
Vaccinia virus: a model system for actin-membrane interactions

ACTIBIND, an Actin-binding Fungal T2-RNase withAntiangiogenic and Anticarcinogenic Characteristics
Levava Roiz, Ph.D.1
Patricia Smirnoff, Ph.D.2
Menashe Bar-Eli, Ph.D.3
Betty Schwartz, Ph.D.2
Oded Shoseyov, Ph.D.1
1 Institute of Plant Science and Genetics in Agri-culture, Faculty of Agricultural, Food and Environ-mental Quality Sciences, The Hebrew University ofJerusalem, Rehovot, Israel.
2 Institute of Biochemistry, Food Science and Nu-trition, Faculty of Agricultural, Food and Environ-mental Quality Sciences, The Hebrew University ofJerusalem, Rehovot, Israel.
3 Department of Cancer Biology, The University ofTexas M. D. Anderson Cancer Center, Houston,Texas.
Address for reprints: Oded Shoseyov, Ph.D., In-stitute of Plant Science and Genetics in Agricul-ture, Faculty of Agricultural, Food and Environ-mental Quality Sciences, The Hebrew Universityof Jerusalem, P.O. Box 12, Rehovot 76100,Israel; Fax: (011) 972-8-9462283; E-mail:[email protected]
Address for reprints: Betty Schwartz, Ph.D., In-stitute of Biochemistry, Food Science and Nutri-tion, Faculty of Agricultural, Food and Environ-mental Quality Sciences, The Hebrew Universityof Jerusalem, P.O. Box 12, Rehovot 76100,Israel; Fax: (011) 972-8-9363208; E-mail:[email protected]
Received July 25, 2005; revision received Decem-ber 11, 2005; accepted January 3, 2006.
BACKGROUND. ACTIBIND is an Aspergillus niger extracellular ribonuclease (T2-
ribonuclease [RNase]) that possesses actin-binding activity. In plants, ACTIBIND
inhibits the elongation and alters the orientation of pollen tubes by interfering with
the intracellular actin network. The question rose whether ACTIBIND can also
affect mammalian cancer development.
METHODS. Cell colony formation was performed in human colon (HT-29, Caco-2,
RSB), breast (ZR-75-1), and ovarian (2780) cancer cells in the presence or absence
of 1 �M ACTIBIND. In HT-29 and ZR-75-1 cells, the effect of ACTIBIND on cell
migration was studied by microscopic observations and by invasion assay through
Matrigel. Tube formation was assessed in human umbilical vein endothelial cells
(HUVEC) in the presence of angiogenin or basic fibroblast growth factor (bFGF) (1
�g/mL each) following overnight incubation with 1 or 10 �M ACTIBIND. In an
athymic mouse xenograft model, HT-29 cells were injected subcutaneously, fol-
lowed by subcutaneous (0.4-8 mg/mouse/injection) or intraperitoneal (0.001-1
mg/mouse/injection) injections of ACTIBIND. In a rat dimethylhydrazine (DMH)-
colorectal carcinogenesis model, ACTIBIND was released directly into the colon via
osmotic micropumps (250 �g/rat/day) or given orally via microcapsules (1.6 mg/
rat/day). Aberrant crypt foci, tumors in the distal colon, and tumor blood vessels
were examined.
RESULTS. ACTIBIND had an anticlonogenic effect unrelated to its ribonuclease
activity. It also inhibited angiogenin-induced HUVEC tube formation in a dose-
responsive manner. ACTIBIND was found to bind actin in vitro. It also bound to
cancer cell surfaces, leading to disruption of the internal actin network and
inhibiting cell motility and invasiveness through Matrigel-coated filters. In mice,
ACTIBIND inhibited HT-29 xenograft tumor development, given either as a sub-
cutaneous or intraperitoneal treatment. In rats, ACTIBIND exerted preventive and
therapeutic effects on developing colonic tumors induced by DMH. It also reduced
the degree of tumor observation.
CONCLUSIONS. This study indicated that ACTIBIND is an effective antiangiogenic
and anticarcinogenic factor. Cancer 2006;106:2295–308.
© 2006 American Cancer Society.
KEYWORDS: ACTIBIND, actin, antiangiogenic, anticarcinogenic, ribonuclease.
In addition to their ability to degrade RNA, certain ribonucleases(RNases) display a variety of biological activities. A great deal of
interest has been directed towards the members of the RNase Afamily. For example, human angiogenin and eosinophil cationic pro-tein (ECP) exhibit angiogenic and cytostatic activities, respectively,1–3
and bovine seminal RNase has antispermatogenic, transplanted-bone-marrow-stimulating, immunosuppressive, and antitumor activ-ities.4,5 Onconase, an RNase isolated from Northern Leopard Frog(Rana pipiens) oocytes and early embryos, has been shown to possess
2295
© 2006 American Cancer SocietyDOI 10.1002/cncr.21878Published online 3 April 2006 in Wiley InterScience (www.interscience.wiley.com).

antitumor properties and is currently being investi-gated in Phase III trials for use in cancer therapy.6,7
Members of the RNase T2 family are widely dis-tributed in living organisms, from viruses to mam-mals.8 Some T2-RNases in microorganisms and plantsare capable of digesting extracellular polyribonucle-otides, thereby accelerating phosphate uptake; othersmay protect against possible pathogens.8,9 In self-in-compatible plants, specific T2-RNases encoded by theS-locus are responsible for rejecting self-pollen, thuspreventing self-fertilization.10,11 In the human ge-nome, deletion of a region of chromosome 6 (6q27) isknown to be associated with several malignancies.12,13
This region contains the putative tumor suppressorgene RNase6PL which shares homology with theRNase T2 family.14,15
ACTIBIND is a T2-RNase produced by Aspergillusniger B1 (CMI CC 324626). Our previous studies haveshown that ACTIBIND is an extracellular glycoproteincontaining 32- and 36-kD isoforms, which share acommon 29-kDa protein moiety.16 –18 Unrelated to itsRNase activity, ACTIBIND inhibits the growth and ori-entation of pollen tubes, thus decreasing fruit set indifferent plant species.16 –18 ACTIBIND binds F-actinin vitro at a molar ratio of 1:2. Consequently, ACTIBINDinduces cross-linkage between actin filaments in pol-len tubes and arrests their elongation, interfering withnormal cytoplasmic streaming at the leading edge(Roiz and Shoseyov, in preparation).
The elongation of plant pollen tubes as well as themotility of mammalian cells require the formation ofactin-rich cell protrusions such as phyllopodia andlamellipodia.19,20 Furthermore, the presence of actin-rich pseudopods has been described as a prerequisitefor cancer-cell function.21 The ability of ACTIBIND tointerfere with the intracellular actin network in plantcells raised the question of whether it can also affectmammalian cancer development. The aim of thepresent research was to study the anticarcinogenicand antiangiogenic effects of ACTIBIND.
