
Aptamers are an innovative and promising tool for phytoplankton taxonomy and biodiversity research
14
This article was downloaded by: [gianluca santamaria] On: 16 January 2015, At: 05:06 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Click for updates Chemistry and Ecology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/gche20 Aptamers are an innovative and promising tool for phytoplankton taxonomy and biodiversity research Gianluca Santamaria abc , Carla Lucia Esposito d , Laura Cerchia d , Giovanna Benvenuto a , Deepak Nanjappa a , Diana Sarno a , Adriana Zingone a , Vittorio De Franciscis d & Maurizio Ribera d'Alcalà a a Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy b PhD Program in Bioinformatics, Sannio University, Via Port'Arsa, 11, 82100 Benevento, Italy c Laboratorio di Medicina Molecolare e Genomica dell'Universitá degli Studi di Salerno, Via S. Allende, 1, 84081 Baronissi (Sa), Italy d Istituto per l'Endocrinologia e Oncologia Sperimentale “G. Salvatore” — CNR, Via Pansini 5, 80131 Napoli, Italy Published online: 13 Jan 2015. To cite this article: Gianluca Santamaria, Carla Lucia Esposito, Laura Cerchia, Giovanna Benvenuto, Deepak Nanjappa, Diana Sarno, Adriana Zingone, Vittorio De Franciscis & Maurizio Ribera d'Alcalà (2015): Aptamers are an innovative and promising tool for phytoplankton taxonomy and biodiversity research, Chemistry and Ecology, DOI: 10.1080/02757540.2014.966700 To link to this article: http://dx.doi.org/10.1080/02757540.2014.966700 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Aptamers are an innovative and promising tool for phytoplankton taxonomy and biodiversity research

This article was downloaded by: [gianluca santamaria]On: 16 January 2015, At: 05:06Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Click for updates
Chemistry and EcologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/gche20
Aptamers are an innovative andpromising tool for phytoplanktontaxonomy and biodiversity researchGianluca Santamariaabc, Carla Lucia Espositod, Laura Cerchiad,Giovanna Benvenutoa, Deepak Nanjappaa, Diana Sarnoa, AdrianaZingonea, Vittorio De Franciscisd & Maurizio Ribera d'Alcalàa
a Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli,Italyb PhD Program in Bioinformatics, Sannio University, Via Port'Arsa,11, 82100 Benevento, Italyc Laboratorio di Medicina Molecolare e Genomica dell'Universitádegli Studi di Salerno, Via S. Allende, 1, 84081 Baronissi (Sa), Italyd Istituto per l'Endocrinologia e Oncologia Sperimentale “G.Salvatore” — CNR, Via Pansini 5, 80131 Napoli, ItalyPublished online: 13 Jan 2015.
To cite this article: Gianluca Santamaria, Carla Lucia Esposito, Laura Cerchia, Giovanna Benvenuto,Deepak Nanjappa, Diana Sarno, Adriana Zingone, Vittorio De Franciscis & Maurizio Ribera d'Alcalà(2015): Aptamers are an innovative and promising tool for phytoplankton taxonomy and biodiversityresearch, Chemistry and Ecology, DOI: 10.1080/02757540.2014.966700
To link to this article: http://dx.doi.org/10.1080/02757540.2014.966700
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.

This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

Chemistry and Ecology, 2015http://dx.doi.org/10.1080/02757540.2014.966700
Aptamers are an innovative and promising tool forphytoplankton taxonomy and biodiversity research
Gianluca Santamariaa,b,c,†, Carla Lucia Espositod, Laura Cerchiad, Giovanna Benvenutoa,Deepak Nanjappaa, Diana Sarnoa, Adriana Zingonea, Vittorio De Franciscisd and
Maurizio Ribera d’Alcalàa∗
aStazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy; bPhD Program in Bioinformatics,Sannio University, Via Port’Arsa, 11, 82100 Benevento, Italy; cLaboratorio di Medicina Molecolare e
Genomica dell’Universitá degli Studi di Salerno, Via S. Allende, 1, 84081 Baronissi (Sa), Italy d Istituto perl’Endocrinologia e Oncologia Sperimentale “G. Salvatore” — CNR, Via Pansini 5, 80131 Napoli, Italy
(Received 28 April 2014; final version received 3 September 2014)
Nucleic acid aptamers are small-size ligands that selectively bind to molecular segments even when theyprotrude from cell surfaces. Due to their high specificity, aptamers are widely used in biomedical researchand as probes for different applications. Here, we tested whether aptamers can also discriminate amongphytoplankton cells. As a proof of concept we focused on the widespread centric diatom Leptocylindrusdanicus and generated two aptamers that selectively bind to its cell surface. The aptamers did not bindto other diatoms tested, which included both pennate (Pseudo-nitzschia multistriata) and centric (Skele-tonema marinoi, Chaetoceros socialis) species. They also showed negative binding to closely relatedspecies (Tenuicylindrus belgicus, Leptocylindrus aporus, Leptocylindrus convexus), which are hardly rec-ognizable with microscopy techniques. In addition, aptamers discriminated also among cells of the sameclone, suggesting a potential use of aptamers as clone-specific/stage-specific probes to track phytoplank-ton life stages in their natural environment. While the method still needs to be tested with natural algalsamples, it can complement in a unique way the existing approaches to discriminate among species andpossibly life stages of marine phytoplankton. The method can find useful application in taxonomic andecological studies as well as in environmental monitoring including early warning strategies.
Keywords: diatoms; detection methods; aptamers
1. Introduction
Identification of phytoplankton is historically based on observations with a microscope to detectcell shape and size, along with color and other morphological details concurring to character-ize a taxon, either as vegetative or resting stage. In many cases, only expert taxonomists areable to recognize differences that are often subtle and identify species correctly. This is oneof the reasons why data on taxonomic composition of phytoplankton communities are so rare
*Corresponding author. Email: [email protected]†Current address: Medicina Sperimentale e Clinica, Universita’ degli studi ‘Magna Graecia’, Viale Europa, 88100,Catanzaro, Italy.
