
CD1d-dependent natural killer T cells attenuate angiotensin II ...

1 De Bundel, D.
Angiotensin IV and LVV-Haemorphin 7 Enhance Spatial
Working Memory in Rats: Effects on Hippocampal Glucose
Levels and Blood Flow.
Dimitri De Bundel1, Ilse Smolders1, Rui Yang1, Anthony L. Albiston2, Yvette
Michotte1, Siew Yeen Chai2,3
1 Research Group Experimental Pharmacology: Department of Pharmaceutical
Chemistry, Drug Analysis and Drug Information, Vrije Universiteit Brussel,
Laarbeeklaan 103, 1090 Brussels, Belgium
2Neuropeptides Group, Howard Florey Institute, 3Centre for Neuroscience, University of
Melbourne, Parkville, Victoria 3010 Australia
Corresponding author:
Siew Yeen Chai
Fax : +61 3 9348 1707
Phone: +61 3 8344 7782
E-mail: [email protected]

2 De Bundel, D.
Abstract
The IRAP ligands Angiotensin IV (Ang IV) and LVV-haemorphin 7 (LVV-H7) enhance
performance in a range of memory paradigms in normal rats and ameliorate memory
deficits in rat models for amnesia. The mechanism by which these peptides facilitate
memory remains to be elucidated. In recent in vitro experiments, we demonstrated that
Ang IV and LVV-H7 potentiate activity-evoked glucose uptake into hippocampal
neurons. This raises the possibility that IRAP ligands may facilitate memory in
hippocampus-dependent tasks through enhancement of hippocampal glucose uptake.
Acute intracerebroventricular (i.c.v.) administration of 1 nmol Ang IV or 0.1 nmol LVV-
H7 in 3 months-old Sprague Dawley rats enhanced spatial working memory in the plus
maze spontaneous alternation task. Extracellular hippocampal glucose levels were
monitored before, during and after behavioral testing using in vivo microdialysis.
Extracellular hippocampal glucose levels decreased significantly to about 70% of baseline
when the animals explored the plus maze, but remained constant when the animals were
placed into a novel control chamber. Ang IV and LVV-H7 did not significantly alter
hippocampal glucose levels compared to control animals in the plus maze or control
chamber. Both peptides had no effect on hippocampal blood flow as determined by laser
Doppler flowmetry, excluding that either peptide increased the hippocampal supply of
glucose. We demonstrated for the first time that Ang IV and LVV-H7 enhance spatial
working memory in the plus maze spontaneous alternation task but no in vivo evidence
was found for enhanced hippocampal glucose uptake or blood flow.

3 De Bundel, D.
Key Words: Angiotensin IV; LVV-haemorphin 7; glucose; memory; hippocampus;
IRAP

4 De Bundel, D.
Introduction
Insulin-regulated aminopeptidase (IRAP) was initially characterized in adipocytes as a
marker protein of specialized vesicles containing the insulin-regulated glucose
transporter-4 (GLUT4) (Keller, Scott, Mastick, Aebersold & Lienhard, 1995; Ross, Scott,
Morris, Leung, Mao, Lienhard & Keller, 1996). IRAP is a membrane bound
aminopeptidase with a large extracellular catalytic domain, a single transmembrane
domain and an extended cytoplasmic domain containing two dileucine motifs preceded by
acidic clusters which are associated with trafficking (Johnson, Lampson & McGraw,
2001; Keller et al., 1995). In adipocytes, IRAP redistributes with GLUT4 to the plasma
membrane in response to insulin. IRAP is also found in other tissues such as the brain
(Keller et al., 1995), where it is predominantly expressed in neurons, with high
concentrations in areas involved in cognitive functions such as the hippocampus, the
amygdala, the basal forebrain and the cerebral cortex (Fernando, Larm, Albiston & Chai,
2005). Intriguingly, in these brain areas, IRAP and GLUT4 are co-localized in secretory
vesicles, similarly to the insulin-regulated vesicles of adipocytes, suggesting that IRAP
may be involved in the regulation of GLUT4 dependent neuronal glucose uptake
(Fernando, Luff, Albiston & Chai, 2007).
The role of IRAP in the brain remained elusive until it was identified as a high affinity
binding site for two endogenous peptides, angiotensin IV (Ang IV) and LVV-haemorphin
7 (LVV-H7) (Albiston, McDowall, Matsacos, Sim, Clune, Mustafa, Lee, Mendelsohn,
Simpson, Connolly & Chai, 2001). Acute or chronic administration of these IRAP ligands
into the cerebral ventricles of rats enhances performance in a range of memory paradigms
including the passive avoidance (Braszko, Kupryszewski, Witczuk & Wisniewski, 1988;

5 De Bundel, D.
Wright, Miller-Wing, Shaffer, Higginson, Wright, Hanesworth & Harding, 1993), Barnes
circular maze (Lee, Albiston, Allen, Mendelsohn, Ping, Barrett, Murphy, Morris,
McDowall & Chai, 2004) and Morris water maze (Wright, Stubley, Pederson, Kramar,
Hanesworth & Harding, 1999) tasks. Moreover, IRAP ligands ameliorate memory deficits
induced by chronic alcohol exposure (Wisniewski, Borawska & Car, 1993), global
ischemia (Wright, Clemens, Panetta, Smalstig, Weatherly, Kramar, Pederson, Mungall &
Harding, 1996), bilateral perforant pathway lesion (Wright et al., 1999), or chemical
perturbations of the septo-hippocampal cholinergic pathways (Albiston, Pederson, Burns,
Purcell, Wright, Harding, Mendelsohn, Weisinger & Chai, 2004; Olson, Olson, Qualls,
Stratton, Harding & Wright, 2004; Pederson, Krishnan, Harding & Wright, 2001).
Whereas several brain areas may be involved in the different memory effects of IRAP
ligands, many studies have focused on the hippocampus because of the robust effects of
these peptides on the hippocampal-dependent spatial learning. The observation that i.c.v.
administration of Ang IV predominantly evokes c-FOS expression in the hippocampus
(Roberts, Krebs, Kramar, Shaffer, Harding & Wright, 1995) indeed suggests that this
brain area is a major target site for the memory effects of IRAP ligands. In support of
their spatial memory enhancing effects, IRAP ligands were demonstrated to facilitate
long-term potentiation in the CA1 region of the hippocampus in vitro (Kramar,
Armstrong, Ikeda, Wayner, Harding & Wright, 2001) and in the dentate gyrus in vivo
(Wayner, Armstrong, Phelix, Wright & Harding, 2001). Furthermore, Ang IV and LVV-
H7 facilitated potassium-evoked acetylcholine release in rat hippocampal slices (Lee,
Chai, Mendelsohn, Morris & Allen, 2001).

