
Antiretroviral therapy with protease inhibitors in human immunodeficiency virus type 1- and human...
8
Anti-retroviral therapy with protease inhibitors decreases virulence enzyme expression in vivo by Candida albicans without selection of avirulent fungus strains or decreasing their anti-mycotic susceptibility Flavia De Bernardis a, * , Evelina Tacconelli b , Francesca Mondello a , Adriana Cataldo b , Silvia Arancia a , Roberto Cauda b , Antonio Cassone a a Department of Infectious, Parasitic and Immune-mediated Diseases, Istituto Superiore di Sanit a, Viale Regina Elena 299, Rome 00161, Italy b Department of Infectious Diseases, Catholic University, Rome, Italy Received 1 September 2003; received in revised form 22 December 2003; accepted 24 December 2003 First published online 15 January 2004 Abstract Highly active anti-retroviral therapies (HAART) with human immunodeficiency virus (HIV) protease inhibitors (PIs) or non- nucleoside reverse-transcriptase inhibitors (NNRTI) were compared for their effect on prevalence, aspartyl proteinase (Sap) pro- duction and the biotypes and anti-mycotic sequential susceptibility of Candida spp. isolates from the oral cavity in a longitudinal prospective study. HAART-PI, but not HAART-NNRTI strongly inhibited Sap expression in the oral cavity without exerting any consistent effect on the role of Candida spp. isolation or selection of low virulence or anti-mycotic resistant fungus biotype. More importantly, the sequential isolates of Candida albicans from HAART-PI, but not those from suspended HAART-NNRTI, showed an increased Sap production in vitro. While further demonstrating that HIV-PI inhibit Sap expressions, our results do not support the view that the mentioned inhibition could eliminate Candida or its selection of the oral cavity. Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Candida; HIV; Proteinase inhibitors; Proteinase secretion; Biotypes 1. Introduction Oropharyngeal candidiasis (OC) was originally a frequent opportunistic infection in human immunode- ficiency virus (HIV)-positive subjects before the ad- vent of highly active anti-retroviral therapy (HAART) [1,2]. The most common agent causing mucosal candi- diasis is Candida albicans, which has the ability to se- crete aspartyl proteinases (Sap), a principal family of virulence enzymes, facilitating the invasion of mucosal tissue [3,4]. This is a relevant aspect of the oral pathol- ogy caused by this fungus that has been demon- strated by the repeated observations of the strains of C. albicans of particularly high Sap production. These were more frequently isolated from the oral and vaginal cavities of HIV-positve rather than from HIV-negative subjects [5,6]. A distinctive set of Sap genes has been found to express the fungus in the oral and vaginal cavity [7]. As mentioned above, HAART including HIV pro- teinase inhibitors (HAART-PI) have caused a marked decline of OC and other opportunistic infections [8–11]. We and others have demonstrated that this therapy exerts an early immune reconstitution-independent ben- eficial effect against OC, which is entirely due to the PI capacity to inhibit Sap enzymes of Candida secretory aspartyl proteinase (Sap) family that belongs to the same family as HIV-proteinase [12–15]. There is now * Corresponding author. Tel.: +39-06-49902809; fax: +39-06- 49387112. E-mail address: [email protected] (F. De Bernardis). 0928-8244/$22.00 Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsim.2003.12.006 FEMS Immunology and Medical Microbiology 41 (2004) 27–34 www.fems-microbiology.org
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Antiretroviral therapy with protease inhibitors in human immunodeficiency virus type 1- and human...

FEMS Immunology and Medical Microbiology 41 (2004) 27–34
www.fems-microbiology.org
Anti-retroviral therapy with protease inhibitors decreasesvirulence enzyme expression in vivo by Candida albicanswithout selection of avirulent fungus strains or decreasing
their anti-mycotic susceptibility
Flavia De Bernardis a,*, Evelina Tacconelli b, Francesca Mondello a, Adriana Cataldo b,Silvia Arancia a, Roberto Cauda b, Antonio Cassone a
a Department of Infectious, Parasitic and Immune-mediated Diseases, Istituto Superiore di Sanit�a, Viale Regina Elena 299, Rome 00161, Italyb Department of Infectious Diseases, Catholic University, Rome, Italy
Received 1 September 2003; received in revised form 22 December 2003; accepted 24 December 2003
First published online 15 January 2004
Abstract
Highly active anti-retroviral therapies (HAART) with human immunodeficiency virus (HIV) protease inhibitors (PIs) or non-
nucleoside reverse-transcriptase inhibitors (NNRTI) were compared for their effect on prevalence, aspartyl proteinase (Sap) pro-
duction and the biotypes and anti-mycotic sequential susceptibility of Candida spp. isolates from the oral cavity in a longitudinal
prospective study. HAART-PI, but not HAART-NNRTI strongly inhibited Sap expression in the oral cavity without exerting any
consistent effect on the role of Candida spp. isolation or selection of low virulence or anti-mycotic resistant fungus biotype. More
importantly, the sequential isolates of Candida albicans from HAART-PI, but not those from suspended HAART-NNRTI, showed
an increased Sap production in vitro. While further demonstrating that HIV-PI inhibit Sap expressions, our results do not support
the view that the mentioned inhibition could eliminate Candida or its selection of the oral cavity.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Candida; HIV; Proteinase inhibitors; Proteinase secretion; Biotypes
1. Introduction
Oropharyngeal candidiasis (OC) was originally a
frequent opportunistic infection in human immunode-
ficiency virus (HIV)-positive subjects before the ad-
vent of highly active anti-retroviral therapy (HAART)
[1,2]. The most common agent causing mucosal candi-
diasis is Candida albicans, which has the ability to se-crete aspartyl proteinases (Sap), a principal family of
virulence enzymes, facilitating the invasion of mucosal
tissue [3,4]. This is a relevant aspect of the oral pathol-
ogy caused by this fungus that has been demon-
* Corresponding author. Tel.: +39-06-49902809; fax: +39-06-
49387112.