MATERIALS AND METHODSChemicals, Drugs, and SuppliesEosin was from Acros Organics (Geel, Belgium); E. coliRNase I was from Ambion (Austin, TX); polyclonalrabbit-anti-ACTIBIND was prepared at Anilab (Reho-vot, Israel); Matrigel was from Becton-Dickinson (Bed-ford, MA); Dulbecco minimum Eagle medium(DMEM), fetal calf serum (FCS), glutamine and anti-biotic–antimycotic solution were from Biological In-dustries (Bet Haemek, Israel); ECGF was from Bio-medical Technologies Inc. (Stoughton, MA); Diff-Quikstain was from Hamilton Thorne Research (Beverly,MA ); bFGF was from Promega (Madison, MI); recom-
binant human angiogenin was from R&D Systems, Inc(Minneapolis, MN); Tetramethyl-rhodamine B iso-thiocyanate (TRITC)-labeled phalloidin, rabbit antiac-tin, fluorescein isothiocyanate (FITC)-conjugated goatantirabbit, Mayer hematoxylin and G-actin were fromSigma-Aldrich Co. (St Louis, MO); Dimethylhydrazine(DMH) was from Fluka (St. Gallen, Switzerland);monoclonal mouse antiactin IgM followed by FITC-conjugated goat antimouse IgM (Ab-1 kit) was fromOncogene (Cambridge, MA). The enhanced chemilu-minescence (ECL) Western blot detection system wasfrom Amersham Pharmacia Biotech, Ltd., (Bucking-hamshire, UK); terminal deoxynucleotidyl transferase-mediated dUTP-nick end labeling (TUNEL) Klenow-Frag-El kit was from Oncogene, (Cambridge, MA);CD31 antibody was from Pharmingen (San Diego, CA);PECAM-1 (H-300), sc-8306 antibody was from SantaCruz Biotechnology Inc. (Santa Cruz, CA).
Colony-formation AssayThe colony-formation assay was performed essentiallyas described7 with minor modifications. Human colon(HT-29, Caco-2, RSB), breast (ZR-75-1), and ovarian(2780) cancer cells were grown in 50-mL flasks (105
cells per flask). The medium contained 7 mL of DMEMsupplemented with 10% fetal calf serum (FCS), 1%glutamine, and 1% antibiotic-antimycotic solution inthe presence or absence of 1 �M ACTIBIND. The coloncancer cells were also exposed to the same concentra-tion of enzymatically inactivated (EI)-ACTIBIND,which was preheated for 30 minutes in an autoclave(120°C, 120 kPa) until no RNase activity was detected.The cells were incubated at 37°C in a humidified at-mosphere containing 5% CO2. After 48 hours, 1000cells/well were seeded in 96-well plates in 200 �Lmedium, in the presence or absence of 1 �M ACTIB-IND. HT-29, Caco-2, and ZR-75-1 cell lines weregrown for 7 days; 2780 and RSB cell lines were grownfor 10 and 14 days, respectively. The cells were thenfixed in 4% formaldehyde and stained with methyleneblue. The number of colonies was counted (N � 3).
Human Umbilical Vein Endothelial Cell (HUVEC)Angiogenesis AssayAs previously described,22 freshly isolated HUVECwere maintained in M199 medium supplemented with20% FCS, 1% glutamine, 1% antibiotic–antimycoticsolution, 0.02% ECGF, and 50 units/100 mL heparin.They were then plated in a 96-well plate (14 � 103
cells/well) previously coated with growth factor-de-pleted Matrigel, in M199 medium containing 5% FCSand 0.005% ECGF supplemented with angiogenin orbFGF (1 �g/mL each), ACTIBIND (1 or 10 �M), orphosphate-buffered saline (PBS) were added, and the
2296 CANCER May 15, 2006 / Volume 106 / Number 10

final volume in each well was 120 �L. After overnightincubation at 37°C, the plates were photographed, andthe extent of tube formation was assessed (N � 3).
Membrane BlottingActin was bound to ACTIBIND or to other proteins asdescribed.23 ACTIBIND, angiogenin and E. coli RNaseI (1 �g of each protein) were loaded on 10% SDS-PAGEand transferred onto a Hybond ECL nitrocellulosemembrane. G-actin (1 �g) was used as a positive con-trol. The membrane was blocked at 4°C by overnightincubation in rinse buffer (10 mM Tris-HCl pH 7.5; 150mM NaCl; 1 mM EDTA; 0.02% Triton X-100) contain-ing 5% bovine serum albumin (BSA), under constantshaking, and then washed twice for 10 minutes withrinse buffer. The membrane was incubated in buffer G(2 mM Tris pH 8; 0.2 mM CaCl2; 0.2 mM ATP), fol-lowed by overnight incubation at 4°C with 5 �g/mlG-actin in buffer G under constant stirring. Following3 washes in rinse buffer containing 0.2 mM CaCl2, themembrane was immunolabeld with antiactin IgM fol-lowing by FITC-conjugated goat antimouse IgM (Ab-1kit). Signals were detected with the ECL Western BlotDetection System.
Microscopic ObservationsFor actin observation, HT-29 cells were cultured for 3days in DMEM in the presence or absence of 1 �MACTIBIND or EI-ACTIBIND and fixed for 10 minutesin 4% formaldehyde in PBS on ice. Cell membrane waspermeabilized with PBS containing 0.05% Tween 20(PBST) (1 hour, room temperature), and then stained for1 hour with TRITC-labeled phalloidin. For ACTIBINDimmunostaining assay, we used rabbit anti-ACTIBINDfollowed by FITC-conjugated goat antirabbit. Cellswere observed with a confocal laser scanning micro-scope, model 510 (Zeiss, Oberkochen, Baden-Wuttem-berg, Germany).
For cytologic observations, HT-29, ZR-75-1, and2780 cells were cultured as described above. Cellswere fixed, stained with Mayer hematoxylin and eosin(H & E) and observed by light microscope (BX-40,Olympus, Hamburg, Germany).
Invasion Assay Through MatrigelHT-29 and ZR-75-1 cells were cultured with 1 or 10 �MACTIBIND or in PBS for 4 days as described above.Wells and Matrigel-coated inserts of a commerciallyavailable 24-well invasion chamber were rehydrated in500 �L of serum-free medium overnight and pro-cessed according to the manufacturer’s instructions;500 �l of HT-29 and ZR-75-1 control or ACTIBIND-treated cell suspensions containing 2.5 � 104 cellseach were added to the top of the chambers, and 750
�l of DMEM media containing 10% FCS was added tothe lower chamber. The invasion chambers were in-cubated for 22 hours in a 37°C cell-culture incubator.The noninvading cells on the upper surface of themembrane of the insert were removed by scrubbing.The cells on the lower surface of the membrane werestained with Diff-Quik stain. The membranes werefixed, and the cells were counted under a light micro-scope (N � 3).