6 De Bundel, D.
The ability of IRAP ligands to facilitate memory in both normal animals and a range of
memory deficit models makes IRAP a potential therapeutic target. However, the
mechanism by which IRAP ligands facilitate memory is not fully understood. Several
candidate mechanisms have been proposed including facilitation of neuronal glucose
uptake (Albiston, Mustafa, McDowall, Mendelsohn, Lee & Chai, 2003; De Bundel,
Smolders, Vanderheyden & Michotte, 2008). In recent in vitro experiments, we
demonstrated that Ang IV and LVV-H7 potentiate activity-evoked glucose uptake into
mice hippocampal neurons (Fernando, Albiston & Chai, 2008). This effect was absent in
IRAP knockout mice, strongly suggesting a role for IRAP in hippocampal glucose uptake
(Fernando et al., 2008). IRAP ligands may therefore enhance spatial memory through
potentiation of glucose uptake into hippocampal neurons. Indeed, it is well established
that glucose is a potent modulator of memory. Systemic or intrahippocampal glucose
administration facilitates spatial memory in rats (Dash, Orsi & Moore, 2006; McNay &
Gold, 1999). Furthermore, a previous microdialysis study elegantly demonstrated a
decrease in extracellular glucose levels in the hippocampus of rats performing a plus
maze spontaneous alternation task (McNay, Fries & Gold, 2000). This decrease in
extracellular glucose is indicative of enhanced glucose uptake by hippocampal cells
during spatial memory processing. We hypothesized that under these conditions, Ang IV
and LVV-H7 may facilitate hippocampal glucose uptake similarly to our observations in
vitro.
The present study uses the in vivo microdialysis technique to test this hypothesis. We
investigated the effects of Ang IV and LVV-H7 on hippocampal glucose levels during
performance of a spatial working memory task. Since the hippocampal extracellular

7 De Bundel, D.
glucose concentration not only depends on the local usage but also on the supply from the
blood, we further determined the effects of Ang IV and LVV-H7 on hippocampal blood
flow. Identification of the mechanism by which IRAP ligands facilitate memory may
provide a novel strategy for the treatment of cognitive impairment.
Materials and methods
Subjects
Male Sprague-Dawley rats (Animal Resource Centre, Canning Vale, WA, Australia and
Charles-River Laboratories, L’Arbresle, Cedex, France ), 3 months old at the time of
surgery, were housed individually in an animal housing facility maintained at a 12/12
hours light/dark cycle initiated at 7:00 h. All animals were given free access to water and
standard rat chow. The experiments were carried out in accordance with the Prevention of
Cruelty to Animals Act and the National Health and Medical Research Council Code of
Practice for the Use of Animals for Scientific Purposes in Australia and according to the
European directive 86/609/EEC with the corresponding national guidelines as approved
by the Ethical Committee for Animal Experimentation of the Faculty of Medicine and
Pharmacy of the Vrije Universiteit Brussel.
Surgery
Behavior and microdialysis experiments
Rats were anesthetized with 5% (v/v) isoflurane (Fluothane; ICI, Melbourne, Australia),
placed on a stereotaxic frame and maintained on 2% (v/v). Rectal temperature was
maintained at 37.5°C using a heating pad. The rats were stereotaxically implanted with a
microdialysis guide cannula (CMA/12; CMA Microdialysis, Solna, Sweden) above the

8 De Bundel, D.
left or right hippocampus and an injection guide cannula (22 g; Plastics One, Roanoke,
USA) above the ipsilateral ventricle. The location of both the microdialysis guide cannula
and injection guide cannula were randomized between left and right hemisphere for
different animals within each experimental group. The flat skull coordinates were 4.6 mm
lateral from midline, 5.6 mm posterior from bregma and 3.6 mm ventral from dura for the
microdialysis guide cannula aimed at the hippocampus and 1.5 mm lateral from midline,
0.8 mm posterior to bregma and 3.5 mm ventral from dura for the injection guide cannula
aimed at the lateral ventricle. After stereotaxic positioning, the guide cannulae were
secured to the skull with stainless steel screws and dental cement. Rats received an
intramuscular injection of 0.05 mg/kg buprenorphine (Temgesic; Reckitt Benckiser, West
Ryde, Australia) and a subcutaneous injection of 10 mg/kg enrofloxacin (Baytril; Bayer,
Pymble, Australia). Dummy cannulae were introduced into the guide cannulae to prevent
occlusion. Rats were allowed to recover for a minimum of 6 days and were handled daily
for at least 1 minute during this time. On completion of testing, the rats received an
overdose of Lethobarb (Virbac, Peakhurst, Australia) and the brains were removed for
verification of the guide cannula localizations.
Hippocampal blood flow experiments
Rats were anesthetized with an intraperitoneal injection of 60 mg/kg pentobarbital
(Nembutal; Ceva Sante Animal, Brussels, Belgium) and maintained with an
intraperitoneal infusion of 0.06 mg/kg pentobarbital at a flow rate of 1 µl/min. Tracheal
intubation was performed to facilitate breathing throughout the experiment and the right
jugular vein of some of the rats was cannulated for supplementary drug administration.
The animals were placed on a stereotaxic frame and rectal temperature was maintained at

9 De Bundel, D.
37.5°C using a heating pad. The rats were stereotaxically implanted with an injection
guide cannula (22 g; Plastics One, Roanoke, USA) above the left or right ventricle and a
circular craniotomy with 2 mm diameter was performed above the ipsilateral
hippocampus. The location of both the injection guide cannula and the craniotomy were
randomized between left and right hemisphere for different animals within each
experimental group. The flat skull coordinates were 1.5 mm lateral from midline, 0.8 mm
posterior to bregma and 3.5 mm ventral from dura for the injection guide cannula aimed
at the lateral ventricle. The injection guide cannula was secured to the scull with a screw
and dental cement. The craniotomy above the hippocampus was performed at flat skull
coordinates 4.6 mm lateral from midline and 5.6 mm posterior from bregma. After
careful removal of the dura, a laser Doppler recording probe (0.8 mm diameter; Vasamed,
Eden Prairie, USA) was lowered 3.6 mm ventral from dura to measure blood flow in the
hippocampus. On completion of testing, the rats received an overdose of pentobarbital
and the brains were removed for verification of the guide cannula and laser Doppler
recording probe localizations.
Peptide administration
In all experiments, the peptides were administered into the lateral ventricle with a 10 µl
Hamilton syringe (Scientific Glass Engineering, Ringwood, Australia) via a FEP tubing
(CMA Microdialysis, Sweden), which was connected to the injection needle (30 g;
Plastics One, Roanoke, USA). The injection needle protruded 0.5 mm beyond the tip of
the guide cannula, thus perforating the wall of the lateral ventricle. The injector was left
in the lateral ventricle for 1 min after the injection to avoid reflux. The animals were not
restrained during the i.c.v. injections and showed no signs of stress or discomfort. All

10 De Bundel, D.
peptides were dissolved in artificial cerebrospinal fluid (aCSF; 128 mM NaCl, 3.0 mM
KCl, 1.3 mM CaCl2, 1.0 mM MgCl2, 21.0 mM NaHCO3, 1.3 mM NaH2PO4, 1.0 mM D-
glucose, pH 7.0) and injected in a volume of 2 µl at a rate of 2 µl/min. Ang IV and LVV-
H7 were obtained from Auspep (Parkville, Australia). All other chemicals were obtained
from Sigma-Aldrich (Castle-Hill, Australia).
The effects of IRAP ligands on spatial working memory and hippocampal glucose levels
were investigated by placing the rats in a plus maze with four closed arms, 5 min after
receiving an injection of either aCSF (n = 12), 1 nmol Ang IV (n = 7), 10 nmol Ang IV (n
= 6), 0.1 nmol LVV-H7 (n = 7) or 1 nmol LVV-H7 (n = 6). The timing of peptide
administration was based on previous results (Lee et al., 2004). The effects of IRAP
ligands on hippocampal glucose levels under control conditions were tested by placing
the animals in the control chamber, 5 min after receiving an injection of either aCSF, 1
nmol Ang IV, or 0.1 nmol LVV-H7 (n = 6 for each experimental group). The effects of
IRAP ligands on hippocampal bloodflow was monitored in anaesthetized rats after
injection of aCSF, 1 nmol Ang IV, or 0.1 nmol LVV-H7 (n = 4 for each experimental
group).
Behavioral testing
All rats were habituated overnight to the testing room, where a 12/12 hours light/dark
cycle was maintained. The effects of IRAP ligands in spatial working memory and the
associated hippocampal glucose concentration was assessed by placing the rats into a
novel control chamber or a plus maze with closed arms for 20 mins, after injection of
either aCSF, Ang IV or LVV-H7. The plus maze and control chamber were thoroughly
cleansed before each experiment with 70% ethanol. The control chamber was a black

