

E-SPONDER System: A new Communication Infrastructure for Future Emergency Networks.
Upload
khangminh22Category
view
0download
0
P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
ii
P1: IYP0521809592agg.xml CB777B/Carrington 0 521 80959 2 April 9, 2005 15:59
This page intentionally left blank

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
Animal Communication Networks
Most animal communication has evolved and now takes place in the context of a
communication network: several signallers and receivers within communication
range of each other. This idea follows naturally from the observation that many
signals travel further than the average spacing between animals. This is
self-evidently true for long-range signals, but at a high density the same is true
for short-range signals (e.g. begging calls of nestling birds). This book provides a
current summary of research on communication networks and appraises future
prospects. It combines information from studies of several taxonomic groups
(insects to people via fiddler crabs, fish, frogs, birds and mammals) and several
signalling modalities (visual, acoustic and chemical signals). It also specifically
addresses the many areas of interface between communication networks and
other disciplines (from the evolution of human charitable behaviour to the
psychophysics of signal perception, via social behaviour, physiology and
mathematical models).
P. K. McGregor was Head of the Department of Animal Behaviour at
Copenhagen University; he is now Reader in Applied Zoology at Cornwall
College, Newquay, UK. He is editor of the journal Bioacoustics and on the editorial
board of several other academic journals.
i

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
ii

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
AnimalCommunicationNetworks
Edited by
P. K. McGregorUniversity of Copenhagen andCornwall College, Newquay, UK
iii

Cambridge, New York, Melbourne, Madrid, Cape Town, Singapore, São Paulo
Cambridge University PressThe Edinburgh Building, Cambridge , UK
First published in print format
- ----
- ----
© Cambridge University Press 2005
2005
Information on this title: www.cambridg e.org /9780521823616
This book is in copyright. Subject to statutory exception and to the provision ofrelevant collective licensing agreements, no reproduction of any part may take placewithout the written permission of Cambridge University Press.
- ---
- ---
Cambridge University Press has no responsibility for the persistence or accuracy ofs for external or third-party internet websites referred to in this book, and does notguarantee that any content on such websites is, or will remain, accurate or appropriate.
Published in the United States of America by Cambridge University Press, New York
www.cambridge.org
hardback
eBook (EBL)
eBook (EBL)
hardback

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
Contents
List of contributors viii
Preface xiii
1 Introduction 1Peter K. McGregor
Part I Behaviours specific to communication networks
Introduction 9
2 Eavesdropping in communication networks 13Tom M. Peake
3 Public, private or anonymous? Facilitating and countering
eavesdropping 38Torben Dabelsteen
4 Performing in front of an audience: signallers and the social
environment 63Ricardo J. Matos & Ingo Schlupp
5 Fighting, mating and networking: pillars of poeciliid
sociality 84Ryan L. Earley & Lee Alan Dugatkin
6 The occurrence and function of victory displays within
communication networks 114John L. Bower
Part II The effects of particular contexts
Introduction 129
v

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
vi Contents
7 Enlightened decisions: female assessment and
communication networks 133Ken A. Otter & Laurene Ratcliffe
8 Predation and noise in communication networks of
neotropical katydids 152Alexander B. Lang, Ingeborg Teppner, Manfred Hartbauer &Heiner Römer
9 Nestling begging as a communication network 170Andrew G. Horn & Marty L. Leonard
10 Redirection of aggression: multiparty signalling within a
network? 191Anahita J. N. Kazem & Filippo Aureli
11 Scent marking and social communication 219Jane L. Hurst
Part III Communication networks in different taxa
Introduction 247
12 Waving in a crowd: fiddler crabs signal in networks 252Denise S. Pope
13 Anuran choruses as communication networks 277T. Ulmar Grafe
14 Singing interactions in songbirds: implications for social
relations and territorial settlement 300Marc Naguib
15 Dawn chorus as an interactive communication network
320John M. Burt & Sandra L. Vehrencamp
16 Eavesdropping and scent over-marking 344Robert E. Johnston
17 Vocal communication networks in large terrestrial
mammals 372Karen McComb & David Reby
18 Underwater acoustic communication networks in marine
mammals 390Vincent M. Janik
19 Looking for, looking at: social control, honest signals and
intimate experience in human evolution and history 416John L. Locke

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
Contents vii
Part IV Interfaces with other disciplines
Introduction 445
20 Perception and acoustic communication networks 451Ulrike Langemann & Georg M. Klump
21 Hormones, social context and animal communication 481Rui F. Oliveira
22 Cooperation in communication networks: indirect
reciprocity in interactions between cleaner fish and client
reef fish 521Redouan Bshary & Arun D’Souza
23 Fish semiochemicals and the evolution of communication
networks 540Brian D. Wisenden & Norman E. Stacey
24 Cognitive aspects of networks and avian capacities 568Irene M. Pepperberg
25 Social complexity and the information acquired during
eavesdropping by primates and other animals 583Dorothy L. Cheney & Robert M. Seyfarth
26 Communication networks in a virtual world 604Andrew M. R. Terry & Robert Lachlan
Index 628

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
Contributors
Filippo AureliSchool of Biological and Earth Sciences, Liverpool John Moores University, Byrom St,Liverpool L3 3AF, UK
John L. BowerFairhaven Office 348, Fairhaven College, Western Washington University,Bellingham, Washington 98225-9118, USA
Redouan BsharyDepartment of Zoology, University of Cambridge, Downing St, Cambridge CB2 3EJ,UK. Present address: Evolutionary Psychology and Behavioural Ecology ResearchGroup, School of Biological Sciences, Crown St, University of Liverpool, LiverpoolL69 7ZB, UK
John M. BurtCornell Laboratory of Ornithology, 159 Sapsucker Woods Rd, Ithaca, NY 14850, USA.Present address: Department of Psychology, Box 351525, University of Washington,Seattle, WA 98195-1525, USA
Dorothy L. CheneyDepartment of Biology, University of Pennsylvania, Philadelphia, PA 19104, USA
Torben DabelsteenDepartment of Animal Behaviour, Copenhagen University Zoological Institute,Tagensvej 16, DK-2200 Copenhagen N, Denmark
Arun D’SouzaDepartment of Animal Ecology and Tropical Biology, University of Wurzburg, 97074Wurzburg, Germany
Lee Alan DugatkinDepartment of Biology, University of Louisville, Louisville, KY 40292, USA
viii

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
List of contributors ix
Ryan L. EarleyDepartment of Biology, Georgia State University, 20 Peachtree Center Ave NE, 402Kell Hall, Atlanta GA 30303, USA
T. Ulmar GrafeDepartment of Animal Ecology and Tropical Biology, University of Wurzburg, 97074Wurzburg, Germany
Manfred HartbauerInstitute of Zoology, Karl-Franzens-University, Universitatsplatz 2, A-8010 Graz,Austria
Andrew G. HornDepartment of Biology, Life Science Centre, Dalhousie University, 1355 Oxford St,Halifax, Nova Scotia, Canada B3H 4J1
Jane L. HurstFaculty of Veterinary Science, University of Liverpool, Leahurst Veterinary FieldStation, Neston, South Wirral L64 7TE, UK
Vincent M. JanikCentre for Social Learning and Cognitive Evolution and the Sea Mammal ResearchUnit, Gatty Marine Laboratory, University of St Andrews, Fife KY16 8LB, UK
Robert E. JohnstonDepartment of Psychology, Uris Hall, Cornell University, Ithaca, NY 14853, USA
Anahita J. N. KazemSchool of Biological Sciences, University of Wales Bangor, Brambell Building, DeiniolRd, Bangor LL57 2UW, UK
Georg M. KlumpCarl von Ossietzky Universitat Oldenburg, AG Zoophysiologie and Verhalten, FB 7,26111 Oldenburg, Germany
Robert LachlanDepartment of Biology, Coker Hall, University of North Carolina at Chapel Hill, NorthCarolina 27599, USA
Alexander B. LangInstitute of Zoology, Karl-Franzens-University, Universitatsplatz 2, A-8010 Graz,Austria
Ulrike LangemannCarl von Ossietzky Universitat Oldenburg, AG Zoophysiologie and Verhalten, FB 7,26111 Oldenburg, Germany
Marty L. LeonardDepartment of Biology, Life Science Centre, Dalhousie University, 1355 Oxford St,Halifax, Nova Scotia, Canada B3H 4J1

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
x List of contributors
John L. LockeNew York University, 719 Broadway (Suite 200), New York, NY 10003, USA. Presentaddress: Department of Speech-Language-Hearing Sciences, Lehman College, CityUniversity of New York, 250 Bedford Park Boulevard West, Bronx, New York 10468,USA
Ricardo J. MatosDepartment of Animal Behaviour, Copenhagen University Zoological Institute,Tagensvej 16, DK-2200 Copenhagen N, Denmark
Karen E. McCombExperimental Psychology, School of Biological Sciences, University of Sussex, Falmer,Brighton BN1 9QG, UK
Peter K. McGregorDepartment of Animal Behaviour, Copenhagen University Zoological Institute,Tagensvej 16, DK-2200 Copenhagen N, Denmark. Present address: Centre for AppliedZoology, Cornwall College Newquay, Trenance Gardens, Newquay Cornwall TR7 2LZ,UK
Marc NaguibDepartment of Animal Behavior, University Bielefeld, PO Box 10 01 31, 33501Bielefeld, Germany
Rui F. OliveiraUnidade de Investigacao em Eco-Etologia, Instituto Superior de Psicologia Aplicada,Rua Jardim do Tabaco 34, 1149–041 Lisbon, Portugal
Ken A. OtterEcosystem Science and Management Program, University of Northern BritishColumbia, 3333 University Way, Prince George, British Columbia, Canada V2N 4Z9
Tom M. PeakeDepartment of Animal Behaviour, Copenhagen University Zoological Institute,Tagensvej 16, DK-2200 Copenhagen N, Denmark
Irene M. PepperbergMIT School of Architecture and Planning, Brandeis University and Department ofPsychology, Waltham, MA 02454, USA
Denise S. PopeDepartment of Animal Behaviour, Copenhagen University Zoological Institute,Tagensvej 16, DK-2200 Copenhagen N, Denmark. Present address: Department ofBiology, Trinity University, 1 Trinity Place, San Antonio, TX 78212-7200, USA
Laurene RatcliffeDepartment of Biology, Queen’s University, Kingston, Ontario, Canada K7L 3N6
David RebyExperimental Psychology, School of Biological Sciences, University of Sussex, Falmer,Brighton BN1 9QG, UK

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
List of contributors xi
Heiner RomerInstitute for Zoology, Karl-Franzens-University, Universitatsplatz 2, A-8010 Graz,Austria
Ingo SchluppZoologisches Institut, Universitat Zurich, Winterthurerstrasse 190, CH-8057 Zurich,Switzerland and Section of Integrative Biology C0930, University of Texas, Austin,TX 78712, USA. Present address: Biozentrum Grindel, Universitat Hamburg,Martin-Luther-King Pl. 3, D-20146 Hamburg, Germany
Robert M. SeyfarthDepartment of Psychology, University of Pennsylvania, Philadelphia, PA 19104-6196,USA
Norman E. StaceyDepartment of Biological Sciences, University of Alberta, Edmonton, Alberta, CanadaT6G 2E9
Ingeborg TeppnerInstitute of Zoology, Karl-Franzens-University, Universitatsplatz 2, A-8010 Graz,Austria
Andrew M. R. TerryDepartment of Animal Behaviour, Copenhagen University Zoological Institute,Tagensvej 16, DK-2200 Copenhagen N, Denmark. Present address: IUCN – The WorldConservation Union, Regional Office for Europe, Rue Vergote 15, 1030 Bruxelles,Belgium
Sandra L. VehrencampCornell Laboratory of Ornithology, 159 Sapsucker Woods Rd, Ithaca, NY 14850, USA
Brian D. WisendenDepartment of Biology, Minnesota State University Moorhead, 1104 7th St S,Moorhead, MN 56563, USA

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
xii

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
Preface
This book attempts to reflect the state of current research on communica-
tion networks: groupings of several individuals that constitute the social context
in which communication takes place. In my view, a structured collection of chap-
ters by active researchers best conveys the excitement of the research findings as
well as the underlying expertise of the authors, especially when a wide range of
taxa and signalling modalities are addressed.
The motivation to edit such a book came from the interest in the topic that
was evident after seminars and conference presentations. However, it was the
symposium on communication networks at the XXVIIth International Ethological
Conference held in Tubingen that converted motivation into action. The symposium
showed (at least to my satisfaction) how well the topic integrated research on
different taxa and signalling modalities. It was also the opportunity to meet Shana
Coates of Cambridge University Press and to appreciate her enthusiasm for a ‘book
of the symposium’.
The book has turned out to be much more than a collection of symposium
papers. First, it covers considerably more ground in its 26 chapters than was pos-
sible in a symposium of nine spoken papers. Second, some of the stimulating
informal discussions that characterize a good conference have contributed to the
section introductions. However, the main ‘added value’ comes from the willing-
ness of the authors to comment on the chapters of others, to incorporate comments
and cross-references into their own chapters and, above all, to look at communi-
cation from a network perspective. In many instances, this has led to insights that
are likely to have a major effect on the direction of research on animal communi-
cation – real Eureka moments. It has been a privilege to share in these moments.
Many people deserve my thanks for the role they have played in the cre-
ation of this book. Marc Naguib suggested that we submit the symposium topic
xiii

P1: JZZ/JZZ P2: JZZ0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:42
xiv Preface
‘communication networks’ to the IEC scientific committee. This committee and
the conference main organizer, Raimund Appfelbach, were kind enough to accept
the topic and in doing so set the ball rolling. Shana Coates of Cambridge University
Press gently, but firmly, ensured that my timescale for editing a book on communi-
cation networks was advanced from ‘some time in the future’to ‘in the next couple
of months’. Considerable credit is also due to Tom Peake and Andrew Terry, who
applied their own particular brand of pressure (accompanied by several cappucci-
nos) to stimulate me to draft the book proposal on the flight back from Tubingen.
Shana gave excellent advice in the early stages of the book, since when Tracey
Sanderson and Martin Griffiths have overseen production. Of course, there would
be no book without the authors. I am very grateful to all of them for finding time
in overcrowded schedules to write their own chapters and to comment on those
of others. Denmark’s Statens Naturvidenskabelige Forskningsrad has supported
my research for the last 5 years. København Universitet supported me during the
initial stages of the book, but the bulk of the work was done with support from
the EU and Cornwall College via a Marie Curie Category 40 Fellowship. Last, but by
no means least, I thank Leonie and Tom McGregor – for sustaining me throughout
the project with their love, good humour and flexibility at all times, particularly
when the going got tough.

P1: IKB/... P2: KOD0521823617c01.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 15:52
1
Introduction
p e t e r k . m c g r e g o r
University of Copenhagen, Denmarkand Cornwall College, Newquay, UK
Some of the most conspicuous behaviours performed by an animal are
related to communication – communication that mediates reproduction and sur-
vival. As explained below, a knowledge of animal communication is important in
more respects than simply its role in understanding such fundamental aspects of
animals’lives. This book is about a perspective that can increase our understanding
of animal communication.
One way in which animal communication is important is that it interfaces
with and links several fields of study. In the field of behaviour, for example, com-
munication is often used to illustrate Niko Tinbergen’s four types of question
(function, mechanism, development and evolution) and how the answers comple-
ment each other (e.g. Krebs & Davies, 1993). Communication has interfaces with
many other areas of biology including evolution, ecology, population genetics,
neurobiology and physiology. For example, it can be a window into the cognitive
worlds of animals (e.g. Ch. 24). Links with other sciences are shown by the use
of ideas and techniques from psychology to understand how communication is
perceived (Ch. 20), and using information from physics and chemistry to explain
how communication is achieved (e.g. Bradbury & Vehrencamp, 1998).
Communication cannot occur in isolation; it is an inherently social behaviour.
This makes it even more surprising that the wider social context in which commu-
nication takes place is rarely considered explicitly. As explained in the next para-
graph, it is likely that communication commonly occurs in the context of a net-
work of several animals. This chapter is both a brief introduction to this context –
animal communication networks – and an explanation of this book’s structure.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
1

P1: IKB/... P2: KOD0521823617c01.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 15:52
2 P. K. McGregor
About communication networks
A communication network is a group of several animals within signalling
and receiving range of each other. If signals travel further than the average spacing
between individuals, then there is potential for a communication network to exist.
This is as true for the ocean-spanning songs of whales as it is for the begging calls
of songbird nestlings crammed into a nest cavity, and it is why networks can be
considered to be the commonest context in which communication occurs (e.g.
McGregor & Peake, 2000).
This would seem to be stating the obvious, especially to those new to the field of
animal communication. Indeed, those studying chorusing animals, particularly
insects and anuran amphibians, have long adopted a network perspective and rec-
ognized the importance of doing so (e.g. Otte, 1974). However, it is only relatively
recently that other types of communication have been considered explicitly in a
network context. Communication was, and still is in many instances, treated as oc-
curring between two individuals – the signaller–receiver dyad – perhaps because
this is the simplest relationship possible between the three basic components
found in communication (the signaller, the signal and the receiver). In this sense,
a dyadic view of communication follows from the stricture of Occam’s razor (also
known as the law of parsimony) to employ the simplest explanation consistent
with the facts. While agreeing wholeheartedly with this standard scientific prac-
tice, it is clear that a dyadic view of communication is often not consistent with
the facts. One example is the high signal level used in close-range aggressive en-
counters – human antagonists nose to nose, yet shouting at each other – surely
high signal levels are not needed to achieve signalling at such close range? In a
network context, such high levels make more sense, because there may be more
distant intended receivers (the gathering crowd in the human example) in addi-
tion to the opponent (Zahavi, 1979). Many further examples of communication
that are best considered in the context of a communication network are found
throughout this book.
Another reason for explicitly considering communication in a network context
is that it identifies communication behaviours that cannot occur in a dyad. A good
example is eavesdropping, particularly social eavesdropping (Ch. 2) in which the
eavesdropper extracts information from a signalling interaction between others.
Social eavesdropping requires a minimum of three individuals (one eavesdrop-
ping, two more interacting) and, therefore, falls outside a dyadic view of com-
munication. The evidence for eavesdropping and its wider implications (e.g. for
comparative cognition) is presented in many of the chapters of this book. Eaves-
dropping and similar network behaviours discussed in this book are considered
by many to be a compelling reason to adopt a network perspective.

P1: IKB/... P2: KOD0521823617c01.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 15:52
Introduction 3
Communication networks and eavesdropping
It is perhaps worth emphasizing that, while eavesdropping is a good ex-
ample of communication network behaviour, it is not the only one, and the value
of the communication network perspective does not depend on a demonstration
of eavesdropping. The reason for its current prominence is that it was considered
first and, therefore, at the moment it is more prevalent in the literature. There is
no merit in shoe-horning a natural example into a definition of eavesdropping,
nor in judging the value of any natural communication behaviour by how well it
fits this (or any other) definition. As several chapters demonstrate (e.g. Chs. 9 and
23), such examples from the real world can probe and challenge our definitions
(e.g. of interactions and of communication more generally) and the thinking that
follows from them. The result can be considerable insight and lead to progress for
the whole field of communication.
A note on definitions
Clear and workable definitions are the essential basis for meaningful
discussion. I have tried to ensure that terms are used clearly and consistently
within a chapter, but there may be good reasons why chapters differ in the detail
of their definitions (e.g. for reasons discussed in the previous paragraph). There
are no instances in this book where the same term is used in a markedly different
way in different chapters, but readers should bear in mind that the detail of the
definition may be important to the topics discussed by the chapter.
There are two nice illustrations of the problems that definitions can create. The
first concerns eavesdropping. Alan Grafen pointed out a problem with the term
eavesdropping after I had used it when presenting ideas on communication net-
works at the Royal Society Meeting on Signalling in 1992 (McGregor, 1993). The prob-
lem he foresaw was that in everyday use the term means secret information gath-
ering, and it was clear to him that there may be advantages to the signallers in pro-
viding information (i.e. promoting eavesdropping), especially if the signaller had
won the agonistic contest (see also Zahavi, 1979). The everyday meaning of eaves-
dropping and its implicit association with acoustic signals have been at the root
of several misunderstandings that could perhaps have been avoided if a more neu-
tral term had been used (at the time Grafen suggested type II receivers). Tom Peake
has sorted out this and other problems to do with definitions of eavesdropping
with admirable clarity in Ch. 2. Nevertheless, information gathered without the
source’s knowledge may have particular value, as John Locke discusses in Ch. 19. I
think this demonstrates that identifying the secrecy or otherwise of information
gathering is the route to progress, rather than rigidly applying a definition.

P1: IKB/... P2: KOD0521823617c01.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 15:52
4 P. K. McGregor
The second example concerns the relationship between information and com-
munication. In my view, the terms are clearly not synonymous; rather signals
are a subset of information because they are specialized to transmit information
(more details in McGregor & Peake (2000)). This could have created a problem with
semiochemicals: if they are not signals (i.e. they contain information but are not
specialized to transmit it) then the behaviour involving them is not communica-
tion and the concept of communication networks would not apply. Fortunately
for the book, Brian Wisenden and Norm Stacey thought carefully about the is-
sue and realised that there were many important similarities that gave them an
opportunity to discuss the functional and evolutionary relationships between in-
formation, signals and networks (Ch. 23). So a problem arising from definitions
has given real insight, rather than the acrimonious defence of definitions that is
all too common in the literature.
About this book
Coverage
There are several types of book on animal communication. Some are syn-
optic treatments of the whole topic (e.g. Hauser, 1996; Bradbury & Vehrencamp,
1998) whereas others concentrate on particular types of signal such as pheromones
(Wyatt, 2003) or on a group of animals such as arthropods (Greenfield, 2002).
Many books do both, for example dealing with acoustic communication in insects
(Gerhardt & Huber, 2002) or birds (Kroodsma & Miller, 1996). This book is rather
different in that it looks at a specific topic in communication and covers several
modalities and taxonomic groups.
Organization
Each chapter has been written so that it can be read alone, since this
is a common way for edited volumes to be read. Inevitably, this has led to some
similarity between chapters in their opening remarks, but I think this is more than
offset by each chapter having its own reference section. The many cross-references
to other chapters in the book also show the extent to which authors have taken
account of material in other chapters and made links between them.
A second way in which the book has been given overall coherence is to group the
chapters into four parts that reflect major aspects of communication networks.
Each of these parts is prefaced by a short overview that identifies chapter themes
and highlights some of the issues that remain to be tackled. The fact that many
chapters could have been put into any of the four parts further demonstrates
the extent of overall coherence of the book and the wide-ranging nature of the

P1: IKB/... P2: KOD0521823617c01.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 15:52
Introduction 5
chapters. Within each part, there is no particular order of chapters, although in
Part III the order is loosely phylogenetic.
The chapters grouped into Part I deal with communication behaviours, such
as eavesdropping and audience effects, that involve three or more individuals (i.e.
a communication network) and as such fall outside the ‘classical’ or traditional
dyadic (one signaller and one receiver) approach to communication.
Part II groups particular contexts that are fruitful to consider from a communi-
cation network perspective: mate choice, predation, begging, aggression and scent
marking.
The reason for grouping chapters in Part III is taxonomic: from fiddler crabs
to humans via most groups of vertebrate. While communication networks may
be more or less ubiquitous, features of different taxa (e.g. main senses, social
organization) can have a major effect on the details of communication networks
and provide insight into the topic as a whole.
The final part contains chapters that, to a greater or lesser degree, link com-
munication and other disciplines in biology and more widely in science. From the
evidence of these chapters, a network perspective seems to be particularly valuable
at such subject interfaces.
Summary
There are several reasons for considering that the natural context in which
communication occurs (and in which it has evolved) is a network of several animals
in signalling and receiving range of each other. However, this context has not been
considered explicitly in many studies of animal communication. The chapters in
this book apply a communication network perspective to a variety of taxa using a
number of signal modalities in several circumstances. The results are illuminating.
To modify a marketing phrase used for mobile phones: the future is bright; the
future is a network view of communication.
References
Bradbury, J. W. & Vehrencamp, S. L. 1998. The Principles of Animal Communication.
Sunderland, MA: Sinauer.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans: Common
Problems and Diverse Solutions. Chicago, IL: Chicago University Press.
Greenfield, M. D. 2002. Signalers and Receivers: Mechanisms and Evolution of Arthropod
Communication. Oxford: Oxford University Press.
Hauser, M. D. 1996. The Evolution of Communication. Cambridge, MA: MIT Press.
Krebs, J. R. & Davies, N. B. 1993. An Introduction to Behavioural Ecology, 3rd edn. Oxford:
Blackwell Scientific.

P1: IKB/... P2: KOD0521823617c01.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 15:52
6 P. K. McGregor
Kroodsma, D. E. & Miller, E. H. 1996. Ecology and Evolution of Acoustic Communication in
Birds. Ithaca, NY: Cornell University Press.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
Otte, D. 1974. Effects and functions in the evolution of signaling systems. Annual
Review of Ecology and Systematics, 5, 385–417.
Wyatt, T. D. 2003. Pheromones and Animal Behaviour: Communication by Smell and Taste.
Cambridge, UK: Cambridge University Press.
Zahavi, A. 1979. Why shouting? American Nauralist, 113, 155–156.

P1: JZZ/IKB P2: JZZ/...0521840716agg.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:0
Part I B E H A V I O U R S S P E C I F I C T O
C O M M U N I C A T I O N N E T W O R K S
7

P1: JZZ/IKB P2: JZZ/...0521840716agg.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:0
8

P1: JZZ/IKB P2: JZZ/...0521840716agg.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:0
Introduction
The reason for grouping together the chapters that appear in this part
of the book is that each of them concerns communication behaviours that are
best viewed from a communication network perspective, rather than from the
more common dyadic (one signaller to one receiver) standpoint. It is a fact that,
with the exception of choruses, most studies to date have implicitly or explicitly
considered communication between a dyad. Although the communication net-
work perspective of several signalling and receiving individuals seems to follow
logically from what we know of natural communication, the dyadic viewpoint has
historical precedence and considerable inertia. A network perspective will become
more commonly adopted only if it is clearly better able to explain communication
behaviours than a dyadic approach. It is for this reason that a network perspective
has long been adopted in studies of choruses; the effect on an individual’s signal
timing of the signals of nearby conspecifics can be striking patterns, such as signal
synchrony in the chorus (e.g. Greenfield, 2002; Ch. 13). Such patterns cannot be
explained by considering communication as a dyad. All of the chapters in this
book demonstrate the value of adopting a network perspective; however, it gives
this demonstration more emphasis to begin with a section covering communi-
cation behaviours that are particularly suited to, or associated with, a network
perspective.
Eavesdropping
In Ch. 1, eavesdropping is identified as a receiving behaviour that has been
particularly identified with, and is only possible in, a communication network.
The first two chapters of Part I look at eavesdropping in more detail.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
9

P1: JZZ/IKB P2: JZZ/...0521840716agg.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:0
10 Part I
In Ch. 2, Tom Peake summarizes the evidence for eavesdropping in different
contexts and also discusses the diverse use of the term in the literature. His di-
vision of eavesdropping into interceptive eavesdropping (e.g. predators locating
prey from prey signals) and social eavesdropping (extracting information from a
signalling interaction) is an important clarification. However, as Tom points out,
clarifying definitions is more important as means of moving arguments on from
the question of whether a given behaviour can be called eavesdropping or not and
towards a more fruitful and general approach based on the nature of information
transfer.
Torben Dabelsteen deals mainly with social eavesdropping on the acoustic sig-
nals of birds in Ch. 3. He identifies the potential costs and benefits of eavesdrop-
ping and uses information from studies of how bird song transmits through the
habitat to explore how eavesdropping is best achieved. The overall balance of costs
and benefits of being eavesdropped upon will determine whether signallers pro-
mote eavesdropping on their signals or whether they try to avoid it. One intriguing
possibility that Torben discusses is whether the costs of being eavesdropped upon
could be avoided if signallers made their signals anonymous by removing infor-
mation on signaller identity.
Audience effects
In communication networks, several receivers are likely to be present
during signalling interactions between others; these receivers do not take part in
the interaction and have been referred to as an audience. The effects they can have
on signalling behaviour are the subject of Ch. 4, in which Ricardo Matos and Ingo
Schlupp draw the distinction between an apparent audience and an evolutionary
audience. The distinction is important because selection pressures imposed by
the presence of audiences in the evolutionary past of the animals may result in
features of the signalling interactions despite the absence of an audience during
any particular interaction. Also, whether an audience is apparent to the signallers
involved in interactions may depend on signal modality: individuals have to be
in the line of sight of visual signals to receive them and, therefore, an audience is
likely to be apparent; however, the same is not true of widely broadcast acoustic
signals.
Bystanders
Being a bystander (i.e. present, but not directly involved) during an agonis-
tic or mating interaction can affect subsequent aggressive and mating behaviour
and is explored in Ch. 5. Ryan Earley and Lee Dugatkin focus on social eaves-
dropping (a subset of bystanding) by two species of poeciliid fishes that are likely

P1: JZZ/IKB P2: JZZ/...0521840716agg.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:0
Behaviours specific to communication networks 11
to be familiar to many – green swordtails Xiphophorus helleri and guppies Poecilia
reticulata – and that communicate largely with visual signals. Their chapter shows
how a network view can encompass and organize diverse aspects of fighting and
mating behaviour (including mate copying); it also identifies the many conditions
that favour eavesdroppers and how the effects of eavesdropping are manifested.
Victory displays
In the final chapter of this section, John Bower examines victory displays:
signals produced by the winner (but not the loser) after an aggressive interaction.
There has been surprisingly little work specifically on this topic, despite the wealth
of studies of signalling before and during aggressive displays, and such informa-
tion is widely scattered. Chapter 6 collates the information on victory displays
and then interprets its functional significance, first from a dyadic perspective and
then from a network perspective. It may have been premature to include victory
displays in this section, because on current evidence it is not clear that victory dis-
plays always function in a network context rather than in the winner–loser dyad.
However, even if their main function is dyadic, their conspicuous nature makes
it likely that other individuals could gain useful information by paying attention
to victory displays.
Future directions
The authors dealing with eavesdropping make several suggestions for the
directions future research should take: incorporating eavesdropping into theoret-
ical models to derive testable predictions that can contribute to understanding
signal evolution (Ch. 2); finding evidence of eavesdropping in non-experimental
natural contexts (perhaps by using a combination of tracking and acoustic location
technologies to follow the individuals in a network), and continuing such studies
long enough to identify differences in reproductive success (Ch. 3); unravelling the
complex interrelationships between features of individuals, their social and wider
environment and the role of bystanders in order to understand communication
fully (Ch. 5).
The authors dealing with eavesdropping clearly consider that the phenomenon
is now well characterized. In contrast, victory displays clearly need more detailed
study in order to establish the phenomenon and to elucidate its function and
whether it is network phenomenon. It is likely that controlled laboratory experi-
ments are the best way to investigate what effect, if any, victory displays have on
other members of the communication network (Ch. 6).
Progress in understanding audience effects seems likely to come from a differ-
ent type of approach. In addition to modelling and controlled experiments, there is

P1: JZZ/IKB P2: JZZ/...0521840716agg.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:0
12 Part I
the potential to integrate information on audience effects with underlying mech-
anisms. Suitable candidate mechanisms exist in the literature (e.g. the hormonal
basis of priming effects) and deserve to be investigated more fully (Ch. 4).
References
Greenfield, M. D. 2002. Signalers and Receivers: Mechanisms and Evolution of Arthropod
Communication. Oxford: Oxford University Press.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
2
Eavesdropping in communicationnetworks
t o m m . p e a k e
University of Copenhagen, Denmark
Introduction
All communication occurs in a network environment with the exception
of a subset of systems that unequivocally meet both of the following criteria: (a) a
signal can never be received by more than one receiver; (b) a receiver can never
receive more than one signal simultaneously. In other words, all communication
networks have at least one of two defining properties: (a) signals can be, at least
potentially, received by several receivers; and (b) receivers can, at least potentially,
receive signals from several signallers at any one time. Consequently, in moving
from a dyadic consideration of communication to a network view, signallers and
receivers both take on a range of costs and benefits, which are the theme of this
book. In this chapter, I will consider the implications of a particular type of re-
ceiving behaviour that becomes possible in a network, namely eavesdropping. I
will begin by reviewing different definitions of eavesdropping that are found in
the literature and the evidence for different types of eavesdropping, distinguish-
ing between eavesdropping on signals and eavesdropping on signal interactions. I
will then examine the costs, benefits and implications of eavesdropping on inter-
actions, as recognition of this phenomenon emerged from considerations of
qualitative differences between dyadic and network views of communication
(McGregor, 1993; McGregor & Dabelsteen, 1996).
Defining eavesdropping
The verb eavesdrop is defined by the New Oxford Shorter English Dictionary as
‘Listen secretly to (a person, private conversation), orig, by standing beneath the
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
13

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
14 T. M. Peake
eaves of a house. Formerly also, stand beneath the eaves of (a building) in order to
overhear conversation within.’ This word, at least to native English speakers, has
an evocative quality that makes it appealing to authors in a variety of often quite
different contexts. The use of the word in everyday language also has connotations
that it may be useful to discard at this stage. First, in everyday use the term applies
only to the acoustic modality; as a technical term in animal communication there
is no good reason why this should be, although undoubtedly this has contributed
to the term not being used by authors working in some modalities. Second, the
idea of secrecy contained in the above definition need not necessarily be carried
over to its use in animal communication.
In the context of animal communication, the term has been used in a number of
different ways that can be summed up by a general definition: the use of information
in signals by individuals other than the primary target. This definition excludes the
use of eavesdropping to describe behaviours such as detecting prey by cues that
are not designed to enhance information transfer (e.g. extraneous noise caused
by movement); in this sense the definition differs from that given by Bradbury &
Vehrencamp (1998, p. 3). Eavesdroppers have been called ‘illegitimate’ (Otte, 1974),
‘unintended’ (Wiley, 1994) or ‘third party’ (Zahavi, 1979) receivers or ‘bystanders’
(e.g. Dugatkin, 2001) according to the context in which they were defined. In the
general definition above, I use the phrase ‘individuals other than the primary
target’ on the grounds that, as I shall outline below, eavesdropping individuals do
not under all circumstances impose a cost on signallers as is implied by some of the
alternative terms above. In situations where the presence of eavesdroppers benefits
signallers, there may be selection pressure to allow information transmission to
eavesdroppers, while the major selective force remains the more apparent (or
primary) receiver.
Within this general definition, there are two classes of use of the term eaves-
dropping that are sufficiently different, yet sufficiently commonly used, to war-
rant discussion and clarification. Here I call these classes interceptive and social
eavesdropping. Interceptive eavesdroppers benefit by intercepting signals in-
tended (in an evolutionary sense) for another individual, usually to the cost of
the signaller. Social eavesdroppers gather information on other individuals by
attending to their signalling interactions with conspecifics. At first glance, these
two types of behaviour may seem very similar; however, as I shall argue in the
remainder of the chapter, the nature of information transfer and resulting selec-
tion pressures differ markedly between them. These terms were chosen carefully
to indicate the source from which eavesdroppers gather information (i.e. inter-
cepting signals versus attending to social interactions), without undue prejudice
towards certain aspects more commonly (but rarely exclusively) associated with
either behaviour. The aim of this distinction is to move arguments or points of
confusion away from the question of whether a given behaviour can be called

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 15
Table 2.1. Generalizations concerning the two types of eavesdropping behaviour
defined in the text
Type of eavesdropping Interceptive Social
Source of information Signals Signal interactions
Type of signal Usually broadcast Always directed
Eavesdropper–signaller
relationship
Usually heterospecific Usually conspecific
Payoff to signaller Usually negative or zero Positive, negative or zero
Information gathered Absolute Relative information also available
eavesdropping or not and towards a more fruitful approach based on the nature
of information transfer. While these definitions of eavesdropping require that sig-
nals be transmitted to more than one receiver (network property (a) above), social
eavesdropping further requires that receivers can detect more than one signal at
the same time (network property (b)). One fact is unavoidable: eavesdropping is,
by definition, a behaviour that can only occur in a network as it requires at least
three individuals: a signaller, a target receiver and an eavesdropper.
A number of generalizations may be made that show the distinctions between
the two types of eavesdropping (summarized in Table 2.1). (a) Interceptive eaves-
dropping usually involves the reception of broadcast signals (i.e. those that have
a class of targets such as females of the signaller’s species rather than a spe-
cific target); social eavesdropping, by definition, involves the exchange of sig-
nals directed towards specific receiving individuals. (b) Interceptive eavesdrop-
ping is most commonly identified in situations where eavesdroppers are a differ-
ent species from the signaller; social eavesdropping is usually identified within
a species. (c) Interceptive eavesdropping usually has a negative or zero effect on
signallers; the payoff to a signaller resulting from social eavesdropping is much
less clear, as will be discussed below. (d) Interceptive eavesdropping focuses on
the absolute signalling behaviour of the signaller (in many cases this may be sim-
ple presence/absence of information); social eavesdropping additionally allows
information to be gathered on the relative performance of interacting signallers
(allowing both direct comparison of interactants and assessment of relationships
between them).
Interceptive eavesdropping
Wiley (1983) defined eavesdropping as the behaviour by which ‘signals
intended for one receiver are intercepted by another’; this definition is explic-
itly given as an example of a receiver ‘obtaining information about the signaller

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
16 T. M. Peake
against its own best interests’. Bradbury & Vehrencamp (1998) adapted this defi-
nition to include as eavesdropping situations where the signaller obtains a zero
benefit, terming as ‘exploitation’ cases in which eavesdroppers are detrimental to
signallers. In defining interceptive eavesdropping above, I make no assumptions
about the nature of the signaller payoff. The definition is intended to be simply
descriptive and one could imagine many subdivisions that could be made. One
clear distinction is in the taxonomic relationship between signaller and eaves-
dropper. Where signaller and eavesdropper are of different species, as is the case
in most examples of signal interception found in the literature, the payoff to the
signaller is almost certainly negative. Information obtained by eavesdropping in
this case may be something as simple as the location of a suitable prey item. The
effect of eavesdropping within a species is likely to be more difficult to determine
and the kinds of information gathered may well be more related to features of the
signaller.
As a final point, it is suggested by some authors (e.g. Bradbury & Vehrencamp,
1998; Greenfield, 2002) that signals intercepted by another species, particularly
in such cases as predators locating prey, should be considered cues as they are not
designed to enhance information transfer to those receivers. This is certainly true
if one considers predator and prey in isolation; however, when considering the
structure of the source of information and factors associated with production it
is important that the wider context is included.
Interspecific examples
The most commonly cited examples of eavesdropping, and those that have
been best studied, are those that occur between trophic levels, i.e. predators and
parasites detecting the signals of prey or hosts or prey detecting predator presence
by their signals. Selection pressures imposed by these kinds of eavesdropper are un-
derstandably high and have been shown to lead to a range of counter-adaptations
that aim to ameliorate or avoid such pressures (e.g. Greenfield, 1994, 2002; Heller,
1995; Stoddard, 1999; Gerhardt & Huber, 2002; Ch. 8). Examples are particu-
larly prevalent in acoustic (e.g. Cade, 1975; Ryan et al., 1982; Sakaluk & Belwood,
1984; Belwood & Morris, 1987) and chemical (Aldrich, 1995; Roberts et al., 2001)
signalling and there are good reviews available (e.g. Stowe et al., 1995; Zuk &
Kolluru, 1998). While many visual signals are conspicuous and may be used by
predators to find prey (e.g. Lloyd & Wing, 1983), it is rare to find such examples
called eavesdropping (see Bruce et al. (2001) for such an example).
Such interceptive eavesdropping reflects communication networks working on
a community level. The selection pressures on communication between trophic
levels are widely acknowledged in most considerations of the evolution of commu-
nication. Much less widely studied and appreciated is the importance of networks

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 17
operating within a species, and there is evidence that interceptive eavesdropping
occurs at this level.
Intraspecific examples
Within a species, individuals may eavesdrop on the signalling behaviour
of others for a variety of reasons. In some cases, animals of one sex eavesdrop
on signals intended for the opposite sex (Ch. 12). For example, Kiflawi & Gray
(2000) looked at eavesdropping by male crickets Acheta domesticus on competing
males’ mating calls. Smaller males showed a phonotactic response towards speak-
ers broadcasting calls preferred by females in a two-speaker choice design, while
larger males varied in their phonotaxis. The suggestion here is that males with
unattractive calls can potentially intercept females as they move towards attrac-
tive males.
A recent example concerns the use of female signals, apparently intended for
mates, as a means of detecting fertile females for extra-pair copulations. Female
robins Erithacus rubecula produce ‘seep’ calls to obtain provisioning from their
mate (East, 1981). Mate removal experiments show that females may attract other
males, which provide courtship feeding that may result in copulation (Tobias &
Seddon, 2000). Tobias & Seddon (2002) found that neighbouring males approached
‘seep’ calls when they were played back at a high rate near the territory boundary,
on occasion bearing provisions. They suggest that, if the female call is a hunger
signal directed towards the mate, neighbouring males might be eavesdropping
(in the interceptive sense). In this case, females may derive a benefit from the
presence of eavesdroppers while the primary benefit comes from the response of
the mate. Tobias & Seddon (2002) also acknowledged the possibility that the ‘seep’
call is directed towards extra-pair males as a means of ‘blackmailing’the mate into
providing food. In either case, the results highlight the influence of operating in
a social network.
Individuals may eavesdrop on signals designed to warn others of the presence of
predators. For example, Shennan et al. (1994) describe the behaviour of group mem-
bers paying attention to the vigilance activities of others in order to avoid preda-
tors. Convict cichlids Cichlasoma nigrofasciatum fin-flick in order to warn young; fish
not guarding young do not fin-flick. Parents were shown to fin-flick in response to
fin-flicking models, suggesting that they are capable of monitoring the vigilance
activities of others in order to warn their own young sooner. Here, the primary tar-
gets of the signal are likely to be relatives of the signaller, while non-relatives may
also benefit by paying attention to the signals at no obvious cost to the signaller.
This early warning feature of the signalling behaviour of others has been sug-
gested as a possible function of territoriality (e.g. Eason & Stamps, 1993). Obser-
vations of red-capped cardinals Paroaria gularis showed that territorial males had

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
18 T. M. Peake
a high chance of detecting intrusions where the intruder had been recently re-
pelled from a neighbour’s territory as a consequence of the conspicuousness of
behaviours involved in eviction. Intruders that had not been detected by neigh-
bours were unlikely to be detected by territorial subjects.
In all of these examples, individuals use signals produced by conspecifics for
their own benefit, as fits the definition of interceptive eavesdropping. Less clear is
the effect of such eavesdropping on the signaller. In some cases (e.g. Kiflawi & Gray,
2000), the signaller may suffer a cost because of the presence of eavesdroppers,
while in some there is no obvious benefit or cost to the signaller (e.g. Shennan et al.,
1994). In some cases (e.g. Tobias & Seddon, 2002), the presence of eavesdroppers
may actually benefit the signaller. These examples show how definitions of eaves-
dropping and communication based on costs to the signaller (e.g. Wiley, 1983;
Bradbury & Vehrencamp, 1998) may not apply to all circumstances. It is for this
reason that I prefer the descriptive definition of eavesdropping in general as the
use of signals by receivers other than the primary target (see above).
Autocommunication and eavesdropping
Eavesdropping has also been used to describe the interception of informa-
tion contained in sounds produced by animals in order to investigate their environ-
ment. Although not strictly within the general definition of eavesdropping given
above, because these sounds are not designed to transmit information to others,
the examples are interesting enough to be worth mentioning. Little brown bats
Myotis lucifugus gather information by paying attention to the echolocation calls
of foraging conspecifics (Barclay, 1982). Bats approached speakers broadcasting
echolocation calls of conspecifics and a heterospecific Eptesicus fuscus, suggesting
that eavesdropping could substantially increase potential prey detection distance.
Balcombe & Fenton (1988) found similar results in M. lucifugus, a congener Myotis
yumanensis and another species (Lasiurus borealis), in which individuals apparently
used others’ echolocation calls to identify ‘vulnerable’ prey in order to steal them
from the eavesdropped bat. Similarly, Xitco & Roitblat (1996) use the term eaves-
dropping to describe the ability of a bottlenose dolphin Tursiops truncatus to identify
objects inspected by another dolphin, via the reception of the inspector’s echolo-
cation clicks.
Social eavesdropping
In one of the first explicit considerations of territorial systems as a commu-
nication network, McGregor (1993) suggested that the term eavesdropping could
be applied within species, particularly in the context of paying attention to inter-
actions between neighbours and rivals. While initially McGregor (1993) considered

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 19
this analogous to interceptive eavesdropping, McGregor & Dabelsteen (1996) later
made the distinction, refining their definition of eavesdropping to ‘extracting in-
formation from an [signalling] interaction between other individuals’. They con-
sidered it ‘a prerequisite of eavesdropping that a third party (the eavesdropper)
gains information from an interaction that could not be gained from a signal
alone’. This, they suggested, was a different level of information transfer than
simply locating a prey item by its signals. McGregor & Peake (2000) attempted to
clarify the hierarchical relationship between information, signals and signalling
interactions, concluding that interactions may be under additional selection pres-
sures to those acting on the signals themselves. The most obvious source of these
selection pressures is those with an interest in the outcome: rivals and mates.
Experimental evidence for social eavesdropping has recently increased dra-
matically as clear experimental paradigms have emerged. Studies that explicitly
address eavesdropping in this context have thus far been carried out exclusively
on acoustic interactions in territorial songbirds and visual interactions in teleost
fish. However, evidence from other experiments not designed to test eavesdrop-
ping per se are strongly supportive of the existence of eavesdropping as a means
of gaining information on the qualities of and/or social relationships between
conspecifics.
Acoustic interactions in songbirds
McGregor et al. (1997) addressed the issue in songbirds using interactive
playback (Dabelsteen et al., 1996) to simulate intrusion upon the neighbours of
subject male great tits Parus major (Fig. 2.1ai). Neighbours were presented with
one of two types of intruder. One type indicated its willingness to escalate by
beginning each song immediately following the onset of neighbour song (over-
lapping: Hulsch & Todt, 1982; Dabelsteen et al., 1996, 1997) and increasing song
length. The other type of intruder playback signalled a lower level of willingness
to escalate by beginning songs only after the neighbour songs had been completed
(alternating) and reducing song length. After a short amount of time, an intru-
sion by the same intruder was simulated in the subject male’s territory (Fig. 2.1aii)
singing an alternating pattern with matched song length. Subjects responded to
previously aggressive intruders by keeping their distance and overlapping song,
while less-aggressive intruders were approached quickly.
A similar experiment carried out on great tits looked at the behaviour of fe-
males in response to intruders interacting with their mates and neighbours (Otter
et al., 1999). In this case, experiments were carried out on dyads of neighbouring
territories each defended by a mated pair. Playback was used to intrude on each
territory on successive days (Fig. 2.1bi, bii) such that the same intruder showed a
high willingness to escalate to one male and a low willingness to escalate to the

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
20 T. M. Peake
(a) (b)
(c)
(d)
(e)
KEY
(i)
(i) (i)
(i)
(i)
(ii) (ii)
(ii)
(ii)
(ii) (iii)
(iii)
Loudspeaker
Loudspeakerbroadcastingplayback
Interaction between two loudspeakers
Interaction between male and loudspeaker
Fig. 2.1. Schematic representations of experiments described in the text investigating
social eavesdropping on acoustic interactions in songbirds. (a) Representation of
design used by McGregor et al. (1997) showing (i) interaction between loudspeaker and
neighbouring male and (ii) subsequent playback intrusion in subject’s territory.
(b) Design used by Otter et al. (1999) and Mennill et al. (2002) showing (i) interaction
between one male and a loudspeaker, (ii) subsequent interaction between
neighbouring male and loudspeaker and (iii) observation of female behaviour.
(c) Design used by Naguib & Todt (1997) and Naguib et al. (1999) showing (i) interaction
between two loudspeakers inside the subject’s territory and (ii) subsequent playback
from loudspeaker not approached initially by the subject. (d) Design used by

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 21
neighbouring male. Otter et al. (1999) then followed the females for some time
subsequent to the treatments (Fig. 2.1biii) and showed that females whose mates
had suffered from an intruder that a neighbour dealt with easily were more likely
to trespass onto neighbouring territories, particularly that of the neighbour who
had performed well during playback. These female forays were not converted into
offspring, however (Otter et al., 2001), suggesting that the short-term nature of
the information was not enough to convince females of the poor quality of their
mates. Mennill et al. (2002), however, did find such an effect in female black-capped
chickadees Poecile atricapillus using a similar experimental paradigm. In this case,
Mennill et al. (2002) had information on males’ dominance ranks during winter
feeding flocks so that dyads of neighbouring territories each consisted of one
mated pair in which the male was high ranking and the other in which the male
was low ranking. Playback was carried out to these dyads in a similar way to that
used by Otter et al. (1999) and was followed by microsatellite paternity analysis
in order to assess female reproductive decisions. The results showed that high-
ranking males that had lost to playback showed a much greater incidence of lost
paternity with extra-pair young in 12 of 23 nests, compared with 2 of 20 in control
nests. Low-ranking males that did well against intruders lost paternity to the same
extent as controls.
In all of the above experiments, the conclusion is that the responses of subjects
are a result of information gained by paying attention to the interactions between
intruders and known males. This interpretation is, however, somewhat limited by
the fact that, in each case, the response of the subject may be affected by the
response of the known male: because the interaction of interest was between
a male simulated by playback and a live male, the subsequent response of the
subject may be affected by changes in behaviour of the live male. For example,
in McGregor et al. (1997), the response of the subject may result from changed
behaviour of the neighbour following different levels of intrusion; similarly, in
the latter two studies, females may have changed their behaviour in response to
changed behaviour of their mates. Mennill et al. (2002) did, in fact, examine the
Fig. 2.1 (cont.) Peake et al. (2004) showing (i) interaction between two loudspeakers
outside the territory boundary and (ii) subsequent intrusion by one of the
loudspeakers. (e) Design used by Peake et al. (2002) showing (i) intrusion by
loudspeaker, (ii) subsequent interaction between that and another loudspeaker
outside the territory boundary and (iii) intrusion by the second loudspeaker. Rounded
rectangles represent territory boundaries; male and female symbols represent
approximate positions of resident males and females; arrows represent movements by
loudspeakers; arrows with curved lines represent monitoring of female movements.
See text and cited references for more details of each experiment.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
22 T. M. Peake
behaviour of males following each playback treatment and could find no effect
on subsequent behaviour.
The problem of lack of control over the signalling behaviour of interactants
and subsequent changes in behaviour can be avoided in songbirds by replacing
males with playback; dyadic encounters can then be simulated using two loud-
speakers. The first study to use this approach was carried out on nightingales
Luscinia megarhynchos by Naguib & Todt (1997), who examined the effect of asym-
metric interactions on the responses of territorial males. The asymmetry in this
case was achieved by having one loudspeaker producing songs that overlapped
the other. Loudspeakers were placed inside the territory boundary of the subject
and interactions lasted for two minutes (Fig. 2.1ci). Males responded by spending
more time near, spending more time singing near, and singing more songs near
the overlapping speaker. Ten minutes after the ‘interaction’ had finished, play-
back for one minute was broadcast from the speaker that was not approached
first during the interaction (Fig. 2.1cii). Males sang more at the location of the
formerly overlapping speaker regardless of whether that speaker was producing
song. Naguib et al. (1999) repeated this experiment with a different kind of inter-
action in which songs did not overlap but were still asymmetrical as one speaker
(the follower) always directly followed the output of another (the leader). In this
case, males showed a stronger response to the speaker that ‘led’; once again the
subjects responded differently to the two types of apparent opponent (Naguib &
Todt, 1997).
In these two experiments, the design meant that subjects could associate roles
during an interaction with the location of a singing intruder. Peake et al. (2001)
looked at whether similar associations could be made between roles and song
features using a similar experimental paradigm in great tits. In this case inter-
actions were carried out between two loudspeakers situated outside the territory
boundary and thus in an area that subjects would be less willing to approach di-
rectly (Fig. 2.1di). Information extracted by subjects was then assayed by means of
a third speaker placed well inside the subject’s territory, which broadcast songs
of one of the interactants 15 minutes after the interaction (Fig. 2.1dii). Three
types of interaction were used, again based on song timing between the speakers:
overlapping, alternating and random. Intruders were then one of four types: over-
lappers, alternators, random (i.e. no consistent role) or males that had been over-
lapped by the opponent. In response to the assay intrusion, subjects responded
with equally high song output to overlappers, alternators and random interactants
but showed a twofold reduction in song towards males that had been overlapped.
In all three of these experiments (Naguib & Todt, 1997; Naguib et al., 1999;
Peake et al., 2001), the only difference between intruders was in relative song
timing during the interaction, i.e. there was no absolute information available

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 23
upon which subjects could base their responses. The results then clearly showed
that subjects had eavesdropped on the interaction as a whole and associated the
roles of interactants with either the location of the singer (Naguib & Todt 1997;
Naguib et al., 1999) or features of his song (Peake et al., 2001).
These three experiments also share the feature that, in each case, the subjects
had no prior experience of either interactant; therefore, decisions must have been
made purely on the basis of the interaction. In reality, territorial songbirds are
likely to have knowledge of the relative strengths of neighbouring males as a result
of direct interactions during territory establishment and maintenance and indi-
rectly from hearing them interact with others. Therefore, individuals may be able
to use these known individuals as ‘yardsticks’against which to measure previously
unencountered individuals. The first three studies mentioned in this section at-
tempted to address this issue by looking at eavesdropping on encounters between
intruders and neighbouring males (McGregor et al., 1997) and/or mates (Otter et al.,
1999; Mennill et al., 2002). In these studies, however, there was little control over
eavesdroppers’prior experience with these yardsticks and the possibility that they
may have themselves contributed to the responses shown (see above).
Peake et al. (2002) attempted to address these problems by carefully control-
ling prior experience with an individual. This experiment was similar to the two-
speaker experiment mentioned above (Peake et al., 2001). The difference was that
one of the interactants (A) was introduced to the subject prior to the interaction by
means of a territorial intrusion simulated by interactive playback (Fig. 2.1ei). The
initial intruder either played an aggressive role, overlapping the subject’s song, or
a much less-aggressive role, beginning a song one second after the subject had fin-
ished each song, allowing the subject to overlap playback. Following this intrusion,
an interaction was simulated outside the territory between the recent intruder
and a male (B) unknown to the subject (Fig. 2.1eii); here either A or B played the
aggressive role by overlapping the song of the other. By combining the outcomes
of the two interactions, four treatment types were carried out that provided in-
formation on the status of B relative to the subject. In two cases the information
available did not clearly show the status of B relative to the subject: either A was
aggressive to both B and the subject or A received aggression from both. In the
other two cases, the information available was clear: A showed low aggression to
the subject but high aggression to B, indicating B to be of low status, or A was
highly aggressive to the subject but received aggression from B, indicating that
B was of high status relative to the subject. In response to subsequent intrusion
by B (Fig. 2.1eiii), males showed a threefold reduction in song towards males that
were of low relative status (as indicated by the information available from the
treatment type), compared with the response to high-status males or males about
which information did not reliably determine status. This result shows that males

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
24 T. M. Peake
combined information from the two interactions, one they took part in and one
they heard, in deciding how to respond to subsequent intrusion.
In all of these experiments, great care was taken to ensure that the informa-
tion available to eavesdroppers was purely relative. This is important in order to
demonstrate social eavesdropping, i.e. that individuals pay attention to the inter-
action rather than simply the absolute outputs of either male. However, during
real interactions it is likely that both absolute and relative information is avail-
able to, and indeed used by, social eavesdroppers. A recent experiment on great tits
(Peake et al., 2004) attempted to address this issue. Male great tits have a repertoire
of one to six song types, many of which are shared by neighbouring individuals
and used during song interactions (matched counter-singing; Krebs et al., 1981;
Falls et al., 1982). Peake et al. (2004) used the two-loudspeaker design of Peake et al.
(2001; Fig. 2.1d) to simulate interactions in which interactants differed in their
use of song types. In each interaction, one speaker (A) produced the same song
type throughout the interaction. The other speaker (B) began producing a differ-
ent song type from A and then switched song types halfway through. On half of
the occasions, B switched to the same song type as A (matching); on the other
half of occasions B switched to a song type that was different from A. Thus, there
were four possible intruders during the assay intrusion: males that switched to
match (matchers), males whose opponent switched to match (matched), males
who switched but did not match their opponent (switchers) and males whose op-
ponents switched but did not match (switched). In both types of interaction there
is a clear absolute difference between males in signalling behaviour, i.e. singing
one song type versus two song types. Between the two types of interaction there is
also relative information, available only in the interaction as a whole, in whether
the switching individual matched his opponent. Subjects responded to simulated
intruders by singing much shorter songs to those individuals that used two song
types compared with those singing one song type. In addition, subjects did not
approach or spend time near switched intruders, compared with no difference
in approach response to the other types of intruder. Therefore, it seems that in
this case the response of males to simulated intruders used both relative informa-
tion in the interaction (switching and matching) and absolute information in the
signalling behaviour of the individual interactants (one or two song types).
Visual interactions in fish
Eavesdropping on visual displays given by male Siamese fighting fish Betta
splendens during male–male interactions has been shown by both males (Oliveira
et al., 1998) and females (Doutrelant & McGregor, 2000). In these experiments, sub-
jects (who could see other males without themselves being seen) were allowed
to witness interactions between males displaying across a transparent barrier

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 25
(Fig. 2.2ai). At the same time, two other males were taking part in a similar in-
teraction that could not be seen by the subject, allowing a control for changes
in the male opponents providing information on the outcome rather than (or as
well as) information from the dynamics of the interaction. Male subjects (Oliveira
et al., 1998) were then introduced to each of the four interactants (two seen and
two unseen) in turn (Fig. 2.2aii) and the response measured. Males responded
to individuals that they had seen lose by approaching and displaying sooner
than with males that they had seen win. No such differences were seen in re-
sponse to the winners and losers of displays that had not been witnessed. Features
of the subjects’ behaviour during the interaction strongly suggested that the
information used by subjects in responding was gathered by eavesdropping (see
discussion in McGregor & Peake, 2000). In experiments with females (Doutrelant
& McGregor, 2000), the seen and unseen interactions were temporally separated
rather than concurrent (Fig. 2.2bi, bii). Following interactions, female subjects
were allowed to move freely so as to exhibit a proximity preference for either
male (Fig. 2.2biii). Females visited seen winners first, more often and spent more
time near and displaying to seen winners than seen losers. Unseen losers were
visited first more often than unseen winners, with no difference in the time spent
near or displaying towards either male.
Similar results have been found in green swordtails Xiphophorus helleri by
Earley & Dugatkin (2002). Males allowed to view contests between other males
without themselves being seen (Fig. 2.2ci) responded more cautiously to perceived
winners, being less willing to initiate contests. Males that had witnessed contests
were much less willing to escalate contests than males that had not seen contests.
Males allowed to interact with contesting males during the contest (Fig. 2.2cii),
and hence assess contestants more directly, were as likely to win contests as those
that had not seen contests, suggesting that individual differences between the fish
settled those contests. Males that had not witnessed contests (Fig. 2.2ciii) tended
to escalate, whereas males that had interacted with contestants were unlikely to
escalate, suggesting that these individual differences were assessed previously.
These results also suggested that, where information on an opponent’s fighting
ability was available from direct interaction, a presumably more reliable source,
males tended to ignore the less-direct information gathered by eavesdropping.
The difficulties of providing visual stimuli in the absence of live animals makes
it much less straightforward to achieve the level of control over interactions
afforded to songbird studies by acoustic playback. The use of models (e.g. Shennan
et al., 1994) or video playback (Oliveira et al., 2000) potentially allows control over
stimuli at a comparable level to acoustic playback, but neither has yet been used to
simulate interactions. In the absence of such an approach, one way to delve deeper
into the relationship between interactions and eavesdroppers is to decouple the

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
26 T. M. Peake
(a)
(d)
(c)
(b)
KEY
(i)
(ii)
(ii)
(i)
(iii)
(i) (ii) (iii)
(i) (ii)
Opaque partition
Transparent partition
One-way glass (arrow shows visible direction)
Fig. 2.2. Schematic representations of experiments described in the text investigating
social eavesdropping on visual interactions in fish. All fish were physically isolated by
partitions; opaque, solid line; transparent, dotted line; one-way glass, dashed line
with arrow showing direction in which visual contact was possible. (a) Design used by
Oliveira et al. (1998) showing (i) two visual signalling interactions across transparent
partitions, one witnessed by the central male, the other not; and (ii) subsequent
presentation (indicated by arrow) of each male to the subject. (b) Design used by
Doutrelant & McGregor (2000) showing (i) interaction between two males witnessed by
a female, (ii) not witnessed and (iii) subsequent observation of female movements
(indicated by arrow). (c) Treatments used by Earley & Dugatkin (2002) in which
(i) interacting males were witnessed by a male that could not be seen by the

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 27
experience of interactants and eavesdroppers, i.e. both parties view the interaction
differently. McGregor et al. (2001) attempted to do this using male Siamese fighting
fish. Subject males were allowed to view two conspecific males apparently inter-
acting across a small gap into which the eavesdropper could not see. The gap was
in reality filled by a small aquarium that either was empty (allowing males to inter-
act across the gap: the ‘real’ interaction; Fig. 2.2di) or contained two fish separated
by an opaque partition such that each of the males viewed by the eavesdropper
was, in fact, interacting with a fish that could not be seen (the ‘apparent’ interac-
tion; Fig. 2.2dii). In the apparent interaction, the eavesdropper’s interpretation of
the aggressive signals given by each visible male was decoupled from that male’s
actual experience in his own interaction with a hidden fish. The results showed
that eavesdroppers responded more aggressively to individuals that had displayed
more during apparent interactions, with no such differentiation between males
involved in real interactions. McGregor et al. (2001) suggested that the proximity
of interactants to (hidden) opponents during the apparent interaction (compared
with the relatively large distance between males (about one fish length) in the
real interaction) resulted in an increase in aggressive displays (tail beating) and
behaviour (attempted biting) in these interactions. Therefore, males either paid
more attention to the information in these particularly aggressive encounters or
viewed the winner of a highly aggressive encounter as a different level of threat
to the winner of an encounter of lower general aggression.
The evidence presented above demonstrates that fish pay attention to informa-
tion available in interactions between conspecifics; however, the source of that
information has not been conclusively shown to be the signals exchanged during
interactions. While the results are consistent with the idea that fish eavesdrop,
the difficulties in presenting subjects with fully controlled visual signalling inter-
actions (cf. acoustic playback) means that using the term ‘social eavesdropping’
for this behaviour may be premature.
Other evidence for social eavesdropping
There is, in addition to the examples given above, a variety of evidence
supporting the importance of social eavesdropping as a means of information
gathering. In some cases, there may be clear physiological effects; adult male
Fig. 2.2 (cont.) interactants, (ii) were witnessed by a male that could be seen (and
interacted with) or (iii) were not seen. (d) Treatments used by McGregor et al. (2001) in
which subjects witnessed two males (i) interacting across an empty divide into which
the subject could not see (the real interaction) or (ii) apparently interacting but in fact
interacting with males hidden from the subject (the apparent interaction). See text
and cited references for more details of each experiment.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
28 T. M. Peake
cichlid fish Oreochromis mossambicus that witnessed fights between conspecifics
showed elevated androgen levels (testosterone and 11-ketotestosterone) compared
with controls (Oliveira et al., 2001).
Female domestic fowl Gallus gallus that witness a known dominant being de-
feated by a stranger readily submit to that stranger in subsequent interactions
(Hogue et al., 1996). When the stranger lost to a known dominant, the witnessing
individual was able to dominate this stranger subsequently on 50% of occasions. A
similar situation was found in juvenile rainbow trout Onchorhynchus mykiss, when
subjects were allowed to interact with fish that had been seen to be dominant in a
previous encounter or who had been dominant but had not been seen (Johnsson &
Akerman, 1998). Individuals that lost to either dominant (seen or unseen) reduced
aggression more rapidly to seen dominants. Individuals that won over these domi-
nants increased aggression more rapidly to seen dominants. These results suggest
that, while the final outcome may be a result of individual differences, informa-
tion obtained before direct interactions occurred enabled individuals to make
decisions about how to respond to these individuals more quickly. In these stud-
ies, the extent to which social eavesdropping occurs is difficult to assess, as it
is not clear whether information extracted by observers is contained in signal
interactions between participants or in other aggressive behaviours.
Knowledge of the social rank relationships of others through observation of
social interactions is also an important part of forming strategic alliances in some
primate species (Seyfarth & Cheney, 2002; Ch. 25). The required amount of knowl-
edge of this kind quickly becomes enormous as group sizes increase and has been
suggested as one selection pressure driving large brain sizes in primates (Seyfarth &
Cheney, 2002).
Identifying types of eavesdropping
So far I have considered examples that clearly fall into one of the two
classes of eavesdropping; interception of signals or of signal interactions. Situ-
ations could be imagined that are not so clearly placed in one category or the
other. Some of these problems may be caused by the difficulties of defining signal
interactions (Chs. 9 and 14). All the examples considered so far have dealt with in-
teractions in which the signallers use the same modality. Many behaviour patterns
that can be viewed as signalling interactions may occur in different modalities or
switch between modalities as the interaction proceeds. If an acoustic signal given
by a male is responded to by a visual signal from the female, then we can clearly
say there has been an interaction involving signals, but does this constitute a sig-
nalling interaction? There is no good reason why signalling interactions, at least in
terms of information transfer, have to occur in the same modality for each party.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 29
However, there may be logistic difficulties in demonstrating social eavesdropping
in such cases; one would have to show that both signals are involved in producing
a response by the eavesdropper, i.e. that neither signal alone produced the same
response.
An example in which the level of information transfer is currently unclear con-
cerns the use of courtship signals by neighbouring males to detect reproductive
attempts in the whitethroat Sylvia communis (Balsby & Dabelsteen, 2003). In this
case, the response of neighbouring males was recorded during experiments that
simulated a territorial male interacting with an intruding male (via playback) or a
receptive female (via a remotely controlled dummy and playback of female calls).
The simulation of a receptive female resulted in greater song flight activity by
neighbours than simulation of an intruding male, and intrusions by the neigh-
bour (and subsequent evictions) were only seen during simulated courtship events.
Song output by the experimental male could not explain the incidence of intru-
sion during courtship interactions by neighbours. As Balsby & Dabelsteen (2003)
acknowledged, their experiment does not rule out the possibility that either sig-
nal alone provides sufficient information to explain the pattern of intrusion, thus
interceptive eavesdropping may be an appropriate description of this behaviour.
However, the detection of a courtship attempt in which the female is receptive
is much facilitated by the presence of both male and female signals; therefore,
it may be the interaction that is important, making this an example of social
eavesdropping.
Information gathering and implications of eavesdropping
So far I have discussed the different ways in which animals may be con-
sidered to be eavesdropping and some of the contexts in which this behaviour has
been shown to occur. As yet I have considered the kinds of information that eaves-
droppers may gain in only the broadest terms. As discussed by McGregor & Peake
(2000), the information available in signalling interactions that is not available in
signals alone is the important distinction between social and interceptive eaves-
dropping. Just as signals are a subset of the information available in an animal’s
environment, signal interactions are a further subset of signalled information.
The way in which animals use signals during interactions may in many cases be
more revealing than the underlying content of the signals themselves, and in any
case the use and content of signals need not necessarily be directly related.
A number of features of signalling interactions may be particularly important
in this respect; many of these features are particularly obvious in agonistic sig-
nalling encounters. Signal exchanges during agonistic encounters are generally
thought to function so as to reduce the likelihood of direct physical aggression

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
30 T. M. Peake
(and its ensuing costs) by allowing assessment of the likely outcome of a direct
fight (e.g. Enquist, 1985). Reliability of signals used in this context may be high
because of the threat of having one’s bluff called by one’s opponent. To this end,
and given the immediacy of the potential punishment for cheating, signals given
during these kinds of exchange may be particularly reliable. It is likely that the
most accurate picture of an opponent’s fighting ability (short of actually fighting
them) is gained by becoming involved in an aggressive signal exchange. The relia-
bility of signals given in this context may also make social eavesdropping a good
alternative in terms of obtaining accurate information while avoiding the risk of
escalation. Similarly, while information on the underlying quality of an individual
may be available in signals, immediate quality (e.g. condition, motivation) may be
assessed more accurately when an opponent calls those factors directly into ques-
tion. Thus signal interactions may provide reliable and up-to-date information on
the current quality of participants.
Second, interactions enable a direct comparison to be made between partici-
pants on a relative scale. Simply knowing the outcome of an interaction provides
the information that A is stronger than B. By paying attention to an interaction it
may be possible to extract information on relative quality, e.g. A is much stronger
than B. If selection favours individuals that ‘just do enough’ to win an interaction,
the available information will underestimate the relative difference in quality
between the opponents. However, the nature of the interaction may provide such
information. For example, one might expect interactions involving highly asym-
metrical opponents to be shorter and less intense than those involving closely
matched opponents. Of course, this assumes that the relative quality of oppo-
nents is the only influence on the information contained in the interaction; the
presence of eavesdroppers may well affect the dynamics of interactions (Ch. 4).
In most situations, the presence of eavesdroppers imposes selection pressures
on signallers. In the case of interceptive eavesdropping, the selection pressures
may be severe, especially in the case of predatory eavesdropping. In this case, the
signaller must accept or avoid the costs of signalling; such avoidance mechanisms
are particularly well understood in insects and anurans (e.g. Gerhardt & Huber,
2002; Ch. 8). In the case of selection pressures imposed by social eavesdroppers, the
situation may be much less clear. In some situations, it is apparent that eavesdrop-
pers have imposed strong selection pressures on signalling interactions by the na-
ture of those interactions; for example, intense song duels in birds often involve
switching to ‘quiet’ song when they approach physical aggression (Dabelsteen
et al., 1998). In some cases, however, the opposite seems to be true: signalling
interactions seem to be much more conspicuous than necessary to transmit infor-
mation between participants (Zahavi, 1979). In these cases, it may be reasonable
to assume that signals used here are, at least partly, ‘intended’ for eavesdroppers,

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 31
i.e. that communication is no longer restricted to the interaction. This view is
supported by work on the effects of audiences on the dynamics of interactions
(Ch. 4) and recent work on altruism (see below).
In cases where asymmetries become clear to interactants, there may be different
pressures acting on each party: for ‘winning’ individuals to advertise the fact and
‘losing’ individuals to hide it. Here eavesdroppers, as part of the selection regime
in which communication systems have evolved, provide individuals with different
payoffs depending on the current social context. In this case, one would expect
a variety of adaptations in signalling behaviour to allow individuals dynamically
to advertise or privatize information in interactions; many such adaptations have
been suggested (e.g. Chs. 3 and 10).
While considerations of social eavesdropping have focused on aggressive in-
teractions, there are a number of similarities between the considerations of in-
formation gathering in this context and in the context of acts of apparent altru-
ism (Johnstone, 2001). Suggestions that altruists may benefit by being perceived
as such assume that observers are able to associate those acts with the individ-
ual performing them and subsequently use that information (Nowak & Sigmund,
1998). Studies of eavesdropping provide clear evidence that this level of association
occurs, albeit in a different context. Many other aspects of cooperative and non-
cooperative behaviour occurring between individuals may similarly be explained
by the passage of information outside the apparent dyad (Ch. 22).
Costs of eavesdropping
Social eavesdropping has been suggested as a relatively cost-free means
of gathering reliable information on rivals or potential mates. However, the costs
have not been explicitly discussed in the literature aside from a sentence by Mc-
Gregor & Dabelsteen (1996) that ‘listening at a distance only involves forgoing
other behaviours such as feeding’. While gathering information in this way is
undoubtedly less costly than becoming involved in aggressive, possibly physical,
interactions, the costs of listening may be greater than commonly assumed.
Evidence from studies of prey detection strongly suggests that animals have lim-
its to the attention they can give to competing tasks (Dukas, 1998; Dukas & Kamil,
2001). Dukas (1998) suggested that the brain has finite capacity to process informa-
tion such that animals can only process a limited amount of information at any
one time. While studies have so far concentrated on visual attention, presumably
a more demanding process than listening, it may be that similar reasoning ap-
plies to the acoustic sense. In this case, eavesdropping may limit the attention
available for other important tasks such as predator vigilance, particularly if so-
cial eavesdropping is a more cognitively demanding task than simply attending

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
32 T. M. Peake
to signals. A study on humans provided support for this view (Pendry, 1998). Hu-
man subjects were asked to form an impression of a target person based upon
information provided to them one item at a time on a computer screen. Without
prior warning and with no specific instruction, participants were simultaneously
played a tape recording of a conversation that was either relevant to them or
not (the contents of the conversations were identical; relevance differences were
achieved by changing the object of the conversation). Subsequent tests showed
that participants extracted more information than expected by chance from the
relevant conversation but not from the irrelevant one. Participants hearing the
relevant conversation were much more likely to obtain stereotypical impressions
of the target and recalled many fewer items of information related to the target
than those hearing a conversation of little relevance.
These sorts of cost may still be comparatively low, particularly if eavesdrop-
ping, or otherwise monitoring the social environment, is relatively rare. However,
social monitoring may represent a large proportion of some animals’ lives. A study
of brown capuchin monkeys Cebus apella suggested that monitoring the social en-
vironment was the main function of vigilance behaviour in this species, and that
the amount of time spent in such activity was highly correlated with the number
of neighbouring individuals (Hirsch, 2002). Individuals spent, on average, 12.7%
of their time vigilant, of which nearly 30% could be directly attributed to social
monitoring and less than 10% to predator vigilance (see also Chs. 19 and 25).
Summary and future possibilities
A defining property of communication networks is that more than one
receiver may detect signals. In many cases, at least some of the receivers are not the
primary target of the signal and in some the majority of potential receivers may
fall into this category. Cases where individuals other than the primary target use
information obtained in signals have been termed eavesdropping by a number of
authors. In this chapter, I have distinguished between examples of eavesdropping
that involve the interception of signals (interceptive eavesdropping) and a more
recently suggested phenomenon of gathering information from signalling inter-
actions between conspecifics (social eavesdropping). Interceptive eavesdropping
can have significant effects on signal design and signalling behaviour when eaves-
droppers impose large costs on signallers (e.g. where such eavesdropping occurs
between trophic levels). The effects of interceptive eavesdroppers within species
and social eavesdroppers in general are less well understood.
The focus of studies on social eavesdropping has so far been territorial species,
reflecting the biases of researchers active in this area. However, many different
social situations would seem ideally suited to promoting eavesdropping as a social

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 33
behaviour. Group and/or colonial living species presumably have an even greater
opportunity to extract information from interactions because of the close proxim-
ity of individuals to one another. Similarly, the focus of most studies of eavesdrop-
ping so far has been on male–male aggressive interactions. Interactions between
males and females (Ch. 7), between group members (Chs. 10 and 25) and between
parents and offspring (Ch. 9) are just some of the areas that potentially offer an
important source of information to eavesdroppers and would benefit from fur-
ther research. Equally as interesting would be the possibility of taking studies of
eavesdropping beyond the trio of two interacting signallers and an eavesdropper
and in so doing place the effect of eavesdroppers in a more extensive and natural
network environment.
The study of social eavesdropping is still in its infancy, yet the number of stud-
ies showing that such eavesdropping occurs has increased dramatically since the
idea was mooted by McGregor & Dabelsteen in 1996. As the prevalence of studies
demonstrating eavesdropping in communication networks increases, the impor-
tance of eavesdropping as a selective force on signalling and social structure will be
better understood. We currently lack a clear theoretical framework within which
to place the importance of social eavesdroppers in the evolution of signalling
systems. As empirical studies continue to provide evidence that animals clearly
have these capabilities, we eagerly await the emergence of models including eaves-
droppers, such as those of Johnstone (2001) and Terry & Lachlan (Ch. 26); models
that make clear predictions about where, how and when eavesdropping should
occur.
Acknowledgements
I would like to thank the following people for providing discussion and comments
that greatly improved both this chapter and my thoughts on communication networks: Pete
McGregor, Thorsten Balsby, Torben Dabelsteen, Giuliano Matessi, Ricardo Matos, Denise Pope
and Andy Terry. While writing this chapter, I was funded by the Zoological Institute, University
of Copenhagen.
References
Aldrich, J. R. 1995. Chemical communication in true bugs and exploitation by
parasitoids and commensals. In Chemical Ecology of Insects II, ed. R. T. Carde & W. J.
Bell. New York: Chapman & Hall, pp. 318–363.
Balcombe, J. P. & Fenton, M. B. 1988. Eavesdropping by bats: the influence of
echolocation call design and foraging strategy. Ethology, 79, 158–166.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
34 T. M. Peake
Balsby, T. J. S. & Dabelsteen, T. 2003. Male calling between courtship sequences in
whitethroats: a way to counter intrusions from neighbouring rivals. Behavioural
Processes, 63, 149–157.
Barclay, R. M. R. 1982. Interindividual use of echolocation calls: eavesdropping by bats.
Behavioral Ecology and Sociobiology, 10, 271–275.
Belwood, J. J. & Morris, G. K. 1987. Bat predation and its influence on calling behavior
in neotropical katydids. Science, 238, 64–67.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland MA: Sinauer.
Bruce, M. J., Herberstein, M. E. & Elgar, M. A. 2001. Signalling conflict between prey
and predator attraction. Journal of Evolutionary Biology, 14, 786–794.
Cade, W. 1975. Acoustically orienting parasitoids: fly phonotaxis to cricket song.
Science, 190, 1312–1313.
Dabelsteen, T., McGregor, P. K., Shepherd, M., Whittaker, X. & Pedersen, S. B. 1996. Is
the signal value of overlapping singing different from that of alternating singing
during matching in great tits? Journal of Avian Biology, 27, 189–194.
Dabelsteen, T., McGregor, P. K., Holland, J., Tobias, J. A. & Pedersen, S. B. 1997. The
signal function of overlapping singing in male robins (Erithacus rubecula). Animal
Behaviour, 53, 249–256.
Dabelsteen, T., McGregor, P. K., Lampe, H. M., Langmore, N. E. & Holland, J. 1998. Quiet
song in songbirds: an overlooked phenomenon. Bioacoustics, 9, 89–106.
Doutrelant, C. & McGregor, P. K. 2000. Eavesdropping and mate choice in female
fighting fish. Behaviour, 137, 1655–1659.
Dugatkin, L. A. 2001. Bystander effects and the structure of dominance hierarchies.
Behavioral Ecology, 12, 348–352.
Dukas, R. 1998. Constraints on information processing and their effects on behavior.
In Cognitive Ecology, ed. R. Dukas. Chicago, IL: University of Chicago Press,
pp. 89–127.
Dukas, R. and Kamil, A. 2001. Limited attention: the constraint underlining search
image. Behavioral Ecology, 12, 192–199.
Earley, R. L. & Dugatkin, L. A. 2002. Eavesdropping on visual cues in green swordtail
(Xiphophorus helleri) fights: a case for networking. Proceedings of the Royal Society of
London, Series B, 269, 943–952.
Eason, P. K. & Stamps, J. A. 1993. An early warning system for detecting intruders in a
territorial animal. Animal Behaviour, 46, 1105–1109.
East, M. 1981. Aspects of courtship and parental care of the European robin Erithacus
rubecula. Ornis Scandinavica, 12, 230–239.
Enquist, M. 1985. Communication during aggressive interactions with particular
reference to variation in choice of behaviour. Animal Behaviour, 33, 1152–1611.
Falls, J. B., Krebs, J. R. & McGregor, P. K. 1982. Song matching in the great tit (Parus
major): the effect of similarity and familiarity. Animal Behaviour, 30, 977–1009.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans. Chicago,
IL: University of Chicago Press.
Greenfield, M. D. 1994. Cooperation and conflict in the evolution of signal
interactions. Annual Review of Ecology and Systematics, 25, 97–126

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 35
2002. Signalers and Receivers. New York: Oxford University Press.
Heller, K. G. 1995. Acoustic signaling in palaeotropical bushcrickets (Orthoptera:
Tettigonioidea: Pseudophyllidae): does predation pressure by eavesdropping
enemies differ in the Palaeo- and Neotropics? Journal of Zoology, 237, 469–485.
Hirsch, B. T. 2002. Social monitoring and vigilance behavior in brown capuchin
monkeys (Cebus apella). Behavioral Ecology and Sociobiology, 52, 458–464.
Hogue, M. E., Beaugrand, J. P. & Lague, P. C. 1996. Coherent use of information by hens
observing their former dominant defeating or being defeated by a stranger.
Behavioral Processes, 38, 241–252.
Hulsch, H. & Todt, D. 1982. Temporal performance roles during vocal interactions in
nightingales (Luscinia megarhynchos B.). Behavioral Ecology and Sociobiology, 11,
253–260.
Johnsson, J. I. & Akerman, A. 1998. Watch and learn: preview of the fighting ability of
opponents alters contest behaviour in the rainbow trout. Animal Behaviour, 56,
771–776.
Johnstone, R. A. 2001. Eavesdropping and animal conflict. Proceedings of the National
Academy of Sciences, USA, 98, 9177–9180.
Kiflawi, M. & Gray, D. A. 2000. Size-dependent response to conspecifics mating calls by
male crickets. Proceedings of the Royal Society of London, Series B, 267, 2157–2161.
Krebs, J. R., Ashcroft, R. & van Orsdol, K. 1981. Song matching in the great tit, Parus
major L. Animal Behaviour, 29, 918–923.
Lloyd, J. E. & Wing, S. R. 1983. Nocturnal aerial predation of fireflies by light-seeking
fireflies. Science, 222, 634–635.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McGregor, P. K., Dabelsteen, T. & Holland, J. 1997. Eavesdropping in a territorial
songbird communication network: preliminary results. Bioacoustics, 8, 253–254.
McGregor, P. K., Peake, T. M. & Lampe, H. M. 2001. Fighting fish Betta splendens extract
relative information from apparent interactions: what happens when what you
see is not what you get. Animal Behaviour, 62, 1059–1065.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Naguib, M. & Todt, D. 1997. Effects of dyadic interactions on other conspecifics
receivers in nightingales. Animal Behaviour, 54, 1535–1543.
Naguib, M., Fichtel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Nowak, M. A. & Sigmund, K. 1998. Evolution of indirect reciprocity by image scoring.
Nature, 393, 573–577.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
36 T. M. Peake
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecifics interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Oliveira, R. F., Rosenthal, G. G., Schlupp, I. et al. 2000. Considerations on the use of
video playbacks as visual stimuli: the Lisbon workshop consensus. Acta Ethologica,
3, 61–65.
Oliveira, R. F., Lopes, M., Carneiro, L. A. & Canario, A. V. M. 2001. Watching fights raises
fish hormone levels. Nature, 409, 475.
Otte, D. 1974. Effects and functions in the evolution of signaling systems. Annual
Review of Ecology and Systematics, 5, 385–417.
Otter, K. A., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits Parus major
assess extra-pair males by eavesdropping? A field study using interactive song
playback. Proceedings of the Royal Society of London, Series B, 266, 1305–1309.
Otter, K., Stewart, I. K., Terry, A. M., McGregor, P. K. & Burke, T. 2001. Extra-pair
paternity in great tits in relation to manipulation of male signals. Journal of Avian
Biology, 32, 338–344.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Peake, T. M., Terry, A. M. R., Dabelsteen, T. & McGregor, P. K. 2002. Do great tits assess
rivals by combining direct experience with information gathered by
eavesdropping? Proceedings of the Royal Society of London, Series B, 269, 1925–1929.
Peake, T. M., Matessi, G., Dabelsteen, T. & McGregor, P. K. 2004. Song type matching,
song type switching and eavesdropping in male great tits. Animal Behaviour,
in press.
Pendry, L. 1998. When the mind is otherwise engaged: resource depletion and social
stereotyping. European Journal of Social Psychology, 28, 293–299.
Roberts, S. C., Gosling, L. M., Thornton, E. A. & McClung, J. 2001. Scent-marking by
male mice under the risk of predation. Behavioral Ecology, 12, 698–705.
Ryan, M. J., Tuttle, M. D. & Rand, A. S. 1982. Bat predation and sexual advertisment in a
neotropical anuran. American Naturalist, 119, 136–139.
Sakaluk, S. K. & Belwood, J. J. 1984. Gecko phonotaxis to cricket calling song: a case of
satellite predation. Animal Behaviour, 32, 659–662.
Seyfarth, R. M. & Cheney, D. L. 2002. What are big brains for? Proceedings of the National
Academy of Sciences, USA, 99, 4141–4142.
Shennan, M. G. C., Waas, J. R. & Lavery, R. J. 1994. The warning signals of parental
convict cichlids are socially facilitated. Animal Behaviour, 47, 974–976.
Stoddard, P. K. 1999. Predation enhances complexity in the evolution of electric fish
signals. Nature, 400, 254–256.
Stowe, M. K., Turlings, T. C. J., Loughrin, J. H., Lewis, W. J. & Tumlinson, J. H. 1995. The
chemistry of eavesdropping, alarm, and deceit. Proceedings of the National Academy
of Sciences, USA, 92, 23–28.
Tobias, J. A. & Seddon, N. 2000. Territoriality as a paternity guard in the European
robin Erithacus rubecula. Animal Behaviour, 60, 165–173.

P1: JZZ/... P2: JZZ/...0521823617c02.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 16:16
Eavesdropping in communication networks 37
2002. Female begging in European robins: do neighbours eavesdrop for extra-pair
copulations? Behavioral Ecology, 5, 637–642.
Wiley, R. H. 1983. The evolution of communication. In Animal Behaviour, Vol. 2:
Communication, ed. P. J. B. Slater & T. R. Halliday. Oxford: Blackwell, pp. 156–189.
1994. Errors, exaggeration and deception in animal communication. In Behavioral
Mechanisms in Evolutionary Ecology, ed. L. Real. Chicago, IL: University of Chicago
Press, pp. 157–189.
Xitco, M. J., Jr & Roitblat, H. L. 1996. Object recognition through eavesdropping: passive
echolocation in bottlenose dolphins. Animal Learning and Behavior, 24, 355–365.
Zahavi, A. 1979. Why shouting? American Nauralist, 113, 155–156.
Zuk, M. & Kolluru, G. R. 1998. Exploitation of sexual signals by predators and
parasitoids. Quarterly Review of Biology, 73, 415–438.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
3
Public, private or anonymous?Facilitating and counteringeavesdropping
t o r b e n da b e l s t e e n
University of Copenhagen, Denmark
Introduction
Animals often live in environments where several conspecifics are within
signalling range of each other simultaneously. This is obvious for lekking species,
but it also applies to territorial species. In theory, this allows complicated patterns
of information flow between the individuals, which can be considered members
of a communication network (Dabelsteen, 1992; McGregor, 1993). A special case of
a network is when dyads of signalling individuals are temporarily or, in the case
of a sparse population, permanently isolated from other signallers or receivers.
Within a network, an individual may gather information about another individ-
ual from simple reception of its signals, but it may also be in a position that allows
simultaneous reception of the signals from two individuals engaged in a dyadic
signalling interaction. Such a position provides a special option for gathering rel-
ative information about the two interacting individuals, for example about their
state, strength or quality (McGregor & Dabelsteen, 1996). The relative information
results from how the two individuals use their signals in the interaction; the in-
formation is relative in the sense that it expresses the relative state or ‘value’ of
the two individuals without necessarily giving information about their absolute
states or values. The extraction of information from a signalling interaction is,
therefore, fundamentally different from simple receiving and so deserves its own
term, eavesdropping (McGregor & Dabelsteen, 1996) or, more specifically, social
eavesdropping (Ch. 2). In theory, relative information about different individu-
als could also be obtained by receiving the signals from each individual in turn,
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
38

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 39
followed by a process in which the absolute information about different individ-
uals is compared.
Do animals eavesdrop?
There are observations of how natural signalling interactions between
two animals may cause conspecifics to approach the interactants and/or signal in
immediate or delayed response to the interaction or directly to interfere with the
interaction (e.g. McGregor & Dabelsteen, 1996). Such observations, whether anec-
dotal (e.g. Snow, 1958) or the result of planned studies (e.g. Bower, 2000), suggest
that animals eavesdrop. There is also experimental evidence that a conspecific
may interfere with a dyadic interaction by signalling predominantly towards the
apparent superior individual of the interaction, suggesting that eavesdropping
took place before interference (Naguib & Todt, 1997; Naguib et al., 1999; Ch. 14).
The ability to gather relative information by eavesdropping and utilize this in
later dyadic encounters with one of the previously interacting individuals has been
demonstrated experimentally in field studies with male birds (e.g. McGregor et al.,
1997), female birds (e.g. Otter et al., 1999; Mennill et al., 2002), captive male fish
(e.g. Oliveira et al., 1998; McGregor et al., 2001; Ch. 5) and captive female fish (e.g.
Doutrelant & McGregor, 2000). Some of the most convincing evidence comes from
field experiments with territorial male great tits, Parus major, that were allowed
to eavesdrop on male–male interactions simulated by means of playback from
two different loudspeakers. The experiments with simulated interactions, which
allowed the best possible control over the relative information made available to
the test subjects, demonstrated that males have the ability to extract relative in-
formation about rivals engaged in a song dual and utilize this in later encounters
with them (Peake et al., 2001). They also demonstrated the ability to combine such
information with the eavesdropper’s own previous direct experience with one of
interacting males (Peake et al., 2002). Given the potential advantages to eavesdrop-
pers of gaining such relative information (see below), this ability is likely to be
used also in non-experimental natural contexts, but we still lack firm evidence
that this really happens.
Potential gains and costs of eavesdropping
Gaining at least relative information about the state, quality or strength
of rivals or potential mates when absolute information is not available, or
when the capacity to compare such information is lacking, must constitute the
aim of any assessment process preceding decision making (e.g. in mate choice).
Eavesdropping was, therefore, predicted to be a widespread phenomenon in both
sexes and because it may constitute a low-cost and low-risk alternative to gathering

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
40 T. Dabelsteen
the same relative information through direct interactions with the individuals
eavesdropped upon (McGregor & Dabelsteen, 1996). For instance, a male that is
eavesdropping on male–male interactions probably uses less energy and runs a
lower risk of injury than it would through a direct interaction, which might es-
calate to actual fighting. Escalated interactions, whether hostile or collaborative
(as in courtship), may also increase predation risk because vigilance is reduced
(e.g. Dabelsteen & Pedersen, 1990; Jakobsson et al., 1995). As eavesdroppers must
divide their attention between interactants, they may have to reduce their vig-
ilance more than simple receivers, which can focus their attention on a single
signaller (e.g. Dukas & Kamil, 2001). It is difficult to identify other potential costs
that are specific to eavesdropping rather than costs that are common to any sort
of information gathering. Overall, the advantages of eavesdropping seem obvious
and predict the evolution of eavesdropping strategies that increase the possibility
of gaining relative information about the participants of an interaction.
Whereas the advantages of eavesdropping are clear, it is not necessarily advan-
tageous to give away information to eavesdroppers. For instance, an individual
that ends up losing a hostile interaction is unlikely to benefit from having its
loss advertised, whereas a winner would have a clear interest in advertising its
superiority (see also discussions in Chs. 2, 4 and 10). An individual should only
start an agonistic interaction if it has no prior knowledge that it will lose to its op-
ponent (e.g. Dabelsteen, 1985). Therefore, participants in an agonistic interaction
should not attempt to withhold information at the start of the interaction, but
do so later should the outcome become uncertain and the interaction escalate.
When the outcome is uncertain, both of the interactants would have an interest
in keeping the interaction private until the interaction has been settled. I know
of no experiments in which subjects were presented with an interaction that had
no difference in relative information, but I predict that a possible response of sub-
jects to such playback would be territorial intrusions and extra-pair behaviour in
relation to the interactants.
Courtship interactions may also be sensitive to eavesdropping. During early
stages of courtship, the two sexes may have conflicting interests. Males have an
interest in preventing rivals from discovering, and perhaps interfering with, their
courtship (Balsby & Dabelsteen, 2003a,b). Females may wish to attract more males
(e.g. Wiley & Poston, 1996), for instance to provoke an interaction between males
upon which they could eavesdrop. When the female has chosen a mate, both the fe-
male and the chosen male have an interest in preventing rivals from eavesdropping
on the intensive courtship interaction that often precedes copulation, because
information that copulation is imminent could lead to attempts to prevent or
even interrupt copulation by rivals. Intrusion by neighbours and subsequent

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 41
interference with courtship (including interruption of copulation) have been ob-
served in a number of different species, for example the robin Erithacus rubecula
(Lack, 1940), the dunnock Prunella modularis (Davies, 1992) and the blackbird Turdus
merula (Snow, 1958). As Snow (1958, p. 86) wrote:
The sight of a pair copulating or about to copulate has an immediate
and powerful effect on neighbouring males. In nearly every case that I
observed, copulations were interfered with by the sudden arrival of one
or two males, who either knocked the copulating male off the female or
prevented him from mounting. And these attacks have been directed
against a territory-holder in the middle of his own territory, where the
neighbours normally never go or, if they do, only with every sign of
nervousness.
Snow (1958) also noticed that courtship of an impassive blackbird female did not
result in such interference. Only when the female responded with copulation
solicitation behaviour (i.e. when there was a real courtship interaction) did the
rival males intrude.
In the whitethroat Sylvia communis, the presence of a female in a territory leads
to significantly more intrusions from neighbouring males than when no female
is present, and territory owners always respond by chasing intruders out of their
territories even when they have to interrupt their courtship of the female (Balsby &
Dabelsteen, 2003a,b). A recent experiment suggested that it is the courtship inter-
actions that make neighbours intrude. The experiment compared the intrusion
rate of male subjects that could eavesdrop on either their neighbours interacting
with a loudspeaker playing normal full whitethroat song (song duel treatment)
or courtship interactions between their neighbours and Jumping Sylvia (Balsby &
Dabelsteen, 2002), a remotely controlled stuffed female that could jump and vocal-
ize (courtship treatment). Whereas the song duel treatment never elicited intru-
sions into the neighbour’s territory by the male subjects, the courtship treatment
did so in 56% of trials, and 44% of intrusions led to direct interference with the
neighbour’s courtship of, or copulation with, Jumping Sylvia (Balsby & Dabelsteen,
2004).
The different potential gains and costs of eavesdropping to interactants in
agonistic and courtship contexts predict the evolution of strategies that counter,
or reduce, the negative consequences and strategies that ignore, or even facilitate,
it. In the rest of this chapter, I discuss how communication behaviour can be made
public, private or anonymous and how eavesdroppers should behave. The focus is
on vocal interactions of territorial songbirds.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
42 T. Dabelsteen
How best to eavesdrop
An eavesdropper should attempt to achieve the best conditions for receiv-
ing the signals of both interactants simultaneously and at the same time reduce
the potential costs of missing more important interactions or being detected by
predators. Since eavesdropping always involves increased risk of predation be-
cause of divided attention, the duration of an eavesdropping session should be
restricted to the time necessary for the ‘intended’ gathering of information. In
addition, a male eavesdropper should attempt to stay undetected by the interac-
tants because at least one (e.g. the loser) will always have an interest in concealing
the interaction irrespective of its nature, i.e. whether it is agonistic or sexual. A
detected female eavesdropper, by comparison, may benefit from the intensified
interaction between male interactants. Such intensification of males’displays and
fights is often observed in lekking species when females pass or arrive on a lek
(e.g. Lack, 1939; Hovi et al., 1995).
Predictions on how best to eavesdrop
Simply approaching the interactants or moving to a position where the
signals from both individuals can be received simultaneously and equally well will,
of course, enhance eavesdropping. When the information gathered from an inter-
action depends on the timing of airborne sound signals from two interactants,
eavesdropping may be complicated by the relatively slow speed of sound trans-
mission in air. In such cases, eavesdroppers should approach to positions where
the distances to the two interactants are equal (e.g. Dabelsteen, 1992). Like simple
receivers, eavesdroppers may also improve the conditions for receiving sound sig-
nals by ascending to a high perch. Depending on the nature of the surrounding
vegetation, this may improve the possibilities for observing visual displays. The
evidence for the improvement of sound reception comes from sound transmission
experiments that quantified the degradation of natural sound signals transmit-
ted over natural communication distances using natural signaller and receiver
positions in the appropriate habitat for the study species (Box 3.1).
Sound transmission experiments indicate that simply leaving the ground and
flying a few metres up to the undergrowth of a forest will not necessarily im-
prove the receiving conditions if the signaller is already located higher up in the
vegetation. This is perhaps not surprising given the ‘ground’effect (attenuation, es-
pecially of low-frequency sounds, when sounds are transmitted along the ground)
when both the signaller and the receiver are close to the ground (e.g. Wempen,
1986; Embleton, 1996; Nemeth et al., 2001). However, ascent by the receiver to
perches above the undergrowth may improve receiving conditions considerably
depending on the type of habitat. For instance, a whitethroat that moves from

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 43
4 to 9 m above ground level in an open whitethroat habitat will not improve
sound receiving conditions further (Balsby & Dabelsteen, 2003c). In whitethroats,
high perches mainly seem to help visual surveying of the surroundings. There is
a very different result in a closed forest habitat before leaf burst. Experiments
with songs of three different species, the blackbird (Dabelsteen et al., 1993), the
wren Troglodytes troglodytes (Holland et al., 1998, 2001) and the blackcap Sylvia atri-
capilla (Schmitz et al., 2000; Mathevon et al., 2004), show that receivers may obtain a
considerable improvement by ascending to high perches. For instance, a blackcap
receiver that moves from 4 to 9 m above ground level obtains improvements that
would correspond to a horizontal approach towards the singer of up to 23 m, i.e.
almost half an average territory diameter (Fig. 3.1 and Box 3.1) (Schmitz et al., 2000;
Mathevon et al., 2004).
Box 3.1
Transmission-caused sound degradation has at least four aspects. (a) Sound
signals are attenuated because of spherical spreading (6 dB per doubling of
distance) and excess attenuation (EA) caused by absorption and multiple
scattering (e.g. Michelsen, 1978). (b) This attenuation will, together with the
addition of background noise, reduce the signal-to-noise ratio (SNR).
(c) Selective frequency filtering, atmospheric turbulence and reverberation
will result in a distortion or blurring within the sounds of their frequency
and amplitude patterns over time (e.g. Wiley & Richards, 1982), which can be
quantified by a blur ratio (BR; Dabelsteen et al., 1993). (d) Reverberation will
cause an elongation of the sounds with tails of echoes, which can be
quantified by a tail-to-signal ratio (TSR; Holland et al., 2001). Sound
transmission experiments indicate that all these aspects of sound
degradation change with distance: EA, BR and TSR increase and SNR
decreases with distance. The experiments also indicate that sound
degradation decreases with increasing height above ground level of signaller
(loudspeaker) and receiver (microphone). This means that both signaller and
receiver may improve communication by sound signals by moving up to
high perches. Exactly how large these improvements are is perhaps best
understood when the regressions of the values of each of the four
degradation measures against the logarithm of the distance are used to
translate the improvements obtained by moving upwards into virtual
horizontal distances that a signaller or receiver would have to approach a
receiver or signaller, respectively, to obtain the same improvements.
Figure 3.1 shows an example.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
44 T. Dabelsteen
ReceiverTSR EABRSNR
Sender
BR TSREASNR
Fig. 3.1. Virtual horizontal distances (see text) that a blackcap receiver or signaller
would have to approach a signaller or a receiver, respectively, to obtain
improvements similar to those obtained by moving in the vegetation from 4 to 9 m
above ground level. In this example, the distance between signaller and receiver is
50 m and the calculations are based on average degradation values for 10 different
blackcap song elements. For a signaller, the average improvements for each of four
degradation measures (indicated by silhouettes of flying birds for SNR, BR, EA and
TSR; defined in Box 3.1 text) correspond to virtual approaches only slightly longer
than the 5 m ascent. For a receiver, the virtual approaches are considerably longer,
corresponding to almost half the average territory diameter of 25 m, indicated by
the horizontal line along which the bird silhouettes are flying. Movements and
vegetation are to scale, whereas bird silhouettes are enlarged. (Modified from
Schmitz et al., 2000.)
These sound transmission experiments predicted that eavesdroppers on vocal
interactions should ascend to high perches in a forest habitat before leaf burst. By
ascending to high perches instead of approaching interactants, an eavesdropper
would save energy and also avoid moving too far away from locations where future
interactions of potential interest might take place. By staying somewhere in the
middle of its territory during periods with high-singing activity, for instance at
dawn or during most of the morning, a perching bird would also potentially be
able to monitor a number of simultaneously occurring interactions and switch its
attention between different interactions depending on where the most interest-
ing developments happen. Staying at some distance from the interactants would

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 45
also help to conceal eavesdropping activity. Exactly where and how high above
ground level an eavesdropper should position itself will, of course, depend on the
local vegetation and the availability of cover to avoid detection by predators or
interactants.
Female eavesdroppers have the same interest as males with respect to predators,
but not necessarily with respect to the interactants (see above). If the predation risk
constrains them to stay hidden in the vegetation, they could, in theory, announce
their presence vocally.
For how long should individuals eavesdrop? Eavesdropping should be as brief
as possible to reduce the risk of predation caused by divided attention, but suf-
ficiently long to allow the extraction of relative information about the interac-
tants. What exactly ‘sufficient’ means seems to depend heavily on the context.
For instance, intensive courtship interactions will almost immediately inform an
eavesdropper that both members of a pair are ready to copulate, whereas agonistic
interactions between males may sometimes progress very slowly and the signalling
may only reveal the superior male after some time has passed. In such cases, we
cannot predict the duration of eavesdropping because this will be controlled by
the interactants.
In some species, song repertoire size is correlated with morphometric measures
of males, suggesting that repertoire size is capable of providing absolute informa-
tion about male quality. In such species, relative information about the quality of
different individuals could be deduced by receiving the songs from individuals in
turn. However, in some of these species, studies have failed to show that repertoire
size has signal value. In experimental studies where the design has only allowed
simple receiving, the negative results could reflect the fact that the species do not
have the capacity for determining absolute repertoire size. It would be interesting
to investigate if such species eavesdrop on song duels to gather relative informa-
tion about repertoire sizes. If so, the minimum time needed to obtain the relative
information by eavesdropping could be predicted from cumulative plots of the
number of new songs sung as a function of the total number of songs sung by
each of the interactants. The time taken to get a reliable indication of the relative
repertoire sizes of two interactants would, of course, depend on who is interacting
with whom since the cumulative plots of some pairs of individuals become dif-
ferent sooner than other pairs. Male whitethroats, for instance, apparently do not
vary their responses to playback of different repertoire sizes (Balsby & Dabelsteen,
2001): maybe because they do not consider repertoire size in agonistic contexts;
maybe because they do not have the capacity for determination of absolute reper-
toire size. If they could perceive relative repertoire sizes, such males would have
to eavesdrop for between two minutes (on an interaction between males A and B

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
0
100
200
300
400
500
600
700
040
00
8000
1200
016
000
Tot
al n
umbe
r of
ele
men
ts
A
B
C
Number of different elements
Fig.
3.2.
Cu
mu
lati
vep
lots
ofn
ewso
ng
elem
ents
fou
nd
asa
fun
ctio
nof
son
gel
emen
tssu
ng
by18
dif
fere
nt
wh
itet
hro
atm
ales
.In
10m
inu
tes,
app
roxi
mat
ely
1000
elem
ents
wer
esu
ng.
The
lett
ers
refe
rto
dif
fere
nt
mal
esd
iscu
ssed
inth
ete
xt.(
Mod
ifie
dfr
om
Bal
sby,
2000
.)
46

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 47
in Fig. 3.2) and more than 20 minutes (when males B and C are interacting) to
detect which male has the largest repertoire (Balsby, 2000).
Do eavesdroppers fit the predictions?
The experiments that sought to establish eavesdropping behaviour in
songbirds did not monitor the locations of subjects in a way that allows the above
predictions to be tested. However, there is evidence (personal observation) that
male great tits may approach and/or be perched during experimentally induced op-
portunities for eavesdropping. There are also observations of neighbouring males
approaching during interactive playback to territorial blackbirds, and male black-
birds may interrupt ongoing behaviour and fly to high perches and stay silent in
response to other males starting to sing (personal observation). Although anecdo-
tal, such observations support the idea that eavesdroppers may attempt to achieve
the best conditions for receiving the signals of interactants. It is also possible that
some of the variation in the results obtained in the field experiments with great
tits (Otter et al., 1999; Peake et al., 2001, 2002) was due to the ‘uncontrollable’
eavesdroppers (i.e. subjects) having been in positions that varied in how well the
interactants could be heard.
Evidence suggesting that birds probably engaging in eavesdropping behave in
a way that would best receive signals of interactants comes from a radio-tracking
study of 11 unmated female great reed warblers Acrocephalus arundinaceus (Bensch
& Hasselquist, 1992). Their routes were taken as evidence for female assessment
preceding mate choice (Bensch & Hasselquist, 1992; but see Ch. 7). Figure 3.3 shows
the route followed by one such female. During the eight hours the female was fol-
lowed, it had four relatively long stays within a limited area, indicated by clusters
of black dots in Fig. 3.3. The first cluster indicates a stay of 40 minutes at the
location of the female early in the morning before it starts to move. The other
three clusters, each corresponding to stays of about 60 minutes’ duration, are at
locations with almost exactly equal, relatively short, distances to two males at a
time: perfect positions for eavesdropping on vocal interactions between males.
One cluster would allow eavesdropping on interactions between males VII and III,
the next on males III and II, and finally on males V and VI. These results suggest
that about 60 minutes was needed to gather information from two males at a time,
including relative information from the occasional singing interactions between
the males.
It has been hypothesized that females of some bird species use loud fertility
advertisement calls to incite male–male interactions (e.g. Montgomerie & Thorn-
hill, 1989; Hoi, 1997). Although the actual fertility advertisement function of the
loud female calls is doubtful, it is possible that such calls may initiate or even
intensify ongoing male–male interactions and hence act as an aid in mate choice
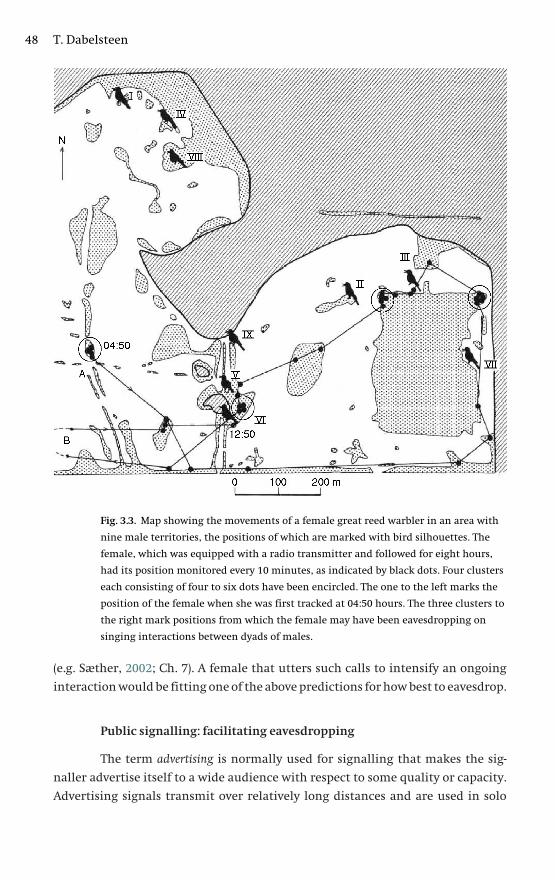
P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
48 T. Dabelsteen
Fig. 3.3. Map showing the movements of a female great reed warbler in an area with
nine male territories, the positions of which are marked with bird silhouettes. The
female, which was equipped with a radio transmitter and followed for eight hours,
had its position monitored every 10 minutes, as indicated by black dots. Four clusters
each consisting of four to six dots have been encircled. The one to the left marks the
position of the female when she was first tracked at 04:50 hours. The three clusters to
the right mark positions from which the female may have been eavesdropping on
singing interactions between dyads of males.
(e.g. Sæther, 2002; Ch. 7). A female that utters such calls to intensify an ongoing
interaction would be fitting one of the above predictions for how best to eavesdrop.
Public signalling: facilitating eavesdropping
The term advertising is normally used for signalling that makes the sig-
naller advertise itself to a wide audience with respect to some quality or capacity.
Advertising signals transmit over relatively long distances and are used in solo

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 49
signalling as well as in signalling interactions. A good example of such an ad-
vertising signal is the full song of songbirds, which often has a dual function:
to attract potential mates and repel rivals. Interactants using advertising signals
will, of course, expose themselves to eavesdropping; however, if such eavesdrop-
ping has no immediate or subsequent adverse consequences, then it should not
change the signalling behaviour of interactants. Yet there could also be situations
where one of the interactants might benefit from the presence of eavesdroppers,
for instance by making its superiority relative to the opponent widely known.
Here the superior individual should continue signalling or even attempt to make
it more effective, whereas the inferior individual should stop using advertising sig-
nals and ultimately stop interacting. It could be argued that an individual should
always make advertising signalling as effective as possible. However, when effec-
tiveness depends on energy used or some other cost, animals may wish to limit
the costs during solo signalling more than during the usually shorter signalling
interactions, perhaps to avoid exhaustion.
Predictions for public signalling
Signallers could allow eavesdropping simply by choosing signals that
transmit effectively in their physical environment and by signalling from posi-
tions and at times of the day that would maximize signal transmission. The ev-
idence for such choices in use of sound signals comes from sound-transmission
experiments. All of the experiments with bird song mentioned above show that
some sound types transmit better than others, even among functionally equivalent
types. For instance, low-frequency, narrowband and unmodulated sounds seem to
transmit best in a forest habitat. In the full song of the blackbird (Fig. 3.4a), the
introductory low-frequency whistle or motif sounds transmit much better than the
terminating highly modulated and broadband twitter sounds; among the whistle
sounds, the relatively unmodulated and constant frequency ‘CF-sounds’ transmit
better than sounds that are frequency modulated (‘FM-sounds’)and/or have energy
rich overtones (‘MIX-sounds’) (Dabelsteen et al., 1993). Also in great tit song, rela-
tively ‘pure-toned’sound elements seem to transmit better than highly modulated
‘buzz’ elements (Blumenrath et al., 2004). In blackcap song the terminating rela-
tively low frequency, pure tone and narrowband motif sounds are less attenuated
than the introductory highly modulated broadband twitter sounds (Dabelsteen &
Mathevon, 2002). In wren song, low frequency sounds are least attenuated
(Holland et al., 1998), and in antbird (Thamnophilidae) song, the narrowband and
low-frequency sounds transmit best (Nemeth et al., 2001).
Low-frequency sounds also have another property that make them suited for
advertising: they usually radiate from the vocalizing individual more or less
equally well in all directions (i.e. they are omnidirectional). This has been shown

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
50 T. Dabelsteen
0.5 1.0 1.5 2.0 2.5 3.0
2
4
6
8
10
0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
2
4
6
8
10kHz
s
s
(a)
(b)
kHz
Fig. 3.4. Sound spectrograms of blackbird song. (a) A male full song recorded in the
field; the introductory whistle part is underlined. (b) A male full song, which is
framed, has the highest frequencies of the terminating twitter part overlapped by a
female copulation trill, the duration of which is indicated by a horizontal line. This
was recorded in an anechoic chamber and shows the part of a playback trial where
playback of full song to an oestradiol-treated female elicits a copulation solicitation
posture and a copulation trill (see Dabelsteen, 1988). The movements associated with
the female posturing are indicated by broadband white noise, especially over the
whistle part of the male’s full song but also over the last third of the copulation trill.
Spectrograms were produced in Avisoft (FFT 512 points, flat top, overlap 75%,
frequency resolution 43 Hz, time resolution 5.805 ms).
in laboratory studies with blackbirds (Larsen & Dabelsteen, 1990). Species using
sounds with a more directional radiation pattern may facilitate advertising by
moving their head from side to side during singing (Brumm & Todt, 2003).
Sound transmission is normally believed to be most effective from high perches.
The sound transmission experiments with bird song also indicate that a signaller
should at least ascend to a few metres above ground level to make sound transmis-
sion effective (e.g. Nemeth et al., 2001); transmission may sometimes be improved
further by ascending to high perches, but only slightly. In a forest, the improve-
ment that a signaller obtains by moving to higher perches sometimes seems very
small relative to that a receiver might obtain by the same movement (e.g. Fig. 3.1);
consequently, high perches should perhaps be called listening posts rather than
song posts in these cases (Schmitz et al., 2000; Mathevon et al., 2004). Sound trans-
mission is also believed to be most effective at dawn (e.g. Henwood & Fabric, 1979).

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 51
However, very few studies have investigated this with sound transmission experi-
ments using natural sound signals. A recent study with blackcap song, at the peak
season of their singing immediately after their return to the breeding sites in
the spring, failed to show that their songs propagate most effectively at dawn
(Dabelsteen & Mathevon, 2002).
Overall, the sound transmission experiments suggest that the most widely prop-
agating sound signals would be relatively loud, low frequency, narrowband and
unmodulated; they would be emitted from at least a few metres above the ground.
The effect of higher song perches is small and the optimal time of day is uncer-
tain and probably depends strongly on the weather. Windy and rainy conditions
will constrain advertising for different reasons (e.g. Lengagne & Slater, 2002), al-
though birds, in theory, might compensate for a high level of background noise
by increasing the output level of songs (Lombard effect; Lombard, 1911). For in-
stance, nightingales Luscinia megarhynchos may use a higher song output level in
noisy environments than at less-noisy locations (Brumm, 2004).
Do interactants use public signals to facilitate eavesdropping?
Observations suggest that birds sometimes increase the loudness of their
singing when they shift from solo singing to counter-singing (i.e. interacting).
Unfortunately, such observations have rarely been verified with sound pressure
level (SPL) measurements. An exception is a study by Brumm & Todt (2002), which
showed that male nightingales singing full songs increase the SPL by more than
5 dB when they shifted from solo singing to playback-induced counter-singing.
However, it is a question of whether the increase in SPL has evolved to facilitate
eavesdropping or whether it represents an increased arousal of the singers, which
as a side effect inevitably facilitates eavesdropping on the vocal interaction. Play-
back experiments support the arousal hypothesis because a higher SPL elicits a
stronger response (e.g. Dabelsteen & Pedersen, 1992).
There is, as yet, no evidence that interactants facilitate eavesdropping dur-
ing interactions by using signals from their repertoire of functionally equivalent
advertising signals that transmit more effectively than those used during solo
signalling. For song types, this would imply that blackbirds engaged in a song
duel across the border between their territories should use more CF-sounds and
fewer FM- and MIX-sounds than during solo singing. However, the opposite seems
to happen, perhaps because the sound types are not functionally equivalent, with
the FM-sounds expressing the highest degree of arousal (Dabelsteen & Pedersen,
1992). Neither is there any evidence that higher perches are used during interac-
tive singing than during solo singing. In both contexts, the use of relatively high
perches is probably mainly to improve sound reception, either of the songs of the
opponent or of vocal responses to the solo songs.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
52 T. Dabelsteen
When singing interactions using advertising signals escalate, the interactants
often shift to special types of close range, private singing (see below). However,
after agonistic encounters, one or both of them may shift back to loud advertising
songs. When both individuals do this and continue to interact, the shift back to
advertising songs can be said to facilitate eavesdropping on their interaction. The
advertising singing of the eventual winner of the escalated interaction is often
different from that of the loser and, therefore, is referred to as an ‘acoustic victory
display’ (e.g. Bradbury & Vehrencamp, 1998; Ch. 6). When only one of the birds
shifts back to advertising songs, it is usually the winner of the interaction. In this
case, the winner’s singing cannot be said to facilitate eavesdropping since there is
only one signaller left.
Private signalling: countering eavesdropping
An interactant should use private signals in an interaction whenever pub-
lic signals would incur potential risks. This is true when the risks are immediate,
for example from predators because of reduced vigilance during interactions com-
pared with solo signalling, or from male eavesdroppers, which may take advan-
tage of an interactant’s involvement with a rival to pay its mate a visit during the
interaction. It is also true when the risks are less immediate, for example eaves-
droppers that can extract information about an interactant and utilize this in
future encounters with it. Whatever the risks, they seem likely to increase with
the duration of the interaction. An important step to make an interaction private
would, therefore, be to make it as brief as possible, because this would reduce the
risk of detecting the signalling interaction and of obtaining useful information.
However, as explained in the next section, there are other options.
Predictions for private signalling
Anything that limits signal transmission and reception may, of course,
help to make signal interactions private, in effect the opposite of advertising.
Private signalling should, therefore, employ signals, signaller positions and sig-
nalling times that are the opposite of those used for advertising. Vocally interact-
ing birds that wish to signal privately should use sound signals that are relatively
high frequency, broadband and highly modulated, and emit them with a low SPL.
This would reduce the number of potential receivers for two reasons: such sounds
attenuate and degrade relatively fast with distance (e.g. Dabelsteen et al., 1993;
Holland et al., 1998) and are directional in the sense of being beamed away from
the sender in one direction (e.g. Larsen & Dabelsteen, 1990). Sound transmission
experiments also show that privatizing would be most effective if the interactions
take place close to the ground or in the undergrowth of a forest.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 53
Background noise, constant as well as transient, may mask communication
sounds and hence contribute to private signalling. This may be especially impor-
tant when eavesdropping involves information gathered from the timing of songs.
For instance, the degree to which songbirds delay their songs relative to each other
or overlap each other’ssongs seem to be important indicators of social dominance
(e.g. Dabelsteen et al., 1997, 1998; Langemann et al., 2000) that are utilized by
eavesdroppers (e.g. McGregor et al., 1997; Peake et al., 2001). Receivers in general
may have internal representations of songs which would help them to reconstruct
songs which are partially masked by transient background noise, for instance the
vocalizations of other birds. However, such representations would probably not
be of much help to eavesdroppers in reconstructing how interactants delay their
songs relative to each other or overlap each other’s songs (e.g. Poesel et al., 2001).
Therefore, private signalling should take place at times of the day when the level
of background noise is high.
Do interactants use private signals to counter eavesdropping?
So-called quiet song in songbirds, sometimes referred to as soft or whis-
per song, seems to be a good candidate for a private signal (e.g. Dabelsteen et al.,
1998). Quiet singing is still relatively unexplored, but wherever it has been discov-
ered it has been accompanied by an active behaviour rather than perching, for
instance posturing and/or escorting or chasing another individual, and it is often
sung more or less continuously without the intersong pauses characteristic of full
singing. Quiet singing usually occurs at close range during escalated interactions,
collaborative as well as competitive, i.e. in contexts of eavesdropping (e.g. Titus,
1998; Balsby, 2000; Balsby & Dabelsteen, 2003a,b).
Good examples of quiet singing occur in the blackbird, the redwing Turdus ili-
acus, the robin, the dunnock and the alpine accentor Prunella collaris (Dabelsteen
et al., 1998). The blackbird has at least three types of quiet singing. Counter-singing
males switch from loud full songs with whistle sounds to quiet twittering without
whistles when the interaction escalates and the interactants approach each other
to within about 10 m. A male also twitters quietly during intense courtship of a
soliciting female immediately before copulation, and the soliciting female sings
a very quiet copulation trill (Figs. 3.4b and 3.5). Until recently, the aggressive and
sexual twitter were believed to be identical because that is how they sound to the
human observer (e.g. Dabelsteen et al., 1998). However, recent spectrographic anal-
yses have suggested that sexual twitter may also include specific sounds (Fleron,
2003) and hence have the potential to communicate the sexual arousal of the
singer in the same way as the accompanying posturing of the male (e.g. Snow,
1958).

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
54 T. Dabelsteen
0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5
2
4
6
8
10kHz
5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5
2
4
6
8
10kHz
s
Fig. 3.5. Sound spectrogram of the quiet vocalizations uttered during 10 seconds of a
natural blackbird courtship interaction early in the morning soon after the dawn
chorus. The courtship vocalizations dominate among the sounds that are visible on
the spectrogram above 4 kHz, most of them being male sexual twitter. A horizontal
line indicates a trill that is likely to be the female copulation trill. Two full songs of a
robin that is singing close by are indicated by dashed horizontal lines: the first has
visible energy up to about 5 kHz; the second starts with a high-frequency part
(approximately 8 kHz) and ends with a more low-frequency part (approximately
4 kHz). Most of the sounds between 1.8 and 3 kHz are whistle parts of more distant
singing blackbird males. Spectrograms were produced as in Fig. 3.4.
Quiet singing in the blackbird seems to fit nearly all of the predictions for a
private signal, with respect to sound type, variability and SPL, and with respect to
where, when and for how long it is used. Relative to the whistle sounds of full song,
all three types of quiet song consist of relatively high-frequency, broadband and
modulated sounds (Figs. 3.4, and 3.5). Twitters are at least 10 dB(A) quieter than
whistles (Dabelsteen, 1984) and sound transmission experiments have shown that
the twitters degrade and attenuate much faster than whistles (Dabelsteen et al.,
1993). Whereas whistles seem capable of transmitting over at least two to four
territory diameters, twitters usually transmit less than one diameter (Dabelsteen
et al., 1993). Twitters are also more directional than whistles (Larsen & Dabelsteen,
1990). Female copulation trills sound even quieter than male twitters (personal
observations) and probably have a radiation pattern and transmission capability

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 55
resembling that of twitters. Both aggressive twitter interactions and courtship
interactions are most frequently performed under cover close to the ground or in
the undergrowth (personal observation) and early in the morning after the males’
dawn chorus at a time where the level of background noise from the vocalizations
of other species is very high (Fig. 3.5) (e.g. Messmer & Messmer, 1956). Finally,
the duration of aggressive twitter interactions can vary quite a lot, but courtship
interactions are always very brief (e.g. Snow, 1958).
The term quiet singing should not be taken too literally since some of the bird
vocalizations referred to as calls seem to be used in the same contexts and to
have the same acoustic structure as quiet singing. Good examples are the dscharp-
and ze-calls of the female whitethroat, which are used in courtship interactions
where the male performs a special diving song display (Balsby & Dabelsteen, 2002,
2003a,b). The two calls fit the requirements for private signals: they have sound
structures and low SPL (measured for dscharp-call, too low to be measured for
ze-call), which make them short-range signals, and they are uttered from low
positions in the vegetation (Balsby & Dabelsteen, 2003a,b).
The eavesdropping contexts with quiet vocalizing are characterized by risks to
the interactants from predation and eavesdropping. This probably applies to all
eavesdropping contexts, and both these risks seem strong enough to cause the
evolution of private signals. It will, therefore, be very difficulty to disentangle the
relative influence of the two risks in the evolution of quiet singing and calling.
Quiet vocalizing might also have evolved simply to save energy during close-range
communication. However, this seems unlikely given that quiet vocalizations are
always accompanied by movements and sometimes even by posturing. This, to-
gether with the continuous nature of quiet vocalizing, suggests that birds do
not necessarily save energy by switching from loud advertising singing to quiet
singing. If energy saving was the main purpose, the bird could simply lower the
output level of singing as in the non-social subsong (e.g. Thorpe & Pilcher, 1959).
They do not have to switch to another song type and otherwise behave in a way
that would counter eavesdropping. However, there are contexts where predation
risk may be the main factor responsible for private signals, but these contexts
do not, as far as I know, involve a signalling interaction and, therefore, do not
constitute eavesdropping in the sense used in this chapter. A good example is
the so-called nest-relief song or calling-out song produced by males of many song-
bird species (e.g. Gompertz, 1961; Stork, 1971; Ficken et al., 1978; Lind et al., 1996).
Structurally, these songs seem identical to the full songs used during advertis-
ing singing, but they are usually sung with a very low SPL (although higher than
in subsong; personal observations) and they are also relatively short or few in
number.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
56 T. Dabelsteen
Anonymity: another way to counter eavesdropping
Private signals counter eavesdropping but do not necessarily prevent it.
However, if interactants can remain anonymous, eavesdroppers cannot attribute
information gained from the interaction to any particular individual and, there-
fore, this cost of eavesdropping to interactants is removed. For instance, eaves-
droppers on an interaction between unknown individuals could not make use of
information in subsequent encounters with them.
Predictions for anonymity
Anything that makes individual identification of a signaller based on its
signalling activity more difficult would help anonymity. One way to constrain
individual recognition of vocalizing birds might be to increase the variability of
their vocal output. For instance, experiments suggest that song repertoires of fewer
than about 25 song types do not interfere with song-based neighbour recognition,
whereas repertoires of more than about 100 do (e.g. Stoddard, 1996; Molles &
Vehrencamp, 2001). Birds that want to make their singing anonymous should,
therefore, change the way they use their repertoire or somehow increase it during
interactions, for instance by switching to another type of singing. If a male can
be identified by the repertoire being sung from particular song posts, switching
to unusual posts combined with frequent post shifts might also help to achieve
anonymity.
Do animals make themselves anonymous in eavesdropping contexts?
Quiet singing in songbirds may aid anonymity because, unlike full song,
it is used almost everywhere and not from preferred posts, and most importantly it
seems a lot more variable than full song (e.g. Dabelsteen et al., 1998). For instance,
when a rival intrudes, a male blackbird uses the aggressive twitter everywhere in
its territory rather than from its usual song posts. Also its repertoire of different
aggressive twitter motifs far exceeds the threshold value (> 100; Fleron, 2003)
that constrains individual identification (e.g. Stoddard, 1996). The repertoire size
of full-song whistles averages 44 (Rasmussen & Dabelsteen, 2001).
The idea of anonymity in signals is novel and, therefore, not really studied yet,
but it seems clear that, unlike private signals, anonymity is difficult to explain
as a response to predation risk. Unlike the aspects of private signals mentioned
above, which can reduce the risk of detection by predators, increased variation
per se does not seem to have such an effect. Relative to full song, quiet song
endowed with a variation similar to that of full song would still be very difficult
for a predator to detect. The evolution of the relatively larger variation in quiet
singing could have resulted from sexual selection, which is believed to play a role

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 57
in the evolution of avian repertoire sizes (e.g. Searcy & Yasukawa, 1996). However,
this would require sexual selection to have acted more strongly on quiet song
than on full song, even though both types of song seem to play important roles in
mate choice and deterrence of rivals. Finally, the relatively large variation of quiet
song could be a side effect of a sound production mechanism, coupling low SPL
or high frequencies with large variation. However, this also seems unlikely given
that full song is sometimes uttered in non-social contexts as very quiet so-called
subsong (Thorpe & Pilcher, 1959) and that high-frequency quiet singing contains
the same type of fixed combinations of sound elements as full song (e.g. Rasmussen
& Dabelsteen, 2001; Fleron, 2003). At the moment, it seems likely that the large
variation of quiet singing relative to full singing in some species reflects the need
for singer anonymity to counter negative consequences of eavesdropping.
Summary
There is now experimental evidence that animals have the ability to gather
relative information about interactants by eavesdropping and utilize this informa-
tion in subsequent decision making about how to behave towards the interactants.
We still lack good observational evidence that this happens in non-experimental
natural contexts, but this seems likely given the obvious advantages to the eaves-
dropper. It is also likely that eavesdroppers behave in ways that enhance their
ability to eavesdrop. The potential gains of being eavesdropped upon are more dif-
ficult to identify but may exist in special situations and, therefore, have led to the
facilitation of eavesdropping, including enhancing advertising signals. The poten-
tial costs of being eavesdropped upon are much more obvious and set the scene for
an evolutionary arms race between eavesdroppers and interactants, with private
signals and anonymity reducing the costs of being subjected to eavesdropping.
In this chapter, I have concentrated on vocal interactions between songbirds
and made predictions for what eavesdroppers and interactants should do in terms
of their relative positioning and the type of sound signals that interactants should
use. Some of the predictions may seem trivial or speculative, while others are
more substantial because they are derived from the results of sound-transmission
experiments. At the moment, there is anecdotal evidence for all of the predictions
except aspects of public signals, where there is stronger evidence. There is also
strong evidence for private signals. Quiet singing in songbirds fulfils most of the
predictions for private signalling and seems to do so for anonymity as well. Other
selection pressures are also important; for instance predation risk is likely to have
played an important role in the evolution of private signals.
An interesting challenge for future research on these matters will be to investi-
gate eavesdropping in natural non-experimental contexts. In territorial songbirds,

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
58 T. Dabelsteen
this could be done by quantifying how territorial subject males behave during vo-
cal interactions between other males and in subsequent vocal interactions with
them. Such a study would need recordings of the vocal activity in a local network
of males using acoustic location systems, as done by, for instance, Bower (2000)
and Burt & Vehrencamp (Ch. 15), combined with monitoring the movements of
silent individuals by radio-tracking. If the study located birds in three dimensions,
it might also test the predictions on positioning and signal use discussed above. If
the study was long term and included collection of data on predation, survival and
reproductive success, it might help to disentangle the influence of the different
selection pressures on the evolution of communicating in a network.
Acknowledgements
Thorsten Balsby kindly commented on the manuscript. Sandra Blumenrath drew Fig. 3.1
and Thorsten Balsby assisted with the production of the remaining figures. Henrik Brumm gave
me access to unpublished manuscripts. The main part of my own research on communication
network activities forming the basis for this chapter has been funded by the Danish National
Science Foundation.
References
Balsby, T. J. S. 2000. Song activity and variability in relation to male quality and female
choice in whitethroats Sylvia communis. Journal of Avian Biology, 31, 56–62.
Balsby, T. J. S. & Dabelsteen, T. 2001. The meaning of song repertoire size and song
length to male whitethroats Sylvia communis. Behavioural Processes, 56, 75–84.
2002. Female behaviour affects male courtship in whitethroats, Sylvia communis. An
interactive experiment using visual and acoustic cues. Animal Behaviour, 63,
251–257.
2003a. Male singing behaviour and female presence in the territory in whitethroats.
Acta Ethologica, 5, 81–88.
2003b. Male calling between courtship sequences in whitethroats: a way to counter
intrusions from neighbouring rivals. Behavioural Processes, 63, 149–157.
2003c. Degradation of whitethroat Sylvia communis song: implications for song flight
and communication network activities. Behaviour, 140, 695–719.
2004. Simulated courtship interactions elicit neighbour intrusions in the
whitethroat Sylvia communis. Animal Behaviour, in press.
Bensch, S. & Hasselquist, D. 1992. Evidence for active female choice in a polygynous
warbler. Animal Behaviour, 44, 301–311.
Blumenrath, S. H., Dabelsteen, T. & Pedersen, S. B. 2004. Being inside nest boxes: does
it complicate the receiving conditions for great tit Parus major females?
Bioacoustics, in press.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 59
Bower, J. L. 2000. Acoustic interactions during naturally occurring territorial conflict
in a song sparrow neighborhood. Ph.D. Thesis, Cornell University, Ithaca,
New York.
Brumm, H. 2004. The impact of environmental noise on song amplitude in a
territorial bird. Journal of Animal Ecology, 73, 434–440.
Brumm, H. & Todt, D. 2002. Noise-dependent song amplitude regulation in a
territorial songbird. Animal Behaviour, 63, 891–897.
2003. Facing the rival: directional singing behaviour in nightingales. Behaviour, 140,
43–53.
Bradbury, J. W. & Vehrencamp, S. L. 1998. The Principles of Animal Communication.
Sunderland, MA: Sinauer.
Dabelsteen, T. 1984. An analysis of the full song of the blackbird Turdus merula with
respect to message coding and adaptations for acoustic communication. Ornis
Scandinavica, 15, 227–239.
1985. Messages and meanings of bird song with special reference to the blackbird
(Turdus merula) and some methodology problems. Det Kongelige Danske
Videnskabernes Selskab Biologiske Skrifter, 25, 173–208.
1988. The meaning of the full song of the blackbird, Turdus merula, to untreated and
oestradiol-treated females. Ornis Scandinavica, 19, 7–16.
1992. Interactive playback: a finely tuned response. In: Playback and Studies of Animal
Communication, ed. P. K. McGregor. New York: Plenum Press, pp. 92–109.
Dabelsteen, T. & Mathevon, N. 2002. Why do songbirds sing intensively at dawn? A test
of the acoustic transmission hypothesis. Acta Ethologica, 4, 65–72.
Dabelsteen, T. & Pedersen, S. B. 1990. Song and information about aggressive
responses of blackbirds Turdus merula: evidence from interactive playback
experiments with territory owners. Animal Behaviour, 40, 1158–1168.
1992. Song features essential for species discrimination and behaviour assessment
by male blackbirds (Turdus merula). Behaviour, 121, 259–287.
Dabelsteen, T., Larsen, O. N. & Pedersen, S. B. 1993. Habitat-induced degradation of
sound signals: quantifying the effects of communication sounds and bird
location on blur ratio, excess attenuation, and signal-to-noise ratio in blackbird
song. Journal of the Acoustical Society of America, 93, 2206–2220.
Dabelsteen, T., McGregor, P. K., Holland, J., Tobias, J. & Pedersen, S. B. 1997. The signal
value of overlapping singing in male robins (Erithacus rubecula). Animal Behaviour,
53, 249–256.
Dabelsteen, T., McGregor, P. K., Lampe, H. M., Langmore, N. E. & Holland, J. 1998.
Quiet singing in song birds: an overlooked phenomenon. Bioacoustics, 9,
89–105.
Davies, N. B. 1992. Dunnock Behaviour and Social Evolution. Oxford: Oxford University
Press.
Doutrelant, C. & McGregor, P. K. 2000. Eavesdropping and mate choice in female
fighting fish. Behaviour, 137, 1655–1659.
Dukas, R. & Kamil, A. C. 2001. Limited attention: the constraint underlying search
image. Behavioral Ecology, 12, 192–199.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
60 T. Dabelsteen
Embleton, T. F. W. 1996. Tutorial on sound propagation outdoors. Journal of the
Acoustical Society of America, 100, 31–48.
Ficken, M. S., Ficken, R. W. & Witkin, S. R. 1978. Vocal repertoire of the black-capped
chickadee. The Auk, 95, 34–48.
Fleron, G. M. W. 2003. Stille sang hos solsorten, Turdus merula. M.Sc. Thesis, University
of Copenhagen.
Gompertz, T. 1961. The vocabulary of the great tit. British Birds, 54, 369–394,
409–418.
Henwood, K. & Fabric, A. 1979. A quantitative analysis of the dawn chorus: temporal
selection for communicatory optimisation. American Naturalist, 114,
260–274.
Hoi, H. 1997. Assessment of the quality of copulation partners in the monogamous
bearded tit. Animal Behaviour, 53, 277–286.
Holland, J., Dabelsteen, T., Pedersen, S. B. & Larsen, O. N. 1998. Degradation of wren
Troglodytes troglodytes song: implications for information transfer and ranging.
Journal of the Acoustical Society of America, 103, 2154–2166.
Holland, J., Dabelsteen, T., Pedersen, S. B. & Paris, A. L. 2001. Potential ranging cues
contained within the energetic pauses of transmitted wren song. Bioacoustics, 12,
3–20.
Hovi, M., Alatalo, R. V. & Siikamaki, P. 1995. Black grouse leks on ice: female mate
sampling by incitation of male competition? Behavioral Ecology and Sociobiology, 37,
283–288.
Jakobsson, S., Brick, O. & Kullberg, C. 1995. Escalated fighting behaviour incurs
increased predation risk. Animal Behaviour, 49, 137–142.
Lack, D. 1939. The display of the blackcock. British Birds, 32, 290–303.
1940. Observations on captive robins. British Birds, 33, 262–270.
Langemann, U., Tavares, J. P., Peake, T. M. & McGregor, P. K. 2000. Responses of great
tits to escalating patterns of playback. Behaviour, 137, 451–471.
Larsen, O. N. & Dabelsteen, T. 1990. Directionality of blackbird vocalization.
Implications for vocal communication and its further study. Ornis Scandinavica,
21, 37–45.
Lengagne, T. & Slater, P. J. B. 2002. The effects of rain on acoustic communication:
tawny owls have good reason for calling less in wet weather. Proceedings of the
Royal Society of London, Series B, 269, 2121–2125.
Lind, H., Dabelsteen, T. & McGregor, P. K. 1996. Female great tits can identify mates by
song. Animal Behaviour, 25, 667–671.
Lombard, E. 1911. Le signe de l’elevation de la voix. Annales des Maladies de l’Oreille et du
Larynx, 37, 101–119.
Mathevon, N., Dabelsteen, T. & Blumenrath, S. H. 2004. Are high perches in the
blackcap Sylvia atricapilla song or listening posts? A sound transmission study.
Journal of the Acoustical Society of America, in press.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the. Royal
Society of London, Series B, 340, 237–244.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
Facilitating and countering eavesdropping 61
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K., Dabelsteen, T. & Holland, J. 1997. Eavesdropping in a territorial
songbird communication network: preliminary results. Bioacoustics, 8, 253–254.
McGregor, P. K., Peake, T. M. & Lampe, H. M. 2001. Fighting fish Betta splendens extract
relative information from apparent interactions: what happens when what you
see isn’t what you get? Animal Behaviour, 62, 1059–1065.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Messmer, E. & Messmer, I. 1956. Die Entwicklung der Lautausserungen und einiger
Verhaltensweisen der Amsel (Turdus m. merula L.) unter naturlichen Bedingungen
und nach Einzelaufzucht in schalldichten Raumen. Zeitschrift fur Tierpsychologie,
13, 341–441.
Michelsen, A. 1978. Sound reception in different environments. In: Sensory Ecology, ed.
M. A. Ali. New York: Plenum Press, pp. 345–373.
Molles, L. E. & Vehrencamp, S. L. 2001. Neighbor recognition by resident males in the
banded wren, Thryothorus pleurostictus, a tropical songbird with high song type
sharing. Animal Behaviour, 61, 119–127.
Montgomerie, R. & Thornhill, R. 1989. Fertility advertisement in birds: a means of
inciting male–male competition? Ethology, 81, 209–220.
Naguib, M. & Todt, D. 1997. Effects of dyadic vocal interactions on other conspecific
receivers in nightingales. Animal Behaviour, 54, 1535–1543.
Naguib, M., Fichtel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Nemeth, E., Winkler, H. & Dabelsteen, T. 2001. Differential degradation of antbird
songs in a Neotropical rainforest: adaptation to perch height? Journal of the
Acoustical Society of America, 110, 3263–3274.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecifics interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Otter, K. A., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits Parus major
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society of London, Series B, 266, 1305–1309
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
2002. Do great tits assess rivals by combining direct experience with information
gathered by eavesdropping? Proceedings of the Royal Society of London, Series B, 269,
1925–1929.
Poesel, A., Dabelsteen, T. & Pedersen, S. B. 2001. Making yourself heard: a study of
masking effects on blue tit Parus caeruleus singing interactions. In Proceedings of the
XVIII International BioAcoustics Conference, p. 12.

P1: GFZ/... P2: JZZ/...0521823617c03.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:1
62 T. Dabelsteen
Rasmussen, R. & Dabelsteen, T. 2001. Song repertoires and repertoire sharing in a local
group of blackbirds. Bioacoustics, 13, 63–76.
Sæther, S. A. 2002. Female calls in lek-mating birds: indirect mate choice, female
competition for mates, or direct mate choice? Behavioral Ecology, 13, 344–352.
Schmitz, S., Dabelsteen, T. & Mathevon, N. 2000. High perches in the blackcap Sylvia
atricapilla: song posts or listening posts? In Proceedings of the 8th International
Behavioral Ecology Congress, p. 175.
Searcy, W. A. & Yasukawa, K. 1996. Song and female choice. In: Ecology and Evolution of
Acoustic Communication in Birds, ed. D. E. Kroodsma and E. H. Miller. Ithaca, NY:
Cornell University Press, pp. 454–473.
Snow, D. W. 1958. A Study of Blackbirds. London: Allen and Unwin.
Stoddard, P. K. 1996. Vocal recognition of neighbours by territorial passerines. In:
Ecology and Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma and E. H.
Miller. Ithaca, NY: Cornell University Press, pp. 356–374.
Stork, H.-J. 1971. Zur socialen Funktion des Gesanges der Amsel Turdus merula L.
Zeitschrift fur Tierpsychologie, 28, 54–58.
Thorpe, W. H. & Pilcher, P. M. 1959. The nature and characteristics of sub-song. British
Birds, 51, 509–514.
Titus, R. C. 1998. Short-range and long-range songs: use of two acoustically distinct
song classes by dark-eyed juncos. The Auk, 115, 386–393.
Wempen, J. 1986. Outdoor sound propagation close to the ground. In: Sound
Propagation in Forested Areas and Shelterbeds, ed. M. J. M. Martens. Nijmegen: Faculty
of Sciences, Catholic University, Nijmegen, pp. 83–106.
Wiley, R. H. & Poston, J. 1996. Indirect mate choice, competition for mates, and
coevolution of the sexes. Evolution, 50, 1371–1380.
Wiley, R. H. & Richards, D. G. 1982. Adaptations for acoustic communication in birds:
transmission and signal detection. In: Acoustic Communication in Birds, Vol. 1, ed.
D. E. Kroodsma & E. H. Miller. New York: Academic Press, pp. 131–181.

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
4
Performing in front of an audience:signallers and the social environment
r i c a r d o j . m a t o s 1 & i ng o s c h l u p p 2
1University of Copenhagen, Denmark2University of Zurich, Switzerland and University of Texas, Austin, USA
Introduction
Several signallers and receivers sharing the same active signalling space
constitute a communication network. This type of environment imposes addi-
tional selection pressures on both signallers and receivers other than those clas-
sically considered in signaller–receiver dyads. In this chapter, we shall discuss
how communication networks influence the behaviour of a signaller and, more
specifically, the effect of an audience (defined below) on signalling behaviour.
An individual signaller has to cope with two main issues when signalling in a
network: (a) it has to compete or cooperate with other signallers, and (b) it has to
deal with the presence of several receivers. Signalling at the same time as other
individuals poses a problem for the signaller: how does it ensure that its specific
signal is detected by a receiver when other conspecifics are signalling? Signallers
solve or minimize this problem by either cooperating or competing for the signal
broadcast space. For example, in frog and insect choruses, individuals time their
signals to avoid acoustic interference (e.g. alternating their calls) or compete for
call order in the chorus (Gerhardt & Huber, 2002; Ch. 13). At the community level,
different species with similar signals may broadcast their signals at different times
of the day (Endler, 1992).
The presence of several receivers presents two additional problems for the sig-
naller. The first is how to direct the signal to a specific receiver. For example, bird
song often has a range that encompasses several neighbouring territories. When a
bird sings, the song could potentially reach all the neighbours in surrounding ter-
ritories. During interactions with neighbours, individuals may need to direct the
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
63

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
64 R. J. Matos & I. Schlupp
signal to a specific individual, for example because that neighbour starts to sing
close to the territory boundary. McGregor & Peake (2000) discussed several ways
in which songbirds can direct the signal to a specific rival neighbour or intruder.
For example, matched counter-singing (Stoddard et al., 1992; Beecher et al., 1996)
is a good candidate for directing the signal to a specific individual bird.
The second issue that arises from the presence of several receivers, and one that
this chapter covers in more detail, is how signallers communicate in the presence
of additional receivers other than the primary target receiver. We will concentrate
on conspecific receivers because heterospecific receivers, especially predators and
parasites, have received considerable attention and are known to be important
in shaping signals and signalling interactions (Bradbury & Vehrencamp, 1998;
Chs. 2 and 8). The term audience has been used to describe conspecific receivers
in the context of a communication network (McGregor & Peake, 2000; Doutrelant
et al., 2001). In this chapter, we shall begin by discussing this term and its use in
the context of communication networks. We shall then discuss how the presence
of several receivers may affect signalling behaviour and the choice and evolution
of signalling strategies.
Definitions of audience and audience effects
Audiences
We define audiences as individuals that are present during, but do not
take part in, signalling interactions between others. We distinguish two types of
audience: evolutionary audiences and apparent audiences.
Evolutionary audiences
By evolutionary audiences we mean individuals that were historically
common in the environment of the signaller and that may have generated selec-
tion on the form and content of signalling behaviour. For example, it is widely
accepted that bird song has a dual function, both as a signal to attract females
and as a signal used in male–male competition (Berglund et al., 1996; Searcy &
Nowicki, 2000). The evolution of this dual function has been widely discussed in
the literature (e.g. Searcy & Nowicki, 2000). One hypothesis suggests that song first
evolved as a male–female signal with males acting as eavesdroppers (see Ch. 12 for
similar discussion on fiddler crabs). This eavesdropping pressure caused by male
audiences may have induced new selective forces on the form and content of the
signal, resulting in the appearance of a dual function signal. If this hypothesis is
correct, then males have acted as an evolutionary audience in bird song evolu-
tion. An evolutionary audience does not need to be present or apparent to affect

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 65
signalling behaviour at any instant in time, because selection has acted in the past
(and presumably continues to act) on the signal (e.g. introducing or emphasizing
features in the design of songs that males use in male–male competition). For
more information on the effects and importance of evolutionary audiences, we
refer the reader to Chs. 2 and 14.
Apparent audiences
Apparent audiences are individuals that affect the behaviour of the sig-
naller only when they are present and detected. For example, in the presence of
females, interacting male Siamese fighting fishes Betta splendens decrease highly
aggressive behaviours (attempted bites) and increase the intensity of conspicuous
displays (tail beats and gill cover display) (Doutrelant et al., 2001). Unlike evolution-
ary audiences, the effects produced by this type of audience are triggered when
the audience is present; males show no such effects on the different displays when
the female is absent.
In this chapter, we are mainly concerned with the study of apparent audiences,
as their effects can be studied experimentally and, unlike studies of evolutionary
audiences, they do not rely on historical inference.
Audience effects
We define an audience effect as changes in the signalling behaviour during
an interaction between individuals caused by the mere presence of an audience.
Matos & McGregor (2002) found that male fighting fish engaged in visual signalling
interactions changed their signalling (i.e. the visual displays directed towards the
rival male) when a male audience was present. It is important to emphasize that
the change in signalling behaviour occurred between the two individuals involved
in the signalling interaction and not directly towards the audience. Whether the
information content of signalling changes will depend on the balance of cost and
benefit to the signallers (see below). This type of effect is specific to a commu-
nication network as it can only occur in situations where a minimum of three
individuals is present: two individuals engaged in a signalling interaction and
one individual making up the audience.
Why audience?
Different authors have used different terms to designate extra potential
receivers in a communication network, such as bystanders (Dugatkin, 2001), unin-
tended receivers (Endler, 1993) or illegitimate receivers (Otte, 1974). Most of these
terms, however, have been used in an interspecific context (with the exception
of bystander) to describe predator detection of prey signalling behaviour (Otte,
1974; Endler, 1993; Bradbury & Vehrencamp, 1998; Ch. 2). Because we restrict our

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
66 R. J. Matos & I. Schlupp
definition of an audience to conspecifics, we exclude predators or parasites re-
sponding to the signal (Ch. 2). We chose the term audience because it is more
descriptive of the role of the individual during the signalling interaction in two
ways. First, it implies that individuals are present but do not take part in the
interaction, although they are clearly able to. Second, it implies that the individ-
uals may pay attention to the signalling interaction and thus potentially extract
information.
We think that it is important to link the term audience to other network be-
haviours such eavesdropping in this way because the presence of eavesdroppers
can impose costs and benefits on signallers and to link these costs–benefits to the
information content of the interaction. For example, the finding that eavesdrop-
pers behave more aggressively to individuals that behave as losers in an aggres-
sive signalling interaction (e.g. Chs. 2 and 14) identifies an immediate cost of an
audience on the losers. It is worth noting though that we do not have to show
that individuals are able to extract information to cause an audience effect. For
example, audiences may be costly just because there is a high risk of the audi-
ence disrupting the signalling interaction (e.g. intervention behaviour of semi-
captive zebras Equus quagga; Schilder, 1990). By comparison, non-apparent eaves-
droppers do not promote an audience effect because signallers are unaware of their
presence.
Other uses of audience and audience effect in the literature
The terms audience and audience effects have been used in the communi-
cation network literature to describe the effects on signalling interactions of the
presence of additional potential receivers that do not take part in the interaction
(Doutrelant et al., 2001; Matos & McGregor, 2002). However, these terms have also
been used in other studies in animal communication. In the following paragraphs
we shall talk about these studies and underline the differences between the two
uses of the term audience.
The first studies to use the terms audience and audience effects looked at the
effect of the presence of a conspecific on the incidence of alarm and food calls
in birds (Gyger et al., 1986; Marler et al., 1986; Gyger, 1990; Evans & Marler, 1994).
These authors were interested in whether these calls were elicited by and directed
to a specific class of individuals or audiences, namely conspecifics (e.g. conspecific
versus predator; male versus female). In these studies, an audience is defined as
any individual that is present in the same location as the subject (an apparent
audience), and the audience effect is the change in signalling behaviour (e.g. in-
crease in food call rate: Marler et al., 1986) caused by the presence of the audience.
In both cases, the signal was assumed to be directed towards the audience; for
example, Gyger et al. (1986) performed two experiments to investigate whether

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 67
male cockerels Gallus domesticus modulated their alarm calls in the presence of
an audience when a model of a predator was presented. The protocol of both
experiments was the same; the birds were placed in a cage above which a model
of a predator was ‘flown’. The audience was housed in another cage next to the
male’s cage; both individuals could see the predator. In the first experiment, the
audience was either their own mate or a female that was mated to another male,
with an empty cage as a control. The second experiment was similar to the first
one with the difference that instead of another male’s female the authors used an
unfamiliar male. The authors found that in both experiments males increased the
rate of alarm calls when a conspecific was present compared with when alone. No
significant difference was found between the presence of the male’s mate com-
pared with another male’s mate, or between the male’s mate compared with an
unfamiliar male. The authors concluded from these results that the presence of
a conspecific audience has an effect on alarm calling and that these calls may be
primarily directed towards conspecifics and not towards the predator. Because
there was no significant effect of the type of conspecific (own mate, other’s mate,
unfamiliar male), one can rule out the hypothesis that the observed increase in
call rate is a result of sharing the risk with the other prey (Gyger et al., 1986).
There are two main differences between the use of the terms audience and
audience effects in these studies and our own use. First, we restrict audience effects
to the signalling interaction between the two individuals; the audience is not the
primary receiver of the signals but acts as a potential non-targeted receiver. In the
predator/food call studies, the distinction between the audience and a primary
receiver of the signal is blurred as the target receiver is the audience (Fig. 4.1). The
second difference is that our definition is specific to communication networks. In
the predator/food call studies, this was not necessarily true; only two conspecifics
were necessary to produce the audience effect: the signaller and the audience. For
example, in a similar study to the one described above, Marler et al. (1986) showed
that male cockerels increased their food calls in response to the presence of one
hen; such a situation is a signaller–receiver dyad.
We point out these differences in use of the terms to ensure that different
phenomena are not confused by the use of a common term and suggest that the
terms should be clearly defined when used.
Audience effects
Relatively few studies have addressed directly the question of whether
audience effects occur. In this section, we summarize these studies and discuss
other systems in which audience effects appear to have an important influence
on signalling behaviour.

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
68 R. J. Matos & I. Schlupp
Fig. 4.1. The audience effect in different types of study. (a) In predator/food call
studies (Gyger, et al., 1986), the change in behaviour (dotted arrows) is triggered by the
presence of the audience (the hen) and directed towards the audience. (b) In the
audience effects described in this chapter, the change in behaviour (dotted arrows) is
triggered by the presence of the audience (non-target receiver, the hen) and directed
towards the target receiver (the other male).
Male–male aggressive signalling interactions
Individuals often use signals to compete for resources such as territories,
food or mates. These displays are used to assess the opponents’ fighting ability
and motivation (Huntingford & Turner, 1987; Bradbury & Vehrencamp, 1998). In
a communication network, this information is available not only to the opponent
but also to other individuals that are within signal range. This audience of non-
targeted receivers may introduce extra costs or benefits to signallers; as explained
above, some studies show that eavesdropping fish are more likely to initiate ag-
gressive interactions with a loser than with a winner (Oliveira et al., 1998; Earley &
Dugatkin, 2002; Chs. 2 and 5). If an audience has high costs or benefits to signallers,
then signallers should adjust their behaviour towards the opponent in order to
conceal or enhance information, respectively (McGregor & Peake, 2000).
Siamese fighting fish
Siamese fighting fish often use signals to mediate competition over re-
sources such as territories, food or mates, and such visual displays have been

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 69
(a) (b)
Fig. 4.2. Representation of the experimental design used in Matos (2002) to study the
effect of a male audience on male–male interactions in Betta splendens. (a) In the first
10 minutes, both males were allowed to interact in the absence of an audience. (b) In
the second 10 minute period, either an audience or an empty tank was revealed
(removal of the opaque partition) to the males. Ma and Mb are the interacting males;
A is the audience tank; o.p. is an opaque partition; arrows represent the direction in
which visual contact was possible.
used as a model system to address different questions related to communication
networks (e.g. eavesdropping: Oliveira et al., 1998; McGregor et al., 2001). One of
the first experiments to address specifically whether male Siamese fighting fish
were affected by the presence of an audience during an aggressive interaction was
performed by Matos (2002). Two males were allowed to interact through a clear
partition (tank walls), and a third male (the audience) was placed at a small dis-
tance from these males (Fig. 4.2). This small distance prevented the audience from
taking part in the interaction yet, at the same time, allowed both males to see the
audience. Each trial of the experiment was divided into two 10 minute periods: in
the first period the two individuals were allowed to interact without the audience
being present; the second period started when an opaque partition that separated
the audience from the two males was removed, allowing the males to see the au-
dience while interacting. Previous studies have shown that one can predict the
winner of a fight between two male fighting fish from display difference at the
beginning of the interaction (Simpson, 1968). In this experiment, the winner of
the signalling interaction was defined as the individual that displayed most during
the first 10 minutes of the interaction (the other male was the loser). It is important
to note that the barriers between males prevented actual fighting and none of the
interactions reached an outcome (e.g. displaying submissive colouration). No dis-
plays directed towards the audience were observed. Matos (2002) found that ‘win-
ners’ did not change their signalling behaviour in the presence of an audience. In

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
70 R. J. Matos & I. Schlupp
contrast, when an audience was present ‘losers’ reduced the time they spent in
gill cover display (a purely visual display) and the time spent near the opponent
compared with when there was no audience. However, there was no significant
change in the more aggressive displays that had both tactile and visual compo-
nents (i.e. attempted bites and tail beats). This change in behaviour may be viewed
as an attempt by the loser to restrict the information available to the eavesdropper
while at the same time providing adequate information for assessment by the op-
ponent. Another hypothesis is that by reducing the less-aggressive displays whilst
maintaining the more aggressive forms, ‘losers’ may seem more aggressive to the
audience. Thus even though the audience may have seen that individual lose, it
would be more reluctant to interact with it because of its aggressiveness (‘good
loser’ hypothesis: Peake & McGregor, 2004).
This study (Matos, 2002) suggests that there is an audience effect when a male
audience is present during male–male interactions and that the presence of the
audience can be more costly for the individual that is losing the interaction than
for the winner. The finding that the audience effects in this situation involved
a change of signalling behaviour by the loser fits both observations that losers
are more rapidly approached by males that saw them lose (Siamese fighting fish:
Oliveira et al., 1998; McGregor et al., 2001; swordtail fish Xiphophorus helleri: Earley &
Dugatkin, 2002) and that this effect disappears in combats where both individuals
escalated (Earley & Dugatkin, 2002).
In an earlier study, Doutrelant et al. (2001) also found that female audiences
affected male–male B. splendens aggressive displays. In this experiment, a female
audience was presented to a pair of males that interacted through a clear partition.
The effect of the presence of an audience was then compared with a treatment
where males were allowed to interact with no audience present. Males increased
the amount of conspicuous displays (e.g. tail beats and time with gill cover erect)
and decreased the more aggressive displays (e.g. attempted bites) towards oppo-
nents when a female was present. The authors interpreted this result as males try-
ing to compromise between having to interact with an opponent and at the same
time provide information to the audience by using more conspicuous displays,
which are more often used in both aggressive and courtship contexts. Doutrelant
et al. (2001) also performed a second experiment to examine whether male audi-
ences affected signalling interactions but did not find an audience effect (except
for a tendency for males to spend less time near the opponent). However, the re-
sult of these two experiments cannot be compared directly because of differences
in the experimental design and procedure (i.e. the audience was closer to the
males and the males were pre-exposed to the audience in the female experiment,
while in the male experiment the audiences were further away and there was no

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 71
(a) (b)
Fig. 4.3. Schematic representation of the experimental design used in both Matos &
McGregor (2002) and Matos et al. (2003). (a) In a five minute pre-exposure period, both
males could see the audience tank. (b) In the 10 minute interaction period, the opaque
partition was removed and both males were allowed to interact with each other in
front of or in the absence of the audience. Ma and Mb are the interacting males; A is
the audience tank; o.p. is an opaque partition; arrows represent the direction in
which visual contact was possible.
pre-exposure period). Both distance and pre-exposure to another individual have
been shown to have a strong effect on male aggressive display (Bronstein, 1989;
Halperin et al., 1998; also see below).
In a more recent experiment, Matos & McGregor (2002) looked directly at the
effect of the sex of the audience. Three different types of audience were used:
male, female B. splendens and female Xiphophorus spp. (to control for responses not
specific to conspecifics). A control with no audience present was also used. The
design and procedure of the experiment was similar to that in Matos (2002), ex-
cept that the males were first pre-exposed to the audience and then were allowed
to see and interact with the opponent (Fig. 4.3). The audience was visible for the
entire trial. No distinction was made between winners and losers as data were only
collected from one of the individuals involved in the interaction. No differences
were observed between the female Xiphophorus spp. treatment and no audience;
therefore the Xiphophorus spp. treatment was used as the control. Males behaved
more aggressively (i.e. attempted more bites and spent less time near the oppo-
nent) when a male audience was present than with a female audience (Matos &
McGregor, 2002). To explain this difference, the authors suggested that the pres-
ence of a female might confront the males with a trade-off between expelling
their male opponent and not driving away a potential mate. Males of this species
often bite when courting a female and highly aggressive males may cause females
to flee because of the high risk of injury (Bronstein, 1984). The results of these

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
72 R. J. Matos & I. Schlupp
experiments suggest that the sex of the audience is important in determining
how males should behave during aggressive signalling interactions.
Field crickets
Tachon et al. (1999) studied male–male competition for resources in the
field cricket Gryllus bimaculatus. They tested whether the presence of a female
influences the aggressive behaviour between males. In each test, a group of five
males in an arena under three different treatments was observed. Besides the two
obvious treatments, presence and absence of females, they used a third condition
where a paper impregnated with female scent was introduced into the arena.
Previous studies had shown that this scent elicited behavioural responses from
males of this species (Otte & Cade, 1976; Hardy & Shaw, 1983).
Tachon et al. (1999) found that males increased their level of aggressive displays
(e.g. aggressive stridulation and mandible flaring) towards other males in the
treatment where the females were present. Interestingly, there was no evidence
that the female scent produced the same effect as the actual presence of a female.
Female scent alone in this system may be a poor predictor of female presence and
the cost of escalating increases when there is a high probability that the female
is not present. However, in this example, it is not clear what effect direct female–
male interactions had on male–male competition, as opposed to the effect of the
mere presence of the female. Further studies are needed to attempt to distinguish
these effects and thus to confirm whether this is an example of an audience effect.
Parental behaviour
Male parental care is common in many species. If there is a direct link
between the care provided to the young and their survival until reproductive age,
it might be of advantage for the females to choose a good father as a potential
mate. One way of assessing paternal care is to observe male interactions with
young (e.g. affiliate signalling behaviour). If females do choose a good father for
their future mate, then it should be to the advantage of the male to try to perform
as a better ‘parent’ when a female is present.
Vervet monkeys
Vervet monkeys Cercopithecus aethiops have a complex social system where
individuals influence their own or other group members’ dominance rank by
socializing with individuals of different rank. In such a system, female mate choice
or preference to associate with a male can influence the male’s future position in
the hierarchy (Ch. 25). Interactions between males and infant are quite common
and males often form strong protective relationships with the females and their
young. These relationships may reduce the harassment that females and infants

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 73
receive from other group members. Therefore, females may prefer to associate
with males that perform more affiliatively towards their infant.
Hector et al. (1989) investigated whether male vervet monkeys changed their
interaction with an infant in the presence versus ‘absence’ of the mother. In this
experiment, the females were placed (a) behind a one-way mirror, where they could
see both male and infant but not vice versa; (b) behind a Plexiglas partition, where
male, female and infant could see each other; and (c) behind a metal partition,
where the female could not see the dyad and the male and infant could not see
the female. The results of this experiment showed that males are sensitive to
the presence of the mother and engaged in more affiliative and less-antagonistic
behaviour toward the infant when the male was able to see the mother. However,
it is not clear whether the effect is simply caused by the presence of the female or
occurs because the females could still potentially signal to the dyad through the
Plexiglas, affecting the behaviour of both infant and male. A further treatment
would be needed to address this question, where the female is placed behind a
one-way mirror and the dyad can see the female but not vice versa.
The authors further studied if females varied their behaviour towards males
that they saw performing more affiliative behaviours towards their infants and
found that females tolerated the males more and also performed more affiliative
behaviours towards them. In spite of the lack of an appropriate control, this study
showed that potentially individuals may adjust their behaviour when an audience
is present and that there are direct consequences to the individual.
Budgerigars
Female birds may assess male parental care behaviour by the male’sextra-
pair behaviour during the period prior to egg laying. In species with obligate
biparental care, males that provide more care to the young should be preferred as
a mate, as less-committed males increase the female’scosts of feeding and spending
more time with the young. Extra-pair activity by the male (e.g. displaying to another
female) may provide information to the female on the male’s attentiveness to-
wards the female and the nest.
Budgerigars Melopsittacus undulatus are socially monogamous birds where both
members of the pair provide parental care. The males of this species provide most
of the food to the nest, both at the start of the nesting period and through brooding.
As a consequence, male commitment to the female and brood is very important to
the female and survival of the brood, and females may use cues of male commit-
ment when they are choosing a potential mate. Baltz & Clark (1994) investigated
whether male budgerigars were less likely to court another female when their own
mate was present. In other words, they tested whether there is an effect of an audi-
ence (their mate) on the male’s extra-pair behaviour. The study was conducted on

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
74 R. J. Matos & I. Schlupp
a captive population housed in an outdoor aviary. Nestboxes were provided, sim-
ulating the nests in natural cavities observed in the wild. The authors assumed
that the females lost visual contact with the male when inside the nestbox. The
behaviour of each male and its mate was recorded in the periods where the female
was inside (no audience) and outside (audience) the nest. Males significantly in-
creased extra-pair courtship behaviour when out of view of the female (i.e. when
the female was inside the nestbox) relative to when the female was in view. How-
ever, the results of this experiment can also be explained by an alternative hypoth-
esis. Males may reduce the time courting other females because with their mate
outside the nest they are more vulnerable to extra-pair courtship and copulations
by other males in the flock. Therefore, the reduction of courtship may be a re-
sult of mate guarding (Baltz & Clark, 1994). In another study, Baltz & Clark (1997)
showed that the necessity for mate guarding did not change the males’ response
to the extra-pair female. The authors used the same experimental design as before
but this time the mate was separated from the rest of the flock in both treat-
ments. This procedure prevented other males from interacting with the female
(subject’s mate) and thus reduced the necessity for mate guarding. Once again,
males reduced courtship behaviour towards extra-pair females when their mate
was visible. Although this study suggests that there maybe an audience effect, we
consider it poor evidence for audience effects as we define them in this chapter.
The main problem with the experimental design of both studies is that the audi-
ence effect is not caused by the mere presence of the audience, the male–female
pair are only separated visually by an opaque partition, and, as the authors state,
both individuals could still contact each other through calls even when they could
not see each other. We suggest that further studies would be required to confirm
the presence of an audience effect in such system.
Human behaviour
Social psychologists have long recognized that audiences have an im-
portant effect on human behaviour (e.g. Zajonc, 1965; Blumstein, 1973; Felson,
1982; Ch. 19). These effects extend from a change in the performance of sim-
ple motor tasks, when compared with apparently ‘non-social’ contexts (Zajonc,
1965), to changes in more complex forms of social behaviour such as interper-
sonal strategies used during social interactions (Blumstein, 1973). One interesting
area of study with regard to audience effects in humans is impression manage-
ment theory. This theory focuses on the principle that a person is aware of being
characterized or typified by others when performing a behaviour and responds
by trying to make these characterizations favourable. As a consequence, most
human behaviour is designed to obtain ‘favourable’ reactions from an audience
(Felson, 1978, 1982). For example, Felson (1982) found an effect of third-party

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 75
presence on aggressive interactions between humans. The study was based on in-
terviews with patients with previous mental health problems, with ex-criminals
and with a sample of the general population. All groups answered a questionnaire
asking them to describe in detail four aggressive incidents. The replies showed
that the outcome of an interaction between individuals of the same sex was more
severe when an audience was present (when allowing for third-party instigation
or mediation of the fight). There was a higher probability that individuals would
escalate from verbal insults to actual physical contact. However, the authors also
found that the same was not true in conflicts between the sexes; the cause of
such a difference may be that the audience is more likely to disapprove of severe
aggression in between-sex conflicts (Felson, 1982).
The general idea that individuals may try to manipulate their characterization
by others has recently been used to explain altruistic behaviour in humans and
non-human animals (Zahavi & Zahavi, 1997; Wedekind & Milinski, 2000; Milinski
et al., 2001; Bshary, 2002). This idea is discussed by Bshary & D’Souza in Ch. 22.
Priming: a mechanism of audience effects or a functional alternative?
In the experiments discussed above showing that male Siamese fighting
fish behaved more aggressively towards an opponent when a male audience was
present (Matos & McGregor, 2002), the trial procedure allowed males to see the
audience before they started interacting. This procedure was used to ensure that
the males were aware of the presence of the audience during the interaction. In
a further series of experiments, Matos et al. (2003) found that the presence of an
audience before an interaction affected how male B. splendens behaved during the
interaction. Using a similar design to that described by Matos & McGregor (2002),
the authors divided each trial into two continuous periods: a pre-exposure period
(when males could either see an empty tank or a tank containing an audience) and
an interaction period (when both males where allowed to interact with each other
in the presence or absence of an audience). In the first experiment, four different
treatments where used in which the audience was (1) present in the pre-exposure
period, (2) present during the interaction period, (3) present in both periods or (4)
absent in both periods. The authors then separated the behaviours overt aggression
(i.e. attempted bites and latency to first bite) and a display score (combined measure
of the other displays, i.e. time spent flaring the gill cover, number of tail beats and
time spent near the opponent); for details on the method see Matos et al. (2003).
Overall, males behaved more aggressively (i.e. shorter latency to attempt to bite
the opponent) during the interaction in the treatments where the males were pre-
exposed to the audience (treatments 1 and 3). This effect is similar to aggressive
priming. The presence of the audience before the interaction may have increased

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
76 R. J. Matos & I. Schlupp
(a) (b)
(c) (d)
Fig. 4.4. Schematic representation of the second experimental design used in Matos
et al. (2003). (a–c) The five minute pre-exposure period when both males were
pre-exposed to an empty tank (no pre-exposure) (a); both males were pre-exposed to an
audience (b); and only one of the males (Ma) was pre-exposed to the audience (c).
(d) The 10 minute interaction period following all treatments, where both males were
allowed to interact in front of an audience. Ma and Mb are the interacting males; A is
the audience; thick lines between the tanks represent opaque partitions; arrows
represent the direction in which visual contact was possible.
the motivation to behave aggressively. As a result, individuals escalated more
rapidly into more aggressive forms of behaviour when they interacted with the
opponent.
The authors also found that priming effects overrode any effect of presenting
the audience only during the interaction. The levels of aggression between the two
treatments where males were pre-exposed (treatments 1 and 3) were similar, in-
dependent of audience presence during the interaction period, while much lower
levels of aggression were seen in treatments 2 and 4. In fact, there was no signifi-
cant difference between the treatments with the audience absent in both periods
(treatment 4) and with the audience present during the interaction (treatment 2).
These results may suggest that audiences do not affect male–male fighting fish
interactions, as the audience affected only treatments with pre-exposure. In this
respect the results matched those of Doutrelant et al. (2001), in which male au-
diences did not have an effect on male–male signalling interactions (see above).
However, we should also note that in both studies the authors did not look at
losers and winners separately.
Matos et al. (2003) performed a second experiment to look at the interac-
tion between audience effects and pre-exposure to the audiences; the design al-
lowed independent pre-expose of the two opponent males (Fig. 4.4). As in the first

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 77
experiment, the trials were divided in two periods: five minutes of pre-exposure
and a 10 minute period in which the two opponents were allowed to interact.
There were three treatments in the pre-exposure period: both males pre-exposed
to an empty tank (no pre-exposure; Fig. 4.4a), both males pre-exposed to the audi-
ence (symmetric pre-exposure; Fig. 4.4b), and one of the opponents pre-exposed to
the audience while the other male was pre-exposed to an empty tank (asymmetric
pre-exposure; Fig. 4.4c). The audience was always present in the interaction period
(Fig. 4.4d). The results confirmed that pre-exposed males tend to behave more ag-
gressively (higher display scores and overt aggression); both the no pre-exposure
and the symmetric treatments showed the same tendencies. In the asymmetric
treatment, pre-exposed males also tended to display more than the ones not pre-
exposed with one exception: non-pre-exposed males matched the number of at-
tempted bites of the pre-exposed males. A possible explanation is that it may be
costly for individuals not to retaliate when its opponent escalates, because of the
high risk of injury, especially in a confined space such as the experimental tanks
(Maan et al., 2001). However, this cost may be enhanced by the presence of the
audience. By matching the opponent in more aggressive behaviour, males may be
either decreasing the ability of an audience to discriminate loser from winner or
manipulating the information to seem more aggressive. These results support the
previously discussed idea that males, particularly losers, may gain by performing
more aggressively during an interaction in the presence of an audience, as it may
decrease the chances of future harassment by that individual (Earley & Dugatkin,
2002; Matos et al., 2003).
These two studies taken together support the idea that previous studies on
audience effects (i.e. Doutrelant et al., 2001; Matos & McGregor, 2002) have under-
estimated the effect of pre-exposure on male aggression. Nevertheless, they also
suggest that the social environment (i.e. audiences) is important in determining
the dynamics of signalling interaction.
Previous studies have shown that priming is an important mechanism mediat-
ing aggressive interactions (e.g. Potegal & Popken, 1984; Bronstein, 1989; Halperin
et al., 1992) as it affects the individual’s aggressive motivation. For example, prim-
ing may decrease the time to initiate aggression or increase the attack behaviour
of individuals (e.g. Potegal & ten Brink, 1984; Halperin et al., 1998). However, the
effect on the outcome of interactions is not always clear. It seems that priming
may have a more pronounced effect during the initial stage of the fight, either
causing the individual to display more actively at the beginning of the inter-
action or to escalate and initiate aggression more quickly (Potegal & Popken,
1984; Bronstein, 1989; Halperin et al., 1998). In several species, individuals that
display more intensively and escalate earlier during an interaction usually gain a

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
78 R. J. Matos & I. Schlupp
competitive advantage over their opponents (Huntingford & Turner, 1987). In such
a case, priming may produce a positive effect as it increases the probability that
the individual will win the fight. In some cases, however, priming can have a
negative effect, male B. splendens that have been isolated and then primed with
a conspecific image behaved more aggressively towards their opponents but lost
most of the interactions (Halperin et al., 1998). These individuals could have been
manipulated by priming into aggressive levels that they were not able to sustain
during the entire fight, causing them to tire faster than the opponents and sub-
sequently lose the interaction. We conclude that priming may have an important
impact on the outcome of the interaction, but whether this impact is positive or
negative may depend on whether the initial stages of the interaction determine
the outcome and on the length of the interaction.
One potential mechanism behind such aggressive priming is the production
of hormones caused by the presentation of a social stimulus. In a recent study,
Oliveira et al. (2000) showed that watching a fight raises the androgen levels of
adult male cichlid fish Oreochromis mossambicus. Priming may involve a similar
mechanism, and the facilitation of aggressive behaviour through pre-exposure
may be caused by an increase in androgen levels initiated by the pre-exposure to
the audience. Oliveira et al. (2000) suggested that these hormones mediate changes
in the perceptual abilities and readiness to interact of males, which, in turn, would
enhance their success in social interactions.
Further studies are needed to comprehend fully the relation between the adap-
tive value of priming and the presence of an audience. Advances in the under-
standing of the effects of the social environment on the neuroendocrinological
system may be an important contribution in this area (Ch. 21).
Summary and future directions
One important question in the general context of communication net-
works is how narrowly or broadly we wish to define the social context of sig-
nalling. Recent studies have shown that mate preferences can be altered by view-
ing sexual interactions (Westneat et al., 2000). In several species, seeing a male
mate enhances this male’s attractiveness to females (Dugatkin, 1992; Ch. 5). Such
choosing females would be eavesdropping on the signalling–mating interaction
of two other individuals and responding accordingly. The same reasoning might
apply for other interactions as well. It appears that most dyadic interactions are
actually embedded in a social context or network. This raises the question of
how common the well-studied dyadic interactions actually are, as these studies
have only considered them in a social void. This situation might be more of an
exception than the rule. In this context, more knowledge on sensory ecology and

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 79
especially the role of private channels would be very helpful. Communication via
private channels uses sensory channels not available to the audience. This has
been documented for swordtails (Xiphophorus spp.): males signal in the ultraviolet,
a part of the spectrum that cannot be detected by a predator, the Mexican tetra
Astyanax mexicanus (Cummings et al., 2003). True dyadic interactions may be brief
and limited to signals transmitted in close contact. A potential example might be
nipping in poeciliid fishes; here, males nibble a female’s genital region and chem-
ical signals are transmitted (Parzefall, 1973). Such signals are not available to any
other individual, although the male’s response to the signal might be (Parzefall,
1973).
Another aspect to consider is that many social interactions relevant to several
aspects of an individual’s life may happen simultaneously and influence each
other. Any given individual will have to include this into its signalling decisions.
For instance, a singing bird may simultaneously be faced with the problems of at-
tracting a female, discouraging a neighbour from entering its territory and avoid-
ing predators. This leads to a more complicated network of social interactions,
the components of which may influence each other to shape a ‘social interac-
tion network’. Our singing bird example also illustrates that each context alone
would select for a different signal or signalling strategy. Signals have to be effective
enough to transmit accurate information to target receivers but private enough to
prevent this information from being detected by ‘unwanted’untargeted receivers.
Any signal that is under such conflicting demands will be a compromise, depend-
ing on the associated costs and benefits. Only recently has formal modelling been
used to address this problem (Johnstone, 2001; Ch. 26).
We have attempted to show that the presence and type of audience can have
important effects on the signalling strategies of individuals. The nature of infor-
mation and the extent to which it is broadcast may depend on the type of audience
and on the role of each signaller during an interaction. Audiences may also in-
fluence the evolution of new types of signal. During signal evolution, different
pressures may arise in signal design depending on whether it is specialized to ad-
vertize or privatize information (e.g. ‘normal’ song versus quiet song: Dabelsteen
et al., 1998; Ch. 3). Audience effects may be closely linked with mechanisms such as
priming effects, which may influence motivation of signallers and consequently
their signalling strategy. In natural systems, the social environment affects how
animals make behavioural decisions. Individuals can use signalling interactions
between others as a source of information; this can in many ways have important
consequences for the fitness of individuals. In order to improve our understanding
of the evolution of signals and signalling strategies, we must take into account
the individuals’ social environment and the costs and benefits associated with the
presence of audiences and eavesdroppers.

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
80 R. J. Matos & I. Schlupp
Acknowledgements
We would like to express our sincere thanks to Peter McGregor for providing us with the
opportunity to write this chapter and to Tom Peake, Ryan Earley, Denise Pope, Giuliano Matessi
and Andrew Terry for their valuable comments and discussion of the manuscript. We thank the
Fundacao para a Ciencia e Tecnologia (Portugal) for funding R. M., whose Ph.D. provided data and
ideas included in this chapter. I. S. was supported by a Heisenberg Fellowship of the Deutsche
Forschungogemeinschaft.
References
Baltz, A. P. & Clark, A. B. 1994. Limited evidence for an audience effect in budgerigars,
Melopsittacus undulatus. Animal Behaviour, 47, 460–462.
Baltz, A. P. & Clark, A. B. 1997. Extra-pair courtship behaviour of male budgerigars and
the effect of an audience. Animal Behaviour, 53, 1017–1024.
Beecher, M. D., Stoddard, P. K., Campbell, S. E. & Horning, C. L. 1996. Repertoire
matching between neighboring song sparrows. Animal Behaviour, 51, 917–923.
Berglund, A., Bisazza, A. & Pilastro, A. 1996. Armaments and ornaments: an
evolutionary explanation of traits of dual utility. Biological Journal of the Linnean
Society, 58, 385–399.
Blumstein, P. W. 1973. Audience, Machiavellianism, and tactics of identity bargaining.
Sociometry, 36, 346–365.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.
Bronstein, P. M. 1984. Agonistic and reproductive interactions in Betta splendens.
Journal of Comparative Psychology, 98, 421–431.
Bronstein, P. M. 1989. The priming and retention of agonistic motivation in male
Siamese fighting fish, Betta splendens. Animal Behaviour, 37, 165–166.
Bshary, R. 2002. Biting cleaner fish use altruism to deceive image-scoring client reef
fish. Proceedings of the Royal Society of London, Series B, 269, 2087–2093.
Cummings, M. E, Rosenthal, G. G. & Ryan, M. J. 2003. A private ultraviolet channel in
visual communication. Proceedings of the Royal Society of London, Series B, 270,
897–904.
Dabelsteen, T., McGregor, P. K., Lampe, H. M., Langmore N. E. & Holland J. 1998.
Quiet song in song birds: an overlooked phenomenon. Bioacoustics, 9,
89–105.
Doutrelant, C., McGregor, P. K. & Oliveira, R. F. 2001. The effect of an audience on
intra-sexual communication in male Siamese fighting fish, Betta splendens.
Behavioral Ecology, 12, 283–286.
Dugatkin, L. A. 1992. Sexual selection and imitation females copy the mate choice of
others. American Naturalist, 139, 1384–1389.
Dugatkin, L. A. 2001. Bystander effects and the structure of dominance hierarchies.
Behavioral Ecology, 12, 348–352.

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 81
Earley, R. L. & Dugatkin, L. A. 2002. Eavesdropping on visual cues in green swordtail
(Xiphophorus helleri) fights: a case for networking. Proceedings of the Royal Society of
London, Series B, 269, 943–952.
Endler, J. A. 1992. Signals, signal conditions, and the direction of evolution. American
Naturalist, 139, S125–S153.
Endler, J. A. 1993. Some general comments on the evolution and design of animal
communication systems. Philosophical Transactions of the Royal Society of London,
Series B, 350, 215–225.
Evans, C. S. & Marler, P. 1994. Food calling and audience effects in male chickens,
Gallus gallus: their relationship to food availability, courtship and social
facilitation. Animal Behaviour, 47, 1159–1170.
Felson, R. B. 1978. Aggression as impression management. Social Psychology, 41,
94–105.
Felson, R. B. 1982. Impression management and the escalation of aggression and
violence. Social Psychology Quarterly, 45, 245–254.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans: Common
Problems and Diverse Solutions. Chicago, IL: University of Chicago Press.
Gyger, M. 1990. Audience effects on alarm calling. Ethology Ecology and Evolution, 2,
227–232.
Gyger, M., Karakashian, S. J. & Marler, P. 1986. Avian alarm calling: is there an
audience effect? Animal Behaviour, 34, 1570–1572.
Halperin, J. R. P., Dunham, D. W. & Ye, S. 1992. Social isolation increases social display
after priming in Betta splendens but decreases aggressive readiness. Behavioural
Processes, 28, 13–32.
Halperin, J. R. P., Giri, T., Elliott, J. & Dunham, D. W. 1998. Consequences of
hyper-aggressiveness in Siamese fighting fish: cheaters seldom prospered. Animal
Behaviour, 55, 87–96.
Hardy, T. N. & Shaw, K. C. 1983. The role of chemo-reception in sex recognition by
male crickets: Acheta domesticus and Teleogryllus oceanicus. Physiological Entomology, 8,
151–166.
Hector, A. C. K., Seyfarth, R. M. & Raleigh, M. J. 1989. Male parental care, female
choice and the effect of an audience in vervet monkeys. Animal Behaviour, 38,
262–271.
Huntingford, F. A. Turner, A. K. 1987. Animal Conflict. London: Chapman & Hall.
Johnstone, A. R. 2001. Eavesdropping and animal conflict. Proceedings of the National
Academy of Sciences, USA, 98, 9177–9180.
Maan, M. E., Groothuis T. G. G. & Wittenberg, J. 2001. Escalated fighting despite
predictors of conflict outcome: solving the paradox in a South American cichlid
fish. Animal Behaviour, 62, 623–634.
Marler, P., Dufty, A. & Pickert, R. 1986. Vocal communication in the domestic chicken:
II. Is a sender sensitive to the presence and nature of a receiver? Animal Behaviour,
34, 194–198.
Matos, R. J. 2002. Social influences on signalling: audience effects and communication
networks. Ph.D. Thesis, University of Copenhagen.

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
82 R. J. Matos & I. Schlupp
Matos, R. J. & McGregor, P. K. 2002. The effect of the sex of an audience on
male–male displays of Siamese fighting fish (Betta splendens). Behaviour, 139,
1211–1221.
Matos, R. J., Peake, T. M. & McGregor, P. K. 2003. Timing of presentation of an
audience: aggressive priming and audience effects in male Siamese fighting fish
(Betta splendens). Behavioural Processes, 63, 53–61.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McGregor, P. K., Peake, T. M. & Lampe, H. M. 2001. Fighting fish Betta splendens extract
relative information from apparent interactions: what happens when what you
see isn’t what you get. Animal Behaviour, 62, 1059–1065.
Milinski, M., Semmann, D., Bakker, T. C. M. & Krambeck, H.-J. 2001. Cooperation
through indirect reciprocity: image scoring or standing strategy? Proceedings of the
Royal Society of London, Series B, 268, 2495–2501.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Oliveira, R. F., Lopes, M., Carneiro, L. A. & Canario, A. V. M. 2000. Watching fights raises
fish hormone levels. Nature, 409, 475–475.
Otte, D. 1974. Effects and functions in the evolution of signalling systems. Annual
Review of Ecology and Systematics, 5, 385–417.
Otte, D. & Cade, W. 1976. On the role of olfaction in sexual and interspecies
recognition in crickets (Acheta and Gryllus). Animal Behaviour, 24, 1–6.
Parzefall, J. 1973. Attraction and sexual cycle of poeciliids. In: Genetics and Mutagenesis
of Fish, ed. J. H. Schroder. Berlin: Springer, pp. 177–183.
Peake, T. M. & McGregor, P. K. 2004. Information and aggression in fishes. Learning and
Behavior, in press.
Potegal, M. & ten Brink, L. 1984. The behavior of attack-primed and attack satiated
female golden hamsters, Mesocricetus auratus. Journal of Comparative Psychology, 98,
66–75.
Potegal, M. & Popken, J. 1984. The time course of attack priming effects in female
hamsters. Behavioural Processes, 11, 199–208.
Schilder, M. B. J. 1990. Interventions in a herd of semi-captive plains zebras. Behaviour,
112, 53–83.
Searcy, W. S. & Nowicki, S. 2000. Male–male competition and female choice in the
evolution of vocal signalling In: Animal Signals: Signalling and Signal Design in
Animal Communication, ed. Y. Espmark, T. Amundsen & G. Roseqvist. Trondheim:
Tapir Academic Press, pp. 301–315.
Simpson, M. J. A. 1968. The display of the Siamese fighting fish Betta splendens. Animal
Behaviour Monographs, 1, 1–73.
Stoddard, P. K., Beecher, M. D., Campbell, S. E. & Horning, C. L. 1992. Song-type
matching in the song sparrow. Canadian Journal of Zoology, 70, 1440–1444.
Tachon, G., Murray, A.-M., Gray, D. A. & Cade, W. H. 1999. Agonistic displays and the
benefits of fighting in the field cricket, Gryllus bimaculatus. Journal of Insect
Behaviour, 12, 533–543.