E-mail address: [email protected] (F. De Bernardis).
0928-8244/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsim.2003.12.006
strated by the repeated observations of the strains of
C. albicans of particularly high Sap production. These
were more frequently isolated from the oral and vaginal
cavities of HIV-positve rather than from HIV-negative
subjects [5,6]. A distinctive set of Sap genes has been
found to express the fungus in the oral and vaginal
cavity [7].
As mentioned above, HAART including HIV pro-teinase inhibitors (HAART-PI) have caused a marked
decline of OC and other opportunistic infections [8–11].
We and others have demonstrated that this therapy
exerts an early immune reconstitution-independent ben-
eficial effect against OC, which is entirely due to the PI
capacity to inhibit Sap enzymes of Candida secretory
aspartyl proteinase (Sap) family that belongs to the
same family as HIV-proteinase [12–15]. There is now
. Published by Elsevier B.V. All rights reserved.

28 F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34
further evidence that PI could directly affect some other
opportunistic agents of disease common in HIV-positive
subjects [16]. However, previous studies did not clarify
whether the anti-retroviral therapy with PI had a per-
sistent effect on the fungus infection or carriage in theoral cavity. They did not confirm whether the therapy
caused the selection of different fungal strains with dif-
ferent Sap-producing abilities or had any impact on
fungus susceptibility to triazole anti-mycotics. In this
study, we took advantage of comparing a longitudinal,
comparative study of subjects who received HAART-PI
with another group who were under a therapeutic regi-
men with a nonnucleoside reverse-transcriptase inhibi-tor (NNRTI) utilized as a control. Thus, we have
longitudinally assessed the prevalence of fungal isolates
as well as the biotypes of C. albicans isolated from each
patient, together with the Sap production ability of each
isolate from each subject. In specific reports on the as-
sociation between Sap production level and anti-mycotic
susceptibility [5,17], we also tested the susceptibility of
all C. albicans isolates to fluconazole and itraconazole.The results clearly demonstrate that PI-containing
HAART regimens strongly inhibit the expression of Sap
activity in the oral cavity without selecting the low-Sap
producer fungus biotypes or affecting Candida preva-
lence and anti-mycotic susceptibility.
2. Subjects and methods
2.1. Study setting
The clinical investigation was performed at the
Catholic University Hospital, a 1700-bed tertiary care
centre located in Rome, Italy, as detailed elsewhere
[15,18]. Seventy HIV-infected subjects were considered
that were naive for anti-retroviral therapy and whowere admitted as outpatients from July 1st through
December 31st 1999. Among this group, 35 subjects
began HAART-PI (Group 1) and 35 received HAART-
NNRTI (Group 2). Therapy start time was designated
as 0 and all subjects were monitored for a period of 180
days. Clinical details and general clinical parameters
recorded from these patients are given elsewhere
[12,15,18].
2.2. Parameters evaluated
With the consent of each enrolled subject in the
study, the following data were obtained: the age, sex,
HIV-risk behavior; any active intravenous drug abuse;
the stage of HIV infection; previous manifestations of
HIV infection; date of AIDS-defining illness; presence ofOC and history of previous OC episodes; concurrent
opportunistic infections; risk factors for OC anti-
microbial therapy; type and use of prophylaxis for
Pneumocystis carinii pneumonia and toxoplasmosis;
and any other medications (anti-tuberculous agents,
anti-virals, corticosteroids, anti-acids); the number of
circulating CD4þT (/mm3) cells and of peripheral poly-
morphonuclear (PMN) cells (/mm3) and HIV-viremiawith a detection limit below 50 copies/ml. The analyzed
risk factors for OC included neutropenia (PMNs <1000/
mm), alcoholism, cirrhosis, diabetes, neoplastic diseases
and chronic renal failure.
The first episode of microbiologically documented
diagnosis of OC was routinely made of the typical signs
and symptoms. All OC were clinically categorized into
three types: pseudomembranous, erythematous and hy-perplastic forms, as detailed elsewhere [18].
2.3. Yeast isolation and identification
Oropharyngeal samples were taken with cotton-
tipped swabs and kept in sterile physiological saline,
before transferring to the laboratory where they were
streaked on plates of Sabouraud–dextrose agar (BBL,Baltimore, MD) with added chloramphenicol (50
lgml�1), and Mycosel agar, before immersion into en-
richment broth. All cultures were incubated for 48–72 h
at 30 �C. All isolates were identified on the basis of their
morphology and profiles in the API 20C system (Bio-
Merieux, Marcy l�Etoile, France).
2.4. Sap detection in vitro and in salivar fluids
All isolates were tested for their ability to grow and
produce a clear zone of hydrolysis in bovine serum al-
bumin (BSA) agar, as detailed elsewhere [4,15].