© 2015 Taylor & Francis
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

2 G. Santamaria et al.
as compared with data on their bulk properties such as biomass, pigment composition and pri-mary production. Over the time, interest has grown in understanding not only the dynamics ofspecific algal blooms, because they are crucial events in biogeochemical cycles, but also theirpossible toxicity and their effects on marine organisms and sea food contamination. Hence aquick and reliable identification of phytoplankton species, or even clones and stages, has becomeincreasingly indispensable.
The phylogenetic and evolutionary distance among phytoplankton groups is conspicuous.[1,2]Relevant differences at the biomolecular level are then expected, despite the evident convergencetoward being unicellular and at times colonial. Therefore, without abandoning the insight pro-vided by morphological characterization which hints at possible adaptive traits linked to cellshapes, an increasing effort to better assess and quantify phytoplankton biodiversity is currentlydevoted to methods based on molecular differences.[3,4] Several methods have been developedfor molecular identification of phytoplankton. Non-coding nuclear (18S, ITS and 28S rDNA) orchloroplastic (16 rDNA, rbcL) DNA sequences [5–7] have been used as common markers formolecular taxonomy and phylogeny, whereas more variable markers such as microsatellites [8]are used to address diversity at the population level. Thanks to these methods it has been possibleto identify many cases of taxa that display subtle morphological differences with their siblingsbut show clear differences at the molecular level [9,10] and are often sexually incompatible [11]thus extending the concept of cryptic species to phytoplankton. The problem of phytoplank-ton identification in natural samples, already relevant in the case of morphologically distinctspecies, is obviously more serious in these cases of species with small or no morphologicaldifferences.
Fluorescent in situ hybridization probes (FISH),[12,13] microarrays [14] and real-time PCR(RT-PCR),[15] which imply the binding of species-specific molecular probes to nucleic acids inwhole cells (FISH) or cell extracts (RT-PCR and Microarrays) have also been developed.
Among those methods, FISH is the only one allowing the observation of the whole cellsafter treatment, but problems are often encountered with the size of the probe and the needof its diffusion toward the cell interior to reach the target molecules. In addition, molecular dif-ferences may be limited to few base pairs, whereas intraspecific molecular diversity may behigh and basically unknown, which leads to several drawbacks in terms of effectiveness andprecision of the method. Finally, none of the methods developed so far can be used on livecells.[13–15]
To overcome all the above-mentioned constraints we decided to explore another class ofprobes, highly successful in biomedical sciences, the aptamers.[16–18] These are oligomers of,preferentially, RNA, DNA or modified RNA, with considerable properties (low size, no immuno-genicity, chemical structure that can be easily modified to improve their in vivo applications).In addition, they display high specificity in binding to different classes of molecules (proteins,polysaccharides, etc.) even when they protrude from cell walls [19] making them useful probesalso to tag whole cells.[20–22]
Since their discovery in the late 1990s (RNA aptamers [23] and DNA aptamers [24]), therehave been constant efforts in harnessing the potential of aptamers as biosensors for biotoxinsand biological pathogens. As biosensors, aptamers are used to detect infectious bacteria [25] andviruses.[26] However, to the best of our knowledge no exploration has been conducted so faron the potential of using aptamers to tag selectively unicellular marine microorganisms, eitherprokaryotic or eukaryotic, while operating in their salty natural medium.
Here we describe a method that allowed the efficient generation of specific 2′-fluoro pyrim-idine aptamers for the unequivocal identification of the marine diatom Leptocylindrus danicusagainst congeneric species or other diatoms, and discuss future steps and perspectives of thisapproach.
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15
Chemistry and Ecology 3
2. Materials and methods
2.1. Overview of the method
The first step is the generation of a nucleic acid library (RNA, DNA or modified RNA) of ∼ 4Ndifferent molecules (N ≥ 24) by randomization of a synthetic sequence stretch from 22 up to100 nucleotides in length. The sequences obtained have a vast array of different conformationswith different binding properties. Subsequently, sequences are submitted to a procedure namedSELEX (Systematic Evolution of Ligands by Exponential Enrichment),[23,27] which allowsisolation of aptamers that specifically bind to a selected target. The target (molecule, cell, tis-sue, etc.) to be tagged is called ‘selection’, whereas the other entities of the same kind to bediscriminated against are called ‘counter-selection’.
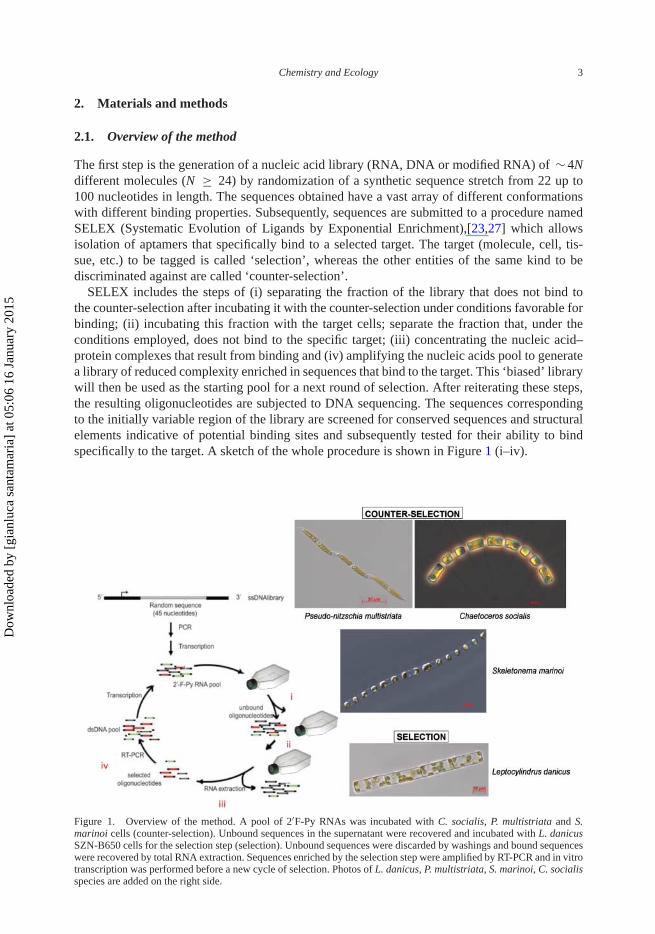
SELEX includes the steps of (i) separating the fraction of the library that does not bind tothe counter-selection after incubating it with the counter-selection under conditions favorable forbinding; (ii) incubating this fraction with the target cells; separate the fraction that, under theconditions employed, does not bind to the specific target; (iii) concentrating the nucleic acid–protein complexes that result from binding and (iv) amplifying the nucleic acids pool to generatea library of reduced complexity enriched in sequences that bind to the target. This ‘biased’ librarywill then be used as the starting pool for a next round of selection. After reiterating these steps,the resulting oligonucleotides are subjected to DNA sequencing. The sequences correspondingto the initially variable region of the library are screened for conserved sequences and structuralelements indicative of potential binding sites and subsequently tested for their ability to bindspecifically to the target. A sketch of the whole procedure is shown in Figure 1 (i–iv).