Animal StudiesXenograftsThe in vivo preventive and therapeutic effects ofACTIBIND on HT-29 – derived xenografts were studiedin athymic mice (CD-1 nu/nu; Charles River, Wilming-ton, MA) by using 2 modes of administration. In thesubcutaneous/subcutaneous (sc/sc, N � 10) model,viable HT-29 cells (2 � 105/100 �l) were injected sub-cutaneously into the left hip, and ACTIBIND (0.4, 2, 4,8 mg/100 �l) or PBS was injected subcutaneously intothe area of the cell injection. ACTIBIND was adminis-trated daily for 42 days, starting from the day of cellinjection. In the subcutaneous/intraperitoneal (sc/ip,N � 6) model, HT-29 cells (106 cell/100 �L) were in-jected as described above. ACTIBIND (0.001, 0.01, 0.1,0.5, 1.0 mg/mouse) or PBS was injected every otherday for 30 days starting from 24 hours after cell injec-tion (preventive treatment) or from when tumors werepalpable (therapeutic treatment). At the end of thisexperiment, the tumors were excised for size measure-ments and for histopathological examinations. Eachtreatment was repeated twice.
Carcinogen-treated ratsThe in vivo preventive and therapeutic effects ofACTIBIND on colorectal carcinogenesis inducedwith dimethylhydrazine (DMH)24 –26 were studied inCharles River-derived outbred male rats. ACTIBIND orEI-ACTIBIND was given via subcutaneously implantedosmotic micropumps (ALZET, Cupertino, CA), allow-ing directed release of ACTIBIND (250 �g/rat/day)into the colon via a catheter.
ACTIBIND was also given orally, using entero-coated cellulose acetate-phthalate (CAP) microcap-sules27 (1.6 mg/rat/day). During the experiment, feceswere collected weekly to monitor ACTIBIND uptake bymeans of RNase activity assays.18 In the preventiveregime, rats were sacrificed 8 or 11 weeks after director oral application of ACTIBIND. Aberrant crypt foci(ACF) in the distal 5 cm of the colons28 or tumors wereexamined. In the therapeutic regime, 11 weeks afterthe first DMH injection, ACTIBIND was administered(as described above) for an additional 6 weeks, andtumors were examined. Tumors were fixed, and
ACTIBIND: a T2-RNase with Anticancer Effect/Roiz et al. 2297

10-�m paraffin sections were stained with H & E forhistology and with the TUNEL procedure for apoptosisaccording to the supplier’s instructions. Blood vesselsin median tumor cross-sections were localized usingCD31 antibody followed by PECAM-1 (H-300), sc-8306antibody. In each median tumor cross-section, theblood vessels were counted, their diameters weremeasured, and the total area was calculated. Relativearea (percentage) was calculated as the ratio betweentotal blood-vessel area and tumor-section area. Theexperiments were repeated twice, and each treatmentwas applied to 6 to 10 rats.
All animal experiments were approved by the Eth-ics Committee for Animal Experimentation, Faculty ofAgricultural, Food and Environmental Quality Sci-ences, the Hebrew University of Jerusalem, Israel.
Statistical AnalysisMeans were compared by analysis of variance(ANOVA) or Student t test. Differences were consid-ered statistically significantly at P �.05. All statisticaltests were two-sided.
RESULTSActibind Effects In VitroACTIBIND solutions were tested for the presence ofbacterial endotoxin by the limulus amebocyte lysatedetection assay (Associates of Cape Cod Inc, East Fal-mouth, MA). The value measured in ACTIBIND solu-tions was below the detection limit of the assay (0.05EU/mL) as obtained in standard curves using endo-toxin as a control standard (from E. coli 0113:H10,Associates of Cape Cod Inc., East Falmouth, MA.).
We assessed the effect of ACTIBIND on the colony-forming ability of human cancer cell lines originatingfrom colon, breast, and ovarian tissues. ACTIBINDsignificantly inhibited colony formation of all cancercells tested (Fig. 1). The anticlonogenic effect was sig-nificant only in ACTIBIND-pretreated cells and was
much less effective in cells not previously exposed toACTIBIND (not shown). In HT-29, Caco-2, and RSBcolon cancer cells (Fig. 1A), continuous exposure toACTIBIND resulted in a substantial reduction in theirclonogenic ability (56%, 71%, and 58%, respectivelycompared with controls P �.01). EI-ACTIBINDshowed a similar effect in HT-29, Caco-2, and RSBcells (53%, 78%, and 80% of inhibition, respectively; P�.01). Similarly, ACTIBIND inhibited the clonogenicability in breast ZR-75-1 and ovarian 2780 cancer cells(Fig 1B; 33% and 81%, respectively, compared withcontrols; P �.05). Colonies of HT-29 and ZR-75-1 cellswere stained with H & E for microscopic observation.The peripheral cells of the control colonies displayednumerous cytoplasmic extensions (Fig. 2A,C), whereasin the ACTIBIND-treated colonies, the peripheral cellextensions were arrested (Fig. 2B,D). These resultssuggest that ACTIBIND’s treatment effects on themorphology of the cells may impinge upon their mo-tility.
To assess the ability of ACTIBIND to inhibit an-giogenesis, we examined its effect on tube formationof vascular structures in an in vitro HUVEC assay.ACTIBIND significantly inhibited angiogenin as wellas bFGF-induced tube formation in a dose– depen-dent manner in comparison with controls (Fig. 3).ACTIBIND did not affect HUVEC proliferation orviability as assessed by MTT assay, either after expo-sure to angiogenin or bFGF (not shown).
The Western blot analysis using G-actin to bind todifferent proteins demonstrated that ACTIBIND, sim-ilar to angiogenin and to E. coli RNase I which is alsoa T2-RNase,29 binds actin (Fig. 4).
Actibind Binds to the Cancer Cell Surface and Affectsthe Intracellular Actin NetworkHT-29 cells grown in the presence or absence ofACTIBIND exhibited different patterns of specific an-ti-ACTIBIND immunostaining. Control cells exhibited
FIGURE 1. The effect of ACTIBIND
on clonogenicity in A, colon cancer cell
lines (HT-29, Caco-2 and RSB) and in
B, breast (ZR-75-1) and ovarian (2780)
cell lines (N � 3 for each cell line).
The number of colonies in ACTIBIND-
treated cells (open square, �) was
significantly lower (P �.01) than in
controls (solid square, ■). In colon
cancer cells, EI-ACTIBIND (hatched
square, o) also showed anticlono-
genic effect (P �.01).
