts with the repression domain of TGA2
to negate its function. Plant Cell 21: 3700–3713
Murmu et al.
1502 Plant Physiol. Vol. 154, 2010

Buchanan BB, Balmer Y (2005) Redox regulation: a broadening horizon.
Annu Rev Plant Biol 56: 187–220
Canales C, Bhatt AM, Scott R, Dickinson H (2002) EXS, a putative LRR
receptor kinase, regulates male germline cell number and tapetal
identity and promotes seed development in Arabidopsis. Curr Biol 12:
1718–1727
Cecchetti V, Altamura MM, Falasca G, Costantino P, Cardarelli M (2008)
Auxin regulates Arabidopsis anther dehiscence, pollen maturation, and
filament elongation. Plant Cell 20: 1760–1774
Chen W, Singh KB (1999) The auxin, hydrogen peroxide and salicylic acid
induced expression of the Arabidopsis GST6 promoter is mediated in
part by an ocs element. Plant J 19: 667–677
Chitwood DH, Guo M, Nogueira FTS, Timmermans MCP (2007) Estab-
lishing leaf polarity: the role of small RNAs and positional signals in the
shoot apex. Development 134: 813–823
Chuang CF, Running MP, Williams RW, Meyerowitz EM (1999) The
PERIANTHIA gene encodes a bZIP protein involved in the determina-
tion of floral organ number in Arabidopsis thaliana. Genes Dev 13:
334–344
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agro-
bacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:
735–743
Despres C, Chubak C, Rochon A, Clark R, Bethune T, Desveaux D, Fobert
PR (2003) The Arabidopsis NPR1 disease resistance protein is a novel
cofactor that confers redox regulation of DNA binding activity to the
basic domain/leucine zipper transcription factor TGA1. Plant Cell 15:
2181–2191
Despres C, DeLong C, Glaze S, Liu E, Fobert PR (2000) The Arabidopsis
NPR1/NIM1 protein enhances the DNA binding activity of a subgroup
of the TGA family of bZIP transcription factors. Plant Cell 12: 279–290
Dietz KJ (2008) Redox signal integration: from stimulus to networks and
genes. Physiol Plant 133: 459–468
Dinneny JR, Weigel D, Yanofsky MF (2006) NUBBIN and JAGGED define
stamen and carpel shape in Arabidopsis. Development 133: 1645–1655
Dong X, Hong Z, Sivaramakrishnan M, Mahfouz M, Verma DPS (2005)
Callose synthase (CalS5) is required for exine formation during micro-
gametogenesis and for pollen viability in Arabidopsis. Plant J 42:
315–328
Fan LM, Wang YF, Wang H, Wu WH (2001) In vitro Arabidopsis pollen
germination and characterization of the inward potassium currents in
Arabidopsis pollen grain protoplasts. J Exp Bot 52: 1603–1614
Fode B, Siemsen T, Thurow C, Weigel R, Gatz C (2008) The Arabidopsis
GRAS protein SCL14 interacts with class II TGA transcription factors
and is essential for the activation of stress-inducible promoters. Plant
Cell 20: 3122–3135
Gapper C, Dolan L (2006) Control of plant development by reactive oxygen
species. Plant Physiol 141: 341–345
Gechev TS, Van Breusegem F, Stone JM, Denev I, Laloi C (2006) Reactive
oxygen species as signals that modulate plant stress responses and
programmed cell death. Bioessays 28: 1091–1101
Hepworth SR, Valverde F, Ravenscroft D, Mouradov A, Coupland G
(2002) Antagonistic regulation of flowering-time gene SOC1 by CON-
STANS and FLC via separate promoter motifs. EMBO J 21: 4327–4337
Hepworth SR, Zhang Y, McKim S, Li X, Haughn GW (2005) BLADE-ON-
PETIOLE-dependent signaling controls leaf and floral patterning in
Arabidopsis. Plant Cell 17: 1434–1448
Higginson T, Li SF, Parish RW (2003) AtMYB103 regulates tapetum and
trichome development in Arabidopsis thaliana. Plant J 35: 177–192
Huffaker A, Ryan CA (2007) Endogenous peptide defense signals in
Arabidopsis differentially amplify signaling for the innate immune
response. Proc Natl Acad Sci USA 104: 10732–10736
Ishiguro S, Kawai-Oda A, Ueda J, Nishida I, Okada K (2001) The
DEFECTIVE IN ANTHER DEHISCIENCE gene encodes a novel phos-
pholipase A1 catalyzing the initial step of jasmonic acid biosynthesis,
which synchronizes pollen maturation, anther dehiscence, and flower
opening in Arabidopsis. Plant Cell 13: 2191–2209
Ito T, Nagata N, Yoshiba Y, Ohme-Takagi M, Ma H, Shinozaki K (2007)
Arabidopsis MALE STERILITY1 encodes a PHD-type transcription fac-
tor and regulates pollen and tapetum development. Plant Cell 19:
3549–3562
Ito T, Wellmer F, Yu H, Das P, Ito N, Alves-Ferreira M, Riechmann JL,
Meyerowitz EM (2004) The homeotic protein AGAMOUS controls
microsporogenesis by regulation of SPOROCYTELESS. Nature 430:
356–360
Jakoby M, Weisshaar B, Droge-Laser W, Vicente-Carbajosa J, Tiedemann
J, Kroj T, Parcy F; bZIP Research Group (2002) bZIP transcription
factors in Arabidopsis. Trends Plant Sci 7: 106–111
Johnson-Brousseau SA, McCormick S (2004) A compendium of methods
useful for characterizing Arabidopsis pollen mutants and gametophyti-
cally-expressed genes. Plant J 39: 761–775
Kesarwani M, Yoo J, Dong X (2007) Genetic interactions of TGA transcrip-
tion factors in the regulation of pathogenesis-related genes and disease
resistance in Arabidopsis. Plant Physiol 144: 336–346
Kohalmi SE, Reader LJW, Samach A, Nowak J, Haughn GW, Crosby WL
(1998) Identification and characterization of protein interactions
using the yeast 2-hybrid system. In SB Gelvin, RA Schilperoot, eds,
Plant Molecular Biology Manual M1. Kluwer Academic Publishers,
Dordrecht, The Netherlands, pp 1–30
Koncz C, Schell J (1986) The promoter of T1-DNA gene 5 controls the
tissue-specific expression of chimeric genes carried by a novel type of
Agrobacterium binary vector. Mol Gen Genet 204: 383–396
Lamb P, McKnight SL (1991) Diversity and specificity in transcriptional
regulation: the benefits of heterotypic dimerization. Trends Biochem Sci
16: 417–422
Li S, Lauri A, Ziemann M, Busch A, Bhave M, Zachgo S (2009) Nuclear
activity of ROXY1, a glutaredoxin interacting with TGA factors, is
required for petal development in Arabidopsis thaliana. Plant Cell 21:
429–441
Ma H (2005) Molecular genetic analyses of microsporogenesis and micro-
gametogenesis in flowering plants. Annu Rev Plant Biol 56: 393–434
Mandaokar A, Browse J (2009) MYB108 acts together with MYB24 to
regulate jasmonate-mediated stamen maturation in Arabidopsis. Plant
Physiol 149: 851–862
Millar AA, Gubler F (2005) The Arabidopsis GAMYB-like genes, MYB33
and MYB65, are microRNA-regulated genes that redundantly facilitate
anther development. Plant Cell 17: 705–721
Mitsuda N, Seki M, Shinozaki K, Ohme-Takagi M (2005) The NAC
transcription factors NST1 and NST2 of Arabidopsis regulate secondary
wall thickenings and are required for anther dehiscence. Plant Cell 17:
2993–3006
Mizuno S, Osakabe Y, Maruyama K, Ito T, Osakabe K, Sato T, Shinozaki
K, Yamaguchi-Shinozaki K (2007) Receptor-like protein kinase 2 (RPK
2) is a novel factor controlling anther development in Arabidopsis
thaliana. Plant J 50: 751–766
Mueller S, Hilbert B, Dueckershoff K, Roitsch T, Krischke M, Mueller
MJ, Berger S (2008) General detoxification and stress responses are
mediated by oxidized lipids through TGA transcription factors in
Arabidopsis. Plant Cell 20: 768–785
Nagpal P, Ellis CM, Weber H, Ploense SE, Barkawi LS, Guilfoyle TJ,
Hagen G, Alonso JM, Cohen JD, Farmer EE, et al (2005) Auxin response
factors ARF6 and ARF8 promote jasmonic acid production and flower
maturation. Development 132: 4107–4118
Ndamukong I, Abdallat AA, Thurow C, Fode B, Zander M, Weigel R,
Gatz C (2007) SA-inducible Arabidopsis glutaredoxin interacts with
TGA factors and suppresses JA-responsive PDF1.2 transcription. Plant J
50: 128–139
Ogawa M, Kay P, Wilson S, Swain SM (2009) ARABIDOPSIS DEHIS-
CENCE ZONE POLYGALACTURONASE1 (ADPG1), ADPG2, and
QUARTET2 are polygalacturonases required for cell separation during
reproductive development in Arabidopsis. Plant Cell 21: 216–233
Paulsen CE, Carroll KS (2010) Orchestrating redox signaling networks
through regulatory cysteine switches. ACS Chem Biol 5: 47–62
Paxson-Sowders DM, Dodrill CH, Owen HA, Makaroff CA (2001) DEX1,
a novel plant protein, is required for exine pattern formation during
pollen development in Arabidopsis. Plant Physiol 127: 1739–1749
Pekker I, Alvarez JP, Eshed Y (2005) Auxin response factors mediate
Arabidopsis organ asymmetry via modulation of KANADI activity. Plant
Cell 17: 2899–2910
Rochon A, Boyle P, Wignes T, Fobert PR, Despres C (2006) The coactivator
function of Arabidopsis NPR1 requires the core of its BTB/POZ domain
and the oxidation of C-terminal cysteines. Plant Cell 18: 3670–3685
Running MP, Meyerowitz EM (1996) Mutations in the PERIANTHIA gene
of Arabidopsis specifically alter floral organ number and initiation
pattern. Development 122: 1261–1269
Sanders PM, Bui AQ, Le BH, Goldberg RB (2005) Differentiation and
TGA9 and TGA10 Are Required for Anther Development
Plant Physiol. Vol. 154, 2010 1503

degeneration of cells that play a major role in tobacco anther dehiscence.
