
Antigen Binding and Solubility Effects upon the Veneering of a Camel VHH in Framework-2 to Mimic a...
14
Antigen Binding and Solubility Effects upon the Veneering of a Camel VHH in Framework-2 to Mimic a VH Katja Conrath 1,2 * , Ce ´ cile Vincke 1,2 , Benoı ˆt Stijlemans 1,2 Joost Schymkowitz 3 , Klaas Decanniere 2,4 , Lode Wyns 2,4 Serge Muyldermans 1,2 and Remy Loris 2,4 1 Laboratorium voor Cellulaire en Moleculaire Immunologie Vrije Universiteit Brussel Pleinlaan 2, B-1050 Brussel Belgium 2 Department of Cellular and Molecular Interactions, Vlaams Interuniversitair Instituut voor Biotechnologie, Pleinlaan 2 B-1050 Brussel, Belgium 3 SWITCH laboratory, Vlaams Interuniversitair Instituut voor Biotechnologie, Pleinlaan 2 B-1050 Brussel, Belgium 4 Laboratorium voor Ultrastructuur, Vrije Universiteit Brussel, Pleinlaan 2 B-1050 Brussel, Belgium Heavy chain only antibodies of camelids bind their antigens with a single domain, the VHH, which acquired adaptations relative to classical VHs to function in the absence of a VL partner. Additional CDR loop confor- mations, outside the canonical loop structures of VHs, broaden the repertoire of the antigen-binding site. The combined effects of part of the CDR3 that folds over the “former” VL binding site and framework-2 mutations to more hydrophilic amino acids, enhance the solubility of VHH domains and prevent VL pairing. cAbAn33, a VHH domain specific for the carbohydrate moiety of the variant surface glycoprotein of trypanosomes, has a short CDR3 loop that does not cover the former VL binding site as well as a VH-specific Trp47 instead of the VHH-specific Gly47. Resurfacing its framework-2 region (mutations Tyr37Val, Glu44Gly and Arg45Leu) to mimic that of a human VH restores the VL binding capacity. In solution, the humanised VHH behaves as a soluble, monomeric entity, albeit with reduced thermodynamic stability and affinity for its antigen. Comparison of the crystal structures of cAbAn33 and its humanised derivative reveals steric hindrance exerted by VHH-specific residues Tyr37 and Arg45 that prevent the VL domain pairing, whereas Glu44 and Arg45 are key elements to avoid insolubility of the domain. q 2005 Elsevier Ltd. All rights reserved. Keywords: camel; VHH; single-domain antibody; VL binding site *Corresponding author Introduction The serum of Camelidae contains a high proportion of functional antibodies devoid of light chains. 1 The antigen-binding site of such a heavy- chain antibody is located in a single variable domain with a mass of only 15,000 Da. This dedicated variable domain, referred to as VHH, is the smallest intact antigen-binding fragment derived from heavy-chain antibodies. 2 Recombi- nant VHHs that recognise specifically and with high affinity their antigens can be retrieved easily by phage display from a bank of VHHs cloned from the heavy-chain antibodies of an immunised camelid. Such VHHs are promising molecular recognition units to generate more complex bivalent, 3 pentavalent 4 and bispecific constructs or immuno-fusions for tumour targeting and drug delivery. 5 In the latter study, the VHH was named Nanobodye for its problate shape, with a diameter of 2.5 nm and height of 4 nm. 5 The capacity to frequently recognise regions that are less antigenic for conventional antibodies, like the catalytic site of enzymes, 6–8 is an asset for the immuno-modulation of enzymes (e.g. gene knock-down in plants 9 ), or preventing retroviral infections. 10 In contrast to human VHs, the VHHs are generally well expressed in Escherichia coli, highly soluble and stable at high concentration in aqueous solutions. 11 The solubility and monomeric behaviour of VHHs is attributed to adaptations in their amino acid sequence, which were introduced into the VHH germline genes during evolution of artiodactyls. 12 Compared to conventional VHs, four residues in otherwise conserved sequences are typically substituted in the framework-2 of VHHs. These VH-VHH hallmarks are Val37Phe, Gly44Glu, Leu45Arg and Trp47Gly (the latter substitution 0022-2836/$ - see front matter q 2005 Elsevier Ltd. All rights reserved. Abbreviations used: GdmCl, guanidinium chloride; PDB, RCSB Protein Data Bank. E-mail address of the corresponding author: [email protected] doi:10.1016/j.jmb.2005.04.050 J. Mol. Biol. (2005) 350, 112–125
Transcript of Antigen Binding and Solubility Effects upon the Veneering of a Camel VHH in Framework-2 to Mimic a...

