ANHUAL - CORE
350
ANHUAL ND .P)CDENT rig94 COLD SPRING HARBOR LABORATORY
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of ANHUAL - CORE

ANHUALND .P)CDENT
rig94
COLD SPRING HARBOR LABORATORY

ANNUALREPORT
1984
COLD SPRING HARBOR LABORATORY
r--
. _
-7-- -/
- !--,
_-
t
*-V-r-J"441,727t110

ANNUAL REPORT 1984
Cold Spring Harbor LaboratoryBox 100Cold Spring Harbor, New York 11724
Editors Elizabeth Ritcey, Dorothy Brown, Annette KirkPhotography Susan Zehl, Herb Parsons, Rosie ChanCover photo by David Mick los
Recto: Weathervane atop Jones laboratory, which was erected in 1893 by the Laboratory'sfounder, John D. Jones.

ContentsOfficers of the Corporation/Board of Trustees v
Governance and Major Affiliations vi
Committees vii
DIRECTOR'S REPORT
DEPARTMENTAL REPORTS 15
Administration 17Buildings and Grounds 18Information Services 21
Library 22Publications 23
RESEARCH 27
Tumor Viruses 29Molecular Genetics of Eukaryotic Cells 65Molecular Genetics 101Cell Biology 129Neurobiology 159
COLD SPRING HARBOR MEETINGS
Symposium on Quantitative Biology 171Meetings 179
169
BANBURY CENTER 253
Director's Report 255Meetings 259
EDUCATIONAL ACTIVITIES 277
Postgraduate Courses 279Seminars 297Undergraduate Research 299Curriculum Study 301Nature Study 305
FINANCIAL STATEMENT 307
GRANTS AND CONTRIBUTIONS 315
Grants 317Corporate Sponsors 322Long Island Business Cancer Campaign 323Contributors 324Financial Support 326
LABORATORY STAFF 327
LONG ISLAND BIOLOGICAL ASSOCIATION 333

(Front row:) E. Pulling, T.J. Knight, Mrs. C. Dolan, W.S. Page, Mrs. H. Harris, Jr.,
Mrs. S. Hatch; (back row:) B. Magasanik, R. Cummings, N.D. Zinder, M. Scharff,
B. Clarkson, J. Klingenstein, T. Whipple, J.D. Watson, R. Webster, W.S. Robertson,
S. Strickland

Officers of the CorporationWalter H. Page, ChairmanDr. Bayard Clarkson, Vice-ChairmanDr. Norton D. Zinder, SecretaryRobert L. Cummings, TreasurerDr. James D. Watson, Director
Board of TrusteesINSTITUTIONAL TRUSTEES
Albert Einstein College of MedicineDr. Matthew Scharff
Columbia UniversityDr. Eric R. Kandel
Duke UniversityDr. Robert Webster
Long Island Biological AssociationEdward Pulling
Massachusetts Institute of TechnologyDr. Boris Magasanik
Memorial Sloan-Kettering Cancer CenterDr. Bayard Clarkson
New York University Medical CenterDr. Claudio Basilico
Princeton UniversityDr. Thomas E. Shenk
The Rockefeller UniversityDr. Norton D. Zinder
State University of New York,Stony Brook
Dr. Sidney Strickland
Wawepex SocietyTownsend J. Knight
Yale UniversityDr. Charles F. Stevens
INDIVIDUAL TRUSTEES
Robert L. CummingsCold Spring Harbor, New York
Mrs. Charles DolanOyster Bay, New York
Oliver R. GraceOyster Bay, New York
Mrs. Henry Harris, Jr.Glen Head, New York
Mrs. Sinclair HatchOyster Bay, New York
Ambassador John P HumesMill Neck, New York
John KlingensteinNew York, New York
Dr. Ralph LandauNew York, New York
Walter S. PageCold Spring Harbor, New York
William S. RobertsonHuntington, New York
Harvey SampsonCold Spring Harbor, New York
Dr. James D. WatsonCold Spring Harbor, New York
Taggart WhippleOyster Bay, New York
HONORARY TRUSTEES
Dr. Harry EagleAlbert Einstein College of Medicine
Dr. H. Bentley GlassEast Setauket, New York
Dr. Alexander HollaenderAssociated University, Inc.

Governance and Major AffiliationsThe Laboratory is governed by a 25-member Board of Trustees which meets three or fourtimes a year. Authority to act for the Board of Trustees between meetings is vested in theExecutive Committee of the Board of Trustees. The Executive Committee is composed ofthe Officers of the Board plus any other members who may be elected to the ExecutiveCommittee by the Board of Trustees. Additional standing and ad hoc committees are ap-pointed by the Board of Trustees to provide guidance and advice in specific areas of theLaboratory's operations.
Representation on the Board of Trustees itself is divided between community represen-tatives and scientists from major research institutions. Ten such institutions are presentlyrepresented on the Board of Trustees: Albert Einstein College of Medicine, Columbia Uni-versity, Duke University, Princeton University, Massachusetts Institute of Technology, Mem-orial Sloan-Kettering Cancer Center, New York University, The Rockefeller University, TheState University of New York at Stony Brook, and Yale University.
Institutional Trustees were brought onto the Board during the 1960s-a difficult periodwhen the Laboratory was undergoing reorganization as an independent entity. In additionto supplying scientific leadership to the governing body, participating institutions also pro-vided emergency funds to help keep the Laboratory afloat during this crucial phase of de-velopment. Although participating institutions now give only token financial support, theirTrustees continue to help steer the course of the Laboratory's scientific and administrativepolicies.
Also represented as participating institutions are the Wawepex Society and the LongIsland Biological Association (LIBA). The Wawepex Society was formed in the mid-nine-teenth century as a philanthropic arm of the Jones family, who supported the formation ofthe Laboratory. LIBA's origins began in 1890 when members of the local community be-came involved as friends and with membership on the Board of Managers of the BiologicalStation in Cold Spring Harbor, under the aegis of the Brooklyn Academy of Arts and Sci-ences. When the Brooklyn Academy withdrew its support of the Biological Station, commu-nity leaders, in 1924, organized their own association that actually administered the Labo-ratory until the Laboratory's reorganization as an independent unit in 1962. Today, LIBAremains a nonprofit organization organized under the Department of Social Welfare of theState of New York and represents a growing constituency of "friends of the Laboratory." Its450 member families support the Laboratory through annual contributions and participa-tion in fund drives to raise money for major construction projects.
The Laboratory is chartered as an educational and research institution by the Board ofRegents of the Education Department of the State of New York. It is designated as a "publiccharity" by the Internal Revenue Service and claims special status as a school under theInternal Revenue Code.

Committees*Audit Executive (Elected) Nominating
Townsend J. Knight, ChairmanOliver R. GraceAmbassador John P Humes
Banbury Program
Dr. Norton D. Zinder, ChairmanDr. Bayard ClarksonRobert L. CummingsWendy Hatch
Building
Edward Pulling, ChairmanHelen DolanOliver R. GraceMary Jeanne HarrisMary LindsayElizabeth Watson
Commercial Relations
Townsend J. Knight, ChairmanDr. James HicksDr. Ralph LandauDr. Richard RobertsDr. Matthew ScharffDr. Robert WebsterTaggart Whipple
Development Planning
Edward Pulling, ChairmanDavid MicklosHarvey SampsonTaggart Whipple
Walter H. Page, ChairmanDr. Bayard ClarksonRobert L. CummingsOliver R. GraceHarvey SampsonDr. Charles F. StevensDr. James D. WatsonDr. Norton D. Zinder
Edward Pulling, ChairmanDr. Bayard ClarksonRobert L. CummingsHarvey SampsonDr. Matthew Scharff
Robertson House
William S. Robertson, Chairman
Finance and Investment Helen DolanMary Jeanne Harris
Robert L. Cummings, Chairman John MaroneyOliver R. Grace Anne MeierWendy Hatch Dr. Michael ShodellAmbassador John P HumesTownsend J. KnightWilliam S. Robertson Robertson Research FundHarvey Sampson TrusteesTaggart Whipple (Laboratory Representatives)
Neurobiology
Dr. Charles F. Stevens, ChairmanDr. Eric R. KandelWilliam S. RobertsonJohn Klingenstein
*Committees and membership as of November 10, 1984.
Robert L. CummingsAmbassador John P. HumesWalter H. PageDr. James D. Watson
Tenure and Appointments
Dr. Norton D. Zinder, ChairmanDr. Claudio BasilicoDr. Boris MagasanikDr. Matthew Scharff

DIRECTOR'SREPORT
Much more than it realizes, our nation now faces the choice of how todeal with the great scientific revolutions whose consequences increasingly domi-nate the quality of human life. Whether we use or misuse this new science willdetermine whether our now old and venerable Western Civilization can be main-tained or whether it will collapse into a form of modern feudalism marked by con-formity to arbitrary ideals and a reluctance to question ideas and customs al-ready in use. Past history provides no example of a civilization that did noteventually dig its own grave and great achievements like the Taj Mahal have oc-curred when the tide was already turning.
In particular, we must face the fact that the human brain is probably no moreeffective than it was at the time of the American Revolution, or for that matterduring the reign of the Pharaohs. Yet the world's affairs move incredibly faster,with the implications of single human acts having infinitely greater long-term con-sequences. So we have to develop institutions that allow us to react better to therapid changes of the moment without stifling our capacity for self-expression andinnovation. Today, our most perplexing concerns arise from the consequences ofmodern physics and the development of nuclear weapons. Their use on even amodest scale would destroy the essence of Western Civilization, leaving what-ever population remained so mesmerized by fear of a recurrence that it would belikely to trade off freedom as we know it today in the West for easy assurances,no matter how hollow, that the holocaust would never be repeated.
It is just 40 years since Hiroshima entered our vocabularies, and yet the imme-diate horror of the apocalypse that it epitomized is now, more often than not, re-placed with an uneasy satisfaction that the threat of mutual annihilation will keepthe peace. Initially I would have thought that the possession of just a few dozenatomic bombs and perhaps only one or two hydrogen bombs would scare offeven the most fearless. Today, however, we find ourselves and our putative en-emy, the Russians, with tens of thousands of nuclear weapons, all too many ofwhich are encased in delivery vehicles that have reduced the time between deci-sion to strike and indescribable destruction to mere minutes. So as each yearpasses, the chance of our respective military establishments knowing how to actand react during any nuclear exchange becomes more and more remote. Butlike all too many cancers, we know not how to remove the nuclear triggers fromour lives.
It is thus natural to want to believe that modern science, having first given usnuclear weapons, would someday be able to provide the magic bullets thatwould flick them off course to explode harmlessly in the desolation of outerspace. Today, in fact, we are told that given some 50-1000 billion dollars and theactive participation of our nation's brightest, we might some 15-20 years from

now throw up a shield of powerful laser beams that would knock down the major-ity of intruders and let us retain the capacity to get back at any power so insaneas to believe that we can be defeated. Conceivably, this modern form of MaginotLine might actually work as planned, giving us the capacity to retaliate againstany evil force that would enslave us.
But before devoting such a large fraction of our national creative talent to thispurpose, we must ask what level of contingency planning does this Star Warsresponse represent. In my opinion, we are reacting to a very, very low probabilityevent, merely because if it were to happen, it would be so catastrophic. That thecollective Russian leadership would actually ever believe that it could win a nu-clear exchange, no matter how one-sided, strikes me as a most improbableevent. Much more likely, though also a very improbable event, would be the fail-ure of our nation's soundest bank, the Morgan Bank, followed by a complete col-lapse of the world's financial institutions. Yet today I suspect that not one treas-urer of a major world corporation is devoting any of his real resources to theconsequences of such a total banking collapse. Assuming the worst possibleoutcome is not the way successful individuals, banks, or nations run their dailylives. If they have the money, they take out just enough insurance (weapons) tosurvive a finitely calculable misfortune. Most certainly I would be thought silly, ifnot insane, to spend most of my funds on life insurance, leaving virtually nomoney to feed, house, educate, or medically protect my family.
The counter arguments, of course, can be made that as biologists we are tooinexperienced to think in a military way and, even more important, that we aretoo small a lobby ever to influence major national priorities. So, no matter howwe feel, we should sit tight, not unnecessarily make enemies in the wrong places,and leave to others the task of controlling the arms race. This is a position I can-not subscribe to as long as I feel that Star Wars money could be spent muchmore wisely to prevent tragedies that we know will actually happen.
Now that we at last have the means to finally understand the molecular natureof cancer, it would be at best perverse for us not to find the added monies neces-sary to deploy this knowledge for the human good within the lifetimes of thosewe love. Today, not 20 years distant, is the time to find out, say, the moleculardefects that make a breast cell cancerous. Yet at this moment, even though can-cer research is still better funded than all other parts of biology (but not phys-ics! ), there is not enough money for even our major laboratories to push aheadfull speed. We are now half-throttled back, often spending more time worryingabout money than our science. Instead, our recent great successes in the isola-tion and molecular dissection of oncogenes should be followed up by a nationalcommitment, expressed by our President himself, to go forward with cancer re-search even faster. By so behaving as a nation, we would be living up to ourgreatest potential as humans, the capacity for accumulating the self-knowledgethat will let us live even more meaningful lives.
Until such a turnabout in national priorities occurs, I fear that an ever-increas-ing malaise will spread through our lives as biologists. Up till now, our scientificsystem has worked well because we could treat our younger scientists with therespect they deserve. It is they who do most of the experiments that our moresenior scientists talk about. So when they succeed, they should know that they,like their bosses at the moment, will soon be able to set up their own labs andtest out their own ideas. Today, however, even our most talented younger scien-tists live with the fear that they will not get research grants of a size sufficient tolet them do effective science. This situation cannot go on too much longer with-
2

out destroying the civility of the peer review system that up till now has workedso well to support our country's science. Without some form of major upward fi-nancial readjustment, it will be all too clear that one's success in getting a grantwill be at the expense of another equally talented scientist's potential for doingscience. So, hopefully, the greatly enlarged funds available to the HowardHughes Medical Research Institute can very soon be used to help the essentialfabric of biology and medical research temporarily survive while our nation reas-sesses our national priorities.
Even the Howard Hughes monies, however, will not be enough to let today'sbiologists do all the jobs of which they are now capable. I would guess that weshall need at least a fifty percent increase in total funds (toward 10 billion dollars)to generate the new facts that will let us not only handle current, immediate chal-lenges like AIDS, but also respond to the fact that as our nation ages, we mustproduce the medical advances that will allow many more of our over-65 popula-tion to continue to work productively. What thus represents the self-interests oftoday's biologists and medical research communities should serve equally wellthe long-term interests of our nation as a whole.
We thus must not be alarmed that an ever-growing part of our national wealthmust go not only to biological research, but also to the other forms of sciencewhose futures look equally bright. Instead, we would have every reason to worryif we thought that our national scientific effort could stay constant while the grow-ing impact of the world's vast population increase continues to present us withdilemmas, both practical and ethical, which at a minimum must be called daunt-ing. In the long run, the effective limit for our scientific efforts is likely to be theeducational aspirations of our population. With the world changing so fast, wehave no reason to believe that our nation will remain a leading power if the peo-ples of other countries have a greater desire to learn the facts necessary to takeup the new technologies of the future. There is no reason at all to believe that ifwe lose the will to move ahead, then other people will drop out at the same time.
Our only sensible course is to look as far into the future as possible, make intel-ligent guesses as to what will and will not happen, and act accordingly. Thosenations who so arrange their affairs will have the best chance to have long-termdestinies for the civilizations they promote.
HIGHLIGHTS OF THE YEAR
Using Yeast Cells to Understand Human Cancer
Over the past several years, Mike Wig ler and his research group have focusedmuch of their research on a group of three human ras genes, mutations in whichlead to the cancerous phenotype. Each of these ras genes codes for one of aclosely related set of small (189 amino acids) proteins that associate with the in-ner surfaces of the outer cytoplasmic membrane of cells and that bind GTP(GDP). These two characteristics suggested that the ras proteins might be func-tionally related to the so-called "G proteins" that mediate the transmission of sig-nals from cell-surface receptors to the membrane-bound enzyme adenyl cyclasewhich converts ATP to cyclic AMP (cAMP), the key intracellular regulator. Be-cause the ras proteins are present in relatively small amounts in cells, the effec-tive purification and characterization of these proteins from cancer cells haveproved very difficult, and the working out of how they function at the molecularlevel has not been straightforward.
3

William R. Udry
Wigler's group has thus recently directed much of its attention to the two RASproteins of yeast whose structures are very similar to those of mammalian cellsand whose functioning is required for yeast cell growth. Most importantly, thepowerful new recombinant DNA procedures for introducing genetic informationinto yeast allow a genetic analysis far more subtle than would ever be possiblewith mammalian cells. Through such experiments they have shown that the sameamino acid replacements that generate cancer-producing ras proteins in mam-malian cells lead to metabolically out-of-control yeast cells that overproducecAMP. Further evidence favoring regulation of cAMP production by the RAS pro-teins comes from analysis of a special class of yeast mutants that compensate forthe absence of functional RAS genes. These genes that effectively suppress theRAS phenotype have been shown by Matsumoto in Japan to code for the regula-tory subunit of the protein kinase normally controlled by cAMP. Thus, mutantsthat lead to the absence of the regulatory subunit need not require cAMP for ki-nase activation. Hence, they have the ability to survive in the absence of the RASproteins whose presence is normally required to make cAMP. These very impor-tant experiments confirm the now almost forgotten mania of some 12 years agowhen the cAMP levels of many cancer cells were found to be abnormally highand the essence of cancer was proclaimed to be an excess of cAMP. Though thishypothesis clearly understates the true complexity of cancer, it is now clear thatone must think about cAMP levels if we are to have a serious understanding ofthe metabolic uniqueness of many forms of cancer.
The Loss of a Superb Administrator
It was with sadness that we bid farewell to Administrative Director Bill Udry at theend of the year. Bill came to us in late 1970, after serving as Chief ExecutiveOfficer of the Eye Research Institute of Bethesda, Maryland. A skillful administra-tor, he introduced professional management to our then financially uinsophisti-cated institution. His advice at many levels, from the receipt of the first CancerCenter grant in 1971, through the renovation of our once highly decrepit physicalplant, to initiating construction on the Grace Auditorium, was a crucial factor inthe development of this Laboratory over the past 15 years.
Bill's departure created a large gap in our administrative infrastructure, whichwas exacerbated by my own slow recovery from injuries sustained in a wretchedcar accident in December. It is a tribute to Bill that the senior administrative staffselected and trained during his tenure pitched in admirably to close the breachduring this difficult period.
Changes in Our Scientific Staff
The Laboratory's great success in recent years has been built upon a steady in-flux of young scientists, to whom we give virtually unlimited intellectual freedomand few administrative duties. Under this system, the Laboratory has nurturedthe early careers of many molecular biologists who have since risen to promi-nence. Although we benefit greatly from the intellectual fervor of youth, it is a sadfact that we cannot permanently absorb a large number of mature scientists.
Due to a lack of endowed research appointments, we often cannot offer thesecurity-and, increasingly, the salaries-offered by major universities and com-panies. Thus, we are forced to contend with a high turnover in our staff. Thoughwe have come to accept this fact of life, we are always saddened to face the
4

departure of long-valued staff members. This is tempered by the knowledge thatthey have accepted positions at some of the foremost universities and compa-nies in the country.
Tom Broker and Louise Chow left their Senior Scientist positions in July to es-tablish a new electron microscopy unit at the University of Rochester. Tom andLouise arrived in 1975, after serving post- and predoctoral periods, respectively,in Norm Davidson's lab at Caltech. Even before coming here, they had estab-lished themselves as leaders in the electron microscopy of nucleic acids. In1977, their superb photos did much to first establish the phenomenon of RNAsplicing.
Jeff Strathern left his Senior Scientist position in June to head up a researchgroup at the Frederick Cancer Research Facility. Jeff arrived in 1977 from IraHerskowitz's lab to organize, with Jim Hicks, the Debi-Lick (then Davenport) YeastGroup. In 1979, Jeff, Jim Hicks, Amar Klar, and Jim Broach firmly establishedyeast as a major system for studying development when they showed at the mo-lecular level that mating-type switches in Saccharomyces cerevisiae are achievedthrough the movement of genetic cassettes.
Also leaving for the Frederick Cancer Research Facility was Steve Hughes.After serving a postdoctoral period with the Bishop-Varmus group in San Fran-cisco, Steve came to the Laboratory in 1979 as a Senior Staff Investigator. Ourfirst neurobiology appointment in 1978, Birgit Zipser left in December from herposition as a Senior Staff Investigator for a visiting professorship at the NationalInstitutes of Health. Senior Staff Investigator Ron McKay, a tumor virologistturned neurobiologist, left in July to accept an associate professorship at MIT. In1980, Ron and Birgit helped revolutionize neurobiology when they developedcell fusion techniques to produce monoclonal antibodies against leech neural an-tigens.
After ten years at the Laboratory, Bill Topp accepted a position as President ofOtisville Biotech, Inc., in upstate New York. Bill came to us in 1974 as a postdoc-toral fellow in Bob Pollack's mammalian cell genetics unit and advanced in 1978to Senior Staff Investigator working in James lab. Leaving in October for an as-sistant professorship at Princeton was Lee Silver, who came to us as a SeniorStaff Investigator in 1981 to begin our effort in mouse embryology. After scoringa major coup by showing that two oncogenic functions are required to transformprimary cells, Earl Ruley left his Senior Staff Investigator position for an assistantprofessorship appointment at MIT.
Accepting a position at the College of Physicians & Surgeons of Columbia Uni-versity was Staff Investigator Mitch Goldfarb, who worked with Mike Wigler overthe last four years and played a key role in isolating the ras oncogenes. Jim Lin,a Staff Investigator in McClintock who developed many of the first monoclonalantibodies against cytoskeletal proteins, left in February for an assistant profes-sorship at Iowa State University. A native of Italy, Staff Investigator Mara Rossiniaccepted a position with the biotechnology firm SCLAVO in Siena. Returning toExxon Research and Engineering Company in Clinton, New Jersey, followingjoint appointments at Cold Spring Harbor, were Susan Bonitz and Dan Lundell.
Visiting scientists Pei-Mao Lin and Guang-Yun Cai returned to the People's Re-public of China, while Jin-Zhao Li took a position at Downstate Medical Center inBrooklyn. Kristen Frederiksen relocated to MIT with Ron McKay's researchgroup.
Leaving after their postdoctoral periods were Bill Huse to become an assistantprofessor at Yale in Chuck Stevens' neurobiology program; Prasad Koka to the
Louise Chow and Tom Broker
Jeff Strathern

Center for Cancer Research at MIT; Nita Maihle to work with Steve Hughes at theFrederick Cancer Research Facility; Paul Mains to the Department of Molecular,Cellular and Developmental Biology at the University of Colorado; Kazuo Maruy-ama to MIT with Earl Ruley; Olof Sundin to the MRC Laboratory in Cambridge;Elizabeth (B.J.) Taparowski to continue postdoctoral work at the University of Vir-ginia; Stuart Weisbrod to pursue a MBA degree at Columbia Business School;and Bob Weiss to the University of Utah.
Joining the Laboratory in 1984 as Senior Staff Investigator was Clive Slaughter.Clive came to us from the University of Texas Health Science Center at Dallas toreestablish a major focus in protein chemistry in the new Demerec extension. Pab-lo Scolnick, formerly a research associate at the University of Chicago, also setup a laboratory in the Demerec extension. Pablo studies gene expression andcarotenoid biosynthesis in Rhodopseudomonas and tomatoes.
During the year, both Russell Malmberg and David Beach were promoted toSenior Staff Investigator from Staff Investigator positions. Russell studies the roleof polyamines in tobacco flower development, and David's studies focus on cell-cycle control in fission yeast. Receiving promotions from Postdoctoral Fellow toStaff Investigator were Steve Dellaporta, Bob Franza, Doug Hanahan, WinshipHerr, and Bill Welch. After completing his dissertation under our joint Ph.D. pro-gram in genetics with Stony Brook, Rich Kostriken began a postdoctoral fellow-ship with Mark Zoller.
Jim Feramisco Appointed to Rolling Five Position
In April, the Trustees of the Laboratory approved the promotion of Jim Feramiscoto Senior Scientist in recognition of his work in focusing the efforts of McClintocklab on the cell biochemistry of oncogenic proteins. Jim came to Cold Spring Har-bor in 1978 as a postdoctoral fellow in our Cell Biology Group. With Guenter Al-brecht-Buehler's departure in 1982, Jim assumed leadership of the cell biologyeffort.
Our Senior Scientist rank carries with it a "rolling" five-year appointment, whichmeans that the position will be funded continuously for a minimum of five yearsfrom any point in time. Our closest approximation to tenure for almost all our sen-ior staff, the rolling five falls short of the job security offered by universities andindustry. Thus, over the next several years, we must work hard to establish sev-eral tenured research professorships funded by hard money if we are to remaincompetitive for the top echelon of mature scientists.
Joe Sambrook F.R.S.
In March of this year, the Fellows of the Royal Society, meeting in London,elected into their Fellowship Joe Sambrook for his distinguished contributions tothe understanding of tumor viruses at the molecular level. That Joe would re-ceive this most coveted honor for a British scientist was long expected since hisrecord of involvement in important discoveries about DNA tumor viruses datesback to the late 1960s when he was at the Salk Institute. Since coming here, hehas attracted an extraordinarily talented group of co-workers who, with him, havemade this Laboratory into a major, internationally recognized site for research onDNA tumor viruses. In doing so, they have given James lab a reputation for intel-lectual intensity of which we are most proud.
6

Joint Agreement with Monsanto to Study Mouse Genetics
In October, we signed a five-year, $2.1 million agreement with Monsanto Com-pany to conduct a cooperative research program on the use of gene transfer inthe study of gene expression during mammalian development. Under the termsof the agreement, Monsanto has the option for an exclusive license to developfor commercial use inventions arising from research covered by the agreement.Cold Spring Harbor will retain ownership of any patents and will receive royaltieson sales resulting from technology developed here. Also, up to two Monsantoscientists will be trained for three-month periods at Cold Spring Harbor.
The program's main goal is to shed light on one of the great unanswered ques-tions in modern biology: How does an organism develop from a single, undiffer-entiated cell into a complex system of many different types of cells, tissues, andorgans? Current theory holds that differentiation is achieved by "tissue-specific"expression of genes-the selective turning on and off of specific genes in spe-cific tissues during different phases of development.
One major approach to this problem is the microinjection of a solution of for-eign DNA directly into fertilized mouse eggs. The eggs are then reinserted intothe oviducts of pseudopregnant female mice and allowed to develop. Approxi-mately six percent of all microinjected eggs give rise to live "transgenic" mice inwhich the injected DNA has become incorporated into the host DNA. Becausethe injected DNA can code for one or more novel proteins not found in normalmice, expression of the foreign DNA in various tissues of transgenic mice can bedetected by looking for the telltale proteins.
Early in 1985, Doug Hanahan obtained a remarkable research result using thistechnique. Following microinjection of a fusion gene that linked the insulin pro-moter to the SV40 large-T antigen, he found that the T antigen is expressed onlyin the insulin-producing beta cells of the pancreas. Now that he has achievedcorrect tissue-specific expression, regulatory DNA sequences within the insulinpromoters which are responsible for turning the transplanted genes on and off atthe proper stages in development can be isolated.
Our current foray into the world of mouse genetics dates to the appointment ofLee Silver in 1981, who came to study the T locus. With the completion of theHarris building in 1982, we had a first-class facility for rearing mice, and in 1983we introduced a course on the Molecular Embryology of the Mouse. The Labora-tory's involvement in mouse genetics actually dates to the 1920s, when Drs. Clar-ence Little and Carlton Mac Dowell of the Carnegie Institution of Washington staffbegan to develop pure strains of inbred mice that have unusually high inci-dences of cancer. Little went on to found the Jackson Laboratory in Bar Harbor,Maine, now a major supplier of genetically pure strains of mice, while Mac Dowellcontinued work here, developing the important high leukemic mouse strain C58black.
New Building Projects Continue Apace
Throughout this decade we have completed an average of one or more majorconstruction projects per year: the Davenport restoration in 1980, DelbrOck labo-ratory and Sammis Hall in 1981, the Harris Animal Facility in 1982, and the De-merec protein chemistry-nucleic acid addition in 1983. This trend has continuedwith the completion, in late fall, of the monoclonal antibody extension to Jameslab.
7

Construction costs for the extension were underwritten by grants of $425,000from the National Cancer Institute, $400,000 from the Pew Memorial Trust, and$30,000 from the William Randolf Hearst Foundation. An additional grant of$30,000 from the Hearst Foundation, a $100,000 grant from the Fannie E. RippelFoundation, $15,000 from the Samuel Freeman Charitable Trust, and $50,000from various Long Island businesses went toward purchasing equipment.
The 6400-square-foot addition to the north end of James contains offices andtwo very spacious laboratories-one for Joe Sambrook's and Mary-Jane Ge-thing's group and another for Ed Harlow-a cell fusion facility, and a largekitchen. Combined with the Harris animal house, we now have state-of-the-artfacilities for the production of monoclonal antibodies needed to isolate cellularproteins and probe their structures. Already the James extension is fulfilling itsrole as a clearinghouse of monoclonal antibody technology for the entire cam-pus; Ed Harlow provides expert technical advice, with Carol Schley available toperform cell fusions and screening procedures.
In May, we began the long-awaited construction of the Oliver and LorraineGrace Auditorium. When completed in late fall 1985, this 360-seat facility will alle-viate the long-standing problem of inadequate seating at our major meetings.With its massive dormers, richly textured brick and stucco exterior, and airy pub-lic spaces, the Grace Auditorium will be perhaps the finest meeting facility onLong Island.
Now planned as a multifunction building, the upper floor of the auditorium, be-sides containing the Lecture Hall, will house our Information Services Depart-ment, while on the lower floor will be our Meetings Office and a new computercenter that will be the hub of an integrated network serving the entire scientificstaff.
Delays in the Construction of Our Squash Court
Two years ago we thought we would soon be starting construction on both theGrace Auditorium and a squash court, to be located near the tennis court, whichwould give us a year-round facility in which our scientific staff, steadily growing innumber, could benefit from short intervals of intense exercise. But as planningfor the Grace Auditorium proceeded, we expanded its scope greatly through thedecision to build a fully functional basement so as to give us the freedom tomove within it the headquarters of our ever-expanding computer system. Withthis decision, the cost of the building rose to nearly $3.6 million, a sum far inexcess of the earlier anticipated $2 million expenditure. In making the decision tostart construction on the auditorium, we have had to commit virtually all the freefunds in our treasury, leaving no monies to cover the estimated $200,000 cost ofthe squash court facility. We thus have had no choice but to postpone construc-tion of the squash court until we find funds specifically donated for this effort.This may not be easy to do, since our neighbors know by now how much we alsoneed new monies to expand our facilities for research and the holding of meet-ings and courses. Hopefully, we can persuade one of the now growing numberof Cold Spring Harbor alumni who have helped form the biotechnology industryto donate his name along with the necessary number of stock certificates tobring this very much needed building into existence.
8

Revised Plans for New Facilities for Plant Molecular Biology
Initially, the most sensible way for us to expand our efforts in plant molecular biol-ogy appeared to be the construction of a joint laboratory-field station complex atUplands Farm, the nearby former dairy farm that was owned by Mrs. GeorgeNichols and whose land and buildings now belong, through her donation, to theNature Conservancy. We thus entered into a contract with the Conservancy inthe fall of 1984 to purchase from them some 10 acres of this land, on which aresituated two houses and a 6000-square-foot garage/apartment building, for aprice of $705,000. As our planning for this complex grew more precise, we real-ized that instead of saving money by using the existing garage for a new lab, therenovations required would actually cost more money and provide less suitablefacilities than a new laboratory facility on our main Bungtown Road site. We thusnow plan to use the Uplands Farm land and buildings exclusively for a Field Sta-tion on which we shall grow corn and erect a major new greenhouse specificallydesigned for corn. At the same time, the garage will be extensively renovated toprovide areas for corn seed storage and examination, microscope facilities forchromosome analysis, plant growth chambers, and the housing of the necessaryfarm equipment. We are simultaneously making detailed plans for a 6000 -square -foot addition to Delbruck laboratory, made possible by moving thenearby Firehouse (which now contains three apartments) to a site 80 feet to thenorth. Relocation of the Firehouse is now planned for late August 1985, with theconstruction of the Delbruck north addition scheduled to start in September andhopefully to be completed in the early summer of 1986. The projected cost ofthese various efforts now totals $2.3 million. Toward this sum, we have received agrant of some $700,000 from the National Science Foundation, $350,000 offoundation support, and have hope of another major donation being receivedbefore construction is to begin.
Already we have initiated work on the Uplands Farm garage, with occupancyscheduled by the end of summer 1985. Bids will soon be asked for the newgreenhouse, with its completion date hopefully in time for a winter planting of thecorn seeds that will result from the summer 1985 crop of Steve Dellaporta andhis associates.
2-D Gel Technology Attracts Major Support
After years of hard work, Jim Garrel's computerized system for analyzing pro-teins separated by two-dimensional gel electrophoresis is attracting the attentionit deserves. In June, the Laboratory granted an exclusive license to Protein Data-bases, Inc. (PDI), to develop this technology for commercial use. In return for thelicense, the Laboratory will receive $400,000 in research support from PDI,which has opened a facility on Oakwood Road in Huntington Station, N.Y.
Later in the year, we received a $2 million Biotechnology Resource grant fromthe National Institutes of Health to develop Cold Spring Harbor as a major centerto train scientists in the uses of 2-D gel analysis in biomedical research. The Bio-technology Resource Center will be set up as a separate unit on the lower floor ofthe Grace Auditorium and will contain three Mass Comp computer workstationspowerful enough to generate the complex graphics that are part of the analysis.Serving on the NIH advisory committee to the Center are Matthew Scharff, Bar-ney Clarkson, Rich Roberts, Sidney Strickland, and James Schwartz.
9

Two-dimensional gel electrophoresis, first developed in 1975 by Patrick O'Far-rell at the University of Colorado, can resolve more than 2000 different proteinsin a single trial. Because of this exquisite sensitivity, 2-D gels contain far moreinformation than can possibly be analyzed by visual observation alone. Jim Car-rels, who came to the Laboratory in 1978 from the Salk Institute, was among thefirst to work out a practical, computerized method for analyzing the complex spotpatterns generated by 2-D gels. One major goal of the Biotechnology ResourceCenter will be to establish complete protein databases for normal vs. transformedrat cell lines, yeast cell lines, and mouse embryo cell lines.
Our Summer Teaching Program Flourishes
Educational outreach-in the form of meetings and courses-has been a ColdSpring Harbor hallmark since the founding of the Biological Laboratory here in1890. At that time, the harbor, mill ponds, and Long Island Sound were used asa natural laboratory in which to test Darwin's notions about how species evolve toexploit various environmental niches. Although our bucolic surroundings are nolonger the object of experimentation, they do continue to inspire a serious,though informal, approach to science.
Our specialized courses in molecular genetics and neurobiology continue toattract three times as many applicants as there are available spaces. More than400 applicants vied for 89 slots in six genetics courses, while 185 researchersapplied for 91 positions in six neurobiology courses. Far and away the most pop-ular courses, Molecular Cloning and Advanced Cloning drew a record 200 and64 applicants, respectively.
The Neurobiology Teaching Program, which has functioned exceptionallysmoothly under the direction of Sue Hockfield, was substantially boosted by thereceipt in December of a $110,000 equipment grant from the McKnight Founda-tion. The grant will allow us to purchase centrifuges, incubators, and other equip-ment necessary to bring recombinant DNA techniques into our Drosophila, im-munoglobin probes, and single channel methods courses. A Banbury meetingon Computational Neuroscience-sponsored by the Sloan Foundation-was sowell received that in 1985 we are creating a new lecture course at Banbury toexplore the topic.
A High-level Symposium on Genetic Recombination
The 49th Symposia on Quantitative Biology had as its major theme recombina-tion at the DNA level, a topic that brought the return of many old friends to theLaboratory as well as first-time visits by many whom we hope will also comeback many times. Serving as its major organizers were Amar Klar and Jeff Strath-ern who brought together a most distinguished and varied collection of speak-ers. Starting off the meeting on a high intellectual level was Bruce Alberts, whogave his view of how proteins work during DNA replication. Equally satisfyingwas Allen Campbell's final summary. Greatly assisting our standing-room-only au-dience was the use of a very large TV screen which often provided more than lifesize views of speakers faces to those watching the proceedings in the adjacentroom of Bush Lecture Hall. As usual, our more than competent Meetings Officestaff received much praise from our visitors for the kindly way in which they wereintroduced to the way of life here, a feature we must always maintain if we are toexpect to keep our position as the leading meetings center for molecular biology.
10

The Robertson Research Fund Provides Our Main Source of DiscretionaryResearch Funds
The gift from Mr. Charles S. Robertson in 1973 of an $8 million fund whose in-come should go exclusively to the support of research changed radically the na-ture of this Laboratory. In 1984, we used some $600,000 of this income to furtherhelp our newly created Mouse Embryology Group, to provide major items ofequipment for the new north extension to James lab, to provide carryover fundsneeded for the temporary maintenance of the late Ahmad Bukhari's research ef-forts, to support our meeting programs, to help buy a cell sorter for James lab,and to support the stipends of several postdoctoral fellows and visiting scientists.
The Banbury Center Now Functions with a Balanced Budget
With each year we have made increasing use of our Banbury Center located onthe site of Mr. Charles S. Robertson's former estate in Lloyd Harbor. During thefive-month-long meeting and summer courses period its facilities are now in full-time use. Particularly important is its conference facility, which serves as the siteof our summer neurobiology lecture courses. During the rest of the year, it servesas the perfect site for a series of small meetings arranged by Mike Shodell, theDirector of Banbury Center. While Mr. Robertson provided a generous endow-ment fund for the upkeep of his marvelous house and the grounds of this beauti-ful 40-acre estate, each of the meetings we hold there demands additional out-side funding. Toward this end, applications for grants are constantly being madeto potentially interested governmental agencies and requests made to major in-dustrial sources and private foundations for core-type support. The raising of thismoney has never been easy, and so it is with much satisfaction that I can reportthat all of last year's $550,000 in operating expenses were raised, leaving us, infact, with a minor surplus. This coming year I fear may not be so satisfactory.Grant support for meetings is now very hard to get, even with high priorityscores, due to the proposed reductions anticipated in the number of grantsawarded. We shall thus have to work even harder to see that Banbury Centerremains an important site of incisive small meetings, many devoted specifically tothe interaction between the latest biological research and human society.
LIBA Continues to Provide Us with Critically Needed Building Funds
Our neighbors who work so effectively on our behalf under the auspices of theLong Island Biological Association (LIBA) came again to our rescue in a momentof great need by providing an additional $100,000 toward the costs of the Oliverand Lorraine Grace Auditorium. This project, initiated with the enthusiastic sup-port of LIBA, will radically upgrade our facilities for the holding of our majormeetings, allowing us to respond effectively to the enormous expansion in thenumber of molecular biologists that is accompanying the recombinant DNA revo-lution. So we remain deeply indebted to all of our LIBA members for their contin-ued assistance which allows us to remain a major center of DNA research for theentire world.
Over the next year the officers and directors of LIBA will be engaged in discus-sions with our Trustees as to how we should prepare for the 100th anniversary ofour founding in 1890. The five years still to pass before this major anniversaryprovide an obvious interval in which to reassess where we want to be on our
11

100th birthday and hopefully to plan out a major program for achieving theseobjectives.
Formation of an Ad Hoc Committee of Our Board of Trustees forNeurobiology
While we offer the most comprehensive series of summer courses in neurobiol-ogy in the world and are now generally recognized worldwide as a major assetfor the promotion of neurobiology, we still do not possess a laboratory in whichneurobiologists can work throughout the year. Jones laboratory, our sole facilityspecifically equipped and renovated to do neurobiology, must also function eachyear as the site of our summer courses. During the summer our neurobiologistsmust go away or crowd together in a modernized mini-lab in the old MouseHouse, where effectively they must stop most of their research. This is clearly anintolerable situation in the long-term. This situation recalls the similar circumstan-ces of our Yeast Group when their booming research on mating-type switchingled us to build for them an addition to Davenport (now DelbrOck) laboratory.There is no way, however, that a major addition can be added to the 1893-builtJones lab without destroying the intrinsically beautiful way our complex of oldbuildings appears when seen from both the Lab grounds and from across theinner harbor.
It was thus virtually inevitable that we would witness the premature breakup ofthe small Neurobiology Group that has worked in Jones lab over the past severalyears. This is inherently very distressing in view of their seminal role in showingthe great potential of monoclonal antibodies not only for probing the anatomy ofthe nervous system, but also for potentially revealing the identity of the key mole-cules whose interactions lead to linking of nerve cells to their correct cellularpartners during embryonic development. First to leave us for the prospect of amore stable long-term future was Ron McKay, who moved in the summer of 1984to the Whittaker School of MIT. Soon afterwards, Birgit Zipser left to continue herresearch on the leech at the National Institutes of Health. And earlier this year wehad to accept the fact that Sue Hockfield would be leaving in mid-1985 for theNeuroanatomy Department of Yale Medical School.
The prospective long-term absence of any year-round efforts in neurobiologyis clearly incompatible with the running of a major summer program. We mustthus design, and then find the funds for, a new laboratory specifically designedfor neurobiology. Toward this end, a committee of our Board of Trustees hasbeen created whose members are Charles Stevens, Eric Kandel, John Klingen-stein, and William Robertson. Hopefully, semiprecise plans as to how we shouldproceed will be available by the end of the summer of 1985.
A Strong New Set of Additions to Our Board of Trustees
That our Board of Trustees combines science and the public sector at its bestremains one of the strongest assets of the Laboratory. I consider myself most for-tunate that from our trustees I can call on the collective wisdom of many verytalented scientists as well as the financial and legal savvy necessary for quick,experienced reactions to a multiplicity of problems that need answers. It is thuswith much regret that the statutory six-year terms of three very devoted trusteesexpired this past November. Roderick Cushman, Mary Lindsay, and AlexanderTomlinson have all served us with distinction. In particular, I wish to note that this
12

is the second time Mary Lindsay has completed a six-year term of office. Happily,all our departing trustees live near to us, and we are sure that their repeated helpcan be relied upon. Following his move to Princeton in the fall, Tom Shenk nowbecomes our trustee from Princeton University, with the Stony Brook slot nowably filled by Sidney Strickland. Newly elected as individual trustees are Mrs.Charles Dolan of Cove Neck, who, with her husband, has been a major imagina-tive force in the development of cable TV in the United States; Mrs. SinclairHatch of Oyster Bay, a longtime activist for a variety of medically important goals;and Mr. Harvey Sampson of Cold Spring Harbor, a distinguished leader of busi-ness and Chairman of the Harvey Electronic Corporation.
We Are Going Through a Major Transition in Leadership
That I have been the director for 17 years and still enjoy the many tasks that gowith my position owes much to the fact that I have for almost all this time effec-tively shared most of my burdens with Joe Sambrook, who has headed Jameslab since his arrival here in 1969 and who for the last seven years has functionedalso as our Assistant Director for Research. It was thus with both unease andregret that last August I learned of Joe's decision to resign from his position herein September of 1985 to join the University of Texas Medical School in Dallas asHead of its Department of Biochemistry. Filling Joe's shoes will be no easy task,since Joe has a forceful, innovative mind that consistently and wisely has workedfor the good of this institution. In doing so, he has played a far more major rolethan generally perceived by the outside world in preserving Cold Spring Harboras a major site for teaching and research in molecular biology.
To help me now with many of the tasks that Joe carried out, I have asked TerriGrodzicker to serve as Assistant Director with the particular charge of overseeingour summer teaching and meetings programs. Still to be filled is the position akinto that of the Scientific Director in many other research institutions, where oneindividual has the major role of overseeing the recruitment and subsequent pro-motions of the scientific staff. This is a task that Joe ably performed for manyyears, since as the Laboratory grew larger, I no longer had the time to effectivelyknow the world of younger scientists. The recruitment of a high-level scientistversed in all aspects of DNA research, if not of biology itself, thus stands out asthe most important goal for me to accomplish over the next year.
In this period when I have more tasks than one individual can handle, I takerelief that in the recent appointment of G. Morgan Browne, we now have a newAdministrative Director in whom we already have great confidence. A 1957 grad-uate of Yale, he is an experienced business executive, with a strong financialbackground, particularly in the high technology area. A member of LIBA for sev-eral years, and with a keen liking for and interest in science, Morgan knowsmany of the Lab's longtime supporters and needs no introduction either into theunique roles of the Laboratory or to our community that so long has been anindispensable asset.
The Grace Auditorium Will Transform the Way We Are Seen by IncomingVisitors
With the completion of the Grace Auditorium, our Laboratory will be ending a 23-year era that commenced in 1953 with the completion of Bush Auditorium andDemerec lab. With these new buildings in hand, Cold Spring Harbor had the re-
13

sources to participate in the biological revolution that took off with the discoveryof the double helix. Though we have continued to make many important newadditions to our physical plant, the Laboratory has looked much the same to un-initiated visitors. The hill overlooking the Blackford entrance hid much of thewestern afternoon sun and was never a site of beauty, remaining permanentlyscarred from the removal of the sand used in the 1906 building of Blackford it-self. Thanks to the powers of modern earth-moving equipment, Blackford Hallnow looks out onto a newly made open area created so that our auditoriumwould be graced with an adjacent area into which our meeting participants canmove freely during coffee breaks and on the way to meals. Soon the nowpushed-back hill will be landscaped and on its top will be placed an ancienteight-sided Gazebo given to us when a recently sold estate was broken up. Backof the gazebo we shall be creating a brand new parking area for some 70 cars tohandle the increased number of visitors that shall come to the new lecture hall. Inopening up this area, we shall be losing a few more trees than we would like, butto compensate for this, we shall plant even more to eventually allow a forestlikefeeling to return. As we rework this upper space, we will face the facts that the26-bed Page Motel built in 1953 functions too often like a 1930s New Hampshiremotel and that guests chosen to live in our four 1930s-type cabins often remem-ber the experience more than they wish. So we are already thinking about whattheir replacements should look like and tentatively have decided to have our ar-chitects come forth with the "Adirondack Style" revisited. And next to their poten-tial site is an area that could be perfect for a new neurobiology building.
Hopefully, all of these dreams will be reality by our 100th birthday. If so, what aglorious next 100 years we can share with the world of biology.
June 26, 1985 J.D. Watson
14

DEPARTMENTALREPORTS
21
20=EXPENDITURES
19 ED ADJUSTED FOR INFLATION
18
17
16
15
14
13
6
5
4
3
2
68 69 70 71 72 73 74 75 76 77 76 79 80 81 82 83 84YEARS
320-310 -300 -290-280 -270 -260 -250 -240 -230 -220.-210-200
9080 -70 -60 -50 -40 -30 -20 -10-00 -9080-70-60-50-40-30-20-IX
-70-60
STAFF
fEz] SCIENTISTS
=SUPPORT
Total Staff
4000
3000
2000
1000-
MEETINGS 8 COURSE PARTICIPANTS
8 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84YEARS
r7 7
8 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84YEARS
*Consists of Technical Support, Core Services, Publications, Library, Buildings and Grounds,Information Services, Administrative Personnel, Banbury Center
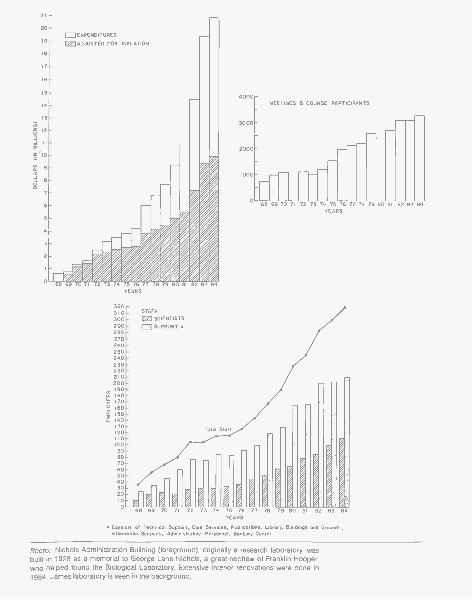
Recto: Nichols Administration Building (foreground), originally a research laboratory, wasbuilt in 1928 as a memorial to George Lane Nichols, a great-nephew of Franklin Hooper,who helped found the Biological Laboratory. Extensive interior renovations were done in1984. James laboratory is seen in the background.
ADMINISTRATION
The continued growth of the Laboratory over the past years has had a dramaticimpact on the Administrative staff. Since 1968, the Laboratory's expenditureshave increased from $580,000 to over $20 million, almost a 3500% increase.The number of research grants has grown from less than 10 to over 200 (notincluding subprojects within a grant), and the Laboratory staff has increasedfrom approximately 35 to more than 325. Meetings and course participants wentfrom approximately 700 in 1968 to 3300 in 1984. Although the challenges havebeen considerable throughout this period, they have been met through in-creased efficiency in virtually all areas.
From a small, in some ways "patchwork," operation, Administration hasevolved into a modern, computerized organization built around a core of experi-enced professional managers and highly qualified clerical personnel. In stream-lining our operations to provide greater support and assistance for the Labora-tory's scientific endeavors, we have not, however, lost sight of the Laboratory'ssmall-town, spontaneous, and scientifically intensive atmosphere that has servedscience so well for so long.
Installation of the Administrative Computer
Prior to the installation of the administrative computer, records and data werespread throughout the campus in separate offices located in separate buildings.The flow of paper from building to building and office to office precluded timelyand coordinated information exchange. Now, with direct access to our MAI BasicFour computer, the various administrative segments can instantly and accuratelyreflect and report their daily activities to related departments.
In addition to greatly assisting clerical functions, the computer has eased theburden of processing purchase orders, maintaining personnel records, and man-aging finances. Up-to-date financial and grant reports are now readily availableto senior management and scientific staff. It is difficult to imagine how the presentvolume and variety of transactions would have been managed without the accu-racy, simplicity, and consolidation afforded by a computer.
Formation of the Commerical Relations Committee
The advance of biotechnology at the Laboratory has drawn the interest of sev-eral commercial organizations. To safeguard the Laboratory's basic researchmission and to avoid potential conflicts of interest, the Commercial RelationsCommittee was established by the Board of Trustees to review such relationshipsand to generate policies and guidelines to govern staff interactions with commer-cial organizations. Townsend Knight, Chairman of the Committee, has contrib-uted enormously toward reviewing the frequently complex and somewhat un-precedented new partnerships between commerce and basic researchinstitutions.
Nichols Renovation
After years of neglect and overcrowding, the Nichols Administration building hadbecome inefficient and no longer suited to the increased administrative tasks. It
17

had also become structurally unsound and could no longer support the numberof people using the facility. Renovation plans were drawn up and Jack Richardsturned the attention of his staff toward making Nichols into a more efficient andbetter place to work. The results are dramatic from both an aesthetic and func-tional point of view. Offices are now arranged on the basis of operating efficiencyand functional considerations, and the staff has a much more productive andpleasant environment.
Tax Exempt Bond Issue
What began over 15 years ago as a critical program to restore the Laboratory'sdeteriorated physical plant has evolved into a substantial program of capital im-provements and expansion which include large additions to Demerec and Jameslabs, construction of the dramatic Grace Auditorium, modernization of BlackfordHall, and the addition of a much needed new parking area. To provide the fundsnecessary for these ambitious projects, we applied for and received an $8 millionNassau County Tax Exempt Industrial Bond Issue. With Morgan Guaranty Trustas bond purchaser, we can proceed with all of these capital projects without limit-ing the ongoing research program. Special recognition is due William R. Udrywho, working together with our former Treasurer and Trustee, Clarence Galston,provided long hours of effort and invaluable financial savvy, which enabled thisbond issue to succeed.
Consolidation of Core Services
Some of the most direct and important services provided in support of the scien-tific staff are the operations referred to as core services. These include the safetydepartment, machine shop, electronics repair shop, equipment repair shop, Har-ris animal care facility, photography and technical illustrating, and the scientificcomputer staff. In 1984 these services were centralized under the direction of ourhighly capable Safety Director, Art Brings. With Art's well-established leadership,we expect that these vital services will prove even more useful to the Laboratory.
The Laboratory continues to be fortunate to have a high-quality administrativestaff. We welcome to the staff Fred Lukas, who has already proven to be an ableassistant to our Comptroller, Bill Keen. Roberta Salant continues to provide inva-luable assistance to me and to the members of the Board of Trustees. Guy Cozzaand Bob Pace deserve specific mention, for without them our computer installa-tion could never have succeeded.
John Maroney
BUILDINGS AND GROUNDS
The overall expansion of the Laboratory since 1969 has resulted in an expandingworkload for the Buildings and Grounds Department and an increase in person-nel from 10 to 42 (including Banbury personnel). Maintenance of an ever-ex-
18

panding physical plant and the attendant landscaping has become increasingly more sophisticated and time consuming. In sharp contrast to the one part-time handyman of years gone by, who'maintained five or six buildings using a ham- mer and screwdriver, our maintenihft! personnel todayBptide men aknowledge of air conditioning and heating, plumbers, and electricians, f i e advanced planning of future buildings, done in conjunction with architects, engineers, and contractors, and the supervision of ongoing construction and renovations also increase the demands on the department.
Jam es Lab Extension for Monoclonal Antibody Research Completed
Th^major highlight of 1984 was the completion; in mid-summer of th i new James laboratory extension by the contracting figjj of Robins and Cowan of Huntington, Long Island. The Buildings and Grounds staff soon after fifti|hed-the-offtces, in- stalled the kitchen, and coordinated the long awaited move of the scientists and their equipment from their previous quarters in James lab. In addition, the park- ing area behind James was extended, and the site around the building and lot was landscaped.
First Underground Conduits in Place for Utility L ines
James Lab extension
A goal ffethe Labo ra to ry to bury all telephone, electrical', and comput:#r:Hiihes for all facilities by late 1986 or early 1987. Through 1984, the lines from Harris to the James extension, including extra computer conduits for future use, have been buried. This leg was quite an undertaking and only with Owen Stewart’s excellent planning were we able to proceed, despite the inevitable complications associated with dealings with major utility companies.
19

Oliver and LorraineGrace Auditorium
Work Is Begun on Much Needed Auditorium
Construction of the new Grace Auditorium began early in 1984, and although thejob is being done by an outside contractor, A.D. Herman, much time has beenspent by members of the department in helping the contractor along. Hopes tocomplete construction in time for the 50th Symposium in June 1985 faded aslabor strikes halted work during the summer. Completion is now expected in fall1985.
Facilities Must Keep Pace as Needs Change
Interior renovations in 1984 were numerous. With a laboratory vacancy comesthe planning for renovation of the facility, and it always seems that it was only"last year" that we were reworking the same rooms. The Slaughter and Tamanoi-Stillman labs in Demerec were completed and the entire first floor of Demerec Awas reworked, including a new kitchen, halls, hall floors, and media room. InMcClintock, the first floor south office and laboratory were redone as a full lab.Refurbishing of James Annex Library included the removal, refinishing, and rein-stallation of all book shelves on foam-insulated walls. The James Annex coffeekitchen received a facelift with refinished cabinets and a ceramic tile countertop,while a laser printer room was constructed in lower James Annex. To correct awater problem caused by our hills and springs, it was necessary to install alongthe west wall of the James Annex a drainage grate and pipe leading to a sumppump pit. Now that the James (North) extension is completed and occupied, acomplete remodeling of the original James (Middle) lab is planned for 1985.
Work was begun on the first floor of the Nichols administration building to mod-ernize the facility and to correct problems that have developed over the years asthe result of rushed expansions to meet critical needs. Duplicate equipment wasinstalled in the wastewater treatment plant, increasing the capacity by at leastone third. None of these renovations could have been accomplished without theadvance sketching done by Charlie Schneider, as well as the talents of DiamondScarduzio in carpentry and overall planning and coordination.
20

General Maintenance and Repairs Are Ever Present
Over and above the capital improvements and renovations, Buildings andGrounds works closely with our scientists and their families to assist them in boththeir work and personal lives. Rarely a day goes by without a maintenance orrepair request from the scientific laboratories and tenants in our housing, both onand off grounds. With off-grounds rentals, the Banbury site, and Uplands Farm,the workload can become quite staggering. The summer courses and meetingsincrease the workload in all departments, and so too in Buildings and Groundswhich must respond to the additional strain placed on all facilities through in-creased use.
Department Saddened by Three Losses
Our department was saddened early in the year by the deaths of Fred Pfeiffer inMarch, Mary Hill in April, and Jim Stanley (who retired in 1980) in June. We hopethat in the future we will be fortunate to have people of their caliber on our team.
Jack Richards
INFORMATION SERVICES
Two Programs Aimed at Nonscientists
One major goal of the Information Services Department is to help explain theLaboratory and its research to nonscientists. Toward this end, "The BiologicalRevolution," a six-week series of public seminars, was held last spring in con-junction with Hutton House Lectures of Long Island University. Dave Mick los,Joe Sambrook, Mary-Jane Gething, Steve Dellaporta, Sue Hockfield, and MikeShodell presented talks on recombinant DNA and its applications to biomedicine,agriculture, and industry.
Also in 1984, the Laboratory's outreach to local schools was formalized as theCold Spring Harbor Curriculum Study. In conjunction with Superintendent Fran-cis Roberts from Cold Spring Harbor High School, a cooperative program wasdeveloped to improve teacher competence and to bring state-of-the-art researchconcepts into school biology classes. Building upon an initial $10,000 grant fromCitibank N.A., we were able to attract pledges of $10,000 apiece from eightneighboring school districts who are participating for the 1984-1985 schoolyear: Cold Spring Harbor, East Williston, Great Neck, Herricks, Jericho, North-port-East Northport, Oyster Bay, and Syosset. It is hoped that the CurriculumStudy will serve as a model for similar efforts throughout the United States.
Support from Foundations Increases
The department set out to match a $100,000 grant from the Fannie E. RippelFoundation for the purchase of equipment for the new Monoclonal Antibody Ex-tension to James lab. The first attempt to involve local companies in the Labora-
21

tory's research-the Long Island Business Cancer Campaign-yielded $50,000in cash and donated equipment. Grants of $30,000 and $15,000 were securedfrom the William Randolf Hearst Foundation and the Samuel Freeman CharitableTrust.
The department also coordinated efforts to augment a $691,000 National Sci-ence Foundation grant to expand the Laboratory's research on the molecular ba-sis of plant development. A reception held at the home of Ambassador JohnHumes led to grants from the William and Maude Pritchard Charitable Trust($60,000) and the Griggs and Burke Foundation ($10,000) and numerous indi-vidual gifts. Two other major grants were secured: $75,000 from the Charles E.Culpeper Foundation and $50,000 from the Surdna Foundation.
Another highlight was the receipt of a $110,000 grant from the McKnight Foun-dation to purchase equipment to teach molecular genetic techniques in our sum-mer neurobiology courses. Also heartening was news of a four-year, $74,000grant in support of the Undergraduate Research Program from the Alfred PSloan Foundation.
Inauguration of the Corporate Sponsor Program
The 1984 Corporate Sponsor Program achieved its two goals: to provide a stablebase of support for our meetings program and to increase dialog between aca-demic and industrial researchers. Last year, 260 industrial scientists attendedCold Spring Harbor Laboratory and Banbury Center meetings. A new series ofBanbury conferences dealing with technical aspects of genetic engineering wasespecially well received. So enthusiastic was the response to the inaugural yearthat 1985 membership has been expanded from 15 to 22 companies. Repre-sented among the Laboratory's sponsors are prestigious Fortune-500 pharma-ceutical and chemical companies and specialty biotechnology firms.
LIBRARY SERVICES
David Mick los
Reference Services Increase and Archives Expand
In 1984, patron/reference services became the focal point for the Library staff.Following the resignation of Audrey Powers as Librarian in July 1983, we werefortunate to have Genemary Falvey, a staff member for three years, step in asActing Librarian while she completed her Master's Degree in Library and Infor-mation Science. In December 1984, Ms. Falvey graduated with honors and waspromoted to Librarian. Genemary's expertise is in reference services, with em-phasis on computer-based literature searches tailored to the needs of individualusers. The impact of these new services on library activity is clear: Scientificsearches increased from a total of 60 in 1983 to 105 in 1984 and reference andinformation questions increased by 41% (from 5000 in 1983 to 8500 in 1984).
Several major objectives were accomplished in the reference and archivalareas this year. The reference collection has been weeded and updated, and the
22

foundation has been laid for upgrading the textbook collection. A DEC personalcomputer was purchased so that extensive on-line searches would no longer sapthe campus-wide system. The preservation and cataloging of 32 maps related tothe history of Cold Spring Harbor Laboratory and its environs were completed.Organization of the historical records of the Laboratory, including completion ofthe registry of the Carnegie correspondence from 1930 to 1950 and the stream-lining of the portrait files, continued. On the recommendation of Trustee Ed Pull-ing, the library also established a Long Island Cultural Arts/Activities File for theLaboratory community. This service provides announcements from all entertain-ment facilities on Long Island.
Permanent Collection and Patron Services Grow Amid Staff Economies
The growth of library resources included the addition of 14 periodical titles, while14 others were withdrawn, keeping our journal subscription count stable at 433.The book collection grew by a net total of 1,404 bound volumes, bringing thenumber of bound volumes to 27,299. Patron services increased an overall 355/ofrom 1983 to 1984.
I would like to publicly acknowledge the contribution made by the entire Li-brary staff in 1984. All individuals contributed beyond their usually high perfor-mance, allowing a smooth operation and reducing salary expenditures by theequivalent of one full-time staff member.
PUBLICATIONS
Susan Gensel
New Titles Published and a New Series Launched
The Publications Department continued to grow in 1984, with a total of 11 newtitles published in addition to 18 reprintings. A new series, Cancer Cells, was in-augurated with the publication of "The Transformed Phenotype" and "Oncogenesand Viral Genes." This series supersedes Cold Spring Harbor Conferences onCell Proliferation, and its journal format provides a forum for scientists from di-verse disciplines. Two new monographs were published this year: Microbial De-velopment, issued simultaneously in cloth and paperback, and Gene Function inProkaryotes, which was later released in paperback in early 1985.
The practice of paperbacking selected titles to extend readership particularlyto the student market was continued with the publication of RNA Tumor Viruses/1,Text. This volume and RNA Tumor Viruses/2, Supplements and Appendixes(due in 1985), with a combined total of more than 2500 pages, constitute an up-dated and expanded edition of the RNA Tumor Viruses volume originally pub-lished in 1982.
Experiments with Gene Fusions, a manual based on the Bacterial Geneticscourses held at Cold Spring Harbor in 1981 and 1982 was well received by re-viewers and by our teaching and research colleagues. A Strain Kit for use withthis manual will be available in 1985.
23

Other titles produced include Modern Approaches to Vaccines, which was anoutgrowth of a 1983 Cold Spring Harbor meeting. So successful was the meetingthat its organizers, Robert Chanock and Richard Lerner, were able to contract forfive more annual meetings. From these meetings will be published a five-yearseries, beginning with Vaccines 85. Another timely publication was Human T-CellLeukemia/Lymphoma Virus, which studies the role of this virus family in malig-nancies and in AIDS. A meeting in April 1984 gave impetus to the publication inDecember of Molecular Biology of the Cytoskeleton.
In 1984, we assumed publication of a biennial series entitled Genetic Maps: ACompilation of Linkage and Restriction Maps of Genetically Studied Organisms.Originally published and distributed as a service in 1980 and 1982 by SteveO'Brien of the National Cancer Institute, the growing demand for, and complexityof, the volume required the expertise of a publisher and we were happy to addthis series to our list.
In recent years, we have strived to publish me results of our meetings as rap-idly as possible, even though most of our books are multi-authored and inevita-bly turn out to be at least 100 pages longer than initially planned. In spite of this,it has been our goal to publish our annual Symposium volume in the same yearin which the meeting takes place. Although the proceedings of the 1983 Sympo-sium on Molecular Neurobiology was at the printers in 1983, bound books werenot available until 1984. For the first time, the Symposium was published simulta-neously in cloth and paperback editions. When the subject matter warrants thepublication of a paperback edition for the student market, this practice will becarried out in the future. In addition to these new titles, we published abstracts ofthe 11 meetings held here in 1984 and the Annual Report.
Use of In-house Computer Saves Time and Money
In-house computer typesetting was utilized in producing seven of the 1984 titles.This technology saves time and cuts composition costs by approximately 30%.We achieved even greater efficiency and savings by initiating the practice ofsending authors letter-quality computer printouts instead of typeset galleys. Thisallows all author corrections to be inserted before the manuscript is transmittedto the typesetter.
Move to Paperbacks Increases Volume
Although unit sales increased from approximately 35,000 units in 1983 to morethan 37,000 in 1984, net sales declined to slightly more than $1,500,000. Thisdecline in the wake of increasing volume is directly related to our major moveinto offering lower-priced paperback editions of many titles. To continue this pro-gram of providing paperbacks priced within reach of the economically hard-pressed student market, every means of increasing sales volume while control-ling costs must be explored.
Marketing Pushes for Greater Foreign Sales
With an eye toward increasing volume, the Marketing Department has been vig-orously pursuing new channels for foreign distribution, including finalizing anagreement with a new distributor for India, Panima Educational Book Agency,and opening negotiations for a new Japanese distributor with the prospect of in-
24

creasing sales to Japan over the next three years from $100,000 to $250,000annually.
A consistent program of follow-up by Marketing on all books sent to journalsfor review has yielded an impressive 40-43% response with a review and/or list-ing within 18-24 months, well above the industry norm of a 25-30% responsewithin the same time frame.
Fulfillment Moves Toward Consolidating AllWarehousing and Shipping
Equally responsive to the need for greater volume, the Fulfillment Departmentincreased t, le ease of placing orders by offering customers the convenience ofan 800 'oll-free number. To speed delivery, 48% of 1984 orders were shippeddirectly from the storage facility at Cold Spring Harbor. With this increase in di-rect shipments it became imperative that sufficient warehousing space be ac-quired locally not only to solve the on-grounds storage problem, but also to con-solidate all warehousing and shipping under our direct control. Havingcompleted negotiations with the Board of Education to lease a portion of the no-longer-needed East Side School in Cold Spring Harbor, it is expected that all in-ventory will be transferred to this new warehouse by mid-summer of 1985.
Publications Completes First Full Year in Urey Cottage
The start of 1984 saw the Publications group happily installed in our attractivenew quarters. Specifically renovated to meet our growing needs, the function-oriented planning has more than lived up to expectations in providing the quie-tude and ease of communication needed for efficiently handling the myriad de-tails involved in editing and producing high-quality scientific books. This combi-nation of an environment conducive to productivity and a superb staff, bothknowledgeable and dedicated, should enable us to continue to increase thenumber of titles published over the next few years while maintaining present stafflevels.
Nancy Ford
25

RESEARCH


TUMOR VIRUSES
Thirteen years ago, the National Cancer Institute funded a major Program Proj-ect Grant to support work on tumor viruses at Cold Spring Harbor Laboratory.This grant is now in its third cycle and has allowed us to focus the efforts of amultidisciplinary team toward the study of adenoviruses, SV40, and papilloma vi-ruses. Because this work has been funded by the Program Project mechanism, ithas been characterized by extensive collaboration both within and between thevarious sections whose activities are described in the following pages.
In addition to the inherent challenge of understanding how they work, theseviruses have provided exciting model systems and tools for exploring the molec-ular biology of eukaryotic organisms in general. Thus, viral vectors have beendesigned that enable the overproduction of both viral and host proteins. Studiesof viral transformation are already leading into the more general phenomena ofgene regulation and cellular growth control. The adenovirus El A region and itsgene products are the subjects of intensive research efforts. Studies of RNAsplicing and viral enhancers continue while the role of the adenovirus VA RNAsfinally seems to be unraveling. In vitro systems that replicate adenovirus andSV40 DNAs have led to the identification of essential viral and host proteins.
Our long-time colleagues, Tom Broker and Louise Chow, have left Cold SpringHarbor Laboratory to take up appointments at the University of Rochester. Theyhave played key roles in many of our past projects and will be sorely missed.Likewise, Earl Ruley, who has moved to the Massachusetts Institute of Technol-ogy, has been a valued and productive colleague.
MOLECULAR BIOLOGY OF TUMOR VIRUSES
Y. Gluzman T. Adams N. Beck M. McKeeverT. Grodzicker R. Gerard N. Cap lin M. MerleE. Harlow J. Harper D. Chao J. MoomawW. Herr M. Manos R. Chisum S. RookH.E. Ruley K. Maruyama J. Clarke L. Sansome
M. Quinlan J. Cosetti C. SchleyK. Van Doren M. DeLuca L. van der WalP. Whyte M. Goodwin J. WigginsM. Yamada R. McGuirk N. WilliamsonB. Ahrens
The major emphasis of our research continues tobe the analysis of DNA tumor viruses, especiallySV40 and adenoviruses. A major effort is di-rected toward the analysis of the transforminggenes of these viruses. A wide variety of mutantsin these genes, viral vectors that overproducetransforming proteins as well as a large set ofmonoclonal antibodies directed against trans-
forming proteins, provide reagents used in a va-riety of studies. Thus, many projects are con-cerned with the role of viral transforming genesin the regulation of cellular and viral gene expres-sion, in the establishment and transformation ofprimary cells, in the interaction of viral trans-forming genes with each other and with other on-cogenes, and with the interaction of virus-en-
29

coded transforming proteins with other cellularproteins. Experiments on the cellular localiza-tion and modification of transforming proteinsare also being carried out. We are continuingstudies of controlling elements of viral early genesthat are involved in the regulation of transcrip-tion, mRNA processing and translation, and theinteraction of viral and cellular proteins withthese controlling elements. Work has continuedon the construction and analysis of viral vectorsthat has allowed us to produce, in mammaliancells, large amounts of eukaryotic proteins, in-cluding transforming proteins that can be used forpurification and biochemical studies.
Analysis of Wild-type and MutantAd2 E1A cDNAsH.E. Ruley, T. Grodzicker [in collaboration withE. Moran, B. Zerler, and M.B. Mathews]
The adenovirus early region IA (E1A) gene is re-quired for adenovirus-induced cell transforma-tion and is involved in the regulation of expres-sion of other adenovirus genes. To distinguish theindividual roles of the 13S, 12S, and 9S E1A geneproducts, we have isolated the correspondingcDNA clones and have recombined them intoboth plasmids and viruses (see Protein Synthesisin this section).
The 13S virus plaques equally well in human293 cells, He La cells, and A549 cells, a humanlung-oat cell carcinoma line that does not con-tain adenovirus DNA. Plaque titers of the 12Svirus are much reduced in He La and A549 cellsas compared with that in 293 cells. The 12S virusis multiplicity-dependent leaky in both He La andA549 cells. At low multiplicities of infection, theyield of 12S virus in He La cells is low; however,in A549 cells, even at low multiplicities of infec-tion, the yield of 12S virus increases with time toapproach the maximum yield from 293 cells.These results suggest that the A549 tumor cellline, even though it contains no adenovirus DNA,can partially complement the E1A defect of the12S virus so that transcription of other early ad-enovirus genes is activated. Immunofluorescencestudies indicate that the adenovirus DNA-bind-ing protein is made during infection of He La cells
30
with the 13S virus but is not detectable during in-fection with the 12S virus. Primary baby rat kid-ney (BRK) cells are immortalized by the 12S virusat very high efficiency, whereas infection of thesecells with 13S virus, as with wild-type El A virus,results mainly in cell death. Transfection with12S or 13S plasmids shows that both the 12S and13S plasmids can, like genomic El A, establishBRK cells or cooperate with plasmids carrying theras gene to transform these cells. In a transientexpression assay in HeLa cells, the 13S plasmidstimulates expression of the chloramphenicolacetyltransferase (cat) gene from the adenovirusE3 promoter, but the 12S plasmid does not. The12S plasmid appears to interfere with the stimu-lation of E3-cat expression by the 13S plasmid.To differentiate the functions of the E1A prod-ucts further, we have used oligonucleotide-di-rected mutagenesis to obtain several point muta-tions in the genomic E1A or cDNA clones. Apoint mutation at position 961, which converts aglutamic acid residue to lysine, appears to impairthe transformation function seen in the transfec-tion assay while not interfering with the gene-reg-ulating functions studied in the E3-cat transientexpression assay. We are presently transferringthe mutant E1A 12S and/or 13S regions into ad-enovirus so that their lytic and transformingproperties can be assessed. Several other pointmutations at nearby sites have also been made andare being analyzed.
Analysis of the Transforming Functionsof Adenovirus E1AP. Whyte, K. Maruyama, T. Grodzicker, H.E. Ruley[in collaboration with E. Moran and B. Zerler]
The polyoma virus middle T antigen, T24 Ha-ras-1, and adenovirus E1B genes are all individuallyunable to transform cultured primary BRK cells.Adenovirus E1A expresses functions that enablethese genes to transform. We have speculatedthat the oncogene cooperativity in transforma-tion is linked to the E1A functions which pro-mote the in vitro establishment of cultured pri-mary cells. To probe the function of E1A intransformation, plasmids containing the E1A re-gions from Ad5hrl and Ad5hr440, as well as

cDNAs for the 12S and 13S E1A mRNAs, weretransfected into primary BRK cells alone and to-gether with the polyoma middle T antigen andT24 Ha-ras-1 genes. The results obtained indi-cate that the establishment functions of E1A arelocated in the aminoterminal segment of the pro-tein.
To determine the minimal aminoterminal se-quences required for oncogene cooperation, a se-ries of deletion mutants have been constructed.A plasmid containing the E1A region was linear-ized at the Smal site, located within the 13Sunique coding region, and deletions were gener-ated by digesting with the exonuclease BAL-31.A linker encoding a termination codon was in-serted and the plasmids were recircularized, re-sulting in plasmids coding for truncated E1Aproteins. The extent of the deletions was deter-mined by sequencing using the dideoxy method.
The ability of these E1A mutants to cooperatein transformation was assayed by cotransfectingplasmids containing the E1A mutants and the T24Ha-ras-1 gene into primary BRK cells and scoringfor transformed foci.
We have determined the minimal aminoter-minal sequence of E1A necessary for oncogenecooperation. This region is common to both 12Sand 13S mRNAs and is different from the E1Afunctions necessary for transcriptional activationof the viral early-region genes. These mutants arecurrently being used to investigate the relation-ship between functions required for oncogene co-operation and in vitro establishment of primarycells.
Adenovirus E1A Blocks In VitroDifferentiation of Rat PC12 CellsK. Maruyama, H.E. Ruiey
Adenovirus E1A and genes related to the myc on-cogene of avian myelocytomatosis virus expressestablishment functions that circumvent the com-mitment of cultured primary cells to growth ar-rest and senescence. To investigate the role of es-tablishment functions in the regulation of cellproliferation further, we have introduced E1Aand myc genes into cell lines that can be induced
to differentiate in vitro and that experiencegrowth arrest in the process.
PC12 is a rat adrenal pheochromocytoma cellline that differentiates in response to both nerve-growth factor (NGF) and dibutyryl cyclic AMP(dcAMP) by extending neuronal processes and bycell growth arrest. Plasmids, containing E1A andmyc genes, linked to the bacterial neomycin phos-photransferase (neo) gene, were introduced intoPC12 cells, and G418-resistant clones were iso-lated. Clones were characterized for E1A andmyc expression and for the ability to differentiatein response to NGF and dcAMP. In the case ofE1A, clones expressing moderate-to-high levels ofE1A-related RNA failed to differentiate in re-sponse to even high levels of NGF and dcAMP,whereas clones expressing lower levels of E1A -re-lated RNA responded only weakly to NGF. Var-iation in the ability of myc-neo transfectants todifferentiate was also observed, and we are pres-ently measuring the levels of myc-related RNA tocorrelate myc expression with the inability to dif-ferentiate in vitro.
Cellular Immortality Is Not Sufficientfor Oncogenic Transformationby the T24 Ha-ras-1 GeneH.E. Ruley, R. Franza, J. Garrels
Although the functions required by the Ha-ras-1gene to transform primary cells have not beenseparated from functions that promote in vitroestablishment, we have recently demonstratedthat the REF52 cell line behaves like primary cellswith regard to transformation by the T24 Ha-ras-1 oncogene. Thus, the T24 Ha-ras-1 gene aloneis unable to transform REF52 cells, but the ad-enovirus E1A gene enables the T24 oncogene totransform. These results indicate that in vitro es-tablishment is not a sufficient prerequisite fortransformation of cultured cells by the T24 Ha-ras-1 gene and suggest that the role of E1A in pri-mary cell transformation extends beyond in vitroestablishment.
Initial evidence that the T24 Ha-ras gene is un-able to transform REF52 cells came from mi-croinjection studies. Microinjection of the T24oncogene plasmid induced neither transient nor
31

permanent morphological transformants. Incontrast, 1-2% of the cells microinjected by acombination of the T24 Ha-ras-1 and adenovirusE1A genes transiently acquired the rounded mor-phology of highly transformed cells, and approx-imately 0.1% of the microinjected cells grew pro-gressively as morphological transformants.
To probe the requirements for transformationof REF52 further, the T24 Ha-ras-1 and adeno-virus E1A genes were linked to the bacterial ami-noglycoside transferase gene (apt) from the Tn5transposon expressed from the SV40 early pro-moter (SVapt). Plasmids were transfected intoREF52 cells by the calcium phosphate coprecipi-tation method. Phenotypic transformation wasscored either by focus formation on cell mono-layers or following selection in G418. The follow-ing results were obtained: (1) The T24 Ha-ras andAd5E1A genes individually are unable to trans-form REF52 cells. Indeed, REF cells expressingthe T24 Ha-ras p21 protein (T24REF) have beenisolated. They are indistinguishable from REF52cells expressing the SVapt gene (REFneo) with re-gard to growth rate, saturation density, failure togrow in 1% serum, and failure to form tumors insyngeneic rats. Cell lines expressing the E1A gene(1AREF) have also been isolated. These cells aremorphologically altered but remain flat and aresimilar to REFneo cells with regard to growth rateand saturation densities. (2) Transformation ofREF52 cells, like that of primary cells, can re-quire at least two collaborating oncogenes. Thus,transformants arose only from cultures cotrans-fected with E1A and T24 Ha-ras-1 genes. Thetransformants grew to high saturation density(106/cm') and formed tumors in syngeneic rats;however, the tumors regressed within 5 weeks. (3)E1A does not collaborate with the T24 Ha-rasgene in transformation by enabling the cell toovercome some toxic consequence of introducingthe T24 Ha-ras-1 gene. Thus, neither E1A norT24 Ha-ras-1 was toxic to REF52 cells, as judgedby the fact that pKOneo (the SVapt gene alone),p 1 Aneo, and pT24neo plasmids induced G418-resistant colonies with similar frequencies.
T24REF cells differ from REFneo cells in atleast two respects. First, T24REF cells can betransformed by plasmids containing the Ad5E1Agene linked to the bacterial guanine phosphori-bosyl transferase gene expressed from the SV40early promoter (SV gpt), whereas REFneo cells
32
cannot be transformed by E1A. This result indi-cates that the T24 Ha-ras-1 gene and Ad5E1A cantransform REF52 cells in a stepwise manner.Second, T24REF cells require much lower serumconcentrations to escape growth arrest induced by1% serum. Over three times more serum was re-quired to induce 50% of the REFneo cells to en-ter S phase (within the labeling period) as com-pared with T24REF cells (9-2.5%); 1AREF cellswere intermediate in their responsiveness, requir-ing 5% serum. Experiments are in progress todetermine the physiological mechanisms under-lying this responsiveness. It is possible that con-stitutive expression of the E1A and T24 Ha-ras-1genes complements specific serum growth factorsin the mitogenic response of serum-arrested cells.Alternatively, the oncogene proteins may playsome role in growth-factor signal transductionand thereby enhance the responsiveness to certainserum growth factors.
Monoclonal Antibodies to theAdenovirus E1A ProteinsE. Harlow, C. Schley
Following infection of mammalian cells with ad-enovirus, the first virus-coded proteins that aresynthesized are the products of the E1A region.To help in the studies of the E1A proteins, wehave constructed a number of monoclonal anti-bodies specific for these polypeptides. A bacte-rial expression plasmid (kindly supplied by K.Spindler and A. Berk, University of California,Los Angeles), which will direct the synthesis of afusion protein between the bacterial tryptophanE gene product and a cDNA copy of the 13SmRNA, was used as a source to purify an E1A-related protein. This polypeptide was purified bySDS-PAGE, electroeluted, and used to immunizeBALB/c mice. Hybridomas were prepared andtissue-culture supernatants were screened for theproduction of antibodies that would bind to thefusion protein in a solid substrate radioimmu-noassay. Eighty-three hybridomas secreting anti-bodies specific for the fusion protein have beenisolated and single-cell-cloned. As demonstratedby indirect immunofluorescence, 29 of these hy-

bridomas secrete antibodies that will bind to E1Aproteins synthesized in He La cells infected withAd5 virus. However, of these 29 antibodies, only12 will efficiently immunoprecipitate the E1Aproteins from lytically infected or transformedhuman cells. Analysis of the E1A polypeptidesby immunoprecipitation and two-dimensionalIEF/PAGE has shown that the E1A products canbe resolved into approximately 60 polypeptidespecies (Fig. 1). We have used mutant viruses thatcan only synthesize portions of the E1A region tolocalize the binding sites of these antibodies. Allof the monoclonal antibodies will precipitate theproducts of the 13S mRNA, but two of the hybri-domas secrete antibodies that will not bind to the12S polypeptides. In addition to the authenticE1A proteins present in the immunoprecipitatesprepared from an adenovirus-transformed hu-man cell line (293 cells), there are several copre-cipitating proteins. These proteins are not im-
13S
12S
30 KFIGURE 1 Two-dimensional analysis of the adenovirus El Aproteins. The adenovirus E1A proteins from Ad5-infectedHeLa cells were immunoprecipitated with M73 hybridomatissue-culture supernatant and separated by two-dimen-sional IEF/PAGE. The region of the autoradiogram that con-tains the E1A polypeptides is shown. The autoradiogram hasbeen annotated to show the polypeptides that are encodedby the 12S and 13S mRNAs. The mRNAs that direct the syn-thesis of the 30K and 35K-40K polypeptides have not beenidentified.
Actin
munochemically related to the E1A polypeptidesand are precipitated from these cells because theyare stably bound to the E1A proteins. We arecurrently investigating the possible roles of thesevirus-host complexes.
Mutations in the Nuclear-envelope-associated Adenovirus E1B 19KTumor Antigen Cause the Degradationof Chromosomal DNAT. Grodzicker [in collaboration with B. Stillmanand E. White]
We reported last year that the adenovirus mutantAd2ts111 contains a mutation in the early region2A (E2A) gene that encodes the single-strandedDNA-binding protein (which results in thermo-labile replication of virus DNA) and a mutationin early-region 1 (El) that causes the degradationof intracellular DNA. A recombinant virus, Ad2-cyt106 has been constructed that contains theAd2ts111 El mutation and the wild-type E2A genefrom Ad5. This virus, like its parent Ad2ts111,has two phenotypes; first, it has the ability tocause enhanced and unusual cytopathic effects onthe host cell (cytocidal phenotype, Cyt) and, sec-ond, it induces degradation of cellular DNA(DNA degradation phenotype, Deg). We havemapped the mutation responsible for these phe-notypes into the early region 1B (E1B) by mark-er rescue, transferring the left-end adenovirusDNA fragments from ts111 to wild-type DNA andsequencing the relevant portion of ts111 E1BDNA.
The mutation responsible for these phenotypesis a single point mutation in the gene encodingthe adenovirus E1B 19K tumor antigen. Thismutation causes a change in the twentieth aminoacid from the amino terminus of the protein froma serine to an asparagine. Three other mutantsthat affect the ElB 19K protein function have alsobeen examined. The mutants Ad2/p5 and Ad5 -d/337 (from G. Chinnadurai [St. Louis UniversityMedical Center] and T. Shenk [SUNY, StonyBrook]) have both the cytocidal and DNA deg-radation phenotypes (Cyt and Deg), whereasAd2/p3 only has the cytocidal phenotype and doesnot induce degradation of cellular DNA (Cyt and
33

Deg - ). Thus, the DNA degradation is not causedby the altered cell morphology. Furthermore, themutant Ad5d1337 does not make any detectableE1B 19K protein product, suggesting that the ab-sence of E1B 19K protein function is responsiblefor the mutant phenotypes. A fully functionalE1B 19K protein is not absolutely required forlytic growth of Ad2 in HeLa cells. Although theseE1B mutants clearly play a role in affecting cellmorphology, the ts111 mutant, which has reducedcytotoxicity because of the mutation in the DNA-binding protein gene, can transform BRK cells atthe nonpermissive temperature.
Molecular Cloning and Expression ofthe Human p53 Tumor AntigenE. Harlow, N. Williamson, T. Adams
To study the biochemistry of the cellular tumorantigen p53, we have begun experiments designedto purify p53. Analysis of a number of cell linesand animal tissues that might serve as goodsources for purification has shown that these cellsdo not express high enough levels of p53 to alloweasy purification. Therefore, we have cloned acDNA copy of the p53 gene and plan to overex-press the gene for p53 in mammalian expressionvectors. A cDNA copy of the mRNA for the hu-man p53 from A431 cells was cloned in a bacterialplasmid vector, and the sequence of the largestinsert was determined by the dideoxy-sequencingmethod. Comparison of the predicted aminoacid sequence of this clone and the amino acidsequence predicted from similar clones preparedfrom mouse mRNA confirms the authenticity ofthis clone. This comparison has also shown thatthese two proteins have long regions of amino acidhomology, particularly in the aminoterminal 20amino acids and in the carboxyterminal 200amino acids. Preliminary results have shown thatthis cDNA sequence may be difficult to expressin mammalian cells in its present form. There-fore, we have isolated a number of subclones thathave deletions in both the 5'- and 3'-untranslatedregions, and we are studying the ability of thesesequences to direct the correct synthesis of hu-man p53.
34
Expression of p53-specific Transcriptsduring Mouse DevelopmentT.E. Adams
We have established the pattern of expression ofp53 during mouse embryogenesis and in adult tis-sues. Polyadenylated mRNA was prepared fromtotal RNA isolated from a variety of adult mousetissues and from embryos at various stages of de-velopment. p53-specific transcripts were identi-fied by Northern blot hybridization using a cDNAprobe derived from 3'-coding sequences of themouse gene. The pattern of expression obtainedwas in keeping with the concept of a role for p53in cellular proliferation; highest levels of p53transcripts were observed in embryos taken earlyin development and postnatally in hematopoieticand lymphopoietic tissues and in skin. Addition-al studies on chemically transformed cell lines,using both Northern hybridization and "sand-wich" radioimmunoassays, have indicated post-transcriptional regulation of the level of p53 inthese cell lines.
Transcriptional Modulation of GeneExpression by the SV40 Enhancer:Duplications in a Mutated EnhancerRestore Its ActivityW. Herr, Y. GIL _man, J. Clarke
The SV40 enhancer contains two 8-bp stretchesof alternating purines and pyrimidines that havebeen implicated in transcriptional activation(Nordheim and Rich, Nature 303: 674 [1983]).We investigated the role of these sequences in thefunction of the enhancer by mutating two nucleo-tides in each 8-bp segment such that the alterna-tion of purines and pyrimidines was destroyed.The relative potential of the wild-type and mu-tant enhancers to activate transcription wasmeasured by placing them upstream of the hu-man 6-globin gene, followed by measurement ofthe levels of properly initiated 6-globin mRNAafter transient expression in HeLa cells. In thisassay, the mutated enhancer is four- to sixfold lessactive than the wild-type enhancer. Furthermore,

SV40 carrying these four point mutations growsvery poorly in CV-1 cells.
Revertants with improved growth potentialcould be obtained after passage of mutant virusstocks in CV-1 cells. Analysis of the DNA from18 plaque-purified revertants showed that the en-hancer regions had rearranged. Each of these
251 178
Revertant
rd135
rd127
rd126
rd115
rdno
rd 97
rd 88
rd85
rd 71
rc166 . J
rd 65
r d 64 -1
rd 64 -2
rc162c1
rd62b
rd 53
rct51
rd45
721,p
ATGTG7GT GCATGC,
Duplicated Region
0/5243
=MCI 12111
E
L-
L -
L
TGTGGAAAGTCCCCA
FIGURE 2 Structure of revertants of an SV40 enhancermutant. At the top of the figure is a diagram of an SV40 con-trol region that contains only one copy of the 72-bp element.Shown from right to left are the early transcriptional start site(zigzag arrow), the origin of replication (on), the AT-rich TATA-like element (A/T), the two GC-rich 21-bp repeats, and thesingle 72-bp element. Hatched boxes indicate the locationof the two 8-bp stretches of alternating purines and pyrimi-dines; directly below these boxes are the sequences of eachof these stretches and the base substitutions found in eachof these segments in the dpm12 mutant. The series of rec-tangular boxes represents the extents of the tandemly dupli-cated region in each of the 18 independent revertants of theenhancer mutant. They are ordered from top to bottom bysize. The two series of double Xs identify the positions of theoriginal point mutations where duplicated. The stippled areawithin each rectangular box corresponds to the 15-bp re-gion that is common to all of the duplications; the sequenceof this region is shown at the bottom of the figure with abracket outlining the "core" element (Weiher et al., Science219: 626 [1983]).
rearranged enhancer elements was studied in theenhancer assay and exhibited increased transcrip-tional activation of the 0-globin promoter inHeLa cells. The nucleotide sequence of these re-vertants showed that each rearrangement was dueto tandem duplication of the enhancer region inwhich the mutations were preserved (see Fig. 2).Although the duplicated regions range from 45by to 135 by in length and the sequences con-tained in each duplication vary, there is a 15-bpsequence, TGTGGAAAGTCCCCA, that is con-sistently duplicated in each reactivated enhancer.This region contains the "core" sequence,GTGGAAAG, that was initially suggested (Laim-ins et al., Proc. Natl. Acad. Sci. 79: 6453 [1982])to be functionally significant, since related se-quences were identified in a variety of viral en-hancers. Our results suggest that the core andsurrounding sequences are a cis-acting elementthat can function independently of and compen-sate for the alternating purine and pyrimidine se-quences.
We have tested the hypothesis that the core ele-ment is important for enhancer function by mak-ing point mutations within this sequence and havefound that indeed these mutations are deleteriousto both SV40 growth and enhancer function. Weare presently analyzing the structure of revertantsof these core mutations to determine what alter-ations of the SV40 genome can compensate forthese new defects. Preliminary results indicatethat duplication of either one or the other of theregions containing the alternating purine and py-rimidine sequences is able to compensate for themutations in the core element. These results sug-gest that the SV40 enhancer is composed of mul-tiple independent cis-acting elements that are ableto compensate for one another. At present, wedo not know whether these independent cis-act-ing elements are interacting with the same or dif-ferent complementary trans-acting factors.
Diethylpyrocarbonate as a ChemicalProbe for DNA Secondary StructureW. Herr
Last year, Y. Gluzman and I reported a study inwhich we analyzed the significance of two 8-bp
35

alternating purine and pyrimidine sequences thatlie within the SV40 enhancer. These two se-quences had been suggested by Nordheim andRich (Nature 303: 674 [1983]) to have the potentialto form left-handed Z-DNA, and these authorsproposed that Z-DNA may be functionally im-portant for enhancer activity. We found thattransition mutations, which maintain the alter-nating patterns of purines and pyrimidines, hadlittle effect on the potential of the SV40 enhancerto activate transcription, whereas transversionmutations, which destroy the alternation of pur-ines and pyrimidines, had much greater deleteri-ous effects. These results were consistent with thepossibility that Z-DNA is involved in enhancerfunction, because alternating purine and pyrimi-dine sequences favor the formation of Z-DNA.However, analysis (with A. Nordheim and A.Rich) of the ability of the mutant DNAs to formZ-DNA using anti-Z-DNA antibodies has beeninconclusive because of the difficulty in identify-ing the precise DNA-binding site of the anti-bodies.
To identify more precisely sites of Z-DNA for-mation, I searched for a chemical probe thatwould discriminate between the B-DNA and Z-DNA secondary structures. As a test substrate, Iused the plasmid pLP32 (Peck et al., Proc. Natl.Acad. Sci. 79: 4560 [1982]) which carries a 32-bpstretch of alternating GC residues. This 32-bp se-quence is in the B conformation when the plas-mid DNA is relaxed, but upon negative supercoil-ing, it will flip to the Z conformation. I foundthat diethylpyrocarbonate (DEPC), which modi-fies the N7 position of adenines and gu'anines, ishyperreactive to the 32-bp alternating GC se-quence in the Z conformation, compared with thesame sequence in the B structure. The compara-tive reactivity of DEPC to specific nucleotideswas displayed as follows: Linear and negativelysupercoiled pLP32 DNAs were partially modifiedwith DEPC, digested with a restriction enzyme,and end-labeled, and the strands of singly end-labeled molecules were cleaved as described byMaxam and Gilbert (Proc. Natl. Acad. Sci. 74:560 [1977]).
Reactivity of DEPC to natural sequences wasanalyzed by examining the modification patternsof a region of the plasmid pBR322 that has beenshown to bind anti-Z-DNA antibodies when theplasmid is negatively supercoiled and contains a
36
9-bp stretch of alternating purines and pyrimi-dines. pBR322 plasmid DNAs carrying increasingdensities of negative supercoils were modifiedwith DEPC. As the supercoil density increased,DEPC exhibited hyperreactivity to the purineswithin the 9-bp alternating purine and pyrimidinesequence. Purines flanking the alternating purineand pyrimidine sequence became hyperreactive athigher supercoil densities. These results suggestthat Z-DNA is favored within the alternating pur-ine and pyrimidine sequence and then spreads tothe adjacent nonalternating purine and pyrimi-dine sequences as the supercoil density increases.
I am currently applying this technique to studythe Z-DNA potential of the SV40 enhancer re-gion. In preliminary experiments, I have not de-tected any differences in the reactivity of DEPCto the enhancer and early promoter regions of re-laxed and negatively supercoiled SV40 DNAs.These results indicate a potential lack of correla-tion between the binding to DNA of anti-Z-DNAantibodies and the hyperreactivity of DEPC. Thisenigma will hopefully be explained by further in-vestigation.
SV40 Mutants That Differentiate theLytic and Transforming Functionsof Large T AntigenM. Manos, Y. Gluzman
We have continued the analysis of DNA replica-tion-defective, but transformation-competent,SV40 mutants such as C2, C6, and C11 and haveisolated and analyzed two more such mutants(C8/SV40 and T22/SV40). These mutants wereisolated from the C8 and T22 simian cell linesthat were transformed with UV-irradiated SV40.Viral DNA was rescued by fusion of these lineswith COS-1 cells, which provide a functional Tantigen in trans and therefore allow mutant inser-tions to excise and replicate in the heterokaryons.The rescued SV40 inserts were cloned into theplasmid vector pK1, restoring a complete SV40genome containing the early region (BglI- BamHI)from the mutants and a wild-type late region.These DNAs were analyzed by marker-rescue andDNA sequencing. The mutants contain lesions indifferent parts of T antigen that render the pro-

tein defective for viral DNA replication but effi-cient in transforming cells in vitro. The muta-tions responsible for the defects in viral DNAreplication are His->Glu at residue 203 (T22) andLys->G1u at residue 224 (C8). There is an addi-tional mutation in the C8 mutant that convertsglutamine (amino acid 660) to a stop codon;therefore, C8 T antigen lacks the 49 carboxyter-minal amino acids. These two mutations were sep-arated to form mutants pC8A and pC8B. Allmutants, pC8, pC8A, pC8B, and pT22, are com-parable to the recombinant wild-type SV40 DNAin transforming primary mouse embryo fibro-blasts or primary BRK cells. Whereas pC8, pC-8A, and pT22 are completely defective in viralDNA replication, pC8B is able to replicate viralDNA but only at 10% wild-type efficiency. Al-though C8B DNA replication occurs with thesame efficiency in both CV-1 and BSC-1 cells, vi-rus only grows on BSC-1 cells. Thus, truncationof 49 amino acids from the carboxyl terminus ofT antigen results in a host-range mutant pheno-type. Analysis of the origin-binding activity ofmutant T antigen reveals that C8, C8A and C8BT antigens bind to SV40 origin as efficiently aswild-type T antigen, but T22 T antigen is com-pletely defective in the origin-binding assay. Allmutants are positive in ATPase assay.
All of the replication-defective T antigen mu-tants we have so far analyzed have been clonedinto an adenovirus expression vector, and largequantities of mutant T antigen have been pro-duced. In collaboration with C. Prives (Colum-bia University), the biochemical properties ofthese mutants are being further analyzed withgreater scrutiny.
Permissive Cell Lines That InduciblyExpress SV40 AntigenR.D. Gerard, Y. Gluzman
We wanted to generate cell lines in which the rep-lication of transfected SV40 origin sequencescould be controlled. One way to accomplish thisis to replace the SV40 early promoter with themouse methallothionein promoter to renderexpression of T antigen inducible by heavy met-als. Transformation of permissive monkey cells
with this chimeric gene has yielded cell lines(CVMT and BSCMT) that synthesize three- totenfold more T antigen upon induction. Unin-duced levels of T antigen are comparable to thosepresent in COS cells. Withdrawal of these celllines from inducer results in a rapid decay in therate of T-antigen synthesis, with estimates of thehalf-time ranging from 3 hours to 5 hours, de-pending on the clone. In addition, pulse-chaseexperiments indicate that T antigen is stable, witha half-life (20-30 hr) comparable to that in COScells.
The rate of replication of transfected SV40 onsequences rapidly slows after heavy metals arewithdrawn even though T antigen is stable. Therate of DNA replication decays with a half-timeof 5-8 hours and parallels the decay of the rate ofsynthesis of T antigen. One explanation of thisphenomenon is that continual synthesis of T an-tigen is required for DNA synthesis because onlynewly synthesized T antigen will support DNAreplication. Further analysis of SV40 on repli-cation in CVMT and BSCMT cell lines indicatesthat induction of T-antigen synthesis by heavymetals results in a 20-100-fold greater accumula-tion of replicated DNA than obtained under un-induced conditions after 48 hours. COS cellsreplicate DNA to levels comparable to those pres-ent in uninduced CVMT and BSCMT cells.
To examine the ability of these cell lines tomaintain SV40 on sequences in a episomal state,we have transformed them with the neo gene car-ried on an SV40 on plasmid (pON3). Low-mo-lecular-weight DNA was isolated from G418-re-sistant cultures and analyzed by both Southernblotting and plasmid rescue. The results demon-strate the persistence of pON3 DNA as an epi-some and an increase of 20-100-fold in the copynumber of these sequences upon induction of T-antigen synthesis by heavy metals. Although sig-nificant rearrangement of the DNA sequencessometimes occurs, the majority of these altera-tions occur within plasmid sequences presumablybecause alterations in the SV40 on and neo struc-tural genes are selected against.
Transformation of CVMT and BSCMT cellswith a similar DNA construct containing the neogene under SV40 early promoter control, theSV40 ori, and the influenza virus hemagglutinin(HA) gene under SV40 late promoter control(provided by M.-J. Gething, Molecular Genetics
37

of Eukaryotic Cells Section) (neoHA DNA) hasalso given rise to G418-resistant clones. Analysisof low-molecular-weight DNA by Southern blot-ting demonstrates the persistence of intact neoHADNA and a 10-100-fold amplification of these se-quences after induction of T-antigen synthesis byheavy metals for 48 hours. Synthesis of the HAprotein is easily assayed by immunoprecipitationand gel electrophoresis and shows a 3-15-fold in-crease above control levels upon heavy metal in-duction.
Although we do not know if chromosomal in-tegration of these sequences has occurred, theseobservations suggest that CVMT and BSCMT celllines may be a useful system for maintainingcloned genes in an amplifiable episomal state inmammalian cells.
Monoclonal Antibodies Specific forSV40 Nucleoprotein ComplexesR. Gerard, E. Harlow
During the lytic cycle of SV40 infection, the viralDNA can be recovered from the nucleus as a nu-cleoprotein complex. These complexes are het-erogeneous in nature and include DNA that isbeing actively replicated and transcribed. Con-sequently, these complexes should contain a vari-ety of eukaryotic proteins that are involved inreplication and transcription. We have begun ananalysis of the immunogenic proteins in thesecomplexes.
SV40 nucleoprotein complexes (NPCs) wereisolated from virus-infected CV-1 monkey cellsand were purified by sucrose gradient centrifu-gation. After the complexes were fixed by formal-dehyde treatment, they were used to immunizea BALB/c mouse. Hybridomas were constructed,and tissue-culture supernatants were screened forantibodies specific for NPCs by indirect immu-nofluorescence on SV40-infected CV-1 monolay-ers. Of the 384 cultures examined, 36 hybrido-mas produced antibodies recognizing antigenspresent in SV40-infected cells. Among these wereantibodies specific for the SV40-coded large Tantigen and viral capsid proteins. Another hybri-doma in this series produces antibodies that pre-cipitate NPCs that have been pulse-labeled with
38
[3II]thymidine. This antibody will immunopre-cipitate a 70K phosphoprotein from infected ormock-infected cells, and we are currently at-tempting to identify this antigen.
An Adenovirus Vector That Expressesthe Protein Product of a NonselectedForeign Gene at a High Level:Synthesis of HSV-1 Thymidine Kinaseand Human a-Chorionic GonadotropinM. Yamada, T. Grodzicker
We have been developing an adenovirus vectorsystem that can express foreign gene products toa high level in mammalian cells. We have shownpreviously that SV40 T antigen is produced at thehighest levels when its coding sequences areplaced adjacent to the third segment of the aden-ovirus tripartite leader. We have also shown thatdefective adenovirus-SV40 recombinants express-ing SV40 large T antigen can be selected by virtueof the large T helper function that allows adeno-virus to grow on normally nonpermissive monkeycells. To insert other foreign genes into the thirdleader of the adenovirus genome, we used a com-bination of in vivo and in vitro recombination,followed by the selection of hybrid viruses thatalso express the helper function of SV40 T anti-gen (Thummel et al., Cell 33: 455 [1983]). We in-serted the thymidine kinase (tk) gene of herpessimplex virus I (HSV), the genomic sequencescoding for human a-chorionic gonadotropin (a-HCG), or its cDNA form at the third segment ofthe tripartite leader. These viruses also containthe SV40 early promoter and T-antigen-codingregion downstream from the tk or a-HCG genes.
Human or monkey cells infected by one of theadeno-tk hybrid viruses (AdTkSVR591) producedin the late stage of infection (1) an abundant TkmRNA with the length expected for a moleculecontaining both the tripartite leader sequence andtk sequences, (2) a 45K protein that reacted withanti-Tk antisera and with the same molecularweight as that found in HSV-infected cells (seeFig. 3), (3) a Tk protein that was synthesized tohigh levels and represented the third most heavilylabeled protein in the cells, after hexon and fiber,and (4) severalfold higher Tk activity than isfound in HSV-infected cells.
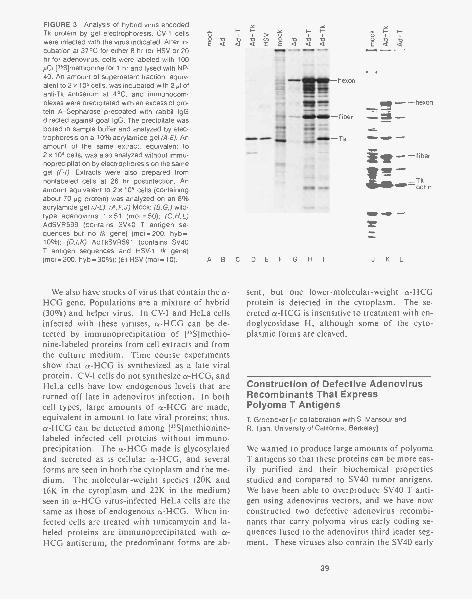
FIGURE 3 Analysis of hybrid-virus-encodedTk protein by gel electrophoresis. CV-1 cellswere infected with the virus indicated. After in-cubation at 37°C for either 8 hr for HSV or 20hr for adenovirus, cells were labeled with 100
[35S]methionine for 1 hr and lysed with NP-40. An amount of supernatant fraction, equiv-alent to 2 x 105 cells, was incubated with 2 pl ofanti-Tk antiserum at 4°C, and immunocom-plexes were precipitated with an excess of pro-tein A-Sepharose precoated with rabbit IgGdirected against goat IgG. The precipitate wasboiled in sample buffer and analyzed by elec-trophoresis on a 10% acrylamide gel (A-E). Anamount of the same extract, equivalent to2 x 104 cells, was also analyzed without immu-noprecipitation by electrophoresis on the samegel (F-1). Extracts were also prepared fromnonlabeled cells at 26 hr postinfection. Anamount equivalent to 2 x 105 cells (containingabout 70 mg protein) was analyzed on an 8%acrylamide gel (J-L). (A,F,J) Mock; (B,G,) wild-type adenovirus 1 x51 (moi = 50); (C,H,L)AdSVR599 (contains SV40 T antigen se-quences but no tk gene) (moi= 200, hyb =10%); (0,1,K) AdTkSVR591 (contains SV40T antigen sequences and HSV-1 tk gene),(moi = 200, hyb = 30%); (E) HSV (moi = 10).
.<3
ABC DEF G HI
We also have stocks of virus that contain the a-HCG gene. Populations are a mixture of hybrid(30°7o) and helper virus. In CV-1 and HeLa cellsinfected with these viruses, a-HCG can be de-tected by immunoprecipitation of ["S]methio-nine-labeled proteins from cell extracts and fromthe culture medium. Time course experimentsshow that a-HCG is synthesized as a late viralprotein. CV-1 cells do not synthesize cx-HCG, andHeLa cells have low endogenous levels that areturned off late in adenovirus infection. In bothcell types, large amounts of a-HCG are made,equivalent in amount to late viral proteins; thus,a-HCG can be detected among ['SS]methionine-labeled infected cell proteins without immuno-precipitation. The a-HCG made is glycosylatedand secreted as is cellular a-HCG, and severalforms are seen in both the cytoplasm and the me-dium. The molecular-weight species (20K and16K in the cytoplasm and 22K in the medium)seen in a-HCG virus-infected HeLa cells are thesame as those of endogenous a-HCG. When in-fected cells are treated with tunicamycin and la-beled proteins are immunoprecipitated with a-HCG antiserum, the predominant forms are ab-
J K L
sent, but one lower-molecular-weight a-HCGprotein is detected in the cytoplasm. The se-creted a-HCG is insensitive to treatment with en-doglycosidase H, although some of the cyto-plasmic forms are cleaved.
Construction of Defective AdenovirusRecombinants That ExpressPolyoma T AntigensT. Grodzicker [in collaboration with S. Mansour andR. Tjian, University of California, Berkeley]
We wanted to produce large amounts of polyomaT antigens so that these proteins can be more eas-ily purified and their biochemical propertiesstudied and compared to SV40 tumor antigens.We have been able to overproduce SV40 T anti-gen using adenovirus vectors, and we have nowconstructed two defective adenovirus recombi-nants that carry polyoma virus early coding se-quences fused to the adenovirus third leader seg-ment. These viruses also contain the SV40 early
39

promoter and T-antigen-coding region to allowhelper function selection for recombinants.
One virus, RAdPySVR498, carries the entirepolyoma early coding region. This virus expres-ses proteins that are immunoprecipitable with anti-polyoma tumor serum and that comigrate withthe authentic polyoma T antigens on SDS-poly-acrylamide gels. The hybrid virus, AdPySVR-498, overproduces both middle T antigen (5-fold)and small T antigen (20-fold) in infected cells.Small T antigen can be detected among total[35S]methionine- labeled infected cell proteinswithout immunoprecipitation.
S1- nuclease analysis of the hybrid-virus-en-coded mRNAs suggests that the polyoma splicingsignals are being correctly utilized and that themRNA levels reflect the levels of the three T-an-tigen proteins. In addition, at least the third ad-enovirus leader segment is present at the 5' end ofthe mRNAs. It should be noted that, althoughall three polyoma T antigens are synthesized inAdPySVR498-infected human or monkey cells,the ratios between the proteins are different fromthose found in polyoma-infected mouse cells.However, these lower levels of large T antigen rel-ative to levels of middle and small T antigens cor-respond to the amounts of the respective mRNAsproduced by the hybrid virus. This suggests thatthe donor splice sites for large T antigen versusmiddle- and small-T-antigen mRNAs are usedwith different efficiencies in adenovirus-infectedcells. A similar situation is found with adenovi-rus-5V40 hybrid virus-infected monkey cells,where the donor splice sites for SV40 large andsmall T antigens are differentially utilized (Thum-mel et al., Cell 23: 825 [1981]).
The second virus, RAdLTSVR545, was con-structed to carry a cDNA coding for the large Tantigen of polyoma virus rather than the entireearly coding region. Immunoprecipitation of pro-teins late in an RAdLTSVP infection shows afivefold overproduction of 100K polyoma large Tantigen. S1- nuclease analysis of RAdLTSVR545-encoded mRNA shows that the large T messagecontains the adenovirus third leader at its 5' end.Using a fragment immunoprecipitation assay, weshowed that the RAdLTSVP large T antigen bindsspecifically to the polyoma noncoding region, i.e.,to the region around the origin of viral DNA rep-lication and the early and late promoters. Inde-
40
pendent binding to fragments on both sides of theBgll site was observed.
We are currently attempting to purify suffi-cient amounts of polyoma large T antigen fromRAdLTSVR545-infected HeLa spinner cells tocarry out detailed DNA-binding studies and todetermine whether this protein will repress tran-scription from the polyoma early promoter in vi-tro. The hybrid virus, AdLTSVR545, containspBR322 sequences downstream from the poly-oma insert and represents less than 5% of the vi-ral population. Thus, it is difficult to assess themaximum amount of polyoma large T antigenthat could be produced. We do know that recom-binant viruses containing the genes encodingHSV-1 Tk or cx-HCG that constitute 10-20% ofthe viral population make very high levels of theseproteins (Yamada and Grodzicker, see above).
Helper-free Ad5 Vectors: Expressionof Complete or Truncated SV40 TAntigens by Recombinant Virusesduring Lytic InfectionJ. Harper, Y. Gluzman
As we reported last year, large amounts of SV40T antigen are synthesized late after infection ofhuman 293 cells with recombinant viruses car-rying SV40-T-coding sequences. The largestamount is produced in cells infected with the vi-rus containing the Ad2 MLP with 2-1/2 leadersegments. Threefold less T antigen is producedin cells infected by viruses containing the Ad2MLP with only half of the first late leader seg-ment. Analysis of T-antigen-specific mRNAs ex-tracted late after infection of 293 cells by the re-combinant viruses revealed that the amount ofthese messages correlated with the amount ofprotein produced in these cells. These data dem-onstrate that under certain (as yet unknown) con-ditions, the mRNA with 2-1/2 leaders is nottranslated more efficiently than mRNA with onlyhalf of the first late leader segment. In additionto authentic T antigen, smaller polypeptides rec-ognized by the large-T-specific monoclonal anti-body PAb416 were seen in cells infected by virusescarrying the SV40 insert in the left-to-right ori-

entation. The truncated T antigens produced byviruses containing the Ad2 MLP + 2-1/2 leadersegments appear to be 26K, and T antigens pro-duced by viruses containing the Ad2 MLP + 1/2leader segment appear to be 20K. The correla-tion between the orientation of the SV40 insert inthe vector and synthesis of truncated T-antigenpolypeptides suggested that the mRNA for theseproteins might be produced by aberrant splicingof the SV40 RNA. Several examples of aberrantsplicing of the RNA encoded by genes insertedinto adenovirus have been reported (Thummel etal., J. Mol. Appl. Genet. 1: 435 [1982]; Berknerand Sharp, Nucleic Acids Res. 10: 1925 [1984];Westphal, J. Virol. 40: 526 [1981]). We have ana-lyzed the T-antigen-related polypeptides synthe-sized in rabbit reticulocyte lysates primed withRNA from cells infected by various recombinantviruses. The occurrence, size, and relativeamounts of truncated T antigens synthesized invitro were similar to those of the in vivo products.These data suggest that these proteins are pri-mary translation products of RNA molecules spe-cific to viruses that carry the SV40 insert in a left-to-right orientation. Both the 20K and 26K trun-cated T antigens contain the aminoterminal re-gion of T antigen, since they are immunoprecipi-tated by monoclonal antibodies PAb430 (whichrecognizes both small T and large T antigens) andPAb416 (which is specific for a large-T-antigendeterminant located within the aminoterminal20% of the molecule), but are not immunoprecip-itated by a mixture of carboxyterminal-specific T-antigen monoclonal antibodies (PAb414, PAb431,and PAb405).
S1- nuclease analysis of corresponding mRNAsindicates that truncated RNA is produced by atleast two splicing reactions: removal of the nor-mal large T intron and an additional splice usinga donor in the second exon approximately 150 nu-cleotides downstream from the normal splice ac-ceptor. A good splice donor consensus sequenceappears in the SV40 DNA sequence at approxi-mately this position (nucleotide 4424). Use of asplice donor in this region has been reported forone of the Ad2 'ND, RNA species (Westphal, J.Virol. 40: 526 [1981]).
Our data show no obvious differences betweenRNAs encoding the 26K and 20K T antigens inthe SV40 HindIIIB region. Differences in the
protein-coding capacity of these RNAs may bedue to small differences in splice sites that are notdetected by this method, the use of differentsplice sites in the SV40 sequences 3' to the regionprobed, or the use of different splice acceptors inthe adenovirus E1B sequences. SI-nuclease ex-periments are in progress to examine the latterpossibilities.
Integration and Stable Expression ofForeign Genes Introduced by HelperIndependent Adenovirus Vectorsin a Variety of Mammalian CellsK. Van Doren
Last year, we reported that recombinant adeno-viruses containing a selectable marker for neo-mycin resistance in El will integrate into the host-cell genome. The data obtained from analysis ofthe integrated viral DNA indicate that, in gen-eral, the viral DNA is present as almost full-lengthlinear genomes. These observations suggest eitherthat the viral DNA is preferentially integratedfrom the ends of the genome or that this occursdue to a positive selective pressure created by hav-ing the selectable gene at one end of the viralchromosome. Recombinant viruses in which theselectable marker for neomycin resistance re-places early region 3 (E3) of the adenovirus vec-tor were used to address this point. These recom-binant viruses contain the selectable marker farfrom the ends of the viral genome. Because theselectable marker is not located at the end of theviral chromosome, there should be no positive se-lective pressure exerted for the integration of theends of the viral genome into the host-cell chro-mosomes. Recombinant viruses containing theneomycin resistance gene in E3 transformed CV-1 cells to G418 resistance as efficiently as recom-binant viruses containing the neomycin resistancegene in El. Preliminary analysis of the viral DNAcontained in cell lines derived from these virusessuggests that the pattern of integration is verysimilar to that found for El replacement viruses.This indicates that integration occurs preferen-tially from the ends of the adenovirus genome.
Previously, we reported that a recombinant vi-
41

rus containing the SV40 early region with a defec-tive origin of replication, located in E1, would ef-ficiently transform human fibroblasts and thatthe pattern of integrated DNA was identical tothat reported for the neomycin recombinant vi-ruses. Because human fibroblasts are semiper-missive for SV40 DNA replication, we were un-able to assess the state of the integrated DNA incells transformed by recombinant viruses con-taining an SV40 functional origin of replicationdue to the presence of large amounts of freelyreplicating DNA. Upon subcloning of the paren-tal transformed cell lines, we observed that two ofthe three subcloned cell lines had lost the abilityto produce freely replicating SV40 DNA mole-cules. The subclones contained T antigen as de-tected by immunofluorescence, indicating thatthey had not lost the SV40 DNA. It was possiblethat a mutation occurred in a cellular permissiv-ity factor or in the viral T antigen so that it couldno longer function in DNA replication. We fusedthe subclones to either COS-1 or CV-1 cells andfound that the T antigen in the human cells hadmutated so that it no longer supported SV40 rep-lication, which suggests that there is a strong se-lective pressure against having a functional SV40replication system in human cells. The pattern ofintegrated viral DNA in these cells was found tobe identical to that observed for human cellstransformed by recombinant viruses containing adefective SV40 origin of replication.
We are currently characterizing the use of adouble-replacement vector in which foreign genescan be cloned into both El and E3. This vectorwill make it possible to introduce nonselectablegenes into mammalian cells by using a selectablemarker in the second replacement site. Doublerecombinant viruses containing the human )3 -glo-bin gene with a selectable marker for either neo-mycin resistance or methotrexate resistance(DHFR) are being constructed and analyzed forthe ability of the viruses to introduce the globingene into a variety of cells. The feasibility of us-ing the DHFR gene as a method to amplify theintegrated viral DNA will also be explored.
PUBLICATIONS
Hanahan, D. and Y. Gluzman. 1984. Rescue of functionalreplication origins from embedded configurations in a
42
plasmid carrying the adenovirus genome. Mol. Cell. Biol.4: 302-309.
Herr, W. and Y. Gluzman. 1985. Duplications of a mutatedSV40 enhancer restore its activity. Nature 313: 711-714.
Karlsson, S., R.K. Humphries, Y. Gluzman, and A.W. Nien-huis. 1985. Transfer of genes into hematopoietic cellsusing recombinant DNA viruses. Proc. Natl. Acad. Sci.82: 158-162.
Klessig, D.F., T. Grodzicker, and V. Cleghon. 1984. Con-struction of human cell lines that contain and express theadenovirus DNA binding protein gene by cotransforma-tion with the HSV-1 tk gene. Virus Res. 1: 169-188.
Manos, M. and Y. Gluzman. 1984. Simian virus 40 large Tantigen point mutants that are defective in viral DNA rep-lication, but competent in oncogenic transformation. Mol.Cell. Biol. 4: 1125-1133.
. 1985. Genetic and biochemical analysis of transfor-mation-competent, replication-defective SV40 large Tantigen. J. Virol. 53: 120-127.
Mansour, S., T. Grodzicker, and R. Tijian. 1985. An adeno-virus vector system used to express polyoma virus tumorantigens. Proc. Natl. Acad. Sci. 82: 1359-1363.
Ruley, H.E., J.F. Moomaw, and K. Maruyama. 1984. Avianmyelocytomatosis virus myc and adenovirus early region1 A promote the in vitro establishment of cultured primarycells. Cancer Cells 2: 481-486.
Ruley, H.E., J. Moomaw, C. Chang, J.I. Carrels, M. Furth,and B.R. Franza. 1985. Multistep transformation of an es-tablished cell line by the adenovirus E1A and T24 Ha-res -1 genes. Cancer Cells 3: 257-264.
Stillman, B., F. White, and T. Grodzicker. 1984. Independentmutations in Ad2ts111 cause degradation of cellular DNAand defect viral DNA synthesis. J. Virol. 50: 598-605.
Van Doren, K. and Y. Gluzman. 1984. Efficient transforma-tion of human fibroblasts by adenovirus-SV40 recombi-nant viruses. Mol. Cell. Biol. 4: 1653-1656.
Van Doren, K., D. Hanahan, and Y. Gluzman. 1984. Infectionof eukaryotic cells by helper-independent recombinantadenoviruses: Early region 1 is not obligatory for integra-tion of viral DNA. J. Virol. 50: 606-614.
White, E., T Grodzicker, and B. Stillman. 1984. Mutations inthe gene encoding the adenovirus early region 1B19,000-molecular-weight tumor antigen cause the deg-radation of chromosomal DNA. J. Virol. 52: 410-419.
In Press, Submitted, and In Preparation
Yamada, M., J. Lewis, and T Grodzicker. 1985. Overpro-duction of the protein product of a nonselected foreigngene using an adenovirus vector. Proc. Natl. Acad. Sci.(in press).

DNA SYNTHESIS
S.W. Stillman P. Cotton G. PrelichJ. Diffley B. FahaE. White S. Penzi
Over the past 5 years, we have concentrated ourstudies on the biochemical analysis of eukaryoticDNA replication using adenovirus as a model sys-tem. This work is now at a stage where it is pos-sible to replicate completely adenovirus DNA invitro with purified proteins. Three of these pro-teins are encoded by the virus and two are pro-duced by the host cell. Thus, despite the atypicalmechanism of adenovirus DNA replication, twocellular proteins required for adenovirus DNAreplication have been identified, and it is possiblethat these proteins function in important cellularprocesses such as DNA replication. However, be-cause adenovirus replicates its DNA differentlyfrom the way a cell replicates its chromosomalDNA, we have turned to the biochemical analysisof DNA replication of another DNA tumor virus,SV40, which provides an excellent model forchromOsomal replication in cells. During thisyear, we have developed a cell-free system for thereplication of SV40 DNA. Since the replicationof SV40 DNA and that of cellular DNA appearto be similar, we expect that characterization ofthis system will provide valuable insight into themechanism of initiation, elongation, and termi-nation of eukaryotic DNA replication and ulti-mately enable a molecular analysis of the controlof these processes.
In addition to our studies of DNA replication,we have continued to probe the biochemical func-tions of the adenovirus tumor antigens. One ofthese proteins has been shown to be localized tothe nuclear envelope of infected and transformedcells, and mutations in the protein induce the deg-radation of cellular DNA. We have now foundthat this DNA degradation is specific for the typeof human cell that is infected by the mutants.
Adenovirus DNA ReplicationJ. Diffley, G. Prelich, B. Stillman
Two cellular proteins are required for the com-plete replication of adenovirus DNA in vitro.
One of these proteins, nuclear factor II, containsa type I topoisomerase activity and is required forcomplete elongation of nascent DNA strands.The other protein, nuclear factor I, is a site-spe-cific DNA-binding protein that binds to se-quences within the origin of adenovirus DNAreplication, and this binding is required for effi-cient initiation of replication. It is known thatthe protein can bind specifically to cellular DNAsequences that are present at approximately 1 inevery 100,000 base pairs in the human cell ge-nome (Gronostajski et al., Proc. Natl. Acad. Sci.81: 4013 [19801), but the function of the proteinin uninfected cells is not known. At present, theonly assay for this protein for purification pur-poses is its ability to stimulate adenovirus DNAreplication in the presence of the three virus-en-coded proteins, the single-stranded DNA-bindingprotein (DBP), the adenovirus DNA polymerase(Adpol), and the adenovirus precursor terminalprotein (pTP). However, this assay requires thatthese three proteins be substantially pure prior topurification of nuclear factor I. We have there-fore developed a simple, more rapid assay for nu-clear factor I that can detect the functional pro-tein in crude extracts of human cells and isindependent of its role in initiation of adenovirusDNA synthesis.
The assay for nuclear factor I utilizes its spe-cific DNA-binding properties. Briefly, DNAcontaining the binding site for nuclear factor I islabeled at each terminus with 32P and mixed withcrude extracts derived from the nuclei of HeLacells and a mixture of synthetic homopolymers ascompetitor DNA. The binding of nuclear factorI to the "P-labeled DNA is assayed by a simplenitrocellulose-filter-binding method, and the re-tention of labeled-DNA-protein complexes on thenitrocellulose is detected by autoradiography orscintillation counting. The assay is dependent ontime and temperature, the presence of Mgr ,
and the concentration of NaC1 in the reactionmixture. The competitor DNA is an equal mix-ture of the synthetic homopolymers poly(dA):
43

oligo(dT) and poly(dC):oligo(dG) and must beadded to the nuclear extract prior to the additionof labeled DNA. It is preferable that the labeledDNA be as short as possible, and we routinely usefragments in the 35-200-bp range. As a controlfor nonspecific binding to DNA of proteins pres-ent in the crude extract, a labeled DNA fragmentthat does not contain the binding site for nuclearfactor I is used, usually a DNA fragment contain-ing a mutated binding site. The presence of nu-clear factor I in the extract is observed when DNAcontaining the binding site is preferentially re-tained on nitrocellulose, but the control DNAfragment fails to bind. Using this assay, we havepurified nuclear factor I from He La cells, and theactivity coelutes at every purification step with thereconstitution of adenovirus DNA replication ac-tivity of the protein. We are currently concen-trating our efforts on determining the normal cel-lular function of the nuclear-factor-I protein.
This assay for site-specific DNA-binding pro-teins in extracts prepared from eukaryotic cells isgenerally applicable to other proteins. Site-spe-cific DNA-binding proteins have been shown toregulate transcription of certain genes in eukary-otes (Dynan and Tjian, Cell 35: 79 [1983]; Parkerand Topol, Cell 36: 357 [1984]) and may be im-portant in other cellular processes such as DNAreplication and cellular differentiation. Using therapid filter-binding assay, we have detected incrude extracts from HeLa cells a protein thatbinds specifically to the 21-bp repeat region of theSV40 early promoter, and it is most likely thatthis protein corresponds to the transcription fac-tor SP1 required for transcription of the SV40early promoter in vitro (Dynan and Tjian, Cell35: 79 [1983]). This will provide a rapid assay forthe purification of this protein and identificationof its molecular weight and any associated enzy-matic activities.
In addition to biochemical analysis of thesecellular encoded proteins, structural and func-tional studies on the adenovirus DBP are contin-uing. This protein is required for elongation ofadenovirus DNA replication, and, in addition, itis involved in the control of early mRNA synthe-sis and late gene expression. We have now se-quenced the mutation in the DBP from the mu-tant Ad2ts111 and have found that it lies in themiddle of the protein sequence, away from theother well-characterized DBP temperature-sensi-
44
tive mutation in Ad5ts125. The Ad2ts111 muta-tion lies adjacent to another mutation present inAd5ts23 (J. Williams, pers. comm.); we have ob-tained this virus and are currently comparing thein vitro biochemical properties of the mutant andwild-type proteins. These studies should comple-ment the structural studies on DBP by Tsern-oglou et al. (J. Mol. Biol. 172: 237 [1984]), whohave crystallized a fragment of the DBP and arecurrently determining the three-dimensionalstructure of the protein.
SV40 DNA ReplicationB. Stillman
The tumor virus SV40 contains a double-stranded, circular DNA genome of 5243 by andreplicates efficiently in monkey cells. DNA rep-lication can also be detected in some, but not all,human cell lines but does not occur in mouse cells.The viral DNA is part of a nucleoprotein complexthat is very similar in structure to the cell's chro-mosome, and replication of SV40 DNA in vivo isfollowed by duplication of the nucleosome struc-ture. Replication proceeds from a single originof replication and is dependent on a single virus-encoded protein, the SV40 tumor (T) antigen.Because of these properties, it is most likely thatSV40 replicates its DNA in a similar way to cellchromosomal DNA replication. We therefore setout to prepare cell-free extracts that would sup-port complete SV40 DNA replication and even-tually study the mechanism and control of eu-karyote DNA synthesis in some detail.
Recently, Li and Kelly (Proc. Natl. Acad. Sci.81: 6973 [1984]) have developed a cell-free systemfor the replication of SV40 DNA, using extractsfrom monkey cells, and we have modified thissystem for the use of human cells. We decided touse human cells for SV40 DNA replication stud-ies, since monkey cells proved to contain high lev-els of protease activities and were difficult tomaintain in suspension cultures, which would benecessary for obtaining large amounts of cellularprotein for purification purposes. It was shownpreviously that human 293 cells could support ef-ficient SV40 DNA replication in vivo but thatreplication in human HeLa cells was very ineffi-
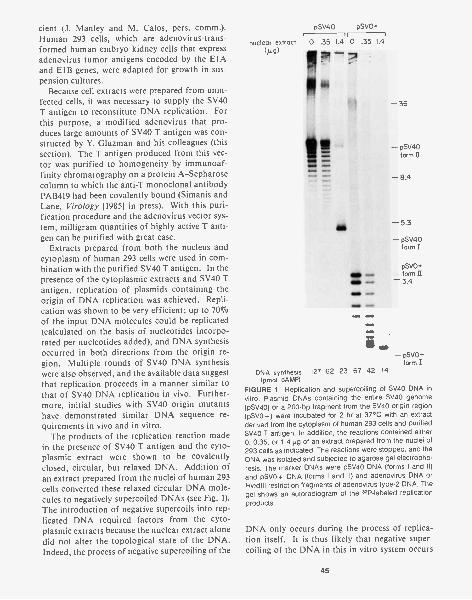
cient (J. Manley and M. Ca los, pers. comm.).Human 293 cells, which are adenovirus-trans-formed human embryo kidney cells that expressadenovirus tumor antigens encoded by the E1Aand E1B genes, were adapted for growth in sus-pension cultures.
Because cell extracts were prepared from unin-fected cells, it was necessary to supply the SV40T antigen to reconstitute DNA replication. Forthis purpose, a modified adenovirus that pro-duces large amounts of SV40 T antigen was con-structed by Y. Gluzman and his colleagues (thissection). The T antigen produced from this vec-tor was purified to homogeneity by immunoaf-finity chromatography on a protein A-Sepharosecolumn to which the anti-T monoclonal antibodyPAB419 had been covalently bound (Simanis andLane, Virology [1985] in press). With this puri-fication procedure and the adenovirus vector sys-tem, milligram quantities of highly active T anti-gen can be purified with great ease.
Extracts prepared from both the nucleus andcytoplasm of human 293 cells were used in com-bination with the purified SV40 T antigen. In thepresence of the cytoplasmic extracts and SV40 Tantigen, replication of plasmids containing theorigin of DNA replication was achieved. Repli-cation was shown to be very efficient; up to 70%of the input DNA molecules could be replicated(calculated on the basis of nucleotides incorpo-rated per nucleotides added), and DNA synthesisoccurred in both directions from the origin re-gion. Multiple rounds of SV40 DNA synthesiswere also observed, and the available data suggestthat replication proceeds in a manner similar tothat of SV40 DNA replication in vivo. Further-more, initial studies with SV40 origin mutantshave demonstrated similar DNA sequence re-quirements in vivo and in vitro.
The products of the replication reaction madein the presence of SV40 T antigen and the cyto-plasmic extract were shown to be covalentlyclosed, circular, but relaxed DNA. Addition ofan extract prepared from the nuclei of human 293cells converted these relaxed circular DNA mole-cules to negatively supercoiled DNAs (see Fig. 1).The introduction of negative supercoils into rep-licated DNA required factors from the cyto-plasmic extracts because the nuclear extract alonedid not alter the topological state of the DNA.Indeed, the process of negative supercoiling of the
pSV40
nuclear extract 0 .35 1.4 0 .35 1.4
(fug)
DNA synthesis(pmol dAMP)
FIGURE 1 Replication and supercoiling of SV40 DNA invitro. Plasmid DNAs containing the entire SV40 genome(pSV40) or a 200-bp fragment from the SV40 origin region(pSVO+) were incubated for 2 hr at 37°C with an extractderived from the cytoplasm of human 293 cells and purifiedSV40 T antigen. In addition, the reactions contained either0, 0.35, or 1.4 yg of an extract prepared from the nuclei of293 cells as indicated. The reactions were stopped, and theDNA was isolated and subjected to agarose gel electropho-resis. The marker DNAs were pSV40 DNA (forms I and II)and pSVO+ DNA (forms I and II) and adenovirus DNA orHindlIl restriction fragments of adenovirus type-2 DNA. Thegel shows an autoradiogram of the 32P-labeled replicationproducts.
- 36
- pSV40form II
- 8.4
- 5.3
- pSV40form I
pSVO+- form II- 3.4
albSNP sowto
,opt, 40,40.
- pSVO+form I
127 62 23 67 42 14
DNA only occurs during the process of replica-tion itself. It is thus likely that negative super-coiling of the DNA in this in vitro system occurs
45

concomitantly with DNA synthesis. This resultwas of some interest because the negatively super-coiled DNA was stable, even in the presence ofhigh levels of the type I topoisomerase in the ex-tracts. This latter activity normally relaxes neg-atively supercoiled DNA. It therefore seems thata novel regulation of the higher order of structureof DNA is occurring in these extracts. It will beof interest to determine whether the DNA is su-percoiled by a DNA gyrase activity that is presentin the extracts or whether the DNA is associatedwith protein that causes the DNA to alter its to-pological structure.
The SV40 replication system should prove use-ful for the elucidation of the mechanism of DNAsynthesis in eukaryotic cells and eventually leadto the identification of replicative proteins. It isexpected that a better understanding of the con-trol of DNA replication and cell proliferation willbe obtained.
Adenovirus Tumor AntigensE. White, P. Cotton, B. Faha, B. Stillman
In addition to our studies on DNA replication,we have continued with the biochemical analysisof the adenovirus tumor antigens that are pro-duced from the E1B genes. Two tumor antigensare synthesized from this early gene region andboth proteins appear to be required for completetransformation of cells. In last year's Annual Re-port, we described our studies on the El B 19,000-dalton (19K) tumor antigen. Single point muta-
tions in, or a deletion within, the coding regionof the E1B 19K gene in the adenovirus genomecaused a drastic phenotype in infected cells. Thecellular morphology was severely altered, andcellular DNA was degraded into low-molecular-weight fragments during the course of infectionof human He La cells with these viral mutants.We have characterized these phenotypes, whichare the result of two separable functions of theE1B 19K protein: The cyt mutations caused analtered cytopathic effect in mutant infected cellsand the deg mutations caused the degradation ofthe cell's chromosomal DNA. All of the deg mu-tants demonstrated the Cyt phenotype, but the cytmutant 1p3 did not induce degradation of cellularDNA. The phenotype of these El B 19K mutantsis summarized in Table 1.
We had previously demonstrated by indirectimmunofluorescence at the light microscopiclevel, and by biochemical fractionation, that theE1B 19K protein in wild-type adenovirus-infectedHe La cells was localized to the nuclear envelope;50% of the protein was tightly associated with thenuclear envelope lamina and the remaining frac-tion was less tightly associated with the laminaand was probably localized in (or on) the nu-clear envelope membranes. In adenovirus-trans-formed cells, the E1B 19K protein localized to thenuclear envelope and also to a cytoplasmic mem-brane structure. Nuclear envelope localization ofthe tumor antigen in infected cells was confirmedby immunoelectron microscopy, which was donein collaboration with S. Blose (Cell Biology Sec-tion). The protein localized to both the inner andouter nuclear envelope membranes but did not lo-calize in the interior of the nucleus (see Fig. 2).
TABLE 1 Phenotypes of E1B 19K Gene Mutants
Virus Cyt' Deg,19K
protein Transformation.
Ad2 wild type Cyt * Deg - + Tra*Ad2cyt106 Cyt Deg + Trod
Ad51p3 Cyt Deg* + Tra
Ad5/p5 Cyt Deg + Tra
Ad5dl337 Cyt Deg Tra
'The Cyt phenotype is an enhanced and abnormal cytopathic effect in infected cells. Wild-type phe-notype is Cyt*.
,The Deg phenotype is the production of degraded cellular DNA during lytic infection. Wild-typephenotype is Deg*.
'Tra' indicates that the virus has transforming potential.dOnly when the mutation is in a temperature-sensitive background.
46

A
Nu7.v
B
Nu
-vJp
FIGURE 2 Immunoperoxidase electron microscopic staining of the nuclear-envelope-associated El B 19K tu-mor antigen. He La cells were infected with wild-type adenovirus for 41 hr or mock-infected, and the cells werethen fixed and stained with antibodies against either the El B 19K protein or the nuclear lamina. Electron micro-graphs of thin sections through nuclei of Ad2 (A-D) or uninfected He La cells (E,F) stained with anti-19K protein(A) or anti-lamin (C,E) antibodies. For comparison, the control panels represent reactions with goat anti-rabbit(8) or rabbit anti-human (D,F) horseradish-peroxidase-conjugated second antibodies alone. The nucleus (Nu),pore complex (p), virion (V), inner (in) and outer (out) nuclear membranes, and lamina (L) are indicated. Magni-fication, 83,915 x .
These results confirmed our previous data andcombined suggest that the E1B 19K protein is as-sociated with the nuclear lamina on the mem-brane side of this structure, as well as with thenuclear envelope membranes.
In addition to the E1B 19K tumor antigen, weobserved that late in infection, several adenovirusstructural proteins were also found associatedwith the lamina. The precursor (but not the pro-cessed forms of virion proteins VI and VII) andprotein IX were selectively enriched in the nuclearlamina preparations. Furthermore, the majornonstructural protein produced late in infection,the 100K phosphoprotein, was shown to be en-riched in nuclear envelope membrane prepara-tions but not in the nuclear lamina preparations.It is therefore likely that this protein is associatedwith the nuclear envelope membranes. These ob-servations suggest that certain aspects of virionmorphogenesis occur on either the nuclear enve-
lope or an internal nuclear structure that is partof a nuclear matrix.
Of particular interest was the association of theE1B 19K tumor antigen with the nuclear envelopeand the fact that mutations in the E1B 19K genecaused degradation of cellular DNA. This cor-relation was observed in virus-infected HeLa cells,and in these cells, the yield of virus was only re-duced about tenfold compared with the wild-typevirus. As part of a search to find a human cellline in which the E1B 19K mutants were more de-fective than wild type, it was noticed that in somecells, the mutants did not induce degradation ofcellular DNA. When primary human embryokidney (HEK) cells, or WI38 human diploid fi-broblast cells, were infected with the E1B 19Kmutants, the mutant viruses did not induce deg-radation of cellular DNA and surprisingly grewto significantly higher titers than the wild-typevirus. On the other hand, the E1B 19K mutants
47

were extremely defective for virus production inhuman KB cells and caused enhanced DNA deg-radation, whereas wild-type virus grew efficientlyin these cells.
We have thus identified three classes of humancells that respond in different ways to the E1B19K mutants and that clearly demonstrate thehost-range phenotype of these mutants. The firstclass, represented by HEK and WI38 cells, allowthe E1B 19K mutants to grow efficiently and es-cape the events that lead to the degradation ofDNA seen in the other cells. In the second class,represented by He La cells and 293 cells, the mu-tants cause degradation of cellular DNA, but thevirus is able to grow at nearly wild-type levels. Inthe third class of cells, represented by KB cells,the mutant viruses are severely defective forgrowth and cause enhanced destruction of thecellular DNA. We are currently examining atwhich stage of the lytic cycle the mutant virusesare blocked in infected KB cells.
The identification of a cell line in which thevirus is defective should enable revertants of theE1B 19K mutants to be isolated and character-ized. The E1B 19K mutant, d1337, which does notproduce any 19K protein due to a large deletionof coding sequences for this protein, can still in-duce DNA degradation upon infection into He Laand KB cells. Thus, since the 19K protein is ab-sent in ti1337-infected cells, another virus protein(or proteins) must be responsible for causing theCyt and Deg phenotypes. It is hoped that the re-vertants will identify this gene product. We arealso creating double mutants in vitro, in whichselected early gene products are mutated in thed1337 or cyt106 viral genomes.
The other protein produced from the E1B re-gion is the 57,000-dalton (57K) tumor antigen.We have previously demonstrated that mutationsin this protein cause a delay in the onset of DNAsynthesis during lytic infection but that the pro-tein is not required for DNA replication directly.The E1B 57K protein also has been shown to as-sociate with the p53 cellular tumor antigen, asdoes the SV40 T antigen, so that it is possible thatSV40 T antigen and the E 1 B 57K protein share acommon biochemical activity. In an attempt toidentify a role for this protein in lytic infectionand cell transformation, we are currently exam-ining methods of overexpressing the protein andeventually purifying the protein. To this end, a
48
number of monoclonal antibodies directedagainst this protein have been isolated and arecurrently being characterized.
PUBLICATIONS
Guggenheimer, R.A., B.W. Stillman, K. Nagata, F. Tamanoi,and J. Hurwitz. 1984. DNA sequences required for thein vitro replication of adenovirus DNA. Proc. Natl. Acad.Sci. 81: 3069-3073.
Stillman, B.W., E. White, and T. Grodzicker. 1984. Independ-ent mutations in Ad2ts111 cause degradation of cellularDNA and defective viral DNA replication. J. Virol.50: 598-605.
White, E., S.H. Blose, and B. W. Stillman. 1984. Nuclear en-velope localization of an adenovirus tumour antigenmaintains the integrity of cellular DNA. Mo/. Cell. Biol.4: 2865-2875.
White, E., T. Grodzicker, and B.W. Stillman. 1984. Mutationsin the gene encoding the adenovirus early region 1B19,000-molecular-weight tumor antigen cause degrada-tion of chromosomal DNA. J. Virol. 52: 410-419.
In Press, Submitted, and In Preparation
Stillman, B.W. 1985. Biochemical and genetic analysis of ad-enovirus DNA replication in vitro. In Genetic engineering:Principles and methods. (ed. A. Hollaender and J.K. Set-low), vol. 7. Plenum Press, New York. (In press.)
Stillman, B.W. and Y. Gluzman. 1985. Replication and super-coiling of SV40 DNA in cell free extracts from human cells.Mo/. Cell. Biol. (Submitted.)
White, E. and B. W. Stillman. 1984. A role for a nuclear en-velope localized adenovirus tumor antigen in virus me-diated transformation and lytic infection. UCLA Symp.Mol. Cell. Biol. (in press).