Sap antigen secretion in BSA broth was also assayed
by ELISA [4,15]. Briefly, the isolates were precultured in
YPD broth (1% yeast extract; 2% peptone and 2% glu-
cose), the cultures were centrifuged and the cellularpellet was washed and incubated (106 cells in 25 ml of
YPD broth plus 0.2% BSA) at 30 �C with slight agita-
tion. After 40 h of incubation, the samples were centri-
fuged at 3400g for 10 min and the supernatants were
treated with 0.5 ml of 50% trichloroacetic acid (TCA),
incubated for 30 min in ice and centrifuged at 3400g for
10 min. The pellets were washed twice with 95% ethanol,
dissolved in 1% (w/v) sodium dodecyl sulfate (SDS),diluted in 0.2 M sodium carbonate buffer (pH 9.6) and
applied to the MicroTest plates. Purified rabbit anti-Sap
serum [12,19] was added at a 1:100 dilution and incu-
bated for 2 h at 37 �C. The second antibody was phos-
phatase-conjugated goat anti-rabbit immunoglobulin G
(1:1000 dilution, Sigma. St. Louis, MO.) The reaction
was detected with phosphatase substrate nitrophenol
phosphate (Sigma). Sap amount was calculated from astandard curve (1–110 ng) determined with a highly
purified Sap preparation [12,19] as coating antigen. The
plates were read with a Titer K Multiscan (Labsystem

F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34 29
Helsinki, Finland) at 405 nm blanked against air. The
specificity of the ELISA for proteinase detection was
confirmed by Western blotting [19].
ELISA was also used for Sap detection in the saliva
of patients, according to a procedure fully describedelsewhere [15].
2.5. Anti-mycotic susceptibility test
A recently modified, improved method of anti-
mycotic susceptibility assays in vitro, derived from the
standardized NCCLS method [20,21] was used
throughout this study for the determination of Candidaisolate susceptibility to fluconazole and itraconazole.
Stock solution of fluconazole (Pfizer Inc., New York,
USA) was prepared in sterile distilled water. Stock so-
lution of itraconazole (Janssens Pharmaceutica, Beerse,
Belgium) was prepared by heating at 75 �C for 45 min in
polyethylene glycol 400.
The yeast isolates were grown on Sabouraud dextrose
agar for 48 h at 35 �C. The inoculum suspension wasprepared by picking five colonies of at least 1 mm in
diameter and suspending them in 5 ml of sterile distilled
water. The cell density of the suspension was adjusted
spectrophotometrically to a final transmission of 85% at
530 nm. The working suspension was made by a 1:50
dilution in sterile distilled water followed by a 1:20
dilution in media to obtain a 2� final suspension. In-
oculum sizes were confirmed by the enumeration ofcolony-forming units (CFU) on Sabouraud dextrose
agar. The medium for susceptibility testing was RPMI
1640 media (American Biorganics, Inc., Niagara Falls,
NJ) added with LL-glutamine and buffered at pH 7.0 with
0.165 M morpholinepropanesulphonic acid. Testing was
performed in sterile, flat-bottom 96-well microtiter
plates. Drugs were prepared at 10� the strength of the
final concentration, and these mother solutions werediluted 1:5 with RPMI 1640 to obtain 2� the final
concentrations. Volumes of 50 ll of the 2� drug dilu-
tions were dispensed into wells; two wells in each row
were used, one as a growth control and the other as a
sterility check. The microtiter plates were stored at )70�C until use. On the day of the test, 50 ll of the yeast
suspension were added to each well.
The microtiter plates were incubated at 35 �C andread at 24 and 48 h with the aid of a reading mirror after
shaking the growth samples in each well and compared
these with that of the growth control (drug-free) well.
The MIC was defined according to previously estab-
lished criteria [19,20].
2.6. Karyotype analysis by pulsed field gel electrophoresis
Cells of C. albicans were grown to a stationary phase
overnight at 28 �C in YPD medium (glucose 2%; yeast
extract 1% and Bacto-Peptone 2%; Difco, Detroit, MI).
The cells were packed using centrifugation (3000 rpm
for 5 min) and washed in 1.2 M sorbitol solution con-
taining 20 mM EDTA, pH 8.0. As described above, in a
solution of EDTA–sorbitol, the pellet was re-suspended
in a cell concentration of 109 ml�1, but this time con-taining 20 mM mercaptoethanol and incubated for 15
min at 37 �C. The samples were embedded in a low-
melting-point agar (L.M.) (BioRad, Richmond, VA).
The spheroplast lysis method, as described by Vollrath
and Davis [22], was used to prepare the chromosomal
DNA. The cell/agarose L.M. mixture was transferred to
plug molds. Solidified pellets were removed from the
molds and placed in 1.2 M sorbitol solution containing20 mM EDTA, 10 mM Tris–HCl, pH 7.5, 100 ll zy-moliasi 100 T (100,000 Ug�1) (1 mgml�1) (Seikagaku
Corporation, Tokyo, Japan). After a 2-h incubation at
37 �C, the plugs were incubated in 1% SDS solution
containing 10 mM EDTA, 10 mM Tris–HCl, pH 7.5,
and left overnight at 37 �C. The pellets were stored in
1% sarcosyl solution containing 100 mM EDTA, 10 mM
Tris–HCl, pH 7.5, at 4 �C.The DNA samples in agarose inserts were resolved by
contour clamped homogenous field electrophoresis
(CHEF). CHEF analysis was performed with the
CHEF-DRII apparatus (BioRad). The operating con-
ditions included three consecutive runs on each gel (14.5
by 20.5 cm, 1-cm thick 1% agarose; BioRad) containing
the agarose inserts of DNA immersed in a running
buffer (50 mM Tris–HCl, 50 mM boric acid and 1.5 mMEDTA, pH 8.2). Saccharomyces cerevisiae (BioRad) was
used as a standard. The parameters for each run were
150 V, 14 �C, 24 h with 90-s, 120-s and 180-s switches.
Gels were then stained with ethidium bromide (0.5
lgml�1; 30 min), de-stained and photographed under
ultraviolet light.