P1: GFZ/... P2: JZZ/...0521823617c04.xml CU1917B/McGregor 0 521 582361 7 April 10, 2005 17:49
Signallers and the social environment 83
Wedekind, C. & Milinski, M. 2000. Cooperation through image scoring in humans.
Science, 288, 850–852.
Westneat, D. F., Walters, A., McCarthy, T. M., Hatch, M. I. & Hein, W. K. 2000.
Alternative mechanisms of nonindependent mate choice. Animal Behaviour, 59,
467–476.
Zahavi, A. & Zahavi, A. 1997. The Handicap Principle. Oxford: Oxford University Press.
Zajonc, R. B. 1965. Social facilitation. Science, 149, 269–274.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
5
Fighting, mating and networking:pillars of poeciliid sociality
r y a n l . e a r l e y 1 & l e e a l a n d u g a t k i n 2
1Georgia State University, Atlanta, USA2University of Louisville, USA
We are both spectators and actors in this great drama of existence
Niels Bohr
Introduction
Poeciliid fishes such as green swordtails Xiphophorus helleri and guppies
Poecilia reticulata aggregate in social groups called shoals. In addition to reducing
predation risk and increasing foraging efficiency (e.g. Magurran & Pitcher, 1987;
Ranta & Juvonen, 1993), fish shoals promote the transfer of social information
within the group. For instance, information about foraging routes is transmit-
ted from trained individuals to naive fish in guppy shoals (Laland & Williams,
1997; Swaney et al., 2001; Brown & Laland, 2002). The type of information transfer
demonstrated in the social learning and foraging literature involves the trans-
mission of signals from one or more individuals to the remaining group mem-
bers. Investigations of social foraging and anti-predator behaviour have demon-
strated that poeciliids attend to a variety of cues emitted by both conspecifics and
heterospecifics (e.g. predators: Brown & Godin, 1999; Mirza et al., 2001; Brosnan
et al., 2003). Although social learning and anti-predator responses constitute
important aspects of group living in poeciliids, this chapter focuses more on how
individuals gain information from observing interactions that occur in their social
environment. Indeed, the concept of communication networks was founded on
the premise that the information exchanged during social interactions (e.g.
agonistic or courtship displays) may be available not only to the participants but
also to bystanders within signal detection range (McGregor, 1993; McGregor et al.,
2000).
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
84

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 85
Throughout this chapter, we make a clear distinction between bystanders and
eavesdroppers, though the two have been used synonymously in the past (Ch. 2).
Bystanders are any individuals within detection range of signalling interactions
while eavesdroppers represent a subset of bystanders that extract information
from these interactions. The primary aim of this chapter is to examine how
social eavesdropping – extracting information from signalling interactions be-
tween others (Ch. 2) – influences aggressive contest behaviour and female mate
choice in male X. helleri and female P. reticulata, respectively. Therefore, we focus
on how eavesdropping affects the subsequent behaviour of poeciliid bystanders
rather than on how the behaviour of participants in an interaction is modified
in the presence of an audience (Ch. 4). Swordtails and guppies are well suited to
investigations of networking phenomena because they are highly social, exhibit
stereotypical agonistic and courtship displays and are especially responsive to a
host of stimuli (e.g. visual, chemical) in their social surroundings.
Social eavesdropping and contest behaviour
Although sociality confers fitness-related benefits to individuals within
the group, competition for social status and limiting resources (e.g. food, mates)
often increases with group size (Pulliam & Caraco, 1984). In many animals, overt
aggressive interactions are most common during hierarchy or territory establish-
ment. Among fishes, rank-order fights involve a series of gradually escalating dis-
plays that convey information about strength, size or willingness to persist in the
encounter. The intensity and/or duration of aggressive contests depend largely on
differences in fighting ability between adversaries. When substantial differences
in fighting ability exist, the interaction may terminate following a bout of non-
contact displays. Interactions between well-matched opponents, however, may
intensify to physical combat, where behavioural tactics such as mouthwrestling
are used to settle the dispute (Enquist et al., 1990). Escalated contests often yield
unambiguous dominant–subordinate relationships among closely matched com-
petitors but the costs associated with such interactions can also be quite high. For
instance, Neat et al. (1998) revealed that prolonged fights result in the accumula-
tion of anaerobic metabolites and depletion of sugar reserves in the muscle tissue
of cichlid fish Tilapia zillii. Other potential contest costs include physical injury,
increased susceptibility to predation, lost mating or foraging opportunities and
increased stress hormone levels, which may impede future reproductive activity
(Haller, 1995; Jakobsson et al., 1995; Schuett, 1997; Halperin et al., 1998; Neat et al.,
1998).
Most theoretical and empirical work on aggressive contest behaviour assumes
that information about fighting ability is available only via direct interactions

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
86 R. L. Earley & L. A. Dugatkin
(Enquist & Leimar, 1983; Payne & Pagel, 1997; Mesterton-Gibbons & Adams, 1998;
Payne, 1998). Although this may be the case for solitary species, the social environ-
ment of group-living animals is ripe with opportunities for indirect assessment
of fighting ability via eavesdropping. Since observing fights may provide infor-
mation about fighting ability without the associated costs of physical combat,
eavesdropping should be an advantageous assessment strategy when both contest
costs and the opportunities for watching interactions are high. In this section, we
review a series of experiments that elucidate how eavesdropping modulates the
agonistic behaviour of male X. helleri. Specifically, we focus on how a bystander’s
behaviour changes after observing fights and the levels at which these behavioural
modifications are manifest.
Spectators in swordtail networks: empirical work
Cast of characters
Green swordtail fish are an excellent system in which to examine visually
based network effects. Although there are few studies on the costs of combat in X.
helleri, corticosteroid hormone levels are elevated above control levels for at least
six hours after contest settlement, particularly in subordinate fish (Hannes et al.,
1984). This finding, together with the data from a number of other studies on fish
(e.g. Neat et al., 1998), indicates that fighting is likely to be costly for male green
swordtails. Moreover, in both laboratory and field settings, male X. helleri establish
social hierarchies where rank-order fights and/or attack–retreat sequences are
common (Beaugrand et al., 1984; Franck et al., 1998). Thus, bystanders probably
have ample opportunity to observe aggressive interactions that occur within their
social environment and, given the potential costs of fighting, may benefit from
doing so. Figure 5.1 depicts a simplified version of the swordtail social network as
being composed of fighters engaged in aggressive signalling interactions, solitary
individuals not involved in dominance interactions, and bystanders. Furthermore,
the solitary individuals and fighters can either be observed or not observed by a
bystander within range to extract information relevant to fighting ability (e.g.
signals exchanged by the fighters or subtle behavioural/morphological cues of
solitary individuals). Of course, each individual within the swordtail groups can
assume the solitary, fighter or bystander position depending on the social circum-
stances they are exposed to at any given moment.
Basic paradigm
Within the swordtail network, bystanders may extract information from
fighters or solitary fish and integrate this information for use in future encounters
with the observed individual(s). Furthermore, eavesdropping may elicit changes
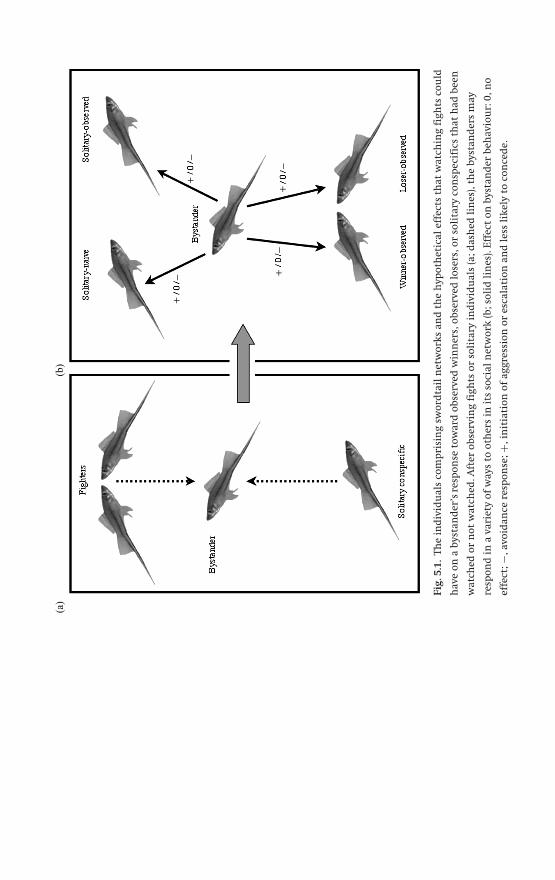
P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
(a)
(b)
+−
+−
+−
+−
Fig.
5.1.
The
ind
ivid
ual
sco
mp
risi
ng
swor
dta
iln
etw
orks
and
the
hyp
oth
etic
alef
fect
sth
atw
atch
ing
figh
tsco
uld
hav
eon
aby
stan
der
’sre
spon
seto
war
dob
serv
edw
inn
ers,
obse
rved
lose
rs,o
rso
lita
ryco
nsp
ecif
ics
that
had
been
wat
ched
orn
otw
atch
ed.A
fter
obse
rvin
gfi
ghts
orso
lita
ryin
div
idu
als
(a;d
ash
edli
nes
),th
eby
stan
der
sm
ay
resp
ond
ina
vari
ety
ofw
ays
toot
her
sin
its
soci
aln
etw
ork
(b;s
olid
lin
es).
Effe
cton
byst
and
erbe
hav
iou
r:0,
no
effe
ct;−
,avo
idan
cere
spon
se;+
,in
itia
tion
ofag
gres
sion
ores
cala
tion
and
less
like
lyto
con
ced
e.
87

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
88 R. L. Earley & L. A. Dugatkin
in the motivational state of the bystander that could influence its interactions
with individuals that had not been observed (Fig. 5.1). To address these issues in
the laboratory, we established a protocol where a bystander was visually isolated
from, or able to observe, fighters or solitary conspecifics. This was accomplished
by placing either an opaque plastic partition (opaque treatment) or a one-way
mirror (mirror treatment) between the bystander and either a pair of interacting
individuals or a solitary individual in laboratory aquaria (Earley & Dugatkin, 2002;
Earley et al., 2003, 2004). In all of the experiments, the bystander and the fighters
(or solitary conspecific) were matched for lateral surface area, a composite mea-
surement of body length, body depth and sword length that corresponds better
with fighting ability than standard length alone (Beaugrand et al., 1996). Following
opportunity or no opportunity to observe, the bystander was pitted against one
of the following individuals: the observed solitary individual, the winner of the
observed fight, the loser of the observed fight, or a solitary fish that had not been
observed (solitary-naive).
In the context of aggressive interactions, eavesdropping could influence a
bystander’s behaviour toward the observed individual(s) in many hypothetical
ways. First, observation could have no effect (0) on the behaviour of the bystander.
In this case, the behaviour of bystanders that had observed should be similar to
the behaviour of naive bystanders. Second, the bystander may exhibit an avoid-
ance response (−) if, through eavesdropping, it assessed the fighting ability of the
observed individual(s) to exceed its own. Avoidance responses include refraining
from initiating aggression or escalation and withdrawing from the contest. Third,
if the bystander assessed the fighting ability of the observed individual(s) to be less
than its own, it may be more inclined to initiate aggression or escalate and less
likely to concede to its opponent (+). It is also possible that eavesdropping could
affect the aggressive behaviour of the bystander outside the context of interacting
with the individuals that had been observed, for example when interacting with
solitary conspecifics that were not observed (Fig. 5.1).
Eavesdropping on fights: confronting winners and losers
Our work on communication networks in X. helleri began with a rela-
tively simple question: does watching a fight influence the agonistic response of
a bystander toward the observed contestants? To determine precisely how eaves-
dropping influences a bystander’s interactions with the observed contestants, one
must also recognize that the fighters enter the interaction with previous domi-
nance or subordination experience. Previous winning or losing experiences are
known to influence a host of contest characteristics in many fish species (e.g.
probability of initiating aggression or winning against future opponents: Bakker

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 89
& Sevenster, 1983; Bakker et al., 1989; Chase et al., 1994; Hsu & Wolf, 1999, 2001).
Prior winning experiences tend to increase the probability of future contest suc-
cess, while prior losing experiences decrease the likelihood of winning in the
future; these experiential effects have been dubbed the ‘winner effect’ and ‘loser
effect’, respectively. The opaque treatment can be seen as a control for the effects
of previous fighting experience. Because the bystander is not allowed to observe
the interaction, the dynamics of bystander versus winner or bystander versus loser
contests are influenced primarily by the fighters’previous experience. In contrast,
both eavesdropping and prior experience effects can mediate bystander contest
dynamics in the mirror treatment. Thus, when the dynamics of contests involv-
ing the bystander are compared between the opaque and mirror treatments, the
contribution of eavesdropping can be determined explicitly.
When the effects of the fighters’ prior experience are controlled, we found
that watching fights had a considerable influence on bystander behaviour, partic-
ularly when confronted with the winner of the observed interaction (Earley &
Dugatkin, 2002). Bystanders that observed the contest exhibited a more
pronounced avoidance response toward winners than bystanders that had not
observed the interaction (Fig. 5.2; Winner). However, the intensity of the observed
contest had no bearing on the eavesdroppers’response toward winners (see caption
to Fig. 5.3). Thus, eavesdroppers avoided observed winners regardless of whether
they defeated their opponent by escalated or non-escalated means. A radically dif-
ferent scenario emerged in the bystander versus loser contests. Here, bystanders
responded in a similar way to all losers, regardless of whether their defeat was
witnessed (Fig. 5.2; Loser). Nevertheless, our data revealed that eavesdroppers were
less likely to initiate aggressive behaviour and win against losers that persisted
versus losers that retreated immediately in the observed contest (Fig. 5.3).
These results demonstrate that swordtails not only make the dichotomous
assessment of ‘winner versus loser’ but also calculate the fighting ability of
each contestant, particularly the losers, independent of final outcome. The ca-
pacity to tease apart fighting ability from final status (i.e. winner or loser) may
be of particular benefit during the initial establishment of dominance hierar-
chies, where the rank of each group member relative to the others, including
the bystander, remains unclear. For instance, if eavesdroppers base their future
agonistic decisions on status assessment alone (winner versus loser), challenging
a relatively strong loser may prove costly. However, eavesdroppers that assess the
fighting ability of the observed contestants based on, for example, persistence
or willingness to escalate may be better equipped to adjust their agonistic be-
haviour in a manner consistent with the actual fighting ability of others in the
network.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Bys
tand
er in
itiat
es a
ggre
ssio
n (p
ropo
rtio
n of
con
test
s)
observed naive
(a)
Prop
ortio
n of
esc
alat
ed c
onte
sts
(b)
observed naive
Fig. 5.2. The effect of watching fights on a swordtail bystander’s propensity to (a)
initiate aggression, (b) participate in escalated contests and (c) win contests against
observed winners, observed losers, solitary conspecifics that had been observed, or
previously unknown individuals (solitary-naive). Grey bars represent the mirror
treatment, where a bystander could see either the fighters or the solitary individual
without being seen; white bars represent the opaque treatment, where a bystander
remained naive to the presence of the fighters or the solitary individual prior to
confrontation. An asterisk indicates significant differences (p < 0.05) between the
mirror and opaque treatments.
90

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 91
Bys
tand
er w
ins
(pro
port
ion
of c
onte
sts)
(c)
observed naive
Fig. 5.2. (cont.)
Eavesdropping on fights: observing solitary individuals
Given that male swordtails modify their agonistic behaviour based on
what appears to be an independent assessment of each contestant, bystanders
may be getting more information from the individual than from the interaction
itself. To address this possibility, we allowed some bystanders to observe a solitary
fish through a one-way mirror; the remaining bystanders were visually isolated
from the solitary individual using an opaque plastic partition. After the obser-
vation period (or lack thereof), the bystander was confronted with its opponent.
In this experiment, we allowed small variation in body length, body depth and
sword length between the two contestants. Asymmetries in any of the three size
measurements did not lead to substantial mismatches in lateral surface area.
When asymmetries in body size between the bystander and the watched individ-
ual were not considered, observation did not appear to elicit modifications in
the bystanders’ behaviour (Earley et al., 2003; Fig. 5.2, Solitary-observed). However,
when small differences in body size were examined as potentially informative
cues, an interesting result emerged. The observers’ propensity to initiate attack
increased as a function of relative body length. Somewhat surprisingly, observers
were more prone to initiate against larger opponents, a trend evident only in the
mirror treatment. Rapid escalation was a key predictor of contest success, with
initiators of attack winning 81% of the interactions (Earley et al., 2003). Therefore,
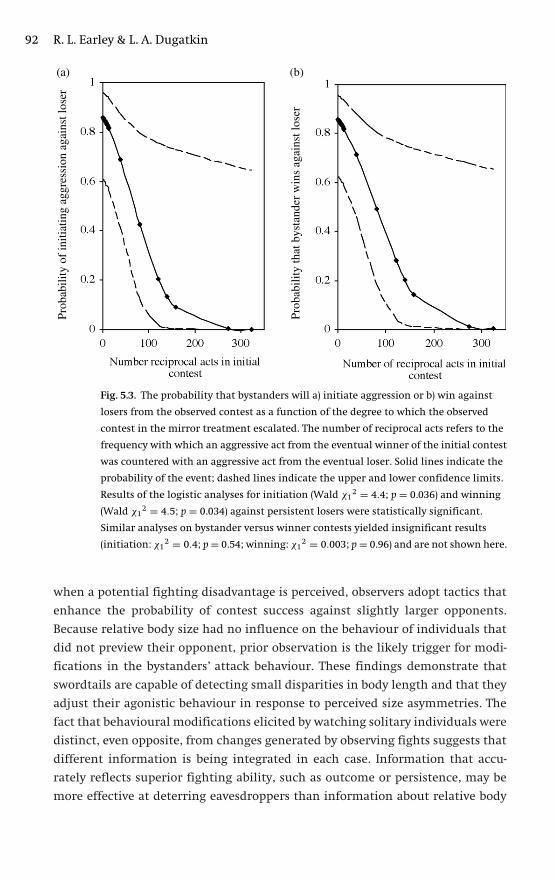
P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
92 R. L. Earley & L. A. Dugatkin
Prob
abili
ty o
f in
itiat
ing
aggr
essi
on a
gain
st lo
ser
Prob
abili
ty th
at b
ysta
nder
win
s ag
ains
t los
er
(a) (b)
Fig. 5.3. The probability that bystanders will a) initiate aggression or b) win against
losers from the observed contest as a function of the degree to which the observed
contest in the mirror treatment escalated. The number of reciprocal acts refers to the
frequency with which an aggressive act from the eventual winner of the initial contest
was countered with an aggressive act from the eventual loser. Solid lines indicate the
probability of the event; dashed lines indicate the upper and lower confidence limits.
Results of the logistic analyses for initiation (Wald χ12 = 4.4; p = 0.036) and winning
(Wald χ12 = 4.5; p = 0.034) against persistent losers were statistically significant.
Similar analyses on bystander versus winner contests yielded insignificant results
(initiation: χ12 = 0.4; p = 0.54; winning: χ1
2 = 0.003; p = 0.96) and are not shown here.
when a potential fighting disadvantage is perceived, observers adopt tactics that
enhance the probability of contest success against slightly larger opponents.
Because relative body size had no influence on the behaviour of individuals that
did not preview their opponent, prior observation is the likely trigger for modi-
fications in the bystanders’ attack behaviour. These findings demonstrate that
swordtails are capable of detecting small disparities in body length and that they
adjust their agonistic behaviour in response to perceived size asymmetries. The
fact that behavioural modifications elicited by watching solitary individuals were
distinct, even opposite, from changes generated by observing fights suggests that
different information is being integrated in each case. Information that accu-
rately reflects superior fighting ability, such as outcome or persistence, may be
more effective at deterring eavesdroppers than information about relative body

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 93
size alone. In systems where prior experience effects, or any other social factor,
have considerable influences on contest behaviour, information about size alone
may not be sufficient to deter eavesdroppers, particularly when asymmetries are
small.
An interesting point that was not addressed by the above experiment is whether
observing individuals exhibiting contest-type behaviour, without actually witness-
ing the interaction itself, would modify a bystander’s agonistic response. The
importance of this question lies in partitioning how the fighting tactics of each
contestant versus the dynamics of the actual interaction influence bystander
behaviour. McGregor et al. (2001) allowed a bystander to observe either ‘real’ or
‘apparent’interactions between two male Betta splendens. The ‘real’ interaction pro-
vided the bystander with information about two contestants that were actually
fighting with one another. In the ‘apparent’interaction, the bystander was exposed
to two males that appeared to be interacting with each other but were actually
fighting against different opponents (Fig. 5.3 and Fig. 2.2d, p. 26). In the ‘apparent’
interactions, bystanders responded to winners (i.e. the individual of a pair that
displayed longest) more strongly than to losers. It is important to note that, in
the ‘apparent’ interactions, an individual that was perceived to have won/lost by
the bystander may have actually obtained a different experience. Therefore, these
results demonstrate that bystanders utilize information about individual contest
behaviour (e.g. display duration), in addition to interaction dynamics, to gauge
their future agonistic decisions. Another way to test this idea would be to allow a
bystander to observe an individual exhibiting aggressive behaviour toward a stim-
ulus that is out of view of the bystander (e.g. mirror image; conspecific opponent).
Following observation, the bystander could be exposed to the individual it had
observed; as potential controls, the bystander could be pitted against individu-
als that were observed not interacting with the stimulus and/or individuals that
were not seen interacting with the stimulus. Provided that the watched individuals
show substantial variation in aggressive behaviour, this type of experiment could
elucidate whether watching individuals exhibiting contest-typical behaviour is
sufficient to elicit modifications in bystander behaviour.
Eavesdropping on fights: confronting naive conspecifics
From the experiments described above, it is clear that observing aggres-
sive interactions prompts an agonistic response in swordtail bystanders. Initially,
we interpreted this as evidence that bystanders extract information about the
fighting ability of each contestant and respond accordingly when confronted
with the observed individuals. Although our results are consistent with the
existence of eavesdropping in swordtails, it is possible that observing fights elicits
general changes in a bystander’s aggressive motivation that could affect future

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
94 R. L. Earley & L. A. Dugatkin
contest behaviour. This alternative hypothesis does not require that bystanders
extract information from signalling interactions between others. Studies on
Mozambique tilapia Oreochromis mossambicus (Oliveira et al., 2001) and Siamese
fighting fish B. splendens (Clotfelter & Paolino, 2003) have shown that observing
fights increases urinary androgen levels and aggressive behaviour/contest success,
respectively. This type of response to social stimuli is best labelled as ‘priming’
(Hollis et al., 1995). However, the motivational changes experienced by swordtail
bystanders, if any, may be quite different from priming. Recall that swordtail spec-
tators responded with increased avoidance behaviour toward winners and losers
that had persisted in the observed contest. Based on these data, any changes in the
motivational state of swordtail bystanders should be manifest as decreases, rather
than increases, in aggressive behaviour, possibly as a consequence of elevated
corticosteroid stress hormones.
To address the ‘motivational’ hypothesis from a behavioural perspective, some
bystanders were exposed to aggressive interactions while others were visually iso-
lated from a pair of fighters. Following the observation period, or the lack thereof,
the bystander was confronted with an inexperienced fish that was not seen.
Bystanders that observed conflict were equally likely to initiate aggression, es-
calate and win against the inexperienced fish as bystanders that were not exposed
to the fight (Earley et al., 2004; Fig. 5.2: Solitary-naive). Therefore, observing fights
does not appear to precipitate general increases or decreases in the aggressive
motivation of swordtail bystanders. Given the lack of support for the ‘motiva-
tional’ hypothesis, it is reasonably clear that swordtail behaviour is modulated
by more sophisticated mechanisms than observation-induced priming or stress.
Namely, the agonistic response of swordtail eavesdroppers is influenced by the
acquisition, integration and retention of information that accurately reflects the
fighting ability of others in the network.
Communication networks and fighting: future considerations
Eavesdropping in aggressive contexts: influence of individual differences
Most investigations of networking phenomena in fishes employ a ‘sym-
metrical’design; that is, all participants (e.g. fighters and bystanders) are matched
for attributes such as size or previous social experience. This type of design mini-
mizes the effect of extraneous variables on the dynamics of contests involving the
bystander and has helped to pinpoint how eavesdropping modulates bystander
behaviour in X. helleri (Earley & Dugatkin, 2002) and B. splendens (Oliveira et al.,
1998). However, the symmetrical design is probably not representative of com-
munication networks in nature where substantial individual variation in size,
previous fighting experience, physiological condition and social status are likely

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 95
to exist. Individual variation in characteristics related to physical prowess may
have important consequences for how the benefits and costs of fighting and/or
observing contests are perceived and, thus, how the effects of eavesdropping are
manifest. For instance, dominant and subordinate members of a social hierarchy
may respond differently to the winners and losers of observed interactions. Main-
tenance of dominance status is likely a priority for dominant individuals while
increasing status may benefit subordinates. Therefore, dominants should respond
more vigorously to individuals that pose the greatest threat of rank usurpation
(i.e. winners), while subordinates should respond more aggressively to losers so as
to take advantage of opportunities to increase rank. Similarly, territorial defence
is essential for resident individuals while non-territorial individuals may be most
interested in seizing a territory of their own. In this case, residents may respond
most aggressively toward upstart winners (Naguib & Todt, 1997; Naguib et al., 1999;
Peake et al., 2001) while intruders should exploit recent losers.
In addition to mediating the ‘direction’ of a bystander’s agonistic response,
differences in individual perceptions of the costs and benefits of fighting may
influence the degree to which eavesdropping is utilized as an assessment strategy
(Johnstone, 2001). The relationship between the benefits and costs of fighting may
be perceived as high for consistent winners, intermediate for inexperienced ani-
mals and low for consistent losers. Given that eavesdropping is most advantageous
under circumstances where combat bears a relatively high cost (low benefit to cost
ratio), inexperienced animals and consistent losers may benefit most by observ-
ing fights. However, as an individual’s perception of combat costs increases past
a certain threshold, it may refrain from aggressive interactions and eavesdrop-
ping altogether (e.g. playing ‘dove’; Johnstone, 2001). An individual’s perception
of the costs and benefits of fighting can be influenced by a host of additional fac-
tors including size, status, ownership, physiological or immunological condition
(e.g. hungry versus satiated; healthy versus weak), reproductive state and resource
value. Whether animals eavesdrop or how they respond to observing fights may
be integrally related to each of these factors. Though the state dependency of
eavesdropping effects has not yet gained empirical attention, this is likely to be
an important avenue of future research in the area of animal communication
networks.
Eavesdropping in aggressive contexts: environmental influences
Just as factors associated with physical prowess can affect eavesdropping,
so too can environmental or population-based variables. The presence of preda-
tors, or cues indicative of predator presence, influences the frequency of aggres-
sive interactions in fishes (e.g. Martel & Dill, 1993; Wisenden & Sargent, 1997;
Ch. 23). Individuals engaging in aggressive contests become more conspicuous to

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
96 R. L. Earley & L. A. Dugatkin
predators; that is, the costs of fighting are increased considerably when predators
are around. Predation risk may also have a negative impact on the frequency of
eavesdropping via at least three potential mechanisms: (a) if fights are less com-
mon in the presence of a predator, (b) if observing fights makes the bystander more
conspicuous to predators (e.g. by association with the fighters), or (c) if the capacity
to dedicate simultaneous attention to fights and predators is limited (see Dukas
(2002) for review on selective attention). Although the frequency of eavesdropping
may decline with predation risk, the efficacy of eavesdropping as an assessment
strategy would not necessarily be compromised. In fact, when aggressive contests
are rare and information flow through the network is reduced, bystanders may
take every opportunity to extract information about the fighting abilities of oth-
ers. In this sense, predation risk decreases the frequency and intensity of aggres-
sive encounters and, by necessity, the opportunities to eavesdrop and the absolute
amount of information available. However, the net benefit of eavesdropping under
these circumstances may remain unaffected (or may even be increased).
To test whether the frequency of eavesdropping is modified under different pre-
dation pressures would entail an analysis of how the proportion of eavesdroppers
in a population changes across predation regimes or how predation risk affects
an individual’s propensity to eavesdrop. Mathematical models may be best suited
to address population-level questions (e.g. how the proportion of eavesdroppers
changes with predation risk). Questions more amenable to empirical study include
whether predation risk affects the extent to which bystanders gather information
or whether bystanders compromise information acquisition in order to remain
hidden from predators. For instance, one could compare whether bystanders
derived from high-predation and low-predation sites differ in their response to
the watched contestants. In addition, one could vary the quality or availability of
refuges, the degree of habitat heterogeneity or the distance of refuges from the
focal fight and subsequently quantify how the bystander’sresponse to the observed
contestants changes with environmental condition.
Besides predation risk, the social and/or mating system of the species in ques-
tion may have a significant impact on how, and to what extent, exposure to fights
alters a bystander’s agonistic decisions. The challenge hypothesis postulated that
individuals should respond to social instability with increased testosterone levels
and, presumably aggression levels, to deter rival males and secure reproductive
opportunity (Wingfield et al., 1990; Ch. 21). In the broad sense, ‘social instability’
could include instances where individuals are being challenged directly (e.g. in
territorial disputes) or where individuals are exposed to but not directly engag-
ing in aggressive interactions. Mozambique tilapia and Siamese fighting fish fit
nicely within this broad scope, as males of these species respond to observing fights
with increased 11-ketotestosterone levels or increased aggression levels (Oliveira

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 97
et al., 2001; Clotfelter & Paolino, 2003). However, we failed to uncover evidence
that general changes in agonistic motivation accompany eavesdropping in green
swordtail fish (Earley et al., 2004). We argued that differences in the mating system
(e.g. breeding seasonality; monogamous versus polygamous) and/or social system
(e.g. territorial versus hierarchical; stable versus unstable) could explain appar-
ent species-specific differences in the response to observing contests (Earley et al.,
2004; Ch. 21 has a more in-depth treatment).
A piece of the hierarchical puzzle?
Eavesdropping is inherently a social phenomenon because it requires at
least two individuals actively engaging with one another and a third party that
extracts information from the signalling interchange. Nevertheless, there have
been no controlled studies in fishes that examine aspects of networking above
and beyond its effects at the dyadic level (for birds see Peake et al., 2002; Ch. 15).
For example, in our work with X. helleri, we exposed a previously inexperienced
bystander to an aggressive interaction and then assessed its response toward one of
the contestants by staging dyadic contests (bystander versus winner or bystander
versus loser). However, is it possible that the effects of eavesdropping are manifest
differently when bystanders interact with a previously observed individual in the
presence of other network members (e.g. observed winners or losers, unknown
conspecifics, previous opponents that defeated or were defeated by the bystander;
see Ch. 4)? Also, since social groups provide the opportunity for a wide range of
interactions, each individual may have a different blend of prior social experience
(e.g. several winning, losing or eavesdropping experiences or any combination
thereof). Could the ways in which several social experiences are integrated over
time, and the mere presence or absence of winner, loser and eavesdropping effects,
have implications for the structure of animal social systems? From an empirical
standpoint, these questions remain unanswered. However, the role of bystanders
in the establishment of linear dominance hierarchies has been a question of con-
ceptual interest for quite some time and, more recently, has attracted theoretical
attention.
Chase (1980, 1982) developed a conceptual model of linear hierarchy formation,
the ‘jigsaw model’,that involved two interacting individuals and a bystander. Once
a dominance relationship was established between the initial contestants, four
possible interaction sequences could follow: initial dominant defeats bystander
(double dominance), initial subordinate submits to the bystander (double subor-
dination), bystander defeats initial dominant, or bystander submits to the initial
subordinate. Double dominance and double subordination most often led to the
establishment of linear hierarchies while the remaining two interaction sequences
generated intransitive dominance orders.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
98 R. L. Earley & L. A. Dugatkin
Chase (1980, 1982) did not address explicitly the behavioural mechanisms
responsible for the double dominance and double subordination sequences and,
thus, the genesis of linear hierarchies. There are at least two mechanisms that
could give rise to such sequences. On the one hand, the initial contestants may
update their estimation of their own fighting ability after having won or lost, i.e.
winners increase and losers decrease their perceived fighting ability (Hsu & Wolf,
2001). As a consequence, previous winners should be more likely to defeat a
bystander with average fighting ability and losers should be more likely to defer to
the same bystander. In this instance, winner and loser effects are the behavioural
mechanisms responsible for the double dominance and double subordination
sequences and, in turn, the formation of linear hierarchies. On the other hand,
the bystander may update its perception of the fighting ability of the initial con-
testants after observing the fight. For instance, the bystander may increase its
estimate of the winner’s fighting ability and decrease its perception of the loser’s
prowess. As a result, the bystander may be liable to attack the loser and submit
to the winner. In this case, eavesdropping is the behavioural mechanism that
generates the double dominance and double subordination sequences and linear
hierarchies. These two scenarios need not be mutually exclusive. That is, winner,
loser and eavesdropping effects may act in concert to promote linear hierarchy
formation in animal groups.
Dugatkin (2001) developed a simulation model to illustrate the potential inter-
actions between eavesdropping and prior-experience effects in shaping dominance
hierarchies. In this model, individuals could increase or decrease their own fight-
ing ability and that of others in the network. The model made two assumptions:
the first was that winner, loser and eavesdropping effects change individual per-
ceptions with equal magnitude; the second was that all bystanders were privy to
every interaction that occurred in their social environment. When eavesdropping
and prior experience were considered separately, only winner effects produced
a linear hierarchy. Conversely, when eavesdropping and prior experience effects
operated simultaneously, a linear hierarchy always emerged. Therefore, linear
hierarchies are most likely to occur when some combination of winner, loser and
eavesdropping effects operate.
In order to improve understanding of how these effects operate in natural sys-
tems, some of the assumptions employed by Dugatkin (2001) need to be relaxed. For
at least two reasons, all bystanders are probably not capable of observing every
interaction that occurs in their social network: first, time spent observing one
interaction interferes with a bystander’sability to observe additional contests and,
second, bystanders are unlikely to be within signal detection range of all aggres-
sive encounters. Moreover, eavesdroppers probably have less information about
the fighting ability of each observed contestant than the contestants themselves.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 99
Therefore, updates of others’ fighting ability may be of lesser magnitude than up-
dates of one’s own fighting ability. R. L. Earley, S. Brosnan & J. Bragg (unpublished
data) developed a simulation model of hierarchy formation in animals, in part to
examine the consequences of relaxing these assumptions (Ch. 26).
Several signalling modalities: seeing is not everything
Our work on eavesdropping and aggression examined the influence of
visual signals on a bystander’s future behaviour. Nevertheless, visual signals may
convey only part of the story in piscine duels. Acoustic (Lugli, 1997; Ladich, 1998;
Amorim & Hawkins, 2000; Thorson & Fine, 2002), chemical (Waas & Colgan, 1992;
Giaquinto & Volpato, 1997) and electrical (McGregor & Westby, 1992) stimuli have
all been implicated as potential modes of communication in aggressive contests
in fishes. Whether these signals elicit similar changes in bystander behaviour as
visual signals has yet to be tested. Thorson & Fine (2002) demonstrated that male
gulf toadfish Opsanus beta emit acoustic signals during the calls of neighbouring
males, a phenomenon they called ‘acoustic tagging’ and interpreted as an aggres-
sive display. If overlapping versus non-overlapping acoustic signals in toadfish pro-
vide information about willingness to escalate (or putative status), then playback
experiments such as those used in territorial bird systems (e.g. Peake et al., 2001;
Ch. 2) may be worth conducting, provided an anechoic aquatic chamber can be
developed. Insights into multimodal signalling, the transmission of these signals
within the network and the availability of such signals to bystanders will surely
weave a more comprehensive story of how communication networks operate in
nature.
Social eavesdropping and female mate choice
As a general rule, females invest more time and energy in the reproduc-
tive process than males (e.g. production of viable eggs, gestation, maternal care,
etc.) and, therefore, should be the choosier of the two sexes. Over the past several
decades, an abundance of conceptual, theoretical and empirical work has focused
on the factors that mediate female mate choice or male success in attracting
females (Ryan, 1997). Most of the female mate-choice models have investigated
how exaggerated male secondary sex characters and female preferences for these
characters evolve, through either direct or indirect selection. For instance, Fisher’s
runaway selection hypothesis postulated that, over evolutionary time, the alleles
responsible for the male trait and the female’spreference for the male trait become
genetically correlated (Fisher, 1958). This genetic linkage initiates a positive feed-
back loop whereby male traits can become more exaggerated as the preference for

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
100 R. L. Earley & L. A. Dugatkin
such traits strengthens and, in turn, further exaggeration of male traits intensifies
female preference.
Fisher’s genetic model, together with alternative models that address direct
(e.g. sensory bias) and indirect (e.g. good genes) selection on female preferences
(Ryan, 1997), have helped to elucidate how exaggerated male traits exist when they
appear to have a negative impact on survival and how strong female preferences
for these traits arise and persist. However, almost all sexual selection models
assume that females choose mates independent of the choices made by other
females (an exception is Kirkpatrick & Dugatkin, 1994; also see below). Is it possible
that female choice depends not only on intrinsic preferences but also on the
preferences of other females in the network? Two lines of evidence suggest that
the decisions a female makes with respect to choosing mates is influenced in large
part by observing interactions. First, monitoring apparent male–male interactions
alters female mating decisions, for example initial mate choice (Doutrelant &
McGregor, 2000) or loyalty to partner (Otter et al., 1999; Mennill et al., 2002; Ch. 7).
Second, observing male–female courtship and/or mating interactions influences
the subsequent mate choice decisions of a female peripheral to the interaction
(e.g. Dugatkin, 1992, 1996a; Dugatkin & Godin, 1992; Grant & Green, 1996; Witte &
Ryan, 1998; Witte & Noltemeier, 2002).
In this section, we focus on the latter aspect of networking in poeciliid fishes:
namely, how intersexual courtship rituals mediate the mating decisions of female
P. reticulata that are not directly involved in the interaction. The principal con-
cept linking communication networks to courtship interactions and female mate
choice is mate copying. Mate copying occurs when “the conditional probability
of choice of a given male by a female is either greater or less than the absolute
probability of choice depending on whether that male mated previously or was
avoided, respectively” (Pruett-Jones, 1992, p. 1001). Furthermore, the female must
obtain information about a male’s mating history (or some part of it) by observa-
tion (Dugatkin, 1996b). In other words, the information gained by eavesdropping
on mating interactions may sway a female’sdecision toward or away from mating
with the observed male. We confine our discussion to unambiguous cases of mate
copying, that is, where a shift in the mating decisions of females is based solely
on observing interactions between males and females other than oneself.
Spectators in guppy networks: empirical work
Cast of characters
Guppies are an ideal species for examining networking phenomena such
as mate copying for a number of reasons. First and foremost, guppies live in mixed-
sex shoals, within which females likely have opportunities to view (and potentially

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 101
copy) the mate choice of nearby conspecifics. In addition, ample evidence suggests
that social information is utilized by guppies in the context of mate choice (see
below), foraging (Laland & Williams, 1997; Laland & Reader, 1999) and in the dy-
namics of shoal motion (Lachlan et al., 1998). Mate choice has been studied exten-
sively in this species (for reviews see Kodric-Brown, 1990; Endler & Houde, 1995;
Houde, 1997) and guppies exhibit normal courtship behaviour when placed in
small aquaria; therefore, they are ideal for manipulative laboratory experiments.
Typical courtship interactions involve the male directing sigmoid displays at the
female; receptive females respond to male displays with a ‘gliding’ motion (for
a more complete description, see Liley (1966)). Lastly, female guppies from the
Paria river (the population that was used in the experiments described below) ex-
hibit heritable preferences for certain male traits (e.g. orange colour: Houde, 1987,
1988; Endler & Houde, 1995). Experiments on such populations provide a unique
opportunity to examine the interaction between genetic-based preferences and
those resulting from interactions within the guppy social network. The cast of
characters in the guppy network resembles that described for the swordtails. The
principal difference is that the network is partitioned into (a) males that are either
quite similar with respect to exaggerated colour patterns or that differ by varying
degrees; (b) at least one female being courted by, or exhibiting a preference for,
a focal male; and (c) a female bystander within range to detect courtship and/or
mating signals (Fig. 5.4). In theory, any of the females within the guppy network
can assume a bystander role or a courtship role at any given time; males however,
are restricted to a courtship role.
Basic paradigm
In the guppy network, we are concerned primarily with how a female
bystander responds to a male that was recently preferred by another female (here,
the focal female). To address this question, one of the authors (Dugatkin) devel-
oped a protocol where a female was either exposed or not exposed to a courtship
interaction between a male and a focal female. Following the observation period,
the female bystander was given the opportunity to make a choice between the
male that was preferred by the focal female and the male that was not preferred
(Dugatkin, 1992). The preference of the focal female was ‘staged’ because she was
restricted to the side of the aquarium occupied by one of the two males; therefore,
the bystander female observed an apparent choice by the focal female. Eavesdrop-
ping on the mate choice of others could have three potential effects on the future
behaviour of the female bystander (Fig. 5.4). If observing courtship interactions
has no effect (0) on the female bystander then she should choose both males with
equal frequency. If females increase their assessment of a male’s quality (+) after
observing him successfully court, then the bystander should choose the male that

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
102 R. L. Earley & L. A. Dugatkin
Brightly coloured male Relatively drab male
Brightly coloured male Model female Relatively drab male
Bystander
+ / 0 /− + / 0 /−
Fig. 5.4. The individuals comprising guppy networks and the hypothetical effects that
watching mating interactions could have on a female bystander’s response toward the
observed, apparently successful male. After witnessing a courtship interaction
(dashed lines), the female bystander can respond in a variety of ways toward males in
her social network (solid lines). Effect on bystander behaviour: 0, no effect; −,
decreased assessment of male quality; +, increased assessment of male quality.
was preferred by the focal female a significant majority of the time, i.e. mate copy-
ing. It is also possible that females decrease their assessment of a male’squality (−)
after observing a courtship interaction, for instance if a recent mating depletes
the male’s sperm supply (Nakatsuru & Kramer, 1982). In this case, the bystander
female should avoid the male that was recently preferred by the focal female. In
the following sections, we provide a general overview of the empirical work on
mate copying that has been conducted in P. reticulata, with special emphasis an
how eavesdropping mediates a female’s subsequent mating decisions.
Female mate copying: does it occur?
Dugatkin’s (1992) research on guppies provided the first controlled study
of female mate copying. In this study, the bystander female chose the male that

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 103
was preferred by the focal female in a significant proportion of the trials. In a
series of five additional experiments, Dugatkin (1992) ruled out a host of alterna-
tive explanations, including female biases toward one side of the aquarium over
the other, female preference for areas of the tank recently occupied by the largest
group of fish (‘schooling preference’) and female choice for males that courted
most recently and thus were more active. These findings provided substantial evi-
dence that females do copy the mate choice of others and that female bystanders
assess the quality of recently preferred males to be higher than those that were
not preferred. Moreover, Dugatkin (1992) demonstrated that courtship inter-
actions between the focal female and a male are crucial for eliciting mate copying
behaviour. These results support the notion that mate choice decisions in P. reticu-
lata are, in part, socially modulated. Two other studies (Brooks, 1996; Lafleur et al.,
1997) found no evidence of mate copying in guppies. It is critical to note, how-
ever, that neither of these studies used guppies from natural streams in Trinidad.
Nonetheless, support for mate copying has also been reported in other poeciliid
species (Poecilia latipinna; Witte & Ryan, 1998) and the Japanese medaka Oryzias
laticeps (Grant & Green, 1996). Given the pivotal role of male–female interactions
in mediating a female bystander’s future mating decisions, the social system of
guppies and other species in which there is unambiguous evidence for mate copy-
ing are clearly amenable to interpretation from the perspective of communication
networks.
Socially modulated versus intrinsic preferences
In the absence of mate copying opportunities, female guppies distinguish
between males based on a number of phenotypic traits, for example tail size
(Bischoff et al., 1985) or colouration patterns (Houde, 1988; Houde & Endler, 1990;
Endler & Houde, 1995). Furthermore, female preferences for male traits such as
orange colouration have a significant heritable component (Houde, 1988; Houde
& Endler, 1990). Because female preferences in guppies are shaped by both ge-
netic and social factors, an interesting question is whether and how these factors
interact. Dugatkin & Godin (1992) conducted an experiment where a female was
initially allowed to choose between two males that differed only in their colour pat-
tern. Following this choice, the female was either exposed or not exposed to a focal
female placed beside the male not chosen in the initial preference test. After the
observation period, the female was again allowed to choose between the same two
males. Interestingly, the female reversed her choice in 75% of the trials, suggesting
that social factors (i.e. eavesdropping on the apparent preference of another female
for the less preferred male) are capable of overriding intrinsic preferences.
Dugatkin (1996a) employed a similar protocol but systematically altered the
asymmetries in orange colouration between the two males provided to the female

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
104 R. L. Earley & L. A. Dugatkin
Prop
ortio
n of
tim
es f
emal
e pr
efer
red
mor
e-or
ange
mal
e
Mean difference in total orange body colour between males
Fig. 5.5. The proportion of times female bystanders chose the more orange of the two
males in the presence (solid squares) or absence (open circles) of a focal female.
(Adapted from Dugatkin, 1996a.)
bystander. When there was no opportunity for mate copying, females exhibited a
more pronounced preference for males with more orange as the asymmetries in
colouration increased (Fig. 5.5). However, when a focal female was placed beside
the male with less orange, so as to simulate an apparent preference for drab
males, the female bystander chose the less-orange male significantly more often
in all cases except when the asymmetry was most drastic (40% difference in total
orange body colouration). Even when substantial asymmetries in male colouration
existed, a female bystander could be coerced into choosing the less-orange male by
increasing his perceived attractiveness via social manipulation (Dugatkin, 1998).
This was accomplished by increasing the number of focal females that exhibited
an apparent preference for the drab male or by increasing the amount of time
a single focal female spent near the less-orange male. Witte & Noltemeier (2002)
obtained strikingly similar results in female sailfin mollies P. latipinna, a related
poeciliid species where females presumably exhibit a genetically based preference
for large males (Marler & Ryan, 1997). Witte & Noltemeier (2002) also demonstrated
that reversals in mate choice incited by a copying bout persisted for long periods

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 105
of time (five weeks), even when a female was allowed to choose between previously
unknown large and small males. This marks one potential direction for the guppy
work: to determine whether observing courtship bouts influences female choice
even in the absence of the observed, previously chosen male.
In all, these results provide compelling evidence that observing courtship inter-
actions is sufficient to overturn female poeciliid’s genetically predisposed choice.
Furthermore, as the amount of social information available to the female by-
stander about a male’s potential quality increases, the more apt she is to rely on
social signals in lieu of the preference algorithm engrained in her genes. It is
important to note that in all of the studies on mate choice copying the observed
female was placed near a male of lesser quality (e.g. smaller or with less orange).
The low-quality male was then considered to be a suitable mate by virtue of his
being chosen earlier. This type of design is necessary to decouple mate choice copy-
ing from established, genetically based preferences. In nature, however, female
bystanders likely observe other females choosing relatively high-quality mates.
Therefore, mate choice copying is likely to reinforce rather than contradict pre-
existing preferences (Brooks, 1996).
Mate copying: a theoretical perspective
Relative to eavesdropping in an aggressive context, mate choice copying
has received an abundance of theoretical attention. Mathematical treatments of
mate copying have addressed two principal evolutionary questions: first, how es-
tablished mate copying strategies influence the evolution of female preferences
and male secondary sex characteristics or the variance in male mating success
(Wade & Pruett-Jones, 1990; Kirkpatrick & Dugatkin, 1994; Laland, 1994; Agrawal,
2001) and, second, the emergence and persistence of copying strategies (Losey
et al., 1986; Pruett-Jones, 1992; Dugatkin & Hoglund, 1995; Servedio & Kirkpatrick,
1996; Stohr, 1998). An exhaustive comparison of these models is beyond the scope
of this chapter. However, mate copying theory has revealed that network phe-
nomena, in particular the behavioural modifications that result from observing
social interactions, can have both short-term effects on individual decision mak-
ing and substantial evolutionary consequences. For instance, established mate
copying strategies can increase the variance in male mating success and, thus,
the opportunity for selection on male phenotypic traits that correlate well with
attractiveness (Wade & Pruett-Jones, 1990). When bystanders most often observe
interactions between females and males that exhibit the most common pheno-
type and when the effects of observing courtship interactions are independent
of male phenotype, mate copying impedes the spread of rare (or novel) male
traits (Kirkpatrick & Dugatkin, 1994; Laland, 1994; Agrawal, 2001). Interestingly,
when eavesdropping has a graded effect on bystander behaviour (e.g. depending

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
106 R. L. Earley & L. A. Dugatkin
on the phenotype of the observed male), a number of additional scenarios
emerge, including the potential for the spread of rare male phenotypes (Agrawal,
2001).
Given that mate copying can have dramatic effects on the distribution of male
traits in a population, it is important to understand how mate copying strategies
could emerge in the first place. A recurrent theme in the literature in this area,
particularly in models that assume female choice is under direct selection (but see
Servedio & Kirkpatrick, 1996), is that copying strategies will thrive when female
choice is costly (e.g. sampling costs, search time, predation risk: Pruett-Jones 1992)
or when substantial differences exist in females’ability to discriminate low- versus
high-quality males (Stohr, 1998). In the next section, we elaborate on how the costs
and benefits of female mate choice (or copying) potentially influence the degree
to which copying is used as a mate-assessment strategy.
Communication networks and mate choice: future directions
Eavesdropping and mate choice: influence of individual differences
In most studies on mate choice copying, the bystander and focal female
are matched for characteristics such as size, age and previous mating experience
(but see Dugatkin & Godin, 1993; Witte & Ryan, 1998). Under natural circum-
stances, females involved in the interaction are likely to differ in some respect.
Individual differences may influence how observation of courtship interactions
modulates bystander behaviour by adjusting the expected costs and benefits of
mate copying or mate choice itself. Dugatkin & Godin (1993) demonstrated that
small (young) females copy the mate choice of large (old) females, but not vice
versa. If older females have more mate choice experience and if experience de-
creases the rate at which errors in mate choice are committed (e.g. choosing a
poor-quality mate), then older females are liable to be better at discriminating
low- from high-quality mates than younger females. Therefore, the fitness benefit
of copying older females, namely having a higher probability of mating with supe-
rior males, may be quite high for young females. Conversely, the costs of copying a
younger, error-prone female may be sufficiently high to discourage mate copying
in older females. A host of other variables, including gravidity, previous mating
experience independent of age and physiological condition may affect how the
benefits and costs of mate choice and/or mate copying are perceived. For instance,
if female mate choice entails substantial sampling or search costs, individuals
who are under significant time constraints (e.g. hungry individuals whose time
would better be spent foraging) should be more likely to rely on the choices of
others. As with eavesdropping in an aggressive context, the state dependency of

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 107
mate copying represents an understudied and potentially important aspect of this
field.
Eavesdropping and mate choice: environmental influences
Environmental factors may also modify an individual’spropensity to copy
the choice of other females. Predation risk is known to influence female choosiness
and this is likely because females encounter a trade-off between the benefits of
remaining vigilant toward predators and the costs of spending time searching for,
or assessing, potential mates (Magnhagen, 1991; Pocklington & Dill, 1995). As the
costs of female choice intensify, mate copying should become increasingly bene-
ficial; therefore, under high predation risk, we might expect females to increase
their propensity to copy other females. However, this also assumes that bystanders
are less conspicuous to predators than individuals involved in active mate choice.
Briggs et al. (1996) found little support for this hypothesis in guppies derived from
streams with relatively high predation risk; apparent predation risk did not influ-
ence the proportion of females reversing their choice in the presence of a focal
female. However, as Briggs et al. (1996) acknowledged, females from populations
derived from streams with a high predation risk may not exhibit differential mate
copying responses under different predation regimes because, given the high cost
to female mate choice under natural circumstances, they may already exhibit the
maximal propensity to copy. Although Briggs et al. (1996) discarded this thesis,
they did not test whether female guppies derived from low-predation streams ex-
hibit divergent responses in the presence or absence of a predator. Nevertheless,
their work marks one of the first attempts to broach a largely unexplored area of
communication networks: namely how predation risk can modulate individual
tendencies to eavesdrop on mating interactions. The probability or frequency for
females to copy the choices of others may also be influenced by temporal factors
(e.g. early vs. late in the breeding season: Dugatkin & Hoglund, 1995), the quality
of one’s own mate, the availability of refuge or the opportunity to observe mating
interactions. Identifying influential environmental factors may aid in formulat-
ing a comparative communication network concept and should certainly trigger
empirical work dedicated to distinguishing the relative effects of individual- and
environment-based factors on decision-making processes in animals.
Summary
The aim of this chapter was to illustrate the importance of extending re-
search on mating and aggressive behaviour beyond the dyad and into a broader
social milieu. We have demonstrated that poeciliid fishes are capable of extracting

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
108 R. L. Earley & L. A. Dugatkin
information from various types of social interaction and integrating this infor-
mation for use in future encounters with the observed individuals. A recurrent
theme in this chapter is how the actual (or perceived) costs and benefits of fighting,
mating or eavesdropping can influence the short-term effects of observing social
interactions and the evolutionary viability of such strategies. This is where mate
copying theory and current, albeit sparse, theory on eavesdropping in an aggres-
sive context intersect. Irrespective of a bystander’s focus, when social interactions
bear a high cost, it pays to be attentive to signalling exchanges between others.
Whether the same selection pressures act on eavesdropping regardless of the con-
text in which it is used remains to be explored theoretically. Nevertheless, we have
attempted to highlight a wealth of factors that could affect the circumstances un-
der which eavesdropping is favoured and how the effects of eavesdropping are
manifest behaviourally. Daunting as the list of candidate influences may be, a
comprehensive understanding of communication networks relies, in part, on our
ability to partition the relative effects of each of these factors using comparative,
theoretical or empirical approaches. Unveiling the complex interactions between
individual (e.g. size, status, age), social (e.g. mating systems) and ecological (e.g.
predation risk, seasonality) variation and bystander behaviour marks a compelling
future direction for the field of communication networks.
Acknowledgements
We express our sincere thanks to Matthew Druen, Trish Sevene-Adams, Meredith McGee,
Michael Boles, Megan Tinsley and Blair Gilliland for their assistance in the laboratory. We are
also grateful to Peter McGregor for extending the invitation to contribute to this book and to
Matthew Grober, Cathleen Drilling and Ed Rodgers for insightful discussion on earlier versions
of this chapter. The work described in this chapter was funded in part by the National Science
Foundation, Sigma Xi, Kentucky Academy of Science, Animal Behavior Society and the American
Livebearers Association.
References
Agrawal, A. 2001. The evolutionary consequences of mate copying on male traits.
Behavioral Ecology and Sociobiology, 51, 33–40.
Amorim, M. & Hawkins, A. 2000. Growling for food: acoustic emissions during
competitive feeding of the streaked gurnard. Journal of Fish Biology, 57, 895–907.
Bakker, T. C. M. & Sevenster, P. 1983. Determinants of dominance in male sticklebacks
(Gasterosteus aculeatus L.). Behaviour, 86, 55–71.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 109
Bakker, T. C. M., Bruijn, E. F. D. & Sevenster, P. 1989. Asymmetrical effects of prior
winning and losing on dominance in sticklebacks (Gasterosteus aculeatus). Ethology,
82, 224–229.
Beaugrand, J. P., Caron, J. & Comeau, L. 1984. Social organization of small
heterosexual groups of green swordtails (Xiphophorus helleri, Pisces, Poeciliidae)
under conditions of captivity. Behaviour, 91, 24–60.
Beaugrand, J. P., Payette, D. & Goulet, C. 1996. Conflict outcome in male green
swordtail fish dyads (Xiphophorus helleri): interaction of body size, prior
dominance/subordination experience and prior residency. Behaviour, 133,
303–319.
Bischoff, R., Gould, J. & Rubenstein, D. 1985. Tail size and female choice in the guppy
(Poecilia reticulata). Behavioral Ecology and Sociobiology, 17, 252–255.
Briggs, S., Godin, J. G. J., Dugatkin, L. A. 1996. Mate-choice copying under predation
risk in the Trinidadian guppy (Poecilia reticulata). Behavioral Ecology, 7, 151–157.
Brooks, R. 1996. Copying and the repeatability of mate choice. Behavioral Ecology and
Sociobiology, 39, 323–329.
Brosnan, S. F., Earley, R. L., Dugatkin, L. A. 2003. Observational learning and predator
inspection in guppies (Poecilia reticulata). Ethology, 109, 823–833.
Brown, C. & Laland, K. N. 2002. Social learning of a novel avoidance task in the guppy:
conformity and social release. Animal Behaviour, 64, 41–47.
Brown, G. & Godin, J. G. J. 1999. Chemical alarm signals in wild Trinidadian guppies
(Poecilia reticulata). Canadian Journal of Zoology, 77, 562–570.
Chase, I. D. 1980. Social process and hierarchy formation in small groups: a
comparative perspective. American Sociological Review, 45, 905–924.
1982. Dynamics of hierarchy formation: the sequential development of dominance
relationships. Behaviour, 80, 218–239.
Chase, I. D., Bartolomeo, C. & Dugatkin, L. A. 1994. Aggressive interactions and
inter-contest interval: how long do winners keep winning? Animal Behaviour, 48,
393–400.
Clotfelter, E. & Paolino, A. 2003. Winner effect is induced in bystanders to aggressive
contests between male fighting fish. Animal Behaviour, 66, 343–347.
Doutrelant, C. & McGregor, P. K. 2000. Eavesdropping and mate choice in female
fighting fish. Behaviour, 137, 1655–1669.
Dukas, R. 2002. Behavioural and ecological consequences of limited attention.
Philosophical Transactions of the Royal Society of London, Series B, 357, 1539–1547.
Dugatkin, L. A. 1992. Sexual selection and imitation: females copy the mate choice of
others. American Naturalist, 139, 1384–1389.
1996a. Interface between culturally based preferences and genetic preferences:
female mate choice in Poecilia reticulata. Proceedings of the National Academy of
Sciences, USA, 93, 2770–2773.
1996b. Copying and mate choice. In: Social Learning in Animals: the Roots of Culture ed.
C. Heyes & G. Galef, New York: Academic Press, pp. 85–105.
1998. Genes, copying, and female mate choice: shifting thresholds. Behavioral
Ecology, 9, 323–327.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
110 R. L. Earley & L. A. Dugatkin
2001. Bystander effects and the structure of dominance hierarchies. Behavioral
Ecology, 12, 348–352.
Dugatkin, L. A. & Godin, J. G. J. 1992. Reversal of female mate choice by copying in the
guppy (Poecilia reticulata). Proceedings of the Royal Society of London, Series B, 249,
179–184.
1993. Female mate copying in the guppy (Poecilia reticulata): age-dependent effects.
Behavioral Ecology, 4, 289–292.
Dugatkin, L. A. & Hoglund, J. 1995. Delayed breeding and the evolution of mate
copying in lekking species. Journal of Theoretical Biology, 174, 261–267.
Earley, R. L. & Dugatkin, L. A. 2002. Eavesdropping on visual cues in green swordtail
(Xiphophorus helleri) fights: a case for networking. Proceedings of the Royal Society of
London, Series B, 269, 943–952.
Earley, R. L., Tinsley, M. & Dugatkin, L. A. 2003. To see or not to see: does previewing a
future opponent affect the contest behavior of green swordtail males (Xiphophorus
helleri)? Naturwissenschaften, 90, 226–230.
Earley, R. L., Druen, M. & Dugatkin, L. A. 2004. Watching fights does not alter a
bystander’s response towards naıve conspecifics in male green swordtail fish
(Xiphophorus helleri). Animal Behaviour, in press.
Endler, J. A. & Houde, A. E. 1995. Geographic variation in female preferences for male
traits in Poecilia reticulata. Evolution, 49, 456–468.
Enquist, M. & Leimar, O. 1983. Evolution of fighting behaviour: decision rules and
assessment of relative strength. Journal of Theoretical Biology, 102, 387–410.
Enquist, M., Leimar, O., Ljungberg, T., Mallner, Y. & Segerdahl, N. 1990. A test of the
sequential assessment game: fighting in the cichlid fish, Nannacara anomala.
Animal Behaviour, 40, 1–14.
Fisher, R. A. 1958. The Genetical Theory of Natural Selection, 2nd edn. New York: Dover.
Franck, D., Klamroth, B., Taebel-Hellwig, A. & Schartl, M. 1998. Home ranges and
satellite tactics of male green swordtails (Xiphophorus helleri) in nature. Behavioural
Processes, 43, 115–123.
Giaquinto, P. C. & Volpato, G. L. 1997. Chemical communication, aggression, and
conspecific recognition in the fish Nile tilapia. Physiology and Behavior, 62,
1333–1338.
Grant, J. & Green, L. 1996. Mate copying versus preference for actively courting males
by female Japanese medaka (Oryzias latipes). Behavioral Ecology, 7, 165–167.
Haller, J. 1995. Biochemical background for an analysis of cost–benefit interrelations
in aggression. Neuroscience and Biobehavioral Reviews, 19, 599–604.
Halperin, J., Giri, T., Elliot, I. & Dunham, D. 1998. Consequences of hyper-
aggressiveness in Siamese fighting fish: cheaters seldom prosper. Animal
Behaviour, 55, 87–96.
Hannes, R. P., Franck, D. & Leimann, F. 1984. Effects of rank-order fights on whole-body
and blood concentrations of androgens and corticosteroids in the male swordtail
(Xiphophorus helleri). Ethology, 65, 53–65.
Hollis, K. L., Dumas, M. J., Singh, P. & Fackelman, P. 1995. Pavlovian conditioning of
aggressive behavior in blue gourami fish (Trichogaster trichopterus): winners become
winners and losers stay losers. Journal of Comparative Psychology, 109, 125–133.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 111
Houde, A. E. 1987. Mate choice based on naturally occurring colour-pattern variation
in a guppy population. Evolution, 41, 1–10.
1988. Genetic differences in female choice between two guppy populations. Animal
Behaviour, 36, 510–516.
1997. Sex, Colour and Mate Choice in Guppies. Princeton, CT: Princeton University Press.
Houde, A. E. & Endler, J. A. 1990. Correlated evolution of female mating preference
and male colour pattern in the guppy, Poecilia reticulata. Science, 248, 1405–1408.
Hsu, Y. & Wolf, L. L. 1999. The winner and loser effect: integrating multiple
experiences. Animal Behaviour, 57, 903–910.
2001. The winner and loser effect: what fighting behaviours are influenced? Animal
Behaviour, 61, 777–786.
Jakobsson, S., Brick, O. & Kullberg, C. 1995. Escalated fighting behaviour incurs
increased predation risk. Animal Behaviour, 49, 235–239.
Johnstone, R. 2001. Eavesdropping and animal conflict. Proceedings of the National
Academy of Sciences, USA, 98, 9177–9180.
Kirkpatrick, M. & Dugatkin, L. A. 1994. Sexual selection and the evolutionary effects of
copying mate choice. Behavioral Ecology and Sociobiology, 34, 443–449.
Kodric-Brown, A. 1990. Mechanisms of sexual selection: insights from fishes. Annales
Zoologici Fennici, 27, 87–100.
Lachlan, R. F., Crooks, L. & Laland, K. N. 1998. Who follows whom? Shoaling
preferences and social learning of foraging information in guppies. Animal
Behaviour, 56, 181–190.
Ladich, F. 1998. Sound characteristics and outcome of contests in male croaking
gouramis (Teleostei). Ethology, 104, 517–529.
Lafleur, D. L, Lozano, G. A. & Sclafani, M. 1997. Female mate-choice copying in guppies,
Poecilia reticulata: a re-evaluation. Animal Behaviour, 54, 579–586.
Laland, K. N. 1994. Sexual selection with a culturally transmitted mating preference.
Theoretical Population Biology, 45, 1–15.
Laland, K. N. & Reader, S. M. 1999. Foraging innovation in the guppy. Animal Behaviour,
57, 331–340.
Laland, K. N. & Williams, K. 1997. Shoaling generates social learning of foraging
information in guppies. Animal Behaviour, 53, 1161–1169.
Liley, N. R. 1966. Ethological isolating mechanisms in four sympatric species of
poeciliid fishes. Behaviour (Supplement), 31, 1–197.
Losey, G., Jr Stanton, F., Telecky, T., Tyler III, W. and the Zoology 691 Graduate
Seminar Class 1986. Copying others, an evolutionary stable strategy for mate
choice: a model. American Naturalist, 128, 653–664.
Lugli, M. 1997. Response of male goby, Padogobius martensii, to aggressive sound
playback following pre-experimental visual stimulation. Behaviour, 134,
1175–1188.
Magnhagen, C. 1991. Predation risk as a cost of reproduction. Trends in Ecology and
Evolution, 6, 183–186.
Magurran, A. E. & Pitcher, T. J. 1987. Provenance, shoal size, and the socio-biology of
predator evasion behaviour in minnow shoals. Proceedings of the Royal Society of
London, Series B, 229, 439–465.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
112 R. L. Earley & L. A. Dugatkin
Marler, C. A. & Ryan, M. J. 1997. Origin and maintenance of a female mating
preference. Evolution, 51, 1244–1248.
Martel, G. & Dill, L. M. 1993. Feeding and aggressive behaviours in juvenile coho
salmon (Oncorhynchus kisutch) under chemically mediated risk of predation.
Behavioral Ecology and Sociobiology, 32, 365–370.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.
McGregor, P. K. & Westby, G. W. M. 1992. Discrimination of individually characteristic
electric organ discharges by a weakly electric fish. Animal Behaviour, 43, 977–986.
McGregor, P. K., Otter, K. & Peake, T. M. 2000. Communication networks: receiver and
signaller perspectives. In: Animal Signals: Signalling and Signal Design in Animal
Communication ed. Y. Espmark, T. Amundsen & G. Rosenqvist. Trondheim: Tapir
Academic Press, pp. 405–416.
McGregor, P. K., Peake, T. M. & Lampe, H. M. 2001. Fighting fish Betta splendens extract
relative information from apparent interactions: what happens when what you
see isn’t what you get. Animal Behaviour, 62, 1059–1065.
Mennill, D., Ratcliffe, L. & Boag, P. 2002. Female eavesdropping on male song contests
in songbirds. Science, 296, 873.
Mesterton-Gibbons, M. & Adams, E. 1998. Animal contests as evolutionary games.
American Scientist, 86, 334–341.
Mirza, R., Scott, J. & Chivers, D. 2001. Differential responses of male and female red
swordtails to chemical alarm cues. Journal of Fish Biology, 59, 716–728.
Naguib, M. & Todt, D. 1997. Effects of dyadic vocal interactions on other conspecific
receivers in nightingales. Animal Behaviour, 54, 1535–1543.
Naguib, M., Fichtel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Nakatsuru, K. & Kramer, D. L. 1982. Is sperm cheap? Limited male fertility and female
choice in the lemon tetra (Pisces, Characidae). Science, 216, 753–755.
Neat, F., Taylor, A. & Huntingford, F. 1998. Proximate costs of fighting in male cichlid
fish: the role of injuries and energy metabolism. Animal Behaviour, 55, 875–882.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Oliveira, R. F., Lopes, M., Carneiro, L. A. & Canario, A. V. M. 2001. Watching fights raises
fish hormone levels. Nature, 409, 475.
Otter, K., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits (Parus major)
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society of London, Series B, 266, 1305–1309.
Payne, R. 1998. Gradually escalating fights and displays: the cumulative assessment
model. Animal Behaviour, 56, 651–662.
Payne, R. & Pagel, M. 1997. Why do animals repeat displays? Animal Behaviour, 54,
109–119.

P1: JZZ/... P2: JZZ/...0521823617c05.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:24
Poeciliid sociality: fighting, mating and networking 113
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
2002. Do great tits assess rivals by combining direct experience with information
gathered by eavesdropping? Proceedings of the Royal Society of London, Series B, 269,
1925–1929.
Pocklington, R. & Dill, L. M. 1995. Predation on females or males: who pays for bright
male traits? Animal Behaviour, 49, 1122–1124.
Pruett-Jones, S. 1992. Independent versus nonindependent mate choice: do females
copy each other. American Naturalist, 140, 1000–1009.
Pulliam, H. R. & Caraco, T. 1984. Living in groups: is there an optimal group size? In:
Behavioural Ecology. An Evolutionary Approach, 2nd edn. ed. J. R. Krebs & N. B. Davies.
pp. Oxford: Blackwell Scientific, pp. 122–147.
Ranta, E. & Juvonen, S. K. 1993. Interference affects food-finding rate in schooling fish.
Journal of Fish Biology, 43, 531–535.
Ryan, M. J. 1997. Sexual selection and mate choice. In: Behavioural Ecology. An
Evolutionary Approach, 4th edn. ed. J. R. Krebs & N. B. Davies. Oxford: Blackwell
Scientific, pp. 179–202.
Schuett, G. 1997. Body size and agonistic experience affect dominance and mating
success in male copperheads. Animal Behaviour, 54, 213–224.
Servedio, M. & Kirkpatrick, M. 1996. The evolution of mate choice copying by indirect
selection. American Naturalist, 148, 848–867.
Stohr, S. 1998. Evolution of mate-choice copying: a dynamic model. Animal Behaviour,
55, 893–903.
Swaney, W., Kendal, J., Capon, H., Brown, C. & Laland, K. N. 2001. Familiarity facilitates
social learning of foraging behaviour in the guppy. Animal Behaviour, 62, 591–598.
Thorson, R. & Fine, M. 2002. Acoustic competition in the gulf toadfish Opsanus beta:
acoustic tagging. Journal of the Acoustical Society of America, 111, 2302–2307.
Waas, J. R. & Colgan, P. W. 1992. Chemical cues associated with the visually elaborate
aggressive displays of threespine sticklebacks. Journal of Chemical Ecology, 18,
2277–2284.
Wade, M. J. & Pruett-Jones, S. G. 1990. Female copying increases the variance in male
mating success. Proceedings of the National Academy of Sciences, USA, 87, 5749–5753.
Wingfield, J. C., Hegner, R. E., Dufty, A. M. & Ball, G. F. 1990. The challenge hypothesis:
theoretical implications for patterns of testosterone secretion, mating systems,
and breeding strategies. American Naturalist, 136, 829–846.
Wisenden, B. D. & Sargent, R. C. 1997. Anti-predator behaviour and suppressed
aggression by convict cichlids in response to injury-released chemical cues of
conspecifics but not to those of an allopatric heterospecific. Ethology, 103, 283–291.
Witte, K. & Noltemeier, B. 2002. The role of information in mate-choice copying in
female sailfin mollies (Poecilia latipinna). Behavioral Ecology and Sociobiology, 52,
194–202.
Witte, K. & Ryan, M. J. 1998. Male body length influences mate-choice copying in the
sailfin molly Poecilia latipinna. Behavioral Ecology, 9, 534–539.

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
6
The occurrence and functionof victory displays withincommunication networks
j o h n l . b ow e r
Fairhaven College, Western Washington University, Washington, USA
Introduction
Much recent research has focused on communication that occurs prior
to and during agonistic interactions in animals, leading to theoretical and em-
pirical advances in our understanding of the evolution of signalling before and
during agonistic contests (Maynard Smith, 1982; Huntingford & Turner, 1987;
Bradbury & Vehrencamp, 1998; Johnstone, 2001). However, very little research
has focused on the signalling that occurs at the conclusion of agonistic con-
tests (but see Ch. 10) despite the fact that such signalling may have important
consequences for animals involved in such contests and nearby conspecifics.
Post-contest signals may be given by the winner or loser of a contest or may
occur when there is no clear winner or loser. Such signalling by a winner or
loser may be directed towards the other combatant or may be directed to oth-
ers, such as potential rivals and mates. One type of post-contest signal has been
called a ‘victory display’ (e.g. Bradbury & Vehrencamp, 1998). Here, I define a vic-
tory display as a display performed by the winner of a contest but not by the
loser. In this chapter, I review the known occurrence of victory displays and then
use those examples to explore the functional significance of victory displays. I
first consider functions within the combatant dyad and then expand the view
to consider functions within a communication network. Along the way, I illus-
trate some difficulties in studying victory displays and suggest areas for further
research.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
114

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
The occurrence and function of victory displays 115
Occurrence of victory displays
It is not known how common victory displays are because few researchers
have studied them or have looked for them, and I am unaware of any published
review of these behaviours. Here, I present examples of displays that are plausi-
bly victory displays. These examples come from a literature survey, enquiries to
researchers and requests for information on victory displays posted to listservers
that focus on behavioural ecology or taxonomic groups. Most potential victory
displays discovered were either side notes within papers devoted to other aspects
of behaviour or were anecdotes from researchers. I will then explore the displays
described here to begin the discussion of how one identifies a victory display.
Because of the inefficiency of the methods I was forced to use, I expect that my list
of potential victory displays does not represent all the occurrences in the litera-
ture, and most certainly the list does not reflect the frequency of its occurrence in
the natural world. In the following paragraphs, I begin with the earliest described
victory display (in waterfowl), then consider other bird examples before travelling
a conventional phylogenetic route from arthropods to humans.
Waterfowl
The earliest described victory display is the ceremonie der triumphe in the
greylag goose Anser anser (Heinroth, 1910). This display occurs in several contexts,
but its use as a victory display occurs when males return to their mates and/or fam-
ilies after ritualized or actual contests. Returning males cackle loudly with their
necks extended and wings half-raised. Their mates sometimes join the display,
creating what Lorenz (1965a) considered to be ‘the most impressive vocal display
of the greylag goose’. Similar displays appear to be common in swans and geese.
Examples from the literature include Canada geese Branta canadensis (Radesater,
1974), barnacle geese Branta leucopsis (Bigot et al., 1995) and black swans Cygnus
atratus (Kraaijeveld & Mulder, 2002). In each of these four examples, the displays
are characterized by acoustic (raucous ‘cackling’ vocalizations) and visual (wing
flapping and water splashing) signals that carry far beyond the area in which the
contest took place. In particular, exaggerated rolling of the neck and cackling
occur simultaneously after a male has won an agonistic contest in the greylag
goose (Lorenz, 1965b). In Canada geese, male ‘high intensity cackling’,occurs after
contests (Radesater, 1974). Raveling (1967) found that triumph ceremonies were
performed by both males following some contests, but that only victors performed
ceremonies following severe attacks. In addition, victors gave more prolonged dis-
plays after severe attacks than they did after less-severe attacks. Thus, while graded
expressions of the triumph ceremony may occur in a variety of behavioural con-
texts, the exaggerated and simultaneous head rolling and cackling may be the

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
116 J. L. Bower
identifying characteristics of a victory display. Furthermore, it may be only af-
ter particularly serious contests that the ceremony can be considered a victory
display.
Other bird groups
In other birds, there are several examples of victory displays. These include
parrots of the genus Trichoglossus, which show post-contest behaviours analogous
to the triumph display in geese (Serpell, 1981) and the ‘bow flipper spread’ given
by winners of contests in the little blue penguin Eudyptula minor (Waas, 1990).
In little blue penguins, aggression commonly occurs between unpaired males
gathered on non-breeding sites within caves that contain a nesting colony. About
10% of agonistic interactions between males escalate to physical fights, which
sometimes caused serious flesh wounds. At the conclusion of such a contest, the
winner typically bows forward with his flippers spread and vocalizes while the
loser remains stationary or retreats.
Some examples involve mainly acoustic signals. For example, a victory display
has recently been described in duetting tropical boubou Laniarius aethiopicus (Grafe
& Bitz, 2004). In playback experiments involving 26 pairs of boubous, pairs sang
one of their 12 shared song types much more often than any other song type
following the cessation of playback. This song type was only rarely sung prior to
playback or during playback, suggesting that the song type functioned as a post-
conflict display. Because only presumptive winners sang the duet and not losers,
the song appears to be a victory display. This song type had unique signal design
features (longer song, higher frequencies, more overlap of male and female notes:
Grafe & Bitz, 2004) and was sung from higher perches and carried farther than
other song types (T. U. Grafe, unpublished data). This is a particularly striking
example of a bird species using a specific song type in a specific behavioural
context.
A second acoustic example of a victory display occurs in song sparrows Melospiza
melodia, in which the winner of a naturally occurring territorial contest (defined
as the bird who remains in the contest area after the contest) increases his song
rate to match the highest song rates sung prior to territorial contests (Table 6.1;
Bower, 2000). During the minute following the conclusion of a contest, the win-
ning bird’ssong rate almost always exceeds those of the other dozen or so males in
the song sparrow neighbourhood (Table 6.2). Like the tropical boubous, winning
song sparrow males typically sang from higher perches after contests ( J. Bower,
unpublished data), making them highly conspicuous to their neighbours and sug-
gesting that neighbours may be intended receivers of the victory display.
A third example of a victory display with a striking acoustic component is the
yodel call of black-throated divers Gavia immer. In a low-density Scottish popula-
tion, yodels were produced by males that had just successfully defended their loch

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
The occurrence and function of victory displays 117
Table 6.1. Song rates of winners, losers, neighbours and non-neighbours during the
minute before and the minute after the end of territorial contests
Songs/min (mean ± SE)
Category Before After No.a p valueb
Winner 3.08 ± 0.99 6.08 ± 0.45 12 0.02∗
Loser 1.17 ± 0.66 2.25 ± 0.63 12 0.26
Unpaired neighbour 4.06 ± 0.59 3.25 ± 2.14 8 0.18
Paired neighbour 1.07 ± 0.41 1.86 ± 0.83 7 0.40
Unpaired non-neighbour 2.32 ± 0.44 2.36 ± 0.40 12 0.86
Paired non-neighbour 0.78 ± 0.25 1.05 ± 0.39 9 0.40
SE, standard erroraSample sizes vary between tests because not all contests included birds of every category,
but males do not appear more than once in the data.bDifferences were compared with Wilcoxon matched-pairs signed-ranks test. Statistically
significant results are marked with an asterisk.
Table 6.2. Song rates of winners, losers, neighbours and non-neighbours during the
minute following the end of territorial contests
Category Songs/min Category Songs/min No.a p valueb
Winner 6.1 ± 0.5 Loser 2.3 ± 0.6 12 0.002∗
Winner 6.3 ± 0.7 Unpaired neighbour 3.3 ± 0.8 8 0.03∗
Winner 6.0 ± 0.4 Paired neighbour 1.9 ± 0.8 7 0.03∗
Winner 6.1 ± 0.5 Unpaired non-neighbour 2.4 ± 0.4 12 0.02∗
Winner 5.7 ± 0.4 Paired non-neighbour 1.1 ± 0.4 9 0.01∗
aSample sizes vary between tests because not all contests included birds of every category.bDifferences were compared with Wilcoxon matched-pairs signed-ranks test. Statistically
significant results are marked with an asterisk.
against another male, and playback of yodels elicits searching rather than yodels
(Gilbert, 1993).
Arthropods to humans
Victory displays have been described in several arthropod species. Reichert
(1978) described one such display in female funnel-web spiders Agelenopsis aptera,
in which the winners and losers of contests over web ownership showed striking
differences in stereotyped behaviours after the contest. Females who take over
a web, in particular, display behaviours such as biting the web manipulating
prey, circling the web or laying new silk. The performance of these behaviours is

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
118 J. L. Bower
exaggerated during post-contest periods beyond their non-contest performance.
For instance, the spiders exaggerate their abdomen movements when laying silk
and walk particularly slowly when circling the web. In crickets and their allies,
post-contest stridulation by winners of male–male contest appears to be common
in several species, including Gryllus bimaculatus (Alexander, 1961; Simmons, 1986;
Adamo & Hoy, 1995), Teleogryllus oceanicus (Burk, 1983) and Acheta domesticus (Hack,
1997). In three species of Australian tree wetas (Hemideina crassidens, H. femorata and
H. ricta), winners of male–male contests stridulate after winning contests while
losers do not (Field & Rind, 1992; Field, 2001).
One species of reptile and one amphibian show what appear to be victory dis-
plays. In some lizards, the end of a contest is marked by ritualized positions, which
may communicate the status of winners and losers to others. For instance, in the
pygmy Mulga monitor Varanus gilleni, individuals who win contests end up atop
the loser and attempt to ride the loser until either a new contest occurs or the
two separate and the loser leaves the contest area (Carpenter, 1976). In green frogs
Rana clamitans, territorial males engage in splashing displays after expelling an
intruding male from a territory (Wells, 1978).
In mammals, potential victory displays have been recorded for a number of
species. Natoli & de Vito (1991) and Natoli et al. (2000) reported that some feral
domestic cats Felis catus roll on their back on the ground, exposing their under-
sides, in front of the contest loser. Males that engaged in this behaviour were
highly ranked within the dominance hierarchy of the feral cat groups studied. In
wolves Canis lupus and coyotes Canis latrans, winners of contests often run about
with their tails held high in the air after winning contests ( J. Way, personal com-
munication). In observations of eastern coyotes at the Stoneham Zoo (Stoneham,
MA), Way reported that winners of pinning contests emerge from the contests
with a high and bouncing gait while losers stay low to the ground in a submissive
pose. Antelope territorial males sometimes engage in a scent-marking behaviour
after expelling an intruder from their territory. For instance, hartebeest Alcelaphus
buselaphus sometimes add to a boundary dung pile after contests (L. M. Gosling,
personal communication).
In marine mammals, the term ‘victory squeal’ is used for the vocalization given
within a second of a fish being seized by a bottlenose dolphin Tursiops truncatus
(S. H. Ridgeway, personal communication) or by a white whale Delphinapterus leucas
(Ridgeway & Carder, 1998). Whether this is a victory display by my definition
depends on whether the fish was caught in a competitive situation: analogous to
goal scoring in human examples.
Lastly, it is worth noting that various human behaviours may be considered vic-
tory displays. For instance, in ritualized sporting contests, winners often engage in
conspicuous displays after goals are scored or victories occur: ice hockey players

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
The occurrence and function of victory displays 119
raise their sticks high after a goal has been scored and football (soccer) players
sometimes take off their shirts and run around the field after scoring a goal. Many
American football players engage in stereotyped displays from ‘spiking’ the ball
against the ground to displays involving dances that are unique to the individual
performing it. At the conclusion of many important team sports events, winning
teams climb on top of each other in ways that would seemingly leave them vulner-
able to aggression from the other team. On a more serious note, victory parades
following armed conflict can be considered a group victory display. It would be
fascinating to know of other examples of victory displays following individual or
group conflict in either adult or juvenile humans.
Functional significance of victory displays
Distinctive design features of victory displays
Before addressing the functional significance of victory displays, I must
address the problem of determining if a victory display has a unique function and
meaning that can be separated from that of signals occurring prior to and dur-
ing contests. One difficulty in answering this question is that many post-contest
signals have similar design features to signals used in other behavioural contexts.
For example, wetas and crickets stridulate and birds sing prior to and during con-
tests as well as after contests, leading one to question whether the meaning of a
post-contest signal is to communicate the end of a contest or whether it is simply
performed in anticipation of the continuation of aggression.
One solution to this problem is to examine whether the context for the post-
contest signalling differs markedly from contexts in which signals with similar
design features are produced. If the sender and potential receivers can determine
that the signal is given in the context of the conclusion of a contest, then one can
surmise that the signal’s meaning in the post-contest context may be specific to
that context. For instance, in song sparrows, the victory display of singing at a
high song rate is similar to the high song rate typical of aggressors before they
initiate contests (Bower, 2000). However, the context in which this post-contest
signal occurs differs from the pre-contest singing in important ways. A major
difference is that at the termination of song sparrow contests, the loser leaves the
area, ending the almost continual chasing and physical contact that marks a song
sparrow territorial contest. Following contests, losers typically remain hidden
and quiet for 30 minutes or more after losing (J. Bower, unpublished data). Thus,
the contest winner and surrounding individuals are likely to be aware very soon
after the contest ends that he has entered a post-contest period in which further
aggression is unlikely to occur in the near future. During most pre-contest periods,
both males sit on perches separated by only a few metres and sing. Consequently,

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
120 J. L. Bower
the context for the victory display is very different from the context for singing
in other stages of interactions. Further research on victory displays may further
our understanding of how such signalling differs from signals with similar design
features given in other contexts.
A second solution to this problem is to test whether the design features of post-
signal displays differ, even subtly, from similar signals given in other contexts. For
instance, one difficulty in considering the triumph ceremony in geese as a victory
display is that the triumph display is composed of a variable and complex mix
of discrete displays, which occur in a variety of social contexts. For instance, the
rolling of the neck and some degree of cackling occurs even when male greylag
geese only feign attacks on other geese and do not actually engage in a contest.
However, both the rolling and cackling are most exaggerated and occur simul-
taneously only after a male has won a real contest (Lorenz, 1965b). Similarly, in
Canada geese, male ‘high intensity cackling’ is the part of the multifaceted tri-
umph display that occurs most commonly after territorial aggression (Radesater,
1974). Therefore, while graded expressions of the triumph ceremony may occur in
a variety of behavioural contexts, the exaggerated and simultaneous head rolling
and cackling may be specific design features of a victory signal.
In other species, behaviours or signal design features may also differ from sig-
nals used in different contexts. For instance, winners in song sparrow contests
often rise quickly above the thick shrubby vegetation to sing from higher perches
than they typically sing from, increasing the active space of their song and the
number of potential receivers ( J. Bower, unpublished data). High perches may
also facilitate listening (Ch. 3). In playback experiments with the duetting tropical
boubou, pairs typically sang one of their 12 shared song-types following the cessa-
tion of playback (Grafe & Bitz, 2004). This song type is rarely sung outside of the
post-contest context, suggesting that it functions as a victory display. Design fea-
tures of this song type differ significantly from other song types in several ways.
Male and female notes overlap more; male notes reach higher frequencies and
the songs are longer in duration than other song types. So, while victory displays
may at first appear to be very similar to other signals, with closer inspection one
may find that they have specific and unique design features that identify them as
victory displays.
Functions of victory displays
Victory displays could function within the winner–loser dyad or more
widely in the communication network. For example, victory displays could make
the victory more memorable (in the sense of Guilford & Dawkins (1991)) to the
loser of the dyad, to other receivers in a network or both. It is also possible that
the displays have no function and are consequences of mechanisms driving

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
The occurrence and function of victory displays 121
contests, such as the hormone changes underlying such interactions (e.g. Ch. 21).
While recognizing that function and mechanism (ultimate and proximate) levels
of explanation are complementary rather than alternatives, the detailed features
of victory displays discussed below make it unlikely that they are functionless
by-products of recent aggression.
Functions within the dyad
If victory displays function within the winner–loser dyad, the winner
would be directing the signal to the loser, most likely in an attempt to decrease
the probability that the loser will initiate a new contest. By discouraging the loser
from starting a new contest, the winner might achieve a lasting victory, thus
avoiding the costs of further contests and creating the opportunity to resume
other activities, such as searching for a mate, maintaining a pair bond, vigilance
against other intruders and predators, and feeding and other maintenance tasks.
The examples described above seem to fit into one of two categories.
First, there are victory displays that seem to invite an extension of the contest.
For instance, in the bow flipper spread display in little blue penguins the winner
bows low the ground and spreads his flippers apart. This appears to put the winner
in a position where he is vulnerable to attack from the loser. Likewise, feral cats
that roll over on their back and expose their undersides would seem to be choosing
a position that is vulnerable to further attacks. It is possible that by providing the
loser with a stimulus for attacking the winner just after the loser has retreated
from the contest, the winner helps to crystallize dominance over the loser. At
a mechanistic level, it is possible that future displays by the winner similar to
the victory display used after a contest victory may result in a change of mental
state in the loser. Such associative learning (e.g. Staddon, 1983), in which the
loser associates losing the contest with the signal used during the winner’svictory
display, may function to discourage a contest loser from initiating future contests
with the winner.
Second, there are victory displays in which the winner may give a display that is
energetically or otherwise costly. For instance, in my song sparrow study (Bower,
2000), winners almost always sang at very high rates in the minutes following a
contest. Since singing is a moderately energetically costly behaviour (Oberweger
& Goller, 2001), the ability to sing at high rates immediately following a contest
may advertise the winner’s vigour or quality. This should be especially true since
song sparrow contests are energetically costly, often characterized by almost con-
tinuous chasing and occasional physical fighting for an hour or more, with no
breaks for feeding or resting (Bower, 2000). By singing at high rates just after a
contest ends, a male may be sending an honest signal to the loser that the winner
has sufficient endurance to defend the area he has just secured from subsequent

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
122 J. L. Bower
challenges, despite the energy costs of the recent contest. Other examples of pos-
sible victory displays that may be metabolically or otherwise costly include post-
victory stridulation in crickets and wetas, circling the web and laying new silk in
funnel-web spiders, splashing displays in frogs and the bouncing gait of eastern
coyotes. As with song sparrows, while victory displays may be relatively short in
duration, and thus not likely to be more than moderately energetically costly, their
occurrence at the end of often long and energetically costly contests may make
them difficult to perform. Consequently, they may function as honest indicators
of aspects of the winner’s quality (for further discussion of handicaps and honest
signalling, see Dawkins (1995)). The ability to perform such a display following a
contest may reduce the chances that the contest loser will re-engage the winner
in a later contest.
Functions within the network
Theory suggests that animals are likely to gain fitness benefits by assess-
ing potential mates and rivals in contest situations and altering their behaviour
according to their assessments (e.g. Cox & Le Boeuf, 1977; McGregor & Dabelsteen,
1996; McGregor & Peake, 2000). If this were so, then one would expect selection
pressure for winners to advertise their victory to other members of a communica-
tion network. Victory displays may inform social eavesdroppers (Ch. 2) and other
members of the network that did not pay attention to the interaction that the
winner has just won a contest. As discussed above, the displays may also provide
further information about the winner’s vigour and/or other measures of quality
by displaying after energetically costly contests.
This possible function of victory displays is distinct from audience effects (Ch. 4)
and considerations of the nature of signals and signalling during interactions (e.g.
private signals (Ch. 3) and the ‘good loser’ hypothesis (Peake & McGregor, 2004)).
However, no studies have been attempted that test whether performing a victory
display modifies the behaviour of network members. At present, investigating the
network function of victory displays has to rely on indirect evidence. One obvious
criterion, of course, is whether conspecifics other than (or as well as) the loser
are able to receive the signal (for similar discussion on the intended receivers for
post-copulatory displays in ducks, see Johnson et al. (2000)). This depends both on
the signal features and the spatial arrangement of the conspecifics. For instance,
of the examples given above, it is least likely that funnel-web spider post-victory
behaviour has a communication network function, since the widely spaced webs
(Reichert, 1978) and reliance on vibration for communication means that rivals
are unlikely to be aware of contests on other webs. Likewise, since varanid lizard
territories tend to be large (R. Earley, personal communication) and the possible
victory display is a visual signal, it is unlikely that rivals in adjoining territories

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
The occurrence and function of victory displays 123
will witness a contest between two lizards. Therefore, the pygmy Mulga monitor’s
‘back riding’post-contest behaviour is more likely directed to the loser rather than
for another potential rival. However, it is not known how often females might
witness such contests, nor how often other males encroaching on the territory
witness the contest. Therefore, even in territorial species in which territories are
large and signals have relatively short ranges, there may be a network function if
other animals are present on territories.
In contrast to these two examples, the other victory displays described above
occur in species in which potential mates and rivals are likely to witness the
display. Territorial songbirds, for instance, often reside in neighbourhoods, with
several territories constituting a neighbourhood. During the song sparrow territo-
rial contests I studied, I noticed that neighbourhood males and females often flew
up to perches from which they could see the contest ( J. Bower, unpublished data).
Such behaviour seemed to indicate that members of the communication network
were paying close attention to contests in their neighbourhood. Bird song is a
moderately long-range signal, typically carrying into and beyond neighbouring
territories (e.g. Brenowitz, 1982; Ch. 20). Therefore, the post-contest vocalizations
of a winning song sparrow, tropical boubou pair or goose would likely be heard
by other conspecifics in the bird’s neighbourhood. Similarly, stridulating crickets
and wetas, as well as splashing green frogs, all produce acoustic signals that are
likely to be accessible to rivals and potential mates. Visual signals made within
open habitats or by species with close spacing may also be candidates for victory
displays directed at the network. Thus, post-contest scent marking by hartebeest,
the movements of geese and the bouncing gait of eastern coyotes all are accessible
to conspecific receivers in the communication network.
Summary
Victory displays are post-conflict signals given by the winner (but not the
loser) of an agonistic contest. There has been little work specifically addressing
such displays and most of the evidence for their existence comes from incidental
descriptions or asides and footnotes. Much remains to be done to characterize
victory displays and to identify their function, including whether they are network
phenomena or are directed at the loser of the interaction. The difficult but exciting
work that lies ahead is to demonstrate that such displays, in conjunction with or
independent from the social eavesdropping that may occur during a contest, alter
the behaviour of other members of the communication network. It is likely that
such a test could be developed more easily in a laboratory setting than in situ;
one approach would be to prevent observers from watching the contest but allow
them to observe the following victory display.

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
124 J. L. Bower
Acknowledgements
I thank the many researchers who responded to my call for potential examples of
victory displays, Ulmar Grafe for sharing unpublished data, and three anonymous referees and
Peter McGregor for comments on previous drafts of this chapter.
References
Adamo, S. & Hoy, R. 1995. Agonistic behaviour in male and female field crickets,
Gryllus bimaculatus, and how behavioural context influences its expression. Animal
Behaviour, 49, 1491–1501.
Alexander, R. 1961. Aggressiveness, territoriality, and sexual behaviour in field
crickets (Orthoptera gryllidae). Behaviour, 17, 130–223.
Bigot, E., Hausberger, M. & Black, J. 1995. Exuberant youth: The example of triumph
ceremonies in barnacle geese (Branta leucopsis). Ethology, Ecology and Evolution, 7,
79–85.
Bower, J. L. 2000. Acoustic interactions during naturally occurring territorial conflict
in a song sparrow (Melospiza melodia) neighborhood. Ph.D. Thesis, Cornell
University, Ithaca, New York.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.
Brenowitz, E. A. 1982. Long-range communication of species identity by song in the
red-winged blackbird. Behavioral Ecology and Sociobiology, 10, 29–38.
Burk, T. 1983. Male aggression and female choice in a field cricket (Teleogryllus
oceanicus): the importance of courtship song. In: Orthopteran Mating Systems: Sexual
Competition in a Diverse Group of Insects, ed. D. T. Gwynne & G. K. Morris. Boulder,
CO: Westview Press, pp. 97–119.
Carpenter, C. C. 1976. A further analysis of the combat ritual of the pygmy mulga
monitor, Varanus gilleni (Reptilia: Varanidae). Herpetologica, 32, 35–40.
Cox, C. R. & Le Boeuf, B. J. 1977. Female incitation of male competition: a mechanism
for mate selection. American Naturalist, 111, 317–335.
Dawkins, M. S. 1995. Unravelling Animal Behaviour, 2nd edn. Harlow, UK: Longman.
Field, L. H. 2001. Aggression behaviour in New Zealand tree wetas. In: The Biology of
Wetas, King Crickets, and Their Allies, ed. L. H. Field & T. H. Jarman. Oxford: CAB
International, pp. 333–350.
Field, L. H. & Rind, F. 1992. Stridulatory behaviour in a New Zealand weta, Hemideina
crassidens. Journal of Zoology, 228, 371–394.
Gilbert, G. 1993. Vocal individuality as a census and monitoring tool: practical
considerations illustrated by a study of the bittern Botaurus stellaris and the
black-throated Diver Gavia arctica. Ph.D. Thesis, University of Nottingham, UK.
Grafe, T. U. & Bitz, J. H. 2004. An acoustic victory display in the duetting tropical
boubou (Laniarius aethiopicus): a signal of victory. BMC Ecology, 4, 1.
http://www.biomedcentral.com/bmcecol/.
Guilford, T. & Dawkins, M. S. 1991. Receiver psychology and the evolution of animal
signals. Animal Behaviour, 42, 1–14.

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
The occurrence and function of victory displays 125
Hack, M. A. 1997. The energetic costs of fighting in the house cricket, Acheta domesticus
L. Behavioral Ecology, 8, 28–36.
Heinroth, O. 1910. Beitrage zur Biologie, namentlich Ethologie und Psychologie der
Anatiden. In: Verhandlungen der V Internationalis Ornithologen Kongressus in Berlin,
pp. 589–702.
Huntingford, F. & Turner, A. K. 1987. Animal Conflict. Cambridge, UK: Cambridge
University Press.
Johnson, K. P., McKinney, F., Wilson, R. & Sorenson, M. D. 2000. The evolution of
postcopulatory displays in dabbling ducks (Anatini): a phylogenetic perspective.
Animal Behaviour, 59, 953–963.
Johnstone, R. A. 2001 Eavesdropping and animal conflict. Proceedings of the National
Academy of Sciences, USA, 98, 9177–9180.
Kraaijeveld, K. & Mulder, R. 2002. The function of triumph ceremonies in the black
swan. Behaviour, 139, 45–54.
Lorenz, K. 1965a. Here I Am – Where Are You?: The Behavior of the Greyleg Goose. New York:
Harcourt Brace.
1965b. The triumph ceremony of the greylag goose. Philosophical Transactions of the
Royal Society of London, Series B, 251, 477–478.
Maynard Smith, J. 1982. Evolution and the Theory of Games. Cambridge, UK: Cambridge
University Press.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
Natoli, E. & de Vito, E. 1991. Agonistic behaviour, dominance rank and copulatory
success in a large multi-male feral cat, Felis catus L., colony in central Rome.
Animal Behaviour, 42, 227–241.
Natoli, E., De Vito, E. & Pontier, D. 2000. Mate Choice in the domestic cat (Felis silvestris
catus L.). Aggressive Behavior, 26, 455–465.
Oberweger, K. & Goller, F. 2001. The metabolic cost of bird song production. Journal of
Experimental Biology, 204, 3379–3388.
Peake, T. M. & McGregor, P. K. 2004. Information and aggression in fishes. Learning and
Behavior, 32, 114–121.
Radesater, T. 1974. Form and sequential associations between the triumph ceremony
and other behaviour patterns in the Canada Goose Branta canadensis L. Ornis
Scandinavica, 5, 87–101.
Raveling, D. G. 1967. Sociobiology and ecology of Canada Geese in winter. Ph.D. Thesis,
Southern Illinois University, Carbondale, USA.
Reichert, S. E. 1978. Games spiders play: behavioral variability in territorial disputes.
Behavioral Ecology and Sociobiology, 3, 135–162.
Ridgeway, S. H. & Carder, D. A. 1998. Net-aided foraging by two white whales. Marine
Mammal Science, 14, 332–334.
Serpell, J. 1981. Duets, greetings and triumph ceremonies: analogous displays in the
parrot genus Trichoglossus. Zeitschrift fur Tierpsychologie, 55, 268–283.

P1: JZZ/... P2: JZZ/...0521823617c06.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:41
126 J. L. Bower
Simmons, L. 1986. Inter-male competition and mating success in the field cricket,
Gryllus bimaculatus (De Geer). Animal Behaviour, 34, 567–579.
Staddon, J. E. R. 1983. Adaptive Behavior and Learning. Cambridge, UK: Cambridge
University Press.
Waas, J. R. 1990. An analysis of communication during the aggressive interactions of
little blue penguins (Eudyptula minor). In: Penguin Biology, ed. LS Davis & JT Darby.
San Diego, CA: Academic Press, pp. 345–376.
Wells, K. D. 1978. Territoriality in the green frog (Rana clamitans): vocalizations and
agonistic behaviour. Animal Behaviour, 26, 1051–1063.

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 10:51
Part II T H E E F F E C T S O F P A R T I C U L A R
C O N T E X T S
127

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 10:51
128

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 10:51
Introduction
The rationale behind the grouping of chapters into this section is to
facilitate comparisons between communication networks found in very different
contexts: mate choice, predation, nestling begging, redirection and scent mark-
ing. One of the attractions of communication networks is that the idea applies to
any context in which the signals used travel far enough to encompass several other
individuals. However, each context will have distinctive features affecting the na-
ture of the information transmitted, the signals used and their travelling power;
therefore, the nature of the communication network may differ. Comparison of
networks found in different contexts could, therefore, advance our understanding
of the topic.
Mate choice
It is probably a fair generalization to say that in recent years the most
widely considered, modelled and experimented upon context for communication
has been the simplest mate choice situation, i.e. that involving a male signaller and
a female receiver. However, Ken Otter and Laurene Ratcliffe point out in Ch. 7 that
a communication network is a more likely context because females have access
to the widely broadcast mate attraction signals of several males. This chapter
discusses which traits females in a communication network use when choosing
between males: both as pair mates and as extra-pair partners. It also discusses the
way choice is achieved (e.g. simultaneous versus sequential assessment) and how
sampling by females can be inferred from the pattern of movement through a
network of signalling males.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
129

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 10:51
130 Part II
Predation
Communication networks are not restricted to individuals of the same
species; predators have long been recognized to be the unwanted guests at com-
munication feasts, with long-range signals advertising the location of potential
prey. The commonest response of choruses of insects and anurans to the detec-
tion of a predator is to cease calling (Gerhardt & Huber, 2002). This may be an
efficient way to avoid predators, but it is an inefficient way to communicate with
the intended receivers of the signals, such as potential mates. In Ch. 8, Alexander
Lang, Ingeborg Teppner, Manfred Hartbauer & Heiner Romer explain how pseu-
dophylline katydids can to some extent overcome this problem by signalling with
vegetation-borne vibrations (tremulations) rather than airborne sounds, because
tremulations cannot be detected by passive listening bat predators. While such bat
predators are an important selection pressure on katydids, they are not the only
ones. This chapter uses neurophysiological preparations in the field and decision-
tree learning algorithms to investigate how katydids communicate in a noisy rain-
forest environment.
Nestling begging
In birds, the begging of nestlings has come to rival mate choice as a model
system for the study of the evolution of biological signalling (Wright & Leonard,
2002). However, in parallel with most of the work on mate choice, begging is
considered as a dyad, with a nestling (or the brood collectively) as the signaller and
one parent as the receiver. This seems odd, perverse even, given the close proximity
of nestlings (both to nestmates and to their parents) and the conspicuous vocal and
visual signals that nestlings produce. In Ch. 9, Andy Horn and Marty Leonard show
how considering begging as a communication network can yield new insights into
begging behaviour. Furthermore, issues for communication networks, in general,
are raised by the close proximity of individuals and the possibility of direct physical
action in a crowded nest. Such issues will be particularly relevant to many highly
social species such as social hymenoptera.
Redirection of aggression
The nature of communication in aggressive encounters has a long history
of study, from Darwin’s antithesis principle (Darwin, 1872) and Lorenz’s classics
King Solomon’s Ring and On Aggression (Lorenz, 1952, 1966) onwards. Most attention
has focused on signal exchanges before and during aggressive encounters. How-
ever, redirection is a puzzling behaviour performed by losers after an aggres-
sive contest. As the target of redirection is an individual other than the winner

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 10:51
The effects of particular contexts 131
(commonly a lower-ranking individual in social groups), it extends even dyadic
interactions into a network. Ani Kazem and Filippo Aureli discuss explanations
for redirection in Ch. 10, focusing on primates where it has been best described.
They conclude that redirection is best interpreted from a network viewpoint, in
terms of how it can influence the behaviour of bystanders.
Scent marking
Scent marking is a rather different context from the others considered in
this section. Whereas the other contexts could be loosely considered to be aspects
of social behaviour, scent marking is a particularly distinct aspect of chemical
communication and can be involved in several social contexts. Scent marking is
also rather different from most other signals because scent marks persist, often for
considerable periods, in the absence of the signaller. Any conspecific visiting the
scent mark can obtain information from it; in this respect, such marks could be
considered analogous to public noticeboards. In Ch. 11, Jane Hurst considers the
selection pressures that result from such undirected and long-lasting signals. Most
of her examples come from studies of mice, where many studies have examined
the behavioural and biochemical basis of scent communication.
Future directions
A common theme of the chapters in this section (and indeed throughout
this book) is that communication needs to be considered in a more complex way in
order to make progress. Far from being a standard recourse to complexities of the
real world when simple explanations fail (and being even further from a counsel
of despair), these chapters demonstrate how adopting a network perspective can
explain troublesome aspects of communication and also indicate directions for
future research.
One future challenge in mate choice is to characterize female assessment be-
haviour, because in many instances potential mates can be assessed at long range,
with close approach possibly representing the outcome of choice rather than
assessment in action. Scent marks seem to offer an opportunity for such char-
acterization because close approach and perhaps contact are required to gather
information from scent marks. Video tracking individuals in a naturalistic enclo-
sure may relatively easily provide data on patterns of visits to scent marks and, by
extension, on information gathering.
Many communication behaviours including mate choice are likely to be con-
strained by the presence of predators, as the chapter on katydids demonstrates. In
such circumstances, information gathering could be a costly exercise if proximity
to signallers increases the risk of being preyed upon. Similarly, being a bystander

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 10:51
132 Part II
at an aggressive interaction could be costly if the bystander becomes the target of
redirected aggression. These examples indicate that animals may have to trade-off
several somewhat disparate costs and benefits when gathering information in a
communication network. As the costs and benefits are likely to change diurnally
and seasonally as well as on much shorter timescales, these trade-offs may be best
explored by modelling.
Many of the chapters have suggested or implied that more detailed empirical
studies, both experimental and observational, are needed to further our under-
standing. In some sense, this is always going to be true because of the enormous
variation in biological systems; however, one value of a communication network
perspective is that it suggests what types of information (e.g. mate choice assess-
ment patterns, variation in the form of begging calls) would allow the field to
develop.
The chapters in this part demonstrate the advantages of a broader view of
communication over and above the advantages of a network view: understand-
ing of communication in any particular context can come from contexts other
than that under immediate consideration. For example, nestling begging would
seem to have little to offer contexts such as mate choice, aggression and resource
defence, because the detailed circumstances of the contexts are so different. How-
ever, begging behaviour focuses attention on issues that are fundamental to all
three contexts and indeed to communication in general, such as the distinction
between signals and physical action. Similarly, several explanations for redirection
behaviour operate over a timescale encompassing a sequence of several contests,
suggesting that some aspects of audience effects and eavesdropping would benefit
from consideration over such longer timescales. Difficult as it may be with an ever-
expanding communication literature, it would seem a good idea to keep an eye on
developments in several communication contexts. Because communication net-
works can be seen to apply to several contexts, adopting a network perspective
helps to promote such a breadth of interest.
References
Darwin, C. 1872. The Expression of the Emotions in Man and Animals. London: John Murray.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans: Common
Problems and Diverse Solutions. Chicago, IL: Chicago University Press.
Lorenz, K. Z. 1952. King Solomon’s Ring; New Light on Animal Ways. New York: Crowell.
1966. On Aggression. New York: Harcourt, Brace and World.
Wright, J. & Leonard, M. L. 2002. The Evolution of Begging: Competition, Cooperation, and
Communication. Dordrecht: Kluwer.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
7
Enlightened decisions: femaleassessment and communicationnetworks
k e n a . o t t e r 1 & l au r e n e r a t c l i f f e 2
1 University of Northern British Columbia, Prince George, Canada2 Queen’s University, Kingston, Canada
Introduction
Asymmetry in parental investment often predicts that females should
be choosier about prospective mates than males. It is commonly assumed that
females assess male characteristics during mate choice, but which traits are
assessed, and how they influence female decision making, is not well understood.
Current models of mate choice suggest females may sequentially sample a pool
of males, memorizing levels of trait expression among comparison males, or else
accept the first male that exceeds some minimum threshold value of mate qual-
ity. Recent tests of communication network theory suggest that these models may
have to be revised because females can tap into advertising signals broadcast in
a network fashion. Such behaviour could reduce costs of mate searching, as sig-
nals are perceived simultaneously, allowing instantaneous relative comparisons.
In this chapter, we explore the potential of females to extract comparative infor-
mation on the relative quality of males for use in reproductive decision making.
We focus primarily on primary mate choice decisions (i.e. initial selection of a
mating partner) and secondary mate choice decisions (i.e. mating decisions that
arise after social pairing, which may include extra-pair copulations or ‘divorce’ of
the current mate to pair with another male) based on acoustic signals in territorial
passerines; however, the ideas that we present should be applicable to other taxa
and other sensory modalities. Finally, we discuss the potential impacts of habitat
alteration on females’ abilities to use network assessments for mate choice.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
133

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
134 K. A. Otter & L. Ratcliffe
In the early 1990s, it was realised that bird song and other animal commu-
nication took place in a more extensive (network) context than male–female
or male–male dyads; several receivers may perceive each signaller simultane-
ously and, conversely, signallers may direct their signal towards several receivers
(McGregor, 1993; McGregor & Dabelsteen, 1996). The idea that females seeking
mates may be able to assess several males simultaneously, not only on leks but
also in situations where males defend more dispersed territories, complicates the
study of female mate choice. For example, we need to rethink how females may
be sampling males, especially during initial territorial settlement and any subse-
quent pursuit of extra-pair copulations.
Communication network theory challenges us to reconsider traditional models
of mate assessment. Constraints on signal transmission and reception may limit
the spread of information to receiving parties. Such constraints should bias assess-
ment to particular kinds of signal and may also help to explain why some signals
important in initial mate choice need not necessarily correlate with secondary
mate choice (e.g. red plumage in house finches, Carpodacus mexicanus, is selected
by females in initial mate choice but does not seem to affect extra-pair success:
Hill, 2002). We must also consider the kinds of signal that are received by females;
how habitat alteration might influence communication in a network, and how,
in turn, that might influence reproductive success.
Deciphering which attributes of males are of most interest to females has
proved to be a challenging task; a common approach has been to ask whether
females pick superior males as partners, where superiority is defined by the expres-
sion of condition-dependent signals. In birds, considerable evidence from dyadic
mate choice studies suggests that colour and vocal cues believed to be energeti-
cally expensive are correlated with mate selection (reviewed by Gil & Gahr, 2002).
Whether such dyadic studies accurately reflect female assessment in natural cir-
cumstances is still unclear. Considerations of communication in a network con-
text have stimulated new experimental approaches that seek to determine how
males evaluate potential rivals (e.g. Naguib et al., 1999; Todt & Naguib, 2000; Ch. 2)
and similar types of study may help researchers to decipher how females evaluate
potential mates.
This chapter reviews data on which traits appear to be important in female
choice in songbirds and then suggests ways to model and test female choice using
communication network theory. Although work in this field is still limited, we
review published and in-progress studies that discuss how signals might spread in
networks and how females might assess such information. Finally, we discuss how
habitat alteration can affect the propagation of signals, and how this influences
the ability of females to assess males in communication networks.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 135
How do females assess males?
Traditional models of primary female mate choice assume that females
assess males sequentially ( Janetos, 1980; Wittenberger, 1983). In choosing social
mates, females move from one male to the next and select mates via either sequen-
tial comparison of the current male versus the last male visited, or via a best-of-n
model, where females sample all males and then return to the individual of highest
quality. Although less-formally modelled, most studies on secondary mate choice
assume similar kinds of decision-making strategy; females select a social partner
then assess this male relative to other available males to determine whether to en-
gage in extra-pair copulations or divorce (e.g. the ‘better options’ model of divorce
(Ens et al., 1993), in which birds select a social mate then assess opportunities to
desert and pair with a male of better quality, or similar strategies in the ‘genetic
benefits’ models of extra-pair copulation (Kempenaers & Dhondt, 1993)). Recent
models of primary mate choice are more realistic because they incorporate costs
of mate searching (Real, 1990; Wiegmann et al., 1996) but still assume that males
are assessed sequentially (e.g. Fig. 7.1a). However, the signals used by females to
assess males may propagate sufficient distance in some circumstances for males
to be assessed simultaneously (Gibson & Langen, 1996).
In communication networks, female receivers can simultaneously detect the
signals of several territorial males. For example, a female moving from one male
to the next may still be able to detect the signals of males she has visited previ-
ously. Thus, females may be able to assess the relative expression of several males’
signals simultaneously, without having to rely on memory of absolute expression.
Females could continue to search in such a fashion until no new male exceeds
a preceding male. However, female searching may not be even this constrained.
If females use a best-of-n strategy, they need not rely solely on memory of trait
expression of each male. All they need do is remember the territory locations of
males of perceived high quality and position themselves in a manner that allows
simultaneous comparison. If signal transmission is sufficiently long range, fe-
males may be able to assess and eliminate a number of males without even closely
approaching them, as described in anuran mating aggregations (e.g. Murphy &
Gerhardt, 2002; see below). Females positioning themselves strategically within
networks and making choices on relative trait expression could decrease search
time, maintain safe distances from territory owners and reduce the chance of
mistaken decisions (Fig. 7.1b).
Field studies typically use female movement patterns to assess the number
of males that are sampled by females. Some studies have used radio telemetry
to track female movement and have assumed that close approach is evidence
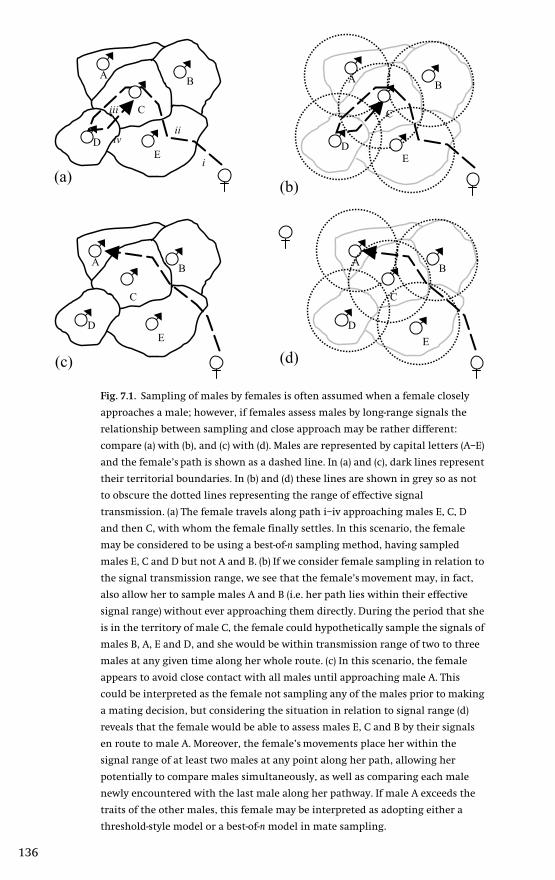
P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
AB
C
DE
i
ii
iii
iv
(d)
(a)
(c)
(b)
AB
C
DE
AB
C
DE
AB
C
D
E
Fig. 7.1. Sampling of males by females is often assumed when a female closely
approaches a male; however, if females assess males by long-range signals the
relationship between sampling and close approach may be rather different:
compare (a) with (b), and (c) with (d). Males are represented by capital letters (A–E)
and the female’s path is shown as a dashed line. In (a) and (c), dark lines represent
their territorial boundaries. In (b) and (d) these lines are shown in grey so as not
to obscure the dotted lines representing the range of effective signal
transmission. (a) The female travels along path i–iv approaching males E, C, D
and then C, with whom the female finally settles. In this scenario, the female
may be considered to be using a best-of-n sampling method, having sampled
males E, C and D but not A and B. (b) If we consider female sampling in relation to
the signal transmission range, we see that the female’s movement may, in fact,
also allow her to sample males A and B (i.e. her path lies within their effective
signal range) without ever approaching them directly. During the period that she
is in the territory of male C, the female could hypothetically sample the signals of
males B, A, E and D, and she would be within transmission range of two to three
males at any given time along her whole route. (c) In this scenario, the female
appears to avoid close contact with all males until approaching male A. This
could be interpreted as the female not sampling any of the males prior to making
a mating decision, but considering the situation in relation to signal range (d)
reveals that the female would be able to assess males E, C and B by their signals
en route to male A. Moreover, the female’s movements place her within the
signal range of at least two males at any point along her path, allowing her
potentially to compare males simultaneously, as well as comparing each male
newly encountered with the last male along her pathway. If male A exceeds the
traits of the other males, this female may be interpreted as adopting either a
threshold-style model or a best-of-n model in mate sampling.
136

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 137
of assessment of a signalling male (Bensch & Hasselquist, 1992; Neudorf et al.,
1997). The pied flycatcher Ficedula hypoleuca offers an extreme example of such
movement, as the nestbox is an assessed resource (Slagsvold et al., 1988; Dale
et al., 1990) and assessment requires close inspection, in much the same way
that a short-range signal would. However, movement patterns can be difficult
to interpret. For example, a female moving along the boundary between terri-
torial males, apparently undetected by them, may still be sampling these males
(Fig. 7.1c). If the males are producing long-range signals, then the female could
sample males based on this signalling network (Fig. 7.1d). Her movement pattern
allows at least two males to be assessed at almost all points along her route. By
such surreptitious sampling (i.e. moving silently and apparently remaining unde-
tected by males along the route: Neudorf et al., 1997), females may avoid some of
the costs associated with close approach to males. These costs could be harassment
from males or aggression from mated females (e.g. Dale & Slagsvold, 1995). After
surreptitious sampling, the female may then closely approach the male she has
selected. In such a scenario, close approach indicates choice rather than sampling
and the amount of sampling is underestimated (in Fig. 7.1d four males have been
sampled rather than one). Murphy & Gerhardt (2002) described such a scenario
in female barking treefrogs Hyla gratiosa in the field; females approach only a
single male in a chorus, suggesting no sampling has occurred and that the first
male encountered is selected. However, further anecdotal evidence suggested that
females may be assessing several males at a distance and then approaching only
the selected male (Murphy & Gerhardt, 2002).
Therefore, we urge caution when patterns of female movement are used to
infer sampling behaviour. We suggest that it may be more appropriate to deter-
mine female movement in relation to the transmission distance of signals used
in mate choice. Such considerations will allow us to determine whether females
are strategically placing themselves in areas that maximize the number of males
that can be simultaneously assessed while concurrently minimizing search costs.
What are females looking for?
Research on mate choice since the early 1990s has focused on females
selecting males based on perceived quality. But how are such distinctions made?
Females are presumably unable to assess male genetic quality directly but can
infer this through assessment of traits that tightly correlate to resource-holding
potential of the male (Grafen, 1990).
Visual signals from plumage are known to be associated with male condition
and ability to acquire resources (Hill, 2002). Dominance status and aggressive
behaviour also reflect relative male condition, as they predict access to limited

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
138 K. A. Otter & L. Ratcliffe
resources (Ekman & Askenmo, 1984; Hogstad, 1988; Desrochers, 1989; Ficken
et al., 1990; Ekman & Lilliendahl, 1992). Singing behaviour is also indicative of
male condition. Repertoire size in several species is related to age (Yasukawa et al.,
1980; Lampe & Espmark, 1994; Birkhead et al., 1997; Eens, 1997), providing infor-
mation on individual survival. Singing behaviour may also reflect a male’s ability
to secure access to limited resources (Reid, 1987; Alatalo et al., 1990; Thomas,
1999a,b; Thomas & Cuthill, 2002). Song output is known to be associated with a
male’s dominance rank (Otter et al., 1997), level of parasite infestation (Møller,
1991) and immune response (Saino et al., 1997a). Even the fine structure of song
may give cues to the survivorship (Forstmeier et al., 2002) or rank (Christie et al.,
2004) of males. Many studies have shown that these behavioural and morpho-
logical characteristics are important in female choice, both for primary mates
(Radesater et al., 1987; Alatalo et al., 1990; Andersson, 1994; Hoi-Leitner et al.,
1995; Buchanan & Catchpole, 1997) and for extra-pair paternity (Smith, 1988;
Morton et al., 1990; Houtman, 1992; Wetton et al., 1995; Hasselquist et al., 1996;
Kempenaers et al., 1997; Saino et al., 1997b; Møller et al., 1998; Otter et al., 1998;
Forstmeier et al., 2002).
Which trait is the best indicator of quality and what is meant by a good indicator
of quality? Strong correlations between male quality and expression of the trait
are assumed for the trait to be ‘reliable’ (Grafen, 1990) and several condition-
dependent traits will often be intercorrelated. To determine which of these signals
are likely to be assessed by females, however, we should focus on the perception
of signals by females rather than the production of signals by males. It does not
necessarily follow that a female will be able to discriminate amongst males even
if male quality is correlated with the expression of the trait. The traits may all
potentially be reliable, but the important question for females in networks is
at what distance are they detectable and discriminable (Ch. 20)? The debate on the
evolution of multiple signals (Møller & Pomiankowski, 1993; Pomiankowski &
Iwasa, 1993; Iwasa & Pomiankowski, 1994; Johnstone, 1996) has focused largely on
whether multiple signals are of use to the female in an additive way, or whether
their apparent redundancy is used by females to confirm their assessment. The
debate assumes that females are able to assess all traits simultaneously. It seems
more likely that, in a natural network context, signals that target different sensory
modalities may be assessed sequentially in relation to their transmission distance.
Distance can have a profound effect on both detection and discrimination
(Wiley & Richards, 1982). Many morphological and behavioural traits that require
visual inspection can only be discriminated at close range, particularly in habi-
tats with dense vegetation. Tactile signals may be similarly restricted. In contrast,
olfactory or auditory signals, particularly song, have evolved to transmit at least
the average interterritory spacing within a species (Brenowitz, 1982; Calder, 1990);

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 139
thus, detection and discrimination are possible at greater range. Long-range sig-
nals may form the basis of initial assessment, which can then be confirmed on
closer inspection by assessment of short-range signals. The apparent redundancy
of intercorrelated signals may thus reflect the hierarchical way in which they are
assessed (Bradbury & Vehrencamp, 1998).
Imagine a situation where a female is assessing a prospective male, either as
a mate or as an extra-pair partner. In many cases, female movement may be con-
strained by such factors as mate guarding, aggression of nearby mated females
(Slagsvold & Lifjeld, 1994; Dale & Slagsvold, 1995) or risk of predation. Under these
circumstances, any mechanism that enables females to narrow down the pool of
acceptable males from a position of relative safety would be favoured (Gowaty,
1996). In songbirds, male advertising song provides an ideal signal for assessment
at a distance (Fig. 7.2a–c), because of its long transmission range. Initial decisions
about males can be made via this single cue; females can then directly approach
subsets of males deemed to be of the best quality among the available pool. Further
discrimination may then occur by shifting assessment to short-range signals, such
as plumage, the expression of which we would expect to correlate with quality
indicated by long-range signals. This sequential assessment of signals may increase
the certainty of assessment.
Females may employ information derived from networks not only to narrow
the pool of potential mates; females may also instigate network communication
to evaluate male quality during close approach. For example, during intrusions
across territorial boundaries, the attraction of neighbouring males may incite
competitive interactions between a female’smate and his neighbours (Fig. 7.2d–f).
Montgomerie & Thornhill (1989) suggested that females might incite interactions
between males as a mechanism for sperm competition, but it is also possible
that such behaviour provides females with more information about the general
quality of available males (Sæther, 2002). This possibility is also supported by
demonstrations that females use information from male–male singing interac-
tions (i.e. they eavesdrop; Ch. 2) in extra-pair behaviour decisions (Otter et al., 1999;
Mennill et al., 2002). Future work on this topic should target species where females
readily engage in secondary mate choice, for example the pursuit of extra-pair
copulations.
Recently, Sæther (2002) demonstrated that such incitement occurs in the great
snipe Gallinago media, a species in which females call from the edges of males’
territories within leks. Using playbacks, he showed that female calls from such
boundaries increase the competitive interactions between neighbouring males,
providing a potential source of information to prospecting females. Similarly,
in territorial songbirds, females may exhibit behaviour that draws their mates
and neighbours into interactions on territorial boundaries. Ramsay et al. (1999)

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
140 K. A. Otter & L. Ratcliffe
(a) (c)(b)
(d) (e) (f)
Fig. 7.2. Secondary mate choice, such as the decision to engage in extra-pair
copulations or divorce a current mate for another male, may be constrained in some
territorial species. Unlike primary (i.e. initial) mate choice, females may not be able to
move freely through the territories of various males because of the presence of
resident females (a). However, long-range signals of males, such as song, transmit
beyond the boundaries of the territories and effectively signal presence to
neighbouring territories (b). In (a, b, d and e), dark lines represent territorial
boundaries with males and females shown as symbols. Dotted lines represent the
range of effective signal transmission and some territory boundaries are show in grey
in (c) and (f ) to prevent obscuring signal ranges. (c) A female may be able to assess all
neighbouring males as well as her mate without having to leave the territory. (d) A
female may incite interactions among males by moving (arrow) towards a boundary.
(e) Her movement may draw neighbouring males to that boundary (arrows) and the
resulting interactions may allow her to assess other signals, including short-range
types of display (plumage, direct dominance interactions or fights). (f ) Thus the
female could assess a subset of the original males using multiple signals in
succession, possibly leading to increased certainty of her assessment.
found that female black-capped chickadees Poecile atricapillus place their nests close
to territorial edges, despite evidence that these areas offer no better resources
or nesting opportunities than central nest locations. One explanation for this
pattern is that females can more easily monitor neighbours in relation to their
mates and capitalize on opportunities for secondary mate choice. One observed
outcome of this nesting pattern by females is increased numbers of territorial
disputes between the resident male and his neighbours (Ramsay et al., 1999), which

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 141
may also make it easier for females to make these relative distinctions. Female
hooded warblers Wilsonia citrina at the peak of their fertility give characteristic
chip calls, which result in increased intrusions by neighbouring males (Neudorf,
1996); female European robins Erithacus rubecula show similar patterns in their
rates of seep calls (Tobias & Seddon, 2002). While such ‘fertility announcements’
are often interpreted as mechanisms that increase female choice through potential
sperm competition, they may also function more directly in active assessment by
females if they increase interactions between mates and other males.
Evidence for female assessment of network information
Few studies have asked whether female mate choice incorporates informa-
tion derived from signals in a communication network, although the potential for
females to use such information seems considerable (e.g. Otter et al., 1999; Mennill
et al., 2002). K. A. Otter, T. M. Peake, A. M. R. Terry and P. K. McGregor (unpublished
data) found that dawn chorus singing of neighbouring male great tits Parus major is
clearly recorded by microphones placed within the nestboxes of roosting females;
therefore, it is likely that females could assess a network of singing males without
leaving the nestbox. Relative song output among males during the dawn chorus is
known to correlate with male condition in a number of species (Reid, 1987; Alatalo
et al., 1990; Otter et al., 1997; Thomas, 1999a,b; Thomas & Cuthill, 2002), and it is,
therefore, a useful cue of quality (Hutchinson et al., 1993). However, surprisingly
little work has investigated whether females attend to variation in male dawn
song. Otter & Ratcliffe (1993, 1996) suggested that changes in dawn singing of
males who have lost their mates might function as useful cues for neighbouring
females seeking better mates, and anecdotal evidence in black-capped chickadees
suggests that divorces occur soon after the dawn chorus ends. Further studies
should be conducted to determine whether the generally higher song output at
dawn is used in assessment by females. This could be done by elevating male song
output by supplementary feeding (e.g. Reid, 1987; Alatalo et al., 1990; Thomas,
1999a,b) and seeing whether radio-tracked females appear attentive to increased
song output of neighbours.
To date, the few studies that have investigated female assessment in commu-
nication networks have focused on eavesdropping upon dyadic male aggressive
singing interactions (Otter et al., 1999; Mennill et al., 2002). These eavesdrop-
ping experiments have used interactive playback (Dabelsteen & McGregor, 1996)
to manipulate the outcome of aggressive singing interactions between males.
Otter et al. (1999) showed that female great tits appear to be aware of the relative
ease with which males interact with a ‘strange intruder’ (the interactive play-
back). The mates of males who lost interactions were more likely to intrude into

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
142 K. A. Otter & L. Ratcliffe
neighbouring territories in following days than were those whose mates won
against the same intruder. As the only difference in singing between the two male
treatments was in their song rates relative to the playback, females appear to base
their movement patterns on the perceived interaction. Moreover, females visited
more frequently the neighbouring male heard to win against the same intruder to
which their mate had lost. Our results, however, found no evidence that females
produced more young with these males; there was no pattern of extra-pair cop-
ulation associated with the playback treatment, although males who had been
cuckolded were of lower genetic heterozygosity than males who were not cuck-
olded (Otter et al., 2001). As genetic heterozygosity appears to be associated with
survival and fecundity (Coulson et al., 1998; Hansson et al., 2001), this result might
indicate that females drawn to males via song may have found other signals (e.g.
colour patterns, which were not assessed during the study) that contradicted the
assessment via song.
By contrast, Mennill et al. (2002) recently showed that female black-capped
chickadees exposed to similar eavesdropping opportunities did modify extra-pair
behaviour, although there was no apparent effect on observed intrusions. Males
of high and low social rank were exposed to challenges simulated by interactive
playback. The challenges either reinforced their rank disparity (e.g. de-escalating
playback to a dominant male, escalating to the subordinate), or countered rank
disparity (escalate to dominant male, de-escalate to subordinate). While females
mated to low-ranking males showed no influence of playback on their decisions
to engage in extra-pair copulation, females mated to high-ranking males that had
lost against the playback ‘intruder’ were more likely to have extra-pair young in
their broods. As females in this species mated to high-ranking males usually forego
extra-pair copulation (Otter et al., 1998), this result suggests that the protocols used
by Mennill et al. (2002) had a profound impact on female decisions. Normally, if
females mated to high-ranking males do engage in extra-pair copulation, they se-
lect males of similar or higher rank than their mate (Otter et al., 1998). Yet, D. J.
Mennill and colleagues (unpublished data) found that extra-pair males selected by
these high-ranking females were nearly random with respect to the relative rank
of their mate; further evidence that assessment by eavesdropping in a network
can have dramatic influences on behavioural decisions.
There is also experimental evidence that female Siamese fighting fish Betta
splendens eavesdrop on male–male aggressive visual displays and are more will-
ing to mate with males that they have seen win such interactions (Doutrelant &
McGregor, 2000; see also Ch. 2) These initial studies provide impetus for future
work. However, a number of fundamental questions still need to be addressed.
For example, the nature and accuracy of the information on relative male quality
available to females in interactions remains to be determined. Another major is-
sue is the extent to which features of signals used for individual identification are

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 143
affected by transmission over long distances. Many ideas related to communica-
tion networks assume that individual signallers can be identified; for example, in
the context of mate choice in birds, females are assumed to be able to distinguish
different males by voice. This has been shown in a number of species (e.g. Davis,
1986; Weary & Krebs, 1992; Lind et al., 1996); however, these studies have not re-
quired females to make these discriminations at long distance. Although song is a
long-distance signal, all acoustic signals are subject to degradation over distance
(Bradbury & Vehrencamp, 1998), which could negatively affect features females
use in discrimination and assessment.
Implications for networks of environmental alteration
Song transmission is affected not only by distance but also by the medium
through which it must travel. Reverberation, differential attenuation and other
effects on sound are imposed by habitat characteristics and may shape the songs of
species inhabiting different areas (Catchpole & Slater, 1996). Habitat alteration can
result in a change in the characteristics of signals to maintain maximum transmis-
sion range within new habitats, for example in rufous-collared sparrows Zonotrichia
capensis inhabiting forested versus grassland habitats (Tubaro et al., 1993; Tubaro &
Segura, 1994). However, it is unknown how long it takes for changes in song to oc-
cur in response to habitat change and how females respond to such changes. Most
habitat alterations occur over very short timeframes, and unless reproductive iso-
lation occurs between undisturbed and disturbed habitats, selection in response
to the altered landscape may be slow (e.g. Dhondt et al., 1992; Dias & Blondel, 1996).
Therefore, changes in the structure of male song may not keep pace with changes
in the habitats, leading to song structure that is mismatched for transmission in
the present environment.
In many species, habitat alteration may change sound transmission conditions
and also decrease resource availability or breeding success (Blondel, 1985; Blondel
et al., 1993; Fort & Otter, 2004). If habitat alteration simultaneously reduces signal
transmission and enlarges territory size in response to lowered resources, the
extent of communication networks and the information to be gained from them
may be seriously reduced. For example, the size of male song networks in black-
capped chickadees appears to be constrained by habitat change. While recording
focal males during the dawn chorus, we conducted standard avian point counts
at three-minute intervals to determine the number and direction of other males
audible at the location of the focal male. The result was that fewer males were
audible to male chickadees that occupy early successional forests (characterized
by a low canopy and dense understorey) than to those occupying nearby mature,
mixed forests (I.-J. Hansen, K. A. Otter & H. van Oort, unpublished data). This is likely
a consequence of decreased transmission of song (I.-J. Hansen, K. A. Otter & H. van

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
144 K. A. Otter & L. Ratcliffe
Oort, unpublished data) and increased territory size (Fort & Otter, 2004) of males
occupying this disturbed habitat.
Such changes to the transmission of signals following habitat alteration can
potentially diminish assessment ability of females. Not only could the networks
decline, reducing the number of males a female could assess for secondary mate
choice, but, in addition, females in such circumstances could also fail to locate
primary mates, or become polygynous. Alternatively, females may mate monog-
amously with males of lesser quality during primary mate selection, because as-
sessment of neighbouring males is constrained (M. Kasumovic, L. M. Ratcliffe &
P. T. Boag, unpublished data). If such assessment is critical in female mating tac-
tics (e.g. Wagner, 1991), females may fail to settle in such altered habitats, even if
the resources would support a breeding effort.
Another impact of habitat alteration that could affect female assessment in
networks is the close relationship between resource access and the ability of males
to produce condition-dependent traits. If habitat quality is poor, the absolute
expression of traits may be diminished (Hill, 1995); moreover Qvarnstrom and
Forsgren (1998) also predict that dominant males may suffer disproportionately
in poor habitat. The costs of achieving dominance status are normally countered by
the benefit of access to rich resources, but if the habitat is unable to produce these
benefits, the high costs paid by dominants may put them in a net metabolic deficit.
In support of this, H. van Oort, K. A. Otter, F. Fort & C. I. Holschuh (unpublished
data) found that song output in the dawn chorus of black-capped chickadees varies
across habitats. As predicted by Otter et al. (1997), birds occupying mature forests in
northern British Columbia, Canada had song output that reflected their relative
rank: high-ranked birds tended to have higher song output than lower-ranked
birds. By comparison, birds settling in neighbouring young, regenerating forests
did not show this same trend. Overall, the birds in the disturbed forests had lower
song output than birds in the undisturbed forests, as predicted by Hill (1995),
but this relationship was driven by abnormally low song output by high-ranking
males in the disturbed woods, as predicted by Qvarnstrom and Forsgren (1998).
Males of either high or low rank in the disturbed forest could not be differentiated
based on song output. Therefore, the transmission of signals may not be the only
impact of landscape alterations; the reliability of signals may also be influenced
by habitat context and may diminish the ability of females to use long-range
signalling networks in assessment.
Summary and future directions
The role of communication networks in female mate choice is ripe for
study using the techniques that simulate signalling interactions. In territorial

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 145
(a) (b)
(c)
Fig. 7.3. A proposed experiment to test females’ use of information in communication
networks in male assessment. The figure shows an aviary in plan view with release
points at the corners (solid squares). (a) The aviary contains four cells (squares) each
containing a choice stimulus (represented by a loudspeaker symbol here, but they
could contain live males. (b) Signals are designed so that, when broadcast, only a
position in the middle of the aviary would allow the female to assess all males
simultaneously (i.e. the point of overlap of the effective signal ranges, shown as dotted
lines). (c) The pattern of female movement observed (dashed lines), for example
consistently moving towards the central areas prior to entering a cell, would indicate
that several males were being compared.
songbirds, studies of eavesdropping (Ch. 2) may help us to understand female
secondary mating tactics. By manipulating the relative signals emanating from
neighbouring males, and using radio-tracking and genetics to measure female
preferences, we should be able to obtain a clearer idea of how (and perhaps why)
socially monogamous females choose secondary partners.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
146 K. A. Otter & L. Ratcliffe
Considering communication in a network context should also aid our under-
standing of primary mate choice, an area of particular interest. It is particularly
important to determine at what range females can discriminate between signals.
As we argued earlier, measures of close approach to males may be insufficient
to determine whether males have been ‘sampled’ by females. This is particularly
the case if females are assessing males at long distance, approaching only chosen
males. Such cryptic choice is potentially difficult to monitor. One way of address-
ing this problem is to plot female movements in relation to known signal trans-
mission range. Speaker replacement studies, such as those used with flycatchers
(Eriksson & Wallin, 1986) or starlings Sturnus vulgaris (Mountjoy & Lemon, 1991),
could simulate clusters of signalling males to determine the effects of altered
singing patterns on female assessment routes and strategies. Aviary studies may
also be informative in this regard. Rather than the traditional dyadic choices pre-
sented to females, aviaries with several males could be presented (Fig. 7.3). The
idea that females position themselves to assess males in a network could be inves-
tigated by manipulating the transmission range of auditory or visual signals and
observing female movements (e.g. Fig. 7.3).
Acknowledgements
We thank Peter McGregor, Bart Kempenaers, Dan Mennill, Harry van Oort, Carmen
Holschuh, Tania Tripp and David Nordstrom for discussion on the topics in this chapter. Ingebjørg-
Jean Hansen, Kevin Fort, Harry van Oort, Peter Christie, Dan Mennill, Mike Kasumovic, Peter
McGregor, Tom Peake and Andrew Terry kindly allowed us to cite the results of unpublished,
co-authored data. Peter McGregor, Dan Mennill, Marc Naguib, Harry van Oort and an anonymous
reviewer also provided useful suggestions on early drafts of the manuscript. Both authors were
funded by NSERC (Canada) research grants during the preparation of this work.
References
Alatalo, R. V., Glynn, C. & Lundberg, A. 1990. Singing rate and female attraction in the
pied flycatcher: an experiment. Animal Behaviour, 39, 601–602.
Andersson, M. 1994. Sexual Selection. Princeton, NJ: Princeton University Press.
Bensch, S. & Hasselquist, D. 1992. Evidence for active female choice in a polygynous
warbler. Animal Behaviour, 44, 301–311.
Birkhead, T. R., Buchanon, K. L., de Voogd, T. J. et al. 1997. Song, sperm quality and
testes asymmetry in the sedge warbler. Animal Behaviour, 53, 965–971.
Blondel, J. 1985. Breeding strategies of the blue tit and coal tit (Parus) in mainland and
island Mediterranean habitats: a comparison. Journal of Animal Ecology, 54,
531–556.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 147
Blondel, J., Dias, P. C., Maistre, M. & Perret, P. 1993. Habitat heterogeneity and
life-history variation of Mediterranean blue tits (Parus caeruleus). The Auk, 110,
511–520.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.
Brenowitz, E. A. 1982. The active space of red-winged blackbird song. Journal of
Comparative Physiology, A, 147, 511–22.
Buchanan, K. L. & Catchpole, C. K. 1997. Female choice in the sedge warbler,
Acrocephalus schoenobaenus: multiple cues from song and territory quality.
Proceedings of the Royal Society of London, Series B, 264, 521–526.
Calder, W. A. 1990. The scaling of sound output and territory size: are they matched?
Ecology, 71, 1810–1816.
Catchpole, C. K. & Slater, P. J. B. 1996. Bird Song. Cambridge, UK: Cambridge University
Press.
Christie, P. J., Mennill, D. J. & Ratcliffe, L. M. 2004. Pitch shifts and song structure
indicate male quality in the dawn chorus of black-capped chickadees. Behavioral
Ecology and Sociobiology, 55, 341–348.
Coulson, T. N., Pemberton, J. M., Albon, S. D. et al. 1998. Microsatellites reveal
heterosis in red deer. Proceedings of the Royal Society of London, Series B, 256, 489–495.
Dabelsteen, T. & McGregor, P. K. 1996. Dynamic acoustic communication and
interactive playback. In: Ecology and Evolution of Acoustic Communication in Birds, ed.
D. E. Kroodsma & E. H. Miller. Ithaca, NY: Cornell University Press, pp. 398–408.
Dale, S. & Slagsvold, T. 1995. Female contests for nest sites and mates in the pied
flycatcher, Ficedula hypoleuca. Ethology, 99, 209–222.
Dale, S., Amundsen, T, Lifjeld, J. T. & Slagsvold, T. 1990. Mate sampling behaviour of
female pied flycatchers: evidence for active mate choice. Animal Behaviour, 27,
87–91.
Davis, W. J. 1986. Acoustic recognition in the belted kingfisher: cardiac response to
playback vocalizations. The Condor, 88, 505–512.
Desrochers, A. 1989. Sex, dominance, and microhabitat use in wintering black-capped
chickadees: a field experiment. Ecology, 70, 636–645.
Dhondt, A. A., Kempenaers, B. & Adriaensen, F. 1992. Density-dependent clutch size
caused by habitat heterogeneity. Journal of Animal Ecology, 61, 643–648.
Dias, P. C. & Blondel, J. 1996. Local specialization and maladaptation in the
Mediterranean blue tit (Parus caeruleus). Oecologia, 107, 79–86.
Doutrelant, C. & McGregor, P. K. 2000. Eavesdropping and mate choice in female
fighting fish. Behaviour, 137, 1655–1669.
Eens, M. 1997. Understanding the complex song of the European starling: an
integrated ethological approach. Advances in the Study of Behaviour, 26, 355–434.
Ekman, J. & Askenmo, C. 1984. Social rank and habitat use in willow tit groups. Animal
Behaviour, 32, 508–514.
Ekman, J. B. & Lilliendahl, K. 1992. Using priority to food access: fattening strategies in
dominance-structured willow tit (Parus montanus) flocks. Behavioral Ecology, 4,
232–2.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
148 K. A. Otter & L. Ratcliffe
Ens, B. J., Safriel, U. N. & Harris, M. P. 1993. Divorce in the long-lived and monogamous
oystercatcher, Haematophagus ostralegus: incompatibility or choosing a better
option? Animal Behaviour, 45, 1199–1217.
Eriksson, D. & Wallin, L. 1986. Male bird song attracts females: a field experiment.
Behavioral Ecology and Sociobiology, 19, 297–299.
Ficken, M. S., Weise, C. M. & Popp, J. W. 1990. Dominance rank and resource access in
winter flocks of black-capped chickadees. Wilson Bulletin, 102, 623–633.
Forstmeier, W., Kempenaers, B., Meyer, A. & Leisler, B. 2002. A novel song parameter
correlates with extra-pair paternity and reflects male longevity. Proceedings of the
Royal Society of London, Series B, 269, 1479–1485.
Fort, K. & Otter, K. A. 2004. The impact of habitat disturbance on reproduction of
chickadees. The Auk, 121, in press.
Gibson, R. M. & Langen, T. A. 1996. How do animals choose their mates? Trends in
Ecology and Evolution, 11, 468–470.
Gil, D. & Gahr, M. 2002. The honesty of bird song: multiple constraints for multiple
traits. Trends in Ecology and Evolution, 17, 133–141.
Gowaty, P. A. 1996. Multiple mating by females selects for males that stay: another
hypothesis for social monogamy in passerine birds. Animal Behaviour, 51,
482–484.
Grafen, A. 1990. Biological signals as handicaps. Journal of Theoretical Biology, 144,
517–546.
Hansson, B., Bensch, S., Hasselquist, D., & Åkesson, M. 2001. Microsatellite diversity
predicts recruitment of sibling great reed warblers. Proceedings of the Royal Society
of London, Series B, 268, 1287–1291.
Hasselquist, D., Bensch, S. & von Schantz, T. 1996. Correlation between male song
repertoire, extra-pair paternity and offspring survival in the great reed warbler.
Nature, 381, 229–232.
Hill, G. E. 1995. Ornamental traits as indicators of environmental health. BioScience, 45,
25–31.
2002. A Red Bird in a Brown Bag: The Function and Evolution of Colourful Plumage in the
House Finch. Oxford: Oxford University Press.
Hogstad, O. 1988. Rank-related resource access in winter flocks of willow tit Parus
montanus. Ornis Scandinavica, 19, 169–174.
Hoi-Leitner, M., Nechtelberger, H. & Hoi, H. 1995. Song rate as a signal for nest site
quality in blackcaps (Sylvia atricapilla). Behavioral Ecology and Sociobiology, 37,
399–405.
Houtman, A. M. 1992. Female zebra finches choose extra-pair copulation with
genetically attractive males. Proceedings of the Royal Society of London, Series B, 249,
3–6.
Hutchinson, J. M. C., McNamara J. M. & Cuthill, I. C. 1993. Song, sexual selection,
starvation, and strategic handicaps. Animal Behaviour, 45, 1153–1177.
Iwasa, Y. & Pomiankowski, A. 1994. The evolution of mate preferences for multiple
sexual ornaments. Evolution, 48, 853–867.
Janetos, A. C. 1980. Strategies of female mate choice: a theoretical analysis. Behavioral
Ecology and Sociobiology, 7, 107–112.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 149
Johnstone, R. A. 1996. Multiple displays in animal communication: ‘backup signals’
and ‘multiple messages’. Philosophical Transactions of the Royal Society of London, Series
B, 351, 329–338.
Kempenaers, B. & Dhondt, A. A. 1993. Why do females engage in extra-pair
copulations? A review of the hypotheses and their predictions. Belgian Journal of
Zoology, 123, 93–103
Kempenaers, B., Verheyen, G. R. & Dhondt, A. A. 1997. Extrapair paternity in the blue
tit (Parus caeruleus): female choice, male characteristics and offspring quality.
Behavioral Ecology, 8, 481–492.
Lampe, H. M. & Espmark, Y. O. 1994. Song structure reflects male quality in pied
flycatchers, Ficedula hypoleuca. Animal Behaviour, 47, 869–876.
Lind, H., Dabelsteen, T. & McGregor, P. K. 1996. Female great tits can identify mates by
song. Animal Behaviour, 52, 667–671.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging, and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–211.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Møller, A. P. 1991. Parasite load reduces song output in a passerine bird. Animal
Behaviour, 41, 723–730.
Møller, A. P. & Pomiankowski, A. 1993. Why have birds got multiple sexual
ornaments? Behavioral Ecology and Sociobiology, 32, 167–176.
Møller, A. P., Saino, N., Taramino, G., Galeotti, P. & Ferrario, S. 1998. Paternity and
multiple signalling: effects of a secondary sexual character and song on paternity
in the barn swallow. American Naturalist, 151, 236–242.
Montgomerie, R. & Thornhill, R. 1989. Fertility advertisment in birds: a means of
inciting male–male competition? Ethology, 81, 209–220.
Morton, E. S., Forman, L. & Braun, M. 1990. Extra-pair fertilisations and the evolution
of colonial breeding in purple martins. The Auk, 107, 275–283.
Mountjoy, D. J. & Lemon, R. E. 1991. Song as an attractant for male and female
European starlings, and the influence of song complexity on their response.
Behavioral Ecology and Sociobiology, 28, 97–100.
Murphy, C. G. & Gerhardt, H. C. 2002. Mate sampling by female barking treefrogs (Hyla
gratiosa). Behavioral Ecology, 13, 472–480.
Naguib, M., Fitchel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Neudorf, D. L. 1996. A dual system of female control of extra-pair copulations in the
hooded warbler (Wilsonia citrina). Ph.D.; Thesis. York University, North York,
Ontario.
Neudorf, D. L., Stutchbury, B. J. M. & Piper, W. H. 1997. Covert extraterritorial behavior
of female hooded warblers. Behavioral Ecology, 8, 595–600.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
150 K. A. Otter & L. Ratcliffe
Otter, K. & Ratcliffe, L. 1993. Changes in the singing behaviour of male black-capped
chickadees (Parus atricapillus) following mate removal. Behavioral Ecology and
Sociobiology, 33, 409–414.
1996. Female initiated divorce in a monogamous songbird: abandoning mates for
males of higher quality. Proceedings of the Royal Society of London, Series B, 263,
351–354.
Otter, K., Chruszcz, B. & Ratcliffe, L. 1997. Honest advertisement and singing during
the dawn chorus of black-capped chickadees, Parus atricapillus. Behavioral Ecology,
8, 167–173.
Otter, K., Ratcliffe, L., Michaud, D. & Boag, P. T. 1998. Do female black-capped
chickadees prefer high ranking males as extra-pair partners? Behavioral Ecology and
Sociobiology, 43, 25–36.
Otter, K., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits (Parus major)
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society of London, Series B, 266, 1305–1310.
Otter, K., Stewart, I. K., Terry, A. M. R., McGregor, P. K. & Burke, T. 2001. Extra-pair
paternity in great tits in relation to manipulation of male signals. Journal of Avian
Biology, 32, 338–344.
Pomiankowski, A & Iwasa, Y. 1993. Evolution of multiple sexual preferences by Fisher’s
runaway process of sexual selection. Proceedings of the Royal Society of London, Series
B, 253, 173–181.
Qvarnstrom, A. & Forsgren, E. 1998. Should females prefer dominant males? Trends in
Ecology and Evolution, 13, 498–501.
Radesater, T., Jakobsson, S., Andbjer, N., Bylin, A. & Nystrom, K. 1987. Song rate and
pair formation in the willow warbler, Phylloscopus trichilus. Animal Behaviour, 35,
1645–1651.
Ramsay, S. M, Otter, K. & Ratcliffe, L. M. 1999. Nest-site selection by female
black-capped chickadees: settlement based on conspecific attraction? The Auk,
116, 604–617.
Real, L. 1990. Search theory and mate choice I: models of single-sex discrimination.
American Naturalist, 136, 376–405.
Reid, M. L. 1987. Costliness and reliability in the singing behaviour of Ipswich
sparrows. Animal Behaviour, 35, 1735–1743.
Sæther, S. A. 2002. Female calls in lek-mating birds: indirect mate choice, female
competition for mates or direct mate choice? Behavioral Ecology, 13, 344–352.
Saino, N., Galeotti, P., Sacchi, R. & Møller, A. P. 1997a. Song and immunological
condition in male swallows (Hirundo rustica). Behavioral Ecology, 8, 364–371.
Saino, N., Primmer, C. R., Ellegren, H. & Møller, A. P. 1997b. An experimental study of
paternity and tail ornamentation in the barn swallow (Hirundo rustica). Evolution,
51, 562–570.
Slagsvold, T. & Lifjeld, J. T. 1994. Polygyny in birds: the role of competition between
females for male parental care. American Naturalist, 143, 59–94.
Slagsvold, T., Lifjeld, J. T., Stenmark, G. & Breiehagen, T. 1988. On the cost of searching
for a mate in female pied flycatchers Ficedula hypoleuca. Animal Behaviour, 36,
433–442.

P1: IYP/... P2: JZZ/...0521823617c07.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 11:49
Enlightened decisions: female assessment 151
Smith, S. M. 1988. Extra-pair copulations in black-capped chickadees: the role of the
female. Behaviour, 107, 15–23.
Thomas, R. J. 1999a. Two tests of a stochastic dynamic programming model of daily
singing routines in birds. Animal Behaviour, 57, 277–284.
1999b. The effect of variability in the food supply on the daily singing routines of
European robins: a test of a stochastic dynamic programming model. Animal
Behaviour, 57, 365–369.
Thomas, R. J. & Cuthill, I. C. 2002. Body mass regulation and the daily singing routines
of European robins. Animal Behaviour, 63, 285–292.
Tobias, J. A. & Seddon, N. 2002. Female begging in European robins: do neighbors
eavesdrop for extrapair copulations? Behavioral Ecology, 13, 637–642.
Todt, D. & Naguib, M. 2000. Vocal interactions in birds: the use of song as a model in
communication. Advances in the Study of Behaviour, 29, 247–295.
Tubaro, P. L. & Segura, E. T. 1994. Dialect differences in the song of Zonotrichia capensis
in the Southern Pampas: a test of the acoustic adaptation hypothesis. The Condor,
96, 1084–1088.
Tubaro, P. L., Segura, E. T & Handford, P. 1993. Geographic variation in the song of the
rufous-collared sparrow in eastern Argentina. The Condor, 95, 588–595.
Wagner, R. H. 1991. Evidence that female razorbills control extra-pair copulations.
Behaviour, 118, 157–169.
Weary, D. M. & Krebs, J. R. 1992. Great tits classify songs by individual voice
characteristics. Animal Behaviour, 43, 283–287.
Wetton, J. H., Burke, T., Parkin, D. T. & Cairns, E. 1995. Single-locus DNA fingerprinting
reveals that male reproductive success increases with age through extra-pair
paternity in the house sparrow (Passer domesticus). Proceedings of the Royal Society of
London, Series B, 260, 91–98.
Wiegmann, D. D., Real, L. A., Capone, T. A. & Ellner, S. 1996. Some distinguishing
features of models of search behavior and mate choice. American Naturalist, 147,
188–204.
Wiley, R. H. & Richards, D. G. 1982. Adaptations for acoustic communication in birds:
sound transmission and signal detection. In: Acoustic Communication in Birds, Vol. 1,
ed. D. E. Kroodsma & E. H. Miller. New York: Academic Press, pp. 131–181.
Wittenberger, J. F. 1983. Tactics of mate choice. In: Mate Choice, ed. P. Bateson.
Cambridge, UK: Cambridge University Press, pp. 435–447.
Yasukawa, K., Blank, J. L. & Patterson, C. B. 1980. Song repertoires and sexual selection
in the red-winged blackbird. Behavioral Ecology and Sociobiology, 7, 233–238.

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
8
Predation and noise in communicationnetworks of neotropical katydids
a l e x a n d e r b . l a ng , i ng e b o rg t e p p n e r , m a n f r e dh a r t b au e r & h e i n e r r Om e r
Karl-Franzen University, Graz, Austria
Introduction
Intraspecific acoustic communication in grasshoppers or katydids ap-
pears to be a very simple and straight forward behaviour: one sex – usually the
male – produces an acoustic signal, and the female, once perceiving and recogniz-
ing the signal as species specific, shows some kind of response, either an acoustic
reply or a phonotactic movement to the male. However, the system is far from
being that simple and involves more than just a sender and receiver. First, com-
munication usually takes place in a physically complex environment, where sound
signals are subject to attenuation and degradation, depending on the carrier fre-
quencies, which are often in the high-sonic or ultrasonic range because of the
small size of the sound radiating structures (Wiley & Richards, 1978; Michelsen,
1992). In addition, the physical conditions of the transmission channel for the
sound may vary strongly during day or night and with weather conditions; con-
sequently, the ability to detect and localize a signal undergoes strong variations.
Second, insects often aggregate and communicate in areas rich in resources or
at periods of the day or night favouring mate attraction. As a result of many sig-
nallers calling in close proximity, masking interference will take place at the site
of receivers, depending on the spacing, as well as the kind and extent of signal tim-
ing. Since such favourable areas and times for signalling are similar for different
species, heterospecific choruses may be formed with impressive sound pressure
levels of biological background noise, which further complicates the detection
of a signal (reviewed for katydids by Schatral (1990)). Third, a female might gain
fitness benefits (directly or indirectly) by choosing a male based on variation of
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
152

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 153
particular properties of the calling song. Such a female preference may drive the
evolution of the signal in a particular direction. For example, in insects, patterns of
preferences based on call rate or the duration of long calls are usually highly direc-
tional, whereas those based on pulse rate or frequency are stabilizing (reviewed by
Gerhardt & Huber, 2002). Female preferences can also result from a sensory bias in
the sensory or nervous system of receivers (Ryan, 1990; Ryan & Keddy-Hector, 1992;
Ryan & Rand, 1993; Endler & Basolo, 1998). Thus sexual selection by female choice
can result in signal traits that enhance the mating success of males. Fourth, traits
preferred by females may also decrease male survivorship by increasing exposure
to predators. Acoustically orienting predators or parasitoids can use the same sig-
nals produced for mate attraction to identify, localize and home in on the signaller
(Cade, 1975; Belwood & Morris, 1987; Lehmann & Heller, 1998; Zuk & Kolluru, 1998;
Allen, 2000). Male fitness can also be decreased by increased competition resulting
from the signal attracting conspecific rivals. Both are cases of interceptive eaves-
dropping in the sense of Peake (Ch. 2). Fifth, signalling at the long duration and
high rate preferred by females may be limited by energetic constraints, as sound
production for small animals is rather inefficient and probably costly (Bailey et al.,
1993; Wagner & Hoback, 1999). Finally, as ectothermic animals, the motor output
of insects depends on the ambient temperature, and in consequence, the temporal
properties of calling songs are influenced by environmental temperature. Vertical
temperature gradients in a grasshopper’s habitat can be 10 ◦C in 30 cm (Romer,
2001); therefore, senders and receivers can differ strongly in body temperature.
As a consequence, the signaller’s temporal patterning of song may not match the
preference function of a receiver.
From this short summary, it is clear that some of the factors contributing to
the evolution of acoustic communication systems could interact in a complex
way. For example, if high predation risk forces a species to communicate acous-
tically at a time of day or night when acoustic competition with other species is
high, the consequence is a high degree of song interference and masking, and the
calling activity of one species can inhibit that of other species (Greenfield, 1988;
Romer et al., 1989). In this chapter we emphasize the importance of an ecological
(integrated) approach to communication networks. By focusing on the intraspe-
cific communication of a subfamily of neotropical katydids, we demonstrate the
complex dependency of predation and signalling, nocturnal ambient light levels,
masking noise levels and alternative signalling strategies.
Predation and antipredator defences in rainforest katydids
A key paper by Belwood & Morris (1987) (see also Belwood, 1990; Morris
et al., 1994) suggested that the evolution of specific anti-predator defences in a

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
154 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
family of neotropical katydids (Pseudophyllinae) is strongly influenced by an-
tipredator defences. Members of this subfamily differ strongly in appearance, be-
haviour and hearing from those of the other larger taxon, the Phaneropterinae.
The latter live in the canopy, have in general a green, leaf-like appearance and
are good flyers. Their ears are about 15 dB more sensitive than the ears of pseudo-
phyllines (unpublished results). By contrast, pseudophyllines live in the rainforest
understorey, they have a long, slender, fusiform body and are bad flyers.
Katydids are a major source of protein for diurnal predators such as birds (e.g.
Formicariidae, Furnariidae and others), rodents and small primates (Nickle &
Heymann, 1996; Martins & Setz, 2000). Some of these birds feed almost exclu-
sively on arthropods by searching curled dead leaves that hang from vegetation
in the lower understorey (Gradwohl & Greenberg, 1980, 1982, 1984; Remsen &
Parker, 1984), thereby counteracting one of the katydids’ primary defence strate-
gies, namely crypsis by general appearance and behaviour (Nickle & Castner, 1995).
During the night, foliage-gleaning bats (Micronycteris hirsuta, Lophostoma silvicolum)
eat large numbers of Pseudophylline katydids (Belwood, 1988). These bats are at-
tracted by calling songs or other sounds involved in phonotactic activities of their
prey. Forest-living katydids exhibit a range of behaviours and signal characters
that appear to be adaptations to avoid predation by these bats: a reduction in call
redundancy (duty cycles of 3% and less), high carrier frequencies over 20 kHz and
the partial (or in one species, complete) replacement of airborne sound signals by
substrate-borne vibrations (tremulation) (Belwood & Morris, 1987).
In this chapter, we present data about the antipredator behaviour of a neotrop-
ical katydid, and its consequences for signal detection in noisy rainforest.
Predation pressure and roost site selection
The study was conducted on Barro Colorado Island (BCI), Panama and
on nearby peninsulas and small islands. The 1500 ha island is located in central
Panama (09◦10′N, 79◦51′W) in Gatun Lake, part of the Panama Canal. BCI is almost
totally covered with secondary and primary semideciduous lowland tropical forest
(Foster & Brokaw, 1982). The study took place in February/March (dry season) and
June/July 2002 (beginning of the rainy season). We studied Docidocercus gigliotosi,
a Pseudophylline katydid with a medium-sized, long and slender brown body.
Its natural history is only poorly known, although it is one of the most common
katydids on the island (Belwood, 1988). M. hirsuta and Micronycteris megalotis are two
insectivorous bats that glean highly cluttered spaces (Kalko et al., 1996) and feed on
D. gigliotosi (established by identification of remains at bat roosts). This katydid con-
stitutes about 20% of the diet of M. hirsuta (Belwood, 1988; personal observations).

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 155
(a) (b)
(c)
20m
Fig. 8.1. Roosting of Docidocercus gigliotosi (Pseudophyllinae) in Aechmea magdalenae.
(a) The A. magdalenae plant; (b) the 2.5 cm leaf edge spines; (c) the location of A.
magdalenae plants in part of field ‘Zetek 15’ on Barro Colorado Island (Panama),
mapped with a geographical information system. Each plant is marked with a circle;
those occupied with one or more D. gigliotosi are shown with a filled circle.
We regularly found D. gigliotosi roosting during the day in Aechmea magdalenae,
a terrestrial bromeliad of the pineapple family that can grow to a height of 2.5 m
(Fig. 8.1a). A striking characteristic of these plants are numerous, inch-long spines
along the leaf edges (Fig. 8.1b). It is abundant throughout BCI, forming dense
stands of sometimes more than 1000 plants. The part of the field ‘Zetek 15’ shown
in Fig. 8.1 comprises about 480 plants covering an area of 2600 m2. The leaves of
the plant form a long tube in the centre and that is where most katydids were
found roosting.
We observed several individuals shortly before sunset and during the night
using infrared video cameras. Sunset occurred around 18:30 h and katydids usually
became active (climbing and cleaning themselves within the plant) between 19:00
and 19:30 h. Three males were observed tremulating for several seconds. Between
20:00 and 21:00 h they used nearby lianas or trees to climb up into the lower
canopy, where they could no longer be observed. We presume that they are active

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
156 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
in the canopy throughout the night, because they returned to their host plants
around 04:00 h and climbed back into the central tube by 04:30 h, which is about
one hour before sunrise. With a mark-and-recapture study, we confirmed this
general scheme of daytime inactivity within the roost plant, nocturnal activity in
the canopy approximately between 21.00 h and 04.00 h and the return to the plant.
We recaptured 35 (out of 65) adults within 17 days; only three recaptures were
more distant than 2 m from the marking site. The maximum recapture distance
was 10 m and we never found marked individuals in different A. magdalenae fields.
Some individuals were found in the same plant for a period of more than two
weeks and 66% of katydids were recaptured in the same plant.
D. gigliotosi were not randomly distributed among A. magdalenae plants but
were found to roost in taller plants in above average condition with leaf-litter-free
central tubes close to canopy access ‘walkways’ (A. B. Lang & H. Romer, unpublished
data). Figure 8.1c shows a field of such plants (Zetek 15), in which plants occupied
by one or more individuals are marked with filled dots. D. gigliotosi roosted in
plants that were significantly taller (mean height (± standard deviation) 1.68 ±0.3 m (n = 32)) than unoccupied plants (1.36 ± 0.36 m (n = 320); (two-tailed Mann-
Whitney U test, p < 0.0001). Similar results were obtained for two study periods in
February/March and May/June 2002. We attempted to quantify the quality of roost
plants by ranking the condition of each plant on a subjective scale from bad (0) to
excellent (3). This ranking included the state of desiccation, number of damaged
leaves, and number of fresh, fleshy leaves, in particular those in the centre.
A survey performed in July 2002 found that the average condition of A. mag-
dalenae plants occupied by katydids was significantly better than that of unoccu-
pied plants (occupied plants, mean rank (± STD) 2.55 ± 0.69 (n = 36); unoccupied
plants, mean rank (± STD) 1.72 ± 0.96 (n = 248); two-tailed Mann–Whitney U test
p< 0.0001). Similar results were found for a survey carried out in March 2002. Most
(81%) of the plants in which adult D. gigliotosi roosted had direct contact with, or
grew within 1 m of a tree or liana reaching at least to the lower canopy.
These data indicate that the life history of the Pseudophylline katydid D.
gigliotosi is strongly influenced by predators. Two pieces of evidence are consis-
tent with D. gigliotosi attempting to avoid predation. First, by roosting during the
day in the spiny bromeliad A. magdalenae they are protected from predatory birds
and mammals. Insectivorous birds can have a pronounced effect on populations
of their arthropod prey (e.g. Lepidoptera larvae: Holmes et al., 1979) and on BCI
Myrmotherula fulviventris (Formicariidae) spends 98% of its foraging time search-
ing aerial leaf litter for arthropods and about 20% of its prey items are crickets
and katydids (Gradwohl & Greenberg, 1982). Therefore, the usual pseudophyllines
habit of roosting in curled leaves is potentially risky. Heavy predation by birds and

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 157
other visually hunting predators selects for various patterns of crypsis in their
katydid prey (Belwood, 1990; Castner & Nickle, 1995) and also for their choice of
backgrounds on which to hide, and on other life-history traits. Second, D. gigliotosi
is not active when gleaning bats show most flight activity (which is likely to be
related to foraging activity). I. silvicolum has a peak in flight activity about one hour
after sunset, when it flies to its hanging perch (C. D. Weise, E. K. V. Kalko, personal
communication), and one hour before sunrise, when it flies back. Radio telemetry
showed a similar pattern of activity in M. hirsuta (S. Spehn, personal communica-
tion). This correlates with the finding that D. gigliotosi, one of the bat’s common
prey species, does not exhibit night-time activity until the major period of bat
flight activity is over, and they also return to their bromeliad roost before the bats
return to their roosts. Although one may argue that flight activity of bats does not
necessarily reflect the time of highest predation pressure for their prey, it is reason-
able to assume that bats would home in on katydid song and the noises caused by
prey flight or landing activity during this time.
The costs of nocturnal communication: masking interference
Many species of insect and anuran communicate acoustically at night
and the resulting multispecies choruses have high sound pressure levels (SPL)
and complex spectral properties. Figure 8.2a shows measurements of the SPL in
the rainforest on BCI over a period of 24 hours. During the day, the SPL was
rather low, measuring 40–50 dB. It rapidly increased shortly after sunset by some
20 dB as a result of calling activity of insects and frogs. SPL declined throughout
the night (depending on the moon cycle; see below), until it reached daytime
levels after sunrise. In such chorus noise, different species occupy different fre-
quency bands. In our recording, the most prominent frequencies were below 8 kHz
(calling songs of crickets) (Fig. 8.2b), but frequencies well above 20 kHz were also
obvious.
We used a ‘biological microphone’ to ‘listen’ through the ears of the biological
receiver to analyse the challenging problem of signal detection after sunset for
D. gigliotosi at the position of potential receivers. The biological microphone is a
small, portable outdoor neurophysiological set-up that records the action poten-
tial activity of a single, identified auditory interneuron of a katydid (Rheinlaender
& Romer, 1986; Romer & Lewald, 1992). Rather than analysing the properties of
signals and noise at the position of potential receivers with conventional micro-
phones, such a method allows one to listen through the ears of the biological
receivers and to draw conclusions from the analysis of afferent nervous activity
under these natural conditions.
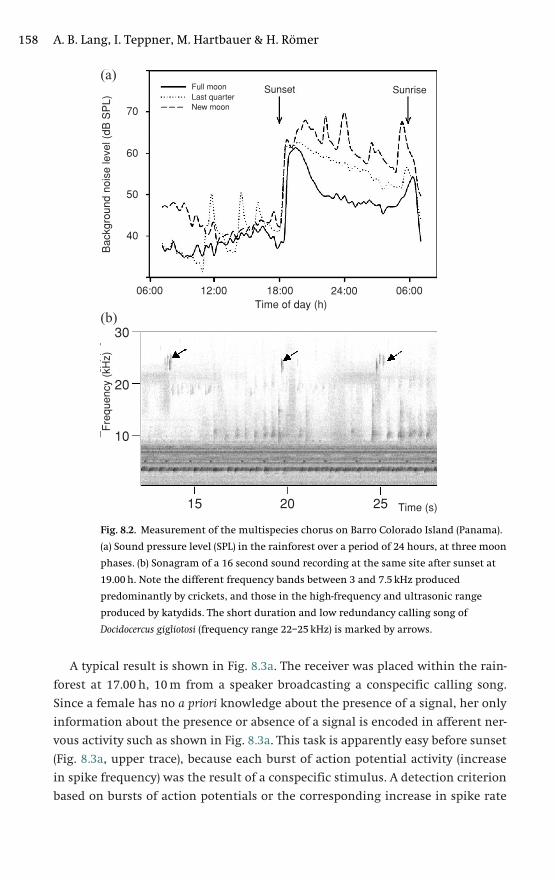
P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
158 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
Time of day (h)
Time (s)
Bac
kgro
und
nois
e le
vel (
dB S
PL)
Fre
quen
cy (
kHz)
(a)
(b)
70
60
50
40
06:00 12:00 18:00 24:00 06:00
Full moon
New moonLast quarter
Sunset Sunrise
30
20
10
15 20 25
Fig. 8.2. Measurement of the multispecies chorus on Barro Colorado Island (Panama).
(a) Sound pressure level (SPL) in the rainforest over a period of 24 hours, at three moon
phases. (b) Sonagram of a 16 second sound recording at the same site after sunset at
19.00 h. Note the different frequency bands between 3 and 7.5 kHz produced
predominantly by crickets, and those in the high-frequency and ultrasonic range
produced by katydids. The short duration and low redundancy calling song of
Docidocercus gigliotosi (frequency range 22–25 kHz) is marked by arrows.
A typical result is shown in Fig. 8.3a. The receiver was placed within the rain-
forest at 17.00 h, 10 m from a speaker broadcasting a conspecific calling song.
Since a female has no a priori knowledge about the presence of a signal, her only
information about the presence or absence of a signal is encoded in afferent ner-
vous activity such as shown in Fig. 8.3a. This task is apparently easy before sunset
(Fig. 8.3a, upper trace), because each burst of action potential activity (increase
in spike frequency) was the result of a conspecific stimulus. A detection criterion
based on bursts of action potentials or the corresponding increase in spike rate

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 159
(a)
(b)Spike frequency
17.00 h
18.30 h
150
100
50
5 s
Fig. 8.3. Outdoor action potential recording of an identified nerve cell of the afferent
auditory pathway of Docidocercus gigliotosi (see text). (a) Recording obtained within the
rainforest at 17.00 h (upper trace), and at 18.30 h (lower trace). Before sunset, masking
noise level is low and each conspecific stimulus elicits bursts of action potentials (hits
shown by open dots). After sunset, many bursts of action potentials are elicited by
noise (false alarms shown by stars). (b) Longer recording of action potentials after
sunset (lower trace) and the corresponding spike rate (upper trace). Note that a signal
presented at five second intervals (arrows on upper trace, points between traces)
elicits an increase in the spike rate, and noise may result in a stronger increase in the
spike rate than the signal.
would give ‘hits’ in terms of signal detection (Green & Swets, 1966). Indeed, in all
cases when there was an acoustic signal during the experiment at 17.00 h, there
was bursting activity in the nerve cell and there was no such activity when a signal
was absent; therefore, there were no ‘misses’ or ‘false alarms’, respectively.
However, this ideal situation for signal detection changed completely after sun-
set, when most katydids and other insects started to communicate acoustically.
The same preparation at exactly the same position in the rainforest now exhibited
high action potential activity (Fig. 8.3a, lower trace) and only an a priori knowledge
of the time of signalling allows correct detection of the stimuli. Using the same
detection criterion as in the situation before sunset would result in many false
alarms (i.e. identifying background noise as signals: stars in Fig. 8.3a, lower trace).
We measured a false alarm rate of more than 1400 in only five minutes, thus ex-
ceeding the hit rate dramatically, rendering communication between conspecifics

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
160 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
Male signal
Representation of signalwithin CNS of receiver
Fig. 8.4. The amplitude-modulated calling song of a male Thamnobates subfalcata
katydid (lower trace) and the corresponding representation of this song in the spike
discharge of a nerve cell (omega-neuron) in the afferent auditory pathway of the
central nervous system (CNS) in a female Docidocercus gigliotosi receiver.
rather ineffective. Figure 8.3b illustrates this point over a longer time scale and
also shows that a signal presented at regular intervals of five seconds elicited a
corresponding increase in the spiking rate, but that noise pulses may result in a
stronger increase in the spike rate than the signal.
Signal detection would be improved by increasing either the duration or the
rate of signalling, and indeed we found such an effect with our preparation. Sim-
ilar experiments to those shown in Fig. 8.3 clearly indicate that the rate of hits
increases and the rate of false alarms decreases with increased signal redundancy
and duration. However, as pointed out by Belwood & Morris (1987), eavesdropping
(interceptive eavesdropping in the sense of Peake (Ch. 2)) by gleaning bats excludes
such a solution and illustrates the opposing selection pressures of avoiding pre-
dation and signalling effectively.
How do the insects solve the problem? A closer look at the signals used by males
offers a possible solution. Figure 8.4 shows a typical, short amplitude-modulated
signal of a male katydid Thamnobates subfalcata and the corresponding represen-
tation of this signal in the spike discharge of a nerve cell (omega-neuron) in the
afferent pathway of a receiver. If the parameters of the spike discharge in response
to a species-specific call differ from those in bursts elicited by background noise,
this difference could be used by the nervous system to discriminate signals from
noise.
To investigate this further we used the biological microphone to record action
potentials in the rainforest at about 21.00 h, when the level of the background
noise was still high (Fig. 8.2). A male signal was broadcast every two seconds
at 10 m from the preparation. The SPL of the signal was adjusted to a value
of approximately 10 dB above the neuron’s masked threshold. We then used a
self-developed Delphi-application (Delphi 6, Borland Software Corporation, Scotts
Valley, CA 95066-3249, USA) to extract bursts from afferent spike recordings. We

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 161
compared several features of bursts (spike variables: the number of spikes in the
sequence; the mean, maximum, minimum; variance; average deviation; standard
deviation; skewness; kurtosis of the interval between two spikes) between hits (i.e.
bursts induced by the playback call) and false alarms (i.e. bursts elicited by back-
ground noise). One half of the recording (30 minutes) was used as the training set to
compute the decision tree: that is, the program learned the specific spike variables
that characterize hits. This tree was then evaluated for the remaining 30 minutes of
the recording, the validation set, to see whether it could detect such responses in
a noisy background. Machine learning on the basis of decision-tree learning is
one of the most widely used and practical data-mining methods to classify very
large amounts of data. It is a method for approximating discrete-valued functions,
in which the learned function is represented by a decision tree that is robust to
noisy data and capable of learning disjunctive expressions (Mitchell, 1997). We
used the algorithms J48 (pruned, unpruned) (Quinlan, 1993) and PART (Frank &
Witten, 1998) for classification of bursts within spike trains, calculating the re-
sults with the Java application WEKA (Trigg et al., 1999). Methods of decision-tree
learning such as J48 and PART search a completely expressive hypothesis space
and thus avoid the difficulties of restricted hypothesis spaces (Mitchell, 1997).
To avoid so-called ‘over fitting’ (see also Mitchell, 1997), we conducted a tenfold
cross-validation.
The unpruned J48 decision tree classified 95.4% of the bursts correctly, meaning
that it was able to distinguish between the double pulse signal and background
noise with an error of only 4.6% (Table 8.1). The effect of varying the duration
of the playback signal (7, 70 and 700 milliseconds) at a constant signal rate of
0.5/second is shown in Table 8.1. The most obvious effect is the lower detectability
of the 7 millisecond signal by all three algorithms; however, there is little increase
in detectability between the 70 and 700 millisecond signals. A similar result was
found for the grasshopper Chorthippus biguttulus, where signal detection improved
with increasing signal duration up to 450 milliseconds but did not improve further
with longer signals (Ronacher et al., 2000).
Although such results do not tell us that the insect’s nervous system makes
use of this information, it does show how bursts of action potentials in response
to conspecific song can, in principle, be discriminated from bursts produced by
other sound sources. The most important spike variables used by the decision trees
in discriminating signals from noise were the kurtosis, number of spikes, mean
and variance. The minimum and maximum spike interval was often used in final
decisions of the trees. Because some of these parameters (e.g. number of action
potentials or the minimum or maximum spike intervals) are also relevant in real
nervous systems for processing and discriminating sensory information, one can
assume that the insect nervous system can also solve this discrimination task.

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
162 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
Table 8.1. The effects of duration (7, 70, 700 milliseconds) of the
playback signal (presented at 0.5/second) on signal detection in
afferent spike trains of a receiver based on decision tree learning
with three different algorithms: J48, unpruned J48, and PARTa
Algorithm Correctly classified bursts (%)
Stimulus 7 ms (n = 609)
J48 80.13
J48 (unpruned) 79.64
PART 85.22
Stimulus 70 ms (n = 616)
J48 94.77
J48 (unpruned) 95.15
PART 94.77
Stimulus 700 ms (n = 108)
J48 95.37
J48 (unpruned) 95.37
PART 95.37
aSee text for further explanation.
Using machine-learning procedures to evaluate afferent spike patterns in sen-
sory systems may also enable us to look more closely at the strategic design of
signals (Guilford & Dawkins, 1991). For example, given the advantage of short-
duration and low-redundancy signalling in the presence of interceptive eaves-
dropping predators, what degree of amplitude modulation in a signal is necessary
to make its representation in afferent channels reliably different from heterospe-
cific signals? This is part of ongoing research on a variety of katydid species on BCI,
some of which pose a real challenge for signal detection by using signals of only a
few milliseconds duration. Additional behavioural experiments under noisy con-
ditions with the same species are urgently needed in order to show whether the
insects’ nervous systems can solve the task.
Variation in ambient light and noise levels, and the use
of a conditional communication strategy
Two features of neotropical rainforest at night vary with the lunar cycle:
insect abundance and noise level. We shall argue that they are related to predation
pressure on communication. We quantified the effect of lunar cycle on katydid
abundance on BCI by collecting at mercury vapour lights in December 1999 and
April 2001 at 21:00 and 24:00 h.

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 163
Day in moonphase
Col
lect
ed k
atyd
ids
per
nigh
t
New
moo
n
Ful
l moo
n
40
30
20
10
0
1 4 7 10 13 16 19 22 25 28
Fig. 8.5. The number of katydids collected at mercury vapour lights on Barro Colorado
Island (Panama) in relation to moon phase. Note that the higher variation at new
moon may be because of rainfall during these nights, when many katydids hide under
leaves and do not fly.
There was a significant relationship between moon phase and the number
of katydids collected (Fig. 8.5): at full moon the number of katydids collected
approached zero and it reached a maximum at new moon. As these data are similar
to results obtained with other insects (e.g. Hardwick, 1972) and are also similar
to collections made with suction traps, these cycles in abundance reflect natural
activity patterns. Light intensity may vary by three to four orders of magnitude
between full moon and new moon (Erkert, 1974). Full-moon ambient light levels
are high enough for humans to orient easily in the forest understorey with their
dark-adapted eyes. It is, therefore, likely that these light conditions allow a variety
of predators to hunt visually and, consequently, their potential prey must adopt
a cryptic lifestyle.
We quantified the effect of lunar cycle on background noise level on BCI with a
continuous recording system. The system consisted of a sound level meter (CEL 414
plus attached CEL-296 digital filter with settings A- weighting and slow time con-
stant) with a condenser microphone (LD 2540, type 4133, range 4–40 kHz). The
set-up was protected from humidity and rainfall and heated to 2 ◦C above am-
bient temperature with an infrared bulb to prevent fogging of the microphone
membrane. The DC output of the sound level meter was monitored at intervals of
five seconds with a Maclab/Powerlab 4e data acquisition system (AD Instruments
Pty Ltd) connected to a portable computer (Sony PCG-F707). Recordings were made
from the end of October to early December 2001, as well as in February, May and
June 2002.
Figure 8.2a shows representative examples of noise measurements over
24 hours at full moon, new moon and the last quarter of a lunar cycle. The increase

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
164 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
Tremulation
Stridulation 2 s
Fig. 8.6. The two types of mating signal produced by a male Docidocercus gigliotosi.
Tremulations (vibratory signals, upper trace) were recorded without mechanical
contact by laser doppler vibrometry, simultaneously with airborne sound signals
produced by elytral stridulation (lower trace). The photograph shows a male on a large
leaf.
in noise at sunset and the decrease at sunrise are common to all recordings. How-
ever, at full moon, noise levels decrease after sunset and for the rest of the night
the noise level is only 10 dB above the daytime level. As a result, the masking noise
level between 09:00 and approximately 05:00 h varies cyclically with the moon
phase; the amplitude of variation is about 10 dB.
Given the fact that the masking noise is the result of acoustic signalling, pre-
dominantly by insects, the drop in noise level at full moon is best explained by
species and/or individuals reducing or abandoning signalling with airborne sound.
We have argued above that the nocturnal lifestyle of many insects avoids predation
from visually hunting predators (e.g. rodents) and that light intensity at full moon
may allow increased visual predation. Therefore, a cryptic lifestyle may include
cryptic signalling.
Direct evidence for this hypothesis comes from the signalling behaviour of
D. gigliotosi. Males produce airborne sound with the usual elytral stridulation and
also tremulations, when the insect rapidly shakes its body up and down or drums
with the abdomen on the substratum (Morris et al., 1994). Tremulations are trans-
mitted through the substratum and females respond with tremulations of their
own. Although the active space of a tremulation signal is limited to the plant
where the tremulation is produced, it may travel 2–3 m (Michelsen et al., 1982;
Markl, 1983). Tremulation is an effective way to communicate in the presence of
acoustic interceptive eavesdroppers such as gleaning bats because only receivers
equipped with sensitive vibration receptors (e.g. spiders) can intercept the signal.
Figure 8.6 demonstrates that the duration and redundancy of the tremulation

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in networks 165
New moon
Time of day
Num
ber
of e
vent
s/5
min
Num
ber
of e
vent
s/5
min
Full moon
60
50
40
30
20
10
0
60
50
40
30
20
10
0
18:0
0
19:1
2
21:3
6
00:0
0
01:1
2
02:2
4
03:3
6
22:4
8
20:2
4
(a)
(b)
Fig. 8.7. The pattern of production of the two types of mating signal (tremulation,
open squares; airborne sound, filled squares and grey area) produced by a male
Docidocercus gigliotosi at new moon (a) and full moon (b). Males were separated from
females a day before data were collected, caged and placed in the rainforest
understorey. An infrared camera recorded both signals over a period of about
nine hours, from sunset to 03.00 h.
signal of D. gigliotosi is higher than the stridulatory airborne sound signal by orders
of magnitude.
As expected if male D. gigliotosi vary the relative proportion of these two modes
of signalling in relation to the chance of predation from gleaning bats, at new
moon, or in the laboratory in complete darkness, males become active about half
an hour after sunset and begin signalling with a period of high-rate tremulation,
followed by a prolonged period of airborne sound production, often for many
hours (Fig. 8.7a). At full moon the onset of signalling after sunset is often delayed,
airborne sound signalling is strongly reduced and tremulation is more common.
It should also be noted that D. gigliotosi reduces acoustic signalling at full moon
despite the fact that the lower noise levels would allow better detection of conspe-
cific signals. The observations are consistent with D. gigliotosi having a conditional
strategy of signalling, where fairly cryptic (i.e. short duration, low redundancy)
airborne sound production is replaced by the even more private mode of commu-
nication with tremulations.

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
166 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
Summary
In this chapter we have emphasized the importance of predation on the
ecology and acoustic communication of a subfamily of neotropical katydids. Pseu-
dophylline katydids switch from airborne sound signalling to tremulation be-
cause of high predation by passive listening bats. This results in either decreased
active space of their signals or decreased detectability of airborne sound signals
in high levels of background nocturnal noise. Neurophysiological experiments
and decision-tree learning algorithms nevertheless indicate that information for
discrimination between signals and noise is still preserved within afferent spike
discharges. Daytime and night-time predation also appears to restrict individuals
to roost sites within bromelid plants. D. gigliotosi exhibits a strong site dependency
for a particular plant in a field over several weeks; consequently, there is very
little horizontal movement of males and females in a population. In conjunction
with a reduced active space of acoustic and/or vibrational signalling, and reduced
flight capability, this situation may strongly reduce the chances of matings with
individuals of neighbouring populations. Current population genetic analysis will
show whether genetic exchange between populations is suppressed, despite the
lack of a geographical barrier between populations.
Acknowledgements
Inspiring discussion with Elisabeth K. V. Kalko, Christa D. Weise, Dina K. N. Dechmann
and Sabine Spehn played a major role in maturing the ideas presented here. We acknowledge the
logistic support of the Smithsonian Tropical Research Institute, Panama. Barbara Bliem, Franz
Kainz, Birgit Roehnfeld and Iris Strauss assisted in the field and in the laboratory. We also thank
Peter McGregor and two anonymous reviewers for many suggestions to improve the manuscript.
Research was supported by the Austrian Science Fund (FWF P14257-BIO to H. R.) and a Ph.D.
scholarship (Austrian Academy of Sciences to A. B. L.).
References
Allen, G. R. 2000. Call structure variability and field survival among bushcrickets
exposed to phonotactic parasitoids. Ethology, 106, 409–423.
Bailey, W. J., Withers, P. C., Endersby, M. & Gaull, K. 1993. The energetic costs of calling
in the bushcricket Requena verticalis (Orthoptera: Tettigoniidae: Listroscelidinae).
Journal of Experimental Biology, 178, 21–37.
Belwood, J. J. 1988. The influence of bat predation on calling behaviour in Neotropical
forest katydids (Insecta: Orthoptera: Tettigoniidae). Ph.D. Thesis, University of
Florida, Gainesville, FL.

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in communication networks 167
1990. Anti-predator defences and ecology of Neotropical forest katydids,
especially the Pseudophyllinae. In: The Tettigoniidae: Behaviour, Systematics,
Evolution, ed. W. J. Bailey & D. C. F. Renz. Bathurst, Australia: Crawford House
Press, pp. 6–26.
Belwood, J. J. & Morris, G. K. 1987. Bat predation and its influence on calling behaviour
in Neotropical katydids. Science, 238, 64–67.
Cade, W. 1975. Acoustically orienting parasitoids: fly phonotaxis to cricket song.
Science, 190, 1312–1313.
Castner, J. L. & Nickle, D. A. 1995. Observations on the behaviour and biology of
leaf-mimicking katydids (Orthoptera: Tettigoniidae: Pseudophyllinae:
Pterochrozini). Journal of Orthopteran Research, 4, 93–98.
Endler, J. A. & Basolo, A. L. 1998. Sensory ecology, receiver biases and sexual selection.
Trends in Ecology and Evolution, 13, 415–420.
Erkert, H. G. 1974. Der Einfluß des Mondlichtes auf die Aktivitatsperiodik
nachtaktiver Saugetiere. Oecologia, 14, 269–287.
Foster, R. B. & Brokaw, N. V. L. 1982. Structure and history of the vegetation of Barro
Colorado Island. In: The Ecology of a Tropical Forest, ed. E. G. Leigh, A. S. Rand & D. M.
Windsor. Washington, DC: Smithsonian Institute Press, pp. 151–172.
Frank, E. & Witten, I. H. 1998. Generating accurate rule sets without global
optimization. In: Proceedings of the 15th International Conference on Machine
Learning, Madison, WI, ed. J. Shavlik. San Francisco, CA: Morgan Kaufmann,
pp. 144–151.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans. Chicago,
IL: University of Chicago Press.
Gradwohl, J. A. & Greenberg, R. 1980. The formation of antwren flocks on Barro
Colorado Island, Panama. The Auk, 97, 395.
1982. The effect of a single species of avian predators on the arthropods of aerial
leaf litter. Ecology, 63, 581–583.
1984. Search behaviour of the checker-throated antwren foraging in aerial leaf
litter. Behavioral Ecology and Sociobiology, 15, 281–285.
Green, D. M. & Swets, J. A. 1966. Signal Detection Theory and Psychophysics. New York:
Wiley.
Greenfield, M. D. 1988. Interspecific acoustic interactions among katydids
Neoconocephalus: inhibition-induced shifts in diel activity. Animal Behaviour, 36,
684–695.
Guilford, T. & Dawkins, M. S. 1991. Receiver psychology and the evolution of animal
signals. Animal Behaviour, 42, 1–14.
Hardwick, D. F. 1972. The influence of temperature and moon phase on the activity of
noctuid moths. Canadian Entomologist, 104, 1767–1770.
Holmes, R. T., Schultz, J. C. & Nothnagle, P. 1979. Bird predation on forest insects: an
exclosure experiment. Science, 206, 462–463.
Kalko, E. K. V., Handley, C. O., Jr & Handley, D. 1996. Organization, diversity, and
long-term dynamics of a Neotropical bat community. In: Long-term Studies of

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
168 A. B. Lang, I. Teppner, M. Hartbauer & H. Romer
Vertebrate Communities, ed. M. L. Cody & J. A. Smallwood. New York: Academic
Press, pp. 503–553.
Lehmann, G. U. C. & Heller, K. G. 1998. Bushcricket song structure and predation by
the acoustically orienting parasitoid fly Therobia leonidei (Diptera: Tachinidae:
Ormiini). Behavioral Ecology and Sociobiology, 43, 239–245.
Markl, H. 1983. Vibrational communication. In: Neuroethology and Behavioural
Physiology, ed. F. Huber & H. Markl. Berlin: Springer, pp. 332–353.
Martins, M. M. & Setz, E. F. Z. 2000. Diet of buffy tufted-eared marmosets (Callithrix
aurita) in a forest fragment in southeastern Brazil. International Journal of
Primatology, 21, 467–476.
Michelsen, A. 1992. Hearing and sound communication in small animals:
evolutionary adaptations to the law of physics. In: The Evolutionary Biology of
Hearing, ed. E. D. Webster, R. R. Fay & A. N. Popper. New York: Springer, pp. 61–77.
Michelsen, A., Fink, F., Gogala, M. & Traue, D. 1982. Plants as transmission channels
for insect vibrational songs. Behavioral Ecology and Sociobiology, 11, 269–281.
Mitchell, T. M. 1997. Machine Learning. New York: McGraw-Hill.
Morris, G. K., Mason, A. C., Wall, P. & Belwood, J. J. 1994. High ultrasonic and
tremulation signals in Neotropical katydids (Orthoptera: Tettigoniidae). Journal of
Zoology (London), 233, 129–163.
Nickle, D. A. & Castner, J. L. 1995. Strategies utilized by katydids (Orthoptera:
Tettigoniidae) against diurnal predators in rainforests of northeastern Peru.
Journal of Orthopteran Research, 4, 93–98.
Nickle, D. A. & Heymann, E. W. 1996. Predation on orthoptera and related orders of
insects by tamarin monkey, Saguinus mystax and Saquinus fuscicollis (Primates:
Callitrichidae), in northeastern Peru. Journal of Zoology, (London), 239, 799–819.
Quinlan, J. R. 1993. C.4.5: Programs for Machine Learning. San Francisco, CA: Morgan
Kaufmann.
Remsen, J. V. & Parker, T. A. 1984. Arboreal dead-leaf-searching birds of the neotropics.
The Condor, 86, 36–41.
Rheinlaender, J. & Romer, H. 1986. Insect hearing in the field: the use of identified
nerve cells as biological microphones. Journal of Comparative Physiology, A, 158,
647–651.
Ronacher, B., Krahe, R. & Hennig, R. M. 2000. Effects of signal duration on the
recognition of masked communication signals by the grasshopper Chorthippus
biguttulus. Journal of Comparative Physiology, A, 186, 1065–1072.
Romer H. 2001. Ecological constraints for sound communication: from grasshoppers
to elephants. In: Ecology of Sensing, ed. F. G. Barth & A. Schmid. Berlin: Springer,
pp. 59–77.
Romer, H. & Lewald, J. 1992. High-frequency sound transmission in natural habitats:
implications for the evolution of insect acoustic communication. Behavioral
Ecology and Sociobiology, 157, 631–642.
Romer, H., Bailey, W. & Dadour, I. 1989. Insect hearing in the field. III. Masking by
noise. Journal of Comparative Physiology, A, 164, 609–620.