NUCLEIC ACID CHEMISTRY
R.J. Roberts A. Bhagwat A. Kiss K. TohillJ. Dellaporta P.-M. Lin M. WallaceG.A. Freyer P.A. Myers B. ZerlerC. Keller K. O'Neill
The BsuRI Restriction-ModificationSystemA. Kiss
We hive continued the characterization of theGGCC-specific BsuRI restriction-modificationsystem of Bacillus subtilis that we have cloned andexpressed in Escherichia coli. The BsuRI genes,cloned originally on a 9.5-kb SphI-Sall fragment,were localized to a 4.3-kb Hpal-Sall fragment.Since the sequencing of the methylase gene, usinga clone that codes only for methylase (Kiss andBaldauf, Gene 21: Ill [1983]), was already in pro-gress in P. Venetianer's laboratory in Hungary,we concentrated our efforts on the endonucleasegene.
A detailed restriction map of the region hasbeen established, and specific fragments havebeen cloned in the M13 phage vectors mp 18 andmp 19 and sequenced by the chain-terminationmethod. The region we have sequenced is 2904by in length and codes for the endonuclease andthe aminoterminal 32 amino acids of the methy-lase. The endonuclease is coded by a 1728-bpopen reading frame specifying 576 amino acids.The two genes are oriented in the same direction,with the methylase gene being downstream fromthe endonuclease gene. The coding regions of thetwo genes are separated by 780 bp, which is longerthan any other intergenic region found so far inrestriction-modification systems. This regioncontains an open reading frame coding for 146amino acids. A Shine-Dalgarno sequence pre-ceding this reading frame suggests that it may di-rect the synthesis of a small protein, but experi-ments with derivatives in which this reading framehas been disrupted indicate that even if this pro-tein is produced, it probably has no role in thefunctioning of the BsuRI system. The open read-ing frame coding for the methylase is preceded bya strong Shine-Dalgarno sequence, whereas thesequence preceding the start codon of the endo-
nuclease gene allows for only weak Shine-Dal-garno interactions. This difference may be im-portant in regulating the expression of the BsuRIsystem.
We have not been able to clone the endonu-clease in the absence of methylase, indicating thatthe expression of the restriction enzyme is not de-pendent on the presence of active methylase. Thetwo genes are expressed when cloned on two dif-ferent plasmids in the same cell, suggesting thatthey do not constitute an operon. Transcriptionof the BsuRI genes is currently being studied byanalysis of the cDNA products synthesized on invivo RNA using specific oligonucleotide primers.These experiments confirm that the two genes aretranscribed separately and indicate that the meth-ylase-specific mRNA is more abundant in the B.subtilis cell than the endonuclease-specific tran-script.
Our main purpose in studying restriction andmodification enzymes is to find out how theseproteins recognize specific DNA sequences.Comparison of different proteins interacting withthe same DNA sequence may be helpful in thisrespect. A search of the amino acid sequencesderived from the DNA sequence revealed no ho-mology between the BsuRI endonuclease andmethylase. However, regions of homology werefound between the BsuRI methylase and twoother GGCC-specific DNA methylases, the BspRIand SPR methylases. We wish to use mutagenesisof the cloned enzymes to determine which partsof the proteins are responsible for sequence-spe-cific DNA recognition.
The EcoRII Restriction-ModificationSystem and the dcm GeneA Bhagwat
The EcoRII endonuclease, the EcoRII methylase,and the product of the Escherichia coli dcm gene
49

all recognize the sequence 5'- CC(A /T)GG -3'. TheEcoRII genes were originally found to reside on alow-copy-number E. coli plasmid N3, and weresubsequently cloned onto multicopy plasmids.The dcm gene, which resides at 43 minutes on theE. coli chromosome, codes for a methylase withthe same specificity as EcoRII methylase. We havefound that despite this known identity in theirspecificities, in vivo, the dcm gene is unable tocomplement endonuclease methylase- (R "M -)mutants of EcoRII.
To understand better the relationship betweenthese genes, we have cloned the dcm locus of wild-type E. coli on to pBR322. From a Sau3A partiallibrary of E. coli DNA in pBR322, the clone car-rying the dcm locus was selected by treating theDNA from a mixture of clones with the EcoRIIendonuclease, followed by transformation intoa dcm deletion (Ddcm) strain. Of the 18 trans-formants thus obtained, 1 carried a plasmid thatwas found to be resistant to the EcoRII endonu-clease. Restriction analysis showed that this plas-mid, designated pDCM1, has an 11.3-kbp insertat the BamHI site.
When used as a probe in Southern blots, thisplasmid hybridizes to several EcoRV fragmentsfrom the chromosomal DNA of a dm' strain butto only one EcoRV fragment from the DNA ofthe Ddcm strain. We thus conclude that the in-sert in pDCM1 is derived from the same region inwhich the Ddcm strain has a deletion. On thebasis of genetic evidence, E. coli codes for onlyone cytosine methylase, dcm; hence, we concludethat pDCM1 must carry the dcm locus.
A probe containing the EcoRII methylase genedoes not hybridize to the insert within pDCM1,nor does it hybridize to any EcoRV fragments ofE. coli chromosomal DNA. However, pDCM1 isable to complement an R+M- mutant of EcoRII.Thus, dcm is able to complement the EcoRIImethylase from a multicopy plasmid location butnot from the single-copy chromosomal location.This is probably due to the greater production ofthe enzyme from the plasmid. Finally, a host car-rying pDCM1 does not restrict incoming phage,which indicates the lack of any endonuclease. Insummary, the products of the dcm gene and theEcoRII methylase gene share the same sequencespecificity, but the genes are not homologous.From its chromosomal location, the dcm geneproduct is not able to methylate all of its sub-
50
strate sites, but it is able to do so from the plas-mid location. There may be no endonuclease genelinked with dcm.
We have constructed protein fusions betweenthe EcoRII endonuclease and 0-galactosidase ofE. coli. The endonuclease gene was cloned, inboth orientations, in front of the 0-galactosidasegene lacking its promoter, initiator methioninecodon, and ribosome-binding site. The resultingphenotype was R"Lac- . E. coli strains (Dlac)carrying one of these two plasmids and a compat-ible plasmid carrying the methylase gene werestreaked out on MacConkey-lactose plates. Sev-eral red (Lac') colonies spontaneously grew upfor one of the orientations of the endonucleasegene but not for the other. These Lac' colonieswere picked and analyzed. Seven independentcolonies contained plasmids with deletions at theendonuclease-0-galactosidase junction ranging insize from 50 by to 1.3 kb. Strains carrying theseplasmids gave rise to white colonies on tetrazo-Hum plates, suggesting the equivalent of 400 ormore units of 13-galactosidase activity as analyzedby the Miller assay. The two smallest deletions,which may have up to 30 amino acids missingfrom the carboxyl terminus of the endonuclease,restrict incoming phage as well as the wild-typegene. A deletion removing up to 66 amino acidsreduces the ability of the cell to restrict incomingphage by a factor of 2 x 102 to 5 x 102. The plat-ing efficiency of the phage is still 20-50-fold loweron this strain compared with that on an R+ strain.Larger deletions completely destroy the ability ofthe cell to restrict phage. These protein fusionsdefine the direction of transcription of the en-donuclease gene. They also suggest that the car-boxyl terminus of the endonuclease may not beessential for its action and argue that a hybridprotein carrying both the endonucleolytic activityand lactose metabolizing activity can be con-structed.
Cloned Genes for the Mspl Restriction-Modification SystemP.M.- Lin, R.J. Roberts
The Mspl restriction-modification system recog-nizes the sequence 5'-CCGG-3' and is found in anorganism originally characterized as a Moraxella

species, although there is now reason to believethat this identification was incorrect. DNA fromthis organism has been cloned in a random fash-ion into the plasmid vector pUC9, and a clone hasbeen isolated that expresses both the methylaseand restriction endonuclease activities of theMspl restriction system. This original clone con-tains a 15-kb insert. A series of subclones havebeen prepared, and one of these, containing a 3-kb insert, still expresses both activities. The en-tire insert of this clone has been sequenced. Threeopen reading frames are present, of which one,encoding a polypeptide of 418 amino acids(M, = 47,664), has been shown to encode Mspmethylase. A second open reading frame en-codes a polypeptide of 262 amino acids (M, =29,833) and is the restriction endonuclease gene.A third open reading frame, of unknown function, partially overlaps with the carboxyl termi-nus of the restriction endonuclease gene and con-tinues uninterrupted into the flanking pBR322sequences.
The genes for the restriction enzyme and themodification methylase are transcribed in oppo-site directions, and the first methionines presentin each reading frame are separated by 110 nu-cleotides. Upstream of both AUGs are sequencesthat closely resemble the Pribnow box found inEscherichia coli promoters. The two genes areextremely AT-rich and show no obvious homol-ogy with each other at either the nucleic acid levelor protein sequence level. Furthermore, theyshow no clear homologies with any other restric-tion enzyme or modification enzyme genes thathave been sequenced to date.
Restriction EndonucleasesP.A. Myers, B. Zerler, R.J. Roberts
The collection of restriction enzymes continues togrow and more than 500 such enzymes are nowknown; 116 different specificities have now beencharacterized, including eight in the last year.Among these new specificities, the recognition se-quences for Bsml (GAATGC), Nhel (GTCTAGC),Sspl (AATTATT), and Spel (ATCTAGT) havebeen characterized, as part of a collaborative pro-gram with I. Schildkraut and D. Comb (NewEngland BioLabs). In addition, we have shown
TABLE 1 Simple Hexanucleotide Palindromes
2nd A1st
C G 3rd
A
CGT
A
C
T
AC
G
T
ACG
T
NcolEcoRl Sphl
AatllSnaBI
Hind IIIPvullSacl
SsplNdelEcoRV
Sma1
Nael
M/ulSac II
BselP1
Nrul
SpelAvr11
NhelXbal
Bg III
PvulBarnH1Bcll
Eco4711I
NarlMstl
StulXmalllApalBall
ScaI
Kpnl
SnaI
ClalXholSallAsull
AvalllPstl
AfIIIHpalAhalll
A
C
G
T
that Mjal is an isoschizomer of Mael (CTAG),Mjall is an isoschizomer of Asul (GGNCC), andNflal is an isoschizomer of EcoRV (GATATC).As shown in Table 1, 44 of the possible 64 en-zymes that could recognize hexanucleotide palin-dromes have now been discovered.
In Vitro Splicing of Ad2 Late RNAG.A. Freyer, K. Tohill
Extracts made from He La cell nuclei have beenused to study RNA splicing in vitro. The sub-strate, which contains the first and second exonand the first intron of the tripartite leader of Ad2major late RNA, is synthesized in vitro using aHe La whole-cell extract. Additionally, we haveconstructed recombinant plasmids that can pro-duce similar precursor RNAs when driven by thebacteriophage X promoter P, or the Salmonellaphage promoter SP6. All of these transcriptscontain a modified version of the first intron, inwhich 934 of the 1020 nucleotides have been de-leted. These substrates have all been shown tosplice efficiently and correctly.
When a "P-labeled runoff transcript of 168 nu-cleotides is incubated in nuclear extracts, the cor-rect joining of the first and second exons takesplace to generate the final splice product; in ad-dition, three potential intermediates are detected.On a 10% polyacrylamide-urea gel, these inter-
51

mediates migrate with apparent lengths of 41, 68,and 118 bases. Fingerprint analysis has shownthat the 41 species is the free first leader. The 68species has not yet been completely character-ized, although it appears to come from the 3' endof the RNA. On the basis of electrophoretic mo-bility and fingerprint analysis of the 118 band, itappears to be a circular molecule, 63 bases inlength, which contains sequences from the 5' endof the intron. It is almost certainly the circularportion of the lariat molecules described by otherinvestigators (Krainer et al., Cell 36: 993 [1985];Grabowski et al. Cell 37: 415 [1985]). These lar-iats contain sequences from the complete intronand the adjacent exon but have an unusual struc-ture in that the 5' end of the intron has formed a2'-5' linkage to an A residue close to the 3' end ofthe intron; thus, they contain a circular RNAstructure with a tail. It is the tail that appears tobe missing in the 118 band. The lariat is believedto be the intermediate that combines with the firstexon to form the final splice product. Its absencein our system suggests either that it turns over ex-tremely rapidly or that some other intermediatemay be involved. The 68-nucleotide-long RNA,which contains the 3' end of the precursor, is themost likely candidate for this role. Experimentsare under way to test this possibility.
To investigate the necessity of lariat formation,we have synthesized a mutant template, in whichboth A residues at and close to the lariat attach-ment site were changed to G residues. When the"P-labeled transcript, made from this plasmid,was incubated in our splicing extract, the onlyvisible product was the 41-nucleotide band thatresults from cleavage at the donor splice site forthe first leader RNA. Although this accumulatedto high levels, the remaining RNA was degradedextensively. This result suggests that at least onefunction of the lariat may be to protect the RNAfrom degradation. It also argues that lariat for-mation is essential for splicing.
Fate of Ad2 Introns In VivoG.A. Freyer, K. O'Neill, R.J. Roberts
Our initial interest in examining introns from invivo RNA was to test a hypothesis concerning themechanism of splicing. This model proposed a
52
simple reciprocal cleavage and ligation event oc-curring at the intron/exon boundary, leading to alinear final splice product and a circular intron.The system that we chose to examine was the tri-partite leader from the major late RNA of Ad2.Two oligonucleotides were synthesized that wouldhybridize across the predicted junctions of circu-lar introns corresponding to either the intron be-tween leaders 1 and 2 or the intron between lead-ers 2 and 3. Nuclear and cytoplasmic RNAs wereprepared separately from Ad2-infected HeLacells. The RNA was electrophoresed on denatur-ing agarose gels, blotted, and probed with these"P-labeled oligonucleotides. The oligonucleo-tide, designed to probe the first intron, failed tohybridize to any RNA species. This result is con-sistent with the circularization of this intron at aninternal position as indicated above. However,the oligonucleotide, designed to probe the secondintron, did hybridize to several RNA species. Incontrol experiments, it was shown that some ofthese species were present in uninfected HeLacells. The structures of the Ad2-specific speciesare under investigation.
To investigate the fate of the first intron se-quences, two oligomers were synthesized, with se-quences complementary to linear portions of thefirst intron. These oligomers gave an interestingand surprising result. They detected RNA spe-cies that were present in abundance in the cyto-plasm of Ad2-infected cells but were undetect-able in the nucleus. Two species of 1000 and 1100nucleotides were found, which is the approxi-mate size of the linear intron. Although the pre-cise structures of these RNAs are still being ex-amined, one behaves like a linear molecule, andthe other migrates anomalously on gels and maywell be a circular molecule.
Both of these oligomers have been used asprimers in dideoxynucleotide sequence analysis ofthese RNAs. A unique sequence was obtainedthat begins near the 3' end of the oligomers andextends up to the 5' penultimate nucleotide of theintron. These results show that the oligomers arehybridizing specifically and also indicate that thestructure of the intron at its 5' boundary is suchthat it prevents further extension with reversetranscriptase. Such a block could be a free 5' endas might be found in a linear molecule cleaved atthe donor splice site or could be an unusual link-age, such as the 2' -5' linkage that characterizes

lariat attachment sites. Additionally, it is clearfrom these results that the oligomer is priming ona processed intron, rather than merely detectingthe linear sequences from the precursor RNA.
Several other oligomers have been synthesizedand used to probe specific regions of these RNAs.First, an oligomer complementary to the 23 nu-cleotides between the lariat attachment site andthe 3' end of the RNA was used as a probe. Al-though this oligomer hybridized to a high-molec-ular-weight RNA in the nucleus, there was no de-tectable hybridization to the cytoplasmic RNA.An oligonucleotide complementary to the 3' re-gion of the intron that spans the lariat attach-ment site was used as a probe at low stringencyand was found to hybridize to both species, sug-gesting that the boundary of these RNAs is at ornear the lariat attachment site. Finally, a thirdoligomer was made that spans the lariat attach-ment site by 13 nucleotides on either side. Underlow stringency, this oligomer was shown to hybri-dize to only one RNA species.
From these results, we conclude that the twointron species contain similar, if not identical, se-quences and that the difference in mobilities islikely due to a difference in structure, i.e., a lin-ear and possibly circular RNA. Neither containsthe 3'-terminal fragment comprising the "handle"of the lariat. However, both hybridize to a probenear the 2' -5' junction. This is the first report ofthe presence of free intron sequences in the cyto-plasm of cells.
Ad2 E1A Gene ProductsB. Zerler
The adenovirus E1A gene is required for adeno-virus-induced cell transformation and is involvedin the regulation of other adenovirus gene prod-ucts. Early in infection, two mRNAs are pro-duced from E1A, the 12S and 13S products. Latein infection, a 9S mRNA product is made. Thenucleotide sequence of the 12S and 13S productsindicate that they code for proteins 243 and 289amino acids long, respectively. These proteinsdiffer only by the presence of an additional 46amino acids unique to the 13S product. The 9Sproduct is predicted to encode a protein 55 aminoacids long, which as a result of splicing enters a
different reading frame from that of the 12S and13S products. Therefore, only the aminoterminal26 amino acids of the 9S protein are common tothe 12S and 13S proteins. Our aim is to distin-guish the roles of the individual E1A products.Last year, we separated the 12S and 13S productsby isolating the corresponding cDNAs. We alsoconstructed a 9S cDNA clone by using syntheticoligomers that span the splice junction. ThesecDNAs were subcloned into plasmids and also re-built into the entire viral genome in place of thegenomic E1A region. For a detailed descriptionof our results on the biological roles of the indi-vidual E1A gene products using these cDNAplasmids and viruses, see Mathews et al. (this sec-tion).
We are presently concentrating on expressingE1A in other eukaryotic systems. We have re-cently expressed the 12S and 13S E1A cDNA prod-ucts in yeast in order to take advantage of theextensive potential for genetic analysis of wild-type and mutant E1A functions in this organism.The E1A cDNAs were cloned into a yeast expres-sion vector under transcriptional control of theinducible GAL10 promoter. Expression of bothE1A gene products has been detected in yeast col-onies by an immunological method described byLyons and Nelson (Proc. Natl. Acad. Sci. 81:7426 [1984]). In addition, immunoprecipitationand Western blot analysis show the presence oftwo 13S-specific proteins with molecular weightssimilar to those of the two major 13S proteinspresent in HeLa cells during Ad2 infection. Sim-ilar studies with the 125-E IA-encoded product inSaccharomyces cerevisiae are in progress.
When grown under inducing conditions, S.cerevisiae containing either of the E1A geneproducts does not exhibit any observable pheno-type. Generation time and morphology appearsimilar to those of controls. We are currently in-vestigating the ability of the E1A 13S and 12Sproducts to complement various yeast tempera-ture-sensitive cdc mutants.
In addition to the above studies, we are cloningE1A into mammalian high-expression vectors toenable purification of the proteins. A BPV vectorcontaining genomic E1A has been constructedand transfected into NIH-3T3 cells. Transform-ants have been isolated and are currently beingscreened by immunofluorescence and immuno-precipitation for the presence of E1A proteins. In
53

addition, we have completed construction of aplasmid containing E1A and the amplifiable di-hydrofolate reductase (DHFR) gene. This plas-mid will be transfected into DHFR- Chinesehamster ovary cells.
PUBLICATIONS
Broker, T.R., C.C. Keller, and R.J. Roberts. 1984. Humanadenovirus serotypes 2,4,8,40 and 41 and adenovirus-simian virus 40 hybrids. Maps of RNA transcripts, pro-tein-coding regions, restriction endonuclease cleavagesites, and DNA structures. In Genetic maps: A compila-tion of linkage and restriction maps of genetically studiedorganisms (ed. S.J. O'Brien), vol. 3, pp 99-110. ColdSpring Harbor Laboratory, Cold Spring Harbor, New York.
Freyer, G.A., Y. Katoh, and R.J. Roberts. 1984. Character-ization of the major mRNAs from adenovirus 2 early re-gion 4 by cDNA cloning and sequencing. Nucleic AcidsRes. 12: 3503-3519.
Keller, C., M. Corcoran, and R.J. Roberts. 1984. Computerprograms for handling nucleic acid sequences. NucleicAcids Res. 12: 379-386.
Roberts, R.J. 1984. Restriction and modification enzymesand their recognition sequences. Nucleic Acids Res.12: r167-r204.
. 1984. Restriction endonucleases, DNA sequencingand computers. In Physics and contemporary needs (ed.A.M. Khan et al.), vol. 6, pp 305-316. Plenum Press, NewYork.
PROTEIN SYNTHESIS
M.B. Mathews G.P. Thomas E. MoranT.M. Harrison M.R. SadaieC.V. Dery C.C. BunnJ.M. Langstaff R. O'Malley
Roberts, R.J. and D. Soll, eds. 1984. The applications ofcomputers to research nucleic acids 11. Part 1 and 2. IRLPress, Oxford and Washington, D.C.
Roberts, R.J., K.E. O'Neill, and C. Yen. 1984. DNA se-quences from the adenovirus-2 genome. J. Biol. Chem.259: 13968-13975.
In Press, Submitted, and In Preparation
Akusjarvi, G., U. Pettersson, and R.J. Roberts. 1985. Struc-ture and function of the adenovirus-2 genome. In Devel-opments in molecular virology (ed. W. Doerfler), vol. 5.(In press.)
Roberts, R.J. 1985. Restriction and modification enzymesand their recognition sequences. In CRC handbook ofmicrobiology. CRC Press, Boca Raton, Florida. (In press.)
. 1985. Restriction enzymes. In Hybridization-Apractical approach (ed. B.D. Names). IRL Press, Wash-ington, D.C. (In press.)
1985. Restriction endonuclease. McGraw-Hill Ency-clopaedia of Science and Technology. McGraw-Hill, NewYork. (In press.)
. 1985. Restriction and modification enzymes and theirrecognition sequences. Nucleic Acids Res. (in press).
Roberts, R.J., G. Akusjarvi, P Alestrom, R.E. Gelinas, T R.Gingeras, D. Sciaky, and U. Pettersson. 1985. A consen-sus sequence for the adenovirus-2 genome. In Develop-ments in molecular virology (ed. W. Doerfler), vol. 5. (Inpress.)
M. FreedmanC.H. HerrmannD.P. PascucciP.A. Reichel
We have continued to work on the regulation ofadenovirus infection and on the heat-shock(stress) response of human cells. In addition, ourstudies of autoantibodies in rheumatic diseasehave expanded in depth and scope during the pastyear.
Functional Analysis of the E1A GeneE. Moran, C.V. Dery, T.M. Harrison, M.B. Mathews
The adenovirus E1A gene is required for virus-induced cell transformation and for the transcrip-
54
tion of other adenovirus genes. Early in infec-tion, two mRNAs are made from E1A, the 12Sand 13S RNAs (see Fig. 1). Their nucleotide se-quences indicate that they encode proteins 243and 289 amino acids long, respectively, differingonly in that the larger protein contains an addi-tional 46 amino acids between positions 140 and185. Our aim is to examine the roles of the sepa-rate E1A products. This goal is being pursued incollaboration with B. Zerler, R. Roberts, and T.Grodzicker (this section). Last year, we took thefirst step, the isolation of cDNA clones corre-sponding to the 12S and 13S cDNAs and the con-struction of a cDNA clone corresponding to the

FIGURE 1 The adenovirus E1A gene. TheE1A gene is located at the left end of the ad-enovirus genome, and some landmarks are in-dicated on the top line. The structures of thethree spliced E1A mRNA5 are depicted by ar-rows and their coding regions are shaded. Thecoding regions of the 12S and 13S mRNA5(hatched) are identical except that the first exonof the 12S RNA is shorter; in both cases, thecodon for one amino acid is split between thefirst and second exons. The amino terminus ofthe 9S protein is identical with that of the 12Sand 13S products, but after the splice junction,the 9S product enters a different reading frame(stippled). Mutations of individual nucleotides(numbered) and a deletion of the Cral/Xmalfragment (shown as a bar) have been used tostudy the functions of the 12S and 13S prod-ucts (see text).
acidic as region 13S unique region
1
ATG ClaI XmaI1 1
poly A
0 559
499 6369S
I2S
I 3S
MEI:3
916 1006
973
138aa
1630
1225Imp104a
928 946 961 deletionCYS PRO GLU Cla/Xma
GLY ALA LYS
ACTIVATE E3-col + + +
TRANSFORM WITH ras V I I
9S mRNA synthesized by the E1A gene later ininfection. To allow further analysis, the cDNAswere inserted into the viral genome in place of thegenomic E1A region, giving so-called 9S, 12S, and13S viruses.
The simplest test of E1A gene function avail-able at present is the transient expression assay,
to regulate transcription from other adenoviruspromoters. In one version of this assay, He La(human) cells are transfected with an E1A deriv-ative, together with the E3-cat plasmid in whichthe adenovirus E3 promoter governs the expres-sion of the readily assayed bacterial enzymechloramphenicol acetyltransferase (cat). As de-scribed last year, plasmids carrying either the en-tire E1A gene or the 13S cDNA stimulate E3-catexpression, whereas the 12S and 9S cDNA plas-mids do not. These experiments have now beenrepeated with the E2-cat plasmid in which en-zyme production is governed by a different ad-enovirus early promoter. Again the results indi-cate that transcriptional activation is a propertyof the 13S product only. We are presently con-structing a set of additional promoter-cat fusionsin order to assess the generality of this conclusionand to measure the sensitivity of other promotersto E1A stimulation.
The functions of the E1A gene were also stud-ied during infection of human cells with thecDNA viruses. The 13S virus is as competent aswild-type virus for lytic growth in He La cells,
55 as
243 as
289 as
whereas the 12S and 9S viruses are defective. In-direct immunofluorescence studies indicate thatthe 72K protein made by the adenovirus E2A geneis present at comparable levels in cells infectedwith 13S or wild-type virus, but the protein is notdetected after infection with 12S virus. As ex-pected, corresponding differences are seen whenviral mRNA levels are compared. These resultsconfirm that the stimulation of expression ofother adenovirus early gene products is a func-tion of the E1A 13S product but not of the 12S or9S products.
The transforming activities of the cDNA plas-mids and viruses were investigated in primarybaby rat kidney (BRK) cells. Both the 12S and13S plasmids can establish BRK cells in culture ina manner similar to that of the intact E1A gene,and all three can cooperate with the ras oncogeneto generate stable transformants. Similar resultsare obtained when the E1A promoter is replacedby the mouse metallothionein promoter, implyingthat the genetic element responsible for the trans-forming function lies within the 12S and 13S cod-ing sequences. Remarkably, the 12S virus trans-forms BRK cells at an efficiency 50-100-foldhigher than wild-type virus. However, the estab-lishment activity observed with the 13S plasmid isnot seen after infection with the 13S virus, prob-ably because it is obscured by the cytopathic ef-fect of this virus. No establishment activity hasbeen detected in the 9S cDNA plasmid or virusconstructs.
55

From these results, we conclude that the estab-lishment function, exhibited by both the 12S and13S products, is separate from the function thatstimulates expression of other adenovirus earlygenes and resides exclusively in the 13S product.Presumably, the 13S unique region is required forefficient stimulation of adenovirus early genes.On the other hand, plasmids carrying the E1A re-gion of an existing E1A mutant virus, hr-A, areable to establish BRK cells and to cooperate withras to transform these cells stably. The hr-A mu-tant fails to produce the 12S mRNA, and its 13SmRNA encodes a truncated E1A product com-prising the first 140 amino acids of the 289-amino-acid protein. Thus, it appears that the first 140amino acids of the 13S product (139 of which areshared by the 12S product) are sufficient for thisactivity and that the activity persists in the trun-cated protein.
To dissect further the functions of E1A, wehave begun a mutational analysis of the gene. Wefirst excised the Clal to Xmal fragment of E1A,producing an in-frame deletion mutant that lacksthe 12S splice donor site and amino acids 121-150of the 289-amino-acid protein (Fig. 1). The mu-tant plasmid fails to stimulate transient expres-sion of E3-cat in He La cells even though it retainscoding information for 35 of the 46 amino acidsof the 13S unique region, including the five cys-teine residues that are a distinctive feature of thisregion. It also fails to cooperate with ras intransformation of BRK cells. These results sug-gest that amino acids 121-150 are required forboth functions and in particular that the sectionfrom residue 121 to residue 140 is required for theestablishment activity.
The most striking feature of this sequence is astring of six acidic amino acids from residue 133to residue 138. The unusual concentration ofnegative charges might form part of an active site.Interestingly, the myc oncogene, which like E1Acan cooperate with ras to transform BRK cells,has a similar run of acidic amino acids. We haveused oligonucleotide-directed, site-specific mu-tagenesis to substitute lysine for glutamic acid atposition 135 (nucleotide 961) to discover whetherintroduction of a positive charge in this nega-tively charged region will disrupt activity. Themutation was constructed in both E1A genomicand 12S cDNA plasmids. The E1A-961 mutationretains the ability to stimulate E3-cat activity, but
56
both the E1A-961 plasmid and the 12S-961 plas-mid are defective in the ability to cooperate withras to transform BRK cells. In addition, whenrebuilt into virus, the 12S-961 construct is also de-fective in the ability to establish foci of immor-talized BRK cells, compared with the wild-type12S virus, although it does appear to give thesecells some extended growth potential. Immuno-precipitation from infected He La and BRK cellsdemonstrates that the mutant protein is producedand is reasonably stable. Thus, the acidic-to-basic amino acid change at position 135 has amarked effect on the establishment function butlittle or no effect on stimulation of E3-cat expres-sion. The activity that stimulates adenovirusearly gene expression can be dissociated from theestablishment function, just as the establishmentfunction can be uncoupled from the early genestimulating activity. We are currently rebuildingthe E1A-961 construct into whole virus to dis-cover whether the mutation affects the growthproperties of the virus.
We have also used oligonucleotides to changethe cysteine residue at position 124 (nucleotide928) to glycine and the proline residue at position130 (nucleotide 946) to alanine. These aminoacids are conserved in the adenovirus serotypes,Ad2, Ad7, and Ad12, suggesting that they mayhave functional significance. The E1A-928 plas-mid construct has a phenotype similar to that ofE1A-961: It stimulates expression of E3-cat but isdefective in cooperating with ras to transformBRK cells, again indicating that the ability tostimulate early gene expression is not dependenton the normal establishment activity. The 13S-946 plasmid is able to stimulate expression of E3-cat, but its establishment potential has not yetbeen determined. Both mutations are currentlybeing incorporated into viruses to assess their lyticand transforming functions.
We are continuing to use oligonucleotide-di-rected mutagenesis as well as random mutagene-sis procedures to isolate additional changes in theregion between the Clal and Xmal sites of E1A.These mutants will be assayed for their lytic andtransforming functions and for their ability toregulate expression of adenovirus genes, cotrans-fected genes, and host-cell products. Further-more, we have recently expressed the E1A cDNAproducts in yeast in order to exploit the potentialfor genetic analysis of wild-type and mutant E1A

functions in this organism. We also hope to ob-tain a sufficient amount of E1A proteins for bio-chemical analysis both from yeast and frommammalian expression vectors. This will permitanalysis of the effects of E1A mutations on thechemical modification and function of the E1Aproteins.
VA RNA and Translational ControlP.A. Reichel, R. O'Malley, M.B. Mathews
The "virus-associated" RNAs, VA RNA, and VARNA, are small, abundant adenovirus tran-scripts that do not code for proteins. They aretranscribed by RNA polymerase III from twoclosely spaced genes located between map units29 and 31 on the viral chromosome. Both speciesare about 160 nucleotides in length, exhibit exten-sive secondary structure, and terminate with a runof U residues that interacts with a cellular proteinknown as the La antigen. Until recently, the roleof the VA RNAs was a matter for speculation,but, as described in last year's report, work withthe Ad5 mutant d1331 (Thimmappaya et al., Cell31: 543 [1982]) pointed to a role in protein syn-thesis. Cells infected with the mutant virus lackVA RNA,, the major VA RNA species, and at latetimes of infection, they are defective in their abil-ity to translate mRNAs of either viral or cellularorigin.
We have now pinpointed the level at which VARNA, functions in translation. Cell-free protein-synthesizing extracts of d1331-infected He La cells,like the intact cells from which they are derived,suffer from a greatly reduced ability to initiatetranslation of mRNA. Surprisingly, the defectcannot be overcome by addition of VA RNA orof VA RNA together with the La antigen, imply-ing that the small RNA acts only indirectly. Fur-thermore, mixing experiments indicated thatd1331 extracts contain an inhibitor capable ofblocking initiation in other cell extracts. Themixing experiments also disclosed that proteinsynthesis can be effectively restored by variousfractions from a rabbit reticulocyte extract, lead-ing us to examine several highly purified reticu-locyte initiation factors for their ability to rescueprotein synthesis in the d1331 extracts. These fac-tors were kindly provided by W Merrick (CaseWestern Reserve University) and J. Siekierka
(Roche Institute). The activity proved to residesolely in eukaryotic initiation factor 2 (eIF-2) andin an ancillary enzyme, the guanosine nucleotideexchange factor (GEF), which enables "used" eIF-2 to function catalytically in initiation.
Figure 2 summarizes the relationships between
GDP Interferon
eIF -2 GTPMet - tRNAF
Ternary complex
40SreIF -2 GTP
Met - TRNAF
eIF -2 :GTPMet - tRNAF 40SmRNA
Met-tRNAF 80S mRNA
LeIF-2(001GEF
Trapped GEF
IBlocked Initiation
Chain elongation
FIGURE 2 Function and control of initiation factor elF-2.During the initiation process, this factor, composed of threepolypeptide subunits, forms a ternary complex with GTP andthe initiating tRNA (methionyl-tRNAF). The complex bindsmRNA, the 40S ribosomal subunit, and the 60S ribosomalsubunit to complete the initiation sequence. In the last step,elF-2 is released bound to GDP, which must be replaced byGTP before elF-2 can recycle. The replacement reaction iscatalyzed by GEE. If elF-2 is phosphorylated on its a-subunitby the double-stranded RNA-activated inhibitor (DAI) or theheme-controlled inhibitor (NCI), it sequesters GEE. The GTP/GDP exchange reaction is then blocked, preventing othermolecules of elF-2 from recycling. VA RNA, prevents phos-phorylation of elF-2 in adenovirus-infected cells. (Diagramfrom Reichel et al., Nature 313: 196 [1985].)
57

these two factors in the initiation process and il-lustrates a mechanism whereby their activity canbe modulated through phosphorylation of eIF -2.Two protein kinases are known to catalyze thisreaction and thereby block protein synthesis: DAIis activated by double-stranded RNA, whereasHCI is triggered by a multiplicity of effectors.Working with T. Mariano and J. Siekierka (RocheInstitute), we have demonstrated the induction ofsuch an activity in d/331-infected cells and haveshown that it is able to block initiation. Al-though the nature of the kinase is not yet certain,in view of the extensively base-paired structure ofVA RNA and its ability to interact with viralmRNA, our working hypothesis is that VA RNAprevents activation of DAI during adenovirus in-fection. We have also tested the possibility thatVA RNA, interacts directly with eIF-2 or GEF buthave failed to detect significant binding.
Studies begun last year of the interaction of VARNAs with specific viral mRNAs in vivo havecontinued. A transient expression system was de-veloped that allows us to detect changes in theamount of the E2A-72K DNA-binding protein(DBP) synthesized in response to VA RNA. Aplasmid containing the complete gene for theDBP is cotransfected into human 293 cells to-gether with plasmids encoding VA RNA, and thesynthesis of DBP is assayed by immunoblottingor immunoprecipitation. In this assay, VA RNAhas little or no effect on DBP synthesis, whereasVA RNA, increases the amount of DBP signifi-cantly. Deletion mutants have allowed prelimi-nary mapping of the VA RNA, sequences re-quired for enhancement of DBP synthesis. Theregion between nucleotides 79 and 157 in the 3'half of the molecule appears to be especially im-portant. More detailed analysis with new mu-tants is projected for the near future, and rapidscreening and assay methods are currently underdevelopment for this purpose.
Ribonucleoprotein Particles andAutoimmunityM.R. Sadaie, C.C. Bunn, M.B. Mathews
Because of our long-standing interest in VA RNAand the La antigen that binds to it, the study ofautoantibodies has developed into a research
58
project in its own right. Our work in this areabenefits from a close association with R. Bern-stein and G.R.V. Hughes (Rheumatology Unit,Hammersmith Hospital, London). Autoantibod-ies circulate in the blood of patients sufferingfrom certain rheumatic and connective-tissue dis-eases, including systemic lupus erythematosus(SLE) and myositis. In these disorders, the built-in safeguards of the immune system are breached,permitting the production of normally "forbid-den" antibodies that react with the body's owncellular components. Examination of these anti-bodies not only sheds light on the nature of au-toimmune disease, but also affords valuable re-agents for molecular and cytological research.Current work is focused on the proliferating cellnuclear antigen (PCNA) and antibodies charac-teristic of the muscle-wasting diseases polymyos-itis and dermatomyositis.
Last year, we reported that PCNA is a proteinwith a molecular weight of 35,000, identical withthe protein "cyclin" that accumulates in dividingand transformed cells. PCNA reacts with auto-antibodies from a group of patients with SLE. Itslevel and localization within the nucleus of cul-tured Wil-2 cells and mitogen-stimulated humanB lymphocytes vary with the phases of the cellcycle (Takasaki et al., J. Exp. Med. 154: 1899[1981]). This protein is therefore of potential sig-nificance in the regulation of cell division as wellas in autoimmunity.
We have begun to study the metabolism ofPCNA in He La cells. It is mainly, if not exclu-sively, nuclear and can be released in large meas-ure by digestion of DNA, which may point to anassociation with chromatin. Indirect immuno-fluorescence using anti-PCNA serum shows thatPCNA is distributed heterogeneously in asyn-chronous cells, but in synchronized cells, severaldistinct patterns are seen (Fig. 3). Particularly in-triguing is its apparent migration into and out ofthe nucleoli. This peripatetic behavior remainsto be explained, and the interpretation is furtherclouded by the discovery that PCNA antibodiesare not entirely monospecific. A group of ninenuclear proteins with molecular weights of12,000-110,000 are detected by immunoblotting,whereas PCNA alone is immunoprecipitated fromour cell extracts. Immunoprecipitation of pro-teins synthesized in vitro gives a single proteinwith the same electrophoretic mobility as PCNA.
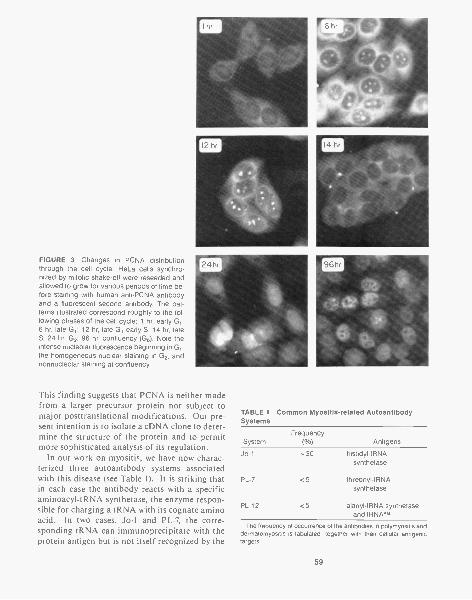
FIGURE 3 Changes in PCNA distributionthrough the cell cycle. He La cells synchro-nized by mitotic shake-off were reseeded andallowed to grow for various periods of time be-fore staining with human anti-PCNA antibodyand a fluorescent second antibody. The pat-terns illustrated correspond roughly to the fol-lowing phases of the cell cycle: 1 hr, early G,;6 hr, late Gl; 12 hr, late G, early S; 14 hr, lateS; 24 hr, G2; 96 hr, confluency (Go). Note theintense nucleolar fluorescence beginning in GI,the homogeneous nuclear staining in G2, andnonnucleolar staining at confluency.
4h
This finding suggests that PCNA is neither madefrom a larger precursor protein nor subject tomajor posttranslational modifications. Our pre-sent intention is to isolate a cDNA clone to deter-mine the structure of the protein and to permitmore sophisticated analysis of its regulation.
In our work on myositis, we have now charac-terized three autoantibody systems associatedwith this disease (see Table 1). It is striking thatin each case the antibody reacts with a specificaminoacyl-tRNA synthetase, the enzyme respon-sible for charging a tRNA with its cognate aminoacid. In two cases, Jo-1 and PL-7, the corre-sponding tRNA can immunoprecipitate with theprotein antigen but is not itself recognized by the
TABLE 1 Common Myositis-related AutoantibodySystems
FrequencySystem ( %) Antigens
Jo-1 -30 histidyl-tRNAsynthetase
PL-7 <5 threonyl-tRNAsynthetase
PL-12 <5 alanyl-tRNA synthetaseand tRNAAl.
The frequency of occurrence of the antibodies in polymyositis anddermatomyositis is tabulated, together with their cellular antigenictargets.
59

antibody. PL-12 sera, on the other hand, recog-nize alanine-accepting tRNA independently of thesynthetase protein. In these patients, the twoseparate antibodies to tRNA and protein coexist,suggesting that the autoimmune response wasoriginally mounted against the enzyme-tRNAcomplex. An alternative theory proposing thatthe initial immunogen is some unidentified cross-reacting macromolecule seems much less likely.With PL -12 as a possible exception, our data alsoargue against the anti-idiotype model of autoim-munity and favor the notion that the antigens areseen as foreign (rather than "self") constituentsbecause they become associated with some for-eign molecule. In the case of the myositis anti-gens, the likeliest candidate for this role wouldseem to be viral RNA, although firm evidence islacking.
The Mammalian Stress ResponseJ.M. Langstaff, D.P. Pascucci, M.B. Mathews, G.P. Thomas
As in past years, our interests in the mammalianstress (or heat-shock) response have centered onthe mechanisms of translational control operat-ing during stress and the analysis of genes encod-ing the human stress-response proteins. Becauseof the relative intractability of the former topic,more emphasis has been placed this year on thelatter.
Probes for the gene encoding the 90,000-dalton(90K) human stress-response protein have beenisolated through a "plus-minus" approach andhave been used to isolate bacteriophage clonesfrom a library of genomic DNA, as indicated inprevious Annual Reports. One such clone, con-taining what appears to be the entire protein-cod-ing sequence for the inducible 90K protein, in ad-dition to about 10 kb of upstream sequence, hasbeen studied intensively. The protein-coding se-quences have been identified through S1- nucleaseprotection experiments coupled with nucleotidesequencing. The sequence of a stretch of about4.5 kb, including the coding regions and an ap-proximately 1 -kb region upstream of the likely in-itiator methionine, has been determined and islargely complete.
Comparing this sequence with the recentlypublished sequences of its counterparts fromDrosophila (fruitfly) and Saccharomyces (yeast),
60
we have identified the putative initiator methio-nine (Fig. 4). The derived protein sequences arevery similar, and in some stretches more than 90%conservation is observed. In Drosophila, the 90Kgene equivalent (HSP83K) is unique in that it isthe only heat-shock gene with an intervening se-quence. Its mRNA initiates some thousand basesupstream of the initiating ATG, but most of thissequence is removed through splicing duringmRNA formation. In yeast, the transcript is notspliced, and the protein-coding region followsabout 50 bases from the start of the mRNA. Ourupstream sequence does not contain likelymatches with the so-called "consensus" (CT-GAA-TTC-AG) sequence implicated in the in-duction of this type of gene, suggesting that the90K gene may be activated by a different mecha-nism. The alternative possibility, that our se-quence may not extend far enough upstream tocontain this region, remains to be excluded, al-though Si- nuclease protection has failed to revealits existence. We hope to resolve this issue in thenear future by primer extension and protectionexperiments using fragments located even furtherupstream than those used so far.
The 90K stress-response gene differs from manyother stress-response genes in that it is expressedat fairly high levels under normal circumstances.The 72K/73K family of stress proteins is subjectto subtle and varied controls: Some members areexpressed normally and repressed under stress,others are expressed normally and seemingly un-affected by stress, and yet others are greatly in-duced during stress. The latter class has drawnattention for two reasons. First, it is a favorablemodel gene to investigate the mechanisms of in-duction since the magnitude of induction exceeds50-fold. Second, its protein product is the majorstress protein of many organisms, has been con-served throughout evolution, and is required forgrowth at elevated temperatures. The "nonindu-cible" members of the family have yet to enjoysuch detailed study, yet are of no less interest, alsobeing well conserved and expressed at substantiallevels under normal circumstances. We have iso-lated cDNA probes for both types of genes, theinduced form (72K) from He La cells treated withthe amino acid analog and, more recently, the un-induced form (73K) from normal rat fibroblasts.
Using the 72K cDNA clone, we have isolatedgenomic clones, and these are currently being
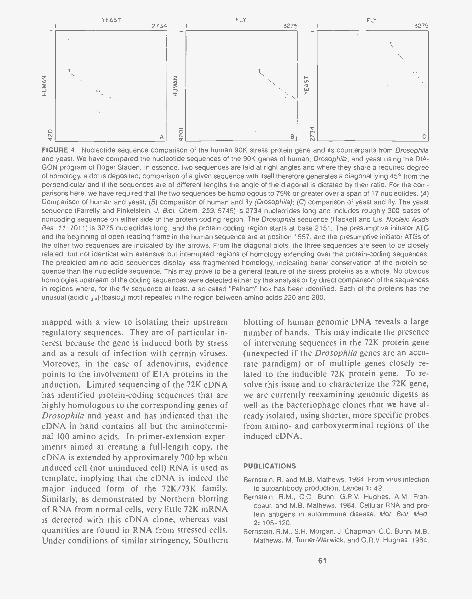
YEAST FLY FLY2734 3275 I 3275
j;-C
FIGURE 4 Nucleotide sequence comparison of the human 90K stress protein gene and its counterparts from Drosophilaand yeast. We have compared the nucleotide sequences of the 90K genes of human, Drosophila, and yeast using the DIA-GON program of Roger Staden. In essence, two sequences are laid at right angles and where they share a required degreeof homology, a dot is deposited; comparison of a given sequence with itself therefore generates a diagonal lying 45° from theperpendicular and if the sequences are of different lengths the angle of the diagonal is dictated by their ratio. For the com-parisons here, we have required that the two sequences be homologous to 75% or greater over a span of 17 nucleotides. (A)Comparison of human and yeast; (8) comparison of human and fly (Drosophila); (C) comparison of yeast and fly. The yeastsequence (Farrel ly and Finkelstein, J. Biol. Chem. 259: 5745) is 2734 nucleotides long and includes roughly 300 bases ofnoncoding sequence on either side of the protein coding region. The Drosophila sequence (Hackett and Lis, Nucleic AcidsRes. 11: 7011) is 3275 nucleotides long, and the protein coding region starts at base 2151. The presumptive initiator ATGand the beginning of open reading frame in the human sequence are at position 1557, and the presumptive initiator ATGs ofthe other two sequences are indicated by the arrows. From the diagonal plots, the three sequences are seen to be closelyrelated, but not identical with extensive but interrupted regions of homology extending over the protein-coding sequences.The predicted amino acid sequences display less fragmented homology, indicating better conservation of the protein se-quence than the nucleotide sequence. This may prove to be a general feature of the stress proteins as a whole. No obvioushomologies upstream of the coding sequences were detected either by this analysis or by direct comparison of the sequencesin regions where, for the fly sequence at least, a so-called "Pelham" box has been identified. Each of the proteins has theunusual (acidic ,)-(basic,) motif repeated in the region between amino acids 220 and 280.
mapped with a view to isolating their upstreamregulatory sequences. They are of particular in-terest because the gene is induced both by stressand as a result of infection with certain viruses.Moreover, in the case of adenovirus, evidencepoints to the involvement of E1A proteins in theinduction. Limited sequencing of the 72K cDNAhas identified protein-coding sequences that arehighly homologous to the corresponding genes ofDrosophila and yeast and has indicated that thecDNA in hand contains all but the aminotermi-nal 100 amino acids. In primer-extension exper-iments aimed at creating a full-length copy, thecDNA is extended by approximately 700 by wheninduced cell (not uninduced cell) RNA is used astemplate, implying that the cDNA is indeed themajor induced form of the 72K/73K family.Similarly, as demonstrated by Northern blottingof RNA from normal cells, very little 72K mRNAis detected with this cDNA clone, whereas vastquantities are found in RNA from stressed cells.Under conditions of similar stringency, Southern
blotting of human genomic DNA reveals a largenumber of bands. This may indicate the presenceof intervening sequences in the 72K protein gene(unexpected if the Drosophila genes are an accu-rate paradigm) or of multiple genes closely re-lated to the inducible 72K protein gene. To re-solve this issue and to characterize the 72K gene,we are currently reexamining genomic digests aswell as the bacteriophage clones that we have al-ready isolated, using shorter, more specific probesfrom amino- and carboxyterminal regions of theinduced cDNA.
PUBLICATIONS
Bernstein, R. and M.B. Mathews. 1984. From virus infectionto autoantibody production. Lancet 1:42.
Bernstein, R.M., C.C. Bunn, G.R.V. Hughes, A.M. Fran-coeur, and M.B. Mathews. 1984. Cellular RNA and pro-tein antigens in autoimmune disease. Mol. Biol. Med.2: 105-120.
Bernstein, R.M., S.H. Morgan, J. Chapman, C.C. Bunn, M.B.Mathews, M. Turner-Warwick, and G.R.V. Hughes. 1984.
61

Anti-Jo-1 antibody: A marker for myositis with interstitiallung disease. Br. Med. J. 289: 151-152.
Francoeur, A.M., E.K.L. Chan, J.I. Garrets, and M.B. Ma-thews. 1984. Characterization and purification of lupusantigen La, an RNA binding protein. Mol. Cell. Biol.5: 586-590.
Mathews, M.B. and A.M. Francoeur. 1984. La antigen rec-ognizes and binds to the 3' oligouridylate tail of a smallRNA. Mol. Cell. Biol. 4: 1134-1140.
Mathews, M.B., R.M. Bernstein, R. Franza, and J.I. Carrels.1984. Identity of the proliferating cell nuclear antigen andcyclin. Nature 309: 374-376.
Mathews, M.B., M. Reichlin, G.R.V. Hughes, and R.M. Bern-stein. 1984. Anti-threonyl-tRNA synthetase, a secondmyositis-related autoantibody. J. Exp. Med. 160:420-434.
Reichel, P.A., W.C. Merrick, J. Siekierka, and M.B. Mathews.1985. Regulation of a protein synthesis initiation factor byadenovirus virus associated RNA,. Nature 313: 196-200.
Thomas, G.P. and M.B. Mathews. 1984. Alterations of tran-scription and translation in HeLa cells exposed to aminoacid analogs. Mol. Cell. Biol. 4: 1063-1072.
In Press, Submitted, and In Preparation
Asselbergs, PAM., A.M. Francoeur, and M.B. Mathews.Expression of adenovirus VA RNA in Xenopus oocytes,and its binding to the La antigen. (In preparation.)
PROTEIN CHEMISTRY
C. Slaughter T FischerC. Moomaw
A Protein Chemistry laboratory has been estab-lished for the isolation and chemical character-ization of the protein products of normal andtransformed cells. The facility has been equippedboth with conventional chromatography systemsto isolate proteins available in quantities largeenough for X-ray crystallography (100 mg ormore) and with high-performance liquid chro-matography (HPLC) and electrophoresis systemsto isolate minor cell proteins in microgram quan-tities for chemical analysis. Two automatedamino acid sequencers have been brought intooperation. The first, a Beckman model 890C,which was on site previously, has been upgradedto model 890M by the addition of a microproces-sor controller and an autoconverter system. Thisinstrument is now being used to perform amino-terminal amino acid sequence analyses on protein
62
Bernstein, R.M., S.H. Morgan, C.C. Bunn, R.C. Gainey,G.R.V. Hughes, and M.B. Mathews. The SL autoanti-body-antigen system: Clinical and biochemical studies.(Submitted.)
Bunn, C.C., R.M. Bernstein, and M.B. Mathews. Autoanti-bodies to alanyl-tRNA synthetase and tRNAAIa coexist andare associated with myositis. (Submitted.)
Moran, E., M.B. Mathews, R.J. Roberts, T. Grodzicker, andB.R. Zerler. Lytic and transforming properties of adeno-viruses expressing the individual E1A products. (Submit-ted.)
Sadaie, M.R. and M.B. Mathews. Immunochemical and bio-chemical analysis of the proliferating cell nuclear antigen(PCNA) in human HeLa cells. (Submitted.)
Siekierka, J., T.M. Mariano, P.A. Reichel, and M.B. Mathews.1985. Translational control by adenovirus: Lack of VARNA, adenovirus infection results in the phosphorylationof initiation factor elF-2 and inhibition of protein synthe-sis. Proc. Natl. Acad. Sci. (in press).
Thomas, G.P. and M.B. Mathews. Structure of the HeLa cellgene encoding the 90,000 dalton heat-shock protein. (Inpreparation.)
Thomas, G.P, J.I. Carrels, and M.B. Mathews. Induction ofthe stress response in HeLa cells and chick fibroblasts.(In preparation.)
Zerler, B.R., E. Moran, and H.E. Ruley. Establishment oftransformation by adenovirus E1A mutants and cDNAs.(In preparation.)
and peptide samples of between 500 pmoles and5 nmoles. The second sequencer, an AppliedBiosystems model 470A, has been newly installedand has been used for protein and peptide sam-ples of between 10 pmoles and 20 nmoles. It hasbeen possible to identify up to 45 amino acids inaminoterminal runs on unknown samples usingboth instruments.
Phenylthiohydantoin (PTH)-amino acids gen-erated by the sequencers are being identified on anewly installed Waters automated HPLC systemby reverse-phase chromatography on a WatersNova Pak C18 column. The procedure has provedto be capable of quantitating 10 pmoles of eachPTH amino acid on a routine basis and detectsconsiderably less. Since the turnaround time foreach analysis is 23 minutes, and the cycle time oneach sequencer is approximately 1 hour, it is

possible for the PTH-amino acid identificationsystem to keep pace with the simultaneous outputof both amino acid sequencers indefinitely.Amino acid sequencing and PTH-amino acididentification are being conducted by T. Fischer.
Estimation of amino acid composition of pep-tides and proteins is being carried out on a newlyinstalled Waters automated HPLC system em-ploying the PICO.TAG procedure. In this pro-cedure, phenylthiocarbamate (PTC)-amino acidderivatives are synthesized in a one-step manualoperation, and the derivatives are separated by re-verse-phase chromatography on a Waters NovaPak C18 column. The PTC derivatives can bequantitated in the low picomole range by their in-trinsic absorption at 254 nm. The turnaroundtime for the separation is 20 minutes.
The synthesis of peptides for the production ofsequence-specific antibodies has been initiatedusing a Beckman model 990 solid-phase peptidesynthesizer. The peptides are subject to verifica-tion prior to uncoupling from their solid supportby sequence analysis on the Applied Biosystemsmodel 470A. After hydrogen fluoride cleavage,purity of the uncoupled peptides is subject tofurther checking by reverse-phase HPLC andby amino acid analysis. Amino acid analysisand peptide synthesis are being conducted byC. Moomaw.
We are currently exploring procedures for ex-tended carboxyterminal amino acid sequencing ofboth proteins and peptides using reverse-phaseHPLC to identify dipeptides cleaved from thecarboxyl terminus by dipeptidyl carboxypeptid-ase digestion. We hope to be able to carry outhigh-sensitivity sequence analyses at both aminoand carboxyl termini of all protein and peptidesamples studied in the laboratory.
Peptide-mapping procedures employing bothgel permeation and reverse-phase HPLC separa-tions in volatile buffer systems have been estab-lished. HPLC peptide mapping has the advan-tage over traditional gel electrophoresis proce-dures that the peptides are readily collected in aform suitable for further chemical analysis. Thecoupling of high-resolution peptide mapping withhigh-sensitivity analytical procedures will provideus with a powerful means of investigating thestructural alterations involved in the processingof the protein products of oncogenes and tumorviruses.
Our laboratory has performed 2566 cycles ofEdman degradation during its first 10 months ofoperation and has been involved with nine proj-ects, seven of which were performed in collabo-ration with scientists on the grounds of ColdSpring Harbor Laboratory. Work on the pro-teins of interest in the field of cell growth has beenequally divided between the human alkalinephosphatases, a family of cell-surface enzymesthat are used extensively as tumor markers inclinical practice, and rat brain protein kinase C,an intracellular enzyme of the plasma membranethat is a receptor for phorbol ester tumor pro-moters. The latter project is being undertaken incollaboration with J. Feramisco's group (Cell Bi-ology Section).
Structural Diversity of HumanAlkaline PhosphatasesC. Slaughter, S. Cheley, T. Fischer, C. Moomaw
The research on alkaline phosphatases (APs) rep-resents the principal focus of the laboratory's in-terest. APs are a family of membrane-associatedenzyme glycoproteins that hydrolyze phosphateesters with a high pH optimum. APs from var-ious tissues differ in biochemical, immunochem-ical, and genetic characteristics, suggesting thatthey are encoded by a family of related structuralgenes. Some of these genes are expressed in ahighly tissue-specific manner, whereas others havea more general pattern of expression. In addi-tion, some members of the gene family are ex-pressed ectopically in tumors and in cell lines de-rived from tumors. In many respects, the APsprovide a useful model system for exploring thegeneral characteristics of multigene families. Theoverall aim of the work is to determine the extentof structural and genetic diversity in both normaland ectopically expressed APs. This problem isbeing approached at both the nucleic acid andprotein levels.
Aminoterminal amino acid sequences havebeen obtained for the products of the principalgenes expressed in human placenta, intestine, andliver. The results confirm the conclusion drawnpreviously from genetic and biochemical evi-
63

dence that different genes are involved in codingfor these tissue-specific forms of AP. The ami-noterminal amino acid sequences of adult and fe-tal intestinal APs have also been compared, andalthough these proteins display distinct antigenicproperties and peptide-mapping patterns, theirsequences thus far appear identical at the aminoterminus. Placental AP has been subjected todigestion with cyanogen bromide, and peptideshave been isolated by reverse-phase HPLC. Inthis way, a variety of amino acid sequences from
64
regions of the molecule other than the amino ter-minus have been deduced. This information hasbeen used by M. Zoller (Molecular Genetics Sec-tion) to synthesize oligonucleotides with which S.Cheley, in collaboration with D. Helfman (CellBiology Section), is screening suitable cDNA li-braries. The clones identified in this way willprovide the means of deducing the completeamino acid sequences of human placental AP andof investigating the genetic and structural com-plexity of a major part of the AP family.

MOLECULAR GENETICS OF EUKARYOTIC CELLS
Included in this section are a diverse range of systems, spanning from yeast toman, and topics, spanning from the control of protein localization to develop-mental biology. The common thread is the application of new methods of molec-ular genetics to long standing biological problems. J. Sambrook and M.-J. Ge-thing (Protein Traffic) describe investigations of protein modification, localization,secretion, and protein assembly in animal cells. Using an influenza hemagglu-tinin gene expression system and mutants constructed in vitro, they are outliningthe pathways for these processes. J. Lewis (Cell-cycle-specific Expression) hascloned the hamster thymidine kinase gene and is examining the mechanisms bywhich expression of this gene is regulated during the cell cycle. D. Kurtz (Hor-monal and Developmental Control of Gene Expression) describes the study ofhormonal and tissue-specific expression of a family of genes encoding closelyrelated proteins, the rat a2,, serum globulins. M. Wig ler (Genetics of Cell Prolifera-tion) details studies on human oncogenes, their isolation, structure, and function.The RAS oncogenes are highly conserved in evolution, and the function of horn-clogs in yeast are also being explored. This work has been carried out in collabo-ration with F. Tamanoi (Biochemistry of Yeast RAS Proteins). L. Silver (Mouse Ge-netics) has been investigating the molecular basis of a group of closely linkeddevelopmental mutants of mice, which together comprise the T locus. These mu-tants are characterized by early developmental death in homozygotes, segrega-tion distortion, and locus-specific inhibition of meiotic recombination. Finally, D.Hanahan (Transgenic Mice) and his group are redesigning the mouse by placinggenes under new controls into the mouse germ line. This experimental systempromises to be one of the most powerful approaches to understanding geneexpression in the developing animal.
PROTEIN TRAFFIC
J. SambrookM.-J. Gething
E. HunterP BirdP GallagherJ. Hearing
M. Roth L. RodgersC. Doyle S. SharmaJ. Henneberry D. York
The general aims of our laboratory are to map themajor routes of protein traffic within mammal-ian cells and to understand how proteins are di-rected to specific destinations.
THE EXOCYTIC PATHWAY
Most of our work on the exocytic pathway in-volves the hemagglutinin (HA) of influenza virus,which is now the best studied of a large class of
integral membrane proteins. Until a few yearsago, work on HA depended almost entirely on thetechniques of protein chemistry (Waterfield et al.,Br. Med. Bull. 35: 57 [1979]), but since the adventof recombinant DNA techniques, the genes en-coding the HAs of many strains of influenza vi-rus have been cloned and sequenced, and this hasallowed the complete amino acid sequence ofmany serotypes of the protein to be deduced (forreview, see Lamb, in Genetics of Influenza Vi-
65
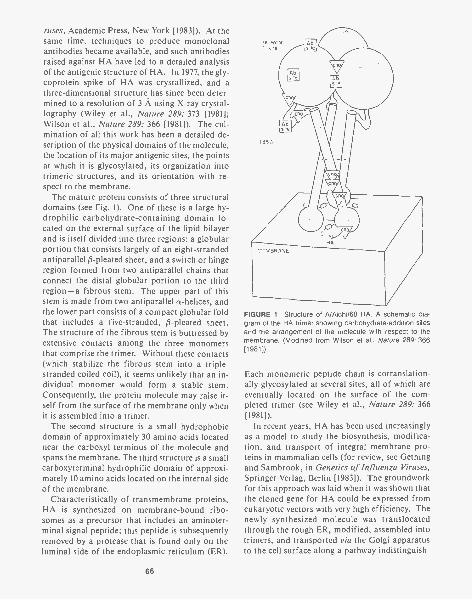
ruses, Academic Press, New York [1983]). At thesame time, techniques to produce monoclonalantibodies became available, and such antibodiesraised against HA have led to a detailed analysisof the antigenic structure of HA. In 1977, the gly-coprotein spike of HA was crystallized, and athree-dimensional structure has since been deter-mined to a resolution of 3 A using X-ray crystal-lography (Wiley et al., Nature 289: 373 [1981];Wilson et al., Nature 289: 366 [1981]). The cul-mination of all this work has been a detailed de-scription of the physical domains of the molecule,the location of its major antigenic sites, the pointsat which it is glycosylated, its organization intotrimeric structures, and its orientation with re-spect to the membrane.
The mature protein consists of three structuraldomains (see Fig. 1). One of these is a large hy-drophilic carbohydrate-containing domain lo-cated on the external surface of the lipid bilayerand is itself divided into three regions: a globularportion that consists largely of an eight-strandedantiparallel 0-pleated sheet, and a switch or hingeregion formed from two antiparallel chains thatconnect the distal globular portion to the thirdregion- a fibrous stem. The upper part of thisstem is made from two antiparallel a-helices, andthe lower part consists of a compact globular foldthat includes a five-stranded, 0-pleated sheet.The structure of the fibrous stem is buttressed byextensive contacts among the three monomersthat comprise the trimer. Without these contacts(which stabilize the fibrous stem into a triple-stranded coiled coil), it seems unlikely that an in-dividual monomer would form a stable stem.Consequently, the protein molecule may raise it-self from the surface of the membrane only whenit is assembled into a trimer.
The second structure is a small hydrophobicdomain of approximately 30 amino acids locatednear the carboxyl terminus of the molecule andspans the membrane. The third structure is a smallcarboxyterminal hydrophilic domain of approxi-mately 10 amino acids located on the internal sideof the membrane.
Characteristically of transmembrane proteins,HA is synthesized on membrane-bound ribo-somes as a precursor that includes an aminoter-minal signal peptide; this peptide is subsequentlyremoved by a protease that is found only on theluminal side of the endoplasmic reticulum (ER).
66
FIGURE 1 Structure of A/Aichi/68 HA. A schematic dia-gram of the HA trimer showing carbohydrate-addition sitesand the arrangement of the molecule with respect to themembrane. (Modified from Wilson et al., Nature 289: 366[1981]).
Each monomeric peptide chain is cotranslation-ally glycosylated at several sites, all of which areeventually located on the surface of the com-pleted trimer (see Wiley et al., Nature 289: 366[1981]).
In recent years, HA has been used increasinglyas a model to study the biosynthesis, modifica-tion, and transport of integral membrane pro-teins in mammalian cells (for review, see Gethingand Sambrook, in Genetics of Influenza Viruses,Springer-Verlag, Berlin [1983]). The groundworkfor this approach was laid when it was shown thatthe cloned gene for HA could be expressed fromeukaryotic vectors with very high efficiency. Thenewly synthesized molecule was translocatedthrough the rough ER, modified, assembled intotrimers, and transported via the Golgi apparatusto the cell surface along a pathway indistinguish-

able from that taken by a large number of endog-enous cellular proteins.
These initial findings were extended to includea number of mutant forms of HA, in which DNAsequences coding for the putative hydrophobicsignal and anchor regions of the molecule weredeleted. Analysis of the expression of the result-ing constructs led to the conclusions (1) that the 2.hydrophobic aminoterminal sequences are neces-sary to cause translocation of the newly synthe-sized molecule across the membrane of the roughER and (2) that the hydrophobic carboxyterminalsequences are sufficient to anchor the completedmolecule in the plasma membrane. Deletion ofthese sequences results in secretion of a truncatedHA from the cell (Gething and Sambrook, Na-ture 300: 598 [1982]).
ALTERATIONS TO THE AMINO TERMINUS ANDCYTOPLASMIC TAIL OF HA CAN AFFECTTRANSPORT OF THE PROTEIN ALONGTHE EXOCYTIC PATHWAY
During the last year, we have constructed andanalyzed a number of additional mutant and chi-meric forms of HA in which the DNA sequencescoding for the signal sequence, the transmem-brane region, and the cytoplasmic domain weremutated at specific sites or replaced by sequencesencoding analogous regions from other proteins.The results of these experiments are summarizedin Table 1 and discussed briefly below:
1. Substitution of the signal sequence of HA withthose from other glycoproteins (the env gly-coprotein of Rous sarcoma virus [RSV] or bo-vine preprochymosin) yields chimeric proteinsthat are translocated into the lumen of the ER,where they undergo core glycosylation. Inboth cases, the signal sequences are cleavedfrom the precursor, leaving an HA in whichthe first two amino acids have been replacedby either 9 (in the case of RSV env) or 12 (pre-prochymosin) novel residues. Neither of thesechimeric proteins is folded correctly (see be-low) and neither is transported further alongthe exocytic pathway to the Golgi apparatus.However, by deleting the DNA sequences cod-ing for the additional aminoterminal aminoacids, we have been able to restore the abilityof the env-HA protein to fold into a form thatis efficiently transported to the cell surface.
3.
We conclude that a signal sequence obtainedfrom a different eukaryotic protein is suffi-cient to guarantee translocation of HA into thelumen of the rough ER. Further movement ofHA along the exocytic pathway is highly de-pendent on the configuration of the amino ter-minus of the protein.Replacement of the transmembrane region andcytoplasmic tail of HA with analogous se-quences from the G glycoprotein of vesicularstomatitis virus (VSV) or the C glycoprotein ofherpes simplex virus yields chimeric glycopro-teins that are transported to the cell surface,where they are displayed in an antigenicallyand biologically active state. The HA-G chi-mera is transported to the surface as effi-ciently as wild-type HA, whereas HA-gC trav-erses the early part of the pathway (from theER to the Golgi apparatus) more slowly andasynchronously. We conclude that the an-choring function of the transmembrane re-gions of HA can be provided by hydrophobicsequences derived from different viral glyco-proteins, without crippling the ability of thechimeric molecule to move through the exocy-tic pathway.Certain alterations to the cytoplasmic tail ofHA drastically lower the rate at which the mol-ecule travels from its site of synthesis to theGolgi apparatus; other alterations affecttransport of HA from the Golgi apparatus tothe cell surface (see Table 1). These results canbe interpreted two ways: Either the cyto-plasmic tail of HA carries signals that deter-mine the rate or efficiency with which the en-tire molecule progresses along the exocyticpathway or the tail is important for the struc-tural integrity of the molecule, and changes inrates of transport are a consequence of an in-ability to fold into or maintain a stable struc-ture.
To distinguish between these possibilities, wehave used a number of approaches to comparethe three-dimensional structure and folding ofwild-type and mutant proteins. An antiserumraised against denatured HA that recognizes anepitope(s) cryptic in the external region of maturewild-type HA reacts strongly with mutant pro-teins that are blocked in the ER (env-HA andHAxpBR). Second, these mutants, unlike wild-
67
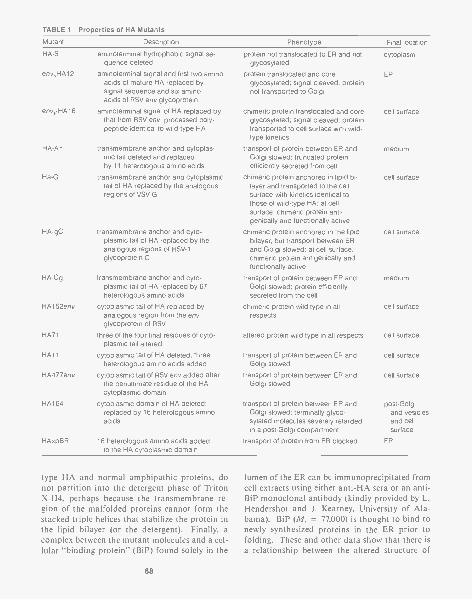
TABLE 1 Properties of HA Mutants
Mutant Description Phenotype Final location
HA-S- aminoterminal hydrophobic signal se-quence deleted
env,HAl2 aminoterminal signal and first two aminoacids of mature HA replaced bysignal sequence and six aminoacids of RSV env glycoprotein
env,-HA16 aminoterminal signal of HA replaced bythat from RSV env; processed poly-peptide identical to wild-type HA
HA-A- transmembrane anchor and cytoplas-mic tail deleted and replacedby 11 heterologous amino acids
Ha-G transmembrane anchor and cytoplasmictail of HA replaced by the analogousregions of VSV G
HA gC transmembrane anchor and cyto-plasmic tail of HA replaced by theanalogous regions of HSV-1glycoprotein C
HA-Cg transmembrane anchor and cyto-plasmic tail of HA replaced by 67heterologous amino acids
HA152env cytoplasmic tail of HA replaced byanalogous region from the envglycoprotein of RSV
HA71 three of the four final residues of cyto-plasmic tail altered
HA11 cytoplasmic tail of HA deleted; threeheterologous amino acids added
HA477env cytoplasmic tail of RSV env added afterthe penultimate residue of the HAcytoplasmic domain
HA164 cytoplasmic domain of HA deleted;replaced by 16 heterologous aminoacids
HAxpBR 16 heterologous amino acids addedto the HA cytoplasmic domain
protein not translocated to ER and not cytoplasmglycosylated
protein translocated and core ER
glycosylated; signal cleaved; proteinnot transported to Golgi
chimeric protein translocated and core cell surfaceglycosylated; signal cleaved; proteintransported to cell surface with wild-type kinetics
transport of protein between ER and mediumGolgi slowed; truncated proteinefficiently secreted from cell
chimeric protein anchored in lipid bi- cell surfacelayer and transported to the cellsurface with kinetics identical tothose of wild-type HA; at cellsurface, chimeric protein anti-genically and functionally active
chimeric protein anchored in the lipid cell surfacebilayer, but transport between ERand Golgi slowed; at cell surface,chimeric protein antigenically andfunctionally active
transport of protein between ER and mediumGolgi slowed; protein efficientlysecreted from the cell
chimeric protein wild type in all cell surfacerespects
altered protein wild type in all respects cell surface
transport of protein between ER andGolgi slowed
transport of protein between ER andGolgi slowed
cell surface
cell surface
transport of protein between ER and post-GolgiGolgi slowed; terminally glyco- and vesiclessylated molecules severely retarded and cellin a post-Golgi compartment surface
transport of protein from ER blocked ER
type HA and normal amphipathic proteins, donot partition into the detergent phase of TritonX-114, perhaps because the transmembrane re-gion of the malfolded proteins cannot form thestacked triple helices that stabilize the protein inthe lipid bilayer (or the detergent). Finally, acomplex between the mutant molecules and a cel-lular "binding protein" (BiP) found solely in the
68
lumen of the ER can be immunoprecipitated fromcell extracts using either anti-HA sera or an anti-BiP monoclonal antibody (kindly provided by L.Hendershot and J. Kearney, University of Ala-bama). BiP (M, = 77,000) is thought to bind tonewly synthesized proteins in the ER prior tofolding. These and other data show that there isa relationship between the altered structure of

mutant forms of HA and their ability to be trans-ported to the cell surface. Apparently, HA needsto be correctly folded in order to move efficientlyalong the exocytic pathway.
FOLDING PATHWAY OF HA
How a protein that is synthesized on one side ofa membrane folds into a specific three-dimen-sional shape on the other is not known. The fol-lowing is our current working model (see Fig. 2):The monomeric polypeptide is translocated es-sentially as a linear string of amino acids. It re-mains attached to the luminal face of the mem-brane not only by the carboxyterminal anchor,but also by the aminoterminal hydrophobic sig-nal. After primary glycosylation is completed,but before trimerization has occurred, the distalregion of the molecule folds into its globularshape and disulfide bonds form. The carboxy-terminal hydrophobic domains of three mono-mers then associate to form a triple-stranded hel-ical coil within the membrane. The three stemregions are then able to twist around each otherto form the coiled coil that comprises the bulk ofthe external fibrous spike. After cleavage of the
-
signal peptide, the completed trimer is free tomove in the plane of the membrane of the ERtoward the Golgi apparatus. Trimerization istherefore a prerequisite for movement. Thismodel is consistent with the structural analysis ofwild-type HA (Wilson et al., Nature 289: 366[1981]; Wiley et al., Nature 289: 373 [1981]) andprovides a plausible explanation for the phe-notype of the mutants. Thus, alterations to theamino and carboxyl termini, although in regionsof the protein that reside on opposite sides of therough ER, might be expected to prevent or delayfolding and oligomerization of the molecule. Inthe most severely affected mutants (HAxpBR andenv-HA), the molecule may be completely unableto form trimers; in other cases (see Table 1) a dis-ordered trimeric structure may retard movementof the protein in the plane of the membrane orobstruct its incorporation into transport vesiclesthat ferry it from the ER to the Golgi apparatus.
These ideas raise many questions: How does thecellular transport machinery distinguish betweenmolecules that are correctly folded and those thatare not? Does malfolding of HA affect its inter-action with hypothetical sets of cellular receptorsthat may be involved in movement of proteins
AAAA
FIGURE 2 A model for the folding of HA and assembly into monomers
69

along the exocytic pathway? If so, are these re-ceptors located in the lumen of the ER, within thelipid bilayer, or on the cytoplasmic face of themembrane? Are there distinct sets of receptorsthat transport proteins through different stagesof the exocytic pathway? We hope that thesequestions can be answered at least in part by fur-ther mutational analysis of HA and the elucida-tion of the mechanism by which the protein foldsand trimerizes.
ROLE OF CARBOHYDRATE IN THE STRUCTUREAND BIOLOGICAL FUNCTION OF HA
Glycosylation of proteins is universal in eukar-yotic organisms. The fact that such a complexand expensive process has been conserved oversuch a long period of evolutionary time suggestsstrongly that glycosylation must be of considera-ble advantage to higher organisms. However,there is no evidence for a single function for gly-cosylation. Instead, it appears (for review, seePollack and Atkinson, J. Cell Biol. 97: 293 [1983])that N-linked oligosaccharides may play a varietyof roles including (1) the maintenance of thestructure and stability of glycoproteins, (2) thedirection of glycoproteins to various intracellularorganelles, and (3) the specific interaction be-tween cells that occurs during tissue organiza-tion.
The oligosaccharide groups attached to HA arethought to serve two functions -to modulate an-tigenicity by shielding potential antigenic sites(Knossow et al., Nature 311: 678 [1984]) and tostabilize the protein structure and thus facilitatethe transport of the protein along the exocyticpathway. The magnitude of this latter effect var-ies widely between HAs of different strains. Thus,the nonglycosylated protein from the A/Aichi/68strain is stable but does not reach the cell surface,whereas that from the A/Japan/57 strain is trans-ported to the cell membrane, albeit at a reducedlevel. Most likely, lack of glycosylation preventsAichi HA from folding into its correct configu-ration for transport; in the case of Japan HA, themolecule is transported to the cell surface, but itis more susceptible than wild-type HA to proteo-lysis.
To study these problems further, we have usedsite-directed mutagenesis to alter the canonicalsequences specifying each of the seven carbohy-
70
drate attachment signal sites on Aichi HA. Thealtered genes have been inserted into SV40 vec-tors and expressed in CV-1 cells. The rate of bio-synthesis, transport, cellular localization, and bi-ological activity of these altered gene products isbeing analyzed in terms of the contribution of theindividual side chains. We are also using site-di-rected mutagenesis to create sites for oligosac-charide addition in areas of the HA surface thatdo not normally carry carbohydrate groups. Wehope to use the novel glycan side chains to shieldparticular surfaces of the molecule, e.g., those in-volved in transport signals or in the recognitionof HA on the membrane of histocompatible tar-get cells by cytotoxic lymphocytes.
EXPRESSION OF HA IN POLARIZEDEPITHELIAL CELLS
The problem of protein traffic is particularlyacute in polarized epithelial cells, in which se-creted and membrane-bound proteins are di-rected to either of two distinct domains of theplasmalemma. These two domains, the apical andthe basolateral, are separated by the zonula oc-cludans, a circumferential belt of tight junctionsthat serves as a barrier to lateral diffusion of pro-teins and lipids in the plane of the plasma mem-brane. Polarized epithelial cells direct both theirown endogenous surface proteins and the glyco-proteins of the viruses that infect them to specificdomains of the plasma membrane (Rodriguez-Boulan, in Modern Cell Biology, A.R. Liss, NewYork [1983]). Thus, during the early stages of in-fection of kidney epithelial cells by influenza vi-rus, HA is expressed almost exclusively at the ap-ical surface of the cell, through which the nascentviral particles will later bud. The infected cellrecognizes HA as belonging to the "apical" set ofproteins, and this recognition has been shown tobe independent of either glycosylation of HA orof the presence of other viral proteins (Roth etal., Cell 33: 435 [1983]). The features of the HApolypeptide recognized by the cellular sortingmachinery could reside either in the large ecto-domain exposed to the luminal environment ofthe exocytic pathway, in the transmembrane re-gion, or in the small carboxyterminal cytoplasmicdomain. We have carried out two types of exper-iments to investigate which regions of HA are im-portant for sorting. First, we have determined the

pattern of secretion in polarized cells of a formof HA lacking the transmembrane and cyto-plasmic domains. This form of HA (HAwc) haspreviously been shown to form trimers and to dis-play all of the biological activities that reside ex-clusively in the ectodomain (Gething and Sam-brook, Nature 300: 598 [1982]). Second, in col-laboration with E. Rodriguez-Boulan (CornellMedical School), we are investigating the distri-bution on the surface of polarized cells of formsof HA in which the transmembrane or cyto-plasmic regions have been replaced with those ofanother glycoprotein. During the early phase ofinfection of MA104 rhesus monkey kidney cellswith a recombinant SV40 vector expressingHFs."c, secretion of HAs'c is almost entirely fromthe apical surface. Thus, the transmembrane andcytoplasmic domains of HA are not needed forthe directional transport of HA in this system.Preliminary results indicate that chimeric glyco-proteins that contain transmembrane and cyto-plasmic domains from viral glycoproteins nor-mally expressed on the basolateral surface areconcentrated on the apical surface of infectedMA104 cells. A simple interpretation of theseobservations is that the large ectodomain of HAcontains dominant features recognized by the cel-lular sorting machinery. We conclude that dis-crimination between apical and basolateral pro-teins probably occurs within the lumen of theorganelle(s) of the exocytic pathway.
While analyzing the biosynthesis of these chi-meric glycoproteins, we realized that they dif-fered from wild-type HA in certain interesting re-spects. In particular, a chimera consisting of theectodomain of HA and the transmembrane andcytoplasmic regions of VSV G protein becamecleaved by protease, generating a protein consist-ing of two subunits (HA, and HA,G) joined by adisulfide bond. The transport of HA-G musttherefore at some stage diverge from that of wild-type HA, causing the chimeric protein to enter acellular compartment in which the relevant pro-tease(s) is active. The kinetics of cleavage are con-sistent with the idea that the chimeric protein en-ters this compartment at about the same rate as itreaches the cell surface, raising the possibility thatHA-G is transported along the conventional exo-cytic pathway and is then removed rapidly fromthe cell surface by endocytosis: Proteolytic cleav-age would therefore occur in endosomes. We are
currently trying to test aspects of this hypothesisand to determine whether the signals that divertHA-G into this novel compartment are carried onthe transmembrane region or the cytoplasmic tailof the protein.
SV40 T ANTIGEN AND THE EXOCYTIC PATHWAY
Proteins tend to have singular locations in cells.Thus, proteins present in one site (e.g., the cellsurface) are by and large not detected in signifi-cant quantities elsewhere. Because specific loca-tions are served by specific transport pathways,proteins generally need to carry only one set oftransport signals. For example, proteins that aredestined for the cell surface almost always con-tain hydrophobic regions that are readily identi-fiable as signal sequences. The large T antigen ofSV40 does not fit easily into this simple pattern.Newly synthesized T antigen is rapidly trans-ported from the cytoplasm into the nucleus of in-fected or transformed cells where it is believed toassociate with cellular p53 and with specific setsof DNA sequences. However, a small proportionof T antigen is also displayed on the surfaces oftransformed cells, where it can be detected in aspecific configuration by a variety of immuno-logical and biochemical techniques (for review,see Rigby and Lane, Adv. Viral Oncol. 3: 31[1983]). However, in contrast to typical trans-membrane proteins, T antigen carries no hydro-phobic signal sequence at or near its amino ter-minus and in fact it lacks any sequence ofhydrophobic or uncharged amino acids longenough to span a membrane. There is no obviousway to explain how considerable portions of Tantigen are translocated across a membrane orwhere in the cell this process may occur.
To approach this problem, we decided to testwhether it would be possible to increase the effi-ciency with which T antigen could be transportedto the cell surface by equipping it with a conven-tional signal peptide at its amino terminus. Wetherefore constructed a chimeric gene consistingof the DNA sequences coding for the aminoter-minal hydrophobic signal peptide of HA fused in-frame to those coding for SV40 T antigen. Thegene was linked to the murine metallothioneinpromoter and inserted into a plasmid containinga transforming fragment of bovine papilloma vi-rus (BPV) DNA. The resulting vector was used
71
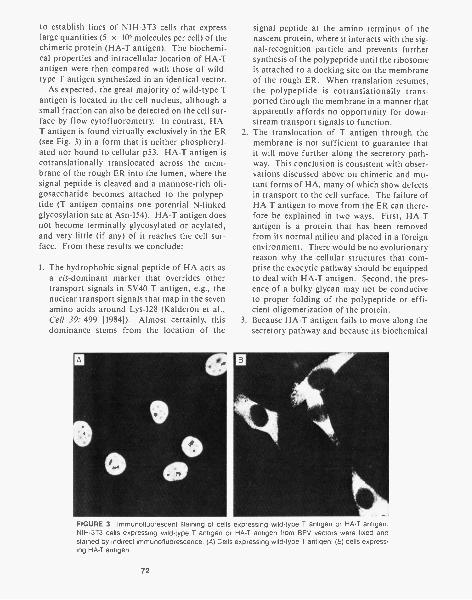
to establish lines of NIH-3T3 cells that expresslarge quantities (5 x 106 molecules per cell) of thechimeric protein (HA-T antigen). The biochemi-cal properties and intracellular location of HA-Tantigen were then compared with those of wild-type T antigen synthesized in an identical vector.
As expected, the great majority of wild-type Tantigen is located in the cell nucleus, although asmall fraction can also be detected on the cell sur-face by flow cytofluorometry. In contrast, HA-T antigen is found virtually exclusively in the ER 2.(see Fig. 3) in a form that is neither phosphoryl-ated nor bound to cellular p53. HA-T antigen iscotranslationally translocated across the mem-brane of the rough ER into the lumen, where thesignal peptide is cleaved and a mannose-rich oli-gosaccharide becomes attached to the polypep-tide (T antigen contains one potential N-linkedglycosylation site at Asn-154). HA-T antigen doesnot become terminally glycosylated or acylated,and very little (if any) of it reaches the cell sur-face. From these results we conclude:
1. The hydrophobic signal peptide of HA acts asa cis-dominant marker that overrides othertransport signals in SV40 T antigen, e.g., thenuclear transport signals that map in the sevenamino acids around Lys-128 (Kalderon et al.,Cell 39: 499 [1984]). Almost certainly, this 3.
dominance stems from the location of the
signal peptide at the amino terminus of thenascent protein, where it interacts with the sig-nal-recognition particle and prevents furthersynthesis of the polypeptide until the ribosomeis attached to a docking site on the membraneof the rough ER. When translation resumes,the polypeptide is cotranslationally trans-ported through the membrane in a manner thatapparently affords no opportunity for down-stream transport signals to function.The translocation of T antigen through themembrane is not sufficient to guarantee thatit will move further along the secretory path-way. This conclusion is consistent with obser-vations discussed above on chimeric and mu-tant forms of HA, many of which show defectsin transport to the cell surface. The failure ofHA-T antigen to move from the ER can there-fore be explained in two ways. First, HA-Tantigen is a protein that has been removedfrom its normal milieu and placed in a foreignenvironment. There would be no evolutionaryreason why the cellular structures that com-prise the exocytic pathway should be equippedto deal with HA-T antigen. Second, the pres-ence of a bulky glycan may not be conduciveto proper folding of the polypeptide or effi-cient oligomerization of the protein.Because HA-T antigen fails to move along thesecretory pathway and because its biochemical
FIGURE 3 lmmunofluorescent staining of cells expressing wild-type T antigen or HA-T antigen.NIH-313 cells expressing wild-type T antigen or HA-T antigen from BPV vectors were fixed andstained by indirect immunofluorescence. (A) Cells expressing wild-type T antigen; (8) cells express-ing HA-T antigen.
72

properties are very different from those ofwild-type T antigen, it seems unlikely to us thatwild-type T antigen reaches the cell surface bytraveling along the conventional exocytic route.We are therefore left with a paradox. At somestage in its life, surface T antigen must cross amembrane in order to be presented on the out-side of the cell: Yet, it has none of the hall-marks of an integral or transmembrane pro-tein. We therefore postulate (see Fig. 4) thatin SV40-transformed cells, all newly synthe-sized T antigen enters the nucleus, where it be-comes associated either directly or via inter-mediate molecules with the inner wall of thenuclear membrane. At cell division, thismembrane is known to break down into smallvesicles that become dispersed throughout thecytoplasm. If after cytokinesis, a minority ofthese vesicles fuses directly with the plasmamembrane or with other vesicles that arebound for the surface, their contents could beeverted onto the outer face of the plasmamembrane. If this hypothesis stands up to ex-perimental test, it will be interesting to deter-mine whether normal cellular proteins travelalong this novel pathway from the nucleus tothe cell surface.
MUTANTS OF MAMMALIAN CELLS THAT AREDEFECTIVE IN TRANSPORT OF PROTEINS
A complete understanding of the exocytic path-way will require not only mutational analysis of
A
GOLGI
HA-T
VaENDOPLASMIC
RETICULUM 4111111110ANINIMIEW11
MEMBRANEBOUND
NUCLEUS RIBOSOMES
proteins like HA, but also the identification andanalysis of cellular components involved in theestablishment and maintenance of the pathway.A good way to identify such components wouldbe to isolate and characterize mutants that specif-ically affect particular steps in the cell's exocyticmachinery. This approach, which has been ex-traordinarily successful in studies of the secretorypathway of Saccharomyces cerevisiae (for review,see Schekman and Novick, in The Molecular Bi-ology of the Yeast Saccharomyces cerevisiae,Cold Spring Harbor Laboratory, Cold SpringHarbor, New York [1982]), has not yet been fullyexploited in mammalian systems, partly becauseof formidable technical difficulties and partly be-cause of the lack of a good selective system toisolate mutants of the correct type.
In an effort to solve some of these problems,we have recently isolated and characterized setsof murine and hamster cell lines that constitu-tively express large quantities of HA on the cellsurface. An important feature of these cell linesis that they carry multiple copies of the HA gene(in BPV vectors) in either an integrated state oran episomal state. Because it seems unlikely thatall of these copies could be mutated simultane-ously, we should be able to avoid the problem ofisolating mutants that fail to express HA on theirsurface merely because of mutations in HA itself.
These cell lines were originally selected for theirability to express large quantities of HA on theirsurface (approximately 107 molecules/cell in thebest case). This amount is more than 100 times
7
0FREE
POLYRIBOSOMES
NUCLEARPORE
SURFACE T ANTIGEN- ---->
NUCLEAR NUCLEUST ANTIGEN
GOLGI
FIGURE 4 A model to illustrate the intracellula location of HA-T antigen (A) and a possible schemefor the transport of wild-type T antigen through the nucleus to the cell surface (B).
73

the threshold required for detection of HA bysurface immunofluorescence and 10 times morethan the amount needed to bind erythrocytes.However, a puzzling characteristic of the originalcell lines was the variation in expression of HAfrom cell to cell, as detected by immunofluores-cence. This problem was solved in September,1984, when we obtained a Coulter Epics C fluo-rescence-activated cell sorter (Fig. 5). This ma-chine has been invaluable in allowing us to obtainderivatives of the original cell lines that homoge-neously express very high levels of HA on theirsurfaces. Examples of typical data are shown inFigure 6. We are at present analyzing these cellpopulations to determine why they express HA soefficiently. Among the more obvious possibili-ties are (1) an increased number of vector mole-cules per cell, (2) an increased amount of mRNAcoding for HA, and (3) an increase in the half-lifeof HA on the cell surface. Whatever the mecha-nism, the elevated expression of HA seems to bea stable characteristic of these cell lines: Some ofthe earlier isolates have now been maintained incontinuous culture for several months without anoticeable decrease in HA production and with-out a return to heterologous expression of HA.Surprisingly, many of these cell lines bind red cellsvery poorly. Apparently, they secrete a glycoca-lyx containing a large amount of neuraminic acidthat shields HA expressed on the cell surface.After the glycocalyx is removed by mild digestion
FIGURE 5 Eric Hunter, Linda Rodgers, and Janet Hearingworking with the Coulter Epics C fluorescence-activated cellsorter.
74
FIGURE 6 Analysis by flow cytometry of cell lines that ex-press large amounts of HA on their surfaces. Each panelshows the number of cells (vertical axis) that fluoresce witha given intensity (horizontal axis). The intensity is displayedin arbitrary units on a logarithmic scale stretching over threelog units. (A) Autofluorescence of a line of hamster cells ex-pressing HA from a BPV vector. (8) Surface fluorescence ofthe same cells stained with fluorescein-conjugated anti-HAserum. (C) Binding of fluorescent erythrocytes to a line ofmurine cells that show heterogeneous expression of HA onits surface. The peak at the left consists of cells that have notbound any fluorescent red cells; the complex series of peaksat the right represents cells that have bound increasing num-bers of fluorescent erythrocytes (1, 2, 3,...etc). (D) Same cellsafter six rounds of selection and growth, in which cells thatbind erythrocytes efficiently were isolated and grown to masspopulation. The proportion of cells that fail to bind red cellsis greatly reduced; the vast majority of the population bindmore than seven red cells.

with trypsin and/or neuraminidase, virtually allof the cells in the population bind erythrocytesvery efficiently. In all of the murine and hamsterlines, HA is transported to the cell surface moreslowly than in simian cells infected with SV40-HA recombinant vectors. There is also an ap-proximately threefold difference in the rate oftransport of HA in different cell lines. The rea-sons for these variations are under investigation.
We are currently using these cell lines to isolateand identify mutant cells that display HA on thecell surface in a temperature-conditional fashion.Cells are mutagenized with ethylmethanesulfon-ate, and after a period of growth at 32°C, theyare shifted to 39°C. Several of the selection pro-cedures that we are using will work only if HAdisplayed on the surface at the time of the tem-perature shift is removed. This is done by incu-bating the cells in the presence of excess anti-HAantibody and allowing the cells to remove thecross-linked antigen during a period of brief in-cubation at 39°C. Preliminary evidence obtainedwith one of the HA-producing cell lines indicatesthat the HA-antibody complexes rapidly become"capped" and then disappear from the cell sur-face. Cells that are unable to reexpress HA dur-ing further incubation at 39°C may then be iden-tified and physically separated from the rest ofthe population by flow cytofluorometry. Thesecells are then grown into mass culture, and theprocess is repeated to enrich for mutants in thesecretory pathway.
A second method for the isolation of cellularsecretory mutants takes advantage of the hemag-glutinating and membrane-fusing activities ofHA. In collaboration with J. White and co-work-ers (Yale University), we have shown that humanerythrocytes loaded with small proteins in vitrocan be made to fuse with lines of hamster andmurine cells expressing HA. The contents of thered cells are then delivered into the cytoplasm.We are using this method to introduce the toxicA chain of the lectin ricin into cells that expressHA on their cell surface. Mutant cells that sur-vive this selection should be either resistant to theaction of the toxin or defective in the biosynthesisand transport of HA.
Finally, we have devised several schemes to iso-late transport mutants of mammalian cells basedon the action of aminoglycoside-3'-phospho-transferase, an enzyme that renders eukaryotic
cells resistant to the antibiotic G418. Under nor-mal circumstances, this antibiotic kills cells bybinding to ribosomes and inhibiting protein syn-thesis. Aminoglycoside-3'-phosphotransferase isa cytoplasmic enzyme that modifies the antibioticand appears to block its entry into the cell. Wepostulate that if the enzyme is directed to a dif-ferent cellular compartment, entry of the anti-biotic will not be blocked and the cell will onceagain be sensitive to G418. By linking specificdominant signal sequences (e.g., the hydrophobicsignal sequences of HA) to the amino terminus ofthe near protein, we expect to synthesize a chi-mera that (like HA-T antigen) crosses the ER.However, cellular mutants that fail to compart-mentalize the chimera in this way should be resis-tant to the antibiotic. We are at present charac-terizing a series of HA-near protein fusions todetermine their localization and activity withincells. This approach can, in principle, be appliedto select mutants in other cellular transport path-ways, by coupling to near, signals that direct pro-teins to other cellular locations like the nucleusor the mitochondria.
MECHANISM OF HA-MEDIATED CELL FUSION
Influenza viruses, like other enveloped animal vi-ruses, enter and infect cells by a process involvingfusion of the viral membrane with a cellularmembrane. Following binding to sialic-acid-con-taining receptors on the cell surface, the virionsare taken into the cell by endocytosis; entry intothe cytoplasm occurs through the membranes ofintracellular vesicles (like endosomes) where theendogenous pH is low. Experimentally, this ac-tivity can be manifested as cell-cell fusion whenmonolayers of cells displaying HA on theirplasma membranes are transiently exposed to lowpH. Alternatively, as described above, fusion in-duced by low pH can be assayed by red-cell-me-diated delivery of horseradish peroxidase (HRP)into cells expressing surface HA. We have previ-ously shown, using cells expressing HA eithertransiently from a recombinant SV40-HA viralvector or constitutively from a recombinant BPV-HA vector, that the HA molecule displays fusionactivity in the absence of any other influenza-virus-coded components (White et al., Nature300: 658 [1982]; Sambrook et al., EMBO J. 3:[1985] in press). This fusion activity requires a
75
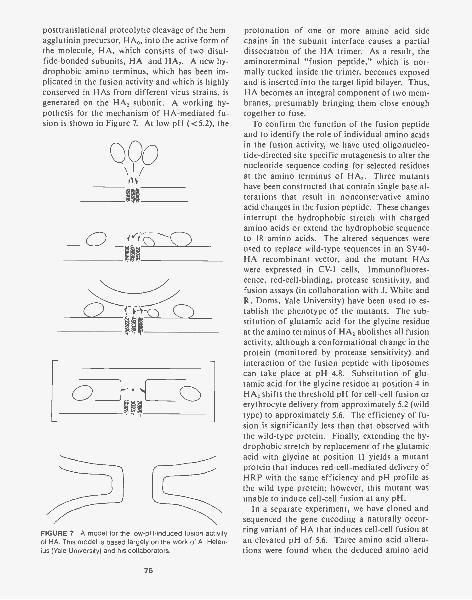
posttranslational proteolytic cleavage of the hem-agglutinin precursor, HA, into the active form ofthe molecule, HA, which consists of two disul-fide-bonded subunits, HA, and HA,. A new hy-drophobic amino terminus, which has been im-plicated in the fusion activity and which is highlyconserved in HAs from different virus strains, isgenerated on the HA, subunit. A working hy-pothesis for the mechanism of HA-mediated fu-sion is shown in Figure 7. At low pH (< 5.2), the
QyQo
0
FIGURE 7 A model for the low-pH-induced fusion activityof HA. This model is based largely on the work of A. Helen-ius (Yale University) and his collaborators.
76
protonation of one or more amino acid sidechains in the subunit interface causes a partialdissociation of the HA trimer. As a result, theaminoterminal "fusion peptide," which is nor-mally tucked inside the trimer, becomes exposedand is inserted into the target lipid bilayer. Thus,HA becomes an integral component of two mem-branes, presumably bringing them close enoughtogether to fuse.
To confirm the function of the fusion peptideand to identify the role of individual amino acidsin the fusion activity, we have used oligonucleo-tide-directed site-specific mutagenesis to alter thenucleotide sequence coding for selected residuesat the amino terminus of HA,. Three mutantshave been constructed that contain single base al-terations that result in nonconservative aminoacid changes in the fusion peptide. These changesinterrupt the hydrophobic stretch with chargedamino acids or extend the hydrophobic sequenceto 18 amino acids. The altered sequences wereused to replace wild-type sequences in an SV40-HA recombinant vector, and the mutant HAswere expressed in CV-1 cells. Immunofluores-cence, red-cell-binding, protease sensitivity, andfusion assays (in collaboration with J. White andR. Doms, Yale University) have been used to es-tablish the phenotype of the mutants. The sub-stitution of glutamic acid for the glycine residueat the amino terminus of HA, abolishes all fusionactivity, although a conformational change in theprotein (monitored by protease sensitivity) andinteraction of the fusion peptide with liposomescan take place at pH 4.8. Substitution of glu-tamic acid for the glycine residue at position 4 inHA, shifts the threshold pH for cell-cell fusion orerythrocyte delivery from approximately 5.2 (wildtype) to approximately 5.6. The efficiency of fu-sion is significantly less than that observed withthe wild-type protein. Finally, extending the hy-drophobic stretch by replacement of the glutamicacid with glycine at position 11 yields a mutantprotein that induces red-cell-mediated delivery ofHRP with the same efficiency and pH profile asthe wild-type protein; however, this mutant wasunable to induce cell-cell fusion at any pH.
In a separate experiment, we have cloned andsequenced the gene encoding a naturally occur-ring variant of HA that induces cell-cell fusion atan elevated pH of 5.6. Three amino acid altera-tions were found when the deduced amino acid

sequence of the variant was compared with thatof wild-type HA. Two of the altered amino acidsare located on the external surface of HA, andare unlikely to be involved in the altered fusionactivity. The third change involves the substitu-tion of an uncharged glutamine residue for a glu-tamic acid residue at position 132 in HA,. Ex-change of restriction fragments between thecloned genes encoding the wild-type and mutantproteins, and expression of the reassorted genesusing SV40-HA recombinant vectors, confirmedthat this substitution was indeed the cause of fu-sion at elevated pH. In the wild-type protein, thisglutamic acid residue is linked by a salt bridge toan arginine at position 185 in the HA, of an ad-jacent subunit of the trimer. The fact that aboli-tion of this salt bridge allows the conformationalchange to occur at pH 5.6 suggests that it mayplay a crucial role in stabilizing the HA trimer.
These results suggest that fusion takes place inat least three stages -a conformational change inHA induced by low pH, which exposes the fusionpeptide; binding or insertion of this peptide intothe target membrane; and, finally, destabilizationand coalescence of the closely apposed lipid bi-layers. Partial destabilization resulting in smallareas of membrane coalescence may be sufficientfor delivery of red cell contents; destabilizationand coalescence of more extended lengths ofmembrane are required for cell-cell fusion andpolykaryon formation.
PUBLICATIONS
Braciale, T.J., V.J. Braciale, T.J. Henkel, J. Sambrook, andM.-J. Gething. 1984. Cytotoxic T lymphocyte recognition
of the influenza hemagglutinin gene product expressedby DNA-mediated gene transfer. J. Exp. Med. 159:341-354.
Doyle, C., M.G. Roth, J. Sambrook, and M.-J. Gething. 1985.Mutations in the cytoplasmic domain of the influenza vi-rus haemagglutinin affect different stages of intracellulartransport. J. Cell Biol. 100: 704-714.
Gething, M.-J., T.J. Braciale, C.M. Brand, and J. Sambrook.1984. Comparison of different eukaryotic vectors for theexpression of hemagglutinin glycoprotein of influenza vi-rus. In Modern approaches to vaccines (ed. R.M. Chan-ock and R.A. Lerner), pp. 263-268. Cold Spring HarborLaboratory, Cold Spring Harbor, New York.
Gething, M.-J., C. Doyle, M. Roth, amd J. Sambrook. 1985.Mutational analysis of the structure and function of theinfluenza virus haemagglutinin. Curr. Top. Membr.Transp. 23: 17-41.
Gething, M.-J., C. Doyle, M. Roth, E. Hunter, and J. Sam-brook. 1985. Site-directed mutagenesis of the hemag-glutinin of influenza virus: Effects on transport throughthe cell. In Protein transport and secretion (ed. M.-J. Ge-thing), pp. 90-96. Cold Spring Harbor Laboratory, ColdSpring Harbor, New York.
Sambrook, J., L. Rodgers, J. White, and M.J. Gething. 1985.Lines of BPV-transformed murine cells that constitutivelyexpress influenza virus haemagglutinin. EMBO J.4: 91-103.
Sharma, S., J. Brandsma, L. Rodgers, M.-J. Gething, and J.Sambrook. 1985. A heterologous signal sequence candivert SV40 T antigen into the exocytic pathway. In Pro-tein transport and secretion (ed. M.-J. Gething), pp.73-78. Cold Spring Harbor Laboratory, Cold Spring Har-bor, New York.
Talmadge, K., W.R. Boorstein, N.C. Vamvakopoulos, M.J.Gething, and J.C. Fiddes. 1984. Only three of the sevenhuman chorionic gonadotropin (3 subunit genes can beexpressed in the placenta. Nucleic Acids Res.12: 8415-8436.
In Press, Submitted, and In Preparation
Sharma, S., J. Brandsma, L. Rodgers, M.J. Gething, and J.Sambrook. 1985. SV40 T antigen and the exocytoticpathway. EMBO J. (in press).
CELL-CYCLE-SPECIFIC EXPRESSION
J.A. Lewis D.A. Matkovich
Genetics of Growth-phase-dependentThymidine Kinase Gene ExpressionJ.A. Lewis, D.A. Matkovich
Our laboratory is fundamentally interested in de-fining the genetic determinants that govern the S-
phase specificity of thymidine kinase (tk) geneexpression in higher eukaryotic cells. To this end,we have (1) cloned the Chinese hamster tk gene ina recombinant phage vector, (2) cloned a nearlyfull-length cDNA to the hamster tk mRNA in aXGT10 vector and characterized this clone by di-deoxynucleotide sequencing, and (3) sequenced
77

selected restriction fragments of the hamster tk-gene clone. This progression of experiments hasled us to a nearly complete resolution of thestructure of the Chinese hamster tk-gene pre-sented in Figure 1. In brief, the hamster tkmRNA sequences are distributed through the 11.2kb of genomic DNA in at least seven exon seg-ments that range in size from the 32-bp Exon IIto the 680-bp Exon VII. These exons are segre-gated by intervening sequences that range in sizefrom the 5.4-kb intron which segregates Exons IVand V to the 92-bp intervening sequence whichsegregates Exons I and II. The nucleotide se-quences that comprise the exon-intron bounda-ries show extended homology with the consensussequences described for splice-donor and splice-acceptor nucleotides.
In an attempt to resolve the structural detailsat the extreme 5' end of the hamster tk gene andto propose a site for tk-gene transcription initia-tion, we have constructed a tk minigene, desig-nated pHaTK2, that efficiently transforms a va-riety of Tk - cells to Tic' and have prepared ExoIII/S, deletion mutants of pHaTK2 through thepromoter region. The endpoints of the deletionsare shown in Figure 1.
We find that mutant derivatives of pHaTK2that retain at least 121 by 5' to the translation in-itiator AUG transform LTk - cells as efficientlyas the pHaTK2 gene, whereas those that retainonly 79 by transform with a nearly 100-fold re-duction in efficiency. It seems clear by this anal-ysis that elements important to tk-gene transcrip-tion are contained within the 42 by between - 79
6353Exon I AGTTAACAAGCTAAACCTGCCCTGAAGG GAGAAAATGAGGAAACCAGGCACGTGGICTCCCGGAGGGGTGGTCGAGTGCCUCCTAACATTCCGGCCCG
L1262 A221GGTTCTGGGCAGCTGCACCC CACCTACCTCTGCAGACAGGGICTTCCAGGGCGGCTCGCGC GGGGAGCGCAGAGGCTGCCATAGCCGGGTCCAGCTCA
A121GAGGGCG
AI77AGCAGCTCGCTTTGCAGGGACAGGGGGAGAGGGGCGGIGGCCCACATIGCGCCCTGGC CTIGGCACGCCTCCGTCTTCGGCTGCGATTGGT
679 142CGGTGAGIT TTTAAA1GGAAAGCGCGAACCTGAGGACGCTTCCCACT CACCGACCCCGGACCTTGGCGCTCAGGCTCGCACAGCCGCCATGAATTACA
GGGGTGAGT
TCAATCTGCCCACCGTGCTGCCCGGCTCCCCCAGCAAGACCCGGGGCCAGATCCAGgtgoggggtccgggctgc
Exon II acttetctoctagGTGATCCTCGGACCCATGTTCTCAGGGAAAAGgtootgootgggcttggg
Exon III tgetgoctottgcogCACCGAGCTCATGAGGAGAGTCCGGCGCTICCAGATCGCCCAGAACAAATGCCTGGICATCAAGTATGCCAAAGACACGCGCTATAGCAGGAGCTTCTCCACACATGACCGgteogtcccaccacccc
Exon IV totttottottoccagGAACACCATGOACGCCCTGCCAGCCTGCCTGCTCCGGGATGTGGCCCAGGAGGCCCTGGGTGCGGCTGICATTGGCATCGATGAAGGACAGTTTgtoagttggettgoo
Exon V tecoccoocttteccogTTCCCTGACATCGTGGAATTCTGTGAAGTGATGGCCAATGCAGGCAAGACAGTGATCGTGGCAGCATTAGACGGAACTITCCAGAGAAAGytaaggtgtctcacc
Exon VI ecceetgtototatogGCTITCGOCAGCATCTTGAACCTGGTGCCCCTGOCTGAGAGTGTGGTGAAGCTGACTGCCGTGIGTATGGAGTGCTICCGAGAAGCCGCCTACACCAAGAGGCTGGGCCTGGAGAAGGAGgtoccttettgtget
Exon VII tggeectgtococogGIGGAGGTGATTGGTGGAGGAGACAAGTACCACTCGGIGTGCCGCGTGIGTTACTTTAAGAAGTCCICGGTACAGCCTGCTGGGCCAGACAACAAAGAGAACTGCCCGGTGCTGGGACAGCCTGGAGAGGCCTCAGCTGTCAGGAAGCTCTTCGCCCCTCAGCAAGTCCTACAGC
ACAACTCCACCAACTGAGAGGACCTGGGGCCTGCCAGCTCCTACCCCAGGTTGGATTCTCAGAGAGCAGAGGACGGGCTGGGACTGCCGTGCCTGATGGA
CAATGTGCCCTGGAGAGGCTCACCCGCTICTCACAGCCTIITTTAGTTCCCTICTIGGITGCTGAGATGCTTTAGCCCAGACTAGAGCCAGCGCCTGCCT
GGTGGITAGGGTTTGACATCCAGCCAGAGGTAGGACAAAGCCACAGGGTGTTGTGACACAGAGGTGCTGGCTICTTCCTICTTGGIGGCTICCAGTCTCA
AGGGCCCCGCCCCCGAGCAAGGCTICACAACCCTCACTITGTGCTGAAGCTTGACCCACAATGGCCCCTAGCGGIGICTTTACAAAGTGGTGCTGTTCTT
GCCCTACTCAGAGCCCCAAGACTCAGGACTCTIGGTGAGAGCCTGTGCTTCTTGTGCTATAGTGTAAAATGAATAATTAATAATATTAAAGTTTCTACTTGACAG
FIGURE 1 The nucleotide sequence of the 380 by 5' to the translation initiator AUG of the hamster tk gene, the seven tk-gene exons, and bordering intron sequences. The endpoints of the Exo III/S, 5' deletions described in the text are indicatedby a triangle, designated to reflect the number of nucleotides remaining 5' to the initiator AUG codon.
78
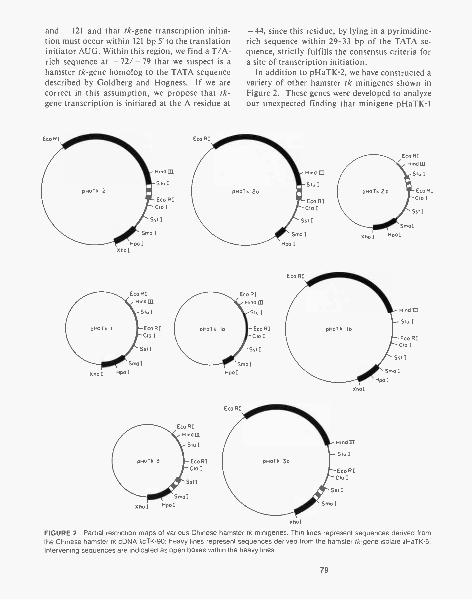
and - 121 and that tk-gene transcription initia-tion must occur within 121 by 5' to the translationinitiator AUG. Within this region, we find a T/A-rich sequence at - 72/ - 79 that we suspect is ahamster tk-gene homolog to the TATA sequencedescribed by Goldberg and Hogness. If we arecorrect in this assumption, we propose that tk-gene transcription is initiated at the A residue at
Eco RI
Hind III
Stu I
Eco RICla I
Sst I
Salo I
Hoc IXho I
Eco RIHind III
Stu I
Eco RIClo I
Sal I
Sm. ISpa IXho I
Eco RI
- 44, since this residue, by lying in a pyrimidine-rich sequence within 29-33 by of the TATA se-quence, strictly fulfills the consensus criteria fora site of transcription initiation.
In addition to pHaTK2, we have constructed avariety of other hamster tk minigenes shown inFigure 2. These genes were developed to analyzeour unexpected finding that minigene pHaTK1
pHoTk 20
Eco RIHind 01
St. I
Eco RICla I
Ssi I
Sma I
Hpa I
Eco RIHind CQ
Stu I
Eco RICla I
Sst I
SrnaI
HpoIXho I
Hind III
Stu I
Eco RIClo I
Sot I
Sala I
Spa I
Eco RIInd III
Stu I
Eco RIClo I
Sat I
SmaI
Spa IXho I
xhol
Xho I
Hind III
Stu I
Eco RICla I
Sat I
Sma I
Hind LEt
Stu I
Coo RICa I
Sat I
Smc
Hpo I
FIGURE 2 Partial restriction maps of various Chinese hamster tk minigenes. Thin lines represent sequences derived fromthe Chinese hamster tk cDNA AoTK90; heavy lines represent sequences derived from the hamster tk-gene isolate AHaTK5.Intervening sequences are indicated as open boxes within the heavy lines.
79

transforms LTk- cells 25-fold less efficiently thanpHaTK2. Since pHaTK1 is composed only ofhamster tk cDNA sequences fused to the hamstertk-gene promoter, it differs significantly in struc-ture from pHaTIC2, which contains all of the se-quences of pHaTK1, introns e and f, as well as3.5 kb of genomic DNA derived from the 3' endof the hamster tk gene. To consider the contri-bution of each of these variables to the transfor-mation efficiency of pHaTK2, we introduced in-trons e and f into pHaTK1 to create pHaTK2band in so doing developed a tk minigene thattransforms LTk- cells to Tic' with a 15-foldgreater efficiency than pHaTK1. At the sametime, we introduced introns a and b of the ham-ster tk gene into pHaTK1 to create pHaTI(3,which transforms LTk- cells nearly 8-fold moreefficiently than pHaTK1 and thereby demon-strated the generality of the intervening sequenceTk transformation-enhancement phenomenon.Both pHaTK2b and pHaTK3 genes, however,were measurably less efficient (2-3-fold) thanpHaTK2, a difference we attributed to the 3.5 kbof genomic DNA contained in pHaTK2. To testthe effects of this DNA segment in other tk-gene constructions, we introduced this DNA intopHaTK1 to create pHaTKlc and into pHaTK3to create pHaTK3a and found that through thesemanipulations, we developed derivatives thattransformed 3-fold and 1.9-fold more efficientlythan the parental tk minigenes.
We do not yet understand the mechanistic basisfor the intervening sequence enhancement phe-nomenon that makes the major contribution topHaTK2 minigene transformation efficiency.Curiously, this phenomenon has been describedpreviously in transformation studies with dihy-drofolate reductase (dhfr) minigenes, in whichcontext the introduction of dhfr intervening se-quences into dhfr minigene constructions im-proved transformation efficiencies as much as 30-fold. We imagine that intervening sequenceseither promote or facilitate the integration of thetrans fected DNAs into host genomic DNA,thereby promoting the number of stable Tk*transformants, or alternatively, intervening se-quences in primary gene transcripts may increasethe efficiency with which these transcripts matureto cytoplasmic mRNA.
We are just as ignorant about the basis of therelatively minor 3.5-kb enhancement phenone-
80
mon. We propose that sequences in this fragmentimprove the efficiency with which the polyaden-ylation signals contained in these minigenes arerecognized, which leads to an improvement in theefficiency of primary transcript maturation.
Our compelling interest in the construction oftk minigenes is of course to exploit them to definethe location and function of the genetic determi-nants governing the growth-phase dependence oftk-gene expression. Since the genetic determi-nants of interest may be contained in 5'-flankingDNA, 3'-flanking DNA, noncoding, coding, orintervening sequences, our experimental plan isto construct a minimal chimeric tk gene, contain-ing only Chinese hamster coding and 3' noncod-ing sequences, and to analyze the pattern of tk-gene expression in Rat 4 Tk* cells stably trans-formed with this gene. If necessary, we intend toelaborate this minimal gene systematically, sup-plying it with structural elements of the hamstertk gene, until we develop a tk minigene whoseexpression is growth-phase-dependent.
As a Tk recipient cell for the analysis of tk-gene expression from our various tk-gene con-structions, we have adopted the Rat 4 Tk cellline derived here at Cold Spring Harbor Labora-tory by W. Topp. This cell line can be readilytransformed to TIC , grows rapidly with a dou-bling time of 14-16 hours, is strictly growth-ar-rested at high density, is stable as a confluentmonolayer for 96-120 hours without serum re-freshment, and can be induced to resume loga-rithmic cell division if passed through fresh me-dia at lower densities. We have generated Rat 4Tk *- transformed lines using either the hamstertk-gene clone, XHaTK5, the tk gene of herpessimplex virus I, or a hamster/herpes chimericgene designated pHa06He (Fig. 3). This chimericgene consists of the herpes tk-gene promoterfused to the coding and 3' noncoding nucleotidesof the Chinese hamster tk cDNA. As we ex-pected, tk-gene expression in Rat 4 lines trans-formed with the hamster Tk genomic cloneXHaTK5 is growth-phase-dependent by the fol-lowing criteria. (1) Specific Tk activity in con-fluent growth-arrested monolayers is only 5-10,7othe specific Tk activity of logarithmically grow-ing cultures; (2) the specific Tk activity of growth-arrested cultures can be increased five- to tenfoldby serum refreshment or by replating at lowerdensity; (3) the specific Tk activity of growth-ar-

B. GCGCCTTGTAGAAGCG CCTCGAGG CCGCCATG500 510
He Xho I Ha
FIGURE 3 Structure of the chimeric tk minigene pHaA6He.This gene was constructed by combining a 700-bp BamH I-Xhol fragment from the HSV tk-gene mutant Tk x 81 con-taining the HSV tk-gene promoter and cap site with an Exo111/S, deletion mutant of pHaTK1 that contains the completecoding and 3' noncoding nucleotides of the hamster tkmRNA and six nucleotides 5' to the translation initiator AUGcodon.
rested cultures can be increased five- to tenfoldby infection with adenovirus type 5. These find-
ings are characteristic of the pattern of endoge-nous tk-gene expression that has been docu-mented throughout a literature of nearly 20 years.In contrast, the specific Tk activity in Rat 4 celllines transformed to Tk' with the tk gene of HSV-1 is growth-phase-independent, being largely in-variant under the variety of culture conditions de-scribed above, results consistent with an extensiveliterature as well.
We were intrigued to find that tk-gene expres-sion in Rat 4 Ti' lines transformed with the chi-meric pHaA6He gene is growth-phase-dependent,by all the criteria established for the XHaTk-5-transformed lines above. These findings consti-tute the first evidence that the determinants oftk-gene growth-phase-dependent expression arecontained exclusively within sequences of the ma-ture tk mRNA. We have yet to establish preciselythe location of these determinants and the man-ner in which they function, although experimentsto consider these fundamental issues are pres-ently under way.
In Press, Submitted, and In Preparation
Lewis, J.A. The structure and function of the Chinese ham-ster thymidine kinase gene. (Submitted.)
Lewis, J.A. and D.A. Matkovich. Tk gene expression in Rat4 cells transformed with TK mini-genes is growth phasedependent and responsive to adenovirus infection. (Sub-mitted.)
HORMONAL AND DEVELOPMENTAL CONTROLOF GENE EXPRESSION
D.T. Kurtz W. Addison J.-Z. LiJ. Maclnnes E. NozikD. Danna A. Louis
For the past several years, we have been studyingthe hormonal and developmental control of theexpression of a rat gene family that codes for aprotein called an globulin. This protein was firstdescribed as a very abundant protein (1-2% oftotal protein synthesis) in adult male rats. Thesynthesis in male rat liver is under complex hor-monal control: Androgens, glucocorticoids,growth hormone, and insulin all induce hepatica2u, whereas estrogens repress the synthesis of this
protein. We have previously shown that an is en-coded by a multigene family (18-25 genes perhaploid complement) clustered on chromosome 5.
TISSUE-SPECIFIC EXPRESSION OF THEa. GENE FAMILY
We have found that 02, is expressed in severalother tissues in rats besides the liver. The an pro-
81

teins that are produced in the various tissues aredistinguishable by differences in isoelectricpoints. We have shown that this is not due toposttranslational modification of the same pri-mary translation product by translating mRNAfrom different a2u-producing tissues, immuno-precipitating the ce2u made in vitro, and subjectingit to isoelectric focusing. The same protein iso-forms found in vivo are found in ce,,, produced inthe cell-free translational systems. It appears thatdifferent ce2, genes are being transcribed in differ-ent tissues.
Liver a, represents a family of four to five dif-ferent proteins (pl 4.8-6.0). The hormonal con-trol of hepatic oz2u is well documented by our lab-oratory and several others. All of the differenta2u genes expressed in the liver rise and fall coor-dinately; i.e., the multihormonal control of ratliver a2, synthesis is apparently not due to differ-ent hormones inducing different genes.
Salivary gland a, consists of four to five pro-tein species with distinctly more acidic pls thanliver a,. The salivary gland proteins run as acluster between pH 4.0 and pH 4.8 on isoelectricfocusing gels. a2, represents about 0.03010 of totalprotein synthesis in salivary gland, i.e., about1/20 to 1/30 the level found in liver. The synthe-sis of salivary gland a2u seems constitutive: Iden-tical levels are found in both males and females,and the levels do not seem to change during sev-eral endocrine manipulations in intact animals.
Lachrymal gland 0,2, comprises a set of pro-teins that look identical to those of salivary glanda,. ce represents about 0.2010 of total proteinsynthesis in lachrymal gland, i.e., about 1/5 thelevel in liver. Male rats produce about three tofive times more lachrymal a, than do female rats.This difference is apparently due to androgen in-duction: Female rats given androgens producelachrymal a2, at the same rate as do males. Allisoelectric forms are induced coordinately.
Mammary glands in pregnant female rats at day18 or 19 of gestation produce a2, at a very lowlevel (about 1/50 the level in male liver). Mam-mary gland a2, is represented by a very largenumber of protein isoforms: 15-18 different a2,proteins can be distinguished (pI 4.0-8.0). Thehormonal modulation of mammary gland ce2,, iscurrently under investigation. The synthesispeaks, sharply at day 18 or 19 of gestation and
82
then falls rapidly until, by day 21 and followingparturition, no mammary gland ce,u is detectable.This timing is consistent with a model in whichprogesterone is inducing Ol2u and then a surge inprolactin, which is known to occur about day 17or 18, represses a2u synthesis.
The preputial gland is the most abundantsource of ce2, proteins that we have found. It rep-resents about 5-10010 of total protein synthesis inthis tissue, and the synthesis seems to be consti-tutive. Preputial gland a2u proteins separate intothe same complex protein isoforms that are foundin mammary glands. The number of differentprotein isoforms distinguishable in the preputialgland is close to the number of ci2u genes that wehave estimated to be in the genome. It thus ap-pears that the preputial gland (and perhaps mam-mary gland) transcribes nearly the entire comple-ment of ce,,, genes present in the genome, whereasthe liver, salivary gland, and lachrymal glandtranscribe much smaller and distinct subsets.
TISSUE-SPECIFIC EXPRESSION OF CLONEDa20 GENES
We have cloned 18 different ce, genes from a li-brary of the rat genome. Identifying which geneis being transcribed in which tissue has proved tobe difficult: The protein-coding regions of thedifferent ce2, genes are for the most part veryhighly conserved. Thus, one cloned gene will hy-bridize to mRNA from all a2u-producing tissues;raising the stringency of hybridization has no dis-cernible effect. Surprisingly, the introns are alsohighly conserved. Intron-specific probes madefrom different cloned genes cannot be used todistinguish different az, transcripts in nuclearRNAs from various tissues. The 5' and 3' non-coding regions of the different mRNAs are iden-tical in length, as assayed by 51-nuclease and byprimer-extension analyses.
We have used two approaches to identify thetissue of expression of specific cloned (y2,, genes:We have sequenced the extreme 5' ends of 12 ofthe 18 cloned genes. In a stretch of approxi-mately 200 nucleotides from the cap site (essen-tially the entire first exon), no two genes are ex-actly identical, although the sequences are verysimilar. We are currently sequencing the corre-

sponding regions of several tissue cDNAs tomatch up the sequences in this region. Sequencedifferences in exon III may also prove to be di-agnostic.
Another approach is to use the pI differencesin the a, proteins that are made in the varioustissues. We have used COS-2 cells as a linkedtranscription-translation system to produce 02,,protein from the individual ce2u genes. COS cellsconstitutively produce SV40 T antigen and willthus effect the replication of any cloned segmentof DNA containing the SV40 origin of replica-tion. We have constructed several SV40-02u fu-sion genes (driven by the SV40 early promoter)and have produced a, protein in COS cells. Inthe course of this research, we made the surpris-ing discovery that adenovirus VA I and VA IIRNAs greatly increase the efficiency of transla-tion of transfected genes in COS cells. If theSV40-a2 hybrid construct is cotransfected intoCOS cells with an adenovirus fragment contain-ing the VA genes, the level of a2u protein pro-duced is increased 15-20-fold, although there isno discernible difference in the extent of replica-tion of, or transcription from, the SV40-a2u hy-brid gene. This finding that adenovirus VA RNAcan have a nonspecific enhancing effect on trans-lation has now been seen in other systems.
IN VIVO TRANSFECTION OF CLONED a2, GENES
Once we have identified specific genes as beingtranscribed in different tissues, we will attempt todetermine the molecular basis of this tissue-spe-cific expression, i.e., to identify the specific DNAsequences responsible for the tissue specificity.There are many recent examples of "tissue-spe-cific enhancers" that greatly increase transcrip-tion in a given tissue but are not part of the pro-moter region per se.
To study this, we would need analogs of thevarious tissues in which a2u is produced. This isnot a trivial problem, since "liver cells" that havebeen put into culture very seldom retain theirdifferentiated properties. It was recently shownthat calcium-phosphate-precipitated polyomaDNA, when resuspended in a buffer containingcollagenase and hyaluronidase, can be injecteddirectly into the liver and spleen of mice. The
polyoma DNA is able to replicate and remainsapparently extrachromosomal for several days.We have managed to duplicate this finding (inboth rats and mice) and, further, we have foundthat if a cloned segment of DNA containing thepolyoma origin is coprecipitated with the entirepolyoma genome, the T antigen produced by thepolyoma DNA can act in trans and effect the rep-lication of the coinjected plasmid.
We have also had success injecting bovine pap-illoma virus (BPV)-containing plasmids directlyinto mouse liver. We have used the "69%"BamHI-HindIII fragment of BPV, which isknown to be sufficient for BPV replication in cul-tured mouse cells. When cloned DNA containingthis fragment is injected into mouse liver, extra-chromosomal BPV DNA is detectable on South-ern blots 2 days after injection. This should be avery direct and simple method by which to studythe tissue-specific expression of cloned liver 0211genes. We can further use this to study whichDNA sequences are required for the hormonalmodulation of 02 transcription in liver. Ourstudies of the hormonal modulation of a2u syn-thesis following transfection into tissue-culturecells have been limited by the lack of cell linesthat contain various hormone receptors. If 02,,genes injected into liver retain their hormonal in-ducibility, we should be able to use this system tostudy DNA sequences that are required for theresponse of liver a2u genes to many different hor-mones.
CONTROL OF RAT TROPOMYOSIN GENEEXPRESSION
In collaboration with D. Helfman (Cell BiologySection), we are studying the mechanism by whichadenovirus infection shuts down the synthesis oftropomyosin I expression, while not affecting theexpression of other tropomyosin forms. We haveintroduced a cloned rat tropomyosin I gene intoHe La cells using DNA-mediated gene transfer,and we are in the process of determining whetherthis transfected gene is negatively controlled fol-lowing adenovirus infection. If so, it should bepossible to determine which DNA regions of thistropomyosin gene are responsible for its negativeregulation.
83

GENETICS OF CELL PROLIFERATION
M. Wig ler C. BirchmeierD. BirnbaumD. BroekS. CameronL. Edlund
S. Efrat P. Sass0. Fasano E. TaparowskyM. Goldfarb T TodaT. Kataoka G. WaitchesS. Powers D. Young
Scientists in our section study oncogenes, genesthat drastically alter the proliferative capacity ofnormal cells. In particular, we have concentratedour efforts on the RAS oncogenes, which wereamong the first genes implicated in human can-cer. The study of RAS has been facilitated by therecent discovery that close homologs exist in theyeast Saccharomyces cerevisiae, an organismreadily subjected to thorough genetic analysis.Studies of the RAS pathway in yeast have led usto examine the cAMP pathway in fine detail. Inaddition to the above, we have recently isolatedand are now studying three new human onco-genes. Much of the work on the yeast RAS wasdone in collaboration with F. Tamanoi (this sec-tion), J. Broach (Princeton University), K. Mat-sumoto (Tottori University, Japan), and I. Unoand T. Ishikawa (University of Tokyo, Japan);some of the work with associated yeast genes wasdone in collaboration with M. Zoller (MolecularGenetics Section); and the work on other verte-brate oncogenes was performed in collaborationwith J. Fogh (Sloan Kettering Institute, Rye, NewYork).
Yeast RAS Genes
D. Broek, S. Cameron, T. Kataoka, S. Powers,T. Toda, M. Wig ler
In the yeast S. cerevisiae, there are two genes,RASI and RAS2, that are closely homologous tothe mammalian RAS genes (Defeo-Jones et al.,Nature 306: 707 [1983]; Dhar et al., Nucleic AcidsRes. 12: 3611 [1984]; Powers et al., Cell 36: 607[1984]). We have cloned both genes (RASI andRAS2) from plasmid libraries and have deter-mined the complete nucleotide sequence of theircoding regions. They encode proteins that havenearly 90% homology with the first 80 positionsof the mammalian RAS proteins, and nearly 50%
84
homology with the next 80 amino acids. YeastRASI and RAS2 proteins are more homologousto each other, with about 90% homology for thefirst 180 positions. After this, at nearly the sameposition that the mammalian RAS proteins beginto diverge from each other, the two yeast RASproteins diverge radically. The yeast RAS pro-teins, like the proteins encoded by the mammal-ian genes, terminate with the sequence CysAAX,where A is an aliphatic amino acid. Thus, theyeast RAS proteins have the same overall struc-ture and interrelationship as the family of mam-malian RAS proteins. The domains of diver-gence may correspond to functional domains ofthe RAS proteins. Monoclonal antibody directedagainst mammalian RAS proteins immunopre-cipitates protein in yeast cells containing highcopy numbers of the yeast RAS2 gene.
We and other investigators have previouslyshown that at least one functional RAS gene isessential for the germination of haploid yeastspores (Kataoka et al., Cell 37: 437 [1984]; Tatch-ell et al., Nature 309: 523 [1984]). This was dem-onstrated by constructing doubly heterozygousdiploid yeast cells containing wild-type RASI andRAS2 alleles and RASI and RAS2 alleles eachdisrupted by a different auxotrophic marker.These diploid cells were then sporulated, and theresulting tetrads were analyzed. Only spores con-taining at least one functional RAS gene couldgerminate. We then introduced into these doublyheterozygous diploid cells a RAS2 gene under thetranscriptional control of the galactose inducibleGAL10 promoter (see Kataoka et al., Cell 40: 19[1985]) linked to a third auxotrophic marker.These diploid cells were then sporulated, and te-trads were germinated on either glucose-contain-ing medium (YPD) or galactose-containing me-dium (YPGa1). The results confirm that at leastone functional RAS gene is required for germi-nation. rasl - ras2- GAL 10-RAS2 spores ger-minate only in the presence of galactose, the in-

ducer for the GAL10 promoter. Cells with thegenotype rasl- ras2- GALIO-RAS2 were grownin YPGa1 and then shifted to YPD. The growthof these cells was then monitored, and cultureswere plated on to YPGaI agar to measure cell vi-ability. Within several cell generations, cellgrowth ceased and cells lost viability. These ex-periments indicated that the RAS genes areneeded not only for spore germination, but alsofor the continued growth and viability of yeastcells.
Certain missense mutations drastically alter thebiological activity of mammalian RAS genes. Inparticular, the Ha-nova"' gene, which encodes va-line instead of glycine at the twelfth codon of thehuman Ha-ras gene, can induce the tumorigenictransformation of NIH-3T3 cells. To test the con-sequences of a similar mutation of RAS2 on theproperties of yeast cells, we constructed theRAS2va19 gene using site-directed mutagenesis(Kataoka et al., Cell 37: 437 [1984]). RAS2va19thus encodes valine instead of glycine at position19, which corresponds to position 12 of the mam-malian Ha-ras protein. We then examined the ef-fects of introducing this gene into yeast cells. Ourfirst observation was that diploid cells containingRAS2va"9 could not be induced to sporulate byincubation under conditions of nutritional depri-vation (Kataoka et al., Ce1137: 437 [1984]). Next,we observed that haploid cells carrying RAS2va"9lost viability if starved for nitrogen, sulfur, orphosphorus and failed to arrest in G, under thoseconditions. Wild-type cells normally arrest in G,and retain viability for long periods when nutri-tionally deprived. In addition to these pheno-types, we also noted that cells carrying RAS2van9failed to accumulate carbohydrate stores as cellsentered the stationary growth phase (Toda et al.,Cell 40: 27 [1985]). Thus, in general, RAS2'119cells appeared to show a defective response to nu-tritional stress.
The cluster of cellular properties just describedclosely resemble the phenotype of cells carryingthe bcyl mutation (Matsumoto et al., Proc. Natl.Acad. Sci. 79: 2355 [1982], Cell 32: 417 [1983],Exp. Cell Res. 146: 151 [1983]; Uno et al., J. Biol.Chem. 258: 10867 [1983]). The bcyl mutationwas first isolated by Matsumoto and co-workersas a mutation that suppresses the lethality thatotherwise results from the disruption of adenyl-
ate cyclase (Matsumoto et al., Proc. Natl. Acad.Sci. 79: 2355 [1982]). Cells carrying bcyl appearto lack the regulatory subunit of the cAMP-de-pendent protein kinase and hence have lost therequirement for cAMP (Uno et al., J. Biol. Chem.257: 14110 [1982]). These observations suggestthat the RAS genes might be participating in thecAMP pathway. In support of this idea, we foundthat bcyl suppressed the lethality that otherwiseresults from disruption of both RAS genes.
The adenylate cyclase activity of the yeast S.cerevisiae is stimulated by guanine nucleotides inthe presence of magnesium (Casperson et al., J.Biol. Chem. 258: 7911 [1983]). In this respect,yeast adenylate cyclase resembles the adenylatecyclase of mammalian cells, which can be stimu-lated by a guanine-nucleotide-binding complexcalled G, (Gilman, Cell 36: 577 [1984]). Since theyeast RAS proteins also bind guanine nucleotides(Tamanoi et al., Proc. Natl. Acad. Sci. 81: 6924[1984]), we reasoned that they might also modu-late adenylate cyclase. This was tested directly bythe assay of membranes from wild-type yeast cellsand rasl- ras2- cells. Membranes from eithercontained appreciable adenylate cyclase activitywhen assayed in the presence of manganese ion,but rasl- ras2- membranes displayed negligiblelevels of activity when assayed in the presence ofmagnesium and a nonhydrolyzable guanine nu-cleotide analog. These results were confirmed ina striking manner by membrane-mixing experi-ments. We prepared membranes from RAS1RAS2 yeast carrying the cyrl-1 mutation andmembranes from rasl- ras2- yeast. The cyrl-1mutation disrupts the catalytic subunit of aden-ylate cyclase. Membranes from these two sourceswere assayed separately and together after mem-brane mixing and fusion. The data indicate thatmembrane mixing and fusion regenerate a guan-ine-nucleotide-stimulated adenylate cyclase activ-ity (Toda et al., Cell 40: 27 [1985]).
The addition of purified yeast RAS2 protein tomembranes from bcyl rasl- ras2- cells restoresadenylate cyclase activity to about 50-fold abovebackground level (Broek et al., Cell [1985] inpress). Adenylate cyclase activity in the bcylrasl- ras2- membranes can also be increaseddramatically by addition of yeast RAS1 protein.Restoration of adenylate cyclase activity by RASproteins is dependent on the presence of guanine
85

nucleotide, with the nonhydrolyzable GTP ana-log, GppNp, yielding twice the activity of thatobserved in the presence of GDP. Incubation ofthe RAS2 protein with GTP prior to mixing withthe bcyl rasl- ras2- membranes results in aden-ylate cyclase activity comparable to that restoredwith RAS2 bound to GDP. In contrast, preincu-bation of RAS2va19 with GTP results in activationof adenylate cyclase to levels identical to that in-duced by RAS2 proteins bound to GppNp. Theseresults confirm our genetic studies and indicatethat there are proteins in membranes that can dis-tinguish between RAS proteins complexed withGTP and RAS proteins complexed with GDP;furthermore, they provide for the first time an invitro bioassay for the effector function of RAS.
Yeast Genes in the RAS/cAMP PathwayT. Kataoka, S. Powers, P. Sass, T. Toda, M. Wig ler
In our effort to understand thoroughly the func-tion of RAS in yeast, we have begun cloning geneswhich encode proteins that operate along theRAS/cAMP pathway. To do this, we have usedstandard genetic strategies.
BCYI was cloned from a centromere-linked li-brary (constructed by M. Rose, Massachusetts In-stitute of Technology, and generously provided byhim) by transforming bcyl cells with the libraryand selecting cells capable of resistance to heatshock. bcyl cells, like RAS21a19 cells, are heat-shock-sensitive (55°C for 30 min), presumablybecause they cannot enter G5. We obtained oneclone that displays all of the genetic features ex-pected of a BCYI clone. Gene disruptions ofBCYI were constructed and used to transformhaploid yeast cells by gene replacement. Thetransformants display the same phenotype of cellscontaining the spontaneously occurring bcyl mu-tation.
CYRI, the gene encoding adenylate cyclase,was cloned in a similar manner by transformingcyrl -2 mutants (temperature-sensitive for aden-ylate cyclase) with centromere-linked libraries andselecting cells that grew at the nonpermissive tem-perature. The nucleotide sequence of this genehas been determined ( >6000 bp), and an aminoacid sequence of the product has been deduced.
86
We are not yet certain which methionine repre-sents the start codon. Aminoterminal deletionmutants of the adenylate cyclase gene give rise toproteins that are catalytically active but do notappear to be regulated by RAS. High-level ex-pression of such genes suppresses the lethality ofthe double rasl- ras2- mutation. We concludefrom this that, in yeast, all of the lethal effectsdue to loss of RAS function are mediated throughadenylate cyclase. We have succeeded in express-ing enzymatically active yeast adenylate cyclase inEscherichia coli.
CAKI, a cAMP-dependent protein kinase cat-alytic subunit, was cloned by complementation ofcdc25, a cell-division control mutant that arrestsin GI at the nonpermissive temperature. cdc25 issuppressed by RAS2'19 and by high-level expres-sion of adenylate cyclase. We therefore reasonedthat CAKI would suppress cdc25, which it does.The nucleotide sequencing of this gene is nearlycomplete, and it shows extensive homology withthe sequence of the bovine cAMP-dependentprotein kinase catalytic subunit. Gene-disruptionexperiments indicate that cells lacking CAKI areviable. We therefore conclude that cells mustcontain a second cAMP-dependent protein ki-nase. Indeed, we observe by hybridization anal-ysis another yeast gene weakly homologous toCAKI, which we are now cloning. High-copyplasmids of CAK1 suppress lethality due to lossof endogenous RAS function or loss of adenylatecyclase. These results are formal proof that theeffects of adenylate cyclase and hence cAMP aremediated through the cAMP-dependent proteinkinase system.
CDC25 was cloned together with CAKI bytransforming a cdc25 strain with a centromere-linked library. CDC25 is not one of the knowngenes in the adenylate cyclase pathway. We arenearly finished with its nucleotide sequence, andthis work and the work on CAKI are proceedingin collaboration with M. Zoller (Molecular Ge-netics Section).
In addition to the genes described above, fourrecessive mutations, supA, B, C, and D, have beenisolated that suppress the phenotype of RAS2va19.One gene, PS1, has been isolated that, in highcopy, suppresses RAS2'"9. The relationship ofthese genes to the classic cAMP pathway has yetto be determined.

