
Anniversary of the Pander Society
19
Conodont Studies Commemorating the 150 th Anniversary of the First Conodont Paper (Pander, 1856) and the 40 th Anniversary of the Pander Society D. Jeffrey Over, editor Palaeontographica Americana Number 62, March 2009
Transcript of Anniversary of the Pander Society

1
Conodont Studies Commemorating the 150th
Anniversary of the First Conodont Paper (Pander,
1856) and the 40th Anniversary of the Pander Society
D. Jeff rey Over, editor
Palaeontographica AmericanaNumber 62, March 2009

2 Palaeontographica Americana, No.
ISSN 0078-8546ISBN 978-0-87710-483-4
Library of Congress Catalog Card Number 2009922563© 2009, Paleontological Research Institution, 1259 Trumansburg Road, Ithaca, New York 14850, U. S. A.
Printed in the United States of AmericaAllen Press, Inc.
Lawrence, Kansas 66044, U. S. A.

3
CONTENTS
Contributors and Reviewers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4-5
Preface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
Klapper, GilbertWilliam M. Furnish (1912-2007) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9-10
Aldridge, Richard J., & Peter von BitterTh e Pander Society: a Brief History at Forty (1967-2007) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11-21
Leslie, Stephen A.Relationships between Upper Ordovician (Sandbian, Mohawkian) lithofacies and conodont biofacies distribution patterns using K- Bentonite Beds as time planes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23-40
Barrick, James E., Mark A. Kleff ner, and Haraldur R. KarlssonConodont faunas and stable isotopes across the mulde event (late Wenlock; Silurian) in southwestern Laurentia (south-central Oklahoma and subsurface west Texas) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41-56
Kleff ner, Mark A., James E. Barrick, James R. Ebert, Damon K. Matteson, and Haraldur R. KarlssonConodont biostratigraphy, δ13C chemostratigraphy, and recognition of Silurian/Devonian boundary in the Cherry Valley, New York region of the Appalachian Basin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57-73
Suttner, Th omas J.Lower Devonian conodonts of the "Baron von Kottwitz" quarry (southern Burgenland, Austria) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75-87
Over, D. Jeff rey, Sarah de la Rue, Peter Isaacson, and Brooks EllwoodUpper Devonian conodonts from black shales of the high latitude Tomachi Formation, Madre de Dios Basin, northern Bolivia . . . . . . 89-99
May, Michael T., Carl B. Rexroad, and Lewis M. BrownCarboniferous conodont biostratigraphy along the southeastern periphery of the Eastern Interior Basin, Kentucky, U. S. A. . . . . . . 101-113
Rosscoe, Steven J., and James E. BarrickRevision of Idiognathodus species from the Desmoinesian-Missourian (~Moscovian-Kasimovian) boundary interval in the Midcontinent Basin, North America . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 115-147
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 148-150

41
CONODONT FAUNAS AND STABLE ISOTOPES ACROSS THE MULDE EVENT LATE WENLOCK; SILURIAN IN SOUTHWESTERN LAURENTIA SOUTHCENTRAL OKLAHOMA
AND SUBSURFACE WEST TEXAS
James E. BarrickDepartment of Geosciences, Texas Tech University, Lubbock, Texas 79409-1053, U. S. A., email [email protected].
Mark A. KleffnerSchool of Earth Sciences, Division of Geological Sciences, Th e Ohio State University at Lima, Lima, Ohio 45804, U. S. A.
and Haraldur R. KarlssonDepartment of Geosciences, Texas Tech University, Lubbock, Texas 79409-1053, U. S. A.
ABSTRACTTh e mid-Homerian Mulde Event coincides with a sequence boundary in distal carbonate ramp settings in southwestern Laurentia (west Texas and Oklahoma). Th e position of the Mulde Event interval and the associated unconformity could only be located by detailed collections of conodonts, which display clearly the Mulde extinction level. Dapsilodus praecipuus Barrick, 1977, D. sparsus Barrick, 1977, Pseudooneotodus linguicornis Jeppsson, 2003, and Panderodus unicostatus (Branson & Mehl ,1933), species characteristic of the Ozarkodina sagitta sagitta Walliser 1964, Zone, disappear abruptly and are replaced by Panderodus equicostatus (Rhodes, 1953) and Belodella silurica Barrick, 1977, species characteristic of the O. bohemica longa Jeppsson, 2003, and Kockelella ortus absidata Barrick & Klapper, 1976, zones. Much of the time represented by the Mulde Event appears to be absent because the positive δ13C excursion associated with the event is incompletely preserved. Strata equivalent to much of the time of the Mulde Event were either removed or not deposited in southwestern Laurentia during the brief eustatic sea level fall that accompanied the Mulde Event. Based on these results, more consideration of the eff ect of eustatic events in interpreting Silurian oceanic events is merited.
Chapter 4, in Conodont Studies Commemorating the 150th Anniversary of the First Conodont Paper (Pander, 1856) and the 40th Anniversary of the Pander Society, edited by D. Jeff rey Over, Palaeontographica Americana, 2009, (62): 41-56.
INTRODUCTIONJeppsson (1990; 1998) developed his model of oceanic events and episodes to explain the observed patterns in the ranges of Silurian conodont and graptolite species and sedimentary shelf deposition, primarily on Gotland, and the potential eff ects of global climate and oceanic circulation on primary productiv-ity. Th e early Wenlock Ireviken Event, the most signifi cant of the Silurian oceanic events, has been recognized globally by diagnostic biostratigraphic and chemostratigraphic signatures (see Jeppsson, 1998; Cramer et al., 2006b). Th e early Home-rian (late Wenlock) Mulde Event is another prominent Silu-rian oceanic event, and the subject of recent scrutiny (Calner et al., 2006; Cramer et al., 2006a; Lenz et al., 2006). Jepps-son (1998) considered the severity of the Mulde Event to be equivalent to that of the Ireviken Event. Unlike most oceanic events that mark the transition between alternate primo and secundo oceanic states, the Mulde Event is a secundo-secundo event (Jeppsson & Calner, 2003). Th e Mulde Event includes the “lundgreni event,” one of the most signifi cant global grap-toloid extinction events, during which the diverse and spe-cialized Wenlock graptoloid fauna was reduced to only a few
taxa (Melchin et al., 1998; Porebska et al., 2005; Lenz et al., 2006). A moderate extinction occurs in conodonts (Jeppsson et al., 1995; Jeppsson & Calner, 2003) and diversity drops in marine palynomorphs (Kaljo et al., 1995). No corresponding decrease in diversity in the marine benthos has been dem-onstrated (Kaljo et al., 1995), although Jeppsson & Calner (2003) suggested that many “shelly” taxa became extinct at that time based on a review of the literature. A signifi cant positive δ 13C excursion associated with the event has been identifi ed across Europe (Samtleben et al., 1996; Kaljo et al., 2003; Jeppsson & Calner, 2003; Porebska et al., 2004; Calner et al., 2006) and in North America (Saltzman, 2001; Cramer et al., 2006a; Lenz et al., 2006).
In this paper we report on the lithological succession, conodont species ranges and biofacies, and carbon isotopes through the Mulde Event interval in three carbonate sections from southwestern Laurentia (Text-fi gs 1-2; Oklahoma and west Texas), and make comparisons to the well-characterized record of the Mulde Event in the shelf successions in the Baltic region (Jeppsson & Calner, 2003; Calner & Jeppsson, 2003; Calner et al., 2006; Kaljo & Martma, 2006).
WENLOCK STRATA IN SOUTHWESTERN LAURENTIA

42 Palaeontographica Americana, No.
Reconstructions of continental positions during the Silurian (Text-fi g. 1) place the southwestern margin of Laurentia ca. 20-30° of latitude south of the equator, facing to the south to southwest (Cocks & Scotese, 1991; Golonka et al., 1994; Cocks & Torsvik, 2002). Wilde et al. (1991) showed that dur-ing the Silurian the southwestern margin of Laurentia would have received relatively cool to cold oceanic waters from a west-erly subpolar current that originated adjacent to Gondwana. Moore et al. (1994: fi g. 5), however, indicated that this part of the world lay in a tropical to subtropical climatic regime, where sea surface temperatures remained at or above 25°C throughout the year. Other recent work on continental posi-tions during the Ordovician and Early Silurian (e. g., Dalziel, 1997; Niocaill et al., 1997) shows Laurentia rotated in such a way that at the beginning of the Silurian, the modern south-ern margin of North America faced west to southwest and Gondwana lay a greater distance away from Laurentia than in the older reconstructions. All of these reconstructions show that southwestern Laurentia was situated thousands of kilo-meters from Baltica and in a position to be aff ected directly by global oceanic circulation.
Barrick (1997) recognized three depositional sequences in Wenlock strata in southern Laurentia, based on sections in subsurface west Texas, south-central Oklahoma, north-ern Arkansas, the Illinois Basin, and central Tennessee: a Sheinwoodian sequence, a lower Homerian sequence, and an upper Homerian to lower Gorstian (Ludlow) sequence. Recent publications on conodont and graptolite biostratigraphy (e. g., Jeppsson et al., 2006) indicate that age assignments of these sequences need revision. Th e lower Wenlock “Sheinwoodian” sequence ranges in age from the ear-ly Sheinwoodian Lower procerus Zone through the early
Homerian sagitta sagitta Zone, and is herein called the “low-er Wenlock sequence.” Across southern Laurentia, variable amounts of latest Sheinwoodian to earliest Homerian strata appear to be missing at the unconformity at the base of the upper Wenlock sequence, the lower Homerian sequence of Barrick (1997), which coincides with the Mulde Event. Th e base of the upper Wenlock sequence lies at, or just above, the Mulde Event interval. Its upper boundary lies in the crassa Zone, which corresponds to the base of the Ludlow.
Recognition of latest Sheinwoodian to earliest Homerian
Text-fi g. 1. Paleogeographic reconstruction for the Silurian. Study area indicated by cross in southern Laurentia. Star marks Gotland. Map modifi ed from Scotese (2002).
Text-fi g. 2. Map of the southern United States showing locations of areas discussed in the text. SOK, southern Oklahoma; TN, Tennes-see; WTX, west Texas. Solid black areas indicate outcrop exposures; lined areas indicate subsurface extent of Silurian strata. Modifi ed from Berry & Boucot (1970).
Text-fi g. 3. Correlation of graptolite zones, condoont zones, and da-tums of the Mulde Event. Modifi ed after Cramer et al. (2006a: fi g. 5) and Jeppsson et al. (2006: fi g. 1).