2.7. Statistics
The comparisons of the mean values were assessed by
ANOVA and Student�s t-test with Bonferroni�s correc-
tion. Nonparametric tests were used for frequency as-
sessment (v2 or Fisher method, as appropriate).
3. Results
3.1. Prevalence and distribution of Candida species
isolated from HIVþ subjects receiving HAART-PI or
HAART-NNRTI
Seventy patients were monitored for a presence of
fungus in the oral cavity for a total of 180 days follow-
up. As shown in Table 1, Candida spp. were isolatedfrom 95 (64%) and 70 (52%) samples from 30 out of 35–
27 out of 35 HAART-PI and HAART-NNRTI patients,
respectively. In both subject groups, C. albicans was

Table 1
Prevalence and distribution of yeast species isolated from HIV-positive subjects receiving HAART-PI or HAART-NNRTI
Treatment Subjects no. Positive no. Samples no. Positive no. No. Isolates species
HAART-PI 35 30 150 95 86 C. albicans�
4 C. glabrata
1 C. tropicalis��
1 C.guilliermondii
1 C. krusei
1 Saccharomyces spp.
1 C. famata
HAART-NNRTI 35 27 135 70 54 C. albicans
7 C. tropicalis
1 C. lipolytica
1 C. guilliermondii
5 C. famata
2 C. kefyr
* Significantly more prevalent in HAART-PI patients.** Significantly more prevalent in HAART-NNRTI patients.
P < 0:05v2 two tailed; P < 0:05 Fisher�s exact test.
30 F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34
largely the most prevalent species, however, it was more
frequently isolated from samples of subjects under
HAART-PI (P < 0:05, two-tailed). The second most
frequent isolate was C. tropicalis (Table 1). Overall, thetwo Sap-producing species (C. albicans and C. tropicalis)
accounted for about 90% of all Candida spp. isolates in
our patients.
3.2. Sap secretion
All isolates of C. albicans and C. tropicalis were
qualitatively confirmed to be proteolytic on BSA-agar[4,19]. This essentially qualitative test did not reveal
gross differences in BSA degradation between the iso-
lates from HAART-PI and those from HAART-
NNRTI subjects (data not shown). All isolates had se-
creted the Sap antigen in the culture medium (tested
only for C. albicans). By this latter quantitative method,
however, a clear trend of increasing gradient of Sap
production among the sequential isolates from the samepatient under treatment with HAART-PI was noticed.
In some subjects, the increased Sap production ability
was already apparent with the initial isolates, in partic-
ular at 7 or 14 days from the initiation of therapy. At the
end of follow-up (day 180), Sap production by the iso-
lates from HAART-PI subjects was significantly higher
than Sap production by the initial isolates [351� 202
versus 100� 66, P < 0:05]. This trend was common inall the subjects examined, which included all whose
isolates were also examined for karyotype (see below)
but not those with a positive Candida isolation (Table 2).
In contrast, the increase in Sap production was not so
evident in HAART-NNRTI subjects, since the consec-
utive isolates of half of these did not show an increase
and in all others the increase was not very significant as
in the HAART-PI subjects (Table 2). Overall, therewas no significant statistical difference between Sap
production of the isolates on day 0 and those on day 90
or 180 for this category of subjects.
The amount of Sap in the saliva of each subject of
both groups was redetermined see also [15]. On day 0,data confirmed that Sap was present in the saliva of all
subjects from whom C. albicans was isolated, and that
quantity significantly declined becoming undetectable in
HAART-PI subjects from the second week of treatment
(as average, from >300 ngml�1 at day 0–100 ngml�1 on
day 14 and <20 ngml�1 on day 30, respectively). In
contrast, the amount of proteinase was not markedly
changed in the saliva samples taken from the patientswith HAART-NNRTI therapy (data not shown, [22]).