doi:10.1016/j.jmb.2005.04.050 J. Mol. Biol. (2005) 350, 112–125
Antigen Binding and Solubility Effects upon theVeneering of a Camel VHH in Framework-2 to Mimic a VH
Katja Conrath1,2*, Cecile Vincke1,2, Benoıt Stijlemans1,2
Joost Schymkowitz3, Klaas Decanniere2,4, Lode Wyns2,4
Serge Muyldermans1,2 and Remy Loris2,4
1Laboratorium voor Cellulaireen Moleculaire ImmunologieVrije Universiteit BrusselPleinlaan 2, B-1050 BrusselBelgium
2Department of Cellular andMolecular Interactions, VlaamsInteruniversitair Instituut voorBiotechnologie, Pleinlaan 2B-1050 Brussel, Belgium
3SWITCH laboratory, VlaamsInteruniversitair Instituut voorBiotechnologie, Pleinlaan 2B-1050 Brussel, Belgium
4Laboratorium voorUltrastructuur, VrijeUniversiteit Brussel, Pleinlaan 2B-1050 Brussel, Belgium
0022-2836/$ - see front matter q 2005 E
Abbreviations used: GdmCl, guanPDB, RCSB Protein Data Bank.
E-mail address of the [email protected]
Heavy chain only antibodies of camelids bind their antigens with a singledomain, the VHH, which acquired adaptations relative to classical VHs tofunction in the absence of a VL partner. Additional CDR loop confor-mations, outside the canonical loop structures of VHs, broaden therepertoire of the antigen-binding site. The combined effects of part of theCDR3 that folds over the “former” VL binding site and framework-2mutations to more hydrophilic amino acids, enhance the solubility of VHHdomains and prevent VL pairing. cAbAn33, a VHH domain specific for thecarbohydrate moiety of the variant surface glycoprotein of trypanosomes,has a short CDR3 loop that does not cover the former VL binding site aswell as a VH-specific Trp47 instead of the VHH-specific Gly47. Resurfacingits framework-2 region (mutations Tyr37Val, Glu44Gly and Arg45Leu) tomimic that of a human VH restores the VL binding capacity. In solution, thehumanised VHH behaves as a soluble, monomeric entity, albeit withreduced thermodynamic stability and affinity for its antigen. Comparisonof the crystal structures of cAbAn33 and its humanised derivative revealssteric hindrance exerted by VHH-specific residues Tyr37 and Arg45 thatprevent the VL domain pairing, whereas Glu44 and Arg45 are key elementsto avoid insolubility of the domain.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: camel; VHH; single-domain antibody; VL binding site
*Corresponding authorIntroduction
The serum of Camelidae contains a highproportion of functional antibodies devoid of lightchains.1 The antigen-binding site of such a heavy-chain antibody is located in a single variabledomain with a mass of only 15,000 Da. Thisdedicated variable domain, referred to as VHH, isthe smallest intact antigen-binding fragmentderived from heavy-chain antibodies.2 Recombi-nant VHHs that recognise specifically and withhigh affinity their antigens can be retrieved easilyby phage display from a bank of VHHs cloned fromthe heavy-chain antibodies of an immunisedcamelid. Such VHHs are promising molecularrecognition units to generate more complex
lsevier Ltd. All rights reserve
idinium chloride;
ing author:
bivalent,3 pentavalent4 and bispecific constructs orimmuno-fusions for tumour targeting and drugdelivery.5 In the latter study, the VHH was namedNanobodye for its problate shape, with a diameterof 2.5 nm and height of 4 nm.5 The capacity tofrequently recognise regions that are less antigenicfor conventional antibodies, like the catalytic site ofenzymes,6–8 is an asset for the immuno-modulationof enzymes (e.g. gene knock-down in plants9), orpreventing retroviral infections.10
In contrast to human VHs, the VHHs aregenerally well expressed in Escherichia coli, highlysoluble and stable at high concentration inaqueous solutions.11 The solubility and monomericbehaviour of VHHs is attributed to adaptations intheir amino acid sequence, which were introducedinto the VHH germline genes during evolution ofartiodactyls.12 Compared to conventional VHs, fourresidues in otherwise conserved sequences aretypically substituted in the framework-2 of VHHs.These VH-VHH hallmarks are Val37Phe, Gly44Glu,Leu45Arg and Trp47Gly (the latter substitution
d.