Figure 1. Overview of the method. A pool of 2′F-Py RNAs was incubated with C. socialis, P. multistriata and S.marinoi cells (counter-selection). Unbound sequences in the supernatant were recovered and incubated with L. danicusSZN-B650 cells for the selection step (selection). Unbound sequences were discarded by washings and bound sequenceswere recovered by total RNA extraction. Sequences enriched by the selection step were amplified by RT-PCR and in vitrotranscription was performed before a new cycle of selection. Photos of L. danicus, P. multistriata, S. marinoi, C. socialisspecies are added on the right side.
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

4 G. Santamaria et al.
2.2. Cell cultures
Type or epitype strains of Leptocylindrus aporus (French III & Hargraves) Nanjappa & Zingone(La, strain SZN-B651), L. convexus Nanjappa & Zingone (Lc, strain SZN-B778), L. danicusCleve (Ld, strain SZN-B650) and Tenuicylindrus belgicus (Meunier) Nanjappa & Zingone (Tb,strain SZN-B739) isolated in previous studies from the Gulf of Naples (GoN) [28] were usedin the current study. A culture of Skeletonema marinoi Sarno & Zingone (Sm) from the Adri-atic Sea was kindly provided by Alessandra Gallina (Bioengineering Department, University ofCalifornia San Diego, La Jolla, CA, USA, formerly at SZN). Other diatom cultures (Chaeto-ceros socialis, H.S. Lauder, Cs and Pseudo-nitzschia multistriata, Takano, Pm) were previouslyobtained from net samples collected at the LTER MareChiara (LTER-MC, 40.80°N, 14.25°E) inthe GoN and maintained in the culture collection of the Laboratory of Ecology and Evolution ofPlankton, Stazione Zoologica Anton Dohrn. During the experiments, individual strain cells wereinoculated from exponentially growing cultures into 300 mL polystyrene culture flasks contain-ing 200 mL of seawater K medium (salinity = 36.0) with silica.[29] The cultures were allowedto grow at 20°C, 100 µmol photons m−2 s−1 and a 12:12 light–dark cycle, until they reached acell density of about 106 cells mL−1.
2.3. SELEX and aptamers
Differential cell SELEX cycle was performed as previously described.[30] The starting RNApool was prepared by the transcription of a DNA library with a variable internal region of45 bp. In vitro transcription was performed in the presence of 1 mM 2′-fluoro-pyrimidine (2′F-Py) in place of natural pyrimidines by using a mutant variant of the T7 RNA polymerase (T7R&DNA polymerase, Epicentre Biotechonologies, Madison, WI) that allows the incorporationof the modified pyrimidines. Incorporation of 2′F-Py in the RNAs was used because of theirhigh resistance to nucleases degradation. The complexity of the starting pool was in the orderof 1014 sequences. The same procedure was used at each SELEX round to obtain the 2′F-PyRNA pool used for selection. In brief, 2′ F-Py RNAs (700 pmol) were heated at 85°C for 5min in 500 µL of distilled water, snap-cooled on ice for 2 min, and allowed to warm up toroom temperature before incubation with the cells. At each SELEX round, the selection stepwas preceded by a counter-selection step in order to deplete the RNA pool of sequences thatmay unspecifically bind the cell surface of L. danicus cells. To this end (counter-selection step),the starting pool (700 pmol) was first incubated for 60 min at 13°C on a rotating wheel with amix of Sm, Cs and Pn cells (6.6 × 105 cells for each diatom species) in a final volume of 15mL. For the selection step, the unbound sequences (supernatant) were recovered by centrifu-gation (2000 rpm, 15 min) and incubated with 106 L. danicus cells with the same volume andprocedure as above, in order to select sequences specifically recognizing this species. After theincubation, cells were centrifuged again (2000 rpm, 10 min), the supernatant was removed andthe pellet was washed (see below for the number of washes) with 1 mL of PBS 1 × . The suspen-sion was then centrifuged, the supernatant removed and the bound sequences were recovered bytotal RNA extraction using the TRIzol Reagent (Life Technologies) following the supplier usermanual.
Reverse transcription was performed with M-MLV Reverse Transcriptase (Roche) followingthe supplier’s instructions. Newly synthesized cDNA was amplified by PCR and will constitutethe DNA library to be transcribed for the following SELEX round (see above).
During the SELEX process, the selective pressure was progressively augmented by increas-ing the number of washing steps (one at the first round, two for washes at the third roundand up to three for the last three rounds); further, Polyinosine (Sigma, St. Louis, MO) wasadded during the incubation step at 100 µg mL−1 as the nonspecific competitor in the last three
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

Chemistry and Ecology 5
rounds; and the incubation time was decreased from 60 at the first round to 30 min in the lastround.
After six rounds of selection, the sequences recovered from each round were sequenced fol-lowing the Illumina MySeq sequence preparation with MySeq Illumina sequencer and analyzed.The two most abundant identified sequences were then custom synthesized by ChemGenesCorporation (Wilmington, MA USA, 01887).