43
strata and scaling of the hiatus at the mid-Wenlock unconfor-mity in southern Laurentia using conodonts has been hindered by the rarity of zonal indices. Th e revised conodont zonation of Calner & Jeppsson (2003) across the Sheinwooodian-Homerian boundary provides additional ways by which latest Sheinwoodian and earliest Homerian strata can be identifi ed in our study area (Text-fi g. 3). Kockelella ortus ortus ranges from the latest Sheinwoodian K. ortus ortus Zone through the succeeding earliest Homerian sagitta sagitta Zone, disap-pearing before the Mulde Event (Jeppsson et al., 2006). Some Kockelella elements from the Clarita Formation in Oklahoma that occur with, or just above, K. patula Walliser, 1964 (samples C1-15 and Haragan Creek-5; Barrick & Klapper, 1976), should be assigned to K. ortus ortus instead of K. ortus absidata, which appears after the Mulde Event. Th e species Pseudooneotodus linguicornis, which appears near the base of the sagitta sagitta Zone and ranges up to the Mulde Event (Calner & Jeppsson, 2003), serves as a secondary means by which to recognize the earliest Homerian sagitta sagitta Zone and strata of the Hellvi Secundo Episode (Calner & Jeppsson,
2003). Th is species is moderately common in samples from the top of the lower Wenlock sequence in Oklahoma and west Texas, and strata bearing it are assigned to the sagitta sagitta Zone. Th e only known occurrence of Ozarkodina sagitta sagitta in southern Laurentia is in the Dougherty West section (DW) in Oklahoma, which is described in this paper. Th ere, O. sag-itta sagitta is present in one sample (DW-3B1) that lies just above the FAD (fi rst appearance datum) of Pseudooneotodus linguicornis and K. ortus ortus (DW-3B; Table 1).
Th e Ozarkodina bohemica longa Zone characterizes the strata including the Mulde Event interval and the base of the succeeding Kockelella ortus absidata Zone lies near the end of the Mulde Event (Calner & Jeppsson, 2003; Jeppsson et al., 2006). Th ese zones are recognized by the FADs of the nomi-nate species, both of which range above their zones. Both spe-cies occur in southern Laurentia, but their zones are diffi cult to identify. Th e next zone, the Ctenognathodus murchisoni (Pander, 1856) Zone, which appears in strata of the Klinte Secundo Episode (Calner & Jeppsson, 2003; Jeppsson et al., 2006), does not occur in southern Laurentia, making it ex-
Table 1. Distribution of conodonts in the Dougherty West section, Murray County, Oklahoma (NE1/4 SW1/4, sec. 3, T-2-S, R-2-E). *, num-bers of elements of Dapsilodus obliquicostatus estimated from number of Sa elements counted (Amsden et al., 1980), except for sample 3F.
Sample 3B 3B1 3C 3D 3E 3F 4 4B 4BB 4C 5 5A
Base (m) 2.50 2.40 2.20 2.10 2.00 1.90 1.80 1.70 1.60 1.50 1.45 1.30
Top (m) 2.40 2.30 2.10 2.00 1.90 1.80 1.70 1.60 1.50 1.45 1.30 1.25
Weight (kg) 2.60 2.53 0.65 1.30 1.15 2.22 1.30 1.30 0.65 1.30 1.30 1.30
Dapsilodus obliquicostatus* (Branson & Mehl, 1933)
X X 3340 5490 2690 2562 1850 1870 1470 1990 2830 2820
Decoriconus fragilis (Branson & Mehl, 1933) X X 235 278 163 19 224 254 123 86 36 170
Pseudooneotodus bicornis Drygant, 1974 X X 30 106 135 156 164 93 22 13 23 77
Ozarkodina excavata (Branson & Mehl, 1933) X X 14 25 6 28 47 137 38 161 23 25
Dapsilodus praecipuus Barrick, 1977 X X 32 93 492 55 - - - - - -
Dapsilodus sparsus Barrick, 1977 X X 2 9 29 19 - - - - - -
Pseudooneotodus linguicornis Jeppsson, 2003 X X 10 20 15 6 - - - - - -
Panderodus recurvatus (Rhodes, 1953) X X 50 65 27 4 - - - - - -
Panderodus unicostatus (Branson & Mehl, 1933) X X 169 241 217 64 - - - - - -
Walliserodus sp. X X 35 76 60 - - - - - - -
Kockelella ortus ortus (Walliser, 1964) X X - - - - - - - - - -
Ozarkodina sagitta sagitta Walliser, 1964 - 2 - - - - - - - - - -
Panderodus equicostatus (Rhodes, 1953) - - - - - - 91 65 - - - -
Belodella silurica Barrick, 1977 - - - - - - 17 270 133 94 40 -
Ozarkodina bohemica longa Jeppsson, 2003 - - - - - - - 9 2 11 - -
Kockelella ortus absidata Barrick & Klapper, 1976 - - - - - - - 12 - 13 - -
?Panderodus greenlandensis Armstrong, 1990 - - - - - - - - 17 29 10 -
Other elements - - 4 1 - - 6 2 12 - 4 2
Barrick ET AL.: Conodont Faunas and Stable Isotopes

44 Palaeontographica Americana, No.
tremely diffi cult to determine precisely the age of the base of the upper Wenlock sequence.
METHODSSamples for conodont processing and isotope analysis were collected at the same time. Th e smallest reasonable sample in-terval for conodont samples is ca. 10 cm, but isotope samples could be taken more closely spaced where potentially short-lived events appeared to be present. Th e mixing eff ects of sedi-ment advection on both conodont biofacies and geochemical signals by burrowing organisms can occur over a thickness of 30-40 cm in slowly deposited sediments today (see discussion by Martin, 1999). It is unlikely that a fi ner-scale time reso-lution of events would be possible with more closely spaced samples because of the averaging eff ects of sediment advec-tion.
Conodont samples were on the order of 1-2 kg. Th e size of samples was limited in the west Texas cores by the material available, but larger sample sizes were possible from the one outcrop section in Oklahoma. Jeppsson (2005) discussed the biases in recovery of conodont elements during sampling and processing. He recommended sample sizes on the order of 50 kg where possible to ensure complete recovery of conodont species at each level. It is well known that larger sample sizes
allow recovery of more species, especially extremely rare taxa, but unless all levels are of equal size, then comparisons among samples becomes problematic. Our samples yielded hundreds to thousands of conodont elements per kilogram (Tables 1-3), which should allow resolution of the probable ranges of the most species. Because this study focuses on extinctions and originations of conodont species relative to the Mulde Event, we are uncertain whether the disappearance or appearance of extremely rare species in the faunas (> 0.1%) will assist with evaluation of the magnitude of environmental change associ-ated with the event. All samples were processed with buff -ered formic acid using the procedure of Jeppsson & Anehus (1995). Faunal counts are based on elements collected on a 125-μm (120 mesh) sieve.
Isotope samples were obtained by drilling out of powder from sawed slabs of the carbonates. Th e report by Cramer et al. (2006b) that micrites and fi ne-grained carbonates are reliable for stable isotope chemostratigraphy has been well demonstrated. Samples were either carbonate mudstones or sparse skeletal wackestones bearing few skeletal grains that were avoided wherever possible. Powders were roasted for 30 min at ca. 480°C under a stream of hot, 99.99% purifi ed he-lium to remove water and organic material. Samples weighing 20-25 mg were reacted with 100% phosphoric acid for 24
Table 2. Distribution of conodonts in the Socony Mobil Pegaus Unit 3 #7-20 core, Midland County, Texas (sec. 30, Blk. 40, T-4-S, T&P RR Survey).
Sample 22 20A 20 19A 19 18 17A 17
Base (m) 3636.20 3635.80 3635.70 3635.60 3635.50 3635.30 3635.10 3634.85
Top (m) 3636.00 3635.70 3635.60 3635.50 3635.40 3635.15 3635.00 3634.75
Weight (kg) 2.50 1.54 0.85 1.15 1.35 1.20 1.30 1.00
Dapsilodus obliquicostatus 656 593 9 64 56 654 875 2135
Decoriconus fragilis 3 24 3 1 11 146 305 173
Pseudooneotodus bicornis 100 64 115 33 16 211 187 59
Ozarkodina excavata - - - 1 3 9 44 87
Dapsilodus praecipuus 593 890 312 214 150 - - -
Dapsilodus sparsus - 7 3 2 - - - -
Pseudooneotodus linguicornis 28 18 2 3 1 - - -
Panderodus recurvatus - 16 - - 7 - - -
Panderodus unicostatus 116 218 115 74 84 - - -
Walliserodus sp. 12 62 47 23 - - - -
Panderodus equicostatus - - - - - 4 134 135
Belodella silurica - - - - - - 6 115
Ozarkodina bohemica longa - - - - - - - 3
Kockelella ortus absidata - - - - - 1 - -
Other elements - - - - - - 2 11