11 De Bundel, D.
Perspex microdialysis bowl with a 40 cm diameter and 40 cm high walls which was
placed on top of a table in the testing room. The plus maze was composed of four closed
arms with each arm measuring 75 x 10 x 20 cm. The floor and the walls of the central
platform and the arms were made of black Perspex. The plus maze was elevated 1 m
above the floor to discourage the rats from leaving the maze. Visual cues were located
around the room, including a door and posters with black on white geometric patterns.
Spontaneous alternation testing was conducted by placing the rats in the centre of the plus
maze while allowing for unimpeded exploration for 20 min. The number and the
sequence of arm entries were recorded for calculation of an alternation score. An
alternation was counted when a rat visited 4 different arms during 5 consecutive arm
entries. Immediate re-entries were not counted. A 4/5 alternation score was calculated by
dividing the number of observed alternations in overlapping quintuplets by the number of
possible alternations and expressed as a percentage.
Microdialysis procedures
Microdialysis probes (CMA/12; CMA Microdialysis, Solna, Sweden), were inserted into
the microdialysis guide cannulae 16-18 hours before the start of the experiment. The
microdialysis probes were 3 mm long with an outer diameter of 0.5 mm and sampled
across the CA1 to CA3 regions of the hippocampal formation. Each animal was used
only once. Probe insertion was timed to give optimal measurement conditions and to
avoid glial scarring at the probe site. The microdialysis probes were perfused via FEP
tubings (1.2 µl/100 mm; CMA Microdialysis, Sweden) with an aCSF at a flow rate of 1.0
µl/min . The aCSF contained 1.0 mM D-glucose to avoid excessive supply or drainage of
glucose from the extracellular fluid (McNay et al., 2000). To establish a dynamic

12 De Bundel, D.
equilibrium between the extracellular fluid and the perfusion liquid, the perfusion
commenced 30 min prior to the experiment. Stable glucose levels in the baseline samples
indicate that this equilibration period was sufficient. Hippocampal dialysate samples
were collected every 5 min for 90 min before during and after behavioral testing.
Dialysate samples were then stored at -20°C for glucose analysis.
The microdialysis tubings were connected to the microdialysis probe 30 min before
collecting baseline samples. A swivel system (Instech, Playmouth, USA) was used to
provide optimal freedom of movement of the rats while avoiding coiling of the
microdialysis tubings. The swivel system was attached to a pivoting arm for
microdialysis sampling in the home cage or the control chamber and was supported by
hand using a clear Perspex rod for microdialysis sampling in the plus maze. The
experimenter always remained at a fixed location to support the swivel system following
the movements of the rats during plus maze testing. The microdialysis tubings were
further supported to the ceiling using nylon wires and swiveling loopholes (Fig. 1).
Glucose analysis
Microdialysis samples were analyzed for glucose content according to the method of
Lowry and Passoneau (1972). Briefly, 5 µl samples were diluted to 400 µl in a 1.0 mM
Tris buffer (pH 7.4) containing 1.0 mM MgCl2, 4 U/ml glucose-6-phosphate
dehydrogenase (EC 1.1.1.49), 0.03 mM ATP and 0.03 mM NADP+. All samples were
analysed in duplicate. Fluorescence was measured with a multi-well plate reader at
excitation wavelength of 355 nm and emission wavelength of 460 nm. A blank reading
was taken, 10 µl of a hexokinase (EC 2.7.1.1.) solution (200 U/ml) was added to each
well and fluorescence was measured after 20 min reaction at 30°C. Hexokinase and

13 De Bundel, D.
glucose-6-phosphate dehydrogenase convert glucose to 6-phosphogluconolactone with
simultaneous reduction of NADP+ to NADPH, which is detected fluorimetrically. All
chemicals were obtained from Sigma-Aldrich (Castle-Hill, Australia).
Blood flow monitoring
The laser Doppler probe was connected to a flow meter (MD-1001; Bioanalytical
Systems, West Lafayette, Indiana, USA). The hippocampal blood flow was monitored
continuously at a scan rate of 1 Hz starting 30 min after implantation of the probe. The
basal hippocampal blood flow was monitored for 30 min. After i.c.v. injection of either
aCSF, Ang IV or LVV-H7, the hippocampal blood flow was further monitored for
another 30 min. The blood flow was averaged over 1 min intervals. The average basal
hippocampal blood flow was set as 100% and the readout after cardiac arrest at the end of
the experiment was set as 0%. The sensitivity of the blood flow measurements at the
surface of the hippocampus were evaluated by measuring the vascular response injection
of 300 µg nicotine/kg in 0.3 ml saline over 1 min into the jugular vein as previously
described (Kagitani, Uchida, Hotta & Sato, 2000). Nicotine was obtained from Sigma-
Aldrich (Bornem, Belgium).
Statistical analysis
All data are presented as the mean ± S.E.M. for replicate measurements. The effects of
peptide treatment on the spontaneous alternation score and the number of arm entries
were determined by Mann-Whitney. Glucose measurements were corrected for probe
recovery. Basal glucose levels in the different experimental groups were compared by
one way analysis of variance. The glucose concentrations in the dialysate samples were

14 De Bundel, D.
expressed as the percentages of the basal levels of glucose and further analyzed by two
way analysis of variance for repeated measures followed by Bonferroni’s post-hoc test.
The hippocampal blood flow was expressed as the percentage of the basal relative blood
flow. Hippocampal blood flow measurements were analyzed by two way analysis of
variance for repeated measures followed by Bonferroni’s post-hoc test. All statistical tests
were performed with GraphPad Prism 4.0 (GraphPad Prism Software Inc, San Diego,
USA) at the 5% level of significance.
Results
Effects of Ang IV and LVV-H7 on spatial working memory
A significant increase of the alternation score was observed after i.c.v. injection of either
1 nmol Ang IV (Mann-Whitney, p=0.005) or 0.1 nmol LVV-H7 (Mann-Whitney,
p=0.005) compared to vehicle-treated control rats (Fig. 2A and 2B). The mean alternation
score (± S.E.M.) for vehicle-treated controls was 62.2 ± 2.2% compared to 78.0 ± 4.0%
and 78.4 ± 3.8% for 1 nmol Ang IV and 0.1 nmol LVV-H7 respectively. The higher
doses of IRAP inhibitors (10 nmol Ang IV, 1 nmol LVV-H7) exhibited mean alternation
scores of 70.1 ± 5.3% and 69.3 ± 7.4% respectively which did not differ significantly
(Mann-Whitney, p > 0.05) from vehicle-treated control (Fig. 2A and 2B). This is in
keeping with previous studies demonstrating the lack of efficacy of IRAP inhibitors at
higher concentrations (Lee et al., 2004; Lee et al., 2001). Analysis of the number of arm
choices did not reveal a significant difference (Mann-Whitney, p > 0.05) between rats
treated with Ang IV and LVV-H7 compared to controls, confirming that the observed