2.4. Binding assay by quantitative reverse transcription-PCR (RT-qPCR)
We adopted the quantitative RT-qPCR reaction to evaluate the binding specificity and affinityof individual RNA sequences to the target cells. L. danicus cells and derivatives cells wereincubated with 100 and 200 nM of either Seq16, Seq20 or with starting RNA pool, used asthe reference control for nonspecific binding, for 30 min at 13°C in the presence of 100 µgmL−1 polyinosine as a nonspecific competitor in a final volume of 1 mL. After centrifugation,the unbound RNA (supernatant) was removed by washings with PBS (3 × 500 µL) and boundsequences (pellet) were recovered by extraction of the total RNA with the TRIzol reagent con-taining 0.5 pmole mL−1 of the unrelated CL4 aptamer [30] used as the internal reference fornormalization. The amount of recovered RNA from all the pellets was determined by perform-ing a two-step RT-qPCR protocol. In step one, recovered RNA was reverse transcribed usingM-MLV Reverse Transcriptase (Roche Diagnostics, Indianapolis, IN). The primers for the RTstep were either the Seq16 and Seq20 3′ primer (5′-GCCTGTTGTGAGCCTCCTGTCGAA-3′)or CL4 aptamer reference control 3′ primer (5′-GCCTCCTGTCGAATCGA-3′). The RT proto-col is as follows. The recovered RNA and the primers were heated at 65°C for 5 min, annealedat 22°C for 5 min and extended at 42°C for 30 min followed by and extension at 48°C for 30min. In step two, the product from the RT reaction was PCR amplified with iQ SYBR GreenSupermix (Bio-Rad, Hercules, CA) using the CFX96-C1000 thermal cycler sequence detectionsystem (Bio-Rad, Hercules, CA). The qPCR protocol is as follows. The RT product was heatedat 95°C for 2 min, followed by 45 cycles of heating at 95°C for 30 s, annealing at 55°C for 30 sand extending at 72°C for 30 s. Reactions were all done in 25 µL volume in triplicate. Data werenormalized to the CL4 reference control and to cell number determined by counting cells cul-tured in conjunction with each experiment. The ratio between the normalized aptamer amountbound to the cells was further normalized dividing that amount by the normalized amount of theoriginal pool bound to the same target (G0 in the following).
Normalized values where then used to determine the specificity of the aptamers for the targetspecies (selection) vs. other species (counter-selection or congeneric species).
The significance of binding affinities and specificities was assessed by a Student t-test on thetwo vectors of unpaired replicates.
Apparent Kd values and the values of the affinity constant of the specific aptamer for thebinding to the cell can be defined by the equation:
[C] = [Cmax][Apt]
Kd + [Apt],
with [C] being the concentration of the bound aptamer and [Apt] being the concentration of theaptamer in the medium (10, 25, 50, 100, 200 and 500 nM) determined for each aptamer by theLineweaver–Burk analysis according to the equation:
1
complex= Kd
[Cmax]× 1
[aptamer]+ 1
[Cmax].
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15
6 G. Santamaria et al.
2.5. Imaging of cell–aptamers complexes
Fluorescence labeling was performed in Eppendorf tubes in triplicate. Seq16 and Seq20 werelabeled with AlexaFluor-488 by using the Ulysis labeling kit (Molecular Probes) according tothe supplier’s instructions. In total, 2 × 105 cells were incubated with 50 nM of labeled Seq16or Seq20 in 500 µL of cultures media for 30 min at 18°C. After three washings with 300 µL ofthe culture medium, the cells were placed on glass bottom dishes (MatTek Corporation). Theimages were taken with a Zeiss Laser Confocal microscope LSM 510 META (Zeiss, Germany).
3. Results
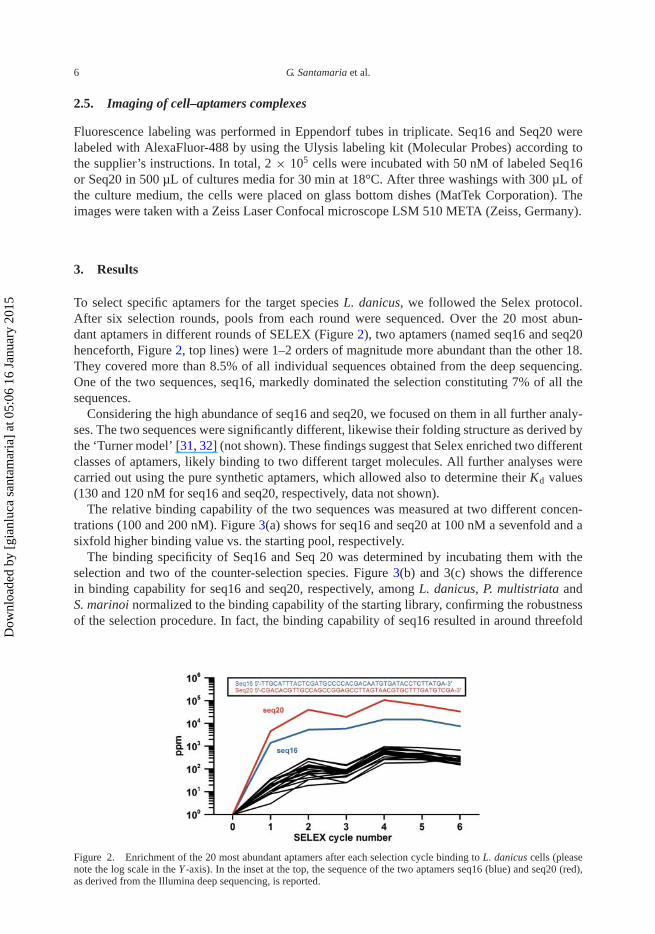
To select specific aptamers for the target species L. danicus, we followed the Selex protocol.After six selection rounds, pools from each round were sequenced. Over the 20 most abun-dant aptamers in different rounds of SELEX (Figure 2), two aptamers (named seq16 and seq20henceforth, Figure 2, top lines) were 1–2 orders of magnitude more abundant than the other 18.They covered more than 8.5% of all individual sequences obtained from the deep sequencing.One of the two sequences, seq16, markedly dominated the selection constituting 7% of all thesequences.