45
hr in an agitated water bath maintained at 25.2°C. Evolved CO2 was extracted and purifi ed following the method of Mc-Crea (1950). Th e relative yield is the percent of measured versus theoretical yield of pure calcite (9.99 μmol CO2/mg). Full yield is rarely obtained for these samples for several rea-sons. First, the highest measured yields from pure calcite are typically 90-95% of the theoretical value for samples reacted at 25°C. Second, acid-resistant phases such as silicates will contribute insignifi cant amounts of gas and lower the overall sample yield. Hence, variable yields are expected for diff erent samples depending on the amount of noncarbonate phases. Th is variation in yield, however, does not imply incomplete reaction of calcite and has no eff ect on the isotopic values.
Gas samples were analyzed on a VG SIRA-12 isotope ratio mass spectrometer equipped with a microinlet system (moved). All values were obtained using an internal standard TTU-2 calcite, calibrated versus international standard NBS-19. All of our isotope results are reported using the conven-tional delta notation (Craig, 1957). Results are reported rela-tive to V-PDB (‰). Oxygen isotope values are adjusted using an acid-fractionation factor of 1.01025. Th e precision of our analysis is determined by overall method and is ± 0.1‰ and ± 0.2‰ for δ 13C and δ 18O, respectively. Jacobi (2004) exam-ined the variation in isotopic values of rock powders for simi-lar Late Silurian carbonate mudstones by making replicate
samples at two levels. He found that in these rocks, the four replicate values at each level diff ered by as much as 0.15‰ for δ 13C and 0.40‰ for δ18O, a product of the natural heteroge-neity of the rocks. Th erefore, we are cautious in making inter-pretations for changes of less than 0.5‰ for either isotope.
THE MULDE EVENT IN SOUTH-WESTERN LAURENTIA
SOUTHERN OKLAHOMA
Dougherty West SectionIn south-central Oklahoma, Wenlock strata are included in the Clarita Formation (Text-fi g. 4). Th e Clarita Formation comprises 4-15 m of carbonate mudstone and wackestone in the central Arbuckle Mountains that grades into pelmatozoan grainstones to the north (Amsden et al., 1980). Th e Clarita Formation in outcrop represents deposition on the deeper part of a carbonate ramp that shallowed northward into cen-tral Oklahoma. Th e Wenlock conodont fauna of the Clarita Formation was described by Barrick & Klapper (1976) and Barrick (1977) and the carbonate lithofacies and biofacies of the formation were discussed by Amsden et al. (1980). Subse-quent work shows that the Clarita spans the entire Wenlock only in the central Arbuckle Mountains and that Clarita sec-
Table 3. Distribution of conodonts in the Cobra Frazier #1 core, Howard County, Texas (sec. 3, Blk. 30, T-1-S, T&P RR Survey).
Sample 78 77B 77A 77 76A 76 75B 75A 75 74A 74
Base (m) 3073.94 3073.80 3073.67 3073.58 3073.45 3073.30 3073.20 3073.05 3072.95 3072.80 3072.70
Top (m) 3073.80 3073.67 3073.58 3073.45 3073.35 3073.20 3073.05 3072.95 3072.85 3072.70 3072.40
Weight (kg) 1.80 1.70 1.10 1.37 1.20 1.80 1.20 1.20 1.00 1.67 1.50
Dapsilodus obliquicostatus 12 11 26 93 15 31 16 57 168 372 24
Decoriconus fragilis - 2 1 - - 16 22 94 54 114 3
Pseudooneotodus bicornis - - 1 1 3 8 14 86 31 36 8
Ozarkodina excavata 2 3 - - - 5 - - 1 16 5
Dapsilodus praecipuus 52 42 1 1 - 6 - - 1? - -
Dapsilodus sparsus - - - - - - - 1? - - -
Pseudooneotodus linguicornis 2 2 1 - - - - - - - -
Panderodus recurvatus 3 4 - 2 1 - - - - - -
Panderodus unicostatus 156 70 34 19 8 149 - - - - -
Walliserodus sp. 5 - - - - 19 - - - - -
Panderodus equicostatus - - - - - - - 32 60 202 6
Belodella silurica - - - - - - - - 2 13 3
Ozarkodina bohemica longa - - - - - - - - - - 1
Kockelella ortus absidata - - - - - - - - - - 3
Other elements - - 1 - - - - 9 - 3 -
Barrick ET AL.: Conodont Faunas and Stable Isotopes

46 Palaeontographica Americana, No.
tions on the Lawrence Uplift to the north are only Sheinwoo-dian in age (sections C1 and P1; Barrick & Klapper, 1976; Amsden et al., 1980; Barrick et al., 1990). In most Clarita sec-tions in the central Arbuckles, argillaceous middle Homerian (post-Mulde Event) strata rest unconformably on a truncated Sheinwoodian section (e. g., sections Ca2 and Highway 77 of Barrick & Klapper, 1976). Latest Sheinwoodian to earli-est Homerian strata overlain by middle Homerian strata ap-pear to occur in only a small area in the Arbuckle Mountains and can be documented in only two sections, Haragan Creek (Barrick & Klapper, 1976), and the Dougherty West section.
Th e Dougherty West section (DW; Text-fi g. 5) is a natu-ral exposure of the Hunton Group along a small stream that drains a small natural lake dammed by Hunton carbonates that dip steeply, 70-80°, to the east. Outfl ow from the lake in times of high water has scoured a small channel. Along the sides and in the bottom of this channel the Hunton units are exposed. Th e lower 5 m of the Clarita Formation comprise the typical dense arthropod micrite facies that is characteristic in the lower part in the Arbuckle Mountains (Amsden et al., 1980). Th e basal unit grades into ca. 2 m of slightly more ar-gillaceous arthropod micrites that are less resistant to weather-ing. Th e upper 0.85 m of the Clarita comprises the ostracode silty marlstone facies typical of the uppermost Clarita in the Arbuckle Mountain region (Amsden et al., 1980). Th e 8 m of Clarita Formation at DW is atypically thick for the central
Arbuckles as compared to the nearby Highway 77 section, where it is only 4 m thick. At the Highway 77 section, the ostracode silty marlstone facies rests directly on the lower ar-thropod micrite facies and late Sheinwoodian to early Home-rian strata appear to be absent. Th e increase in thickness at DW seems to be the result of better preservation of the upper part of the lower Wenlock sequence and perhaps the base of the upper Wenlock sequence. Th e level of the Mulde Event can be identifi ed best by conodont faunas, and less well by a δ13C excursion (Text-fi g. 5). Because the Clarita-Henryhouse contact is well exposed, but the base of the Clarita is covered, all measurements are given in meters below the top of the Clarita. Th e level of the Mulde Event lies at a bedding plane 1.80 m below the top of the Clarita.
Conodont Faunas.–Kockelella patula occurs 3.95-3.85 m below the top of the Clarita in the upper part of the lower arthropod micrites. Th e lowest occurrence of K. ortus ortus is substantially higher, at 2.50 m, within the middle interval of slightly argillaceous arthropod micrites (sample 3B; Table 1). Th e FAD of Pseudooneotodus linguicornis occurs at 2.40 m, which indicates that the FAD of K. ortus ortus lies near or in the sagitta sagitta Zone in this section. Th e only occurrence of Ozarkodina sagitta sagitta in southwestern Laurentia known to date is in the next bed (2.40-2.30 m, sample 3B1; Table 1), which confi rms the presence of the sagitta sagitta Zone in the section.
Conodont abundance is high, over 1,000 elements/kg. Th e sagitta sagitta Zone is strongly dominated by Dapsilo-dus obliquicostatus (> 80% of elements; Text-fi g. 5; Table 1). Panderodus unicostatus, Decoriconus fragilis, Pseudooneotodus bicornis, and Dapsilodus praecipuus are moderately common. Ozarkodina excavata, Walliserodus sp., Panderodus recurvatus, Pseudooneotodus linguicornis, and Dapsilodus sparsus are less abundant, usually fewer than 50 elements/kg. Th e LAD (last appearance datum) of Walliserodus sp. occurs at 1.90 m, which coincides with a slight decrease in abundance of several spe-cies, and a slight increase in abundance of P. bicornis. At 1.80 m, several species disappear: D. praecipuus, D. sparsus, P. lin-guicornis, P. recurvatus, and P. unicostatus. In the next sample, 1.80-1.70 m, Panderodus equicostatus and Belodella silurica ap-pear in moderate abundance. Th e fauna above 1.80 m contin-ues to be dominated by Dapsilodus obliquicostatus, Ps. bicornis remains common, and De. fragilis increases in abundance. Th e FADs of both O. bohemica longa and Kockelella ortus absidata occur at 1.70, the latter species indicating a time near the end of the Mulde Event. At 1.6 m, P. equicostatus disappears and a robust form of Panderodus, tentatively referred here to P. greenlandensis, appears.
Stable Isotopes.–Values of δ13C below the sagitta sagitta Zone
Text-fi g. 4. Stratigraphic columns for Wenlock-Prídolí strata for southern Laurentia, including west Texas, southern Oklahoma, and western Tennessee.