15 De Bundel, D.
effect of the IRAP inhibitors is due to improved working memory and not to differences
in locomotor activity (Fig. 2C and 2D).
Effect of spatial working memory testing on hippocampal glucose levels
The in vivo microdialysis probe recovery was previously determined under identical
experimental conditions as 46% using the zero net-flux technique (McNay & Gold,
1999). In the present study, the mean ± S.E.M. basal hippocampal glucose concentration
from 3 month-old Sprague Dawley rats was 1.36 ± 0.04 mM (n = 56). This is in
agreement with the previously reported extracellular hippocampal glucose concentrations
of 1.24 ± 0.04 in 3 month-old Fisher 344 rats (n = 15) (McNay & Gold, 1999). The basal
hippocampal glucose concentrations of the different experimental groups were not
significantly different (F7,48 = 1,134, p = 0,36). The effect of spontaneous alternation
testing was compared to the effect of exposure to a novel environment by placing the rats
in either the plus maze or the control chamber for 20 minutes, after receiving a sham
i.c.v. injection. The animals placed in the control chamber actively explored the novel
environment but despite their increased locomotor activity, no effect was observed on
hippocampal glucose. In animals placed in the plus maze, hippocampal glucose levels
dropped to approximately 70% of basal levels. This observation corresponds to previous
findings (McNay et al., 2000). A significant time versus treatment interaction was
observed between the hippocampal glucose levels of both groups, F(16,256) = 2.00, p =
0.01 (Fig. 3). Post-hoc analysis revealed that 5 min after the transfer, the hippocampal
glucose concentration of animals placed in the plus maze was significantly lower (p <
0.05) compared to animals placed in the control chamber.
Effects of Ang IV and LVV-H7 on hippocampal glucose levels

16 De Bundel, D.
Administration of Ang IV or LVV-H7 into the lateral ventricle did not further enhance
the decrease of hippocampal glucose in animals performing the plus maze task. There
was no significant time versus treatment effect of Ang IV or LVV-H7 on the
hippocampal glucose concentration in animals performing the plus maze task compared
to sham treated controls, F(32,368) = 0.63 and 0.53, p = 0.94 and 0.98 for Ang IV (Fig. 4)
and LVV-H7 (Fig. 5) respectively. Similarly, in the control environment, i.c.v.
administration of memory-enhancing doses of Ang IV (1 nmol) or LVV-H7 (0.1 nmol)
had no effect on hippocampal glucose concentrations compared to sham-treated controls,
F(32,240) = 0.57, p = 0.97 (Fig. 6).
Effects of Ang IV and LVV-H7 on hippocampal blood flow
The extracellular hippocampal concentration of glucose is dependent on local usage and
supply from the blood. Since a further decrease in extracellular glucose may be masked
by an increased glucose flux from the bloodstream to the brain we investigated whether
i.c.v. administration of IRAP ligands enhances hippocampal blood flow. However,
hippocampal bloodflow remained unchanged after administration of aCSF, 1 nmol Ang
IV or 0.1 nmol LVV-H7 into the lateral ventricle (Fig. 7A). No time versus treatment
interaction was observed (F(118,531) = 0.75, p = 0.97). The sensitivity of the system for
hippocampal blood flow measurement was assessed by measuring the response to
administration of 300 µg/kg into the carotid vein. Corresponding to previous findings
(Kagitani et al., 2000), we observed an increase hippocampal blood flow to
approximately 125% of basal levels (Fig. 7B).

17 De Bundel, D.
Discussion
The present manuscript investigated a potential mechanism underlying the memory
enhancing effects of Ang IV and LVV-H7 in the hippocampus during spatial memory
testing. Acute or chronic i.c.v. administration of IRAP ligands enhances spatial learning
in Morris water maze (Wright et al., 1999) and Barnes maze (Lee et al., 2004) tasks. The
detection of high levels of IRAP expression in the hippocampal formation (Fernando et
al., 2005) provides anatomical support for a role for this enzyme in spatial memory.
However, the mechanism by which IRAP ligands facilitate spatial memory remains
unknown. Our previous anatomical studies revealed that in certain regions of the brain,
including the hippocampus, there is an almost complete co-localization of IRAP with
GLUT4 in intracellular compartments in neurons (Fernando et al., 2007). This finding
strongly suggests that in these neurons, IRAP plays a role in regulating the trafficking of
GLUT4, as is the case in insulin-responsive tissues. We recently found in hippocampal
slices that IRAP inhibitors facilitated activity-evoked glucose uptake into pyramidal
neurons through a GLUT4 mediated mechanism (Fernando et al., 2008). Therefore, the
hypothesis that Ang IV and LVV-H7 facilitate spatial working memory through
facilitation of hippocampal glucose uptake was investigated in the present in vivo study.
Ang IV and LVV-H7 facilitate spatial working memory in the plus maze
spontaneous alternation task
In this study, we demonstrated that acute i.c.v. administration of Ang IV or LVV-H7
enhanced spatial working memory as detected by increased spontaneous alternation in the
plus maze, in addition to the known effects of these peptides on hippocampus-dependent
spatial reference memory. The observed effects of these IRAP ligands on spatial working

18 De Bundel, D.
memory performance were not secondary to changes in locomotor activity or exploratory
behavior since no significant effects between the treatment groups were observed in the
number of arm entries during spontaneous alternation testing.
The percent alternation score is equal to the ratio of actual alternations/possible
alternations multiplied by 100. Chance performance on this task is 44.4% (Lennartz,
2008). The mean ± S.E.M. alternation score for vehicle-treated controls was 62.5 ± 2.4%
compared to 78.0 ± 4.0% and 78.4 ± 3.9% respectively for 1 nmol Ang IV or 0.1 nmol
LVV-H7 treated animals. In a previous study, we demonstrated the facilitatory effects of
Nle1-Ang IV and LVV-H7 on spatial reference memory in Barnes circular maze
experiments (Lee et al., 2004). A rapid, significant effect on the acquisition of spatial
reference memory was observed after a single injection of Nle1-Ang IV or LVV-H7 into
the lateral ventricle in rats 5 min before the first trial on the first day (Lee et al., 2004).
Both spatial working and reference memory are hippocampus-dependent, but require
distinct processing mechanisms and molecular pathways (Bannerman, Niewoehner,
Lyon, Romberg, Schmitt, Taylor, Sanderson, Cottam, Sprengel, Seeburg, Kohr &
Rawlins, 2008). Our findings in this study provide further evidence that IRAP ligands are
effective at improving performance in a wide range of memory paradigms, affecting both
short and longer term memories.
Spatial working memory testing decreases hippocampal extracellular glucose levels
The present data confirm previous observations of a decrease in hippocampal
extracellular glucose during spatial working memory testing in rats (McNay et al., 2000).
Spatial working memory testing leads to functional activation of hippocampal neurons
(Puryear, King & Mizumori, 2006) which correlates strongly with increases in local

19 De Bundel, D.
cerebral blood flow (Cox, Woolsey & Rovainen, 1993) and in energy metabolism
(Shulman, Hyder & Rothman, 2002). The observed decrease in extracellular glucose to
30% below baseline is therefore indicative of an increase in local glucose uptake and
utilization.
Both neurons and astroglia are capable of taking up glucose under basal conditions
through the glucose transporters, GLUT3 for neurons and GLUT1 for astroglia (Abe,
Takahashi & Suzuki, 2006; Itoh, Abe, Takaoka & Tanahashi, 2004). A recent model of
cerebral glucose utilization predicts approximately 30% glucose utlilization by neurons
and 70% glucose utilization by astroglia under basal conditions (Hyder, Patel, Gjedde,
Rothman, Behar & Shulman, 2006). However, neurons metabolize a combination of
glucose and lactate (Erlichman, Hewitt, Damon, Hart, Kurascz, Li & Leiter, 2008).
Whether glucose or lactate is the major source of energy for neurons during synaptic
activity is currently subject to debate (Simpson, Carruthers & Vannucci, 2007).
According to the lactate-shunt hypothesis, glucose is predominantly metabolized by glia
into lactate, which is subsequently transported to neurons, forming their major energy
source (Magistretti, 2006). In contrast, recent studies demonstrated that enhanced activity
of neurons of the brain stem and the cerebellum is associated with an increase in neuronal
glucose utilization rather than lactate metabolism (Bak, Schousboe, Sonnewald &
Waagepetersen, 2006).
We previously demonstrated that activation of mouse hippocampal slices by K+
depolarization or cAMP treatment, enhanced 3H-2-deoxyglucose uptake into the
pyramidal neurons (Fernando et al., 2008). This activity-dependent glucose uptake was
attenuated by indinavir, which preferentially blocks the inducible glucose transporter