2298 CANCER May 15, 2006 / Volume 106 / Number 10

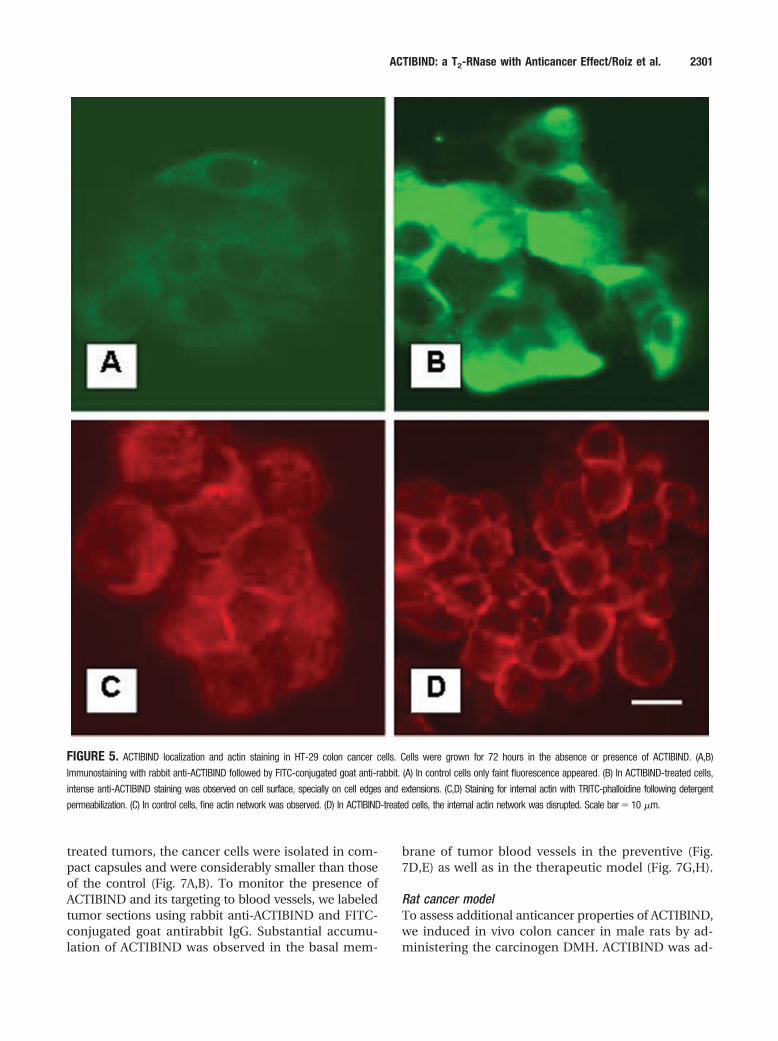
very faint fluorescence (Fig. 5A), whereas ACTIBIND-treated cells showed intense fluorescence, particularlyin the cell edges and extensions (Fig. 5B). Treatmentwith rabbit preimmune serum resulted in a very faintsignal, demonstrating the specificity of anti-ACTIBIND(not shown). The effect of ACTIBIND on the organi-zation of intracellular actin network was tested usingTRITC-phalloidin staining. Control cells showed a fineactin network filling the cell cytoplasm (Fig. 5C). Incontrast, ACTIBIND-treated cells demonstrated in-tense staining in the peripheral zone of the cytoplasm,whereas the inner area remained unstained (Fig. 5D).Similar results were obtained in cells treated with EI-ACTIBIND (not shown).
Inhibition of Tumor Cell Invasion by ACTIBINDBecause ACTIBIND was shown to affect cancer cellmorphology and actin organization, we examinedwhether cell motility was also affected. Both HT-29and ZR-75-1 cells were able to penetrate the Matrigel-coated filters (Table 1). ACTIBIND significantly anddose-dependently inhibited invasiveness of both cell
lines. ACTIBIND (1 �M) reduced the invasiveness ofHT-29 and ZR-75-1 cells by 40% (P�.05 and P�.01,respectively). At a concentration of 10 �M, the inhib-itory effect of ACTIBIND was increased to 80% (P�.01and P�.001, respectively). These findings further em-phasize that the ability of ACTIBIND to inhibit cyto-plasmic extensions (Fig. 2) impinges on the cancercell’s migration and invasion capabilities.
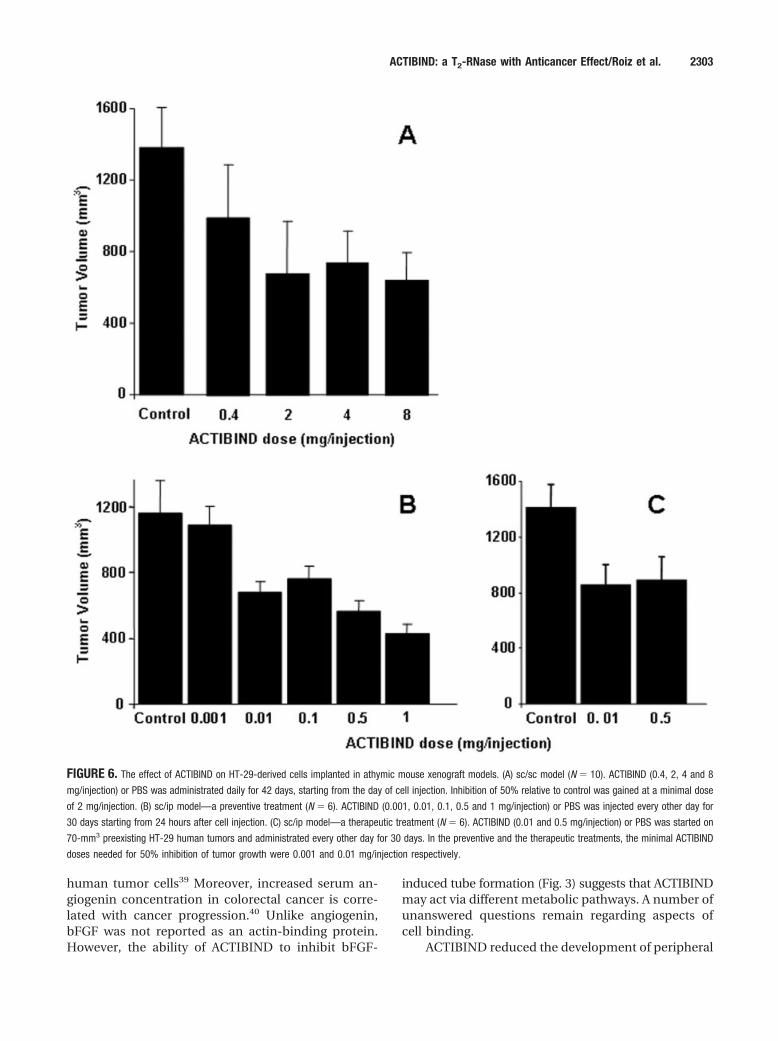
Actibind Inhibits Colon Cancer Growth In VivoMouse xenograft cancer modelBy using the xenograft athymic mouse model, wefound that ACTIBIND caused a significant reductionin the growth rate of HT-29-derived carcinoma in asaturation mode (Fig. 6). In the sc/sc model, inhibitionof 50% relative to control was gained at a minimaldose of 2 mg/injection ACTIBIND (Fig. 6A). In thesc/ip model, the minimal ACTIBIND doses needed for50% inhibition of tumor growth were 0.001 and 0.01mg/injection in the preventive and therapeutic treat-ments, respectively (Fig. 6B,C).