47
are generally a little greater than +1.0‰ (Table 4). In the sag-itta sagitta Zone, there is a gradual increase from +1.1‰ to +1.6‰ which is terminated by a abrupt drop to +0.7‰ at 1.87 m that is followed by a sharp increase to +2.6‰ at 1.82 m, within the bed just below the conodont extinction level at 1.80 m. In the beds just above the extinction level (1.8-1.45 m), δ13C values fall from +1.9‰ to +1.4‰, and then rise from +1.6‰ to +2.3‰ at 1.35 m. Above this peak, δ13C val-ues decrease gradually to less than +2.0‰ over the next me-ter. Values of δ18O are remarkably uniform below and above the extinction level, but display a distinct downward shift of 1.5‰ to -5.3‰ just below the extinction level.
SUBSURFACE WEST TEXAS
Silurian strata of Wenlock to Ludlow age in the subsur-face of west Texas are assigned to the Wristen Formation, which includes carbonate shelf, slope, and basinal facies in the west and a ramp facies to the east (Text-fi g. 4; Ruppel & Holtz, 1994). On the northern shelf area, Wenlock and younger strata are included in the Fasken Formation, whereas
age-equivalent slope to basin strata to the south of Andrews County, Texas, are placed into the older Wink Formation (Wenlock to Ludlow) and the younger Frame Formation [Prídolí-Lochkovian (Ruppel & Holtz, 1994; Barrick, 1995; Barrick et al., 2004)]. Th e Wink Formation is relatively mo-notonous in lithology and is composed of stylolitic slightly argillaceous nodular-bedded mudstones to ostracode wacke-stones that probably represent deposition in a deeper water, outer platform to ramp setting (Ruppel & Holtz, 1994). It thins from less than 7 m in the south to over 90 m in Andrews County, Texas, where it merges with the Fasken Formation. Two cores of the Wink Formation are included in this study, the Socony Pegasus 7-20 core in Midland County (Barrick et al., 1993) and the Cobra Frasier core 50 km northeast in Howard County (Kohl, 2004).
Socony Pegasus 7-20 CoreTh e Socony Pegasus 7-20 core in Midland County, Texas (Text-fi g. 6; Table 2) includes approximately 9 m of monoto-nous gray slightly argillaceous carbonate mudstone to ostra-
Barrick ET AL.: Conodont Faunas and Stable Isotopes
Text-fi g. 5. Distribution and abundance of conodont elements and stable isotope values across the Mulde Event level in the Dougherty West section, southern Okahoma. Sample intervals and tabulations of conodont elements are given in Table 1. Sample intervals and values of stable isotopes are given in Table 4. BS, Belodella silurica; DF, Decoriconus fragilis; DO, Dapsilodus obliquicostatus; DP, Dapsilodus praecipuus; DS, Dapsilodus sparsus; KA, Kockelella ortus absidata; OB, Ozarkodina bohemica longa; OE, Ozarkodina excavata; PE, Panderodus equicostatus; PG, P. greenlandensis?; PR, P. recurvatus; PU, P. unicostatus; PsB, Pseudooneotodus bicornis; PsL, Pseudooneotodus linguicornis; W, Walliserodus sp.

48 Palaeontographica Americana, No.
code wackestone that is assigned to the Wink Formation. Th e Wink becomes slightly more siliceous and argillaceous near the top, but otherwise the only signifi cant features are stylo-lites and thin clay seams distorted by stylolites. Barrick et al. (1993) described the lithological succession and conodont biostratigraphy of the core. Th e Mulde Event level coincides with one of these clay/styolitic seams at ca. 3,635.35 m (sub-sea depth). Sampling of the core for conodonts and isotopes was constrained by the availability of core segments, but a 1.5 m interval across the Mulde Event interval was complete.
Conodont Faunas.–Conodont abundance in the Pegasus core ranges from several hundred to a few thousand elements per kilogram (Text-fi g. 6; Table 2). Pseudooneotodus linguicornis appears at 3,636.20 m, which indicates the sagitta sagitta Zone. Th e sagitta sagitta Zone conodont fauna is domi-nated by elements of Dapsilodus praecipuus (ca. 40%) and elements of D. obliquicostatus, Panderodus unicostatus, and Pseudooneotodus bicornis are common. Decoriconus fragilis, P. recurvatus, and Walliserodus sp. occur in moderate numbers,
and Ozarkodina excavata and P. linguicornis are uncommon. Th e LAD of Walliserodus sp. occurs at 3,635.50 m, and the LADs of D. praecipuus, P. unicostatus, P. recurvatus, and P. lin-guicornis occur at 3,635.40 m. Kockelella ortus absidata, which marks the end of the Mulde Event, appears at 3,635.25 m, just above the extinction horizon. Panderodus equicostatus ap-pears at the same level, and Belodella silurica in the next sam-ple. Ozarkodina bohemica longa appears higher at 3,634.85 m. Th e post-extinction conodont fauna is strongly dominated by D. obliquicostatus. Pseudooneotodus bicornis and D. fragilis are the only other species common just above the extinction level, but in the next sample up, P. equicostatus becomes com-mon. By the level at 3,634.85 m, Ozarkodina excavata and B. silurica are more abundant.
Stable Isotopes.–Th e values of δ13C decrease slightly from +1.4‰ near the base of the sagitta sagitta Zone to +1.0‰ just below the extinction level (Table 4). Just above the clay seam that lies at the extinction level, δ13C jumps to +3.7‰ and falls to +2.0‰ over just 0.30 m. Th rough the next 0.5 m,
Text-fi g. 6. Distribution and abundance of conodont elements and stable isotope values across the Mulde Event level in the Socony Mobil Pegaus Unit 3 #7-20 core, Midland County, Texas. Sample intervals and tabulations of conodont elements are given in Table 2. Sample intervals and values of stable isotopes are given in Table 4. See Text-fi g. 5 for explanation of symbols.
49
δ13C remains at ca. +2.1‰. Th e values of δ18O parallel those of δ13C. Th ey display an abrupt shift down to -6.2‰ below and then up to -3.0‰ just above the extinction level before declining to values between -5.0‰ and -4.0‰.
COBRA FRAZIER # 1 CORE
Th e Cobra Frazier #1 core in Howard County, Texas (Text-fi g. 7), includes 18.5 m of carbonate strata assigned to the Wink Formation. Th e lower 4.20 m (3,080.20-3,076.00 m, subsea depth) comprise light to dark gray, slightly argillaceous skel-etal dolostone and calcitic dolostone (Kohl, 2004). Dark gray calcitic dolostone and dolomitic limestone extend for another 6.50 m, up to 3,069.50 m, and dark gray argillaceous limestone dominates to the top of the core at 3,062.00 m. Th e Wink in the Cobra Frasier core bears numerous karst fractures that are fi lled with sediments that contain Late Devonian conodonts (Meyer & Barrick, 2000). Conodont samples often yielded a mixture of Silurian and Devonian forms. Extreme care was taken in sampling for isotopes to avoid karst-aff ected areas. Th e Mulde Event level lies at 3,073.20 m within a thin (10 cm) interval of deformed clayey seams based on the level of conodont extinctions.
Conodont Faunas.–Pseudooneotodus linguicornis appears at 3,075.00 m in the Cobra Frazier core and is used to in-
dicate the base of the sagitta sagitta Zone. Conodont abun-dance is moderate in the sagitta sagitta Zone; usually just 50-100 elements/kg (Text-fi g. 7; Table 3). Th e lower part of the sagitta sagitta Zone is dominated by elements of Dapsi-lodus praecipuus and Panderodus unicostatus. Higher in the zone, D. obliquicostatus dominates briefl y, but P. unicosta-tus becomes more abundant just before the extinction level. Other species are relatively uncommon to rare: Walliserodus sp., P. linguicornis, Ps. bicornus, Decoriconus fragilis, Ozarko-dina excavata, and P. recurvatus. Th e LADs of P. unicostatus, D. praecipuus, and Walliserodus sp. at 3,073.20 m defi ne the
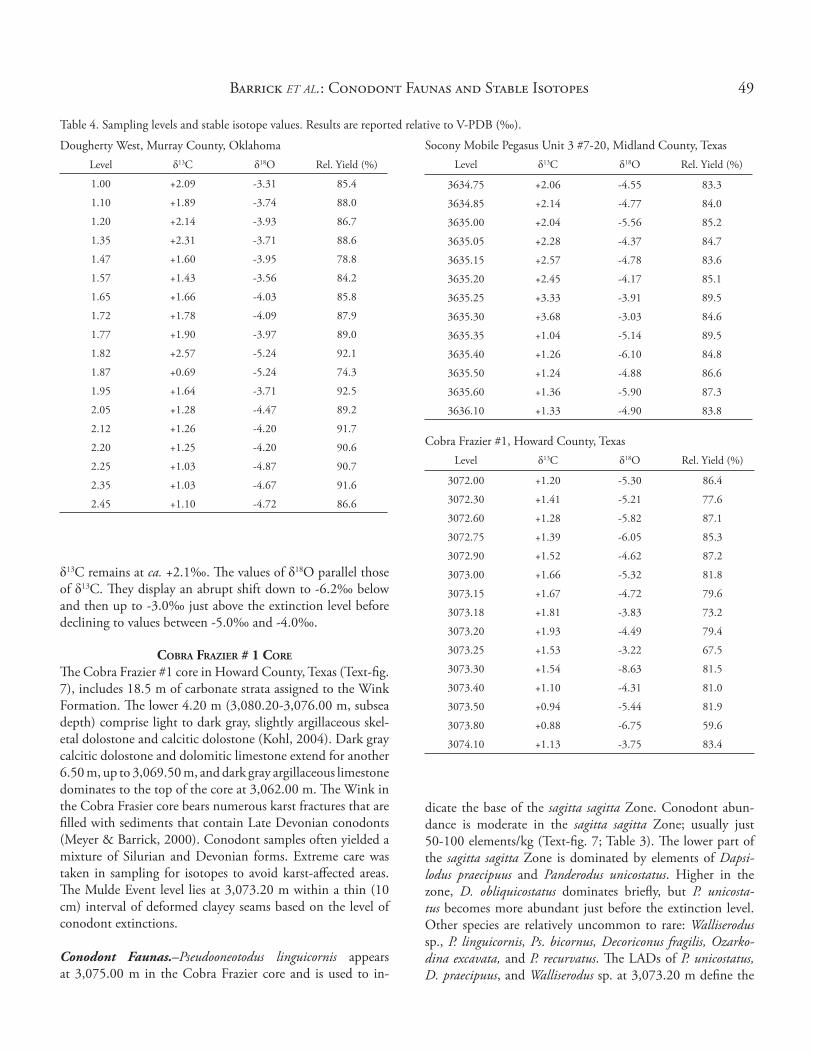
Dougherty West, Murray County, OklahomaLevel δ13C δ18O Rel. Yield (%)
1.00 +2.09 -3.31 85.4
1.10 +1.89 -3.74 88.0
1.20 +2.14 -3.93 86.7
1.35 +2.31 -3.71 88.6
1.47 +1.60 -3.95 78.8
1.57 +1.43 -3.56 84.2
1.65 +1.66 -4.03 85.8
1.72 +1.78 -4.09 87.9
1.77 +1.90 -3.97 89.0
1.82 +2.57 -5.24 92.1
1.87 +0.69 -5.24 74.3
1.95 +1.64 -3.71 92.5
2.05 +1.28 -4.47 89.2
2.12 +1.26 -4.20 91.7
2.20 +1.25 -4.20 90.6
2.25 +1.03 -4.87 90.7
2.35 +1.03 -4.67 91.6
2.45 +1.10 -4.72 86.6
Barrick ET AL.: Conodont Faunas and Stable Isotopes
Table 4. Sampling levels and stable isotope values. Results are reported relative to V-PDB (‰).
Socony Mobile Pegasus Unit 3 #7-20, Midland County, TexasLevel δ13C δ18O Rel. Yield (%)
3634.75 +2.06 -4.55 83.3
3634.85 +2.14 -4.77 84.0
3635.00 +2.04 -5.56 85.2
3635.05 +2.28 -4.37 84.7
3635.15 +2.57 -4.78 83.6
3635.20 +2.45 -4.17 85.1
3635.25 +3.33 -3.91 89.5
3635.30 +3.68 -3.03 84.6
3635.35 +1.04 -5.14 89.5
3635.40 +1.26 -6.10 84.8
3635.50 +1.24 -4.88 86.6
3635.60 +1.36 -5.90 87.3
3636.10 +1.33 -4.90 83.8
Cobra Frazier #1, Howard County, TexasLevel δ13C δ18O Rel. Yield (%)
3072.00 +1.20 -5.30 86.4
3072.30 +1.41 -5.21 77.6
3072.60 +1.28 -5.82 87.1
3072.75 +1.39 -6.05 85.3
3072.90 +1.52 -4.62 87.2
3073.00 +1.66 -5.32 81.8
3073.15 +1.67 -4.72 79.6
3073.18 +1.81 -3.83 73.2
3073.20 +1.93 -4.49 79.4
3073.25 +1.53 -3.22 67.5
3073.30 +1.54 -8.63 81.5
3073.40 +1.10 -4.31 81.0
3073.50 +0.94 -5.44 81.9
3073.80 +0.88 -6.75 59.6
3074.10 +1.13 -3.75 83.4