Humanised Framework-2 of a Camelid VHH 113
being less well conserved among VHHs).12 Within aVH, these key-residues are hydrophobic and crucialfor proper interaction with the VL domain, whereasin VHHs these hallmark amino acid residues aresolvent-exposed, more hydrophilic and areassumed to impede the association with a possibleVL domain. A longer CDR3 size is another featurethat favours the solubility of VHHs13 and a highaffinity for its antigen. From crystallographic data, itbecame clear that part of this CDR3 loop typicallyfolds over the former VL-side,14–18 thereby increas-ing the antigen interaction surface and compensat-ing for the absence of the three CDR loops of the VLdomain of classical antibodies. In addition, thispreferred CDR3 folding covers residual hydro-phobic patches of the framework-2 region andfurther prevents any possible interaction with aVL domain.
Apart from the CDR3 loop, the CDR1 and CDR2loop structures differ nearly always from thecanonical loop structures defined for conventionalVHs despite good preservation of the key residues.This broader structural repertoire of loops mightcontribute to VHH recognising alternative epitopeson an antigen. However, the exact reason for theabsence of a restricted set of structures for theH1/H2 loops in VHHs remains unclear.19 The lackof constraints by VL association might allow alarger variety of loop architectures, and the occur-rence of the VHH hallmark amino acid residues inframework-2 may be involved. Indeed, it has beenreported that the (partial) camelisation of a humanVH by mutating three out of the four hallmarkamino acid residues (positions 44, 45 and 47)resulted in backbone deformations and in abolitionof antigen-binding activity.20,21
In the present study, an antigen-specific VHHwith a CDR3 loop of ten amino acid residues andthe Trp47 (VH-like features) was used to start aveneering of the VHH framework-2 region to mimicthe amino acid residues of a human VH in that area.This might be a first step towards a strategy tohumanise a camelid VHH, while maintainingoptimal solution properties, for subsequent appli-cation in tumour targeting, therapy or in vivodiagnostics.5,18,22,23 The effects of this domainresurfacing on protein stability and antigen affinitywere evaluated. Structural data on the CDR loopconformations, before and after VHH humanisa-tion, were obtained from crystallography, and thelikelihood of the fragment to pair with itself or witha VL was assessed.
Results
A VHH, referred to as cAbAn33 (GenBankaccession no. AAP22644) with specificity for theconserved carbohydrate moiety on variant surfaceglycoprotein of the trypanosome coat23 was chosento assess the effect of the camelid VHH hallmarkamino acid residues on stability, solubility, andantigen-binding loop conformation of the VHH
fragment. The cAbAn33 encodes Tyr37, Glu44,Arg45 in its framework-2 region, which are hall-marks of VHHs (numbering is according to Kabatet al.24). Gly47, the commonest amino acid residue atthe fourth hallmark position in framework-2 ofVHHs is occupied by the bulky aromatic Trp47 incAbAn33, as it is in human or mouse VHs(Figure 1(a)). This deviation from the VHH con-sensus sequence has no effect on the monomericstate of the protein as judged by its elution profileon gel-filtration chromatography (a single peak atthe expected molecular mass; Figure 2). Further-more, cAbAn33 is purified in standard yields fromthe bacterial periplasm. The cAbAn33 has an H3loop (i.e. the third antigen-binding loop) of only tenamino acid residues (Figure 1(a)), which is close tothe average H3 length of a mouse VH (nineresidues) or human VH (12 residues).25 Moreover,the H3 loop length of cAbAn33 is probably too shortto cover the entire “former VL-binding site”, asnoticed in other VHHs,26 so that the hallmarkamino acid residues within this area of the domainremain solvent-accessible. This assumption wasfirst verified by crystallography.