Histologic analysis showed that in ACTIBIND-
FIGURE 2. The effect of ACTIBIND
on the morphology and motility of
HT-29 colon (A,B) and ZR-75-1 breast
cancer cells (C,D). (A,C) In the control
cells, fully developed cell extensions
were observed (arrows). (B,D) In ACT-
IBIND-treated cells, the cell extensions
were inhibited (arrowheads). Scale
bar � 10 �m.
ACTIBIND: a T2-RNase with Anticancer Effect/Roiz et al. 2299

FIGURE 3. HUVEC tube formation on Matrigel induced by angiogenin (A–C) and bFGF (D–F). (A,D) control; (B,E) 1 �M ACTIBIND; (C,F) 10 �M ACTIBIND. ACTIBIND
inhibits both angiogenin and bFGF-induced tube formation (N � 3 for each treatment).
FIGURE 4. Actin binding activity on SDS-PAGE. Proteins were blotted on a nitrocellulose membrane, and actin-binding capability was accomplished after overnight
incubation with 5 �g/mL actin. Proteins bound to actin were in evidence subsequent to reaction with antiactin followed by HRP-conjugated secondary antibody.
Lanes 1-6: SDS-PAGE protein staining. Lanes 7-11: Western blot analysis. Lane 1: Molecular weight markers. Lanes 2,7: actin (positive control); Lanes 3,8:
ACTIBIND; Lanes 4,9: RNase I; Lanes 5,10: Angiogenin; Lanes 6,11: BSA (negative control).
2300 CANCER May 15, 2006 / Volume 106 / Number 10
treated tumors, the cancer cells were isolated in com-pact capsules and were considerably smaller than thoseof the control (Fig. 7A,B). To monitor the presence ofACTIBIND and its targeting to blood vessels, we labeledtumor sections using rabbit anti-ACTIBIND and FITC-conjugated goat antirabbit IgG. Substantial accumu-lation of ACTIBIND was observed in the basal mem-
brane of tumor blood vessels in the preventive (Fig.7D,E) as well as in the therapeutic model (Fig. 7G,H).
Rat cancer modelTo assess additional anticancer properties of ACTIBIND,we induced in vivo colon cancer in male rats by ad-ministering the carcinogen DMH. ACTIBIND was ad-
FIGURE 5. ACTIBIND localization and actin staining in HT-29 colon cancer cells. Cells were grown for 72 hours in the absence or presence of ACTIBIND. (A,B)
Immunostaining with rabbit anti-ACTIBIND followed by FITC-conjugated goat anti-rabbit. (A) In control cells only faint fluorescence appeared. (B) In ACTIBIND-treated cells,
intense anti-ACTIBIND staining was observed on cell surface, specially on cell edges and extensions. (C,D) Staining for internal actin with TRITC-phalloidine following detergent
permeabilization. (C) In control cells, fine actin network was observed. (D) In ACTIBIND-treated cells, the internal actin network was disrupted. Scale bar � 10 �m.
ACTIBIND: a T2-RNase with Anticancer Effect/Roiz et al. 2301

ministered orally in CAP-coated microcapsules or bydirect intestinal application via microosmotic pumps.The latter mode of application was used to supplyACTIBIND as well as EI-ACTIBIND. We applied 2 dif-ferent protocols: 1) a preventive treatment in whichACTIBIND or EI-ACTIBIND was administered con-comitantly with the carcinogenic treatment, and 2) atherapeutic treatment in which we administeredACTIBIND or EI-ACTIBIND subsequent to adenocar-cinoma formation. RNase activity and Western blotanalysis were employed to monitor the concentrationof ACTIBIND in feces samples. Fivefold higher RNaseactivity was measured in feces of ACTIBIND-treatedrats than in the controls, and intact ACTIBIND wasobserved in Western blot analyses of all modes ofapplication (not shown).
ACTIBIND significantly inhibited tumor develop-ment in both preventive and therapeutic treatmentregimes (Table 2). The preventive treatment in whichACTIBIND was continuously administered for 8 weeksby osmotic pump, induced a 50% reduction in thenumber of ACF (P �.01), which are known as earlymarkers of colon carcinogenesis. ACTIBIND adminis-tered orally in microcapsules led to a reduction ofabout 50% in the number (P �.05) and size (P �.05) oftumors as compared with the control following 11 weeksof treatment. In the same manner, ACTIBIND signifi-cantly reduced the number of adenocarcinomas com-pared with the control (P �.01). In the therapeutic treat-ment, we applied ACTIBIND directly via osmotic pumps,from Weeks 12 to 17, after the first DMH injection.ACTIBIND treatments resulted in a 50% reduction in thenumber of tumors per colon (P �.01). In all the abovetreatments, EI-ACTIBIND, like ACTIBIND, showed asimilar inhibitory effect on tumor development.
The effect of ACTIBIND on angiogenesis was mea-sured when the therapeutic treatment with osmoticpumps was completed (Table 3). ACTIBIND signifi-cantly reduced the number of blood vessels per tumor
by 40% relative to the control (P �.01). In addition, a70% reduction in total blood-vessel area per tumorsection (P�.05) was observed. Tumor-associated neo-vascularization, as indicated by microvessel density(MVD), was determined by means of identifying bloodvessels within the tumor (Fig. 8A,B). Concomitant withthe decrease in MVD, a significant 16.5-fold increase(P�.0001) in the number of apoptotic cells was ob-served in ACTIBIND-treated relative to control tumors(Table 3, Fig. 8 C,D).
In none of the described treatments did ACTIBINDaffect body weight, behavior, or liver histology. All ofthese parameters were comparable to those of healthyanimals.
DISCUSSIONIn this study, we show that ACTIBIND directly affectscancer cells by altering their clonogenic ability as wellas their morphology and intracellular actin organiza-tion. We also demonstrate that ACTIBIND is an effec-tive antiangiogenic agent in vivo and in vitro, becauseit significantly affects neovascularization of tumorsand HUVEC tube formation. In the above experi-ments, ACTIBIND and EI-ACTIBIND show similar ef-fects, indicating that the antitumorigenic and antian-giogenic activities may not related to RNase activity.
Previous studies have shown that Northern Leop-ard Frog-derived RNase (onconase) inhibits clonoge-nicity and protein synthesis of 9L glioma cancer cellswithout affecting cell density. These studies claim thatonconase activity depends on its ability to bind to acell-surface receptor.30,31 Our findings demonstratethat the novel RNase described herein, ACTIBIND,binds actin in vitro (Fig. 4), as well as on the cancercell’s surface (Fig.5 A,B). Surface actin has been shownto act as a receptor for angiogenin in endothelialcells.23,32–34 The actin-angiogenin complex activatesplasminogen and plasmin, bearing essential proteo-lytic activities involved in angiogenesis.35–37 Our invivo experiments demonstrate that ACTIBIND signif-icantly inhibits angiogenesis and apoptosis (Fig. 8,Table 3) processes in DMH-induced colonic tumors inrats. Cumulatively, the evidence leads us to proposethat cell-surface actin may be a target for ACTIBIND incancer cells as well as in endothelial cells. Therefore,ACTIBIND may compete with angiogenin for cell-sur-face actin and, in this manner, blocks the formation ofthe actin-angiogenin complex required for cancer cellorganization and angiogenesis in developing neoplas-tic tissue.