Studies of the Vertebrate RAS ProteinsC. Birchmeier, D. Broek, 0. Fasano, T. Kataoka, M. Wig ler
Some tumor cells contain mutant RAS genes thatare capable of transforming NIH-3T3 cells.Those genes that have been analyzed arise fromthe wild-type nontransforming RAS genes by mu-tations producing single amino acid substitutionsat position 12 or 61 of the encoded protein. Wehave performed random bisulfite-induced muta-genesis on the cloned wild-type human Ha-rasgene to determine whether mutations at other po-sitions can activate the transforming potential ofthat gene (Fasano et al., Proc. Natl. Acad. Sci.81: 4008 [1984]). Most mutations are not activat-ing, but mutations that specify single amino acidsubstitutions at position 12, 13, 59, or 63 of theencoded protein do activate the transforming po-tential of the Ha-ras gene. Some, but not all,mutant RAS proteins show an altered electropho-retic mobility in SDS-polyacrylamide gels.
Using the approach described in the previoussection, we have been able to test if expression ofthe normal mammalian Ha-ras protein is suffi-cient for viability in yeast cells lacking their ownendogenous RAS genes. To this end, we con-structed a GAL10-Ha-ras transcription unit thatutilized a full-length cDNA clone of the humanHa-ras mRNA under the control of the galactose-inducible GALIO promoter. This unit, closelylinked to a LEU2 marker, was inserted into dip-loid yeast cells that were doubly heterozyous fortheir endogenous RAS genes. Cells were inducedto sporulate, and tetrads were examined after ger-mination on YPD or YPGa1 plates. Approxi-mately 40% of spores with the genotype rasl-ras2- GAL10-Ha-ras were capable of germina-tion when plated on YPGa1, from which we con-clude that the human Ha-ras protein can supplyessential RAS function to yeast. Direct biochem-ical experiments yielded the same result. Aden-ylate cyclase activity in bcyl rasl- ras2- mem-branes was dramatically increased by the additionof purified human Ha-ras protein. These resultsindicate that the effector function of yeast andmammalian RAS proteins has been conserved inevolution. In particular, these results suggest thatRAS proteins may be involved in regulating aden-ylate cyclase in vertebrates.
To test our ideas about RAS function derivedfrom studies in yeast, we have developed a frogoocyte microinjection system. We have foundthat purified mammalian Ha-ras proteins (Grosset al., Mol. Cell. Biol. 5: 1015 [1985]) can induceimmature oocytes to progress from prophase tometaphase. The Ha-rasv.12 is 200-fold more po-tent than Ha-rasGw2 in inducing oocyte matura-tion. This effect is blocked by cholera toxin,which increases cAMP production in oocytes. Wehave observed no significant changes in cAMPconcentrations due to injection of Ha-ras proteinand conclude that the yeast RAS system is notentirely functionally analogous to the mammal-ian system. Nevertheless, it is clear that the oo-cyte system provides an excellent model systemfor testing ideas concerning RAS protein func-tion.
Isolation and Characterization ofOther OncogenesC. Birchmeier, D. Birnbaum, S. Efrat, 0. FasanoM. Wig ler, D. Young
The transfer of genomic DNA into NIH-3T3 cellshas led to the discovery that genes present in sometumor cells are capable of inducing foci of mor-phologically transformed NIH-3T3 cells. Most ofthe transforming genes detected in this way havenow been identified as members of the RAS genefamily, either Ha -ras, Ki -ras, or N-ras. The trans-forming RAS genes detected by the NIH-3T3 fo-cus assay have structural gene mutations that ac-count for their transforming activity. Since thegreat majority of tumor DNAs tested fail to in-duce transformed foci, we have speculated thatthe focus assay has a bias for RAS genes contain-ing structural mutations. Therefore, we have be-gun a series of experiments to explore alternativeassays for transforming genes present in NIH-3T3cells after DNA transfer. The system is a modi-fication of the one described by D. Blair and co-workers (National Institutes of Health). Liketheirs, our assay also relies on the ability of trans-formed NIH-3T3 cells to form tumors in nudemice, but it incorporates methods of cotransfec-tion to heighten sensitivity. Using this assay, wehave detected three human transforming genesfrom the DNA of MCF-7, a human breast carci-
87

noma cell line. One of these is N-ras, which isamplified in MCF-7 cells but does not appear tocontain structural mutations. We have not yet es-tablished whether the other two genes, which wehave called mcf2 and mcf3, are associated withany genetic abnormality in MCF-7.
We are still in the process of characterizingmcf2 and mcf3. mcf3 appears to be the humanhomolog of the v-ros gene and has the structurepredicted for a transmembrane receptor. It hasbeen activated by a gene rearrangement resultingin truncation of the region encoding the putativeextracellular domain. In addition, we have iso-lated and are characterizing a gene called masl,which was isolated from a human mastoid epider-mal carcinoma.
PUBLICATIONS
Fasano, 0., D. Birnbaum, L. Edlund, J. Fogh, and M. Wig ler.1984. New human transforming genes detected by a tu-morigenicity assay. Mol. Cell. Biol. 4: 1695-1705.
Fasano, 0., T. Aldrich, F. Tamanoi, E. Taparowsky, M. Furth,and M. Wig ler. 1984. Analysis of the transforming poten-tial of the human H-ras genes by random mutagenesis.Proc. Natl. Acad. Sci. 81: 4008-4012.
Gross, M., R. Sweet, G. Sathe, S. Yokoyama, 0. Fasano, M.Goldfarb, M. Wigler, and M. Rosenberg. 1985. Charac-terization of human H-ras proteins produced in E. coli.Mol. Cell. Biol. 5: 1015-1024.
Kataoka, T., S. Powers, S. Cameron, 0. Fasano, M. Gold-farb, J. Broach, and M. Wigler. 1985. Functional homol-ogy of mammalian and yeast RAS genes. Cell 40: 19-26.
Kataoka, T., S. Powers, C. McGill, 0. Fasano, J. Strathern, J.Broach, and M. Wigler. 1984. Genetic analysis of yeastRAS1 and RAS2 genes. Cell 37: 437-445.
Powers, S., T. Kataoka, 0. Fasano, M. Goldfarb, J. Strathern,J. Broach, and M. Wigler. 1984. Genes in S. cerevisiaeencoding proteins with domains homologous to themammalian ras proteins. Cell 36: 607-612.
Tamanoi, F., M. Walsh, T. Kataoka, and M. Wigler. 1984. Aproduct of yeast RAS2 is a guanine nucleotide bindingprotein. Proc. Natl. Acad. Sci. 81: 6924-6928.
Toda, T., I. Uno, T. Ishikawa, S. Powers, T. Kataoka, D. Broek,S. Cameron, J. Broach, K. Matsumoto, and M. Wigler.1985. In yeast, RAS proteins are controlling elements ofthe adenylate cyclase. Cell 40: 27-36.
In Press, Submitted, and In Preparation
Broek, D., N. Samiy, 0. Fasano, A. Fujiyama, F. Tamanoi, J.Northup, and M. Wigler. 1985. Differential activation ofyeast adenylate cyclase by wild type and mutant RASproteins. Cell (in press).
BIOCHEMISTRY OF YEAST RAS PROTEINS
F. Tamanoi A. Fujiyama N. SamiyM. Rao M. Walsh
Structural and Functional Analyses ofYeast RAS ProteinsF. Tamanoi, A. Fujiyama, M. Rao, N. Samiy, M. Walsh
A product of the ras oncogene continues to be asource of intellectual excitement from severalstandpoints. First, it is a membrane-bound pro-tein that contains covalently bound fatty acids,which raises the question of how it travels to andinteracts with the membrane. Second, it is a GTP-binding protein, as well as a GTPase, and is amember of the GTP-binding proteins that regu-late various biochemical reactions within a cell.Finally, a single amino acid alteration is knownto convert a normal protein to a form active intransformation. Why such a small change results
88
in dramatic effects on the physiology of cells hasbeen a tantalizing question to us. Our researcheffort is focused on carrying out physical andchemical studies of the ras protein.
In last year's Annual Report, we describedexpression of the ras gene in Escherichia coliaimed at preparing a large amount of the pureprotein. During the past year, our emphasis hasbeen to analyze products of ras-oncogene homo-logs in the yeast, Saccharomyces cerevisiae. Theyeast homologs, RAS1 and RAS2, code for pro-teins that have at their amino termini more than90% homology with the mammalian ras protein.Furthermore, alteration of a 19th amino acid onthe yeast RAS2 protein from glycine to valine, amutation analogous to the one found with thetransforming ras protein, results in dramatic

changes in the growth properties of yeast cells.Thus, mechanistic aspects of the action of rasproteins could be elucidated using the yeast hom-ologs. Two advances have recently been made.First, we have shown that the yeast RAS proteinsexhibit GTP binding and GTP hydrolysis activi-ties similar to those seen with the mammalianprotein, which establishes similarities in bio-chemical activities between the yeast and mam-malian proteins. Second, in collaboration withD. Broek and M. Wig ler (this section), we havedeveloped an adenylate cyclase system from yeastmembranes that is dependent on the RAS pro-teins.
OVERPRODUCTION AND PURIFICATION OFYEAST RAS PROTEINS
Since the amount of RAS proteins produced inwild-type yeast cells is very low, we decided to over-produce these proteins. To accomplish this, theRAS2 gene was cloned into a YEp51 plasmid thatcontained a galactose-inducible GALIO pro-moter. The construct directed the synthesis of aprotein with an apparent molecular weight of41,000 that was immunoprecipitable by a mono-clonal antibody raised against theprotein. The 41,000-molecular-weight proteinappears to be a primary translation product, sincecell-free translation of mRNA extracted fromyeast cells expressing the cloned RAS2 gene re-sulted in the production of a protein that comi-grated with the 41,000-molecular-weight proteinon SDS-polyacrylamide gels. Overproducing theRASI protein, on the other hand, required themanipulation of the gene, since no suitable re-striction sites were available close to the start ofthe gene. By using synthetic oligonucleotides, wecreated a Sphl restriction site at the start of thegene. After recloning into Ml3mpl8 to add a Sallsite, the gene was cloned into the YEp51 plasmidDNA. The construct directed the synthesis of aprotein with an apparent molecular weight of34,000.
Expression in E. coli provides an alternativeand powerful means to produce the RAS proteinsin large amounts. Furthermore, it is relativelyeasy to purify a protein from E. coli. Our firstconstruction utilized a lac promoter that is induc-ible by the addition of isopropyl-O-o-thiogalac-toside. The yeast RAS2 gene was inserted into
the EcoRI site of pUC8 plasmid, which directedthe synthesis of a protein containing 20 extraamino acids added to its amino terminus. A sim-ilar construct was made with the RASI gene,which directed the synthesis of a protein contain-ing five extra amino acids added to its amino ter-minus. We also constructed an expression vectorthat produced intact RASI or RAS2 protein in E.coli. The expression is under the control of arecA promoter that can be induced by the addi-tion of nalidixic acid.
Extracts prepared from cells carrying the aboveconstructs exhibited a high level of GDP-bindingactivity. This provided an assay to purify theRAS proteins. Several column chromatographymethods, such as phosphocellulose, DEAE-cel-lulose, and gel filtration, are being employed forthe purification. The fusion RAS2 protein hasbeen purified to approximately 70% purity asjudged from densitometric scanning of a stainedgel. Further attempts to purify the proteins us-ing high-performance liquid chromatography(HPLC) and GTP agarose are being made.
MODIFICATION OF RAS PROTEINS
The mammalian ras proteinsbound palmitic acid. The fatty acylation mostprobably takes place on a cysteine residue locatedclose to the carboxy terminus, since mutation ofthis cysteine destroys the modification. In con-trast to the ras protein, a product of another on-cogene (src) contains myristic acid. In this case,the fatty acylation occurs on a glycine residueclose to its amino terminus. Whether there existtwo distinct types of fatty acylations and whetherthey are related to the difference in their func-tions are two questions we would like to address.
As a first step toward investigating these points,we established that fatty acylation also occurs inyeast RAS proteins. First, both RASI and RAS2proteins could be labeled with [3H]palmitic acid.Furthermore, we were able to recover the radio-activity on the protein as palmitic acid after mildalkali treatment. Currently, we are investigatingthe site of modification as well as the nature oflinkage. We are also making use of secretory mu-tants of yeast with the hope of gaining insight intothe transport of the protein.
It has recently been reported that yeast RASIprotein is produced as a 34K protein that is later
89

modified to a 39K protein (Temeles et al., Mol.Cell. Biol. 4: 2298 [1984]). This suggests that an-other type of modification is taking place. Inagreement with this observation, we found a sim-ilar 39K protein using our construction. How-ever, we only detected a minor portion of theRAS1 protein modified, and the majority of theprotein remains as a 34K protein even after a longchase. The cause for this discrepancy is currentlybeing investigated.
BIOCHEMICAL ACTIVITIES OF RAS PROTEINS
Using the RAS2 protein purified from E. coli, wehave carried out a detailed characterization of itsenzymatic activities. First, the RAS2 proteinbinds GDP and GTP. The binding was depen-dent on incubation temperature; very little bind-ing was observed at 0°C, whereas maximum bind-ing was observed at 45°C. The pH optimum was7.6. Optimum binding was observed in the pres-ence of 0.5 mm Mg", but some binding was alsoobserved in the presence of EDTA. A Scatchardplot of GDP binding was consistent with single-site binding kinetics, which gave a calculated Kdvalue of 4.7 x 10-8 wt. Kd for a nonhydrolyzableGTP analog, Gpp(NH)p, was 8.7 x 10-8 M. Thus,the RAS2 protein appears to bind GDP and GTPwith a similar affinity. This is in contrast to otherGTP-binding proteins, such as elongation factorEF-Tu, which binds GDP much more tightly thanGTP. Another biochemical activity that the RAS2protein exhibits is a GTPase. The hydrolysis wasspecific to GTP or dGTP and no hydrolysis ofATP, CTP, dATP, dCTP, or dTTP was observed.The products of hydrolysis were GDP and inor-ganic phosphate. The maximum level of GTPaseactivity was obtained at 45°C and in the presenceof 0.5 mm Mg**. The rate of GTPase activitywas calculated to be 60 mmoles/min/mole ofprotein.
By incubating at low temperature and usingSephadex column chromatography, we were ableto isolate a stable complex between the RAS2protein and GTE. Subsequent incubation at 37°Cresulted in the conversion of the bound GTP toGDR. The GDP was still bound to the RAS2 pro-tein as revealed by nitrocellulose assay. We fur-ther showed that a GDP bound to the RAS2 pro-tein could be exchanged with a fresh GTP. Thus,
90
the RAS2 protein is capable of shuttling betweentwo forms: the GTP-bound form and the GDP-bound form. We are currently interested in de-termining whether there are any differences, suchas protein conformation, between the two forms.
The shuttling between the GTP-bound formand the GDP-bound form appears to be an im-portant aspect of RAS protein action, since thisis precisely the process that is impaired with mu-tant RAS proteins containing an amino acid al-teration at either the 19th (Gly -Val) or 66th(Ala --Thr) residues. Expression of these mutantproteins results in a dramatic change in thegrowth properties of yeast cells, including de-creased sporulation efficiency, increased sensitiv-ity to nutrient starvation, and failure to accumu-late carbohydrate reserves. The mutant proteinspurified after their expression in E. coli exhibitedGDP-binding activity similar to that seen with thewild-type protein. In addition, GDP bound tothe mutant proteins could be exchanged with freeGTP with an efficiency comparable to that seenwith the wild-type protein. However, a signifi-cant difference was observed in their GTPase ac-tivities. The activity of the valine mutant wasabout 1501s of that of the wild-type protein, andthe threonine mutant displayed further reducedGTPase activity.
EFFECT OF RAS PROTEINS ON YEASTADENYLATE CYCLASE
Genetic and biochemical studies carried out byM. Wigler's group (this section) have indicatedthat the yeast RAS genes are involved in the con-trol of adenylate cyclase. Thus, membranes ofyeast cells lacking both RAS1 and RAS2 genesfail to show any detectable level of adenylate cy-clase (Toda et al., Cell 40: 27 [1985]). In collab-oration with D. Broek and M. Wigler, we wereable to demonstrate that the purified RAS2 pro-teins are capable of revitalizing this adenylate-cyclase-defective membrane. RAS2 protein pu-rified from E. coli, as well as the protein purifiedfrom yeast, was capable of complementing theadenylate-cyclase-defective membrane. Thecomplementing activity absolutely required thepresence of the nonhydrolyzable GTP analog,Gpp(NH)p. Replacing Gpp(NH)p with GDP re-duced the stimulation to 50010 of that obtained

with Gpp(NH)p. Thus, RAS2-GTP is more ef-fective in stimulating the adenylate cyclase thanRAS2-GDP. Wild-type RAS2 protein preincu-bated with GTP stimulated adenylate cyclaseonly to the level obtained with GDP, presumablybecause GTP hydrolysis took place during thepreincubation. The valine mutant protein prein-cubated with GTP, on the other hand, gave thesame level of stimulation obtained when Gpp-(NH)p was used. Thus, the mutant proteins over-activate adenylate cyclase, which might be re-sponsible for causing changes in the physiologyof yeast cells. A large amount of purified RASproteins provide an invaluable tool to investigatetheir function within yeast membranes.
MOUSE GENETICS
L. Silver V. BautchJ. BahrenbergL. CisekH. FoxC. Jackson
D. LukralleP. MainsS. PanzerN. Sarvetnick
Mouse t haplotypes are naturally occurring,highly variant forms of a region of mouse chro-mosome 17 that has multiple and interacting lociwith profound effects on embryogenesis andsperm differentiation. The t haplotypes main-tain themselves as discrete genomic entitiesthrough recombination suppression, and thesechromosomal units are propagated through wildpopulations by a male-specific transmission ratiodistortion in their favor. Our understanding of thaplotypes has undergone radical changes re-cently, and it is now possible to focus researchefforts in attempts to answer specific questionsthrough the combined use of molecular and ge-netic technitues. Although we have chosen toconcentrate our efforts on the t complex, we ex-pect many of our results to be indicative of gen-eral principles of mammalian genome structureand function.
PUBLICATIONS
Tamanoi, F., N. Samiy, M. Rao, and M. Walsh. 1985. Enzy-matic properties of yeast RAS2 protein. Cancer Cells3: 251-256.
Tamanoi, F., M. Walsh, T. Kataoka, and M. Wig ler. 1984. Aproduct of yeast RAS2 gene is a guanine nucleotidebinding protein. Proc. Natl. Acad. Sci. 81: 6924-6928.
In Press, Submitted, and In Preparation
Broek, D., N. Samiy, 0. Fasano, A. Fujiyama, F. Tamanoi, J.Northup, and M. Wigler. 1985. Differential activation ofadenylate cyclase by wild type and mutant RAS proteins.Cell (in press).
Tamanoi, F., N. Samiy, M. Rao, and A. Fujiyama. 1985. Shut-tling of yeast RAS2 protein between a GTP bound formand a GDP bound form. (Submitted.)
Integration of Retroviral Sequences intothe DNA of Early Mouse EmbryosV. Bautch, L. Silver
One approach to the study of mammalian devel-opment is to identify mutations that disrupt thenormal program of embryogenesis. Insertionalmutations also provide a molecular marker forthe recovery of genes disrupted by the mutation.However, in order to be able to clone the preinte-gration site and analyze the disrupted gene, theintegration event must preserve the linear orderof cellular sequences in a recognizable way. Re-troviral genomes are particularly useful as poten-tial agents of insertional mutation because retro-viruses integrate with only a small (4 to 9 bp)duplication of cellular DNA. We are generatinginsertions in mice using a breeding scheme (N.
91

Jenkins and N. Copeland, pers. comm.) that al-lows for the de novo integration of endogenousretroviral genomes into the DNA of early em-bryos at a relatively high frequency. A subset ofretroviral genomes called ecotropic proviruseshas been followed because most inbred strains ofmice contain a few (0-6) copies that can be easilyfollowed using molecular techniques.
Mice of the RF/J strain contain three ecotropicproviruses. Two of these loci, Emv-16 and Emv-17, are genetically linked and have been impli-cated in integration events in early embryos. Wehave crossed male RF/J mice to females of twodifferent strains (CBA/CaJ and SWR/J) that arenegative for endogenous ecotropic proviruses.The F, progeny were backcrossed to the ecotropicvirus-negative strains. The proviral sequences ofthe N, offspring were analyzed by hybridizinggenomic DNAs prepared from tail clips to an eco-tropic virus-specific probe. N, females carryingEmv-16 and Emv-17 were then backcrossed toproduce N, progeny that were analyzed for newlyintegrated proviruses.
We have scored a total of 29 ecotropic provirus(EP) fragments not found in parental DNAs from184 mice (15.8%). Analysis of the results led tothe following conclusions: (1) New proviral inte-gration events occur when Emv-16 and Emv -17are carried in the maternal genome but not whencarried in the paternal genome. (2) CrossingEmv-16 and Emv-17 onto the SWR/J strain forthree generations results in a higher frequency ofintegration events (15.8%) than the equivalentcrosses onto the CBA/CaJ strain (0%). (3) N,integration events appear to be distributed non-randomly with respect to breeding pairs and lit-ters. (4) The presence or absence of Emv-16 andEmv-17 in the embryonic genome does not affectthe probability of a de novo integration in thatgenome. These findings suggest that both ge-netic and epigenetic events contribute to this phe-nomenon. The results are consistent with amechanism of reintegration involving infection ofthe egg or early embryo by maternal tissues shed-ding virus, although other possibilities exist.
The EP fragments were visualized at 5-50% theintensity of homozygous bands run on the samegel, with an average at approximately 20%. Thisheterogeneity shows that there is variation in thestage of embryonic development during which the
92
integration event occurs, with those fragments at50% intensity of homozygous bands most prob-ably integrating at the one-cell stage. In practicalterms, we can predict, assuming that genomicDNA from tails is representative of germ-lineDNA, that we will see germ-line transmission ofthe new loci at these levels. We have been able topass 12 of the EP fragments through the germ lineto date, and preliminary results suggest that infact germ-line transmission parallels the repre-sentation of these fragments in tail DNA. There-fore, we should be able to pass most of the scoredEP fragments through the germ line.
We are currently establishing mouse lines con-taining these new proviral loci and plan to ana-lyze them for insertional mutations. Unfortu-nately, mutations cannot be easily selected for inmice; however, we can check several parameters.First, visible structural or behavioral mutationscosegregating with new loci can be identified asdominant mutations if found in heterozygotesand as recessive mutations if found in homozy-gotes. We can also check for recessive lethal mu-tations by using molecular and genetic techniquesto score a homozygous class of offspring. Theabsence of this class in the appropriate cross willbe indicative of a recessive lethal mutation. Fi-nally, we can score any X-chromosome-linked le-thal mutations by deviations in the sex ratios ofoffspring.
A Partial t Haplotype ContainsBoth Copies of a PolymorphicGenomic DNA SequenceV. Bautch, L. Silver
We have been using short low-copy genomic DNAsequence clones (microclones) to dissect thestructure of the partial t haplotypes. Partial thaplotypes are the result of rare recombinationevents between complete t haplotypes and wild-type chromosome 17. Because t chromosomescontain an inversion in the distal region of thecomplex and a hypothesized inversion in theproximal region, it is proposed that these rare re-combination events result in duplication of some

DNA sequences and deletion of other sequences.One of these partial t haplotypes, t"8, containsthe proximal portion of the t complex and carriesa recessive lethal mutation that maps in the vicin-ity of the breakpoint between t DNA and wild-type DNA. Recently, one of the microclones,designated 89S, was shown to hybridize to gen-omic sequences that also map near the break-point in this chromosome. This was done usinga restriction-fragment-length polymorphism(RFLP) between t DNA and DNA from mice ofthe 129 inbred strain, which carries the twm hap-lotype. Genomic DNAs from mice with the t'18chromosome and either the wild-type chromo-some 17 ( + 129/tw'8) or a complete t haplotype(t'/r/8) were analyzed with a probe made fromthe 89S microclone. As expected, the ( + 129/tw'8)DNA contained fragments specific for both thewild-type chromosome 17 and t-haplotype DNA;however, the (t'it'8) DNA also contained bothsets of fragments. Since ( + 129/ + 129) DNA con-tains only wild-type fragments hybridizing tothe 89S probe, and since DNA of mice with twocomplete t haplotypes (t'/t°) contains only the t-DNA-specific fragments, the wild-type bands inthe (t"8/t"'") DNA must have been contributed bythe tw'8 chromosome. Therefore, the recombina-tion event generating this partial 1 haplotype wasprobably an unequal crossing-over resulting in thepresence of both sets of these fragments on the1'8 chromosome. Subsequently, several other in-vestigators (H. Fox, this section, and H. Lehrach,European Molecular Biology Laboratory, Heidel-berg) have shown that three other independentlyderived microclones are also duplicated in the r"chromosome, suggesting that this chromosomecontains a substantial amount of duplicatedDNA. Current studies are aimed at determiningthe extent of the duplication. Preliminary resultsfrom Lehrach's laboratory indicate that the 89Smicroclone maps to the TLA region of the H-2complex on wild-type chromosome 17. We arecurrently analyzing the r" chromosome with H-2 DNA probes to determine whether any of thesesequences are also duplicated. Because the reces-sive lethality carried by the t'18 chromosome is as-sociated with the recombination breakpoint, it ispossible that either the duplication or the pro-posed corresponding deletion of sequences couldbe responsible for the lethal phenotype.
Characterization of t Haplotypes witht Complex ClonesH. Fox, L.M. Silver [in collaboration with G.R. Martin,University of California, San Francisco]
Last year, we described the mapping of DNAclones obtained by chromosome microdissection(by H. Lehrach et al., European Molecular Biol-ogy Laboratory, Heidelberg) to the t complex byidentifying restriction fragment length poly-morphisms with these clones in the DNA of micecongenic for t haplotypes. Specific restrictionfragments identified by these clones were foundto be t-specific, since they were present in allcomplete t haplotypes but replaced by other re-striction fragments in all of the wild-type inbredstrains tested. However, there were no restrictionfragments present in wild-type or t that were notfound in the other, and the level of polymorphismobserved between t haplotypes and wild type isroughly comparable to that observed betweeninbred strains. Thus, the t haplotypes appear tohave a similar origin and have not diverged greatlyfrom other mice.
These clones have been used to classify partialt haplotypes, which are derived from rare re-combinant events between complete t haplotypesand wild-type chromosomes. Partial 1 haplotypeswere found to contain subsets of the t-specific re-striction fragments, and each could be classifiedaccording to the t-specific fragments it contains.This is the first molecular evidence that inde-pendent partial t haplotypes contain differentlengths of t haplotype DNA and confirms the ge-netic studies of M. Lyon (Harwell), who has usedpartial t haplotypes to elucidate the genetic basisof the t haplotype phenotypes.
In a recent survey of wild mice, J. Klein (Tub-ingen) has identified t haplotypes at a frequencyof approximately 25% in mouse populationsthroughout the world. We have analyzed thesenew t haplotypes with our molecular probes andfound that all of the mice resemble the previouslycharacterized t haplotypes except one. This thaplotype, originally isolated in Haifa, is uniquein that it only carries a subset of the t-specificmarkers. Genetic studies indicate that it similarlyonly expresses a subset of the t-specific pheno-types. Several lines of evidence support the the-
93

ory that this haplotype may be an intermediate inthe formation of t haplotypes in the wild.
Evidence for a Proximal t HaplotypeInversionN. Sarvetnick, L.M. Silver
Recently, the product of one rare recombinationevent between a t haplotype and wild type (namedtwLub2) was characterized and found to express twounusual phenotypes not associated with either ofthe parental chromosomes from which it was de-rived. First, the twLub2 chromosome suppresses thedominant, short-tail phenotype expressed by mu-tations at the Brachyury (T) locus so that animalsdoubly heterozygous for T and twLub2 are bornwith tails of normal length. Second, the twLub2chromosome expresses a dominant lethal effect(Tme) in all embryos that inherit the mutantchromosome from their mother, so that hetero-zygous females cannot transmit twLub2 to theiroffspring. Embryos that receive either mutantchromosome from their mother develop in an ap-parently normal fashion until late in gestationwhen death inevitably occurs. At the time ofdeath, mutant embryos are edematous and showan increase in weight relative to their normallittermates. In contrast, when embryos receiveeither mutant chromosome from their father, de-velopment proceeds normally to the birth ofhealthy and fertile individuals.
To gain a better understanding of the geneticbasis for the various phenotypes expressed by thetwLub2 chromosome, we have used several molecu-lar probes to analyze the structure of this unusualchromosome. Our genetic and biochemical anal-yses of twLub2-containing mice have generated evi-dence that the chromosome contains a deletion.First, twLub2 expresses a new lethality not ex-pressed by its parental t haplotype. Second, thechromosome fails to complement the lethality as-sociated with another t haplotype, tw", that mapsin the proximal region. Finally, the chromosomefails to express either known form of the TCP-1protein. Clustered mutations are most simply ex-plained by a deletion.
We have further investigated the structure ofthe twLub2 chromosome by performing a genetic
94
analysis with a cDNA clone (pMK174) that hy-bridizes to sequences located in the proximal por-tion of chromosome 17. The results indicate thatthe twLub2 chromosome must contain both t-spe-cific and wild-type alleles at this locus. Thesedata demonstrate that the twLub2 chromosomecarries a duplication of genetic material, withboth wild-type and t-specific forms of thepMK174 locus.
The accumulated data indicate that a recombi-nation event between a t haplotype and a wild-type chromosome caused the duplication of cer-tain genetic loci (T and pMK174 sequences), aswell as the deletion of other loci (Tme, Tcp-1, andthe tw" lethal locus) in the generation of the twLub2chromosome. However, a simple, unequal re-combination event between homologous chro-mosomes would cause either a duplication or adeletion but not both. To explain the generationof twLub2, we postulate the existence of an inver-sion over the proximal region of t haplotypes, ex-tending from the T locus and pMK174 sequencesto the loci of Tcp-1 and Tme. A correctly ori-ented recombination event between these two setsof inverted loci would produce a chromosomewith all of the genetic properties of twLub2 (see Fig.1 below).
Studies performed in a similar manner indicatethat the Tt°" haplotype appears to have a struc-ture reciprocal to that of twLub2. Our model for thegeneration of both twLub2 and Tt°" predicts thatthe crossover event occurred while the two chro-mosomes were paired in this type of linear, non-homologous fashion. The demonstration of non-homologous chromosomal pairing in t heter-
0-11
0-1
twLub2
TtOrl
48 (T 174) (Tme Tcp- I) 122I I
48
48
(Tcp-I Tme) (174 T)
(T 174) (174 T)
48 (Tcp-I Tme) (Tme Tcp-I)
122
122
I I
122
Figure 1 Representation of the proximal region of mousechromosome 17. The loci inverted in t haplotypes relativeto wild type are indicated, with the postulated recombinationevent that generated twLub2 and Tt°".

ozygotes makes it possible to explain the mecha-nism by which rare recombination occurs in thisregion. The accumulated molecular data suggestthat many rare recombination events have oc-curred within the well-characterized inverted re-gions, with the production of partial t haplotypesthat have both duplicated and deleted loci. Theseevents can be explained by the occasional, chancepairing and recombination of nonallelic membersof a repetitive DNA family. It is important to em-phasize that such events will occur only rarely,and only a fraction will produce viable recombi-nant products.
Genetic Analysis of a Mouse DominantMaternal EffectN. Sarvetnick
The detailed structural analysis of t""" and an-other partial t haplotype, Tt'', has implied thatthe two chromosomes represent reciprocal recom-bination products. Thus, a duplication of a func-tional Tine locus may be present on the Tt°'chromosome. If the lack of maternal transmis-sion of t""" were purely a dosage problem, thenthe duplication could be utilized to complementthe defect. We designed a series of breeding pro-tocols to explore the potential of the Tt °" chro-mosome to alter the penetrance of the t'L"b2 ma-ternal effect.
When Ttf/rL"2 females were mated with wild-type males, 94% of the 188 animals born had theshort-tail phenotype indicative of the T-carryingchromosome. The remaining 6% of the animalshad normal tails and appeared healthy; all thatcould be analyzed were found to have a recombi-nant chromosome that carried neither I " 1"' norT. Hence, with this mating, there is no evidencefor the survival of any twLuw-carrying individualsto the time of birth.
When Ttf/twLu" females were mated to Tt°"tf/+ tf males, a third phenotypic class of neonatal
mice was observed. Of 195 offspring, 16% had atail of normal length, but were much larger thantheir littermates and appeared edematous; all diedwithin 48 hours after birth. This phenotype isreminiscent of that observed in utero for mater-nally derived twL"' embryos from other crosses.
Another 5% of the offspring had tails of normallength, but were healthy, and when tested, theywere found not to carry the t"'" chromosome.The total fraction of mice born with normal tailsfrom this cross (21%) is too large to be accountedfor by crossing-over alone, and the majority ofthese mice express the characteristic r"b2 phe-notype. It appears that the presence of a Tt °"chromosome in the father allows a percentage ofthe maternally derived t"'"" individuals to sur-vive to birth.
To determine whether the presence of the Tt°"chromosome in the mother's genotype has any ef-fect on the survival of t'ub2 individuals, Tt°' /t"L °b2 females were mated to wild-type males. Noanimals were born with the twl-"b2 phenotype.Only 1 (2%) of the 45 offspring born had a nor-mal tail, and when tested, this animal was foundto have a recombinant chromosome without thet'u" haplotype. Hence, the presence of Tt°" inthe primary oocyte is not sufficient to rescue em-bryos that receive the alternative t''Lub2 homologfrom a doubly heterozygous mother.
There are two possible mechanisms by whichthe t"'""2 maternal effect could be expressed. Thefirst mechanism is dependent on an absolute re-quirement for a functional Tine locus during thehaploid phase of oogenesis; this condition is notmet when tv-ub2 is inherited maternally. The sec-ond mechanism requires the differential func-tioning of the maternally and paternally inheritedforms of Tine at some critical stage of embryoge-nesis; e.g., if the paternal locus is inactive, em-bryos having no maternal form will die.
Several facts argue against the first mechanismdescribed above. First, it seems unlikely that amolecule produced (or not produced) during oo-genesis could begin to have a dramatic effect onan embryo that is 10,000 times larger at 15-19 daysafter conception. Second, nuclear transplant re-sults obtained by J. McGrath and D. Solter (Wis-tar) indicate that a wild-type cytoplasm andmembrane are not sufficient to rescue a mutantnucleus. Finally, our results indicate that a spe-cific paternal genome can alter the penetrance ofthe t""" phenotype. This experiment providessupport for the model of a differential function-ing of the Tme locus dependent on the parentfrom which it is inherited. If the maternal alleleis thought of as fully activated and the paternalallele as relatively inactive, then the embryo hav-
95

ing a maternally derived t"'"" chromosome mustsurvive solely on the paternal allele, resulting inlethality. In our experiment, the double dose ofthe relatively inactive Tme paternal locus par-tially complements the deficiency and increasesthe mean life span, so that a large percentage ofthe P4-.2-containing embryos are born.
Cis-Trans Analysis of t HaplotypeLethal GenesP. Mains
It has long been noted that the viability of miceheterozygous for complementing t haplotypesgenerally ranges from 18% to 85%. A previousstudy tested whether this viability could be im-proved by placing two lethal mutations togetheron one chromosome (in cis) rather than on sepa-rate chromosomes (in trans). It was found thatthe complementation between two different le-thals was increased to 100% if they were on thesame chromosome when they are heterozygouswith a fully wild-type chromosome. This wouldindicate that the wild-type alleles work more ef-ficiently when they are on one chromosome (incis) than when they are on separate chromosomes(inirans). Other investigators interpreted this cis-trans effect as indicating that the two loci werepart of a single functional unit, even though theymap 10-20-cM apart. They proposed that the ne-cessity for the alleles at widely separated loca-tions to be on the same chromosome in order tofunction efficiently may be indicative of a gen-omic rearrangement.
An alternative interpretation of the cis-transtest is that t haplotypes may carry additional "sub-optimal" alleles at loci other than their uncondi-tional lethal mutations. The incomplete comple-mentation of two lethal haplotypes could resultfrom a requirement for a cis interaction of wild-type alleles or from an effect of other deleteriousgenes associated with the t haplotypes. To con-trol for this, a t chromosome lacking either lethalis required to provide both "wild-type" alleles ina cis arrangement. This chromosome would bethe reciprocal crossover of the t chromosome car-
96
rying the two lethals in cis. However, because ofthe limited genetic resolution in the mouse, pre-cise, or even nearly precise, reciprocal crossoverscannot be easily isolated. Any two reciprocalpairs will contain overlapping regions that cancomplicate the interpretation of the data. We re-peated the cis-trans test using several independentrecombinant chromosomes in different pairwisecombinations. The different combinations arelikely to share different regions of overlap. Wefound that some combinations improve the com-plementation of two t lethal alleles, whereas oth-ers did not. We believe that the data can be ex-plained simply by the presence of a slightly del-eterious gene(s) within the overlap, or they mayreflect the effect of genetic background.
The minor differences in the viability of the cisand trans heterozygotes are unlikely to be causedby a rigorous requirement that they be carried onthe same chromosome to function efficiently.Instead, it may be due to additional deleteriousgenes carried by t haplotypes. t haplotypes areeffectively isolated from the gene pool of mice dueto the suppression of recombination with thewild-type chromosome 17. This may allow the t-bearing chromosomes to acquire deleterious mu-tations at loci in addition to their unconditionallethal mutations. In fact, some t chromosomeshave been found to carry multiple lethal alleles.These other deleterious alleles may be unique tocertain t haplotypes, or they could be shared byall, and could account for the decrease in the vi-ability of the trans heterozygote compared withthe cis.
PUBLICATIONS
Fox, H.S., L.M. Silver, and G.R. Martin. 1984. An alphaglobin pseudogene is located within the mouse t com-plex. lmmunogenetics 19: 125-130.
Fox, H.S., G.R. Martin, M.F. Lyon, B. Herrmann, A.-M. Fri-schauf, H. Lehrach, and L.M. Silver. 1985. Molecularprobes define different regions of the mouse t complex.Cell 40: 63-69.
Rohme, D., H. Fox, B. Herrmann, A.-M. Frischauf, J.-E. Ed-strom, P. Mains, L.M. Silver, and H. Lehrach. 1984. Mo-lecular clones of the mouse t complex derived from mi-crodissected metaphase chromosomes. Cell 36:783-788.
Silver, L.M., J.I. Carrels, and H. Lehrach. 1984. Molecularstudies of mouse chromosome 17 and the t complex. In

Genetic engineering-Principles and methods, vol. 6, pp.141-156. Plenum Press, New York.
Silver, L.M., D. Lukralle, J.I. Garrels, and K.R. Willison. 1984.Persistence of a lethal t haplotype in a laboratory stockof outbred mice. Genet. Res. 43: 21-25.
Winking, H. and L.M. Silver. 1984. Characterization of arecombinant mouse t haplotype that expresses a domi-nant lethal maternal effect. Genetics 108: 1013-1020.
TRANSGENIC MICE
D. Hanahan S. Alpert M. LacyS. Efrat V. Bautch
Cold Spring Harbor laboratory has establishedand is expanding a group that is studying variousaspects of gene expression and its consequencesusing the mouse as an experimental organism.Currently, the major approach of this group in-volves transferring cloned and often-altered genesback into the mouse germ line, so as to examinetheir effects in that context. This is being accom-plished by injecting a solution of DNA into apronucleus of a one-cell embryo, which is thenreimplanted into a foster mouse and allowed todevelop. Mice that heritably carry the new ge-netic information (so-called transgenic mice) arisefrom about 10% of those born from embryos in-jected with the DNA. Transgenic mice are beingused to examine control of gene expression, theconsequences of oncogene expression, and theprospects for using insertional mutagenesis byDNA microinjection as a genetic tool.
Recombinant Insulin SV40 Oncogenesin Transgenic MiceD. Hanahan, S. Alpert, S. Efrat, M. Lacy
Hybrid oncogenes composed of regulatory infor-mation flanking the rat insulin (II) gene linked toprotein-coding information for SV40 T antigenhave been transferred into the mouse germ line.The rationale for employing such combinations is(1) to identify elements associated with the insulingene that mediate its correct tissue-specific ex-pression in the insulin-producing 3 cells of the
In Press, Submitted, and In Preparation
Mains, P and L.M. Silver. Cis-trans analysis of t haplotypes.Genetics (Submitted.)
Sarvetnick, N., E. Mann, P Mains, R. Elliott, and L.M. Silver.Nonhomologous crossing-over in t haplotype heterozy-gotes produces recombinant progeny with unbalancedgenotypes. Cell (Submitted.)
endocrine pancreas, (2) to examine the conse-quences of oncogene expression in /3 cells, and (3)to address the prospect that oncogene expressioncan be employed to facilitate the establishment ofcultured 0 cell lines.
Transgenic mice were produced that harbor twodifferent insulin/SV40 T-antigen genes. One(called RIPI-Tag) carries the insulin gene pro-moter aligned so as to transcribe the T-antigengene. The second (called RIR-Tag) has the insu-lin promoter inverted with respect to the SV40-coding information. Both recombinant geneselicit the heritable formation of solid 3 -cell tu-mors in transgenic mice that harbor them, sug-gesting that the tissue specificity element(s) canact bidirectionally. Expression of T antigen is de-tected only in the insulin-producing f3 cells. Thehybrid genes are expressing in virtually all (3 cellsprior to tumor formation, which occurs in only afew of the several hundred islets of Langerhansthat comprise the endocrine pancreas. This indi-cates that secondary events are necessary to con-vert a collection of islet cells expressing SV40 Tantigen into a solid, well-vascularized tumor. Itis possible that a cellular oncogene is involved.The other major cell types normally found in is-lets - glucagon-producing a cells and somatosta-tin-producing S cells -are rare and disordered inthese transgenic mice.
In summary, sequences located within 660 by 5'to the transcribed portion of the rat insulin geneare sufficient to elicit cell-type-specific expres-sion in transgenic mice, and the synthesis of Tantigen produces islet disruption and tumor for-
97

mation. These transgenic mice should providefurther insight into the development of the en-docrine pancreas and on the mechanisms of on-cogenesis.
Analysis of DNA Microinjection intoMouse Embryos to CreateInsertional MutantsV. Bautch, D. Hanahan
It is known that insertion of DNA into the mousegerm line via microinjection can result in inser-tional mutations. Microinjected DNA is usuallyfound as 5-50 copies in a head-to-tail tandemarray at a single integration site, and the 5' and3' ends of the array are sometimes missing se-quences. Limited analyses by several investiga-tors indicate that approximately 10% of trans-genic mice carry a recessive-lethal mutation thatcosegregates with the inserted DNA. However, inno line has the integrity of the flanking cellularsequences been adequately analyzed. This pa-rameter is important with respect to insertionalmutagenesis, because only if integration of DNApreserves the linear order of cellular sequences orgives rise to small deletions or rearrangements(e.g., no larger than cosmid size) will it be possi-ble to clone the preintegration site easily and ana-lyze the disrupted gene(s). Therefore, we plan toinvestigate both the rate of generating recessive-lethal mutations and the integrity of flanking se-quences after integration of microinjected DNA.
We have therefore constructed a plasmid vectorthat can be used to rescue, by transformation ofEscherichia coli, the inserted DNA and flankingcellular sequences. Although this vector is de-rived from pBR322, it has been altered so thatdeletion of some plasmid sequences from the endof a tandem array by "nibbling" prior to integra-tion should not affect the ability to rescue thisplasmid. The vector contains three different drugresistance genes, and each gene contains severalrestriction enzyme sites unique to the plasmid. Inaddition, a suppressor tRNA gene has been in-serted that can complement bacteriophage X mu-tations and provide an additional selection. Apolylinker containing unique restriction sites hasbeen inserted between two of the drug resistancemarkers in order to linearize the plasmid prior
98
to microinjection. It is expected that the DNAwill integrate in a head-to-tail tandem repeat sothat the linear order after integration will be-kank-ori-sup'-ampR-tetR- . The genomic DNAwill be digested with a restriction enzyme whosesite is in a drug resistance gene and religated. Anyinternal copies of the tandem DNA should recir-cularize to yield plasmids resistant to all threedrugs upon transformation; however, each endshould contain some cellular DNA and also beuniquely sensitive to the drug whose resistancegene was originally cleaved. For example, digest-ing with an enzyme that cleaves the tee geneshould yield a DNA fragment containing a trun-cated tetR gene at one end and flanking cellularDNA at the other end. This fragment will ligateto form a plasmid with cellular DNA that is amp",kanR, and tets. Moreover, in this scheme, a sec-ond rescue using an enzyme that cleaves the kanRgene should yield a plasmid containing cellularDNA from the opposite end of the tandem array.In the event that plasmid sequences are altered bythe integration so as to make plasmid rescue im-possible, it should be possible to retrieve theflanking DNA by cloning into phage containinga nonsense mutation and selecting for the sup-
of sup-pressor tRNA sequences makes phage rescue im-possible, the flanking DNA can be retrieved bymaking a library of genomic DNA from thetransgenic mouse and screening with the plasmidas a probe.
We plan to generate and analyze 10-20 lines oftransgenic mice containing this vector. Theflanking sequences will be cloned and their coli-nearity with the preintegration site will be as-sessed. We will also use breeding and molecularanalysis to determine the efficiency of generatinginsertional mutants in mice using this technique.
Insulin/SV40 T-antigen Hybrid GeneExpression in the DevelopingMouse PancreasS. Alpert, D. Hanahan
Transgenic mice containing an insulin/SV40 T-antigen hybrid gene will be used to investigate thedevelopmental control of insulin. In mammals,

there are two developmental phases of insulin se-cretion. The first stage is a low level of insulinproduction detected at about day 14 of gestationin mice. This correlates with the early develop-ment of the pancreas. The second phase is astriking increase in insulin synthesis at approxi-mately the 16th day of gestation; cells are firstapparent at this time. During the last 4 days ofgestation, insulin synthesis remains high. Themolecular mechanism controlling this develop-mental sequence has yet to be explained.
Our experimental approach is to compare thedevelopmental expression of the endogenous in-sulin genes to that of the recombinant insulingene. This hybrid gene contains 660 by of the 5'-flanking region of the insulin gene joined to thecoding region for SV40 large T antigen. Analysisof transgenic mice carrying this gene has shownthat the sequences required for tissue-specificregulation are present in the 5'-flanking region ofthe insulin gene (see above report by D. Hanahanet al). The mouse pancreata will be excised andexamined throughout pre- and postnatal de-velopment. Immunohistology to sections of thepancreata will be used to characterize the expres-sion of insulin and other hormone products ofthe endocrine pancreas. If the sequences thatelicit proper developmental expression of the in-sulin gene are contained in this recombinant gene,we would expect to see T antigen expressed in thesame temporal manner as insulin. Various con-structions of the hybrid gene containing differentpieces of the insulin gene and its flanking regionsshould allow us to identify the sequences requiredfor the developmental regulation of insulin. Theinfluence of the altered cells on the develop-ment of the islets of Langerhans will be assessed.This will provide more information of the bioge-nesis of the disordered islets found in the trans-genic mice.
Establishment of (3 Cell Lines fromTransgenic MiceS Alpert, D. Hanahan
Permanent lines of the 3 cells of the pancreas, aswell as biochemical studies on insulin geneexpression, can provide a useful tool for diabetes
research. Islet-cell cultures have been maintainedfor fairly long periods of time in culture (Farkasand Joo, Diabetes 33: 1165 [1984]), but estab-lished cell lines do not arise from these cultures.Rat insulinomas have been used to create insulin-secreting (3 cell lines. However, these cell lines donot appear to be glucose-inducible, which is animportant attribute of authentic cells (Gazdaret al., Proc. Natl. Acad. Sci. 77(6): 3519 [1980]).The use of a hybrid gene containing a tissue-spe-cific promoter to regulate an oncogene could bea tool for expanding different primary cell typesinto established cell lines.
As transgenic mice containing the insulin/SV40T-antigen hybrid gene express T antigen specifi-cally in the ,8 cells of the pancreas, we plan toculture the islets of Langerhans in an attempt toestablish authentic glucose-inducible 13 cell lines.This will involve verification of cultured cell iden-tity by determining (1) that the endogenous insu-lin gene is glucose-inducible, (2) that the trans-gene is glucose-inducible, and (3) that glucagonand somatostatin are not secreted by the cells.Further RNA and protein analyses will be per-formed to quantitate the basal- and glucose-chal-langed levels of insulin and T antigen.
Bovine Papilloma Virus inTransgenic MiceS. Alpert, D. Hanahan
Bovine papilloma virus (BPV) is an 8-kb double-stranded DNA virus. In the natural host, BPVinduces epithelial or fibroepithelial tumors byabortively infecting the epidermal germinal cells.BPV usually remains a free circular molecule, butit can integrate into the genome in certain cases.Although there is no in vitro system for virus pro-duction, C127, an epithelial mouse cell line, cansupport BPV replication. This replication re-quires both cis- and trans-acting factors.
We have injected a BPV plasmid containing apartial tandem repeat of the BPV genome (BPV1.69; a gift from D. DiMiao, Yale University) intomouse embryos. The injections produced twomice positive for the BPV molecule. Only oneof these mice, containing approximately five cop-ies of BPV 1.69 integrated into high-molecular-
99

weight DNA, survived to adulthood. We haveperformed analyses on the offspring of thismouse, and have observed no episomal BPV mol-ecules in total DNA preparations, or low-molec-ular-weight (Hirt) DNA extractions, from 16mouse tissues. We are now assessing the expres-sion of the BPV genome in this transgenic mouse
100
line, and preliminary results indicate that BPVtranscripts are present in the testes of BPV-posi-tive mice. We also plan to try to superinject cir-cular BPV plasmids into this transgenic mouseline to maintain extrachromosomal BPV mole-cules by complementation with the integratedcopies.

MOLECULAR GENETICS
Classic genetics experiments conducted at Cold Spring Harbor Laboratory in theearly 1940s on bacteriophage by DelbrOck, Luria, and Hershey and on maize byMcClintock have paved the way for the field of molecular biology. In recentyears, the tools of genetic engineering have allowed the analysis of genes at thenucleotide level. These tools consist of DNA-mediated transformation, gene clon-ing by complementation, site-directed DNA mutagenesis, transposon tagging,and phenotypic analysis of in-vitro-produced mutations. They have been appliedat Cold Spring Harbor Laboratory to dissect the genetic mechanisms, in molecu-lar terms, of various biological systems. We summarize here the results of thepast year, involving various eukaryotic organisms (the yeast mating types andcell-cycle genes, maize transposable elements, and somatic-cell genetics of to-bacco) and the prokaryotic organisms Clostridium sp., Rhodopseudomonas, E.co/i, and Azotobactor sp.
YEAST GENETICS
A. KlarJ. HicksD. Beach
J. IvyM. McLeodG. LiviR. BooherJ. Gould
S. KakarM. KellyC.-1.13. LinL. MiglioC. Stevens
Research by the Delbruck Laboratory YeastGroup during 1984 continued to focus on themechanism of mating-type switching and its re-lationship to the cell-division cycle, but changeswere developing in the profile of the group andthe building. In the Spring, Jeff Strathern andhis family left Cold Spring Harbor Laboratoryfor the Frederick Cancer Research Center inFrederick, Maryland. Jeff was appointed Direc-tor of Eukaryotic Genetics and is charged withdeveloping a large research effort on eukaryoticmicroorganisms. Our group and the Laboratoryas a whole will miss Jeff scientifically and so-cially. He was constantly acting as a catalyst forinterdisciplinary discussions and used his broadbackground in biology to see the relationshipsamong seemingly disparate observations. We wishhim well in his challenging new job.
Another development occurred when the plansfor the Plant Group to move to nearby Uplands
Farm were altered and it was realized that a newaddition to Delbruck Laboratory might be re-quired, similar to that which allowed the YeastGroup to have permanent quarters in 1982. Planshave been drawn for a north wing that would pro-vide laboratory space for the Plant Group to dou-ble in size while retaining the Uplands Farm landand buildings as a working field station. The nextstep is to develop funding for this planned expan-sion in addition to the support for Uplands Farmthat was provided by the National Science Foun-dation last year.
Finally, 1984 was the last full year that the re-search group studying the the cell-division cycleof fission yeast occupied the Delbruck Labora-tory. This group, headed by D. Beach, will moveto the newly renovated quarters in the DemerecLaboratory in 1985. The growth of this researchprogram, along with the cell-cycle studies of yeast
oncogenes in M. Wigler's laboratory, will keep
101

Cold Spring Harbor Laboratory at the forefrontof efforts to use these simple microorganisms touncover the reasons behind the unchecked growthof human cancer cells.
SUM/: Regulator of the HML and HMRLoci of Saccharomyces cerevisiaeA. Klar, J. Hicks, S. Kakar, J. Ivy, G. Lim.0,-I.P. Lin, L. Miglio
In previous years' reports, we have shown thatboth MATa and MAT code for two transcriptsdivergently transcribed from within the Y region(Fig. 1), suggesting that their promoters proba-bly lie within Y. The structure of the silent coun-terparts of MAT, HML, and HMR is, but for afew bases, identical to that of the constitutivelyexpressed MAT locus, yet the HM loci are unex-pressed. The task of keeping the HM loci unex-
pressed is assigned to at least four trans-actingMAR/SIR loci and cis-acting E and I sites (seeFig. 1).
To determine possible interactions between theMAR/SIR gene functions and to search for newloci required for regulating the HM loci, wesought extragenic suppressors of the marl-I mu-tation. The following selection scheme was used:The HMLa MATa HMRa marl-1 strain exhibitsa sterile phenotype because of the simultaneousexpression of a and a information. A mutantthat exhibits the a phenotype, and therefore hasrepressed the HM loci, was isolated. The muta-tion is unlinked to MAT, to the HM loci, or tothe known MAR/SIR genes. We designate thenew locus as defined by the mutation as SUM/(SUpressor of Mar). The mutation sum/4 is al-lele nonspecific because it also suppresses a marldeletion mutation. Even more interesting, themart (sir3) and sir4 mutations are also sup-pressed. To account for these data, we suggestthat the MAR/SIR functions negatively regulate
MAR-SIRREPRESSION
ZI Z2
E W X YO< ZI Z2 I W X Y°A<A,* E X Ya ZI I
HMLoc MATo<
kwot Yo ZI Z2
W X AM
HMRa
MATaFIGURE 1 Structure of chromosome III showing the mating-type cassettes. The HML and the HMRcopies of the mating-type genes are normally kept silent by the MAR /SIR genes and the cis-acting Eand I sites. Interconversion of MATa and MATa involves a unidirectional transposition-substitutionevent from HML or HMR to MAT.
102

the SUM1 gene function, which in turn is re-quired to "turn on" the HM loci by a positivecontrol. Similarly, we have been successful inisolating suppressors of the mar2 (sir3) mutation.Future genetic and molecular studies will allow usto understand more fully the regulation of theHM loci.
Analysis of MAR/SIR Gene ClonesJ. Ivy, G. Livi, C. Stephens, M. Kelly, J. Hicks, A. Klar
As described in previous Annual Reports, we haveisolated and identified the MAR/SIR genes of S.cerevisiae from within a cloned genomic libraryof DNA fragments by their ability to complementMAR/SIR mutations in vivo. MAR/SIR mutantstrains, which are normally nonmaters due toexpression of HML and HMR, become matingproficient when they contain an extrachromo-somal, independently replicating plasmid bearinga wild-type MAR/SIR gene. Using the fourMAR/SIR gene clones, we have examined thepossibility of a cascade of regulation amongthem. RNA blot analysis, however, has ruled outa cascade of transcriptional regulation.
One observation that multiple copies of SIR3will suppress SIR4 mutations has suggested thepossibility that Sir4 combines with Sir3 to alter itsterically and to enhance its activity. Overprod-uction of Sir3 in the high-copy-number SIR3transformants might obviate the need for thesteric enhancement of Sir3 activity. In support ofthe idea that Sir4 combines with other SIR geneproducts in a dimer or multimer is the observa-tion that a cloned, truncated SIR4 gene has anAntimar activity; transformation of wild-typeSIR strains with Antimar results in expression ofthe silent cassettes.
We have constructed null mutations in the fourMAR/SIR genes by altering and transplacingsuch disruptions into the genome. Null muta-tions in all four genes are viable, and marl (sir2),mar2(sir3), and sir4 null mutations have a non-mating phenotype. Cells with a null mutation insirl, however, still retain partial regulation of thesilent cassettes. We conjecture that a regulatorycomplex still forms in the absence of Sir I but thatit is not stable.
MAR1(SIR2) and MAR2(SIR3) have beenmapped by standard genetic analysis, taking ad-vantage of the fact that crosses can be made withmar-mutant spores if the spores came from a het-erozygous MAR/mar diploid. The cloned geneshave provided us with an alternate means of map-ping these loci. By "tagging" the SIRI and SIR4loci by chromosomal integration of plasmids andmapping the plasmids' selectable marker, we havemade unnecessary the mapping of mutations thatnormally render the strain a nonmater. SIR1 hasbeen assigned to the tip of the right arm of chro-mosome XI and SIR4 has been assigned to themiddle of the right arm of chromosome IV;MAR1 is on the left arm of chromosome IV andMAR2 is located distal to URA4 on chromosomeXII.
To understand more fully the mechanisms in-volved in regulating silent mating-type geneexpression, we have begun experiments to iden-tify and purify each of the MAR/SIR gene prod-ucts. DNA fragments containing functional se-quences of SIR], SIR2, and SIR3 have beensubcloned into the open-reading-frame vectorsconstructed by M. Berman and colleagues (Wein-stock et al., Proc. Natl. Acad. Sci. 80: 4432[1983]), which allow the expression (in Esche-richia colt) of open-reading-frame DNA as a tri-hybrid protein containing sequences from the E.coli proteins ompF and 0-galactosidase. Anti-sera raised against purified fusion proteins will beused to purify the yeast MAR/SIR gene productsand to study MAR/SIR protein synthesis, locali-zation, and DNA-binding properties.
In our screen of the gene bank for sequencesable to suppress MAR/SIR mutations in vivo, wefound one other cloned gene that could do so.Disruption in this locus is lethal to the cell, indi-cating that it encodes an essential function. Itsuppresses mutation in all four MAR/SIR loci,but it does not correspond to any of them. Ge-netic evidence indicates that the suppression ofMAR/SIR mutations is not caused by restorationof MAR/SIR regulation. Rather, it appears thatregulation of mating functions by MAT is dis-rupted. Since all transformants have displayed aweak a mating type, this sequence formally be-haves as an antagonist of MATa1 function;hence, our working designation for the locus isantagonist of al (ANA). We are presently tryingto distinguish between the possibilities that either
103

a truncated gene product or the cloned sequencesthemselves disrupt regulation of mating func-tions.
To determine if the E and I sites are able toregulate expression of genes other than the mat-ing-type genes, we replaced the mating-type a in-formation normally found at HMR with the HIS3gene. We found that in one orientation, E and Icould repress the expression of the HIS3 gene.These results indicate that promoters other thanthe mating-type promoters can come under thecontrol of E and I.
Behavior of the Double-strandedDNA Breaks at MAT inSchizosaccharomyces pombeA. Klar, D. Beach, L. Miglio
During the past year, we reported three classes ofmutants in fission yeast that are defective in mat-ing-type switching: (1) those that lacked the abil-ity to make double-stranded breaks at MAT, (2)those that had the break but apparently did notswitch, and (3) those that were defective in reso-lution. The second class is quite paradoxical-either they switch only homologously or this yeastcan heal the broken ends without obligatelyswitching. To distinguish between these possibil-ities, we deleted both MAT2 and MAT3 in Swi -(switching-deficient mutants) and Swi' strains.
The deletions were made by first cloning a bitof sequence proximal to MAT2 and a small se-quence distal to MAT3. Between these se-quences, a selectable marker (the Saccharomycescerevisiae LEU2 gene, which complements theleul defect of Schizosaccharomyces pombe) wasinserted. The in-vitro-made construct was thencut out of the vector and transformed into appro-priate strains by selecting for the Leu+ pheno-type. As shown in Figure 2, MAT2 and MAT3were deleted, and a new fusion band of the con-struct was found. A quite interesting result is thatSwi' strains with the deletion maintain a stablemating type, still generate the wild-type level ofDNA cut at MAT, and do not exhibit lethality.The presence of the swi mutation in these strainshas no additional phenotype. Thus, this yeast caneasily heal the broken ends without switching. We
104
smt -S h90 11-5
A BC D EF GH I J
FIGURE 2 Southern blot analysis demonstrating deletionsof MAT2 and MAT3 loci. smt-s is a cis-acting mutation thatreduces the level of the double-stranded break at MAT. hs°is a wild-type strain and is shown to contain a double-stranded break at MAT1 in about 200/0 of its DNA (markedby SMT, lane G). h-s is a strain that contains a fusion be-tween MAT2 and MAT3. DNA displayed in B-F, H, and Jshow deletions of MAT2 and MAT3.
therefore propose that mutations in the secondclass (which comprise three complementationgroups) are defective in using the double-strandbroken ends for switching.
In both S. cerevisiae and S. pombe, we haveshown that double-stranded breaks are highly re-combinogenic during mitotic growth (see pre-vious years' reports). Most of the recombinationstudies in fungi, however, are conducted inmeiosis. The question arises as to whether thedouble-stranded DNA breaks are also recombi-nogenic in meiosis. To address this question, weused the MAT2, MAT3 deletion strains of S.pombe. In these strains, the double-strandedbreaks at MAT cannot be healed by switching be-cause of the lack of the donor loci. We testedthese breaks to determine whether they can beused for recombination in meiosis. Tetrad dissec-tion of appropriate hybrids of these deletionstrains has shown that MAT gives gene conver-sion of the 3:1 and 1:3 types in about 20% of theasci, whereas standard wild-type strains hardlygenerate such events. If we reduce the amount ofthe in-vivo-cut DNA, either by using the appro-priate swi mutations or by using the cis-actingspeckled mutation, the gene conversion fre-

quency is correspondingly reduced. Thus, thedouble-stranded breaks can also promote effi-cient gene conversion in meiosis. Similar obser-vations have been made recently in our colla-borative work with A. Kolodkin and E Stahl(University of Oregon) in the S. cerevisiae system.
Control of the Mitotic Cell Cycle andMeiosis in Fission YeastD. Beach, M. McLeod, R. Booher, J. Gould
This year's work concentrated on two key genesof fission yeast, cdc2 and rani. cdc2' is requiredfor progression through both the G, and G,phases of the mitotic cell cycle, and rani' inhib-its the transition from mitotic cell division to
0 hrs
3 hrs
5 hrs
9 hrs
A.
h
B.
meiosis. In the absence of ran]', a haploid cellundergoes premeiotic S phase, partial meiosis,and sporulation, despite the lack of all the nor-mal meiotic requirements (e.g., diploidy, hetero-zygosity at the mating-type locus, and nutritionalstarvation).
The cdc2 gene, which is homologous to thecdc28 gene of S. cerevisiae, is predicted to encodea protein kinase on the basis of homology withknown mammalian kinases. An intron-less formof the cdc2 gene has been constructed in an E.coli expression vector, and a new kinase activityis currently being sought in bacterial extracts.
In vitro mutagenesis has been used to createnonphosphorylatable residues at two potentialsites of phosphorylation in the cdc2 protein. Onesite (Tyr-166-,-Phe; identified by homology withthe major site of phosphorylation in src and fps)was mutated with no effect. Mutation of the
h+i h
c.
h rnei2 /h mei2
2c 4c 2c 4c 2c 4c
FIGURE 3 Analysis of propidium-iodide-stained cells on the EPICS cell sorter. Three strains, h-/h-, h./h-,and h+mei21h-mei2, were starved of a nitrogen source and analyzed at intervals. In h-/h-, cells accumulate inG,, and in h +/h-, cells accumulate in G, but then undergo premeiotic DNA synthesis. In the mei2 mutant (C),premeiotic DNA synthesis does not occur.
105

ranl
GDSLRFVSIIiikAY2VVYKAEDIYDQ,TL WEAL CKDG LNEK
cdc2
NH2MEN Y Q KTE KO(DE@TO000000R H K L SE)13 I V ®M(DK I R L EDESgG
CADPK
LC)Q FE II I K T L@T(DS F®li®M L V K H ME TgN HCAMTI@DTCrT 000®ATP
FIGURE 4 Sequence homologies between fission yeast rant, cdc2, and bovine cAMP-dependent protein kinase. The regionof homology contains the known site of ATP binding in the mammalian kinase.
neighboring site (Thr-167-+Val; identified by ho-mology with the major site of phosphorylation inbovine cAMP-dependent protein kinase) gave aphenotype of cell-cycle arrest. It is unknown atpresent whether Thr-167 is indeed a site of phos-phorylation in cdc2, but the inability of cells car-rying a mutation at this site to undergo DNA syn-thesis raises the interesting possibility that
of cdc2 may be required for cell-cycle progression.
The EPICS cell sorter (James Laboratory) hasbeen used to characterize the transition of cellsfrom the mitotic cell cycle to meiosis (see Fig. 3).It has been confirmed that initiation of meiosis issignaled by a round of DNA replication (pre-meiotic S phase) that does not take place in non-sporulating cells. We have found that the pre-meiotic S phase and meiosis I, induced in atemperature-sensitive rani- strain, do not requirethe activity of the cdc2' gene. This feature dis-tinguishes the premeiotic S phase and meiosis Ifrom the mitotic S phase and mitotic cell divi-sion, both of which require cdc2' function. Therani' gene has been shown to produce a 4.3-kbRNA transcript. The gene has been partially se-quenced, and a small region of homology withthe cdc2 gene has been observed (Fig. 4). Thisregion is particularly significant as it representsthe site of ATP binding in a family of protein kin-ases. The homology suggests that rani' proba-bly encodes a protein with ATPase activity.
A mass of genetic evidence suggests that
106
meiosis is initiated by turning off the rani' prod-uct. We have shown that this inhibition does notoccur at the transcriptional level. Future workwill be directed at identifying the rank' proteinproduct and attempting to understand its bio-chemical activity.
PUBLICATIONS
Abraham, J., K.A. Nasmyth, J.N. Strathern, A.J.S. Klar, andJ.B. Hicks. 1984. Regulation of mating-type informationin yeast: Negative control requiring sequences both 5'and 3' to the regulated region. J. Mol. Biol. 176: 307-331.
Beach, D.H. and A.J.S. Klar. 1984. Rearrangements of thetransposable mating-type cassettes of fission yeast.EMBO J. 3: 603-610.
Egel, R., D.H. Beach, and A.J.S. Klar. 1984. Genes requiredfor initiation and resolution steps of mating-type switch-ing in fission yeast. Proc. Natl. Acad. Sci. 81: 3481-3485.
Feldman, J.B., J.B. Hicks, and J.R. Broach. 1984. Identifi-cation of sites required for repression of a silent mating-type locus in yeast. J. Mol. Biol. 178: 815-834.
Hicks, J.B., J. Strathern, A. Klar, S. Ismail, and J. Broach.1984. Structure of SAD mutation and the location of con-trol sites at silent mating-type genes in Saccharomycescerevisiae. Mol. Cell. Biol. 4: 1278-1285.
Klar, A.J.S. and J.N. Strathern. 1984. Resolution of recom-bination intermediates generated during yeast matingtype switching. Nature 310: 744-748.
Klar, A.J.S., J.N. Strathern, and J.A. Abraham. 1984. Theinvolvement of double-strand chromosomal breaks formating-type switching in Saccharomyces cerevisiae. ColdSpring Harbor Symp. Quant. Biol. 49: 77-88.
Klar, A.J.S., J.N. Strathern, and J.B. Hicks. 1984. Develop-mental pathways in yeast. In Microbial development (ed.R. Losick and L. Shapiro), pp. 151-195. Cold SpringHarbor Laboratory, Cold Spring Harbor, New York.

PLANT GENETICS
S.L. Dellaporta A.C. Hiatt J. MclndooJ.B. Hicks P.S. Chomet J. WoodR.L. Malmberg
Maize controlling elements have been the subjectof genetic studies for more than 40 years. How-ever, work on the molecular biology of maizecontrolling elements is relatively recent. In afew short years, several controlling elements, in-cluding the first controlling element system de-scribed, Activator-Dissociation (Ac/ Ds), byMcClintock (Cold Spring Harbor Symp. Quant.Biol. 16: 13[1951]), have been isolated and char-acterized at the nucleotide level. Several studieshave shown that autonomous controlling ele-ments, such as Ac, are transposable elements thatshare homology with dispersed middle repetitiveDNA sequences. Autonomous elements such asAc can transactivate nonautonomous elementssuch as Ds that are genetically inactive in the ab-sence of the autonomous Ac element. There ap-pear to be two classes of Ds elements: Ds ele-ments that are structural derivatives of Ac (mostappear to be deletion derivatives) and a family ofDs elements that shares little homology with AcDNA, except for the 11-bp inverted terminal re-peats characteristic of all Ac/Ds elements. Thistype of Ds element has been named Dsl, since itrepresents the first Ds element cloned (Peacock etal., Cold Spring Harbor Symp. Quant. Biol. 49:347[1984]). Interestingly, the Ds1 element hy-bridizes to approximately 40 conserved DNA ele-ments in the maize genome.
Ds1 Element at the waxy Locus
S.L. Dellaporta [in collaboration with S. Wessler,University of Georgia]
We have isolated another Dsl element which wasfound as an insertion in the waxy mutation, wx-ml, first isolated by McClintock (Carnegie Inst.Wash. Year Book 47: 155[1948]). This insertionis approximately 400 bp, contains the expected 11-bp inverted terminal repeats of Ac/Ds elements,and is flanked by 8-bp direct repeats. The directrepeats represent a target-site duplication causedduring integration of Ds.
In the presence of an active Ac element, the wx-ml allele is somatically and germinally unstable.This germinal instability reflects the Ac-mediatedexcision of Ds1 from the waxy locus. We havecloned and sequenced several germinal Wx mu-tations from plants containing an Ac element andthe wx -ml allele. These mutations were selectedfor partial, rather than full, restoration of the Wxphenotype. Sequence analysis showed that theseexcision events are not precise, leaving the 8-bpdirect repeats in modified form (Fig. 1). In eachcase, however, the modifications represent theaddition of nucleotides in multiples of three. Wepredict that the insertion lies within exon se-quences and that the partial restoration of the Wx
Wx TAR GET SITEwx-m Iwx-m l(G M1)wx-m l(G M2)
GCATCACCGCATCACC tagggatgaaa...Ds...tttcatcccta GCATCACCG CATCACG GCCATCACCGCATCACC ATCACC
FIGURE 1 Sequence analysis of the wx-m 1 mutation and germinal mutations to partial Wx activity. Partial sequence is shownfor the insertion termini of the Ds element found in the wx-m/ mutation. The 11-bp inverted terminal repeats (lowercase) areflanked by 8-bp direct repeats (uppercase). Ac-mediated germinal mutations of the wx-ml allele can be recovered that showpartial Wx activity. Sequence analysis of the excision sites of two partial Wx alleles (GM1 and GM2) are shown. Both mutationshave lost Ds sequences but leave behind the 8-bp direct repeats in modified form. The modifications result in the addition ofnucleotides in multiples of three. This may affect Wx gene action by introducing additional amino acids into the primarysequence of the Wx protein.
107

phenotype is due to the addition of amino acidresidues to the Wx protein. We are presently test-ing this hypothesis by examining a number of wx-ml germinal mutations and by cDNA analysis ofthe region of the waxy gene near the Ds1 inser-tion.
Gene Cloning by Transposon TaggingS.L. Dellaporta [in collaboration with S. Wessler,University of Georgia, and I. Greenblatt,University of Connecticut, Storrs]
The autonomous controlling element Ac containsinternal DNA sequences that are present in as few
0 R-nj
mR-nj
YFG
as 8-10 copies per maize genome, even if the plantdoes not contain an Ac element. The active Acelement can be distinguished from these related,but inactive, sequences by structural features suchas the presence of unique restriction fragmentsand the state of DNA methylation (see below).The presence of relatively few sequences relatedto internal Ac DNA and the ability to distinguishthe active form of Ac from the inactive crypticAc -like sequences are criteria that can be used toclone genes containing Ac insertions.
We have used Ac tagging to clone the R locus ,of maize. This locus is one of several epistaticgenes involved in the anthocyanin biosyntheticpathway. Our interest in R stems from previousgenetic analysis of this locus which suggests thatR conditions tissue-specific expression of antho-
+ Ac
\ /clone Ac -like DNA
specificprobes
Identify Ac
FIGURE 2 Gene cloning strategy using Ac tagging. A functional allele of the R locus is coupledwith an active Ac element by using maize stocks containing reciprocal translocations between adonor allele (P-vv locus located on chromosome 1) and the recipient locus (R-nj locus located onchromosome 10). Transposition of Ac from donor to recipient site results in an unstable R-nj pheno-type that shows the genetic characteristics of Ac-induced mutations. (1) The mutation shows acharacteristic dosage effect; with increasing copies of Ac-induced mutation, the apparent frequencyand developmental timing of somatic and germinal mutation is delayed; (2) the Ac-induced mutationis capable of mediating Ds transposition in trans. Once an Ac-induced R mutation is geneticallyidentified, recombinant clones are isolated from genomic libraries with low-copy internal Ac DNAprobes. The recombinant clone containing the R allele is identified by screening recombinants forthe canonical Ac sequences by restriction site analysis. DNA flanking the Ac element is used as anR-specific probe to isolate a functional R allele.
108

cyanin in both seed and plant organs. The locuscan be fractionated by meiotic recombination intodistinct seed and plant components, suggestingthat these regions may represent tandem dis-placements of homologous sequences with di-vergent function. Hence, the availability of amolecular probe to R DNA may prove to be avaluable tool to examine the tissue-specific com-ponents and regulation of gene expression in de-veloping plant tissues.
Because the product(s) of the R locus is un-known and may be expressed in trace amounts inplant tissues, conventional cloning by immuno-logical cDNA screening seemed inappropriate.Therefore, we used the strategy outlined in Figure2 to clone the R locus. This strategy requires aninsertion of the Ac element at the R locus. Thisevent was dictated by (1) the transposition pat-terns of an Ac element, (2) the availability of thetarget site to Ac insertion, and (3) the ability toidentify an Ac- induced mutation at the R locus.Previous studies demonstrated that the majorityof Ac transpositions from the P locus on chro-mosome 1 are intrachromosomal events that ex-hibit a preference for short-range distal events. I.Greenblatt (unpubl.) obtained an Ac insertion atR by using strains carrying a reciprocal translo-cation involving breakpoints proximal to the longarm of chromosome 10 (containing the R locus)and distal to the P locus on chromosome 1 (con-taining an Ac- induced unstable P mutation).Several putative transpositions of Ac from theP locus to the R locus were obtained. An au-tonomous unstable allele of R was geneticallycharacterized as Ac- induced by the ability of theelement located at R to transactivate the standardDs element located on chromosome 9. This mu-tation also exhibited a dosage pattern character-istic of Ac- induced mutations.
Several recombinant clones were isolated fromgenomic libraries, constructed with DNA fromplants homozygous for the Ac-induced mutableR mutation, and screened with a low-copy inter-nal Ac probe. One recombinant contained a can-onical Ac element as judged from restriction en-zyme analysis. DNA flanking the Ac element wasdetermined to be R-specific on the basis of astructural comparison of various transposon-in-duced R mutations and their respective rever-tants. We are now in the process of determining
the structure and are cloning the tissue-specificcomponents of the standard R locus.
Active and Inactive Ac ElementsP.S. Chomet, S.L. Dellaporta
Changes in phase refers to the ability of an Acelement to cycle from an active state to an inac-tive state. In the active state, the Ac element iscapable of transposition and transactivation of Dselements (Fig. 3). This ability is thought to bedue to the production of an unidentified transpos-ase referred to as the mutator component of Ac.Ac can cycle from an active phase to an inactivephase where no mutator function is geneticallydetectable. We examined an Ac element at thewaxy locus, isolated by McClintock (CarnegieInst. Wash. Year Book 63: 59211964]), in both theactive and inactive phases. The active and inac-tive Ac elements were identical in both size andposition at the waxy locus. However, the Ac inthe inactive phase has undergone DNA modifi-cation. This modification was detected with re-striction enzymes sensitive to 5-methylcytosine.The modified Ac DNA was found in much re-duced amounts during the active phase at waxy.Examination of a number of active Ac elementsat other loci shows that extensive modificationwas specific for the inactive phase of Ac.
Tz Elements of MaizeS.L. Dellaporta, P.S. Chomet, J.B. Hicks [in collaborationwith J. Mottinger, University of Rhode Island]
Last year, we described the molecular character-ization of several unstable mutations at theshrunken locus in maize. These mutations wereselected in maize lines descendant from plantssystemically infected with barley stripe mosaic vi-rus (BSMV). BSMV infection in maize was as-sociated with high rates of genetic instability, in-cluding spontaneous mutations. Three shrunkenmutations were caused by insertion elements thatare structurally unrelated. The insertion sites andelements are shown in Figure 4.
Genomic clones of the mutant sh-5586 allele
109

AFIGURE 3 Changes in phase of Ac activity at the waxy locus. An Ac element insertion at thewaxy locus [(Ac)wx-m7] has been identified by McClintock (Carnegie Inst. Wash. Year Book 62:486[1964]) and shown to undergo changes in phase of activity. The cycling activity of Ac canbe seen in kernels A-C. Each kernel has an endosperm genotype: al- m3 /al -m3 /al wxlwxl(Ac)wx-m7. In kernel A, the Ac at waxy remained active throughout endosperm development.The activity 'of Ac can be monitored by the instability of Ds at the al-m3 mutation. In the pres-ence of an active Ac element, the Ds at al-m3 is somatically unstable, giving the kernel avariegated colored phenotype. When the Ac cycles from active to inactive, the Ds at al-m3 isstable, inhibiting At gene action. This can be seen as a colorless sector in kernel B. Kernel Crepresents an allele of (Ac)wx-m7 that is both somatically and germinally inactive. This kernelshows no Ac activity and is fully colorless. Molecular analyses (see text) of Ac at wx-m7 indicatethat DNA modification of Ac DNA occurs in the inactive phase. This modification is detectablewith restriction enzymes that are sensitive to 5-methylcytosine.
containing the Tz86 element were characterizedby restriction mapping, genomic blot analysis,and M13 dideoxy-sequencing. We have shownthat Tz86 is a 3.6-kb element flanked by 10-bpdirect repeats. This 10-bp sequence was foundonce in the progenitor Sh allele, indicating thatTz86 causes a 10-bp duplication of target Sh DNAupon insertion. Unlike other maize transposableelements, Tz86 does not contain terminal in-verted or direct repeats. When subclones of Tz86
B EI I
5 2KbTz84 Bc B2 H3
I I I
2 0 KbTz82
were hybridized to genomic blots of maize DNA,we found that Tz86 DNA represents dispersedmiddle repetitive DNA. Internal subclones ofTz86 hybridize to relatively fewer copies, perhaps10 fragments or less, depending on the strain ex-amined. This situation is similar to that in othermaize controlling elements and suggests that viralinfection may destabilize these components in themaize genome and contribute to high rates of ge-netic instability. Moreover, Tz86 continues to
; 7 T3K X H3I I t
3.6 KbTz 86
Illy AllObi r AlEr AMIE
Mexon 0 intronFIGURE 4 Relative position of the Tz insertion element at the shrunken-1 locus. (The intron/exonborders of the Sh gene have been previously determined by W. Werr et al., pers. comm.) We positionseveral insertion elements at Sh by restriction mapping and dideoxy-sequence analysis of mutant shalleles (sh-5582, sh-5584, sh-5586) recovered from virus-infected progeny (see text). The size ofTz84 is a minimum value and has not been precisely determined. On the basis of restriction mappingand hybridization analysis, we find no significant homology among these elements. Yet each elementhybridizes to multiple genomic sequences of maize DNA.
110

show germinal instability in advanced progeny.This instability often restores the Sh phenotype(plump kernels) and is usually accompanied byexcision of Tz86 from the shrunken locus. Se-quence analysis of one such event showed a res-toration of the DNA sequence of the progenitorSh DNA, except for a single-base substitution inthe 10-bp target sequence.
Cell and Developmental Geneticsof TobaccoA.0 Hiatt, R.L. Malmberg, J. Mclndoo
For the past few years, we have been studying thepolyamine synthesis pathway in tobacco. Ourapproach was to focus on genetics: the isolationof mutants that are in some manner altered in thepathway. As documented in the 1982 and 1983Annual Reports, we have developed successfulmutagenesis and selection protocols. These makeuse of some powerful features of the tobacco so-matic-cell system and some useful features of thepolyamine pathway. Tobacco is relatively easy to
manipulate in somatic-cell culture and can besubsequently regenerated into a whole plant fordevelopmental and genetic analyses. The poly-amine pathway has been intensively studied bio-chemically in mammalian systems, and there existspecific inhibitors for four of the six enzymes inthe pathway. Figure 5 shows a simplified dia-gram of the pathway and the inhibitors. Our re-search during the past year has been concernedwith the molecular and biochemical characteriza-tion of the mutants we have isolated previouslyand with the developmental genetics of the regen-erated mutant plants.
DEVELOPMENTAL REGULATION OF ORNITHINEAND ARGININE DECARBOXYLASES
One interesting feature of the pathway is thattobacco synthesizes putrescine in two ways: viaornithine decarboxylase (OrnDC) or arginine de-carboxylase (ArgDC). We have analyzed the dis-tribution of these two enzymes in different tissuesof the whole plant and in cell cultures. The re-sults, shown in Figure 6, reveal that OrnDC ispreferentially found in flowers and in cell cul-
Putrescine
Saim2 DC, C) M6B6
SpSyn 10 DCHA
Spermidine
Spermine
Mgr3301
FIGURE 5 Polyamine synthesis pathway. Abbreviations: (OrnDC) ornithine decar-boxylase; (ArgDC) arginine decarboxylase; (SamDC) S-adenosylmethionine decar-boxylase; (DFMO) difluoromethylornithine; (DFMA) difluoromethylarginine; (MGBG)methylglyoxal-bis(guanyihydrazone).
111

ENZYME SPECIFIC ACTIVITIES
RATIO OF OrnDC TO ArgDC
flowers 1,81
floral buds 1,75
shoots 0.04
roots 0.04
cell cultures 1.70
FIGURE 6 Developmental regulation of OrnDC and ArgDC in tobacco.
tures but not in leaves, stems, or roots. ArgDC isfound in all tissues. Two independently isolatedmutants (ts4 and Dfrl) each have near-zero levelsof OrnDC in cell cultures. These two mutants re-generate into very similar plants (dwarf, smallleaves), and they have not flowered under normalgrowth conditions. This suggests that OrnDC isnot only a flower-specific enzyme but may ac-tually be required for flowering. We also exam-ined the effects of the inhibitors of these enzymeson cell cultures. DFMO inhibits the growth ofcultures after a delay of several generations, butDFMA does not. Thus, even though both en-zymes are found in culture, OrnDC is required,but ArgDC is normally not.
We have used the known tissue distribution ofOrnDC (and the existence of two mutants withlow levels of OrnDC) as an aid to molecular clon-ing of the gene encoding the enzyme. L. Mc-Conlogue and P. Coffino (University of Califor-nia, San Francisco) have previously obtainedcDNA clones for the mouse OrnDC. We haveused their clone as a probe against RNA isolatedfrom different tissues of the plant and againstRNA from wild-type and mutant cell cultures.When washed at moderate stringency, the mouse
112
OrnDC hybridized in a pattern identical to thatpredicted by the distribution of the enzyme activ-ities. This suggested that developmental regula-tion was occurring at a pretranslational level andthat we could use the mouse OrnDC cDNA as aninterkingdom probe to isolate the tobacco gene.With this strategy, we have found two tobaccogenomic clones that hybridize to the mouse probeand to tobacco RNA in the pattern predicted bymeasurements of enzyme activity. We are cur-rently in the process of sequencing the tobaccogenomic clones for OrnDC.
EFFECTS OF MGBG ON ENZYME TURNOVER
We had previously found that wild-type cellsgrown on sublethal levels of MGBG overprod-uced a polypeptide of about 35,000 daltons. Toexamine this increase in abundance, we carriedout a series of protein-labeling experiments, withand without MGBG. These experiments demon-strated that the increase in abundance of the 35-kD polypeptide was due not to new synthesis butto an accumulation of the previously made poly-peptide. Apparently, the MGBG was stabilizingthe polypeptide. This mode of regulation sug-
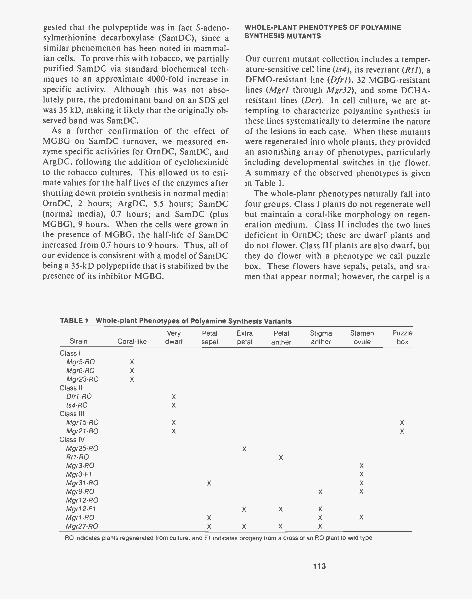
gested that the polypeptide was in fact S-adeno-sylmethionine decarboxylase (SamDC), since asimilar phenomenon has been noted in mammal-ian cells. To prove this with tobacco, we partiallypurified SamDC via standard biochemical tech-niques to an approximate 4000-fold increase inspecific activity. Although this was not abso-lutely pure, the predominant band on an SDS gelwas 35 kD, making it likely that the originally ob-served band was SamDC.
As a further confirmation of the effect ofMGBG on SamDC turnover, we measured en-zyme specific activities for OrnDC, SamDC, andArgDC, following the addition of cycloheximideto the tobacco cultures. This allowed us to esti-mate values for the half lives of the enzymes aftershutting down protein synthesis in normal media:OrnDC, 2 hours; ArgDC, 5.5 hours; SamDC(normal media), 0.7 hours; and SamDC (plusMGBG), 9 hours. When the cells were grown inthe presence of MGBG, the half-life of SamDCincreased from 0.7 hours to 9 hours. Thus, all ofour evidence is consistent with a model of SamDCbeing a 35-kD polypeptide that is stabilized by thepresence of its inhibitor MGBG.
WHOLE-PLANT PHENOTYPES OF POLYAMINESYNTHESIS MUTANTS
Our current mutant collection includes a temper-ature-sensitive cell line (ts4), its revertant (Rtl), aDFMO-resistant line (Dfr1), 32 MGBG-resistantlines (Mgrl through Mgr32), and some DCHA-resistant lines (Dcr). In cell culture, we are at-tempting to characterize polyamine synthesis inthese lines systematically to determine the natureof the lesions in each case. When these mutantswere regenerated into whole plants, they providedan astonishing array of phenotypes, particularlyincluding developmental switches in the flower.A summary of the observed phenotypes is givenin Table 1.
The whole-plant phenotypes naturally fall intofour groups. Class I plants do not regenerate wellbut maintain a coral-like morphology on regen-eration medium. Class II includes the two linesdeficient in OrnDC; these are dwarf plants anddo not flower. Class III plants are also dwarf, butthey do flower with a phenotype we call puzzlebox. These flowers have sepals, petals, and sta-men that appear normal; however, the carpel is a
TABLE 1 Whole-plant Phenotypes of Polyamine Synthesis Variants
StrainVery Petal
Coral-like dwarf sepalExtra Petal Stigmapetal anther anther
Stamen Puzzleovule box
Class IMgr5 -ROMgr6 -ROMgr23 -RO
Class IIDfr1-R0ts4-RO
Class IIIMgr15-R0Mgr21-R0
Class IVMgr25 -RORt1-R0Mgr3 -ROMgr3-F1Mgr31-R0Mgr9-R0Mgr12-R0Mgr12-F1Mgr1-R0Mgr27 -RO
X
RO indicates plants regenerated from culture, and Fl indicates progeny from a cross of an RO plant to wild type.
113

hollow cylinder that does not appear to havefused properly during development. Inside thehollow carpel is a second, and sometimes a third,row of stamen. Apparently, these mutants can-not finish carpel development correctly and thusrepeat the previous stage of stamen.
Class IV phenotypes represent a range of de-velopmental switches, including stamenoidovules, stigmoid anthers, petaloid anthers, extra-petals, petaloid sepals, and simple sterilities.There is overlap within this group; e.g., one plantmay have petaloid anthers, another may havestigmoid anthers, and yet another may have bothphenotypes, as well as others. Thus, the class IVphenotypes mutually overlap in a way that sug-gests they are related. The sterilities generatedhave hampered sexual crossing and conventionalgenetic analysis. However, in two cases, Mgr3and Mgr12, we have generated a small F, popu-lation from crosses to wild type. In these cases,the polyamine phenotype and the abnormalflower cosegregate as nuclear dominant muta-tions.
The possible role of polyamines in altering theexpression of floral meristems is indicated by thefact that all of the polyamine mutants that doflower have some sort of abnormal flower. Var-ious artifacts are possible: double mutants, cell-culture-induced variation, and nongenetic regen-eration abnormalities. Thus, the most critical ex-periments to be performed in the future will in-volve further meiotic genetic analysis of themutants to test the nature of the suggested link-age between polyamine synthesis and floral de-velopment.
SITE-SPECIFIC MUTAGENESIS
M. Zoller K. JohnsonR. Kostriken
DNA/Protein Interactions: Recognitionand Cleavage of MATa DNA byHO EndonucleaseM. Zoller, K. Johnson, R. Kostriken
Our research is aimed at the problem of how aprotein recognizes a specific sequence of DNA.
114
FLORAL MONOCLONAL ANTIBODIES
Our mutant collection should allow us to dissectfloral development genetically, regardless of thephysiological connection to the polyamine syn-thesis pathway. As an aid to analysis of the ab-normal structures seen in the mutant flowers,we are making monoclonal antibodies against flo-ral antigens, in collaboration with S. Hockfield(Neurobiology Section). The strategy is the fa-miliar one of immunizing mice with floral ex-tract, and then screening the resulting hybrido-mas against extracts from individual parts of theflower. In our initial experiment, we were able tofind an antibody that recognizes sepal and petalextracts, but not other tissues; we also found twoantibodies that reacted with only carpel extracts.We are currently using these antibodies to optim-ize histoimmunological techniques with sectionsof tobacco flowers.
PUBLICATIONS
Dellaporta, S.L. and P.S. Chomet. 1984. The activation ofmaize controlling elements. In Plant gene research (ed.B. Hohn and E. Dennis), Chap. 10, pp. 169-216. Spring-er-Verlag, Berlin.
Dellaporta, S.L., P.S. Chomet, J.P. Mottinger, J.A. Wood, S.M.Yu, and J.B. Hicks. 1984. Endogenous transposable ele-ments associated with virus infection in maize. ColdSpring Harbor Symp. Quant. Biol. 49: 321-328.
Malmberg, R.L. and J. Mcindoo. 1984. Ultraviolet mutage-nesis and genetic analysis of resistance to methylglyoxal-bis (guanylhydrazone) in tobacco. Mol. Gen. Genet. 196:28-34.
Mottinger, J.P., S.L. Dellaporta, and P Keller. 1984. Stableand unstable mutations in aberrant ratio stocks of maize.Genetics 106: 751-767.
Much has been learned from the study of prokar-yotic proteins such as the K repressor and EcoRIendonuclease. The interaction between a eukar-yotic protein and a specific sequence of DNA hasyet to be defined to the same extent. Toward thisgoal, we are investigating the manner in which theHO endonuclease from the yeast Saccharomycescerevisiae binds to and cleaves MAT DNA. HO

endonuclease is one of a number of elements re-quired for mating-type switching in S. cerevisiae.A model has been proposed in which the forma-tion of a specific double-strand break in the chro-mosome at MAT is an initiating event in theprocess. The formation of this break is thoughtto be catalyzed by HO endonuclease. R. Kostri-ken and F. Heffron (Research Institute of ScrippsClinic) demonstrated that HO endonucleasecleaved MAT DNA in vitro. Through biochemi-cal as well as genetic experiments, the specific siteof the HO cut has been localized.
The biochemical events involved in HO-de-pendent DNA transposition are not fully under-stood. The problem is being approached fromtwo directions. (1) What are the determinantswithin the HO cut site that are important for rec-ognition and cleavage? (2) What are the specificamino acid residues within HO endonuclease thatare responsible for recognition and cleavage?
MUTATIONS IN MATa THAT AFFECT HOCLEAVAGE IN VITRO
Since genetic approaches yielded only two pointmutations within the HO cut site that affectedswitching and cutting, we used oligonucleotide-directed mutagenesis to saturate this region withsingle base changes. The ability of each mutantto serve as a substrate for HO endonuclease wasassessed in vitro using a yeast extract from an HOstrain and an Escherichia coli extract from astrain engineered to express the HO-gene product(see below). Qualitatively, both the yeast- and theE. coli- produced enzymes cleaved the mutants ina similar manner. Figure 1 shows the analysis ofall 17 mutant substrates after incubation with HOendonuclease produced in E. coll. The mutantsubstrates can be grouped into three categories onthe basis of the rate with which each is cleavedcompared with wild type. Four mutations com-pleted inhibited cleavage, nine mutations resultedin a reduced rate of cleavage, and the remainingfour mutations had no effect on the rate of cleav-age as compared with wild type. These data re-veal that the core of the recognition sequence isCGCNNC, since alteration of any one of the fourbases completely inhibited cleavage. Further-more, the sequence TTTCCGCAACAGT repre-sents the region in close contact with the enzyme,
volZ1WWCT TTCCGCAACAIGTAAAAT
MT TGA AAGGCGITTGTCATITT A
_uncut-11.5 kb- 3.4 kb
FIGURE 1 Agarose gel analysis of MAT substrates cleavedin vitro with HO endonuclease expressed in E. coli. (1) DNAcleaved with Hindlll for markers; (2,3) BamHI-cleaved wild-type MATa DNA without and with HO endonuclease, re-spectively; (4-22) mutant MATa substrates cleaved with HOendonuclease. The corresponding wild-type MATa se-quence is shown above each lane.
since alteration of noncore positions in this re-gion affected the rate of cleavage. It is signifi-cant to note that the extent of the contact regionspans the so-called Y/Z boundary into the allele-specific Ya region. Does the enzyme recognizeMATa in the same manner? A parallel set of mu-tations have not been made in MATa to answerthis question. However, inspection of the allele-specific DNA sequences of MATa and MARX inthe region of the Y/Z boundary revealed the samesequence (CTT) at positions - 5, - 4, and -3(with respect to the Y/Z boundary). As shown inFigure 1 only the mutations at -4 and -3 af-fected cleavage.
Future studies will assess the effect of eachpoint mutation on the frequency of mating-typeswitching in vivo. Currently, we are integratingeach of the mutant MATa substrates into thechromosome, thereby replacing the wild-typeMATa sequences. We predict that the frequencyof switching will correlate with the efficiency ofcleavage in vitro.
CHARACTERIZATION OF THE HO ENDONUCLEASE
The concentration of HO endonuclease in yeastis too low to serve as a source for purified pro-tein. Therefore, we have engineered the HO geneto be expressed in E. coll. The gene was placedunder the isopropyl-O-D-thiogalactoside (IPTG)-inducible lac operator/trp promoter of the vectorpKK233-2 (courtesy of J. Brosius, Columbia Uni-versity). This construction yielded approxi-mately 50-100 times higher expression per gram
115

of wet cells compared with yeast. Expressionfrom this vector was 5 times better than that ofan earlier plasmid utilizing the XP, promoter.Currently, we are purifying HO endonuclease.Future experiments will involve (1) determinationof native stoichiometry and molecular size andshape, (2) alkylation protection studies using thewild-type and mutant recognition sequences, (3)quantitative binding studies with mutant sub-strates, (4) investigation of domains by limitedproteolysis, and (5) active-site affinity labelingusing synthetic recognition sequences.
Oligonucleotide-directed MutagenesisM. Zoller
In vitro mutagenesis of cloned DNA has becomea standard tool in the functional analysis ofnucleic acids and proteins. A number of ap-proaches exist that can be broadly grouped intorandom and site-directed methods. The reper-toire of procedures to mutagenize a fragment ofDNA randomly continues to grow each year.These random techniques are used to identify thelocation and boundaries of a particular function.Once an important region has been identified,site-directed mutagenesis can be employed to de-termine the role of specific nucleotides (or aminoacids). The construction of cloned DNA bearinga specific alteration is best accomplished by eithergene synthesis or oligonucleotide-directed muta-genesis.
The availability of reliable procedures for man-ual oligonucleotide synthesis, along with an in-creasing access to DNA synthesizers, has madegene synthesis a reasonable method to producespecific mutations. The gene is constructed usingsegments of oligonucleotides that are ligated to-gether, and thus an entire gene can be synthesizedthat contains a desired change. In addition, asimple construct can be produced that allows forthe replacement of segments by a number of mu-tated "oligonucleotide cartridges" positioned be-tween restriction endonuclease sites.
Oligonucleotide-directed mutagenesis is similarin principle to mutagenesis by gene synthesis in
116
that an oligonucleotide that bears the desiredmutant sequence is inserted into a cloned gene.However, oligonucleotide-directed mutagenesisdiffers from gene synthesis in the manner in whichthe cloning is accomplished. In this case, an oli-gonucleotide consisting of the mutant sequence ishybridized to its complementary sequence in aclone of wild-type DNA, thereby forming a mu-tant-wild-type heteroduplex. The oligonucleo-tide serves as a primer for in vitro enzymatic DNAsynthesis of regions that are to remain genotypi-cally wild type. A double-stranded heteroduplexis formed, which is subsequently segregated invivo into separate mutant and wild-type clones.The two can be distinguished by a number ofscreening procedures.
The basic strategy of oligonucleotide-directedmutagenesis was developed using the single-stranded phage OX174. The technique has beensuccessfully applied to genes cloned into eitherplasmid or phage vectors. Previously, I developedan efficient procedure for oligonucleotide-di-rected mutagenesis using M13-derived vectors.The major features of this method were the useof single-stranded clone DNA as template, thepurification of in-vitro-synthesized covalentlyclosed, circular DNA by alkaline sucrose gradientcentrifugation, and the use of the mutagenic oli-gonucleotide to screen for mutants by hybridiza-tion.
I have developed a variation of the originalmethod that is simpler, faster, and of equal effi-ciency (Zoller and Smith, DNA 3: 479 [1984]). Itis based on the use of two primers, one of whichis the mutagenic oligonucleotide and the other isa standard sequencing primer (see Fig. 2). Bothprimers are simultaneously annealed to single-stranded template DNA, extended by DNA poly-merase I (large fragment), and ligated together toform a mutant/wild-type gapped heteroduplex.E. coli is transformed directly with this DNA.Mutants are identified by plaque-lift hybridiza-tion using the mutagenic oligonucleotide as aprobe. The same procedure can be used withoutmodification to create site-specific insertions aswell as deletions. This method obviates the needto isolate covalently closed, circular DNA to ob-tain mutants in high yield and retains the conven-ience of the M13 system for template isolation,sequencing, and screening.

HO Sequencing
P MutagenicOligonucleotide
Anneal(55 °C /5 mint
OH
Extend / Ligase (15°C/6-I2 hrs)Klenow, DNA LigasedNTPs, rATP
TransformE coli
Screen plaquesfor mutants
FIGURE 2 Scheme for oligonucleotide-directed mutage-nesis using two primers and a single-stranded template.
DNA Synthesis FacilityM. Zoller
Synthetic oligonucleotides have expanded theability of the molecular biologist to producecloned DNA with a precisely defined sequence.Applications include the construction of pointmutations in a gene, custom-designed linkers, andthe precise fusion of promoter and coding se-quences. In addition, oligonucleotides have beento used to detect a specific gene within a library.Although procedures exist for the manual synthe-sis of oligonucleotides, the operations have beenautomated, thereby freeing the scientist for moreimportant problems. In December of 1983, I be-gan operating an Applied Biosystems DNA Syn-thesizer, which initially served my laboratoryprimarily. However, during 1984, about 300oligonucleotides were synthesized for more than30 individual scientists at the Laboratory. Oli-gonucleotides were produced at about one-tenth
the cost charged by commercial sources and wereobtained much faster, usually within 1 week. In-itially, the machine synthesized only a single oli-gomer at a time; however, the capability was ex-panded so that three separate oligonucleotidescould be synthesized at once. This has decreasedthe waiting time for an individual oligonucleotideand has made usage of solvents more efficient.
The chemistry of the Applied Biosystems DNASynthesizer is the solid-phase phosphite triestermethod based on procedures developed by M.Caruthers (University of Colorado). Currently,the scale is 1 /mole of starting material and syn-thesis proceeds at 20 minutes per cycle; oligo-mers as long as 100 bases have been successfullyproduced on this machine. In the future, a 0.1-Amo le scale synthesis will be offered in conjunc-tion with a 6-minute cycle, which will result infaster syntheses and a 50% reduction in the costper oligonucleotide. The small-scale synthesiswill be especially useful for producing short oli-gonucleotides for sequencing projects. We havefound that the efficiency of synthesis is highenough that the crude (nonpurified) sample canbe used as a primer for M13 dideoxy sequencingexperiments. In addition to the small-scale syn-thesis, a 10-Amo le scale synthesis is available forproducing oligonucleotides to be used for struc-tural experiments on nucleic acids.
PUBLICATIONS
Zoller, M.J. and M. Smith. 1984. Oligonucleotide-directedmutagenesis: A simple method using two oligonucleotideprimers and a single-stranded template. DNA 3:479-488.
In Press, Submitted, and In Preparation
Zoller, M.J., K.E. Johnson, M. Smith, and R.G. Kostriken.Point mutations in the yeast MA Ta gene that affect cleav-age by HO endonuclease. (Submitted).
117

PROKARYOTIC GENETICS
F. DaldalS. HintonP.A. ScolnikC. SlaughterD.C. Youvan
G. FreyerJ. ApplebaumE.J. BylinaS. ChengE. Davidson
W. EisnerT. FischerS. IsmailS. JohannJ. Schumann
Molecular Genetics of Cytochrome c2of Rhodopseudomonas capsulateF. Daldal, J. Applebaum, S. Cheng.
Cytochrome c2 is an electron carrier common tothe photosynthetic and respiratory machinery ofRhodopseudomonas capsulata. It is structurallyhomologous and functionally analogous to thecytochrome c of mitochrondria and to the plas-tocyanin of chloroplasts. During photosynthesis,c2 is the primary electron donor to the reactioncenter, and during respiration, it is an importantcomponent of the c-type cytochrome-containingrespiratory chain. This implies that c2, an evolu-tionarily well-conserved heme-containing pro-tein, must be evolved in such a way that, whenrequired, it can interact with the components ofthe reaction center, of the ubiquinol:cytochromec2 oxidoreductase (bc, complex), and of the cyto-chrome oxidase. Even though cytochrome c2 ofphotosynthetic bacteria has been well studiedbiochemically, biophysically, and immunological-ly, genetic data are still scarce. Until recently, nomutant deficient solely in cytochrome c2 wasavailable, and schemes devised to select c,- mu-tants often yielded pleiotropic mutants devoid ofall known c-type cytochromes. Therefore, the invivo role of c2 on photosynthesis has remaineduntested.
To isolate a c2- structural mutant, we havetaken a "reverse genetic" approach. First, the c2structural gene of R. capsulata was cloned by us-ing synthetic mixed oligonucleotide probes andthen its nucleotide sequence was determined. Thededuced amino acid sequence was found to be inperfect agreement with the known amino acid se-quence obtained from the purified c2 protein.Moreover, the nucleotide sequence of the amino-terminal part of the c2 gene indicated the presenceof a 21-amino-acid "signal sequence." Therefore,in agreement with its known periplasmic loca-tion, the c2 protein is synthesized in the cyto-
118
plasm as a precursor and is then processed duringits secretion to the periplasm.
Using the cloned gene, we constructed in vitroc2 genes containing insertion and insertion-dele-tion mutations, and we used these to replace thechromosomal wild-type allele by genetic crossesmediated by the "gene-transfer agents" (defectivephage-like particles specific to R. capsulata).This led us to the isolation of the first known c2structural mutants of a photosynthetic bacte-rium. In these mutants, the absence of the c2 genewas confirmed by Southern analysis using eitherthe cloned c2 gene or the inserted kanamycinmarker as probes. Furthermore, the absence ofthe c2 gene product was proved by immunoprecip-itation using polyclonal antibodies raised againstthe purified c2 protein, by c-type cytochromestaining of SDS-polyacrylamide gels of total cellextracts, and by reduced oxidized spectrum anal-ysis.
c2- mutants are stable and can grow well byphotosynthesis and by cytochrome-c-dependentrespiration. Considering the fact that during pho-tosynthesis, cytochrome c2 is the primary electrondonor to the reaction center, these mutants showthat in the absence of c2, R. capsulate may havealternative ways unknown until now to reduce theoxidized photosynthetic reaction center. To un-derstand this interesting observation, several hy-potheses, such as the presence of other cyto-chromes functionally analogous to c2 or thenonessentiality of a mobile carrier in photosyn-thesis, are currently under investigation.
Clostridium pasteurianumGalactokinaseF Daldal, J. Applebaum
Until now, no gene from a bacterium like Clostri-dium pasteurianum, which has a very high A + T

content (70%), has been isolated. To study thestructure of a clostridial gene, to assess the pos-sibility of its expression in other hosts, and even-tually to use it as a genetic tool, we have clonedthe C. pasteurianum galactokinase by comple-mentation of an Escherichia coli mutant deficientin galactokinase. Restriction enzyme analysis,subcloning, and Tn5 mutagenesis indicated thatthe gene was located on a 1.8-kb Clal-Sau3Afragment that encoded a polypeptide with an ap-parent molecular weight of approximately 40,000.Although they have similar subunit molecularweights, Southern hybridization analysis indi-cated no strong homology between the C. pasteu-rianum and the E. coli galactokinases. In E. coli,the Gal' phenotype provided by this clostridialgalactokinase was found to be unstable, and wehave shown that spontaneous Gal- derivativeswere originated by insertion sequences (IS/ andIS5) inactivating the clostridial galactokinase.Even though this clone was able to complement agalK mutant of E. coli, it expressed the clostridialgalactokinase at a low level in this host. Using asa probe a DNA fragment internal to the clostri-dial galactokinase, we isolated by hybridizationanother clone carrying (presumably) the wild-typegene. This latter clone was not able to grow ongalactose-containing media due to the overprod-uction of the clostridial galactokinase and theconcomitant inhibition of the E. coli host by"galactosemia."
Comparison of these two plasmids producinglow and high amounts of galactokinase revealedthat the low-level producer carries a deletion ofapproximately 300 by located in the 5'-proximalpart of the gene. Operon fusions constructed be-tween this region where the deletion is localizedand a promoterless E. coli galactokinase gene in-dicated that these clostridial DNA sequences havepromoter activity in E. coli and that the over-expression of the clostridia] galactokinase is atleast partly transcriptional in this latter clone.
The nucleotide sequences of the 5'-proximalpart of the galactokinase gene from several clonesproducing high, low, or undetectable amounts ofgalactokinase were determined. Their compari-son revealed that the clone producing highamounts of galactokinase has clostridial nucleo-tide sequences very homologous to the E. coliconsensus promoter sequences. The role of thesesequences in Clostridium is not yet known, but
fusion studies demonstrated that they mediate theexpression of the clostridial galactokinase in E.coli. Therefore, considering that the transcrip-tion of a gene is an important barrier in hetero-logous expression systems, the A + T richness ofthe clostridial genome appears to be a lucky situ-ation when the chosen host is E. coll. This workshows that the complementation of E. coli mu-tants by the clostridial genes is feasible and is agood way to isolate clostridial genes whenever ap-propriate mutants are available. It remains to beseen whether the C. pasteurianum galactokinasegene will turn out to be a useful genetic markerfor the study, by gene fusions, of the structureand the regulation of other clostridial genes inClostridium using the recently developed trans-formation technique.
Finally, the cloning and characterization of theC. pasteurianum galactokinase indicate that inthis organism, galactose is used through the "Le-Loir pathway" via galactose-l-phosphate.
Characterization of MT113, a Ps-Mutant of R. capsulata ThatLacks a Functional bc, Complexand c-type CytochromesE. Davidson, F. Daldal
MT113, a nonphotosynthetic mutant of Rhodo-pseudomonas capsulata, not mapping in theknown cluster of photosynthetic-machinery-re-lated genes, was isolated previously by Marrs etal. (FEBS Lett. 113: 289 [1980]) and was shownby spectroscopic analysis to lack cytochrome c2.However, with the later establishment that cyto-chrome c,, a component of the bc, complex, alsoabsorbs at the same wavelength (550 nm) as c2, itbecame clear that MT113 also lacks cytochromec,. Therefore, we decided to ascertain, in collab-oration with R. Prince and B. Marrs (Exxon Re-search and Engineering Co.), whether other com-ponents of the bc, complex were also missing inthis mutant.
Electron paramagnetic resonance (EPR) spec-troscopy indicated that membranes derived fromMT113 contained the Rieske Fe-S center at a muchlower amount (< 10%) than the wild-type mem-branes. This lack of the Rieske Fe-S center can-
119

not be attributed to a general defect in the syn-thesis of Fe-S clusters, since the same mutantcontains an almost normal amount of total Fe-Sclusters. Similarly, on the basis of EPR spectro-scopy, an antimycin-sensitive signal characteristicof the anionic semiquinone Qc - was not detecta-ble in MT113 membranes. Absorption spectros-copy showed that, even though an almost normalamount of b-type cytochromes was present in thismutant, the antimycin-induced red shift of thecytochrome b560 of the bc, complex was absent.
Analysis of the MT113 membrane proteins bynondenaturing polyacrylamide gels (LDS-PAGE)coupled to a heme-specific staining procedure in-dicated a general lack of c-type cytochromes. Todetermine whether the lack of a functional bc,complex was related to the absence of specific ap-oproteins corresponding to the bc,-complex com-ponents, cell extracts were submitted to Westernanalysis using antibodies raised against purifiedcytochrome c2 of R. capsulata and cytochrome b,cytochrome c and Rieske Fe-S proteins of Rho-dopseudomonas sphaeroides. The apoprotein ofc2 was not detected, but those of the bc,-complexcomponents were present, suggesting that MT113was able to synthesize the analyzed bc,-complexcomponents but was not able to assemble them asa functional complex. Genetic analysis using thegene-transfer agent of R. capsulata and severalplasmids carrying the structural genes for cyto-chrome c2 and for at least some of the bc,-com-plex components (Rieske Fe-S gene) clearly indi-cated that the primary effect of the singlepleiotropic mutation in MT113 is unlikely to berelated to the structural genes of the cytochromec2 and of the bc,-complex components. The na-ture of this mutation is still unknown, but a gen-eral lack of c-type (but not b-type) cytochrome inMT113 suggests that it could be related to the latesteps of c-type cytochrome biosynthesis.
Cartridge Mutagenesis and Deletion ofthe Photosynthetic Reaction-centerand Light-harvesting Genesfrom R. capsulataD.C. Youvan, S. Ismail
Our purpose in constructing photosynthetic ap-paratus deletion strains is to provide back-
120
grounds for the assay of in-vitro-mutagenized re-action-center and light-harvesting genes carriedby plasmid vectors (Fig. 1). The nucleotide se-quence and deduced polypeptide sequences for allof the reaction-center and light-harvesting struc-tural polypeptides have been determined (Youvanet al., Cell 37: 949 [1984]; Youvan and Ismail,Proc. Natl. Acad. Sci. 82: 58 [1985]). On the ba-sis of primary sequence data, models have al-ready been proposed for reaction-center quinonebinding and light-harvesting bacteriochlorophyllbinding. The three-dimensional structure of the
ReturnWild-typeOperon
))-Sp
Clone genes:rxcilLI- /_7I
operon
Wild - type
IReplace structuralgenes withspectinomycin cartridge
Deletion Strain
ReturnMutagenizedOperon
Wild - type Mutant
FIGURE 1 Construction and complementation of photo-synthetic deletion strains. Deletion backgrounds con-structed by cartridge mutagenesis may be complementedby the wild-type reaction-center or light-harvesting operonsin trans. Alternatively, in-vitro-mutagenized plasmids may bereturned to the deletion strain in order to assay the affect ofengineered point mutations.

reaction center is being solved by X-ray crystal-lography. We anticipate that it will be essentialto test reaction-center and light-harvesting struc-ture-function models by in vitro mutagenesis ofthe structural genes. Progress in this rapidlyadvancing multidisciplinary field has been re-viewed recently (Youvan and Marrs, Cell 39: 1[1984]; Youvan and Marrs, in Molecular Biologyof the Photosyntheic Apparatus, Cold SpringHarbor Laboratory, Cold Spring Harbor, NewYork [1985] in press).
Deletion strains were constructed using the car-tridge mutagenesis technique of P. Scolnik (thissection). This procedure employs the gene-trans-fer agent (GTA) that is specific to Rhodopseudo-monas capsulata. The GTA is similar to a gen-eralized transducing phage, randomly packaging4.6-kb DNA segments from the chromosome.Plasmids carrying R. capsulata genes are muta-genized in vitro by the insertion of small (2-4 kb)restriction fragments specifying resistance tokanamycin or spectinomycin (from Tn5 or Tn 7,respectively). These plasmids are then transferredvia conjugation from Escherichia coli to a GTAoverproducer strain. GTA from the overproduceris crossed with a suitable recipient, and transduc-tants are selected for the appropriate antibioticmarker. Antibiotic-resistant transductants haveincorporated the mutagenized copy of the geneinto their chromosome by homologous recombi-nation, and the corresponding wild-type allelesare lost. Cartridge mutagenesis need not be lim-ited to simple insertional interruption of genes:Deletions may be engineered through ligation ofthe cartridge in place of a target restriction frag-ment.
Strains have been constructed wherein all orpart of the reaction-center (RC), light-harvestingI (LHI), and light-harvesting II (LHII) structuralgenes have been deleted. In one series of strains,the 2778-bp Apal fragment bearing more than90% of the rxcA operon (promoter and structuralgenes for LHI 6, LHI a, RC L, and RC M) hasbeen deleted from the chromosome. When therxcA operon is deleted from the wild type, result-ant strains possess only LHII and are photosyn-thetically defective. The rxcA deletion in a LHII-background results in a strain lacking all light-harvesting antennae and reaction-center sub-units. As expected, this strain has no near-in-frared absorption characteristic of light-harvest-
ing or reaction-center bacteriochlorophyll. TherxcA deletion may be complemented by a pBR322derivative (pU21) carrying the entire rxcA ope-ron. In a second series of deletion strains, the2500-bp BstEll-Stul fragment, including the 6and a structural genes for LHII, has been deletedfrom the chromosome. In the wild-type back-ground, the functional reaction center and LHIare expressed. LHII may be restored by return-ing the plasmid pU2 that carries the LHII ope-ron.
Characterization of the PhotosyntheticReaction CenterD.C. Youvan
Photosynthetic reaction centers from the purplenonsulfur bacterium Rhodopseudomonas cap-sulata have been purified to near homogeneityin amounts (100 mg) suitable for biophysicalanalysis. Photosynthetic membranes (chromato-phores) can be prepared by differential centrifu-gation using a cartridge mutant (U34) lacking theLHII antenna. Otherwise, reaction centers andLHI are wild type. Membrane proteins are solu-bilized in the anionic detergent lauryl-dimethy-lamine-N-oxide (LDAO), and reaction-centerproteins may be purified by affinity chromatog-raphy on horse heart cytochrome c2 coupled toSepharose 4B. This procedure was originally de-veloped by Brudvig et al. (Proc. Natl. Acad. Sci.80: 683 [1983]) for reaction centers from Rhodo-pseudomonas sphaeroides.
Reaction centers purified by affinity chroma-tography are very pure and photochemically ac-tive. No contaminating light-harvesting polypep-tides or bacteriochlorophyll is observed by SDS-PAGE or absorption spectroscopy. The purifiedreaction centers are capable of the initial photo-synthesis reaction involving charge separation:The 870-nm absorption band assigned to the spe-cial pair is photobleached by actinic light.
Reaction centers have been crystallized fromtwo species, Rhodopseudomonas viridis and R.sphaeroides. The detailed biophysics of the pho-toelectron cycle is best characterized in the latterspecies. This is somewhat of an unfortunatesituation since genetics is best developed in R.
121

capsulata. The amino acid sequence of the pho-tosynthetic reaction-center polypeptides is com-pleted for only R. capsulata. We believe thatgenetics is an essential feature for future struc-ture-function problems; hence, X-ray crystallog-raphy should be repeated using R. capsulatareaction centers. The ability to prepare largequantities of purified R. capsulata reaction cen-ters should help to remedy the problem.
In Vitro Mutagenesis of thePhotosynthetic Reaction-centerand Light-harvesting I GenesE.J. Bylina, D.C. Youvan
We are using in vitro mutagenesis to addressthe fundamental question: How are proteins in-volved in the primary light reactions of photosyn-thesis? In the light reactions, membrane proteinsand cofactors (chlorophyll, quinone, and caro-tenoids) interact in a highly efficient way to har-vest light energy and to transduce this energy in-to a biochemically useful reductant. Presumably,the exact placement of the cofactors by the mem-brane proteins facilitates the unprecedented pho-tochemical efficiency of the photosynthetic ap-paratus (quantum yield approaching unity).Some of the amino acid residues within the mem-brane proteins may play more than a structuralrole and participate directly in the photochemicalreactions, facilitating proton and electron con-duction.
We have recently developed in vitro mutagene-sis procedures and bacterial strains to assay thephenotype of plasmid-borne rxcA operons. This3000-bp operon carries the structural genes en-coding four membrane polypeptides: the photo-chemical core of the reaction center (L and Msubunits) and LHI (a and f3 subunits). The clonedrxcA operon may be returned to rxcA deletionstrains in order to assay the phenotype of theplasmid genes in trans.
The plasmid bearing the rxcA operon (pU21)has been mutagenized in vitro by oligonucleo-tide-mediated site-directed mutagenesis. Plas-mid pU21 is nicked with EcoRI in the presence ofethidium bromide, and exonuclease III is then
122
used to convert the nicked DNA into gappedDNA. The mutagenic oligonucleotide is an-nealed to this gapped region, and the gap is thenrepaired with Klenow and ligase. The mutagen-ized plasmids are transformed, and the trans-formants are screened by colony hybridizationusing the mutagenic oligonucleotide as the probe.Direct mutagenesis of the plasmid avoids shut-tling, since pU21 may be conjugated from an E.coli donor to the deletion background with thehelp of a mobilizing plasmid. Hence, the pheno-type of a light-harvesting or reaction-center mu-tation may be rapidly assayed. Additionally, re-vertants may be selected and the sequence of pU21redetermined.
Experiments have been initiated to test the light-harvesting bacteriochlorophyll-binding model ofTheiler and Zuber (Hoppe-Seyler's Z. Physiol.Chem. 365: 721 [1984]). The proposed bindingsite Ala-X-X-X-His has been targeted by in vitromutagenesis of pU21, assayed in the deletionbackground. A histidine-to-aspartic acid changein the LHI ca- subunit results in the loss of thisantenna. This is consistent with the model whichproposed that the a and polypeptides interactthrough a chlorophyll dimer, each bound to a his-tidine lone pair on both polypeptides. The lossof LHI also causes the pleiotropic derangementof the reaction center and a photosynthesis-de-fective (PS -) phenotype. PS revertants havebeen subsequently isolated by direct selection.We plan to sequence the site of reversion in orderto determine whether histidine is the only func-tional residue for chlorophyll binding.
Future in vitro mutagenesis targets include thereaction-center putative quinone-binding site Pro-Phe-His-Met-Leu, which is conserved with higherplants. Mutations in this sequence may affectherbicide resistance, since these chemicals arecompetitive inhibitors of quinone binding. An-other possible target of particular interest tospectroscopists is a possible carotenoid bindingsite in LHII: Tyr-Arg. The arginine residue is be-lieved to be the point charge responsible for thecarotenoid electrochromic band shift that is
sensitive to membrane potential. As crystallo-graphic data become available, mutagenesisshould play an increasingly important role in test-ing light-harvesting and reaction-center struc-ture-function models.

Cartridge Mutagenesis in R. capsulataP.A. Scolnik
Modern genetic approaches require the capabilityof selectively inactivating genes by insertions ordeletions. I have developed a system for intro-ducing gene cartridges into the chromosome ofRhodopseudomonas capsulata by homologousrecombination (Scolnik and Haselkorn, Nature307: 289 [1984]). This method takes advantageof a defective transducing phage (gene-transferagent, GTA) produced only by this bacterium.GTA transduces linear double-stranded segmentsof DNA of 4.5 kbp, which by homologous recom-bination are integrated into the chromosome ofthe recipient cell. I determined that DNA fromrecombinant plasmids, introduced into the cell byconjugation, are also packaged into GTA heads.Consequently, the following method (Fig. 2) wasdeveloped: (1) A gene cartridge is introduced invitro into a cloned sequence by standard recom-binant DNA techniques, generating either an in-sertion or a deletion. (2) The resulting plasmid istransformed into Escherichia colt and then intro-duced into an R. capsulata GTA overproducer byconjugation. (3) GTA is then prepared and usedto treat wild-type recipient cells, selecting for theantibiotic resistance of the gene cartridge. (4)Since the incoming molecule is not a replicon, an-tibiotic resistance cells can only be the product ofhomologous recombination with the host chro-mosome.
This method has two main advantages over
A. B.
GTA
transposon mutagenesis. First, the insertions ordeletions can be engineered in vitro using restric-tion sites. Second, since this method does not in-volve the introduction of plasmids into the cell,the mutants obtained can be used for comple-mentation studies with recombinant plasmids. I
have successfully used kanamycin resistance fromTn5 and spectinomycin resistance from Tn7 asgene cartridges.
Maximum efficiency (10-3 with respect to theefficiency of transduction of a point mutation) isobtained when both ends of the molecule containR. capsulata rather than vector DNA, probablyreflecting the fact that the free ends are recombi-nogenic. However, insertions have been obtainedwith as little as 250 by of homologous DNAflanking the gene cartridge, indicating that eitherinternal recombination occurs or the linear mol-ecule is attacked by exonucleases, generating freeends.
Cryptic Genes and Gene Rearrangementsin Response of a Bacterial Genometo Stress: Precise Excisionof a Gene CartridgeP.A. Scolnik, J. Schumann
Rhodopseudomonas capsulata can fix atmospher-ic nitrogen through the enzyme nitrogenase. Wehave cloned the structural genes (nil HDK) andhave determined that they are organized in a tran-scriptional unit (Scolnik and Haselkorn, Nature
C. D.
I Plasmid Vector R. capsulata DNA i Gene CartridgeFIGURE 2 Strategy for site-directed gene cartridge mutagenesis in R. capsulata (see text).
123

307: 289 [1984]; J. Bacteriol. 156: 251 [1983]),entirely contained within a 12-kbp HindIII frag-ment. Insertions and deletions within this frag-ment (produced by the cartridge mutagenesismethod using kanamycin as the selection marker)give rise to a Nif- phenotype and introduce intothe region an additional HindIII site that is pres-ent in the cartridge.
These mutants are stable in their phenotypewhen grown in medium supplemented with fixednitrogen. However, Nit.* pseudorevertants can beisolated by challenging these cells to grow in am-monia-free medium. A time-course experimentstarting with an insertional mutation grown inammonia-free medium showed that the first classof pseudorevertants appears within the first fewhours of selection. These cells are resistant tokanamycin, and the gene cartridge remains in theoriginal place of insertion, as shown by Southernhybridization; however, a low (10% of wild-typelevels) nitrogenase activity can be detected. Kan-amycin-sensitive cells, with full nitrogenase activ-ity, are observed after at least 2 days of growth.Southern hybridization shows that the gene car-tridge has been deleted from the genome, and twonew HindIII bands (12 and 16 kbp) are observed.The 16-kbp fragment is a transient form, whichresolves in the 12-kbp band upon further incuba-tion. Cells containing this 12-kbp band are indis-tinct from wild types.
We interpret these results as follows: When thenif genes are inactivated by cartridge insertion,R. capsulata will respond to the need for nitrogenby bringing a cryptic nif gene(s) into operation,and this event is responsible for the low level ofnitrogenase activity initially observed. That thisresponse does not involve some type of activationof the inactivated nif genes is evident because itwill occur whenever these genes are deleted. Sub-sequently, the gene cartridge is excised from thegenome, probably by translocation of a 4-kbpelement into the site of kanamycin insertion.This event generates a 16-kbp band, which in turnresolves (probably by excision of the 4-kbp ele-ment) into a 12-kbp HindIII band, and the fullnitrogenase activity phenotype is restored. Theactivation of the cryptic gene provides the neces-sary energy for the process of gene rearrangementto take place.
Since nitrogenase activity is fully restored, theexcision of this cartridge must be precise or near
124
precise. The fact that this process occurs indi-cates that gene cartridges as well as transposonscan be excised from the genome, and thereforetransposon activity is not required. We are cur-rently characterizing the excision process, and wehave sequenced the nifH and nifD genes in orderto be able to determine the nature of the rear-rangement observed.
The nature of the cryptic gene remains unclear.Southern hybrydization of nif structural genes tototal genomic DNA shows several (at least eight)HindIII bands that contain homology with allthree nifH, nifD, and nifK genes. We have clonedone of them and have determined that, althoughit contains 65-80% homology with nifH at thenucleotide level, it does not have the capability ofcoding for a nitrogenase gene. There are fourcopies of this element in the genome, and cosmidcloning shows that they are dispersed in the chro-mosome. Insertions in this region have no phe-notype (P. Schumann and P. Scolnik, in prep.). It
is possible that this element constitutes a nif ho-mologous insertion sequence, a possibility cur-rently being explored.
Cloning Prokaryotic Genes byComplementation with aCosmid LibraryP.A. Scolnik
A situation often exists in which no physical probeto clone a certain gene is available, but a mutantwith the desired phenotype has been obtained. Agene complementing such a mutation can be eas-ily isolated from a genomic library, provided amethod exists to introduce this library massivelyinto the mutant cells. In most gram-negative bac-teria, transformation, at least at high efficiency,is not available. However, wide-host-range plas-mids (developed by D. Helinski's group, Univer-sity of California, San Diego) can be mobilizedby conjugation from E. colt. I have constructeda genomic library from R. capsulata in a cosmidderivative of these vectors and designed a proto-col that allows cloning of the mutants by comple-mentation. In conjunction with cartridge muta-genesis, this method can be used to clone and toidentify R. capsulata genes unequivocally. Morethan ten different sequences, affecting both

photosynthesis and nitrogen fixation, have beencloned using this approach.
A Study of Bacterioferritin fromAzotobacter vinelandiiS. Hinton, C. Slaughter, T. Fischer
Bacterioferritin is a b-type cytochrome, but it alsoclosely resembles mammalian ferritin because ofits quaternary structure (24 identical subunits)and contains an average of 1600 Fe atoms in anoxide-hydroxide-phosphate core. Bacterioferritindiffers from mammalian ferritin in that it con-tains heme and has the ability to take up hundredsof electrons at a low redox potential, which sug-gests that it may function as an electron storageprotein for use in respiration, biosynthesis, andnitrogen fixation. It has been proposed that bac-terioferritin may also be an iron storage proteinfor Azotobacter vinelandii.
We are initiating structure-function studies ofbacterioferritin to understand how it resemblesand differs from ferritin, as well as biochemicalstudies to establish its physiological role. Bacter-ioferritin was purified and crystallized. The ami-noterminal amino acid sequence was determined,and antibodies were raised against the purifiedprotein. Oligonucleotides were synthesized basedon the aminoterminal amino acid sequence of theprotein, and antibodies recognizing bacteriofer-ritin are being used to screen various genomic li-braries of A. vinelandii. The complete primarystructure of bacterioferritin will be determinedby amino acid sequencing as well as by genesequencing. Future experiments will includesite-directed mutagenesis for structure-functionstudies to determine the effect mutations have onnitrogen fixation, respiration, and iron metabo-lism.
Molybdenum Metabolism inClostridium pasteurianumS. Hinton, C. Slaughter, G. Freyer, W. Eisner, T. Fischer
There are two distinct cofactors for various redoxenzymes. The molybdenum-iron protein of ni-
trogenase contains a unique iron-molybdenumcofactor, FeMo-co, whereas all other known mol-ybdoenzymes contain a common molybdenumcofactor, Mo-co, a molybdenum-pterin species.We have proposed a hypothesis that during thebiogenesis of Mo cofactors, precursors are pro-tein-bound. We have identified four molybdo-proteins in crude extracts of Clostridium pasteu-rianum that have characteristics which suggestthat they may play a role in Mo metabolism.
The major Mo-binding protein (Mop) in C.pasteurianum, suspected to be involved in Mometabolism, was purified and biochemicallycharacterized, and the mop gene was cloned andsequenced. Our studies have shown that the iso-lated Mo protein contains Mo as well as a pterin-like fluorescent chromophore. Spectroscopicanalysis indicates that the Mo environment andthe pterin-derivative associated with Mop aresimiliar to the components of the Mo-co isolatedfrom clostridial formate dehydrogenase. TheMo-co dissociated from formate dehydrogenasewas used to reconstitute the activity of Mo-co-deficient nitrate reductase (biological assay forcatalytically active Mo-co). The molybdopterinspecies dissociated from Mop was not active inthe Mo-co reconstitution assay. The results sug-gest that the catalytically inactive molybdopterinspecies associated with Mop might be a precursorto Mo-co.
We identified an expression clone in Esche-richia coli cross-reacting with Mop antibodies byimmunoscreening genomic libraries of C. pasteu-rianum. Western blot analysis showed that theclone produced a protein recognized by the anti-body with the same effective electrophoretic mo-bility as purified Mop. DNA sequence analysisof the expression clone revealed an open readingframe coding for 67 amino acids (m.w. ^-,8000).The aminoterminal amino acid sequence of theprotein showed perfect alignment with the first 12codons in the open reading frame. Amino acidsequence analysis also showed that there appearto be two amino acid residues at six different po-sitions. The DNA sequence predicts only one ofthe amino acid residues identified at each of thesepositions. This ambiguity suggests that there aretwo or more related proteins in the purified pro-tein preparation. Southern blot analysis of C.pasteurianum chromosomal DNA showed thatthe original (Mop) clone hybridized to two DNA
125

fragments. The results indicate that the mop genehas been duplicated and the two variant Mo pro-teins copurify. We have cloned and are presentlysequencing what appears to be the second copyof the mop gene.
Amino acid sequence analysis of Mop main-tained under anaerobic conditions showed that atpositions where the DNA sequence predicted aglutamic acid residue, an amino acid phenylthio-hydantoin (PTH) derivative was identified thatdid not correspond to any known standard. Themodified glutamic acid appears to be oxygen-sen-sitive because glutamic acid PTH was identifiedin the positions predicted by the gene sequencewhen the protein was not handled under anaero-bic conditions. It is interesting to note that Mobinding to Mop was also oxygen-sensitive. Cur-rently, we are determining the structure of theglutamic acid derivative by fast-atom bombard-ment mass spectroscopy and nuclear magneticresonance analysis.
Biogenesis of the MolybdenumCofactor in Escherichia coliS. Hinton, S. Johann
The molybdenum cofactor, Mo-co, common to avariety of molybdoenzymes, contains an as yetundefined pterin derivative associated with a mo-lybdenum (Mo) atom. The products of at leastfive genes (chi A, B, D, E, and G) in Escherichiacoli are needed for Mo-co biogenesis. It has beenproposed that the gene products of chlA and chlEare involved early in Mo-co synthesis, possiblymodifying the pterin. The ch1D, ch1G, and chlBgene products are thought to be involved in sta-bilizing the pterin-derivative in the absence of Mo,inserting Mo into the cofactor, and inserting theMo-co into the apoenzyme, respectively. Ourgoal is to understand the biology and bioinor-ganic chemistry in the transformation of molyb-date, MoO, , into Mo-co. To achieve this end,we are studying the role the chi gene productshave in Mo metabolism by identifying the geneproducts, determining the DNA sequence of thegenes, and studying the effect each chi mutant hason Mo accumulation in E. coli.
Genomic libraries of E. coli DNA were con-
126
structed in a pUC expression vector, several clonesthat complemented the chi (Mu insertion) mu-tants were identified, and complementation wasshown to be dependent on the inserts of E. coliDNA. Southern hybridization analysis showedthat the clones contained fragments of E. coliDNA that have sequence homology with chro-mosomal DNA flanking the Mu insertion in theirrespective chi mutant. Maxicell experimentsshowed that the plasmids which conferred com-plementation encoded proteins with apparentmolecular weights of 24,000 and 10,000 with thechlB clone, 26,000 with the chlD clone, 44,000 and23,000 with the chlE clone, and 25,000 and 6,500with the ch1G clone. Restriction analysis, sub-cloning, and site-specific mutagenesis have beenperformed to localize the chi genes further for fu-ture maxicell and DNA-sequencing experiments.DNA-sequencing analyses of the chlD and ch1Ggenes are in progress.
Radioisotope "Mo-labeling experiments inconjunction with native (anaerobic) multiphasicgel electrophoresis are being used to identify themolybdoproteins in crude extracts of E. coli. Weare interested in determining whether any of thechi gene products are molybdoproteins possiblybinding a precursor of Mo-co.
PUBLICATIONS
Daldal, F 1984. Nucleotide sequence of gene pfkB encod-ing the minor phosphofructokinase of Escherichia coli K-12. Gene 28: 337-342.
Scolnick, P.A. and R. Haselkorn. 1984. Activation of extracopies of genes coding for nitrogenase in Rhodopseu-domonas capsulate. Nature 307: 289-292.
Scolnik, P., P. Autges, and R. Haselkorn. 1984. Multiple cop-ies of genes coding for nitrogenase in the phototrophicbacterium. In Advances in nitrogen fixation research (ed.C. Veeger and Newton), p. 745.
Sedivy, J., F. Daldal, and D.G. Fraenkel. 1984. Fructose bis-phosphatase of Escherichia coli: Cloning of the structuralgene (ibp) and preparation of a chromosomal deletion.J. Bacteriol. 158: 1048-1053.
Youvan, D.C. and S. Ismail. 1985. Light-harvesting II (B800-B850 complex) structural genes from Rhodopseudomo-nas capsulata. Proc. Natl. Acad. Sci. 82: 58-62.
Youvan, D.C. and B.L. Marrs. 1984. Molecular genetics andthe light reactions of photosynthesis. Cell 39: 1-3.
Youvan, D.C., M. Alberti, H. Begusch, E.J. Byline, and J.E.Hearst. 1984. Reaction center and light-harvesting I
genes from Rhodopseudomonas capsulata. Proc. Natl.Acad. Sci. 81: 189-192.
Youvan, D.C., E.J. Byline, M. Alberti, H. Begusch, and J.E.Hearst. 1984. Nucleotide and deduced polypeptide se-

quences of the photosynthetic reaction -center, B870 an-tenna, and flanking polypeptides from R. capsulata. Cell37: 949-957.
In Press, Submitted, and In Preparation
Daldal, F. 1985. Isolation of mutants of Clostridium pasteu-rianum ATCC 6013 and formation and regeneration ofprotoplasts. Arch. Microbiol. (in press),
Daldal, F. and J. Applebaum. 1985. Cloning and expressionof Clostridium pasteurianum galactokinase gene in Esch-erichia co/i and nucleotide sequence analysis of a regionaffecting the amount of the enzyme. J. Mol. Biol. (inpress).
Davidson, E., R.C. Prince, F. Daldal, G. Hauska, and B.L.Marrs. Characterization of nonphotosynthetic mutant
lacking ubiquitol, cytochrome c2 oxidoreductase (bc,complex) and c-type cytochromes. J. Biol. Chem. (Sub-mitted.)
Youvan, D.C. and B.L. Marrs. 1985. Photosynthetic appa-ratus genes from Rhodopseudomonas capsulata. In Mo-lecular biology of the photosynthetic apparatus. ColdSpring Harbor Laboratory, Cold Spring Harbor, New York.(In press.)
. Molecular genetics and the light reactions of photo-synthesis. Sci. Am. (Submitted.)
Youvan, D.C., S. Ismail, and E.J. Bylina. Chromosomal dele-tion and plasmid complementation of the photosyntheticreaction center and light-harvesting genes from Rhodo-pseudomonas capsulata. Gene (in press).
127


CELL BIOLOGY
The focus of the Cell Biology Group has slowly evolved from solely the analysisof cell structure to the analysis of cell growth control. During the past year, vir-tually all of our work involved some aspect of normal- or transformed-cell growth-control pathways. The biochemical and cell biological roles of the ras proteinshave been studied in great detail. Analysis of the effects of adenovirus El A pro-teins and genes and those of the human oncogene, c-myc, in living cells has justbegun. Analyses of these proteins and their effects on cells are also being stud-ied by computerized two-dimensional gels. The role and properties of protein ki-nase C (and of the inositol phospholipids) in cellular functions are also areas ofactive research by the Group. Investigations into the molecular components ofthe heat shock or stress response of cells, which characteristically leads to a tem-porary shutoff of growth, are continuing at a fast pace. In addition, cloning, ge-netic organizational studies, and protein biochemical analysis of the actin-bind-ing proteins, the tropomyosins, are being pursued with vigor. The expression ofthe tropomyosins is rapidly and dramatically changed during the transformationprocess. Progress is being made, but the complexity of the molecular events thatmust be involved in mammalian cell-growth control is so great that the field willcontinue at a high pitch here and elsewhere for some time.
GENES FOR MAJOR STRUCTURAL PROTEINS
D.M. Helfman Y. Yamawaki-KataokaL. Finn
Isolation and Characterization ofcDNA Clones EncodingNonmuscle TropomyosinY. Yamawaki-Kataoka, L. Finn, D.M. Helfman
Rat embryonic fibroblasts contain five forms oftropomyosin, TM-1 (M, = 40,000), TM-2 (M, =36,500), TM-3 (M, = 35,000), TM-4 (M, = 32,400),and TM-5 (Mr= 32,000). The structure and func-tional significance of the different forms oftropomyosin in nonmuscle cells remain to be de-termined. In addition, recent studies have indi-cated significant alterations in the pattern oftropomyosins expressed in cells transformed byvarious DNA and RNA tumor viruses. These al-terations in tropomyosin expression may in partbe responsible for the reduction of microfilamentbundles (stress fibers) and alterations of cell shape
characteristic of transformed cells. The mecha-nism(s) responsible for the altered patterns oftropomyosin synthesis in transformed cells is un-known. With these questions in mind, we havebeen studying tropomyosins in nonmuscle cells(rat embryonic fibroblasts) and the genes that en-code these proteins. To isolate cDNA clones en-coding nonmuscle tropomyosins, we constructeda rat embryonic fibroblast cDNA expression li-brary using the plasmid expression vectors pUC8and pUC9. Using both an immunological screen-ing procedure and "P-labeled cDNA probes, wehave identified and isolated cDNA clones en-coding three of the five tropomyosins found inrat embryonic fibroblasts. By hybrid-selectiontranslation and analysis of the translation prod-ucts by one- and two-dimensional gel electropho-resis, we have identified clones that contain se-quences complementary to TM-I, TM-2, and TM-
129

4. Northern blot analysis indicated transcripts of1.1, 1.8, and 2.2 kb for TM-1, TM-2, and TM-4,respectively. We have identified overlappingclones for the entire coding region of rat TM-1and have determined the complete amino acid se-quence of the protein deduced from nucleic acidsequence analysis (see below). In addition, wehave sequenced cDNA clones encoding a portionof TM-2 and TM-4. Comparison of the nucleicacid sequences of clones for TM-1, TM-2, andTM-4 indicates differences in their coding re-gions, although their predicted amino acid se-quences show more than 85% homology. South-ern blot analysis of rat DNA showed severalunique fragments hybridizing to each probe madefrom cDNA clones encoding TM-1, TM-2, or TM-4. These results indicate that these proteins areencoded by separate but related genes and thattropomyosin species present in rat embryonic fi-broblasts are encoded by at least three separategenes. We are currently analyzing genomic clonesthat encode nonmuscle tropomyosins (see below).
Rat Embryonic Fibroblast Tropomyosin1: cDNA and Complete PrimaryAmino Acid SequenceY. Yamawaki-Kataoka, D.M. Helfman
A first step in understanding the molecular basisfor the differences in the various forms of tropo-myosin in rat embryonic fibroblasts will be a de-termination of their primary structure. Usingoverlapping cDNA clones for TM-1, we have de-termined the sequence of the entire coding regionof 852 bp, 61 by of 5'-untranslated sequences, and73 by of the 3'-untranslated region (Yamawaki-Kataoka and Helfman, J. Biol. Chem. [1985]submitted). Since the 3'-untranslated sequencecontains a poly(A) track and the polyadenyla-tion signal AATAAA is found 14 by from thepoly(A) track, we conclude that our clones con-tain the complete 3'-untranslated sequence. Thus,TM-1 has a relatively short (73 bp) 3'-untrans-lated region. Comparison of the deduced aminoacid sequence of the rat TM-1 with those oftropomyosins isolated from chicken smooth mus-
130
cle, chicken and rabbit skeletal muscle, andequine platelet indicates extensive homologyamong the various tropomyosin isoforms. TM-1,like smooth- and skeletal-muscle tropomyosins,was found to contain 284 amino acids. On theother hand, platelet tropomyosin has been re-ported to contain only 247 amino acids. Thetropomyosin most homologous to rat TM-1 ischicken smooth-muscle a-tropomyosin (95% ho-mology); in contrast, rat TM-1 shows only 77%and 78% homology with skeletal-muscle andplatelet tropomyosins, respectively. Interestingly,TM-1, smooth-muscle, and platelet tropomyosinswere found to be highly homologous at their car-boxyterminal domains, compared with skeletal-muscle tropomyosins. Whether all nonmuscleand smooth-muscle tropomyosins share this fea-ture remains to be determined. In this respect,we have also determined the sequence of the car-boxyterminal end of rat embryonic fibroblastTM-4. This sequence is also highly homologousto TM-1, smooth-muscle, and platelet tropo-myosins. These differences in sequence correlatewell with work from other laboratories showingthat skeletal-muscle troponin T binds stronglyto the carboxyterminal end of skeletal-muscletropomyosin. In contrast, troponin T interactspoorly with smooth-muscle and nonmuscletropomyosins. Work is currently under way toisolate and characterize cDNA clones encodingthe other nonmuscle tropomyosins.
A Single Rat Tropomyosin GeneSpecifies Two DifferentTissue-specific mRNAsD.M. Helfman. L. Finn
To isolate the gene encoding rat embryonic fibro-blast tropomyosin 1 (TM-1), we screened a ratgenomic library for sequences complementary toour cloned tropomyosin cDNAs. Analysis ofgenomic clones indicates there are two separateloci in the rat genome that contain sequencescomplementary to TM-1. One appears to containa functional gene and the other represents a pro-cessed pseudogene. The functional gene is ap-proximately 10 kb long. Northern blot analysis

using "P-labeled cDNA probes encoding TM-1revealed that rat embryonic fibroblasts andsmooth muscle (stomach or uterus) contain a sin-gle transcript of 1.1 kb. In contrast, skeletal mus-cle contains a single transcript of 1.3 kb. Si pro-tection experiments indicate that the skeletal-muscle transcript differs from both the fibroblastand smooth-muscle messages only at the 3' end(corresponding to a region from approximatelyamino acid 190 to 284); This is supported by se-quence comparisons demonstrating that skeletal-muscle I3- tropomyosin is virtually identical withrat TM-1 from amino acid 1 to 190, but differssubstantially at the carboxyterminal end. Wehave now identified two distinct exons at the 3'end of the gene; one is specific for the 1.1-kb tran-script in rat embryonic fibroblasts and smoothmuscle and the other is specific for the 1.3-kbtranscript in skeletal muscle. In addition, thereare at least two exons common to the 1.1-kb and1.3-kb transcripts. Thus, a single gene encodestwo different transcripts, whose expression is reg-ulated by a tissue-specific mechanism. We arecontinuing to characterize the genomic clonesfully and plan to study cis- and trans-acting ele-ments involved in the regulation of this gene.
PUBLICATIONS
Heitman, D.M., J.R. Feramisco, W.R. Ricci, and S.H. Hughes.1984. Isolation and sequence of a cDNA clone that con-tains the entire coding region for chicken smooth musclea-tropomyosin. J. Biol. Chem. 259: 14136-14143.
Helfman, D.M., Y. Yamawaki-Kataoka, W.M. Ricci, S.H.Hughes, and J.R. Feramisco. 1984. Isolation and char-acterization of cDNA clones encoding smooth-muscleand nonmuscle tropomyosins. In Molecular biology of thecytoskeleton (ed. G. Borisy et al.), pp. 479-491. ColdSpring Harbor Laboratory, Cold Spring Harbor, New York.
Lin, J.J.-C., D.M. Helfman, S.H. Hughes, and C.-S. Chou.1985. Tropomyosin isoforms in chicken embryo fibro-blasts. Purification, characterization and changes in Roussarcoma virus-transformed cells. J. Cell Biol.
100: 692-703.
In Press, Submitted, and In Preparation
Harlow, E., N.M. Williamson, R. Ralston, D.M. Helfman, andT.E. Adams. 1985. Molecular cloning and in vitro expres-sion of a cDNA clone for the human cellular tumor anti-gen p53. MO/. Cell. Biol. (in press).
Helfman, D.M., J.R. Feramisco, J.C. Fiddes, J.P. Thomas,and S.H. Hughes. 1985. Identification and isolation ofclones by immunological screening of cDNA expressionlibraries. In Genetic engineering (ed. J.K. Setlow and A.Hollaender), vol. 7. Plenum Press, New York. (In press.)
Yamawaki-Kataoka, Y. and D.M. Helfman. Rat embryonic fi-broblast tropomyosin 1: cDNA and complete primaryamino acid sequence. J. Biol. Chem. (Submitted.)
BIOLOGY OF THE CYTOSKELETON
F. Matsumura S. Yamashiro-Matsumura
Physical and Chemical Properties ofNormal- and Transformed-cellTropomyosinF. Matsumura, S. Yamashiro-Matsumura
We have previously shown that multiple forms oftropomyosin are differentially expressed uponmany types of oncogenic transformation (Matsu-mura et al., J. Biol. Chem. 258: 13954 [1983]; Linet al., Cancer Cells 1: 57 [1984]). Generally, thelevels of tropomyosin with higher apparent mo-lecular weights (prominent in normal cells) aredecreased in transformed cells, whereas the levelsof tropomyosin with lower apparent molecularweights (minor tropomyosins in normal cells) be-
come predominant. Of the five forms of rat cul-tured-cell tropomyosin (TM-1-TM-5) (Matsu-mura et al., J. Biol. Chem. 258: 6636 [1983]),TM-1 (M, = 40,000) and TM-2 (M, = 36,000) arehigh-molecular-weight tropomyosins (prominentin normal cells) and TM-4 (M, = 32,400) and TM-5 (M, = 32,000) are low-molecular-weight tro-pomyosins (prominent in transformed cells). Toelucidate how the changes in tropomyosin expres-sion affect the altered organization of microfila-ments observed in transformed cells, we have ex-amined the physical and chemical properties ofthese normal- and transformed-cell tropomyos-ins. We have also tested whether the changes intropomyosin pattern in microfilaments alter theinteractions between actin and other actin-bind-
131
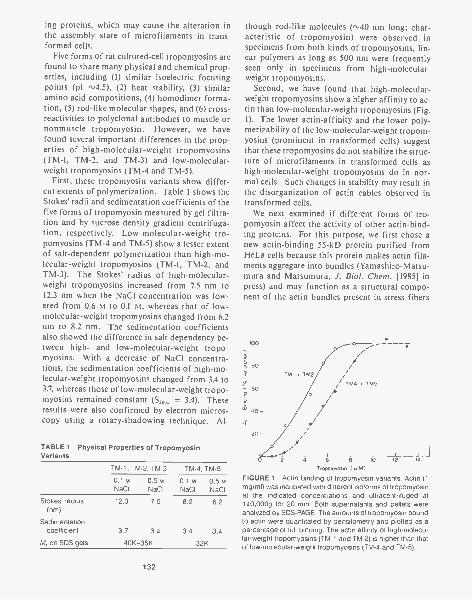
ing proteins, which may cause the alteration inthe assembly state of microfilaments in trans-formed cells.
Five forms of rat cultured-cell tropomyosins arefound to share many physical and chemical prop-erties, including (1) similar isoelectric focusingpoints (pI (2) heat stability, (3) similaramino acid compositions, (4) homodimer forma-tion, (5) rod-like molecular shapes, and (6) cross-reactivities to polyclonal antibodies to muscle ornonmuscle tropomyosin. However, we havefound several important differences in the prop-erties of high-molecular-weight tropomyosins(TM-1, TM-2, and TM-3) and low-molecular-weight tropomyosins (TM-4 and TM-5).
First, these tropomyosin variants show differ-ent extents of polymerization. Table 1 shows theStokes' radii and sedimentation coefficients of thefive forms of tropomyosin measured by gel filtra-tion and by sucrose density gradient centrifuga-tion, respectively. Low-molecular-weight tro-pomyosins (TM-4 and TM-5) show a lesser extentof salt-dependent polymerization than high-mo-lecular-weight tropomyosins (TM-1, TM-2, andTM-3). The Stokes' radius of high-molecular-weight tropomyosins increased from 7.5 nm to12.3 nm when the NaC1 concentration was low-ered from 0.6 to 0.1 rvi, whereas that of low-molecular-weight tropomyosins changed from 6.2nm to 8.2 nm. The sedimentation coefficientsalso showed the difference in salt dependency be-tween high- and low-molecular-weight tropo-myosins. With a decrease of NaCI concentra-tions, the sedimentation Coefficients of high-mo-lecular-weight tropomyosins changed from 3.4 to3.7, whereas those of low-molecular-weight tropo-myosins remained constant = 3.4). Theseresults were also confirmed by electron micros-copy using a rotary-shadowing technique. Al-
TABLE 1 Physical Properties of TropomyosinVariants
TM-1, TM-2, TM-3 TM-4, TM-5
Stokes' radius(nm)
Sedimentationcoefficient
M, on SDS gels
0.1 M 0.5 M 0.1 M 0.5 MNaCI NaCI NaCI NaCI
12.3 7.5 8.2 6.2
3.7 3.4 3.4 3.4
40K-35K 32K
132
though rod-like molecules (,,40 nm long; char-acteristic of tropomyosin) were observed inspecimens from both kinds of tropomyosins, lin-ear polymers as long as 500 nm were frequentlyseen only in specimens from high-molecular-weight tropomyosins.
Second, we have found that high-molecular-weight tropomyosins show a higher affinity to ac-tin than low-molecular-weight tropomyosins (Fig.1). The lower actin -affinity and the lower poly-merizability of the low-molecular-weight tropom-yosins (prominent in transformed cells) suggestthat these tropomyosins do not stabilize the struc-ture of microfilaments in transformed cells ashigh-molecular-weight tropomyosins do in nor-mal cells. Such changes in stability may result inthe disorganization of actin cables observed intransformed cells.
We next examined if different forms of tro-pomyosin affect the activity of other actin-bind-ing proteins. For this purpose, we first chose anew actin-binding 55-kD protein purified fromHe La cells because this protein makes actin fila-ments aggregate into bundles (Yamashiro-Matsu-mura and Matsumura, J. Biol. Chem. [1985] inpress) and may function as a structural compo-nent of the actin bundles present in stress fibers
100-
80--
- 60
'c 40 -
20
TM' + TM2
TM4 + TM5
4 6 8 It 12
Tropomyosin ( p.M)
FIGURE 1 Actin binding of tropomyosin variants. Actin (1mg/ml) was incubated with different isoforms of tropomyosinat the indicated concentrations and ultracentrifuged at140,000g for 20 min. Both supernatants and pellets wereanalyzed by SDS-PAGE. The amounts of tropomyosin boundto actin were quantitated by densitometry and plotted as apercentage of full binding. The actin affinity of high-molecu-lar-weight tropomyosins (TM-1 and TM-2) is higher than thatof low-molecular-weight tropomyosins (TM-4 and TM-5).

