
Anti-ganglioside anti-idiotypic monoclonal antibody-based cancer vaccine induces apoptosis and...
12
Cancer Immunol Immunother (2009) 58:1117–1128 DOI 10.1007/s00262-008-0634-y 123 SHORT COMMUNICATION Anti-ganglioside anti-idiotypic monoclonal antibody-based cancer vaccine induces apoptosis and antiangiogenic eVect in a metastatic lung carcinoma Y. Diaz · A. Gonzalez · A. Lopez · R. Perez · A. M. Vazquez · E. Montero Received: 5 April 2008 / Accepted: 17 November 2008 / Published online: 10 December 2008 © Springer-Verlag 2008 Abstract Anti-idiotype monoclonal antibody (mAb) 1E10 was generated by immunizing BALB/c mice with an Ab1 mAb which recognizes NeuGc-containing ganglio- sides, sulfatides and some tumor antigens. 1E10 mAb induces therapeutic eVects in a primary breast carcinoma and a melanoma model. However, the tumor immunity mechanisms have not been elucidated. Here we show that aluminum hydroxide-precipitated 1E10 mAb immunization induced anti-metastatic eVect in the 3LL-D122 Lewis Lung carcinoma, a poorly immunogenic and highly metastatic model in C57BL/6 mice. The therapeutic eVect was associ- ated to the increment of T cells inWltrating metastases, the reduction of new blood vessels formation and the increase of apoptotic tumor cells in lung nodules. Interestingly, active immunization does not induce measurable antibodies to the 1E10 mAb, the NeuGc-GM3 or tumor cells, which may suggest a diVerent mechanism which has to be eluci- dated. These Wndings may support the relevance of this target for cancer biotherapy. Keywords Cancer vaccine · Lung carcinoma metastases · Anti-idiotype monoclonal antibody · NeuGc-containing ganglioside · Apoptosis · Angiogenesis Abbreviations Ag Antigen Ab(s) Antibodies ELISA Enzyme-linked immunosorbent assay Id Idiotype mAb(s) Monoclonal antibody(ies) NeuGc Neu-glycolyl sc Subcutaneous Introduction Active immunotherapy is one of the most promising Welds in cancer research. One strategy to generate an eVective immune response against tumor-associated antigens involves the use of anti-idiotypic antibodies (Ab2), bearing the “inter- nal image” of the antigen. The use of Ab2 as vaccines is based on Jerne’s idiotypic network theory, which postulate that the immunoglobulin idiotype repertoire must mimic the universe of non-self and self-antigens [1]. DiVerent Ab2s have been generated and some of them have demonstrated to inhibit tumor growth in preclinical studies [2, 3]. Anti-idiotypic vaccines directed to tumor-associated anti- gens have shown their capability to induce antitumor spe- ciWc humoral and cellular responses, and in some cases a delayed tumor progression and an improved survival [4–8]. We have previously reported the generation of an Ab2 monoclonal antibody (mAb) to a murine Ab1, named P3, which binds NeuGc-containing gangliosides, sulfated gly- colipids and antigens expressed in diVerent murine and human tumor cells [9–12]. This IgG1 Ab2 mAb, designated Y. Diaz · A. Gonzalez · A. Lopez · E. Montero (&) Experimental Immunotherapy Department, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba e-mail: [email protected] R. Perez Research and Development, Center of Molecular Immunology, Havana, Cuba A. M. Vazquez Antibody Engineering Department, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba
Transcript of Anti-ganglioside anti-idiotypic monoclonal antibody-based cancer vaccine induces apoptosis and...

Cancer Immunol Immunother (2009) 58:1117–1128
DOI 10.1007/s00262-008-0634-ySHORT COMMUNICATION
Anti-ganglioside anti-idiotypic monoclonal antibody-based cancer vaccine induces apoptosis and antiangiogenic eVect in a metastatic lung carcinoma
Y. Diaz · A. Gonzalez · A. Lopez · R. Perez · A. M. Vazquez · E. Montero
Received: 5 April 2008 / Accepted: 17 November 2008 / Published online: 10 December 2008© Springer-Verlag 2008
Abstract Anti-idiotype monoclonal antibody (mAb)1E10 was generated by immunizing BALB/c mice with anAb1 mAb which recognizes NeuGc-containing ganglio-sides, sulfatides and some tumor antigens. 1E10 mAbinduces therapeutic eVects in a primary breast carcinomaand a melanoma model. However, the tumor immunitymechanisms have not been elucidated. Here we show thataluminum hydroxide-precipitated 1E10 mAb immunizationinduced anti-metastatic eVect in the 3LL-D122 Lewis Lungcarcinoma, a poorly immunogenic and highly metastaticmodel in C57BL/6 mice. The therapeutic eVect was associ-ated to the increment of T cells inWltrating metastases, thereduction of new blood vessels formation and the increaseof apoptotic tumor cells in lung nodules. Interestingly,active immunization does not induce measurable antibodiesto the 1E10 mAb, the NeuGc-GM3 or tumor cells, whichmay suggest a diVerent mechanism which has to be eluci-dated. These Wndings may support the relevance of thistarget for cancer biotherapy.
Keywords Cancer vaccine · Lung carcinoma metastases · Anti-idiotype monoclonal antibody · NeuGc-containing ganglioside · Apoptosis · Angiogenesis
AbbreviationsAg AntigenAb(s) AntibodiesELISA Enzyme-linked immunosorbent assayId IdiotypemAb(s) Monoclonal antibody(ies)NeuGc Neu-glycolylsc Subcutaneous
Introduction
Active immunotherapy is one of the most promising Welds incancer research. One strategy to generate an eVectiveimmune response against tumor-associated antigens involvesthe use of anti-idiotypic antibodies (Ab2), bearing the “inter-nal image” of the antigen. The use of Ab2 as vaccines isbased on Jerne’s idiotypic network theory, which postulatethat the immunoglobulin idiotype repertoire must mimic theuniverse of non-self and self-antigens [1]. DiVerent Ab2shave been generated and some of them have demonstrated toinhibit tumor growth in preclinical studies [2, 3].
Anti-idiotypic vaccines directed to tumor-associated anti-gens have shown their capability to induce antitumor spe-ciWc humoral and cellular responses, and in some cases adelayed tumor progression and an improved survival [4–8].
We have previously reported the generation of an Ab2monoclonal antibody (mAb) to a murine Ab1, named P3,which binds NeuGc-containing gangliosides, sulfated gly-colipids and antigens expressed in diVerent murine andhuman tumor cells [9–12]. This IgG1 Ab2 mAb, designated
Y. Diaz · A. Gonzalez · A. Lopez · E. Montero (&)Experimental Immunotherapy Department, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cubae-mail: [email protected]
R. PerezResearch and Development, Center of Molecular Immunology, Havana, Cuba
A. M. VazquezAntibody Engineering Department, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba
123

1118 Cancer Immunol Immunother (2009) 58:1117–1128
1E10, was able to induce in mice and monkeys anti-anti-idiotypic antibodies (Ab3) bearing P3 mAb idiotopes, but itwas unable to generate Ab3 antibodies with the same anti-gen speciWcity as the P3 mAb [10, 13, 14]. In contrast,1E10 mAb was capable to generate a very strong and spe-ciWc antibody response against NeuGc-containing ganglio-sides in chickens [14]. Similar results have been obtained incancer patients immunized with aluminum hydroxide(Alum)-precipitated 1E10 mAb [11, 12, 15]. In these twospecies, humans and chickens, N-glycolilated gangliosidesare non-self antigens [16–18]. Previous results showed thattreatment with 1E10 Ab2 mAb induced a strong antitumoractivity in mice bearing melanoma or a breast carcinoma[13], but so far the mechanism by which this mAb exertsthis antitumor eVect has not been elucidated. These resultsin syngeneic and allogeneic models of diVerent origin dem-onstrate the potential of 1E10 mAb to activate mechanismsleading to an antitumor immunity, potentially beyond thedemonstrated anti-idiotypic antibody response [10].