Sex Plant Reprod 17: 219–241
Sanders PM, Bui AQ, Weterings K, McIntire KN, Hsu YC, Lee PY, Truong
MT, Beals TP, Goldberg RB (1999) Anther developmental defects in
Arabidopsis thaliana male-sterile mutants. Sex Plant Reprod 11: 297–322
Sanders PM, Lee PY, Biesgen C, Boone JD, Beals TP, Weiler EW, Goldberg
RB (2000) The Arabidopsis DELAYED DEHISCENCE1 gene encodes an
enzyme in the jasmonic acid synthesis pathway. Plant Cell 12: 1041–1061
Schiefthaler U, Balasubramanian S, Sieber P, Chevalier D, Wisman E,
Schneitz K (1999) Molecular analysis of NOZZLE, a gene involved in
pattern formation and early sporogenesis during sex organ develop-
ment in Arabidopsis thaliana. Proc Natl Acad Sci USA 96: 11664–11669
Schiermeyer A, Thurow C, Gatz C (2003) Tobacco bZIP factor TGA10 is a
novel member of the TGA family of transcription factors. Plant Mol Biol
51: 817–829
Scott RJ, Spielman M, Dickinson HG (2004) Stamen structure and func-
tion. Plant Cell (Suppl) 16: S46–S60
Sessions A, Nemhauser JL, McColl A, Roe JL, Feldmann KA, Zambryski
PC (1997) ETTIN patterns the Arabidopsis floral meristem and repro-
ductive organs. Development 124: 4481–4491
Sieburth LE, Meyerowitz EM (1997) Molecular dissection of the AGA-
MOUS control region shows that cis elements for spatial regulation are
located intragenically. Plant Cell 9: 355–365
Smyth DR, Bowman JL, Meyerowitz EM (1990) Early flower development
in Arabidopsis. Plant Cell 2: 755–767
Sorensen AM, Krober S, Unte US, Huijser P, Dekker K, Saedler H (2003)
The Arabidopsis ABORTED MICROSPORES (AMS) gene encodes a
MYC class transcription factor. Plant J 33: 413–423
Steiner-Lange S, Unte US, Eckstein L, Yang C, Wilson ZA, Schmelzer E,
Dekker K, Saedler H (2003) Disruption of Arabidopsis thaliana MYB26
results in male sterility due to non-dehiscent anthers. Plant J 34: 519–528
Tabata R, Ikezaki M, Fujibe T, Aida M, Tian CE, Ueno Y, Yamamoto KT,
Machida Y, Nakamura K, Ishiguro S (2010) Arabidopsis auxin response
factor6 and 8 regulate jasmonic acid biosynthesis and floral organ
development via repression of class 1 KNOX genes. Plant Cell Physiol
51: 164–175
Toriba T, Suzaki T, Yamaguchi T, Ohmori Y, Tsukaya H, Hirano HY (2010)
Distinct regulation of adaxial-abaxial polarity in anther patterning in
rice. Plant Cell 22: 1452–1462
Unte US, Sorensen AM, Pesaresi P, Gandikota M, Leister D, Saedler H,
Huijser P (2003) SPL8, an SBP-box gene that affects pollen sac devel-
opment in Arabidopsis. Plant Cell 15: 1009–1019
Walsh J, Freeling M (1999) The liguleless2 gene of maize functions during
the transition from the vegetative to the reproductive shoot apex. Plant J
19: 489–495
Walsh J, Waters CA, Freeling M (1998) The maize gene liguleless2 encodes a
basic leucine zipper protein involved in the establishment of the leaf
blade-sheath boundary. Genes Dev 12: 208–218
Walter M, Chaban C, Schutze K, Batistic O, Weckermann K, Nake C,
Blazevic D, Grefen C, Schumacher K, Oecking C, et al (2004) Visual-
ization of protein interactions in living plant cells using bimolecular
fluorescence complementation. Plant J 40: 428–438
Wasternack C (2007) Jasmonates: an update on biosynthesis, signal trans-
duction and action in plant stress response, growth and development.