or philopodia. The bundling activity of the 55-kD protein was measured by low-speed centrifu-gation (Fig. 2, lane 1), where bundles of actin fil-aments are pelleted (lane lb) while free actin re-mains in the supernatant (Fig. 2, lane la). Actinwas first complexed with either high- or low-mo-lecular-weight tropomyosins and then the 55-kDprotein was added. When the 55-kD protein wasmixed with the actin/TM-4/TM-5 complex, itchased out TM-4 and TM-5 from actin and madebundles of actin filaments free of tropomyosin(Fig. 2, lane 2). In contrast, when the 55-kD pro-tein was mixed with the actin/TM-1/TM-2/TM-3complex, it made bundles of actin filaments con-taining these tropomyosins (Fig. 2, lane 3). Wehave further examined the effect of skeletal-mus-cle tropomyosin on bundling activity. Interest-ingly, skeletal-muscle tropomyosin inhibited thebundling activity of the 55-kD protein. As lane 4
1 2 3 4 5ab ab ab ab
-"`" 55K
-1110 -Ow -111111rar-.Actin- ATM.40a -.TM
FIGURE 2 The interaction between the 55-kD protein anddifferent forms of tropomyosin. Actin (1 mg/ml) was firstcomplexed with different forms of tropomyosin (0.5 mg/ml)and then the 55-kD protein (0.2 mg/ml) was added. The ac-tivity of actin bundling was examined by low-speed centrif-ugation (12,000g for 15 min). Both pellets (lane a) and su-pernatants (lane b) were analyzed on SDS-polyacrylamidegels. (1) Control without tropomyosin (actin bundles wereprecipitated with the 55-kD protein [lane bp; (2) actin plusTM-4 and TM-5; (3) actin plus TM-3, TM-4, and TM-5; (4)actin plus TM-1 and TM -2; (5) actin plus rabbit skeletal-mus-cle tropomyosin. Note that TM-4 and TM-5 were chased outby the 55-kD protein, whereas TM-1, TM-2, and TM-3 bindto actin bundles made with the 55-kD protein. Skeletal-mus-cle tropomyosin, in contrast, inhibits the bundling activity of55-kD protein.
shows, most actin remains in the supernatant(lane 4a).
The modification of the interaction betweenactin and the 55-kD protein by the multiple formsof tropomyosin may be involved in the regulationof microfilament assembly. We are now studyingwhether multiple forms of tropomyosin changethe interaction of actin with other types of actin-binding protein (actin-filament-severing proteinsuch as gelsolin).
Intracellular Localization of a NewActin-bundling 55-kD Proteinin Cultured CellsS. Yamashiro-Matsumura, F. Matsumura
Last year, we reported the purification and char-acterization of a new actin-bundling 55-kD pro-tein from HeLa cells (Yamashiro-Matsumura andMatsumura, J. Biol. Chem. [1985] in press). Todetermine the intracellular localization of thisprotein in cultured cells, we have made both po-lyclonal and monoclonal antibodies to the 55-kDprotein. Indirect immunofluorescence using bothpolyclonal and monoclonal antibodies shows thepredominant localization of this protein in phil-opodia and retraction fibers, as well as in stressfibers (Fig. 3B).
The 55-kD protein is very similar to fimbrin inthat both proteins are actin-bundling proteins andshow similar localization in cultured cells. Al-though biochemical characterization suggests thatthey are different proteins, we have tested the 55-kD protein by immunoblot analysis to determineif it is immunologically related to fimbrin. Anti-55-kD protein antibody was reactive with the 55-kD protein but not with fimbrin. Likewise, anti-fimbrin antibody reacted with fimbrin but notwith the 55-kD protein. Furthermore, indirectimmunofluorescence of chemically transformedhuman fibroblasts double-stained with poly-clonal anti-fimbrin and with monoclonal anti-55-kD protein showed similar but different localiza-tion. Although both antibodies stained micro-spikes, the anti-55-kD protein antibody stainedthe stress fibers, but the anti-fimbrin staining wasvery weak. These results indicate that the 55-kDprotein and fimbrin are not related to each other
133
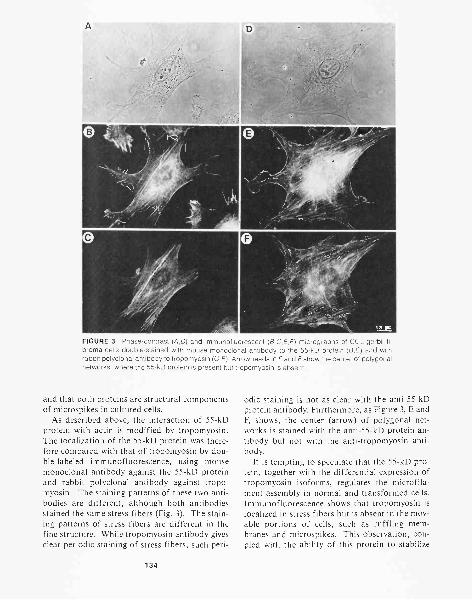
FIGURE 3 Phase-contrast (A,D) and immunofluorescent (B,C,E,F) micrographs of CCL-gerbil fi-broma cells double-stained with mouse monoclonal antibody to the 55-kD protein (B,E) and withrabbit polyclonal antibody to tropomyosin (C,F). Arrowheads in E and F show the center of polygonalnetworks, where the 55-kD protein is present but tropomyosin is absent.
and that both proteins are structural componentsof microspikes in cultured cells.
As described above, the interaction of 55-kDprotein with actin is modified by tropomyosin.The localization of the 55-kD protein was there-fore compared with that of tropomyosin by dou-ble-labeled immunofluorescence, using mousemonoclonal antibody against the 55-kD proteinand rabbit polyclonal antibody against tropo-myosin. The staining patterns of these two anti-bodies are different, although both antibodiesstained the same stress fibers (Fig. 3). The stain-ing patterns of stress fibers are different in thefine structure. While tropomyosin antibody givesclear periodic staining of stress fibers, such peri-
134
odic staining is not as clear with the anti-55-kDprotein antibody. Furthermore, as Figure 3, E andF, shows, the center (arrow) of polygonal net-works is stained with the anti-55-kD protein an-tibody but not with the anti-tropomyosin anti-body.
It is tempting to speculate that the 55-kD pro-tein, together with the differential expression oftropomyosin isoforms, regulates the microfila-ment assembly in normal and transformed cells.Immunofluorescence shows that tropomyosin islocalized in stress fibers but is absent in the mov-able portions of cells, such as ruffling mem-branes and microspikes. This observation, cou-pled with the ability of this protein to stabilize

actin filaments, suggests that tropomyosin mayregulate the organization of microfilaments intostress fibers. In normal cells with well-spreadmorphology, where high-molecular-weight tro-pomyosins are predominant, the 55-kD proteinprobably makes bundles of actin filaments con-taining high-molecular-weight tropomyosins.These tropomyosin-containing bundles may bestable enough to form stress fibers. In contrast,in transformed cells where high-molecular-weighttropomyosins are substituted by low-molecular-weight tropomyosins, the 55-kD protein chasesout the tropomyosin from actin and makes actinbundles without tropomyosin. Such tropomyo-sin-free bundles may not be stable enough to formstress fibers but may be used for the formation ofmicrospikes that are prominent in many trans-formed cells. We are currently examining this hy-pothesis through the combined use of biochemi-cal, cell biological, and immunological methods.
PUBLICATIONS
Lin, J.J.-C., F. Matsumura, and S. Yamashiro-Matsumura.1984. Tropomyosin-enriched and alpha-actinin-enrichedmicrofilaments isolated from chicken embryo fibroblastsby monoclonal antibodies. J. Cell Biol. 98: 116-127.
Lin, J.J.-C., S. Yamashiro-Matsumura, and F Matsumura.1984. Microfilaments in normal and transformed cells:Changes in the multiple forms of tropomyosin. CancerCells 1:57 -65.
Lin, J.J.-C., J.R. Feramisco, S.H. Blose, and F. Matsumura.1984. Monoclonal antibodies to cytoskeleton. In Mono-clonal antibodies and functional cell lines (ed. R.H. Ken-nett et al.), pp. 119-151. Plenum Press, New York.
Yamashiro-Matsumura, S. and F. Matsumura. 1985. Purifi-cation and characterization of actin-bundling 55K proteinfrom He La cells. J. Biol. Chem. 260: 5087-5097.
In Press, Submitted, and In Preparation
Garrels, J.I., S. Yamashiro-Matsumura, J.J.-C. Lin, and F.Matsumura. 1985. A set of actin-filament associated pro-teins characterized by quantitative two-dimensional gelelectrophoresis. In Gene expression in muscle. MuscularDystrophy Association. (In press.)
Matsumura, F. and S. Yamashiro-Matsumura. 1985. Molec-ular analysis of microfilament components in culturedcells: The role of tropomyosin in cell transformation. InCell motility 2. Mechanism and regulation (ed. S. Hatanoet al.), University of Tokyo Press. (In press.)
. Purification and characterization of multiple isoformsof tropomyosin from rat cultured cells. J. Biol. Chem.(Submitted.)
. Interactions of multiple isoforms of tropomyosin with55 kDa actin-bundling protein. (In preparation.)
Yamashiro-Matsumura, S. and F. Matsumura. Intracellular lo-calization of 55 kDa actin-bundling protein in culturedcells. (In preparation.)
BIOCHEMISTRY OF TRANSFORMING PROTEINS
J . R. Feramisco D. Bar-SagiC. Slaughter D.M. HellmanW.J. Welch T. KamataM. Zoller M. Wooten
G. Binns
T. FischerC. HallaranN. KronenbergS. Smith
Intrinsic GTPase Activity of the rasProteins: A Biochemical Analogyto the "G" Protein Family
T. Kamata, G. Binns, J.R. Feramisco [in collaboration withR.W. Sweet, S. Yokoyama, M. Rosenberg, and M. Gross,Smith, Kline and French Laboratories, Pennsylvania]
Ha-ras is a member of a multigene family in manthat encodes highly related proteins of 189 aminoacids (p21). In vitro, ras proteins bind GTP andp21 mutants with threonine at position 59 and
autophosphorylate at that residue. Mutation (atamino acids 12 or 61) and elevated expression ofras genes result in cell transformation in cultureand are also observed in many types of humantumors. Normal and mutant transforming ras
proteins show no differences in localization, lipi-dation, or GTP binding. However, mutations atposition 12 in recombinant (Thr-59) p21 mole-cules were observed to affect autophosphoryla-tion. We have expressed the full-length normaland T24 transforming (Gly --Val at position 12)Ha-ras proteins in Escherichia coli and have pu-
135

rified them to homogeneity; these proteins boundGTP with approximately molar stoichiometry andwith an affinity comparable to that of partiallypurified mammalian proteins. Microinjection ofthe T24 protein into quiescent rodent fibroblastsresulted in a rapid alteration in cell morphology,stimulation of DNA synthesis, and cell division;in contrast, little response was observed with thenormal protein. We now report that the normalras protein has an intrinsic GTPase activity, yield-ing GDP and P whereas the T24 transformingprotein is reduced tenfold in this activity (Fig. 1).We suggest that this deficiency in GTPase is theprobable cause for the transforming phenotype ofthe T24 protein.
The tenfold reduction in GTPase activity of theT24 protein represents the first major biochemi-cal differentiation between the products of a nor-mal and a transforming mutant ras gene. It istempting to speculate on the consequence of thisaltered activity by drawing a functional analogyto transducin and the G stimulatory protein -both of these proteins are activators of enzymesaffecting cyclic nucleotide pools. This activationrequires GTP binding (or exchange of GTP forbound GDP) and appears to terminate on hydro-lysis to GDP. If the ras proteins have an analo-gous regulatory function in whichever prolifera-tive control pathway they participate, then thedeficiency in GTPase activity of the T24 mutantcould result in prolonged stimulation of an oth-erwise regulatable activity. As shown for the Gstimulatory protein, perhaps modulation of rasactivity itself occurs through interactions with
other proteins, and these interactions may be sen-sitive to whether GTP and GDP (or neither) oc-cupies the nucleotide binding site.
Molecular Characterization of Ha-rasProteins: Proteolytic SubstructureD. Bar-Sagi, J.R. Feramisco
Oncogenic ras-encoded proteins differ from theirnormal homologs by an amino acid substitutionfor Gly-12, Ala-59, or Gln -61. To provide insightinto the structural requirements involved in theoncogenic activation of ras proteins, we have ini-tiated studies concerning the characterization oftheir structural and functional domains. Fromthis analysis, we hope to build a framework tostudy the function of ras proteins.
We have purified to homogeneity the protoon-cogenic and oncogenic (T24) forms of Ha-rasproteins produced in Escherichia coli. Both pro-teins were subjected to digestion by trypsin undermild conditions. Figure 2A displays the productsof limited tryptic digestion of the proteins atdifferent times during proteolysis. The resultsindicate a progressive disappearance of the intactHa-ras proteins (24 kD) and the appearance of23.5-kD and 21-kD polypeptides. The 23.5-kDpolypeptide is likely to be an intermediate in thehydrolysis of Ha-ras protein as its appearanceprecedes that of the 21-kD polypeptide. It thendeclines in amount, and by 2 hours it has vir-
h
Gaq
G-111 III III III
1 2 3 1 2 3 2 3 1 2 3
20 00
136
120
FIGURE 1 GTP hydrolysis catalyzed by the ras pro-teins. PEI-cellulose thin-layer analyses of incubations ofla-32PIGTP with the normal ras protein (lane 1), with theactivated oncogene ras protein (lane 2), or with BSA(lane 3) are shown, Notice the low GTPase activity ofthe oncogene protein.
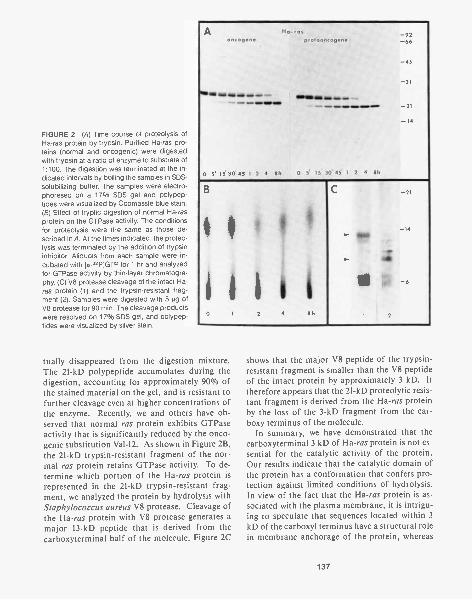
FIGURE 2 (A) Time course of proteolysis ofHa-ras protein by trypsin. Purified Ha-ras pro-teins (normal and oncogenic) were digestedwith trypsin at a ratio of enzyme to substrate of1:100. The digestion was terminated at the in-dicated intervals by boiling the samples in SDS-solubilizing buffer. The samples were electro-phoresed on a 17% SDS gel and polypep-tides were visualized by Coomassie blue stain.(8) Effect of tryptic digestion of normal Ha-rasprotein on the GTPase activity. The conditionsfor proteolysis were the same as those de-scribed in A. At the times indicated, the proteo-lysis was terminated by the addition of trypsininhibitor. Aliquots from each sample were in-cubated with [0,-32P]GTP for 1 hr and analyzedfor GTPase activity by thin-layer chromatogra-phy. (C) V8 protease cleavage of the intact Ha-ras protein (1) and the trypsin-resistant frag-ment (2). Samples were digested with 5µg ofV8 protease for 90 min. The cleavage productswere resolved on 17% SDS gel, and polypep-tides were visualized by silver stain.
Aoncogene
Ho-, osprotooncogene
-92-68
-45
-31
411. 0/1111 III 11 MS-21
-14
0 5 15 30 5' 1 2 4 8h 0 5 15 30 45 1 2 4 8h
B -21
-14
2 4 8h 2
tually disappeared from the digestion mixture.The 21-kD polypeptide accumulates during thedigestion, accounting for approximately 90% ofthe stained material on the gel, and is resistant tofurther cleavage even at higher concentrations ofthe enzyme. Recently, we and others have ob-served that normal ras protein exhibits GTPaseactivity that is significantly reduced by the onco-genic substitution Val-12. As shown in Figure 2B,the 21-kD trypsin-resistant fragment of the nor-mal ras protein retains GTPase activity. To de-termine which portion of the Ha-ras protein isrepresented in the 21-kD trypsin-resistant frag-ment, we analyzed the protein by hydrolysis withStaphylococcus aureus V8 protease. Cleavage ofthe Ha-ras protein with V8 protease generates amajor 13-kD peptide that is derived from thecarboxyterminal half of the molecule. Figure 2C
shows that the major V8 peptide of the trypsin-resistant fragment is smaller than the V8 peptideof the intact protein by approximately 3 kD. Ittherefore appears that the 21-kD proteolytic resis-tant fragment is derived from the Ha-ras proteinby the loss of the 3-kD fragment from the car-boxy terminus of the molecule.
In summary, we have demonstrated that thecarboxyterminal 3 kD of Ha-ras protein is not es-sential for the catalytic activity of the protein.Our results indicate that the catalytic domain ofthe protein has a conformation that confers pro-tection against limited conditions of hydrolysis.In view of the fact that the Ha-ras protein is as-sociated with the plasma membrane, it is intrigu-ing to speculate that sequences located within 3kD of the carboxyl terminus have a structural rolein membrane anchorage of the protein, whereas
137

the GTPase activity is carried by a portion of theamino terminus.
Microinjection of the ras OncogeneProtein into Quiescent CellsResults in Rapid ProliferationJ.R. Feramisco, N. Kronenberg, C. Hallaran, G. Binns,T. Kamata [in collaboration with M. Gross,M. Rosenberg, and R.W. Sweet, Smith, Klineand French Laboratories, Pennsylvania]
Although the function of the ras proteins is notyet known, several observations concerning theirpossible role in cell proliferation have been de-scribed. Epidermal growth factor or insulin stim-ulates the GTP-dependent phosphorylation of themurine v-ras protein and the guanine-nucleotide-'binding activity of the ras proteins in ras-trans-formed cells or membranes derived thereof. Asecond observation is that (with only limited in-formation available) there is some structuralhomology between the ras proteins and the Gprotein, which is known to regulate the hor-mone-sensitive adenylate cyclase activity. Theseobservations suggest that the ras proteins may beinvolved in growth-factor receptor activity, be itat the level of the receptor(s) itself or in the sig-nal transduction that may follow receptor acti-vation with a mechanism that may employ a G-like activity, and/or may be directly regulating theadenylate cyclase system as deduced by M. Wig lerand his colleagues (Molecular Genetics of Eukar-yotic Cells Section).
To study the biochemical and biological activi-ties of the human protooncogenic and oncogenicforms of the Ha-ras proteins, we have begun toexamine the effects of the purified Ha-ras pro-teins in living, normal cells by needle microinjec-tion. Here, we show that the oncogenic form ofthe human Ha-ras protein (T24) produced in andpurified from Escherichia coli causes rapid mor-phological changes and induces cell division wheninjected into quiescent, contact-inhibited cells(see Fig. 3). In certain established cell lines, theprotein completely obviates the need for growthfactors in the proliferation process. In contrast,microinjection of the wild-type protooncogenicform of Ha-ras exhibits little, if any, effect onthese cells.
138
Cooperation of the ras OncogeneProtein with the Adenovirus El AOncogene Protein followingMicroinjection into QuiescentNonestablished CellsJ.R. Feramisco, D. Bar-Sagi, W.J. Welch, T. Kamata[in collaboration with N.F. Sullivan, R.W. Sweet,T.L. Chao, B.Q. Ferguson, M.S. Gross, andM. Rosenberg, Smith, Kline and FrenchLaboratories, Pennsylvania]
We have shown that early-passage nonestablishedrat embryo fibroblasts respond to microinjectionof Escherichia coli-made ras oncogene protein bysynthesizing DNA but not undergoing division.This effect is specific to ras oncogene proteinsince no response is observed upon injection ofcomparable levels of the ras protooncogene pro-tein. In contrast, microinjection of ras oncogeneprotein into late-passage REF cells caused bothDNA synthesis and cell division. Early-passageREF cells could be stimulated to divide in re-sponse to oncogenic ras by either preincubationin conditioned medium from late-passage cells orcoinjection experiments using the adenovirus E IA13S mRNA gene product in combination with rasoncogene protein. The localizationproteins in single cells after microinjection isshown in Figure 4.
Of the two known growth-factor pathways, i.e.,competence as induced by platelet-derived growthfactor (PDGF) and progression as induced by ep-idermal growth factor (EGF), insulin, and soma-tomadin C, only the progression pathway can beblocked by cycloheximide. Since cycloheximideblocks DNA synthesis in response to microin-jected oncogenic ras, we believe that the ras pro-tein must be involved in the progression pathwayof cell growth. In addition, ras must act beforethe proposed restriction point between G, and S,since after this point DNA synthesis cannot beblocked by cycloheximide. Furthermore, sinceoncogenic ras stimulates DNA synthesis with onlytrace amounts of serum, it can apparently bypassthe need for exogenous competence and progres-sion factors. Previous studies have indicated thatEGF and insulin treatment of cell membranesleads to effects on the guanine-nucleotide-bind-ing activity of the ras protein, observations thattend to support some role for ras in these growth-factor pathways.
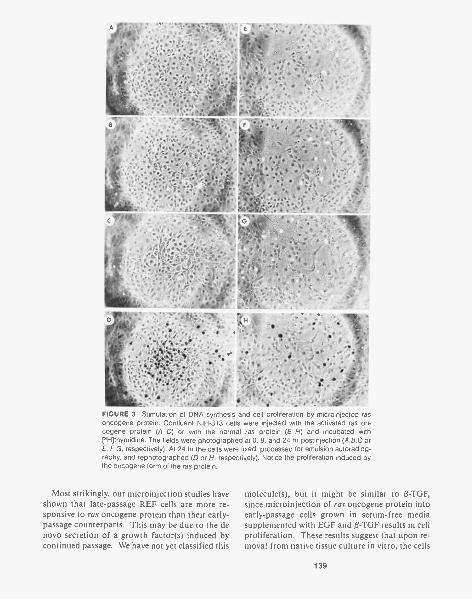
- '
FIGURE 3 Stimulation of DNA synthesis and cell proliferation by microinjected rasoncogene protein. Confluent NIH-3T3 cells were injected with the activated ras on-cogene protein (A-D) or with the normal ras protein (E-H) and incubated with[3H]thymidine. The fields were photographed at 0, 8, and 24 hr postinjection (A,B,C or
F,G, respectively). At 24 hr the cells were fixed, processed for emulsion autoradiog-raphy, and rephotographed (D or H, respectively). Notice the proliferation induced bythe oncogene form of the ras protein.
Most strikingly, our microinjection studies haveshown that late-passage REF cells are more re-sponsive to ras oncogene protein than their early-passage counterparts. This may be due to the denovo secretion of a growth factor(s) induced bycontinued passage. We have not yet classified this
molecule(s), but it might be similar to 0-TGF,since microinjection of ras oncogene protein intoearly-passage cells grown in serum-free mediasupplemented with EGF and 13-TGF results in cellproliferation. These results suggest that upon re-moval from native tissue culture in vitro, the cells
139
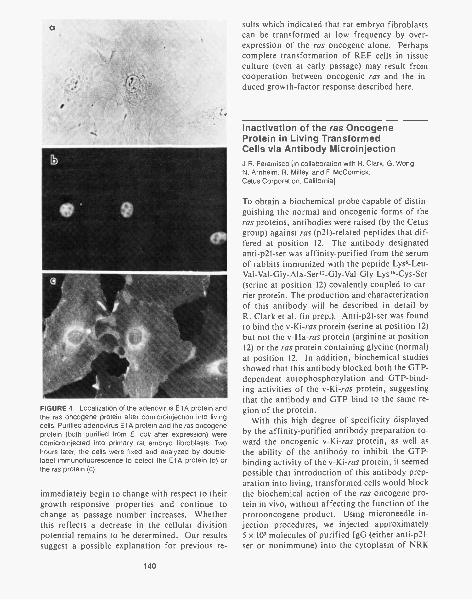
a
I
FIGURE 4 Localization of the adenovirus E1A protein andthe ras oncogene protein after comicroinjection into livingcells. Purified adenovirus El A protein and the ras oncogeneprotein (both purified from E. coil after expression) werecomicroinjected into primary rat embryo fibroblasts. Twohours later, the cells were fixed and analyzed by double-label immunofluorescence to detect the E1A protein (b) orthe ras protein (c).
immediately begin to change with respect to theirgrowth-responsive properties and continue tochange as passage number increases. Whetherthis reflects a decrease in the cellular divisionpotential remains to be determined. Our resultssuggest a possible explanation for previous re-
140
sults which indicated that rat embryo fibroblastscan be transformed at low frequency by over-expression of the ras oncogene alone. Perhapscomplete transformation of REF cells in tissueculture (even at early passage) may result fromcooperation between oncogenic ras and the in-duced growth-factor response described here.
Inactivation of the ras OncogeneProtein in Living TransformedCells via Antibody MicroinjectionJ.R. Feramisco [in collaboration with R. Clark, G. Wong,N. Arnheim, R. Mil ley, and F. McCormick,Cetus Corporation, California]
To obtain a biochemical probe capable of distin-guishing the normal and oncogenic forms of theras proteins, antibodies were raised (by the Cetusgroup) against ras (p21)-related peptides that dif-fered at position 12. The antibody designatedanti-p21-ser was affinity-purified from the serumof rabbits immunized with the peptide Lys6-Leu-Val-Val-Gly-Ala-Ser12-Gly-Val -Gly-Lys I6-Cys-Ser(serine at position 12) covalently coupled to car-rier protein. The production and characterizationof this antibody will be described in detail byR. Clark et al. (in prep.). Anti-p21-ser was foundto bind the v-Ki-ras protein (serine at position 12)but not the v-Ha-ras protein (arginine at position12) or the ras protein containing glycine (normal)at position 12. In addition, biochemical studiesshowed that this antibody blocked both the GTP-dependent autophosphorylation and GTP-bind-ing activities of the v-Ki-ras protein, suggestingthat the antibody and GTP bind to the same re-gion of the protein.
With this high degree of specificity displayedby the affinity-purified antibody preparation to-ward the oncogenic v-Ki-ras protein, as well asthe ability of the antibody to inhibit the GTP-binding activity of the v-Ki-ras protein, it seemedpossible that introduction of this antibody prep-aration into living, transformed cells would blockthe biochemical action of the ras oncogene pro-tein in vivo, without affecting the function of theprotooncogene product. Using microneedle in-jection procedures, we injected approximately5 x 105 molecules of purified IgG (either anti-p21-ser or nonimmune) into the cytoplasm of NRK

cells transformed by the v-Ki-ras oncogene (calledKi-NRK cells). This represents a five- to tenfoldexcess of antibody over the estimated amount ofthe ras protein in these cells. Within 15 hours,and persisting for 48 hours after injection of theanti-p21-ser antibodies, the Ki-NRK cells showeda flattened, normal morphology, and after 48hours, the cells eventually returned to theirrounded morphology. In contrast, cells on thesame dish injected with control IgG showed noobvious change in morphology. (Control Ig Gsincluded goat anti-mouse IgG and rabbit nonim-mune IgG.) Inspection of the photographs takenfrom these types of experiments indicated that inaddition to the reversion of the transformed phe-notype, the flattened cells appeared to grow at aslower rate than cells injected with control IgG orin surrounding uninjected cells. Considering thedata from the genetic analysis of the yeast rasgenes which indicated that complete elimination
of ras gene expression leads to nonviable cells (seeM. Wig ler et al., Molecular Genetics of Eukar-yotic Cells Section), we might predict that anti-bodies that block the function of both the normaland oncogenic forms of the ras proteins wouldkill or prevent growth of microinjected, trans-formed mammalian cells. However, the cells in-jected with the anti-p21-ser preparation are capa-ble of growth, a result that is in keeping with thebiochemical experiments which showed that thisantibody only binds to the oncogenic form of theras protein with serine at position 12.
By utilizing immunofluorescence microscopy,we determined exactly which cells were inocu-lated with the two antibody preparations. An ex-act correlation between the cells inoculated withthe anti-p21-ser preparation and the morphologi-cally normal, flattened cells was seen (Fig. 5).Again, cells inoculated with the control IgG re-tained their rounded, transformed appearance.
(.1.4FIGURE 5 Injection of antibodies specific for the ras oncogene protein causes reversion of the transformed phenotype ofras-transformed cells. Ki-ras-transformed NRK cells were injected either with nonimmune IgG (A,B) or with IgG specific for theKi-ras oncogene protein (C,D). Twenty-four hours later, the cells were fixed and analyzed by immunofluorescence to detectthe injected IgGs. Notice the flat, normal appearance of the cells injected with the anti-ras IgG.
141

Protein Kinase CK. Matlack, T Kamata, M. Wooten, D. Bar-Sagi,D.M. Helfman, S. Smith, J.R. Feramisco
As our interest in the molecular mechanisms ofgrowth control widens, we have undertaken stud-ies to examine the role of phospholipids and pro-tein kinase C in the cell. A wide variety of exper-iments have suggested regulatory roles for proteinkinase C, phospholipids, and their breakdownproducts in growth-factor action and thus havecaptured our attention. Here and in the follow-ing report, we describe the purification, charac-terization, and cloning of protein kinase C.
One of the major problems one faces in work-ing with protein kinase C is a lack of purified,stable enzyme. To overcome this problem, wedesigned a protocol applicable to many tissuesand sources that enabled us to generate relativelylarge quantities of purified protein kinase C.The principal feature of the purification proce-dure is an affinity column utilizing protamine at-tached to a solid support. Protamine is the oneknown substrate that is utilized by protein kin-ase C in the absence of Ca' or phospholipids.Thus, protein kinase C binds tightly to the affin-ity support in the absence of any added activatorsof the enzyme. It is eluted from the column whenMgATP is added, which most likely reflects alower affinity of the enzyme to protamine afterphosphorylation (i.e., product release). By com-bining this step with several steps taken from theoriginal, lengthy procedure of Y. Nishizuka (KobeUniversity, Okazaki, Japan), we can easily purifyrelatively large quantities (,,500 jug) in less than 1week (Fig. 6).
Isolation of cDNA Clones EncodingProtein Kinase CD.M. Helfman, T. Kamata, M. Wooten, S. Smith, M. Zoller,C. Slaughter, T. Fischer, J.R. Feramisco [in collaborationwith J.F. Kuo, P Girard, and I.C. Gonsalus, Department ofPharmacology, Emory University School of Medicine]
To isolate cDNA clones encoding protein kinaseC, we constructed a cDNA expression library of
142
FIGURE 6 Purified protein kinase C. Approxirnately 5µg ofrat brain protein kinase C was applied to an SDS-polyacryl-amide gel; the resultant gel was stained with Coomassie blue.
approximately 80,000 members from adult ratbrain using the plasmid expression vectors pUC8and pUC9. Using antisera for brain protein ki-nase C, we have identified and isolated six bacte-rial colonies producing protein products antigen-ically related to protein kinase C. The plasmidswere isolated, and the cDNA inserts ranged in sizefrom 0.6 kb to 2.7 kb. The fusion proteins pro-duced in bacteria have a molecular weight rang-ing from 17,000 to 80,000. The size of the cDNAinserts correlated with the size of the fusion pro-teins. Three independent colonies were found toproduce a fusion protein with a molecular weightof about 80,000, approximately the size of au-thentic protein kinase C. Comparison by peptidemapping of the 80,000-molecular-weight bacte-rial fusion proteins with authentic protein kinaseC isolated from rat brain indicated that the twoproteins were indeed related. In addition, thecDNA clones encoding the large fusion proteinwere found to cross-hybridize to a synthetic oli-gonucleotide probe prepared from direct aminoacid sequence analysis of purified brain proteinkinase C. We are continuing to characterize theseputative cDNA clones encoding protein kinase C.

PUBLICATIONS
Blose, S.H., D.I. Meltzer, and J.R. Feramisco. 1984. 10-nmfilaments are induced to collapse in living cells microin-jected with monoclonal and polyclonal antibodies againsttubulin. J. Cell Biol. 98: 847-858.
Feramisco, J.R., M. Gross, T. Kamata, M. Rosenberg, andR.W. Sweet. 1984. Microinjection of the oncogene formof the human H-ras (T-24) protein results in rapid prolif-eration of quiescent cells. Cell 38: 109-117.
Hellman, D.M., J.R. Feramisco, W. Ricci, and S.H. Hughes.1984. Isolation and sequence of a cDNA clone that con-tains the entire coding region for chicken smooth musclea-tropomyosin. J. Biol. Chem. 259: 14136-14143.
Kamata, T. and J.R. Feramisco. 1984. Is the ras oncogeneprotein a component of the epidermal growth factor re-ceptor system? Cancer Cells 1: 11-16.
. 1984. Epidermal growth factor stimulates guaninenucleotide binding activity and phosphorylation of rasoncogene proteins. Nature 310: 147-150.
Lin, J.J.C., J.R. Feramisco, S.H. Blose, and F. Matsumura.1984. Monoclonal antibodies to cytoskeletal proteins. InMonoclonal antibodies and functional cell lines (ed. R.H.Kennett et al.), pp. 119-151. Plenum Press, New York.
Sullivan, N., W. Welch, R. Watt, D. Bar-Sagi, B. Ferguson, T.Chao, T. Kamata, R. Sweet, M. Gross, M. Rosenberg, andJ. Feramisco. 1985. Microinjection of oncogene proteinscauses DNA synthesis and proliferation in nonestablished cells. Cancer Cells 3: 243-249.
Sweet, R.W., S. Yokoyama, T. Kamata, J.R. Feramisco, M.Rosenberg, and M. Gross. 1984. The product of ras is a
MAMMALIAN STRESS RESPONSE
W.J. Welch G. Binns N. KronenbergJ.R. Feramisco C. Hallaran J.P. Suhan
Cell Biology of the MammalianStress Response
W.J. Welch, J.R. Feramisco, G. Binns,N. Kronenberg, C. Hallaran
Mammalian cells, when confronted with adverseenvironmental conditions, undergo what is re-ferred to as "stress response." This universal re-sponse is characterized by the rapid, increasedtranscription and translation of a small numberof genes, resulting in the increased synthesis andaccumulation of a group of proteins, the stress
GTPase and the T24 oncogenic mutant is deficient in thisactivity. Nature 311: 273-275.
Welch, W.J. and J.R. Feramisco. 1984. Nuclear and nudes-lar localization of the 72,000-dalton heat shock protein inheat-shocked mammalian cells. J. Biol. Chem. 259:4501-4513.
In Press, Submitted, and In Preparation
Catelli, M., N. Binart, I. Jung-Testas, E. Baulieu, J.R. Feram-isco, and W.J. Welch. The 90kDa stress protein is a com-ponent of the avian progesterone 8S receptor complex.(Submitted.)
Feramisco, J.R. and W.J. Welch. 1985. Modulation of cellu-lar activities via microinjection. In Microinjection and or-ganelle transplantation techniques: Methods and appli-cations (ed. J.E. Celis at al.). Academic Press, London.(In press.)
Feramisco, J.R., R. Clark, G. Wong, N. Arnheim, R. Milley,and F. McCormick. 1985. Microinjection of antibodiesspecific for amino acid 12 of the ras oncogene proteininto ras transformed cells causes a transient reversion ofthe transformed phenotype. Nature (in press).
Welch, W.J. and J.R. Feramisco. 1985. Rapid purification ofthe mammalian 70,000 dalton stress proteins: Affinity ofthe proteins for nucleotides. Mol. Cell. Biol. (in press).
. 1985. Disruption of the three cytoskeletal networksin mammalian cells does not affect transcription, transla-tion or protein translocation changes induced by heatshock. Mo/. Cell. Biol. (in press).
proteins. Although the function of any one ofthe individual stress proteins is unknown, collec-tively, these proteins appear to serve some pro-tective capacity both during and after recoveryof a particular environmental insult. We havecontinued to study and define many of the basicchanges occurring in the stressed cell. In addi-tion, using both biochemical and immunologicalapproaches, we have paid considerable attentionto the structure, properties, and intracellular lo-cation of the individual stress proteins with theeventual aim to determine the function these pro-teins serve in the cell.
143

Morphological StudiesW.J. Welch, J.P. Suhan
To get a better feeling of the many events occur-ring in the stressed cell, we have been examiningbasic morphological changes that accompany in-duction of the stress response, using both lightand electron microscopy. We have characterizeda number of interesting alterations occurring inboth the cytoplasm and nucleus of cells followingheat-shock treatment. For example, we have ob-served that incubation of rat fibroblasts at 42°Cfor 3 hours results in a fragmentation and vesi-cularization of intracellular membranous organ-elles, including the endoplasmic reticulum andGolgi complex. The disruption in these organ-elles is of interest since two of the mammalianstress proteins, the 80-kD and 100-kD proteins,are associated with the endoplasmic reticulumand Golgi complex, respectively (Welch et al., J.Biol. Chem. 258: 7102 [1983]). We have also ob-served changes that occur in the mitochondria ofcells following heat-shock treatment, includinga general swelling of the mitochondria, an in-creased electron density of the individual cristae,and an accompanying increase in the intramem-brane space between individual cristae. Thesesubtle changes in the structure of the mitochon-dria may turn out to be significant, since it hasbeen suggested that changes in mitochondrialfunction per se may be involved in the trigger-ing of the stress response. We are continuing toinvestigate this possibility by analyzing changesin mitochondrial activity using various biochem-ical techniques. Numerous changes with respectto various cytoskeletal components in stressedmammalian cells have also been found, includingan increase in the number of actin-containingstress fibers that span the cytoplasm of thestressed cell, indicative of the growth-arrestedstate of the cells (Thomas et al., Cold SpringHarbor Symp. Quant. Biol. 46: 985 [1982]). Inaddition, there is a complete reorganization in thedistribution of the vimentin-containing interme-diate filaments following physiological stress. Thefilaments are observed to collapse around the nu-cleus, with large aggregated bundles of filamentsvery near the nuclear membrane. Removal of thestress agent and incubation of the cells under nor-
144
mal conditions result in a gradual restoration ofnormal intermediate filament distribution.
Considerable alterations also occur within thenucleus of the cells following heat-shock treat-ment. For example, we have observed unusual nu-clear inclusion bodies that are composed of bun-dles of parallel filaments. Using biochemical andimmunological techniques, we have determinedthat these structures consist of finely packed ac-tin-containing filaments (see Fig. 1). These bun-dles of actin filaments were never observed to ex-tend through the nuclear membrane into thecytoplasm. Suffice it to say, we as yet do not un-derstand the significance of these structures withrespect to the other changes occurring within thestressed cell. We have also found that dramaticchanges occur within the nucleoli of cells follow-ing physiological stress, including a decondensa-tion of the nucleoli and an accompanying swell-ing and/or unraveling of the nucleoli, a dimin-ishment in the number of granular components,and changes in the organization of the fibrillarreticulum. These nucleolar alterations may besignificant, since there is an accompanying inhi-bition of ribosomal RNA synthesis and ribo-somal assembly following physiological stress.Moreover, these nucleolar alterations are of con-siderable interest since two of the major mam-malian stress proteins, the 72-kD and 110-kD pro-teins, localize within the nucleolus in cells bothduring and after recovery from physiologicalstress (Welch and Suhan, J. Cell Biol. [1985] sub-mitted).
Purification, Characterization, andIntracellular Localization of theMajor Mammalian Stress ProteinW.J. Welch, J.R. Feramisco, G. Binns
Mammalian cells placed under stress synthesize(and accumulate) proteins with apparent molec-ular masses of 28, 72, 73, 80, 90, 100, and 110 kD.All of these proteins, with the exception of the 72-kD species, are synthesized at appreciable levelsin cells grown under "normal" conditions. Theirpresence in normal cells suggests that many of thestress proteins probably serve some function(s) in
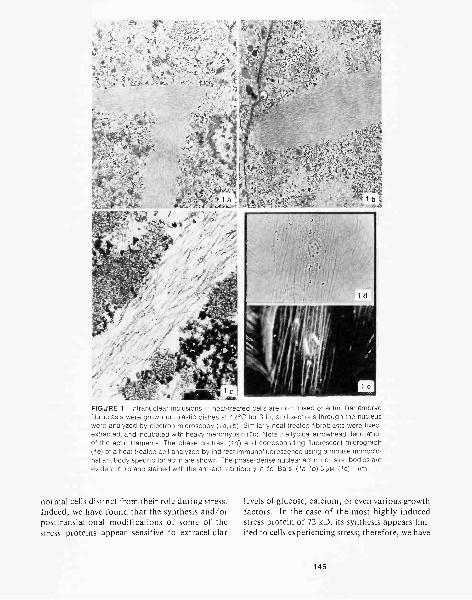
FIGURE 1 Intranuclear inclusions in heat-treated cells are composed of actin. Rat embryofibroblasts were grown on plastic dishes at 42°C for 3 hr, and sections through the nucleuswere analyzed by electron microscopy (1a,lb). Similarly heat-treated fibroblasts were fixed,extracted, and incubated with heavy meromyosin (/c). Note the typical arrowhead decorationof the actin filaments. The phase-contrast (1d) and corresponding fluorescent micrograph(/e) of a heat-treated cell analyzed by indirect immunofluorescence using a mouse monoclo-nal antibody specific for actin are shown. The phase-dense nuclear actin inclusion bodies areevident in ld and stained with the anti-actin antibody in le. Bars: (1a,lb) 5 pm; (1c) 1 pm.
normal cells distinct from their role during stress. levels of glucose, calcium, or even various growthIndeed, we have found that the synthesis and/or factors. In the case of the most highly inducedposttranslational modifications of some of the stress protein of 72 kD, its synthesis appears lim-stress proteins appear sensitive to extracellular ited to cells experiencing stress; therefore, we have
145

paid considerable attention to the properties ofthis protein (Welch and Feramisco, J. Biol. Chem.257: 14949 [1982]). We and others have foundthat the 72-kD protein appears to interact in vivowith nucleic acids, specifically RNA. In addition,we have recently observed that the 72-kD proteinand its partially related counterpart (73 kD) inter-act with various nucleotides in vitro. We havetaken advantage of their affinity for nucleotidesand have now developed a rapid two-step purifi-cation of the 72-kD and 73-kD proteins, utilizingaffinity chromatography on ATP agarose. In ad-dition, this purification procedure has allowed usto purify one of the other major mammalianstress proteins, the 80-kD protein, as well as anoninduced protein, the 75-kD protein, which in-terestingly appears to be related to both the 72-kD and 73-kD proteins (Welch and Feramisco,Mol. Cell. Biol. [1985] in press).
We have raised both polyclonal and monoclo-nal antibodies to several of the mammalian stressproteins, including the major-induced 72-kD spe-cies. Using these antibodies, we have shown thatthe 72-kD protein localizes to the nucleolus instressed cells and also during the recovery periodfollowing stress (Welch and Feramisco, J. Biol.Chem. 259: 4501 [1984]). More recently, we havefound a number of significant differences withrespect to the localization of the 72-kD protein incells recovering from stress after prior exposureto either heat shock, amino acid analogs, or so-dium arsenite (W.J. Welch, in prep.). We are con-tinuing to examine the nuclear, nucleolar, and cy-toplasmic distribution of the 72-kD protein andhave defined some of the basic parameters gov-erning its intracellular localization. First, we havefound that the nucleolar deposition of the 72-kDprotein is a stress-specific phenomenon. For ex-ample, microinjection of purified and fluores-cently labeled 72-kD protein into the cytoplasmof cells incubated at 37°C resulted in the proteinbeing distributed throughout the cytoplasm andthe nucleus, but not the nucleolus. Followingheat treatment of the cells, however, the fluores-cently labeled 72-kD protein migrated into thenucleolus (see Fig. 2). This migration appears tobe independent of the synthesis of the other stressproteins, since prior treatment with cyclohexa-mide and subsequent heat-shock treatment stillresulted in the 72-kD protein being deposited
146
within the nucleolus. We are currently assessingthe effects of transcriptional inhibitors on the in-tranucleolar distribution of the 72-kD protein andare determining whether its distribution is af-fected by RNase and/or DNase treatment of thecells. In addition, using immunoelectron micros-copy, we are very carefully examining where inthe nucleolus the 72-kD protein localizes. Specif-ically, is the protein located within the granularor fibrillar regions? If in the granular region, thiswould be indicative of the protein migrating tothe nucleolus to be assembled into some sort ofribonucleoprotein component. We find this pos-sibility most plausible considering that the pro-tein does show an affinity for RNA and that thenucleolus is the site of assembly of many differentribonucleoprotein complexes. Preliminary im-munoelectron microscopy studies, as well as bio-chemical studies analyzing stabilized UV cross-linked ribonucleoprotein complexes, are consis-tent with this idea.
Changes in the Small 28-kD StressProtein in Response to Mitogensand Growth FactorsW.J. Welch
We are paying considerable attention to the small28-kD stress protein of mammalian cells. Thisprotein, which contains no methionine, is com-posed of at least three highly related isoforms, twoof which are heavily phosphorylated. During in-duction and recovery from stress, synthesis of the28-kD protein increases approximately tenfold,yet there appears to be little change in the netphosphorylation of the protein. Interestingly, wehave found that the phosphorylation of this pro-tein, but not its synthesis, increases in quiescentcells following refeeding with fresh serum or af-ter exposure of the cells to either phorbol estersor calcium ionophores (Welch, J. Biol. Chem.260: 3058 [1985]). In addition, preliminary datain conjunction with J.R. Feramisco (this section)have shown an increased phosphorylation of the28-kD protein in cells induced to proliferate fol-lowing microinjection of the oncogenic form ofthe ras protein. Our excitement regarding these
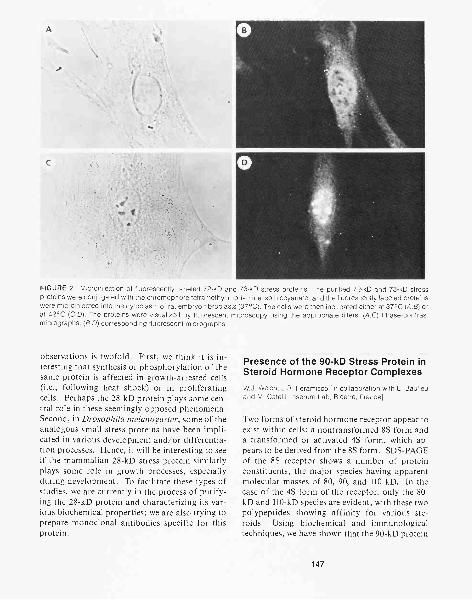
A
C
FIGURE 2 Microinjection of fluorescently labeled 72-kD and 73-kD stress proteins. The purified 72-kD and 73-kD stressproteins were conjugated with the chromophore tetramethyl rhodamine isothiocyanate, and the fluorescently labeled proteinswere microinjected into the cytoplasm of rat embryo fibroblasts (37°C). The cells were then incubated either at 37°C (A,B) orat 42°C (C,D). The proteins were visualized by fluorescent microscopy using the appropriate filters. (A,C) Phase-contrastmicrographs; (B,D) corresponding fluorescent micrographs.
observations is twofold. First, we think it is in-teresting that synthesis or phosphorylation of thesame protein is affected in growth-arrested cells(i.e., following heat shock) or in proliferatingcells. Perhaps the 28-kD protein plays some cen-tral role in these seemingly opposed phenomena.Second, in Drosophila melanogaster, some of theanalogous small stress proteins have been impli-cated in various development and/or differentia-tion processes. Hence, it will be interesting to seeif the mammalian 28-kD stress protein similarlyplays some role in growth processes, especiallyduring development. To facilitate these types ofstudies, we are currently in the process of purify-ing the 28-kD protein and characterizing its var-ious biochemical properties; we are also trying toprepare monoclonal antibodies specific for thisprotein.
Presence of the 90-kD Stress Protein inSteroid Hormone Receptor Complexes
W.J. Welch, J.R. Feramisco [in collaboration with E. Baulieuand M. Catelli, Inserum Lab, Bicetre, France]
Two forms of steroid hormone receptor appear toexist within cells: a nontransformed 8S form anda transformed or activated 4S form, which ap-pears to be derived from the 8S form. SDS-PAGEof the 8S receptor shows a number of proteinconstituents, the major species having apparentmolecular masses of 80, 90, and 110 kD. In thecase of the 4S form of the receptor, only the 80-kD and 110-kD species are evident, with these twopolypeptides showing affinity for various ste-roids. Using biochemical and immunologicaltechniques, we have shown that the 90-kD protein
147

component present in the 8S progesterone recep-tor complex is in fact the 90-kD stress protein(Cate lli et al., Nature [1985] submitted). It wouldappear that the 90-kD protein, which itself doesnot show an affinity for steroids, is lost from the8S complex following binding of the steroid andthe accompanying conversion to the 4S form ofthe receptor. Precedence for a transient associa-tion of the 90-kD stress protein with other ma-cromolecules has been demonstrated. For exam-ple, investigators in many laboratories have shownthat a number of retrovirus transforming pro-teins, all of which are tyrosine-specific kinases,are transiently associated with the 90-kD proteinfollowing their synthesis on free polysomes in thecytoplasm. This complex, containing the partic-ular tyrosine kinase, the 90-kD stress protein, andanother cellular protein of 50 kD, appears to beinvolved in the translocation of the oncogene pro-tein from its site of synthesis to its final destina-tion in the plasma membrane. Interestingly, whenassociated with the 90-kD and 50-kD proteins, theparticular tyrosine kinase shows diminished ac-tivity. Thus, we now have a second example inwhich the 90-kD stress protein displays a tran-sient association with other biologically impor-tant macromolecules, that being the steroid re-ceptor. We are currently determining whether the8S form of the steroid receptor also contains anassociated tyrosine kinase activity.
Production of Monoclonal Antibodiesto the Mammalian Stress ProteinsW.J. Welch, N. Kronenberg
To facilitate our studies on their structure, func-tion, and intracellular location, we are preparingmonoclonal antibodies to the various stress pro-teins. These efforts are now beginning to showsuccess. Currently, we have obtained at least 20different monoclonal antibodies to the major in-duced 72-kD stress protein. We are in the processof characterizing these antibodies with respect totheir antigenic site within the molecule. In addi-tion, we are in the process of making monoclonalantibodies specific to the 73-, 80-, and 90-kDstress proteins.
148
Comparison of the Stress Responsein Normal and Transformed CellsW.J. Welch, J.R. Feramisco
As we discussed in last year's report, transformedcells, in general, appear much more thermosensi-tive than do their nontransformed counterparts.However, the basis for this difference in thermalsensitivity is not at all clear. Athough synthesisof the stress proteins appears to occur in thetransformed cells placed under stress, there doappear to be some differences with respect to theintracellular location of at least the 72-kD and110-kD stress proteins. For example, whereasmost all of the normal rat fibroblasts show a nu-cleolar locale of 72 kD during and after recoveryfrom heat-shock treatment, only about 5% ofeither SV40- or adenovirus-transformed rat fi-broblasts show a nucleolar distribution of theprotein. Similar differences in the intranucleolardistribution of the 110-kD stress protein in nor-mal and transformed cells have also been ob-served. Interestingly, the few transformed cellsthat did show the 72-kD nucleolar staining are al-ways found in close association with one another,indicative perhaps of there being some correla-tion between the cell cycle and the ability of thecells to exhibit the nucleolar staining. Therefore,we are in the process of examining more carefullythe possible correlation between induction and/or proper localization of the stress proteins as afunction of the cell cycle in both normal andtransformed cells. In addition, we should pointout, with regard to cell cycle, that physiologicalstress itself seems to somehow arrest cells at aparticular point in the cell cycle (W.J. Welch andJ.R. Feramisco, in prep.).
We are also pursuing the possibility that basicdifferences in certain metabolic pathways of thetransformed cells may be responsible for their ob-served high thermosensitivity. For example, it hasbeen well established that transformed cells ap-pear more dependent on glycolytic rather thanrespiratory metabolism, and therefore the cellsutilize high levels of extracellular carbon sourcessuch as glucose. Due to their high glycolytic ac-tivity (and consequent lactic acid production), thetransformed cells tend to acidify their extracellu-lar medium. Hence, we have been asking whether
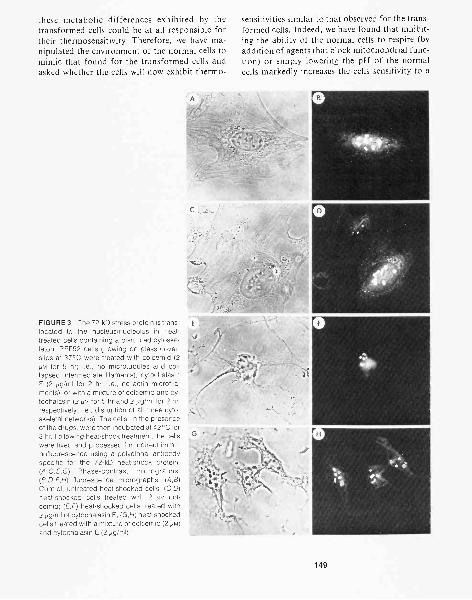
these metabolic differences exhibited by thetransformed cells could be at all responsible fortheir thermosensitivity. Therefore, we have ma-nipulated the environment of the normal cells tomimic that found for the transformed cells andasked whether the cells will now exhibit thermo-
FIGURE 3 The 72-kD stress protein is trans-located to the nucleus/nucleolus in heat-treated cells containing a disrupted cytoske-leton. REF52 cells growing on glass cover-slips at 37°C were treated with colcemid (2/1M for 5 hr; i.e., no microtubules and col-lapsed intermediate filaments), cytochalasinE (2 pg/m1 for 2 hr; i.e., no actin microfila-ments), or with a mixture of colcemid and cy-tochalasin (2 pm for 5 hr and 2 pg/m1 for 2 hr,respectively; i.e., disruption of all three cyto-skeletal networks). The cells, in the presenceof the drugs, were then incubated at 42°C for3 hr. Following heat-shock treatment, the cellswere fixed and processed for indirect immu-nofluorescence using a polyclonal antibodyspecific for the 72-kD heat-shock protein.(A,C,E,G) Phase-contrast micrographs;(B,D,F,H) fluorescence micrographs. (A,B)Control, untreated heat-shocked cells; (C,D)heat-shocked cells treated with 2 pm col-cemid; (E,F) heat-shocked cells treated with2 µg /ml of cytochalasin E; (G,H) heat-shockedcells treated with a mixture of colcemid (2 pilvI)and cytochalasin E (2 ',gimp.
sensitivities similar to that observed for the trans-formed cells. Indeed, we have found that inhibit-ing the ability of the normal cells to respire (byaddition of agents that block mitochondrial func-tion) or simply lowering the pH of the normalcells markedly increases the cells sensitivity to a
149

heat-shock challenge. We are continuing thesetypes of studies in hope of defining the basic pa-rameters responsible for the high thermal sensi-tivities exhibited by the transformed cells.
Use of the Heat-shock Response toStudy the CytoskeletonW.J. Welch, J.R. Feramisco
As mentioned above, physiological stress resultsin a collapse of the vimentin-containing inter-mediate filaments. We therefore asked whetherother agents that similarly result in collapse of theintermediate filaments would also result in an in-duction of the stress response. Others have shownthat treatment of the cells with various drugs thatdepolymerize microtubules (colcemid, colchicine,etc.) also cause a collapse of the intermediate fil-aments. However, we found that treatment of ratfibroblasts with these drugs did not result in aninduction of the stress response. Moreover, thereappeared to be little or no change in the overallpattern of protein synthesis in those cells treatedwith these cytoskeletal disrupting drugs. This wassomewhat surprising since investigators in otherlaboratories have suggested that translationalmachinery in eukaryotic cells functions when inassociation with cytoskeletal elements. Using theheat-shock response to induce de novo transcrip-tion and translation events, we asked whether cellscontaining a disrupted cytoskeleton could faith-fully make and translate new mRNAs even whenthe cells contain no integral and intact macro-cytoskeleton. For these experiments, cells weretreated with drugs that disrupt the actin-contain-ing microfilaments, vimentin-containing inter-mediate filaments, and microtubules; they wereplaced under heat shock and then labeled with ra-dioactive amino acids. Despite the absence of anintact cytoskeleton, the cells were still able to rec-ognize and respond to the heat treatment as as-sayed by the vigorous production of the stressproteins. In addition, the drug-treated cells, afterheat-shock treatment, were still able to translo-cate the 72-kD stress protein from the cytoplasminto the nucleus and nucleolus (see Fig. 3). These
150
results thus raise doubts with regard to previoussuggestions implicating a dependence of newtranslational events and the association of trans-lational machinery with the cytoskeleton. Theyalso demonstrate that some protein translocationevents, specifically, migration of proteins fromtheir site of synthesis in the cytoplasm into thenucleus, can occur in the absence of any integralcytoskeletal networks (Welch and Feramisco,Mol. Cell. Biol. [1985] in press).
PUBLICATIONS
Sullivan, N., W. Welch, R. Watt, D. Bar-Sagi, B. Ferguson, T.Chao, T. Kamata, R. Sweet, M. Gross, M. Rosenberg, andJ. Feramisco. 1985. Microinjection of oncogene proteinscauses DNA synthesis and proliferation in nonesta-blished cells. Cancer Cells 3: 243-249.
Welch, W.J. 1985. Phorbol ester, calcium ionophore, orserum added to quiescent rat embryo fibroblast cells allresult in the elevated phosphorylation of two 28,000 dal-ton mammalian stress proteins. J. Biol. Chem. 260:3058-3062.
Welch, W.J. and J.R. Feramisco. 1985. Nuclear and nucleo-lar localization of the 72,000-dalton heat shock protein inheat-shocked mammalian cells. J. Biol. Chem. 259:4501-4513.
In Press, Submitted, and In Preparation
Catelli, M., N. Binart, I. Jung-Testas, E. Baulieu, J.R. Fera-misco, and W.J. Welch. The 90kDa stress protein is acomponent of the avian progesterone 8S receptor com-plex. Nature (Submitted.)
Feramisco, J.R. and W.J. Welch. 1985. Modulation of cellu-lar activities via microinjection. In Microinjection and or-ganelle transplantation techniques: Methods and appli-cations (ed. J.E. Cells et al.). Academic Press, London.(In press.)
Welch, W.J. and J.R. Feramisco. 1985. Rapid purification ofthe mammalian 70,000 dalton stress proteins: Affinity ofthe proteins for nucleotides. Mo/. Cell. Biol. (in press).
. 1985. Disruption of the three cytoskeletal networksin mammalian cells does not affect transcription, transla-tion or protein translocation changes induced by heatshock. Mo/. Cell. Biol. (in press).
Welch, W.J. and J. Suhan. Morphological study of the mam-malian stress response: Alteration in the cytoskeleton,mitochondria, Golgi, and nucleoli in cells following heatshock. J. Ce// Biol. (Submitted.)
Welch, W.J., J.R. Feramisco, and S.H. Blose. 1985. Themammalian stress response and the cytoskeleton: Alter-ations in intermediate filaments. Ann. N.Y. Acad. Sci. (inpress).

QUEST TWO-DIMENSIONAL GEL LABORATORY
J.I. Garrels C. Chang P. SmithB.R. Franza, Jr. L. Cascio H. Wenzel
The Quest System for Two-dimensionalGel AnalysisJ.I. Garrels, B.R. Franza, Jr., P. Smith,L. Cascio, H. Wenzel
DEVELOPMENT OF PROTEIN DATABASES
During 1984, our program for development ofprotein databases derived from two-dimensionalgel analysis has turned from software develop-ment to problems of data management, distribu-tion, and presentation. In our laboratory, theproduction of standardized two-dimensional gelshas been routine, and the computer software forspot quantitation and matching has been highlyrefined. Although our computer system has beenmuch too small for the size of the programs re-quired and for the amount of data to be ana-lyzed, the hardware limitations will be overcomein 1985 (see below). Even with the relatively smalldatabase that we now have, it has been clear thatthe next major area of technical developmentmust be in the handling of the databases them-selves. Here, we present several forms of analysesof our rat fibroblast protein database, illustratingsome of the ways we have found to present largeamounts of two-dimensional gel data reduced toan easily understood form. In addition, we dis-cuss the hierarchical strategy we have developedfor the management of much larger databases.
Most of the data in our rat fibroblast databasecome from an intensive study of the REF52 cellline and its transformed derivatives. The lines aredescribed in our previous reports and in the fol-lowing report. A number of experiments on theselines have been analyzed and some of the data arepresented in Figure 1. This example points outthe magnitude of the data we must manage. Theoutput, of which only a small portion is shown,presents the quantitative changes for more than900 proteins in 16 different experiments. Eachsmall graph shows how the relative incorporationinto a given protein changed throughout thecourse of an experiment. We find that the graphs,each scaled to put the highest point at full scale,
are easily scanned visually for interesting regula-tory patterns. The large array of graphs, with adifferent spot on each row and a different exper-iment on each column, can be quickly scanned ineither dimension for changes of interest. Al-though the amount of data presented in this wayis still large, it is vastly reduced from the originalimage data represented on hundreds of films.The fact that data in this form is usable and in-teresting can be seen by examination of the graphsfor any particular spot.
The protein PCNA (proliferating cell nuclearantigen), spot 388, has been studied previously inour system. We find that its rate of synthesis de-creases at confluence in normal cells (graph incolumn 1) but that its rate of synthesis does notdecrease in proportion to density in transformedcells (next nine columns). The last two columnsshow the relative rates of synthesis of PCNA innormal and in nine transformed cells duringgrowth phase on day 3 (column 15) or day 4 (col-umn 16) after plating. These two graphs show al-most identical patterns because little change inrates of protein synthesis occurs from one day tothe next during the proliferative phase. The otherexperiments presented here are described in Fig-ure 1.
When viewed on the graphics screen, the usercan point to graphs of interest and have the com-puter display the appropriate regions of the gelimages. Having found a protein of particular in-terest, the user can ask for more informationabout the protein, or having found an interestingpattern of change, the user can ask the computerto highlight other proteins that change in similarways. The reader can see that other proteins inFigure 1 show interesting regulatory patterns, in-cluding the tropomyosins (spots 448, 350, 398,328, 320, 420 for tropomyosins 1 through 6, re-spectively).
In another look at the data from REF52 andthe nine viral transformants, we asked the com-puter to count the number of proteins that re-spond to fresh serum (after being deprived ofserum) in normal REF52 cells and how many of
151

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16309rnmmmEzEiE0Effiffnein318 EEIEIEEEEEEI0©nIfIE1171732017=171171=7=7117PIMM328 EIMEIEnEMEEMEMENN350 ©=I© NEIEIE111E1IFIDE370 El Ellfl EIM 17= 71388 0EE1=17107M1170171a121398 [0air70ni IE02MURI400 EIZIgM1711711171 ILUMILIEPE1409ununma0rm Di El n4202[2]a1u ELIEIDU422 I1:0 El r1r 111
448 ITIZEIErnElE 1711ULEZEJ450zmaziEarivi emF9Eruutu470 EEMMEMEEIEEMEIFIEVIFIGURE 1 Spot graphs showing patterns of incorporation into 15 proteinsthroughout the course of 16 experiments. Numbers at the left are the standard spotnumber, and each column (1-16) represents a different experiment. In each exper-iment, six to ten samples were radiolabeled and analyzed by quantitative two-di-
The graphs are scaled to put the highest point at fullscale. These small graphs are designed to present visually recognizable regulatorypatterns that can be compared among experiments and among proteins. The ex-periments are as follows: (1 -10) Growth curves for REF52, REF52-WT2, REF52 -AG2, REF52-NU2, REF52-WT6, REF52-AG6, REF52-NU6, REF52-Ki-MSV, REF52-Ad5D.4A, and REF52-Ad5W4A; (11,12) comparisons of REF52 with six SV40 trans-formants in normal and defined medium, respectively; (13) comparison of REF52cells in normal and defined medium, followed by REF52-SV1A cells in normal anddefined medium, followed by REF52-SV2A cells in normal and defined medium; (14)REF52 cells labeled for 30 min (two samples), 2 hr (two samples), 24 hr (two sam-ples), and 24 hr followed by a 48-hr chase (two samples); (15, 16) comparisons ofREF52 cells and the nine transformed lines mentioned above on days 3 and 4,respectively, after plating. For proteins not detected in a given sample, the datapoint is plotted at the base line. When a protein was not detected in any of thesamples of the experiment, no graph is plotted.
these proteins are still serum-sensitive in each ofthe transformants. We find that analysis of dy-namic differences (changes in the response of aprotein to a stimulus) is often more rewardingthan just the analysis of static differences(changes in the level of a protein). In the analysisof response to serum stimulation, we reduced thedata from about 60 films to yield the resultsshown in Table 1. Of the 77 proteins that in-crease in rate of synthesis after addition of fresh
152
serum to REF52 cells, only about 50% are in-duced in cells minimally transformed by SV40(WT2 and WT6), only 35% are induced in SV40cells selected for growth in soft agar (AG2 andAG6), and only 25-30% are induced in SV40-transformed cells selected for tumorigenicity(NU2 and NU6). The adenovirus-transformedREF52 cells are even less responsive to serumstimulation, and in one adenovirus transformant(a mutant lacking the 57K E I B protein), only 1 of
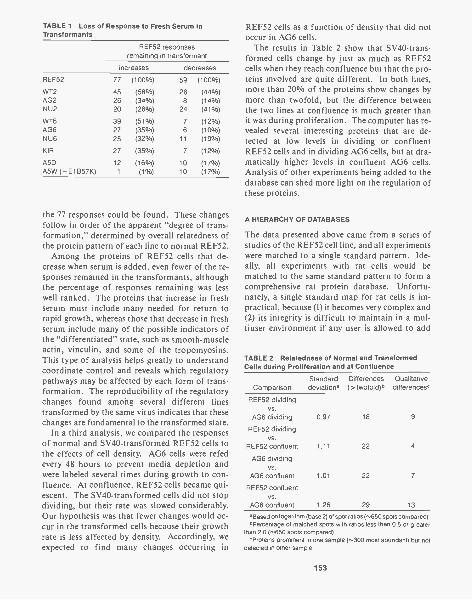
TABLE 1 Loss of Response to Fresh Serum inTransformants
REF52 responsesremaining in transformant
increases decreases
REF52 77 (100%) 59 (100%)
WT2 45 (58%) 26 (44%)AG2 26 (34%) 8 (14%)NU2 20 (26%) 24 (41%)
WT6 39 (51%) 7 (12%)AG6 27 (35%) 6 (10%)NU6 25 (32%) 11 (19%)
KIR 27 (35%) 7 (12%)
A5D 12 (16%) 10 (17%)A5W (- E1B57K) 1 (1%) 10 (17%)
the 77 responses could be found. These changesfollow in order of the apparent "degree of trans-formation," determined by overall relatedness ofthe protein pattern of each line to normal REF52.
Among the proteins of REF52 cells that de-crease when serum is added, even fewer of the re-sponses remained in the transformants, althoughthe percentage of responses remaining was lesswell ranked. The proteins that increase in freshserum must include many needed for return torapid growth, whereas those that decrease in freshserum include many of the possible indicators ofthe "differentiated" state, such as smooth-muscleactin, vinculin, and some of the tropomyosins.This type of analysis helps greatly to understandcoordinate control and reveals which regulatorypathways may be affected by each form of trans-formation. The reproducibility of the regulatorychanges found among several different linestransformed by the same virus indicates that thesechanges are fundamental to the transformed state.
In a third analysis, we compared the responsesof normal and SV40-transformed REF52 cells tothe effects of cell density. AG6 cells were refedevery 48 hours to prevent media depletion andwere labeled several times during growth to con-fluence. At confluence, REF52 cells became qui-escent. The SV40-transformed cells did not stopdividing, but their rate was slowed considerably.Our hypothesis was that fewer changes would oc-cur in the transformed cells because their growthrate is less affected by density. Accordingly, weexpected to find many changes occurring in
REF52 cells as a function of density that did notoccur in AG6 cells.
The results in Table 2 show that SV40-trans-formed cells change by just as much as REF52cells when they reach confluence but that the pro-teins involved are quite different. In both lines,more than 20% of the proteins show changes bymore than twofold, but the difference betweenthe two lines at confluence is much greater thanit was during proliferation. The computer has re-vealed several interesting proteins that are de-tected at low levels in dividing or confluentREF52 cells and in dividing AG6 cells, but at dra-matically higher levels in confluent AG6 cells.Analysis of other experiments being added to thedatabase can shed more light on the regulation ofthese proteins.
A HIERARCHY OF DATABASES
The data presented above came from a series ofstudies of the REF52 cell line, and all experimentswere matched to a single standard pattern. Ide-ally, all experiments with rat cells would bematched to the same standard pattern to form acomprehensive rat protein database. Unfortu-nately, a single standard map for rat cells is im-practical, because (1) it becomes very complex and(2) its integrity is difficult to maintain in a mul-tiuser environment if any user is allowed to add
TABLE 2 Relatedness of Normal and TransformedCells during Proliferation and at Confluence
ComparisonStandarddeviation.
Differences(>twofold)°
Qualitativedifferences°
REF52 dividingvs.
AG6 dividing 0.97 18 9
REF52 dividingvs.
REF52 confluent 1.11 22 4
AG6 dividingvs.
AG6 confluent 1.01 22 7
REF52 confluentvs.
AG6 confluent 1.26 29 13
Based on logarithm (base 2) of spot ratios (",650 spots compared).bPercentage of matched spots with ratios less than 0.5 or greater
than 2.0 (,,650 spots compared).aProteins prominent in one sample (300 most abundant) but not
detected in other sample.
153

new spots to it. Most users first need their ownstandard pattern so that they can compare the gelsof one experiment among themselves, and onlysecondarily do they need to match their data tothe standard patterns used for the higher-level da-tabases. Therefore, we have developed our sys-tem to support an unlimited number of standardpatterns. Using programs that allow standardpatterns to be matched to one another, a hierar-chical database structure becomes supportable.
The lowest level of the hierarchical databasestructure is the individual experiment. The typi-cal experiment involves a related group of sam-ples that can be easily compared with one an-other. A standard pattern representing all thespots detected in the experiment is automaticallygenerated, and a mini-database is generated bymatching each of the gels of the experiment tothis standard. At this level, users can plot thechanges of each protein throughout the course ofthe experiment and can find interesting differ-ences. When users have highly related experi-ments and have used the same standard for each,they can plot the changes of each protein in sev-eral experiments. The procedures for analysis ofa single experiment are simple enough that mostusers can build their own standards and analyzetheir own experiments with minimal help from thecomputer staff. Such a database should be pri-vate, accessible only to the scientists involved inthe project.
At the second level, users may want to comparetheir data with other data in the system. Thesemay be data from other experiments that weredone using different standard patterns, as well asdata made available from other investigators. Toserve these needs, larger databases must be builtup from individual experiments. These will becalled public databases, because they contain dataaccessible to all investigators. Again, these data-bases must not be too general but should be fo-cused in targeted areas, such as rat fibroblasts,mouse embryo cells, and human diploid fibro-blasts. Microorganisms such as yeast or Esche-richia coli can probably be handled as a singledatabase, but a single database encompassing allproteins of a multicellular organism is still toocomplex. When data from a private database aremade available to the public, the data can be en-tered into the public database by matching thestandard patterns. This must be done by a trained
154
staff person who can use special features of thesystem to check the quality of each spot and thecorrectness of the matches. Whenever possible,additional information about each spot, such ascharacteristic regulatory patterns, subcellular lo-calization, or amino acid composition will be usedto confirm the match to the appropriate spot inthe higher database. Users of the private data-bases can perform preliminary matches of theirstandards to the public standards in order tocompare their data to the public databases beforereleasing their data to the public.
At the third level, a species database is needed.This should encompass as many of the proteinsas can be unambiguously identified in the diversecell types and tissues studied within a single spe-cies. Most of the major proteins and many of theless abundant proteins will fall into this category.Proteins that are under intensive investigationoften can be identified by antibodies, by compo-sition, or by characteristic regulatory propertiesin many different cell types. The species data-base would contain the names of most identifiedproteins and would provide, as much as is possi-ble, a cross-referencing between the lower-levelpublic databases. Proteins in the species data-base would be identified on the species standardmap as well as on the standard maps for one ormore of the lower databases. Through the spe-cies database, a researcher working with NIH-3T3cells could cross-reference from that database intothe databases for mouse embryos, mouse eryth-roleukemia cells, or mouse muscle cells to ask howthe proteins of interest are regulated in other celltypes.
A NEW RESOURCE FACILITY
Our present computer facility has served well forprogram development, but falls far short of thecapacity needed to analyze whole experimentsroutinely. Furthermore, scientists from other in-stitutions have expressed strong interest in the useof our system and our databases. We felt that wewere ready for an expanded program devoted pri-marily to database development and that if thetwo-dimensional gel databases were to find gen-eral utility, they would need to be made morereadily available. During 1984, we made a pro-posal to become a Biotechnology Resource andwere awarded 5 years of funding from the Na-

tional Institutes of Health for such a facility, be-ginning in January, 1985. The new facility willcontain three computer workstation systems, eachtenfold more powerful than our present computersystem. A staff will be hired to operate the facil-ity, to serve the outside users, and to manage thepublic databases. Core research funds will beused to build up the public databases and to putmany more identified proteins on the maps. Theplanning for this facility and the work we havedone on database methods during this year shouldhelp us to build large multiuser databases in sev-eral areas of key scientific interest during the nextyear.
Response of Cellular Proteins toOncogene ExpressionB.R. Franza, Jr., H.E. Ruley, J. Moomaw, C. Chang,H. Wenzel, J.I. Garrets
One of the major goals of our research is to studythe effects of expression of different oncogeneson cellular proteins. The REF52 cell line has beenused in much of this work because of its stabilityin tissue culture, because individual oncogenessuch as T24 Ha-ras-1 or Ad5E1A will not onco-genically transform these cells, and because com-binations of oncogenes result in a variety oftransformed phenotypes, some of which are on-cogenic. We have now isolated single cell clonesof REF52 that contain either the T24 Ha-ras4 orAd5E1A oncogene. We have also isolated a num-ber of transformants that result from coexpres-sion of these two oncogenes. For the first time,we are able to compare systematically the effectsof each oncogene, expressed alone or together, oncellular proteins. Here, we present a brief de-scription of a few of the interesting observationswe have already been able to make.
The motivation for comparing the cells undernormal as well as heat-shock conditions primarilywas the previously reported adenovirus effect onexpression of the "HSP7OK" gene. Work done inthe past in collaboration with W.J. Welch (thissection) had already revealed that one protein("73K") in REF52 cells, though expressed at 37°C,is substantially increased in response to heatshock, and another protein("72K") is synthesized
at detectable levels only when the cells arestressed. We therefore wanted to know if eitherof these two proteins would respond to constitu-tive expression of Ad5E1A, and if so, would thisin any way alter their synthesis during heat shock.
Analysis of the experiment described in Figure2 revealed that expression of Ad5E1A alone re-sulted in changes in protein synthesis that were insome cases exactly the opposite of those induced
FIGURE 2 Graphic analysis of certain cellular proteins al-tered by oncogene expression. A display of database infor-mation for five cellular proteins is presented. A region of thestandard REF52 protein map was displayed on the graphicsmonitor, the five spots were individually selected, and theircorresponding graphs were displayed. After displaying allfive graphs the photograph shown in this figure was made.Each graph shows the relative intensity for that spot in eachof the samples analyzed, with the highest intensity set at fullscale. The experiment reported herein involves metaboli-cally labeling of 70% confluent, dividing cells with [31-1]-1eucine for 2 hr. Cells exposed to elevated temperature for 4hr were labeled for the final 2 hr of the heat shock. (A) Nor-mal REF52 cells, 37°C; (8) normal REF52 cells, 42°C; (C)REF52 cells expressing Ad5E1A, 37°C; (D) REF52 cells ex-pressing Ad5E1A, 42°C; (E) REF52 cells expressing T24 Ha-ras-1, 37°C; (F) REF52 cells expressing T24 Ha-ras-1, 42°C;(G) REF52-transformed cells expressing both Ad5E1A andT24 Ha-ras-1, 37°C; (H) REF52-transformed cells express-ing both Ad5E1A and T24 Ha-ras-1, 42°C. The response ofthe following proteins are shown in the graphs: tropomyosin1 (858), stress-response protein, 73K (3708), stress-re-sponse protein, 72K (4678), stress-response protein, 28K(4208), and previously unidentified transformation-sensitiveprotein, 50K (9618).
155

by T24 Ha-ras-1 expression. Spot 858 (shown inFig. 2) demonstrates the reduction in synthesis ofthe cytoskeleton protein, tropomyosin 1, in cellsexpressing Ad5E1A, and an increase in synthesisin cells expressing T24 Ha-ras-1. The same resultwas observed with the sm-Actin (smooth muscleform) synthesized in REF52 cells. We have al-ready demonstrated that these two proteins arecoordinately regulated in growth-curve experi-ments, in serum-stimulation experiments, andin SV40 and adenovirus DNA transformants ofREF52. Interestingly, in the AD5E1A + T24 Ha-ras-1 transformant, the prevailing effect on tro-pomyosin 1 is that seen in the Ad5E1A-expressingcells, i.e., decreased synthesis. In the case of sm-Actin, no detectable synthesis occurs in the trans-formant. Therefore, expression of each onco-gene alone can exert effects that are exactly theopposite of the other. In addition, their co-expression is not the simple summation of ef-fects. The interaction of two oncogenes in atransformed cell should therefore be thought ofin terms of comodulation of cellular protein syn-thesis. The predominant effect, of one oncogeneor the other when coexpressed in the transformedcell, is discernible as a result of having each on-cogene individually expressed in cell lines derivedfrom the same parent as the transformant.
Spot 3708 presents data on the synthesis of 73Kin each of the cell lines under study. As shownin Figure 2, cells expressing Ad5E1A synthesizeslightly more 73K than do normal cells at 37°C.In addition, the induced expression at 42°C ishigher than in normal cells. The transformant(expressing both oncogenes) shows even greatersynthesis of 73K at 37°C, but coexpression of T24Ha-ras-1 may account for the reduction in thelevel synthesized during heat shock. Comparisonof the results shown in the graph of spot 4678 for72K clearly shows that these two proteins are reg-ulated differently. Ad5E1A-containing cells showhigher induction of 72K than normal cells whenboth are exposed to 42°C, and T24 Ha-ras-1 cellsshow an even greater induction of 72K when heat-shocked. In all the T24 Ha -ras -1 + Ad5E1A-trans-formed REF52 clones, the level of 72K inducedby heat shock is less than that seen in clones con-taining one or the other gene, and in the case ofthis particular transformant, it is not detectable.In this particular transformant, the level of theheat-shock 110K proteins (graph not shown) is
156
substantial even at 37°C, although it is not de-tectable in the other lines shown in this study un-less they are heat-shocked. It may be that consti-tutive production of 110K at 37°C is the result ofcoexpression of the two oncogenes and affects theinducibility of 72K in these cells.
The graph of spot 9618 represents a previouslyunidentified transformation-sensitive protein,"50K." Interestingly, this protein is synthesizedat 37°C in the transformed cell line (much like110K mentioned above) but not in other lines. Inthe transformant, 50K is also inducible by ele-vated temperature to even higher levels of synthe-sis. 50K and 110K therefore represent proteinswhose regulation is sensitive to both stress andcoexpression of two oncogenes.
The final protein presented here is known as"28K" and its response is shown in graph 4208.The two spots immediately to the left of 28K (seeFig. 2) are its phosphorylated forms. The identi-fication of these phosphate-modified forms wasdone in collaboration with W.J. Welch (this sec-tion). He has shown that serum, stress, and 12-0-tetradecanoylphorbol-13-acetate (TPA) can in-duce increased phosphorylation of 28K. Indeed,in experiments in which we have compared the ef-fects of serum, platelet-derived growth factor(PDGF), and TPA, within 3 minutes of exposureto any of these agents, a significant increase inthe density of the spots representing the phospho-rylated forms of 28K occurs. It was thereforevery interesting to see that the synthesis of thenonphosophorylated 28K was substantially re-duced in cells expressing Ad5E1A but was in-duced in the cells expressing T24 Ha-ras-1 alone.The prevailing influence in the transformantseems to be Ad5E1A. The response of this pro-tein to 42°C in each line is also shown and indi-cates again the complexity of coexpression versusindividual expression of the two oncogenes understudy.
Normal REF52 cells require fetal calf serum forgrowth. W. Topp (Otisville Biotech, Inc.) and D.McClure (W. Alton Jones Cell Science Center)have shown that in defined media, PDGF will notinduce REF52 cells to leave density-arrest. Theyhave also shown that in defined media, dividingREF52 cells do not require PDGF for continuedgrowth and cell division. Cells expressing the T24Ha-ras-1 oncogene, either stably incorporated orafter microinjection, demonstrate a substantial

increase in serum dependence. Serum depriva-tion causes the cells to flatten out, to develop sig-nificant cytoplasmic vacuolization, and to accu-mulate multiple nuclei in individual cells. Theserum deprivation response is induced if the me-dia containing 10% fetal calf serum is notchanged every other day. Cells expressing theAd5E1A gene alone also show increased serumdependence, but their response to fresh serum aswell as serum deprivation is different. Whenplated in fresh serum, these cells are often small,more refractile, and show a greater number ofmitotic forms. One to two days after serum isadded they begin to flatten out and divide moreslowly. Unless the cells are refed with fresh serumthey become round, detach from the plastic, anddie. Thus, the effects of the Ad5E1A gene andthe T24 Ha-ras-1 gene are distinctly different withrespect to morphology and survival in serum-de-pleted medium. Cells coexpressing both genes arefully transformed and serum-independent.
Analysis of proteins such as "28K" may help ussort out the differences in growth control exhib-ited by cells expressing either Ad5E1A or T24 Ha-ras-1. We are currently studying the response of28K in experiments in which PDGF is added tothe cells to see if the phosphorylation that occursin the cells expressing Ad5E1A is substantiallydifferent from that seen in the cells expressing T24Ha-ras-1. When these results are placed in thecontext of the other induced changes in phospho-rylation/dephosphorylation of cellular proteinsacutely exposed to different growth-promoting orretarding agents, we expect that sets of proteinsspecifically altered in cells expressing each onco-gene will be identified. Such knowledge will al-low us to begin defining the proteins involved inthe growth-control pathways of the cell.
Among the applications available once theabove analysis is complete is a strategy for moreefficient and alrected isolation of the genes en-coding proteins involved in cellular growth con-
trol. It will employ two-dimensional analysis ofin vitro translates of cellular mRNA preparedfrom each of the different cell types exposed tovarious conditions to determine quantitativelywhen the message for a particular protein is mostabundant. From this information, protocols fordifferential hybridization can be optimized, andthe resultant hybrid-selected mRNA can be trans-lated and analyzed on two-dimensional gels toensure that the proteins of interest are indeed en-coded by the selected cDNA.
PUBLICATIONS
Francoeur, A.M., E.K.L. Chan, J.I. Garrels, and M.B. Ma-thews. 1985. Characterization and purification of Lupusantigen La, an RNA-binding protein. Mol. Cell. Biol.
5: 586-590.Franza, B.R. and J.I. Garrels. 1984. Transformation-sensi-
tive proteins of REF52 cells detected by computer-ana-lyzed two-dimensional gel electrophoresis. Cancer Cells1: 137-146.
Garrels, J.I. and B.R. Franza. 1984. Identification of proteinmarkers for transformed cells by two-dimensional gelelectrophoresis. In Genetic variability in responses tochemical exposure, pp. 275-284. Banbury Reports, ColdSpring Harbor Laboratory, Cold Spring Harbor, New York.
Garrels, J.I., J.T. Farrar, and C.B. Burwell IV. 1984. TheQUEST system for computer-analyzed two-dimensionalelectrophoresis of proteins. In Two-dimensional gel elec-trophoresis of proteins: Methods and applications (ed.J.E. Celis and R. Bravo), pp. 37-91. Academic Press,New York.
Kurkinen, M., A. Taylor, J.I. Garrels, and B.L.M. Hogan.1984. Cell surface-associated proteins which bind na-tive Type IV collagen or gelatin. J. Biol. Chem. 259:5915-5922.
Mathews, M.B., R.M. Bernstein, B.R. Franza, and J.I. Gar-rels. 1984. Identity of the proliferating cell nuclear anti-gen and cyclin. Nature 309: 374-376.
Ruley, H.E., J. Moomaw, C. Chang, J.I. Garrels, M. Furth,and B.R. Franza. 1985. Multi-step transformation of anestablished cell line by the adenovirus E1A and T24 Ha-ras-1 genes. Cancer Cells 3: 257-264.
Silver, L.M., D. Lukralle, J.I. Garrels, and K. Willison. 1984.Persistence of a lethal t haplotype in a laboratory stockof outbred mice. Genet. Res. 43: 21-25.
157


NEUROBIOLOGY
During mammalian neurogenesis, a plate of approximately 105 morphologicallyhomogeneous cells gives rise to 1012 morphologically and physiologically hetero-geneous neurons in the adult brain. Estimates of mRNA complexity indicate thatthe adult mammalian brain contains more than 30,000 different brain-specificproteins. The studies of the Neurobiology Group continue to explore several fea-tures of this enormous molecular diversity using both vertebrate and, simpler, in-vertebrate nervous systems. Our previous studies showed that in contrast to theimmune system, where molecular diversity among lymphocytes is expressedwithin a single class of molecule (immunoglobulin), diversity in the nervous sys-tem is not confined to one class of molecule (such as neurotransmitter), as anti-genic heterogeneity is manifest in many different subcellular structures.
One of the potential uses of cell-specific markers is to trace lineage relation-ships through development. Somewhat surprising was our finding that all of ourmarkers that differentiate between adult mammalian neurons recognize antigensthat are first expressed relatively late in development. Some of these antigensfirst appear long after the apparent establishment of adult structure, In the lastyear, we have focused increasingly on developmental issues. Our developmentalstudies suggest that the temporal control of antigen expression and the transientpresence of some classes of cells are importantly related to the normal develop-ment of the adult nervous system. One of the most exciting observations in thelast year was the demonstration that the expression of an antigen recognized byone of our monoclonal antibodies is controlled by early visual experience. Thissuggests that the mechanisms of experience-induced negronal developmentmay be accessible to molecular analysis.
Our studies using invertebrate nervous systems have addressed issues thatare not easily accessible in the mammalian brain. Using the moth, we are explor-ing the basis of sexual dimorphism in the central nervous system (CNS). Ourstudies on the leech CNS continue to examine features of the segmental organi-zation of nervous systems.
VERTEBRATES
S. Hockfield B. Friedman C. BautistaR. McKay S. Zaremba E. Waldvogel
A MARKER FOR EARLY AXONS ALLOWS USTO VISUALIZE THE FIRST EVENTS INAXON OUTGROWTH
One of the great difficulties in studying how con-nections are made in the developing mammaliannervous system has been the inability to identifythe earliest axons reproducibly and unambigu-ously. Available methods have severe limitations.
Using electron microscopy at very early stages ofdevelopment, one cannot reliably discriminatebetween axons and other profiles. Neither neu-rofibrillary stains nor intra-axonal tracers can beused to detect axons at their earliest stages ofgrowth. We have generated a monoclonal anti-body, Rat-202, that identifies axons at the earlieststages of axon growth. The antibody binds to an
159

antigen distributed throughout growing axons,including the growth cone and filopodia, thestructures believed to be responsible for axonnavigation.
Studies in tissue culture of growing axons havedescribed the morphology and behavior of growthcones and filopodia. These studies have demon-strated that growing axons show a preference forparticular substrates. Due to the vastly greatercomplexity of the intact nervous system, similarstudies in tissue sections were not possible. Usingantibody Rat-202, we have been able to identifyand study growth cones and filopodia in sections.One of the advantages of antibody identificationis that histological analyses can be carried to theultrastructural level. Electron microscopic exam-ination of Rat-202-stained material shows thatthe filopodia of growth cones in the developingspinal cord contact nonneuronal profiles. Usingdouble-labeling techniques, we are now trying toidentify simultaneously these nonneuronal pro-files and early axons to determine the structuresfilopodia contact to guide axon growth in theCNS.
A NONNEURONAL CELL MAY SUPPORT INITIALAXON GROWTH AND AXON REGENERATIONIN THE PERIPHERY
Axon regrowth after injury occurs in the periph-eral nervous system, but not in the CNS, of adultmammals. Accumulating evidence indicates thataxon growth depends on environmental factorssuch as nonneuronal cells and extracellular ma-trix present in peripheral nerve. However, themechanism of axon interaction with the periph-eral nerve environment is not well understood.One unsettled question is whether axons contactone specific class of nonneuronal cells duringgrowth in immature or regenerating peripheralnerves and/or whether they preferentially contactnoncellular, extracellular matrix components se-creted by these nonneuronal cells. In the past,these questions have been inaccessible to studybecause early nonneuronal cell types cannot bereliably distinguished on morphological groundsand there was no ready means to generate mark-ers for peripheral nerve nonneuronal cells or ex-tracellular matrix components. The developmentof hybridoma technology has provided a usefultool to generate anatomical probes for the study
160
of early events during peripheral nerve develop-ment and regeneration.
Monoclonal antibody Rat-401, generated in ourlaboratory by immunizing mice with paraformal-dehyde-fixed spinal cord from rats at day 15 ofgestation, recognizes nonneuronal cells in boththe developing peripheral nervous system and theCNS. Rat-401-stained cells are present in periph-eral nerve roots as early as day 11 of gestation.Rat-401-positive nonneuronal cells are present be-fore axons grow into the peripheral nerve. Inter-estingly, as the early axons enter the nerve roots,they contact Rat-401-stained cells. Thus, thesenonneuronal cells are in a position where theymay guide or support initial peripheral axonalgrowth. Embryonic optic nerve and other CNSaxon tracts contain Rat-401-stained cells, an in-dication that Rat-401-positive cells may also assistaxon growth in the CNS.
Rat-401-stained cells in the developing CNS arepresent in the early neural tube and are morpho-logically similar to the radial glial cells. The ra-dial glial cells act as guides for neuronal migra-tion in the CNS, suggesting that the cells thatguide axon outgrowth and neuronal migration arerelated. Our earlier studies in the leech suggestedthat axons may selectively fasciculate with oneanother through diverse surface-recognition mol-ecules. The observations with Rat -40l raise thepossibility that initial axon outgrowth and navi-gation occur in relation to a nonneuronal cell, andsubsequent navigation may be mediated by inter-axonal interactions. We have now establishedcultures of Rat-401 cells from the embryonic CNSso that the role of these cells in cell migration andneurite outgrowth can be studied and manipu-lated in vitro.
The adult CNS lacks Rat-401-stained cells,which is consistent with the absence of neuronalcell migration and axon regeneration followinginjury in the mature CNS. In contrast, the ma-ture peripheral nerve (where axon regenerationdoes occur) contains Rat-401-positive process.Rat-401-stained cells in the sciatic nerve are lo-cated external to myelin sheathes in a configu-ration similar to that of Schwann cells, a non-neuronal cell type that ensheathes axons. Thissuggests that a set of the embryonic Rat-401 cellsdevelop into myelin-related Schwann cells. Wehave now shown that cultured Schwann cells, butnot fibroblasts, are Rat-401-positive, increasing
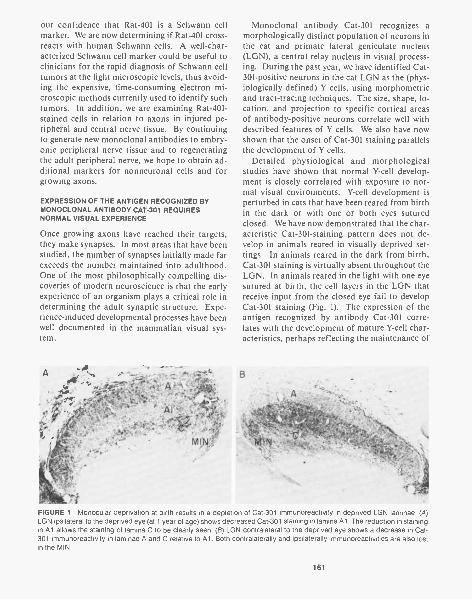
our confidence that Rat-401 is a Schwann cellmarker. We are now determining if Rat-401 cross-reacts with human Schwann cells. A well-char-acterized Schwann cell marker could be useful toclinicians for the rapid diagnosis of Schwann celltumors at the light microscopic levels, thus avoid-ing the expensive, time-consuming electron mi-croscopic methods currently used to identify suchtumors. In addition, we are examining Rat -401-stained cells in relation to axons in injured pe-ripheral and central nerve tissue. By continuingto generate new monoclonal antibodies to embry-onic peripheral nerve tissue and to regeneratingthe adult peripheral nerve, we hope to obtain ad-ditional markers for nonneuronal cells and forgrowing axons.
EXPRESSION OF THE ANTIGEN RECOGNIZED BYMONOCLONAL ANTIBODY CAT-301 REQUIRESNORMAL VISUAL EXPERIENCE
Once growing axons have reached their targets,they make synapses. In most areas that have beenstudied, the number of synapses initially made farexceeds the number maintained into adulthood.One of the most philosophically compelling dis-coveries of modern neuroscience is that the early
of an organism plays a critical role indetermining the adult synaptic structure. Expe-rience-induced developmental processes have beenwell documented in the mammalian visual sys-tem.
A
Monoclonal antibody Cat-301 recognizes amorphologically distinct population of neurons inthe cat and primate lateral geniculate nucleus(LGN), a central relay nucleus in visual process-ing. During the past year, we have identified Cat-301-positive neurons in the cat LGN as the (phys-iologically defined) Y cells, using morphometricand tract-tracing techniques. The size, shape, lo-cation, and projection to specific cortical areasof antibody-positive neurons correlate well withdescribed features of Y cells. We also have nowshown that the onset of Cat-301 staining parallelsthe development of Y cells.
Detailed physiological and morphologicalstudies have shown that normal Y-cell develop-ment is closely correlated with exposure to nor-mal visual environments. Y-cell development isperturbed in cats that have been reared from birthin the dark or with one or both eyes suturedclosed. We have now demonstrated that the char-acteristic Cat-301-staining pattern does not de-velop in animals reared in visually deprived set-tings. In animals reared in the dark from birth,Cat-301 staining is virtually absent throughout theLGN. In animals reared in the light with one eyesutured at birth, the cell layers in the LGN thatreceive input from the closed eye fail to developCat-301 staining (Fig. 1). The expression of theantigen recognized by antibody Cat-301 corre-lates with the development of mature Y-cell char-acteristics, perhaps reflecting the maintenance of
B
MIN
FIGURE 1 Monocular deprivation at birth results in a depletion of Cat-301 immunoreactivity in deprived LGN laminae. (A)LGN ipsilateral to the deprived eye (at 1 year of age) shows decreased Cat-301 staining in lamina Al. The reduction in stainingin Al allows the staining of lamina C to be clearly seen. (B) LGN contralateral to the deprived eye shows a decrease in Cat-301 immunoreactivity in laminae A and C relative to Al. Both contralaterally and ipsilaterally immunoreactivities are also lostin the MIN.
161
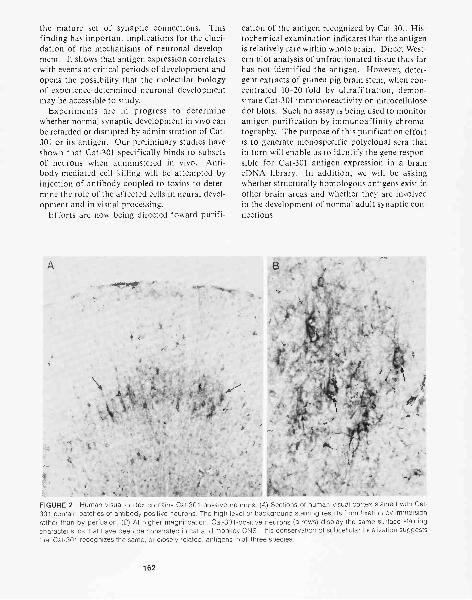
the mature set of synaptic connections. Thisfinding has important implications for the eluci-dation of the mechanisms of neuronal develop-ment. It shows that antigen expression correlateswith events at critical periods of development andopens the possibility that the molecular biologyof experience-determined neuronal developmentmay be accessible to study.
Experiments are in progress to determinewhether normal synaptic development in vivo canbe retarded or disrupted by administration of Cat-301 or its antigen. Our preliminary studies haveshown that Cat-301 specifically binds to subsetsof neurons when administered in vivo. Anti-body-mediated cell killing will be attempted byinjection of antibody coupled to toxins to deter-mine the role of the affected cells in neural devel-opment and in visual processing.
Efforts are now being directed toward purifi-
cation of the antigen recognized by Cat-301. His-tochemical examination indicates that the antigenis relatively rare within whole brain. Direct West-ern blot analysis of unfractionated tissue thus farhas not identified the antigen. However, deter-gent extracts of guinea pig brain stem, when con-centrated 10-20-fold by ultrafiltration, demon-strate Cat-301 immunoreactivity on nitrocellulosedot blots. Such an assay is being used to monitorantigen purification by immunoaffinity chroma-tography. The purpose of this purification effortis to generate monospecific polyclonal sera thatin turn will enable us to identify the gene respon-sible for Cat-301 antigen expression in a braincDNA library. In addition, we will be askingwhether structurally homologous antigens exist inother brain areas and whether they are involvedin the development of normal adult synaptic con-nections.
FIGURE 2 Human visual cortex contains Cat-301-positive neurons. (A) Sections of human visual cortex stained with Cat-301 contain patches of antibody-positive neurons. The high level of background staining results from fixation by immersionrather than by perfusion. (8) At higher magnification, Cat-301-positive neurons (arrows) display the same surface stainingcharacteristics that have been demonstrated in cat and monkey CNS. This conservation of subcellular localization suggeststhat Cat-301 recognizes the same, or closely related, antigens in all three species.
162

MARKING AREAL BOUNDARIES WITHMONOCLONAL ANTIBODIES
Classical neurobiology has provided maps of thevertebrate cerebral cortex that divide the braininto areas with specific functions and body rep-resentations. Nonspecific cell stains have illus-trated different neuronal cell types in the cortexand their organization. Hybridoma technologyprovides a means of selectively staining identifiedneuronal types and establishing a molecular basisfor their anatomical and physiological diversity.Monoclonal antibodies have shown differencesamong cell populations that were heretofore un-recognized.
Previously, we have reported that in the pri-mate visual cortex, Cat-301 identifies neurons ina pattern that reflects the ocular dominance or-ganization of the primary visual cortical area(VI). A dramatic shift in staining patterns marksthe boundary between VI and the adjacent sec-ond visual cortical area (V2). This boundary canbe resolved using more classical techniques, butelsewhere in the primate cortex, Cat-301 stain-ing marks boundaries between less well resolvedareas. For example, the boundaries between areasMT and MST and between V3 and V4 appear tobe marked by changes in Cat-301-staining pat-terns. Currently, electrophysiological and tract-tracing techniques are being used in conjunctionwith Cat-301 immunohistochemistry to confirmthese areal borders.
One of the difficulties in understanding humancortical function has been the inability to set nor-mal ranges for the size and location of corticalareas. In experimental animals, areal boundarieshave been determined by invasive methods, suchas electrophysiological recording and tract-trac-ing studies. The conservation of antigens throughevolution may allow us to use markers like Cat-301 to study the organization of the human cor-tex in postmortem material. We have alreadyshown that Cat-301 recognizes an antigen in hu-man cortex (Fig. 2) that is related to the antigenrecognized in cat and monkey. The cells that ex-press the Cat-301 antigen in human area VI havea patchy distribution similar to that seen in themacaque monkey. Our current studies are aimedat establishing rapid, reproducible criteria formapping areal boundaries in the human cortex.
Another recently generated monoclonal anti-
body, Rat-203, recognizes subsets of neurons inthe rat CNS and shows sharply defined corticalboundaries in rat cortex. The antibody, whichidentifies a 180,000-molecular-weight polypep-tide in Western blot experiments, recognizes pro-jection neurons in rat neocortex, but not in pa-leocortical areas such as the olfactory cortex anddentate gyrus. In the cerebellar cortex, Rat-203recognizes a small group of interneurons in thepaleocerebellar area, but not in the archicerebel-lar area, whereas projection neurons are Rat-203-positive throughout the cerebellum.
PUBLICATIONS
Hendry, S.H.C., S. Hockfield, E.G. Jones, and R.D.G.McKay. 1984. A monoclonal antibody that identifies sub-sets of neurons in the central visual system of the monkeyand cat. Nature 307: 267-269.
Johansen, J., R. McKay, and S. Hockfield. 1984. Distributionand morphology of nociceptive cells in the central nerv-ous system of the leech. J. Comp. Neurol. 226: 255-262.
. 1984. Axonal projections of mechanosensory neu-rons in the connective and peripheral nerves of the leech.J. Comp. Neurol. 226: 255-262.
McKay, R.D.G., J. Johansen, and S. Hockfield. 1984. Mono-clonal antibody identifies a 63,000 dalton antigen foundin all central neuronal cell bodies but in only a subset ofaxons in the leech. J. Comp. Neurol. 226: 448-455.
Zaremba, S. and R. Hogue-Angeletti. 1985. A reliable meth-od for assessing topographical arrangement of proteinsin the chromaffin granule membrane. Neurochem. Res.10: 19-32.
In Press, Submitted, and In Preparation
Friedman, B. and A. Aguayo. 1985. Injured neurons in theolfactory bulb of the adult rat grow axons along grafts ofperipheral nerve. J. Neurosci. 5: (in press).
Hockfield, S. and R. McKay. 1985. Identification of major cellclasses in the developing mammalian nervous system. J.Neurosci. (in press).
Sur, M., D. Frost, and S. Hockfield. 1985. Expression of asurface antigen on Y-cells in cat lateral geniculate nu-cleus is regulated by visual experience. Nature (Submit-ted.)
Zaremba, S. and J.H. Keen. 1985. Limited proteolytic diges-tion of coated vesicle assembly polypeptides abolishesreassembly activity. J. Cell. Biochem. (in press).
163

INVERTEBRATES
S. Hockfield A. Hishinuma T. FlanaganR. McKay G. Bablanian C. SchleyB. Zipser
Monoclonal Antibodies DemonstrateSexually Dimorphic Structuresin the Moth CNSA. Hishinuma, S. Hockfield, R. McKay
The relative simplicity of the invertebrate CNScompared with the vertebrate CNS often makesbasic problems in neurobiology more accessibleto study. The existence of sexual dimorphism inthe CNS is a highly charged issue, but severalstudies have shown differences between sexes inboth invertebrate and vertebrate systems. In themoth, Manduca sexta, male-specific sensory neu-rons in the antennae, which respond to the femalesex pheromone, connect to male-specific neuronsin the antennal lobes of the CNS at the male-spe-cific synaptic region, the macroglomerular com-plex (MGC). Previous studies showed that sur-gical transplantation of the imaginal disks of themale antennae into female hosts gives rise toMGC-like structures in the gynandromorphicfemale antennal lobe. To obtain reagents withwhich to study the mechanisms that govern thedevelopment of sexually dimorphic structures,monoclonal antibodies were generated to the malemoth CNS.
The olfactory-specific antibody (OSA) stainsthe primary olfactory neurons both in males andfemales. Cell bodies, proximal parts of den-drites, and axons are recognized by this antibody.The intracranial portion of the antennal nervecontains sensory axons of two different modali-ties. Here, the olfactory axons are stained by theOSA but the mechanosensory axons are not. Inthe antennal lobe, the olfactory axons make syn-aptic contacts in structures called glomeruli. TheOSA recognizes every glomerulus (Fig. 1A). Cellbodies of another set of primary olfactory neu-rons are located in a part of the mouth, the labialpulp organ, which also sends axons into the an-tennal lobe. The tracts from the labial pulp or-gan to the antennal lobes are also stained by theOSA antibody. To confirm the specificity of OSA
164
for olfactory structures, the whole brain and thethoracic ganglion were tested. OSA staining wasconfined to the primary olfactory neurons. Atthe electron microscopic level, OSA staining isassociated with the external side of the plasmamembrane of cell bodies and axons. As in ourprevious studies in the leech, axons with surfaceOSA staining travel together in fascicles. We havepreviously suggested that such axonal surfacemolecules may be chemoaffinity molecules bywhich axons navigate along specific routes totheir targets. The synaptic terminals of the pri-mary olfactory neurons in the glomeruli of theantennal lobe, which cannot be resolved at thelight microscopic level, can be identified by thepresence of antibody staining at the electron mi-croscopic level.
The male-specific antibody (MSA) stains theprimary olfactory neurons in males but not infemales. The MSA recognizes cell bodies, den-drites, and axons. There are two types of hair-like sensillae in the antennae; longer ones arefound only in males and shorter ones in bothsexes. The dendrites of the primary olfactoryneurons that lie in long sensillae are stained byMSA. In females, primary olfactory neurons arenot stained by MSA. In the antennal lobe, onlythe male-specific glomerulus is stained by MSA(Fig. 1B). The ordinary glomeruli, which arecommon to males and females, are unstained.Another cell type, associated with the limitingmembrane of the antennae, is stained in bothmales and females. The position and develop-mental appearance of this second type of MSA-positive cell suggest that these cells are develop-mentally related to the primary olfactory neu-rons, perhaps being an undifferentiated progeni-tor cell.
At present, we are characterizing the antigensrecognized by these antibodies by Western blotanalysis and studying the expression of their an-tigens in the development of the moth. We hopeto be able to describe developmental events thatgive rise to male-specific structures.
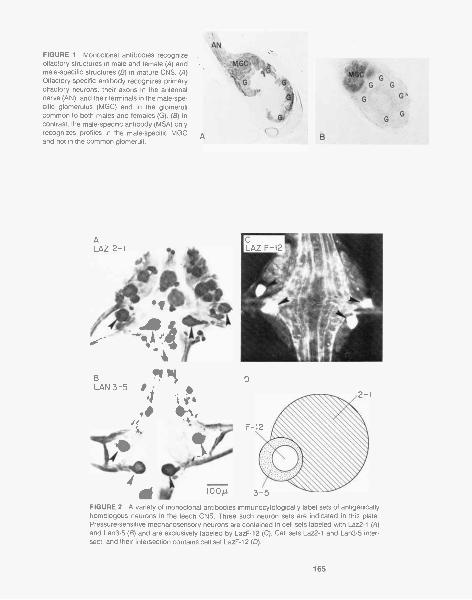
FIGURE 1 Monoclonal antibodies recognizeolfactory structures in male and female (A) andmale-specific structures (B) in mature CNS. (A)Olfactory-specific antibody recognizes primaryolfactory neurons, their axons in the antennalnerve (AN), and their terminals in the male-spe-cific glomerulus (MGC) and in the glomerulicommon to both males and females (G). (8) Incontrast, the male-specific antibody (MSA) onlyrecognizes profiles in the male-specific MGCand not in the common glomeruli.
BLAN 3-5
D
FIGURE 2 A variety of monoclonal antibodies immunocytologically label sets of antigenicallyhomologous neurons in the leech CNS. Three such neuron sets are indicated in this plate.Pressure-sensitive mechanosensory neurons are contained in cell sets labeled with Laz2-1 (A)and Lan3-5 (8) and are exclusively labeled by LazF-12 (C). Cell sets Laz2-1 and Lan3-5 inter-sect, and their intersection contains cell set LazF-12 (D).
165

Cellular Organizing Systems in theLeech: Are Nervous Systems Built byInteractions among AntigenicallyRelated, Functionally HeterogeneousSets of Neurons?G. Bablanian, T. Flanagan, B. Zipser
An extension of classic approaches to the studyof nervous system networks has been the appli-cation of immunocytological probes to define setsof immunochemically related cells. In the leech,we have identified 11 different antigenically re-lated neuron sets. Such sets vary in size, contain-ing from 2 to 50 neurons. We have identified in-dividual neurons within these sets and we realizethat antigenically related neurons can serve di-verse physiological functions. Some identifiedneurons belong to more than one set of antigeni-cally related cells. In the leech, antigenically re-lated cell sets are thus either intersected by orcontained within other antigenically related cell
COUNTING BY TWOS
0©0©000©0©1 2 3 4 5 6 7 8 9 10
COUNTING BY SEVEN
0000VOC)005 6 7 8 12 13 14 15 16
COUNTING FROM AN ORIGIN
CC:00 0©00 0 01 2 3 4 I 2 3 4 5 6
I I
Cephalic Segmental Series
FIGURE 3 Immunocytologically labeled neurons that ap-pear in nonadjacent ganglia along the axis of the leech CNSsuggest the existence of segmental counting mechanisms.One such mechanism assigns immunoreactive neurons toeven-numbered ganglia, counting through the CNS in two-segment units (top). Another such mechanism assigns im-munoreactive neurons to seven-segment units within the mid-body ganglionic chain (middle). A third mechanism counts offtwo-segment units from the origin of the cephalic and mid-body ganglionic series (bottom).
166
sets (see Fig. 2). We have recently identified apair of neurons that are common to a dispropor-tionately large number of antigenically related cellsets. Why do these cells (which we call a cells)carry such a rich array of set-specific antigens?Do these antigens correlate with or specify syn-aptic connections? If such a correlation exists,then these a cells function in a variety of neu-ronal networks. Perhaps they play a pivotal rolein activating one of several mutually exclusive be-havioral networks. Electrophysiological analysiswill be able to address this hypothesis in futurestudies of the leech nervous system. At present,the broader question of the significance of di-verse and intersecting antigenically related cellsets remains unanswered. As we expand our cat-alog of antigenically related cell sets, we are con-tinually searching for clues (such as a cells) thatmight suggest the biological basis for this phe-nomenon.
Supersegmental Organizing Systemsin the Leech: ImmunocytologicallyIdentified Neurons That Appear inSerially Repeated MultiganglionUnits Along the Segmental Axis of theLeech CNS Suggest the Existence ofSupersegmental Counting MechanismsT. Flanagan, C. Schley, B. Zipser
The leech central nervous system (CNS) is com-posed of a series of 32 homologous ganglia thatare each derived from one of the 32 embryonicsegments that form the leech body axis. Eachganglion consists of a core population of about400 individually identifiable neurons and an ad-ditional set of specialized cells restricted to se-lected ganglia. We have used immunocytologicalmethods to identify three sets of specialized cellsthat are serially expressed in nonadjacent gangliathroughout the leech CNS. The distribution ofthese cells define multisegmental units within theleech CNS and suggest the existence of superseg-mentation counting mechanisms that individuallylabel each leech ganglion (Fig. 3). We have begunelectrophysiological analysis of these specializedcounting neurons, and we now realize that theyare interganglionic interneurons. Some of these
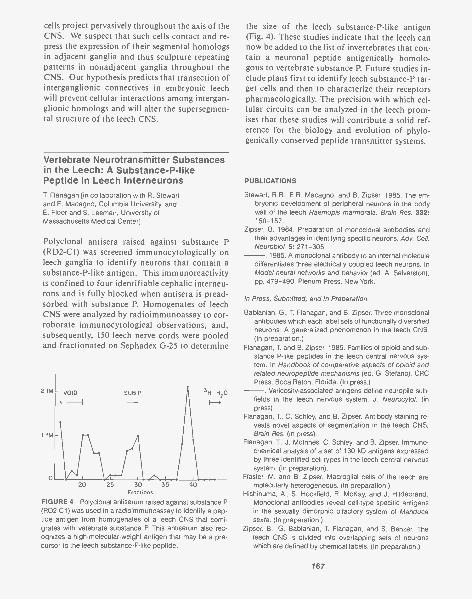
cells project pervasively throughout the axis of theCNS. We suspect that such cells contact and re-press the expression of their segmental homologsin adjacent ganglia and thus sculpture repeatingpatterns in nonadjacent ganglia throughout theCNS. Our hypothesis predicts that transection ofinterganglionic connectives in embryonic leechwill prevent cellular interactions among intergan-glionic homologs and will alter the supersegmen-tal structure of the leech CNS.
Vertebrate Neurotransmitter Substancesin the Leech: A Substance-P-likePeptide in Leech InterneuronsT. Flanagan [in collaboration with R. Stewartand E. Macagno, Columbia University, andE. Floor and S. Leeman, University ofMassachusetts Medical Center]
Polyclonal antisera raised against substance P(RD2-C1) was screened immunocytologically onleech ganglia to identify neurons that contain asubstance-P-like antigen. This immunoreactivityis confined to four identifiable cephalic interneu-rons and is fully blocked when antisera is pread-sorbed with substance P. Homogenates of leechCNS were analyzed by radioimmunoassay to cor-roborate immunocytological observations, and,subsequently, 150 leech nerve cords were pooledand fractionated on Sephadex G-25 to determine
20 25 30 35Fractions
FIGURE 4 Polyclonal antiserum raised against substance P(RD2-C1) was used in a radioimmunoassay to identify a pep-tide antigen from homogenates of a leech CNS that comi-grates with vetebrate substance P This antiserum also rec-ognizes a high-molecular-weight antigen that may be a pre-cursor to the leech substance-P-like peptide.
40
the size of the leech substance-P-like antigen(Fig. 4). These studies indicate that the leech cannow be added to the list of invertebrates that con-tain a neuronal peptide antigenically homolo-gous to vertebrate substance P. Future studies in-clude plans first to identify leech substance-P tar-get cells and then to characterize their receptorspharmacologically. The precision with which cel-lular circuits can be analyzed in the leech prom-ises that these studies will contribute a solid ref-erence for the biology and evolution of phylo-genically conserved peptide transmitter systems.
PUBLICATIONS
Stewart, R.R., E.R. Macagno, and B. Zipser. 1985. The em-bryonic development of peripheral neurons in the bodywall of the leech Haemopis marmorata. Brain Res. 332:150-157.
Zipser, B. 1984. Preparation of monoclonal antibodies andtheir advantages in identifying specific neurons. Adv. Cell.Neurobiol. 5: 271-305.
. 1985. A monoclonal antibody to an internal moleculedifferentiates three electrically coupled leech neurons. InModel neural networks and behavior (ed. A. Selverston),pp. 479-490. Plenum Press, New York.
In Press, Submitted, and In Preparation
Bablanian, G., T. Flanagan, and B. Zipser. Three monoclonalantibodies which each label sets of functionally diversifiedneurons: A generalized phenomenon in the leech CNS.(In preparation.)
Flanagan, T. and B. Zipser. 1985. Families of opioid and sub-stance P-like peptides in the leech central nervous sys-tem. In Handbook of comparative aspects of opioid andrelated neuropeptide mechanisms (ed. G. Stefano). CRCPress, Boca Raton, Florida. (In press.)
. Varicosity-associated antigens define neuropile sub-fields in the leech nervous system. J. Neurocytol. (inpress).
Flanagan, T., C. Schley, and B. Zipser. Antibody staining re-veals novel aspects of segmentation in the leech CNS.Brain Res. (in press).
Flanagan, T, J. McInnes, C. Schley, and B. Zipser. Immuno-chemical analysis of a set of 130 kD antigens expressedby three identified cell types in the leech central nervoussystem. (In preparation).
Flaster, M. and B. Zipser. Macroglial cells of the leech aremolecularly heterogeneous. (In preparation.)
Hishinuma, A., S. Hockfield, R. McKay, and J. Hildebrand.Monoclonal antibodies reveal cell-type specific antigensin the sexually dimorphic olfactory system of Manducasexta. (In preparation.)
Zipser, B., G. Bablanian, T. Flanagan, and S. Benzer. Theleech CNS is divided into overlapping sets of neuronswhich are defined by chemical labels. (In preparation.)
167



Recto: Vannevar Bush Lecture Hall, since its construction in 1953, has been the site of theannual Symposium and other meetings. Vannevar Bush was president of the Carnegie Insti-tution of Washington when this building was erected.

49TH COLD SPRING HARBOR SYMPOSIUMON QUANTITATIVE BIOLOGY
Recombination at the DNA LevelMay 30-June 6
248 participants
The principle of genetic linkage in Mendelian inheritance was established by T.H.Morgan in 1911. That genetic recombination occurs by chromosomal crossing-over events was first shown in two independent studies, both published in1931 -one by H.S. Creighton and B. McClintock working with maize and theother by C. Stern working with Drosophilia. Since that time a small, elite group ofgeneticists have thought about and experimentally analyzed genetic recombina-tion with such profound consequences that introductory genetics was, and still is,learned by studying chromosome recombination.
Publication by R. Holliday in 1964 of the first conceptual framework to explaingenetic recombination in molecular terms spurred an ever-increasing number ofscientists to work in this area. However, the main tool of investigation remainedstrictly transmission genetics until the dawn of recombination in molecular detail.Our choice of the topic "Recombination at the DNA Level" for the 49th annualSymposium underscores these recent achievements as well as the future pros-pects in this field. In planning this Symposium, we emphasized primarily the bio-chemical aspects of recombination, including systems ranging from bacterio-phage to mammalian cells.
To make this meeting most exciting and informative, we called upon the ad-vice of our colleagues in the field. In particular we wish to acknowledge the won-derful counsel given by Charles Radding, Nick Cozzarelli, Barbara McClintock,and Bob Weisberg. The formal meeting program contained 94 excellent presen-tations, with a high proportion of attendees staying through to the very end ofthis rather lengthy meeting. The introductory remarks by Bruce Alberts and thesummary delivered by Allan Campbell were most appropriate; we thank them fortheir efforts.
Financial support for this year's meeting was provided by the National CancerInstitute, Fogarty International Center, the Department of Energy, and the Na-tional Science Foundation.
Introduction B. Alberts, University of California, San Francisco
SESSION 1 CHROMOSOME MECHANICS
Chairperson: F. Stahl, University of Oregon, Eugene, Oregon
Rossignol, J.-L., Nicolas, A., Hamza,H., Kalogeropoulos, A., Langin, T,Laboratoire d'Etude des Interac-tions Moleculaires Genomiques,Universite Paris, France: Origins ofgene conversion and reciprocalexchange in Ascobolus.
Koh li, J., Munz, P., Aebi, R., Amstutz,H., Gysler, C., Heyer, W., Schuch-ert, P., Szankasi, P., Leupold, U.,Institute of General Microbiology,University of Bern, Switzerland: In-tergenic conversion betweentRNA genes and the influence of
specific mutations on allelic re-combination.
Carpenter, A.T.C., Dept. of Biology,University of California, SanDiego, La Jolla: Meiotic coconver-sion length in Drosophila recombi-nation-defective mutants.
171

A. Klar, B. McClintock
Esposito, M.S.,' Hosoda, J.,1 Go lin,J.,2 Moise, H.,' Bjornstad, K.,' Me-lees, D.,"Lawrence BerkeleyLaboratory, University of Califor-nia, Berkeley; 2Central Research
SESSION 2 YEAST SYSTEMS
and Development, E.I. Dupont deNemours, Wilmington, Delaware:Recombination in S. cerevis-iae-rec-gene mutants and ssDNA-binding proteins.
Chairperson: J. Strathern, Cold Spring Harbor Laboratory, New York
Fogel, S., Welch, J.W., Louis, E.,Dept. of Genetics, University ofCalifornia, Berkeley: Meiotic geneconversion mediates gene amplifi-cation in yeast.
Haber, J., Borts, R., Lichten, M.,
Hearn, M., Rosenstiel Center,Brandeis University, Waltham,Massachusetts: Initiation and tim-ing of meiotic recombination inyeast.
Klar, A., Strathern, J., Hicks, J., Cold
A. Campbell, N. Cozzarelli
172
J. Strathern
Hastings, P.J., Dept. of Genetics, Uni-versity of Alberta, Edmonton, Can-ada: Conversion by mismatch re-pair in yeast.
Spring Harbor Laboratory, NewYork: Initiation and resolution stepsof yeast mating-type gene trans-position.
Kostriken, R.,' Heffron, F.,2 'ColdSpring Harbor Laboratory, NewYork; 2Dept. of Molecular Biology,Salk Institute, La Jolla, California:The product of the HO gene is asite-specific, double-strand endo-nuclease that initiates mating-typeswitching.
Jensen, R., Stern, M., Herskowitz, I.,Dept. of Biochemistry and Bio-physics, University of California,San Francisco: Multifaceted con-trol of HO-gene expression-Pat-tern of mating-type interconver-sion.
Miller, A.M., Nasmyth, K.A., MRCLaboratory of Molecular Biology,Cambridge, England: Role forDNA replication in the control ofgene expression at a distance.
Butow, R.A., Hudson, A.P., Zinn, A.,Dept. of Biochemistry, Universityof Texas Health Science Center,Dallas: Gene conversion at theyeast mitochondrial 21S rDNA lo-cus.

SESSION 3 MAMMALIAN HOMOLOGUS RECOMBINATION
Chairperson: 0. Smithies, University
Folger, K., Thomas, K., Capecchi,M.R., Dept. of Biology, Universityof Utah, Salt Lake City: Analysis ofhomologous recombination in cul-tured mammalian cells.
Lin, F. -L., Sperle, K., Sternberg, N.,NCI, Frederick Cancer ResearchFacility, Frederick, Maryland: Ho-mologous recombination in cul-tured mouse L cells.
Brenner, D.A., Smigocki, A.C., Ca-merini-Otero, R.D., NIADDK, Na-tional Institutes of Health, Be-thesda, Maryland: Effect ofheterology on homologous recom-bination between mutant tk genesin mouse Ltk- cells.
Smithies, 0.,' Koralewski, M.A.,'Song, K.-Y.,2 Kucherlapati, R.S.,2'Laboratory of Genetics, Univer-sity of Wisconsin, Madison;2Center for Genetics, University ofIllinois College of Medicine, Chi-cago: Homologous recombinationwith DNA introduced into mam-malian cells.
Smith, A.J.H., Berg, R, Dept. of Bio-chemistry, Stanford University,California: Measurements of thefrequencies of homologous re-combination events between de-fective neo genes in mouse 3T6cells.
SESSION 4 TRANSPOSONS
of Wisconsin, Madison, Wisconsin
Liskay, R.M., Stachelek, J.L., Letsou,A., Depts. of Therapeutic Radiol-ogy and Human Genetics, YaleUniversity School of Medicine,New Haven, Connecticut: Homol-ogous recombination between tan-demly duplicated genes in cul-tured mouse cells.
Kucherlapati, R., Hannaken, A.,Spenser, J., Moore, P., Dept. of Mi-crobiology and Immunology, Uni-versity of Illinois College of Medi-cine, Chicago: Study ofhomologous recombination inmonkey cells and human cell-freeextracts.
B. Alberts, 0. Smithies, P. Starlinger
Chairperson: N. Kleckner, Harvard University, Cambridge, Massachusetts
Redfield, R., Campbell, A., Dept. ofBiological Sciences, Stanford Uni-versity, California: Generation ofnontandem duplications by homol,ogous recombination with a A pro-phage.
Ohtsubo, E.,' Ishizaki, K.,2 'Instituteof Applied Microbiology, Univer-sity of Tokyo; 2Radiation BiologyCenter, Kyoto University, Japan:Functional difference of the twoends of insertion element IS/ intransposition and cointegration.
Berg, D.E., Lodge, J., Weston-Hafer,K., Lowe, J.B., Carle, G.F., Sasak-awa, C., Depts. of Microbiologyand Immunology, and Genetics,Washington University MedicalSchool, St. Louis, Missouri: Speci-
ficity determinants in Tn5 transpo-sition.
Sherratt, D.,' Dyson, R,2 Brown, L.,'Summers, D.,' Stewart, G.,"Dept.of Genetics, University of Glas-gow, Scotland; 2Dept. of Microbi-ology, Dalhousie University, Hali-fax, Canada: Site-specificrecombination in transposition andplasmid stability.
Morisato, D., Bender, J., Kleckner, N.,Dept. of Biochemistry and Molecu-lar Biology, Harvard University,Cambridge, Massachusetts: Physi-cal and genetic analysis of theTn 10 transposition.
Reed, R.R., Moser, C.D., Dept. of Mo-lecular Biology and Genetics,Johns Hopkins School of Medi-
cine, Baltimore, Maryland: Map-ping regions of the transposase im-portant in the recognition of Tn3-family terminal inverted repeats.
Weinert, T.A., Grindley, N.D.F., Dept.of Molecular Biophysics and Bio-chemistry, Yale University, NewHaven, Connecticut: Transpositionof Tn903- Evidence for both repli-cative and conservative modes oftranspositional recombination.
Newman, B., Falvey, E., Wiater, L.,Grindley, N., Dept. of MolecularBiophysics and Biochemistry, YaleUniversity, New Haven, Connecti-cut: Site-specific recombination bythe y d resolvase-Analysis of theprotein and its interaction with theres site.
173

SESSION 5 Mu
Chairperson: N. Symonds, University of Sussex, Sussex, England
Akroyd, J., Barton, B., Lund, P., Chaconas, G., Gloor, G., Miller, J.L.,Smith, S.M., Sultana, K., Symonds, Giddens, E.B., Depts. of Biochem-N., University of Sussex, School of istry and Microbiology, and Immu-Biological Sciences, England: Two nology, University of Western On-aspects of Mu recombination. tario, London, Canada: Studies on
Patterson, TA., Martin, K.A., Weiss, bacteriophage Mu DNA transposi-R., Gould, J., Bukhari, A.I., Cold tion.Spring Harbor Laboratory, New Kahmann, R., Mertens, G., Max-York: Studies of the expression Planck-Institut, Berlin, Federal Re-and regulation of bacteriophage public of Germany: Substrate andMu genes using Mu -IacZ fusions, enzyme requirements for in vitro
Harshey, R.M., Dept. of Molecular site-specific recombination in bac-Biology, Scripps Clinic and Re- teriophage Mu.search Foundation, La Jolla, Cali- Plasterk, R.H.A., van de Putte, P.,fornia: One mode of bacterio- Laboratory of Molecular Genetics,phage Mu transposition is not State University of Leiden, Thecoupled to replication. Netherlands: Inversion of the G-
SESSION 6 PLANT TRANSPOSONS/T4 RECOMBINATION
DNA segment in bacteriophageMu and related systems in pro-karyotes.
Kamp, D., Kardas, E., Ritthaler, W.,Max-Planck-Institut, Martinsried,Federal Republic of Germany:Comparative analysis of invertibleDNA in phage genomes.
Shapiro, J.A., Dept. of Microbiology,University of Chicago, Illinois: Pro-gramming of DNA rearrangementsinvolving Mu prophages.
Chairperson: P. Star linger, University of Cologne, Cologne, Federal Republic of Germany
Dellaporta, S.L.,' Chomet, P.S.,1 Mot- gie Institution of Washington, Balti-tinger, J.P.,2 Hicks, J.B.,"Cold more, Maryland; 2Dept. of Bio-Spring Harbor Laboratory, New chemistry, University ofYork; 2University of Rhode Island, Minnesota, St. Paul; 3Dept. of Ge-Kingston: Spontaneous mutation netics, University of Wisconsin,caused by insertion elements in Madison: Molecular analysis ofmaize. transposable controlling elements
Courage, U., D6ring, H.-P., Kunze, R., in maize.Laird, A., Merckelbach, A., Willer- Peacock, W.J.,' Dennis, E.S.,' Ger-Neumann, M., Star linger, P., Till- lack, W.L.,1 Schwartz, D.,2mann, E., Weck, E., Werr, W., 'Division of Plant Industry, CSIRO,Yoder, J., Institut fOr Genetik, Univ- Canberra, Australia; 2Dept. of Biol-ersitat Kan, Federal Republic of ogy, Indiana University, Blooming-Germany: Transposable elements ton: Insertion and excision of Ds-Ac and Ds in Z. mays. elements in maize.
Fedoroff, N.,' Kelly, S.,' Pohlman, R.,2 Saedler, H., Bonas, U., Harrison, B.,Messing, J.,2 Furtek, D.,3 Nelson, Krebbers, E., Nevers, P., Piotrow-0.,"Dept. of Embryology, Carne- iak, R., Sommer, H., Upadhyaya,
SESSION 7 TOPOISOMERASE, RESOLVASE, AND GYRASE
Chairperson: N. Cozzarelli, University of California, Berkeley
Benjamin, H., Wasserman, S., Ger-rard, S., Dungan, J., Matzuk, M.,Krasnow, M., Cozzarelli, N., Dept.of Molecular Biology, University ofCalifornia, Berkeley: Topologicaland structural analysis of the re-combination by Tn3 resolvase.
Ikeda, H., Miura, A., Shiozaki, M., In-stitute of Medical Science, Univer-sity of Tokyo, Japan: Nonhomolo-gous recombination mediated byE. coli DNA gyrase- Possible in-volvement of DNA replication.
174
Wang, J., Dept. of Biochemistry/Mo-lecular Biology, Harvard Univer-sity, Cambridge, Massachusetts:Breakage and rejoining of DNAstrands by DNA topoisomerases.
O'Connor, M.B., Malamy, M.H., TuftsUniversity School of Medicine,Boston, Massachusetts: Mappingof in vivo DNA-gyrase cleavagesites in pBR322 and their role insite-specific recombination.
Menzel, R., Gellert, M., NIADDK, Na-tional Institutes of Health, Be-
K., Gierl, A., KlOsgen, R.B., Peter-son, PA., Schwartz-Sommer, Z.,Wienand, U., Max-Planck-Institute,Cologne, Federal Republic of Ger-many: Transposable elements inA. majus and Z. mays.
Formosa, T., Alberts, B.M., Dept. ofBiochemistry and Biophysics, Uni-versity of California, San Fran-cisco: Studies on genetic recombi-nation proteins of bacteriophageT4.
Mosig, G., Dept. of Molecular Biology,Vanderbilt University, Nashville,Tennessee: Processing of recom-binational intermediates in bacte-riophage T4.
thesda, Maryland: Control of theexpression of DNA gyrase in E.coll.
Champoux, J.J., McCoubrey, W.K.,Been, M.D., Dept. of Microbiologyand Immunology, University ofWashington School of Medicine,Seattle: Specificity of eukaryotictype-1 DNA topoisomerase.

SESSION 8 E. coli GENERAL RECOMBINATION
Chairperson: A. Clark, University of California, Berkeley
Mahajan, S.K., Pandit, N.N., Sarkari,J.F., Biology and Agriculture Divi-sion, Bhabha Atomic ResearchCentre, Bombay, India: Host func-tions in TN9 amplification-A newmodel for DNA amplification.
Clark, A.J., Sandler, S.J., Blanar,M.A., Willis, D.K., Chu, C., Univer-sity of California, Berkeley: Genesof the RecF pathway of recombi-nation in E, coll.
Kushner, S.R., Dykstra, C.C., Dept. ofGenetics, University of Georgia,Athens: Purification and character-ization of exonuclease V from E.coil K-12.
Lloyd, R., Picksley, S., Attfield, P.,Benson, F, Buckman, C., Dept. ofGenetics, University of Not-tingham, England: Genetic analy-sis and regulation of inducible re-combination in E. co/i.
Hays, J.B., Smith, A.T., Friedman,S.F., Lee, E., Coffman, G.L., Dept.of Chemistry, University of Mary-land-Baltimore County, Catons-ville, Maryland: Recombination ofnonreplicating UV-irradiated bac-teriophage A DNA.
Smith, G.R., Amundsen, S.K., Chau-dhury, A.M., Cheng, K.C., Ponti-celli, A.S., Roberts, C.M., Schultz,D.W., Taylor, A.F., Fred Hutchinson
SESSION 9 recA
Cancer Research Center, Seattle,Washington: Roles of RecBC en-zyme and Chi sites in homologousrecombination.
Kobayashi, I., Stahl, M.M., Stahl, F.W.,Institute of Molecular Biology, Uni-
versity of Oregon, Eugene: Spe-cial sites in homologous recombi-nation-Mechanism of the Chi-cosinteraction in RecA-RecBC-me-diated recombination in bacterio-phage 1.
Top: S. Shaner, R. Kahn, S. Honigberg, R.P. Cunningham, D. Gonda, J. Flory,C.M. Radding, T. Shibata, M. LeahyBottom: M. Bianchi, C. DasGupta, S. Chow
Chairperson: C. Radding, Yale University, New Haven, Connecticut
Muniyappa, K., Tsang, S.S., Shaner,S., Radding, C., Depts. of HumanGenetics and Molecular Biophys-ics and Biochemistry, Yale Univer-sity School of Medicine, New Ha-ven, Connecticut: Relationship ofsecondary structure, aggregation,and conjunction of DNA moleculesto the mechanism of homologouspairing promoted by recA protein.
Morrical, S., Neuendorf, S., Cox, M.,Dept. of Biochemistry, Universityof Wisconsin, Madison: Light-scat-tering studies of recA protein fila-ments in the presence and ab-sence of ssDNA.
Bryant, F.R., Riddles, P.W., Lehman,I.R., Dept. of Biochemistry, Stan-ford University School of Medi-
cine, California: Studies of themechanism of strand exchangepromoted by the recA protein of E.co /i.
Shibata, T.,1 Makino, O.,' Ohtani, T.,'Iwabuchi, M.,1 lkawa, S.,' Ando,T.,' Shibata, Y,2 Maeda, H.,''Dept. of Microbiology, Riken Insti-tute, Saitama; 2Blood TransfusionService, Tokyo University Hospital,Japan: Roles of processive un-winding by E. coli recA protein inhomologous recombination.
Griffith, J., Chrysogelos, S., Register,J. III., Welsh, G., Lineberger Can-cer Research Center, University ofNorth Carolina, Chapel Hill: A hy-brid complex involving SSB andRecA proteins bound to ssDNA in-
175

A. Skalka, A. Hershey
SESSION 10 REPAIR
Chairperson: S. Linn, University of California, Berkeley
Linn, S., Samson, L., Brooks, P, Mos-baugh, D., Evans, D., Dept. of Bio-chemistry, University of California,Berkeley: Enzymology of DNA ex-cision repair in mammalian cells.
Lu, A.-L., Welsh, K., Su, M., Clark, S.,Modrich, P., Dept. of Biochemistry,Duke University Medical Center,Durham, North Carolina: Repair ofDNA base-pair mismatches in vitro.
Walker, G.C., Pang, RP., Tsen, S.D.,Dept. of Biology, MassachusettsInstitute of Technology, Cam-
bridge: Mismatch repair genes ofS. typhimurium.
Fishel, R.A., Kolodner, R., Laboratoryof Molecular Genetics, Dana Far-ber Cancer Institute, and Dept. ofBiological Chemistry, HarvardMedical School, Boston, Massa-chusetts: An E. coil cell-free sys-tem that catalyzes the repair ofsymmetrically methylated hetero-duplex DNA.
Radman, M., Mutagenes Group, Insti-tut Jacques Nomod, RBM, Paris,
itiates the proper assembly ofRecA-DNA filaments.
Stasiak, A., Stasiak, A.Z., Koller, T., In-stitute for Cell Biology, Zurich,Switzerland: Complexes of recAwith DNA under condition of in vi-tro recombination.
Howard-Flanders, P., West, S.C.,Rusche, J.R., Dept. of MolecularBiophysics and Biochemistry, andDept. of Therapeutic Radiology,Yale University, New Haven, Con-necticut: recA protein can promotestrand-exchange reactions pastdouble-strand breaks in certainpositions, indicating that eachrecA monomer may have two non-equivalent DNA-binding sites.
France: Genetic consequences ofmismatch repair in E. coli.
Kunes, S., Botstein, D., Fox, M.S.,Dept. of Biology, MassachusettsInstitute of Technology, Cam-bridge: Inverted dimer formationand interplasmid recombination inthe repair of linearized plasmidsduring yeast transformation.
Rothstein, R., Dept. of Microbiology,New Jersey Medical School, New-ark: Double-strand-break repairand recombination.
SESSION 11 EUCARYOTIC ENZYMES
Chairperson: R. Holliday, National Institute for Medical Research, London, England
Resnick, M., Chow, T., Nitiss, J., Na-tional Institute of EnvironmentalHealth Sciences, Research Trian-gle Park, North Carolina: Changesin chromosomal DNA duringmeiosis in repair mutants and thepossible role of a deoxyribonucle-ase.
Skalka, A.M.,' Duyk, G.,2 Longiaru,M.,' DeHaseth, F,2 Terry, R.,' Leis,J.,2 'Dept. of Molecular Genetics,Hoffmann-La Roche Inc., RocheResearch Center, Nutley, New Jer-sey; 2Dept. of Biochemistry, CaseWestern Reserve University,
176
Cleveland, Ohio: Integrative re-combination-A role for the retro-viral reverse transcriptase.
Honjo, T, Kataoka, T, Konda, S.,Nishi, M., Kodaira, M., Dept. ofGenetics, Osaka University Medi-cal School, Japan: In vitro studieson immunoglobulin gene rear-rangement.
Holliday, R.,' Taylor, S.Y.,' Kmiec, E.,2Holloman, W.K.,2 'National Insti-tute for Medical Research, Lon-don, England; 2Dept. of Immunol-ogy and Medical Microbiology,University of Florida, Gainesville:
Characterization of red mutantsand recl protein of U. maydis.
Kmiec, E., Kroeger, F, Holloman, W.,University of Florida, Gainesville:Interaction between Ustilago reclprotein and Z-DNA.
Ferro, A.M., McElwain, M.C., Reid,E.J., Olivera, B.M., Dept. of Biol-ogy, University of Utah, Salt LakeCity: Poly ADP-ribose synthetase,DNA topoisomerase I and thesuppression of strand-exchangeevents in somatic cells.

SESSION 12 INTEGRATION AND EXCISION OF BACTERIOPHAGE
Chairperson: H. Nash, National Institutes of Health, Bethesda, Maryland
Miller, H.I., Dept. of Molecular Biol- nism of recombination by A Int andogy, Genentech Inc., South San Tn3 resolvase.Francisco, California: himA gene Spengler, S.,1 Stasiak, A.,2 Cozzarelli,of E. coli-Gene structure and reg- N.,"Dept. of Molecular Biology,ulation, homology with DNA-bind- University of California, Berkeley;ing proteins, and role in site-spe-cific recombination and generegulation.
Leong, J.,' Nunes-Duby, S.,' Oser,A.,1 Lesser, C.,' Youderian, P.,2Susskind, M.,2 Landy, A.,"Dept.of Molecular and Cell Biology,Brown University, Providence,Rhode Island; 2Dept. of Microbiol-ogy, University of Massachusetts,Worcester: Site-specific recombi-nation systems of +80, P22, and A.
de Massy, B.,1 Studier, F.W.,2 Dorgai,L.,' Appelbaum, E.,' Weisberg,R.,' 'NICHD, National Institutes ofHealth, Bethesda, Maryland;'Dept. of Biology, Brookhaven Na-tional Laboratory, Upton, NewYork: Role of Holliday structures inrecombination.
Echols, H.,1 Dodson, M.,' Roberts,J.D.,2 McMacken, R.,2 'Dept. ofMolecular Biology, University ofCalifornia, Berkeley; 2Dept. of Bio-chemistry, Johns Hopkins Schoolof Hygiene and Public Health, Bal-timore, Maryland: Role of special-ized nucleoprotein structures insite-specific recombination and ini-tiation of DNA replication.
Nash, H.A., Kitts, P., Richet, E., Na-tional Institute of Mental Health,Bethesda, Maryland: A Integrativerecombination-Approaches tothe synaptic mechanism.
Spengler, S.,' Stasiak, A.,2 Benjamin,H.,' Wasserman, S.,' Gerrard, S.,'Dungan, J.,' Matzuk, M.,' Kras-now, M.,' Cozzarelli, N.,"Dept. ofMolecular Biology, University ofCalifornia, Berkeley; 2lnstitute forCell Biology, Zurich, Switzerland:Topological tests of the mecha-
2Institute for Cell Biology, Zurich,Switzerland: Structure of knotsand catenanes produced by AInt-Implications for mechanismand DNA structure.
Wine and Cheese Party
SESSION 13 SITE-SPECIFIC RECOMBINATION
Chairperson: J. Broach, State University of New York, Stony Brook, New York
Johnson, R., Bruist, M., Glaccum, M.,Simon, M., Biology Division, Cali-fornia Institute of Technology, Pas-
adena: Molecular mechanism ofphase variation.
Hoess, R., Abremski, K., Sternberg,
177
N., NCI, Frederick Cancer Re-search Facility, Frederick, Mary-land: Studies on the Cre-/ox site-

specific recombination system ofbacteriophage
lida, S., Huber, H., Hiestand-Nauer,R., Meyer, J., Bickle, T., Arber, W.,Biozentrum, University of Basel,Switzerland: Sequences used incrossover by the site-specific cinrecombinase of bacteriophage P1.
McLeod, M., Volkert, F., Broach, J.,
Dept. of Microbiology, State Uni-versity of New York, Stony Brook:Analysis of the FLP recombinationsystem of the yeast plasmid 2-pmcircle.
Sadowski, R, Lee, D., Beatty, L., An-drews, B., Babineau, D., Protean,G., Vetter, D., Dept. of Medical Ge-netics, University of Toronto, Can-
SESSION 14 RECOMBINATION IN VITRO
Chairperson: M. Gellert, National Institutes of Health, Bethesda, Maryland
Kolodner, R., Symington, L., Howard,M., Muster-Nassal, C., Dana Far-ber Cancer Institute, Dept. of Bio-logical Chemistry, Harvard Medi-cal School, Boston,Massachusetts: Plasmid recombi-nation catalyzed by cell-free ex-tracts of yeast.
Kemper, B., Depka, M.V., Hoffmann,H., Jensch, F., Institute of Ge-netics, University of Cologne, Fed-eral Republic of Germany: Resolu-tion of Holliday structures in
cruciform DNA by endonucleaseVII.
Higgins, N.P.," Krause, H.M.,1 Oliv-era, B.M.,2 'Dept.of Biochemistry,University of Alabama, Birming-ham; 2Dept. of Biology, Universityof Utah, Salt Lake City: Definitionof the Mu operator and the use ofrepressor in analyzing in vivo andin vitro intermediates of transposi-tion.
Mizuuchi, K., Mizuuchi, M., Craigie,R., NIADDK, National Institutes of
Summary: A. Campbell, Stanford University, Stanford, California
178
ada: Genetic recombination ofbacteriophage T7 and yeast 2-pmcircle DNA in vitro.
Cox, M., Bruckner, R., Meyer-Leon,L., Senecoff, J., Dept. of Biochem-istry, University of Wisconsin,Madison: Purification and proper-ties of the FLP protein of the yeast2-pm plasmid.
Health, Bethesda, Maryland:Mechanism of transposition ofbacteriophage Mu-Site-specificrecognition of Mu ends by the MuA-gene product and polarity of thestrand-transfer reaction.
Rosenberg, J., Dept. of Biological Sci-ences, University of Pittsburgh,Pennsylvania: Kinked DNA incrystalline complex with EcoRI en-donuclease.