50 Palaeontographica Americana, No.
extinction level, but P. linguicornis disappears well below this level. Th e sample immediately above the extinction horizon contains only elements of D. obliquicostatus, D. fragilis, and Ps. bicornis. Slightly higher, Panderodus equicostatus appears and increases in abundance, Belodella silurica appears, and D. obliquicostatus rises to dominance. Ozarkodina bohemica longa and Kockelella ortus absidata appear well above the extinction level at 3,072.70 m. One element each of D. praecipuus and Dapsilodus sparsus was recovered well above the extinction lev-el. Th eir presence might be true range extensions, but could also be a product of reworking or sample contamination.
Stable Isotopes.–Th e values of δ13C through most of the sag-itta sagitta Zone vary slightly ca. +1.0‰, then increase to just above +1.5‰ just below the extinction level (Table 4). δ13C increases sharply to +1.9‰ just above the extinction level, drops to 1.7‰, and then declines gradually to ca. 1.2‰. Val-ues of δ18O fl uctuate more erratically, with a pronounced fall to -8.3‰ and sharp rise to -3.5 ‰ just below the extinction level.
DISCUSSION
Th e DW section in southern Oklahoma and the two sections from the Pegasus and Cobra-Frazier cores in west Texas pro-vide three somewhat diff erent views of conodont faunas and stable isotopes across the Mulde Event in southwestern Lau-rentia. Th e three sections were deposited in approximately the same off shore depositional setting, on the distal portion of a carbonate ramp, and all are characterized by slightly argil-laceous carbonate mudstones to arthropod wackestones. Th e lithological succession remains unchanged through the Mulde Event interval in each section. At the DW section the extinc-tion level lies at a bedding plane, in the Pegasus core it lies at a clay seam/stylolite, and in the Cobra Frazier core the ex-tinction level corresponds to a thin interval of deformed clay seams. None of these features is so obvious that they would have been identifi ed as the Mulde Event level without faunal and isotopic data.
Th e Mulde Event interval is best identifi ed in the three sections using the extinction level in the conodont faunas. Th e fi nal extinctions of Dapsilodus praecipuus, D. sparsus, and
Text-fi g. 7. Distribution and abundance of conodont elements and stable isotope values across the Mulde Event level in the Cobra Frazier #1 core, Howard County, Texas. Sample intervals and tabulations of conodont elements are given in Table 3. Sample intervals and values of stable isotopes are given in Table 4. See Text-fi g. 5 for explanation of symbols.

51
Pseudooneotodus linguicornis and the temporary disappearance of Panderodus unicostatus, P. recurvatus, and Walliserodus sp. occur at the same level or over a very short stratigraphic inter-val. Th e appearance of Panderodus equicostatus and Belodella silurica, the fi rst of which ranges into older strata, are the most obvious features of the post-event fauna. Th e sample immedi-ately above the extinction level generally has low diversity and low abundance of the same species, both of which increase upward. Th e occurrence of Ozarkodina bohemica longa and Kockelella ortus absidata above the extinction level provides confi rmation that the Mulde Event interval lies below.
Th e composition of conodont biofacies diff ers consider-ably from section to section both above and below the ex-tinction level. At DW, in southern Oklahoma, Dapsilodus obliquicostatus, which strongly dominates all samples, and the common forms Decoriconus fragilis, Pseudooneotodus bicornis, and Ozarkodina excavata persist through the Mulde Event in-terval with little change in abundance. Th ese species comprise over 90% of the elements recovered. Th e pre-event fauna in-cludes relatively abundant Dapsilodus praecipuus, Panderodus unicostatus, and Walliserodus sp., which are replaced by rela-tively abundant Panderodus equicostatus and Belodella silurica in post-event strata. In contrast, pre-event strata in the Pegasus core are characterized by near equal numbers of D. praecipuus and D. obliquicostatus, relatively abundant P. unicostatus and Ps. bicornis, and few O. excavata. Post-event strata are strongly dominated by D. obliquicostatus, Decoriconus fragilis, Ps. bi-cornis, and O. excavata are more abundant than below, and P. equicostatus is common. In the lower abundance faunas of the Cobra Frazier core, pre-event strata are dominated by P. unicostatus, D. obliquicostatus, and D. praecipuus up to just before the event, where D. obliquicostatus and P. unicostatus dominate. Other taxa are uncommon before the event, but after the event, faunas are dominated by D. obliquicostatus. D. fragilis, Ps. bicornis, and P. equicostatus are common. No generalized pattern of biofacies shifts in the conodont fau-nas before the Mulde Event interval can be resolved. Th e fi rst sample above the extinction level appears to be less diverse and abundant than those at higher levels, but no other gen-eralization can be made. Th e observed changes in conodont biofacies, especially relative abundance of species, appears to be more a response to local environmental conditions than to global oceanic eff ects.
Th e patterns of values of δ13C through the Mulde Event interval allow recognition of the event, although not as reliably as by the conodont faunas. Each section displays a somewhat diff erent pattern of values. Pre-event values of δ13C are ca. 1.0 ± 0.5‰ at each section and increase slightly up section. Just below the extinction event, δ13C increases slightly (0.5‰) in the Cobra Frazier core. But in the DW section, δ13C drops 1‰ before rising to +2.6‰, and in the Pegasus core only a
slight decrease of 0.5‰ occurs. Th e position and maximum value of δ13C also varies. At DW, the peak of +2.6‰ lies at the top of the bed just below the extinction level, but in the Pegasus core, the peak of +3.5‰ lies at the base of the bed overlying the extinction level. In the Cobra Frazier core, the small peak of +2.0‰ also lies just above the extinction hori-zon. In all three sections, the values of δ13C are slightly higher, ca. 0.5-1.0‰ higher, in strata above the Mulde Event interval than in the beds below.
COMPARISON WITH OTHER MULDE EVENT SUCCESSIONS
Calner & Jeppsson (2003) and Jeppsson & Calner (2003) provided details of sequence stratigraphy, facies patterns, and conodont faunas through the Mulde Event interval on the Silurian carbonate platform of Gotland. Th ese authors recognized three conodont datum levels through the fi rst 60-65 thousand years of the 350-380 thousand years of the Mulde Event (Text-fi g. 3). Datum 1 lies at the extinction of Ozarkodina sagitta sagitta and strongly reduced abundance of Pseudooneotodus linguicornis; this is the top of the sagitta sagit-ta Zone and the base of the O. bohemica longa Zone, as well as the top of the Cyrtograptus lundgreni Carruthers, 1867, Zone in the graptolite succession (Jeppsson et al., 2006). Th is level is interpreted to coincide with the extinction of most grapto-lite lineages. Th e O. bohemica longa Zone is divided into fi ve subzones, based largely on relative abundances of ramiform and coniform elements. Th e basal Subzone 0 represents an in-terregnum because neither O. sagitta sagitta nor O. bohemica longa is present. Walliserodus, Panderodus panderi (Stauff er, 1940) (= P. recurvatus herein), and P. linguicornis persist from the sagitta sagitta Zone through this subzone, up to Datum 1.5. Datum 1.5 represents the local extinction of Walliserodus. Subzone 1, like Subzone 0, is strongly dominated by ramiform elements, mostly those of O. excavata. Datum 2 lies at the base of Subzone 2 and at the top of the lundgreni Zone, at the moment in time where all graptoloids of the lundgreni Zone, except Gothograptus nassa (Holm, 1890) and Pristiograptus dubius (Suess, 1851), became extinct. Th e conodont fauna of Subzone 2 is dominated by coniforms, especially Panderodus equicostatus. In Subzone 3, ramiform elements return to domi-nance in conodont faunas, and in Subzone 4, a more balanced conodont fauna is present. Not until the Kockelella ortus absi-data Zone and Colonograptus praedeubeli (Jaeger, 1990) Zone, near the end of the Mulde Event, does conodont diversity return to near typical levels.
Th e pattern of conodont faunas across the Mulde Event in-terval in southwestern Laurentia makes only a general match with the Gotland conodont faunas. Th e extinction event in southwestern Laurentia is like that on Gotland. However, it is confi ned to a single horizon, for all species aff ected disap-
Barrick ET AL.: Conodont Faunas and Stable Isotopes