Considering the high abundance of seq16 and seq20, we focused on them in all further analy-ses. The two sequences were significantly different, likewise their folding structure as derived bythe ‘Turner model’ [31, 32] (not shown). These findings suggest that Selex enriched two differentclasses of aptamers, likely binding to two different target molecules. All further analyses werecarried out using the pure synthetic aptamers, which allowed also to determine their Kd values(130 and 120 nM for seq16 and seq20, respectively, data not shown).
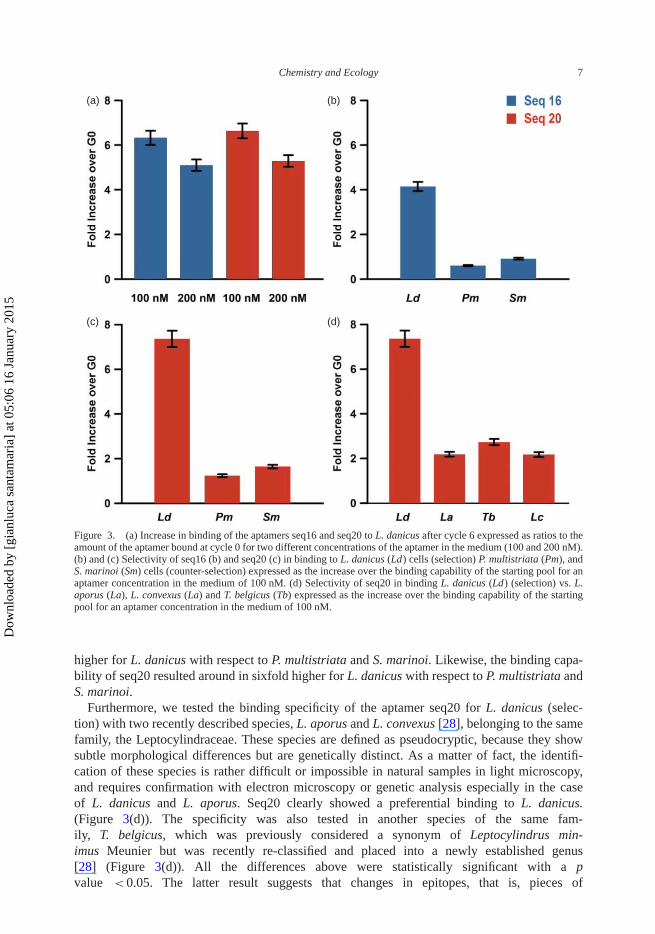
The relative binding capability of the two sequences was measured at two different concen-trations (100 and 200 nM). Figure 3(a) shows for seq16 and seq20 at 100 nM a sevenfold and asixfold higher binding value vs. the starting pool, respectively.
The binding specificity of Seq16 and Seq 20 was determined by incubating them with theselection and two of the counter-selection species. Figure 3(b) and 3(c) shows the differencein binding capability for seq16 and seq20, respectively, among L. danicus, P. multistriata andS. marinoi normalized to the binding capability of the starting library, confirming the robustnessof the selection procedure. In fact, the binding capability of seq16 resulted in around threefold
Figure 2. Enrichment of the 20 most abundant aptamers after each selection cycle binding to L. danicus cells (pleasenote the log scale in the Y -axis). In the inset at the top, the sequence of the two aptamers seq16 (blue) and seq20 (red),as derived from the Illumina deep sequencing, is reported.
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15
Chemistry and Ecology 7
(a) (b)
(c) (d)
Figure 3. (a) Increase in binding of the aptamers seq16 and seq20 to L. danicus after cycle 6 expressed as ratios to theamount of the aptamer bound at cycle 0 for two different concentrations of the aptamer in the medium (100 and 200 nM).(b) and (c) Selectivity of seq16 (b) and seq20 (c) in binding to L. danicus (Ld) cells (selection) P. multistriata (Pm), andS. marinoi (Sm) cells (counter-selection) expressed as the increase over the binding capability of the starting pool for anaptamer concentration in the medium of 100 nM. (d) Selectivity of seq20 in binding L. danicus (Ld) (selection) vs. L.aporus (La), L. convexus (La) and T. belgicus (Tb) expressed as the increase over the binding capability of the startingpool for an aptamer concentration in the medium of 100 nM.
higher for L. danicus with respect to P. multistriata and S. marinoi. Likewise, the binding capa-bility of seq20 resulted around in sixfold higher for L. danicus with respect to P. multistriata andS. marinoi.
Furthermore, we tested the binding specificity of the aptamer seq20 for L. danicus (selec-tion) with two recently described species, L. aporus and L. convexus [28], belonging to the samefamily, the Leptocylindraceae. These species are defined as pseudocryptic, because they showsubtle morphological differences but are genetically distinct. As a matter of fact, the identifi-cation of these species is rather difficult or impossible in natural samples in light microscopy,and requires confirmation with electron microscopy or genetic analysis especially in the caseof L. danicus and L. aporus. Seq20 clearly showed a preferential binding to L. danicus.(Figure 3(d)). The specificity was also tested in another species of the same fam-ily, T. belgicus, which was previously considered a synonym of Leptocylindrus min-imus Meunier but was recently re-classified and placed into a newly established genus[28] (Figure 3(d)). All the differences above were statistically significant with a pvalue < 0.05. The latter result suggests that changes in epitopes, that is, pieces of