In this work, we evaluated the anti-metastatic eVect of1E10 mAb in Alum adjuvant (1E10/Alum) in the 3LL-D122Lewis Lung carcinoma model. 1E10/Alum immunizationwas tested in two diVerent settings distinguished by the fre-quency of the immunizations, the amount of vaccine and theinitiation of the vaccine schedule related to the tumor chal-lenge. Independently to the immunization schedule, 1E10/Alum promoted a signiWcant reduction of spontaneous lungmetastases. The therapeutic eVect was associated to the incre-ment in the number of T cells inWltrating metastases, a reduc-tion of new blood vessels formation and the increment ofapoptotic tumor cells in lung nodules. Interestingly, activeimmunization does not induce measurable antibodies to the1E10 mAb, the NeuGc-GM3 or tumor cells, which may sug-gest a diVerent mechanism which has to be elucidated.
Materials and methods
Animals
Female C57BL/6 inbred mice, 8–10 weeks old, were pur-chased from the Center for Laboratory Animal Production(CENPALAB, Havana, Cuba). Animals were housed underconventional conditions with free access to water and foodand maintained in accordance with the guidelines stipulatedby Animal Subject Committee Review Board of the Centerof Molecular Immunology (CIM) and CIM’s InstitutionalAnimal Care and Use Committees.
Gangliosides
Gangliosides are named according to the nomenclature ofSvennerholm (1964) NeuAcGM3 and NeuGcGM3 were
kindly provided by Dr L.E. Fernández, Vaccine Depart-ment, Center of Molecular Immunology (Havana, Cuba).NeuAcGM3 and NeuGc-GM3 gangliosides were puriWedfrom dog and horse erythrocytes, respectively, as described[19]. Homogeneity and purity of gangliosides is more than95% as determined by thin layer chromatography and den-sitometry [17].
Aluminum hydroxide-precipitated 1E10 mAb (1E10/Alum)
1E10 mAb (IgG1, �) was puriWed from mouse ascites byaYnity chromatography on Protein A-CL Sepharose 4 Bcolumn (Amersham Pharmacia Biotech, Uppsala,Sweden). The purity of the isolated immunoglobulin wasmore than 97% as determined by SDS-PAGE, high-pres-sure liquid chromatography, and isoelectric focusing. Forthis study puriWed 1E10 mAb was aseptically mixed at aWnal concentration of 1 mg/ml with 5 mg/ml of aluminumhydroxide as adjuvant (Superfos Biosector, Frederikssund,Denmark). The mixture was gently stirred for 3 h at roomtemperature. The aluminum hydroxide-precipitated mAbwas aliquot and stored at 4°C until use. Aluminum hydrox-ide-precipitated iorC5 (iorC5/Alum), a murine mAb thatrecognizes an antigen preferentially expressed in humancolorectal cells [9], was used as isotypic and allotypiccontrol.
Cell line
Lewis Lung carcinoma clone D122 (3LL-D122) is low-immunogenic and high-metastatic in syngeneic C57BL/6mice [20]. 3LL-D122 cells were cultured in DMEM (GibcoLife Technologies, Grand Island, NY, USA) supplementedwith 10% heat inactivated fetal bovine serum (HyClone,UT, USA), 2-mM L-glutamine, 100 U/ml penicillin,100 �g/ml streptomycin and 50 �M 2-mercaptoethanol. Forinjection into mice, cell suspensions were prepared bydetaching cells from culture Xasks with trypsin-EDTA(0.05%), followed by washing and resuspension in phos-phate-buVered saline (PBS), pH 7.4. Tumor cells werecounted and viability was greater than 95%, as assayed bytrypan blue exclusion test.
Antitumor experiments
C57BL/6 mice (six to eight mice per group) were chal-lenged subcutaneous (sc) into one hind footpad with2 £ 105 of 3LL-D122 cells (designated as day 0 of theexperiment). We tested two immunization schedules: (1)mixed setting; animals were immunized before and afterthe challenge with tumor, on days ¡28, ¡14, 7 and 21, with50 �g of 1E10/Alum, iorC5/Alum or only with the adju-vant, in 0.2 ml of PBS sc; (2) therapeutic setting; mice were
123

Cancer Immunol Immunother (2009) 58:1117–1128 1119
treated after tumor challenge, on days 3, 10, 17, 24, 31 and38, sc with 100 �g of 1E10/Alum or only with the adjuvant.Primary tumors were surgically removed when reached adiameter of 8–9 mm (around days 25–29). Mice were sacri-Wced 21 days later and their lungs were removed, weighted,Wxed in Bouin’s solution and the number of surface lungmetastases was counted under a dissecting microscope.Sera were collected pretreatment and 14 days after the Wrstthree doses and 21 days after the last dose. Mice treatedwith iorC5/Alum or the adjuvant alone in PBS (PBS/Alum)were used as controls. Experiments were repeated at leastthree times.
Antibody binding assay
Reactivity to 1E10 (Ab3) in sera from immunized mice wasdetermined by ELISA as described previously [21]. BrieXy,96-well microtiter plates (high binding; Costar, Cambridge,MA, USA) coated with 10 �g/ml of F(ab�) 1E10 mAb wereincubated with animal sera. The second antibody, alkaline-phosphatase-conjugated goat anti-mouse IgG + IgM (Fcfragment speciWc; Sigma, St Louis, MO, USA), was addedand the reaction was developed with p-nitrophenylphos-phate substrate (Sigma) in diethanolamine buVer, pH 9.8.Absorbance was measured at 405 nm in an ELISA reader(Organon Teknika, Salsburg, Austria). iorC5 F(ab�)2 frag-ments were used as controls. The highest serum dilutiongiving optical density values ¸ 0.2 and being at least twotimes the value corresponding to the pre-immune serum atthe same dilution was considered as the antibody titer.
Binding of mice Ab3 to puriWed gangliosides wasdetermined using an ELISA assay described previously[21]. BrieXy, gangliosides (4 �g/ml) in 50 �l of methanolwere dried in 96-well microtiter plates (Polysorp, Nunc,Denmark). Plates were blocked with 1% BSA in 0.05-MTris–HCl buVer, pH 7.8, and incubated with serum sam-ples. The biotin-goat anti-mouse IgG + IgM (Fc fragmentspeciWc; Sigma) was added and the streptavidin conju-gate was developed with p-nitrophenylphosphate sub-strate (Sigma) in diethanolamine buVer, pH 9.8. Toconsider that a serum sample had a positive reaction to aparticular ganglioside, values of absorbance had tobe ¸ 0.2 and at least two times the absorbance valuegiven by incubating the serum in wells containing nogangliosides.
Flow cytometry analysis
The recognition of 3LL cells by the sera of tumor-free ortumor-bearing mice, treated with four doses of 50 �g of1E10/Alum, iorC5/Alum or PBS/Alum, were analyzed byXow cytometry. BrieXy, 3LL-D122 cells were incubated at4°C for 25 min with diVerent mice sera dilutions, then
washed in PBS and subsequently incubated with PE-conju-gated goat anti-mouse immunoglobulins (Dako, Denmark)for 25 min at 4°C. Cells were resuspended and expressionof molecules of interest was monitored by FACScan andanalyzed using the WinMDI 2.8 Program.
Histological and immunohistochemical analyses
For histological and immunohistochemical studies, frag-ments of metastatic lungs were formalin-Wxed andparaYn-embedded. Five-micrometer tissue sections werecut, air-dried on glass slides, deparaYnized and rehy-drated. Tissue sections were stained with hematoxylin andeosin under standard procedures for detection of mitoticbodies.
For endothelial cell detection by CD31 (platelet-endo-thelial cell adhesion molecule-1) staining, endogenousperoxidase activity was blocked by hydrogen peroxidase(0.3%) for 30 min at room temperature. Later, tissue sec-tions were treated with pepsin for 30 min at 37°C andblocked with PBS containing 5% of BSA for 15 min.Sections were incubated with an anti-CD31 mAb(Pharmingen, San Diego, CA, USA), for 90 min and thenwashed with Tris buVered saline, pH 7.4. Binding of theprimary antibody was detected using a biotinylated goatanti-mouse immunoglobulin secondary antiserum(Jackson Laboratories Bar Harbor, ME, USA), followedby the avidin–biotin–peroxidase system (Dako, Denmark)with 3,3-diaminobenzidine (Sigma, St Louis, MO, USA)as the chromogen. Sections were counterstained withhematoxylin.
Apoptosis was performed using Apoptag™ Plus Kit(Oncor, Canada), according to manufacturer’s instructions.Methyl green was used as counterstaining.