Angiogenin was first isolated from HT-29 cellsconditioned medium.38 It was shown to be a key factorfor tumor growth because antagonists of its activityinhibited the establishment and metastatic spread of
TABLE 1Effect of ACTIBIND on HT-29-Colon and ZR-75-1-Breast Cancer CellInvasiveness Through Matrigel-coated Filters (N � 3)
Cell Lines
Cell Counts, Mean � SE
Control
ACTIBIND
1 �M 10 �M
HT-29 963.7 � 95.7 *623.3 � 104.2 †223.7 � 13.1ZR-75-1 1297.7 � 62.5 †784.0 � 51.5 ‡271.7 � 16.6
* P �.05† P �.01‡ P �.001
2302 CANCER May 15, 2006 / Volume 106 / Number 10
human tumor cells39 Moreover, increased serum an-giogenin concentration in colorectal cancer is corre-lated with cancer progression.40 Unlike angiogenin,bFGF was not reported as an actin-binding protein.However, the ability of ACTIBIND to inhibit bFGF-
induced tube formation (Fig. 3) suggests that ACTIBINDmay act via different metabolic pathways. A number ofunanswered questions remain regarding aspects ofcell binding.
ACTIBIND reduced the development of peripheral
FIGURE 6. The effect of ACTIBIND on HT-29-derived cells implanted in athymic mouse xenograft models. (A) sc/sc model (N � 10). ACTIBIND (0.4, 2, 4 and 8
mg/injection) or PBS was administrated daily for 42 days, starting from the day of cell injection. Inhibition of 50% relative to control was gained at a minimal dose
of 2 mg/injection. (B) sc/ip model—a preventive treatment (N � 6). ACTIBIND (0.001, 0.01, 0.1, 0.5 and 1 mg/injection) or PBS was injected every other day for
30 days starting from 24 hours after cell injection. (C) sc/ip model—a therapeutic treatment (N � 6). ACTIBIND (0.01 and 0.5 mg/injection) or PBS was started on
70-mm3 preexisting HT-29 human tumors and administrated every other day for 30 days. In the preventive and the therapeutic treatments, the minimal ACTIBIND
doses needed for 50% inhibition of tumor growth were 0.001 and 0.01 mg/injection respectively.
ACTIBIND: a T2-RNase with Anticancer Effect/Roiz et al. 2303

FIGURE 7. Paraffin sections of HT-29 derived xenografts in nude mice. (A) Contol xenografts. (B) ACTIBIND-treated xenografts. (C-E) Blood vessels from tumors
treated with sc injections. (F-H) Blood vessels from tumors treated with ip injections. (C,F) In controls, H & E staining showed the basal membrane (arrow). (D,G)
Immunostaining resulted in only faded image. (E,H) In ACTIBIND-treated tumors, ACTIBIND accumulated in the arterial basal membrane. H & E staining: A-C, F, H;
FITC-labeled immunostaining with anti-ACTIBIND: D,E,G,H. Scale bars � 5 �m (E,H); 20 �m (B).
2304 CANCER May 15, 2006 / Volume 106 / Number 10

cytoplasmic extensions in different cancer cell lines(Fig. 2). These results correlated with ACTIBIND’sability to disrupt the intracellular cytoskeletal networkand to generate F-actin accumulation towards thecell-membrane boundaries. In addition, ACTIBINDsignificantly and dose-dependently affected the inva-siveness of HT-29 and ZR-75-1 cells through Matrigel(Table 1). The effect of ACTIBIND on the morphologyof cancer cells mimicked our previous observations inlily pollen tubes where ACTIBIND bound F-actin in-duced massive F-actin accumulation at the growingtip, which led to growth arrest (Roiz and Shoseyov, inpreparation). In migrating cells, the extension of lead-ing edges depends upon actin assembly in response tosignal-transduction pathways.20,41 Furthermore, incancer cells, the structure of the actin network is di-rectly related to malignant potential, because it deter-mines cell motility.21
We, therefore, suggest that ACTIBIND interfereswith cancer cells’ ability to communicate, organizeinto colonies, and motilize through extracellular ma-trix substrates by binding to cell-surface actin anddisrupting the intracellular actin network.
The anticarcinogenic and antiangiogenic effectsof ACTIBIND were demonstrated in vivo in differentanimal models. In athymic mice, treatment withACTIBIND inhibited HT-29-derived xenograft devel-opment in both sc/sc and sc/ip models (Fig. 6). Wealso demonstrated that ACTIBIND accumulates in themouse artery basement membrane (Fig. 7). Endothe-lial cells, pericytes, and their respective supportingextracellular matrix, all contain considerable concen-trations of �-smooth–muscle actin.34,42,44 – 47 For thisreason, targeting extracellular actin seems to be anappropriate approach to arresting angiogenesis. Wecan claim that actin residues provide epitopes forACTIBIND binding in a fashion similar to the specificactin binding observed in vitro (Fig. 4). These resultsare supported by previous studies in which antibodiesagainst either angiogenin or actin delayed or pre-vented the appearance of HT-29-inoculated tumors inathymic mice.39,43
In rats, ACTIBIND inhibited DMH-induced coloncancer. ACTIBIND applied directly to the colon viaosmotic pump was effective from the early stages ofACF-induced carcinogenesis to later stages of malig-nant tumor development for all parameters examined.Furthermore, a reduction in the number of blood ves-sels per tumor as well as blood-vessel size was ob-served at the end of the therapeutic treatment withosmotic pumps (Table 3). Therapeutic treatment withACTIBIND administered via osmotic pumps not onlydiminished MVD, but concomitantly induced cell ap-optosis in the colonic tumors (Fig. 8). Apoptosis couldbe affected directly by interaction between ACTIBINDand tumor cells, or, alternatively, it could be the resultof hypoxia due to significant reduction in the numberof blood vessels.48,49 In this regard, cortical actin lo-calization has been observed in cells undergoing ap-optosis. The specific mechanism linking these twophenomena is still unclear.50 –52 These results impli-cate ACTIBIND as a potential antiangiogenic sub-stance.
ACTIBIND given orally exerted a clear preventive,but not therapeutic effect (Table 2). We followed theroute of fluorescent, protein-loaded, CAP microcap-sules along the rat gut and found that only traces ofthe initial fluorescence reached the colon. Encapsu-lated ACTIBIND most likely meets the same fate.These traces may be sufficient to prevent cancer ini-tiation but are not sufficient to treat fully developedtumors. We believe that improving the encapsula-tion coating would allow for improved delivery ofACTIBIND to the colon.
