
ADHD Comorbidity Findings From the MTA Study: Comparing Comorbid Subgroups
Upload
independentCategory
view
0download
0
Analysis of IgVH gene mutations in B cell chronic lymphocyticleukaemia according to antigen-driven selection identifiessubgroups with different prognosis and usage of the canonicalsomatic hypermutation machinery
B-cell chronic lymphocytic leukaemia (B-CLL) cells were
originally considered to be the neoplastic counterpart of B
lymphocytes unable to undergo somatic hypermutation (SHM;
Hamblin, 2002; Chiorazzi & Ferrarini, 2003). The demonstra-
tion that approximately 50% of B-CLL cases express a mutated
configuration of immunoglobulin (Ig)VH genes (Schroeder &
Dighiero, 1994; Oscier et al, 1997; Fais et al, 1998) changed
this view suggesting the existence of at least two distinct B-CLL
subsets that probably originated from pre- or post-germinal
centre (GC) B cells (Naylor & Capra, 1999). This different
histogenesis has important prognostic implications, as B-CLL
cases with a mutated (M) configuration of IgVH genes have a
better prognosis than those with unmutated (UM) B-CLL
(Damle et al, 1999; Hamblin et al, 1999; Maloum et al, 2000;
Krober et al, 2002; Lin et al, 2002). More recently, the
expression of a homogeneous phenotype and gene profile
related to experienced B cells virtually in all B-CLL cases (Klein
et al, 2001; Rosenwald et al, 2001; Damle et al, 2002) have
strongly questioned such a pathogenetic model, rather favour-
ing a common transformation mechanism for both M and UM
B-CLLs.
The SHM process, as it occurs during a T-cell-dependent
GC differentiation of normal B cells, introduces mutations in a
non-random manner generating several biased nucleotide
substitutions. Among them, the skewing of replacement (R)
mutations from framework (FR) to complementarity-deter-
mining regions (CDR) of IgVH genes is the result of a positive
selection operated by the antigen, aimed at preserving the
Massimo Degan,1 Riccardo Bomben,1
Michele Dal Bo,1 Antonella Zucchetto,1
Paola Nanni,1 Maurizio Rupolo,2
Agostino Steffan,3 Vincenza Attadia,1
Pier Ferruccio Ballerini,4 Daniela
Damiani,5 Carlo Pucillo,6 Giovanni Del
Poeta,7 Alfonso Colombatti8 and Valter
Gattei1
1Clinical and Experimental Haematology
Research Unit, 2Medical Oncology, 3Blood Bank,
IRCCS, Aviano (PN), 4Division of Internal
Medicine, De Gironcoli Hospital, Conegliano
(TV), 5Division of Hematology, and 6Laboratory
of Immunology, University of Udine, Udine,7S. Eugenio Hospital, University of Tor Vergata,
Rome, and 8Experimental Oncology 2 and Centro
di Riferimento Oncologico, IRCCS, Aviano (PN),
Italy
Received 23 January 2004; accepted for
publication 22 March 2004
Correspondence: Valter Gattei MD, Clinical and
Experimental Hematology Research Unit,
Centro di Riferimento Oncologico, IRCCS, Via
Pedemontana Occidentale, 12, Aviano (PN),
Italy. E-mail: [email protected]
Summary
Cases of B-cell chronic lymphocytic leukaemia (B-CLL) with mutated (M)
IgVH genes have a better prognosis than unmutated (UM) cases. We analysed
the IgVH mutational status of B-CLL according to the features of a canonical
somatic hypermutation (SHM) process, correlating this data with survival. In
a series of 141 B-CLLs, 124 cases were examined for IgVH gene per cent
mutations and skewing of replacement/silent mutations in the framework/
complementarity-determining regions as evidence of antigen-driven
selection; this identified three B-CLL subsets: significantly mutated (sM),
with evidence of antigen-driven selection, not significantly mutated (nsM)
and UM, without such evidence and IgVH gene per cent mutations above or
below the 2% cut-off. sM B-CLL patients had longer survival within the good
prognosis subgroup that had more than 2% mutations of IgVH genes. sM,
nsM and UM B-CLL were also characterized for the biased usage of IgVH
families, intraclonal IgVH gene diversification, preference of mutations to
target-specific nucleotides or hotspots, and for the expression of enzymes
involved in SHM (translesion DNA polymerase f and g and activation-
induced cytidine deaminase). These findings indicate the activation of a
canonical SHM process in nsM and sM B-CLLs and underscore the role of
the antigen in defining the specific clinical and biological features of B-CLL.
Keywords: chronic lymphocytic leukaemia, somatic hypermutation, antigen
selection, translesion DNA polymerases, prognosis.
research paper
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 doi:10.1111/j.1365-2141.2004.04985.x

structural integrity of Igs by concomitantly providing a high
mutational rate in antigen-binding regions (Chang & Casali,
1994; Dorner et al, 1998a; Lossos et al, 2000). Moreover, the
excess of transitions over transversions, as well as of mutations
targeting specific nucleotides or nucleotide motifs, may be
explained by the activity of a specific mutational machinery
(Smith et al, 1996; Dorner et al, 1998b; Neuberger et al, 1998).