To determine the inWltrated lymphocyte population inlung metastasis, frozen sections were Wxed in acetone andincubated with biotinylated rat anti-mouse CD4 and CD8antisera (Pharmingen). The binding of biotinylated antibod-ies was detected as described above.
For all the techniques, 12 independent lung metastasesWelds from two mice from each group were counted.
Statistical analysis
The mean and the error of the values obtained in each groupwere calculated. Each experiment was repeated at leasttwice. Statistical diVerences between groups were deter-mined. Kruskal–Wallis with Dunn’s multiple comparisonstest was used to compare three groups and Mann–Whitneytest for the analysis of two groups. Chi-square was per-formed to compare the incidence of lung metastasisbetween the treatment groups. Analyses were performedwith Graph Pad Prism, version 4.0.
123

1120 Cancer Immunol Immunother (2009) 58:1117–1128
Results
1E10/Al immunization reduces the incidence of spontaneous lung metastases in the murine 3LL-D122 Lewis Lung carcinoma model
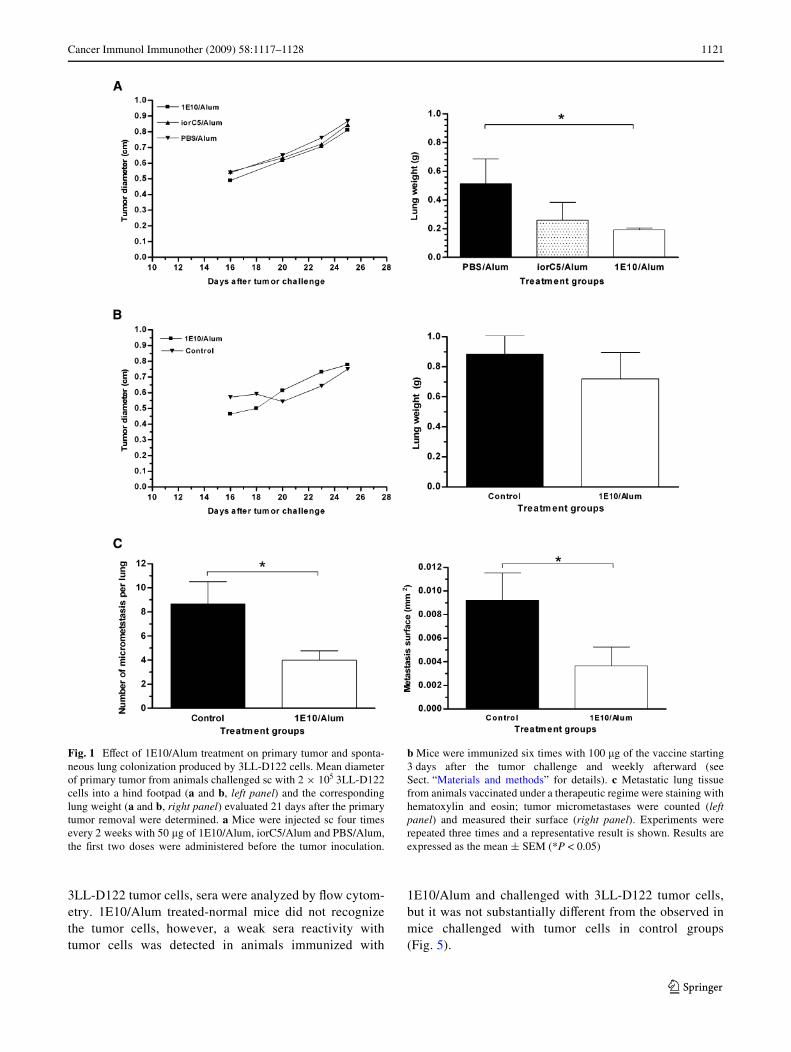
To investigate if 1E10/Alum had the capacity to inhibit thespontaneous lung colonization of 3LL-D122 cells, micewere injected sc with this vaccine preparation. Mice treatedwith four doses of 50 �g of 1E10/Alum based on the sched-ule 1 (see Sect. “Materials and methods” for details),showed a signiWcant reduction of both lung weight and thenumber of macrometastases (mean: 1E10/Alum: 1.8, iorC5/Alum: 14.3 an PBS/Alum: 18.2); compared to controlstreated with an irrelevant isotype matched mAb or Alumalone (P < 0.05, Kruskal–Wallis test; Fig. 1a and Table 1).However, that treatment did not have an incidence on theprimary tumor growth (Fig. 1a).
Moreover following a therapeutic setting, animals weretreated six times with 100 �g of 1E10/Alum (schedule 2),beginning 3 days after tumor challenge. Despite that treat-ment did not modify the incidence the primary tumor vol-ume or lung weight (Fig. 1b), there was a signiWcantreduction in the number of micrometastasis and the micro-metastasis surface in the lung tissue (Fig. 1c) of vaccinatedanimals compared to the control group (P < 0.05, Mann–Whitney test).
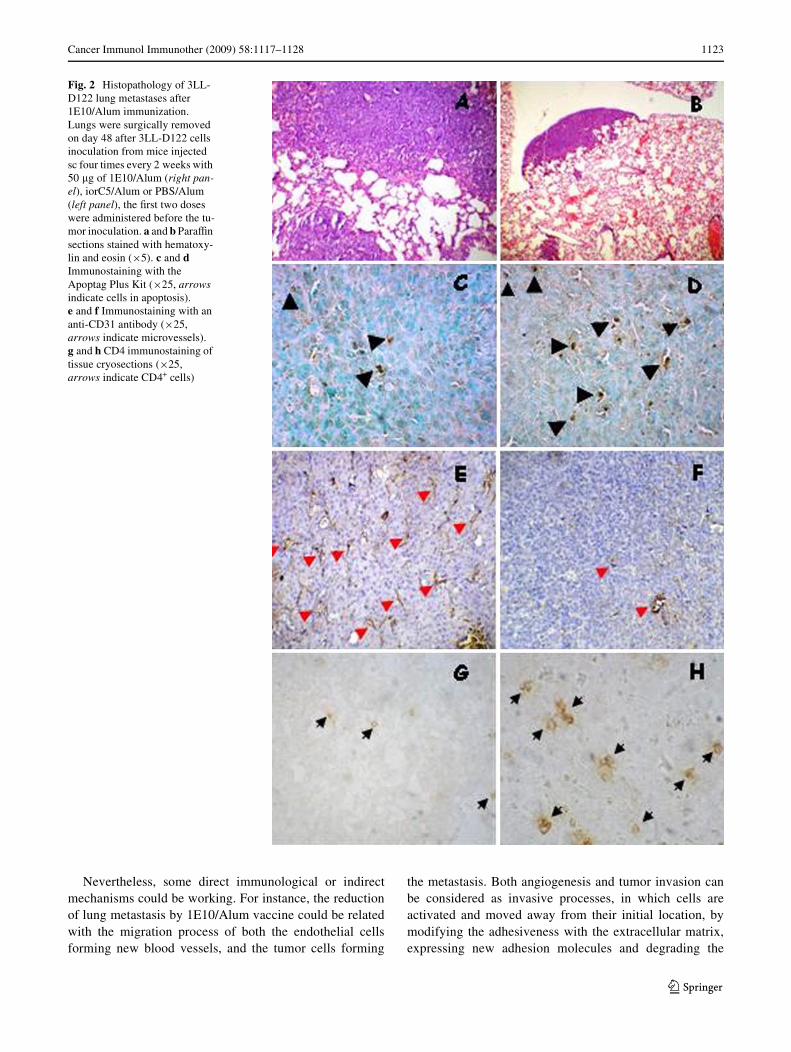
The histopathological study of the lungs from controlanimals (PBS/Alum or iorC5/Alum) revealed a completeloss of lung parenchyma due to a massive invasion bytumor cells. There is a high incidence of lung metastaticnodules associated to a lack of visible alveolar spaces with-out a particular localization (Fig. 2a). In contrast, in tissuesfrom mice treated with 1E10/Alum, a reduced number oflung metastases and a smaller size of metastatic nodules inlung sections were observed. The alveolar spaces were sim-ilar to normal lung histology and small tumor nodules weredetected mainly in the proximity of the pleura (Fig. 2b).
In summary the vaccination with 1E10/Alum induced asigniWcant anti-metastatic eVect in the 3LL-D122, a poorlyimmunogenic and highly metastatic version of the LewisLung carcinoma, evidenced by a reduction in lung weightand/or a reduction in the number of spontaneous macro- ormicrometastasis.