52 Palaeontographica Americana, No.
pear at the same level or in random order in lower abundance samples. Datum 1 cannot be distinguished from Datum 1.5. Th e appearance of Kockelella ortus absidata immediately above the extinction level at DW (10 cm), in the Pegasus core (5-10 cm) or a sample or two higher in the Cobra Frazier core (50 cm) suggests that strata of the Mulde Event are partially missing in this region. Th e low diversity interval just above the extinction level in southwestern Laurentia could be inter-preted as a “recovery” fauna, but one unlike what is seen on Gotland. Even so, the variation in conodont biofacies among the three sections in Oklahoma and Texas is as great or greater than that used by Calner & Jeppsson (2003) to designate the series of subzones for the Ozarkodina bohemica longa Zone. Th ese Baltic subzones could represent only the local faunal changes after the extinction event and might not be correlat-able outside of the Baltic region.
Calner & Jeppsson (2003) and Calner et al. (2006) showed that the Mulde Event on Gotland corresponds to the top of a highstand systems tract, a fi fth-order sequence boundary be-tween a forced regressive systems tract and a transgressive sys-tems tract (Bara Oolite Member, Halla Formation), and the succeeding highstand systems tract. Th e strong positive δ 13C excursion in the Gotland section possesses a major peak that lies within the Ozarkodina bohemica longa Zone, just below Datum 2 (Calner & Jeppsson, 2003: fi g. 13) in the regres-sive systems tract. A δ 13C low point lies near the base of the overlying Kockelella ortus absidata Zone, which corresponds to a sequence boundary and the low point in sea level. A sec-ond peak is restricted to the Ctenognathodus murchisoni Zone (Calner et al., 2005). In the graptolite succession (Text-fi g. 3), the lower peak lies near the FAD of Gothograptus nassa, the low point near the boundary of the Pristograptus dubius-G. nassa and Colonograptus praedeubeli zones. Th e second peak lies within the C. praedeubeli Zone (Porebska, et al., 2004; Cramer et al., 2006a: fi g. 5; Kaljo & Martma, 2006; Lenz et al., 2006).
Cramer et al. (2006a) described the positive carbon iso-tope excursion associated with the Mulde Event interval in central Tennessee, ca. 1,000 km east of the Oklahoma study area. In the McCrory Lane section near Nashville, the excur-sion occurs over an interval of ca. 4 m, starting in the upper strata of the Maddox Member of the Wayne Formation, peak-ing in the Waldron Member, and ending in the lower Lego Member. Th e top of the Maddox Member constitutes the up-per part of the highstand systems tract at the top of the lower Wenlock sequence in southern Laurentia and shows lithologi-cal and biofacies evidence of shallowing (Barrick, 1983). Th e Waldron Shale lies at the base of the upper Wenlock sequence and might represent the transgressive system tract of that se-quence and the overlying Lego Member is the highstand sys-tems tract of the upper Wenlock sequence. Th e values of δ13C
start near +1‰ in the upper Maddox Member and rise gently though the upper Maddox before dipping slightly. Above the Maddox, values increase sharply to attain a peak of +2.8‰ in the lower shale of the Waldron Member. Th is fi rst peak occurs in strata of the transgressive system tract, unlike in the Baltic region (Calner et al., 2006). A low point in values (+1.5‰) lies in the middle limestone of the Waldron followed by a sec-ond peak of +2.4‰ in the upper shale of the Waldron. In the base of the overlying Lego Member, a steady decline occurs in δ13C to a baseline level of ca. +1.0‰. Little biostratigraph-ic information is available for the McCrory Lane section. Pseudooneotodus linguicornis occurs just below the Waldron in western Tennessee, but has not been recovered from central Tennessee. Th e Lego sections in western Tennessee have yield-ed few examples of Ozarkodina bohemica longa and Kockelella ortus absidata, the former appearing ca. 2 m above the base of the Lego and latter appearing 6 m or more above the base of the Lego (Barrick, 1983).
Th e double-peak isotopic excursion reported for the Mulde Event interval cannot be recognized in the three Oklahoma-Texas sections, probably because the critical strata are missing. As can be demonstrated in the Tennessee sections, the Mulde Event interval in southern Laurentia coincides with a sequence boundary that extends through the Oklahoma and West Texas sections (Barrick, 1997). Th e sharp increase in δ13C before the extinction level at DW could represent the fi rst rise in δ13C that occurs in Gotland in the regressive system tract (Calner et al., 2006), as does the more gradual increase in δ13C in the Cobra Frazier core. If so, the top of the highstand system tracts in these two sections might preserve strata leading up to the fi rst stages of the Mulde Event. In contrast, the absence of an increase in δ13C in the Pegasus core below the extinction level suggests that some strata are missing from the top of the lower Wenlock highstand systems tract. Th e major peak above the extinction level in the Pegasus core could correspond to the descending limb of the second, younger Mulde peak that lies within the transgressive system tract in the Baltic (Calner et al., 2006). If so, then one can estimate the magnitude of the hiatus that lies in the Oklahoma-west Texas sections stud-ied here. Th e hiatus should correspond in time to the dura-tion of the Mulde Event, the interval from approximately the top of the Ozarkodina sagitta sagitta Zone (lundgreni Zone) to within the Ctenognathodus murchisoni Zone (upper praedeu-beli Zone), which Calner & Jeppsson (2003) estimated to be ca. 350-380 thousand years.
DISCUSSIONFrom comparisons of the succession of conodont faunas and the stable isotope record, it is clear that only a fragmentary record of the Mulde Event interval is preserved in southwest-ern Laurentia because it lies at an unconformity associated