3.3. Strain biotyping
The data above demonstrated a preserved carriage of
the fungus in the oral cavity and, in the absence of the
disease [22], a decreased amount of saliva Sap in the
HAART-PI subjects. Interestingly, for these subjects, C.albicans isolates tended to produce increased Sap
quantities in vitro. One possible explanation of the latter
data could be intra therapy replacement of the initial
strain of C. albicans by other strains with increased
potential for Sap production. To obtain an insight into
this aspect, we analyzed the electrophoretic karyotype of
C. albicans isolates from HAART-PI subjects which
showed the best definite trend to increased Sap pro-duction (37 isolates from 11 subjects). As expected, the
complete chromosomal pattern of C. albicans isolates
was unique for each initial (day 0) isolate, consisting of
7–9 distinct bands ranging from more than 3.0 to less
than 1.0 Mbp. Nevertheless, in all but two patients (both
belonging to the HAART-NNRTI) all sequential iso-
lates showed the same electrophoretic karyotype sug-
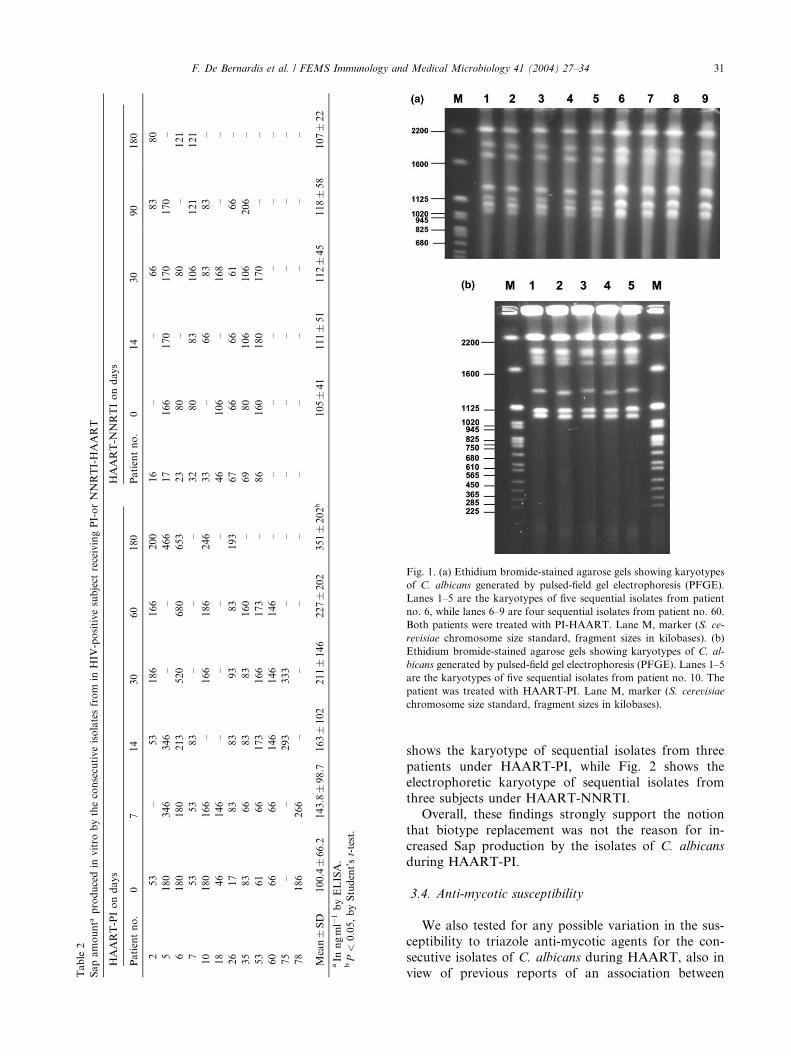
gesting there was persistent oral colonization by thesame initial strain. As an example, Fig. 1(a) and (b)
Table
2
Sapamounta
producedin
vitro
bytheconsecutiveisolatesfrom
inHIV
-positivesubject
receivingPI-orNNRTI-HAART
HAART-PIondays
HAART-N
NRTIondays
Patientno.
07
14
30
60
180
Patientno.
014
30
90
180
253
–53
186
166
200
16
––
66
83
80
5180
346
346
––
466
17
166
170
170
170
–
6180
180
213
520
680
653
23
80
–80
–121
753
53
83
––
–32
80
83
106
121
121
10
180
166
–166
186
246
33
–66
83
83
–
18
46
146
––
––
46
106
–168
––
26
17
83
83
93
83
193
67
66
66
61
66
–
35
83
66
83
83
160
–69
80
106
106
206
–
53
61
66
173
166
173
–86
160
180
170
––
60
66
66
146
146
146
––
––
––
–
75
––
293
333
––
––
––
––
78
186
266
––
––
––
––
––
Mean�SD
100.4�66.2
143.8�98.7
163�102
211�146
227�202
351�202b
105�41
111�51
112�45
118�58
107�22
aIn
ngml�
1byELISA.
bP<
0:05,byStudent�s
t-test.
Fig. 1. (a) Ethidium bromide-stained agarose gels showing karyotypes
of C. albicans generated by pulsed-field gel electrophoresis (PFGE).
Lanes 1–5 are the karyotypes of five sequential isolates from patient
no. 6, while lanes 6–9 are four sequential isolates from patient no. 60.
Both patients were treated with PI-HAART. Lane M, marker (S. ce-
revisiae chromosome size standard, fragment sizes in kilobases). (b)
Ethidium bromide-stained agarose gels showing karyotypes of C. al-
bicans generated by pulsed-field gel electrophoresis (PFGE). Lanes 1–5
are the karyotypes of five sequential isolates from patient no. 10. The
patient was treated with HAART-PI. Lane M, marker (S. cerevisiae
chromosome size standard, fragment sizes in kilobases).
F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34 31
shows the karyotype of sequential isolates from three
patients under HAART-PI, while Fig. 2 shows the
electrophoretic karyotype of sequential isolates from
three subjects under HAART-NNRTI.
Overall, these findings strongly support the notion
that biotype replacement was not the reason for in-
creased Sap production by the isolates of C. albicans
during HAART-PI.
3.4. Anti-mycotic susceptibility
We also tested for any possible variation in the sus-
ceptibility to triazole anti-mycotic agents for the con-
secutive isolates of C. albicans during HAART, also in
view of previous reports of an association between

Fig. 2. Ethidium bromide-stained agarose gels showing karyotypes of
C. albicans generated by pulsed-field gel electrophoresis (PFGE).
Lanes 1–4 are the karyotypes of four sequential isolates from patient
no. 32, lanes 5 and 6 are two sequential isolates from patient no. 23,
while lanes 7 and 8 are two sequential isolates from patient no. 46. All
patients were treated with HAART-NNRTI. Lane M, marker (S. ce-
revisiae chromosome size standard, fragment sizes in kilobases).