Ann Bot (Lond) 100: 681–697
Wilson ZA, Morroll SM, Dawson J, Swarup R, Tighe PJ (2001) The
Arabidopsis MALE STERILITY1 (MS1) gene is a transcriptional regu-
lator of male gametogenesis, with homology to the PHD-finger family of
transcription factors. Plant J 28: 27–39
Wilson ZA, Zhang DB (2009) From Arabidopsis to rice: pathways in pollen
development. J Exp Bot 60: 1479–1492
Wu HM, Cheung AY (2000) Programmed cell death in plant reproduction.
Plant Mol Biol 44: 267–281
Xiang C, Miao Z, Lam E (1997) DNA-binding properties, genomic organi-
zation and expression pattern of TGA6, a new member of the TGA
family of bZIP transcription factors in Arabidopsis thaliana. Plant Mol
Biol 34: 403–415
Xiang C, Miao ZH, Lam E (1996) Coordinated activation of as-1-type
elements and a tobacco glutathione S-transferase gene by auxins,
salicylic acid, methyl-jasmonate and hydrogen peroxide. Plant Mol
Biol 32: 415–426
Xie DX, Feys BF, James S, Nieto-Rostro M, Turner JG (1998) COI1: an
Arabidopsis gene required for jasmonate-regulated defense and fertil-
ity. Science 280: 1091–1094
Xing S, Rosso MG, Zachgo S (2005) ROXY1, a member of the plant
glutaredoxin family, is required for petal development in Arabidopsis
thaliana. Development 132: 1555–1565
Xing S, Zachgo S (2008) ROXY1 and ROXY2, two Arabidopsis glutaredoxin
genes, are required for anther development. Plant J 53: 790–801
Xu M, Hu T, McKim SM, Murmu J, Haughn GW, Hepworth SR (2010)
Arabidopsis BLADE-ON-PETIOLE1 and 2 promote floral meristem
fate and determinacy in a previously undefined pathway targeting
APETALA1 and AGAMOUS-LIKE24. Plant J 63: 974–989
Yang C, Vizcay-Barrena G, Conner K, Wilson ZA (2007a) MALE STERIL-
ITY1 is required for tapetal development and pollen wall biosynthesis.
Plant Cell 19: 3530–3548
Yang C, Xu Z, Song J, Conner K, Vizcay Barrena G, Wilson ZA (2007b)
Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening
in the endothecium and is essential for anther dehiscence. Plant Cell 19:
534–548
Yang SL, Xie LF, Mao HZ, Puah CS, YangWC, Jiang L, Sundaresan V, Ye D
(2003) Tapetum determinant1 is required for cell specialization in the
Arabidopsis anther. Plant Cell 15: 2792–2804
Yang WC, Ye D, Xu J, Sundaresan V (1999) The SPOROCYTELESS gene of
Arabidopsis is required for initiation of sporogenesis and encodes a
novel nuclear protein. Genes Dev 13: 2108–2117
Zhang W, Sun Y, Timofejeva L, Chen C, Grossniklaus U, Ma H (2006)
Regulation of Arabidopsis tapetum development and function by
DYSFUNCTIONAL TAPETUM1 (DYT1) encoding a putative bHLH
transcription factor. Development 133: 3085–3095
Zhang Y, Tessaro MJ, Lassner M, Li X (2003) Knockout analysis of
Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their
redundant and essential roles in systemic acquired resistance. Plant Cell
15: 2647–2653
Zhang ZB, Zhu J, Gao JF, Wang C, Li H, Li H, Zhang HQ, Zhang S, Wang
DM, Wang QX, et al (2007) Transcription factor AtMYB103 is required
for anther development by regulating tapetum development, callose
dissolution and exine formation in Arabidopsis. Plant J 52: 528–538
Zhao DZ, Wang GF, Speal B, Ma H (2002) The excess microsporocytes1
gene encodes a putative leucine-rich repeat receptor protein kinase that
controls somatic and reproductive cell fates in the Arabidopsis anther.
Genes Dev 16: 2021–2031
Zhu J, Chen H, Li H, Gao JF, Jiang H, Wang C, Guan YF, Yang ZN (2008)
Defective in tapetal development and function 1 is essential for anther
development and tapetal function for microspore maturation in Arabi-
dopsis. Plant J 55: 266–277
Murmu et al.
1504 Plant Physiol. Vol. 154, 2010
Copyright © 2022 FDOKUMEN




