The inactivated ACTIBIND maintained its actin-binding, anticlonogenic, and anticancer activities invitro as well as in vivo. Unlike ACTIBIND, RNase ac-
TABLE 2Effect of ACTIBIND on DMH-induced Colon Cancer in Rats: PooledData From 2 Subsequent Experiments (N � 16)
Treatment Parameter Control ACTIBIND
Preventive *ACF from osmotic pumps 117.8 � 13.2 55.5 � 3.5†Tumor no. from microcapsules 8.3 � 2.0 4.5 � 0.6*Tumor malignancy from
microcapsulesAdenoma 3.6 � 1.1 3.1 � 0.8Adenocarcinoma 5.9 � 0.8 2.4 � 0.6
Therapeutic †Tumor no. from osmoticpumps
11.3 � 6.0 5.5 � 1.5
DMH: dimethylhydrazine; ACF: aberrant crypt foci.
* P �.01† P �.05
TABLE 3Effect of ACTIBIND on Angiogenesis (Control N � 12, ACTIBIND N� 10) and Apoptosis (N � 3, 10 Microscopic Fields of Each) in DMH-induced Tumors in Rats
Parameter Control ACTIBIND
Angiogenesis Blood vessels per tumor *56.8 � 5.0 31.9 � 3.6Relative area, % †1.0 � 0.3 0.34 � 0.1
Apoptosis Apoptotic cells permicroscopic field
†2.0 � 0.2 37.0 � 5.0
DMH: dimethylhydrazine.
* P �.01† P �.05
ACTIBIND: a T2-RNase with Anticancer Effect/Roiz et al. 2305

tivity is essential for onconase anticancer activity.30 We,thus, assume that the anticancer effects of ACTIBINDand onconase are mediated by different mecha-nisms.
The resistance of many tumors to conventionaltherapy has led to attempts to develop novel and moreeffective strategies to improve anticancer agents. Mo-bile cells (e.g., angiogenic and cancer cells) may bemore sensitive to ACTIBIND because they display sig-nificantly more cell-surface actin than stationarycells.2,45 RNases are now the focus of a great deal ofattention because of their newly discovered activitiesand applications, particularly in cancer biology.1
Therefore, a better understanding of the mecha-nism(s) through which ACTIBIND, the fungal T2-
RNase, exerts its antiangiogenic and anticarcinogenicproperties is required.
REFERENCES1. Schein CH. From housekeeper to microsurgeon: the diag-
nostic and therapeutic potential of ribonucleases. NatureBiotech. 1997;15;529 –536.
2. Maeda T, Kitazoe M, Tada H, de Llorens R, Salomon DS,Ueda M, Yamada H, Seno M. Growth inhibition of mamma-lian cells by eosinophil cationic protein. Eur J Biochem.2002;269:307–316.
3. Strydom D J. The angiogenins. Cell Mol Life Sci. 1998;54:811– 824.
4. Antignani A, Naddeo M, Cubellis MV, Russo A D’Alessio G.Antitumor action of seminal ribonuclease, its dimeric struc-ture, and its resistance to the cytosolic ribonuclease inhib-itor. Biochemistry. 2001;40:3492–3496.
FIGURE 8. The effect of ACTIBIND on blood vessels and on apoptosis rate in DMH-induced colonic tumors. (A,C) Controls; (B,D) ACTIBIND-treated tumors. (A,B)
Immunostaining for CD31. Red areas represent the numerous blood vessels in control tumor. (C,D) TUNEL assay. Brown nuclei represent the numerous apoptotic
cells in ACTIBIND-treated tumors. Scale bar � 100 �m.
2306 CANCER May 15, 2006 / Volume 106 / Number 10

5. Matousek J. Ribonucleases and their antitumor activity.Comp Biochem Physiol C Toxicol Pharmacol. 2001;129:175–191.
6. Lin JJ, Newton DL, Mikulski SM, Kung HF, Youle RJ, RybakSM. Characterization of the mechanism of cellular and cell-free protein-synthesis inhibition by an antitumor ribonucle-ase. Biochem Biophys Res Commun. 1994;204:156 –162.
7. Lee I, Lee YH, Mikulski SM, Lee J, Covone K, Shogen K.Tumoricidal effects of onconase on various tumors. J SurgOncol. 2000;73:164 –171.
8. Irie M, Ohgi K. Ribonuclease T2. Meth Enzymol. 2001;341:42–55.
9. Nurnberger T, Abel S, Jost W, Glund K. Induction of extra-cellular ribonucleases in cultured tomato cells upon phos-phate starvation. Plant Physiol. 1990;92:970 –976.
10. Clarke AE, Newbigin E. Molecular aspects of self-incompat-ibility in flowering plants. Annu Rev Genet. 1993;27:257–279.
11. Silva NF Goring DR. Mechanisms of self-incompatibility inflowering plants. Cell Mol Life Sci. 1997;58:1988 –2007.
12. Cooke IE, Shelling AN, Le Meuth VG, Charnock ML, Gane-san TS. Allele loss on chromosome arm 6q and fine mappingof the region at 6q27 in epithelial ovarian cancer. GenesChromosomes Cancer. 1996;15:223–233.
13. Saito S, Saito H, Ko S, Sagae S, Kudo R, Saito J, Noda K,Nakamura Y. Fine-scale deletion mapping of the distal longarm of chromosome 6 in 70 human ovarian cancers. CancerRes. 1992;52:5815–5817.
14. Trubia M., Sessa L. Taramelli R. Mammalian Rh/T2/S-gly-coprotein ribonuclease family genes: cloning of a humanmember located in a region of chromosome 6 (6q27) fre-quently deleted in human malignancies. Genomics 1997;42:342–344.
15. Acquati F, Nucci C, Bianchi MG, Gorletta T, Taramelli R.Molecular cloning, tissue distribution, and chromosomallocalization of the human homolog of the R2/Th/Stylar ri-bonuclease gene family. Meth Mol Biol. 2001;160:87–101.
16. Roiz L, Goren R, Shoseyov O. Stigmatic RNase in ca-lamondin (Citrus reticulata var. Austera x Fortunella sp.).Physiol Plant. 1995;94:585–590.
17. Roiz L, Shoseyov O. Stigmatic RNase in self-compatiblepeach (Prunus persica). Int J Plant Sci. 1995;156:37–341.
18. Roiz L, Ozeri U, Goren R, Shoseyov O. Characterization ofAspergillus niger B-1 RNase and its inhibitory effect on pol-len germination and pollen tube growth in selected treefruit. J Amer Soc Hort Sci. 2000;125:9 –14.
19. Palanivelu R, Preuss D. Pollen tube targeting and axon guid-ance: parallels in tip growth mechanisms. Trends Cell Biol.2000;10:517–524.
20. Sheetz MP, Felsenfeld D, Galbraith CG, Choquet D. Cellmigration as a five-step cycle. Biochem Soc Symp. 1999;65:233–243.