P1: IRK/KAA P2: IYP-KOD0521823617c08.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 13:38
Neotropical katydids: predation and noise in communication networks 169
Ryan, M. J. 1990. Sensory systems, sexual selection, and sensory exploitation. Oxford
Surveys in Evolutionary Biology, 7, 157–195.
Ryan, M. J. & Keddy-Hector, A. 1992. Directional patterns of female mate choice and
the role of sensory biases. American Naturalist, 139, S4–S35.
Ryan, M. J. & Rand, S. A. 1993. Sexual selection and signal evolution: the ghost of biases
past. Philosophical Transactions of the Royal Society of London, Series B, 340, 187–195.
Schatral, A. 1990. Interspecific acoustic behaviour among bushcrickets. In: The
Tettigoniidae: Behaviour, Systematics, Evolution, ed. W. J. Bailey & D. C. F. Renz.
Bathurst, Australia: Crawford House Press, pp. 150–165.
Trigg, L., Hall, M. & Frank, E. 1999. WEKA (Java collection of machine learning
algorithms). Hamilton, New Zealand: University of Waikato;
http://www.cs.waikato.ac.nz/ml/weka/.
Wagner, W. E., Jr & Hoback, W. W. 1999. Nutritional effects on male calling behaviour
in the variable field cricket. Animal Behaviour, 57, 89–95.
Wiley, R. H. & Richards, D. G. 1978. Physical constraints on acoustic communication in
the atmosphere: implications for the evolution of animal vocalizations. Behavioral
Ecology and Sociobiology 3, 69–94.
Zuk, M. & Kolluru, G. R. 1998. Exploitation of sexual signals by predators and
parasitoids. Quarterly Review of Biology, 73, 415–438.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
9
Nestling begging as acommunication network
a n d r e w g . h o r n & m a r t y l . l e o n a r d
Dalhousie University, Halifax, Canada
Introduction
In many bird species, young beg for care from their parents. A parent
arriving at the nest with food is met by begging nestlings, which are waving their
wings, calling and stretching to expose brightly coloured gapes, all within the
confines of a nest that may contain several other begging nestlings. This mode
of parent–offspring communication has become a model for the study of the
evolution of biological signalling.
Hungrier nestlings beg more intensely, so the parent can use the display to
decide which nestling to feed and to decide how soon it should return to the nest
with food (reviewed by Budden & Wright, 2001). The fact that the parent can extract
information on nestling hunger from such a confusing burst of signalling raises
numerous questions. How does each nestling ensure that its own signal of need
is received above the din of its nestmates’ displays? How do parents differentiate
among these displays to choose which nestling to feed? How much do the displays,
as opposed to the physical jostling toward the parent that also goes on in the nest,
determine which nestlings are fed?
To answer such questions we need to understand how the begging behaviours
of whole broods function together. Concepts derived from the new field of com-
munication networks seem well suited to this task but have not yet been explicitly
applied to begging. As currently defined (McGregor & Dabelsteen, 1996; McGregor
& Peake, 2000), a communication network forms whenever several individuals
communicate within transmission range of each other’s signals. Nestlings noisily
begging within the confines of a nest clearly fit this definition, since most or all of
the nestlings within a brood are within transmission range of each other’ssignals.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
170

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 171
In this chapter, we hope to show that considering begging as a communication
network yields new insights, not only into begging behaviour but also into commu-
nication networks in general. We begin by briefly summarizing previous research
on begging, most of which has treated the display as dyadic communication: that
is, as signalling from one individual, the nestling (or the brood considered as send-
ing one joint signal), to one receiver, the parent. We then apply ideas from studies
of communication networks to nestling begging, identifying several conceptual
issues that we think studies of begging can help to clarify. We also discuss aspects
of the design of begging and parental behaviour that may have evolved in re-
sponse to the network environment and finally we make some suggestions for
future work.
Begging as dyadic communication
In this section, we summarize the theoretical and empirical work on beg-
ging to date, most of which has treated parent–offspring communication as a
dyadic communication system. This summary provides background information
for the discussion of communication networks that follows, while also illustrat-
ing some of the strengths and weaknesses of the dyadic approach to begging
behaviour.
Theoretical work
Begging has attracted considerable attention from evolutionary biologists
largely because of its apparently needless conspicuousness. Because parents are
only a few centimetres away from their young, it is not clear why offspring signal
for food with such an elaborate display. Perhaps the best-known explanation for
this apparent extravagance stems from parent–offspring conflict theory (reviewed
by Godfray, 1995; Parker et al., 2002). Natural selection favours parents that dis-
tribute resources optimally amongst both their current and future offspring. Each
of these offspring, however, is selected to solicit resources so as to benefit its own
fitness, rather than the inclusive fitness of its siblings. Therefore, offspring might
signal for resources that parents would do better giving to siblings or reserving
for future broods. To overcome parental reluctance, offspring may have to send
exaggerated signals of need (Trivers, 1974; Godfray, 1995).
This basic explanation has been revised or extended in various ways, making
the parent–offspring dyad one of the most thoroughly modelled animal communi-
cation systems. Some of the most influential models, both for begging and for ani-
mal signals in general, have asked how reliable signalling can evolve in the face of
conflict between signallers and receivers (reviewed by Godfray & Johnstone, 2000;
Johnstone & Godfray, 2002). Specifically, if young are prone to exaggerate, then

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
172 A. G. Horn & M. L. Leonard
why would parents respond at all to begging signals? The answer is that, whereas
parents might easily be able to assess some aspects of their nestlings, like their
size, parents might not be able to assess important aspects of their nestlings’needs,
for example their immediate need for food. If begging provides information on
these aspects of nestling need, then parents should provision nestlings accord-
ing to variation in the begging signal. This situation can be evolutionarily stable,
however, only if the signal is costly for nestlings to produce. Therefore, in effect,
nestlings might have to put on a costly begging display to prove that they really
are hungry (Godfray, 1991).
These results have been largely responsible for the general acceptance of the
idea that reliable signals must be costly if they are to evolve. Some of the com-
plexities of this story are less widely known, however. For instance, recent models
have suggested that, in some situations, nestlings might signal their needs ac-
curately without large costs, for example if exaggeration draws so much care
away from siblings that the cost to the signaller’s inclusive fitness outweighs
the direct benefits of the extra signalling (Maynard Smith, 1994; Bergstrom &
Lachmann, 1998; Johnstone, 1999; Price et al., 2002).
For the purposes of this chapter, two features of theoretical work on begging
particularly stand out. First, these models have focused on fundamental issues
in dyadic communication, such as how signalling can evolve despite conflicts of
interest between signallers and receivers. Thus they are relevant to our under-
standing of a wide range of communication systems. Second, the emphasis these
models have placed on particular aspects of signalling, such as its honesty and
costliness, has led empirical studies to focus on these aspects of begging to the
neglect of others (see below). One of these neglected aspects is the communication
network in which begging occurs; although recent attempts to model the effects
of signalling on nestmates (reviewed by Royle et al., 2002; Johnstone & Godfray,
2002), which we discuss further below, are steps in that direction.
Empirical work
The theoretical possibility that begging might be exaggerated led many
researchers to test whether begging is indeed a reliable signal of need. Studies in
a wide range of species confirm that the intensity of both the visual and vocal
aspects of the display increase with food deprivation (Budden & Wright, 2001). In
turn, parents use the begging signal in two ways to make provisioning decisions.
First, the more intense the begging of the brood as whole, the more often parents
return to the nest with food. This level of response has been shown most clearly
in experiments in which playback of nestling begging calls stimulates higher
provisioning rates (Budden & Wright, 2001). Second, once parents arrive at the
nest, nestlings that beg more intensely than their nestmates are more likely to be

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 173
fed. Experiments again provide the clearest demonstrations of this effect: parents
are more likely to direct feedings to nestlings with brighter gapes (Gotmark &
Ahlstrom, 1997; Kilner, 1997; Saino et al., 2000; Saino and Møller, 2002) or to
nestlings placed next to speakers playing higher call rates (Leonard & Horn, 2001a;
Kilner, 2002a; but see Glassey & Forbes, 2002a). Therefore, begging appears to
communicate to parents the requirements both of the brood as a whole and of
individual nestlings.
Begging is more than a simple cry for food, however, for two reasons. First, food
deprivation is not the only aspect of nestling need that the begging display adver-
tises. For example, in some species begging may signal long-term nutritional need
as opposed to the short-term hunger described above, with nestlings in poorer
condition (e.g. having lower mass than nestmates) begging more than their nest-
mates (Price et al., 2002). Additionally, some aspects of begging, especially begging
calls, can change when nestlings lose heat, thus signalling the need for brooding
(Evans, 1994; Leonard & Horn, 2001b; Clotfelter et al., 2003; B. Glassey, personal
communication). Finally, gape colour in some species may advertise a nestling’s
immunocompetence (Saino & Møller, 2002). Clearly, the message that begging is
sending may be more complex than just short-term hunger.
A second complicating factor is the effect of siblings on nestling begging. Beg-
ging intensity, whether measured by the intensity of the postural display or overall
call rate, increases with brood size in many species (Budden & Wright, 2001) and
may also increase when nestmates beg (e.g. Leonard & Horn, 1998). Also, nestlings
compete physically for access to parents (see below) and their display and its effect
on parents may vary according to the nature and intensity of this physical com-
petition (e.g. Price et al., 1996; Cotton et al., 1999). Interest in the effects of both
signalling interactions and physical competition among nestmates has mainly fo-
cused on how they complicate honest signalling of need (e.g. Rodrıguez-Girones et
al., 2001; Price et al., 2002). We will be discussing them further below because they
are clearly central to any discussion of begging as a communication network.
Summary
This brief review shows that the main emphasis of work on begging has
been on how it functions as a signal of need from nestlings to parents. Begging has
been treated mainly as a dyadic signalling system: that is involving one signaller
(the nestling or the brood considered as sending one joint signal) and one re-
ceiver (the parent). Siblings have been included in the picture, but mainly because
they might affect the dyadic signalling of need. Only recently have researchers
started to consider the effects of competing signalling by nestmates in any de-
tail, an important step toward treating the begging brood as a communication
network.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
174 A. G. Horn & M. L. Leonard
Begging as a communication network
If we are to broaden our view of begging to include the communication
network in which it occurs, we must first characterize that communication net-
work. By definition, a brood of begging young is a communication network because
nestmates are all within range of each other’s signals (McGregor & Dabelsteen,
1996; McGregor & Peake, 2000). Going beyond this definition, however, to charac-
terize the network and explore its implications, raises more conceptual challenges
than this simple definition might suggest.
In this section, we discuss three of these issues. First, to apply the definition of
communication networks at all, we must distinguish signalling from other acts.
This can be especially problematic in the case of begging, in which signalling and
direct physical competition are tightly linked. Second, to examine some of the
more interesting implications of the network, we must carefully consider the na-
ture of signals and signalling interactions – again, a challenging distinction when
applied to begging. Third, there are factors, such as the genetic relatedness of
nestlings, which are at least as important for characterizing this communication
network as the overlapping transmission ranges of signals that define it. While all
three of these areas present challenges for studies of begging networks, they also
provide opportunities for testing some key concepts in the study of communica-
tion networks.
Physical competition versus signalling
Nestlings form a communication network because they are within sig-
nalling range of each other. Unlike members of many other communication net-
works, however, nestlings are also in direct physical contact with each other. This
tight proximity highlights difficulties that can arise when we try to distinguish
between signalling and other acts, in this case physical competition. Since a com-
munication network, by definition, consists of signalling (i.e. of behaviours spe-
cialized to communicate information (McGregor & Peake, 2000)), this distinction
is fundamental for understanding any communication network.
Nestlings jostle with one another for access to parental feeding locations
within the nest and their success at reaching the parent strongly affects which
nestlings are fed (Budden & Wright, 2001). Nestlings can physically compete in
several ways, for example by usurping positions close to where parents arrive
at the nest, by blocking parents’ access to other nestlings or, particularly in non-
passerine species, by directly pushing or pecking one another (Mock & Parker, 1997;
Budden & Wright, 2001; Drummond, 2002).
Much of this physical competition is hard to distinguish from signalling.
Jostling for position and direct aggression seem to be non-signalling acts by which

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 175
nestlings get better access to parents. Parents may nonetheless get information
about nestling need and quality from these physical interactions, which they then
use to choose which nestling to feed (Rodrıguez-Girones, 1996; Lotem et al., 1999).
This informativeness alone does not make them signals. If, however, the interac-
tions are designed to affect that choice, rather than merely to thrust a nestling
forward to rob the parent of its choice, then they are signals, by the above defini-
tion, despite their outward appearance. Conversely, some features of begging that
appear to have been designed partly to convey information and thus are signals by
definition (McGregor & Peake, 2000), such as posturing (Kilner, 2002b), seem just as
clearly designed for effective jostling toward the parent. Even the design features
of begging displays that are adaptations for overcoming interference from nest-
mates (reviewed below) may be seen either as ways to signal information on need
more effectively to parents (Horn & Leonard, 2002) or as scrambles for parental
attention (Rodrıguez-Girones et al., 2001; Royle et al., 2002). In the latter case, their
ultimate function would differ little from that of physical competition, since by
dominating the parents’ visual and acoustic fields they too would not inform
parents so much as reduce the parents’ opportunity to choose which nestling to
feed.
Therefore, a nest full of begging nestlings is part communication network,
part scrum toward the parent. Which view of begging is more accurate depends
largely on how parents interpret begging signals and physical competition, a topic
we discuss further below. Given that display behaviours ultimately evolve from
non-signalling acts, however, we can at least conclude that begging offers an
interesting system for studying how social behaviours besides signalling affect
communication networks.
Signalling interactions versus just signalling
One of the aspects of communication that has become more prominent
as a result of the communication network approach is the information content of
signalling interactions: the give and take of signals among members of the network
(McGregor & Peake, 2000). It is from the interactions between signallers, rather
than the signals themselves, that some particularly interesting consequences of
communication networks arise, such as signalling to avoid interference (Ch. 13)
and eavesdropping (Peake et al., 2002; Ch. 2).
Distinguishing signals (directed at the parent) from signal interactions (di-
rected at nestmates) in the case of nestling begging is difficult, however. On the
one hand, several lines of evidence show that nestmates’ signals influence how
a nestling signals. In many studies, nestlings beg more intensely when in bigger
broods or when with nestmates than when alone (Budden & Wright, 2001; but see
Cotton et al., 1996). More direct evidence comes from studies in which nestlings

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
176 A. G. Horn & M. L. Leonard
increase their postural display when their nestmates do (e.g. Leonard & Horn, 1998)
or call more when they hear nestmates calling (e.g. Leonard & Horn, 2001c).
On the other hand, it is not clear that these changes in signalling constitute
signalling interactions in the sense implied by current discussions of commu-
nication networks, especially work on social eavesdropping (McGregor & Peake,
2000; Ch. 2). According to this work, a signalling interaction consists of a sender
directing a signal at a receiver, which then responds. To the degree that begging
is directed at the parent, then competitive interactions among nestlings to catch
the parents’ attention are not signalling interactions in this sense (Royle et al.,
2002). By extension, parents that choose to feed nestlings that beg more than their
nestmates (Budden & Wright, 2001), like the predators that are attracted to nests
whose calling is increased by competition (Haskell, 2002), are interceptive rather
than social eavesdroppers, because social eavesdroppers must base their response
on signalling interactions not just on signals (Ch. 2).
This conclusion may partly reflect our still sketchy understanding of nestling
interactions. For example, Roulin (2002) has recently suggested that at least some
signalling by nestlings may be directed at nestmates. Nestling barn owls Tyto alba,
for example, appear to have calling contests between parental visits, in which
nestlings negotiate which of them will receive a feeding when the parent next
returns (Roulin, 2002). If nestlings do direct signals to each other in this way, then
parents that extract information from these interactions would fit the definition
of social eavesdroppers (Ch. 2).
In the particular case of barn owls, nestling negotiations occur when the par-
ent is absent and so cannot be overheard by parents. In principle, however, there
is no reason why similar interactions between nestlings could not also occur in
the parent’s presence, especially in species in which parents spend enough time
transferring food to their young that the young have time to interact (e.g. par-
rots (Psittaciformes); Krebs, 2002). Certainly, if nestlings do direct their signals to
each other, the importance of considering nestling begging as a communication
network is considerably strengthened.
Functional relationships among nestlings and network structure
Communication networks were first defined in the context of communi-
cation among territorial songbirds, which are widely separated on different terri-
tories but are interconnected by the overlapping transmission ranges of their songs
(McGregor & Dabelsteen, 1996). Song is, thus, the main way in which these birds
interact; consequently, characterizing interacting songbirds as a communication
network captures much of how they affect each other’s signalling behaviour.
Nestlings packed together within a nest, however, are interconnected in many
ways besides the overlapping ranges of their signals. We have already discussed

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 177
how they interact through physical competition and how that may have strong
effects on their signalling behaviour. In this section, we briefly list three other
interconnections that are integral to any explanation of how nestmates affect
each other’s signalling behaviour.
Unlike physical competition, these effects do not present difficulties for defin-
ing signals and hence applying the definition of communication networks to
begging. They do, however, illustrate that, in some communication networks, sig-
nallers are so mutually dependent on one another that the overlapping transmis-
sion ranges of their signals are only one way in which their signalling behaviours
are interconnected.
We will list three such relationships among nestlings: genetic relatedness,
shared fate and heat transfer. For each category, we touch briefly on their pos-
sible implications for signalling. We then discuss perhaps their most interesting
implication, which is how all these relationships might combine to give a structure
to the communication network within the brood.
Genetic relatedness
Genetic relatedness is perhaps the most important of the relationships
among nestlings, because it so heavily influences the fitness consequences of all
the other types of relationship. Since nestlings tend to be highly related to one
another, relatedness probably affects signalling in this communication network
more than in most of the other networks described in this volume. Indeed, for
most theoretical models of begging, the main route of sibling effects on begging is
through a nestling’s inclusive fitness. In general, theory predicts less-exaggerated
or less-costly begging the higher the relatedness among nestmates ( Johnstone &
Godfray, 2002; Price et al., 2002). Consistent with such predictions, interspecific
brood parasites, whose relatedness with their host nestmates is zero, such as Euro-
pean cuckoos Cuculus canorus, great spotted cuckoos Clamator glandarius and brown-
headed cowbirds Molothrus ater, call more loudly and more frequently than their
nestmates (Dearborn & Lichtenstein, 2002; Redondo & Zuniga, 2002).
Evidence for non-parasitic species, however, is scant. In one comparison across
species for which data on genetic parentage were available, begging calls were
louder in species with more frequent mixed parentage (Briskie et al., 1994). This
result suggests that a species’ average level of relatedness within broods might set
its average level of begging. A more relevant result for communication networks,
however, would be if nestlings within a species could assess their relatedness to
broodmates and adjust their levels of competitive signalling accordingly. Nestlings
are generally thought to lack the cues by which their nestmates could assess
their relatedness (e.g. Whittingham & Dunn, 2001); indeed there may be selection
against such cues ( Johnstone, 1997). As for kin recognition in birds in general

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
178 A. G. Horn & M. L. Leonard
(Komdeur & Hatchwell, 1999), addressing this issue directly will require more
sophisticated experiments than have been applied to date.
Shared fate
Along with relatedness, a fundamental feature underlying nestling inter-
actions is that, like the proverbial eggs in one basket, nestmates often share the
same fate. For better or worse, they have the same adults feeding them, share the
same local environmental conditions around the nest and, therefore share their
chances of survival to a greater degree than participants in most other types of
communication network.
This shared fate has inevitable consequences for signalling behaviour; if one
nestling begs more loudly, for example, the parents might return more often to
feed all the nestlings or a predator might be more likely to find the nest and eat
all the nestlings. Thus, both the benefits and the costs of begging by any given
nestling are at least partially visited on the whole brood. Indeed, Wilson & Clark
(2002) went still further and suggested that broods are subject to a form of group
selection which may lead nestlings to signal cooperatively. Aspects of begging that
are usually presented as competitive, such as signal characteristics that ostensibly
serve to circumvent interference (see below), might instead function cooperatively
to coordinate nestmates’ signals (Wilson & Clark, 2002). How individual signals
fit together to form aggregate brood signals has not been studied yet, but we can
safely expect that the shared fate of nestlings will make signalling interactions
within their networks differ in interesting ways from those of signallers with more
independent fates, such as chorusing frogs.
Heat transfer
Nestling birds cannot thermoregulate until partway through the nesting
period. Before that point, they rely not only on brooding by parents but also on
heat from their nestmates. Nests where young hatch asynchronously may consist
of older, heat-producing nestlings and younger, heat-consuming nestlings (e.g.
Hill & Beaver, 1982). Such thermal relationships among nestlings may increase the
variety of their signals and signalling interactions. Specifically, in several species,
some aspects of begging, especially begging calls, change when nestlings lose heat
and may signal their need for brooding (see above). Nestlings might, therefore, have
to compete for attention from nestmates that are sometimes signalling for food
and sometimes for warmth, and they might adopt different signalling strategies
for each situation. Thermal relationships might also affect signalling through
more direct effects on individual signallers. For example, some evidence suggests
that house sparrow Passer domesticus nestlings lose heat when the stretching and
gaping of begging increases their surface area (Ovadia et al., 2002). They might,

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 179
therefore, be able to beg more when next to larger nestmates, since any thermal
loss during begging would be reduced. Thermal relationships among nestmates
are still poorly understood, but, like physical competition and signalling, they
are probably readily perceived by nestlings and thus may have immediate and
dynamic effects on patterns of signalling within the nest.
Network structure
The net result of all the relationships listed above, including the physical
competition also discussed, is that they may lend structure to the communica-
tion network within the nest. By ‘structure,’ we mean a pattern in which not all
nestlings have the same sorts of relationship with one another. Most obviously,
physical competition can lead to dominance hierarchies, with larger or stronger
nestlings suppressing the begging signals of smaller nestlings or displacing them
from positions near the parent where their begging signals would attract the
parent’s attention more effectively (Mock & Parker, 1997).
Hierarchies, however, are only one of a variety of network architectures
that might arise. Speaking more generally, Glassey & Forbes (2002b) noted that
nestlings can often be divided into ‘core’ and ‘marginal’ nestlings (Mock & Forbes,
1995). Survival of core nestlings is usually predictable, whereas marginal nestlings,
which may be smaller, in poorer condition, younger, subordinate and/or less able
to thermoregulate, survive only if ecological conditions are favourable. This ‘struc-
tured sibship’ (Glassey & Forbes, 2002b) may yield three different sorts of nestling
relationships within the brood: core to core, marginal to marginal, and core to
marginal (Glassey & Forbes, 2002b).
Variation among species in this underlying structure will affect physical com-
petition and signalling interactions within the nest. For example, one core and
one marginal nestling might yield a simple dominance hierarchy, whereas three
nestlings in each category might yield two ‘cliques’ of nestlings, between which
there is a dominance hierarchy but within which signalling behaviours are simi-
lar and physical competition is equitable. In any case, the underlying structure of
relationships within the brood, even though they do not consist of signalling rela-
tionships, nevertheless may strongly affect the structure of the overlying commu-
nication network – no doubt a recurring theme for most communication networks
(e.g. Chs. 10 and 25).
Summary
We have raised three complexities in applying the concept of communi-
cation networks to nestling begging. First, characterizing the communication net-
work entails a difficult distinction between signalling and physical interactions.
Second, demonstrating some of the more interesting effects of communication

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
180 A. G. Horn & M. L. Leonard
networks entails another difficult distinction: between signalling to the parent
and signalling interactions with nestmates. Third, any realistic description of the
communication network must include interrelationships among nestmates that
do not involve signalling but nevertheless may shape the structure of the network.
These particular issues, of course, have less of an impact on communication in
some other kinds of network. Territorial birds singing from their song posts, for ex-
ample, are far beyond the range of physical interaction, are clearly directing their
signals at each other (but see Ch. 14), and are generally unrelated to one another.
Nonetheless, the issues we have raised are not unique to begging nestlings. Even
territorial birds, for example, can engage in close-range interactions that combine
signals with direct aggression, sing in ways that can be seen either as signalling
interactions or as attempts to overcome interfering signals, and have dominance
relationships that structure their communication network. If communication net-
works are indeed ‘the commonest social environment in which communication
occurs’(McGregor & Peake, 2000), then network concepts will inevitably be applied
to other systems that do present some of the complications we have discussed to
varying degrees. If we are to understand how these networks function, we need
to clarify these issues and begging should prove to be a particularly useful system
for doing so.
Consequences of the network for begging
We now turn from attempting to characterize the communication net-
work within the nest to exploring how it might affect communication, from both
signallers’ and receivers’ perspectives. Most discussions of communication net-
works have emphasized two consequences in particular (e.g. McGregor & Peake,
2000; McGregor et al., 2000; see also other chapters in this volume) and we begin
with these. First, from the signaller’s point of view, signals must be designed to
catch the receiver’s attention in the face of interference from other signals in the
network. Second, receivers, for their part, can more readily compare signallers in
a network because they are in transmission range of several signallers at once. A
third possible consequence has received less attention: communication networks
might reduce error in the information that signals convey. Specifically, as we
explain below, nestlings are particularly error prone in deciding when and how
intensely to beg. When nestlings partly base these decisions on the behaviour of
other nestlings, as they can when signalling within a network, these errors might
have less effect on their signals of need.
Design to catch receiver attention
McGregor and Peake (2000) suggested that the main effect of networks on
signal design arises through competition for receiver attention, as each signaller

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 181
attempts to circumvent the interference caused by competing signals in the net-
work. Perhaps no other communication system is more obviously a competition
for receiver attention than a brood of noisy nestlings. Given the interest in the
exaggeration of this signal and its role in nestmate competition, however, there
are surprisingly few studies that specifically address how begging signals are de-
signed to overcome interference from nestmates. Our understanding of begging
and nestling competition might be considerably enhanced by thinking of begging
nestlings as a communication network.
In particular, we suspect that many of the most striking characteristics of the
begging display may be designed for overcoming interference. If so, then the con-
spicuousness and complexity of the display, which seems unnecessarily extrava-
gant for such a short-range signal, may, in fact, be a proportionate response to
signal interference (Dawkins & Guilford, 1997; Horn & Leonard, 2002). Here we
briefly discuss how selection for overcoming interference might account for a few
of the more obvious features of begging (see also Horn & Leonard, 2002).
High output
The most straightforward way to overcome any background noise is to
increase the amplitude or duty cycle of one’s signal. There is ample evidence that
nestlings respond in this way to signalling by nestmates (Budden & Wright, 2001;
but see Cotton et al., 1996). For example, nestlings in some species beg more in-
tensely when placed near a begging nestmate (Leonard & Horn, 1998) and call at
higher rates when they can hear a nestmate calling (Leonard & Horn, 2001c).
Locatable signals
Surprisingly small apparent angular separation between stimuli can sig-
nificantly enhance a receiver’s ability to tell them apart (Ch. 20). Thus design
features that enhance the locatability of nestlings are likely to enhance how well
they stand out from competing signals and so focus parental attention on an in-
dividual nestling. The visual components of begging, brightly coloured gapes in
particular, seem designed to be readily locatable targets for parental attention.
These gapes have particularly bright outlines in species that nest in cavities, most
likely so that the location of each nestling’s gape is distinct despite the darkness
(Kilner & Davies, 1998; Heeb et al., 2003).
Begging calls, in contrast, do not seem as obviously suited for locating nestlings
because they are broadcast noisily throughout the nest. Also, there is little evidence
so far that their structures are individually distinct in ways that would make
them easy for parents to distinguish (Leonard & Horn, 2001c; but see Popp &
Ficken, 1991). Indeed, some theoretical models suggest that they should not be
individually distinct because that would risk rejection by the parent (Beecher,
1991; Johnstone, 1997).

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
182 A. G. Horn & M. L. Leonard
Fig. 9.1. Spectrograms of three nestling begging calls: tree swallow Tachycineta bicolor,
hairy woodpecker Picoides pubescens and white-browed scrubwren Sericornis frontalis.
Vertical bar is 10 kHz, horizontal bar is 500 milliseconds and filter bandwidth is
700 Hz.
Nonetheless, many calls do display features thought to enhance locatability,
including abrupt onsets and offsets, broad frequency ranges and use of frequencies
to which parents are most acutely tuned (Horn & Leonard, 2002; Fig. 9.1). Whether
these features really do enhance locatability within the confines of a nest has
not been tested directly. Comparative evidence, however, suggests that begging
calls do display some of these features, except when subject to counteracting
selective pressure from predators that use locatable calls to find and depredate
nests (Haskell, 2002; Horn & Leonard, 2002).
Multiple components
Which features of signals stand out from the noise of competing signals
will depend on the situation, and the multiple components of the begging display
may allow nestlings to signal effectively in each of these different situations. For
example, a nestling competing with a nestmate in the front of a cavity nest might
gain more from gaping wider and posturing more intensely than a nestling stuck
in the back of the nest, because the nestling in the front is in plain sight of the
parent. In contrast, a nestling in the back of the box cannot be clearly seen by the
parent and would probably gain more from large increases in call rate than from
any changes in the visual signal (Leonard et al., 2003). Therefore, in addition to
the numerous other psychological advantages of multimodal signal components
(Rowe, 1999), they may provide nestlings with a toolbox of ways to make their
signal stand out despite changing conditions.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 183
Precedence
Precedence effects, the tendency of receivers to take more notice of signals
that occur first, may favour signallers that signal before their competitors do
(McGregor et al., 2000). Note that such effects, as shown in insect and frog choruses,
for example, may (Greenfield, 2002) or may not (Gerhardt & Huber, 2002) be the
result of certain psychological effects also known as precedence (for which Dent &
Dooling (2003a,b) provide an avian example). Begging may provide a particularly
good example of this effect on signalling. Parents in a wide range of species are
more likely to feed nestlings that beg before their nestmates (Budden & Wright,
2001) and nestlings appear to have been selected for hair-trigger responses to
the first sign of the parent’s arrival (Leonard & Horn, 2001d). The importance of
precedence effects may vary considerably among species, providing interesting
opportunities for comparative tests of their effects on signalling. For example,
they may be less important in species in which parents spend more time assessing
begging signals at each visit (e.g. Krebs, 2002) or in which hasty responses by
nestlings might waste energy or attract predators (Leonard & Horn, 2001d).
Signal suppression
All the aspects of signal design we have outlined so far can overcome sig-
nal competition by enhancing the signaller’s own signal. Signallers might also,
however, overcome competition by suppressing the signals of competitors. For ex-
ample, nestling whydahs Vidua spp. spread their wings to block their parents’view
of nestmate signals (B. Mines, personal communication) and dominant nestlings
of many non-passerine species aggressively punish subordinate nestmates that
beg in their presence (Drummond, 2002; Roulin, 2002). Subtler versions of such
direct approaches to signal competition may be widespread and should be looked
for in other species.
Comparison among signals
A second consequence of communication networks is that they allow re-
ceivers to compare information from several signallers at once. Social eavesdrop-
ping, extracting information from a signalling interaction (Ch. 2), is a particularly
interesting special case of such comparisons. However, receivers might also ben-
efit from the network simply by being able to compare signals simultaneously
rather than having to assess each signaller in succession (Chs. 7 and 14).
Surprisingly, how or even whether parents compare begging signals to decide
which nestling to feed is still poorly understood. Many studies, using various mea-
sures of begging intensity, have shown that more intensely begging nestlings are
more likely to be chosen, but such evidence is only correlational. Only a few re-
cent studies have experimented on parental choice in sufficient detail to separate

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
184 A. G. Horn & M. L. Leonard
the roles of non-signalling and signalling components of begging, or to demon-
strate preferences based on individual components of the begging display (Horn &
Leonard, 2002; Kilner, 2002a,b). Demonstrating whether parents use information
from signalling interactions among nestlings will require still more refined ex-
periments (see above).
Interestingly, recent models suggest that parents must assess interactions
among nestlings if begging is to evolve as a signal at all (Rodrıguez-Girones et al.,
2001; Royle et al., 2002). Specifically, if parents simply select the most obvious
signal, then the information content of begging becomes irrelevant and begging
consists merely of a scramble for the parents’ attention. If, however, parents can
calibrate the information in the signals to correct for competitive differences
among nestlings, whether those are expressed via signalling (e.g. Roulin, 2002)
or physical competition, then begging can indeed convey information on need
(Rodrıguez-Girones et al., 2001; Royle et al., 2002). Under this scenario, a network
environment may have been of central importance in the evolution of begging.
Error reduction
The last possible consequence of communication networks that we will
discuss has received little attention, although it seems simple in principle and
broad in implications. Specifically, because information in a network is transferred
via not just one but several signals, the impact of error from any given signal might
be reduced. To explain this possibility, we first outline some possible sources of
error in begging displays and then discuss how the communication network may
reduce this error.
Begging by individual nestlings may be considerably error prone for at least
two reasons (Clark, 2002; Horn & Leonard, 2002). First, nestlings may be poor at
assessing their own needs, especially since doing so requires integrating their cur-
rent condition with their future requirements and their likely returns from beg-
ging, both of which are partly under control of their parents, their nestmates and
the vagaries of the environment (Clark, 2002). Second, nestlings are often poor
at distinguishing the parent’s arrival at the nest from other sights and sounds
and, therefore, often beg in response to irrelevant stimuli. In older tree swallow
Tachycineta bicolor nestlings, for example, while nestlings often simply start beg-
ging after their nestmates do, many of the initial begging responses are to events
other than the parent, like the wind blowing through the trees or the bump of
another bird species landing by the nest (Leonard & Horn, 2001d; Horn & Leonard,
2002). Conversely, nestlings apparently hold back on begging when they are un-
sure whether the parent actually has arrived and so may miss the parent’s arrival
or may send an inappropriately weak signal (Clark, 2002). From the nestling’spoint
of view, these are errors in how they deliver the begging signal. From the parent’s

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 185
point of view, however, such errors corrupt any information that the parent might
obtain from the begging signal.
Begging in a network may buffer such errors, because each nestling bases its
decision of when and how intensely to beg partly on the begging of its nestmates
(e.g. Leonard & Horn, 1998, 2001c). This influence of nestmates should reduce the
impact of the errors that each nestling would make if it were begging on its own;
from the parent’spoint of view, it would provide a more reliable signal of offspring
need (Clark, 2002).
This argument could, of course, be reversed. Specifically, one might argue that
the more links in the information chain from nestlings to parents, the less accurate
and reliable information will be (Royle et al., 2002). Determining whether networks
reduce or increase error requires modelling of information flow through the net-
work. A nestling’s decision of when to beg, to take the first step in the chain, may
be seen as a game of signal detection, in which the nestling can either try to be the
first to detect the parent’s arrival, at the risk of more false alarms (as shown above
for tree swallows), or can free-ride by eavesdropping on the responses of nestmates,
at the risk of begging later than its nestmates (Erev et al., 1995). Notwithstanding
the promise of such models, probably the most pressing need for understand-
ing information flow through networks, indeed for all the possible consequences
of the begging network surveyed above, is for more empirical research on how
parents assess begging signals.
Summary and future directions
In this chapter we have tried to show that begging by nestling birds is a
promising system for clarifying fundamental aspects of communication networks,
particularly the grey but conceptually fruitful areas between physical acts and
signals, between signalling competitively and interacting, and between commu-
nication and other functional relationships among signallers. Theoretical work
on the evolution of begging has already started exploring each of these areas,
but it has been inspired more by field workers’ insistence that begging behaviour
is complex than by any attempt to treat begging as a communication network.
In the future, theoretical work would likely benefit from a more explicit appli-
cation of network concepts, much as studies of economics and cooperation in
humans have benefited from models of social networks (e.g. Slikker & van den
Nouweland, 2001). Conversely, those studying other communication networks
will likely benefit from staying abreast of theoretical developments in the study of
begging.
Perhaps the greatest opportunities for future work, however, are in empirical
studies that focus on signalling and nestmate interactions in more detail. Despite

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
186 A. G. Horn & M. L. Leonard
enormous variation in the form of begging calls within and across species, for
example (Popp & Ficken, 1991), only a handful of studies have addressed the func-
tion of this variation in any detail (Horn & Leonard, 2002; see also Kilner (2002b) for
the display as a whole). Similarly, despite a long history of interest in intrabrood
competition in birds (Mock & Parker, 1997), few studies have tried to identify the
specific functions of the various behaviours that nestlings use in competition,
especially what information they might convey to both parents and nestmates
(Clark, 2002; Roulin, 2002). Perhaps most importantly, how, or even whether, par-
ents choose which nestling to feed remains largely unknown because the requisite
experiments have not been done (Royle et al., 2002). Hopefully, greater appreciation
that nestlings communicate within a network of signallers, with all its attendant
challenges and opportunities, will inspire more research on all of these funda-
mental questions.
Acknowledgements
We thank Pete McGregor for the chance to contribute to this volume and for his pa-
tience and constructive advice during the preparation of this chapter. We also thank John Bower
and an anonymous reviewer for their helpful comments on an earlier draft. Conversations with
participants at the Gregynog 2000 Begging Workshop and with authors of the resultant book have
been invaluable in developing our ideas about begging, as has our collaboration with Rob Magrath
and his students at the Australian National University and with the many students and assistants
who have worked on tree swallows with us. We also thank the Coldwell, Hines and Minor families
for allowing us to work on their land, and NSERC for financial support.
References
Beecher, M. D. 1991. Successes and failures of kin recognition in animals. In: Kin
Recognition, ed. P. G. Hepper. Cambridge, UK: Cambridge University Press,
pp. 94–124.
Bergstrom, C. T. & Lachmann, M. 1998. Signaling among relatives. III. Talk is cheap.
Proceedings of the National Academy of Sciences, USA, 95, 5100–5105.
Briskie, J. V., Naugler, C. T. & Leech, S. M. 1994. Begging intensity of nestling birds
varies with sibling relatedness. Proceedings of the Royal Society of London, Series B, 258,
73–78.
Budden, A. E. & Wright, J. 2001. Begging in nestling birds. Current Ornithology, 16,
83–118.
Clark, A. B. 2002. Appetite and the subjectivity of hunger. In: The Evolution of Begging:
Competition, Cooperation, and Communication, ed. J. Wright & M. L. Leonard.
Dordrecht: Kluwer, pp. 173–198.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 187
Clotfelter, E. D., Shubert, K. A., Nolana, V. Jr & Ketterson, E. D. 2003. Mouth color
signals thermal state of nestling dark-eyed juncos ( Junco hyemalis). Ethology, 109,
171–182.
Cotton, P. A., Kacelnik, A. & Wright, J. 1996. Chick begging as a signal: are nestlings
honest? Behavioral Ecology, 7, 178–182.
Cotton, P. A., Wright, J. & Kacelnik, A. 1999. Chick begging strategies in relation to
brood hierarchies and hatching asynchrony. American Naturalist, 153, 412–420.
Dawkins, M. S. & Guilford, T. 1997. Conspicuousness and diversity in animal signals.
Perspectives in Ethology, 12, 55–72.
Dearborn, D. C. & Lichtenstein, G. 2002. Begging behaviour and host exploitation in
parasitic cowbirds. In: The Evolution of Begging: Competition, Cooperation, and
Communication, ed. J. Wright & M. L. Leonard. Dordrecht: Kluwer, pp. 361–
387.
Dent, M. K. & Dooling, R. J. 2003a. Investigations of the precedence effect in
budgerigars: effects of stimulus type, intensity, duration, and location. Journal of
the Acoustical Society of America, 113, 2146–2158.
2003b. Investigations of the precedence effect in budgerigars: the perceived location
of auditory images. Journal of the Acoustical Society of America, 113, 2159–2169.
Drummond, H. 2002. Begging versus aggression in avian broodmate competition. In:
The Evolution of Begging: Competition, Cooperation, and Communication, ed. J. Wright &
M. L. Leonard. Dordrecht: Kluwer, pp. 337–360.
Erev, I., Gopher, D., Itkin, R. & Greenshpan, Y. 1995. Toward a generalization of signal
detection theory to n-person games: the example of two person safety problem.
Journal of Mathematical Psychology, 39, 360–375.
Evans, R. M. 1994. Cold-induced calling and shivering in young American white
pelicans: honest signalling of offspring need for warmth in a functionally
integrated thermoregulatory system. Behaviour, 129, 14–34.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans. Chicago,
IL: University of Chicago Press.
Glassey, B. & Forbes, S. 2002a. Muting individual nestlings reduces parental feeding
for the brood. Animal Behaviour, 63, 779–786.
2002b. Begging and asymmetric nestling competition. In: The Evolution of Begging:
Competition, Cooperation, and Communication, ed. J. Wright & M. L. Leonard.
Dordrecht: Kluwer, pp. 269–282.
Godfray, H. C. J. 1991. The signalling of need by offspring to their parents. Nature, 353,
328–330.
1995. Evolutionary theory of parent–offspring conflict. Nature, 376, 133–138.
Godfray, H. C. J. & Johnstone, R. A. 2000. Begging and bleating: the evolution of
parent–offspring conflict and sibling rivalry. Philosophical Transactions of the Royal
Society of London, Series B, 355, 1581–1591.
Gotmark, F. & Ahlstrom, M. 1997. Parental preferences for red mouth of chicks in a
songbird. Proceedings of the Royal Society of London, Series B, 264, 959–962.
Greenfield, M. D. 2002. Signalers and Receivers: Mechanisms and Evolution of Arthropod
Communication. Oxford: Oxford University Press.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
188 A. G. Horn & M. L. Leonard
Haskell, D. G. 2002. Begging behaviour and nest predation. In: The Evolution of Begging:
Competition, Cooperation, and Communication, ed. J. Wright & M. L. Leonard.
Dordrecht: Kluwer, pp. 163–172.
Heeb, P., Schwander, T. & Faoro, S. 2003. Nestling detectability affects parental feeding
preferences in a cavity-nesting bird. Animal Behaviour, 66, 637–642.
Hill, R. W. & Beaver, D. L. 1982. Inertial thermostability and thermoregulation in
broods of redwing blackbirds. Physiological Zoology, 55, 250–260.
Horn, A. G. & Leonard, M. L. 2002. Efficacy and the design of begging signals. In: The
Evolution of Begging: Competition, Cooperation, and Communication, ed. J. Wright &
M. L. Leonard. Dordrecht: Kluwer, pp. 127–142.
Johnstone, R. A. 1997. Recognition and the evolution of distinctive signatures: when
does it pay to reveal identity? Proceedings of the Royal Society of London, Series B, 264,
1547–1553.
1999. Signaling of need, sibling competition, and the cost of honesty. Proceedings of
the National Academy of Sciences, USA, 96, 12644–12649.
Johnstone, R. A. & Godfray, H. C. J. 2002. Models of begging as a signal of need. In: The
Evolution of Begging: Competition, Cooperation, and Communication, ed. J. Wright &
M. L. Leonard. Dordrecht: Kluwer, pp. 1–20.
Kilner, R. M. 1997. Mouth colour is a reliable signal of need in begging canary
nestlings. Proceedings of the Royal Society of London, Series B, 264, 963–968.
2002a. The evolution of complex begging displays. In: The Evolution of Begging:
Competition, Cooperation, and Communication, ed. J. Wright & M. L. Leonard.
Dordrecht: Kluwer, pp. 87–106.
2002b. Sex differences in canary (Serinus canaria) provisioning rules. Behavioral
Ecology and Sociobiology, 52, 400–407.
Kilner, R. M. & Davies, N. B. 1998. Nestling mouth colour: ecological correlates of a
begging signal. Animal Behaviour, 56, 705–712.
Komdeur, J. & Hatchwell, B. J. 1999. Kin recognition: function and mechanism in avian
societies. Trends in Ecology and Evolution, 14, 237–241.
Krebs, E. A. 2002. Sibling competition and parental control: patterns of begging in
parrots. In: The Evolution of Begging: Competition, Cooperation, and Communication, ed.
J. Wright & M. L. Leonard. Dordrecht: Kluwer, pp. 319–336.
Leonard, M. L. & Horn, A. G. 1998. Need and nestmates affect begging in tree swallows.
Behavioral Ecology and Sociobiology, 42, 431–436.
2001a. Begging calls and parental feeding decisions in tree swallows (Tachycineta
bicolor). Behavioral Ecology and Sociobiology, 49, 170–175.
2001b. Acoustic signalling of hunger and thermal state by nestling tree swallows.
Animal Behaviour, 61, 87–93.
2001c. Dynamics of calling by tree swallow (Tachycineta bicolor) nestmates. Behavioral
Ecology and Sociobiology, 49, 170–175.
2001d. Begging in the absence of parents by nestling tree swallows. Behavioral
Ecology, 12, 501–505.
Leonard, M. L., Horn, A. G. & Parks, E. 2003. The role of calling and posturing in the
begging behavior of nestling birds. Behavioral Ecology and Sociobiology, 54, 188–
193.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
Nestling begging as a communication network 189
Lotem, A., Wagner, R. H. & Balshine-Earn, S. 1999. The overlooked signaling
component of nonsignaling behavior. Behavioral Ecology, 10, 209–212.
Maynard Smith, J. 1994. Must reliable signals always be costly? Animal Behaviour, 42,
1034–1035.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McGregor, P. K., Otter, K. & Peake, T. M. 2000. Communication networks: receiver and
signaller perspectives. In: Signalling and Signal Design in Animal Communication, ed.
Y. Espmark, T. Amundsen & G. Rosenqvist. Trondheim: Tapir Academic Press,
pp. 329–340.
Mock, D. W. & Forbes, L. S. 1995. The evolution of parental optimism. Trends in Ecology
and Evolution, 10, 130–134.
Mock, D. W. & Parker, G. A. 1997. The Evolution of Sibling Rivalry. Oxford: Oxford
University Press.
Ovadia, O., Pinshow, B. & Lotem, A. 2002. Thermal imaging of house sparrow nestlings:
the effect of begging behavior and nestling rank. The Condor, 104, 837–842.
Parker, G. A., Royle, N. J. & Hartley, I. R. 2002. Intrafamilial conflict and parental
investment: a synthesis. Philosophical Transactions of the Royal Society of London, Series
B, 357, 295–307.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2002. Do great tits assess
rivals by combining direct experience with information gathered by
eavesdropping? Proceedings of the Royal Society of London, Series B, 269, 1925–1929.
Popp, J. & Ficken, M. S. 1991. Comparative analysis of acoustic structure of passerine
and woodpecker nestling calls. Bioacoustics, 3, 255–274.
Price, K., Harvey, H. & Ydenberg, R. 1996. Begging tactics of nestling yellow-headed
blackbirds, Xanthocephalus xanthocephalus, in relation to need. Animal Behaviour, 51,
421–435.
Price, K., Ydenberg, R. & Daust, D. 2002. State-dependent begging with asymmetries
and costs: a genetic algorithm approach. In: The Evolution of Begging: Competition,
Cooperation, and Communication, ed. J. Wright & M. L. Leonard. Dordrecht: Kluwer,
pp. 21–42.
Redondo, T. & Zuniga, J. M. 2002. Dishonest begging and host manipulation by
Clamator cuckoos. In: The Evolution of Begging: Competition, Cooperation, and
Communication, ed. J. Wright & M. L. Leonard. Dordrecht: Kluwer, pp. 389–412.
Rodrıguez-Girones, M. A. 1996. Siblicide: the evolutionary blackmail. American
Naturalist, 148, 101–122.
Rodrıguez-Girones, M. A., Enquist, M. & Lachmann, M. 2001. Role of begging and
sibling competition in foraging strategies of nestlings. Animal Behaviour, 61,
733–745.
Roulin, A. 2002. The sibling negotiation hypothesis. In: The Evolution of Begging:
Competition, Cooperation, and Communication, ed. J. Wright & M. L. Leonard.
Dordrecht: Kluwer, pp. 107–126.

P1: JZZ/... P2: JZZ/...0521823617c09.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 12:20
190 A. G. Horn & M. L. Leonard
Rowe, C. 1999. Receiver psychology and the evolution of multicomponent signals.
Animal Behaviour, 58, 921–931.
Royle, N. J., Hartley, I. R. & Parker, G. A. 2002. Begging for control: when are offspring
solicitation behaviours honest? Trends in Ecology and Evolution, 17, 434–440.
Saino, N. & Møller, A. P. 2002. Immunity and begging. In: The Evolution of Begging:
Competition, Cooperation, and Communication, ed. J. Wright & M. L. Leonard.
Dordrecht: Kluwer, pp. 245–267.
Saino, N., Ninni, P., Calza, S. et al. 2000. Better red than dead: carotenoid based mouth
coloration reveals infection in barn swallow nestlings. Proceedings of the Royal
Society of London, Series B, 267, 57–61.
Slikker, M. & van den Nouweland, A. 2001. Social and Economic Networks in Cooperative
Game Theory. Dordrecht: Kluwer.
Trivers, R. L. 1974. Parent–offspring conflict. American Zoologist, 14, 249–264.
Whittingham, L. A. & Dunn, P. 2001. Male parental care and paternity. Current
Ornithology, 16, 257–298.
Wilson, D. S. & Clark, A. B. 2002. Begging and cooperation: an exploratory flight. In:
The Evolution of Begging: Competition, Cooperation, and Communication, ed. J. Wright &
M. L. Leonard. Dordrecht: Kluwer, pp. 107–126.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
10
Redirection of aggression: multipartysignalling within a network?
a na h i t a j . n . k a z e m 1 & f i l i p p o au r e l i 2
1University of Wales, Bangor, UK2Liverpool John Moores University, UK
Introduction
In many species, an individual that finds itself in a losing position may
interrupt a contest to harass a bystander (i.e. an apparently uninvolved third party)
aggressively or may do so immediately after hostilities with the original oppo-
nent have ceased. Such ‘redirection’ of aggression (Bastock et al., 1953) is often
interspecific; for example, rollers and chasseur-type kingfishers (Coraciiformes)
are reported to dash away frequently during disputes to attack small passerines,
doves and plovers (Moynihan, 1998). The scapegoats are typically not ecological
competitors but do tend to be smaller and inoffensive individuals, both literally
and figuratively, and thus relatively safe targets. In socially living taxa, however,
redirection is most commonly directed towards a lower-ranking group member
(where available) and, therefore, is usually intraspecific. Both aspects of redirec-
tion are conveyed by the description of tensions between spotted hyaenas Crocuta
crocuta at a kill, producing a cascade of aggression, in which ‘A chases B, B chases
C, C chases D, and D chases vultures’ (Zabel et al., 1992, p. 129).
Redirection of aggression has traditionally been explained as a means of reduc-
ing the physiological arousal associated with participation in a conflict. The neuro-
endocrine responses underlying the preparation for ‘fight or flight’, whilst
essential in the immediate context, can be detrimental if they remain activated
over prolonged periods. Chronically elevated secretion of glucocorticoids, for
example, is associated with a range of cardiovascular pathology, depressed im-
mune function and compromised digestion, growth and reproduction (reviewed
by Sapolsky, 1998). Any action that prompts the endocrine stress response to
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
191

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
192 A. J. N. Kazem & F. Aureli
attenuate to baseline levels more rapidly can, therefore, reduce the physiological
costs of being victimized. When an animal is unable to retaliate directly against
an aggressor (for example because it is a higher-ranking or otherwise superior
competitor), redirecting offers an outlet for ‘frustration’ and allows the actor to
perceive a measure of ‘control’over the social situation. In experimental studies on
rodents and primates, both psychological effects modulate the elevation in heart
rate and glucocorticoid levels that repeated exposure to an unavoidable physical
stressor usually produces (reviewed by Levine et al., 1989; Sapolsky, 1998). Further-
more, rats Rattus norvegicus given the opportunity to attack a conspecific when
subjected to a mild electric shock subsequently developed fewer gastric lesions
than yoked controls that were not provided with this outlet (Weiss et al., 1976).
Amongst wild olive baboons Papio anubis, males that readily initiate aggression in
appropriate contexts and that frequently respond to losing fights by redirecting
aggression against others have significantly lower basal glucocorticoid levels (and
a better response to acute challenge) than similarly ranked individuals that do
not exhibit these behaviour patterns as frequently (Sapolsky & Ray, 1989; Virgin &
Sapolsky, 1997). However, the extent to which this can be attributed to beneficial
consequences of redirection per se is not clear, as it was only one of a suite of
traits concerning temperamental style in handling male–male competition that
characterized these individuals. Nevertheless, recent experimental work has con-
firmed that redirection does have an inhibitory effect on neuroendocrine stress
responses in rainbow trout Oncorhynchus mykiss (Øverli et al., 2004).
Here we summarize evidence suggesting there may, in some species, be more
to redirection than this. The behaviour is particularly prevalent and has been
best demonstrated in primates, notably several members of the genus Macaca
(Table 10.1). In these species, harassing a conspecific would seem a rather costly
way of gaining stress relief. Aggression carries the risk of injury, attracting preda-
tors and, despite a careful choice of target, may still provoke retaliation from its
allies. Why harass bystanders rather than quietly chew on wood (which has bene-
ficial effects in rats (Weiss et al., 1976))? Naturally, redirection may confer benefits
in terms of resource acquisition or reinforcing one’s status over the target; yet
this would not explain why individuals are particularly likely to instigate such
aggression immediately after losing a conflict: a time when their energy reserves
may be depleted and the former opponent likely to join in coalition against them.
Instead, the answer may lie in how redirection influences the behaviour of by-
standers, rather than the target. A number of additional hypotheses have been put
forward to explain redirection behaviour in macaques: for example that it diverts
the aggressor’s attention (Itani, 1963), provides an opportunity for the two oppo-
nents to resolve their differences by joining forces against a common foe (de Waal &
Yoshihara, 1983) or encourages the former aggressor to participate in a conciliatory

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 193
Table 10.1. Non-human primate studies of intraspecific redirected aggression within
intact social groups
Species Holding conditions Redirection Evidencea Source
Barbary macaque, Captive Yes 1a Aureli et al., 1994
Macaca sylvanus
Japanese macaque, Captive Yes 1c Aureli et al., 1992b
Macaca fuscata Captive Yes 1a Aureli et al., 1993
Captive Yes 2 Eaton, 1984; Scucchi et al.,
1988
Longtailed
macaque,
Wild and captive Yes 1a Aureli & van Schaik, 1991a;
Aureli, 1992
Macaca fascicularis
Pigtail macaque, Captive Yes 1b Judge, 1982b
Macaca nemestrina
Rhesus macaque,
Macaca mulatta
Free ranging and
captive
Yes 1a de Waal & Yoshihara,
1983;c Kazem, 1999
Captive Yes 2 Gore, 1994
Stumptail macaque, Captive Yes 2 Walker Leonard, 1979
Macaca arctoides
Hamadryas baboon,
Papio hamadryas
Captive Yes 1a F. Zaragoza &
F. Colmenares,
unpublished data
Captive (females) No 2 Gore, 1994
Olive baboon, Wild (females) No 1a Castles & Whiten, 1998a
Papio anubis Wild (males) Yes 2 Sapolsky & Ray, 1989;
Virgin & Sapolsky, 1997
Sooty mangabey, Captive Yes 1a Gust & Gordon, 1993
Cercocebus
torquatus atys
Vervet monkey, Wild Yes 1b,c Cheney & Seyfarth, 1986,
Cercopithecus 1989b,c
aethiops
Spectacled langur, Captive No 1a Arnold & Barton, 2001
Trachypithecus
obscurus
Mountain gorilla,
Gorilla gorilla
Wild (males and
immatures)
Yes 1a Watts, 1995bd
beringei Wild (females) No 1a Watts, 1995b
(cont.)
P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
194 A. J. N. Kazem & F. Aureli
Table 10.1. (cont.)
Species Holding conditions Redirection Evidencea Source
Common Captive Yes 1a Fuentes et al., 2002e
chimpanzee
Pan troglodytes
White-faced Wild Yes 2 S. E. Perry,
capuchin, unpublished data
Cebus capucinus
Black lemur, Semi free ranging No 1a Roeder et al., 2002
Eulemur macaco
Brown lemur,
Eulemur fulvus
Captive No 1a Roeder et al., 2002
aThe nature of quantitative evidence varies: 1, comparison of post-conflict period with matched-
controls (1a), with baseline focal observations or pre-conflict period (1b) or with other data (1c); 2,
inference from quantitative but uncontrolled data. Type 2 data do not necessarily demonstrate
that a significant post-conflict increase in aggression against bystanders exists. For Japanese,
longtailed and rhesus macaques, additional anecdotal citations can be found in several other
studies.bAnalysis of kin-oriented redirection, not overall incidence against all targets.cSubject’s role in previous conflict not distinguished; therefore, analyses potentially include for-
mer aggressors.dResult for adult males was trend only, owing to sample size (n = 4).eConspecific and human targets were not distinguished in analyses.
reunion with the actor (Aureli & van Schaik, 1991a). Although not explicitly
couched in signalling terms, all rely on the principle that the former aggressor
perceives the act and responds in ways that indirectly reduce the likelihood of
further aggression against the redirecting individual. We develop this notion by
proposing that redirection functions as a signal aimed at both the former oppo-
nent and other bystanders, which conveys information about the perpetrator’s
competitive ability and current state and thus directly reduces challenges from
these receivers. Such pre-emptive strikes offer a novel interpretation of the maxim
that ‘offence is the best defence’.
The macaque system
The results we describe are primarily drawn from three studies examin-
ing post-conflict behaviour in both captive and wild longtailed macaques Macaca
fascicularis (Aureli & van Schaik, 1991a,b; Aureli, 1992), and in juveniles from two
social groups within a free-ranging colony of rhesus macaques Macaca mulatta

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 195
(Kazem, 1999, A. J. N. Kazem, unpublished data). Both species form permanent
multimale–multifemale groups and within the genus are regarded as possessing
a relatively ‘despotic’ dominance style (de Waal & Luttrell, 1989; Thierry, 2000).
Clear-cut dominance relationships are apparent in both sexes, and aggressive dis-
putes are frequent, often injurious and overwhelmingly directed down the social
hierarchy. Although individuals will defend relatives and close associates against
attack, the most common pattern of intervention is support for the aggressor, with
dominant group members receiving assistance from both kin and non-kin against
subordinates that attempt to contravene the established hierarchy (Chapais, 1995).
Unsurprisingly, direct retaliation by targets against aggressors is uncommon (e.g.
less than 9% of conflicts involve counter-aggression in rhesus macaques (de Waal &
Luttrell, 1989)).
Anecdotal reports of redirected aggression abound in the literature on non-
human primates, but the phenomenon has been explored and statistically con-
firmed in rather fewer species (Table 10.1). Demonstrating that a defeat influences
the likelihood of initiating aggression against bystanders requires a comparison
with the victim’s behaviour during an equivalent period not preceded by a con-
test. In naturalistic studies within intact social groups, this is typically achieved
by comparing the immediate post-conflict period with a ‘matched-control’ obser-
vation collected on the next possible day (PC–MC method (de Waal & Yoshihara,
1983)) or, less frequently, with periods selected from a distribution of baseline
focal observations on the same individual. Current best-practice protocols entail
matching the conditions at the start of these paired samples with respect to fac-
tors likely to influence rates of aggression or other social interactions between
relevant parties. These include the time of day, prevailing climatic conditions,
predominant activity of both the subject and the wider group (if different) and,
in analyses of interactions between former adversaries, inter-opponent distance
(e.g. Aureli, 1992; Kazem, 1999). Studies differ in whether the demonstration of
redirection emphasizes the occurrence of a single critical early post-conflict event
(as in the PC–MC method), or simply compares overall rates of aggression initiated
within a defined timeframe. Discussion of the relative merits of methods that can
be used to demonstrate a significant post-conflict increase and/or operationally
identify particular bouts of aggression as ‘redirection’ events, and the statistical
issues involved, can be found in Veenema (2000), Das et al. (1997) and Kazem (1999).
Characteristic post-conflict aggressive phenomena
Together our three studies provide a database of more than 2550 post-
conflict samples from 137 individuals, in which the subject had been the victim of
unidirectional aggression with a clear outcome (i.e. the initial recipient was always
the loser). Despite differences in species, holding conditions, group composition,

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
196 A. J. N. Kazem & F. Aureli
age of subjects and methods of analysis, the patterns documented were strikingly
similar. Subjects threatened or attacked third parties significantly earlier and
more frequently in the aftermath of a defeat than under control conditions. In
both species, the increase was most pronounced within the first two minutes,
quickly declining to baseline levels thereafter (Aureli & van Schaik, 1991a; Kazem,
1999). On average, rhesus victims harassed at least one bystander within the first
minute after 22% of contests, corresponding to a 10-fold increase over control lev-
els (an underestimate, as the methodology used excludes bouts initiated during
the original conflict and individuals may redirect multiple times). The scapegoats
were almost always lower ranking, but lack of a suitable conspecific did not neces-
sarily stop individuals. For example, a young female rhesus victim, finding herself
surrounded by members of the alpha matriline, proceeded to energetically and
noisily pursue several lizards and a rat before peering intently into and repeatedly
threatening a small bush (completely devoid of vertebrates), whilst continually at-
tempting to solicit assistance from her former foe situated some distance away (A.
J. N. Kazem, personal observation). Such scenes are not uncommon but were not
operationally classified as redirection.
By comparison, the post-conflict behaviour of former aggressors has received
little attention. In rhesus macaques, victors also exhibit a post-conflict elevation
in attacks against bystanders (and disproportionately target relatives of their for-
mer victim (Kazem, 1999)), while in longtailed macaques they do not (Das, 1998).
Nevertheless, rhesus subjects were significantly more likely to harass bystanders
after losing a conflict than after winning one, identifying redirection by former
victims as the more distinctive and pervasive phenomenon.
Another notable feature is that, having lost one contest, individuals are also
liable to receive further aggression. In the aftermath of conflicts where the victim
had neither reconciled with its opponent (i.e. engaged in a post-conflict affiliative
reunion (Aureli et al., 2002)) nor redirected against a third party, defeated individu-
als were subjected to significantly elevated rates of threats and attacks in all three
studies. The levels received were particularly high for the first three to four min-
utes (and at least 10 minutes in longtailed macaques), gradually waning toward
baseline incidence over time. Interestingly, the renewal of hostilities by the former
adversary was not the only cause; in many cases, the aggressor was a previously
uninvolved bystander. Such increased receipt of aggression is a common find-
ing, being the predominant sequence of events in triadic interactions between
Japanese macaques Macaca fuscata (‘mobbing’; Eaton, 1984) and documented in
controlled post-conflict studies of both macaques (e.g. de Waal & Yoshihara, 1983;
Cords, 1992; Kutsukake & Castles, 2001) and other cercopithecines (e.g. mountain
gorillas Gorilla gorilla beringei (Watts, 1995a) and olive baboons (Castles & Whiten,
1998a)). The effect is specific to former victims; neither the victor nor participants

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 197
in bidirectional contests suffer an enhanced risk (Castles & Whiten, 1998a; Das,
1998; Kazem, 1999). In the rhesus study, the number of aggressive incidents be-
tween other group members recorded within a 10 m radius of the focal animal did
not differ consistently between post-conflict and control samples (A. J. N. Kazem
unpublished data). This rules out the possibility that the increases observed in
both initiation and receipt of aggression by victims could have been caused by
the post-conflict samples being obtained during periods of generally heightened
group aggressivity, and it confirms that control samples effectively matched post-
conflict conditions even in the more variable free-ranging situation.
A consequence of loser effects and eavesdropping?
The most likely explanation for the temporal patterning of challenges
from bystanders is that victims undergo some form of ‘loser experience’(cf. Scott &
Fredericson, 1951), which renders them more easily beaten than under other
circumstances. Research in a broad range of vertebrates has documented a ten-
dency for animals who have suffered a defeat to lose in subsequent interactions
against randomly selected and otherwise equally matched individuals (reviewed by
Chase et al., 1994; Hsu & Wolf, 1999). In some cases, prior losers are even at a
disadvantage against considerably smaller opponents – ones that they would nor-
mally be expected to defeat easily in any other encounter. To our knowledge, the
standard protocols used to demonstrate this behavioural pattern (staging succes-
sive dyadic contests between unfamiliar individuals, with all factors other than
the competitors’ prior social experiences held constant) have not been applied
in a primate. However, in experiments in which novel triads (trios) of unfamil-
iar rhesus macaques were convened, the sequences of agonistic interactions
observed were consistent with the operation of a loser (and indeed a winner) effect
(Mendoza, 1993). Triad members had been matched for size, age, sex and activity
level, making it unlikely that the predominance of consecutive losses (or wins)
against different opponents was a result of pre-existing differences in intrinsic
attributes.
These patterns may arise from physiological changes precipitated by an indi-
vidual’sexperiences in a prior contest. During the initial minutes of an encounter,
both contestants typically exhibit rapid, and often qualitatively similar, changes
in central neurotransmitter activity (serotonergic, dopaminergic and noradren-
ergic) as well as increased secretion of adrenal axis hormones such as gluco-
corticoids and testosterone (e.g. van Erp & Miczek, 2000; Summers et al., 2003).
However, as a contest progresses and the outcome becomes perceived, the neu-
roendocrine profiles of winner and loser diverge. Notably, in many vertebrates,
central serotonin activity and peripheral glucocorticoid concentrations return to
baseline levels relatively rapidly in victors, while greater initial increases and more

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
198 A. J. N. Kazem & F. Aureli
prolonged elevation are characteristic of defeated individuals (Schuurman, 1980;
Hannes et al., 1984; Øverli et al., 1999; Summers et al., 2003). These differences
are compounded if the protagonists remain confined together in the longterm,
with subordinate individuals often exhibiting chronic elevation of these para-
meters (Blanchard et al., 1993; Gust et al., 1993; Winberg & Lepage, 1998). Winners
are also reported to undergo increases in levels of circulating androgens such as
testosterone, while those in losers appear temporarily suppressed (Bernstein et al.,
1974; Rose et al., 1975; Hannes et al., 1984; Booth et al., 1989). In keeping with this,
dominant animals generally possess higher basal testosterone levels than do sub-
ordinates (see Ch. 21), although neither the causal direction nor the cumulative
influence of successive contests can be distinguished in such data.
High levels of circulating testosterone are known to sharpen concentration
and enhance social attention and memory processes (e.g. Andrews, 1991; Cynx &
Nottebohm, 1992), and they are associated with both greater risk taking (Kavaliers
et al., 2001) and expression of offensive aggression (e.g. Delville et al., 1996; Higley
et al., 1996; Ch. 21). In contrast, serotonin generally exerts an inhibitory effect on
aggressive behaviour. Experimentally enhancing central serotonin levels reduces
an animal’s readiness to initiate aggressive acts (Olivier et al., 1995; Ferris et al.,
1997; Perreault et al., 2003), while primates with chronically low serotoninergic
functioning exhibit greater impulsivity, perseverance and use of severe unre-
strained aggression (i.e. engage in aggression without regard for its consequences
(Mehlman et al., 1994; Higley et al., 1996; Fairbanks et al., 2001)). It is, therefore,
possible that the physiological changes typical of winners and losers produce tran-
sient alterations in factors that affect actual fighting ability. Alternatively, recent
evidence in fish suggests that it may simply be an individual’s perception of its
own relative ability that is modified (Hsu & Wolf, 2001), hence affecting subse-
quent decisions to initiate, escalate or withdraw. Either way, bystanders can take
advantage of these changes to gain a temporary competitive edge over recent
losers, offering an opportunity to reverse or reinforce an existing dominance rela-
tionship at relatively low cost. However, the behavioural consequences of a single
prior loss are short lived (and species specific). They persist for only a matter of min-
utes or hours in many taxa, even after the prolonged and intense fights often char-
acteristic of experimentally staged encounters between unfamiliar competitors
(Chase et al., 1994; Hsu & Wolf, 1999). Timing may assume additional importance
in species where coalitions are common, because a challenger’scosts will be further
reduced if, by choosing this moment to attack, its actions are also likely to receive
support from the victim’sformer opponent (an event that may produce additional
dividends by strengthening the assailant’s bonds with the latter individual).
One means of being alerted to a possible loser effect is by attending to the out-
come of conflicts between other group members. In some cases, simple cues might

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 199
betray a recent defeat (e.g. the dorsal darkening and flank patterns that appear
in newly subordinate individuals in many fish), but in primates the only possibil-
ity would seem to be specific postural or behavioural changes only apparent after
particularly severe losses. Instead, experimental evidence from a growing number
of species suggests that third parties ‘eavesdrop’ (McGregor, 1993), extracting in-
formation on the relative fighting abilities of conspecifics from interactions they
have witnessed (Ch. 2). Eavesdroppers subsequently treat perceived winners and
losers differently, in ways consistent with having identified those individuals as
relatively strong or weak competitors, respectively (e.g. Hogue et al., 1996; Oliveira
et al., 1998; McGregor et al., 2001; Peake et al., 2001; Earley & Dugatkin, 2002). A mul-
titude of evidence suggests that cercopithecines are similarly aware of the nature
and outcome of contests in which they are not themselves involved. Macaques and
baboons commonly exhibit apparent knowledge of the dominance relations be-
tween their groupmates (e.g. Silk, 1999; Ch. 25). In these species, the relative rank
of third parties typically cannot be deduced directly from cues such as relative
body size (especially in females) and is most likely derived by scrutinizing ago-
nistic interactions between the individuals concerned. As an illustration, female
chacma baboons Papio ursinus pay greater attention when presented with manipu-
lated sequences of calls in which the affiliative grunt of a subordinate individual
is closely followed by a scream from a higher-ranking female – a situation incon-
sistent with the existing dominance relationship between those particular group
members – than they do to a control (and causally consistent) sequence (Cheney
et al., 1995). Eavesdropping may also have contributed, in part, to the pattern of
double-wins and double-losses observed in the rhesus experiment described above
(Mendoza, 1993), given that the interactions occurred within a triad setting (see
Chase et al. (2002) for similar logic).
Eavesdropping is often regarded as a means of gathering information on an un-
known competitor’s abilities without incurring the costs of directly confronting
the party concerned. For example, a bystander might integrate information on
how animal A fares against B with prior knowledge (gained via direct interaction)
of its own standing relative to A, to extrapolate how it too might fare against
the unknown B. Alternatively, even if informed only about the relative prowess
of two strangers, this may still provide a probabilistic indication of how the de-
feated individual might rate relative to oneself and thus be useful in guiding
behaviour. We suggest that another function of eavesdropping is simply to detect
that an individual (known or unknown) has suffered a loss and, therefore, may
be undergoing a loser experience. After all, in the cercopithecine systems being
described, individuals are generally well aware of who outranks whom within the
group; therefore, when contest outcome is in the expected direction, it may be
the information about temporary changes in an individual’s current state that

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
200 A. J. N. Kazem & F. Aureli
is more important in the decision to challenge. Note that while eavesdropping
can account for why known losers are more readily challenged than their victorious
counterparts (as in the experiments cited above), it does not in itself explain why an
individual should be challenged more frequently following a defeat than at other
times (as in our data). Acquiring information on the relative abilities of potential
competitors does not necessitate that it be put to use immediately; in contrast,
the transient nature of a putative loser effect would require immediate use of the
information.
Redirection influences the behaviour of bystanders
In both longtailed and rhesus macaques, the amount of aggression a vic-
tim received in the minutes following a redirection event was significantly lower
than that during comparable periods on occasions when it had lost but had not
redirected (Aureli & van Schaik, 1991b; Kazem, 1999; A. J. N. Kazem, unpublished
data). In other words, targeting third parties after a defeat appears to confer a
protective effect. It is always possible that some feature of the preceding conflict
influenced both the likelihood of redirection and of receiving subsequent aggres-
sion. Two results counsel against this view. First, in the rhesus study the likelihood
of receiving post-conflict harassment in the period before redirection took place
did not differ systematically between contests in which the victim did, or did not,
go on to redirect (the individual’s mean latency to redirect was used to define the
relevant timeframe in the latter). Second, rhesus youngsters redirected aggression
more frequently after low-level contests (threats or minor lunges) than after more
intense confrontations (involving prolonged pursuits or physical contact) – a point
to which we shall return. However, less-intense disputes were not in themselves
associated with receipt of low levels of subsequent harassment; victims incurred
virtually identical rates of aggression following mild or severe incidents. There-
fore, the reduction in harassment documented appears to be associated with the
act of redirecting itself.
It is not yet known whether the beneficial effect is achieved primarily via an
alteration in the behaviour of the former opponent or in that of opportunistic
bystanders. A change in the disposition of the former aggressor, at least, seems
likely because redirection apparently influences its behaviour in other respects.
Among longtailed macaques, redirecting is associated with an increased likelihood
that the former aggressor will later participate in an affiliative reunion with the
perpetrator (Aureli & van Schaik, 1991a), although a causal connection remains
to be demonstrated. Reconciliation between former adversaries is known to have
positive consequences in terms of restoring tolerance and reducing subsequent
aggression between the protagonists (e.g. Cords, 1992); although note that this
effect cannot have been indirectly responsible for the results we report above,

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 201
because post-conflict periods in which reconciliation had occurred were excluded
from analyses.
An honest indicator of post-conflict condition and motivation?
The evidence that redirection events may influence the disposition and
behaviour of individuals other than the target of aggression suggests there is a sig-
nalling advantage to be gained by behaving antagonistically in post-conflict con-
texts. In essence, the primary benefit of the behaviour might derive from it being
witnessed by third (or often, fourth) parties. It is often claimed that redirection
serves its purpose by focusing attention upon an alternative target, thus cutting
short the original contest and/or persuading potential challengers to look else-
where (Itani, 1963; Gust & Gordon, 1993). Data supporting such an outcome are
rarely, if ever, presented. This tactic may be successful in spotted hyaenas, social
carnivores that exhibit a high degree of within-group coordination in activities
and are liable to join in against any animal that is currently losing – to the point
where supporters of an aggressor will often switch sides simply because their target
counter-attacked its original opponent (Zabel et al., 1992). It seems less plausible
in macaques, where there is little compelling reason why the former aggressor
(at least) should so readily divert to a different target. Nor would it account for
why aggressors seem more willing to reconcile with victims that have redirected
against others.
We propose that redirecting does more than merely draw attention to a
new stimulus; the act itself may provide bystanders with useful information. As
outlined above, the physiological and psychological consequences of a defeat gen-
erally reduce the likelihood that an individual will initiate aggression in the
ensuing minutes, and they may render it less likely to persist and be less effective
in combat when challenged. However, a recent victim that is nevertheless suffi-
ciently confident and capable of rapidly redirecting against a third party thereby
demonstrates (to the scapegoat, and more importantly to others) that it has not
been unduly compromised by the preceding experience. The act may serve as an
unfakeable marker of the perpetrator’spost-conflict state, indicating that it would
be ready and/or able to defend itself, thus dissuading renewed or opportunistic
challenges from bystanders. It might also make individuals that redirect appear
more formidable rivals within the group, both to the former aggressors and their
kin (see below; Aureli & van Schaik, 1991a), which could explain why former aggres-
sors become more willing to reconcile. This hypothesis assumes that the neuroen-
docrine response to a perceived loss is not an all-or-nothing affair: that features of
the prior contest (primarily its intensity and duration, and perhaps the opponent’s
identity) can modulate the magnitude and/or type of changes undergone and the
time course of recovery. The fact that rhesus macaques were much more likely to

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
202 A. J. N. Kazem & F. Aureli
redirect following receipt of low-intensity as opposed to high-intensity harassment
(see above) is consistent with this argument; comparable results are also available
in rainbow trout (Øverli et al., 2004). Furthermore, in many taxa, individuals differ
consistently not only in their baseline levels of physiological parameters but also
in the magnitude and nature of neuroendocrine response (‘reactivity’) produced
by stressors such as received aggression (reviewed by Koolhaas et al., 1999). Conse-
quently, the degree to which a victim has been compromised following any specific
defeat is not straightforward for bystanders to surmise. Variation in post-conflict
state provides both the impetus for conveying this information to potential chal-
lengers and the means (motivation or ability) to do so.
Ensuring others notice the event
Redirection events possess several features likely to draw these acts to
the attention of bystanders. For example, reports often emphasize that victims
redirect ‘in front of ’ their former assailant (e.g. de Waal & Yoshihara, 1983; Aureli
et al., 1992; but see Watts, 1995b). Bouts of redirection by young rhesus macaques
in a free-ranging situation were also more likely to take place within 5 m of (and
hence within view and earshot of ) their former opponents than were equivalent
bouts instigated under control conditions, even when inter-opponent distance at
the start of paired observation periods was statistically controlled (Kazem, 1999).
As in other studies, it was not uncommon to observe subjects glancing back at
their previous adversary both prior to and while threatening the target, imply-
ing that the aggressor’s presence (and perhaps even line of gaze: Emery et al.,
1997; Tomasello et al., 1998) was actively taken into account. Experiments have
demonstrated that vervet monkeys Cercopithecus aethiops and macaques often take
the presence and composition of bystanders into account before behaving an-
tagonistically toward others (Keddy Hector et al., 1989; Cheney & Seyfarth, 1990).
Equally, a rule of thumb simply prompting victims to act quickly could ensure
that their former adversary was likely to have remained nearby (the mean latency
to redirect was 12 and 28 seconds in longtailed and rhesus macaques, respec-
tively, excluding those bouts which occurred within the original conflict (Aureli
& van Schaik, 1991b; Kazem, 1999)). In some cases, victims even approached and
attempted to enlist their former opponent’s support in the venture with con-
spicuous head-flagging (a recruitment gesture; see also Cords (1988) for simi-
lar behaviour in young longtailed macaques). It has been speculated that such
solicitation may be aimed at using partnership in a coalition to achieve some
form of ‘reconciliation’ with the former adversary (de Waal & Yoshihara, 1983;
Aureli & van Schaik, 1991b), as well as more directly reducing the likelihood
of renewed aggression from that quarter. Again, aggression against the target

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 203
appears an almost incidental by-product of communication with one’s former
opponent.
Redirection is also a particularly noisy affair. These interactions disproportion-
ately often incorporate vocal threats in comparison with equivalent bouts initiated
during control observations (Kazem, 1999). Increased incidence of vocal forms of
aggression might simply reflect greater arousal in animals that have themselves
recently been subjected to attacks, but whatever the proximate mechanism the
resulting events will be more effective at alerting bystanders. This is reminiscent
of Zahavi’s (1979) observation that many aggressive signals are far louder than
actually required for effective information transfer between the two parties di-
rectly concerned. He interpreted the probable interception of these signals by
several more distant receivers as imposing greater costs upon the signaller, there-
fore ensuring the reliability of the degree of threat conveyed to the opponent. We
suggest that one ‘shouts’ precisely in order to advertise the signal to those more
distant receivers, because one gains a benefit by doing so. Furthermore, evidence
for individual discrimination by voice exists in macaques, although the extent of
individual signatures varies according to call type (rhesus: Gouzoules et al., 1986;
Rendall et al., 1996, 1998). Perception of identity in threat calls does not appear to
have been tested but, if present, would allow the signaller’sactions to be identified
even if the redirection event was not observed.
Kin-oriented redirection: a special case?
An intriguing variant is that victims appear specifically to target ma-
ternal relatives of their former assailant. Examples have been reported in some
cercopithecines: vervet monkeys (Cheney & Seyfarth (1989), although they did not
specify whether the actor was victim or aggressor in the original conflict), juvenile
longtailed macaques (Aureli & van Schaik, 1991a), Japanese macaques (Aureli et al.,
1992) and pigtail macaques Macaca nemestrina ( Judge, 1982). As maternal relatives
share the (typically higher) familial rank of the original aggressor, the strategy
is not without its risks. Japanese macaques circumvent this issue by selecting
younger – and therefore often lower-ranking and more vulnerable – relatives of
their former opponent and take advantage of ‘safe’ opportunities to join ongoing
coalitions against the target, making it difficult for the target (or the former ag-
gressor) to retaliate (Aureli et al., 1992). Unsurprisingly, kin-oriented threats and
attacks typically account for only a small fraction of redirection events and may
take place over a longer timescale (minutes or even hours) because of the need
to encounter appropriate conditions. In Japanese macaques the majority (74%) of
these incidents occurred within view of the former aggressor, leading Aureli et al.
(1992) to propose that inflicting indirect fitness costs might serve as a form of so-
cial leverage, to deter further aggression from the same individual over the longer

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
204 A. J. N. Kazem & F. Aureli
term (assuming the latter associates the two events). The suggestion, therefore,
relies on a signalling argument, in this case restricted to a particular third-party
receiver. It has recently been demonstrated that apparently ‘spiteful’ acts (such as
these) can be evolutionarily stable in systems where observers accord each other
status on the basis of aggression witnessed, at least under certain simplified social
conditions ( Johnstone & Bshary, 2004).
Kin-oriented redirection need not be restricted to primates. Appropriate con-
ditions are provided in any system where individuals are often constrained from
retaliating directly against aggressors, reside in groups composed of a mixture of
related and unrelated conspecifics, and close kin are preferred associates or coali-
tion partners. Macaques and baboons can discriminate kin relationships between
third parties (e.g. Dasser, 1988; Cheney & Seyfarth, 1999), possibly by observing the
association patterns of other group members, and are often assumed to act on
this basis. However, use of simpler proximity-based rules may suffice. If relatives
of protagonists tend to cluster at the scene of conflicts (as is often the case in taxa
where individuals preferentially support their kin in coalitions), they will be over-
represented among the bystanders present. Therefore, a tendency to strike at any
vulnerable individual nearby (other than one’sown close associates) could have the
effect of disproportionately targeting the opponent’s kin under post-conflict con-
ditions. Spotted hyaenas and greylag geese Anser anser, both of which are reported
to redirect aggression (Table 10.2), might be promising candidates. Spotted hyae-
nas form large multimale–multifemale clans with high variance in within-group
relatedness and a matrilineal structure similar to macaques (Frank, 1986; Mills,
1990). Many geese form cohesive family units (pairs and their immature offspring)
that aggregate in large feeding flocks during winter. Both are highly competitive
societies with pronounced dominance relationships and a high propensity to in-
tervene aggressively on behalf of relatives in disputes; consequently, rank is highly
dependent on social support (hyaenas: Zabel et al., 1992; Engh et al., 2000; geese:
Lamprecht, 1986; Black & Owen, 1987; K. Kotrschal, personal communication). Kin
are valuable allies and often in spatial proximity, affording competitors the op-
portunity to learn (or otherwise locate) the habitual associates of others, as well
as ‘safe’ opportunities to redirect in coalition.
Intraspecific redirection in other taxa
A number of predictions can be made regarding systems in which redi-
rection might operate as a signal. First and foremost, conditions should facilitate
eavesdropping: the communication modality, social structure and typical habitat
should be such that agonistic signals often transmit further than the average spac-
ing between conspecifics and can, therefore, be detected by several receivers. The

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 205
extent of the communication network will often constrain whether bystanders are
likely to be aware of a contestant’s defeat, providing both the impetus to redirect
and the means by which its occurrence is detected.
Where individuals reside in permanent and cohesive social groups with de-
fined dominance relationships between group members (e.g. many primates, so-
cial carnivores, some ungulates), the majority of interactions witnessed are likely
to involve animals whose capabilities are already known to the individual and
aggression in the expected direction within a dyad (the exceptions being con-
tests involving immigrants and rank reversals occurring as individuals mature
or decline in ability). As we have argued for macaques, much of the utility of
eavesdropping may then lie in detecting transient loser effects (determining the
optimum timing of challenges), and redirection may primarily convey updated
information on a victim’spost-conflict state (as well as reaffirming the ‘status quo’
to both target and bystanders). If so, redirection should occur in conjunction with
a physiological/behavioural loser effect (at least in the wake of intense contests)
in systems where victims often receive several attacks from different individuals
in quick succession (coyotes Canis latrans may be an example; Table 10.2).
However, the redirection principle may be more broadly applicable. In many
taxa, individuals have knowledge of the abilities of a consistent subset of con-
specifics yet frequently encounter others whose status is not yet known. Examples
include species where kin units coalesce into larger but spatially structured aggre-
gations (e.g. winter feeding flocks in geese and some corvids), or individuals form
dominance relationships within a temporary display aggregation (e.g. male ducks).
Other possibilities include species where individuals defend nest sites within a
breeding colony or form closely spaced territories during the breeding season (as
in many birds and fish), thus forming relationships with their immediate neigh-
bours. Where eavesdropping is used mainly to estimate the fighting ability of un-
familiar competitors, redirecting could limit the negative impression conveyed
by a loss. Observers should be wary of challenging those losers that have neverthe-
less demonstrated the motivation and ability still to dominate some opponents
( just as eavesdroppers in some species respond more cautiously toward losers
that have exhibited persistent counter-aggression during their previous contest
(Earley & Dugatkin, 2002)). In this case, loser effects need not be present for redi-
rection to be worthwhile (although where they are, the incentive for redirecting
may be further enhanced).
There are also many systems where a signalling aspect to redirection seems
unlikely. For example, both eavesdropping and redirection would seem to be of less
use when individuals reside in large and continually shifting aggregations (either
year-round or during certain seasons), in which they possess little information on
conspecifics’ identities. In such cases, ‘badges of status’ may be used to mediate

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Tabl
e10
.2.
Repo
rts
ofin
tras
peci
fic
redi
rect
ion
ofag
gres
sion
inot
her
spec
ies
Spec
ies
Con
text
inw
hic
hSy
stem
aC
oun
ter-
Lose
rV
icti
ms
Mai
nso
urc
ee
red
irec
tion
obse
rved
aggr
essi
onb
effe
ctc
chal
len
ged
d
Rai
nbo
wtr
out,
Onc
orhy
nchu
s
myk
iss
Juve
nil
etr
iad
s(u
nse
xed
),m
eeti
ng
insu
cces
sive
pai
rwis
e
exp
erim
enta
lco
nte
sts
(cap
tive
)
BV
aria
ble
1Ye
si–
Øve
rli
etal
.,20
04f
Atl
anti
csa
lmon
,Sal
mo
sala
rPa
rrh
eld
inm
ixed
-sex
quin
tets
and
sext
ets
infl
um
e(s
emin
atu
ral
stre
am)j
BYe
sH
.C.S
ute
r,p
erso
nal
com
mu
nic
atio
ng
Blu
egil
lsu
nfi
sh,L
epom
is
mac
roch
irus
Nes
tin
gm
ales
wit
hin
bree
din
g
colo
ny
(wil
d)
AYe
s?k
M.R
.Gro
ss,p
erso
nal
com
mu
nic
atio
ng
Am
aril
lofi
sh,G
irar
dini
chth
ys
mul
tira
diat
us
Mal
esin
mix
ed-s
exex
per
imen
tal
grou
ps
ofse
ven
(cap
tive
)
AV
aria
ble
1C
.Mac
ıas
Gar
cia,
per
son
al
com
mu
nic
atio
ng
Gre
ylag
goos
e,A
nser
anse
rG
and
ers
inw
inte
rfe
edin
gfl
ocks
(fre
era
ngi
ng)
CLo
wYe
sK
.Kot
rsch
alet
al.,
un
pu
blis
hed
dat
ah
Mal
lard
,Ana
spl
atyr
hync
hos
Dra
kes
wit
hin
pre
-pai
rin
gd
isp
lay
aggr
egat
ion
s(w
ild
and
outd
oor
pen
s)
CLo
wYe
sE.
J.A
.Cu
nn
ingh
am,
per
son
alco
mm
un
icat
ion
g
Gre
atti
t,Pa
rus
maj
orA
du
lts,
typ
ical
lym
ales
,in
smal
l
floc
ks(a
viar
ygr
oup
san
dw
inte
r
fora
gin
gfl
ocks
inw
ild
)
DV
aria
ble
2Ye
slYe
slP.
J.D
ren
t,p
erso
nal
com
mu
nic
atio
n;D
ren
t,
1983
g
Labo
rato
ryra
t,Ra
ttus
norv
egic
us
Dom
inan
tm
ales
inm
ixed
-sex
colo
nie
s(t
riad
s),i
nst
aged
resi
den
t–in
tru
der
test
s(c
apti
ve)n
BYe
smS.
Pell
is,p
erso
nal
com
mu
nic
atio
ng
Spot
ted
hya
ena,
Croc
uta
croc
uta
Ad
ult
san
dju
ven
iles
ofbo
thse
xes,
ofte
nfe
mal
es(w
ild
clan
san
d
cap
tive
coh
orts
)
ELo
wYe
soZ
abel
etal
.,19
92;M
.L.E
ast
&
H.H
ofer
,per
son
al
com
mu
nic
atio
ng
206

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Coy
ote,
Cani
sla
tran
sB
oth
sexe
s,cu
bsw
ith
insi
blin
g
sext
ets
(wil
dan
dca
pti
veli
tter
s)
ELo
wYe
spYe
sM
.Bek
off,
per
son
al
com
mu
nic
atio
ng
Hu
man
,Hom
osa
pien
sB
oth
sexe
s,ad
ult
ssu
bjec
ted
to
verb
alp
rovo
cati
on
(exp
erim
enta
l)
E−
Mar
cus-
New
hal
let
al.,
2000
f
aSo
cial
syst
emty
pic
alof
this
con
text
inth
ew
ild
:A–D
,in
div
idu
als
fam
ilia
rw
ith
only
asu
bset
ofco
nsp
ecif
ics
regu
larl
yen
cou
nte
red
;A
,def
ence
ofn
est
or
cou
rtin
gsi
tein
seas
onal
aggr
egat
ion
byon
ese
x;B
,def
ence
ofte
rrit
ory
orfe
edin
gsi
te,d
omin
ance
rela
tion
ship
sbe
twee
nin
div
idu
als
resi
den
tin
loca
lare
a;C
,
def
ence
ofsh
ifti
ng
feed
ing
zon
ear
oun
din
div
idu
alor
fam
ily,
dom
inan
cere
lati
onsh
ips
betw
een
subs
ets
ofin
div
idu
als
ofon
eor
both
sexe
s;D
,mem
bers
hip
of
non
-ter
rito
rial
floc
kor
terr
itor
ialp
air,
dom
inan
cere
lati
onsh
ips
betw
een
subs
ets
ofco
nsp
ecif
ics;
E,m
embe
rsh
ipof
coh
esiv
egr
oup
,dom
inan
cere
lati
onsh
ips
betw
een
all
grou
pm
embe
rs.
bLo
w,b
etw
een
fam
ilia
rin
div
idu
als
occu
rsin
≤10
%of
dya
dic
con
test
s;va
riab
le1,
typ
ical
lylo
wbe
twee
nfa
mil
iar
ind
ivid
ual
s,bu
th
igh
erin
this
inst
ance
,
e.g.
beca
use
con
test
ants
un
fam
ilia
rat
star
tof
pro
toco
l;va
riab
le2,
typ
ical
lylo
wbe
twee
nfa
mil
iar
ind
ivid
ual
s;−
,pre
ven
ted
byex
per
imen
talp
roto
col.
The
like
lyfr
equ
ency
ofco
un
ter-
aggr
essi
ond
ecre
ases
from
low
tova
riab
le2
tova
riab
le1
toex
per
imen
tall
yp
reve
nte
d.
c Exp
erim
enta
lly
dem
onst
rate
du
sin
gsu
cces
sive
pai
rwis
eco
nte
sts,
orin
ferr
edfr
omse
quen
ces
ofin
tera
ctio
nin
inta
ctgr
oup
s.dR
epor
tsth
atvi
ctim
sre
ceiv
esu
bseq
uen
th
aras
smen
tfr
omby
stan
der
sin
the
min
ute
sfo
llow
ing
anin
itia
ld
efea
t.e Ev
iden
cefo
rre
dir
ecti
ond
emon
stra
ted
exp
erim
enta
lly.
f inco
mp
aris
onw
ith
con
trol
dat
a;gba
sed
onan
ced
otal
rep
orts
from
auth
ors
coll
ecti
ng
quan
tita
tive
dat
aon
aggr
essi
vein
tera
ctio
ns;
hor
infe
rred
from
quan
tita
tive
but
un
con
trol
led
dat
a.i A
bbot
tet
al.,
1985
.j Se
min
atu
ral
grou
ps
hel
dat
rela
tive
lyh
igh
den
sity
;red
irec
tion
not
obse
rved
inw
ild
.k B
ehav
iou
ral
lose
ref
fect
has
been
doc
um
ente
din
Lepo
mis
gibb
osus
(Bea
cham
&N
ewm
an,1
987;
Ch
ase
etal
.,19
94)a
nd
L.cy
anel
lus
(McD
onal
det
al.,
1968
).l V
erbe
ek,1
998;
Ver
beek
etal
.,19
99.
mSe
war
d,1
946;
van
der
Poll
etal
.,19
82.
nN
otkn
own
wh
eth
erth
esu
bjec
tw
aslo
ser
orw
inn
erof
the
init
ial
con
test
.o O
nly
occa
sion
ally
rece
ived
from
byst
and
ers,
but
Wah
ajet
al.,
2001
dem
onst
rate
rece
ipt
from
form
erop
pon
ent.
pB
ekof
f&
Du
gatk
in,2
000.
207

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
208 A. J. N. Kazem & F. Aureli
access to resources such as food, where individual items are of relatively low
value (e.g. certain finches: Rohwer, 1975; Johnstone & Norris, 1993). Displaying a
badge means an individual’s relative aggressive propensity and/or ability should
be immediately apparent to those it meets and to others who witness its social
interactions, reducing the utility of eavesdropping. However, if dominance is not
mediated solely via plumage badges (e.g. individuals may still form relationships
with a subset of familiar conspecifics), and as the magnitude of any loser effect
may still vary, a signalling role for redirection is not ruled out.
Finally, a low probability of counter-aggression during contests is thought to
promote redirection (e.g. Thierry, 1985), because this factor influences both the
necessity and the costs of the behaviour. As mentioned above, redirection offers an
outlet for arousal and frustration in situations where animals cannot retaliate ag-
gressively against an initiator, either because they are physically prevented from
doing so (in experiments) or because the adversary is perceived as too powerful.
A signalling interpretation can also predict a negative association between the
two forms of response; they might be alternative methods of ‘proving oneself ’ to
pre-empt strikes from conspecifics that had witnessed the defeat (although indi-
viduals might sometimes do both to reinforce the message; cf. a multicomponent
signal). Second, if there is a high risk of reprisal from the target, redirection may
become an excessively costly option. Both arguments have been used to explain
the absence of operationally defined redirection in adult female olive baboons
and mountain gorillas (Watts, 1995b; Castles & Whiten, 1998b); bidirectional ag-
gression is common between adult females in these species. Potential retaliation
from targets might also explain why the majority of ‘redirection’ in a small cap-
tive group of chimpanzees Pan troglodytes was directed toward human caretakers,
rather than conspecifics (Malone et al., 2000). Suitable targets are expected to be
those likely to capitulate immediately. Selection may be based on familiarity:
targeting a known subordinate makes sense, especially in species with a ‘strict’
dominance style. However, certain classes of unfamiliar conspecific can also be
identified as unlikely to dispute the outcome, for example targeting first-year in-
dividuals in birds (identifiable by their size and plumage) or smaller floater and
satellite males in fish.
Reports of intraspecific redirection
When placed in situations where they are unable to retaliate directly to
verbal provocation, humans will readily redirect hostility by verbally or physically
punishing substitute targets, both animate and inanimate (reviewed by Marcus-
Newhall et al., 2000). Experimentally induced ‘displaced aggression’ has also been
observed in rodents. Although females are not ordinarily subjected to aggression
by males, mates may become the target of ‘redirected’ attacks in the seconds

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 209
following staged contests between resident males and same-sex intruders (mice
Peromyscus spp.: Eisenberg, 1962; Simmel & Walker, 1970; montane and prairie
voles Microtus montanus and Microtus ochrogaster: S. Pellis, personal communication),
especially if continued access to the opponent(s) is suddenly barred. Unfortunately,
in rodent studies it is often unclear whether the redirecting individual was, or
would eventually have become, the loser in the original contest. A rather artificial
situation is also imposed by the research foci of these experiments; the subject
may not have the option of retaliating against the original aggressor because
s/he is no longer present (often the case in human studies) or may be physically
prevented from pursuing the original contest to its conclusion (e.g. because of
partitions separating opponents in work on rodents). Consequently the subject is
generally presented with a limited range of options and stimuli against which to
direct any response. However, apparent redirection of aggression by victims has
been reported under more naturalistic social contexts in a wide range of species
(a selection, by no means a comprehensive survey, is presented in Table 10.2).
In some species, the behaviour may be performed purely for the physiologi-
cal effects: the benefits of rapidly reducing arousal and perhaps of experiencing
victory. However, the occurrence of redirection is likely to be used as a cue by
bystanders, because it inevitably carries information about an unknown competi-
tor’s relative ability within a population and, where winner or loser effects are
in operation, its current state. If this means bystander behaviour is influenced to
the redirecting individual’sadvantage, one would expect redirection to have been
selected as a signal that is performed more than required for strictly physiological
reasons (or even in the absence of any physiological benefits) and adjusted so as
to be effectively publicized. The challenge is to identify where particular taxa fall
within this spectrum of possibilities.
Testing the occurrence and function of redirection
An essential preliminary is verifying whether an identifiable phe-
nomenon exists – i.e. a pronounced increase in initiation of aggression after
being victimized in comparison with the incidence under appropriate control
conditions. As the primate data illustrate (Table 10.1), redirection may fail to be
demonstrated operationally in species (or age–sex classes) where, on the basis of
uncontrolled data, it had been assumed to be present. It would also be useful to clar-
ify whether counter-aggression and redirection are dissociated, and what factors
favour expression of one tactic over the other. In observational work to date, there
is often insufficient data per individual to allow analyses that compare the inci-
dence of redirection after unidirectional versus bidirectional contests, whilst also
controlling for systematic differences in aggression intensity, duration and num-
ber of participants in the contest. Crucially, the existence of a causal relationship

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
210 A. J. N. Kazem & F. Aureli
between redirecting and a reduction in the number of challenges subsequently
received by the actor requires experimental verification. This could be addressed
by experimentally manipulating the access of the former aggressor and other
bystanders to visual/acoustic information concerning the initial conflict and its
aftermath – how do counter-aggression and redirection compare in terms of their
influence upon the behaviour of witnesses? Furthermore, demonstrating that vic-
tims are sensitive to the perceived presence and composition of bystanders when
redirecting (and not just the availability of a suitable target) would clearly sup-
port a signalling interpretation. Given the likely costs of engaging in aggression,
such facultative use of redirection is predicted. Although absence of an ‘audience
effect’ (i.e. a change in behaviour when in the perceived presence of conspecifics;
see Ch. 4) does not preclude a signalling function, one would expect that use of an
unconditional rule could evolve only if, under natural circumstances, a relevant
audience is almost invariably present. Finally, in taxa that exhibit a loser effect,
it should be possible to confirm whether conflicts concluded after differing dura-
tions and intensity of aggression produce different physiological changes in the
loser. This would provide a basis for exploring how neuroendocrine profiles in the
immediate post-conflict period, and the time course of recovery, might differ in
cases where the animal did or did not redirect.
Summary
We propose that redirection of aggression may function, in part, as a sig-
nal, used to pre-empt subsequent challenges from the former aggressor and/or
other bystanders that witness the act. In systems where behavioural loser effects
exist, such as macaques, this might be achieved because redirection serves as an
honest indicator of post-conflict condition and motivation, demonstrating that
the perpetrator has not been unduly compromised by its preceding defeat. Al-
ternatively, redirection might simply limit the extent of the negative impression
usually conveyed by a loss, reaffirming one’s position within a status hierarchy to
bystanders or, where the bystander is unfamiliar with the protagonist, conveying
an ability to defeat at least some individuals within the population.
This suggestion has a number of broader implications. Being observed to lose a
contest incurs social penalties, affecting not only the likelihood of being chal-
lenged by others but also the prospects of acquiring or retaining mates (e.g.
Doutrelant & McGregor, 2000; Mennill et al., 2002). So far, attention has focused
exclusively on how contestants counter these pressures by modifying their dis-
plays during the original agonistic exchange if in the perceived presence of an
audience (e.g. Doutrelant et al., 2001; Matos & McGregor, 2002; A. J. N. Kazem,
R. J. Motos & P. K. McGregor, unpublished data). In reality, there may be several

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 211
points at which the eventual loser can influence others’ perceptions of its ability:
in the decision to counter-attack rather than defer, to fight harder (or differ-
ently) in the current exchange, or even by redirecting after the contest has con-
cluded. First, this suggests it would be profitable to consider audience effects and
eavesdropping over a longer timescale, encompassing sequences of contests. Sec-
ond, individuals who approach conflicts between others risk becoming targets
of redirection (this may explain why, in our study species, low-ranking individ-
uals often flee the scene of incipient or ongoing aggression). Therefore, depend-
ing on the signal modality used, the costs of eavesdropping may not be as low
as previously assumed – particularly for low-ranking or otherwise poor competi-
tors, which stand to gain the most from acquiring information via this route.
Finally, redirection may constitute a rare example of a signal aimed primar-
ily at ‘secondary’ receivers. The communication networks perspective has high-
lighted how many dyadic displays evolve within the context of, and may even
be designed to advertise to, several receivers. However, redirection may be an in-
stance of a signal performed almost entirely for its effect on third parties rather
than upon the apparent receiver. There has been substantial theoretical inter-
est in the possibility that apparently altruistic behaviour may be maintained via
this route (e.g. Zahavi, 1977; Nowak & Sigmund, 1998; Roberts, 1998; Leimar &
Hammerstein, 2001), but few empirical examples have been discovered outside
humans. Redirection has the potential to prove a taxonomically widely distributed
case, but experimental verification and analyses to explore the evolutionary sta-
bility of such a signalling system in aggression are still required.
Acknowledgements
We are very grateful to the many correspondents who provided access to unpublished
data and detailed observations on their study taxa. We would also like to thank Peter McGregor
for inviting us to contribute to this volume, and his encouragement and support of the first
author’s ongoing work on redirection. The latter’s research on rhesus macaques was financially
supported by awards from the H. F. Guggenheim Foundation, L. S. B. Leakey Foundation, Wenner
Gren Foundation for Anthropological Research, Zunz Foundation and a University of Durham
Research Studentship; the generosity of these organizations is greatly appreciated.
References
Abbott, J. C., Dunbrack, R. L. & Orr, C. D. 1985. The interaction of size and experience
in dominance relationships of juvenile steelhead trout (Salmo gairdneri). Behaviour,
92, 241–253.
Andrews, R. J. 1991. Testosterone, attention and memory. In: The Development and
Integration of Behaviour, ed. P. P. G. Bateson. Cambridge, UK: Cambridge University
Press, pp. 171–190.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
212 A. J. N. Kazem & F. Aureli
Arnold, K. & Barton, R. A. 2001. Postconflict behavior of spectacled leaf monkeys
(Trachypithecus obscurus). II. Contact with third parties. International Journal of
Primatology, 22, 267–286.
Aureli, F. 1992. Post-conflict behaviour among wild long-tailed macaques (Macaca
fascicularis). Behavioral Ecology and Sociobiology, 31, 329–337.
Aureli, F. & van Schaik, C. P. 1991a. Post-conflict behaviour in long-tailed macaques
(Macaca fascicularis): I. The social events. Ethology, 89, 89–100.
1991b. Post-conflict behaviour in long-tailed macaques (Macaca fascicularis): II.
Coping with the uncertainty. Ethology, 89, 101–114.
Aureli, F., Cozzolino, R., Cordischi, C. & Scucchi, S. 1992. Kin-oriented redirection
among Japanese macaques: an expression of a revenge system? Animal Behaviour,
44, 283–291.
Aureli, F., Veenema, H. C., van Panthaleon van Eck, C. J. & van Hooff, J. A. R. A. M. 1993.
Reconciliation, consolation, and redirection in Japanese macaques (Macaca
fuscata). Behaviour, 124, 1–21.
Aureli, F., Das, M., Verleur, D. & van Hooff, J. A. R. A. M. 1994. Postconflict social
interactions among Barbary macaques (Macaca sylvanus). International Journal of
Primatology, 15, 471–484.
Aureli, F., Cords, M. & van Schaik, C. P. 2002. Conflict resolution following aggression
in gregarious animals: a predictive framework. Animal Behaviour, 64, 325–343.
Bastock, M., Morris, D. & Moynihan, M. 1953. Some comments on conflict and
thwarting in animals. Behaviour, 6, 66–84.
Beacham, J. L. & Newman, J. A. 1987. Social experience and the formation of
dominance relationships in the pumpkinseed sunfish, Lepomis gibbosus. Animal
Behaviour, 35, 1560–1563.
Bekoff, M. & Dugatkin, L. A. 2000. Winner and loser effects and the development of
dominance relationships in young coyotes: an integration of data and theory.
Evolutionary Ecology Research, 2, 871–883.
Bernstein, I. S., Rose, R. M. & Gordon, T. P. 1974. Behavioral and environmental
events influencing primate testosterone levels. Journal of Human Evolution, 3,
517–525.
Black, J. M. & Owen, M. 1987. Determinants of social rank in goose flocks: acquisition
of social rank in young geese. Behaviour, 102, 129–146.
Blanchard, D. C., Sakai, R. R., McEwen, B. & Blanchard, R. J. 1993. Subordination stress:
behavioral, brain and neuroendocrine correlates. Behavioural Brain Research, 58,
113–121.
Booth, A., Shelley, G., Mazur, A., Tharp, G. & Kittok, R. 1989. Testosterone, and winning
and losing in human competition. Hormones and Behavior, 23, 556–571.
Castles, D. L. & Whiten, A. 1998a. Post-conflict behaviour of wild olive baboons. II.
Stress and self-directed behaviour. Ethology, 104, 148–160.
1998b. Post-conflict behaviour of wild olive baboons. I. Reconciliation, redirection
and consolation. Ethology, 104, 126–147.
Chapais, B. 1995. Alliances as a means of competition in primates: evolutionary,
developmental and cognitive aspects. Yearbook of Physical Anthropology, 38, 115–
136.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 213
Chase, I. D., Bartolomeo, C. & Dugatkin, L. 1994. Aggressive interactions and
inter-contest interval: how long do winners keep winning? Animal Behaviour, 48,
393–400.
Chase, I. D., Tovey, C., Spangler-Martin, D. & Manfredonia, M. 2002. Individual
differences versus social dynamics in the formation of animal dominance
hierarchies. Proceedings of the National Academy of Sciences, USA, 99, 5744–5749.
Cheney, D. L. & Seyfarth, R. M. 1986. The recognition of social alliances by vervet
monkeys. Animal Behaviour, 34, 1722–1731.
1989. Redirected aggression and reconciliation among vervet monkeys, Cercopithecus
aethiops. Behaviour, 110, 258–275.
1990. Attending to behaviour versus attending to knowledge: examining monkeys’
attribution of mental states. Animal Behaviour, 40, 742–753.
1999. Recognition of other individuals’ social relationships by female baboons.
Animal Behaviour, 58, 67–75.
Cheney, D. L., Seyfarth, R. M. & Silk, J. B. 1995. The responses of female baboons (Papio
cynocephalus ursinus) to anomalous social interactions: evidence for causal
reasoning? Journal of Comparative Psychology, 109, 134–141.
Cords, M. 1988. Resolution of aggressive conflicts by immature long-tailed macaques,
Macaca fascicularis. Animal Behaviour, 36, 1124–1135.
1992. Post-conflict reunions and reconciliation in long-tailed macaques. Animal
Behaviour, 44, 57–61.
Cynx, J. & Nottebohm, F. 1992. Testosterone facilitates some conspecific song
discriminations in castrated zebra finches Taeniopygia guttata. Proceedings of the
National Academy of Sciences, USA, 89, 1376–1378.
Das, M. 1998. Conflict management and social stress in long-tailed macaques. Ph.D.
Thesis, University of Utrecht, the Netherlands.
Das, M., Penke, Z. & van Hooff, J. A. R. A. M. 1997. Affiliation between aggressors and
third parties following conflicts in long-tailed macaques (Macaca fascicularis).
International Journal of Primatology, 18, 157–179.
Dasser, V. 1988. A social concept in Java monkeys. Animal Behaviour, 36, 225–230.
Delville, Y., Mansour, K. M., Ferris, C. F. 1996. Testosterone facilitates aggression by
modulating vasopressin receptors in the hypothalamus. Physiology and Behavior,
60, 25–29.
de Waal, F. B. M. & Luttrell, L. M. 1989. Toward a comparative socioecology of the
genus Macaca: different dominance styles in rhesus and stumptail macaques.
American Journal of Primatology, 19, 83–109.
de Waal, F. B. M. & Yoshihara, D. 1983. Reconciliation and redirected affection in
rhesus monkeys. Behaviour, 85, 224–241.
Doutrelant, C. & McGregor, P. K. 2000. Eavesdropping and mate choice in female
fighting fish. Behaviour, 137, 1655–1669.
Doutrelant, C., McGregor, P. K. & Oliveira, R. F. 2001. The effect of an audience on
intra-sexual communication in male Siamese fighting fish, Betta splendens.
Behavioral Ecology, 12, 283–286.
Drent, P. J. 1983. The functional ethology of territoriality in the great tit (Parus major
L.). Ph.D. Thesis, University of Groningen, the Netherlands.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
214 A. J. N. Kazem & F. Aureli
Earley, R. L. & Dugatkin, L. A. 2002. Eavesdropping on visual cues in green swordtail
(Xiphophorus helleri) fights: a case for networking. Proceedings of the Royal Society of
London, Series B, 269, 943–952.
Eaton, G. G. 1984. Aggression in adult male primates: a comparison of confined
Japanese macaques and free-ranging olive baboons. International Journal of
Primatology, 5, 145–160.
Eisenberg, J. F. 1962. Studies on the behavior of Peromyscus maniculatus gambelli and
Peromyscus californicus parasiticus. Behaviour, 19, 177–207.
Emery, N. J., Lorincz, E. N., Perret, D. I., Oram, M. W. & Backer, C. I. 1997. Gaze
following and joint attention in rhesus monkeys (Macaca mulatta). Journal of
Comparative Psychology, 111, 286–293.
Engh, A. L., Esch, K., Smale, L. & Holekamp, K. E. 2000. Mechanisms of maternal rank
‘inheritance’ in the spotted hyaena, Crocuta crocuta. Animal Behaviour, 60,
323–332.
Fairbanks, L. A., Melega, W. P., Jorgensen, M. J., Kaplan, J. R. & McGuire, M. T. 2001.
Social impulsivity inversely associated with CSF 5-HIAA and fluoxetine exposure
in vervet monkeys. Neuropsychopharmacology, 24, 370–378.
Ferris, C. F., Melloni, R. H., Jr, Koppel, G. et al. 1997. Vasopressin/serotonin interactions
in the anterior hypothalamus control aggressive behavior in golden hamsters.
Journal of Neuroscience, 17, 4331–4340.
Frank, L. G. 1986. Social organization of the spotted hyena (Crocuta crocuta). 2.
Dominance and reproduction. Animal Behaviour, 34, 1510–1527.
Fuentes, A., Malone, N., Sanz, C., Matheson, M. & Vaughan, L. 2002. Conflict and
post-conflict behavior in a small group of chimpanzees. Primates, 43, 223–235.
Gore, M. A. 1994. Dyadic and triadic aggression and assertiveness in adult female
rhesus monkeys, Macaca mulatta, and hamadryas baboons, Papio hamadryas. Animal
Behaviour, 48, 385–392.
Gouzoules, H., Gouzoules, S. & Marler, P. 1986. Vocal communication: a vehicle for the
study of social relationships. In: The Cayo Santiago Macaques: History, Behavior and
Biology, ed. R. G. Rawlins & M. J. Kessler. Albany, NY: State University of New York
Press, pp. 111–129.
Gust, D. A. & Gordon, T. P. 1993. Conflict resolution in sooty mangabeys. Animal
Behaviour, 46, 685–694.
Gust, D. A., Gordon, T. P., Hambright, M. K. & Wilson, M. E. 1993. Relationship between
social factors and pituitary-adrenocortical activity in female rhesus monkeys
(Macaca mulatta). Hormones and Behavior, 27, 318–331.
Hannes, R. P., Franck, D. & Liemann, F. 1984. Effects of rank order fights on body and
blood concentrations of androgens and corticosteroids in the male swordtail
(Xiphophorus helleri). Zeitschrift fur Tierpsychologie, 65, 53–65.
Higley, J. D., Mehlman, P. T., Poland, R. E. et al. 1996. CSF testosterone and 5-HIAA
correlate with different types of aggressive behaviors. Biological Psychiatry, 40,
1067–1082.
Hogue, M.-E., Beaugrand, J. P. & Lague, P. C. 1996. Coherent use of information by hens
observing their former dominant defeating or being defeated by a stranger.
Behavioural Processes, 38, 241–252.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 215
Hsu, Y. & Wolf, L. L. 1999. The winner and loser effect: integrating multiple
experiences. Animal Behaviour, 57, 903–910.
2001. The winner and loser effect: what fighting behaviours are influenced? Animal
Behaviour, 61, 777–786.
Itani, J. 1963. Vocal communication of the wild Japanese monkeys. Primates, 4, 11–66.
Johnstone, R. A. & Bshary, R. 2004. Evolution of spite through indirect reciprocity.
Proceedings of the Royal Society of London, Series B, 271, 1917–1922.
Johnstone, R. A. & Norris, K. 1993. Badges of status and the cost of aggression.
Behavioral Ecology and Sociobiology, 32, 127–134.
Judge, P. G. 1982. Redirection of aggression based on kinship in a captive group of
pigtail macaques. International Journal of Primatology, 3, 301.
Kavaliers, M., Choleris, E. & Colwell, D. D. 2001. Brief exposure to female odors
‘emboldens’ male mice by reducing predator-induced behavioral and hormonal
responses. Hormones and Behavior, 40, 497–509.
Kazem, A. J. N. 1999. Conflict management in free-ranging immature rhesus
macaques, Macaca mulatta. Ph.D. Thesis, University of Durham, UK.
Keddy Hector, A., Seyfarth, R. M. & Raleigh, M. J. 1989. Male parental care, female
choice, and the effect of an audience in vervet monkeys. Animal Behaviour, 38,
262–271.
Koolhaas, J. M., Korte, S. M., de Boer, S. F. et al. 1999. Coping styles in animals: current
status in behavior and stress-physiology. Neuroscience and Biobehavioral Reviews, 23,
925–935.
Kutsukake, N. & Castles, D. L. 2001. Reconciliation and variation in post-conflict stress
in Japanese macaques (Macaca fuscata fuscata): testing the integrated hypothesis.
Animal Cognition, 4, 259–268.
Lamprecht, J. 1986. Structure and causation of the dominance hierarchy in a flock of
bar-headed geese (Anser indicus). Behaviour, 96, 28–48.
Leimar, O. & Hammerstein, P. 2001. Evolution of cooperation through indirect
reciprocity. Proceedings of the Royal Society of London, Series B, 268, 745–753.
Levine, S., Coe, C. & Wiener, S. G. 1989. Psychoneuroendocrinology of stress: a
psychobiological perspective. In: Psychoendocrinology, ed. S. Levine & R. Brush.
London: Academic Press, pp. 341–377.
Malone, N., Vaughan, L. & Fuentes, A. 2000. The role of human caregivers in the
post-conflict interactions of captive chimpanzees (Pan troglodytes). Laboratory
Primate Newsletter, 39, 1–3.
Marcus-Newhall, A., Pedersen, W. C., Carlson, M. & Miller, N. 2000. Displaced
aggression is alive and well: a meta-analytic review. Journal of Personality and Social
Psychology, 78, 670–689.
Matos, R. J. & McGregor, P. K. 2002. The effect of the sex of an audience on male–male
displays of Siamese fighting fish (Betta splendens). Behaviour, 139, 1211–1221.
McDonald, A. L., Heimstra, N. W. & Damkot, D. K. 1968. Social modification of
agonistic behaviour in fish. Animal Behaviour, 16, 437–441.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
216 A. J. N. Kazem & F. Aureli
McGregor, P. K., Peake, T. M. & Lampe, H. M. 2001. Fighting fish, Betta splendens, extract
relative information from apparent interactions: what happens when what you
see is not what you get. Animal Behaviour, 62, 1059–1065.
Mehlman, P. T., Higley, J. D., Faucher, I. et al. 1994. Low CSF 5-HIAA concentrations and
severe aggression and impaired impulse control in nonhuman primates. American
Journal of Psychiatry, 151, 1485–1491.
Mendoza, S. P. 1993. Social conflict on first encounters. In: Primate Social Conflict, ed.
W. A. Mason & S. P. Mendoza. Albany, NY: State University of New York Press,
pp. 85–110.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Mills, M. G. L. 1990. Kalahari hyaenas: Comparative Behavioural Ecology of Two Species.
London: Unwin Hyman.
Moynihan, M. H. 1998. The social regulation of competition and aggression in animals.
Washington, DC: Smithsonian Institution Press.
Nowak, M. A. & Sigmund, K. 1998. Evolution of indirect reciprocity by image scoring.
Nature, 393, 573–577.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Olivier, B., Mos, J., van Oorschot, R. & Hen, R. 1995. Serotonin receptors and animal
models of aggressive behavior. Pharmacopsychiatry, 28, 80–90.
Øverli, Ø., Harris, C. A. & Winberg, S. 1999. Short-term effects of fights for social
dominance and the establishment of dominant-subordinate relationship on brain
monoamines and cortisol in rainbow trout. Brain Behavior and Evolution, 54,
263–275.
Øverli, Ø., Korzan, W. J., Larson, E. T. et al. 2004. Behavioural and neuroendocrine
correlates of displaced aggression in trout: cycle of violence is evolutionarily
conserved. Hormones and Behavior, 45, 3224–3349.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Perreault, H. A. N., Semsar, K. & Godwin, J. 2003. Fluoxetine treatment decreases
territorial aggression in a coral reef fish. Physiology and Behavior, 79,
719–724.
Rendall, D., Rodman, P. S. & Emond, R. E. 1996. Vocal recognition of individuals and
kin in free-ranging rhesus monkeys. Animal Behaviour, 51, 1007–1015.
Rendall, D., Owren, M. J. & Rodman, P. S. 1998. The role of vocal tract filtering in
identity cueing in rhesus monkey (Macaca mulatta) vocalizations. Journal of the
Acoustical Society of America, 103, 602–614.
Roberts, G. 1998. Competitive altruism: from reciprocity to the handicap principle.
Proceedings of the Royal Society of London, Series B, 265, 427–431.
Roeder, J.-J., Fornasieri, I. & Gosset, D. 2002. Conflict and postconflict behaviour in two
lemur species with different social organizations (Eulemur fulvus and Eulemur
macaco): a study on captive groups. Aggressive Behavior, 28, 62–74.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
Redirection of aggression: multiparty signalling 217
Rohwer, S. 1975. The social significance of avian plumage variability. Evolution, 29,
593–610.
Rose, R. M., Bernstein, I. S. & Gordon, T. 1975. Consequences of social conflict on
plasma testosterone levels in rhesus monkeys. Psychosomatic Medicine, 37, 50–61.
Sapolsky, R. M. 1998. Why Zebras Don’t get Ulcers: An Updated Guide to Stress, Stress-related
Diseases, and Coping, 2nd edn. New York: Freeman.
Sapolsky, R. M. & Ray, J. C. 1989. Styles of dominance and their endocrine correlates
among wild olive baboons (Papio anubis). American Journal of Primatology, 18, 1–13.
Schuurman, T. 1980. Hormonal correlates of agonistic behaviour in adult male rats.
Progress in Brain Research, 53, 415–520.
Scott, J. P. & Fredericson, E. 1951. The causes of fighting in mice and rats. Physiological
Zoology, 24, 273–309.
Scucchi, S., Cordischi, C., Aureli, F. & Cozzolino, R. 1988. The use of redirection in a
captive group of Japanese monkeys. Primates, 29, 229–236.
Seward, J. P. 1946. Aggressive behavior in the rat. IV. Submission as determined by
conditioning, extinction and disuse. Journal of Comparative Psychology, 39,
51–57.
Silk, J. B. 1999. Male bonnet macaques use information about third-party rank
relationships to recruit allies. Animal Behaviour, 58, 45–51.
Simmel, E. C. & Walker, D. A. 1970. Social priming for agonistic behavior in a ‘docile’
mouse strain. American Zoologist, 10, 486–487.
Summers, C. H., Summers, T. R., Moore, M. C. et al. 2003. Temporal patterns of limbic
monoamine and plasma corticosterone response during social stress. Neuroscience,
116, 553–563.
Thierry, B. 1985. Patterns of agonistic interactions in three species of macaque (Macaca
mulatta, M. fascicularis, M. tonkeana). Aggressive Behavior, 11, 223–233.
2000. Covariation of conflict management patterns in macaque societies. In: Natural
Conflict Resolution, ed. F. Aureli & F. B. M. de Waal. Berkeley, CA: University of
California Press, pp. 106–128.
Tomasello, M., Call, J. & Hare, B. 1998. Five primate species follow the visual gaze of
conspecifics. Animal Behaviour, 55, 1063–1069.
van de Poll, N. E., de Jonge, F., van Oyen, H. G. & van Pett, J. 1982. Aggressive behaviour
in rats: effects of winning and losing on subsequent aggressive interactions.
Behavioural Processes, 7, 143–155.
van Erp, A. M. M. & Miczek, K. A. 2000. Aggressive behavior, increased accumbal
dopamine and decreased cortical serotonin in rats. Journal of Neuroscience, 15,
9320–9325.
Veenema, H. C. 2000. Methodological progress in post-conflict research. In: Natural
Conflict Resolution, ed. F. Aureli & F. B. M. de Waal. Berkeley, CA: University of
California Press, pp. 21–23.
Verbeek, M. 1998. Bold or cautious: behavioural characteristics and dominance in
great tits. Ph.D. Thesis, University of Wageningen, the Netherlands.
Verbeek, M. E. M., de Goede, P., Drent, P. J. & Wiepkema, P. R. 1999. Individual
behavioural characteristics and dominance in aviary groups of great tits.
Behaviour, 136, 23–48.

P1: GFZ/... P2: JZZ/...0521823617c10.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 14:27
218 A. J. N. Kazem & F. Aureli
Virgin, C. E., Jr & Sapolsky, R. M. 1997. Styles of male social behavior and their
endocrine correlates among low-ranking baboons. American Journal of Primatology,
42, 25–39.
Wahaj, S. A., Guse, K. R. & Holekamp, K. E. 2001. Reconciliation in the spotted hyena
(Crocuta crocuta). Ethology, 107, 1057–1074.
Walker Leonard, J. 1979. A strategy approach to the study of primate dominance
behaviour. Behavioural Processes, 4, 155–172.
Watts, D. P. 1995a. Post-conflict social events in wild mountain gorillas (Mammalia,
Hominoidea). I. Social interactions between opponents. Ethology, 100, 139–157.
1995b. Post-conflict social events in wild mountain gorillas. II. Redirection, side
direction, and consolation. Ethology, 100, 158–174.
Weiss, J. M., Pohorecky, L. A., Salman, S. & Gruenthal, M. 1976. Attenuation of gastric
lesions by psychological aspects of aggression in rats. Journal of Comparative and
Physiological Psychology, 90, 252–259.
Winberg, S. & Lepage, O. 1998. Elevation of brain 5-HT activity, POMC expression, and
plasma cortisol in socially subordinate rainbow trout. American Journal of
Physiology, 274, R645–R654.
Zabel, C. J., Glickman, S. E., Frank, L. G., Woodmansee, K. B. & Keppel, G. 1992.
Coalition formation in a colony of prepubertal spotted hyenas. In: Coalitions and
Alliances in Humans and Other Animals, ed. A. H. Harcourt & F. B. M. de Waal. Oxford:
Oxford University Press, pp. 113–135.
Zahavi, A. 1977. Reliability in communication systems and the evolution of altruism.
In: Evolutionary Ecology, ed. B. Stonehouse & C. M. Perrins. London: Macmillan,
pp. 253–259.
1979. Why shouting? American Naturalist, 113, 155–156.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
11
Scent marking and socialcommunication
j a n e l . h u r s t
University of Liverpool, Liverpool, UK
Introduction
The use of chemical scents for communication between individuals is
widespread among both vertebrate and invertebrate animals. Scent signals ema-
nating from an animal’s body can be used for intimate and immediate commu-
nication when two or more individuals interact at close quarters, but scents can
also be deposited in the environment in the form of scent marks. Unlike most
visual or acoustic signals used by animals, scent marks persist in the absence of
the signaller, often over extended periods. The prolonged duration of signals in
deposited scent marks makes them particularly suited for broadcasting informa-
tion to all conspecifics that visit a scent-marked site. Further, scent marks might be
deposited to signal to certain individuals, such as when animals are attempting to
attract potential mates or to indicate a territorial boundary to neighbours. Once
deposited in the environment, however, the scent is not physically directed to-
wards specific recipients and the information will be available to any other ani-
mals in the locality. This ready availability of scent marks to third parties is likely
to provide strong selection pressure to ensure that the information deposited
in scent marks is appropriate for communication to any individual likely to en-
counter the scent. Consequently, scent marks are likely to have evolved to be used
for network communication rather than as signals between specific individuals.
While volatile components of the scent may be detected at some distance from
a scent mark, alerting animals to the presence and location of scent signals, non-
volatile components can be detected only by close contact investigation. Typically,
animals approach and sniff scent marks very closely; many species will also lick the
scent. Scent marks may thus communicate information via both volatile odorants
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
219

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
220 J. L. Hurst
and non-volatile components or those of low volatility, and perception may involve
more than one chemosensory system. For example, mammals have both a main
olfactory system linked to the olfactory epithelium (reviewed by Nibu, 2002) and an
accessory olfactory system linked to the vomeronasal organ (reviewed by Brennan,
2001; Takami, 2002; Zufall et al., 2002).
Scent marking is particularly common among terrestrial, arboreal and subter-
ranean mammals and plays an important role in a number of social contexts,
including recognition of group members and kin, the advertisement of territory
ownership and social dominance, assessment of the quality of competitors and
potential mates, and the advertisement and control of reproductive status. In
each of these contexts, communication characteristically involves all individuals
that deposit and detect scent marks within a particular area rather than private
communication between two individuals. To illustrate this, I will review the use
of scent marks for communication among house mice Mus musculus domesticus,
in which both the behavioural and biochemical mechanisms underlying scent
communication have been well studied, both in the laboratory and among wild
mice in more naturalistic enclosure experiments. In the concluding part of the
chapter, I will discuss the advantages of using scent marks as broadcast signals to
communicate to a network of conspecifics rather than directing signals to specific
individuals.
Scent marking among house mice
Urine is the primary source of social odours among house mice and con-
tains ‘fixed’ (genomic) information about the species, sex, individual identity, ma-
jor histocompatibility complex (MHC) type and other genetic background of the
owner, as well as ‘variable’ (metabolic) information on the owner’s current social,
reproductive and health status, and its food resources (reviewed by Brown, 1985a,
1995; Hurst et al., 2001a; Malone et al., 2001). Once old enough to leave the nest,
mice of both sexes scent mark by depositing urine in small spots and streaks on
the substrate in a deliberate pattern of deposition as they move around their home
area (Fig. 11.1). All surfaces are rapidly covered with this background scent mark-
ing and scent mark density patterns generally correspond with spatial patterns of
activity, most marks being deposited around physical edges such as walls, feeding
sites and near nest sites (Fig. 11.2). Mice do not appear to urine mark within their
nest site. Once mice have scent marked a patch of substrate thoroughly, the rate of
further background marking decreases but it persists at a low level so the animals
are continually refreshing their scent. Because mice generally live in territorial so-
cial groups consisting of one dominant male together with one or more breeding
females and their offspring, and variable numbers of non-breeding adult females

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 221
(c) Urine scent posts
(a) Water stimulus (b) Intruder urine stimulus
Fig. 11.1. Scent marks of wild house mice. Urine scent marks deposited by a resident
male territory owner on a patch of substrate (15 cm × 15 cm of absorbent paper)
introduced into the territory for 30 min. The patch was first treated with 10µl of
either water (a) or urine from an intruder male (b). The white box indicates location of
the stimulus). Urinary scent marks (white) were visualized under ultraviolet light
using a FluorS scanner (Bio-Rad Laboratories Ltd, Hemel Hempsted, UK). (c) Urine scent
posts (approx 2 cm tall, see white box) on a wooden batten where mice frequently
rested in an infested poultry house.
and subordinate males (reviewed by Barnard et al., 1991), the substrate becomes
smothered in urinary scent marks from all mice that share the same home area
(Hurst, 1989). Not all mice contribute equally to this background scent marking;
the dominant male territory owner, responsible for most defence of the territory,
marks at a much higher rate than other individuals. In situations where males

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
222 J. L. Hurst
1.2 m
Male mouse
Male mouse
(a)
(b)
Fig. 11.2. Correspondence between activity and scent deposition when an adult male
mouse explored a clean enclosure neighbouring his territory for 15 min. Arrow
indicates the hole through which the male entered from his own territory.
(a) Location sampled three times per second. (b) Urine marks were sampled by
swabbing each 15 cm × 15 cm floor section. The darkness of spots shows the amount
of urine recovered, visualized by immunoassay of the major urinary proteins. The
nestbox (circle) and food station (rectangle) were not swabbed. (Data collected by
Karen Sanders.)
have to defend their territory from other males, dominant male territory owners
deposit large streaks as well as smaller spots of urine wherever they go (Desjardins
et al., 1973; Hurst, 1990), aided by hairs on the end of the adult male prepuce
(Maruniak et al., 1975).
Adult mice excrete a number of species- and sex-specific volatiles in their urine
that are under hormonal control, in addition to a large number of non-specific
volatile metabolites (Novotny et al., 1984, 1990; Schwende et al., 1986; Harvey
et al., 1989). Male-specific signalling volatiles include two sesquiterpenes, (E , E )
α-farnesene and (E ) β -farnesene, which are secreted into the urine by the preputial
glands, and 2-sec-butyl-4,5-dihydrothiazole and 2,3-dehydro-exo-brevicomin, which

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 223
are present in the bladder before any additions from accessory glands (Harvey et al.,
1989; Novotny et al., 1990). A large number of volatiles in female urine vary over the
oestrous cycle (Andreolini et al., 1987) and during pregnancy and lactation ( Jemiolo
et al., 1987), although it is not yet known which (if any) of these are chemosignals or
metabolic by-products. One urinary constituent, 2,5-dimethylpyrazine, produced
under adrenal control by group-housed females is known to inhibit reproduction
in other females (Novotny et al., 1986; Jemiolo & Novotny, 1993, 1994).
In addition to these sex-specific volatiles, mouse urine is characterized by the
presence of a high concentration of protein, over 99% of which is contributed
by a group of 18–20 kDa lipocalins known as the major urinary proteins (MUPs).
The lipocalins are synthesized in the liver and secreted into the plasma; they
subsequently pass through the glomerular filter into the urine (Beynon et al., 2001).
Although urine of both sexes contains a substantial quantity of these proteins,
adult male urine typically contains 20–30 mg/ml protein, approximately three
times as much as female urine (Beynon & Hurst, 2003). This sex difference occurs
at puberty when there is an increase in excretion among males (Payne et al., 2001).
These lipocalin proteins have a central cavity that binds lipophilic molecules.
In males, MUPs bind a number of ligands but principally the two male-specific
signalling volatiles 2-sec-butyl-4,5-dihydrothiazole and 2,3-dehydro-exo-brevicomin
(Bacchini et al., 1992; Robertson et al., 1993; Novotny et al., 1999a). Once urine is
deposited as a scent mark, the binding of signalling volatiles to MUPs greatly slows
down their evaporation from the scent mark (Hurst et al., 1998; Robertson et al.,
2001), extending the duration over which volatiles can be detected (Hurst et al.,
1998; Humphries et al., 1999). No MUP ligands have yet been identified in female
urine.
Although volatiles may be detected from a distance of several centimetres or
more, depending on the amount of scent deposited, the response of mice is almost
always to approach and contact the scent mark, investigating it at very close
quarters. Consequently, it is likely that mice gain information from both the
volatile and non-volatile components in scents (Humphries et al., 1999). Recent
evidence suggests that the vomeronasal system only detects scents when these
are pumped to the vomeronasal organ after contact with a stimulus (Luo et al.,
2003).
Advertising territory ownership and competitive ability
The network of scent marks deposited and investigated by all individuals
within the local population provides a mechanism by which animals can advertise
their competitive ability to both competitors and to potential mates in a manner
that makes it difficult, if not impossible, to cheat. This reliable mechanism involves

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
224 J. L. Hurst
both the spatial and the temporal pattern of competitive scent marking as well as
the quality of an individual’s scent.
Spatial and temporal distribution of scent
Like many other mammals, dominant male house mice advertise their
territory ownership by scent marking throughout their defended area, marking
at a much higher rate than other individuals (Ralls, 1971; Desjardins et al., 1973;
Hurst, 1990). Territory owners continually refresh these scents at a high rate so
that, in frequently used sites, small posts of dried urine can build up like small
stalagmites (Fig. 11.1c). Because only animals that dominate a territory can ensure
that their marks predominate in that area, the spatial pattern and density of scent
marks bearing the owner’s individual identity signature provide physical proof of
an individual’s territory ownership (Gosling, 1982; Hurst, 1993). Further, the spa-
tial and temporal pattern of scent marks from other males indicates the success
with which an owner dominates its scent-marked territory. Only males that de-
fend their territory effectively can ensure that no other males deposit competing
signals that might attract mates (Hurst, 1993; Hurst & Rich, 1999). Conversely,
the presence of any competing signals that are as fresh or fresher than those of
the owner will indicate that the owner is not stopping competitors from deposit-
ing competing marks and, therefore, is not being very successful in dominating
the area, even if the area is suffused with the owner’s scent. Accordingly, domi-
nant male mice rapidly counter-mark if they encounter competing scent signals
from other males in their territory, as well as attacking and excluding from the
territory any competitors that deposit such competing scent marks (Ralls, 1971;
Hurst & Rich, 1999). In house mice, counter-marking consists of a rapid increase
in the rate of urine scent marking in the vicinity of, but not specifically on top
of, a competitor male’s scent (Fig. 11.1b; Hurst, 1989; Humphries et al., 1999). In
other species, counter-marking may take the form of over-marking the competi-
tor’sscent to prove which scent was deposited most recently ( Johnston et al., 1997;
Johnston, 1999; Ch. 16). House mice do not attempt to over-mark the scent marks
of a competitor, perhaps because their urinary scent marks are scattered so widely
in numerous spots and streaks. Instead, mice assess the difference in age of nearby
scent marks from competing males to determine which male’sscent was deposited
most recently (Rich & Hurst, 1999). Because scent marks remain in the environ-
ment and are long lived (see p. 232), the spatial and temporal pattern of scents
from different individuals provides a continuous record of any challenges for dom-
inance over the area and, crucially, the outcome of those challenges. This record is
available for investigation by any other animal in the area. Animals do not need to
witness, or to eavesdrop on, individual challenges for dominance while these are
occurring, because the outcome will be readily apparent from scent marks for as

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 225
long as the marks retain the individual signatures of the depositors (but see Ch. 2
for a discussion of extra information available from interactions). Scent marks
provide the ‘minutes’ of a meeting that are made public to all interested parties,
although they may not include a transcript of all of the information involved in
the detailed arguments expressed during the meeting.
Experimental manipulation of the scent marks within male mouse territories
has shown that this information is used by third parties to assess the competitive
ability of territory owners: (a) by females when selecting high-quality mates, and
(b) by other males when deciding whether to avoid a territory owner or to challenge
the owner for dominance themselves. These are discussed separately below.
Assessment by females
Although female house mice generally nest and raise their offspring
within one male’s territory, they often visit or range over several neighbouring
male territories (Hurst, 1987) and extraterritorial matings occur frequently (e.g.
43% of all matings in large captive populations occurred when a female travelled
to, and mated with, a male owning a nearby territory (Potts et al., 1991)). Inter-
estingly, among wild house mice, resident male territory owners appear to show
little or no discrimination against offspring sired by other males that are reared
within their territory (Hurst & Barnard, 1992). Therefore, females usually have a
choice between several high-quality male territory owners as potential mates in
the local population, regardless of where they choose to nest, because there is
little risk to their offspring.
Most simply, females can compare the scent marks left by two competing males
to assess which male’s scent was deposited most recently. Since only a male that
is successfully preventing other males from depositing competing scents can en-
sure that his marks are the freshest in that location, this male must have been the
winner of the conflict. Subsequently given a choice between the two signalling
males, females generally prefer the owner of the most recently deposited scent
(e.g. Johnston et al., 1997; Fig.11.3a). However, females can also use the presence
of scent marks from any males in an owner’s territory to make a much more
general assessment of each territory owner’s competitive ability when choosing
between territory owners. By manipulating the scent marks in equivalent male
territories, Rich & Hurst (1998) showed that female house mice prefer the owners
of exclusively scent-marked territories (those containing no scents from competi-
tor males) over neighbouring males whose territories contain some competing
counter-marks from an intruder male (Fig. 11.3b). After exploring the males’ ter-
ritories, females spent more time sniffing and chewing a barrier to gain access
to the owner of the exclusively scent-marked territory and were more affiliative
and invited attempted mounts from this territory owner when they were allowed

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
226 J. L. Hurst
Fig. 11.3. Effects of counter-marking between males on female preferences in
different types of test. (a) Female encounters a single scent mark from male A that
is deposited on top of scent from male B (e.g. Johnston et al., 1997). (b) Females
explore neighbouring male scent-marked territories, one of which contains
patches of intruder (i) scent counter-marking the owners scent (Rich & Hurst, 1998).
(c) Both neighbouring male territories contain patches of intruder (i) scent (Rich &
Hurst, 1999).
to interact. When given a choice between two males whose territories both con-
tained some intruder scent marks, but one owner had counter-marked the intruder
scents while some of the other owner’s scent had been counter-marked by the in-
truder, females preferred the owner that had counter-marked the intruder’s scent
(Fig. 11.3c; Rich & Hurst, 1999). In both cases, intruder scent marks came from
unfamiliar males; therefore, females were not making a simple choice between
two interacting males. Neither were females simply responding to the freshest
scents encountered because both territories contained fresh scent marks from the

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 227
territory owners. Female preferences resulted from their assessment of the pattern
of male scent marks and counter-marks as an indicator of each male’scompetitive
ability. Scent marks were manipulated in such a way that only the females were
exposed to the manipulation, while the territory-owning males were temporar-
ily removed from their territories, so responses were not a result of any changes
in male behaviour in response to the intruder scent marks introduced into each
male’s territory.
Female mice appear to distinguish the most recent scent (i.e. the counter-mark)
by the age difference between the male’s scents (Rich & Hurst, 1999). When both
territory owners’ and intruders’ scents were of very similar age, females failed to
show a preference in favour of an owner that had counter-marked intruder scents.
This could be because females were unable to discriminate between scent marks
and counter-marks without a substantial age difference in the scents (24 hours
in these experiments), or because the similar age of the scents indicated that the
competition between the males had yet to be resolved.
Males can thus gain a reproductive advantage from scent marking their territo-
ries and from counter-marking the scents of any competitors to ensure that their
own scent marks are those most recently deposited. Although it is often assumed
that the very high rates of marking at borders between neighbouring territory
owners are signals to warn neighbours to keep out, the main function of frequent
scent marking at shared borders may be the need of both territory owners to en-
sure that their own scent is as fresh as their neighbour’s wherever their scents are
in close proximity. This would require both animals continually to refresh their
scent at a shared border as an advertisement to females in the locality (including
females resident in the male’s territory and those living in neighbouring areas
(Hurst & Rich, 1999)). Male territory owners can thus compete with each other
simply through their scent-marking behaviour because competitive scent marks
are used by females when selecting a mate.
Assessment by males
Third-party competitors also appear to use the record of competitive scent-
mark signals to identify and avoid challenging owners that are defending their
territory effectively against other males. Unfamiliar intruders use the scent marks
deposited around a territory to identify the territory owner and are much less
likely to challenge a male whose individual scent signature matches the local
scent marks than a male whose scent does not match (Gosling & McKay, 1990).
Adding a small drop of fresh urine from the territory owner onto one of the own-
ers’ scent marking posts increases the frequency with which intruders and resi-
dent subordinates spontaneously flee when they encounter the owner, without
any attack or pursuit. In contrast, addition of urine from a neighbouring territory
owner reduces their evasion and increases challenges against the territory owner,

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
228 J. L. Hurst
regardless of their own previous experience of the high competitive ability of
the territory owner during direct interactions (Hurst, 1993; Hurst & Rich, 1999).
Thus, territory owners also appear to gain a strong advantage in competitive in-
teractions with other males through broadcasting their scent around the territory
and through advertising their ability to overcome challenges from other males,
reducing their need to invest in direct aggression. Other third-party males gain
by avoiding challenging a male that is successfully defending his territory against
other competitors but will also rapidly detect when the owner is struggling to
maintain dominance (for example, if his competitive ability is reduced by ageing,
injury or disease). This may be particularly important to resident subordinates,
which are likely to be highly familiar with the greater competitive ability of the
dominant male and reluctant to challenge if this might result in their exclusion
from the territory.
Deposition patterns and scent age
In contrast to isolated or subordinate (non-competitive) mice, males that
are advertising territory ownership or competing to establish a territory change
their pattern of scent marking by scattering their urine in a much larger number
of streaks and small spots. While this helps to ensure that their scent is distributed
throughout the territory, numerous scent marks are deposited close together in
the same local area (see Fig. 11.1b). When counter-marking another male’s scent,
mice do not attempt to deposit a bigger scent mark than that of the competitor,
which would contain a greater intensity of volatile signalling molecules. Instead,
they deposit many small marks in the vicinity, returning repeatedly to add more
scent marks usually over a period of several hours (Humphries et al., 1999). By drib-
bling out their urine rather than depositing it all in one go, they are maximizing
the freshness of their scent marks by increasing the rate of replenishment (Hurst
et al., 2001a). Thus, each time they deposit a new scent mark, they are increas-
ing the age difference between their own scent and that of the competitor, while
volatiles in their own fresh scent attract the attention of others to the aged scents
of a competitor. Notably, males counter-mark both fresh and aged scents from
competitors but deposit most marks near to the aged competitor scent where the
contrast will be greatest (Humphries et al., 1999). Most male signalling volatiles
are lost from scents within a few hours (Hurst et al., 1998; Humphries et al., 1999;
Robertson et al., 2001), but non-volatile components of scent marks continue to be
detected for at least seven days if males are aware of the presence of scent marks
in the area (Humphries et al., 1999, 2001).
Both volatile and non-volatile components of a scent mark are likely to be in-
volved in providing a reliable signal of scent-mark age. While volatile components

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 229
will be lost as a scent mark ages, the intensity of a volatile signal at any point
in time will depend on the amount initially deposited as well as the time since
deposition. Receivers will not be able to assess the age of the mark from a volatile
signal alone without knowing the amount that was deposited (Hurst et al., 2001b).
In contrast, non-volatile components are not lost through time and may provide a
measure of the amount of scent deposited. As yet, we do not know the molecular
mechanism used to assess scent-mark age, but MUPs that bind and slowly release
volatile ligands in mouse urine have the capacity to provide a very reliable indi-
cator of scent-mark age. Because each protein molecule can only bind one ligand
molecule, and ligands are slowly released and evaporate from the scent mark,
the proportion of protein molecules that contain ligands will decrease with time
since deposition. Making one component the ligand of the other defines implic-
itly the relationship between them. By contrast, the ratio between two unrelated
volatile components that have different rates of evaporation requires that the re-
ceiving animal knows the ratio between them at the time of deposition. Although
MUPs are not odorants, these non-volatile proteins appear to be detected through
the vomeronasal system (Brennan et al., 1999; Krieger et al., 1999). Direct contact
with the scent source appears to be essential for activation of this system, sug-
gesting that non-volatile components like MUPs are necessary to deliver volatile
pheromones to the vomeronasal organ (Luo et al., 2003). The release of volatiles
from MUPs also plays an important function in alerting mice to the presence of a
scent mark (Hurst et al., 1998; Humphries et al., 1999).
Advertising subordinate status
Females are attracted by sex-specific volatiles in the urine of adult
male mice ( Jemiolo et al., 1985, 1991), which also act as reproductive priming
pheromones to stimulate female oestrus cycling (see below). These same male sig-
nalling volatiles elicit aggression from other competitive males (Novotny et al.,
1985), or avoidance by subordinates ( Jones & Nowell, 1989; Novotny et al., 1990;
Gosling et al., 1996; Mucignat-Caretta et al., 1998). However, subordinate males
that live within the territory of another male and are defeated repeatedly by the
dominant territory owner reduce their production of these male-specific volatiles
( Jones & Nowell, 1989; Harvey et al., 1989); their preputial glands are smaller than
those of dominant males (Hucklebridge et al., 1972; Bronson & Marsden, 1973) and
they are much less likely to initiate competitive interactions (Crowcroft & Rowe,
1963; Hurst, 1987). As a consequence of these changes in scent quality, subordinate
male urine is no longer attractive to females (Bronson & Caroom, 1971; Jones &
Nowell, 1974; Jemiolo et al., 1991) and females will not mate with subordinate
males (Wolff, 1980; Hurst, 1987; Potts et al., 1991). In compensation, subordinate

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
230 J. L. Hurst
male odours elicit less aggression from other males (Mugford & Nowell, 1970; Jones
& Nowell, 1973, 1975; Novotny et al., 1985). Subordinate male mice also show a dra-
matic and immediate reduction in scent-marking behaviour, although this is not
completely suppressed and they continue to deposit scent marks around their
home area in larger spots and pools (Desjardins et al., 1973; Sandnabba, 1986).
Because their urinary scent differs in quality from that of dominant males, these
scent marks advertise their subordinate status within the territory to all other
animals in the area, including females.
Why should subordinates advertise their low quality in such a public manner?
Experimental manipulations of these substrate scents indicate that they are criti-
cal in determining tolerance of the subordinate by other resident males. Male mice
generally attempt to exclude other adult males from their scent-marked territo-
ries (relatives or non-relatives) and they are highly aggressive towards unfamiliar
mice or familiar neighbours that intrude into the territory even if intruders are
of subordinate status (Barnard et al., 1991). However, complete exclusion can be
extremely difficult to achieve in complex habitats where persistent males can hide
(Crowcroft, 1966; Poole & Morgan, 1976). Familiar males living in the same terri-
tory establish a social structure in which one male becomes dominant and main-
tains dominance over familiar subordinates through brief attacks and aggressive
postures, rather than attempting to evict the subordinates from the territory. If
these familiar males are housed in separate cages but their soiled cage substrate is
regularly mixed to maintain their contact with group scent cues, males continue
to be relatively tolerant of each other. However, if a familiar subordinate male
is suddenly prevented from contributing fresh scent to the mixed group-marked
substrate, although the subordinate itself continues to encounter group substrate
scents as if it were still a group member, within 24–48 hours both the resident
dominant and other subordinate males in the group start to investigate and to
attack and chase this male as if he was no longer a tolerated group member (Hurst
et al., 1993). In contrast, scent marks deposited to compete with the signals of the
dominant territory owner, for example on territorial scent-marking posts, induce
attack against the familiar subordinate (Hurst, 1993).
Competitive pressure from dominant male territory owners thus appears to
force subordinate males both to change the quality of their urinary scent and to
deposit urinary scent marks that advertise their subordinate status to any animals
using or visiting the area. Because the scent marks remain in the environment once
deposited, and carry the subordinate’s individual identity signature (see below), a
subordinate would be unable to cheat by altering the quality of his scent during
direct interactions with females. In order to be tolerated and allowed to remain
within another male’s territory, subordinates appear to be forced to broadcast
honest signals of their low competitive ability at the cost of their reproductive

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 231
success, because their scent marks will be encountered by females. The outcome
of this is to reduce considerably the risk that subordinate males might compete
for females through sneaky matings (mating with subordinate males is very rarely
observed in seminaturalistic studies of mouse populations), making it safe for the
territory owner to tolerate subordinates within the territory that contribute to
group substrate scents. Notably, experiments have revealed that animals of low
competitive ability only suppress the production of competitive male scents when
they live in very close proximity to a dominant individual; they do not suppress
scent signals simply in response to defeat by another male. Jones and Nowell (1989)
confirmed that, if males are repeatedly defeated by a higher quality competitor
and are kept in continuous olfactory contact with the scent of their victor (as if
they lived within his territory), the defeated subordinate male’s scent loses the
aversive effect on other males caused by male signalling volatiles. However, if
males experience the same frequency of defeat but are housed in separate cages
from their victor (as if they could escape to another territory), the defeated male’s
scent retains the high levels of male signalling volatiles that are aversive to other
males. Not surprisingly, males do not advertise their low quality unless they are
forced to do so by the constant threat of attack and displacement from higher
quality competitors.
Female reproductive priming
Female reproductive physiology is strongly influenced by a number of
reproductive priming pheromones in the urine of male or female mice (reviewed
by Brown, 1985b; Novotny et al., 1999a). Volatile priming pheromones in the urine
of adult male mice have stimulatory effects on female physiology, accelerating
puberty in young females (Vandenberg, 1969; Novotny et al., 1999b) and stimulat-
ing oestrous cycling (Whitten, 1956; Jemiolo et al., 1986), while urinary odours
from pregnant or lactating females have similar though not identical effects
(Drickamer & Hoover, 1979; Hoover & Drickamer, 1979). The stimulatory effects of
urine from pregnant or lactating females may reflect the preference of house mice
for communal nesting, because females raise more offspring when cooperating
with another female than they can when breeding alone (Konig, 1994a). In contrast,
nestling survival is greatly reduced in overcrowded nest sites, particularly among
females of low social status (Southwick, 1955; Hurst, 1987). Accordingly, when
non-breeding females live in groups with several other non-breeding females, or
have frequent contact with the urine of other non-breeding females, they produce
a priming pheromone in their own urine that inhibits oestrous cycling in adults
(Champlin, 1971) and delays puberty in prepubertal females (Drickamer, 1977;
Jemiolo & Novotny, 1994). However, females of high social status are not affected

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
232 J. L. Hurst
and continue to breed (Lloyd & Christian, 1969). Lastly, if a female encounters the
scent of a novel male within four days of mating, implantation fails and she will
abort if she is not protected by continued exposure to urine from the familiar
stud male, a phenomenon known as the Bruce effect (Bruce, 1959; Brennan, 1999).
This is likely to provide females with the opportunity to mate with a new territory
owner if their previous mating partner is displaced. Since male territory owners
do not appear to discriminate against the offspring of other males born within
their territory (Hurst & Barnard, 1992), this may be a tactic to increase offspring
quality.
Although priming pheromones are volatile, these are detected through the
vomeronasal system and animals must make contact with the scent source to
allow chemical stimuli to be pumped to the vomeronasal organ (Brown, 1985b;
Luo et al., 2003). Under natural conditions, females are surrounded by urinary scent
marks from all individuals using the same sites, exposing them to both stimulatory
and inhibitory priming pheromones. Because these urinary scents are very widely
distributed and females do not appear deliberately to control their exposure to
these cues (Hurst & Nevison, 1994), this network of scent signals from all animals
using the same area appears to provide a mechanism for females to adjust their
own reproductive physiology appropriately, according to the current local social
conditions and to the individual’s own age and social status.
Individual scent signatures
Scent marks are deposited in the environment to provide information in
the absence of the signaller (unlike other types of signal), so it is essential that they
provide stable and persistent information about the donor’s individual identity.
Ideally, individuality scents should be ‘hard-coded’ in the individual’s genome,
exhibit a high degree of individual polymorphism to uniquely identify the donor
and be expressed by all individuals regardless of social status or sex (Beynon et
al., 2001). Attention has focused largely on the volatile components of scents as
sources of individuality signatures, particularly those associated with the highly
polymorphic major histocompatibility complex (MHC) odortypes, although many
other genetic loci also influence individual differences in urinary volatile profiles
(Boyse et al., 1987; Beauchamp et al., 1990; Eggert et al., 1996). The MHC encodes
for glycoproteins involved in individual (self versus non-self) recognition at the
cellular level but has also been shown to affect the volatile scent signals produced
by animals such as mice, rats, fish and humans (reviewed by Jordan & Bruford,
1998; Singh, 2001; see also Olsen et al., 1998; Reusch et al., 2001; Jacob et al., 2002).
Laboratory studies using MHC congenic strains of mice and rats have confirmed
that rodents are able to discriminate differences in urinary odours from donors

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 233
that differ genetically at alleles within the MHC region, even when donors differ
only at a single MHC locus (Yamazaki et al., 1999; Singh, 2001; Carroll et al., 2002).
Although the molecular basis of MHC-associated odours is not known, it appears to
involve a complex mixture of volatile metabolites bound and released by urinary
proteins (Singer et al., 1993, 1997). One hypothesis is that soluble fragments of MHC
class I and class II molecules in urine differentially bind volatile metabolites in
the antigen-binding groove once the peptide normally bound in this groove is lost
(Singh, 2001). Alternatively, MHC haplotype may affect the volatile metabolites
that are released into the urine (Singer et al., 1997; Yamazaki et al., 1999). These
MHC-dependent volatiles might then be bound and released by the MUPs, which
are present at protein concentrations up to a million times higher than MHC
class I molecule fragments and possess a large flexible binding pocket for small
lipophilic (and thus potentially volatile) molecules (Beynon et al., 2001).
Although MHC polymorphism results in differences in scents that are discrim-
inable by mice, these volatile signals appear to be easily disrupted by environ-
mental factors that affect an individual’s metabolite profile, such as changes in
food type, bacterial gut flora or social status (reviewed by Brown, 1995; Nevison
et al., 2000). This presents a problem for an individual recognition signature, sug-
gesting that MHC-associated odours may not provide sufficient stability or per-
sistence to act as individuality signals in scent marks. Indeed, mice do not use
MHC-associated odours to discriminate their own scent marks from those of other
males, despite their clear ability to detect differences in their own MHC type and
those of other individuals ( J. L. Hurst, unpublished data). Instead, they use the
different patterns of MUPs that individual mice express in their urine (Hurst et al.,
2001b).
MUPs are coded by a multigene family on chromosome 4 and are expressed
at high concentration by adult house mice of both sexes, although males invest
more than females in both scent marking and MUP production (Beynon et al.,
2001; Payne et al., 2001; Beynon & Hurst, 2003). MUPs exhibit a very high level
of genetic polymorphism and individual mice express a combination of MUPs
(typically at least 7–12) such that the combinatorial diversity of individual MUP
profiles among wild mice may be as great as for MHC (Robertson et al., 1997;
Beynon et al., 2001, 2002; Payne et al., 2001). These individual-specific patterns of
urinary MUPs in the scent marks of wild mice appear to be essential in allowing
outbred wild mice to distinguish another individual’s scent marks from their
own, regardless of differences at many other genetic loci such as MHC (Hurst et al.,
2001b). If a male territory owner encounters a competing scent mark from another
male in his territory, he normally investigates closely and then counter-marks
and spends time in the vicinity of the scent mark. However, males only recognize
an intruder’s scent mark if it carries a different MUP pattern from their own.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
234 J. L. Hurst
Scent marks with their own MUP pattern draw initial investigation but no further
response (Hurst et al., 2001b; J. L. Hurst, unpublished data). This is not because
males avoid competing with a relative (i.e. kin discrimination) as they respond
strongly to urinary scents of different MUP type whether from a close relative
or an unrelated male. In contrast, if their own urinary MUP profile is altered by
adding a recombinant MUP to their urine, males counter-mark as if the scent was
from an intruder (Hurst et al., 2001b). There is a high degree of individual variability
in MUP patterns expressed by males captured from the same population (Payne
et al., 2001) and MUPs show little or no degradation in scent marks (Hurst et al.,
2001a). These patterns, therefore, provide a stable and persistent individual scent
signature that is hard-coded in the individual’s genome and remains constant
throughout the individual’s lifetime.
Kin and group member recongnition
Inherited scent signatures are also important in allowing animals to rec-
ognize whether others are likely to be close relatives. Many genetic differences
appear to contribute to inherited scents used for kin recognition, including MHC-
associated odours. For example, female mice prefer to rear their offspring com-
munally with close relatives rather than with unrelated females, and offspring
survival is greater when cooperating with a familiar sister (Konig, 1994b). When
unfamiliar female mice are mixed together in seminatural populations, they ap-
pear to recognize the similar scents of other females of the same MHC type as
themselves and are more likely to share nests with these females (Manning et al.,
1992).
Recognition of close relatives is particularly important to avoid inbreeding.
There is considerable evidence that MHC type affects female preference between
male scents, with females generally preferring the scents of males of different MHC
type to themselves or their parents (e.g. Egid & Brown, 1989; Penn & Potts, 1998).
By crossing wild mice with laboratory strains to create wild-type mice of known
homozygous MHC type, Potts et al. (1991) confirmed that females in seminatural
populations showed a significant preference for MHC-disassortative mating. Inter-
estingly, females showed no MHC bias when mating with the owner of the territory
in which they nested (suggesting that MHC type did not influence territory prefer-
ence despite the genetic information in the owner’s scent marks). However, when
females went outside their territory to mate, they preferred owners of neighbour-
ing territories that had a different MHC type to their own familiar MHC-associated
odours.
In mice, the mechanism of kin recognition appears to be largely through im-
printing on scents experienced during early life rather than on cues inherited by

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 235
the female herself (D’Udine & Alleva, 1983). Mating preferences can be reversed
by cross-fostering. Females fostered onto a different strain will later avoid mating
with males related to their foster parents rather than with those related to them-
selves (e.g. Penn & Potts, 1998; though see Eklund, 1997). Because relatives will
carry a much greater range of alleles than the female herself, imprinting on the
scents of relatives experienced in their early environment may be more effective
than phenotype matching to self for recognizing potential relatives or those from
a similar genetic background.
In addition to inherited scents, mice acquire scents on their bodies from other
group members that influence recognition when mice interact (Aldhous, 1989).
All group members are likely to become tainted with the scents of the resident
territory owner from the sticky scent marks deposited throughout the area. This
acquired group scent may make an important contribution to the ability of mice
to recognize their own group members regardless of their inherited scents (e.g.
Hurst & Barnard, 1992, 1995).
Scent marking as broadcast signals
Animals spontaneously scent mark their territories in the absence of in-
teraction with others, although scent marking is usually significantly enhanced
by competitive and sexual interaction with others or with their scent marks. In
addition to the chemical information in an individual’s scent, the spatial and
temporal patterning of scent marks from all individuals in a locality provides
information about their social and genetic relationships. Scent marks are partic-
ularly suitable for network communication between many individuals. By their
very nature, they are long lasting and readily available for inspection by any indi-
viduals that visit a scent-marked site. Conversely, scent marks are not appropriate
for private communication (see Ch. 3) unless individual access to the scent marked
site is restricted.
There are two ways in which scent marking might be used in a signalling net-
work. First, scent marks may be deposited as broadcast signals, designed to com-
municate information to all other animals in the area. Alternatively, scents may
be deposited as signals to specific individuals, with third parties making use of
information in scent marks and counter-marks by eavesdropping on the commu-
nication between others. Eavesdropping has been defined as ‘extracting informa-
tion from signalling interactions between others’ (McGregor & Dabelsteen, 1996;
McGregor & Peake, 2000; Ch. 2). The implication here is that signals are designed
to provide information to one or more interacting individuals (e.g. during the in-
teraction between two competitors), not to provide information to eavesdroppers.
However, eavesdroppers make use of this information to their own advantage. It

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
236 J. L. Hurst
is, therefore, important to establish who gains an advantage from the use of these
signals.
With respect to competitive counter-marking by territory owners in response
to aggressive challenges and scent marking from other males, the main fitness ad-
vantage to signallers appears to be the response of females to these scent marks,
although females may be viewed as ‘third parties’ responding to signalling inter-
actions between males (see Ch. 7 for parallels with bird song). Evidence from our
scent-manipulation studies in house mice indicates that these scent signals have
highly significant effects on female preferences, in accordance with our hypothe-
sis that this is a very reliable way to assess the competitive ability of different po-
tential mates. Similarly, territorial scent marking and the counter-marking of any
intruder scents by the owner increases avoidance responses and decreases compet-
itive challenges from third-party males (i.e. males that are not the owners of either
the scent marks or counter-marks). As this affects the responses of all males in the
vicinity, this is likely to have a big impact on the ease with which males defend
their territories. Therefore, a successful territory owner gains clear advantages
from advertising his territory ownership and competitive ability widely, includ-
ing his ability to overcome the challenges of competitors. While increased scent
marking to counter-mark a competitor’s scent might, at first sight, appear to be a
signalling interaction between two competitors, the main selective advantage to
successful competitors comes from broadcasting this information to all animals in
the vicinity. There is no evidence that counter-marking the scent of an aggressive
competitor reduces challenges from the competitor itself without direct aggres-
sion, although once defeated, competitors will show a generalized avoidance of
competitor scents (e.g. Jones & Nowell, 1989; Hurst et al., 1997). McGregor & Peake
(2000) pointed out that there seem to be few demonstrated advantages to signallers
of communicating in the social environment of a network. However, competitive
scent marking provides an excellent example of the advantages that successful
territory owners can gain from depositing competitive scent marks within the
network of signals from other males, because this provides a mechanism for the
reliable advertisement of their own competitive ability. These scent marks are
clearly broadcast signals, designed to communicate information to all other ani-
mals in the area. As such, the concept of eavesdropping on signals aimed at others
does not seem to be appropriate.
The deposition of scent signals that advertise an individual’s subordinate sta-
tus appears to be enforced by the resident territory owner and other resident
subordinate males, and thus subordinate scent marking might be viewed as a
signal aimed principally at local competitors to reduce aggression against the
subordinate. However, the selective advantage to local competitors of reducing
their aggression is that such scent marks broadcast the subordinate status and

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 237
identity of a competitor to females. Subordinates that signal their subordinate
status through their body scent but not through substrate scent marks are not
tolerated. From an evolutionary viewpoint, it again seems more appropriate to
view scent marks deposited to advertise subordinate status as broadcast signals,
aimed at publicising this information to others despite the immediate reduction
in reproductive opportunities to the signaller from doing so. This at least allows
unsuccessful males to remain in a suitable habitat with the potential to become
a successful territory owner, and gain reproductive success, in the future.
Summary
In conclusion, since scent marks persist in the environment and cannot be
directed towards specific recipients (unlike most visual and acoustic signals), scent
marks are only likely to be used as broadcast signals and are used in social contexts
where the signaller can gain an advantage from communicating information to
a public audience.
Acknowledgements
I am grateful to Rob Beynon for providing Fig. 11.2, for invaluable discussion and for
comments on the manuscript. Research carried out in my laboratory was supported by grants
from the Biotechnology and Biological Sciences Research Council.
References
Aldhous, P. 1989. The effects of individual cross-fostering on the development of
intrasexual kin discrimination in male laboratory mice. Animal Behaviour, 37,
741–750.
Andreolini, F., Jemiolo, B. & Novotny, M. 1987. Dynamics of excretion of urinary
chemosignals in the house mouse (Mus musculus) during the natural estrous cycle.
Experientia, 43, 998–1002.
Bacchini, A., Gaetani, E. & Cavaggioni, A. 1992. Pheromone binding proteins of the
mouse, Mus musculus. Experientia, 48, 419–421.
Barnard, C. J., Hurst, J. L. & Aldhous, P. 1991. Of mice and kin: the functional
significance of kin bias in social behaviour. Biological Reviews of the Cambridge
Philosophical Society, 66, 379–430.
Beauchamp, G. K., Yamazaki, K., Duncan, H., Bard, J. & Boyse, E. A. 1990. Genetic
determination of individual mouse odour. In: Chemical Signals in Vertebrates 5, ed.
D. W. Macdonald, S. Natyncuk & D. Muller-Schwarze. Oxford: Oxford University
Press, pp. 244–255.
Beynon, R. J. & Hurst, J. L. 2003. Multiple roles of major urinary proteins in the house
mouse, Mus domesticus. Biochemical Society Transactions, 31, 142–146.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
238 J. L. Hurst
Beynon, R. J., Hurst, J. L., Gaskell, S. J. et al. 2001. Mice, MUPs and myths:
structure-function relationships of the major urinary proteins. In: Chemical Signals
in Vertebrates, ed. A. Marchelewska-Koj, D. Muller-Schwarze & J. Lepri. New York:
Plenum Press, pp. 149–156.
Beynon, R. J., Veggerby, C., Payne, C. E. et al. 2002. Polymorphism in major urinary
proteins: molecular heterogeneity in a wild mouse population. Journal of Chemical
Ecology, 28, 1429–1446.
Boyse, E. A., Beauchamp, G. K. & Yamazaki, K. 1987. The genetics of body scent. Trends
in Genetics, 3, 97–102.
Brennan, P. A. 1999. Bruce effect. In: Encyclopedia of Reproduction, ed. E. Knobil & J. Neill.
San Diego, CA: Academic Press, pp. 433–438.
2001. The vomeronasal system. Cellular and Molecular Life Science, 58, 546–555.
Brennan, P. A., Schellinck, H. M. & Keverne, E. B. 1999. Patterns of expression of the
immediate-early gene egr-1 in the accessory olfactory bulb of female mice exposed
to pheromonal constituents of male urine. Neuroscience, 90, 1463–1470.
Bronson, F. H. & Caroom, D. 1971. Preputial gland of the male mouse; attractant
function. Journal of Reproductive Fertility, 25, 279–282.
Bronson, F. H. & Marsden, H. M. 1973. The preputial gland as an indicator of social
dominance in male mice. Behavioral Biology, 9.
Brown, R. E. 1985a. The rodents II: suborder Myomorpha. In: Social Odours in Mammals,
ed. R. E. Brown & D. W. Macdonald. Oxford: Clarendon Press, pp. 345–457.
1985b. The rodents I: effects of odours on reproductive physiology (primer effects).
In: Social Odours in Mammals, ed. R. E. Brown & D. W. Macdonald. Oxford:
Clarendon Press, pp. 245–344.
1995. What is the role of the immune system in determining individually distinct
body odours? International Journal of Immunopharmacology, 17, 655–661.
Bruce, H. M. 1959. An exteroceptive block to pregnancy in the mouse. Nature, 184, 105.
Carroll, L. S., Penn, D. J. & Potts, W. K. 2002. Discrimination of MHC-derived odors by
untrained mice is consistent with divergence in peptide-binding region residues.
Proceedings of the National Academy of Sciences, USA, 99, 2187–2192.
Champlin, A. K. 1971. Suppression of oestrus in grouped mice: the effects of various
densities and the possible nature of the stimulus. Journal of Reproductive Fertility,
27, 233–241.
Crowcroft, P. 1966. Mice All Over. London: Foulis.
Crowcroft, P. & Rowe, F. P. 1963. Social organization and territorial behaviour in the
wild house mouse (Mus musculus L.). Proceedings of the Zoological Society of London,
140, 517–531.
Desjardins, C., Maruniak, J. A. & Bronson, F. H. 1973. Social rank in the house mouse:
differentiation revealed by ultraviolet visualisation of urinary marking patterns.
Science, 182, 939–941.
Drickamer, L. C. 1977. Delay of sexual maturation in female house mice by exposure
to grouped females or urine from grouped females. Journal of Reproductive Fertility,
51, 77–81.
Drickamer, L. C. & Hoover, J. E. 1979. Effects of urine from pregnant and lactating
female house mice on sexual maturation of juvenile females. Developmental
Psychobiology, 12, 545–551.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 239
D’Udine, B. & Alleva, E. 1983. Early experience and sexual preferences in rodents. In:
Mate Choice, ed. P. Bateson. Cambridge, UK: Cambridge University Press,
pp. 311–327.
Eggert, F., Holler, C., Luszyk, D., Muller-Ruchholtz, W. & Ferstl, R. 1996.
MHC-associated and MHC-independent urinary chemosignals in mice. Physiology
and Behavior, 59, 57–62.
Egid, K. & Brown, J. L. 1989. The major histocompatibility complex and female mating
preferences in mice. Animal Behaviour, 38, 548–550.
Eklund, A. 1997. The major histocompatibility complex and mating preferences in
wild house mice. Behavioral Ecology, 8, 630–634.
Gosling, L. M. 1982. A reassessment of the function of scent marking in territories.
Zeitschrift fur Tierpsychologie, 60, 89–118.
Gosling, L. M. & McKay, H. V. 1990. Competitor assessment by scent matching: an
experimental test. Behavioral Ecology and Sociobiology, 26, 415–420.
Gosling, L. M., Atkinson, N. W., Dunn, S. & Collins, S. A. 1996. The response of
subordinate male mice to scent marks varies in relation to their own competitive
ability. Animal Behaviour, 52, 1185–1191.
Harvey, S., Jemiolo, B. & Novotny, M. 1989. Pattern of volatile compounds in dominant
and subordinate male-mouse urine. Journal of Chemical Ecology, 15, 2061–2072.
Hoover, J. E. & Drickamer, L. C. 1979. Effects of urine from pregnant and lactating
female house mice on oestrous cycles of adult females. Journal of Reproductive
Fertility, 55, 297–301.
Hucklebridge, F. H., Nowell, N. W. & Wouters, A. 1972. A relationship between social
experience and preputial gland function in the albino mouse. Journal of
Endocrinology, 55, 449–450.
Humphries, R. E., Robertson, D. H. L., Beynon, R. J. & Hurst, J. L. 1999. Unravelling the
chemical basis of competitive scent marking in house mice. Animal Behaviour, 58,
1177–1190.
Humphries, R. E., Robertson, D. H. L., Nevison, C. M., Beynon, R. J. & Hurst, J. L. 2001.
The role of urinary proteins and volatiles in competitive scent marking among
male house mice. In: Chemical Signals in Vertebrates, ed. A. Marchelewska-Koj, D.
Muller-Schwarze & J. Lepri. New York: Plenum Press, pp. 353–360.
Hurst, J. L. 1987. Behavioural variation in wild house mice Mus domesticus Rutty: a
quantitative assessment of female social organization. Animal Behaviour, 35,
1846–1857.
1989. The complex network of olfactory communication in populations of wild
house mice Mus domesticus Rutty: urine marking and investigation within family
groups. Animal Behaviour, 37, 705–725.
1990. Urine marking in populations of wild house mice Mus domesticus Rutty. 1.
Communication Between Males. Animal Behaviour, 40, 209–222.
1993. The priming effects of urine substrate marks on interactions between male
house mice, Mus musculus domesticus Schwarz and Schwarz. Animal Behaviour, 45,
55–81.
Hurst, J. L. & Barnard, C. J. 1992. Kinship and social behavior in wild house mice:
effects of social group membership and relatedness on the responses of dominant
males toward juveniles. Behavioral Ecology, 3, 196–206.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
240 J. L. Hurst
1995. Kinship and social tolerance among female and juvenile wild house mice: kin
bias but not kin discrimination. Behavioral Ecology and Sociobiology, 36, 333–342.
Hurst, J. L. & Nevison, C. M. 1994. Do female house mice, Mus domesticus, regulate their
exposure to reproductive priming pheromones. Animal Behaviour, 48, 945–959.
Hurst, J. L. & Rich, T. J. 1999. Scent marks as competitive signals of mate quality. In:
Advances in Chemical Communication in Vertebrates, ed. R. E. Johnson, D.
Muller-Schwarze & P. W. Sorensen. New York: Plenum Press, pp. 209–226.
Hurst, J. L., Fang, J. M. & Barnard, C. J. 1993. The role of substrate odours in
maintaining social tolerance between male house mice, Mus musculus domesticus.
Animal Behaviour, 45, 997–1006.
Hurst, J. L., Gray, S. J., Davey, P. et al. 1997. Social interaction alters attraction to
competitor’s odour in the mouse Mus spretus Lataste. Animal Behaviour, 54, 941–953.
Hurst, J. L., Robertson, D. H. L., Tolladay, U. & Beynon, R. J. 1998. Proteins in urine scent
marks of male house mice extend the longevity of olfactory signals. Animal
Behaviour, 55, 1289–1297.
Hurst, J. L., Beynon, R. J., Humphries, R. E. et al. 2001a. Information in scent signals of
competitive social status: the interface between behaviour and chemistry. In:
Chemical Signals in Vertebrates, ed. A. Marchelewska-Koj, D. Muller-Schwarze & J.
Lepri. New York: Plenum Press, pp. 43–52.
Hurst, J. L., Payne, C. E., Nevison, C. M. et al. 2001b. Individual recognition in mice
mediated by major urinary proteins. Nature, 414, 631–634.
Jacob, S., McClintock, M. K., Zelano, B. & Ober, C. 2002. Paternally inherited HLA alleles
are associated with women’s choice of male odour. Nature Genetics, 30, 175–179.
Jemiolo, B. & Novotny, M. 1993. Long-term effect of a urinary chemosignal on
reproductive fitness in female mice. Biology of Reproduction, 48, 926–
929.
1994. Inhibition of sexual maturation in juvenile female and male mice by a
chemosignal of female origin. Physiology and Behavior, 55, 519–522.
Jemiolo, D., Alberts, J., Sochinski-Wiggins, S., Harvey, S. & Novotny, M. 1985.
Behavioural and endocrine responses of female mice to synthetic analogs of
volatile compounds in male urine. Animal Behaviour, 33, 1114–1118.
Jemiolo, B., Harvey, S. & Novotny, M. 1986. Promotion of the Whitten effect in female
mice by synthetic analogs of male urinary constituents. Proceedings of the National
Academy of Sciences, USA, 83, 4576–4579.
Jemiolo, B., Andreolini, F., Wiesler, D. & Novotny, M. 1987. Variations in mouse (Mus
musculus) urinary volatiles during different periods of pregnancy and lactation.
Journal of Chemical Ecology, 13, 1941–1956.
Jemiolo, B., Xie, T. M. & Novotny, M. 1991. Socio-sexual olfactory preference in female
mice: attractiveness of synthetic chemosignals. Physiology and Behavior, 50,
1119–1122.
Johnston, R. E. 1999. Scent over-marking. How do hamsters know whose scent is on
top and why should it matter? In: Advances in Chemical Signals in Vertebrates, ed.
R. E. Johnson, D. Muller-Schwarze & P. W. Sorensen. New York: Plenum Press,
pp. 227–238.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 241
Johnston, R. E., Sorokin, E. S. & Ferkin, M. H. 1997. Female voles discriminate males’
over-marks and prefer top-scent males. Animal Behaviour, 54, 679–690.
Jones, R. B. & Nowell, N. W. 1973. Aversive and aggression-promoting properties of
urine from dominant and subordinate male mice. Animal Learning and Behavior, 1,
207–210.
1974. A comparison of the aversive and female attractant properties of urine from
dominant and subordinate male mice. Animal Learning and Behavior, 2,
141–144.
1975. Effects of clean and soiled sawdust substrates and of different urine types
upon aggressive behaviour in male mice. Aggressive Behavior, 1, 111–121.
1989. Aversive potency of urine from dominant and subordinate male laboratory
mice (Mus musculus): resolution of a conflict. Aggressive Behavior, 15, 291–296.
Jordan, W. C. & Bruford, M. W. 1998. New perspectives on mate choice and the MHC.
Heredity, 81, 239–245.
Konig, B. 1994a. Components of lifetime reproductive success in communally and
solitarily nursing house mice: a laboratory study. Behavioral Ecology and
Sociobiology, 34, 275–283.
1994b. Fitness effects of communal rearing in house mice: the role of relatedness
versus familiarity. Animal Behaviour, 48, 1449–1457.
Krieger, J., Schmitt, A., Lobell, D. et al. 1999. Selective activation of G protein subtypes
in the vomeronasal organ upon stimulation with urine-derived compounds.
Journal of Biological Chemistry, 274, 4655–4662.
Lloyd, J. A. & Christian, J. J. 1969. Reproductive activity of individual females in three
experimental freely growing populations of house mice (Mus musculus L.). Journal
of Mammalogy, 50, 49–59.
Luo, M., Fee, M. S. & Katz, L. C. 2003. Encoding pheromonal signals in the accessory
olfactory bulb of behaving mice. Science, 299, 1196–1201.
Malone, N., Payne, C. E., Beynon, R. J. & Hurst, J. L. 2001. Social status, odour
communication and mate choice in wild house mice. In: Chemical Signals in
Vertebrates, ed. A. Marchelewska-Koj, D. Muller-Schwarze & J. Lepri. New York:
Plenum Press, pp. 217–224.
Manning, C. J., Wakeland, E. K. & Potts, W. K. 1992. Communal nesting patterns in
mice implicate MHC genes in kin recognition. Nature, 360, 581–583.
Maruniak, J. A., Desjardins, C. & Bronson, F. H. 1975. Adpatations for urinary marking
in rodents: prepuce length and morphology. Journal of Reproductive Fertility, 44,
567–570.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
Mucignat-Caretta, C., Caretta, A. & Baldini, E. 1998. Protein-bound male urinary
pheromones: differential responses according to age and gender. Chemical Senses,
23, 67–70.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
242 J. L. Hurst
Mugford, R. A. & Nowell, N. W. 1970. Pheromones and their effect on aggression in
mice. Nature, 226, 967–968.
Nevison, C. M., Barnard, C. J., Beynon, R. J. & Hurst, J. L. 2000. The consequences of
inbreeding for recognising competitors. Proceedings of the Royal Society of London,
Series B, 267, 687–694.
Nibu, K. 2002. Introduction to olfactory neuroepithelium. Microscopy Research and
Technique, 58, 133–134.
Novotny, M., Schwende, F. J., Wiesler, D. Jorgenson, J. W. & Carmack, M. 1984.
Identification of a testosterone-dependent unique volatile constituent of male
mouse urine: 7-exo-ethyl-5-methyl-6,8- dioxabicyclo(3.2.1)-3-octene. Experientia, 40,
217–219.
Novotny, M., Harvey, S., Jemiolo, B. & Alberts, J. 1985. Synthetic pheromones that
promote inter-male aggression in mice. Proceedings of the National Academy of
Sciences, USA, 82, 2059–61.
Novotny, M., Jemiolo, B., Harvey, S., Wiesler, D. & Marchlewska-Koj, A. 1986.
Adrenal-mediated endogenous metabolites inhibit puberty in female mice.
Science, 231, 722–725.
Novotny, M., Harvey, S. & Jemiolo, B. 1990. Chemistry of male dominance in the house
mouse, Mus domesticus. Experientia, 46, 109–113.
Novotny, M. V., Ma, W., Zidek, L. & Daev, E. 1999a. Recent biochemical insights into
puberty acceleration, estrus induction and puberty delay in the house mouse. In:
Advances in Chemical Communication in Vertebrates, ed. R. E. Johnson, D.
Muller-Schwarze & P. W. Sorensen. New York: Plenum Press, pp. 99–116.
Novotny, M. V., Ma, W., Wiesler, D. & Zidek, L. 1999b. Positive identification of the
puberty-accelerating pheromone of the house mouse: the volatile ligands
associating with the major urinary protein. Proceedings of the Royal Society of London,
Series B, 266, 2017–2022.
Olsen, K. H., Grahn, M., Lohm, J. & Langefors, A. 1998. MHC and kin discrimination in
juvenile Arctic charr, Salvelinus alpinus (L.). Animal Behaviour, 56, 319–327.
Payne, C. E., Malone, N., Humphries, R. E. et al. 2001. Heterogeneity of major urinary
proteins in house mice: population and sex differences. In: Chemical Signals in
Vertebrates, ed. A. Marchelewska-Koj, D. Muller-Schwarze & J. Lepria. New York:
Plenum Press, pp. 233–240.
Penn, D. & Potts, W. 1998. MHC-disassortative mating preferences reversed by
cross-fostering. Proceedings of the Royal Society of London, Series B, 265, 1299–
1306.
Poole, T. B. & Morgan, H. D. R. 1976. Social and territorial behaviour in mice (Mus
musculus L.) in small complex areas. Animal Behaviour, 24, 476–480.
Potts, W. K., Manning, C. J. & Wakeland, E. K. 1991. Mating patterns in seminatural
populations of mice influenced by MHC genotype. Nature, 352, 619–621.
Ralls, K. 1971. Mammalian scent marking. Science, 171, 443–449.
Reusch, T. B., Haberli, M. A., Aeschlimann, P. B. & Milinski, M. 2001. Female
sticklebacks count alleles in a strategy of sexual selection explaining MHC
polymorphism. Nature, 414, 300–302.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
Scent marking and social communication 243
Rich, T. J. & Hurst, J. L. 1998. Scent marks as reliable signals of the competitive ability
of mates. Animal Behaviour, 56, 727–735.
1999. The competing counter-marks hypothesis: reliable assessment of competitive
ability by potential mates. Animal Behaviour, 58, 1027–1037.
Robertson, D. H. L., Beynon, R. J. & Evershed, R. P. 1993. Extraction, characterization
and binding analysis of two pheromonally active ligands associated with major
urinary protein of house mouse (Mus musculus). Journal of Chemical Ecology, 19,
1405–1416.
Robertson, D. H., Hurst, J. L., Bolgar, M. S., Gaskell, S. J. & Beynon, R. J. 1997. Molecular
heterogeneity of urinary proteins in wild house mouse populations. Rapid
Communications in Mass Spectrometry, 11, 786–790.
Robertson, D. H. L., Marie, A. D., Veggerby, C., Hurst, J. L. & Beynon, R. J. 2001.
Characteristics of ligand binding and release by major urinary proteins. In:
Chemical Signals in Vertebrates, ed. A. Marchelewska-Koj, D. Muller-Schwarze & J.
Lepri. New York: Plenum Press, pp. 169–176.
Sandnabba, N. K. 1986. Changes in male odors and urinary marking patterns due to
inhibition of aggression in male mice. Behavioral Processes, 12, 349–361.
Schwende, F. J., Wiesler, D., Jorgenson, J. W., Carmack, M. & Novotny, M. 1986. Urinary
volatile constituents of the house mouse, Mus musculus, and their endocrine
dependency. Journal of Chemical Ecology, 12, 277–296.
Singer, A. G., Tsuchiya, H., Wellington, J. L., Beauchamp, G. K. & Yamazaki, K. 1993.
Chemistry of odortypes in mice: fractionation and bioassay. Journal of Chemical
Ecology, 19, 569–579.
Singer, A. G., Beauchamp, G. K. & Yamazaki, K. 1997. Volatile signals of the major
histocompatibility complex in male mouse urine. Proceedings of the National
Academy of Sciences, USA, 94, 2210–2214.
Singh, P. B. 2001. Chemosensation and genetic individuality. Reproduction, 121,
529–539.
Southwick, C. H. 1955. Regulatory mechanisms of house mouse populations: social
behavior affecting litter survival. Ecology, 36, 627–634.
Takami, S. 2002. Recent progress in the neurobiology of the vomeronasal organ.
Microscopy Research and Technique, 58, 228–250.
Vandenberg, J. G. 1969. Male odor accelerates female sexual maturation in mice.
Endocrinology, 84, 658–660.
Whitten, W. K. 1956. Modification of the oestrous cycle of the mouse by external
stimuli associated with the male. Journal of Endocrinology, 13, 399–404.
Wolff, R. J. 1980. Mating behaviour and female choice: their relation to social
structure in wild caught house mice (Mus musculus) housed in a semi-natural
environment. Journal of Zoology (London), 207, 43–51.
Yamazaki, K., Singer, A. & Beauchamp, G. K. 1999. Origin, functions and chemistry of
H-2 regulated odorants. Genetica, 104, 235–240.
Zufall, F., Kelliher, K. R. & Leinders-Zufall, T. 2002. Pheromone detection by
mammalian vomeronasal neurons. Microscopy Research and Techniques, 58, 251–
260.

P1: JZZ/... P2: JZZ/...0521823617c11.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:56
244

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
Part III C O M M U N I C A T I O N N E T W O R K S
I N D I F F E R E N T T A X A
245

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
246

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
Introduction
Communication networks can be found in any taxonomic group of an-
imals, all that is required is that their signals travel further than the average
distance between individuals. This potential for taxonomically widespread oc-
currence is one of the reasons that communication networks are likely to be an
important concept for the understanding of communication in general. However,
taxa vary considerably in several aspects that could affect communication net-
works, including the senses used by receivers (signal modality), processing power
and social organization. The potential insights gained from such taxon-related
differences are the reason for grouping chapters into this section.
Not all taxa are covered in Part III: for example, fish do not appear, but they do
in Parts I and IV (Chs. 4, 5, 21, 22 and 23). Also some taxa are underrepresented:
there is a preponderance of endothermic vertebrate groups, which is recognized
to be a general feature of the literature (Bonnet et al., 2002), and invertebrates
have many fewer chapters than their species richness would seem to require. The
invertebrate balance is redressed slightly by the fact that insects are the focus of
a chapter elsewhere in this book (Ch. 8) and by recent books on insect communi-
cation that deal extensively with chorus behaviour (e.g. Gerhardt & Huber, 2002;
Greenfield, 2002). Nevertheless, this part does have chapters ranging from fiddler
crabs to humans and that is a sufficiently broad taxonomic coverage to demon-
strate common themes and illuminating differences.
Fiddler crabs
To the casual observer, fiddler crabs on a mudflat would seem to be a
clear example of a communication network because of the density of crabs and
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
247

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
248 Part III
the male behaviour of conspicuously waving an enlarged claw. However, as Denise
Pope points out in Ch. 12, that is a human perception; we need to know whether
the crabs consider themselves part of a network. After carefully weighing up the
evidence, particularly on their visual abilities, she concludes that it is likely that
most fiddler crab populations do function as networks. Careful review also in-
dicates that the claw-waving display functions predominantly in male–female
contexts, focusing attention on the intriguing receiver behaviour of conspecific
interceptive eavesdropping and types of competitive signalling interaction.
Anuran amphibians
The frogs and toads (anurans) were one of the first taxonomic groups to
be considered from a communication network perspective. This early interest was
a result of their habit of communicating in striking choruses: groups of calling
individuals that can number several thousand. The calls of male anurans incor-
porate adaptations to enhance the effectiveness of mate attraction in the noisy
environment of a breeding pool. However, calling males also have to repel male
competitors and avoid the unwanted attentions of predators and parasites. In
Ch. 13, Ulmar Grafe considers how such compromises affect the design of acoustic
signals. He also points out that in natural circumstances the precise timing of calls
(e.g. whether calls are synchronized or alternated with the calls of neighbouring
males) may be as important in determining a male’s reproductive success as the
acoustic properties of the calls themselves.
Songbirds
Bird song has long excited the interest and admiration of humans, a fact
reflected in the large literature on songbirds. Song is a long-range advertising
vocalization, so although males defending territories may be widely separated (in
contrast to birds on leks, in flocks or at roosts) they can function as a network
because of song. In many parts of the world, there is also a distinct dawn chorus,
when most individuals of most species are singing at much the same time. Such
characteristics explain why this section has two chapters on songbirds and may
explain why studies of songbirds from a communication network perspective are
becoming more common.
Marc Naguib concentrates on vocal interactions between territorial (usually
male) songbirds in Ch. 14, integrating such information with aspects of territo-
rial behaviour such as settlement patterns. The spatial and social relationships
that define territorial neighbours are mediated by vocal interactions and have
implications for general spacing behaviour.

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
Communication networks in different taxa 249
John Burt and Sandy Vehrencamp (Ch. 15) tackle the dawn chorus of songbirds.
It is a striking acoustic phenomenon when song rate, singing diversity and song
complexity reach their peak. However, the function of the dawn chorus is not
readily explained by a single hypothesis. In Ch. 15, the dawn chorus is considered
from a network perspective, which seems more likely to reveal its function.
Terrestrial mammals
Terrestrial mammals are the second taxonomic grouping represented by
two chapters in this section. This reflects the diversity of terrestrial mammals;
for example, they span a size range from shrews to elephants, with obvious con-
sequences for the feasibility of laboratory studies, and employ signal modalities
such as scent and sound.
Bob Johnston deals with scent communication in small terrestrial mammals
in Ch. 16. Scent marks may contain information for several weeks and during that
time the original scent mark may be over-marked by several individuals. As some
species can determine the order of over-marking, this will create a sort of scent
bulletin board.
Scent marking cannot be detected unaided by human observers; this is in con-
trast to the loud calls of many large terrestrial mammals but similar to the infra-
sonic signals of very large mammals. Karen McComb and David Reby, in Ch. 17,
consider the loud calls of large mammals that can and cannot be heard by humans.
They also point out the implications of social organization for communication net-
works, particularly how the fluid fission–fusion nature of many large mammal
groups is likely to increase opportunities for contact.
Marine mammals
Far-carrying acoustic signals and fluid social systems are also characteris-
tics of marine mammals. In Ch. 18, Vincent Janik summarizes the effect of these
factors on communication networks of pinnipeds and cetaceans and explains how
the nature of sound transmission in water means that sounds can potentially travel
much further than in air. He also discusses whether the communication networks
of marine mammals have been reduced in size in recent decades as oceans have
become noisier (Andrew et al., 2002).
Humans
Many of the terms used to discuss communication network behaviours
(e.g. eavesdropping, audience effects) have their origin in our everyday human

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
250 Part III
experiences. It is, therefore, a surprise to find that studies of human language
generally consider dyads, in close parallel to other animal communication. John
Locke in Ch. 19 argues that a network perspective is more realistic and that the laws
forbidding eavesdropping found in some of the earliest known legal codes show
that such behaviour has always been common. He also argues that information
gained surreptitiously by eavesdropping may be particularly reliable and hence
valuable, repaying the considerable effort and ingenuity often expended by human
eavesdroppers.
Future directions
There is consensus between the chapters in this section that a communica-
tion network perspective is an advance in understanding, but most of the chapters
also point out that more information is necessary in order to evaluate fully the
utility of this perspective by establishing the costs and benefits of communicating
in a network. For this reason, it is premature to contemplate a formal comparative
study of features of communication networks at the level of taxa represented by
chapters in this section. However, it should be possible to attempt comparative
analysis considerably sooner in species-rich groups with diverse features likely to
affect networks (e.g. density and habitat) such as fiddler crabs and anurans.
A key feature of any communication network addressed by most of the chapters
in Part III is the extent of the network: how many individuals are encompassed by a
signal? Theoretical estimates of maximum signal transmission distance combined
with average separation distances between individuals are a very useful first ap-
proximation (for the role of perceptual abilities, see Ch. 20). However, it should be
remembered that such maximum estimates of network size might be considerably
larger than the actual size to which individuals respond. For example, in many
anuran choruses, males adjust their call timing only to immediate neighbours.
Similarly, it would be interesting to know the effect of fission–fusion societies on
network size. Resolving such issues requires detailed study of actual networks,
sometimes involving relatively new techniques (such as passive acoustic location)
or features of signalling modalities that have been recognised as important rela-
tively recently (e.g. scent over-marking).
Several chapters raise the issue of the cognitive requirements for operating
in a communication networks; for example, the extent to which an ability to
identify individuals constrains social eavesdropping. Similarly, the nature of the
information contained in signals will help to determine information flow through
the network: from our human viewpoint, it would seem obvious that human
language is a far richer source of information than the long-range advertising
signals of other animals. Cognitive aspects of communication networks are also

P1: JZZ/... P2: JZZ/...0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 15:10
Communication networks in different taxa 251
dealt with by several chapters in Part IV, indicating that such questions are of
interest to many.
References
Andrew, R. K., Howe, B. M., Mercer, J. A. & Dzieciuch, M. A. 2002. Ocean ambient sound:
comparing the 1960s with the 1990s for a receiver off the California coast. Acoustic
Research Letters Online, 3, 65–70.
Bonnet, X., Shine, R. & Lordais, O. 2002. Taxonomic chauvinism. Trends in Ecology and
Evolution, 17, 1–3.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans: Common
Problems and Diverse Solutions. Chicago, IL: Chicago University Press.
Greenfield, M. D. 2002. Signalers and Receivers: Mechanisms and Evolution of Arthropod
Communication. Oxford: Oxford University Press.