1E10/Alum immunization induces apoptosis without aVecting cell mitosis of spontaneous lung metastases in the murine 3LL-D122 Lewis Lung carcinoma model
To determine whether the therapeutic eVect was associatedto the tumor cell apoptosis in 3LL-D122 pulmonary metas-tases, sections of lung tissues from C57BL/6 mice immu-nized with 1E10/Alum, iorC5/Alum or adjuvant were
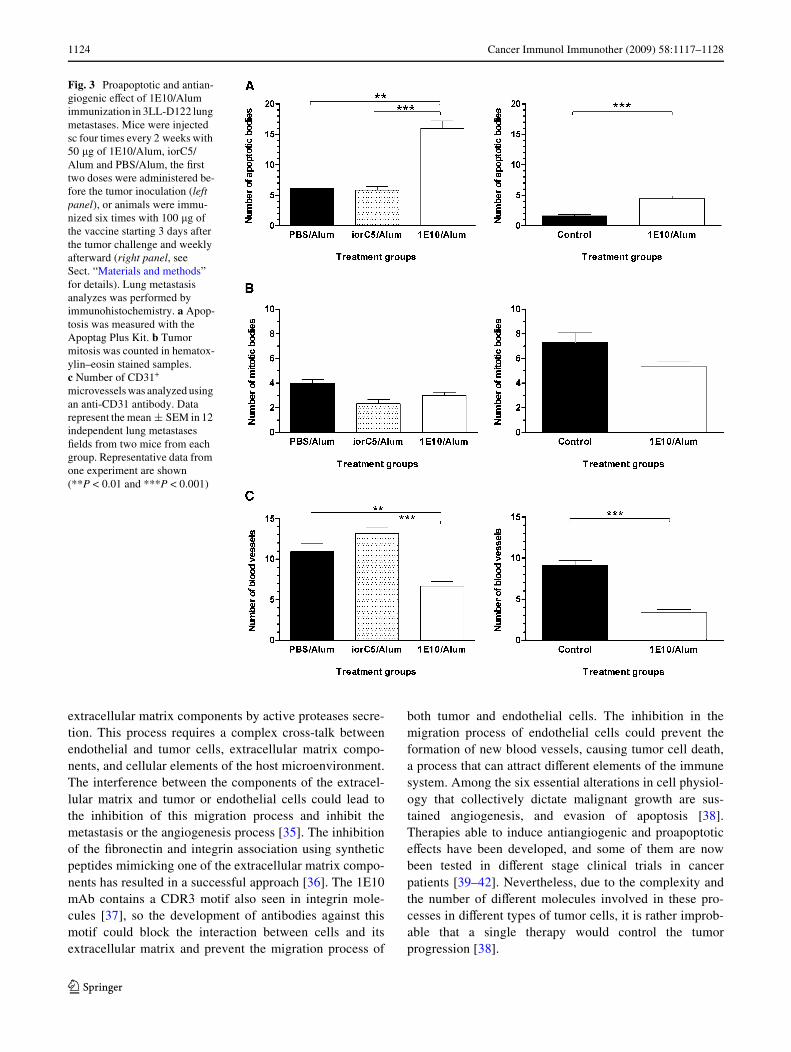
examined using the Apoptag Plus Kit. As shown inFigs. 2c, d and 3a, tumors from mice treated with 1E10/Alum displayed a signiWcant increment of apoptotic cellscompared to the control groups (P < 0.05, Kruskal–Wallisor Mann–Whitney test).
The inXuence of 1E10/Alum treatment in tumor mitosiswas also examined. The analysis of mitotic cell counts inlung metastatic tissues did not show diVerences betweenthe control groups and the animals that received 1E10/Alum (Fig. 3b). Therefore, 1E10/Alum treatment promotedapoptosis of lung metastatic cells without interfering withthe normal mitosis process of tumor cells.
1E10/Alum immunization promotes an antiangiogenic eVect in spontaneous lung metastases of the murine 3LL-D122 Lewis Lung carcinoma model
Immunohistochemical analysis by CD31 staining of meta-static lungs from mice injected with 1E10/Alum, in bothtested schedules, revealed a signiWcantly lower mean ofmicrovessel counts compared with those from animalstreated with the irrelevant isotype mAb precipitated inAlum or only with the adjuvant alone (Figs. 2e, f, 3c;P < 0.05, Kruskal–Wallis test).
1E10/Al immunization increases the T cell inWltration in spontaneous lung metastases of the murine 3LL-D122 Lewis Lung carcinoma model
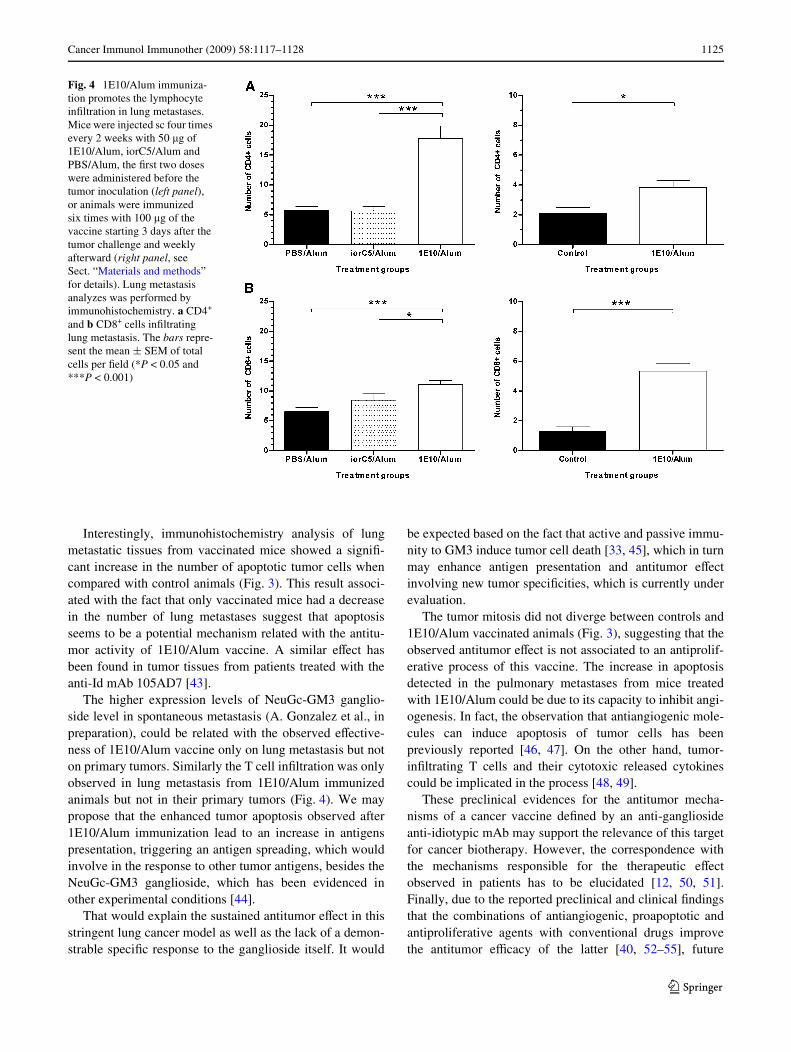
Tumor tissues from 3LL-D122 Lewis Lung carcinomabearing mice displayed a limited leukocyte inWltration.However, lung sections from 1E10/Alum treated miceshowed higher intra-tumor inXammatory inWltrates thancontrols. To measure the lymphocyte inWltration in metas-tases, lung cryostat sections from animals immunizedwith 1E10/Alum, iorC5/Alum or Alum alone, werelabeled with anti-CD4 and anti-CD8 antibodies and ana-lyzed by immunohystochemistry. The examinationrevealed a signiWcant increase in the number of CD4+ andCD8+ cells inWltrating lung metastases from mice immu-nized with 1E10/Alum (Figs. 2g, h, 4; P < 0.05, Kruskal–Wallis test).