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15
8 G. Santamaria et al.
(a) (b)
(c) (d)
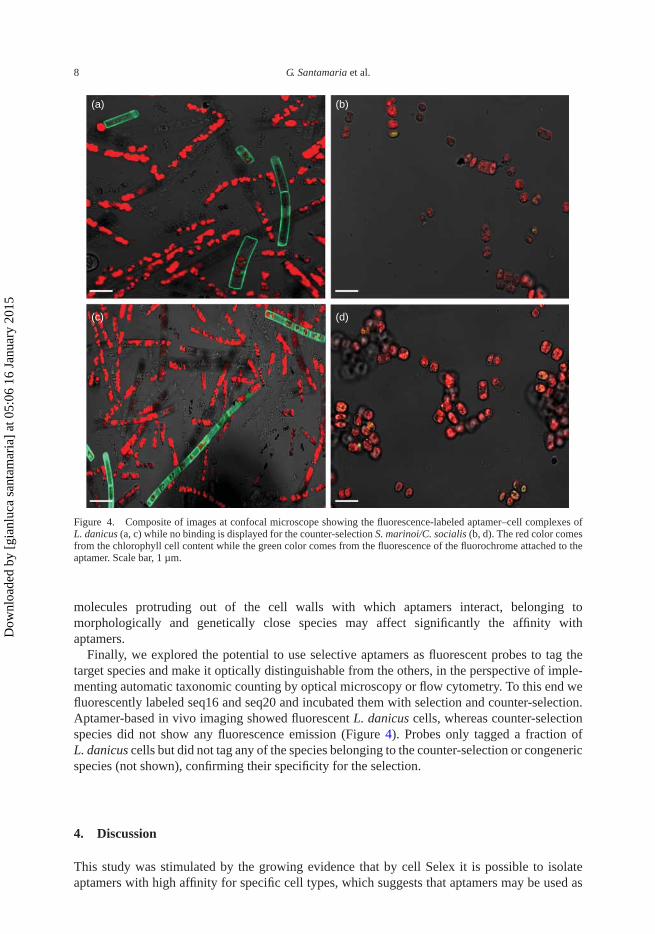
Figure 4. Composite of images at confocal microscope showing the fluorescence-labeled aptamer–cell complexes ofL. danicus (a, c) while no binding is displayed for the counter-selection S. marinoi/C. socialis (b, d). The red color comesfrom the chlorophyll cell content while the green color comes from the fluorescence of the fluorochrome attached to theaptamer. Scale bar, 1 µm.
molecules protruding out of the cell walls with which aptamers interact, belonging tomorphologically and genetically close species may affect significantly the affinity withaptamers.
Finally, we explored the potential to use selective aptamers as fluorescent probes to tag thetarget species and make it optically distinguishable from the others, in the perspective of imple-menting automatic taxonomic counting by optical microscopy or flow cytometry. To this end wefluorescently labeled seq16 and seq20 and incubated them with selection and counter-selection.Aptamer-based in vivo imaging showed fluorescent L. danicus cells, whereas counter-selectionspecies did not show any fluorescence emission (Figure 4). Probes only tagged a fraction ofL. danicus cells but did not tag any of the species belonging to the counter-selection or congenericspecies (not shown), confirming their specificity for the selection.
4. Discussion
This study was stimulated by the growing evidence that by cell Selex it is possible to isolateaptamers with high affinity for specific cell types, which suggests that aptamers may be used as
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

Chemistry and Ecology 9
effective molecular probes also for the discrimination among unicellular organisms belonging todifferent species, for example, phytoplankton.
Aptamers have several features that make them a very promising tool in the field of phyto-plankton identification. They are relatively cheap, once selected, and handy to manage becausetheir size (8–15 kDa) ranges between those of single chain antibody fragments and smallpeptides. They can be entirely chemically synthesized, thus avoiding complex manufacturingprocesses using cell-based (eukaryotic or prokaryotic) expression systems that are requiredfor antibodies production. They are low-immunogenic and low-toxic molecule, likely becausenucleic acids are not typically recognized by the human immune system as foreign agents.[33]They can support chemical modifications whilst maintaining their structure and function.
This first attempt targeted diatoms because of their global relevance in marine ecosystemdynamics.[34, 35] As the first test case we chose L. danicus because it is a widely distributedcoastal marine diatom species, which often co-occurs with the species we chose for the counter-selection.[36, 37] An additional challenge was the growing evidence that what it is normallyclassified as L. danicus is indeed a cluster of different species that co-exist in the same area show-ing an intriguing seasonal cycle.[28] This offered the opportunity for testing the discriminatingcapability of aptamers vs. congeneric, hardly distinguishable species.
The finding of specific aptamers binding to marine diatom cell surface was not an obviousresult, considering the particular structure of the diatom cell wall, with a thick siliceous theca,the frustule, perforated by minute pores (c. 20 nm), surrounding the cell membrane, and oftenfurther covered with mucopolymers. Indeed the presence of a frustule and of a mucous coveringhas been considered a reason why diatom viruses have been identified only for a few diatomspecies.[38,39]
In our first experiments, aptamers selectively bound the target organism but not all the cellsof the culture were labeled. This can be caused by problems with the protocol, which certainlydeserves more efforts toward optimization. However, it is also possible that the aptamer(s) testedwere specific for target molecules not always present on the cell surface. Indeed within a cultureindividual cells may differ in their cell cycle phase, physiological status and life cycle phase,which may be reflected at the cell surface level. In fact aptamers bind to epitopes, that is, piecesof molecules protruding out of the cell walls with which aptamers interact, which may varywith the physiological status of the cell even in the same species. Especially cells undergoinggametogenesis may produce specific substances that can favor cell contact and recognition ofthe opposite mating type.[40, 41] While this allows us to speculate on the high potential ofaptamers as powerful biomarkers for phytoplankton biology, this may hamper their use as probesfor quantitative determination of species abundance in a natural sample. However, the selectedaptamers did not attach to any cells of the counter-selection, which confirmed the robustness ofthe selection procedure.
The results presented in this contribution should be considered still preliminary, since wehave not tested yet the two sequences on a mixed population. This will be done in the nearfuture having synthetized an aptamer permanently bound to a fluorophore. In our case, withtwo different aptamers showing high affinity for the target cells, the plan is to use two differentfluorophores. To develop a robust protocol, more tests are needed on the optimal conditions forbinding, starting from synchronized cultures, with likely low intraspecific variability, to move onto explore more complex suspension.
Despite this preliminary stage, a few general comments can be made: (i) the aptamers appeara promising molecular probe for phytoplankton ecological and biological studies, deserving fur-ther investigation; (ii) they differ from all the other probes because they are the only ones, sofar, that can be used on live cells. Even though the selection of a highly specific aptamer is moreexpensive and time consuming than the generation of a FISH, the possibility of their use in vivomakes them worth the initial effort. In addition, once the sequence has been characterized, and
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

10 G. Santamaria et al.
the length optimized, which we have not yet done, it can be synthetized in great quantities atrelatively low cost and it is very stable; and (iii) the rapid advances in deep sequencing technolo-gies will certainly promote the design of molecular probes based on species-specific nucleotidesequences. However, the problem of discriminating among cells sharing the same, or very simi-lar, genome will be hardly solved with those probes. By contrast, the aptamers we isolated werecapable to do it for the three strains of Leptocylindrus.