32 F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34
increased Sap production and refractoriness to some
anti-mycotics as for instance, fluconazole [17].All the 85 isolates of C. albicans from the 35 patients
receiving HAART-PI were tested. They were all found
susceptible to fluconazole (MIC range 0.125–32 lgml�1
and itraconazole (MIC range 0.078–0.5 lgml�1). In
particular, no evidence of susceptibility changes in
consecutive fungus isolates were found, and no relation
was either found between MIC level and Sap production
in vitro (data not shown).
4. Discussion
We have previously demonstrated that HIV protein-
ase inhibitors (PIs) exert an early, immune reconstitu-
tion-independent clinical benefit against oral candidiasis
in HIV-positive subjects. This effect is mediated by theinhibition of C. albicans secretory aspartyl proteinase
(Sap), a family of a critical virulence factor for mucosal
candidiasis [4,6,7,12,14,23,24]. From these initial ob-
servations, a wealth of data has been generated con-
firming and extending these PI unexpected effects on
other agents of opportunistic infections [16], also pro-
voking the active search of anti-fungal PI [25]. The anti-
candidal benefit of HAART-PI was entirely due to PIhas recently been ascertained by the comparison with a
controlled group of subjects under HAART with PI-
sparing regimens [15].
We have now extended these studies in the same co-
horts of previously examined HIV-positive subjects [15]
with the specific aim of examining whether HAART-PIfavored the elimination of the fungus from the oral
cavity or it was substituted with less virulent specie or
strain. Sap appears to play a role in fungus adherence to
epithelial cells [26], an effect which should in turn create
a serious obstacle to Candida colonization. Moreover,
we speculated whether the therapy caused variations in
the anti-mycotic susceptibility of the oral fungus iso-
lates, which were shown by others to be affected in theirsusceptibility to some anti-fungals from an elevated Sap
production [17,27,28].
In order to solve these questions, we identified all
sequential oral yeast isolates during the whole follow-up
period of 180 days and determined their capacity to
produce Sap in vitro. We also determined their suscep-
tibility to some of the most common anti-fungal agents,
such as fluconazole and itraconazole, from which es-tablished standards of susceptibility testing exist [21].
Finally we re-determined the amount of oral Sap during
the whole follow-up period to confirm the clear-cut
difference of this parameter for subjects under PI- and
those under HAART-NNRTI [15,18].
We isolated Candida spp. from the oral cavity of the
large majority (from 80% to 90%) of all HAART-treated
subjects, irrespective of treatment regimen. Althoughthere were differences in the prevalence of Candida spp.
between the two groups of patients, the two Sap-
producing species (C. albicans and C. tropicalis) ac-
counted overall for more than 90% of all isolates and
were consistently isolated throughout the follow-up
period. More importantly, without exception, all se-
quential isolates from each positive subject under HA-
ART-PI had identical electrophoretic karyotype andidentical susceptibility to fluconazole and itraconazole.
Conversely, the data clearly confirmed, from the early
initiation of HAART-PI, but not HAART-NNRTI,
that no Sap could be detected in the oral cavity.
Consequently this data strongly suggests that PI may
have a quantitative yet certainly not a qualitative effect
on colonization of the oral cavity. It is also clear that PI
do not induce a replacement of strains incapable ofcausing disease or more susceptible to triazole anti-
mycotics. Although not ruling out completely the pres-
ence of subtle genetic changes in the sequential strains,
the electrophoretic karyotype is rather generally taken
as a method which can reliably define Candida biotype
[29]. (In fact, only two isolates showed an apparent
strain replacement: both belonged to NNRTI-group and
did not show significant increase in Sap production.This is in keeping with an observation made by Taylor
et al. [29] who showed that C. albicans isolates from the
oral cavity of HIV-positive patients under HAART

F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34 33
regimen had no decrease in virulence, compared to ref-
erence strains, for experimental mucosal infection. Un-
fortunately however, Sap quantities or activity by these
isolates were not determined in the above study [29].
Besides host immunereconstitution, the beneficialclinical effect of PI appears to be entirely due to their
capacity to inhibit Sap production in vivo, as they po-
tently do with in vitro [12,23,24]. The above interpre-
tation appears still more plausible if we consider an
unexpected and intriguing finding of this investigation.
That is, the fact that the sequential C. albicans isolates
from HAART-PI subjects we examined do not only
produce Sap in vitro but have a clear trend to producemore Sap in vitro as long as the subject is treated. At the
end of treatment, the average Sap production by the
isolates on day 180 (351� 87 ngml�1; means� SE) was
significantly higher than Sap production first day (day 0)
isolates (100� 21 ngml�1; means� SE). Although this
did not occur in the isolates from HAART-NNRTI
subjects, it is unlikely that PI are the indirect cause of
this phenomenon. In several studies it was shown thatoral and vaginal isolates of C. albicans from HIV-posi-
tive subjects to be higher Sap producers in vitro than the
isolates of HIV-negative subjects, in the pre-HAART
era [4–6].