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
12
Waving in a crowd: fiddler crabs signalin networks
d e n i s e s . p o p e
University of Copenhagen, Denmark
Introduction
A communication network is formed when more than one receiver can in-
tercept the signal produced by a signaller, and when more than one signal reaches
a receiver at the same time (McGregor & Dabelsteen, 1996). Communication net-
work theory broadens the consideration of selection pressures on signallers and
receivers to include selection on signallers by receivers other than the primary or
target receiver, and selection on receivers when they receive more than one signal
simultaneously or intercept a signal that was not targeted at them (McGregor &
Dabelsteen, 1996; McGregor & Peake, 2000).
A mudflat full of male fiddler crabs (genus Uca, family Ocypodidae), all rhyth-
mically waving their enlarged claw, seems a perfect example of a communication
network: there are several signallers and receivers in close proximity, and many
signals are being produced simultaneously. However, this human perception of a
coordinated network may be partly a product of our excellent visual ability and
large size in relation to these small crabs. What about the crabs themselves: how
many receivers does a signal reach, and how many signals can individuals receive
simultaneously? Is our impression that they form signalling networks simply an il-
lusion caused by our extreme size and high visual acuity? Most importantly, what
can we learn about the communication system of fiddler crabs by considering
networks of signallers and receivers rather than simple sender–receiver dyads?
In this chapter, I will first introduce the biology of fiddler crabs, then review
the evidence that groups of displaying males of these species form communica-
tion networks; finally, I will examine the implications that such networks may
have for our understanding of the fiddler crab communication system and suggest
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
252

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 253
possible routes for future investigation. Although I am focusing on a single taxon,
my hope is that these ideas may stimulate similar lines of investigation into com-
parable signalling systems in other taxa. In particular, much of the attention in
communication network theory has, to date, focused on the phenomena of social
eavesdropping and audience effects (see Chs. 2 and 4). Such social eavesdropping
is unlikely in fiddler crabs, for reasons explained below, but in this review I aim to
illustrate the utility of the network approach for identifying other consequences
for signallers and receivers that are not predicted from the dyadic view of animal
communication. In addition, much of the previous work on networks has focused
on agonistic interactions between males, while in this chapter I focus on signals
used by males to attract and court females.
The biology of fiddler crabs
Fiddler crabs are small, deposit-feeding semiterrestrial crabs that inhabit
protected shores worldwide in tropical and some warm temperate regions (Crane,
1975). There are 97 recognized species and subspecies in the genus (Rosenberg,
2001). Their intertidal and semiterrestrial existence governs their life in many
ways: their activities are confined to low-tide periods and their lives are cen-
tred around their individual burrows, which they defend against conspecifics
and which serve as shelters during tidal inundation and as refuges from heat,
desiccation, and predation during low tide periods. Their reproductive lives are
also constrained by an obligate pelagic larval stage. This larval stage often results
in lunar or semilunar cycles of reproductive activity (Christy, 1978; Zucker, 1978;
Greenspan, 1982; Yamaguchi, 2001a), as egg hatching and larval release are timed
to coincide with optimal times for larval transport (Morgan & Christy, 1995). The
timing of peak mating activity is, therefore, set by the timing of larval release and
the duration of egg incubation (approximately two weeks depending on temper-
ature and species; reviewed in Yamaguchi, 2001b).
Among behavioural biologists, fiddler crabs are perhaps best known for their
striking sexual dimorphism: males have highly asymmetrical claws, with the ma-
jor claw greatly enlarged (up to five times in length) relative to both the male’s
own minor claw and the female’s two symmetrical small claws (Rosenberg, 2002).
Since the minor claw is used for scooping up the sediment for deposit feeding,
females have the advantage of two feeding appendages while males have only one
and hence have lower intake rates (Weissburg, 1992). The male’smajor claw is used
primarily in two types of activity: fighting and signalling. Both males and females
defend their burrows and the surrounding area from intruders, and the major
claw is a very effective weapon in these disputes. Male–male aggression generally
progresses through a series of stereotyped threat postures with the major claw to

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
254 D. S. Pope
eventual pushing and grappling with interlocked claws, which sometimes results
in one opponent being thrown (Crane, 1975; Hyatt & Salmon, 1977; Jennions &
Backwell, 1996). When males are displaced from burrows they ‘wander’, either
searching for an empty burrow or attempting to take over a burrow from another
male (Crane, 1975; Jennions & Backwell, 1996; Backwell et al., 2000). Much of the
variation in major claw morphology between species may be related to differences
in fighting techniques (Crane, 1975).
Fights between males for burrow ownership may be common because male bur-
rows serve as a breeding resource in many species of fiddler crab (Christy, 1982;
Backwell & Passmore, 1996). In what has sometimes been regarded as the ‘typical’
fiddler crab mating system, females leave their own burrows to ‘wander’ and sam-
ple courting males and their burrows, eventually choosing to stay in one burrow
to mate with the male and lay her eggs; she then usually remains there for the
duration of egg incubation until the larvae are released (Christy, 1983; Backwell
& Passmore, 1996). A second mating system involves copulation on the surface
close to the female’s own burrow (Crane, 1975; Salmon, 1984; Christy & Salmon,
1984); in this case, a male may defend his burrow as a base from which to court
neighbouring females (Salmon, 1984). Recently, it has been recognized that many
species exhibit both of these modes of mating in the same population; hence, they
might best be thought of as alternative mating tactics (Koga et al., 1998; de Rivera
& Vehrencamp, 2001; de Rivera et al., 2003). Several factors probably contribute
to the opportunity for species to engage in surface mating in addition to, or in-
stead of, burrow mating: small clutch size (Christy & Salmon, 1984; de Rivera &
Vehrencamp, 2001), anatomical receptivity of females (if gonopore opercula are
decalcified to allow copulation throughout the lunar cycle (Salmon, 1984)), preda-
tion level (Koga et al., 1998), density (de Rivera et al., 2003) and the spatial overlap
between feeding areas where females burrow and breeding areas where males
court (Christy, 1982, 1983), because in species where males and females gener-
ally do not inhabit adjacent burrows, the opportunity for surface mating is very
limited.
In addition to fighting, males use their major claw in a variety of movement
signals and signalling postures, most conspicuously in the claw-waving display.
This display is a species specific (Crane, 1975) and relatively stereotyped (Hyatt,
1977; Doherty, 1982) pattern of claw elevation (and in some species, unflexing),
sometimes accompanied by movements of the minor claw, legs and body, and
occasionally combined with a stereotyped pattern of locomotion (Crane, 1975).
The display is performed only during the breeding season (Salmon, 1965; Crane,
1975; Wolfrath, 1993) and generally only by territorial, burrow-holding males.
This context of the display led to suggestions that it may function to attract
receptive females to the burrow, repel rival males from it or serve the dual

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 255
function of signalling to both types of receiver (reviewed by Crane, 1975; Moriito &
Wada, 2000).
Recent experimental work has attempted to distinguish between these pro-
posed functions of claw waving by identifying the target receiver of claw wav-
ing in several species of fiddler crabs (U. pugilator (Pope, 2000); U. beebei and U.
terpsichores (formerly U. musica; Rosenberg, 2001; D. S. Pope, unpublished data);
U. annulipes (P. R. Y. Backwell, unpublished data); U. tangeri (D. S. Pope & P. K.
McGregor, unpublished data)) and one other species of ocypodid crab, Scopimera
globosa (Moriito & Wada, 2000). The details of the experimental design differed
between studies, but the general experimental approach involved isolating males
either in cages or with temporary fences to control their visual environment,
and then exposing them to different categories of potential receiver: neighbour-
ing and/or introduced males, and neighbouring and/or introduced females. The
results of these studies are best understood by first explaining that waving can
be classified into at least two categories based on the intensity of the display,
as has also been pointed out by other authors (von Hagen, 1962; Salmon, 1965;
Crane, 1975; Doherty, 1982). High-intensity waving can be differentiated from low-
intensity waving both by an increased rate and, in some species, by the addition
or deletion of display components. In all six species studied, high-intensity wav-
ing was evoked only by the introduction of females, simulating the presence of a
mate-searching female in the male’svicinity, strongly implying that high-intensity
waving is directed exclusively to wandering females in these species.
High-intensity waving is, therefore, clearly part of the courtship sequence in
burrow-mating species. During times of peak mating, males in good condition
generally wave at the low intensity, or background level, more or less continuously.
von Hagen (1962) suggested that low-intensity waving functions to orient mate-
searching females towards the male from relatively long distances. When a male
detects a wandering female near his burrow, he switches to high-intensity waving
by increasing the wave rate and adding or subtracting display components. When
females approach closely, males often switch to yet another courtship signal (e.g.
rapping the major claw against the substrate in U. pugilator (Salmon, 1965) and the
raised carpus display in U. beebei (Christy, 1988a)). The rate of waving also varies
with temperature (Hyatt, 1977; Doherty, 1982) and male size (Hyatt, 1977; Jennions
& Backwell, 1998).
While high-intensity waving seems clearly directed to a particular receiver, low-
intensity waving may function more as a broadcast signal, in the sense that it is
not directed to any particular individual; however, as von Hagen (1962) suggested,
it may be targeted at a general class of receiver: the mate-searching female. This
function may imply that the presence of other individuals should have no effect
on the ‘background’ level of low-intensity waving. The six species studied so far

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
256 D. S. Pope
differed in the effect of neighbours’ presence on low-intensity waving: in other
words, whether the rate of waving differed when males were completely visually
isolated and when they could see their near neighbours. S. globosa, U. pugilator and
U. annulipes showed no background, or low-intensity, waving in the experiments
(Moriito & Wada, 2000; Pope, 2000; P. R. Y. Backwell, unpublished data); U.
terpsichores waved at the same rate when alone as when surrounded by neighbours
(D. S. Pope, unpublished data), and in both U. beebei and U. tangeri the presence of
neighbours increased the wave rate above the background level when visually iso-
lated (D. S. Pope, unpublished data; D. S. Pope & P. K. McGregor, unpublished data).
The lack of background waving in the S. globosa and U. pugilator studies may have
been experimental artefacts as the males in these cases were translocated to caged
areas, so the natural level of background waving in those species remains to be
clarified. The U. annulipes study also involved caged males, although in this case the
males were not displaced; the fact that males waved only when females approached
accords with observations of natural interactions in this species (Backwell et al.,
1998). The reasons for lack of low-intensity or background waving in this species
deserve further investigation.
The different effects of the presence of neighbours on the waving rate in the
remaining three species may be attributable to differences among the species in
the spatial overlap between the sexes. In U. terpsichores, males and females are
generally spatially segregated while in U. beebei and U. tangeri, males and females
intermingle in the same microhabitat (personal observation); therefore, U. terpsi-
chores males have only male neighbours while the neighbours of the other two
species would include both males and females. This potential correspondence be-
tween the presence of female neighbours and an increased rate of low-intensity
waving may indicate either that low-intensity waving is simply stimulated by the
presence of females in the vicinity, whether they are neighbours or wandering
females, or that female neighbours themselves are part of the target receivers of
the low-intensity display. Another difference between U. terpsichores and the other
two species is that the other species engage in surface mating in addition to bur-
row mating. In these species, female neighbours are potential mates, and if these
females assess the quality of neighbouring males, low-intensity claw waving may
be targeted at them (see discussion on p. 268). However, a comparative study of
four Panamanian species that exhibit different combinations of mating tactics,
including U. terpsichores and U. beebei (D. S. Pope, unpublished data) found no in-
dication that males of any species faced female neighbours while waving, and in
every species, males waved most often not facing any individual in the vicinity,
suggesting that they may actively avoid facing neighbours while waving. If female
neighbours do prove to be part of the target receivers of low-intensity waving, it
is likely still more accurate to think of this level of waving as a broadcast display,

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 257
directed not at specific individuals but rather targeted at females in general as po-
tential receivers. Despite suggestions based on behavioural observation that claw
waving appears to be directed sometimes to males as a territorial or threat display
(reviewed by Moriito & Wada, 2000), the experimental evidence indicates that this
is not the case in the six species studied so far.
From current knowledge of fiddler crab mating systems, it appears that the
majority of species engage in burrow mating, either alone or in combination with
surface mating and other less-common tactics (Pope, 1998). All of the experimen-
tal investigations into the targeted receivers of the display involved species with
burrow mating only or mixed tactics; therefore, in these cases, the importance
of attracting females for burrow mating is clear. No studies have investigated
the function of waving in species that copulate only on the surface, but several
authors have noted that surface mating is not preceded immediately by waving
(Salmon, 1984; Yamaguchi, 2001c), implying that it does not serve as a courtship
signal in this context in the same sense that it does in burrow mating. The fact
that the waving display has been retained in these species implies that it continues
to serve some function as a communication signal, although phylogenetic com-
parative evidence suggests that the complexity of the display may be reduced in
these species (Pope, 1998; de Rivera & Vehrencamp, 2001). Further investigations
are warranted into the possibilities that neighbouring females assess males for
surface mating by their waving display (see discussion below) or that waving is
used more in male–male interactions in these species.
In addition to claw waving, fiddler crabs have a rich repertoire of other signals,
not all of which involve the major claw. Using the major claw held outstretched,
males produce threat signals to other males (similar to the threat displays of
many other brachyuran crabs (Wright, 1968)). By rapping the claw against the
substrate or through stridulation of body parts, males also produce vibration sig-
nals (Salmon & Horch, 1972; Popper et al., 2001). These vibration signals are most
commonly produced at night by species that are nocturnally active, although they
may also play a role in the final sequence of diurnal courtship, as described above.
The other major class of signals in fiddler crabs are structures constructed from
the sediment (sand or mud), generally in close proximity to the signaller’sburrow.
Courting males of 16 species build structures that range from the small semidomes
of U. pugilator to the elaborate pillars of U. beebei and hoods of U. terpsichores (re-
viewed by Christy, 1988b; Christy et al., 2002). Hoods and pillars increase the attrac-
tiveness of the males that build them to mate-searching females (Christy, 1988a;
Christy et al., 2002). While I will focus on claw waving in this review, some of
the conclusions may also be applicable to the other potentially long-range signals
of fiddler crabs, specifically vibration signals and structures such as pillars and
hoods.

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
258 D. S. Pope
Much of the preceding descriptions of the crabs’ biology, as well as many of
the suggestions to come in this chapter, may also apply to other members of the
family Ocypodidae. There are five subfamilies (Kitaura et al., 1998) and fiddler crabs
belong to the subfamily Ocypodinae, along with their closest relatives, the ghost
crabs (genus Ocypode). This close relationship with ghost crabs can be misleading
as ghost crabs are unusual within the family (many ghost crabs are specialized
predators; many are nocturnal; they often inhabit exposed shorelines; and they
are, on average, larger than other members of the family), and many of the other
species more closely resemble fiddler crabs in ecology and behaviour than do ghost
crabs. Crabs in the genera Macrophthalmus, Ilyoplax and Scopimera, in particular,
resemble fiddler crabs in that they are small, deposit-feeding crabs inhabiting
protected shores, which show a rich repertoire of signalling behaviour including
claw waving (e.g. Wada, 1991; Kosuge et al., 1994; Moriito & Wada, 1997) and
structure building (e.g. Wada, 1994, Kitaura et al., 1998). They also show both
surface- and burrow-mating tactics, with many species exhibiting both tactics
(e.g. Wada, 1984; Henmi et al., 1993).
Do fiddler crabs signal in networks?
All signals produced by fiddler crabs have four classes of potential con-
specific adult receiver: wandering females, burrow-holding females, wandering
males and burrow-holding males. At least in species with substantial overlap in
microhabitat use by males and females, all four classes of individual are potentially
within receiving range of the display at any given time, creating the conditions
necessary for the formation of communication networks: that is, the active space
of a signal exceeds the average spacing between individuals (McGregor, 1993;
McGregor & Dabelsteen, 1996). What is the evidence that these conditions are met
within fiddler crabs?
Interindividual spacing
Fiddler crab colonies are often described as dense, but this is of course
from the human observer’s point of view. In fact, average densities vary both be-
tween and within species. Larger species are in general less densely distributed
than smaller species (de Rivera & Vehrencamp, 2001). The density of fiddler crab
colonies can be estimated by counting surface-active individuals, by counting the
number of open burrows or by excavating the sediment and counting all crabs un-
covered in a given area; the last gives the best estimate of true population density
(Macia et al., 2001). Counts of surfacing individuals underestimate the true popula-
tion density in U. annulipes while burrow counts tend to overestimate the number
of excavated crabs because some burrows are empty (Macia et al., 2001). However,

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 259
when considering the social environment available to waving male crabs, the
density of individuals on the surface at any given time is the best estimate of
the potential density of interactants in the putative communication network. We
can get an estimate of the range of interindividual distances by considering two
well-studied species at opposite ends of the size spectrum: U. beebei and U. tangeri
(with average adult male body sizes, measured as carapace width or the distance
between the outer edge of the eye sockets, of approximately 0.9 cm and 3.0 cm,
respectively; D. S. Pope, unpublished data). Density measures can be transformed
into estimates of interindividual distances by taking the reciprocal of the density
to yield the average space per individual; if it is then assumed that the area is circu-
lar, the radius of that circle can be calculated from the area. Multiplying the radius
by two gives the average distance between two individuals. U. beebei is found at high
densities of 49 active individuals per m2 on average (D. S. Pope, unpublished data),
which translates into 16 cm between individuals. The larger species U. tangeri is
more widely spaced, at an average of 4.6 active individuals per m2, giving an aver-
age interindividual distance of 53 cm (D. S. Pope, unpublished data). Each of these
densities was estimated from areas of high crab activity, but crab distribution is
patchy within the colonies of both species (U. beebei (de Rivera et al., 2003), U. tangeri
(D. S. Pope, unpublished data)), so there will be areas with larger spacing between
individuals. These estimates may, therefore, be regarded as the optimal conditions
for communication networks in these species. In addition, other species are found
at lower densities. For example, U. terpsichores is in the same size class as U. beebei
but is often found in sandier areas with densities of 17.5 individuals per m2, or
27 cm between neighbours. U. stylifera is a larger crab (approximately 2.4 cm body
size (Crane, 1975)) and is found at 2.6 individuals per m2 (70 cm spacing) at high
local densities (D. S. Pope, unpublished data).
These estimated interindividual distances represent the average spacing of
burrow-holding individuals, but the distance of a wandering individual from con-
specifics will be approximately half this distance, assuming that a wanderer main-
tains an equal distance between conspecific burrows. Therefore, wandering males
or wandering mate-searching females of U. beebei would be, on average, 8 cm from
the closest burrow-holding conspecifics and U. tangeri wanderers would be, on
average, 26.5 cm from the closest burrow-holding conspecifics.
Detection distance of conspecifics
Given that we know the average spacing of individuals, we now need to
know at what distance fiddler crabs are likely to perceive a waving male conspeci-
fic. On an absolute level, the vision of fiddler crabs is constrained by the size
of the ommatidia in the crabs’ eyes, as its size determines the resolution of the
eye. The ability of the eye to resolve an object depends on both the angular size

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
260 D. S. Pope
of the object and the object’s contrast with the background. Therefore, a con-
servative estimate of the limit of detectability is that an object with an angular
size less than the size of a single ommatidium is unlikely to be resolved from
the background unless it contrasts strongly with it (Land & Nilsson, 2001; Zeil &
Hofmann, 2001). However, ommatidial size varies throughout the eye. Fiddler
crabs have eyes well adapted to their primarily flat visual environment: like all
‘flat world’ crabs, they have a band of high vertical resolution around the hori-
zon of the eye, which they align with the visual horizon (Zeil et al., 1986; Land
& Layne, 1995a; Zeil & Al-Mutairi, 1996). Since fiddler crabs carry their eyes on
long stalks, objects below the height of their eyes, including the bodies of most
conspecifics, will be seen below the horizon line. Several authors have argued that
this visual horizon can allow crabs easily to categorize stimuli into either ‘con-
specific’ or ‘predator’, as predators, being larger than the crab, would be imaged
above the horizon line (Land & Layne, 1995a; Layne, 1998). In this high-resolution
zone close to the horizon, the theoretical resolution threshold is approximately
0.5–1◦. Given this resolution and assuming an eye height of 2.5 cm for crabs of
1.0 cm average body size, a 1 cm conspecific should be easily resolvable at 57 cm;
this distance doubles to 114 cm for a 2 cm conspecific ( J. Zeil, personal communi-
cation). Larger species, such as U. tangeri, should theoretically be able to detect con-
specifics at greater distances, both because of the larger stimulus size and because
of the increased height of the eyes above the ground (estimated to be 4 cm in U.
tangeri (D. S. Pope, unpublished data)), which expands the range over which dis-
tance judgements can be made based on retinal elevation (J. Zeil, personal com-
munication). Therefore, the likely detection distance for conspecifics exceeds the
average interindividual spacing in both large and small species of fiddler crab.
Unfortunately, we are technically constrained in our measurements of what a
crab can see by what its behaviour tells us. In other words, a field measurement of
the distance at which a crab reacts to a specific stimulus is a combined measure
of both the crab’s ability to detect it and the relevance of the stimulus. A given
stimulus may well be detectable at much greater distances but it only elicits a re-
sponse once it enters a specific zone around the individual’s burrow. Given these
caveats, what reaction distances have been reported for fiddler crabs? Reaction
distances to conspecifics and conspecific-sized stimuli have been tested in only a
few species. Land & Layne (1995a) reported 30 cm as the distance at which court-
ing male U. pugilator (large male carapace width 1.6 cm (de Rivera & Vehrencamp,
2001)) seemed to notice a wandering conspecific, based on an increase in wave rate.
They reported 10–15 cm as the distance at which males apparently discriminated
the sex of an approaching individual (presumably by the presence or absence of a
major claw), based on the approach distance at which males switched from wav-
ing to threat behaviour to males. The 30 cm distance corresponds to an angular

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 261
size of two to three interommatidial angles, which is two to three times larger
than the theoretical detection threshold. These distances should be interpreted
as minimum estimates of detection distance, however, because they are probably
modulated by the size of a male’s territory, as illustrated by the fact that these
authors also measured responses to predator-like stimuli above the horizon (to
which the motivation to respond is presumably higher) at much smaller angular
sizes. Hemmi & Zeil (2003) performed experiments on burrow surveillance by male
U. vomeris by moving crab-sized dummies (2.25 cm wide and 1.2 cm high) across the
surface towards the male’s burrow and measuring the distance at which the crabs
responded by returning to their burrow to defend it. Male response was better
predicted by the distance of the dummy to the burrow than by the distance of the
dummy to the crab: they consistently responded when the dummy was an average
of 23.8 cm from the burrow, suggesting that this distance might represent the
radius of their defended territory. However, the distance between the dummy and
the crab when the crab reacted varied greatly, since this depended on the male’s
distance to his burrow and the orientation of approach of the dummy towards the
burrow. Crabs responded to dummies at distances of up to 80 cm (J. Zeil, personal
communication), indicating that these conspecific-sized objects were clearly re-
solvable at that distance. Finally, in U. tangeri (3 cm, as above), response distances
to wandering females have been estimated as 150–200 cm (von Hagen, 1962). In
a preliminary experiment on the same species, I attempted to determine a max-
imum reaction distance by controlling a male’s visual environment with fences
and then providing him with a stimulus to which he should be highly motivated
to respond, a tethered but realistically moving female. In these preliminary mea-
surements, I estimated a reaction threshold distance of 100–150 cm. More field
experiments and observations, in addition to neurobiological work on crab vi-
sion, will help us to understand better how fiddler crabs process and respond to
their visual environment.
All of these measured reaction distances were reactions to non-waving con-
specifics, or conspecific-sized objects. There are several reasons to suspect that the
claw-waving display makes a male more detectable to conspecifics than a motion-
less male. First, at adult male body sizes, the claw length exceeds the carapace
width of the male (Crane, 1975), increasing the size of the visual stimulus. Second,
the claw itself is probably the most detectable part of the male: in displaying males
it is often bleached white to contrast strongly with the body (Crane, 1975) and it
is the part of the body that contrasts most strongly with the substrate in terms of
spectral reflectance and polarization (Zeil & Hofmann, 2001). There is, as yet, no
definitive evidence that fiddler crabs have colour vision, but recent findings are
suggestive of a dual-pigment system (Horch et al., 2002), and the smooth wet cuti-
cle of fiddler crabs generate ample specular and ultraviolet reflectance contrasts

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
262 D. S. Pope
with the mudflat background (Zeil & Hofmann, 2001). The movement of the claw
provides motion signatures that should also make the crab more detectable (Zeil &
Zanker, 1997). In addition, during claw waving, the claw itself is elevated above the
level of the crabs’ eyestalks and crosses the visual horizon of viewing conspecifics,
hence entering the zone above the horizon that is used to detect predators (Land
& Layne, 1995a; Zeil & Zanker, 1997). Some authors have suggested that claw wav-
ing thus taps into the predator-escape responses of conspecifics (e.g. Land & Layne,
1995a). Clearly, even if the waving display did initially exploit the female’sreceiver
biases for detection of moving objects in the zone above the horizon, selection has
since modified the response from a generalized negative or inhibitory response
towards threatening stimuli to a more specialized positive or attractive response
towards preferred male conspecifics (see discussion of exploitation of antipredator
receiver biases in Greenfield (2002)). In summary, it is likely that several aspects of
signal design work together to increase the detectability of displaying male fiddler
crabs to conspecifics, although we do not yet know the absolute ‘signal space’ of a
male’s claw-waving display.
Inferences and assumptions
The evidence reviewed above suggests that both the distances at which
crabs can theoretically detect conspecific-sized objects and the distances at which
they have been shown to react to conspecifics exceed the average spacing between
individuals, especially the distances between a wandering individual and the clos-
est burrow owner, thus setting the stage for potential communication networks to
exist, at least in species or populations with relatively high densities of individuals.
Clearly more research is needed on the perceptual ability of crabs, including poten-
tial differences between species resulting from factors such as phylogeny (fiddler
crabs are traditionally divided into broad-front and narrow-front species, based on
the space between their eyes and hence a difference in relative eyestalk length be-
tween species – what effect does this have on their vision and behaviour?), crab size
(does ommatidial size and number scale with body size?) and visual environment
(do mangrove-dwelling fiddler crabs have any special adaptations to deal with
their more complex visual environment?). All of these considerations are likely
to mean that the extent of communication networks will vary both between and
within species of fiddler crabs, and this variation could be a fruitful avenue for
further research.
Implications of network signalling for fiddler crab communication
I will now examine the consequences of assuming that fiddler crabs signal
in networks. What are the possible behavioural effects this will have on how fiddler

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 263
crabs produce and respond to signals, as distinct from what would be predicted
from a traditional ‘dyadic’ signalling scenario?
Production of signals by fiddler crabs in a network: strategies for signal competition
In a communication network, signallers are faced with a more complex
problem than in a simple sender–receiver dyad in the sense that they must neces-
sarily compete with other signallers for the attention of the targeted receiver. This
is self-evident in many cases of sexual signalling, where males are competing to
attract females. In most situations in fiddler crabs, as in many chorusing species of
insects and anurans, females will be in a position to receive more than one signal
simultaneously, so the environment is by default a network (Ch. 2). Such signal
competition probably has effects on at least two timescales: in terms of gross sig-
nal timing (whether to signal or not), and in terms of the fine-scale patterns of
signal timing among neighbouring males. I will term these two timescales bout
timing and signal timing, respectively (Gerhardt & Huber, 2002; Greenfield, 2002).
Bout timing
At the level of bout timing, males undoubtedly use other males as cues for
when to start signalling. Presumably because of energetic constraints, males do not
constantly wave their claws, so they should time their signalling to coincide with
the maximum likelihood of attracting receptive females, assuming that, at least
in burrow-mating species, the display functions primarily in attracting females
to the male’s burrow and persuading them to mate. As noted above, male fiddler
crabs track the timing of receptive females on the scale of the breeding season
(Salmon, 1965; Crane, 1975; Wolfrath, 1993), the lunar cycle (Christy, 1978; Zucker,
1978) and also the daily cycle (Christy et al., 2001). In addition, males are probably
selected to signal whenever their neighbours are signalling lest they miss mating
opportunities; hence males are probably likely to begin signalling if another male
does so. There is some evidence that males do respond this way in U. pugilator,
both to waving neighbours and acoustically signalling neighbours (Salmon, 1965;
Pope, 1998). This effect may also be inferred in U. beebei and U. tangeri, as males of
these species show an increase in waving rate with an increasing number of male
neighbours (D. S. Pope, unpublished data). There is good evidence that males use
other males as cues to begin signalling bouts in acoustic insects (Greenfield, 2002)
and so this should be a fruitful avenue for research in fiddler crabs as well.
Signal timing
Choruses of acoustically signalling anurans and insects often exhibit
group coordination of signals such as synchronous or alternating calling, which
must be accomplished through fine-scale timing adjustments of individual males

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
264 D. S. Pope
to the calls of other calling males (Gerhardt & Huber, 2002; Greenfield, 2002;
Ch. 13). These chorusing interactions are thought to be epiphenomena result-
ing from the preference of females for leading signals (Greenfield, 1994, 2002).
Recent experiments using acoustic playback have revealed in many anuran and
insect species that if two signals overlap in time but are otherwise equal, females
will prefer the leading of the two signals (reviewed by Gerhardt & Huber, 2002;
Greenfield, 2002). Such a preference is thought to result from psychoacoustic con-
straints such as masking (Gerhardt & Huber, 2002) or the precedence effect (a
phenomenon by which two signals, overlapping in time, are perceived as a single
acoustic object by the receiver (Greenfield et al., 1997; Greenfield, 2002)). Males
have thus been selected to avoid being the following male and hence have evolved
timing mechanisms by which they delay their calling if a rival male calls within a
certain critical interval following the male’s own call (Greenfield et al., 1997). As a
consequence, males subtly adjust their timing in an effort always to be the first to
call of a pair of calling males. Such mutual adjustment leads to either synchrony
or call alternation, depending on the call timing of the species. There is no evi-
dence that either synchronous or alternating patterns of calling are cooperative
in the sense that there is any benefit owing to greater attraction of females per
capita by grouped versus solitary signallers (Gerhardt & Huber, 2002; Greenfield,
2002); hence signal competition is the most parsimonious explanation for these
chorusing phenomena.
In a few species of fiddler crabs and a related ocypodid, there is some very good
evidence that males adjust the timing of their signals in relation to their neigh-
bours, resulting in the production of synchronous signals: U. annulipes (Gordon,
1958; Backwell et al., 1998, 1999), U. perplexa and U. saltitanta (P. R. Y. Backwell, M. D.
Jennions, K. Wada, M. Murai & J. H. Christy, unpublished data), and Ilyoplax pusilla
(Aizawa, 1998). The phenomenon of synchronous waving has been most thor-
oughly documented in U. annulipes (Backwell et al., 1998, 1999), an Indo-West Pacific
broad-fronted species. U. annulipes is unusual in that males apparently do not pro-
duce low-intensity, or background, waves but only wave in the presence of a female
(Backwell et al., 1998). When a mate-searching female approaches, males cluster
around her, and males within this cluster synchronize their waves with each other.
As in most synchronously calling insects and anurans, Backwell et al. (1998) also
found a female preference for the leading male of a group of synchronous wa-
vers. These males also signal at a faster rate than their neighbours (Backwell et al.,
1999), producing some waves that are not overlapped by other males. It is not clear,
therefore, whether the female preference for the leading male is the result of a per-
ceptual constraint as it appears to be in insects and anurans, or whether it is simply
a consequence of females preferring males that signal at the fastest rate, which
is condition-dependent in this species (Jennions & Backwell, 1998). The fact that

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 265
synchronous waving results, however, would imply that perhaps the overriding
preference is for the leader (Backwell et al., 1999), and the fastest waving male
is simply the one who least needs to adjust his signal to those of his neighbours
(Greenfield, 2002). In both U. perplexa and U. saltitanta, males produce synchronous
waves in both the presence and absence of mate-searching females and increase
their wave rate in the presence of females (P. R. Y. Backwell, M. D. Jennions, K. Wada,
M. Murai & J. H. Christy, unpublished data), as expected if high intensity waving
serves to court females and persuade them to mate, as discussed above. The waves
of U. perplexa become less synchronous in the presence of females, perhaps because
the increased rate of high-intensity waving means that some males are not able to
produce waves at such a high rate and fall out of synchrony with their neighbours
(P. R. Y. Backwell, M. D. Jennions, K. Wada, M. Murai & J. H. Christy, unpublished
data). In these two species, females also prefer the male waving at the fastest rate,
but only in U. perplexa is that male also most often the leading male. This would
suggest that the overwhelming preference is for high-quality males, waving at the
fastest rate. The fact that U. perplexa and U. saltitanta continue to wave in synchrony
even when mate-searching females are not nearby suggests other aspects of signal
competition: perhaps it results from a female preference for males with the high-
est wave rate even at a distance, a preference for groups of synchronous wavers, or
because such signal competition allows males to compete better with their male
neighbours in other contexts such as territory acquisition and maintenance (P. R. Y.
Backwell, M. D. Jennions, K. Wada, M. Murai & J. H. Christy, unpublished data). The
results of these studies clearly demonstrate that the network phenomenon of sig-
nal competition for receivers’ responses, resulting in signal synchrony, occurs in
these species. If a high rate of waving indicates male quality in other species of
fiddler crabs, females may be expected to prefer males that wave at the highest
rate in these species as well, which leads to the question of why signal competition
resulting in synchronous signalling is not more widespread in fiddler crabs. Fu-
ture research should be directed at investigating what factors may have promoted
the evolution synchronous waving in some species of Uca and not others.
While synchronous waving has been observed in a few other Uca species (P. R. Y.
Backwell, personal communication), bouts of synchrony do not appear to be as
sustained or as tightly timed as they are in U. annulipes, U. perplexa and U. saltitanta.
In the vast majority of species (i.e. those that do not signal synchronously), it is not
clear whether males adjust the timing of their signals in relation to each other,
other than at the gross level of initiating bouts when neighbours do. However,
results from the ocypodid I. pusilla suggest that males may adjust their signal
timing in more subtle ways. Aizawa (1998) found that males delayed the timing of
their waves so that they overlapped with both live and videotaped male neighbours
in a laboratory setting, agreeing with the observations that neighbouring males

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
266 D. S. Pope
of this species signal synchronously. More surprisingly, field measurements also
demonstrated that male I. pusilla adjusted the timing of their signals to match that
of the much larger fiddler crab U. lactea, whereas there was no evidence that U.
lactea adjusted their timing reciprocally (Aizawa, 2000). These results suggest that
males of other species of fiddler crab (and other waving ocypodids) may make fine-
scale adjustments to their signal timing in ways that are not immediately obvious
to the naked eye and will only be uncovered by careful analysis. Burford et al. (1998)
found some suggestion of an adjustment of wave rate by large males of U. tangeri
when signalling in the presence of smaller neighbours, but the comparisons were
made only at the level of wave rate, and not the males’ timing of signal initiation
in relation to each other’ssignals. The considerations above lead to the conclusion
that males in many fiddler crab species are easily able to perceive the signals of
at least their immediate neighbours, implying that such timing adjustments are
likely.
Reception of signals by fiddler crabs in a network:strategies for information gathering
Some of the most intriguing possible consequences of fiddler crab commu-
nication networks relate to how receivers may use signals to gather information in
ways not traditionally considered in a dyadic framework. This section is somewhat
speculative as, to date, there is little evidence for these effects. However, given the
clear existence of network effects on the production of signals by male fiddler
crabs (reviewed above), the network environment is likely to have consequences
for receivers as well.
As outlined in the introduction, the experimental evidence from the five fiddler
crab species so far studied points to wandering, receptive females as the primary
receivers to which claw waving is directed. In this section, I will work under the
assumption that wandering females are the target receivers. As all of these five
species engage in burrow mating, and since less research to date has been focused
on species that only mate on the surface, the discussion is biased towards species
that mate in burrows. Our understanding of communication networks in fiddler
crabs will benefit from more in-depth study into the function and use of the claw-
waving display and other signals in surface-mating species, and such study will
provide a useful test of the generality of the following inferences about fiddler
crab networks.
While I assume here that wandering females are the primary receivers of the
claw-waving display, I reiterate that by ‘receivers’ here I mean not only the ‘in-
tended’ receivers, or primary targets, of the signal but also other individuals that
may be paying attention to the signal. As described above, in most species there
are at least two other classes of adult conspecifics within receiving range of the

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 267
display: neighbouring males and wandering males. In addition, in species with ex-
tensive spatial overlap between males and females, neighbouring females will also
be within receiving range. Such potential receivers may be considered ‘conspecific
interceptive eavesdroppers’,that is, conspecifics that benefit from intercepting sig-
nals targeted at another individual (see Ch. 2). In addition, other receivers such
as bird predators, juvenile conspecifics and heterospecific crabs may also bene-
fit from intercepting the signals of males (by better locating or avoiding waving
males, depending on the context), but I will not consider these additional receivers
further. I will consider each of the four categories of adult conspecific receivers in
separate sections.
Wandering females: the target receivers
Females may be attracted to groups of signalling males because the group
as a whole is more detectable or because it provides the female with an enhanced
opportunity to compare mates. Such opportunities might benefit a female because
they reduce search time (presumably also reducing search risk) and perhaps result
in a higher overall mate quality if signalling in some way allows her to assess male
quality. There is mixed evidence from acoustically signalling insects and anurans
that females are differentially attracted to groups of signalling males. Studies test-
ing whether choruses themselves attract females better than lone signallers have
found primarily negative results (reviewed by Gerhardt & Huber, 2002; Greenfield,
2002). Yet, there is fairly good evidence that females may be attracted to some
groups of males over others within choruses, based on the size or density of the
group (reviewed by Gerhardt & Huber, 2002). As resources in fiddler crabs are not
as clumped as in these acoustic species, the situation in fiddler crabs may be more
analogous to the second case, of females moving within choruses; as such, the
potential for similar effects exist: females may be more attracted to areas with a
higher density of signalling males. In the eastern Pacific species U. beebei, higher
density increased the likelihood of mate searching by females in experimentally
manipulated areas, and wandering females more frequently entered areas of nat-
urally higher density than low-density areas (de Rivera et al., 2003). If females are
directly attracted to higher densities of waving males, this can be tested by exper-
imentally offering females a choice of patches of males that differ in density of
signallers.
Factors affecting the attractiveness of claw waving to females have been
best studied in synchronously waving species (Backwell et al., 1998, 1999; P. R. Y.
Backwell, M. D. Jennions, K. Wada, M. Murai & J. H. Christy, unpublished data).
Further work is needed on other species to determine what aspects of waving are
most attractive to females and how these factors correlate with male quality and re-
source (burrow) quality. More information on what attracts females to claw waving

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
268 D. S. Pope
may provide insight into the potential exploitation of receiver biases and how
males may use waving to enhance their detectability and attractiveness to fe-
males, as Christy and colleagues have done for male courtship structures (Christy
et al., 2001, 2002).
Neighbouring females
As reviewed above, there is as yet no evidence that claw waving functions
as a direct prelude to surface copulation. However, in species that have both mixed-
sex colonies and either mixed mating tactics (both surface and burrow mating)
or surface mating only, the possibility exists that females may assess and com-
pare their male neighbours (who are their most likely surface copulation partners
(Yamaguchi, 2001c)) for potential future copulations. In an unpublished study of
four Uca species in Panama, I found no evidence that males directed their displays
to female neighbours, but this does not preclude the possibility that females may
still use the displays to gain information. A recent study by Murai et al. (2002) found
that in U. paradussimieri, a species with a unique mating tactic in which males enter
female’s burrows for mating, territorial males showed evidence of being able to
assess the reproductive state of their female neighbours and directed non-waving
courtship at ones that were close to being receptive, mating with them up to three
days later. Therefore, if males are able to assess and integrate such information
over such time periods, females may in a parallel fashion be able to assess and com-
pare male neighbours and integrate such information into their decisions about
surface-mating partners. The potential for longer-term assessment by neighbour-
ing females in species that surface mate only should be investigated in concert
with experimental manipulations to assess the target receivers of waving in these
species; this would improve our understanding of how waving functions in these
species. Such knowledge will also help us to assess how waving has evolved across
the genus and family as a whole.
Wandering males
The traditional view of claw waving as a dual function territorial signal
holds that claw waving would repel a wandering male from approaching the bur-
row of a waving male, because the display would in this case signal the male’s
ownership of the burrow and his willingness to defend it. Given that there is no
evidence that males address their waving to wandering males, and the fact that
they switch to distinct threat displays when another male intrudes (Salmon &
Stout, 1962; Land & Layne, 1995a; D. S. Pope & P. K. McGregor, unpublished data),
claw waving may not have this function in fiddler crabs. Conversely, the network
view suggests that the display might have the opposite effect: that of attracting a
wandering male to the vicinity. If claw waving is directed at females, wandering
males might use it to gather information about potential areas to establish a new

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 269
burrow or attempt a take over of an occupied one. If the goal of burrow ownership
is to attract females for mating (over and above the necessity of a burrow as a
shelter and refuge), then areas of high waving activity might indicate to the male
an area of high likelihood of mate attraction. This effect may be magnified in the
case of high-intensity waving, as it indicates the actual presence of a female. A
wandering male’s response to a waving male may be modulated by the wanderers’
size relative to the resident, since larger crabs usually win fights, although there
is also a resident advantage ( Jennions & Backwell, 1996). Thus, waving may actu-
ally repel smaller males, whereas equally sized or larger males may be attracted
by it.
Male attraction to rival males’ advertisement signals has been suggested in
a few taxa. Stamps (1988) found that juvenile Anolis aeneus lizards showed ‘con-
specific attraction’ when settling in new habitat and suggested that they might
use the territorial advertisement displays of head-bobbing to assess conspecific
density. Alatalo et al. (1982) demonstrated that broadcasting the song of the pied
flycatcher Ficedula hypoleuca attracted settlement by conspecifics in nearby nest
boxes. Playback of male advertisement calls in laboratory phonotaxis experiments
attracted conspecific males in the spadefoot toad Spea multiplicata (Pfennig et al.,
2000) and the house cricket Acheta domesticus (Kiflawi & Gray, 2000). In both of
these cases, smaller males in particular differentially approached calls that were
most attractive to females. Such behaviour can result in male aggregations that
are independent of resource distribution, and it may also be a prerequisite for
satellite male-calling behaviour (reviewed by Gerhardt & Huber, 2002). Conspe-
cific interceptive eavesdropping of this type, involving male attraction to signals
of their rivals, may be much more common than is widely recognized, and the
dearth of examples may simply result from an absence of studies. Observational
studies of the movements of wandering males and experimental studies testing
whether males respond aversively or positively to other males’ signals would help
to clarify the existence of this phenomenon in fiddler crabs.
Neighbouring males
One final possible consequence of fiddler crab communication networks
is another form of interceptive eavesdropping, in this case by neighbouring terri-
torial males. The fact that males are more likely to switch to high-intensity waving
when wandering females approach (D. S. Pope & P. K. McGregor, unpublished data)
suggests the possibility that other males could use their neighbours as ‘female
detectors’. By monitoring the signalling of neighbouring males, in particular not-
ing when they switch to high-intensity waving, they might effectively expand the
distance at which they can detect a wandering female. This monitoring would
presumably need to be upregulated by the actual detection of the female her-
self, so that males do not end up wasting energy unnecessarily. The number of

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
270 D. S. Pope
neighbours that are high-intensity waving may also have a synergistic effect, such
that a male is more likely to pay attention if more than one neighbour switches to
high intensity, because this is a more reliable indicator of female presence. Recent
experiments on U. tangeri suggest that this effect may indeed be occurring (D. S.
Pope & P. K. McGregor, unpublished data). The high-intensity waving display of
this species is easily differentiated from the low-intensity display by the addition
of an introductory curtsey, and males reliably switch to high-intensity waving
when a female approaches (D. S. Pope & P. K. McGregor, unpublished data). The vi-
sual environment of neighbouring pairs of males was controlled with an opaque
fence so that the males could view each other but females could be introduced
on one side of the fence in such a way that one male and not the other could see
them. Males that could view the introduced females significantly increased their
rate of high-intensity waving, as expected. Males that could see their neighbour,
but not the introduced female, significantly increased their rate of low-intensity
waving; the rate of high intensity waving also increased, but not significantly.
Thus these ‘interceptor’ males showed an intermediate level of waving between
the lower background level and the higher level in the presence of a female. These
results suggest that males do attend to the waves of their neighbours and use
the information to compete to attract females, but they are more responsive to
the actual detection of the female nearby. A similar result was found in the syn-
chronously waving species U. annulipes (M. D. Jennions, unpublished data): males
began to wave in synchrony with a neighbouring male even when their view of the
female was blocked with a fence. This ‘female detector’effect might be common in
fiddler crab species but may not normally be noted by a human observer because
it is often not clear whether a male is responding to seeing the female herself or
to his neighbour.
The possibility of males using rival males’ signals as ‘female detectors’ may be
extended to other taxa as well. Such interceptive eavesdropping by rival males
is a type of socially acquired information (Giraldeau et al., 2002) in which ani-
mals collect information from other conspecifics on resource location and quality.
When the probability of acting on socially acquired information goes up when the
same information is obtained from more than one individual, information cas-
cades can result in which many individuals make the same behavioural decision
without obtaining direct information themselves (Giraldeau et al., 2002; Watts,
2002). Such information cascades can result in information being transmitted
faster than the rate of direct information acquisition, for example when escape
responses are transmitted throughout a group faster than the approach of a model
predator (Treherne & Foster, 1981), which the authors termed the Trafalgar effect.
These information cascades can sometimes lead to suboptimal behaviour if the
behavioural decisions of the initiators of the cascades were erroneous (e.g. escape
response to a sudden movement of vegetation rather than a predator (Giraldeau

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 271
et al., 2002)). The experiments described above on U. tangeri and U. annulipes involved
only exposure to the behavioural decision (in this case, high-intensity waving) of a
single conspecific; it remains to be tested if the effect is stronger if males witness
the waving of more than one individual, as would be predicted if the situation is
a true information cascade (Giraldeau et al., 2002). The potential also exists that
males may track the movement of a female through a group of males by monitor-
ing the signals of other males, as suggested by McGregor & Dabelsteen (1996) for
territorial intrusions in songbirds.
Summary and future directions
The ecology of fiddler crabs (typically high densities and open habitat)
as well as their sensory physiology (good visual resolution) argue that most fid-
dler crab colonies operate as communication networks. The fact that their most
common and conspicuous signal, the claw-waving display, does not appear to be
used in male–male interactions makes it unlikely that the network effect of so-
cial eavesdropping (i.e. eavesdropping on signalling interactions; Ch. 2) occurs in
these crabs. However, many other consequences of communication networks sug-
gest themselves, particularly involving competitive signalling interactions and
novel information-gathering strategies, including forms of conspecific intercep-
tive eavesdropping. Such possibilities should provide fruitful avenues for future
investigation.
The best-studied cases of interceptive eavesdropping, which involve intercep-
tion by predators or parasites, clearly harm the signallers, but the situation is
less clear for conspecific interceptive eavesdropping (reviewed in Ch. 2). In fiddler
crabs, males are likely to benefit from any interception of their signals by females,
whether or not a given individual female was the target of the signal. Even in cases
where a male’s signal compares unfavourably with that of a neighbour because it
is given at a lower rate or is not a leading signal in a synchronous species, males are
still likely to benefit more from waving than from not signalling at all. However,
males may be more likely to suffer costs, in terms of missed mating opportuni-
ties, from the interception of their signals by other males. Are there any strategies
that males can use to minimize these potential costs? The broadcast nature of the
claw-waving display (at least at low-intensity, or background, levels) make it un-
likely that males can eliminate the potential for eavesdropping (cf. in songbirds;
Ch. 3); however, they may be able to target specific individuals when signalling
by orienting either their front or back sides towards the approaching individual
(female). This would present the largest visual stimulus to the female while reduc-
ing the view for neighbouring males situated off-axis to the signalling male. There
is some suggestion that males orient in this way towards approaching females in
U. pugilator (Land & Layne, 1995b), although such orientation was not in evidence

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
272 D. S. Pope
in U. beebei (Christy, 1988a). The effectiveness of such a strategy for minimizing the
information available to neighbours is unknown and more information is needed
on how widespread this orientation strategy is in fiddler crabs. Future research
should be directed towards investigating not only the existence of the potential
conspecific interceptive eavesdropping described here but also whether males suf-
fer substantial costs from such eavesdropping, and if so, what counter-strategies
they might employ for minimizing these costs. Only further work can illuminate
the costs and benefits to fiddler crab signallers and receivers of operating in a
network environment and uncover behavioural strategies each party might use
to exploit the situation to their best advantage. The diversity among fiddler crab
species, in terms of habitats (mudflat versus mangrove), mating tactics and dis-
play form and function will create variation in the extent to which the network
perspective is applicable to fiddler crabs and could serve as useful tests of the
predicted consequences to these crabs of signalling in a network environment.
Acknowledgements
I would like to thank Peter McGregor and the ‘communication crew’at the University of
Copenhagen for welcoming me into their network and stimulating and clarifying my thinking on
the communication networks of fiddler crabs. Michael Jennions, Giuliano Matessi, Ricardo Matos,
Peter McGregor, Tom Peake, Andrew Terry and an anonymous reviewer all provided very helpful
feedback on earlier versions of this chapter. I would like to thank Jochen Zeil in particular for
his comments and insight into the visual world of fiddler crabs. Patricia Backwell and Catherine
de Rivera generously shared their results with me before they were published. My own research
described here was funded by the US National Science Foundation and the Danish Natural Science
Research Council, which also supported me during the writing of this chapter.
References
Aizawa, N. 1998. Synchronous waving in an ocypodid crab, Ilyoplax pusilla: analyses of
response patterns to video and real crabs. Marine Biology, 131, 523–532.
2000. Heterospecific timing of waving display in ocypodid crabs Ilyoplax pusilla and
Uca lactea. Journal of the Marine Biological Association of the UK, 80, 947–948.
Alatalo, R. V., Lundberg, A. & Bjorkland, M. 1982. Can the songs of male birds attract
other males? An experiment with the Pied Flycatcher Ficedula hypoleuca. Bird
Behavior, 4, 42–45.
Backwell, P. R. Y. & Passmore, N. I. 1996. Time constraints and multiple choice criteria
in the sampling behavior and mate choice of the fiddler crab, Uca annulipes.
Behavioral Ecology and Sociobiology, 38, 407–416.
Backwell, P., Jennions, M., Passmore, N. & Christy, J. 1998. Synchronized courtship in
fiddler crabs. Nature, 391, 31–32.

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 273
Backwell, P. R. Y., Jennions, M. D., Christy, J. H. & Passmore, N. 1999. Female choice in
the synchronously waving fiddler crab, Uca annulipes. Ethology, 105, 415–421.
Backwell, P. R. Y., Christy, J. H., Telford, S. R., Jennions, M. D. & Passmore, N. I. 2000.
Dishonest signalling in a fiddler crab. Proceedings of the Royal Society of London, Series
B, 267, 719–724.
Burford, F. R. L., McGregor, P. K. & Oliveira, R. F. 1998. Chorusing by male European
fiddler crabs, Uca tangeri: a study of visual communication networks. Acta
Ethologica, 1, 33–41.
Christy, J. H. 1978. Adaptive significance of reproductive cycles in the fiddler crab Uca
pugilator: a hypothesis. Science, 199, 453–455.
1982. Burrow structure and use in the sand fiddler crab, Uca pugilator (Bosc). Animal
Behaviour, 30, 687–694.
1983. Female choice in the resource-defense mating system of the sand fiddler crab,
Uca pugilator. Behavioral Ecology and Sociobiology, 12, 169–180.
1988a. Pillar function in the fiddler crab Uca beebei (II): competitive courtship
signaling. Ethology, 78, 113–128.
1988b. Pillar function in the fiddler crab Uca beebei (I): effects on male spacing and
aggression. Ethology, 78, 53–71.
Christy, J. H. & Salmon, M. 1984. Ecology and evolution of mating systems of fiddler
crabs (genus Uca). Biological Reviews 59, 483–509.
Christy, J. H., Backwell, P. R. Y. & Goshima, S. 2001. The design and production of a
sexual signal: hoods and hood building by male fiddler crabs Uca musica.
Behaviour, 138, 1065–1083.
Christy, J. H., Backwell, P. R. Y., Goshima, S. & Kreuter, T. 2002. Sexual selection for
structure building by courting male fiddler crabs: an experimental study of
behavioral mechanisms. Behavioral Ecology, 13, 366–374.
Crane, J. 1975. Fiddler Crabs of the World (Ocypodidae: Genus Uca). Princeton, CT: Princeton
University Press.
de Rivera, C. E. & Vehrencamp, S. L. 2001. Male versus female mate searching in fiddler
crabs: a comparative analysis. Behavioral Ecology 12, 182–191.
de Rivera, C. E., Backwell, P. R. Y., Christy, J. H. & Vehrencamp, S. L. 2003. Density
affects female and male mate searching in the fiddler crab, Uca beebei. Behavioral
Ecology and Sociobiology, 53, 72–83.
Doherty, J. A. 1982. Stereotypy and the effects of temperature on some spatio-temporal
subcomponents of the ‘courtship wave’ in the fiddler crabs Uca minax (Le Conte)
and Uca pugnax (Smith) (Brachyura, Ocypodidae). Animal Behaviour, 30, 352–363.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans: Common
Problems and Diverse Solutions. Chicago, IL: University of Chicago Press.
Giraldeau, L.-A., Valone, T. J. & Templeton, J. J. 2002. Potential disadvantages of using
socially acquired information. Philosophical Transactions of the Royal Society of London,
Series B, 357, 1559–1566.
Gordon, H. R. S. 1958. Synchronous claw-waving of fiddler crabs. Animal Behaviour, 6,
238–241.
Greenfield, M. D. 1994. Cooperation and conflict in the evolution of signal
interactions. Annual Review of Ecology and Systematics, 25, 97–126.

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
274 D. S. Pope
2002. Signalers and Receivers: Mechanisms and Evolution of Arthropod Communication.
Oxford: Oxford University Press.
Greenfield, M. D., Tourtellot, M. K. & Snedden, W. A. 1997. Precedence effects and the
evolution of chorusing. Proceedings of the Royal Society of London, Series B, 264,
1355–1361.
Greenspan, B. N. 1982. Semi-monthly reproductive cycles in male and female fiddler
crabs, Uca pugnax. Animal Behaviour, 30, 1084–1092.
Hemmi, J. M. & Zeil, J. 2003. Robust judgement of inter-object distance by an
arthropod. Nature, 421, 160–163.
Henmi, Y., Koga, T. & Murai, M. 1993. Mating behavior of the sand bubbler crab
Scopimera globosa. Journal of Crustacean Biology, 13, 736–744.
Horch, K., Salmon, M. & Forward, R. 2002. Evidence for a two pigment visual system in
the fiddler crab, Uca thayeri. Journal of Comparative Physiology A, 188, 493–499.
Hyatt, G. W. 1977. Quantitative analysis of size-dependent variation in the fiddler crab
wave display (Uca pugilator, Brachyura, Ocypodidae). Marine Behavior and Physiology,
5, 19–36.
Hyatt, G. W. & Salmon, M. 1977. Combat in the fiddler crabs Uca pugilator and U.
pugnax: a quantitative analysis. Behaviour, 65, 182–211.
Jennions, M. D. & Backwell, P. R. Y. 1996. Residency and size affect fight duration and
outcome in the fiddler crab Uca annulipes. Biological Journal of the Linnean Society, 57,
293–306.
1998. Variation in courtship rate in the fiddler crab Uca annulipes: is it related to
male attractiveness? Behavioral Ecology, 9, 605–611.
Kiflawi, M. & Gray, D. A. 2000. Size-dependent response to conspecific mating calls by
male crickets. Proceedings of the Royal Society of London, Series B, 267, 2157–2161.
Kitaura, J., Wada, K. & Nishida, M. 1998. Molecular phylogeny and evolution of unique
mud-using territorial behavior in ocypodid crabs (Crustacea: Brachyura:
Ocypodidae). Molecular Biology and Evolution, 15, 626–637.
Koga, T., Backwell, P. R. Y., Jennions, M. D. & Christy, J. H. 1998. Elevated predation risk
changes mating behaviour and courtship in a fiddler crab. Proceedings of the Royal
Society of London, Series B, 265, 1385–1390.
Kosuge, T., Poovachiranon, S. & Murai, M. 1994. Male courtship cycles in three species
of tropical Ilyoplax crabs (Decapoda, Brachyura, Ocypodidae). Hydrobiolgia 285,
93–100.
Land, M. & Layne, J. 1995a. The visual control of behavior in fiddler crabs: I.
Resolution, thresholds and the role of the horizon. Journal of Comparative Physiology
A, 177, 81–90.
1995b. The visual control of behavior in fiddler crabs: I1. Tracking control systems
in courtship and defence. Journal of Comparative Physiology A, 177, 91–103.
Land, M. F. & Nilsson, D.-E. 2001. Animal Eyes. Oxford: Oxford University Press.
Layne, J. E. 1998. Retinal location is the key to identifying predators in fiddler crabs
(Uca pugilator). Journal of Experimental Biology, 201, 2253–2261.
Macia, A., Quincardete, I. & Paula, J. 2001. A comparison of alternative methods for
estimating population density of the fiddler crab Uca annulipes at Saco Mangrove,
Inhaca Island (Mozambique). Hydrobiologia, 449, 213–219.

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
Fiddler crabs signal in networks 275
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society, Series B, 340, 237–244.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
Morgan, S. G. & Christy, J. H. 1995. Adaptive significance of the timing of larval release
by crabs. American Naturalist 145, 457–479.
Moriito, M. & Wada, K. 1997. When is waving performed in the ocypodid crab
Scopimera globosa? Crustacean Research, 26, 47–55.
2000. The presence of neighbours affects waving display frequency by Scopimera
globosa (Decapoda, Ocypodidae). Journal of Ethology, 18, 43–45.
Murai, M., Koga, T. & Yong, H.-S. 2002. The assessment of female reproductive state
during courtship and scramble competition in the fiddler crab, Uca
paradussumieri. Behavioral Ecology and Sociobiology, 52, 137–142.
Pfennig, K. S., Rapa, K. & McNatt, R. 2000. Evolution of male mating behavior: male
spadefoot toads preferentially associate with conspecific males. Behavioral Ecology
and Sociobiology 48, 69–74.
Pope, D. S. 1998. The fiddler crab claw waving display: function and evolution of a
sexually selected signal. Ph.D. thesis, Duke University, USA.
2000. Testing function of fiddler crab claw waving by manipulating social context.
Behavioral Ecology and Sociobiology, 47, 432–437.
Popper, A. N., Salmon, M. & Horch, K. W. 2001. Acoustic detection and communication
by decapod crustaceans. Journal of Comparative Physiology A, 187, 83–89.
Rosenberg, M. S. 2001. The systematics and taxonomy of fiddler crabs: a phylogeny of
the genus Uca. Journal of Crustacean Biology, 21, 839–869.
2002. Fiddler crab claw shape variation: a geometric morphometric analysis across
the genus Uca (Crustacea: Brachyura: Ocypodidae). Biological Journal of the Linnean
Society, 75, 147–162.
Salmon, M. 1965. Waving display and sound production in the courtship behavior of
Uca pugilator, with comparisons to U. minax and U. pugnax. Zoologica, 50,
123–150.
1984. The courtship, aggression and mating system of a ‘primitive’ fiddler crab (Uca
vocans: Ocypodidae). Transactions of the Zoological Society of London, 37, 1–50.
Salmon, M. & Horch, K. 1972. Acoustic signaling and detection by semi-terrestrial
crabs of the family Ocypodidae. In: Behavior of Marine Animals, vol. 1, ed. H. E. Winn
& B. L. Olla, New York: Plenum Press, pp. 60–96.
Salmon, M. & Stout, J. F. 1962. Sexual discrimination and sound production in Uca
pugilator Bosc. Zoologica 47: 15–19.
Stamps, J. A. 1988. Conspecific attraction and aggregation in territorial species.
American Naturalist 131, 329–347.
Treherne, J. E. & Foster, W. A. 1981. Group transmission of predator avoidance
behaviour in a marine insect: the Trafalgar effect. Animal Behaviour 29, 911–917.

P1: IYP/KAB P2: KPB-KOD0521823617c12.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:36
276 D. S. Pope
von Hagen, H.-O. 1962. Frielandstudien zur sexual- und forpflanzungs-biologie von Uca
tangeri in Andalusien. Zietschrift fur Morphologie und Okologie der Tiere, 51, 611–725.
Wada, K. 1984. Pair formation in the two forms of Macrophthalmus japonicus De Haan
(Crustacea: Brachyura) at a co-occurring area. Journal of Ethology, 2, 7–10.
1991. Biogeographic patterns in waving display, and body size and proportions of
Macrophthalmus japonicus species complex (Crustacea: Brachyura: Ocypodidae).
Zoological Science, 8, 135–146.
1994. Earthen structures built by Ilyoplax dentimerosa (Crustacea, Brachyura,
Ocypodidae). Ethology, 96, 270–282.
Watts, D. J. 2002. A simple model of global cascades on random networks. Proceedings of
the National Academy of Sciences, USA, 99, 5766–5771.
Weissburg, M. 1992. Functional analysis of fiddler crab foraging: sex-specific
mechanics and constraints in Uca pugnax (Smith). Journal of Experimental Marine
Biology and Ecology, 156, 105–124.
Wolfrath, B. 1993. Observations on the behaviour of the European fiddler crab Uca
tangeri. Marine Ecology Progress Series, 100, 111–118.
Wright, H. O. 1968. Visual displays in brachyuran crabs: field and laboratory studies.
American Zoologist 8, 655–665.
Yamaguchi, T. 2001a. The breeding period of the fiddler crab, Uca lactea (Decapoda,
Brachyura, Ocypodidae) in Japan. Crustaceana 74, 285–293.
2001b. Incubation of eggs and embryonic development of the fiddler crab, Uca
lactea (Decapoda, Brachyura, Ocypodidae). Crustaceana 74, 449–458.
2001c. The mating system of the fiddler crab, Uca lactea (Decapoda, Brachyura,
Ocypodidae). Crustaceana 74, 389–399.
Zeil, J. & Al-Mutairi, M. M. 1996. Variation of resolution and of ommatidial dimensions
in the compound eyes of the fiddler crab Uca lactea annulipes (Ocypodidae,
Brachyura, Decapoda). Journal of Experimental Biology, 199, 1569–1577.
Zeil, J. & Hofmann, M. 2001. Signals from ‘crabworld’: cuticular reflections in a fiddler
crab colony. Journal of Experimental Biology, 204, 2561–2569.
Zeil, J. & Zanker, J. M. 1997. A glimpse into crabworld. Vision Research, 37, 3417–3426.
Zeil, J., Nalbach, G. & Nalbach, H.-O. 1986. Eyes, eye stalks and the visual world of
semi-terrestrial crabs. Journal of Comparative Physiology A, 159, 801–811.
Zucker, N. 1978. Monthly reproductive cycles in three sympatric hood-building
tropical fiddler crabs (genus Uca). Biological Bulletin, 155, 410–424.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
13
Anuran choruses as communicationnetworks
t. u l m a r g r a f e
University of Wurzburg, Germany
Introduction
It is becoming more and more apparent that communication often takes
place in a network of several signallers and receivers (as shown by most of the chap-
ters in this volume and reviewed by McGregor & Peake (2000)). The network view
of communication stresses that signallers and receivers have additional costs and
benefits to those usually found in dyadic interactions. For example, in communica-
tion networks signallers often face the problem of intense intra- and interspecific
competition whereas receivers must discriminate information from individuals
under conditions of high background noise.
In many frogs and toads, males aggregate in large choruses to advertise for
females. The signals they use are conspicuous and long range; therefore, choruses
constitute a classic example of a communication network. The challenge of com-
municating in such large choruses is to balance the costs and benefits of attracting
a mate, repelling rivals and avoiding predators and/or parasites. Consequently, ad-
vertising in choruses will have far-reaching effects on vocal behaviour. If we want
to understand signal design and signalling behaviour in such aggregations, we
need to look at communication in the network context in which these different
selective pressures operate.
In this chapter, I will review why it is important to investigate communication
in chorusing anurans within the network environment. I will focus on the be-
haviour of both signallers and receivers. First, I will discuss patterns of male–male
vocal competition that can best be understood within the network environment.
In particular, I will discuss how the timing of signals within the chorus determines
mating success. The variation in signal timing between anurans suggests that they
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
277

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
278 T. U. Grafe
are fine-tuned to both the level of competition and the receiver biases. Further-
more, high chorus density necessitates that males interact with only a subset of
males. Determining the degree of selective attention (i.e. the number of individu-
als males interact with) is, therefore, an important parameter of connectivity in
the network. Aggregations of chorusing males allow females to monitor male–
male interactions prior to mate choice. What evidence from anurans is there that
females eavesdrop? Similarly, both calling and satellite males may benefit from
monitoring male–male interactions and position themselves within the chorus to
increase their active space, associate with attractive callers or gain information
about the location of gravid females.
Signaller behaviour
Patterns of male–male vocal competition
Many anurans form large and conspicuous aggregations in which high lev-
els of background noise reduce their effectiveness in attracting females (Gerhardt
& Klump, 1988; Narins & Zelick, 1988; Gerhardt & Schwartz, 1995). Similar effects
are found in insects (e.g. Romer et al., 1989) and birds (Klump, 1996). Chorusing
intensifies competition between males over females or over resources of interest
to females. Temporal segregation of calling activity or the partitioning of calling
frequencies are solutions available to some species in some situations. However,
in many cases, males cannot avoid calling in dense aggregations and are subject to
high levels of intra- and interspecific acoustic interference. This is especially evi-
dent in species that breed in temporary ponds where reproductive success depends
on laying eggs as early as possible to ensure that larval development is completed
before the breeding sites dry out.
Males that vocalize in dense aggregations deal with high levels of acoustic
competition in several ways (reviewed by Wells, 1988; Gerhardt & Schwartz, 1995;
Gerhardt & Huber, 2002). They may increase their call repetition rate, increase the
complexity of their calls or defend calling sites and acoustic space against other
males. Many species show all of these adaptations. The classic example is that of the
tungara frog Physalaemus pustulosus where males add chuck notes to their whines
to form complex advertisement calls and increase their call rate depending on
the social milieu (Ryan, 1985). Some frogs even increase call intensity in response
to playbacks of conspecific advertisement calls (Lopez et al., 1988) or lower the
dominant frequency of their advertisement calls during aggressive interactions
with other males (reviewed by Bee & Bowling, 2002). In addition, chorusing males
alternate or synchronize their calls with neighbouring males (e.g. Zelick & Narins,
1983; Forester & Harrison, 1987; Klump & Gerhardt, 1992; Greenfield & Roizen,
1993; Grafe, 1999; Bosch & Marquez, 2002). The precise timing of calls may, in

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 279
fact, be as important in determining a male’s mating success as the acoustic call
properties that, traditionally, are investigated. I discuss this in more detail in
following sections.
Repertoire sizes can be large in some anurans, reflecting the potential to mod-
ulate social interactions on a fine scale. Repertoires appear especially impressive
in Boophis madagascariensis, with 28 distinct calls reported (Narins et al., 2000) and
Amolops tormotus, with an amazing variety of calls that have defied categorization
(Feng et al., 2002). Such call variety may reflect the need to signal to males and
females simultaneously (see below).
Signals used during aggressive interactions
Aggressive calls play an important role in mediating the spacing between
male anurans within a chorus and there is tremendous variation in their use
(Schwartz, 2001; Gerhardt & Huber, 2002). In fact, many species never produce
distinct aggressive calls. In the genus Kassina, for example, aggressive calls have
only rarely been reported (Fleischack & Small, 1978), despite extensive playback
experiments in most species (T. U. Grafe, unpublished data). In some species, the
advertisement call grades into the aggressive call (e.g. Wells, 1989; Grafe, 1995a);
in others the advertisement call is distinct from the aggressive call, with the ag-
gressive call being graded (e.g. Schwartz, 1989). Some species have several kinds
of aggressive call (e.g. Given, 1987). Much discussion revolves around the func-
tional significance of graded versus discrete signalling systems and how cheat-
ing can be prevented. Handicap models predict that aggressive signals should be
graded to convey best information about the probability that the signaller will
attack (Grafen, 1990). In contrast, conventional signalling (Enquist et al., 1998) and
discrete handicap models (Johnstone, 1994) both predict that signals should be
discrete. This discussion has not considered the necessity for chorusing males si-
multaneously to repel rivals and attract females. Here, signals need to reach two
different classes of receiver (males and females) and may need to be designed dif-
ferently. In general in anurans, aggressive calls are less attractive to females than
advertisement calls (e.g. Brenowitz & Rose, 1999). Therefore, signals that grade be-
tween advertisement and aggressive calls or discrete aggressive call variants may
allow males to increase the aggressive content of a signal gradually or discretely,
while only partially reducing a male’s attractiveness to females. This may be a
general solution to signalling when trying to reach different classes of receiver
and a resolution to a signalling conflict that is typical of a network environment.
The variation in the use of aggressive calls in anurans may depend on the degree
of flexibility needed in dealing with the conflicting demands of signalling to
males and females simultaneously (Brenowitz et al., 2000; Marshall et al., 2003). An
elegant solution to this problem is the use of two-part advertisement calls, in which

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
280 T. U. Grafe
one part is directed to females and the other to males. In Geocrinia victoriana, for
example, males produce an introductory note that is directed towards males and
a series of shorter repeated notes that are directed towards females (Littlejohn &
Harrison, 1985). A similar case can be made for the diphasic call of Eleutherodactylus
coqui (Narins & Capranica, 1978). Viewed from an energetic perspective, such a
solution appears wasteful because only every second note is directed to females.
This may explain why such a solution is not very common.
Fine-scale patterns of signal timing
Signal timing can be an important feature influencing mating success
in chorusing anurans and, therefore, males need to monitor the call timing of
competitors (reviewed by Greenfield, 1994a; Gerhardt & Huber, 2002). The fine-
scale patterns of signal timing are often described by taking the calling period
of one male as a reference point and relating the timing of the second individ-
ual to this reference (reviewed by Klump & Gerhardt, 1992; Greenfield, 1994a). At
one end of the continuum of call-timing patterns, individuals signal in perfect
synchrony (relative phase of 0◦); at the other end signals are spaced with equal
intervals between them resulting in perfect alternation (relative phase of 180◦).
Most anurans do not show either of these extreme patterns on a regular basis.
Synchrony can, therefore, refer to signalling patterns in which signals overlap,
whereas alternation characterizes patterns in which signals regularly are more or
less evenly spaced in time (Gerhardt & Huber, 2002). Examples of different call-
timing patterns are shown in Fig. 13.1. I use the term entrainment to refer to
call-timing patterns with relative phase angles below 45◦ but not overlapping
(a similar classification is used in insects (Greenfield, 1994a)). This represents
an operational definition that helps to classify a continuously varying param-
eter. Alternating would then refer to call-timing patterns with relative phase
angles above 45◦. In dyadic interactions of entrained calling, the calls of one
male will lead and the calls of the other male will follow. Leading and lagging
roles often switch between individuals (Fig. 13.1; Gerhardt & Huber, 2002; Grafe,
2003) are less discernible in alternating species (Klump & Gerhurdt 1992; Bosch &
Marquez, 2002).
Levels of overlap rise dramatically in aggregations of three and four males (e.g.
Schneider et al., 1988; Schwartz, 1993; Grafe, 1996a). However, overlap remains
lower than expected if frogs were calling at random. This suggests that males
are interacting in ways that prevent high levels of overlap. In most cases, males
will attempt to place their calls in relatively silent gaps, thus avoiding masking
interference by other males. Zelick & Narins (1985), in their pioneering work with
the Puerto Rican treefrog E. coqui, documented experimentally that males were
able to place their calls in unpredictable gaps of silence. Males were triggered to

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 281
Fig. 13.1. Spectrograms of call-timing patterns (defined in text) in five different
species of running frogs (Kassina). K. cassinoides shows alternation, K. schioetzi
entrainment, K. senegalensis entrainment with occasional overlap, K. fusca synchronous
calling and K. kuvangensis synchronous call groups with alternating calls. The letters
(A, B) designate individual males. Note the switching of leader and follower roles
between males in the top four spectrograms. These species call syntopically (i.e. in the
same pond and at the same time) and were recorded in the Guinea savannah region of
the Comoe National Park, Ivory Coast. K. kuvangensis was recorded at Hillwood Farm,
northwestern Zambia.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
282 T. U. Grafe
call by the rapid offset of background sound. Similar studies on other anurans
have supported these basic results and the underlying mechanisms of call ini-
tiation (Schwartz, 1993; Grafe, 1996a). Interestingly, constant high noise levels
do not prevent calling, suggesting that call inhibition subsides with time. Like
E. coqui, Broadley’s painted reed frogs Hyperolius marmoratus broadleyi initiate their
calls in response to drops in background noise levels, showing an ‘off-response’
(Grafe, 1996a). The modal response latencies in the reaction of males were be-
tween 40 and 80 milliseconds (in some bushcrickets it is less than 20 milliseconds
(Robinson, 1990)), suggesting that higher auditory centres such as the thalamus
are not involved in processing such a fast response (Walkowiak, 1992). Although
modal response latencies are very short, most calls are given with much longer
latencies. Males avoid call overlap by calling within windows of low noise levels
(‘silence’) by selectively attending to near and thus loud neighbours.
The flexibility with which males can adjust their calls even on a note-by-note
basis is remarkable. Schwartz (1993), studying Hyla microcephala, found that males
increased the spacing between their calls when interrupted, thus avoiding further
overlap of subsequent notes in their call. I found a similar response in the Central
African frog Kassina kuvangensis (Grafe, 2003). Calling in this species is characterized
by synchronizing call groups while at the same time alternating advertisement
calls with those of neighbouring males. As in H. microcephala, males readjusted their
inter-call intervals within milliseconds in response to the playback stimulus.
In contrast to alternating or entrained calling, synchronous calling is unusual
in anurans. It has been reported for only a handful of species: the neotropical hylids
Smilisca sila (Ryan, 1986), Hyla ebraccata (Wells & Schwartz, 1984) and Centrolenella
granulosa (Ibanez, 1993) as well as the African running frogs Kassina fusca and
Kassina senegalensis (Grafe, 1999; T. U. Grafe & H. Lussow, unpublished data). In
pairwise interactions of the savannah running frog K. fusca, 81.5% (overall median)
of calls overlapped (relative phase of 8.6 ± 4.4◦) with a median degree of overlap
of 20.8%. In the Senegal running frog K. senegalensis, only 21.6% of calls between
neighbouring males overlapped on average (T. U. Grafe & H. Lussow, unpublished
data).
Synchrony in anurans, in contrast to many insects (reviewed by Greenfield,
1994a), is achieved by a rapid acoustic response to the onset of a concurrent signal
produced by a neighbour. This ‘on-response’ mechanism is thus fundamentally
different from the ‘off-response’ found in the alternating and entrained response
type. It should be stressed, however, that at least one anuran, K. fusca, can vary
its response type. When presented with playbacks of conspecific advertisement
calls, males showed a synchronous response whereas they entrained calls to het-
erospecific calls or white noise stimuli, thus showing both on- and off-responses
(Grafe, 1999).

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 283
There is considerable variation in the patterns of call timing between species. In
insects and anurans, call alternation is found predominantly in species that show
low call rates, whereas synchrony is more common in species that call at high rates
(Narins, 1982; Greenfield, 1994a). An exception to this rule is the genus Kassina.
Males show synchronous or entrained calling at both low and high call rates inde-
pendent of density, suggesting that call rate alone is not the prime determinant of
signal timing (Grafe, 1999). Moreover, some species show both synchrony and al-
ternation (e.g. Moore et al., 1989). In K. kuvangensis, males synchronize call groups
while at the same time alternating calls within call groups. Such synchronized
interdigitated calling may serve to reduce predation while maintaining species-
specific temporal information important to females (Grafe, 2003). This may be a
solution to reducing the costs associated with synchrony and alternation.
Adaptive significance of call timing
Both cooperative and competitive hypotheses can explain the evolution
and maintenance of call timing in anurans and insects. Cooperative explanations
for synchrony include (a) confusion of predators by decreasing the locatability of
signals; (b) enhancement of detection by females by increasing the peak ampli-
tude of signals; and (c) improving the detection of female acoustic responses. Sev-
eral authors have noted that there is little support for these cooperative explana-
tions (Greenfield, 1994a; McGregor & Peake, 2000). Only in one species, S. sila, may
synchronous calling provide protection against frog-eating bats Trachops cirrhosus,
since bats were shown to be more attracted to alternating than to synchronous
playback of calls (Tuttle & Ryan, 1982). There is no evidence for enhanced detection
of overlapping calls by females. Peak amplitude of synchronous calls or choruses of
males is not much higher than that of individual signallers (Bradbury, 1981), thus
providing only a marginal increase in the active space of males. Although females
are attracted to larger choruses in a number of species, the mating success per
male often declines as lek size increases (Deutsch, 1994; Widemo & Owens, 1995).
Similarly, playback experiments with the grasshopper Ligurotettix coquilletti and
the treefrog H. microcephala showed that females were attracted to larger arrays of
speakers but the attractiveness per speaker was not higher for larger arrays (Shelly
& Greenfield, 1991; Schwartz, 1993). Finally, acoustic responses to male advertise-
ment calls are given in only a few species, most notably in midwife toads Alytes spp.
(Bosch, 2001). Since calling effort (duty cycle) is generally not very high in midwife
toads, there is little gained by synchronizing calls to improve the detection of
female acoustic responses.
There is strong evidence that competition to produce leading calls often ex-
plains call timing in anurans (and insects) because there is a strong preference
by females for leading signals (e.g. Grafe, 1996a; Greenfield et al., 1997; Bosch &

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
284 T. U. Grafe
Marquez, 2002). This preference for leading calls has been termed the precedence
effect (reviewed by Gerhardt & Huber, 2002). In species in which females prefer
leading calls, males that produce trailing calls will benefit by delaying the onset
of their calls to avoid overlap or by speeding up calls to produce leading calls.
Selective attention
In the previous section, I have outlined why chorusing anurans should
generally avoid interference with other signalling males. The problem in a large
aggregation is that if a male is to avoid interference with all males in the chorus,
he would have to stop calling. The solution is to attend only to a subset of nearby
males. Brush & Narins (1989) were the first to investigate systematically the ques-
tion of to how many chorus members an individual male attends. They found that
E. coqui males typically avoided overlap with just two neighbours, only rarely with
three individuals. Monitoring small choruses of four to six male H. microcephala,
Schwartz (1993) found that males generally responded to their loudest rivals. In
addition, more centrally located males typically attended to more males (one to
four) than those at the periphery of the chorus (one). Greenfield & Rand (2000)
have further demonstrated the plasticity involved in selective attention. Male P.
pustulosus responded to two to three neighbouring males depending on the chorus
structure and intensity of those males’ calls. These results suggest that selective
attention is a dynamic process that will vary as males enter the chorus and move
within it.
Selective attention to nearest neighbours may occur even in anurans that do not
avoid call overlap. In K. fusca, call overlap itself is a measure of the attention paid
to other males. Preliminary work (T. U. Grafe, unpublished data) suggests that the
spatial distribution of males in the chorus is an important factor in determining
the number of males that are paid attention.
Another interesting case is the gray treefrog Hyla versicolor (Schwartz, 2001;
Schwartz et al., 2001). Although females discriminate against overlapping calls,
neighbouring males in small choruses do not avoid overlap. Avoidance of overlap
appears to be overridden by female preference for longer calls and a step-like
decrease in attractiveness of short calls even if they are unmasked. Given the
preferences of females, the best strategy for male gray treefrogs appears to be the
production of long calls that partly overlap with those of neighbours. The plasticity
in response to acoustic competition and the differences in auditory perception
of receivers suggest the absence of a unifying general rule governing selective
attention.
Energetics of calling
Important determinants of the interactions of calling male anurans are
the energetic constraints of calling. Calling is the most energetically expensive

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 285
behaviour of ectothermic vertebrates (reviewed by Wells, 2001). In the European
treefrog Hyla arborea, for example, instantaneous rates of oxygen consumption
can reach 41 times the resting rate (Grafe & Thein, 2001). Such high levels of
expenditure cannot be maintained for long and they set an upper limit to the
rate and complexity of calling. Female anurans prefer to mate with males calling
at high rates in all species tested (Ryan & Keddy-Hector, 1992; Gerhardt & Huber,
2002). Consequently, males need to adjust the rate and complexity of calls to the
levels of competition or risk having to drop out of the chorus prematurely because
they have run out of energy. In species that vary the duration of calls, such as
H. versicolor, the best predictor of energy expenditure is the product of call rate
and call duration (calling effort or duty cycle; reviewed by Wells, 2001).
In most anurans, calling effort increases with chorus density (e.g. Taigen et al.,
1985; Grafe, 1996b). In H. versicolor, however, males reduce the rate of calling while
increasing call duration as chorus density increases, thus maintaining a constant
level of energy expenditure (Wells & Taigen, 1986; Grafe, 1997a). Females prefer
long calls at low rates to short calls at high rates (Klump & Gerhardt, 1987). These
studies show that the energetic constraints of calling require males to monitor
the behaviour of others to maintain their attractiveness towards females and that
the way males partition their energy depends on female preferences.
Receiver behaviour
Mechanisms of female preferences
Important selection pressures on signal design and signalling behaviour
are the sensory and neuronal abilities of females. In anurans, acoustic commu-
nication plays a central role in mate choice. The wide range in the threshold of
auditory neurons and the sensitivity for narrow frequency bands in the peripheral
auditory system are important in allowing females to choose between conspecific
males in the presence of background noise (reviewed by Narins & Zelick, 1988).
Numerous studies have shown that females prefer males that produce loud and
conspicuous signals with large active space (Ryan & Keddy-Hector, 1992). Conse-
quently, receivers generally exert strong selection for loud and ritualized signals.
In recent years, it has become clear that the fine-scale patterns of signal timing
have a large influence on female choice. Females of many taxonomic groups show
a preference for leading, but not necessarily overlapping, signals in the olfactory
(voles: Johnston et al., 1997), visual (fiddler crabs: Backwell et al., 1998; fireflies:
Vencl & Carlson, 1998) and auditory modalities (field crickets: Wyttenbach &
Hoy, 1993; katydids: Greenfield, 1994b; Greenfield et al., 1997; frogs: Gerhardt &
Huber, 2002; rats: Kelly, 1974; cats: Cranford & Oberholzer, 1976). This preference
for leading signals has entered the literature under the term precedence effect. It
was originally described by Wallach et al. (1949) in humans and describes the

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
286 T. U. Grafe
observation that when two spatially separated sounds are presented with a brief
delay in onset the leading sound dominates localization. However, apart from
the work on mammals, the experimental designs of the studies listed above do
not distinguish between masking of the trailing call and the inability to locate
the trailing call even though it is easily heard (i.e. it is not masked). To distin-
guish between these alternatives, psychoacoustic studies will be necessary. From
an evolutionary perspective, however, the selection pressures on males to produce
leading calls will be strong irrespective of the underlying mechanisms.
I was able to demonstrate a strong preference for leading calls in Broadley’s
painted reed frog H. m. broadleyi (Grafe, 1996a) and the savannah running frog
K. fusca (Grafe, 1999). In reed frogs, the preference by females for the leading call
was largely independent of sound pressure, underscoring the robustness of this
preference (see also Dyson & Passmore, 1988a). In the synchronously calling run-
ning frog, preference for both leading and trailing calls was observed depending
on the degree of overlap (Grafe, 1999). Females preferred leading calls when calls
overlapped by 75% and 90% but switched their preference to trailing calls at 10%
and 25% of overlap. Thus, males are selected to overlap the calls of neighbours;
however, they should not do so with high degrees of overlap. Interestingly, play-
back experiments also showed that males were able to initiate their calls sooner
than they actually do, suggesting that special mechanisms are involved that in-
hibit males from calling with high degrees of overlap. For the savannah running
frog, the adaptive significance of synchronous calling is explained, at least in part,
by the auditory preferences of females.
Whereas the preference for leading signals appears to be a basic design feature
of nervous systems and thus a constraining feature of receivers that males need
to attend to, the preference of females for trailing signals is likely to be a more
fine-tuned adaptation by receivers to specific signalling environments. It remains
unclear why female savannah running frogs prefer trailing calls at low degrees of
overlap.
A comparative analysis within the genus Kassina may provide some answers as
to how a species’environment, in particular the communication network in which
a population finds itself, influences signal design and signalling behaviour. Pre-
liminary female choice experiments suggest that the call-timing pattern of males
is also tuned to the respective preference functions of females in K. senegalensis
(T. U. Grafe & H. Lussow, unpublished data). However, call-timing patterns do not
correlate with habitat characteristics such as degree of cover or calling site. Fur-
ther comparative work needs to be done to elucidate the environmental correlates
of call timing in anurans.
It should be noted that the physical characteristics of the communication chan-
nel, the transmission properties of the environment and the network structure

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 287
(i.e. distance between network members) might select for different signals (Wiley
& Richards, 1978; Ryan et al., 1990; Staaden & Romer, 1997). Signal attenuation and
degradation through reverberations or irregular amplitude fluctuations are fac-
tors that limit the active space of signals (Wiley & Richards, 1978; see also Ch. 20).
These effects vary between habitats, with distance between sender and receivers
and with height above ground. Spectral components of the call are degraded least,
for example, if either sender or receivers are elevated (Wiley & Richards, 1978).
Ryan & Wilczynski (1991) demonstrated that differences in habitat characteris-
tics explained a large part of the variance in the frequency of advertisement calls
between populations of the chorus frog Acris crepitans (see Wiley (1991) for bird
examples). In several recent studies with anurans, however, none of the predicted
differences in call features was found between species from different habitats
(Penna & Solis, 1998; Kime et al., 2000). The evidence for the influence of transmis-
sion properties of the environment and network structure on signal design and
signalling behaviour in anurans is equivocal at best.
Comparing female choice in two-choice trials and in natural choruses
To identify which acoustic parameters are important in determining male
mating success, researchers traditionally use two-choice trials in which female
anurans are given the choice between two acoustic stimuli. Gravid female frogs
and toads readily phonotactically approach one of the speakers and will search for
the male on or in the speaker. Typically population-wide preferences of females
are then noted and inferences drawn about the importance of male acoustic traits
(Gerhardt, 1994). Regarded from a network perspective, such experiments should
be viewed with caution because females are being tested in very simplified envi-
ronments in which background noise is reduced to a minimum (see also Sullivan
et al., 1995). To illustrate this point, I will review three examples of how prefer-
ences demonstrated in simple arena trials may not translate into sexual selection
in natural populations.
The first example is from the detailed studies on the South African painted reed
frog Hyperolius marmoratus marmoratus by Neville Passmore and his colleagues. They
showed that females preferred lower frequency calls, suggesting that larger males
should have a mating advantage. This, however, was only the case when comparing
the mating success and body size of males in small choruses (Telford et al., 1989). In
large choruses, large males no longer had an advantage. Instead of preferring large
males, females tested in the field preferred males calling at high rates (Passmore
et al., 1992). Therefore, under noisy field conditions, female preference for calls of
lower frequency were overridden by preferences for call rate.
I made a similar observation when studying mate choice in Broadley’s painted
reed frog H. m. broadleyi. To determine female preferences in the small choruses

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
288 T. U. Grafe
in the field, I monitored male calling behaviour using an array of microphones
set around the periphery of small natural ponds (Grafe, 1997b). This technique of
passive sound localization (McGregor & Dabelsteen, 1996) enabled me to match
precisely each call on the recording to an individual male in the chorus. I moni-
tored both the spectral and the temporal components of each male’s calls while a
female, released from the edge of the pond, was choosing among males. The analy-
sis showed that call rate and proximity to the female release site were the best
predictors of male mating success. These preferences were corroborated in tradi-
tional two-choice trials. Interestingly, females also preferred medium-frequency
calls to high or low frequencies in two-choice trials, a preference not mani-
fested under noisy field conditions. Furthermore, call parameters of interest to
females are often intensity dependent: that is, preference for a call trait can be
reversed by increasing the intensity of an alternative stimulus (e.g. Arak, 1988;
Gerhardt, 1988).
The third example showing the importance of testing females under natural
conditions is the work on H. versicolor by Schwartz (2001). As mentioned above, fe-
males tested in arena choice trials prefer males producing longer calls even if they
call at a lower rate as long as calling effort remains the same. The preference for
long calls is non-linear, with strong discrimination against very short calls. Under
quiet conditions, the discrimination was remarkable. On average, females discrim-
inated in favour of calls on the basis of 1.5 pulses (out of 20). Background noise
played to females over an additional speaker reduced the ability to discriminate
to 2.3 pulses. Field experiments supported the importance of call duration in mat-
ing success; however, it was limited. An array of eight speakers was placed along
the perimeter of a pond and calls of varying duration and call rate were broad-
cast over many nights. Naturally arriving females were trapped at the speaker of
their choice. The extent of the preference for call duration was quite restricted,
with only the shortest call being discriminated against. Overall, the preference
for long calls explained less than 10% of the variation in male mating success in
the field.
These examples do not argue against the utility of two-choice trials but suggest
that they should not stand alone. In chorusing anurans, females have to make
decisions under acoustically unfavourable environments, often under the risk of
predation, and must, therefore, limit their choosiness. The utility of two-choice
trials comes into play when testing hypotheses generated by field observations or
experiments, as recently demonstrated by Murphy & Gerhardt (2002). They com-
bined field observations of mate sampling by female barking treefrogs Hyla gratiosa
with two- and three-speaker choice trials to determine the influence of distance
of calling males on female choice. As in Broadley’s painted reed frog, most female
barking treefrogs approached the closest male and mated with him. Evidence sug-
gests that these species simultaneously sample males. Such sampling is especially

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 289
vulnerable to background noise and will be limited by the perceptual abilities of
females (Murphy & Gerhardt, 2002). It should be noted, however, that females of
other anurans probably use sequential sampling techniques and, therefore, are
less influenced by background noise because they closely approach several males
(reviewed by Murphy & Gerhardt, 2002).
Eavesdropping by females
Do females extract information from male–male interactions that influ-
ences their mating decisions? Rephrasing this question in communication net-
work terminology: are females eavesdropping on male–male interactions (i.e. in-
dulging in social eavesdropping; Ch. 2)? The evidence that they are comes from
observing female choice for the relative timing of male advertisement calls. Like
bird song, the advertisement call of anurans is directed to both males and females
(i.e. it is not just a mating call). Females can potentially extract information from
how males interact. Chorusing male anurans adjust the timing of their advertise-
ment calls to that of neighbouring males in a competitive way (Klump & Gerhardt,
1992; Schwartz, 2001). As outlined above, in most species, females prefer the calls
of leading males, thereby often overriding their preferences for other call param-
eters (e.g. Dyson & Passmore, 1988b; Grafe, 1996a). In a few cases, females prefer
follower calls (Schwartz & Wells, 1984; Grafe, 1999).
Evidence that females eavesdrop on male–male interactions requires simulta-
neous monitoring of males, i.e. that they show simultaneous mate choice. Good
evidence for simultaneous mate choice comes from species that approach males
only after spending some time, often several minutes, at the edge of breeding
ponds and from the relative preferences of females tested in two-choice trials
(Grafe, 1997b; Murphy & Gerhardt, 2002). However, a convincing study of social
eavesdropping would need to show that females are not just approaching the first
male they can distinguish from the background noise, in most cases this would be
the nearest male. In H. m. broadleyi, females based their choice not only on nearby
males but also on male call rate (Grafe, 1997b). Likewise, female Hyla gratiosa did
not just approach the first male they could distinguish from the background noise
(Murphy & Gerhardt, 2002). Such observations and experimental evidence suggests
that females monitor the calling behaviour of more than one male. Two-choice
trials have shown that females prefer males that jam the calls of other males.
For social eavesdropping to occur, one need not assume high cognitive abilities.
In fact, the proximate mechanisms underlying female preference for leading or
follower calls may not even require the involvement of higher auditory centres.
Eavesdropping by males
Potentially, anuran choruses provide ample opportunities for males to
eavesdrop on the interactions between other males or between males and females.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
290 T. U. Grafe
0
5
10
15
20
25
30
0 2.5 5 7.5 10 12.5 15 17.5 20
Cal
l rat
e (c
alls
/0.5
min
)
Time (min)
Fig. 13.2. Call rates of seven males (open and closed symbols) in a small chorus over
20 minutes. Note that the ‘opportunistic male’ (closed symbol) is not calling until
1.25 minutes before he went into amplexus (at 19.8 minutes). Since call rates were
high at other times during the recording, other cues, such as seeing the female, in
addition to high call rates are likely to have directed the attention of the
‘opportunistic male’ to the female.
Males of many species increase their call rate substantially when approached by
a female and this can lead to local interactions with heightened activity between
males. Other silent or satellite males in the vicinity may make use of such infor-
mation by approaching this chorus area and attempting to intercept the female
or attract females by vigorously starting to call themselves. I recorded three cases
of such ‘opportunistic’ calling in Broadley’s painted reed frog using a microphone
array (Grafe, 1995b). These males were silent throughout most of the recording
period (3, 19, 20 minutes) and started calling vigorously only after a female had
been introduced into the chorus and was moving towards other calling males. In
all three cases, males approached the area of heightened activity and were success-
ful in attracting the female’s attention and mating with her. One case is shown in
Fig. 13.2. Documenting the behaviour of non-signalling individuals is particularly
difficult and may be one of the reasons why little is known about ‘silent’ (eaves-
dropping) strategies. This underscores the utility of using microphone arrays to
record chorusing activity because the absence of calling can be documented using
this technique.
Interceptive eavesdropping (Ch. 2) is known to occur in some anurans. In spade-
foot toads and green frogs, for example, satellite males associate with speak-
ers that produce attractive advertisement calls (Pfennig et al., 2000; Gerhardt &
Huber, 2002), suggesting that these males are monitoring the activity of other

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 291
males and making adaptive decisions. Such behaviour is probably more common
than reported.
In anurans, in which females give acoustic responses to male advertisement
calls (e.g. midwife toads, see above), it would be interesting to see if either sex
uses these male–female interactions to interfere with the courting pair. For exam-
ple, female Australian bushcrickets Elephantodeta nobilis give acoustic responses to
male advertisement calls (Bailey & Field, 2000). Males that are probably satellites
are attracted by these duets and produce advertisement calls, thereby occasionally
attracting these females themselves. Eavesdropping on male–female vocal interac-
tions may occur in Alytes spp. as well. Two studies report eavesdropping in captive
Alytes obstreticans in which females competed for male parental care by approach-
ing vocalizing pairs, attempting to block the path of other females and displacing
amplectant males (Verrell & Brown, 1993; Grafe et al., 1999).
Indirect evidence for eavesdropping comes from observing the signal type and
signalling intensity used during communication in a network environment. Priva-
tizing an interaction is a likely consequence of eavesdropping (Ch. 3). Many male
anurans have distinct courtship calls that are quieter than their advertisement
calls (reviewed by Wells, 1988). In E. coqui, for example, males use these quiet
courtship calls to lead females to oviposition sites on the forest floor (Townsend &
Stewart, 1986). Since females are not being mate guarded (i.e. are not in amplexus),
it is important for the male to prevent interference by other males.
Another point of interest is that anuran advertisement signals are generally
omnidirectional (e.g. Passmore, 1981), probably because males cannot predict from
where females approach. Spherical spreading, however, facilitates eavesdropping.
One would predict that courtship calls should be more directional; however, sound
fields of courtship calls have not been measured in anurans.
Summary and future directions
Aggregations of calling frogs and toads are characterized by high levels of
background noise. The common problems of communicating in a noisy environ-
ment, with its variety of conflicting selection pressures that act on both signallers
and receives, have led to a diversity of solutions. Masking interference, for exam-
ple, is reduced in most species by adjusting the timing of signals to avoid overlap.
In some species, however, overlap increases as males compete to become more
attractive to females.
Many features of communicating in networks, such as signal timing interac-
tions, selective attention and simultaneous mate choice, have been relatively well
studied in anurans. Other specific signalling behaviours, predicted from commu-
nication network theory, such as social eavesdropping (Ch. 14), audience effects

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
292 T. U. Grafe
(Ch. 4) and victory displays (Ch. 6), have generally not been considered or investi-
gated explicitly. Such behaviours would be predicted to occur in species that have
several encounters with the same individuals, such as in many territorial ranid
frogs that show individual recognition (e.g. Bee & Gerhardt, 2002) or in dendro-
batid frogs with year-round territorial behaviour and complex patterns of parental
care (e.g. Summers, 1992).
Social eavesdropping is likely to be a general phenomenon of anuran choruses.
Identifying additional cases of social eavesdropping in anurans, as outlined above,
would provide a fruitful avenue for future studies and would further highlight that
anuran choruses are complex communication networks. An open question in this
context is the functional significance of female preferences for leading (or lagging)
advertisement calls. Determining any indirect and direct benefits females may
obtain from their choice of leading males would be highly desirable. In addition,
studies that demonstrate social eavesdropping through aggressive interactions
would be of interest.
The relative ease with which phonotaxis can be induced in females in the lab-
oratory and the availability of sound synthesis software for the production of
synthetic signals that can be constructed with signal parameters varying indepen-
dently of each other have diverted attention from testing female preferences in
the chorus. New techniques, such as multiple channel recordings as well as mi-
crophone and speaker arrays, will undoubtedly contribute to our understanding
of patterns of male vocal competition and female choice. More observational data
on female sampling behaviour would also contribute to revealing how receivers
deal with complex signalling networks (e.g. Murphy & Gerhardt, 2002).
The perceptual basis of communication in noisy environments remains largely
unexplored (Ch. 20). It seems likely that receivers group sounds into auditory
streams in order to improve recognition and to assign them to individual signallers
(i.e. auditory scene analysis: Feng & Ratnam 2000; Hulse 2002). A recent study
by Farris et al. (2002) demonstrated auditory grouping in female tungara frogs
in which the whine and chuck are grouped together even when presented from
widely different directions. Understanding how auditory systems group incoming
signals or signal components will help to explain how animals communicate in
noisy networks and how they achieve selective attention; it also has the potential to
provide a mechanistic basis for understanding the evolution of multicomponent
signals.
Anurans offer unique opportunities to study communication networks. Males
of many species aggregate in choruses of varying size and complexity. Advertise-
ment calls of anurans are long-range signals used to attract females and repel rival
males. Investigations are facilitated by the species-specific and highly stereotyped
signals as well as by the generally small signal repertoires. The consequences of

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 293
strategic decisions can readily be observed because fertilization is generally
external, making paternity analysis unnecessary. Consequently, the evolutionary
consequences of communicating in network environments can be assessed with
relative ease. In addition, anurans occupy a wide variety of habitats and use a
variety of reproductive strategies. This diversity provides numerous opportunities
for comparative analyses.
Acknowledgements
I thank Peter McGregor and two anonymous reviewers for their helpful comments on
a previous version of this chapter. My work on running frogs in West Africa was supported by the
Deutsche Forschungsgemeinschaft (Gr 1584).
References
Arak, A. 1988. Female mate selection in the natterjack toad: active choice or passive
attraction? Behavioral Ecology and Sociobiology, 22, 317–327.
Backwell, P., Jennions, M., Passmore, N. & Christy, J. 1998. Synchronized courtship in
fiddler crabs. Nature, 391, 31–32.
Bailey, W. J. & Field, G. 2000. Acoustic satellite behaviour in the Australian bushcricket
Elephantodeta nobilis (Phaneropterinae, Tettigonidae, Orthoptera). Animal
Behaviour, 59, 361–369.
Bee, M. A. & Bowling, A. C. 2002. Socially mediated pitch alteration by territorial male
bullfrogs, Rana catesbeiana. Journal of Herpetology, 36, 140–143.
Bee, M A. & Gerhardt, H. C. 2002. Individual voice recognition in a territorial frog
(Rana catesbeiana). Proceedings of the Royal Society of London, Series B, 269, 1443–1448.
Bosch, J. 2001. Female reciprocal calling in the Iberian midwife toad (Alytes cisternasii)
varies with male call rate and dominant frequency: implications for sexual
selection. Naturwissenschaften, 88, 434–437.
Bosch, J. & Marquez, R. 2002. Female preference function related to precedence effect
in an amphibian anuran (Alytes cisternasii): tests with non-overlapping calls.
Behavioral Ecology, 13, 149–153.
Bradbury, J. W. 1981. The evolution of leks. In: Natural Selection and Social Behavior, ed. R.
D. Alexander & D. W. Tinkle. New York: Chiron Press, pp. 138–169.
Brenowitz, E. A. & Rose, G. J. 1999. Female choice and plasticity of male calling
behaviour in the Pacific treefrog. Animal Behaviour, 57, 1337–1342.
Brenowitz, E. A., Rose, G. J. & Alder, T. 2000. The neuroethology of acoustic
communication in Pacific treefrogs. In: Anuran Communication, ed. M. J. Ryan.
Washington, DC: Smithsonian Institute Press, pp. 145–155.
Brush, J. S. & Narins, P. M. 1989. Chorus dynamics of a Neotropical amphibian
assemblage: comparison of computer simulation and natural behaviour. Animal
Behaviour, 37, 33–44.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
294 T. U. Grafe
Cranford, J. & Oberholzer, M. 1976. Role of neocortex in binaural hearing in the cat. II:
The ‘precedence effect’ in sound localization. Brain Research, 111, 225–239.
Deutsch, J. C. 1994. Uganda kob mating success does not increase on larger leks.
Behavioral Ecology and Sociobiology, 34, 451–459.
Dyson, M. L. & Passmore, N. I. 1988a. Two-choice phonotaxis in Hyperolius marmoratus
(Anura: Hyperoliidae): the effect of temporal variation in presented stimuli.
Animal Behaviour, 36, 648–652.
1988b. The combined effect of intensity and the temporal relationship of stimuli on
phonotaxis in female painted reed frogs Hyperolius marmoratus. Animal Behaviour,
36, 1555–1556.
Enquist, M., Ghirlanda, S. & Hurd, P. L. 1998. Discrete conventional signalling of a
continuous variable. Animal Behaviour, 56, 749–754.
Farris, H. E., Rand, A. S. & Ryan, M. J. 2002. The effects of spatially separated call
components on phonotaxis in tungara frogs: evidence for auditory grouping.
Brain, Behavior and Evolution, 60, 181–188.
Feng, A. S. & Ratnam, R. 2000. Neural basis of hearing in real-world situations. Annual
Review of Psychology, 51, 699–725.
Feng, A. S., Narins, P. M. & Xu, C.-H. 2002. Vocal acrobatics in a Chinese frog, Amolops
tormotus. Naturwissenschaften, 89, 352–356.
Fleischack, P. C. & Small, C. P. 1978. The vocalizations and breeding behaviour of
Kassina senegalensis (Anura, Rhacophoridae) in summer breeding aggregations.
Koedoe, 21, 91–99.
Forester, D. C. & Harrison, W. K. 1987. The significance of antiphonal vocalisation by
the spring peeper, Pseudacris crucifer (Amphibia, Anura). Behaviour, 103, 1–15.
Gerhardt, H. C. 1988. Acoustic properties used in call recognition by frogs and toads.
In: The Evolution of the Amphibian Auditory System, ed. B. Fritzsch, M. J. Ryan, W.
Wilczynski, T. E. Hetherington & W. Walkowiak. New York: John Wiley,
pp. 455–483.
1994. The evolution of vocalization in frogs and toads. Annual Review of Ecology and
Systematics, 25, 293–324.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans: Common
Problems and Diverse Solutions. Chicago, IL: Chicago University Press.
Gerhardt, H. C. & Klump, G. M. 1988. Masking of acoustic signals by the chorus
background noise in the green treefrog: a limitation on mate choice. Animal
Behaviour, 36, 1247–1249.
Gerhardt, H. C. & Schwartz, J. J. 1995. Interspecific interactions in anuran courtship.
In: Amphibian Biology: Social Behaviour, ed. H. Heatwole & B. K. Sullivan. Chipping
Norton, UK: Surrey Beatty, pp. 601–632.
Given, M. F. 1987. Vocalizations and acoustic interactions of the carpenter frog, Rana
virgatipes. Herpetologica, 43, 467–481.
Grafe, T. U. 1995a. Graded aggressive calls in the African painted reed frog Hyperolius
marmoratus (Hyperoliidae). Ethology, 101, 67–81.
1995b. Sexual selection and communication in the African painted reed frog
(Hyperolius marmoratus). Ph.D. Thesis, Cornell University, Ithaca, NY.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 295
1996a. The function of call alternation in the African reed frog Hyperolius
marmoratus: precise call timing prevents auditory masking. Behavioral Ecology and
Sociobiology, 38, 149–158.
1996b. Energetics of vocalization in the African reed frog (Hyperolius marmoratus).
Comparative Biochemistry and Physiology A, 114, 235–243.
1997a. Use of metabolic substrates in the gray treefrog, Hyla versicolor: implications
for calling behavior. Copeia, 1997, 356–362.
1997b. Costs and benefits of mate choice in the lek-breeding reed frog, Hyperolius
marmoratus. Animal Behaviour, 53, 1103–1117.
1999. A function of synchronous calling and a novel female preference shift in an
anuran. Proceedings of the Royal Society of London, Series B, 266, 2331–2336.
2003. Synchronised interdigitated calling in the Kuvangu running frog (Kassina
kuvangensis). Animal Behaviour, 66, 127–136.
Grafe, T. U. & Thein, J. 2001. Energetics of calling and metabolic substrate use during
prolonged exercise in the European treefrog Hyla arborea. Journal of Comparative
Physiology B, 171, 69–76.
Grafe, T. U., Spieler, M. & Konig, B. 1999. Soziobiologische Erklarungsansatze
des Verhaltens von Amphibien und Reptilien. Zeitschrift fur Feldherpetologie, 6,
1–42.
Grafen, A. 1990. Biological signals as handicaps. Journal of Theoretical Biology, 144,
517–546.
Greenfield, M. D. 1994a. Cooperation and conflict in the evolution of signal
interactions. Annual Review of Ecology and Systematics, 25, 97–126.
1994b. Synchronous and alternating choruses in insects and anurans: common
mechanisms and diverse functions. American Zoologist, 34, 605–615.
Greenfield, M. D. & Rand, A. S. 2000. Frogs have rules: selective attention algorithms
regulate chorusing in Physalaemus pustulosus (Leptodactylidae). Ethology, 106,
331–347.
Greenfield, M. D. & Roizen, I. 1993. Katydid synchronous chorusing is an evolutionary
stable outcome of female choice. Nature, 364, 618–620.
Greenfield, M. D., Tourtellot, M. K. & Snedden, W. A. 1997. Precedence effects and the
evolution of chorusing. Proceedings of the Royal Society of London, Series B, 264,
1355–1361.
Hulse, S. 2002. Auditory scene analysis in animal communication. Advances in the Study
of Behavior, 31, 163–200.
Ibanez, D. R. 1993. Female phonotaxis and call overlap in the Neotropical glassfrog
Centrolenella granulosa. Copeia, 1993, 846–850.
Johnston, R. E., Sorokin, E. S. & Ferkin, M. H. 1997. Female voles discriminate males’
over-marks and prefer top-scent males. Animal Behaviour, 54, 679–690.
Johnstone, R. A. 1994. Honest signalling, perceptual error and the evolution of
‘all-or-nothing’ displays. Proceedings of the Royal Society of London, Series B, 256,
169–175.
Kelly, J. B. 1974. Localization of paired sound sources in the rat: small time differences.
Journal of the Acoustical Society of America, 55, 1277–1284.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
296 T. U. Grafe
Kime, N. M., Turner, W. R. & Ryan, M. J. 2000. The transmission of advertisement calls
in Central American frogs. Behavioral Ecology, 11, 71–83.
Klump, G. M. 1996. Bird communication in the noisy world. In: Ecology and Evolution of
Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY:
Cornell University Press, pp. 321–338.
Klump, G. M. & Gerhardt, H. C. 1987. Use of non-arbitrary acoustic criteria in mate
choice by female gray tree frogs. Nature, 326, 286–288.
1992. Mechanisms and function of call-timing in male–male interactions in frogs.
In: Playback and Studies of Animal Communication, ed. P. K. McGregor. New York:
Plenum Press, pp. 153–174.
Littlejohn, M. J. & Harrison, P. A. 1985. The functional significance of the diphasic
advertisement call of Geocrinia victoriana (Anura: Leptodactylidae). Behavioral
Ecology and Sociobiology, 16, 363–373.
Lopez, P. T., Narins, P. M., Lewis, E. R. & Moore, S. W. 1988. Acoustically induced call
modification in the white-lipped frog, Leptodactylus albilabris. Animal Behaviour, 36,
1295–1308.
Marshall, V. T., Humfeld, S. C. & Bee, M. A. 2003. Plasticity of aggressive signalling in
male spring peepers (Pseudacris crucifer). Animal Behaviour, 65, 1223–1234.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
Moore, S. W., Lewis, E. R., Narins, P. M. & Lopez, P. T. 1989. The call-timing algorithm of
the white-lipped frog, Leptodactylus albilabris. Journal of Comparative Physiology A,
164, 309–319.
Murphy, C. G. & Gerhardt, H. C. 2002. Mate sampling by female barking treefrogs (Hyla
gratiosa). Behavioral Ecology, 13, 472–480.
Narins, P. M. 1982. Behavioral refractory period in neotropical treefrogs. Journal of
Comparative Physiology A, 148, 337–344.
Narins, P. M. & Capranica, R. R. 1978. Communicative significance of the two-note call
of the treefrog Eleutherodactylus coqui. Journal of Comparative Physiology A, 127, 1–9.
Narins, P. M. & Zelick, R. 1988. The effects of noise on auditory processing and
behavior in amphibians. In: The Evolution of the Amphibian Auditory System, ed. B.
Fritzsch, T. Hetherington, M. Ryan, W. Wilczynski & W. Walkowiak. New York:
John Wiley, pp. 455–483.
Narins, P. M., Lewis, E. R. & McClelland, B. E. 2000. Hyperextended call note repertoire
of the endemic Madagascar treefrog Boophis madagascariensis (Rhacophoridae).
Journal of Zoology, 250, 283–298.
Passmore, N. I. 1981. Sound levels of mating calls of some African frogs. Herpetologica,
37, 166–171.
Passmore, N. I., Bishop, P. J. & Caithness, N. 1992. Calling behaviour influences mating
success in male painted reed frogs, Hyperolius marmoratus. Ethology, 92, 227–241.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 297
Penna, M. & Solis, R. 1998. Frog call intensities and sound propagation in the South
American temperate forest region. Behavioral Ecology and Sociobiology, 42, 371–381.
Pfennig, K. S., Rapa, K. & McNatt, R. 2000. Evolution of male mating behavior: male
spadefoot toads preferentially associate with conspecific males. Behavioral Ecology
and Sociobiology, 48, 69–74.
Robinson, D. 1990. Acoustic communication between the sexes in bushcrickets. In: The
Tettigoniidae: Behavior, Systematics, Evolution, ed. W. J. Bailey & D. C. F. Rentz.
Bathurst, Australia: Crawford House Press, pp. 110–129.
Romer, H., Bailey, W. J. & Dadour, I. 1989. Insect hearing in the field. III. Masking by
noise. Journal of Comparative Physiology A, 164, 609–620.
Ryan, M. J. 1985. The Tungara Frog. Chicago, IL: University of Chicago Press.
1986. Synchronized calling in a treefrog (Smilisca sila). Brain, Behavior and Evolution,
29, 196–206.
Ryan, M. J. & Keddy-Hector, A. 1992. Directional patterns of female mate choice and
the role of sensory biases. American Naturalist, 139, S4–S35.
Ryan, M. J. & Wilczynski, W. 1991. Evolution of intraspecific variation in the
advertisement call of a cricket frog (Acris crepitans). Biological Journal of the Linnean
Society, 44, 249–271.
Ryan, M. J., Cocroft, R. B. & Wilczynski, W. 1990. The role of environmental selection in
intraspecific divergence of mate recognition signals in the cricket frog, Acris
crepitans. Evolution, 44, 1869–1872.
Schneider, H., Hoermann, G. & Hodl, W. 1988. Calling and antiphonal calling in four
Neotropical anuran species of the family Leptodactylidae. Zoologische Jahrbucher
Physiologie, 92, 77–103.
Schwartz, J. J. 1989. Graded aggressive calls of the spring peeper, Pseudacris crucifer.
Herpetologica, 45, 172–181.
1993. Male calling behavior, female discrimination and acoustic interference in the
Neotropical treefrog Hyla microcephala under realistic acoustic conditions.
Behavioral Ecology and Sociobiology, 32, 401–414.
2001. Call monitoring and interactive playback systems in the study of acoustic
interactions among male anurans. In: Anuran Communication, ed. M. J. Ryan.
Washington, DC: Smithsonian Institute Press, pp. 183–204.
Schwartz, J. J. & Wells, K. D. 1984. Interspecific acoustic interactions of the neotropical
treefrog Hyla ebraccata. Behavioral Ecology and Sociobiology, 14, 211–224.
Schwartz, J. J., Buchanan, B. W. & Gerhardt, H. C. 2001. Female mate choice in the gray
treefrog (Hyla versicolor) in three experimental environments. Behavioral Ecology
and Sociobiology, 49, 443–455.
Shelly, T. E. & Greenfield, M. D. 1991. Dominions and desert clickers (Orthoptera:
Acrididae): influences of resources and male signaling on female settlement
patterns. Behavioral Ecology and Sociobiology, 28, 133–140.
Staaden, M. J. v. & Romer, H. 1997. Sexual signalling in bladder grasshoppers: tactical
design for maximizing calling range. Journal of Experimental Biology, 200,
2597–2608.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
298 T. U. Grafe
Sullivan, B. K., Ryan, M. J. & Verrell, P. A. 1995. Female choice and mating system
structure. In: Amphibian Biology: Social Behaviour, ed. H. Heatwole & B. K. Sullivan.
Chipping Norton, UK: Surrey Beatty, pp. 469–517.
Summers, K. 1992. Dart-poison frogs and the control of sexual selection. Ethology, 91,
89–107.
Taigen, T. L., Wells, K. D. & Marsh, R. L. 1985. The enzymatic basis of high metabolic
rates in calling frogs. Physiological Zoology, 58, 719–726.
Telford, S. R., Dyson, M. L. & Passmore, N. I. 1989. Mate choice occurs only in small
choruses of painted reed frogs (Hyperolius marmoratus). Bioacoustics, 2, 47–53.
Townsend, D. S. & Stewart, M. M. 1986. Courtship and mating behavior of a Puerto
Rican frog, Eleutherodactylus coqui. Herpetologica, 42, 165–170.
Tuttle, M. D. & Ryan, M. J. 1982. The role of synchronized calling, ambient light, and
ambient noise, in anti-bat-predator behavior of a treefrog. Behavioral Ecology and
Sociobiology, 11, 125–131.
Vencl, F. V. & Carlson, A. D. 1998. Proximate mechanisms of sexual selection in the
firefly Photinus pyralis (Coleoptera: Lampyridae). Journal of Insect Behavior, 11,
191–207.
Verrell, P. A. & Brown, L. E. 1993. Competition among females for mates in a species
with male parental care, the widwife toad Alytes obstetricans. Ethology, 93, 247–257.
Walkowiak, W. 1992. Acoustic communication in the fire-bellied toad: an integrative
neurobiological approach. Ethology, Ecology and Evolution, 4, 63–74.
Wallach, H., Newman, E. B. & Rosenzweig, M. R. 1949. The precedence effect in sound
localization. American Journal of Psychology, 62, 315–336.
Wells, K. D. 1988. The effect of social interactions on anuran vocal behavior. In: The
Evolution of the Amphibian Auditory System, ed. B. Fritzsch, M. J. Ryan, W. Wilczynski,
T. E. Hetherington & W. Walkowiak. New York: John Wiley,
pp. 433–454.
1989. Vocal communication in a Neotropical treefrog, Hyla ebraccata: responses of
males to graded aggressive calls. Copeia, 1989, 461–466.
2001. The energetics of calling in frogs. In: Anuran Communication, ed. M. J. Ryan.
Washington, DC: Smithsonian Institute Press, pp. 45–60.
Wells, K. D. & Schwartz, J. J. 1984. Vocal communication in a Neotropical treefrog, Hyla
ebraccata: advertisement calls. Animal Behaviour, 32, 405–420.
Wells, K. D. & Taigen, T. L. 1986. The effect of social interactions on calling energetics
in the gray treefrog (Hyla versicolor). Behavioral Ecology and Sociobiology, 19, 9–18.
Widemo, F. & Owens, I. P. F. 1995. Lek size, male mating skew and the evolution of
lekking. Nature, 373, 148–151.
Wiley, R. H. 1991. Associations of song properties with habitat for territorial oscine
birds of eastern North America. American Naturalist, 138, 973–993.
Wiley, R. H. & Richards, D. G. 1978. Physical constraints on acoustic communication in
the atmosphere: implications for the evolution of animal vocalizations. Behavioral
Ecology and Sociobiology, 3, 69–94.
Wyttenbach, R. A. & Hoy, R. R. 1993. Demonstration of the precedence effect in an
insect. Journal of the Acoustical Society of America, 94, 777–784.

P1: JZZ/... P2: JZZ/...0521823617c13.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 16:26
Anuran choruses as communication networks 299
Zelick, R. D. & Narins, P. M. 1983. Intensity discrimination and the precision of call
timing in two species of neotropical treefrogs. Journal of Comparative Physiology A,
153, 403–412.
1985. Characterization of the advertisement call oscillator in the frog
Eleutherodactylus coqui. Journal of Comparative Physiology A, 156, 223–229.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
14
Singing interactions in songbirds:implications for social relationsand territorial settlement
m a rc n ag u i b
University of Bielefeld, Germany
Introduction
Interactions between individuals make up a significant part of life in so-
cial animals. They form a crucial behavioural mechanism establishing and main-
taining particular spacing patterns among individuals and groups of individuals
and are inherent in the regulation of social relations. Animals interact with each
other in a broad range of contexts, such as during intersexual competition, mate
choice, or parent–offspring communication, but still many of the underlying prin-
ciples share common ground (Hauser, 1996; Bradbury & Vehrencamp, 1998). It is
well documented that the performance of individuals in interactions has profound
implications for the resolution of conflicts over resources, such as mates, food or
space. Interactions may consist of complex behavioural displays or may be based
exclusively on signals in either one or several signalling modalities. Vocal interac-
tions are among the most conspicuous forms of interactions and have been well
studied in several taxonomic groups, such as insects, anurans and birds (Bradbury
& Vehrencamp, 1998). In birds, vocal interactions are most evident in parent–
offspring communication (Kilner & Johnstone, 1997; Ch. 9), calling and singing in
group-living species (Farabaugh & Dooling, 1996; Zann, 1996), duetting in tropi-
cal songbirds (von Helversen, 1980; Farabaugh, 1982) and in singing interactions
between male territorial songbirds (Todt & Naguib, 2000).
In this review, I will focus on singing interactions in male territorial song-
birds. Their vocal interactions are among the most striking examples of bird
vocal communication and are an established model for studies on territoriality
and communication networks (McGregor, 1993; McGregor & Dabelsteen, 1996;
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
300

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 301
Todt & Naguib, 2000). Male songbirds commonly hold adjacent territories forming
neighbourhoods; so usually several males sing within signalling range of each
other. The typical pattern of settlement and spacing of individuals of the same
species thus sets the framework for the evolution of communication behaviour,
including the evolution of complex patterns of vocal interactions and the strate-
gies for gathering information on conspecifics.
Singing interactions between males take place in a variety of different contexts
and the information exchanged may strongly depend on the singers’ social and
spatial relations. They take place during immediate competition over resources
such as space or mates in addition to being a conspicuous component in the social
relations between established territorial neighbours. In general, vocal interactions
differ from the classical song traits such as singing activity (Kempenaers et al.,
1997; Gil et al., 1999; Amrhein et al., 2002, 2004), singing versatility (Hasselquist
et al., 1996; Searcy & Yasukawa, 1996) or other performance-related traits (Podos,
1996; Forstmeier et al., 2002). These classical traits can be regarded as ‘individual’
traits that are present regardless of the social context in which a male is singing
(Fig. 14.1). Vocal interactions, in contrast, have an additional interactive dimension
as the message conveyed depends strongly on the pattern of song interchange
between the interacting singers (Todt & Naguib, 2000). This interactive dimension
has resulted in considerable current research interest in vocal interactions, as the
performance of singers during an interaction provides immediate information on
relative differences between them: information that also is used by eavesdropping
individuals in a communication network (McGregor & Dabelsteen, 1996).
My principal goal in this chapter is to integrate current knowledge on strategies
of vocal interactions in territorial songbirds with concepts of territorial behaviour
and territorial settlement (Waser & Wiley, 1980; Stamps, 1994; Stamps & Krishnan,
2001) and to explore how recent advances in studies on vocal interactions con-
tribute to our understanding of the social relations among neighbouring territo-
rial songbirds. The social and spatial relationships among neighbouring males can
be mediated by their vocal interactions; consequently, vocal interactions can have
profound implications for the evolution of strategies for territorial settlement
and spacing behaviour in general. I will also evaluate how principles of vocal in-
teractions contribute to our understanding of the evolution of singing strategies
and the evolution of receivers’adaptations to gather information from conspecific
signalling behaviour. A key trait of vocal interactions responsible for much of the
interest is that they are commonly asymmetric in the sense that each of the singers
involved uses its songs differently in relation to those of its counterpart. These
asymmetries can reflect differences in motivation or quality among singers and
consequently provide information about the relationship between them (Todt &
Naguib, 2000). Therefore, this chapter will consider the internal aspects of

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
302 M. Naguib
Pathways of information gathering
Social traits
Vocal interactions (e.g. asymmetries in vocal matching, or in timing of songs)
Song structure
Diurnal singing activity
Singing rate
Individual song traits
Singing versatility
Extracting information from individual signals Extracting information from interactions and their asymmetries (eavesdropping)
Fig. 14.1. Singing traits and pathways of information gathering. Receivers can gather
information from individual traits during and in the absence of interactions.
Information gathering by extracting information from signalling interactions
(eavesdropping) provides additional information that cannot be extracted from
individual song traits. In interactions, additional social factors come into play such as
the relative relation of the songs, which can provide immediate information on
differences between singers.
interactions (i.e. their function in the interaction between the singers) as well as
their external implications (i.e. their wider importance as a source of information
for other listening (eavesdropping) individuals; see also Ch. 2).
After reviewing some recent studies of vocal interactions and by drawing several
examples from our own studies on nightingales Luscinia megarhynchos (reviewed
in more detail by Todt & Naguib, 2000), I will evaluate more closely the general
association of social and spatial relationships between males and their strategies in
vocal interactions, as well as their strategies in gathering information by attending
to others’ vocal interactions.
Nature of interactions
Interactions in communication can be defined as the exchange of in-
formation through signals by at least two individuals where the signals of both
signallers have some direct relation to each other. In acoustic communication,
interactions can be best determined when there are two individuals signalling;
such interactions are referred to commonly as dyadic interactions. When several
individuals are signalling, they may form a communication network with highly
complex modes of interactions (see Chs. 13 and 15) but this need not be the case

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 303
(Shackleton & Ratcliffe, 1994). Although interactions pragmatically can be defined
broadly as instances when two or more singers are singing at the same time, true
interactions are best identified by demonstrating that the singers influence each
other in their choice and timing of song types or in other aspects of their singing
strategy. Such a definition is analogous to our understanding of dialogues in hu-
man speech and allows us to extract and define specific strategies and to study
their causations and evolutionary implications.
Song matching
Most songbirds have song repertoires, allowing them flexibility in deci-
sions on the next song to be sung (Kroodsma, 1982). The most conspicuous way of
using specific song types during an interaction is song matching, a situation in
which a singer replies with a song of the same type as the preceding song sung
by the opponent. Such matching of signals is also found in other taxa (Ch. 18) and
can give insights into the functions of signalling strategies as well as addressing
mechanistic questions such as how animals perceive and categorize signals (Falls
et al., 1982, 1988; Weary et al., 1990; Naguib et al., 2002). Song matching is known to
be used to address a particular rival and often it appears to function as an aggres-
sively directed signal to increase the level of threat towards a specific rival (Krebs
et al., 1981; McGregor et al., 1992; Nielsen & Vehrencamp, 1995; Vehrencamp, 2001).
However, several studies have shown that song matching is not always a strong
aggressive signal and may be used as a graded signal of intent (Searcy et al., 2000;
Burt et al., 2002; Naguib et al., 2002). Moreover, males may sometimes match some,
but not all, features of songs. Males may match only parts of a song or certain song
parameters such as the frequency of the full song (Otter et al., 2002) or specific song
components (Burt et al., 2002; Naguib et al., 2002), duration of songs (Weary et al.,
1990) or categories of song (Wiley et al., 1994; Naguib et al., 2002). Neighbouring
song sparrows Melospiza melodia have been shown to match repertoires by reply-
ing with non-matching songs that are shared with the singing opponent (Beecher
et al., 1996). The meaning of matching also may vary with the distance between
singers, the general context, the song type or even the specific song component
that is matched. In nightingales, we discovered patterns of song matching that
clearly differed from the most widespread principle that song matching increases
with the level of perceived threat (Naguib et al., 2002). In playbacks conducted
on nightingales’ nocturnal song, males increased the precision of matching the
pitch of whistles in so-called whistle songs (Fig. 14.2) with increasing distance
to the simulated unfamiliar opponent but not the overall rate of matching the
song category. Matching whistle songs and specifically the pitch of the whistles
may have a different biological significance than matching ‘normal’ songs. The
narrow spectral bandwidth of the whistles implies that they transmit with less

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
304 M. Naguib
2 4 6 s
2
4
6
8
10kHz
2 4 s
2
4
6
8
10kHz
1 12
21
(a)
(b)
Fig. 14.2. Spectrographic examples of whistle song matching and overlapping in
nightingales. Hatched arrows indicate beginning of songs, numbers indicate different
males. (a) The whistle song in the centre from male 2 is a non-overlapping, full song
type match of the first song of male 1, i.e. the whistle part and terminal sections are
the same in both songs and there is no noticeable overlap of songs. Male 1 replied
immediately to match and clearly overlap his opponent. (b) The two whistle songs
match but also differ in frequency so that the whistle parts do not mask each
other.
spectral degradation over long distances and thus may function particularly in
long-range signalling (Wiley & Richards, 1978; Slabbekoorn et al., 2002). At first
glance, more matching at long distance is puzzling as the social importance and,
therefore, the urgency of addressing a rival is assumed to decrease with interindi-
vidual spacing. However, even distant neighbours that do not share a territorial
boundary are part of the same neighbourhood, in which males have to establish
and maintain social and spatial relations that are likely to be regulated through
long-range vocal interactions. Radio-tracking data have shown that males make
substantial excursions into territories of direct neighbours and even more dis-
tant ones within auditory range from the own territory (Hanski, 1992; Chandler
et al., 1997; Pitcher & Stutchbury, 2000; Naguib et al., 2001), indicating that the
social and spatial relations between males go beyond their immediate neighbours
with whom territorial boundaries are shared. Long-distance matching, therefore,
may be a mechanism involved directly in establishing and maintaining spatial
relations between males.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 305
In summary, the extensive research on song matching (reviewed more fully
by Todt & Naguib, 2000) shows that matching can do more than simply address a
rival; it can have several functions, depending on the social context, the territorial
relationships of the interacting males and the song type or its specific parameters
that are matched. It will be interesting to see how the refinements of playback
design and playback technologies (e.g. Dabelsteen, 1992) will address more subtle
questions on the function of song matching in different contexts and thus provide
more detailed insights into the kinds of message conveyed.
Timing of songs
In a singing interaction, the relative timing of song production by the
interactants can signal specific information on the singer’s state and intention,
such as its readiness to escalate the contest (Todt & Naguib, 2000). The relative
timing of songs during an interaction differs from song matching in two ways.
First, song sharing or detailed knowledge of a rival’s repertoire is not required.
Second, relative song timing can vary continuously whereas song matching is more
categorical (i.e. matching occurs or does not occur) even though recent research
has emphasized that song matching can be subtle with graded components and
that it is not restricted to matching full song types, as discussed above.
Despite the continuous nature of relative song timing, two categories of timing
of songs have been shown to occur to date (Hultsch & Todt, 1982) and to have func-
tionally different signal value: song alternating and song overlapping (Brindley,
1991; Dabelsteen et al., 1996; Naguib et al., 1999; Langemann et al., 2000). Song al-
ternating is a common strategy where males take turns in delivering their songs.
This strategy can also be observed in concurrent interspecific singing where males
avoid acoustic competition (Ficken et al., 1974). During song overlapping, in con-
trast, males begin to sing a song before the opponent has ended its song. Song
overlapping has been shown in several species to function as a directed agonistic
signal (Brindley, 1991; McGregor et al., 1992; Dabelsteen et al., 1996, 1997; Naguib,
1999) whereas song alternating is the seemingly predominating singing strategy
during less-intense contexts. Interestingly, song overlapping is not only treated
as an agonistic signal by the singer whose songs are overlapped but also is used
by eavesdropping males and females to assess differences in the relative quality
or motivation in two interacting males (Naguib & Todt, 1997; Naguib et al., 1999;
Otter et al., 1999; Peake et al., 2001, 2002; Mennill et al., 2002), as discussed below
(see also Ch. 2). Song overlapping can result in considerable masking of part of the
rival’s song, but this effect is not inherent in song overlapping. Masking effects
will depend on the distance between singers (i.e. their relative difference in am-
plitude), the amount of song overlapped in terms of duration and the similarity
in phonology between the overlapping parts of the two songs (Fig. 14.2; see also

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
306 M. Naguib
Ch. 20). In addition, the extent of perceived masking will vary with whose percep-
tion is considered: the singer whose songs are overlapped, the overlapper or other
individuals, which may be at any relative location to the two singers (see below). As
a result, benefits through masking the opponent’ssongs may be limited to certain
conditions and locations and, despite the attractiveness of the argument, it is un-
likely to be the primary consequence that led to the evolution of song overlapping
as a singing strategy in long-distance interactions. Although overlap will affect
detection and recognition of subtle sound features (Wiley, 1983, 1994) by other lis-
tening conspecifics, spatial release from masking (Klump, 1996) is a compensatory
mechanism that can help in coping with problems resulting from masking. Todt
and Naguib (2000) further suggested that a male that is overlapping the songs
of its opponent might benefit by shifting the attention of eavesdroppers to the
overlapper. Finally, despite such a range of basic effects of song overlap on sig-
nal perception, song overlapping may also be a conventional signal of dominance
that is maintained by retaliation costs if overlapping increases the probability to
escalate a contest.
Song overlapping and song alternating have been shown to be of biological
significance, but the issue of how much overlap needs to be achieved in order to
accomplish a certain function remains to be studied. Similarly, the precise timing
of songs during alternating singing and also during song overlapping in natural
conditions may have specific signal value (McGregor et al., 1992), another issue
that deserves to be explored in more detail in future studies. For instance, some
studies suggest that song alternating is not a homogeneous strategy but that the
exact timing during alternating is also of functional significance. Specifically,
leader–follower relationships, in which the follower sings soon after the leader,
have been interpreted as the leader representing the more dominant singer (Smith
& Norman, 1979; Popp, 1989; Naguib et al., 1999). Gathering information on very
fine temporal differences in timing of songs will require knowledge of the dis-
tance to the opponent (Naguib & Wiley, 2001) and eavesdroppers will require to
know the distance to each interactant because of the different time delay of the
songs originating from two sources at unequal distance. However, confusion over
whether or not a song is overlapping can occur only in long-range interactions in
two extremes: when both singers start their song at about the same time (so that
each singer will perceive the opponent’s song as overlapping) or when an overlap-
ping song sets on late during the song that is overlapped (so that the opponent
may receive it as non-overlapping). It remains to be studied if overlapping events
that fall into this ‘confusion range’are interpreted differently from unambiguous
events. In nightingales, overlapping songs most commonly fall outside these con-
fusion ranges (Hultsch & Todt, 1982; Naguib, 1999); consequently, in most cases
overlapping is an unambiguous event.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 307
Matching and timing
In the preceding sections, we have seen that both song matching and
song timing are characteristic of bird vocal interactions and in many cases are
related to a specific meaning of singing. However, the possibility that both may
be dependent on each other (Todt, 1981; Wolffgramm & Todt, 1982) has been little
explored from a functional perspective. There are several possible combinations
of these two aspects. For example, males may match and overlap a rival’s song, as
nightingales frequently do when matching whistle songs (Fig. 14.2), or they may
match a song with varying delays during alternating singing.
Matching songs during boundary disputes often occurs in interactions with
high song rates and thus short delays in responses. Matching with short delays
may be of specific value in signalling the willingness to escalate a contest. When
males interact where no immediate dispute is apparent, such as during the dawn
chorus or with low song rates in long-range interactions between neighbours that
sing at the same time of day or night, matching may be timed differently and,
therefore, may have a different meaning and consequence for the social relations
between singers. One possibility is that frequent matching between established
territory holders acts to repel non-territorial males seeking to establish a territory
(Amrhein et al., 2004) (thus benefiting both territorial males) by signalling long-
term territory tenure and an established spatial and social relationship. There is
evidence that males sharing song types have longer territory tenure (Beecher et al.,
2000) and neighbouring males often share more songs than non-neighbouring
males (Kroodsma, 1974; Hultsch & Todt, 1981; McGregor & Krebs, 1982; Schroeder
& Wiley, 1983; Beecher, 1996; Payne, 1996; Beecher et al., 2000; Griessmann &
Naguib, 2002): both features that make song matching more likely. An increase
in song sharing over the season, as shown for thrush nightingales Luscinia luscinia
(Sorjonen, 1987), may increase the probability of matching during vocal interac-
tions as the season progresses. If so, matching during long-range vocal interactions
such as the thrush nightingales’ nocturnal song may function to strengthen ter-
ritorial residency in neighbouring males rather than being an agonistic signal.
Vocal interactions and social relationships among singers
Songbirds interact with song in at least five different social contexts that
need to be considered when singing strategies and their evolutionary implications
are studied (Fig. 14.3). Territorial males interact by song with; (a) neighbours
over long distances when there is no immediate dispute noticeable (Fig. 14.3a);
(b) neighbours in immediate disputes over territorial boundaries or possibly over
access to females (Fig. 14.3b); (c) neighbours that have crossed the shared territo-
rial boundary (Fig. 14.3c); (d) unfamiliar rivals that have intruded into the territory

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
308 M. Naguib
Fig. 14.3. Five contexts of vocal interactions between male songbirds. Territories are
represented by elliptical shapes. (a) Long-range interaction between established
neighbours (filled and open circles); (b) boundary interaction between established
neighbours with both males singing in their own territory; (c) interaction in which
the resident male of the right-hand territory has intruded; (d) interaction between a
territory holder and a non-territorial stranger (hatched circle) that has intruded into
the territory; (e) interaction between a territory holder and a distant stranger
(hatched circle) that may attempt to establish a territory nearby.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 309
and start vocally claiming part of it (Fig. 14.3d); or (e) unfamiliar rivals that attempt
to establish a territory nearby without directly threatening the resident’sterritory
(Fig. 14.3e). Singing strategies in these different contexts are under different selec-
tion pressures as the social context differs; accordingly, the implications of specific
singing strategies will vary in these situations. In encounters between a resident
male and an intruder, asymmetries in site-specific dominance are inherent, as
the payoff for each male differs because of the prior investment of the resident
male in establishing and maintaining a territory before the contest (Maynard
Smith & Parker, 1976; Waser & Wiley, 1980). Interactions with strangers are often
single and time-limited events and, therefore, males clearly have to signal their
strength and should signal a higher readiness to escalate the contest. Territorial
residents are more likely to win a contest than intruders, so it is adaptive for
residents to invest more in the interaction (Pusey & Packer, 1997). Interactions
among neighbours, in contrast, are repeated and it may pay males to use a differ-
ent singing strategy. Moreover, the asymmetries in site-specific dominance that
is evident in all encounters between residents and intruders does not apply to
neighbour–neighbour interactions, provided that both are singing from within
their territories at locations that are not under direct dispute. The exact ways
males interact with their neighbours will depend, therefore, on their locations
and on previous experience of the dyad (Wiley & Wiley, 1980). Interactions be-
tween neighbours, as between residents and intruders, are still characterized by
asymmetries, presumably as a result of inherent differences among males, such as
differences in age, duration of prior residency or mating status. Remaining asym-
metries in status between territorial neighbours can then well be reflected in the
way they interact with each other vocally. Moreover, males may develop specific
expectations when interacting with specific neighbours because of the specific
ontogenetic trajectory of their relationship; consequently a given singing strat-
egy may have a different functional significance when used with neighbours than
when it is used with strangers. Biologically, the interactions among neighbours
are particularly interesting as they presumably reflect the social relationships be-
tween them and, therefore, provide deeper insights into the territorial and social
system of songbirds.
Experiments that focused explicitly on vocal interactions have simulated a
stranger’s or neighbour’s intrusion into a territory (or appearance near the ter-
ritory), simulating the contexts illustrated in Fig. 14.3d,e (Todt & Naguib, 2000).
Vocal interactions in such high-intensity contexts have served as an important
experimental model to unravel the functions of specific singing strategies during
an interaction. In general, such situations are characterized by high song rates,
song overlap, song matching and song switching (in species in which males usu-
ally repeat the same song type several times before switching to a different one).

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
310 M. Naguib
Depending on locations of simulated intrusions, territorial males distinguish be-
tween neighbours and strangers in their responses (Falls, 1982; Stoddard, 1996).
Such individual recognition is the prerequisite for different response strategies
with specific neighbours and new rivals and raises issues about how interactions
vary in dynamics and functions with the familiarity of the singers and their spatial
and social relationship.
The information on functions and principles of interactions obtained from
these playback experiments simulating intrusions are likely to be of general value
and applicability also to long-range interactions among established neighbours
that are interacting when singing on their own territory. However, it is impor-
tant to consider that neighbours can be expected to exchange much more subtle
information in vocal interactions in the absence of an immediate dispute. Neigh-
bours having prior experience with each other may be better at using nuances
in variation of singing patterns, such as song rate, quality of sound production
or use of specific song variants or song types. Communication among established
males may thus reach a much higher level of complexity with higher cognitive de-
mands than communication among unfamiliar males, where disputes are driven
by more immediate contests in specific contexts over specific resources. More de-
scriptive and experimental studies on long-range vocal interactions in the absence
of immediate disputes will be needed to test these ideas further.
Functions of vocal interactions in territorial defence against intruders
The functions of vocal interactions among residents and intruders are ob-
vious as there is an immediate conflict over space. Vocal interactions here make
up a significant fraction of the behaviour during such conflicts, underlying their
importance in spacing behaviour. Asymmetries in these interactions may be an im-
portant predictor for subsequent behaviour over the spatial conflict and, therefore,
may set the stage for the occurrence of subsequent and intermittent movements
and the probability of physical encounters.
Vocal interactions commonly escalate in intensity in immediate disputes over
territorial boundaries. Consequently, most intense interactions can be observed
early in the season when territories are established, whenever boundaries are vi-
olated by neighbours, and when males attempt to establish a new territory in an
area with males that have been resident for some time. In these situations, terri-
tory holders are highly aroused and attempt to drive the rival from the disputed
area by intense singing and an interactive strategy that signals high readiness
for escalation. Commonly, males then sing at a high song rate and interactions
are characterized by frequent song overlapping and high rates of song matching
or song switching, as discussed above. Most experimental research on the func-
tion of singing during vocal contests has used playback simulating an unfamiliar

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 311
intruder and demonstrated that song matching, song switching (not reviewed
here) and song overlapping are strategies used and perceived as an agonistic sig-
nalling behaviour, as discussed above. Therefore, these singing variables are likely
to determine the outcome of a contest over space. Future playback experiments
that focus on consequences of song overlap and song matching in terms of choice
of song posts by the opponent will be important to address this issue in more
detail. Males may avoid singing at posts where they provoke intense responses,
for example being overlapped (e.g. Todt, 1981) and challenged by high song rates,
and retreat earlier when their singing evokes such responses by resident males.
Functions of vocal interactions among neighbouring males
Songbirds sing extensively in the phase when territories are established
but continue to interact vocally when conflicts over space become less intense,
that is when territories appear to be established. The functions of these contin-
uing vocal interactions must have a different evolutionary significance, as their
outcome is less likely to have drastic effects in conflicts over space. Territorial
neighbours exhibit site-specific dominance; therefore, their vocal interactions are
not associated with spatial asymmetries unless intrusions take place. In these sit-
uations, when vocal interactions are unlikely to function to resolve immediate
conflicts over space, such interactions are more likely to function in maintaining
a spacing pattern and keeping remaining asymmetries of the territorial neigh-
bours at an equilibrium that avoids conflicts in which no clear winner is likely to
emerge. Information on conspecifics will be imperfect; consequently, neighbour-
ing males may need to continuously update their information on neighbours to
refine their assessment. Therefore, after the spatial arrangement in a territorial
system becomes established, vocal interactions occur most frequently between fa-
miliar neighbouring males singing from their own territories in the absence of im-
mediate boundary disputes. However, long-range interactions among neighbours
are much less well studied than interactions in high-intensity contexts (Kramer &
Lemon, 1983; Kramer et al., 1985).
Factors that are important to consider when assessing the function of specific
singing strategies are that basic principles of singing are likely to depend on (a) the
specific prior relationship of the singers, (b) the current singers’ relationship and,
(c) the expected future relationship. Many of the basic principles of singing, such as
song matching and strategies of song timing, may have the same function regard-
less of the singers’ specific relationship as long as they reflect the singers’ internal
states. However, the interpretation of a singing strategy may differ depending on
whether or not the singers are familiar with each other: whether or not they are ter-
ritorial neighbours. Neighbouring males often already have substantial previous
experience with each other and have to expect a long-term relationship. Singing

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
312 M. Naguib
strategies during their interactions may be different as the strategy is an integral
component of the previous relationship and that expected in the future. Thus,
males may be able to code information more subtly in choice of song patterns or
in the timing of songs. Males who have established dominance over rivals through
previous interactions may not escalate substantially during subsequent interac-
tions. Rather a few occurrences of song overlap (for instance) may suffice to signal
alertness or the readiness to escalate a contest. Therefore, the function and the
consequences of singing strategies during vocal interactions among neighbours
may depend not only on the current singing strategy but also on how it relates to
the strategy in previous interactions. By changing song posts during vocal inter-
actions with neighbours, males may probe each other. If males avoid song posts at
which they encounter repeated vocal aggression, not only territory boundaries but
also the choice of song posts may be determined by neighbours’ singing behaviour
during vocal interactions. Overall, more descriptive studies on the nature of vocal
interactions between established males in relation to their spatial relationships
will be needed to answer questions on how vocal interactions reflect and affect
the social relations between males in long-term spatial relationships.
Vocal interactions and territorial settlement
The role of vocal interactions in territorial settlement is particularly inter-
esting when singing strategies during vocal interactions reflect the males’qualities
or their motivation to defend a particular space. Although the function of song
in territorial defence is well established once a male has an established territory
(Krebs et al., 1978; Nowicki et al., 1998; Naguib et al., 2001), there has been little
discussion of how song and vocal interactions determine spatial relations of males
during territorial settlement or when territories shift in the course of the breeding
season. Interpretations that particular singing strategies reflect a winner (Peake
et al., 2001, 2002) can apply to single interactions, but single interactions may
not necessarily reflect the overall relationship between the singers. For example,
the singing behaviour of individuals establishing territories may differ from that
when they are defending an established territory. The terms winner and loser in
competition over resources usually imply that one individual gets all (or has first
access) and the other gets nothing (retreats). Stamps and Krishnan (1997, 1999,
2001) pointed out that during territorial settlement the consequences of winning
and losing contests are much more complex. During territorial establishment, in-
dividuals are dividing up space and such division of space usually involves repeated
interactions from different locations. Therefore, the outcome of vocal interactions
may be determined by the net outcome of repeated interactions at different lo-
cations rather than a single interaction. This situation presumably predominates

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 313
during establishment of territories in areas that are not yet saturated and where
both interacting males can expect to succeed in establishing a territory in a par-
ticular area. In some instances, males may even divide up space without a definite
winner. By shifting song posts, space may be divided up passively when males
avoid song posts in which they elicit high-intensity interactions with their rival.
In this way, losing an interaction in territorial conflicts does not necessarily mean
that losers fully retreat but rather that they shift song posts and they may be the
winner at a different location. The final spatial arrangement of singing territo-
ries may be determined by the pattern of repeated vocal interactions with males
singing at different song posts. Vocal interactions between established neighbours
may reveal information on remaining differences among them that does not re-
sult from asymmetries in site-dependent dominance, where each male may be the
winner at a certain location. In nightingales, males may systematically ‘win’ re-
peated interactions with a particular neighbour, but this pattern is not true for all
neighbouring males (M. Naguib, unpublished data). If recurring asymmetries exist
in the interactions between particular males, this may reflect a stable dominance
relationship that is maintained after space is divided up.
Vocal interactions in communication networks
Vocal interactions have received particular attention in recent years as
their asymmetries have been shown to be used by other individuals as a source
of information. Vocal interactions in songbirds are a clearly defined signalling
context and so have become one of the main models in studies of communica-
tion networks (McGregor & Dabelsteen, 1996; McGregor & Peake, 2000; Whitfield,
2002). To date, several studies have shown that male songbirds eavesdrop on rival
vocal interactions and have expanded the understanding of information gather-
ing when several individuals are within signalling rage of each other (Naguib &
Todt, 1997; Naguib et al., 1999; Otter et al., 1999; Peake et al., 2001, 2002; Mennill
et al., 2002). In our own studies, we showed in two-loudspeaker experiments that
territorial nightingales discriminated between asymmetries in vocal interactions:
subjects responded significantly more strongly to a simulated rival that was over-
lapping the songs of the opponent, i.e. was the more aggressive intruder (Naguib &
Todt, 1997). When songs in the interaction simulated by the loudspeakers did not
overlap each other but were played in an alternating order with songs of one loud-
speaker leading (closely followed by the songs of the other loudspeaker), subjects
responded more strongly to the loudspeakers playing the leading songs (Naguib
et al., 1999). These combined experiments indicate that different proximate cues
were used depending on the kind of asymmetry simulated. The use of oppos-
ing proximate cues by subjects depending on the kind of asymmetry perceived

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
314 M. Naguib
provides evidence that subjects use an adaptive strategy that is independent of
a putative general proximate shift of attention to a first or last heard stimuli.
Peake and coworkers (2001) elegantly expanded this playback design and showed
that great tits Parus major attended to such asymmetries in interactions played
to them from outside their territory and then responded differently to subse-
quent intrusions, depending on which previous singer was simulated as intruder.
They further showed that males varied their song output in responses to intrud-
ers depending on the kind of experience they had with the intruder prior to an
interaction with another male (Peake et al., 2002). This suggests more complex
ways of gathering information than shown in any of the previous experiments
on eavesdropping. Studies by Otter et al. (1999) and Mennill et al. (2002) indicated
that females also use asymmetries in male–male interactions as sources of infor-
mation in their responses and seemingly even in reproductive decisions (see also
Ch. 7). Therefore, it is well documented that songbirds not only attend to vocal in-
teractions between males but also extract information coded in the asymmetries
in singing strategies and use the information adaptively. An interesting question
to be answered in order to understand the further implications of eavesdropping
in communication networks with widely spaced individuals is whether birds are
able accurately to extract meaningful asymmetries in an interaction occurring at
a distance, or whether they are only able to do so when they are close to, or at
equal distance from, the singers. We already have clear evidence that vocal inter-
actions have much wider implications than the exchange of information between
interacting males. Strategies of singing during vocal interactions then are likely
not only to evolve through responses of the opponent but also through effects on
other listening individuals (Ch. 2). Viewing communication from the perspective
of communication networks considerably broadens the view of social implica-
tions of the prevalence of vocal interactions and the functional implications of
certain singing strategies during vocal interactions. Most experimental studies of
the functional significance of different singing strategies during an interaction
have used different playback protocols and there is a need to complement these
studies by more descriptive studies on natural interactions.
Summary and future directions
Vocal interactions in songbirds have many facets that need to be inte-
grated into models of information gathering in communication. Because of the
accessibility of song, vocal interactions are a suitable model to address wider con-
cepts in communication, to address questions in cognitive ecology (Ch. 24) and
to obtain new insights into the social relationships between territorial neigh-
bours. Singing strategies during vocal interactions and strategies for information
gathering from vocal interactions by participants and by eavesdroppers have

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 315
evolved under ecological constraints, such as the spacing patterns of conspecifics
or more specifically the abundance and distribution of signallers in space and
time. As we continue to gain more information on strategies of vocal interactions
in natural settings among established neighbours and during establishment of
territories, we are likely to obtain new insights into how social relations are me-
diated by song and how strategies of singing interactions are related to spatial
ecology.
Acknowledgements
I thank Peter McGregor, Ken Otter and an anonymous referee for helpful comments on
a previous version of the manuscript
References
Amrhein, V., Korner, P. & Naguib, M. 2002. Nocturnal and diurnal singing activity in
the nightingale: correlations with mating status and breeding cycle. Animal
Behaviour, 64, 939–944.
Amrhein, V., Kunc, H. P. & Naguib, M. 2004. Non-territorial nightingales prospect
territories during the dawn chorus. Proceedings of the Royal Society of London, Series B,
(Supplement), 271, S167–S169.
Beecher, M. D. 1996. Bird song learning in the laboratory and field. In: Ecology and
Evolution in Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 61–78.
Beecher, M. D., Stoddard, P. K., Campbell, S. E. & Horning, C. L. 1996. Repertoire
matching between neighbouring song sparrows. Animal Behaviour, 51, 917–923.
Beecher, M. D., Campbell, S. E. & Nordby, J. C. 2000. Territory tenure in song sparrows
is related to song sharing with neighbours, but not to repertoire size. Animal
Behaviour, 59, 29–37.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.
Brindley, E. L. 1991. Response of European robins to playback of song: neighbour
recognition and overlapping. Animal Behaviour, 41, 503–512.
Burt, J. M., Bard, S. C., Campbell, S. E. & Beecher, M. D. 2002. Alternative forms of song
matching in song sparrows. Animal Behaviour, 63, 1143–1151.
Chandler, C. R., Ketterson, E. D. & Nolan, V. 1997. Effects of testosterone on use of
space by male dark-eyed juncos when their mates are fertile. Animal Behaviour, 54,
543–549.
Dabelsteen, T. 1992. Interactive playback: a finely tuned response. In: Playback and
Studies of Animal Communication, ed. P. K. McGregor. New York: Plenum Press,
pp. 97–109.
Dabelsteen, T., McGregor, P. K., Shepherd, M., Whittaker, X. & Pedersen, S. B. 1996. Is
the signal value of overlapping different from that of alternating during matched
singing in great tits? Journal of Avian Biology, 27, 189–194.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
316 M. Naguib
Dabelsteen, T., McGregor, P. K., Holland, J., Tobias, J. A. & Pedersen, S. B. 1997. The
signal function of overlapping singing in male robins. Animal Behaviour, 53,
249–256.
Falls, J. B. 1982. Individual recognition by sounds. In: Acoustic Communication in Birds,
Vol. 2, ed. D. E. Kroodsma & E. H. Miller. New York: Academic Press, pp. 237–278.
Falls, J. B., Krebs, J. R. & McGregor, P. K. 1982. Song matching in the great tit (Parus
major): the effect of similarity and familiarity. Animal Behaviour, 30, 997–1009.
Falls, J. B., Horn, A. G. & Dickinson, T. E. 1988. How western meadowlarks classify their
songs: evidence from song matching. Animal Behaviour, 36, 579–585.
Farabaugh, S. M. 1982. The ecological and social significance of duetting. In: Acoustic
Communication in Birds, Vol. 2, ed. D. E. Kroodsma & E. H. Miller. New York:
Academic Press, pp. 85–124.
Farabaugh, S. M. & Dooling, R. J. 1996. Acoustic communication in parrots: laboratory
and field studies of budgerigars, Melopsittacus undulatus. In: Ecology and Evolution of
Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY:
Cornell University Press, pp. 97–117.
Ficken, R. W., Ficken, M. S. & Hailman, J. P. 1974. Temporal pattern shifts to avoid
acoustic interference in singing birds. Science, 183, 762–763.
Forstmeier, W., Kempenaers, B., Meyer, A. & Leisler, B. 2002. A novel song parameter
correlates with extra-pair paternity and reflects male longevity. Proceedings of the
Royal Society of London, Series B, 269, 1479–1485.
Gil, D., Graves, J. A. & Slater, P. J. B. 1999. Seasonal patterns of singing in the willow
warbler: evidence against the fertility announcement hypothesis. Animal
Behaviour, 58, 995–1000.
Griessmann, B. & Naguib, M. 2002. Song sharing in neighboring and non neighboring
thrush nightingales (Luscinia luscinia) and its implications for communication.
Ethology, 108, 377–387.
Hanski, I. K. 1992. Territorial behaviour and mixed reproductive strategy in the
chaffinch. Ornis Scandinavica, 23, 475–482.
Hasselquist, D., Bensch, S. & von Schantz, T. 1996. Correlation between male song
repertoire, extra-pair paternity and offspring survival in the great reed warbler.
Nature, 381, 229–232.
Hauser, M. D. 1996. The Evolution of Communication. Cambridge, MA: MIT Press.
Hultsch, H. & Todt, D. 1981. Repertoire sharing and song-post distance in nightingales
(Luscinia megarhynchos B.). Behavioral Ecology and Sociobiology, 8, 183–188.
1982. Temporal performance roles during vocal interactions in nightingales
(Luscinia megarhynchos). Behavioral Ecology and Sociobiology, 11, 253–260.
Kempenaers, B., Verheyren, G. R. & Dhondt, A. A. 1997. Extrapair paternity in the blue
tit (Parus caeruleus): female choice, male characteristics, and offspring quality.
Behavioral Ecology, 8, 481–492.
Kilner, R. & Johnstone, R. A. 1997. Begging the question: are offspring solicitation
behaviours signals of need? Trends in Ecology and Evolution, 12, 11–15.
Klump, G. 1996. Bird communication in the noisy world. In: Ecology and Evolution of
Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY:
Cornell University Press, pp. 321–338.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 317
Kramer, H. G. & Lemon, R. E. 1983. Dynamics of territorial singing between
neighboring song sparrows (Melospiza melodia). Behaviour, 85, 198–223.
Kramer, H. G., Lemon, R. E. & Morris, M. J. 1985. Song switching and agonistic
stimulation in the song sparrow (Melospiza melodia). Animal Behaviour, 33, 135–149.
Krebs, J. R., Ashcroft, R. & Webber, M. 1978. Song repertoires and territory defense in
the great tit Parus major L. Nature, 271, 539–542.
Krebs, J. R., Ashcroft, R. & van Orsdol, K. 1981. Song matching in the great tit Parus
major L. Animal Behaviour, 29, 918–921.
Kroodsma, D. E. 1974. Song learning, dialects and dispersal in the Bewick’s wren.
Zeitschrift fur Tierpsychologie, 35, 352–380.
1982. Song repertoires: problems in their definitions and use. In: Acoustic
Communication in Birds, Vol. 1, ed. D. E. Kroodsma & E. H. Miller. New York:
Academic Press, pp. 125–146.
Langemann, U., Tavares, J. P., Peake, T. M. & McGregor, P. K. 2000. Response of great
tits to escalating patterns of playback. Behaviour, 137, 451–471.
Maynard Smith, J. & Parker, G. A. 1976. The logic of asymmetric contests. Animal
Behaviour, 24, 159–175.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Krebs, J. R. 1982. Mating and song types in the great tit. Nature, 297,
60–61.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McGregor, P. K., Dabelsteen, T., Shepherd, M. & Pedersen, S. B. 1992. The signal value of
matched singing in great tits: evidence from interactive playback experiments.
Animal Behaviour, 43, 987–998.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873–873.
Naguib, M. 1999. Effects of song overlapping and alternating on nocturnally singing
nightingales. Animal Behaviour, 58, 1061–1067.
Naguib, M. & Todt, D. 1997. Effects of dyadic vocal interactions on other conspecific
receivers in nightingales. Animal Behaviour, 54, 1535–1543.
Naguib, M. & Wiley, R. H. 2001. Estimating the distance to a source of sound:
mechanisms and adaptations for long-range communication. Animal Behaviour,
62, 825–837.
Naguib, M., Fichtel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders in simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 265, 537–542.
Naguib, M., Altenkamp, R. & Griessmann, B. 2001. Nightingales in space: song and
extra-territorial forays of radio tagged song birds. Journal fur Ornithologie, 142,
306–312.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
318 M. Naguib
Naguib, M., Mundry, R., Hultsch, H. & Todt, D. 2002. Responses to playback of whistle
songs and normal songs in male nightingales: effects of song category, whistle
pitch, and distance. Behavioral Ecology and Sociobiology, 52, 216–223.
Nielsen, B. M. B. & Vehrencamp, S. L. 1995. Responses of songs sparrows to song-type
matching via interactive playback. Behavioral Ecology and Sociobiology, 37, 109–117.
Nowicki, S., Searcy, W. A. & Hughes, M. 1998. The territory defense function of song in
song sparrows: a test with the speaker occupation design. Behaviour, 135, 615–628.
Otter, K., McGregor, P. K., Terry, A. M. R. et al., 1999. Do female great tits (Parus major)
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society of London, Series B, 266, 1305–1309.
Otter, K. A., Ratcliffe, L., Njegovan, M. & Fotheringham, J. 2002. Importance of
frequency and temporal song matching in black-capped chickadees: evidence
from interactive playback. Ethology, 108, 181–191.
Payne, R. B. 1996. Song traditions in indigo buntings: origin, improvisation, dispersal,
and extinction in cultural evolution. In: Ecology and Evolution of Acoustic
Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY: Cornell
University Press, pp. 198–220.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2002. Do great tits assess
rivals by combining direct experience with information gathered by
eavesdropping? Proceedings of the Royal Society of London, Series B, 269, 925–1929.
Pitcher, T. E. & Stutchbury, B. J. M. 2000. Extraterritorial forays and male parental care
in hooded warblers. Animal Behaviour, 59, 1261–1269.
Podos, J. 1996. Motor constraints on vocal development in a songbird. Animal
Behaviour, 51, 1061–1070.
Popp, J. W. 1989. Temporal aspects of singing interactions among territorial ovenbirds
(Seiurus aurocapillus). Ethology, 82, 127–133.
Pusey, A. E. & Packer, C. 1997. The ecology of relationships. In: Behavioural Ecology, an
Evolutionary Approach, ed. J. R. Krebs & N. B. Davies. Oxford: Blackwell Scientific,
pp. 254–283.
Schroeder, D. J. & Wiley, R. H. 1983. Communication with shared song themes in
tufted titmice. The Auk, 100, 414–424.
Searcy, W. A. & Yasukawa, K. 1996. Song and female choice. In: Ecology and Evolution of
Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY:
Cornell University Press, pp. 454–473.
Searcy, W. A., Nowicki, S. & Hogan, C. 2000. Song type variants and aggressive context.
Behavioral Ecology and Sociobiology, 48, 358–363.
Shackleton, S. A. & Ratcliffe, L. 1994. Matched counter-singing signals escalation of
aggression in black-capped chickadees (Parus atricapillus). Ethology, 97, 310–316.
Slabbekoorn, H., Ellers, J. & Smith, T. B. 2002. Bird song and sound transmission: the
benefits of reverberations. The Condor, 104, 564–573.
Smith, D. G. & Norman, D. O. 1979. ‘Leader–follower’ singing in red-winged blackbirds.
The Condor, 81, 83–84.

P1: IRK/KFO P2: IRK-KOD0521823617c14.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:1
Singing interactions in songbirds 319
Sorjonen, J. 1987. Temporal and spatial differences in traditions and repertoires in the
song of the thrush nightingale (Luscinia luscinia) birds. Behaviour, 102, 196–212.
Stamps, J. 1994. Territorial behavior: testing the assumptions. Advances in the Study of
Behavior, 23, 173–232.
Stamps, J. A. & Krishnan, V. V. 1997. Functions of fights in territory establishment.
American Naturalist, 150, 393–405.
1999. A learning-based model of territory establishment. Quarterly Review of Biology,
74, 291–318.
2001. How territorial animals compete for divisible space: a learning-based model
with unequal competitors. American Naturalist, 157, 154–169.
Stoddard, P. K. 1996. Vocal recognition of neighbors by territorial passerines. In:
Ecology and Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H.
Miller. Ithaca, NY: Cornell University Press, pp. 356–376.
Todt, D. 1981. On functions of vocal matching: effects of counter-replies on song post
choice and singing. Zeitschrift fur Tierpsychologie, 57, 73–93.
Todt, D. & Naguib, M. 2000. Vocal interactions in birds: the use of song as a model in
communication. Advances in the Study of Behaviour, 29, 247–296.
Vehrencamp, S. L. 2001. Is song-type matching a conventional signal of aggressive
intentions? Proceedings of the Royal Society of London, Series B, 268, 1637–1642.
von Helversen, D. 1980. Structure and function of antiphonal duets. Acta XVIIIth
International Ornithological Congress, Berlin, 682–688.
Waser, P. M. & Wiley, R. H. 1980. Mechanisms and evolution of spacing in animals. In:
Handbook of Behavioral Neurobiology: Social Behavior and Communication, ed. P. Marler
& J. G. Vandenbergh. New York: Plenum Press, pp. 159–233.
Weary, D., Falls, J. B. & McGregor, P. K. 1990. Song matching and perception of song
types in great tits, Parus major. Behavioral Ecology, 1, 43–47.
Wiley, R. H. 1983. The evolution of communication: information and manipulation.
In: Animal Behaviour : Communication, Vol. 2, ed. T. R. Halliday & P. J. B. Slater.
Oxford: Blackwell Scientific, pp. 156–189.
1994. Errors, exaggeration and deception in animal communication. In: Behavioral
Mechanisms in Ecology, ed. L. Real. Chicago, IL: University of Chicago Press,
pp. 157–189.
Wiley, R. H. & Richards, D. G. 1978. Physical constraints on acoustic communication in
the atmosphere: implications for the evolution of animal vocalizations. Behavioral
Ecology and Sociobiology, 3, 69–94.
Wiley, R. H. & Wiley, M. S. 1980. Territorial behavior of a blackbird: mechanisms of site
dependent dominance. Behaviour, 73, 130–154.
Wiley, R. H., Godard, R. & Thompson, A. D. 1994. Use of two singing modes by hooded
warblers as adaptations for signaling. Behaviour, 129, 243–278.
Whitfield, J. 2002. Nosy neighbours. Nature, 419, 242–243.
Wolffgramm, J. & Todt, D. 1982. Pattern and time specificity in vocal responses of
blackbirds Turdus merula. Behaviour, 81, 264–286.
Zann, R. A. 1996. The Zebra Finch: a Synthesis of Field and Laboratory Studies. Oxford:
Oxford University Press.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
15
Dawn chorus as an interactivecommunication network
j o h n m . b u r t & s a n d r a l . v e h r e nc a m p
Cornell Laboratory of Ornithology, Ithaca, USA
Introduction
Dawn chorus singing is a striking behaviour pattern, performed by some
temperate-zone and tropical songbird species, as well as a few non-passerine and
non-avian species. In a typical chorusing songbird species, all territorial males in
a neighbourhood synchronously start singing 30 to 90 minutes before sunrise.
During the ensuing chorus period, song rate, singing diversity and song complex-
ity reach maximal levels, and often birds do not seem to be interacting with any
one particular neighbour (Hultsch & Todt, 1982). Then, as the light level increases
around sunrise, this mode of singing usually abruptly ends. Soon after dawn cho-
rus is over, birds begin to forage and patrol their borders, and they switch to
courtship singing or dyadic (i.e. paired) counter-singing with nearby neighbours.
Post-chorus singing is typically more sporadic and overall song rates tend to be
lower and much more variable than they are at dawn chorus (for a review of dawn
chorus behaviour, see Staicer et al. (1996)).
Numerous hypotheses have been proposed to explain dawn chorus singing.
In an insightful review, Staicer et al. (1996) outlined 12 non-exclusive hypotheses
and compared their predictions against the existing empirical evidence across
many songbird and non-songbird species. The 12 hypotheses were grouped into
three categories: intrinsic, environmental and social. Intrinsic explanations such
as circadian cycles of testosterone and self-stimulation are likely proximate mecha-
nisms for dawn singing (Wingfield & Farner, 1993; Goodson, 1998). Environmental
explanations, such as low predation risk, good acoustic transmission, inefficient
foraging with low ambient light and unpredictable night-time conditions lead-
ing to excess energy reserves on most mornings, provide reasons why singing at
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
320

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 321
dawn might be less costly than singing at other times (Henwood & Fabrick, 1979;
Kacelnik & Krebs, 1982; Mace, 1987; McNamara et al., 1987; Hutchinson, 2002;
Dabelsteen & Mathevon, 2002). However, none of these intrinsic or environmental
hypotheses provide a functional explanation of the selective advantage for vigor-
ous, continuous, complex vocal displays prior to sunrise, nor do they explain why
only some species exhibit this phenomenon.
The social hypotheses outlined by Staicer et al. (1996) that do attempt to provide
functional explanations for dawn chorus singing include mate attraction and/or
stimulation, territory defence and resolution of social dynamics. Predictions for
these alternatives have been used to support or reject certain hypotheses in several
species. For example, mate attraction and mate stimulation can be ruled out as
primary explanations in species where dawn chorus singing is uniformly high
across the breeding season, rather than being concentrated during periods of mate
attraction and mate fertility as these hypotheses predict (Kroodsma et al., 1989;
Part, 1991; Slagsvold et al., 1994). Moreover, males have generally been observed to
stop dawn chorus singing as soon as their mates emerge from their night roost,
and males of some species drop out of the dawn chorus altogether on days of
peak mate fertility, findings that do not support mate stimulation hypotheses
for those species (Mace, 1986; Cuthill & Macdonald, 1990; Part, 1991; Otter &
Ratcliffe, 1993). The territory defence hypothesis, suggesting that dawn song is an
extra vigorous keep-out signal, is contradicted in some sedentary species because
males cease dawn chorus singing during the non-breeding season even though
they continue to occupy and defend a territory (Staicer et al., 1996). Staicer et al.
concluded that the social hypothesis that best fits the existing evidence is the social
dynamics hypothesis, which proposes that the function of dawn chorus is the
interactive communication and adjustment of social relationships among males.
Their conclusion is based partly on findings in numerous species that dawn chorus
singers use signals and modes of signalling that are specifically associated with
male–male interaction and that dawn chorus singers of some species appear to be
listening to and directing their songs towards particular neighbours (Kroodsma
et al., 1989; Staicer, 1989; Nelson & Croner, 1991; Spector, 1991; Dabelsteen, 1992;
Staicer et al., 1996).
Females of at least some species may acquire information about mate qual-
ity from dawn singing. In several species, individual differences in dawn song
output are correlated with male age and/or dominance and with female laying
date and/or fecundity (Welling et al., 1995; Otter et al., 1997; Poesel et al., 2001;
Ballentine et al., 2003). Females could possibly assess the quality of singing males
at dawn chorus either by attending to song traits that are directly associated with
indicators of fitness, such as stamina, age or dominance, or by eavesdropping to

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
322 J. M. Burt & S. L. Vehrencamp
acquire information about relative quality if males are interacting (Otter et al.,
2001; Mennill et al., 2002; Ch. 7).
Taken as a whole, the pattern of dawn chorus behaviour across species does not
seem to fit any single currently hypothesized function. Furthermore, the social
hypotheses are not mutually exclusive, suggesting that the dawn chorus might
have multiple functions that may differ in relative importance, depending on
species. If that is the case, then it may be necessary to examine more closely (or
re-examine) the singing behaviour of each species that has a dawn chorus, pay-
ing attention to such factors as the intended receiver(s) and whether interactions
are occurring among singers within the chorus. Staicer et al. (1996) provided a set
of predictions for such evidence that could be helpful in distinguishing which
hypotheses might apply to a particular species (cf. Table 24.1 in Staicer et al.,
1996).
Investigation of these target and interaction issues would benefit from an ap-
proach that considers the neighbourhood of singing males as a communication
network. Network communication is broadly defined as the involvement of at least
three individuals, one or more of them signalling and all receiving (McGregor &
Dabelsteen, 1996; see other chapters in this volume). The dawn chorus, with many
simultaneous signallers (possibly interacting) and many potential receivers, cer-
tainly fits the broad definition of a communication network. However, to date,
no study has examined dawn chorus from a network perspective. We suggest
that a study of the characteristics of the communication network that occurs
at dawn chorus for a given species could provide further information about its
function. In this chapter, we discuss what kinds of communication network the
different functional hypotheses might predict for dawn chorus and how one might
go about testing for them. We then test some of these ideas using a dawn chorus
recording of a neighbourhood of banded wrens Thryothorus pleurostictus as an
example.
Communication network structures at dawn chorus
The detailed structure and complexity of communication networks can
vary and depend largely on the degree to which the signalling ‘links’between com-
municating individuals are one way or are interactive (i.e. signals flow both ways
between individuals). Using three hypothetical individuals (Fig. 15.1), it is possible
to define three basic network structures: broadcast networks, in which one sender
broadcasts a one-way omnidirectional signal to two receivers (Fig. 15.1a); eaves-
dropping networks, where two senders interact and a third receiver eavesdrops
on the interaction to obtain information about the interactants (Fig. 15.1b); and in-
teractive networks, with three senders all interacting with each other (Fig. 15.1c).