53
with a sequence boundary. Based on the shelf succession on Gotland, Calner & Jeppsson (2003) indicated that only a brief sea-level event occurred (fi fth-order sequence bound-ary). Johnson (2006) calibrated the short-lived fall/drop in sea level associated with the Mulde Event to be only ca. 16 m based on data from Gotland (Calner & Säll, 1999). Th is is a smaller eustatic event than one might expect to produce the hiatus observed in carbonate sections interpreted to have been deposited in a distal ramp setting. Resolution of this problem will require additional stratigraphic, sedimentologic, and pe-trographic studies of the units bounding the unconformity in southern Laurentia (Text-fi gs 2, 4).
Cramer et al. (2006a) and other authors suggest that short-lived isotope excursions can allow precise correlation of Silurian strata across large regions. In this study, however, the Mulde Event interval was discovered only because the au-thors used the change in conodont faunas to narrow down the interval of strata that might preserve the isotope excursion. Th e coincidence of the Mulde Event with a cryptic regional unconformity reduced the record of the isotope excursion, all traces of which would have been removed with a larger hiatus. Saltzman (2001) sampled the Clarita for stable isotopes at the Highway 77 section, but did not detect the Mulde isotope excursion for this very reason.
As Holland & Patzkowsky (2007) pointed out, many outcrop-based studies have recognized that episodes of eco-logical change frequently occur at sequence boundaries and major hiatal surfaces. Th ey stated that these surfaces are where stratigraphic artifacts that could produce the appear-ance of an abrupt faunal turnover are most likely to occur (Holland, 1995, 1996, 2000). Determining whether abrupt faunal turnover that occurs at a sequence boundary, such as the Mulde extinction event, is wholly or partly an artifact of stratigraphic architecture, has proven to be diffi cult. Th e in-complete conodont and stable isotope record of the Mulde Event in the west Texas and Oklahoma sections is clearly an artifact of the stratigraphic architecture associated with an eu-static sea-level event. Barrick & Männik (2005) questioned how much of the Mulde Event interval on Gotland could be interpreted as an artifact of stratigraphic sequences as opposed to an extinction event in conodonts. Jeppsson et al. (2006: 110) acknowledged that several Silurian oceanic events are re-lated to a “very brief sea-level fall,” but were optimistic about reconstructing a complete stratigraphic record from off shore distal and stratigraphically continuous sections. Th e chance of fi nding sections that are depositionally continuous across a se-quence boundary is extremely low because unconformities are predicted to occur in both shelf and off shore sections for Type 1 stratigraphic sequences (see Emery & Myers, 1996: fi g. 5.2 for a chronostratigraphic representation). Calner et al. (2006) reconstructed a series of sections across the Baltic region that
show a regressive system tract on Gotland and correlative con-formities off shore in the graptolite facies. Regressive system tracts are the result of special conditions, rapid subsidence combined with high sediment supply, and this situation could apply to the sections on Gotland, but few other places. Even in a situation where depositional continuity exists, the rapid shifts in facies and environments will create artifacts in faunal distributions (Holland, 1996, 2000).
Kaljo et al. (2003) and Johnson (2006) considered the sig-nifi cance of eustatic events in interpreting Silurian episodes and events. Kaljo et al. (2003) proposed that Silurian glacial events were the most likely mechanisms for isotope excursions during the Llandovery and Wenlock, because the distribution of stable isotope excursions were a better match to glacial-eus-tatic record than to the pattern of oceanic episodes and events. Johnson (2006) expressed concern over the mismatch of sea-level events predicted by the oceanic episode and event model of Jeppsson (1998: fi gs 1-2) with the sea-level curves gener-ated by other means. Johnson (2006: 119, fi g. 1) noted that many (7 of 8) of the oceanic events agree with low stands of sea level, including the Mulde Event, and suggested that gla-cio-eustacy could have been more than suffi cient to generate climate change that altered large area of the global ocean. We share the concerns of these authors and suggest that as more detailed models of Silurian oceanic history are constructed based on climatic models (e. g., Cramer et al., 2006b), that greater attention be paid to the potential of glacio-eustacy as a primary cause rather than a secondary eff ect. As Kaljo et al. (2003) pointed out, a huge variety of physical and biological mechanisms could have generated the isotopic excursions and all these possibilities should be investigated.
SUMMARYTh e Mulde Event interval is poorly represented in distal car-bonate ramp settings in southwestern Laurentia because it coincides with an unconformity at a sequence boundary. Th e unconformity is diffi cult to recognize in outcrop or in cores because it lies at a bedding plane or as a thin, shaly interval in monotonous, slightly argillaceous, carbonate mudstone and arthropod wackestone. Detailed conodont collections were used to locate the unconformity, which separates a lower pre-Mulde, sagitta sagitta Zone conodont fauna (Dapsilodus praecipuus, D. sparsus, Pseudooneotodus linguicornis, and Pan-derodus unicostatus) from a post-Mulde, Ozarkodina bohem-ica longa to Kockelella ortus absidata zone fauna (Panderodus equicostatus and Belodella silurica). Th e stable isotope excur-sion of the Mulde Event is preserved only as incomplete posi-tive δ13C peaks just below or just above the unconformity sur-face that was discovered only with detailed sampling of cono-dont faunas. Strata equivalent to most of the Mulde Event interval were either removed or not deposited in southwestern
Barrick ET AL.: Conodont Faunas and Stable Isotopes

54 Palaeontographica Americana, No.
Laurentia during the brief fall in eustatic sea level that accom-panied the Mulde Event. Th e faunal and stable isotope record of the Mulde Event in southwestern Laurentia is largely an ar-tifact of sequence stratigraphic architecture, which should also aff ect the preservation of the Mulde Event in most shelf and cratonic settings and lessen the ability for it to be correlated using stable isotopes alone.
ACKNOWLEDGMENTSTh is material is based upon work supported by the National Science Foundation under grant no. NSF EAR-0517976 to J. E. Barrick and M. A. Kleff ner. James Browning assisted with many aspects of sample preparation and isotope analyses at Texas Tech. Graham Butler (Texas Tech) assisted with the fi eld work in Oklahoma. S. M. Bergström, C. Bright, D. J. Over, and A. Simpson provided careful and valuable reviews of the manuscript.
LITERATURE CITEDAmdsen, T. W., D. F. Toomey, & J. E. Barrick. 1980. Paleoenvironment
of Fitzhugh Member of Clarita Formation (Silurian, Wenlockian), southern Oklahoma. Oklahoma Geological Survey Circular 83, University of Oklahoma, Norman, 54 pp.
Barrick, J. E. 1977. Multielement simple-cone conodonts from the Clarita Formation (Silurian), Arbuckle Mountains, Oklahoma. Geologica et Palaeontologica, 11: 47-68.
Barrick, J. E. 1983. Wenlockian (Silurian) conodont biostratigra-phy, biofacies and carbonate lithofacies, Wayne Formation, cen-tral Tennessee. Journal of Paleontology, 57: 208-239.
Barrick, J. E. 1995. Biostratigraphy of uppermost Ordovician through Devonian depositional sequences in the Permian Basin, west Texas and southeastern New Mexico. Pp 207-216, in: Carbonate Facies and Sequence Stratigraphy: Practical Application of Carbonate Models, P. H. Pausé & M. P. Candelaria, eds. Permian Basin Section-SEPM Publication 95-36, Permain Basin Section SEPM, Midland, Texas.
Barrick, J. E. 1997. Wenlock (Silurian) depositional sequences, eustatic events, and biotic change on the southern shelf of North America. Pp 47-65, in: Paleozoic Sequence Stratigraphy, Biostratigraphy, and Biogeography: Studies in Honor of J. Granville (“Jess”) Johnson, G. Klapper, M. A. Murphy, & J. A. Talent, eds. Geological Society of America Special Paper 321, Geological Society of America, Boulder, Colorado.
Barrick, J. E., S. C. Finney, & J. N. Haywa-Branch. 1993. Revision of ages of the Fusselman, Wristen, and Th irtyone Formations (Late Ordovician-Early Devonian) in the subsurface of West Texas based on conodonts and graptolites. Texas Journal of Science, 45: 231-247.
Barrick, J. E., & G. Klapper. 1976. Multielement Silurian (late Llandoverian-Wenlockian) conodonts of the Clarita Formation, Arbuckle Mountains, Oklahoma, and phylogeny of Kockelella. Geologica et Palaeontologica, 10: 59-100.
Barrick, J. E., G. Klapper, & T. W. Amsden. 1990. Late Ordovician-Early Devonian conodont succession in the Hunton Group, Arbuckle Mountains and Anadarko Basin, Oklahoma. Pp 55-92,
in: Early to Middle Paleozoic Conodont Biostratigraphy, S. M. Ritter, ed. Oklahoma Geological Survey Guidebook 27, University of Oklahoma, Norman.
Barrick, J. E., & P. Männik. 2005. Silurian conodont biostratigra-phy and palaeobiology in stratigraphic sequences. Pp 103-116, in: Conodont Biology and Phylogeny: Interpreting the Fossil Record, M. A. Purnell & P. C. J. Donoghue, eds. Th e Palaeontological Association Special Papers in Palaeontology 73, Th e Palaeontological Association, London.
Barrick, J. E., B. D. Meyer, & S. C. Ruppel. 2004. Th e Silurian-Devonian boundary and the Klonk event in the Frame Formation, subsurface West Texas. Pp 105-122, in: A Standing Ovation: Papers in Honor of Gilbert Klapper, J. E. Barrick & H. R. Lane, eds. Bulletins of American Paleontology 369.
Berry, W. B. N., & A. J. Boucot. 1970. Correlation of the North American Silurian Rocks. Geological Society of America Special Paper 102, Geological Society of America, Boulder, Colorado, 289 pp.
Calner, M., & L. Jeppsson. 2003. Carbonate platform evolution and conodont biostratigraphy during the middle Silurian Mulde Event, Gotland, Sweden. Geological Magazine, 140: 173-203.
Calner, M., L. Jeppsson, & M. E. Ericksson. 2005. Th e Baltic ba-sin and the Silurian strata of Gotland, Sweden. Pp 6-20, in: Th e Dynamic Silurian Earth, M. E. Eriksson & M. Calner, eds. Subcommission on Silurian Stratigraphy Field Meeting 2005. Sveriges Geologiska Undersökning, Rapporter och Meddelanden, 121.
Calner, M., A. Kozlowska, M. Masiak, & B. Schmitz. 2006. A shoreline to deep basin correlation chart for the middle Silurian coupled extinction-stable isotope event. GFF, 128: 79-84.
Calner, M., & E. Säll. 1999. Transgressive oolites onlapping a Silurian rocky shoreline unconformity. GFF, 121: 91-100.
Cocks, L. R. M., & C. R. Scotese. 1991. Th e global paleogeography of the Silurian Period. Th e Palaeontological Association Special Papers in Paleontology, 44: 109-122.
Cocks, L. R. M., & T. H. Torsvik. 2002. Earth geography from 500 to 400 million years ago: a faunal and paleomagnetic review. Journal of the Geological Society, London, 159: 631-644.
Craig, H. 1957. Isotopic standards for carbon and oxygen and cor-rection factors for mass-spectrometric analysis of carbon diox-ide. Geochimica et Cosmochimica Acta, 12: 133-149.
Cramer, B. D., M. A. Kleff ner, & M. R. Saltzman. 2006a. Th e Late Wenlock Mulde positive carbon isotope (δ13Ccarb) excursion in North America. GFF, 128: 85-90.
Cramer, B. D., M. R. Saltzman, & M. A. Kleff ner. 2006b. Spatial and temporal variability in organic carbon burial during global positive carbon isotope excursions: new insight from high-res-olution δ13Ccarb stratigraphy from the type area of the Niagaran (Silurian) Provincial Series. Stratigraphy, 2: 327-340.
Dalziel, I. W. D. 1997. Neoproterozoic geography and tectonics: review, hypothesis, environmental speculation. Geological Society of America Bulletin, 109: 16-42.
Emery, D., & K. J. Myers. 1996. Sequence Stratigraphy. Blackwell Scientifi c, Oxford, 297 pp.
Golonka, J. M., M. I. Ross, & C. R. Scotese. 1994. Phanerozoic paleogeographic and paleoclimatic modeling maps. Pp 1-47, in: Pangea: Global Environments and Resources, A. F. Embry, B.