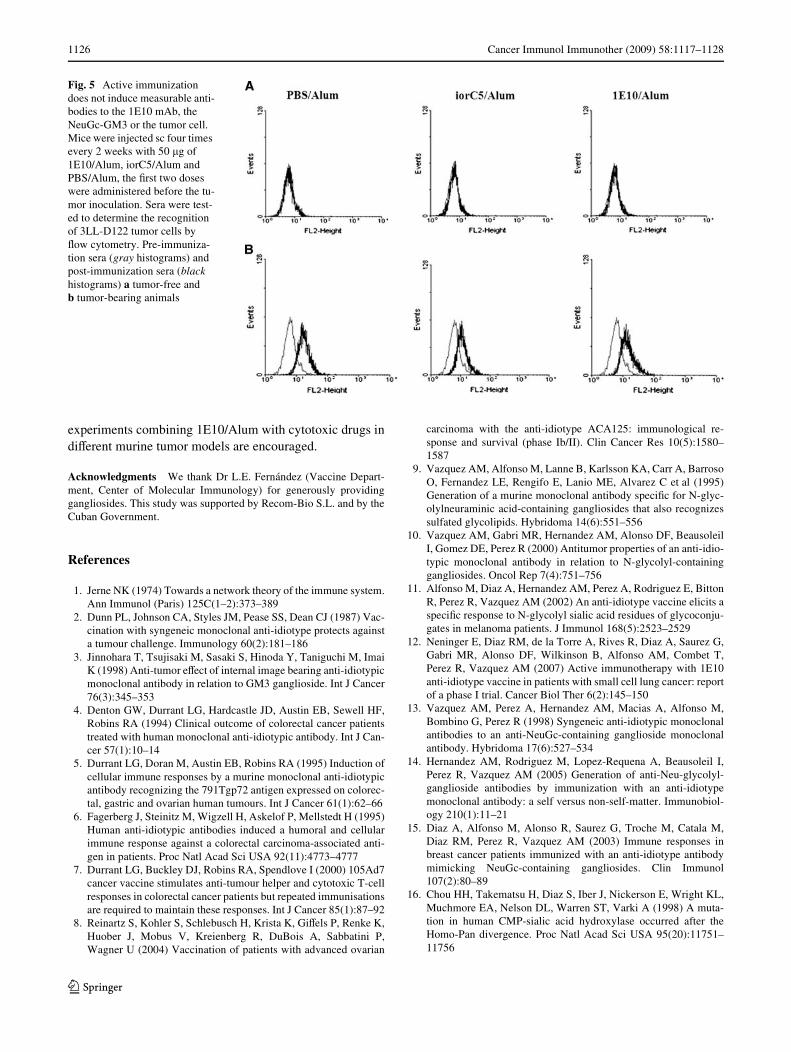
1E10/Al immunization does not induce measurable antibodies to the 1E10 mAb, the NeuGc-GM3 or the 3LL-D122 Lewis Lung carcinoma cells
Sera from mice immunized with four doses of 1E10/Alumwere analyzed by an indirect ELISA against the F(ab�)2
fragments of the anti-Id mAb or against NeuGc-GM3ganglioside. No reactivity of the sera against these anti-gens was detected. To deWne whether 1E10/Alum treat-ment is able to induce antibodies capable to recognize the
123
Cancer Immunol Immunother (2009) 58:1117–1128 1121
3LL-D122 tumor cells, sera were analyzed by Xow cytom-etry. 1E10/Alum treated-normal mice did not recognizethe tumor cells, however, a weak sera reactivity withtumor cells was detected in animals immunized with
1E10/Alum and challenged with 3LL-D122 tumor cells,but it was not substantially diVerent from the observed inmice challenged with tumor cells in control groups(Fig. 5).
Fig. 1 EVect of 1E10/Alum treatment on primary tumor and sponta-neous lung colonization produced by 3LL-D122 cells. Mean diameterof primary tumor from animals challenged sc with 2 £ 105 3LL-D122cells into a hind footpad (a and b, left panel) and the correspondinglung weight (a and b, right panel) evaluated 21 days after the primarytumor removal were determined. a Mice were injected sc four timesevery 2 weeks with 50 �g of 1E10/Alum, iorC5/Alum and PBS/Alum,the Wrst two doses were administered before the tumor inoculation.
b Mice were immunized six times with 100 �g of the vaccine starting3 days after the tumor challenge and weekly afterward (seeSect. “Materials and methods” for details). c Metastatic lung tissuefrom animals vaccinated under a therapeutic regime were staining withhematoxylin and eosin; tumor micrometastases were counted (leftpanel) and measured their surface (right panel). Experiments wererepeated three times and a representative result is shown. Results areexpressed as the mean § SEM (*P < 0.05)
123

1122 Cancer Immunol Immunother (2009) 58:1117–1128
Discussion
Most preclinical studies for the evaluation of antitumoragents are frequently based on the use of rapidly growingtransplantable mouse tumors which usually progress aslocalized solid masses. This kind of experimental scenariolargely diVers from the real situation in patients bearingmalignant tumors, where in most cases are the metastasesresponsible for cancer lethality [22]. Therefore, we focusour present preclinical studies on the anti-metastatic capac-ity of 1E10/Alum in a model of spontaneous lung metasta-ses generated after the surgical removal of the primary3LL-D122 Lewis Lung carcinoma inoculated subcutane-ously in C57BL/6 mice.
The anti-metastatic eVect of 1E10/Alum immunizationwas evaluated in two diVerent settings distinguished by thefrequency of the immunizations, the amount of vaccine andthe initiation of the vaccine schedule related to the tumorchallenge. In a mixed schedule mice were injected sc fourtimes every 2 weeks with 50 �g of 1E10/Alum where theWrst two doses were administered before the tumor inocula-tion. Additionally, in a fully therapeutic setting mice wereimmunized six times with 100 �g of the vaccine starting3 days after the tumor challenge and weekly afterward,resulting closer to the clinical scenario.
Independently to the immunization schedule, 1E10/Alum promoted a signiWcant reduction of spontaneous lungmetastases (Fig. 1). The sc administration of 1E10/Alum,before and after the implantation of 3LL-D122 lung carci-noma cells in mice, produced a dramatic decrease in thenumber of macro- and micrometastases in the animal lungs.These results agree with our previous Wnding that intraperi-toneal vaccination with this anti-idiotypic antibody coupledwith a carrier protein in Freund’s adjuvant, or its intravenous
administration alone, was capable to reduce the spontane-ous lung metastases induced by the murine F3II mammarycarcinoma or the experimental lung metastases produced bythe B16 melanoma cells, respectively [10].
In order to explore the role of the immune response inthe anti-metastatic activity elicited by 1E10/Alum, we Wrstevaluated the association of induced antibodies to this phe-nomenon. Ab3 response was not detected in the sera ofmice treated with 1E10/Alum when analyzed by an indirectELISA against F(ab�)2 fragments of the anti-Id mAb oragainst NeuGc-GM3 ganglioside. Moreover, an enhance-ment of the reactivity of the mouse sera against the 3LL-D122 tumor cells was not demonstrable by Xow cytometry(Fig. 5 and data not shown). These results are in agreementwith those previously reported by our group treating micewith this anti-idiotypic mAb [10, 13]. Taking these resultsinto account, the anti-metastatic eVect induced by 1E10/Alum seems not to be directly linked to a potential antibodyresponse induced by this vaccine.
In contrast, immunohistochemical analyses demon-strated an increase in the number of CD4+ and CD8+ cellsinWltrating lung metastases from 1E10/Alum treated mice.Involvement of CD8+ and CD4+ cells in the antitumoractivity of diVerent vaccines has been documented inmouse tumor models [23, 24]. In addition, the developmentof cellular immune responses in cancer patients due to thevaccination with anti-idiotypic antibodies mimickingtumor-associated antigens has been well demonstrated [7,25–27]. On the other hand, it is important to remark the factthat gangliosides based cancer vaccine (using the intactmolecule or anti-idiotypic Ab) usually induce antibodyresponse but not cellular immunity [28, 29]. The glycolipidnature of gangliosides may limit their presentation in theclassical major histocompatibility complex molecules [30].However, the recent Wnding of CD1-restricted human T cellspeciWc for GM1 gangliosides, which are able to secreteIFN� and TNF� cytokines [31], and the presentation of theGD3 ganglioside by CD1d to mouse NKT cells [32], mayunveil the potential development of T cell response to thiskind of antigens.