20 De Bundel, D.
GLUT4 (Fernando et al., 2008). In the hippocampus, GLUT4 is exclusively expressed in
neurons (Apelt, Mehlhorn & Schliebs, 1999; Fernando et al., 2005) and is co-localized
with IRAP in cytoplasmic vesicles under basal conditions (Fernando et al., 2005;
Fernando et al., 2007). This suggests that the enhanced glucose uptake in hippocampal
slices requires recruitment of GLUT4 to the cell surface. We hypothesized that activation
of hippocampal neurons by increased cognitive demand such as in spatial memory testing
may provide a stimulus for such a mobilization of GLUT4, which could enhance
neuronal glucose uptake above basal levels provided by the constitutively expressed
glucose transporter GLUT3.
Ang IV and LVV-H7 did not alter hippocampal glucose levels or blood flow
The observation that exogenous administration of glucose improves spatial working
memory in rats and attenuates the associated decrease in extracellular glucose in the
hippocampus, suggests that spatial memory processing is limited by the rate of local
glucose uptake and/or usage and is susceptible to improvement (McNay et al., 2000).
Since Ang IV and LVV-H7 enhanced activity-evoked glucose uptake in hippocampal
slices (Fernando et al., 2008), we investigated whether these IRAP inhibitors improved
spatial working memory through facilitation of glucose transport into hippocampal
neurons in vivo. Surprisingly, Ang IV and LVV-H7 did not reduce further the
extracellular glucose concentration during performance of spatial working memory task,
despite eliciting an improvement in the alternation scores.
A potential explanation for present findings could be the competition of neurons and glia
for the available energy substrates and the competition of glucose and lactate for
cytosolic nicotinamide-adenine-dinucleotide, which results in the redox switch

21 De Bundel, D.
phenomenon where utilization of one of the two substrates limits the utilization of the
other (Bouzier-Sore, Voisin, Bouchaud, Bezancon, Franconi & Pellerin, 2006). In this
context it was previously demonstrated that disruption of the lactate shuttle from astroglia
to neurons increases the neuronal glucose uptake (Erlichman et al., 2008). Conversely, a
recent model for glucose uptake predicts that under conditions of increased neuronal
glucose uptake, glia may take up lactate rather than glucose as an energy substrate
(Simpson et al., 2007). Therefore, it is conceivable that an increase of neuronal glucose
uptake, induced by IRAP inhibitors, may result in a decrease of glial glucose uptake.
Under these conditions, the overall extracellular glucose would remain constant.
Alternatively, a further decrease in extracellular glucose could be masked by an increase
in glucose flux from the bloodstream to the brain. In this context, Ang IV was
demonstrated to increase cerebral microcirculation when administered into the carotid
artery of anesthetized rats in a dose of 0.1 nmol (Kramar, Harding & Wright, 1997).
Similarly, administration of 1 nmol Ang IV into the carotid artery protected rats against
ischemic damage by inducing a redistribution of blood flow to the ischemic areas (Faure,
Bureau, Oudart, Javellaud, Fournier & Achard, 2008; Faure, Oudart, Javellaud, Fournier,
Warnock & Achard, 2006). These effects were not associated with an increase in arterial
blood pressure but were blocked by the nitric oxide synthase inhibitor L-NAME (Faure et
al., 2006; Kramar, Krishnan, Harding & Wright, 1998). Ang IV was therefore suggested
to increase cerebral blood flow through stimulation of endothelial nitric oxide production.
In the present setup, acute i.c.v. administration of Ang IV or LVV-H7 had no effect on
hippocampal blood flow in anaesthetized rats. This lack of vasodilatatory effects on the
hippocampal vasculature could be due to the difference in site of administration. Indeed,

22 De Bundel, D.
the present data suggest that the access of Ang IV or LVV-H7 to the endothelial layer of
the hippocampal microvasculature after i.c.v. injection may be limited.
It should also be considered that IRAP is a functional aminopeptidase and that Ang IV
and LVV-H7 bind competitively and with relatively high affinity to the catalytic site of
IRAP (Lew, Mustafa, Ye, McDowall, Chai & Albiston, 2003). IRAP is capable of
cleaving a number of peptides associated with memory in vitro (Matsumoto, Nagasaka,
Hattori, Rogi, Tsuruoka, Mizutani & Tsujimoto, 2001). These substrates include arginine-
vasopressin, oxytocin, met-enkephalin, somatostatin and dynorphin A (1-8). Therefore an
alternative mechanism of these to peptides could be the inhibition of the enzymatic
activity of IRAP resulting in the extension of the half-life of its neuropeptide substrates.
Our recent finding in support of this hypothesis is the demonstration that one of our newly
developed, highly selective, small molecule IRAP inhibitors has memory-enhancing
properties in two distinct memory paradigms (Albiston, Morton, Ng, Pham, Yeatman, Ye,
Fernando, De Bundel, Ascher, Mendelsohn, Parker & Chai, 2008).
Conclusion
The current study shows that a single injection of an IRAP ligand, Ang IV or LVV-H7,
into the lateral ventricle of normal rats enhances spatial working memory in the
spontaneous alternation task. Extracellular hippocampal glucose concentrations and
hippocampal blood flow were unaffected by i.c.v. administration of Ang IV or LVV-H7,
suggesting that facilitation of glucose uptake or enhancement of local blood flow in the
hippocampus were not involved in the observed memory effects. IRAP inhibitors have
been found to facilitate other types of memory such as spatial reference memory, object
recognition memory, passive avoidance memory and fear conditioning. Elucidation of the

23 De Bundel, D.
mechanism underlying these cognitive effects, including possible tests of additional brain
areas, may therefore provide a novel strategy for repair of cognitive function.
Acknowledgements
This research was supported by a project grant from the National Health and Medical
Research Council of Australia (NHMRC) and the Robert J. Kleberg Jr. and Helen C.
Kleberg, and CASS Foundations (Australia), the Queen Elisabeth Medical Foundation
(Geneeskundige Stichting Koningin Elisabeth, Belgium); the Research Foundation
Flanders (Fonds voor Wetenschappelijk Onderzoek Vlaanderen, Belgium) and the Vrije
Universiteit Brussel (GOA-2007 and OZR 895, Belgium). S.Y.C. is an Australian
NHMRC Senior Research Fellow. D.D.B. is a Belgian Doctoral Research Fellow of the
Research Foundation Flanders.
References
Abe, T., Takahashi, S., & Suzuki, N. (2006). Oxidative metabolism in cultured rat
astroglia: effects of reducing the glucose concentration in the culture medium and
of D-aspartate or potassium stimulation. J Cereb Blood Flow Metab, 26, 153-160.
Albiston, A. L., McDowall, S. G., Matsacos, D., Sim, P., Clune, E., Mustafa, T., Lee, J.,
Mendelsohn, F. A., Simpson, R. J., Connolly, L. M., & Chai, S. Y. (2001).
Evidence that the angiotensin IV (AT(4)) receptor is the enzyme insulin-
regulated aminopeptidase. J Biol Chem, 276, 48623-48626.
Albiston, A. L., Morton, C. J., Ng, H. L., Pham, V., Yeatman, H. R., Ye, S., Fernando, R.
N., De Bundel, D., Ascher, D. B., Mendelsohn, F. A., Parker, M. W., & Chai, S.