MEETINGS
Molecular Biology of the CytoskeletonApril 25-April 29
ARRANGED BYGary Borisy, University of WisconsinDon Cleveland, Johns Hopkins Medical SchoolDouglas Murphy, Johns Hopkins Medical School
122 participants
The meeting Molecular Biology of the Cytoskeleton sought to bring together adiverse group of researchers whose principal interests center on some molecularaspect of cytoskeletal elements, but who by virtue of the particular experimentalapproach or organism employed do not have an opportunity for scientific ex-change at other meetings. To this end, 122 investigators were assembled whoseefforts included: genetic approaches to determination of function of cytoskeletalproteins in a diverse number of genetic systems, determination of the dynamicsof assembly of cytoskeletal components both in vivo and in vitro, biochemicalanalysis of various cytoskeletal proteins and their variants, and finally, dissectionof the regulatory pathways that control cytoskeletal gene expression. Overall, thecongregation of investigators using diverse approaches to study cytoskeletalfunction and regulation achieved the original goal of introducing workers in eacharea of cytoskeletal investigation to alternative approaches and concepts.
This meeting was supported in part by the National Science Foundation andthe following divisions of the National Institutes of Health: Fogarty InternationalCenter, National Cancer Institute, and the National Institute of General MedicalSciences.
Welcoming Remarks: D. Murphy, Johns Hopkins Medical School, Baltimore, Maryland
SESSION 1 STRUCTURE AND DYNAMICS OF THE CYTOSKELETON
Chairperson: G. Borisy, University of Wisconsin, Madison, Wisconsin
Borisy, G., Kronebusch, P., Labora-tory of Molecular Biology, Univer-sity of Wisconsin, Madison: Micro-tubule dynamics in vivo.
Kirschner, M., Mitchison, T., Dept. ofBiochemistry, University of Califor-nia, San Francisco: Microtubuledynamics in vitro.
Weber, K., Osborn, M., Geis ler, N.,Max-Planck-Institute, Goettingen,Federal Republic of Germany: In-termediate filaments-Structuraland immunological analysis.
Kreis, TE.,' Schlessinger, J.,2 Geiger,B.,2 'European Molecular BiologyLaboratory, Heidelberg, FederalRepublic of Germany; 2Dept. of
Chemical Immunology, WeizmannInstitute of Science, Rehovot, Is-rael: Dynamic properties of actin,a-actinin, and vinculin in focal con-tact areas.
Salmon, E.D.,' Saxton, W.M.,2 Leslie,R.J.,2 Karow, M.L.,2 Neighbors,B.,2 McIntosh, J.R.,2 'Dept. of Bi-ology, University of North Caro-lina, Chapel Hill; 2Dept. of Molecu-lar, Cellular, and DevelopmentalBiology, University of Colorado,Boulder: Measurements of spindlemicrotubule dynamics by fluores-cence redistribution after photo-bleaching.
S. Ward, R Anderson
179

SESSION 2 MOSTLY MOLDS: MOLECULAR GENETICS
Chairperson: D. Botstein, Massachusetts Institute of Technology, Cambridge,Massachusetts
Botstein, D., Shortie, D., Novick, P.,Thomas, J.H., Huffaker, T., Schatz,P., Dept. of Biology, Massachu-setts Institute of Technology, Cam-bridge: Applying old and newgenetic technology toward under-standing the cytoskeleton of yeast.
Kilmartin, J.V., MRC Laboratory ofMolecular Biology, Cambridge,England: Microtubules and actinfilaments during the yeast cellcycle.
Pringle, J.R., Coleman, K., Lillie, S.,Haarer, B., Adams, A., Jacobs, C.,Robinson, J., Evans, C., Stapleton,A., Division of Biological Sciences,University of Michigan, Ann Arbor:Cellular morphogenesis and thecytoskeleton in yeast.
Thomas, J.H., Botstein, D., Dept. ofBiology, Massachusetts Institute ofTechnology, Cambridge: Yeastmutants affecting spindle function.
Novick, P.J., Osmond, B., Botstein,D., Dept. of Biology, Massachu-setts Institute of Technology, Cam-bridge: Genetic analysis of actinand interacting proteins.
Rose, M., Fink, G.R., Whitehead Insti-tute for Biomedical Research andMassachusetts Institute of Technol-ogy, Cambridge: Mutations affect-ing nuclear fusion in yeast.
Birkett, C.R.,' Boston, R.S.,2 Burland,TG.,2 Dove, W.F.,2 Foster, K.E.,'Green, L.,2 Gull, K.,' Johnston,L.,2 Paul, E.C.A.,' Roobol, A.,'Schedl, T.,2 Wilcox, M.,''Biological Laboratory, Universityof Kent, Canterbury, England;2McArdle Laboratory, University ofWisconsin, Madison: Differentialexpression of the tubulin genefamily in the life cycle of Physarumpolycephalum.
Bur land, TG.,' Schedl, T.,' Gull, K.,2
SESSION 3 MOSTLY MITOSIS: CENTROMERES AND KINETOCHORES
Chairperson: M. Kirschner, University of California, San Francisco, California
Amaya, E., Bloom, K., Dept. of Biol-ogy, University of North Carolina,Chapel Hill: Molecular architectureof a yeast centromere.
Valdivia, M.M.,' Sykes, R.C.,2 Chi-nault, A.C.,2-3 Brinkley, B.R.,'Depts. of 'Cell Biology,2Biochemistry, 3Medicine, BaylorCollege of Medicine, Houston,Texas: Biochemical studies of thekinetochore/centromere of mam-malian chromosomes.
Cox, J.V., Olmsted, J.B., Dept. of Biol-ogy, University of Rochester, NewYork: Kinetochore antigens-Syn-thesis, phosphorylation, and evo-lutionary conservation.
Vandre, D.D.,' Davis, F.M.,2 Borisy,G.G.,"Laboratory of MolecularBiology, University of Wisconsin,Madison; 2University of Texas,M.D. Anderson Hospital and Tu-mor Institute, Houston: Phospho-proteins are components of micro-
SESSION 4 MOSTLY WORMS: MOLECULAR GENETICS IN WORMS AND HIGHEREUKARYOTES
Chairperson: E. Raff, Indiana University, Bloomington, Indiana
Raff, E.C., Fuller, M.T, Caulton, J.H.,Rudolph, J.E., Kimble, M., Hutch-ens, J.A., Dept. of Biology, IndianaUniversity, Bloomington: Tubulingene expression and microtubulefunction in Drosophila.
Cox, K.H., Angerer, L.M., Angerer,R.C., Dept. of Biology, Universityof Rochester, New York: Individual
180
actin gene expression-Markersfor early differentiation of cell lin-eages in developing sea urchinembryos.
Bejsovec, A., Eide, D., Anderson, P.,Dept. of Genetics, University ofWisconsin, Madison: Genetic tech-niques for analysis of nematodemuscle.
Dove, W.F.,"McArdie Laboratory,University of Wisconsin, Madison;2Biological Laboratory, Universityof Kent, Canterbury, England: Ge-netics of the tubulin gene familiesin Physarum.
Morris, N.R., Weatherbee, J.A., Gam-bino, J., Bergen, L.G., Dept. ofPharmacology, UMDNJ-RutgersMedical School, Piscataway, NewJersey: Tubulins of Aspergillus ni-dulans-Genetics, biochemistry,and function.
Lai, E.Y., Remillard, S.P., Fulton, C.,Dept. of Biology, Brandeis Univer-sity, Waltham, Massachusetts: Thea-tubulin gene family of Naegleriaand its expression during differen-tiation.
tubule organizing centers of themitotic spindle in Chinese hamsterovary cells.
Mitchison, T.J., Kirschner, M., Dept. ofBiochemistry, University of Califor-nia, San Francisco: Kinetochoresas in vitro microtubule organizingcenters.
Ward, S., Dept. of Embryology, Car-negie Institution of Washington,Baltimore, Maryland: Acquisitionof cellular asymmetry during C.elegans spermatogenesis.
Epstein, H.F, Ortiz, I., Berliner, G.C.,Miller, D.M. III, Dept. of Neurology,Baylor College of Medicine, Hous-ton, Texas: Functionally distinct

myosins in C. elegans analyzedthrough mutants.
Honda, S., Berman, S.A., Epstein,H.F., Dept. of Neurology, BaylorCollege of Medicine, Houston,
Texas: Regulation of myosinheavy-chain expression in C. ele-gans.
Krause, M.,' Wild, M.,' Hirsh, D.,' Wa-terston, R.,' 'Dept. of Molecular,
SESSION 5 MOSTLY CHO AND CHLAMY: MOLECULAR GENETICS OFCYTOSKELETAL PROTEINS
Chairperson: J. Rosenbaum, Yale University, New Haven, Connecticut
Cabral, F., Schibler, M., Division ofEndocrinology, University of TexasMedical School, Houston: Analysisof the role of microtubules in mito-sis using CHO mutants requiringtaxol for cell growth.
Gottesman, M.M.,1 Abraham, I.,' Kuri-yama, R.,2 Mackensen, S.,' Whit-field, C.,"NCI, National Institutesof Health, Bethesda, Maryland;'Dept. of Molecular Biology, Uni-versity of Wisconsin, Madison: Ge-netic analysis of microtubule func-tion in CHO cells.
Abraham, I., McClurkin, C., Gottes-man, M.M., NCI, National Insti-tutes of Health, Bethesda, Mary-land: Cross-resistance of CHOcells with tubulin mutations.
Whitfield, C., Doherty, P., Gottesman,M.M., NCI, National Institutes ofHealth, Bethesda, Maryland: Clon-ing wild-type and mutant tubulinsfrom CHO cells.
Elliott, E.M.,' Okayama, H.,' Sarangi,F.,' Henderson, G.,' Ling, V.,1
'Dept. of Medical Biophysics, Uni-versity of Toronto and OntarioCancer Institute, Canada;'National Institutes of Health, Be-thesda, Maryland: Expression ofmultiple a-tubulin genes in CHOcells.
Dutcher, S.K.,1 Ramanis, Z.,' Luck,D.J.L.,2 'Dept. of Molecular, Cellu-lar, and Developmental Biology,University of Colorado, Boulder;'Rockefeller University, New York:Flagellar mutants of C. reinhardtiithat show altered molecular phe-notypes in isolated basal bodypreparations.
Youngblom, J.,' Schloss, J.,2 Silflow,C.,"Dept. of Genetics and CellBiology, University of Minnesota,St. Paul; 'Dept. of Biology, YaleUniversity, New Haven, Connecti-cut: Comparison of the two f3-tubu-lin genes in Chlamydomonas.
Brunke, K.,12 Anthony, J.,1,2 Stern-berg, Weeks, D.,2 'Institutefor Cancer Research, Fox Chase
SESSION 6 PROTEIN ISOFORMS
Chairperson: D. Murphy, Johns Hopkins Medical School, Baltimore, Maryland
Murphy, D.B., Grasser, W.A., Wallis,K.T., Dept. of Cell Biology andAnatomy, Johns Hopkins MedicalSchool, Baltimore, Maryland:Expression of a unique /3-tubulinvariant in chicken-red-cell devel-opment.
Craig, S.W.,' Pardo, J.V.,2 Pittenger,M.F.,"Johns Hopkins MedicalSchool, Baltimore, Maryland;'Duke Medical School, Durham,North Carolina: Suellular sorting ofisoactins.
Blose, S.H., Matsumura, F., ColdSpring Harbor Laboratory, NewYork: Molecular analysis of micro-tubules rapidly purified from cells
using monoclonal-antibody-in-duced aggregation.
Vallee, R., Bloom, G., Luca, F., Cell Bi-ology Group, Worcester Founda-tion for Experimental Biology,Shrewsbury, Massachusetts: Dif-ferential cellular and suellular dis-tribution of microtubule-associatedproteins.
Lazarides, E., Nelson, W.J., Kasa-matsu, T., Division of Biology, Cali-fornia Institute of Technology, Pas-adena: Expression, assembly, andtopogenesis of membrane-cyto-skeletal domains in neurons.
Piperno, G., Rockefeller University,New York: Analysis of the mitotic
Cellular, and Developmental Biol-ogy, University of Colorado, Boul-der; 'Dept. of Genetics, Washing-ton School of Medicine, St. Louis,Missouri: C. elegans actin genes.
Cancer Center, Philadelphia,Pennsylvania; 2Zoecon Corp., PaloAlto, California; 3Smith-Kline andFrench Laboratories, Swede land,Pennsylvania: Coordinate expres-sion of the four tubulin genes inChlamydomonas.
Lefebvre, P. Barsel, S., Stuckey, M.,Swartz, L., Dept. of Genetics andCell Biology, University of Minne-sota, St. Paul: Isolation and char-acterization of mutants of Chlamy-domonas defective in theregulation of flagellar gene expres-sion.
Kuchka, M., Jarvik, J., Dept. of Bio-logical Sciences, Carnegie-MellonUniversity, Pittsburgh, Pennsylva-nia: Genetic analysis of flagellarsize control in C. reinhardtii.
Baldwin, D., Chojnacki, B., Jarvik, J.,Dept. of Biological Sciences, Car-negie-Mellon University, Pitts-burgh, Pennsylvania: Defective fla-gellar assembly in stumpy flagellamutants of C. reinhardtii.
apparatus by monoclonal anti-bodies to components of flagellaraxonemes.
Wiche, G.,' Briones, E.,' Koszka, C.,'Leichtfried, FE.,1 Krepler, R.,' Art-lieb, U.,2 'Institute of Biochemistry;'Dept. of Pathology, University ofVienna, Austria: Widespread oc-currence of microtubule-associ-ated proteins MAP 1 and MAP 2 innonneuronal cells/tissues and as-sociation of both polypeptides withmicrotubules polymerized in vitro.
Aebi, U.,' Salvo, H.,' Leavitt, J.,2'Dept. of Cell Biology and Anat-omy, Johns Hopkins MedicalSchool, Baltimore, Maryland;
181

2Linus Pau ling Institute of Scienceand Medicine, Palo Alto, Califor-nia: In vitro characterization of amutant f3 -actin expressed in a ma-lignant human fibroblast cell line.
Cappuccinelli, P., Institute of MedicalMicrobiology, University of Sas-sari, Sardinia, Italy: Immunological
heterogeneity of cytoskeletal pro-teins in eukaryotic microorga-nisms.
Owens, R.J., Totty, N., Waterfield,M.D., Crumpton, M.J., ImperialCancer Research Fund, Lincoln'sInn Fields, London, United King-dom: Structural characterization of
SESSION 7 MOSTLY MAMMALIAN GENE EXPRESSION AND REGULATION
Chairperson: D. Cleveland, Johns Hopkins Medical School, Baltimore, Maryland
Cleveland, D.W.,' Pittenger, M.F,'Feramisco, J.R.,2 Havercroft,J.C.,"Dept. of PhysiologicalChemistry, Johns Hopkins Univer-sity School of Medicine, Baltimore,Maryland; 2Cold Spring HarborLaboratory, New York: Autoregu-lated expression of vertebrate tu-bulins.
Lee, M.G.-S., Cowan, N.J., Dept. ofBiochemistry, New York UniversityMedical Center, New York: Modu-lation of mRNA levels transcribedfrom a human /3- tubulin genecloned in a eukaryotic shuttle vec-tor.
J. Spudich
Baker, E., Bandziulis, R., Keller, L.,Rosenbaum, J., Schloss, J., Dept.of Biology, Yale University, NewHaven, Connecticut: Synthesisand accumulation of mRNAs forflagellar proteins during flagellarregeneration in Chlamydomonas.
Schedl, T,' Burland, T.G.,, Gull, K.,2Dove, W.F.,' McArdle Laboratory,University of Wisconsin, Madison;?Biological Laboratory, Universityof Kent, Canterbury, England:Regulation of tubulin expression inthe Physarum cell cycle.
Landfear, S.M., Wirth, D.F, HarvardSchool of Public Health, Boston,Massachusetts: Structure and de-velopmental regulation of tubulingenes in the parasitic protozoonLeishmania enriettii.
Farmer, S.R., Bond, J.F., Robinson,G.S., Mbangkollo, D., Fenton,M.J., Knight , G.B., Berkowitz,E.M., Dept. of Biochemistry, Bos-ton University Medical School,Massachusetts: Differentialexpression of the p-tubulin multi-gene family during rat brain devel-opment-Identification of neuralspecific and constitutive mRNAs.
Sullivan, K.F., Lau, J.T.Y., Cleveland,D.W., Dept. of PhysiologicalChemistry, Johns Hopkins Univer-sity, School of Medicine, Balti-more, Maryland: Primary structureof a vertebrate p-tubulin gene fam-
SESSION 8 MOSTLY MUSCLE' GENE STRUCTURE AND EXPRESSION
Chairperson: S. Penman, Massachusetts Institute of Technology, Cambridge,Massachusetts
Penman, S., Dept. of Biology, Massa-chusetts Institute of Technology,Cambridge: Morphology and pro-tein composition of the structural
182
elements of cytoplasm and nu-cleus in cells and tissues.
Zimmer, W.E., Jr., Chang, K.S.,Bergsma, D., Schwartz, R., Dept.
p68, a new Ca2+-binding proteinof the cytoskeleton.
Maruta, H., Knoerzer, W., Isenberg,G., Max-Planck-Institute, Munich,Federal Republic of Germany:Nonpolymerizable variants regu-late actin polymerization in Physa-rum.
ily-At least three divergent ( -tu-bulin isotypes are expressed in thechicken.
Havercroft, J.C., Cleveland, D.W.,Dept. of Physiological Chemistry,Johns Hopkins University Schoolof Medicine, Baltimore, Maryland:Programmed expression of (3-tu-bulin genes during developmentand differentiation of the chicken.
Ginzburg, I., Littauer, U.Z., Dept. ofNeurobiology, Weizmann Instituteof Science, Rehovot, Israel:Expression and cellular regulationof microtubule proteins.
Drubin, D.,' Feinstein, 5.,2 Shooter,E.,2 Kirschner, M.,' 'Dept. of Bio-chemistry, University of California,San Francisco; 2Dept. of Neuro-biology, Stanford University, Cali-fornia: Expression of tau proteinand tau mRNA during PC12 differ-entiation.
Holtzer, H., Friedman, T, Forry-Schaudies, S., Pacifici, M., Dept.of Anatomy, University of Pennsyl-vania Medical School, Philadel-phia: Effects of Colcemid andtaxol on the synthesis and assem-bly of tubulin, vimentin, and des-min during chondrogenesis andmyogenesis.
Welch, W.J., Blose, S., Feramisco,J.R., Cold Spring Harbor Labora-tory, New York: Heat-shock re-sponse and the cytoskeleton.
of Cell Biology, Baylor College ofMedicine, Houston, Texas: Organi-zation and expression of thechicken actin multigene family.

Leavitt, J.,1 Ng, S.-Y.,1 Lin, C.-S.,'Gunning, P.,2 Kedes, L.,2 1LinusPau ling Institute of Science andMedicine, Palo Alto, California;'Dept. of Medicine, Stanford Medi-cal School and Veterans Adminis-tration Medical Center, California:Isolation and characterization ofwild-type and mutant human a -ac-tin genes.
Gunning, P.,1 Hickey, R.,2 Ng, 5.-Y,3Ponte, P.,1 Minty, A.,1 Erba, H.,'Leavitt, J.,3 Skoultchi, A.,2 Kedes,L.,"Stanford Medical School andVeterans Administration MedicalCenter, Palo Alto, California;'Albert Einstein School of Medi-cine, Bronx, New York; 3LinusPau ling Institute of Science andMedicine, Palo Alto, California:Human actin-gene structure, evo-lution, and expression in heterolo-gous cells.
Helfman, D.M., Feramisco, J.R., Kost,T.A., Yamawaki-Kataoka, Y., Ricci,W.M., Hughes, S.H., Cold SpringHarbor Laboratory, New York:Cloning by immunological screen-ing of cDNA expression libraries-Analysis of smooth-muscle a- andp-tropomyosins.
MacLeod, A.R., Talbot, K., Ludwig In-stitute for Cancer Research, MRC,Cambridge, England: Human cy-toskeletal tropomyosin genes.
G. Borisy, D. Cleveland, D. Murphy
Cooper, T., Ordahl, C., Dept. of Anat-omy, University of California, SanFrancisco: Genomic structure of atroponin-T isogene expressed ac-cording to two regulatory pro-grams in heart and skeletal mus-cle development.
Hammer, J.A. III, Korn, E.D., Pater-son, B.M., National Institutes ofHealth, Bethesda, Maryland: Clon-ing of the Acanthamoeba myosinheavy-chain genes.
Leinwand, L.,' de Lozanne, A.,2
SESSION 9 ENTIRELY INTERMEDIATE FILAMENTS
Chairperson: E. Fuchs, University of Chicago, Chicago, Illinois
Fuchs, E., Marchuk, D., Tyner, A.,McCrohon, S., Dept. of Biochemis-try, University of Chicago, Illinois:Expression of keratin genes in hu-man epithelia.
Steinert, P.,' Roop, D.,2 'DermatologyBranch; 'NCI, National Institutesof Health, Bethesda, Maryland:Amino acid sequences of epider-mal keratins correlate with theirdevelopmental expression.
Jorcano, J.L., 1,2 Rieger, M.,2 Franz,J.K.,2 Magin,T.M.,2 Schiller, D.L.,2Franke, W.W.,2 'Center of Molecu-lar Biology, University of Heidel-berg; 'Institute of Cell and TumorBiology, German Cancer ResearchCenter, Heidelberg, Federal Re-
public of Germany: Principles ofcytokeratin expression and cyto-keratin relationships.
Zehner, Z.,' Paterson, B.,2 'MedicalCollege of Virginia, Virginia Com-monwealth University, Richmond;'NCI, National Institutes of Health,Bethesda, Maryland: Sequence ofthe chicken vimentin gene.
Lazarides, E., Capetanaki, Y.G., Ngai,J., Division of Biology, CaliforniaInstitute of Technology, Pasadena:Vimentin- From gene to morpho-genesis.
Quax, W., Quax-Jeuken, Y., Egberts,W.V., van den Heuvel, R., Hen-driks, W., Bloemendal, H., Dept. ofBiochemistry, University of Nijme-
Lewis, M.,' Saez, L.,' Spudich, J.,2'Albert Einstein College of Medi-cine, Bronx, New York; 'StanfordUniversity School of Medicine,California: Isolation and character-ization of nonmuscle myosinheavy-chain cDNA clones fromDictyostelium.
Izant, J.G., Dept. of Genetics, FredHutchinson Cancer ResearchCenter, Seattle, Washington: Anti-sense gene transcription as a toolfor molecular analysis.
gen, Netherlands: Genes for vi-mentin and desmin.
Ben-Ze'ev, A., Dept. of Genetics,Weizmann Institute of Science, Re-hovot, Israel: Differential control ofcytokeratins and vimentin synthe-sis by cell-cell contact and cellspreading in cultured epithelialcells.
Roop, D.R.,1 Toftgard, R.,' Kronen-berg, M.S.,' Clark, J.H.,2 Yuspa,S.H.," NCI, National Institutes ofHealth, Bethesda, Maryland;'Dept. of Cell Biology, Baylor Col-lege of Medicine, Houston, Texas:Changes in keratin gene expres-sion during differentiation.
Closing Remarks: D. Cleveland, Johns Hopkins Medical School, Baltimore, Maryland
-183

Ciliate Molecular GeneticsMay 2-May 6
ARRANGED BY
Peter Bruns, Cornell UniversityMartin Gorovsky, University of Rochester
98 participants
Investigators from laboratories in the United States, Europe, and Japan attendedthe Ciliate Molecular Genetics meeting. Among the many high points of the meet-ing were: reports of telomere location for Paramecium surface antigens; indica-tions from three different laboratories that ciliates may have a somewhat uniquegenetic code (unspliced termination codons in coding regions); the description oftwo remarkable cases (one partly characterized at the molecular level) of cyto-plasmic mutants that alter macronuclear development; report of a Tetrahymenagene that is expressed only during conjugation and that appears to be homolo-gous to the zeste locus in Drosophila; demonstration that E. coil CAP binds in asequence-specific manner to the promoter of Tetrahymena rDNA; purification ofgamones responsible for mating in a hypotrichous ciliate; and demonstration of atopoisomerase-like molecule bound to specific sequences in the promoter regionof the Tetrahymena rDNA.
The meeting was reviewed both in Nature and Genetics.Funding for this meeting was supplied in part by Amicon Corporation; J.T.
Baker Chemical Co.; Boehringer Mannheim Biochemicals; Fotodyne, Inc.; H.P.Genenchem; Hans Biologics, Inc.; Kontes Scientific Glassware; Nikon, Inc.; Pro-mega Biotech; Schleicher & Schuell; Society for Developmental Biology, Inc.;and the following divisions of the National Institutes of Health: Fogarty Interna-tional Center and National Institute of General Medical Sciences.
SESSION 1 OVERVIEW
Chairperson: J. Engberg, University of Copenhagen, Copenhagen, Denmark
Gall, J.G., Dept. of Embryology, Car-negie Institution, Baltimore, Mary-land: Ciliate molecular genetics.
Prescott, D.M., Swanton, M.T., Roth,M., Klobutcher, L.A., Dept. of Mo-lecular, Cellular, and Develop-mental Biology, University of Colo-
rado, Boulder: Modification ofDNA in ciliate development.
'Freer, J., Jr., Dept. of Biology, IndianaUniversity, Bloomington: Over-view-Immobilization antigens.
Nanney, D.L., Dept. of Genetics andDevelopment, University of Illinois,
SESSION 2 GENOME ORGANIZATION AND REORGANIZATION
Chairperson: S.L. Allen, University of Michigan, Ann Arbor, Michigan
Klobutcher, L.,' Jahn, C.,2 Prescott,D.,3 'Dept. of Biochemistry, Uni-versity of Connecticut Health Cen-ter, Farmington; 2Dept. of Biologi-cal Sciences, University of Illinois,Chicago; 3Dept. of Molecular, Cel-lular, and Developmental Biology,University of Colorado, Boulder:
184
DNA sequence elimination andaddition are both involved in genematuration during macronucleardevelopment in Oxytricha nova.
Cartinhour, S., Herrick, G., Dept. ofCellular, Viral, and Molecular Biol-ogy, University of Utah, Salt LakeCity: Alternate juxtaposition of ma-
Urbana; Humboldt Fellow, Zoolo-gisches Institut, Universitat Muns-ter, Federal Republic of Germany:Evolutionary genetics of the cil-iates.
cronuclear sequences in Oxytrichafallax.
Lipps, H.J., Kraut, H., Institut fOr Biol-ogie III, Tubingen, Federal Repub-lic of Germany: DNA reorganiza-tion during macronucleardevelopment of Stylonychia.
Yao, M.-C., Altschuler, M., Auster-

berry, C., Dept. of Biology, Wash-ington University, St. Louis, Mis-souri: DNA rearrangement inTetrahymena.
Blackburn, E., Bliska, J., Challoner, P.,Cherry, J.M., Ryan, T., Spangler,E., Dept. of Molecular Biology,University of California, Berkeley:Sequence analysis and synthesisof macronuclear telomeres.
Diamond, C.H., Ohashi, RS., Rose,A., Tsao, N.N.G.,Pearlman, R.E.,Dept. of Biology, York University,Toronto, Canada: Analysis of a mi-cronucleus limited sequence fromTetrahymena thermophila.
SESSION 3 POSTER SESSION
Ammermann, D., Institute of BiologieIII, University of TObingen, FederalRepublic of Germany: Is the chro-mosome elimination in Stylonychialemnae dependent on the micron-uclear DNA content?
Bruns, P.J., Section of Genetics andDevelopment, Cornell University,Ithaca, New York: Genetic maps ofTetrahymena thermophila.
Allen, S.L., Ervin, RR., McLaren,N.C., Division of Biological Sci-ences, University of Michigan,East Lansing: Organization of the5S rRNA gene clusters in thegerm-line and somatic genomes ofTetrahymena thermophila.
Pederson, D.S., Bannon, G.A.,Bowen, J.K., Shupe, K., Gorovsky,M.A., Dept. of Biology, Universityof Rochester, New York: Relation-ship of DNAse I hypersensitivesites to transcription of 5S and hi-stone genes in Tetrahymena ther-mophila.
Hufschmid, J.-D., de Haller, G., Labo-ratory of Protozoology, Universityof Geneva, Switzerland: Studieson the DNA of the heterotrich cil-iate Climacostomum virens.
Dawson, D., Herrick, G., Dept. of Cel-lular, Viral, and Molecular Biology,University of Utah, Salt Lake City:Rare internal C,A4 repeats in themicronuclear genome of Oxytrichafallax.
Tschunko, A., Allen, S., University ofMichigan, Ann Arbor: Structureand genomic organization of an in-verted repeated DNA sequence ofTetrahymena thermophila.
White, T, Allen, S., University of Mich-igan, Ann Arbor: Eliminated DNAsequences and a specific site ofmethylation in the macronucleusof Tetrahymena thermophila.
Yaeger, P,' Orias, E.,' Larson, D.,2Howard, E.A.,2 Blackburn, E.H.,2,Dept. of Biological Sciences, Uni-versity of California, Santa Bar-bara; 2Dept. of Molecular Biology,University of California, Berkeley:Mutations affecting rDNA amplifi-cation in Tetrahymena thermo-phila.
Olins, D.E., Allen, R.L., Cadilla, C.L.,Olins, A.L., University of Tennes-
see-Oak Ridge Graduate Schoolof Biomedical Sciences and Biol-ogy Division, Oak Ridge: Macron-uclear and chromatin structure inEuplotes eurystomus.
Gottschling, D.E., Cech, T.R., Dept. ofChemistry, University of Colorado,Boulder: A complex at the molecu-lar ends of Oxytricha macronu-clear DNA imparts nucleosomephasing.
Poster Session
Haller, B.L., Orias, E., Dept. of Biolog-ical Sciences, University of Califor-nia, Santa Barbara: Aphidicolin af-fects differential amplification ofrDNA molecules in B/C3 heterozy-gotes of Tetrahymena thermo-phila.
Conover, R.K., Brunk, C.F., Dept. ofBiology, University of California,Los Angeles: Isolation of telom-eres from Tetrahymena thermo-phila.
Rogers, M., Karrer, K., Dept. of Biol-ogy, Brandeis University, Waltham,Massachusetts: Adolescence inTetrahymena thermophila.
Suhr-Jessen, P., Dept. of Anatomyand Cytology, University of
Odense, Denmark: Stage-specificchanges in protein synthesis pat-tern during conjugation in Tetrahy-mena thermophila.
Gutierrez, J.C., Orias, E., Dept. of Bi-ological Sciences, University ofCalifornia, Santa Barbara: Doubletcells of Tetrahymena thermophilaoriginated after capsule inductionon conjugating pairs.
Hamilton, E.P., Orias, E., Dept. of Bio-logical Sciences, University of Cal-ifornia, Santa Barbara: Macronu-clear retention and otherdevelopmental effects induced byvinblastine in conjugating Tetrahy-mena.
Eustice, D.C., Wilhelm, J.M. Dept. of
185

Microbiology, University of Roch-ester, New York: Mechanism ofac-tion of aminoglycoside antibioticswith wild-type Tetrahymena ther-mophila and drug-resistant mu-tants.
Skelley, S., Wilhelm, J.M., Dept. of Mi-crobiology, University of Roches-ter, New York: Multiple mutationsin Tetrahymena thermophila con-
SESSION 4 CONJUGATION
ferring resistance to aminoglyco-side antibiotics.
Sripati, C.E., Institut de Biologie phys-ico-chimique, Paris, France: Invivostability of ribosomes in Tetrahy-mena thermophila.
Cleffmann, G., Palissa, H., Institute ofAnimal Physiology, University ofGiessen, Federal Republic of Ger-many: Division mutants as tools to
Chairperson: A. Miyake, University di Camerino, Camerino, Italy
Miyake, A., Dipartimento di BiologiaCellulare, University di Camerino,Italy: Ciliate conjugation-Intro-ductory remarks.
Heckmann, K., Zoological Institute,University of MOnster, Federal Re-public of Germany: Towards themolecular basis of cell recognitionin Euplotes octocarinatus.
Luporini, P., Miceli, C., Dipartimentodi Biologia Cellulare, University diCamerino (MC), Italy: The molecu-lar mechanism of sexual interac-tion in Euplotes.
Miceli, C., Luporini, P., Dipartimentodi Biologia Cellulare, University di
Camerino, Italy: Isogamones inEuplotes raikovi.
Fujishima, M., lwao, Y., Biological In-stitute, Faculty of Science, Yama-guchi University, Japan: Inductionof reinitiation of meiosis in toadoocytes by injection of Parame-cium extracts.
Eckert, W.A., Weiske-Benner, A., Insti-tute of Zoology, University of Hei-delberg, Federal Republic of Ger-many: Differentiation andtranscriptional activity of nucleiduring conjugation in Tetrahy-mena thermophila.
Allis, C.D., Wiggins, J.C., Richman,
SESSION 5 RIBOSOMES, RIBOSOMAL GENES, AND MITOCHONDRIAL GENES
Chairperson: R. Hallberg, Iowa State University, Ames, Iowa
Amin, A.A., Pearlman, R.E., Dept. ofBiology, York University, Toronto,Canada: Autonomously replicating
segments from Tetrahymena ther-mophila rDNA-Sequence, struc-ture, and function.
P.J. Bruns
186
study cell cycle control in Tetrahy-mena.
Smith-Sonneborn, J., Dept. of Zool-ogy and Physiology, University ofWyoming, Laramie: Clonal age-and cell-specific response to low-frequency electromagnetic fields.
Dyer, B.D., Dept. of Biology, BostonUniversity, Massachusetts: Anaer-obic, sulfide -tolerant ciliates.
R., Chicoine, L., Wenkert, D., Bay-lor College of Medicine, TexasMedical Center, Houston: Histonerearrangements accompany nu-clear differentiation and dediffer-entiationin Tetrahymena thermo-phila.
Martindale, D.W.,1.2 Goldberg, M.,'Martindale, H.,1,2 Bruns, P.J.,"Dept. of Genetics and Develop-ment, Cornell University, Ithaca,New York; 2Dept. of Microbiology,McGill University, Quebec, Can-ada: Sequence conservation of aconjugation-specific gene from Te-trahymena.
Engberg, J., Nielsen, H., Jeppesen,C., Biochemical Institute B, Univer-sity of Copenhagen, Denmark:The ribosomal RNA genes of Te-trahymena - Structure and func-tion.
Higashinakagawa, T., Matsuura, T.,Matsumoto, K., Matsuda, R., Dept.of Biology, Tokyo MetropolitanUniversity, Japan: Structure andexpression of Tetrahymena ribo-somal DNA.
Niles, E.G., Matusick, L., Dept. of Bio-chemistry, State University of NewYork, Buffalo: CAP binding to theTetrahymena pyriformis rDNA pro-moter.
Gocke, E., Bonven, B.J., Wester-gaard, 0., Dept. of Molecular Biol-ogy and Plant Physiology, Univer-sity of Aarhus, Denmark: Is rRNAsynthesis in Tetrahymena regu-lated by a sequence-specific topo-isomerase I framing the gene?
K ster, K.-P, Eckert, W.A., Dept. of

Physiology, Institute of Zoology,Heidelberg, Federal Republic ofGermany: In vitro processing ofprecursor rRNA in isolated ma-cronuclei of Tetrahymena thermo-phila.
Zaug, A.J., Kent, J.R., Cech, T.R.,Dept. of Chemistry, University ofColorado, Boulder: Auto-reopen-ing of the circular intervening se-quence of Tetrahymena- lmplica-
Lions for the mechanism of RNAself-splicing.
Wilhelm, J.M., Kelly, E., Skelly, S.,Dept. of Microbiology, Universityof Rochester, New York: Ribo-somal proteins from antibiotic-re-sistant strains of Tetrahymenathermophila- Absence of starva-tion-induced protein phosphoryla-tion.
Suyama, Y, Dept. of Biology, Univer-
SESSION 6 BEHAVIOR, CELL SURFACE, AND CELL ARCHITECTURE
Chairperson: C. Kung, University of Wisconsin, Madison, Wisconsin
Kung, C., Dept. of Genetics, Labora-tory of Molecular Biology, Univer-sity of Wisconsin, Madison: Behav-ioral genetics of Paramecium -Ancient and recent history.
Haga, N.,' Forte, M.,2 Ramanathan,R.,' Saimi, Y.,' Kung, C.,''Laboratory of Molecular Biology,University of Wisconsin, Madison;2Dept. of Biology, Case WesternReserve University, Cleveland,Ohio: Purification of a soluble pro-tein controlling Ca" channel ac-tivity in Paramecium.
Saimi, Y., Hinrichsen, R.D., Labora-tory of Molecular Biology, Univer-sity of Wisconsin, Madison: Ge-
netic dissection of electrogenesisand characterization of mutants inParamecium.
Schultz, J.E., Grunemund, R., Gier-lich, D., Klumpp, S., Pharmaceuti-cal Institute, University of TObin-gen, Federal Republic ofGermany: Ionic regulation of thecyclic nucleotide system in Para-mecium tetraurelia.
Bonini, N.M., Gustin, M.C., Mason, P,Nelson, D.L., Piper, L., Dept. ofBiochemistry, University of Wis-consin, Madison: A possible rolefor cyclic AMP in the regulation ofthe ciliary beat in Paramecium te-traurelia.
SESSION 7 GENE ORGANIZATION AND EXPRESSION
Chairperson: E. Orias, University of California, Santa Barbara, California
Caron, F., Meyer, E., Centre de Gend-tique Moleculaire, CNRS, Gif-sur-Yvette, France: Cloning of the en-tire structural gene of the G sur-face antigen and its expression inE. coli.
Find ly, R.C., Dept. of Biology, YaleUniversity, New Haven, Connecti-cut: Genomic organization of theTetrahymena hsp70 heat-shockgenes.
Hallberg, R., Kraus, K., Dept. of Zool-ogy, Iowa State University, Ames:Regulation of heat-shock proteinmRNA utilization in Tetrahymenathermophila.
Galego, L., Rodriques-Pousada, C.,Dept. of Microbiology, InstitutoGulbenkian de Ciencia, Oeiras,Portugal: Stress response in Tetra-hymena pyriformis- Differences
between starvation and heatshock.
Ness, J.C., Morse, D.E., Dept. of Bio-logical Sciences, University of Cal-ifornia, Santa Barbara: Biochemi-cal analysis of the regulation ofgalactokinase gene expression inTetrahymena thermophila.
Cupples, C., Pearlman, R.E., Dept. ofBiology, York University, Toronto,Canada: Actinlike sequences inTetrahymena.
Helftenbein, E., Institut fur Biologie III,Universitat TObingen, Federal Re-public of Germany: Nucleotide se-quence of a macronuclear gene-sized DNA molecule coding for a-tubulin in Stylonychia lemnae.
Seyfert, H.-M., Institute of AnimalPhysiology, University of Giessen,Federal Republic of Germany:
sity of Pennsylvania, Philadelphia:Genes and genome organizationof Tetrahymena mitochondrialDNA.
Heinonen, T.Y.K., Young, P.G., Dept.of Biology, Queen's University,Kingston, Canada: Transcriptionmap of Tetrahymena mitochon-dria! DNA.
Van Houten, J., Schulz, S., Preston,R., Dept. of Zoology, University ofVermont, Burlington: Role of mem-brane in Paramecium chemore-ception.
Kaneshiro, E.S., Matesic, D.F., Dept.of Biological Sciences, Universityof Cincinnati, Ohio: The sphingo-lipids of the Paramecium baA mu-tant.
Frankel, J., Jenkins, L.M., Nelsen,E.M., Lansing, T.J., Bakowska, J.,Dept. of Biology, University ofIowa, Iowa City: Mutations affect-ing patterning of cell-surface struc-tures in Tetrahymena thermophila.
M.A. Gorovsky
187

Regulation of tubulin synthesis versity of Rochester, New York:throughout ciliary regeneration in Regulation of tubulin synthesisTetrahymena. during cilia regeneration in
Calzone, F.J., Callahan, R.C., Gorov- starved Tetrahymena thermophila.sky, M.A., Dept. of Biology, Uni- Orias, E., Baum, M.P., Dept. of Bio-
SESSION 8 SECRETION AND CYTOPLASMIC PHENOMENA
Chairperson: B.H. Satir, Albert Einstein College of Medicine, Bronx, New York
Adoutte, A.,' Garreau de Loubresse,N.,2 Beisson, J.,"Centre deGenetique Moleculaire du CNRS,Gif-sur-Yvette; 2Centre de Cytolo-gie Experimentale du CNRS, Ivry-sur-Seine, France: Proteolyticcleavage and maturation of thecrystalline secretion products ofParamecium.
Garofalo, R.S., Satir, B.H., Dept. ofAnatomy, Albert Einstein Collegeof Medicine, Bronx, New York:
Quantitative studies on trichocystmatrix expansion in vitro and itsregulation by Ca" and pH.
Aufderheide, K.J., Dept. of Biology,Institute of Developmental Biology,Texas A&M University, CollegeStation: Genetic and cortical phe-notypes associated with earlyclonal death in Paramecium te-traurelia.
Kanabrocki, J.A., Quackenbush, R.L.,Dept. of Microbiology, University
SESSION 9 TRANSFORMATION WORKSHOP
Chairperson: R.E. Pearlman, York University, Toronto, Canada
Killeen, M.T.,1 Duda, E.,2 Pearlman,R.E.,"Dept. of Biology, York Uni-versity, Toronto, Canada; 2Instituteof Biochemistry, Hungarian Acad-emy of Sciences, Szeged, Hun-gary: Attempts to geneticallytransform Tetrahymena.
Duda, E.G.,' Pearlman, R.,2 'Instituteof Biochemistry, Biological Re-search Center, Szeged, Hungary;
188
2Dept. of Biology, York University,Downsview, Canada: Transforma-tion of Tetrahymena thermophila.
Luehrsen, K.R., Orias, E., Dept. of Bi-ological Sciences, University ofCalifornia, Santa Barbara-At-tempts to transform Tetrahymena:A progress report.
Brunk, C.F., Conover, R.K., Dept. ofBiology, University of California,
logical Sciences, University of Cal-ifornia, Santa Barbara: The "firstreplicated, most amplified" hypoth-esis for caryonidal mating type de-termination.
of South Dakota, School of Medi-cine, Vermillion, South Dakota: R-body synthesis in E. coli.
Hufnagel, L.A., Dept. of Microbiology,University of Rhode Island, Kings-ton, and Marine Biological Labora-tory, Woods Hole, Massachus-setts: Ciliary rosettes-A new typeof intramembranous particle arrayin starved and mating Tetrahy-mena thermophila.
Forney, J.D., Epstein, L.M., Dept. ofBiology, lndiania University,Bloomington: Cytoplasmic inherit-ance of a macronuclear deletion inParamecium.
Williams, N.E.,' Ron, A.,2 'Dept. ofZoology, University of Iowa, IowaCity; 2Dept. of Anatomy and Em-bryology, Hebrew University-Had-assah Medical School, Jerusalem,Israel: Synthesis and deploymentof immobilization antigen H3 intransforming Tetrahymena thermo-phila cells.
Meyer, E., Caron, F., Guiard, B.,Centre de Genetique Moleculaire,CNRS, Gif-sur-Yvette, France: Invitro translation of Parameciummessenger RNAs is blocked atprecise sites.
Kaney, A.R., Dept. of Biology, BrynMawr College, Pennsylvania: Unu-sual mating behavior in an ami-cronucleate strain of Tetrahymenathermophila.
Los Angeles: Transformation of Te-trahymena to paromomycin resist-ance with pSV2-neo DNA.
WOnning, I.U., Lipps, H.J., Institut fiirBiologie III, Universitat TObingen,Federal Republic of Germany: Atransformation system for the hy-potrichous ciliate Stylonychia myti-lus.

Molecular Biology of the Photosynthetic ApparatusMay 9-May 13
ARRANGED BY
Charles Arntzen, DOE/Plant Research Laboratory, Michigan State UniversityLawrence Bogorad, The Biological Laboratories, Harvard UniversitySusan Bonitz, Cold Spring Harbor Laboratory
146 participants
The process of photosynthesis involves many reactions-light absorption, elec-tron transfers, energy-coupling processes leading to ATP synthesis, and enzy-matic reactions leading to CO, fixation and substrate and product cycling. Themain purpose of this meeting was to emphasize the advancements that have oc-curred in the study of photosynthesis using new tools of molecular biology. Spe-cial emphasis was placed on organizing sessions in which specialists in the anal-ysis of the functional components of the photosynthetic processes presentedrecent data in parallel with reports by molecular biologists dealing with the ge-netic basis and developmental events that lead to the synthesis and stable inte-gration of these components.
This meeting was supported in part by the Advanced Genetic Sciences, Inc.;Amoco; Arco Solar, Inc.; CIBA-GEIGY; Dow Chemical Co.; Exxon Research andEngineering; National Science Foundation; N.A,T.O.; Rhone-Poulenc, Inc.; ShellDevelopment Co.; Stauffer Chemical Co.; and U.S. Dept. of Agriculture.
SESSION 1 DNA REPLICATION, TRANSCRIPTION, AND PROTEIN SYNTHESISIN PLASMIDS
Chairperson: J. Weil, University of Strasbourg, Strasbourg, France
Gillham, N.,' Palmer, J.,' Boynton,J.,2 Harris, E.,2 Depts. of 'Zoology,2Botany, Duke University, Durham,North Carolina: Organization ofplastid chromosomes.
Bogorad, L., Crossland, L.D., Russell,D., Stirdivant, S., Harvard Univer-sity, Cambridge, Massachusetts:Transcription of chloroplast genes.
Hallick, R.B., Johanningmeier, U.,Karabin, G.D., Nickoloff, J.A., Pas-savant, C., Dept. of Chemistry,University of Colorado, Boulder:Transcription of chloroplast genes.
Orozco, E.M., Jr., Hanley-Bowdoin,L., Poulsen, C., Mullet, J.E., Chua,N.-H., Laboratory of Plant Molecu-lar Biology, Rockefeller University,New York, New York: In vitro tran-scriptional analysis of the rL andatpB genes from spinach andmaize.
Rodermel, S.R., Bogorad, L., Dept. ofCellular and Developmental Biol-ogy, Harvard University, Cam-bridge, Massachusetts: Photore-gulation of transcript levels during
light-induced plastid developmentin maize.
Greenberg, B.,' Narita, J.,' DeLuca-Flaherty, C.,'Gruissem, W.,2 Hal-lick, R.,"Dept. of Chemistry, Uni-
versity of Colorado, Boulder;2Dept. of Botany, University of Cal-ifornia, Berkeley: Properties of twochloroplast RNA polymerase activ-ities.
Coffee break
189

SESSION 2 THE FUNCTIONAL ORGANIZATION OF THE PHOTOSYNTHETIC ENERGYTRANSDUCING APPARATUS
Chairperson: B. Cramer, Purdue University, West Lafayette, Indiana
Arntzen, C.J., DOE/Plant ResearchLaboratory, Michigan State Uni-versity, East Lansing: Chloroplastmembrane structure and function.
Andersson, B., Akerlund, H.-E., Jans-son, C., Dept. of Biochemistry,University of Lund, Sweden: Poly-peptides on the oxidizing side ofPSII.
Herrmann, R., Botanishes Institut,University of Dusseldorf, FederalRepublic of Germany: Plastidgenes for PSII proteins.
Erickson, J.M.,' Rochaix, J.-D.,'Mets, L.,2 'Depts. of Molecular Bi-ology and Plant Biology, Universityof Geneva, Switzerland; 2Dept. ofBiology, University of Chicago, Illi-
B. Andersson, C. Arntzen, J. Bennett
SESSION 3 POSTER SESSION
Crossland, L., Rodermel, S., Bogo-rad, L., Dept. of Cellular and De-velopmental Biology, Harvard Uni-versity, Cambridge,Massachusetts: Single gene forthe large subunit of RuBP carbox-ylase in maize yields two mRNAs.
Gingrich, J.C., Hal lick, R.B., Dept. ofChemistry, University of Colorado,Boulder: The E. gracilis ribulose-1,5-bisphosphate carboxylasegene-Nine introns in a chloro-plast protein gene.
Passavant, C.W., Hal lick, R.B., Dept.of Chemistry, University of Colo-rado, Boulder: Analysis of fourstructural genes encoded in the E.gracilis chloroplast chromosome.
Mullet, J.E., Boyer, S., Dept.of Bio-chemistry and Biophysics, Texas
190
A &M University, College Station: Invitro transcription of the chloro-plast psbA gene.
Hanley-Bowdoin, L., Chua, N.-H.,Laboratory of Plant Molecular Biol-ogy, Rockefeller University, NewYork, New York: A 109-bp chloro-plast sequence contains the pro-moter for the maize rL gene.
Woessner, J.,1 Masson, A.,3 Harris,E.,2 Bennoun, P,3 Gillham, N.,'Boynton, J.,2 Depts. of 'Zoology,2Botany, Duke University, Durham,North Carolina; 3lnstitut de Biolo-gie Physico-Chimique, Paris,France: Molecular and geneticanalysis of the chloroplast ATPaseof Chlamydomonas.
Margulies, M.M., Smithsonian Envi-ronmental Research Center, Rock-
nois: Using mutants to probe thefunctional structure of the 32 kilo-dalton protein of PSII.
Cohen, B.N., Bloom, M.V., Coleman,T, Weissbach, H., Roche Instituteof Molecular Biology, Nutley, NewJersey: Analysis of in vitro chloro-plast gene expression using a sim-plified dipeptide synthesis system.
Golden, S.S.,1 Sherman, L.A.,2 Hasel-korn, R.," Dept. of Biophysics,University of Chicago, Illinois;2Division of Biological Sciences,University of Missouri, Columbia:Analysis of A. nidulans R2 psbAgenes from wild-type and DCMU-resistant strains.
Mattoo, A.K.,' Marder, J.B.,2 Edel-man, M.,' 'University of Maryland,College Park, and Plant HormoneLaboratory, USDA Agricultural Re-search Service, Beltsville, Mary-land; 2Dept. of Plant Genetics,Weizmann Institute of Science, Re-hovot, Israel: Light and the 32Kherbicide binding protein-Metab-olism and function.
Kyle, D.J., DOE/Plant Research Labo-ratory, Michigan State University,East Lansing: Light-induced mem-brane damage and repair; photo-inhibition.
ville, Maryland: The function ofchloroplast membrane-bound ri-bosomes.
Edwards, C.A.,' Bellemare, G.,2Dharm, B.,' Chua, N.-H.,''Laboratory of Plant Molecular Bi-ology, Rockefeller University, NewYork, New York; 2Dept. of Bio-chemistry, Laval University, Que-bec, Canada: Chloroplast DNA-dependent RNA polymerase con-tains subunits that cross-react withthe subunits of E. coli RNA poly-merase.
Muskavitch, K.M.T., Krebbers, E.T.,Bogorad, L., The Biological Labo-ratories, Harvard University, Cam-bridge, Massachusetts: Light-stim-ulated transcription of a region ofthe Z. mays chloroplast genome.

McKown, R.L., Tewari, K.K., Dept. ofMolecular Biology and Biochemis-try, University of California, Irvine:Purification and properties of apea chloroplast single-strandedDNA-binding protein.
Larrinua, I.M., Bogorad, L., The Bio-logical Laboratories, Harvard Uni-versity, Cambridge, Massachu-setts: The characterization oftranslational signals for higherplant plastid genes.
Graan, T., Ort, D.R., Dept. of Plant Bi-ology, USDA/ARS, University of Il-linois, Urbana: Quantitation of therapid electron donors to P700, thefunctional plastoquinone pool, andthe ratio of the photosystems inspinach chloroplasts.
Wolber, P.K., Steinback, K.E., Ad-vanced Genetic Sciences, Oak-land, California: Analysis of a tryp-
tic peptide photoaffinity-labeled byazidoatrazine.
Williams, J.C.,' Steiner, L.A.,2 Simon,M.I.,2 Feher, G.,' 'University ofCalifornia, San Diego;2Massachusetts Institute of Tech-nology, Cambridge; 3Agouron In-stitute, La Jolla, California: Se-quencing of the genes encodingthe subunits of the reaction centerfrom R. sphaeroides.
Worland, S.,' Wilson, K.,2 Hearst, J.,'Sauer, K.,"Division of ChemicalBiodynamics, Lawrence BerkeleyLaboratory, and Dept. of Chemis-try, University of California, Berke-ley; 2Cetus Corporation, Emery-ville, California: Amino-terminalsequences of reaction-center poly-peptides from R. capsulate.
Zsebo, K.M.,' Wu, F.,2 Hearst, J.,2'Group in Comparative Biochem-
SESSION 4 BACTERIAL PHOTOSYNTHESIS
Chairperson: S. Kaplan, University of Illinois, Urbana, Illinois
Crews, G., Peters, J., Dierstein, R., In-stitute of Biology, Albert-LudwigsUniversity, Freiburg, Federal Re-public of Germany: Organizationand synthesis of the photosyn-thetic apparatus of R. capsulate.
Youvan, D.C., Cold Spring HarborLaboratory, New York, and ExxonResearch and Engineering Com-pany, Clinton, New Jersey: Reac-tion-center, B870 and B800 andB850 genes and deduced poly-peptide sequences from R. capsu-late.
Feher, G., Allen, J.R, Dept. of Phys-ics, University of California, SanDiego: Crystallization of reactioncenters from R. sphaeroides; Pre-liminary characterization.
Donohue, T.J.,' Chory, J.,' Varga,A.R.,2Staehelin, A.L.,2 Kaplan,S.,"University of Illinois, Urbana;2University of Colorado, Boulder:Biochemical and morphologicalstudies of the induction of the pho-tosynthetic membrane of R.sphaeroides.
Fuller, R.C., Red linger, T.E., Dept. ofBiochemistry, University of Massa-chusetts, Amherst: The develop-ment and topology of the photo-synthetic apparatus of C.aurantiacus.
Hearst, J.E., University of California,Berkeley: The relationship be-tween amino acid sequences andquinone binding in photosyntheticmembranes.
Gilbert, C.W.,' Brown, A.E.,2Wy-drzynski, T.J.,1 Williams, K.,'
istry; 2Dept..of Chemistry, Univer-sity of California, Berkeley: Ge-netic-physical mapping of aphotosynthetic gene cluster fromR. capsulate.
Kiley, P.J., Kaplan, S., University of Illi-nois, Urbana: Identification oflight-harvesting polypeptideswithin the chromatophores of R.sphaeroides by analysis of light-harvesting mutants.
Salvucci, M.E., Ogren, W.L., USDA/ARS and Agronomy Dept., Univer-sity of Illinois, Urbana: Biochemi-cal characterization of two Chia-mydomonas Calvin cycle mutants.
Chattoo, B.B., Hohn, B., FriedrichMiescher-Institut, Basel, Switzer-land: Molecular basis for an unsta-ble chloroplast phenotype in petu-nia hybrida.
Arntzen, C.J.,"Michigan StateUniversity, East Lansing; 2AuburnUniversity, Alabama: Molecularanalysis of triazine herbicide re-sistance in the photosynthetic bac-terium R. sphaeroides.
Poster Discussion
Vannevar Bush Lecture Hall
191

SESSION 5 NUCLEAR GENE REGULATION
Chairperson: E. Tobin, University of California, Los Angeles, California
Boynton, J.,' Schmidt, R.,' Myers,A.,2 Hosier, J.,' Gil lham, N.,2Depts. of 'Botany, 2Zoology, DukeUniversity, Durham, North Caro-lina: Biogenesis of chloroplast ri-bosomes-Cooperation of chloro-plast and nuclear genes.
Jenkins, G.I.,' Bennett, J.,2 'Dept. ofBiological Sciences, University ofWarwick, Coventry, England;2Dept. of Biology, Brookhaven Na-tional Laboratory, Upton, NewYork: Photoregulation of chloro-
SESSION 6 POSTER SESSION
Chory, J., Muller, E.D., Kaplan, S.,Dept. of Microbiology, Universityof Illinois, Urbana: Cloning,expression, and assembly of theform II ribulose-1,5-bisphosphatecarboxylase from R. sphaeroides.
Andrews, T.J., Bailment, B., Austra-lian Institute of Marine Science,Townsville: Reassembly of Rub-isco from isolated subunits- Car-bamate formation and 2-CABPbinding by isolated large-subunitoctamers.
Gutteridge, S., Sigal, I., Thomas, B.,Arentzen, R., Cordova, A., Lori-mer, G., Central Research and De-velopment Dept., E.I. du Pont deNemours and Company, Wilming-ton, Delaware: Directed mutage-nesis of the carbamate-metal-ion-binding site of RuBP carboxylase.
Pierce, J., Reddy, G.S., Central Re-search and Development Division,E.I. du Pont de Nemours andCompany, Wilmington, Delaware:A structure for the catalytic com-plex of RuBP carboxylase.
Coughlan, S.J., Clark, R.D., Hawkes-ford, M.J., Bennett, J., Hind, G.,Dept. of Biology, Brookhaven Na-tional Laboratory, Upton, NewYork: Identification of ferre-doxin : NADP oxidoreductase as acomponent of a 5-polypeptide cy-tochrome b/f complex.
Sen, A., Dept. of Biophysics, RoswellPark Memorial Institute, Buffalo,New York: Polymorphic phase be-havior of chloroplast thylakoidmembrane lipids.
Chia, C.F., Arntzen, C.J., DOE/PlantResearch Laboratory, Michigan
192
plast proteins-mRNA levels, pro-tein stability, and phosphorylation.
Morelli, G., Coruzzi, G., Broglie, R.,Moses, P., Chua, N.-H., Labora-tory of Plant Molecular Biology,Rockefeller University, New York,New York: The expression of rSgenes in vivo and in transformedplant cells.
Cashmore, A., Timko, M., Hand, M.,Kausch, A., Fassler, J., Laboratoryof Cell Biology, Rockefeller Univer-sity, New York, New York: Nuclear
State University, East Lansing: Ac-celerated membrane protein turn-over in a chloroplast-encoded mu-tant.
Leto, K.J.,1 Bell, E.,2 McIntosh, L.,2'DuPont Company, Central Re-search and Development, Wil-mington, Delaware; 2MichiganState University, East Lansing:Failure to assemble PSII in a nu-clear maize mutant is accom-panied by accelerated turnover ofchloroplast-encoded PSII polypep-tides.
Clark, R.D., Bennett, J., Hind, G.,Dept. of Biology, Brookhaven Na-tional Laboratory, Upton, NewYork: Extraction and purification ofa spinach thylakoid kinase thatcan phosphorylate light-harvestingchlorophyll a/b protein (LHCP) invitro.
Abbott, M.S.,' Czarnecki, J.J.,2 Sel-man, B. R.,' of Biochemis-try, 2lnstitute for Enzyme Re-search, University of Wisconsin,Madison: Localization of the high-affinity binding site for ADP andATP on the (3 subunit of the chloro-plast CF1.
Merchant, S., Selman-Reimer, S., Ab-bott, M.S., Selman, B.R., Dept. ofBiochemistry, University of Wis-consin, Madison: Analysis of thechloroplast CF1 in C. reinhardtii.
Tobin, E.M., Neumann, G.A., Kohorn,B.D., Thornber, J.P., Dept. of Biol-ogy, University of California, LosAngeles: Sequence and structureof an intron containing chlorophylla/b-protein gene expressed in L.gibba.
genes encoding the light-harvest-ing chlorophyll a/b-binding poly-peptides and the small subunit ofribulose-1,5-bisphosphate carbox-ylase.
Herrera-Estrella, L.,1 Van den Broeck,G.,1 Maenhaut, R.,1 Timko, M.,2Cashmore, A.,2 Van Montagu, M.,1Schell, J.,"Laboratorium voorGenetica, Gent, Belgium;2Rockefeller University, New York,New York: Photoregulation of nu-clear gene expression.
Anderson, L.,1 Eiserling, F.,1.2'Molecular Biology Institute,2Dept.of Microbiology, University of Cali-fornia, Los Angeles: Assembly in-teractions in Synechocystis 6701phycobilisomes.
Kaufman, L.S., Watson, J.C., Briggs,W.R., Thompson, W.F., CarnegieInstitution of Washington, Stan-ford, California: Photoregulation ofnuclear genes in developing peabuds.
Wimpee, C.F., Tobin, E.M., Dept. ofBiology, University of California,Los Angeles: Structure andexpression of genes encoding thesmall subunit of RuBP carboxylasefrom L. gibba.
Yen, H.-C.B., Huggins, B.J., Georgen,K.N., Corporate Research, Stand-ard Oil Company, Naperville, Illi-nois: Genetic transformation of for-eign DNA in the photosyntheticbacterium R. sphaeroides.
Ausich, R.L., Murphy, R.M., Corpo-rate Research, Standard Oil Com-pany, Naperville, Illinois: Experi-ments on the transformation of C.reinhardtii.
Gold, B., Bogorad, L., The BiologicalLaboratories, Harvard University,Cambridge, Massachusetts: Maizechloroplast DNA fragments are ca-pable of acting as autonomouslyreplicating sequences in yeast.
Cushman, J.C., Reardon, E.M., Miller,M.E., Price, C.A., Waksman Insti-tute, Rutgers University, Piscata-way, New Jersey: Proline mimicsthe effect of light in the regulationof plastid protein synthesis in Eu-glena.

SESSION 7 RUBISCO STRUCTURE AND FUNCTION
Chairperson: W. Ogren, University of Illinois, Urbana, Illinois
Ellis, R.J., Gallagher, T.F., Robinson,C., Jenkins, G.I., Hemmingsen, S.,Dept. of Biological Sciences, Uni-versity of Warwick, Coventry, Eng-land: Synthesis, processing, andassembly of Rubisco subunits.
Nierzwicki-Bauer, S.A.,' Curtis, S.E.,2Haselkorn, R.,"Dept. of Biophys-ics, University of Chicago, Illinois;2Dept. of Genetics, North Carolina
State University, Raleigh: Cotran-scription of genes coding for thelarge and small subunits of RuBPcarboxylase in the cyanobacter-ium Anabaena 7120.
Poster DiscussionLorimer, G., Pierce, J., Gutteridge, S.,
Central Research and Develop-ment Division, E.I. du Pont deNemours and Company, Wilming-
SESSION 8 CYTOCHROME but AND PHOTOSYSTEM I
Chairperson: W. Krogman, Purdue University, West Lafayette, Indiana
Hauska, G., Dept. of Biology, Univer-sity of Regensburg, Federal Re-public of Germany: Organizationand function of the cytochrome b6fcomplex.
Clark, R.D., Hind, G., Hawkesford,M.J., Bennett, J., Dept. of Biology,Brookhaven National Laboratory,Upton, New York: Ferredoxin :NADP oxidoreductase of chloro-plasts is also a protein kinase.nese.
Ortiz, W., Lam, E., Malkin, R., Divisionof Molecular Plant Biology, Univer-sity of California, Berkeley: Organi-zation of the thylakoid PSI com-plex.
Lundell, D.J.,' Glazer, A.N.,' Melis,A.,2 Malkin, R.,2 'Dept. of Microbi-ology and Immunology; 2Divisionof Molecular Plant Biology, Univer-sity of California, Berkeley: Char-acterization of a cyanobacterialPS1 complex.
Fish, L.E.,1 Kuck, U.,2 Bogorad, L.,1'Dept. of Cellular and Develop-mental Biology, Harvard Univer-sity, Cambridge, Massachusetts;
2Lehrstuhl fur Allegemeine Bo-tanik, Bochum, Federal Republicof Germany: Structural analysis oftwo adjacent light-inducible chlo-roplast genes in maize with partialamino acid homology encodingpolypeptides of the CPI complexof PSI.
Jagendorf, A.T., Anthon, G., Sectionof Plant Biology, Cornell Univer-sity, Ithaca, New York: Unresolvedproblems in photophosphoryla-tion.
Selman-Reimer, S., Selman, B.R.,Dept. of Biochemistry, Universityof Wisconsin, Madison: Reversibleconformational changes of chloro-plast CF1 induced by selective or-ganic solvents.
Bottom ley, W.,' Zurawski, G.,2 Mason,J.G.,' Whitfeld, P.R.,' 'Division ofPlant Industry, CSIRO, Canberra,Australia; 2DNAX Research Insti-tute, Palo Alto, California: Organi-zation and structure of the chloro-plast-encoded subunits of ATPsynthase.
Walker, J.E., Gay, N.J., Hampe, A.,
SESSION 9 LIGHT HARVESTING SYSTEMS IN CYANOBACTERIA
Chairperson: B. Zilinskas, Cook College, New Brunswick, New Jersey
Glazer, A.N., Dept. of Microbiologyand Immunology, University ofCalifornia, Berkeley: Phycobilipro-teins as PBS components and aspurified proteins.
Gantt, E., Red linger, T, Lipschultz,C.A., Smithsonian EnvironmentalResearch Center, Rockville, Mary-land: Phycobilisomes-A compari-son of the terminal acceptor pig-
ment in red algae and cyano-bacteria.
Lemaux, P.G.,12 Grossman, A.R.,''Carnegie Institution of Washing-ton, Stanford, California; 2Dept. ofBiological Sciences, Stanford Uni-versity, California: Isolation ofgenes encoding phycobiliproteinsubunits.
Chang, 5.5.,' Kycia, J.H.,' Ledbetter,
ton, Delaware: RuBP carboxyl-ase-Stereochemical aspects andthe involvement of the carbamate-metal-ion complex in catalysis.
Hanks, J.F., Somerville, C.R., Mc-Intosh, L., DOE/Plant ResearchLaboratory, Michigan State Uni-versity, East Lansing: Site-specificmutagenesis of RuBP carboxyl-ase/oxygenase.
Falk, G., Runswick, M.J., Tybulew-icz, V.L.J., MRC, Laboratory ofMolecular Biology, Cambridge,England: Polypeptides of couplingfactors and their genes.
H. Zuber
M.C.,' Feldmann, R.J.,2 Siegel-man, H.W.,"Dept. of Biology,Brookhaven National Laboratory,Upton, New York; 2Division ofComputer Research and Technol-ogy, National Institutes of Health,Bethesda, Maryland: Cyanobac-terial PBS.
de Lorimier, R.,' Bryant, D.A.,' Porter,R.D.,' Fields, P.I.,' Liu, W.-Y,2 Jay,
193

L. Bogorad, C. Arntzen, S. Bonitz
SESSION 10 TRANSFORMATION SYSTEMS AND DIFFERENTIATION OF THEPHOTOSYNTHETIC APPARATUS
Chairperson: I. Ohad, Hebrew University, Jerusalem, Israel
Kolowsky, K.S., Williams, J.G.K., Sza-lay, A.A., Boyce Thompson Insti-tute for Plant Research, CornellUniversity, Ithaca, New York: Sta-ble integration of foreign DNA intothe chromosome of cyanobac-teria.
Flores, E., Wolk, C.P., DOE/Plant Re-search Laboratory, Michigan StateUniversity, East Lansing: Ap-proach to the genetic analysis of
oxygenic photosynthesis-Trans-fer of plasmid vectors from E. colito facultatively heterotrophic cy-anobacteria.
Taylor, W., Mayfield, S., Martineau, B.,Harpster, M., Nelson, T, Yamagu-chi, J., Dept. of Genetics, Univer-sity of California, Berkeley: Onto-genetically regulated genes.
Watson, J.C., Kaufman, L.S., Thomp-son, W.F., Dept. of Plant Biology,
Concluding Discussion: The Use of Genetic Transformation in UnderstandingPhotosynthetic Complexity
RNA Processing MeetingMay 16-May 20
ARRANGED BY
Michael Mathews, Cold Spring Harbor LaboratoryHugh Robertson, Rockefeller UniversityAaron Shatkin, Roche Institute
258 participants
E.,2 Stevens, S.E., Jr.,''Microbiology Program, Pennsyl-vania State University, UniversityPark; 2Dept. of Chemistry, Univer-sity of New Brunswick, Frederic-ton, Canada: Genes for the a andI3 subunits of PC.
Zuber, H., Sid ler, W., FOglistaller, P.,Brunisholz, R., Theiler, R., Institutfur Molekularbiologie and Biophy-sik, Eidgen Technische Hochs-chule, Zurich, Switzerland: Struc-tural studies on the light-harvesting polypeptides from cy-anobacteria and bacteria.
Loach, R, Parkes, P., Miller, J., Hin-chigeri, S., Callahan, P., Dept. ofBiochemistry, Molecular Biologyand Cell Biology, NorthwesternUniversity, Evanston, Illinois:Structure-function relationships ofthe light-harvesting complexes inphotosynthetic bacteria.
Carnegie Institution of Washing-ton, Stanford, California: Organiza-tion and expression of light-regu-lated nuclear genes in pea andmung bean.
Horsch, R.B., Fraley, R.T., Rogers,S.G., Monsanto Company, St.Louis, Missouri: Inheritance offunctional foreign genes in plants.
This year's meeting, the third of the series, attracted nearly 250 scientists andcreated a great deal of excitement. The field's prosaic-sounding name does lessthan justice to metamorphoses that transform RNA molecules both structurally
194

and functionally. Primary transcripts, simple copies of the nucleotide sequence inthe gene, are modified by chemical substitution, by addition or deletion ofstretches of RNA, and by splicing-the joining of previously distant RNA se-quences.
One high point this year was the recognition of the catalytic role played by theRNA moiety of the RNase P enzyme in transfer RNA maturation. This moleculenow joins what is likely to be a growing list of RNA molecules with catalytic func-tion in cleavage and splicing reactions. The self-splicing reaction of the inauguralmember of this group, Tetrahymena ribosomal RNA, was subjected to closescrutiny, and the candidacy of some further RNAs was advanced. A secondpeak was reached with the development of cell-free extracts capable of efficientlysplicing mammalian messenger RNAs. Intense interest was occasioned by thediscovery of unexpected chemical bonds and lariat-like intermediate structuresformed during the splicing reaction, echoing illustrations of rope-makers' art onthe cover of the abstract book. Progress was seen in areas such as ribonucleo-protein particle assembly and ribosomal RNA processing, and considerable de-bate was stirred by experiments implying the existence of a splicing mechanismin phage-infected bacteria. Particularly promising was the development of an invitro system for the processing of mRNA 3' ends, which will undoubtedly aug-ment the elegant genetic studies of this event in the future.
This conference, the third to be held here in consecutive years, firmly estab-lished the need for an annual meeting on RNA processing, The series has cre-ated a vital forum for discussion of a range of topics related to RNA metabolism,and its success is founded on the dynamism and broad scope of the field. Withsome reluctance on this side of the ocean, it was decided that the 1985 meetingwill be held in Rome. The meeting will return to its original venue in 1986, possi-bly launching an annual alternation between a European site and Cold SpringHarbor.
This meeting was supported in part by funds from the National Science Foun-dation and the following divisions of the National Institutes of Health: Fogarty In-ternational Center and National Institute of General Medical Sciences.
Introduction: T. Cech, University of Colorado, Boulder, Colorado
SESSION 1 RNA CATALYZED REACTIONS
Chairperson: J. Steitz, Yale University, New Haven, Connecticut
Price, J.,' Engberg, J.,3 Kieft, G.,2Sievers, E.,2 Kent, J.,2 Morin, G.,2Been, M.,2 Cech, T.,1,2 Depts. of'Molecular, Cellular, and Develop-mental Biology; 2Chemistry, Uni-versity of Colorado Boulder;3Blochemical Institute, Panum In-stitute, University of Copenhagen,Denmark: The effects of deletionsand insertions on the in vitro self-splicing of T. thermophila pre-rRNA.
Sullivan, F.X., Cech, T.R., Dept. ofChemistry, University of Colorado,Boulder: The covalent addition ofan oligoribonucleotide to the circu-lar IVS of Tetrahymena is a fully re-
versible reaction mediated by theRNA.
Allen, L.C., Haydock, K., Dept. ofChemistry, Princeton University,New Jersey: Molecular mecha-nisms of catalysis by RNA.
Garriga, G., Collins, R., Lambowitz,A., Dept. of Biochemistry, St. LouisUniversity School of Medicine,Missouri: RNA splicing in Neuro-spora mitochondria.
Perlman, P., Haldi, M., Jarrell, R.,Morawiec, A., Dept. of Genetics,MCDB Program, Ohio State Uni-versity, Columbus: Studies of mito-chondrial intron structure.
Guerrier-Takada, C., Baer, M., Law-
rence, N., Altman, S., Dept. of Bi-ology, Yale University, New Haven,Connecticut: Structure and func-tion of the RNA component ofRNase P from E. coil.
Pace, N.R., Gardiner, K., Marsh, TL.,National Jewish Hospital and Re-search Center, and University ofColorado Health Sciences Center,Denver: The B. subtilis RNase P.
Prody, G.A., Bakos, J.T., Buzayan,J.M., Schneider, I.R., Bruening,G., Dept. of Biochemistry and Bio-physics, University of California,Davis: Self-processing of multi-meric forms of satellite tobaccoringspot virus RNA.
195

SESSION 2 STRUCTURE AND MECHANISM
Chairperson: 0. Uhlenbeck, University of Illinois, Urbana, Illinois
Munroe, S.H., Dept. of Biology, Mar-quette University, Milwaukee, Wis-consin: Higher-order structure ofRNA sequences at splice sites ofadenoviral pre-mRNA.
Shelness, G.S., Williams, D.L., Dept.of Pharmacological Sciences,State University of New York,Stony Brook: Structure mapping ofan estrogen-stabilized mRNA.
Lawrence, C., Browner, M., Ramsey,W.J., Dept. of Cell Biology, BaylorCollege of Medicine, Houston,Texas: Conserved secondarystructure features in mRNA andhnRNA.
Moore, P., Kime, M.J., Abdel-Meguid,S., Dept. of Chemistry, Yale Uni-versity, New Haven, Connecticut:Structure and properties of 5SRNA and its complexes with ribo-somal proteins.
SESSION 3 POSTER SESSION
Andersen, J.,' Delihas, N.,' Hanas,J.S.,2 Wu, C.-W.,2 Depts. of 'Mi-crobiology; 'Pharmacological Sci-ences, State University of NewYork, Stony Brook: 5S RNA struc-ture and interaction with transcrip-tion factor A from X. laevis oo-cytes.
Ares, M., Mangin, M., Weiner, A.M.,Dept. of Molecular Biophysics andBiochemistry, Yale University, NewHaven, Connecticut: Expression ofhuman U2 genes in Xenopus oo-cytes.
Arrigo, A.-P., Simon, M., Darlix, J.-L.,Khandjian, E.W., Spahr, P. -F.,Dept. of Molecular Biology, Uni-versity of Geneva, Switzerland:Heat-shock proteins of D. melano-gaster are associated with bothnuclear and cytoplasmic RNP par-ticles.
Bark, C., Hammarstrbm, K.,'Westin,G., Pettersson, U., Dept. of Medi-cal Genetics, University of Upps-ala, Sweden: Pseudogenes for hu-man U4 RNA-Strong support infavor of an RNA-mediated mecha-nism of dispersion.
Barta, A.,' Steiner, G.,' Noller, H.F.,'Kuechler, E.,"Institute of Bio-chemistry, University of Vienna,Austria; 2Thimann Laboratory, U ni-
196
Filipowicz, W.,' Konarska, M.,' Stru-gala, K.,' Shatkin, A.J.,2 'Instituteof Biochemistry and Biophysics,Polish Academy of Sciences, War-saw, Poland; 'Roche Institute ofMolecular Biology, Nutley, NewJersey: Mechanism of action ofRNA 3'-terminal phosphate cy-clase and identification of 2', 3'-cyclic 3'-phosphodiesterase in ex-tracts of HeLa cells.
Furneaux, H., Arenas, J., Perkins,K.K., Reinberg, D., Pick, L., Ad-hya, S., Carleton, S., Hurwitz, J.,Dept. of Developmental Biologyand Cancer, Albert Einstein Col-lege of Medicine, Bronx, NewYork: Purification and properties ofan RNA cyclase and RNA ligasefrom HeLa cells.
Ohshima, Tatei, K.,' Takemura,K.,' Maeda, A.,' Fujiwara, Y.,' Tan-
versity of California, Santa Cruz:Identification of a domain on 23SrRNA located at the peptidyl trans-ferase site.
Bass, B., Cech, T.R., Dept. of Chemis-try, University of Colorado, Boul-der: Specific interaction betweenthe self-splicing RNA of Tetrahy-mena and its guanosine substrate.
Berrios, M., Fisher, PA., Dept. ofPharmacological Sciences, StateUniversity of New York, StonyBrook: Is the major photolabelableATPase associated with the Droso-phila nuclear matrix-pore, com-plex-lamina fraction a form ofmyosin?
Blanchard, J.M., Piechaczyk, M.,Panabieres, F, Dani, C., Riad, S.,Marty, L., Jeanteur, P., Universitydes Sciences et Techniques duLanguedoc, and Centre P. Lam-argue, Montpellier, France: Tissue-specific differences in the steady-state polyadenylation of mRNAcoding for glyceraldehyde-3-P-de-hydrogenase in various chickentissues.
Bonitz, S., Cold Spring Harbor Labo-ratory, New York: Nuclear regula-tion of RNA processing in yeastmitochondria.
Breitenberger, C.A., Alzner-DeWeerd,
aka, H.,' 'Institute of BiologicalSciences, University of Tsukuba;'National Chemical Laboratory forIndustry, Ibaraki, Japan: U1RNA-protein complex preferen-tially binds to both 5' and 3' splicejunction sequences.
Aebi, M., Reiser, J., Weissmann, C.,Institut fOr Molekularbiologie I,Universitat Zurich, Switzerland:Point mutations within the 5' spliceregion of the rabbit p-globin largeintron -Conversion of the "Cham-bon rule" GT to GC or GA doesnot preclude splicing.
Solnick, D., Dept. of Molecular Bio-physics and Biochemistry, YaleUniversity Medical School, NewHaven, Connecticut: Secondarystructure and scanning mecha-nisms in mRNA splicing.
B., Browning, K.S., RajBhandary,U.L., Dept. of Biology, Massachu-setts Institute of Technology, Cam-bridge: tRNA sequences serve assignals in processing of mitochon-dria) rRNAs and mRNAs in Neuro-spore crassa.
Calvet, J.P., Smardo, EL., Jr., Maser,R.L., Dept. of Biochemistry, Uni-versity of Kansas Medical Center,Kansas City: Small RNAs that doand do not interact with rRNA.
Capasso, J.M., Docherty, M.A., Ray,A., Kaplan, E.D., Eliceiri, G.L., St.Louis University Medical School,Missouri: U1 small, nuclear RNA-like sequences in high-molecular-weight hnRNA.
Carrasco, A.E., Muller, M., De Rober-tis, E.M., Dept. of Cell Biology,University of Basel, Switzerland:Homeoticlike genes in verte-brates-Analysis of a gene that isexpressed during early Xenopusdevelopment.
Chae, C.-B., Patton, J.R., Dept. ofBiochemistry, University of NorthCarolina, Chapel Hill: Specific re-gions of U1 snRNP are sensitiveand resistant to nuclease.
Chu, F.K., Belfort, M., Maley, F.,Maley, G., Center for Laboratoriesand Research, New York State De-

partment of Health, Albany: Struc-ture of T4 bacteriophage thymidyl-ate synthase gene and its product.
Crouch, R.J., Itaya, M., Kanaya, S.,NCI, National Institutes of Health,Bethesda, Maryland: rnh mutantsare defective in cell growth.
de Massy, B.,' Fayet, 0.,2 Kogoma,T.,3 'NCI, National Institutes ofHealth, Bethesda, Maryland;2CRBGC, Toulouse, France;3Dept. of Biology, University ofNew Mexico, Albuquerque: Acti-vation of origins of DNA replica-tion in sdrA (rnh) mutants of E. coilK-12.
Dihanich, M.,' Ellis, S.,2 Najarian, D.,2Morales, M.,2 Clark, R.,2 Martin,N.C.,2 Hopper, A.K.,' 'Dept. of Bi-ological Chemistry, Hershey Medi-cal Center, Pennsylvania; 2Dept. ofBiochemistry, University of TexasHealth Science Center, Dallas:Modification of both cytoplasmicand mitochondrial tRNAs are af-fected by single nuclear muta-tions.
Dinter-Gottlieb, G., Cech, T.R., Dept.of Chemistry, University of Colo-rado, Boulder: Viroids containhomologies with the TetrahymenaIVS and may resemble group I in-trons in structure.
Drabkin, H., RajBhandary, U.L., Dept.of Biology, Massachusetts Instituteof Technology, Cambridge: Site-specific mutagenesis on a humaninitiator tRNA gene-Effect of A54to T54 change on RNA processingand on modification of U55 tow55.
Earley, J., Roebuck, K.A., Stumph,W.E., Dept. of Chemistry and Mo-lecular Biology Institute, SanDiego State University, California:Some chicken U1 RNA genes areclustered and have limited regionsof flanking DNA sequence homol-ogy.
Engberg, J., Nielsen, H., Jeppesen,C., Biochemical Institute B, Univer-sity of Copenhagen, Denmark:The ribosomal RNA genes of Te-trahymena- Structure and func-tion.
Falkenthal, S.,' Parker, V.P.,2 David-son, N.,3 'Dept. of Genetics, OhioState University, Columbus;'Amgen, Newberry Park, Califor-nia; 3Dept. of Chemistry, CaliforniaInstitute of Technology, Pasadena:
An unexpected 3' splice site is in-volved in differential splicing ofRNA transcribed from the Droso-phila MLC-ALK gene.
Forbes, D.,1,3 Dahlberg, J.,2 Lund,E.,2 'Dept. of Biochemistry andBiophysics, University of Califor-nia, San Francisco; °Dept. ofPhysiological Chemistry, Univer-sity of Wisconsin, Madison; 3Dept.of Biology, University of California,San Diego, La Jolla: Differentialexpression of multiple U1 RNAspecies in Xenopus oocytes andembryos.
Gupta, K.C., Kingsbury, D.W., Dept.of Virology and Molecular Biology,St. Jude Children's Research Hos-pital, Memphis, Tennessee: Poly-transcripts-An enigma in para-myxoviruses and rhabdoviruses.
Gurevitz, M., Apirion, D., Dept. of Mi-crobiology and Immunology,Washington University School ofMedicine, St. Louis, Missouri: Theribonuclease III processing sitenear the 5' end of an RNA precur-sor of bacteriophage T4 and its ef-fect on termination.
Hashimoto, C., Steitz, J.A., Dept. ofMolecular Biophysics and Bio-chemistry, Yale University, NewHaven, Connecticut: U4 and U6RNAs coexist in a single small nu-clear ribonucleoprotein particle.
Hay, N., Aloni, Y, Dept. of Genetics,Weizmann Institute of Science, Re-hovot, Israel: Efficient and accu-rate in vitro splicing of SV40-asso-ciated, small RNA (SAS-RNA).
Homison, G., Dieckmann, C.L., Ko-erner, T.J., Tzagoloff, A., Dept. ofBiological Sciences, ColumbiaUniversity, New York, New York:Identification, isolation, and char-acterization of a yeast nucleargene (CBPI) that is required forthe proper processing of the 5'end of the cytochrome b mRNA.
Hurt, D.J., Hopper, A.K., HersheyMedical Center, Pennsylvania:Cloning and characterization ofLOST, a gene of S. cerevisiae in-volved in tRNA processing.
Inoue, T., Cech, T.R., Dept. of Chem-istry, University of Colorado, Boul-der: Secondary structure of thecircular form of the TetrahymenarRNA intervening sequence-Anew technique for RNA structureanalysis using chemical probes
141$4017$
3 07'- -
Wine and cheese party
and reverse transcriptase.ltaya, M., Kanaya, S., Crouch, R.J.,
NCI, National Institutes of Health,Bethesda, Maryland: E. coilRNase H (rnh) mutations can besuppressed.
Kaltwasser, G., DiMaria, P, Golden-berg, C.J., Dept. of Pathology,Washington University, St. Louis,Missouri: Characterization andpartial purification of an mRNAsplicing activity.
Klein, B., Staden, A., Schlessinger, D.,Division of Biology and BiomedicalSciences, Washington UniversitySchool of Medicine, St. Louis, Mis-souri: Alternate conformations in16S rRNA.
Klootwijk, J., Kempers-Veenstra, A.E.,Dekker, A.F., van Heerikhuizen,H., Musters, W., Oliemans, J.,Planta, R.J., Biochemisch Labora-torium, Vrije Universiteit, Amster-dam, The Netherlands: Expressionof artificial rRNA genes in yeast.
Konarska, M., Padgett, R.A., Gra-bowski, P.J., Sharp, PA., Dept. ofBiology, Massachusetts Institute ofTechnology, Cambridge: mRNAsplicing in vitro-Requirement fora cap structure.
Krainer, A.R., Ruskin, B., Green,
197

M.R., Maniatis, T., Dept. of Bio- sity School of Medicine, Boston,chemistry and Molecular Biology, Massachusetts: Endogenous
Harvard University, Cambridge, cleavages in the polyribosomal /3-Massachusetts: In vitro splicing of globin mRNA of rabbit reticulo-normal and mutant human 13-glo- cytes.
bin pre-mRNAs. Krowczynska, A., Brawerman, G.,Krowczynska, A., Albrecht, G., Braw- Dept. of Biochemistry and Phar-
erman, G., Dept. of Biochemistry macology, Tufts University Schooland Pharmacology, Tufts Univer- of Medicine, Boston, Massachu-
SESSION 4 tRNA PROCESSING
Chairperson: S. Altman, Yale University, New Haven, Connecticut
Tobian, J.A., Castano, J.G., Zasloff,M.A., NCI, National Institutes ofHealth, Bethesda, Maryland: Pointmutations in a human tRNA7,gene-Effect on transport andprocessing.
Doersen, C., Gaines, G., Shuey, D.,Attardi, G., California Institute ofTechnology, Pasadena: Synthesisand processing of human mRNAin in vitro systems.
Gandini-Attardi, D., Margarit, I., Toc-chini-Valentini, G.P., Institute of CellBiology, Rome, Italy: Role of thestructure of the exon in tRNA splic-ing.
Hollingsworth, M.J., Martin, N.C.,Dept. of Biochemistry, Universityof Texas Health Science Center,
SESSION 5 rRNA PROCESSING
Dallas: A mitochondrial gene is re-quired for the expression of yeastmitochondrial RNase P.
Willis, I., Chisholm, V., Pearson, D.,Sall, D., Dept. of Molecular Bio-physics and Biochemistry, YaleUniversity, New Haven, Connecti-cut: Mutations affecting 5'-end mat-uration and splicing of an S.pombe dimeric tRNA precursor.
Atkinson, N.S., Dunst, R.W., Nolan,S.L., Hopper, A.K., Dept. of Bio-logical Chemistry, Hershey Medi-cal Center, Pennsylvania: Charac-terization of a yeast gene affectingRNA processing.
Furdon, P, Altman, S., Dept. of Biol-ogy, Yale University, New Haven,Connecticut: Site-specific muta-
Chairperson: J. Dunn, Brookhaven National Laboratory, Upton, New York
Dahlberg, A., Zwieb, C., Jemiolo, D.,Jacob, W., Skinner, R., Stark, M.,Section of Biochemistry, Brown
University, Rhode Island: Effects ofsite-directed mutations on pro-cessing of E. coil rRNA.
M. Mathews, A. Shatkin
198
setts: Control of mRNA stability inmouse erythroleukemia cells.
Lahiri, D.K., Thomas, J.0., Dept. ofBiochemistry, New York UniversitySchool of Medicine, New York:What happens to nuclear RNAand its associated proteins duringmitosis?
genesis of the dihydrouridine loopof the tRNAT, su3, gene from E.coil.
Reilly, R.M., RajBhandary, U.L., Dept.of Biology, Massachusetts Instituteof Technology, Cambridge: Bio-synthesis of E. coil Sunl tRNA isdefective in mutants encodingGAUC in place of the universalGTipC sequence.
Deutscher, M.P., Marlor, C.W., Zan-iewski, R., Dept. of Biochemistry,University of Connecticut HealthCenter, Farmington: The exoribo-nuclease family of E. coil- Identifi-cation and characterization of anew enzyme, RNase T
Sirdeshmukh, R., Schlessinger, D., Di-vision of Biology and BiomedicalSciences, Washington UniversitySchool of Medicine, St. Louis, Mis-souri: Maturation of 23S rRNA invitro.
Szeberenyi, J., Apirion, D., Dept. ofMicrobiology, Washington Univer-sity, St. Louis, Missouri: Primaryand secondary processing of 5SrRNA in E. coll.
Cole, J.R., Singer, P., Nomura, M.,Depts. of Genetics and Biochemis-try, University of Wisconsin, Madi-son: Translational regulation of ri-bosomal proteins in E. coli affectsthe half-lives of ribosomal proteinmRNAs.
Bolla, R.,' Shiomi, Y.,2 Braaten, D.,2Herbert, M.B.,2 Schlessinger, D.,2'Dept. of Biology, University ofMissouri, St. Louis; 2Dept. of Mi-crobiology and Immunology,Washington University School of

Medicine, St. Louis, Missouri:Characterization of mouse L-cellcore nucleoli.
Reddy, R., Lischwe, M., Epstein, P.,Henning, D., Tan, E., Reich lin, M.,Busch, H., Dept. of Pharmacology,Baylor College of Medicine, Hous-ton, Texas: Nucleolar RNP anti-gens and U3 genes.
Tabak, Osinga, K.A., Groot
SESSION 6 POSTER SESSION
Lee, M.G.,' Young, R.A.,2 Begs,J.D.,"Dept. of Biochemistry, Im-perial College of Science andTechnology, London, England;2Dept. of Biochemistry, StanfordUniversity School of Medicine,California: Cloning and analysis ofthe RNA2 gene from S. cerevisiae.
Leser, G.P., Martin, T.E., Dept. of Biol-ogy, University of Chicago, Illinois:hnRNP core protein complementsof cells examined with monoclonalantibodies.
Leys, E.J.,' Crouse, G.F.,2 Frayne,E.G.,' Kellems, R.E.,"Dept. ofBiochemistry, Baylor College ofMedicine, Houston, Texas; 2NCI,Frederick Cancer Research Facil-ity, Frederick, Maryland: Identifica-tion of an RNA polymerase II tran-scription unit with an initiation siteless than 200 by upstream fromthe mouse dhfr gene.
Lockard, R.E., Garrett-Wheeler, E.,Kumar, A., Dept. of Biochemistry,George Washington UniversityMedical School, Washington,D.C.: Mapping of psoralen cross-linked nucleotides in RNA.
Mangin, M., Ares, M., Jr., Weiner,A.M., Dept. of Molecular Biophys-ics and Biochemistry, Yale Univer-sity, New Haven, Connecticut:DNA conformation of snRNAgenes-Mapping of hypersensitivesites on the human U1 and U2transcription units.
Maran, A., Kalyanaraman, S., Shan-mugam, G., School of BiologicalSciences, Madurai Kamaraj Uni-versity, India: Double-strandedRNA specific nuclease from germi-nating embryos of Pennisetum ty-phoides.
Mattaj, I.W., Zeller, R., DeRobertis,E.M., Dept. of Cell Biology, Bio-
Koerkamps, M.J.A., van derHorst, G.T.J., Section for Molecu-lar Biology, University of Amster-dam, The Netherlands: Transcrip-tion of yeast mitochondrial DNAand processing of RNA.
Bachellerie, J.P., Michot, B., Has-souna, N., Raynal, F, Centre deRecherche de Biochimie et deGenetique Cellulaires du CNRS,
center, University of Basal, Switz-erland: Xenopus U-snRNA genetranscription units.
Mecklenburg, K.,' Sass, P.,2 Mahler,H.R.,2 Perlman, P.S.,' 1MCDB Pro-gram, Ohio State University, Co-lumbus; 2Dept. of Chemistry, Indi-ana University, Bloomington:Splicing defective mutants in theOxi3 gene of yeast mtDNA.
Miller, K., Zbrzezna, V., Pogo, A.O.,Lindsey F. Kimball Research Insti-tute, New York Blood Center, NewYork: RNA interstrand duplexes inmurine erythroleukemia cells andDrosophila KC cells.
Morris, G.F., Marzluff, W.F., Dept. ofChemistry, Florida State University,Tallahassee: Biosynthesis of seaurchin N1 snRNA.
Noble, J., Prives, C., Manley, J.L.,Dept. of Biological Sciences, Co-lumbia University, New York, NewYork: In vitro splicing and poly-adenylation of an SV40 early pre-mRNA.
Ore liana, 0., Cooley, L., SOil, D.,Dept. of Molecular Biophysics andBiochemistry, Yale University, NewHaven, Connecticut: Transcriptionand processing of a clonedtRNAH's gene from E. coll.
Parker, R., Guthrie, C., Dept. of Bio-chemistry and Biophysics, Univer-sity of California, San Francisco: Agenetic analysis of mRNA splicingin yeast.
Patton, J.R., Chae, C.-B., Dept. ofBiochemistry, University of NorthCarolina, Chapel Hill: Specific pro-tein binding on p-globin RNA inwhole nuclei, the nuclear matrix,and 40S hnRNP of chicken reticu-locyte.
Pedersen, N., Hel lung-Larsen, P.,Engberg, J., Dept. of Biochemis-
Toulouse, France: Structure andprocessing of mouse rRNA pre-cursors-Genuine spacers andspacerlike domains within maturerRNA.
Vance, V., Bowman, L.H., Dept. of Bi-ology, University of South Caro-lina, Columbia: Transfection ofmouse rDNA into rat cells-Faith-ful transcription and processing.
try, Panum Institute, University ofCopenhagen, Denmark: snRNAsin Tetrahymena.
Reinberg, D., Roeder, R.G., Rockefel-ler University, New York, NewYork: RNA polymerase II transcrip-tion system-A biochemical ap-proach.
Reveillaud, I., Lelay-Taha, M.N., Sri-Widada, J., Brunel, C., Jeanteur,P., Laboratoire de Biochimie,Centre P. Lamarque, and Labora-toire de Biologie Moleculaire,USTL, Montpellier, France: Dra-matic effect of Mg ** on snRNPconformation application to isola-tion of individual U1 and U5 sn-RNPs.
Roe, B.A., Ma, D.-P., Wong, J.F.H.,Wilson, R.K., Ahmed, M.A., Dept.of Chemistry, University of Okla-homa, Norman: Structure andexpression of several eukaryote,prokaryote, and organelle tRNAgenes.
Rooney, R.J., Harding, J.D., Dept. ofBiological Sciences, ColumbiaUniversity, New York, New York:Kinetics of in vitro transcriptionand processing of mouse tRNAgenes.
Ryner, L., Manley, J.L., Dept. of Bio-logical Sciences, Columbia Uni-versity, New York, New York: Anovel poly(A) polymerase activityin a cytoplasmic S100 extract ofHe La cells.
Samuels, D.S. Peebles, C.L., Dept. ofBiological Sciences, University ofPittsburgh, Pennsylvania: Over-production of pre-tRNAs.
Sarkar, S., Jayabaskaran, C., Das-gupta, S., Dept. of Muscle Re-search, Boston Biomedical Re-search Institute, Massachusetts: Anovel cytoplasmic, translation-in-
199

hibitory 10S RNP particle of chickembryonic muscle.
Schmidt, F.J., Motamedi, H., Lee, Y.,Dept. of Biochemistry, Universityof Missouri, Columbia: Tandempromoters preceding the gene forE. coli M1 RNA.
Schrier, W.H., Okarma, T.B., StanfordUniversity, California: Induction ofgrowth hormone transcription bytriiodothyronine and dexametha-sone is accompanied by in-creased 4S-8S RNA synthesis.
Shelness, G.S., Williams, D.L., Dept.of Pharmacological Sciences,State University of New York,Stony Brook: Apo lipoprotein IImRNA-Alternate splicing andtranscriptional heterogeneityyields six untranslated 5' leadersequences.
Simon, M., Arrigo, A. -P., Spahr, P. -F.,Dept. of Molecular Biology, Uni-versity of Geneva, Switzerland: Acellular protein phosphorylated byRSV transforming product is asso-ciated with small cytoplasmic RNPparticles.
Smith, D.B., Inglis, S.C., Dept. of Pa-thology, University of Cambridge,England: Influenza virus mRNAsplicing is modulated during infec-tion.
Strobel, M.C.,12 Abelson, J.,2 'Dept.of Biology, University of California,San Diego, La Jolla; 2Division ofBiology, California Institute ofTechnology, Pasadena: The inter-vening sequence directs modifica-tion of the SUP53 (tRNALeu) antico-don.
SESSION 7 mRNA SPLICING
Subbarao, M.N., Watson, N., Apirion,D., Dept. of Microbiology, Wash-ington University, St. Louis, Mis-souri: Processing reactions cata-lyzed by RNA in E. coli.
Talib, S., O'Brien-Vedder, C., Okarma,T.B., Stanford University, Califor-nia: In vivo effects of psoralencross-linking on U-snRNA geneexpression.
Tanner, N.K., Cech, T.R., Dept. ofChemistry, University of Colorado,Boulder: Probing the structure andfunction of RNA with ethidium bro-mide and methidiumpropyl-EDTAFe(11).
Thompson, W.S., Abernathy, E.G.,Gross, R.H., Dept. of Biology,Dartmouth College, Hanover, NewHampshire: Stability and localiza-tion of liposome-introduced sn-RNAs and anti-snRNP antibodiesin Drosophila cells.
Tomcsanyi, T, Apirion, D., Dept. ofMicrobiology and Immunology,Washington University School ofMedicine, St. Louis, Missouri: Asmall RNA processed by RNase Eis involved in plasmid DNA repli-cation.
Tu, C.-P.D., Lai, H.-C.J., Li, N., Weiss,M.J., Reddy, C.C., PennsylvaniaState University, University Park:Tissue-specific expression of a Yaand a Y, subunit of rat glutathioneS-transferases.
Ul lu, E., Weiner, A.M., Dept. of Molec-ular Biophysics and Biochemistry,Yale University, New Haven, Con-necticut: Efficient and accurateexpression of a 7SL hnRNA gene
Chairperson: J. Manley, Columbia University, New York, New York
Belfort, M., Chu, F., Maley, G., Ehren-man, K., Pedersen-Lane, J.,Maley, F, Center for Laboratoriesand Research, New York State De-partment of Health, Albany: Ex-pression of an intron-containingprokaryotic thymidylate synthasegene.
Pikielny, C.W.,' Rodriguez, J.,2 Ros-bash, M.,2 'Dept. of Biochemistry;2Dept. of Biology, Brandeis Uni-versity, Waltham, Massachusetts:Possible intermediate in yeastmRNA splicing.
200
Rossi, J.J., McMahon, J., Cellini, A.,Beckman Research Institute of theCity of Hope, Los Angeles, Califor-nia: Deletion of the ICS from theactin intron reduces but does notabolish splicing.
van Santen, V., Spritz, R.A., Dept. ofMedical Genetics, University ofWisconsin, Madison: Sequence re-quirements for splicing of eukar-yotic pre-mRNA-Deletion analy-sis of an intervening sequence.
Hwang, L., Park, J., Gilboa, E., Dept.of Molecular Biology, Princeton
is strongly dependent upon anelement located in the 5'- flankingsequences.
Vold, B., Green, C., SRI International,Menlo Park, California: Large clus-ters of tRNA genes from B. sub-tills,
Watson, N., Dallmann, G., Dallmann,K., Sonin, A., Apirion, D., Dept. ofMicrobiology and Immunology,Washington University School ofMedicine, St. Louis, Missouri:Cloning of genes for E. coli RNAprocessing enzymes.
Westin, G.,' Hammarstrtim, K.,' Za-bielski, J.,' Bark, C.,' Murphy, J.,2Lund, E.,2 Dahlberg, J.,2 Petters-son, U.,"Dept. of Medical Ge-netics, University of Uppsala, Swe-den; 2Dept. of PhysiologicalChemistry, University of Wiscon-sin, Madison: Isolation andexpression of genes for U2hnRNA.
Willis, I., Frendewey, D., Nichols, M.,Schaack, J., Soli, D., Dept. of Mo-lecular Biophysics and Biochemis-try, Yale University, New Haven,Connecticut: A single basechange in the anticodon of an S.pombe serine tRNA affects RNaseP cleavage and splicing in vitro.
Wollenzien, PL.,' Expert-Bezancon,A.,2 Cantor, C.R.,' 'Dept. of Hu-man Genetics and Development,Columbia University, New York,New York; 2lnstitut de Rechercheen Biologie Moleculaire, Paris,France: Three dimensional ar-rangement of the E. coli 16S rRNAin the 30S subunit.
University, New Jersey: Mecha-nism of splicing of Mo-MLV envmRNA.
DiMaria, P., Kaltwasser, G., Golden-berg, C.J., Dept. of Pathology,Washington University, St. Louis,Missouri: In vitro Ad2 mRNA splic-ing of purified, synthetic RNA pre-cursors.
Kramer, A.,' Hernandez, N.,' Reiss,B.,' Lahr, G.,' Frick, M.,' Keller,W.,1 Rinke, J.,2 Appel, B.,2 LOhr-mann, R.,2 'Institute of Cell andTumor Biology, German Cancer

Research Center, Heidelberg;2Max-Planck-Institute, Berlin, Fed-eral Republic of Germany: In vitrosplicing of mRNA precursors.
Ruskin, B., Krainer, A., Maniatis, T.,Green, MAR., Dept. of Biochernis-
try and Molecular Biology, Har-vard University, Cambridge, Mas-sachusetts: The fate of intronsequences during in vitro splicing.
Grabowski, P.J., Padgett, R.A., Sharp,P.A., Dept. of Biology, Massachu-
SESSION 8 mRNA PROCESSING AND METABOLISM
Chairperson: W. Le Stourgeon, Vanderbilt University, Nashville, Tennessee andNational Science Foundation, Washington, D.C.
Halbreich, A., Grandchamp, C.,Foucher, M., Centre de GenetiqueMoleculaire du CNRS, Gif-sur-Yvette, France: Yeast mitochondriacontain a linear RNA strand com-plementary to the circular, intronicb11 RNA of cytochrome b.
Simon, M., Faye, G., Institut Curie, Or-say, France: Organization and pro-cessing of the OX13/0L12 multi-genic transcript in yeastmitochondria.
Last, R., Woolford, J.L., Dept. of Bio-logical Sciences, Carnegie-MellonUniversity, Pittsburgh, Pennsylva-nia: Evidence for related functionsof the RNA2, RNA3, and RNA4gene products of S. cerevisiae.
Pape, L.K., Tzagoloff, A., Dept. of Bi-ological Sciences, Columbia Uni-versity, New York, New York: A nu-clear gene required for processingof the S. cerevisiae mitochondrialapocytochrome b pre-mRNA.
Anderson, K.P., Klessig, D.F., Dept. ofCellular, Viral and Molecular Biol-ogy, University of Utah School ofMedicine, Salt Lake City: AlteredmRNA splicing in monkey cells
SESSION 9 SMALL RNAs AND RNPs
abortively infected with human ad-enovirus may be responsible forinefficient synthesis of the virion fi-ber polypeptide.
Dreyfuss, G., Choi, Y-D., Adam, S.A.,Dept. of Biochemistry, Molecularand Cell Biology, NorthwesternUniversity, Evanston, Illinois: Char-acterization of the protein com-plexes of hnRNA in vivo withmonoclonal antibodies.
Raziuddin, A.K., Thomas, J.0., Szer,W., Dept. of Biochemistry, NewYork University School of Medi-cine, New York: Functional do-mains of HD40, the major corehnRNP protein of Artemia.
Greenberg, J.R., Carroll, E. III, CellBiology Group, Worcester Foun-dation for Experimental Biology,Shrewsbury, Massachusetts: Re-constitution of functional mRNP-protein complexes.
GOrlach, M., Schulze, G., Sarre, T.,Hilse, K., Institute kir Biology UN-versitat, Freiburg, Federal Repub-lic of Germany: Characterization ofcytoplasmic mRNPs from two dif-ferent erythropoietic systems (rab-
Chairperson: R. Geste land, University of Utah, Salt Lake City, Utah
Patterson, B., Tollervey, D., Swerdlow,H., Wise, WA., Guthrie, C., Dept.of Biochemistry and Biophysics,University of California, San Fran-cisco: A genetic analysis of snRNAfunction in yeast.
Skuzeski, WM.,' Lund, E.,' de Vegvar,H.E.N.,' Murphy, J.T,2 Steinberg,T.,2 Dahlberg, J.E.,"Dept. ofPhysiological Chemistry; 2McArdleLaboratory, University of Wiscon-sin, Madison: Signals for transcrip-tion and maturation of U1 RNA.
Perez-Stable, C., Hess, J., Ayres,TM., Shen, C.-K.J., Dept. of Ge-netics, University of California,Davis: Novel features of in vitrosynthesis of human a/u familysmall RNAs-Distinctive promoterorganization and a new eukaryoticclass III terminator.
Luhrmann, R., Appel, B., Rinke, J.,Reuter, R., Theissen, H., Bring-mann, P., Max-Planck-Institut, Ber-lin, Federal Republic of Germany:Structure of U-snRNPs- Evidence
setts Institute of Technology, Cam-bridge: mRNA splicing in vitro-An excised intervening sequenceand a potential intermediate.
bit reticulocytes/murine erythroleu-kemia cells).
Katze, M.G., Chen, Y. -T., Krug, R.M.,Memorial Sloan-Kettering CancerCenter, New York, New York: Theprocessing and expression ofRNA polymerase II transcripts ininfluenza virus infected cells.
H. Robertson
for the existence of snRNAs U4and U6 in a single RNP complex.
Fisher, D.E., Conner, G.E., Reeves,W.H., Blobel, G., Kunkel, H.G.,Rockefeller University, New York,New York: SnRNP assembly is atwo-step process, accompaniedby posttranslational modification.
Kloetzel, P.M., Schult, C., Bautz,E.K.F, Dept. of Molecular Ge-netics, University of Heidelberg,Federal Republic of Germany:Analysis of specific RNP-com-
201

plexes in heat-shocked Drosophila Sm- and U1- specific snRNP anti-cells. gens at transcription sites, includ-
Martin, T.E., Monsma, S.A., Leser, ing some lacking introns.G.P., Dept. of Biology, University Masukata, H., Tomizawa, J., NIADDK,of Chicago, Illinois: Localization of National Institutes of Health, Be-
SESSION 10 POLY A AND 3' ENDS
Chairperson: R. Perry, Institute for Cancer Research, Philadelphia, Pennsylvania
Mott, J., Galloway, J.L. Farnham, RJ.,Platt, T., Dept. of Molecular Bio-physics and Biochemistry, YaleUniversity, New Haven, Connecti-cut: RNA termination and process-ing at the 3' end of tryptophan op-eron mRNA.
Higgins, C.F., Dept. of Biochemistry,University of Dundee, Scotland: Anovel structure (the "REP" se-quence) present in a large numberof bacterial mRNA species mayregulate mRNA processing anddegradation.
Aloni, Y., Hay, N., Bengal, E., Laub,O., Dept. of Genetics, WeizmannInstitute of Science, Rehovot, Is-rael: Attenuation and transcription-termination in the control of eukar-yotic gene expression -SV40 as amodel system.
Frayne, E.G.,' Leys, E.J.,' Crouse,G.F.,2 Kellems, R.E.,"Dept. of
RNA Tumor VirusesMay 22-May 27, 1984
ARRANGED BY
Biochemistry, Baylor College ofMedicine, Houston, Texas; 2NCI,Frederick, Maryland: Transcriptionof the dhfr gene proceeds uninter-rupted through all seven poly(A)sites and terminates further down-stream.
Wickens, M., Stephenson, P, Dept. ofBiochemistry, University of Wis-consin, Madison: Point mutationsin any one of three positions of theAAUAAA sequence prevent cleav-age of SV40 late pre-mRNAs inXenopus oocytes.
McDevitt, M.A., Imperiale, M.J., Ali,H., Nevins, J.R., Rockefeller Uni-versity, New York, New York: Re-quirement of a downstream se-quence for generation of a poly(A)addition site.
Sadofsky, M., Alwine, J.C., Dept. ofMicrobiology, University of Penn-sylvania, Philadelphia: Deletions of
thesda, Maryland: Alteration inRNA structure affects primer for-mation for ColEl DNA replication.
SV40 DNA downstream from thelate AAUAAA, including a regionof U4 RNA homology, decreasethe efficiency of 3'-end process-ing.
Moore, C., Sharp, PA., Dept. of Biol-ogy, Massachusetts Institute ofTechnology, Cambridge: Site-spe-cific poladenylation in a cell-freereaction.
Manley, J.L., Yu, H., Ryner, L., Dept.of Biological Sciences, ColumbiaUniversity, New York, New York:The hexanucleotide AAUAAAforms part of a "promoter" formRNA polyadenylation in vitro.
Nordstrom, J.L., Kessler, M.M., Wes-thafer, M.A., Dept. of Biochemistryand Biophysics, Texas A&M Uni-versity, College Station: The 3' endof the sea urchin H2A gene servesas a polyadenylation signal in COScells.
Tony Hunter, Salk InstituteSteve Martin, University of California, Berkeley
434 participants
RNA tumor virus research continues to be an exciting and rapidly moving field.This was the ninth RNA tumor virus meeting, and its continuing popularity wasdemonstrated by the large number of attendees and presentations.
More than half of the presentations were concerned with work on viral andcellular oncogenes. Recurring themes included: the different mechanisms thatresult in the activation of proto-oncogenes; the use of in vitro mutagenesis tech-niques to define functional domains within viral transforming proteins; and thediscovery of new, common integration sites for viruses without one genes. Thedemonstration that mutations in the yeast RAS genes have marked phenotypiceffects underscored the advantages of simple eukaryote models for studies onthe cellular functions of oncogenes. Another exciting observation was the finding
202

that the Philadelphia translocation in chronic myelogenous leukemia results in aunique c-ab/ transcript and an altered and apparently activated c-ab/ protein.
Although work on oncogenes received the lion's share of attention, there werenovel findings in other areas: the requirement for po/ endonuclease activity forDNA integration; the role of the MLV po/ protease in the processing of gag pre-cursors; and the tissue specificity of the enhancer elements within viral LTRs as abasis for the tissue tropism of these viruses, Considerable progress was also re-ported in the characterization of the human T-cell lymphotropic viruses (HTLVand LAV) and their roles in T-cell leukemia and AIDS.
Funding was provided in part by the Cold Spring Harbor Laboratory CancerCenter Grant from the National Cancer Institute, National Institutes of Health.
SESSION 1 ONCOGENES (src, rel, fps, abl)
Chairperson: H. Hanafusa, Rockefeller University, New York, New York
lba, H.,' Cross, F.R.,1 Takeya, T.,2Hanafusa, T.,1 Hanafusa, H.,"'Rockefeller University, New York,New York; 2lnstitute for ChemicalResearch, Kyoto University, Ja-pan: Low protein kinase activity invivo may be correlated to the ina-bility of p60c-sre to transform CEF.
Yaciuk, P., Coussens, P.M., Shalloway,D., Molecular and Cell BiologyProgram, Pennsylvania State Uni-versity, University Park: Effects ofmodifying the pp6Osrc carboxyl ter-minus.
Coussens, P.M., Johnson, P.J., Shallo-way, D., Molecular and Cell Biol-ogy Program, Pennsylvania StateUniversity, University Park: Highlyefficient c-src expression plasmidsinduce low-level transformation.
Jakobovits, E.B., Majors, J.E., Var-mus, H.E., Dept. of Microbiologyand Immunology, University ofCalifornia, San Francisco: Hor-monal regulation of the RSV srcgene via the MMTV promoter de-fines a threshold dose for transfor-mation.
Wilhelmsen, K.C., Tarp ley, W.G.,Miller, C.K., Temin, H.M., McArdleLaboratory, University of Wiscon-sin, Madison: Identification ofsome of the parameters governingtransformation by oncogenes inretroviruses.
Yonemoto, W.,' Jarvis-Morar, M.,1Bolen, J.,2 Israel, M.,2 Brugge, J.,''Dept. of Microbiology, State Uni-versity of New York, Stony Brook;2NCI, National Institutes of Health,Bethesda, Maryland: Associationof the cellular src gene product
with the middle T antigen of poly-oma virus results in enhancementof associated tyrosine-specific ki-nase activity and modification ofthe cellular src protein.
lijima, S., Wang, L.-H., RockefellerUniversity, New York, New York:Tissue-specific expression of c-srcmRNA by alternative splicing.
Rohrschneider, L., Reynolds, S.,Rothwell, V., Fred HutchinsonCancer Research Center, Seattle,Washington: Regulation of cellularmorphology by the RSV src geneproduct-Analysis of fusiform mu-tants.
Cross, FR., Garber, E.A., Pellman, D.,Hanafusa, H., Rockefeller Univer-sity, New York, New York: The ex-treme amino terminus of the RSV-transforming protein is requiredfor myristylation, membrane asso-ciation, and transformation.
Garber, E.A., Cross, FR., Pellman, D.,Hanafusa, H., Rockefeller Univer-sity, New York, New York: Myristy-lation of p6Osrc and its role in mem-brane association-Comparison ofwild-type RSV and in vitro con-structed aminoterminal deletionmutants.
Snyder, M.A., Bishop, J.M., GeorgeWilliam Hooper Research Founda-tion, Dept. of Microbiology andImmunology, University of Califor-nia, San Francisco: A mutation atthe major phosphotyrosine site inpp60v-sm alters oncogenic poten-tial.
Stone, J.C.,' Atkinson, T.,2 Smith, M.,2Pawson, T.,' Depts. of'Microbiology, 2Biochemistry, Uni-
versity of British Columbia, Van-couver, Canada: Identification offunctional regions in the transform-ing protein of FuSV by in-phase in-sertion mutagenesis.
Weinmaster, G.,' Zoller, M.,3 Smith,M.,2 Pawson, T.,' Depts. of'Microbiology, 2Biochemistry, Uni-versity of British Columbia, Van-couver, Canada; 3Cold SpringHarbor Laboratory, New York:Site-directed mutagenesis of FuSVreveals that tyrosine phosphoryla-tion of p1309a9 -fPs modulates itsenzymatic and biological activities.
Foster, D.A., Shibuya, M., Hanafusa,H., Rockefeller University, NewYork, New York: The structure andtransformation potential of cellularDNA sequences homologous tothe fps gene of FuSV.
Feldman, R.A.,1,2 Gabrilove, J.L.,3Tam, J.,2 Moore, M.A.S.,3 Hana-fusa, H.,2 'NCI, National Institutesof Health, Bethesda, Maryland;2Rockefeller University; 3MemorialSloan-Kettering Cancer Center,New York, New York: Differentialexpression of the cellular fps/fes-encoded protein in human hema-topoietic tumors.
Mathey-Prevot, B., Baltimore, D.,Whitehead Institute for BiomedicalResearch, Massachusetts Instituteof Technology, Cambridge: Effectof replacing the abl sequence bythe avian sequences src or fps inAb-MLV.
Prywes, R.,' Hoag, J.,1 Rosenberg,N.,2 Baltimore, D.,"Whitehead In-stitute for Biomedical Research,Massachusetts Institute of Technol-
203

ogy, Cambridge; 2Tufts UniversitySchool of Medicine, Boston: Thep15 region of gag is required forlymphoid transformation by Ab-MLV.
Foulkes, J.G.,' Baltimore, D.,' Mailer,
J.L.,2 'Whitehead Institute forBiomedical Research, Massachu-setts Institute of Technology, Cam-bridge; 2Dept. of Pharmacology,University of Colorado, School ofMedicine, Denver: Microinjection
SESSION 2 REPLICATION AND INTEGRATION
Chairperson: H. Varmus, University of California, San Francisco, California
Lobel, L.I., Tanese, N., Goff, S.P.,Dept. of Biochemistry, ColumbiaUniversity College of Physiciansand Surgeons, New York, NewYork: Transfer of mutations fromthe 5' LTR to the 3' LTR during rep-lication of Mo-MLV-Strong-stopDNAs may "jump" prematurely.
Smith, J.K., Cywinski, A., Taylor, J.M.,Institute for Cancer Research, FoxChase Cancer Center, Philadel-phia, Pennsylvania: The specificityof initiation of strong-stop plusDNA by RSV.
Resnick, R.M., Faras, A.J., Dept. ofMicrobiology, University of Minne-sota, Minneapolis: Recognitionbinding sites for reverse transcrip-tase during RNase H-mediated(+) DNA synthesis in vitro.
Panganiban, A., Temin, H.M., Mc-Ardle Laboratory for Cancer Re-search, University of Wisconsin,Madison: Sequences required forintegration of SNV DNA.
Colicelli, J., Goff, S.P., Dept. of Bio-chemistry, Columbia University,College of Physicians and Sur-geons, New York, New York: Mu-tants of Mo-MLV bearing a singlebase pair change at the LTR tipcan still integrate into host DNA.
SESSION 3 POSTER SESSION I
Dolberg, D.S., Bissell, M.J., Dept. ofBiology and Medicine, LawerenceBerkeley Laboratory, University ofCalifornia, Berkeley: The role ofwounding in RSV-induced tumorformation.
Kirschmeier, P.T., Perkins, A.S.,Shane, E., Bilezikian, J.P., Wein-stein, I.B., Columbia UniversityCollege of Physicians and Sur-geons, New York, New York:Suppression of the transformedphenotype by the Salmonella ty-phimurium adenyl cyclase gene.
Laugier, D.,' Poirier, F.,' Marx, M.,2
204
Longiaru, M.,' Duyk, G.,2 Leis, J.,2Skalka, A.M.,"Dept. of MolecularGenetics, Hoffman-La Roche Inc.,Roche Research Center, Nutley,New Jersey; 2Case Western Re-serve University School of Medi-cine, Cleveland, Ohio: Selectivecleavage of wild-type and mutateddouble-strand retroviral LTR DNAby the endonuclease associatedwith AMV reverse transcriptase.
Duyk, G.,' Longiaru, M.,2 DeHaseth,P.,' Terry, R.,2 Shalka, A.,2 Leis,J.,"Case Western Reserve Uni-versity School of Medicine, Cleve-land, Ohio; 3Floche Institute of Mo-lecular Biology, Nutley, NewJersey: Sequence requirementsfor site selective cleavage of theRAV-2 LTR by the reverse tran-scriptase endonuclease.
Donehower, L.A., Varmus, H.E., Dept.of Microbiology and Immunology,University of California, San Fran-cisco: poi mutants of MLV are de-fective for integration.
Hippenmeyer, P.J., Grandgenett, D.P.,Institute for Molecular Virology, St.Louis, Missouri: Amino terminusand mutational analysis of the ASVpp32 protein.
Levin, J.G.,' Hu, S.C.,' Rein, A.,2
Genvrin, P.,1 Dambrine, G.,3 Cal-othy, G.,' 'Institut Curie, Orsay;2CNRS, ICIG, Villejuif; 3Station dePathologie Aviaire de l'INRA Mon-naie, France: A RSV mutant withhost cell-dependent transformingcapacity.
Stoker, A.W., Enrietto, P.J., Wyke,J.A., Dept. of Tumour Virology, Im-perial Cancer Research Fund,London, England: Temperature-sensitive mutations in functionaldomains of pp60v-s.
Hughes, S.,' Mellstrom, K.,2 Kosik,E.,' Tamanoi, F.,' Brugge, J.,2
of Ab-MLV tyrosyl-protein kinaseinto Xenopus oocytes inducesphosphorylation of ribosomal pro-tein S6 on serine residues.
Messer, L.I.,1 Gerwin, B.l.,31NICHD, National Institutes ofHealth, Bethesda; 2Frederick Can-cer Research Facility, Frederick;31\11C, National Institutes of Health,Bethesda, Maryland: Functionalmapping of MLV poi gene by char-acterization of a frameshift mutant.
Crawford, S., Goff, S.P., Dept. of Bio-chemistry, Columbia University,College of Physicians and Sur-geons, New York, New York: Adeletion mutation in the 5' part ofthe poi gene of Mo-MLV preventsprocessing of the Pr659a9 precur-sor polyprotein.
Yoshinaka, Y, Katoh, I., Copeland,T.D., Oroszlan, S., NCI, FrederickCancer Research Facility, Freder-ick, Maryland: Purification, bio-chemical and structural character-ization of Mo-MLV proteaseencoded by the gag -poi gene.
Leis, J.,' Fu, X.,' Traugh, J.,3 'CaseWestern Reserve UniversitySchool of Medicine, Cleveland,Ohio; 2University of California, Riv-erside: The RNA-binding proper-ties of the avian retrovirus pp12are regulated by phosphorylationof a specific serine residue.
'Cold Spring Harbor Laboratory; 2Dept. of Microbiology, State Uni-versity of New York, Stony Brook:Mutation of a termination codonaffects src initiation.
Perez, L., Wills, J.W., Hunter, E., Dept.of Microbiology, University of Ala-bama, Birmingham: Effects of up-stream initiation codons on theexpression of the RSV env gene.
Feuerman, M.,' Davis, B.,' Patten-gale, P.,2 Fan, H.,"Dept. of Mo-lecular Biology and Biochemistry,University of California, Irvine;2University of Southern California,

Los Angeles: Transformation andpathogenicity of in vitro recombi-nant retrovirus.
Tanaka, A.,' Gibbs, C.,2 Anderson,S.K.,' Kung, H.-J.,2 Fujita, D.J.,''Cancer Research Laboratory,Dept. of Biochemistry, Universityof Western Ontario, London, Can-ada; 2Dept. of Biochemistry, Michi-gan State University, East Lansing:Structural analysis and nucleotidesequencing of a human c-srcgene.
Parvin, J.,' lijima, S.,2 Wang, L.-H.,2'Mount Sinai School of Medicine;2Rockefeller University, New York,New York: Mechanisms for thegeneration of src-deletion mutantsand recombination between viraland c-src sequences.
Kotler, M., Roguel, N., Relevy, H.,Dept. of Molecular Genetics, He-brew University-Hadassah Medi-cal School, Jerusalem, Israel: Am-plification of proviruses and theirmethylated state in avian sarcoma-transformed rat cells.
Mannino, R.J., Winters, L., Dept. ofMicrobiology and Immunology, Al-bany Medical College, New York:Regulation of src expression ac-companies growth inhibition ofRSV-transformed cells.
Barnekow, A., Gessler, M., Institut fOrMedical Virologie, Giessen, Fed-eral Republic of Germany: Differ-ences in expression of c-src and c-yes during chicken development.
Maness, P,' Fults, 0.,2 'Dept. of Bio-chemistry, University of North Car-olina School of Medicine, ChapelHill; 2Dept. of Neurosurgery, Bow-man Gray School of Medicine,Winston-Salem: pp60c-s'° is local-ized in processes of developingcerebellar neurons.
Cotton, P.C., Brugge, J.S., Dept. ofMicrobiology, State University ofNew York, Stony Brook: Neural tis-sues express a modified form ofthe cellular src gene productpp60c-s' c.
Gould, K.,' Cooper, J.A.,' Hunter, T.,'Bretscher, A.,2 'Molecular Biologyand Virology Laboratory, Salk In-stitute, San Diego, California;2Section of Biochemistry, Molecu-lar and Cell Biology, Cornell Uni-versity, Ithaca, New York: Tyrosineprotein kinase substrate p81 is ho-mologous to a chicken microvillarcore protein.
Burr, J.G., Lui, J.C., Linder, M.E.,Dept. of Biology, University ofTexas, Dallas: In situ phosphoryla-tion of cytoskeleton-associatedproteins by pp60sr..
Chackalaparampil, I., Poirier, Y., Ba-nerjee, D., Mukherjee, B.B., Dept.of Biology, and Centre for HumanGenetics, McGill University, Mon-treal, Canada: Alteration in the se-cretion of a specific phosphopro-tein in RSV-transformed rat cellswhich had undergone reversibleloss of anchorage-independentgrowth after retinoic acid treat-ment.
Jarosik, G., Morar, M.J., Broach, J.,Brugge, J., Dept. of Microbiology,State University of New York,Stony Brook: Functional expres-sion of the RSV src gene in yeast.
Huang, C.-C., Hammond, C., Bishop,J.M., George William HooperFoundation, Dept. of Microbiology,University of California, San Fran-cisco: The nucleotide sequenceand topography of chicken c-fps.
Goh, W.C., Sodroski, J., Haseltine,W.A., Dana-Farber Cancer Insti-tute, Boston, Massachusetts: Se-quence organization of the humanc-fps/fes gene.
Young, J., Moss, P., Martin, G.S.,Dept. of Zoology, University ofCalifornia, Berkeley: Relationshipbetween the intracellular locationof polypeptides containing fps se-quences and transforming activity.
Bergold, P.J.,1 Wang, J.Y.J.,2 John-son, E.M.,. Littau, V.C.,3 Hardy,W.D., Jr.,' Besmer, P.,' Laborato-ries of 'Molecular Oncology,2University of California, SanDiego; 3Flockefeller University,New York; 'Veterinary Oncology,Memorial Sloan-Kettering CancerCenter, New York, New York;Structural analysis of the abl insertof HZ2-FeSV and an alteration ofc-ab/ in a primary acute lympho-cytic leukemia.
Lederman, L.,' Zuckerman, E.E.,2Hardy, W.D., Jr.,2 Snyder, H.W.,Jr.,3 Pilch, P.F.,"Dept. of Bio-chemistry, Boston University Medi-cal School, Massachusetts;2Laboratory of Veterinary Oncol-ogy, Memorial Sloan-KetteringCancer Center, New York, NewYork; 3lmmune Response Pro-gram, Pacific Northwest ResearchFoundation, Seattle, Washington:
Interaction of HZ2-FeSV P98 gag-abl translation product and associ-ated protein kinase activity withcellular membranes, cytoskeleton,and a possible cytoskeletal sub-strate.
Tidmarsh, G., Pillemer, E., Whitlock,C., Dailey, M., Weissman, I., Labo-ratory of Experimental Oncology,Stanford University School of Med-icine, California: A novel surfaceantigen on Ab-MLV and sponta-neous B-cell lymphomas.
Schiff-Maker, L.,' Konopka, J.B.,2Davis, R.L.,2 Watanabe, S.M.,2Ponticelli, A.S.,2 Witte, 0.N.,2 Ro-senberg, N.,"Cancer ResearchCenter, Immunology GraduateProgram, Tufts University Schoolof Medicine, Boston, Massachu-setts; 2Dept. of Microbiology andMolecular Biology Institute, Uni-versity of California, Los Angeles:Membrane localization of the Abel-son protein.
Lamph, W.W., Arfsten, A., Green, P.L.,Shilling, J., Risser, R., McArdleLaboratory for Cancer Research,University of Wisconsin, Madison:Monoclonal antibodies to a cellularin vitro kinase found in Abelsonlymphoma cells.
Watanabe, S.M., Konopka, J.B.,Witte, 0.N., Molecular Biology In-stitute, University of California, LosAngeles: Antisera specific for se-quences within the Ab-MLV 3'open reading frame.
Neckameyer, W.,' Shibuya, M.,2 Hsu,M.-T,' Wang, L. -H.,' RockefellerUniversity, New York, New York;2University of Tokyo, Japan: Nu-cleotide sequence of the trans-forming gene of ASV UR2 andstructural analysis of its cellularhomolog.
Propst, F., Vande Woude, G., NCI,Frederick Cancer Research Facil-ity, Frederick, Maryland: A clusterof mouse repetitive sequencesdownstream from the c-mos locuscontains a novel repeat inter-rupted by an LTR element.
Dean, M., Vande Woude, G.F., LBI-Basic Research Program, NCI,Frederick Cancer Research Facil-ity, Frederick, Maryland: Multiplesites in front of the v-mos genecan serve as transcription initiationpoints.
Seth, A., Zweig, M., Showalter, S.,Vande Woude, G., NCI, Frederick
205

Cancer Research Facility, Freder-ick, Maryland: Production of anti-sera with v-mos protein synthe-sized in E.coli.
Brizzard, B.L., Nash, M.A., Cizdziel,PE., Murphy, E.C., Jr., Dept. of Tu-mor Biology/Virology Section, Uni-versity of Texas System CancerCenter, M.D. Anderson Hospitaland Tumor Institute, Houston: Or-ganization of viral DNA in MSVts110-infected cell lines.
Singh, B.,' Escobedo, J.,2 Dina, D.,2Arlinghaus, R.B.,1 'Dept. of Molec-ular Biology, Scripps Clinic andResearch Foundation, La Jolla;2Chiron Corp., Emeryville, Califor-nia: Reduced level of a 55,000-dalton cellular protein upon infec-tion by Mo-MSV.
Papkoff, J., Ringold, G., Dept. ofPharmacology, Stanford UniversitySchool of Medicine, California:Use of the MMTV LTR to promotesteroid-inducible expression of v-mos.
Righi, M.,1 Coll, J.,' Dozier, C.,1 Gali-bert, F.,2 Stehelin, D.,1 lOncologieMoleculaire, INSERM, Institut Pas-teur, Lille; 2Centre Hayem, HopitalSt. Louis, Paris, France: Analysisof the mil oncogene of MH2 andits cellular counterpart.
S. Hughes
206
Kan, N.,' Rovigatti, U.,' Duesberg,P.,2 Papas, T.,"NCI, FrederickCancer Research Facility, Freder-ick, Maryland; 2Dept. of MolecularBiology, University of California,Berkeley: Avian carcinoma virusMH2 contains two onc-specificgenes, myc and mht, which areclosely related to the transforminggenes of the avian MC29 and mu-rine sarcoma 3611 viruses.
Walther, N., Jansen, H.W., Greiner, E.,Patschinsky, T, Bister, K., Max-Planck-Institut, Berlin, Federal Re-public of Germany: The myc andmil oncogenes of MC29-subgroupviruses.
Seeley, T.W., Mark, G.E., NCI, Na-tional Institutes of Health, Be-thesda, Maryland: cDNA clonesfrom human fetal liver homologousto the raf oncogene.
Mark, G.E.,' Flordellis, C.S.,2 Rapp,U.R.,2 Harris, C.C.,"NCI, Na-tional Institutes of Health, Be-thesda; 2NCI, Frederick CancerResearch Facility, Frederick, Mary-land: The raf oncogene is tran-scribed in small cell lung carcino-mas.
Bonner, T.,2 Goldsborough, M.,1Kerby, S.,' Gunnell, M.,' Opper-mann, H.,3 Seeburg, P.,. Rapp,U.,1 'NCI, National Institutes ofHealth, Frederick, Maryland;2NIMH, National Institutes ofHealth, Bethesda, Maryland;.Genentech, Inc., South San Fran-cisco, California: The human andmouse cellular homologs of theraf/mil oncogene.
Lewis, W., Raines, M., Wagner, R.,Crittenden, L., Kung, H.J., Dept.of Biochemistry, Michigan StateUniversity, USDA Regional PoultryResearch Laboratory, East Lan-sing: c-erb-B polymorphism -Cloning and characterization of asecond c-erb-B allele.
Akiyama, T,' Yamada, Y.,' Ogawara,H.,' Yamamoto, T,2 Kasuga, M.,3'Second Dept. of Biochemistry,Meiji College of Pharmacy;2lnstitute of Medical Science,3Third Dept. of Internal Medicine,University of Tokyo, Japan: Site-specific antibodies to erb-B onco-gene product immunoprecipitateepidermal growth factor receptor.
Qi, X.D., Guilhot, S., Galibert, F., La-boratoire d'Hematologie, HospitalSaint Louis, Paris, France: Restric-
tion fragment length polymorph-ism of the human c-fms gene.
Bloemers, H.P.J., Verbeek, J.S., vanden Ouweland, A.M.W., Van deVen, W.J.M., Dept. of Biochemis-try, University of Nijmegen, TheNetherlands: Structural analysis ofa hereditary abnormal c-fmsproto-oncogene locus in man.
Najita, L., Rohrschneider, L., FredHutchinson Cancer ResearchCenter, Seattle, Washington:Expression of the v-fms proteinproduct at the cell surface.
Donoghue, D.J., Dept. of Chemistry,University of California, SanDiego, La Jolla: Analysis of v-sisby deletion mutagenesis- ldentifi-cation of regions required for bio-logical activity.
Niman, H.L.,1 Houghten, R.A.,'Bowen-Pope, D.F.,2 'Dept. of Mo-lecular Biology, Scripps Clinic andResearch Foundation, La Jolla,California; 2Dept. of Pathology,University of Washington, Seattle:Widespread expression of high-molecular-weight forms of PDGF.
Thiel, H.-J., Hafenrichter, R., FederalResearch Centre for Virus Dis-eases of Animals, TObingen, Fed-eral Republic of Germany: SSVtransformation-specific glycopep-tide- Immunological relationshipto human PDGF.
Clark, J.,1 Wagner, E.,2 Gilboa, E.,''Dept. of Molecular Biology, Prin-ceton University, New Jersey;2European Molecular Biology Lab-oratory, Heidelberg, Federal Re-public of Germany: The nature ofthe block in Mo-MLV replication inembryonal carcinoma cells.
DesGroseillers, L., Jolicoeur, F, Insti-tut de Recherches Cliniques deMontreal, Universite de Montreal,Quebec, Canada: Delayed replica-tion of a MLV LTR mutant-Con-struction and biological character-ization.
Chen, P. -J., Cywinski, A., Taylor, J.M.,Institute for Cancer Research, Phil-adelphia, Pennsylvania: Endoge-nous reverse transcription of cellu-lar RNA species-An approach tounderstanding the origin of certainpseudogenes and processedgenes.
Van Beveren, C., Verma, I.M., Molec-ular Biology and Virology Labora-tory, Salk Institute, San Diego, Cal-ifornia: Novel eukaryotic initiation

codon (GUG) for murine glycosy-lated gag protein.
Pepinsky, R.B., Potts, W., Steeg, C.M.,Southard, L., Vogt, V.M., Sectionof Biochemistry, Cornell University,Ithaca, New York: Structure andfunction of p19°2° and p19-relatedproteins of ASV and ALV.
Darlix, J.-L., Meric, C., Spahr, P-F,Dept. of Molecular Biology, Uni-versity of Geneva, Switzerland:The functions of RSV protein p12
in reverse transcription, transla-tion, and packaging of the viralgenome.
Katoh, Yoshinaka, Y,1 Oroszlan,S.,1 Coward, J.,2 Luftig, R.B.,2'Frederick Cancer Research Cen-ter, Frederick, Maryland; 2Dept. ofMicrobiology and Immunology,Louisiana State University MedicalCenter, New Orleans: The effect ofcerulenin on MLV assembly.
Ro, J.H.-S., Ghosh, H.P., Dept. of Bio-
SESSION 4 MORE ONCOGENES (erbB, fms, raf/mil, mos, sis)
Chairperson: C. Sherr, St. Jude Childrens Hospital, Memphis, Tennessee
Raines, M.,' Lewis, W.,1 Crittenden,L.B.,2 Kung, H.J.,1 'Dept. of Bio-chemistry, Michigan State Univer-sity; 2USDA Poultry Research Lab-oratory, East Lansing, Michigan:Structure and expression of acti-vated c-erb-B genes in ALV-in-duced erythroblastosis.
Raines, M.,1 Fad ly, A.,2 Crittenden,L.B.2 Kung, H.J.,1 'Dept. of Bio-chemistry, Michigan State Univer-sity; 2USDA Regional Poultry Re-search Laboratory, East Lansing,Michigan: Biological characteriza-tion of ALV-induced erythro-blasto-sis-A leukemia associated with c-erb-B activation.
Xu, Y.-H.,' Ishii, S.,1 Clark, A.,1 Rich-ert, N.,1 Ito, S.,1 Roe, B.,2 Yama-moto, T.,3 Merlino, G.,1 Pastan, I.,''NCI, National Institutes of Health,Bethesda, Maryland; 2Dept. ofChemistry, University of Okla-homa, Norman; 3 Institute of Medi-cal Science, University of Tokyo,Japan: Cloning of cDNA encodingthe human EGF receptor.
Merlino, G.,' Ishii, S.,' Xu, Y.-H.,'Clark, A.,1 Richert, N.,1 Ito, S.,1 Ya-mamoto, T.,2 Pastan, I.,' 'NCI, Na-tional Institutes of Health, Be-thesda, Maryland; 2lnstitute ofMedical Science, University of To-kyo, Japan: Correlation of en-hanced expression and amplifica-tion of the EGF receptor gene inA431 cells.
Gilmore, T., De Clue, J., Dehazya, P.,Martin, G.S., Dept. of Zoology,University of California, Berkeley:Tyrosine phosphorylation inducedby the v-erb-B protein in vivo andin vitro.
Rettenmier, C.W., Roussel, M.F., An-derson, S.J., Look, A.T., Sherr,
C.J., St. Jude Children's ResearchHospital, Memphis, Tennessee:Cell-surface localization of some v-fms-coded glycoproteins is re-quired for transformation.
Rettenmier, C.W., Roussel, M.F.,Sherr, C.J., St. Jude Children's Re-search Hospital, Memphis, Ten-nessee: In vitro kinase activity as-sociated with the v-fmsglycoprotein.
Rapp, U.R.,1 Jansen, H.W.,2 Bister,K.,2 'NCI Frederick Cancer Re-search Fa Cility, Frederick, Mary-land; 2Max-Planck-Institut, Berlin,Federal Republic of Germany:Transforming ability of retrovirusescontaining the v-raf/mil and v-myconcogenes.
Biegalke, B.,' Morrow, M.,' Rapp,U.,2 Linial, M.,1 'Fred HutchinsonCancer Research Center, Seattle,Washington: 2NCI, Frederick Can-cer Research Facility, Frederick,Maryland: The v-mil gene productis not required for transformationby MH2.
Bechade, C.,' Calothy, G.,' Coll, J.,2Saule, 5.,2 Ghysdael, J.,2 Stehelin,D.,2 'Institut Curie, Orsay;2INSERM University, Institut Pas-teur, Lille, France: Expression ofthe v-mi/-encoded p100gag-^,lis re-quired for transforming and mito-genic properties of MH2 in chickembryo neuroretinal cells.
Hannink, M., Dept. of Chemistry, Uni-versity of California, San Diego, LaJolla: Oligonucleotide mutagene-sis of viral oncogenes.
Blair, D.G.,1 Seth, A.,2 Dean, M.,2Zweig, M.,3 Vande Woude, G.,2'NCI; 2LBI; 3PRI, NCI, FrederickCancer Research Facility, Freder-ick Maryland: Expression of MSV
chemistry, McMaster University,Ontario, Canada: RSV reversetranscriptase gene contains pro-moter and ribosomal binding se-quences needed for its expressionin E. co/i.
Deen, K.C., Sweet, R.W., Dept. of Mo-lecular Genetics, Smith Kline andFrench Laboratories, Philadelphia,Pennsylvania: Retroviral-relatedpolymerase sequences in humanDNA.
v-mos human c-mos hybrid re-combinants in transfected NIH-3T3 cells.
McGeady, M.L.,1 Wood, T.G.2 VandeWoude, G.F.,1 'NCI, FrederickCancer Research Facility, Freder-ick, Maryland; 2Dept. of Biochem-istry, Georgetown University,Washington, DC: Inhibition of mosexpression by normal mouse cellsequences.
Hamelin, R.,' Murphy, E.C., Jr., 2 Ar-linghaus, R.B.,"Dept. of Molecu-lar Biology, Scripps Clinic and Re-search Foundation, La Jolla,California; 2Dept. of Tumor Biol-ogy, M.D. Anderson Hospital,Houston, Texas: Transcriptionproducts of the ts110 Mo-MSVand its wild-type revertants.
Maxwell, S.A., Kloetzer, W.S., Arlin-ghaus, R.B., Dept. of Molecular Bi-ology, Scripps Clinic and Re-search Foundation, La Jolla,California: P8509111 .s encoded byts110 Mo-MSV contains a thermol-abile kinase activity associatedwith the mos sequences.
Gazit, A., lgarashi, H., Chiu, Sri-
nivasan, A., Yaniv, A., Tronick, S.,Robbins, K., Aaronson, S.A., NCI,National Institutes of Health, Be-thesda, Maryland: Activation ofthe normal human c-sis/PDGF-2proto-oncogene as a transforminggene.
Chiu, I.-M., Tronick, S.R., lgarashi, H.,Reddy, E.P., Givol, D., Robbins,K.C., Aaronson, S.A., NCI, Na-tional Institutes of Health, Be-thesda, Maryland: Molecular clon-ing and nucleotide sequenceanalysis of the human c-sis PDGFgene.
Westin, E., Clarke, M., Gorse, K. Rat-
207

ner, L., Josephs, S., Reitz, M.,Gallo, R.C., Wong-Staal, F., NCI,National Institutes of Health, Be-thesda Maryland: Characterizationof cells transformed by a cDNAclone of the human c-sis gene.
Josephs, S.,' Ratner, L.,' Westin, E.,'Clarke, M.,2 Reitz, M.S.,' Wong-Staal, F.,' 'NCI, National Institutesof Health; 2NIADDKD, Bethesda,Maryland: Nucleotide sequence
analysis of transforming human c-sis cDNA.
Josephs, S.,' Lautenberger, J.,' Wes-tin, E.,' Vergilio, L.,' Ratner, L.,'Clarke, M.,2 Reitz, M.S.,' Wong-Staal, F.,' Papas, T.S.,"NCI, Na-tional Institutes of Health;2NIADDKD, Bethesda, Maryland:Expression in bacteria of human c-sis protein-Evidence against apolyprotein precursor including
chain A and chain B of humanPDGF.
Deuel, T.F., Huang, S.S., Huang, J.S.,Jewish Hospital, Washington Uni-versity Medical Center, St. Louis,Missouri: Human PDGF-A prod-uct of the human proto-oncogenec-sis which interacts with cellsthrough a PDGF receptor/proteintyrosine kinase.
SESSION 5 YET MORE ONCOGENES (ras, SIMPLE EUKARYOTES AND NEW oncs)
Chairperson: D. Lowy, National Cancer Institute, National Institutes of Health,Bethesda, Maryland
Willumsen, B.M.,' Christensen, A.,'Hubbert, N.L.,2 Papageorge,A.G.,2 Lowy, D.R.,2 'Fibiger Insti-tute, Copenhagen, Denmark;2NCI, National Institutes of Health,Bethesda, Maryland: The carboxylterminus of the Ha-MSV p21 pro-tein is required for three func-tions-Cellular transformation,membrane association, and lipidbinding.
Tambourin, P.E., Chattopadhyay, S.K.,Lowy, D.R., NCI, National Insti-tutes of Health, Bethesda, Mary-land: HaSV-Noncoding and p21-coding sequences affect its onco-genic activity.
Vousden, K.H., Phillips, D.H., Mar-shall, C.J., Chester Beatty Labora-tories, Institute of Cancer Re-search, London, England:Activation of a cellular proto-onco-
gene by in vitro modification with achemical carcinogen.
Santos, E., Barbacid, M., NCI, Freder-ick Cancer Research Facility, Fred-erick, Maryland: Frequent geneticrearrangements of the Ki-ras locusin human tumors.
Gibbs, J.B.,' Ellis, R.W.,' Sigal,Stein, R.B.,' Poe, M.,2 Scolnick,E.M.,"Virus and Cell Biology Re-search, Merck Sharp and DohmeResearch Laboratories, WestPoint, Pennsylvania; 2Dept. of Bio-physics, Merck Sharp and DohmeResearch Laboratories, Rahway,New Jersey: Autophosphorylationof v-Ha-ras p21 is modulated inwhole cells and in vitro by aminoacid residue 12.
Der, C.J.,' Finkel, T,2 Cooper, G.M.,''Dept. of Pathology, Harvard Med-ical School; 2Committee on Bio-
E. Ruley, J. Lautenberger, N. Teich
208
physics, Harvard University, Bos-ton, Massachusetts: Structural andfunctional analyses of normal andactivated cellular ras proteins.
Wigler, M.,' Powers, S.,' Kataoka, T,'Fasano, 0.,' Taparowsky, E.,'Birnbaum, D.,' Goldfarb, M.,'Strathern, J.,' Broach, J.,2 'ColdSpring Harbor Laboratory; 2Dept.of Microbiology, State University ofNew York, Stony Brook: Structure,genetics, and activation of yeastand mammalian ras genes.
Tamanoi, F, Walsh, M., Rao, M., Pow-ers, S., Kataoka, T., Wig ler, M.,Cold Spring Harbor Laboratory,New York: Yeast ras proteins.
MacAuley, A., Auersperg, N., Weeks,G., Pawson, T., Depts. of Microbi-ology and Anatomy, University ofBritish Columbia, Vancouver, Can-ada: Expression and functional ac-tivity of ras proteins are regulatedin developmental systems fromsimple and higher eukaryotes.
Simon, M.A.,' Drees, B.,' Kornberg,T,' Bishop, J.M.,2 'Dept. of Bio-chemistry and Biophysics,2George William Hooper Founda-tion, University of California, SanFrancisco: The expression of threesrc homologs during Drosophiladevelopment.
Mark, G.E.,' Jackman, M.,' Mac-Intyre, R.,2 'NCI, National Insti-tutes of Health, Bethesda, Mary-land; 2Cornell University, Ithaca,New York: Drosophila melanogas-ter contains unique loci related tothe raf oncogene.
Besmer, P.,' Murphy, J.E.,' Lader, E.,'George, PC.,' Snyder, H.W., Jr.,3Zuckerman, E.E.,2 Hardy, W.D.,

Jr.2 Laboratories of 'Molecular On-cology, 2Veterinary Oncology,Memorial Sloan-Kettering CancerCenter, New York; 3Pacific North-west Research Foundation, Seat-tle, Washington: New acute trans-forming feline retroviruses-TheHZ4-FeSV contains unique retro-viral oncogene v-kit---The HZ5-FeSV is a new fms virus.
SESSION 6 POSTER SESSION II
Guerrero, Villasante, A.,' D'Eusta-chio, P.,2Corces, V.,3 Pellicer, A.,'Depts. of 'Pathology,2Biochemistry, New York Univer-sity Medical Center, New York,New York; 3Dept. of Biology,Johns Hopkins University, Balti-more, Maryland: In vivo activationof mouse ras oncogenes.
Tainsky, M.A.,' Cooper, C.S.,2 Blair,D.G.,' Giovanella, B.C.,3 VandeWoude, 'NCI, FrederickCancer Research Facility, Freder-ick, Maryland; 2Dana-Farber Can-cer Institute, Boston, Massachu-setts: 3Stehlin Foundation forCancer Research, Houston, Texas;4Director, LEI Basic Research Pro-gram, Frederick Cancer ResearchFacility, Frederick, Maryland: Anactivated N-ras gene is detected inlate- but not early-passage humanPA1 teratocarcinoma cells.
Westin, E., Wong-Staal, F, Gorse, K.,Gallo, R.C., NCI, National Insti-tutes of Health, Bethesda, Mary-land: Activation of N-ras in humanacute leukemias.
Souyri, M., Furth, M., Fleissner, E.,Memorial Sloan-Kettering CancerCenter, New York, New York: Ac-tive N-ras oncogenes from humanT-cell leukemias-Structural char-acteristics and pathogenic proper-ties.
Gambke, C., Moroni, C., FriedrichMiescher-lnstitut, Basel, Switzer-land: Identification and character-ization of transforming genes fromhuman leukemias.
Padua, R.A., Barrass, N., Currie,G.A., Marie Curie Memorial Foun-dation Research Institute, Oxted,England: Transforming genes inmalignant melanoma.
Kraus, M., Yuasa, Y., Aaronson, S.,NCI, National Institutes of Health,Bethesda, Maryland: A position
Willer, R.,' Willer, D.,' Molders, H.,'Defesche, J.,' Bonner, TI.,2 Rapp,U.R.,2 'European Molecular Biol-ogy Laboratory, Heidelberg, Fed-eral Republic of Germany; 2NCI,National Institutes of Health, Fred-erick Cancer Research Facility,Frederick, Maryland: An alterna-tive strategy for the identificationof potential c-onc genes.
12-activated Ha-ras oncogene inall HS578T mammary carcinosar-coma cells but not normal cells ofthe same patient.
Robins, T., Showalter, S., VandeWoude, G., NCI, Frederick CancerResearch Facility, Frederick, Mary-land: Biological properties of amurine retrovirus expressing thehuman EJ bladder oncogene.
Chang, E.H.,' Morgan, Pl.,' White,E.A.,' Tsichlis, P.N.,2 Patrick,D.H.,1 'Uniformed Services Uni-versity of the Health Sciences, Be-thesda, Maryland; 2Fox ChaseCancer Center, Philadelphia,Pennsylvania: Pathogenicity of re-troviruses containing either humanc-Ha-ras-/ or EJ/T24 ras gene.
Lee, E.J., Chang, E.H., Dept. of Pa-thology, Uniformed Services Uni-versity of the Health Sciences, Be-thesda, Maryland:Characterization and cDNA clon-ing of human c-Ha-ras RNA tran-scripts.
Anderson, G.R., Stoler, D.L., Saa-vedra, R.A., Dept. of Cell and Tu-mor Biology, Roswell Park, Buf-falo, New York: The KiSV, itsanoxic shock genes, and LDH,.
Ulsh, L.S., Shih, T.Y., NCI, FrederickCancer Research Facility, Freder-ick, Maryland: Metabolic turnoverof human c-ras p21 of EJ bladdercarcinoma and its normal cellularhomolog.
Srivastava, S.K., Yuasa, Y., Kraus,M.H., Fijita, J., Aaronson, S.A.,NCI, National Institutes of Health,Bethesda, Maryland: Comparativeanalysis of p21 molecules en-coded by human ras oncogenes.
Gallick, G.E.,' Kurzrock, R.,2 Gutter-man, J.U.,2 Depts. of 'Tumor Biol-ogy, 2Clinical Immunology,UTSCC, M.D. Anderson Hospital,Houston, Texas: Expression of ras
Cooper, C.S.,' Park, M.,2 Blair, D.G.,2Tainsky, M.A.,2 Vande Woude,G.F.,2 'Laboratory of MolecularCarcinogenesis, Dana-FarberCancer Institute, Boston, Massa-chusetts; 2NCI, Frederick CancerResearch Facility, Frederick, Mary-land: Molecular cloning of a newtransforming gene from a chemi-cally transformed human cell line.
gene products in primary humancolon tissues and metastatic le-sions.
Kurzrock, R.,2 Gallick, G.E.,' Liang,J.,3 Gutterman, J,U.,2 Depts. of'Tumor Biology, Section of Virol-ogy, 2Clinical Immunology, 3Dept.of Genetics, UTSCC, M.D. Ander-son Hospital, Houston, Texas:Expression of ras gene productsin primary human lung tumors.
Samid, D.,' Friedman, R.M.,' Schaff,Z.,2 Chang, E.H.,' 'Dept. of Pa-thology, Uniformed Services Uni-versity of the Health Sciences,2Hepatitis Branch, Bureau of Biol-ogics, Bethesda, Maryland: Re-duction of ras expression accom-panies phenotypic reversion ininterferon-treated c-Ha-rasoncogene-transformed mousecells.
Debuire, B.,' Henry, C.,' Benaissa,M.,' Biserte, G.,' Saule, 5.,2 Mar-tin, P.,2Stehelin, D.,2 'Institut deRecherches sur le Cancer de Lille;2INSERM, Institut Pasteur, Lille,France: Sequencing of the erb-Agene of AEV reveals a new type ofoncogene.
Wadsworth, S.C.,' Madhavan, K.,2'Cell Biology Group, WorcesterFoundation for Experimental Biol-ogy, Shrewsbury; 2Dept. of Biol-ogy, College of the Holy Cross,Worcester, Massachusetts:Expression and structure of Droso-phila src genes.
Katzen, A.L.,' Bishop, J.M.,2 'Dept. ofBiochemistry and Biophysics,2George William Hooper Founda-tion, University of California, SanFrancisco: fps-related loci in Dro-sophila melanogaster.
Gilmore, T,' Thorner, J.,2 Martin,G.S.,' Depts. of 'Zoology,2Microbiology and Immunology,University of California, Berkeley:
209