Recently, the development of CD8 T cells response pro-tecting against the melanoma B16 challenge, immunizingwith a GM3-based ganglioside vaccine has been docu-mented. In addition, T cells from immunized animals rec-ognized the dendritic cells pulsed with melanoma extract,secreting IFN� [33]. Altogether, these emerging resultsmay suggest a direct involvement of the cellular immuneresponse to gangliosides in the antitumor immunity. In par-ticular, patients with advanced breast cancer immunizedwith 1E10/Alum induced a NeuGc-speciWc T cell IFN�response [34]. However, the direct mechanism for antitu-mor eVect of tumor-inWltrating T cell promoted by 1E10/Alum requires to be further dissected.
Table 1 EVect of 1E10/Alum immunization on 3LL-D122 spontane-ous metastases
Mice were injected sc four times every 2 weeks with 50 �g of 1E10/Alum, iorC5/Alum and PBS/Alum, the Wrst two doses were adminis-tered before the sc inoculation with 2 £ 105 3LL-D122 cells into a hindfootpad. At day 48, mice were sacriWced and lung macrometastaseswere counted
*** P < 0.001, Chi-squared test
Treatments (number of animals)
Lung macrometastases
Percent of animals per group
Mean § SEM
0–4 ¸5
PBS/Alum (14) 27 73 14.3 § 4.8
iorC5/Alum (6) 33 67 18.2 § 7.4
1E10/Alum (15)*** 93 7 1.8 § 0.4
123
Cancer Immunol Immunother (2009) 58:1117–1128 1123
Nevertheless, some direct immunological or indirectmechanisms could be working. For instance, the reductionof lung metastasis by 1E10/Alum vaccine could be relatedwith the migration process of both the endothelial cellsforming new blood vessels, and the tumor cells forming
the metastasis. Both angiogenesis and tumor invasion canbe considered as invasive processes, in which cells areactivated and moved away from their initial location, bymodifying the adhesiveness with the extracellular matrix,expressing new adhesion molecules and degrading the
Fig. 2 Histopathology of 3LL-D122 lung metastases after 1E10/Alum immunization. Lungs were surgically removed on day 48 after 3LL-D122 cells inoculation from mice injected sc four times every 2 weeks with 50 �g of 1E10/Alum (right pan-el), iorC5/Alum or PBS/Alum (left panel), the Wrst two doses were administered before the tu-mor inoculation. a and b ParaYn sections stained with hematoxy-lin and eosin (£5). c and d Immunostaining with the Apoptag Plus Kit (£25, arrows indicate cells in apoptosis). e and f Immunostaining with an anti-CD31 antibody (£25, arrows indicate microvessels). g and h CD4 immunostaining of tissue cryosections (£25, arrows indicate CD4+ cells)
123
1124 Cancer Immunol Immunother (2009) 58:1117–1128
extracellular matrix components by active proteases secre-tion. This process requires a complex cross-talk betweenendothelial and tumor cells, extracellular matrix compo-nents, and cellular elements of the host microenvironment.The interference between the components of the extracel-lular matrix and tumor or endothelial cells could lead tothe inhibition of this migration process and inhibit themetastasis or the angiogenesis process [35]. The inhibitionof the Wbronectin and integrin association using syntheticpeptides mimicking one of the extracellular matrix compo-nents has resulted in a successful approach [36]. The 1E10mAb contains a CDR3 motif also seen in integrin mole-cules [37], so the development of antibodies against thismotif could block the interaction between cells and itsextracellular matrix and prevent the migration process of
both tumor and endothelial cells. The inhibition in themigration process of endothelial cells could prevent theformation of new blood vessels, causing tumor cell death,a process that can attract diVerent elements of the immunesystem. Among the six essential alterations in cell physiol-ogy that collectively dictate malignant growth are sus-tained angiogenesis, and evasion of apoptosis [38].Therapies able to induce antiangiogenic and proapoptoticeVects have been developed, and some of them are nowbeen tested in diVerent stage clinical trials in cancerpatients [39–42]. Nevertheless, due to the complexity andthe number of diVerent molecules involved in these pro-cesses in diVerent types of tumor cells, it is rather improb-able that a single therapy would control the tumorprogression [38].
Fig. 3 Proapoptotic and antian-giogenic eVect of 1E10/Alum immunization in 3LL-D122 lung metastases. Mice were injected sc four times every 2 weeks with 50 �g of 1E10/Alum, iorC5/Alum and PBS/Alum, the Wrst two doses were administered be-fore the tumor inoculation (left panel), or animals were immu-nized six times with 100 �g of the vaccine starting 3 days after the tumor challenge and weekly afterward (right panel, see Sect. “Materials and methods” for details). Lung metastasis analyzes was performed by immunohistochemistry. a Apop-tosis was measured with the Apoptag Plus Kit. b Tumor mitosis was counted in hematox-ylin–eosin stained samples. c Number of CD31+ microvessels was analyzed using an anti-CD31 antibody. Data represent the mean § SEM in 12 independent lung metastases Welds from two mice from each group. Representative data from one experiment are shown (**P < 0.01 and ***P < 0.001)
123
Cancer Immunol Immunother (2009) 58:1117–1128 1125
Interestingly, immunohistochemistry analysis of lungmetastatic tissues from vaccinated mice showed a signiW-cant increase in the number of apoptotic tumor cells whencompared with control animals (Fig. 3). This result associ-ated with the fact that only vaccinated mice had a decreasein the number of lung metastases suggest that apoptosisseems to be a potential mechanism related with the antitu-mor activity of 1E10/Alum vaccine. A similar eVect hasbeen found in tumor tissues from patients treated with theanti-Id mAb 105AD7 [43].
The higher expression levels of NeuGc-GM3 ganglio-side level in spontaneous metastasis (A. Gonzalez et al., inpreparation), could be related with the observed eVective-ness of 1E10/Alum vaccine only on lung metastasis but noton primary tumors. Similarly the T cell inWltration was onlyobserved in lung metastasis from 1E10/Alum immunizedanimals but not in their primary tumors (Fig. 4). We maypropose that the enhanced tumor apoptosis observed after1E10/Alum immunization lead to an increase in antigenspresentation, triggering an antigen spreading, which wouldinvolve in the response to other tumor antigens, besides theNeuGc-GM3 ganglioside, which has been evidenced inother experimental conditions [44].
That would explain the sustained antitumor eVect in thisstringent lung cancer model as well as the lack of a demon-strable speciWc response to the ganglioside itself. It would
be expected based on the fact that active and passive immu-nity to GM3 induce tumor cell death [33, 45], which in turnmay enhance antigen presentation and antitumor eVectinvolving new tumor speciWcities, which is currently underevaluation.
The tumor mitosis did not diverge between controls and1E10/Alum vaccinated animals (Fig. 3), suggesting that theobserved antitumor eVect is not associated to an antiprolif-erative process of this vaccine. The increase in apoptosisdetected in the pulmonary metastases from mice treatedwith 1E10/Alum could be due to its capacity to inhibit angi-ogenesis. In fact, the observation that antiangiogenic mole-cules can induce apoptosis of tumor cells has beenpreviously reported [46, 47]. On the other hand, tumor-inWltrating T cells and their cytotoxic released cytokinescould be implicated in the process [48, 49].
These preclinical evidences for the antitumor mecha-nisms of a cancer vaccine deWned by an anti-gangliosideanti-idiotypic mAb may support the relevance of this targetfor cancer biotherapy. However, the correspondence withthe mechanisms responsible for the therapeutic eVectobserved in patients has to be elucidated [12, 50, 51].Finally, due to the reported preclinical and clinical Wndingsthat the combinations of antiangiogenic, proapoptotic andantiproliferative agents with conventional drugs improvethe antitumor eYcacy of the latter [40, 52–55], future
Fig. 4 1E10/Alum immuniza-tion promotes the lymphocyte inWltration in lung metastases. Mice were injected sc four times every 2 weeks with 50 �g of 1E10/Alum, iorC5/Alum and PBS/Alum, the Wrst two doses were administered before the tumor inoculation (left panel), or animals were immunized six times with 100 �g of the vaccine starting 3 days after the tumor challenge and weekly afterward (right panel, see Sect. “Materials and methods” for details). Lung metastasis analyzes was performed by immunohistochemistry. a CD4+ and b CD8+ cells inWltrating lung metastasis. The bars repre-sent the mean § SEM of total cells per Weld (*P < 0.05 and ***P < 0.001)
123
1126 Cancer Immunol Immunother (2009) 58:1117–1128
experiments combining 1E10/Alum with cytotoxic drugs indiVerent murine tumor models are encouraged.