Crystal structure of cAbAn33
Two ligand-free molecules of cAbAn33 arepresent within the asymmetric unit of the crystal.These two molecules are essentially identical, with abackbone rmsd value of 0.79 A (0.62 A for theframework residues, defined as Kabat VH aminoacid positions 4–23, 37–48, 66–92, and 103–111).Superposition of the framework residues on otherdromedary and llama VHHs gives rmsd valuesbetween 1.0 A and 1.5 A for all backbone atoms.
The cAbAn33 structure has the expectedimmunoglobulin fold (Figure 1(b)) with nineb-strands spread over two sheets, an architecturethat is observed constitutively in other VHH andVH domains.27 As regularly present in dromedaryVHHs and predicted from the cAbAn33 sequence,an interloop disulphide bond is formed betweenCys32 in CDR1 and Cys96 in CDR3.26 A closer lookat the antigen-binding loops reveals non-canonicalloop conformations for H1 and H2 loops(Figure 3(a) and (b)), as observed for most othercamelid VHH loops.14–16,19
Although the H1 loop is defined clearly in onlyone of the two molecules in the asymmetric unit, itis evident that this loop of cAbAn33 does not adoptany of the three canonical structures described forH1 loops of VHs,28,29 nor the type-4 structureobserved in two VHHs (Figure 3(a)).17 In all VHstructures a seven residue H1 loop fits a canonicalstructure type-1.30 This canonical loop structure isshaped by a sharp turn at Gly26, clustering thehydrophobic side-chains of Ala24, Phe27 andMet34, and the hydrophobic part of the Arg94side-chain. Although the H1 loop of cAbAn33 con-tains seven amino acid residues, its sequence com-position at key sites for a type-1 structure differssubstantially (substitutions Ala24Val, Phe27Ser,
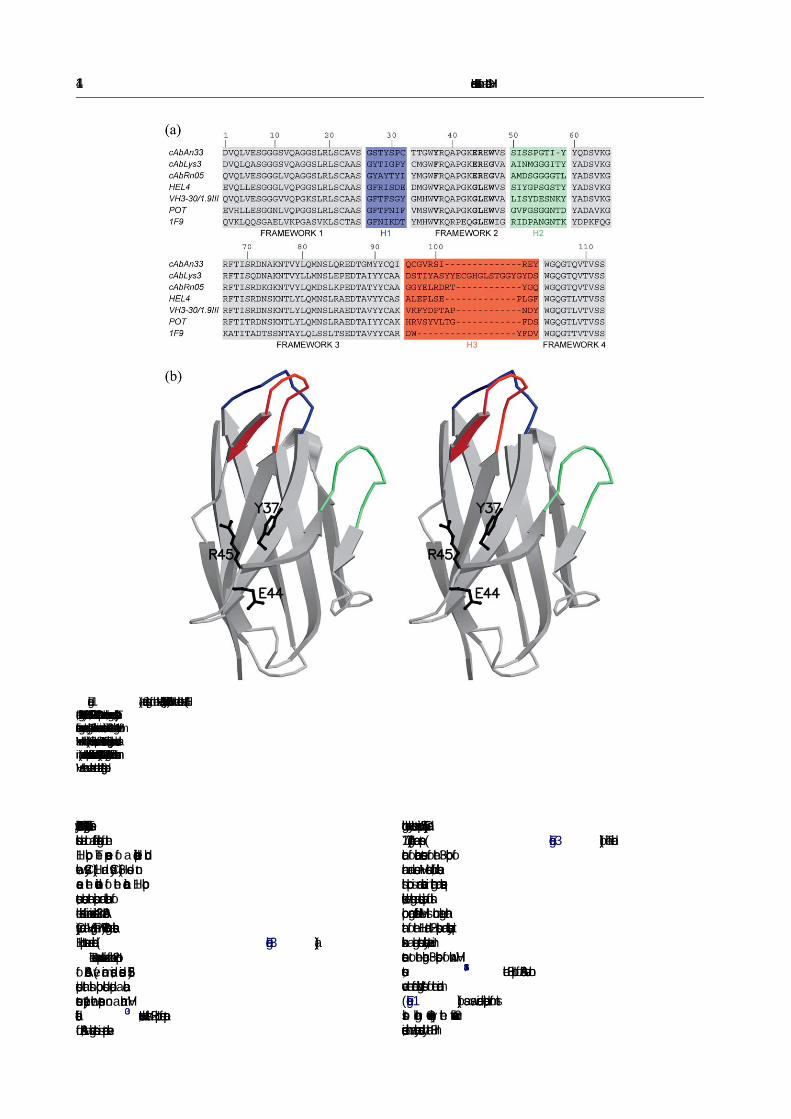
Figure 1. (a) Sequence alignment of the VHHs (cAbAn33, cAbLys3, cAbRN05) with the human/mouse VHs (HEL4,Pot, IgM-Rf-2A2 and 1F9). The CDR1/H1, CDR2/H2 and CDR3 loops are coloured blue, green and red, respectively. Theframework regions are coloured grey. The four hallmark amino acid residues in framework-2 that distinguish VHs fromVHHs are in bold. (b) Stereo ribbon representation of cAbAn33 structure. The antigen-binding loops are colour-coded asin (a), and the VHH-speci�c framework-2 residues in cAbAn33 (Tyr37, Glu44 and Arg45) that differ from human/mouseVH are shown as ball-and-stick and labelled (single-letter code).