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 323
(a)
(b)
(c)
Fig. 15.1. Three basic network components may occur within a communication
network, either singly or in combination: (a) broadcast networks, in which at least one
sender produces undirected one-way signals that are received by potentially many
receivers; (b) eavesdropping networks, in which two signallers interact and
eavesdroppers obtain relative information about each interactant; (c) interactive
networks containing three or more individuals signalling interactively to one another
and eavesdropping on nearby interactions.
Any real-world communication network is likely to include many more than just
three individuals, and networks can theoretically consist of one or any combina-
tion of these basic components, adding more potential variety and complexity
to network structure. For example, it is likely that any bird-song network with
an interactive component probably also has an eavesdropping component in the
form of non-interacting listeners such as females and floater males, as well as
interacting males who are eavesdropping on their neighbours’ interactions.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
324 J. M. Burt & S. L. Vehrencamp
Investigating the structure of a dawn chorus communication network might
provide useful clues about its function, with a key variable being the degree to
which signallers within the network are interactive. For example, omnidirectional
non-interactive signalling at dawn would indicate a broadcast network. A broad-
cast network at dawn chorus would support male keep-out, female attraction or
direct male quality-assessment functions, since these hypotheses do not strictly
require male–male interaction. Conversely, evidence of two-way or multi-way sig-
nalling between singers at dawn would indicate an interactive network, support-
ing the notion that the dawn chorus serves an inter-male communication role
(the social dynamics hypothesis). Evidence of eavesdropping would also support a
relative male quality-assessment function for dawn chorus.
Searching for interactions at dawn
The presence or absence of an interactive network may be a key diagnostic
for the function of dawn chorus, but on a practical level how do we go about
looking for interactions at dawn? One possible first step would be to examine
what we know about how males in a given dawn-chorusing species use their songs
to communicate during the daytime. Indeed, much effort has been devoted to
analysing daytime counter-singing interactions between pairs of adjacent males.
Observational studies of daytime dyadic counter-singing between focal birds and
a neighbour or intruder are relatively easy to conduct, and playback experiments
have been used to test hypotheses for the function of male song interactions. In
territorial species, song has generally been found to function as a keep-out signal to
other males (Krebs et al., 1978; Yasukawa et al., 1982; Nowicki et al., 1998). Studies
of daytime singing have found that song can also be used in complex ways to
mediate aggression between neighbours. For example, in populations with high
levels of song-type sharing between neighbours, birds can match their neighbour’s
song with their own version of that type. Matching is particularly useful as a
directed signal, since by replying with the same type a bird can unambiguously
address a rival. Post-chorus, birds have been shown to use song matching as a
directed threat to indicate subsequent aggressive intentions (Krebs et al., 1981;
McGregor et al., 1992; Burt et al., 2001; Vehrencamp, 2001). The rate of switching
between song types, temporal overlapping of songs, duration matching and pitch
matching are additional potential directed signalling strategies that vary with
the intensity of agonistic interaction and serve to mediate aggression between
neighbours (reviewed by Vehrencamp, 2000).
The directional properties of signals such as song matching make them poten-
tially useful for detecting interactions at dawn since an audio recording of both
singers can determine the singer and target. However, conventional one- or two-
channel recording methods will not usually be adequate for this task for three

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 325
reasons. First, a dawn chorus interactive network would span a large area of ad-
jacent territories and birds could interact with any neighbour, thereby reducing
the effectiveness of focal recording. Second, identification and location of singers
by visual means is very difficult in the twilight before dawn; third, the sheer quan-
tity of vocalizations that occur at dawn in many locations tends to mask easy
identification of specific interacting participants. In fact, these difficulties may be
the reason why other researchers have not detected interactive networks at dawn
chorus, even in species where males clearly interact with their neighbours during
daylight.
New methods for studying dawn chorus
One solution to the problem of detecting interactions at dawn is the use of
distributed microphone arrays, which have been proposed as an ideal method for
monitoring communication networks in territorial neighbourhoods (McGregor &
Dabelsteen, 1996). Such systems are ideal for studying dawn chorus because they
can simultaneously record the songs of many singers in a large area. Another
advantage to using microphone arrays is the ability to determine the location
(and, therefore, in many cases the identity) of each singer using sound arrival
time differences (Watkins & Schevill, 1972; Speisberger & Fristrup, 1990). The
specific details of how acoustic location systems work is reviewed more thor-
oughly in McGregor et al. (1997). Array recording is a particularly good method
for detecting interactive networks in species that are known to use some form
of directed signal, such as immediate matching or overlapping during vocal ex-
changes. Changes in singing behaviour associated with movement to different
parts of the territory may also indicate interaction, most likely of a dyadic na-
ture. Additionally, microphone array recordings can be used to document changes
in the singing behaviour of non-interacting individuals before, during and af-
ter an intense interaction between two other individuals in a neighbourhood
(Eason & Stamps, 1993; Bower, 2000). Evidence that non-vocalizing receivers act
on information gained by eavesdropping (i.e. eavesdropping networks) is best ac-
quired with carefully designed playback experiments (e.g. Naguib & Todt, 1997;
Oliveira et al., 1998; Naguib et al., 1999; McGregor et al., 2001; Peake et al., 2001;
see also Ch. 2).
Banded wren song behaviour
In this chapter, we describe one of the first attempts to study a dawn cho-
rus communication network using a microphone array system (see also Bower,
2000). Our study species is the banded wren, in which males possess repertoires
of discrete, distinctive song types and usually switch to a different type after
each consecutive song. Neighbouring males share many of the same song types

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
326 J. M. Burt & S. L. Vehrencamp
and prior studies have documented the use of type matching and other singing
patterns during aggressive encounters (Molles & Vehrencamp, 1999, 2001). This
species is also a vigorous dawn chorus singer. Here we present an initial analysis
of the singing behaviour and interactions among four neighbouring male wrens
recorded during a single morning encompassing dawn chorus and the subsequent
hour of post-dawn chorus. We look for evidence of network communication in-
teractions involving two, three or more birds by searching for the presence of
directed signals such as matching and overlapping.
The banded wren is a common and vocally active species that inhabits the trop-
ical dry deciduous forest of the Pacific slope of Central America. It breeds only
during the first half of the rainy season (May–August) but remains resident and
paired on the same territory during the rest of the year. The mating system is so-
cially monogamous and each pair defends an all-purpose territory approximately
0.4 ha in area. Although not a true duet, a female occasionally sings short male-
like songs following or overlapping her mate’s songs. Males possess a repertoire
of 15 to 30 discrete song types, which may be delivered with a high rate of switch-
ing between song types (immediate variety mode) or in a more repetitive fashion
(eventual variety mode). Young males tend to copy whole song types from nearby
males and generally do not disperse very far from their natal territory, so estab-
lished adjacent neighbours share between 50 and 90% of their song-type repertoire
(Molles & Vehrencamp, 1999).
In the course of our research, we have identified a variety of song-delivery
patterns that banded wrens use to communicate with their neighbours in the
daytime during bouts of counter-singing. Song matches appear to be a threat sig-
nal; repertoire matches (singing a song shared with but not currently sung by the
neighbour) are used as a low-threat directed signal maintaining the interaction,
and switches to non-shared song types indicate a desire to deescalate (Molles &
Vehrencamp, 2001). Finally, banded wrens also appear to use song overlapping
during escalated interactions. Although the function of overlapping is not well
understood, it is often associated with (and often simultaneously combined with)
high rates of song matching, suggesting it also has a threat function. Overlapping
is apparently avoided during low-intensity counter-singing interactions between
distant males (Molles & Vehrencamp, 1999).
Banded wrens have a pronounced dawn chorus during the breeding season
months and are relatively silent at dawn during the rest of the year, despite remain-
ing on their territories. The chorus starts at twilight (approximately 05:00 h) and
lasts about 30 minutes. During this period, males sing vigorously and loudly. They
initially perch high in an emergent tree (10–20 m) and sing without pause from
one location for several minutes while constantly changing their body orientation.
As light levels increase, dawn chorus singers usually shift to other high perches

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 327
in other parts of their territories. Prior to the array study, we had anecdotally ob-
served numerous type matching events at dawn, but, given the conditions, it was
unknown to what extent, and with whom, the birds were matching. These prelim-
inary observations of song matching suggested to us that dawn chorus might be
an interactive phenomenon.
A male usually abruptly ceases dawn chorusing behaviour when his mate ap-
proaches and interacts with him. She may join him in a brief, uncoordinated duet.
Occasionally a male forgoes the dawn chorus completely to interact with his (pre-
sumably fertile) mate. Unmated males continue to sing at a high rate for another
30 to 60 minutes. After the dawn chorus, males begin foraging, interspersed with
bouts of singing, and patrol the borders of their territories more actively. At this
time, males seem to shift to more focused counter-singing with nearby neighbours.
Each territorial male has three to four adjacent neighbours with whom he
regularly interacts. Males construct bulky covered nests and are constantly ini-
tiating new ones because of high nest-predation rates. Females appear to select
the nest location and sometimes choose a site near a territorial border, which
forces the male to renegotiate that boundary with his neighbour through close-
range counter-singing and fights. The dynamic nature of territory boundaries in
this species could be one source of changes in social status hypothetically being
signalled during the dawn chorus.
Recording methods and subjects
As part of our research project studying the function of banded wren
song, we developed a microphone array recording system as a tool for acousti-
cally monitoring several vocalizing individuals. The technique involves placing
an array of many microphones at strategic locations within and around a small
neighbourhood of banded wren territories and simultaneously recording all song
interactions picked up by the microphones on a central multiple-channel receiving
unit. With these array recordings, we can quantify neighbourhood-wide singing
patterns and also focus on individual birds to gain a more complete picture of
their interactions with all of their neighbours.
The array data presented in this chapter were taken from an analysis of a
recording made on 20 June, 2001 at one of our study sites in Santa Rosa Park,
Costa Rica. This recording was selected from our library of daily recordings made
during the 2000, 2001 and 2002 May–July field seasons as a representative example
of dawn chorus singing in our study population. The chapter dataset runs from
05:02 h (Central American time zone) to 06:42 h, a total of 100 minutes. On that
day, civil twilight occurred at 04:58 h and sunrise at 05:21 h. Behaviourally, the
recording covered the first song sung in the neighbourhood at 05:04 h, the entire
dawn chorus and about one hour of post-chorus singing when dyadic counter-

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
328 J. M. Burt & S. L. Vehrencamp
450
400
LE
BO
N
N
N
N
OH
PU
YO
WBUB
WE
350
300
250
200
150
100
50
50 100 150 200
Distance (m)
Dis
tanc
e (m
)
250 300 350 400
*
*
* *
*
***
**
**
*
Fig. 15.2. Map of banded wren territorial neighbourhood recorded by the array. Solid
lines show the boundaries of the focal neighbours, while dotted lines are the
boundaries of adjacent neighbours. An asterisk indicates a microphone position and
N indicates current nest sites for each focal male. The shaded region in bird OH’s
territory indicates the area recently annexed from bird YO by OH.
singing predominated. The array consisted of 13 microphones, situated among
and surrounding four focal neighbours (males OH, YO, BO and WB; Fig. 15.2 shows
array configuration and territories). Since the focal birds were usually within
the array, nearly all of the songs sung by these four birds could be identified
and located. Songs of four other neighbours adjacent to the central four often
could be detected and identified if the bird was close to the array, but usually
not located (birds WE, PU, UB and LE; Fig. 15.2). Songs of these outlying birds
were included in calculations of overlapping and song matching to create more
accurate song statistics for the central focal birds. During the recording, four
observers were posted near each focal bird to take behavioural notes so that later
we could reconstruct patterns of interaction.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 329
The recording on 20 June 2001 was typical for a mid-breeding season dawn
chorus and post-dawn chorus day of singing. The focal birds varied in age and
stage of nesting. OH had recently built a nest near his border with YO and had
extended his territory into that of YO (shaded area on OH’s territory in Fig. 15.2).
This boundary shift was a source of ongoing aggression between OH and YO, as well
as with BO, another of OH’s neighbours who was also affected by the shift. OH was
a long-term resident of the field site (at least seven years old). YO was banded as an
adult two years earlier (so was three or more years of age) and on the recording day
had just started building a new nest, which was located approximately 10–20 m
from OH’s border and approximately 50 m from OH’s nest. WB was an offspring of
the previous owner of PU’s territory, hatched in 1999 (two years old) and had an
active nest with nestlings. BO was a newly banded bird (probably first year), whose
nest was predated the day before by capuchin monkeys. During the recording, BO
was observed to be building a new nest.
Temporal patterns of singing behaviour
In this section, we provide a quantitative description of changes in several
key singing behaviours over time, based on the four focal birds in our recording. To
help the reader to visualize these dynamic patterns, we have adopted a presenta-
tion format that plots a running mean and standard error of the mean (SEM) of the
four focal birds’scores, starting at the time of the first song of the morning (05:04 h)
over a series of overlapping five minute intervals that move forward in time in
one minute increments (Fig. 15.3). Since the numbers of matches and overlaps are
highly dependent on the number of songs delivered in each interval, percentages
are given for these measures. Table 15.1 shows the correlations among these mea-
sures. For matching and overlapping analyses, all focal birds were considered to be
‘adjacent’neighbours (see Fig. 15.1), because all focal birds could easily hear all the
other focals and we had previously observed matched counter-singing between all
combinations of focal neighbours during the daytime.
Bout structure
Intersong interval (ISI), measured as the time from the end of a song
to the beginning of the next song, was used as an index of bout structure. This
variable is particularly sensitive to shifts between continuous singing and bout
singing (periods of relatively high song rates interspersed with pauses in singing)
and can be used to indicate the point in time that dawn chorus ends and post-
chorus bout singing begins. When all birds sing continuously at high rates, as in
the classic dawn chorus, the mean and variability of ISI will be small. When birds
shift to singing in asynchronous bouts, both the mean and the variability of ISI
will increase. Figure 15.3a shows the running mean and SEM of ISI over the course

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
330 J. M. Burt & S. L. Vehrencamp
200twilight
(a)
(b)
(c)
(d)
(e)
150
100
50
0
20
15
10
5
0
100
50
0
100
50
0
100
50
0
05:00 05:15 05:30 05:45 06:00 06:15 06:30 06:45
05:00 05:15 05:30 05:45 06:00 06:15 06:30 06:45
05:00 05:15 05:30 05:45 06:00 06:15 06:35 06:45
05:00 05:15 05:30 05:45 06:00 06:15 06:30 06:45
ISI (
s)S
ongs
/bird
Mov
emen
t (m
)M
atch
es (
%)
Ove
rlaps
(%
)
Time of day
05:00 05:15 05:30 05:45 06:00 06:15 06:30 06:45
Fig. 15.3. Changes in song behaviour and movement patterns over the course of the
recording, averaged across the four focal birds OH, YO, BO and WB. Values are
measures calculated over a series of overlapping five minute intervals that move
forward in time in one minute increments. Values are plotted at the centre time for
each segment. Means are shown as a solid line and the grey region represents ± SEM.
(a) Intersong interval (ISI); (b) number of songs per bird; (c) distance moved from each
bird’s average position at the previous to the current time segment; (d) percentage of
songs within a segment that were matches; (e) percentage of songs within a segment
that were overlaps.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 331
Table 15.1. Pearson correlation coefficients among the key singing behaviours above the
diagonal, p values below the diagonala
Intersong
Song rate interval Movement Matches (%) Overlaps (%)
Song rate 0.046 −0.222 0.102 0.378
Intersong interval 0.657 0.280 −0.0128 −0.062
Movement 0.029 0.005 −0.149 −0.163
Matches (%) 0.322 0.211 0.147 0.251
Overlaps (%) 0.0001 0.549 0.111 0.013
a Significant correlation coefficients are shown in bold. The time series data were separated into
one minute bins. Each variable was corrected for autocorrelation by regressing it against its
lagged values and the residuals were used for the correlations (n = 97).
of the recording. Song bout structure for these banded wrens clearly differed
between dawn chorus and later. From the start of dawn chorus and for 20 minutes
into the recording, the mean ISI was very brief (around 10 seconds) and SEM was
very low, indicating that all birds were singing more or less continuously. At
around 05:21 h, the ISI measure exhibits a noticeable break from the previous
trend and both the mean and SEM increase and become slightly more variable
over time. At 05:45 h, a major break in ISI occurred, reflecting the fact that birds
had begun to sing in asynchronous bouts with variable interbout pauses. From
that point on, each bird stopped singing at least once for five minutes or longer.
These pauses are shown as spikes in mean ISI in Figure 15.3a, which occurred at a
different time for each bird (WB: 05:44–05:50 and 06:16–06:23 h; BO: 06:06–06:12 h;
YO: 06:23–06:29 h; OH: 06:25–06:32 h).
Song rates
In this recording, birds sang at uniformly high rates throughout dawn cho-
rus (Fig. 15.3b). OH began singing two minutes before the other birds, then YO and
BO began to sing, and two minutes later WB finally joined the chorus. At 05:22 h,
WB began to sing at a lower rate, while OH, BO and YO continued to sing at high but
more variable rates (shown as an increase in SEM in Fig. 15.3b). At about 05:40 h, all
four birds began a bout of intense counter-singing, and OH continued to sing at an
exceptionally high rate. After this synchronized bout, song rates declined again
but were punctuated by four more peaks, which reflect brief bouts of counter-
singing between different sets of neighbours. Individually, the four birds clearly
differed in their overall song output, with WB in particular being consistently
lower in song rate, starting later and quitting earlier compared with the others.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
332 J. M. Burt & S. L. Vehrencamp
Movement patterns
Patterns of movement were analysed using passive acoustic location to
calculate the position of each bird whenever he sang. Figure 15.3c plots distances
between mean locations for successive time segments. This measure has low values
when birds are singing continuously from one position and increases when birds
move to new positions. The focal birds showed an initial spike of movement at the
beginning of dawn chorus (05:04–05:07 h) and then remained relatively stationary
until about 05:22 h. The initial movement spike was caused by several birds (BO,
OH and WB), who appear to have sung for a brief period from positions near their
sleeping nests and then moved to a more centrally located high song post for the
bulk of their dawn chorus singing. Between 05:27 and 05:43 h, birds made two
or three short distance movements to different parts of their territories while
still singing fairly vigorously and continuously. After 05:43 h, movements became
larger and more frequent. At this time, birds moved relatively quickly between
song perches and tended to stay at each location for several minutes before moving
again (these movements are seen as a pattern of brief spikes in Fig. 15.3c).
Song matching
Figure 15.3d plots the mean percentage across birds of songs sung that
were matches. A song was judged to be a match if it was the same type as an
adjacent neighbour’s recent song (either a song the neighbour had just sung,
or the one previous) and occurred less than 30 seconds after the matched song.
Matching was initially lower because OH had sung alone for two minutes, but
as soon as the other birds began to sing, matching rates quickly increased. Song
matching peaked at 70% at 05:14 h (12 minutes into dawn chorus). After the first
peak, matching declined slightly (although it remained quite high at around 40%)
and peaked again at 05:30 h (52%). Matching declined thereafter, but peaked again
at 05:56 (43%), 06:07 (30%) and 06:26 h (25%).
The four focal birds sang an average of 22 different song types during the record-
ing (range, 20–24) and shared 82% of their song types with any given neighbour
(range, 77.3–85.1). With high singing rates, frequent song type switches and high
levels of song type sharing, the possibility of birds matching by chance will be
higher at dawn chorus. We created a model to test whether the observed rates
of matching were higher than that expected through chance. To estimate chance
matching rates, we generated new datasets using the observed singing data for
all males with randomly shuffled song-type assignments within each bird’s reper-
toire. By averaging the matching rates obtained over multiple permutations of
shuffled song types, we could estimate the probability of chance matching if the
birds had chosen their songs without regard to other singers. Figure 15.4 compares
observed matching versus expected chance matching calculated on the basis of

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 333
Mea
n pe
rcen
tage
mat
ches
per
5 m
in
Fig. 15.4. Observed (± SEM) and expected percentage matching rate per five minute
interval. Expected values were calculated by averaging matching outcomes of 100
random permutations of song types within focal birds. Observed matching rates that
were significantly higher than expected are marked with an open circle (two-tailed
binomial tests, Holm corrected for multiple comparisons; criterion p < 0.05).
100 permutations of random song-type shuffling. Observed rates were significantly
higher than expected throughout the first 15 minutes of dawn chorus and during
the four subsequent peaks later on. The peak at 05:28 h was caused by an intense
interaction between the four focal birds, with a three-way matching interaction
between OH, YO and BO, and a separate matching interaction between WB and
BO (conclusions drawn from analysing individual bird data not shown in the fig-
ure). The three later peaks are attributable to further intense bouts of matched
counter-singing between pairs and trios of neighbours (OH, YO and PU at 05:56 h;
WB, UB and WE at 06:07 h; and WB, BO and LE at 06:27 h).
If song matching is an indicator of conflict, knowing who is matching with
whom can provide useful information about what is going on in a neighbourhood.
Bird OH was involved in many of the interactions that morning. We think that his
singing behaviour and his neighbours’ responses to him were related to his recent
annexation of space at the corner of his, YO’sand BO’sboundaries to defend a newly
active nest in that area (Fig. 15.2). Figure 15.5 shows the patterns of matching by

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
334 J. M. Burt & S. L. Vehrencamp
Fig. 15.5. Patterns of matching by and to bird OH throughout the recording period.
Grey squares indicate OH’s song matches to one of his four neighbours. Black squares
indicate that OH sang a non-matching song. Open squares indicate a match by a
particular neighbour to OH.
and to bird OH, giving us a finer picture of the interactions between him and the
other birds that morning. OH’s bout of solo singing two minutes before his other
neighbours accounts for the lower mean song matching at the very beginning of
dawn chorus. During the initial peak of song at 05:12 h, OH was matched by and
was matching the neighbours at the disputed corner (BO and YO). At this time, the
majority of matches were initiated by the neighbours (each matched a different
one of OH’s songs) and OH replied with his own matches to those neighbours (i.e.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 335
Table 15.2. Percentage of matches by and to each focal bird
Target bird Matching bird
BO OH YO WB Other neighbours
BO – 25 38 20 17
OH 32 – 43 16 8
YO 22 49 – 11 17
WB 21 38 24 – 17
Other neighbours 14 8 16 25 –
in Fig. 15.5 the open squares indicating a neighbour match usually preceded OH’s
own match to that neighbour, so the neighbour matched first). OH’s border shift
had particularly affected YO, and YO and OH matched most often throughout the
morning (43% of YO’s matches were to OH, while 49% of OH’s matches were to
YO; Table 15.2), as would be predicted by an agonistic function for dawn chorus
singing. Another trend is visible in Fig. 15.5, primarily during the dawn chorus
(05:07–05:32 h) and briefly later (05:52–05:57 h): OH, as well as the other birds not
shown in this figure, frequently alternately matched multiple neighbours within
a short period of time. Often these multiple matches involved using a different
song to match each neighbour. For example, at the start of dawn chorus, OH had
been switching between two song types. The first song sung by YO was a match
to one of OH’s types, while BO first sang a match to the other type. Matching was
not merely isolated between the OH/YO/BO trio – all of these birds were often
matching other birds at the same time. Based on our observations that morning,
we know that WB spent most of his time interacting with two other neighbours
(WE and UB) but, nevertheless, at various times did match and was matched by
OH, BO and YO.
Overlapping
In prior work, we had noticed many occurrences of what appeared to be
deliberate overlapping during close-range counter-singing interactions between
neighbouring males. Furthermore, overlapping often occurred in conjunction
with a song match, timed to cover the majority of the other bird’s song as if
the matcher were trying to ‘jam’ the other singer. The context of the ‘overlapping
match’ phenomenon suggested that overlapping is used as an aggressive signal.
Figure 15.3e plots the mean percentage across birds of songs that overlap an-
other song. A song was considered to be an overlap if it ‘covered’ an adjacent
neighbour’ssong by 50% or more of its duration. Songs are sufficiently long in this
species (mean duration is 3.4 seconds) and the territories relatively small (centres
120 m apart) that errors in perceived overlapping caused by the slow speed of

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
336 J. M. Burt & S. L. Vehrencamp
sound are small for this high overlap criterion (Dabelsteen, 1992). Overlapping
was strongly correlated with mean song rate (Table 15.1). Our overlapping mea-
sure cannot distinguish between accidental and deliberate overlapping and it is
likely that much of the trend in overlapping resulted from an increase in the prob-
ability of accidental overlapping during times of increased song rates. Overlapping
may still be used deliberately, as we have observed, but much less frequently or
under very specific contexts. We have some evidence for this: overlapping rate was
significantly correlated with matching rate, even after controlling for the effects
of song rate (partial r = 0.215; p = 0.024), a pattern that would occur if birds oc-
casionally deliberately combined overlapping with song matching. There were 27
occurrences of overlapping matches out of 924 songs delivered by the focal birds
during the recording.
Overall patterns of banded wren singing at dawn chorus
The structure of the banded wren dawn chorus follows the pattern com-
monly described for other species: males begin singing at twilight and sing con-
tinuously at high rates. Then, coincident with increasing light levels and female
emergence, males change to a daytime pattern of interacting individually with
nearby neighbours. Banded wrens also show high rates of song matching to neigh-
bouring singers at dawn chorus. Although dawn chorus matching has been noted
in several other species (Todt, 1970; Spector, 1991), it had not been quantified be-
fore our study, making it difficult to know whether high-rate matching at dawn
occurs in many other species. Therefore, to the degree that dawn chorus has been
characterized in other birds, banded wrens appear to behave similarly to other
chorusing species.
We found clear differences among the four focal males recorded on this single
morning in their rate and duration of singing, which could be caused by vari-
ation in male quality, condition, dominance status or territory quality, as de-
scribed for several species (Cuthill & Macdonald, 1990; Otter et al., 1997; Poesel
et al., 2001). However, there was evidence that these differences could be caused
by short-term variation in motivation arising from differences in nesting stage or
territory boundary disputes. The lowest-rate singer in this recording, WB, was a
highly successful male who was feeding nestlings, whereas the other three males
had recently lost their nests and were engaged in boundary disputes resulting
from OH’s incursion into YO’s and BO’s territories. Analysis of additional record-
ings would clearly be needed to determine whether individual differences in song
output are consistent over time or vary with breeding conditions and/or short-term
motivation.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 337
Evidence for an interactive network
Our strongest evidence for an interactive network during the banded wren
dawn chorus is the high level of song matching that takes place during the first
half hour of singing. Matching at rates significantly higher than chance indicates
that song types delivered by one bird are affecting song types delivered by other
birds and that implies, by definition, that they are interacting. The dawn matching
was likely of a competitive nature too, given that bouts of matching also occur
in the daytime during synchronized counter-singing between dyads or trios of
neighbouring males and when males approach each others’ boundaries for closer
interaction. Our simultaneous recordings showed that males alternately matched
different neighbours in rapid succession with different song types, the key type of
evidence for a fully multi-way interactive communication network. Furthermore,
matching was strongly directed toward one male, OH, by his neighbours YO and
BO at a time when OH was expanding his territory into mostly YO’s territory to
accommodate a new nest site. We thus see particularly clear evidence of multi-way
competitive interactions among these three males.
An observational study conducted on the same population in 2000 indirectly
corroborates our claim of male–male interaction at dawn. In that study, we found
that banded wren males use their most vigorous song type renditions during the
most intense period of singing at dawn, for example longer song-type variations,
compound songs (two or more types sung together) and song types with longer
trills, wider bandwidth and rattle and buzz elements, all of which are associated
with intense male–male interactions at other times (S. L. Vehrencamp & A. Trillo,
unpublished data). Similar patterns have been described for the European black-
bird Turdus merula and yellow warbler Dendroica petechia, which use louder, longer
songs of higher intensity during the dawn chorus and when counter-singing from
a distance with other males (Dabelsteen, 1992; Lowther et al., 1999).
Comparing song patterns with aggression
Patterns of matching and switching in this recording could also reveal
short-term changes in motivation and the outcomes of recent interactions. For
example, OH’s prior aggressive behaviour (annexing portions of two of the focal
neighbours’ territories) appears to have had a strong effect on the behaviour ob-
served in this recording. From the start of dawn chorus, the pattern of matching
toward individual birds was strongly asymmetrical, with most of the matching
directed toward OH (Table 15.2). OH’s boundary shift had probably involved pro-
longed bouts of escalated counter-singing and physical fighting with the affected
neighbours (YO and BO), making OH a particularly threatening neighbour. OH’s

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
338 J. M. Burt & S. L. Vehrencamp
early start to dawn chorus, two minutes before the others, may have been intended
to reinforce his tentative ownership of the disputed property. The matching re-
sponse directed toward OH by the affected neighbours may, in turn, have been
retaliatory threats in response to OH’s announcement of continued occupation.
Frequent type switching during a matching interaction may enable a bird to as-
sess more easily which neighbour is feeling most threatened or to challenge each
neighbour with a distinctive signal.
Dawn chorus singing as an indicator of male quality
In addition to revealing information about short-term changes in moti-
vation, dawn singing could also give eavesdropping receivers information about
longer-term or intrinsic differences among males related to dominance, condi-
tion and age. Montgomerie (1985) suggested that energy reserves should be lowest
at dawn, imposing a handicap such that the vigour and amount of singing hon-
estly reflects a male’s condition or territory quality. Food supplementation was
shown to increase the amount of song in blackbirds (Cuthill & Macdonald, 1990).
Peak song rate during the dawn chorus was correlated with winter dominance at
feeders in black-capped chickadees Parus atricapillus (Otter et al., 1997) and with
earlier female laying date in the blue tit Parus caeruleus (Poesel et al., 2001). Banded
wrens attain peak daily song rates during the dawn chorus. In addition, their
songs seem to be especially loud at this time, although this impression could be
caused by the high song perches (Dabelsteen & Mathevon, 2002). One drawback
to array recording is that individuals who do not vocalize are ‘invisible’ to the
analysis and so we have not been able to show any evidence for eavesdropping
with this dataset. However, with more recordings and more detailed analysis,
it may be possible to show some direct effects of eavesdropping on vocalizing
interactants.
The value of matching at dawn chorus
A defining feature of the dawn chorus is a continuous high rate of singing
by all territorial neighbours. To a listener in a forest at dawn chorus, there is a
confusingly high density of song coming from many directions. This unique ‘song
environment’ poses a challenge to singers, who may be trying to direct signals
to specific neighbours and simultaneously listen for signals directed at them
from all of their neighbours. For this reason, highly directional signals may in-
crease in value and birds would be predicted to shift their singing strategies to
favour more directional signals and avoid using signals that might create ambi-
guity as to the singer’s intended target. In particular, dawn chorusing birds may
avoid using signals that rely on song rate and timing such as song overlapping,

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 339
synchronized song rates and synchronized song-type switching, because these
signals are more likely to be masked when many individuals are singing nearby at a
high rate.
Song matching is one of the few signals that retains its usefulness as a direc-
tional signal at dawn chorus, because it is based on song-type selection, rather
than song timing or rate. In addition, the higher rates of singing and song-type
switching often seen at dawn chorus provide birds with more opportunities for
directed song matching than at other times. For these reasons, it is possible that
song matching accompanied by rapid switching rates will be a common occur-
rence in dawn chorusing species that engage in neighbour–neighbour interactive
networks.
Summary
The single array recording presented here gives the reader a glimpse into
the behaviour of banded wrens at dawn chorus. Our observations provide evi-
dence of a highly interactive communication network, which is most consistent
with the social dynamics hypothesis for the function of dawn chorus, as presented
in Staicer et al. (1996). We are currently analysing a number of similar recordings,
made between 2000 and 2002, on the same focal birds, as well as on different
sets of focal neighbours. With more recordings and more birds, we intend to test
more rigorously the trends we saw in the recording presented in this chapter. In
particular, evidence that the bird who is the focus of matching changes on dif-
ferent days, in relation to current patterns of boundary and nest-site movements,
would greatly strengthen the argument for dawn chorus mediating changes in
social status. Repeated observations on the same birds will also be necessary to
determine whether any of the dawn chorus behaviours are related to male age,
repertoire size, sharing level or quality, which would indicate an additional male
quality-assessment role for the banded wren dawn chorus, and the existence of
eavesdropping.
Based on our findings, we think that the microphone array recording tech-
nique is an ideal method for studying the details of dawn chorus song behaviour.
In particular, multimicrophone recording in some form is possibly the only fea-
sible way to detect and study multi-way interactive communication networks,
such as we found in the banded wren dawn chorus. It is our hope that other
species can be studied using techniques similar to ours so that cross-species com-
parisons can be made of dawn chorus communication networks. Until that time,
it will not be known whether the highly interactive dawn chorus network we
have documented in banded wrens is unique or common among dawn chorusing
species.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
340 J. M. Burt & S. L. Vehrencamp
Acknowledgements
Logistical support was provided by the staff of the Area de Conservacion Guanacaste.
We thank Alex Trillo, Liz Campbell, Carlos Botero, Richard Mills, Dan Pendleton and Harold Mills
for helping us set up the array and make observations during recording sessions, and Cary Leung
for analysing much of the array data. This research was funded by NIH grant R01-MH60461.
References
Ballentine, B., Badyaev, A. & Hill, G. 2003. Changes in song complexity correspond to
periods of female fertility in Blue Grosbeaks (Guiraca caerulea). Ethology, 109,
55–66.
Bower, J. L. 2000. Acoustic interactions during naturally occurring territorial conflict
in a song sparrow neighbourhood. Ph.D. Thesis, Cornell University, New York.
Burt, J. M., Campbell, S. E. & Beecher, M. D. 2001. Song type matching as threat: a test
using interactive playback. Animal Behaviour, 62, 1163–1170.
Cuthill, I. C. & MacDonald, W. A. 1990. Experimental manipulation of the dawn and
dusk chorus in the blackbird Turdus merula. Behavioral Ecology and Sociobiology, 26,
209–216.
Dabelsteen, T. 1992. Interactive playback: a finely tuned response. In: Playback and
Studies of Animal Communication, ed. P. K. McGregor. New York: Plenum Press,
pp. 97–109.
Dabelsteen, T. & Mathevon, N. 2002. Why do songbirds sing intensively at dawn? A test
of the acoustic transmission hypothesis. Acta Ethologica, 4, 65–72.
Eason, P. K. & Stamps, J. A. 1993. An early-warning system for detecting intruders in a
territorial animal. Animal Behaviour, 46, 1105–1109.
Goodson, J. L. 1998. Territorial aggression and dawn song are modulated by vasotocin
and vasoactive intestinal polypeptide in male field sparrows (Spizella pusilla).
Hormones and Behavior, 34, 66–77.
Henwood, K. & Fabrick, A. 1979. A quantitative analysis of the dawn chorus: temporal
selection for communicatory optimization. American Naturalist, 114, 260–274.
Hultsch, H. & Todt, D. 1982. Temporal performance roles during vocal interactions in
nightingales (Luscinia megarhynchos). Behavioral Ecology and Sociobiology, 11, 253–260.
Hutchinson, J. M. C. 2002. Two explanations of the dawn chorus compared: how
monotonically changing light levels favour a short break from singing. Animal
Behaviour, 64, 527–539.
Kacelnik, A. & Krebs, J. R. 1982. The dawn chorus in the great tit (Parus major):
proximate and ultimate causes. Behaviour, 83, 287–309.
Krebs, J. R., Ashcroft, R. & Webber, M. 1978. Song repertoires and territory defence in
the great tit. Nature, 271, 539–542.
Krebs, J. R., Ashcroft, R. & van Orsdol, K. 1981. Song matching in the great tit Parus
major. Animal Behaviour, 29, 918–923.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 341
Kroodsma, D. E., Bereson, R. C., Byers, B. E. & Minear, E. 1989. Use of song types by the
chestnut-sided warbler: evidence for both intra- and inter-sexual functions.
Canadian Journal of Zoology, 67, 447–456.
Lowther, P. E., Celada, C., Klain, N. K., Rimmer, C. C. & Spector, D. A. 1999. Yellow
warbler. In: The Birds of North America, no. 454, ed. A. Poole & F. Gill. Philadelphia:
The Birds of North America, Inc.
Mace, R. H. 1986. The importance of female behaviour in the dawn chorus. Animal
Behaviour, 34, 621–622.
1987. Why do birds sing at dawn? Ardea, 75, 123–132.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K., Dabelsteen, T., Shepherd, M. & Pedersen, S. B. 1992. The signal value of
matched singing in great tits: evidence from interactive playback experiments.
Animal Behaviour, 43, 987–998.
McGregor, P. K., Dabelsteen, T., Clark, C. W. et al. 1997. Accuracy of a passive acoustic
location system: empirical studies in terrestrial habitats. Ethology, Ecology and
Evolution, 9, 269–286.
McGregor, P. K., Peake, T. M. & Lampe, H. M. 2001. Fighting fish Betta splendens extract
relative information from apparent interactions: what happens when what you
see is not what you get? Animal Behaviour, 62, 1059–1065.
McNamara, J. M., Mace, R. H. & Houston, A. I. 1987. Optimal daily routines of singing
and foraging in a bird singing to attract a mate. Behavioral Ecology and Sociobiology,
20, 399–405.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Molles, L. E. & Vehrencamp, S. L. 1999. Repertoire size, repertoire overlap, and
singing modes in the banded wren, Thryothorus pleurostictus. The Auk, 116,
677–689.
2001. Songbird cheaters pay a retaliation cost: evidence for auditory conventional
signals. Proceedings of the Royal Society of London, Series B, 268, 2013–2019.
Montgomerie, R. D. 1985. Why do birds sing at dawn? In: Proceedings of the XIX
International Ethology Congress, p. 242.
Naguib, M. & Todt, D. 1997. Effects of dyadic vocal interactions on other conspecific
receivers in nightingales. Animal Behaviour, 54, 1535–1543.
Naguib, M., Fitchel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Nelson, D. A. & Croner, L. J. 1991. Song categories and their functions in the field
sparrow (Spizella pusilla). The Auk, 108, 42–52.
Nowicki, S., Peters, S. & Hughes, M. 1998. The territory defense function of song in
song sparrows: a test with the speaker occupation design. Behaviour, 135,
615–628.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
342 J. M. Burt & S. L. Vehrencamp
Oliveira, R. F., McGregor, P. K. & Latuffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Otter, K. & Ratcliffe, L. 1993. Changes in singing behaviour of male black-capped
chickadees (Parus atricapillus) following mate removal. Behavioral Ecology and
Sociobiology, 33, 409–414.
Otter, K. A., Chruszcz, B. & Ratcliffe, L. 1997. Honest advertisement and song output
during the dawn chorus of black-capped chickadees. Behavioral Ecology, 8, 167–173.
Otter, K. A., Stewart, I. R. K., McGregor, P. K. et al. 2001. Extra-pair paternity among
great tits Parus major following manipulation of male signals. Journal of Avian
Biology, 32, 338–344.
Part, T. 1991. Is dawn singing related to paternity insurance? The case of the collared
flycatcher. Animal Behaviour, 41, 451–456.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Poesel, A., Foerster, K. & Kempenaers, B. 2001. The dawn song of the blue tit Parus
caeruleus and its role in sexual selection. Ethology, 107, 521–531.
Slagsvold, T., Dale, S. & Saetre, G.-P. 1994. Dawn singing in the great tit (Parus major):
mate attracting, mate guarding, or territorial defence? Behaviour, 131, 115–138.
Spector, D. A. 1991. The singing behaviour of yellow warblers. Behaviour, 117, 29–52.
Speisberger, J. L. & Fristrup, K. M. 1990. Passive location of calling animals and sensing
of their acoustic environment using acoustic tomography. American Naturalist,
135, 107–135.
Staicer, C. A. 1989. Characteristics, use, and significance of two singing behaviours in
Grace’s Warbler (Dendroica graciae). The Auk, 106, 49–63.
Staicer, C. A., Spector, D. A. & Horn, A. G. 1996. The dawn chorus and other diel
patterns in acoustic signaling. In: Ecology and Evolution of Acoustic Communication in
Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY: Cornell University Press,
pp. 426–453.
Todt, D. 1970. Gesang und gesangliche Korrespondenz der Amsel. Naturwissenschaften,
57, 61–66.
Vehrencamp, S. L. 2000. Handicap, index, and conventional signal elements of bird
song. In: Signalling and Signal Design in Animal Communication, ed. Y. Espmark,
T. Amundsen & G. Rosenqvist. Trondheim: Tapir Academic Press,
pp. 277–300.
2001. Is song-type matching a conventional signal of aggressive intentions?
Proceedings of the Royal Society of London, Series B, 268, 1637–1642.
Watkins, W. A. & Schevill, W. E. 1972. Sound source location by arrival times on a
non-rigid three-dimensional hydrophone array. Deep-Sea Research, 19,
691–706.
Welling, P., Koivula, K. & Lahti, K. 1995. The dawn chorus is linked with female
fertility in the willow tit Parus montanus. Journal of Avian Biology, 26, 241–246.

P1: JZZ/... P2: JZZ/...0521823617c15.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 17:8
Dawn chorus as an interactive network 343
Wingfield, J. C. & Farner, D. S. 1993. Endocrinology of reproduction in wild species. In:
Avian Biology, Vol. 9, ed. D. S. Farner, J. R. King & K. C. Parkes. London: Academic
Press, pp. 163–327.
Yasukawa, K., Bick, E. I., Wagman, D. W. & Marler, P. 1982. Playback and
speaker-replacement experiments on song-based neighbour, stranger, and self
discrimination in male red-winged blackbirds. Behavioral Ecology and Sociobiology,
10, 211–215.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
16
Eavesdropping and scent over-marking
ro b e r t e . j o h n s t o n
Cornell University, Ithaca, USA
Introduction
Compared with communication in other sensory domains and with scents
that are released into the air, scent marking is unusual because the signal remains
long after the signalling behaviour; for example, the flank gland marks of male
golden hamsters Mesocricetus auratus deposited on glass in the laboratory are de-
tected by other hamsters 40 days later and vaginal secretion marks are detected
at least 100 days after deposition (Johnston & Schmidt, 1979). In the field, the
paste scent marks deposited by brown hyaenas Crocuta crocuta can be detected
by humans for at least 30 days (Gorman, 1990) and klipspringers Oreotragus oreo-
tragus respond to preorbital gland marks that have been exposed to direct sun
for at least seven days by an increase in scent marking (Roberts, 1998). In many
species, especially those that live solitarily, there is often no receiver present
when the marks are deposited. Consequently, scent marks are necessarily gen-
eral broadcast signals that usually have several functions, depending on the age,
sex, reproductive status, social status and individual identities of both senders and
receivers.
One type of marking, scent counter-marking, is directed at the scent marks
of other individuals, but again these individuals are often not present to ob-
serve the signalling behaviour. I consider the term scent counter-marking to
include two different types of behaviour: (a) over-marking, in which the sec-
ond individual’s scent at least partially overlaps that of the first individual; and
(b) adjacent marking, in which the second individual’s scent is close to that of
the first individual but does not overlap it. In the species with which I am most
familiar (golden hamsters and meadow voles Microtus pennsylvanicus), both types of
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
344

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 345
counter-marking are usually intermixed, but it is possible that some species would
engage in one type of counter-marking but not the other, or would vary the type of
counter-marking or the proportions of different types of counter-mark in different
contexts.
Based on the variety of situations in which counter-marking is observed, it no
doubt has a variety of functions (Brown & Macdonald, 1985). Counter-marking of-
ten occurs only between adult males, suggesting a sexually selected trait related
to competition for females (Hurst & Rich, 1999; Johnston, 1999), but in some
species adult males mark over the marks of their mates, suggesting mate guard-
ing (Mertl, 1977; Moore & Byers, 1989; Kappeler, 1998; Roberts, 2000; Powzyk,
2002). Females may counter-mark other females (e.g. golden hamsters (Johnston,
1977), house mice Mus musculus (Hurst, 1990)) or mark over the marks of males
(e.g. moustached tamarin Saguinus mystax (Heymann, 1998)). Among species that
live in social groups, most or all of the members of a group may mark in the same
place sequentially, producing a group counter-mark (e.g. Rasa, 1973; Mills et al.,
1980; Gorman & Mills, 1984); sometimes dominant individuals in the group mark
most often (Peters & Mech, 1975), but sometimes subordinate, subadult individ-
uals mark most (Lazaro-Perea et al., 1999). Groups may also counter-mark during
territorial encounters (Jolly, 1966). There is a great need for more observational
and experimental field studies focused on the functions of over-marking.
Several years ago, I proposed that the functions of over-marking could be ap-
proached from the question of what kinds of information third parties could
obtain about the marking individuals from scents in over-marks (Johnston et al.,
1994). I suggested that there were three different types of effect that might occur:
(a) masking, in which the most recently deposited scent covers previous scent
marks and thereby eliminates access to the information they contain (e.g. individ-
ual identity, sex, reproductive state); (b) mixing, in which the scents of different
individuals become a chemical mixture, thus producing a new odour quality (e.g.
a group odour) and thereby eliminating information about particular individu-
als; and (c) posting, in which each scent mark to some extent remains separate or
distinguishable, thus producing a bulletin board at which information about all
individuals that marked there can be obtained. Subsequent research suggests that
an important fourth possibility is that scent over-marks can also provide informa-
tion about the relationships between the odours of different individuals, perhaps
including the relative freshness, amount of scent, number of marks, top or bottom
position, or geometric layout (Wilcox & Johnston, 1995; Johnston et al., 1997a;
Johnston & Bhorade, 1998; Hurst & Rich, 1999). Although these early experiments
addressed the perception of over-marks and the subsequent memory for individ-
ual odours, they also can be viewed as indicating the kinds of information that
are available to eavesdroppers.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
346 R. E. Johnston
Scent marking, scent over-marking and eavesdropping
Regardless of the different functions that counter-marking might have,
eavesdropping can occur when a third individual investigates a place marked by
two or more other individuals. I use eavesdropping in the broad sense of an animal
witnessing some type of interaction or an exchange of information between two
other individuals (similar to ‘social eavesdropping’ sensu Peake, Ch. 2). This does
not imply conscious intent or secrecy on the part of the animal doing the observ-
ing nor any awareness or attempt to conceal the interaction by the animals whose
interaction is observed. Eavesdropping on scent counter-marking may be particu-
larly common because the marks are so long lasting, allowing many individuals
to investigate them, not just the individuals present when the marking behaviour
was performed. Furthermore, when eavesdropping on scent counter-marks, indi-
viduals can more easily avoid a potential cost of eavesdropping on vocalizations or
visual displays, namely being detected and threatened, chased or attacked. Among
vertebrates, neither scent marking nor other aspects of chemical communication
have been explicitly analysed using the framework of eavesdropping. Among inver-
tebrates, however, numerous cases have been described using these concepts, but
authors studying insects seem to have adopted a different meaning for the term.
All of the examples I have found involve predators, parasites or parasitoids using
the chemical signals of the host or prey as a means of locating them (e.g. Stowe
et al., 1995). This is a different set of phenomena (‘interceptive eavesdropping’; see
Chs. 2 and 23) and will not be considered here.
What evidence is there for eavesdropping based on scent counter-marking
among mammals? I will discuss three different types of evidence, primarily from
our own work: (a) apparent sensory and perceptual specializations for the evalua-
tion of scent over-marks; (b) evidence that the information in over-marks leads to
differential responses by the perceiver towards individuals whose marks are in dif-
ferent positions in over-marks (top versus bottom); and (c) specialized mechanisms
for the production of over-marks.
Scent marking in golden hamsters
There are two types of scent marking in golden hamsters. Flank marking
is carried out by both males and females and scent is deposited from the flank
gland, which is a region of specialized, pigmented sebaceous glands on the poste-
rior flank. This glandular field is larger in males than in females and is testosterone
dependent (Vandenbergh, 1973). Flank marking is performed as a part of general
maintenance activities (e.g. shortly after animals wake up and groom themselves)
and is especially prevalent in potentially agonistic situations (e.g. in the presence
of odours of other individuals). Hamsters do not usually flank mark during actual

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 347
interactions (Johnston, 1975a,b,c, 1977, 1985). In seminatural laboratory environ-
ments, hamsters mark just inside the tunnel to their burrow, in the vicinity of
the burrow entrance and in other locations (Johnston, 1975c). Subordinate males
in these environments have been observed marking in their nest when a dom-
inant male is attempting to enter (Johnston, 1975c). Therefore, it seems likely
that flank marking is involved in defending the burrow and food hoard by both
males and females (Johnston, 1975c). The second type of scent marking is a type
of anogenital marking, called vaginal marking, that deposits vaginal secretions.
Hamsters have a specialized pouch surrounding the distal vagina that produces
and collects this secretion. The frequency of vaginal marking is related to the fe-
male’sreproductive state, peaking during the night 12–24 hours before receptivity
(Johnston, 1977, 1985). This secretion is highly attractive to males; it stimulates
copulatory behaviour, reduces aggressive behaviour and causes increases in cir-
culating luteinizing hormone and testosterone (Johnston, 1985, 1990). In a study
in seminatural enclosures in the laboratory, females attracted males the night
before receptivity, slept with them during the day, mated early the next day and
then drove the male away (Lisk et al., 1983). Therefore, one primary function of this
type of marking is to advertise sexual receptivity and to attract males. In addition,
females may over-mark the vaginal marks of other females (Fischer & McQuiston,
1991), perhaps as a means of competing for the attention of males or as a secondary
aspect of defence of the burrow and food hoard against other females.
Specialized mechanisms for evaluation of scent over-marks
The first evidence suggesting special mechanisms for evaluation of scent
over-marks came from experiments aimed at understanding the information
obtained about individuals from the scents in an over-mark, as described above.
In particular, these experiments were designed to discover what golden hamsters
would remember after investigating an over-mark consisting of the scents of two
individuals (Johnston et al., 1994). Corresponding to the idea that an over-mark by
one individual might mask, mix or remain distinguishable from the underlying
individual’s scent, would hamsters remember just the top scent, neither scent, or
both scents? We used an habituation technique in which male subjects were first
exposed to a newly deposited, flank-gland over-mark from two donor males (male
B always on top of male A) on four or five successive trials and then were tested on
a final trial with the scent of one of these donors and the scent of a novel donor.
In the first experiment, we wanted to be sure that the scent of male B covered the
scent of male A, so we simulated natural scent marks by picking up the animals
and rubbing their flank gland region against the substrate, a glass plate. We placed
a paper cardstock template over the glass so that scent was deposited only in a

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
348 R. E. Johnston
limited area on the plate (the exact size of the area corresponded to the usual size
of flank or vaginal scent marks (Johnston et al., 1994)). Across repeated habituation
trials, the investigation of the scent over-mark decreased, as it would to a single
individual’s scent. On the test trial, subjects should investigate the familiar scent
significantly less than the novel scent, indicating memory for the familiar scent
(e.g. Johnston et al., 1993). The results were quite interesting: subjects treated the
flank gland odour of the top-scent individual as familiar (investigated it less than
that of a novel individual) but investigated the flank odour of the bottom scent
male the same amount as that of the novel individual (Johnston et al., 1994). We
obtained similar results when males were habituated to vaginal secretion over-
marks of females and then tested with each scent individually compared with a
novel vaginal secretion. Because the scent of the second donor was placed on top
of that of the first donor, our interpretation was that the top scent masked the
bottom scent and, therefore, the bottom scent actually was novel to the subjects
(Johnston et al., 1994).
This first experiment was, however, somewhat unrealistic in that the scent of
the second individual completely covered the scent of the first individual; when
hamsters deposit their own scent they are usually not so thorough or precise. In
a second experiment, we placed the top and bottom scents at right angles to one
another such that they formed a cross; that is, there was a region of overlap of
the two individuals’ scent marks, but also regions where each individual’s scent
was by itself. The same results were obtained as in the first experiment: male
subjects treated the vaginal scent of donor B as familiar but treated the scent of the
bottom-scent individual (donor A) the same as that of a novel individual (Fig. 16.1a).
Results using flank-gland secretions showed the same pattern (Johnston et al., 1995;
Johnston, 1995). These results indicate that the subjects had a preferential memory
for the top scent compared with the bottom scent, despite being able to investigate
both scents during the habituation trials. This preferential memory suggests that
the subjects have a mechanism for evaluating over-marks and either preferentially
remember the top scent, selectively forgetting the bottom scent, or tag the memory
of the top scent so that it is more salient than that of the bottom scent. In further
experiments of this type, we found evidence for preferential memory for the top
scent even if, during the habituation trials, there was an additional scent mark of
the bottom-scent donor that was not marked over (Fig. 16.1b; Wilcox & Johnston,
1995). These experiments suggest that hamsters extracted information about the
relative position (top or bottom) or the relative freshness of the two individuals’
scent marks and either selectively remembered just one of them or had placed
a different value on the memory of the top and bottom odours. The existence
of either of these mechanisms suggests that the relative position of an
individual’s scent in an over-mark is important to the perceiver and that this

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 349
0.005
Mea
n in
vest
igat
ing
tim
e (s
)
0.0025
Fig. 16.1. The time that male hamsters spent investigating the vaginal scents of
females on the test trial after habituation to a pattern of vaginal scent-marks as
shown above the graphs. In both (a) and (b), males investigated the scent from the
top-scent female significantly less than the novel female’s scent (n = 9 in (a); n = 10
in (b)) whereas there was no significant difference in time spent investigating the
scent of the bottom-scent male and the novel scent (n = 10 in both groups), thus
indicating a preferential memory for the top scent of the over-mark. The bar indicates
the standard error; p values derived from t-tests. (From Wilcox & Johnston, 1995.)
information may influence subsequent social interactions between the perceiver
and the individuals that deposited the scent marks.
Did the subjects in these experiments actually forget the bottom scent in an
over-mark or did they just attach less value or importance to it? The latter seems
more likely. First, it is difficult to believe that hamsters would not remember one
of two individually distinctive odours after investigating them four or five times,
since a single scent is remembered at least 10 days after such exposures (Johnston,
1993). Second, later experiments showed that hamsters would remember two ad-
jacent scents (see below). Third, in another experiment, we obtained evidence
that males did have some memory of the bottom scent from an over-mark. Male
hamsters were first exposed to experimenter-produced over-marks of male flank
glands in the pattern of a cross during four habituation trials with 15 minutes
between trials. Investigation of the crossed scents decreased significantly in all
three groups (10 per group) across the habituation trials: the results for trial 1 and
trial 4, respectively were 16.8 and 3.8 seconds in group 1 (t = 10.216; p < 0.001),
21.1 and 3.2 seconds in group 2 (t = 14.802; p < 0.001), and 19.3 and 3.7 seconds
in group 3 (t = 7.156; p < 0.001). These groups were then tested 15 minutes after

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
350 R. E. Johnston
0.0007
0.01M
ean
inve
stig
atin
g ti
me
(s)
Fig. 16.2. The time male hamsters spent investigating the flank scent of one male in
the test trial after habituation to a scent over-mark in the pattern of a cross (shown
above) in a series of trials. The top scent from the over-mark was investigated least,
indicating habituation to and memory for this odour compared with the novel odour.
The bottom scent from the scent mark was investigated an intermediate amount,
suggesting some memory for this odour but one that was significantly different than
that for the top scent. The bar indicates the standard error; p values derived from
t-tests; 10 animals in each group. (R. E. Johnston & M. Schiller, unpublished data.)
the last habituation trial with the flank scent from just one male on the test
trial: the flank scent of the top-scent male for group 1, the bottom-scent male for
group 2 and a novel male for group 3. The novel flank scent was investigated most
and the top scent was investigated significantly less, as in the previous tests in
which there were two stimuli present in the test trial (Fig. 16.2; R. E. Johnston &
M. Schiller, unpublished data). The bottom scent, however, was investigated an
intermediate amount, significantly more than the top scent but significantly less
than the novel scent (Fig. 16.2). This experiment suggests that hamsters do re-
member the scent of the bottom-scent male but the memory is not as strong or as
salient, or that the behaviour based on the memory is different from that for the
top-scent male. We do not know exactly in what way it is different, but we suspect
that the bottom scent is less important to the subjects (see p. 359–360). Experiments
on meadow voles support the notion that the odour of the bottom-scent male
is devalued relative to top-scent males or novel males (Woodward et al., 2000).
Before describing our experiments on how hamsters and voles determine which
scent is on top, it is useful to review what we know about the mechanisms under-
lying discrimination between odours from different individuals. As in any other

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 351
sensory domain, individual recognition is accomplished by pattern recognition
mechanisms: perceptual mechanisms by which animals discriminate between sim-
ilar, complex stimuli (e.g. faces, voices, odours). In such processes, specific features
are generally much less important than the relationships between features. In the
case of odours, the pattern is generated by a large number of individual chemical
compounds that occur in differing proportions across individuals (Gorman, 1976;
Bagneres et al., 1991; Gamboa et al., 1996; Singer et al., 1997; Smith et al., 2001). It is
these differences in proportions of chemical compounds that give each individual
its distinctive odour quality. The particular chemicals that differ in proportion
vary across pairs of individuals; that is, there does not seem to be a particular set
of chemical compounds that are used for this purpose (Smith et al., 2001). Conse-
quently, one might expect that a mixture of two scents would be created when
one individual over-marks another’s scent that was different from either of the
two original mixtures and that this new mixture would produce a new odour
quality. The results of the experiments reported in the preceding paragraphs, in
which subjects were habituated to over-marks, argue against this hypothesis be-
cause in the test trial hamsters showed memory for one individual but not the
other (Johnston et al., 1994, 1995; Wilcox & Johnston, 1995).
We have undertaken a series of experiments to try to characterize the mecha-
nisms used to distinguish top and bottom scents in an over-mark. The strategy in
these experiments was to determine if a particular kind of information in over-
marks was sufficient, by itself, to promote differential responses to the odour of
one donor versus the other donor.
One possible cue is the relative freshness of the two scents, because the top scent
of an over-mark is necessarily fresher than the bottom scent. In the experiments
discussed above, the difference in the age of the two scent marks in over-marks
was about 30 seconds (not more than 60 seconds); however, in nature, scent marks
are likely to differ more than this in freshness (e.g. by at least tens of minutes
and often by hours or days). Therefore, a series of experiments were carried out to
see if differences in freshness alone would cause the differential memory effect.
In addition, we wanted to be sure that animals could detect and remember the
scents of two individuals that were in close proximity. In the first experiment, sub-
jects were habituated to vaginal secretion scents from two individuals (A and B)
that were placed adjacent to one another in an open cross pattern, as shown
in Fig. 16.3a, and then were tested for their responses to scent A or B versus a
novel scent. Males investigated both scents (A and B) less than the novel scent,
indicating that they could remember the scents of two individuals from habitu-
ation trials (for flank gland scents, see Cohen et al. (2001)). Then we tested males
to determine if differences in freshness between the two scents would result in
differential memory for the fresher scent. The scents were again presented in the

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Mean investigating time (s)
(a)
Hab
ituat
ion
stim
uli
Scen
t 1
Scen
t 2
0.00
10.
005
0.00
250.
025
0.02
5
Scen
t 1gr
oup
Scen
t 2gr
oup
scen
tnove
l
scen
t
Fig.
16.3
.Th
eti
me
spen
tby
mal
eh
amst
ers
inve
stig
atin
gva
gin
alse
cret
ion
scen
tsfo
llow
ing
hab
itu
atio
nto
vagi
nal
secr
etio
nm
arks
inth
ep
atte
rnsh
own
abov
eth
egr
aph
.(a)
Aft
erh
abit
uat
ion
totw
osc
ent
mar
ksin
the
pat
tern
ofan
open
cros
s,bo
thsc
ents
wer
ein
vest
igat
edsi
gnif
ican
tly
less
than
an
ovel
scen
t,in
dic
atin
gm
emor
yfo
rbo
th.(
b)A
fter
hab
itu
atio
nto
two
scen
tsof
dif
feri
ng
deg
rees
offr
esh
nes
s,bo
thsc
ents
are
inve
stig
ated
sign
ific
antl
yle
ssth
anth
en
ovel
scen
ts,i
nd
icat
ing
that
ad
iffe
ren
cein
fres
hn
ess
doe
sn
otle
adto
dif
fere
nti
alm
emor
yfo
ron
eof
them
.(c)
Aft
er
hab
itu
atio
nto
scen
tsin
an‘a
pp
aren
tcr
oss’
(on
eco
nti
nu
ous
scen
tan
don
ein
terr
up
ted
scen
t),t
he
con
tin
uou
ssc
ent
was
inve
stig
ated
sign
ific
antl
yle
ssth
anth
en
ovel
scen
tbu
tth
ere
was
no
sign
ific
ant
dif
fere
nce
inti
me
spen
tin
vest
igat
ing
the
inte
rru
pte
dsc
ent
and
the
nov
elsc
ent,
sugg
esti
ng
poo
rer
mem
ory
for
the
inte
rru
pte
dsc
ent
than
for
the
con
tin
uou
s
scen
t.Th
eba
rin
dic
ates
the
stan
dar
der
ror;
pva
lues
der
ived
from
t-te
sts;
10an
imal
sin
each
grou
p.(
From
Joh
nst
on&
Bh
orad
e,19
98.)
352

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 353
open cross pattern during habituation trials, but one vaginal secretion scent was
30 seconds old and the other was 24 hours old. On the test trial, male hamsters
investigated novel scents more than familiar scents of the same age, indicating
memory of both of the familiar scents. The investigation of the familiar, 30 second
scent did not differ from that of the familiar 24 hour scent, indicating no preferen-
tial memory for the fresher scent (Fig. 16.3b; Johnston & Bhorade, 1998; for similar
data on male flank gland scent, see Cohen et al. (2001)). Similar experiments with
meadow voles exposed to adjacent anogenital marks differing in age by 60 minutes
yielded no significant difference in response to the donor of the fresh scent versus
the donor of the 60-minute-old scent (Ferkin et al., 1999). Therefore, freshness by
itself did not lead to preferential treatment of the scent of one individual. It is
worth noting that four different types of odour were used in these experiments:
vaginal secretions and flank glands in hamsters (Johnson & Bhorade, 1998), anogen-
ital area scent in voles (Ferkin et al., 1999) and urine in voles (M. H. Ferkin,
J. Dunsavage & R. E. Johnston, unpublished data). This suggests that the lack of
an effect of freshness is not caused by the chemistry of one particular type of
scent (such as the sebaceous scent from flank glands) that might change little over
24 hours. We do not know how long individually specific information lasts, but
hamster flank and vaginal scents deposited on glass in the laboratory are detected
and investigated after 40 and 100 days, respectively (Johnston & Schmidt, 1979).
A second possible cue that might be used as a guide for preferential responses to
one individual over another is the relative amount of scent or number of marks de-
posited by two individuals. Among many species, dominant or high-ranking indi-
viduals mark more often than subordinate or low-ranking individuals (Ralls, 1971;
Eisenberg & Kleiman, 1972; Johnson, 1973; Johnston, 1975a,c; Bronson, 1976; Hurst
& Rich, 1999) and the relative amount of over-marking is probably correlated with
the frequency of marking. Therefore, the number of marks or over-marks could
be an indication of dominance status. Even when there was a single over-mark, it
might be the case that subjects would perceive that the top-scent individual de-
posited more scent if the top scent masked some of the bottom-scent individual’s
mark. The ‘amount of scent’ and ‘number of marks’ hypotheses were, however,
ruled out as explanations of the differential effects in hamsters and meadow
voles. That is, hamsters still remembered the top-scent individual and meadow
voles investigated the top-scent individual more even when, during the expo-
sure to over-marks, there were more marks or more area covered by the bottom-
scent individual (e.g. Fig. 16.1b; Wilcox & Johnston, 1995; Johnston et al., 1997a,b;
Johnston & Bhorade, 1998; Ferkin et al., 1999). In one experiment with meadow
voles, for example, female subjects investigated the home cage of a male that
had been briefly investigated and marked by an ‘intruder’ male. When tested in
the Y-maze, females spent more time investigating the whole-body odour of the

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
354 R. E. Johnston
Mea
n in
vest
igat
ing
time
(s)
0.01
Fig. 16.4. The time that female meadow voles spent investigating whole-body odours
from cotton bedding in a Y-maze after exposure to the home cage of one male that
had been briefly investigated and scent marked by another male, as represented by
the drawing on the left. The bar indicates the standard error; p values derived from
Wilcoxon matched-pairs signed-rank test; n = 12. (From Johnston et al., 1997b.)
intruder male than that of the home-cage male, even though the home-cage male
must have had many more marks and covered more area with his marks than
the intruder did (Fig. 16.4; Johnston et al., 1997b). Therefore, neither hamsters nor
voles seem to use the relative amount of scent or number of marks, by themselves,
as a means of selective responses to other individuals.
Other experiments suggest that it is some type of information from the area of
overlap, or the area of overlap compared with adjacent areas with non-overlapped
scents, that leads to preferential responses to the top-scent individual. One line
of evidence for this conclusion is that among all of the experiments that we have
done with hamsters and meadow voles, using a variety of testing methods, we
have found differential responses in the test phase only when there were scent
marks that overlapped during the exposure phase (with one exception, see below)
(Johnston et al., 1994, 1995, 1997a,b; Johnston & Bhorade, 1998; Wilcox & Johnston,
1995; Ferkin et al., 1999; Cohen et al., 2001). A second line of evidence for the
importance of scent overlap comes from experiments specifically designed to test
whether overlap of two individuals’ scent marks was necessary for differential
responses to the two animals or to their odours. For example, we first habituated

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 355
Fig. 16.5. The time that female meadow voles spent investigating whole-body odours
from cotton bedding in a Y-maze after an exposure to scent marks in the patterns
shown above the graph. (a) After exposure to a large area of scent from one male with
a small spot of scent from another male on top, females (n = 12) spent significantly
more time investigating the top-scent male. (b) When the small spot was placed in a
clean area of the slide, so that there is no overlap, females (n = 10) show no significant
difference in investigation time. Bar indicates the standard error; p values derived
from Wilcoxon test. (From Johnston et al., 1997a.)
male golden hamsters either to two scent marks in the pattern of a cross or to
two scent marks in a pattern of an ‘open cross’ – that is, there was no overlap in
the middle, just an unscented space. On the test trial, males exposed to this latter
pattern showed an equivalent response to the scent of the two donors, treating
them both as familiar (e.g. Fig. 16.3a; Cohen et al., 2001), but males exposed to
the crossed scents showed memory for the top scent but not for the bottom scent
(Fig. 16.1a; Johnston et al., 1995; Johnston & Bhorade, 1998; Cohen et al., 2001). In
another example, one group of female meadow voles were exposed for 15 minutes
to a microscope slide largely covered by the anogenital scent of one male but with
a small spot of scent from a second male placed on top. During the test trial, these
females spent significantly more time investigating the whole-body odours of the
small-spot, top-scent male (Fig. 16.5a). If, however, the second male’s scent was
placed in a clean ‘hole’ surrounded by scent from the first male, female meadow
voles showed no significant difference in response to the odours of the two males
(Fig. 16.5b; Johnston et al., 1997a). In both hamsters and voles, an area of overlap
was necessary to produce a differential response to their odours. We do not know

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
356 R. E. Johnston
exactly how hamsters or voles use the area of overlap to determine which scent is
on top, but one hypothesis is that they compare the odour quality of the overlap
region with that in the adjacent areas. Assuming that there is partial mixing in the
area of overlap, the odour in this region should smell more like the top-scent male
than the bottom-scent male. In the hamster experiment, animals could compare
the area of overlap with the two adjacent, non-overlapped scents and determine
which was the top-scent by which scent was closest in odour quality to the region
of overlap. This mechanism would not work for the vole experiment, however,
because in this situation there is just an area of overlap and an area of scent from
the bottom-scent male.
Finally, there is some evidence that hamsters may be able to use the geomet-
ric relationships between the two scent marks (interrupted versus uninterrupted
streaks) to determine which is the top scent. We reasoned that, by analogy with
depth perception in vision, if one scent occludes another, it must be on top. If,
when investigating scent over-marks, hamsters or other animals develop a repre-
sentation of the geometrical layout of the marks, they might be able to determine
which of two scents was on top by determining which one occluded the other. To
test this possibility while at the same time eliminating cues from a region of over-
lap, we investigated how male hamsters would respond after being habituated
to a pattern of scent marks in which it might appear that one individual’s scent
occluded the other but in fact there was no region of overlap; rather, there was one
continuous scent and, at right angles to it, two scent marks that approached this
scent closely but did not touch it (Fig. 16.3c). Male hamsters showed a preferential
memory for the continuous scent compared with the interrupted scent for vaginal
scent marks (Fig 16.3c; Johnston & Bhorade, 1998) and male flank marks (Cohen
et al., 2001). This is the only case in which a region of overlap was not necessary to
obtain a differential response to one animal or its odours after exposure to scent
marks of two individuals. In contrast, meadow voles show no evidence of using
the same kind of spatial information (Ferkin et al., 1999). Perhaps the primary rea-
son for this species difference is that, whereas hamsters deposit marks in linear
streaks, voles’ marks are more often small spots or larger irregularly shaped areas
(especially urine marks) and it is not obvious how spots and blobs could provide
spatial cues about which scent was on top.
The results summarized above indicate that at least two species of rodents
have evolved specialized mechanisms for the perception and analysis of scent over-
marks. These abilities are quite striking – indeed, amazing – and we do not yet fully
understand them. Many other species have probably evolved similar mechanisms,
but, because of the diversity of functions served by scent marking, it is not likely
that all species have evolved such mechanisms. The existence of these abilities,

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 357
however, should stimulate us to think about other ways in which animals might
extract information from arrays of scent marks.
Specialized mechanisms for production of over-marks
If over-marking has functions that are distinct from the functions of reg-
ular scent marking, specialized mechanisms should have evolved to promote ac-
curate placement of scent on top of that of another individual: that is, mecha-
nisms to target the scent marks of others. Numerous observations of a variety of
mammalian species in nature indicate that such targeted over-marking does oc-
cur (Ralls, 1971; Eisenberg & Kleiman, 1972; Johnson, 1973; Brown & Macdonald,
1985). Several recent field studies have provided quantitative data on the extent of
over-marking in natural environments (Kappeler, 1998; Lazaro-Perea et al., 1999).
In perhaps the most dramatic case, male diademed sifakas Propithecus diadema over-
mark 94% of the scent marks deposited by their mates (Powzyk, 2002). Observations
of over-marking in nature are convincing evidence for a targeting mechanism be-
cause the probability of even one over-mark occurring by chance is extremely low,
given that there are many possible places to mark. In addition, most animals that
over-mark also engage in other related activities when encountering scent marks
from another individual, such as careful investigation, pawing or scratching the
ground, biting the bark of the tree on which the marks are deposited or becoming
visibly aroused or agitated, indicating that they are reacting to this odour and are
focused on it.
Virtually all experimental studies of the mechanisms underlying scent mark-
ing and over-marking, however, have been carried out in captivity or in laboratory
settings, where space is limited. In such circumstances, it is much more difficult
to determine if over-marks occur by targeting mechanisms or occur by chance.
For example, a common (but not universal) observation is that a particular odour
(e.g. urine, flank gland) will increase the frequency of scent marking with the
same scent (Ralls, 1971; Johnston, 1975a, 1977; Hurst, 1990). Some of these scent
marks will be over-marks. In the limited spaces used in the laboratory, however,
it is difficult to know whether the increase in the number of over-marks occurs
just because the rate of marking increased and, by chance, some were deposited
in places that had been marked previously, or because a specific targeting mecha-
nism exists. Similarly, other mechanisms that increase marking frequency, such as
changes in hormone levels with reproductive state, can increase the frequency of
over-marking. Furthermore, many species have preferred types of site for marking,
such as particular topological features (visually prominent rocks or vegetation, in-
tersections of trails, particular types of plant or tree, water holes or other rare

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
358 R. E. Johnston
resources). Hamsters in laboratory environments, for example, prefer to mark in
confined spaces, such as the corners of a rectangular arena and just outside of their
burrow entrance (Johnston, 1975c). An increase in the number and the percentage
of over-marks could occur if the rate of marking is increased by stimulation and
there are a limited number of preferred places in which to mark.
We designed a simple method to determine if individuals have a specific mecha-
nism for targeting another individual’sscent mark to produce an over-mark, which
we have applied to both flank marking and vaginal marking by hamsters (R. E.
Johnston, S. Szmuilowicz, D. J. Mayeaux, S. K. Barot, & N. S. Schwarz, unpublished
data). This involves comparing the number of marks that are deposited over scent
marks to the number of marks deposited over mirror-symmetric locations in the
same arena that have no scent (imaginary marks). Since the types of locations are
identical and are in the same arena, this method controls for both the problem of
preferred locations and the problem of odour-stimulated increase in overall mark-
ing frequency. In one recent experiment of this type (S. K. Barot, N. S. Schwarz &
R. E. Johnston, unpublished data), we found that the mean number of flank marks
by 12 male hamsters that overlapped another male’s flank marks was 6.0, whereas
the number of flank marks that overlapped imaginary marks (symmetric locations
but clean) was 2.8, (t = 3.171; p < 0.01). Females, however, did not selectively flank
mark over the flank scents of other females or males. We are currently replicating
and refining these experiments, but these initial experiments suggest the exis-
tence of a specific mechanism in adult, male golden hamsters that targets the
flank marks of other adult males but not the marks of juvenile males or females.
These results suggest that flank over-marking by males is a sexually selected char-
acteristic related to male–male competition, but that flank marking by females is
a more generalized kind of broadcast signalling.
Functions of over-marking and eavesdropping on over-marks
What is the function of scent over-marking, and why have hamsters and
meadow voles evolved specialized mechanisms for evaluating over-marks? We
have hypothesized that in meadow voles and golden hamsters, both of which live
solitarily, scent over-marking by males may be a type of intrasex competition in
which each male targets its male neighbours (Johnston et al., 1997a,b; Johnston
& Bhorade, 1998; Johnston, 1999; Cohen et al., 2001). This mutual over-marking
presumably reflects a struggle for dominance between males. If each male is, at
least during the breeding season, continuously trying to keep its scent on top of
the scents of its male neighbours, over-marking should be an energetically costly
activity. I am not aware of any data on the energetic costs of over-marking in
natural settings, but the cost of marking for defence of territory is important for

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 359
some species (Mills et al., 1980; Gorman & Mills, 1984; Gorman, 1990; Gosling et al.,
2000; Gosling & Roberts, 2001). The degree to which an individual is successful at
keeping its marks on top of those of its neighbours should be an honest indicator
of phenotypic vigour and quality and, to the extent that these characteristics are
dependent on genotype, genetic quality. Information gathered by third par-
ties from over-marks about ‘whose scent is on top’ should, therefore, be valu-
able information for mate-choice decisions by females and it may influence in-
teractions between like-sex rivals as well. Is there evidence to support these
hypotheses?
The evidence suggests that analysis of scent over-marks does affect preferences
for opposite-sexed individuals and that such preferences are likely to be impor-
tant in mate-choice decisions. Most of the tests we carried out with meadow voles
described above were preference tests rather than habituation tests: female voles
were first exposed to anogenital or urine over-marks from two males for 15 min-
utes; 10 minutes later they were tested for their preferences for the whole-body
odours (cotton bedding material, that contained additional body odours and pos-
sibly urine odours) of donor males in a Y-maze. Females spent more time close
to and investigating the whole-body odour of the top-scent male than that of
the bottom-scent male in a variety of experiments (Figs. 16.4, 16.5a; Johnston et al.,
1997a,b). Males and females were housed in long-day light cycles and were in re-
productive condition, so this preferential behaviour suggests that females would
prefer the top-scent males as mates (Johnston et al., 1997b). In addition, we also
exposed female meadow voles to naturally deposited over-marks of males and
found that females again preferred the whole-body odours (cotton bedding) from
top-scent males over that of bottom-scent males (Fig. 16.4; Johnston et al., 1997b).
It is worth noting that in several of these experiments there was far more of
the bottom-scent male’s odour present during the exposure phase than of the
top-scent male’s odour, but females still preferred the top-scent male (Figs. 16.4
and 16.5; Johnston et al., 1997a,b). We interpret these results as a preference for
the top-scent male because the test stimulus (whole-body odours in bedding ma-
terial) incorporated whole set of body odours and was a different stimulus from
the one odour collected by us and presented during the exposure phase. Further-
more, the test was carried out in a different environment from the one used for
the exposure to the over-marks (Y-maze rather than subject’shome cage). These re-
sults cannot be explained as merely the result of habituation for two reasons. First,
females were exposed to the odours of both stimulus animals during the presenta-
tion of the over-marks. Second, our studies using a habituation paradigm showed
that after habitation to an over-mark subjects spent less time investigating the
scent from the top-scent male than that from the bottom-scent male, which is the
opposite of what we found in our Y-maze experiments.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
360 R. E. Johnston
Pilot experiments with hamsters suggest that after females eavesdrop on males’
over-marks they show preferences for the top-scent over the bottom-scent male.
Female hamsters living in a seminatural enclosure in the laboratory were allowed
to explore regions of the environment that had been explored and marked by two
males, but one male was always first (male A) and the other was always second
(male B); consequently, when the female explored the arena, the marks of male
B should have been on top. (Although we did observe males marking, we did not
have a method of determining the location of the marks with sufficient accuracy
to be certain for all cases that they overlapped or not.) Females were tested in
the seminatural environment approximately 18 hours prior to receptivity (when
they are soliciting males) and also when they were receptive. The stimulus males
(A and B) were confined in small, wire-mesh enclosures. Females spent more time
investigating male B (that had explored and marked the arena second) than male A
(there first): combined investigation time for day before oestrous was 238.1 ± 39.8
seconds and for day of oestrous was 159.1 ± 24.6 seconds (degrees of freedom (df ) =9; t = 2.39; p = 0.04). Females also vaginal marked more in the vicinity of male B (7.4
± 1.4 seconds for male B and 4.9 ± 1.0 seconds for male A; df = 9; t = 2; p = 0.057
(S. K. Barot, N. S. Schwarz & R. E. Johnston, unpublished data)). In other experiments
with hamsters, males explored an arena in which there were six vaginal secretion
marks of female B overlapping those of female A and two marks of female A
by themselves. After 45–60 minutes, males were tested for their preference in a
simultaneous choice apparatus (Steel, 1984). Males spent more time sniffing the
top-scent female B (108 ± 4.8 seconds) than female A (88.7 ± 5.4 seconds; df =15; t = 2.956; p = 0.01 (R. E. Johnston & C. Lee, unpublished data)). These results
suggest that males may prefer as mates females that successfully over-mark other
females with vaginal secretions; such over-marking by females may reflect their
vigour and ability to defend their burrow and food hoard from other females.
Maintaining a safe burrow and a food hoard could lead to greater pup survival and
if so it would be advantageous for males to mate preferentially with such females.
Experiments with other species also indicate that information gained by inves-
tigation of scent marks influences mate preferences. After female house mice ex-
plored areas in which the marks of only one male were present (exclusively marked
territory) and another area in which a second male had also marked (and over-
marked), females preferred the males that had the exclusively marked areas (Rich &
Hurst, 1998). Females also preferred a male that had counter-marked another
male to one that had been counter-marked (Rich & Hurst, 1999; see also Ch. 11).
Similar supporting evidence comes from studies with a primate, the pygmy loris
Nycticebus pygmaeus (Fisher et al., 2003). Male lorises over-mark the urine marks of
other males. Females were exposed to naturally produced male over-marks over
a period of 14–20 weeks, until the females came into oestrous. Several measures

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 361
of female behaviour (proximity and orientation to males, investigation of odours
and socio-sexual behaviour) all indicated a preference for the top-scent male by
oestrous females.
The evidence so far suggests that in several species eavesdropping on scent
over-marks influences mate preferences, perhaps because over-marks usually re-
flect the relative phenotypic and possibly genotypic quality of individuals. Much
more research needs to be done to determine how widespread this phenomenon
is, the degree to which eavesdropping on over-marks influences mate choice in
natural settings, and the factors leading to the evolution of over-marking as a
sexually selected trait that provides useful information about potential mates. It
would also be valuable to determine the degree to which over-marking correlates
with other measures of behaviour or physiology that are related to the geno-
typic or phenotypic quality of individuals within a population. Although I have
stressed the usefulness of over-marks as a means by which third parties might
assess opposite-sexed individuals, analysis of over-marks may also influence com-
petitive interactions between third parties of the same sex as those that deposited
the over-marks (e.g. in competition for territory, food or water resources, nesting
sites or mates) could be seen as more likely to dominate in confrontations. Also yet
to be investigated are the effects of over-marks on the individuals that are engaged
in over-marking contests. No doubt there are many interesting phenomena yet to
be discovered.
Field tests of specific hypotheses about over-marking
Both field and laboratory studies are needed to test specific hypotheses
about the functions of over-marking. If over-marking is a type of advertising con-
test between like-sex individuals for mates for example, one would expect that
over-marking the scent of potential rivals (adults of the same sex in reproductive
condition) would be especially prevalent. There are many studies that have shown
that scent marking in general is stimulated by like-sex adults, but relatively few
studies that have measured over-marking and even fewer that have compared
over-marking towards rivals and non-rivals. Likewise, if over-marking is a means
of mate-guarding, one would expect it to be predominantly done by the sex that is
most actively competing for mates (usually males) and that males would mark over
the scent of their mate, as has been observed in the pronghorn antelope Antilocapra
americana (Byers & Bekoff, 1986). Particularly valuable should be field experiments
in which scent marks are manipulated in ways similar to song playbacks (Sliwa &
Richardson, 1998), for example, by experimentally over-marking some residents
but not others, or by using the scent of one resident to over-mark the scents marks
of rivals.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
362 R. E. Johnston
Audience effect on scent marking?
Audience effects refer to an alteration in behaviour because of the pres-
ence of specific individuals or classes of individuals: the audience (see Ch. 4).
Generally, the presence of other animals is defined as the subject being able to
see or hear these other animals. Scent provides interesting possibilities that have
not been systematically explored. For example, might the behaviour of one indi-
vidual toward a second individual be altered by the presence of scent from some
individuals but not by scent from others? If so, would fresh scent be more effective
than older scent? Although fresh scent is not exactly the same as being observed
or heard by another individual, it could indicate the recent presence of an individ-
ual, and thus it could indicate a high probability of being observed or discovered
by the animal that deposited the scent. One can imagine that very fresh scent of
a dominant individual, for example, could inhibit some types of aggressive be-
haviour of a subordinate or that fresh scent of a male paired with that of a female
might inhibit another male from courting her.
In a more traditional sense, do animals alter their scent marking behaviours
based on the social environment? Both of the species that we have studied in
detail (golden hamsters and meadow voles) live solitarily and in nature one would
predict that they usually mark when alone. In laboratory settings, golden hamsters
do not usually flank mark in the presence of another hamster but often mark
vigorously just after social encounters (Johnston, 1975a). This could be considered
a type of audience effect, but not one that depends on the presence or absence of a
specific audience. The one exception that I have observed to this pattern of marking
when alone is that subordinate males do sometimes mark just inside their burrow
entrance when they defend their burrows from a dominant male (Johnston, 1975c).
This behaviour, however, seems to be readily explained by flank marking as an
agonistic behaviour involved in the defence of the burrow or food hoard: the
notion of an audience effect does not aid our understanding. Female hamsters
are stimulated to vaginal mark in the presence of a male or a male’s odours but
not in the presence of a female or her odours: they mark most frequently in the
period 12–24 hours before sexual receptivity (Johnston, 1977, 1979). Once again,
this pattern seems to be primarily related to the function of sexual advertisement
and is not a modulation of behaviour based on specific relationships between
individuals.
Perhaps the most likely situations in which audience effects might be observed
would be in gregarious species. For example, in the ring-tailed lemur, Lemur catta,
individuals are attentive to the marking behaviour of others and 62% of all marks
are investigated within 30 seconds of deposition; in 89% of these cases the original
mark is over-marked (Kappeler, 1998). In this study, however, no evidence was

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 363
found for an influence of social context on the likelihood of scent marking. In
species that live in groups that have a clear dominance hierarchy, such as wolves
Canis lupus or free-ranging packs of domestic dogs, dominant individuals mark
much more than subordinates (Peters & Mech, 1975; Bekoff, 1979). One could
imagine that subordinates might alter their marking based on the presence or
absence of a more dominant individual, but I am not aware of any data that
explicitly supports this speculation. In some group-living species, such as dwarf
mongooses Helogale undulata, all members of the group may mark in the same
place, especially when first emerging from the burrow in the morning (Rasa, 1973).
Marking by some individuals could be stimulated by observing others mark (social
facilitation) or by the mere presence of others in the group at that time of day (a
possible audience effect). Another situation in which an audience effect might be
observed is in cases in which the scent-marking behaviour serves as a visual signal
as well as a means of depositing a chemical signal. For example, males among
all species of gazelles engage in stereotyped visual displays when marking their
territories. This type of marking has been called ‘demonstrative marking’ because
of its obvious nature and probable value as a simultaneous visual and olfactory
signal (Estes, 1967). Again, one could hypothesize that the occurrence or vigour
of such marking displays might vary depending on the relationships between the
marker and specific males that were present or the presence, or absence, of females.
I am not aware of any data that have been analysed in this context, however. The
alpha male and female dogs of a pack are more likely to mark with a raised leg
urination after observing another dog do so than are lower-ranking individuals
(Bekoff, 1979), but again it is not clear if this is a competitive reaction to observing
the mark or is dependent on the presence of the audience of the other members
of the pack.
Networks, cognition and individual recognition: speculations on
species differences in underlying mechanisms
The notion of a communication network is important because it empha-
sizes that individuals are a part of a community of interacting individuals (for an
early version of this view, see Estes (1969)). This is true even in species in which in-
dividuals spend most of their time by themselves. The concepts of eavesdropping
and audience effects draw attention to two specific ways that the social context
can modulate the behaviour of individuals. Although evidence demonstrating
eavesdropping or audience effects does not imply anything specific about the
mechanisms underlying these effects, most researchers working in this area seem
to assume that the animals they work with treat one another as unique individuals
with unique sets of distinctive characteristics. This is an inference about cognitive

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
364 R. E. Johnston
and neural processes that I will refer to as true individual recognition or having
a concept of other individuals. One important aspect of having such an attribute
is that animals should have memories consisting of several types of information
about other individuals and these memories should be stored as organized units
or representations of individuals (Johnston & Jernigan, 1994; Johnston & Bullock,
2001). This type of representation contrasts with other, simpler mechanisms that
are nonetheless sufficient to explain many of the findings that are referred to
as individual recognition, neighbour recognition, etc. Although the mechanisms
involved in recognition and memory are not directly observable, it is possible
to characterize these mechanisms by appropriate behavioural and physiological
measurements. The fields of cognitive psychology and cognitive neuroscience, for
example, depend on this kind of analysis. With regard to recognition of individ-
uals by non-human animals, for example, it is possible to discover the kinds of
information that one animal knows about another (Johnston & Bullock, 2001). Fur-
ther, this information can help to explain why individuals behave the way they
do. Indeed, the complexity of an individual’s knowledge about other individuals
is likely to have a profound influence on how that individual behaves in social
interactions with them (e.g. Chs. 24 and 25).
I have previously argued that many demonstrations of individual recognition
in the field and in the laboratory do not allow us to discriminate between merely
recognizing familiar versus unfamiliar cues or combinations of cues and recogniz-
ing individuals as unique entities (Johnston & Jernigan, 1994; Johnston & Bullock,
2001). For example, in the literature on neighbour recognition by song in birds,
the majority of studies merely demonstrate that a territorial male responds more
strongly to a novel song than a familiar song, or a novel song–direction combi-
nation than to a familiar song–direction combination. They do not provide proof
that the birds recognize their neighbours as individuals. A slightly more complex
mechanism might be categorization of information into heterogeneous categories
(e.g. a frequently heard song versus a rarely heard song) but again, not a catego-
rization based on individuals as the unit of analysis (Barrows et al.,1975; Caldwell,
1985). Cases in which birds engage in repertoire matching, in contrast, suggest the
existence of true individual recognition. That is, when a song sparrow Melospiza
melodia hears a neighbour sing one song type and then responds with a song
type that he shares with this neighbour but that is different from the one that the
neighbour just sang (Beecher et al., 1996), this suggests that the male knows several
characteristics of his neighbour and that they differ for different neighbours. This
type of observation provides evidence that song sparrows respond to neighbours
as unique individuals. Another example that demonstrates complex, multicom-
ponent representations of individuals comes from work with golden hamsters, in
which we showed that after males had interacted with several females they had

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 365
memories of these females that incorporated at least three different odours. Using
an habituation task, we showed that habituation to one of these odours resulted
in habituation to the other odours as well. That is, when becoming habituated to a
particular stimulus the males also became habituated to the individual and thus
other features of this individual (Johnston & Jernigan, 1994; Johnston & Bullock,
2001). These effects do not occur if the males have not interacted with the stimulus
females and, therefore, have not had an opportunity to learn about the features
of these females; that is, the effects are not a result of inherent similarities across
odours and consequent generalization across odours.
Similarly, an experimental demonstration of eavesdropping may result from
true individual recognition or it could result from a simpler mechanism. For
example, if one male fighting fish observes a fight between two other males of
the species, he could remember that fish A with the purple fringe on this dorsal
fin and red stripes on his tail and a distinctive wiggle in his display is much
more aggressive than fish B, characterized by all red fins and tail but a purple
spot in the middle of the tail fin. The observer fish could have memories of these
two individuals, and each memory would consist of an integrated memory of
that male’s physical and behavioural characteristics. A simpler type of memory
would, however, also be sufficient to explain the effects. The observer fish might
associate ‘purple edge above and red stripes in back’ with fear and ‘all red’ with
lack of fear. He could learn an association between a few specific cues and fear
or danger but not have memories of individuals as such. Both types of memory
would result in differential responses to the two individuals, but the mechanisms
underlying the responses would be different and indicate different levels of neural
and cognitive complexity. Additional experiments could provide evidence for or
against the existence of true individual recognition. For example, does the subject
react differently to two individuals that were observed to have similar experiences
(e.g. lost a fight) but some aspects of the interaction were different (e.g. how quickly
the animal lost)? Does the subject react differently to the two individuals in a
non-aggressive context? Does the subject cross-habituate to different features (e.g.
odours, sounds, other visual information) from the same individuals (Johnston &
Jernigan, 1994; Johnston & Bullock, 2001)?
There is probably a continuum of complexity in the types of representation that
animals have of other individuals, and these differences in complexity should cor-
relate roughly with the complexity of the social behaviour of different species
and taxonomic groups. At higher levels of complexity are species that not only
remember individuals as such but also remember something about the relation-
ships between different individuals (Cheney & Seyfarth, 1990a, b; Chs. 24 and 25).
At the most complex end of the continuum would be species, such as humans and
perhaps some other highly social animals, that have partially or well-developed

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
366 R. E. Johnston
abilities to understand that other individuals have different knowledge or inten-
tions than themselves; that is, they have a so-called ‘theory of mind’ (Premack &
Woodruff, 1978; Wimmer & Perner, 1983; Cheney & Seyfarth, 1990a,b; Ch. 25). One
crucial task is to identify the simplest level of representation that can explain a
particular phenomenon because any phenomena could be explained by a more
complex mechanism. Identifying the simplest mechanism that could be used can
provide a starting place for a taxonomy of the kinds of representation that indi-
viduals have of other individuals, species differences in these representations and
hypotheses about the neural mechanisms underlying them. Ultimately, it may
help us to understand the evolution of social behaviour.
The complexity of representations that individuals have of other individuals is
important in the context of communication networks because these representa-
tions will influence the way in which individuals interact with others, the kinds
of information they extract from observing interactions between others and the
effects that such information has on their own behaviour. This, in turn, will influ-
ence the nature of the networks that develop and the ways that individuals interact
within those networks (Chs. 24 and 25). I suggest that animals with complex, in-
tegrated representations of individuals will have a number of advantages over
those with simpler mechanisms. For example, complex representations contain
more information and should reduce errors in recognition, especially recognition
over long intervals during which changes in relevant cues may have occurred
through age, injury, nutrition or hormonal status. More complex representations
may also facilitate the evaluation of relationships between two or more individuals
obtained via observation of interactions.
Summary and future directions
The observations described in this chapter suggest that individuals in
some species obtain information from over-marks about the relative quality of
the individuals that deposited these marks and that this information influences
subsequent responses to those individuals, thus providing evidence for eavesdrop-
ping (but see Ch. 11). In addition, some species appear to have specialized sensory
mechanisms for the evaluation of scent over-marks and specialized mechanisms
that promote accurate placement of a scent mark over the scent of another in-
dividual (targeting). At present there is little evidence for audience effects on
scent marking, but this may be because few observers have looked for such ef-
fects. More studies are desperately needed on the functions of scent over-marking,
eavesdropping on over-marks and the effects of such eavesdropping, especially
observations and experiments in natural settings. Also important are compara-
tive studies on a set of related species with a rich, diverse repertoire of scent-
marking behaviours. Little is understood about the variety of functions that scent

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 367
over-marking has or about the kinds of information that may be obtained from
such over-marks.
Studies of eavesdropping on singing interactions in birds suggest some inter-
esting possibilities for further experiments with scent over-marking. For exam-
ple, use of information gathered by eavesdropping to direct extra-pair behaviour
(Mennill et al., 2002; Chs. 2 and 7). Among mammals, similar behaviour may be
found in monogamous pairs. In species in which females usually mate with sev-
eral partners, individual females might reduce their interactions with additional
males if the first mating partner was known to be highly successful in over-mark
competitions; in contrast, females that live near males that are less successful in
over-marking competitions might be more vigorous in advertising for, or in seek-
ing out, other males. That is, a female might be influenced by knowledge of the
history of over-marking interactions between numerous males in her vicinity.
Acknowledgements
Thanks to E. Regan, Peter McGregor, Jane Hurst and two anonymous reviewers for
comments on the manuscript. Thanks to Joan Johnston for help with graphics and other technical
help.
References
Bagneres, A.-G., Killian, A., Clement, J.-L. & Lange, C. 1991. Interspecific recogntion
among termites of the genus Reticulitermes: evidence for a role for the cuticular
hydrocarbons. Journal of Chemical Ecology, 17, 2397–2420.
Barrows, E. M., Bell, W. J. & Michner, C. D. 1975. Individual odor differences and their
social functions in insects. Proceedings of the National Academy of Sciences, USA, 72,
2824–2828.
Beecher, M. D., Stoddard, P. K., Campbell, S. E. & Horning, C. L. 1996. Repertoire
matching between neighboring song sparrows. Animal Behaviour, 51, 917–923.
Bekoff, M. 1979. Scent marking by free-ranging domestic dogs: olfactory and visual
components. Biology of Behavior, 4, 123–139.
Bronson, F. H. 1976. Urine marking in mice: causes and effects. In: Mammalian
Olfaction, Reproductive Processes and Behavior, ed. R. L. Doty. New York: Academic
Press, pp. 119–141.
Brown, R. E. & Macdonald, D. W. 1985. Social Odours in Mammals, Vols. 1 & 2. Oxford:
Oxford University Press.
Byers, J. A. & Bekoff, M. 1986. What does ‘kin recognition’ mean? Ethology, 72, 342–345.
Caldwell, R. L. 1985. A test of individual recognition in the stomatopod, Gonodactylus
festae. Animal Behaviour, 33, 101–106.
Cheney, D. L. & Seyfarth, R. M. 1990a. How Monkeys See the World. Chicago, IL: University
of Chicago Press.
1990b. The representation of social relations by monkeys. Cognition, 37, 167–196.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
368 R. E. Johnston
Cohen, A. B., Johnston, R. E. & Kwon, A. 2001. How golden hamsters (Mesocricetus
auratus) discriminate top from bottom flank scents in over-marks. Journal of
Comparative Psychology, 115, 241–247.
Eisenberg, J. F. & Kleiman, D. 1972. Olfactory communication in mammals. Annual
Review of Ecology and Systematics, 3, 1–31.
Estes, R. D. 1967. The comparative behavior of Grant’s and Thomson’s gazelles. Journal
of Mammalogy, 48, 189–209.
1969. Territorial behavior of the wildebeest (Connochaetes taurinus Bruchell, 1823).
Zeitschrift fur Tierpsychologie, 26, 284–370.
Ferkin, M. H., Dunsavage, J. & Johnston, R. E. 1999. What kind of information do
meadow voles, Microtus pennsylvanicus, use to distinguish between the top and
bottom scent of an overmark? Journal of Comparative Psychology, 113, 43–51.
Fischer, R. B. & McQuiston, J. 1991. A possible role for Syrian hamster, Mesocricetus
auratus, vaginal secretion in inter-female competition. Animal Behaviour, 42,
949–954.
Fisher, H. S., Swaisgood, R. R. & Fitch-Snyder, H. 2003. Countermarking by male pygmy
lorises (Nycticebus pygmaeus): do females use odor cues to select mates with high
competitive ability? Behavioral Ecology and Sociobiology, 53, 123–130.
Gamboa, G. J., Grudzien, T. A., Espelie, K. E. & Bura, E. A. 1996. Kin recognition
pheromones in social wasps: combining chemical and behavioural evidence.
Animal Behaviour, 51, 625–629.
Gorman, M. L. 1976. A mechanism for individual recognition by odour in Herpestes
auropunctatus (Carnivora: Viverridae). Animal Behaviour, 24, 141–145.
1990. Scent marking strategies in mammals. Revue suisse de Zoologie, 97, 3–29.
Gorman, M. L. & Mills, M. G. L. 1984. Scent marking strategies in hyaenas (Mammalia).
Journal of Zoology, 202, 535–547.
Gosling, L. M. & Roberts, S. C. 2001. Scent-marking by male mammals: cheat-proof
signals to competitors and mates. Advances in the Study of Behavior, 30, 169–217.
Gosling, L. M., Roberts, S. C., Thorton, E. A. & Andrew, M. J. 2000. Life history costs of
olfactory status signalling in mice. Behavioral Ecology and Sociobiology, 48,
328–332.
Heymann, E. W. 1998. Sex differences in olfactory communication in a primate, the
moustached tamarin, Saguinus mystax (Callitrichinae). Behavioral Ecology and
Sociobiology, 43, 37–45.
Hurst, J. L. 1990. Urine marking in populations of wild house mice Mus domesticus
Rutty. II. Communication between females. Animal Behaviour, 40, 223–232.
Hurst, J. L. & Rich, T. J. 1999. Scent marks as competitive signals of mate quality. In:
Advances in Chemical Signals in Vertebrates, ed. R. E. Johnston, D. Muller-Schwarze &
P. W. Sorensen. New York: Kluwer Academic/Plenum, pp. 209–225.
Johnson, R. P. 1973. Scent marking in mammals. Animal Behaviour, 21, 521–535.
Johnston, R. E. 1975a. Scent marking by male hamsters: I. Effects of odors and social
encounters. Zeitschrift fur Tierpsychologie, 37, 25–98.
1975b. Scent marking by male hamsters II. The role of flank gland odor in the
causation of marking. Zeitschrift fur Tierpsychologie, 37, 138–144.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 369
1975c. Scent marking by male hamsters III. Behavior in a semi-natural environment.
Zeitschrift fur Tierpsychologie, 37, 213–221.
1977. The causation of two scent marking behaviour patterns in female golden
hamsters. Animal Behaviour, 25, 317–327.
1979. Olfactory preferences, scent marking and ‘proceptivity’ in female hamsters.
Hormones and Behavior, 13, 21–39.
1985. Communication. In: The Hamster: Reproduction and Behavior, ed. H. I. Siegel. New
York: Plenum Press, pp. 121–124.
1990. Chemical communication golden hamsters: from behavior to molecules and
neural mechanisms. In: Contemporary Issues in Comparative Psychology, ed. D. A.
Dewsbury. Sunderland, MA: Sinauer, pp. 381–409.
1993. Memory for individual scent in hamsters (Mesocricetus auratus) as assessed by
habituation methods. Journal of Comparative Psychology, 107, 201–207.
1995. Responses to individual signatures in scent counter-marks: a striking new
phenomenon. In: Chemical Signals in Vertebrates 7, ed. R. Apfelbach, D.
Muller-Schwarze, K. Reutter & E. Weiler. Oxford: Elsevier, pp. 361–369.
1999. Scent over-marking: how do hamsters know whose scent is on top and why
should it matter? In: Advances in Chemical Communication in Vertebrates, ed. R. E.
Johnston, D. Muller-Schwarze & P. W. Sorensen. New York: Kluwer
Academic/Plenum, pp. 227–238.
Johnston, R. E. & Bhorade, A. 1998. Perception of scent over-marks: novel mechanisms
for determining which individual’s mark is on top. Journal of Comparative
Psychology, 112, 230–243.
Johnston, R. E. & Bullock, T. A. 2001. Individual recognition by use of odours in golden
hamsters: the nature of individual representations. Animal Behaviour, 61,
545–557.
Johnston, R. E. & Jernigan, P. 1994. Golden hamsters recognize individuals, not just
individual scents. Animal Behaviour, 48, 129–136.
Johnston, R. E. & Schmidt, T. 1979. Responses of hamsters to scent marks of different
ages. Behavioral and Neural Biology, 26, 64–75.
Johnston, R. E., Derzie, A., Chiang, G., Jernigan, P. & Lee, H.-C. 1993. Individual scent
signatures in golden hamsters: evidence for specialization of function. Animal
Behaviour, 45, 1061–1070.
Johnston, R. E., Chiang, G. & Tung, C. 1994. The information in scent over-marks of
golden hamsters. Animal Behaviour, 48, 323–330.
Johnston, R. E., Munver, R. & Tung, C. 1995. Scent counter marks: selective memory for
the top scent by golden hamsters. Animal Behaviour, 49, 1435–1442.
Johnston, R. E., Sorokin, E. S. & Ferkin, M. H. 1997a. Female voles discriminate males’
over-marks and prefer top-scent males. Animal Behaviour, 54, 679–690.
1997b. Scent counter-marking by male meadow voles: females prefer the top-scent
male. Ethology, 103, 443–453.
Jolly, A. 1966. Lemur Behavior. Chicago, IL: University of Chicago Press.
Kappeler, P. M. 1998. To whom it may concern: the transmission and function of
chemical signals in Lemur catta. Behavioral Ecology and Sociobiology, 42, 411–421.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
370 R. E. Johnston
Lazaro-Perea, C., Snowdon, C. T. & Arruda, M. F. 1999. Scent-marking behavior in wild
groups of common marmosets (Callithrix jacchus). Behavioral Ecology and
Sociobiology, 46, 313–324.
Lisk, R. D., Ciaccio, L. A., & Catanzaro, C. 1983. Mating behaviour of the golden
hamster under seminatural conditions. Animal Behaviour, 31, 659–666.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Mertl, A. S. 1977. Habituation to territorial scent marks in the field by Lemur catta.
Behavioral Biology, 21, 500–507.
Mills, M. G. L., Gorman, M. L. & Mills, M. E. J. 1980. The scent marking behaviour of the
brown hyena Hyaena brunnea. South African Journal of Zoology, 15, 240–248.
Moore, J. D. & Byers, J. A. 1989. The function of scent marking by males on female
urine in pronghorns. Journal of Mammalogy, 70, 812–814.
Peters, R. & Mech, L. D. 1975. Scent marking in wolves. American Scientist, 63,
628–637.
Powzyk, J. A. 2002. Scent wars. Natural History, 111, 82–85.
Premack, D. & Woodruff, G. 1978. Does the chimpanzee have a theory of mind?
Behavioral and Brain Sciences, 4, 515–526.
Ralls, K. 1971. Mammalian scent marking. Science, 171, 443–449.
Rasa, O. A. E. 1973. Marking behaviour and its social significance in the African dwarf
mongoose, Helogale undulata rufula. Zeitschrift fur Tierpsychologie, 32, 293–318.
Rich, T. J. & Hurst, J. L. 1998. Scent marks as reliable signals of the competitive ability
of mates. Animal Behaviour, 56, 727–735.
1999. The competing countermarks hypothesis: reliable assessment of competitive
ability by potential mates. Animal Behaviour, 58, 1027–1037.
Roberts, S. C. 1998. Behavioural responses to scent marks of increasing age in
klipspringer Oreotragus oreotragus. Ethology, 104, 585–592.
2000. Female territoriality and the function of scent-marking in a monogamous
antelope (Oreotragus oreotragus). Behavioral Ecology and Sociobiology, 47, 417–423.
Singer, A. G., Beauchamp, G. K. & Yamazaki, K. 1997. Volatile signals of the major
histocompatibility complex in male mouse urine. Proceedings of the Naional
Academy of Sciences, USA, 94, 2210–2214.
Sliwa, A. & Richardson, P. R. K. 1998. Responses of aardwolves, Proteles cristatus,
Sparrman 1783, to translocated scent marks. Animal Behaviour, 86, 137–146.
Smith, T. E., Tomlinson, A. J., Mlotkiewicz, J. A. & Abbott, D. H. 2001. Female marmoset
monkeys (Callithrix jaccus) can be identified from the chemical composition of
their scent marks. Chemical Senses, 26, 449–458.
Steel, E. 1984. Effect of the odour of vaginal secretion on non-copulatory behaviour of
male hamsters (Mesocricetus auratus). Animal Behaviour, 32, 597–608.
Stowe, M. K., Turlings, T. C. J., Loughrin, J. H., Lewis, W. J. & Tumlinson, J. H. 1995. The
chemistry of eavesdropping, alarm and deceit. Proceedings of the National Academy of
Sciences, USA, 92, 23–28.
Vandenbergh, J. G. 1973. Effects of gonadal hormones on the flank gland of the golden
hamster. Hormone Research, 4, 28–33.

P1: GFZ/... P2: JZZ/...0521823617c16.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:57
Eavesdropping and scent over-marking 371
Wilcox, R. M. & Johnston, R. E. 1995. Scent counter-marks: specialized mechanisms of
perception and response to individual odors in golden hamsters, Mesocricetus
auratus. Journal of Comparative Psychology, 109, 349–356.
Wimmer, H. & Perner, J. 1983. Beliefs about beliefs: representation and constraining
function of wrong beliefs in young childrens’s understanding of deception.
Cognition, 13, 103–128.
Woodward, R. L. J., Bartos, K. & Ferkin, M. H. 2000. Meadow voles (Microtus
pennsylvanicus) and prairie voles (M. ochrogaster) differ in their responses to
over-marks from opposite- and same-sex conspecifics. Ethology, 106, 979–992.

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
17
Vocal communication networks in largeterrestrial mammals
k a r e n m c c o m b & dav i d r e b y
University of Sussex, Falmer, Brighton, UK
Introduction
Many mammals give long-range calls that can be received over wide areas,
often containing large numbers of receivers. In the case of mammals with fluid so-
cial systems, opportunities for exposure to the calls of others are further enhanced
by the movement of individuals with respect to one another. In our chapter, we
discuss the relevance of eavesdropping and communication networks in a range
of mammal species, first considering how these concepts apply in cases where
loud calls are used to exchange social information in static territorial and fluid
fission–fusion societies, and then exploring their potential importance where
mammals use loud sexual calls to broadcast information about resource-holding
potential. We also outline the mechanisms by which information in mammalian
calls is encoded, broadcast and acquired, and we consider the possible fitness con-
sequences that attending to calling interactions can confer. Finally, we evaluate
how the vocal communication networks described for non-human mammals differ
from human communication networks and discuss possible explanations for these
differences.
When mammals give loud calls, the area over which the signal can be re-
ceived is potentially extensive. Such calls are typically emitted at high sound-
pressure levels (greater than 100 dB at 1 m) and while spherical spreading and
excess attenuation from the environment eventually result in the signal being
engulfed in background noise, it often remains intelligible over distances of
several kilometres from the source: for example the calls of lions Panthera leo
(Ogutu & Dublin, 1998; Funston, 1999; K. McComb, unpublished data), hyaenas
Crocuta crocuta (Ogutu & Dublin, 1998) and elephants Loxodonta africana (McComb
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
372

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 373
et al., 2003). Many mammals occur at relatively high densities; therefore, the ac-
tive space may contain a large number of potential receivers. In addition, some
mammals live in fluid social systems where there are unusual opportunities for
exposure to vocal signals from others in the population. In such social systems,
where the identity of immediate neighbours constantly changes because of the
movement of individuals and groups in relation to one another, an individual may
broadcast to and receive signals from a larger section of the overall population
than is normally possible in territorial systems. These two important characteris-
tics of mammal signalling systems have the potential to generate a much wider
audience for the signaller.
On the basis of the characteristics outlined above, it seems appropriate to con-
ceptualize the production and perception of mammal loud calls in the context
of an array of several receivers (after McGregor & Dabelsteen, 1996). Indeed we
will argue, based on the examples presented in our review, that this is the best
way to view communication involving loud calls. There is direct evidence that
mammals attend to vocal signals that are not explicitly directed at them. For ex-
ample, elephant contact calls, although directed at family and bond group mem-
bers, are attended to by others in the population, who exhibit knowledge of these
calls and adjust their social behaviour on the basis of them (McComb et al., 2000).
Diana monkeys Cercopithecus diana attend to the alarm calls of another primate
(Campbell’s monkey Cercopithecus campbelli) and appear to obtain functionally rel-
evant information from the detailed combination of different vocalizations used
(Zuberbuhler, 2002, see also below). However, in considering the extent to which
this form of audience effect (Ch. 4) in loud-calling mammals involves ‘eavesdrop-
ping’ or constitutes a ‘communication network’, three important issues need to
be considered.
First, eavesdropping in the context of animal signalling has been defined as
‘extracting information from an interaction between other individuals’(McGregor
& Dabelsteen, 1996; see also Ch. 2). This is a technical definition of a term that
in colloquial usage implies more specifically that receivers ‘secretly listen to a
conversation’(Concise Oxford English Dictionary). While behavioural acts constituting
secrecy or deception are notoriously difficult to identify in mammals (Semple &
McComb, 1996), it seems important to distinguish between cases of eavesdropping
in which transmission of information to receivers other than the main recipient
would be selected for and those where it would not. This can be achieved by
conducting cost–benefit analyses of particular signalling interactions (e.g. Ch. 3).
Second, an important aspect of the McGregor & Dabelsteen (1996) definition
of eavesdropping is that it involves extracting information from ‘an interaction’
rather than simply attending to the call itself. At this stage, evidence that mammals
attend to the signalling interaction (rather than just the signal) is very sparse. An

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
374 K. McComb & D. Reby
isolated example is provided by the studies of Cheney and colleagues (Cheney
et al., 1995; Ch. 25) on baboons Papio cynocephalus ursinus, which demonstrated
that receivers distinguish between appropriate and anomalous vocal exchanges
between dominants and subordinates in their group. However, lack of evidence
for attention to the vocal interaction itself may be more apparent than real. Few
researchers other than Cheney and colleagues have conducted the appropriate
experiments specifically to examine this phenomenon.
Finally, while it is certainly the case that a system of mammal communication
linking signallers to several receivers has some properties of a network, it lacks
others. A network can be viewed simply as ‘a system of interconnected people
or things’. However, advanced networks such as human social networks or the
Internet are generally understood to involve the passage of information from one
remote part of the network to another, via intermediate recipients that pass on
information to other individuals. Non-human vocal communication (including
bird and mammal communication systems) appears to fall short of this and we
will consider possible explanations for this in the course of the review.
Loud calls and social behaviour
Availability of information in a simple territorial system
The typical nature of mammal loud calls that are used to mediate so-
cial behaviour suggests that selection for concealing information from unwanted
receivers has not been paramount. Their high sound-pressure level, abrupt on-
set and broadband (often noisy) nature are properties that would be expected to
make them easy to detect and locate by listeners (e.g. Brown et al., 1979, 1980).
Given that such calls often function to advertise territory ownership, it would
usually be advantageous for them to attract the attention of any conspecifics in
the vicinity. There is evidence from a range of mammals that individuals can
distinguish between familiar and unfamiliar callers, even where receivers are
separated from signallers by large distances relative to the size of the animal:
for example pikas Ochotona princeps (Conner, 1985), cotton-top tamarins Sanguinus
oedipus (Snowdon et al., 1983) mangabeys Cercocebus albigena (Waser, 1977), rhesus
macaques Macaca mulatta (Rendall et al., 1996), wolves Canis lupus (Tooze et al., 1990),
and lions (McComb et al., 1993; Grinnell & McComb, 2001).
Where calls characteristics are adapted for long-distance transmission and are
easy to locate, the only mechanism for withholding information from unwanted
receivers would be to suppress calling altogether. The potential for loud calls to
attract unwanted attention may well be considerable in social mammals that use
loud calls not only in territorial defence but also to maintain contact with widely
spaced social companions: for example wolves (Harrington & Mech, 1979), lions

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 375
(McComb et al., 1994; Grinnell & McComb, 1996), and chimpanzees Pan troglodytes
(Mitani & Nishida, 1993). Here particular classes of individual that cannot afford
to risk escalated encounters with competitors that might hear their calling could
benefit by remaining silent even though, by doing so, they would forfeit the ben-
efits of coordinating their movements with members of their own social group.
The behaviour of free-ranging nomadic male lions in the Serengeti National Park
is consistent with these predictions (Grinnell & McComb, 2001).
Prides of African lions consist of matrilineal kin groups of females, their depen-
dent offspring and a coalition of resident males that enter the pride from outside
(Packer et al., 1988). In the pride, both sexes use loud calls (roaring) to advertise
ownership of a territory and to stay in contact with other members of their so-
cial group (Schaller, 1972; McComb et al., 1994; Grinnell et al., 1995; Grinnell &
McComb, 1996). At any one time, however, a high proportion of male lions in the
population are not in possession of a pride. These ‘nomads’wander widely, passing
through pride ranges singly or in coalitions until they are successful in taking over
a pride of their own (Hanby & Bygott, 1987; Pusey & Packer, 1987). It is crucial for
nomadic males to gain and maintain social bonds with their companions while
they wander because success in competition for prides is strongly dependent on
group size (Bygott et al., 1979; Grinnell et al., 1995). Roaring provides a means by
which nomadic males might coordinate their movements with coalition partners
or recruit new ones (see also McComb et al., 1994; Grinnell et al., 1995). However,
if nomads used this loud, long-distance signal to communicate with social com-
panions, they would also advertise their position to resident males in the area.
Nomadic males are likely to pay high costs if they attract the attention of
resident males in the area. Resident males have been consistently shown to ap-
proach aggressively playbacks of roaring from strange males that are broadcast in
their territories (Grinnell et al., 1995; Grinnell & McComb, 2001) and intercoalition
encounters can be fatal (Schaller, 1972; Grinnell et al., 1995). Given these costs, no-
mads might benefit by reducing their rate of roaring or even abandoning roaring
altogether and concealing their presence – despite the detrimental effects that
this would be likely to have on their ability to maintain contact with coalition
partners and attract potential mates. Grinnell & McComb (2001) found that in the
Serengeti population only male lions that were resident in a pride ever roared.
Nomadic males were never observed roaring when they were followed at night,
even when they became separated from their coalition partners. They also failed
to roar when played recordings of unfamiliar males roaring. In contrast, resident
males maintained a high rate of roaring in both these circumstances.
There are two possible explanations for why nomadic male lions fail to roar:
first, non-resident males gain no benefits from roaring and so never do so; second,
non-resident males could benefit from roaring, particularly by enhancing their

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
376 K. McComb & D. Reby
ability to recruit and maintain contact with coalition partners (McComb et al.,
1994; Grinnell et al., 1995), but the costs of engaging in this behaviour outweigh
the benefits. If the first explanation is true, then nomadic males should never roar
under any circumstances, whereas the second explanation predicts that nomadic
males will only roar when the probability of incurring costs, specifically attracting
the attention of resident males in the area, is low. Observational studies at other
field sites suggest that the second explanation is correct (Grinnell & McComb,
2001). Funston (1999), working on nomadic male lions in Kruger National Park,
South Africa, found that nomadic males do sometimes roar, but at greatly reduced
rates in comparison with resident males. Of the three nomadic coalitions that he
followed, one was explicitly noted to spend most of their time in an area without
resident males and thus where the social costs to roaring would be reduced. In
addition, observations by Grinnell in Pilanesberg National Park, South Africa sug-
gested that here, too, nomadic males roar when local resident males are unlikely to
hear them. Pilanesberg is an ancient volcanic caldera that contains valleys which
are acoustically isolated from each other by mountainous ridges. A non-resident
male coalition was observed roaring in one of these valleys that was not occupied
by resident males or females (Grinnell & McComb, 2001). It is also important to
note that, while nomadic males in the Serengeti study did not roar, males that
had been nomadic were seen to begin roaring as soon as they launched a chal-
lenge for ownership of a pride (Grinnell & McComb, 2001). This emphasizes that
roaring is a flexible behaviour that signallers may have been selected to adjust ac-
cording to the potential costs and benefits of revealing information on location to
listeners.
There are reports from other species of low signalling rates, or suppression of
signals altogether, in situations in which conspicuous signals could attract the
attention of potential aggressors. Chimpanzees have been observed to remain un-
usually quiet during excursions into the territories of other communities (Goodall,
1986) and, when they hunt monkeys (Colobus and Cercopithecus spp.), are reported
to fall silent on hearing the prey’s calls (Boesch & Boesch-Achermann, 2000). Lone
wolves howl less than do territorial pairs and packs (Harrington & Mech, 1979).
Similarly, transient coyotes Canis latrans howl at greatly reduced rates compared
with residents when passing through others’ territories (Gese & Ruff, 1998). It is
interesting that resident male lions may also adopt an apparently stealthy strat-
egy when ranging outside their territory. Grinnell & McComb (2001) noted that
resident males that had ventured well beyond their territory boundaries never
roared even when missing male companions. Long-distance signalling may well
be controlled in similar ways in other social species where eavesdroppers can
impose high costs on signalling (see also Ch. 4). Recent work on transient killer
whales (Orcinus orca) suggests that these animals adjust their calling behaviour to

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 377
minimize the costs of being detected by their acoustically sensitive mammalian
prey (Deecke, 2003).
Availability of information in mammals with fluid social systems
Above we have presented evidence that information on caller character-
istics such as identity is potentially available over quite long distances in species
that use loud calls for social communication within territorial systems. Nonethe-
less, degradation of calls with distance from the source will eventually result in
such information being engulfed in background noise and lost to receivers. In
certain mammal social systems, however, receivers are not limited to learning
only the calls of individuals in their own group or of particular territorial neigh-
bours within their hearing range. Some mammal societies are highly fluid, with
individuals and social units moving freely with respect to each other and ranging
widely. In these fluid societies, individuals pass through the signalling ranges of a
much larger number of conspecifics and are provided with opportunities to learn
to recognize the vocalizations of many more signallers than just their immediate
neighbours (see also discussions in Chs. 20 and 25). If mental capacities for storing
information on the identity of conspecifics’ signals are adequate, these circum-
stances would provide individuals with opportunities to become familiar with
the signals of many different conspecifics that form part of a widespread popula-
tion. Thus in mammals with fluid social systems, the unusually high encounter
rates that individuals have with conspecifics should interact with long-distance
signalling abilities to increase greatly the opportunities that receivers have for
learning to recognize the vocalizations of other individuals in the population. A
number of large mammals, including some primates (e.g. chimpanzees: Boesch
& Boesch-Achermann, 2000), cetaceans (e.g. sperm whales Physeter macrocephalus:
Whitehead et al., 1991) and African elephants use long-distance signals for social
communication and have fluid social systems.
In African elephants, the closest social relationships exist between members
of a family unit, typically composed of adult females that are matrilineal rela-
tives and their immature offspring, and between bond groups of families that
associate frequently and often greet one another when they meet (Moss & Poole,
1983). However, individual family units move freely with respect to one another
and range widely, frequently coalescing with other family units in the popula-
tion as they move and feed, thus forming highly fluid fission–fusion societies
(Moss & Poole, 1983). In a population of elephants in Amboseli National Park,
Kenya, with known life histories and ranging patterns, the extent to which fe-
male subjects were capable of recognizing others in the population through
long-distance contact calls was evaluated from playback experiments (McComb
et al., 2000). These experiments demonstrated that female African elephants not

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
378 K. McComb & D. Reby
only give a characteristic reaction to the contact calls of family or bond group
members but also can discriminate between the calls of less-frequent associates,
distinguishing the calls of individuals in this category with whom they have
higher association indices from those with whom they have lower association
indices. Based on the association indices involved, McComb et al. (2000) estimated
that subjects would have to be familiar with the contact calls of a mean of 14
different families (including about 100 adult females) in order to perform this
discrimination.
Empirical studies of the extent of networks of vocal recognition in cetaceans
and primates, which are currently lacking, may reveal similar patterns. Networks
of vocal recognition are likely to be particularly extensive where individuals
are long lived and social knowledge can be accumulated over considerable time
periods.
Loud calls and sexual behaviour
Many large mammals have loud calls that function to attract individuals
of the opposite sex and advertise resource-holding potential to competitors in
the vicinity (e.g. Clutton-Brock & Albon, 1979; Tyack, 1981 ; McElligott et al., 1999).
These calls are often very conspicuous and seem specifically adapted for attracting
the attention of a wide audience. The loud reproductive calls of polygynous deer,
which typically serve several functions, provide some of the best examples.
Loud mating calls in deer
Male red deer Cervus elaphus roar at high rates during the autumn breeding
season or rut and these loud vocalizations are known not only to affect the outcome
of contests between males (Clutton-Brock & Albon, 1979; Reby & McComb, 2003b)
but also to influence mate attraction (McComb, 1991) and advance ovulation in
females (McComb, 1987). There are consequently several receivers to whom male
roars might be relevant, including other males, the signaller’s own harem of fe-
males and other potential mates within hearing range. Video footage of red deer
stags orientating their responses to the roars of several neighbours with distinct
spatial locations clearly indicates that they take the complex spatial distribution
of callers around them into account (D. Reby & K. McComb, personal observation).
Moreover, it has been shown that female red deer are able to discriminate between
the roars of their own stag and those of surrounding males (Reby et al., 2001).
Finally, there is some evidence that when signallers would benefit by advertising
the outcome of their interactions, they use particularly conspicuous vocalizations.
Roaring bouts given during roaring competitions with rival stags and after chasing
hinds tend to contain a high proportion of ‘harsh roars’,which are unusually loud

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 379
and easy to locate and have an acoustic structure that emphasizes the caller’sbody
size (Reby & McComb, 2003b).
Groaning in fallow deer bucks Dama dama also appears adapted for more than
one category of receiver and each of these must be considered when modelling the
vocal behaviour of callers (McElligott & Hayden, 1999, 2001; McElligott et al., 1999).
McElligott et al. (1999) found that the bucks that achieved most matings were those
who had initiated vocal activity early in the season and who had remained vocal on
most days. This led the authors to conclude that females may discriminate between
males on the basis of long-term cumulative investment in vocal activity. However,
although rates of groaning were higher when females were present, males with
females exhibited higher groaning rates in the presence of nearby vocal males,
suggesting that the signal was also a threat aimed at male rivals (McElligott &
Hayden, 1999).
Given the several functions of deer vocalizations, it is clear that there are situa-
tions in which signals that would be beneficial in one context may be costly in an-
other: for example, when an individual male could gain reproductive advantages
by signalling to attract mates but in doing so would invite escalated contests with
male competitors. We have observed that young red deer stags (four to five year
old) who have gained access to a harem of females while the mature harem holder
is temporarily absent, and who have started to roar, will rapidly fall silent when
the harem holder returns, often dropping their heads to feed as he approaches
(K. McComb & D. Reby, personal observation). Similarly, playback experiments on
fallow deer (Komers et al., 1997) have shown that immature males decrease their
rate of groaning in response to playbacks of groans from mature males, whereas
mature males increase their groaning rates in this situation. Fallow deer bucks
may, therefore, adjust groaning rate in relation to several receivers, responding to
the complex balance between the benefits of deterring other males and displaying
to females and the costs of inviting contests with potentially stronger males in
the vicinity.
Since red and fallow deer rutting calls are individually distinct (McComb, 1988;
Reby, 1998; Reby et al., 1998), females and males may be able to recognize individual
callers from their vocalizations and accumulate knowledge on both a signaller’s
short-term vocal interactions with others and it’s long-term calling behaviour. Re-
search on red deer has revealed that females can discriminate between the roars
of their own stag and those of neighbouring harem holders (Reby et al., 2001). It
is possible that red deer hinds could receive information from roaring exchanges
and move between harems accordingly. Stags may also attend to contests between
other males for information on the body size and motivational state and adjust
their decisions to challenge harem holders on this basis. In this context, all re-
ceivers, whether they are directly involved in an interaction with the caller or not,

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
380 K. McComb & D. Reby
would benefit from attending to cues to resource-holding potential madeavailable
in this way. What is now required is empirical work to investigate directly the ex-
tent to which receivers attend to interactions in which they are not themselves
involved.
Loud mating calls in other mammals
Loud and acoustically complex sexual songs produced by humpback
whales Megaptera novoeangliae (Payne & McVay, 1971) and fin whales Balaenoptera
physalus (Croll et al., 2002) have the potential to travel unprecedented distances
underwater (e.g. Croll et al., 2002). While it is known that the individuals who give
these songs are male, the intended receivers have not yet been unambiguously
identified; they may be rival males, potential mates or both (Tyack, 1983; Mobley
et al., 1988; Noad et al., 2000; Croll et al., 2002). What is clear is that such vocaliza-
tions are detectable over vast tracts of ocean and may reach a much larger audience
than the sexual calls of terrestrial mammals discussed above. Male pinnipeds also
have loud sexual advertisement calls (e.g. Northern elephant seals Mirounga angu-
stirostris (Shipley et al., 1981, and common seals Phoca vitulina (van Parijs et al., 2000))
and calling interactions between males on land or underwater may be attended
to by rival males, potential mates or both. It remains to be seen whether receivers
alter their subsequent behaviour on the basis of which male dominates in a calling
interaction (see fuller discussion in Ch. 18).
Mammal anti-predator calls
In contrast to long-distance social and sexual calls given by large mam-
mals, alarm calls typically have acoustic features that would be expected to make
them difficult to locate. While these calls may be delivered at moderate ampli-
tudes, the information that they contain is likely to be available over shorter dis-
tances. Despite this, they are clearly attended to by a range of receivers, including
members of other mammal species (Schaller, 1967; Oda, 1997; Zuberbuhler, 2002).
In responding to the alarm calls of Campbell’smonkeys, Diana monkeys attend not
only to the referent of the alarm call, responding with their own species-specific
alarm call for the same predator, but also appear sensitive to the detailed composi-
tion of the alarm-calling sequence. In situations where the presence of a predator
is less threatening, Campbell’s monkeys emit a pair of ‘boom’ calls before their
alarm calls. Playbacks of Campbell’s alarm calls with booms did not elicit alarm
calls from Diana monkey subjects (Zuberbuhler, 2002).
Some anti-predator calls may have an even wider audience. A study of roe
deer Capreolus capreolus revealed that barks, previously identified as ‘alarm calls’,
in fact function to elucidate the cause of disturbance (Reby et al., 1999a). In this

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 381
communication system, calls inform any predator that might be present that it has
been detected and simultaneously reveal the caller’sidentity and status to any con-
specific (whether the latter is the cause of the disturbance or not). The likelihood of
barking in response to a predator-like disturbance is independent of the presence
of (related or unrelated) conspecifics in the close vicinity, demonstrating that it is
not an alarm call (Reby et al., 1999a). However, barking is contagious, with one in-
dividual’s barks often being followed by antiphonal calling behaviour from up to
seven neighbouring individuals of both sexes (Reby et al., 1999a). Since the acoustic
structure of the vocalization carries information on the sex, age and identity of
the caller (Reby et al., 1999b), barking may enable roe deer to identify and locate
each other, and possibly assess dominance status (particularly during counter-
barking sessions involving several animals). Playback experiments supported the
hypothesis that although barking may have initially evolved as an anti-predator
strategy it is also a signal attended to by conspecific receivers, in particular other
males during the territorial period (Reby et al., 1999a). Therefore, when a roe deer
barks, irrespective of the stimulus that elicits it (predator or conspecific), it reveals
its location, identity and status to a diverse audience of receivers, the composi-
tion of which will have marked effects on the costs and benefits associated with
calling.
Encoding of information on individuality and size
Within a network, the ability of individuals to determine each other’s
identity, physical status or internal state from signals dramatically increases the
level of functionally relevant information that is potentially exchanged. Whereas
in some cases identity may be inferred from the location of the caller or by using
visual or olfactory signals, acoustic cues are likely to be of primary importance
when individuals range widely. Such cues can provide receivers with instanta-
neous information on the location and attributes of the caller and may represent
the only effective signalling modality in nocturnal or forest-dwelling species. There
is a considerable body of evidence indicating that the vocalizations of terrestrial
mammals contain information on the identity and physical attributes of the caller
(see below).
In principle, individual differences can be present at several levels in the acous-
tic structure of the call. When mammals give voiced calls, the resultant sound
is the product of a source signal, generated in the larynx, that is subsequently
filtered in the cavities of the vocal tract (Fant, 1960). The source–filter theory of
voice production separates the source components, generated by the vibration of
the vocal folds, from the filter components, generated when certain frequencies
in the source spectrum are selectively amplified or filtered as the signal passes

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
382 K. McComb & D. Reby
through the supralaryngeal vocal tract. The characteristics of the source include
the duration of the call, its fundamental frequency, the periodicity of the signal,
its spectral slope and the presence of phenomena associated with non-linear dy-
namics, such as subharmonics, biphonation and deterministic chaos (Wilden et al.,
1998). Differences in these characteristics of call structure arise from variation in
subglottal pressure and in the length and shape of the vocal folds and their stress
and tension. All of these parameters can vary between individuals, either as a re-
sult of differences in the way the larynx is operated or simply because of random
variation in the morphology of callers. In comparison, the key characteristics of
the filter are the position and bandwidths of the formant frequencies, which de-
scribe the shape of the spectral envelope. Formant frequencies are determined by
the length and shape of the cavities of the vocal tract, namely the pharynx, mouth
and nasal cavities. Individual differences in formant frequencies can arise from
differences in vocal tract morphology or from variation in the way the shape of
the vocal tract is actively modified during vocalization (e.g. the extent of mouth
opening, lip rounding and vocal tract extension).
Variation in source and filter characteristics both appear to be important in
encoding individual identity in a range of large mammals. Differences in the
fundamental frequency contour have been identified as important in broadcast-
ing information on identity in wolves (Tooze et al., 1990) and elephants (McComb
et al., 2003), while individuality in formant frequencies has been demonstrated in
fallow deer (Reby et al., 1998), roe deer (Reby et al., 1999b), red deer (McComb, 1988;
Reby, 1998), elephants (McComb et al., 2003) and rhesus macaques (Rendall et al.,
1998). Filter characteristics, in particular the frequency spacing between succes-
sive formants, provide the most reliable cues to body size (Fitch, 1997; Riede &
Fitch, 1999; Reby & McComb, 2003a). In contrast, source characteristics, in partic-
ular fundamental frequency values, provide relatively poor information on size
(Masataka, 1994; Reby & McComb, 2003b) but are better indicators of age and sex
and may, therefore, reflect important variation in vocal fold length between sexes
and throughout the lifetime (Reby & McComb, 2003b).
It is important to appreciate that source and filter characteristics that have the
potential to provide receivers with information on caller identity can be distorted
or lost as distance from the signaller increases. Even where calls can theoretically
be transmitted over long distances because they possess acoustic characteristics
that are well adapted for sound transmission in a particular environment, it is
unsafe to conclude that receivers can extract socially relevant information from
degraded calls at these distances. In female African elephants, playback exper-
iments and re-recordings indicate that abilities for social recognition through
long-distance contact calls become limited when frequency components around
115 Hz become immersed in background noise (McComb et al., 2003). This typically

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 383
occurs at distances of 1–2 km from the caller, which is considerably shorter than
the propagation distances that have been proposed for calls with infrasonic funda-
mental frequencies (McComb et al., 2003). This finding highlights the importance
of considering the distances over which vocal signals within communication net-
works can propagate without losing their intelligibility to receivers, which are
not necessarily equivalent to the distances over which such signals are physically
detectable (see also Ch. 20).
Acquiring and storing information on vocal characteristics
Little is known about the factors that influence how effectively individ-
uals acquire and store information about their social companions, although it is
known that social knowledge, particularly that used in vocal recognition between
mothers and offspring, can be retained for several years even when individuals do
not encounter each other (Insley, 2000; McComb et al., 2000). In African elephants,
where adult females are familiar with the contact calls of a large proportion of
the population around them (see above), the key factor that affects social discrim-
ination abilities is the age of the oldest female in the group (McComb et al., 2001).
Playback experiments revealed that families with older matriarchs were signifi-
cantly better at discriminating the contact calls of genuine strangers from those
of more familiar associates than were families with younger matriarchs (McComb
et al., 2001). While families with older matriarchs were several thousand times
more likely to bunch into defensive formation when played the calls of families
they have encountered only rarely than when played the calls of families they
frequently associate with, families with younger matriarchs were only marginally
more likely to bunch (McComb et al., 2001). Log-linear analysis revealed that vari-
ables such as the number of other females present in the group, and their respec-
tive ages, did not affect vocal discrimination abilities. An additional factor that did
appear to be important was the rate at which subjects encountered other families
in the population. An elephant family unit directly encounters, on average, 25
other families over the course of the year in Amboseli National Park, and passes
within 1–2 km of 35, providing family members with plenty of opportunity to
become familiar with the calls of others. Recent analyses suggest that having a
high encounter rate with others in the population can enhance a family’sability to
identify the calls of genuine strangers, and that this may be particularly beneficial
for families with younger matriarchs (K. McComb, unpublished data).
The above results suggest that the age of one crucial individual, the oldest
female or matriarch, can affect the social knowledge of the group as a whole. Age
and experience are likely to affect abilities to acquire and store information on
vocal signals in other societies where animals are long lived and remain part of

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
384 K. McComb & D. Reby
a social network for many years. The social systems of some whales have strong
parallels with those of elephants (e.g. Pennisi, 2001) and Ford et al. (1994) noted
that in killer whales the death of the oldest female, from whom many of the
individuals are usually descended, may destabilize a pod. The effects of age and
other factors on abilities to recognize the vocalizations of conspecifics has not been
investigated for species that defend individual territories (rather than sharing a
range with matrilineal relatives as in elephants and some whales) and studies
of this sort are now required. Moreover, we as yet know nothing of the extent
to which large mammals develop knowledge of the mating calls of others in the
population and the factors that affect the acquisition of this knowledge.
The fitness consequences of attending to the calls of others
There is some evidence that attending to the exchange of social calls
between other individuals can confer fitness benefits on receivers. In African ele-
phants, where the matriarch appears to act as a repository for information on
the calls of others in the population (see above), families with older matriarchs
have greater reproductive success, at least some of which appears to derive from
superior social knowledge (McComb et al., 2001). The fitness consequences of at-
tending to vocal interactions involving sexual calls have never been quantified
for mammals but are likely to be highly significant. Acquiring information on
resource-holding potential by monitoring the outcome of vocal contests may al-
low receivers to assess rivals and potential mates much more accurately, and to
benefit from better decisions made as a consequence.
Summary and back to definitions
In light of the examples discussed above, the term communication net-
work can be usefully employed to identify sets of links between individuals (not
necessarily contiguous) that are known to each other through vocal signals or
that acquire information about each other’s interactions through vocalizations.
However, animal communication networks appear to be limited in a number of
important respects (see the Introduction to this chapter). In their typical form, they
describe overlapping lattices each consisting of three individuals: the signaller,
the intended receiver and an extraneous listener. In such systems, extraneous lis-
teners do not normally interact – in particular they do not pass on information
that they gain from attending to interactions. This is in stark contrast with hu-
man communication networks, where information can be transferred from one
remote part of the network to another and where intermediate recipients may
not be the ultimate receivers. Several key constraints on vocal communication

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 385
may prevent non-human mammals from sharing information in this more com-
plex way. These include the limited size of mammalian vocal repertoires, the
rarity of fully referential calls and the limited productivity possible in the ab-
sence of duality of patterning – a unique feature of human language whereby
phonemes can be combined into words and words into sentences (Hockett, 1960;
Pinker, 1994). These characteristics are likely to have been selected for in the
course of the massive expansion in sociality and social fluidity that occurred
during human evolution, creating an environment where the ability to use sym-
bolism and syntax to communicate about displaced activities would be of great
importance. It is important to appreciate that once these abilities had evolved,
the fitness benefits of attending to the calls of others would increase by orders of
magnitude.
Acknowledgements
The research described was funded by grants from BBSRC, NERC, the Royal Society (all
to K.M.) and INRA (to D.R.). David Reby was supported by Fyssen and E. U. Marie Curie Fellowships
and the University of Sussex. We also thank Vincent Janik, Peter McGregor and an anonymous
referee for helpful comments on the first draft of the chapter.
References
Boesch, C. & Boesch-Achermann, H., 2000. The Chimpanzees of the Tai Forest. Oxford:
Oxford University Press.
Brown, C. H., Beecher, M. D., Moody, D. B. & Stebbins, W. C. 1979. Locatability of vocal
signals in Old World monkeys: design features for the communication of
position. Journal of Comparative Physiological Psychology, 5, 806–819.
1980. Localization of noise bands by Old World monkeys. Journal of the Acoustical
Society of America, 68, 127–132.
Bygott, J. D., Bertram, B. C. R. & Hanby, J. P. 1979. Male lions in large coalitions gain
reproductive advantages. Nature, 282, 839–841.
Cheney, D. L., Seyfarth, R. M. & Silk, J. B. 1995. The responses of female baboons (Papio
cynocephalus ursinus) to anomalous social interactions: evidence for causal
reasoning? Journal of Comparative Psychology, 109, 134–141.
Clutton-Brock, T. H. & Albon, S. D. 1979. The roaring of red deer and the evolution of
honest advertisement. Behaviour, 69, 145–169.
Conner, D. A. 1985. Analysis of the vocal repertoire of adult pikas: ecological and
evolutionary perspectives. Animal Behaviour, 33, 124–134.
Croll, D. A., Clark C. W., Acevedo, A. et al. 2002. Only male fin whales sing loud songs.
Nature, 417, 809.
Deecke, V. B. 2003. The vocal behaviour of transient killer whales (Orcinus orca):
communicating with costly calls. Ph.D. Thesis, University of St Andrews.

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
386 K. McComb & D. Reby
Fant, G. 1960. Acoustic Theory of Speech Production. The Hague: Mouton.
Fitch, W. T. 1997. Vocal tract length and formant frequency dispersion correlate with
body size in rhesus macaques. Journal of the Acoustical Society of America, 102,
1213–1222.
Ford, J. K. B., Ellis, G. M. & Balcomb, K. C. 1994. Killer Whales. Vancouver: UBC Press;
Seattle, WA: University of Washington Press.
Funston, P. J. 1999. Predator–prey relationships between lions and large ungulates in
the Kruger National Park. Ph.D. Thesis, University of Pretoria.
Gese, E. M. & Ruff, R. L. 1998. Howling by coyotes (Canis latrans): variation among
social classes, seasons, and pack sizes. Canadian Journal of Zoology, 76,
1037–1043.
Goodall, J. 1986. The Chimpanzees of Gombe. Cambridge, MA: Belknap.
Grinnell, J. & McComb, K. 1996. Maternal grouping as a defense against potentially
infanticidal males: evidence from field playback experiments on African lions.
Behavioral Ecology, 7, 55–59.
2001. Roaring and social communication in African lions: the limitations imposed
by listeners. Animal Behaviour, 62, 93–98.
Grinnell, J., Packer, C. & Pusey, A. E. 1995. Co-operation in male lions: kinship,
reciprocity or mutualism? Animal Behaviour, 49, 95–105.
Hanby, J. P. & Bygott, J. D. 1987. Why do subadult lions leave their natal pride? Animal
Behaviour, 35, 161–169.
Harrington, F. H. & Mech, L. D. 1979. Wolf howling and its role in territory
maintenance. Behaviour, 68, 207–249.
Hockett, C. F. 1960. The origin of speech. Scientific American, 203, 88–96.
Insley, S. J. 2000. Long-term vocal recognition in the northern fur seal. Nature, 406,
404–405.
Komers, P. E., Pelabon, C. & Stenstrom, D. 1997. Age at first reproduction in male
fallow deer: age-specific versus dominance-specific behaviors. Behavioral Ecology, 8,
456–462.
Masataka, N. 1994. Lack of correlation between body size and frequency of
vocalisations in young female Japanese macaques (Macaca fuscata). Folia
Primatologica, 63, 115–118.
McComb, K. 1987. Roaring by red deer stags advances the date of oestrus in hinds.
Nature, 330, 648–649.
1988. Roaring and reproduction in red deer, Cervus elaphus. D. Phil. Thesis,
University of Cambridge.
1991. Female choice for high roaring rates in red deer, Cervus elaphus. Animal
Behaviour, 41, 79–88.
McComb, K., Pusey, A., Packer, C. & Grinnell, J. 1993. Female lions can identify
potentially infanticidal males from their roars. Proceedings of the Royal Society of
London, Series B, 252, 59–64.
McComb, K., Packer, C. & Pusey, A. E. 1994. Roaring and numerical assessment in
contests between groups of female lions, Panthera leo. Animal Behaviour, 47,
379–387.

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 387
McComb, K., Moss, C., Sayialel, S. & Baker, L. 2000. Unusually extensive networks of
vocal recognition in African elephants. Animal Behaviour, 59, 1103–1109.
McComb, K., Moss, C., Durant, S. M., Baker, L. & Sayialel, S. 2001. Matriarchs act as
repositories of social knowledge in African elephants. Science, 292, 491–494.
McComb, K., Reby, D., Baker, L., Moss, C. & Sayialel, S. 2003. Long-distance
communication of social identity in African elephants. Animal Behaviour, 65,
317–329.
McElligott, A. G. & Hayden, T. J. 1999. Context-related vocalization rates of fallow
bucks, Dama dama. Animal Behaviour, 58, 1095–1104.
2001. Postcopulatory vocalisations of fallow bucks: who is listening? Behavioral
Ecology, 12, 41–46.
McElligott, A. G., O’Neill, K. P. & Hayden, T. J. 1999. Cumulative long-term investment
in vocalization and mating success of fallow bucks, Dama dama. Animal Behaviour,
57, 1159–1167.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma and E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
Mitani, J. C. & Nishida, T. 1993. Contexts and social correlates of long-distance calling
by male chimpanzees. Animal Behaviour, 45, 735–746.
Mobley, J. R., Herman L. M. & Frankel A. S. 1988. Responses of wintering humpback
whales (Megaptera novaeangliae) to playback of recordings of winter and summer
vocalizations and of synthetic sound. Behavioral Ecology and Sociobiology, 23,
211–223.
Moss, C. J. & Poole, J. H. 1983. Relationships and social structure of African elephants.
In: Primate Social Relationships: An Integrated Approach, ed. R. A. Hinde. Oxford:
Blackwell Scientific, pp. 315–325.
Noad, M. J., Cato, D. H., Bryden, M. M. et al., 2000. Cultural revolution in whale songs.
Nature, 408, 537.
Oda, R. 1997. The response of Verreaux’s sifakas to anti-predator alarm calls given by
sympatric ring tailed lemurs. Folia Primatologica, 69, 357–360.
Ogutu, J. O. & Dublin, H. T. 1998. The response of lions and spotted hyenas to sound
playbacks as a technique for estimating population size. African Journal of Ecology,
36, 83–95.
Packer, C., Herbst, L., Pusey, A. E. et al. 1988. Reproductive success of lions. In
Reproductive Success, ed. T. H. Clutter-Brock. Chicago, IL: University of Chicago
Press, pp. 363–383.
Payne, R. S. & McVay, S. 1971. Songs of humpback whales. Science, 173, 585–597.
Pennisi, E. 2001. Elephant matriarchs tell friend from foe. Science, 292, 417–418.
Pinker, S. 1994. The Language Instinct. New York: William Morrow.
Pusey, A. E. & Packer, C. 1987. The evolution of sex-biased dispersal in lions. Behaviour,
101, 275–310.
Reby, D. 1998. Etude de la communication sonore chez les cervides. Prise en compte de
la dimension individuelle. Ph.D. Thesis, Universite Paul Sabatier, Toulouse,
France.

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
388 K. McComb & D. Reby
Reby, D. & McComb, K. 2003a. Anatomical constraints generate honesty: acoustic
cues to age and weight in the roars of red deer stags. Animal Behaviour, 65,
519–530.
2003b. Vocal communication and reproduction in deer. Advances in the Study of
Behaviour, 33, 231–264.
Reby, D., Joachim, J., Lauga, J., Lek, S. & Aulagnier, S. 1998. Individuality in the groans
of fallow deer (Dama dama) bucks. Journal of Zoology (London), 245, 79–84.
Reby, D., Cargnelutti, B. & Hewison, A. J. M. 1999a. Contexts and possible functions of
barking in roe deer. Animal Behaviour, 57, 1121–1128.
Reby, D., Cargnelutti, B., Joachim, J. & Aulagnier, S. 1999b. Spectral acoustic structure
of barking in roe deer: sex-, age-, and individual-related variations. Comptes Rendus
de l’Academie des Sciences, 322, 271–279.
Reby, D., Izquierdo, M., Hewison, A. J. M. & Pepin, D. 2001. Red deer (Cervus elaphus)
hinds discriminate between the roars of their current harem holder stag and
those of neighbouring stags. Ethology, 107, 951–959.
Rendall, D., Peter S. R. & Emond R. E. 1996. Vocal recognition of individual and kin in
free ranging rhesus monkeys. Animal Behaviour, 51, 1007–1015.
Rendall, D., Owren, M. J. & Rodman, P. S. 1998. The role of vocal tract filtering in
identity cueing in rhesus monkey (Macaca mulatta) vocalizations. Journal of the
Acoustical Society of America, 103, 602–614.
Riede, T. & Fitch, W. T. 1999. Vocal tract length and acoustics of vocalization in the
domestic dog, Canis familiaris. Journal of Experimental Biology, 202, 2859–2869.
Schaller, G. B. 1967. The Deer and the Tiger: A Study of Wildlife in India. Chicago, IL:
University of Chicago Press.
1972. The Serengeti Lion. Chicago, IL: University of Chicago Press.
Semple, S & McComb, K. 1996. Behavioural deception. Trends in Ecology and Evolution,
11, 434–437.
Shipley, C., Hines, M. & Buchwald J. S. 1981. Individual differences in threat calls of
northern elephant seal bulls. Animal Behaviour, 29, 12–18.
Snowdon, C. T. Cleveland, J. & French, J. A. 1983. Responses to context- and
individual-specific cues in cotton top tamarin long-calls. Animal Behaviour, 31,
92–101.
Tooze, Z. J., Harrington, F. H. & Fentress, J. C. 1990. Individually distinct vocalisations
in timber wolves, Canis lupus. Animal Behaviour, 40, 723–730.
Tyack, P. 1981. Interactions between singing Hawaiian humpback whales and
conspecifics nearby. Behavioral Ecology and Sociobiology, 8, 105–116.
1983. Differential response of humpback whales, Megaptera novaeangliae, to playback
of song or social sounds. Behavioral Ecology and Sociobiology, 13, 49–55.
van Parijs, S. M., Hastie, G. D. & Thompson, P. M. 2000. Individual and geographic
variation in display behaviour of male harbour seals in Scotland. Animal Behaviour,
59, 559–568.
Waser, P. M. 1977. Individual recognition, intragroup cohesion and intergroup
spacing: evidence from sound playback to forest monkeys. Behaviour, 60, 28–74.

P1: JZZ/... P2: JZZ/...0521823617c17.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:14
Vocal networks in large terrestrial mammals 389
Whitehead, H., Waters, S. & Lyrholm, T. 1991. Social organisation of female sperm
whales and their offspring: constant companions and casual acquaintances.
Behavioral Ecology and Sociobiology, 29, 385–389.
Wilden, I., Herzel, H., Peters, G. & Tembrock, G. 1998. Subharmonics, biphonation and
deterministic chaos in mammal vocalisation. Bioacoustics, 9, 171–196.
Zuberbuhler, K. 2002. A syntactic rule in forest monkey communication. Animal
Behaviour, 63, 293–299.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
18
Underwater acoustic communicationnetworks in marine mammals
v i nc e n t m . j a n i k
University of St Andrews, St Andrews, UK
Introduction
Communication in networks has received considerable research attention
over the last few years (Naguib & Todt, 1997; Otter et al., 1999; Peake et al., 2001;
Mennill et al., 2002; Ch. 1). This is true for two types of network interaction, one
in which several receivers react to the signal of just one individual and a more
complex one in which receivers eavesdrop on the signal exchange of two or more
individuals and use the information they gained in their own decision making
(McGregor & Dabelsteen, 1996; Ch. 2). If we think about communication, the fact
that there often is more than just one individual receiving any given signal is not
surprising. One reason that many studies on more transient signals concentrated
on only one signaller and one receiver was a methodological problem. It is notori-
ously difficult to sample behaviour from more than one or two individuals at a
time, especially if interactions are rapid and involve movements of individuals.
Recently, the simultaneous tracking of several individuals in a large group has
become feasible. This has led to an increase in studies investigating the effects
of signals on several receivers in rapid communication interactions. Many such
studies have concentrated on the acoustic domain, a modality that is inherently
transient. Signals rarely last more than a few seconds and usually provide a variety
of different messages within a single signal.
While there is ample evidence from terrestrial environments that more than
one individual can receive and use information from a call or a calling interaction,
data on acoustic communication networks in marine environments are sparse.
The marine environment imposes constraints on, and presents opportunities for,
communication in networks that are different from those found in the terrestrial
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
390

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 391
world (Janik, 1999). Light only penetrates a few tens of metres into the ocean from
the surface. This limits the use of colouration or movement signals in the ocean.
Signals that use active light sources are more successful but also cost more to
develop. Sound is a much cheaper option and travels much further than in air.
However, a variety of parameters, especially pressure and temperature gradients,
influence sound propagation at different depths.
The loudest and best-known biological sound sources in the ocean are marine
mammals. Pinnipeds and cetaceans use underwater sound in very similar ways.
Many features of their social lives make them interesting subjects for the study of
communication networks. For example, sound transmission characteristics of the
sea allow individuals to stay in acoustic contact over very long distances (Tyack,
1998). Several species live in large aggregations or fission–fusion societies in which
group composition changes frequently while individuals maintain preferences
for certain associates (Wells et al., 1999). Many marine mammals are capable of
vocal learning, which helps them to produce a variety of different sounds (Janik &
Slater, 1997). Finally, many of their cognitive abilities rival those of the great apes
(Herman, 1987; Kako, 1999; Schusterman & Kastak, 2002). This combination of
environmental conditions and social skills is unique among mammals. Each of
them affects how animals communicate and will have profound effects on the
characteristics of communication networks. In this chapter, I summarize what we
know about acoustic communication networks in marine mammals.
Size and characteristics of marine mammal communication
networks at sea
Payne & Webb (1971) suspected that cetacean communication networks
are among the largest in the world. To identify the potential size of a communica-
tion network, we need information on the density of receivers and the active space
of a signal. The active space is the area in which another individual can perceive
the calls of a conspecific (Brenowitz, 1982). Active space can either be measured
directly by playing back calls of known source level and observing a predicted
reaction of the receiver, or through theoretical calculations using call source lev-
els, perception thresholds of individuals and models of sound propagation. Direct
measurement and theoretical calculation have advantages and disadvantages. For
example, an advantage of direct measurement using playback is that the sound
reached the animal through its actual environment and no assumptions about
propagation loss are necessary. Such assumptions can be a weak point of theoret-
ical calculations since propagation is influenced by several parameters, many of
which can change from one minute to the next. A disadvantage of direct measure-
ment using playback is that the active space is likely to be underestimated because

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
392 V. M. Janik
receivers may extract information from a call but not show an immediate reaction.
An advantage of theoretical calculations of active space is that empirical data on
sound perception thresholds in different noise conditions can be used to predict
when an animal can perceive a sound. However, an ability to perceive a sound of
a particular frequency does not necessarily mean that the signal is recognizable
as a call of a conspecific.
Marine mammal sound propagation in the sea
Sound propagation in the sea differs greatly from that in air. In addition
to normal spreading loss, underwater a sound of 1 kHz loses around 0.04 dB/km
through absorption while the same sound in air loses 4 dB/km (Richardson et al.,
1995). The result is that marine mammal calls have a much larger active space
than those of most terrestrial animals. Given that animals can usually detect sig-
nals at, or greater than, the level of background noise (Ch. 20), as can we using
microphones, it is safe to assume that animals can perceive sounds if we are able
to record them. If such recordings are made with passive acoustic localization
techniques (e.g. Watkins & Schevill, 1972; Clark & Ellison, 2000; Janik et al., 2000),
we are able to determine the distance to the sound source. Studies using such
equipment have confirmed that many marine mammals produce signals that can
be detected from more than 10 km away (Table 18.1). These distances coincide
roughly with those at which reactions to calls have been observed. A fin whale
Balaenoptera physalus has been observed to start swimming towards a vocalizing
group 20–25 km away (Watkins, 1981). Fin whales also refrain from using certain
sound types if there are no other whales within a 20 km radius (Watkins, 1981).
Humpback whales Megaptera novaeangliae respond to sounds 9 km away (Tyack &
Whitehead, 1983). Such responses are indications that the communication net-
work includes animals at such distances. However, the actual network may be
much larger. Recent use of ocean-wide microphone arrays offshore has allowed
researchers to record baleen whales over several hundred kilometres (Table 18.1).
At large distances, it is unlikely that an animal shows an immediate reaction to a
single call. Nevertheless, marine mammals may use distant sounds that indicate
the location of other individuals to find breeding or foraging grounds.
Detailed theoretical calculations of active space have been conducted for bot-
tlenose dolphins Tursiops truncatus, sperm whales Physeter macrocephalus and killer
whales Orcinus orca. Figure 18.1 shows the active space of bottlenose dolphin whis-
tles. Given the fact that most bottlenose dolphin whistles do not have much energy
below 3 kHz, whistles should be detected over distances of 20–25 km at maximum
source levels. The average source level of 158 dB re 1 µPa measured by Janik (2000a)
still gives an active space of 9–16 km in calm seas. However, transmission loss in-
creases with frequency, which means that high-frequency whistles or parts of

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 393
Table 18.1. Maximum distances from which marine mammal calls can be detected
Species Frequency range of Recorded from Source
vocalizations (kHz)a distance (km)
Bearded seal, Erignathus
berbatus
0.02–6 25 Cleator et al., 1989
Harp seal, Phoca
groenlandica
< 0.1–16+ 30 Watkins & Schevill,
1979
Ringed seal, Phoca
hispida
0.4–16 1 Richardson et al., 1995
Delphinids 0.1–27.3 16 Barlow et al., 2001
Peale’s dolphin,
Lagenorhynchus
australis
0.3–12 0.02 Schevill & Watkins,
1971
Sperm whale, Physeter
macrocephalus
0.1–30 37 Barlow & Taylor, 1997
Bowhead whale,
Balaena mysticetus
0.025–3.5 17 Clark et al., 1986
Humpback whale, 0.02–8.2 15 Helweg et al., 1992
Megaptera 160 Clark, 1995
novaeangliae
Fin whale, Balaenoptera
physalus
0.01–0.75 > 20 Watkins, 1981;
Watkins et al., 1987
Blue whale, 0.012–0.39 600 Stafford et al., 1998
Balaenoptera musculus 1600 Clark, 1995
aTaken from overview in Richardson et al., 1995.
whistles would not have the same active space as low-frequency components. Cur-
rently, we know little about how this would affect the information that is available
to the receiver. With experience, bottlenose dolphins can identify individually dis-
tinctive signature whistles of conspecifics even if they only hear parts of the whistle
(Caldwell et al., 1990). However, subtle variations in whistle parameters can carry
additional information (Janik et al., 1994), which could be lost in such cases. Using
similar methods, Miller (2004) found that killer whale calls have an active space of
up to 26 km in calm seas and Madsen et al. (2002) calculated 60 km for slow clicks
and 16 km for usual clicks of sperm whales. Again, the maximum source levels
were used for these calculations and the same restrictions for high frequencies
apply.
While these active spaces seem very large, there are some marine mammal
signals that are much quieter and do not travel nearly as far (Table 18.1). Most
marine mammals produce sounds at various different source levels many of which

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
394 V. M. Janik
0
10
20
30
40
1 2 3 4 5 6 7 8 9 10 11 12
Frequency (kHz)
Ran
ge (
km)
0
10
20
30
40
1 2 3 4 5 6 8 10 12
Frequency (kHz)
Ran
ge (
km)
(b)
(a)
Fig. 18.1. The estimated radius of the active space of dolphin whistles without
frequency modulation at different frequencies for sea state 0 (�, calm seas, no wind)
and sea state 4 (�, moderate breeze of 13–18 miles/h). (a) Whistles produced at
maximum source level of 169 dB re 1 µPa. (b) Whistles produced at mean source level
of 158 dB re 1 µPa. Transmission loss in a habitat of homogeneous temperature and
10 m depth (source and receiver at 5 m depth) was calculated following Marsh &
Schulkin (1962) and Urick (1983). Ambient noise was taken from Knudsen et al. (1948).
Data for auditory thresholds and critical ratios of Tursiops truncatus were taken from
Johnson (1967, 1968). (After Janik, 2000a.)

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 395
would only be audible to conspecifics within a 100 m or less. Furthermore, several
species like harbour porpoises Phocoena phocoena (Busnel & Dziedzic, 1966) and
Hector’s dolphins Cephalorhynchus hectori (Dawson, 1991), rarely use low-frequency
sounds (i.e. <20 kHz) but instead use clicks to communicate. Their signals are
subject to much larger transmission loss. Communicative clicks are very similar to
echolocation clicks and travel only a few hundred metres, making the active space
of these species relatively small. Communication networks in these species are,
therefore, much smaller and more comparable to those found in some terrestrial
species.
The active space calculations for bottlenose dolphins and killer whales used
empirical models of sound propagation to predict transmission loss in shallow
water. While this is a useful method for this estimate, several other factors influ-
ence transmission in different parts of the water column. Shallow water trans-
mission is greatly influenced by reflections off the surface and the bottom. This
leads to reverberation that can make acoustic signals unrecognizable. Perception
experiments using degraded signals would help us to understand how degrada-
tion affects signal detection and recognition in marine mammals. Furthermore,
other parameters like transmitter depth or frequency can have a strong effect on
active space (Mercado & Frazer, 1999). Consequently, the loudest marine mammal
signals are not necessarily the ones that transmit the furthest.
In deep water, temperature and pressure profiles give the propagation path a
unique shape that is very different from those found in terrestrial environments
(Richardson et al., 1995). The speed of sound increases with depth and tempera-
ture. In summer, when the surface layer is warmer than the water below, sound is
refracted downwards, leading to a shadow zone ahead of the sound source. As the
sound travels deeper, temperature does not change much but pressure increases.
This leads to refraction towards the surface. As a result, sound travels up and
down through the water column as it travels away from the source (Fig. 18.2). If
the surface layer is mixed or shows little temperature layering, as is often the case
in winter, sound travels more easily through the upper layers. However, some en-
ergy still leaks into lower layers and travels in the same ray pattern as in summer.
The result of these conditions is that animals at the surface enter and leave con-
vergence zones of the ray paths of a sound produced at great distance. Therefore,
to locate a calling animal, an individual needs to consider the special propagation
path. If it listened at the surface in one of the convergence zones, it may encounter
an area that appears to have a caller in its centre with sound energy decreasing in
all directions from it. However, the centre does not have the calling animal in it. In-
stead, the sound was produced several kilometres away and has travelled through
deep waters before returning to the surface. The result is that animals cannot
use changes in received levels at the surface to locate a distant caller. However,

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
0
500
1000
1500
2000
2500
3000
3500
4000
4500
50001500 1510 1520 1530 1540 1550 1560
Dep
th (
m)
Sound speed (m/s)
(a)
(b)
Fig. 18.2. Changes in the speed of sound and ray paths with depth. (a) Typical profile
of speed of sound versus depth for temperate or tropical seas. (b) Calculated ray paths
for a 20 Hz signal produced at a depth of 50 m in an environment with the speed of
sound profile of (a). Ray paths were calculated using a parabolic equation model.
White blocks indicate attenuation of ≤ 60 dB; black blocks indicate attenuation of
≥100 dB. Note the convergence zone (shown by arrows) near the surface at ranges of
6.5, 130 and 190 km.
396

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 397
following parts of the actual sound path underwater may allow an individual to
recognize that it is listening in a convergence zone rather than being close to the
original caller.
Another interesting aspect of sound transmission at sea is the deep sound
channel or SOFAR channel. This is a layer at approximately 600–1200 m in which
sound is trapped and travels almost horizontally with much less transmission
loss (because of the shorter travel path and no losses from surface or bottom
reflections). It can be found in the layer with minimum sound speed. Little is
known on whether marine mammals use this channel, but it has been suggested
that whales may use it for long-distance communication (Payne & Webb, 1971).
This is only possible for a few species that travel routinely to this depth, for example
elephant seals Mirounga spp. (Le Boeuf et al., 1989; Hindell et al., 1991) or northern
bottlenose whales Hyperoodon ampullatus (Hooker & Baird, 1999). However, in Arctic
waters, where the minimum sound speed (and thus the SOFAR channel) can be at
much shallower depths, it may be within reach of more species.
The conditions described here are idealizations assuming little variation in
other parameters. They describe general patterns but the actual situation faced
by a marine mammal changes with location and time. One conclusion from these
patterns is that it must be difficult to estimate range from a caller using parame-
ters such as sound intensity. However, they may be able to use other parameters
to determine their distance from a caller. Premus & Spiesberger (1997) analysed
fin whale sounds recorded in the Gulf of California. They found that the signal
arrived several times at each hydrophone, which is typical if the sound takes sev-
eral different paths to reach the receiver. Longer paths result in later arrivals, and
at great distances these time delays can be substantial. However, Premus & Spies-
berger (1997) noted that the first arrival of a fin whale call was much sooner than
expected even if it was taking the shortest route available through the water. This
fast sound transmission could only be explained if the first arrival represented
sound energy that entered and travelled in the sediment, where sound speed is
much higher than in water. If such multipath arrivals through different media
are common, whales may be able to tell the distance of the caller by listening to
the differences in the time of arrival of the sound travelling through the sediment
and that travelling through water. This sound path may even allow them to listen
to individuals on the other side of an island. Another way in which distance in-
formation could be extracted is by listening to the extent of sound degradation.
Again, we know little about the abilities of marine mammals to use such features
to judge distance to the caller.
The number of animals in a communication network
The second variable that determines communication network size is the
number of animals within the transmission range of a signal. This varies greatly

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
398 V. M. Janik
Table 18.2. Examples of animal density and average group sizes for selected sites
Species Location Density Average Source
(animals/100 km2) group size
Bottlenose
dolphin
Gulf of Mexico
(US coast)
6–480 2–15 Shane et al., 1986
Harbour
porpoise
Northwest
Europe
10–80 1.49 Hammond et al.,
2002
White-beaked
dolphin
Northwest
Europe
0–5 3.78 Hammond et al.,
2002
Minke whale Northwest
Europe
0–3 1.04 Hammond et al.,
2002
Vocalizing fin
whales
Hawaii 0.0027;
maximum 0.0081
1 McDonald &
Fox, 1999
and depends on the area, species and behaviour of a marine mammal. Bottlenose
dolphins, for example, can be found in groups of hundreds (Saayman et al., 1973)
or even thousands (Scott & Chivers, 1990) offshore, while individuals in coastal
areas may at times find themselves acoustically isolated from all conspecifics if
they enter small inlets in which sound is blocked by land. Furthermore, many ma-
rine mammal species, especially delphinids, live in fission–fusion societies where
group composition and size can change rapidly. Finally, if we consider that in-
dividuals are capable of restricting signal transmission to specific receivers (see
below), it becomes clear that network size is difficult to assess. On an evolutionary
scale, however, it is interesting to look at how many potential receivers there are
for any given signal. This might help us to understand the relationship between
network size and specific strategies to direct or restrict signals. Because of the lack
of information on average transmission distances of marine mammal sounds, we
can currently only look at data on the number of animals in an area rather than
calculate network sizes. Ultimately, to calculate network sizes, population den-
sities and the average active space of a signal from the same area need to be
combined.
The average group size of a marine mammal species is a good indicator of the
most commonly found minimum network size (Table 18.2). If group size is very
large, as in some oceanic dolphin species, the transmission range of a signal can be
limited by masking noise from conspecifics and the actual network would contain
fewer animals than are in the group (for similar considerations in anurans, see
Ch. 13). In most cases, however, the network will be larger than the average group
size because of the large active space of marine mammal calls. Population-density
data can be used to estimate average network sizes for unrestricted signals that

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 399
travel beyond the group’s boundaries. Table 18.2 gives population densities and
average group sizes for a few selected marine mammal species. Some studies have
assessed animal density using acoustic surveys. This means that only vocalizing
animals are registered. While this gives a good density estimate of signallers, it
represents a minimum rather than a representative average estimate of network
size. However, population density studies usually look at very large areas that are
less relevant for estimates of communication network sizes. One tight group of 20
dolphins in 1000 km2, for example, will yield a very low population density but still
represents a communication network of 20 animals. Like sound propagation in the
sea, population density is a highly dynamic variable. Therefore, the size of marine
mammal communication networks is likely to vary greatly on a temporal as well
as spatial scale. We can expect that territorial and, therefore, relatively stationary
species display more stability in network size, but studies on the dynamics of such
network sizes are still lacking.
Directing and restricting signals
In a network, we can expect to find two different kinds of signal: those
that are directed at all receivers within range and others aimed at only one or a few.
Callers directing signals at specific individuals benefit from adding information
that indicates who they are addressing. This is even more important if, like in
marine mammals, the network can be large, locations of individuals are difficult
to predict (e.g. if animals are not territorial) and if only one sensory modality is
available. Concurrently, such conditions render it more difficult for the sender
to identify who is within range as a potential receiver that it is worth calling to.
One way of solving this problem for the caller is to give unequivocal information
about its own identity or group membership. This makes it more likely to be
recognized by other group members or close associates within range. While it
might be disadvantageous to broadcast one’s identity or location if predators use
such cues to find prey, signalling this information can be evolutionarily stable
if it improves information transmission for the sender to the required receiver
(Johnstone, 1997).
Most animal species cannot avoid providing identity information through indi-
vidually specific voice cues. Such cues result from individually specific genetic and
environmental influences on the morphology of the vocal apparatus during devel-
opment. Similarly, genetically related individuals may share a voice feature that
can be used in kin or even group recognition if related individuals stay together.
However, voice cues are relatively subtle and can be difficult to decode over long
distances or in high background noise. There are several ways to improve the en-
coding of information on identity. First, if groups are genetically isolated, genetic

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
400 V. M. Janik
drift can increase group differences. Even between sympatric groups, this is pos-
sible if their members do not interbreed. Another solution is to use call types
that are shared by all animals in a population at higher or lower rates than the
rest of the population. Finally, animals can develop group- or individual-specific
call types. This last solution can usually only be achieved through vocal learning
or invention (Janik & Slater, 2000). Nevertheless, several of these influences can
act together to create individual differences. For example, limited skills in vocal
learning that only allow a slight change in the fundamental frequency of a given
signal may be used to enhance individual differences caused by environmental
influences during development.
Many marine mammal species show pronounced differences between groups of
animals. Weddell seals Leptonychotes weddelli in breeding colonies only 20 km apart
have been found to use colony-specific call types and show differences in usage
of shared call types (Morrice et al., 1994). Similar geographic variation over much
larger distances has been described for leopard seals Hydrurga leptonyx (Thomas &
Golladay, 1995), bearded seals Erignathus berbatus (Cleator et al., 1989), harp seals
Phoca groenlandica (Terhune, 1994) and harbour seals Phoca vitulina (van Parijs et al.,
2000a). However, in these cases it is possible that individuals from different sites
are geographically isolated. Humpback whales in the Atlantic and the Pacific, for
example, sing very different songs (Winn et al., 1981). Since they cannot encounter
each other, these differences are not necessary for group recognition. Thus, the
occurrence of differences between the calls of groups of animals is not evidence
for a specific adaptation for group recognition.
Killer whales (Ford & Fisher, 1983), sperm whales (Rendell & Whitehead, 2003)
and blue whales Balaenoptera musculus (Stafford et al., 2001) also have distinctive
group calls, but here these groups overlap in their geographic ranges. In these
cases, the distinctiveness in the repertoire may be more important for directing
signals than in geographically isolated groups. However, the calls in these exam-
ples are not individually specific.
Individual specificity may not be necessary for animals that live in stable fam-
ily groups like killer whales. If group composition is less stable though, more
unequivocal signals may be required for individual recognition. Bottlenose dol-
phins, for example, associate preferentially with specific individuals, but their
daily ranging behaviour results in regular changes of group composition and
short-term associations (Wells et al., 1987). This organization is often referred to
as a fission–fusion society. Bottlenose dolphins develop individually distinctive
signature whistle types (Fig. 18.3) that are used while animals are out of visual
contact (Caldwell et al., 1990; Janik & Slater, 1998). These signals have a much
larger interindividual variability than isolation calls of other animal species and
thus transmit individual identity more reliably (Tyack, 2000). Signature whistles

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 401
Fig. 18.3. Three randomly chosen spectrograms (columns; FFT size, 1024; time
resolution, 20.5 milliseconds; frequency resolution, 50 Hz; number of FFT steps, 200;
weighting function, Hanning window) of signature whistles from each of four
different individual bottlenose dolphins (rows). Background noise and harmonics
have been removed on all spectrograms to show the pronounced difference in the
shape of the fundamental frequency of signature whistles of the different individuals.
(After Janik & Slater, 1998).

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
402 V. M. Janik
have been reported to be stable over more than 12 years in female bottlenose dol-
phins in the Bay of Sarasota, Florida, USA (Sayigh et al., 1990). Tyack (1997) found
that vocal learning influences whistle development. In fact, learning or innova-
tion may be the only way to develop such signals that have to be different from
those of a large number of conspecifics within a fission–fusion society. Janik &
Slater (1997) suggested that individual or group recognition might have been one
of the main selection pressures on the evolution of vocal learning in cetaceans.
Evidence for similar individually distinctive signature signals exists for common
dolphins Delphinus delphis (Caldwell & Caldwell, 1968), Pacific white-sided dolphins
Lagenorhynchus obscurus (Caldwell & Caldwell, 1971), spotted dolphins Stenella pla-
giodon (Caldwell et al., 1973), Pacific humpback dolphins Sousa chinensis (van Parijs &
Corkeron, 2001) and sperm whales (Watkins & Schevill, 1977).
While such shared calls may facilitate recognition in general, calls can also
be directed at specific individuals through vocal matching. In vocal matching, an
individual responds to the signal of a caller by producing a signal of the same type.
Vocal matching can be used without the existence of individual-specific calls as
long as other individuals can copy calls or if they have a repertoire of shared calls.
Many species of cetaceans have been observed to produce calls of the same kind
in response to a call of a conspecific, but such anecdotal reports cannot exclude
the possibility of matching occurring by chance. If individuals share a repertoire
and produce sounds independently, by chance alone two different individuals
can produce signals of the same type in close succession. However, this does not
necessarily mean they interact vocally. The proportion of such interactions has to
be larger than expected by chance to represent evidence for vocal matching. True
vocal matching has been demonstrated for bottlenose dolphins (Janik, 2000b) and
for killer whales (Miller et al., 2004). In bottlenose dolphins, signature whistles
can be copied by another individual in such matching interactions (Janik & Slater,
1998). Tyack (1991) raised the interesting possibility that bottlenose dolphins may
use signature whistles of other individuals to initiate contact with the ‘owner’ of
the signature whistles. However, in all reported cases in which signature whistle
matching has been observed and the identities of the calling individuals were
known, the ‘owner’ of the signature whistle called first (Janik & Slater, 1998).
A different strategy is changing the directionality of calls. The fundamental
frequencies of most marine mammal calls are usually transmitted in a relatively
omnidirectional pattern (Evans et al., 1964; Lammers & Au, 2003). However, clicks
are highly directional (reviewed by Au, 1993) and have the potential to be used
in addressing specific individuals. Dolphins use clicks in echolocation as well as
communication. Several species (see above) rely on clicks for communication and
do not produce any whistles at all (Dawson, 1991; reviewed by Herman & Tavolga,
1980). It is possible that these species use the directionality of their clicks to direct

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 403
or restrict signals in social interactions. Even in whistles, the high-frequency parts
are highly directional (Lammers & Au, 2003) so that the same signal may carry dif-
ferent types of information with some of it only available to animals ahead of the
caller. This could be achieved by filtering high-frequency components in specific
patterns, similar to formants in human signals. Such filtering would not be dis-
cernible from listening to the low-frequency component alone. Dolphins have con-
trol over the filtering of higher-frequency harmonics (Fig. 1 in Janik et al., 1994), but
the significance of such changes is unclear. Miller (2002) found that killer whales
have call types with high-frequency components that show higher directionality
than the low-frequency parts of the same call or calls without these components.
High- and low-frequency components of the same killer whale call are not har-
monically related. Therefore, the modulation pattern of one component cannot
be discerned from the other one. This makes withholding information even easier.
Miller (2002) suggested that killer whales might use calls with high-frequency
components to indicate their direction of movement. Alternatively, they may
be used to direct signals at specific individuals and withhold information from
others.
Another way of restricting the spread of signals through a communication net-
work is by decreasing the source level so that they do not carry as far. Pinnipeds
and cetaceans produce the same call types at a variety of different source levels
(reviewed by Richardson et al., 1995). Many species of odontocetes also have sig-
nals of very different frequency in their repertoires. High-frequency signals are
attenuated much more rapidly than low-frequency sounds. Odontocetes may be
able to restrict transmission range by choosing high-frequency clicks rather than
lower-frequency whistles even though they are produced with the same source
level. However, calling depths have different optimal frequencies for long-range
signal transmission (Mercado & Frazer, 1999). Higher frequencies can sometimes
travel further than lower ones, especially in relatively shallow water (i.e. less than
100 m deep). Further studies are needed to explore the possible use of source-level
adjustments and frequency selection in directing and restricting signals.
Eavesdropping
Peake (Ch. 2) has distinguished two types of eavesdropping; interceptive
eavesdropping (e.g. predators locating prey by listening to prey vocalizations) and
social eavesdropping (extracting information from a signalling interaction). Brad-
bury & Vehrencamp (1998) also used the term ‘cue’ for prey signals that are used
by predators to locate prey. While it is arguable whether such interactions can be
called communication, effects of calls on predators and their prey are an interest-
ing ecological variable that can influence the design of communication systems.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
404 V. M. Janik
Several studies have looked at the impact of killer whale signals on other ma-
rine mammal species that are potential prey. The diet of killer whales can vary
considerably from one location to another. In British Columbia, Canada, some
killer whales only eat fish while others take marine mammals as prey. Fish eaters
are known as resident killer whales since they have smaller ranging patterns than
the so-called transient killer whales that feed on marine mammals. Deecke et al.
(2002) conducted playback experiments and inferred from diving patterns that
harbour seals in British Columbia avoided sounds made by transient killer whales
but they did not react to sounds of resident killer whales. Transients and residents
use different call types and individual killer whale pods have repertoires of up
to 17 call types (Ford, 1989). Deecke et al. (2002) carefully selected specific sound
types for each comparison to ensure that the discrimination performed by the
seals could not be based on just one or two call types. Harbour seals also avoided
playbacks of sounds from Norwegian killer whales. These whales concentrate on
herring as prey for at least part of the year, which makes it unlikely that fish-
eating killer whales share voice features that identify them as harmless to seals.
It is unclear how harbour seals distinguish between known residents and other
killer whales. There are genetic differences between killer whale populations and
even between sympatric residents and transients of British Columbia (Hoelzel
et al., 1998). Perhaps residents share a voice feature that affects all their calls and
makes them recognizable. Alternatively, the seals may have learned all call types
used by resident killer whales and avoid all other call types.
Grey whales Eschrichtius robustus (Cummings & Thompson, 1971) and beluga
whales Delphinapterus leucas (Fish & Vania, 1971) have been found to avoid loca-
tions from which killer whale sounds had been played. Unfortunately, it is not
clear whether the sounds used in these studies came from mammal-eating or fish-
eating killer whales. Belugas (Schevill 1964; Fish & Vania, 1971) and grey whales
(Cummings & Thompson, 1971) also ceased vocalizing when exposed to killer
whale sounds, another well-known reaction of cetaceans to any unusual stimulus
(Herman & Tavolga, 1980). Other examples are pilot whales Globicephala melaena
falling silent when hunted (Schevill, 1964) and bottlenose dolphins (Caldwell &
Caldwell 1967) and Peale’s dolphins Lagenorhynchus australis (Schevill & Watkins,
1971) falling silent when captured or when approached by a boat. Interestingly,
transient killer whales appear to use fewer echolocation clicks than resident
whales while they forage (Barrett-Lennard et al., 1996). This may be a counter-
strategy to avoid early detection by their prey.
Several aspects of marine mammals make it difficult to establish whether social
eavesdropping occurs in this group (i.e. whether information has been extracted
from a signalling interaction). Individuals often approach callers, for example.
Groups of surface-active humpback whales produce a variety of sounds that can

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 405
attract males that are several kilometres away (Tyack, 1983) and similar results
have been found for southern right whales Eubalaena glacialis (Clark & Clark, 1980).
However, it is difficult to determine who is interacting (i.e. whether individuals in
such groups signal to each other or to individuals outside the group) and, there-
fore, whether there is potential for eavesdropping. Distant individuals may extract
information from the calls of animals interacting in the group and decide to ap-
proach (which would qualify as social eavesdropping), or they may be attracted by
calls that are directed at distant animals (which is a good example of communi-
cation in a network but not for eavesdropping). It will be difficult to distinguish
between these possibilities experimentally; furthermore, these two scenarios are
not mutually exclusive.
One context in which social eavesdropping has been demonstrated is in song
interactions between birds (Naguib & Todt, 1997; Otter et al., 1999; Peake et al.,
2001; Mennill et al., 2002). Many marine mammal species also produce song dur-
ing the mating season and some, like Weddell seals (Bartsh et al., 1992) and har-
bour seals (van Parijs et al., 2000b), establish underwater territories. By analogy
with songbirds, social eavesdropping by marine mammals may be found in such
circumstances. However, other singing species of marine mammals are less sta-
tionary. For example, while singing humpback whales are spaced further apart
than non-singers and singers often avoid each other (Frankel et al., 1995), individ-
uals can rarely be found in the same location from one day to the next (Clapham,
2000). Clapham termed this arrangement a floating lek, in which females are able
to listen to several males but males are not stationary. Given the apparent lack of
direct vocal interactions outside of the surface-active groups that form when sev-
eral males start to escort a female, eavesdropping is less likely to be of importance
here. However, further studies relating vocal displays to movement of individuals
are needed before we can assess the relevance of eavesdropping in this context.
Another very different context in which the term eavesdropping has been used
is echolocation (Xitco & Roitblat, 1996). These authors found that a bottlenose dol-
phin could extract information about the location and shape of an object without
having to produce echolocation sounds itself; it did so by listening to the echoes
of echolocation clicks produced by another individual. This might be a common
feature of echolocating animals. Bats have been found to be attracted by feeding
buzzes of conspecifics (Barclay, 1982; Balcombe & Fenton, 1988). However, the stud-
ies on bats could not determine whether feeding buzzes are generally attractive,
like food calls of non-echolocating animals, or whether they can provide informa-
tion about the exact location and shape of the target to the eavesdropper. In the
study of Xitco & Roitblat (1996), the eavesdropping animal was very close to the
echolocating one and such close proximity may be a prerequisite for gathering
such target-specific information. This form of eavesdropping can be defined as

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
406 V. M. Janik
extracting information from an interaction between another individual’s echolo-
cation signal and an echolocation target. While this might explain some of the
swimming formations that dolphins use during foraging, it is of less relevance to
the topic of communication networks.
Conservation implications
A major concern in marine mammal conservation is the impact of noise
made by human activity in the sea. There has been an increasing amount of indus-
trial, shipping and seismic survey noise over the last century. For example, engine
noise of ships in the busy shipping lanes of the North Atlantic increases the average
ambient noise levels below 500 Hz by 10–40 dB (Urick, 1983). Ross (1976) estimated
that shipping led to a 10 dB increase in ambient noise in these areas from 1950
to 1975. The issue of such noise has been discussed recently in the context of
the Acoustic Thermometry of Ocean Climate (ATOC) study and the low-frequency
active sonar systems deployed by the military (Richardson et al., 1995). These tech-
niques can potentially harm marine mammals because of high source levels and
signals that are similar to those of some marine mammal species.
The main concern in noise exposure has been potential physical damage to the
animals. For example, several Cuvier’s beaked whale Ziphius cavirostris strandings
occurred at the same time as military exercises (e.g. Frantzis, 1998; Balcomb &
Claridge, 2001) and Jepson et al. (2003) reported acute and chronic tissue damage
caused by gas bubbles in whales stranded during such exercises. Weddell seals
exposed to underwater blasts showed severe damage to their inner ears (Bohne
et al., 1986). Another form of impact is a change in the animal’s behaviour. This
can have the same consequences as physical damage since isolation from group
members or the exclusion from feeding grounds can easily lead to the death of an
animal. There are many studies showing short-term avoidance by marine mam-
mals of sound sources (review in Richardson et al., 1995). Examples are killer whales
(Morton & Symonds, 2002) and harbour porpoises (Johnston, 2002) avoiding areas
ensonified by acoustic harassment devices deployed to reduce seal predation on
fish farms; beluga whales avoiding ice-breakers by as much as 80 km for up to
48 hours (Finley et al., 1990; Foote et al., 2004); and bottlenose dolphins in Florida
avoiding specific feeding grounds on weekends when boat activity is highest (Allen
& Read, 2000).
Another response to noise of human origin is a change in calling behaviour.
Such responses can involve a change in temporal or structural parameters of a
call (e.g. Au et al., 1985; Foote at al., 2004) or lead to animals changing call rates
or ceasing to vocalize (Terhune et al., 1979; Bowles et al., 1994). Such changes
could either indicate a direct disruption of communication or be a by-product of a

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 407
general change in behaviour if animals stop activities that involve specific calling
rates or types of signal to avoid a sound source.
A source of human-derived noise may also affect the behaviour of an animal
by masking marine mammal sounds, thereby disrupting communication. This
could be a serious problem for animals in a number of circumstances. First, many
marine mammals use acoustic signals to maintain contact between mothers and
calves (e.g. Renouf, 1984; Smolker et al., 1993) and noise can shorten the range
over which they are able to hear each other. Second, if information is gathered
by eavesdropping on interactions of more distant individuals, noise could mask
such interactions. Erbe (2002) found that the noise of a fast-moving boat can mask
quiet killer whale sounds if the vessel is 14 km from the listening animal. Simi-
lar calculations predict that icebreaking noise can mask quiet beluga sounds if
the icebreaker is up to 71 km from the animal that is listening (Erbe & Farmer,
2000). While we do not know to what extent information gathered through eaves-
dropping is used by marine mammals, masking certainly has an effect on signals
designed to reach more distant receivers, as in marine mammal song. Therefore,
apart from inflicting physical damage, noise could have a severe effect by disrupt-
ing acoustic contact between individuals.
Summary and future directions
While we have data on maximum transmission distances for some marine
mammal sounds, it is still unclear to what extent acoustic signals from distant
animals provide valuable information to a conspecific. If the active space of the
signal is particularly large, as seems to be the case for many marine mammals,
the information from distant animals may not be of much use. For example, it
is of only limited value for a predator to know that an animal is foraging 20 km
away if a long time is needed to travel that distance. For marine mammals, most
aggregations of prey species are very dynamic and either move quickly or only
last for brief periods of time. Consequently, a large active space may just increase
noise for distant receivers and could have contributed to the evolution of redun-
dancy and distinctiveness in communication signals: two features that can help to
improve information transmission and that are pronounced in marine mammal
communication systems.
One way of addressing the question of the value of distant signals would be
to compare reactions to distant marine mammal calls with reactions to artificial
broadband noise at similar levels. If conspecifics only add noise to the commu-
nication channel, responses should be the same. Only at a closer, more relevant
distance should reactions differ. Yet communication over large distances may
help in mate attraction or coordination of behaviour patterns. In that case, the

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
408 V. M. Janik
animals should show specific reactions to distant signals of conspecifics, for exam-
ple specific changes in movement direction. Any studies investigating how marine
mammals react to distant signals would be extremely valuable.
Two related issues are the effect of degradation and how marine mammals
judge the distance to a caller. The distinctiveness of signature whistles in bot-
tlenose dolphins, for example, suggests that at least the identity information
encoded is relatively resistant to degradation. However, what happens to more
subtle cues? The auditory system of marine mammals is adapted to detect and
identify marine mammal signals. Therefore, it would be difficult to make predic-
tions from experiments with artificial test signals. How receivers estimate their
distance from a sound source has been studied extensively in birds and humans
(reviewed by Naguib & Wiley, 2001). The most important parameter appears to
be the degree of reverberation. However, other parameters such as overall and
frequency-dependent attenuation or amplitude fluctuations can also help in the
assessment of distances if the receiver has some experience with the source signal
and the environment. Marine mammals may also use additional cues like time
delays of multiple arrivals via different sound paths (Premus & Spiesberger, 1997)
or changes in signal composition of the same received signal at different receiver
depths (Mercado & Frazer, 1999). Whether and how such information is used by
marine mammals is still unknown.
Eavesdropping on interactions of conspecifics in marine mammals is still vir-
tually unstudied. Territorial seal species would probably be the best starting point
for such studies as interactions between neighbours and intruders are the most
likely source of relevant information that could be obtained through eavesdrop-
ping. However, to simulate such interactions experimentally we need to know
the acoustic parameters that identify a successful or unsuccessful animal in such
contests. Furthermore, we need to investigate whether individuals can recognize
other individuals by general voice features. Without voice recognition, it is diffi-
cult to explain how an animal would recognize an individual that it previously
eavesdropped on. Studying how marine mammals address specific individuals can
also help to understand how relevant eavesdropping is. If marine mammals not
only address specific individuals by matching or the use of signature signals but
also actively exclude potential receivers through the selective use of highly direc-
tional signals, eavesdropping might have been a factor in the evolution of such
strategies.
Theoretical estimates of maximum signal transmission distance and commu-
nication network sizes are useful but they need verification in the real world.
Most likely such extremes are rarely relevant for communicating in everyday life.
Nevertheless, marine mammal communication networks are clearly among the
largest that can be found. As we have seen, this opens up interesting opportunities

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 409
but also imposes further constraints by increasing background noise. Future stud-
ies that investigate the dynamics of marine mammal signalling will improve our
understanding of how underwater sound transmission helped to shape their com-
munication systems and to what extent marine mammals use the extra informa-
tion provided by such large active spaces in their communication networks.
Acknowledgements
I would like to thank Peter McGregor for valuable comments on earlier drafts of this
chapter. The chapter was written with support from a Royal Society University Research Fellow-
ship. Figure 18.1 has been reprinted from Janik (2000a) with permission from Springer Verlag.
Figure 18.3 has been reprinted from Janik & Slater (1998) with permission from Elsevier Science.
References
Allen, M. C. & Read, A. J. 2000. Habitat selection of foraging bottlenose dolphins in
relation to boat density near Clearwater, Florida. Marine Mammal Science, 16,
815–824.
Au, W. W. L. 1993. The Sonar of Dolphins. New York: Springer.
Au, W. W. L., Carder, D. A., Penner, R. H. & Scronce, B. L. 1985. Demonstration of
adaptation in beluga whale echolocation. Journal of the Acoustical Society of America,
77, 726–730.
Balcomb, K. C. & Claridge, D. E. 2001. A mass stranding of cetaceans caused by naval
sonar in the Bahamas. Bahamas Journal of Science, 8, 2–12.
Balcombe, J. P. & Fenton, M. B. 1988. Eavesdropping by bats: the influence of
echolocation call design and foraging strategy. Ethology, 79, 158–166.
Barclay, R. M. R. 1982. Interindividual use of echolocation calls: eavesdropping by bats.
Behavioral Ecology and Sociobiology, 10, 271–275.
Barlow, J. & Taylor, B. 1997. Acoustic census of sperm whales in the eastern temperate
North Pacific. Journal of the Acoustical Society of America, 102, 3213.
Barlow, J., Rankin, S., Oswald, J. N. & Mellinger, D. K. 2001. Long-range detection of
delphinid whistles using a towed hydrophone array. In Proceedings of the 14th
Biennial Conference on the Biology of Marine Mammals, Vancouver, p. 17.
Barrett-Lennard, L. G., Ford, J. K. B. & Heise, K. A. 1996. The mixed blessing of
echolocation: differences in sonar use by fish-eating and mammal-eating killer
whales. Animal Behaviour, 51, 553–565.
Bartsh, S. S., Johnston, S. D. & Siniff, D. B. 1992. Territorial behaviour and breeding
frequency of male Weddell seals (Leptonychotes weddelli) in relation to age, size, and
concentration of serum testosterone and cortisol. Canadian Journal of Zoology, 70,
680–692.
Bohne, B. A., Bozzay, D. G. & Thomas, J. A. 1986. Evaluation of inner ear pathology in
Weddell seals. Antarctic Journal, 21, 208.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
410 V. M. Janik
Bowles, A. E., Smultea, M., Wursig, B., DeMaster, D. P. & Palka, D. 1994. Relative
abundance and behavior of marine mammals exposed to transmissions from the
Heard Island feasibility test. Journal of the Acoustical Society of America, 96,
2469–2482.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.
Brenowitz, E. A. 1982. The active space of red-winged blackbird song. Behavioral Ecology
and Sociobiology, 147, 511–522.
Busnel, R.-G. & Dziedzic, A. 1966. Acoustic signals of the pilot whale Globicephala
melaena and of the porpoises Delphinus delphis and Phocoena phocoena. In: Whales,
Dolphins, and Porpoises, ed. K. S. Norris. Berkeley, CA: University of California Press,
pp. 607–646.
Caldwell, M. C. & Caldwell, D. K. 1967. Intraspecific transfer of information via the
pulsed sound in captive odontocete cetaceans. In: Animal Sonar Systems: Biology and
Bionics, ed. R. G. Bullock. Jouy-en-Josas: Laboratoire Physiologie Acoustique,
pp. 879–936.
1968. Vocalization of naive captive dolphins in small groups. Science, 159, 1121–1123.
1971. Statistical evidence for individual signature whistles in Pacific whitesided
dolphins, Lagenorhynchus obliquidens. Cetology, 3, 1–9.
Caldwell, M. C., Caldwell, D. K. & Miller, J. F. 1973. Statistical evidence for individual
signature whistles in the spotted dolphin, Stenella plagiodon. Cetology, 16, 1–21.
Caldwell, M. C., Caldwell, D. K. & Tyack, P. L. 1990. Review of the signature-
whistle-hypothesis for the Atlantic bottlenose dolphin. In: The Bottlenose Dolphin,
ed. S. Leatherwood & R. R. Reeves. San Diego, CA: Academic Press, pp. 199–234.
Clapham, P. J. 2000. The humpback whale: seasonal feeding and breeding in a baleen
whale. In: Cetacean Societies: Field Studies of Dolphins and Whales, ed. J. Mann, R. C.
Connor, P. L. Tyack & H. Whitehead. Chicago, IL: University of Chicago Press,
pp. 173–196.
Clark, C. W. 1995. Application of US Navy underwater hydrophone arrays for
scientific research on whales. Reports of the International Whaling Commission, 45,
210–212.
Clark, C. W. & Clark, J. M. 1980. Sound playback experiments with southern right
whales (Eubalaena australis). Science, 207, 663–665.
Clark, C. W. & Ellison, W. T. 2000. Calibration and comparison of the acoustic location
methods used during the spring migration of the bowhead whale, Balaena
mysticetus, off Pt Barrow, Alaska, 1984–1993. Journal of the Acoustical Society of
America, 107, 3509–3517.
Clark, C. W., Ellison, W. T. & Beeman, K. 1986. A preliminary account of the acoustic
study conducted during the 1985 spring bowhead whale, Balaena mysticetus,
migration off Point Barrow, Alaska. Reports of the International Whaling Commission,
36, 311–316.
Cleator, H. J., Stirling, I. & Smith, T. G. 1989. Underwater vocalizations of the bearded
seal (Erignathus barbatus). Canadian Journal of Zoology, 67, 1900–1910.
Cummings, W. C. & Thompson, P. O. 1971. Gray whales, Eschrichtius robustus, avoid the
underwater sounds of killer whales, Orcinus orca. Fishery Bulletin, 69, 525–530.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 411
Dawson, S. M. 1991. Clicks and communication: the behavioural and social contexts of
Hector’s dolphin vocalizations. Ethology, 88, 265–276.
Deecke, V. B., Slater, P. J. B., Ford, J. K. B. 2002. Selective habituation shapes predator
recognition in harbor seals. Nature, 420, 171–173.
Erbe, C. 2002. Underwater noise of whale-watching boats and potential effects on
killer whales (Orcinus orca), based on an acoustic impact model. Marine Mammal
Science, 18, 394–418.
Erbe, C. & Farmer, D. M. 2000. Zones of impact around icebreakers affecting beluga
whales in the Beaufort Sea. Journal of the Acoustical Society of America, 108,
1332–1340.
Evans, W. E., Sutherland, W. W. & Beil, R. G. 1964. The directional characteristics of
delphinid sounds. In: Marine Bio-acoustics, ed. W. N. Tavolga. Oxford: Pergamon
Press, pp. 353–370.
Finley, K. J., Miller, G. W., Davis, R. A. & Greene, C. R. 1990. Reactions of belugas,
Delphinapterus leucas, and narwhals, Monodon monoceros, to ice-breaking ships in
the Canadian high Arctic. Canadian Bulletin for Fisheries and Aquatic Sciences, 224,
97–117.
Fish, J. F. & Vania, J. S. 1971. Killer whale, Orcinus orca, sounds repel white whales,
Delphinapterus leucas. Fishery Bulletin, 69, 531–535.
Foote, A. D., Osborne, R. W. & Hoelzel, A. R. 2004. Whale-call response to masking boat
noise. Nature, 428, 910.
Ford, J. K. B. 1989. Acoustic behaviour of resident killer whales (Orcinus orca) off
Vancouver Island, British Columbia. Canadian Journal of Zoology, 67, 727–745.
Ford, J. K. B. & Fisher, H. D. 1983. Group-specific dialects of killer whales (Orcinus orca)
in British Columbia. In: Communication and Behavior of Whales, ed. R. Payne.
Boulder, CO: Westview Press, pp. 129–161.
Frankel, A. S., Clark, C. W., Herman, L. M. & Gabriele, C. M. 1995. Spatial distribution,
habitat utilization, and social interactions of humpback whales, Megaptera
novaeangliae, off Hawai’i, determined using acoustic and visual techniques.
Canadian Journal of Zoology, 73, 1134–1146.
Frantzis, A. 1998. Does acoustic testing strand whales? Nature, 392, 29.
Hammond, P. S., Berggren, P., Benke, H. et al. 2002. Abundance of harbour porpoise
and other cetaceans in the North Sea and adjacent waters. Journal of Applied
Ecology, 39, 361–376.
Helweg, D. A., Frankel, A. S., Mobley, J. R. & Herman, L. M. 1992. Humpback whale
song: our current understanding. In: Marine Mammal Sensory Systems, ed. J. Thomas,
R. Kastelein & S. Supin. New York: Plenum Press, pp. 459–483.
Herman, L. M. 1987. Receptive competencies of language-trained animals. Advances in
the Study of Behavior, 17, 1–60.
Herman, L. M. & Tavolga, W. N. 1980. The communication systems of cetaceans. In:
Cetacean Behavior: Mechanisms and Functions, ed. L. M. Herman. New York: John
Wiley, pp. 149–209.
Hindell, M. A., Slip, D. J. & Burton, H. R. 1991. The diving behaviour of adult male and
female southern elephant seals, Mirounga leonina (Pinnipedia: Phocidae).
Australian Journal of Zoology, 39, 595–619.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
412 V. M. Janik
Hoelzel, A. R., Dahlheim, M. & Stern, S. J. 1998. Low genetic variation among killer
whales (Orcinus orca) in the Eastern Pacific and genetic differentiation between
foraging specialists. Journal of Heredity, 89, 121–128.
Hooker, S. K. & Baird, R. W. 1999. Deep-diving behaviour of the northern bottlenose
whale, Hyperoodon ampullatus (Cetacea: Ziphiidae). Proceedings of the Royal Society of
London, Series B, 266, 671–676.
Janik, V. M. 1999. Origins and implications of vocal learning in bottlenose dolphins.
In: Mammalian Social Learning: Comparative and Ecological Perspectives, ed. H. O. Box &
K. R. Gibson. Cambridge, UK: Cambridge University Press, pp. 308–326.
2000a. Source levels and the estimated active space of bottlenose dolphin (Tursiops
truncatus) whistles in the Moray Firth, Scotland. Journal of Comparative Physiology A,
186, 673–680.
2000b. Whistle matching in wild bottlenose dolphins (Tursiops truncatus). Science, 289,
1355–1357.
Janik, V. M. & Slater, P. J. B. 1997. Vocal learning in mammals. Advances in the Study of
Behavior, 26, 59–99.
1998. Context-specific use suggests that bottlenose dolphin signature whistles are
cohesion calls. Animal Behaviour, 56, 829–838.
2000. The different roles of social learning in vocal communication. Animal
Behaviour, 60, 1–11.
Janik, V. M., Dehnhardt, G. & Todt, D. 1994. Signature whistle variations in a
bottlenosed dolphin, Tursiops truncatus. Behavioral Ecology and Sociobiology, 35,
243–248.
Janik, V. M., van Parijs, S. M. & Thompson, P. M. 2000. A two-dimensional acoustic
localization system for marine mammals. Marine Mammal Science, 16,
437–447.
Jepson, P. D. Arbelo, M., Deaville, R. et al. 2003. Gas-bubble lesions in stranded
cetaceans. Nature, 425, 575.
Johnson, C. S. 1967. Sound detection thresholds in marine mammals. In: Marine
Bio-acoustics, Vol. 2, ed. W. N. Tavolga. Oxford: Pergamon Press, pp. 247–255.
1968. Masked tonal thresholds in the bottlenosed porpoise. Journal of the Acoustical
Society of America, 44, 965–967.
Johnston, D. W. 2002. The effect of acoustic harassment devices on harbour porpoises
(Phocoena phocoena) in the Bay of Fundy, Canada. Biological Conservation, 108,
113–118.
Johnstone, R. A. 1997. Recognition and the evolution of distinctive signatures: when
does it pay to reveal identity? Proceedings of the Royal Society of London, Series B, 264,
1547–1553.
Kako, E. 1999. Elements of syntax in the systems of three language-trained animals.
Animal Learning and Behavior, 27, 1–14.
Knudsen, V. O., Alford, R. S. & Emling, J. W. 1948. Underwater ambient noise. Journal of
Marine Research, 7, 410–429.
Lammers, M. O. & Au, W. W. L. 2003. Directionality in the whistles of Hawaiian
spinner dolphins (Stenella longirostris): a signal feature to cue direction of
movement? Marine Mammal Science, 19, 249–264.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 413
Le Boeuf, B. J., Naito, Y., Huntley, A. C. & Asaga, T. 1989. Prolonged, continuous, deep
diving by northern elephant seals. Canadian Journal of Zoology, 67, 2514–2519.
Madsen, P. T., Wahlberg, M. & Møhl, B. 2002. Male sperm whale (Physeter macrocephalus)
acoustics in a high-latitude habitat: implications for echolocation and
communication. Behavioral Ecology and Sociobiology, 53, 31–41.
Marsh, H. W. & Schulkin, M. 1962. Shallow-water transmission. Journal of the Acoustical
Society of America, 34, 863–864.
McDonald, M. A. & Fox, C. G. 1999. Passive acoustic methods applied to fin whale
population density estimation. Journal of the Acoustical Society of America, 105,
2643–2651.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Comstock, pp. 409–425.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Mercado III, E. & Frazer, L. N. 1999. Environmental constraints on sound transmission
by humpback whales. Journal of the Acoustical Society of America, 106, 3004–3016.
Miller, P. J. O. 2002. Mixed-directionality of killer whale stereotyped calls: a direction
of movement cue? Behavioral Ecology and Sociobiology, 52, 262–270.
Miller, P. J. O. 2004. Estimated active space of killer whale sounds. Journal of
Comparative Physiology A, in press.
Miller, P. J. O., Shapiro, A. D., Tyack, P. L. & Solow, A. R. 2004. Call-type matching in
vocal exchanges of free-ranging resident killer whales (Orcinus orca). Animal
Behaviour, 67, 1099–1107.
Morrice, M. G., Burton, H. R. & Green, K. 1994. Microgeographic variation and songs in
the underwater vocalization repertoire of the Weddell seal (Leptonychotes weddelli)
from the Vestfold Hills, Antarctica. Polar Biology, 14, 441–446.
Morton, A. B. & Symonds, H. K. 2002. Displacement of Orcinus orca (L.) by high amplitude
sound in British Columbia, Canada. ICES Journal of Marine Science, 59, 71–80.
Naguib, M. & Todt, D. 1997. Effects of dyadic interactions on other conspecific
receivers in nightingales. Animal Behaviour, 54, 1535–1543.
Naguib, M. & Wiley, R. H. 2001. Estimating the distance to a source of sound:
mechanisms and adaptations for long-range communication. Animal Behaviour,
62, 825–837.
Otter, K. A., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits (Parus major)
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society of London, Series B, 266, 1305–1309.
Payne, R. & Webb, D. 1971. Orientation by means of long range acoustic signaling in
baleen whales. Annals of the New York Academy of Sciences, 188, 110–140.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Premus, V. & Spiesberger, J. L. 1997. Can acoustic multipath explain finback (B.
physalus) 20-Hz doublets in shallow water? Journal of the Acoustical Society of America,
101, 1127–1138.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
414 V. M. Janik
Rendell, L. E. & Whitehead, H. 2003. Acoustic clans in sperm whales (Physeter
macrocephalus). Proceedings of the Royal Society of London, Series B, 270, 225–231.
Renouf, D. 1984. The vocalization of the harbour seal pup (Phoca vitulina) and its role in
the maintenance of contact with the mother. Journal of Zoology, 202, 583–590.
Richardson, W. J., Greene, C. R., Jr, Malme, C. I. & Thomson, D. H. 1995. Marine
Mammals and Noise. San Diego, CA: Academic Press.
Ross, D. 1976. The mechanics of underwater noise. New York: Pergamon Press.
Saayman, G. S., Taylor, C. K. & Bower, D. 1973. Diurnal activity cycles in captive and
free-ranging Indian Ocean bottlenose dolphins (Tursiops truncatus Ehrenburg).
Behaviour, 44, 212–233.
Sayigh, L. S., Tyack, P. L., Wells, R. S. & Scott, M. D. 1990. Signature whistles of
free-ranging bottlenose dolphins, Tursiops truncatus: mother–offspring
comparisons. Behavioral Ecology and Sociobiology, 26, 247–260.
Schevill, W. E. 1964. Underwater sounds of cetaceans. In: Marine Bio-Acoustics, Vol. 1, ed.
W. N. Tavolga. Oxford: Pergamon Press, pp. 307–316.
Schevill, W. E. & Watkins, W. A. 1971. Pulsed sounds of the porpoise Lagenorhynchus
australis. Breviora, 366, 1–10.
Schusterman, R. J. & Kastak, D. 2002. Problem solving and memory. In: Marine Mammal
Biology: an Evolutionary Approach, ed. A. R. Hoelzel. Oxford: Blackwell, pp. 371–387.
Scott, M. D. & Chivers, S. J. 1990. Distribution and herd structure of bottlenose
dolphins in the eastern tropical Pacific Ocean. In: The Bottlenose Dolphin, ed. S.
Leatherwood & R. R. Reeves. San Diego, CA: Academic Press, pp. 387–402.
Shane, S. H., Wells, R. S. & Wursig, B. 1986. Ecology, behavior and social organization
of the bottlenose dolphin: a review. Marine Mammal Science, 2, 34–63.
Smolker, R. A., Mann, J. & Smuts, B. B. 1993. Use of signature whistles during
separations and reunions by wild bottlenose dolphin mothers and infants.
Behavioral Ecology and Sociobiology, 33, 393–402.
Stafford, K. M., Fox, C. G. & Clark, D. S. 1998. Long-range acoustic detection and
localization of blue whale calls in the northeast Pacific Ocean. Journal of the
Acoustical Society of America, 104, 3616–3635.
Stafford, K. M., Nieukirk, S. L. & Fox, C. G. 2001. Geographic and seasonal variation of
blue whale calls in the North Pacific. Journal for Cetacean Research and Management,
3, 65–76.
Terhune, J. M. 1994. Geographical variation of harp seal underwater vocalizations.
Canadian Journal of Zoology, 72, 892–897.
Terhune, J. M., Stewart, R. E. A. & Ronald, K. 1979. Influence of vessel noises on
underwater vocal activity of harp seals. Canadian Journal of Zoology, 57, 1337–1338.
Thomas, J. A. & Golladay, C. L. 1995. Geographic variation in leopard seal (Hydrurga
leptonyx) underwater vocalizations. In: Sensory Systems of Aquatic Mammals, ed. R. A.
Kastelein, J. A. Thomas & P. E. Nachtigall. Woerden: De Spil, pp. 201–221.
Tyack, P. 1983. Differential response of humpback whales, Megaptera novaeangliae, to
playback of song or social sounds. Behavioral Ecology and Sociobiology, 13, 49–55.
1991. If you need me, whistle. Natural History, 8, 60–61.