It is not clear why the same strain or biotype of C.
albicans could be induced to produce more Sap in vitro
when isolated from subjects undergoing treatment with
PI, in a situation where Sap production is inhibited invivo. In the pre-HAART era, this phenomenon has been
attributed to Sap induction by HIV envelope proteins
[30], a fact that seems to be unlikely with a therapy
causing rapid loss of viremia such as the present-day
HAART. Nor can it be attributed to anti-mycotic usage
before entering HAART and acquired resistance to
them, since, as shown here, all isolates were susceptible
to both fluconazole and itraconazole. It is possible thatduring oral colonization and persistence, the strain of
Candida is induced in vivo. In the presence of PI, it alters
the type of secreted Sap, which could be more reactive
with our anti-serum in the ELISA test. For this, we used
a polyclonal reagent raised against a purified Sap2
preparation. Because of the strong homologies within
Sap family, it is expected that a polyclonal reagent
would contain antibody specificity against the majorityof Saps, but clearly not to the same extent or reactivity.
If so, our data anticipate that during therapy with PI C.
albicans could change the type of secreted Sap. Most of
all, the type of Sap produced on BSA medium in vitro
may be different from that (or those) secreted in the oral
cavity [7]. At any rate, this change should not impact on
the clinical benefit of PI against oral candidosis which is
evident in all subjects regardless Sap production abilityin vitro by the fungus isolate.
There have been interesting reports suggesting a
correlation between high Sap production and decreased
susceptibility to some azoles by C. albicans isolates from
HIV-positive subjects, in the pre-HAART era [5,17]. We
could not confirm these reports as there was no sus-
ceptibility decrease to both anti-fungal during either or
HAART-PI or HAART-NNRTI. Importantly, the iso-lates on day 0 were, as a whole, all susceptible to
fluconazole and itraconazole, in contrast to the isolates
of C. albicans studied by Ollert et al. [5] which were
resistant to these drugs. It appears that a population of
triazole-susceptible isolates, although persisting in the
oral cavity and gaining some increase in Sap production,
is not going to change its pattern of drug susceptibility
during 180 days of PI-HAART.
Acknowledgements
We are grateful to Daniela Adriani and Antonietta
Girolamo for their technical assistance. Thanks are ex-
tended to Dr. Giusi Mandarino for helping in the
preparation of the manuscript.This work was supported in part by National AIDS
Research Program under ISS contract 50E.
References
[1] Holmberg, K. and Meyer, R.D. (1986) Fungal infections in
patients with AIDS and AIDS-related complex. Scand. J. Infect.
Dis. 18, 179–185.
[2] Klein, R.S., Harris, C.A., Butkus Small, C., Moll, B., Lesser, M.
and Friendland, G.H. (1984) Oral candidiasis in high risk patients
as the initial manifestation of the acquired immunodeficiency
syndrome. N. Engl. J. Med. 311, 354–357.
[3] Cutler, J.E. (1991) Putative virulence factors of Candida albicans.
Ann. Rev. Microbiol. 45, 187–218.
[4] De Bernardis, F., Boccanera, M., Rainaldi, L., Guerra, C.E.,
Quinti, I. and Cassone, A. (1992) The secretion of aspartyl
proteinase, a virulence enzyme, by isolates of Candida albicans
from the oral cavity of HIV-infected subjects. Eur. J. Epidemiol. 8,
362–367.
[5] Ollert, M.W., Wende, C., Gorlich, M., McMullan-Vogel, C.G.,
Borg von-Zepelin, M., Vogel, C.W. and Korting, H.C. (1995)
Increased expression of Candida albicans secretory proteinase, a
putative virulence factor in isolates from human immunodefi-
ciency virus-positive patients. J. Clin. Microbiol. 33, 2543–2549.
[6] De Bernardis, F., Mondello, F., Scaravelli, G., Pach�ı, A.,
Girolamo, A., Agatensi, L. and Cassone, A. (1999) High aspartyl
proteinase production and vaginitis in human immunodeficiency
virus-infected women. J. Clin. Microbiol. 37, 1376–1380.
[7] Naglick, J.R., Rodgers, C.A., Shirlaw, P.J., Dobbie, J.L.,
Fernandez-Naglick, l., Greenspan, D., Agabian, N. and Challa-
combe, S.J. (2003) Differential expression of Candida albicans
secreted aspartyl proteinase and phospholipase B genes in human
correlates with active oral and vaginal infections. J. Infect. Dis.
188, 469–479.
[8] Deeks, S.G., Smith, M., Holodniy, I. and Kahn, J.O. (1997) HIV-
1 protease inhibitors. A review for clinicians. JAMA 277, 145–155.
[9] Palella, F.J., Delaney, K.M., Moorman, A.C., Loveless, M.O.,
Fuhrer, J., Satten, G.A., Aschman, D.J. and Holmberg, S.D.
(1998) Declining morbidity and mortality among patients with
advanced human immunodeficiency virus infection. New Eng. J.
Med. 338, 853–860.

34 F. De Bernardis et al. / FEMS Immunology and Medical Microbiology 41 (2004) 27–34
[10] Sepkowitz, K.A. (1998) Effect of HAART on natural history of
AIDS-relate opportunistic disorders. Lancet 351, 228–230.
[11] Brodt, H.R., Kamps, B.S., Gute, P., Knupp, B., Staszewski, S.
and Helm, E.B. (1997) Change incidence of AIDS-defining illness
in the era of antiretroviral therapy. AIDS 11, 1731–1738.
[12] Cassone, A., De Bernardis, F., Torosantucci, A., Tacconelli, E.,
Tumbarello, M. and Cauda, R. (1999) In vitro and vivo
anticandidal activity of human immunodeficiency virus protease
inhibitors. J. Infect. Dis. 180, 448–453.