21. Janmey PA, Chaponnier C. Medical aspects of the actincytoskeleton. Curr Opin Cell Biol. 1995;7:111–117.
22. Ponoe ML. In vitro matrigel angionesis assays. In: MurrayJC. Methods in Molecular Medicine. Vol. 46: Angiogenesisprotocols. New York :Totowa Humana Press; 2001:205–209.
23. Hu GF, Strydom DJ, Fett JW, Riordan JF, Vallee BL. Actin isa binding protein for angiogenin. Proc Natl Acad Sci U S A.1993;90:1217–1221.
24. Schwartz B, Avivi C, Lamprecht SA. Isolation and character-ization of normal and neoplastic colonic epithelial cell pop-ulations. Gastroenterology. 1991;100:692–702.
25. Schwartz B, Smirnoff P, Shany S, Liel Y. Estrogen controls
expression and bioresponse of 1,2,5-dihydroxyvitamin D re-ceptors in the rat colon. Mol Cell Biochem. 2000;203:87–93.
26. Smirnoff P, Liel Y, Gnainsky J, Shany S, Schwartz B. Theprotective effect of estrogen against chemically induced mu-rine colon carcinogenesis is associated with decreased CpGisland methylation and increased mRNA and protein ex-pression of the colonic vitamin D receptor. Oncol Res. 1999;11:255–264.
27. Lin SY, Tzan YL, Weng CN, Lee CJ. Preparation of enteric-coated microspheres of Mycoplasma hyopneumoniae vac-cine with cellulose-acetate phthalate. 2. Effect of tempera-ture and pH on the stability and release behavior ofmicrospheres. J Microencapsul. 1991;8:537–545.
28. Bird RP. Observation and quantification of aberrant cryptsin the murine colon treated with a colon carcinogen: pre-liminary findings. Cancer Lett. 1987;37:147–151.
29. Padmanabhan S, Zhou K, Chu CY, Lim RW Lim LW Over-expression, biophysical characterization and crystallizationof ribonuclease I from Escherichia coli, a broad-specificityenzyme in the RNase T2 family. Arch Biochem Biophys. 2001;390:42–50.
30. Wu Y, Mikulski SM, Ardelt W, Rybak SM, Youle RJ. A cyto-toxic ribonuclease. Study of the mechanism of onconasecytotoxicity. J Biol Chem. 1993;268:10686 –10693.
31. Wu Y, Saxena SK, Ardelt W, Gadina M, Mikulski SM, DeLorenzo C, D’Alessio G, Youle RJ. Study of the intracellularrouting of cytotoxic ribonucleases. J Biol Chem. 1995;270:17476 –17481.
32. Pardridge WM, Nowlin DM, Choi TB, Yang J, Calaycay J,Shively JE. Brain capillary 46,000 dalton protein is cytoplas-mic actin and is localized to endothelial plasma membrane.J Cereb Blood Flow Metab. 1989;9:675– 680.
33. Hu GF, Chang SI, Riordan JF, Vallee BL. An angiogenin-binding protein from endothelial cells. Proc. Natl Acad SciUSA. 1991;88:2227–2231.
34. Moroianu J, Fett JW, Riordan JF, Vallee BL. Actin is a surfacecomponent of calf pulmonary artery endothelial cells inculture. Proc Natl Acad Sci U S A. 1993;90:3815–3819.
35. Hu GF, Riordan JF. Angiogenin enhances actin accelerationof plasminogen activation. Biochem Biophys Res Commun.1993;197:682– 687.
36. Hu G.F, Riordan JF, Vallee BL. Angiogenin promotes inva-siveness of cultured endothelial cells by stimulation of cell-associated photolytic activities. Proc Natl Acad Sci U SA.1994;91:12096 –12100.
37. Gho YS, Yoon WH, Chae CB. Antiplasmin activity of a pep-tide that binds to the receptor-binding site of angiogenin.J Biol Chem. 2002;277:9690 –9694.
38. Yoshimura T, Matsushima K, Tanaka S, Robinson EA, Ap-pella E, Oppenheim JJ. Leonard EJ. Purification of a humanmonocyte-derived neutrophil chemotactic factor that haspeptide sequence similarity to other host defense cytokines.Proc Natl Acad Sci U S A. 1987;84:9233–9237.
39. Olson KA, Fett JW, French TC, Key ME, Vallee BL. Angioge-nin antagonists prevent tumor growth in vivo. Proc NatlAcad Sci U S A. 1995;92:442– 446.
40. Shimoyama S, Yamasaki K., Kawahara M, Kaminishi M.Increased serum angiogenin concentration in colorectalcancer is correlated with cancer progression. Clin CancerRes. 1999;5:1125–1130.
41. Etienne-Manneville S, Hall A. Rho GTPases in cell biology.Nature, 2002;420:629 – 635.
42. Bach PR, Bentley JP. Structural glycoprotein, fact or artifact.Connect Tissue Res. 1980;7:185–196.
ACTIBIND: a T2-RNase with Anticancer Effect/Roiz et al. 2307

43. Olson KA, French TC, Vallee BL, Fett JW. A monoclonalantibody to human angiogenin suppresses tumor growth inathymic mice. Cancer Res. 1994;54:4576 – 4579.
44. Accinni L, Natali PG, Silvestrini M, De Martino C, Actin inthe extracellular matrix of smooth muscle cells. An immu-noelectron microscopic study. Connect Tissue Res.1983;11:69 –77.
45. Morikawa S, Baluk P, Kaidoh T, Haskell A, Jain RK, McDonaldDM. Abnormalities in pericytes on blood vessels and endothe-lial sprouts in tumors. Am J Pathol. 2002;160:985–1000.
46. Frame MC, Brunton VG. Advances in Rho-dependent actinregulation and oncogenic transformation. Curr Opin GenetDev. 2002;12:36 – 43.
47. Shestenko OP, Nikonov SD, Mertvetsov NP. Angiogenin andits function in angiogenesis. Mol Biol. 2001;35:294 –314.
48. Jimenez B, Volpert OV. Mechanistic insights on the inhibi-tion of tumor angiogenesis. J Mol Med. 2001;78:663– 672.
49. Knowles HJ, Harris AL. Hypoxia and oxidative stress inbreast cancer: Hypoxia and tumourigenesis. Breast CancerRes. 2001;3:318 –322.
50. Palladini G, Finardi G, Bellomo G. Disruption of actin mi-crofilament organization by cholesterol oxides in 73/73 en-dothelial cells. Exp Cell Res. 1996;223:72– 82.
51. Guenal I, Risler Y, Ignotte B. Down-regulation of actin genesprecedes microfilament network disruption and actin cleav-age during p53-mediated apoptosis. J Cell Sci. 1997:110:489 –495.
52. Staiger CJ Franklin-Tong VE. The actin cytoskeleton is atarget of the self-incompatibility response in Papaverrhoeas. J Exp Bot. 2002;54:103–113.
2308 CANCER May 15, 2006 / Volume 106 / Number 10
Copyright © 2022 FDOKUMEN




