Along this path, the additional potential of aptamers can be in the capability of discriminatingamong different stages of the same species. We did not test yet this capability but the fact thataptamers can discriminate in mammals among tumoral and healthy cells of the same individualand the same tissue strongly suggests that the potential is there. Therefore, any difference inthe molecules partially protruding from the cell wall associated with a different metabolic stateor life cycle stage could be potentially tagged by a proper aptamer. If our inference will beproved, aptamers can become a crucial tool to study phytoplankton biodiversity, biology andecology.
5. Conclusions
While still being in the stage of a proof-of-concept study we demonstrated that:
• SELEX can be used also in seawater and is effective in selecting aptamers that specificallybind to phytoplankton cells;
• it is possible to select species-specific aptamers for diatoms with a high discriminating power,for example, vs. other species of the same group;
• there is a great potential for aptamers to be able to bind selectively to one among different,cryptic taxa within a supposed unique species, thus offering a unique tool for overcoming thelimits of existing identification methods;
• it could be hypothesized that specific aptamers distinguish among cell phases or mating types,which is basically impossible with probes based on nucleotide sequences; and
• the synthesis of aptamers with an attached fluorochrome may be a complementary method toexisting FISH probes.
Because the use of selective external probes could be more rapid in tagging target cells thanprobes that need internalization, aptamers could also be better suited probes for in situ continuousflow systems. However, they should be tested on a mixed population. To the best of our knowl-edge this is the first time that RNA aptamers generated from cell-SELEX are used for in vivofluorescence imaging on diatoms. As molecular imaging probes, these aptamers showed biosta-bility, specificity and sensitivity for optical imaging. With these advantages, molecular imagingof diatoms can be performed with relative ease in laboratories.
In conclusion, we envision many areas of phytoplankton ecology where aptamers can be suc-cessfully developed and applied. These include discrimination, within a species, of cells that aresexually active from inactive ones, toxin-producing cells, or any other physiologically differentcells for which specific aptamers may be developed. Such applications could also be coupledwith flow cytometry for identification, quantification and enrichment of positive cells throughcell sorting. Thus, aptamer-based functional studies can contribute to solve complex biologi-cal problems, especially in the era of single-cell genomics. We believe that this method, whichresults from a joined effort of two different areas, biomedical science and ecology, while at a verypreliminary stage, is worth to be further explored because it can become a powerful tool pavingthe way to unprecedented opportunities for the study of the biology of phytoplankton species.
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

Chemistry and Ecology 11
Acknowledgements
The authors thank the Italian Flagship Programs RITMARE (grant MRdA) and NanoMax (DESIRED) (grant VdF) forpartially funding this research and the Joint SZN-Open University PhD program for supporting DN and GS. GS and DNwere supported by SZN PhD fellowship.
References
[1] Falkowski PG, Katz ME, Knoll AH, Quigg A, Raven JA, Schofield O, Taylor FJ. The evolution of moderneukaryotic phytoplankton. Science. 2004;305:354–360.
[2] Not F, Siano R, Kooistra WHCF, Simon N, Vaulot D, Probert I. Diversity and ecology of eukaryotic marinephytoplankton. Adv Bot Res. 2012;64:1–53.
[3] de Bruin A, Ibelings BW, Van Donk E. Molecular techniques in phytoplankton research: from allozymeelectrophoresis to genomics. Hydrobiologia. 2003;491:47–63.
[4] Medlin LK, Kooistra WHCF. Methods to estimate the diversity in the marine photosynthetic protist communitywith illustrations from case studies: a review. Diversity. 2010;2:973–1014.
[5] Marande W, López-García P, Moreira D. Eukaryotic diversity and phylogeny using small- and large-subunitribosomal RNA genes from environmental samples. Environ Microbiol. 2009;11:3179–3188.
[6] Hamsher SE, Evans KM, Mann DG, Poulícková A, Saunders GW. Barcoding diatoms: exploring alternatives toCOI-5P. Protist. 2011;162:405–422.
[7] Stern RF, Andersen RA, Jameson I, Kupper FC, Coffroth MA, Vaulot D, Le Gall F, Veron B, Brand JJ, Skel-ton H, Kasai F, Lilly EL, Keeling PJ. Evaluating the ribosomal Internal Transcribed Spacer (ITS) as a candidatedinoflagellate barcode marker. PLoS ONE. 2012;7:e42780.
[8] Tesson SV, Borra M, Kooistra WHCF, Procaccini G. Microsatellite primers in the planktonic diatom Pseudo-nitzschia multistriata (Bacillariophyceae). Am J Bot. 2011;98:e33–35.
[9] Sarno D, Kooistra WCHF, Medlin LK, Percopo I, Zingone A. Diversity in the genus Skeletonema (Bacillario-phyceae). Bacillariophyceae II. An assessment of the taxonomy of S. costatum-like species with the description offour new species. J Phycol. 2005;41:151–176.
[10] Sarno D, Koistra WHCF, Balzano S, Hargraves PE, Zingone A. Diversity in the genus Skeletonema (Bacillar-iophyceae): III. Phylogenetic position and morphological variability of Skeletonema costatum and Skeletonemagrevillei, with the description of Skeletonema ardens sp. nov. J Phycol. 2007;43:156–170.
[11] Amato A, Kooistra WH, Ghiron JH, Mann DG, Pröschold T, Montresor M. Reproductive isolation among sympatriccryptic species in marine diatoms. Protist. 2007;158:193–207.
[12] Miller PE, Scholin CA. Identification and enumeration of cultured and wild Pseudo-nitzschia (Bacillariophyceae)using species-specific LSU rRNA-targeted fluorescent probes and filtered-based whole cell hybridization. J Phycol.1998;34:371–382.
[13] Miller PE, Scholin CA. On detection of Pseudo-nitzschia Bacillariophyceae species using whole cell hybridization:sample fixation and stability. J Phycol. 2000;36:238–250.