Phosphotyrosine in Saccharo-myces cerevisiae and E. coli.
Woodland, E.C., Horowit, T.S.,Reinsch, S., Shank, P.R., Divisionof Biology and Medicine, BrownUniversity, Providence, Rhode Is-land: Characterization of a sponta-neous deletion mutant of B77 RSVwhich has transduced a cellularsequence.
Rhim, J.S., Arnstein, P., Jay, G., San-ford, K.K., Fujita, J., Aaronson,S.A., NCI, National Institutes ofHealth, Bethesda, Maryland: Hu-man epithelial cell viral carcino-genesis-Combined action ofDNA and RNA tumor viruses pro-duces malignant transformation ofhuman epidermal keratinocytes.
O'Hara, B., Dusing-Schwartz, S., Blair,D.G., NCI, Frederick Cancer Re-search Facility, Frederick, Mary-land: Transfection of human cellswith cloned oncogenes.
Hynes, N., Kozma, S., Jaggi, R., Gro-nemeyer, H., Carrozza, M.L., Ball,R., Groner, B., Ludwig Institute forCancer Research, Bern, Switzer-land: Detection of oncogenes acti-vated in human mammary tumorcells.
Gardner, M.J.,' Estival, A.,3 Bassin,R.H.,3 Yang, W.K.,21University ofTennessee, Oak Ridge GraduateSchool of Biomedical Sciences,2Biology Division, Oak Ridge Na-tional Laboratory; 3NCI, NationalInstitutes of Health, Bethesda,Maryland: Chemically transformedNIH -3T3 cells selected from softagar do not contain activated cel-lular ras oncogenes.
Diamond, A., Devine, J.M., Cooper,G.M., Dept. of Pathology, Dana-Farber Cancer Institute, HarvardMedical School, Boston, Massa-chusetts: The nucleotide se-quence of the human Blym-1transforming gene.
Cullen, B.R., Ju, G., Dept. of Molecu-lar Genetics, Hoffmann-La RocheInc., Roche Research Center, Nu-tley, New Jersey: Retroviral infec-tion greatly increases expressionof transfected DNA in avian cells.
Goss, J.A., Sen, A., Ingene Inc.,Santa Monica, California: Osteob-last-specific transforming growthfactor from a metastatic osteosar-coma harboring an active onco-gene.
Cook, W.D., Melbourne Tumour Biol-
210
ogy Branch, Ludwig Institute forCancer Research, Australia: Thy-mocyte subset specificity of Abel-son virus transformation.
Kloetzer, W.,' Smith, L.,' Spiller, M.,'Kurzrock, R.,2 Talpaz, M.,2 Gutter-man, J.,2 Arlinghaus, R.,''Johnson and Johnson Biotech-nology Center Inc., La Jolla, Cali-fornia; 2M.D. Anderson Hospitaland Tumor Institute, Houston,Texas: Detection of a tyrosine pro-tein kinase in the chronic myelo-genous leukemia cell line K562 us-ing antisera to the mouse v-ablgene product.
Balachandran, R.,' Pierce, J.P.,2 Aar-onson, S.A.,2 Reddy, E.P.,2 Swan,D.C.,2 'Laboratory of Biochemis-try; 2NCI, National Institutes ofHealth, Bethesda, Maryland:Characterization of T lymphocytestransformed by Mo-MLV.
Serunian, L.A.' 2 Rosenberg, N.,''Cancer Research Center, TuftsUniversity School of Medicine,Boston; 2Dept. of Cellular and De-velopmental Biology, Harvard Uni-versity, The Biological Laborato-ries, Cambridge, Massachusetts:Modulation of Abelson virus-targetcell interactions by a B-cell mito-gen.
Casey, G., Smith, R., Peters, G., Dick-son, C., Imperial Cancer ResearchFund Laboratories, London, Eng-land: Identification and character-ization of human DNA sequencesrelated to the murine int-2 locus.
Dudley, J.,' Arfsten, A.,3 Kozak, C.,2Risser, R.,3 'Dept. of Microbiology,University of Texas, Austin;2NIAID, National Institutes ofHealth, Bethesda, Maryland;3McArdle Laboratory for CancerResearch, Madison, Wisconsin:Characterization of cloned MMTVproviruses and their intergrationsites from T-cell lymphomas.
Prakash, 0., Sarkar, N.H., MemorialSloan-Kettering Cancer Center,New York, New York: Molecularcloning and characterization of theMMTV proviral unit I of C3H/Hemice.
Pathak, V., Young, L.J.T., Morris,D.W., Cardiff, R.D., Dept. of Pa-thology, University of California,Davis: int-1 and int-2 rearrange-ment in mouse mammary neoplas-tic progression.
Fanning, T.G., Hu, W.-S., Cardiff, R.D.,
Dept. of Pathology, School of Med-icine, University of California,Davis: Methylation patterns ofMMTV DNA in mammary and non-mammary tissues.
Morris, D.W.,' Bradshaw, H.D.,2 Fan-ning, T.G.," Young, L.J.T.,' Cardiff,R.D.,1 'Dept. of Pathology, Univer-sity of California, Davis; 2Dept. ofBiochemistry, Louisiana State Uni-versity Medical Center, New Orle-ans: Isolation and preliminarycharacterization of a tumor-associ-ated MMTV proviral integration lo-cus-GR int-2.
Lee, W.T.-L., Prakash, 0., Etkind, P.R.,Sarkar, N.H., Memorial Sloan-Ket-tering Cancer Center, New York,New York: Identification and mo-lecular cloning of a variant MMTVprovirus in a DBA/2 leukemia.
Michalides, R., Weijers, P., Dept. ofMolecular Biology, The Nether-lands Cancer Institute, Amster-dam; Dept. of Electronmicroscopy,Laboratory of Biochemistry, Uni-versity of Amsterdam, The Nether-lands: Rearrangements in the LTRof the extra MMTV proviruses in T-cell leukemias in GR mice result inan enhancerlike structure.
Graham, D.E.,' Medina, D.,2 Smith,G.H.,"NCI, National Institutes ofHealth, Bethesda, Maryland;2Dept. of Cell Biology, BaylorSchool of Medicine, Houston,Texas: A novel MMTV subgenomictranscript which contains gag-polsequences.
Devaux, B., Lefrancois, D., Crepin,M., Pasteur Institute, Paris,France: Transcription and methyla-tion of MMTV LTR DNA intergratedin fibroblast clones.
Durban, E.M.,' Slagle, B.L.,' Medina,11,2 Butel, J.S.,' Depts. of'Virology and Epidemiology, 2CellBiology, Baylor College of Medi-cine, Houston, Texas: Modulationof endogenous MMTV proteinexpression during normal mam-mary cell differentiation in BALB/cmice.
Racevskis, J., Prakash, 0., MemorialSloan-Kettering Cancer Center,New York, New York: Proteins en-coded by the LTR region ofMMTV- Identification by hybrid-selected translation.
Smith, E.J., Crittenden, L.B., USDARegional Poultry Research Labo-ratory, East Lansing, Michigan:

Novel infectious endogenous vi-ruses in chickens.
Dorner, A., Stoye, J., Coffin, J., TuftsUniversity Medical School, Bos-ton, Massachusetts: Molecular ba-sis of host-range variation in avianretroviruses.
Simon, M.C.,12 Smith, R.E.,3 Necka-meyer, W.S.,2 Hayward, W.S.,"'Memorial Sloan-Kettering CancerCenter; 2Rockefeller University,New York, New York; 3ColoradoState University, Fort Collins:Mechanisms of oncogenesis bysubgroup-F ALV.
Shank, RR.,' Reinsch, S.S.,' Jensen,L.,2 Robinson, H.L.,' 'Division ofBiology and Medicine, Brown Uni-versity, Providence, Rhode Island;2Worcester Foundation for Experi-
mental Biology, Shrewsbury, Mas-sachusetts: Sequences in the gagor po/ genes are responsible forhigh-incidence production of os-teopetrosis by nonacute avian re-troviruses.
Kotler, M.,1 Burstein, H.,1 Gilead, M.,2'Dept. of Molecular Genetics, He-brew University-Hadassah Medi-cal School, Jerusalem; 2PoultryDiseases Laboratory, Beit She-mesh, Israel: Viral etiology of he-mangiosarcoma outbreaks amonglayer hens.
Ewert, D.L., Wistar Institute, Philadel-phia, Pennsylvania: ALV infectionand tumor induction in infection-resistant and susceptible chimericchickens.
Dunwiddie, C., Faras, A., Dept. of Mi-
SESSION 7 CARCINOGENESIS AND LEUKEMOGENESIS
Chairperson: R. Risser, McArdle Laboratory, University of Wisconsin, Madison,Wisconsin
Nusse, R., van Ooyen, A., Dept. ofMolecular Biology, The Nether-lands Cancer Institute, Amster-dam: Structure, nucleotide se-quence and mode of activation ofint-1- Implications for mammarytumorigenesis.
Fung, Y.-K.T., Seeger, C., Shackelford,G., Varmus, H.E., Dept of Microbi-ology and Immunology, Universityof California, San Francisco: Mo-lecular cloning of int-1 cDNA andits expression in E. coll.
Moore, R., Brookes, S., Casey, G.,Dickson, C., Peters, G., ImperialCancer Research Fund Laborato-ries, London, England: The struc-ture and expression of the MMTVint-2 locus in mice.
Gallahan, D., Robbins, J., Callahan,R., NCI, National Institutes ofHealth, Bethesda, Maryland: Evi-dence for a new common regionfor MMTV integration in mammarytumors of feral Mus muscu/usmuscu/us.
Hankins, W.D.,1 Thorgeirsson, S.,' Ta-bin, C.,2 Cones, R.,2 Weinberg,R.,2 Mulligan, R.,2 'NCI, NationalInstitutes of Health, Bethesda,Maryland; 2Massachusetts Insti-tute of Technology, Boston: Trans-formation of hemopoietic precur-sors by replication-defective,helper-free virions carrying the
bladder carcinoma cellular onco-gene, EJ-1.
Rein, A.,1 'hie, J.,' Medicus, R.,'Holmes, K.,2 Schultz, A.,' Keller,J.,1 'NCI, Frederick Cancer Re-search Facility, Frederick; 2NIAID,National Institutes of Health, Be-thesda, Maryland: Immortalizationof a class of hematopoietic cellsby infection with HaSV.
Boettiger, D.,' Dexter, T.M.,2 'Dept. ofMicrobiology, University of Penn-sylvania, Philadelphia; 2PatersonLaboratories, Manchester, Eng-land: Infection of murine hemo-poietic stem cells with a recombi-nant retrovirus containing the srcgene from RSV alters their self-re-newal capacity.
Palmieri, S., NIAID, National Institutesof Health, Hamilton, Montana:Erythroid cell transformation byRSV.
Heisterkamp, N.,1 Stephenson, J.R.,1Grosveld, G.,2 de Klein, A.,2 Grof-fen, J.,"Oncogene Science Inc.,Mineola, New York; 2Erasmus Uni-versity, Rotterdam, The Nether-lands: Cellular (onco)genes in-volved in chronic myelocyticleukemia.
Canaani, E.,' Shtivelman, E.,' Lifshiz,B.,' Klar, A.,' Gale, R.P.,2 'Dept. ofChemical Immunology, WeizmannInstitute of Science, Rehovot, Is-
crobiology, University of Minne-sota, Minneapolis: The presenceof avian retroviral gag-, pot -, andenv-related gene sequences inALV lacking chick embryo fibro-blasts and quail embryo fibro-blasts.
Fung, Y-K.T.,1 Lai, C.L.,2 Todd, D.,2Ganem, D.,' Varmus, H.E.,"Dept.of Microbiology and Immunology,University of California, San Fran-cisco; 2Dept. of Medicine, QueenMary Hospital, University of HongKong, China: Evidence against in-sertion mutagenesis in hepatitis Bvirus-associated hepatoma.
rael: 2Dept. of Medicine, Univer-sity of California School of Medi-cine, Los Angeles: A unique ablRNA transcript in chronic myelo-genous leukemia.
Konopka, J.B., Watanabe, S.M.,Witte, 0.N., Dept. of Microbiologyand Molecular Biology Institute,
S. Martin
211

University of California, Los Ange-les: Activation of the human c-abloncogene.
Wolf, D., Harris, N., Rotter, V., Dept. ofCell Biology, Weizmann Institute ofScience, Rehovot, Israel: Recon-stitution of p53 expression of Ab-
MLV-transformed L12 cells con-taining a nonfunctional gene bytransfection of a genomic p53clone.
Shen-Ong, G.L.C., Reddy, E.P., Mu-shinski, J.F., Potter, M., NCI, Na-tional Institutes of Health, Be-
SESSION 8 ENHANCERS AND VECTORS
Chairperson: G. Vande Woude, National Cancer Institute, National Institutes of Health,Bethesda, Maryland.
Celander, D., Haseltine, W.A., Dana-Farber Cancer Institute, Boston,Massachusetts: The role of thetranscriptional enhancer elementin cell tropism and leukemogenic-ity of murine leukemia viruses.
Lenz, J.,' Cloyd, M.,2 Rosen, C.,3 Ce-lander, D.,3 Haseltine, W.,3 'Dept.of Genetics, Albert Einstein Col-lege of Medicine, Bronx, NewYork; 2 Dept. of Surgery, Duke Uni-versity, Durham, North Carolina;3Dana-Farber Cancer Institute,Harvard Medical School, Boston,Massachusetts: Leukemogenicityand thymotropism of a murine re-trovirus and determined by enhan-cer-related sequences in the viralLTR.
Rosen, C.,' Lenz, J.,2 Cloyd, M.,3Haseltine, W.,1 'Dana-Farber Can-cer Institute, Boston, Massachu-setts; 2Albert Einstein College ofMedicine, Bronx, New York;3NIAID, Hamilton, Montana: Ele-ments that confer tissue tropismreside within the U3 region of theLTR of Fr-MLV.
DesGroseillers, L., Villemur, R., Jo li-coeur, P., Institut de RecherchesCliniques de Montreal, Universitede Montreal, Quebec, Canada:The tandem direct repeats withinthe LTR of MLV are the primary de-terminant of their leukemogenicpotential.
Clark, S.P., Mak, T.W., Ontario CancerInstitute, Toronto, Canada: Com-parison of murine retroviral enhan-cers as transcriptional promoters.
Davis, B.,' Chao, E.,' Overhauser, J.,'Linney, E.,2 Fan, H.,"Depti of Mo-
SESSION 9 POSTER SESSION III
Yoshimura, F., Hedin, K., Fred Hutch-inson Cancer Research Center,Seattle, Washington: Tissue-spe-
212
lecular Biology and Biochemistry,University of California, Irvine; 2LaJolla Cancer Research Founda-tion, California: Deletion and sub-stitution mapping within the Mo-MLV LTR.
Raymond, K.J.,' Berg, P.,2Ju, G.,''Dept. of Molecular Genetics,Roche Research Center, Hoff-mann-La Roche Inc., Nutley, NewJersey: 2National Institutes ofHealth, Bethesda, Maryland: Lo-calization of enhancer sequencesin the RSV LTR.
Norton, PA., Coffin, J.M., Dept. ofMolecular Biology and Microbiol-ogy, Tufts University School ofMedicine, Boston, Massachusetts:
in the RSV LTR.Robins, T, Vande Woude, G., NCI,
Frederick Cancer Research Facil-ity, Frederick, Maryland: Cloningand expression of intronlessgenes using a retrovirus "shuttle"vector.
Jhappan, C.,' Chirikjian, J.,' VandeWoude, G.,2 Robins, T.,2 'Dept. ofBiochemistry, Georgetown Univer-sity, Washington, DC; 2FrederickCancer Research Facility, Freder-ick, Maryland: Direct molecularcloning of G418R "shuttle" vectorproviral DNA mediated by cell fu-sion.
Gilboa, E., Park, J., Hwang, L., Sei-berg, M., Yu, S., Dept. of Molecu-lar Biology, Princeton University,New Jersey: Strategies of genetransfer with retrovirus-derivedvectors.
Hwang, L., Park, J., Gilboa, E., Dept.
cific expression of enhancer activ-ity of an MCF MLV LTR.
Overhauser, J., Fan, H., Dept. of Mo-
thesda, Maryland: Disruption andactivation of the c-myb locus byMo-MLV insertion in plasmacytoidlymphosarcomas induced by pris-tane and Abelson virus.
of Molecular Biology, PrincetonUniversity, New Jersey: Mecha-nism of splicing of Mo-MLV envmRNA-Role of intron-containedsequences.
Reik, W., Wolf, B., Harbers, K., Jaen-isch, R., Heinrich-Pette Institut fOrExperimentelle Virologie and Im-munologie, Hamburg, Federal Re-public of Germany: Vectors for theanalysis of retroviral integration-Construction of replication compe-tent Mo-MLV derivatives selectablein bacteria.
Miller, A.D., Eckner, R.J., Ong, E.S.,Evans, R.M., Verma, I.M., Molecu-lar Biology and Virology Labora-tory, Salk Institute, San Diego, Cal-
into culturedcells and animals using retroviralvectors.
Cone, R.D., Reilly, E., Weber, A., Ei-sen, H.N., Mulligan, R.C., Centerfor Cancer Research and White-head Institute, Massachusetts In-stitute of Technology, Cambridge:Passage- and tissue-specificexpression of small genomic se-quences inserted into murine re-troviral vectors.
Mann, R., Baltimore, D., WhiteheadInstitute of Biomedical Research,Massachusetts Institute of Technol-ogy, Cambridge: The position ofthe Mo-MLV packaging site can bevaried.
Mann, R., Baltimore, D., WhiteheadInstitute for Biomedical Research,Massachusetts Institute of Technol-ogy, Cambridge: Packaging ofspliced retroviral RNAs.
lecular Biology and Biochemistry,University of California, Irvine: In-sertion of DNA fragments into the

Moloney LTR -Effects on pro-moter activity and viral expression.
Herman, S.A., Coffin, J.M., Dept. ofMolecular Biology and Microbiol-ogy, Tufts University School ofMedicine, Boston, Massachusetts:Analysis of downstream transcrip-tion from randomly integrated pro-viruses.
Keshet, E., Rotman, G., ltin, A., Dept.of Virology, Hebrew University-Hadassah Medical School, Jeru-salem, Israel: Diversity of LTR unitsassociated with a murine "retrovi-rus-like" (VL30) gene family.
Khan, A.S.,' Kessel, M.,22NCI, National Institutes of Health,Bethesda, Maryland: Structuraland functional analysis of the LTRsassociated with a cloned endoge-nous retrovirus genome isolatedfrom African green monkey DNA.
Morgan, R.A., Huang, R.C.C., Dept.of Biology, Johns Hopkins Univer-sity, Baltimore, Maryland: Under-methylation of intracisternal A-par-ticle genes is correlated withtranscriptional activity in murineplasmacytomas but not in NIH-3T3embryo fibroblasts.
Honigman, A., Bar-Shira, A., Panet,A., Hebrew University HadassahMedical School, Jerusalem, Israel:Processing of the 3' ends of Mo-MLV RNA.
Ridgway, A.A.,' Swift, R.A.2 Dodgson,J.B.,3 Kung, H.-J.2 Fujita, D.J.,''Cancer Research Laboratory,University of Western Ontario,Canada; Depts. of 2Biochemistry,3Microbiology, Michigan State Uni-versity, East Lansing: Activation ofthe c-myc gene by chicken syncy-tial virus-Transcription in vitro ini-tiates from two sites within the viralLTR.
Stoltzfus, C.M., Lorenzen, S.K.,Chang, L.-J., Dept. of Microbiol-ogy, University of Iowa, Iowa City:Comparison of src mRNA splicingin cells infected with differentstrains and intron deletion mutantsof RSV.
Robinson, H., Worcester Foundationfor Experimental Biology, Shrews-bury, Massachusetts: Retroviraltransduction of proto-oncogenesequences.
Niwa, 0., Dept. of Experimental Ra-diology, Faculty of Medicine,Kyoto University, Japan: Expres-sion of the transfecting Mo-MLV
genome and the neo gene fusedto the LTR of Mo-MLV in teratocar-cinoma cells,
Taketo, E.,3 Sherman,M.I.,"Dept. of Cell Biology,Roche Institute of Molecular Biol-ogy, Nutley, New Jersey; 2JacksonLaboratory, Bar Harbor, Maine;3Dept. of Biochemical Sciences,Princeton University, New Jersey:Expression in embryonal carci-noma cells of neomycin resistancegene placed under the control ofMo-MLV LTR promoter.
Anderson, C., Dept. of Chemistry,University of California, SanDiego, La Jolla: Retroviral vectorsand gene expression.
Weber-Benarous, A., Cone, R., Lon-don, I.M., Mulligan, R., Research;Massachusetts Institute of Technol-ogy, Cambridge: Expression ofK562 p-globin gene in mouseerythroleukemia cells.
Cone, R.D., Mulligan, R., Center forCancer Research and WhiteheadInstitute, Massachusetts Institute ofTechnology, Cambridge: Con-struction of a broad host-rangepackaging line for murine retro-viral vectors.
Armentano, D., Shimamura, A., Gil-boa, E., Dept. of Molecular Biol-ogy, Princeton University, NewJersey: The packaging signal ofMo-MLV.
Olsen, J., Swanstrom, R., Dept. ofBiochemistry, Cancer ResearchCenter, University of North Caro-lina, Chapel Hill: Analysis of plas-mids from a RSV-derived shuttlevector rescued in bacteria.
Lautenberger, J.,' Court, D.,' Jorcyk,C.,' Kan, N.,' Seth, A.,' Vergilio,L.,1 Josephs, 5.,2 Papas, T,"NCI,Frederick Cancer Research Facil-ity, Frederick; 2NCI, National Insti-tutes of Health, Bethesda, Mary-land: High-level expression ofoncogene proteins in E. col' usingplasmid pJL6 and its variants.
Jensen, L.,' Coffin, J.,2 Robinson,H.,"Worcester Foundation for Ex-perimental Biology, Shrewsbury;2Tufts University School of Medi-cine, Boston, Massachusetts: gag-po/ sequences play a key role inthe lymphomogenic potential ofRAV-1.
Yang, W.K.,' Boone, L.R.,12 Ou, C.Y.,'Yang, D.M.,' Myer, F.E.,' Tennant,R.W.,' 'Biology Division, Oak
Ridge National Laboratory, Ten-nessee; 2National Institute of Envi-ronmental Health Sciences, Re-search Triangle Park, NorthCarolina: A distinct viral p309.9-di-rected host restriction of endoge-neous murine type-C retroviruses.
DesGroseillers, L., Barrette, M., Jo li-coeur, P., Institut de RecherchesCliniques de Montreal, Universityde Montreal, Quebec, Canada:Physical mapping of the paralysis-inducing derterminant of wildmouse ecotropic neurotropic re-troviruses.
Yuen, PH., Malehorn, D., Nau, C.,Knupp, C., Wong, P.K.Y., Dept. ofMicrobiology, University of Illinois,Urbana: Localization of the paraly-togenic determinant(s) of ts1, atemperature-sensitive mutant ofMoloney murine leukemia virus.
Ikeda, H., Laigret, F., Repaske, R.,Theodore, T, Kozak, C.A., Martin,M.A., NCI, National Institutes ofHealth, Bethesda, Maryland:Characterization of molecularlycloned retroviral sequence associ-ated with Fv-4 resistance.
Kozak, C., NIAID, National Institutesof Health, Bethesda, Maryland:Susceptibility of cells from wildmice to exogenous xenotropicmouse leukemia virus-Regulationby a single chromosome 1 gene.
Silver, J., NIAID, National Institutes ofHealth, Bethesda, Maryland: AKRgenes on chromosomes 7 and 15affect susceptibility to T-cell lym-phoma.
Dandekar, S., Rositto, P., Gardner, M.,Dept. of Pathology, School of Med-icine, University of California,Davis: Mechanism of an ecotropicMLV restriction gene, Akyr-1., inwild mice.
Levine, K., Yoshimura, F., Fred Hutch-inson Cancer Research Center,Seattle, Washington: Characteriza-tion of molecular clones of AKRthymoma-specific proviruses.
Hays, E.F.,' Levy, J.A.,2 'Laboratoryof Biomedical and EnvironmentalSciences, University of California,Los Angeles; 2Cancer ResearchInstitute, University of California,San Francisco: Differences in lym-phomagenic properties of AKRmouse retroviruses.
Wolff, L., Ruscetti, S., NCI, NationalInstitutes of Health, Bethesda,Maryland: Loss of the U3 region
213

tandem repeat appears to be acommon feature of pathogenicFr-MLV-derived recombinantviruses.
Sitbon, M., Evans, L., Nishio, J.,Wehrly, K., Chesebro, B., NIAID,National Institutes of Health,Rocky Mountain Laboratories,Hamilton, Montana: Difference invirulence of two related Fr-MLV isnot due to difference in their abilityto replicate or to induce MCF vi-ruses.
Evans, L.H.,' Cloyd, M.W.,2 'NIAID,National Institutes of Health,Rocky Mountain Laboratories,Hamilton, Montana; 2Dept. of Sur-gery, Duke University, Durham,North Carolina: Specificity of re-combination of ecotropic MLVswith endogenous murine retrovi-rus sequences to generate MCFviruses.
Villemur, R., DesGroseillers, L., Jo li-coeur, P., Institut de RecherchesCliniques de Montreal, Universitede Montreal, Quebec, Canada: At-tempts to detect specific integra-tion site of ecotropic and MCF-type proviruses in gross passageof a MLV-induced mouse thy-moma.
Thomas, C.Y, Boykin, B.J., Coppola,M.A., University of Virginia Schoolof Medicine, Charlottesville: Re-combinant ecotropic and class-IIpolytropic viruses from CW miceaccelerate the development ofsplenic lymphomas.
Cloyd, M.W.,' Evans, L.H.,2 'Dept. ofSurgery, Duke University, Durham,North Carolina; 2NIAID, NationalInstitutes of Health, Rocky Moun-tain Laboratories, Hamilton, Mon-tana: Primary murine leukemiacells lack xenotropic MLV envproducts but do express MCF-likeproteins-Type of MCF env varieswith mouse strain and ecotropicvirus inoculated.
Buchhagen, D.L., Dept. of Microbiol-ogy and Immunology, State Uni-versity of New York, DownstateMedical Center, Brooklyn, NewYork: Unintegrated MCF provi-ruses are amplified in the thy-muses of young AKR mice.
Asje, B., Palminger, I., Fenye, E.M.,Dept. of Virology and Tumor Biol-ogy, Karolinska Institutet, Stock-holm, Sweden: Leukemogenesisby Mo-MLV involves two subsets
214
of T cells probably through differ-ent mechanisms.
Balachandran, R.,1 Pierce, J.P.,2 Aar-onson, S.A.,2 Reddy, E.P.,2 Swan,D.C.,2 1 Laboratory of Biochemis-try, 2NCI, National Institutes ofHealth, Bethesda, Maryland:Characterization of T lymphocytestransformed by Mo-MLV.
Newcomb, E.W., Bieler, J.G., Binari,R., Fleissner, E., Memorial Sloan-Kettering Cancer Center, Sloan-Kettering Division, GraduateSchool of Medical Sciences, Cor-nell University, New York, NewYork: Evidence against a viral ori-gin of most radiation leukemias inBALE /c mice.
Snyder, H.W., Jr.,' Singhal, M.C.,1Grant, O.K.,' Hardy, W.D., Jr.,2Jones, F.R.,"Pacific NorthwestResearch Foundation, Seattle,Washington; 2Memorial Sloan-Ket-tering Cancer Center, New York,New York: Remission of FeLV-as-sociated lymphosarcoma and per-sistent FeLV infection after extra-corporeal immunoadsorptionusing Staphylococcus aureusCowan I (SAC)-Details of immuneresponse.
Mally, M.I.,' Haas, M.,12 'CancerCenter, 2Dept. of Biology, Univer-sity of California, San Diego:Expression of oncogenes in mu-rine T-cell neoplasms.
Haas, M.,'3 Bogart, M.,2 Patch, J.,3'Dept. of Biology, 2Division of Ge-netics, 3Cancer Center, Universityof California, San Diego: Autosti-mulation of X-ray-induced T-Iym-phoma cells is mediated by anovel growth factor.
Elder, J.H., Johnson, D.A., Dept. ofMolecular Biology, Scripps Clinicand Research Foundation, LaJolla, California: Distribution of 81-TAg thymocyte marker on normaland neoplastic cells.
Fuqua, S.A.,' Saxinger, W.C.,2 Bron-son, D.L.,' 'Southwest Foundationfor Biomedical Research, San An-tonio, Texas; 2NCI, National Insti-tutes of Health, Bethesda, Mary-land: Retrovirus production andoncogene expression as a func-tion of differentiation in human em-bryonal carcinoma cells.
Spodick, D.A.,' Soe, L.H.,2 Roy-Bur-man, Depts. of 'Biochemistry,2Pathology, University of SouthernCalifornia School of Medicine, Los
Angeles: Isolation and analysis ofendogenous RD-114 loci in thegenome of the domestic cat.
Bacheler, L.T., Fels Research Institute,Temple University School of Medi-cine, Philadelphia, Pennsylvania:Molecular clones of endogenousMLV-related DNA sequences fromBALB/c mice-Characterization ofintegration sites.
Boots, L.,' Bacheler, L.T.,2 'Dept. ofMicrobiology and Immunology,2Fels Research Institute, TempleUniversity School of Medicine,Philadelphia, Pennsylvania: Viralgene expression in Mo-MLV-in-fected rat cells.
Horowitz, J.M., Kaehler, D., Risser, R.,McArdle Laboratory for CancerResearch, Madison, Wisconsin:Molecular and biological charac-terization of the endogenous eco-tropic provirus of BALB/c mice.
Horowitz, J.M., Kaehler, D., Risser, R.,McArdle Laboratory for CancerResearch, Madison, Wisconsin:Rescue of endogenous MLV ge-nomes by transfection withsubgenomic viral fragments.
Jolicoeur, P.,' Villeneuve, L.,' Kozak,C.,2 'Institut de Recherches Cli-niques de Montreal, Universite deMontreal, Quebec, Canada;2NIAID, National Institutes ofHealth, Bethesda, Maryland: TheRMoInt-1, a locus rearranged inseveral Mo-MLV-induced rat thy-momas, maps on mouse chromo-some 15.
Economou-Pachnis, A., Lohse, M.,Tsichlis, P.N., Fox Chase CancerCenter, Philadelphia, Pennsylva-nia: Polymorphism in Mlvi-2 andthe immunoglobulin heavy-chainlocus in the rat due to individualmembers of a family of highly re-peated elements.
Merregaert, J.,1 Janowski, M.,' Bon-iver, J.,2 Maisin, J.R.,1 'Dept. ofRadiobiology, CEN/SCK;2Laboratory of Pathological Anat-omy, University of Liege, Belgium:The proviral genome of the murineradiation leukemia virus-Integra-tion in cellular DNA, characteriza-tion of an infectious molecularclone, and nucleotide sequence ofthe LTR.
Pal, B.K., Cooper, R.E., Dept. of Bio-logical Sciences, California StatePolytechnic University, Pomona:Methylation, virogene expression,

and stages of differentiation of Bcells transformed by wild mouseretrovirus.
Sitbon, M., Nishio, J., Wehrly, K.,Chesebro, B., NIAID, National In-stitutes of Health, Rocky MountainLaboratories, Hamilton, Montana:A new sensitive focal immunofluo-rescence assay with monoclonalantibodies allowing detection of in-
dividual strains of MLV in complexvirus mixtures.
Wolfe, J.H., Janssen, W.E., Pozsgay,J., Blank, K.J., Dept. of Pathology,University of Pennsylvania Schoolof Medicine, Philadelphia: Altera-tions in viral protein processing inMLV-induced cell lines from H-2-congenic mice.
Zijlstra, M.,' de Goede, R.E.Y.,'
SESSION 10 MORE CARCINOGENESIS AND LEUKEMOGENESIS
Schoenmakers, H.J.,' Radasz-kiewics, T.,2 Melief, C.J.M.,''Central Laboratory of BloodTransfusion Service, Amsterdam,The Netherlands; 2Institute of Pa-thology, Vienna, Austria: The H-2complex of the mouse influencesthe phenotype of MLV-inducedlymphomas.
Chairperson: E. Fleissner, Memorial Sloan-Kettering Cancer Center, New York, New York
Famulari, N.G.,' Holland, C.A.,2 Hop-kins, N.H.,2 O'Donnell, P.V.,''Memorial Sloan-Kettering CancerCenter, New York, New York;2Dept. of Biology, MassachusettsInstitute of Technology, Boston:The contribution of gp70, p15(E),and LTR sequences of MCF 247virus to thymotropism and patho-genicity in AKR mice.
Holland, C.,' Chatis, P.,' Hartley, J.,2Hopkins, N.,' 'Center for CancerResearch, Massachusetts Instituteof Technology, Cambridge; 2NCI,National Institutes of Health, Be-thesda, Maryland: The 3' end ofnondefective Friend virus conferserythroleukemogenesis on MoLV.
Oliff, A., Memorial Sloan-KetteringCancer Center, New York, NewYork: Both LTR and env gene se-quences contribute to leukemoge-nesis by the helper-independentFriend viruses.
O'Donnell, P.V., Koehne, C., Woller,R., Lonial, H., Fleissner, E., Memo-rial Sloan-Kettering Cancer Center,New York, New York: Identificationof a clonal population of cells withlimited transformation capacityduring the latent period of MCF vi-rus-accelerated leukemia in AKR/Jmice.
SESSION II THE LAST ONCOGENE
Chairperson: R. Eisenman, Fred HWashington
Meijlink, F.C.P.W., Curran, T., Miller,A.D., Verma, I.M., Molecular Biol-ogy and Virology Laboratory, SalkInstitute, San Diego, California:Transformation by the c-fosgene- Intramolecular interaction
Li, Y.,' Holland, C.,' Hopkins, N.,'Hartley, J.,2 'Center for CancerResearch, Massachusetts Instituteof Technology, Cambridge; 2NCI,National Institutes of Health, Be-thesda, Maryland: Viral integrationnear c-myc in about 15./o ofMCF247-induced AKR thymomas.
Selten, G.,' Cuypers, H.T.,1 Quint,W.,' Zijlstra, M.,2 Maandag, E.R.,'Boelens, W.,' van Wezenbeek, P,'Melief, C.,2 Berns, A.,' 'Dept. ofBiochemistry, University of Nijme-gen; 2CLB, Amsterdam, The Neth-erlands: MLV-induced T-cell lym-phomagenesis in mice- Proviralactivation of c-myc and pim-1.
Steffen, D., Worcester Foundation forExperimental Biology, Shrewsbury,Massachusetts: Proviruses adja-cent to c-myc in Mo-MLV-inducedlymphomas.
Tsichlis, P.N., Strauss, PG., Lohse,M., Fox Chase Cancer Center,Philadelphia, Pennsylvania: Mo-MLV-induced rat thymomas con-tain at least three common regionsof provirus integration.
Neil, J.C.,' Stewart, M.,' Wilkie, N.,'Lees, G.,2 Jarrett, 0.,2 Onions,D.,2 'Beatson Institute for CancerResearch; 2Dept. of Veterinary Pa-thology, University of Glasgow,
SHOW (fos, myc, myb, ski)
utchinson Cancer Research Center, Seattle,
in the 3' region regulates expres-sion.
Curran, T.,' Van Beveren, C.,2 Verma,I.M.,2 'Dept. of Molecular Ge-netics, Hoffmann-LaRoche Inc.,Roche Research Center, Nutley,
Scotland: Multiple mechanisms ofmyc activation in feline T-cell leu-kemias.
Mullins, J.I., Doggett, D.L., Broda,D.S., Binari, R.C., Jr., Cotter, S.M.,Dept. of Cancer Biology, HarvardSchool of Public Health, Boston,Massachusetts: c-myc transduc-tion in naturally occurring feline T-cell leukemias.
Bestwick, R., Machida, C., Kabat, C.,Oregon Health Sciences Univer-sity, Portland: Seed oncogenesthat cause progressive leukemo-genesis- Membrane glycoproteingenes of SFFV.
Breindl, M., Harbers, K., Jaenisch, R.,Heinrich-Pelle-Institut for Experi-mental Virologie, Hamburg, Fed-eral Republic of Germany: Retrovi-rus-induced lethal mutation inmice is associated with an alteredchromatin structure.
Levy, D.,1.2 Lerner, R.,2 Wilson, M.,2'Division of Biology, California In-stitute of Technology, Pasadena;2Dept. of Molecular Biology, Re-search Institute of Scripps Clinic,La Jolla: Tissue-specific transcrip-tion of defective endogenous re-troviruses under coordinate ge-netic control.
New Jersey; 2Molecular Biologyand Virology Laboratory, Salk In-stitute, San Diego, California: Viraland cellular fos proteins are corn-plexed with a 39,000-dalton cellu-lar protein.
215

Wiman, K.G.,' Hayday, A.C,.2 Saito,H.,2 Tonegawa, S.,2 Hayward,W.S.,"Memorial Sloan-KetteringCancer Center, New York, NewYork; 'Massachusetts Institute ofTechnology, Boston: Mechanismof activation of a translocated butintact c-myc gene that is not closeto an immunoglobulin enhancerelement.
Vennstrom, B.,' Kahn, P.,' Adkins,B.,' Enrietto, P.,' Hayman, M.,2Graf, T.,' Luciw, P.,3 'EuropeanMolecular Biology Laboratory, Hei-delberg, Federal Republic of Ger-many; 'Imperial Cancer ResearchFund, London, England; 3ChironCorp., Emeryville, California:Transformation capacities of a mu-rine retrovirus encoding an avianmyc oncogene.
Ramsay, G.,' Evan, G.,' Lerner, R.,'Bishop, J.M.,"George WilliamHooper Foundation, University ofCalifornia, San Francisco; 'ScrippsClinic and Research Foundation,La Jolla, California: Identificationof the human c-myc protein usingsera raised against a series of c-mycI'ma31 peptides.
Hann, S., Eisenman, R., Fred Hutch-inson Cancer Research Center,Seattle, Washington: Protein prod-ucts of the human c-myc onco-gene.
SESSION 12 POSTER SESSION IV
Willer, R., Wagner, E.F., EuropeanMolecular Biology Laboratory, Hei-delberg, Federal Republic of Ger-many: Induction of differentiationafter transfer of a cellular onco-gene (c-fos) into F9 teratocarci-noma stem cells.
Shibuya, M.,' Yokota, J.,' Ueyama,Y.,' 'Dept. of Genetics, Institute ofMedical Science, University of To-kyo; 'Dept. of Pathology, TokaiUniversity, Central Institute for Ex-perimental Animals, Japan: Ampli-fication of a cellular oncogene (c-myc) in human gastric adenocarci-noma cell lines.
Rovigatti, U.,' Blair, D.,1 Nelson, L.,2Papas, T.,' Rader, J.,"NCI, Fred-erick Cancer Research Facility,Frederick Maryland; 'MemorialSloan-Kettering Cancer Center,New York, New York: Oncogenealterations in human neuroblas-
216
Heaney, M.L., Parsons, J.T., Dept. ofMicrobiology, University of Vir-ginia, Charlottesville: Site-directedmutagenesis of the v-myc gene-Analysis of regions that share ho-mology with the v-myb and El Agenes.
Morgan, J.H., Parsons, J.T., Dept. ofMicrobiology, University of VirginiaSchool of Medicine, Charlottes-ville: Analysis of the myc proteinfrom normal and transformedcells.
Lee, W., Schwab, M., Westaway, D.,Bishop, J.M., Varmus, H., Depts.of Medicine, Microbiology and Im-munology, University of California,San Francisco: Transformation-promoting activity of c-myc genesderived from normal and tumor tis-sues.
Schwab, M.,1 Busch, M.,2 Rosenau,W.,2 Brodeur, G.,3 Varmus, H.E.,1Bishop, J.M.,' Depts. of'Microbiology, 'Pathology, Univer-sity of California, San Francisco;3Dept. of Pediatrics, St. Louis, Mis-souri: Amplification and over-expression of N-myc may be re-lated to malignant progressionrather than to cellular transforma-tion in human neuroblastoma.
Pelicci, PG., Lanfrancone, L., Brath-waite, M.D., Wolman, S.R., Dalla-Favera, R., Dept. of Pathology, Ka-
toma and primitive neuroectoder-mal tumors.
Sklar, M.,12 Rosson, D.,2 Tereba, A.,2Depts. of 'Radiation Therapy,'Virology and Molecular Biology,St. Jude Children's Research Hos-pital, Memphis, Tennessee: Onco-genic activity of v-myc and a hu-man c-myc-related sequence in amouse bone marrow-derived cellline and primary mouse macro-phages.
Brightman, B.K., Fan, H., Dept. ofMolecular Biology and Biochemis-try, University of California, Irvine:Construction of a recombinant mu-rine leukemia virus carrying MC29v-myc gene.
Matthews, E.A., Bishop, J.M., GeorgeWilliam Hooper Research Founda-tion, Dept. of Microbiology andImmunology, University of Califor-nia, San Francisco: A recombinant
plan Cancer Center, New YorkUniversity School of Medicine,New York: Oncogene (c-myc, c-myb) amplification and acute mye-logenous leukemia-Molecularand cytogenetic studies.
Klempnauer, K.-H.,' Symonds, G.,'Evan, GI,' Bishop, J.M.,1'George William Hooper ResearchFoundation, Dept. of Microbiologyand Immunology, University ofCalifornia Medical Center, SanFrancisco; 'Ludwig Institute forCancer Research, MRC Centre,Cambridge, England: Subcellularlocalization of myb gene productsand the fate of p452 -rrIY, during thedifferentiation of AMV-transformedmyeloblasts.
Lipsick, J.S., Boyle, W.J., Lampert,M.A., Ong, J., Baluda, M.A., Jons-son Comprehensive Cancer Cen-ter, Dept. of Pathology, Universityof California School of Medicine,Los Angeles: Both the E26 andAMV viral oncogene products arenuclear proteins.
Brodeur, D., Barkas, A., Stavnezer, E.,Molecular Biology and VirologyProgram, Memorial Sloan-Ketter-ing Cancer Center, New York, NewYork: v-ski polyproteins are lo-cated in the nuclei of SKV-trans-formed cells.
virus containing cellular and viralmyc sequences.
Van Beneden, R.,' Watson, D.,2 Son-stegard, R.,' Chen, T.,2 'Dept. ofBiology, McMaster University, On-tario, Canada; 'NCI, FrederickCancer Research Facility, Freder-ick, Maryland: Isolation and char-acterization of fish c-myc.
Soe, L.H., Roy-Burman, P, Depts. ofPathology and Biochemistry, Uni-versity of Southern CaliforniaSchool of Medicine, Los Angeles:Possible association between DNApolymorphism at the c-myc locusand tumors in the domestic cat.
Shih, C.-K.,'-' Clurman, B.E.,' Good-enow, M.M.,' Linial, M.,3 Hayward,W.S.," Memorial Sloan-KetteringCancer Center; 'Rockefeller Uni-versity, New York, New York; 3FredHutchinson Cancer Center, Seat-tle, Washington: Characterization

of the 5' region of the chicken c-myc locus.
Dean, M.,' Campisi, J.,2Sonenshein,G.,3 Pardee, A.B.,2 1LBI -Basic Re-search Program, NCI, FrederickCancer Research Facility, Freder-ick, Maryland; 2Dana-Farber Can-cer Institute, Boston, Massachu-setts; 3Boston University MedicalCenter, Massachusetts: Regulationof the c-myc proto-oncogene dur-ing the cell cycle and differentia-tion.
Butnick, N.J., Miyamoto, C., Chizzon-ite, R., Ju, G., Skalka, A.M., Dept.of Molecular Genetics, Hoffmann-La Roche, Inc., Roche ResearchCenter, Nutley, New Jersey: Regu-lation of c-myc expression in HL60cells-Transcriptional and transla-tional control.
Keath, E.J., Kelekar, A., Cole, M.D.,Dept. of Biochemistry, St. LouisUniversity School of Medicine,Missouri: Transcriptional activationof the translocated c-myc onco-gene in murine plasmacytomas.
Chen, T.,' Watson, D.,' Pry, T.,' Orosz-Ian, S,,2 Bader, J.,' Papas, T.,''NCI, National Institutes of Health;2Litton Bionetics Inc., FrederickMaryland: Characterization bytryptic mapping, and determina-tion of the amino terminusof thechicken c-myc protein.
Bader, J.P., Ray, D.A., Laboratory ofMolecular Oncology, NCI, Freder-ick Cancer Research Facility, Fred-erick, Maryland: The proteins ofMC29 virus and the c-myc onco-gene occur as intranuclear di-mers.
Duprey, S.P., Boettiger, D.E., Dept. ofMicrobiology, University of Penn-sylvania School of Medicine, Phila-delphia: Elevated levels of c-mybRNA are associated with earlystages of hemopoietic differentia-tion.
Watson, D.,' Nunn, M.,2 Duesberg,P.,2 O'Brien, S.,' Papas, T.,"NCI,Frederick Cancer Research Facil-ity, Frederick, Maryland; 2Dept. ofMolecular Biology, University ofCalifornia, Berkeley: Isolation,characterization, and chromo-somal localization of cellular se-quences related to v-ets, theunique one gene of the avian re-trovirus, E26.
Graziano, 5.,1.2 Merl, S.,'2.4 Lehr, B.,'Ehrlich, G.,3 Krichbaum, K.,3.4
Hubbell, C.,' Planas, A.,2 Mitter,N.,' Davey, F.,' Moore,Comis, Poiesz,'State University of New York;2Veterans Administration MedicalCenter; 3State University, Syra-cuse; 4Barbara Kopp ResearchCenter, Auburn, New York: Aquantitative assay of HTLV trans-formation.
Krichbaum, K.,'.2 Merl, 5.,1.3.4 Keller,Poiesz, Ruscetti, F.,6
Vournakis, J.,2 Comis, R.,'.3Moore, J.,'.3 'Barbara Kopp Re-search Center, Auburn; 2State Uni-versity, Syracuse; 3State Universityof New York 4Veterans Administra-tion Medical Center, Syracuse;.Merck Sharp and Dohme Re-search Laboratories, West Point,Pennsylvania; 6NCI, FrederickCancer Research Facility, Freder-ick, Maryland: A quantitative flu-orescent assay for adsorption ofHTLV to cellular membranes.
Longo, D.,' Matis, L.,' O'Brien, 5.,2Gelmann, E.," NCI, National Insti-tutes of Health, Bethesda; 2NCI,Frederick Cancer Research Facil-ity, Frederick, Maryland: Transfor-mation of human B and T lympho-cytes by infection with HTLV-IHs, astrain of HTLV-I produced by a B-cell clone.
Merl, 5.,1.2.4 Barcos, M.,5 Han, T.,5Pollard, C.,. Ehrlich,Zamkoff, Moore, J.,'Davey, F.,' Mitter, N.,' Goldberg,J.,' Vournakis, J.,3 Comis, R.,'.4Poiesz, B.,1.2.4 'State University ofNew York, 2Veterans Administra-tion Medical Center, 3State Univer-sity, Syracuse; 4Barbara Kopp Re-search Center, Auburn; .RoswellPark Memorial Institute, Buffalo:HTLV information in non-T-cell he-matologic neoplasia.
Aldovini, A.,' De Rossi, A.,' Franchini,G.,2 Gallo, R.C.,2 Wong-Staal, F.,2'Laboratory of Oncology, Univer-sity of Padova, Italy; 2NCI, Na-tional Institutes of Health, Be-thesda, Maryland: Cell-freetransmission of HTLV-I to normalhuman lymphocytes-Molecularstudies on HTLV integration andexpression.
Guo, H.-G., Reitz, M.S., Saxinger,W.C., Gallo, R.C., Wong-Staal, F.,NCI, National Institutes of Health,Bethesda, Maryland: HTLV-likeproviruses in nonhuman primates.
Tomar, R.V.,' Kloster, B.,' Ehrlich, G.,2Lamberson, H.V.,3 Stockman, J.,3Groth, D.,' Vournakis, J.,2 Merl,S.,' Poiesz, B.,"Upstate MedicalCenter; 2Syracuse University;3Syracuse American Red Cross,New York: Evidence for associa-tion of HTLV in hemophilic pa-tients.
Kalyanaraman, V., Feorino, P, Getch-ell, J., Francis, D., Narayanan, R.,Palmer, E., Ramsey, R., Evatt, B.,Chorba, R., Cabradilla, C., Cen-ters for Disease Control, Atlanta,Georgia: Isolation and character-ization of HTLV-II-related virusfrom cultured T cells of a patientwith hemophilia A and pancyto-penia.
Narayanan, R.,' Kalyanaraman, V.,'Srinivasan, A.,2 Getchell, J.,' Hol-loway, B.,' Feorino, P.,' Francis,D.,' Ramsey, R.,' Evatt, B.,'Chorba, T.,' Cabradilla, C.,'2Centers for Disease Control, At-lanta, Georgia; NCI, National Insti-tutes of Health, Bethesda, Mary-land: HTLV-II-related sequences inA-cell lines derived from a hemo-philic patient with pancytopenia.
Shaw, G.M.,' Gonda, M.A.,2 Flickin-ger, G.H.,2 Hahn, B.H.,' Gallo,R.C.,' Wong-Staal, F.,"NCI, Na-tional Institutes of Health, Be-thesda; 2Frederick Cancer Re-search Facility, Frederick,Maryland: Conservation in theviral genomes of evolutionarily di-vergent members of the HTLVfamily.
Hahn, B.H.,' Shaw, G.M.,' Popovic,M.,' LoMonico, A.,' Leibowitch,J.,2Wong-Staal, F,1 Gallo, R.C.,''NCI, National Institutes of Health,Bethesda, Maryland; 2Dept. of Im-munologie, Hospital RaymondPoincare, Garches, France: Mo-lecular cloning and analysis of anew variant of human T-cell leuke-mia virus (HTLV-Ib) from an Afri-can patient with T-cell lym-phoma.
Drake, A.L.,' Franchini, G.,2 Wilson,L.,3 Faller, D.V.,3 Mullins, J.I.,''Dept. of Cancer Biology, HarvardSchool of Public Health, Boston,Massachusetts; 2NCI, National In-stitutes of Health, Bethesda, Mary-land; 3Dana-Farber Cancer Insti-tute, Boston, Massachusetts:Arrangement and expression ofHTLV-I proviruses derived from a
217

virus-productive adult T-cell leuke-mia cell line.
Eiden, M.,' Fisher, A.,' Newman, M.,2Mann, D.,' Mullins, J,,3 Howley,P,' Reitz, M.,' Gallo, R.C.,"NCI,National Institutes of Health;'Uniformed Services University ofthe Health Sciences, Bethesda,Maryland; 3Harvard School ofPublic Health, Boston, Massachu-setts: Expression of cloned HTLV-Ismall env protein in transfectedmouse cells.
Homma, T.,' Kanki, N.W.,
Jr.,' Hunt, R.D.,2 Essex, M''Dept. of Cancer Biology, HarvardSchool of Public Health, Boston;'New England Regional PrimateResearch Center, Harvard MedicalSchool, Southborough, Massachu-setts: Antibodies to cell membraneantigens associated with HTLV inmacaques with lymphoma.
Newman, M.,' Baker, I.,' Reitz, M.,2Eiden, M.,2 Blattner, W.,2 Harris,C.,2 Mann, D.,2 'Uniformed Serv-ices University of the Health Sci-ences; 'NCI, National Institutes ofHealth, Bethesda, Maryland: De-tection of viral env protein onHTLV-infected cells using mono-clonal antibodies.
Sarngadharan, M.G.,' Popovic, M.,'Salahuddin, Z.,2 Bruch, L.,' Mark-ham, P.D.,' Schuepbach, J.,2Gallo, R.C.,2 'Litton Bionetics,Kensington; 'NCI, National Insti-tutes of Health, Bethesda, Mary-land: Antibodies reactive with ahuman T-Iymphotropic retrovirus(HTLV-III) in the sera of patientswith AIDS.
Narayanan, R.,'Srinivasan, A.,2 Kaly-anaraman, V.,' Getchell, J.,' Fran-cis, D.,' Holloway, B.,' Cabradilla,C.,"Centers for Disease Control,Molecular Virology Branch, At-lanta, Georgia; 'NCI, National In-stitutes of Health, Bethesda, Mary-land: Survey of tissues from AIDSpatients for the expression of cel-lular homologs of retroviral onco-genes.
Mullins, J.I., Riedel, N., Dept. of Can-cer Biology, Harvard School ofPublic Health, Boston, Massachu-setts: Identification of proviral DNAweakly related to HTLV in cells de-rived from a patient with AIDS.
Mullins, J.I.,1 McLane, Essex,M.,' Groopman, J.,2 Gold, J.,4Schooley, R.,3 Ho, D.,3 Hirsch,
218
M.,3 Feorino, P.,' Cabradilla, C.,5'Harvard School of Public Health;'New England Deaconess Hospi-tal; 3Massachusetts General Hos-pital, Boston; eMemorial Sloan-Kettering Cancer Center, NewYork, New York; 'Centers for Dis-ease Control, Atlanta, Georgia: Asearch for HTLV-related DNA se-quences in AIDS.
Ehrlich, G.,1,2 Merl, S.,2,3,4 Han, T.,'Tomar, R.,2 Barcos, M.,' Moore, J.,2,3,4 Blair, D.,2 Zamkoff, K.,2'3 Vour-nakis, J.,' Comis,B.,2.3.4 'State University of NewYork; 'State University of New YorkMedical Center; 3Veterans Admin-istration Medical Center, Syracuse,New York; eBarbara Kopp Re-search Center, Auburn, New York;'Roswell Park Memorial Institute,Buffalo, New York: HTLV-I homolo-gous sequences in fresh and cul-tured cells from AIDS and LAS pa-tients.
Barker, C.,' Willis, J.W.,' Bradac, J.,'Tainsky, M.,2 Hunter, E.,"Dept. ofMicrobiology, University of Ala-bama, Birmingham; 'NCI, Freder-ick Cancer Research Facility, Fred-erick, Maryland: The molecularcloning and characterization of thegenome of Mason-Pfizer monkeyvirus.
Kashmiri, S.V.S., Mehdi, R., Ferrer,J.F., University of Pennsylvania,Philadelphia: Hypermethylation ofbovine leukemia proviral DNA inleukemic cells and infected lym-phocytes.
O'Neill, R.R., Buckler, C.E., Theodore,T, Martin, M.A., Repaske, R.,NIAID, National Institutes ofHealth, Bethesda, Maryland: Thecomplete nucleotide sequence ofenv and LTR regions of an infec-tious xenotropic MLV-Relation-ship to MCF and ecotropic MLVsequences.
Friedrich, R.,' Hackl, B.,' Oliff, A.,2'Institute of Immunobiology, Uni-versity of Freiburg, Federal Re-public of Germany; 'MemorialSloan-Kettering Cancer Center,New York, New York: Structure ofthe env gene of the nonleukemo-genic murine retrovirus Ampho4070.
Steele, P., Rabson, A., Maloy, L., Coli-gan, J., Bryan, T., Martin, M.,NIAID, National Institutes ofHealth, Bethesda, Maryland: Hu-
man endogenous env sequencesare shared by a novel class of pri-mate endogenous retroviral DNAsand are expressed in human tis-sue.
Elder, J.H.,' Mullins, J.I.,2 'Dept. ofMolecular Biology, Scripps Clinicand Research Foundation, LaJolla, California; 'Dept. of CancerBiology, Harvard School of PublicHealth, Boston, Massachusetts:Comparative analysis of the envgenes of FeLV-B.
Katz, R.A., Skalka, A.M., Dept. of Mo-lecular Genetics, Hoffman-LaRoche Inc., Roche Research Cen-ter, Nutley, New Jersey: Biochemi-cal analysis of a defect in themRNA leader region of the endog-enous avian retrovirus ev-1.
Bennett, D.D.,' Wright, S.E.,1.2 'ViralOncology Laboratory, VeteransAdministration Medical Center;'Depts. of Medicine and Cellular,Viral and Molecular Biology, Uni-versity of Utah School of Medi-cine, Salt Lake City: Immunizationwith envelope glycoprotein of anavian RNA tumor virus protectsagainst sarcoma virus tumor in-duction-Role of subgroup.
Nunberg, J.H.,' Gilbert, J.H.,' Rodg-ers, G.,' Snead, R.M.,' Winston,5.,2 'Cetus Corporation, Emery-ville, California; 2Syngene Prod-ucts and Research, Fort Collins,Colorado: Localization of a deter-minant of virus neutralization onFeLV gp70.
Schultz, A.M., Copeland, T.D., Orson-Ian, S., NCI, Frederick Cancer Re-search Facility, Frederick, Mary-land: The env proteins of BLV-Purification and sequence analy-sis.
Pinter, A., Honnen, W.J., MemorialSloan-Kettering Cancer Center,New York, New York: Functionalroles of the carbohydrates of MLVenv proteins-Studies with inhibi-tors of oligosaccharide process-ing.
Soong, M.M.,' Wong, P.,2 Tompkins,W.,' Depts. of 'Pathobiology,'Microbiology, University of Illi-nois, Urbana: Evidence for pro-cessing of Mo-MLV gPr80e" in as-sociation with the cellcytoskeleton.
Suni, Narvanen, A.,' Wahlstrom,T.,2 Vaheri, A.,' Copeland, T.,3Cohen, M.,3 Oroszlan, 5.,3 Depts.

of 'Virology, 'Pathology, Universityof Helsinki, Finland; 3NCI, Freder-ick Cancer Research Facility, Fred-erick, Maryland: Retrovirus-relatedM, 75,000 polypeptide from cul-tured choriocarcinoma cells andrenal adenocarcinoma tissue.
Simonneau, L.,' Crisanti, P.,2 Lorinet,A.M.,' Calothy, G.,3 Courtois, Y.,'Pessac, B.,2 'INSERM; 2INSERM,Hopital Broussais, Paris, 3InstitutCurie-Biologie, Orsay, France:Regulation of crystallins expres-
SESSION 13 HTLV BLV, AND D TYPES
sion and lentoid bodies formationby oncogenic retroviruses in quailembryo neuroretinal cultures.
Rossomando, A., Meruelo, D., Dept.of Pathology, New York UniversityMedical Center, New York, NewYork: A common 300-bp viral/gen-ornic DNA sequence is associatedwith minor and major histocompa-tibility (Ft) genes.
Ou, C.Y.,1 Koh, C.K.,' Boone, L.R.,2Callahan, R.,3 Yang, W.K.,''Biology Division, Oak Ridge Na-
Chairperson: W. Haseltine, Dana-Farber Cancer Institute, Boston, Massachusetts
Sodroski, J.,1 Perkins, D.,' Briggs,D.,' Patarca, R.,'Wong-Staal, F.,2Gallo, R.,2 Haseltine, W.,11Dana-Farber Cancer Institute, Boston,Massachusetts; 'National Institutesof Health, Bethesda, Maryland:Structural differences betweenHTLV-I and HTLV-II.
Sodroski, J., Rosen, C., Haseltine,W.A., Dana-Farber Cancer Insti-tute, Boston, Massachusetts:Functional differences betweenthe LTRs of HTLV types I and II.
Chen, I.S.Y.,1Wachsman, W.,1 Shimo-tohno, K.,2 Golde, D.W.,1 'Dept. ofMedicine, University of California,Los Angeles; 'National CancerCenter, Tokyo, Japan: Moleculargenetic studies of the HTLV-II LTRand the pX region.
Shimotohno, K.,' Takahashi, Y.,'Golde, D.W.,2 Miwa, M.,' Sugi-mura, T.,1 Chen, I.S.Y.,2 'NationalCancer Center Research Institute,Tokyo, Japan; 'Dept. of Medicine,University of California, Los Ange-les: Nucleotide sequence analysisof HTLV typell.
Ratner, L., Josephs, S.F., Shaw, G.M.,Hahn, B.H., Wong-Staal, F., Gallo,R.C., NCI, National Institutes ofHealth, Bethesda, Maryland: pXdeletion in HTLV isolates from anAfrican adult T-cell leukemia pa-tient.
Yoshida, M.,' Kiyokawa, T.,' Yoshik-ura, H.,' Hattori, S.,' Seiki, M.,''Dept. of Viral Oncology, CancerInstitute, Tokyo; 'Dept. of Bacteri-ology, Faculty of Medicine, Univer-sity of Tokyo, Japan: Identificationand production of HTLV env pro-tein and cytotoxic antisera to virus-positive cells.
Hattori, S.,' Imagawa, K.-1.,2 Shimizu,F.,' Hashimura, E.,2 Seiki, M.,'Yoshida, M.,"Dept. of Viral On-cology, Cancer Institute, Tokyo;2Otsuka Assay Laboratory, OtsukaPharmacy Co. Ltd., Tokushima,Japan: Identification of gag andenv gene products of HTLV.
Copeland, T.D., Tsai, W.P., Schultz,A.M., Oroszlan, S., LBI-Basic Re-search Program, NCI, FrederickCancer Research Facility, Mary-land: Human HTLV gag and envgene products-Structural and an-tigenic relatedness to type-C andtype-D retroviruses.
Franchini, G., Popovic, M., Mann, D.,Wong-Staal, F., Gallo, R.C., NCI,National Institutes of Health, Be-thesda, Maryland: HTLV se-quences in the T cells and B cellsof a patient with adult T-cell leuke-mia.
Franchini, G., Wong-Staal, F Gallo,R.C., NCI, National Institutes ofHealth, Bethesda, Maryland:HTLV-I transcripts in fresh and cul-tured cells of patients with adult T-cell leukemia.
Popovic, M.,' Sarngadharan, M.G.,'Read, E.,1 Gallo, R.C.,' 'NCI, Na-tional Institutes of Health, Be-thesda; 'Litton Bionetics, Kensing-ton, Maryland: Rescue andcontinous production of human T-cell lymphotropic retrovirus (HTLV-111) from patients with AIDS.
Popovic, M.,' Flomenberg, N.,3 Volk-man, D.,2 Mann, D.,' Fauci, A.S.,'Dupont, B.,3 Gallo, R.C.,"NCI,National Institutes of Health,2NIAID, Bethesda, Maryland;3Dept. of Human Immunogenetics,Memorial Sloan-Kettering Cancer
tional Laboratory, Tennessee;2NIEHS, Research Triangle Park,North Carolina; 3National Institutesof Health, Bethesda, Maryland: Anapparent novel family of retroviralsequences in the mouse genome.
Rabson, A.B., Steele, P.E., Hama-gishi, Y., Martin, M.A., NIAID, Na-tional Institutes of Health, Be-thesda, Maryland: Structuralanalysis of human endogenous re-troviral mRNAs.
Center, New York, New York: Alter-ation of specific T-cell function byinfection with HTLV-I or HTLV-II.
Barre-Sinoussi, F, Chermann, J.C.,Montagnier, L., Viral OncologyUnit, Institut Pasteur, Paris,France: Characteristics of a newhuman T- lymphotropic retrovirusassociated with acquired immunedeficiency syndrome.
Lee, TH.,1 McLane, M.F.,' Tachibana,N.,1-2-3 Francis, D.P.,2-3 Homma,T.,' Essex, M.,' 'Dept. of CancerBiology, Harvard UniversitySchool of Public Health, Boston,Massachusetts; 'Miyazaki MedicalSchool, Japan; 3Center for Dis-ease Control, Atlanta, Georgia:Relative immunogenicity of HTLV-specific proteins in adult T-cell leu-kemia/lymphoma patients, healthycarriers, asymptomatic hemophil-iacs, and AIDS patients.
Lee, TH.,' Coligan, J.E.,2-3 Homma,T.,' McLane, M.F.,1 Tachibana,N.,1.2,3 Essex, M.,"Dept. of Can-cer Biology, Harvard UniversitySchool of Public Health, Boston,Massachusetts; 2NIAID, NationalInstitutes of Health, Bethesda,Maryland; 3Miyazaki MedicalSchool, Japan: env gene-encodedantigens detected in cells trans-formed by different strains ofHTLV.
Gelmann, E.,' Fleurdelys, B.,' Kett-mann, R.,' Burny, A.,' 'NCI, Na-tional Institutes of Health, Be-thesda, Maryland; 'Dept. ofBiologie Moleculaire, UniversiteLibre de Bruxelles, Belgium: Mo-lecular cloning of integrated BLVproviral DNA from lymphoma tis-sue.
219

Sagata, N.,' Yasunaga, T,2 Ogawa,Y.,' Tsuzuku-Kawamura, J.,'lkawa, Y.,"Laboratory of Molecu-lar Oncology, 2Computation Cen-ter, Institute of Physical and Chem-ical Research (RIKEN), Wako,Japan: BLV-Unique structuralfeatures of its LTRs and evolutionalrelationship to HTLV.
Stromberg, K.,' Arthur, L.O.,' Rabin,H.,' Giddens, W.E.,2 Benveniste,
R.E.," 'NCI, Frederick Cancer Re-search Facility, Frederick, Mary-land; 2Dept. of Pathology, Univer-sity of Washington, Seattle:Characterization of an exogenoustype-D retrovirus isolated from ma-caques with simian AIDS and ret-roperitoneal fibromatosis.
Desrosiers, R.,' Daniel, M.D.,' Butler,C.,' King, N.,' Wills, J.,2 Barker,C.,2 Hunter, E.,2 Hunt, R.,' Letvin,
SV40, Polyoma, and Adenoviruses
N.,' 'New England Regional Pri-mate Research Center, HarvardMedical School, Southboro, Mas-sachusetts; 2Laboratory for Spe-cial Cancer Research, Universityof Alabama, Birmingham: Charac-terization of a new type-D retrovi-rus-ls it the cause of the immuno-deficiency syndrome ofmacaques?
August 15-August 19
ARRANGED BY
Terri Grodzicker, Cold Spring Harbor LaboratoryMichael Botchan, University of California, Berkeley
401 participants
The meeting on the molecular biology of SV40, polyoma, and adenoviruses,which is held in alternate years at the Cold Spring Harbor Laboratory, was at-tended by more than 400 scientists. The small DNA tumor viruses continue to beintensively studied model systems for investigating mechanisms of eukaryoticgene control, replication, and transformation. Many papers were presented onactive areas of research, such as: control of transcription and RNA splicing bothin vivo and in vitro; analysis and purification of viral and cellular transcription fac-tors; properties of viral and cellular enhancers; regulation of gene expression bythe adenoviral E1A protein; control of translation of VA RNAs; replication of viralDNAs in vitro; the role of viral transforming proteins in immortalization, transfor-mation, and cooperation with other oncogenes; and association of tumor anti-gens with cellular proteins such as p53 and c-src. Several viral transforming pro-teins have been overproduced by using a variety of prokaryotic and eukaryoticexpression vectors. The adenovirus E1A protein and SV40 T antigen have beenpurified and their functions in a variety of cells as well as in in vitro transcriptionand replication systems are being intensively investigated.
This meeting was supported in part by the Cancer Center Grant to ColdSpring Harbor Laboratory from the National Cancer Institute, National Institutesof Health.
SESSION 1 ADENOVIRUSES: TRANSCRIPTION
Chairperson: J. Manley, Columbia University, New York, New York
Hearing, P., Shenk, T., Dept. of Micro-biology, State University of NewYork, Stony Brook: Transcription ofAd5 E1A is regulated in cis by twoseparate sequences and in transby the E1A gene products.
Kingston, R.E., Baldwin, A.S., Sharp,PA., Center for Cancer Research,
220
Massachusetts Institute of Technol-ogy, Cambridge: Regulation ofheat-shock-protein-70 geneexpression by the adenoviral El Aregion and by c-myc.
Kumar, R., Gong, S.-S., Subramanian,K.N., Dept. of Microbiology andImmunology, University of Illinois
Health Sciences Center, Chicago:Transcriptional regulatory ele-ments located at the left terminusof Ad5.
Stein, R., Ziff, E.B., Kaplan CancerCenter, New York University Medi-cal Center, New York: He La cell (3-tubulin gene transcription is stimu-

lated by Ad5 in parallel with viral Moore, C.L., Sharp, PA., Center forearly genes by an E1A-dependent Cancer Research, Massachusettsmechanism. Institute of Technology, Cam-
Gaynor, R.B.,1 Yoshinaga, S.,2 Han, bridge: Site-specific utilization ofM.,2 Feldman, L.,3 Berk, A.,2 viral polyadenylation sites in a cell-Depts. of 'Medicine, 2Micro- free reaction.biology, 3Microbiology and Immu- McDevitt, M.A., Hart, R.P., Imperiale,nology, University of California, M.J., Ali, H., Nevins, J.R., Rocke-Los Angeles: Adenoviral E1A pro- feller University, New York, Newtein enhances transcription of York: Requirement of a down-genes transcribed by RNA poly- stream sequence for generation ofmerase III. a poly(A) addition site.
Hoeffler, W.K., Roeder, R.G., Rocke- Falck-Pedersen, E.,1 Logan, J.,3feller University, New York, New Shenk, T.,2 Citron, B.,1 Darnell,York: Extraction and quantitation J.E., Jr.,"Rockefeller University,of pol III transcription factors from New York, New York; 2Dept. of Mi-adenovirally infected HeLa cells. crobiology, State University of
SESSION 2 SV40, POLYOMA: TRANSCRIPTION
Chairperson: W. Folk, University of Michigan, Ann Arbor, Michigan
Gidoni, D.,' Dynan, W.,' Saffer, J.,2Singer, M.,2 Tjian, R.,"Dept. ofBiochemistry, University of Califor-nia, Berkeley; 2NCI, National Insti-tutes of Health, Bethesda, Mary-land: Protein-DNA interactionsbetween transcription factor Sp1and regulatory sequences in viraland cellular promoters.
Mishoe, H.,1 Brady, J.N.,2 Radonov-ich, M.,' Salzman, N.P.,1 'Lab-oratory of Biology of Viruses,2Laboratory of Molecular Virology,NCI, National Institutes of Health,Bethesda, Maryland: SV40 GC-rich sequences function as an in-dependent transcriptional controlelement in vitro.
Keller, J.M., Alwine, J.C., Dept. of Mi-crobiology, University of Pennsyl-vania, Philadelphia: Elements ofthe SV40 late promoter and trans-activation by T antigen.
Brady, J., Khoury, G., NCI, NationalInstitutes of Health, Bethesda,Maryland: SV40 late transcrip-tion- Induction by T antigen.
Chandrasekharappa, S., Hartzell, S.,Byrne, B., Subramanian, K., Dept.of Microbiology and Immunology,University of Illinois Health Sci-ences Center, Chicago: Two non-contiguous domains of the SV40late promoter are required for itsactivation with T antigen.
Michaeli, T., Prives, C., Dept. of Bio-logical Sciences, Columbia Uni-versity, New York, New York: Reg-ulation of SV40 late gene
expression in Xenopus laevis ooc-ytes.
Mertz, J.E., McArdle Laboratory, Uni-versity of Wisconsin, Madison: TheSV40 late leader region encodes aregulated transcriptional termina-tor.
Tseng, R.W., Acheson, N.H., Dept. ofMicrobiology and Immunology,McGill University, Montreal, Can-ada: Transcription is terminated atmultiple sites on the L strand ofpolyoma virus DNA during lytic in-fection.
New York, Stony Brook: The use ofrecombinant mutants of Ad5 d1309for examination of transcriptionalcontrol DNA elements-Termina-tion of the E1A gene by a mouse13-major globin termination se-quence.
Natarajan, V., Madden, M.J., Salz-man, N.P, NIAID, NCI, NationalInstitutes of Health, Bethesda,Maryland: Proximal and distal do-mains that control in vitro tran-scription of the adenoviralgene.
Sadofsky, M., Alwine, J.C., Dept. ofMicrobiology, University of Penn-sylvania, Philadelphia: Deletionsdownstream from the SV40 lateAAUAAA decrease the efficiencyof 3'-end processing.
Kern, F.G., Dailey, L., Basilico, C.,Dept. of Pathology, New York Uni-versity School of Medicine, NewYork: Effects of deletions and poly-adenylation signal substitution ontranscription initiating from the latepromoter in rat cells transfectedwith chimeric plasmids.
F. Sherman, M. Botchan, T. Grodzicker
221

SESSION 3 POSTER SESSION I
Duprey, E., Hearing, P., Shenk, T.,Dept. of Microbiology, State Uni-versity of New York, Stony Brook:The bidirectional nature of theAd5-E1A enhancer is environmentspecific.
Dickerson, I., Jones, N.C., Dept. ofBiological Sciences, Purdue Uni-versity, West Lafayette, Indiana:Sequence requirements for splic-ing of Ad5 El A pre-mRNA.
Tremblay, M.L., Yee, S.-R, Persson,R., Smiley, J.R., Branton, RE.,Dept. of Pathology, McMaster Uni-versity, Hamilton, Canada: Lack ofcomplementation between Ad5mutants and mouse cell lines con-stitutively expressing the ICP4 im-mediate early gene of HSV-1.
Leff, T, Sassone-Corsi, P, Zajch-owski, D., Boeuf, H., Goding, C.,Jalinot, P., Kedinger, C., Cham-bon, P., INSERM, Faculte de Me-decine, Strasbourg, France: Se-quence elements controlling theexpression of adenoviral E2A andE3 transcription units.
Murthy, S., Bhat, G., Thimmappaya,B., Cancer Center, NorthwesternUniversity Medical School, Chi-cago, Illinois: Adenoviral E2Aearly promoter contains two tran-scriptional control regions butlacks nucleotide sequences spe-cific for its transactivation by El A.
Huang, D.-H., Roeder, R.G., Rocke-feller University, New York, NewYork: Localization of upstream ele-ments that modulate transcriptionof the adenoviral E2A early genes.
lmperiale, M.J., Hart, R.P., Nevins,J.R., Rockefeller University, NewYork, New York: Mapping andproperties of Ad5 E2 transcrip-tional regulatory sequences.
Berger, S.L.,1 Folk, 'Cell andMolecular Biology Program,2Dept. of Biological Chemistry,University of Michigan, Ann Arbor:Differential activation of an RNApolymerase II and an RNA poly-merase III-transcribed gene bythe polyoma virus enhancer andthe adenoviral El A gene prod-ucts.
Yoder, S.S., Berget, S.M., Dept. ofBiochemistry, Rice University,Houston, Texas: Control of dhfrgene expression by Ad2.
Khalili, K., Weinmann, R., Wistar Insti-
222
tute, Philadelphia, Pennsylvania:Shutoff of host protein synthesisafter adenoviral infection.
Reich, N.C.,' Babich, A.,' Weinber-ger, C.,2 Darnell, J.,' 'RockefellerUniversity, New York, New York;2State University of New York,Stony Brook: Nuclear-cytoplasmictransport of viral and cellularmRNAs late in adenoviral infec-tion.
Iwamoto, S.,' Eggerding, F.A.,2 Falck-Pedersen, E.,' Darnell, J.E., Jr.,''Rockefeller University, New York,New York; 2Dept. of Pathology,University of California School ofMedicine, Los Angeles: Transcrip-tion of the Ad2 major late tran-script at early and late times afterinfection.
Ko, J.-L., Harter, M.L., Dept. of Micro-biology, University of Medicineand Dentistry of New Jersey, New-ark: In vitro characterization of abacterially produced Ad2 El A-likeprotein-DNA binding and tran-scriptional activation.
Abmayr, S.M., Roeder, R.G., Rocke-feller University, New York, NewYork: The effect of pseudorabiesvirus infection on in vitro transcrip-tion.
Bondopadhyay, S., Spector, D.J.,Dept. of Microbiology, Pennsylva-nia State University College ofMedicine, Hershey: Effect of tem-plate conformation on transcrip-tion of adenoviral promoters in vi-tro.
Samuels, M., Carthew, R., Sharp,RA., Center for Cancer Research,Massachusetts Institute of Technol-ogy, Cambridge: Purification ofspecific RNA polymerase II tran-scription factors.
Freyer, G.A., O'Neill, K.E., Tohill, K.A.,Roberts, R.J., Cold Spring HarborLaboratory, New York: In vitro andin vivo studies of Ad2 major latemRNA splicing.
Lee, R., Concino, M., Weinmann, R.,Wistar Institute, Philadelphia,Pennsylvania: The effect on in vi-tro transcription of single basechanges in the adenoviral majorlate promoter.
Mirza, A., Institut fOr Molekularbiolo-gie, Universitatsklinikum Essen,Federal Republic of Germany:Protein kinase(s) as positive and
negative modulators of adenoviraltranscription.
Mok, M., Maderious, A., Chen-Kiang,S., Memorial Sloan-Kettering Can-cer Center, Sloan-Kettering Divi-sion of Cornell University Gradu-ate School of Medical Sciences,New York, New York: Prematuretermination of transcription occurstemporally and is inducible in ad-enoviral infection.
Fraser, N.W., Owen, J.L., Dressler,G.R., Wistar Institute, Philadelphia,Pennsylvania: Transcriptional ter-mination in Ad2.
Bertuch, A., Hansen, U., Dana-FarberCancer Institute, Boston, Massa-chusetts: Nitrocellulose filter-bind-ing assay for the cellular protein(s)specifically binding the 21-bp re-peat region of the SV40 pro-moters.
Nobel, J., Prives, C., Manley, J.L.,Dept. of Biological Sciences, Co-lumbia University, New York, NewYork: In vitro splicing and poly-adenylation of an SV40 early pre-mRNA.
Hay, N., King, D., Aloni, Y., Dept. ofGenetics, Weizmann Institute ofScience, Rehovot, Israel: Efficientand accurate in vitro excision ofSV40-associated small RNA (SAS-RNA).
Somasekhar, M.B., Mertz, J.E., Mc-Ardle Laboratory for Cancer Re-search, University of Wisconsin,Madison: Factors involved in splic-ing of the SV40 late transcripts.
Laub, 0,, Hay, N., Bengal, E., Aloni,Y., Dept. of Genetics, WeizmannInstitute of Science, Rehovot, Is-rael: Attenuation and transcription-termination in the control of SV40gene expression.
Grass, D.S., Manley, J.L., Dept. of Bi-ological Sciences, Columbia Uni-versity, New York, New York: Inser-tion of the Ad2 late promoter intothe SV40 late leader region and itseffect on late mRNA synthesis.
Lamarche, M.F., Bourgaux-Ramoisy,D., Bourgaux, P, Faculte de Me-decine, Universite de Sherbrooke,Canada: Activity of a late promoterin integrated polyoma DNA.
Bergeron, J.L., Acheson, N.H., Dept.of Microbiology and Immunology,McGill University, Montreal, Can-ada: Viable insertion mutants of

polyoma virus containing a sec-ond copy of the signal for poly-adenylation of RNA.
Skarnes, W.C., Tessier, D.C., Ache-son, N.H., Dept. of Microbiologyand Immunology, McGill Univer-sity, Montreal, Canada: Pausing orpremature termination of RNA po-lymerase II nearby the early andlate promoters of polyoma virusRNA.
Cunningham, T.P., Pipas, J.M., Dept.of Biological Sciences, Universityof Pittsburgh, Pennsylvania: SA12is a BK-like papovavirus whichreplicates in monkey cells.
Delmas, V.,' Bastien, C.,1Scherneck,5.,2 Feunteun, J.,"IRSC, Villejuif,France; 'Academy of Sciences ofthe German Democratic Republic,Berlin: Sequence and genetic or-ganization of HapV, a papovaviruscausing lesions in Syrian ham-sters.
Davies, B.,' Arrand, J.,2 Griffiths, M.,1Fried, M.,' 'Dept. of Tumor VirusGenetics, Imperial Cancer Re-search Fund, London; 2PatersonLaboratories, Christie Hospital,Manchester, England: Sequenceof enhancer and origin regions ofpolyoma virus defectives.
von Hoyningen-Huene, V.,' Ruley, E.,2Davies, B.,' Griffiths, M.,' Fried,M.,1 'Dept. of Tumour Virus Ge-netics, Imperial Cancer ResearchFund, London, England; 2ColdSpring Harbor Laboratory, NewYork: Enhancer activity of duplica-ted sequences in the polyoma vi-rus noncoding region.
Herbomel, F, Bourachot, B., Yaniv,M., Dept. of Molecular Biology,Pasteur Institute, Paris, France:Homologous and heterologoustransactivation and transrepres-sion of early and late transcription.
SESSION 4 POSTER SESSION II
Glenn, G.M., Ricciardi, R.R, Wistar In-stitute, Philadelphia, Pennsylvania:Location of the functional nucleo-tide defects in Ad5 El A host-range mutants.
Smith, D., Velcich, A., Kegler, D., Ziff,E., Kaplan Cancer Center, Dept. ofBiochemistry, New York UniversityMedical Center, New York: Com-plementation of the Ad5 El A mu-tant d/312 by vectors that express
Passananti, C., Maione, R.,. Pozzi, L.,De Simone, V., Amati, F, Dept. diBiopatologia Umana, UniversityLa Sapienza, Roma, Italy: Poly-oma mutants selected in differen-tiated cell lines.
Boehnlein, E., Chowdhury, K., Gruss,P., Institute for Microbiology,ZMBH, Heidelberg, Federal Re-public of Germany: Structural andfunctional analysis of a polyomaenhancer/promoter mutation re-quired for growth in embryonalcarcinoma cells.
Templeton, D.,' Eckhart, W.,' Blangy,D.,' 'Salk Institute, San Diego,California; 2IRSC, Villejuif, France:Mouse sequences can functionallysubstitute polyoma enhancer se-quences.
Dandolo, L., Aghion, J., Blangy, D.,IRSC, Villejuif, France: Regulationof polyoma virus late expression inembryonal carcinoma cells.
Zenke, M., GrundstrOm, T., Wintzerith,M., Matthes, H., Chambon, F, IN-SERM, Faculte de M6decine,Strasbourg, France: Point muta-tion scanning of the SV40 enhan-cer.
Hen, R,. Borelli, E., Chambon, P., IN-SERM, Faculte de Medecine,Strasbourg, France: Blocking ofSV40 and polyoma enhancer ac-tivity by the Ad2 E1A products.
Axelrod, N., Farabaugh, P., Dept. ofMicrobiology, University of Con-necticut Health Center, Farming-ton: Can SV40 enhancer se-quences substitute for yeastenhancerlike sequences (up-stream activation sites)?
Sergeant, A.,' Bohmann, D.,' Zent-graf, H.,2 Keller, W.,' 'Institute ofCell and Tumor Biology, 2lnstituteof Virology, German Cancer Re-search Center, Heidelberg, Fed-
linker insertion mutants of the Ad5E1A gene.
Krippl, B.,' Ferguson, B.,2 Westphal,H.,' Rosenberg, M.,2 Jones, N.,3'NICHD, NCI, National Institutes ofHealth, Bethesda, Maryland;2Dept. of Molecular Genetics,Smith Kline and French Laborato-ries, Philadelphia, Pennsylvania;3Purdue University, West Lafay-ette, Indiana: Functionalcharac-
eral Republic of Germany: The.ef-fects of chromatin assembly andof the SV40 enhancer on tran-scription of homologous and het-erologous genes in vitro.
Kenny, S.,' Natarajan, V.,1 Strike, D.,'Khoury, G.,2 Salzman, N.R,''Laboratory of Biology of Viruses,'Laboratory of Molecular Virology,National Institutes of Health, Be-thesda, Maryland: JC virus enhan-cer active in human brain cells.
Dabrowski, C.E., Blankenhorn, E.R,Alwine, J.C., Dept. of Microbiol-ogy, University of PennsylvaniaMedical School, Philadelphia:Analysis of the efficiency of trans-lational initiation in SV40-contain-ing transient expression vectors.
Litzkas, P., Jha, K.K., Ozer, H.L.,Dept. of Biological Sciences,Hunter College, City University ofNew York, New York: Efficienttransfer of clones SV40 and non-viral DNA into human diploid cells.
Dalie, B., Harter, M.L., Dept. of Micro-biology, University of Medicineand Dentistry of New Jersey, New-ark: Expression of Ad2 E1A andEl B gene products from an SV40vector.
Lebkowski, J., DuBridge, R., Greisen,K., Antell, E., Cabs, M., Dept. ofGenetics, Stanford UniversitySchool of Medicine, California:Studying mutation with viral vec-tors.
Reddy, V.B., Garramone, A.J., Beck,A.K., Vellussi, V., Bernstine, E.G.,Integrated Genetics, Framingham,Massachusetts: Expression of di-meric, biologically active humanchoriogonadotropin (hCG) in CV-1cells, using a single SV40 vector.
terization of the EtA proteins ofhuman adenovirus.
Schmitt, R.C., Fahnestock, M.L.,Lewis, J.B., Fred Hutchinson Can-cer Research Center, Seattle,Washington: Ad2 E1A proteins lo-calized using antipeptide antisera.
Tsukamoto, A., Berk, A.J., MolecularBiology Institute, University of Cali-fornia, Los Angeles: A colony blotmethod for the selection of mono-
223

clonal antibodies to adenoviralEl A proteins and their use in puri-fication.
Harlow, E., Schley, C., Cold SpringHarbor Laboratory, New York:Monoclonal antibodies to the ad-enoviral El A proteins.
Yee, S.-P., Tremblay, M.L., Mc Glade,J., Branton, P.E., Dept. of Pathol-ogy, McMaster University, Hamil-ton, Canada: Analysis of Ad5 Elproteins, using antipeptide sera.
Subramanian, T.,' Kuppuswamy, M.,1Gysbers, J.,2 Mak, 5.,2Chinnadu-rai, G.,' 'Institute for Molecular Vi-rology, St. Louis University Medi-cal Center, Missouri; 2Dept. ofBiology, McMaster University,Hamilton, Canada: 175R T antigencoded by adenoviral El B plays adual role in viral DNA replication.
D'Halluin, J.C., Milleville, M., Labora-toire de Virologie Moleculaire, IN-SERM, Lille, France: An Ad2 El Bgene product is involved in thecontrol of cell endonuclease activ-ity.
Barker, D., Berk, A.J., Molecular Biol-ogy Institute, University of Califor-nia, Los Angeles: The role of Ad2El B gene products during infec-tion and transformation.
Virtanen, A., Pettersson, U., Dept. ofMedical Genetics, University ofUppsala, Sweden: Novel mRNAsfrom the transforming region El Bof human Ad2.
Deutscher, S.L.,' Kapoor, Q.S.,2 Cla-daras, C.,2 Wold, W.S.M.,2'Edward A. Doisy Dept. of Bio-chemistry; 2lnstitute for MolecularVirology, St. Louis University Medi-cal School, Missouri: The 19K gly-coprotein coded by region E3 ofadenovirus-Purification, charac-terization, and structural analysis.
Chatterjee, D., Maize', J.V., Jr.,NICHD, NCI, National Institutes ofHealth, Bethesda, Maryland: Theadenoviral E3 glycoprotein is ho-mologous to HLA-DR heavy chain.
Cutt, J.R., Halbert, D.N., Shenk, T.E.,Dept. of Microbiology, State Uni-versity of New York, Stony Brook:Analysis of adenoviral E4 mutants.
Lewis, J.B.,' Fahnestock, M.L.,'Hardy, M.M.,2 Anderson, C.W.,2'Fred Hutchinson Cancer Re-search Center, Seattle, Washing-ton; 2Dept. of Biology, BrookhavenNational Laboratory, Upton, NewYork: Nonvirion protein products
224
resulting from expression of theAd2 major late promoter at earlytimes after the infection of He Lacells.
Silverman, L.,' Johnston, J.M.,' An-derson, K.P.,1 Deppert, W.,2 Kles-sig, D.F.,"Dept. of Cellular, Viraland Molecular Biology, Universityof Utah, Salt Lake City; 2Dept. ofBiochemistry, University of Ulm,Federal Republic of Germany:Characterization of the block toreplication of human adenovirus inabortively infected monkey cells.
Bhat, R.A., Domer, P., Thimappaya,B., Cancer Center, NorthwesternUniversity Medical School, Chi-cago, Illinois: Structural require-ments of adenoviral VA, RNA forits translation enhancement func-tion.
Rohan, R.M., Ketner, G., Dept. of Bi-ology, Johns Hopkins University,Baltimore, Maryland: Point muta-tions in the adenoviral VA, pro-moter region.
Horvath, J., Weber, J., Dept. de Mi-crobiologie, Faculte de Medecine,University de Sherbrooke, Can-ada: Ad2 complementary DNA se-quences in human B and T lym-phocytes.
Signas, C.,' Akusjarvi, G.,' Petters-son, U.,2 Depts. of 'Microbiology,2Medical Genetics, University ofUppsala, Sweden: The gene forAd3 fiber.
van Loon, A.E., Rozijn, T.H., Sussen-bach, J.S., Laboratory for Physio-logical Chemistry, State Universityof Utrecht, The Netherlands: Gen-eral characterization of DNAs ofthe enteric Ad40 and Ad41.
Belak, 5.,1 Virtanen, A.,3 Berencsi,G.,2 Pettersson, U.,3 'Dept. of Epi-zootiology, University of VeterinaryScience, 2lnstitute of Microbiology,Semmelweis Medical University,Budapest, Hungary; 3Dept. ofMedical Genetics, University ofUppsala, Sweden: Sequence ho-mology between bovine adenovi-ruses with different speciesspecificity-Importance of regionE3.
Becerra, S.P.,' Anderson, C.W.,2Rose, J.A.,"NIAID, NCI, NationalInstitutes of Health, Bethesda,Maryland; 2Dept. of Biology,Brookhaven National Laboratory,Upton, New York: Direct mappingand mechanism of origin of the
adeno-associated viral structuralproteins.
Ko, J.-L., Coles, R.S., Harter, M.L.,Dept. of Microbiology, Universityof Medicine and Dentistry of NewJersey, Newark: Improved meth-ods for maximizing expression ofcloned Ad2 E1A and El B genesin E. coll.
Vasavada, R.,' Barbanti-Brodano,G.,2 Ricciardi, R.,' 'Wistar Insti-tute, Philadelphia, Pennsylvania;2lnstitute of Microbiology, Univer-sity of Ferrara, Italy: Constructionof human cell lines that expressthe Ad12 E1A proteins, using anovel BK-plasmid vector.
VanDoren, K., Solnick, D., Reich!, H.,Gluzman, Y., Cold Spring HarborLaboratory, New York: Helper in-dependent Ad5 vectors.
Green, P.,1 Hearing, P.,2 Fowlkes, D.,'Dept. of Pathology, University of
North Carolina, Chapel Hill; 2Dept.of Microbiology, State University ofNew York, Stony Brook: Expres-sion of a heterologous gene in ad-enovirus.
Haj-Ahmad, Y., Graham, FL., Depts.of Biology and Pathology, Mc-Master University, Hamilton, Can-ada: Packaging constraints in hu-man Ad5.
Hearing, F, Samulski, R.J., Shenk, T.,Dept. of Microbiology, State Uni-versity of New York, Stony Brook:Identification of the sequences re-quired for efficient packaging ofAd5 DNA into virions.
Franza, R.,' Garrels, J.," Furth, M.,2Ruley, E.,"Cold Spring HarborLaboratory, New York; 2MemorialSloan-Kettering Cancer Center,New York, New York: Cooperatingoncogenes are required for trans-formation of established REF52cells.
Senear, A.W., Lewis, J.B., FredHutchinson Cancer ResearchCenter, Seattle, Washington: Effi-cient transformation of rodent celllines by the adenoviral E1A gene.
Kuppuswamy, M., Hurwitz, D.R.,Chinnadurai, G., Institute for Mo-lecular Virology, St. Louis Univer-sity Medical Center, Missouri: Asingle E1A T antigen (243R) canimmortalize and fully transform ratembryo fibroblasts in cooperationwith El B.
Moran, B., Whyte, P., Zerler, B., Ma-ruyama, K., Grodzicker, T., Ruley,

E., Cold Spring Harbor Labora-tory, New York: Analysis of thetransforming functions of adeno-viral E1A.
Haley, K.,' Jones, N.C.,' Babiss, L.,2Ginsberg, H.S.,2 'Dept. of Biologi-cal Sciences, Purdue University,West Lafayette, Indiana; 2Dept. ofMicrobiology, College of Physi-cians & Surgeons, Columbia Uni-versity, New York, New York: Therole of Ad5 El A gene products intransformation of rat cells.
Jochemsen, A.G., Bernards, R., Bos,J.L., van der Eb., A.J., Dept. ofMedical Biochemistry, State Uni-versity of Leiden, The Nether-lands: Oncogenic transformationby adenoviral E1A regions in thepresence of the c-Hras oncogene.
Eager, K.,' Williams, J.,3 Appel la, E.,3Ricciardi, R.,' "Wistar Institute,'Carnegie-Mellon University, Pitts-burgh, Pennsylvania; 3NCI, Na-tional Institutes of Health, Be-thesda, Maryland: Expression ofclass-I cell-surface antigens ofAdl 2-transformed mouse cells ishighly reduced.
Akagi, K.,' Cook, J.L.,3 Patch, C.T.,1Kato, T.,1 Lewis, A.M.,2 Levine,A.S.,' 1NICHD, 2NIAID, NCI, Na-tional Institutes of Health, Be-thesda, Maryland; 3National Jew-ish Hospital and Research Center,Denver, Colorado: Dose-depen-dent correlation of Ad2 geneexpression and sensitivity to lysis
by nonspecific effector cellsamong hybrid cells formed fromSV40- and Ad2-transformed cells.
Yoshida, K., Ren, C.S., Yamashita, T.,Fujinaga, K., Cancer Research In-stitute, Sapporo Medical College,Japan: The transforming gene ofcanine Adl.
Klimkait, T., Doerfler, W., Institute ofGenetics, University of Cologne,Federal Republic of Germany:Ad2 or Ad5 El functions elicit rep-lication and late expression ofAd12 DNA in hamster cells.
Wojcik, B., Dermody, J., LaBella, F.,Radna, R., Ozer, H.L., Dept. of Bi-ological Sciences, Hunter College,City University of New York, NewYork: Viral DNA synthesis in wild-type and mutant Chinese hamstercells.
Pierce, WC., Eggerding, F.A., Dept.of Pathology, University of Califor-nia, Los Angeles: Expression ofAd2 replicative functions in semi-permissively infected cells.
Korn, R.,' Friefeld, B.R.,1 deJong, P.,2Sninsky, J.J.,2.3 Horwitz, M.S.,12Depts. of 'Cell Biology, 2Micro-biology and Immunology, 3Molec-slur Biology, Albert Einstein Col-lege of Medicine, Bronx, NewYork: Expression of a domain ofthe adenoviral DNA polymerase ina prokaryotic vector.
Wang, K., Pearson, G.D., Dept. ofBiochemistry and Biophysics, Ore-gon State University, Corvallis: Ad-
SESSION 5 ADENOVIRUSES: REGULATION OF GENE EXPRESSION
Chairperson: N. Jones, Purdue University, West Lafayette, Indiana
Jones, N.C.,' Andrisani, O.,' Richter,J.,2 Ferguson, B.,3 Rosenberg,M.,3 'Dept. of Biological Sciences,Purdue University, West Lafayette,Indiana; 2Worcester Foundationfor Experimental Biology, Shrews-bury, Massachusetts; 3Dept. ofMolecular Genetics, Smith Klineand French Laboratories, Philadel-phia, Pennsylvania: Functionaland biochemical characterizationof purified Ad5 and Ad12 E1Aproteins.
Spindler, KR., Eng, C.Y., Berk, A.J.,Dept. of Microbiology and Molecu-lar Biology Institute, University ofCalifornia, Los Angeles: Maximal
adenoviral DNA replication ingrowth-arrested cells requires theproduct of the El A 12S mRNA.
Moran, E., Zerler, B., Ruley, E., Ma-thews, M.B., Grodzicker, T., ColdSpring Harbor Laboratory, NewYork: Analysis of wild-type andmutant Ad2 E1A cDNAs.
Pilder, S., Logan, J., Shenk, T., HealthSciences Center, State Universityof New York, Stony Brook: Func-tional analysis of the Ad5 El Bgene products.
White, E., Grodzicker, T., Stillman,BAN., Cold Spring Harbor Labora-tory, New York: Mutations in thenuclear envelope-associated ad-
enoviral sequences required forreplication in vivo.
Munz, P.L.,' Volkert, F.C.,2 Young,C.S.H.,' 'Dept. of Microbiology,Columbia University, New York,New York; 2Dept. of Molecular Bi-ology, Princeton University, NewJersey: The characterization of ad-enoviral overlap recombination.
Kinloch, R., Hay, R., Mautner, V.,Medical Research Council Virol-ogy Unit, Glasgow, Scotland: Re-combination between Ad2 andAd5 hexon-coding regions.
Dery, C.V.,' de Murcia, G.,3 Lamarre,D.,2 Poirier, G.,2 Weber, J.,' Depts.of 'Microbiologie, 2Biologie, Univ-ersite de Sherbrooke, Canada;3lnstitut de Biologie Moleculaire etCellulaire de CNRS, Strasbourg,France: PolyADP-ribosylation ofadenoviral core proteins.
Wang, C.-N.J., Chen-Kiang, S., Mem-orial Sloan-Kettering Cancer Cen-ter, Sloan-Kettering Division ofCornell University GraduateSchool of Medical Sciences, NewYork, New York: Characterizationof transcriptionally active Ad2chromatin during lytic infection.
Silver, L.S., Hafemann, B., Anderson,C.W., Dept. of Biology, Brookha-ven National Laboratory, Dept. ofMicrobiology, State University ofNew York, Stony Brook: Acquisi-tion of Ad2 receptors by normal,peripheral human lymphocytes.
enoviral El B 19K tumor antigencause the degradation of chromo-somal DNA.
Weinberg, D.H., Ketner, G., Dept. ofBiology, Johns Hopkins University,Baltimore, Maryland: Character-ization of an adenoviral E4 mutantdefective in viral late gene expres-sion.
Svensson, C., Larsson, S., Akusjarvi,G., Dept. of Medical Genetics,University of Uppsala, Sweden:Adenoviral VA, RNA-A generalstimulator of mRNA translation.
Schneider, R.J., Weinberger, C.,Shenk, T., Dept. of Microbiology,State University of New York,
225

Stony Brook: Translational regula-tion by adenoviral VA, RNA-Anal-ysis of a human a-globin gene inVA,-"- backgrounds. Analysis of aVA,-'VA- double mutant.
Mathews, M.B.,1 O'Malley, R.,' Rei-chel, P.A.,' Merrick, W.C.,2 Sier-kierka, J.,3 'Cold Spring Harbor
Laboratory, New York; 2CaseWestern Reserve University,Cleveland, Ohio; 3Roche Instituteof Molecular Biology, Nutley, NewJersey: Adenoviral VA, RNA main-tains the activity of protein synthe-sis initiation factor elF -2.
Rice, S., Klessig, D., Dept. of Cellular,
SESSION 6 SV40, POLYOMA: REGULATION OF GENE EXPRESSION
Chairperson: G. Khoury, National Institutes of Health, Bethesda, Maryland
Lewis, D., Manley, J., Dept. of Biologi- nia: Can SV40 replication antago-cal Sciences, Columbia University, nize transcription?New York, New York: Replication-defective mutants restore expres-sion of the SV40 early region inhuman 293 cells.
Robbins, P., Botchan, M., Dept. ofMolecular Biology, University ofCalifornia, Berkeley: T-antigen canstimulate the effects of the SV40enhancer on the HSV TK pro-moter.
Velcich, A., Ziff, E., Kaplan CancerCenter, New York University Medi-cal Center, New York: An Ad2 E1Aexpression vector reduces tran-scription from the early SV40 pro-moter.
Lebkowski, J., Greisen, K., Cabs, M.,Dept. of Genetics, Stanford Uni-versity School of Medicine, Califor-
SESSION 7 POSTER SESSION III
Asselin, C., Bastin, M., Dept. of Micro-biology, University of Sherbrooke,Canada: Immortalization of sec-ondary rat embryo cells by trun-cated forms of the large T genesof SV40 and polyoma virus.
de Ronde, A., Sol, C., MacDonald,M., ter Schegget, J., van Strien,A., van der Noordaa, J., Dept. ofVirology, Academical MedicalCentre, Amsterdam, The Nether-lands: Host range for transforma-tion of BKV and SV40.
Kelly, F.,' Agha, M.E.,2 Livingstone,D.,21Institut Pasteur, Paris,France; 2Dana-Farber Cancer In-stitute, Boston, Massachusetts: Im-mortalization of mouse fibroblastswith small T antigen.
Chiang, L.-C., Silnutzer, J., Spence,S.L., Greenwood, D., Pipas, J.M.,Barnes, D.W., Dept. of BiologicalSciences, University of Pittsburgh,Pennsylvania: Selection of SV40-
226
Hertz, G.Z., Mertz, J.E., McArdle Lab-oratory for Cancer Research, Uni-versity of Wisconsin, Madison:SV40 sequences required for tran-scription and viral DNA replica-tion.
Herr, W., Gluzman, Y., Cold SpringHarbor Laboratory, New York: Du-plications of a mutated SV40 en-hancer restore its activity.
Piette, J., Yaniv, M., Dept. of Molecu-lar Biology, Pasteur Institute, Paris,France: In vivo and in vitro studiesof the interactions between pro-teins of 3T6 cells and the polyomaenhancer.
Wildeman, A.G., Sassone-Corsi, P,Chambon, P, Laboratoire deGenetique Mol4culaire des Eu-
and polyoma virus-transformedcells in a serum-free media.
Hutchinson, N., Chang, L.S., Pater,M.M., di Mayorca, G., Dept. of Mi-crobiology, University of Medicineand Dentistry of New Jersey, New-ark: Discovery of a is mutant lo-cated at 0.455 map units in theSV40 large T antigen that trans-forms F111 rat cells to anchorageindependence at the nonpermis-sive temperature for lytic growth,
Gurney, T.,1;2Gurney, E.,12Blanck,G.,3 'Oncologie Moleculaire, IRSCdu CNRS, Villejuif, France; 2Dept.of Biology, University of Utah, SaltLake City; 3Dept. of Biology, Co-lumbia University, New York, NewYork: Particular rearrangementswithin SV40 DNA accompany re-version and retransformation inclones of SVT2 cells.
O'Neill, F.J., Miller, T.H., Renzetti, L.,Veterans Administration Medical
Viral and Molecular Biology, Uni-versity of Utah, Salt Lake City:Construction and analysis of aden-oviral E2A deletion mutants.
caryotes du CNRS, Faculte de Me-decine, Strasbourg, France: TheSV40 enhancer stimulates tran-scription in vitro from both homol-ogous and heterologous pro-moters by interacting with specificfactor(s).
Schb ler, H.R.,' Sergeant, A.,2 Keller,W.,2 Gruss, P.,"Institute for Micro-biology, ZMBH, Im Neuenheimer;2Division of Molecular Biology,German Cancer Research Center,Heidelberg, Federal Republic ofGermany: Cell-specific transcrip-tional enhancement in vitro.
Me lin, F, Pinon, H., Kress, C., Blangy,D., IRSC, Villejiuf, France: Poly-oma mutants with extended host-range and high lytic potential onEC cells.
Center, University of Utah, SaltLake City: Inhibition of transforma-tion of permissive cells followinginfection with SV40 stocks contain-ing defectives with reiteratedorigins and termini.
Vesco, C.,' Fischer Fantuzzi, L.,2'Instituto di Biologia Cellulare,2Centro Studi Acidi Nucleici, CNR,Roma, Italy: A 43-amino-acid dele-tion in the amino-terminal half ofthe SV40 large T antigen results ina non-karyophilic protein capableof transforming established cells.
Kennett, S., Samad, A., Blum, A., Car-roll, R.B., Dept. of Pathology, NewYork University Medical Center,New York: Characterization of astable aminoterminal fragment ofSV40 large T antigen.
Montenarh, M., Kohler, M., StOrz-becher, H.-W., SchOrmann, C.,Muller, D., Henning, R., Dept. ofBiochemistry, University of Ulm,

Federal Republic of Germany: Dis-section of structural modificationsof SV40 T antigen with regard toviral replication and transforma-tion.
Hand, R., Whittaker, L., Ismail, A.,McGill Cancer Centre, Montreal,Canada: Membrane and nuclearforms of SV40 large T antigen dif-fer in immunological and biochem-ical characteristics.
Jarvis, D.L., Butel, J.S., Dept. of Virol-ogy and Epidemiology, Baylor Col-lege of Medicine, Houston, Texas:Modification of SV40 T antigen byglycosylation.
Tack, L.C.,' Tack, B.F.,2 'Molecular Bi-ology and Virology Laboratory,Salk Institute, 2Dept. of Immunol-ogy, Scripps Clinic and ResearchFoundation, La Jolla, California:Identification of an active esterbond in SV40 T antigen,
Montano, X., Lane, D.P., Dept. of Bio-chemistry, Imperial College, Lon-don, England: Cellular localizationand immunochemistry of SV40small T, using the small-T-specificmonoclonal antibody pAp280.
Levine, S., O'Driscoll, K., Wolowodiuk,V., Maltzman, W., Waksman Insti-tute of Microbiology, Rutgers Uni-versity, New Brunswick, New Jer-sey: Cellular DNA synthesis andgene expression are necessary forthe interaction of SV40 T antigenwith the p53 cellular tumor anti-gen.
Winberry, L., Friderici, K., Fluck, M.,Dept. of Microbiology, MichiganState University, East Lansing:Transformation by polyoma tsAmutants-Characterization of thetemperature-sensitive phenotype.
Clark, K., Folk, W., Dept. of BiologicalChemistry, University of Michigan,Ann Arbor: Transformation of ratcells by a polyoma-gastrin recom-binant.
Mark land, W.,' Cheng, S.H.,' Oostra,B.A.,' Markham, A.,2 Smith, A.E.,''National Institute for Medical Re-search, London; 2ICI Pharmaceuti-cals Division, Macclesfield, Eng-land: In vitro mutagenesis ofpolyoma middle T transformation-associated domains.
Bolen, J.B., Yonemoto, W., Brugge,J.S., Israel, M.A., NCI, National In-stitutes of Health, Bethesda, Mary-land: Increased specific activity ofpp60c-s'° phosphotransferase in
rodent cells following polyoma vi-rus infection and transformation.
Dalianis, T., Ramqvist, T., Klein, G.,Dept. of Tumor Biology, KarolinskaInstitutet, Stockholm, Sweden:Studies on the polyoma virus-induced tumor-specific transplan-tation antigen (TSTA)- Does mid-dle or large T antigen play a role?
Street, A.J., Griffin, B.E., ImperialCancer Research Fund, London,England: Purification and charac-terization of polyoma virus middleT antigen.
Pater, M.M., di Mayorca, G., Pater, A.,Dept. of Microbiology, Universityof Medicine and Dentistry of NewJersey Medical School, Newark:Transformation of primary humanembryonic kidney cells with BKV,a human papovavirus.
Major, E., Miller, A., Mourrain, P.,Traub, R., NINCDS, NCI, NationalInstitutes of Health, Bethesda,Maryland: JCV multiplication in aline of human fetal glial cells, SVG,established with an origin-defec-tive mutant of SV40.
Shaw, P., Keller, W., Institute of Celland Tumor Biology, German Can-cer Research Center, Heidelberg,Federal Republic of Germany:Identification of cell-viral hybridmRNA and proteins expressed bythe SV40-transformed rat cell line14B.
Conrad, S.E.,1 Ito, M.,' Stewart, C.,'Stuart, P.,"Dept. of Microbiology,Michigan State University, EastLansing; 2Dept. of Molecular Biol-ogy, University of California,Berkeley: SV40-induced expres-sion of cellular thymidine kinase.
Sauer, G., Amtmann, E., Dann, M.,Hoss, A., Institute for Virus Re-search, German Cancer ResearchCenter, Heidelberg, Federal Re-public of Germany: Reversion ofthe transformed phenotype in thepresence of the SV40 T antigen byan antiviral xanthate compound.
Matlashewski, G.,' Lamb, P.,1 Pim,D.,' Benchimol, 5.,2 Crawford, L.,''Molecular Virology Laboratory,Imperial Cancer Research Fund,London, England; 2Division of Bio-logical Research, Ontario CancerInstitute, Toronto, Canada: Analy-sis of p53 genomic and cDNAclones from human cells.
Schmieg, F., Simmons, D., School of
Life and Health Sciences, Univer-sity of Delaware, Newark: Intracel-lular location and kinetics of com-plex formation between SV40 Tantigen and p53.
Van Roy, F., Liebaut, G., Fiers, W.,Laboratory of Molecular Biology,State University of Ghent, Bel-gium: Complexes between virallarge T antigen and cellular p53are dispensable for most SV40-in-duced transformation characteris-tics.
Rundell, K., Bossert, A., Dept. of Mi-crobiology and Immunology,Northwestern University, Chicago,Illinois: Recognition of cellular pro-teins by small T antigen producedin bacteria.
de Villiers, J.,' Schaffner, W.,' Tyndall,C.,2 Lupton, 5.,2 Kamen, B.,2'Institut kir Molekularbiologie II,Zurich, Switzerland; 2lmperial Can-cer Research Fund Laboratories,Lincoln's Inn Fields, England: En-hancer function is essential forpolyoma virus DNA replication.
Osterlund, M., Risuleo, G., Magnus-son, G., Dept. of Virology, UppsalaUniversity Biomedical Center,Sweden: Effect of polyoma virusearly gene expression and DNAsynthesis of mutations in the en-hancer region and base se-quences 3' to the poly(A) site.
Garcea, R.L.,' Raptis, L.,2 Liang, J.,2Benjamin, T,2 'Dana-Farber Can-cer Institute, 2Dept. of Pathology,Harvard Medical School, Boston,Massachusetts: Separating thefunctions of polyoma's transform-ing genes.
Cowie, A., Kamen, R., Genetics Insti-tute, Boston, Massachusetts: Pro-tein-DNA interactions betweenpolyoma virus large T antigen andthe viral DNA.
Triezenberg, S.J., Folk, W.R., Dept. ofBiological Chemistry, University ofMichigan, Ann Arbor: Essentialnucleotides in the polyoma virusorigin region.
Manos, M.,2 Gluzman, Y.,"ColdSpring Harbor Laboratory, NewYork; 2Dept. of Molecular Ge-netics, Cetus Corporation, Emery-ville, California: Genetic and bio-chemical analysis of SV40 large Tantigen mutants.
Scheller, A., Covey, L., Prives, C.,Dept. of Biological Sciences, Co-lumbia University, New York, New
227

York: Binding of polyoma andSV40 large T antigens to viral andcellular DNA.
Hinzpeter, M., Deppert, W., Dept. ofBiochemistry, Ulm, Federal Re-public of Germany: Nuclear sub-classes of SV40 large T exhibit dif-ferent SV40 ORI DNA-bindingproperties.
Tack, L.C., Molecular Biology and Vi-rology Laboratory, Salk Institute,La Jolla, California: Loss of T anti-gen from replicating SV40 chro-mosomes is associated with asyn-chronous progression ofreplication forks.
Fields-Berry, S.,' Weaver, D.,2 De-Pamphilis, M.,"Dept. of Biologi-cal Chemistry, Harvard MedicalSchool, 'Dept. of Biology, Massa-chusetts Institute of Technology,Boston: Physiological conditionsand DNA sequence determine thepathway for segregation of SV40DNA replicative intermediates.
Hsu, M.-T Dept. of Molecular Cell Bi-ology, Rockefeller University, NewYork, New York: Study of SV40DNA repair and replication by two-dimensional gel electrophoresisand psorlane cross-link.
Paxton, W.B., Subramanian, K.N.,Dept. of Microbiology and Immu-nology, University of Illinois HealthSciences Center, Chicago: Chro-matin structure of the early SV40origin region.
Innis, J.W., Scott, W.A., Dept. of Bio-chemistry, University of MiamiSchool of Medicine, Florida: DNAreplication and chromatin struc-ture of SV40 insertion mutants.
Milavetz, B., Cyze, C., Payne, C.,Hopkins-Davis, T., Depts. of Bio-chemistry, Microbiology and Im-munology, University of WesternOntario, London, Canada:Changes in chromatin associatedwith the early stages in SV40 en-capsidation.
Ambrose, C., Blasquez, V., Bina, M.,Dept. of Chemistry, Purdue Uni-versity, West Lafayette, Indiana:The virion assembly process playsan active role in the organizationof SV40 chromatin.
Ng, S.-C.,' Mertz, I.E.,' Bina, M.,''Dept. of Chemistry, Purdue Uni-versity, West Lafayette, Indiana;2McArdle Laboratory for CancerResearch, University of Wisconsin,Madison: SV40 assembly in cells
228
infected with mutants deleted inthe agnogene.
Lutter, L.C., Quasney, M., Cellularand Molecular Biology Program,Dept. of Biological Chemistry, Uni-versity of Michigan, Ann Arbor:lmmunoprecipitation of SV40 repli-cation and transcription complex,using monoclonal antibodiesagainst large T antigen.
Gerard, R.D., Harlow, E., Cold SpringHarbor Laboratory, New York:Monoclonal antibodies to SV40nucleoprotein complexes.
Mole, S.E., Lane, D.P., Dept. of Bio-chemistry, Imperial College of Sci-ence and Technology, London,England: Fusion proteins betweenp- galactosidase and SV40 large Tin an immunochemical investiga-tion of T antigen.
Simanis, V., Gannon, J., Lane, D.P.,CRC Eukaryotic Molecular Ge-netics Group, Dept. of Biochemis-try, Imperial College of Scienceand Technology, London, Eng-land: An immunoaffinity purifica-tion procedure for SV40 large Tantigen.
Milavetz, B., Hopkins-Davis, T,Payne, C., Cancer Research Lab-oratory, Depts. of Biochemistry,Microbiology and Immunology,University of Western Ontario,London, Canada: Purification ofSV40 T antigen.
Gerard, R.D., Gluzman, Y., ColdSpring Harbor Laboratory, NewYork: Inducible expression of SV40T antigen.
Kruczek, I., KOhnel, H., Fanning, E.,Institute of Biochemistry, Munich,Federal Republic of Germany:Small rodent RNAs and highermolecular weight RNAs of SV40-infected and SV40-transformedmonkey cells have sequences ho-mologous to the integration site ofAd12 DNA in a transformed ham-ster cell.
Hand, R.,' Zeng, G.C.,' Ozer, H.,2'McGill Cancer Centre, Montreal,Canada; 'Hunter College, CityUniversity of New York, New York:Inhibition of an early step in DNAchain elongation of polyoma DNAin ts20, a DNA's mutant of BALB/3T3 cells, and correlation of thatdefect with a heat-labile topoiso-merase I activity from mutant cells.
Lambert, M.,' Pelligrini, S.,2 Gattoni-Celli, S.,' Weinstein, I.B.,"Institute
of Cancer Research, ColumbiaUniversity; 'Dept. of Pathology,New York University School ofMedicine, New York, New York: Acarcinogen induces asynchronousreplication of polyoma DNA in ratcells via a trans-acting factor.
Bullock, P., Miller, J., Botchan, M.,Dept. of Molecular Biology, Uni-versity of California, Berkeley:Analysis of the sequences usedduring SV40 excision.
Roth, D.B., Wilson, J.H., Verna andMarrs McLean Dept. of Biochem-istry, Baylor College of Medicine,Houston, Texas: Measurement ofthe relative rates of homologousand nonhomologous recombina-tion in transfected SV40 DNA.
Nepveu, A., Wallenburg, J., Gusew,N., Chartrand, P., Dept. de Micro-biologie, Faculte de Medecine,CHUS, Sherbrooke, Canada: Evi-dence that a polyoma virus ge-nome containing an insertion ofhighly repetitive cellular DNA inte-grates via homologous recombina-tion.
Manor, H., Yarom, R., Neer, A., Baran,N., Dept. of Biology, Technion- Is-rael Institute of Technology, Haifa:Recombination between inte-grated viral DNA and flanking cel-lular sequences occurs within "on-ion skin" replication intermediatesin polyoma-transformed cells.
Friderici, K., Priehs, C., Fluck, M.M.,Dept. of Microbiology and PublicHealth, Michigan State University,East Lansing: Superinfection res-cue of integrated polyoma se-quences from Py-6 cells.
Oh, S.Y., Fluck, M.M., Dept. of Micro-biology and Public Health, Michi-gan State University, East Lansing:Differences in integration patternsof polyoma virus genomes be-tween Fischer rat cells and babyhamster kidney cells.
Priehs, C., Fluck, M.M., Dept. of Mi-crobiology, Michigan State Univer-sity, East Lansing: High frequencyof recombination between viral ge-nomes in the integration pathwayof polyoma virus.

SESSION 8 WORKSHOP: VIRAL VECTORS
Chairperson: John Hassell, McGill University, Montreal, Canada
Berkner, Sharp, PA.,' Rob-erts, T.M.,' Schaffhausen, B.,4'Center for Cancer Research,Massachusetts Institute of Technol-ogy, Cambridge; 2ZymoGenetics,Inc., Seattle, Washington; 'Dana-Farber Cancer Institute, 4TuftsNew England Medical Center,Boston, Massachusetts: Abundantexpression of active polyoma mid-dle Tin 293 cells infected with anAd5 (pym T) recombinant.
Massie, B.,' Gluzman, Y.,2 Hassell,J.A.,"Dept. of Microbiology andImmunology, McGill University,Montreal, Canada; 2Cold SpringHarbor Laboratory, New York:Construction of a helper-free ad-enovirus-polyomavirus hybrid thatexpresses polyomavirus large Tantigen from the Ad2 major latepromoter.
Logan, J., Shenk, T., Dept. of Microbi-ology, State University of New
York, Stony Brook: The adenoviraltripartite leader sequence en-hances translation of viral mRNAslate after infection.
Mansour, S.,' Tjian, R.,' Grodzicker,T.,2 'Dept. of Biochemistry, Univer-sity of California, Berkeley; 'ColdSpring Harbor Laboratory, NewYork: Construction of defective ad-enoviral recombinants that ex-press polyoma T antigens.
Yamada, M., Grodzicker, T., ColdSpring Harbor Laboratory, NewYork: An adenoviral vector that ex-presses the protein product of anonselected foreign gene at ahigh level.
Jat, F, Cepko, C., Sharp, P.A., Centerfor Cancer Research, Dept. of Bi-ology, Whitehead Institute forBiomedical Research, Massachu-setts Institute of Technology, Cam-bridge: Construction of recombi-nant retroviruses encoding
SESSION 9 SV40, POLYOMA: REPLICATION
Chairperson: C. Prives, Columbia University, New York, New York
Veldman, G.,' Lupton, 5.,2 Kamen,R.,"Genetics Institute, Boston,Massachusetts; 2Dept. of Microbi-ology, State University of NewYork, Stony Brook: The polyomaenhancer contains multiple, redun-dant sequence elements that acti-vate both viral DNA replicationand gene expression.
Muller, W.J., Hassell, J.A., Dept. ofMicrobiology and Immunology,McGill University, Montreal, Can-ada: Structural requirements forthe function of the polyoma virusorigin for DNA replication.
Lee, G.J., Woodworth-Gutai, M.,Dept. of Cell and Tumor Biology,Roswell Park Memorial Institute,Buffalo, New York: Eukaryotic reg-ulation elements that enhanceDNA replication.
Magnusson, G., Nilsson, S., Sand-berg, G., Dept. of Virology, Upps-ala University Biomedical Center,Sweden: Activities of polyoma vi-rus T antigens in lytic infection.
Vakalopoulou, E., Vogt, B., Schneider,J., Mertz, R., Sperka, S., Huber,B., Winnacker, E.-L., Fanning, E.,
Institute for Biochemistry, Munich,Federal Republic of Germany:Binding of SV40 T antigen to viraland synthetic DNA sequences.
Chalifour, L.E., Wirak, D.O., Wassar-man, P.M., DePamphilis, M.L.,Dept. of Biological Chemistry, Har-vard Medical School, Boston,Massachusetts: Replication andexpression of SV40 and polyomavirus DNA in mouse embryoniccells.
Bradley, M., Hudson, J., Livingston,D., Dana-Farber Cancer Institute,Harvard Medical School, Boston,Massachusetts: Specific in vitroadenylation of SV40 large T anti-gen.
Clertant, P.,' Guizani, I.,' May, E.,2Cuzin, F.,"INSERM, Centre deBiochimie, Universite de Nice;'Institut de Recherche Scientifiquesur le Cancer du CNRS, Villejuif,France: The nucleotide bindingactivity of polyoma and SV40 largeT proteins.
Levitt, A., Chen, S., Blanck, G.,George, D., Pollack, R., Dept. ofBiological Sciences, Columbia
transforming genes from DNA tu-mor viruses.
Cherington, V.,' Mahoney, M.,' Ka-plan, D.,' Corbley, M.,' Morgan,B.,' Schaffhausen, B.,2 Roberts,T,"Dana-Farber Cancer Institute,'Tufts New England Medical Cen-ter, Boston, Massachusetts: Retro-viral shuttle vector for polyomaearly region proteins and neomy-cin resistance.
Roberts, T.M.,' Kaplan, D.,' Pallas,D.,' Mahoney, M.,' Bockus, B.,2Bolen, J.,3 Schaffhausen, B.,2'Dana-Farber Cancer Institute,'Dept. of Biochemistry and Phar-macology, Tufts University, Bos-ton, Massachusetts; 'NCI, Na-tional Institutes of Health,Bethesda, Maryland: Isolation ofpolyoma T antigens from prokar-yotic and eukaryotic sources.
University, New York, New York:Two integrated, partial repeats ofSV40 together code for a super-Tantigen.
Cereghini, S., Yaniv, M., Dept. of Mo-lecular Biology, Pasteur Institute,Paris, France: Chromatin assem-bly of transfected DNA-Structuralchanges in the origin/promoter/en-hancer region of SV40 upon repli-cation.
Bryan, P.,' Folk, W.,2 'Dept. of Molec-ular Genetics, Genex Corporation,Gaithersburg, Maryland; 'Dept. ofBiological Chemistry, University ofMichigan, Ann Arbor: Sequencesrequired for the DNase I hyper-sensitive structure in polyoma.
229

SESSION 10 TRANSFORMATION. I
Chairperson: J. Butel, Baylor College of Medicine, Houston, Texas
Butel, J.S.,1 Lanford, R.E.,2 'Dept. ofVirology and Epidemiology, BaylorCollege of Medicine, Houston;'Dept. of Microbiology, SouthwestFoundation for Biomedical Re-search, San Antonio, Texas: Trans-forming potential of an SV40 mu-tant unable to transport T antigento the nucleus.
Richardson, W.D., Kalderon, D., Rob-erts, B., Paucha, E., Col ledge,W.H., Smith, A.E., National Insti-tute for Medical Research, Lon-don, England: Sequence require-ments for nuclear location of SV40large T
Krieg ler, M.,' Perez, C.F.,' Hardy, C.,'Feinberg, M.,2 Botchan, M.,''Dept. of Molecular Biology, Uni-versity of California, Berkeley;'Dept. of Radiology, Stanford Uni-versity Medical Center, California:Transformation mediated by theSV40 T antigens-Separation ofthe overlapping SV40 early geneswith a retroviral vector.
Ryan, K., Brockman, W., University ofMichigan Medical School, Ann Ar-bor: Isolation of a cell line resistantto transformation by SV40 T anti-gens.
SESSION 11 TRANSFORMATION. II
Sharma, S., Rodgers, L., Cold SpringHarbor Laboratory, New York: Celllines that express exclusively amembrane-bound form of SV40large T antigen.
Raptis, L., Benjamin, T.L., Dept. of Pa-thology, Harvard Medical School,Boston, Massachusetts: Regulatedexpression of middle T antigen ina line of rat fibroblasts.
Gelinas, C., Bouchard, L., Bastin, M.,Dept. of Microbiology, Universityof Sherbrooke, Canada: Stabiliza-tion of polyoma middle T antigenby a cellular factor is required fortransformation.
Dailey, L.,' Brugge, J.S.,2 Basilico,0.," Dept. of Pathology, New YorkUniversity Medical Center, NewYork; 'Dept. of Microbiology, StateUniversity of New York, StonyBrook: The amino terminus ofpolyoma middle T antigen is re-quired for formation of active c-xrc /middle T complexes and poly-oma-mediated transformation.
Schaffhausen, B.,' Parsons, S.,3 Rob-erts, T.,2 Bockus, B.," Dept. ofBiochemistry and Pharmacology,Tufts Medical School, 'Dana-Far-ber Cancer Institute, Boston, Mas-
Chairperson: M. Fried, Imperial Cancer Research Fund, London, England
Kimelman, D.,' Miller, J.,' Cepko, C.,'Mulligan, R.,2 Roberts, B.,"Dept.of Biological Chemistry, HarvardMedical School, 'Center for Can-cer Research, Massachusetts Insti-tute of Technology, Boston: Indi-vidual Ad5 Et A gene productselicit distinct alterations of cellularmorphology and gene expression.
Maruyama, K., Ruley, E., Cold SpringHarbor Laboratory, New York: Ad-enoviral E1A blocks in vitro differ-entiation of rat PC12.
Kao, H.-T, Capasso, 0., Heintz, N.,Nevins, J.R., Rockefeller Univer-sity, New York, New York: Controlof cellular gene expression in ad-enovirally transformed cells.
Vassen, R.T.M.J., Houweling, A., Ber-nards, R., Jochemsen, A.G., Of-fringa, R., van der Eb, A.J., Dept.of Medical Biochemistry, Sylvius
230
Laboratories, State University ofLeiden, The Netherlands: Regula-tion of MHC expression in adeno-virally transformed cells.
Cook, J.L.,' Lewis, A.M., Jr.,''National Jewish Hospital and Re-search Center, Denver, Colorado;2NIAID, NCI, National Institutes ofHealth, Bethesda, Maryland: Dif-ferential hosts' inflammatory-cellkilling of hamster cells infectedwith or transformed by nononco-genic or highly oncogenic adeno-viral species- Implications for Tantigen function.
Simanis, V., Lane, D.P., CRC Eukar-yotic Molecular Genetics Group,Dept. of Biochemistry, ImperialCollege of Science and Technol-ogy, London, England: Cloning ofcellular sequences to which SV40large T antigen is bound in the
sachusetts; 3Dept. of Microbiol-ogy, University of Virginia,Charlottesville: Association ofpolyoma middle T with c-src.
Grussenmeyer, T,' Walter, G.,2 Hutch-inson, M.A.,3 Templeton, D.,3 Eck-hart, W.,3 'Institut fur Immunbiolo-gie, Universitat Freiburg, FederalRepublic of Germany; 'Dept. ofPathology, University of California,San Diego, La Jolla; 3Salk Insti-tute, San Diego, California: Prop-erties of polyoma virus medium Tantigen protein kinase purified byaffinity chromatography using twoantipeptine monoclonal anti-bodies.
Wilson, J.B.,' Courtneidge, S.,2 Grif-fiths, M.,1 Fried, M.,"ImperialCancer Research Fund, 2MedicalResearch Council, London, Eng-land: Analysis of unstable, flat re-vertants of a polyoma-transformedcell line.
chromatin of SV40-transformedcells.
Glaichenhaus, N.,' Masiakowski, P.,2Mougneau, E.,' Lawrence, J.-J.,3Cuzin, F.,"INSERM, Centre deBiochimie, Universite de Nice;2Labortoire de Genetique Molecu-laire des Eucaryotes, Faculte deMedecine, Strasbourg; 3Centred'Etudes Nucleaires, Grenoble,France: Differential expression ofcellular genes in the G2 and 01phases of the cell cycles, modifiedin cells expressing the large T pro-tein of polyoma virus.
Singh, K., Saragosti, S., Carey, M.,Botchan, M., Dept. of MolecularBiology, University of California,Berkeley: Enhanced levels of poly-merase III transcripts derived fromthe B2 repetitive element in SV40transformed mouse cell lines.