P1: IYP/... P2: JZZ/...0521823617c18.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:33
Underwater acoustic communication in marine mammals 415
1998. Acoustic communication under the sea. In: Animal Acoustic Communication:
Sound Analysis and Research Methods, ed. S. L. Hopp, M. J. Owren & C. S. Evans. Berlin:
Springer, pp. 163–220.
1997. Development and social functions of signature whistles in bottlenose
dolphins Tursiops truncatus. Bioacoustics, 8, 21–46.
2000. Dolphins whistle a signature tune. Science, 289, 1310–1311.
Tyack, P. & Whitehead, H. 1983. Male competition in large groups of wintering
humpback whales. Behaviour, 83, 132–154.
Urick, R. J. 1983. Principles of Underwater Sound, 3rd edn. New York: McGraw-Hill.
van Parijs, S. M. & Corkeron, P. J. 2001. Evidence for signature whistle production by a
Pacific humpback dolphin, Sousa chinensis. Marine Mammal Science, 17, 944–949.
van Parijs, S. M., Hastie, G. D. & Thompson, P. M. 2000a. Individual and geographic
variation in display behaviour of male harbour seals in Scotland. Animal Behaviour,
59, 559–568.
van Parijs, S. M., Janik, V. M. & Thompson, P. M. 2000b. Display area size, tenure and
site fidelity in the aquatic mating male harbour seal. Canadian Journal of Zoology,
78, 2209–2217.
Watkins, W. A. 1981. Activities and underwater sounds of fin whales. Scientific Reports
of the Whales Research Institute, 33, 83–117.
Watkins, W. A. & Schevill, W. E. 1972. Sound source location by arrival times on a
non-rigid three-dimensional hydrophone array. Deep-Sea Research, 19, 691–706.
1977. Sperm whale codas. Journal of the Acoustical Society of America, 62, 1485–1490.
1979. Distinctive characteristics of underwater calls of the harp seal, Phoca
groenlandica, during the breeding season. Journal of the Acoustical Society of America,
66, 983–988.
Watkins, W. A., Tyack, P., Moore, K. E. & Bird, J. E. 1987. The 20-Hz signals of finback
whales (Balaenoptera physalus). Journal of the Acoustical Society of America, 82,
1901–1912.
Wells, R. S., Scott, M. D. & Irvine, A. B. 1987. The social structure of free-ranging
bottlenose dolphins. In: Current Mammalogy, ed. H. H. Genoways. New York:
Plenum Press, pp. 247–305.
Wells, R. S., Boness, D. J. & Rathbun, G. B. 1999. Behavior. In: Biology of Marine Mammals,
ed. J. E. Reynolds III & S. A. Rommel. Washington, DC: Smithsonian Institute Press,
pp. 324–422.
Winn, H. E., Thompson, T. J., Cummings, W. C. et al. 1981. Song of the humpback
whale: population comparisons. Behavioral Ecology and Sociobiology, 8, 41–46.
Xitco, M. J. & Roitblat, H. L. 1996. Object recognition through eavesdropping: passive
echolocation in bottlenose dolphins. Animal Learning and Behavior, 24, 355–365.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
19
Looking for, looking at: social control,honest signals and intimate experiencein human evolution and history
j o h n l . l o c k e
City University of New York, USA
Introduction
Recently, Hauser et al. (2002) argued that if we are to understand human
language, several disciplines must work cooperatively. Predictably, these include
linguistics and certain areas within psychology and anthropology as well as some
relative newcomers: biology and animal behaviour. However, if collaboration can
facilitate the investigation of language, long held to be a uniquely human faculty,
it is surely indispensable to the study of human communication, for which a
number of homologous or analogous processes exist in other species.
In the case of language, a behaviour with countless social benefits, researchers
have tended to focus on dyadic interactions. In the typical model, the ‘sender’ is a
rational human being who has information. As a social being, the sender wishes to
share it. The ‘receiver’, equally rational and social, wants to hear it; so the receiver
listens and makes an appropriate response. ‘Communication occurs,’according to
one authoritative source, ‘when one organism (the transmitter) encodes informa-
tion into a signal which passes to another organism (the receiver) which decodes
the signal and is capable of responding appropriately’ (Ellis & Beattie, 1986, p. 3).
Dyadic interactions such as these occur, of course, and deserve linguists’ theo-
retical attention. However, in a gregarious species such as ours – and this is a major
point of divergence between social communication and linguistic interaction –
dyads are often embedded in aggregations of individuals, in various arrange-
ments (communication networks in the sense of this book), and these will usually
include one or more perceptual bystanders. If thought to be unobserved, dyads
tend to behave in an unguarded way, making them unusually interesting, and
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
416

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 417
their behaviour unusually informative, to uninvited viewers and listeners. Of
course, the perceptual target may be alone, acting without recourse to the dis-
plays or material objects that are normally used to project his more public self.
These solitary behaviours will be less veiled than the dyadic interactions, mak-
ing them especially useful to anyone who stands to benefit from prediction-grade
social knowledge.
What I am suggesting is that behaviour which is neither donated by the actor
nor observed with his awareness is likely to be unusually high in reliability. It is
also likely to be intimate: that is, sufficiently personal that the actor might like
to shield it from prying eyes and ears. Reliability and intimacy give prospective
observers two rather compelling reasons to sample such behaviour, but it will
usually be impossible for them to do so overtly. This gives rise to eavesdropping,
a form of information gathering that in humans occurs only by stealth.
In animals, research has addressed two broad areas of observation. One relates
to the information that is obtained when animals look for other animals. When
non-human primates do this type of looking, their focus is typically on the loca-
tion and activities of outsiders, including predators and competitors from other
groups. Typically, this is referred to as vigilance. Other types of information are
obtained when animals look at the constituents of their own groups. This type
of looking, social observation, is addressed later in the chapter and elsewhere in
this volume (e.g. Ch. 25). Predictably, these discriminable functions are associated
with different benefits.
Looking for
Animal vigilance
When animals look for other animals, their tendency is to scan territorial
boundaries in order to detect encroachment of predators or competitors. Early
detection alerts individuals and, through their reactions, other group members
to the need for evasive or defensive action. The perceptual act is performed from
the naturally exposed position of group-living animals. Vigilance appears to be
a form of perceptual alertness that occurs in anticipation of important events,
rather than a form of observation per se, and may even be discontinued when
those events occur. The observing itself is performed as sporadic interruption of
other activities, rather than a circumscribed commitment of looking time (an
exception, ‘sentinels,’ will be discussed below).
Much of the research on vigilance involves non-human primates. They, like
other animals, need to look out for predators. Vigilance thus produces valuable
information, but it comes at a price. Red colobus Procolobus badius tephrosceles and
redtail monkeys Cercopithecus ascanius schmidtii typically spend over 50% of their

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
418 J. L. Locke
time just visually scanning (Treves, 1998). Similar figures have been obtained for
chacma baboons Papio cynocephalus ursinus (Cowlishaw, 1998). This does not count
all the time these primates spend in a vigilant state, a figure that may approach the
totality of free-ranging animals’ waking time.
In several different species of monkey, animals that were physically isolated
spent more time looking than monkeys that were near a group member (Steenbeek
et al., 1999; Treves et al., 2001). This difference may reflect a greater fear of predation
on the part of solitaires. Where attacks are unlikely, however, animals might be
expected to devote more of their attention to members of their group. Research on
brown capuchin monkeys Cebus apella in Iguazu Falls National Park in Argentina,
where the annual rate of predation is extremely low, suggests that this may be so
(Hirsch, 2002).
Predictably, there are also variations in time spent looking among the mem-
bers of a single group. Subordinate animals tend to look more than dominant
ones, largely because they spend a great deal of time watching the dominant
animals themselves. These rank differences were suggested some years ago by
Chance (Chance, 1967; Chance & Jolly, 1970) and have since been confirmed in
a number of species, including long-tailed macaques Macaca fascicularis (Pitcairn,
1976), talapoins Miopithecus talapoin (Dixson et al., 1975; Keverne et al., 1978) and
brown capuchins (Hirsch, 2002). In each group, subordinates more often look at
dominant animals than the reverse arrangement.
In many studies, there have also been sex effects. A male vigilance bias has been
witnessed in various primate groups in at least seven different studies (Cheney &
Seyfarth, 1981; Fragaszy, 1990; Baldellou & Henzi, 1992; Rose, 1994; Rose & Fedigan,
1995; Gould et al., 1997; also see reviews by Quenette, 1990; Steenbeek et al., 1999). A
great deal of male vigilance appeared to be directed outside the group, presumably
to predators or sexual competitors, but perceptual targets are notoriously difficult
to identify in free-ranging animals.
There is one exception to the usual hierarchical pattern. High-ranking males
frequently assume unusual responsibilities for vigilance (Rose & Fedigan, 1995).
In cooperative groups such as vervets and marmosets, as well as baboons, they – or
some other large male – may even take on the role of sentinel and adopt a superior
vantage point. These individuals then become the focus of attention for group
members, who monitor the sentinel instead of looking for predators themselves
(Hall, 1960; Horrocks & Hunte, 1986; Koenig, 1994).
The behaviour of sentinels has been described in detail. The tendency is for the
sentinel to ascend a tree or rock, mainly so foraging animals can achieve visual con-
tact, making auditory warnings unnecessary (Horrocks & Hunte, 1986). In green
monkeys Cercopithecus aethiops sabaeus, this works so well that when a sentinel
detects approaching humans, his quietly visible movements may enable the troop

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 419
to disperse without detection (Poirier, 1972). Therefore, in this species at least, the
sentinel seemed to ‘tip off ’ the foragers with subtle visual behaviours instead of
warning them with loud barks. In the case of baboons, the vigilance itself involves
repeated 180◦ head turns, which take about five seconds (Hall, 1960), and in this
group detection produces barks. In every report I have read, the only ‘predators’of
possible concern to the sentinel were human and the ongoing feeding behaviours
best characterized as ‘raids’ on a plantation (Maples et al., 1976). Consequently the
use of sentinels may be predator and context specific.
Much of the time, the targets of vigilant males are other males, including
interlopers from outside conspecific groups (Rose & Fedigan, 1995). Therefore, the
motivation for male vigilance may be more closely linked to a self-oriented control
function than a contribution to group welfare. This issue will assume a broader
significance below when we see some related sex differences in our own species.
Levels of vigilance vary, affected by a variety of internal factors such as mating
periods, births and infant excursions away from their mothers. In vervet monkeys
Cercopithecus aethiops, males are more vigilant than females, especially during the
breeding season (Baldellou & Henzi, 1992). In a study of black howler monkeys
Alouatta pigra, female vigilance rates increased after the birth of infants (Treves
et al., 2001). In squirrel monkeys Saimiri boliviensis, tape recordings of infant vocal-
izations increased the time that adult females spent looking for predators fivefold
(Biben et al., 1989).
In several species, it has been shown that visual obstruction alters the usual
benefits of herding and flocking. The first to study this effect was Underwood
(1982). He noted that African antelopes frequently interrupted their foraging to
look around, but when grazing in tall grass they spent even more time lifting their
heads to look at distant areas. Metcalfe (1984a,b) observed a similar pattern in two
different species of shorebirds in western Scotland. He found that in both species
the time devoted to vigilance rose with increases in the density of obstructions such
as rocks, boulders and banks of seaweed. He also noticed that obstructions broke
up the usual relationship between flock size and vigilance. Metcalfe reasoned that
obstructed animals were in a vulnerable position, unable to see if predators or
potentially protective neighbours were nearby.
Recapitulating, the primary functions of primate vigilance appear to be defen-
sive when looking is externally directed and the threat of predation is high, and
social when looking is internally directed and the threat of predation is low. In
the latter, vigilance enables animals to evaluate dominance relations – a criti-
cal function in primate societies – and resource-holding potential. In the species
studied, the primary sensory modality has been visual. In non-human primates,
males and subordinates generally devote more time to vigilance than females and
dominants. The target of review is a physical area, such as the perimeter of an

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
420 J. L. Locke
occupied territory, or other animals that are either within or outside the group.
The observer may make no attempt either to conceal or to expose his position.
Human vigilance
Humans have not been reluctant to engage in the ancient and deeply
ingrained behaviour of vigilance. Some of the benefits derived from these activities
resemble those enjoyed by our evolutionary ancestors, broadly understandable as
social knowledge and social control. As we will see, our species critically relies
on information that can only be obtained through these one-way processes, and
yet there is little record of empiricism. To be sure, there have been psychological
studies of vigilance – usually defined as the detection of prespecified perceptual
targets that occur infrequently, irregularly and weakly – in relation to a range of
military and industrial issues. Currently, there is concern with baggage scanning
in relation to airport security screening.
It is difficult, however, to find reports of research conducted within an ethologi-
cal framework. Few investigators have asked how humans exercise vigilance with
respect to strangers or potentially aggressive intruders. Yet, in societies wishing
to guard against crime and terrorism, citizens are concerned with precisely this
issue.
Until about 20 000 years ago, our ancestors spent much of their time follow-
ing herds of large animals from place to place. But when the herds dissipated,
nomads began to hunt smaller game, to fish and to gather. This shift enabled
the new sedentists to spend more time in their resting places before seasonal
changes precipitated the next round of migration. At this stage in history, one
assumes that human and non-human primates behaved rather similarly with re-
spect to vigilance. Since human groups were several times larger, there were more
individuals that had to be monitored, but this was obviously manageable as our
premodern ancestors lived almost as openly as the other primates.
The nature of their encampments is implied both by archaeological evidence
and the behaviour of an existing group whose way of life is thought to replicate
ancient patterns of living (Lee, 1979). This group is the !Kung, a population of
largely egalitarian hunter–gatherers who inhabit the Kalahari Desert of Botswana
and southwest Africa. Although their way of life is changing, in the mid- to late-
twentieth century, when they were studied fairly intensively, most of the !Kung
lived in bands of 50 or 60. These bands periodically dispersed into still smaller
groups or concentrated into larger ones, as suited their needs.
The typical camp was laid out in concentric circles. In the centre was a public
gathering place. Rimming this plaza were the bandsmen’s grass huts, which were
used mainly for storage. These were packed very closely together, enabling bands-
men to perceive and react to the earliest and subtlest acts of an antisocial nature.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 421
‘If a person is angry,’ wrote Draper (1978, p. 47), ‘someone, if not everyone, will
soon know about it.’
Significantly, the !Kung rarely if ever entered their huts to escape scrutiny. The
reason is that it was considered improper for anyone to withdraw from the sociality
of camp life, either physically or psychologically. ‘To seek solitude,’ according to
Lee (1979, p. 32), ‘is regarded as bizarre behaviour.’But this attitude toward privacy
was not unique to the !Kung. There are several other openly living groups.
1. The Baktaman of New Guinea. ‘There are no recognised and respected
ways in which the public gaze can be cut off, no way of separating
oneself out from others present’ (Barth, 1975, p. 24).
2. The Mehinacu of Central Brazil. ‘Wherever a person goes in the village
he can be seen or heard. When he speaks there is a chance that a third
person is listening, and that in a short time everyone else will know
what he said. Even the most intimate details of his sex life often become
a matter of public knowledge’ (Gregor, 1970, p. 238).
3. The Nayaka of southern India. ‘They remain sited by their respective
fire-places, and talk across space from fire to fire . . . they rarely try to
conceal their domestic activities’ (Bird-David, 1994, pp. 590–591).
4. The Samoans. They ‘live most of their lives in a very public arena. The
more private aspects of experience are strongly discouraged by the
absence of walls in a Samoan house, and by powerful norms of social
life, which keep people in almost constant social interaction’ (Shore,
1982, p. 148).
5. The Sakalava people of Madagascar. ‘To stay alone in the house is
considered a sure sign of evil intent.’ (Feeley-Harnik, 1980, p. 568). A
house with curtains on the outside doors, or fences and walls, was also
seen as a threat to normal sociality. Even the house itself could pose
problems, Feeley-Harnik wrote, since it is meant to remove the
occupants from the larger social order. ‘Secrecy and separation,’ she
continued, ‘indicate at best a lack of generosity, a suspiciously
anti-social striving for distinction’ (Feeley-Harnik, 1980, p. 581).
6. Villagers in the mountainous Zinacantan region of southern Mexico.
They too have also been suspicious of too much domestic privacy. The
typical home is fenced in, and village folk are forbidden from passing
through the fence without prior approval. However, staying indoors, or
closing the house door, is considered ‘a gross and open admission of
being up to no good’ (Haviland & Haviland, 1983, p. 347).
Many of these cultures that have opposed privacy and favoured social visibil-
ity were egalitarian; according to Bailey (1971, p. 19), ‘equality is the reward for

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
422 J. L. Locke
constant vigilance’. In parallel, there was also a suspiciousness of structures and
behaviours that reduced visibility, since these would surely foil the only proven
means of keeping the group together and under control.
I consider that there are three benefits of vigilance and eavesdropping: social
control, honest signals and intimate experience. Let us now examine the first of
these, which is closely linked to vigilance. Honest signals and intimate experience
are tied to privacy, thus to eavesdropping, and we will address these benefits in
that section. Since there is little in the way of relevant research, my treatment of
social control will necessarily be historical, discursive and somewhat speculative.
Social control
Some things that occur in private are intended to be secret. They may be
offensive, morally wrong or even criminal. There are good reasons for humans to
observe this activity, too, but in some cases there may be little benefit in doing so
covertly. The reason is that looks, if interpreted as gazes, can also send messages of
their own. Some are confrontations that vary in intensity from ‘I see you’ to ‘I’m
keeping an eye on you’ and, in the extreme case, ‘Back off ’.
On the community level, the protective function of surveillance has always
been clear. ‘If by chance some good-for-nothing appeared in the neighbourhood,’
wrote Yves Castan (1989a, p. 49) in reference to French villages, ‘there were plenty
of eyes to survey his movements’. In Victorian England, the rich and powerful
lived side by side with the poor and powerless. This made it possible for each
group to observe the other and particularly for establishment figures to keep an
eye on potentially troublesome subordinates. ‘The middle-classes desired privacy
for themselves,’ wrote Olsen (1974, pp. 275–276) ‘but wished the lives of the lower
orders to be lived in the full blaze of publicity. Street improvements and slum
clearance schemes were designed to bring the poor out into the open, where they
could be observed, reproved and instructed by their superiors.’
On an individual level, vigilance also enables humans to avoid quarrelsome
or dangerous people, our equivalent of predators. In large cities, one is forced to
acquire ‘street smarts’, an awareness of menacing strangers in relation to oneself,
and the relation of one’s own location to places of safety. Predictably, the best
security – as criminologists have shown – is the presence of some reasonable
number of non-predatory people on the street. For a city to be safe, ‘there must be
eyes upon the street,’ wrote Jacobs (1961, p. 45), ‘eyes belonging to those we might
call the natural proprietors of the street’. Research in the ensuing years has been
supportive of this view (Kelling & Coles, 1996).
Primates in hierarchically organized groups spend more time looking at each
other when they could be looking for predators or food (Caine & Marra, 1988)
and animals do more social looking within mixed than in homogeneous groups
(Treves, 1999). These findings are relevant to Putnam’s (1993) study of provincial

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 423
self-government in Italy. Provinces that lacked trust, he found, spent a great deal
of time keeping an eye on each other.
Watching humans
With vision alone, non-human primates pick up cues to sex, age and rank,
the last inferred from dominance and submissive displays. We humans care about
these things, too, and also transmit much of our information visually, through
physical alterations and adornments. Among the Kayapo of the Amazon forest,
visible affectations include pierced ears, lip plugs, penis sheaths and body painting
(Turner, 1980). These adornments convey messages about status as well as personal
roles and significance and do so just as surely as verbal signals. In modern societies,
hairstyle, cosmetics, jewellery, eyeglasses, tattoos and body rings – to say nothing
of cell phones, water bottles, clothes, shoes, handbags, briefcases, fanny packs,
shopping bags and backpacks – send visual signals about who we are or how we
wish to be perceived.
The desire to enhance personal images goes back at least 28 000 years. Studies
of the ‘Venus’ figurines and burial sites indicate that women many millennia ago
were already wearing hats, dresses and various bodily adornments (Soffer et al.,
2000). This suggests that, before they were securely and privately housed, our
historical ancestors already had some sense of self, a matter to which we will
return shortly.
We are not, of course, merely intelligible through our clothes and other objects
of material culture. Like other species, humans have a number of ritualized action
patterns that presuppose visualization (cf. Smith, 1977). These include the facial
and bodily displays that emerge in infancy, are seemingly universal (Schiefenhovel,
1997) and occur in blind as well as sighted infants (Eibl-Eibesfeldt, 1973). Under
the influence of culture, humans take on additional gestures – some functioning
as salutations, others signalling transition points in verbal engagements (Kendon,
1990) – and learn rules of proxemics that suggest possible ranges of interpersonal
distance (Hall, 1966). Personal status and relational intimacy are also revealed by
touching (Hall, 1996) and, in the case of single women in America, hair flips and
head tosses (Moore, 1985).
The eyes send many different types of social and emotional signal. We saw
earlier that in primates socially dominant individuals receive more gazes than
subordinate ones. This relationship also holds in humans. At any given moment
in time, the person who is being looked at is usually the person who is talking and
that will typically be the person with the highest status (Bales et al., 1951; Fisek &
Ofshe, 1970; Exline et al., 1975; Abramovitch, 1976; Kalma, 1991).
With all these visible signals, it would be surprising indeed if people did not
create opportunities to be looked at and to do so on their own terms. In 1800,
Parisians began to put their public selves on parade. That is when pavements

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
424 J. L. Locke
came to Paris and merchants adaptively repositioned their shops and displays.
In the new pavement cafes, the chairs were ‘always placed towards the street,’
wrote an urbanologist, ‘as the chairs in a theatre are placed towards the stage’
(Oosterman, 1992, p. 161).
Promenades were once expected to achieve an instructive or regulatory func-
tion. When a family went out for an evening stroll, it was assumed that the husband
would ‘see himself as others saw him,’ according to Cranz (1980, p. S80), ‘the head
of a family, wife on arm, children in tow, all in Sunday best. Reformers reasoned
that he would experience this as pleasurable and resolve to make it the mainstay
of his life.’ In 1890, the commissioners of Boston’s parks department saw public
viewings as a course of moral instruction. The mere sight of families was expected
to exert ‘a wholesome influence’on other patrons, and to do so far more effectively
than laws and police ever could (Cranz, 1980, p. 581).
In contemporary America, recreational vigilance is largely carried out in parks
and malls. In a survey conducted in the early 1970s, a fifth or fewer of the patrons
of two parks in Portland, Oregon said they went to the parks to walk, eat, talk,
read, engage in crafts or hobbies, or exercise. Far more patrons, fully 55%, said
they went to the parks in order to watch other people (Love, 1973). In a survey
conducted in the Los Angeles area a decade later, adolescents said that the main
reason they went to a particular mall, after shopping, was to look for members of
the opposite sex (Anthony, 1985).
There are several circumstances in which vigilance is exercised in relation to
intimate relationships. Buss (1988, 1997) surveyed American couples to see how
frequently they reported the use of vigilance in order to control intimate relation-
ships. Items in the survey included unexpectedly calling and dropping by a place
to see if the partner was there and remaining nearby, or at least in visual contact,
during social engagements. Men and women reported equal levels of vigilance,
but there was a significant correlation for men, and not women, between levels
of vigilance and ratings of partner attractiveness.
Control and intimacy are also conjoined in many cases of the crime ‘stalking’.
Since stalking is usually defined as an unwelcome act of ‘perceptual following’
that is overt or blatant, it qualifies as vigilance. In one study, 57% of stalkers had
previously been in an intimate relationship with the victim (Hall, 1998). In another
study, approximately a third of all stalkers were considered intimacy seekers. Most
lived alone and had never had a romantic partner (Mullen et al., 2000).
Ethological studies of human vigilance
In non-human primates, individuals tend to look up less often when a
group member is nearby (Hirsch, 2002). A similar trend has been found in humans.
Observing students in a university snack bar, Barash (1972) found that cumulative

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 425
looking-up frequency was significantly higher in solitaires than individuals in
groups. In a similar study conducted in Germany, Wirtz & Wawra (1986) observed
university students having lunch in a refectory. Each sat alone or with one to four
other students. In these subjects, it was found that the time spent looking away
from the table steadily decreased as the number of number of people at the table
increased, possibly because this increased the proximity of others.
In Wirtz & Wawra (1986), male students spent significantly more time looking
away from their table than females. This fits with primate research, reviewed
earlier, that revealed a male looking bias, particularly for distant areas. It also
agrees with Aiello (1972, 1977), who found that men looked significantly longer
at each other than women did when seated 10 feet (3.2 m) or more apart, a trend
that was reversed for shorter distances.
Paradoxically, most of the work on vigilance in humans involves detection
of signals, whereas in animals, vigilance involves attention to the existence and
behaviour of individuals (also see studies of social monitoring and comparison).
The disposition of females to look longer at near individuals may be linked to a
tendency to rely on the support of group members, while the disposition of males
to look longer at distant individuals may be associated with the need to address the
threats posed by strangers. Stripped to the basics, here are two issues – intimacy
(the network ‘glue’) and control – that concern human women and men. These
issues, as we will see, have been connected to sex differences in social monitoring
for the last six or seven centuries of recorded history.
Looking at
Eavesdropping in animals
Much information is acquired by social observation: looking at con-
specifics. For example, male Mallee dragon lizards Ctenophorous fordi produce sig-
nificantly more ejaculate and spend 60% more time copulating with a female
previously seen copulating with another male than do males not having this prior
perceptual experience (Olsson, 2001). In this example, the source of information
did not involve signals. However, an important subset of observational informa-
tion comes from the signals of others. Such information is gathered by eavesdrop-
ping, a behaviour that is defined in animals as ‘the use of information in signals by
individuals other than the primary target’ (Ch. 2). The context for eavesdropping
is a communication network (Ch. 1); therefore it is not surprising that this volume
discusses at length the evidence for eavesdropping by animals (e.g. Chs. 2 and 5).
There have also been a number of recent reviews of eavesdropping (e.g. McGregor &
Peake, 2000).

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
426 J. L. Locke
Among the primates, maintenance of societies – including kin and power rela-
tions – requires that individuals spend a certain amount of time gathering infor-
mation about other group members. These internal appraisals, which in human
research are usually called social comparison and in non-human primates are con-
sidered a form of social vigilance (Hirsch, 2002), are required if animals are to alter
or maintain their status. Since Cheney & Seyfarth (Ch. 25) describe research in this
area, I will limit my own review to studies that expose links to our own species.
Primates’resources include cooperative relationships, which may involve high-
ranking animals. As Whiten (1993, p. 719, italics his) has pointed out, ‘simply to be
seen by others grooming with high-ranking A, or chatting with high-status B, is worth
something to the individual because of what this advertises with respect to future
coalition.’ An individual that has these kinds of social resource is considered to be
rich in ‘social attention holding potential’ (Gilbert, 1989). Animals may also look
within the group for individuals with valued physical resources. For example, a
perceptual target of so-called ‘scroungers’ is the foraging success of other animals
(Beauchamp, 2001).
There are variations between species in social vigilance, partly because of dif-
ferences in social organization. Consider squirrel monkeys, which live in large
groups that are characterized by cliques, subgroups and dominance hierarchies,
and cotton-top tamarins Saguinus oedipus, who live in more egalitarian family
groups known for cooperation, sharing and relative peacefulness. In a compar-
ison of social looking during foraging, the congenial tamarins devoted 17% of
their time to within-group vigilance. The more competitive squirrel monkeys, by
contrast, devoted 45% of their time attending to group members (Caine & Marra,
1988).
A feature of eavesdropping by animals is that it is usually carried out by isolated
individuals who do not subsequently share their perceptual intake with others,
although in a densely populated area other observers may individually sample
the same activity on their own. The perceptual target of eavesdropping is often a
pair or small group of individuals, which provides the observer with interactive or
relational information connected with fighting or sex, and it does so with minimal
risk or expenditure of effort.
Eavesdropping in humans
In this section, I follow accepted semantic practice and use the term eaves-
dropping only where the act of observation occurs surreptitiously. I also include
cases of social vigilance that do not involve signals or interactions (as do Cheney &
Seyfarth in Ch. 25).
I noted earlier that domestic vigilance has gone largely unstudied ethologi-
cally, but when it comes to human eavesdropping there is no record of empiricism

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 427
whatsoever. To be sure, there are publications that use the word eavesdropping,
but these typically describe government anti-crime programmes that include wire-
tapping and surveillance. What is missing is research on the behaviour of social
eavesdropping in a naturalistic context.
The lack of research on human eavesdropping seems odd, since the practice
is neither rare nor lacking in benefits. When asked, people usually admit that
they have eavesdropped in the past, or even do so habitually. Frequently, the
admissions are offered shyly, occasionally with embarrassment, but I have yet to
find anyone who denies ever having engaged in this practice. This is not to say
that everyone peeks through keyholes. Most of us ‘tune in’ less adventurously.
When in a restaurant or waiting room, for example, we tend to accomplish our
perceptual business in a number of optical stabs, interrupted by bogus glances at
other features of the physical or social landscape. If the subject suddenly looks
up, the invasion may be disguised by a slow and smooth deflection, as though a
continuous sweep was in progress when the ‘interruption’ occurred.
If people are naturally inclined to penetrate the private spaces of others, and
just as naturally resist such intrusions themselves, one might expect historical
evidence of these dispositions, perhaps in art or literature. In fact, there was activ-
ity in both media in the seventeenth century, from the paintings of Dutch artist
Nicolaes Maes to the novels of Le Sage and Hawthorne and the plays of Marivaux.
These depictions suggest that our historical ancestors were acutely aware of eaves-
dropping and may even have approved of it. However, there are also church and
court documents going back three centuries earlier, in several different cultures,
and these tell us something about the relative frequency of eavesdropping as a
behaviour, and a crime.
The !Kung hunter–gatherers, as we have seen above, welcomed round-the-clock
surveillance and intentionally subjected themselves to a panoptical living arrange-
ment (cf. Bentham, 1791). This made vigilance easy, but for the same reason it made
covert eavesdropping impossible (P. Wiessner, personal communication).
One assumes the !Kung’s residential arrangement was somewhat representa-
tive of historically earlier ways of living, when variations in the availability of
food required individuals constantly to relocate. With the advent of agriculture,
however, the new sedentists departed from the hunter–gatherer pattern, build-
ing huts that could be lived in – not just used for storage – and spacing them
more widely. This necessitated an aggressive form of perceptual intrusion. At the
same time, groups began to expand, and strangers grew more numerous. In a brief
space of time, a lifestyle that was two million years old – open living, with visual
monitoring – began to unravel.
Before the development of structural privacy as it is enjoyed in modern soci-
eties, some degree of solitude was achieved behaviourally. Bird (1983) reported that

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
428 J. L. Locke
the Naiken people of India exhibit nachika, literally a ‘shyness’ or ‘reticence’ that
protects them from direct encounters with others. Since the Naiken live openly,
reticence provides relief against what Bird called ‘involuntary intimacy.’
Similar observations were made by Fejos (1943), who studied the Yagua people
of northeastern Peru in the early 1940s. All the families of a clan, which ranged
from 25 to 50 members, lived communally in one large house. Fejos noted that
although there were no partitions, members could achieve privacy at any time
simply by turning away. ‘No one in the house,’ wrote Fejos (1943, p. 87), ‘will look
upon, or observe, one who is in private facing the wall, no matter how urgently
he may wish to talk to him’.
Note that the privacy achieved by these individuals was in each case negotiated
with, and conferred by, others. It began when the privacy-seeking individual gave
an observable sign. The observers, out of respect for the person, then reduced or
suspended evaluation. Goffman (1963) called this ‘civil inattention’.
Perhaps these behavioural means of securing privacy were sufficient, for even
with inclement weather, social competitors and wild animals, little interest was
shown in domestic walls (Carpenter, 1966; Rapoport, 1969). Consequently one is
curious about the residents of more hospitable climes who nonetheless chose
to live behind walls. The reasons for these exceptions to climatic determinism,
Rapoport (1969) pointed out, may have had something to do with religion, status
or some ‘other’ factor. One candidate for the ‘other’ factor, according to Wilson
(1988), would have been the desire to escape constant scrutiny. But there is another
possibility.
We have already seen that the !Kung sat in full view of each other during their
time in camp. If new members continually join such arrangements, eventually
something has to give. Individuals who cut back on their looking time will dis-
cover that the machinations of an increasingly complex, if not Machiavellian,
society have left them frightfully unaware and out of step. Alternatively, those
who continue to crank up their looking time in step with population growth will
soon have no time to do anything but look. Therefore, the critical factor may have
been the need to minimize the time that they, as members of burgeoning groups,
had to spend surveying the social landscape.
Little wonder that the desire for privacy grew as people became accustomed to
domestic life. In the 1960s, the Sarakatsani were a small group of shepherds who
alternately, by season, inhabited the Zagori Mountains and plains of Greece. To
them, a hut was inviolate. ‘Whatever takes place within the sanctuary of its walls
is private and sacred to the members of the family’ wrote Campbell (1964, p. 292).
‘No stranger may invade it without an invitation.’
Occupants could only be safe by assuming that they, like birds (e.g. Metcalfe,
1984a,b), grazing animals (e.g. Underwood, 1982) or isolated monkeys (e.g. Treves

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 429
et al., 2001) were ineligible for assistance. Therefore, the need was for walls
that were not merely visually obstructive but also secure. Eavesdropping became
the only way to restore information that had once been available. However,
eavesdropping was not merely restorative, for with increasing privacy, this very
penetrant means of observation became the only way to obtain the newest
and highest grade of personal information, one that had never been available
previously.
Honest signals
As people spent more time behind walls, direct sensory information about
them became less available. This posed problems for the community, but the
experience of privacy also altered the people themselves. In time, the most honest
and reliable information about individuals was only available to those who were
behind the walls, or in personal relationships, with them. Others were excluded.
Therefore, behaving as trained ethologists, eavesdroppers attempted to conceal
themselves in order to avoid detection, which would alter or discontinue the flow
of desired information.
People behave differently when they believe others are unable to see them.
When shielded from public view, they have the opportunity literally to compose
themselves – to decide who they are and how they would like to be perceived
by others. When they plunge into the social world again, they may then do so
appropriately dressed and ornamented, presenting others with the image they
would most like to convey.
Earlier I referred to a low-grade sense of self that antedated, or occurred early in
the development of, domestic life. While the hominids may have had some level of
self-awareness, along with the other primates (Hauser, 2000), every domestically
living human now has two selves: one public, the other private. The public self is
the way we are in the presence of others. Our private self is on view only when
individuals are alone or with intimate friends. There is a telling fact about the
private self, in connection with the process of perceptual theft. It is, as Baumeister
(1986, p. v, italics mine) said, ‘the way the person really is’.
The dishonest signals that are issued in public are not worthless, of course.
These may provide information as to the way a person really is not. For example, a per-
son who is making a conspicuous display of wealth may be ‘financially strapped’–
not wealthy at all – and also seeking to hide this fact for a reason, one that with
further analysis may be discovered. Still, people in private are likely to act in ways
that are, as Baumeister said, more ‘them’. This fact, by itself, increases the reliabil-
ity of private behaviour. However, private behaviour is also privileged. This gives
others reasons to want it, for as humans they have the inherited dispositions of
evolutionary ancestors whose survival was dependent on the ability to observe

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
430 J. L. Locke
behaviour and infer intentions, and they enjoy vicariously experiences that are
not issued for the benefit of observers.
Intimate experience
After nearly two million years of watchfulness, walls enabled the chroni-
cally wary Homo sapiens to cast its senses inwards for a change. Dwellers could pay
undivided attention to compelling tasks not just for three seconds, but for three
hours. Free from the stares and queries of villagers, the new residents could begin
to examine their own lives and think about how they differed from others. With
shielding, they could create or discover the existence of a deeper and more reflec-
tive form of themselves and begin to contrast this with their public presentation.
The time that domestication liberated from external vigilance could be devoted
to matters that were occurring – or with additional attention could be initiated –
on the inside. These would have included personal and communal activities. Fam-
ily members could, at last, devote their undivided attention to each other. By
creating an ‘outside’, individuals found ways in which more intimate relations
could be developed with members of the family on the ‘inside’. Consequently, to
look inside a house was to get unprecedented glimpses of intimate behaviour.
If someone peeks through a crack or keyhole, how will this come to the attention
of researchers? Eavesdroppers are no more likely to be detected by an ethologist
than by their perceptual prey, nor would they be likely to describe their activities
truthfully to an interviewer. The situation seems hopeless, and yet we do know
something about eavesdropping, especially the kind that occurred many centuries
ago when the threat of privacy was new and attempts to breech it were frequent,
adaptive and perhaps even honourable. In sixteenth century England, there was
a law against adultery and it required eye-witness testimony. Court records have
been preserved, revealing the testimony and identity of witnesses. Frequently, the
lead or sole witness was a woman who had peeked through a door from within
the house, or crack in the wall from an adjoining house. In a case that occurred in
London in 1598, a housewife named Margaret Browne watched a tryst involving
the woman who lived next door, her looking ‘bout’ – like the adulterous activity
itself – lasting for an entire afternoon (Crawford & Gowing, 2000).
I have inspected many cases involving this sort of domestic eavesdropping.
Although I kept no detailed count, it is clear that the typical perpetrator was
female. One might suppose that this is because women were merely home more
often, but in the sixteenth and seventeenth centuries, the husband was often
somewhere about the house, too. When Margaret Browne saw what was happening
next door, she called her husband to the crack to confirm her observations.
Mr Browne took a brief look and left, but Margaret remained at the crack,
taking mental notes. Her courtroom testimony two weeks later was extraordinarily
detailed, down to the exact words and phrases of the lovers and details of their

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 431
various sex acts, as well as the colour of her neighbour’s underwear. Clearly, this
was a memorable experience for Mrs Browne.
In early fourteenth century France, they had something more ominous than
adulterers: heretics. In Montaillou, a small village in the Pyrenees, there was a
group of Cathars that were actively working to oppose the Catholic Church. In
order to get needed evidence, the church requested parishioners to bring in eye-
witness testimony, which was subsequently used in court (Le Roy Ladurie, 1978).
These church records were preserved, testimony revealing that the women of Mon-
taillou, in general, were unusually active in the more subtle form of eavesdropping
that involves listening at keyholes and looking through holes in domestic doors
and walls. The men ‘were inquisitive enough,’ wrote Le Roy Ladurie (1978, p. 257),
‘but their curiosity was nothing beside that of the women’ (italics mine).
Centuries later, in a completely different context, a similar comment was made
about the women of Italy. These women, too, were ‘curious by nature’ according
to Nicole Castan (1989b, p. 417). ‘Women of the lower orders shamelessly admitted
it.’ One confessed that ‘she was “obliged” to follow the movements of a passer-by,
another that she could not help overhearing a conversation or lying in wait for a
neighbor’.
While the courts welcomed eavesdropping as eyewitness testimony, they pun-
ished cases of eavesdropping when it proved to be disruptive to community life. In
England, it was a crime to ‘listen under walls or windows, or the eaves of a house
to hearken after discourse, and thereupon to frame slanderous and mischievous
tales’. Data analysed by McIntosh (1998) revealed that for a good 200 years, begin-
ning in the 1370s, eavesdropping made up about 8% of all social crimes. But here
we find a sex reversal; during this period, about 80% of the courts having some
incidence of eavesdropping happened to hear male cases only.
Why such a high percentage of men? McIntosh (1998) suggested that the men
who were caught listening under eaves were actually attempting to control their
communities by investigating the possibility of domestic misbehaviour. If so, many
of the arrests for eavesdropping may well have been instances of vigilance, an
activity that in other primates also favours males. The irony is that much of the
eavesdropping – a misdemeanour – was undertaken by people who may have been
attempting to prevent domestic misbehaviour.
When the English eavesdroppers witnessed moral transgressions, the obvious
next step was to broadcast what they had seen and this is what many did. However,
recall that the second part of the eavesdropping law involved framing ‘slanderous
and mischievous tales’. The fourteenth and fifteenth century English were still in
the process of privatizing and so felt ambivalent about publicizing the results of
perceptual invasion.
The male eavesdroppers may have been attempting to police their communi-
ties, but they could also have been attempting to control individuals. I base this,

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
432 J. L. Locke
in part, on the fact that stalking, as discussed earlier, is widely understood as a
means of controlling the life of another and it also has a near-identical sex bias to
medieval English eavesdropping. In a large American survey, 87% of the stalkers
were male (Tjaden & Thoennes, 2000) and similar statistics are available for other
cultures (Mullen et al., 2000).
If honest signals, social control and intimate experience are interconnected, it
would not be surprising if visual monitoring sometimes leads to eavesdropping. In
Mineville, a town of 1000 inhabitants in America’s Rocky Mountains, Blumenthal
(1932, p. 103) noted that peoples’attempts to live privately merely inflamed the cu-
riosities of others. Some, he wrote, became ‘more thoroughly known than would
have been the case had they not tried so obviously to guard their privacies, for
in doing so they made themselves mysterious, and thus stimulated the curios-
ity of the people so that more than ordinary attention was given to discovering
something about them’.
Because of such interconnections, it may be difficult to carry out a motivational
analysis on anything but the initial bout of observation. In the daily parade of
public selves, people in search of honest signals have been forced to invade private
spaces, thereby accessing the intimate experience that occurs there, finally finding
themselves in possession of knowledge of the kind that leads to social control.
Looking at and for: a functional comparison
Cheney & Seyfarth (Ch. 25) have described the need of primates to moni-
tor their fellow group members, but do non-human primates actually engage in
dictionary-definition eavesdropping: that is, observe under conditions of stealth?
It is not clear that researchers have asked this question, and yet it appears that an-
imals sometimes secure conditions of perceptual privacy – a circumstance that
favours eavesdropping – before undertaking certain behaviours. For example,
when subordinate males approach females in oestrus, they look around, evidently
to see if they and their intended partners are under review. This is evidently be-
cause the sight of a presenting female is arousing and may produce unwanted
competition (Hall & De Vore, 1965). Females do the same. Kummer (1968, p. 41) de-
scribed the attempt by adult female baboons to copulate with young males ‘behind
the backs of their leaders’ and Smuts (1987) presented photographic evidence of a
rhesus female checking to see if she and an extragroup male were being watched
before they commenced mating activities.
Whether primates ever undertake within-group evaluation from obscure posi-
tions remains to be demonstrated, but some males take measures that make this
unlikely. I refer to consortship, the practice whereby a male browbeats a female
into following his exodus from the group, for mating purposes, sometimes over
considerable distances and for extended periods of time (Goodall, 1986; McGinnis,

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 433
1979; Tutin, 1979). These bouts of absenteeism are of particular interest in rela-
tion to eavesdropping, since they suggest an awareness, on some level, that some
activities are better pursued in perceptual privacy.
As groups enlarge beyond some optimal size, any savings in predator vigilance
may be mitigated by new observational needs within the group. Other things being
equal, the larger the group the more competition there will be for food and other
resources (van Schaik et al., 1983). This increases conspecific threat and, with it,
alliances, which also must be visually monitored (Treves, 2000) and personally
serviced (Dunbar, 1993).
In red colobus and redtail monkeys, Treves (1999) found that if vigilance was
needed for external activity it came out of the time that would otherwise be
devoted to within-group looking. This, he speculated, might explain the fact that
in primates there has been little evidence for the hypothesis that looking time
decreases as group size increases (Pulliam, 1973; Elgar, 1989). For it is difficult to
see how total looking time could decrease if individuals are forced to keep an eye
on individuals in their own group.
Indeed, there is a tension between the time devoted to vigilance outside one’s
group and the time spent looking within it. In white-faced capuchin males, as
indicated above, external competition increases males’attention to outside males,
at the expense of internal vigilance (Rose & Fedigan, 1995), although presumably
reducing cuckoldry (Gould et al., 1997). In one study, redtail monkeys glanced at
associates more often when in the presence of red colobus monkeys than in purely
conspecific groups (Treves, 1999).
In various species, focus of attention is susceptible to rapid and dynamic shifts
from family and alliance members to strangers and predators. Some types of moni-
toring of the physical and social environment are carried out openly – even demon-
stratively – while other types may be effected with stealth. These shifts require a
dynamic model that recognizes the continuous interplay of multiple variables.
Indeed, the optimal paradigm would seem to be one that flexibly admits all types
of observation.
Toward a unified model
Is it possible to achieve a model that accounts for core principles asso-
ciated with vigilance, social observation and eavesdropping? In both human and
non-human primates, individuals appear to spend less time looking for predators
and competitors, and more time looking at each other, if group members are
nearby. In hierarchically organized societies – whether inherently complex squir-
rel monkey groups or highly politicized human societies – a great deal of internal
attention appears to be needed if individuals are to keep or to feel adequately
informed. In Machiavellian societies, inference would seem to play an exaggerated

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
434 J. L. Locke
role. In such societies, there may be greater need of observation, particularly of
the surreptitious kind.
It should be noted that when one looks at fellow group members, information
becomes available not only about them but also about the events to which they
may be reacting. In our own species, for example, when the young enter novel
situations they often monitor caregivers’ facial activity, which serves as a reliable
index of danger. Thus, internal observation offers both within-group information
needed for social comparison and extra-group information about predators and
competitors.
In both human and non-human primates, subordinate individuals spend more
time watching dominant individuals than the reverse arrangement. In Marivaux’s
plays, according to Trapnell (1987), keyholes enabled the young social climber ‘to
distinguish between his ally and his enemies’, to ‘observe the terrain on which he
must manoeuver, assess the efforts his ambition will require, determine the appro-
priate strategy and gauge his chances of success’. Were it not for eavesdropping,
Trapnell (1987, p. 109) wrote, the world would be ‘inaccessible and even unknown’
to people born without special advantages and privileges.
In Victorian England, as we saw above, the upper classes in many instances lived
beside the lower ones. This arrangement gave the ruling classes unobstructed vi-
sion of the individuals they wished to control, but it also gave the lower classes
a regular view of behaviours they had reason to emulate. In nineteenth century
America, upwardly mobile men and women had limited perceptual access to the
upper class behaviours they needed to absorb. To compensate, they used biogra-
phies as ‘handbooks’. They did so, according to Casper (1999), in the belief that
the difference between public success and failure lay in the private habits that
defined one’scharacter, or true self: the stuff of which compelling biographies are
made.
Earlier, we saw that, in non-human primates, males do more looking than fe-
males, presumably in an attempt to detect competition and danger. This vigilance
is very clearly tied to control and defence. In our own species, too, males seem to
have performed in a vigilant capacity more than females, boldly standing under
domestic eaves and then broadcasting the perceptual ‘take’.
Female networks are more extensive and stronger both in non-human primates
(Dunbar, 1988) and in our own species. There is evidence of a strong female ad-
vantage in human grooming (Sugawara, 1984, 1990) as well as touching (Jones &
Yarbrough, 1985) and concerted social action (Motz, 1983). There also is evidence
of a female preference for gossip – the use of speech to discuss mutual acquain-
tances not physically present – that spans cultures and most decades of the twen-
tieth century (Bischoping, 1993). There are indications, additionally, that when
peers offend young women in various cultures, the victims respond by working

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 435
indirectly through female friends (Bjorkqvist et al., 1994; Galen & Underwood,
1997; Crick & Bigbee, 1998).
We saw earlier that vigilance levels in non-human primates are affected by
transient factors such as mating and birth. In humans, too, vigilance levels are
clearly influenced by environmental change, including increases in population,
economic competition and terrorism. Surveillance cameras proliferated in the
USA after 11 September 2001. Although there had been a long-standing fear of
surveillant societies of the type envisaged in Orwell’s book 1984, the terrorist
attacks on New York and Washington seemed to have had the opposite effect. The
lack of objection noted by several newspaper columnists suggests that citizens may
have derived solace from the knowledge that government officials were looking
for and screening out ‘predators’.
In non-human primates, as we have seen, animals may devote as much as
half their waking hours to looking. Dunbar (1993) reported that animals spend as
much as 20% of their time grooming in some primate groups. There are indications
that crowding increases grooming (Nieuwenhuijsen & de Waal, 1982; Novak et al.,
1992; Judge & de Waal, 1997) and grooming has been found to decrease within-
group monitoring (Maestripieri, 1993; Hirsch, 2002). In future work, it would be
interesting to look at within-group vigilance and grooming in the same animals
as a function of density and predational threat.
Merely by comparing species, certain common patterns emerge, but more can
be done, beginning with the resolution of definitional issues. If interdisciplinary
collaboration facilitates the study of processes by which ‘senders’ use language to
donate information, as Hauser et al. (2002) have argued, it will surely affect the
investigation of processes by which ‘receivers’ of widely ranging communicative
abilities use their senses to extract it.
Acknowledgements
This chapter developed from a paper delivered to the Konrad Lorenz Institute in Al-
tenberg, Austria in December of 2001. Portions coevolved with a larger work in progress about
eavesdropping. The author wishes to acknowledge helpful comments by Adrian Treves, Eric
Salzen, Michael Studdert-Kennedy, Polly Wiessner and Ben Hirsch.
References
Abramovitch, R. 1976. The relation of attention and proximity to rank in preschool
children. In: The Social Structure of Attention, ed. M. R. A. Chance & R. R. Larsen.
London: John Wiley, pp. 153–176.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
436 J. L. Locke
Aiello, J. R. 1972. A test of equilibrium theory: visual interaction in relation to
orientation, distance and sex of interactants. Psychonomic Science, 27, 335–336.
1977. Visual interaction at extended distances. Personality and Social Psychology
Bulletin, 3, 83–86.
Anthony, K. H. 1985. The shopping mall: a teenage hangout. Adolescence, 20, 307–312.
Bailey, F. G. 1971. Gifts and poison. In: Gifts and Poison: The Politics of Reputation, ed. F. G.
Bailey. Oxford: Basil Blackwell, pp. 1–25.
Baldellou, M. & Henzi, P. 1992. Vigilance, predator detection and the presence of
supernumerary males in vervet monkey troops. Animal Behaviour, 43, 451–461.
Bales, R. F., Strodtbeck, F. L., Mills, T. M. & Roseborough, M. E. 1951. Channels of
communication in small groups. American Sociological Review, 16, 461–468.
Barash, D. P. 1972. Human ethology: the snack-bar security syndrome. Psychological
Reports, 31, 577–578.
Barth, F. 1975. Ritual and Knowledge Among the Baktaman of New Guinea. New Haven, CT:
Yale University Press.
Baumeister, R. F. 1986. Preface. In: Public Self and Private Self, ed. R. F. Baumeister. New
York: Springer, pp. v–viii.
Beauchamp, G. 2001. Should vigilance always decrease with group size? Behavioral
Ecology and Sociobiology, 51, 41–46.
Bentham, J. 1791. Panopticon: Postscript; Part I: Containing Further Particulars and Alterations
Relative to the Plan of Construction Originally Proposed; Principally Adapted to the Purpose
of a Panopticon Penitentiary-House. London: Printed for T. Payne, at the Mews-Gate.
Biben, M., Symmes, D. & Bernhards, D. 1989. Vigilance during play in squirrel
monkeys. American Journal of Primatology, 17, 41–49.
Bird, N. 1983. Conjugal units and single person: an analysis of the social system of the
Naiken of the Nilgiris (South India). D. Phil. Thesis, University of Cambridge.
Bird-David, N. 1994. Sociality and immediacy: or, past and present conversations on
bands. Man, 29, 583–603.
Bischoping, K. 1993. Gender differences in conversation topics, 1922–1990. Sex Roles,
28, 1–18.
Bjorkqvist, K., Osterman, K. & Lagerspetz, K. M. J. 1994. Sex differences in covert
aggression among adults. Aggressive Behaviour, 20, 27–33.
Blumenthal, A. 1932. Small Town Stuff. Chicago, IL: University of Chicago Press.
Buss, D. M. 1988. From vigilance to violence: tactics of mate retention in American
undergraduates. Ethology and Sociobiology, 9, 291–317.
1997. From vigilance to violence: mate retention tactics in married couples. Journal
of Personality and Social Psychology, 72, 346–361.
Caine, N. G. & Marra, S. L. 1988. Vigilance and social organization in two species of
primates. Animal Behaviour, 36, 897–904.
Campbell, J. K. 1964. Honour, Family and Patronage: A Study of Institutions and Moral Values
in a Greek Mountain Community. Oxford: Clarendon Press.
Carpenter, E. 1966. Image making in Arctic art. In: Sign, Image, Symbol, ed. G. Kepes.
New York: George Braziller, pp. 206–225.
Casper, S. E. 1999. Constructing American Lives: Biography and Culture in Nineteenth-Century
America. Chapel Hill, NC: University of North Carolina Press.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 437
Castan, N. 1989b. The public and the private. In: A History of Private Life. III. Passions of the
Renaissance, ed. R. Chartier. Cambridge, MA: Harvard University Press, pp. 403–446.
Castan, Y. 1989a. Politics and private life. In: A History of Private Life. III. Passions of the
Renaissance, ed. R. Chartier. Cambridge, MA: Harvard University Press, pp. 21–67.
Chance, M. R. A. 1967. Attention-structure as the basis of primate rank orders. Man, 2,
503–518.
Chance, M. R. A. & Jolly, C. J. 1970. Social Groups of Monkeys, Apes and Men. London:
Jonathan Cape.
Cheney, D. L. & Seyfarth, R. M. 1981. Selective forces affecting predator alarm calls of
vervet monkeys. Behaviour, 76, 25–61.
Cowlishaw, G. 1998. The role of vigilance in the survival and reproductive strategies of
desert baboons. Behaviour, 135, 431–452.
Cranz, G. 1980. Women in urban parks. Signs: Journal of Women in Culture and Society, 5,
S79–S95.
Crawford, P. & Gowing, L. 2000. Women’s World in Seventeenth-Century England: A
Sourcebook. London: Routledge.
Crick, N. R. & Bigbee, M. A. 1998. Relational and overt forms of peer victimization: a
multiinformant approach. Journal of Consulting and Clinical Psychology, 66, 337–347.
Dixson, A. F., Scruton, D. & Herbert, J. 1975. Behaviour of the talapoin monkey
(Miopithecus talapoin) studied in groups in the laboratory. Journal of Zoology,
(London), 176, 177–210.
Draper, P. 1978. The learning environment for aggression and anti-social behavior
among the !Kung. In: Learning Non-Aggression: The Experience of Non-Literate Societies,
ed. M. F. A. Montagu. Oxford: Clarendon Press, pp. 31–53.
Dunbar, R. I. M. 1988. Primate Social Systems. Ithaka, NY: Cornell University Press.
1993. On the origins of language: a history of constraints and windows of
opportunity. Behavioral and Brain Sciences, 16, 720–735.
Eibl-Eibesfeldt, I. 1973. The expressive behaviour of the deaf- and -blind-born. In: Social
Communication and Movement: Studies of Interaction and Expression in Man and
Chimpanzee, ed. M. von Cranach & I. Vine. London: Academic Press, pp. 163–194.
Elgar, M. A. 1989. Predator vigilance and group size in mammals and birds: a critical
review of the empirical evidence. Biological Review, 64, 13–33.
Ellis, A. & Beattie, G. 1986. The Psychology of Language and Communication. New York:
Guilford Press.
Exline, R. V., Ellyson, S. L. & Long, B. 1975. Visual behaviour as an aspect of power role
relationships. In: Nonverbal Communication of Aggression 2, ed. P. Pliner, L. Krames &
T. Alloway. New York: Plenum press, pp. 21–52.
Feeley-Harnik, G. 1980. The Sakalava house (Madagascar). Anthropos, 75, 559–585.
Fejos, P. 1943. Viking Fund Publications in Anthropology, No. 1: Ethnography of the Yagua.
New York: Viking Fund.
Fisek, M. H. & Ofshe, R. 1970. The process of status evolution. Sociometry, 33, 327–346.
Fragaszy, D. M. 1990. Age and sex differences in the organization of behavior in
wedge-capped capuchins, Cebus olivaceus. Behavioral Ecology, 1, 81–94.
Galen, B. R. & Underwood, M. K. 1997. A developmental investigation of social
aggression among children. Developmental Psychology, 33, 589–600.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
438 J. L. Locke
Gilbert, P. 1989. Human Nature and Suffering. London: Erlbaum.
Goffman, E. 1963. Behavior in Public Places. New York: Free Press.
Goodall, J. 1986. The Chimpanzees of Gombe: Patterns of Behavior. Cambridge, MA: Harvard
University Press.
Gould, L., Fedigan, L. M. & Rose, L. M. 1997. Why be vigilant? The case of the alpha
animal. International Journal of Primatology, 18, 401–414.
Gregor, T. 1970. Exposure and seclusion: a study of institutionalized isolation among
the Mehinacu Indians of Brazil. Ethnology, 9, 234–250.
Hall, D. M. 1998. The victims of stalking. In: The Psychology of Stalking: Clinical and
Forensic Perspectives, ed. J. R. Meloy. San Diego, CA: Academic Press, pp. 115–137.
Hall, E. T. 1966. The Hidden Dimension. New York: Doubleday.
Hall, J. A. 1996. Touch, status, and gender at professional meetings. Journal of Nonverbal
Behavior, 20, 23–44.
Hall, K. R. L. 1960. Social vigilance, behaviour of the chacma baboon, Papio ursinus.
Behaviour, 16, 261–294.
Hall, K. R. L. & De Vore, I. 1965. Baboon social behavior. In: Primate Behavior: Field Studies
of Monkeys and Apes, ed. I. De Vore. New York: Holt, Rinehart & Winston. pp. 53–110.
Hauser, M. 2000. Wild Minds: What Animals Really Think. London: Allen Lane.
Hauser, M., Chomsky, N. & Fitch, W. T. 2002. The faculty of language: what is it, who
has it, and how did it evolve? Science, 298, 1569–1579.
Haviland, L. K. & Haviland, J. B. 1983. Privacy in a Mexican Indian village. In: Public and
Private in Social Life, ed. S. I. Benn & G. F. Gaus. London: Croom Helm, pp. 341–361.
Hirsch, B. T. 2002. Social monitoring and vigilance behavior in brown capuchin
monkeys (Cebus apella). Behavioral Ecology and Sociobiology, 52, 458–464.
Horrocks, J. A. & Hunte, W. 1986. Sentinel behaviour in vervet monkeys: who sees
whom first. Animal Behaviour, 34, 1566–1567.
Jacobs, J. 1961. The Death and Life of Great American Cities. London: Penguin Books.
Jones, S. E. & Yarbrough, A. E. 1985. A naturalistic study of the meanings of touch.
Communication Monographs, 52, 19–56.
Judge, P. G. & de Waal, F. B. M. 1997. Rhesus monkey behaviour under diverse
population densities: coping with long-term crowding. Animal Behaviour, 54,
643–662.
Kalma, A. 1991. Hierarchisation and dominance assessment at first glance. European
Journal of Social Psychology, 21, 165–181.
Kelling, G. L. & Coles, C. M. 1996. Fixing Broken Windows: Restoring Order and Reducing
Crime in our Communities. New York: The Free Press.
Kendon, A. 1990. Conducting Interaction: Patterns of Behavior in Focused Encounters.
Cambridge: Cambridge University Press.
Keverne, E. B., Leonard, R. A., Scruton, D. M. & Young, S. K. 1978. Visual monitoring in
social groups of talapoin monkeys (Miopithecus talapoin). Animal Behaviour, 26,
933–944.
Koenig, A. 1994. Random scan, sentinels or sentinel system? A study in captive
common marmosets (Callithrix jacchus). In: Current Primatology, Vol. 11: Social
Development, Learning and Behaviour, ed. J. J. Roeder, B. Thierry, J. R. Anderson & N.
Herrenschmidt. Strasbourg: University Louis Pasteur, pp. 69–76.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 439
Kummer, H. 1968. Social Organization of Hamadryas Baboons. Chicago, IL: University of
Chicago Press.
Lee, R. B. 1979. The Dobe !Kung. New York: Holt, Rinehart & Winston.
Le Roy Ladurie, E. 1978. Montaillou: Cathars and Catholics in a French Village. London:
Scolar Press.
Love, R. L. 1973. The fountains of urban life. Urban Life and Culture, 2, 161–209.
Maestripieri, D. 1993. Vigilance costs of allogrooming in macaque mothers. American
Naturalist, 141, 744–753.
Maples, W. R., Maples, M. K., Greenhood, W. F. & Walek, M. L. 1976. Adaptations of
crop-raiding baboons in Kenya. American Journal of Physical Anthropology, 45,
309–316.
McGinnis, P. R. 1979. Sexual behavior in free-living chimpanzees: consort
relationships. In: The Great Apes, ed. D. A. Hamburg & E. R. McCown. Menlo Park,
CA: Benjamin/Cummings, pp. 429–439.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McIntosh, M. K. 1998. Controlling Misbehavior in England, 1370–1600. Cambridge:
Cambridge University Press.
Metcalfe, N. B. 1984a. The effects of mixed-species flocking on the vigilance of
shorebirds: who do they trust? Animal Behaviour, 32, 986–993.
1984b. The effects of mixed-species flocking on the vigilance of shorebirds: is
visibility important? Animal Behaviour, 32, 981–995.
Moore, M. M. 1985. Nonverbal courtship patterns in women: context and
consequences. Ethology and Sociobiology, 6, 237–247.
Motz, M. F. 1983. True Sisterhood: Michigan Women and their Kin 1820–1920. Albany, NY:
State University of New York Press.
Mullen, P. E., Pathe, M. & Purcell, R. 2000. Stalkers and their Victims. Cambridge:
Cambridge University Press.
Nieuwenhuijsen, K. & de Waal, F. B. M. 1982. Effects of spatial crowding on social
behavior in a chimpanzee colony. Zoo Biology, 1, 5–28.
Novak, M. A., O’Neill, P. & Suomi, S. J. 1992. Adjustments and adaptations to indoor
and outdoor environments: continuity and change in young adult rhesus
monkeys. American Journal of Primatology, 28, 125–138.
Olsen, D. J. 1974. Victorian London: specialization, segregation, and privacy. Victorian
Studies, 17, 265–278.
Olsson, M. 2001. ‘Voyeurism’ prolongs copulation in the dragon lizard Ctenophorous
fordi. Behavioral Ecology and Sociobiology, 50, 378–381.
Oosterman, J. 1992. Welcome to the pleasure dome: play and entertainment in urban
public space – the example of the sidewalk cafe. Built Environment, 18, 155–164.
Pitcairn, T. K. 1976. Attention and social structure in M. fascicularis. In: The Social
Structure of Attention, ed. M. R. A. Chance & R. R. Larsen. London: John Wiley
pp. 51–81.
Poirier, F. E. 1972. The St Kitts green monkey (Cercopithecus aethiops sabaeus): ecology,
population dynamics, and selected behavioral traits. Folia primatologica, 17,
20–55.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
440 J. L. Locke
Pulliam, H. R. 1973. On the advantages of flocking. Journal of Theoretical Biology, 38,
419–422.
Putnam, R. D. 1993. Making Democracy Work: Civic Traditions in Modern Italy. Princeton,
NJ: Princeton University Press.
Quenette, P.-Y. 1990. Functions of vigilance behaviour in mammals: a review. Acta
Œcologica, 11, 801–818.
Rapoport, A. 1969. House Form and Culture. Englewood Cliffs, NJ: Prentice-Hall.
Rose, L. M. 1994. Benefits and costs of resident males to females in white-faced
capuchins (Cebus capucinus). American Journal of Primatology, 32, 235–248.
Rose, L. M. & Fedigan, L. M. 1995. Vigilance in white-faced capuchins, Cebus capucinus,
in Costa Rica. Animal Behaviour, 49, 63–70.
Schiefenhovel, W. 1997. Universals in interpersonal interactions. In: Nonverbal
Communication: Where Nature Meets Culture, ed. U. Segerstrale & P. Molnar. Mahwah,
NJ: Lawrence Erlbaum, pp. 61–79.
Shore, B. 1982. Sala ‘Ilua: A Samoan Mystery. New York: Columbia University Press.
Smith, W. J. 1977. The Behavior of Communicating: An Ethological Approach. Cambridge,
MA: Harvard University Press.
Smuts, B. B. 1987. Sexual competition and mate choice. In: Primate Societies, ed. B. B.
Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham & T. T. Struhsaker. Chicago,
IL: University of Chicago Press, pp. 385–309.
Soffer, O., Adovasio, J. M. & Hyland, D. C. 2000. The ‘Venus’ figurines: textiles, basketry,
gender, and status in the Upper Paleolithic. Current Anthropology, 41, 511–537.
Steenbeek, R., Piek, R. C., van Buul, M. & van Hooff, J. A. R. A. M. 1999. Vigilance in
wild Thomas langurs (Presbytis thomasi): the importance of infanticide risk.
Behavioral Ecology and Sociobiology, 45, 137–150.
Sugawara, K. 1984. Spatial proximity and bodily contact among the Central Kalahari
San. African Study Monographs, 3, 1–43.
1990. Interactional aspects of the body in co-presence: observations on the Central
Kalahari San. In: Senri Ethnological Studies 27: ‘Culture Embodied,’ ed. M. Moerman &
M. Nomura. Osaka, Japan: National Museum of Ethnology, pp. 79–122.
Tjaden, P. & Thoennes, N. 2000. Prevalence and consequences of male-to-female and
female-to-male intimate partner violence as measured by the National Violence
Against Women Survey. Violence Against Women, 6, 142–161.
Trapnell, W. H. 1987. Eavesdropping in Marivaux. Geneve: Librairie Droz.
Treves, A. 1998. The influence of group size and neighbors on vigilance in two species
of arboreal monkeys. Behaviour, 135, 453–481.
1999. Within-group vigilance in red colobus and redtail monkeys. American Journal of
Primatology, 48, 113–126.
2000. Theory and method in studies of vigilance and aggregation. Animal Behaviour,
60, 711–722.
Treves, A., Drescher, A. & Ingrisano, N. 2001. Vigilance and aggregation in black
howler monkeys (Alouatta pigra). Behavioral Ecology and Sociobiology, 50, 90–95.
Turner, T. S. 1980. The social skin. In: Not Work Alone: A Cross-Cultural View of Activities
Superfluous to Survival, ed. J. Cherfas & R. Lewin. London: Temple Smith,
pp. 112–140.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
Humans: control, signalling and intimate experience 441
Tutin, C. E. G. 1979. Mating patterns and reproductive strategies in a community of
wild chimpanzees (Pan troglodytes schweinfurthii). Behavioral Ecology and Sociobiology,
6, 29–38.
Underwood, R. 1982. Vigilance behaviour in grazing African antelopes. Behaviour, 79,
81–107.
van Schaik, C. P., van Noordwijk, M. A., Warsono, B. & Sutriono, E. 1983. Party size and
early detection of predators in Sumatran forest primates. Primates, 24, 211–221.
Whiten, A. 1993. Social complexity: the roles of primates’ grooming and people’s
talking. Behavioral and Brain Sciences, 16, 719.
Wilson, P. J. 1988. The Domestication of the Human Species. New Haven, CT: Yale University
Press.
Wirtz, P. & Wawra, M. 1986. Vigilance and group size in Homo sapiens. Ethology, 71,
283–286.

P1: JZZ/... P2: JZZ/...0521823617c19.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 18:41
442

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
Part IV I N T E R F A C E S W I T H O T H E R
D I S C I P L I N E S
443

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
444

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
Introduction
Communication has a history of addressing topics of interest to other
disciplines, both in biology and more generally. The interface between disciplines
has been long recognized to generate paradigm shifts and the same has been
true of interfaces with communication. The interface between communication
and neurobiology provides a good example. The discovery that the brain nuclei
controlling song production varied in size seasonally (Nottebohm, 1981) was a
finding that overturned accepted notions of the stability of brain architecture and
triggered studies of evolutionary plasticity in brain structure (e.g. Sherry, 1998).
An important question then is whether the communication network approach
enhances communication’s interest to, and interfaces with, other disciplines.
This section shows that the answer is an emphatic yes, it does. In part, this is
shown by the wide range of topics addressed: from perception and physiology,
through aspects of cognition to the evolution of altruism. However, it is in the
details of the chapters that the value of the approach becomes apparent, as does
an enthusiasm about the further research possibilities.
Perception
The extent of a communication network is often an important issue and
is discussed by several chapters in this book. Network size is related to the distance
at which signals can be received and this distance is influenced by several factors.
These factors include the distorting and attenuating effects of the environment
through which the signal travels and the level of interference from the signals of
others. A key factor that is often overlooked is the sensory abilities of the receiver.
Such abilities can be extraordinary; for example, some bird species respond to a
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
445

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
446 Part IV
signal even when that signal is embedded in noise that is louder than the signal.
Ulli Langemann and Georg Klump (Ch. 20) discuss such perceptual abilities and
how they relate to aspects of signal structure and transmission in the acoustic
modality. Their chapter, therefore, covers the interface between communication
networks, psychophysics and physics.
Endocrinology
Hormones play an established role in determining when animals com-
municate, by controlling annual and circadian rhythms. They are also known to
be involved in mediating the response to signals; for example, raising the level
of oestradiol can induce female songbirds to perform copulation solicitation dis-
plays in response to song in the absence of the male singer (Searcy & Yasukawa,
1996). In Ch. 21, Rui Oliveira uses examples mainly from bony fishes to show that
androgens can be affected by the social environment as well as modulating be-
haviours that partly create the social environment. Establishing this reciprocal
link between hormones and behaviour (comparable to that for stress hormones)
has obvious consequences for both endocrinology and communication behaviour.
Cooperation and altruism
Altruism between unrelated individuals (that can include cooperation)
has long been considered an evolutionary puzzle. Recent mathematical models
and experiments with humans have shown that altruism can evolve through an
increase in the altruist’s ‘prestige’or ‘image’ in the eyes of others (e.g. Wedekind &
Milinski, 2000). Redouan Bshary and Arun D’Souza point out in Ch. 22 that these
recent advances in cooperation theory are a specific instance of a communication
network because others not directly involved in the altruistic interaction must
observe it. They then investigate the evolution and maintenance of altruistic be-
haviour, tactical deception and spiteful behaviour using data gathered in the field
from interactions between coral reef cleaner fish and their clients.
Semiochemicals
Information is obviously a key concept in communication, but as Brian
Wisenden and Norm Stacey make clear in Ch. 23, communication is a subset of
information. Aquatic animals can obtain information to guide reproductive and
predator-avoidance behaviour from chemicals released as a by-product of other
processes. Information in such semiochemicals can have striking effects; for ex-
ample chemicals released by predators attacking or digesting their prey can change

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
Interfaces with other disciplines 447
the behaviour, life history and morphology of potential prey (e.g. Brancelj et al.,
1996). After considering the use of chemical information by fishes in predator–
prey and sexual interactions, Wisenden and Stacey set out the case for communica-
tion networks as a subset of information networks and the potential evolutionary
routes for the origins of network behaviours such as eavesdropping. Their sug-
gestions include the possibility that a process analogous to eavesdropping can
precede the origin of communication, rather than such information gathering
following the development of communication interactions.
Cognition
The role of communication in cognition has ensured a lively interface
with the cognitive sciences, often concerning the extent to which non-human
communication can be considered a language (e.g. Hauser, 1996). Irene Pepper-
berg considers the cognitive abilities of birds in Ch. 24, particularly the issue of
transitive inference, using information from two different approaches. The first
approach uses communication to explore the cognitive abilities of parrots, with
human speech being used as the tool in much the same way as it is in explorations
of human cognitive abilities. The second approach uses the results of field ex-
periments investigating social eavesdropping to indicate the cognitive abilities of
territorial songbirds.
Several chapters in other sections of this book touch upon the cognitive abilities
of animals communicating in networks (e.g. the extent to which individual identi-
fication is a prerequisite of eavesdropping in Ch. 16). Dorothy Cheney and Robert
Seyfarth expand this theme in Ch. 25. They review the evidence for eavesdropping
in primates, concentrating on species living in large, permanent social groups,
often with complex social relationships. They then suggest a framework for assess-
ing the occurrence in other animal groups of social intelligence (i.e. mechanisms
such as transitive inference used to gather information relevant to social interre-
lationships). This framework, together with the information presented in other
chapters of this book, provides an opportunity for a taxonomically wide-ranging
comparative approach to the issue of social intelligence.
Mathematical models
Mathematical models have provided insights in many areas of biology,
including communication. However, most models of communication have not
dealt with networks, at least partly because of the difficulty in applying tractable
analytical models to networks. In Ch. 26, Andrew Terry and Rob Lachlan describe
models that capture two important aspects of networks by being spatially explicit

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
448 Part IV
and based on individual behaviour. These simulation models of anuran acoustic
choruses and eavesdropping strategies generate different results from more tra-
ditional models applied to the same questions. Incorporating strategic communi-
cation decisions into such spatially realistic individually based models seems an
approach that is likely to generate the kind of testable predictions about commu-
nication in networks that are needed to stimulate further research.
Applied aspects
Communication is central to animals’ lives and as such can be used to
modify their behaviour for human ends, such as in pest control, animal welfare and
conservation. A chapter jointly written with Tom Peake was planned for this book;
it would have explored the possible interfaces between communication networks
and aspects of applied biology. However, the relative newness of the network
perspective means that there are, as yet, few concrete examples. We decided,
therefore, to outline here some of the interfaces that we think are promising
and to draw attention to chapters elsewhere in the book that have mentioned
applications.
One way to judge the welfare of animals is to assess the extent to which ani-
mals in captivity, including those on public display in zoos and aquaria, are able to
display the full range of behaviours shown by free-ranging animals under natural
conditions. Many of the chapters in this book have argued that communication in
a network is such a natural condition and, therefore, the ability to communicate as
part of a network could be regarded as a feature of adequate captive provision. Ad-
verse effects on breeding performance have been noted when group-living species
are kept in small groups, and communication networks that are much smaller
than occur in the wild could underlie this effect. Attempts to increase apparent
group size (e.g. use of mirrors with captive flamingos (Whitfield, 2002)) have met
with mixed success, perhaps because the manipulations did not adequately mimic
communication networks. It may be easier to create an apparent communication
network for species that are widely spaced and possibly territorial, because at long
range it is only signals that are detected and signal playback is straightforward, at
least with acoustic signals. Tom Peake has suggested that interactive acoustic sig-
nals could be provided as a type of environmental enrichment in communication
for captive animals. For example, zoo visitors could interact with vocal species
such as gibbons Hylobates spp. via playback from remotely sited loudspeakers trig-
gered from a control panel in the cage’s viewing area (obviously there would have
to be safeguards to ensure that the nature and extent of interaction did not exceed
natural levels). Communication in a network may be equally difficult if a popula-
tion is held at an unnaturally high density for production reasons (e.g. fish farms),

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
Interfaces with other disciplines 449
because the network is overloaded. From the discussions above, it can be expected
that removing the ability to signal will have adverse effects on welfare (e.g. removal
of rodent scent marks by cage cleaning (Gray & Hurst, 1995)). Signal removal can
also have additional network-wide effects. Removing the major chela of male fid-
dler crabs Uca tangeri clearly prevents individual males from signalling visually in
their usual manner. In addition, it may skew the sex ratio apparent to males and
females because males without a major chela appear female to both sexes, and the
response to the apparent sex ratio could accelerate population declines (Oliveira
et al., 2000).
There are several established applications of communication in conservation,
such as identifying individuals from features of their vocalizations (e.g. McGregor
et al., 2000). The network perspective emphasizes the possibility that anthro-
pogenic noise could have adverse effects by disrupting or restricting the size of
acoustic communication networks (e.g. McGregor & Dabelsteen, 1996). Vincent
Janik considers this applied aspect in more detail in relation to marine mammals
in Ch. 18. In the terrestrial environment, road noise may similarly restrict the
acoustic communication networks of other taxa such as songbirds. Habitat frag-
mentation is another way of disrupting songbird communication networks and
it is discussed by Ken Otter and Laurene Ratcliffe in Ch. 7. In Ch. 8, Alexandra
Lang and colleagues discuss how the need to use signals with limited range and
detectability to predators may have the side effect of reducing the effective popula-
tion size of katydids, with associated increased susceptibility to random extinction
processes.
We think that these brief examples illustrate that communication networks can
have relevance to applied biology and that often the implications are not straight-
forward. We suggest that those researching communication networks have an
obligation to explore the applications of their findings. Arguably the best way to
make applied biologists aware of the relevance of network research is to make
suggestions on how best to modify current practice to incorporate new findings.
Future directions
The chapters in this section clearly show how several areas of research
interface with communication networks. An obvious question is whether this will
also be true for interfaces with disciplines that are not represented in this section. A
number of chapters (e.g. Chs. 12, 14 and 26) mention the possibility of fruitful links
with ecology, or more specifically spatial ecology, suggesting that this would seem
to be a good interface to explore. Also, given that we humans consider ourselves
to be supreme communicators and often do so in a network environment, it is
possible that many aspects of social psychology and sociology could interface

P1: KPB/... P2: KOD0521823617agg.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:11
450 Part IV
fruitfully with a communication network approach. These brief considerations
suggest that the answer to the question posed above is that interfaces between
communication networks and many other disciplines can be sources of insight
and inspiration in the future.
References
Brancelj, A., Celhar, T. & Sisko, M. 1996. Four different head shapes in Daphnia hyalina
(Leydig) induced by the presence of larvae of Chaoborus flavicans (Meigen).
Hydrobiologia, 339, 37–45.
Gray, S. & Hurst, J. L. 1995. The effects of cage cleaning on aggression within groups of
male laboratory mice. Animal Behaviour, 49, 821–826.
Hauser, M. D. 1996. The Evolution of Communication. Cambridge, MA: MIT Press.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K., Peake, T. M. & Gilbert, G. 2000. Communication behaviour and
conservation. In: Behaviour and Conservation, ed. L. M. Gosling & W. J. Sutherland.
Cambridge, UK: Cambridge University Press, pp. 261–280.
Nottebohm, F. 1981. A brain for all seasons: cyclical anatomical changes in song
control nuclei of the canary brain. Science, 214, 1368–1370.
Oliveira, R. F., Machado, J. L., Jordao, J. M. et al. 2000. Human exploitation of male
fiddler crab claws: behavioural consequences and implications for conservation.
Animal Conservation, 3, 1–5.
Searcy, W. A. & Yasukawa, K. 1996. Song and female choice. In: Ecology and Evolution of
Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca, NY:
Cornell University Press, pp. 454–473.
Sherry, D. F. 1998. The ecology and neurobiology of spatial memory. In: Cognitive
Ecology: The Evolutionary Ecology of Information Processing and Decision Making, ed. R.
Dukas. Chicago, IL: Chicago University Press, pp. 261–296.
Wedekind, C. & Milinski, M. 2000. Cooperation through image scoring in humans.
Science, 288, 850–852.
Whitfield, J. 2002. Mirrors to help birds mate. Nature Science Update, 19 March:
http://www.nature.com/news/2002/020318/full/020318-2.html.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
20
Perception and acousticcommunication networks
u l r i k e l a ng e m a n n & g e o rg m . k l u m p
University of Oldenburg, Germany
Introduction
Traditionally, the analysis of acoustic communication has been based on
a model system composed of a sender, the transmission channel and a receiver
(Shannon & Weaver, 1949). Since the early 1990s, this view has been extended to
communication networks, in which several signallers and receivers are involved
(e.g. McGregor & Peake, 2000). Two general approaches have been adopted in order
to investigate communication behaviour. First, measurements of physical modifi-
cations to the signal during transmission (e.g. Wiley & Richards, 1978; Dabelsteen
et al., 1993; Holland et al., 1998) have been used to assess the feasibility of com-
munication (e.g. estimating maximum communication distances) or to evaluate
which features of signals might be adaptive in a certain context. Second, playback
studies have been used to conclude which features may be of importance for signal
discrimination: different behavioural responses can be elicited by playback of sig-
nals that have been modified by physical properties of the environment or by the
experimenter. Often the physical properties of signals are manipulated in ways
that are informed by studies of signal transmission in the animal’s environment.
However, behavioural responses can be understood more fully if the animal’s per-
ceptual abilities are taken into account (Wiley & Richards, 1982; Klump, 1996).
Perception includes the transduction process by the animal’s sensory organs and
the subsequent processing by the nervous system. However, perception can only
be inferred indirectly from the animal’s responses. An animal’s failure to respond
differentially to playback can either mean that the animal was not motivated
to discriminate within the experimental context (e.g. because the modified sig-
nal deviated too far from species-specific signals) or that the animal’s auditory
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
451

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
452 U. Langemann & G. M. Klump
system could not ‘resolve’ modifications of the signal and, therefore, they were
not perceived. In the latter case, perceptual constraints render a behavioural re-
sponse impossible. If more than one signal parameter was modified in playback
experiments, the results might be even more difficult to interpret. Studies in the
laboratory focusing on perceptual mechanisms allow us to control for motivation
of an animal, help to conclude which modifications can be exploited by the ani-
mal and make it possible to determine the perceptual resolution of the animal’s
sensory system.
In this chapter, we will explain how the current knowledge on perceptual
mechanisms offers a better understanding of animal communication, especially
in the context of a communication network. For example, new results from per-
ceptual studies now give us a more complete and accurate picture of how animals
parse signals from noise and from different sources in acoustic scenes (e.g. Feng &
Ratnam, 2000; Hulse, 2002). We will demonstrate this by showing how knowledge
on perceptual masking can aid in explaining signal assessment by the animal. An
understanding of how animals segregate several sources of signals in a commu-
nication network requires knowledge of how accurately signals are localized and
how the spatial arrangement of sources affects masking (e.g. Klump, 2000). Finally,
the behaviour of an animal in a communication network often requires an abil-
ity to range a signal, that is, assess the distance of a sound source (e.g. Naguib &
Wiley, 2001). Explaining results from playback studies of ranging requires an un-
derstanding of the perception of degradation. Providing a comprehensive review
of all the topics mentioned here is beyond the scope of this chapter. Instead, we
will present specific examples from the animal behaviour literature and discuss
them in light of knowledge of the physiology of perception.
Detection and recognition
Spectral aspects of masking and signal detection
Communication in any context requires signal detection. This would
hardly be a problem in a silent world. However, the environment is noisy, perhaps
particularly so in a communication network. Therefore, signal detection needs
to be considered in relation to the level of the background noise. Environmen-
tal noise originates from biotic sources such as calling insects (e.g. grasshoppers
and cicadas (Waser & Waser, 1977; Ryan & Brenowitz, 1985)) and calling frogs
(Wollerman & Wiley, 2002) or singing birds (especially during the dawn chorus,
e.g. Staicer et al. (1996)). Biotic environmental noise constitutes an especially severe
problem in large assemblies of individuals of the same species (e.g. roosts, breed-
ing colonies, choruses) since the masking noise matches the frequency spectrum
of the signal (as the signals of conspecifics constitute most of the masking noise

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 453
experienced by an individual). There are other biotic sources that may contribute
to background noise. For example, the rustling of the foliage and the movement
of twigs and branches in a deciduous wood provide a substantial level of back-
ground noise (e.g. Klump, 1996) that increases with wind speed (Fegeant, 1999).
A similar increase in background noise with wind speed can be observed in open
grassland habitats. For example, wind with a moderate speed of 5 m/s produces
sound pressure levels of more than 60 dB in the one-third-octave band at 20 Hz for
at least 95% of the time (Boersma, 1997). Much lower levels of background noise are
observed at frequencies above 500 Hz. Coastal environments and running waters
may produce a substantial amount of non-biotic noise (Dubois & Martens, 1984;
Douglas & Conner, 1999).
The difference between the frequency spectrum of the signal and the back-
ground noise and their relative amplitudes determine the amount of masking.
The amplitude of sounds can either be specified in terms of their overall sound-
pressure level or in terms of the amplitude contained in the individual compo-
nents. Commonly, the components are resolved into frequency bands 1 Hz wide
and the sound pressure level (which is a measure of the amplitude relative to the
standard reference pressure of 2 × 10−5 Pa) in each 1 Hz band is determined. This
amplitude measure is conventionally called the spectral density or the spectrum
level (e.g. Moore, 2003). The difference between the signal amplitude and the am-
plitude of the background can be described by the signal-to-noise (S/N) ratio. The
S/N ratio can be either expressed as the difference between the overall signal level
and the overall level of the noise or as the difference between the signal spectral
density and the spectral density of the noise. The S/N ratio allows us to estimate
whether a signal can be detected or not (see Box 20.1). The same rules for signal
detection apply for single receivers as well as for individuals in a communication
network. Each ‘node’ in a network (a receiver sitting at a different place), however,
might experience quite different S/N ratios for the same signal. Behavioural ex-
periments in the laboratory can determine how random background noise affects
absolute auditory sensitivity for tonal signals or signals with a distinctive peak in
the spectrum.
The value that denotes the shift in auditory sensitivity when random (wide-
band) noise is present is called the critical masking ratio or critical ratio (CR).
The CR is simply the S/N ratio at detection threshold in random wideband noise
expressed as the difference between the level of a tonal signal (which is identical
to its spectral density) and the spectral density of the noise (N0). The CR is usu-
ally independent of the level of random background noise. However, the CR is
frequency dependent, increasing at about 3 dB per octave (overview in Fay, 1988).
The CR also provides a rough estimate of the bandwidth of auditory analysis filters
(CR filter bandwidth in Hz is given by 10CR/10; e.g. Yost, 1994). There are numerous

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
454 U. Langemann & G. M. Klump
Box 20.1 Determining the signal-to-noise (S/N) ratio forsignal detection
Calculating the S/N ratio for signal detection in animal studies is relatively
straightforward in the laboratory environment with suitable equipment and
all parameters well controlled. Field measurements are more difficult.
Nevertheless, as shown here, it is possible to get an estimate of the S/N ratio
for signal detection in the natural environment.
Which equipment to use
Preferably, a sound-level meter should be used in the field to
determine signal level directly. Often it is possible to record sounds through
the microphone of the sound-level meter (e.g. on a DAT recorder or directly
with a note-book computer) for subsequent analysis, but any microphone
and recording equipment that allows signals to be recorded with an
accurately defined gain is suitable. The directional characteristics of the
microphone should be adapted to the question of interest. For example, an
omnidirectional microphone is the best choice when the general level of
background noise is being measured. If the goal is to measure a signal
originating from a specific source, it is recommended to use a directional
microphone and approach the source as close as possible. The
frequency-transfer function of the microphone should be as flat as possible
to avoid a bias in the later analysis. Recording a calibration signal of known
sound-pressure level with the signal of interest makes it subsequently
possible to compute the absolute power spectra of the recorded signals (the
calibration signal can be recorded at 1 m from the microphone, at the
location from which the animal was recorded or by using a calibrator placed
on the microphone). Even inexpensive sound-level meters can be used for
accurate field measurements if they are calibrated against a high-quality
instrument in the laboratory. Spectral analysis can be carried out with
specialized spectrum analyser hardware or (if the signals are digitized and
stored on a computer) with suitable software, e.g. freeware.
Computing signal-to-noise ratios
First, one needs to decide which of the two common measures of the
S/N ratio should be used: comparing overall sound-pressure levels or
comparing the sound-pressure levels with reference to spectral density (i.e.
compute the ratio between signal and noise spectrum level N0; for further
details, see text). The frequency spectrum and the sound-pressure level of

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 455
both the signal of interest and the background noise have to be known in
order to determine the S/N ratio correctly.
Step 1: measuring sound-pressure levels of signal and background noise
It is important that the sound-pressure level of the signal of interest
is at least 10 dB above the sound pressure level of other sound making up the
background noise. If the difference is less than 10 dB, the sound-pressure
level of the signal cannot be measured independently from the
sound-pressure level of the background noise and, therefore, a S/N ratio
cannot be computed accurately (the ratio would then be the level of signal
plus noise divided by the level of the noise). Strategies to achieve the 10 dB
minimum difference include getting close enough to the signal source,
getting as far away as possible from sources of noise or using a highly
directional microphone. The distance between the microphone and the
sound source(s) should be reported. This allows an estimate of the signal
level at a specified distance from the source (e.g. by using the rule of thumb
that the signal level is reduced by 6 dB for every doubling of the distance
and, if more accurate estimates are required, the estimates should include
effects of excess attenuation).
Sound-level meters frequently offer at least two types of filter setting: the
A and C settings. The A filter has a low-frequency cut-off of about 800 Hz, a
high-frequency cut-off of about 9 kHz and emphasizes the intermediate
frequencies in this range. The C filter has a low-frequency cut-off of
approximately 30 Hz, a high-frequency cut-off of approximately 8 kHz and
has a flat unbiased frequency response. To be sure that an appropriate filter
is used, the signals of interest must fall within the frequency range of the
filter. For example, measuring ambient background noise over a wide
frequency range is only possible with a C filter setting. The integration time
constant of the sound-level meter ideally should match the integration time
of the auditory system of the study species or at least the duration of its
signals. The ‘fast’ integration time is suitable for measuring the
sound-pressure level of animal signals of 125 milliseconds or longer
duration. Using a very long integration time (1 second at the ‘slow’ setting of
the sound-level meter) will underestimate the level of signals that are
composed of brief components. If shorter signals than 125 milliseconds need
to be analysed, the ‘impulse’ or ‘peak’ settings are more suitable. These
provide integration times of 35 and 0.05 milliseconds, respectively, for
fast-rising signal levels and a very slow decay. The integration times used to
determine root mean square sound pressure with a sound-level meter allow
only approximations of the real sound pressure (that can be calculated from

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
456 U. Langemann & G. M. Klump
the calibrated digitized signal) or the perceived sound pressure (for which
one should apply the integration time of the animal’s auditory system).
When digitizing the recordings for further analysis with a computer, the
sampling rate must be at least twice the highest frequency of the signals to
avoid serious sampling errors.
Step 2: calculating signal-to-noise ratios
If the signal and the background noise have the same frequency
spectra (e.g. frog calls in a dense chorus of conspecifics) the S/N ratio is
simply the difference between the overall signal level and the overall level of
the background noise, both measured in decibels. If the signal and the
background noise have different bandwidths and thus different frequency
spectra (e.g. a tonal bird vocalization in a wideband background noise), a S/N
ratio based on measurements of the spectrum level is easier to interpret. The
spectrum level can be calculated from the overall level by subtracting the
bandwidth (in decibels) of the frequency filter used in the measurement
from the overall sound-pressure level measured through this filter, i.e.
spectrum level (dB) = overall level (dB) − 10 × log10(bandwidth)
If the signal has a smaller bandwidth than the frequency filter throughwhich its sound-pressure level was determined, the bandwidth of the signal
must be used instead of the bandwidth of the filter. Then the S/N ratio is
calculated as the difference between the spectrum level of the signal and of
the background noise measured in decibels. It must be noted that this
engineering-type measure of the S/N ratio provides only an approximation of
stimulus characteristics relevant for perception. The filters relevant for the
perception of the animal are the auditory analysis filters. Furthermore, the
measure of the S/N ratio as calculated here does not incorporate temporal
aspects of the structure of the signal and background noise. The physiology
of the auditory system will determine how temporal aspects affect the
perceived S/N ratio. If one wants to know more accurately how the acoustic
environment is perceived by an animal, the measurements should be
interpreted using a model of the physiology of the animal’s auditory system.
studies determining the CR values at various frequencies of the hearing range in
fish, amphibians, birds and mammals (summarized by Fay, 1988). The average CR
in birds is about 23 dB at 1 kHz, 24.5 dB at 2 kHz, 27.5 dB at 4 kHz and 36 dB at 8 kHz
(Klump, 1996). Typical CR values in mammals are 22.5 dB at 1 kHz, 25.5 dB at 2 kHz,
29.5 dB at 4 kHz and 32 dB at 8 kHz (average for cat, rat, chinchilla and human,
respectively (Fay, 1988)). The average slope of 3 dB per octave that is often described
probably does not hold for frequencies below 500 Hz and the CR will decrease much

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 457
less for such frequencies (e.g. Moore & Glasberg, 1983). There are some exceptions
from the usual pattern of an increasing values of CR with increasing frequency.
In mammals, the greater horseshoe bat Rhinolophus ferrumequinum has the lowest
CR values at the frequencies of their ultrasonic echolocation calls (Long, 1977).
In birds, great tits Parus major show relatively little change in CR with frequency.
At high frequencies, CR values for great tits are much lower than those of other
bird species (e.g. 25.9 dB at 8 kHz). This may be an adaptation that makes the high-
pitched communication sounds of great tits much less susceptible to masking
by environmental noise in their deciduous forest habitat (Klump & Curio, 1983;
Langemann et al., 1998). Signal-detection thresholds in background noise, called
masked thresholds, thus depend on the level of background noise and on the
CR. Estimates of masked auditory thresholds (MAT) are commonly calculated by
adding the CR to the spectrum level of the noise (N0 + CR = MAT). Knowing both
CR and the level of the background noise allows us to estimate approximately how
background noise in a specific communication setting will influence the auditory
sensitivity of animals.
We will use an example to demonstrate how such calculations can estimate
the distance over which communication is possible: that is, the extent of the
communication network (see also Box 20.1). The detection distance for a typical
great tit song element with a spectral peak frequency of 2 kHz and signal amplitude
of 90 dB can be estimated to be 331 m (Table 20.1). This estimate takes into account
the great tit’s absolute threshold at 2 kHz (6.5 dB), spherical spreading of the song
(6 dB per doubling of distance for all frequencies) and the habitat-dependent signal
attenuation (excess attenuation, e.g. Marten & Marler, 1977; Dabelsteen et al., 1993).
However, the main acoustic energy of background noise in a deciduous forest
occurs at lower frequencies (Klump, 1996; Fegeant, 1999) and the 2 kHz signal
will be masked by background noise of 10 dB, and the perceptual threshold of a
great tit receiver in noise (masked auditory threshold) will be much worse than
6.5 dB: in fact the masked threshold of a 2 kHz signal would be 35.6 dB (N0+ CR, i.e.
10 + 25.6 dB; Table 20.1). Hence, a great tit can only perceive song elements of 2 kHz
as long as the sound level does not drop below the great tit’s masked threshold,
giving a maximum detection distance of 124 m. At this distance, the sound level
of the element equals the value of the masked threshold (35.6 dB). In contrast, a
‘seeet’ alarm call indicating the presence of an aerial predator (peak frequency
around 8 kHz, mean amplitude 60.1 dB) experiences much less masking by the
forest background noise. At 8 kHz, the background noise spectrum level is −5.2 dB
and since great tits’CR at 8 kHz is similar to the CR at 2 kHz, the masked threshold
is very close to great tits’ absolute threshold. In consequence, the perception of
this alarm call is mainly limited by great tits’ absolute auditory sensitivity.
Infrasound communication in elephants is assumed to be a communica-
tion network extending over several kilometres (Larom et al., 1997; Ch. 17).

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
458 U. Langemann & G. M. Klump
Table 20.1. An example of estimates of masked thresholds (i.e. the signal-detection
threshold in background noise) and perceptual distances (i.e. the maximum distance where
behavioural responses would be expected) of single great tit song elements with spectral
peak frequencies between 2 and 8 kHz or the aerial predator ‘seeet’ alarm call
(of approximately 8 kHz)
Signal frequency (kHz)
2 4 6.3 8 ‘Seeet’ call
Spherical spreading (dB/dd) 6 6 6 6 6
Excess attenuation (dB/100 m) 10.0 13.7 18.0 21.1 21.1
Source level (dB) 90 90 90 90 60.1
Absolute hearing threshold (dB) 6.5 9.1 12.8 18.1 18.1
How far in quiet (m) 331 242 179 137 56
Noise level N0 (dB) 10.0 4.3 −1.7 −5.2 −5.2
Critical ratio (dB) 25.6 23.8 25.9 25.9 25.9
Masked auditory threshold (dB) 35.6 28.1 24.2 20.7 20.7
Detection distance in noise (m) 124 138 130 128 49
Recognition threshold (dB) 38.6 31.1 27.2 23.7 23.7
Perceptual distance in noise (m) 107 124 118 117 41
dd, doubling of distance; CR, critical ratio;
Signal detection is a function of physical signal properties (signal frequency, source level), of
the environment (background noise level N0, intensity loss from excess attenuation), of physics
(intensity loss from spherical spreading) and of physiological constraints set by the auditory
system. The critical ratio denotes the shift in auditory sensitivity from the absolute hearing
threshold (in quiet) to the masked auditory threshold when random wideband noise is present.
For calculating perceptual distances, random noise is used as an approximation of the ambient
background noise. Note that the background noise level N0 is expressed with reference to spectral
density of the noise in order to calculate masked thresholds (N0 + CR) Spectral density is the sound-
pressure level of each 1 Hz wide frequency component relative to the standard reference pressure
of 2 × 10−5 Pa). Detection is only the first step in perception; the second step is recognition (or
discrimination), which is also influenced by noise. Recognition thresholds may be estimated to be
an additional 3 dB higher in signal-to-noise ratio than detection thresholds. Values for thresholds
in great tits are from Langemann et al. (1998), noise level from Klump (1996), source level of
‘seeet’ calls from Klump & Shalter (1984). An excess attenuation of 10 dB/100 m was assumed for
2 kHz; the excess attenuation above 2 kHz was increased by 1.85 dB/100 m for every additional
1 kHz. The amount (A) to which the original sound pressure level of the source (S) can drop to
be just detectable in quiet (i.e. to the absolute threshold AAT), in noise (the masked auditory
threshold AMAT) or recognized in noise (the recognition threshold ART) is one variable used for
estimating the perceptual distance (m). The other components include the amplitude decrease
from spherical spreading (20 log10 m) and the decrease from excess attenuation (EA/100 m). Solving
the following equation for m yields the perceptual distance (see also Marten & Marler, 1977):
A(dB) = S (dB) − 20 log10 m − m(E A/100).

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 459
Communication distances of nearly 10 km are obtained by assuming that absolute
thresholds are limiting and that wind speeds are low at night. However, during the
day, wind speed is much higher and it is likely that wind-induced noise provides
sufficient masking to reduce communication distances. Using measurements of
the sound-pressure level of wind-induced noise in grassland (Boersma, 1997) and
assuming a CR of 10 dB (low-frequency CR values can only be extrapolated from
studies in humans, e.g. Moore & Glasberg, 1983) gives a masked threshold of 73 dB.
This is much higher than the absolute threshold of 50 dB that was used in the
calculation by Larom et al. (1997) and suggests that infrasound communication
networks may be less extensive than previously thought.
Some caveats must be borne in mind when estimating detection distances
or transmission distances from CR values. First, CR is measured by presenting a
narrowband signal in wideband masking noise. If the frequency spectrum of the
signal and the noise are rather similar (e.g. detection of an individual calling frog
in the masking noise provided by a chorus of thousands of conspecific frogs), then
the masked threshold calculated from the CR will be overestimated. With signals
of similar frequency spectra, the S/N ratio is probably close to, or even below, 0 dB,
i.e. signals can be detected when their level is equal to, or even below, the level of
the background noise. In this case, the task resembles an increment detection in
overall sound amplitude when the signal is added to the background noise (e.g.
Miller, 1947). Second, the CR may not provide a good estimate of masked thresholds
when the temporal structure of the background noise has very pronounced slow
envelope fluctuations (see below). In this case, masked thresholds may be up to
20 dB more sensitive than would be expected from the CR. Third, it should be
remembered that communication signals are often broadcast repeatedly, or at
least some signal elements are repeated, whereas in laboratory studies the test
signals for detection will often be presented only once. Detection sensitivity is
known to improve by the square root of the number of independent observations
(e.g. Swets et al., 1950), suggesting that repetitive signals may be detected more
readily.
Separating sounds by exploiting temporal patterns
Environmental background noise will usually not resemble the random
noise with a steady-state envelope that is often employed in the laboratory as
a masker. Animals in communication networks appear to be adapted to exploit
temporal envelope fluctuations in the background noise that are typical of the
natural environment (Klump, 1996; Nelken et al., 1999). Signallers have often been
observed to call or sing during periods of reduced amplitude of the background
noise. For example, many frog species call when the nearest signalling neighbours
are silent (Klump & Gerhardt, 1992; Ch. 13). Some frog species go even further and

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
460 U. Langemann & G. M. Klump
(a)
(b)
(c)
Time (s)
Fig. 20.1. Waveforms illustrating different noise envelopes. (a) Unmodulated
wideband random noise exhibiting a Gaussian distribution of amplitude values. This
type of noise exhibits little variation in the temporal envelope. Unmodulated
wideband noise is normally used in determining critical masking ratios.
(b) Coherently amplitude modulated noise. Note the slow envelope fluctuations. This
noise was synthesized by multiplying random wideband noise by a low-passed noise
with a bandwidth of 12.5 Hz. It has the same overall bandwidth as the unmodulated
noise in (a). This type of noise resembles more closely the structure of environmental
noise. (c) A dawn chorus recorded in a European deciduous forest. It has a waveform
with pronounced slow amplitude fluctuations that are more similar to the
fluctuations in the envelope of coherently modulated noise (b) than of unmodulated
noise (a).
will use call timing strategies to mask their neighbours’ calling (e.g. Gerhardt &
Huber, 2002). Receivers may also benefit from temporal patterns in signals and
in background noise because the separation of sounds from different sources is
improved if their amplitude patterns differ considerably. The separation of sounds
originating from different sources into different ‘auditory streams’ is also known
as sound segregation (Bregman, 1990). The ‘unmasking’ effect associated with
sound segregation is well documented in laboratory studies with humans and
other animals (e.g. Moore, 1990; Klump & Langemann, 1995; Nelken et al., 1999;
Klump & Nieder, 2001; Pressnitzer et al., 2001). It should be noted that unmasking

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 461
Table 20.2. Masking release observed in the laboratory for detection of pure tone signalsa
Species Masking Amplitude Study
release (dB) factor
Human, Homo sapiens 12 4 e.g. Schooneveldt & Moore, 1989
European starling, Sturnus
vulgaris
12 4 Klump & Langemann, 1995
Gerbil, Meriones unguiculatus 17 7 Klump et al., 2001
Chinchilla, Chinchilla laniger 6 2 Niemiec, 2001; A. J. Niemieac,
personal communication
Cat, Felis catus 5 2 Budelis et al., 2002; b B. J. May,
personal communication
aMasking release is the masked threshold in wide-band coherently amplitude-modulated back-
ground noise compared with the masked threshold in unmodulated background noise of the
same overall signal energy and bandwidth. The ‘gain’ in signal detection can be expressed either
in decibels or as an amplitude factor (10dB/20). Similar masker envelopes were used in all species,
i.e. the noise bands had dominant frequencies of envelope fluctuations below 50 Hz.bTaking into account only the masking release across auditory analysis filters.
does not mean that masking is absent, rather it means that a partial release from
masking can be observed.
For example, this unmasking effect is shown by the ability of European starlings
Sturnus vulgaris to detect a tone (i.e. a signal of a particular frequency) in noise
(e.g. Klump & Langemann, 1995). Their tone-detection threshold in the type of
wideband random noise commonly used in studies of the CR (Fig. 20.1a) is up to
20 dB worse than in noise of the same overall signal energy and bandwidth that
has been coherently amplitude modulated (Fig. 20.1b) to resemble more closely
typical environmental noise (e.g. bird song dawn chorus; Fig. 20.1c).
The amount of masking release resulting from the modulation of noise de-
pends on the bandwidth of the masking noise: masking release decreases with
decreasing bandwidth of the masking noise. For bandwidths of the size of an au-
ditory analysis filter (about 10–20% of the centre frequency), the masking release
is reduced. For example, in the starling, the masking release is reduced to about
5 dB if 2 kHz signals have to be detected in 200 Hz wide noise centred at the sig-
nal frequency (Klump et al., 1998). Slow rates of envelope fluctuation result in a
larger masking release than fast rates of fluctuation (Klump & Langemann, 1995).
Masking release in amplitude-modulated background noise has been observed in
all species studied so far, although to a different extent (Table 20.2). In humans, an
unmasking effect is also observed if the signal and the masking background noise
are both amplitude-modulated noise bands (no other species has been tested with

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
462 U. Langemann & G. M. Klump
this paradigm). If the noise-band signal has a pattern of envelope fluctuation that
is different from the masking noise bands, the detection of the noise band sig-
nal is improved in comparison with stimuli that have similar correlated envelope
fluctuations in both signal and masking bands. For signals with durations of more
than 200 milliseconds, detection differences of about 8 dB have been observed (e.g.
McFadden & Wright, 1990).
The unmasking effects described in the previous paragraph are likely to be
relevant in animal communication systems (e.g. Klump, 1996). Signals that are
broadcast from a sender will often be amplitude modulated during transmission
(Richards & Wiley, 1980). The same signal travelling along different paths will ex-
perience different modulation patterns since the modulation results from local
turbulence in the atmosphere (arising from temperature gradients or wind). This
applies also for signals from different sources that travel along different paths
to a receiver. Thus, receivers should be able to exploit the different modulation
patterns imposed on signals by the natural environment to gain an advantage in
signal detection. In addition, signallers themselves create amplitude modulation
patterns when broadcasting calls or songs. For example, king penguins Aptenodytes
patagonicus appear to be able to utilize amplitude modulations in their calls to im-
prove their sensitivity when searching for their mate or chick in an assembly of
hundreds of individuals (e.g. Aubin & Jouventin, 1998). The contact calls of the
emperor penguin Aptenodytes forsteri and of the king penguin exhibit a syllable
structure with pronounced amplitude modulations, and their two-voice mode of
call generation creates amplitude beats (Aubin et al., 2000; Lengagne et al., 2001).
Both species of penguin thus produce calls with a distinctive envelope-modulation
pattern that facilitates unmasking effects. The observation by Aubin & Jouventin
(1998) that king penguin chicks are able to detect their parents’ calls within the
colony background noise with a S/N ratio of about −6 dB can be adequately ex-
plained by masking release in amplitude-modulated background noise.
Spatial release from masking
Signal detection also depends on the spatial arrangement of the sound
sources. If the sources of signal and masking noise are well separated, such as by
territorial songbirds, a considerable masking release may be observed. Hine et al.
(1994) measured the detection thresholds of ferrets Mustela putorius for 500 Hz tones
masked by narrowband noise. Signal and noise were presented either from the
same direction (+90◦, i.e. from the subject’s right side) or the signal was presented
from −90◦ and the noise bilaterally from +90◦ and −90◦. Signal detection was
improved by about 10 dB in the bilateral case. Signal detection did not improve in
animals that were only allowed monaural listening (Hine et al., 1994), indicating

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 463
that the release from masking was a result of binaural processing of signal and
noise. Dent and colleagues (1997) replicated the experiment with budgerigars
Melopsittacus undulatus using wideband noise as the masker and tone signals of
different frequencies. The amount of masking was, on average, 7.5 dB less for
bilaterally presented masking noise versus unilateral masking noise presented
from the same direction as the tone signal (Dent et al., 1997). In an additional
experiment, the authors demonstrated that the directional characteristics of the
birds’ auditory system are sufficient to explain the amount of unmasking. By
keeping the masking noise source constant at 0◦ azimuth and moving the signal
source around the animal in 30◦ steps, they observed a masking release of up to
10 dB (Dent et al., 1997; see also the review by Klump, 1996). Binaural processing
will thus contribute to signal detection in communication networks as well as in
other natural situations. However, such a large masking release has not been found
in all species that have been tested. For example, in the green treefrog Hyla cinerea,
spatial unmasking of up to 3 dB has been observed by phonotaxis experiments
with separate noise and signal sources (Schwartz & Gerhardt, 1989).
Recognition of signals
Detecting a signal is the first step in perception. Individuals in an acoustic
communication network may become alert when they detect a signal, but further
reaction will depend on the specific message: that is, the signal needs to be rec-
ognized. Signal recognition in the sense of statistical separation of signals may be
explained best by an everyday example. When listening to the radio while driving
a car, an individual may tune to a programme and can just detect ‘some signal’
or even ‘human speech’. By turning up the volume, and thus increasing the S/N
ratio between the speech and the engine noise of the car, words and, therefore, the
meaning of the message can be recognized. More theoretically, detecting a signal
means observing the occurrence of a signal, i.e. the addition of some signal to a
(possibly noisy) background. However, recognition implies the ability to classify
a detected signal as a member of a set of many (e.g. Green et al., 1977; Wiley &
Richard, 1982). We have little direct evidence on how the S/N ratio for recognition
compares with the S/N ratio required for detection of a signal.
Lohr et al. (2003) presented budgerigars and zebra finches Taeniopygia guttata
with contact calls of three species (zebra finch, budgerigar, canary Serinus canarius)
in order to determine thresholds for the detection of the calls in noise. They also
determined each species’ ability to discriminate between different call types of
zebra finches or budgerigars in the same masking noise. Birds were thus forced in
noisy background conditions to ‘hear out’ and recognize a deviant call in a series
of repeating reference calls. Thresholds for the discrimination of both conspecific

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
464 U. Langemann & G. M. Klump
and heterospecific calls in noise were about 3 dB worse than thresholds for call
detection: the S/N ratio for recognition had to be, on average, 3 dB higher than the
S/N ratio for mere detection (Lohr et al., 2003).
There is no information from field studies that allows us to compare the S/N
ratio necessary for detection of signals with that for recognition of signals in the
natural environment. This is because field studies use playback to elicit natural
species-specific responses and, therefore, the subjects have both detected and rec-
ognized the signal (e.g. Brenowitz, 1982a,b; Aubin & Jouventin, 1998). Brenowitz
(1982a,b) studied the reaction of territorial male red-winged blackbirds Agelaius
phoeniceus to song signals that were either played alone or with wideband random
noise added to the playback song. Playback elicited more high-intensity song and
visual display when the S/N ratio was increased from 0 to 3 dB (as measured in
the 4 kHz octave band that contained most of the spectral energy of the song).
Since this male response required the recognition of the signal, one can conclude
that 3 dB is a conservative estimate of the S/N ratio necessary for signal recogni-
tion. Using data on auditory signal detection in red-winged blackbirds obtained
in the laboratory (Hienz & Sachs, 1987), Klump (1996) calculated that the S/N ratio
necessary for detection should be about 8 dB less than the S/N ratio necessary for
recognition. In king penguins, the S/N ratio necessary for recognition appears to
be lower than in the red-winged blackbird (Aubin & Jouventin, 1998). However,
as suggested above, this may be because of the distinctive envelope-modulation
pattern of penguin calls.
The sender’s adaptations for maximizing signal transmission
Behavioural observations of signalling birds suggest that senders have
evolved mechanisms to modify signal production in order to improve detection
by the receiver. Holland et al. (1998) concluded from measurements of broadcast
song of the wren Troglodytes troglodytes that higher song posts could optimize sound
transmission. Broadcasting song from high perches where the vegetation is less
dense also possibly improves the ability to detect responses of conspecifics in
wrens (Holland et al., 1998) and blackbirds Turdus merula (Dabelsteen et al., 1993).
However, it should be noted that changing location alone may improve perception
by allowing sequential integration of acoustic information, for example in the
context of sound localization.
Another strategy employed by a sender to increase information transfer within
a communication network is to adapt its sound output to the level of the back-
ground noise. This requires that a sender constantly monitors the level and spec-
tral composition of the background noise interfering with its own vocalization.
The increase in sound-pressure level of vocal output at times of increased levels of

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 465
background noise is called the Lombard effect and it has been well investigated
in humans (e.g. Pick et al., 1989). Sinnott et al. (1975) demonstrated that trained
monkeys Macaca fascicularis and M. nemestrina spontaneously increased their call
amplitude if noise bands of the same fundamental frequencies as their call masked
them, but not if noise bands were of much higher frequencies. The monkeys’voice
amplitude increased by about 2 dB for every 10 dB of masking noise. Also budgeri-
gars significantly increase the level of their vocalization in response to noise of the
same frequency spectrum as their contact calls but not to noise outside this spec-
tral range (Manabe et al., 1998). The Lombard effect has also been reported in zebra
finches (Cynx et al., 1998) and nightingales Luscinia megarhynchos (Brumm & Todt,
2002). The first animal species in which the Lombard effect has been shown un-
der field conditions is the blue-throated hummingbird Lampornis clemenciae (Pytte
et al., 2003). The authors observed that naturally occurring and experimentally
controlled amplitude changes of the ambient noise level induced change in am-
plitude of the birds’ territorial advertisement call.
Localization and distance perception
Perceiving the direction of a sound source
For participants in communication networks, it is advantageous to be
able to identify the location of the signal source. For example, the pattern of
alarm calls in a bird community could provide a good estimate of the path taken
by a predator (e.g. McGregor & Dabelsteen, 1996). Also, in territorial interactions,
birds appear to combine information from the song signal and the direction from
which it is heard to evaluate the potential threat by a competitor (e.g. McGregor &
Avery, 1986). There is considerable variation in the accuracy of sound localization
between species. Furthermore, each species’ ability to localize sound depends on
the physical characteristics of the sound, such as the frequency spectrum of the
sound or its temporal characteristics.
The accuracy and mechanisms of sound localization have been reviewed in
frogs (e.g. Rheinlaender & Klump, 1988), in birds (e.g. Klump, 2000) and in terres-
trial mammals (e.g. Gourevitch, 1987; Brown, 1994). Two cues are used in sound
localization: the difference in the time of arrival (or the phase difference) and the
intensity differences between the spectral components of the sound impinging
on the two ears. Figure 20.2a shows an example of interspecific variation in the
accuracy of the localization of tones in the horizontal plane (azimuth) for two
birds of prey, barn owl Tyto alba (Knudsen & Konishi, 1979) and sparrowhawk Ac-
cipiter nisus (G. M. Klump & E. Kretzschmar, unpublished data), and for four species
of small birds, great tit (Klump et al., 1986), zebra finch, budgerigar and canary

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
466 U. Langemann & G. M. Klump
(a)
(b)
Fig. 20.2. The accuracy of azimuth sound localization. (a) Minimum detectable angles
of pure-tone stimuli in relation to frequency as determined in the laboratory in
two avian predators (open symbols, Knudsen & Konishi, 1979; G. M. Klump &
E. Kretzschmar, unpublished data) and four species with smaller interaural distances
than the raptors (filled symbols; Klump et al., 1986; Park & Dooling, 1991).

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 467
(Park & Dooling, 1991). The superior accuracy of the two birds of prey may be
explained at least partly by the physical properties of their auditory system pro-
viding larger interaural cues and by specializations in processing the interaural
sound differences in the auditory pathway (see Klump, 2000).
When discussing the accuracy of sound localization in the natural environ-
ment, data from field studies are relevant (e.g. Nelson & Stoddard, 1998). Figure
20.2b shows the sound localization accuracy of a trained male sparrowhawk in
the laboratory and in the field (G. M. Klump & E. Kretzschmar, unpublished data).
The data in both sets were obtained with the same stimulus paradigm and operant
procedure and so can be compared directly. The sparrowhawk’ssound localization
accuracy was considerably reduced in the field. Localization accuracy for natural
sounds was similar to the accuracy for tones of comparable frequency. The data
shown in Fig. 20.2 were obtained by forcing the bird to localize a single signal
presented at an unpredictable time. This procedure ensures that the bird is only
using open-loop sound localization (Klump, 1995) and cannot use strategies to in-
tegrate information over several signal presentations or maximize binaural cues
in some other way. It is to be expected that field studies presenting several signals
before the subject responds will result in more accurate sound localization (e.g.
Nelson & Stoddard, 1998).
Perceiving the distance of a sound source
To assess the location of a signal source, knowledge of distance is as impor-
tant as information about the direction from which the signal is heard. Assessing
distance information is often referred to as ranging (Morton, 1982). In the past,
distance assessment has almost exclusively been investigated in birds by simulat-
ing territorial intrusions and studying the behavioural response of the territory
owner (reviewed by Naguib & Wiley, 2001). There are also many other common
contexts in which distance is assessed, for example animals maintaining contact
with mates or flock members when moving through dense vegetation.
Fig. 20.2 (cont.) (b) Minimum angles detectable by a sparrowhawk Accipiter nisus
determined either in an anechoic chamber in the laboratory (open symbols, thick grey
lines) or in a natural deciduous forest (closed symbols, thick black lines) using a
two-alternative forced-choice procedure that had been established for measuring the
accuracy of sound localization in small birds (Klump, 1995). Circles show the
minimum detectable angle for pure tones (open circles are the same data as in (a).
Diamonds indicate the minimum detectable angle for the ‘seeet’ aerial predator call.
Horizontal lines represent the frequency range and the sparrowhawk’s localization
accuracy for great tit mobbing calls (dashed thick lines) and scolding calls (solid thick
lines). (G. M. Klump & E. Kretzschmar, unpublished data.)

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
468 U. Langemann & G. M. Klump
Two recent studies have provided data on the accuracy of distance percep-
tion as revealed by the approach responses of birds in the natural environment.
Nelson & Stoddard (1998) measured the accuracy of the approach by Eastern
towhees Pipilo erythrophthalmus to a loudspeaker playing back the species’ calls.
The error in distance assessment was determined from the birds’ closest approach
to the loudspeaker. The birds’ initial distance to the sound source was approxi-
mately 10, 20 or 30 m and resulted in average distance errors of 2.3, 3.5 and 3.4 m,
respectively. In additional experiments, the actual distance to the sound source
did not match the characteristics of the playback signal. For example, a signal rere-
corded after being transmitted over a distance of 20 m was played from an actual
distance of 10 m resulting in a simulated distance of 30 m. About half of the ex-
perimental birds responded by flying the actual distance towards the loudspeaker
and the other half by flying the simulated distance, suggesting that some cues for
distance assessment are derived from signal characteristics and some from the
actual location of the sound source (Nelson & Stoddard, 1998).
Simulated distances have also been used by Naguib et al. (2000) to study distance
assessment by chaffinches Fringilla coelebs. Unlike previous field studies, Naguib
et al. (2000) manipulated song signals by simulating their transmission in a vir-
tual forest with the help of a computer. This allowed control over the amount of
reverberation imposed and control over frequency-dependent attenuation. Simu-
lated transmission distances ranged from 0 (original source signal) to 120 m. The
approach response to playback of a single song with different virtual distances
showed that chaffinches mainly discriminated between playback signals simu-
lating shorter distances (0, 20 and 40 m) and playback signals simulating longer
distances (80 and 120 m) but did not discriminate within short- or long-distance
categories. This means that chaffinches exhibit a categorical response to simulated
intruders close to, versus more distant from, their territory, rather than gauging
their approach to the virtual distance of the sound source (Naguib et al., 2000).
Laboratory studies of birds’ perception allow us to evaluate the salience of the
cues that may be used for distance assessment. Cues suggested by field studies in-
clude the overall signal amplitude, the frequency-dependent excess attenuation,
the amplitude modulation of the signal envelope imposed by atmospheric turbu-
lence along the transmission path, the addition of noise to the signal, and rever-
beration resulting from echoes overlapping or trailing the signal (e.g. Dabelsteen
et al., 1993; McGregor, 1994). Overall signal amplitude has been identified as
a useful cue for distance assessment in psychoacoustic studies. Phillmore et al.
(1998) trained zebra finches and black-capped chickadees Poecile atricapillus with
operant procedures to distinguish calls and songs from either species that were
recorded in a woodland habitat at distances of 5, 25, 50 and 75 m from the

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 469
loudspeaker. After training with a set of signals from all four distances, the birds
could discriminate between the signals of the set and also between unknown
songs from these recording distances. The stimuli representing various distances
differed in amplitude at least as much as the known intensity-difference limen (the
just-noticeable intensity difference) in birds (Dooling & Saunders, 1975; Klump &
Baur, 1990). Removing amplitude cues made discrimination considerably worse
(Phillmore et al., 1998), indicating its potential role in distance assessment. How-
ever, in the field, amplitude alone may not be a reliable indicator of distance,
since head movements of the singing bird can lead to amplitude differences at the
receiver’s position (e.g. Larsen & Dabelsteen, 1990).
A number of field studies suggest reverberation as an important cue for distance
assessment (e.g. McGregor, 1994; Naguib & Wiley, 2001). Echos that are imposed
on each signal element during transmission ‘degrade’ (i.e. distort) its original
amplitude and time pattern: the study by Holland et al. (2001) suggested that
wrens can extract cues that allow distance assessment from the echo tail trailing
the signal. Wrens responded to songs consisting of undegraded elements with
added trailing echo tails (from degraded elements) in the same way as to degraded
songs (i.e. degraded element and echo tail). Songs consisting of degraded elements
without echo tails elicited a response that was intermediate between that to an
undegraded and that to a degraded song with echo tail, stressing the salience of the
echo tail as a cue. Psychoacoustic studies of humans also indicate that echo tails
provide an important cue for distance assessment; we appear to use the direct-
to-reverberant energy ratio (Zahorik, 2002). This cue may allow us to estimate
sound-source distance independent of the sound level, which in turn may allow
the loudness of the source to be inferred (Zahorik & Wightman, 2001).
SINDSCAL: an analysis method for perceptual distances
Laboratory studies in the great tit also indicate that echoes alone may
provide a sufficient cue for distance assessment. In this final part, we would like
to describe how perceptual differences can be examined in trained animals. We
present a multidimensional scaling procedure (SINDSCAL: symmetric individual
difference scaling; e.g. Arabie et al., 1987) that is especially suited to investigate
which signal modifications are salient to the animals. A virtual forest (Naguib
et al., 2000) was used to impose reverberation on synthetic great tit song signals
equivalent to sound transmission distances of between 5 and 320 m (Fig. 20.3).
The sound pressure of the song signals was then adjusted to the same overall root
mean square amplitude so that the reverberation pattern imposed on the signal
remained the only possible cue to distance. Great tits were then trained in an oper-
ant Go/NoGo procedure with repeating background to discriminate between song

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
470 U. Langemann & G. M. Klump
Fig. 20.3. An example of stimuli used to estimate perceptual differences in laboratory
experiments with great tits. The spectrogram shows a two-element great tit song at a
virtual distance of 5 m and the waveforms show the same song elements at virtual
distances of 5, 80 and 160 m. Further details in text.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 471
elements in which echo patterns alone indicated different virtual distances (e.g.
Dooling & Okanoya, 1995). The response latencies of the great tits to all possible
reference–test differences were recorded (Fig. 20.4a) and analysed by a SINDSCAL
(e.g. Arabie et al., 1987) model with log-transformed data. The result of SINDSCAL is
a three-dimensional object space (Fig. 20.4b), providing a kind of ‘perceptual map’
in which the response latencies are translated into relative distances between
data points. The distance between data points in the perceptual space provides a
measure of the perceived similarity of the acoustic signals. Data points that are
close to each other indicate that large response latencies were observed in the
discrimination (i.e. the difference was not salient to the birds). Short response
latencies indicate a salient differences that will lead to a large spread of the data
points. A three-dimensional SINDSCAL model accounts for 86–90% of the variance
in response latencies. Since the first dimension explained most of the total vari-
ance, the latency data for the different experimental songs were reanalysed with a
one-dimensional SINDSCAL model to allow easier comparison between song types.
Perceptual space coordinates (from the one-dimensional model) of the four differ-
ent two-note songs that were tested are shown in Fig. 20.5 as a function of the
virtual distance of each signal. As in Fig. 20.4b, close perceptual space coordinates
indicate signals that have been perceived as being more similar. There was no sig-
nificant difference between the different song types tested (two-way ANOVA with
distance and song type as factors and subsequent Tukey-tests, F3,24 < 0.001; p = 1).
However, within the factor ‘virtual distance’, significant differences were obvious
(F6,21 = 73.7; p ≤ 0.001). Space coordinates for ‘long’ virtual distances (160 and
320 m) differed significantly from those for the other distances but not from each
other (i.e. ‘long’ virtual distances were similar for all the birds’). Space coordinates
for the virtual distance of 80 m significantly differed from those of the three ‘short’
distances (5, 10 and 20 m). ‘Short’ virtual distances of 5, 10 and 20 m were treated
as similar by the birds (i.e. no significant difference was found). Therefore, great
tits in the laboratory provided with echo patterns as the only available distance
assessment cue showed a categorical response that was similar to the response
observed in field experiments with chaffinches (Naguib et al., 2000).
Auditory scene analysis
So far we have discussed basic perceptual mechanisms involved in rela-
tively simple auditory detection and discrimination tasks. Real-world situations,
however, require receivers to analyse sounds from a mixture of simultaneously
active sources: that is, to perform auditory scene analysis (Bregman, 1990). For ex-
ample, this applies to communication networks of birds during the dawn chorus,
when a receiver has to analyse streams of song elements from each individual

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
472 U. Langemann & G. M. Klump
500
1000
1500
2000
2500
0 50 100 150 200 250 300 350
Virtual distance difference (m)
rs = −0.839 p < 0.001
Song 5
Perceptual space(song 5)
5 m10 m
20 m
80 m
160 m
320 m
40 m
(a)
(b)
Res
po
nse
late
ncy
(m
s)
−
−
−
−
−
−
−
−
−
−
−
−
−
−
−
−
Fig. 20.4.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 473
Fig. 20.5. One-dimensional perceptual space coordinates as a function of the virtual
distance (i.e. the distance simulated by imposing reverberations upon the signal) for
four test songs. Similar perceptual space coordinates indicate that the differences in
the echo pattern of the signals are not very salient to the great tits, and large
differences between data points indicate salient differences have been detected
between the respective echo modifications. There are no salient differences between
signals of ‘short’ virtual distances of 5, 10 and 20 m or between signals of ‘long’ virtual
distances of 160 and 360 m. Differences between signals of ‘short’ virtual distances
and ‘long’ virtual distances are very salient to the birds. Virtual distances of 40 and
80 m lie in a transition range. This pattern occurs for all songs that were tested (see
ANOVA results in the text).
Fig. 20.4 (cont.) Results of great tits scaling differences between songs manipulated to
represent various virtual distances. The signal variants consisted of the same song
elements that differed only in echo pattern, which simulated sound transmission
distances of 5 m to 320 m. (a) Average response latencies of four great tits as a function
of the difference in virtual distance (e.g. the virtual-distance difference between
signals of simulated transmission distances of 320 and 80 m is 240 m). Larger response
latencies indicate less-salient differences in the cues; shorter response latencies
indicate more-salient differences. (b) Three-dimensional object space or ‘perceptual
map’ (SINDSCAL model with log-transformed data, see text; Arabie et al., 1987)
demonstrating the salience of reverberation for great tits. Distances between data
points in perceptual space reflect response-latency differences in discriminating
between the signal variants. Small distances indicate that signals are treated as being
similar, and large distances indicate that salient differences between signals have
been perceived. (U. Langemann, U. Pander & G. M. Klump, unpublished data.)

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
474 U. Langemann & G. M. Klump
singer. Laboratory experiments have shown that animals form auditory streams
and analyse auditory scenes in a similar way to humans (e.g. Hulse et al., 1997;
Feng & Ratnam, 2000; Moss & Surlykke, 2001; Hulse, 2002). However, some of
the basic mechanisms of masking discussed above also contribute to an auditory
scene analysis that is characterized by an improved segregation of overlapping
signals. For example, we have shown that common modulation of components
of sounds aids signal segregation and results in reduced masking of one sound
by another. Similarly, common onsets or offsets of signal components lead to the
formation of auditory objects (e.g. Geissler & Ehret, 2002) that can be analysed
separately from other objects in the same auditory scene. Spatial separation of
sources will also aid auditory object formation, and the spatial release from mask-
ing discussed above may be partly a result of improved signal segregation from the
background.
Summary
In this chapter, we have illustrated how sensory abilities of individuals af-
fect auditory perception and thus acoustic communication. How does this relate
to communication between individuals in networks? In a network, individuals
are distributed in space. The relative position of any ‘node’ in this network, the
distance between individuals, profoundly determines a receiver’s ability to detect
and recognize acoustic signals. Because of the spatial distribution of signallers
and receivers, the same propagated signal may result in quite different percep-
tion at different places in a communication network. Acoustic signals will be
modified along their transmission path and will be masked by acoustical energy
from other sources. On the one hand, masking is certainly the most important fac-
tor severely impairing the detection of acoustic communication signals. Spectral
aspects and the temporal patterns of masking sounds affect the amount of mask-
ing that is exerted and the spatial distribution of concurrent sound sources (or
individuals) contributes to masking efficiency. On the other hand, receivers may
exploit changes imposed on a signal during transmission. For example, reverbera-
tion patterns will allow distance assessment of sound sources and, together with
binaural cues, render it possible to gain insights into the spatial distribution of
the individuals in a network.
Our current knowledge from perceptual studies will provide a better under-
standing of animal behaviour within acoustically complex communication net-
works. In addition, an approach that takes the receiver’s perception into account
will allow a better evaluation of communication behaviour in the field than ap-
proaches that rely mainly on physical properties of signals and their transmission
(McGregor et al., 2000).

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 475
Acknowledgements
The research was supported by grants from the Deutsche Forschungsgemeinschaft (SFB
204, FOR 306). Ulrike Pander provided data from experiments with great tits that were reanalysed
for this study.
References
Arabie, P., Carroll, J. D. & DeSarbo, W. S. 1987. Three-Way Multidimensional Scaling and
Related Techniques. Newbury Park, CA: Sage.
Aubin, T. & Jouventin, P. 1998. Cocktail-party effect in king penguin colonies.
Proceedings of the Royal Society of London, Series B, 265, 1665–1673.
Aubin, T., Jouventin, P. & Hildebrand, C. 2000. Penguins use the two-voice system to
recognize each other. Proceedings of the Royal Society of London, Series B, 267,
1081–1087.
Boersma, H. F. 1997. Characterization of the natural ambient sound environment:
measurements in open agricultural grassland. Journal of the Acoustical Society of
America, 103, 2104–2110.
Bregman, A. S. 1990. Auditory Scene Analysis: The Perceptual Organization of Sound.
Cambridge, MA: MIT Press.
Brenowitz, E. A. 1982a. The active space of red-winged blackbird song. Journal of
Comparative Physiology A, 147, 511–522.
1982b. Long-range communication of species identity by song in the red-winged
blackbird. Behavioral Ecology and Sociobiology, 10, 29–38.
Brown, C. H. 1994. Sound localization. In: Comparative Hearing: Mammals, ed. R. R. Fay &
A. N. Popper. New York: Springer, pp. 57–96.
Brumm, H. & Todt, D. 2002. Noise-dependent song amplitude regulation in a
territorial songbird. Animal Behaviour, 63, 891–897.
Budelis, J., Fishbach, A. & May, B. J. 2002. Behavioral assessment of comodulation
masking release in cats. Abstracts of the Association for Research in Otolaryngology, 25,
211.
Cynx, J., Lewis, R., Tavel, B. & Tse H. 1998. Amplitude regulation of vocalizations in
noise by a songbird, Taeniopygia guttata. Animal Behaviour, 56, 107–113.
Dabelsteen, T., Larsen, O. N. & Pedersen, S. B. 1993. Habitat-induced degradation of
sound signals: quantifying the effects of communication sounds and bird
location on blur ratio, excess attenuation, and signal-to-noise ratio in blackbird
song. Journal of the Acoustical Society of America, 93, 2206–2220.
Dent, M. L., Larsen, O. N. & Dooling, R. J. 1997. Free-field binaural unmasking in
budgerigars (Melopsittacus undulatus). Behavioral Neuroscience, 111, 590–598.
Dooling, R. J. & Okanoya, K. 1995. Psychophysical methods for assessing perceptual
categories. In: Methods in Comparative Psychoacoustics, ed. G. M. Klump, R. J. Dooling,
R. R. Fay & W. C. Stebbins. Basel: Birkhauser, pp. 307–318.
Dooling, R. J. & Saunders, J. C. 1975. Auditory intensity discrimination in the parakeet
(Melopsittacus undulatus). Journal of the Acoustical Society of America, 58, 1308–1310.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
476 U. Langemann & G. M. Klump
Douglas, H. D. & Conner, W. E. 1999. Is there a sound reception window in coastal
environments? Evidence from shorebird communication systems.
Naturwissenschaften, 86, 228–230.
Dubois, A. & Martens, J. 1984. A case of possible vocal convergence between frogs and a
bird in Himalayan torrents. Journal fur Ornithologie, 125, 455–463.
Fay, R. R. 1988. Hearing in Vertebrates: a Psychophysics Databook. Winnetka, IL: Hill-Fay.
Fegeant, O. 1999. Wind-induced vegetation noise. Part II: field measurements.
Acustica/Acta Acustica, 85, 241–249.
Feng, A. S. & Ratnam R. 2000. Neural basis of hearing in real-world situations. Annual
Review of Psychology, 51, 699–725.
Gerhardt, H. C. & Huber, F. 2002. Acoustic Communication in Insects and Anurans. Chicago,
IL: University of Chicago Press.
Geissler, D. B. & Ehret, G. 2002. Time-critical integration of formants for perception of
communication calls in mice. Proceedings of the National Academy of Sciences, USA, 99,
9021–9025.
Gourevitch, G. 1987. Binaural hearing in land mammals. In: Directional Hearing, ed.
W. A. Yost & G. Gourevitch. New York: Springer, pp. 226–246.
Green, D. M., Weber, D. L. & Duncan, J. E. 1977. Detection and recognition of pure
tones in noise. Journal of the Acoustical Society of America, 62, 948–954.
Hienz, R. D. & Sachs, M. B. 1987. Effects of noise on pure-tone thresholds in blackbirds
(Agelaius phoeniceus and Molothrus ater) and pigeons (Columba livia). Journal of
Comparative Psychology, 101, 16–24.
Hine, J. E., Martin, R. L. & Moore, D. R. 1994. Free-field binaural unmasking in ferrets.
Behavioral Neuroscience, 108, 196–205.
Holland, J., Dabelsteen, T., Pedersen, S. B. & Larsen, O. N. 1998. Degradation of wren
Troglodytes troglodytes song: implications for information transfer and ranging.
Journal of the Acoustical Society of America, 103, 2154–2166.
Holland, J., Dabelsteen, T., Bjørn, C. P. & Pedersen, S. B. 2001. The location of ranging
cues in wren song: evidence from calibrated interactive playback experiments.
Behaviour, 138, 189–206.
Hulse, S. H. 2002. Auditory scene analysis in animal communication. Advances in the
Study of Behaviour, 31, 163–200.
Hulse, S. H., MacDougall-Shackleton, S. A. & Wisniewski, A. B. 1997. Auditory scene
analysis by songbirds: stream segregation of birdsong by European starlings
(Sturnus vulgaris). Journal of Comparative Psychology, 111, 3–13.
Klump, G. M. 1995. Sound localization studies in nonspecialized birds. In: Methods in
Comparative Psychoacoustics, ed. G. M. Klump, R. J. Dooling, R. R. Fay & W. C.
Stebbins. Basel: Birkhauser, pp. 171–182.
1996. Bird communication in the noisy world. In: Ecology and Evolution of Acoustic
Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca NY: Cornell
University Press, pp. 321–338.
2000. Sound localization in birds. In: Comparative Hearing: Birds and Reptiles, ed. R. J.
Dooling, R. R. Fay & A. N. Popper. New York: Springer, pp. 247–305.
Klump, G. M. & Baur, A. 1990. Intensity discrimination in the European starling
(Sturnus vulgaris). Naturwissenschaften, 77, 545–548.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 477
Klump, G. M. & Curio, E. 1983. Warum liegen viele Lautaußerungen von Singvogeln
oberhalb des Bereichs besten Horens? Verhandlungen der Deutschen Zoologischen
Gesellschaft, 76, 182.
Klump, G. M. & Gerhardt, H. C. 1992. Mechanisms and function of call-timing in
male–male interactions in frogs. In: Playback and Studies of Animal Communication,
ed. P. K. McGregor. London: Plenum Press, pp. 153–174.
Klump, G. M. & Langemann, U. 1995. Comodulation masking release in a songbird.
Hearing Research, 87, 157–164.
Klump, G. M. & Nieder, A. 2001. Release from masking in fluctuating background
noise is represented in a songbird’s auditory forebrain. Neuroreport, 12,
1825–1829.
Klump, G. M. & Shalter, M. D. 1984. Acoustic behaviour of birds and mammals in the
predator context: I. Factors affecting the structure of alarm calls. II. The
functional significance of alarm signals and their evolution. Zeitschrift fur
Tierpsychologie, 66, 189–226.
Klump, G. M., Windt, W. & Curio E. 1986. The great tit’s (Parus major) auditory
resolution in azimuth. Journal of Comparative Physiology, 158, 383–390.
Klump, G. M., Langemann, U. & Nieder, A. 1998. Mechanisms that improve signal
detection in noise: a study of comodulation masking release in a songbird. In:
Psychophysical and Physiological Advances in Hearing, ed. A. R. Palmer, A. Rees, A. Q.
Summerfield & R. Meddis. London: Whurr, pp. 270–276.
Klump, G. M., Kittel, M., Wagner, E. 2001. Comodulation masking release in the
Mongolian gerbil. Abstracts of the Association for Research in Otolaryngology, 24, 84.
Knudsen, E. I. & Konishi, M. 1979. Mechanisms of sound localization in the barn owl
(Tyto alba). Journal of Comparative Physiology, 133, 13–21.
Langemann, U., Gauger, B. & Klump, G. M. 1998. Auditory sensitivity in the great tit:
perception of signals in the presence and absence of noise. Animal Behaviour, 56,
763–769.
Larom, D., Garstang, M., Lindeque, M. et al. 1997. Meteorology and elephant
infrasound at Etosha National Park, Namibia. Journal of the Acoustical Society of
America, 101, 1710–1717.
Larsen, O. N. & Dabelsteen, T. 1990. Directionality of blackbird vocalization.
Implications for vocal communication and its further study. Ornis Scandinavica,
21, 37–45.
Lengagne, T., Lauga, J. & Aubin, T. 2001. Intra-syllabic acoustic signatures used by the
king penguin in parent-chick recognition: an experimental approach. Journal of
Experimental Biology, 204, 663–672.
Lohr, B., Wright T. F. & Dooling, R. J. 2003. Detection and discrimination of natural
calls in masking noise by birds: estimating the active space of a signal. Animal
Behaviour, 65, 763–777.
Long, G. R. 1977. Masked auditory thresholds from the bat, Rhinolophus ferrumequinum.
Journal of Comparative Physiology A, 116, 247–255.
Manabe, K., Sadr, E. I. & Dooling, R. J. 1998. Control of vocal intensity in budgerigars
(Melopsittacus undulatus): differential reinforcement of vocal intensity and the
Lombard effect. Journal of the Acoustical Society of America, 103, 1190–1198.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
478 U. Langemann & G. M. Klump
Marten, K. & Marler, P. 1977. Sound-transmission and its significance for animal
vocalization. I. Temperate habitats. Behavioral Ecology and Sociobiology, 2,
271–290.
McFadden, D. & Wright, B. A. 1990. Temporal decline of masking and comodulation
detection differences. Journal of the Acoustical Society of America, 88, 711–724.
McGregor, P. K. 1994. Sound cues to distance: the perception of range. In: Perception and
Motor Control in Birds: An Ecological Approach, ed. M. N. O. Davies & P. R. Green.
Berlin: Springer, pp. 74–94.
McGregor, P. K. & Avery, M. I. 1986. The unsung songs of great tits (Parus major):
learning neighbours’ songs for discrimination. Behavioral Ecology and Sociobiology,
18, 311–316.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca
NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McGregor, P. K., Otter, K. A. & Peake, T. M. 2000. Communication networks: receiver
and signaller perspectives. In: Animal Signals. Signalling and Signal Design in Animal
Communication, ed. Y. Espmark, T. Amundsen & G. Rosenqvist. Trondheim: Tapir
Academic Press, pp. 405–416.
Miller, G. A. 1947. Sensitivity to changes in the intensity of white noise and its
relation to masking and loudness. Journal of the Acoustical Society of America, 19,
609–619.
Moore, B. C. J. 1990. Co-modulation masking release: spectro-temporal pattern analysis
in hearing. British Journal of Audiology, 24, 131–137.
2003. An Introduction to the Psychology of Hearing. Amsterdam: Academic Press.
Moore, B. C. J. & Glasberg, B. R. 1983. Suggested formulae for calculating
auditory-filter bandwidths and excitation pattern. Journal of the Acoustical Society of
America, 74, 750–753.
Morton, E. S. 1982. Grading, discreteness, redundancy, and motivation–structural
rules. In: Acoustic Communication in Birds, Vol. I, ed. D. E. Kroodsma, E. H. Miller & H.
Ouellet. New York: Academic Press, pp.183–212.
Moss, C. F. & Surlykke, A. 2001. Auditory scene analysis by echolocation in bats. Journal
of the Acoustical Society of America, 110, 2207–2226.
Naguib, M. & Wiley, R. H. 2001. Estimating the distance to a source of sound:
mechanisms and adaptations for long-range communication. Animal Behaviour,
62, 825–837.
Naguib, M., Klump, G. M., Hillmann, E., Griessman, B. & Teige, T. 2000. Assessment of
auditory distance in a territorial song bird: accurate feat or rule of thumb? Animal
Behaviour, 59, 715–721.
Nelken, I., Rotman, Y. & Bar Yosef, O. 1999. Responses of auditory-cortex neurons to
structural features of natural sounds. Nature, 397, 154–157.
Nelson, B. S. & Stoddard, P. K. 1998. Accuracy of auditory distance and azimuth
perception by a passerine bird in natural habitat. Animal Behaviour, 56, 467–477.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
Perception and acoustic communication networks 479
Niemiec, A. J. 2001. The effects of increasing masker temporal regularity on
co-modulation masking thresholds in chinchillas. Abstracts of the Association for
Research in Otolaryngology, 24, 85.
Park, T. J. & Dooling, R. J. 1991. Sound localization in small birds: absolute localization
in azimuth. Journal of Comparative Psychology, 105, 125–133.
Pick, H. L., Siegel, G. M., Fox, P. W., Garber, S. R. & Kearney, J. K. 1989. Inhibiting the
Lombard effect. Journal of the Acoustical Society of America, 85, 894–900.
Phillmore L. S., Sturdy, C. B., Ramsay, S. M. & Weisman, R. G. 1998 Discrimination of
auditory distance cues by black-capped chickadees (Poecile atricapillus) and zebra
finches (Taeniopygia guttata). Journal of Comparative Psychology, 112, 282–291.
Pressnitzer, D., Meddis, R., Delahaye, R. & Winter, I. M. 2001. Physiological correlates
of comodulation masking release in the mammalian ventral cochlear nucleus.
Journal of Neuroscience, 21, 6377–6386.
Pytte, C. L., Rusch, K. M. & Ficken, M. S. 2003. Regulation of vocal amplitude by the
blue-throated hummingbird, Lampornis clemenciae. Animal Behaviour, 66, 703–710.
Rheinlaender, J. & Klump, G. M. 1988. Behavioural aspects of sound localization. In: The
Evolution of the Amphibian Auditory System, ed. B. Fritsch, M. J. Ryan, W. Wilczynski,
T. E. Hetherington & W. Walkowiak. New York: John Wiley, pp. 297–305.
Richards, D. G. & Wiley, R. H. 1980. Reverberations and amplitude fluctuations in the
propagation of sound in a forest: implications for animal communication.
American Naturalist, 115, 381–399.
Ryan, M. J. & Brenowitz, E. A. 1985. The role of body size, phylogeny and ambient noise
in the evolution of bird song. American Naturalist, 126, 87–100.
Schooneveldt, G. P. & Moore, B. C. J. 1989. Comodulation masking release (CMR) as a
function of masker bandwidth, modulator bandwidth, and signal duration.
Journal of the Acoustical Society of America, 85, 273–281.
Schwartz, J. J. & Gerhardt, H. C. 1989. Spatially mediated release from auditory
masking in an anuran amphibian. Journal of Comparative Physiology A, 166, 37–41.
Shannon, C. E. & Weaver, W. 1949. The Mathematical Theory of Communication. Urbana, IL:
University of Illinois Press.
Sinnott, J. M., Stebbins, W. C. & Moody, D. B. 1975. Regulation of voice amplitude by
the monkey. Journal of the Acoustical Society of America, 58, 412–414.
Staicer, C. A., Spector, D. A. & Horn, A. G. 1996. The dawn chorus and other dial
patterns in acoustic signalling. In: Ecology and Evolution of Acoustic Communication in
Birds, ed. D. E. Kroodsma & E. H. Miller. Ithaca NY: Cornell University Press,
pp.426–453.
Swets, J. A., Shipley, E. F., McKey, M. J. & Green, D. M. 1950. Multiple observations of
signals in noise. Journal of the Acoustical Society of America, 31, 514–521.
Waser, P. M. & Waser, M. S. 1977. Experimental studies of primate vocalization:
specializations for long-distance propagation. Zeitschrift fur Tierpsychologie, 43,
239–263.
Wiley, R. H. & Richards, D. G. 1978. Physical constraints on acoustic communication in
the atmosphere: implications for the evolution of animal vocalizations. Behavioral
Ecology and Sociobiology, 3, 69–94.

P1: KPB/... P2: KOD0521823617c20.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:27
480 U. Langemann & G. M. Klump
1982. Adaptations for acoustic communication in birds: sound transmission and
signal detection. In: Acoustic Communication in Birds, Vol. I, ed. D. E. Kroodsma, E. H.
Miller & H. Ouellet. New York: Academic Press, pp. 131–181.
Wollerman, L. & Wiley, R. H. 2002. Background noise from a natural chorus alters
female discrimination of male calls in a Neotropical frog. Animal Behaviour, 63,
15–22.
Yost, W. A. 1994. Fundamentals of Hearing: An Introduction. San Diego CA: Academic Press.
Zahorik, P. 2002. Direct-to-reverberant energy ratio sensitivity. Journal of the Acoustical
Society of America, 112, 2110–2117.
Zahorik, P. & Wightman, F. L. 2001. Loudness constancy with varying sound source
distance. Nature Neuroscience, 4, 78–83.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
21
Hormones, social contextand animal communication
r u i f. o l i v e i r a
Instituto Superior de Psicologia Aplicada, Lisbon, Portugal
Introduction
The views on the role that hormones play in the control of behaviour
have changed progressively with time. Hormones were classically seen as causal
agents of behaviour, acting directly on the display of a given behaviour. This view
was mainly supported by early studies of castration and hormone-replacement
therapy, which showed that some behaviours were abolished by castration and
restored by exogenous administration of androgens (Nelson, 2001). Later this view
shifted towards a more probabilistic approach and hormones started to be seen
more as facilitators of behaviour than as deterministic factors (Simon, 2002). Ac-
cording to this new view, hormones may increase the probability of the expression
of a given behaviour by acting as modulators of the neural pathways underlying
that behavioural pattern. For example, the effects of androgens on the expres-
sion of aggressive behaviours in mammals are mediated by modulatory effects
on central serotonergic and vasopressin pathways (Simon, 2002). Yet, it is also
known that the social environment (i.e. network of interacting individuals) also
feeds back to influence hormone levels (Wingfield et al., 1990), suggesting a two-
way type of interaction between hormones and behaviour. In this chapter, I will
develop the hypothesis that social modulation of androgens is an adaptive mech-
anism through which individuals adjust their motivation according to the social
context that they are facing. Thus, the social interactions within a given social
network would stimulate the production of androgens in the individuals and the
individual levels of androgens would be a function of the perceived social status
and the stability of the social environment in which the animal is living. According
to this view, androgens may play a key role as endocrine mediators of the effects
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
481

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
482 R. F. Oliveira
Fig. 21.1. Interplay between androgens, social behaviour and social context.
Androgens influence the production of a number of social behaviours involved in
communication interactions between animals. In turn, these social interactions
among a network of individuals will shape the social context in which these animals
live, which subsequently will modulate their androgen levels.
of social context on the expression of social behaviour, allowing the animal to
adjust its social behaviour to the context by modulating sensory, cognitive and
motor neural mechanisms underlying animal communication (Fig. 21.1).
In this chapter, I only consider vertebrates because they have a remarkably
similar endocrine system, whereas that of invertebrates can be very different (e.g.
the androgenic hormone in crustaceans is a peptide not a steroid as in vertebrates
(Hasegawa et al., 2002)). Within the vertebrates, I mainly use examples from bony
fishes, a group with wide diversity in mating and parental care systems that makes
up about half the existing vertebrate species (Nelson, 1994). I have also concen-
trated on androgens and male behaviour because I argue that the social network in
which the individual lives modulates its neuroendocrine system, which, in turn,
adjusts the expression of behaviour according to social context. Stress hormones
are, by definition, affected by the social environment and a number of reviews on
social stress and hypothalamus–pituitary–adrenal axis have been published since
the early 1990s (e.g. Sapolsky, 1992). Consequently, an additional benefit of this
chapter is to claim that, like stress hormones, androgens (and perhaps also other
hormones) respond in an adaptive way to the social context, preparing the animal
for the social interactions that it has to face in its everyday life.
Hormones and communication I: the dyadic view
Conceptually, the neurochemical pathways modulated by hormones can
be part of one of three major functional compartments of the nervous system:
sensory, central processing and effector systems (Nelson, 2001). If we translate
this rationale to the communication paradigm, one can consider that hormones
may affect communication by modulating the production of the signal in the

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 483
Signal
Input/perception
Motoroutput
Centralprocessing
Hormones
Receiver
Input/perception
Motoroutput
Centralprocessing
Hormones
Somatic releasers
Sender
Fig. 21.2. Flow of information in a communication dyad. The arrows indicate the
direction of circulation of the information within and between individuals. In the
sender, sensory information received will influence central processing mechanisms
in the central nervous system (CNS), which control, at a higher level, the behavioural
motor output systems that produce the signals. Hormones may modulate signal
production by the sender by acting on central mechanisms, on motor output
mechanisms or by modifying somatic structures that affect the emission of the signal
(i.e. somatic releasers). In the receiver, the signal will be detected by sensory systems
and after peripheral processing will be forwarded to central processing systems in the
CNS. Hormones may affect signal reception and processing in the receiver by acting
directly on the sensory systems that perceive it and/or by acting at a higher level on
the central processing mechanism of the CNS. The central processing of the signal by
the limbic system (and other structures involved in motivational mechanism) may
feed back on hormone levels. The boxes delimit the two organisms and within the
boxes the grey elliptical areas represent the nervous system.
sender, the perception of the signal by the receiver or the central processing of
the message in both senders and receivers (Fig. 21.2).
Hormonal modulation of effector pathways
In senders, hormones may modulate the effector pathways that are in-
volved in the motor circuits underlying the production of the signal. In this way,
hormones can affect the expression of visual displays, vocalizations or pheromone
production and/or release. From the numerous examples in the literature, I have
selected the following, which are intended to cover different communication chan-
nels in different vertebrate taxa.
Androgens and the production of acoustic signals
In songbirds, circulating levels of testosterone are higher at the peak of
the breeding season when singing behaviour reaches its maximum (e.g. Rost, 1990,

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
484 R. F. Oliveira
1992; Smith et al., 1997). Moreover, song production is substantially reduced after
castration and is restored after androgen-replacement therapy (e.g. Arnold, 1975;
Heid et al., 1985). Finally, both androgen and oestrogen receptors have been local-
ized in the song control nuclei of the bird brain: the former in the high vocal cen-
tre, the robust nucleus of the archistriatum, the lateral part of the magnocellular
nucleus of the anterior neostriatum and the dorsomedial part of the intercollic-
ular nucleus; the latter in the high vocal centre and the intercollicular nucleus
(Balthazart et al., 1992; Brenowitz & Arnold, 1989, 1992; Gahr et al., 1987, 1993). Song
is produced by the coordinated contraction of respiratory, syringeal and cranio-
mandibular muscles (Suthers et al., 1999). The activity of syringeal muscles regu-
lates both the timing and the fundamental frequency of the sound (Suthers et al.,
1999). Therefore, by acting directly on the activity and development of syringeal
muscles, hormones may affect song production. For example, in zebra finches
Taenopygia guttata, androgens inhibit the activity of the enzyme cholinesterase,
which breaks down the neurotransmitter acetylcholine in the neuromuscular
junctions of the syrinx. This results in a longer lifetime for the neurotransmitter
in the synaptic cleft, which will affect the syringeal contraction pattern and, sub-
sequently, song output and/or structure (Luine et al., 1980). Testosterone is also
known to increase both syringeal muscle mass (Luine et al., 1980) and the density
of acetylcholine receptors in syringeal muscles, suggesting that circulating levels
of testosterone may increase the size and number of endplates in neuromuscular
junctions (Bleisch et al., 1984). Also, in non-oscine birds, testosterone is effective in
inducing changes in call structure by acting on the motor vocal structure underly-
ing these calls. In grey partridges Perdix perdix, male mating calls used by females
in mate choice are affected by testosterone treatment, which induces a thickening
of the external tympanic membranes that are known to be the main sound source
in galliforms (Beani et al., 1995).
These effects of androgens on motor systems underlying the production of vocal
signals are not exclusive to birds. Many fish species also use sounds to communi-
cate. Male toadfish are among the most vocal fish, producing loud humming calls
to attract females to their nest site (e.g. plainfin midshipman Porichthys notatus;
Brantley & Bass, 1994). Also in toadfish, the exogenous administration of andro-
gens promotes the development of the sonic muscles, for example the oyster toad-
fish Opsanus tau (Fine & Pennymaker, 1986) and the plainfin midshipman (Brantley
et al., 1993). Another example comes from amphibians, in which vocal behaviour
is sexually dimorphic in most species and thus potentially androgen dependent
(Kelley, 2002). In the African frog Xenopus laevis, males produce mating calls charac-
terized by fast trills that attract females (Wetzel & Kelley, 1983; Kelley, 2002). The
call-production organ of X. laevis is the larynx and all sounds are produced under-
water (Kelley, 2002). The sex differences in vocal behaviour observed in this species

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 485
are mostly a result of sex differences in adult laryngeal synapses. Male larynx motor
neurons release less neurotransmitter, which will produce lower postsynaptic po-
tentials than in females, allowing the fibres to reach a spike threshold (Tobias et al.,
1995). This synaptic facilitation in male motor neurons allows modulation of the
amplitude of the trills, a characteristic of the call that is used by females when
assessing the males as potential mates (Tobias et al., 1995). Contrary to most cases
of sexual dimorphism in which the default situation is female, the sex differences
in postsynaptic response emerge in females under the influence of oestradiol,
with the default being the slow neurotransmitter release typical of males (Tobias
& Kelley, 1995). However, other sex differences in this vocal system are androgen
dependent, namely the differentiation of laryngeal motor neurons, muscles fibres
and laryngeal cartilage (Kelley, 2002).
Androgens and pheromone production and/or release
A very large number of mammals use chemical signals (i.e. pheromones)
in intraspecific communication. These pheromones can be produced by specific
scent glands or are released into the environment in the urine or in other body
fluids (Bradbury & Vehrencamp, 1998). Most mammals use marking behaviour to
release these pheromones, a behaviour that is sexually dimorphic ( Johnson, 1973;
Brown & McDonald, 1985; Chs. 11 and 16). There are classic examples of marking
behaviour, such as the scent marking of reindeer Rangifer tarandus, with preorbital,
caudal and tarsal glands as well as with urine (see Brown & MacDonald, (1985)
for other examples and detailed references). Scent marks are also widespread in
rodents such as mice Mus musculus, hamsters Mesocricetus auratus and rats Ratus spp.
(Hurst, 1990 Chs. 11 and 16).
In general, both pheromone production and its release (i.e. scent marking)
are androgen dependent in males, as shown by castration and testosterone-
replacement therapy experiments for example hamsters Mesocricetus auratus
(Gawienowsky et al., 1976), meadow voles Microtus pennsylvanicus (Ferkin & Johnston,
1993), tree shrews Tupaia belangeri (Holst & Eichman, 1998) and Wistar rats Manzo
et al., 2002); however, see Lepri & Randall (1983) and Randall (1986) for an ex-
ception regarding the endocrine control of sandbathing in male kangaroo rats
Dipodomys spp. The scent-marking behaviour decreases after castration and is
restored after treatment with testosterone (e.g. rats: Brown, 1978; Taylor et al.,
1987; Manzo et al., 2002). Interestingly, in many species, testosterone is the pro-
hormone for this effect, because it needs to be metabolized in specific brain
areas into oestradiol or dihydrotestosterone in order to become biologically
active, for example rabbits Oryctolagus cuniculus (Gonzalez-Mariscal et al., 1993),
gerbils Meriones unguiculatus (Yahr & Stephens, 1987) and Wistar rats (Manzo
et al., 2002).

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
486 R. F. Oliveira
Chemical communication is also widespread in urodeles, playing a major role
in sex recognition and mate attraction (e.g. European newts Triturus spp.: Cedrini
& Fasolo, 1970; Malacarne et al., 1984; Belvedere et al., 1988). One of the best-studied
species is the Japanese red-bellied newt Cynops pyrrhogaster. In this species, males
produce a female-attracting pheromone (sodefrin) with the abdominal glands,
which is released by the cloaca of the male (Kikuyama et al., 1995, 1997). Both
castration and hypophysectomy reduced the sodefrin content of the abdominal
glands and testosterone administration restored it (Yamamoto et al., 1996).
Androgens and visual displays
Many species of vertebrates use complex visual displays in intraspecific
communication, both in the context of conflict resolution (i.e. aggressive displays)
and for mate attraction (i.e. courtship displays) (Bradbury & Vehrencamp, 1998).
The evolution of stereotypic species-specific movements suggests that specific
neuromuscular systems (i.e. motoneurons and their target muscles) may have
evolved specifically for the production of these behaviours.
In some bird species, courtship displays involve coordinated wing and leg move-
ments with the individuals on the ground, on perches or in the air (Schlinger et al.,
2001). These visual displays are usually sexually dimorphic. Because sex steroids,
including androgens, have been shown to play a major role in secondary sex dif-
ferentiation in most vertebrate species studied so far, they are also potential can-
didates for a key role in the control of these displays. In wild golden-collared
manakins Manacus vitellinus, a tropical arena bird, males perform a courtship dis-
play that consists of a sequence of jumps and wing snaps (i.e. upward flips of
the bird’s wings that produce an acoustic signal). The feathers involved in the
production of these wing snaps are the primary and secondary wing feathers
(Schlinger et al., 2001), which are sexually dimorphic (Chapman, 1935). Also the
muscles controlling the wing movements and/or feather position and the jump
often associated with the wing snap are hypertrophied in male manakins (Lowe,
1942). The muscles involved in the wing-snap movement also show sex differ-
ences when examined in more detail (e.g. in fibre diameter, metabolic enzyme
activity and myosin isoform expression), which suggests that they are specialized
for greater force generation and speed of contraction (Schultz et al., 2001). These
sex differences in this neuromuscular system are not present in species in which
males do not use these muscles in courtship displays (e.g. zebra finch), although
they are still functional for other activities (e.g. for raising and lowering of the
wings during flying). These muscles are innervated by motor neurons that accu-
mulate [3H]-testosterone in their soma in the spinal cord, suggesting a role for
androgens in the control of these behavioural mechanisms (Schultz & Schlinger,
1999).

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 487
Another example of an androgen-dependent display is the amplexus behaviour
displayed by amphibian males to clasp females during mating. The forelimb mus-
cle involved in this behaviour (i.e. the flexor carpi radiali), is androgen sensi-
tive (Dorlochter et al., 1994). Castration induces atrophy and testosterone treat-
ment of castrated males causes hypertrophy of some regions of this muscle;
immunocytochemistry techniques have identified the presence of androgen re-
ceptors (Dorlochter et al., 1994). Adult males have slower acetylcholine recep-
tor kinetics than females, which facilitates slow and tonic muscle contractions
appropriate for the function of this behaviour (Brennan & Henderson, 1995).
Moreover, testosterone has been shown to act both at the pre- and postsynap-
tic level in these neuromuscular junctions, which may be viewed as an adap-
tation for a more flexible modulation of this behaviour (Nagaya & Herrera,
1995).
Finally in fish, androgens induce the development of somatic structures used in
visual signalling such as the elongation of the dorsal and anal fins used in lateral
displays and the thickening of the jaw used in mouthfighting (e.g. Mozambique
tilapia Oreochromis mossambicus: Oliveira & Almada, 1998).
Androgens and electrocommunication signals
There are two orders of fish that produce weak electric signals with an
electric organ located in their tails: the Gymnotiformes from South America and
the Mormyriformes from Africa (Zakon & Smith, 2002). The evolution of these
weak electric signals most probably occurred independently in the two orders
because they are phylogenetically distant (Alves-Gomes, 1999). Nevertheless, in
both orders, this electric sense is used for the same two functions: electrolocation
(i.e. locating objects in the environment) and intraspecific communication (Zakon
& Smith, 2002). Electrical signals are perceive by the receivers with specialized
electroreceptors mainly located in the midline of the fish (Zakon & Smith, 2002).
There are two types of electric organ discharges: pulse type and wave type. Each
species only produces one or the other (Zakon & Smith, 2002). Within species, there
are marked sex differences in the electric organ discharge. In most Gymnotiform
species that generate wave-type discharges, the males produce signals of lower
frequency than females. For example, in Sternopygus macrurus males produce an
electric discharge of 50–90 Hz while female signals range from 100 to 150 Hz
(Hopkins, 1972). Sex steroids, in particular androgens, seem to be important in
the determination of electric organ discharge frequency. In male S. macrurus, cir-
culating levels of androgens are negatively correlated with frequency (Zakon et al.,
1991) and when their reproductive axis was challenged with human chorionic go-
nadotrophin, they responded with an increase in circulating 11-ketotestosterone
levels and a decrease in the frequency of the discharge from their electric organs

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
488 R. F. Oliveira
(Zakon et al., 1990). Moreover, treatment of wave gymnotiforms with androgens
induces a masculinization of the waveform (i.e. higher wave frequency and in-
creased duration (Meyer, 1983; Mills & Zakon, 1987; Dunlap & Zakon, 1998)). Inter-
estingly, 11-ketotestosterone increased the frequency of electric organ discharge
(Meyer et al., 1987) in species in which the discharge pattern is sex reversed, that
is males generate higher-frequency discharges than females (e.g. brown ghost,
Apternotus leptorhynchus, Hagedorn & Heiligenberg, 1985). In all pulse-type species,
both mormyriforms and gymnotiforms, the treatment of juveniles, females, cas-
trated males or non-reproductive males with androgens masculinizes the pulse
form (Bass & Hopkins, 1983, 1985; Hagedorn & Carr, 1985; Bass & Volman, 1987;
Landsman & Moller, 1988; Freedman et al., 1989; Landsman et al., 1990; Herfeld
& Moller, 1998). The effects of androgens on the frequency and/or duration of
electric organ discharges may be mediated by their effects on the morphology of
the electric organ (i.e. size and/or shape of electrocytes) or by an influence on the
ionic currents of the electromotor system (e.g. Bass et al., 1986; Bass & Volman,
1987; Mills & Zakon, 1991). Apart from its influence on electric organ discharge
parameters, testosterone also activates the onset of electric signalling in weakly
electric fish (Landsman & Moller, 1988).
Hormonal effects on signal reception
A literature search revealed fewer studies of androgen modulation of
sensory perception than of the effects of androgens on effector mechanisms. The
four studies below are examples of effects on perception.
In many cyprinid fishes, females produce a sex pheromone that elicits male
courtship behaviour. The response of males to the female pheromones can be
measured either behaviourally or electrophysiologically, by placing electrodes
in the olfactory epithelium and measuring the potentials evoked by the expo-
sure of the epithelium to different odorants (i.e. electroolfactograms: Stacey &
Sorensen, 2002). In the tinfoil barb Puntius schwanenfeldi, females release a sex
pheromone (15-ketoprostaglandin-2α) that stimulates male courtship behaviour
(Cardwell et al., 1995). This response is greatest during the breeding season in sexu-
ally mature males; such males have visible breeding tubercules, dermal structures
that are known to be androgen dependent (Smith, 1974). Moreover, juveniles im-
planted with androgens (either 11-ketotestosterone or methyltestosterone) show
both an increased electroolfactogram response to 15-ketoprostaglandin-2α and
increased sexual behaviours directed towards stimuli fish (i.e. juveniles injected
with 15-ketoprostaglandin-2α (Cardwell et al., 1995)). These results clearly demon-
strate a peripheral effect of androgens on olfactory sensitivity. Other species also
show increased olfactory sensitivity to such stimuli during the breeding season
when androgen levels are also higher (e.g. electroolfactogram responsiveness to

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 489
testosterone in the Atlantic salmon Salmo salar (Moore & Scott, 1991)), which
suggests that the effect described above may be a general phenomenon in fish
olfaction.
In addition, electroreception in weakly electric fish seems to be modulated
by androgens (Keller et al., 1986; Sisneros & Tricas, 2000). Testosterone not only
affects the frequency of discharge from electric organs (as described above) but
also shifts the maximum receptivity of the electroreceptor to the new frequency
produced (Meyer & Zakon, 1982; Bass & Hopkins, 1984). Thus, androgens keep
the electroreceptors of a given individual fine-tuned to its own electric organ
discharge, which might be viewed as an adaptation for electrolocation.
A third example comes from studies of auditory sensitivity in the plainfin mid-
shipman. As mentioned above, in this species type I males produce a humming
call during the breeding season that is used to attract spawning females to their
nests (Ibara et al., 1983; Brantley & Bass, 1994). Male reproductive success must
depend heavily on their calling behaviour because females are choosy regarding
call parameters of the ‘hum’ signal (McKibben & Bass, 1998). Female reproductive
success is also expected to depend on their ability to locate and choose males
based on their acoustic signals. Recently, it has been demonstrated that, during
the summer when females need to exert their mate choice preferences based on
the male call, the auditory saccular units in the females increase their temporal
encoding capacity up to 340 Hz, compared with only 100 Hz in winter females
(Sisneros & Bass, 2003). This seasonal plasticity of the peripheral auditory system
is most probably driven by sex steroids, because it follows the seasonal variation
in steroid profiles (Forlano et al., 2003) and because expression of the oestrogen
receptor β has been identified recently in auditory hair cells (P. M. Forlano & A. H.
Bass, unpublished data). Therefore, an increase in sex steroids at the beginning of
the breeding season may induce changes in the frequency sensitivity of these hair
cells in a similar way to androgen-dependent changes in electroreceptor tuning
described above.
Finally, there are suggestions that sex steroids may also be involved in the
modulation of visual perception in teleost fish. In the three-spined stickleback Gas-
terosteus aculeatus, sexually active females prefer to mate with males with redder
bellies (e.g. Milinski & Bakker, 1990). Using optomotor responses, Cronley-Dillon &
Sharma (1968) have demonstrated that the sensitivity of the female visual system
to red wavelengths increases during the breeding season, suggesting a potential
role for female sex hormones. In this example, it can be argued that the effect
found could be acting either at the level of the sensory organ or at the level of
visual information processing by the central nervous system (i.e. optic tectum). In-
terestingly, aromatase activity has been found in fish retina, indicating that these
cells are actively metabolizing sex steroids (Callard et al., 1993) and supporting

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
490 R. F. Oliveira
the idea that the steroid modulation of visual sensitivity to key colours may occur
in the periphery.
These four studies taken together suggest that sex steroid modulation of sen-
sory perception is a common phenomenon in different sensory modalities.
Hormonal modulation of motivational and memory mechanisms
Androgens can also affect central mechanisms of information processing
both in senders and receivers. At this level, the modulatory action of hormones
may affect signalling behaviour by acting either on motivational neural circuits
underlying decision-making mechanisms, or on learning and memory systems
(Schulkin, 2002; Dohanich, 2002). By acting on central mechanisms, androgens
may set up the subject to perceive stimuli and to behave in particular ways,
for example by increasing the likelihood of the expression of a given behaviour,
ranging from food ingestion to maternal behaviour or aggression. For example,
androgens modulate central mechanisms of chemical perception in male ham-
sters. In this species, vaginal secretions stimulate male sexual behaviour after
male anogenital investigation of the female ( Johnston, 1975; Ch. 16). These se-
cretions are detected by two different sensory systems, the olfactory mucosa and
the vomeronasal organ, that use different neural pathways converging in three
central areas: the medial nucleus of the amygdala, the bed nucleus of the stria
terminalis and the medial preoptic area (Scalia & Winans, 1975). Androgen re-
ceptors are found in all these three areas (Wood et al., 1992) and direct androgen
implantation here restores sexual behaviour in castrated males (Lisk & Bezier,
1980).
Usually the effects of steroids, including androgens, on motivational mecha-
nisms involves the regulation of neuropeptide gene expression in the limbic sys-
tem, namely of arginine-vasopressin (or its homologue arginine-vasotocin in non-
mammalian vertebrates), which subsequently influence central states that con-
trol the behavioural output (Herbert, 1993). There are numerous examples of this
principle. In hamsters, testosterone enhances the effects of arginine-vasopressin
infused in the bed nucleus of the stria terminalis on scent-marking behaviour
(Albers et al., 1988). In male prairie voles, testosterone also promotes the expres-
sion of parental behaviour by increasing arginine-vasopressin synthesis and by
preventing the apoptosis of responsive neurons (De Vries, 1995). Finally, in amphib-
ians, sex steroids control both female egg-laying behaviour and male courtship via
arginine-vasotocin modulation (Moore et al., 1992).
The potential effects of sex steroids on learning, memory and other cognitive
functions have been addressed using two main approaches: (a) by documenting
the distribution of androgen and oestrogen receptors in brain areas known to
be involved in these functions, and (b) by testing hormone-treated subjects in

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 491
cognitive tasks. There is a much larger body of literature on oestrogens than on
androgens regarding this topic. The available data on androgens will be summa-
rized below.
Androgen receptors are found in the hippocampus of mammals and birds (Kerr
et al., 1995; Saldanha et al., 1999) and in the homologue dorsolateral telencephalon
of fish (Northcutt & Davis, 1983; Gelinas & Callard, 1997). These brain areas are
involved in relational memory processes, namely in spatial memory (Eichenbaum
et al., 1992; Squire, 1992). Androgen receptors have also been found in pyramidal
cells of the cortex in rats, monkeys and humans (Pomerantz & Sholl, 1987; Kerr
et al., 1995; Tohgi et al., 1995). These results set the stage for a potential functional
direct effect of androgens on memory mechanisms. The occurrence of oestrogen
receptors together with aromatase (an enzyme that metabolizes androgens into
oestrogens) also suggests a potential alternative route for aromatizable androgens
to affect cognitive function (e.g. Gelinas & Callard, 1997).
There are numerous examples of sex differences in spatial memory tasks, with
males outperforming females, which suggests a role for sex steroids in spatial
memory mechanisms (reviewed by Dohanich, 2002). Early androgen exposure ap-
parently has organizational effects on adult spatial abilities, and the masculiniza-
tion of spatial learning involves the aromatization of androgens into oestrogens
in rodents (Williams et al., 1990; Roof & Havens, 1992; Roof, 1993). In humans,
early exposure to androgens masculinizes spatial function, as is suggested by
data on girls suffering from congenital adrenal hyperplasia. These girls are ex-
posed to androgens in utero as a result of hypertrophy of the adrenal glands and
are born with virilized genitalia. When compared with their unaffected sisters,
girls with congenital adrenal hyperplasia have better performances in mental
object-rotation tests designed to measure spatial ability (Resnick et al., 1986; see
Kimura (1996) for further references). In adults, the relationship between circu-
lating androgen levels and spatial ability is not linear. Lower testosterone levels
in males, and higher testosterone levels in females, are associated with better
performances in an object-rotation task, which suggests an optimum circulating
level of testosterone to excel in this task (Moffat & Hampson, 1996). As regards
other cognitive mechanisms, in general the administration of androgens to birds
and mammals outside the critical period of development fails to affect learn-
ing and memory tasks (Dohanich, 2002). However, social memory is an exception
to this rule in rats and zebra finches (Sawyer et al., 1984; Cynx & Nottebohm,
1992).
Hormones and somatic releasers
There are a number of somatic structures that act as sign stimuli (sensu
Tinbergen, 1951) evoking a behavioural response in conspecifics. The classic

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
492 R. F. Oliveira
example of these releasers is the red belly of the male three-spined stickleback,
which elicits aggressive responses in other male sticklebacks (Tinbergen, 1951).
Since initially proposed by Tinbergen, these social releasers have been described
in many other species and can range from nuptial colouration patterns in
fish and birds to dermal appendages in fish (e.g. dermal tubercules), birds (e.g.
combs, elongated tail feathers) and reptiles (e.g. dewlap membrane in Anolis
spp.). The development of at least some of these somatic structures with a re-
leaser function is under hormonal control. There are various examples in the
teleosts. First, male nuptial colouration in African cichlids is suppressed in cas-
trated males and restored in castrates and females by exogenous administration
of testosterone (Levy & Aronson, 1955; Reinboth & Rixner, 1972; Wapler-Leong &
Reinboth, 1974; Fernald, 1976). Also in male sticklebacks, the nuptial colouration
can be suppressed by castration (Ikeda, 1933) or by the exogenous administration
of an anti-androgen (cyproterone acetate) (Rouse et al., 1977). Finally, in the sex-
role-reversed peacock blenny Salaria pavo, in which some ‘sneaker’ males mimic
female nuptial colouration, androgens (i.e. 11-ketotestosterone) inhibit the ex-
pression of female nuptial colouration in these sneaker males (Oliveira et al.,
2001a).
However, nuptial colouration is not the only releaser to be androgen depen-
dent in fish. The development of the sword as an extension of the caudal fin in
male swordtail fish Xiphophorus helleri and the development of the dermal breed-
ing tubercules in male cyprinids are both also induced by testosterone (Baldwin
& Goldin, 1939; Smith, 1974). Therefore, another way for hormones to affect com-
munication is by affecting the expression of somatic releasers in senders.
Social modulation of androgen levels
As shown above, androgens can be viewed, on the one hand, as causal
agents of behaviour, including signalling behaviour among animals in a commu-
nication network. On the other hand, the endocrine system is responsive to the
network of social relationships in which the animal is involved. Several studies
have shown the effects of social interactions on the short-term modulation of
androgen levels. In the early 1940s, it was established that male mice that lost
an agonistic interaction had lower levels of androgens than winners (Ginsberg &
Allee, 1942). This pattern has been found repeatedly in other vertebrate taxa from
fish (e.g. Hannes, 1984, 1986) to primates, including humans (e.g. Rose et al., 1971,
1975; Bernstein et al., 1974; Booth et al., 1989; see Mazur & Booth, 1998 for more
references). This set of results led to the proposal of the ‘challenge hypothesis’ by
John Wingfield and co-workers (Wingfield, 1984; Wingfield et al., 1987, 1990), ac-
cording to which the social interactions involving the subject determine androgen
levels. This hypothesis gives a conceptual framework for the study of the interplay

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 493
between social factors and endocrine responses and generates a number of testable
predictions.
1. Androgen levels should be higher during periods of social instability
when social interactions are more frequent and more intense. In fact, in
bird species in which a clear breeding cycle can be recognized,
testosterone levels are higher during the period of territory
establishment than when territories are established (Hegner &
Wingfield, 1987a; see Wingfield et al., 1999, 2000 for more examples).
2. Territorial and dominant males are expected to show higher androgen
levels than non-territorial or subordinate males because territorial males
have to defend their territories from intruders and dominant males
have actively to maintain their status. Again the available evidence
supports this hypothesis (e.g. see Oliveira et al. (2002) for a review of
teleost fish and Wingfield et al. (1999, 2000) for reviews of birds).
3. Populations of the same species breeding under different
population-density regimes should also show differences in the average
androgen levels of breeding males as a result of a different probability of
territory intrusions. This prediction should be taken with caution
because in a population with increased density, physiological and/or
behavioural mechanisms may be present to avoid aggression.
Nevertheless, positive correlations have been found between density of
breeding territories and androgen levels both in fish and in birds (e.g.
Ball & Wingfield, 1987; Beletsky et al., 1990, 1992; Pankhurst & Barnett,
1993).
Interestingly, during periods of social inertia, the levels of social interaction fall
to a baseline and androgen levels become decoupled from social behaviour. These
results have been interpreted as an adaptation (or an exaptation sensu Gould &
Vrba (1982), depending on the underlying historical evolutionary pathway) for
the individuals to adjust their behaviour (motivation) to the social milieu that
they are currently experiencing. Thus, social interactions would stimulate the
production of androgens and androgen levels would be a function of the stability
of the social environment in which the animal is living (Wingfield et al., 1990,
1999, 2000; Oliveira et al., 2002).
It is interesting to note here that it is the perception that the individual has
of the interaction in which it is involved or which it is observing that activates
the endocrine response and not the objective structure of the situation per se. To
investigate this idea we have recently tested the effect of mirror-elicited aggres-
sion on androgen levels in a cichlid fish (L. Carneiro & R. F. Oliveira, unpublished
data). The mirror image stimulation test is widely used in fish ethology to assess

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
494 R. F. Oliveira
aggressiveness (Rowland, 1999), but some inconsistencies have been found in the
relationship between social status and the aggressive score of an individual in
this test (Ruzzante, 1992). In Mozambique tilapia, we showed that androgen levels
before the fish were grouped were not good predictors of social status, but andro-
gen levels at the end of the time spent in a group were highly correlated with the
social status of each individual, suggesting that androgens are being modulated
by the social interactions experienced by the grouped individuals (Oliveira et al.,
1996). In the mirror image stimulation test, the individual is placed in a very pe-
culiar situation. Because fish do not recognize as themselves the image reflected
by the mirror, they respond to it as an intruder and attack. In our experiment,
males reacted aggressively to their own images in the mirror and escalated the
interaction using more overt aggressive behaviours (e.g. biting) as time went by.
However, because the mirror reflects exactly the same behaviours that the experi-
mental fish is displaying, the interaction has no outcome (winning versus losing).
Therefore, if the endocrine response to the social interaction is triggered by the
behavioural output during the interaction (e.g. number of displays or time spent
displaying), a variation in androgen levels is predicted. However, if it depends
on behavioural feedback received from the opponent, then no androgen varia-
tion is predicted in the test. In our mirror image stimulation experiment with
Mozambique tilapia, we found a strong behavioural response but a complete lack
of an androgen response (L. Carneiro & R. F. Oliveira, unpublished data), which
suggests that the endocrine system responds to a clear perception of the outcome
of the social interaction. This result is also interesting because it shows that it is
the communication component of the social interaction that may affect hormone
levels.
Hormones and communication II: the network view
In the previous section, the interrelationship between hormones (i.e. an-
drogens) and social behaviour was considered at the dyadic level. First, the mech-
anisms through which androgens may affect communication between a pair of
individuals were described. Second, the ways in which these androgen levels might
be modulated by the social environment (network) in which the animal is living
were described.
If we now consider an interaction that occurs within a social network (or a
communication network sensu (McGregor, 1993)), with possibilities for other in-
dividuals in the network to eavesdrop on the interaction (e.g. Ch. 2) and for the
interacting pair to adjust their behaviour according to the presence of an audi-
ence (Ch. 4), the complexity of the interrelationship between hormones and be-
haviour/communication mechanisms could increase substantially. The presence

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 495
Input/perception
Motoroutput
Centralprocessing
Hormones
Receiver
Signal
Input/perception
Motoroutput
Centralprocessing
Hormones
Somatic releasers
Sender
Input/perception
Motoroutput
Centralprocessing
Hormones
Bystander
Fig. 21.3. Flow of information in a communication network. This figure is similar to
Fig. 21.2 with the following extra elements. The presence of a third individual (the
bystander) may be perceived by both the sender and the receiver and affect their signal
production and signal reception mechanisms, respectively. The perception of the
presence of the bystander may also affect central (motivational) mechanisms in the
central nervous system in both senders and receivers, which may, in turn, modulate
hormone levels in both individuals. The perception of the signal by the bystander
would also affect its central processing of information at the level of motivational
mechanisms, which could affect its hormone levels.
of a bystander that could act both as an eavesdropper and as an audience (Fig. 21.3),
may affect androgen levels in both the sender and the receiver and subsequently
affect their androgen-modulated communication behaviour in the same ways as
described above (see also Fig. 21.2). The androgen levels of the bystander itself may
respond to the observed interaction, which, in turn, will affect its own subsequent
social behaviour.
Consequently, androgens may play a key role as physiological mediators of the
modulation of behaviour by the social context. A number of social phenomena (e.g.
winner–loser effects, bystander effect, dear enemy effect) that have been described
in social networks may then be physiologically mediated by changes in hormone
levels, especially androgens.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
496 R. F. Oliveira
Adjusting behaviour to the social context: a role for androgens?
Winner–loser effects
There is an extensive literature (including most of the chapters of this
book) that clearly shows that animals use information on relative competitive
abilities in the social network in which they are placed to adjust their behaviour
accordingly. They may obtain this information by direct assessment of their peers
by interacting with one another in a dyadic fashion and then adjusting their
behaviour in subsequent interactions depending on the outcome of previous in-
teractions. For example, individuals that win an interaction increase their prob-
ability of winning a subsequent interaction and vice versa for losers. In this case,
although only two individuals have to be present during the initial interaction,
unless there were other individuals with whom the interactants subsequently
interacted, there would be no winner–loser effect. Therefore, this effect is better
understood within the framework of social networks than with a dyadic approach.
This winner–loser effect may last from a few minutes up to several hours or even
days and has been reported for several taxa, for example invertebrates (Alexander,
1961; Otronen, 1990; Whitehouse, 1997), fish (McDonald et al., 1968; Frey & Miller,
1972; Bakker & Sevenster, 1983; Francis, 1983, 1987; Abbott et al., 1985; Beaugrand
& Zayan, 1985; Beacham & Newman, 1987; Franck & Ribowski, 1987; Beacham,
1988; Bakker et al., 1989; Beaugrand et al., 1991, 1996; Chase et al., 1994; Hsu &
Wolf, 1999), reptiles (Schuett, 1997) and birds (Drummond & Osorio, 1992). The
winner effect is usually of shorter duration than the loser effect (e.g. Chase et al.,
1994), and when integrating prior social experiences more recent outcomes are
more effective in predicting the probability of winning a subsequent interaction
than previous ones (Hsu & Wolf, 1999). Another interesting characteristic of this
effect is that it is more effective when winning or losing against a well-matched
opponent than when there is a large asymmetry in resource-holding potential
(sensu Parker, 1974) between the two individuals (Beaugrand & Goulet, 2000).
The behavioural mechanism proposed to explain the winner effect is based on
the fact that initiators of interactions have higher probabilities of winning and that
winners of recent encounters become more likely to initiate future interactions
( Jackson, 1991). This is especially true for the initiators of attacks (Hsu & Wolf,
2001).
It is conceivable that by winning an interaction an individual raises its androgen
levels, which, in turn, increases its willingness to initiate future interactions and
the probability of winning the next interaction in which it participates. The reverse
would be predicted for losers.
We are conducting an ongoing literature survey to collect data on the two
steps of this endocrine hypothesis for the winner–loser effect: (a) that winners

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 497
Table 21.1. Literature survey of reported differences in male androgen levels among
vertebrates according to social status and the phase of the sexual cycle and of the effects of
androgen treatment on aggressive behaviour
Taxa Androgens and Androgens and Effect of androgen
social status phase of treatment on
sexual cycle aggressive behaviour
D = S or MP < PP or No
No. D > S D < S No. MP > PP MP = PP No. Effect effect
Fish 12 9 3 9 8 1 12 7 5
Amphibians 1 0 1 2 2 0 0 – –
Reptiles 3 2 1 0 – – 3 3 0
Birds 10 4 6 48 47 1 10 6 4
Mammals 18 15 3 4 3 1 9 5 4
Total 44 30 14 63 58 5 34 21 13
Male androgen levels in; D, dominant; S, subordinate; MP, mating phase; PP, parental care phase.
K. Hirschenhauser & R. F. Oliveira, unpublished data.
have higher androgen levels than losers; (b) that androgens increase aggressive
behaviour and hence the probability of victory in a subsequent interaction. As
there are not enough studies that we can find that measured the androgen vari-
ations in response to a social interaction to address the first step, it was decided
to search for correlational data in the form of reported androgen differences be-
tween dominant and subordinate individuals. We found 44 published studies,
68% of which confirmed that androgen levels were higher in dominants than in
subordinates (Table 21.1). Our literature survey revealed that 62% of the studies
confirmed that administration of androgens increased aggressive behaviour in
different taxa (Table 21.1), thereby supporting the second step of the endocrine
hypothesis. Although the majority of the studies supported the assumptions of the
proposed hypothesis, the percentages do not provide overwhelming support and
so we decided to test this hypothesis experimentally with the Mozambique tilapia
(A. Silva & R. F. Oliveira, unpublished data). After staging a first fight between two
males, the winner and the loser fought two independent, naive individuals (i.e.
males that have not been involved in social interactions recently) (Fig. 21.4a). As ex-
pected, our preliminary data showed that winners of the first encounter won the
majority of the interactions with the naive fish and vice versa for losers (Fig. 21.4b).
When winners were treated with an anti-androgen (cyproterone acetate) between
the two interactions (which were two hours apart), the winner effect was no longer
detectable in the second fight with the neutral fish, suggesting an involvement

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
498 R. F. Oliveira
0102030405060708090
100
Winner Loser
control
Per
cent
age
treated
(a) (b)
Winner LoserNeutralmale
Neutralmale
t1
t2t2
Fig. 21.4. Androgens and the winner–loser effect. (a) Experimental-set up: four
Mozambique tilapia Oreochromis mossambicus males were introduced to individual
compartments separated by opaque partitions in an aquarium. After a period of
acclimation (t1), the central partition was removed and the two individuals in the
central compartments were allowed to interact until a winner and a loser could be
recognized. Then the partition was put back in place. Two hours later (t2), the two
lateral partitions were simultaneously removed and the winner and the loser of the
previous interaction were allowed to interact with the neutral males that were placed
in the end compartments. This second interaction went on until a winner and a loser
could be recognized. (b) Percentage of second interactions won by winners or losers of
the first interaction. Four groups of experimental animals were compared: t1 winners
treated with an androgen inhibitor (cyproterone acetate); control t1 winners (treated
with a placebo saline solution); t1 losers treated with an androgen
(11-ketotestosterone); control t1 losers (treated with a placebo saline solution). Twelve
replicates of the experiment were run. (A. Silva & R. F. Oliveira, unpublished data.)
of androgens in the winner effect (Fig. 21.4b). However, the loser effect was not
inhibited in the second interaction by treating losers with exogenous androgens,
which suggests that, although a fall in androgens is observed in losers, it is not
the underlying causal mechanism for the loser effect. Although this result may
seem paradoxical at first sight, it makes some sense; androgen variations induced
by social interactions occur in the short term and so do winner effects; however,
the loser effect may last up to several days depending on a number of factors.
Consequently, other neuroendocrine mechanisms must be involved in the loser
effect. One of the best candidates for this role is the serotonergic system. The fol-
lowing evidence from studies using different fish species seems to support this
hypothesis: (a) losers experience increased brain levels of serotonin and subordi-
nate individuals have chronically elevated brain levels of serotonin (Winberg &

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 499
Nilsson, 1993a,b; Winberg et al., 1997; Winberg & Lepage, 1998); and (b) serotonin
appears to be inhibitory to behavioural responsiveness in general and to inhibit
aggressive behaviour in particular (Winberg & Nilsson, 1993a,b; Adams et al., 1996;
Edwards & Kravitz, 1997). Therefore, losers would display a marked behavioural in-
hibition, with increased attack latencies in subsequent interactions, which would
prevent them from winning these interactions and would reinforce their subordi-
nate role. Interestingly, the administration of a precursor of dopamine (L-dopa) to
individuals that had lost an interaction two days before induced lower serotoner-
gic activity and reduced the attack latency in subsequent interactions, suggesting
that the dopaminergic system counteracts the serotonin-mediated effects of social
subordination (Hoglund et al., 2001).
Bystander effects
Information on the relative competitive ability of conspecifics within a
social network can also be gathered using indirect methods, namely by extract-
ing information from watching conspecific interactions that the subject uses in
subsequent interactions with the observed individuals (eavesdropping (McGre-
gor, 1993; McGregor & Peake, 2000) and social eavesdropping sensu Peake, Ch. 2).
This sort of information gathering on the relative ability of conspecifics has been
demonstrated in a number of species (see Ch. 2), for example fish ( Johnsson &
Åkerman, 1998; Oliveira et al., 1998; Earley & Dugatkin, 2002; Ch. 5) and birds
(Hogue et al., 1996; McGregor et al., 1997; Naguib et al., 1999; Peake et al., 2001),
and has the advantage of avoiding the costs associated with fighting (e.g. McGre-
gor, 1993; McGregor & Peake, 2000; Dugatkin, 2001). Some authors consider that
there is a difference between bystander and eavesdropping effects: eavesdropping
implies an active gathering of information by bystander individuals that will be
used in future interactions within the social network (McGregor, 1993; McGregor
& Peake, 2000), while the bystander effect was originally described as a priming
of aggressive motivation in bystanders of agonistic interactions (Hogan & Bols,
1980; Bronstein, 1989). Therefore, from the point of view of the required cognitive
abilities, eavesdropping is expected to be more demanding than a mere priming
response. However, both phenomena are adaptive because they might increase the
probability of eavesdroppers/bystanders of winning their next social interaction
(Clotfelter & Paolino, 2003; Hollis et al., 1995; Peake & McGregor, 2004).
The priming response associated with the bystander effect is another phe-
nomenon that could be mediated by androgens. To investigate if bystanders
experience an increase in their androgen levels, we conducted an experiment
with Mozambique tilapia in which a bystander fish had visual access through a
one-way mirror to two conspecific neighbours separated by an opaque partition

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
500 R. F. Oliveira
Male A
Male C
One way mirror
−12−10−8−6−4−2
0246
30 min 2 h 6 h
Time after exposure to stimuli
11-K
eto
test
ost
ero
ne
vari
atio
n
(ng
/ml)
p < 0.05
p < 0.01p < 0.01
Male B
(a)
(b)
Fig. 21.5. Social modulation of androgen levels in bystander male Mozambique tilapia
Oreochromis mossambicus. (a) Experimental-set up: three males were introduced to
individual compartments in an aquarium. Males A and B were separated by an opaque
partition and male C was separated from the other two males by a one-way mirror
that allowed it to observe the other two males without being observed. After a period
of acclimation, two conditions were created. (i) In the experimental group, the opaque
partition separating males A and B was removed (grey arrow) and the two individuals
were allowed to interact for 20 minutes while male C observed the interaction
(bystander). (ii) In the control group, the opaque partition separating males A and B
was not removed and the bystander individual observed its two neighbours resting or
swimming around for 20 minutes. Urine samples were collected from male C at
regular intervals (just before the start of the test, 30 minutes after, two hours after
and six hours after the experiential situation) and assayed for androgens using
radioimmunoassays. (b) Androgen (11-ketotestosterone) variation (i.e. urine
concentrations after the experiential situation minus the urine concentrations just
before the experiential situation) in bystander males of the experimental (black bars)
and control (white bars) groups. (Adapted from Oliveira et al., 2001b.)
(Fig. 21.5; Oliveira et al., 2001b). After a period of familiarization, in the exper-
imental treatment the opaque partition between neighbours was removed and
the bystander was allowed to observe the agonistic interaction between its neigh-
bours. In the control group after the same period of familiarization, the opaque
partition between neighbours remained in place and the bystander could see its

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 501
two neighbours resting or swimming around in their respective compartments. As
predicted, androgen levels (both 11-ketotestosterone and testosterone) increased
significantly in the experimental group of bystanders after watching their neigh-
bours fighting and no effect was detected in the control group (Oliveira et al.,
2001b). This result has an interesting parallel in humans. It has been demon-
strated that sports fans experience variations in testosterone levels depending on
the outcome of the game they have attended, both for college basketball and for
soccer. Fans of the winning team display an increase in salivary testosterone levels
and there is a decrease in testosterone levels in fans of the losing team (Bernhardt
et al., 1998).
Audience effects
The term audience effect was first used in the ethological literature to de-
scribe the facilitation effect of the presence of other individuals on the production
of food calls or alarm calls in response to food items or a predator, respectively
(Gyger et al., 1986; Marler et al., 1986; Evans & Marler, 1994). Here the term will be
used in a more restricted way, following the definitions provided by McGregor &
Peake (2000) and by Matos (2002) (see also Ch. 4): individuals participating in an in-
teraction may also manipulate the information available to others and adjust their
signalling behaviour according to the presence and composition of an audience
of conspecifics. These audience effects have been demonstrated in different ver-
tebrate taxa, including fish (e.g. Doutrelant et al., 2001; Matos & McGregor, 2002),
birds (e.g. Searcy et al., 1991; Baltz & Clark, 1997) and mammals (e.g. Hector et al.,
1989), and have involved different social contexts, from agonistic interactions in
Siamese fighting fish Betta splendens (Doutrelant et al., 2001; Matos & McGregor,
2002) to extra-pair copulations in male budgerigars Melopsittacus undulatus (Baltz &
Clark, 1997).
The audience effect has been interpreted as a way for the individual to ma-
nipulate the information broadcast to its social network, which may influence
subsequent social interactions in which it will have to participate. Therefore, it
can be predicted that subjects behave more promptly and aggressively towards
an intruder when a male audience is present. Again it is predicted that this ef-
fect may be mediated by increased androgen levels in the interacting individuals
induced by the presence of the audience. This aggressive priming effect of an au-
dience has already been established in Siamese fighting fish (Matos, 2002) but its
androgen-mediation remains to be tested.
Dear enemy effects
In territorial systems, residents react less aggressively towards famil-
iar opponents than to intrusions by strangers, a phenomenon called the dear