[14] Barra L, Ruggiero MV, Sarno D, Montresor M, Kooistra WH. Strengths and weaknesses of microarray approachesto detect Pseudo-nitzschia species in the field. Environ Sci Pollut R. 2013;20:6705–6718.
[15] Perini F, Casabianca A, Battocchi C, Accoroni S, Totti C, Penna A. New approach using the real-time PCRmethod for estimation of the toxic marine dinoflagellate Ostreopsis cf. ovata in marine environment. PLoS ONE.2011;6:e17699.
[16] Jayasena SD. Aptamers: an emerging class of molecules that rival antibodies in diagnostics. Clin Chem1999;45:1628–1650.
[17] Bunka DH, Stockley PG. Aptamers come of age – at last. Nat Rev Microbiol. 2006;4:588–596.[18] Cerchia L, de Franciscis V. Targeting cancer cells with nucleic acid aptamers. Trends Biotechnol. 2010;28:517–525.[19] Klussmann S, editor. The aptamers handbook: functional oligonucleotides and their application. Weinheim: Wiley-
vch Verlag; 2006.[20] Homann M, Göringer HU. Combinatorial selection of high affinity RNA ligands to live African trypanosomes.
Nucleic Acids Res. 1999;27:2006–2014.[21] Ulrich H, Magdesian MH, Alves MJ, Colli W. In vitro selection of RNA aptamers that bind to cell adhesion
receptors of Trypanosoma cruzi and inhibit cell invasion. J Biol Chem. 2002;277:20756–20762.[22] Shangguan D, Meng L, Cao ZC, Xiao Z, Fang X, Li Y, Cardona D, Witek RP, Liu C, Tan W. Identification of liver
cancer-specific aptamers using whole live cells. Anal Chem. 2008;80:721–728.[23] Tuerk C, Gold L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4
DNA polymerase. Science. 1990;249:505–510.[24] Bock LC, Griffin LC, Latham JA, Vermaas EH, Toole JJ. Selection of single-stranded DNA molecules that bind
and inhibit human thrombin. Nature. 1992;355:564–566.[25] Dwivedi HP, Smiley RD, Jaykus LA. Selection and characterization of DNA aptamers with binding selectivity to
Campylobacter jejuni using whole-cell SELEX. Appl Microbiol Biotechnol. 2010;87:2323–2334.[26] Pavski V, Le XC. Detection of human immunodeficiency virus type 1 reverse transcriptase using aptamers as probes
in affinity capillary electrophoresis. Anal Chem. 2001;73:6070–6076.[27] Fitzwater T, Polisky B. A SELEX primer. Methods Enzymol. 1996;267:275–301.
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15

12 G. Santamaria et al.
[28] Nanjappa D, Kooistra WHCF, Zingone A. A reappraisal of the genus Leptocylindrus (Bacillariophyta), with theaddition of three species and the erection of Tenuicylindrus gen. nov. J Phycol. 2013;49:917–936.
[29] Keller MD, Selvin RC, Claus W, Guillard RRL. Media for the culture of oceanic ultraphytoplankton. J Phycol.1987;23:633–638.
[30] Esposito CL, Passaro D, Longobardo I, Condorelli G, Marotta P, Affuso A, deFranciscis V, Cerchia L. A neu-tralizing RNA aptamer against EGFR causes selective apoptotic cell death. PLoS One. 2011;6(9):e24071. doi:10.1371/journal.pone.0024071
[31] Freier SM, Kierzek R, Jaeger JA, Sugimoto N, Caruthers MH, Neilson T, Turner DH. Improved free-energyparameters for predictions of RNA duplex stability. Proc Natl Acad Sci USA. 1986;83:9373–9377.
[32] Turner DH, Sugimoto N, Freier SM. RNA structure prediction. Annu Rev of Biophys BiophysChem. 1988;17:167–192.
[33] Song K-S, Lee S, Changill B. Aptamers and their biological applications. Sensors. 2012;12:612–631.doi:10.3390/s120100612
[34] Smetacek VS. Role of sinking in diatom life-history cycles: ecological, evolutionary and geological significance.Mar Biol. 1985;84:239–251.
[35] Kooistra WHCF, Gersonde R, Medlin LK, Mann DG. The origin and evolution of the diatoms: their adaptation to aplanktonic existence. In: Falkowski PG, Knoll AH, editors. Evolution of primary producers in the sea. Burlington:Elsevier Academic Press; 2007. p. 207–249.
[36] Jasprica N, Caric M, Kršinic F, Kapetanovic T, Batistic M, Njire J. Planktonic diatoms and their environment in thelower Neretva River estuary (Eastern Adriatic Sea, NE Mediterranean). Nova Hedwigia. 2012;141:405–430.
[37] Ribera d’Alcalà M, Conversano F, Corato F, Licandro P, Mangoni O, Marino D, Mazzocchi MG, Modigh M,Montresor M, Nardella M, Saggiomo V, Sarno D, Zingone A. Seasonal patterns in plankton communities in apluriannual time series at a coastal Mediterranean site (Gulf of Naples): an attempt to discern recurrences andtrends. Scientia Marina. 2004;68:65–83.
[38] Murray AG. Phytoplankton exudation: exploitation of the microbial loop as a defence against algal viruses.J Plankton Res. 1995;17:1079–1094.
[39] Losic D, Rosengarten G, Mitchell JG, Voelcker NH. Pore architecture of diatom frustules: potential nanostructuredmembranes for molecular and particle separations. J Nanosci Nanotechnol. 2006;6:982–989.
[40] Sato S, Beakes G, Idei M, Nagumo T, Mann DG. Novel sex cells and evidence for sex pheromones in diatoms. PloSone. 2011;6:e26923
[41] Koester JA, Swanson WJ, Armbrust EV. Positive selection within a diatom species acts on putative proteininteractions and transcriptional regulation. Mol Biol Evol. 2013;30:422–434.
Dow
nloa
ded
by [
gian
luca
san
tam
aria
] at
05:
06 1
6 Ja
nuar
y 20
15




