[13] Hube, B. (1996) Candida albicans secreted aspartyl proteinase.
Curr. Top. Med. Mycol. 7, 55–69.
[14] De Bernardis, F., Arancia, S., Morelli, L., Hube, B., Sanglard, D.
and Schaefer, W. (1999) Evidence that members of the secretory
aspartyl proteinases gene family (SAP), in particolar SAP2, are
virulence factors for Candida vaginitis. J. Infect. Dis. 179, 201–208.
[15] Cassone, A., Tacconelli, E., De Bernardis, F., Tumbarello, M.,
Torosantucci, A. and Chiani, P. (2002) Antiretroviral therapy
with protease inhibitors has an early, immune reconstitution-
independent beneficial effect on Candida virulence and oral
candidiasis in human immunodeficiency virus-infected subjects.
J. Infect. Dis. 185, 188–195.
[16] Atzori, C., Angeli, E., Mainini, A., Agostoni, F., Micheli, V. and
Cargnel, A. (2000) In vitro activity of human immunodeficiency
virus protease inhibitors against Pneumocystis carinii. J. Infect.
Dis. 181, 1629–1634.
[17] Wu, T., Wright, K., Hurst, S.F. and Morrison, C.J. (2000)
Enhanced extracellular production of aspartyl proteinase, a
virulence factor by Candida albicans isolates following growth in
subinhibitory concentration of fluconazole. Antimicrob. Agents
Chemother. 44, 1200–1208.
[18] Cauda, R., Tacconelli, E., Tumbarello, M., Morace, G., De
Bernardis, F., Torosantucci, A. and Cassone, A. (1999) Role of
protease inhibitors in preventing recurrent oral candidosis in
patients with HIV infection: a prospective case-control study.
J. Acquir. Immun. Defic. Syndr. Hum. Retrovirol. 21, 20–25.
[19] De Bernardis, F., Agatensi, L., Ross, I.K., Emerson, G.W.,
Lorenzini, R., Sullivan, P.A. and Cassone, A. (1990) Evidence for
a role for secreted aspartate proteinase of Candida albicans in
vulvovaginal candidiasis. J. Infect. Dis. 161, 1276–1283.
[20] Barchiesi, F.A., Colombo, L., McGough, D.A., Fothergill, A.W.
and Rinaldi, M.G. (1994) In vitro activity of itraconazole
against fluconazole-susceptible and -resistant Candida albicans
isolates from oral cavities of patients infected with human
immunodeficiency virus. Antimicrob. Agents Chemother. 38,
1530–1533.
[21] National Committee for Clinical Laboratory Standard (1997).
Reference method for broth dilution antifungal susceptibility
testing of yeasts; Approved Standard M27-A, NCCLS, Wayne,
PA.
[22] Vollrath, D. and Davis, R.W. (1987) Resolution of DNA
molecules greater than 5 megabases by contour clamped homog-
enous electric fields. Nucleic Acids Res. 15, 7865–7876.
[23] Korting, H.C., Schaller, M., Eder, G., Hamm, G., Bohmer, U.
and Hube, B. (1999) Effects of the human immunodeficiency virus
(HIV) proteinase inhibitors saquinavir and indinavir on in vitro
activities of secreted aspartyl proteinases of Candida albicans
isolates from HIV-infected patients. Antimicrob. Agents Che-
moth. Aug., 2038–2042.
[24] Munro, C.A. and Hube, B. (2002) Antifungal therapy at the
HAART of viral therapy. Trends Microb. Opin. 10, 173–177.
[25] Skrbec, D. and Romeo, D. (2002) Inhibition of Candida albicans
secreted aspartic protease by a novel series of peptidomimetics,
also active on the HIV-1 protease. Biochem. Biophys. Res.
Commun. 297, 1350–1353.
[26] Bektic, J., Lell, C.P., Fuchs, A., Stoiber, H., Speth, C., Lass-Florl,
C., Borg-von Zepelin, M., Dierich, M.P. and Wurzner, R. (2001)
HIV protease inhibitors attenuate adherence of Candida albicans
to epithelial cells in vitro. FEMS, Immun. Med. Microb. 31,
65–71.
[27] Schaller, M., Krnjac, N., Niewerth, M., Hamm, G., Hube, B. and
Korting, H. (2003) Effect of antimycotic agents on the activity of
aspartyl proteinases secreted by Candida albicans. J. Med. Microb.
52, 247–249.
[28] Stewart, K. and Abad-Zapatero, C. (2001) Candida proteases and
their inhibition: prospects for antifungal therapy. Curr. Med.
Chem. 8, 941–948.
[29] Taylor, B.N., Fischenbaum, C., Saavedra, M., Slaviinsky III, J.,
Swoboda, R., Wozniak, K., Arribas, A., Powderly, W. and Fidel
Jr., P.L. (2000) In vivo virulence of Candida albicans isolates
causing mucosal infections in people infected with the human
immunodeficiency virus. J. Inf. Dis. 182, 955–959.
[30] Gruber, A., Speth, C., Lukasser-Vogl, E., Zangerle, R., Borg-von
Zepelin, M., Dierich, M.P. and Wurzner, R. (1999) Human
immunodeficiency virus type 1 protease inhibitor attenuates
Candida albicans virulence properties in vitro. Immunopharma-
cology 41, 227–234.




