24 De Bundel, D.
Y. (2008). Identification and characterization of a new cognitive enhancer based
on inhibition of insulin-regulated aminopeptidase. Faseb J.
Albiston, A. L., Mustafa, T., McDowall, S. G., Mendelsohn, F. A., Lee, J., & Chai, S. Y.
(2003). AT4 receptor is insulin-regulated membrane aminopeptidase: potential
mechanisms of memory enhancement. Trends Endocrinol Metab, 14, 72-77.
Albiston, A. L., Pederson, E. S., Burns, P., Purcell, B., Wright, J. W., Harding, J. W.,
Mendelsohn, F. A., Weisinger, R. S., & Chai, S. Y. (2004). Attenuation of
scopolamine-induced learning deficits by LVV-hemorphin-7 in rats in the passive
avoidance and water maze paradigms. Behav Brain Res, 154, 239-243.
Apelt, J., Mehlhorn, G., & Schliebs, R. (1999). Insulin-sensitive GLUT4 glucose
transporters are colocalized with GLUT3- expressing cells and demonstrate a
chemically distinct neuron-specific localization in rat brain. J Neurosci Res, 57,
693-705.
Bak, L. K., Schousboe, A., Sonnewald, U., & Waagepetersen, H. S. (2006). Glucose is
necessary to maintain neurotransmitter homeostasis during synaptic activity in
cultured glutamatergic neurons. J Cereb Blood Flow Metab, 26, 1285-1297.
Bannerman, D. M., Niewoehner, B., Lyon, L., Romberg, C., Schmitt, W. B., Taylor, A.,
Sanderson, D. J., Cottam, J., Sprengel, R., Seeburg, P. H., Kohr, G., & Rawlins, J.
N. (2008). NMDA receptor subunit NR2A is required for rapidly acquired spatial
working memory but not incremental spatial reference memory. J Neurosci, 28,
3623-3630.
Bouzier-Sore, A. K., Voisin, P., Bouchaud, V., Bezancon, E., Franconi, J. M., & Pellerin,
L. (2006). Competition between glucose and lactate as oxidative energy substrates

25 De Bundel, D.
in both neurons and astrocytes: a comparative NMR study. Eur J Neurosci, 24,
1687-1694.
Braszko, J. J., Kupryszewski, G., Witczuk, B., & Wisniewski, K. (1988). Angiotensin II-
(3-8)-hexapeptide affects motor activity, performance of passive avoidance and a
conditioned avoidance response in rats. Neuroscience, 27, 777-783.
Cox, S. B., Woolsey, T. A., & Rovainen, C. M. (1993). Localized dynamic changes in
cortical blood flow with whisker stimulation corresponds to matched vascular and
neuronal architecture of rat barrels. J Cereb Blood Flow Metab, 13, 899-913.
Dash, P. K., Orsi, S. A., & Moore, A. N. (2006). Spatial memory formation and memory-
enhancing effect of glucose involves activation of the tuberous sclerosis complex-
Mammalian target of rapamycin pathway. J Neurosci, 26, 8048-8056.
De Bundel, D., Smolders, I., Vanderheyden, P., & Michotte, Y. (2008). Ang II and Ang
IV: Unraveling the Mechanism of Action on Synaptic Plasticity, Memory, and
Epilepsy. CNS Neurosci Ther, 14, 315-339.
Erlichman, J. S., Hewitt, A., Damon, T. L., Hart, M., Kurascz, J., Li, A., & Leiter, J. C.
(2008). Inhibition of monocarboxylate transporter 2 in the retrotrapezoid nucleus
in rats: a test of the astrocyte-neuron lactate-shuttle hypothesis. J Neurosci, 28,
4888-4896.
Faure, S., Bureau, A., Oudart, N., Javellaud, J., Fournier, A., & Achard, J. M. (2008).
Protective effect of candesartan in experimental ischemic stroke in the rat
mediated by AT2 and AT4 receptors. J Hypertens, 26, 2008-2015.
Faure, S., Oudart, N., Javellaud, J., Fournier, A., Warnock, D. G., & Achard, J. M.
(2006). Synergistic protective effects of erythropoietin and olmesartan on

26 De Bundel, D.
ischemic stroke survival and post-stroke memory dysfunctions in the gerbil. J
Hypertens, 24, 2255-2261.
Fernando, R. N., Albiston, A. L., & Chai, S. Y. (2008). The insulin-regulated
aminopeptidase IRAP is colocalised with GLUT4 in the mouse hippocampus--
potential role in modulation of glucose uptake in neurones? Eur J Neurosci, 28,
588-598.
Fernando, R. N., Larm, J., Albiston, A. L., & Chai, S. Y. (2005). Distribution and cellular
localization of insulin-regulated aminopeptidase in the rat central nervous system.
J Comp Neurol, 487, 372-390.
Fernando, R. N., Luff, S. E., Albiston, A. L., & Chai, S. Y. (2007). Sub-cellular
localization of insulin-regulated membrane aminopeptidase, IRAP to vesicles in
neurons. J Neurochem.
Hyder, F., Patel, A. B., Gjedde, A., Rothman, D. L., Behar, K. L., & Shulman, R. G.
(2006). Neuronal-glial glucose oxidation and glutamatergic-GABAergic function.
J Cereb Blood Flow Metab, 26, 865-877.
Itoh, Y., Abe, T., Takaoka, R., & Tanahashi, N. (2004). Fluorometric determination of
glucose utilization in neurons in vitro and in vivo. J Cereb Blood Flow Metab, 24,
993-1003.
Johnson, A. O., Lampson, M. A., & McGraw, T. E. (2001). A di-leucine sequence and a
cluster of acidic amino acids are required for dynamic retention in the endosomal
recycling compartment of fibroblasts. Mol Biol Cell, 12, 367-381.

27 De Bundel, D.
Kagitani, F., Uchida, S., Hotta, H., & Sato, A. (2000). Effects of nicotine on blood flow
and delayed neuronal death following intermittent transient ischemia in rat
hippocampus. Jpn J Physiol, 50, 585-595.
Keller, S. R., Scott, H. M., Mastick, C. C., Aebersold, R., & Lienhard, G. E. (1995).
Cloning and characterization of a novel insulin-regulated membrane
aminopeptidase from Glut4 vesicles. J Biol Chem, 270, 23612-23618.
Kramar, E. A., Armstrong, D. L., Ikeda, S., Wayner, M. J., Harding, J. W., & Wright, J.
W. (2001). The effects of angiotensin IV analogs on long-term potentiation within
the CA1 region of the hippocampus in vitro. Brain Res, 897, 114-121.
Kramar, E. A., Harding, J. W., & Wright, J. W. (1997). Angiotensin II- and IV-induced
changes in cerebral blood flow. Roles of AT1, AT2, and AT4 receptor subtypes.
Regul Pept, 68, 131-138.
Kramar, E. A., Krishnan, R., Harding, J. W., & Wright, J. W. (1998). Role of nitric oxide
in angiotensin IV-induced increases in cerebral blood flow. Regul Pept, 74, 185-
192.
Lee, J., Albiston, A. L., Allen, A. M., Mendelsohn, F. A., Ping, S. E., Barrett, G. L.,
Murphy, M., Morris, M. J., McDowall, S. G., & Chai, S. Y. (2004). Effect of
I.C.V. injection of AT(4) receptor ligands, NLE(1)-angiotensin IV and LVV-
hemorphin 7, on spatial learning in rats. Neuroscience, 124, 341-349.
Lee, J., Chai, S. Y., Mendelsohn, F. A., Morris, M. J., & Allen, A. M. (2001).
Potentiation of cholinergic transmission in the rat hippocampus by angiotensin IV
and LVV-hemorphin-7. Neuropharmacology, 40, 618-623.