Van Dyke, T.,' Levine, A.J.,' Brinster,R.,2 Chen, H.Y.,2 Messing, A.,2 Pal-miter, R.D.,3 'Dept. of Microbiol-ogy, State University of New York,Stony Brook; 'School of VeterinaryMedicine, University of Pennsylva-
SESSION 12
Chairperson:
nia, Philadelphia; 3HowardHughes Medical Institute, Univer-sity of Washington, Seattle: Trans-genic mice containing the SV40early region genes develop tu-mors of the choroid plexus.
ADENOVIRUSES: REPLICATION
Garrels, J., Franza, B.R., Jr., ColdSpring Harbor Laboratory, NewYork: A two-dimensional gel data-base for proteins of normal andvirally transformed rat cells.
B. Stillman, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
Nagata, K., Guggenheimer, R.A.,Kenny, M., Field, J., Lindenbaum,J., Murakami, Y., Gronostajski,R.M., Hurwitz, J., Dept. of Devel-opmental Biology, Albert EinsteinCollege of Medicine, Bronx, NewYork: Enzymatic DNA replicationof adenoviral DNA.
Rawlins, D.R.,' Rosenfeld, P.J.,'Wides, R.J.,' Challberg, M.D.,'Kelly, T.J., Jr.,"Dept. of MolecularBiology and Genetics, Johns Hop-kins University School of Medi-cine, Baltimore, Maryland; 'Dept.of Microbiology and Molecular Bi-ology, Tufts University School ofMedicine, Boston, Massachusetts:Sequence analysis of the adeno-viral origin of replication.
Gronostajski, R.M., Nagata, K., Ad-hya, S., Guggenheimer, R.A., Hur-witz, J., Dept. of DevelopmentalBiology and Cancer, Albert Ein-stein College of Medicine, Bronx,
New York: Characterization of theHe La cell DNA sequences homol-ogous to the replication origin ofAd5.
Leegwater, P.A.J., de Vries, E., vanDriel, W., van Wijnen, A., Kwant,M.M., van der Vliet, P.C., Labora-tory for Physiological Chemistry,University of Utrecht, The Nether-lands: Replication of adenoviralDNA with purified proteins-Originsequences and stimulation byRNA.
Schneider, R., Dorper, T., Groger, W.,Winnacker, E.-L., Institut fOr Bioch-emie, Universitat Munchen, Fed-eral Republic of Germany: Struc-ture and function of the adenoviralreplicon.
Hay, R.T., Medical Research CouncilVirology Unit, Glasgow, Scotland:The initiation of adenoviral DNAreplication in vivo.
Bernstein, J., Porter, J. Challberg,
Phage and Bacterial Regulatory Mechanisms
M.D., Dept. of Molecular Biologyand Microbiology, Tufts MedicalSchool, Boston, Massachusetts:Nucleotide sequence require-ments for the replication of plas-mid DNA in Ad5-infected He Lacells.
Rekosh, D., Mertz, L., Brewster, J.,Prestine, L., Dept. of Biochemistry,State University of New York, Buf-falo: Cloning and expression in E.coli of the gene encoding the ad-enoviral DNA polymerase.
Chow, K.-C., Pearson, G.D., Dept. ofBiochemistry and Biophysics, Ore-gon State University, Corvallis: Ad-enoviral infection elevates He Latopoisomerase activity.
Graham, FL., Depts. of Biology andPathology, McMaster University,Hamilton, Canada: Covalent lyclosed circles of human adeno-viral DNA are infectious.
August 21-August 26
ARRANGED BY
Sankar Adhya, National Cancer InstituteHarrison Echols, University of California, Berkeley
289 participants
The study of bacteriophage stands as a cornerstone of our present knowledge ofthe biological universe, and investigating gene structure and behavior in phagecontinues to be enormously satisfying and rewarding. What emerged at thisyear's phage meeting was testament to this fact. It was apparent from the pres-entations that both genetic and biochemical problems are being studied at a mo-lecular detail that was only a dream in the last decade. Because the regulatorymechanisms of phage are now known to be so intricately related to host regula-tory machinery and because host genes and proteins are also being studied at a
231

similar level of sophistication, it was thus proper that the meeting had wider par-ticipation.
In the new format the sessions were arranged not according to organism, ashas been the usual practice, but by biochemical processes; e.g., recombination,replication, and transcription initiation and termination. Of course, it was difficultto draw a line between these processes, which are so interdependent. Each ofthe sessions was chaired by one of the veterans, whose direct participation in thediscussions, instant analysis of the situation, and summary of the take-home les-sons showed that a chairperson can make a significant difference.
Finally, the meeting brought back the "nonphage" prokaryotic molecular biolo-gists to Cold Spring Harbor and provided a forum in which they and the phagebiologists can interact, sharing problems as well as excitement with each other.
This meeting was supported in part by the Robertson Research Fund.
SESSION 1 BACTERIOPHAGE Mu
Goosen, N., van Heuvel, M., Moo le-naar, G.F., van de Putte, P., Labo-ratory of Molecular Genetics, StateUniversity of Leiden, The Nether-lands: The regulation of early andrepressor transcriptions of bacte-riophage Mu.
Schumann, W., Institut fur OrganChemie and Biochemie, Tech-nische Hochschule Darnstadt,Federal Republic of Germany:Regulation of the early functions oftemperate E. coli phage
Priess, H., Schmidt, C., Li, G.-Di.,Kamp, D., Max-Planck-Institut,Martinsried, Federal Republic ofGermany: Structure and some as-pects of regulation of the early re-gion of phage Mu.
Hattman, S., Ives, J., Dept. of Biology,University of Rochester, New York:Regulation of the methylation-sen-sitive DNA modification function ofbacteriophage Mu.
Kahmann, R., Heisig, P., Seiler, A.,Max-Planck-Institut, Berlin, Fed-eral Republic of Germany: Themom gene of bacteriophage Mu-A complex regulatory cascade tocontrol a lethal function.
Kahmann, R., Rudt, F, Mertens, G.,
Max-Planck-Institut, Berlin, Fed-eral Republic of Germany: Site-specific recombination in bacterio-phage Mu-G inversion in vitro.
Grundy, F.J., Howe, M.M., Dept. ofBacteriology, University of Wiscon-sin, Madison: The role of invertibleG-segment genes in the determi-nation of phage Mu tail fiber struc-ture and host specificity.
Castilho, B., Casadaban, M., Dept. ofMolecular Genetics and Cell Biol-ogy, University of Chicago, Illinois:The specificity of bacteriophageMu insertion.
Craigie, R., Mizuuchi, M., Mizuuchi,K., NCI, National Institutes ofHealth, Bethesda, Maryland: Site-specific interactions between thebacteriophage Mu A protein andthe Mu ends.
Desmet, L.,' Toussaint, A.,' Pato, M.,2Waggoner, B.,2 'Universite libre deBruxelles, Laboratoire de GOne-tique, Belgium; 2National JewishHospital and Research Center,Denver, Colorado: Requirement ofprimase for phage Mu replicationand transposition.
Pato, M., Reich, C., Waggoner, B.,National Jewish Hospital and Re-
SESSION 2 DNA REPLICATION AND REPAIR
Dodson, M.,' Echols, H.,' Roberts,J.D.,2 McMacken, R.,2 'Dept. ofMolecular Biology, University ofCalifornia, Berkeley; 2Dept. of Bio-chemistry, Johns Hopkins Schoolof Hygiene and Public Health, Bal-timore, Maryland: Specialized nu-
232
cleoprotein structures at the repli-cation origin of bacteriophage x.
Zylicz, M., Yamamoto, T, Sell, S.,Georgopoulos, C., Dept. of Cellu-lar, Viral, and Molecular Biology,University of Utah Medical Center,Salt Lake City: In vitro Adv DNA
search Center, Denver, Colorado:Replicative transposition of bacte-riophage Mu DNA.
Harshey, R.M., Scripps Clinic and Re-search Foundation, Dept. of Mo-lecular Biology, La Jolla, Califor-nia: Nonreplicative DNAtransposition-Integration of in-fecting bacteriophage Mu.
Burlingame, R., Lynn, D.L., Obukow-icz, M.G., Howe, M.M., Propertiesof phages with cis-dominant point
Mu replication.Glasgow, A.C., Miller, J.L., Howe,
M.M., Dept. of Bacteriology, Uni-versity of Wisconsin, Madison: Mu-specific sites and functions in-volved in growth inhibition ofA.::mini-Mu.
Shore, S.H., Ross, W., Howe, M.M.,Dept. of Bacteriology, University ofWisconsin, Madison: E. coli DNAgyrase mutations that inhibit thegrowth of bacteriophage Mu.
Yoshida, R.K., Howe, M.M., Dept. ofBacteriology, University of Wiscon-sin, Madison: Analysis of the blockto phage Mu development in E.coli himA mutant strains.
replication dependent on the E.coli dnaJ and dnaK proteins.
Wickner, S., NCI, National Institutes ofHealth, Bethesda, Maryland: Lim-its of A origin DNA required for theinitiation of replication in vitro.
Macdonald, P., Mosig, G., Dept. of

Molecular Biology, Vanderbilt Uni-versity, Nashville, Tennessee: Sig-nals in a DNA replication origin ofphage T4.
Hinton, D., Nossal, N., NCI, NationalInstitutes of Health, Bethesda,Maryland: Cloning of T4 DNA en-coding the T4 primase proteins,genes 41 and 61, and a DNA-binding protein of 46 kD.
Se lick, H.E., Nakanishi, M., Kreuzer,K.N., Alberts, B.M., Dept. of Bio-chemistry and Biophysics, Univer-sity of California, San Francisco:Molecular dissection of potentialT4 replication genes.
Kreuzer, K.N., Alberts, B.M., Dept. ofBiochemistry and Biophysics, Uni-versity of California, San Fran-cisco: Molecular cloning and anal-ysis of phage T4 replicationorigins.
Fulford, W., Model, F, RockefellerUniversity, New York, New York:Gene x of bacteriophage fl is re-quired for phage DNA synthesis-Mutagenesis of in-frame overlap-ping genes.
Horiuchi, K., Rockefeller University,New York, New York: Dissection offunctional domains of DNA repli-cation origin in bacteriophage fl.
Scheuermann, R.H., Young, K.,Echols, H., Dept. of Molecular Bi-ology, University of California,Berkeley: The role of the c-subunit
SESSION 3 PHAGE MORPHOGENESIS
Feiss, M., Sippy, J., Miller, G., Dept. ofMicrobiology, University of Iowa,Iowa City: Terminase readthroughduring sequential packaging ofbacteriophage A chromosomes.
Gold, M., Parris, W., Dept. of MedicalGenetics, University of Toronto,Canada: A bacterial protein re-quirement for bacteriophage A ter-minase activity in vitro.
Rosenberg, S.M., Stahl, F.W., Instituteof Molecular Biology, University ofOregon, Eugene: A monomerscannot be packaged in vitro.
Stahl, F.W.,' Stahl, M.M.,' Kobayashi,I.,2 Thaler, D.," Institute of Molec-ular Biology, University of Oregon,Eugene; 2Dept. of Bacteriology,University of Tokyo Faculty ofMedicine, Japan: Recombinationand packaging of phage A.
of E. colt DNA polymerase III inthe fidelity of DNA replication.
Panayotatos, N., Fontaine, A., Dept.of Molecular Biology, Bingen S.A.,Switzerland: Expression of an en-donuclease in E. colt damages theDNA and induces the SOS re-sponse.
Ennis, D.G.,' Fisher, B.L.,' Mount,D.W.,1.2 Depts. of 'Molecular andCellular Biology, 2Biochemistry,University of Arizona, Tucson: Di-rect involvement of recA protein inumuCD or pkm101-mediated in-ducible mutagenesis.
Bhatia, K., Martin, S., Lee, E., Hays,J., Dept. of Chemistry, Universityof Maryland Baltimore County, Ca-tonsville: uvr, phr, and sos func-tion in the dark repair of nonrepli-cating uv-irradiated phage A DNA.
Wong, A., Lemmermann, L., Maurer,R., Dept. of Molecular Biology andMicrobiology, Case Western Re-serve University, Cleveland, Ohio:Requirement for dnaC proteins inS. typhimurium harboring a strongsuppressor mutation in dnaB.
Engstrom, J., Maurer, R., Dept. ofMolecular Biology and Microbiol-ogy, Case Western Reserve Uni-versity School of Medicine, Cleve-land, Ohio: Mutations in the dnaA-dnaN region in S. typhimuriumsuppress dnaZ(ts).
Pruss, G.J., Drlica, K., Dept. of Biol-
Russel, M., Model, P., Rockefeller Uni-versity, New York, New York: Thio-redoxin is required for filamentousphage assembly.
Smith, G.P., Division of Biological Sci-ences, University of Missouri, Co-lumbia: Filamentous phage mor-phogenesis-Which genes arereally essential?
Wyckoff, E., Casjens, S., Dept. of Cel-lular, Viral, and Molecular Biology,University of Utah Medical Center,Salt Lake City: Expression of hy-brid scaffolding protein-p-Galac-tosidase gene is regulated by un-assembled scaffolding protein.
Berget, P.B., Kylberg, R.W., Rang-wala, S.H., Plishker, M.F., Dept. ofBiochemistry and Molecular Biol-ogy, University of Texas MedicalSchool, Houston: In vitro analysis
S. Adhya
ogy, Unviersity of Rochester, NewYork: Extremely high levels of neg-ative supercoiling in supX mu-tants.
GOnthert, U., Max-Planck-Institut, Ber-lin, Federal Republic of Germany:The DNA methyltransferase geneof the B. subtilis phage SPR.
Kellman, S.I., Margolin, P., PublicHealth Research Institute of theCity of New York, New York: Toler-ance of S. typhimurium to the ab-sence of DNA topoisomerase Iand the effect on prophage induc-tion.
of the sequential assembly of pro-teins of the bacteriophage T4 ba-seplate.
Davis, N., Model, P., Rockefeller Uni-versity, New York, New York: Anal-ysis of the membrane anchoringrequirements of a model integralmembrane protein.
Kurnit, D.,' Neve, R.,' Lagos, R.,2Lee, S.-J.,2 Goldstein, R.,2'Genetics Division, Children's Hos-pital, °Dept. of Microbiology andMolecular Genetics, Harvard Med-ical School, Boston, Massachu-setts: Plasmid cloning vector P4vir1 sid amt -supF dosage me-diates capsid size.
233

SESSION 4 TRANSPOSITION
Dalyrmple, B., Mollet, B., Ida, S., Ar-ber, W., Dept. of Microbiology,Biozentrum of the University ofBasel, Switzerland: Structure andfunctions of the prokaryotic is ele-ments IS26 and IS30.
Shen, M., Raleigh, E., Kleckner, N.,Dept. of Biochemistry and Molecu-lar Biology, Harvard University,Cambridge, Massachusetts: Anal-ysis of unselected IS10 rearrange-ments.
Roberts, D., Kleckner, N., Dept. ofBiochemistry and Molecular Biol-ogy, Harvard University, Cam-bridge, Massachusetts: Effects ofadenine methylation on activity ofIS10.
Raleigh, E.A., Kleckner, N., Dept. ofBiochemistry and Molecular Biol-ogy, Harvard University, Cam-bridge, Massachusetts: Rate oftranscription and translation ofIS10- right.
Kittle, J.D., Jr., Kleckner, N., Dept. ofBiochemistry and Molecular Biol-ogy, Harvard University, Cam-bridge, Massachusetts: Pairing be-tween IS/O's PIN and Po,transcripts in vitro.
Morisato, D., Kleckner, N., Dept. ofBiochemistry and Molecular Biol-
SESSION 5 REGULATION I
Ebright, R.H.,1 Miller, J.,3 'HarvardMedical School, Boston, Massa-chusetts; 3University of California,Los Angeles: Putative contact be-tween GIn18 of lac repressor andbase pair 7 of /ac operator.
Majumdar, A., Polymeropoulos, M.,Adhya, S., NCI, National Institutesof Health, Bethesda, Maryland: In-teraction between repressor andoperators of gal operon of E. coli.
Garges, S., Adhya, S., NCI, NationalInstitutes of Health, Bethesda,Maryland: Analysis of mutations incrp gene of E. coll.
Ebright, R.H.,12 Krakow, J.,3 Kunkel,T.,4 Cossart, P.,2 Gicquel-Sanzey,B.,2 Beckwith, J.,' Buc, H.,2'Harvard Medical School, Boston,Massachusetts; 2lnstitute Pasteur,Paris, France; 3Hunter College,New York, New York; 4National In-stitutes of Environmental HealthSciences, Research Triangle Park,North Carolina: Ligand binding,
234
ogy, Harvard University, Cam-bridge, Massachusetts: Physicalanalysis of Tn10 transposition.
Davis, M.A., Simons, R.W., Kleckner,N., Dept. of Biochemistry and Mo-lecular Biology, Harvard Univer-sity, Cambridge, Massachusetts:ISIO protects itself at two levelsfrom fortuitous activation by exter-nal promoters.
Bender, J., Kleckner, N., Dept. of Bio-chemistry and Molecular Biology,Harvard University, Cambridge,Massachusetts: Tn10 transpositionproducts contain information fromboth strands of donor transposonmolecule.
Mazodier, P., Giraud, E., Gasser, F.,Dept. de Biochimie et Gen4tiqueMoleculaire, Institut Pasteur, Paris,France: Genetic organization,DNA sequence, and expression ofthe three resistance genes of thecentral region of Tn5.
Nag, D.K., Dasgupta, U., Berg, D.E.,Dept. of Microbiology, WashingtonUniversity, St. Louis, Missouri:IS50-mediated inverse transposi-tion-Specificity and precision.
Amundsen, S.K.,1 Ennis, D.G.,2Smith, G.R.,"Fred HutchinsonCancer Research Center, Seattle,
conformation, and altered DNA-sequence recognition propertiesof CAP(G1u181-.Leu) andCAP(Glu181-.Val).
Okamoto, K., Pondo, C., Freundlich,M., Dept. of Biochemistry, StateUniversity of New York, StonyBrook: LexA protein repressestranscription of the crp gene.
Wertman, K.F., Mount, D.W., Dept. ofMolecular and Cellular Biology,University of Arizona, Tucson:Identification of the nucleotide se-quence determinants of site-spe-cific binding of lexA protein toDNA.
Martin, K., Schleif, R., Dept. of Bio-chemistry, Brandeis University,Waltham, Massachusetts: Repres-sion-negative mutations of thearaBAD operon lie in multiple re-gions.
Schleif, R., Dept. of Biochemistry,Brandeis University, Waltham,Massachusetts: Addition of DNA
Washington; 3University of Ari-zona, Tucson: Analysis of factorsinfluencing inversion of a A DNAsegment flanked by inverted re-peats.
Lichens, A., Syvanen, M., Dept. of Mi-crobiology and Molecular Ge-netics, Harvard Medical School,Boston, Massachusetts: Isolationof a novel bacteriophage l plas-mid that contains the right stem ofTn5.
Selwin Thomas, P., Shanmugasun-daram, S., Lakshmanan, M.,School of Biological Sciences,Madurai Kamaraj University, India:Transposon mutagenesis in A.chroococcum using Mud(Ap.lac).
Cheng, K.C.,12 Smith, G.R.,' 'FredHutchinson Cancer ResearchCenter, 3Dept. of Pathology, Uni-versity of Washington, Seattle: Re-combinational hotspot activity ofChi-like seqeunces.
Murphy, K.,' Casey, L.,' Yannoutsos,N.,' Poteete, T.,' Hendrix, R.,2'University of Massachusetts Med-ical School, Worcester; 2Universityof Pittsburgh, Pennsylvania: Func-tional domains of P22 Erf protein.
helical turns between the araO2operator and araBAD promotercyclically hinders repression.
Hendrickson, W., Brunelle, A., Schleif,R., Dept. of Biochemistry, Bran-deis University, Waltham, Massa-chusetts: Are C protein of E. co/icontacts three turns of the DNAhelix.
Peterson, M.L., Reznikoff, W.S., Dept.of Biochemistry, University of Wis-consin, Madison: Overlapping andpolymerase-binding sites withinthe lac promoter.
Gussin, G.N., Matz, K., Hwang, J.J.,Zuber, M., Court, D.L., Universityof Iowa, Iowa City, and NCI, Fred-erick Cancer Research Facility,Frederick, Maryland: Alteredspacing in the PRE promoter re-verses the effect of Acll protein.
Hoopes, B.C., McClure, W.R., Dept.of Biological Sciences, Carnegie-Mellon University, Pittsburgh,Pennsylvania: Activation of bacte-

riophage A promoters by cll pro-tein.
Wulff, D., Bogorad, L., Mahoney, M.,Dept. of Biological Sciences, StateUniversity of New York, Albany:Mutations in phage A partially res-toring activity to a NE-promotermutant.
Keilty, S., Ho, Y.-S., Sathe, G., Rosen-berg, M., Dept. of Molecular Ge-netics, Smith, Kline and FrenchLaboratories, Swede land, Penn-sylvania: Transcription of the APan. promoter-Absence of " -35"region sequence specificity.
Basu, S., Maitra, U., Albert EinsteinCollege of Medicine, Bronx, NewYork: Studies on the interactions ofT3 RNA polymerase with its cog-nate promoter.
SESSION 6 POSTER SESSION
Hasan, N., Szybalski, W., McArdleLaboratory for Cancer Research,University of Wisconsin, Madison:Structure of the coliphage A null_antiterminator module.
Mac Hattie, L.A., Dept. of MolecularGenetics and Cell Biology, Univer-sity of Chicago, Illinois: Expressionof IS-interrupted coding sequencerestored by plasmid dimerization.
Daniels, D., Isackson, P, Bertrand, K.,Dept. of Microbiology and Molecu-lar Genetics, University of Califor-nia, Irvine: Interactions betweenthe divergent overlapping pro-moters for Tn10 tetA and tetR.
Miller G., Christiansen, S., Feiss, M.,Dept. of Microbiology, Universityof Iowa, Iowa City: Spacing andsequence changes in the Xmnlsymmetry segment of cosA.
Weisemann, J., Weinstock, G., NCI,Frederick Cancer Research Facil-ity, Frederick, Maryland: A positiveselection for recA mutations usinglacZ protein fusions- Isolation ofmutations affecting recA expres-sion.
Chan, PT.,' Ohmori, H.,2 Tomizawa,J.-I.,2 Lebowitz, J.,"Dept. of Mi-crobiology, University of Alabama,Birmingham; 2NCI, National Insti-tutes of Health, Bethesda, Mary-land: Nucleotide sequence andgene organization of ColE1 DNA.
Lambert, P.F., Reznikoff, W.S., Dept.of Biochemistry, University of Wis-consin, Madison: DNA require-
D. Ennis, H. Echols
ments at the bacteriophage G4 or-igin of complementary strand DNAreplication,
Sisk, W., Court, D., NCI, FrederickCancer Research Facility, Freder-ick, Maryland: A plasmid vectorfor identifying and expressingopen reading frames utilizing the Api. promoter.
Harbrecht, D.F., Newman, D., Ander-son, D., Salstrom, J.S., MolecularGenetics, Incorporated, Minne-tonka, Minnesota: E. coil vectorsfor the expression of cloned genesunder the control of PL promoterand cro ribosome-binding site ofphage A.
Yin, J.C.P., Krebs, M.P., Reznikoff,W.S., Dept. of Biochemistry, Uni-versity of Wisconsin, Madison:Identification of the translationalinitiation sites for the right repeatproteins of Tn5.
Yu, X.M., Reznikoff, W.S., Dept. ofBiochemistry, University of Wis-consin, Madison: Deletion analysisof the CAP-cAMP-binding site ofthe E. coil lactose promoter.
Watson, L.A., Cheung, A., SOII, D.,Dept. of Molecular Biophysics andBiochemistry, Yale University, NewHaven, Connecticut: An unlinkedgene gInR enhances expression ofthe glutaminyl-tRNA synthetasegene, gInS, in E. coli.
Twitchell, D., Poteete, A., Dept. of Mo-lecular Genetics and Microbiology,University of Massachusetts Medi-
cal School, Worcester: Relation-ships among the lysis functions ofP22, A, and T4.
Trojanowska, M., Alford, C., Dawson,M., Karam, J., Dept. of Biochemis-try, Medical University of SouthCarolina, Charleston: Control ofexpression of the T4 regA gene.
Shanblatt, S.H., Revzin, A., Dept. ofBiochemistry, Michigan State Uni-versity, East Lansing: Activation oftranscription at the E. coli galac-tose operon by two CAP mole-cules.
Robinson, A.C., Dept. of MolecularBiology, University of Edinburgh,Scotland: DNA sequence of ftsQand ftsA -Essential cell divisiongenes of E. coif.
Cram, D., Ray, A., Skurray, R., Dept.of Microbiology, Monash Univer-sity, Clayton, Australia: Cloningand analysis of F plasmid pif re-gion specifying abortive infectionof phage T7.
Podhajska, A.J., Hasan, N., Szybal-ski, W., McArdle Laboratory forCancer Research, University ofWisconsin, Madison: Regulation ofcloned gene expression by in vivostructural rearrangements in theplasmid.
Podhajska, A.J., Szybalski, W., Mc-Ardle Laboratory for Cancer Re-search, University of Wisconsin,Madison: An N-unresponsive left-ward terminator region in coli-phage A.
235

Peltz, S., Brown, A.L., Szybalski, W.,McArdle Laboratory for CancerResearch, University of Wisconsin,Madison: Loss of boxA results inthe terminator-specific thermosen-sitivity of the cloned nutL antitermi-nator of coliphage A.
Parker, R., Dept. of Chemistry, Indi-ana University, Bloomington: Reg-ulation of transcription from dual-promoter regions in ColE1 DNA.
Newman, E.B., Ahmed, D., Dumont,D., Miller, B., Ramotar, D., Walker,C., Dept. of Biological Sciences,Concordia University, Montreal,Canada: Multiple controls on Ir.sense deaminase activity in E. coltK-12 -Why so much fuss for anonessential activity?
McFall, E.,' Palchaudhuri, S.,2 Born-stein, S.,1 'Dept. of Microbiology,New York University School ofMedicine, New York; 2Dept. of Mi-crobiology, Wayne State UniversitySchool of Medicine, Detroit, Michi-gan: Organization and minimumsize of the E. colt K-12 p-serine de-aminase control region.
McCormick, J.R., Zengel, J.M., Lin-dahl, L., Dept. of Biology, Univer-sity of Rochester, New York: Fur-ther definition of ribosome-bindingsites in E. coil.
Marsh, L., Walker, G.C., Dept.of Biol-ogy, Massachusetts Institute ofTechnology, Cambridge: UmuDC-induced cold sensitivity in E. colt.
K. Horiuchi
236
Marinus, M.G.,' Carraway, M.,' Frey,A.,' Youderian, P.,2 1University ofMassachusetts Medical School,Worcester; 2University of SouthernCalifornia, Los Angeles: Mutationspectrum in a dam mutant of E.coil.
Ludtke, D.,' Silhavy, T,' Torriani, A.,2'NCI, Frederick Cancer ResearchFacility, Frederick, Maryland;2Dept. of Biology, MassachusettsInstitute of Technology, Cam-bridge: Regulation of phoM andthe identification of its gene prod-uct in E. colt.
Liao, S. M., McClure, W.R., Dept. ofBiological Sciences, Carnegie-Mellon University, Pittsburgh,Pennsylvania: Properties of thesmall antisense RNA promoter(Psar) of bacteriophage P22.
LaRossa, R., Smulski, D., Central Re-search and Development Depart-ment, E.I. du Pont de Nemoursand Company, Wilmington, Dela-ware: Inverse escape synthesismediated by Mud(Aplac)-1 pro-phage induction.
Koop, A.H., Hartley, M., Bourgeois,S., Regulatory Biology Laboratory,Salk Institute, San Diego, Califor-nia: Construction of single-copyampicillin-resistance plasmids forcloning and promoter analysis.
Kest ler, H., Zengel, J.M., Lindahl, L.,Dept. of Biology, University ofRochester, New York: A novel reg-ulation of r-protein synthesis fromthe a operon of E. coil.
Jakubowski, H., Goldman, E., Dept.of Microbiology, University of Med-icine and Dentistry of New JerseyMedical School, Newark: Quan-tities of individual aminoacyl-tRNAfamilies and their turnover in E.colt.
Hoopes, R.R., Jr., Mink ley, E.G., Jr.,Dept. of Biological Sciences, Car-negie-Mellon University, Pitts-burgh, Pennsylvania: Activation ofthe transfer operon of E. coil Fplasmid.
Hinton, D.,' Musso, R.,2 'NCI, Na-tional Institutes of Health, Be-thesda, Maryland; 'Dept. of Biol-ogy, University of South Carolina,Columbia: In vitro transcriptionfrom a promoter in the insertionsequence IS2 correlates withexpression of a 14-kD IS2 protein.
Kalty, K., Goldman, E., Dept. of Mi-crobiology, New Jersey Medical
School, Newark: Pleiotropicsuppression obtained by cloningin pBR322 restriction fragmentsfrom total DNA of Su +6 E. coil.
Friedman, S.,' Hays, J.,2Zagursky,R.,' 'NCI, Frederick Cancer Re-search Facility, Frederick; 2Dept.of Chemistry, University of Mary-land, Baltimore County, Catons-ville: Specific inhibition of E. coltrecBC nuclease activity by Agamclones.
Garciarrubio, A., Le 6n, P., Rocha, M.,Vazquez, M., Covarrubias, A.A.,Nitrogen Fixation Research Cen-ter, National University of Mexico,Cuernavaca, Morelos: Two func-tional promoters for the expressionof the glutamine synthetase geneof E. colt.
Colasanti, J., Denhardt, D.T., CancerResearch Laboratory, University ofWestern Ontario, London, Can-ada: Effects of the cloned Ox174A* gene on E. coil metabolism.
Christie, G.E.,' Ljungquist, E.,2 Calen-dar, R.,' 'Dept. of Molecular Biol-ogy, University of California,Berkeley; 2Dept. of Microbial Ge-netics, Karolinska Institute, Stock-holm, Sweden: Sequence of theP2 ogr gene, a regulator of P2 lategene transcription.
Cohen, G., Dept. of Microbiology, TelAviv University, Ramat-Aviv, Israel:Early lytic replication of bacterio-phage P1.
Brooks, J.E.,1 Howard, K.A.,1 Ther-iault, G.,2 Roy, P.,2 Gingeras, T.R.,3'New England Biological Labora-tories, Beverly, Massachusetts;2Dept. of Biochemistry, UniversiteLaval, Quebec, Canada; 3SaIk In-stitute for Biotechnology and In-dustrial Associates, La Jolla, Cali-fornia: Characterization of PaeR7,a restriction-modification systemcloned from P. aeruginosa.
Bialkowska-Hobrzanska, H., Den-hardt, D.T., Cancer Research Lab-oratory, University of Western On-tario, London, Canada: Use of invitro gene fusions to study the repgene of E. coil K -12: Analysis oftranscription regulation.
Belfort, M., Pedersen-Lane, J., Centerfor Laboratories and Research,New York State Department ofHealth, Albany: A genetic systemfor studying the thyA gene andanalyzing E. colt thymidylate syn-thase.

Zylicz, M., Sell, S., Yamamoto, T.,Georgopoulos, C., Dept. of Cellu-lar, Viral, and Molecular Biology,University of Utah Medical Center,Salt Lake City: Host and phageprotein interactions involved in Areplication.
SESSION 7 REGULATION II
Hayes, C., Tuer, R., Hayes, S., Dept.of Microbiology, University of Sas-katchewan, Saskatoon, Canada:Nonimmune exclusion phenotypeexhibited by defective a lysogens.
Christie, G.E.,1 Dale E.,' Calendar,R.,' Lin, C.,2 Six, E.,2 'Dept. of Mo-lecular Biology, University of Cali-fornia, Berkeley; 'Dept. of Microbi-ology, University of Iowa, IowaCity: Comparison of the late genepromoters of coliphage P2 andsatellite phage P4.
Christie, G.E., Pfaff, S., Calendar, R.,Dept. of Molecular Biology, Uni-versity of California, Berkeley: Invivo induction of P2 late geneexpression from galK fusion plas-mids.
Alano, P., Dehd, G., Ghisotti, D., Si-roni, G., Zangrossi, S., Dept. diBiologia, Universita de Milano, It-aly: Alternative regulatory statesfor the intracellular propagation ofthe genetic element P4.
Kim, S.K., Lagos, R., Goldstein, R.,Dept. of Microbiology and Molecu-lar Genetics, Harvard MedicalSchool, Boston, Massachusetts:Plasmid P4-An integrated mas-
SESSION 8 REGULATION III
Arps, P.J.,1 Marvel, C.C.,' Rubin,B.C.,' Winkler, M.E.,"North-western University MedicalSchool, Chicago, Illinois; 'NavalBiomedical Research Laboratoryof the University of California,Berkeley: The hisT gene is part ofa multigene operon in E. coil K-12.
Nieuwkoop, A.J., Bender, RA., Divi-sion of Biological Sciences, Uni-versity of Michigan, Ann Arbor:"Double negative" control of thehuhUH operon in K. aerogenes byCAP-cAMP.
Reitzer, L., Magasanik, B., Dept. of Bi-ology, Massachusetts Institute of
Thylen, C., Dept. of PharmaceuticalMicrobiology, Biomedical Center,Uppsala, Sweden: Expression ofthe T4 gene for fl-glucosyltransfer-ase, cloned on a plasmid vector, isdependent on the allelic state ofthe rho gene of the host.
tercopy is required for stable plas-mid maintenance.
Lozano, P, Lagos, R., Agarwal, M.,Lee, S.-J., Kim, S., Goldstein, R.,Dept. of Microbiology and Molecu-lar Genetics, Harvard MedicalSchool, Boston, Massachusetts:Differential gene expression byphasmid P4 in the plasmid andphage stages.
Horabin, J.I., Webster, R.E., Dept. ofBiochemistry, Duke University,Durham, North Carolina: Regula-tion of gene I expression of the fi-lamentous bacteriophage fl.
Greene, J.R., Geiduschek, E.R, Dept.of Biology, University of California,San Diego: A bacteriophageSP01-coded type-II DNA-bindingprotein shows site-selectivity.
Markiewicz, P.,' Haynes, L.,' Malone,C.,1 Chase, J.,2 Rothman-Denes,L.B.,"Dept. of Molecular Ge-netics and Cell Biology, Universityof Chicago, Illinois; 'Dept. of Mo-lecular Biology, Albert EinsteinCollege of Medicine, Bronx, NewYork: N4 virion RNA polymerase-Template and host-factor require-ments for transcription.
Technology, Cambridge: The threepromoters of the g/nALG operon.
Lupski, J., Nesin, M., Svec, P, God-son, G.N., Dept. of Biochemistry,New York University Medical Cen-ter, New York: Regulation of therpsU-dnaG-rpoD macromolecularsynthesis operon of E. col/K-12.
Haughn, G.W.,1 Squires, C.H.,2 De-Felice, M.,3 Calvo, J.M.,' 'CornellUniversity, Ithaca, New York;'Synergen, Boulder, Colorado; 3CNR, Naples, Italy: Transcriptionalinitiation of the ilvIH operon of E.coli.
Springer, M.,' Plumbridge, J.A.,'
Rothstein, D.M., Bertonis, Bio-gen Research Corp., Cambridge,Massachusetts: Efficiency of tran-scription of the plasminogen acti-vator gene in E. coli.
Jin, D.J., Gross, C.A., Dept. of Bacte-riology, University of Wisconsin,Madison: Search for structural andfunctional domains in the (3-subunitof RNA polymerase in E. coli bystudying rifampicin-resistant muta-tions.
Guidi-Rontani, C.,' Spassky, A.,2Depts. of 'Biochimie et GenetiqueMaleculaire, 'Biologie Moleculaire,Institut Pasteur, Paris, France: Mu-tation of the 0-subunit of E. coliRNA polymerase that affectsexpression of the catabolite-sensi-tive operons.
Hu, J.C., Gross, C.A., Dept. of Bacte-riology, University of Wisconsin,Madison: Mutations of the a-sub-unit of E. coli RNA polymerase thataffect positive control of geneexpression.
Franco, R., Drlica, K., Dept. of Biol-ogy, University of Rochester, NewYork: Supercoiling and gyrasegene expression in E. coll.
Daniel, ,J., Danchin, A., Dept. deBiochimie et Genetique Molecu-laire, Institut Pasteur, Paris,France: On the mediators of cata-bolite repression in E. coli.
Fayat, G.,2 Mayaux, J.F.,2 Blan-quet, S.,2 Grunberg-Manago, M.,''Institut de Biologie Physico-chi-mique, Paris, 2Laboratoire deBiochimie, Ecole Polytechnique,Paris, France: Different controlmechanisms for the expression oftwo aminoacyl-tRNA synthetasesin E. coll.
Gourse, R., Takebe, Y., Nomura, M.,Institute for Enzyme Research,University of Wisconsin, Madison:Feedback regulation of rRNA andtRNA transcription in E. coll.
Imamoto, F., Ishii, S., Maekawa, T.,Nagase, T, Dept. of Molecular Ge-
237

netics, Riken, Wako, Japan: Struc-ture and expression of the nusAand nuuB genes in E. coll.
Albin, R., Silverman, P., Albert Ein-stein College of Medicine, Bronx,New York: The cpxA locus of E.coil K-12 is a single gene.
Guiso, N., Roy, A., Danchin, A., Ull-mann, A., Institut Pasteur, Paris,France: Structure and regulationof adenylate cyclase in E. coll.
LaRossa, R.A., Smulski, D.R., VanDyk, T.K., Central Research andDevelopment Department, E.I. duPont de Nemours and Company,
SESSION 9
Wilmington, Delaware: Sulfometu-ron methyl-A new probe of the i/vregulon.
Kusta, S., Hirschman, J., Meeks, J.C.,Dept. of Bacteriology, University ofCalifornia, Davis: Covalent modifi-cation of bacterial glutamine syn-thetase-Physiological signifi-cance.
Chapon, C.,' Michaelis, S.,12 Wille-mot, K.,3 Cornelis, P.,, Raibaud,O.,' Schwartz, M.,"Institut Pas-teur, Paris, France; 2University ofCalifornia, San Francisco;2Universite de Louvain, Belgium:
REGULATION IV AND TRICKS
Bardwell, J.C.A., Craig, E.A., Dept. ofPhysiological Chemistry, Univer-sity of Wisconsin, Madison: Isola-tion of an E. coil heat-induciblegene homologous to the Droso-phila 83,000-m.w. heat-shock pro-tein gene.
Grossman, A.D.,' Zhou, Y.,' Heilig,J.,2 Christie, G.,2 Calendar, R.,'Gross, C.,"Dept. of Bacteriology,University of Wisconsin, Madison;2Dept. of Molecular Biology, Uni-versity of California, Berkeley: Ex-tragenic suppressors of a is muta-tion in the o subunit of E. coli RNApolymerase.
Grossman, A.D., Erickson, J.W.,Gross, C.A., Dept. of Bacteriology,University of Wisconsin, Madison:The htpR-gene product of E. coil isa ci factor for heat-shock pro-moters.
Paek, K.-H., Walker, G.C., Dept. of Bi-ology, Massachusetts Institute ofTechnology, Cambridge: A defectin induction of heat-shock proteinsat high temperature in xthA mu-tants.
Winter, R.B., Gold, L., Dept. of Molec-ular, Cellular, and DevelopmentalBiology, University of Colorado,Boulder: The maturation (A2) pro-tein from the RNA bacteriophageQB induces the synthesis of someE. coil heat-shock proteins.
Zengel, J.M., Dept. of Biology, Uni-
SESSION 10 REGULATION V
Schauer, A.T., Eades, L.J., Bigelow,B.D., Friedman, D.I., Dept. of Mi-crobiology and Immunology, Uni-versity of Michigan Medical
238
versity of Rochester, New York:The synthesis of ribosomal pro-teins in E. coil is inhibited duringthe heat-shock response.
Christman, M.F., Morgan, R.W., Ja-cobson, ES., Ames, B.N., Dept. ofBiochemistry, University of Califor-nia, Berkeley: Regulation of cellu-lar defenses against oxidativestress.
Gaitanaris, G.A., Gottesman, M.E.,McCormick, M., Howard, B.H.,NCI, National Institutes of Health,Bethesda, Maryland: Reconstitu-tion of an operon from overlap-ping fragments.
Backman, K., O'Connor, M.J., Ma-ruya, A., Erfle, M., BioTechnica In-ternational, Inc., Cambridge, Mas-sachusetts: Regulation of geneexpression by in vivo rearrange-ment of DNA sequences.
Refolo, L.M., Clarke, M.A., Conley,M.P., Jacobsen, J.S., Humayun,M.Z., Dept. of Microbiology, Uni-versity of Medicine and Dentistryof New Jersey Medical School,Newark: Nucleotide sequencecontext effects in spontaneousframeshift mutagenesis of M13cloning vectors.
Mott, J.E.,' Grant, R.A.,' Ho, Y-S.,2Platt, T.,"Dept. of Molecular Bio-physics and Biochemistry, YaleUniversity, New Haven, Connecti-cut; 2Smith, Kline and French Lab-
School, Ann Arbor: Host elementsof the A N antitermination reac-tion-Studies of the nusA andnusE genes.
Pullulanase, an extracellular prod-uct of a gene in the maltose regu-Ion of K. pneumoniae.
Gilson, E., Bedouelle, H., Charbit, A.,Dassa, E., Duplay, P., Rousset,J.P., Hofnung, Institut Pasteur, IN-SERM, CNRS, Paris, France: ThemalB region in E. coil K-12 -Struc-ture and expression.
Stern, M., Ferro-Luzzi Ames, G.,Dept. of Biochemistry, Universityof California, Berkeley: Repetitiveextragenic palindromic (REP) se-quences-A major component ofthe bacterial chromosome.
oratories, Philadelphia, Pennsylva-nia: Maximizing gene expressionfrom plasmid vectors containingthe A pi_ promoter-Strategies foroverproducing transcription termi-nation factor rho.
Raibaud, 0., Mock, M., Schwartz, M.,Institut Pasteur, Paris, France: Atechnique for integrating any DNAfragment into the chromosome ofE. coif.
Shub, D.A., Casna, N.J., Dept. of Bio-logical Sciences, State Universityof New York, Albany: Expressionof 13-galactosidase fusion proteinsin bacteriophage T4.
Groisman, E., Casadaban, M., Dept.of Molecular Genetics and Cell Bi-ology, University of Chicago, Illi-nois: In vivo cloning of DNA fromenterobacteriaceae with mini-Mubacteriophage containing plasmidreplicons.
Weinstock, G., Bremer, E., NCI, Fred-erick Cancer Research Facility,Frederick, Maryland: x placMuphages.
Van Gijsegem, F., Toussaint, A.,Schoonejans, E., Piecq, M., La-boratoire de Genetique, Universitelibre de Bruxelles, Belgium: In vivocloning of pectinase and cellulasegenes of E. chrysanthemi usingRP4::mini-Mu plasmids.
Franklin, N.C., Howard Hughes Medi-cal Institute, Dept. of Biology, Uni-versity of Utah, Salt Lake City: Re-lationships deduced from

completed DNA sequences ofphages A, 021, and P22 in earlyexpressed genome regions rele-vant to the N transcription-antiter-mination function.
Zuber, M., Court, D., NCI, FrederickCancer Research Facility, Freder-ick, Maryland: Transcription termi-nation and antitermination at thet51 terminator of bacteriophage x.
Brown, A.L., Szybalski, W., McArdleLaboratory for Cancer Research,University of Wisconsin, Madison:Effect of site-specific mutations onthe antitermination activity of theboxA-nutR sequence of coliphageA.
Hasan, N., Szybalski, W., McArdleLaboratory for Cancer Research,University of Wisconsin, Madison:Translation of the nutL site and itseffect on the N mediated antitermi-nation function of phage A.
Roberts, J.W., Grayhack, E.J., Yang,X., Goliger, J.A., Section of Bio-chemistry, Molecular, and Cell Bi-ology, Cornell University, Ithaca,New York: Mechanism of late generegulation by phage A gene Q-Activity of an antiterminator in vi-tro.
K., University of Connecticut,
SESSION 11 POST-TRANSCRIPTION
Shimamoto, N., Ikushima, N., Horie,K., Utiyama, H., Life ScienceGroup, Hiroshima University, Ja-pan: Regulation of free form of sin-gle-stranded DNA-binding proteinin E. coll.
Wu, T,' Liao, S.,3 McClure, W.R.,2Susskind, M.M.,1 'Dept. of Molec-ular Genetics and Microbiology,University of Massachusetts Medi-cal School, Worcester; 2Dept. ofBiological Sciences, Carnegie-Mellon University, Pittsburgh,Pennsylvania: Negative control ofP22 antirepressor synthesis by asmall antisense RNA.
Young, F.S., Reid, J., Genentech, Inc.,South San Francisco, California:Stabilization of cloned mRNAs bysequences from the E. coli lipopro-tein gene.
Berkhout, B., Kastelein, R.A., vanDuin, J., Dept. of Biochemistry,University of Leiden, The Nether-lands: Translational interference at
Farmington: An analysis of E. col'"nus" function by in vitro comple-mentation.
Ghosh, B., University of Connecticut,Farmington: Formation of the anti-termination apparatus-Role ofnusA and nusB proteins.
Sparkowski, J., University of Connect-icut, Farmington: The "ran" allelesof RNA polymerase-A class ofrpoB mutations that restore antiter-mination in the nusAl strain.
Somasekhar, G., Szybalski, W., Mc-Ardle Laboratory for Cancer Re-search, University of Wisconsin,Madison: Structure and function ofdeletion mutants in the Q-utiliza-tion (gut) site required for tran-scription antitermination in coli-phage A.
Holben, W.E., Morgan, E.A., Dept. ofBiology, State University of NewYork, Buffalo: Antitermination oftranscription from an E. coli rRNApromoter.
Kuroda, M.I., Yanofsky, C., Dept. ofBiological Sciences, Stanford Uni-versity, California: Nuclease cleav-age analysis of the transcript sec-ondary structures that form in thetrp leader transcript.
Engelberg-Kulka, H., Kopelowitz, J.,
overlapping reading frames inmRNA.
Davidson, A., Gold, M., Dept. of Med-ical Genetics, University of To-ronto, Canada: Regulation andstructure of the bacteriophageterminase genes.
Sampson, L., Casjens, S., Dept. ofCellular, Viral, and Molecular Biol-ogy, University of Utah MedicalCenter, Salt Lake City: Translationinitiation controls the relativerates of expression of the x lategenes.
Belfort, M., Chu, F, Maley, G., Ehren-man, K., Pedersen-Lane, J.,Maley, F., Center for Laboratoriesand Research, New York State De-partment of Health, Albany:Expression of the interrupted T4thymidylate synthase gene.
Gottesman, S., Torres-Cabassa, A.,Maurizi, M., Trisler, P., NCI, Na-tional Institutes of Health, Be-thesda, Maryland: Genetics of pro-
Schoulaker-Schwarz, R., Dept. ofMolecular Biology, Hebrew Uni-versity Hadassah Medical School,Jerusalem, Israel: UGA read-through -A new regulatory mech-anism involved in attenuation ofthe tryptophan operon of E. coli.
Lynn, S.P., Donohue, T.J., Kasper,L.M., Gardner, J.F., Dept. of Mi-crobiology, University of Illinois,Urbana: Effects of mutations in theRNA stem and polyuridine tract ontranscription termination andpausing in the threonine operon ofE. coli.
Post le, K., Godd, R.F., Dept. of Micro-biology and Molecular Genetics,University of California, Irvine: ThetonB terminator also terminatestranscription from an opposinggene.
Mc Kenney, K., NCI, National Institutesof Health, Bethesda, Maryland:Analysis of the "simple" A to tran-scription-termination site.
Galloway, J.L., Platt, T., Dept. of Mo-lecular Biophysics and Biochemis-try, Yale University, New Haven,Connecticut: p-Dependent tran-scription termination of the E. colitryptophan operon.
tein degradation in E. coli -Ionmutations.
Gimble, FS., Sauer, R.T., Dept. of Bi-ology, Massachusetts Institute ofTechnology, Cambridge: repres-sor mutants having a noninducible(ind-) phenotype.
Froshauer, S., Beckwith, J., Dept. ofMicrobiology and Molecular Ge-netics, Harvard Medical School,Boston, Massachusetts: Analysisof the localization and the regula-tion of the malF-gene product.
Cheng, H., MacFarlane, L., Hoyt,M.A., Echols, H., Dept. of Molecu-lar Biology, University of Califor-nia, Berkeley: The regulatory func-tion of hflA -Controls on the cllprotein of phage A and on bacte-rial proteins.
Manning, F.,' Rosas, D.,' Christian-sen, G.,2 Svanborg -Eden, C.,3Wilkinson, B.,4 Rubin, B.,5 Han-son, P.,6 O'Brien, T.,4 Goldstein,R., 'Harvard University, Cam-
239

bridge, Massachusetts; 2Universityof Aarhus, Denmark; 3GoteborgUniversity, Sweden; Brigham andWomen's Hospital, Boston, Massa-
SESSION 12 PLASMIDS AND EXCLUSION
Chattoraj, D., Abe les, A., NCI, Freder-ick Cancer Research Facility, Fred-erick, Maryland: Control of P1plasmid replication.
Austin, S., Abe les, A., Frederick Can-cer Research Facility, Frederick,Maryland: The partition functionsof P1, P7, and F mini-plasmids.
Fowler, T., Thompson, R., Institute ofVirology, University of Glasgow,Scotland: Control points in F plas-mid conjugation.
Lupski, J., Ozaki, L., Projan, S., God-son, G.N., Dept. of Biochemistry,New York University Medical Cen-ter, New York: A temperature-de-
chusetts; 5Massachusetts GeneralHospital, Boston; 6Cambridge Re-search Laboratory, Massachu-setts: The pap operon of urogenic
pendent pBR322 copy-numbermutant due to a Tn5 position ef-fect,
Novick, R.P., Highlander, S.K., Projan,S.J., Kumar, C.C., Carleton, S.,Dept. of Plasmid Biology, PublicHealth Research Institute of theCity of New York, New York: Repli-cation of plasmid pT181 is regu-lated by two countertranscripts.
Sun, T.P., Webster, R.E., Dept. of Bio-chemistry, Duke University, Dur-ham, North Carolina: Bacterialmutants defective in phage fl in-fection.
Molineux, I.J., Schmitt, M., Kearney,
Modern Approaches to Vaccines
E. coli-Cloning, expression, func-tion, and epidemiology.
C., Condreay, J.P., Dept. of Micro-biology, University of Texas, Aus-tin: A basis for F-mediated restric-tion of phage T7.
Bross, P., Burger, B., Bussmann, K.,Rasched, I., Fakultat fOr Biologie,Universitat Konstanz, Federal Re-public of Germany: Analysis of theeffects of sequences of the fd-phage gene III on the host conju-gative system.
September 12-September 16
ARRANGED BY
Robert Chanock, National Institutes of HealthRichard Lerner, Research Institute of Scripps Clinic
229 participants
This second meeting on Modern Approaches to Vaccines included presentationsthat described the identification, molecular cloning, and sequence analysis ofgenes coding for antigens that are specific for each stage of the life cycle of themalaria parasite. Many problems remain to be solved in the development of amalaria vaccine, but it should be noted that several encouraging findings weredescribed. Progress was also reported in the construction of viable bacterialdeletion mutants that lack the sequences for exotoxin. Other encouraging eventsin the development of modern bacterial vaccines included the synthesis of anti-genic peptides that correspond to active regions of bacterial toxins or adhesionproteins.
The meeting was also concerned with the rapidly moving area of viral vac-cines. Significant progress was reported for candidate vaccinia-viral gene recom-binant vaccines as well as viral host range and reassortant vaccines. In addition,a number of presentations described advances in the current understanding ofprotective antigenic sites on viral surface proteins. Finally, the molecular basis forattenuation of one of the major licensed live-virus vaccines (i.e., poliovirus vac-cine) now appears to be coming into focus.
At the conclusion of the conference the organizers and members of the ColdSpring Harbor Laboratory staff decided that further annual meetings would be
240

held during the next 5 years in order to bring together scientists from differentdisciplines who might interact to accelerate progress in vaccine development.
Funding for this meeting was provided in part by Johnson & Johnson Biotech-nology Center, Inc.; Wellcome Biotechnology Ltd.; and the following divisions ofthe National Institutes of Health: Fogarty International Center and the NationalInstitute of Allergy and Infectious Diseases.
SESSION 1 CHEMISTRY OF MALARIA SURFACE ANTIGENS
Chairperson: V. Nussenzweig, New York University Medical Center, New York
Miller, L.H., Laboratory of ParasiticDiseases, National Institutes ofHealth, Bethesda, Maryland: Anoverview on research toward avaccine against malaria.
Nussenzweig, V., New York UniversityMedical Center, New York: Pres-ence of multiple epitopes on theCS protein of the monkey malariaparasite P. knowlesi.
Dame, J.B.,1 McCutchan, Hock-meyer, W.T.,2 Miller, L.H.,"NIAID,National Institutes of Health, Be-thesda, Maryland; 2Dept. of Immu-nology, Walter Reed Army Instituteof Research, Washington, D.C.:Structure of the gene encoding theimmunodominant surface antigen
of the sporozoite of the humanmalaria parasite P. falciparum.
Cowman, A.F, Saint, R.B., Coppel,R.L., Brown, G.V., Favaloro Dahl,J., Langford, C.J., Crewther, P.Stahl, H.-D., Bianco, E.A., Mitch-ell, G.F., Kemp, D.J., Anders, R.F.,Walter and Eliza Hall Institute ofMedical Research, Melbourne,Australia: Repeat structures in pro-tein antigens of asexual erythro-cyte stages of P. falciparum.
Cohen, S.,' Deans, J.A.,' Mitchell,G.H.,1 Thomas, A.W.,' Waters,A.P.,' Alderson, T, 'Dept. ofChemical Pathology, Guy's Hospi-tal Medical School, London;2Laboratory of Molecular Biology,
SESSION 2 SURFACE ANTIGENS OF MALARIA, TRYPANOSOMES, ANDSCHISTOSOMES
Chairperson: L.H. Miller, National Institutes of Health, Bethesda, Maryland
Kumar, N., Graves, PM., Rener, J.,Kaushal, D.C., Grotendorst, C.,Miller, L.H., Carter, R., Laboratoryof Parasitic Diseases, National In-stitutes of Health, Bethesda, Mary-land: Target antigens of transmis-sion-blocking immunity of malariaparasites.
Langsley, G.,' Mercereau, O.,' Ko-nen, M.,2 Scherf, A.,2 Pereira daSilva, L.,' Muller-Hill, B.,2 'InstitutPasteur, Paris, France; 2Institut fOrGenetik, Cologne, Federal Repub-lic of Germany: Identification of P.falciparum antigens by differentialantibody screening of a genomicexpression library.
Wahlgren, Perlmann, H.,' Wah-lin, B.,' Berzins, K.,' Carlsson, J.,'BjOrkman, A.,2 McNicols,Dame, J.B.,3 McCutchan,Perlmann, P.,' 'Dept. of Immunol-ogy, University of Stockholm;2Dept. of Infectious Diseases, Ka-rolinska Institutet, Roslagstulls
Hospital, Stockholm, Sweden;3NIAID, National Institutes ofHealth, Bethesda, Maryland: Pf155, a putative vaccine candidatefor protection against asexualstages of P. falciparum malaria.
Cross, G.A.M., Rockefeller University,New York, New York: Variant sur-face antigens of T. brucei.
Boothroyd, J.C., Campbell, D.A., Sut-ton, R.E., Dept. of Medical Micro-biology, Stanford UniversitySchool of Medicine, California:Expression of surface-antigengenes in T. brucei involves a novelsystem of discontinous transcrip-tion.
Nogueira, N., Lizardi, P, Lerner, T, DiGiovanni, L., Gonzalez, A., Rocke-feller University, New York, NewYork: Expression in E. coif ofcDNA clones encoding surfaceantigens of T. cruzi- Insect andmammalian stages.
Snary, D., Wellcome Research Labo-
Cambridge, England: Putativeprotective antigens of the erythro-cyte stage of P. knowlesi.
McGarvey, M.J., Perrin. L., Mach, B.,Dept. of Microbiology, Universityof Geneva Medical School, Switz-erland: Cloning and expression inE. coli of genes corresponding tospecific merozoite proteins ex-pressed late in the asexual bloodcycle of P. falciparum.
Holder, A.A., Odink, K.G., Lockyer,M.J., Freeman, R.R., Dept. of Mo-lecular Biology, Wellcome Re-search Laboratories, Beckenham,England: A major surface antigenof P. falciparum merozoites-Stud-ies on the protein and its gene.
ratories, Beckenham, England:Cell-surface glycoproteins on T.cruzi.
L. Miller
241

Sher, A.,' James, S.,2 Correa-Oliveria,R.,' Lanar, D.,' Pearce, E.,''Laboratory of Parasitic Diseases,NIAID, Bethesda, Maryland;
°Dept. of Medicine, George Wash-ington University School of Medi-cine, Washington, D.C.: Immuno-logic and immunochemical
SESSION 3 NEW DIRECTIONS AND PRIORITIES; BACTERIAL VACCINES
Chairperson: M. So, Research Institute of Scripps Clinic, La Jolla, California
Warren, K., Rockefeller University, tory of Bacteriology and Immunol-New York, New York: The great ogy, Rockefeller University, Newneglected diseases of mankind. York, New York: Structural and
Beale, A.J., Wellcome Biotechnology functional properties of gonococ-Ltd., Beckenham, England: Per- cal porins.spective of a traditional manufac- So, M., Hagblom, P., Segal, E., Deal,surer. C., Getzoff, E., Trainer, J., Dept. of
Petricciani, J.C., Center for Drugs and Molecular Biology, Research Insti-Biologics, Food and Drug Admin- Lute of Scripps Clinic, La Jolla,istration, Bethesda, Maryland: California: Genetic and structuralSummary of a workshop on the studies of N. gonorrheae pilus an-use of abnormal cells to produce tigenic variation.new products. Rothbard, J.B., Schoolnik, G.K.,
Gotschlich, E.C., Blake, M.S., Labora- Depts. of Medicine and Medical
SESSION 4 BACTERIAL VACCINES AND IMMUNOLOGY
Chairperson: M. Oldstone, Research Institute of Scripps Clinic, La Jolla, California
Miller, T.,' Auerbach, J.,2 Peetz, R.,' Fields, R, Heffron, F, Dept. of Molec-Reed, P.,2 Kost, T.,' Brown, A.,' ular Biology, Research Institute ofRosenberg, M.,2 'Norden Labora- Scripps Clinic, La Jolla, California:tories, Lincoln, Nebraska; 2Dept. Genetic analysis of intracellularof Molecular Genetics, Smith, parasitism and virulence in S. ty-Kline and French Laboratories, phimurium.Philadelphia, Pennsylvania: High- Mekalanos, J.,' Goldberg, Miller,level expression of E. coli heat-la- V.,' Pearson, G.,' Swartz, D.,' Tay-bile toxin by a A lysogen. lor, R.,' Harford, N.,2 Groyne, F.,2
242
analysis of an attenuated vaccineagainst experimental schistosom-iasis.
Microbiology, Stanford UniversitySchool of Medicine, California: Ap-proaches toward a gonococcalvaccine.
Houghten, R.A.,' Klipstein, FA.,2 Ler-ner, R.A.,' Wright, RE.,"Dept. ofMolecular Biology, Scripps Clinicand Research Foundation, LaJolla, California; 2Dept. of Medi-cine, University of Rochester, NewYork: Synthetic vaccine against E.coil heat stable enterotoxin.
Simoen, E.,2 deWilde, M.,2 'Dept.of Microbiology and Molecular Ge-netics, Harvard Medical School,Boston, Massachusetts;2Molecular Genetics Division,Smith Kline-RIT, Rixensart, Bel-gium: Genetic construction ofcholera vaccine prototypes.
Kaper, J.B., Levine, M.M., Lockman,H.A., Baldini, M.M., Black, R.E.,Clements, M.L., Center for Vac-cine Development, University ofMaryland School of Medicine, Bal-timore: Development and testingof a recombinant live oral coleravaccine.
Reiser, J., Friedman, R.L., Germanier,R., Swiss Serum and Vaccine Insti-tute, Berne, Switzerland: Molecu-lar cloning of B. pertussis toxinand filamentous HA genes.
Ladant, D.,' Brezin, C.,2 Guiso, N.,'Michelson, S.,3 Alonso, J.M.,2Barzu, O.,' Ullmann, A.,"Unitede Biochimie des Regulations Cel-lulaires, 2Unite d'Ecologie Bacter-ienne, 3Unite de Virologie Medi-cale, Institut Pasteur, Paris,France: Purification and character-ization of extracellular adenylatecyclase of B. pertussis.
Mallett, C.P., Everhart, D.L., Dept. ofMicrobiology, New York UniversityDental Center, New York: Mono-

clonal antibody analysis of a ribo-some-derived immunogen from S.mutans.
Ahmed, R., Southern, P., Oldstone,M.B.A., Dept. of Immunology, Re-search Institute of Scripps Clinic,
SESSION 5 LESSONS FROM SMALLPDX ERADICATION AND USE OF VACCINIARECOMBINANTS
Chairperson: F. Fenner, John Curtin School of Medical Research, Australian NationalUniversity, Canberra, Australia
Fenner, F., John Curtin School ofMedical Research, Australian Na-tional University, Canberra: Les-sons from the smallpox eradica-tion campaign.
Paoletti, E., Perkus, M., Piccini, A.,Wos, S., Lipinskas, B.R., Centerfor Laboratories and Research,New York State Department ofHealth, Albany, New York: Geneti-cally engineered poxviruses.
Koprowski, H., Wiktor, T.J., Reagan,K., Macfarlan, R., Dietzschold, B.,Wistar Institute, Philadelphia,Pennsylvania: New generation ofrabies vaccines-Rabies glycopro-tein gene recombinants, anti-idi-otypic antibodies, and syntheticpeptides.
Buller, R.M.L.,' Smith, G.L.,' Cremer,K.,2 Notkins, A.L.,3 Moss, B.,''NIAID, 2NIDR, National Institutes
SESSION 6 POSTER SESSION
Amann, E., Br Oker, M., Wurm, F., Re-search Laboratories of Behring-werke, Marburg, Federal Republicof Germany: Expression of HSV-1glycoprotein-C antigens in E. coli.
Audibert, F.,1 Jolivet, M.,' Gras-Masse, H.,2 Tartar, A.,2 Chedid,L.,"CNRS, Institut Pasteur, Paris;2Chimie Organique, Faculte dePharmacie, Lille, France: Con-struction of antimicrobial polyva-lent synthetic vaccines active insaline.
Britton, P., Garwes, D.J., Bountiff, L.,Millson, G.C., AFRC Institute forResearch on Animal Diseases,Compton, England: Identificationof the immunogenic antigen ofporcine transmissible gastroenteri-tis virus.
Crainic, R., Blondel, B., Coste, M.,Horaud, F., Virologie Medica le, In-stitut Pasteur, Paris, France: In-vi-tro-derived neurovirulant rever-tants of attenuated Sabin type-1poliovirus.
of Health, Bethesda, Maryland: In-fectious vaccinia virus TK- hybridrecombinants that express foreigngenes are less virulent than wild-type virus in BALB/cByJ mice.
Cremer, K.,' Mackett, M.,2 Wohlen-berg, C.,' Notkins, Al.,' Moss,B.,21NIDR, 2NIAID, National Insti-tutes of Health, Bethesda, Mary-land: Vaccinia virus recombinantthat expresses the HSV-1 glyco-protein-D gene protects miceagainst lethal challenge by HSV-1or HSV-2 and the establishment ofa latent ganglionic infection byHSV-1.
Small, RA., Jr.,2 Smith, G.L.,' Moss,B.,3 'Dept. of Immunology andMedical Microbiology, Universityof Florida, Gainesville; 2NIAID, Na-tional Institutes of Health, Be-thesda, Maryland: Intranasal vac-
Deufel, T., Vissing, H., Kofod, H.,Lernmark, A., Hagedorn ResearchLaboratory, Gentofte, Denmark:Antibodies against synthetic pep-tides recognize an HLA-class-II-positive subpopulation of humanperipheral blood cells.
Fox, G.M., Hu, S., Bruszewski, J.,Langley, D., Amgen, Inc., Thou-sand Oaks, California: Immunolog-ical response to porcine parvovi-rus proteins cloned and expressedin E. coli.
Guesdon, J.L.,' Bouges-Bocquet, B.,2Debarbouille, M.,3 Hofnung, M.,2'Unite dlmmunocytochimie,2Unite de Programmation Molecu-laire et Toxicologie Genetique,3Unite de Virologie Moleculaire, In-stitute Pasteur, Paris, France: Anenzymoimmunoassay allowing de-tection of E. coil cells expressing agiven antigenic determinant attheir surfaces.
Hackett, C.J., Hurwitz, J.L., Moller,C., Dietzschold, B., Heber-Katz,
La Jolla, California: Viral genes,cytotoxic T lymphocytes and im-munity.
cination with recombinant vacciniacontaining influenza HA preventsboth influenza viral pneumoniaand nasal infection-Intradermalvaccination prevents only viralpneumonia.
Geysen, H.M.,' Mason, T.J.,' Rodda,S.,' Meloen, R.H.,2 Barteling,S.J.,2 'Commonwealth SerumLaboratories, Parkville, Australia;2Central Veterinary Institute, Lelys-tad, The Netherlands: Amino acidcomposition of antigenic determi-nants-Implications for antigenprocessing by the immune systemof animals,
Shih, M.-F.,' Tiollas, R,2 Roizman, B.,'1KovIer Viral Oncology Laborato-ries, University of Chicago, Illinois;2INSERM, Institut Pasteur, Paris,France: Herpes simplex virus as avector for eukaryotic viral genes.
E., Gerhard, W., Wistar Institute,Philadelphia, Pennsylvania:Helper-T-cell (TH) determinants ofinfluenza HA-Immunization ofmice with a synthetic decapeptide
F. Brown
243

M.A. Epstein, F. Prince
of an immunodominant region elic-its TH cells that recognize whole vi-rus.
Hu, S.-L., Potts, D.E., Molecular Ge-netics, Inc., Minnetonka, Minne-sota: Expression of antigenic de-terminants of bovine adenovirushexon in E. colt.
Hu, S., Bruszewski, J., Smalling, R.,Browne, J.K., Amgen, Inc., Thou-sand Oaks, California: Studies ofTGEV spike protein gp195 ex-pressed in E. colt and by a TGE-vaccinia virus.
Hudecz, GOnczOl, E.,' Pereira,L.,2 Dietzschold, B.,' Plotkin, S.,'1Wistar Institute, Philadelphia,Pennsylvania; 2Viral and Rickett-sia) Disease Laboratory, Dept. ofHealth Services, Berkeley, Califor-nia: Immune responses to enve-lope proteins of HCMV.
Keller, P.M., Ellis, R.W., Lowe, R.S.,Zivin, R.A., Merck, Sharp &Dohme Research Laboratories,West Point, Pennsylvania: Geneticmapping of the major glycoproteingene gC of varicella-zoster virus.
Kennedy, R.C., Henkel, R.D., Fren-chiek, P.J., Eichberg, J.W., Drees-man, G.R., Dept. of Virology andImmunology, Southwest Founda-tion for Biomedical Research, SanAntonio, Texas: Use of anti-idi-otype antibodies to modulate theimmune response in HBV chroniccarrier chimpanzees.
Kieny, M.-P.,' Lathe, R.,' Drillien, R.,2Wiktor, T.,3 Koprowski, H.,3 Le-cocq, J.-P,' Transgene S.A.;2Institut de Virologie, Strasbourg,France; 3Wistar Institute, Philadel-
244
phia, Pennsylvania: Effective im-munization against rabies using avaccinia-rabies recombinant virusexpressing the surface glycopro-tein.
Lenstra, J.A., van der Zeijst, B.A.M.,Niesters, H.G.M., de Groot, R.J.,ter Haar, R.J., Jacobs, C.E., Rot-tier, P.J.M., Spaan, W.J.M., Horzi-nek, M.C., Institute of Virology,State University, Utrecht, TheNetherlands: Toward syntheticvaccines against coronaviruses.
Levings, R.L., National VeterinaryServices Laboratories, Ames,Iowa: Identification of a neutraliza-tion epitope and a complementarydiagnostic antigen of some inacti-vated bovine herpesvirus vac-cines.
Manavathu, E., Hasnain, S.E., Liu,A.P., Bouma, C., Zwaagstra, J.,Leung, W.C., Dept. of Medicine,University of Alberta, Edmonton,Canada: Construction of HSV-1and HSV-2 glycoproteins, their hy-brids, and subfragments in yeastexpression vector.
Mazigh, D.A., Chalvignac, M.A., Quil-icci, M.L., Mollaret, H.H., InstitutPasteur, Ecologie Bacterienne,Paris, France: A 36,000-D proteincoded by the 47-MD virulence-as-sociated plasmid of Y. enterocoli-tica implicated in the immunoge-nicity against Y. pertis.
Minor, PD.,' Evans, D.M.A.,' Schild,G.C.,' Almond, J.W.,2 'NIBSC,London; 2University of Leicester,England: Antigenic sites in theneutralization of poliovirus.
Morrissey, P.,' Russel, R.,2 Novotny,
P.,' Dougan, G.,"Wellcome Bio-technology Ltd., Beckenham,2Royal College of Surgeons,Downe, England: Molecular ap-proaches to the development ofsafe, acellular bacterial vaccines.
Neurath, A.R.,' Kent, S.B.H.,2 Strick,N.,' 1Lindsley F. Kimball ResearchInstitute of the New York BloodCenter, New York; 2California Insti-tute of Technology, Pasadena: Im-mune response to pre-S-gene-coded determinants of HBV.
Parry, N.R.,' Ouldridge, E.J.,' Bar-nett, P.V.,' Rowlands, D.J.,'Brown, F.,' Bittle, J.L.,2 Houghten,R.A.,2 Lerner, R.A.,2 'WellcomeBiotechnology Ltd., Pirbright, Eng-land; 2Research Institute ofScripps Clinic, La Jolla, California:Identification of neutralizing epi-topes of FMDV.
Petteway, S.R., Jr.,' Ivanoff, L.A.,'Ray, J.,2 Korant, B.D.,"Dept. ofCentral Research & Development,E.I. du Pont de Nemours, Wilming-ton; 2Dept. of Biomedical Prod-ucts, E.I. du Pont de Nemours,Glasgow, Delaware: Expression ofrubella and poliovirus proteins inE. co/i.
Robertson, J.S.,' Naeve, C.W.,2Webster, R.G.,2 Bootman, J.S.,'Newman, R.,' Schild,'National Institute for BiologicalStandards and Control, London,England; 2St. Jude Children's Re-search Hospital, Memphis, Ten-nessee: Molecular changes in theHA of influenza virus associatedwith adaptation of the virus togrowth in eggs.
Siddiqui, A., Bulla, G., Dept. of Micro-biology and Immunology, Univer-sity of Colorado Health SciencesCenter, Denver: Expression ofhepatitis B surface antigen inmammalian cells.
Southern, P, Singh, M., Oldstone,M.B.A., Buchmeier, M.J., Dept. ofImmunology, Scripps Clinic andResearch Foundation, La Jolla,California: Application of syntheticpeptides to gene mapping andprotein characterization with lym-phocytic choriomeningitis virus.
Timmins, J.G., Rea, T.J., Post, L.E.,Molecular Biology Unit, UpjohnCompany, Kalamazoo, Michigan:Cloning, sequencing, and bacte-rial expression of the excreted gly-coprotein of PRV.

Weijer, W.J.,' Kerling, K.E.T.,2 Hoog-erhout, P.,2 Bloemhoff, W.,2 Geer-ligs, H.,' Wilterdink, J.B.,' Popken-Boer, T,' Welling-Wester, S.,' Well-ing, G.W.,"Laboratorium voorMedische Microbiologie, Gronin-gen; 2Laboratorium voor Organ-ische Chemie, Leiden, The Neth-
erlands: Synthetic peptides asbasis for a herpesvirus vaccine.
Whetstone, C.A., National VeterinaryServices Laboratories, Ames,Iowa: Mechanisms of adenovirusneutralization as defined withmonoclonal antibodies.
Wright, Bennett, D.,"Viral On-
SESSION 7 MAJOR ANTIGENIC SITES ON PROTECTIVE VIRAL ANTIGENS
Chairperson: R. Lerner, Research Institute of Scripps Clinic, La Jolla, California
Kent, S.B.H.,' Neurath, A.R.,' Strick,N.,2 'Division of Biology, CaliforniaInstitute of Technology, Pasadena;2Lindsley F. Kimball Research In-stitute of the New York Blood Cen-ter, New York: Location and chem-ical synthesis of a pre-S-gene-coded immunodominant epitopeof HBV.
Jameson, B.A.,' Bonin, J.,' Murray,M.,' Kew, O.,' Wimmer, E.,1 'Dept.of Microbiology, State University ofNew York, Stony Brook;'Molecular Virology Branch, Cen-ter for Infectious Diseases, Atlanta,Georgia: Studies of poliovirus neu-tralization.
Francis, M.J.,' Fry, C.M.,1 Rowlands,D.J.,' Brown, F.,' Bittle, J.L.,'Houghten, R.A.,2 Lerner, R.A.,2'Wellcome Biotechnology Ltd.,Beckenham, England; 'ScrippsClinic, La Jolla, California: Primingwith peptides of FMDV.
Emini, E.A.,' Boger, J.,2 Hughs, J.V.,'Mitra, S.W.,3 Linemayer, D.L.,3Depts. of 'Virus and Cell Biology,'Medicinal Chemistry, Merck,Sharp & Dohme Research Labora-tories, West Point, Pennsylvania;3Dept. of Biochemical Genetics,Merck, Sharp & Dohme Laborato-ries, Rahway, New Jersey: Primingof an anti-HAV antibody responseby poliovirus-specific syntheticpeptides-Localization of potentialHAV-neutralizing antigenic sites.
Nunberg, J.H.,' Gilbert, J.H.,' Rodg-ers, G.,' Snead, R.M.,' Nitecki,D.,' Winston, S.,2 'Cetus Corpora-tion, Emeryville, California;2Syngene Products and Research,Fort Collins, Colorado: Localiza-tion of a determinant of virus neu-tralization of FeLV envelope pro-tein gp70.
Dietzschold, B.,' Heber-Katz, E.,1 Hu-decz, F.,' Hollosi, M.,2 Fasman,
SESSION 8 MAJOR ANTIGENIC SITES ON PROTECTIVE VIRAL ANTIGENS ANDMOLECULAR BASIS FOR ATTENUATION OF POLIOVIRUSES
Chairperson: F. Brown, Animal Virus Research Institute, Pirbright, England.
Diamond, D.C., Jameson, B.A., Wim-mer, E., Dept. of Microbiology,State University of New York,Stony Brook: Antibody-resistant'variants of poliovirus type 1 -Cap-sid structure, antibody binding,and neutralization are complexphenomena.
Baroudy, B.M.,' Ticehurst, J.R.,'Miele, T.,' Maizel, J.V.,2 Purcell,R.H.,' Feinstone, S.M.,' 1NIAID,2NICHD, National Institutes ofHealth, Bethesda, Maryland: Se-quence analysis of HAV cDNAcoding for the capsid region andits application to vaccine develop-ment.
Hughes, J. Bennett, C. Stanton, L.,
Linemeyer, D., Mitra, S., Merck,Sharp & Dohme Research Labora-tories, West Point, Pennsylvania:HAV structural proteins-Sequenc-ing and ability to induce virus-neu-tralizing antibody responses.
Patzer, E.,' Gregory, T,2 Nakamura,G.,' Simonsen, C.,3 Hershberg,R.,' Levinson, A.,3 Depts. of'Vaccine Development, 'ProcessDevelopment, 3Molecular Biology,Genetech, Inc., South San Fran-cisco, California: Immunogenicpotency of recombinant HBV fromcell culture.
Valenzuela, F, Coit, D., Kuo, G., Med-ina-Selby, M.A., Graves, P.V., Chi-ron Research Laboratories, Chiron
cology Laboratory, VA MedicalCenter; 'Depts. of Medicine andCellular, Viral, and Molecular Biol-ogy, University of Utah School ofMedicine, Salt Lake City, Utah: Po-tential avian RNA tumor virus vac-cine.
G.,2 Eisenberg, R.J.,3 Cohen,G.H.,3 'Wistar Institute of Anatomyand Biology, Philadelphia, Penn-sylvania; 'Dept. of Biochemistry,Brandeis University, Waltham,Massachusetts; 3University ofPennsylvania School of DentalMedicine, Philadelphia, Pennsyl-vania: Structure-function relation-ship in immunogenic syntheticHSV peptides.
Gerin, J.L.,' Purcell, R.H.,2 Lerner,R.A.,3 'Georgetown University,Rockville; 2NIAID, National Insti-tutes of Health, Bethesda, Mary-land; 3Research Institute ofScripps Clinic, La Jolla, California:A search for protective epitopes ofthe hepatitis B surface antigen.
Corporation, Emeryville, Califor-nia: Synthesis and assembly inyeast of modified HBsAg particles.
Almond, J.W.,' Westrop, G.D.,' Stan-way, G.,' Cann, A.J.,' Minor, P.D.,'Evans, D.M.A.,' Schild, G.C.,2'Dept. of Microbiology, Universityof Leicester, 2NIBSC, London,England: Molecular basis of atten-uation in the Sabin type-3 polio-virus vaccine.
Omata, T.,' Kohara, M.,2 Abe, S.,2Itoh, H.,2 Komatsu, T.,3 Arita, M.,3Semler, B.L.,5 Wimmer, E.,5 Kuge,S.,2 Kameda, A.,4 Nomoto, A.,4'Dept.of Public Health, School ofPharmaceutical Sciences, KitasatoUniversity, 'Japan Poliomyelitis
245

B. Chanock, R.M. Chanock, M.B.A. Oldstone
SESSION 9 GLYCOPROTEIN ANTIGENS AND VIRULENCE OF ENVELOPED VIRUSES
Chairperson: R. Chanock, National Institutes of Health, Bethesda, Maryland
Epstein, J., Qi, Y., Quinnan, G., Cen-ter for Drugs and Biologics, Foodand Drug Administration, Be-thesda, Maryland: Immunogenicstructural proteins of CMV.
Collins, PL.,' Wertz, G.W.,2'Laboratory of Infectious Dis-eases, National Institutes ofHealth, Bethesda, Maryland;2University of North CarolinaSchool of Medicine, Chapel Hill:Nucleotide sequences of sixmRNAs of human RS virus.
Paterson, R.G., Hiebert, S.W., Harris,T.J.R., Lamb, R.A., Dept. of Bio-chemistry, Northwestern Univer-
sity, Evanston, Illinois: Fusionprotein and hemagglutinin-neu-raminidase protein of the para-myxovirus SV5-Nucleotide se-quence derived from mRNAs.
Shaw, M.W., Choppin, P.W., Rockefel-ler University, New York, NewYork: Studies on the synthesis andantigenicity of the NB glycoproteinof influenza-B virus.
Epstein, M.A., Dept. of Pathology,University of Bristol MedicalSchool, England: Prevention ofEBV-associated malignant dis-eases.
Miller, G.,1,2,3 Heston, L.,2 Rabson,
SESSION 10 MOLECULAR BASIS FOR ATTENUATION OF VIRUSES
Chairperson: R. Purcell, National Institutes of Health, Bethesda, Maryland
Cox, N.J.,' Naeve, C.W.,2 Kitame, F.,3Kendal, A.R.,' WHO CollaboratingCenter for Influenza, Centers forDisease Control, Atlanta, Georgia;2Dept. of Virology, St. Jude Chil-dren's Hospital for Research,Memphis, Tennessee; 3Dept. ofBacteriology, Yamagata University,Japan: Sequence changes ingenes of the attenuated influenzaA vaccine donor strain, A/Ann Ar-bor/6/60.
Buckler-White, A.,' Naeve, C.,2 Mur-phy, B.,' 1NIAID, National Insti-tutes of Health, Bethesda, Mary-land; 2St. Jude Children's Hospital,Memphis, Tennessee: Character-ization of the genes involved inhost range restriction of avian in-fluenza A viruses in primates.
Thompson, R.L.,1.2 Stevens, J.G.,'Wagner, E.K.,2 Devi, G.B.,''University of California School ofMedicine, Los Angeles; 2Dept. ofMolecular Biology and Biochemis-try, University of California, Irvine:Genetic control of herpes simplexvirus neurovirulence.
Kapikian, A.Z.,' Midthun, K.,' Levine,M.M.,2 Clements, M.L.,2 Do lin, R.,3Wright, P.F.,4 Belshe, R.B.,6 Ander-son, E.L.,6 Hoshino, Y.,' Flores,J.,' Wyatt, R.G.,1 Askaa, J.,'Glass, Nakagomi, O.,' Nako-gomi, T.,' Potash, L.,6 Chanock,R.M.,"NIAID, National Institutesof Health, Bethesda, Maryland;2University of Maryland, Balti-more; 3University of Rochester,New York; 4Vanderbilt University,
Summary: E. Norrby, Karolinska Institute, Stockholm, Sweden
246
Research Institute, 3Dept. of Enter-oviruses, National Institute ofHealth, °Dept. of Microbiology,Faculty of Medicine, University ofTokyo, Japan; 5Dept. of Microbiol-ogy, School of Medicine, StateUniversity of New York, StonyBrook: Construction of recombi-nant viruses between Mahoneyand Sabin strains of type-1 polio-virus and their biological charac-teristics.
M.,2 Countryman, J.,3 Jenson, H.,'Depts. of 'Pediatrics, 2Epidem-iology and Public Health, °Molec-ular Biophysics and Biochemistry,Yale University School of Medi-cine, New Haven, Connecticut:EBVs that do not immortalize lym-phocytes nor remain latent.tent.
Maassab, H.F., DeBorde, D.C., Dona-bedian, A.M., Smitka, C.W.,School of Public Health, Universityof Michigan, Ann Arbor: Develop-ment of cold-adapted "master"strains for type-B influenza virusvaccines.
Nashville, Tennessee; qvlarshallUniversity, Huntington, West Vir-ginia; 6FIow Labs Inc., McLean,Virginia: Rhesus rotavirus strainMMU 18006-Candidate vaccinefor humans.
Vesikari, T.,' lsolauri, E.,1 Delem, A.,2D'Hondt, E.,2 Andre, F.E.,2 'Dept.of Clinical Sciences, University ofTampere, Finland; 2Biological Divi-sion, Smith Kline-RIT, Rixensart,Belgium: Protection of infantsagainst human rotavirus diarrheaafter vaccination with live atten-uated bovine (NCDV) rotavirusstrain RIT 4237.

Growth Factors and TransformationSeptember 19-September 23
ARRANGED BY
James Feramisco, Cold Spring Harbor LaboratoryBrad Ozanne, University of Dallas, TexasCharles Stiles, Dana-Farber Cancer Institute
208 participants
The rapidly accumulating evidence of the intimate connection between cellulargrowth factors and transformation provided the impetus as well as the focus fororganizing the second in the series of meetings devoted to the cancer cell. Inves-tigators from various disciplines, including cell biologists, molecular biologists,and tumor virologists, came together to present their latest findings in these con-verging fields. Topics covered at the meeting included oncogenes, kinases, lym-phokines, PDGF, EGF, TGF, insulin, and a concluding session that exploredpromising lines of future research. Among the highlights of the conference were:(1) the correlation of the expression of the c-fos and c-myc proto-oncogenes withthe stimulation of cells by growth factors; (2) the involvement of the ras oncogenewith adenylate cyclase in yeast; and (3) the demonstration that microinjection ofthe oncogenic form of ras protein into normal cells causes rapid proliferation ofthese cells without the need for growth factors.
This second Cancer Cells meeting was an exciting demonstration of the factthat this area of research has reached a detailed molecular level. The first pro-posals of the molecular mechanisms of the oncogenes sis, erb-B, and ras in cellproliferation have been stated, and it is clear that further research in the areas ofgrowth factors and oncogenes will yield tremendous new information in the mo-lecular biology of cancer. This series of meetings will certainly serve as a main-stay for the presentation of this work.
Funding for this meeting was supplied in part by the Cold Spring Harbor Can-cer Center Grant from the National Cancer Institute, National Institutes of Health.
Introduction: C. Stiles, Dana-Farber Cancer Institute, Boston, Massachusetts
SESSION 1 ONCOGENES
Chairperson: S. Martin, University of California, Berkeley, California
Foulkes, J.G., Baltimore, D., White-head Institute for Biomedical Re-search, Cambridge, Massachu-setts: Characterization of a tyrosyl-protein kinase encoded by Ab-MLV.
Feramisco, J.R.,1 Fasano, O.,' Gold-farb, M.,' Wig ler, M.,' Katama, T.,1Gross, M.,2 Yokoyama, S.,2 Rosen-berg, M.,2 Sweet, R.W.,2 'ColdSpring Harbor Laboratory, NewYork; 2Dept. of Molecular Ge-netics, Smith, Kline and FrenchLaboratories, Swede land, Penn-sylvania: Microinjection of the on-
cogene form of the human Ha-ras(T24) protein results in rapid prolif-eration of quiescent cells.
Cooper, G.M., Dana-Farber CancerInstitute, Harvard Medical School,Boston, Massachusetts: Structureand function of ras and Blym on-cogenes.
Wig ler, M.,' Kataoka, T.,1 Powers, S.,1Toda, T.,1 Fasano, O.,' Strathern,J.,2 Broach, J.,3 'Cold Spring Har-bor Laboratory, New York;2Frederick Cancer Research Facil-ity, Frederick, Maryland; 3Dept. ofMolecular Biology, Princeton Uni-
versity, New Jersey: Structure, ge-netics, and activation of yeast andmammalian ras genes.
Ruley, E.,' Moomaw, J.,' Garrels, J.,'Furth, M.,2 Franza, R.,"ColdSpring Harbor Laboratory, NewYork; 2Memorial Sloan-KetteringCancer Center, New York, NewYork: Cellular immortality is notsufficient for oncogenic transfor-mation by the T24 Ha-ras onco-gene.
Verma, I.M.,1 Meijlink, F.,' Mitchell,R.,' Zokas, L.,' Schreiber, R.,2Curran, T.,' Miller, A.D.,' Van Bev-
247

C.D. Stiles
SESSION 2 KINASES
eren, C.,"Molecular Biology andVirology Laboratory, Salk Institute,San Diego; 2Scripps Clinic andResearch Foundation, La Jolla,California: Oncogene los-Struc-ture and expression.
Ferguson, B.,' Krippl, B.,2 Westphal,H.,2 Jones, N.,3 Rosenberg, M.,''Dept. of Molecular Genetics,Smith Kline and French Laborato-ries, Philadelphia, Pennsylvania;2NICHD, National Institutes ofHealth, Bethesda, Maryland;
Chairperson: J. Feramisco, Cold Spring Harbor Laboratory, New York
Cooper, J.A., Hunter, T., Molecular Bi-ology and Virology Laboratory,Salk Institute, San Diego, Califor-nia: Endogenous substrates for ty-rosine protein kinases in growthfactor-treated cells
J. Feramisco
SESSION 3 POSTER SESSION
Bonin, P.D., Tebaldi, A.F., D'Alessan-dro, J.S., Singh, J.P., CollaborativeResearch Incorporated, Lexing-ton, Massachusetts: Isolation ofthree physicochemically distinctIL-2 activities from human lympho-cyte conditioned media.
Chen, M.-J., Weiss, R., Sathe, G.,Henner, J., Young, P., Rosenberg,M., Dept. of Molecular Genetics,Smith Kline and French Laborato-ries, Philadelphia, Pennsylvania:Expression of human c-sis gene innormal and tumor cells.
Dani, C., Blanchard, J.M., Piechac-
248
Frackelton, A.R., Jr., Roger WilliamsGeneral Hospital and Brown Uni-versity, Providence, Rhode Island:Use of a monoclonal antibody tophosphotyrosine to characterizephosphotyrosyl proteins in cellstransformed by Ab-MLV.
Rosner, M.R., McCaffrey, PG., Fried-man, B.-A., Massachusetts Insti-tute of Technology, Cambridge:Modulation of growth factor actionby tumor promoters and C kinase.
Parker, P.,' Downward, J.,' Earp,H.S.,' Gullick, W.,' Waterfield, M.,'Seeberg, P.,2 Ullrich, A.,. 'ImperialCancer Research Fund Laborato-ries, London, England;2Genentech, Incorporated, SouthSan Francisco, California: Thestructure and function of EGF re-ceptor and its interaction with pro-tein kinases A and C.
Macara, Marinetti, G.,. Livings-ton, J.N.,. Balduzzi, P.C.,4 Depts.of 'Radiation Biology and Bio-
zyk, M., El Sabouty, S., Marty, L.,Jeanteur, P., Universite de Sci-ences et Techniques de Langue-doc, Centre Paul Lamarque,Montpellier, France: Unstability ofc-myc mRNA in normal and trans-formed human cells.
Dunn, A.R.,' Metcalf, 11,2 Burgess,A.W.,' Gough, N.,"Ludwig Insti-tute for Cancer Research; 2Walterand Eliza Hall Institute for MedicalResearch, Victoria, Australia: Mo-lecular characterization of murinehematopoietic growth regulators.
Fine, L.G., Badie-Dezfooly, B., Lowe,
.Purdue University, West Lafay-ette, Indiana: Functional charac-terization of human adenovirusEl A proteins synthesized and pu-rified from E. coli.
Lavu, S., Mushinski, J.F., Shen-ong,G.L.C., Potter, M., Reddy, E.P.,NCI, National Institutes of Health,Bethesda, Maryland: Structural or-ganization of the mouse c-myb lo-cus and the mechanism of its rear-rangement in mouse plasmacytoidlymphosarcomas.
physics, 2Biochemistry, .Medicine,4MicrobioIogy, University of Roch-ester School of Medicine and Den-tistry, New York: Lipid phosphoryl-ating activity associated withtyrosine kinases-A possible rolefor phosphatidylinositol turnover intransformation.
Goldberg, A.R.,' Wong, R.W.,' Tse-Dinh, Y-C.,. 'Rockefeller Univer-sity, New York, New York; 2Dept. ofCentral Research and Develop-ment, E.I. du Pont de Nemoursand Company, Wilmington, Dela-ware: Properties of the major spe-cies of tyrosine protein kinase inrat liver-Effects of DNA topoiso-merase activity.
Blenis, J., Sugimoto, Y., Biemann, H.-P., Erikson, R.L., Dept. of Cellularand Developmental Biology, Har-vard University, Cambridge, Mas-sachusetts: Control of cell prolifer-ation and transformation by theRSV transforming gene product.
A., Dept. of Medicine, Universityof California School of Medicine,Los Angeles: Induction of hyper-trophy in a primary culture of renalproximal tubular cells by stimula-tion of Na*-H* antiport.
Frick, K.K.,1 Doherty, P.,2 Scher,C.D.,' Liau, G.,2 Ganguly, K.,'Gottesman, M.M.,2 'Children'sHospital of Philadelphia, Pennsyl-vania; 2NCI, National Institutes ofHealth, Bethesda, Maryland:PDGF, TPA, and retroviruses in-crease the accumulation of MEPRNA transcripts.

Gross, M.,' Yokoyama, S.,' Sathe,G.,' Rosenberg, M.,' Sweet, R.,'Fasano, O.,' Goldfarb, M.,2 Wig ler,M.,2 Kamata, T.,2 Feramisco, J.,2'Smith Kline and French Laborato-ries, Philadelphia, Pennsylvania;'Cold Spring Harbor Laboratory,New York: Purification and charac-terization of mammalian ras pro-teins produced in E. coll.
Haskell, J., Nissley, P, Rechler, M.,Sasaki, N., Greenstein, L., Lee, L.,National Institutes of Health, Be-thesda, Maryland: Phosphoryla-tion of the type-II insulinlike growthfactor receptor in cultured cells.
Heath, WK.,' Isacke, C.M.,2 'Dept. ofZoology, University of Oxford,England; 'Molecular Biology andVirology Laboratory, Salk Institute,San Diego, California: PC13 em-bryonal/carcinoma-derived growthfactor.
Holmgren, A., Dept. of Chemistry, Ka-rolinska Institutet, Stockholm, Swe-den: Functions of thioredoxin in in-sulin and IGFs degradation,receptor modulation, and growthfactor action.
Jonak, G.J., Anton, E.D., Fahey, D.,Friedland, B., Cheng, Y.-S.E.,Knight, E., Jr., Dept. of Central Re-search and Development, E.I. duPont de Nemours and Company,Wilmington, Delaware: Interferon-mediated regulation of the c-myconcogene expression in humancells and its relation to the inhibi-tion of cell growth.
Kruijer, W., Cooper, J., Verma, I.M.,Molecular Biology and VirologyLaboratory, Salk Institute, SanDiego, California: Induction of thec-fos gene by PDGF and FGF inquiescent fibroblasts.
Lanahan, A.,' Strazdis, J.,2 Johnson,D.E.,' Kucherlapati, R.,3 Bothwell,M.,"Dept. of Molecular Biology,Princeton University, New Jersey;'Dept. of Biological Sciences,Oakland University, Rochester,Michigan; .Center for Genetics,University of Illinois, College ofMedicine, Chicago: Transfer of thehuman EGF receptor gene inmouse L cells.
Lehrman, S.R.,' Felix, A.M.,' Heimer,E.,' Boublik, M.,2 'Hoffman-LaRoche Incorporated; 'Roche Insti-tute of Molecular Biology, Nutley,New Jersey: Synthesis and confor-mational analysis of a 37-amino-
Poster Session
acid peptide form the amino termi-nus of p21.
Linzer, D., Lee, S.-J., Nathans, D.,Howard Hughes Medical Institute,Johns Hopkins University Schoolof Medicine, Baltimore, Maryland:Expression of a growth-relatedmember of the prolactin-growthhormone family.
Macdonald-Bravo, H., Bravo, R., Eu-ropean Molecular Biology Labora-tory, Heidelberg, Federal Repubicof Germany: Growth factors in-duce the expression of the nuclearprotein "cyclin" in quiescent 3T3cells.
Magun, B.E., Finch, J.S., Dept. ofAnatomy and Cell Biology, OregonHealth Sciences University, Port-land: Effect of methylamine on in-duction of VL30 gene activity byEGF.
Marchildon, G.A.,' Casnellie, J.E.,2Walsh, K.A.,' Krebs, E.G.,''University of Washington, Seattle;'University of Rochester, NewYork: The LSTRA tyrosine proteinkinase is myristilated.
Moolenaar, W., Defize, L., van derSaag, R, de Last, S., HubrechtLaboratory, Utrecht, The Nether-lands: Cytosolic free Ca++, Na+/H+ exchange and tyrosine phos-phorylation in the action of growthfactors.
Mowat, M., Bernstein, A., Mak, TW.,Benchimol, S., Ontario Cancer Re-search Institute and University ofToronto, Canada: Heterogeneity incellular p53 antigen expressionand tumorigenicity in F-MuLV in-duced cell lines.
Newman, M.J.,' Lane, E.A.,' Racker,E.,2 'Dept. of Biochemistry, Bran-
deis University, Waltham, Massa-chusetts; 'Section of Biochemistry,Cornell University, Ithaca, NewYork: Isolation of a partially trans-formed derivative of normal ratkidney cells-Anchorage-inde-pendent growth induced bygrowth factors.
Norris, J.S.,' Syms, A.J.,2 Smith,R.G.,2 'Depts. of Medicine andPhysiology, University of ArkansasSchool of Medical Sciences, LittleRock; 'Depts. of Urology and CellBiology, Baylor College of Medi-cine, Houston, Texas: Glucocorti-coids inhibit production of c-sis, c-myc poly(A)+ RNA and inhibitgrowth.
Patch, C.T., Akagi, K., Lewis, A.M.,Levine, A.S., National Institutes ofHealth, Bethesda, Maryland: Dif-ferences in the mitogenic activityof hamster cells transformed byadenovirus 2 or SV40.
P asecki, A., Paul, D., Institute of Toxi-cology, Hamburg University Medi-cal School, Hamburg, Federal Re-public of Germany: Growth controlof rat hepatocytes by hormones +rat platelet mitogens + selectionof phenotypic hepatocyte variantsfrom DEN-treated rats in primarycultures.
Quarless, S.A., Dept. of BiologicalSciences, Massachusett8 Instituteof Technology, Cambridge: Prop-erties of a novel coupled deoxyri-bonuclease-protein kinase activityfrom HeLa cells.
Radding, W., Dept. of Cell Biology,New York University MedicalSchool, New York, New York: Ahypothesis about the purpose ofautophosphorylation.
249

Ritt ling, S.R., Hirschhorn, R.R., Gib-son, C.W., Al ler, P, Yuan, Z.-A.,Calabretta, B., Kaczmarek, L.,Baserga, R., Fells Research Insti-tute, Temple University School ofMedicine, Philadelphia, Pennsyl-vania: Cell cycle-specific genesmay be essential for cell cyclecontrol.
Ross, A.H.,' Bothwell, M.,2 Grob, P.,2Marano, N.,2 Chao, M.,3 Ernst,C.S.,4 Elder, D.E.,4 Koprowski,H.,' 'Wistar Institute, Philadelphia,Pennsylvania; 2Dept. of Biochem-istry, Princeton University, NewJersey; 3 Dept. of Cell Biology,Cornell Medical School, New York,New York; 4Dept. of Pathology,Hospital of the University of Penn-sylvania, Philadelphia: Character-ization of the nerve growth factorreceptor using monoclonal anti-bodies.
Scheurich, P, Ucer, U., Pfizenmaier,K., Max-Planck-Society, Gottingen,Federal Republic of Germany: In-fluence of y-IFN on human T cellproliferation via regulation of IL-2receptor expression.
Seifert, B.A.,' Ross, R.,'.2 Bowen-Po-per, D.F,' Depts. of 'Pathology,2Biochemistry, University of Wash-ington, Seattle: Demonstration of"cryptic" receptors for PDGF intransformed cells.
SESSION 4 LYMPHOKINES
Simmen, FA.,' Gope, M.L.,' Schultz,TZ.,' Carpenter, G.,2 Wright,D.A.,2 O'Malley, B.W.,"Dept. ofCell Biology, Baylor College ofMedicine, Houston, Texas; 'Dept.of Biochemistry, Vanderbilt Univer-sity School of Medicine, Nashville,Tennessee; 2Dept. of Genetics,M.D. Anderson Hospital and Tu-mor Institute, Houston, Texas: Hu-man EGF receptor proteins-Anal-ysis of mRNA by translation inXenopus oocytes and cDNA clon-ing.
Stacey, D.W.,' Kung, H.-F.,2 'RocheInstitute of Molecular Biology,2Dept. of Molecular Genetics,Roche Research Center, Nutley,New Jersey: Translation of NIH-3T3 cells by microinjected Ha-rasp21 protein.
Stoppelli, M.P., Verde, P., Locatelli, E.,Blasi, F., International Institute Ge-netics and Biophysics, Naples, It-aly: Induction of urokinase mRNAgrowth factors-Similarities with c-myc induction.
Tamanoi, F, Rao, M., Samiy, S.,Walsh, M., Cold Spring HarborLaboratory, New York: Enzymaticproperties of yeast RAS proteins.
Tempst, P.,' Barth, R.,' Fors, L.,' Win-oto, A.,1 Aebersold, R.,' Woo, D.,2Antoniades, H.,3 Hood, L.,''California Institute of Technology,
Chairperson: K.A. Smith, Dartmouth Medical School, Hanover, New Hampshire
Smith K.A., Dartmouth Medical stage-specific transforming geneSchool, Hanover, New Hampshire: from T-cell lymphomas.Interleukin 2-receptor regulation of Ihle, J.N., Keller, J., Rein, A., NCI,T-cell proliferation. Frederick Cancer Research Facil-
Lane, M.A., Stephens, H.A.F., Doh- ity, Frederick, Maryland: Regula-erty, K.M., Tobin, M.B., Dept. of tion of the proliferation of normalPathology, Dana-Farber Cancer In- and transformed cells by IL-3.stitute, Harvard Medical School, Waldmann, T.A., Leonard, W.J., Dep-Boston, Massachusetts: TLym-I, a per, J.M., Greene, W.C., NCI, Na-
SESSION 5 PDGF
Pasadena; 2University of Califor-nia, Los Angeles; 3 HarvardSchool of Public Health, Boston,Massachusetts: Structural studiesof the PDGF (c-sis) gene, thePDGF-receptor protein, and theirpossible involvement in the forma-tion of specific tumors.
Wang, J.L., Hsu, Y-M., Dept. of Bio-chemistry, Michigan State Univer-sity, East Lansing: Isolation andcharacterization of a growth regu-latory factor from 3T3 cells.
Willey, J.,' Moser, C.,' Moody, T,2Harris, C.,"NCI, National Insti-tutes of Health, Bethesda, Mary-land; 2Dept. of Biochemistry,George Washington University,Washington, D.C.: Characteriza-tion of receptors for the carboxy-terminal tetradecapeptide of gas-trin-releasing peptide in normalhuman bronchial epithelial cells.
Yoakum, G.,' Lechner, J.,' Gabriel-son, E.,' Korba, B.,' Malan-Shi-bley, L.,' Willey, J.,' Valerio, M.,2Shamsuddin, A.,3 Trump, B.,3 Har-ris, C.,"NCI, National Institutes ofHealth, Bethesda; 2Litton Bionet-ics, Rockville; 2Dept. of Pathology,University of Maryland School ofMedicine, Baltimore: Transforma-tion of human bronchial epithelialcells transfected by H-ras onco-gene.
tional Institutes of Health, Be-thesda, Maryland: The structure,function, and expression of thelymphocyte receptor for IL-2.
Wong, G.G., Wang, E.A., Kaufman,R.J., Clark, S.C., Genetics Insti-tute, Boston, Massachusetts: Mo-lecular cloning of a human GM-CSF cDNA.
Chairperson: H. Antoniades, Harvard School of Public Health, Boston, Massachusetts
Antoniades, H.N., Pantazis, P., Owen,A.J., Center for Blood Research,Harvard School of Public Health,Boston, Massachusetts: PDGFand cell transformation.
Pantazis, P.,' Morgan, D.A.,2 Brodsky,1.,2 Antoniades, H.N.,' Center forBlood Research, Harvard School
250
of Public Health, Boston, Massa-chusetts; 2Dept. of Hematologyand Oncology, Hahnemann Uni-versity, Philadelphia, Pennsylva-nia: PDGF polypeptides in humanmegakaryocytelike cell lines.
Aaronson, S.A., Gazit, A., Igarashi,H., Chiu, I.-M., Tronick, S.R., Rob-
bins, K.C., NCI, National Institutesof Health, Bethesda, Maryland:Activation of a normal humangene coding for a growth factor toone with transforming properties.
Deuel, T.F., Tong, B.D. Huang, J.S.,Jewish Hospital at WashingtonUniverstiy Medical Center, St.

Louis, Missouri: PDGF-Roles ingrowth of normal and SSV-trans-formed cells.
Williams, LT.,' Garrett, J.S.,' Cough-lin, &R.: Wang, J.Y.H.,2 Frackel-ton, A.R.,' Daniel, TO.,' 'HowardHughes Medical Institute, Univer-sity of California, San Francisco;2Dept. of Biology, University ofCalifornia, San Diego: Blockade ofautocrine stimulation in SSV-trans-formed cells reverses downregula-tion of PDGF receptors.
Wang, J.Y.J.,' Williams, LT.,' 'Dept.of Biology, University of California,San Diego; 2Howard HughesMedical Institute, University of Cal-ifornia, San Francisco: Biologicalactivity of v-sis-encoded proteinproduced in bacteria.
Bowen-Pope, D.F., DiCorleto, P.E.,Harlan, J.M., Reidy, M.A., Ross,R., Schwartz, S.M., Seifert, R.A.,Vogel, A., Walker, L.N., Dept. ofPathology, University of Washing.
SESSION 6 EGF
Chairperson: B. Ozanne, University
Ullrich, A.,' Coussens, L.,' Hayflick,J.S.,' Dull, T.J.,' Gray, A.,' Tam,A.W.,' Lee, J.,' Yarden, Y,2 Liber-mann, T.A.,2 Schlessinger, J.,2Downward, J.,3 Mayes, E.L.V.,3Whittle, N.,3 Waterfield, M.D.,3Seeburg, RH.,' 'Dept. of Molecu-lar Biology, Genentech, Incorpo-rated, South San Francisco, Cali-fornia; 2Dept. of ChemicalImmunology, Weizmann Institute ofScience, Rehovot, Israel; 3ProteinChemistry Laboratory, ImperialCancer Research Fund, London,England: Structure and expressionof the human EGF receptor.
Gill, G.N., Dept. of Medicine, Univer-sity of California School of Medi-
SESSION 7 TGF
Chairperson: G. Todaro, Oncogen,
Assoian, R., Roberts, A., Anzano, M.,Frolik, C., Wakefield, L., Sporn, M.,NCI, National Institutes of Health,Bethesda, Maryland: Structureand action of transforming growthfactor-p.
Moses, H.L., Shipley, G.D., Tucker,R.F., Dept. of Cell Biology, Mayo
B. Ozanne, C.S. Richards
ton, Seattle: Nonplatelet sourcesof PDGF-Possible roles in trans-formation, normal development,and response to injury.
Pledger, W.J., Herman, B., Universityof North Carolina, Chapel Hill:PDGF-stimulated cellular eventsinitiate proliferation.
of Texas, Dallas, Texas
cine San Diego, La Jolla: Charac-
Merlino, G.T,' Xu, Y. -H.,' Ishii, S.,'Clark, A.,' Wilson, R.,2 Ma, D.P.,2Roe, B.,2 Knutsen, T,' Whang-Peng, J.,' Pastan, I.,"NCI, Na-tional Institutes of Health, Be-thesda, Maryland; 2Dept. ofChemistry, University of Okla-homa, Norman: Cloning and char-acterization of EGF receptor genesequences in A431 human carci-noma cells.
Gilmore, T., DeClue, J., Martin, G.S.,Dept. of Zoology, University ofCalifornia, Berkeley: Tyrosinephosphorylation induced by the v-erb-B protein in vivo and in vitro.
Seattle, Washington
Foundation and Medical School,Rochester, Minnesota: Type-ptransforming growth factor is agrowth stimulator and a growth in-hibitor.
Massague, J., Dept. of Biochemistry,University of Massachusetts Medi-cal Center, Worcester: Identifica-
Armelin, M.C.S.,' Armelin, H.,' Coch-ran, B.H.,2 Stiles, C.D.,2 'Dept. ofBioquimica, Universidade de SaoPaulo, Brazil; 2Dana-Farber Can-cer Institute, Harvard MedicalSchool, Boston, Massachusetts:Intracellular mediators of the mito-genic response to PDGF
Novak-Hofer, I., Martin Perez, J.,Thomas, G., Fried-
rich Miescher-Institut, Basel, Switz-erland: EGF-mediated phosphoryl-ation of 40S ribosomal protein S6phosphorylation in Swiss mouse3T3 cells.
Hendler, F.,' Shum, A.,' Richards,Cassells, D.,' Gusterson,
B.,2 Ozanne, B.,"Depts. of Inter-nal Medicine and Microbiology,Universiy of Texas Health ScienceCenter, Dallas; 2Ludwig Institute,Sutton, England: Evidence for in-creased EGF receptor in epider-moid malignancies.
Lion of membrane receptors fortype-p transforming growth factorin target cells.
Derynck, R.,' Roberts, A.B.,3 Winkler,M.E.,2 Chen, E.Y.,' Goeddel,D.V.,' Depts. of 'Molecular Biol-ogy, 2Protein Biochemistry, Ge-nentech, Incorporated, South San
251

Francisco, California; 'NCI, Na-tional Institutes of Health, Be-thesda, Maryland: Human trans-forming growth factor -a-Precursor structure and heterolo-gous expression.
Adkins, B.,',2 Leutz, A.,' Graf, T,''European Molecular Biology Lab-oratory, Heidelberg, Federal Re-
SESSION 8 INSULIN
public of Germany; 'Dept. of Pa-thology, Stanford University,California: Viruses carrying v-src-related oncogenes induce auto-crine growth in transformedchicken myeloid cells.
Sager, R., Craig, R., Dana-FarberCancer Institute, Harvard MedicalSchool, Boston, Massachusetts:
Chairperson: 0. Rosen, Albert Einstein College of Medicine, Bronx, New York
Czech, M.P., Mottola, C., Oka, Y., Yu,K.-T., Davis, R., MacDonald, R.G.,Dept. of Biochemistry, Universityof Massachusetts Medical School,Worcester: Mechanisms that regu-late the receptors for insulin andthe insulinlike growth factors.
Roth, R.A.,' Morgan, D.O.,' Masara-chia, R.A.,2 'Dept. of Pharmacol-ogy, Stanford University School ofMedicine, California; 'Dept. of Bio-chemistry, North Texas State Uni-versity, Denton: Monoclonal anti-bodies as probes of the structureand function of the insulin recep-tor.
SESSION 9 FUTURES
Rosen, 0.M., Ganguly, S., Herrera,R., Petruzzelli, L.M., Stadtmauer,L., Memorial Sloan-Kettering Can-cer Center, New York, New York:The insulin-dependent tyrosineprotein kinase of human placenta.
Fujita-Yamaguchi, Y., Kathuria, S.,Beckman Research Institute of theCity of Hope, Duarte, California:Relationship between insulin re-ceptor structure and its protein ki-nase activity.
Bruni, C.B.,' Frunzio, R.,' Whitfield,H.J.,2 Rechler, M.M.,2 'Centro diEndocrinologia ed OncologiaSperimentale, CNR, Naples, Italy;
Chairperson: A. Pardee, Dana-Farber Cancer Institute, Boston, Massachusetts
Pardee, A.B., Dana-Farber Cancer In-stitute, Harvard Medical School,Boston, Massachusetts: New di-
252
rections-Molecular. basis ofgrowth regulation.
Cunningham, D., Thompson, J., Hira-moto, S., Van Nostrand, K., Dept.of Microbiology and Molecular Ge-netics, University of California, Ir-vine: Thrombin-stimulated cell divi-sion.
Pouyssegur, J., Chambard, J.-C.,Franchi, A., L'Allemain, G., Paris,S., Van Obberghen-Schilling, E.,Centre de Biochimie, CNRS, Nice,France: Growth factor activation ofthe Na' /H' antiporter controlsgrowth of fibroblasts by regulatingintracellular pH.
Jakobovits, A., Martin, G.R., Dept. ofAnatomy, University of California,San Francisco: Embryonal carci-noma-derived growth factors-Specific growth-promoting and dif-ferentiation-inhibiting activities.
Villereal, M., Owen, N., Vic.entini, L.,Mix-Muldoon, L., Jamieson, G.,
Genetic suppression of tumori-genicity in hybrids from fusions ofnormal x EJ-transformed CHEF/18 cells.
2NIADDKD, National Institutes ofHealth, Bethesda, Maryland:Structure and expression of rat in-sulinlike growth factor-II gene.
Jacobs, S., Wellcome Research Labo-ratories, Research Triangle Park,North Carolina: Phorbol ester-stim-ulated phosphorylation of recep-tors for insulin and IGF-I.
Dept. of Pharmacology and Phys-iological Science, University ofChicago, Illinois: Mechanism forgrowth factor-induced increase ofNa+/H + exchange and in Ca+ +activity in cultured human fibro-blasts.
O'Brian, C., Columbia University Col-lege of Physicians & Surgeons,New York, New York: Studies onPKC and their relevance to tumorpromotion.
Maness, P.F., Dept. of Biochemistry,University of North CarolinaSchool of Medicine, Chapel Hill:Expression of pp60c-src inthe de-veloping nervous system.
Chen, L.B., Weiss, M.J., Davis, S.,Shepherd, E.L., Walker, E.S.,Dana-Farber Cancer Institute, Har-vard Medical School, Boston,Massachusetts: The use of rhoda-mine-123 to probe mitochondria!bioenergetics in living cells.

BANBURYCENTER

Recto: Detail of Sammis Hall guest house at Banbury Center.

BANBURY CENTERDIRECTOR'S REPORT
This was the seventh consecutive year of operation of the Banbury Center ofCold Spring Harbor Laboratory. Throughout this period, there has been a contin-ual expansion of the Center's conference programs and publications, particularlyin the realms of health risk assessment and carcinogenesis, as well as in the im-pact upon these areas of recent developments in molecular biology. That thisprogram has been able to flourish during a period when governmental supporthas become increasingly scarce and problematic is due, to a large extent, to theresponse the Center has been fortunate in eliciting from private sector sources. Aparticularly important step in 1984 was the partial consolidation of such supportunder the Cold Spring Harbor Laboratory Corporate Sponsor Program. This andthe ongoing essential contributions from Core Supporters of the Banbury Centerand specific corporate contributions toward particular programs remain crucialfor the continuance and further development of the Banbury program.
It is perhaps fitting that the 1984 schedule began with the first meeting of thenewly initiated Corporate Sponsor Program. This, the first of three such Sponsormeetings, was on Site-directed Mutagenesis and Protein Structure and Function.The meeting was almost prototypic of those to be held within this program, whichenables the development of small scientific conferences in the rapidly develop-ing field of molecular biology, especially in the area of recombinant DNA method-ologies and their applications. These emphases were reflected in the remainingtwo Sponsor meetings of the year, a June conference on Yeast Expression Vec-tors and a November meeting on Transport and Secretion of Proteins.
Robertson House provideshousing and diningaccommodations atBanbury Center
255

Sammis Hall, guest house
The Sponsor Program thus acted as a rich complement to the ongoing Ban-bury concern with human health risk assessment in addition to the Center's in-creasing interest in newly emerging areas in the biological sciences that mayparticularly bear social or regulatory implications. The first major conference ofthe year in these areas was an April conference on the Biological Mechanisms ofDioxin Action. Organized jointly by Dr. Alan Poland (McArdle Laboratory of theUniversity of Wisconsin) and Dr. Renate Kimbrough (Centers for Disease Con-trol), this conference focused on the specific mechanisms by which the dioxinsand closely related halogenated aromatic hydrocarbons bring about their broadrange of biological effects. These compounds represent potentially far-reachinghuman health consequences and are among the most highly publicized and inci-tive of environmental pollutants. Bringing epidemiological and public healthpoints of view together with major cellular and molecular research approaches,in the nonadversarial yet highly interactive setting of a small Banbury confer-ence, was the impetus for this important conference. The papers and discussionsof this meeting will appear early in 1985 as volume 18 in the Banbury Reportsseries.
The second major risk assessment conference of the year, to be published in1985 as Banbury Report 19, was a mid-May conference on Risk Quantitationand Regulatory Policy. Organized by Dean Richard Merrill (University of VirginiaSchool of Law) and Dr. David Hoel (Director of the Biometry and Risk Assess-ment Program of the National Institute of Environmental Health Sciences), thisconference brought together legal, regulatory, and research representatives toconsider the most efficacious ways to make the evolving science of risk assess-ment accessible and amenable to incorporation into regulatory policy formula-tions. The papers and discussions of this meeting proved a most useful compen-dium of centralized information and ideas in an area that has otherwise becomea frequent source of public consternation and institutional conflict.
The final 1984 conference, also to be published in the Banbury series, was anOctober meeting organized by Dr. Frank Costantini (Columbia University) and Dr.Rudolph Jaenisch (now of the Whitehead Institute) on the Genetic Manipulation
256

of the Mammalian Ovum and Early Embryo. This relatively new and rapidly ad-vancing research area has already received considerable public attention. Yetthis attention has rarely focused on the new abilities that such advances havebeen bringing to current approaches toward an understanding of gene regula-tion and the genetic control of development. Given the nature and potentially far-reaching impact of this field, a present-state assessment and consideration of itslikely future developments seemed especially appropriate at this particular time.
The 1984 Banbury program was rounded out by a full complement of summercourses and a series of smaller workshops held throughout the year. Among
these smaller workshops was one held in early spring on the large T antigen of
the SV40 oncogenic virus, followed by a National Institute of Mental Health Work-
shop on Single Unit Activity and Behavior, an Alfred P Sloan Foundation Compu-
tational Neuroscience Workshop, and a late October workshop on the Structure
and Function of Gap Junctions. The meeting year concluded at the end of No-
vember with a further Journalists' Workshop in the series held under the auspices
of an ongoing grant from the Alfred P Sloan Foundation. As previously men-
tioned, public confusion and concern often arises over issues of en-
vironmental health risk assessment. This is especially true in areas of occupa-
tional and environmental carcinogenesis. This year's journalists' workshop topic
was thus "Assessing Risk Assessment". The workshop again proved to be both
an intense and enjoyable few days, leading to considerably greater journalistic
sophistication concerning the strengths and weaknesses in the quantitation of
risk.
1984 Support
As noted earlier, the ability to carry on such an overall program is due largely to
support that the Banbury Center has been fortunate to receive from private sec-
tor sources. The donation to Cold Spring Harbor Laboratory of the Banbury es-
tate by Charles S. Robertson generously included an endowment for the mainte-
nance of the grounds and of Robertson House. Funding of actual programs,
257
Robertson House Library
Robertson House hall

Banbury Center Meeting House
however, as well as maintenance and operation of the Meeting House and ofSammis Hall remain the responsibility of the Banbury Center itself. It is thus withconsiderable gratitude that I take this opportunity to thank the Corporate Spon-sors and Core Supporters of the Center. The names of these companies arelisted separately in this report. It is also with great thanks that I acknowledge thesupport of specific programs from the following sources: Biological Mechanismsof Dioxin Action was supported by contributions from Hoffmann-La Roche Inc.,Monsanto Company, the Dow Chemical Company, and Diamond Shamrock Cor-poration; Risk Quantitation and Regulatory Policy was supported in part by agrant from the U.S. Department of Energy, together with contributions from theMonsanto Company and National Distillers and Chemical Corporation; and Ge-netic Manipulation of the Mammalian Ovum and Early Embryo was held undergrants from the March of Dimes Birth Defects Foundation, the Fogarty Interna-tional Center, and the National Institute of Child Health and Human Develop-
ment.
Michael Shodell
258

MEETINGS
SV40 Large T AntigenMarch 8-March 11
ARRANGED BY
E. Harlow, Cold Spring Harbor Laboratory, New York
SESSION 1 MUTANTS
Chairperson: P. Tegtmeyer, State University of New York, Stony Brook
C. Cole, Dartmouth Medical School,Hanover, New Hampshire
D. Kalderon, MRC National Institutefor Medical Research, London,England
R. Lanford, Baylor College of Medi-cine, Houston, Texas
M. Manor, Cold Spring Harbor Labo-ratory, New York
K. Peden, Johns Hopkins University,Baltimore, Maryland
T. Shenk, State University of NewYork, Stony Brook
A.E. Smith, MRC National Institute forMedical Research, London, Eng-land
SESSION 2 TRANSFORMATION
Chairperson: C. Prives, Columbia University, New York, New York
R. Baserga, Temple University Medi- juif, Francecal School, Philadelphia, Pennsyl- J. Pipas, University of Pittsburgh,vania Pennsylvania
F Birg, INSERM, Marseille, France R. Pollack, Columbia University, NewM. Botchan, University of California, York, New York
BerkeleyJ. Feunteun, Institut de Recherches
Scientifiques sur le Cancer, Ville-
SESSION 3 SURFACE T
Chairperson: E. Harlow, Cold Spring Harbor Laboratory, New York
J. Butel, Baylor College of Medicine,Houston, Texas
W. Deppert, Universitat Ulm, FederalRepublic of Germany
L. Gooding, Emory University, At-lanta, Georgia
SESSION 4 EXPRESSION SYSTEMS
B. Knowles, Wistar Institute, Philadel-phia, Pennsylvania
S. Tevethia, Pennsylvania State Uni-versity, Hershey
Chairperson: E. Harlow, Cold Spring Harbor Laboratory, New York
Y. Gluzman, Cold Spring Harbor Lab-oratory, New York Laboratory, New York ley
P. Rigby, Imperial College, London,England
L. Sompayrac, University of Colo-rado, Boulder
T. Grodzicker, Cold Spring Harbor D. Rio, University of California, Berke-
259

SESSION 5 DNA BINDING AND REGULATION
Chairperson: T. Shenk, State University of New York, Stony Brook
J.C. Alwine, University of Pennsylva-nia, Philadelphia
J. Brady, Laboratory of Molecular Vi-rology, National Institutes ofHealth, Bethesda, Maryland
R. Dixon, Merck Sharp & Dohme,West Point, Pennsylvania
E. Fanning, Ludwig Maximillian Uni-versity, Munich, Federal Republicof Germany
D.P. Lane, Imperial College, London,England
D.M. Livingston, Sidney Farber Can-cer Institute, Boston, Massachu-setts
J.L. Manley, Columbia University,New York, New York
D. Rio, University of California, Berke-ley
J. Stringer, University of Cincinnati,Ohio
P. Tegtmeyer, State University of NewYork, Stony Brook
SESSION 6 MODIFICATIONS WORKSHOP
Chairperson: D.M. Livingston, Sidney Farber Cancer Institute, Boston, Massachusetts
R.B. Carroll, New York University E. May, Institut de Recherches Scien-Medical Center, New York tifiques sur le Cancer, Villejuif,
W. Deppert, Universitat Ulm, Federal FranceRepublic of Germany D. Simmons, University of Delaware,
R. Henning, Universitat Ulm, Federal NewarkRepublic of Germany
SESSION 7 ANTIBODIES WORKSHOP
Chairperson: D.P. Lane, Imperial College, London, England
L. Gooding, Emory University, At- E. Paucha, MRC National Institute forlanta, Georgia Medical Research, London, Eng-
E. Gurney, Institut de Recherches landScientifiques sur le Cancer, Ville-juif, France
SESSION 8 SEQUENCES WORKSHOP
Chairperson: S. Weissman, Yale University, New Haven, Connecticut
G. Walter, University of California, SanDiego, La Jolla
Site-directed Mutagenesis and Protein Structure and FunctionMarch 18-March 21
ARRANGED BY
R.T. Sauer, Massachusetts Institute of Technology, Cambridge
260

SESSION 1 ALTERATIONS OF ENZYMATIC PROPERTIES I
Chairperson: M. Smith, University of British Columbia, Vancouver, Canada
M. Smith, University of British Colum-bia, Vancouver, Canada: Oligonu-cleotide mutagenesis and thestudy of heme proteins.
S.J. Benkovic, Pennsylvania StateUniversity, Hershey: Strategies formutagenesis: Preliminary resultson dihydrofolate reductase.
S.A. Benner, Harvard University,Cambridge, Massachusetts: Syn-
thetic genes for fundamental stud-ies in protein chemistry.
G.A. Petsko, Massachusetts Instituteof Technology, Cambridge: Site-di-rected alteration of enzymaticproperties.
M.L. Sinnott, University of Bristol,England: The catalytic conse-quences of experimental evalua-tion of the ebg gene of E. coli.
SESSION 2 ALTERATIONS OF ENZYMATIC PROPERTIES II
Chairperson: J.R. Knowles, Harvard University, Cambridge, Massachusetts
J.R. Knowles, Harvard University,Cambridge, Massachusetts: Re-gional mutagenesis in the signalcodons of the fl-lactamase gene.
A.R. Fersht, Imperial College of Sci-ence and Technology, London,England: Structure and activity oftyrosyl-tRNA synthetase.
M. Zoller, Cold Spring Harbor Labora-tory, New York: Specific DNA/pro-tein interactions in yeast. Point mu-tations in the DNA recognitionsequence of the "HO" endonu-clease.
P.R. Schimmel, Massachusetts Insti-tute of Technology, Cambridge:Domain structure of amino acyltRNA synthetase.
R. Wetzel, Genentech, Inc., South SanFrancisco, California: Thermosta-bility of phage T4 lysozyme.
J.H. Richards, California Institute ofTechnology, Pasadena: /3-lacta-mase: Mutagenic strategies andproperties of mutants-Process-ing, secretion, stability, catalysis.
J.A. Ger lt, Yale University, New Ha-'ven, Connecticut: Genetic studiesof nucleotidyl transferases.
D. Shortie, State University of NewYork, Stony Brook: Physical chem-ical genetics of staphylococcal nu-clease.
SESSION 3 NONENZYME STRUCTURE/ACTIVITY CORRELATIONS
J.N. Abelson, California Institute ofTechnology, Pasadena.
J. Kraut, University of California, SanDiego, La Jolla: Site-directed mu-tagenesis of E. col/ dihydrofolatereductase.
W.J. Rutter, University of California,San Francisco: Altering trypsinproteolytic specificity.
Chairperson: H.O. Smith, Johns Hopkins University School of Medicine, Baltimore,Maryland
P.B. Berget, University of Texas Medi- Technology, Cambridge: Usingcal School, Houston: Structure/ mutants to probe the stability andfunction relationships in the bacte- activities of DNA-binding proteins.riophage P22 tail protein. R.H. Ebright, Harvard Medical
J.M. Pipas, University of Pittsburgh, School, Boston, Massachusetts:Pennsylvania: The large tumor an- Mutations that alter the DNA se-tigen of simian virus 40-Genetic quence specificity of the cataboliteanalysis of a viral oncogene. gene activator protein of E. col /.
G.K. Ackers, Johns Hopkins Univer- M. Ptashne, Harvard University, Cam-sity, Baltimore, Maryland: Hemo- bridge, Massachusetts: Repres-globin mutants and cooperative sors and cro proteins-Structureenergies. and function.
P.A. Youderian, University of Southern S.C. Harrison, Harvard University,California, Los Angeles: Altering Cambridge, Massachusetts: Struc-DNA-binding specificities of re- tural organization of sphericalpressor proteins. plant viruses.
R.T. Sauer, Massachusetts Institute of
261

SESSION 4 PROTEIN FOLDING DYNAMICS AND STABILITY
Chairperson: R.L. Baldwin, Stanford University Medical Center, California
R. Matthews, Stanford University M. Karplus, Harvard University, Cam-Medical Center, California: Protein bridge, Massachusetts: Proteinalteration and folding of dihydro- folding: Some model studies.folate reductase. M. Levitt, Weizmann Institute of Sci-
B.T. Nall, University of Texas Medical ence, Rehovot, Israel: ComputerSchool, Houston: Folding of yeast modeling of mutant proteins.cytochrome c. J.B. Matthew, Genex Corporation,
I.B. Kingston, MRC Laboratory of Mo- Gaithersburg, Maryland: Electro-lecular Biology, Cambridge, Eng- static contribution to protein stabil-land. ity and macromolecular assembly.
SESSION 5 NOVEL PEPTIDES
G. Rose, Pennsylvania State Univer-sity, Hershey: Toward a taxonomyof protein structure.
H.A. Scheraga, Cornell University, Ith-aca, New York: Molecular recogni-tion in proteins.
Chairperson: M.F. Perutz, MRC Laboratory of Molecular Biology, Cambridge, England
M. Eigen, Max-Planck-Institut fur bio-physikalische Chemie, Gottingen,Federal Republic of Germany:Evolving systems.
E.T. Kaiser, Rockefeller University,New York, New York: Amphiphilic
secondary structures-The designof biologically active peptides.
C.O. Pabo, Johns Hopkins MedicalSchool, Baltimore, Maryland: De-signing novel peptides.
SESSION 6 HEMOGLOBIN AND PROTEIN EVOLUTION
J.S. Richardson, Duke University,Durham, North Carolina: Designcriteria for inventing a protein.
Chairperson: M.F. Perutz, MRC Laboratory of Molecular Biology, Cambridge, England
M.F. Perutz, MRC Laboratory of Mo-lecular Biology, Cambridge, Eng-land: The structural basis of pro-tein evolution.
R.F. Doolittle, University of California,San Diego, La Jolla: Evolving pro-teins with novel functions.
Biological Mechanisms of Dioxin ActionApril 1-April 4
ARRANGED BY
A. Poland, University of Wisconsin, MadisonR.D. Kimbrough, Centers for Disease Control, Atlanta, Georgia
SESSION 1 CHEMISTRY AND GENERAL PATHOLOGY I
Chairperson: R.D. Kimbrough, Centers for Disease Control, Atlanta, Georgia
D. Firestone, U.S. Food and DrugAdministration, Washington, D.C.:Chlorinated aromatic compoundsand related dioxins and furans-Production, uses, and environ-mental exposure.
C
E.
. Rappe, University of Umea, Swe-den: Chemistry and analysis ofpolychlorinated dioxins and diben-zofurans in biological samples.
E. McConnell, National Institute ofEnvironmental Health Sciences,
SESSION 2 CHEMISTRY AND GENERAL PATHOLOGY II
Chairperson: J.A. Moore, Environmental Protection Agency, Washington, D.C.
H. Poiger, Federal Institute of Technol- Switzerland: The metabolism ofogy and the University of Zurich, TCDD in the dog and rat.
262
Research Triangle Park, NorthCarolina: Clinicopathologic con-cepts of dibenzo-p-dioxin intoxica-tion.
R.A. Neal, Chemical Industry Instituteof Toxicology, Research Triangle

Park, North Carolina: Metabolismof TCCD intoxication.
R.M. Pratt, National Institute of Envi-ronmental Health Sciences, Re-
SESSION 3 RECEPTOR BINDING
search Triangle Park, North Caro-lina: Mechanism of TCDD-inducedcleft palate in the mouse.
R.J. Kociba, Dow Chemical USA,
Chairperson: A. Poland, University of Wisconsin, Madison
D.R. Koop, University of MichiganMedical School, Ann Arbor: Iden-tity of cytochromes P-450 inducedby diverse xenobiotics.
B.A. Taylor, Jackson Laboratory, BarHarbor, Maine: The aryl hydrocar-bon hydroxylase inducibility locus(Ah) of the mouse, a genetic per-spective.
A. Poland, University of Wisconsin,Madison: Reflections on the mech-anisms of action of halogenatedaromatic hydrocarbons.
J.A. Goldstein, National Institute ofEnvironmental Health Sciences,Research Triangle Park, NorthCarolina: Regulation of a multi-gene family of P-450 isozymes byTCDD and related compounds.
S.H. Safe, Texas A&M University, Col-lege Station: Binding to the TCDDreceptor and AHH/EROD induc-tion-In vitro QSAR.
SESSION 4 ENZYME INDUCTION
Midland, Michigan: Evaluation ofthe carcinogenic and mutagenicpotential of TODD and other chlor-inated dioxins.
Chairperson: J.A. Goldstein, National Institute of Environmental Health Sciences,Research Triangle Park, North Carolina
J.-A. Gustafsson, Huddinge Univer-sity Hospital, Sweden: Physico-chemical characteristics of theTCDD receptor.
T.A. Gasiewicz, University of Roches-ter Medical Center, New York: Evi-dence for a homologous nature ofAh receptors among various mam-malian species.
A. Schecter, State University of NewYork, Binghamton: Ultrastructuralalterations of liver mitochondria inresponse to dioxins, furans, PCBs,and biphenylenes.
J.P. Whitlock, Jr., Stanford UniversitySchool of Medicine, California:TODD regulates cytochrome P,-450 gene expression.
SESSION 5 BIOCHEMICAL CHANGES IN LIVER
Chairperson: J.B. Greig, MRC Toxicology Unit, Carshalton, England
S. Sassa, Rockefeller University, NewYork, New York: Inhibition of uro-porphyrinogen decarboxylase ac-tivity in polyhalogenated aromatedhydrocarbon poisoning.
G.D. Sweeney, McMaster University,Hamilton, Canada: Mechanismsunderlying the hepatotoxicity ofTCDD.
S.J. Stohs, University of NebraskaMedical Center, Omaha: Inductionof lipid peroxidation and inhibitionof glutathione peroxidase byTCDD.
A.B. Rifkind, Cornell University Medi-cal Center, New York, New York:The chick embryo as a model forPCB and dioxin toxicity-Evidence
H.J. Eisen, National Institute of ChildHealth and Human Development,Bethesda, Maryland: The nuclearTODD Ah receptor complex andthe induction of cytochrome P,-450 mRNA.
of cardiotoxicity and increasedprostaglandin synthesis.
F. Matsumura, Michigan State Univer-sity, East Lansing: Toxicologicalsignificance of pleiotropic changesof plasma membrane functions,particularly that of EGF receptorcaused by TCDD.
263

SESSION 6 LIPID METABOLISM AND WASTING DISEASE
Chairperson: R.E. Peterson, University of Wisconsin, Madison
R.E. Peterson, University of Wiscon-sin, Madison: The wasting syn-drome in TCDD toxicity-Basicfeatures and their interpretation.
S.D. Aust, Michigan State University,East Lansing: On the mechanismof anorexia and toxicity of TODDand related compounds.
C.M. Schiller, National Institute of En-vironmental Health Sciences, Re-search Triangle Park, North Caro-lina: Metabolic impairmentassociated with a low dose ofTCDD in adult male Fischer rats.
T. Thunberg, Karolinska Institute,Stockholm, Sweden: Effect of
SESSION 7 SKIN AND IN VITRO RESPONSES
Chairperson: E.E. McConnell, National Insitute of Environmental Health Sciences,Research Triangle Park, North Carolina
R.D. Kimbrough, Centers for DiseaseControl, Atlanta, Georgia: Skin le-sions in animals and humans: A R.
brief overview.W.F. Greenlee, Chemical Industry In-
stitute of Toxicology, Research Tri-angle Park, North Carolina: Stud- J.
ies on the mechanisms of toxicityof TCDD to human epidermis.
H. Rice, Harvard School of PublicHealth, Boston, Massachusetts:Response of malignant epidermalkeratinocytes to TCDD.
Knutson, University of Wisconsin,
SESSION 8 IMMUNOLOGICAL MECHANISMS
TCDD on vitamin A and its relationto TCDD toxicity.
K.K. Rozman, University of KansasMedical Center, Kansas City: Roleof thyroid hormones and brownadipose tissue in the toxicity ofTCDD.
Madison: XB cells-An in vitromodel for the differentiation andproliferation response to TCDD.
J.B. Greig, MRC Toxicology Unit, Car-shalton, England: Differences be-tween skin and liver toxicity ofTCDD in mice.
Chairperson: J.G. Vos, National Institute of Public Health, Bilthoven, The Netherlands
J.G. Vos, National Institute of PublicHealth, Bilthoven, The Nether-lands: Dioxin-induced thymic atro-phy and suppression of thymus-dependent immunity.
M.I. Luster, National Institute of Envi-ronmental Health Sciences, Re-search Triangle Park, North Caro-
SESSION 9 EPIDEMIOLOGY
lina: In vivo and in vitro effects ofTCDD on stem cell and B cell dif-ferentiation.
G.D. Sweeney, McMaster University,Hamilton, Canada: Dose re-sponse, time-course, and mecha-nism for suppression of cytotoxic Tcell generation by TCDD.
Chairperson: N. Nelson, New York University Medical Center, New York
H. Falk, Centers for Disease Control,Atlanta, Georgia: A pilot epidemio-logic study of health effects due toTCDD contamination in Missouri.
M.A. Fingerhut, National Institute for
Occupational Safety and Health,Cincinnati, Ohio: An evaluation ofreports of dioxin exposure andsoft tissue sarcoma pathology inU.S. chemical workers.
Risk Quantitation and Regulatory Policy
W.F. Greenlee, Chemical Industry In-stitute of Toxicology, Research Tri-angle Park, North Carolina: A pro-posed model for the actions ofTCDD on epidermal and thymicepithelial target cells.
G.D. Lathrop, Brooks Air Force Base,San Antonio, Texas: An epidemio-logic investigation of health effectsin Air Force personnel followingexposure to herbicides.
May 13-May 16
ARRANGED BY
D. Hoel, National Institute of Environmental Health Sciences, Research Triangle Park, North CarolinaR.A. Merrill, University of Virginia, CharlottesvilleM.C. Pike, Imperial Cancer Research Fund, Oxford, England
264

SESSION 1 REGULATORY PROGRAMS UTILIZING RISK ASSESSMENT
Chairperson: L.B. Lave, Carnegie-Mellon University, Pittsburgh, Pennsylvania
L.B. Lave, Carnegie-Mellon Univer-sity, Pittsburgh, Pennsylvania:Quantitative risk assessment inregulation.
P.B. Hutt, Covington & Burling, Wash-ington, D.C.: Use of quantitativerisk assessment in regulatory deci-sion making under federal healthand safety statutes.
M.R. Taylor, King & Spalding, Wash-
ington, D.C.: The use of risk as-sessment in food safety decisionmaking: The scope of FDA's dis-cretion and the safeguards againstabuse.
R.A. Merrill, University of VirginiaSchool of Law, Charlottesville: Thesignificance of risk quantitation inhealth and environmental regula-tion.
SESSION 2 EPIDEMIOLOGY IN RISK ESTIMATION I
Chairperson: M.C. Pike, Imperial Cancer Research Fund, Oxford, England
M.C. Pike, Imperial Cancer ResearchFund, Oxford, England: Epide-miology and risk assessment: Esti-mation of GI cancer risk from as-bestos in drinking water.
P. Landrigan, National Institute for Oc-cupational Safety and Health, Cin-cinnati, Ohio: Approaches to theevaluation of dose in occupationalepidemiology.
SESSION 3 EPIDEMIOLOGY IN RISK ESTIMATION II
Chairperson: M.C. Pike, Imperial Cancer Research Fund, Oxford, England
J.M. Kaldor, International Agency forResearch on Cancer, Lyon,France: The use of epidemiologi-cal data for the assessment of hu-man cancer risk.
J. Peto, Institute of Cancer Research,Sutton, England: Limitations in thequantitation of carcinogenic risk-The asbestos data base.
SESSION 4 MODELING AND EXTRAPOLATION
Chairperson: R. Wilson, Harvard University, Cambridge, Massachusetts
M.D. Hogan, National Institute of En-vironmental Health Sciences, Re-search Triangle Park, North Caro-lina: Risk estimation models inepidemiology studies.
J. Van Ryzin, Columbia UniversitySchool of Public Health, New York,New York: Consequences of non-
linear kinetic dose responsemodels on carcinogenic risk as-sessment.
R. Wilson, Harvard University, Cam-bridge, Massachusetts: Expres-sion of uncertainty in risk assess-ment.
SESSION 5 MUTAGENIC RISK IN HUMAN POPULATIONS
Chairperson: F.P. Perera, Columbia University School of Public Health, New York,New York
M.F. Lyon, MRC Radiobiology Unit,Harwell, England: Attempts to esti-mate genetic risks caused by mu-tagens to later generations.
J.V. Neel, University of Michigan
Medical School, Ann Arbor: Howdo we get better human geneticrisk data, and how then do we usethe data?
265

SESSION 6 TOXICOLOGY AND BIOLOGICAL MECHANISMS I
Chairperson: I.B. Weinstein, CollegeNew York, New York
I.F.H. Purchase, Imperial Chemical Industries PLC, Macclesfield, Eng-land: The toxicologist's contribu-tion to risk quantitation.
R.L. Dedrick, National Institutes ofHealth, Bethesda, Maryland: Ap-plication of model systems in phar.macokinetics.
of Physicians & Surgeons, Columbia University,
J.R. Gillette, National Institutes ofHealth, Bethesda, Maryland: Bio-logical variation: The unsolvableproblem in quantitative extrapola-tions from laboratory animals andother surrogate systems to humanpopulations.
SESSION 7 TOXICOLOGY AND BIOLOGICAL MECHANISMS II
Chairperson: B.D. Goldstein, Environmental Protection Agency, Washington, D.C.
I.B. Weinstein, College of Physicians& Surgeons, Columbia University,New York, New York: Relevance of
SESSION 8 SPECIFIC CASE HISTORIES
mechanisms of action of tumorpromoters to risk assessment.
R.H. Reitz, Dow Chemical Company,
F.P. Perera, Columbia UniversitySchool of Public Health, New York,New York: Methods of measuringbiologically effective doses of car-cinogenic substances-Monitoringusing DNA adducts.
Midland, Michigan: Mechanisticconsiderations in the formulationof carcinogenic risk estimations.
Chairperson: P.W. Preuss, Consumer Product Safety Commission, Bethesda, Maryland
J.A. Swenberg, Chemical Industry In-stitute of Toxicology, Research Tri-angle Park, North Carolina: A sci-entific approach to formaldehyderisk assessment.
M. Cohn, Consumer Product SafetyCommission, Bethesda, Maryland:
Risk assessment and the formal-dehyde data base.
M. Corn, Johns Hopkins University,Baltimore, Maryland: Human ex-posure estimates in hazardouswaste site risk assessment.
B.D. Goldstein, Environmental Protec-tion Agency, Washington, D.C.:Risk assessment and risk manage-ment of benzene by the EPA.
SESSION 9 RISK QUANTITATION AND THE DYNAMICS OF POLICY FORMULATION
Chairperson: R.S. Merrill, University of Virginia School of Law, Charlottesville
R.E. Albert, New York University Med- EPA's guidelines for carcinogenical Center, New York: Issues of risk assessment.concern in the revision of the U.S. P.W. Preuss, Consumer Product
Yeast Expression Vectors
Safety Commission, Bethesda,Maryland: The changing role of riskassessment in Federal regulation.
June 21-June 24
ARRANGED BY
J. Strathern, Cold Spring Harbor Laboratory, New YorkJ.R. Broach, State University of New York, Stony Brook
SESSION 1 VECTORS
M.V. Olson, Washington UniversitySchool of Medicine, St. Louis, Mis-souri
R. Rothstein, New Jersey MedicalSchool, Newark
S. Weisbrod, Cold Spring HarborLaboratory, New York
266
K. Bloom, University of North Caro- V.A. Zakian, Fred Hutchinson Cancerlina, Chapel Hill Research Center, Seattle, Wash-
R.W. Davis, Stanford University Medi- ingtoncal Center, California
C.P. Hollenberg, Universitat Dussel-dorf, Federal Republic of Germany

SESSION 2 PROMOTERS
L.P. Guarente, Massachusetts Insti-tute of Technology, Cambridge
A. Hinnen, Ciba-Geigy AG, Basel,Switzerland
F Sherman, University of RochesterSchool of Medicine, New York
J. Hopper, Pennsylvania State Univer-sity, Hershey
J.R. Broach, State University of NewYork, Stony Brook
S. Fields, University of California, SanFrancisco, School of Medicine
SESSION 3 PROMOTERS II
P. Silver, Harvard University, Cam-bridge, Massachusetts
A. Kingsman, University of Oxford,England
G.R. Fink, Massachusetts Institute ofTechnology, Cambridge
K. Nasmyth, Laboratory of MolecularBiology, Cambridge, England
SESSION 4 PROTEIN LOCALIZATION AND SECRETION
J. Thorner, University of California,Berkeley
R.W. Schekman, University of Califor-nia, Berkeley
SESSION 5 EXPRESSION SYSTEMS
R.A. Hitzeman, Genentech, Inc.,South San Francisco, California
G.A. Bitter, Applied Molecular Ge-
S. Emr, California Institute of Technol-ogy, Pasadena
G. Sprague, Jr., University of Oregon,Eugene
netics, Inc., Thousand Oaks, Cali-fornia
M. Duncan, Collaborative Research,
J. Abraham, California Biotechnology,Inc., Palo Alto
L. Hereford, Dana-Farber Cancer In-stitute, Boston, Massachusetts
K. Bostian, Brown University, Provi-dence, Rhode Island
Inc., Lexington, MassachusettsG. Ammerer, ZymoGenetics, Inc., Se-
attle, Washington
U.S. Environmental Protection Agency Workshop on "PossibleShort-term Evolutionary Consequences of Biotechnology"August 28-August 31
ARRANGED BY
J.R. Fowle III, U.S. Environmental Protection Agency, Washington, D.C.
Chairperson: P. Regal, University of Minnesota, Minneapolis
PARTICIPANTS
D. Archer, Food and Drug Administra-tion, Washington, D.C.
F. Betz, U.S. Environmental Protec-tion Agency, Washington, D.C.
R. Bierbaum, Office of TechnologyAssessment, Washington,D.C.
A.W. Bourquin, U.S. EnvironmentalProtection Agency, Gulf Breeze,Florida
J. Brown, University of Arizona, Tuc-son
E. Carlson, State University of NewYork, Stony Brook
R. Colwell, University of California,Berkeley
M.A. Dane llo, Food and Drug Admin-istration, Washington, D.C.
J.R. Fowle III, U.S. EnvironmentalProtection Agency, Washington,D.C.
267

M. Gough, Office of Technology As-sessment, Washington, D.C.
L.L. Greenlee, Agrigenetics Corp.,Boulder, Colorado
F Harris, National Science Founda-tion, Washington, D.C.
A. Hollander, U.S. Environmental Pro-tection Agency, Washington, D.C.
D. Kamely, U.S. Environmental Pro-tection Agency, Washington, D.C.
D. Kaplan, Dow Chemical Company,Midland, Michigan
National Institute of Mental Health Workshopon "Single Unit Activity and Behavior"
R. Lensky, University of Massachu-setts, Amherst
B.R. Levin, University of Massachu-setts, Amherst
M. Levin, House Subcommittee on In-vestigations and Oversight, U.S.Congress, Washington, D.C.
M. Lloyd, University of Chicago, Illi-nois
D. MacKenzie, Louisiana State Uni-versity, Baton Rouge
B. McClintock, Cold Spring HarborLaboratory, New York
A. Mc Daniels, U.S. EnvironmentalProtection Agency, Washington,D.C.
E Milewski, National Institutes ofHealth, Washington, D.C.
S. Panem, U.S. Environmental Protec-tion Agency, Washington, D.C.
J. Rissler, U.S. Environmental Protec-tion Agency, Washington, D.C.
A. Rose, Department of Health andHuman Services, Washington,D.C.
D Simberloff, Florida State University,Tallahassee
Z. Vaituzis, U.S. Environmental Pro-tection Agency, Washington, D.C.
September 9-September 11
ARRANGED BY
R. Schoenfeld, National Institute of Mental Health, Rockville, Maryland
Chairpersons: B.S. Bunney, Yale University School of Medicine, New Haven,ConnecticutB.L. Jacobs, Princeton University, New Jersey
PARTICIPANTS
N. Bernick, National Institute of Men-tal Health, Rockville, Maryland
M.R. De Long, Johns Hopkins Hospi-tal, Baltimore, Maryland
P.S. Goldman-Rakic, Yale UniversitySchool of Medicine, New Haven,Connecticut
M. Konishi, California Institute ofTechnology, Pasadena
268
S.H. Koslow, National Institute ofMental Health, Rockville, Mary-land
I. Kupfermann, College of Physicians& Surgeons, Columbia University,New York, New York
J.B. Ranck, State University of NewYork, Downstate Medical Center,Brooklyn
R. Schoenfeld, National Institute ofMental Health, Rockville, Mary-land
R.F. Thompson, Stanford University,California
T.N. Wiesel, Rockfeller University,New York, New York
D.J. Woodward, University of TexasHealth Science Center, Dallas

Alfred P. Sloan Foundation ComputationalNeuroscience WorkshopSeptember 28-September 30
ARRANGED BY
E. Wanner, Alfred P. Sloan Foundation, New York, New York
SESSION 1 VISION
Organizers: T. Poggio, Massachusetts Institute of Technology, CambridgeT. Sejnowski, Johns Hopkins University, Baltimore, Maryland
PARTICIPANTS
D. Ballard, University of Rochester,New York
H.B. Barlow, Cambridge University,England
C. Gilbert, Rockefeller University,New York, New York
D.A. Glaser, University of California,Berkeley
E. Hildreth, Massachusetts Institute ofTechnology, Cambridge
A.J. Movshon, New York University,New York
G.F. Poggio, Johns Hopkins Univer-sity School of Medicine, Baltimore,Maryland
T. Poggio, Massachusetts Institute ofTechnology, Cambridge
T Sejnowski, Johns Hopkins Univer-sity, Baltimore, Maryland
C.F. Stevens, Yale University School
SESSION 2 VESTIBULO-OCULAR PROCESSING
Organizer: D.A. Robinson, Johns Hopkins Hospital, Baltimore, Maryland
PARTICIPANTS
A.F. Fuchs, University of Washington,Seattle
H. Galiana, McGill University, Mon-treal, Canada
E.L. Keller, Medical Research Instituteof San Francisco at Pacific Medi-cal Center, California
G.E. Loeb, National Institute of Neu-
SESSION 3 MOTOR CONTROL
rological and Communicative Dis-orders and Stroke, Bethesda,Maryland
L.M. Optican, National Eye Institute,Bethesda, Maryland
D.A. Robinson, Johns Hopkins Hospi-tal, Baltimore, Maryland
Organizers: E. Bizzi, Massachusetts Institute of Technology, CambridgeJ.M. Hollerbach, Massachusetts Institute of Technology, Cambridge
PARTICIPANTS
E. Bizzi, Massachusetts Institute ofTechnology, Cambridge
E.V. Evarts, National Institutes ofHealth, Bethesda, Maryland
L. Finkel, Rockefeller University, NewYork, New York
C. Ghez, College of Physicians & Sur-geons, Columbia University, NewYork, New York
N. Hogan, Massachusetts Institute ofTechnology, Cambridge
J.M. Hollerbach, Massachusetts Insti-tute of Technology, Cambridge
J.C. Houk, Northwestern University,Chicago, Illinois
R.W. Mann, Massachusetts Institute ofTechnology, Cambridge
M.H. Raibert, Carnegie-Mellon Uni-versity, Pittsburgh, Pennsylvania
of Medicine, New Haven, Con-necticut
V. Torre, Universita di Genova, ItalyS. Ullman, Massachusetts Institute of
Technology, Cambridge
269

Genetic Manipulation of the Mammalian Ovum and Early EmbryoOctober 7-October 10
ARRANGED BY
F. Costantini, Columbia University, New York, New YorkR. Jaenisch, Heinrich-Pette-Institut, Hamburg, Federal Republic of Germany
SESSION 1 DEVELOPMENTAL GENETICS
Chairperson: L. Silver, Princeton University, New Jersey
L. Silver, Princeton University, NewJersey: The origin and evolution ofmouse t haplotypes.
V.E. Chapman, Roswell Park Memo-rial Institute, Buffalo, New York: X-chromosome regulation in femalemammals.
S.M. Tilghman, Fox Chase CancerCenter, Philadelphia, Pennsylva-nia: Tissue-specific expression ofcloned AFP genes in cells andmice,
M.J. Evans, University of Cambridge,England: EK cell contribution tochimaeric mice: From tissue cul-ture to sperm.
D. Solter, The Wistar Institute of Anat-omy and Biology, Philadelphia,Pennsylvania: Capacity of nucleifrom preimplantation mouse em-bryos to support normal develop-ment.
A. Surani, Institute of Animal Physiol-
ogy, Cambridge, England: Regula-tion of embryogenesis by maternaland paternal genomes in themouse.
SESSION 2 ENDOGENOUS VIRUSES AND VIRAL VECTORS
Chairperson: P.W.J. Rigby, Imperial College of Science and Technology, London,England
N.A. Jenkins, University of CincinnatiCollege of Medicine, Ohio: Insta-bility of ecotropic proviruses in RF/J-derived hybrid mice.
N.G. Copeland, University of Cincin-nati College of Medicine, Ohio:Molecular genetic approaches to
the study of murine lymphoid cellneoplasms.
R. Jaenisch, Heinrich -Pette- Institut,Hamburg, Federal Republic ofGermany: Recombinant viralprobes of gene regulation andmethylation patterns in the mouse.
SESSION 3 INTRODUCTION OF CLONED GENES INTO THE MAMMALIANGERM LINE I
Chairperson: F.H. Rudd le, Yale University, New Haven, Connecticut
R.L. Brinster, University of Pennsylva-nia, Philadelphia: Introduction ofSV40 genes into mice.
R.D. Palmiter, University of Washing-ton, Seattle: Expression of growth-hormone genes in mice.
D. Hanahan, Cold Spring Harbor
270
Laboratory, New York: Expressionof insulin-SV40 T-antigen hybridgenes in transgenic mice.
T.A. Stewart, Genentech, Inc., SouthSan Francisco, California: Trans-genic mice carrying MTV/myc fu-
K. Willison, Chester Beatty Labora-tory, London, England: Haploidgene expression and the mouse tcomplex.
E. F. Wagner, European Molecular Bi-ology Laboratory, Heidelberg,Federal Republic of Germany: In-troducing and expressing genes instem cells and mice.
sion genes develop mammary ad-enocarcinomas.
R. Evans, Salk Institute, San Diego,California: Novel tissue-specificexpression of growth-hormonegenes in transgenic animals.

SESSION 4 INTRODUCTION OF CLONED GENES INTO THE MAMMALIANGERM LINE II
Chairperson: A.J. Levine, Princeton University, New Jersey
F Costantini, Columbia University, expression of human HPRT in
New York, New York: Regulated CNS of transgenic mice.
expression of a foreign (3-globin D.C. Kraemer, Texas A&M University,
gene in transgenic mice. College Station: Gene transfer in
D. Baltimore, Massachusetts Institute cattle and sheep.
of Technology, Cambridge: Tissue- C. Polge, Institute of Animal Physiol-
specific expression of immuno- ogy, Cambridge, England: Gene
globulin genes in transgenic mice transfer approaches in farm ani-
and cultured cells. mall.U. Storb, University of Washington,
Seattle: Expression of a microin-jected immunoglobulin x gene intransgenic mice.
C.T. Caskey, Baylor College of Medi-cine, Houston, Texas: Elevated
SESSION 5 NONMAMMALIAN SYSTEMS
Chairperson: E. Davidson, California Institute of Technology, Pasadena
E. Davidson, California Institute ofTechnology, Pasadena: Transferand expression of cloned se-quences in the sea urchin embryo.
J. Newport, University of California,San Diego: Nuclear formation inthe Xenopus embryo.
D.T. Stinchcomb, Harvard University,Cambridge, Massachusetts: DNAtransformation of C. elegans; ex-trachromosomal arrays are repli-cated, segregated, and ex-pressed.
T Maniatis, Harvard University, Cam-bridge, Massachusetts: Analysis ofDrosophila alcohol dehydrogen-ase gene expression by P-elementtransformation.
Gap JunctionsOctober 21-October 24
ARRANGED BY
M.V.L. Bennett, Kennedy Center, Bronx, New York
SESSION 1 STRUCTURE
Chairperson: M.V.L. Bennett, Kennedy Center, Bronx, New York
L. Makowski, College of Physicians & domains in gap junctions and im-
Surgeons, Columbia University, plications for gating.
New York, New York: Structural G. Zampighi, University of California,
Los Angeles, School of Medicine:Structure of lens and liver junc-tional proteins.
271

R. Hanna, State University of NewYork College of Environmental Sci-ence and Forestry, Syracuse, NewYork: Fast-frozen gap junctions.
S.B. Yancey, and J. -P. Revel, Califor-
SESSION 2 BIOCHEMISTRY
Chairperson: J.-P. Revel, California
E.L. Hertzberg, Baylor College ofMedicine, Houston, Texas: Tissuesand species specificity of gapjunctional proteins.
K. Willecke, Universitat Essen, Fed-eral Republic of Germany: Immu-nochemical characterization ofgap junction protein from differentmammalian tissues.
SESSION 3 BIOPHYSICS
nia Institute of Technology, Pasa-dena: Organization of junctionalproteins in the membrane as de-duced from sequence data.
Institute of Technology, Pasadena
M. Finbow, Beatson Institute for Can-cer Research, Glasgow, Scotland:Tissue, species, and phylogeneticvariation of gap junctional pro-teins.
R.G. Johnson, University of Minne-sota, St. Paul: Lens junctions-Antibodies to and phosphorylationof MP26.
Chairperson: R. Llinas, New York University Medical Center, New York
P Brink, State University of New York,Stony Brook: Solvent effects onjunctional permeability.
D.C. Spray, Kennedy Center, Bronx,New York: Gating by voltage andchemical agents.
F. Ramon, Centro de Investlgacion yde Estudios Avanzados Del IPN,
SESSION 4 CONTROL OF FORMATION
J.
J.
Mexico City, Mexico: Physiologicalcontrol mechanisms.
Wojtchak, Rockefeller University,New York, New York: Electrical un-coupling induced by general an-esthetics-A calcium independentprocess?
E. Hall, University of California, Ir-
Chairperson A. Warner, University College London, England
J. Sheridan, University of Minnesota,Minneapolis: Altered junctionalpermeability between virally trans-formed cells.
J.D. Pitts, Beatson Institute for Can-cer Research, Glasgow, Scotland:Tissue specificity.
W. Coles, McMaster University, Hamil-
ton, Canada: Alterations in cou-pling in uterine muscle.
J. Kessler, Kennedy Center, Bronx,New York: Coupling between cul-tured vertebrate neurons.
S.B. Kater, University of Iowa, IowaCity: Specificity of coupling at mol-luscan neurons.
SESSION 5 ROLE IN INTERCELLULAR COMMUNICATION AND DEVELOPMENT
E. Page, University of Chicago, Illi-nois: Structure and protein com-position of cardiac gap junctions.
D. Paul, Harvard University MedicalSchool, Boston, Massachusetts:Proteolytic cleavage of junctionalproteins from lens and liver.
vine: Channel reconstitution fromjunctional proteins.
C. Peracchia, University of RochesterMedical School, New York: Phos-phorylation and reconstitution oflens junctions.
Chairperson: J.D. Pitts, Beatson Institute for Cancer Research, Glasgow, Scotland
C.W. Lo, University of Pennsylvania,Philadelphia: Compartmentaliza-tion in embryos.
S. Caveney, University of Western On-tario, London, Canada: Control ofmolecular movement within andbetween developmental compart-ments.
272
A. Warner, University College Lon-don, England: Antibodies to gapjunction proteins-Probes forstudying development.
W.J. Larsen, University of CincinnatiCollege of Medicine, Ohio: Rela-tionships of gap junction modula-tion to cell and tissue function.
W. Beers, New York University, NewYork: Gap junctions and ovulation.
O.H. Petersen, University of Liver-pool, England: Communication insecretory epithelia.