114 Humanised Framework-2 of a Camelid VHH
Phe29Tyr, Met34Thr and Arg94Ile). Therefore, thesesubstitutions lead to a different organisation of theH1 loop. The presence of a disulphide bondbetween Cys32 (H1) and Cys96 (H3) does notcause the deviation of the canonical H1 loopstructure, because the position and orientation ofthe side-chain of amino acid residue 32 in cAbAn33(Cys) and a VH (e.g. Phe in POT) having a classicalH1 loop structure are identical ( Figure 3(a)).
The sequence contents and the size of the H2 loopof cAbAn33 (�ve amino acids, residues 52–56)predicts that this loop should adopt a canonicalstructure type-1 when present on a human VHscaffold.30 However, when this H2 loop forms partof cAbAn33, a deviating structure is adopted even
though the key elements at positions 55 (Gly55) and71 (Arg71) are present (Figure 3(b)). The identi�-cation of canonical structures for the H3 loop ofhuman and mouse VH remains dif�cult, becausethis loop is more variable in length and sequence.Likewise, the length and sequence variation of thisloop among different VHHs is much larger thanthat of the H1 and H2 loops and consequently itexhibits a large structural variability. However, incontrast to the longer H3 loops of known VHHstructures,14,15,18the H3 loop of cAbAn33 does notcover the “former VL-binding site” of the domain(Figure 1(b)), as was indeed anticipated from itsshort length. Consequently, the framework-2residues, which are normally covered by the H3 in











Humanised Framework-2 of a Camelid VHH 125
anti-Z-DNA antibody variable domains. The role ofheavy and light chains measured by surface plasmonresonance. J. Immunol. 154, 2198–2208.
32. Fan, Z. C., Shan, L., Guddat, L. W., He, X. M., Gray,W. R., Raison, R. L. & Edmundson, A. B. (1992). Three-dimensional structure of an Fv from a human IgMimmunoglobulin. J. Mol. Biol. 228, 188–207.