28 De Bundel, D.
Lennartz, R. C. (2008). The role of extramaze cues in spontaneous alternation in a plus-
maze. Learn Behav, 36, 138-144.
Lew, R. A., Mustafa, T., Ye, S., McDowall, S. G., Chai, S. Y., & Albiston, A. L. (2003).
Angiotensin AT4 ligands are potent, competitive inhibitors of insulin regulated
aminopeptidase (IRAP). J Neurochem, 86, 344-350.
Magistretti, P. J. (2006). Neuron-glia metabolic coupling and plasticity. J Exp Biol, 209,
2304-2311.
Matsumoto, H., Nagasaka, T., Hattori, A., Rogi, T., Tsuruoka, N., Mizutani, S., &
Tsujimoto, M. (2001). Expression of placental leucine
aminopeptidase/oxytocinase in neuronal cells and its action on neuronal peptides.
Eur J Biochem, 268, 3259-3266.
McNay, E. C., Fries, T. M., & Gold, P. E. (2000). Decreases in rat extracellular
hippocampal glucose concentration associated with cognitive demand during a
spatial task. Proc Natl Acad Sci U S A, 97, 2881-2885.
McNay, E. C., & Gold, P. E. (1999). Extracellular glucose concentrations in the rat
hippocampus measured by zero-net-flux: effects of microdialysis flow rate, strain,
and age. J Neurochem, 72, 785-790.
Olson, M. L., Olson, E. A., Qualls, J. H., Stratton, J. J., Harding, J. W., & Wright, J. W.
(2004). Norleucine1-angiotensin IV alleviates mecamylamine-induced spatial
memory deficits. Peptides, 25, 233-241.
Pederson, E. S., Krishnan, R., Harding, J. W., & Wright, J. W. (2001). A role for the
angiotensin AT4 receptor subtype in overcoming scopolamine-induced spatial
memory deficits. Regul Pept, 102, 147-156.

29 De Bundel, D.
Puryear, C. B., King, M., & Mizumori, S. J. (2006). Specific changes in hippocampal
spatial codes predict spatial working memory performance. Behav Brain Res, 169,
168-175.
Roberts, K. A., Krebs, L. T., Kramar, E. A., Shaffer, M. J., Harding, J. W., & Wright, J.
W. (1995). Autoradiographic identification of brain angiotensin IV binding sites
and differential c-Fos expression following intracerebroventricular injection of
angiotensin II and IV in rats. Brain Res, 682, 13-21.
Ross, S. A., Scott, H. M., Morris, N. J., Leung, W. Y., Mao, F., Lienhard, G. E., &
Keller, S. R. (1996). Characterization of the insulin-regulated membrane
aminopeptidase in 3T3-L1 adipocytes. J Biol Chem, 271, 3328-3332.
Shulman, R. G., Hyder, F., & Rothman, D. L. (2002). Biophysical basis of brain activity:
implications for neuroimaging. Q Rev Biophys, 35, 287-325.
Simpson, I. A., Carruthers, A., & Vannucci, S. J. (2007). Supply and demand in cerebral
energy metabolism: the role of nutrient transporters. J Cereb Blood Flow Metab,
27, 1766-1791.
Wayner, M. J., Armstrong, D. L., Phelix, C. F., Wright, J. W., & Harding, J. W. (2001).
Angiotensin IV enhances LTP in rat dentate gyrus in vivo. Peptides, 22, 1403-
1414.
Wisniewski, K., Borawska, M., & Car, H. (1993). The effect of angiotensin II and its
fragments on post-alcohol impairment of learning and memory. Pol J Pharmacol,
45, 23-29.
Wright, J. W., Clemens, J. A., Panetta, J. A., Smalstig, E. B., Weatherly, L. A., Kramar,
E. A., Pederson, E. S., Mungall, B. H., & Harding, J. W. (1996). Effects of

30 De Bundel, D.
LY231617 and angiotensin IV on ischemia-induced deficits in circular water
maze and passive avoidance performance in rats. Brain Res, 717, 1-11.
Wright, J. W., Miller-Wing, A. V., Shaffer, M. J., Higginson, C., Wright, D. E.,
Hanesworth, J. M., & Harding, J. W. (1993). Angiotensin II(3-8) (ANG IV)
hippocampal binding: potential role in the facilitation of memory. Brain Res Bull,
32, 497-502.
Wright, J. W., Stubley, L., Pederson, E. S., Kramar, E. A., Hanesworth, J. M., & Harding,
J. W. (1999). Contributions of the brain angiotensin IV-AT4 receptor subtype
system to spatial learning. J Neurosci, 19, 3952-3961.

31 De Bundel, D.
Figure Legends
Figure 1. Overview of the setup for microdialysis sampling during plus maze
spontaneous alternation testing. The microdialysis tubings were supported through
loopholes which were attached to the ceiling with nylon wire. The microdialysis swivel
system could easily be moved from a static arm above the home cage or control chamber
to a loophole above the plus maze which was attached to the ceiling with an elastic band.
A transparent rod was used to manually guide the movement of the microdialysis swivel
following the movements of the rat in the plus maze, while the experimenter stayed at a
fixed position relative to the maze.
Figure 2. (A and C) Spatial working memory performance and the corresponding
locomotor activity after i.c.v. injection of aCSF (n=12), 1 nmol Ang IV (n = 7) or 10
nmol Ang IV (n = 6). (B and D) Spatial working memory performance and the
corresponding locomotor activity after i.c.v. injection of aCSF (n=12), 0.1 nmol LVV-H7
(n = 7) or 1 nmol LVV-H7 (n = 6). The mean ± S.E.M. 4/5 alternation score on the plus
maze spontaneous alternation task is expressed as the percentage of possible
alternations.** p < 0.01 compared to aCSF
Figure 3. Mean ± S.E.M. extracellular hippocampal glucose levels during control
chamber ● (n = 6) and plus maze ○ (n = 12) experiments after i.c.v. injection of aCSF,
expressed as the percentage of the baseline concentration (A). Baseline samples were
collected with the rats in the homecage from 0-30 min. The rats received the i.c.v.