SESSION 6 ELECTROTONIC SYNAPSES
Chairperson: D.C. Spray, Kennedy Center, Bronx, New York
F.E. Dudek, Tulane University Schoolof Medicine, New Orleans, Louisi-ana: Electrical interactions andsynchronization of hippocampalneurons-Electrotonic couplingvs. field effects.
R. Llinas, New York University Medi-cal Center, New York: Functionalsignificance of coupling in theinferior olive.
M.V.L. Bennett, Kennedy Center,Bronx, New York: Interactionsamong chemical and electrotonicsynapses.
C. Giaume, Institut Pasteur, Paris,France: The rectifying electrotonicsynapse of crayfish.
H.M. Gerschenfeld and J. Neyton,Eco le Norma le Superieure, Paris,France: Dopaminergic control ofcoupling between horizontal cells.
E. Lasater and J. Dowling, HarvardUniversity, Cambridge, Massachu-setts: Characteristics of couplingbetween pairs of cultured horizon-tal cells.
Transport and Secretion of ProteinsNovember 7-November 10
ARRANGED BY
M.-J. Gething, Cold Spring Harbor Laboratory, New York
SESSION 1 TRANSLOCATION OF PROTEINS ACROSS THE LIPID BILAYER
Chairperson: G. Blobel, Rockefeller University, New York, New York
P. Walter, University of California, SanFrancisco: Targeting of nascentsecretory proteins to the endo-plasmic reticulum membrane.
D. Meyer, European Molecular Biol-ogy Laboratory, Heidelberg, Fed-eral Republic of Germany: Factorsmediating protein translocation inthe endoplasmic reticulum -Thedocking protein and beyond.
R. Gilmore, Rockefeller University,New York, New York: Evidence forthe existence of a signal receptorin the microsomal membrane.
S. Benson, Princeton University, NewJersey: Intragenic information re-quired for the export of LamB tothe outer membrane of E. coli.
W. Wickner, University of California,Los Angeles, School of Medi-cine-Mechanisms of bacterialcell-surface assembly.
S. Ferro-Novick, Harvard MedicalSchool, Boston, Massachusetts:Genetic evidence for the couplingof the synthesis and secretion ofproteins in E. coli.
M. Inouye, State University of New
York, Stony Brook: Functional andstructural analysis of the signalpeptide.
C. Kaiser, Massachusetts Institute ofTechnology, Cambridge: Muta-tions in the signal sequence affect-ing the localization of invertase.
273

SESSION 2 TRANSPORT FROM THE ENDOPLASMIC RETICULUM TO THE PLASMAMEMBRANE
Chairperson: H.F. Lodish, Whitehead Institute for Biomedical Research, Cambridge,Massachusetts
I. Schauer, University of California,Berkeley: Invertase sequence sub-stitutions interfere with transport ofactive enzyme from the ER.
J. Sambrook, Cold Spring HarborLaboratory, New York: How to getSV40 T antigen into membranes.
J. Rose, Salk Institute, San Diego,California: Redesigning transportsignals in the VSV glycoprotein.
P Berman, Genentech, Inc., South
San Francisco, California: Pro-cessing and export of membrane-bound and secreted forms ofcloned HSV-1 glycoprotein D incontinuous cell lines.
M.-J. Gething, Cold Spring HarborLaboratory, New York: Site-di-rected mutagenesis of the hemag-glutinin of influenza virus-Effectson transport through the cell.
A. Brake, Chiron Corporation, Emery-
SESSION 3 STRUCTURES INVOLVED IN TRANSPORT AND ENDOCYTOSIS
Chairperson: G.E. Pa lade, Yale University School of Medicine, New Haven,Connecticut
P.W. Robbins, Massachusetts Institute I. Mellman, Yale University School ofof Technology, Cambridge: Protein Medicine, New Haven, Connecti-glycosylation. cut: Fc receptor transport, proton
M.G. Farquahar, Yale University transport, and the control of intra-School of Medicine, New Haven, cellular membrane traffic.Connecticut: Receptor traffic to G. Warren, European Molecular Biol-Golgi subcompartments. ogy Laboratory, Heidelberg, Fed-
W.S. Sly, St. Louis University School eral Republic of Germany: Recon-of Medicine, Missouri: Mannose-6- struction of an endocytic fusionP receptor-mediated sorting and event in a cell-free system.transport of lysosomal enzymes. S. Schmid, Stanford University School
SESSION 4 DIRECTED TRANSPORT I
Chairperson: D. Sabatini, New York University Medical Center, New York
D. Sabatini, New York University Med-ical Center, New York: Intracellularsorting and distinct recycling pat-terns of viral glycoproteins in po-larized epithelial cells.
K. Mostov, Whitehead Institute forBiomedical Research, Cambridge,Massachusetts: Transepithelialtransport of IgA and IgM.
H.-P. Moore, University of California,San Francisco: Sorting and trans-port of proteins in regulated secre-tory cells.
J.-P. Kraehenbuhl, University of Lau-sanne, Switzerland: Biogenesis of(Na,,K+)-ATPase in ion-transport-ing epithelia.
D. Louvard, Institut Pasteur, Paris,France: Basolateral membraneprotein markers are expressed atthe surface of undifferentiated pre-cursors of enterocytes in vitro,whereas transport of apical mark-ers is abortive.
274
ville, California: Sequence of twogenes encoding precursors to theyeast peptide mating pheromonea factor.
J. Thorner, University of California,Berkeley: Cell biology and enzy-mology of the specific endopepti-dase required for processing ofyeast precursor proteins at pairsof basic residues.
of Medicine, California: Enzymaticrecycling of clathrin from coatedvesicles.
P.C. Tai, Boston Biomedical ResearchInstitute, Massachusetts: Clathrinin B.subtilis? Bacterial proteintranslocation.
E. Rodriguez-Boulan, Cornell Univer-sity Medical College, New York,New York: Vectorial exocytosis ofplasma membrane glycoproteinsin epithelial cells.
K. Simons, European Molecular Biol-ogy Laboratory, Heidelberg, Fed-eral Republic of Germany: Sortingof apical and basolateral proteinsin MDCK cells,

SESSION 5 DIRECTED TRANSPORT II
Chairperson: D. Botstein, Massachusetts Institute of Technology, Cambridge
S.D. Emr, California Institute of Tech-nology, Pasadena: Use of gene fu-sions to study protein traffic inyeast.
T.H. Stevens, University of Oregon,Eugene: Yeast mutants defectivein the sorting of vacuolar proteins.
M.N. Hall, University of California,
San Francisco: Studies on themechanism of nuclear protein lo-calization in yeast.
E.C. Hurt, University of Basel, Switz-erland: Import of proteins into mi-tochondria.
J. Kaput, Rockefeller University, NewYork, New York: Trans location of
nuclearly encoded mitochrondrialproteins translated from SP6-pro-moted transcripts.
M.G. Douglas, University of TexasHealth Sciences Center, San Anto-nio: The biochemistry and ge-netics of protein import into mito-chondria.
Journalists' Workshop on "Assessing Health Risk Assessment"November 28-November 30
ARRANGED BY
M. Shodell, Banbury Center, Cold Spring Harbor Laboratory, New York
SESSION 1
D.P. Rall, National Institute of Environ-mental Health Sciences, ResearchTriangle Park, North Carolina: Anintroduction to the principles ofrisk assessment.
J.A. Swenberg, Chemical Industry In-stitute of Toxicology, Research Tri-angle Park, North Carolina:Models in biological risk assess-ment.
SESSION 2
I.C.T. Nisbet, Clement Associates,Inc., Arlington, Virginia: Assessingthe models used for assessingrisk.
F.P. Perera, Columbia UniversitySchool of Public Health, New York,New York: New approaches in riskassesssment.
SESSION 3
R.E. Albert, New York University Med-ical Center, New York, New York:Risk assessment in the regulationof carcinogens.
275



Recto: James laboratory, erected in 1929, houses the Laboratory's tumor virology programand a number of summer postgraduate research courses. The Annex (shown here), built in1971, contains offices, a seminar room, and a library.

Postgraduate Courses
The summer program of Postgraduate Courses at Cold Spring Harbor Labora-tory is aimed at meeting the special need for training in interdisciplinary subjectsthat are either so new or so specialized that they are not adequately treated byuniversities. Our aim is to provide intensive study in the most recent develop-ments and techniques in these subjects and to prepare students to enter directlyinto research in a particular area. To ensure up-to-date coverage of current re-search work, we bring together a workshop staff from many laboratories aroundthe world and supplement this staff with a series of seminar speakers.
Immunoglobulins: Molecular Probes of the Nervous SystemJune 8-June 28
INSTRUCTORS
Hockfield, Susan, Ph.D., Cold Spring Harbor Laboratory, New YorkKintner, Chris, Ph.D., Medical Research Council, Kings College London, EnglandSilbertstein, Laura, Ph.D., Stanford Medical Center, CaliforniaWeber Eckard, M.D., Stanford Medical School, California
ASSISTANT
Evans, Chris, Ph.D., Stanford University, California
Over the last decade, antibodies have become powerful and popular reagents inneurobiological research. This course, intended for research scientists of all lev-els, was designed to provide an advanced understanding of the power and limi-tations of immunoglobulins both as biochemical and anatomical reagents.Through laboratory work and lectures, we explored immunochemical and immu-nohistochemical techniques in detail and the application of these techniques tocurrent issues in neurobiology.
The laboratory work included: generation of monoclonal antibodies; prepara-tion and characterization of antibodies to synthetic peptides and to complex anti-gens; solid-phase immunoassays; antibody characterization and purification; an-tigen purification using antibody affinity methods; Western blotting; antibodyconjugation; biochemical characterization of neuropeptides using R IA; light andelectron microscopic immunohistochemistry using peroxidase, fluorescence, andbiotin-avidin techniques; and methods for the colocalization of multiple anti-bodies.
A series of lectures by invited speakers covered: structure and function of im-munoglobulins; molecular genetics of antibody diversity; cellular regulation of theimmune response; hybridoma technology; studies using synthetic peptides; im-munological characterization of the acetylcholine receptor and the neuromuscu-lar junction; and immunological characterization of cellular diversity in vertebrateand invertebrate nervous systems and in cell culture.
279

PARTICIPANTS
Drysdale, Rachel A., B.A., Universityof Wisconsin, Madison
Flanagan, Thomas R., Ph.D., ColdSpring Harbor Laboratory, NewYork
Kalil, Katherine, Ph.D., University ofWisconsin, Madison
Messing, Albee, Ph.D., University ofPennsylvania School of Medicine,Philadelphia
Milner, Teresa Ann, Ph.D., CornellUniversity School of Medicine,New York, New York
Ridgway, Richard L., Ph.D., Washing-ton State University, Pullman
Simonds, William F., M.D., National In-stitutes of Health, Bethesda, Mary-land
Thompson, Wesley J., Ph.D., Univer-sity of Texas, Austin
Treiman, Marek, M.D., RockefellerUniversity, New York, New York
Wahle, Petra, Msc., Max-Planck-Insti-tut fOr Biophysikalische Chemie,Frankfurt, Federal Republic ofGermany
SEMINARS
Wilson, D., University of Pennsylva-nia. Cellular immunology.
lmmanishi-Karl, T., Massachusetts In-stitute of Technology. Molecularimmunology.
Davis, D., National Institutes of
Health. 3-D structure of immuno-globulins.
Lindstrom, J., Salk Institute. Usingmonoclonal antibodies to studyacetylcholine receptors.
Burden, S., Massachusetts Institute ofTechnology. Subsynaptic proteinat nerve-muscle synapses.
Matthews, B., Harvard MedicalSchool. Monoclonal antibodiesused to study neurite-promotingfactors.
Hendry, S., University of California, Ir-
Molecular Biology of Plants
vine. Immunocytochemical studiesof the mammalian cerebral cortex.
Jessel, T., Harvard University MedicalSchool. Probes for mammaliandorsal root ganglion cells.
Pickel, V., Cornell University School ofMedicine. Use of combined immu-nocytochemical techniques for thelocalization of more than one anti-gen.
Karten, H., State University of NewYork, Stony Brook. Immunochemi-cal analysis of the retina.
June 8-June 28
INSTRUCTORS
Malmberg, Russell, Ph.D., Cold Spring Harbor Laboratory, New YorkMessing, Joachim, Ph.D., University of Minnesota, St. PaulSussex, Ian, Ph.D., Yale University, New Haven, Connecticut
ASSISTANTS
Boylan, Margaret, Ph.D., Yale University, New Haven, ConnecticutMclndoo, Jean, B.S., Cold Spring Harbor Laboratory, New YorkZarowitz, Michael, Ph.D., University of Minnesota, St. Paul
This course provided an introduction to current techniques for the manipulationof plant material as applied to experiments in molecular biology and genetics. Itwas designed primarily for scientists working in other areas who wish to pursue
280

research in plants. It was assumed that applicants had a general working knowl-edge of molecular biology, but no previous experience with plants was required.Experiments were designed to emphasize aspects unique to plant systems, in-cluding mutagenesis and analysis of mutants, tissue-culture techniques, proto-plast isolation and culture, regeneration of plants from culture, DNA and RNAisolation and cloning procedures, crown gall tumorigenesis, chloroplast genetics,and nitrogen fixation. Guest lecturers provided a background in plant morphoge-nesis, physiology, pathology, genetics, and cytology. Different plant species wereused to illustrate particular experimental techniques, but the course emphasizedtobacco and maize as model systems.
PARTICIPANTS
Avivi, Aaron A., Ph.D., Weizmann In-stitute of Science, Rehovot, Israel
Choi, Cha Yong, Ph.D., Seoul Na-tional University, Korea
DasSarma, Shiladitya, B.S., Massa-chusetts Institute of Technology,Cambridge
De Block, M., Ph.D., RijksuniversiteitGent, The Netherlands
Dilworth, Machi F., U.S. Departmentof Agriculture, Washington, D.C.
Dynan, William S., Ph.D., University ofCalifornia, Berkeley
Hille, Jacques, Ph.D., University ofWageningen, The Netherlands
Jones, Davy, Ph.D., University of Ken-tucky, Lexington
Kakidani, Hitoshi, Ph.D., SagamiChemical Research Center, Japan
Lammers, Peter ,J., Ph.D., Universityof Chicago, Illinois
Lee, Nancy, Ph.D., Interferon Sci-ences, Inc., New Brunswick, NewJersey
Mindrinos, Michael N., Ph.D., BostonCollege, Massachusetts
Peterson, Thomas A., B.S., Universityof California, Davis
Sugar, Anurag, M.S., Rockefeller Uni-versity, New York, New York
Tavantzis, Stellos M., Ph.D., Univer-sity of Maine, Orono
Thirion, J.-P, Ph.D., Centre Hospital-ier Universitaire, Sherbrooke, Can-ada
SEMINARS
van Montagu, M., RijksuniversiteitGent. Agrobacterium and crowngall disease.
Sims, T, University of California, LosAngeles. Soybean embryonicgenes.
Gengenbach, B., University of Minne-sota. Embryogenesis in Zea tissuecultures.
. Mutant selection in Zea tissuecultures.
Broglie, R., L. Hanley-Bowdoin, Rock-efeller University. Nuclear andchloroplast gene regulation.
Walbot, V., Stanford University. Devel-opmental genetics of maize.
Long, S., Stanford University. Rhizo-bium-legume developmental ge-netics.
Quail, P, University of Wisconsin.Phytochrome gene expression.
Lonsdale, D., Plant Breeding Institute.The mitochondrial genome.
Crouch, M., University of Indiana.Compatibility/incompatibility sys-tems.
Phillips, R., University of Minnesota.Somaclonal variation.
. Cytogenetics of plant cells inculture.
Ellingboe, A., University of Wisconsin,Genetics of plant-pathogen inter-actions.
Hanson, M., University of Virginia. Or-ganelle genetics.
Ho, D., Washington University. Hor-mone response mutants.
Palukaitis, P, Cornell University. Plantviruses.
Miles, D., University of Missouri. Pho-tosynthesis.
Donner, H., Advanced Genetic Sci-ences. Controlling elements ofmaize.
Binns, A., University of Pennsylvania.Biology of transformed plants.
281

Molecular Cloning of Eukaryotic GenesJune 8-June 28
INSTRUCTORS
Bothwell, Al, Ph.D., Yale Medical School, New Haven, ConnecticutAlt, Fred, Ph.D., Columbia University College of Physicians & Surgeons, New York, New
YorkMullins, James, Ph.D., Harvard School of Public Health, Boston, Massachusetts
ASSISTANTS
LeClair, Ken, B.A., Yale Medical School, New Haven, ConnecticutYancopoulos, George, B.A., Columbia University, New York, New YorkBruck, Claudine, Ph.D., Harvard Medical School, Boston, Massachusetts
This laboratory and lecture course covered the principles of recombinant DNAtechnology and the application of these procedures to the study of eukaryoticgenes. The isolation and characterization of lymphocyte-specific genes were em-phasized. Among the topics covered were: construction of cDNA libraries inplasmid or bacteriophage A vectors, construction of bacteriophage A and cosmidlibraries of high-molecular-weight eukaryotic DNA, screening DNA libraries withgene-specific hybridization probes and by recombination, purification, and char-acterization of recombinant clones using restriction endonuclease and blot hy-bridization analyses, and reintroduction and expression of cloned genes in heter-ologous systems. Strategies for isolating genes that encode rare mRNAsequences were discussed. Guest lecturers discussed the application of molecu-lar cloning procedures to the study of specific eukaryotic gene systems.
PARTICIPANTS
Bignami, Margherita, Ph.D., IstitutoSuperiore Di Sanita, Rome, Italy
Binninger, Dorothea, M.S., Paul-Ehr-lich-Institut, Frankfurt, Federal Re-public of Germany
Cox, Diane W., Ph.D., Hospital forSick Children, Toronto, Canada
Crittenden, Lyman B., Ph.D., USDA
282
Regional Poultry Research Labo-ratory, East Lansing, Michigan
Foster, Patricia L., Ph.D., Boston Uni-versity School of Public Health,Massachusetts
Iscove, Norman N., Basel Institute forImmunology, Switzerland
Jalkanen, Markku T., Ph.D., StanfordUniversity Medical Center, Califor-nia
Kasuga, Masato, M.D., University ofTokyo, Japan
Lever, Julia E., Ph.D., University ofTexas Medical School, Houston
Nelson, David L., M.D., National Can-cer Institute, Bethesda, Maryland
ole-MoiYoi, Onesmo, M.D., Interna-tional Laboratories for Researchon Animal Diseases, Nairobi,Kenya

Prusiner, Stanley B., M.D., Universityof California, San Francisco
Remold, Heinz G., Ph.D., HarvardMedical School, Boston, Massa-chusetts
Schuurmann, R.K.B., Ph.D., Univer-sity Hospital Leiden, The Nether-lands
Sidman, Charles L., Ph.D., JacksonLaboratory, Bar Harbor, Maine
Zingales, Bianca, D.Sc., Institute DeQuimica, Brazil
SEMINARS
Lehrach, H., EMBL. Molecular clon-ing of the T Locus.
Botstein, D., Massachusetts Instituteof Technology. Reverse genetics:cloning of cytoskeletal genes.
Goff, S., Columbia University. In vitromutagenesis of retroviral genes.
Reth, M., Columbia University. Anti-bodies and Western blotting.
Baltimore, D., Massachusetts Institute
of Technology. Expression of Iggenes introduced into the germ-line.
Miyada, G., City of Hope Hospital.Use of synthetic oligonucleotidesfor cloning and expression of mu-rine MHC genes.
Lacy, E., Memorial Sloan-KetteringCancer Center. Gene transfer intomice.
Rubin, J., University of California,Berkeley. P- element -mediatedgene transfer in Drosophila.
Schimke, B., Stanford University. Mo-lecular mechanisms of gene am-plification.
Maniatis, T., Harvard University. Glo-bin gene transcription and RNAsplicing.
Schwarzbauer, J., Massachusetts In-stitute of Technology.Transfer ofgenes into hematopoeitic stemcells.
Seed, B., Massachusetts GeneralHospital. Recovery and manipula-tion of recombinant DNA in vivo.
Neurobiology of Human Disease
Gluzman, Y., Cold Spring Harbor Lab-oratory. Eukaryotic cloning vec-tors.
Hughes, S., Frederick Cancer Re-search Facility. Oncogenes.
Messing, J., University of Minnesota.M13 vectors.
McKnight, S., Hutchinson Cancer Re-search Institute. Regulation oftranscription.
Wahl, G., Salk Institute. Gene transfertechnology.
Hamyln, J., University of Virginia. Iso-lation of eukaryotic origins of repli-cation.
Mullins, J., Harvard School of PublicHealth. FeLV, AIDS, and HTLV.
Ruley, E., Cold Spring Harbor Labora-tory. Transformation and oncoge-nesis.
Wig ler, M., Cold Spring Harbor Labo-ratory. Yeast RAS genes.
Szostak, J., Dana-Farber Cancer Insti-tute. Replication and segregationof linear chromosomes in yeast.
June 8-June 17
INSTRUCTORS
Breakfield, Xandra 0., Ph.D., E.K. Shriver Center and Harvard Medical School, Boston,Massachusetts
Black, Ira B., M.D., Cornell University School of Medicine, New York, New YorkGusella, James, Ph.D., Massachusetts General Hospital, Boston
This intensive seminar course explored the molecular and cellular basis of abnor-mal neural function. It focused on basic scientific studies that have provided in-sight into the etiology and pathogenesis and neurologic and psychiatric dis-eases. Topics included: (1) molecular pathology of neurotransmitterderangement; (2) developmental plasticity and choice of neurotransmitter pheno-type; (3) synthesis and regulation of neuropeptides; (4) cellular events in neuralregeneration and brain transplantation; (5) neural pathways involved in pain syn-dromes; (6) genetic linkage analysis using DNA polymorphisms; (7) defects inDNA repair and activation of one genes; (8) biochemistry of the lipidoses; (9) au-toimmune diseases; (10) brain imaging and metabolism; (11) epilepsy and sei-zure disorders; (12) cell death in degenerative disorders; (13) viral infections ofthe nervous system; and (14) experimental models of learning and memory.
PARTICIPANTS
Arias, Clorinda, M.D., UniversidadNacional Autonoma de Mexico,Mexico City
Bellini, Fabrizio, Ph.D., Fidia Re-search Laboratories, Terme, Italy
Cambi, Franca, M.D., E.K. ShriverCenter, Harvard Medical School,Boston, Massachusetts
Desan, Paul, Ph.D., Harvard MedicalSchool, Boston, Massachusetts
Foxman, Brett T, M.D., Cornell Medi-cal Center, New York, New York
Kagan, Bruce L., Ph.D., University of
283

California Neuropsychiatric Insti-tute, Los Angeles
Kremer, Norbert E., Ph.D., State Uni-versity of New York, Stony Brook
Kurlan, Roger M., M.D., University ofRochester Medical Center, NewYork
Maehlen, Jan, M.D., Institute of Neu-rophysiology, Oslo, Norway
Mangun, George R., B.S., Universityof California, San Diego
Moroi, Sayoko E., B.A., MedicalAdministration Center, Columbus,Ohio
Nakajima, Gene A., M.D., Johns Hop-kins Medical School, Baltimore,Maryland
Oliver, Eugene J., Ph.D., National In-stitutes of Health, Bethesda, Mary-land
Sanes, Dan H., M.A., Princeton Uni-versity, New Jersey
Segal, Rosalind A., B.A., RockefellerUniversity, New York, New York
Sims, Stephen J., B.A., DownstateMedical Center, New York, NewYork
Singh, Dyal N.P., Ph.D., University ofAlberta, Canada
SEMINARS
Gage, F., Lund University. Brain trans-plantation.
Aguayo, A., Montreal General Hospi-tal. Neural regeneration.
Norwich, A., Yale University School ofMedicine. Molecular genetic ap-proaches to organic acidemias.
Sokoloff, L., National Institute of Men-tal Health. Brain metabolism andimaging.
Advanced Bacterial Genetics
Phelps, M., University of CaliforniaMedical School, Los Angeles.Brain metabolism and imaging.
Coyle, J., Johns Hopkins UniversitySchool of Medicine. Movementdisorders and degenerative dis-eases: experimental models.
Wexler, N., Hereditary Disease Foun-dation. Huntington's disease inVenezuela.
Ginns, E., National Institutes ofHealth. Lipidoses.
Basbaume, A., University of Califor-nia, San Francisco. Pain syn-dromes and neuropeptides.
Jessell, T, Harvard Medical School.Pain syndromes and neuropep-tides.
Herbert, E., University of Oregon.Neuroendocrinology.
Lindstrom, J., Salk Institute. Autoim-mune diseases: myastheniagravis.
McNamara, J., Duke University Medi-cal Center. Epilepsy and seizuredisorders,
Crill, W., University of WashingtonSchool of Medicine. Epilepsy andseizure disorders.
Hawkins, R., Columbia University Col-lege of Physicians and Surgeons.Learning and memory: experi-mental models.
Mishkin, M., National Institute of Men-tal Health. Learning and memory:primate models.
Gazziniga, M., Cornell UniversityMedical College. Cognitive disor-ders.
Haase, A., Veterans AdministrationMedical Center, San Francisco.Viral infections: latency and defec-tive particles.
Fields, B., Harvard Medical School.Neurotropic viruses.
July 1-July 21
INSTRUCTORS
Bear, Susan, Ph.D., National Institutes of Health, Bethesda, MarylandBerman, Michael, Ph.D., Frederick Cancer Research Facility, Frederick, MarylandEnquist, Lynn, Ph.D., National Institutes of Health, Bethesda, MarylandSilhavy, Thomas, Ph.D., Frederick Cancer Research Facility, Frederick, Maryland
284

ASSISTANTS
Trun, Nancy, Frederick Cancer Research Facility, Frederick, MarylandZagursky, Bob Frederick Cancer Research Facility, Frederick Maryland
This course demonstrated the use of gene fusions, transposable elements, andrecombinant DNA for genetic analysis in Escherichia coli. Students learned toconstruct gene fusions both in vivo and in vitro. Subsequent experimentsstressed the use of these fusions for monitoring gene expression and, in conjunc-tion with transposable elements, for obtaining defined mutations (nonsense, dele-tion, and insertion) either in the target gene or in regulatory genes. RecombinantDNA was applied to clone and define physically the target gene, the regulatorygenes, and the sites of action of the regulatory proteins at the target gene. Inaddition, the cloned DNA and the defined mutations were used to analyze thegenes and the regulatory system genetically. For the sake of clarity, the coursefocused on a particular region; however, the experimental techniques presentedare sufficiently general to be applicable to any gene in E. coli for which thereexists a mutation conferring a recognizable phenotype.
PARTICIPANTS
Bardwell, James, B.S., University ofWisconsin, Madison
Booth, Ian R., Ph.D., University of Ab-erdeen, Scotland
Edwards, Mary Francis, B.S., Stan-ford University, California
Gardiol, Alicia E., B.S., MichiganState University, East Lansing
Hsiung, Hansen M., Ph.D., Eli Lillyand Co., Indianapolis, Indiana
Jones, Helen M., B.S., University ofCalifornia, Los Angeles
Kirschner, Richard J., Ph.D., UpjohnCo., Kalamazoo, Michigan
Niesel, David W., Ph.D., University ofTexas Medical Branch, Galveston
Olmedo, Gabriela, B.S., Centro DeEstudios Avanzados del I.P.N.,Mexico City, Mexico
Sanfacon, Helene, B.Sc., Laval Uni-versity, Quebec, Canada
Selvaraj, Gopalan, Ph.D., CarletonUniversity, Ottawa, Canada
Seto-Young, Donna L.T., Ph.D., Na-tional Research Council of Can-ada, Ottawa
Skvirsky, Rachel C., Ph.D., SimmonsCollege, Boston, Massachusetts
Stout, Valerie G., B.S., Kansas StateUniversity, Manhattan
Vary, Patricia S., Ph.D., Northern Illi-nois University, De Kalb
Welkos, Susan L., Ph.D., U.S. ArmyMedical Research Institute, Fred-erick, Maryland
SEMINARS
Austin, S., Frederick Cancer Re-search Facility. Stability and main-tenance of bacterial plasmids.
Howe, M., University of Wisconsin.Bacteriophage Mu: properties anduses.
Friedman, D., University of Michigan.Protein and nucleic acid signals intranscription termination and anti-termination.
Wolf, R., Jr., University of Maryland.
Growth-rate-dependent regulationof an E. coli central metabolismgene, gnd.
Youdarian, R, University of SouthernCalifornia. Genetic analysis of pro-tein-DNA interactions.
Beckwith, J., Harvard MedicalSchool. Genetic studies on the ba-sis of the specificity of a DNAbinding protein, CAP
Roth, J., University of Utah. Selectionand analysis of bacterial chromo-some inversions.
285

Neurobiology of DrosophilaJuly 1-July 21
INSTRUCTORS
Jan, Lily Yeh, Ph.D., University of California, San FranciscoJan, Yuh Nung, Ph.D., University of California, San FranciscoO'Farrell, Patrick H., Ph.D., University of California, San FranciscoGreenspan, Ralph J., Ph.D., Princeton University, New Jersey
ASSISTANT
Royden, Stanzi, University of California, San Francisco
This laboratory/lecture course was designed for people who may want to useDrosophila as an experimental system for studying function or development ofthe nervous system. One major aim was to introduce students to the various ge-netic, molecular, and physiological techniques that are currently available in Dro-sophila research and that make it distinctive.
The course began with a crash course on Drosophila genetics (a series of lec-tures supplemented with laboratory demonstrations and exercises) to familiarizethe students with classical genetics of Drosophila, strategies of mutant isolation,methods used to analyze newly isolated mutations, and mosaic analysis. Thiswas followed by workshops and laboratory projects on molecular genetics and
286

electrophysiology. In the area of molecular genetics topics covered were strate-gies and techniques for cloning genes, in situ hybridization to polytene chromo-somes, PM hybrid dysgenesis, and DNA-mediated transformation, while theelectrophysiology section covered neuromuscular transmission and voltageclamp analysis, as applied to mutants with altered ion channels. Additionally, aseries of lectures by invited speakers illustrated the application of these tech-niques to current research. The research topics included: molecular and func-tional analysis of mutants with distinctive ion channels, establishment of neuronalpathways in the developing embryo as well as in imaginal discs, analysis of be-havioral mutants, maternal and zygotic mutations that alter polarity and segmen-tation, homeotic mutations, and the use of DNA transformation for the study ofgene expression.
PARTICIPANTS
Bredesen, Dale, M.D., San FranciscoGeneral Hospital, California
Burnell, Anne, M.Sc., St. Patrick's Col-lege, Kildare, Ireland
Caudy, Michael, B.S., University ofCalifornia, Berkeley
DiNardo, Stephen, Ph.D., Universityof California, San Francisco
Hasan, Gaiti, Ph.D., Tata Institute ofFundamental Research, Bombay,India
Nash, Howard, Ph.D., National Insti-tutes of Mental Health, Bethesda,Maryland
Pollock,versity, New York
Roof, Dorothy, Ph.D., Purdue Univer-sity, Lafayette, Indiana
Shelton, David, B.A., University ofCalifornia, San Francisco
Taylor, Barbara, B.A., University ofCalifornia, San Diego
SEMINARS
Hall, J., Brandeis University. Muta-tions affecting sexual behaviorand circadian rhythm.
Timpe, L., University of California,San Francisco. Voltage-clampstudies of Drosophila mutants.
Quinn, W., Princeton University.Learning mutants.
Pak, W., Purdue University. Visual mu-tants.
Spradling, A., Carnegie Institution ofWashington. Hybrid dysgenesisand P-element transformation.
Use of transformation to dis-
amplification.Cherbas, P., R. Schultz, Harvard Uni-
versity. Complexities of eukaryotictranscription units and techniquesfor studying transcription.
Ecdysone-inducible transcrip-tion.
Molecular Biology of the Nervous SystemJuly 5-July 18
INSTRUCTORS
Kelly, Regis B., Ph.D., University of California, San FranciscoMcKay, Ronald D., Ph.D., Cold Spring Harbor Laboratory, New York
Levine, M., University of California,Berkeley. In situ localization oftranscripts.
. Engrailed and homeotic loci.Pardue, M.L., Massachusetts Institute
of Technology. Cytogenetics.Bender, W., Harvard Medical School.
Molecular biology of mutation.. Combining classical genetics
with molecular biology, homeoticmutations.
Ghysen, A., University Libre de Brux-elles. Sensory projections in hom-eotic mutants.
Wyman, R., Yale University. Early em-bryogenesis, segmentation mutants.
Thomas, J., Stanford University. Neu-rogenesis in embryos.
Horvitz, R., Massachusetts Institute ofTechnology. Cell lineage in nema-tode.
Goodman, C., Stanford University.Pathway formation in grasshopperembryos.
The technologies of patch-clamping and immunochemistry have revolutionizedmolecular neurobiology. Increasingly, the tools of recombinant DNA technologyand molecular genetics are being used to identify and characterize moleculesinvolved in neuronal function and development. The aim of this lecture coursewas to familiarize the participants with recent developments in molecular biologyand to discuss their application to neurobiological problems.
287

The following topics were discussed: patch-clamping living cells and channelsin bilayers, gene cloning, DNA sequencing, regulation of gene expression, in vi-tro mutagenesis, expressing foreign genes in heterologous hosts, in situ hybridi-zation, transposable elements and hybrid dysgenesis, in vitro immunization, anti-idiotype antibodies, T cell/B cell interaction, new optical and electron microscopetechniques, the cytoskeleton, organelle sorting and movement, regulation oftransmitter release, growth factors, cell lineages in development, assaying syn-aptogenesis in vitro, surface molecules and the extracellular matrix.
PARTICIPANTS
Amador, Roberto, M.D., RockefellerUniversity, New York, New York
Audigier, Yves, Ph.D., CNRS-INSERMde Pharmacologie/Endocrinolo-gie, Montpellier, France
Baldwin, Timothy, B.Sc., UniversityCollege London, England
Breer, Heinza, Ph.D., University Os-nabruck, Federal Republic of Ger-many
Calof, Anne, M.S., University of Cali-fornia School of Medicine, SanFrancisco
Cohen-Cory, Susana, B.S., Centro deInvestigation sobre Ingenieria Ge-netica y Biotecnologia, UNAM,Mexico City, Mexico
Earnest, Thomas, M.A., Boston Uni-versity, Massachusetts
Estacion, Mark, B.S., University ofCalifornia, Irvine
Hescheler, Jurgen, Universitat desSaarlandes, Hamburg, FederalRepublic of Germany
Hsiao, Karen Ke-jui, Ph.D., Universityof California, San Francisco
Jennings, Charles, B.A., UniversityCollege of London, England
Kadan, Michael, B.S., Johns HopkinsUniversity, Baltimore, Maryland
La Gamma, Edmund, M.D., CornellUniversity, Ithaca, New York
Lai, Yvonne, M.S., Rockefeller Univer-sity, New York, New York
Maelicke, Alfred, Ph.D., Max-PlanckInstitut, Dortmund, Federal Re-public of Germany
McCabe, Joseph, Ph.D., RockefellerUniversity, New York, New York
Michelsohn, Arie, B.A., California
288
Institute of Technology, PasadenaMiller, Stephen, B.S., California Insti-
tute of Technology, PasadenaSarlieve, Louis, Ph.D., INSERM et
Centre de Neurochimie du CNRS,Strasbourg, France
Silverstein, Faye, M.D., University ofMichigan, Ann Arbor
Tipnis, Ulka, Ph.D., University ofTexas Health Science Center, Dal-las
SEMINARS
Claudio, T., Columbia University, R.Scheller, Stanford University. Char-acterization of DNA. Cloning.
. Northerns. Southerns. Re-striction maps.
. Mapping. Synthetic DNA.cDNA and genomic libraries.
. Sequencing DNA. Electronmicroscopy.
. Hybridization section.Angerer, R., University of Rochester.
In situ hybridization.Claudio, T., Columbia University. In-
troducing foreign genes into cells.Costantini, F., E. Lacy, Columbia Uni-
versity. Introducing foreign genesinto the germ line.
Sharp, P., Massachusetts Institute ofTechnology. Introduction to eukar-yotic gene expression: processingon the mRNA level.
Maniatis, T., Harvard University. Roleof DNA sequences in regulatinggene expression.
Evans, R., Salk Institute. Genomicexpression and regulation in bac-teria.
Scheller, R., Stanford University, R.Evans, Salk Institute. Current mo-lecular research on the nervoussystem.
Kennedy, M., California Institute ofTechnology. Protein phosphoryla-tion.
Solomon, F., Massachusetts Institute

of Technology. Cytoskeleton modi-fication.
Siegelbau, S., Columbia University.Phosphorylation and learning.
Getter, M., Massachusetts Institute ofTechnology. Nature of the immuneresponse.
. Cell interactions in the im-mune response.
Schinnick, T, Scripps, Clinic and Re-search Center. Synthetic peptidesas antigens.
Alt, F., Columbia University. Molecularbasis of antibody diversity.
Lindstrom, J., Salk Institute. Func-tional domains of the AChR.
Aldrich, R., Yale University. Introduc-tion to patch-clamping.
Miller, C., Brandeis University. Recon-stitution of channels in mem-branes.
White, M., California Institute of Tech-nology. Synthesizing and injectingmRNA.
Khorana, G., Massachusetts Instituteof Technology. Genetic modifica-tion of a proton pump.
Bender, W., Harvard University. Ge-
Molecular Embryology of the Mouse
netic approaches to developmentusing Drosophila.
Jan, L., University of California. Iden-tifying the K" channel in Droso-phila.
Gusella, J., Massachusetts GeneralHospital. Mapping human geneticdefects.
Horvitz, R., Massachusetts Institute ofTechnology. Genetics of nematodedevelopment.
July 8-July 21
INSTRUCTORS
Hogan, Brigid, Ph.D., ICRF, Mill Hill, EnglandCostantini, Frank, Ph.D., Columbia University College of Physicians & Surgeons, New
York, New YorkLacy, Liz, Ph.D., Memorial Sloan-Kettering Cancer Center, New York, New York
ASSISTANTS
Robertson, Liz, University of Cambridge, EnglandChada, Kiran, Columbia University, New York, New York
This course was designed for biochemists, molecular biologists, and cell biolo-gists interested in applying their expertise to the study of mouse development. Inparticular, the genetic manipulation of the mouse through the introduction of for-eign genes and cells in early embryos was stressed. Through laboratory exer-cises and lectures, the participants were introduced to the following proceduresand their possible applications: the isolation and in vitro culture of germ cells andpreimplantation and early postimplantation embryos, the microinjection of DNAinto fertilized eggs, nuclear transplantation, the formation of chimeras, dissectionof germ layers, the localization of antigens and mRNAs in embryonic tissue sec-tions, and basic mouse handling and breeding techniques. Guest speakers dis-cussed current research in related fields.
PARTICIPANTS
Brulet, Philippe, Ph.D., Institut Pas-teur, Paris, France
Emerson, Beverly, Ph.D., National In-stitutes of Health, Bethesda, Mary-land
Gimlich, Robert, B.A., University ofCalifornia, Berkeley
Gross, Kenneth, Ph.D., Roswell ParkMemorial Institute, Buffalo, NewYork
Linney, Elwood, Ph.D., La Jolla Can-cer Research Foundation, Califor-nia
Mendel, Jane, B.A., MassachusettsInstitute of Technology, Cambridge
Murray, Ben, Ph.D., Rockefeller Uni-versity, New York, New York
Nepveu, Alain, Ph.D., Centre Hospi-talier Universitaire, Sherbrooke,Canada
Seising, Erik, Ph.D., Brandeis Univer-sity, Waltham, Massachusetts
Sorge, Joe, M.D., Scripps ResearchInstitute, La Jolla, California
Vogt, Thomas, B.S., Fox Chase Can-cer Institute, Philadelphia, Penn-sylvania
Weaver, David, Ph.D., MassachusettsInstitute of Technology, Cambridge
Kollias, George, National Institute ofMedical Research, London, Eng-land
289

SEMINARS
Wasserman, P.M., Harvard MedicalSchool. Nature and expression ofthe egg's receptor for sperm.
Damsky, C., Wistar Institute. Celladhesion molecules in embryonictissues.
Chapman, V., Roswell Park MemorialInstitute. Cell adhesion moleculesin embryonic tissues.
Silvery, L., Cold Spring Harbor Labo-ratory. Mouse genetics.
Stuhlman, H., Whitehead Institute. Mi-croinjection.
Copeland, N., University of Cincin-
Developmental Neurobiology
nati. Demonstration of blastocystinjection.
Jenkins, N., University of Cincinnati.Uterine transfer.
Solter, D., Wistar Institute. Post-im-plantation mouse development.
. Transnuclear mice.Silver, L., Cold Spring Harbor Labora-
tory. The T complex.McLaren, A., MRC Mammalian Devel-
opment Unit. Origin and differen-tiation of germ cells.
. Sex determination.Mulligan, R., Whitehead Institute. Re-
troviral vectors.Adamson, E., La Jolla Cancer Center.
Expression of growth factors, theirreceptors, and oncogenes inmouse embryos and teratocarci-noma cells,
Rigby, Imperial College. Microinjec-tion.
Posakony, J., Harvard University.Gene insertion in Drosophila.
July 20-August 2
INSTRUCTORS
Purees, Dale, M.D., Washington University School of Medicine, St. Louis, MissouriPatterson, Paul H., Ph.D., California Institute of Technology, Pasadena
The aim of this intensive two-week course is to expose a group of approximately20 students to the history of ideas and classical literature on neural development,with special emphasis on recent advances and controversies in this rapidly grow-ing field. The course is offered every other year, and the session in 1984 repre-sented its fourth cycle. In general, the information covered is not readily availablein a coherent form to students (especially those from abroad) because (1) manyimportant results are quite new, and (2) results both old and new rely on a widevariety of technical approaches and are therefore not encompassed in any sys-tematic way in courses arranged along conventional lines (e.g., courses on neu-rocytology and neurophysiology). The course on developmental neurobiologyseeks to fill this gap inviting about 15 guest lecturers, distinguished by their di-dactic skills as well as by their scientific accomplishments, to present their area ofinterest to students in a relatively informal setting where student-faculty inter-change and discussion are much encouraged. The lecturers are required toteach their material as broadly as possible so the students will take away someidea of the overall direction of research in neural development rather than spe-cific accounts of the endeavors of a few individuals.
290

This year's class of 23 students (a somewhat larger number of students wereaccepted in 1984 because of the quantity of outstanding applicants) was com-posed of 7 women and 16 men from 7 countries. The students were aboutequally divided between postdoctoral fellows and advanced graduate studentsalready involved in some phase of developmental neurobiology. The research in-terests of the students were extremely varied, ranging from behavior to molecularbiology; the only common denominator was an interest in pursuing research re-lated to neural development.
PARTICIPANTS
Bovolenta, Paola, Ph.D., New YorkUniversity, New York
Buxbaum, Joseph, M.S., WeizmannInstitute, Rehovot, Israel
Camp, Ann, B.A., Vanderbilt Univer-sity, Nashville, Tennessee
Coopersmith, Robert, B.S., Universityof California, Irvine
DeGennaro, Louis, Ph.D., Max-Planck-Institut, Martinsried, Fed-eral Republic of Germany
Ferrari, Giovanna, Ph.D. New YorkUniversity, New York
Fladby, Tormod, University of Oslo,Norway
Geijo, Emilio, M.D., University of Ali-cante, Spain
Hitchcock, Peter, Ph.D., Salk Institute,San Diego, California
Janowsky, Jeri, B.A., Cornell MedicalCenter, New York, New York
LaMantia, Anthony-Samuel, B.A., YaleUniversity, New Haven, Connecti-cut
Lefcort, Francis, B.A., University ofCalifornia, Berkeley
Levy, Daniel, Ph.D., Harvard Univer-sity, Cambridge, Massachusetts
Lo, Donald, B.S., Yale University, NewHaven, Connecticut
Mack ler, Scott, B.A., University ofPennsylvania, Philadelphia
Mikkelsen, Bente, M.D., University ofOslo, Norway
New, Helen, B.S., University CollegeLondon, England
Ross, Jenny, B.S., University ofOtago, New Zealand
Schrank, Ethan, B.S., University ofNorth Carolina, Chapel Hill
Stretavan, David, B.S., Stanford Uni-versity, California
Tessier-Levigne, Marc, B.S., Univer-sity College London, England
Voyvodic, James, B.S., WashingtonUniversity, St. Louis, Missouri
Wolf, Werner, M.D., Max-Planck-Insti-tut, G6ttingen, Federal Republic ofGermany
SEMINARS
Jacobson, M., University of Utah.Early events in embryogenesis.
Rakic, P., Yale University. Cell prolifer-ation and neuronal migration.
Bate, M., Cambridge University, Gra-dients, compartments, and posi-tional information in morphogene-sis.
Horvitz, R., Massachusetts Institute ofTechnology. Studies of cell lineagein C. elegans.
Goodman, C., Stanford University.Development of the grasshoppernervous system.
Stent, G., University of California,Berkeley. Development of theleech nervous system.
Bray, D., Kings College, The growthcone and mechanisms of axonelongation.
Advanced Electrophysiological Methods
Landmesser, L.T, University of Con-necticut. Axon outgrowth in verte-brates.
Goodman, C., Stanford University.Axon outgrowth in invertebrates.
Aguayo, A., McGill University. Axonelongation in the central and pe-ripheral nervous systems of mam-mals: the role of glial cells andother factors.
Greene, L., New York University Me-dial Center. Studies of the mecha-nism of action of nerve growth fac-tor.
Sanes, J.R., Washington UniversitySchool of Medicine. Formation ofsynapses in the central and pe-ripheral nervous system.
Easter, S., University of Michigan.Specificity in the retinotectal sys-tem.
Edelman, G., Rockefeller University.CAM and its role in development.
Nirenberg, M., National Heart andLung Institute. A possible molecu-lar basis of positional information.
Shatz, C., Stanford University Schoolof Medicine. Development of themammalian visual system.
Hildebrand, J., Columbia University.The development of behavior in in-sects.
Nottebohm, F, Rockefeller University.The development of birdsong andits relevance to learning.
July 24-August 13
INSTRUCTORS
Ascher, Phillippe, D. Sc., Ecole Norma le Superieure, Paris, FranceSchuetze, Stephen, Ph.D., Columbia University, New York, New YorkSiegelbaum, Steven, Ph.D., Columbia University, New York, New York
291

In this experimental course, students were introduced to the various electrophys-iological techniques that are currently applied to the study of transmitter actionon ion channels in skeletal muscle and invertebrate neurons. They used bothbasic voltage-clamp techniques (including a single microelectrode voltageclamp) as well as the patch-clamp technique for single-channel recording. As-pects of the theory of the techniques as well as some basic biochemical and bio-physical principles of transmitter actions were also covered.
Advanced Techniques in Molecular CloningJuly 24-August 13
INSTRUCTORS
Atkinson, Thomas, B.S., University of British Columbia, Vancouver, CanadaFiddes, John, Ph.D., California Biotechnology, Inc., Palo AltoShort le, David,, Ph.D., State University of New York, Stony BrookSmith, Michael, Ph.D., University of British Columbia, Vancouver, CanadaZoller, Mark, Ph.D., Cold Spring Harbor Laboratory, New York
ASSISTANT
Pielak, Gary, University of British Columbia, Vancouver
PARTICIPANTS
Chuman, Mary, Ph.D., Rush Presby-terian St. Luke's Medical Center,Chicago, Illinois
Frederiksen, Kristen, M.S., ColdSpring Harbor Laboratory, NewYork
Hay, Bruce, B.A., University of Califor-nia, San Francisco
Madison, Daniel, Ph.D., University ofCalifornia, San Francisco
Marshall, Cameron, B.S., Brown Uni-versity, Providence, Rhode Island
Pitchford, Simon, B.S., University ofBristol, England
Sigihara, Izumi, M.D., Tokyo MedicalSchool, Japan
Tobin, Thomas, Ph.D., State Univer-sity of New York Downstate Medi-cal Center, New York
Worley, Jennings, B.S., University ofMaryland, College Park
Zeitoun, Irene, M.S., Tel Aviv Univer-sity, Israel
This was a laboratory and lecture course on advanced aspects of molecularcloning designed for scientists who are familiar with basic recombinant DNAtechniques. It included the chemical synthesis of oligodeoxyribonucleotides ofdefined sequence and their characterization by the Maxam-Gilbert sequencing
292

method. These oligonucleotides were used in various ways, including their useas probes for the isolation of specific genes from libraries and as reagents fordirecting site-specific in vitro mutagenesis. Specific mutants were isolated, andthe sequence of the altered genes was determined. Other methods of in vitro mu-tagenesis such as nucleotide misincorporation into DNA, sodium bisulfate treat-ment of DNA, and linker insertion were covered in both laboratory exercises andlectures.
Guest lectures and demonstrations covered the application of these tech-niques to analysis of various cloned genes, as well as the use of alternate meth-ods of mutagenesis, expression, and analysis of cloned genes by immunologicalscreening.
PARTICIPANTS
Cane, Jerome, B.S., Indiana Univer-sity, Bloomington
Chaudhuri, Bhabatosh, Ph.D., Fried-rich Miescher Institut, Basel, Switz-erland
Dean, Carolyn, Ph.D., Advanced Ge-netic Sciences, Oakland, Califor-nia
Geller, Ruth, Ph.D., Walter Reed ArmyInstitute of Research, Washington,D.C.
Giroux, Craig N., Ph.D., NIEHS, Re-search Triangle Park, North Caro-lina
Goff, Stephen, Ph.D., Harvard Medi-cal School, Boston, Massachusetts
Hilt, Dana, M.D., National Institutes ofHealth, Bethesda, Maryland
Hopper, Peggy, B.S., Tufts University,Medford, Massachusetts
Mostov, Keith, Ph.D., Rockefeller Uni-versity, New York, New York
Mroczkowski, Barbara, Ph.D., Van-derbilt University, Nashville, Ten-nessee
Rdest, Ursula, Ph.D., University ofWurzberg, Federal Republic ofGermany
Robinson, Mary Ann, Ph.D., NationalInstitutes of Health, Bethesda,Maryland
SEMINARS
Gumport, R., University of Illinois. Therole of homology in bacteriophage
site-specific recombination.
Goff, S., Columbia University. Muta-tions in the gag and po/ genes ofMoloney murine leukemia virus.
Bogenhagen, Daniel, State Universityof New York, Stony Brook. Tran-scription of vertebrate mitochon-dria! DNA.
Russell, D., University of Texas, Dal-las. The structure of human low-density lipoprotein receptor.
Davis, M., Stanford University. Isola-tion and characterization of the T-cell receptor gene.
Essigmann, J., Massachusetts Insti-tute of Technology. Extrachromo-somal probes for mutagenesis bychemical carcinogens.
Meyers, R., Harvard University. Satu-ration mutagenesis of eukaryoticregulatory sequences.
293

Yeast GeneticsJuly 24-August 13
INSTRUCTORS
Sherman, Fred, Ph.D., University of Rochester, New YorkFink, Gerald, Ph.D., Massachusetts Institute of Technology, CambridgeHicks, James, Ph.D., Cold Spring Harbor Laboratory, New York
ASSISTANT
Chiu, Isabel, Massachusetts Institute of Technology, Cambridge
The major laboratory techniques used in the genetic analysis of yeast were stud-ied, including the isolation and characterization of chromosomal and mitochon-dria) mutants, tetrad analysis, chromosomal mapping, mitotic recombination, andtest of allelism and complementation. Micromanipulation used in tetrad analysiswas carried out by all students. Recombinant DNA techniques, including yeasttransformation, filter hybridization, and gel electrophoresis, were applied to clon-ing and genetic analysis of yeast DNA. Lectures on fundamental aspects of yeastgenetics were presented along with seminars given by outside speakers on top-ics of current interest.
PARTICIPANTS
Bibus, Claudia, B.A., University ofBasel, Switzerland
Butt, Tauseef, Ph.D., Smith Kline &French Laboratories, Philadelphia,Pennsylvania
Chin, Jean, Ph.D., Harvard Univer-sity, Cambridge, Massachusetts
Eisenberg, Shlomo, Ph.D., WeizmannInstitute, Rehovot, Israel
Folger, Kim, Ph.D., University of Utah,Salt Lake City
Harmony, Judith, PH.D., University ofCincinnati, Ohio
LaPorte, David, Ph.D., University ofMinnesota, Minneapolis
Lautenberger, Jams, Ph.D., National
294
Cancer Institute, Bethesda, Mary-land
Martin, Charles, Ph.D., Rutgers Uni-versity, New Brunswick, New Jer-sey
Nelson, Daniel, Ph.D., University ofHouston, Texas
Nolan, Catherine, Ph.D., NationalCancer Institute, Bethesda, Mary-land
Pines, Ophrey, Ph.D., State Universityof New York, Stony Brook
Rosendahl, Mary, Ph.D., RockefellerUniversity, New York, New York
Sawadogo, Michele, Ph.D., Rockefel-ler University, New York, New York
Varshaysky, Alexander, Ph.D., Massa-chusetts Institute of Technology,Cambridge
SEMINARS
Fox, T, Cornell University. Mitochon-dria( genetics.
Warner, J., Albert Einstein College ofMedicine. Ribosomal genes: struc-ture and regulation.
Kolodner, R., Dana Farber Cancer In-stitute. Genetic recombination cat-alyzed by cell-free extracts.
Wig ler, M., Cold Spring Harbor Labo-ratory. RAS oncogenes in yeast.

Cantor, C., Columbia University. Sep-aration of chromosome-sized DNAmolecules.
Fitzgerald-Hayes, M., University ofMassachusetts. Base-specific mu-tations affecting centromere func-tions.
Carlson, M., Columbia University.Regulation and structure of yeastinvertase genes.
Lindquist, S., University of Chicago.
Yeast sporulation. heat shock pro-teins and a whole lot more.
Tye, B.-K., Cornell University. Func-tional components of yeast chro-mosomes.
Broach, J., State University of NewYork, Stony Brook. Control of plas-mid replication in yeast.
Zoller, M., Cold Spring Harbor Labo-ratory. Mutagenesis of the HO cutsite.
Botstein, D., Massachusetts Instituteof Technology. Genetics of the cy-toskeleton and cell cycle.
Szostak, J., Dana Farber Cancer Insti-tute. Yeast telomeres and artificialchromosomes.
Silver, F, Harvard University. Nuclearlocalization of proteins in yeast.
Prakash, S., University of Rochester.Structure and regulation of DNArepair genes.
295


Seminars
Cold Spring Harbor Seminars were initiated to provide a semiformal avenue forcommunication between the various research groups at the Laboratory. They areparticularly useful for research personnel who have joined the Laboratory duringthe summer. The seminars also afford a necessary opportunity for the graduatestudents and postgraduate staff to develop their skills in defending, organizing,and presenting their research. In addition to those listed below, seminars weregiven by many others involved in research at this Laboratory.
1983 -1984
September
Tom Curran, Salk Institute, San Diego, California: A com-parative analysis of the viral and cellular fos oncogenes.
Richard Treisman, Harvard University, Cambridge, Massa-chusetts: Expression of cloned globin genes in culturedcells.
October
James Alwine, University of Pennsylvania, Philadelphia:Control of Simian virus 40 late gene expression.
Edward Trifonov, Weisman Institute, Rehovot, Israel: Typicalsecondary structures around splicing junctions.
Pat Hearing, State University of New York, Stony Brook:Analysis of the Ad5 E1A-transcriptional control regionand sequences required for packaging of viral DNA.
Rob Jackson, Albert Einstein College of Medicine, Bronx,New York: A genetic analysis of voltage sensitive so-dium channels in Drosophila.
Pat Levitt, Medical College of Pennsylvania, Philadelphia:Molecular specificity in stem cells in functional systemsof the brain.
Sayeeda Zain, University of Rochester School of Medicineand Dentistry, New York:I. Two different human liver mRNAs encode serum al-
bumin.II. Development of a simple and efficient in vitro mRNA
splicing system.
November
Eric Hunter, University of Alabama, Birmingham: Mutationsthat alter the biosynthesis of the Rous sarcoma virus en-velope glycoprotein.
Saran Narang, National Research Council of Canada, Ot-tawa: Total synthesis, cloning, and expression of humanpreproinsulin.
Nils Loneberg, Harvard University, Cambridge, Massachu-setts: Pyruvate kinase genes.
Elizabeth Blackburn, University of California, Berkeley: Mo-lecular structures of telomeres in lower eukaryotes.
Paul Ludden, University of Wisconsin, Madison: Regulationof nitrogenase in photosynthetic bacteria.
David Lowe, University of Toronto, Canada: The mouse hsp70 gene family: Cloning of cDNAs from inducible andconstitutive mRNAs.
Rob Jackson, Albert Einstein College of Medicine, Bronx,New York: A genetic analysis of voltage sensitive so-dium channels in Drosophila.
Claude Dery, University of Sherbrooke, Quebec, Canada:Structure of adenovirus chromatin in lytic infection.
December
Michael Young, Rockefeller University, New York, NewYork: Transposable elements and mutations of a com-plex locus in Drosophila.
Dirk Elseviers, New York University Medical College, NewYork: Readthrough of nonsense codons: A tool for ge-netics of proteins.
Neil Copeland, University of Cincinnati, Ohio: Endogenousmurine retroviruses: Molecular probes for dissecting themammalian genome.
Nancy Jenkins, University of Cincinnati, Ohio: Retrovirusesas insertional mutagens of mice.
Fred Alt, Columbia University, New York: Rearrangement ofendogenous and introduced heavy chain variable re-gion gene segments in pre-B cell lines,
David Grumwald, University of Oregon, Eugene: Geneticsof zebra fish: A new approach to the study of vertebratedevelopment.
January
Hans Lehrach, European Molecular Biology Laboratory,Heidelburg, Federal Republic of Germany: Molecularstudies of mouse chromosome 17.
Joe Moscal, National Institutes of Health, Bethesda, Mary-land: Characterization and purification of a topographi-cally distributed molecule in chick retina.
Richard Novick, Public Health Research Institute of the Cityof New York, New York: Counter transcription regulationof plasmid replication.
Bertrand Jordan, Center of Immunology, Marseille, France:Structure and expression of human histocompatibilitygenes.
James McCubrey, Basel Institute for Immunology, Sweden:Gene transfer experiments with the functionally rear-ranged immunoglobulin genes.
297

February
Joanne Williams, University of California, San Diego: Reac-tion center genes from Rhodopseudomonas sphae-roides.
Walter Keller, German Cancer Research Center, Heidel-berg, Federal Republic of Germany: In vitro splicing ofmRNA precursors.
Malcolm Casadaban, University of Chicago, Illinois: In vivogenetics manipulations with bacteria phage Mu,
March
Marcel lo Siniscalco, Memorial Sloan-Kettering Cancer Cen-ter, New York, New York: Fragile sites of human chromo-somes and meiotic recombination.
Michael A. Lischwe, Baylor College of Medicine, Houston,Texas: Nucleolar proteins: Phosphorylation, argininemethylation, immunofluorescence, and scleroderma an-tigen.
Charles Craik, University of California, San Francisco:Expression and site-specific mutagenesis of rat pan-creatic trypsin.
April
Kirsten Fischer Lindahl, Basel Institute for Immunology,Sweden: Mitochondria and the mouse MHC.
Adrian Heyday, Massachusetts Institute of Technology Can-cer Center, Cambridge: Activation of c-myc in human B-cell lymphoma.
298
Alan Senior, University of Rochester School of Medicine,New York: The use of mutants in studies of E. coli pro-ton-ATPase.
Masayori Inouye, State University of New York, StonyBrook: A novel regulatory mechanism using an RNAtranscript complementary to a specific mRNA (micRNA).
Rick Myers, Harvard University, Cambridge, Massachu-setts: Saturation mutagenesis of eukaryotic regulatorysequences.
David Housman, Massachusetts Institute of TechnologyCancer Center, Cambridge: Gene mapping in mouseand man.
Paul Bingham, State University of New York, Stony Brook:Molecular Genetics of Drosophila transposons.
Tom Braciole, Washington University, St. Louis, Missouri:Major histocompatability complex restricted cytotoxic T-cells: What do they see?
May
Steve Cheley, University of Western Ontario, London, Can-ada: Intracellular mouse hepatitis virus polypeptidesand RNAs.
Leone llo Bossi, University of Utah, Salt Lake City: Unusualconformation of the DNA in a region upstream from abacterial promoter is associated with maximal levels ofexpression.
June
Dave Pauza, MRC University Medical School, Cambridge,England: Early events in human lymphocyte activation.

Undergraduate Research
An important aspect of the summer program at the Laboratory is the participation ofcollege undergraduate students in active research projects under the supervision offull-time laboratory staff members. The program was initiated in 1959. Since thatyear, 260 students have completed the course, and many have gone on to creativecareers in biological science.
The objectives of the program are to provide (1) a greater understanding of thefundamental principles of biology, (2) an increased awareness of major problemareas under investigation, (3) better understanding of the physical and intellectualtools for modern research and the pertinence of this information to future training,and (4) a personal acquaintance with research, research workers, and centers forstudy.
The following students, selected from a large number of applicants, took part inthe program, which was supported by the J.M. Foundation, Burroughs WellcomeFund, and Bayer Aspirin (Glen Brook Labs). Students are listed below with theirLaboratory sponsors and topics of research.
Mark Alfenito, Cornell UniversityResearch Advisor: S. DellaportaGenetic and molecular study of maize controlling elements.
Catherine Chen, Massachusetts Institute of TechnologyResearch Advisor: B. StillmanAnalysis of the deg phenotype associated with mutations in
the gene encoding the adenovirus E1B 19K tumor anti-gen.
Shinta Cheng, Yale UniversityResearch Advisor: F. DaldalMolecular genetics of cytochrome C2 of Rhodopseudomonas
capsulata.
Susan Euling, Columbia UniversityResearch Advisor: D. KurtzDetermination of tissue-specific alpha 2,1 globulin.
William Howell, University of WisconsinResearch Advisor: P. ScolnikCharacterization of the end-sites of the DNA of the gene trans-
fer agent of Rhodopseudomonas capsulata.
Phyllis Kristal, Massachusetts Institute of TechnologyResearch Advisor: L. SilverDevelopmental analysis of lethal mutations on mouse chromo-
some 17.
Mark Montgomery, Princeton UniversityResearch Advisor: J. LewisTranscription studies with the cell-cycle-regulated Chinese
hamster thymidine kinase gene.
Ramona Morfeld, Wheaton CollegeResearch Advisor: J. HicksInvestigation of a transformation system in maize.
Jon Rubin, Harvard UniversityResearch Advisor: D. HelfmanIsolation and characterization of cDNA clones encoding non-
muscle tropomyosin.
David Southern, University of GlasgowResearch Advisor: R. SadaiePCNA: The need for research.
Olney FellowNasrollah Samiy, Harvard UniversityResearch Advisor: F. TamanoiExpression in two oncogenes.
299


Cold Spring Harbor Curriculum Study
The lag time between new discoveries in the laboratory-the basic stuff of sci-ence-and their effective integration into secondary science teaching can be asmuch as 10-15 years. Unfortunately, during this interval there has occurred arevolution in biology whose vast social and economic implications promise to in-fluence the lives of the current generation of youth profoundly.
The explosive advent of powerful techniques to probe the molecular structureof DNA and proteins has fundamentally changed the focus of modern biology.Unfortunately, present school curricula do not reflect this significant revolution orits product- biotechnology. We are faced with an untenable situation: The biol-ogy now being taught in high schools is of a different era and of a separate mind-set than that which is currently being practiced at the laboratory bench.
In an effort to help close this gap, Cold Spring Harbor Laboratory has joinedwith eight Long Island school districts in forming the Cold Spring Harbor Curricu-lum Study-a cooperative project to improve teacher competence and to de-velop model curricula on molecular biology, recombinant DNA, and biotechnol-ogy. The program is funded by a $10,000 grant from Citibank, N.A., andappropriations from the participating school districts: Cold Spring Harbor, EastWilliston, Great Neck, Herricks, Jericho, Northport-East Northport, Oyster Bay,and Syosset. Included in the $90,000 budget for the Curriculum Study's initialyear is over $40,000 in equipment needed to perform state-of-the-art experi-ments that culminate in the production of recombinant DNA molecules.
For many years, the Laboratory was considered an ivory-tower institution,aloof in an esoteric cloud from the average person. Now, through the CurriculumStudy, the institution is reaching out to share its knowledge and expertise withthe local community. By acting as a resource for public schools, we hope to helpestablish Long Island as a nationwide leader in advanced science education.The largest coordinated effort in the country to teach the "new biology," the Cur-riculum Study will serve as a model for similar efforts, and the materials devel-oped will find use in schools throughout the United States. The program is de-signed to take a broad approach to the problems of biology education byimproving teacher competence and by developing new classroom materials andmethods. It is hoped that by effecting change at the top of the education pyra-mid-at the level of administrators, science chairpersons, and teachers-benefitswill filter down to the largest number of students.
Teachers from participating districts take part in seminars presented by ColdSpring Harbor Laboratory staff members and meet regularly to develop laborato-ries and instructional materials. Enrichment activities for students include "honorscolloquia" at the Laboratory, a site visit to each school district by a Cold SpringHarbor staff scientist, and an intensive 6-day workshop to be held during thesummer.
Science and Society
Few would argue the extent to which scientific advancements and new technolo-gies affect our socioeconomic environment. On a personal level, technologyreorders our life-styles at a bewildering rate. On a public level, science- and tech-nology-related issues play an increasingly important role in public policy. Al-
301

though our involvement in technology is increasing at an exponential rate, publicunderstanding of the scientific factors that shape our lives lags far behind. Thus,another goal of the Curriculum Study is to prepare students to evaluate more ac-curately the social and ethical issues generated by the new biology."
Recent survey data indicate that Americans attribute our global prestige andinfluence to our technological know-how. However, there is mounting evidencethat the United States is beginning to lose its advantage in international techno-logical competition. Clearly, maintaining our technological edge demands astrong commitment to developing new scientific talent. Yet, it appears that we arefailing to stimulate expansion of our scientific base at a grass-roots level in ourelementary and secondary schools. We seem to have forgotten that increasedemphasis on science education is the only way to keep up with the technologicalrealities of our rapidly changing world. Our technological competitors have longrealized this fact and have taken pains to expand their science curricula. Yet, wehave been too smug or too blind to see that we are in the same race with tech-nology; science curricula in our secondary schools are steadily shrinking.
American students are required to take far fewer science and math coursesthan are their peers in other countries. In Japan; the Soviet Union, and East Ger-many, the vast majority of high school graduates have taken chemistry, physics,and biology; the majority of American students take only 1 year of biology. Spec-ialized science study in these countries begins in the 6th grade, with separatecourses in biology, chemistry, and physics.
Science Education Coming Full Circle
The scenario is not new. The launch of the Soviet satellite Sputnik in 1957 trig-gered similar fears that the United States was losing its position as world leaderin technology. A public outcry demanded increased support of science educa-tion. During the 1960s, an infusion of federal funds and leadership from the Na-tional Science Foundation (NSF) stimulated a renaissance in secondary scienceeducation-new curricular materials were developed and science teaching wasbrought up-to-date.
By the mid-1970s, concern was replaced by complacency. The spectacular re-sults of the space program and great advances in computer technology seemedto indicate that we were, once again, ahead in the technology race. In an era ofsocial unrest, NSF-developed curricula were increasingly attacked as a source ofbreakdown in traditional values. The NSF precollege program declined and wasessentially dismantled between 1980-82; curriculum development ground to ahalt.
Now we have come full circle. We stand at another watershed in science edu-cation in this country. Over the past several years, numerous major assessmentsof precollege science education have pointed to the urgent need for new effortsto update science teaching. The government appears to have heeded the call;the NSF budget for precollege education rose from $15 million in 1983 to over$54 million in 1984. The Cold Spring Harbor Curriculum Study will stand at theheadwaters of this new surge in American science education.
Why Johnny Doesn't Learn Science
Children start out as "natural scientists." They routinely carry out investigations tounderstand the world around them. Although science teaching should build on
302

this natural inquisitiveness, recent surveys indicate that children acquire a dislikefor formal science at an early age and that these negative attitudes increase withschooling. For the majority of students, there appears to be little growth in inter-est or knowledge of science during the high school years.
The lack of student interest in science may stem, in large part, from inadequa-cies in the way it is taught. The vast majority of science teachers rely heavily onstandard texts that tend to stress terminology and definitions. Thus, rote learningof terms, which emphasizes only the academic aspects of science, is the majorcomponent of precollege biology education. Little attention is paid to the real-world applications of biology or its underlying unity with the chemical and physi-cal sciences.
Although learning by doing has long been a goal of effective science teaching,the sad fact is that our schools place too little emphasis on discovery learning.Too often, teachers faced with overambitious syllabi and large classes opt to skipthe laboratory work that could make science more relevant to students. Short-ages in supplies and equipment have drastically reduced laboratory time in thesecondary schools. Even when laboratories are used, they tend to be demon-strations of information already presented, rather than true discovery learning. Itseems clear that although they may impart some formal knowledge of science,current secondary biology curricula are failing miserably to prepare individuals tounderstand better or participate in an increasingly technological world.
Why Teach Molecular Biology?
Despite the fact that DNA is the starting point of all life, its seminal importance isnot emphasized in current science curricula. Intuitively, it makes great sense tofocus biology teaching more closely on DNA. It provides a molecular basis uponwhich can be built the major concepts of biology: the difference between livingand nonliving, development and the differentiation of tissues, reproduction, therelationship between structure and function, and the diversity of life.
One criticism of science education is that it has become too aligned with dis-crete academic disciplines. Survey data indicate that high school students arefrightfully ignorant of chemistry as it relates to life. This is a shame, because, inpractice, science is becoming less and less compartmentalized. A synthesis ofseveral disciplines, including genetics, microbiology, biochemistry, physicalchemistry, and physics, molecular biology highlights the unity of science. Sincemany high school students' only introduction to science is in a general biologycourse, it makes sense to stress chemistry and physics as it relates to life. Thus,emphasizing a molecular biological approach would be a step toward teaching"integrated" science, By providing an entrée to biotechnology, such a molecularapproach would feed students' interest in technology and demand for practicalrelevance.
In addition to increasing knowledge of the pivotal role of DNA in life processes,a major aim of the Curriculum Study is to stimulate awareness of career oppor-tunities in biotechnology. Molecular biology is a science that will figure largely inthe future economic development of this nation. Like the explosive growth ofcomputer technology, biotechnology will provide unprecedented professionalopportunities in research and development. Students who participate in this proj-ect will have a clear advantage in the competition for acceptance in science pro-grams at prestigious universities. These students will be well prepared to pursue
303

other enrichment activities, such as individualized research, projects for sciencecompetitions, and part-time work at research institutions. This experience will beespecially crucial for students planning careers in biological research.
304

Nature Study Program
The Nature Study Program gives elementary and secondary school students the opportunityto acquire a greater knowledge and understanding of their environment. Through a series ofspecialized field courses, students can engage in such introductory programs as Nature De-tectives, Seashore Life, and Pebble Pups, as well as more advanced programs such as MarineBiology, Nature Photography, and the Geology of Long Island.
During the summer of 1984 a new high of 525 students participated in the Nature StudyProgram. Most classes were held outdoors, when weather permitted, or at the Uplands FarmNature Preserve of the Long Island Chapter of the Nature Conservancy. The Laboratory hasequipped and maintains a darkroom and classroom/laboratories at Uplands Farm. This facilityis used as a base for the student's exploration of the local environment. Field classes are heldon Laboratory grounds, St. John's Preserve, Shu Swamp Preserve, Montauk Point State Park,the Long Island Pine Barrens, Caumsett Park, and in other area parks.
In addition to the four-week courses, a series of one-day marine biology workshops wasoffered to students. Studies on the marine ecology of L.I. Sound were conducted aboard the100 foot Sloop Clearwater chartered from Hudson River Sloop Clearwater, Inc. of Poughkeep-sie, New York. Students were able to study the Sound chemically, physically, and biologicallyusing the ship's instrumentation. The three-day Adventure Education class took children on an18 mile bike hike to Caumsett State Park, a six-mile canoe trip on the Nissequogue River, anda day of sailing on the Sloop Clearwater.
PROGRAM DIRECTOR
Edward Tronolone, M.S., RD., Science Curriculum Associate,East Williston Public Schools
INSTRUCTORS
Kathryn Bott, M.S., science teacher, Friends AcademyRuth Burgess, B.A., naturalist, Nassau County BOCESEric Knuffke, M.S., science teacher, Mineola High SchoolFred Maasch, M.S., science teacher, Islip High SchoolPaul Paino, M.S., science teacher, The Wheatley High SchoolBill Payoski, M.S., science instructor, Nassau Community CollegeLinda Payoski, B.A., naturalist, Nassau County BOCES
COURSES
Nature BugsNature DetectivesAdvanced Nature StudyIntroduction to EcologyFrogs, Flippers, and Fins
Pebble PupsBird StudyFresh Water LifeSeashore LifeThey Swim, Walk, and Crawl
Geology of Long IslandMarine BiologyNature PhotographyAdventure EducationMarine Biology Workshop
305

Facing page: A replica of an adenovirus particle decorates the gazebo built on top of theLaboratory's waste water treatment plant.

FINANCIALSTATEMENT
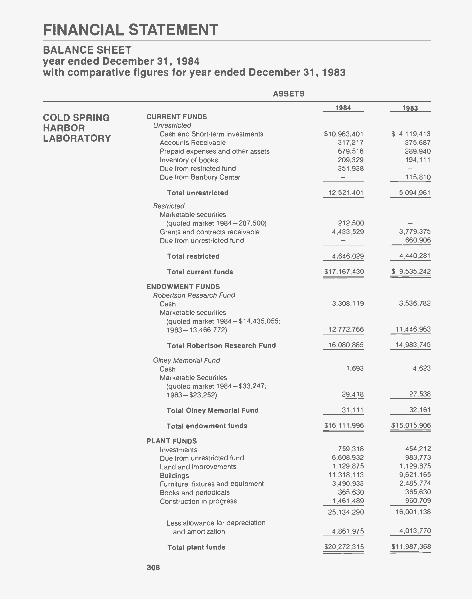
FINANCIAL STATEMENTBALANCE SHEETyear ended December 31, 1984with comparative figures for year ended December 31, 1983
ASSETS
1984 1983
COLD SPRING CURRENT FUNDS
HARBOR Unrestricted
LABORATORY Cash and Short-term investmentsAccounts Receivable
$10,963,401317,217
$ 4,119,413375,687
Prepaid expenses and other assets 679,516 289,940Inventory of books 209,329 194,111Due from restricted fund 351,938Due from Banbury Center 115,810
Total unrestricted 12,521,401 5,094,961
RestrictedMarketable securities
(quoted market 1984-287,500) 212,500Grants and contracts receivable 4,433,529 3,779,375Due from unrestricted fund 660,906
Total restricted 4,646,029 4,440,281
Total current funds $17,167,430 $ 9,535,242
ENDOWMENT FUNDSRobertson Research Fund
Cash 3,308,119 3,536,782Marketable securities
(quoted market 1984-$14,435,055;1983-13,466,772) 12,772,766 11,446,963
Total Robertson Research Fund 16,080,885 14,983,745
Olney Memorial FundCash 1,693 4,623Marketable Securities
(quoted market 1984-$33,247;1983-$23,252) 29,418 27,538
Total Olney Memorial Fund 31,111 32,161
Total endowment funds $16,111,996 $15,015,906
PLANT FUNDSInvestments 759,318 454,212Due from unrestricted fund 6,608,932 983,773Land and improvements 1,129,875 1,129,875Buildings 11,318,113 9,621,165Furniture, fixtures and equipment 3,490,933 2,485,774Books and periodicals 365,630 365,630Construction in progress 1,461,489 960,709
25,134,290 16,001,138
Less allowance for depreciationand amortization 4,861,975 4,013,770
Total plant funds $20,272,315 $11,987,368
308

LIABILITIES AND FUND BALANCES
1984 1983
CURRENT FUNDSUnrestricted
Accounts payable $ 747,261 $ 835,166Deferred income 120,000 140,000Due to Banbury Center 74,580Due to plant fund 6,608,932 983,773Due to restricted fund 660,906Fund balance 4,970,628 2,475,116
Total unrestricted 12,521,401 5,094,961
RestrictedAccounts payable 65,792Due to Banbury Center 8,952Due to unrestricted fund 351,938 -Fund balance 4,250,458 4,440,281
Total restricted 4,646,029 4,440,281
Total current funds $17,167,430 $ 9,535,242
ENDOWMENT FUNDS
Fund balance $16,111,996 $15,015,906
PLANT FUNDSAccounts payable 121,055Loan payable 8,000,000Fund balance 12,151,260 11,987,368
Total Plant Funds $20,272,315 $11,987,368
309

ASSETS
BANBURY CENTER1984 1983
CURRENT FUNDSUnrestricted
Cash 700 700Prepaid and deferred expenses 19,184 15,025Inventory of books 46,763 36,857Due from Banbury restricted fund 95,505Due from CSHL unrestricted fund 16,497
Total unrestricted 83,144 148,087
RestrictedGrants and contracts receivable 111,450 142,988Due from CSHL unrestricted fund 58,083Due from CSHL restricted fund 8,952
Total restricted 178,485 142,988
Total current funds 261,629 291,075
ENDOWMENT FUNDSRobertson Maintenance Fund
Cash 597,661 494,054Marketable securities
(quoted market 1984 -$2,347,644;1983 -$2,314,259) 2,077,778 1,978,853
Total endowment funds 2,675,439 2,472,907
PLANT FUNDSLand 772,500 772,500Buildings 846,028 845,967Furniture, fixtures and equipment 180,383 176,318Construction in progress 1,385 1,386
1,800,296 1,796,171Less allowance for depreciation 357,019 320,450
Total plant funds 1,443,277 1,475,721
Total Banbury Center $ 4,380,345 $ 4,239,703
Total-All funds $57,932,086 $40,778,219
310

LIABILITIES AND FUND BALANCES
1984 1983
CURRENT FUNDSUnrestricted
Accounts payable $ 18,466 $ 32,277Due to CSHL unrestricted fund 115,810Fund balance 64,678
Total unrestricted 83,144 148,087
RestrictedAccounts payable 1,394Due to Banbury unrestricted 95,505Fund balance 177,091 47,483
Total restricted 178,485 142,988
Total current funds 261,629 291,075
ENDOWMENT FUNDS
Fund balance 2,675,439 2,472,907
PLANT FUNDS
Fund balance 1,443,277 1,475,721
Total Banbury Center $ 4,380,345 $ 4,239,703
Total -All funds $57,932,086 $40,778,219
311
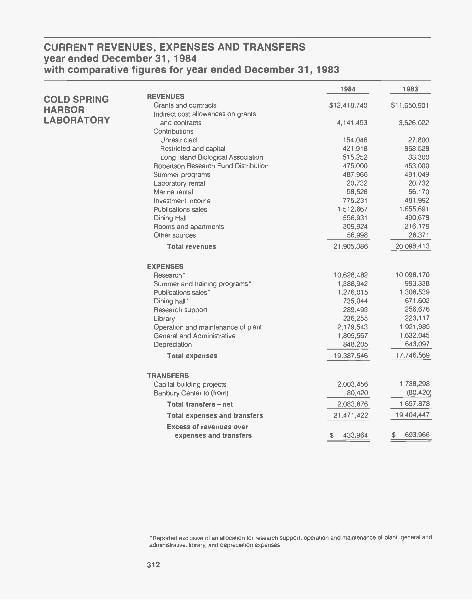
CURRENT REVENUES, EXPENSES AND TRANSFERSyear ended December 31, 1984with comparative figures for year ended December 31, 1983
1984 1983
COLD SPRING REVENUES
HARBOR Grants and contractsIndirect cost allowances on grants
$12,418,740 $11,650,901
LABORATORY and contracts 4,141,453 3,626,022Contributions
Unrestricted 154,046 27,800Restricted and capital 421,918 868,528Long Island Biological Association 515,252 33,300
Robertson Research Fund Distribution 475,000 453,000Summer programs 487,966 491,049Laboratory rental 20,732 20,732Marina rental 58,528 56,170Investment income 775,231 481,992Publications sales 1,512,667 1,655,691Dining Hall 556,931 490,678Rooms and apartments 309,924 216,179Other sources 56,998 26,371
Total revenues 21,905,386 20,098,413
EXPENSESResearch* 10,628,482 10,098,170Summer and training programs* 1,388,942 993,338Publications sales* 1,276,015 1,306,539Dining hall* 735,044 671,602Research support 289,493 256,676Library 236,255 223,117Operation and maintenance of plant 2,179,543 1,921,985General and Administrative 1,805,567 1,632,045Depreciation 848,205 643,097
Total expenses 19,387,546 17,746,569
TRANSFERSCapital building projects 2,003,456 1,738,298Banbury Center to (from) 80,420 (80,420)
Total transfers- net 2,083,876 1,657,878
Total expenses and transfers 21,471,422 19,404,447
Excess of revenues overexpenses and transfers $ 433,964 $ 693,966
'Reported exclusive of an allocation for research support, operation and maintenance of plant, general andadministrative, library, and depreciation expenses.
312

1984 1983
BANBURY CENTER REVENUESEndowment income $106,000 $100,000Grants & contributions 407,806 261,436Indirect cost allowances on
grants and contracts 2,060 13,576Rooms and apartments 65,757 76,522Publications 102,410 191,993Conference fees 14,697 54,096Dining Hall 34,884 4,708Transfer from (to) Cold
Spring Harbor Laboratory 80,420 (80,420)
Total revenues 814,034 621,911
EXPENSESConferences 171,095 206,659Publications 128,181 126,618Operation and maintenance
of plant 120,103 93,548Programiiadministration 196,244 202,651Depreciation 36,569 45,107Capital plant 4,125
Total expenses 656,317 674,583
Excess (deficit) of revenuesover expenses $157,717 $(52,672)
NOTE: Copies of our complete, audited financial statements, certified by our independentauditors, Peat, Marwick, Mitchell & Co., are available upon request from the Comptroller, ColdSpring Harbor Laboratory.
313
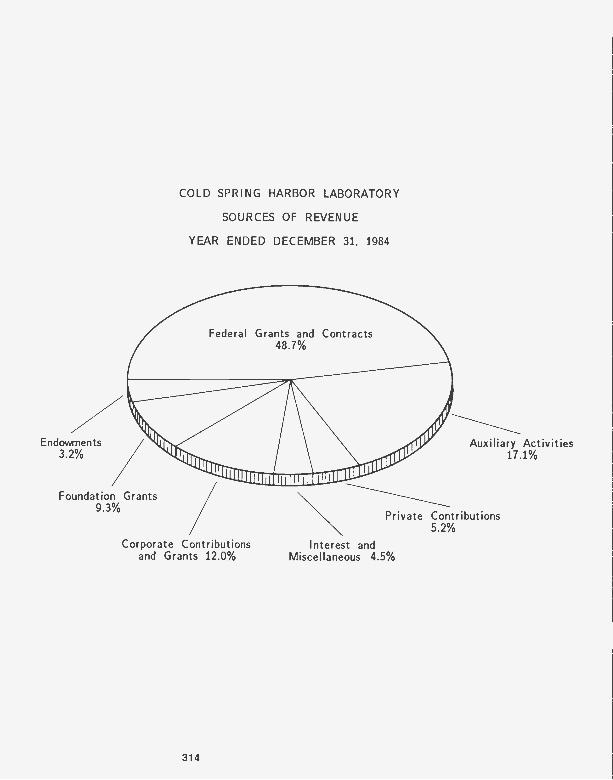
COLD SPRING HARBOR LABORATORY
SOURCES OF REVENUE
YEAR ENDED DECEMBER 31, 1984
Federal Grants and Contracts48.7%
Endowments3.2%
Foundation Grants9.3%
Corporate Contributionsand Grants 12.0%
314
Auxiliary Activities17.1%
Private Contributions5.2%
Interest andMiscellaneous 4.5%


Recto: Detail of Carnegie Library, built in 1905 by the Carnegie Institution of Washington.Originally designed as a laboratory, the building was refitted as a library in 1953 and partiallyrenovated in 1980 to house the Marketing offices.
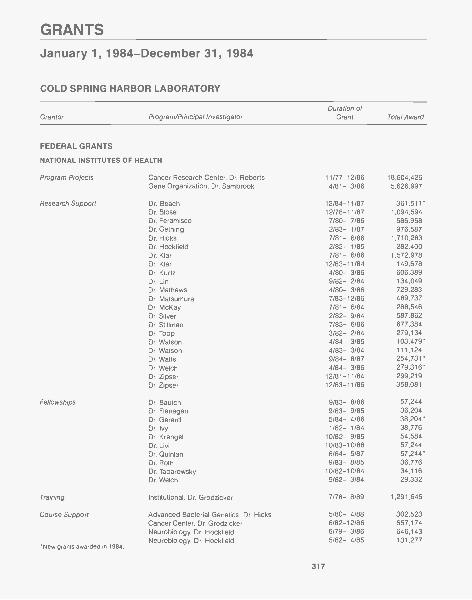
GRANTS
January 1, 1984-December 31, 1984
COLD SPRING HARBOR LABORATORY
Duration of
Grantor Program/Principal Investigator Grant Total Award
FEDERAL GRANTS
NATIONAL INSTITUTES OF HEALTH
Program Projects
Research Support
Cancer Research Center, Dr. RobertsGene Organization, Dr. Sambrook
Dr. BeachDr. Blose
11/77-12/864/81- 3/86
12/84-11/8712/78-11/87
18,604,4265,626,997
361,511"1,094,594
Dr. Feramisco 7/80- 7/86 686,968Dr. Gething 2/83- 1/87 976,587Dr. Hicks 7/81- 6/86 1,710,283Dr. Hockfield 2/82- 1/85 282,400Dr. Klar 7/81- 6/86 1,572,978Dr. Klar 12/83-11/84 149,678Dr. Kurtz 4/80- 3/86 606,389Dr. Lin 9/82- 2/84 134,049Dr. Mathews 4/80- 3/86 729,283Dr. Matsumura 7/83-12/86 469,737Dr. McKay 7/81- 6/84 288,546Dr. Silver 2/82- 9/84 587,862Dr. Stillman 7/83- 6/86 677,384Dr. Topp 3/82- 2/84 279,134Dr. Watson 4/84- 3/85 103,479*Dr. Watson 4/83- 3/84 111,124Dr. Watts 9/84- 8/87 254,731Dr. Welch 4/84- 3/86 279,316*Dr. Zipser 12/81-11/84 299,219Dr. Zipser 12/83-11/86 358,081
Fellowships Dr. Bautch 9/83- 8/86 57,244Dr. Flanagan 9/83- 9/85 36,204Dr. Gerard 5/84- 4/86 38,204*Dr. Ivy 1/82- 1/84 38,776Dr. Krangel 10/82- 9/85 54,584Dr. Livi 10/83-10/86 57,244Dr. Quinlan 6/84- 5/87 57,244*Dr. Roth 9/83- 8/85 36,776Dr. Taparowsky 10/82-10/84 34,116Dr. Welch 9/82- 3/84 29,332
Training Institutional, Dr. Grodzicker 7/78- 8/89 1,291,646
Course Support Advanced Bacterial Genetics, Dr. Hicks 5/80- 4/88 302,523Cancer Center, Dr. Grodzicker 6/82-12/86 657,174Neurobiology, Dr. Hockfield 6/79- 3/86 646,143Neurobiology, Dr. Hockfield 5/82- 4/85 131,277
*New grants awarded in 1984.
317
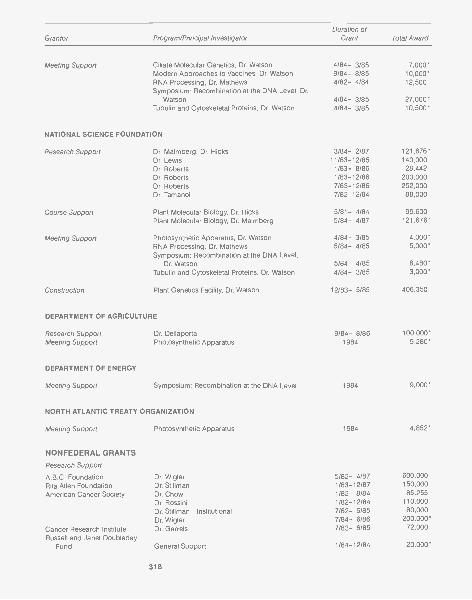
Grantor Program/Principal InvestigatorDuration of
Grant Total Award
Meeting Support Ciliate Molecular Genetics, Dr. Watson 4/84- 3/85 7,000*Modern Approaches to Vaccines, Dr. Watson 9/84- 8/85 10,000*RNA Processing, Dr. Mathews 4/82- 4/84 12,500Symposium: Recombination at the DNA Level, Dr.
Watson 4/84- 3/85 27,000*Tubulin and Cytoskeletal Proteins, Dr. Watson 4/84- 3/85 10,500*
NATIONAL SCIENCE FOUNDATION
Research Support Dr. Malmberg, Dr. Hicks 3/84- 2/87 121,676*Dr. Lewis 11/83-12/85 140,000Dr. Roberts 1/83- 8/86 28,442Dr. Roberts 1/83-12/86 200,000Dr. Roberts 7/83-12/86 252,000Dr. Tamanoi 7/82-12/84 88,000
Course Support Plant Molecular Biology, Dr. Hicks 5/81- 4/84 99,600Plant Molecular Biology, Dr. Malmberg 5/84- 4/87 121,676*
Meeting Support Photosynthetic Apparatus, Dr. Watson 4/84- 3/85 4,000*RNA Processing, Dr. Mathews 5/84- 4/85 5,000"Symposium: Recombination at the DNA Level,
Dr. Watson 5/84- 4/85 6,480*Tubulin and Cytoskeletal Proteins, Dr. Watson 4/84- 3/85 3,000*
Construction Plant Genetics Facility, Dr. Watson 12/83- 5/85 406,350
DEPARTMENT OF AGRICULTURE
Research Support Dr. Dellaporta 9/84- 8/86 100,000*Meeting Support Photosynthetic Apparatus 1984 9,280*
DEPARTMENT OF ENERGY
Meeting Support Symposium: Recombination at the DNA Level 1984 9,000*
NORTH ATLANTIC TREATY ORGANIZATION
Meeting Support Photosynthetic Apparatus 1984 4,852*
NONFEDERAL GRANTS
Research Support
A.B.C. Foundation Dr. Wigler 5/82- 4/87 600,000
Rita Allen Foundation Dr. Stillman 1/83-12/87 150,000
American Cancer Society Dr. Chow 1/82- 8/84 85,255
Dr. Rossini 1/82-12/84 110,000
Dr. Stillman-Institutional 7/82- 6/85 80,000
Dr. Wigler 7/84- 6/86 200,000*
Cancer Research Institute Dr. Garrels 7/83- 6/85 72,000
Russell and Janet DoubledayFund General Support 1/84-12/84 20,000*
318
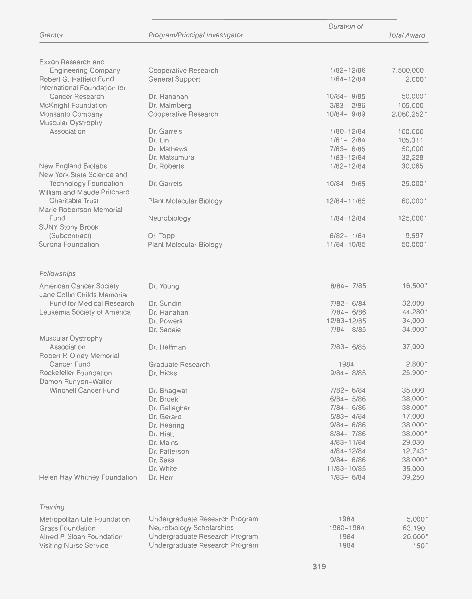
Grantor Program/Principal InvestigatorDuration of
Total Award
Exxon Research andEngineering Company Cooperative Research 1/82-12/86 7,500,000
Robert G. Hatfield Fund General Support 1/84-12/84 2,000*International Foundation for
Cancer Research Dr. Hanahan 10/84- 9/85 50,000*McKnight Foundation Dr. Malmberg 3/83- 2/86 105,000Monsanto Company Cooperative Research 10/84- 9/89 2,060,252*Muscular Dystrophy
Association Dr. Garrels 1/80-12/84 100,000Dr. Lin 1/81- 2/84 105,311Dr. Mathews 7/83- 6/85 50,000Dr. Matsumura 1/83-12/84 32,228
New England Biolabs Dr. Roberts 1/82-12/84 30,065New York State Science and
Technology Foundation Dr. Garrels 10/84- 9/85 25,000*William and Maude Pritchard
Charitable Trust Plant Molecular Biology 12/84-11/85 60,000*Marie Robertson Memorial
Fund Neurobiology 1/84-12/84 125,000*SUNY Stony Brook
(Subcontract) Dr. Topp 8/82- 1/84 9,597Surdna Foundation Plant Molecular Biology 11/84-10/85 50,000*
Fellowships
American Cancer Society Dr. Young 8/84- 7/85 16,500*Jane Coffin Childs Memorial
Fund for Medical Research Dr. Sundin 7/82- 6/84 32,000Leukemia Society of America Dr. Hanahan 7/84- 6/86 44,280*
Dr. Powers 12/83-12/85 34,000Dr. Sadaie 7/84- 8/85 34,000*
Muscular DystrophyAssociation Dr. Hellman 7/83- 6/85 37,000
Robert P Olney MemorialCancer Fund Graduate Research 1984 2,800*
Rockefeller Foundation Dr. Hicks 9/84- 8/85 25,900*Damon Runyon-Walter
Winchell Cancer Fund Dr. Bhagwat 7/82- 6/84 35,000Dr. Broek 6/84- 5/86 38,000*Dr. Gallagher 7/84- 6/86 38,000*Dr. Gerard 5/83- 4/84 17,000Dr. Hearing 9/84- 8/86 38,000*Dr. Hiatt 8/84- 7/86 38,000*Dr. Mains 4/83-11/84 29,030Dr. Patterson 4/84-12/84 12,743*Dr. Sass 9/84- 8/86 38,000*Dr. White 11/83-10/85 35,000
Helen Hay Whitney Foundation Dr. Herr 1/83- 6/84 39,250
Training
Metropolitan Life Foundation Undergraduate Research Program 1984 5,000"Grass Foundation Neurobiology Scholarships 1980-1984 63,190Alfred P Sloan Foundation Undergraduate Research Program 1984 20,000*Visiting Nurse Service Undergraduate Research Program 1984 150"
319
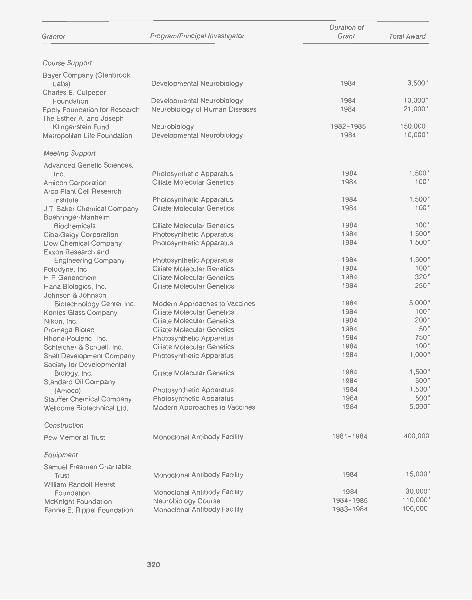
Grantor Program/Principal InvestigatorDuration of
Grant Total Award
Course Support
Bayer Company (GlenbrookLabs) Developmental Neurobiology 1984 3,500*
Charles E. CulpeperFoundation Developmental Neurobiology 1984 10,000*
Epp ly Foundation for Research Neurobiology of Human Diseases 1984 21,000*The Esther A. and Joseph
Klingenstein Fund Neurobiology 1982-1985 150,000
Metropolitan Life Foundation Developmental Neurobiology 1984 10,000*
Meeting Support
Advanced Genetic Sciences,Inc. Photosynthetic Apparatus 1984 1,500*
Amicon Corporation Ciliate Molecular Genetics 1984 100*
Arco Plant Cell ResearchInstitute Photosynthetic Apparatus 1984 1,500*
J.T. Baker Chemical Company Ciliate Molecular Genetics 1984 100*
Boehringer-ManheimBiochemicals Ciliate Molecular Genetics 1984 100*
Ciba-Geigy Corporation Photosynthetic Apparatus 1984 1,500*
Dow Chemical Company Photosynthetic Apparatus 1984 1,500*
Exxon Research andEngineering Company Photosynthetic Apparatus 1984 1,500*
Fotodyne, Inc. Ciliate Molecular Genetics 1984 100*
H.P. Genenchem Ciliate Molecular Genetics 1984 320*
Hans Biologics, Inc. Ciliate Molecular Genetics 1984 250*
Johnson & JohnsonBiotechnology Center Inc. Modern Approaches to Vaccines 1984 5,000*
Kontes Glass Company Ciliate Molecular Genetics 1984 100*
Nikon, Inc. Ciliate Molecular Genetics 1984 200*
Promega Biotec Ciliate Molecular Genetics 1984 50*
Rhone-Poulenc, Inc. Photosynthetic Apparatus 1984 750*
Schleicher & Schuell, Inc. Ciliate Molecular Genetics 1984 100*
Shell Development Company Photosynthetic Apparatus 1984 1,000*
Society for DevelopmentalBiology, Inc. Ciliate Molecular Genetics 1984 1,500*
Standard Oil Company 1984 500*
(Amoco) Photosynthetic Apparatus 1984 1,500*
Stauffer Chemical Company Photosynthetic Apparatus 1984 500*
Wellcome Biotechnical Ltd. Modern Approaches to Vaccines 1984 5,000*
Construction
Pew Memorial Trust Monoclonal Antibody Facility 1981-1984 400,000
Equipment
Samuel Freeman CharitableTrust Monoclonal Antibody Facility 1984 15,000*
William Randolf HearstFoundation Monoclonal Antibody Facility 1984 30,000*
McKnight Foundation Neurobiology Course 1984-1985 110,000*
Fannie E. Rippel Foundation Monoclonal Antibody Facility 1983-1984 100,000
320

BANBURY CENTER
Grantor Program/Principal InvestigatorDuration of
Grant Total Award
FEDERAL GRANTS
NATIONAL INSTITUTES OF HEALTH
Meeting Support Genetic Manipulation of Mammalian Ovum and Early 9/84- 8/85 9,550*Embryo, M. Shodell
DEPARTMENT OF ENERGY
Meeting Support Risk Quantitation 4/84- 3/85 8,000*
OFFICE OF NAVALRESEARCH
Meeting Support Gap Junctions 1984 4,700*
NONFEDERAL GRANTS
Meeting Support
Council for Research Planningin Biological Sciences, Inc. EPA Workshop 8/84-12/84 4,000*
Diamond Shamrock Biological Mechanisms of Dioxin Action 4/84- 3/85 2,000*Dow Chemical, U.S.A. Biological Mechanisms of Dioxin Action 4/84- 3/85 2,000*Hoffmann-La Roche, Inc. Biological Mechanisms of Dioxin Action 4/84- 3/85 5,000*March of Dimes Genetic Manipulation of Mammalian Ovum and Early
Embryo 10/84- 9/85 7,500*Monsanto Company Biological Mechanisms of Dioxin Action 4/84- 3/85 5,000*
Risk Quantitation 4/84- 3/85 5,000*National Distillers & Chemical
Corporation Risk Quantitation 4/84- 3/85 1,000*Alfred P. Sloan Foundation Journalists' Workshop 1984-1986 162,000*
Computational Neuroscience Workshop 7/84- 6/85 21,000*
321
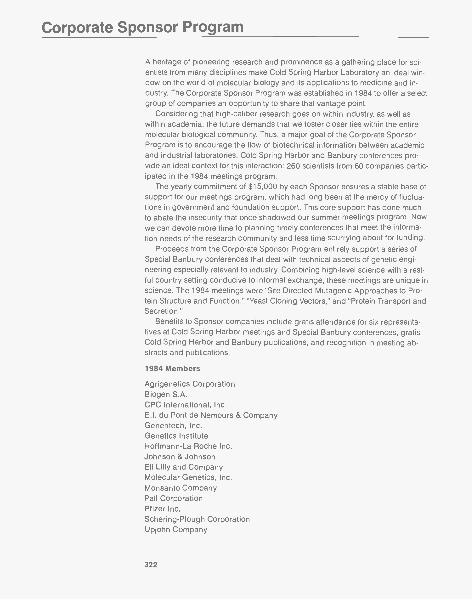
Corporate Sponsor Program
A heritage of pioneering research and prominence as a gathering place for sci-entists from many disciplines make Cold Spring Harbor Laboratory an ideal win-dow on the world of molecular biology and its applications to medicine and in-dustry. The Corporate Sponsor Program was established in 1984 to offer a selectgroup of companies an opportunity to share that vantage point.
Considering that high-caliber research goes on within industry, as well aswithin academia, the future demands that we foster closer ties within the entiremolecular biological community. Thus, a major goal of the Corporate SponsorProgram is to encourage the flow of biotechnical information between academicand industrial laboratories. Cold Spring Harbor and Banbury conferences pro-vide an ideal context for this interaction: 260 scientists from 60 companies partic-ipated in the 1984 meetings program.
The yearly commitment of $15,000 by each Sponsor ensures a stable base ofsupport for our meetings program, which had long been at the mercy of fluctua-tions in government and foundation support. This core support has done muchto abate the insecurity that once shadowed our summer meetings program. Nowwe can devote more time to planning timely conferences that meet the informa-tion needs of the research community and less time scurrying about for funding.
Proceeds from the Corporate Sponsor Program entirely support a series ofSpecial Banbury conferences that deal with technical aspects of genetic engi-neering especially relevant to industry. Combining high-level science with a rest-ful country setting conducive to informal exchange, these meetings are unique inscience. The 1984 meetings were "Site Directed Mutagenic Approaches to Pro-tein Structure and Function," "Yeast Cloning Vectors," and "Protein Transport andSecretion."
Benefits to Sponsor companies include gratis attendance for six representa-tives at Cold Spring Harbor meetings and Special Banbury conferences, gratisCold Spring Harbor and Banbury publications, and recognition in meeting ab-stracts and publications.
1984 Members
Agrigenetics CorporationBiogen S.A.CPC International, Inc.E.I. du Pont de Nemours & CompanyGenentech, Inc.Genetics InstituteHoffmann-La Roche Inc.Johnson & JohnsonEli Lilly and CompanyMolecular Genetics, Inc.Monsanto CompanyPall CorporationPfizer Inc.Schering-Plough CorporationUpjohn Company
322

Long Island Business Cancer Campaign
For many years, the international scientific community has known what a greatasset Cold Spring Harbor Laboratory is to science. It is a sad irony that the labo-ratory is not well known to many local people and, thus, remains one of LongIsland's best-kept "secrets." However, just as the world belatedly recognized theachievements of CSH researcher Dr. Barbara McClintock with the 1983 NobelPrize, so, too, are Long Islanders now becoming aware of the great asset thathas quietly been at work in their own backyard since 1890.
The Long Island Association, the official chamber of commerce for Nassauand Suffolk Counties, voted Cold Spring Harbor Laboratory as recipient of its1984 Medal of Honor. This award was given "in recognition of the Laboratory'svital work for the good of Long Islanders and all mankind."
In an era of shrinking government funding the private sector must assume alarger responsibility for supporting biomedical research. The Long Island Busi-ness Cancer Campaign offers businesses an opportunity to participate in theLaboratory's research on the molecular basis of cancer. We hope that this localcancer initiative will help build a greater sense of community on Long Island.
The 1984 campaign netted $50,000 in cash and donated equipment, whichwas used in the development of the new monoclonal antibody laboratory. Wethank the following companies who joined in saying "Long Island is doing some-thing about cancer."
Benefactor Contributors
Chemap, Inc. Castro Convertibles
PatronsExecutoneGovernment Employees Insurance
Brinkmann Instruments CompanyEntenmann's Bakery Hartman SystemsGrumman Corporation Hazeltine Corporation
SponsorsJameco IndustriesMichelin Tire Corporation
Fortunoff's NEC America, Inc.Long Island Savings Bank Olympus Camera CorporationLong Island Trust Company Travelers Companies FoundationLumexManufacturers Hanover Trust
Other Donations
Newsday Analytab Products, Inc.Sperry Corporation Fairchild Weston SystemsTampax, Inc. Health Extension Services
IJT LimitedLawrence Aviation IndustriesPhotocircuitsPinkerton's Inc.R.J. Rudden Associates, Inc.
323

CONTRIBUTORS
Mr. Christopher C. AbbotACTION for the Preservation and Conservation
of the North Shore of L.I., Inc.Rita Allen FoundationMs. Judy AltmanMrs. Charles E. AmesMs. Rosamond R. ArthurMr. George AvazisDr. & Mrs. Alfred AzzoniMr. & Mrs. William N. BannardMr. & Mrs. Daniel BarsantiMrs. Robert E. BelknapMr. & Mrs. Thomas H. BelknapDr. Paul BergMs. Barbara BermanMr. Bruno BichEdith Blum FoundationMrs. Shirley F BowringMr. & Mrs. William BremerMrs. Louise McDowell BrightonMr. & Mrs. James T Bryan, Jr.Mr. & Mrs. Sydney G. ButlerMr. Thorne ButlerMr. Jeremiah J. CallahanMr. & Mrs. Samuel R. CallawayMiss Patricia CasentiniMr. & Mrs. ChaikowskiDr. Lan Bo ChenMr. & Mrs. Beverly C. ChewCitibank, N.A.Mr. John P. ClearyColumbia UniversityDr. & Mrs. Crispin CookeMr. & Mrs. Robert CunninghamMr. Roderick CushmanMr. & Mrs. George W. Cutting, Jr.Mr. Lawrence L. DavisMr. & Mrs. John W. DaytonMr. Howard W. Dippel, Jr.Mr. & Mrs. Lawrence DrennanMr. & Mrs. Richard T. du MoulinDr. & Mrs. Lester DubnickDuke UniversityMr. & Mrs. Edgar P. DunlaevyMr. William T Dunn, Jr.Mrs. Emily EarleyAlbert Einstein College of MedicineMr. & Mrs. Harold L. Fates
Mr. & Mrs. E.E. FessendenFolk Music Society of HuntingtonWalter N. Frank & CompanyMs. Jamie GagarinMr. Clarence E. GalstonMr. & Mrs. John W. GalstonMr. & Mrs. John W. GatesMr. & Mrs. Henry A. GerryMr. & Mrs. Bentley GlassMr. & Mrs. Lawrence R. GlennMr. & Mrs. Robert G. GoeletDr. & Mrs. Richard L. GoldenMs. Gail W. GoltraMr. & Mrs. Oliver R. GraceMr. Frank GravenMr. Edward R. GreefMr. & Mrs. Richard HaarMs. Rochelle HabibMr. & Mrs. Hamilton HaddenHarley Avenue P.T.A.Mr. & Mrs. Henry U. HarrisMs. Louise HarrisonMs. Maxine HarrisonMr. & Mrs. George HegemanDr. William W. HeroyDr. Ira HerskowitzDr. Bernhard HirtMs. Sandy HoffmanDr. Alexander HollaenderMr. Charles S. HoppinMr. & Mrs. Abraham HsieThe Hon. John P HumesMrs. David IngrahamMr. & Mrs. Richard M. JacksonI. Janvey & Sons, Inc.Mr. Abraham A. JanveyMr. David J. KalalMs. Tae KamiharaMr. & Mrs. John P. KippMrs. T.W. LaphamMr. & Mrs. Ernest V. Lazio, Jr.Mr. & Mrs. Richard E. LeckerlingMrs. Louise Woods LevensonDr. & Mrs. Monroe LevinMr. & Mrs. George N. LindsayMr. & Mrs. Robert V. LindsayMs. Debbie LipmanMr. & Mrs. Russell W. London
324

Long Island League to Abolish CancerLong Island UniversityMr. & Mrs. William R. LucasMr. & Mrs. Michael LuckeyMktg. Inc.Mr. John B. MarshMassachusetts Institute of TechnologyMr. & Mrs. Dick McAdooMiss Leila McKnightMr. Jo Zach Miller IVMr. William H. MillerMr. & Mrs. Daniel C. Mil lettMrs. Stella MonchickMr. & Mrs. F. Franklin MoonMr. & Mrs. John P. MorganMs. Alice MorgansternMr. George MurnaneMr. & Mrs. Francis W. Murray IIIMr. F Wisner MurrayMr. & Mrs. Rem V. MyersNew York UniversityThe Hon. Eugene H. & Marie-Louise NickersonMr. & Mrs. Basil NorthernMr. Charles NoyesMr. & Mrs. George D. O'NeillMr. & Mrs. Arthur W. PageMr. & Mrs. John PageMr. & Mrs. Walter H. PageMrs. Marcel PalmaroMr. Nicholas R. ParksParkway SchoolMs. Irene PelledRev. Linda PeytonMs. Pamela Peyton PostMr. J. Gardiner PieperMs. Claire PlatkinMr. & Mrs. Graham L. PlattMr. & Mrs. Richardson Pratt, Jr.William and Maude Pritchard Charitable TrustMs. Mar len C. RobbMrs. Donald W. RoseMr. & Mrs. Jack RosenbaumMr. & Mrs. Walter RothschildMr. & Mrs. J.A. RousmaniereMs. Rosina B. RueMr. Quentin B. SammisMrs. John K. SandsMrs. Norman Schaff, Jr.Schiff FoundationDr. & Mrs. Carl SchmidlappMrs. Franz Schneider
325
Dr. & Mrs. Roland SebbaneMr. Willard SektbergDr. & Mrs. W. Shropshire, Jr.Memorial Sloan-Kettering Cancer CenterDr. & Mrs. Robert SmirnowMr. & Mrs. William SmootState University of New York, Stony BrookMr. & Mrs. Thomas SteitzDr. Gunther StentMr. Norman W. StewartMr. & Mrs. Richard S. StorrsMrs. Jeannette Jennings TaylorMr. & Mrs. Evan W. ThomasMs. Joan P TilneyMr. & Mrs. E.T. TurnerMr. & Mrs. Baxter VenableMr. & Mrs. Colton P WagnerMr. & Mrs. Sherwood WaldronWaldron Memorial FundMr. James C. WangMr. & Mrs. Charles W.B. WardellMrs. David E. WardenMrs. Ethelbert WarfieldMr. Samuel WarrenDr. James D. WatsonMs. Phyllis WeekesMr.
Mr. & Mrs. Edward C.R. WhitcraftWhitehall FoundationMrs. Fred WilliamsUniversity of WisconsinYale University
In memory of . .
Mr. Robert CavanaghMr. Aldo EspositoMr. Michael FoleyMs. Mildred Sally GalanMs. Phyllis KilkennyMrs. G. KirsteinMrs. Beatrice LevinMr. Salvatore ManninaMrs. Janet PsipsikasMrs. Sara Delano RedmondMr. Stanley RossMrs. Mildred SmithMr. James StaisMr. Chester StilesMr. Walter G. UhlMr. Harry D. Williams

FINANCIAL SUPPORT OF THE LABORATORY
Cold Spring Harbor Laboratory is a nonprofit research and educational institu-tion chartered by the University of the State of New York. Contributions are taxexempt under the provisions of Section 501(c)(3) of the Internal Revenue Code.In addition, the Laboratory has been designated a "public charity" and, there-fore, may receive funds resulting from the termination of "private" foundations.
Because its endowment is limited and the uses of research grants are formallyrestricted, the Laboratory depends on generous contributions from private foun-dations, sponsors, and friends for central institutional needs and capital improve-ments.
Over the years, the Laboratory has won a reputation for innovative researchand high-level science education. By training young scientists in the latest exper-imental techniques, it has helped spur the development of many research fields,including tumor virology, cancer genes, gene regulation, moveable genetic ele-ments, yeast genetics, and molecular neurobiology. However, the continued de-velopment of new research programs and training courses requires substantialsupport from private sources.
METHODS OF CONTRIBUTING TO COLD SPRING HARBOR LABORATORY
Gifts of money can be made directly to Cold Spring Harbor Laboratory.Securities You can generally deduct the full amount of the gift on your income tax return,
and, of course, you need pay no capital gains tax on the stock's appreciation.We recommend any of the following methods:(1) Have your broker sell the stock and remit the proceeds to Cold Spring Harbor Labo-
ratory.(2) Deliver the stock certificates to your broker with instructions to him to open an ac-
count for Cold Spring Harbor Laboratory and hold the securities in that accountpending instructions from Cold Spring Harbor Laboratory.
(3) Send the unendorsed stock certificates directly to the Laboratory: Comptroller, ColdSpring Harbor Laboratory, Cold Spring Harbor, New York 11724. In a separate enve-lope send an executed stock power.
Bequests Probably most wills need to be updated. Designating Cold Spring Harbor Lab-oratory as beneficiary ensures that a bequest will be utilized as specified for continuinggood.
Appreciated real estate or personal property Sizable tax benefits can result from suchdonations; the Laboratory can use some in its program and can sell others.
Life insurance and charitable remainder trusts can be structured to suit the donor'sspecific desires as to extent, timing, and tax needs.
Conversion of privation foundation to "public" status on termination This may bedone by creating a separate fund within Cold Spring Harbor Laboratory whereby theassets of the private foundation are accounted for as a separate fund and used for thepurposes specified by the donor. Alternatively, the private foundation can be establishedas a "supporting organization of Cold Spring Harbor Laboratory."
For additional information, please contact the Director of Information Services, Cold SpringHarbor Laboratory, Box 100, Cold Spring Harbor, N.Y. 11724, or call 516-367-8397.
326

LABORATORYSTAFF

Recto: Demerec laboratory extension, erected in 1983.

Laboratory Staff
Research Staff
David BeachStephen BloseSusan BonitzFevzi DaldalSteven DellaportaJames FeramiscoRobert FranzaJames GarrelsMary-Jane GethingYakov GluzmanTerri GrodzickerDouglas HanahanEdward HarlowWinship HerrJames HicksSteven HintonSusan HockfieldAmar KlarDavid KurtzJohn LewisDaniel LundellRussell MalmbergMichael MathewsFumio MatsumuraRichard RobertsMara RossiniPablo ScolnikClive SlaughterBruce StillmanFuyuhiko TamanoiPaul ThomasWilliam WelchMichael Wig lerDouglas YouvanMark Zoller
PostdoctoralResearch Fellows
Timothy AdamsWilliam AddisonDafna Bar-SagiVictoria BautchAshok BhagwatCarmen Birchmeier
Philip BirdDaniel BroekSteven CheleyPatricia CottonClaude DeryJohn DiffleyShimon EfratThomas FlanaganGregory FreyerBeth FriedmanPatricia GallagherRobert GerardJoan HarperJanet HearingDavid HelfmanAndrew HiattJohn IvyTohru KamataYuriko KataokaAntal KissKrystyna Slaska-KissRichard KostrikenMichael KrangelJohn LangstaffPatsy LinGeorge LiviJanet MaclnnesKathy MartinShigeko MatsumuraMaureen McLeodElizabeth MoranScott PowersMargaret QuinlanMichael RothReza SadaiePhilip SassPetra SchumannToda TakashiSusan WattsEileen WhiteMasao YamadaDalian YoungSam ZarembaBradley Zerler
James D. Watson, DirectorJoseph Sambrook, Assistant Director for ResearchWilliam R. Udry, Administrative DirectorJohn Maroney, Assistant Administrative DirectorWilliam Keen, ComptrollerJack Richards, Director Buildings & GroundsMichael Shodell, Banbury Center Director
Genetics Research UnitCarnegie Institutionof Washington
Alfred D. HersheyBarbara McClintock
Visiting Scientists
Susan AlpertDaniel BirnbaumOttavio FasanoAsao FujiyamaNickolas Gillhamtimothy HarrisonMark HomonoffEric HunterTohru Kataoka
Graduate Students
Robert BooherEdward BylinaScott CameronPaul ChometWilliam CostelloCarolyn DoyleMargaret KellyLonny LevinRobert O'MalleyGregory PrelichMadan RaoNora SarvetnickSreenath SharmaCarmel la StephensKevin Van DorenPeter Whyte
Support Staff
Patricia BirdLouisa DalessandroJoanne DellaportaMarilyn GoodwinChristopher KellerPhilip Renna
329

Regina Schwarz Rodney Chisum Elizabeth Ritcey
Joseph Suhan Mary Freedman Pauline Tannenholz
Madeline Szadkowski Connie Hallaran Michaela Taylor
Barbara Weinkauff Brian KleeEleanor MathewsSandra Penzi Library/Marketing
Laboratory Technicians Maureen RiceSusan Rook
Susan Gensel
Barbara Ahrens Douglas Sankar LibraryJoy Applebaum Leonard Sansone Genemary FalveyCarmelita Bautista Suzanne Vavrinec Laura HymanGeorgia Binns Albert Yan Margaret MainGladys Blose Michela McBrideChristopher BunnCecile Chang Medium Makers Marketing
Dolly Chao Ellen DeWeerdt
Jennifer Clarke William Eisner Monica Ewing
Joan Cossetti Margaret Falkowski Elaine Gaveglia
Debra Danna Marie Hallaran Amy Gibson
Kathleen Dinota Monika Stein Maureen Spirn
Louise Edlund Margaret Wallace
Barbara FahaLinda Finn AccountingThomas Fischer Safety and CoreJane Gould Facilities Services Accounting
Eric GrubmanGuy Cozza
Arthur Brings Fran GerardJean Henneberry Mary Ellen GoldsteinSajida Ismail Safety Carolyn KempfStephen Johann Marlene Rubino Dorrine LambertKaren JohnsonNancy Kronenberg
Computing Center Fred Lukas
Michael Balamuth Carlos MendezMark Lacey Robert PaceAlan Louis Harris Animal Facility June PolistinaRobert McGuirk Deborah LukralleJeanne Mclndoo Fulfillment
Richard ShubertLisa Miglio Charlaine Apsel
Carolyn MoomawConnie Parra
Ann FeltonPhyllis Myers Art Department Barbara SkellyKathy O'Neill Michael Ockler Barbara TerryDiane PascucciDaniel Pollock
David Greene GrantsD
Michael RiggsDebra Lindner Susan SchultzJean Michaelis
Linda Rodgers Patricia KurfessNasrollah Samiy Technical Services Patricia MaroneyCarol Schley Robert BorrusoPatricia Scolnik August DulisScott Silbiger Raymond Mickus Information ServicesSusan Smith Clifford SutkevichLinda Vanderwal David Mick losGayle Waitches Jenny FlanaganElizabeth Waldvogel Publications Herb ParsonsPatricia WendelHeidi Wenzel Nancy Ford
Jeanne Wiggins Annette Kirk Personnel and MeetingsJonathan Wood
Dorothy Brown William PutnamJudith Cuddihy
Laboratory Aides Joan Ebert PersonnelAdrienne Guerra Rita Duffy
Jennifer Brooks Martha O'Gorman Catherine ErbAngela Ca lasso Douglas Owen Jacqueline Terrence
330

MeetingsMaureen BerejkaChris BretGladys KistMerilyn SimkinsBarbara Ward
Purchasing
Peter Rice
Deon BakerSandra D'ArcangeloBruce FahlbushSteven HorlyGraceAnn LovellThomas MaloneyJames SabinElissa Troisi
Central Administration
Roberta SalantAndrea Stephenson
Buildings and Grounds
AdministrationCharles Schneider
Barbara Cuff
Carpentry and PaintingDiamond Scarduzio
Joseph BrodawchukJohn ChilcottLawrence DawJohn LarsenThomas LydenJohn MeyerJoseph Pirnak
Custodial Banbury CenterCharles Marshall
John BerejkaJoseph CannerWilliam GardnerDaniel JusinoAlfred NickelRichard O'LearyFrank RussellSteven TangPeter Tierney
Electrical and MechanicalMaintenanceRobert CollinsFrank MessinaJerry ReichertOwen Stewart
GroundsHans Trede
Vincent CareyWarren EddyJoseph EllisJohn Robert FuchsChristopher McEvoyAnthony Zanoni
HousekeepingDorothy Youngs
Rita AbramsCarol CaldarelliTurid FaretBarbara RobinsonCherie Thurn
PlumbingLane Smith
Douglas Haskett
SecurityPaul Anzalone
Banbury Grounds and MaintenancePeter Stahl
Daniel Miller
Judith BlumDessie CarterKatya DaveyBeatrice To liver
Scientific StaffDepartures during 1984
Senior Staff ScientistsThomas BrokerLouise ChowJeffrey Strathern
Senior Staff InvestigatorsStephen HughesRonald McKayEarl RuleyLee SilverWilliam Topp
Staff InvestigatorsMitchel GoldfarbJames Lin
Visiting ScientistsGuang-Yun CaiKristen FrederiksenJin-Zhao Li
Postdoctoral FellowsRichard GuilfoyleWilliam HusePrasad KokaNita MaihlePaul MainsKazuo MaruyamaOlof SundinElizabeth TaparowskiStuart WeisbrodRobert Weiss
Graduate StudentsChin Sheng ChouMichele ManosKent Matlack
331


"2',r"'.,
..

Recto: Williams House (ca. 1850), rebuilt in 1977 with funds raised by LISA, provides fiveapartments for staff scientists. It is named for Col. Timothy S. Williams, first president of LISA.

Officers Executive CommitteeMr. Edward Pulling, ChairmanMr. George J. Hossfeld, Jr., Vice-ChairmanMr. James A. Eisenman, TreasurerMrs. James J. Pirtle, SecretaryMr. William R. Udry, Asst. Secretary-Treasurer
Directors
The Officers, alsoMr. Samuel R. CallawayMrs. Henry U. Harris, Jr.Mrs. Richardson Pratt
Mrs. Donald ArthurDr. Alfred A. AzzoniMrs. Gilbert A. BallMr. Edmund BartlettMr. Samuel R. CallawayMrs. John P CampbellMrs. Miner D. Crary, Jr.Mr. Arthur M. CrockerMr. Roderick H. CushmanMr. George W. Cutting, Jr.
Mr. Lawrence L. DavisMr. Joseph C. Dey, Jr.Mr. Charles DolanMr. James A. EisenmanMrs. R. Duncan ElderMrs. Henry U. Harris, Jr.Mr. George J. Hossfeld, Jr.Mr. Edward M. LamontMr. Arthur C. MerrillMr. Grinnell Morris
335
Mrs. James J. Pirtle, Jr.Mrs. Richardson PrattMr. Edward PullingMr. John R. ReeseMr. Harvey E. SampsonMr. Stanley S. Trotman, Jr.Dr. James D. WatsonMrs. Alex M. White

The LONG ISLAND BIOLOGICAL ASSOCIATION (LIBA)and Its Relation to the Cold Spring Harbor Laboratory
Biological research at Cold Spring Harbor began in 1890 when the Brooklyn Institute of Artsand Sciences was looking for a place to set up a summer laboratory as the headquarters of itsDepartment of Zoology. Representatives of the Institute were invited by Eugene S. Blackford,at that time the Fish Commissioner of the State of New York, to inspect a site on BungtownRoad across Northern Boulevard (25A) from the Fish Hatchery. The site was found to be ideal,and so the original Laboratory was organized with Mr. Blackford as president of the Board ofTrustees. The land was leased from Mr. John D. Jones, whose family since 1819 had operatedvarious industries including shipbuilding, coopering, and textile manufacture (later whaling,also) at the head of Cold Spring Harbor. Bungtown Road, which runs through the Lab prop-erty, got its name from the factory that specialized in making bungs-or stoppers-for barrels.
In 1892 the Laboratory's land was leased for a dollar a year from the Wawepex Society,which Mr. Jones had organized as a corporation for holding real estate and for investing fundsfor the propagation of fishes and for scientific research. In 1904 the Wawepex Society leasedadditional land to the Carnegie Institution of Washington, which wanted to locate a Departmentof Experimental Evolution in the Cold Spring Harbor area. Charles B. Davenport, who hadbeen directing the Laboratory since 1896, assumed the additional duties of director of the Car-negie Institution's experimental station. It is interesting to note that Mr. Davenport lived in whatcame to be known later as the Carnegie Dormitory, the Victorian house on 25A built by JohnD. Jones before the turn of the century, and recently repainted in its original colors,
The Long Island Biological Association was established in 1924 when the Brooklyn Institutedecided to discontinue its research at Cold Spring Harbor and offered its laboratory to twouniversities. Fortunately, a local group of interested neighbors decided to assume responsibilityfor the Lab, and thus LIBA came into being. For 38 years LIBA actually operated the Labora-tory in conjunction with the Carnegie Institution, but in 1962 it seemed advisable for the Labo-ratory to be reorganized as an independent unit. Therefore, the property on which it nowstands was conveyed to it by LIBA, which, however, still retains reversionary rights. TodayLIBA is one of twelve institutions participating in the support of the Laboratory, each institutionbeing represented on the Laboratory's Board of Trustees.
What has happened, in effect, is that LIBA has become an expanding group of local"Friends of the Laboratory" who help support it through annual contributions. Also, from timeto time, the Association undertakes campaigns to finance special important projects for whichthe Lab cannot obtain funds from the Federal Government or from other sources. For instance,in recent years, building the James Laboratory Annex and the renovation of Blackford Hall, therebuilding of Williams House, the acquisition of the land formerly belonging to the CarnegieInstitution, and (in part) the construction of the new Grace Auditorium.
The affairs of LIBA are handled by a board of 28 directors who are elected to office by themembership at an annual meeting, At least twice a year LIBA members are invited to bringtheir friends to a lecture or an open house at the Lab.
Membership in LIBA requires a minimum annual contribution (tax deductible) of $25 fora husband and wife, $15 for a single adult, $5 for a junior member (under 21). Furtherinformation can be obtained from the Long Island Biological Association, Box 100, ColdSpring Harbor, N.Y. 11724, or by telephoning the Laboratory at (516) 367-8300.
336

CHAIRMAN'S REPORT
At the Autumn meeting of the Board of Directors I was pleased to announce a sub-stantial growth in the number of LIBA members during the year, also that the gener-ous response to our campaign to raise at least $500,000 towards the cost of thenew auditorium indicated that we would be able to increase our contribution to$600,000. At the annual meeting of LIBA members in December it was voted to dothis. Approval was given also to the directors' recommendation that our next projectshould be the financing of a much needed documentary movie (or tape) depictingthe history and current research of the Laboratory.
As usual, LIBA members and their guests were invited to two lectures during theyear. At the first, the Dorcas Cummings Memorial Lecture, in May, our speaker wasProfessor Ian Sussex of the Department of Biology at Yale University. Because of therecently established Department of Plant Genetics at the Lab, Dr. Sussex appropri-ately spoke on "The new plant genetics and its implications for biotechnology". Helucidly explained how independent lines of research had come together in the pastfew years to allow us to get a whole new approach to understanding how plantsdevelop and are regulated.
At the annual meeting in December we were most fortunate to secure as ourspeaker Captain Eugene A. Cernan, U.S.N., who made three space flights forNASA and, as commander of Apollo 17, was the last man to leave his footprints onthe surface of the moon. Captain Cernan described his fascinating experiences asan astronaut and then went on to explain the advantages of outer space for scien-tific research. As the overflow audience was leaving the auditorium we could hearon all sides the comment that Captain Cernan's lecture was exceptionally interestingand also inspiring.
The custom of giving dinner parties for visiting scientists during the June Sympo-sium was successfully continued in 1984. It is a custom enjoyed equally by the sci-entists and by their hosts and hostesses. This year's parties were given by
Mrs. Donald ArthurMrs. Gilbert A. BallMr. & Mrs. John P. CampbellMr. & Mrs. Miner D. CraryMr. & Mrs. James A. EisenmanMr. & Mrs. Henry U. Harris, Jr.Mr. & Mrs. Kennedy B. Middendorf
Mr. & Mrs. Grinnell MorrisMr. & Mrs. Walter H. PageMrs. H. Irving PrattMr. & Mrs. John K. SandsMr. & Mrs. Franz SchneiderMr. & Mrs. Richard S. StorrsMr. & Mrs. Martin B. Travis
At the Annual Meeting regret was expressed that the terms of office as directorsfor Messrs. Charles S. Gay and William F. Parsons, Jr. had expired. Their placeswere filled by the election of Mrs. R. Duncan Elder and Mr. Roderick H. Cushman.
At a subsequent meeting of the Board of Directors the previous officers and
members of the Executive Committee were re-elected.
Edward Pulling, ChairmanLong Island Biological Association
337
LIBA audience listens to . .
Capt. Eugene A. Cernan, U.S.N.,describing life in outer space.

Membersof the Long Island Biological AssociationMr. & Mrs. Thomas Ackerman Mr. & Mrs. T.C. Cattrall, Jr. Mr. & Mrs. James C. Dudley
Mr. & Mrs. Amyas Ames Mr. & Mrs. Lionel Chaikin Dr. & Mrs. John L. Duffy
Mrs. Charles E. Ames Dr. Carton Chen Mr. Anthony Drexel Duke
Mr. & Mrs. Hoyt Ammidon Mrs. Helen Chenery Mr. & Mrs. Edgar P Dunlaevy
Miss Barbara Amott Mr. & Mrs. Beverly C. Chew Mr. & Mrs. William T Dunn, Jr.
Drs. Harold & Shirley Andersen Mr. Jan S. Chock Dr. & Mrs. Gerard L. Eastman
Mr. Robert W. Anthony Mr. & Mrs. Charles M. Clark Dr. & Mrs. James D. Ebert
Mr. & Mrs. J.S. Armentrout Mr. & Mrs. David C. Clark Mr. & Mrs. Paul S. Eckhoff
Mrs. Donald Arthur Dr. & Mrs. Bayard Clarkson Mr. & Mrs. James A. Eisenman
Dr. & Mrs. Joseph T August Mrs. Robert L. Clarkson, Jr. Mrs. Fred J. Eissler
Mr. & Mrs. Henry Austin III Mr. & Mrs. John P. Cleary Mr. & Mrs. Duncan Elder
Mrs. Robert W. Ayer Mr. & Mrs. Thomas N. Cochran Mr. & Mrs. Ashton G. EldredgeDr. & Mrs. Alfred Azzoni Dr. & Mrs. Peter Cohn Mr. & Mrs. Martin Elias
Mr. & Mrs. Henry D. Babcock, Jr. Mrs. C. Payson Coleman Mr. & Mrs. John Evans
Mr. & Mrs. Benjamin H. Balkind Mr. & Mrs. Francis X. Coleman, Jr. Mr. Henri Eyl
Mrs. Gilbert Ball Mr. & Mrs. John H. Coleman Mr. & Mrs. Raymond Z. Fahs
Mr. & Mrs. William M. Bannard Mrs. John K. Colgate Mr. & Mrs. Joel M. Fairman
Mrs. George C. Barclay Mr. & Mrs. Emilio G. Collado Mr. & Mrs. Harold L. Fates
Dr. & Mrs. Henry H. Bard Mr. & Mrs. Bradley Collins Mr. John R. Fell, Jr.
Mrs. Lila Barnes Mr. & Mrs. Patrick Collins Mrs. Allston Flagg
Mr. & Mrs. Edmund Bartlett Mr. & Mrs. Kingsley Colton Mr. & Mrs. Mario Fog
Mrs. Robert E. Belknap, Jr. Mrs. Ralph C. Colyer Mr. & Mrs. Joseph G. Fogg Ill
Dr. & Mrs. Stefan Belman Dr. & Mrs. Joseph B. Conolly, Jr. Mr. & Mrs. Henry L. Fox
Miss Linda Bennetta Dr. & Mrs. Crispin Cooke Mr. Walter N. Frank, Jr.
Mr. & Mrs. Loren C. Berry Mrs. Howard Corning, Jr. Mr. & Mrs. George S. Franklin
Mr. & Mrs. Robert P. Beuerlein Mr. Duncan B. Cox Ms. Mary Freedman
Mr. & Mrs. Nicholas Biddle, Jr. Mr. & Mrs. Charles L. Craig Mr. & Mrs. Jack B. Friedman
Mrs. William Binnian Mr. & Mrs. Miner D. Crary, Jr. Mr. Louis M. Fuccio
Mr. F Roberts Blair Mr. & Mrs. Arthur M. Crocker Mr. D. Kent Gale
Mrs. Mary Lenore Blair Mr. & Mrs. Robert Cuddeback Mr. & Mrs. Clarence E. Galston
Mr. & Mrs. Bache Bleecker Mr. Robert L. Cummings Mr. & Mrs. John W. Galston
Mrs. Margery Blumenthal Mr. Robert L. Cummings Ill Mrs. James E. Gardner, Jr.
Mr. & Mrs. Elito Bongarzone Mr. & Mrs. Richard T. Cunniff Mr. & Mrs. Robert Gardner, Jr.
Mr. & Mrs. A.L. Boorstein Dr. & Mrs. Paul Cushman, Jr. Mr. & Mrs. Charles S. Gay
Mr. & Mrs. Murray Borson Mr. & Mrs. Roderick H. Cushman Mr. & Mrs. Robert A. Gay
Dr. & Mrs. George Bower Mr. & Mrs. George W. Cutting, Jr. Mrs. Louis F Geissler, Jr.
Mr. & Mrs. William Braden Miss Lucy Cutting Mr. & Mrs. Harry Giffords
Dr. & Mrs. Arik Brissenden Mrs. Howard Dana Mr. & Mrs. Stephen E. Gilhuley
Mrs. Horace Brock Mr. Theodore H. Danforth Mrs. Ruth Reed Gillespie
Mr. & Mrs. F. Sedgwick Browne Mr. & Mrs. Norris Darrell, Jr. Mr. & Mrs. Robert Gilmore, Jr.
Mr. & Mrs. G. Morgan Browne, Jr. Mr. & Mrs. A. George Dartt Dr. & Mrs. H. Bentley Glass
Mr. & Mrs. Frank Bruder Miss Katya Davey Mrs. J. Wooderson Glenn
Mr. & Mrs. James Bryan, Jr. Mr. & Mrs. Lawrence Davis Mr. & Mrs. Bernard Gloisten
Mr. & Mrs. Julian Buckley Mrs. F Trubee Davison Mr. & Mrs. E. Rawson Godfrey
Mrs. R.P. Burr Mr. & Mrs. Donald C. Death Mr. C.F. Gordon
Mr. & Mrs. John Busby Mr. & Mrs. Jay De Bow Mr. & Mrs. Kilbourn Gordon, Jr.
Mr. & Mrs. Albert Bush-Brown Mr. & Mrs. Raymond DeClairville Mr. & Mrs. Hank Grabowski
Mr. & Mrs. Sidney G. Butler Mr. & Mrs. Donald L. Deming Mr. & Mrs. Oliver R. Grace
Miss Grace T Caidin Mr. & Mrs. Douglas Despard, Jr. Mr. & Mrs. William R. Grant
Mr. & Mrs. T.J. Calabrese, Jr. Mr. & Mrs. Joseph C. Dey, Jr. Mr. & Mrs. Craig P Greason
Miss Monique Calabro Mr. & Mrs. Charles Dolan Mr. & Mrs. Alfred T Gregory
Mr. & Mrs. Samuel Callaway Russell & Janet Doubleday Fund Mr. & Mrs. George Haberer
Mr. & Mrs. John P Campbell Mr. & Mrs. Clark M. Drasher, Jr. Mr. & Mrs. Sidney Hack
Mr. & Mrs. Ward C. Campbell Dr. & Mrs. Arthur Drazan Mrs. Joanna Hadden
Miss Martha Worth Carder Dr. & Mrs. Lester Dubnick Mr. & Mrs. John W.B. Hadley
Mr. & Mrs. John Carr Mrs. Eugene DuBois Mr. & Mrs. Lawrence Hahn
338

The Rev. Linda Peyton HancockMrs. Beverly HansonMr. & Mrs. Gordon HargravesMr. & Mrs. Henry U. HarrisMr. & Mrs. Henry U. Harris, Jr.Miss Maxine HarrisonDr. & Mrs. C. HartensteinMr. & Mrs. Rolf D. HartmannMrs. Sinclair HatchMr. & Mrs. Horace Havemeyer, Jr.Mr. & Mrs. T.M. HearnMr. Huy ler C. HeldMr. & Mrs. James E. HellierMr. & Mrs. W.P. HendersonDr. & Mrs. William W. HeroyMr. & Mrs. Charles L. HewittMrs. Herman A. HeydtMrs. Thomas L. HigginsonMr. & Mrs. Charles R. HolcombMr. & Mrs. Karl HoltzschueMr. & Mrs. George J. HossfeldMr. & Mrs. George J. Hossfeld, Jr.Mrs. Margaret H. HoweMrs. J. Taylor HowellMr. & Mrs. Charles J. HubbardMr. & Mrs. Philip G. HullHon. & Mrs. John P HumesMr. & Mrs. J.A.V. HyattMrs. Elmira S. IngersollMrs. David IngrahamMr. & Mrs. Frederic B. IngrahamMr. & Mrs. Fred IrwinMrs. Elizabeth W. IvinsMr. & Mrs. Richard M. JacksonMr. & Mrs. Valdemar F. JacobsenMr. & Mrs. Irving D. JakobsonWalter B. James FundMr. & Mrs. Robert D. JayMrs. B. Brewster JenningsMr. & Mrs. L.S. JohnsonMr. & Mrs. John J. JudaMrs. Ruth KarpMr. & Mrs. Morris I. KarpenDr. & Mrs. Francis C. KeilMr. & Mrs. Francis E. Kelly, Jr.Mrs. Walter A. KernanMr. & Mrs. Martin S. KimmelMiss Louise M. KingMr. & Mrs. Francis S. KinneyMr. & Mrs. Richard L. KlineDr. & Mrs. Rudolf W. KnoepfelMrs. Robert P. KoenigMr. & Mrs. Jack KolodnyMr. & Mrs. Edward W. KozlikMr. & Mrs. Paul KramerMr. & Mrs. Morton Kunst lerMrs. Carol LambMr. & Mrs. Gordon LambMr. & Mrs. Edward M. LamontMs. Laurie J. LandeauMr. Charles Lassen
Mrs. Orin T LeachMr. & Mrs. Leslie S. LearnedMrs. Randall J. LeBoeuf, Jr.Mr. & Mrs. Henry C. LeeRev. & Mrs. T Carleton LeeMr. & Mrs. Thomas LeonDr. & Mrs. R.V. LewisMrs. Elizabeth A. LindsayMr. & Mrs. George N. Lindsay, Jr.Mr. & Mrs. Robert V. LindsayMr. & Mrs. Bernard LippmanMr. & Mrs. V.S. LittauerMr. & Mrs. Angus LittlejohnMr. & Mrs. John H. LivingstonMr. & Mrs. Candido V. LloveraMr. & Mrs. Alan J. LongMr. Thomas P Losee, Jr.Mrs. Grace E. LotowyczMr. & Mrs. John F MacKayMrs. Malcolm MacKayMr. & Mrs. Sayre MacLeodMrs. Henry R. MacyMrs. John R. MahoneyMr. & Mrs. John MarianiMrs. John B. MarshMr. & Mrs. John M. MartinMr. & Mrs. William H. MathersMrs. John D. MaxwellDr. Ernst MayrMr. & Mrs. Edward F McCannMr. & Mrs. James A. McCurdy IIMr. & Mrs. Richard McDanielMr. & Mrs. Victor McElhenyMiss Diana McllvaineMr. & Mrs. Angus P McIntyreMr. & Mrs. Randall P McIntyreMr. & Mrs. Lawrence M. MeadDr. & Mrs. Walter MeierMr. Arthur C. MerrillMrs. Frances MeurerDr. & Mrs. Leo M. MeyerMr. & Mrs. Philip MichelmanMr. & Mrs. Kennedy B. MiddendorfMiss Patricia C. MillerMr. & Mrs. William H. MillerMr. Dudley H. MillsMrs. A.M. MiltonMr. & Mrs. George G. Montgomery, Jr.Mrs. F Franklin MoonMr. & Mrs. James D. Mooney, Jr.Mr. Louis deB MooreMr. & Mrs. Richard K. MooreMr. & Mrs. George MorgeseMr. & Mrs. Grinnell MorrisMrs. Robert C. MuhlhausenMr. & Mrs. Gardner MundyMr. & Mrs. Allan F MunroMr. & Mrs. Francis W. Murray IIIDr. & Mrs. Larry NathansonMr. & Mrs. John S. NicholsMr. & Mrs. Adams Nickerson
Mrs. John W. NieldsMr. & Mrs. William S. NivenMr. & Mrs. Albert C. Nolte, Jr.Mr. Lawrence W. NorthamMr. & Mrs. Charles P NoyesMr. & Mrs. David C. Noyes, Jr.Mr. & Mrs. George D. O'NeillMr. & Mrs. Richard B. OpsahlMr. & Mrs. Robert OsterhusMr. & Mrs. Arthur W. Page, Jr.Dr. & Mrs. David A. PageMr. & Mrs. John H. PageMr. & Mrs. Walter H. PageMr. & Mrs. Fred W. PainMr. & Mrs. Samuel D. ParkinsonMr. & Mrs. Joseph M. ParriottMr. & Mrs. William ParsonsMr. & Mrs. William Parsons, Jr.Mr. & Mrs. Fremont C. PeckMr. & Mrs. William H. PeckMr. & Mrs. Paul Pennoyer, Jr.Mr. & Mrs. John M. PerkinsMr. & Mrs. Bernard Peyton IIIMr. & Mrs. Howard Phipps, Jr.Mr. John PickettMrs. Richard N. PiersonMr. & Mrs. Robert PilkingtonMr. & Mrs. James J. PirtleMr. & Mrs. Collier PlattMr. & Mrs. Graham L. PlattMrs. Francis T.P. PlimptonMr. & Mrs. Martin PollakMr. & Mrs. Edward Everett PostMrs. William S. PostMs. Helen W. PowersMrs. H. Irving PrattMr. & Mrs. Richardson PrattDr. William PrestonMr. Edward PullingMr. Thomas L. PullingMr. & Mrs. Preston V. PumphreyMr. & Mrs. Tarrant PutnamMr. & Mrs. James T. PyleMr. & Mrs. William RandMr. & Mrs. John R. ReeseMr. & Mrs. Willis L.M. ReeseMrs. Cornelius J. ReidDr. Robert M. ReissMr. Bernard J. ReveredinMr. & Mrs. John T. RicksMr. & Mrs. J. Gregory RileyMr. & Mrs. George RobertsMr. & Mrs. William S. RobertsonMr. & Mrs. Samuel B. RogersMiss Elizabeth E. RooseveltMrs. Phillip J. RooseveltMrs. Quentin RooseveltMrs. Donald W. RoseMr. & Mrs. Arthur W. Rossiter, Jr.Mr. & Mrs. Walter N. Rothschild, Jr.Mr. & Mrs. Francis J. Rue, Jr.
339

Mrs. F.E. RulandMr. Robert SabinMr. & Mrs. Harold P SalembierMr. & Mrs. Bernard SalzbergMr. & Mrs. Harvey E. SampsonMr. & Mrs. John K. SandsMr. Rudolph J. SantoroMr. John M. SchiffMr. Peter G. SchiffMr. & Mrs. Carl SchlangerDr. & Mrs. Carl J. Schmidlapp IIMr. & Mrs. Frederick P SchmittMr. & Mrs. Franz SchneiderDr. Irving M. SchneiderMr. & Mrs. Hermann C. SchwabMr. & Mrs. David R. SchwarzMr. & Mrs. Frederick E. SeymourMr. & Mrs. Marvin SharksDr. & Mrs. Edward M. ShepardMr. & Mrs. Douglas A. Shepardson, Jr.Mr. & Mrs. Robert E. SheridanMr. & Mrs. Oscar ShoenfeldMr. Paul W. SeigertDr. & Mrs. Paul SiminoffMr. & Mrs. William C. SimpsonMrs. James C. SkiffMr. & Mrs. Rowland G. SkinnerMr. & Mrs. H. Turner SlocumMr. & Mrs. Robert SmirnowMr. & Mrs. William G. SmithMr. & Mrs. William S. SmootMr. & Mrs. James SommermeyerMrs. Julian C. StanleyMr. W.O. StantonMr. David SteinMr. & Mrs. James F StebbinsMrs. Theodore E. Stebbins
340
Mr. & Mrs. John C. StevensonMr. & Mrs. Norman W. StewartMr. Dudley W. StoddardMr. & Mrs. Richard S. StorrsMr. & Mrs. Joseph S. StoutMr. & Mrs. Joseph S. Stout, Jr.Mr. Theodore C. StreibertMr. Scott SutcliffeMr. & Mrs. Robert SwiggertDr. Basil TangrediMr. & Mrs. Benjamin TaylorMr. & Mrs. Frederick TaylorMr. & Mrs. John W. TaylorMiss Jeanie TengelsonMr. & Mrs. Daniel G. Tenney, Jr.Mr. & Mrs. D.B. TenneyMr. & Mrs. Fred ThielkingMr. Charles R. ThompsonMrs. Denyse D. ThorsDr. & Mrs. Robert TilneyMr. & Mrs. Warren I. Titus, Jr.Mr. & Mrs. Alexander C. TomlinsonDr. & Mrs. M.B. TravisMr. & Mrs. Bronson Trevor, Jr.Mr. & Mrs. Stanley TrotmanMr. & Mrs. A.M. TrotterMr. & Mrs. E.T. TurnerMr. & Mrs. William R. UdryMr. & Mrs. Stephen Van R. UlmanDr. & Mrs. Thornton VandersallMr. & Mrs. Martin VictorMr. Eric Von RaitsMr. & Mrs. John WagnerMr. & Mrs. Colton P WagnerMs. Christine M. WalkerMrs. George WalkerMr. & Mrs. R.W. Walker
Mrs. Robert P WaltonDr. & Mrs. David E. WardenMrs. Ethelbert WarfieldMr. & Mrs. Bradford A. WarnerMr. & Mrs. Harold L. Warner, Jr.Dr. & Mrs. James D. WatsonEdwin S. Webster FoundationMrs. Arthur D. Weekes, Jr.Mrs. Bradford G. WeekesMr. & Mrs. Bradford G. Weekes IIIMr. & Mrs. R. Ray WeeksMrs. F Carrington WeemsMr. John C. WeghornMr. & Mrs. Richard J. WeghornMr. & Mrs. L.B. Wehle, Jr.Mr. & Mrs. Taggart WhippleMrs. Alexander M. WhiteMr. & Mrs. Warren D. WhiteMr. & Mrs. Hendricks WhitmanMr. Theodore S. WickershamMr. Malcolm D. WidenorMr. & Mrs. Douglas WilliamsMr. & Mrs. Duane N. WilliamsMr. Henry S. WilliamsMr. & Mrs. Ichabod WilliamsMr. & Mrs. Thorndike WilliamsMrs. John C. WilmerdingMrs. Henry S. WingateMrs. Scudder WinslowMr. & Mrs. William A. WoodcockMr. & Mrs. James A. WoodsMrs. Ford WrightMiss Flavia Helen WyethMr. & Mrs. Woodhull YoungMr. Robert Zakary