Acknowledgments We thank Dr L.E. Fernández (Vaccine Depart-ment, Center of Molecular Immunology) for generously providinggangliosides. This study was supported by Recom-Bio S.L. and by theCuban Government.
References
1. Jerne NK (1974) Towards a network theory of the immune system.Ann Immunol (Paris) 125C(1–2):373–389
2. Dunn PL, Johnson CA, Styles JM, Pease SS, Dean CJ (1987) Vac-cination with syngeneic monoclonal anti-idiotype protects againsta tumour challenge. Immunology 60(2):181–186
3. Jinnohara T, Tsujisaki M, Sasaki S, Hinoda Y, Taniguchi M, ImaiK (1998) Anti-tumor eVect of internal image bearing anti-idiotypicmonoclonal antibody in relation to GM3 ganglioside. Int J Cancer76(3):345–353
4. Denton GW, Durrant LG, Hardcastle JD, Austin EB, Sewell HF,Robins RA (1994) Clinical outcome of colorectal cancer patientstreated with human monoclonal anti-idiotypic antibody. Int J Can-cer 57(1):10–14
5. Durrant LG, Doran M, Austin EB, Robins RA (1995) Induction ofcellular immune responses by a murine monoclonal anti-idiotypicantibody recognizing the 791Tgp72 antigen expressed on colorec-tal, gastric and ovarian human tumours. Int J Cancer 61(1):62–66
6. Fagerberg J, Steinitz M, Wigzell H, Askelof P, Mellstedt H (1995)Human anti-idiotypic antibodies induced a humoral and cellularimmune response against a colorectal carcinoma-associated anti-gen in patients. Proc Natl Acad Sci USA 92(11):4773–4777
7. Durrant LG, Buckley DJ, Robins RA, Spendlove I (2000) 105Ad7cancer vaccine stimulates anti-tumour helper and cytotoxic T-cellresponses in colorectal cancer patients but repeated immunisationsare required to maintain these responses. Int J Cancer 85(1):87–92
8. Reinartz S, Kohler S, Schlebusch H, Krista K, GiVels P, Renke K,Huober J, Mobus V, Kreienberg R, DuBois A, Sabbatini P,Wagner U (2004) Vaccination of patients with advanced ovarian
carcinoma with the anti-idiotype ACA125: immunological re-sponse and survival (phase Ib/II). Clin Cancer Res 10(5):1580–1587
9. Vazquez AM, Alfonso M, Lanne B, Karlsson KA, Carr A, BarrosoO, Fernandez LE, Rengifo E, Lanio ME, Alvarez C et al (1995)Generation of a murine monoclonal antibody speciWc for N-glyc-olylneuraminic acid-containing gangliosides that also recognizessulfated glycolipids. Hybridoma 14(6):551–556
10. Vazquez AM, Gabri MR, Hernandez AM, Alonso DF, BeausoleilI, Gomez DE, Perez R (2000) Antitumor properties of an anti-idio-typic monoclonal antibody in relation to N-glycolyl-containinggangliosides. Oncol Rep 7(4):751–756
11. Alfonso M, Diaz A, Hernandez AM, Perez A, Rodriguez E, BittonR, Perez R, Vazquez AM (2002) An anti-idiotype vaccine elicits aspeciWc response to N-glycolyl sialic acid residues of glycoconju-gates in melanoma patients. J Immunol 168(5):2523–2529
12. Neninger E, Diaz RM, de la Torre A, Rives R, Diaz A, Saurez G,Gabri MR, Alonso DF, Wilkinson B, Alfonso AM, Combet T,Perez R, Vazquez AM (2007) Active immunotherapy with 1E10anti-idiotype vaccine in patients with small cell lung cancer: reportof a phase I trial. Cancer Biol Ther 6(2):145–150
13. Vazquez AM, Perez A, Hernandez AM, Macias A, Alfonso M,Bombino G, Perez R (1998) Syngeneic anti-idiotypic monoclonalantibodies to an anti-NeuGc-containing ganglioside monoclonalantibody. Hybridoma 17(6):527–534
14. Hernandez AM, Rodriguez M, Lopez-Requena A, Beausoleil I,Perez R, Vazquez AM (2005) Generation of anti-Neu-glycolyl-ganglioside antibodies by immunization with an anti-idiotypemonoclonal antibody: a self versus non-self-matter. Immunobiol-ogy 210(1):11–21
15. Diaz A, Alfonso M, Alonso R, Saurez G, Troche M, Catala M,Diaz RM, Perez R, Vazquez AM (2003) Immune responses inbreast cancer patients immunized with an anti-idiotype antibodymimicking NeuGc-containing gangliosides. Clin Immunol107(2):80–89
16. Chou HH, Takematsu H, Diaz S, Iber J, Nickerson E, Wright KL,Muchmore EA, Nelson DL, Warren ST, Varki A (1998) A muta-tion in human CMP-sialic acid hydroxylase occurred after theHomo-Pan divergence. Proc Natl Acad Sci USA 95(20):11751–11756
Fig. 5 Active immunization does not induce measurable anti-bodies to the 1E10 mAb, the NeuGc-GM3 or the tumor cell. Mice were injected sc four times every 2 weeks with 50 �g of 1E10/Alum, iorC5/Alum and PBS/Alum, the Wrst two doses were administered before the tu-mor inoculation. Sera were test-ed to determine the recognition of 3LL-D122 tumor cells by Xow cytometry. Pre-immuniza-tion sera (gray histograms) and post-immunization sera (black histograms) a tumor-free and b tumor-bearing animals
123

Cancer Immunol Immunother (2009) 58:1117–1128 1127
17. Ledeen RW, Yu RK (1982) Gangliosides: structure, isolation, andanalysis. Methods Enzymol 83:139–191
18. Olson MV, Varki A (2003) Sequencing the chimpanzee genome:insights into human evolution and disease. Nat Rev Genet4(1):20–28
19. Stults CL, Sweeley CC, Macher BA (1989) Glycosphingolipids:structure, biological source, and properties. Methods Enzymol179:167–214
20. Eisenbach L, Hollander N, Greenfeld L, Yakor H, Segal S,Feldman M (1984) The diVerential expression of H-2 K versusH-2D antigens, distinguishing high-metastatic from low-meta-static clones, is correlated with the immunogenic properties of thetumor cells. Int J Cancer 34(4):567–573
21. Alfonso M, Vazquez AM, Carr A, Haerslev T, Fernandez LE,Lanio ME, Alvarez C, Zeuthen J, Perez R (1995) T cell-indepen-dent B cell response to self-monosialogangliosides: primaryresponse monoclonal antibodies. Hybridoma 14(3):209–216
22. Kerbel RS (2000) Tumor angiogenesis: past, present and the nearfuture. Carcinogenesis 21(3):505–515
23. Kohlgraf KG, Gawron AJ, Higashi M, VanLith ML, Shen X,CaVrey TC, Anderson JM, Hollingsworth MA (2004) Tumor-spe-ciWc immunity in MUC1.Tg mice induced by immunization withpeptide vaccines from the cytoplasmic tail of CD227 (MUC1).Cancer Immunol Immunother 53(12):1068–1084
24. Liu JY, Wei YQ, Yang L, Zhao X, Tian L, Hou JM, Niu T, Liu F,Jiang Y, Hu B, Wu Y, Su JM, Lou YY, He QM, Wen YJ, Yang JL,Kan B, Mao YQ, Luo F, Peng F (2003) Immunotherapy of tumorswith vaccine based on quail homologous vascular endothelialgrowth factor receptor-2. Blood 102(5):1815–1823
25. Birebent B, Mitchell E, Akis N, Li W, Somasundaram R, Purev E,Hoey D, Mastrangelo M, Maguire H, Harris DT, Nair S, Cai D,Zhang T, Herlyn DM (2003) Monoclonal anti-idiotypic antibodymimicking the gastrointestinal carcinoma-associated epitopeCO17–1A elicits antigen-speciWc humoral and cellular immune re-sponses in colorectal cancer patients. Vaccine 21(15):1601–1612