32 De Bundel, D.
injection of aCSF at 30 min (arrow) and were placed in the control box or the plus maze
(shaded area) from 35-55 min. Between 55-85 min the rats were placed back in the
homecage. * p < 0.05 compared to control chamber.
Figure 4. Mean ± S.E.M. extracellular hippocampal glucose levels during plus maze
spontaneous alternation experiments after i.c.v. injection of aCSF ○ (n = 12), 1 nmol Ang
IV ▲ (n = 7) or 10 nmol Ang IV ▲ (n = 6) expressed as the percentage of the baseline
concentration. Baseline samples were collected with the rats in the homecage from 0-30
min. The rats received the i.c.v. injection of aCSF, Ang IV or LVV-H7 at 30 min (arrow)
and were placed in the plus maze (shaded area) between 35-55 min. From 55-85 min the
rats were placed back in the homecage.
Figure 5. Mean ± S.E.M. extracellular hippocampal glucose levels during plus maze
spontaneous alternation experiments after i.c.v. injection of aCSF ○ (n = 12), 0.1 nmol
LVV-H7 (n = 7) or 1 nmol LVV-H7 (n = 6) expressed as the percentage of the
baseline concentration. Baseline samples were collected with the rats in the homecage
from 0-30 min. The rats received the i.c.v. injection of aCSF, Ang IV or LVV-H7 at 30
min (arrow) and were placed in the plus maze (shaded area) between 35-55 min. From
55-85 min the rats were placed back in the homecage.
Figure 6. Mean ± S.E.M. extracellular hippocampal glucose levels during control box
experiments after i.c.v. injection of aCSF ● (n = 6), 1 nmol Ang IV ▲ (n = 6) or 0.1 nmol
LVV-H7 (n = 6) expressed as the percentage of the baseline concentration. Baseline

33 De Bundel, D.
samples were collected with the rats in the homecage from 0-30 min. The rats received
the i.c.v. injection of aCSF, Ang IV or LVV-H7 at 30 min (arrow) and were placed in the
control chamber (shaded area) between 35-55 min. From 55-85 min the rats were placed
back in the homecage.
Figure 7 Mean ± S.E.M. hippocampal bloodflow after (A) i.c.v. injection (arrow) of
aCSF ● (n = 4), 1 nmol Ang IV ▲ (n = 4) or 1 nmol LVV-H7 (n = 4) and (B) after i.v.
infusion (arrow) of 300 µg/kg nicotine as a positive control (n = 4). The relative blood
flow is expressed as the percentage of the baseline blood flow.

Figure 1

Figure 2
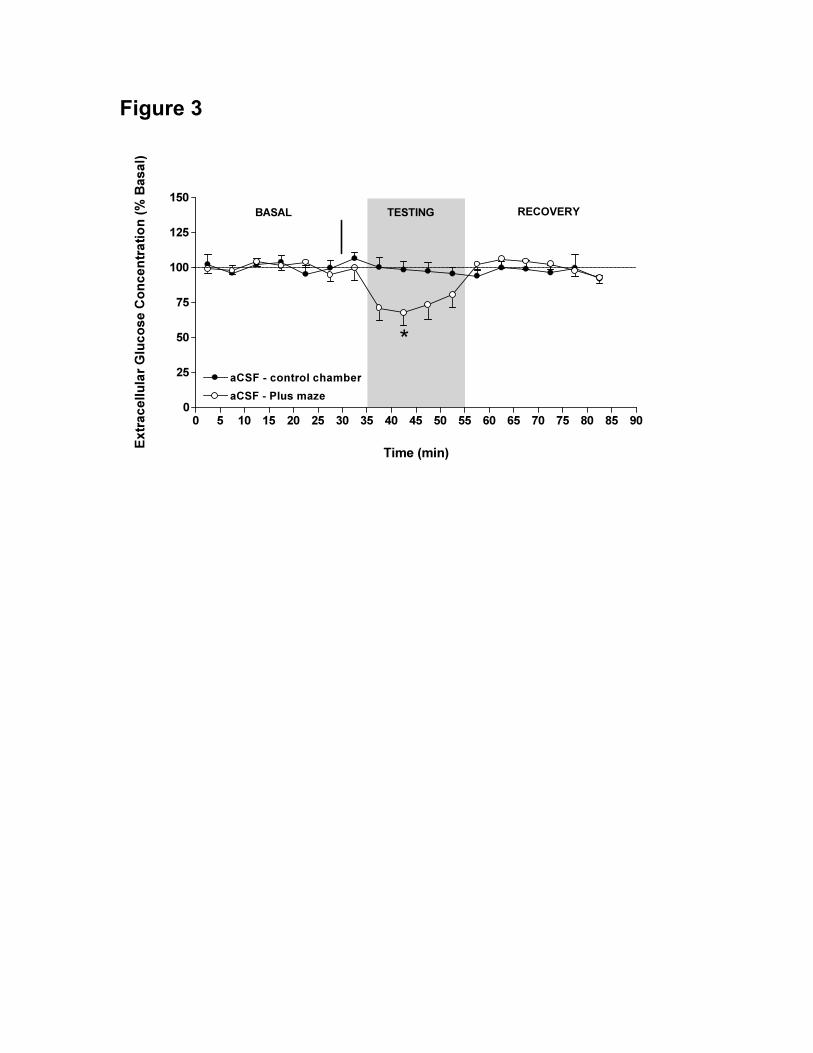
Figure 3

Figure 4

Figure 5
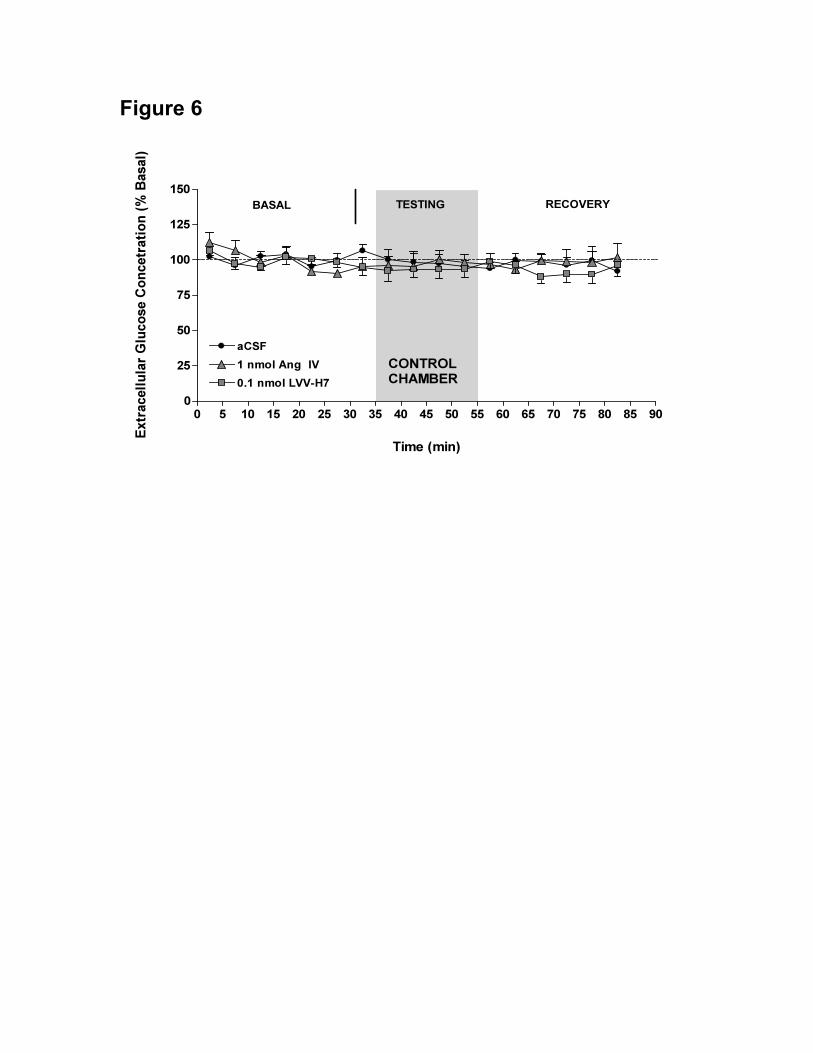
Figure 6

Figure 7
Copyright © 2022 FDOKUMEN




