33. Graille, M., Stura, E. A., Corper, A. L., Sutton, B. J.,Taussig, M. J., Charbonnier, J. B. & Silverman, G. J.(2000). Crystal structure of a Staphylococcus aureusprotein A domain complexed with the Fab fragmentof a human IgM antibody: structural basis forrecognition of B-cell receptors and superantigenactivity. Proc. Natl Acad. Sci. USA, 97, 399–404.
34. Ay, J., Keitel, T., Kuttner, G., Wessner, H., Scholz, C.,Hahn, M. & Hohne, W. (2000). Crystal structure of aphage library-derived single-chain Fv Fragmentcomplexed with turkey egg-white lysozyme at 2.0 Aresolution. J. Mol. Biol. 301, 239–246.
35. Jespers, L., Schon, O., James, L. C., Veprintsev, D. &Winter, G. (2004). Crystal structure of HEL4, a soluble,refoldable human V(H) single domain with a germ-line scaffold. J. Mol. Biol. 337, 893–903.
36. Nymalm, Y., Kravchuk, Z., Salminen, T.,Chumanevich, A., Dubnovitsky, A., Kankare, J. et al.(2002). Antiferritin VL homodimer binds humanspleen ferritin with high specificity. J. Struct. Biol.138, 171–186.
37. Nguyen, V. K., Desmyter, A. & Muyldermans, S.(2001). Functional heavy-chain antibodies inCamelidae. Advan. Immunol. 79, 261–296.
38. Dottorini, T., Vaughan, C. K., Walsh, M. A., LoSurdo,P. & Sollazzo, M. (2004). Crystal structure of a humanVH: requirements for maintaining a monomericfragment. Biochemistry. 43, 622–628.
39. Worn, A. & Pluckthun, A. (1998). Mutual stabilizationof VL and VH in single-chain antibody fragments,investigated with mutants engineered for stability.Biochemistry, 37, 13120–13127.
40. Cai, X. & Garen, A. (1996). A melanoma-specific VHantibody cloned from a fusion phage library of avaccinated melanoma patient. Proc. Natl Acad. Sci.USA, 93, 6280–6285.
41. Skerra, A. & Pluckthun, A. (1988). Assembly offunctional immunoglobulin Fv fragments inEscherichia coli. Science, 240, 1038–1041.
42. Gill, S. & von Hippel, P. (1989). Calculation of proteinextinction coefficients from amino acid sequence data.Anal. Biochem. 182, 319–326.
43. Dumoulin, M., Conrath, K., Van Meirhaeghe, A.,Meersman, F., Heremans, K., Frenken, L. G. J. et al.(2002). Single-domain antibody fragments with highconformational stabilities. Protein Sci. 11, 500–515.
44. Otwinowski, Z. & Minor, W. (1997). Processing ofX-ray diffraction data collected in oscillation mode.Methods Enzymol. 276, 307–326.
45. Collaborative Computational Project Number 4.(1994). The CCP4 suite: programs for protein crystal-lography. Acta Crystallog. sect. D, 50, 760–763.
46. Navaza, J. (1994). AMORE: an automated package formolecular replacement. Acta Crystallog. sect. A, 50,157–163.
47. Brunger, A. T., Adams, P. D., Clore, G. M., DeLano,W. L., Gros, P., Grosse-Kunstleve, R. W. et al. (1998).Crystallography & NMR system: a new software suitefor macromolecular structure determination. ActaCrystallog. sect. D, 54, 905–921.
48. Roussel, A. & Cambillau, C. (1989). TURBO-FRODOSilicon Graphics Geometry Partner Directory, SiliconGraphics, Mountain View, CA pp. 71–78.
49. Kraulis, P. J. (1991). MOLSCRIPT: a program toproduce both detailed and schematic plots of proteinstructures. J. Appl. Crystallog. 24, 946–950.
50. Merritt, E. A. & Bacon, D. J. (1997). Raster3D:photorealistic molecular graphics. Methods Enzymol.277, 505–524.
Edited by I. Wilson
(Received 24 December 2004; received in revised form 14 April 2005; accepted 22 April 2005)




