26. Pride MW, Shuey S, Grillo-Lopez A, Braslawsky G, Ross M,Legha SS, Eton O, Buzaid A, Ioannides C, Murray JL (1998)Enhancement of cell-mediated immunity in melanoma patientsimmunized with murine anti-idiotypic monoclonal antibodies(MELIMMUNE) that mimic the high molecular weight proteogly-can antigen. Clin Cancer Res 4(10):2363–2370
27. Wagner U, Kohler S, Reinartz S, GiVels P, Huober J, Renke K,Schlebusch H, Biersack HJ, Mobus V, Kreienberg R, BauknechtT, Krebs D, Wallwiener D (2001) Immunological consolidation ofovarian carcinoma recurrences with monoclonal anti-idiotypeantibody ACA125: immune responses and survival in palliativetreatment. Clin Cancer Res 7(5):1154–1162
28. Livingston PO, Ragupathi G (1997) Carbohydrate vaccines thatinduce antibodies against cancer. 2. Previous experience andfuture plans. Cancer Immunol Immunother 45(1):10–19
29. Chapman PB (2003) Vaccinating against GD3 ganglioside usingBEC2 anti-idiotypic monoclonal antibody. Curr Opin InvestigDrugs 4(6):710–715
30. Ishioka GY, Lamont AG, Thomson D, Bulbow N, Gaeta FC, SetteA, Grey HM (1992) MHC interaction and T cell recognition ofcarbohydrates and glycopeptides. J Immunol 148(8):2446–2451
31. Shamshiev A, Donda A, Carena I, Mori L, Kappos L, De Libero G(1999) Self glycolipids as T-cell autoantigens. Eur J Immunol29(5):1667–1675
32. Wu DY, Segal NH, Sidobre S, Kronenberg M, Chapman PB(2003) Cross-presentation of disialoganglioside GD3 to naturalkiller T cells. J Exp Med 198(1):173–181
33. Mazorra Z, Mesa C, Fernandez A, Fernandez LE (2008) Immuni-zation with a GM3 ganglioside nanoparticulated vaccine confersan eVector CD8(+) T cells-mediated protection against melanomaB16 challenge. Cancer Immunol Immunother 57(12):1771–1780
34. Guthmann MD, Castro MA, Cinat G, Venier C, Koliren L, BittonRJ, Vazquez AM, Fainboim L (2006) Cellular and humoralimmune response to N-Glycolyl-GM3 elicited by prolongedimmunotherapy with an anti-idiotypic vaccine in high-risk andmetastatic breast cancer patients. J Immunother 29(2):215–223
35. Sasisekharan R, Shriver Z, Venkataraman G, Narayanasami U(2002) Roles of heparan-sulphate glycosaminoglycans in cancer.Nat Rev Cancer 2(7):521–528
36. Rajotte D, Arap W, Hagedorn M, Koivunen E, Pasqualini R,Ruoslahti E (1998) Molecular heterogeneity of the vascular endo-thelium revealed by in vivo phage display. J Clin Invest102(2):430–437
37. Perez A, Mier ES, Vispo NS, Vazquez AM, Perez Rodriguez R(2002) A monoclonal antibody against NeuGc-containing ganglio-sides contains a regulatory idiotope involved in the interactionwith B and T cells. Mol Immunol 39(1–2):103–112
38. Hanahan D, Weinberg RA (2000) The hallmarks of cancer. Cell100(1):57–70
39. Atkins MB, Hidalgo M, Stadler WM, Logan TF, Dutcher JP,Hudes GR, Park Y, Liou SH, Marshall B, Boni JP, Dukart G,Sherman ML (2004) Randomized phase II study of multiple doselevels of CCI-779, a novel mammalian target of rapamycin kinaseinhibitor, in patients with advanced refractory renal cellcarcinoma. J Clin Oncol 22(5):909–918
40. Hicklin DJ, Ellis LM (2005) Role of the vascular endothelialgrowth factor pathway in tumor growth and angiogenesis. J ClinOncol 23(5):1011–1027
41. Klasa RJ, Gillum AM, Klem RE, Frankel SR (2002) OblimersenBcl-2 antisense: facilitating apoptosis in anticancer treatment.Antisense Nucleic Acid Drug Dev 12(3):193–213
42. Rini BI, Small EJ (2005) Biology and clinical development ofvascular endothelial growth factor-targeted therapy in renal cellcarcinoma. J Clin Oncol 23(5):1028–1043
43. Amin S, Robins RA, Maxwell-Armstrong CA, ScholeWeld JH,Durrant LG (2000) Vaccine-induced apoptosis: a novel clinicaltrial end point? Cancer Res 60(12):3132–3136
44. Nowak AK, Lake RA, Marzo AL, Scott B, Heath WR, Collins EJ,Frelinger JA, Robinson BW (2003) Induction of tumor cellapoptosis in vivo increases tumor antigen cross-presentation,cross-priming rather than cross-tolerizing host tumor-speciWc CD8T cells. J Immunol 170(10):4905–4913
45. Roque-Navarro L, Chakrabandhu K, de Leon J, Rodriguez S,Toledo C, Carr A, de Acosta CM, Hueber AO, Perez R (2008)Anti-ganglioside antibody-induced tumor cell death by loss ofmembrane integrity. Mol Cancer Ther 7(7):2033–2041
46. Crombet-Ramos T, Rak J, Perez R, Viloria-Petit A (2002) Anti-proliferative, antiangiogenic and proapoptotic activity of h-R3: ahumanized anti-EGFR antibody. Int J Cancer 101(6):567–575
47. Jung YD, MansWeld PF, Akagi M, Takeda A, Liu W, Bucana CD,Hicklin DJ, Ellis LM (2002) EVects of combination anti-vascularendothelial growth factor receptor and anti-epidermal growth fac-tor receptor therapies on the growth of gastric cancer in a nudemouse model. Eur J Cancer 38(8):1133–1140
48. Komita H, Homma S, Saotome H, Zeniya M, Ohno T, Toda G(2006) Interferon-gamma produced by interleukin-12-activatedtumor inWltrating CD8+ T cells directly induces apoptosis ofmouse hepatocellular carcinoma. J Hepatol 45(5):662–672
49. Dorothee G, Vergnon I, Menez J, Echchakir H, Grunenwald D,Kubin M, Chouaib S, Mami-Chouaib F (2002) Tumor-inWltratingCD4+ T lymphocytes express APO2 ligand (APO2L)/TRAILupon speciWc stimulation with autologous lung carcinoma cells:role of IFN-alpha on APO2L/TRAIL expression and -mediatedcytotoxicity. J Immunol 169(2):809–817
50. Manjili MH (2007) Come forth 1E10 anti-idiotype vaccine: deliv-ering the promise to immunotherapy of small cell lung cancer.Cancer Biol Ther 6(2):151–152
123

1128 Cancer Immunol Immunother (2009) 58:1117–1128
51. Alfonso S, Diaz RM, de la Torre A, Santiesteban E, Aguirre F,Perez K, Rodriguez JL, Barroso MD, Hernandez AM, Toledo D,Gabri MR, Alonso DF, Viada C, Gomez RE, Suarez E, VazquezAM, Perez R, Macias AE (2007) 1E10 anti-idiotype vaccine innon-small cell lung cancer: experience in stage IIIb/IV patients.Cancer Biol Ther 6(12):1847–1852
52. Gasparini G, Longo R, Fanelli M, Teicher BA (2005) Combina-tion of antiangiogenic therapy with other anticancer therapies:results, challenges, and open questions. J Clin Oncol 23(6):1295–1311
53. Pegram MD, Lipton A, Hayes DF, Weber BL, Baselga JM, Tripa-thy D, Baly D, Baughman SA, Twaddell T, Glaspy JA, Slamon DJ
(1998) Phase II study of receptor-enhanced chemosensitivity usingrecombinant humanized anti-p185HER2/neu monoclonal anti-body plus cisplatin in patients with HER2/neu-overexpressingmetastatic breast cancer refractory to chemotherapy treatment.J Clin Oncol 16(8):2659–2671
54. Teicher BA (1999) Potentiation of cytotoxic cancer therapies byantiangiogenic agents. Humana Press Inc, Totowa
55. Woude GF, KelloV GJ, Ruddon RW, Koo HM, Sigman CC, Bar-rett JC, Day RW, Dicker AP, Kerbel RS, Parkinson DR, Slichenm-yer WJ (2004) Reanalysis of cancer drugs: old drugs, new tricks.Clin Cancer Res 10(11):3897–3907
123




