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
502 R. F. Oliveira
enemy effect (Ydenberg et al., 1988; Temeles, 1994). In evolutionary terms, this
phenomenon can be viewed as an adaptation for the individual to adjust its ter-
ritorial behaviour according to the threat posed by the intruder (Temeles, 1994):
having a dear enemy neighbour allows the resident individual to defend its ter-
ritory against unfamiliar intruders with the same efficiency as if they were the
only competitors in the area, which reduces the costs of territory defence (Leiser
& Itzkowitz, 1999; Whiting, 1999).
The dear enemy phenomenon can be explained in terms of proximate mech-
anisms by an ability of the resident male to discriminate between familiar and
unfamiliar intruders together with a habituation to the neighbours, which would
explain the lower response that they elicit, for example visual habituation to neigh-
bours in Siamese fighting fish (Bronstein, 1994) and habituation to neighbours’
calls in frogs (Owen & Perrill, 1998). Therefore, it can be predicted that resident
males will react more aggressively towards strangers than towards familiar intrud-
ers and that the increase in androgen levels expected from the social challenge
experienced will be higher in the case of intrusions by strangers. Moreover, it is
also predicted that, for repeated intrusions by neighbouring males, the androgen
response should be higher in the first trials and decrease with the number of trials
(i.e. habituation). These two predictions remain to be tested.
The adaptive value of social modulation of hormones: a cost–benefit
analysis of androgen levels
As stated above, the main adaptive reason for androgens to respond to
the social environment is to allow individuals to fine-tune the expression of their
behaviours in a context-dependent fashion. For example, this mechanism would
allow subordinate individuals to downregulate the expression of their aggressive
behaviour and thus avoid the initiation of agonistic encounters that they have
low probabilities of winning. In the long run, this mechanism can be seen as
an opportunity for individuals to adopt a behavioural tactic that suits best their
relative competitive ability. As a result, androgen-mediated behavioural tuning
to the social environment may result in either a continuous or a discrete vari-
ation of behavioural phenotypes. For example, even small changes in androgen
levels induced by social interactions in electric fish, can affect the pulse duration,
resulting in dominant males with more masculinized discharges from their elec-
tric organs than subordinates (e.g. Brienomyrus brachyistius: Carlson et al., 2000).
Also, in the Mozambique tilapia, the acquisition of dominant status induces the
exaggeration of male morphological traits, an effect that has been shown to be
mediated by androgens (Oliveira & Almada, 1998). By comparison, in a number

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 503
of teleost species, individuals of lower competitive ability can adopt frequency-
or condition-dependent alternative reproductive tactics (Taborsky, 1994) or even
change sex (Grober, 1998).
However, it can be argued that, instead of having their androgen levels open to
social influences, selection could have favoured animals that permanently keep
their androgen levels at an optimum high value in order to optimize their social
behaviour at all times. It follows that there must be costs associated with maintain-
ing high levels of androgens that counteract the social benefits of high androgen
levels. Therefore, a cost–benefit analysis is needed to establish the adaptive value
of the social modulation of androgens.
Potential benefits of high androgen levels
Among the potential benefits of increasing androgen levels at periods of
social challenge, one can think of androgen effects both on aggressive motivation
and on cognitive tasks that would promote the success of the animals in social
interactions.
The available data on the effects of androgens on aggressive motivation has
already been review above, and in most studies an effect has been found (Ta-
ble 21.1). Sex steroids, including androgens, are known to play a major role in
cognitive processes such as social attention, learning and memory in a variety
of vertebrate taxa (e.g. Andrew, 1991; Cynx & Nottebohm, 1992) and so they may
help the animal to be prepared for a competitive context (see text above for more
references).
We have recently tested the effects of androgens on social attention in Siamese
fighting fish. Eavesdropping has already been demonstrated in this species and
male Siamese fighting fish are known to spend time observing conspecific inter-
actions (Oliveira et al., 1998). So we designed an experiment to assess the effect of
the administration of exogenous androgens on the time males spend observing
social interactions between conspecific males. Not surprisingly, androgen-treated
males spent more time observing social interactions than controls, suggesting an
effect of androgens on selective attention to the social environment (R. F. Oliveira
& L. Carneiro, unpublished data).
Another potential benefit that androgens may convey in a competitive situa-
tion is an increased probability of the expression of risk-taking behaviours, which
might be adaptive in a competitive situation. A nice example of this phenomenon
has recently been published (Kavaliers et al., 2001). Male mice were pre-exposed
to the odour of an oestrous female and subsequently exposed to the odours of
predators (cat and weasel). Mice that were only exposed to the predator odour,
simulating a situation of increased predation risk, showed increased circulating

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
504 R. F. Oliveira
levels of corticosterone and decreased levels of testosterone. The pre-exposure to
the female odour attenuated this response to predator odour, which might reflect
a greater tendency for risk taking in the presence of predators (Kavaliers et al.,
2001).
Potential costs of high androgen levels
Elevated androgen levels have been shown to have associated costs;
consequently, one would expect high circulating levels to be restricted to
periods of social challenge. The following potential costs have been dis-
cussed in the literature: (a) increased energy consumption; (b) impairment of
immunocompetence; (c) higher rates of injuries and reduced survival; (d) inter-
ference with parental care; and (e) potential oncogenic effects (Wingfield et al.,
1999). Of all these potential costs, two will be analysed in more detail below:
the potential negative effects of androgens on metabolic rate and on parental
behaviour.
Metabolic costs of high androgen levels
Studies of the metabolic effects of androgens have produced contradic-
tory results. In bird species, testosterone treatment increased the basal metabolic
rate in house sparrows Passer domesticus (Buchanan et al., 2001), reduced it in white-
crowned sparrows Zonotrichia leucophrys (Wikelsky et al., 1999) and had no effect in
dark-eyed juncos Junco hyemalis (Deviche, 1992). However, in juncos, an indepen-
dent study found an association between high testosterone levels and increased
lipid catabolism and nocturnal body temperature (Vezina & Thomas, 2000). In the
lizard Sceloporus jarrovi, testosterone treatment increased the maximal metabolic
rate but had no effect on basal metabolic rate (Marler et al., 1995) and male tilapia
treated with 11-ketotestosterone showed an increase in the resting metabolic rate
and in metabolic scope but a non-significant increase in the basal metabolic rate
(Ros et al., 2004). This discrepancy in the results can be attributed to methodolog-
ical variations among studies, including the choice of the measures taken, the
timespan of the experiment, the season, etc. Nevertheless, androgens failed to
affect the metabolic measure used in only two studies, and in one case the data
are contradicted by a subsequent study on the same species. In the other four
studies, androgens affected different metabolic measures. Consequently, it can be
said that androgens may affect metabolism in a non-linear way and a metabolic
cost associated with higher levels of androgens should not be excluded.
Parental care trade-off with androgens
One of the predictions of the challenge hypothesis is that male andro-
gen levels above a breeding baseline are incompatible with male parental care

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 505
(Wingfield et al., 1990). If androgen levels increase as a result of social challenges,
males will invest less time in paternal activities, and thus a trade-off between
social interactions and paternal care, mediated by androgens emerges. In many
bird species with male parental care, the experimental increase of circulating
testosterone in parental males suppressed paternal behaviour and promoted ago-
nist interactions (Silverin, 1980; Hegner & Wingfield, 1987b; Ketterson et al., 1992;
Beletsky et al., 1995). Moreover, several studies on the seasonal variation of andro-
gen levels in birds show that during the breeding season male androgen levels
are higher during the mating phase than during the parental phase (Wingfield et
al., 1987). To document this trade-off further, we have gathered published data on
androgen levels in vertebrate species with respect to paternal care: out of the 63
species of vertebrates for which data are available, 92% show the expected pattern
of lower circulating androgen concentrations during the parental phase (Table
21.1). In summary, keeping high levels of androgens at all times is detrimental to
the individual in many ways and so the stage is for the evolution of a flexible
system modulated by the social environment.
Summary and future directions
Androgen modulation by social context and the subsequent role of an-
drogens in the activation of expression of social behaviour have been proposed in
this chapter to explain the mechanisms underlying experiential effects. However,
this hypothesis does not exclude explanations of the phenomena described, in
terms of associative learning mechanisms. Cognitive abilities such as individual
recognition and discrimination would explain some of the described behavioural
responses to social context (e.g. McDonald et al., 1968) and winning or losing can
be seen as having reinforcing properties. For instance, male Siamese fighting fish
will perform an operant response to have access to an opponent that they can
subsequently fight (Hogan, 1967; Bols, 1977). Similar results have been reported
for mice (Tellegen et al., 1969), suggesting that the opportunity to interact with
an opponent may be a universal positive reinforcer in vertebrates. However, the
two explanations (i.e. endocrine modulation and associative learning) should not
be seen as mutually exclusive but as complementary, and it is even possible that
they represent two levels of analysis that are tightly interconnected. Condition-
ing of the endocrine response by social stimuli is a possibility that remains to
be tested, and there are already examples of androgen modulation of learning
mechanisms (e.g. in castrated zebra finches testosterone facilitates conspecific
song discrimination (Cynx & Nottebohm, 1992)). Therefore, the interrelationship
between androgens and associative learning mechanisms is certainly a key topic
for future research in this area.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
506 R. F. Oliveira
Acknowledgements
I wish to thank all the present and past members of the ‘Mackerel Academy’ (what
other name could a fish ethology group have in a Psychology School?) for all the reading club
discussions and the Monday seminars that generated hypotheses, shaped experimental designs
and dissected the results of some of the research reported here. They are in alphabetical order: R.
Andrade, K. Becker, L. Carneiro, N. Castro, T. Fagundes, D. Goncalves, K. Hirschenhauser, J. Jordao,
M. Lopes, T. Oliveira, A. Ros, J. Saraiva and A. Silva. I also thank P. McGregor, R. Bshary and two
anonymous reviewers for providing helpful comments on an earlier version of the manuscript.
I would like to dedicate this chapter, as a posthumous expression of thanks, to the memory of
the late Luis Carneiro (b. 18 January 1969, d. 12 September 2002). More than a PhD student with
a promising career, Luis was a beloved friend and his humour and attitude towards life made
him an example of how intensely life can, and should, be lived. The unpublished studies reported
here were funded by two ongoing research grants from Fundacao para a Ciencia e a Tecnologia –
FCT (PRAXIS XXI/P/BIA/10251/1998 and POCTI/BSE/38395/2001). The writing of this chapter was
partially funded by the Plurianual Program from FCT (UI&D 331/94).
References
Abbott, J. C., Dunbrack, R. L. & Orr, C. D. 1985. The interaction of size and experience
in dominance relationships of juvenile steelhead trout (Salmo gairdneri). Behaviour,
92, 241–253.
Adams, C. F., Liley, N. R. & Gorzalka, B. B. 1996. PCPA increases aggression in male
firemouth cichlids. Pharmacology, 53, 328–330.
Albers, H. E., Liou, S. Y. & Ferris, C. F. 1988. Testosterone alters the behavioural
response of the medial preoptic–anterior hypothalamus to microinjection of
arginine vasopressin in the hamster. Brain Research, 456, 382–386.
Alexander, R. D. 1961. Aggressivness, territoriality and sexual behaviour in field
crickets. Behaviour, 17, 130–223.
Alves-Gomes, J. 1999. Systematic biology of gymnotiform and mormyriform electric
fishes: phylogenetic relationships, molecular clocks and rates of evolution in the
mitochondrial rRNA genes. Journal of Experimental Biology, 202, 1167–1183.
Andrew, R. J. 1991. Testosterone, attention and memory. In: The Development and
Integration of Behaviour, ed. P. Bateson. Cambridge, UK: Cambridge University Press,
pp. 171–190.
Arnold, A. P. 1975. The effects of castration and androgen replacement on song,
courtship and aggression in zebra finches (Poephilia guttata). Journal of Experimental
Zoology, 191, 309–326.
Bakker, T. C. M. & Sevenster, P 1983. Determinants of dominance in male sticklebacks
(Gasterosteus aculeatus L.). Behaviour, 86, 55–71.
Bakker, T. C. M., Bruijn, E. & Sevenster, P. 1989. Asymmetrical effects of prior winning
and losing on dominance in sticklebacks (Gasterosteus aculeatus). Ethology, 82,
224–229.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 507
Baldwin, F. M., Goldin, H. S. 1939. Effects of testosterone propionate on the female
viviparous teleost, Xiphophorus helleri Heckel. Proceedings of the Society for
Experimental Biology and Medicine, 42, 813–819.
Ball, G. F. & Wingfield, J. C. 1987. Changes in plasma luteinizing hormone and sex
steroid hormones in relation to multiple-broodness and nest-site density in male
starlings. Physiological Zoology, 602, 191–199.
Balthazart, J., Foidart, A., Wilson, E. M. & Ball, G. F. 1992. Immunocythochemical
localization of androgen receptors in the male songbird and quail brain. Journal of
Comparative Neurology, 317, 407–420.
Baltz, A. P. & Clark, A. B. 1997. Extra-pair courtship behaviour of male budgerigars and
the effect of an audience. Animal Behaviour, 53, 1017–1024.
Bass, A. H. & Hopkins, C. D. 1983. Hormonal control of sexual differentiation: changes
in electric organ discharge wave-form. Science, 220, 971–974.
1984. Shifts in frequency tuning in electroreceptors in androgen-treated mormyrid
fish. Journal of Comparative Physiology A, 155, 713–724.
1985. Hormonal control of sex differences in the electric organ discharge (EOD) of
mormyrid fishes. Journal of Comparative Physiology A, 156, 587–604.
Bass, A. H. & Volman, S. F. 1987. From behavior to membranes: testosterone-induced
changes in action potential duration in electric organs. Proceedings of the National
Academy of Sciences, USA, 84, 9295–9298.
Bass, A. H., Denizot, J. P. & Marchaterre, M. A. 1986. Ultrastructural features and
hormone-dependent sex differences of mormyrid electric organs. Journal of
Comparative Neurology, 254, 511–528.
Beacham, J. L. 1988. The relative importance of body size and aggressive experience as
determinants of dominance in pumpkinseed sunfish, Lepomis gibbosus. Animal
Behaviour, 36, 621–623.
Beacham, J. L. & Newman, J. A. 1987. Social experience and the formation of
dominance relationships in the pumpkinseed sunfish, Lepomis gibbosus. Animal
Behaviour, 35, 1560–1562.
Beani, L., Panzica, G., Briganti, F., Persichella, P. & Dessı-Fulgheri, F. 1995.
Testosterone-induced changes of call structure, midbrain and syrinx anatomy in
partridges. Physiology and Behavior, 58, 1149–1157.
Beaugrand, J. P. & Goulet, C. 2000. Distinguishing kinds of prior dominance and
subordination experiences in males of green swordtail fish (Xiphophorus helleri).
Behavioural Processes, 50, 131–142.
Beaugrand, J. P. & Zayan, R. 1985. An experimental model of aggressive dominance in
Xiphophorus helleri (Pisces, Poeciliidae). Behavioural Processes, 10, 1–52.
Beaugrand, J. P., Goulet, C. & Payette D 1991. Outcome of dyadic conflict in male green
swordtail fish, Xiphophorus helleri: effects of body size and prior dominance. Animal
Behaviour, 41, 417–424.
Beaugrand, J. P., Payette, D. & Goulet, C. 1996. Conflict outcome in male green
swordtail fish dyads (Xiphophorus helleri): interaction of body size, prior
dominance/subordination experience, and prior residency. Behaviour, 133,
303–319.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
508 R. F. Oliveira
Beletsky, L. D., Orians, G. H. & Wingfield, J. C. 1990. Steroid hormones in relation to
territoriality, breeding density, and parental behavior in male yellow-headed
blackbirds. The Auk, 107, 60–68.
1992. Year-to-year patterns of circulating levels of testosterone and
corticosterone in relation to breeding density, experience, and reproductive
success of the polygynous red-winged blackbird. Hormones and Behavior, 26,
420–432.
Beletsky, L. D., Gori D. F., Freeman, S. & Wingfield, J. C. 1995. Testosterone and
polygyny in birds. In: Current Ornithology, Vol. 12, ed. D. M. Power. New York:
Plenum Press, pp. 1–41.
Belvedere, P., Colombo, L., Giacoma, C., Malacarne G. & Andreoletti, G. E. 1988.
Comparative ethological and biochemical aspects of courtship pheromones in
European newts. Monitore Zoologico Italiano, 22, 397–403.
Bernhardt, P. C., Dabbs, J. M., Fielden, J. A. & Lutter, C. D. 1998. Testosterone changes
during vicarious experiences of winning and losing among fans at sports events.
Physiology and Behavior, 65, 59–62.
Bernstein, I. S., Rose, R. M. & Gordon T. P. 1974. Behavioral and environmental events
influencing primate testosterone levels. Journal of Human Evolution, 3,
517–525.
Bleisch, W., Luine, V. N. & Nottebohm, F. 1984. Modification of synapses in
androgen-sensitive muscle. I. Hormonal regulation of acetylcholine receptor
number in the songbird syrinx. Journal of Neuroscience, 4, 786–792.
Bols, R. J. 1977. Display reinforcement in the Siamese fighting fish, Betta splendens:
aggressive motivation or curiosity? Journal of Comparative and Physiological
Psychology, 91, 233–244.
Booth, A., Shelley, G., Mazur, A., Tharp, G. & Kittok, R. 1989. Testosterone, and winning
and losing in human competition. Hormones and Behavior, 23, 556–571.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.
Brantley, R. K. & Bass, A. H. 1994. Alternative male spawning tactics and acoustic
signals in the plainfin midshipman fish, Porichthys notatus (Teleostei,
Batrachoididae). Ethology, 96, 213–232.
Brantley, R. K., Marchaterre, M. A. & Bass, A. H. 1993. Androgen effects on vocal muscle
structure in a teleost fish with inter- and intra-sexual dimorphisms. Journal of
Morphology, 216, 305–318.
Brenowitz, E. A. & Arnold, A. P. 1989. Accumulation of estrogen in a vocal control
brain region of a duetting songbird. Brain Research, 359, 364–367.
1992. Hormone accumulation in song regions of the canary brain. Journal of
Neurobiology, 23, 871–880.
Brennan, C. & Henderson, L. P. 1995. Androgen regulation of neuromuscular junction
structure and function in a sexually dimorphic muscle of the frog Xenopus laevis.
Journal of Neurobiology, 27, 172–188.
Bronstein, P. M. 1989. The priming and retention of agonistic motivation in male
Siamese fighting fish, Betta splendens. Animal Behaviour, 37, 165–166.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 509
1994. On the predictability, sensitisation, and habituation of aggression in male
bettas (Betta splendens). Journal of Comparative Psychology, 98, 421–431.
Brown, R. E. 1978. Hormonal control of odor preferences and urine-marking in male
and female rats. Physiology and Behavior, 20, 21–24.
Brown, R. E. & MacDonald, D. W. 1985. Social Odours in Mammals. Oxford: Clarendon
Press.
Buchanan, K. L., Evans, M. R., Goldsmith, A. R., Bryant, D. M. & Rowe, L. V. 2001.
Testosterone influences basal metabolic rate in male house sparrows: a new cost
of dominance signalling? Proceedings of the Royal Society of London, Series B, 268,
1337–1344.
Callard, G. V., Drygas, M. & Gelinas, D. 1993. Molecular and cellular physiology of
aromatase in the brain and retina. Journal of Steroid Biochemistry and Molecular
Biology, 44, 541–547.
Cardwell, J. R., Stacey N. E., Tan, E. S. P., McAdam, D. S. O. & Lang, S. L. C. 1995.
Androgen increases olfactory receptor response to a vertebrate sex pheromone.
Journal of Comparative Physiology A, 176, 55–61.
Carlson, B. A., Hopkins, C. D. & Thomas, P. 2000. Androgen correlates of socially
induced changes in the electric organ discharge waveform of a mormyrid fish.
Hormones and Behavior, 38, 177–186.
Cedrini, L. & Fasolo, A. 1970. Olfactory attractants in sex recognition of the crested
newt: an electrophysiological research. Monitore Zoologico Italiano, 5, 223–229.
Chapman, F. M. 1935. The courtship of Gould’s manakin (Manacus vitellinus vitellinus) on
Barrado Colorado Island, Canal Zone. American Museum of Natural History Bulletin,
68, 472–521.
Chase, I. D., Bartolomeo, C. & Dugatkin, L. A. 1994. Aggressive interactions and
inter-contest interval: how long do winners keep winning? Animal Behaviour, 48,
393–400.
Clotfelter, E. & Paolino, A. 2003. Winner effect is induced in bystanders to aggressive
contests between male fighting fish. Animal Behaviour, 66, 343–347.
Cronley-Dillon, J. & Sharma, S. C. 1968. Effect of season and sex on the photopic
spectral sensitivity of the three-spined stickleback. Journal of Experimental Biology,
49, 679–687.
Cynx, J. & Nottebohm, F. 1992. Testosterone facilitates some conspecific song
discriminations in castrated zebra finches Taeniopygia guttata. Proceedings of the
National Academy of Sciences, USA, 89, 1376–1378.
De Vries, G. J. 1995. Studying neurotransmitter systems to understand the
development and function of sex differences in the brain: the case of vasopressin.
In: Neurobiological Effects of Sex Steroid Hormones, ed. P. E. Micevych & R. P. Hamer Jr.
Cambridge, UK: Cambridge University Press, pp. 254–278.
Deviche, P. 1992. Testosterone and opioids interact to regulate feeding in a male
migratory songbird. Hormones and Behavior, 26, 394–405.
Dohanich, G. 2002. Gonadal steroids, learning and memory. In: Hormones, Brain and
Behavior, Vol. 2, ed. D. W. Pfaff, A. P. Arnold, A. M. Etgen, S. E. Fahrbach & R. T.
Rubin. New York: Academic Press, pp. 265–327.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
510 R. F. Oliveira
Dorlochter, M., Astrow, S. H. & Herrera, A. A. 1994. Effects of testosterone on a sexually
dimorphic frog muscle: repeated in vivo observations and androgen receptor
distribution. Journal of Neurobiology, 25, 897–916.
Doutrelant, C., McGregor, P. K. & Oliveira, R. F. 2001. The effect of an audience on
intra-sexual communication in male Siamese fighting fish, Betta splendens.
Behavioral Ecology, 12, 283–286.
Drummond, H. & Osorio, J. L. 1992. Training siblings to be submissive losers:
dominance between booby nestlings. Animal Behaviour, 44, 881–893.
Dugatkin, L. A. 2001. Bystander effects and the structure of dominance hierarchies.
Behavioral Ecology, 12, 348–352.
Dunlap, K. D. & Zakon, H. H. 1998. Behavioral actions of androgens and androgen
receptor expression in the electrocommunication system of an electric fish,
Eigenmannia virescens. Hormones and Behavior, 34, 30–38.
Earley, R. L. & Dugatkin, L. A. 2002. Eavesdropping on visual cues in green swordtail
(Xiphophorus helleri) fights: a case for networking. Proceedings of the Royal Society of
London, Series B, 269, 943–952.
Edwards, D. H. & Kravitz, E. A. 1997. Serotonin, social status and aggression. Current
Opinion in Neurobiology, 7, 812–819.
Eichenbaum, H., Otto, T. & Cohen, N. J. 1992. The hippocampus: what does it do?
Behavioral and Neural Biology, 57, 2–36.
Evans, C. S. & Marler, P. 1994. Food calling and audience effects in male chickens,
Gallus gallus: their relationships to food availability, courtship and social
facilitation. Animal Behaviour, 47, 1159–1170.
Ferkin, M. H. & Johnston, R. E. 1993. Roles of gonadal hormones in control of five
sexually attractive odors in meadow voles (Microtus pennsylvanicus). Hormones and
Behavior, 27, 523–538.
Fernald, R. D. 1976. The effect of testosterone on the behavior and coloration of adult
male cichlid fish, Haplochromis burtoni. Hormone Research, 7, 172–178.
Fine, M. L. & Pennymaker, K. R. 1986. Hormonal basis for sexual dimorphism of the
sound-producing apparatus of the oyster toadfish. Experimental Neurology, 92,
289–298.
Forlano, P. M., Sisneros, J. A., Knapp, R. & Bass, A. H. 2003. Reproductive periodicity of
gonadal steroid hormone levels in the vocal plainfin midshipman, Porichthys
notatus. Hormones and Behavior, 44: 49.
Francis, R. C. 1983. Experiential effects on agonistic behavior in the paradise fish,
Macropodus opercularis. Behaviour, 85, 292–313.
1987. The interaction of genotype and experience in the dominance success of
paradise fish (Macropodus opercularis). Biology of Behaviour, 12, 1–11.
Franck, D. & Ribowski, A. 1987. Influences of prior agonistic experiences on
aggression measures in the male swordtail (Xiphophorus helleri). Behaviour, 103,
217–240.
Freedman, E. G., Olyarchuk, J., Marchaterre, M. A. & Bass, A. H. 1989. A temporal
analysis of testosterone-induced changes in electric organs and electric organ
discharges of mormyrid fishes. Journal of Neurobiology, 20, 619–634.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 511
Frey, D. F. & Miller, R. J. 1972. The establishment of dominance relationships in the
blue gourami Trichogaster trichopterus (Pallas). Behaviour, 42, 8–62.
Gahr, M., Flugge, G. & Guttinger, H. R. 1987. Immunocytochemical localization of
estrogen-binding neurons in the songbird brain. Brain Research, 402, 173–177.
Gahr, M., Guttinger, H. R. & Kroodsma, D. A. 1993 Estrogen receptors in the avian
brain: survey reveals general distribution and forebrain areas unique to
songbirds. Journal of Comparative Neurology, 327, 112–122.
Gawienowsky, A. M., De Nicola, D. B. & Stacewicz-Sapuntzakis, M. 1976. Androgen
dependence of a marking pheromone in rat urine. Hormones and Behavior, 7,
401–405.
Gelinas, D. & Callard, G. V. 1997 Immunolocalization of aromatase and androgen
receptor-positive neurons in the goldfish brain. General and Comparative
Endocrinology, 106, 155–168.
Ginsberg, B. & Allee, W. C. 1942. Some effects of conditioning of social dominance and
subordination in inbred strains of mice. Physiological Zoology, 15, 485–506.
Gonzalez-Mariscal, G., Melo, A. E., Zavala, A. & Beyer, C. 1993. Sex steroid regulation of
chin-marking behavior in male New Zealand rabbits. Physiology and Behavior, 54,
1035–1040.
Gould, S. J. & Vrba, E. S. 1982. Exaptation: a missing term in the science of form.
Paleobiology, 8, 4–15.
Grober, M. S. 1998. Socially controlled sex-change: integrating ultimate and proximate
levels of analysis. Acta Ethologica, 1, 3–17.
Gyger, M., Karakashian, S. J. & Marler, P. 1986. Avian alarm calling: is there an
audience effect? Animal Behaviour, 34, 1570–1572.
Hagedorn, M. & Carr, C. 1985. Single electrocytes produce a sexually dimorphic signal
in South American electric fish Hypopomus occidentalis (Gymnotiformes,
Hypopomidae). Journal of Comparative Physiology A, 156, 511–523.
Hagedorn, M. & Heiligenberg, W. 1985. Court and spark: electric signals in the
courtship and mating of gymnotid fish. Animal Behaviour, 32, 254–265.
Hannes, R. P. 1984. Androgen and corticoid levels in blood and body extracts of high
and low ranking swordtail males (Xiphophorus helleri) before and after social
isolation. Zeitschrift fur Tierpsychologie, 66, 70–76.
1986. Blood and whole-body androgen levels of male swordtails correlated with
aggression measures in a standard-opponent test. Aggressive Behavior, 12, 249–254.
Hasegawa, Y., Okuno, A. & Nagasawa, H. 2002. Immunohistochemical study of
androgenic gland hormone: localization in the male reproductive system and
species specificity in the terrestrial isopods. General and Comparative Endocrinology,
125, 218–225.
Hector, A. C. K., Seyfarth, R. M. & Raleigh, M. 1989. Male parental care, female choice
and the effect of an audience in vervet monkeys. Animal Behaviour, 38, 262–271.
Hegner, R. E. & Wingfield, J. C. 1987a. Social status and circulating levels of hormones
in flocks of house sparrows, Passer domesticus. Ethology, 76, 1–14.
1987b. Effects of experimental manipulations of testosterone levels on parental
investment and breeding success in male house sparrows. The Auk, 104, 462–469.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
512 R. F. Oliveira
Heid, P., Guttinger, H. R. & Prove, E. 1985. The influence of castration and testosterone
replacement on the song architecture of canaries (Serinus canarius). Zeitschrift fur
Tierpsychologie, 69, 224–236.
Herbert, J. 1993. Peptides in the limbic system: neurochemical codes for coordinated
adaptive responses to behavioural and physiological demand. Neurobiology, 41,
723–791.
Herfeld, S. & Moller, P. 1998. Effects of 17-alfa-methyltestosterone on sexually
dimorphic characters in the weakly electric fish Brienomyrus niger (Gunther, 1866)
(Mormyridae): electric organ discharge, ventral body wall indentation, and
anal-fin ray bone expansion. Hormones and Behavior, 34, 303–319.
Hogan, J. A. 1967. Fighting and reinforcement in Siamese fighting fish Betta splendens.
Journal of Comparative and Physiological Psychology, 64, 356–359.
Hogan, J. A. & Bols, R. J. 1980. Priming of aggressive motivation in Betta splendens.
Animal Behaviour, 28, 135–142.
Hoglund, E., Kolm, N. & Winberg, S. 2001. Stress-induced changes in brain
serotonergic activity, plasma cortisol and aggressive behavior in Arctic charr
(Salvelinus alpinus) is counteracted by L-DOPA. Physiology and Behavior, 74,
381–389.
Hogue, M.-E., Beaugrand, J. P. & Lague, P. C. 1996. Coherent use of information by hens
observing their former dominant defeating or being defeated by a stranger.
Behavioural Processes, 38, 241–252.
Hollis, K. L., Dumas, M. J., Singh, P. & Fackelman, P. 1995. Pavlovian conditioning of
aggressive behavior in blue gourami fish (Trichogaster trichopterus): winners become
winners and losers stay losers. Journal of Comparative Psychology, 109, 123–133.
Holst, D. V. & Eichmann, F. 1998. Sex-specific regulation of marking behavior by sex
hormones and conspecifics scent in tree shrews (Tupaia belangeri). Physiology and
Behavior, 63, 157–164.
Hopkins, C. D. 1972. Sex differences in signalling in an electric fish. Science, 176,
1035–1037.
Hsu, Y. & Wolf, L. L. 1999. The winner and loser effect: integrating multiple
experiences. Animal Behaviour, 57, 903–910.
2001. The winner and loser effect: what fighting behaviours are influenced? Animal
Behaviour, 61, 777–786.
Hurst, J. L. 1990. Urine marking in populations of wild mice Mus domesticus Rutty. III.
Communication between the sexes. Animal Behaviour, 40, 233–243.
Ibara, R. M., Penny, L. T., Ebeling, A. W., van Dykhuizen, G. & Cailliet, G. 1983. The
mating call of the plainfin midshipman fish, Porichthys notatus. In: Predators and
Prey in Fishes, ed. D. G. L. Noakes, D. G. Lindquist, G. S. Helfman & J. A. Ward. The
Hague: Junk Press, pp. 205–212.
Ikeda, K. 1933. Effect of castration on the sexual characters of anadromous
three-spined stickleback, Gasterosteus aculeatus aculeatus (L.). Japanese Journal of
Zoology, 5, 135–157.
Jackson, W. 1991. Why do winners keep winning? Behavioral Ecology and Sociobiology, 28,
271–276.
Johnson, R. P. 1973. Scent marking in mammals. Animal Behaviour, 21, 521–535.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 513
Johnsson, J. I. & Åkerman, A. 1998. Watch and learn: preview of the fighting ability of
opponents alters contest behaviour in rainbow trout. Animal Behaviour, 56,
771–776.
Johnston, R. E. 1975. Sexual excitation function of hamster vaginal secretions Animal
Learning and Behavior, 3, 161–166.
Kavaliers, M., Choleris, E. & Colwell, D. D. 2001. Brief exposure to female odors
‘emboldens’ male mice by reducing predator-induced behavioural and hormonal
responses. Hormones and Behavior, 40, 497–509.
Keller, C. H., Zakon, H. H. & Sanchez, D. Y. 1986. Evidence for a direct effect of
androgens on electroreceptor tuning. Journal of Comparative Physiology A, 158,
301–310.
Kelley, D. C. 2002. Hormonal regulation of motor output in amphibians: Xenopus laevis
vocalizations as a model system. In: Hormones, Brain and Behavior, Vol. 2, ed. D. W.
Pfaff, A. P. Arnold, A. M. Etgen, S. E. Fahrbach & R. T. Rubin. New York: Academic
Press, pp. 445–468.
Kerr, J. E., Allore, R. J., Beck, S. G. & Handa, R. J. 1995. Distribution and hormonal
regulation of androgen receptors (AR) and AR messenger ribonucleic acid in the
rat hippocampus. Endocrinology, 136, 3213–3221.
Ketterson, E. D., Nolan, V., Jr, Wolf, L. & Ziegenfus, C. 1992. Testosterone and avian life
histories: effects of experimentally elevated testosterone on behavior and
correlates of fitness in the dark-eyed junco, Junco hyemalis. American Naturalist, 140,
980–999.
Kikuyama, S., Toyoda, F., Ohmiya, Y. et al. 1995. Sodefrin: a female-attracting
pheromone in newt cloacal glands. Science, 267, 1643–1645.
Kikuyama, S., Toyoda, F., Yamamoto, K., Tanaka, S. & Hayashi, H. 1997. Female-
attracting pheromone in newt cloacal glands. Brain Research Bulletin, 44, 415–422.
Kimura, D. 1996. Sex, sexual orientation and sex hormones influence human
cognitive function. Current Opinion in Neurobiology, 6, 259–263.
Landsman, R. E. & Moller, P. 1988. Testosterone changes the electric organ discharge
and external morphology of the mormyrid fish Gnathonemus petersii
(Mormyriformes). Experientia, 44, 900–903.
Landsman, R. E., Harding, C. F., Moller, P. & Thomas, P. 1990. The effects of androgens
and estrogen on the external morphology and electric organ discharge waveform
of Gnathonemus petersii (Mormyriformes, Teleostei). Hormones and Behavior, 24,
532–553.
Leiser, J. K. & Itzkowitz, M. 1999. The benefits of dear enemy recognition in
three-contender convict cichlid (Cichlasoma nigrofasciatum) contests. Behaviour, 136,
983–1003.
Lepri, J. & Randall, J. A. 1983. Hormonal regulation of sandbathing in male kangaroo
rats (Dipodomys merriami). Behavioral and Neural Biology, 37, 256–264.
Levy, M. & Aronson, L. R. 1955. Morphological effects of castration and hormone
administration in the male cichlid fish Tilapia macrocephala. Anatomical Records,
122, 450–451.
Lisk, R. D. & Bezier, J. L. 1980. Intrahypothalamic hormone implantation and
activation of sexual behavior in the male hamster. Neuroendocrinology, 30, 220–222.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
514 R. F. Oliveira
Lowe, P. 1942. The anatomy of Gould’s manakin (Manacus vitellinus) in relation to its
display. Ibis, 6, 50–83.
Luine, V., Nottebohm, F., Harding, C. & McEwen, B. S. 1980. Androgen affects
cholinergic enzymes in syringeal motor neurons and muscle. Brain Research, 192,
89–107.
Malacarne, G., Bottoni, L., Massa, R. & Vellano, C. 1984. The abdominal gland of the
crested newt: a possible source of courtship pheromones. Monitore Zoologico
Italiano, 18, 33–39.
Manzo, J., Garcia, L. I., Hernandez, M. E., Carrillo, P. & Pacheco, P. 2002.
Neuroendocrine control of urine-marking behavior in male rats. Physiology and
Behavior, 75, 25–32.
Marler, C. A., Walsberg, G., White, M. I. & Moore, M. 1995. Increased energy
expenditure due to increased territorial defense in male lizards after phenotypic
manipulation. Behavioral Ecology and Sociobiology, 37, 225–231.
Marler, P., Dufty, A. & Pickert, R. 1986. Vocal communication in the domestic chicken.
II. Is a sender sensitive to the presence and nature of a receiver? Animal Behaviour,
34, 194–198.
Matos, R. J. 2002. Social influences on signalling: audience effects and communication
networks. Ph.D. Thesis, University of Copenhagen.
Matos, R. J. & McGregor, P. K. 2002. The effect of the sex of an audience on male–male
displays of Siamese fighting fish (Betta splendens). Behaviour, 139, 1211–1221.
Mazur, A. & Booth, A. 1998. Testosterone and dominance in men. Behavioral and Brain
Sciences, 21, 353–397.
McDonald, N. W., Heimstra, A. L. & Damkot, D. K. 1968. Social modification of
agonistic behaviour in fish. Animal Behaviour, 16, 437–441.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.
McGregor, P. K. & Peake, T. 2000. Communication networks: social environments for
receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
McGregor, P. K., Dabelsteen, T. & Holland, J. 1997. Eavesdropping in a territorial
songbird communication network: preliminary results. Bioacoustics, 8, 253–254.
McKibben, J. R. & Bass, A. H. 1998. Behavioral assessment of acoustic parameters
relevant to signal recognition and preference in a vocal fish. Journal of the Acoustic
Society of America, 104, 3520–3533.
Meyer, J. H. 1983. Steroid influences upon the discharge frequency of a weakly electric
fish. Journal of Comparative Physiology A, 153, 29–38.
Meyer, J. H. & Zakon, H. H. 1982. Androgens alter the tuning of electroreceptors.
Science, 217, 635–637.
Meyer, J. H., Leong, M. & Keller, C. H. 1987. Hormone-induced and ontogenetic changes
in electric organ discharge and electroreceptor tuning in the weakly electric fish
Apteronotus. Journal of Comparative Physiology A, 160, 385–394.
Millinski, M. & Bakker, T. C. M. 1990. Female sticklebacks use male coloration in mate
choice and hence avoid parasitised males. Nature, 344, 330–333.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 515
Mills, A. C. & Zakon, H. H. 1987. Coordination of EOD frequency and pulse duration in
a weakly electric wave fish: the influence of androgens. Journal of Comparative
Physiology A, 161, 417–430.
1991. Chronic androgen treatment increases action potential duration in the
electric organ of Sternopygus. Journal of Neuroscience, 11, 2349–2361.
Moofat, S. D. & Hampson, E. 1996. A curvilinear relationship between testosterone and
spatial cognition in humans: possible influence of hand preferences.
Psychoneuroendocrinology, 21, 323–337.
Moore, A. & Scott, A. P. 1991. Testosterone is a potent odorant in precocious male
Atlantic salmon (Salmo salar L.) parr. Philosophical Transactions of the Royal Society of
London, Series B, 332, 241–244.
Moore, F. L., Wood, R. E. & Boyd, S. K. 1992. Sex steroids and vasotocin interact in a
female amphibian (Taricha granulosa) to elicit female-like egg-laying behavior or
male-like courtship. Hormones and Behavior, 26, 156–166.
Nagaya, N. & Herrera, A. A. 1995. Effects of testosterone on synaptic efficacy at
neuromuscular junctions in a sexually dimorphic muscle of male frogs. Journal of
Physiology, 483, 141–153.
Naguib, M., Fichtel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Nelson, J. S. 1994. Fishes of the World, 3rd edn. New York: John Wiley.
Nelson, R. 2001. An Introduction to Behavioural Endocrinology, 2nd edn. Sunderland, MA:
Sinauer.
Northcutt, R. G. & Davis, R. E. 1983. Telencephalic organization in ray-finned fishes. In:
Fish Neurobiology, Vol. 2: Higher Brain Areas and Functions, ed. R. E. Davis & R. G.
Northcutt. Ann Arbor, MI: University of Michigan Press, pp. 203–236.
Oliveira, R. F. & Almada, V. C. 1998. Androgenization of dominant males in a cichlid
fish: androgens mediate the social modulation of sexually dimorphic traits.
Ethology, 104, 841–858.
Oliveira, R. F., Almada, V. C. & Canario, A. V. M. 1996. Social modulation of sex steroid
concentrations in the urine of male cichlid fish Oreochromis mossambicus. Hormones
and Behavior, 30, 2–12.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecifics interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Oliveira, R. F., Carneiro, L. A., Goncalves, D. M., Canario, A. V. M. & Grober, M. S.
2001a. Effects of 11-ketotestosterone in sneaker males of the peacock blenny, a
species with male alternative mating tactics. Brain, Behavior and Evolution, 58,
28–37.
Oliveira, R. F., Lopes, M., Carneiro, L. A. & Canario, A. V. M. 2001b. Watching fights
raises fish hormone levels. Nature, 409, 475.
Oliveira, R. F., Hirschenhauser, K., Carneiro, L. & Canario, A. V. M. 2002. Social
modulation of androgen levels in male teleost fish. Comparative Biochemistry and
Physiology B, 132, 203–215.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
516 R. F. Oliveira
Otronen, M. 1990. The effect of prior experience on the outcome of fights in the
burying beetle, Nicrophorus humatio. Animal Behaviour, 40, 980–1004.
Owen, P. C. & Perrill, S. A. 1998. Habituation in the green frog, Rana clamitans.
Behavioral Ecology and Sociobiology, 44, 209–213.
Pankhurst, N. W. & Barnett, C. W. 1993. Relationship of population density, territorial
interaction, and plasma levels of gonadal steroids in spawning male demoiselles
Chromis dispilus (Pisces, Pomacentridae). General and Comparative Endocrinology, 90,
168–176.
Parker, G. A. 1974. The assessment strategy and the evolution of fighting behaviour.
Journal of Theoretical Biology, 47, 223–243.
Peake, T. M. & McGregor, P. K. 2004. Information and aggression in fishes. Learning and
Behavior 32, 114–121.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Pomerantz, S. M. & Sholl, S. A. 1987. Analysis of sex and regional differences in
androgen receptors in fetal rhesus monkey brain. Brain Research, 433, 151–154.
Randall, J. A. 1986. Lack of gonadal control of the dorsal gland and sandbathing in
male and female Bannertail kangaroo rats (Dipodomys spectabilis). Hormones and
Behavior, 20, 95–105.
Reinboth, R. & Rixner, W. 1972. Verhalten des Kleinen Maulbruters Hemihaplochromis
multicolor nach Kastration und Behandlung mit Testosteron. [Film C 1019/, 1970].
Gottingen: Institut fur den Wissenschaftlichen Film.
Resnick, S. M., Berenbaum, S. A., Gottesman, I. J. & Bouchard, T. J. 1986. Early
hormonal influences on cognitive functioning in congenital adrenal hyperplasia.
Developmental Psychology, 22, 191–198.
Roof, R. L. 1993. Neonatal exogenous testosterone modifies sex difference in radial
arm and Morris water maze performance in prepubescent and adult rats.
Behavioral and Brain Research, 53, 1–10.
Roof, R. L. & Havens, M. D. 1992. Testosterone improves maze performance and induces
development of a male hippocampus in females. Brain Research, 572, 310–313.
Ros, A. F. H., Becker, K., Canario, A. V. M. & Oliveira, R. F. 2004. Androgen levels and
energy metabolism in Oreochromis mossambicus. Journal of Fish Biology, 65,
895–905.
Rose, R. M., Bernstein, I. S. & Holaday, J. W. 1971. Plasma testosterone, dominance
rank and aggressive behavior in a group of male rhesus monkeys. Nature, 231,
366.
Rose, R. M., Berstein, I. S. & Gordon, T. 1975. Consequences of social conflict on plasma
testosterone levels in rhesus monkeys. Psychosomatic Medicine, 37, 50–61.
Rost, R. 1990. Hormones and behavior: a joint examination of studies on seasonal
variation in song production and plasma levels of testosterone in the great tit
Parus major. Journal of Ornithology, 131, 403–411.
1992. Hormones and behavior: a comparison of studies on seasonal changes in song
production and testosterone plasma levels in the Willow tit Parus montanus. Ornis
Fennica, 69, 1–6.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 517
Rouse, E. F., Coppinger, C. J. & Barnes, P. R. 1977. The effect of an androgen inhibitor
on behavior and testicular morphology in the stickleback Gasterosteus aculeatus.
Hormones and Behavior, 9, 8–18.
Rowland, W. J. 1999. Studying visual cues in fish behavior: a review of ethological
techniques. Environmental Biology of Fishes, 56, 285–305.
Ruzzante, D. E. 1992. Mirror image stimulation, social hierarchies, and population
differences in agonistic behavior: a reappraisal. Canadian Journal of Fisheries and
Aquatic Sciences, 49, 1966–1968.
Saldanha, C. J., Clayton, N. S. & Schlinger, B. A. 1999. Androgen metabolism in the
juvenile oscine forebrain: a cross-species analysis at neural sites implicated in
memory function. Journal of Neurobiology, 40, 397–406.
Sapolsky, R. M. 1992. Neuroendocrinology of the stress-response. In: Behavioral
Endocrinology, ed. J. B. Becker, S. M. Breedlove & D. Crews. Cambridge, MA: MIT
Press, pp. 287–324.
Sawyer, T. F., Hengehold, A. K. & Perez, W. A. 1984. Chemosensory and hormonal
mediation of social memory in male rats. Behavioral Neuroscience, 98, 908–913.
Scalia, F. & Winans, S. S. 1975. The differential projections of the olfactory bulb and
accessory olfactory bulb in mammals. Journal of Comparative Neurology, 161, 31–56.
Schlinger, B. A., Schultz, J. D. & Hertel, F. 2001. Neuromuscular and endocrine control
of an avian courtship behavior. Hormones and Behavior, 40, 276–280.
Schuett, G. W. 1997. Body size and agonistic experience affect dominance and mating
success in male copperheads. Animal Behaviour, 54, 213–224.
Schulkin, J. 2002. Hormonal modulation of central motivational states. In: Hormones,
Brain and Behavior, Vol. 1, ed. D. W. Pfaff, A. P. Arnold, A. M. Etgen, S. E. Fahrbach &
R. T. Rubin. New York: Academic Press, pp. 663–657.
Schultz, J. D. & Schlinger, B. A. 1999. Widespread accumulation of [3H]testosterone in
the spinal cord of a wild bird with an elaborate courtship display. Proceedings of the
National Academy of Sciences, USA, 96, 10428–10432.
Schultz, J. D., Hertel, F., Bauch, M. & Schlinger, B. A. 2001. Adaptations for rapid and
forceful contraction in wing muscles of the male golden-collared manakin: sex
and species comparisons. Journal of Comparative Physiology A, 187, 677–684.
Searcy, W. A., Eriksson, D. & Lundberg, A. 1991. Deceptive behavior in pied flycatchers.
Behavioral Ecology and Sociobiology, 29, 167–175.
Silverin, B. 1980. Effects of long-acting testosterone treatment on free-living pied
flycatchers, Ficedula hypoleuca, during the breeding period. Animal Behaviour, 28,
906–912.
Sisneros, J. A. & Bass, A. H. 2003. Seasonal plasticity of peripheral auditory frequency
sensitivity. Journal of Neuroscience, 23, 1049–1058.
Sisneros, J. A. & Tricas, T. C. 2000. Androgen-induced changes in the response
dynamics of ampullary electrosensory primary afferent neurons. Journal of
Neuroscience, 20, 8586–8595.
Simon, N. G. 2002. Hormonal processes in the development and expression of
aggressive behavior. In: Hormones, Brain and Behavior, Vol. 1, ed. D. W. Pfaff, A. P.
Arnold, A. M. Etgen, S. E. Farbach & R. T. Rubin. New York: Academic Press,
pp. 339–392.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
518 R. F. Oliveira
Smith, G. T., Brenowitz, E. A., Beecher, M. D. & Wingfield, J. C. 1997. Seasonal changes
in testosterone, neural attributes of song control nuclei, and song structure in
wild songbirds. Journal of Neuroscience, 17, 6001–6010.
Smith, R. J. F. 1974. Effects of 17-methyltestosterone on the dorsal pad and tubercles of
fathead minnows (Pimephales promelas). Canadian Journal of Zoology, 52, 1031–1038.
Squire, L. R. 1992. Memory and the hippocampus: a synthesis from findings with rats,
monkeys, and humans. Psychological Reviews, 9, 195–231.
Stacey, N. & Sorensen, P. 2002. Hormonal pheromones in fish. In: Hormones, Brain and
Behavior, Vol. 2, ed. D. W. Pfaff, A. P. Arnold, A. M. Etgen, S. E. Fahrbach & R. T.
Rubin. New York: Academic Press, pp. 375–434.
Suthers, R. A., Goller, F. & Pytte, C. 1999. The neuromuscular control of bird song.
Philosophical Transactions of the Royal Society of London, Series B, 354, 927–939.
Taborsky, M. 1994. Sneakers, satellites, and helpers: parasitic and cooperative
behaviour in fish reproduction. Advances in the Study of Behaviour, 23, 1–100.
Taylor, G. T., Bartko, G. & Farr, S. 1987. Gonadal hormones and conspecific marking in
male rats. Hormones and Behavior, 21, 234–244.
Tellegen, A., Horn, J. M. & Legrand, R. G. 1969. Opportunity for aggression as a
reinforcer for mice. Psychonomic Science, 14, 104–105.
Temeles, E. J. 1994. The role of neighbours in territorial systems: when are they ‘dear
enemies’? Animal Behaviour, 47, 339–350.
Tinbergen, N. 1951. The Study of Instinct. Oxford: Clarendon Press.
Tobias, M. L. & Kelley, D. B. 1995. Sexual differentiation and hormonal regulation of
the laryngeal synapse in Xenopus laevis. Journal of Neurobiology, 28, 515–526.
Tobias, M. L., Kelley, D. B. & Ellisman, M. 1995. A sex-difference in synaptic efficacy at
the laryngeal neuromuscular junction of Xenopus laevis. Journal of Neuroscience, 15,
1660–1668.
Tohgi, H., Utsugisawa, K., Yamagata, M. & Yoshimura, M. 1995. Effects of age on mRNA
expression of glucocorticoid, thyroid hormone, androgen, and estrogen receptors
in postmortem human hippocampus. Brain Research, 700, 245–253.
Vezina, M. & Thomas, D. W. 2000. Social status does not affect resting metabolic rate
in wintering dark-eyed junco (Junco hyemalis). Physiological and Biochemical Zoology,
73, 231–236.
Wapler-Leong, D. C. Y. & Reinboth, R. 1974. The influence of androgenic hormone on
the behaviour of Haplochromis burtoni (Cichlidae). Fortschritte der Zoologie, 22,
334–339.
Wetzel, D. M. & Kelley, D. B. 1983. Androgen and gonadotrophin effects on male mate
calls in South African clawed frogs, Xenopus laevis. Hormones and Behaviour, 17,
388–404.
Whitehouse, M. A. 1997. Experience influences male–male contests in the spider
Argyrodes antipodiana (Theridiidae: Aranea). Animal Behaviour, 53, 913–923.
Whiting, M. J. 1999. When to be neighbourly: differential agonistic responses in the
lizard Platysaurus broadleyi. Behavioral Ecology and Sociobiology, 46, 210–214.
Wikelsky, M., Lynn, S., Breumer, C., Wingfield, J. C. & Kenagy, G. J. 1999. Energy
metabolism, testosterone and corticosterone in white-crowned sparrows. Journal
of Comparative Physiology A, 185, 463–470.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
Hormones, social context and animal communication 519
Williams, C. L., Barnett, A. M. & Meck, W. H. 1990. Organizational effects of early
gonadal secretions on sexual differentiation in spatial memory. Behavioral
Neuroscience, 104, 84–97.
Winberg, S. & Lepage, O. 1998. Elevation of brain 5-HT activity, POMC expression, and
plasma cortisol in socially subordinate rainbow trout. American Journal of
Physiology, 274, R645–R654.
Winberg, S. & Nilsson, G. E. 1993a. Roles of brain monoamine neurotransmitters in
agonistic behaviour and stress reactions, with particular reference to fish.
Comparative Biochemistry and Physiology C, 10, 597–614.
1993b. Time course of changes in brains serotonergic activity and brain tryptophan
levels in dominant and subordinate juvenile Arctic charr. Journal of Experimental
Biology, 179, 181–195.
Winberg, S., Winberg, Y. & Fernald, R. D. 1997. Effect of social rank on brain
monoaminergic activity in a cichlid fish. Brain, Behavior and Evolution, 49,
230–236.
Wingfield, J. C. 1984 Environmental and endocrine control of reproduction in the
song sparrow, Melospiza melodia. II. Agonistic interactions as environmental
information stimulating secretion of testosterone. General and Comparative
Endocrinology, 56, 417–424.
Wingfield, J. C., Ball, G. F., Dufty, A. M., Hegner, R. E. & Ramenofsky, M 1987.
Testosterone and aggression in birds: tests of the challenge hypothesis. American
Scientist, 75, 602–608.
Wingfield, J. C., Hegner, R. E., Dufty, A. M. & Ball, G. F. 1990. The ‘challenge hypothesis’:
theoretical implications for patterns of testosterone secretion, mating systems,
and breeding strategies. American Naturalist, 136, 829–846.
Wingfield, J. C., Jacobs, J. D., Soma, K. et al. 1999. Testosterone, aggression, and
communication: ecological bases of endocrine phenomena. In: The Design of
Animal Communication, ed. M. D. Hauser & M. Konishi. Cambridge, MA: MIT Press,
pp. 257–283.
Wingfield, J. C., Jacobs, J. D., Tramontin, A. D. et al. 2000. Toward an ecological
basis of hormone–behavior interactions in reproduction of birds. In: Reproduction
in Context, ed. K. Wallen & J. E. Schneider. Cambridge, MA: MIT Press,
pp. 85–128.
Wood, R., Brabec, R. K., Swann, J. M. & Newman, S. W. 1992. Androgen and estrogen
concentrating neurons in chemosensory pathways in the male Syrian hamster
brain. Brain Research, 596, 89–98.
Yahr, P. & Stephens, D. R. 1987. Hormonal control of sexual and scent marking
behaviors of male gerbils in relation to the sexually dimorphic area of the
hypothalamus. Hormones and Behavior, 21, 331–346.
Yamamoto, K., Toyoda, F., Tanaka, S., Hayashi, H. & Kikuyama, S. 1996.
Radioimmunoassay of a newt sex pheromone, sodefrin, and the influence of
hormones on its level in the abdominal gland. General and Comparative
Endocrinology, 104, 356–363.
Ydenberg, R. C., Giraldeau, L. A. & Falls, J. B. 1988. Neighbours, strangers and the
asymmetric war of attrition. Animal Behaviour, 36, 343–347.

P1: GFZ-IYP/... P2: KOD0521823617c21.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:12
520 R. F. Oliveira
Zakon, H. H. & Smith, G. T. 2002. Weak electric fish: behavior, neurobiology and
neuroendocrinology. In: Hormones, Brain and Behavior, Vol. 2, ed. D. W. Pfaff, A. P.
Arnold, A. M. Etgen, S. E. Fahrbach & R. T. Rubin. New York: Academic Press,
pp. 349–374.
Zakon, H. H., Yan, H. Y. & Thomas, P. 1990. Human chorionic gonadotropin-induced
shifts in the electrosensory system of the weakly electric fish, Sternopygus. Journal
of Neurobiology, 21, 826–833.
Zakon, H. H., Thomas, P. & Yan, H. Y. 1991. Electric organ discharge frequency and
plasma sex steroid levels during gonadal recrudescence in a natural population
of the weakly electric fish Sternopygus macrurus. Journal of Comparative Physiology A,
169, 493–499.

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
22
Cooperation in communicationnetworks: indirect reciprocity ininteractions between cleaner fishand client reef fish
r e d o ua n b s h a r y 1 & a r u n d ’s o u z a 2
1University of Cambridge, Cambridge, UK2University of Wurzburg, Wurzburg, Germany
Introduction
The aim of this chapter is twofold. First, to outline how recent develop-
ments in cooperation theory are so similar to the communication network concept
(McGregor, 1993) that a unified terminology would be useful to facilitate exchange
of ideas. Second, we argue that the communication network concept provides an
evolutionary framework to predict the widespread occurrence of phenomena that
until now have been discussed in the context of highly developed cognitive abili-
ties. This creates a problem: as it stands, there appear to be no words in cooperation
theory that were not developed in the human context and hence do not include
a cognitive component. We have to use definitions that only constitute the func-
tional aspects of phenomena (like tactical deception and indirect reciprocity) and
ignore the mechanistic aspects (i.e. theory of mind, intentionality) that are com-
monly part of the definitions. We ask readers always to keep in mind that our
definitions never imply any specific cognitive abilities. We illustrate our ideas
with data on the mutualism between the cleaner wrasse Labroides dimidiatus and
its ‘client’ reef fish, which visit cleaners to have ectoparasites removed from their
surface, gills and mouth (reviewed by Losey et al., 1999; Cote, 2000).
Cooperation provides a challenge to evolutionary theory because it often in-
volves apparently altruistic behaviour. Hamilton (1964) provided a framework to
understand why altruism between kin can be evolutionary stable; specifically, an
altruist gains indirect fitness benefits from its action. However, there are plenty of
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
521

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
522 R. Bshary & A. D’Souza
examples where recipients of altruistic acts are unrelated to the helping individual
(Dugatkin, 1997). Trivers (1971) pointed out that such altruism might be evolu-
tionarily stable if the recipient later reciprocates, an idea that was formalized
by Axelrod & Hamilton (1981). They used an iterated version of the prisoner’s
dilemma game, a two-player game in which opponents can either cooperate or
defect. Defection yields a higher payoff than cooperation independently of the
partner’s action, but if both players cooperate they receive a higher payoff than
if both defect, hence the dilemma. In a computer tournament in which several
strategies competed with each other, a simple strategy called ‘tit-for-tat’ emerged
as a cooperative solution to the game. Tit-for-tat players start by being cooperative;
then in the next round they play what their opponent did in the previous round.
Thus, tit-for-tat players can reap the benefits of mutual cooperation while avoid-
ing exploitation by an uncooperative opponent (except for the first occasion on
which an opponent defected). Several new strategies that are similar but appar-
ently superior to tit for tat have been tested since the first computer tournament
(reviewed by Dugatkin, 1997).
Despite the intuitive appeal of reciprocal altruism and behavioural strategies
similar to tit for tat to ensure cooperative behaviour, few empirical examples have
been reported and some that have are contentious (Dugatkin, 1997). In addition,
Alexander (1987) pointed out that many examples of human altruistic behaviour
do not fit an iterated prisoner’s dilemma game: humans often help individuals
who are highly unlikely to ever reciprocate. He proposed that humans might help
others simply to increase their own image within the society. Nowak & Sigmund
(1998) explored this idea by developing a game theory model in which direct
reciprocity on altruistic acts was excluded. Instead, an altruist gained an increase
in his image score. If an individual’s image score was linked to the probability that
others were willing to provide help when needed, cooperation readily emerged
and was evolutionarily stable (Nowak & Sigmund, 1998). Individuals helped in
order to be helped themselves during future interactions with current bystanders
(for further theoretical developments see Lotem et al. (1999, 2003) and Leimar &
Hammerstein (2001)). An experiment with first-year students confirmed a crucial
prediction of the models: students that helped more than average received more
help and, therefore, a final payoff that was above the average (Wedekind & Milinski,
2000). This new approach to the evolution of cooperation is a specific instance of
a communication network: the interactions between individuals do not happen
in a social vacuum but in the presence of other individuals who eavesdrop and
thereby extract relevant information for own future interactions with the actors
(McGregor, 1993). As a consequence of eavesdropping, it pays individuals to alter
their behaviour, either as a general unconditional response (Johnstone, 2001) or
specifically in situations where bystanders are present (audience effects; see Ch. 4).

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 523
Nowak & Sigmund (1998) proposed that altruism based on indirect reciprocity
is a hallmark of human evolution. Although they do not specify why this should
be so, their proposal implies that non-human animals either do not live in so-
cial environments that would favour the evolution of altruism through indirect
reciprocity or lack some of the cognitive abilities required (but see Zahavi, 1995;
Roberts, 1998). In contrast, we suggest that the communication network concept
has the advantage of coming from a purely functional perspective rather than
trying to explain apparently maladapted human behaviour. Early discussions of
communication networks (McGregor, 1993) argued that behaviours such as eaves-
dropping and audience effects should evolve in the context of a network, with-
out detailed consideration of the underlying cognitive mechanisms. By ignoring
mechanisms, we use the communication network concept to predict eavesdrop-
ping and audience effects in potentially cooperative contexts without worrying
about cognitive constraints. Experimental evidence for the existence of eaves-
dropping has been provided for a wide variety of taxa (Ch. 2). Whereas human
subjects can be asked about their behaviour, eavesdropping in other animals has
to be inferred from the eavesdroppers’ subsequent behaviour towards individu-
als observed interacting. Differences in individuals’ roles must elicit differences
in subsequent eavesdropper behaviour towards them. It was thus an implicit as-
sumption of communication-network studies that eavesdroppers attribute some
sort of image score to observed individuals and that this score governs their own
future behaviour towards those individuals. Scoring an individual’s tendency to
help is just one type of image score. Fighting ability, aggressiveness and mating
success with regard to female choice are the image scores typically studied in
communication networks (e.g. Ch. 5).
That eavesdroppers adjust their own behaviour to what they have witnessed has
important implications for the behaviour of individuals that are observed. While a
classical approach would suggest that individuals maximize payoffs in each single
interaction (with the exception of reciprocal altruism and punishment, where ben-
efits are delayed), any occurrence of eavesdropping implies that selection favours
individuals that optimize current actions by integrating both immediate payoffs
and future consequences of their behaviour. Within the framework of coopera-
tion theory, it may pay individuals to be altruistic if this increases the probability
of meeting more cooperative eavesdroppers in the future; in contest theory, it
may pay to be more aggressive in the presence of potential challengers if winning
a fight results in fewer attacks from eavesdroppers. Individuals can respond to
eavesdropping in two ways. First, they can alter their behaviour in any interaction
in relation to the average probability that eavesdroppers are present. In this case,
all individuals behave in the same way (with respect to eavesdroppers) in all inter-
actions. Second, individuals can pay attention to specific cues that eavesdroppers

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
524 R. Bshary & A. D’Souza
are present for a particular interaction and alter their behaviour accordingly. In
the latter, individuals show a flexible behavioural pattern. Communication net-
work models have until now dealt with the first scenario (Nowak & Sigmund, 1998;
Johnstone, 2001; Leimar & Hammerstein, 2001). There is, however, increasing ev-
idence that animals adjust their behaviour in a particular interaction according
to the presence or absence of eavesdroppers (Doutrelant et al., 2001; Bshary, 2002;
Ch. 4).
Altruism towards unrelated individuals has been linked to positive reciprocity,
be it direct (Trivers, 1971) or indirect (Nowak & Sigmund, 1998): individuals help
because they will receive help in return. However, this is not necessarily the case.
An alternative is that an individual helps in order to raise its image and uses
its image to exploit recipients or eavesdroppers, which will behave cooperatively
because of this high image. If all individuals in a population exploited eavesdrop-
pers, then image scoring would break down. However, as long as image scoring
yields an overall benefit, either because most altruistic acts are honest in that
an individual’s willingness to cooperate is revealed or because the benefits of co-
operation exceed the costs of being exploited, altruism may be used both as an
honest and as a deceptive signal. Therefore, altruism may sometimes serve as a
signal out of context, causing other individuals to react in the signaller’s favour
and to their own disadvantage. This is the functional definition of tactical decep-
tion (Hauser, 1998). In communication-network terms, it may pay individuals to
be altruistic if this signal is misinterpreted by eavesdroppers in a way that allows
future exploitation of them.
Such a functional approach to tactical deception is in strong contrast to the
traditional cognitive approach. Though such behaviour has been described, for
example in birds (Munn, 1986), tactical deception is often seen as a hallmark
of primate ‘Machiavellian intelligence’ (Byrne & Whiten, 1988): the notion that
most primate species have been strongly selected for the cognitive abilities to
cope with their social environment (see references in Byrne & Whiten (1988) and
Ch. 25). The ability to use tactical deception has, therefore, been linked to the
concept of theory of mind (Premack & Woodruff, 1978): the ability to speculate
how another individual might perceive a certain situation. However, Heyes (1998)
cautioned that any observations of tactical deception do not imply the existence of
particularly high cognitive abilities. Instead, originally animals might have made
an error (i.e. produced a signal out of context) but it may have had a favourable
outcome for the signaller. As a result the signaller may associate this error with
a reward and consequently would be more likely to produce the signal again in
this context. The notion that simple associative learning might suffice to produce
signals that fit the functional definition of tactical deception offers the possibility
of using a much more functional, rather than mechanistic, approach to the topic

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 525
(see also Hauser, 1998) – and the appropriate framework for the study of tactical
deception is communication networks.
Interactions between cleaner fish and clients
In the remainder of this chapter, we present data on interactions between
the cleaner wrasse and client reef fish to illustrate the arguments outlined above.
Data were collected in the Red Sea, at Ras Mohammed National Park, Egypt. Meth-
ods of data collection are described in detail elsewhere (Bshary, 2001, 2002), so here
we will keep this kind of information to a minimum. All data are field observa-
tions; therefore, experimental proof is still lacking. However, these data illustrate
that it is worthwhile searching for potential examples of positive indirect reci-
procity and tactical deception with a functional perspective rather than worrying
about cognitive constraints.
Clients regularly visit the cleaners at their small territories called ‘cleaning
stations’ (cleaning mutualism reviewed by Losey et al., 1999; Cote, 2000). As in-
dividual cleaner wrasse may have more than 2000 interactions per day (Grutter,
1995), interactions often take place in the presence of other potential visitors.
Such bystanders can eavesdrop and evaluate the cleaner’s service quality. While
the cleaner fish eat parasites, in particular gnathiid isopods (Grutter, 1996), they
also feed on client mucus and scales (Randall, 1958; Grutter, 1997). Feeding on
healthy client tissue is correlated with the occurrence of client ‘jolts’, an observ-
able short shake of the client’s body, in response to mouth contact by the cleaner
fish (Bshary & Grutter, 2002a). The frequency of client jolts correlates negatively
with parasite load; therefore, client jolts are not a byproduct of parasite removal.
Rather, jolts are an easily observable correlate of cleaner fish cheating (Bshary, &
Grutter, 2002a). Note that only non-predatory clients (i.e. species that could not eat
cleaner fish) jolt on a regular basis, while jolts of predatory clients are infrequent
(Bshary, 2001). Therefore, we will only present data on non-predatory clients. In
response to a jolt, clients often dart off or chase the cleaner, depending on their
strategic options. Client species with large home ranges that cover several cleaning
stations (‘choosy clients’) usually make use of their choice options and swim off
and visit a different cleaning station for their next inspection (Bshary & Schaffer,
2002), as predicted by biological market theory (Noe et al., 1991; reviewed by Noe,
2001). In contrast, client species with small territories or home ranges, and hence
with access to only one cleaning station (‘resident clients’),tend to punish cleaners
by chasing them (Bshary & Grutter, 2002a). Both darting off and punishment could
readily provide bystanders with the information that a cleaning service was bad.
In contrast, if an observed interaction ends without apparent conflict, then the
service had probably been good. Therefore, clients could easily attribute an image

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
526 R. Bshary & A. D’Souza
score to a cleaner fish, and cleaners could adjust their behaviour to the presence
of eavesdropping potential clients.
Why should clients attribute an image score to cleaners?
Attributing an image score to an individual and basing behavioural de-
cisions during interactions with that individual on that score only makes sense
if the score has some predictive power about how that individual will behave. In
potentially cooperative interactions, a positive image score should be attributed
to an individual only if cooperation on one occasion is usually followed by coop-
eration on the next occasion. In the context of cleaning mutualism, this implies
that there must be consistent variation in cheating rates either between indi-
vidual cleaners or within individual cleaners. Indeed, Bshary (2002) found that a
minority of cleaners cheated more frequently than the rest. These ‘biting cleaners’,
compared with normal cleaners, specifically targeted larger non-predatory clients,
both residents (median client jolt rate was 12/100 seconds in interactions with bit-
ing cleaners compared with 2/100 seconds in interactions with normal cleaners)
and choosy clients (18/100 seconds compared with 3/100 seconds), while there was
no evidence for increased cheating of predatory clients (0/100 seconds compared
with 0/100 seconds) or small resident clients (6/100 seconds compared with 6/100
seconds) (Bshary, 2002). These data suggest that it would pay larger non-predatory
clients to avoid interactions with such biting cleaners. One way they could do this
is to extract information from ongoing interactions and attribute an image to a
cleaner.
There is another reason why constant image scoring of cleaner behaviour is
advantageous for clients. Data from one biting cleaner fish revealed that cheating
rates changed considerably over a period of six weeks. Some of the 99% confi-
dence intervals around observed daily jolt rates of choosy clients did not overlap,
suggesting that the variation is significant (Fig. 22.1). This individual was a female,
as were all other biting cleaners that have been observed (n = 7). Cheating of non-
predatory choosy clients peaked at the two periods of full moon that occurred
during the observation period, and full moon coincided with repeated spawning
with her male partner. After the second spawning period, the male disappeared
and cheating rates fell to very low values. The cleaner wrasse is a protogynous
hermaphrodite; that is, individuals start their reproductive career as females and
eventually switch sex to become males (Robertson, 1972). Males have a larger repro-
ductive output because they often have a harem. Therefore, females face a trade-off
between investing in current reproductive effort through the production of eggs
and investing in growth to become a male. If the energy requirements for egg
production are maximal close to spawning, females needed extra energy in order
to avoid compromising growth too much. We suggest that the females’ switch to a

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 527
Cho
osy
clie
nt jo
lts/1
00 s
Fig. 22.1. Jolt rates (with 99% confidence intervals) of choosy clients (species with large
home ranges that cover several cleaning stations) when interacting with one
particular female cleaner on nine different days, based on one hour of observations
on each day.
temporarily deceptive strategy yields short-term energetic advantages. In aquaria,
clients jolt more frequently when interacting with hungry cleaners that when in-
teracting with satiated cleaners (A. S. Grutter, unpublished data). The benefits of
cheating, therefore, seem to be condition dependent, and the client control mech-
anisms like punishment (Bshary & Grutter, 2002a) and partner switching (Bshary
& Schaffer, 2002) only work most of the time.
It even appears that the same individual can switch back and forth between a
cooperative and a biting strategy within seconds. Another biting female, observed
over a six-week period, cheated clients frequently during the spawning period
but client jolt levels remained high after that. Her male partner tolerated her
presence at his cleaning station only during spawning but not thereafter and
chased her off repeatedly. The female spent about equal amounts of time at her
own cleaning station on the other side of the reef patch and on excursions to
the male’s cleaning station. When the female was at the male’s cleaning station,
her resident and choosy clients jolted significantly more frequently than when
interacting with the female at her own cleaning station (residents (n = 8): t = 1.5,
p = 0.021; choosy clients (n = 7): t = 0, p = 0.016; Wilcoxon matched-pair signed-
ranks tests; Fig. 22.2). The variation both within and between cleaners apparent
from these examples means that client image scoring is a profitable strategy to
avoid (temporarily) cheating cleaners.
Do clients attribute image scores to cleaners?
As shown elsewhere (Bshary, 2002), clients use information about the out-
come of ongoing interactions when visiting a cleaning station. To appreciate fully
what is happening, it is important to note that clients usually do not ‘hang out’ at
cleaning stations but visit them only when they seek an inspection by a cleaner.

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
528 R. Bshary & A. D’Souza
Jolts
/100
s
C C
p p
Fig. 22.2. Jolt rates (medians and interquartile ranges) of eight resident species (clients
without choice; n = 8) and seven client species with choice (defined in text; n = 7)
during interactions with one particular female cleaner depending on the location of
the interaction (at her own station (•) or at the male’s station (�); see text for further
details).
Therefore, clients can gather information on how cleaners treat other clients only
when they visit the station themselves and only if a cleaner is busy inspecting
another client while they approach. So visiting clients can base their decision to
invite inspection on current information only if they can observe another client
being inspected. The newly arrived individual can attribute a positive image score
to the cleaner if the current interaction ends without apparent conflict and a
negative image score if the current interaction ends with the client darting off
or chasing the cleaner. If another client is not present when the client arrives,
no current information is available and the image score might be neutral. This is
what the data suggest. If an ongoing interaction ended without apparent conflict,
clients that had arrived during the interaction invited inspection by the cleaner in
almost 100% of observed interactions. In contrast, if the interaction ended with an
apparent conflict, clients hardly ever invited inspection (Bshary, 2002). When no
information about a cleaner’s previous interaction was available, clients invited
inspection with intermediate probability and the actual outcome of the previous
interaction (that was unobserved by the client) had no significant effect. When
clients do not invite inspection, they often exhibit an ambiguous response; they
let the cleaner approach and inspect but do not stop coordinated swimming move-
ments before the interaction starts (they may stop afterwards). They may also flee
from the approaching cleaner. Fleeing most often results in no inspection and fre-
quently happens when clients are approached immediately after an interaction
has ended with a conflict. In contrast, fleeing hardly ever occurs after a positive
interaction had just ended or if the previous interaction had ended a while ago
(Fig. 22.3). These observations are consistent with the statement above that clients
only visit cleaning stations to seek an inspection. The decision to invite inspection
is only altered if they observe a negative interaction.

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 529
P
P N
C
Fig. 22.3. The frequency of fleeing (accelerating away from approaching cleaners) by
resident client species (•) and choosy client species (defined in text, �) arriving at a
cleaning station in four different situations: the previous interaction ended either ≤5 s or ≤ 5 s ago and had positive (without conflict) or negative (with conflict)
outcomes. Values are median and interquartile ranges. The letters b and b′ above the
values for fleeing in the situation where the previous interaction had ended
negatively ≤ 5 s ago indicate a significant difference to the other three situations,
which are not statistically significant between each other, as indicated by using the
same letters a and a′.
Response of ‘normal’ cleaner fish to image-scoring clients
If clients attribute image scores to cleaners, one would expect that clean-
ers adjust their behaviour and cheat current clients less frequently if bystanders
are present than when no bystanders are present. Such audience effects should
be particularly common if bystanders have access to several cleaning stations, as
these species (see above) might not only delay their interaction with the cleaner
but also swim to another cleaning station. Resident bystanders can only delay
their interaction or avoid interactions altogether and remain uncleaned. To look
for such effects, we assumed that all individuals within 50 cm of cleaner–client
interactions at the beginning and at the end of each interaction were able to
collect information about the ongoing interaction and all individuals ≥ 10 cm
total length were potential next clients. We quantified the number of all such
individuals and their species identity for 12 cleaners. For each client species and
cleaner station, we calculated correlations between the frequency of jolts and the
number of bystanders. We analysed four (partly overlapping) classes of bystander;
conspecific, heterospecific, resident species and choosy species. We only calculated
correlations for observations where only one of these classes of bystander was
present. For each client species and bystander category, we compared the number

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
530 R. Bshary & A. D’Souza
No.
spe
cies
C Heterospecifics R C
pp
Fig. 22.4. The influence of the presence (within 50 cm at the beginning and the end of
an interaction) of four bystander classes on the jolt rate of clients during interactions
with cleaners. The histograms show the number of client species for which the
correlation between jolt rate and the number of bystanders of a category was either
negative (black) or positive (white).
of positive and negative correlations and scored a plus if the majority was positive
and a minus if the majority was negative. Thus, we had one data point for each
client species and bystander category and evaluated any significant impacts of by-
stander categories on client jolt rates using sign tests. We did not find a significant
effect of conspecific bystanders on client jolt rates (n = 13; x = 6; NS) while the pres-
ence of heterospecific bystanders had a significantly negative effect (n = 23; x = 6;
p = 0.034; Fig. 22.4). The effect of heterospecific bystanders was mainly owing to
choosy bystanders (n = 17; x = 1; p < 0.001) while resident bystanders did not have
a significant effect on client jolt rates (n = 15; x = 6; NS; Fig. 22.4).
While the data presented above are in line with the hypothesis that client image
scoring influences cleaner fish behaviour, there is an alternative explanation. It
could be that when more clients are present it is easier for cleaners to pick the few
obvious parasites from each of them and the reduction in client jolt rate is a result
of an optimal foraging decision of cleaners rather than caused by bystander image
scoring. In favour of the optimal foraging interpretation, it is known that choosy
clients are, on average, larger than resident clients (Bshary 2001), which could
explain the stronger effect of their presence on the current clients’ jolt rates than
the smaller residents. However, optimal foraging cannot explain our observation
that choosy bystanders have different effects on cleaner fish behaviour, depending
on whether they are the same species as the interacting client or whether they are
a different species. We can explore this effect further by considering only inter-
actions in which individuals of one choosy species, the sergeant major Abudefduf
vaigiensis, were bystanders. We picked 13 cleaning stations for data collection on
the basis that these clients were frequent visitors. Sergeant majors may visit as
single individuals or as large shoals of 20–50 individuals. For 11 out of 12 client
species, we found more negative than positive correlations between the number
of sergeant majors present and client jolt rates (sign test: n = 12; x = 1; p < 0.01).
This result is the opposite of the effects of sergeant major bystanders on the jolt

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 531
1–40Bystanders
p
p
Clie
nt (
jolts
/100
s)
≥ 5
Fig. 22.5. The influence of the number of sergeant major Abudefduf vaigiensis
bystanders present (within 50 cm at the beginning and the end of an interaction) on
the jolt rate of the sergeant major client.
rates of sergeant majors being cleaned: in comparison to interactions in which no
bystanders were present, small numbers of bystanders did not have any detectable
effect and the presence of large numbers led to an increase in jolt rates (Friedman
test: n = 12; χ2 = 9.9; df = 2; p < 0.01; Fig. 22.5).
Response of biting cleaners to image scoring clients
Biting cleaners have more interactions that end with a conflict and clients
approaching their cleaning station more often avoid them than normal cleaners
(Bshary, 2002). As explained above, it seems likely that the latter observation is the
result of client image scoring rather than previous direct experience of clients.
Do biting cleaners still have some means to improve their image? In this respect,
it is important to note that biting cleaners behave very differently from normal
cleaners, not only with respect to jolt rates of large clients but also with respect to
their behaviour towards small resident clients. Biting cleaners often ride above the
small residents’dorsal area and provide tactile stimulation with their pectoral and
pelvic fins. While this behaviour is part of every cleaner’s repertoire, about 50% of
the interactions between biting cleaners and small residents consisted of tactile
stimulation only (Bshary, 2002). Providing tactile stimulation is incompatible with
foraging; hence interactions that consist of tactile stimulation only are clearly
costly to cleaners. Usually, cleaners provide tactile stimulation in response to the
behaviour of the client; for example, manipulating clients that are unwilling to
interact. The manipulation serves to slow down the clients, allowing the cleaners
to forage on the clients’ surface (Bshary & Wurth, 2001). As tactile stimulation of
small residents did not appear to provide the cleaners with any direct benefits from
the recipients, Bshary (2002) proposed that it may serve as a signal to attract image-
scoring clients, which can then be exploited. In line with this argument, it was
found that interactions that consisted of tactile stimulation only were followed

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
532 R. Bshary & A. D’Souza
C
B N
p
Fig. 22.6. Frequencies with which biting and normal cleaners ignored the invitations
for inspection of small resident clients. Values are the median and interquartile
ranges for five biting and eleven normal cleaners. (The p value is derived from
Mann–Whitney U-test.)
by interactions ending with a conflict immediately after a client jolt more often
than expected. It appears that tactile stimulation of small residents is a signal out
of context that attracts image-scoring clients to their own disadvantage (they will
be cheated) and to the cleaners’ advantage, fulfilling the functional definition of
tactical deception (Hauser, 1998).
The presence or absence of bystanders was not noted, so it remains unclear
whether biting cleaners seek small residents in particular when larger clients are
nearby or whether they start such interactions independently of the presence of
bystanders. The latter scenario is more plausible, as larger clients, in particular the
choosy ones, are not willing to queue for inspection (Bshary & Schaffer, 2002) and
would, therefore, swim off despite the cleaner’spositive image. So, cleaners appear
unable to time interactions with small residents for maximal effects. However,
some evidence suggests that tactile stimulation of small residents is part of biting
cleaners’ strategies to improve their image. Cleaner fish sometimes ignore clients
that invite inspection, in particular small resident clients (Bshary & Wurth, 2001).
These clients do not offer a large food source and do not have the option to visit
another cleaner if ignored. When approached by small residents, biting cleaners
ignore them significantly less frequently than normal cleaners (Mann-Whitney
U-test: n = 11; m = 5; U = 2; p = 0.004; Fig. 22.6).
Discussion
We have provided a description of behavioural patterns in interactions
between cleaner fish and client reef fish that emphasizes the importance of the
communication-network framework in understanding the dynamics of coopera-
tive interactions and the occurrence of tactical deception. Cleaning interactions
often occur in the presence of other potential clients of cleaners. These bystanders
eavesdrop on ongoing interactions, and the information that they collect appears

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 533
to be crucial for their decision to invite inspection or to avoid the cleaner. In
response, it appears that normal cleaners reduce cheating frequencies in the pres-
ence of eavesdroppers, in particular if these eavesdroppers have access to several
cleaning stations. Data of this kind are still missing for biting cleaners. Biting clean-
ers frequently engage in costly (or at least non-profit) interactions with small resi-
dents that appear to serve to attract larger image-scoring clients, which can then
be exploited. The results have important implications for theoretical approaches
to indirect reciprocity. Existing models predict that image scoring drives altruistic
behaviour towards fixation (Nowak & Sigmund, 1998; Lotem et al., 1999; Leimar
& Hammerstein, 2001). Cheating individuals can only reinvade an image-scoring
population after genetic drift has led to an increase in non-discriminatory altru-
ists. This scenario does not fit the cleaner fish mutualism very well. Image scoring
of clients mediates cooperative behaviour of cleaners, but this cooperative be-
haviour may be an honest or a deceptive signal. Cheating individual cleaners use
one class of clients for altruistic behaviour to produce a signal that allows them
to exploit, through image scoring, another class of clients. Image scoring thus
works for the receiver of altruistic behaviour but it does not always work for the
eavesdropper.
The major difference between the cleaner fish system and the models might
concern the payoff matrix. While it is assumed in the models that payoffs are the
same in every interaction, payoffs are variable for cleaners. First, an advantage
of cheating clients with access to several cleaning stations rather than resident
clients is that the former just swim off after being cheated, while the latter chase
the cleaner fish around (Bshary & Grutter, 2002a), so the cleaner loses some of the
energy it has just gained. Second, cleaners can probably gain very little from in-
teractions with small clients anyway, no matter whether they cooperate or cheat.
This contrasts with interactions with large clients, which have more parasites but
also more mucus and a larger surface for the cleaner to scrape along with its lower
jaw. This gives the opportunity for cleaners to behave altruistically when payoffs
are low and to be exploitative when payoffs are high, as long as the altruism in-
creases the frequency of high payoff interactions. Image scoring would not persist
if it did not yield a benefit to its performer, but it also provides an opportunity
for individuals to perform altruistic acts in order to gain access to and exploit
image-scoring individuals. Therefore, image scoring in communication networks
may explain both the evolution of altruistic behaviour and the occurrence of
tactical deception. The commonness of dishonest signals that nevertheless still
fool observers has yet to be evaluated. While verbal arguments predicted low fre-
quencies (Dawkins & Krebs, 1978), game theoretic models indicate that this is
not necessarily the case (Johnstone & Grafen, 1993; Szamado, 2000). In particu-
lar, if the benefits of finding a cooperative partner largely outweigh the cost of

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
534 R. Bshary & A. D’Souza
interacting with a cheating partner, tactical deception may occur at quite high
frequencies.
Future work with the cleaner system
Several important points of the cleaner fish mutualism still have to be
clarified. First, we need experimental evidence for both client image scoring and
cleaner fish audience effects. There is increasing evidence that a reduction in client
jolt rates in the presence of bystanders reflects a more cooperative behaviour by
cleaners (Bshary & Grutter, 2002b) and that such behaviour is indeed more altru-
istic. Grutter & Bshary (2003) offered cleaners the choice between equal amounts
of mucus, gnathiid isopods and monogeneans attached to plexiglas plates and
found that cleaners ate mucus more often than parasites, in particular gnathiids.
Assuming that the results reflect the items’quality as a food source, cleaners profit
even more from feeding on mucus when interacting with real clients as mucus is
abundantly spread over the clients’ surface whereas parasites have to be searched
for. In conclusion, while the experiment did not quantify energy intake, it makes
it very plausible that feeding on mucus yields a higher energy gain than feeding
on parasites. Another point that needs to be addressed is the biting cleaners’ be-
haviour with respect to small and large clients. Is it really true that interactions
with small clients generally offer low payoffs compared with interactions with
larger clients, and that the margin between the benefits from cooperation and
cheating increase with client size? Does image scoring of clients indeed inhibit
the cleaners’ tendency to cheat in low-payoff interactions but not in high-payoff
interactions? Currently, no data are available to evaluate these questions. Finally,
one might expect that clients should respond to the biting cleaners’ behaviour
by fine-tuning their image scoring, paying less attention to the outcome of inter-
actions between cleaners and small residents. Pooling of existing data indicate
that this is indeed the case. Invitation for inspection (i.e. spreading the pectoral
fins and stopping coordinated swimming movements) occurred more frequently
if cleaners interacted with choosy clients than with small residents. This prelim-
inary result has to be tested with a larger data set that allows statistical analysis
based on behaviour of individual client species.
Cognitive aspects
While indirect reciprocity and tactical deception were considered to be
a hallmark of human evolution (Nowak & Sigmund, 1998) and primate Machi-
avellian intelligence (Byrne & Whiten, 1988), the data on cleaner–client interac-
tions suggest that, on a purely descriptive level ignoring underlying mechanisms,
these phenomena are more widespread. We have argued that they should occur

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 535
frequently in social networks. These phenomena should occur if it pays to alter
the optimal behaviour in a situation in order to alter one’s image, which will, in
turn, produce benefits during future interactions with bystanders that exceed the
momentary costs. With respect to aggression, a game theoretic model shows that
it may even pay individuals to act spitefully towards a partner (in the sense that
the spiteful act will not lead to any benefits gained from future behaviour of
the recipient) if this spiteful act reduces, for example, the threat of attack from
bystanders (Johnstone & Bshary, 2004; Ch. 10).
With respect to cognition, the data generally support the view of Heyes (1998)
that we need to establish what kind of information animals use for their decision
making to find out what cognitive abilities are involved in a given phenomenon. In-
direct reciprocity and tactical deception may be something ‘smart’in some species
and simple conditioning in others. Cleaners certainly have ideal conditions to de-
velop their behaviour through conditioning. They have more than 2000 interac-
tions per day (Grutter, 1995), making it easy to connect altruistic behaviour with
reward (i.e. the invitation from bystanders to inspect) and cheating with punish-
ment (i.e. evasive actions of bystanders when approached by the cleaner). In the
absence of decisive experiments, it could even be possible that parts of cleaner
fish and client behaviour may be governed by endocrine responses rather than
through learning (Ch. 21). A good candidate for an endocrine-mediated behaviour
might be the good service that cleaners provide to predatory clients: there might
be an innate programme to recognize predators, and the presence of a predator
might trigger a stress response that, in turn, may inhibit cheating behaviour. Al-
ternatively, one could also generalize the Machiavellian intelligence hypothesis
and predict that a complex social network should have similar effects on cognitive
abilities in all species (Byrne & Whiten, 1988). It may turn out that cleaners have
relatively high cognitive abilities, as their large interspecific social network is at
least in part based on individual recognition (Tebbich et al., 2002) and demands
the solving of a variety of problems (Bshary et al., 2002). In this context, it is worth
pointing out that both the biting females which were observed over longer time
periods showed considerable variation in their behaviour, as so did their male part-
ners. One male often prevented his female from interacting with clients while the
other did not (Fisher test: n = 34; p = 0.003; Table 22.1). The preventive male was
also almost significantly more likely to chase his female when their client darted
off after a jolt, while the other male often followed the client to provide tactile
stimulation (Fisher test: n = 12; p = 0.053; Table 22.1). This observation of flexi-
bility of both males and females is important as it was the careful description of
individual-specific strategies in primates that eventually led to a cognitive, rather
than genetic, approach towards behaviour (Strum et al., 1998).

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
536 R. Bshary & A. D’Souza
Table 22.1. The behaviour of two males that were partners of biting
females
Male responses to female biting
Tactile Preventative
stimulation Chasing No obvious chasing of
of clienta femalea reactiona femaleb
First male 5 2 3 1
Second male 0 5 2 16
a Reaction to clients darting off following cheating by female.b Keeping female away from clients.
Summary
In summary, we think that the concept of communication networks has
major implications for our understanding of the evolution and maintenance of
altruistic behaviour, tactical deception and spiteful behaviour. Because of its func-
tional approach, the communication-network framework may help to demystify
phenomena that are often considered to demand high cognitive abilities, opening
the way to focus on the underlying mechanisms and the complexity of information
processing and decision rules in order to illuminate cognitive differences between
species (for a parallel discussion, see Ch. 24). Game theory models should help to
generate testable predictions of the circumstances in which altruism, tactical de-
ception and spiteful behaviour may yield fitness benefits within communication
networks. In particular, it is time to develop cognitive models rather than genetic
models, allowing individuals to process information about their social environ-
ment before making a behavioural decision (see Stephens & Clements (1998) for a
first approach towards cognitive game theory).
Acknowledgements
We thank Peter McGregor for inviting us to write this chapter. We are grateful to the
EEAA in Cairo for the permit to work in the Park and to Alain de Grissac, the Park rangers and Ingo
Riepl for their support at the Park. The study was supported by the Deutsche Forschungsgemein-
schaft (grants BS 2/2-1 to BS 2/2-4) and written while RB was on a Marie Curie Fellowship of the EU.
We want to thank Wolfgang Wickler, Karin Bergmann and Barbara Knauer for additional support.
The chapter was greatly improved by comments from Peter McGregor, Rui Oliveira, Alexandre
Roulin, Sabine Tebbich and an anonymous referee.

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Bibliography 537
References
Alexander, R. D. 1987. The Biology of Moral Systems. New York: Aldine de Gruyter.
Axelrod, R. & Hamilton, W. D. 1981. On the evolution of co-operation. Science, 211,
1390–1396.
Bshary, R. 2001. The cleaner fish market. In: Economics in Nature, ed. R. Noe, J. A. R. A. M.
van Hooff & P. Hammerstein. Cambridge, UK: Cambridge University Press,
pp. 146–172.
2002. Biting cleaner fish use altruism to deceive image scoring clients. Proceedings of
the Royal Society of London, Series B, 269, 2087–2093.
Bshary, R. & Grutter, A. S. 2002a. Experimental evidence that partner choice is the
driving force in the payoff distribution among cooperators or mutualists: the
cleaner fish case. Ecology Letters, 5, 130–136.
2002b. Asymmetric cheating opportunities and partner control in a cleaner fish
mutualism. Animal Behaviour, 63, 547–555.
Bshary, R. & Schaffer, D. 2002. Choosy reef fish select cleaner fish that provide high
service quality. Animal Behaviour, 63, 557–564.
Bshary, R. & Wurth, M. 2001. Cleaner fish Labroides dimidiatus manipulate client reef
fish by providing tactile stimulation. Proceedings of the Royal Society of London, Series
B, 268, 1495–1501.
Bshary, R., Wickler, W. & Fricke, H. 2002. Fish cognition: a primate’s eye view. Animal
Cognition, 5, 1–13.
Byrne, R. W. & Whiten, A. 1988. Machiavellian Intelligence. Oxford: Clarendon Press.
Cote, I. M. 2000. Evolution and ecology of cleaning symbioses in the sea. Oceanography
and Marine Biology Annual Review, 38, 311–355.
Dawkins, R. & Krebs, J. R. 1978. Animal signals: information or manipulation? In:
Behavioural Ecology: An Evolutionary Approach, ed. J. R. Krebs & N. B. Davies. Oxford:
Blackwell, pp. 282–309.
Doutrelant, C., McGregor, P. K. & Oliveira, R. F. 2001. The effect of an audience on
intrasexual communication in male Siamese fighting fish, Betta splendens.
Behavioral Ecology, 12, 283–286.
Dugatkin, L. A. 1997. Cooperation among Animals: A Modern Perspective. Oxford: Oxford
University Press.
Grutter, A. S. 1995. Relationship between cleaning rates and ectoparasite loads in
coral reef fishes. Marine Ecology Progress Series, 118, 51–58.
1996. Parasite removal rates by the cleaner wrasse Labroides dimidiatus. Marine Ecology
Progress Series, 130, 61–70.
1997. Spatio-temporal variation and feeding selectivity in the diet of the cleaner fish
Labroides dimidiatus. Copeia, 1997, 346–355.
Grutter, A. S. & Bshary, R. 2003. Cleaner wrasse prefer client mucus: support for
partner control mechanisms in cleaning interactions. Proceedings of the Royal
Society of London, Series B, Biology Letters Supplement, 2, 242–244.
Hamilton, W. D. 1964. The genetical evolution of social behaviour. Journal of Theoretical
Biology, 7, 1–52.

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
538 R. Bshary & A. D’Souza
Hauser, M. D. 1998. Minding the behaviour of deception. In: Machiavellian Intelligence II,
ed. A. Whiten & D. W. Byrne. Cambridge, UK: Cambridge University Press,
pp. 112–143.
Heyes, C. M. 1998. Theory of mind in non human primates. Behavioral and Brain Sciences,
21, 101–148.
Johnstone, R. A. 2001. Eavesdropping and animal conflict. Proceedings of the National
Academy of Sciences, USA, 98, 9177–9180.
Johnstone, R. A. & Bshary, R. 2004. The evolution of spite through indirect reciprocity.
Proceedings of the Royal Society of London, Series B, 271, 1917–1922.
Johnstone, R. A. & Grafen, A. 1993. Dishonesty and the handicap principle. Animal
Behaviour, 46, 759–764.
Leimar, O. & Hammerstein, P. 2001. Evolution of cooperation through indirect
reciprocity. Proceedings of the Royal Society of London, Series B, 268, 745–753.
Losey, G. C., Grutter, A. S., Rosenquist, G., Mahon, J. L. & Zamzow, J. P. 1999. Cleaning
symbiosis: a review. In: Behaviour and Conservation of Littoral Fishes, ed. V. C. Almada,
R. F. Oliveira & E. J. Goncalves. Lisbon: Instituto Superior de Psicologia Aplicada,
pp. 379–395.
Lotem, A., Fishman, M. A. & Stone, L. 1999. Evolution of cooperation between
individuals. Nature, 400, 226–227.
2003. From reciprocity to unconditional altruism through signaling benefit.
Proceedings of the Royal Society of London, Series B, 270, 199–205.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society of London, Series B, 340, 237–244.
Munn, C. A. 1986. Birds that ‘cry wolf’. Nature, 319, 143–145.
Noe, R. 2001. Biological markets: partner choice as the driving force behind the
evolution of cooperation. In: Economics in Nature, ed. R. Noe, J. A. R. A. M. van Hooff
& P. Hammerstein. Cambridge, UK: Cambridge University Press, pp. 92–118.
Noe, R., van Schaik, C. P. & van Hooff, J. A. R. A. M. 1991. The market effect: an
explanation for pay-off asymmetries among collaborating animals. Ethology, 87,
97–118.
Nowak, M. A. & Sigmund, K. 1998. Evolution of indirect reciprocity by image scoring.
Nature, 393, 573–577.
Premack, D. & Woodruff, G. 1978. Does the chimpanzee have a theory of mind?
Behavioral and Brain Sciences, 4, 515–526.
Randall, J. E. 1958. A review of labrid fish genus Labroides, with descriptions of two
new species and notes on ecology. Pacific Scientist, 12, 327–347.
Roberts, G. 1998. Competitive altruism: from reciprocity to the handicap principle.
Proceedings of the Royal Society of London, Series B, 265, 427–431.
Robertson, D. R. 1972. Social control of sex reversal in a coral-reef fish. Science, 177,
1007–1009.
Stephens, D. W. & Clements, K. C. 1998. Game theory and learning. In: Game theory and
Animal Behaviour, ed. L. A. Dugatkin & H. K. Reeve. Oxford: Oxford University Press,
pp. 239–260.

P1: JZZ/... P2: JZZ/...0521823617c22.xml CU1917B/McGregor 0 521 582361 7 April 11, 2005 19:30
Indirect reciprocity in interactions in fish 539
Strum, S. C., Forster, D. & Hutchins, E. 1998. Why machiavellian intelligence may not
be machiavellian. In: Machiavellian Intelligence II, ed. A. Whiten & D. W. Byrne.
Cambridge, UK: Cambridge University Press, pp. 50–85.
Szamado, S. 2000. Cheating as a mixed strategy in a simple model of aggressive
communication. Animal Behaviour, 59, 221–230.
Tebbich, S., Bshary, R. & Grutter, A. S. 2002. Cleaner fish Labroides dimidiatus recognise
familiar clients. Animal Cognition, 5, 139–145.
Trivers, R. L. 1971. The evolution of reciprocal altruism. Quarterly Review of Biology, 46,
35–57.
Wedekind, C. & Milinski, M. 2000. Cooperation through image scoring in humans.
Science, 288, 850–852.
Zahavi, A. 1995. Altruism as a handicap : the limitations of kin selection and
reciprocity. Journal of Avian Biology, 26, 1–3.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
23
Fish semiochemicals and the evolutionof communication networks
b r i a n d . w i s e n d e n 1 & no r m a n e . s t ac e y 2
1Minnesota State University, Moorhead, USA2University of Alberta, Edmonton, Canada
Introduction
The concept that animals typically communicate in networks (involving
at least one signaller and more than one receiver) derives from the active space of
signals and social spacing of conspecific and heterospecific receivers (McGregor
& Peake, 2000; Ch. 1). The ecological and evolutionary consequences of such net-
works have been explored most thoroughly for visual (e.g. Ch. 12) and acoustic
signals (e.g. Otter et al., 1999; Ch. 2), although it is clear that chemical signalling
also can involve networks (Chs. 11 and 16). Research on aquatic communication
networks has so far been limited to the context of visual and acoustic signalling
(e.g. Oliveira et al., 1998; Chs. 5 and 18). Semiochemicals (i.e. chemicals that trans-
fer information within and/or between species) exert important and diverse ef-
fects on the behaviour and physiology of aquatic animals (Liley, 1982; Chivers &
Smith, 1998; Kats & Dill, 1998; Sorensen & Stacey, 1999; Stacey & Sorensen, 2002;
Wisenden, 2003). Studies of two key aspects of fish chemical ecology (predator–
prey and reproductive interactions) have revealed great differences in the sources
and nature of the semiochemicals released, their active spaces and their biologi-
cal functions. These studies also provide sufficient information to assess, in fish,
the existence and function of semiochemical information networks, which we de-
fine more fully below as a general category of network that includes not only
communication networks employing specialized signals but also other networks
employing unspecialized cues. Here, we briefly describe well-studied examples of
predator–prey and reproductive semiochemicals to explore the applicability of cur-
rent communication network theory to aquatic chemical information networks
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
540

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 541
and consider how their function and evolution might differ from those employing
other sensory modalities and information-transmission media.
Research on intra- and interspecific transfer of chemical information in ter-
restrial species has generated a bewildering terminology related not only to the
nature, actions and functions of the chemicals but also to the concept of com-
munication (Hauser, 1996; Beauchamp, 2000; Hasson, 2000; McClintock, 2002).
Although we do not presume to clarify such a complex terminological problem in
this brief paper, it is imperative that we begin by clearly defining key terms, par-
ticularly as much of our subject matter appears to be distinct from that typically
discussed in the context of communication networks.
Definition of terminology
Semiochemicals include allomones and pheromones that, respectively, trans-
mit interspecific and intraspecific information. We will consider fish pheromones
and allomones involved in predator–prey interactions (p. 544) separately from
those involved in reproduction (p. 549). We define a pheromone as ‘a substance, or
mixture of substances, which is released by an individual and that evokes a specific
and adaptive response in conspecifics’ (Stacey & Sorensen, 2002). This definition is
more inclusive than the original definition of pheromone (Karlson & Luscher, 1959)
because, for reasons explained below, it omits any requirement that pheromones
be involved in communication. We use the terms releaser and primer not to classify
pheromones but only to describe their rapid behavioural and slower physiological
actions, respectively, for the simple reason that ‘it is quite possible for the same
pheromone to be both a releaser and a primer’ (Wilson & Bossert, 1963), as is the
case for sex pheromones of goldfish Carassius auratus (p. 549).
Central to our terminological schema is the concept (Sorensen & Stacey, 1999;
Stacey & Sorensen, 2002) that evolution of chemical communication progresses
through a series of three functional phases: ancestral, spying and communication. In
the ancestral phase, individuals (originators) release a chemical(s) that does not in-
fluence receivers (Fig. 23.1). This primitive, prepheromonal condition progresses
to spying if receivers evolve the ability to detect and respond adaptively to the
originator’s released chemical(s), now termed a pheromonal or allomonal cue(s).
In spying, originators may or may not benefit from the receiver’s response but,
importantly, remain in an unspecialized state with respect to production and re-
lease of pheromonal cues. Finally, spying progresses to communication if there is a
mechanism for receiver responses to select for specialization in production and/or
release of the detected cue(s), now termed a pheromonal or allomonal signal(s) and
released by a signaller. Signals evolve through natural selection because of fitness
benefits the signaller receives by manipulating the behaviour or physiology of
receivers. In many cases, signal senders and receivers form a mutualism in which

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
542 B. D. Wisenden & N. E. Stacey
Fig. 23.1. The evolution of communication from the ancestral state, where the
originator does not possess specializations for synthesis and release of
semiochemicals, to spying, where receivers possess specializations for detecting
semiochemicals but originators do not possess specializations, to true
communication, in which both originator (now signaller) and receiver possess
specializations for semiochemical exchange of information.
signals coevolve with the sensory biology of receivers. Once established, however,
mutually beneficial communicative relationships could be susceptible to deceit-
ful signal manipulations by signallers, which reduce the receiver fitness, as seen
in visual, acoustic and chemical signals (e.g. Lloyd, 1965; Møller, 1989; Paxton &
Tengo, 2001).
The ancestral state applies to released chemicals not currently functioning in
spying or communication. We restrict the terms signal and communication to
those situations in which there is clear evidence for signal specialization, such
as tissue hypertrophy or discrete structures for signal production; in contrast to
the situation in terrestrial insects and vertebrates, where pheromone-producing

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 543
glandular structures are common, such specializations in fish appear to be the ex-
ception rather than the rule (e.g. Laumen et al., 1974; Colombo et al., 1980; van den
Hurk & Resink, 1992). Consequently, we regard the great majority of fish predator–
prey and reproductive semiochemicals to function in spying, which is, in effect,
the default condition for cases where there is no evidence for specialization in
semiochemical production or release and/or where the social system apparently
precludes selection for signal specialization. It is to be expected that future re-
search may reveal some putative examples of spying to be true communication
because they involve previously undetected signal specialization.
Although we believe the distinction between cues and signals is fundamental
to an understanding of the function and evolution of semiochemical systems,
fish olfactory systems evidently do not make this distinction and process cues
and signals through similar mechanisms, which differ considerably from those
processing food odours (amino acids). Therefore, in comparison with food odours,
semiochemical cues and signals are detected by more sensitive and specific ol-
factory receptor mechanisms and generate neuronal activity that is processed in
distinct arrays (glomeruli) in the olfactory bulbs, is conducted to the brain by
distinct nerve bundlets (olfactory tracts), and is projected to distinct brain areas
(Sorensen et al., 1998; Hamdani et al., 2000, 2001; Brown et al., 2001; Stacey &
Sorensen, 2002). The distinction between chemical spying (via cues) and chemical
communication (via signals) highlights a dichotomy relevant not only to our un-
derstanding of semiochemicals (the functional relationships among originators,
signallers and receivers; evolutionary origins of species-specific cues and signals:
Sorensen & Stacey (1999)) but also to the concept of communication networks
(McGregor & Peake, 2000). In particular, first, how might networks involving com-
munication differ from those involving spying and, second, can the concept of
eavesdropping, defined as ‘extracting information from signalling interactions
between others’ (McGregor and Peake, 2000) be applied to information networks
that do not involve signalling?
Transfer of chemical information
Propagation of chemical information differs fundamentally from propa-
gation of visual and acoustic information. In general, visual and acoustic sig-
nals are propagated with predictable speed and direction, and they generate pre-
dictable active spaces throughout which much of the temporal information con-
tained in the signal’s initial pattern can be retained. In contrast, semiochemi-
cals are released into fluid media (air or water) in which local variation in flow
typically creates turbulent odour plumes, which not only distort or destroy tem-
poral pattern but also make the position, shape and size of the chemical’s active

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
544 B. D. Wisenden & N. E. Stacey
space highly unpredictable (Weissburg, 2000). Moreover, semiochemicals can per-
sist in the environment for considerable time (e.g. Wisenden et al., 1995; Sorensen
et al., 2000; Polkinghorne et al., 2001), and thus can become disassociated from
originators/signallers either when currents carry away a transiently released
semiochemical or when the originator/signaller moves to a new location. Although
there is considerable information on the mechanisms by which some invertebrates
(e.g. crustaceans and moths) navigate in physically characterized odour plumes,
this complex issue is poorly understood in fish (Vickers, 2000). Finally, it is im-
portant to realise that semiochemical function in water also can be influenced
by additional solutes that affect olfactory response, such as heavy metals (Hansen
et al., 1999) and organics (Hubbard et al., 2002).
The olfactory system is similar to other sensory systems in being functionally
delimited by the sensitivity and specificity of its sensory neurons, but it differs
in the nature of the information it processes. Visual and acoustic systems process
linear arrays of light and sound frequencies in spectra common to many species,
particularly if they are related; olfactory systems process information from odor-
ants that cannot be arranged in a linear dimension by means of receptors that
are sensitive to one or a few chemicals. These differences have two important im-
plications for the nature and evolution of semiochemicals. First, whereas visual
and acoustic signals usually encode species-typical information in frequency and
temporal pattern, semiochemicals encode this information through the presence,
absence or ratio of specific odorants. Second, whereas visual and acoustic signals
are potentially detectable by all individuals and species sensitive to the emitted
spectra, semiochemical detection will be restricted to individuals with olfactory
receptors sensitive to the odorant(s). Thus, large differences in semiochemical
production and detection can occur with only small changes either in chemical
metabolism and release or in olfactory receptor specificity.
Assessment of predation risk
Natural selection strongly promotes attendance to cues that reduce the
probability of predation. Consequently, temporal and spatial variation in pre-
dation risk governs much of animal behaviour. Chemicals reliably inform about
predation risk because they are carried well in water, persist for ecologically appro-
priate amounts of time, transmit information through turbid or highly structured
habitat and darkness, and provide types of information not contained in visual
and acoustic modalities.
To apply communication-network theory to chemical assessment of predation
risk, we must first determine whether use of chemical information for the pur-
poses of risk assessment involves spying (via cues) or communication (via signals).

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 545
Fig. 23.2. Semiochemicals associated with predation. Predation escalates from initial
detection (top) to attack (middle) and, finally, ingestion (bottom). Chemical cues (solid
arrows) released at each stage inform nearby prey (conspecific and heterospecific) and
predators of the presence and extent of the interaction between predator and prey.
Known and hypothesized benefits are indicated by dotted arrows.
We conclude in the discussion below that, despite a plethora of semiochemically
mediated mechanisms for predator avoidance, evidence for signals is not com-
pelling. Although these information networks may not be communication networks
per se, there is evolutionary opportunity for communication networks and eaves-
dropping to evolve because receivers have evolved the ability to detect and respond
to many types of semiochemical (see below).
Chemicals correlated with predation
The literature concerning chemicals linked to predation has been re-
viewed elsewhere (Smith, 1992; Chivers & Smith, 1998; Kats & Dill, 1998; Wisenden,
2000, 2003; Chivers & Mirza, 2001) and only a brief overview will be presented here.
Several classes of chemical compound inform prey about predation risk. Gener-
ally, these cues are released passively before, during and after a predation event
(Fig. 23.2). Before an attack is initiated, prey can detect and respond to three types
of chemical cues: (a) odour of disturbed (startled but uninjured) prey (Chivers &
Smith, 1998; Wisenden, 2003), (b) species-specific kairomones (a predator’s natural
odour) (Kats & Dill, 1998) and (c) injury-released alarm cues of prey that leak from

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
546 B. D. Wisenden & N. E. Stacey
the gut of the predator (Chivers & Mirza, 2001). When a predator attacks and in-
jures a prey organism, damaged prey tissues release chemical compounds that are
released only in this context; consequently, these cues reliably indicate risk and
elicit intense anti-predator behaviour (Chivers & Smith, 1998). These are alarm
cues. Most aquatic taxa exhibit anti-predator behaviour in response to alarm cues
(Chivers & Smith, 1998; Wisenden, 2003) in ways that reduce the probability of
predation (Hews, 1988; Mathis & Smith, 1993; Wisenden et al., 1999; Gazdewich &
Chivers, 2002). Ingested prey release alarm cues, or their metabolites, from the gut
of their predators (Chivers & Mirza, 2001). The ecological reality is undoubtedly
more complex than the interactions depicted in Fig. 23.2. Additional interactions
arise from variation in (a) diet breadth and overlap among predators, (b) relative
threat from each predator species over time and space, (c) interacting ontogenies
of prey and predator species, and (d) learned behavioural responses to correlates
of alarm cues.
The vast majority of chemical information used by aquatic prey to assess preda-
tion risk appears to be opportunistic use of chemical information mediated by un-
specialized chemical cues. This information is of great fitness benefit to receivers,
but receiver response, with one notable exception discussed below, generally
has not been shown to accrue benefit to the originator/signaller. Chemically me-
diated predation risk might be described most parsimoniously as an information
network, where a suite of prey species spy on the foraging activities of a suite of
predator species.
Ostariophysan alarm substance cells
For passively released chemical cues to qualify as signals, specializations
for their synthesis and/or release must occur that plausibly have been selected
for by benefits accruing to the originator/signaller. This condition appears to be
met in fishes of the superorder Ostariophysi (minnows, tetras, catfishes, suckers
and sundry others). This large group of vertebrates (> 5500 species) makes up ap-
proximately 27% of the global ichthyofauna and 64% of all freshwater fish species
(Nelson, 1994). In addition to successful occupation of a diverse array of habitats,
they are often the numerically dominant vertebrates in aquatic ecosystems.
Ostariophysans possess specialized epidermal cells that contain a potent alarm
chemical(s), termed schreckstoff or alarm substance (von Frisch, 1941; Pfeiffer, 1977;
Smith, 1992); this appears to activate components of the olfactory system that also
are activated by sex pheromones (Hamdani et al., 2000). It is not known how much
skin area is typically damaged during a predatory attack, but homogenates of
1 cm2 skin can create active spaces of 10 000 litres (zebrafish Danio rerio; Gandolfi
et al., 1968) to 58 000 litres (fathead minnow Pimephales promelas; Lawrence &
Smith, 1989), equivalent to spheres 2.6–4.8 m in diameter. The active ingredient in

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 547
ostariophysan alarm cells is likely, at least in part, to be hypoxanthine 3N-oxide,
a compound first isolated from European minnows Phoxinus phoxinus (Argentini,
1976; after Smith, 1999). Subsequent work demonstrated that hypoxanthine 3N-
oxide elicits anti-predator behaviour from a characin (Pfeiffer et al., 1985) and from
fathead minnows at concentrations as low as 0.4 nmol/l (Brown et al., 2001). How-
ever, efforts to detect hypoxanthine 3N-oxide in fathead minnow skin with high
performance liquid chromatography have not been successful (Smith, 1999) and
fractionation of skin extract indicates the biologically active component is found
with the polypeptides with molecular weights greater than 1100 (Kasumyan &
Ponomarev, 1987) rather than with the small molecules such as hypoxanthine 3N-
oxide. It is possible that hypoxanthine 3N-oxide is associated with protein while
within skin cells and remains associated with protein once released. Heat-treated
skin extract of fathead minnows loses 70% of its protein and its ability to elicit
alarm (N. L. Korpi, L. D. Louisiana, J. J. Provost & B. D. Wisenden, unpublished data).
A protein–hypoxanthine association would be consistent with cross-species alarm
reactions that decline with phylogenetic distance (Schutz, 1956). Whatever the
active ingredient(s) of alarm cue might be, their biological potency (Lawrence &
Smith, 1989; Brown et al., 2001) suggests selection for olfactory sensitivity similar
to that seen with sex pheromones (Stacey & Sorensen, 2002).
Is ostariophysan alarm substance a passively released cue or a specialized sig-
nal? Although the epidermal cells appear to be structures specialized for infor-
mation transfer of alarm, selection for signal specialization via benefits to the
originator (i.e. the individual that released the substance) is not immediately ap-
parent. There has been much speculation over the historical and current selection
benefits to individuals that invest in these cells (Smith, 1992, 1997; Williams,
1992; Magurran et al., 1996; Henderson et al., 1997). Smith (1992) summarized 16
hypotheses by which signallers may benefit from alarm signalling. One of these,
attraction of secondary predators (Fig. 23.2), has empirical support. Laboratory
and field experiments have demonstrated that predators are attracted to minnow
skin extract containing alarm substance cells over minnow skin lacking alarm
substance cells or skin extract from non–ostariophysan species (Mathis et al.,
1995; Wisenden & Thiel, 2002). Interruption of a predation event by the arrival of
a second predator allows prey an opportunity to escape (Chivers et al., 1996), a ben-
efit that elevates passively released cues to signal status. In this context, however,
this signal is not an alarm signal, but an attractant signal because the signaller
benefits because the from the responses of secondary predators, rather than from
responses of conspecific and heterospecific members of the prey community.
Is this a communication network? Are members of the prey community eaves-
droppers on the predator-attractant signal? Assessment of predation risk via chem-
ical cues does not depend on receiver (secondary predators) response (i.e. not social

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
548 B. D. Wisenden & N. E. Stacey
eavesdropping) but interceptive eavesdropping (defined in Ch. 2) on this attrac-
tant signal provides highly salient temporal and contextual information about
predation risk. Therefore, from the perspective of the general non-ostariophysan
prey community, detection of injury-released chemical compounds may be con-
sidered as (interceptive) spying; however for the ostariophysan fishes, spying on
the predator-attractant signal might best be considered as a case of interceptive
eavesdropping (Ch. 2).
Evolutionary opportunities for communication networking
Several lines of evidence suggest potential for communication networks
in the ostariophysan alarm cue system. First, fishes frequently survive predatory
attacks (Smith & Lemly, 1986). An originator/signaller that survives an attack may
benefit from group behavioural responses of the prey community (Smith, 1992; Fig.
23.2) increased shoal cohesion and dashing or skittering behaviour that confuse
predators and reduce attack efficiency. Second, minnows associate alarm cues
with correlates of predation such as predator appearance and odour (reviewed
by Chivers & Smith, 1998). Alarm cues enable conspecifics and heterospecifics
to acquire predator recognition after a single simultaneous or non-simultaneous
encounter with a novel indicator of risk (Suboski, 1990; Suboski et al., 1990; Chivers
& Smith, 1994; Hall & Suboski, 1995; Korpi & Wisenden, 2001). Therefore, a second
benefit to the signaller could be providing shoalmates with an opportunity to learn
predator identity, as a shoalmate trained in this way may detect that predator in
the future and alert the signaller. If a shoal contains individuals related to the
signaller, then a third benefit might accrue to the signaller’s inclusive fitness
through kin selection.
In this context, eavesdropping on signaller–group communication in the os-
tariophysan system could occur when minnows observe the visual stimulus of
anti-predator behaviour of an alarmed shoal (Verheijen, 1956; Magurran, 1989;
Suboski et al., 1990; Brown et al., 1999).
Although evolutionary ecologists have focused on cells producing alarm sub-
stances in ostariophysans, these fishes are not unique in possessing specialized
epidermal cells (Smith, 1992). The epidermal layer of freshwater perch, wall-
eye and darters (superorder Acanthopterygii, order Perciformes, family Percidae:
Smith, 1979, 1982; Wisenden, 2003), Australian bullies (order Perciformes, family
Eleotridae: Kristensen & Closs, 2004) and poeciliids (superorder Acanthopterygii,
Order Cyprinodontiformes, family Poeciliidae: Bryant, 1987) all possess epidermal
club cells with similar histological properties. The tropical marine and freshwater
fishes in the Gobiidae (order Perciformes, 1875 species) possess epidermal vacuo-
late cells but have an inconsistent behavioural response to skin extract (Smith,

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 549
1992). This leaves open the possibility that analogous communication networks
for assessment of predation risk occur among other fish taxa.
In summary, it is parsimonious, based on current knowledge of chemical alarm
cues, to conclude that for aquatic taxa, including most fishes, chemically mediated
risk assessment does not constitute a true communication network because it is
not based on specialized signals. However, the ostariophysan alarm semiochemical
system appears to be a good candidate for an incipient communication system (see
p. 558). Future research may reveal signaller–group communication of alarm and
potentially uncover communication networks and eavesdropping.
Sex pheromones in information networks
In addition to alarm responses discussed in the previous section,
pheromonal cues and signals of teleost fish influence many diverse non-
reproductive (migration, parent–young interactions, schooling and related social
behaviours: Liley, 1982) and reproductive (Stacey et al., 1986; Stacey & Sorensen,
2002) phenomena. Best understood are those cases (the great majority being re-
productive) in which chemical identification has allowed study of pheromone
production, detection and biological effects under controlled and repeatable
conditions. Since Colombo et al. (1980) first proposed that the male black goby,
Gobius niger, releases a conjugated steroid (etiocholanolone glucuronide) to func-
tion as a pheromone that attracts the female to his nest for spawning, many studies
have reported putative pheromonal roles for steroid and prostaglandin hormones,
and their precursors and metabolites (hereafter termed hormonal pheromones) in a
variety of fish (reviewed by Sorensen & Stacey, 1999; Stacey & Sorensen, 2002).
Indeed, we expect the use of hormonal pheromones might be universal among
fish, given that information-rich hormones and hormonal metabolites are neces-
sarily released into the same water medium bathing the olfactory systems of con-
specifics. Here, we briefly discuss two species in which identification of distinctly
different reproductive pheromones has led to an understanding of pheromone
function germane to concepts of chemically mediated information networks.
Goldfish
The hormonal pheromones of goldfish are currently the best under-
stood of any fish and have recently been reviewed in detail (Sorensen & Stacey,
1999; Kobayashi et al., 2002; Stacey & Sorensen, 2002); therefore, we provide a
brief summary before considering aspects that appear directly related to con-
cepts of semiochemical information networks. Goldfish live in mixed-sex, appar-
ently unstructured, groups, undergoing gonadal growth during the winter and
spawning a number of times in spring and summer. At ovulation, which occurs

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fig. 23.3. Nature and actions of goldfish hormonal pheromones released by
periovulatory females (see Stacey & Sorensen (2002) for additional details and original
sources). (a) Female periovulatory events. An afternoon surge of pituitary (P)
gonadotrophin II (GTH-II) release induces follicular synthesis of
17α,20ß-dihydroxy-4-pregnen-3-one (17,20ß-P), which induces final maturation
(completion of arrested meiosis) in mature oocytes. When ovulation occurs
approximately 12 hours later, oocytes in the oviduct stimulate synthesis of
prostaglandin F2α (PGF2α), which remains at high concentrations in the blood until
ovulated oocytes are shed. (b) Preovulatory pheromone. During the GTH-II surge,
females release a changing mixture of three steroids: 17,20ß-P and androstenedione
(AD), which are released together across the gills, and a sulphated 17,20ß-P metabolite
(17,20ß-P-S), which is released in urine pulses. Peak release of AD (which inhibits
endocrine response to 17,20ß-P) occurs early in the GTH-II surge, followed by peaks of
17,20ß-P and 17,20ß-P-S release. The preovulatory steroid acts on specific and sensitive
(picomolar detection threshold) olfactory receptors, both inducing male behavioural
responses and, by the time of ovulation, increasing the quantity and quality of sperm
550

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 551
near dawn, groups of males vigorously compete for spawning access as females
repeatedly enter aquatic vegetation to oviposit adhesive, undefended eggs over
a period of several hours. In such a promiscuous mating system, where male
reproductive success likely depends only on the number of eggs fertilized, we
believe sperm competition has been a major selective force in the evolution of
male reproductive tactics.
The cascade of events leading to spawning begins when exogenous factors (in-
creased water temperature and aquatic vegetation) trigger an afternoon surge
release of pituitary gonadotrophin II (GTH-II), which stimulates follicular syn-
thesis of the oocyte maturation-inducing steroid 17α,20ß-dihydroxy-4-pregnen-
3-one (17,20ß-P) (Fig. 23.3a). Ovulation occurs approximately 12 hours later; at
which point females become sexually active for the several hours that eggs in the
oviduct stimulate synthesis of prostaglandin F2α (PGF2α), a behavioural hormone
(Fig. 23.3c). During the approximately 15 hours between the onset of the
GTH-II surge and completion of spawning, females sequentially release a preovula-
tory steroid pheromone (Fig. 23.3b) and a postovulatory prostaglandin pheromone
(Fig. 23.3c), which dramatically affect male physiology and behaviour.
The preovulatory steroid pheromone (Fig. 23.3b) is a dynamic mixture in which
the primary components appear to be 17,20ß-P, its sulphated metabolite (17,20ß-
P-S) and androstenedione (a testosterone precursor). Although the nature and ac-
tions of the preovulatory pheromone are complex (Stacey & Sorensen, 2002), it
induces in males both releaser effects on socio-sexual behaviours (e.g. Poling et al.,
2001) and a dramatic primer effect: a rapid increase in blood GTH-II that increases
both the quantity and quality of releasable stores of milt (sperm and seminal
fluids) in the sperm ducts prior to ovulation and spawning (e.g. Zheng et al., 1997).
At ovulation, females terminate release of the preovulatory steroid pheromone
and begin to release the postovulatory prostaglandin pheromone (PGF2α and its
more potent metabolite 15-keto-PGF2α) (Fig. 23.3c). The prostaglandin pheromone
not only triggers male courtship and attracts the male to the ovulated female
(anosmic males do not spawn) but also activates non-endocrine and endocrine
Fig. 23.3 (cont.) stores in the ducts (inducing GTH-II release, which stimulates
testicular 17,20ß-P synthesis). (c) Postovulatory pheromone. Entry of ovulated oocytes
to the oviduct stimulates synthesis of PGF2α, which acts in the brain (B) to stimulate
female sexual behaviours. PGF2α and its more potent metabolite 15-keto-PGF2α are
released in urinary pulses and act on olfactory receptors to trigger male sexual
behaviours. Sexual interactions then stimulate movement of sperm to the ducts by
two mechanisms: an endocrine mechanism distinct from that mediating testicular
response to the preovulatory pheromone; and a rapid and apparently non-endocrine
mechanism that begins to increase sperm stores within 15 minutes.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
552 B. D. Wisenden & N. E. Stacey
mechanisms (different from those mediating responses to the preovulatory
pheromone) that further increase the volume of releasable milt.
In summary, male goldfish first increase their potential fertility through en-
docrine responses to reliable chemical indicators of imminent ovulation (17,20ß-P
and 17,20ß-P-S) and then use reliable indicators that ovulation has occurred (PGF2α
and 15-keto-PGF2α) to locate the female and maintain sperm stores. For a number
of reasons (Stacey & Sorensen, 2002), most notably a lack of evidence for specialized
pheromone production and release, we regard these components of the goldfish
hormonal pheromone system as an example of male spying on female chemical
cues. Although it is difficult to exclude the possibility that domestication has in-
fluenced the goldfish pheromone system, it appears remarkably similar to those
of the closely related Crucian carp Carassius carassius and common carp Cyprinus
carpio (Irvine & Sorensen, 1993; Stacey et al., 1994; Bjerselius et al., 1995). Further-
more, it is likely that other cyprinids (Family Cyprinidae; > 2000 species) possess
similar pheromone systems given that olfactory detection of 17,20ß-P-like steroids
and prostaglandins is widespread among this taxon (Stacey & Sorensen, 2002).
The effects of goldfish pheromones described above have been studied in
the context of dyadic interactions between female originators of hormonal
pheromone cues and their male receivers (Fig. 23.3b,c). However, given the prox-
imity of individuals in aggregations, and the size of pheromonal active spaces
estimated from release rates and olfactory detection threshold (Sorensen et al.,
2000), it is obvious that these ovulatory cues normally operate in an information
network, where a female’s preovulatory steroids can potentially be detected by
many males and her postovulatory prostaglandins are the proximate trigger pro-
moting sperm competition at spawning. Moreover, the network activated by the
preovulatory pheromone evidently includes not only the ovulatory female and her
potential spawning partners but also additional females and males not directly
exposed to her preovulatory cues (Fig 23.4.).
The evidence for female interactions is based on the finding that low concen-
trations of water-borne 17,20ß-P induced ovulation in goldfish (Kobayashi et al.,
2002), suggesting a mechanism for the ovulatory synchrony observed in the field
and laboratory. The female benefit(s) of ovulatory synchrony is not known but
may involve predator swamping, amplification of preovulatory cues that stimu-
late male fertility, or (perhaps counter-intuitively) reduction of male to female
ratios at spawning (high ratios can result both in ‘forced’ egg release away from
suitable spawning substrate and skin damage through abrasion by the male’s
breeding tubercles or ‘pearl organs’).
Interactions among males appear more complex because they both decrease
(Fig. 23.4.a) and increase sperm stores in response to unidentified cues from other
males (Stacey et al., 2001; Fraser & Stacey, 2002). For example, males isolated from

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
m
fm
m
f
2
1
2
Tem
pera
ture
+su
bstr
ate
Fii
i
ii
(a)
(b)
iii
3
?
?
Fig.
23.4
.N
etw
ork
acti
vati
onby
ph
erom
ones
.(a)
Inth
ep
rese
nce
ofm
atu
refe
mal
esth
atar
en
otov
ula
tory
(f),
mat
ure
mal
es
rem
ain
ina
‘bas
al’e
nd
ocri
ne
stat
e(m
)an
dre
leas
eu
nkn
own
cues
that
sup
pre
sssp
erm
stor
es.(
b)If
exog
enou
sst
imu
li(i
ncr
ease
d
wat
erte
mp
erat
ure
and
aqu
atic
vege
tati
on)i
nd
uce
anov
ula
tory
gon
adot
rop
hin
IIsu
rge
inon
efe
mal
e(s
had
edgr
ey),
rele
ase
of
the
pre
ovu
lato
ryp
her
omon
eca
nd
irec
tly
stim
ula
teen
doc
rin
ech
ange
sin
one
mal
e(1
)th
atin
crea
seh
issp
erm
stor
e;h
eth
an
rele
ases
un
know
ncu
esin
du
cin
gsi
mil
arch
ange
sin
add
itio
nal
mal
es(2
).In
add
itio
n,t
he
pre
ovu
lato
ryp
her
omon
eca
nin
du
ce
ovu
lati
onin
add
itio
nal
fem
ales
(F),
wh
ose
pre
ovu
lato
ryp
her
omon
ere
leas
ew
ill
acti
vate
add
itio
nal
net
wor
ksof
dir
ectl
y(i
)an
d
ind
irec
tly
(ii)
stim
ula
ted
mal
es.I
tis
not
know
nif
mal
es(2
and
ii)t
hat
rece
ive
ind
irec
tin
form
atio
nth
atov
ula
tion
isim
min
ent
intu
rnin
flu
ence
add
itio
nal
mal
es(3
and
iii)
.
553

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
554 B. D. Wisenden & N. E. Stacey
a male group dramatically increase sperm stores within 24 hours, indicating they
normally suppress their potential for milt production in response to an inhibitory
male cue(s). However, if sperm stores of one of a group of males are increased (either
by gonadotrophin injection or exposure to 17,20ß-P), untreated males in the group
also increase their stores. Therefore, it appears that, in the absence of cues from
preovulatory or ovulated females, a mature male goldfish is both originator and
receiver of unknown cues that suppress sperm stores in other males by maintain-
ing basal GTH-II and steroids (Fig. 23.4.a). This stable, negative-feedback situation is
rapidly and transiently perturbed, however, when exogenous stimuli trigger a pre-
ovulatory GTH-II surge in females, resulting in release of the preovulatory steroid
pheromone. Males and non-ovulatory females encountering this stimulatory cue
in turn increase their GTH-II, amplifying and disseminating the original cue(s) and
promoting synchronous final maturation (ovulation and increased sperm stores)
of individuals within the network (Fig. 23.4.b).
Numerous unresolved questions make it difficult to compare the complex re-
productive interactions of goldfish with the classical visual and acoustic commu-
nication networks that have been studied in terrestrial species. Perhaps the key
issue is whether the pheromonal interactions known among goldfish involve only
responses of receivers to unspecialized cues, or whether some are mediated by
specialized signals. There is no evidence that female preovulatory and postovu-
latory pheromones are specialized signals to males or other females (Sorensen
& Stacey, 1999; Stacey & Sorensen, 2002). Nor is it obvious how the male’s adap-
tive endocrine–testicular response to female preovulatory cues (Fig. 23.3b) would
also be shaped by selection to include the release of a specialized signal that ev-
idently increases the fertility of his competitors. Indeed, it seems more probable
that the indirect responses of males (2 and ii in Fig. 23.4.b) to female preovula-
tory cues are mediated by spying on unspecialized cues released as by-products of
the endocrine responses of males (1 and i in Fig. 23.4.b) directly stimulated by a
preovulatory female. In a species where males are territorial and where females
mate with several males on their territories, the interactions depicted in Fig. 23.4.b
might be expected to have arisen from female tactics to promote sperm competi-
tion. However, given that the female goldfish cannot control the number of males
competing for fertilization attempts, that virtually all her eggs can be fertilized
by a single male (Zheng et al., 1997) and that, as noted above, additional males may
disrupt spawning activity, we feel it most probable that the interactions depicted
in Fig. 23.4.b result solely from male competition.
We hope that our proposal that the hormonal pheromones of goldfish function
in spying interactions will stimulate discussion of how this and similar systems
can be integrated into current theoretical concepts of information networks based

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 555
on true communicative interactions. To open such a discussion, can we speak of
eavesdropping in spying networks (e.g. males 2 and ii in Fig 23.4.), given that
eavesdropping appears to be restricted to interactions mediated by specialized
signals (McGregor & Peake, 2000; Ch. 2) or do we require new terms and/or new
definitions?
Sea lamprey Petromyzon marinus
Anadromous sea lamprey Petromyzon marinus spend most of their life as
stream-dwelling, filter-feeding ammocoete larvae before undergoing a dramatic
metamorphosis, migrating to the ocean or large lakes and feeding parasitically
on large fish, whose unpredictable movements can carry the lamprey far from
their natal streams. After approximately a year, the parasites cease feeding, be-
gin to mature sexually and search for a spawning stream, guided by a potent
pheromone that serves as a reliable indicator of suitable larval habitat. Since gain-
ing access to the American Great Lakes from the Atlantic Ocean about a century
ago, the sea lamprey has seriously depleted many of these lakes’ fisheries. Based
on preliminary evidence (Teeter, 1980) that larvae release a pheromone attracting
migrating adults and that spawning adults employ sex pheromones, sea lamprey
pheromones have been extensively studied in the hope of identifying semiochemi-
cals for use in biological control, as has successfully been achieved for many insects
(Chapman, 2000).
Larval pheromone attracting migratory adults
Both field and laboratory studies provide compelling evidence that mi-
gratory adult lamprey do not return preferentially to natal streams but instead
locate suitable spawning habitat by responding to a pheromone released by stream-
dwelling larvae. Historical capture records show that estimated numbers of mi-
grating adults fall by up to 50% following application of larvicides to remove
larval populations (reviewed by Sorensen & Vrieze, 2003). Evidence that such re-
ductions in migrant numbers result from removal of larval odour comes from
studies of captive migrants in large two-choice mazes (Vrieze & Sorensen, 2001).
Water from streams without larvae is much less attractive to adults than is water
from larva-bearing streams, but water from streams without larvae becomes at-
tractive following addition of low concentrations of larval odour. The potency of
larval odour is such that a single larva (weighing only several grams) creates an
active space of 400–4000 l/h, sufficient to account for the attractive properties of
streams with larvae. These studies also reveal that spawning-stream selection is
based on more than larval odour alone: migratory adults prefer stream water (even

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
556 B. D. Wisenden & N. E. Stacey
without larval odour) to lake water, suggesting the presence of unknown stream
odorants that act synergistically with larval odour (Vrieze & Sorensen, 2001).
The larval pheromone attracting adult migrants has been fully character-
ized and shown to be a mixture of chemicals (Vrieze & Sorensen, 2001; P. W.
Sorensen, personal communication); two of the primary components are the novel
bile acids, allocholic acid (ACA; 3α,7α,12α-trihydroxy-5α-cholan-24-oic acid) and
petromyzonol sulphate (PS; 3α,12α,24-trihydroxy-5α-cholan-24-sulphate). PS may
be a unique lamprey product and is synthesized by the liver of larvae but not by the
parasitic or adult phases (Polkinghorne et al., 2001). Because larvae undergo gall
bladder and bile duct atrophy at metamorphosis and also cease synthesis of PS and
ACA (Polkinghorne et al., 2001), these compounds should be specific indicators of
streams containing favourable spawning and nursery habitat. PS and ACA, which
are released primarily in larval faeces (Polkinghorne et al., 2001), are detected by
the olfactory organ of migratory adults (Li & Sorensen, 1997) not only with great
specificity, but also with a sensitivity (1 pmol/l olfactory detection threshold) that
would account for behavioural responsiveness at the low concentrations estimated
to occur in spawning streams (Polkinghorne et al., 2001). Furthermore, these bile
acids attract migratory adults (but not parasites) in maze tests (Bjerselius et al.,
2000; Vrieze & Sorensen, 2001).
Taken together, the results indicate that a suite of conspecific cues regulate
stream selection and upstream migration of maturing adult lamprey, and that
response to larval odour is adaptive in so far as it increases the likelihood of lo-
cating habitat suitable for larval growth. Moreover, because there is no evidence
at this time that larval production and release of PS and ACA are specialized for
functions other than digestion (Polkinghorne et al., 2001), and no evident mecha-
nism whereby adult response could select for specialized signalling functions for
these compounds, we regard these components of the migratory pheromone as
cues involved in chemical spying. Unlike the transient pheromonal steroid and
prostaglandin cues of goldfish, which are released only at specific stages of repro-
duction, however, the bile acid cues of lamprey appear to be released not only
during the period of peak adult migration in May but throughout the extended
period (April–August) of larval feeding (Sutton & Bowen, 1994; Polkinghorne et al.,
2001). In addition, whereas the transient pheromonal cues of female goldfish
are estimated to generate only small active spaces (Sorensen et al., 2000), PS and
ACA released by larval lamprey are estimated to create very large active spaces
sufficient to serve effectively as an upstream attractant given that larval popu-
lations can contain hundreds of thousands of individuals (Polkinghorne et al.,
2001). Perhaps the greatest departure from the goldfish situation, however, is
that, whereas goldfish pheromonal cues promote interactions of originators and
receivers within a small social unit, the lamprey larval pheromone functions in

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 557
a vast network of dispersed originators and receivers that do not interact be-
haviourally.
Sex pheromones
During upstream migration, adult male and female lamprey undergo
final maturation (spermiation and ovulation), lose behavioural responsiveness
to the larval pheromone and develop behavioural responsiveness to the odour
of mature conspecifics of the opposite sex (Bjerselius et al., 2000; Li et al., 2002).
Although the described behavioural responses of mature adults (positive rheotaxis,
increased locomotory behaviours) are rather non-specific, they are appropriate to
mediate upstream movement to spawning grounds and facilitate male–female
interactions, although this has not been demonstrated experimentally. However,
the traditional use of mature males to trap females (Fontaine, 1938; discussed in
Teeter, 1980) supports the existence of a potent male attractant, which is the only
lamprey sex pheromone to be studied intensively. This pheromone, estimated to
have a large active space (> 106 l/h per adult male (Li et al., 2002)), is proposed to
function in attracting females to mature males, which are reported to precede
females to the spawning grounds.
Major components of the pheromone released by spermiated male lam-
prey are proposed to be 3-keto-petromyzonol-sulphate (3-keto-PS; 7α,12α,24-
trihydroxy-3-one-5α-cholan-24-sulphate) and 3-keto-allocholic acid (3-keto-ACA;
7α,12α-dihydroxy-5α-cholan-3-one-24-oic acid) (Li et al., 2002; Yun et al., 2003). Al-
though both these compounds are detected by the lamprey olfactory system, only
3-keto-PS has been investigated for pheromonal activity. As with the odour of sper-
miated males, 3-keto-PS when added to a two-choice maze both attracts ovulated
females (but not preovulatory females or males) and stimulates their searching
behaviours (Li et al., 2002). Moreover, whereas non-spermiated males (whose odour
does not attract ovulated females in the maze) do not release appreciable quanti-
ties of 3-keto-PS, spermiated males release large quantities of 3-keto-PS (approxi-
mately 500 g/h) (Li et al., 2002; Yun et al., 2002).
As with the bile acid pheromone of larval lamprey (Polkinghorne et al., 2001),
3-keto-PS has been found in the liver of spermiated males (Li et al., 2002). However,
unlike the larval pheromone, which is released primarily in faeces, the pheromone
from spermiated males appears to be released by the gills, which in mature males
(but not females) develop glandular cells (Pickering, 1977) that evidently are spe-
cialized for pheromone release (Siefkes et al., 2003).
The current information on male lamprey pheromone suggests its synthesis
occurs through a subtle shift in bile acid metabolism that results in the larval
pattern of PS and ACA production changing to 3-keto-PS and 3-keto-ACA in spermi-
ating males (presence of the 3-keto acids in livers of ovulating females appears not

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
558 B. D. Wisenden & N. E. Stacey
to have been examined). Furthermore, because fully mature adults are exposed to
larval and adult bile acids in spawning streams, it is expected that the lamprey ol-
factory system has been selected to discriminate larval (3-hydroxy acid) and adult
(3-keto acid) odours, although this remains to be examined.
The identified sex pheromones of lamprey and goldfish are similar in that
they operate within a complex network of originators/signallers and receivers,
although they differ fundamentally both in the interactions between genders
and in the ancestral (prepheromonal) functions of the cues and signals. Moreover,
the evidence for signal specialization in production and release of male lamprey
pheromone suggests a true communicatory interaction, which is unlikely in
goldfish.
Synthesis
Current theory about the function of animal communication networks
(e.g. McGregor & Peake, 2000) has been heavily influenced by studies of acoustic and
visual systems, where it seems clear that true communication between specialized
signallers and receivers has arisen through the bilateral benefits resulting from
their reciprocal interactions. Although studies of fish semiochemicals also provide
evidence of specializations indicative of communication, the specific functions of
such specialized semiochemicals within networks are not well understood.
In sea lamprey, for example, both the large active space of the proposed male
sex pheromone 3-keto-PS and apparent male-specific gill structure facilitating its
release (Li et al., 2002) suggest specializations for increased amplitude of a spe-
cialized tonic signal. The proposed function of this male lamprey signal appears
analogous to the aggregate signal produced by chorusing male anurans (Ch. 13),
in so far as the combined odour of many males induces the upstream movement
of many females. However, it remains to be determined if attracted female lam-
prey also use the male pheromone in mate choice and if this might have been the
pheromone’s original function.
Also, in the black goby, non-spermatogenic portions of the testes appear spe-
cialized for synthesis of a steroid pheromone, etiocholanolone glucuronide, orig-
inally proposed simply to attract ovulated females to the male’s nest (Colombo
et al., 1980). In the round goby Neogobius melanostomus, however, both males and fe-
males respond behaviourally to etiocholanolone glucuronide (Murphy et al., 2001),
suggesting that the pheromone functions in a more complex network involving
both intra- and intersexual communication.
Given that semiochemical communication appears to have evolved in sea lam-
prey and gobies, and perhaps in some other fish such as blennies (Laumen et al.,
1974; Goncalves et al., 2002) and African catfish Clarias gariepinus (van den Hurk &

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 559
Resink, 1992), these species may communicate in semiochemical networks analo-
gous to those seen in terrestrial systems involving acoustic and visual signals. How-
ever, other fish semiochemicals, such as the alarm cues of ostariophysans and the
sex pheromones of goldfish, appear to function not in communication but rather
in spying, where specialization for information transfer evidently is restricted to
receivers. Nonetheless, these semiochemical cues also operate in complex infor-
mation networks in which semiochemicals can influence several conspecifics both
directly (through exposure) and indirectly (through changes induced in exposed
individuals) (e.g. Figs. 23.2 and 23.4.b)
Because such fish semiochemical networks based on unspecialized cues have
the potential to give rise to true communication networks, they should not only
extend the scope of current network theory but also raise important issues rele-
vant to the evolutionary processes by which such communicatory networks evolve.
To cite just one example, when discussion of information networks is restricted
to those that involve communication, it might seem reasonable to assume that
eavesdropping arises only after communicative interaction has been established.
However, the ability of male goldfish to derive information indirectly about female
cues by spying on the responses of exposed males (e.g. Figs. 23.2 and 23.4.b) demon-
strates that a process analogous (and possibly homologous) to eavesdropping can
precede the origin of communication.
To promote discussion of the functional and evolutionary relationships among
spying, eavesdropping and communication, we propose two hypothetical schemes.
One is based on the intraspecific interactions induced by the goldfish preovulatory
steroid pheromone (Fig. 23.5a); the second involves both intra- and interspecific
predator–prey interactions in ostariophysan fishes (Fig. 23.5b), and both are de-
rived from our general model for the evolution of communication (Fig. 23.1).
In goldfish, spying by male receivers (R) on an unspecialized steroid cue released
by female originators (O; Fig. 23.5a1) could lead to communication (Fig. 23.5a2)
if male response to heritable variation in cue production leads to differential
female fitness. If this occurs, females would then be signallers (S) releasing a
specialized pheromonal signal and the male’s role would change (R1), as he now
influences, and is influenced by, signal evolution. As we emphasize in this chapter,
however, the goldfish preovulatory pheromone mediates more than the simple
dyadic spying event depicted in Fig. 23.5a1. The pheromone directly stimulates
behavioural and endocrine–testicular responses in more than one male (R) and
also induces a distinct response (ovulation) in females (Fig. 23.5a3). In addition,
the pheromone indirectly stimulates males (R2) via cues released by pheromone-
exposed males (Fig. 23.5a4).
In the ancestral condition of predator-induced prey chemical alarm cues, preda-
tor (P) attack releases general cues from the originator (O) that can be received both

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
560 B. D. Wisenden & N. E. Stacey
spying
communication
R1
2
S
3
R
R
OO R
1
OO
R2
R
4
OOO
IE
R1
5
S
SE
R1
6
S
(a)
1
spying
communication
(b)R
OO
R
P2
Rr
3 4
OO P2
OC+
OO R
2 6 SE
S P3
IE
S P3
IE
R1
OC+
5
PPP
P P P
S
Fig. 23.5. Theoretical evolutionary pathways of the transition between spying and
communication networks involving semiochemicals used in reproductive (a) and
predator–prey (b) interactions. Thin solid and dashed arrows indicate spying
functions; thick, opposed, black and white arrows indicate communicative functions,
and large white arrows indicate transitions between proposed stable states. O,
originator; S, signaller; R, receiver (r, heterospecific receiver); IE, interceptive
eavesdropper; SE, social eavesdropper; P, predator; C, alarm cue. See text for further
explanation.
as an alarm cue by conspecific prey (R) and as a feeding cue by secondary preda-
tors (P2; Fig. 23.5b1). If interference by secondary predators benefits originators and
leads to alarm cue specialization, originators become signallers (S), the secondary
predator’s role changes (P3), and receiving conspecific prey become interceptive
eavesdroppers (IE) in a communication network (Fig. 23.5b2). As with the goldfish
pheromone (Fig. 23.5a3,4), predator-induced alarm cues can exert complex effects
prior to the evolution of communication. For example, alarm cues are used to

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 561
associate risk with stimuli (C) correlated with predation, which later serve as in-
dicators of predation risk (Fig. 23.5b3). This latter system may become elevated to
that of a communication network without involvement of a secondary predator
if an originator’s shoalmates learn to recognize a novel indicator of risk and later
alert the surviving originator to the presence of risk through early response to dan-
ger (Fig. 23.5b5). In direct relevance to the evolution of eavesdropping, alarm cues
can also affect predator–prey interactions indirectly through social facilitation
(social spying?) of alarm behaviour both in conspecifics (R2) and in heterospecifics
(r; Fig. 23.5b4).
If it is reasonable to assume that sex and alarm pheromone communication
evolves from spying, as depicted in Figs. 23.1, 23.5a1,2 and 23.5b1,2, then it also
seems reasonable to ask whether and how communication networks evolve from
spying networks. We, therefore, propose two general scenarios, which differ pri-
marily in the evolutionary origins of eavesdropping. In the first scenario, a simple
dyadic communication (Figs. 23.5a2 and 23.5b2) could lead to the evolution of
interceptive or social eavesdropping (Ch. 2) if receivers evolve adaptive responses
either to the signalling behaviour per se (interceptive eavesdropper (IE): Figs. 23.5a5
and 23.5b6) or to the signalling interaction (social eavesdropper (SE): Figs. 23.5a6
and 23.5b6). In this scenario, where the evolution of communication precedes that
of eavesdropping, eavesdropper functions (interceptive and social) are analogous
to the various receiver functions in spying networks (Figs. 23.5a3,4 and 23.5b3,4).
In the second scenario, incipient eavesdropping arises in spying networks, either
as direct (Figs. 23.5a3 and 23.5b4) or indirect (Figs. 23.5a4 and 23.5b4) spying by
receivers on originators and is retained as interceptive and social eavesdropping,
respectively, following the evolution of communication. In this scenario, receivers
in spying networks are homologous to eavesdroppers in communication networks.
In all the scenarios shown in Fig. 23.5, we depict eavesdropping in its proposed
initial state: that is, spying via a cue that is not specialized for transmission to
eavesdroppers, despite being a signal specialized for information transfer to the
primary target (Ch. 2). At this early stage, the network functions of eavesdropper
and primary target differ in kind. However, if subsequent selection by eavesdrop-
pers leads to signal specialization specific to the eavesdropping interaction, and
thus forming a communicative relationship between eavesdropper and signaller,
functions of eavesdroppers and receivers will come to differ only in degree. Studied
examples of eavesdropping (Ch. 2) typically appear to involve costs or benefits to
signallers that would be expected to modify signal function; consequently, it will
be important to determine whether, as has been suggested for sex pheromone
function in fish (Fig. 23.5a1), various forms of eavesdropping in communication
networks (Figs. 23.5a5,6 and 23.5b2,5,6) can persist as spying. Moreover, it will be
important to document covariance in the relative proportions of spying versus

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
562 B. D. Wisenden & N. E. Stacey
communicative eavesdropping and the ecological and social factors that lead to
the spying–communication transition.
Acknowledgements
The authors gratefully acknowledge support from MSUM College of Social and Nat-
ural Sciences, MSUM Dille Fund for Excellence, MSUM Alumni Foundation, MnSCU Learning
by Doing (B. Wisenden) and the Natural Sciences and Engineering Research Council of Canada
(N. Stacey).
References
Argentini, M. 1976. Isoleirung des Schreckstoffes aus der Haut der Erlitz Phoxinus
phoxinus (L.). Ph.D. Thesis, University of Zurich.
Beauchamp, G. K. 2000. Defining pheromones. In: The Monell Connection, Fall 2000, ed.
L. J. Stein. Philadelphia, PA: Monell Chemical Senses Center, p. 2.
Bjerselius, R., Olsen, K. H. & Zheng, W. 1995. Endocrine, gonadal and behavioral
responses of male crucian carp (Carassius carassius) to the hormonal pheromone
17α,20β -dihydroxy-4-pregnen-3-one. Chemical Senses, 20, 221–230.
Bjerselius, R., Li, W., Teeter, J. H. et al. 2000. Direct behavioral evidence that unique
bile acids released by larval sea lamprey (Petromyzon marinus) function as a
migratory pheromone. Canadian Journal of Fisheries and Aquatic Sciences, 57,
557–569.
Brown, G. E., Godin, J.-G. J. & Pedersen, J. 1999. Fin-flicking behaviour: a visual
anti-predator alarm signal in a characin fish, Hemigrammus erythrozonus. Animal
Behaviour, 58, 469–475.
Brown, G. E., Adrian, J. C., Jr & Shih, M. L. 2001. Behavioural responses of fathead
minnows to hypoxanthine-3N-oxide at varying concentrations. Journal of Fish
Biology, 58, 1465–1470.
Bryant, P. B. 1987. A study of the alarm system in selected fishes of Northern
Mississippi. Ph.D. Thesis, University of Mississippi.
Chapman, R. F. 2000. Entomology in the twentieth century. Annual Reviews of
Entomology, 45, 261–285.
Chivers, D. P. & Mirza, R. S. 2001. Predator diet cues and the assessment of predation
risk by aquatic vertebrates: a review and prospectus. In Chemical Signals in
Vertebrates, ed. A. Marchlewska-Koj, J. Lepri & D. Muller-Schwarze. New York:
Plenum Press, pp. 277–284.
Chivers, D. P. & Smith, R. J. F. 1994. The role of experience and chemical alarm
signalling in predator recognition by fathead minnows, Pimephales promelas
Rafinesque. Journal of Fish Biology, 44, 273–285.
1998. Chemical alarm signalling in aquatic predator–prey systems: a review and
prospectus. Ecoscience, 5, 338–352.
Chivers, D. P., Brown, G. E. & Smith, R. J. F. 1996. The evolution of chemical alarm
signals: attracting predators benefits alarm signal senders. American Naturalist,
148, 649–659.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 563
Colombo, L., Marconato, A., Belvedere, P. C. & Frisco, C. 1980. Endocrinology of teleost
reproduction: a testicular steroid pheromone in the black goby, Gobius jozo L.
Bolletino di Zoologia, 47, 355–364.
Fontaine, M. 1938. La lamproie marine. Sa peche et son importance economique.
Bulletin de la Societe Oceanographique de France, 17, 1681–1687.
Fraser, E. J. & Stacey, N. 2002. Isolation increases milt production in goldfish. Journal of
Experimental Zoology, 293, 511–524.
Gandolfi, G., Classon, L. J. & Rossi, A. C. 1968. The fright reaction of zebra fish. Atti
Societa Italiana di Scienze Naturali, 107, 74–88.
Gazdewich, K. J. & Chivers, D. P. 2002. Acquired recognition by fathead minnows:
influence of habitat characteristics on survival. Journal of Chemical Ecology, 28,
439–445.
Goncalves, D. M., Barata, E. N., Oliveira, R. F. & Canario, A. V. M. 2002. The role of male
visual and chemical cues on the activation of female courtship behaviour in the
sex-role reversed peacock blenny. Journal of Fish Biology, 61, 90–105.
Hall, D. & Suboski, M. D. 1995. Visual and olfactory stimuli in learned release of alarm
reactions by zebra danio fish (Brachydanio rerio). Neurobiology of Learning and
Memory, 63, 229–240.
Hamdani, E. H., Stabell, O. B., Alexander, G. & Døving, K. B. 2000. Alarm reaction in the
crucian carp is mediated by the medial bundle of the medial olfactory tract.
Chemical Senses, 25, 103–109.
Hamdani, E. H., Kasumyan, A. & Døving, K. B. 2001. Is feeding behaviour in crucian
carp mediated by the lateral olfactory tract? Chemical Senses, 26, 1133–1138.
Hansen, J. A., Rose, J. D., Jenkins, R. A., Gerow, K. G. & Bergman, H. L. 1999. Chinook
salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss) exposed
to copper: neurophysiological and histological effects on the olfactory system.
Environmental Toxicology and Chemistry, 18, 1979–1991.
Hasson, O. 2000. Knowledge, information, biases and signal assemblages. In:
Animal Signals: Signalling and Signal Design in Animal Communication, ed.
Y. Espmark, T. Amundsen & G. Rosenqvist. Trondheim: Tapir Academic Press,
pp. 119–139.
Hauser, M. 1996. The Evolution of Communication. Cambridge, MA: MIT Press.
Henderson, P. A., Irving, P. W. & Magurran, A. E. 1997. Fish pheromones and
evolutionary enigmas: a reply to Smith. Proceedings of the Royal Society of London,
Series B, 264, 451–453.
Hews, D. K. 1988. Alarm response in larval western toads, Bufo boreas: release of larval
chemicals by a natural predator and its effect on predator capture efficiency.
Animal Behaviour, 36, 125–133.
Hubbard, P. C., Barata, E. N. & Canario, A. V. M. 2002. Possible disruption of
pheromonal communication by humic acid in the goldfish, Carassius auratus.
Aquatic Toxicology, 60, 169–183.
Irvine, I. A. S. & Sorensen, P. W. 1993. Acute olfactory sensitivity of wild common carp,
Cyprinus carpio, to goldfish sex pheromones is influenced by gonadal maturity.
Canadian Journal of Zoology, 71, 2199–2210.
Karlson, P. & Luscher, M. 1959. ‘Pheromones’: a new term for a class of biologically
active substances. Nature, 183, 55–56.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
564 B. D. Wisenden & N. E. Stacey
Kasumyan, A. O. & Ponomarev V. Y. 1987. Biochemical features of alarm pheromone in
fish of the order Cypriniformes. Journal of Evolutionary Biochemistry and Physiology,
23, 20–24.
Kats, L. B. & Dill, L. M. 1998. The scent of death: chemosensory assessment of predation
risk by prey animals. Ecoscience, 5, 361–394.
Kobayashi, M., Sorensen, P. W. & Stacey, N. E. 2002. Hormonal and pheromonal control
of spawning behaviour in the goldfish. Fish Physiology and Biochemistry, 26, 71–84.
Korpi, N. L. & Wisenden, B. D. 2001. Learned recognition of novel predator odour by
zebra danios, Danio rerio, following time-shifted presentation of alarm cue and
predator odour. Environmental Biology of Fishes, 61, 205–211.
Kristensen, E. A. & Closs, G. P. 2004. Anti-predator response of naive and experienced
common bully to chemical alarm cues. Journal of Fish Biology, 64. 643–652.
Laumen, T. J., Pern, U. & Blum, V. 1974. Investigations on the function and hormonal
regulation of the anal appendices in Blennius pavo (Risso). Journal of Experimental
Zoology, 190, 47–56.
Lawrence, B. J. & Smith, R. J. F. 1989. Behavioral response of solitary fathead minnows,
Pimephales promelas, to alarm substance. Journal of Chemical Ecology, 15, 209–219.
Li, W. & Sorensen, P. W. 1997. Highly independent olfactory receptor sites for
conspecific bile acids in the sea lamprey, Petromyzon marinus, Journal of Comparative
Physiology A, 180, 429–438.
Li, W., Scott, A. P., Siefkes, M. J. et al. 2002. Bile acid secreted by male sea lamprey that
acts as a sex pheromone. Science, 296, 138–141.
Liley, N. R. 1982. Chemical communication in fish. Canadian Journal of Fisheries and
Aquatic Sciences, 39, 22–35.
Lloyd, J. E. 1965. Aggressive mimicry in Photuris: firefly femme fatales. Science, 149,
653–654.
Magurran, A. E. 1989. Acquired recognition of predator odour in the European
minnow (Phoxinus phoxinus). Ethology, 82, 216–223.
Magurran, A. E., Irving, P. W & Henderson, P. A. 1996. Is there a fish alarm pheromone?
A wild study and critique. Proceedings of the Royal Society of London, Series B, 263,
1551–1556.
Mathis, A. & Smith, R. J. F. 1993. Chemical alarm signals increase the survival time of
fathead minnows (Pimephales promelas) during encounters with northern pike
(Esox lucius). Behavioral Ecology, 4, 260–265.
Mathis, A., Chivers, D. P. & Smith, R. J. F. 1995. Chemical alarm signals: predator
deterrents or attractants? American Naturalist, 146, 994–1005.
1996. Cultural transmission of predator recognition in fishes: intraspecific and
interspecific learning. Animal Behaviour, 51, 185–201.
McClintock, M. 2002. Pheromones, odors, and vasanas: the neuroendocrinology of
social chemosignals in humans and animals. In: Hormones, Brain, and Behavior,
Vol. 2, ed. D. Pfaff, D. Arnold, A. Etgen, S. Fahrbach & R. Rubin. New York:
Academic Press, pp. 797–870.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 565
Møller, A. P. 1989. Deceptive use of alarm calls by male swallows, Hirundo rustica: a new
paternity guard. Behavioral Ecology, 1, 1–6.
Murphy, C. A., Stacey, N. E. & Corkum, L. D. 2001. Putative steroidal pheromones in the
round goby, Neogobius melanostomus: olfactory and behavioural responses. Journal
of Chemical Ecology, 27, 443–470.
Nelson, J. S. 1994. Fishes of the World, 4th edn. New York: Wiley-Interscience.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 254, 1045–1049.
Otter, K. A., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits (Parus major)
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society of London, Series B, 266, 1305–1310.
Paxton, R. J. & Tengo, J. 2001. Double duped males: the sweet and sour of the orchid’s
bouquet. Trends in Ecology and Evolution, 16, 167–169.
Pfeiffer, W. 1977. The distribution of fright reaction and alarm substance cells in
fishes. Copeia, 1977, 653–665.
Pfeiffer, W., Riegelbauer, G., Meir, G. & Scheibler, B. 1985. Effect of
hypoxanthine-3(N)-oxide and hypoxanthine-1(N)-oxide on central nervous
excitation of the black tetra Gymnocorymbus ternetzi (Characidae, Ostariophysi,
Pisces) indicated by dorsal light response. Journal of Chemical Ecology, 11, 507–524.
Pickering, A. D. 1977. Sexual dimorphism in the gills of the spawning river lamprey,
Lampetra fluviatilis L. Cell and Tissue Research, 180, 1–10.
Poling, K. R., Fraser, E. J. & Sorensen, P. W. 2001. The three steroidal components of the
goldfish preovulatory pheromone signal evoke different behaviors in males.
Comparative Biochemistry and Physiology B, 129, 645–651.
Polkinghorne, C. N., Olson, J. N., Gallaher, D. G. & Sorensen, P. W. 2001. Larval sea
lamprey release two unique bile acids to the water at a rate sufficient to produce
detectable riverine pheromone plumes. Fish Physiology and Biochemistry, 24, 15–30.
Schutz, F. 1956. Vergleichende Untersuchungen uber die Schreckreacktion bei Fischen
und Verbreitung. Zeitschrift fur Vergleichende Physiologie, 38, 84–135.
Siefkes, M. J., Scott, A. P., Zielinski, B. et al. 2003. Male sea lampreys Petromyzon marinus,
excrete a sex pheromone from gill epithelia. Biology of Reproduction, 69, 125–132.
Smith, R. J. F. 1979. Alarm reaction of Iowa and johnny darters (Etheostoma, Percidae) to
chemicals from injured conspecifics. Canadian Journal of Zoology, 57, 1278–1282.
1982. Reaction of Percina nigrofasciata, Ammocrypta beani, and Etheostoma swaini
(Percidae, Pisces) to conspecific and intergeneric skin extracts. Canadian Journal of
Zoology, 17, 2253–2259.
1992. Alarm signals in fishes. Reviews in Fish Biology and Fisheries, 2, 33–63.
1997. Does one result trump all others? A response to Magurran, Irving and
Henderson. Proceedings of the Royal Society of London, Series B, 264, 445–450.
1999. What good is smelly stuff in the skin? Cross function and cross taxa effects in
fish ‘alarm substances’. In: Advances in Chemical Signals in Vertebrates, ed. R. E.
Johnston, D. Muller-Schwarze & P. W. Sorensen. New York: Kluwer
Academic/Plenum Press, pp. 66–84.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
566 B. D. Wisenden & N. E. Stacey
Smith, R. J. F. & Lemly, A. D. 1986. Survival of fathead minnows after injury by
predators: the possible role in the evolution of alarm signals. Environmental Biology
of Fishes, 15, 147–149.
Sorensen, P. W. & Stacey, N. E. 1999. Evolution and specialization of fish hormonal
pheromones. In: Advances in Chemical Signals in Vertebrates, ed. R. E. Johnston, D.
Muller-Schwarze & P. W. Sorensen. New York: Kluwer Academic/Plenum Press,
pp. 15–47.
Sorensen, P. W. & Vrieze, L. A. 2003. The chemical ecology and potential application of
the sea lamprey migratory pheromone. Journal of Great Lakes Research, 29 (Suppl. 1),
66–84.
Sorensen, P. W., Christensen, T. A. & Stacey, N. E. 1998. Discrimination of pheromonal
cues in fish: emerging parallels with insects. Current Opinion in Neurobiology, 8,
458–467.
Sorensen, P. W., Scott, A. P. & Kihslinger, R. L. 2000. How common hormonal
metabolites function as relatively specific pheromonal signals in goldfish. In:
Proceedings of the Sixth International Symposium on the Reproductive Physiology of Fish,
ed. B. Norberg, O. S. Kjesbu, G. L. Taranger, E. Andersson & S. O. Stefansson.
Bergen: John Grieg, pp. 125–128.
Stacey, N. E. & Sorensen, P. W. 2002. Hormonal pheromones in fish. In: Hormones, Brain,
and Behavior, Vol. 2, ed. D. Pfaff, D. Arnold, A. Etgen, S. Fahrbach & R. Rubin. New
York: Academic Press, pp. 375–435.
Stacey, N. E., Kyle, A. L. & Liley. N. R. 1986. Fish reproductive pheromones. In: Chemical
Signals in Vertebrates, Vol. 4, ed. D. Duvall, D. Muller-Schwarze & R. M. Silverstein.
New York: Plenum Press, pp. 117–133.
Stacey, N. E., Zheng, W. B. & Cardwell, J. R. 1994. Milt production in common carp
(Cyprinus carpio): stimulation by a goldfish steroid pheromone. Aquaculture, 127,
265–276.
Stacey, N. E., Fraser, E. J., Sorensen, P. W. & van der Kraak, G. J. 2001. Milt production
in goldfish: regulation by multiple social stimuli. Comparative Biochemistry and
Physiology C, 130, 467–476.
Suboski, M. D. 1990. Releaser-induced recognition learning. Psychology Review, 97,
271–284.
Suboski, M. D., Brian, S., Carty, A. E. et al. 1990. Alarm reaction in acquisition and
social transmission of simulated predator recognition by zebra danio fish
(Brachydanio rerio). Journal of Comparative Physiology, 104, 101–112.
Sutton, T. M. & Bowen, S. H. 1994. Significance of organic detritus in the diet of larval
lampreys in the Great Lakes basin. Canadian Journal of Fisheries and Aquatic Sciences,
51, 2380–2387.
Teeter, J. 1980. Pheromone communication in sea lampreys (Petromyzon marinus):
implications for population management. Canadian Journal of Fisheries and Aquatic
Sciences, 37, 2123–2132.
van den Hurk, R. & Resink, J. W. 1992. Male reproductive system as sex pheromone
producer in teleost fish. Journal of Experimental Zoology, 261, 204–213.
Verheijen, F. J. 1956 Transmission of fright reaction amongst a school of fish and the
underlying sensory mechanisms. Experientia, 12, 202–204.

P1: IYP/... P2: IYP0521823617c23.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 12:49
Fish semiochemicals 567
Vickers, N. J. 2000. Mechanisms of animal navigation in odor plumes. Biological Bulletin
198, 203–212.
von Frisch, K. 1941. Uber einen Schreckstoff der Fischaut and seine biologische
Bedeutung. Zeitschrift Fur Vergleichende Physiologie, 29, 46–145.
Vrieze, L. A. & Sorensen, P. W. 2001. Laboratory assessment of the role of a larval
pheromone and natural stream odor in spawning stream localization by
migratory sea lamprey (Petromyzon marinus). Canadian Journal of Fisheries and Aquatic
Sciences, 58, 2374–2385.
Weissburg, M. J. 2000. The fluid dynamical context of chemosensory behavior.
Biological Bulletin, 198, 188–202.
Williams, G. C. 1992. Natural Selection: Domains, Levels and Challenges. Oxford: Oxford
University Press.
Wilson, E. O. & Bossert, W. H. 1963. Chemical communication among animals. In:
Recent Progress in Hormone Research, Vol. 19, ed. G. Pincus. New York: Academic
Press, pp. 673–716.
Wisenden, B. D. 2000. Scents of danger: the evolution of olfactory ornamentation in
chemically mediated predator–prey interactions. In: Animal Signals: Signalling and
Signal Design in Animal Communication, ed. Y. Espmark, T. Amundsen & G.
Rosenqvist. Trondheim: Tapir Academic Press, pp. 365–386.
2003. Chemically mediated strategies to counter predation. In: Sensory Assessment of
the Aquatic Environment, ed. S. P. Collin & N. J. Marshall. New York: Springer,
pp. 236–251.
Wisenden, B. D. & Thiel, T. A. 2002. Field verification of predator attraction to minnow
alarm substance. Journal of Chemical Ecology, 28, 433–438.
Wisenden, B. D., Chivers, D. P., Brown, G. E. & Smith, R. J. F. 1995. The role of experience
in risk assessment: avoidance of areas chemically labelled with fathead minnow
alarm pheromone by conspecifics and heterospecifics. Ecoscience, 2, 115–122.
Wisenden, B. D., Cline, A. & Sparkes, T. C. 1999. Survival benefit to anti-predator
behavior in the amphipod Gammarus minus in response to injury-released
chemical cues from conspecifics and heterospecifics. Ethology, 105, 407–414.
Yun, S.-S., Scott, A. P., Siefkes, M. & Li, W. 2002. Development and application of an
ELISA for a sex pheromone released by the male sea lamprey (Petromyzon marinus
L.). General and Comparative Endocrinology, 129, 163–170.
Yun, S.-S., Scott, A. P. & Li, W. 2003. Pheromones of the male sea lamprey, Petromyzon
marinus L: structural studies on a new compound, 3-keto allocholic acid, and
3-keto petromyzonol sulfate. Steroids, 68, 297–304.
Zheng, W., Strobeck, C. & Stacey, N. E. 1997. The steroid pheromone
17α,20β -dihydroxy-4-pregnen-3-one increases fertility and paternity in goldfish.
Journal of Experimental Biology, 200, 2833–2840.

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
24
Cognitive aspects of networks andavian capacities
i r e n e m . p e p p e r b e rg
Brandeis University, Waltham, USA
Introduction
The natural world is an extremely complicated place of myriad interac-
tions – some obvious, some hidden – but all of critical importance if one is to
understand its workings. Information must be processed, sorted, ignored or acted
upon by all creatures, even though the levels of processing ability vary across
species. Scientists, although well aware of these complexities and eager to make
sense of them, often begin by reducing interactions to their simplest form, un-
der the assumption that one can gain an understanding of more complex issues
by first gaining full knowledge of the simplest. Consequently, in most scientific
endeavours, initial studies examine the effect of a single stimulus on an entity:
in physics, how light waves interact with a single atom, or how two atoms might
interact; in child psychology, the reaction of an infant to a caretaker’ssmile or to a
novel toy; in animal behaviour laboratories, the effect of a shock on the behaviour
of a rat’smovement in a simple laboratory maze or the effect of a tape loop of song
on a bird in a sound isolation box. In each instance, however, the data obtained
provide only a small glimmer of the complexity that exists in the real world, and
in many cases inferences drawn from data in such experiments truly explain only
the specific laboratory situation being studied. To expand to a larger system and a
broader base often requires – and triggers – the development of more sophisticated
tools, be they mathematical theories (e.g. the Nash equilibrium), more powerful
computers for handling data or more sophisticated equipment for gathering data
(e.g. complex recording arrays). Sometimes, however, what is first needed is simply
the capacity to think outside of traditional forms of experimentation.
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
568

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 569
The study of animal (and particularly avian) cognition was one such paradigm
shift, and the consequent realization that animals needed to – and could – process
several sets of information (e.g. Pepperberg, 1990) was a logical outcome of such a
shift. The story begins with the so-called cognitive revolution (Hulse et al., 1968),
when researchers began to accept that levels and types of intelligence in non-
humans formed a continuum with those of humans, and to investigate a wide
range of behaviour and its development through many techniques in various
species. Most resulting studies, however, simply adapted material from human
cognitive experiments, often focused on a single set of tasks (i.e. not forcing sub-
jects to choose an appropriate set of responses from several possibilities, as they
must do in nature), continued to use a small number of species (predominantly
monkeys, rats, and pigeons) and made fairly sweeping conclusions as to the rel-
ative intelligence of all animals on the basis of these data (review in Pepperberg,
2001). Researchers, mostly in the laboratory but even in the field, initially failed
to examine species’ innate predispositions, evolutionary histories or ecological
constraints and, possibly most important of all, focused on how mostly social ani-
mals reacted in situations of social isolation. Although laboratory tasks presented
to animals may indeed have been cognitively complex, many animals failed to
demonstrate advanced capacities because of the specific nature of the task and
the situation in which the task was presented (for examples see Menzel & Juno,
1982).
Moreover, researchers often allowed their prejudices about animal capacities
to influence their hypotheses. The phylogenetic closeness of primates to humans
(e.g. Sarich & Cronin, 1977) and the large brains of cetaceans (e.g. Russell, 1979)
led scientists to anticipate and accept that their communicative and cognitive
capacities would be comparable to those of humans. (It should be noted however,
that Morgane et al. (1986) expressed concern that the dolphin brain, although large
even with respect to body size, may lack some of the complexity found in primates.)
Yet experimenters rarely expected analogous abilities in birds and failed to search
for such capacities. For many years, researchers argued that cognitive capacity
was likely a consequence of relative cortical size, and that birds, lacking much in
the way of cortical development, had to be inferior to mammals and primates (e.g.
Premack, 1978). My own research on the cognitive and communicative capacities
of the African grey parrots Psittacus erithacus is a particularly striking example of
these issues (Pepperberg, 1999): birds once thought to be capable merely of mind-
less mimicry have demonstrated, under appropriate experimental conditions,
referential use of elements of English speech and cognitive abilities (e.g. concepts
of category, number, bigger/smaller, same/different, absence) comparable to
those of a human child aged four to six years. Similarly, songbirds once thought
merely to be emitting sets of innately predisposed vocal patterns acquired during

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
570 I. M. Pepperberg
a brief stage in their lives, have demonstrated vocal behaviour indicating various
levels of cognitive processing and extensive memory. There are a number of good
examples. White-crowned sparrows Zonotrichia leucophrys, assumed to acquire only
their species-specific song during a limited sensitive period (Marler, 1970), show
flexibility in learning elements of other species’songs when living in a complex so-
cial environment, not only in the laboratory but also in areas of sympatry (Baptista
& Catchpole, 1989). Marsh wrens Cistothorus palustris actively choose which song in
their 100–400 song repertoire to use in competitive countersinging so as to match,
anticipate and possibly ‘jam’ the next song in their neighbours’ 100–400 song
series (Kroodsma, 1979; Kroodsma & Byers, 1998). Nightingales Luscinia megarhyn-
chos learn their 100 or so songs in chunks, much like humans learning long lists
(Todt & Hultsch, 1998). Numerous avian species recognize subtle variations that
differentiate their neighbours’ songs from those of strangers (Stoddard, 1996) and
some even remember neighbours’ songs from one year to the next (Godard, 1991).
Finally, if a song sparrow Melospiza melodia does not have an appropriate song type
in its repertoire for an exact match in a countersinging bout, it selects one that
is most similar (e.g. with the same introductory section,showing some level of
same/different comprehension (Burt et al., 2002)). However, even these examples
generally have focused on a single individual or one-on-one interactions and have,
therefore, to some extent ignored the real world: that these birds are actually part
of a larger network and that a countersinging bird, for example, would interact
over time with usually at least two or three individuals (i.e. all its territorial
neighbours), processing and storing all that information. Interestingly, advances
in field techniques, both in recording and playback (e.g. McGregor et al., 1992;
Naguib & Todt, 1997; Burt 2000), have not only allowed researchers to examine all
the information available to their subjects but have also led these researchers to
appreciate the complex cognitive processes that birds must be using to make sense
of this information. A specific avian case involves the relationship between the
complex cognitive task called ‘transitive inference’ and the natural situational
behaviour of ‘eavesdropping’ among networks of songbirds (Dabelsteen et al.,
1997; McGregor et al., 1997; Naguib & Todt, 1997; Naguib et al., 1999; Otter et al.,
1999; Peake et al., 2001, 2002; Mennill et al., 2002). A discussion of the complexity of
transitive inference, including the advances and pitfalls of laboratory work on the
topic and a brief review of data on eavesdropping (for details, see Ch. 2 and several
other chapters in this volume) will demonstrate some extent of avian cognitive
capacities.
Transitive inference
Transitive inference is one of several psychological tasks that engender
incredible amounts of discussion both as to the actual mechanism by which it is

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 571
performed and as to whether animals (particularly birds) are indeed capable of its
performance (reviewed by Zentall, 2001). The task, as originally stated for human
children, goes something like ‘Sam is taller than Bob. Bob is taller than Jack. Is
Jack shorter than Sam?’ (e.g. McGonigle & Chalmers, 1984a); sometimes the prob-
lem is given as ‘Bob is taller than Jack. Sam is taller than Bob. Who is shortest?’.
No training or rewards are involved when the task is given to children and the
child is assumed to understand the concept of taller (or bigger, stronger, etc.).
The child is thought to succeed by being able to integrate the two different vocal
pieces of information, including the reversal from taller to shorter, in a conscious,
cognitive manner; the complexity of the task derives from this integration and
reversal and the strong likelihood that the process requires a mental represen-
tation of the integrated pieces of information for success. In one instance, adult
subjects were given only five seconds to solve each of a series of transitive infer-
ence problems of various forms (‘Triangle is above circle. Square is below circle. Is
triangle above square?’ ‘Circle is darker than square. Circle is lighter than triangle.
Is triangle darker than square?’)and were then asked to report their reasoning pat-
terns (Egan, 1983). Subjects who used different reasoning strategies (e.g. ordering
the objects on a linear scale, which they then ‘scanned’, versus making individual
images of the objects, which they then compared) made different amounts and dif-
ferent types of reasoning error (the linear thinkers had approximately 10% errors
whereas the imaging group had approximately 38% errors). Use of a particular rea-
soning strategy was affected by aptitude for visualizing spatial transformations
of figures and the context in which reasoning problems were posed, but each
strategy involved some form of representation and integration: that is, cognitive
processing. Whatever the strategy, the connection is evident between such a task
and real-world knowledge of dominance hierarchies for any species living in a
network of individuals; therefore, researchers assumed that demonstrating this
understanding in non-human animals would be straightforward. Such, however,
has not been the case.
The task, as presented to non-humans, usually differs in a number of ways. First,
the number of contrasting pairs involved usually is at least five (note that some
studies such as those of McGonigle & Chalmers (1984b) use comparable numbers
for children). Second, a hungry animal undergoes extensive training on pairwise
comparisons where one of the pair is reinforced by a food reward (designated by +);
the other is not reinforced (designated by −); the amount and type of food re-
ward never varies. So the animal is trained to criterion on one pair (A+/B−),
then to criterion on the next pair (B+/C−), likewise for subsequent pairs (C+/D−,
D+/E−), and finally tested on an internal novel pair such as B/D to see which it
will choose. The elements of the pair to be tested have never individually been
shown to be the best or the worst and their relative worth has never been trained.
Third, the animal is not specifically cued, as humans generally are, that the task

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
572 I. M. Pepperberg
involves relative judgements. Remember, humans are given specific verbal cues
that could be seen as the equivalent of ‘four chocolates if you chose A, three if
you choose B, two if you choose C, one if you choose D and none if you choose
E; now, do you prefer B over D?’ So in some sense the animals are given a task
that is more difficult than that initially given to children: the animal has not only
to learn and remember a series of comparisons but also to understand the point
of the query when presented with a novel pair, each member of which had ac-
tually led to reward in some instances. Rules that might have initially assisted
during training (e.g. ‘choose the familiar item in a new pair even if it was not
previously rewarded’) are of no use during testing, and additional rules developed
during pairwise training (‘choose what was most recently rewarded’) would be
misleading. Moreover, because the tests are not rewarded, the subject may not
even be able to learn through successive iterations. Note, however, that the ani-
mal does not need to engage in reversal (i.e. the taller-to-shorter change mentioned
above) and whether the animal is indeed cognitively engaged is unclear. Several
researchers argue that non-cognitive mechanisms based simply on reward might
be sufficient to explain the results of the pigeon subjects tested, which received
only a reward/no-reward condition within pairs of items (i.e. given no reason to
expect explicit relative relationships; see Couvillon & Bitterman, 1992; Wynne,
1997). At issue is the fact that the speed of acquisition and thus the number of
trials would differ somewhat for each of the pairs, and consequently more errors
would be made to some elements than others (see Zentall, 2001). Other researchers
argue for cognitive processes based on spatial mapping (e.g. Weaver et al., 1997),
such that the animals form some kind of linear set or mental representation to
which they can retrospectively refer during testing. Interestingly, pigeons given
explicit size cues to assist in forming a linear hierarchy did not learn any faster
nor were they more accurate than those without such cues (von Ferson, 1989),
suggesting that a linear model was not necessary for success. Possibly a form of
‘value transfer’ is involved, in that B, although never rewarded when given with A,
accrues some of A’s value of 100% reward, whereas D, although always rewarded
with E, loses some value by being connected to C, which accrues only 50% reward,
and E, which is never rewarded (Weaver et al., 1997). Although a number of differ-
ent species, from pigeons (Zentall, 2001) to chimpanzees (Gillan, 1981; Boysen et
al., 1993), appear to succeed, the mechanisms used by animals and humans might
differ.
The transitive inference studies my students and I are attempting with African
grey parrots, although not involving problems faced in the natural world, should
avoid these issues; I present the material to clarify how an experiment on transi-
tive inference could be performed in a laboratory to test whether a bird is using
a representational, cognitive mechanism. The oldest parrot, Alex, already vocally

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 573
designates the bigger/smaller of an object pair with respect to mass (Pepperberg &
Brezinsky, 1991) and can quantify collections of up to six objects with vocal
English number labels (Pepperberg, 1994); he is learning to label Arabic numerals
so that we can determine if he can combine these abilities to rank order number
symbols using transitive inference. Will he understand, without specific training
to associate Arabic numerals directly with their physical values, that the symbol
‘5’ is greater than the symbol ‘3’? Only a task using equivalence relations and
transitive inference can test this ability: he must use the commonality of English
to correlate (form equivalence relations between) quantity and Arabic numerals,
then use a form of transitive inference to identify the colour of one of a pair of
Arabic numbers that is bigger or smaller (e.g. a blue 3, red 5). To succeed, he must
base choice of ‘Arabic numeral X bigger/smaller than Arabic numeral Y’ on de-
ductions and on inferences: deduce that an Arabic numeral has the same value
as a vocal label, compare representations of quantity (mass) for which the numeral
stands, infer rank ordering based on these representations (transitive inference)
and then vocally report the result. Specific stimuli within pairs are not associated
with reward (Wynne, 1997), and by requiring colour, not number label responses,
rote replies cannot be used for a given pair. He has had no explicit training on
‘more/less than’rankings of individual elements (Arabic numerals) to be tested. The
task involves use of both working and long-term memory (Geary et al., 2000). Note,
too, that Alex was not trained to associate numbers with quantities sequentially:
He first learned 3 and 4, then 2 and 5, then 6 (Pepperberg, 1987), and he does
not produce vocal strings of number labels (i.e. does not say ‘one, two, three . . . ’);
therefore, he has had no training in rank ordering numbers whatsoever. Initial
trials are encouraging; he scored 15/18 in probes.
Although Alex’s task will provide some intriguing information on transitive
inference, the procedure still does not allow us to equate animal and human
studies. Consequently, given the differences between the standard animal and
human tasks, of particular interest are data collected when tasks more like those
given to the animals were given to adult humans. When adult humans were given
a non-vocal transitive task based on a computer game and not told that the game
involved transitive inference, only 70% succeeded (Siemann, 1993). In a different
experiment, when adult college students were given the exact same task as the
animals (Werner et al., 1992), their accuracy for the transitive pair was impressive
(approximately 95%), but only about two-thirds could explicitly state how they
solved the problem. When two groups of humans were given the non-vocal animal
task, with only one having been told that the task was inferential (Greene et al.,
2001), both groups succeeded, again suggesting that the processing need not be
conscious. Nevertheless, the informed group performed slightly better than those
in the uninformed group, many of whom, after testing, indicated that they had

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
574 I. M. Pepperberg
inferred the hierarchical relationship. Finally, when human subjects are given the
vocal human task with five elements even without reversal (‘John is taller than
Bob. Bob is taller than Jim. Jim is taller than Richard. Richard is taller than David.
Who is taller, Bob or Richard?’), humans often fail because the information given
in such a task exceeds their memory capacity; many repetitions of the information
are necessary before they succeed (Delius & Siemann, 1998; see also Woocher
et al., 1978). Therefore, one can argue that the emphasis on overt hierarchical
presentation in the standard human vocal task (A > B, B > C, A ? C) provides strong
cues for its solution, and that in the longer case (A > B, B > C, C > D, D > E, B ? D)
some form of learning is likely involved. When the task involves specific sequential
training on abstract pairs and trial-and-error learning, however, subjects – be they
pigeons (and by inference birds in general) or humans – do not truly demonstrate
transitive inference; rather their results appear to be merely an artefact of the
training and reward situation. Furthermore, transitive inference involving groups
larger than three, even with explicit instruction as to the hierarchical nature of
the task, appears to be more difficult than expected.
The real issue, then, is not whether a pigeon can be taught something that has
the surface appearance of transitive inference, but whether birds (a) are indeed
capable of a task that has the same cognitive complexity of the human task and,
probably more importantly, (b) are faced with, and able to solve, such a task in the
real world. Although my research on the former issue is only in the earliest stages,
my findings suggest that a parrot, with vocal and cognitive capacities that resemble
those of very young children, is a good candidate for such a task (Pepperberg, 1999;
see comment in Delius et al., 2000). Some data on chickens (Hogue et al., 1996) and
recent field research on avian song (e.g. Peake et al., 2002), however, have provided
intriguing information that birds use a network of information to solve at least
simple transitive inference problems in their daily lives. Such data demonstrate
a level of cognitive processing unexpected in a creature with a brain not much
bigger than the size of a pea, or at most a shelled walnut, and a brain that is
organized so differently from that of humans (cf. Jarvis & Mello, 2000). Excluding
the chickens because they watched actual physical interactions, I will concentrate
on songbirds, where the data involve decisions based on vocalizations.
Songbirds and transitive inference
Songbirds, as noted above, live in a noisy environment of vocalizations
and other sounds of numerous species. Even if we make simplifying assumptions
that they rarely need to attend to the songs of other species or that they may not
need to learn to ignore such songs, they still need to learn the repertoires of their
neighbours. By so doing, they can determine, for example, whether, a territorial

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 575
encroachment is by a neighbour with whom they can resolve the boundary dispute
fairly quickly (e.g. Kroodsma, 1979; Beecher et al., 1996; Stoddard, 1996) or by a
stranger who may pose a serious threat (Stoddard, 1996). Recent research suggests
that songbirds also process and remember how their neighbours fare in territorial
disputes with other neighbours and strangers and, by eavesdropping, they become
aware of the relative dominance hierarchy of these birds and react with respect
to that information, a case of transitive inference. Note that, for example, an
unfamiliar floater male passing through a given area is quite likely to challenge
several residents; knowing one’s relationship to one’s neighbours and how one’s
neighbours have fared in such interactions could be advantageous.
Now, eavesdropping by itself does not provide direct evidence for transitive
inference, but it clearly sets the stage. Female black-capped chickadees Poecile
atricapillus, for example, attend to the vocal duels between males and make their
reproductive choices based on the outcomes (Mennill et al., 2002); they actively seek
extra-pair copulations when their high-ranking mate has lost an interaction with
a simulated intruder, and rarely if he has won. Data from such experiments (Ch. 7)
show that the females are capable of processing information from at least two
sources and making comparisons: A has beaten B, so B is less appealing. However,
such data merely suggest that rankings can be made on the basis of several com-
parisons; for transitive inference, the question is whether females rank a number of
different males and, if so, must they use overt interactions or can they interpolate
(true transitive inference)? Note, too, that several females will be competing for
the winning male in nature and female quality must also be taken into account.
An interesting study would be to see how females judge their relative quality and
whether they use some form of transitive inference.
At a different level are results from nightingales using a simulated playback
between two rivals; the target male noted which rival was overlapping the other
(a sign of dominance) and proceeded to respond more strongly to the overlapper
(Naguib & Todt, 1997; Naguib et al., 1999). The targeted male processed the inter-
action it heard, apparently viewed the overlapper as the greater threat and chose
to react as though he needed to establish dominance over the overlapper: that is,
given that A has beaten B, I had better beat A because he poses the greater threat;
the untested inference is that B will be attending and will not have to be dealt with
independently. Additional compelling data come from a study on male great tits
Parus major (Peake et al., 2002; Ch. 2), which also appear to base their interactions
with a simulated stranger on how that stranger has fared with a neighbour of
known rank with respect to themselves.
Another interesting case involves female great tits, who appear to decide
whether to enter a male neighbour’s territory based on eavesdropping upon ex-
perimentally manipulated interactions between a stranger and her mate and the

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
576 I. M. Pepperberg
same stranger and said neighbouring male. The female makes her decision by
inferring the ranking of the two resident males based on their respective abilities
in dealing with the same intruder and is much more likely to enter the terri-
tory of the neighbour if he is inferred to be dominant to her mate (Otter et al.,
1999; Chs. 2 and 7; see also Fig. 2.1b, p. 20). Interestingly, because the relative
ranking of males chosen for the experiment was unknown and the choice was
random as to whether a given playback would simulate a dominant or a subordi-
nate interaction, the information might counter what she knows about previous
interactions between her mate and her neighbour. Even if her response was merely
to obtain more information, the experiment shows how much attention is paid
to such interactions. Of course, her mate might act differently overall after los-
ing a simulated encounter, and no one has yet observed natural interactions of
this type, although experiments are underway to examine this possibility (K. A.
Otter, personal communication; note that Mennill et al. (2002) did not observe any
post-playback behavioural differences in chickadees).
However, how do these interactions demonstrate cognitive complexity? Is the
level of complexity as great as it is in the human case? Is cognition involved at all,
or are some other mechanisms at play, as in the case of the trained pigeons?
Discussion
These field studies did not test several levels of inference as did occur in
the laboratory studies: that is, the birds were not exposed to a large number of
different interactions among a simulated intruder and several different neighbours
whose rankings were known and asked to rank the simulated intruder with respect
to an untested comparison with these birds. Yet the field studies did demonstrate
an interesting level of cognitive complexity. The male birds were asked to place
themselves inside the rankings and to determine how they were likely to fare in
a previously untested situation; the female birds appeared to act on an inference
based on their observations. To determine the specific complexity of the situation,
let us deconstruct two of the field tasks.
In one task involving male responses, the bird judges the relative worth of the
intruders in order to decide how to respond (Peake et al., 2002; see also Fig. 2.1e,
p. 20). First, the bird must recognize that stranger A is in its territory, duel with
it, then store its own rank with respect to that stranger. It must then attend to
an interaction outside its territory between stranger A and a second stranger, B,
and determine which has the higher rank. Subsequently, it must listen to one of
those strangers, determine that it was B and not A (with whom it had previously
interacted), remember this stranger’s rank with respect to the bird A with whom

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 577
it did interact, remember its rank with respect to A, and then infer whether it has
a chance against B or whether it should avoid B.
In the task involving female responses, a bird judges the relative worth of her
mate and neighbours based on the males’ interactions with a simulated intruder
in order to decide how to respond (Otter et al., 1999; see also Fig. 2.1b, p. 20). First,
the female has to distinguish her various neighbours from her mate and has likely
stored the relative worth of her mate M, and each neighbour N. She must identify
a new male, S, listen and determine his rank in a contest versus M, and then in
another contest versus N. She must store and compare these two rankings and
then infer the relative ranking of M and N based on their rankings with S, possibly
updating her stored original memory. Although not examined, of interest would
be whether she would try to search out S if he beat both M and N.
Conceivably, birds, like humans (Duchaine et al., 2001), have an easier time
making decisions that involve a social, familiar setting than they do if the same
decision is required in an abstract context (Greene et al., 2001). That is, these tasks
are somewhat simplified by being explicitly important to the bird’s survival and
its reproductive success. Nevertheless, both situations may involve reversal and
neither involves specific pairwise rewards; consequently, the likelihood of the
results being merely some experimental artefact as in the case of, for example,
the pigeon studies is unlikely.
Clearly, of future interest would be the addition of simulated interactions by
more intruders, C and D with A or B, and with the subjects’ other neighbours to
determine how many dominance relationships a bird might encode. Chickadees
may present an interesting case: their sense of overall ranking at winter feeding
stations may be settled well in advance of their daily interactions, because the
signals between any two birds landing at a feeding station are somewhat cursory
(Popp et al., 1990). Given that such flocks involve approximately a dozen birds, the
data suggest that individual birds may have some general understanding of their
rank on a global basis, that is, via transitive relationships (K. A. Otter, personal
communication). Could an experiment be designed to test whether (or how) a
subject could rank others independent of their relationships to himself, or would
a bird be able to rank others only in relationship to its need to avoid or engage in a
direct confrontation? Possibly the rankings with respect to self actually complicate
the issue, in that the bird must demonstrate some level of self-awareness as to
where it fits into the hierarchy.
Self-awareness is a separate but related issue in terms of animal abilities (e.g.
Griffin, 1998) and merits some discussion in the present case, at least for clar-
ification. Self-awareness, as used here, is distinguishable from ‘consciousness’:
the full-blown central monitoring of sensory inputs and mental states, executive

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
578 I. M. Pepperberg
control of decision making and voluntary action, awareness of one’sown thoughts
(being aware that one is aware (Carruthers, 1992)) and attribution of mental states
to others (see discussion in Pepperberg & Lynn, 2000). Here, awareness describes
a state of higher-order cognition in which information is represented, processed
and used to control behaviour (e.g. Pepperberg, 1992). Thus the male tit is aware of
its relationship to A, aware of A’s relationship to B, and makes a decision based on
processing of these pieces of information; likewise, the female tit is aware of its re-
lationships to M and N, the relationships between these males and S, and makes a
decision based on the processing of these pieces of information. The point of evok-
ing awareness is that the tits will probably have a mental representation of these
pieces of information (e.g. Saidel, 2002) and use this representation in a ‘mind’s
eye’ view to make a decision; a researcher would be hard pressed to character-
ize the tits’ processing in any of the non-cognitive, non-representational manners
used to characterize the laboratory-based pigeon studies described above. The tits,
however, may not be consciously aware of their use of these representations (e.g.
be reacting to the specific situation in which, for example, its relationship to B
is unclear by consciously weighing all the possible risks and future benefits on a
personal basis and imputing the same to B, rather than reacting by chance based
on lack of information). Devising a test to uncover conscious processing would be
difficult.
Summary
In sum, at least some birds appear capable of solving transitive inference
tasks when dealing with a network of information, thus demonstrating complex
cognitive processing requiring the formation of several representations, exten-
sive memory for these various representations, and the ability to make inferences
based on a hierarchical organization of these representations. The situations pre-
sented to great tits (Otter et al., 1999; Peake et al., 2002) are at least as complicated as
those presented vocally to young children, and the results are not likely artefacts
of experimental manipulation. Should Alex’s preliminary data hold, African grey
parrots will also have succeeded in transitive inference. Although many objections
exist to evaluating animal intelligence and cognition based on human tasks (see
Pepperberg, 2001), the issue of importance here is that animals in nature are often
faced with the same types of task as their human counterparts (both at present and
historically) and so have been faced with the same evolutionary pressures on cog-
nitive development. In such circumstances, evaluating their competence on what
at first appears to be a human task does not ignore their natural behaviour, their
motivations, their ecological niches or their sensorimotor competence. Rather,
when animals are given tasks that are fully comparable to those given to humans

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 579
with respect both to ecological validity and experimental control, we can make
clear comparisons of animal and human abilities.
Acknowledgements
Writing of this manuscript was supported by the MIT School of Architecture and Plan-
ning and a grant from the American Foundation. Research on African grey parrots was supported by
NSF (IBN 96–03803) and REU supplements, the John Simon Guggenheim Foundation, the Kenneth
A. Scott Charitable Trust, the Pet Care Trust, the University of Arizona Undergraduate Biology
Research Program and many donors to the Alex Foundation.
References
Baptista, L. F. & Catchpole, C. K. 1989. Vocal mimicry and interspecific aggression in
songbirds: experiments using white-crowned sparrow imitation of song sparrow
song. Behaviour, 109, 247–257.
Beecher, M. D., Stoddard, P. K., Campbell, S. E. & Horning, C. L. 1996. Repertoire
matching between neighbouring song sparrows. Animal Behaviour, 51, 917–923.
Boysen, S. T., Berntson, G. G., Shreyer, T. A. & Quigley, K. S. 1993. Processing of
ordinality and transitivity by chimpanzees (Pan troglodytes). Journal of Comparative
Psychology, 107, 208–215.
Burt, J. M. 2000. Use of a radio microphone array to study banded wren song
interactions at the neighborhood level. Journal of the Acoustical Society of America,
108, 2583.
Burt, J. M., Bard, S. C., Campbell, S. E. & Beecher, M. D. 2002. Alternative forms of song
matching in song sparrows. Animal Behaviour, 63, 1143–1151.
Carruthers, P. 1992. The Animals Issue: In Search of a Fundamental Theory. New York:
Oxford University Press.
Couvillon, P. A. & Bitterman, M. E. 1992. A conventional conditioning analysis of
‘transitive inference’ in pigeons. Journal of Experimental Psychology: Animal Behavior
Processes, 18, 308–310.
Dabelsteen, T., McGregor, P. K., Holland, J., Tobais, J. & Pedersen, S. B. 1997. The signal
value of overlapping singing in male robins (Erithacus rubecula). Animal Behaviour,
53, 249–256.
Delius, J. D. & Siemann, M. 1998. Transitive responding in animals and humans:
exaptation rather than adaptation? Behavioural Processes, 42, 107–137.
Delius, J. D., Jitsumori, J. & Siemann, M. 2000. Stimulus equivalencies through
discrimination reversals. In: The Evolution of Cognition, ed. C. Heyes & L. Haber.
Cambridge, MA: MIT Press, pp. 103–122.
Duchaine, B., Cosmides, L. & Tooby, J. 2001. Evolutionary psychology and the brain.
Current Opinion in Neurobiology, 11, 225–230.
Egan, D. E. 1983. Retrospective reports reveal differences in people’s reasoning. Bell
System Technical Journal, 62, 1675–1697.