55
Beauchamp, & D. J. Glass, eds. Canadian Society of Petroleum Geologists Memoir 17, Canadian Society of Petroleum Geologists, Calgary, Alberta.
Holland, S. M. 1995. Sequence stratigraphy, facies control, and their eff ects on the stratigraphic distribution of fossils. Pp 1-13, in: Sequence Stratigraphy and Depositional Response to Eustatic, Tectonic, and Climatic Forcing, B. U. Haq, ed. Kluwer Academic, Dordrecht.
Holland, S. M. 1996. Guidelines for interpreting the stratigraphic record of extinctions: distinguishing pattern from artifact. P. 174, in: Sixth North American Paleontological Convention, Abstracts of Papers, J. E. Repetski, ed. Th e Paleontological Society Special Publication 8, Th e Paleontological Society, Lawrence, Kansas.
Holland, S. M. 2000. Th e quality of the fossil record: a sequence stratigraphic approach. Pp 148-168, in: Deep Time, D. H. Erwin & S. L. Wing, eds. Th e Paleontological Society, Lawrence, Kansas.
Holland, S. M., & M. E. Patzkowsky. 2007. Gradient ecology of a biotic invasion: biofacies of the type Cincinnatian Series (Upper Ordovician), Cincinnati, Ohio region, USA. Palaios, 22: 392-407.
Jacobi, D. J. 2004. Stable Isotope Chemostratigraphy Across the Silurian-Devonian Boundary in Oklahoma and West Texas. Unpublished Masters thesis, Texas Tech University, Lubbock, 97 pp.
Jeppsson, L. 1990. An oceanic model for lithological and faunal changes tested on the Silurian record. Journal of the Geological Society, London, 147: 663-674.
Jeppsson, L. 1998. Silurian oceanic events: summary of general char-acteristics. Pp 239-257, in: Silurian Cycles: Linkages of Dynamic Stratigraphy with Atmospheric, Oceanic, and Tectonic Changes, E. Landing & M. E. Johnson, eds. New York State Museum Bul-letin 491, New York State Education Department, Albany, New York.
Jeppsson, L. 2005. Bias in the recovery and interpretation of mi-cropalaeontological data. Pp 57-71, in: Conodont Biology and Phylogeny: Interpreting the Fossil Record, M. A. Purnell & P. C. J. Donoghue, eds. Th e Palaeontological Association Special Papers in Palaeontology 73, Th e Palaeontological Association, London.
Jeppsson, L., R. J. Aldridge, & K. J. Dorning. 1995. Wenlock (Silurian) oceanic episodes and events. Journal of the Geological Society, London, 152: 487-498.
Jeppsson, L., & R. Anehus. 1995. A buff ered formic acid technique for conodont extraction. Journal of Paleontology, 69: 790-794.
Jeppsson, L., & M. Calner. 2003. Th e Silurian Mulde event and a scenario for secundo-secundo events. Transactions of the Royal Society of Edinburgh, Earth Sciences, 93: 135-154.
Jeppsson, L., M. E. Eriksson, & M. Calner. 2006. A latest Llandovery to latest Ludlow high-resolution biostratigraphy based on the Silurian of Gotland – a summary. GFF, 128: 109-114.
Johnson, M. E. 2006. Relationship of Silurian sea-level fl ucuations to oceanic episodes and events. GFF, 128: 115-121.
Kaljo, D., A. J. Boucot, R. M. Corfi eld, A. LeHéRisse, T. N. Koren, J. Kriz, P. Männik, T. Marss, V. Nestor, R. H. Shaver, D. Siveter, & V. Viira. 1995. Silurian bioevents. Pp 173-224, in: Global Events and Event Stratigraphy in the Phanerozoic: Results of International Interdisciplinary Cooperation in the IGCP Project
216 “Global Biological Events in Earth History,” O. H. Walliser, ed., Springer-Verlag, Berlin.
Kaljo, D., & T. Martma. 2006. Application of carbon isotope stra-tigraphy to dating the Baltic Silurian rocks. GFF, 128: 123-129.
Kaljo, D., T. Martma, P. Männik, & V. Viira. 2003. Implications of Gondwana glaciations in the Baltic late Ordovician and Silurian and a carbon isotopic test of environmental cyclicity. Bulletin de la Société Géologique de France, 174: 59-66.
Kohl, M. L. 2004. Integrated Stratigraphy and Petrology of the Silurian Wink Formation (Wenlock-Ludlow), Howard County, Texas. Unpublished Masters thesis, Texas Tech University, Lubbock, 118 pp.
Lenz, A. C., P. J. Noble, M. Masiak, S. R. Poulson, & A. Kozlowska. 2006. Th e lundgreni extinction event: integration of paleonto-logical and geochemical data from Arctic Canada. GFF, 128: 153-158.
Martin, R. E. 1999. Taphonomy: A Process Approach. Cambridge University Press, Cambridge, U. K., 508 pp.
McCrea, J. 1950. Th e isotope chemistry of carbonates and a pale-otemperature scale. Journal of Chemical Physics, 18: 849-857.
Melchin, M. J., T. N. Koren, & P. Storch. 1998. Global diversity and survivorship patterns of Silurian graptoloids. Pp 165-182, in: Silurian Cycles: Linkages of Dynamic Stratigraphy with Atmospheric and Oceanic, and Tectonic Changes -- James Hall Centennial Volume, E. Landing & M. E. Johnson, eds. New York State Museum Bulletin 491, New York State Education Department, Albany, New York.
Meyer, B. D., & J. E. Barrick. 2000. Conodonts from the Woodford Formation (Late Devonian) and adjacent units, subsurface West Texas and eastern New Mexico. Pp 229-237, in: Th e Permian Basin: Proving Ground for Tomorrow’s Technologies, W. D. DeMis, M. K. Nelis, & R. C. Trentham, eds. West Texas Geological Society Publication 00-19, West Texas Geological Society, Midland.
Moore, G. T., S. R. Jacobson, C. A. Ross, & D. N. Hayashida. 1994. A paleoclimate simulation of the Wenlockian (Late Early Silurian) world using a general circulation model with implications for early land plant paleoecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 110: 115-144.
Niocaill, C. B. M., B. A. van der Pluijm, & R. van der Voo. 1997. Ordovician paleogeography and the evolution of the Iapetus Ocean. Geology, 25: 159-162.
Porebska, E. A., A. Kozlowska-Dawidziuk, & M. Masiak. 2004. Th e lundgreni event in the Silurian of the East European platform, Poland. Palaeogeography, Palaeoclimatology, Palaeoecology, 213: 271-294.
Ruppel, S. C., & M. H. Holtz. 1994. Depositional and diage-netic facies patterns and reservoir development in Silurian and Devonian rocks in the Permian Basin. Th e University of Texas, Bureau of Economic Geology, Report of Investigations 216, Texas Bureau of Economic Geology, Austin, 89 pp.
Saltzman, M. R. 2001. Silurian δ13C stratigraphy: a view from North America. Geology, 29: 671-674.
Samtleben, C. A., A. Munnecke, T. Bickert, & J. Päzold. 1996. Th e Silurian of Gotland: facies interpretation based on stable iso-topes in brachiopod shells. Geologische Rundschau, 85: 278-292.
Scotese, C. R. 2002. PALEOMAP website. http://www.scotese.com,
Barrick ET AL.: Conodont Faunas and Stable Isotopes

56 Palaeontographica Americana, No.
last accessed July 2007.Wilde, P., W. B. N. Berry, & M. S. Quinby-Hunt. 1991. Silurian
oceanic and atmospheric circulation and chemistry. Pp 123-143, in: Th e Murchinson Symposium: Proceedings of an International
Conference on the Silurian System, M. G. Bassett et al., eds. Th e Palaeontological Association Special Papers in Palaeontology 44, Th e Palaeontological Association, London.




