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
580 I. M. Pepperberg
Geary, D. C., Hamson, C. O. & Hoard, M. K. 2000. Numerical and arithmetical
cognition: a longitudinal study of process and concept deficits in children with
learning disability. Journal of Experimental Child Psychology, 77, 236–263.
Gillan, D. J. 1981. Reasoning in the chimpanzee: II. Transitive inference. Journal of
Experimental Psychology: Animal Behavior Processes, 7, 150–164.
Godard, R. 1991. Long-term memory of individual neighbours in a migratory songbird.
Nature, 350, 228–229.
Greene, A. J., Spellman, B., Dusek, J. A., Eichenbaum, H. B. & Levy, W. B. 2001.
Relational learning with and without awareness: transitive inference using
nonverbal stimuli in humans. Memory & Cognition, 29, 893–902.
Griffin, D. R. 1998. From cognition to consciousness. Animal Cognition, 1, 3–16.
Hogue, M.-E., Beaugrand, J. P. & Lague, P. C. 1996. Coherent use of information by hens
observing their former dominant defeating or being defeated by a stranger.
Behavioural Processes, 38, 241–252.
Hulse, S. H., Fowler, H. & Honig, W. K. (eds.) 1968. Cognitive Processes in Animal Behavior.
Hillsdale, NJ: Erlbaum.
Jarvis, E. D. & Mello, C. V. 2000. Molecular mapping of brain areas involved in parrot
vocal communication. Journal of Comparative Neurology, 419, 1–31.
Kroodsma, D. E. 1979. Vocal dueling among male marsh wrens: evidence for ritualized
expressions of dominance/subordinance. The Auk, 96, 506–515.
Kroodsma, D. E. & Byers, B. E. 1998. Songbird song repertoires: an ethological approach
to studying cognition. In: Animal Cognition in Nature, ed. R. P. Balda, I. M.
Pepperberg & A. C. Kamil. London: Academic Press, pp. 305–336.
Marler, P. 1970. A comparative approach to vocal learning: song development in
white-crowned sparrows. Journal of Comparative and Physiological Psychology, 71,
1–25.
McGonigle, B. & Chalmers, M. 1984a. The selective impact of question form and input
mode on the symbolic distance effect in children. Journal of Experimental Child
Psychology, 37, 525–554.
1984b. Are children any more logical than monkeys on the five-term series
problem? Journal of Experimental Child Psychology, 37, 355–377.
McGregor, P. K., Dabelsteen, T., Shepherd, M. & Pedersen, S. B. 1992. The signal value of
matched singing in great tits: evidence from interactive playback experiments.
Animal Behaviour, 43, 987–998.
McGregor, P. K., Dabelsteen, T. & Holland, J. 1997. Eavesdropping in a territorial
songbird communication network: preliminary results. Bioacoustics, 8, 253–254.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Menzel, E. W., Jr & Juno, C. 1982. Marmosets (Saguinus fuscicollis): are learning sets
learned? Science, 217, 750–752.
Morgane, P. J., Jacobs, M. S. & Galaburda, A. 1986. Evolutionary morphology of the
dolphin brain. In: Dolphin Cognition and Behavior: A Comparative Approach, ed. R. J.
Schusterman, J. A. Thomas & F. G. Wood. Hillsdale, NJ: Erlbaum, pp. 5–29.
Naguib, M. & Todt, D. 1997. Effects of dyadic vocal interactions on other conspecific
receivers in nightingales. Animal Behaviour, 54, 1535–1543.

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
Cognitive aspects of networks and avian capacities 581
Naguib, M., Fitchel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society London,
Series B, 266, 537–542.
Otter, K. A., McGregor, P. K., Terry, A. M. R. et al. 1999. Do female great tits Parus major
assess males by eavesdropping? A field study using interactive song playback.
Proceedings of the Royal Society London, Series B, 265, 1045–1049.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male interaction. Proceedings of the Royal Society
London, Series B, 268, 1183–1187.
2002. Do great tits assess rivals by combining direct experience with information
gathered by eavesdropping? Proceedings of the Royal Society London, Series B, 269,
1925–1929.
Pepperberg, I. M. 1987. Evidence for conceptual quantitative abilities in the African
grey parrot: labeling of cardinal sets. Ethology, 75, 37–61.
1990. Cognition in an African grey parrot (Psittacus erithacus): further evidence for
comprehension of categories and labels. Journal of Comparative Psychology, 104,
41–52.
1992. Proficient performance of a conjunctive, recursive task by an African grey
parrot (Psittacus erithacus). Journal of Comparative Psychology, 106, 295–305.
1994. Numerical competence in an African grey parrot. Journal of Comparative
Psychology, 108, 36–44.
1999. The Alex Studies: The Cognitive and Communicative Abilities of Grey Parrots.
Cambridge, MA: Harvard University Press.
2001. Evolution of avian intelligence. In: The Evolution of Intelligence, ed. R. Sternberg &
J. Kaufman. Mahwah, NJ: Erlbaum, pp. 315–337.
Pepperberg, I. M. & Brezinsky, M. V. 1991. Relational learning by an African grey parrot
(Psittacus erithacus): discriminations based on relative size. Journal of Comparative
Psychology, 105, 286–294.
Pepperberg, I. M. & Lynn, S. K. 2000. Possible levels of animal consciousness with
reference to grey parrots (Psittacus erithacus). American Zoologist, 40, 893–901.
Popp, J. W., Ficken, M. S. & Weise, C. M. 1990. How are agonistic encounters among
black-capped chickadees resolved? Animal Behaviour, 39, 980–986.
Premack, D. 1978. On the abstractness of human concepts: why it would be difficult to
talk to a pigeon. In: Cognitive Processes in Animal Behavior, ed. S. H. Hulse, H. Fowler &
W. K. Honig. Hillsdale, NJ: Erlbaum, pp. 421–451.
Russell, I. S. 1979. Brain size and intelligence: a comparative perspective. In: Brain,
Behavior and Evolution, ed. D. A. Oakley & H. C. Plotkin. London: Methuen,
pp. 126–153.
Saidel, E. 2002. Animal minds, human minds. In: The Cognitive Animal, ed. M. Bekoff,
C. Allen & G. M. Burghardt. Cambridge, MA: Bradford Books and MIT Press,
pp. 53–57.
Sarich, V. M & Cronin, J. E. 1977. Generation length and rates of hominid evolution.
Nature, 269, 354–355.
Siemann, M. 1993. Transitive Inferenz: Experimentelle Untersuchung einer
kognitiven Leistund. [Transitive inference: experimental investigation of a

P1: IYP/... P2: IYP0521823617c24.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 13:31
582 I. M. Pepperberg
cognitive performance.] Ph.D. Thesis, University of Konstanz. [Cited in Delius &
Siemann, 1998.]
Stoddard, P. K. 1996. Vocal recognition of neighbors by territorial passerines. In:
Ecology and Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H.
Miller. Ithaca, NY: Cornell University Press, pp. 356–374.
Todt, D. & Hultsch, H. 1998. Hierarchical learning, development and representation of
song. In: Animal Cognition in Nature, ed. R. Balda, I. M. Pepperberg & A. C. Kamil.
London: Academic Press, pp. 275–303.
von Ferson, L. 1989. Kognitive Prozesse bei Tauben (Columba livia). [Cognitive processes in
pigeons (Columba livia).] Pfaffenweiler: Centaurus.
Weaver, J. E., Steirn, J. N. & Zentall, T. R. 1997. Transitive inference in pigeons: control
for differential value transfer. Psychonomic Bulletin & Review, 4, 113–117.
Werner, U. B., Koeppl, U. & Delius, J. D. 1992. Transitive Inferenz bei nicht-verbaler
Aufgabendarbietung. [Transitive inference in nonverbal task presentation.]
Zeitschrift fur Experimentelle und Angewandte Psychologie, 39, 662–683.
Woocher, F. D., Glass, A. L. & Holyoak, K. J. 1978. Positional discriminability in linear
orderings. Memory & Cognition, 6, 165–173.
Wynne, C. D. L. 1997. Pigeon transitive inference: tests of simple accounts of a
complex performance. Behavioural Processes, 39, 95–112.
Zentall, T. R. 2001. The case for a cognitive approach to animal learning and behavior.
Behavioural Processes, 54, 65–78.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
25
Social complexity and the informationacquired during eavesdropping byprimates and other animals
d o ro t h y l . c h e n e y & ro b e r t m . s e y fa r t h
University of Pennsylvania, Philadelphia, USA
Introduction
In many of the studies reviewed in this book, eavesdropping takes the
following form: a subject has the opportunity to monitor, or eavesdrop upon, an
interaction between two other animals, A and B. The subject then uses the informa-
tion obtained through these observations to assess A’s and B’s relative dominance
or attractiveness as a mate (e.g. Mennill et al., 2002; Ch. 2). For example, Oliveira
et al. (1998) found that male fighting fish Betta splendens that had witnessed two
other males involved in an aggressive interaction subsequently responded more
strongly to the loser of that interaction than the winner. Subjects’behaviour could
not have been influenced by any inherent differences between the two males, be-
cause subjects responded equally strongly to the winner and the loser of compet-
itive interactions they had not observed. Similarly, Peake et al. (2001) presented
male great tits Parus major with the opportunity to monitor an apparent competi-
tive interaction between two strangers by simulating a singing contest using two
loudspeakers. The relative timing of the singing bouts (as measured by the degree
of overlap between the two songs) provided information about each ‘contestant’s’
relative status. Following the singing interaction, one of the ‘contestants’ was
introduced into the male’s territory. Males responded significantly less strongly
to singers that had apparently just ‘lost’ the interaction (see also McGregor &
Dabelsteen, 1996; Naguib et al., 1999; Ch. 2).
What information does an individual acquire when it eavesdrops on others?
In theory, an eavesdropper could acquire information of many different sorts:
about A, about B, about the relationship between A and B, or about the place of
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
583

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
584 D. L. Cheney & R. M. Seyfarth
A’s and B’s relationship in a larger social framework. The exact information ac-
quired will probably reflect the particular species’ social structure. For example,
songbirds like great tits live in communities in which six or seven neighbours
surround each territory-holding male. Males appear to benefit from the knowl-
edge that certain individuals occupy specific areas (e.g. Brooks & Falls, 1975), that
competitive interactions between two different neighbours have particular out-
comes, and that these outcomes are stable over time. We would, therefore, expect
an eavesdropping great tit not only to learn that neighbour A was dominant to
neighbour B, for example, but also to form the expectation that A was likely to
defeat B in all future encounters. More speculatively, because the outcome of ter-
ritorial interactions are often site specific (reviewed by Bradbury & Vehrencamp,
1998), we would expect eavesdropping tits to learn further that A dominates B
in some areas but B dominates A in others. In contrast, the information gained
from monitoring neighbours’ interactions would unlikely be sufficient to allow
the eavesdropper to rank all of its neighbours in a linear dominance hierarchy,
because not all neighbouring males would come into contact with one another.
Such information would be difficult if not impossible to acquire; it might also be
of little functional value.
In contrast, species that live in large, permanent social groups have a much
greater opportunity to monitor the social interactions of many different indi-
viduals simultaneously. Monkey species such as baboons Papio cynocephalus, for
example, typically live in groups of 80 or more individuals, which include several
matrilineal families arranged in a stable, linear dominance rank order (Silk et al.,
1999). Offspring assume ranks similar to those of their mothers, and females main-
tain close bonds with their matrilineal kin throughout their lives. Cutting across
these stable long-term relationships based on rank and kinship are more tran-
sient bonds: for example, the temporary associations formed between unrelated
females whose infants are of similar ages, and the ‘friendships’ formed between
adult males and lactating females as an apparent adaptation against infanticide
(Palombit et al., 1997, 2001). In order to compete successfully within such groups, it
would seem advantageous for individuals to recognize who outranks whom, who
is closely bonded to whom, and who is likely to be allied to whom (Harcourt, 1988,
1992; Cheney & Seyfarth, 1990; see below). The ability to adopt a third party’s per-
spective and discriminate among the social relationships that exist among others
would seem to be of great selective benefit.
In this chapter, we review evidence for eavesdropping in selected primate
species and we consider what sort of information is acquired when one individual
observes or listens in on the interactions of others. We then compare eavesdrop-
ping by primates with eavesdropping in other animal species, focusing on both
potential differences and directions for further research.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 585
B1 threat-grunt + E1 scream B and E
B1 threat-grunt + D scream B and E
A threat-grunt + C scream B and E
Playback sequence Subjects
Test
Control 1(dominant’s kin)
Control 2(no kin)
Fig. 25.1. The protocol for playback experiments testing baboon females’ recognition
of other individuals’ kin. B, the more dominant of the subjects; E, the more
subordinate; B1 and E1, the subjects’ close kin; A, C and D, signallers unrelated to
either subject.
Knowledge about other animals’ kin
Some of the first evidence that monkeys recognize other individuals’ so-
cial relationships emerged as part of a relatively simple playback experiment de-
signed to document individual vocal recognition in vervet monkeys Cercopithecus
aethiops (Cheney & Seyfarth, 1980). We had noticed that mothers often ran to sup-
port their juvenile offspring when these individuals screamed during aggressive
interactions. This observation, like many others (e.g. Hansen, 1976; Gouzoules
et al., 1984), suggested that mothers recognized the calls of their offspring. To
test this hypothesis, we designed a playback experiment in which we played the
distress scream of a juvenile to a group of three adult females, one of whom
was the juvenile’s mother. As expected, mothers consistently looked toward the
loudspeaker for longer durations than did control females. Even before she had re-
sponded, however, a significant number of control females looked at the mother.
In so doing, they behaved as if they recognized not only the identity of signallers
unrelated to themselves but also associated those individuals with specific adult
females (Cheney & Seyfarth, 1980, 1982).
In an attempt to replicate these results, we carried out a similar set of ex-
periments on free-ranging baboons in the Okavango Delta of Botswana. In these
experiments, two unrelated female subjects were played a sequence of calls that
mimicked a fight between their close relatives (Fig. 25.1). The females’ immediate
responses to the playback were videotaped and both subjects were followed for
15 minutes after the playback to determine whether their behaviour was affected
by the calls they had heard. In separate trials, the same two subjects also heard two
control sequences of calls (Fig. 25.1). The first sequence mimicked a fight involving
the dominant subject’s relative and an individual unrelated to either female; the
second mimicked a fight involving two individuals who were both unrelated to
either female (for details see Cheney & Seyfarth, 1999).
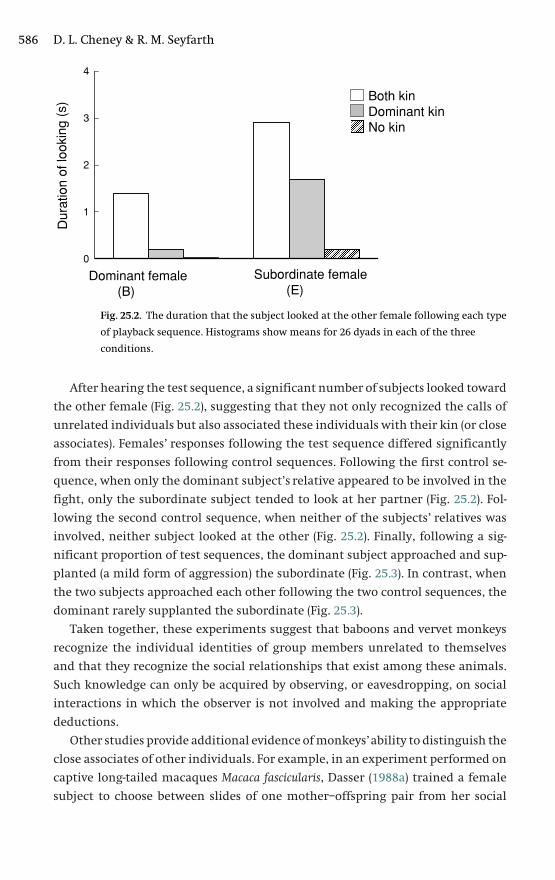
P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
586 D. L. Cheney & R. M. Seyfarth
0
1
2
3
4D
urat
ion
of lo
okin
g (s
) Both kinDominant kinNo kin
Dominant female(B)
Subordinate female(E)
Fig. 25.2. The duration that the subject looked at the other female following each type
of playback sequence. Histograms show means for 26 dyads in each of the three
conditions.
After hearing the test sequence, a significant number of subjects looked toward
the other female (Fig. 25.2), suggesting that they not only recognized the calls of
unrelated individuals but also associated these individuals with their kin (or close
associates). Females’ responses following the test sequence differed significantly
from their responses following control sequences. Following the first control se-
quence, when only the dominant subject’s relative appeared to be involved in the
fight, only the subordinate subject tended to look at her partner (Fig. 25.2). Fol-
lowing the second control sequence, when neither of the subjects’ relatives was
involved, neither subject looked at the other (Fig. 25.2). Finally, following a sig-
nificant proportion of test sequences, the dominant subject approached and sup-
planted (a mild form of aggression) the subordinate (Fig. 25.3). In contrast, when
the two subjects approached each other following the two control sequences, the
dominant rarely supplanted the subordinate (Fig. 25.3).
Taken together, these experiments suggest that baboons and vervet monkeys
recognize the individual identities of group members unrelated to themselves
and that they recognize the social relationships that exist among these animals.
Such knowledge can only be acquired by observing, or eavesdropping, on social
interactions in which the observer is not involved and making the appropriate
deductions.
Other studies provide additional evidence of monkeys’ability to distinguish the
close associates of other individuals. For example, in an experiment performed on
captive long-tailed macaques Macaca fascicularis, Dasser (1988a) trained a female
subject to choose between slides of one mother–offspring pair from her social

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 587
0
10
20
30
40
50
Dominantsupplants
Per
cent
age
of tr
ials
Both kinDominant kinNo kin
Dominantapproaches
Subordinateapproaches
Fig. 25.3. The percentage of subjects’ first interactions with each other that took
various forms following each playback sequence. Histograms show means for 26 dyads
in each condition. Dominant supplants indicates that the dominant subject
approached and supplanted the more subordinate subject. Dominant approaches
indicates that the dominant subject approached the subordinate subject without
supplanting her and/or interacted with her in a friendly manner. Subordinate
approaches indicates that the subordinate subject approached the dominant subject
and/or interacted with her in a friendly manner.
group and slides of two unrelated individuals from her group. Having learned to
respond to one mother–offspring pair, the subject was then tested with 14 novel
slides of familiar mothers and offspring paired with an equal number of novel
slides of familiar unrelated animals matched for age and sex. In all tests, she
correctly selected the mother–offspring pair. In so doing, she appeared to use an
abstract category to classify pairs of individuals that was analogous to our concept
of ‘mother–child affiliation’. Dasser (1988a) was able to exclude the possibility that
mothers and offspring were matched according to physical resemblance, because
subjects were unable to match unfamiliar mothers and offspring. Instead, indi-
viduals appeared to be classified according to their degree of association. Again,
such knowledge of other individuals’ close associates can only be obtained by
monitoring, or eavesdropping upon, their social interactions.
Under natural conditions, it is difficult to determine whether animals distin-
guish between different categories of social relationships. Do monkeys recognize,
for example, that mother–offspring bonds are distinct from sibling bonds or friend-
ships even when all are characterized by high rates of interaction? In perhaps the
only test of monkeys’ ability to recognize different categories of social affiliation,
Dasser (1988b) trained a long-tailed macaque to identify a pair of siblings from

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
588 D. L. Cheney & R. M. Seyfarth
her social group and then tested her ability to distinguish novel slides of familiar
sibling pairs from familiar mother–offspring pairs, familiar pairs of less-closely-
related matrilineal kin and familiar unrelated pairs. Although the subject did
distinguish siblings from unrelated pairs and pairs of less-closely-related individ-
uals, she was unable to discriminate between siblings and mothers and offspring.
This failure may have occurred because the same female had previously been re-
warded for picking the mother–offspring pair. It is also possible, however, that
she did not distinguish between different kinship categories and simply chose the
pair that was more closely affiliated.
Natural patterns of aggression also reflect the knowledge that monkeys have
of their group’s social network. In many monkey species, an individual who has
just threatened or been threatened by another animal will often ‘redirect aggres-
sion’ by threatening a third, previously uninvolved, individual. Judge (1982) was
the first to note that redirected aggression in rhesus macaques Macaca mulatta
does not always occur at random. Rather than simply threatening any nearby
individual, animals will instead specifically target a close matrilineal relative of
their recent opponent. Similar kin-biased redirected aggression occurs in Japanese
macaques Macaca fuscata (Aureli et al., 1992) and vervets (Cheney & Seyfarth, 1986,
1989). Kazem & Aureli (Ch. 10) further discuss the relationship between redirected
aggression and communication networks.
Knowledge about other animals’ dominance ranks
Dominance ranks offer another opportunity to test whether non-human
primates gain information about other animals’relationships by eavesdropping on
their social interactions. Like matrilineal kinship, linear, transitive dominance re-
lations are a pervasive feature of social behaviour in groups of Old World monkeys.
A linear, transitive rank order might emerge because individuals simply recognize
who is dominant or subordinate to themselves. In this case, a linear hierarchy
would occur as an incidental outcome of paired interactions and there would be
no evidence to suggest that animals eavesdropped on others’interactions. Alterna-
tively, a linear hierarchy might emerge because individuals genuinely recognize
the transitive dominance relations that exist among others: a middle-ranking in-
dividual, for example, might know that A is dominant to B and B is dominant
to C and, therefore, conclude that A must be dominant to C. Like knowledge of
matrilineal kin, such knowledge could only be acquired through eavesdropping
on the interactions of others.
In many species of Old World monkeys, female dominance ranks are deter-
mined by the rank of an individual’smatriline (Walters & Seyfarth, 1987; Chapais,
1988). Knowledge of another female’s rank cannot, therefore, be obtained by

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 589
attending to absolute attributes such as age or size; instead, it demands the mon-
itoring of other individuals’ interactions. Several observations and experiments
suggest that monkeys do recognize the rank relations that exist among other
females in their group. For example, dominant female baboons often grunt to
mothers with infants as they approach the mothers and attempt to handle or
touch their infants. Grunts seem to function to facilitate social interactions by ap-
peasing anxious mothers, because an approach accompanied by a grunt is signifi-
cantly more likely to lead to subsequent friendly interaction than is an approach
without a grunt (Cheney et al., 1995a). Occasionally, however, a mother will utter a
submissive call, or ‘fear bark’, as a dominant female approaches. Fear barks are an
unambiguous indicator of subordination; they are never given to lower-ranking
females.
To test whether baboons recognize that only a more dominant animal can
cause another individual to give a fear bark, we designed a playback experiment
in which adult female subjects were played a causally inconsistent call sequence
in which a low-ranking female apparently grunted to a high-ranking female and
the higher-ranking female apparently responded with fear barks. As a control, the
same subjects heard the same sequence of grunts and fear barks made causally
consistent by the inclusion of additional grunts from a third female who was dom-
inant to both of the other signallers. For example, if the inconsistent sequence was
composed of female 6’sgrunts followed by female 2’sfear barks, the corresponding
consistent sequence might begin with female 1’s grunts, followed by female 6’s
grunts and ending with female 2’s fear barks. Some subjects were higher-ranking
than the signallers; others were lower ranking. Regardless of their own relative
ranks, subjects responded significantly more strongly to the causally inconsistent
sequences, suggesting that they recognize not only the identities of different sig-
nallers but also the rank relations that exist among others in their group (Cheney
et al., 1995b).
Further suggestion that monkeys recognize other individuals’ ranks comes
from observations on competition among adult female vervet monkeys for access
to a grooming partner (Seyfarth, 1980). Such competition occurs when one fe-
male approaches two that are grooming, supplants one of them and then grooms
with the female that remains. Interestingly, in those cases when a female ap-
proaches two groomers who are both subordinate to her, the lower ranking
of the two groomers typically moves away, while the higher ranking remains
(Cheney & Seyfarth, 1990). By remaining seated, the higher ranking of the two
groomers acts as if she recognizes that, although they are both lower ranking
than the approaching female, she is the higher ranking. Though not definitive,
these observations suggest that females recognize not only their own status rela-
tive to other individuals but also other individuals’ status relative to each other.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
590 D. L. Cheney & R. M. Seyfarth
In other words, they appear to recognize a rank hierarchy (Cheney & Seyfarth,
1990).
The ability to rank other group members is perhaps not surprising, given the
evidence that captive monkeys and apes can be taught to rank objects according to
an arbitrary sequential order (D’Amato & Colombo, 1989; Treichler & van Tilberg,
1996), the amount of food contained within a container (Gillan, 1981), their size
or the number of objects contained within an array (e.g. Matsuzawa, 1985; Hauser
et al., 1996; Brannon & Terrace, 1998). What distinguishes the social example, how-
ever, is the fact that, even in the absence of human training, female monkeys seem
able to construct a rank hierarchy and then place themselves at the appropriate
location within it.
Knowledge about more transient social relationships
All of the studies discussed so far focus on interactions among females
in groups where matrilineal kin usually retain close bonds and similar ranks
throughout their lives. It might seem, therefore, that an individual could simply
memorize the close associates and relative ranks of other females and thereafter
navigate easily through a predictable network of social relationships. Not all social
and rank relationships, however, are as stable as those among matrilineal kin.
Some types of social bond are relatively transient, and some rank relationships –
particularly among adult males – change often. Nonetheless, there is evidence
that non-human primates also recognize these more transient associations.
For example, under natural conditions, male and female hamadryas baboons
Papio hamadryas form close, long-term bonds that can last for a number of years.
Potential rivals appear to recognize the ‘ownership’ of specific females by other
males and refrain from challenging those males for their females (Kummer et al.,
1974). Experiments conducted in captivity have shown that rival males assess the
strength of other males’ relationships with their females before attempting to
challenge them. They do not attempt to take over a male’s female if the pair ap-
pears to have a close social bond (Bachmann & Kummer, 1980). Although similar
experiments have not yet been conducted with savannah baboons, observational
data suggest that these baboons, too, recognize the temporary bonds, or ‘friend-
ships’, that are formed between males and lactating females (Palombit et al., 1997).
For example, Smuts (1985) observed that males who had recently been threatened
by another male often redirected aggression toward the female friends of their op-
ponent (see Dunbar (1983) for similar observations on gelada baboons Theropithecus
gelada).
Monkeys also seem to recognize the bonds that exist between males and particu-
lar infants. In Tibetan macaques Macaca thibetana, males are often closely affiliated

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 591
with a particular infant in the group. Competitive interactions between males are
mediated by the carrying of infants and a male will frequently carry an infant and
present it to another male. In a study of such carrying (or ‘bridging’) behaviour,
Ogawa (1995) observed that males more frequently provided other males with
those males’ affiliated infants than with other, non-affiliated infants.
Finally, there is evidence that monkeys recognize even very transient dom-
inance relations among others. Dominance among male vervets, baboons and
macaques is determined primarily by age, fighting ability, and, in some popula-
tions, the presence of alliance partners. As a result, rank relations among males are
considerably less stable than they are among females (Walters & Seyfarth, 1987).
In a study of a large social group of captive bonnet macaques Macaca radiata, Silk
(1993, 1999) found that males formed linear, transitive dominance hierarchies that
remained stable for only short periods of time. As in other primate species, males
occasionally attempted to recruit alliance support during aggressive interactions
(approximately 12% of all aggressive encounters). Significantly, males consistently
solicited allies that outranked both themselves and their opponents. Males did not
simply solicit the highest-ranking individual in the group or choose allies that out-
ranked only themselves. Instead, soliciting males seemed to recognize not only
their own rank relative to a potential ally but also the rank relation between the
ally and their opponent. If dominance ranks remained stable, this might not have
been a difficult task. However, over the course of one year, approximately half
of the 16 males changed dominance rank each month (data from Table 3 in Silk,
1993). The males’ apparent ability to keep track of such highly transient rank re-
lations suggests that they carefully monitored all aggressive interactions among
other males, constantly updated their list of relative ranks and placed themselves
accurately into each new list.
Eavesdropping by other mammals
Data from dolphins Tursiops truncatus and hyaenas Crocuta crocuta suggest
that non-human primates are not the only mammals in which individuals acquire
information about many different individuals’social relationships (for other mam-
mals see Chs. 17 and 18). When competing over access to females, male dolphins
form dyadic and triadic alliances with selected other males, and allies with the
greatest degree of partner fidelity are most successful in acquiring access to fe-
males (Connor et al., 1992, 1999, 2001). The greater success of high-fidelity alliances
raises the possibility that males in newly formed alliances, or in alliances that have
been less stable in the past, recognize the strong bonds that exist among others
and are more likely to retreat when they encounter rivals with a long history of
cooperative interaction.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
592 D. L. Cheney & R. M. Seyfarth
Like many species of Old World monkeys, hyaenas live in social groups compris-
ing matrilines in which offspring inherit their mothers’ dominance ranks (Smale
et al., 1993; Engh et al., 2000). Holekamp et al. (1999) played recordings of cubs’
‘whoop’ calls to mothers and other breeding females. As with vervet monkeys
and baboons, hyaena females responded more strongly to the calls of their own
offspring and those of close relatives than to the calls of unrelated cubs. In con-
trast to vervets and baboons, however, unrelated females did not look at the cubs’
mothers. One explanation for these negative results is that hyaenas are unable to
recognize third-party relationships, despite living in social groups that are super-
ficially similar to those of many primates. It also remains possible, however, that
hyaenas are simply uninterested in the calls of unrelated cubs.
In fact, hyaenas’ patterns of alliance formation suggest that they do monitor
other individuals’ interactions and extrapolate information about other animals’
relative ranks from their observations. During competitive interactions over meat,
hyaenas often solicit alliance support from other, uninvolved individuals. When
choosing to join ongoing skirmishes, hyaenas that are dominant to both of the
contestants almost always support the more dominant of the two individuals
(Engh et al., 2004). Similarly, when the ally is intermediate in rank between the
two opponents, it inevitably supports the dominant individual. These data provide
the first evidence in a non-primate species that alliance partners may be chosen on
the basis of both the allies’and the opponents’relative ranks (Harcourt, 1988, 1992).
They are consistent with the hypothesis that hyaenas are able to infer transitive
rank relations among other group members.
Possible differences between primates and other animals
Do primates differ from other animals in their ability to infer third-party
social relationships through eavesdropping? We can identify at least three com-
peting hypotheses.
The first hypothesis argues that primates are in fact more intelligent than non-
primates. This intelligence is reflected not only in tests of captive animals but also
in primates’ superior ability to keep track of complex social relationships. The
difference between primates and non-primates is qualitative and fundamental
and will be corroborated by future research.
The second hypothesis maintains that selection has favoured the ability to rec-
ognize other individuals’ relationships in all species that live in large, complex
social groups. According to this hypothesis, monkeys only appear to have a greater
capacity to recognize third-party social relationships because they have received
more attention than non-primates living in similarly large groups. Once this im-
balance in research has been redressed, differences between primates and other

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 593
animals will disappear, to be replaced by a difference that depends primarily on
group size and composition.
The third hypothesis claims that neither phylogeny nor group size and composi-
tion have influenced animals’ ability to gain information about other individuals’
social relationships. It argues, in effect, that there are no species differences in
‘social intelligence’. Monkeys and hyaenas, for example, only appear to excel in
their ability to recognize the relative ranks of allies and opponents because their
large social groups allow them to display this knowledge. In contrast, studies of
species that live in small social groups have to date focused primarily on observers’
ability to assess the dominance of only two individuals. Once monogamous and
even solitary species have been given the opportunity to reveal what they know
about the social relationships of many different individuals, they will be shown
to possess a level of social intelligence that is no different from that found among
animals living in large social groups.
At present, it is difficult to test these alternative hypotheses; below we review
some information that may be relevant.
Hypothesis 1: primates have greater social intelligence than other species
Primates have larger brains for their body size than other vertebrates
(Martin, 1983). Dunbar (2000) argued that this arises because primate social groups
are not only larger but also more complex than those of other taxa. Primate groups
are typically composed of many reproductively active males and females, and
individuals interact regularly with both kin and non-kin, with whom they must
simultaneously cooperate and compete for resources. Such social complexity may
place strong selective pressure on the ability to recognize close associates of other
individuals.
To date, only monkeys and possibly dolphins have been shown to recognize
the affiliative relationships that exist among other group members. In monkey
groups, closely bonded individuals are usually matrilineal kin, but this is not
always the case. The ability to classify other individuals into matrilineal or closely
bonded subgroups is likely to be relatively complex, for several reasons.
Matrilineal kin groups vary in size and not all individuals within a kin group
interact at the same rate or in the same way. Moreover, no single behavioural mea-
sure underlies the associations between individuals and there is no threshold or
defining criterion for a ‘close’ social bond. For example, females in many monkey
species form the majority of their alliances with matrilineal kin, and high-ranking
kin usually form alliances at higher rates than low-ranking kin (reviewed by Silk,
1987; Walters & Seyfarth, 1987). There is no evidence, however, that other group
members more easily recognize the kin (or close associates) of high-ranking in-
dividuals than the kin of low-ranking individuals. Similarly, female kin usually

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
594 D. L. Cheney & R. M. Seyfarth
occupy adjacent dominance ranks. This rule of thumb, however, cannot reliably be
used to classify females into kin groups, because not all adjacently ranked females
are kin. We do not yet know whether monkeys discriminate among different types
of social bond: whether they distinguish, for example, among the bonds formed
by mothers and offspring, sisters, or friends. Moreover, the degree to which there
is a quantitative or qualitative threshold for learning to recognize that two other
individuals share a close bond is not known.
Furthermore, some social relationships among monkeys are transitive, while
others are not. For example, if infant A1 and juvenile A2 both associate at high
rates with a particular adult female A, it is usually correct to infer that the ju-
venile and infant are also closely bonded. Similarly, if A is dominant to B and B
is dominant to C, it is usually true that A is dominant to C. In other cases, how-
ever, transitivity cannot be assumed. If infant baboon A1 and juvenile baboon A2
both associate at high rates with the same adult female and she associates with
an adult male ‘friend’, we can infer that the male is probably also closely allied
to the infant. However, it would incorrect to assume that he is equally closely
allied to the juvenile, who may instead be more closely allied to another male
who was previously the mother’s friend (Seyfarth, 1978; Smuts, 1985; Palombit
et al., 1997). Baboon females from the same matriline often form friendships with
different males; conversely, the same male may form simultaneous friendships
with females from two different matrilines. In the latter case, the existence of a
close bond between a male and two females does not predict a close bond between
the two females. In fact, their relationship is likely to be as competitive as it is
friendly (Palombit et al., 2001).
Finally, as group size increases, the challenge of monitoring other individuals’
social relationships and dominance ranks increases exponentially. In a group of
80 animals (not an unusual size for many monkey species), each individual con-
fronts 3160 different possible dyadic combinations and 82 160 different triadic
combinations of individuals: numbers that may place considerable demands on
the observer’s memory and inferential abilities.
Preliminary evidence suggests that monkeys are able to monitor and remember
the social ranks and relationships of many individuals simultaneously. Despite
the lack of a consistent criterion for determining which individual is likely to be
closely bonded with which others, monkeys appear to be able to distinguish the
close associates of other group members. They appear to view their social groups
not just in terms of the individuals that constitute them but also in terms of a
web of social relationships in which certain individuals are linked with several
others.
Some learning experiments with captive animals support the view that pri-
mates are generally more adept than non-primates at classifying items according

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 595
to their relative relations. In oddity tests, for example, a subject is presented with
three objects, two of which are the same and one of which is different, and asked
to choose the object that is different. Monkeys and apes achieve high levels of ac-
curacy in such tests even when tested with novel stimuli (Harlow, 1949; D’Amato
et al., 1985; see also reviews by Tomasello & Call, 1997; Shettleworth, 1998). Baboons
and chimpanzees can also learn to make abstract discriminations about relations
between relations, matching patterns containing repeated samples of the same
item with similar ‘same’ patterns (Premack, 1983; Oden et al., 1988; Fagot et al.,
2001). In all cases, subjects’ performances suggest the use of an abstract hypoth-
esis, because concepts like ‘odd’ specify a relation between objects independent
of their physical features. In a similar manner, the concept ‘closely bonded’ can
be applied to any two individuals and need not be restricted to specific pairs that
look alike.
Judgements based on relations among items have been demonstrated more
often in non-human primates than in other taxa, and primates seem to recognize
abstract relations more readily than at least some other animals. Although it is
possible, for example, to train pigeons to recognize relations such as ‘same’, the
procedural details of the test appear more critical for pigeons than they are for
monkeys, and relational distinctions can easily be disrupted (Herrnstein, 1985;
Wright et al., 1988; Wasserman et al., 1995). Rather than attending to the relations
among stimuli, pigeons seem predisposed to focus on absolute stimulus properties
and to form item-specific associations (reviewed by Shettleworth, 1998). Similarly,
in tests of transitive inference, monkeys and apes appear to acquire a representa-
tion of series order that allows them to rank items even when some items in the
list are missing. In contrast, pigeons seem to attend primarily to the association
between adjacent pairs, which limits their ability to add or delete items from a list
(D’Amato & Colombo, 1989; von Fersen et al., 1991; Treichler & van Tilberg, 1996;
Zentall et al., 1996).
Hypothesis 2: differences in ‘social intelligence’ are related to group sizeand complexity
If, as has been hypothesized, the recognition of third-party relationships
confers a selective advantage because it allows individuals to remember who as-
sociates with whom, who outranks whom and who is allied to whom, we should
expect to find evidence for this ability not just in non-human primates but also
in any animal species that lives in large social groups composed of individuals of
varying degrees of dominance rank and genetic relatedness. We would also pre-
dict that selection should have acted less strongly on this ability in solitary species
and species living in small, egalitarian groups that are composed primarily either
of close kin or of unrelated individuals. Thus, the ability to recognize the close

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
596 D. L. Cheney & R. M. Seyfarth
associates of others should be evident in non-primate species such as hyaenas
and lacking or less evident in some ape species, including gorillas Gorilla gorilla
and orangutans Pongo pygmaeus. Although recent evidence that hyaenas recognize
other individuals’ relative ranks lends support to this hypothesis, other compara-
tive data are lacking. For example, no study has yet attempted to determine the
extent to which any ape species is able to recognize the social relationships of
other group members.
Within the Primate order, species that live in large groups have a relatively
larger neocortex than those that are solitary or live in small groups (Barton &
Dunbar, 1997). A similar relation is found in carnivores (Barton & Dunbar, 1997)
and toothed whales (Connor et al., 1998a,b; Marino, 1998), supporting the hypoth-
esis that sociality has favoured the evolution of large brains (see also Jolly, 1966;
Humphrey, 1976; Cheney & Seyfarth, 1990). Indeed, differences in social complex-
ity may exert their effect even in species that lack a cortex entirely. In paper wasps
Polistes dominulus, for example, there is a significant increase in the size of the
antennal lobes and collar (a substructure of the calyx of the mushroom body) in
females that nest colonially – with other queens – as opposed to solitary breeders
(Ehmer et al., 2001). This increase in neural volume may be favoured because so-
ciality places increased demand on the need to discriminate between familiar and
unfamiliar individuals and to monitor other females’ dominance and breeding
status. Clearly, therefore, neural correlates of sociality need not be restricted to
higher mammals.
Further supporting this argument are data from some other laboratory studies
suggesting fewer differences between primates and other animals in the ability
to make relational distinctions. For example, Alex, an African grey parrot Psitta-
cus erithacus, is reported to make explicit same/different judgements about sets
of objects (Pepperberg, 1992, Ch. 24). Similarly, sea lions Zalophus californianus
(Schusterman & Krieger, 1986; Schusterman & Gisiner 1988) and dolphins (Herman
et al. 1993; Mercado et al. 2000) have been taught to respond to terms such as ‘left’
and ‘bright’, which require the animals to assess relations among a variety of dif-
ferent objects. Finally, a number of species, including parrots (Pepperberg, 1994)
and rats (Church & Meck, 1984; Capaldi, 1993), are able to assess quantities, sug-
gesting that relatively abstract concepts of numerosity and transitivity may be
pervasive among animals (reviewed by Shettleworth, 1998).
Hypothesis 3: there are few differences in ‘social intelligence’ across species
Recent research on social eavesdropping (Ch. 2) by birds and fish indi-
cates that even animals living in small social groups are capable of acquiring
detailed information about other individuals’ relative dominance or attractive-
ness as a mate. Often, this information is of necessity restricted to a few other

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 597
individuals. For territorial species living in small family groups, questions about
the ability to track social relationships among many other individuals are largely
moot, because the opportunity to monitor interactions among all possible neigh-
bours rarely arises. Eavesdropping on the competitive singing duets of strangers,
for example, allows territorial songbirds to extract information about the two
contestants’ relative dominance. Whether these birds would also be capable of
recognizing a dominance hierarchy involving numerous individuals remains un-
clear. Although many species of songbirds form flocks during the winter, little
is known about the social interactions that take place within such flocks, or the
degree to which flock members recognize other individuals’ relative ranks (but
see Popp, 1987).
Recently, Bond et al. (2003) tested the prediction that socially living birds will
display enhanced abilities to make transitive inferences by comparing the per-
formance of highly social pinyon jays Gymnorhinus cyanocephalus with relatively
non-social western scrub jays Aphelocoma californica. Using operant procedures,
subjects were required to order a set of arbitrary stimuli by inference from a se-
ries of dyadic comparisons. Subjects of both species learned the sequence order,
but pinyon jays did so more rapidly and more accurately than scrub jays. Although
not conclusive, these results lend support to the hypothesis that social complexity
may be correlated with superior performance in tasks involving the ranking of
multiple stimuli (see also Hogue et al. (1996) for experiments with flock-dwelling
domestic chickens Gallus domesticus).
As yet, very little is known about the ability of non-primate mammals or birds
to recognize social relationships of other individuals. Colonial white-fronted bee-
eaters Merops bullockoides offer one example of an avian society in which there
would appear to be strong selective pressure for the recognition of the kin groups
of other individuals. Observational evidence suggests that bee-eaters may recog-
nize other individuals and kin groups and associate these groups with specific
feeding territories (Emlen et al., 1995), although this has not yet been tested
experimentally.
Clearly, more data are needed from both natural and laboratory studies before
we can draw any definitive conclusions about cognitive differences between pri-
mates and other animals, or between species living in large as opposed to small
groups. It remains entirely possible that apparent species differences between
primates and other animals in the recognition of third-party social relationships
result more from differences in the social context in which eavesdropping occurs
than from any cognitive differences in the ability to monitor social interactions.
Given the opportunity to evaluate the social relationships of many different indi-
viduals, species living in small family groups and even primarily solitary species
may well be shown to have similar abilities to those living in large social groups. It

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
598 D. L. Cheney & R. M. Seyfarth
is to be hoped that future research will attempt to investigate the extent to which
gregarious species in taxa other than primates are capable of recognizing the
close associates and allies of other group members, and to determine the neural
correlates, if any, of this ability.
Summary
Non-human primates are skilled voyeurs. By observing or listening to the
interactions of others, they acquire information about the social relationships
of other individuals and learn to place these relationships within a larger social
framework, such as a group of ranked, matrilineal families. Given the large, com-
plex societies in which monkeys cooperate and compete, the adaptive value of
such eavesdropping seems clear. At present, however, we do not know whether
the information acquired by eavesdropping in primates differs significantly from
the information acquired by individuals in other species. Primates (and a few
other mammals) may be qualitatively different from other species in their ability
to monitor the social relationships of many other individuals. Alternatively, the
societies of birds, fish and other non-primate species – often superficially simpler
than those of primates – may have led us to underestimate the information that
individuals acquire about others. Finally, both hypotheses may have some valid-
ity. There may be qualitative differences in social intelligence between different
taxonomic groups, but within each group the information acquired from eaves-
dropping may increase in sophistication with increasing social complexity. The
chapters in this volume demonstrate that eavesdropping is widespread among
animals. They set the stage for comparative research that examines differences
between species in the information acquired about others.
References
Aureli, F., Cozzolino, R., Cordischi, C. & Scucchi, S. 1992. Kin-oriented redirection
among Japanese macaques: an expression of a revenge system? Animal Behaviour,
44, 283–291.
Bachmann, C. & Kummer, H. 1980. Male assessment of female choice in hamadryas
baboons. Behavioral Ecology and Sociobiology, 6, 315–321.
Barton, R. A. & Dunbar, R. 1997. Evolution of the social brain. In: Machiavellian
Intelligence II: Extensions and Evaluations, ed. A. Whiten & R. W. Byrne. Cambridge,
UK: Cambridge University Press, pp. 240–263.
Bond, A. B., Kamil, A. C. & Balda, R. P. 2003. Social complexity and transitive inference
in corvids. Animal Behaviour, 65, 479–487.
Bradbury, J. W. & Vehrencamp, S. L. 1998. Principles of Animal Communication.
Sunderland, MA: Sinauer.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 599
Brannon, E. & Terrace, H. 1998. Ordering of the numerosities 1 to 9 by monkeys.
Science, 282, 746–749.
Brooks, R. J. & Falls, J. B. 1975. Individual recognition by song in white-throated
sparrows. III. Song features used in individual recognition. Canadian Journal of
Zoology, 53, 1749–1761.
Capaldi, E. J. 1993. Animal number abilities: implications for a hierarchical approach
to instrumental learning. In: The Development of Numerical Competence, ed. S. T.
Boysen & E. J. Capaldi. Hillsdale, NJ: Lawrence Erlbaum, pp. 191–209.
Chapais, B. 1988. Rank maintenance in female Japanese macaques: experimental
evidence for social dependency. Behaviour, 104, 41–59.
Cheney, D. L. & Seyfarth, R. M. 1980. Vocal recognition in free-ranging vervet monkeys.
Animal Behaviour, 28, 362–367.
1982. Recognition of individuals within and between groups of free-ranging vervet
monkeys. American Zoologist, 22, 519–529.
1986. The recognition of social alliances among vervet monkeys. Animal Behaviour,
34, 1722–1731.
1989. Reconciliation and redirected aggression in vervet monkeys. Behaviour, 110,
258–275.
1990. How Monkeys See the World: Inside the Mind of Another Species. Chicago, IL:
University of Chicago Press.
1999. Recognition of other individuals’ social relationships by female baboons.
Animal Behaviour, 58, 67–75.
Cheney, D. L., Seyfarth, R. M. & Silk, J. B. 1995a. The role of grunts in reconciling
opponents and facilitating interactions among adult female baboons. Animal
Behaviour, 50, 249–257.
1995b. The responses of female baboons (Papio cynocephalus ursinus) to anomalous
social interactions: evidence for causal reasoning? Journal of Comparative
Psychology, 109, 134–141.
Church, R. M. & Meck, W. H. 1984. The numerical attribute of stimuli. In: Animal
Cognition, ed. H. L. Roitblat, T. G. Bever & H. S. Terrace. Hillsdale, NJ: Lawrence
Erlbaum, pp. 445–464.
Connor, R. C., Smolker, R. A. & Richards, A. F. 1992. Dolphin alliances and coalitions.
In: Coalitions and Alliances in Humans and other Animals, ed. A. H. Harcourt & F. de
Waal. Oxford: Oxford University Press, pp. 415–443.
Connor, R. C., Mann, J., Tyack, P. L. & Whitehead, H. 1998a. Social evolution in toothed
whales. Trends in Ecology and Evolution, 13, 228–232.
1998b. Reply. Trends in Ecology and Evolution, 13, 408.
Connor, R. C., Heithaus, R. M. & Barre, L. M. 1999. Superalliance of bottlenose
dolphins. Nature, 371, 571–572.
2001. Complex social structure, alliance stability, and mating access in a bottlenose
dolphin ‘super-alliance’. Proceedings of the Royal Society of London, Series B, 268,
263–267.
D’Amato, M. & Colombo, M. 1989. Serial learning with wild card items by monkeys
(Cebus apella): implications for knowledge of ordinal rank. Journal of Comparative
Psychology, 103, 252–261.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
600 D. L. Cheney & R. M. Seyfarth
D’Amato, M., Salmon, P. & Colombo, M. 1985. Extent and limits of the matching
concept in monkeys (Cebus apella). Journal of Experimental Psychology: Animal Behavior
Processes, 11, 35–51.
Dasser, V. 1988a. A social concept in Java monkeys. Animal Behaviour, 36, 225–230.
1988b. Mapping social concepts in monkeys. In: Machiavellian Intelligence: Social
Expertise and the Evolution of Intellect in Monkeys, Apes, and Humans, ed. R. W. Byrne
and A. Whiten. Oxford: Oxford University Press, pp. 85–93.
Dunbar, R. 1983. Structure of gelada baboon reproductive units. III. The male’s
relationship with his females. Animal Behaviour, 31, 565–575.
2000. Causal reasoning, mental rehearsal, and the evolution of primate cognition.
In: Vienna Series in Theoretical Biology: The Evolution of Cognition, ed. C. Heyes & L.
Huber. Cambridge, MA: MIT Press, pp. 205–219.
Ehmer, B., Reeve, H. K. & Hoy, R. R. 2001. Comparison of brain volumes between single
and multiple foundresses in the paper wasp Polistes dominulus. Brain, Behavior and
Evolution, 57, 161–168.
Emlen, S. T., Wrege, P. H. & Demong, N. J. 1995. Making decisions in the family: an
evolutionary perspective. American Scientist, 83, 148–157.
Engh, A. L., Esch, K., Smale, L. & Holekamp, K. E. 2000. Mechanisms of maternal rank
‘inheritance’ in the spotted hyaena, Crocuta crocuta. Animal Behaviour, 60, 323–332.
Engh, A. L., Siebert, E., Greenberg, D. & Holekamp, K. 2004. Primate-like cognition in a
social carnivore. Animal Behaviour, in press.
Fagot, J., Wasserman, E. A. & Young, M. 2001. Discriminating the relation between
relations: the role of entropy in abstract conceptualization by baboons (Papio
papio) and humans (Homo sapiens). Journal of Experimental Psychology: Animal Behavior
Processes, 27, 316–328.
Gillan, D. J. 1981. Reasoning in the chimpanzee, II. Transitive inference. Journal of
Experimental Psychology: Animal Behavior Processes, 7, 150–164.
Gouzoules, S., Gouzoules, H. & Marler, P. 1984. Rhesus monkey (Macaca mulatta)
screams: representational signalling in the recruitment of agonistic aid. Animal
Behaviour, 32, 182–193.
Harlow, H. F. 1949. The formation of learning sets. Psychological Review, 56, 51–65.
Hansen, E. W. 1976. Selective responding by recently separated juvenile rhesus
monkeys to the calls of their mothers. Developmental Psychobiology, 9, 83–88.
Harcourt, A. H. 1988. Alliances in contests and social intelligence. In: Machiavellian
Intelligence: Social Expertise and the Evolution of Intellect in Monkeys, Apes, and Humans,
ed. R. W. Byrne & A. Whiten. Oxford: Oxford University Press, pp. 132–152.
1992. Coalitions and alliances: are primates more complex than non-primates? In:
Coalitions and Alliances in Humans and Other Animals, ed. A. H. Harcourt & F. de Waal.
New York: Oxford University Press, pp. 86–99.
Hauser, M. D., MacNeilage, P. & Ware, M. 1996. Numerical representations in primates.
Proceedings of the National Academy of Sciences, USA, 93, 1514–1517.
Herman, L. M, Pack, A. A & Morrel-Samuels, P. 1993. Representational and conceptual
skills of dolphins. In: Comparative Cognition and Neuroscience, ed. H. L. Roitblat, L. M.
Herman & P. E. Nachtigall. Hillsdale, NJ: Lawrence Erlbaum, pp. 403–442.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 601
Herrnstein, R. J. 1985. Riddles of natural categorization. Philosophical Transactions of the
Royal Society of London, Series B, 308, 129–144.
Hogue, M. E., Beaugrand, J. P. & Laugue, P. C. 1996. Coherent use of information by
hens observing their former dominant defeating or being defeated by a stranger.
Behavioural Processes, 38, 241–252.
Holekamp, K. E., Boydston, E. E., Szykman, M. et al. 1999. Vocal recognition in the
spotted hyaena and its possible implications regarding the evolution of
intelligence. Animal Behaviour, 58, 383–395.
Humphrey, N. 1976. The social function of the intellect. In Machiavellian Intelligence:
Social Expertise and the Evolution of Intellect in Monkeys, Apes, and Humans, ed. R. W.
Byrne and A. Whiten. Oxford: Oxford University Press, pp. 13–26.
Jolly, A. 1966. Lemur Behavior. A Madagascar Field Study. Chicago, IL: University of
Chicago Press.
Judge, P. 1982. Redirection of aggression based on kinship in a captive group of pigtail
macaques. International Journal of Primatology, 3, 301.
Kummer, H., Goetz, W. & Angst, W. 1974. Triadic differentiation: an inhibitory process
protecting pair bonds in baboons. Behaviour, 49, 62–87.
Marino, L. 1998. Quantifying brain–behavior relations in cetaceans and primates.
Trends in Ecology and Evolution, 13, 408.
Martin, R. D. 1983. Primate Origins and Evolution: A Phylogenetic Reconstruction. Princeton:
Princeton University Press.
Matsuzawa, T. 1985. Use of numbers by a chimpanzee. Nature, 315, 57–59.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, eds. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
Mennill, D. J., Ratcliffe, L. M. & Boag, P. T. 2002. Female eavesdropping on male song
contests in songbirds. Science, 296, 873.
Mercado, E., Killebrew, D. A., Pack, A. A., Macha, I. & Herman, L. M. 2000.
Generalization of ‘same-different’ classification abilities in bottlenosed dolphins.
Behavioural Processes, 50, 79–94.
Naguib, M., Fichtel, C. & Todt, D. 1999. Nightingales respond more strongly to vocal
leaders of simulated dyadic interactions. Proceedings of the Royal Society of London,
Series B, 266, 537–542.
Oden, D. Thompson, R. & Premack, D. 1988. Spontaneous transfer of matching by
infant chimpanzees (Pan troglodytes). Journal of Experimental Psychology: Animal
Behavior Processes, 14, 140–145.
Ogawa, H. 1995. Recognition of social relationships in bridging behavior among
Tibetan macaques (Macaca thibetana). American Journal of Primatology, 35, 305–310.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Palombit, R. A., Seyfarth, R. M. & Cheney, D. L. 1997. The adaptive value of ‘friendships’
to female baboons: experimental and observational evidence. Animal Behaviour,
54, 599–614.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
602 D. L. Cheney & R. M. Seyfarth
Palombit, R. A., Cheney, D. L. & Seyfarth, R. M. 2001. Female–female competition for
male ‘friends’ in wild chacma baboons (Papio cynocephalus ursinus). Animal
Behaviour, 61, 1159–1171.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male–male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.
Pepperberg, I. M. 1992. Proficient performance of a conjunctive, recursive task by an
African grey parrot (Psittacus erithacus). Journal of Comparative Psychology, 106,
295–305.
1994. Numerical competence in an African gray parrot (Psittacus erithacus). Journal of
Comparative Psychology, 108, 36–44.
Popp, J. W. 1987. Choice of opponents during competition for food among American
goldfinches. Ethology, 75, 31–36.
Premack, D. 1983. The codes of man and beast. Behavioral and Brain Sciences, 6, 125–167.
Schusterman, R. J. & Gisiner, R. 1988. Artificial language comprehension in dolphins
and sea lions: the essential cognitive skills. Psychological Record, 38, 311–348.
Schusterman, R. J. & Krieger, K. 1986. Artificial language comprehension and size
transposition by a California sea lion (Zalophus californianus). Journal of Comparative
Psychology, 100, 348–355.
Seyfarth, R. M. 1978. Social relationships among adult male and female baboons. II.
Behavior throughout the female reproductive cycle. Behaviour, 64, 227–247.
Seyfarth, R. M. 1980. The distribution of grooming and related behaviors among adult
female vervet monkeys. Animal Behaviour, 28, 798–813.
Shettleworth, S. 1998. Cognition, Evolution, and Behaviour. Oxford: Oxford University
Press.
Silk, J. B. 1987. Social behavior in evolutionary perspective. In: Primate Societies, ed. B. B.
Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham & T. Struhsaker. Chicago, IL:
University of Chicago Press, pp. 318–329.
1993. Does participation in coalitions influence dominance relationships among
male bonnet macaques? Behaviour, 126, 171–189.
1999. Male bonnet macaques use information about third-party rank relationships
to recruit allies. Animal Behaviour, 58, 45–51.
Silk, J. B., Seyfarth, R. M. & Cheney, D. L. 1999. The structure of social relationships
among female savanna baboons. Behaviour, 136, 679–703.
Smale, L., Frank, L. G. & Holekamp, K. E. 1993. Ontogeny of dominance in free-living
spotted hyaenas: juvenile rank relations with adult females and immigrant
males. Animal Behaviour, 46, 467–477.
Smuts, B. 1985. Sex and Friendship in Baboons. Chicago, IL: Aldine.
Tomasello, M. & Call, J. 1997. Primate Cognition. Oxford: Oxford University Press.
Treichler, F. & van Tilburg, D. 1996. Concurrent conditional discrimination tests of
transitive inference by macaque monkeys: list linking. Journal of Experimental
Psychology, Animal Behavior Processes, 22, 105–117.
von Fersen, L., Wynne, C., Delius, J. & Staddon, J. 1991. Transitive inference formation
in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 17, 334–341.

P1: JZZ/... P2: JZZ/...0521823617c25.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:36
Social complexity and eavesdropping 603
Walters, J. R. & Seyfarth, R. M. 1987. Conflict and cooperation. In: Primate Societies, ed.
B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham & T. Struhsaker.
Chicago, IL: University of Chicago Press, pp. 306–317.
Wasserman, E. A., Hugart, J. A. & Kirkpatrick-Steger, K. 1995. Pigeons show
same-different conceptualization after training with complex visual stimuli.
Journal of Experimental Psychology: Animal Behavior Processes, 21, 248–252.
Wright, A., Cook, R. & Rivera, J. 1988. Concept learning by pigeons: matching to sample
with trial-unique video picture stimuli. Animal Learning and Behavior, 16, 436–444.
Zentall, T., Weaver, J. & Sherburne, L. 1996. Value transfer in concurrent-schedule
discriminations by pigeons. Animal Learning and Behavior, 24, 401–409.

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
26
Communication networks ina virtual world
a n d r e w m . r . t e r r y 1 & ro b e r t l ac h l a n 2
1University of Copenhagen, Denmark2University of North Carolina, Chapel Hill, USA
Introduction
When one individual is signalling, or two individuals are interacting, they
do so within a network of potential receivers (see McGregor, 1993; McGregor &
Dabelsteen, 1996; Ch. 1). As the other chapters in this book show, the decisions
that both signallers and receivers make about their future behaviour are thus
contingent not only on each other’s behaviour but also on a wider network of
individuals (McGregor & Peake, 2000). This view is finding support in empirical
studies showing that individuals use information that could only be extracted
from network interactions (e.g. Oliveira et al., 1998; Peake et al., 2001, 2002; Ch. 2).
These empirical findings also have implications for the theoretical study of sig-
nalling strategies (e.g. Nowak & Sigmund, 1998; Johnstone, 2001). For example,
an individual’s signalling strategy may no longer be predicted solely from the re-
sponses of an opponent. We consider that the signalling strategies of individuals
will only be explored realistically by models that include the potential responses
of signallers to other individuals. In this chapter, we ask whether current mod-
elling approaches can be adapted to include networks or whether new modelling
techniques need to be considered.
The aim of creating a model is to advance our conceptual understanding of a
system and create empirically testable hypotheses (Wilson, 2000; Hemelrijk, 2002)
by simplifying the real world using words or mathematical expressions. Most hy-
potheses start with a verbal model and develop into mathematical models, which
more precisely specify limiting conditions and assumptions and often provide
a deeper understanding of the logic underlying the hypothesis. Models vary in
Animal Communication Networks, ed. Peter K. McGregor. Published by Cambridge University Press.c© Cambridge University Press 2005.
604

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 605
their complexity and the level of explanatory power they provide: often, simple
models provide a deeper and general understanding of underlying dynamics but
also contain more restrictive (or unrealistic) assumptions. For example, simple
biological models, such as the Lotka–Volterra equations of predator–prey interac-
tions, provided a powerful heuristic insight into the cyclic nature of population
abundances but contained highly restrictive assumptions, primarily concerning
the heterogeneity of populations and environments, that limited their applica-
tion to specific cases (Maynard Smith, 1982; Begon et al., 1990; Wilson, 2000). In
population genetics models, assumptions are typically made about population
sizes and the pattern of distribution of traits. Nevertheless, such assumptions of-
ten do not qualitatively affect the conclusion (e.g. Turelli & Barton, 1994). Simpler
models are likely to be mathematically tractable, allowing a more complete analy-
sis and understanding of the processes underlying the system being investigated.
Complexity increases as more realistic assumptions are incorporated and the pos-
sibility of mathematical analysis becomes more remote. However, in recent years,
the rapid rise in computer power has allowed theoretical methods to acquire new
levels of complexity, mostly through the use of simulation modelling (Grimm,
1999). Simulation models allow as many variables to be included as the investiga-
tors have imagination, programming skills and time. The downside to this is that
the models are harder to generalize (it is harder to ensure that all the variables
are realistic for a wide range of conditions) and that it is more difficult to iso-
late precisely the factors that are causing an effect of interest (Wilson, 2000). An
attractive solution to this dilemma is to use a variety of modelling techniques, dif-
fering in how many assumptions are required (Dieckman, 1997), with the specific
aim of identifying the parameters, variables and assumptions that are critical in
explaining the behaviour of the system under investigation (Wilson, 2000).
In this chapter, we discuss the role modelling has played in the conceptual
development of communication networks. In doing this, we also examine which
features of communication network models are especially important, often by
identifying which unrealistic assumptions are likely to change qualitatively the
conclusions reached about behaviour in networks. Finally, we examine how theo-
ries of communication that include networks are likely to differ from those that
do not.
Conceptualizing networks
Network structure in existing models
A network is an association of nodes connected to each other by some
means. In animal communication, the nodes are individual animals and the con-
nections (or links) are patterns of communication between them. For every signal,

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
606 A. M. R. Terry & R. Lachlan
there are likely to be several possible receivers, leading to the concept of the
communication network. This concept modifies our understanding of the costs
and benefits of signalling (e.g. McGregor & Peake, 2000). For example, individuals
winning an interaction may be more willing to publicize the interaction than
those losing it. How does this view of communication fit with standard methods
used to model animal signalling and do different methods need to be developed?
Most current models of animal communication consider only dyadic encoun-
ters between individuals. For example, game theory models of communication
normally study the evolution of strategies through the responses of two interac-
tants to each other (examples in Maynard Smith, 1982). When analytical models
have been used to study interactions within large groups, they typically make
restrictive assumptions that may limit their ability to assess the evolutionary
pressures of communicating within networks. For example, individuals may be
drawn at random from the population to interact or they may have perfect knowl-
edge about the behaviour of all other individuals (e.g. Nowak & Sigmund, 1998;
Johnstone, 2001). It seems that, whereas traditional evolutionary game theory
models are well suited to the study of dyadic encounters and contests, a network
of individuals all gathering information from each other and using this in sig-
nalling interactions, which may or may not be directed at other receivers as well
as the primary receiver, may prove too complicated to be tractable (although see
p. 617). It is with this in mind that we consider the role of simulation modelling
as a tool for studying communication in networks.
Simulation models are widely used in ecology as they allow the user to incor-
porate an unlimited number of variables and parameters in the model, and they
are being increasingly used in behavioural studies. A recent trend has favoured
individually based and spatially explicit models, which contain a discrete popula-
tion of individual animals within a defined spatial environment. Individuals are
governed by a series of movement and behavioural rules (Grimm, 1999). These
models, therefore, allow an increase in biological realism (Wilson, 2000) by re-
placing the inaccurate assumptions about the random or structured patterns of
interaction within a population found in simpler models with assumptions that
better capture spatial structuring. These types of model have been used to show
how complex collective behaviours can arise from the interaction of individu-
als obeying simple behavioural rules (Hemelrijk, 2002). For example, Hemelrijk
(2000) created an individually based model of dominance interactions within a
social group to show that increased aggression caused the emergence of ‘selfish
herd’ organization in the group. Previous theoretical studies of selfish herding
had difficulties equating the complex movement rules needed to make individ-
uals aggregate in the models (i.e. with tight clustering of dominants surrounded
by subordinate individuals (Hamilton, 1971)) with observations of this herding

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 607
behaviour in the field (Morton et al., 1994; Viscido et al., 2002). Individuals in the
model of Hemelrijk (2000) were governed by two rules; the tendency to aggregate
and the tendency to enter into dominance interactions. When an individual lost
a dominance interaction, it would flee and the winner would move to occupy its
space. In the model, it became possible for weak individuals to get ‘caught in a rut’
and lose several interactions in a row causing them to move to the periphery of
the group, whereas the strongest individuals would remain in the centre. This is
an example of a model that made no assumptions about the way in which individ-
uals chose to aggregate and yet patterns of social organization that were similar
to those observed in the wild emerged as products of aggressive interactions and
individual differences.
Small-world network analysis
As explained above, most models of communication have not considered
the role of individuals other than the immediate interactants; when they have,
they placed unrealistic assumptions on the information gathered. A further con-
sideration in the theoretical analysis of communication and information gather-
ing in a network is whether the way in which the network is organized affects
its function. In recent years, considerable research has focused on the structure
and organization of networks. Most networks associated with social interactions
may be physically limited. For example, the interactions in a territorial system are
mostly restricted to neighbouring individuals. However, a consideration of com-
munication networks means that a larger group of individuals must be considered
in the analysis as the long-range signals most commonly used for advertising or ag-
gressive interactions usually travel much further than an individual’s immediate
neighbours (McGregor & Peake, 2000).
The analysis of networks in fields as diverse as metabolic pathways in eukaryote
cells (Jeong et al., 2000), food webs (Williams et al., 2000; but see Dunne et al., 2002)
and links in the World Wide Web (Albert et al., 1999) has found that they show a
number of similar structural properties. As a result of these similarities, such net-
works are referred to as ‘small-worlds’ or ‘scale-free’ networks (Watts & Strogatz,
1998; Barabasi & Albert, 1999). Fundamentally, these networks are dynamic: a
new node joining the network is likely to attach preferentially to certain exist-
ing nodes. As a result, networks arise that contain tight clustering around some
highly connected nodes, called ‘hubs’.There is no one centralized dominant node;
consequently, organization is spread between the few highly connected hubs. This
organization is both the network’s main asset and its Achilles heel, as it means
that the behaviour of most nodes has little impact on the network at large, but
removing one of the hubs can have a critical impact on the flow of information
through the network (Albert & Barabasi, 2002).

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
608 A. M. R. Terry & R. Lachlan
A key consequence of this structure is that these networks have short path
lengths between any two nodes (i.e. any two individuals within the population
can be linked by a small number of connections). For example, Kudo & Dunbar
(2001) found that primate groups remained socially interconnected despite be-
ing fragmented into small cliques, with males possibly serving the role as hubs,
connecting female groups. The implication of this phenomenon is that informa-
tion exchanged between individuals may spread widely and rapidly throughout
the network. Watts & Strogatz (1998) considered a simple model of a contagion
that examined how a disease might spread through a small-world network. They
found, rather frighteningly, that only a few hubs were required for the disease to
spread rapidly through the population. This model can be directly compared to
a communication situation, where the probability of an individual producing a
signal is affected by the number of others it perceives signalling. The conclusion
would be that, in a small-world network, a bout of signalling could spread very
rapidly throughout an entire population (see Ch. 12 on information cascades).
In summary, small-world analyses suggest that, when studying the pattern of
communication within a population, it may be important to identify how asym-
metric the communication networks of individuals are. In this context, asymme-
try means the number of connections each individual has with other individuals.
In a random network, all individuals would have, on average, the same num-
ber of connections. However, in a small-world network, the few hub individuals
have the majority of connections, while the rest of the population has very few.
Asymmetry could be imposed by environmental features (a hub individual could
occupy a more central position within a forest or a more open spot from which his
signal could propagate further) or by social roles (e.g. male primates connecting
female groups; or possibly ‘floating’juveniles connecting territorial adults in bird
species).
Models of communication network dynamics from
signallers’ perspectives
There are a number of potential costs and benefits associated with sig-
nalling in a network. Signallers must compete with each other to make their sig-
nals detectable by receivers. They must also balance the benefits of the intended
receiver perceiving the signal with the costs of other receivers doing the same.
These costs can range from heterospecific predators or parasites to competing
conspecifics. Signalling within a network may also coordinate behaviour among
individuals within larger social groups. Although many experimental studies have
shown the effects of signalling within networks of several individuals, there have
been far fewer studies modelling the effects of networks on signals and signalling
dynamics (but see Chs. 2, 5 and 13). Here we highlight some of the benefits of

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 609
modelling signalling within groups with three case studies: the chorus behaviour
of acoustic insects and frogs, the social coordination of ant foraging, and territory
establishment. The case studies share similarities in the methods they employ and
the conclusions they draw. In each case, the examples show that complex patterns
of behaviour can arise from simple individual decision patterns.
Signal dynamics in acoustic choruses
Choruses are of interest when studying behaviour in communication
networks from both experimental and theoretical perspectives as they can show
tightly coordinated patterns of signalling within large groups (Chs. 12 and 13). Indi-
viduals within an acoustic chorus must deal with a complex acoustic background
generated by other signallers within which they must maximize the efficacy
of their signals in terms of transmission and female attraction. Females can show
preferences for specific temporal features of male signals; for example, they may
respond less to calls that are overlapped (e.g. gray tree frogs Hyla versicolor: Schwartz
et al., 2001) or may prefer leading calls (Snedden & Greenfield, 1998; Greenfield,
2002). Thus, female preferences for certain temporal features may have led to pat-
terns of synchronous and alternating choruses in anuran and insect species. In
general, when a species calls at a rapid rate, choruses tend towards synchrony; as
the call period becomes longer, choruses are more likely to alternate (Grafe, 1999).
Anuran and insect choruses are amenable to experimental studies of net-
work behaviour because the whole network can be controlled and manipulated
(Schwartz et al., 2002) and individual behaviour within the network can be mea-
sured. This level of experimental control allows the predictions of models to be
tested. However, to date, there have been few theoretical considerations of sig-
nalling dynamics within choruses. We discuss two different approaches that have
been used to model signalling within choruses (Brush & Narins, 1989; Greenfield et
al., 1997). Both models consider mechanisms individuals may use to control their
call timing and hence avoid interference in a chorus. The mechanism used in each
case is a form of inhibitory resetting. Each individual has an internal mechanism
that increases from a basal state to a peak where it initiates a call. If, before call-
ing, the individual perceives another individual’s call, the mechanism is reset to
its basal level and it begins to increase again. Such mechanisms have been shown
to exist for many anuran and insect species (Zelick & Narins, 1985; Greenfield
et al., 1997).
Brush & Narins (1989) adapted models of computer networks to study whether
choruses in the Puerto Rican treefrog Eleutherodactylus coqui were controlled by this
inhibitory resetting mechanism. Computer-network models simulate the flow of
data between interconnected computers and study how a shared resource (i.e.
bandwidth) is partitioned between them. Individual computers are linked to each
other via data lines and send packets of information through the network. Before

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
610 A. M. R. Terry & R. Lachlan
sending a packet, each computer checks if the data line is clear. If it is not, the
computer waits for a random time period before checking the line again (Brush &
Narins, 1989). However, even when the line is free, computers have a probability of
deferring the transfer. Also in some cases, the terminals must transmit informa-
tion even if the lines are busy. In the model, frogs represented the terminals; the
data lines represented the communication network between them, and their calls
were the packets of information. Although their model was an individually based
simulation, it was not spatially explicit and, therefore, distance between individ-
uals was not a factor in the analysis. The frogs in this model used their inhibitory
resetting to avoid being jammed by other individuals; if they detected another frog
calling during their refractory period, they delayed the next call by a randomly
chosen period and then returned to the standard refractory period for the next
call (Brush & Narins, 1989). Using this model, Brush & Narins (1989) showed that
this mechanism would lead to fewer calls being overlapped and that there was an
optimal chorus size of between three and four individuals at which information
transfer was maximized (i.e. overlap was minimized). The results of the model
were also corroborated by field data collected on the treefrog, which showed that
choruses occurred in small groups and when group size was large (five or six
individuals) males showed selective attention to one or two neighbours. These
results are similar to those of Greenfield & Rand (2000) who show that tungara
frogs Physalaemus pustulosus paid attention to a subset of the potential signallers
in artificially generated choruses.
Greenfield et al. (1997) developed a linear model of chorus signalling in the
katydid Neoconocephalus spiza, which initially was based on dyadic interactions.
Individuals in their model used an inhibitory resetting mechanism similar to that
of Brush & Narins (1989). They modelled two individuals signalling at the same
time and measured signal overlap from a receiving female’s perspective. They
showed that individuals overlapped far less and avoided producing following calls
(i.e. calls that were initiated after the onset of another male’s call) when using an
inhibitory resetting mechanism, and that signals tended towards synchrony or
alternation depending on the speed at which the mechanism reached its peak
level (i.e. at call initiation). If males within a given call period could return to
their peak level quickly from inhibition (‘rebound’), alternation would arise as
males could quickly begin calling again. However, if the rebound took almost as
long as the call cycle, males would fall into bouts of synchrony (Greenfield et al.,
1997). They also considered the case of a larger number of calling males. This
model was an individually based and spatially explicit simulation, with males
randomly placed on a 20 m × 20 m grid. Simulated females were also randomly
located around this grid; the model assessed the level of call overlapping from the
female’s perspective. Male attractiveness was assessed as the number of their calls

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 611
that were overlapped. When they ran simulations for a number of males (2–10),
Greenfield et al. (1997) found that males had to pay attention to a subset of the
chorus for inhibitory resetting to function as a chorus mechanism, but when they
did, patterns of synchrony and alternation emerged.
The theoretical analyses of call timing in choruses of signalling anurans and
acoustic insects have shown that simple mechanisms that control signal timing,
which may have arisen in response to female preferences for leading signals, can
generate the complex patterns of alternation and synchrony observed in nature.
An emergent feature of these models was that, given the existence of such a re-
setting mechanism, males could only pay attention to a subset of the individuals
in their chorus when determining their call timing. Choruses, in general, would
seem to represent a tractable means of studying signalling dynamics in networks.
Although the individuals within the chorus are signalling to females, males in-
directly compete with each other for acoustic space and directly compete over
actual resources, all within the scope of the chorus. Therefore, it is surprising that
chorus dynamics have received such limited theoretical attention given that they
represent such an amenable study system for simulation models.
Swarm intelligence and self-organization in social insects
Social insects are not noted for their individual cognitive abilities, yet
they are famous for their ability collectively to ‘solve’ problems of how best to ex-
ploit food resources in their environment. Such coordination requires communi-
cation between individuals; for example, the honeybee Apis mellifera uses a waggle
dance that indicates the location of food sources to other individuals (von Frisch,
1967). Similarly, ants use pheromone trails to lead colony members to food. The
collectively adaptive processes that arise out of these interactions are examples
of self-organization. Key ‘ingredients’ of self-organization (Bonabeau et al., 1999,
p. 9) are positive feedback (e.g. one forager recruits more bees to a food source by
dancing) and multiple communicative interactions.
The field of foraging strategies in social insects has a rich empirical background.
Moreover, several recent models of such behaviour (Deneubourg & Goss, 1989;
Deneubourg et al., 1990; Camazine & Sneyd, 1991; Seeley et al., 1991; reviewed by
Bonabeau et al., 1999) have shed light on the much simpler underlying individual
behaviour patterns. In these models, the type of foraging problem that is faced
typically structures the social and communication networks of the population.
For example, individuals that are laying a pheromone trail for a food source that
is nearby will overlay that trail more frequently than those laying a trail for a food
source further away, simply because they move along it more frequently. Such a
model can explain the ability of ants preferentially to use the nearest food source
first (Resnick, 1994).

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
612 A. M. R. Terry & R. Lachlan
Most of these models have implicitly incorporated communication networks.
For example, Deneubourg et al. (1990) investigated the ability of ants to choose
one of two paths to a food source. Experimental evidence suggests that Argentine
ants Linepithema humile quickly decide to use one of the two paths. Deneubourg
et al. (1990) fitted this behaviour to a simple positive-feedback model in which
the more ants had travelled along a path, the more likely it was to be chosen by
another ant. Interestingly, the best fit between model parameters and empirical
data occurred when each extra ant travelling along a path had a greater impact on
others’ behaviour (e.g. the twentieth ant had a disproportionately larger impact
than the first). This is yet another example of asymmetry in networks, where the
most recent information has a greater impact than old, and possibly outdated,
information. A similar situation is found in the chemical over-marking signals of
mice (e.g. Rich & Hurst 1999; Ch. 11).
Territory establishment
To compare and contrast the different approaches to modelling behaviour
and communication, we use an example from models of territory establishment.
Models of territory establishment are predominantly based on game theory, where
a series of dyadic encounters occur over indivisible areas. These models contain
‘winner-takes-all’ assumptions, in that whoever wins the contest takes the re-
source, and contests cannot end in draws or with division of the area.
Stamps & Krishnan (1999) developed an individually based spatial simulation
model of territory establishment. In their model, individuals moved around a spa-
tially heterogeneous area containing patches of different size. At each time step,
they assessed the attractiveness of the patches around them and moved into the
one with the highest attractiveness. The attractiveness of an area was based on two
key parameters: positive and aggressive experiences. Positive experiences occurred
when an individual entered a patch and did not become involved in an aggres-
sive interaction. Positive experiences increased the attractiveness of the patch and
thus increased the likelihood that the individual would return to the patch in the
future. Aggressive experiences occurred when two individuals entered the same
patch. They would then enter into a costly interaction, which, for this model,
would end in a draw (i.e. there would be no clear winner). Aggressive encounters
would discourage individuals from returning to that patch. The model predicted
that individuals would gradually build territories by incorporating novel patches
into their home range. Individuals would show periods of sustained aggression
when fighting over familiar (i.e. repeatedly visited) sites and they could also take
over sites by repeatedly entering into aggressive interactions. The net result of
the model was that a pattern of stable territories would be generated through
repeated interactions where there was no clear winner taking a resource.

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 613
Game theoretic models make similar predictions to those of Stamps & Krishnan
(1999, 2001), albeit for different reasons (Sih & Mateo, 2001). Similar predictions
include the increased benefits of being a resident, prolonged encounters between
two residents and the more desperate attempts of newcomers to claim territorial
space when they have few remaining options. In game theory models, the territory
has an intrinsic value, which is greater for residents than newcomers, and the
stable strategy that evolves is one of territory choice (i.e. when to stay and when
to move on).
As with the previous case studies, the models of Stamps & Krishnan (1999, 2001)
do not explicitly study networks; however, they contain network-like features, as
the behaviour of individuals are affected by their previous encounters with several
other individuals. The model could be extended to include some explicit network
effects such as eavesdropping (e.g. Ch. 2). For example, an individual’s decision
to enter an area could, in part, be based on previous encounters it had observed
take place there. This would allow it to discriminate between two novel areas, one
hotly contested by other individuals and one that was not. This form of modelling
shows that individually based simulation models can create predictions similar
to those of game models but without the same restrictive assumptions. As in this
case, they can also extend the predictions that the models can make.
Summary
In this section, we have chosen examples of communication within net-
works that are very different. While these studies have not explicitly used the
concept of communication networks, they would not be possible without such a
viewpoint. They also show how the network may influence a signaller’s behaviour
and show that complex patterns of organization and behaviour within networks
may be possible through the implementation of simple rules. The territory estab-
lishment models also show how the simulation models can be compared with
game theoretic models and generate similar predictions while making fewer re-
strictive assumptions.
Models of communication networks from receivers’ perspectives
In the previous section, we emphasized the consequences of communi-
cation networks for signalling behaviour and how it influenced population level
patterns of signalling and organization. In this section, we examine how receivers
can use the network environment to extract information about others and modify
their own behaviour. We deal primarily with the role of eavesdropping (Ch. 2) as a
means of information gathering. Eavesdropping is one source of information that
becomes available to receivers within a network and it is the one that has received

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
614 A. M. R. Terry & R. Lachlan
the most empirical study. Here we describe two case studies that reflect the new ap-
proaches being taken to the theoretical analysis of communication networks; the
game theory simulation model (R. Lachlan & A. M. R. Terry, unpublished data) stud-
ies the evolution of eavesdropping as a strategy and the spatial simulation model
(R. L. Earley, S. Brosnan & J. Bragg, unpublished data) determines how eavesdrop-
ping may shape the formation of linear dominance hierarchies. Both models are
spatial simulations and emphasize the point made throughout this chapter: that
consideration of the spatial nature of communication is required for the success-
ful modelling of communication networks. In this section, we also discuss how
game theory has been used to model communication in groups and why tradi-
tional game theory methods may not be best suited to the study of networks. We
conclude that significant advances may be made through the combined use of
both proximate-based simulation models and strategic decision making through
game theory models.
Game theory and eavesdropping
Evolutionary game theory encompasses a well-established set of tech-
niques for determining which strategies prove most effective in interactions be-
tween individuals. The aim is to establish whether a given strategy can be invaded
by any other strategy; if not, it can be called an evolutionarily stable strategy (ESS,
Maynard Smith, 1982). There have been several studies using traditional game the-
ory in which individuals use communication networks to predict the behaviour
of others. The studies adapted well-established game scenarios and investigated
how a given eavesdropping strategy would fare in the game. Johnstone (1998)
developed a model of signal detection that aimed to determine whether ‘con-
spiratorial whispers’ (low-cost and inconspicuous signals (Krebs & Dawkins, 1984))
could be evolutionarily stable in cooperative communication systems. The idea of
conspiratorial whispers implicitly recognizes the role of eavesdroppers and hence
communication networks. Here eavesdroppers were modelled in the most general
sense of the word (i.e. both conspecific and heterospecifics receivers, see Ch. 2) and
it was assumed that it was generally costly to be overheard by an eavesdropper.
Johnstone’s (1998) model maintained that even when signalling was cooperative,
expenditure was required to make the signal detectable for a receiver and that
this creates a conflict of interest between signallers and receivers in the face of
the costly eavesdroppers. However, in many situations it may not be costly to be
overheard. Pollock & Dugatkin (1992) investigated eavesdropping in the famous
Prisoner’s Dilemma game. The Prisoner’s Dilemma is an extreme abstraction of
many cooperative situations where individuals have the option of either cooper-
ating or defecting in any given round of the game (for a review, see Dugatkin,
1998). Cooperators benefit if they play one another, compared with two defectors

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 615
playing one another. However, if a cooperator plays a defector, the former does
badly, while the latter does well (i.e. the defector exploits the cooperator’s benefi-
cence). Therefore, in a single round of the game, individuals should always defect
as they will benefit the most; if, however, the game is repeated over a number
of rounds, a strategy of reciprocation or ‘tit-for-tat’ becomes most beneficial and
stable (Axelrod & Hamilton, 1981; Stephens et al., 2002).
Since the publication of the classic paper by Axelrod and Hamilton (1981), many
revisions have been published, but versions of tit-for-tat are still regarded as the
most successful strategy. Although individuals playing tit-for-tat copy the strategy
their opponent played in the previous round, in reality this information may not
be present (if they only play each other once) or might be unreliable (if individ-
uals update their strategy frequently). In these situations, individuals could use
eavesdropping as a way of obtaining up-to-date information about their opponents’
strategies. Pollock & Dugatkin (1992) found that their so-called ‘observer tit-for-tat’
strategy was sometimes successful when tit-for-tat itself was not an evolutionarily
stable strategy (although observer tit-for-tat was out-competed by tit-for-tat un-
der many conditions). Sigmund & Nowak (1998) examined a similar situation and
found that indirect reciprocity through ‘image scoring’ (i.e. cooperating with in-
dividuals that had a record of cooperation) was a successful strategy. In this case,
individuals gain an increase to their image score each time they cooperate and a
decrease when they do not (for more details, see Ch. 22).
One of the main criticisms of image scoring was that observing an individual’s
image score did not take into account the behaviour of that individual’sopponent.
Thus, an observer would react the same way to an individual that defected against a
notoriously uncooperative opponent as to one that defected against a good cooper-
ator (Leimar & Hammerstein, 2001). To remedy this, Leimar & Hammerstein (2001)
investigated the evolutionary stability of the ‘good standing’ strategy (Sugden,
1986), in which individuals strive to maintain their good social standing. Under
this strategy, individuals could improve their standing by cooperating and could
have it damaged by defecting, but defecting against a player that was uncooper-
ative was not punished. Leimar & Hammerstein (2001) found that this strategy
was very successful. Milinski et al. (2001) investigated whether humans used good
standing in cooperative games but found that the simpler image scoring tended to
be used. They concluded that a strategy using good standing might ask too much
of working memory as individuals would have to remember each opponent’s pre-
vious interactions and when there were errors in perception of the roles adopted
in the interaction, image scoring predominated (Milinski et al., 2001).
The models discussed above studied the evolution of cooperation through
indirect reciprocity, something which takes place within communication net-
works (e.g. Ch. 22); however, they do not really examine how individuals interpret

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
616 A. M. R. Terry & R. Lachlan
interactions between others, a prerequisite for eavesdropping. In a more explicit
model of eavesdropping, Johnstone (2001) adapted the hawk–dove game to assess
the use of eavesdropping within such a framework of contests. The hawk–dove
model is a simple game that models some of the essential features of animal con-
tests (Maynard Smith, 1982; Riechert, 1998). In this game, individuals can play
one of two strategies; aggressive hawks escalate contests into fights while peaceful
doves rely on non-aggressive displays. Hawks, therefore, always beat doves but risk
damage if they face another hawk. Doves always lose to hawks but suffer no cost to
meeting another dove. With a high risk of damage from fights, the evolutionar-
ily stable strategy of this game is a mixture of hawks and doves (Maynard Smith,
1982).
In Johnstone’s (2001) model, the success of a third strategy, eavesdropper was
investigated in comparison to the pure strategies. The eavesdropper chose to play
dove if its opponent had won its previous encounter, and hawk otherwise. To repli-
cate ‘error’ in an eavesdropper’s assessment of the outcome of an interaction,
there was a chance that individuals could misinterpret their eavesdropping, for
example, by playing hawk against a winner. The model showed that eavesdropper
was most common when there was a high cost to fighting. When eavesdropping
errors were more frequent, eavesdroppers reached their peak frequency at a lower
fighting cost. However, eavesdropping never spread to fixation within the popu-
lation. Johnstone (2001) suggests that this is because eavesdroppers are unable to
assess the strategy that other eavesdroppers will adopt, because it may change each
round. Therefore, eavesdroppers are reducing the fitness benefit upon which they
are based. When rare, they have the advantage of predicting the correct role in
most cases. Consequently, a mixed equilibrium evolves with eavesdroppers at low
frequencies. A surprising result of the model was that eavesdropping promoted
increased aggression. This is because the model includes a form of ‘winner–loser’
effect: an individual that won in one round is more likely to win in the next as
eavesdroppers will chose a submissive role to play.
In the models described above, individuals were restricted to obtaining infor-
mation about only a small part of the interaction between two others. For example,
Johnstone’s (2001) model focused on the outcomes of interactions (who won and
who lost) and not on the roles adopted in the interactions themselves. This repre-
sents one form of eavesdropping (interceptive eavesdropping, see Ch. 2); however, it
is likely that real eavesdroppers also consider the roles that the interactants play
(who played hawk or dove). Finally, except for the simple viscosity factor of Pollock
& Dugatkin (1992), which defined a probability that an individual would have
eavesdropped on his opponent’s last contest (Nowak & Sigmund (1998), include
a similar factor), the models did not consider how structuring the communica-
tion networks might be important. However, as stressed throughout this chapter,

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 617
spatial structuring of populations is likely to have a significant effect on the pre-
dictions of theoretical models. A spatial simulation model of the hawk–dove game,
in which strategies could evolve in response to all aspects of the interaction, has
been developed by R. Lachlan and A. M. R. Terry (unpublished data) to analyse both
these factors. In a hawk–dove game between two individuals, A and B, player A
could be involved in one of six different types of interaction:
1. A plays hawk, defeats B, which also plays hawk
2. A plays hawk, loses to B, which also plays hawk
3. A plays dove, defeats B, which also plays dove
4. A plays dove, loses to B, which also plays dove
5. A plays hawk, defeats B, which plays dove
6. A plays dove, loses to B, which plays hawk.
An eavesdropper on the interaction, who would eventually play A, could, therefore,
obtain one of six pieces of information about A.
The spatial simulation model (R. Lachlan & A. M. R. Terry, unpublished data)
examines how eavesdroppers should respond to obtaining one of these pieces of
information. The model is a spatially explicit game theoretic simulation: individ-
uals within the population were placed in a 40 × 40 two-dimensional grid (i.e.
population size of 1600). In each round of the simulation, individuals engaged
in one contest with each of their four neighbours. The strategy adopted by each
individual was determined by a vector containing six values (varying between 0
and 1). Each value represented the probability of playing either hawk or dove hav-
ing just witnessed the opponent in one of the six situations mentioned above.
For example an individual with the vector {0.9 0.8 0.2 0 0.7 0.2} would follow an
image-scoring strategy as it would tend to play hawk, having observed its oppo-
nent play hawk in a previous round (refer to the six types of information listed
above).
Mortality and reproduction were modelled by having individuals periodically
update their strategy by choosing one of their neighbours’ strategies. This choice
was determined by the success of individuals: that is, their total payoff after play-
ing the game during the previous round. We investigated two types of inheritance;
either the strategy of the most successful neighbour was inherited, or the probabil-
ity of inheritance was directly proportional to the neighbours’relative success. The
difference between these conditions was that only the most successful strategies
were rewarded in the first, whereas in the second, moderately successful strategies
could also be inherited. A mutation rate was also included, which would create
novel strategies in new individuals.
Over a range of conditions, a variety of eavesdropping strategies evolved, but
only two groups of strategies were found to arise regularly. The first strategy is

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
618 A. M. R. Terry & R. Lachlan
Fig. 26.1. The pattern of aggression in a simulated population of 1600 individuals.
Each square represents an individual and the square’s shade of grey represents how
many times the individual was attacked over the previous 20 rounds of interaction:
attacked every time (black); never attacked (white). (a) The pattern for an image-scoring
strategy (individuals copy the hawk or dove behaviour of their opponents) is a similar
level of aggression to that of neighbours (the pattern is a fairly even shade of grey) but
darker areas show local waves of aggression. (b) The pattern for a reputation-scoring
strategy, in which individuals copied the behaviour of their opponent’s opponent (i.e.
if an individual was witnessed being attacked, it was subsequently more likely to be
attacked), shows that squares are usually either black or white.
somewhat similar to the image-scoring (or observer tit-for-tat) strategy: play hawk if
you witness your opponent playing hawk, and dove if he played dove. However, under
a wider range of conditions, especially when only the most successful individuals
could pass their strategies on, a second group of strategies was most successful.
This group consisted of two extreme strategies and a range of intermediates be-
tween them. The first extreme corresponded to Johnstone’s (2001) eavesdropper
strategy (i.e. play hawk if you eavesdrop on situations 2, 4 or 6; play dove otherwise,
see Fig. 26.1a). The more common extreme (which we call reputation scoring),
however, consisted of a novel strategy in which individuals essentially copied the
behaviour of their opponent’sopponent (i.e. play hawk in situations 1, 2 and 6; play
dove otherwise). An anthropomorphism of this strategy would be that individuals
paid attention to an individual’s reputation rather than its deeds. The reputation-
scoring strategy was successful because it tended to lead to neighbours ‘ganging
up’ on the same individuals (Fig. 26.1b); as a result, some individuals were highly
successful, and others were very unsuccessful (Fig. 26.2). The overall mean level of
success for the image-scoring and reputation-scoring strategies was actually rather
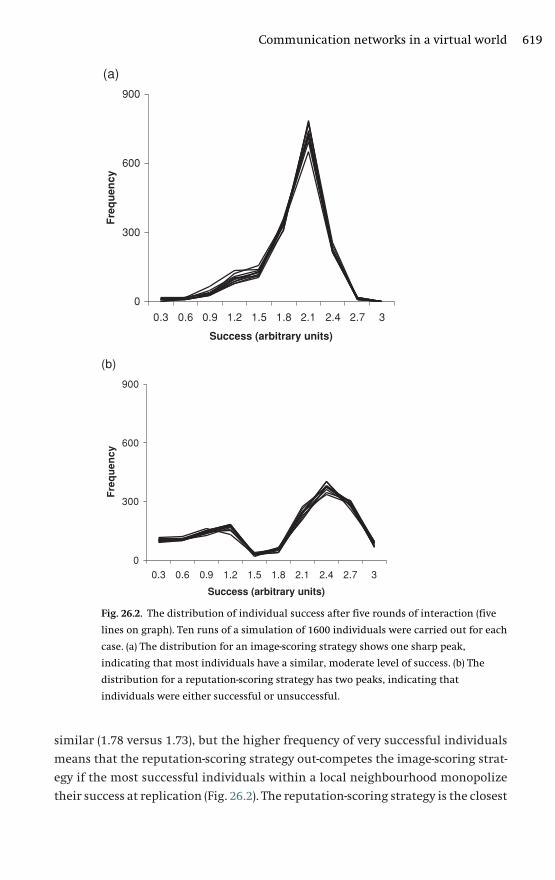
P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 619
(b)
0
300
600
900
0.3 0.6 0.9 1.2 1.5 1.8 2.1 2.4 2.7 3
Success (arbitrary units)
Fre
qu
ency
(a)
0
300
600
900
0.3 0.6 0.9 1.2 1.5 1.8 2.1 2.4 2.7 3
Success (arbitrary units)
Fre
qu
ency
Fig. 26.2. The distribution of individual success after five rounds of interaction (five
lines on graph). Ten runs of a simulation of 1600 individuals were carried out for each
case. (a) The distribution for an image-scoring strategy shows one sharp peak,
indicating that most individuals have a similar, moderate level of success. (b) The
distribution for a reputation-scoring strategy has two peaks, indicating that
individuals were either successful or unsuccessful.
similar (1.78 versus 1.73), but the higher frequency of very successful individuals
means that the reputation-scoring strategy out-competes the image-scoring strat-
egy if the most successful individuals within a local neighbourhood monopolize
their success at replication (Fig. 26.2). The reputation-scoring strategy is the closest

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
620 A. M. R. Terry & R. Lachlan
approximation to social eavesdropping (Ch. 2), because it requires eavesdroppers
to pay attention to the interaction and the role of each individual. The model’s
combination of a spatial simulation limiting individuals to information gathered
from the interactions going on around them, with a cost–benefit approach of de-
termining the optimal strategy to follow should provide a deeper understanding
of the evolution of eavesdropping as a strategy for gathering information.
Network effects on the formation of linear dominance hierarchies
The formation of dominance hierarchies represents another area where
information from interactions between individuals can be used by observers to
influence future encounters (e.g. Dugatkin, 2001). A linear dominance hierarchy
is defined by the number of component triads (trios) within the group that form
transitive relationships (i.e. if A beats B, and B beats C, then A also beats C); as the
number of transitive relationships increases, linearity increases. Here we consider
a simulation model of the effects of communication networks on the formation
of linear dominance hierarchies (R. L. Earley, S. Brosnan & J. Bragg, unpublished
data).
Linear dominance hierarchies are established via overt aggressive interactions
and their establishment leads to the unequal distribution of resources among
dominant and subordinate individuals. Hierarchies also cause a general decrease
in the overall aggression levels within the group. Dominance hierarchy forma-
tion has been studied in a wide range of taxa; however, the factors involved in
their formation remain contentious. Conceptual models attribute the formation
of linear hierarchies either to some aspect related to fighting ability (e.g. Slater,
1986; Jackson & Winnegrad, 1988) or to social effects such as winner effects, loser
effects and eavesdropping (e.g. Chase, 1980; Bonabeau et al., 1996; Dugatkin, 1997,
2001). The individual-based spatially explicit simulation model is being developed
(R. L. Earley, S. Brosnan & J. Bragg, unpublished data) to study how social eaves-
dropping may influence the dynamics of hierarchy formation in groups of virtual
animals.
In each simulation, a group of 10 individuals are allowed to interact for a
predetermined period of time. At each time step, one individual can initiate an
interaction with another and, if the other individual responds an aggressive in-
teraction begins. Individuals can interact through displays or they can escalate
the contest to fighting. At the conclusion of the contest, individuals update their
estimate of their own fighting ability. These updates mimic the winner and loser
effects where dominant animals increase and subordinate animals decrease their
perception of their own fighting ability (Hsu & Wolf, 2001). A certain proportion
of individuals close to the interaction (eavesdroppers) can observe it and, in conse-
quence, update their estimates of the interactants’ fighting abilities. The estimate

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 621
of an opponent’sfighting ability determines whether an individual will initiate an
interaction and to what extent it will be pursued. The outcome of the simulation
is a dominance matrix and the degree of linearity is determined using Landau’s
index (0 < h < 1; Landau, 1951). If Landau’s h > 0.9, the hierarchy is considered to
be linear (Chase, 1974).
In an initial assessment of the model, the social effects were kept symmetrical
(i.e. winner effects = loser effects); there was no initial variation in fighting abili-
ties, and all individuals could eavesdrop on each interaction within their network.
When social factors were excluded from the model, display and escalated inter-
actions occurred with equal frequency and non-linear hierarchies emerged. Also
when winner or loser effects operated alone, the linearity index remained low
and did not increase greatly when the magnitude of the winner/loser effects was
increased. However, the model showed that eavesdropping, when it was included,
acted to increase the estimates of the fighting abilities of winning individuals
and to decrease those of losers. When included with winner–loser effects, eaves-
dropping caused the formation of strongly linear hierarchies. The most important
factor was the inflation of the winner’s estimated fighting ability (decreasing the
loser’s estimate had less effect). As with our model of a spatial hawk–dove game,
this model provided a simulation of social eavesdropping (Ch. 2) as individuals
paid attention to both the interactants. The resulting modification of the esti-
mates of fighting ability depended not only on each of the interactants but also
on the relative differences in fighting ability when the two individuals met in the
interaction. This model is a first attempt to study the implications of networks
in the formation of dominance hierarchies. Future studies could investigate the
relationship between asymmetries in both fighting abilities and eavesdropping in
promoting or hindering linear dominance hierarchies.
Summary
The models detailed here have provided the first theoretical studies of the
role of eavesdropping in communication networks. Eavesdropping is one of the
potential sources of information available to receivers and has received the most
experimental attention (McGregor & Peake, 2000, Ch. 2). Experimental data from
different taxa have shown that individuals can and do pay attention to the inter-
actions of conspecifics and that these interactions will modify the behaviour of
individuals in future encounters (e.g. Oliveira et al., 1998; Peake et al., 2001, 2002). It
is likely that eavesdropping is a common behaviour. Although originally thought
to be a cost-free source of information (McGregor, 1993), it is likely that eavesdrop-
ping has costs associated with the partitioning of cognitive processes required to
follow interactions and the fact that individuals may have to abstain from per-
forming other behaviours to witness interactions. Initial theoretical treatments

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
622 A. M. R. Terry & R. Lachlan
of eavesdropping showed that it was only stable as a minority strategy because
eavesdroppers could not predict how other eavesdroppers would behave and, as
a result, it would lead to more aggressive encounters (Johnstone, 2001). However,
when eavesdroppers were able to follow the interactions of individuals around
them, as opposed to being randomly drawn into interactions, it became apparent
that eavesdropping was a viable strategy (R. Lachlan & A. M. R. Terry, unpublished
data). Further analyses will address how individuals balance the costs and benefits
of eavesdropping against other sources of information or other behaviours.
Summary
When communication is considered to occur within a network, new pos-
sibilities emerge for individuals to broadcast and receive information that will
affect their behaviour in future interactions. Networks also extend the considera-
tion of the costs and benefits of signalling to include other signallers and receivers
that may or may not be apparent to the respective interactants. Current models
of animal communication have been dominated by game theory, which is well
suited to the analysis of strategies used by individuals in dyadic encounters. How-
ever, when applied to networks, pure game theory models allow only a superficial
analysis of the costs and benefits of signalling. While consideration of the strategic
nature of communication in networks remains important, we feel that it must
be combined with more process-based approaches that place fewer restrictions on
individual behaviour. Individually based simulation models are becoming increas-
ingly common in behavioural ecology and a combination of these more proximate
level models with game theory approaches will give a greater understanding of the
evolution of communication networks. In particular, in the examples we have de-
scribed (anuran acoustic choruses and eavesdropping strategies), spatially realistic
individually based models have generated different results from more traditional
techniques applied to the same question. In this chapter, we have emphasized the
importance of studying realistically structured populations and have identified
the spatial nature of communication in networks as an important feature of any
theoretical consideration. The importance of the spatial nature of networks may
also extend to network organization. There are likely to be asymmetries in the
extent to which individuals contribute to the flow of information in a network:
some individuals providing more information than others. One future avenue
for both theoretical and experimental research may be to determine how com-
munication networks are organized and whether hub individuals act as routers
through which most information flows. The nature of the flow of information
through networks, whether it is a signal spreading through a chorus or an alarm

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 623
call spreading through a population, will undoubtedly have implications for the
evolution of signalling within that system.
Although there is a wide literature on the theoretical aspects of strategic deci-
sions when communicating and cooperating in groups, there is little experimen-
tal evidence to support these models (for exceptions see Wedekind & Milinski,
2000; Milinski, et al., 2001; Stephens et al., 2002). Work on cooperation in groups
has shown that modelled strategies may not consider the complex combination
of assumptions, working rules and limitations that individuals face when decid-
ing how to respond in interactions with known individuals (Milinski et al., 2001;
Stephens et al., 2002). We suggest that future models incorporating both strategic
and simulation aspects will be able to model more closely the dynamics involved
in observing and taking part in repeated interactions, and this, in turn, will lead to
a better understanding of the strategies and behaviours that individuals employ
when communicating in networks.
Acknowledgements
We are very grateful to Ryan Earley for allowing us to discuss and present his model. We
would also like to thank several people whose comments helped make this chapter more readable;
Ryan Earley, Ricardo Matos, Tom Peake and Denise Pope. A. T. was funded by the Zoological
Institute at the University of Copenhagen during the writing of this chapter.
References
Albert, R. & Barabasi, A. L. 2002. Statistical mechanics of complex networks. Reviews of
Modern Physics, 74, 47–97.
Albert, R., Jeong, H. & Barabasi, A. L. 1999. Diameter of the World-Wide Web. Nature,
401, 130–131.
Axelrod, R. & Hamilton, W. D. 1981. The evolution of cooperation. Science, 211,
1390–1396.
Barabasi, A. L. & Albert, R. 1999. Emergence of scaling in random networks. Science,
286, 509–512.
Begon, M., Harper, J. L. & Townsend, C. R. 1990. Ecology: Individuals, Populations and
Communities, 2nd edn. Oxford: Blackwell.
Bonabeau, E., Theraulaz, G. & Deneubourg, J.-L. 1996. Mathematical model of
self-organizing hierarchies in animal societies. Bulletin of Mathematical Biology, 58,
661–717.
Bonabeau, E., Dorigo, M. & Theraulaz, G. 1999. Swarm Intelligence: From Natural to
Artificial Systems. Oxford: Oxford University Press.

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
624 A. M. R. Terry & R. Lachlan
Brush, J. S. & Narins, P. M. 1989. Chorus dynamics of a neotropical amphibian
assemblage: comparison of computer-simulation and natural behavior. Animal
Behaviour, 37, 33–44.
Camazine, S. & Sneyd, J. 1991. A model of collective nectar source selection by
honey-bees: self-organization through simple rules. Journal of Theoretical Biology,
149, 547–571.
Chase, I. D. 1974. Models of hierarchy formation in animal societies. Behavioral Science,
19, 374–382.
1980. Social process and hierarchy formation in small groups: a comparative
perspective. American Sociology Review, 45, 905–924.
Deneubourg, J. L. & Goss, S. 1989. Collective patterns and decision making. Ethology,
Ecology and Evolution, 1, 295–311.
Deneubourg, J. L., Aron, S., Goss, S. & Pasteels, J. M. 1990. The self-organizing
exploratory patterns of the Argentine ant. Journal of Insect Behaviour, 3, 159–168.
Dieckman, U. C. 1997. Can adaptive dynamics invade? Trends in Ecology and Evolution,
12, 128–131.
Dugatkin, L. A. 1997. Winner and loser effects and the structure of dominance
hierarchies. Behavioral Ecology, 8, 583–587.
1998. Game theory and cooperation. In: Game Theory and Animal Behavior, ed. L. A.
Dugatkin & H. K. Reeve. Oxford: Oxford University Press, pp. 38–63.
2001. Bystander effects and the structure of dominance hierarchies. Behavioral
Ecology, 12, 348–352.
Dunne, J. A., Williams, R. J. & Martinez, N. D. 2002. Food-web structure and network
theory: the role of connectance and size. Proceedings of the National Academy of
Sciences, USA, 99, 12917–12922.
Grafe, T. U. 1999. A function of synchronous chorusing and a novel female preference
shift in an anuran. Proceedings of the Royal Society of London, Series B, 266, 2331–2336.
Greenfield, M. D. 2002. Signalers and Receivers: Mechanisms and Evolution of Arthropod
Communication. Oxford: Oxford University Press.
Greenfield, M. D. & Rand, A. S. 2000. Frogs have rules: selective attention algorithms
regulate chorusing in Physalaemus pustulosus (Leptodactylidae). Ethology, 106,
331–347.
Greenfield, M. D., Tourtellot, M. K. & Snedden, W. A. 1997. Precedence effects and the
evolution of chorusing. Proceedings of the Royal Society of London, Series B, 264,
1355–1361.
Grimm, V. 1999. Ten years of individual-based modelling in ecology: what have we
learned and what could we learn in the future? Ecological Modelling, 115, 129–148.
Hamilton, W. D. 1971. Geometry of the selfish herd. Journal of Theoretical Biology, 31,
295–311.
Hemelrijk, C. K. 2000. Towards the integration of social dominance and spatial
structure. Animal Behaviour, 59, 1035–1048.
2002. Understanding social behaviour with the help of complexity science. [Invited
article.] Ethology, 108, 655–671.

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 625
Hsu, Y. Y. & Wolf, L. L. 2001. The winner and loser effect: what fighting behaviours are
influenced? Animal Behaviour, 61, 777–786.
Jackson, W. M. & Winnegrad, R. L. 1988. Linearity in dominance hierarchies: a second
look at the individual attributes model. Animal Behaviour, 36, 1237–1240.
Jeong, H., Tombor, B., Albert, R., Oltval, Z. N. & Barabasi, A. L. 2000. The large-scale
organization of metabolic networks. Nature, 407, 651–654.
Johnstone, R. A. 1998. Conspiratorial whispers and conspicuous displays: games of
signal detection. Evolution, 52, 1554–1563.
2001. Eavesdropping and animal conflict. Proceedings of the National Academy of
Sciences, USA, 98, 9177–9180.
Krebs, J. R. & Dawkins, R. 1984. Animal signals: mind-reading and manipulation. In:
Behavioural Ecology: An Evolutionary Approach, 2nd edn, ed. J. R. Krebs & N. B. Davies.
Oxford: Blackwell, pp. 380–402.
Kudo, H. & Dunbar, R. I. M. 2001. Neocortex size and social network size in primates.
Animal Behaviour, 62, 711–722.
Landau, H. G. 1951. On dominance relations and the structure of animal societies: I.
Effect of inherent characteristics. Bulletin of Mathematical Biophysics, 13, 1–19.
Leimar, O. & Hammerstein, P. 2001 Evolution of cooperation through indirect
reciprocity. Proceedings of the Royal Society of London, Series B, 268, 745–753.
Maynard Smith, J. 1982. Evolution and the Theory of Games. Cambridge, UK: Cambridge
University Press.
McGregor, P. K. 1993. Signalling in territorial systems: a context for individual
identification, ranging and eavesdropping. Philosophical Transactions of the Royal
Society, Series B, 340, 237–244.
McGregor, P. K. & Dabelsteen, T. 1996. Communication networks. In: Ecology and
Evolution of Acoustic Communication in Birds, ed. D. E. Kroodsma & E. H. Miller.
Ithaca, NY: Cornell University Press, pp. 409–425.
McGregor, P. K. & Peake, T. M. 2000. Communication networks: social environments
for receiving and signalling behaviour. Acta Ethologica, 2, 71–81.
Milinski, M., Semmann, D., Bakker, T. C. M. & Krambeck, H.-J. 2001. Cooperation
through indirect reciprocity: image scoring or standing strategy? Proceedings of the
Royal Society of London, Series B, 268, 2495–2501.
Morton, T. L., Haefner, J. W., Nugala, V. & Decino, R. D. 1994. The selfish herd revisited:
do simple movement rules reduce relative predation risk? Journal of Theoretical
Biology, 167, 73–79.
Nowak, M. A. & Sigmund, K. 1998. Evolution of indirect reciprocity by image scoring.
Nature, 393, 573–577.
Oliveira, R. F., McGregor, P. K. & Latruffe, C. 1998. Know thine enemy: fighting fish
gather information from observing conspecific interactions. Proceedings of the
Royal Society of London, Series B, 265, 1045–1049.
Peake, T. M., Terry, A. M. R., McGregor, P. K. & Dabelsteen, T. 2001. Male great tits
eavesdrop on simulated male-to-male vocal interactions. Proceedings of the Royal
Society of London, Series B, 268, 1183–1187.

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
626 A. M. R. Terry & R. Lachlan
2002. Do great tits assess rivals by combining information gathered by
eavesdropping? Proceedings of the Royal Society of London, Series B, 269, 1925–1929.
Pollock, G. & Dugatkin, L. A. 1992. Reciprocity and the evolution of reputation. Journal
of Theoretical Biology, 159, 25–37.
Resnick, M. 1994. Turtles, Termites and Traffic Jams. Cambridge, MA: MIT Press.
Rich, T. J. & Hurst, J. L. 1999. The competing countermarks hypothesis: reliable
assessment of competitive ability by potential mates. Animal Behaviour, 58,
1027–1037.
Riechert, S. E. 1998. Game theory and animal contests. In: Game Theory and Animal
Behaviour, ed. L. A. Dugatkin & H. K. Reeve. Oxford: Oxford University Press,
pp. 64–93.
Schwartz, J. J., Buchanan, B. W. & Gerhardt, H. C. 2001. Female mate choice in the gray
treefrog (Hyla versicolor) in three experimental environments. Behavioral Ecology
and Sociobiology, 49, 443–455.
2002. Acoustic interactions among male gray treefrogs, Hyla versicolor, in a chorus
setting. Behavioral Ecology and Sociobiology, 53, 9–19.
Seeley, T. D., Camazine, S. & Sneyd, J. 1991. Collective decision making in honey-bees:
how colonies choose among nectar sources. Behavioral Ecology and Sociobiology, 28,
277–290.
Sigmund, K. & Nowak, M. A. 1998. The complexity of cooperation: agent-based models
of competition and collaboration. Nature, 392, 457–457.
Sih, A. & Mateo, J. 2001. Punishment and persistence pay: a new model of territory
establishment and space use. Trends in Ecology and Evolution, 16, 477–479.
Slater, P. J. B. 1986. Individual differences and dominance hierarchies. Animal
Behaviour, 34, 1264–1265.
Snedden, W. A. & Greenfield, M. D. 1998. Females prefer leading males: relative call
timing and sexual selection in katydid choruses. Animal Behaviour, 56, 1091–1098.
Stamps, J. A. & Krishnan, V. V. 1999. A learning-based model of territory establishment.
Quarterly Review of Biology, 74, 291–318.
2001. How territorial animals compete for divisible space: a learning-based model
with unequal competitors. American Naturalist, 157, 154–169.
Stephens, D. W., McLinn, C. M. & Stevens J. R. 2002. Discounting and reciprocity in an
iterated prisoner’s dilemma. Science, 298, 2216–2218.
Sugden, R. 1986. The Economics of Rights, Co-operation and Welfare. Oxford: Blackwell.
Turelli, M. & Barton, N. H. 1994. Genetic and statistical analyses of strong selection
and polygenic traits: what, me normal? Genetics, 138, 913–941.
Viscido, S. V. & Wethey, D. S. 2002. Quantitative analysis of fiddler crab flock
movement: evidence for ‘selfish herd’ behaviour. Animal Behaviour, 63, 735–741.
von Frisch, K. 1967. The Dance Language and Orientation of Bees. Cambridge, MA: Harvard
University Press.
Watts, D. J. & Strogatz, S. H. 1998. Collective dynamics of ‘small-world’ networks.
Nature, 393, 440–442.
Wedekind, C. & Milinski, M. 2000. Cooperation through image scoring in humans.
Science, 288, 850–852.

P1: JZZ/... P2: JZZ/...0521823617c26.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 10:42
Communication networks in a virtual world 627
Williams, R. J. & Martinez, N. D. 2000. Simple rules yield complex food webs. Nature,
404, 180–183.
Wilson, W. 2000. Simulating Ecological and Evolutionary Systems in C. Cambridge, UK:
Cambridge University Press.
Zelick, R. & Narins, P. M. 1985. Characterization of the advertisement call oscillator in
the frog Eleutherodactylus coqui. Journal of Comparative Physiology A: Sensory Neural and
Behavioral Physiology, 156, 223–229.

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index
Abudefduf vaigiensis see sergeant major fish
Acheta domesticus see house cricket
Accipiter nisus see sparrowhawk
acoustic choruses, models of signalling
dynamics 609–611
see also anuran choruses; insect choruses
acoustic communication
caller identity and status information 381
factors affecting evolution of 152–153
problem of masking interference 157–162
see also signal production; sound
transmission
Acrocephalus arundinaceus see great reed
warbler
advertising signals, to facilitate
eavesdropping 48–49, 52
African catfish (Clarias gariepinus),
semiochemicals 558–559
African elephant (Loxodonta africana)
benefits of attending to others’ social calls
384
contact call discrimination and memory
383–384
fluid social systems and long-distance
signalling 377–378
group social knowledge and age of
matriarch 383–384
individuality in fundamental frequency
contour in vocalizations 382
infrasound communication 457–459
intelligible distance of calls 372, 382–383
social knowledge related to age and
experience 383–384
African grey parrot (Psittacus erithacus),
cognitive and communicative capacities
569, 572–573
Agelaius phoeniceus see red-winged blackbird
Agelenopsis aptera see funnel-web spider
aggression 2
and social instability in fishes 96
androgens and 481–482
in models of territory establishment
612–613
modelling linear dominance hierarchy
formation 620–621
physiological costs 191–192
song overlapping in songbirds 304,
305–306
victory displays 11
see also redirected aggression
aggressive calls, variety in male anurans
279–280
aggressive priming, audience effects
75–78
aggressive signal exchanges see
eavesdropping; signalling interactions
agonistic contests see aggression
Alcelaphus buselaphus see hartebeeste
allomones see fish semiochemicals
Alouatta pigra see black howler monkey
alpine accentor (Prunella collaris), quiet singing
53
628

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 629
altruism
and cooperation in communication
networks 446
audience effects 30–31
dishonest signals used for tactical
deception 524–525
eavesdropping and indirect reciprocity
522–523
evolution and maintenance 536
functional rather than cognitive approach
522–523
image scoring and evolution of 533–534
reciprocal altruism and behavioural
strategies 521–522
Alytes spp. see midwife toads
Amolops tormotus see Chinese frog
androgens
adaptive value of social modulation 497,
502–505
and aggressive behaviour 481–482, 496–498,
499
and bystander priming response 499–500,
501
and electrocommunication signals
487–488
and pheromone production and/or release
485–486
and sex differences in spatial memory
490–491
brain receptors 490–491
costs and benefits of high levels 497,
502–505
effects of early exposure (critical period)
490–491
effects of high levels on male parental care
497, 504–505
effects of population density 492–494
effects on cognitive functions 490–491
effects on expression of somatic releasers
491–492
effects on pheromone production in
urodeles 485–486
effects on singing behaviour of songbirds
483–485
effects on visual displays in vertebrates
486–487
effects on vocal structures of amphibians
484–485
effects on vocal structures of birds 483–484
effects on vocal structures of toadfish 484
interactive effects with social environment
482
interrelationship with associative learning
mechanisms 505
levels during social instability 492–494
levels in dominant and subordinate males
492–494, 496–498, 499
modulation of behaviour in
communication networks 494–502
modulation of central mechanisms
affecting motivation 490
modulation of sensory perception 488–489
possible mediation of audience effects 501
possible mediation of dear enemy effects
501–502
role in winner–loser effects 198, 492–494,
496–498, 499
social modulation of androgen levels
492–494
social modulation of behavioural effects
481–482
stimulation by social interactions 492–494
Anolis aeneus see lizard
anonymity, used to counter eavesdropping
56–57
Anser anser see greylag goose
antbirds (Thamnophilidae), sound
characteristics 49
Antilocapra americana see pronghorn antelope
anuran amphibians
communication networks 2, 248
effects of androgens 484–485, 486–487
use of private signalling 291
victory displays 118, 122
anuran choruses 263–264
adaptations for acoustic competition
278–279
alternation of signalling 280–282

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
630 Index
anuran choruses (cont.)
auditory systems and background noise 292
call-timing, evolution and maintenance
282–284
conflicting demands of signalling 279–280
entrained calling 280–282
features of network communications
291–292
female acoustic responses 283
female attraction to larger choruses 283
female call preferences 264, 283–284,
285–286, 292
female choices in trials and natural
choruses 287–289
female cognitive requirements for social
eavesdropping 289
female eavesdropping on male–male
interactions 289
female mate sampling techniques 288–289
female sensory abilities and signal
discrimination 285
fine-scale patterns of signal timing 280–283
graded or discrete calls 279
leading and lagging roles 280–282
male aggressive calls 279–280
male calling energy costs 284–285
male interceptive eavesdropping 289–291
male–male vocal competition 278–279
males spatial distribution and selective
attention 284
models of signalling dynamics 609–610, 611
network view 277–278
‘off response’ call initiation 280–282
repertoire size of signallers 279
selection pressures on signals 277, 283–284
signal overlap avoidance in males 280–282
suitability for studying communications
networks 292–293
synchronous calling 280, 281, 282
two-part advertising call 279–280
use of both ‘on-response’ and ‘off-response’
calling 282–283
Aphelocoma californica see western scrub jay
Apis mellifera see honeybee
Aptenodytes forsteri see emperor penguin
Aptenodytes patagonicus see king penguin
Apternotus leptorhynchus see brown ghost
Argentine ant (Linepithema humile), foraging
strategies 611–612
arthropods, victory displays 117–118,
122
associative learning
and tactical deception 524–525
interrelationship with androgen effects
505
Atlantic salmon (Salmo salar), androgen effects
on olfactory sensitivity 488–489
audience effects 10, 30–31
and altruism 522–523
and eavesdropping 66
and victory displays 122–123
clarification of terminology 66–67, 68
definition 65–66
human behaviour 74–75
in redirected aggression 209–210
male–male aggressive signalling 68–72
male parental behaviour 72–74
on cheating behaviour 529–530, 531
on signalling 65–66
possible influences on scent marking
362–363
possible mediation by androgens 501
pre-exposure and male aggression 75–78
review of evidence for 67–75
audiences
and signal evolution 79
apparent 65
definition of 64
evolutionary 64–65
terminology 66–67, 68
see also eavesdropping
auditory systems
analysis by receivers 471–474
coping with background noise 292
hormonal modulation of sensitivity 489
Australian bushcricket (Elephantodeta nobilis),
evidence for eavesdropping 291
autocommunication, and eavesdropping 18

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 631
avian cognition, research history and
prejudices 568–570
baboon (Papio cynocephalus)
knowledge of other animals 585–586, 587,
589
social structure and relationship
recognition 584
background noise
and functioning of auditory systems 292
effects of lunar phase 158, 163–164
see also masking interference; masking
release
Baktaman people (New Guinea) 421
Balaenoptera musculus see blue whale
Balaenoptera physalus see fin whale
banded wren (Thryothorus pleurostictus)
behaviour after dawn chorus 327
bout structure during and after dawn
chorus 329–331
dawn chorus as an interactive network 322,
337
dawn chorus features 326–327
dawn chorus singing and male quality 338
dawn chorus structure 336
daytime song-delivery patterns 326
male interactions at dawn chorus 335,
337–338
movement patterns around dawn chorus
330, 332
multiple song matching 335
overlapping and matching songs 331, 334,
335–336
recording methods 327–329, 330, 339–340
singing behaviour during and after dawn
chorus 323, 329–336
song behaviour 325–327
song matching as indicator of conflict
333–335
song matching during and after dawn
chorus 330, 332–335
song overlapping as an aggressive signal
331, 334, 335–336
song rates during dawn chorus 330, 331
song repertoire 326
territorial conflict and song matching
333–335
territorial behaviour 325–326, 327
variation in dawn chorus singing of
individual males 336
barking treefrog (Hyla gratiosa), female
assessment of males 137, 288–289
barn owl (Tyto alba)
nestling interactions 176
sound localization ability 465–467
barnacle goose (Branta leucopsis), victory
display 115–116
bats (various species)
attraction to echolocation signals 405
predation on katydids 154, 156–157
bearded seal (Erignathus berbatus), distinctive
group calls 399–400
behaviour, reciprocal link with hormones
481–482
beluga whale (Delphinapterus leucas)
avoidance of ice-breakers 406–407
reactions to killer whale sounds 403–404
victory display 118
Betta splendens see Siamese fighting fish
binaural processing, and acoustic signal
masking release 462–463
birds
effects of androgens on vocal structures
483–484
equating human and avian cognitive
studies 573–574
signal distance assessment 467–468
victory displays 115–117
visual displays and effects of androgens 486
see also songbirds; individual species
black-capped chickadee (Poecile atricapillus)
dawn chorus and male condition 141, 338
eavesdropping and transitive inference (TI)
574–576
female mate choice 140–141, 142
habitat change and song networks 143–144
habitat quality and song output 144
ranking process 577

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
632 Index
black-capped chickadee (cont.)
signal amplitude as a distance cue 468–469
use of social eavesdropping 21–22
black goby (Gobius niger)
male sex pheromone 549–558
semiochemical communication 558–559
black howler monkey (Alouatta pigra), factors
affecting levels of vigilance 419
black swan (Cygnus atratus), victory display
115–116
black-throated diver (Gavia immer), victory
display 116–117
blackbird (Turdus merula)
courtship interruption by neighbours 41
dawn chorus singing 337
male quality and dawn chorus singing 338
sound characteristics and attenuation
49–50
use of high perches 43, 44, 464–465
use of quiet song 50, 53–55, 56
blackcap (Sylvia atricapilla)
sound characteristics and attenuation 49
use of high perches 43, 44
blenny semiochemical communication
558–559
blue-throated humming bird (Lampornis
clemenciae), Lombard effect 464–465
blue tit (Parus caeruleus), dawn chorus singing
and male quality 338
blue whale (Balaenoptera musculus), distinctive
group calls 400
bonnet macaque see macaques
Boophis madagascariensis see Madagascar
treefrog
bottlenose dolphin (Tursiops truncatus)
active space of signals 392–394, 395
avoidance of feeding grounds with boat
noise 406–407
eavesdropping on other’s echolocation
clicks 18, 405–406
fission–fusion societies 400–402
individual signature whistle types 392–395,
400–402
numbers of animals in networks 397–399
recognition of others’ social relationships
591
silence when captured or near a boat
403–404
victory display 118
vocal learning and individual whistle
development 400–402
vocal matching to signal a specific
individual 402
boubou (Laniarius aethiopicus), victory display
116
Branta canadensis see Canada goose
Branta leucopsis see barnacle goose
Brienomyrus brachyistius see electric fish
Broadley’s painted reed frog (Hyperolius
marmoratus broadleyi)
female call preferences 286, 287–288
‘off-response’ call initiation 280–282
simultaneous mate choice in females
289
brown capuchin monkey (Cebus apella)
demands of social monitoring
32
predation levels and vigilance 417–418
time spent looking by subordinates
417–418
brown ghost (Apternotus leptorhynchus),
androgen effects on electric signals
487–488
brown-headed cowbird (Molothrus ater),
nestling begging 177
budgerigar (Melopsittacus undulatus)
male parental behaviour 73–74
masking release and binaural processing
462–463
signal-to-noise ratios for recognition and for
detection 463–464
sound localization ability 465–467
bystanders
and redirected aggression 192–194, 200,
201–203
costs and benefits of information gathering
131–132
distinction from eavesdroppers 84–85

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 633
effects on dominance hierarchies 97–99
influence of eavesdropping on behaviour
86–91, 93
influence of previous experience on
behaviour 88–89
postconflict attacks on losers 196–197
priming response and androgens 499–500,
501
social eavesdropping 10–11
taking advantage of ‘loser effects’ 198–200
see also eavesdroppers
Canis latrans see coyote
Canis lupus see wolf
Campbell’s monkey (Cercopithecus campbelli),
attention to alarm calls by another
species 373, 380
Canada goose (Branta canadensis), victory
display 115–116
canary (Serinus canarius), sound localization
ability 465–467
Capreolus capreolus see roe deer
Carassius auratus see goldfish
Carassius carassius see Crucian carp
cat, feral ( Felis catus), victory display 118–119,
121–122
Cebus apella see brown capuchin monkey
Cebus capucinus see white-faced capuchin
monkey
Cephalorhyncus hectori see Hector’s dolphin
Cercocebus albigena see mangabey
Cercopithecus aethiops see vervet monkey
Cercopithecus ascanius schmidtii see redtail
monkey
Cercopithecus campbelli see Campbell’s monkey
Cercopithecus diana see Diana monkey
Cervus elaphus see red deer
cetaceans, vocal matching to signal a specific
individual 402
chacma baboon (Papio cynocephalus ursinus)
attending to signal interactions 373–374
awareness of dominance relationships
199
time spent visually scanning 417–418
chaffinch (Fringilla coelebs), signal distance
assessment 467–468
chemical communication
correlation with predation 545–546
evolution through different functional
phases 541–542, 543
predation risk assessment 544–549
scent marking 131
signal propagation 543–544
see also fish semiochemicals; scent marking
chimpanzee (Pan troglodytes)
choice of targets for aggression 208
fluid social systems and long-distance
signalling 377
situations where calling is suppressed
374–375, 376–377
Chinese frog (Amolops tormotus) acoustic signal
repertoire 279
chorusing interactions
and female preference for leading signals
264
and signal competition 263–264
in anurans and insects 263–264
precedence effect 264
to avoid signal masking 264
see also anuran choruses
Cichlasoma nigrofasciatum see convict cichlid
cichlid see Mozambique tilapia
Cistothorus palustris see marsh wren
Clamator glandarius see great spotted cuckoo
Clarias gariepinus see African catfish
claw waving see fiddler crabs
cleaner wrasse (Labroides dimidiatus)
behaviour towards clients 534
cheating behaviour 525–526
client image scoring and wrasse behaviour
526–528, 529–530, 531
cognitive abilities used in tactical deception
535, 536
effects of bystander types on cheating
behaviour 529–530, 531
interactions with client reef fish 525–526
mutualism with client reef fish 521
possible endocrine-mediated response 535

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
634 Index
cleaner wrasse (cont.)
preferred food 534
response of biting cleaners to image scoring
clients 531–532
tactile stimulation 531–532
variable payoffs from different clients
533–534
variations in cooperative and cheating
behaviour 526–527, 528
cognition in animals, research history and
prejudices 568–570
cognitive aspects of communication networks
250–251
cognitive capacity
and cortical size 569
of birds 568–570
requirements for tactical deception 524–525
cognitive sciences, interface with
communication networks 447
common carp (Cyprinus carpio), pheromone
system 552
common dolphin (Delphinus delphis), individual
signature signals 400–402
common seal (Phoca vitulina)
avoidance of killer whale sounds 403
distinctive group calls 399–400
loud sexual advertisement calls 380
territorial behaviour 404–406
communication, dyadic view 2
and hormones 482–483, 494
limitations of 9
nestling begging 171–173
social context 78–79
communication networks 2, 9
androgen-modulated behaviour in 494–502
animal versus human 384–385
application to welfare of captive animals
assessing social intelligence 447
banded wren dawn chorus 337
chemical assessment of predation risk
544–549
cognitive requirements for participants
250–251, 447
concept of true individual recognition
363–366
context effects 1, 78–79, 129–132
cooperation and altruism 446, 536
defining properties 13
effects on signalling and receiving 180–185
evolution of spiteful behaviour 536
evolution of tactical deception 536
evolutionary process 558–559
game theory models 536
habitat alteration effects 143–144
hormones and communication 494–502
impact on signalling errors 184–185
implications for theoretical study of
signalling strategies 604
individual recognition mechanism 363–366
information cascades 270, 607–608
interfaces with other disciplines 1, 445–450
mathematical modelling 447–448
models of dominance hierarchy formation
620–621
models of eavesdropping by receivers
613–622
models of effects on signallers and
signalling dynamics 608–613
models of structure and organization
607–608
ostariophysan alarm system 546, 547–549
possible links with applied biology 448–450
receiver’s perception 445–446, 451–452, 474
semiochemicals 446–447, 540–541
size and extent 250
spatial distribution effects on acoustic
signals 474
structure influences signals and signalling
286–287
structure within a nest 179
use of nestling begging to study 179–180
see also modelling communication
contest behaviour see aggression; redirected
aggression; victory displays
contest behaviour in fishes 85
effects of eavesdropping 85–86, 94–95
environmental influences 95–97

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 635
‘loser effect’ 88–89
opportunities to eavesdrop 86, 87
physical effects of observing fights 90–91,
93–94
potential costs 86
‘winner effect’ 88–89
convict cichlid (Cichlasoma nigrofasciatum),
eavesdropping 17
cooperation and altruism, in communication
networks 446, 521–525
cooperation theory 521
cotton-top tamarin (Saguinus oedipus)
ability to distinguish unfamiliar callers 374
courtship interactions, interruption by
eavesdroppers 40–41
‘social organization and vigilance 426
coyote (Canis latrans)
potential for redirected aggression 205,
206
situations where calling is suppressed
376–377
victory display 118–119, 122
crabs (various species), similarities to fiddler
crabs 258
crickets (various species), victory displays 118,
122
critical (masking) ratio 459
Crocuta crocuta see hyaena
Crucian carp (Carassius carassius), pheromone
system 552
Ctenophorous fordii see Mallee dragon lizard
Cuculus canorus (European cuckoo), nestling
begging 177
Cuvier’s beaked whale (Ziphius cavirostris),
strandings and noise pollution 406–407
Cygnus atratus see black swan
cyprinid fishes
androgen effects on olfactory sensitivity
488–489
androgen effects on somatic releasers
491–492
pheromone systems 552
Cynops pyrrhogaster see Japanese red-bellied
newt
Cyprinus carpio see common carp system
552
Dama dama see fallow deer
Danio rerio see zebrafish
dark-eyed junco (Junco hyemalis), metabolic
effects of high androgen levels 504
dawn chorus
characteristics 320
communication network view 322
comparison with known daytime singing
interactions 324
directed song matching 338–339
environmental explanations for 320–321
female eavesdropping to assess males
321–322
hypotheses to explain 320–322
indicator of male condition 141
interactions in relation to functions 324–325
meaning of song matching and timing 307
mediating changes in social status 339
multi-way male interaction 335, 337–338
possible network structures 322–323
recording methods 324–325, 327–329
singing and male quality 338
social dynamics hypothesis 321, 339
structure in relation to functions 324
temporal patterns of singing behaviour 323,
329–336
see also banded wren
dear enemy effects, possible androgen
mediation 501–502
Delphinapterus leucas see beluga whale
Delphinus delphis see common dolphin
diademed sifaka (Propithecus diadema), scent
over-marking 357
Diana monkey (Cercopithecus diana), attention
to alarm calls of another species 373,
380
Dipodomys spp. see kangaroo rats
Docidocercus gigliotosi see katydid
dolphins
filtering of high–frequency signal
components 402–403

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
636 Index
dolphins (cont.)
use of directionality of clicks to target
signals 402–403
see also bottlenose dolphin
domestic fowl (Gallus domesticus)
audience effects on calls 66–67, 68
eavesdropping and dominance 28
dominance hierarchies
among nestlings 179
and social eavesdropping 28
eavesdropping effects on 97–99, 620–621
effects of prior experience (winner/loser
effects) 97–99
in non-human primates 588–589
modelling formation of 620–621
dominance interactions, simulation
modelling 606–607
dominance status, problems in poor habitats
144
dunnock (Prunella modularis)
courtship interruption by neighbours 41
quiet singing 53
dwarf mongoose (Helogale undulata), group
scent marking 363
dyadic view see communication, dyadic view
Eastern towhee (Pipilo erythrophthalmus), signal
distance assessment 467–468
eavesdroppers
alternative terms for 14
and mate choice 142–143
attending to outcomes of conflicts 198–200
attention to asymmetries in songbird vocal
interactions 313–314
awareness of dominance relationships
198–200
behavioural responses of those observed
523–524
costs and benefits 31–32, 39–40, 45–47, 211
distinction from bystanders 84–85
image scoring allows exploitation by cheats
532–533
image scoring of observed individuals
523
information from scent counter-marking
345, 346
interruption of courtship interactions
40–41
selection pressures caused by 30–31,
523–524
strategies for effective eavesdropping 42
use of song overlapping information 304,
305–306
see also bystanders, eavesdropping
eavesdropping
and altruism 522–523
and audience effects 66
and autocommunication 18
and dominance hierarchies 97–99
and predation risk 45
and secrecy 13–14
and transitive inference 574–576
as a type of bystander effect 499
at dawn chorus 338
bystander behaviour and social instability
96
comparison of primates with other animals
592–598
costs and benefits for signallers 30–31,
40–41, 48–49
countering with private signalling 52–53, 55
definitions of 3, 10, 13–15
effects on bystanders’ behaviour 85, 86–91,
93, 94–95
effects on female mate choice 100–103
effects on interactions 30–31
environmental influences on 95–97, 107
evidence for 39
evolution in semiochemical
communication 542, 559–560, 562
facilitation by interactants 51–52
factors influencing 97, 107–108
for song repertoire information 45–47, 76
identifying different types 28–29
image scoring and dishonest signals
524–525
image scoring in client reef fish 526–528,
529

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 637
implications and future research 57–58
in anuran choruses 289
in communication networks 2–3
in non-primate mammals 591, 592
in relation to scent marking 235–237
indicators of dominance 52–53
individual differences 106–107
information not shared in animals 426
knowledge about other animals’ dominance
ranks 588–590
knowledge about transient social
relationships 590–591
knowledge of other animals’ kin 585–587,
588
models of how receivers use networks
613–622
ostariophysan alarm system 545, 547–549
physical effects of observing fights 90–91,
93–94
predation pressures 95–96
quiet song as response to risks 55
reliability and intimacy of information
416–417
risks for interactants 52
scent over-marking and mate choice 359,
360–361
signalling in different modalities 14, 28–29
social observation in animals 425
social structure and types of information
available 87, 583–584
sound transmission in natural habitats
42–43, 44
state dependency of bystander effects
95
strategies for effective eavesdropping
44–48
strategies for private signalling 52–53
to assess potential mates 141–142
to take advantage of ‘loser effects’ 198–200
true recognition or simple association
363–366
use of advertising to facilitate 48–49
use of high perches to improve reception
43, 44
used to assess fighting ability 85–86
using anonymity to counter 56–57
see also audiences; interceptive
eavesdropping; social eavesdropping;
mate copying
electric fish (Brienomyrus brachyistius),
androgen levels and dominance signals
502–505
electrocommunication signals, effects of
androgens 487–488, 489
elephant see African elephant
elephant seals (Mirounga spp.)
loud sexual advertisement calls 380
use of deep sound channel 397
Elephantodeta nobilis see Australian bushcricket
Eleutherodactylus coqui see Puerto Rican treefrog
emperor penguin (Aptenodytes forsteri),
amplitude modification in calls 462
endocrine response, importance of
individual’s perception of event 493–494
endocrine system
interface with communication networks
446
vertebrates 482
environment, influences on signals and
signalling 286–287 see also habitat
alteration
Erignathus berbatus see bearded seal
Erithacus rubecula see robin
Eschrichtius robustus see grey whale
Eubalaena glacialis see southern right whale
Eudyptula minor see little blue penguin
European cuckoo (Cuculus canorus), nestling
begging 177
European minnow (Phoxinus phoxinus), alarm
substance 546–548
European newts (Triturus spp.), chemical
communication and mate attraction
485–486
European starling (Sturnus vulgaris),
unmasking effect of sound segregation
460, 461
European treefrog (Hyla arborea), energy costs
of male calling 284–285

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
638 Index
fallow deer (Dama dama)
costs and benefits of groaning 379
discrimination of individual male callers
379–380
functions of groaning in males 379
individuality in formant frequencies in
vocalizations 382
fathead minnow (Pimephales promelas), active
space of alarm substance 546–548
Felis catus see cat, feral
female mate choice
assessment by eavesdropping 141–142
call preferences in anurans 264, 283–284,
285–286, 292
desirable male attributes 134
effects of observing interactions 78, 100–103
effects on male trait distribution 99–100,
101, 105–106
emergence of strategies 106
environmental influences 107
future work on 144–145, 146
genetic-based preferences 99–100,
101
in communication networks 129, 133–134
inferences from female movement patterns
135–136, 137
influence of differences in females 106–107
influence of particular signals 134
influence of predation risk 107
information from dawn chorus 141
instigation of male–male interactions
139–140, 141
male trait preference versus mate copying
103–105
measures of male quality 137–141
movement patterns infer assessment of
males 135–136, 137
preference for leading signals 264, 285–286
scent mark assessment in house mice
225–227
secondary mate choice 133, 135
simultaneous assessment 135, 136
song preferences, costs and benefits for
males 152–153
state-dependent influences 106–107
strategies for assessing males 135–136, 137
transmission distance and multiple signals
138–140
use of dawn chorus to assess males 321–322
use of network information 141–143
see also mate choice; mate copying
ferret (Mustela putorius), masking release and
binaural processing 462–463
Ficedula hypoleuca see pied flycatcher
fiddler crabs (Ocypodidae, Uca spp.)
biology 253–256, 257, 258
claw waving display 254–256, 261–262
communication networks 247–248,
252–253, 258
competition for burrow ownership 268–269
conspecific interceptive eavesdroppers
266–267, 268–271
costs and benefits of interceptive
eavesdropping 271–272
costs and benefits of signalling 271–272
courtship displays 254–256
detection distance for conspecifics
259–260
estimating density 258–259
female assessment of male quality 264–266,
267
gross signal timing among males 263
male response to rivals’ waving 268–271
neighbouring eavesdroppers 268, 269–271
primary and secondary receivers 266–271
range and functioning of visual system
259–260
reaction distances for conspecifics 260–261
signals other than claw waving 257
strategies for information gathering
266–271
strategies for signal competition 263–266
synchronous waving in males 264–266
wandering females as target receivers 266,
267
wandering males as eavesdroppers 268–269
waving rate as indication of male quality
264–266

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 639
field cricket (Gryllus bimaculatus), audience
effects on males 72
fin whale (Balaenoptera physalus)
call response distances 392–395
loud sexual songs 380
use of multipath signal arrivals to locate
callers 397
fish
androgen-induced development of somatic
display structure 487
eavesdropping on visual interactions 24–27,
28
information transfer within shoals 84
vocal sounds 484
fish semiochemicals
ancestral phase 541–542, 543
correlation with predation 545–546
distinction between cues and signals
541–542, 543
eavesdropping 543
evolution into communication networks
541–542, 543, 558–559
evolution into spying 541–542, 543
evolution through different functional
phases 541–542, 543
fish taxa with specialized epidermal cells
548–549
fitness benefits for alarm signallers 545,
548–549
hormonal pheromones in information
networks 549–558
hypoxanthine N-oxide 540–541
occurrence of true communication
networks 545, 547–549
ostariophysan alarm substance 546–548
processing by fish olfactory system 543–544
signal propagation 543–544
spying and communication in information
networks 542, 545, 553, 558–560, 562
terminology 541–543
Fringilla coelebs see chaffinch
frogs, habitat influence on signalling 286–287
funnel-web spider (Agelenopsis aptera), victory
display 117–118, 122
Gallinago media see great snipe
Gallus domesticus see domestic fowl
game theory models 536, 606, 612–613,
614–618, 619, 620
Gasterosteus aculeatus see three-spined
stickleback
Gavia immer see black-throated diver
genetic relatedness and nestling begging
177–178
gerbil (Meriones unguiculatus), effects of
androgens on scent marking behaviour
485
ghost crabs (Ocypode spp.) 258
Globicephala melaena see pilot whale
Gobius niger see black goby
golden-collared manakins (Manacus vitellinus),
effects of androgens on visual display 486
golden hamster (Mesocricetus auratus)
androgen effects on central motivational
mechanisms 490
androgen effects on scent marking
behaviour 485
female counter-marking
female preference for top-scent males 360
flank marking 346–347
male preference for top-scent females 360
mechanisms to distinguish top and bottom
scents 347–357
over-marking and territory defence 358–359
persistence of scent marks 344
preferential memory for top scents 347–350
process of counter-marking 344–345
scent marking and social environment 362
targeted over-marking in males 358
true individual recognition 364–365
use of geometric relationships to determine
top scent 352, 356
vaginal marking 347
goldfish (Carassius auratus)
hormonal pheromones and spawning
549–551, 552
hormonal pheromones in an information
network 550–551, 552–553, 555
Gorilla gorilla beringei see mountain gorilla

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
640 Index
grasshopper (Ligurotettix coquilletti), female
attraction to larger choruses 283
great reed warbler (Acrocephalus arundinaceus),
female mate assessment 47–48
great snipe (Gallinago media), female
assessment of males 139
great spotted cuckoo (Clamator glandarius),
nestling begging 177
great tit (Parus major)
attention to asymmetries in vocal
interactions 314
calculation of maximum detection distance
454–456, 457, 458, 459
critical ratios 457
dawn chorus and male condition 141
eavesdropping and transitive inference
575–576
female mate choice and eavesdropping
141–142
information acquired from eavesdropping
19–21, 22–24, 583–584
signal reverberation as a distance cue
469–471, 472–473
sound localization ability 465–467
use of unmodulated sounds 49
greater horseshoe bat (Rhinolophus
ferrumequinum), critical ratios 457
green frog (Rana clamitans), victory display 118,
122
green swordtail (Xiphophorus helleri)
eavesdropping on visual displays 25,
26–27
social interactions 84, 85
influence of eavesdropping on bystander
behaviour 86–91, 93
opportunities to eavesdrop 86, 87
potential costs of combat 86
green treefrog (Hyla cinerea), spatial
unmasking of signals 462–463
grey partridge (Perdix perdix), effects of
androgens on vocal structures 483–484
grey treefrog (Hyla versicolor)
energy costs of male calling 284
female call preferences 284, 288
grey whale (Eschrichtius robustus), reactions to
killer whale sounds 403–404
greylag goose (Anser anser)
kin-oriented redirected aggression 204
victory display 115–116
Gryllus bimaculatus see field cricket
gulf toadfish (Opsanus beta), acoustic signalling
99
guppy (Poecilia reticulata)
eavesdropping and mate choice 100–103
mate copying versus male trait preference
103–105
social interactions 84, 85
Gymnorhinus cyanocephalus see pinyon jay
Gymnotiformes, androgen effects on weak
electric signals 487–488
habitat alteration
and female mate assessment 144
effects on song transmission 143–144
see also environment
habitat quality, and dominance effects
144
hamadryas baboon (Papio hamadryas),
knowledge about transient social
relationships 590
hamster see golden hamster
harbour porpoise (Phocoena phocoena)
avoidance of noise of human activity
406–407
use of clicks for communication 395
harbour seal see common seal
harp seal (Phoca groenlandica), distinctive
group calls 399–400
hartebeeste (Alcelaphus buselaphus), victory
display 118
Hector’s dolphin (Cephalorhyncus hectori), use of
clicks for communication 395
Helogale undulata see dwarf mongoose
Hemideina spp. see wetas
honeybee (Apis mellifera), communication and
social coordination 611–612
hooded warbler (Wilsonia citrina), female
assessment of males 141

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 641
hormones and communication
adaptive value of social modulation 497,
502–505
dyadic view 482–483, 494
effects on cognitive functions 490–491
effects on communication 482–483
effects on learning and memory 490–491
effects on signal reception 488–490
expression of somatic releasers 491–492
modulation of central mechanisms
affecting motivation 490
modulation of effector pathways 483–488
network view 483, 494–495, 502
pheromones 549–558
reciprocal link with behaviour 481–482
role in control of behaviour 481–482
social modulation of androgen levels
492–494
house cricket (Acheta domesticus)
eavesdropping 17
male attraction to rivals’ signals 269
house mouse (Mus musculus domesticus)
effect of androgens on scent marking 485
female reproductive priming through scent
231–232
genetic sources of individual scent
signatures 232–234
information from age of scent marks
228–229
information in urine about owner 220–222
kin and group member recognition by scent
234–235
major urinary proteins present in urine
222–223, 233–234
male dominance structures 229–231
male territorial scent marking 221,
223–225, 227–228
MHC odour types and individual scent
signatures 232–233, 234–235
pheromones in urine 231–232
reproductive priming and the Bruce effect
232
scent mark assessment of males by females
225–227
scent mark assessment of males by other
males 227–228
scent mark detection and the vomeronasal
organ 219–220, 223, 229, 232
scent marking patterns 220–222, 228
scent marks of subordinate males 229–231
scent over-marking and mate choice 360
urine scent marking 220–237
volatile and non-volatile components of
urine 222–223, 228–229
see also mice
house sparrow (Passer domesticus), metabolic
effects of high androgen levels 504
human communication
dyadic view 416–417
network view 416–417
human eavesdropping 426–432
achieving privacy by behavioural means
427–428
and female curiosity 430–431
as a result of increasing privacy 428–429
caused by the need for privacy 428–430, 432
everyday occurrence of 427
historical evidence for 427, 430–432
honest signals and private behaviour
416–417, 429–430
in communication networks 249–250
lack of research on 426–427
male attempts to control others 431–432
privacy and intimate experience 430
social observation time costs in large
groups 428–429
stalking as a means of control 431–432
humans
altruism and indirect reciprocity 522–523
ancestral way of life 420
androgen effects on spatial memory
490–491
animal networks compared with human
networks 384–385
audience effects on behaviour 74–75
benefits from vigilance 420
common behaviour patterns with other
primates 433–435

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
642 Index
humans (cont.)
community benefits of vigilance 422–423
congenital adrenal hyperplasia 490–491
creating opportunities to be looked at
423–424
demands of social monitoring 31–32
domestication and freedom from external
vigilance 430
domestication and intimate behaviour 430
enhancing personal image with visual cues
423
equating human and non-human cognitive
studies 573–574
ethological studies of vigilance 424–425
factors affecting vigilance 435
individual benefits of vigilance 422
information gained from observation 422,
433–434
Lombard effect 464–465
male vigilance and control 434
parading in front of other people 423–424
privacy and self-awareness 429–430
redirected aggression 208–209
sex differences in social monitoring
424–425
sex differences in spatial memory 490–491
signal echo tail as a distance cue 469–471,
472–473
social comparisons 425–426
social control and vigilance 421–423, 424
stalking 424, 431–432
strength and features of female networks
434–435
suspicion of private behaviour in openly
living groups 420–422
transitive inference task 570–571
unifying model for vigilance, social
observation and eavesdropping 433–435
unmasking effects of amplitude-modulated
background noise 461–462
using vigilance to control 424
victory displays 118–119
vigilance 420–425
visual cues in movements and gestures 423
humpback whale (Megaptera novaeangliae)
call response distances 392–395
distinctive group calls 399–400
loud sexual songs 380
possible social eavesdropping 404–406
hyaena (Crocuta crocuta)
ability to infer rank among other group
members 592
intelligible distance of loud calls 372
kin-oriented redirected aggression 204
persistence of scent marks 344
redirected aggression 191
redirected aggression and target diversion
201
Hydrurga leptonyx see leopard seal
Hyla arborea see European treefrog
Hyla cinerea see green treefrog
Hyla gratiosa see barking treefrog
Hyla microcephala see neotropical treefrog
Hyla versicolor see grey treefrog
Hyperolius marmoratus broadleyi see Broadley’s
painted reed frog
Hyperolius marmoratus marmoratus see South
African painted reed frog
Hyperoodon ampullatus see northern bottlenose
whale
image scoring
and tactical deception 531–532, 533–534
and evolution of altruistic behaviour
533–534
benefits for client reef fish 526–527, 528
cheats exploit eavesdroppers 532–533
evidence in client reef fish 527–528, 529
indirect reciprocity
and cheating behaviour 532–534
cognitive abilities involved 535
occurrence in social networks 534–535
information cascades 270, 607–608
information gathering see bystanders;
eavesdroppers
information networks
and sex pheromones 549–558
fish semiochemicals 540–541

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 643
information transfer
hierarchies 18–19
within fish shoals 84
infrasound communication, in African
elephants 457–459
insect choruses 2, 263–264
female preference for leading signals 264
models of signalling dynamics 609, 610–611
insects
foraging strategies 611–612
self-organization among social insects
611–612
interceptive eavesdropping
among invertebrates 346
among marine mammals 403
compared with social eavesdropping 14–15
interspecific 16
intraspecific 17–18
ostariophysan alarm system 547–548
signaller payoff 15–16, 18
see also eavesdropping
invertebrates, interceptive eavesdropping
346
Japanese macaque see macaques
Japanese medaka (Oryzias laticeps), mate
copying 103
Japanese red-bellied newt (Cynops pyrrhogaster),
androgens and pheromone production
485–486
Junco hyemalis see dark-eyed junco
kangaroo rats (Dipodomys spp.), endocrine
control of sandbathing in males 485–486
Kassina fusca see savannah running frog
Kassina kuvangensis see Kuvangu running frog
Kassina senegalensis see Senegal running frog
katydids (various species)
acoustic communication 152–153
activity patterns and predator avoidance
153–154
chorus signalling dynamics 610–611
cryptic signalling mode 164–165
lunar phase effects 158, 162–164
masking interference in acoustic
communication 157–162
predation of 154, 156–157
predator avoidance 154–157, 164–165
Kayapo people physical adornments 423
killer whale (Orcinus orca)
active space of signals 392–395
avoidance of human noise 406–407
call suppression near prey 376–377
distinctive group calls 400
filtering of high-frequency signal
components 403
importance of oldest female 384
mammal- and fish-eating groups 403
resident and transient groups 403
sound avoidance by potential prey 403
vocal matching to signal a specific
individual 402
king penguin (Aptenodytes patagonicus)
amplitude modifications in calls 462
signal-to-noise ratio for recognition
463–464
kingfishers (Coraciiformes), interspecific
aggression 191
kin-oriented redirected aggression 203–204
klipspringer (Oreotragus oreotragus), persistence
of scent marks 344
!Kung people, openly living groups 420–421,
427
Kuvanga running frog (Kassina kuvangensis)
adjustment of male call 282
call response types 282–283
Labroides dimidiatus see cleaner wrasse
Lagenorhynchus australis see Peale’s dolphin
Lagenorhynchus obscurus see Pacific white-sided
dolphin
Lampornis clemenciae see blue-throated
humming bird
Laniarius aethiopicus see boubou
Lemur catta see ring-tailed lemur
Leptonychotes weddelli see Weddell seal
leopard seal (Hydrurga leptonyx), distinctive
group calls 399–400

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
644 Index
Ligurotettix coquilletti see grasshopper
Linepithema humile see Argentine ant
lion (Panthera leo)
ability to distinguish unfamiliar callers
374
benefits and costs of loud calling 374–376
intelligible distance of loud calls 372
male suppression of loud calling 374–376,
376–377
little blue penguin (Eudyptula minor), victory
display 116, 121–122
lizards (various species)
male attraction to rivals’ signals 269
metabolic effects of high androgen levels
504
Lombard effect 464–465
longtailed macaque (Macaca fascicularis)
ability to distinguish social relationships
586–588
kin-oriented redirected aggression 203–204
Lombard effect 464–465
postconflict behaviour 194–204
time spent looking by subordinates
417–418
losers see also winner–loser effects
increased receipt of aggression 196–197
loser effects in victims 197–198
physiological changes from conflict
197–198
postconflict changes 195–196, 201–202
role of androgens 496–498, 499
serotonin-related behavioural inhibition
498–499
Luscinia megarhynchos see nightingale
Loxodonta africana see African elephant
Macaca fascicularis see longtailed macaque
Macaca fuscata see macaques
Macaca mulatta see rhesus macaque
Macaca nemestrina see macaques
Macaca radiata see macaques
Macaca thibetana see macaques
macaques (various species)
dominance relationships 194–195
kin-orientated redirected aggression
203–204, 588
knowledge about transient social
relationships 586–588, 590–591
Lombard effect 464–465
postconflict behaviour 194–204
see also longtailed and rhesus macaques
Madagascar treefrog (Boophis madagascariensis),
acoustic signal repertoire 279
male parental behaviour, audience effects
72–74
male signal traits
and exposure to predators 152–153
and female preferences 152–153
male traits
distribution effects of mate copying
105–106
effects of female mate choice 99–100,
101
preference versus mate copying 103–105
male-male aggressive signalling, audience
effects 68–72
Mallee dragon lizard (Ctenophorous fordii),
social observation affects sexual
behaviour 425
mammals
acquiring and storing social knowledge
383–384
animal versus human networks 384–385
anti-predator calls 380–381
benefits of attending to others’ social calls
384
caller identity and status information in
acoustic signals 381
communication network view of loud
calling 373–374
contact call discrimination and memory
383–384
distinguishing unfamiliar callers 374
effects of androgens on scent marking
behaviour 485
filter characteristics of vocalizations 382
fluid social systems increase receivers of
loud calls 373

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 645
high densities increase receivers of loud
calls 373
high encounter rates and long-distance
signalling 377–378
information availability in fluid social
systems 377–378
information availability in territorial
systems 374–377
intelligible distance of calls 372, 382–383
loud calls 249, 372, 374–376, 378–380
recognition of vocalizations from
conspecifics 377–378
situations where calling is suppressed
374–377
scent marking in small terrestrial
mammals 249
source characteristics of vocalizations 382
source-filter theory of voice production
381–382
victory displays 118–119
see also marine mammals
Manacus vitellinus see golden-collared manakins
mangabey (Cercocebus albigena), ability to
distinguish unfamiliar callers 374
marine environment and acoustic
communications 390–391
marine mammals
acoustic communication networks 249
active space of signals 391–395
caller identity information in signals
399–400
determining the distance of a caller
395–396, 397
disruption by human noise 406–407
directional high-frequency signal
components 402–403
eavesdropping 403–406, 408
features of communications 391
fission–fusion societies 397–399
group identity information in signals
399–400
maximum call detection distances 393
methods of restricting and directing signals
402–403
numbers of animals in networks 397–399
population density and network size
397–399
predator–prey interceptive eavesdropping
403
restricting range by selecting
high-frequency signals 403
restricting signal by decreasing source level
403
size of communication networks 391–394,
399, 408–409
sound propagation in the sea 392–394, 396,
397, 408
value of distant signals 407–408
vocal learning in cetaceans 400–402
vocal matching to signal a specific
individual 402
see also mammals
marsh wren (Cistothorus palustris), cognitive
abilities 569–570
masking interference 157–162
masking release
and binaural processing 462–463
and spatial separation of sound sources
462–463
in amplitude-modulated background noise
459–461, 462
mate choice see also female mate choice
and eavesdropping 141–143
and scent over-marking 359, 360–361
in communication networks 129
mate copying 100–103
costs and benefits for different females
106–107
effects on male trait distribution 105–106
emergence of strategies 106
environmental influences 107
evolutionary consequences 105–106
influence of predation risk 107
state-dependent influences 106–107
versus male trait preference 103–105
mathematical modelling, interface with
communication networks 447–448 see also
modelling communication

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
646 Index
meadow vole (Microtus pennsylvanicus)
effects of androgens on scent marking
behaviour 485
female preference for top-scent males 354,
355, 359
mechanism to distinguish top and bottom
scents 351–356
over-marking and territory defence
358–359
overlap of scent marks to determine top
scent 349, 352, 354–356
preferential memory for top scents 350
process of counter-marking 344–345
Megaptera novaeangliae see humpback whale
Mehicacu people, openly living groups
421
Melopsittacus undulatus see budgerigar
Melospiza melodia see song sparrow
Meriones unguiculatus see gerbil
Mesocricetus auratus see golden hamster
mice (Peromyscus spp.), studies of redirected
aggression 208–209
Micronycteris spp. see bats
Microtus montanus see montane vole
Microtus ochrogaster see prairie vole
Microtus pennsylvanicus see meadow vole
midwife toads (Alytes spp.)
evidence for eavesdropping 291
female acoustic responses 283
Miopithecus talapoin see talapoin 417–418
Mirounga spp. see elephant seals
modelling communication
comparing eavesdropping strategies
614–618, 619, 620
complex behaviour from simple rules
606–607
conceptualizing networks 605–608
cooperation strategies 614–615
development and assumptions 604–605
eavesdropping by receivers in networks
613–622
effects of female preferences on signalling
in choruses 611
effects of hubs in a network 607–608
effects of networks on signal and signalling
dynamics 608–613
emergence of ‘selfish herd’ organization
606–607
game theory and eavesdropping 614–618,
619, 620
game theory compared with simulations
612–613
game theory models of dyadic encounters
606
hawk–dove game 615, 618, 619
implications of communication networks
604
individually based spatially explicit
simulations 606–607, 612–613
information cascades in networks
607–608
mechanisms to control call timing in
choruses 611
network effects on linear dominance
hierarchy formation 620–621
network structure in existing models
605–607
new possibilities with network approach
622–623
signal dynamics in acoustic choruses
609–611
simulation modelling 605
small-world (scale-free) network analysis
607–608
swarm intelligence and self-organization in
social insects 611–612
territory establishment 612–613
Molothrus ater see brown-headed cowbird
montane vole (Microtus montanus), studies of
redirected aggression 208–209
Morymyriformes, androgen effects on weak
electric signals 487–488
mountain gorilla (Gorilla gorilla beringei)
absence of redirected aggression in females
208
postconflict attacks on losers
196–197
mouse see house mouse

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 647
Mozambique tilapia (Oreochromis mossambicus)
androgen effects 487, 491–492, 499–500,
501, 502–505
effects of eavesdropping 27–28
endocrine response to social interaction
493–494
hormonal effects of aggressive priming
78
metabolic effects of high androgen levels
504
winner–loser effects 496–498, 499
Mus musculus see house mouse
Mustela putorius see ferret
Myotis lucifugus see brown bat
Naiken people, achieving privacy by
behavioural means 427–428
Nayaka people, openly living groups
421
Neoconocephalus spiza see katydids
Neogobius melanostomus see round goby
neotropical tree frog (Hyla microcephala)
female attraction to larger choruses 283
fine adjustment of male calls 282
male selective attention to neighbours
284
nestling begging
and genetic relatedness 177–178
as communication network 179–180,
185–186
costs and benefits to the brood 178
distinguishing signalling from physical
competition 174–175
dyadic communication approach 171–173
evolution of 171–172, 184
future work 185–186
heat loss and signalling behaviour 178–179
importance of signalling first 183
in interspecific brood parasites 177
influence of nestmates 173, 175–176, 178
locatability of calls 181–182
nestling signalling interactions 175–176
nestlings as a communication network 130,
170–171, 174–179, 180
parents’ responses to 172–173, 188
physical competition and dominance
hierarchies 179
reliability as a signal of need 172–173
signal costs and reliability 171–172
signalling errors 184–185
signalling to catch receiver attention
180–183
suppressing competitors’ signals 183
use of locatable signals 181–182
nestling gape 181
network view of communication 2
nightingale (Luscinia megarhynchos)
background noise and song output level
51
cognitive abilities 569–570
eavesdropping and transitive inference (TI)
574–576
Lombard effect 464–465
male vocal interactions 302
solo and interactive singing 51
song matching 303–304, 307
use of social eavesdropping 20–21, 22
non-human primates
common behaviour patterns with humans
433–435
consortship behaviour 432–433
defensive and social functions of vigilance
417, 419–420, 425–426
factors affect levels of vigilance 419
male bias for vigilance outside the group
417–418
male vigilance and social control 419
proportion of time spent in vigilance
417–418
securing perceptual privacy for some
interactions 432–433
sentinel behaviour among high-ranking
males 418–419
time spent looking 417–418, 433
unifying model for vigilance, social
observation and eavesdropping 433–435
vigilance 417
see also primates

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
648 Index
northern bottlenose whale (Hyperoodon
ampullatus), use of deep sound channel
397
Nycticebus pygmaeus see pygmy loris
Ochotona princeps see pika
odontocetes, restricting range by selecting
high-frequency signals 403
oestrogens, receptors in the brain 490–491
olfactory system
hormonal modulation of sensitivity
488–489
signal detection and encoding 543–544
olive baboon (Papio anubis)
absence of redirected aggression in females
208
effects of redirected aggression 191–192
postconflict attacks on losers 196–197
Onchorhynchus mykiss see rainbow trout
Opsanus beta see gulf toadfish
Opsanus tau see oyster toadfish
Orcinus orca see killer whale
Oreochromis mossambicus see Mozambique
tilapia
Oreotragus oreotragus see klipspringer
Oryctolagus cuniculus see rabbit
Oryzias laticeps see Japanese medaka
oyster toadfish (Opsanus tau), effects of
androgens on vocal structures 484
Pacific humpback dolphin (Sousa chinensis),
individual signature signals 400–402
Pacific white-sided dolphin (Lagenorhynchus
obscurus), individual signature signals
400–402
Pan troglodytes see chimpanzee
Panthera leo see lion
paper wasp (Polistes dominulus), neural
development in colonial females
595–596
Papio anubis see olive baboon
Papio cynocephalus see baboon
Papio cynocephalus ursinus see chacma
baboon
Papio hamadryas see hamadryas baboon
parent birds
comparison of nestlings’ signals 183
information from behaviour of nestlings
174–175
parental care, effects of high androgen levels
in males 497, 504–505
Paroaria gularis see red-capped cardinal
parrots (Trichoglossus spp.), victory displays
116
Parus atricapillus see black-capped chickadee
Parus caeruleus see blue tit
Parus major see great tit
Passer domesticus see house sparrow
peacock blenny (Salaria pavo), androgen effects
on somatic releasers 491–492
Peale’s dolphin (Lagenorhynchus australis),
silence when captured or near a boat
403
Perdix perdix (grey partridge), effects of
androgens on vocal structures
483–484
Petromyzon marinus see sea lamprey
pheromones
fish semiochemicals 541–543
hormonal pheromones 549–558
releaser and primer effects 541
reproductive priming effects of mouse
urine 231–232
Phoca groenlandica see harp seal
Phoca vitulina see common seal
Phocoena phocoena see harbour porpoise
Phoxinus phoxinus see European minnow
Physalaemus pustulosus see tungara frog
Physeter macrocephalus see sperm whale
physics, interface with communication
networks 445–446
pied flycatcher (Ficedula hypoleuca)
female assessment of males 135–136,
137
male attraction to rivals’ signals 269
pigtail macaque see macaques
pika (Ochotona princeps), ability to distinguish
unfamiliar callers 374

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 649
pilot whale (Globicephala melaena), falling
silent when hunted 403
Pimephales promelas see fathead minnow
pinyon jay (Gymnorhinus cyanocephalus), ability
to rank multiple stimuli 596–598
Pipilo erythrophthalmus see Eastern towhee
plainfin midshipman (Porichthys notatus)
auditory sensitivity modulation by sex
steroids 489
effects of androgens on vocal structures
484
Poecile atricapillus see black-capped chickadee
Poecilia latipinna see sailfin molly
Poecilia reticulata see guppy
Polistes dominulus see paper wasp
Porichthys notatus see plainfin midshipman
prairie vole (Microtus ochrogaster)
androgen effects on central motivational
mechanisms 490
studies of redirected aggression 208–209
precedence effect, female preference for
leading signals 285–286
predation risk
and eavesdropping 14, 16, 17, 45, 95–96
and katydid activity patterns 154–157
and katydid roost site selection
154–157
and male signal traits 152–153
and signal detection 159, 160
avoidance in communication networks
130
chemical assessment of 544–549
influence on female mate choice 107
influence on mate copying 107
interceptive eavesdropping among marine
mammals 403
mammal anti-predator calls 380–381
quiet song as response to 55
predators
information from prey alarm calls 380–381
use of locatable calls to find prey 181–182
Procolobus badius tephrosceles see red colobus
monkey
Propithecus diadema see diademed sifaka
primates
ability to classify objects based on abstract
concepts 594–595
ability to rank objects 590
awareness of social rank relationships 28,
588–591
benefits of associations with high-ranking
animals 426
complexities of recognizing affiliative
relationships 593–594
eavesdropping abilities versus that in other
animals 592–598
effects of social organization on vigilance
426
‘greater intelligence’ hypothesis for
eavesdropping abilities 592, 593–595, 598
interconnected groups with males as ‘hubs’
608
knowledge of other animals’ kin 585–587,
588
‘large social groups’ hypothesis for
eavesdropping abilities 592–593,
595–596, 598
monitoring social relationships as group
size increases 594
‘no species difference’ hypothesis about
eavesdropping abilities 593, 596–598
social resources 426
types of information acquired from
eavesdropping 584
see also non-human primates
priming response
and male aggression 75–78
androgen effects on bystanders 499–500,
501
private signalling, in male anurans 291
see also quiet song
pronghorn antelope (Antilocapra americana),
scent over-marking 361
Prunella collaris see alpine accentor
Prunella modularis see dunnock
Psittacus erithacus see African grey parrot
psychophysics, interface with communication
networks 445–446

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
650 Index
Puerto Rican treefrog (Eleutherodactylus coqui)
diphasic advertisement call 280
male selective attention to neighbours
284
models of chorus signalling dynamics
609–610
‘off response’ call initiation 280–282
Puntius schwanenfeldi see tinfoil barb
pygmy loris (Nycticebus pygmaeus), scent
over-marking and mate choice 360–361
pygmy Mulga monitor lizard (Varanus gilleni),
victory display 118
quiet song
as aid to anonymity 56–57
as response to predation risks 55
reasons for variability 56–57
used for private signalling 53–55
rabbit (Oryctolagus cuniculus), effects of
androgens on scent marking behaviour
485
rainbow trout (Onchorhynchus mykiss)
eavesdropping and dominance 28
effects of redirected aggression 191–192
Rana clamitans see green frog
Rangifer tarandus see reindeer
rank-order fights see contest behaviour
rat (Rattus norvegicus), effects of redirected
aggression 191–192
receivers
auditory scene analysis 471–474
comparison of signals in a network 183–184
precedence effects 183
signals designed to catch attention 180–183
see also signal detection
reciprocal altruism, and behavioural
strategies 521–522
red-capped cardinal (Paroaria gularis),
territorial behaviour 17–18
red colobus monkey (Procolobus badius
tephrosceles)
time spent looking and group size 433
time spent looking 417–418
red deer (Cervus elaphus)
discrimination of individual male callers
379–380
individuality in formant frequencies in
vocalizations 382
loud mating calls 378–379
red-winged blackbird (Agelaius phoeniceus),
signal-to-noise ratios for recognition and
for detection 463–464
redirected aggression
and postconflict attacks on losers 196–197,
200
and reconciliation with aggressor 192–194,
200, 202–203
and the ‘fight or flight’ response 191–192
as an outlet for ‘frustration’ 191–192
as possible target diversion 201
as audience effect 202–203, 209–211
benefits for losers 200, 201–202
in communication networks 130–131
in non-human primates 192–194, 195
in species other than primates 204–205, 206
in winners 196
influence on bystanders’ behaviour
192–194, 200
intraspecific aggression 206, 208–209
kin-oriented 203–204, 588
‘loser effects’ 195–196, 198–200
possible use a signal 201–202, 204–205, 208
summary of functions 210–211
testing occurrence and function 193,
209–210
to attenuate endocrine stress response
191–192
to signal postconflict condition to
bystanders 201–202
redtail monkey (Cercopithecus ascanius schmidtii)
time spent looking 417–418, 433
redwing (Turdus iliacus), quiet singing 53
reindeer (Rangifer tarandus), androgen effects
on scent marking behaviour 485
relational distinction
abilities of non-primates 596–598
abilities of primates 594–595

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 651
reptiles, victory displays 118
rhesus macaque (Macaca mulatta)
ability to distinguish unfamiliar callers
374
kin-biased redirected aggression 588
individuality of frequencies in vocalizations
382
postconflict behaviour 194–204
see also macaques
Rhinolophus ferrumequinum see greater
horseshoe bat
ring-tailed lemur (Lemur catta), scent
over-marking 362–363
robin (Erithacus rubecula)
courtship interruption by neighbours 41
eavesdropping 17
female assessment of males 141
quiet singing 53
rodents, redirected aggression 208–209
roe deer (Capreolus capreolus)
barking call 380–381
individuality of vocal frequencies 382
round goby (Neogobius melanostomus),
semiochemical communication 558–559
rufous-collared sparrow (Zonotrichia capensis),
song changes with habitat 143–144
Saguinus oedipus see cotton-top tamarin
sailfin molly (Poecilia latipinna), mate copying
103, 104–105
Saimiri boliviensis see squirrel monkey
Sakalava people, openly living groups 421
Salaria pavo see peacock blenny
Salmo salar see Atlantic salmon
Samoan people, openly living groups 421
Sarakatsani people, privacy of the hut 428
savannah baboon, knowledge about transient
social relationships 590
savannah running frog (Kassina fusca)
female call preference 286
variation in call response types 282–283
Sceloporus jarrovi see lizards
scent marking
adjacent marking 344–345
amount of scent and top scent
discrimination 349, 353–354
and communication networks 131
and mate choice 359, 360–361
androgen effects 485
as a visual signal 362–363
as broadcast signals 235–237, 344
discriminating individual odours
350–351
functions of 345, 354, 355, 358–361
for network communication 219, 362–363
in communication networks 366–367
information available to eavesdroppers 345,
346
information in spatial and temporal
distributions 221, 223–225
in social contexts 220
mechanisms for targeted over-marking
357–358
mechanisms to distinguish top and bottom
scents 347–357
olfactory detection 219–220, 223
persistence of signals 219, 344
possible olfactory consequences 345
relative freshness and top scent
discrimination 351–353
reliability of signals 221, 223–225
to advertise competitive ability 221,
223–225, 227–228
to advertise territory ownership 221,
223–225, 227–228
use of geometric relationships to determine
top scent 352, 356
volatile and non-volatile components
219–220
sea lamprey (Petromyzon marinus)
larval pheromone attracts migrating adults
555–557
life cycle 555
male sex pheromone in communication
networks 557–558
possible specialization in signal production
and release 557–559
search for biological control 555

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
652 Index
selection pressures
female song preferences 152–153
from eavesdroppers 14, 16, 30–31
imposed by audiences 10, 64–65, 79
on interactive singing strategies 309
on primate brain size 28
on signalling interactions 18–19, 79
on victory displays 122–123
self-awareness in animals 577–578
semiochemicals
information from 446–447
see also fish semiochemicals
Senegal running frog (Kassina senegalensis),
synchronous calling 282
sergeant major fish (Abudefduf vaigiensis),
bystander effects on cleaner fish
behaviour 530–531
Serinus canarius see canary
serotonin
and aggressive behaviour 197–198
and behavioural inhibition in losers
498–499
sex steroids, modulation of sensory
perception 488–490
Siamese fighting fish (Betta splendens)
androgen effects on eavesdropping 503–504
audience effects 65, 68–72
eavesdropping on visual displays 24–27
female mate choice and eavesdropping 142
priming and male aggression 75–78
signal detection
amplitude as a distance cue 468–469
auditory scene analysis by receivers 471–474
biological background noise 152
calculation of maximum detection distance
454–456, 457, 458, 459
compared with signal recognition 463–464
critical (masking) ratio for various animals
453–457, 459
decision tree learning to discriminate
signals 158, 160–162
distance assessment 467–468, 470, 471,
472–473
female song preferences 152–153
high background noise 157–160
hormonal effects on 488–490
in a complex environment 152
perceptual mechanisms of receivers
451–452
precedence effect 183, 285–286
reverberation as a distance cue 469–471,
472–473
signal ranging 467–471, 472–473
signal-to-noise ratio 453–456, 457
simultaneous comparison by receiver 183
spatial separation of sound sources 462–463
use of ‘biological microphone’ 157–160
see also receivers
signal production
amplitude modification 462
anonymous signalling 56–57
audience effects 64, 65–66
broadcasting from high perches 464–465
catching receiver attention 180–183
competition and cooperation with other
signallers 63
competition in chorusing interactions
263–264
cryptic signalling 164–165
environmental temperature effects on
signallers 153
high output to overcome interference 181
impact of signal errors in communication
networks 184–185
Lombard effect 464–465
predation risk 159, 160
sender adaptations to maximize
transmission 464–465
signal evolution and selection pressures
79
suppressing competitors’ signals 183
targeting a specific receiver 63–64
use of multimodal components 182
using locatable signals 181–182
signal transmission
background noise sources and levels 52–53,
452–453
environmental influences on 286–287

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 653
frequency and amplitude masking effects
49–50, 453
influence of communication network
structure 286–287
information transfer hierarchies 29
in natural habitats 42–43, 44
masked auditory thresholds 457
masking release in amplitude-modulated
background noise 459–460, 461, 462
private signalling 32–53
signaller proximity and masking
interference 152
sound segregation unmasking effect
460–461
sound types 49–51
use of high perches 44, 50
using background noise fluctuations to
reduce masking 459–460, 461, 462
signal recognition, compared with signal
detection 463–464
signal-to-noise ratio
and signal detection 453–456, 457
determination 454–456
for detection and for recognition 463–464
signalling interactions
between nestlings 175–176
influence of shared fate of nestlings
178
information gathered from 29–30, 38–39
models of effects of networks on signalling
dynamics 608–613
selection pressures on 18–19
see also eavesdropping
signalling modalities
and audience effects 10
acoustic signalling in 99
multimodal signalling 99, 182
use of different modalities 28–29
signals, distinguishing from physical
competition in nestlings 174–175
simulation models see modelling
communication
SINDSCAL multidimensional scaling analysis
469–471, 472–473
small-world (scale-free) network analysis
607–608
Smilisca sila see treefrog
social complexity, possible selection pressure
on brain development 595–596
social context
influence on eavesdropping effects 97
influence on hormone levels 481–482,
492–494
of communication 1
of communication networks 78–79
of dyadic interactions 78–79
social eavesdropping 10, 18–32
and dominance hierarchies 28
and victory displays 122–123
cognitive requirements in female anurans
289
compared with interceptive eavesdropping
14–15
form of bystander behaviour 10–11
in communication networks 2–3
in territorial songbirds 19–21, 24
in territorial systems 18–19
information gathered from 38–39
visual interactions in fish 24–27, 28
see also bystanders; eavesdropping
social instability, and bystander decisions 96
social intelligence, assessment of 447
social modulation of androgen levels 492–494
song matching
during and after dawn chorus 330,
332–335
value as directional signal at dawn chorus
338–339
song output, and habitat quality 144
song overlapping, as an aggressive signal 331,
334, 335–336
song repertoire size, and individual
identification 56–57
song sparrow (Melospiza melodia)
cognitive abilities 569–570
individual recognition of neighbours 364
song repertoire matching 303
victory display 116, 117, 121–122

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
654 Index
song transmission, effects of habitat on
143–144 see also sound transmission
songbirds eavesdropping
and female mate assessment 19–21, 22,
47–48, 314, 577
attention to interaction asymmetries
313–314
cognitive complexity of male ranking
judgements 576–577
courtship interruptions by eavesdroppers
40–41
for repertoire size information 45–47
male eavesdropping 313–314
use of quiet singing 53–55
songbirds vocal interactions
among neighbours 309–310, 311–312
among territorial males 301–302
and territorial behaviour 301–302, 312–313
and transitive inference 574–576
androgen effects 483–485
asymmetries in vocal interactions 301, 309,
310–311, 313–314
cognitive processes in a communication
network 569–570, 574–576
evolution of vocal interaction strategies
314–315
function of the dawn chorus 248–249
functions of specific singing strategies 308,
309–310
in dialogues 302–303
in various contexts 301
interactive dimension of vocal interactions
301, 302–303
memory capacity 569–570
maintenance of territorial spacing pattern
311–312
precise timing and interpretation of
interactions 306
resident–intruder vocal interactions
309–311
selection pressures on singing strategies
309
self-awareness 577–578
site-specific dominance in neighbours
311–312
social contexts of vocal interactions
307–308, 310
solo versus interactive singing 51–52
song alternating 305, 306
song matching 303–305, 307
song overlapping 304, 305–306
song production relative timing 304,
305–306
territorial settlement by multiple
interactions 312–313
territories as communication networks
248–249
sound localization
cues used for 465
variation in ability between species
465–467
sound transmission see signal transmission
source-filter theory of voice production
381–382
Sousa chinensis see Pacific humpback dolphin
South African painted reed frog (Hyperolius
marmoratus marmoratus), female call
preferences 287
southern right whale (Eubalaena glacialis),
possible social eavesdropping
404–406
spadefoot toad (Spea multiplicata), male
attraction to rivals’ signals 269
sparrowhawk (Accipiter nisus), sound
localization ability 465–467
spatial memory, sex differences and effects of
androgens 490–491
Spea multiplicata see spadefoot toad
sperm whale (Physeter macrocephalus)
active space of signals 392–395
distinctive group calls 400
fluid social systems and long-distance
signalling 377
individual signature signals 400–402
spiteful behaviour, evolution and
maintenance 534–535, 536
spotted dolphin (Stenella plagiodon), individual
signature signals 400–402
spotted hyaena see hyaena
spying see fish semiochemicals

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 655
squirrel monkey (Saimiri boliviensis)
factors affect levels of vigilance 419
social organization and vigilance 426
Stenella plagiodon see spotted dolphin
Sternopygus macrurus see weakly electric fish
stress hormones, effects of social environment
482
Sturnus vulgaris see European starling
swordtail (Xiphophorus helleri)
androgen effects on sword development
491–492
audience effects on males 70
communication via private channels 79
Sylvia atricapilla see blackcap
Sylvia communis see whitethroat
Tachycineta bicolor see tree swallow
tactical deception
cognitive abilities required 524–525, 535
concept of theory of mind 524–525
evolution and maintenance 536
exploitation of eavesdroppers 524–525
image scoring and evolution of 533–534
occurrence in social networks 534–535
Taeniopygia guttata see zebra finch
talapoin (Miopithecus talapoin), time spent
looking by subordinates 417–418
terrestrial mammals see mammals
territorial behaviour
and eavesdropping 17–18
and transitive inference 574–576
relative timing of song production 305–306
resident–intruder vocal interactions
309–311
scent marking in house mice 220–222,
223–225, 227–228
song matching 303–305, 307
vocal interactions among neighbours
309–310, 311–312
victory displays 122–123
territorial systems
and communication networks 143–144
availability of information from loud calls
374–377
dear enemy effects 501–502
modelling of territory establishment
612–613
social eavesdropping in 18–19
testosterone levels, increase in winners 198
three-spined stickleback (Gasterosteus aculeatus)
androgen effects on somatic releasers
491–492
visual perception modulation by sex
steroids 489–490
Thryothorus pleurostictus see banded wren
Tibetan macaque see macaques
tilapia see Mozambique tilapia
Tinbergen, Niko 1
tinfoil barb (Puntius schwanenfeldi), androgen
effects on olfactory sensitivity 488–489
toadfish, effects of androgens on vocal
structures 484
Tonatia silvicola see bats
transitive inference
cognitive abilities of birds 578–579
equating human and avian studies 573–574,
578–579
mechanisms used by animals 572
presentation to humans 570–571
presentation to non-humans 571–572
songbirds in communication networks
574–576
studies with parrots 572–573
tree shrew (Tupaia belangeri), effects of
androgens on scent marking behaviour
485
tree swallow (Tachycineta bicolor), nestling
signalling errors 184–185
treefrog (Smilisca sila), synchronous calling
282, 283–284
tremulation, as cryptic signalling mode
164–165
Trichoglossus spp. see parrots
Triturus spp. see European newts
triumph ceremonies see victory displays
Troglodytes troglodytes see wren
tungara frog (Physalaemus pustulosus)
adaptations for acoustic competition 278
male selective attention to neighbours 284
Tupaia belangeri see tree shrew

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
656 Index
Turdus iliacus see redwing
Turdus merula see blackbird
Tursiops truncatus see bottlenose dolphin
Tyto alba see barn owl
urodeles, androgen effects on pheromone
production 485–486
Uca spp. see fiddler crabs
Varanus gilleni see pygmy Mulga monitor
lizard
vertebrate endocrine systems 482
vervet monkey (Cercopithecus aethiops)
bystander effects on aggression 202–203
factors affecting levels of vigilance
419
kin-oriented redirected aggression 203–204
knowledge of other animals 585, 589–590
male parental behaviour 72–73
victory displays 11, 114
amphibians 118
androgen effects on vertebrates 486–487
arthropods 117–118, 122
birds 115–117
categories of 121–122
distinctive features of 120
distinguished by context 119–120
effects within the communication network
120–121, 122–123
functions within the winner–loser dyad
120–122
humans 118–119
mammals 118–119
occurrences of 119
reptiles 118
Vidua spp. see whydahs
vigilance
in animals 417–420
in humans 420–425
visual interactions in fish, eavesdropping on
24–27, 28
visual system, hormonal modulation
489–490
vocal learning in cetaceans 400–402
vocal matching, by cetaceans to signal a
specific individual 402
vomeronasal organ 219–220, 223, 229,
232
weakly electric fish (Sternopygus macrurus),
androgen effects on electroreception 489
Weddell seal (Leptonychotes weddelli)
colony-specific call types 399–400
ear damage from underwater noise 406–407
territorial behaviour 404–406
western scrub jay (Aphelocoma californica),
ability to rank multiple stimuli 596–598
wetas (Hemideina spp.), victory displays 118,
122
white-crowned sparrow (Zonotrichia leucophrys)
cognitive abilities 569–570
metabolic effects of high androgen levels
504
white-faced capuchin monkey
(Cebus capucinus)
male vigilance and social control 419
time spent looking as group size increases
433
white whale see beluga whale
whitethroat (Sylvia communis)
courtship interruption by neighbours 41
private signalling 55
signalling in different modalities 29
use of high perches to improve reception
42–43
whydahs (Vidua spp.), nestmate signal
suppression 183
Wilsonia citrina see hooded warbler
winner–loser effects
role of androgens 496–498, 499
serotonin-related behavioural inhibition in
losers 498–499
see also losers; winners
winners
increase in androgen levels 198
physiological changes following victory 198
postconflict aggression 196
‘winner effects’ following victory 198

P1: IRK/KFO P2: KOD0521823617ind.xml CU1917B/McGregor 0 521 582361 7 April 12, 2005 11:14
Index 657
Wistar rat, effects of androgens on scent
marking behaviour 485
wolf (Canis lupus)
ability to distinguish unfamiliar callers
374
individuality in vocal frequency 382
scent marking 363
situations where calling is suppressed
376–377
victory display 118–119
wren (Troglodytes troglodytes)
signal echo tail as distance cue 469–471,
472–473
use of high perches 43, 44, 464–465
use of low-frequency sounds 49
Xenopus laevis see African frog
Xiphophorus helleri see green swordtail;
swordtail
Yagua people, achieving privacy by
behavioural means 427–428
zebra finch (Taeniopygia guttata)
effects of androgens on vocal structures
483–484
Lombard effect 464–465
signal-to-noise ratios 463–464
signal amplitude as a distance cue 468–469
sound localization ability 465–467
zebrafish (Danio rerio), active space of alarm
substance 546–548
Zinacantan people, suspicion of private
behaviour 421–422
Ziphius cavirostris see Cuvier’s beaked whale
Zonotrichia capensis see rufous-collared
sparrow
Zonotrichia leucophrys see white-crowned
sparrow
Copyright © 2022 FDOKUMEN




