This includes the DNA polymerases (pol) f and g, which are
efficient translesion and error-prone DNA polymerases, i.e.
capable of carrying out lesion bypassing DNA synthesis by
copying DNA with a high error rate (Zan et al, 2001; Zeng
et al, 2001; Storb & Stavnezer, 2002). Moreover, the activation-
induced cytidine deaminase (AID), a member of the cytidine
deaminase family exclusively expressed by GC B cells, is
another essential enzyme for both SHM and class switch
recombination of normal B cells (Muramatsu et al, 1999, 2000;
Cerutti et al, 2002; Storb & Stavnezer, 2002).
While some reports indicate a broad expression of AID in
B-CLL (Albesiano et al, 2003; Oppezzo et al, 2003), with the
highest levels being confined to UM CD38high subsets
(McCarthy et al, 2003; Degan et al, 2004), expression of
translesion DNA polymerases has not been yet investigated in
this disease. However, the finding of an intraclonal VHDJH
gene diversification in most B-CLL cases (Gurrieri et al, 2002)
suggests the presence of mutational machinery that is still
functionally active following neoplastic transformation. Finally,
evidence of antigen-driven selection in B-CLL, as evaluated by
computing the unbalanced distribution of R/silent (S) muta-
tions in CDR/FR, although reported by some studies (Hamblin
et al, 1999; Gurrieri et al, 2002; Messmer et al, 2004), has
never been correlated with survival.
In the present study, the IgVH mutational status was
investigated in a large cohort of B-CLLs by using a strategy of
cloning and sequencing of at least 5–10 transcripts for each
patient. By extending some of our preliminary observations
(Gattei et al, 2003), we demonstrated that the application of
statistical models aimed at evaluating the impact of positive
selection operated by the antigen (Chang & Casali, 1994;
Lossos et al, 2000) enabled the identification of three B-CLL
subsets, each with a different prognosis and usage of the
canonical SHM machinery.
Patients, materials and methods
B-CLL patients and cell samples and immunophenotypicalanalysis
The study was performed on peripheral blood (PB) samples,
collected after informed consent for routine diagnostic and
follow-up procedures, from 141 patients affected by typical B-
CLL. Diagnosis was made according to standard criteria, inclu-
ding expression of CD19, CD5, CD23 and low levels of clonally
restricted surface Igs (Matutes et al, 1994). The median age at
diagnosis was 64 years (range 32–97 years) and the male to female
ratio was 1Æ1 (75 males, 66 females). The clinical stage at diagnosis
according to Rai et al (1975) was available for 127 of 141 patients
as follows: stage 0, 33% (n ¼ 42); stage I, 39Æ4% (n ¼ 50);
stage II, 23Æ6% (n ¼ 30); stage III, 1Æ6% (n ¼ 2); stage IV, 2Æ4%
(n ¼ 3). Mononuclear cells were isolated by centrifugation on
Ficoll–Hypaque (Pharmacia, Uppsala, Sweden) gradient and
either used directly or cryopreserved in liquid nitrogen until use.
Expression of phenotypic markers, including CD38, was inves-
tigated by flow cytometry, as previously described (Gattei et al,
1997; Damle et al, 1999; Del Poeta et al, 2001). Cases in which
CD38 was detected in <30% of cells were identified as CD38low,
while cases with ‡30% CD38 B-CLL cells were considered
CD38high (Damle et al, 1999; Del Poeta et al, 2001).
Amplification, cloning and sequencing of VHDJH
transcripts
Total RNA was extracted and reverse-transcribed as described
(Gattei et al, 1997). The resulting cDNAs, checked for first-
strand synthesis (Degan et al, 2000), were amplified using a
mixture of sense primers annealing either to the VH1 through
VH6 leader sequences or to the 5¢-end of VH1–VH6 FR1
utilized in conjunction with a mixture of antisense primers
complementary to the germ line JH regions, as reported (Fais
et al, 1998; Damle et al, 1999; Hamblin et al, 1999; Gurrieri
et al, 2002; Krober et al, 2002). The purified amplified
products, inserted into the PCR2.1-TOPO vector (Invitrogen
S.R.L., San Giuliano Milanese, Milan, Italy), were expanded in
TOP10 One ShotTM competent cells (Invitrogen) and cloned.
Plasmid DNAs were isolated from overnight cultures of
randomly selected colonies and sequenced by using an
automatic DNA sequencer (ABI PRISM 3100, Applied
Biosystem, Foster City, CA, USA; Beckman CEQ2000XL,
Fullerton, CA, USA). When tested for the DNA polymerase
error rate according to Pasqualucci et al (1998), this strategy
yielded an incorporating error rate of 2Æ8 · 10)4/bp. In
additional experiments, plasmid DNA from two distinct
colonies from two different B-CLL patients (B176 and B211)
were amplified and cloned by exactly reproducing the proce-
dures described above. Among 20 sequenced colonies
(6732 bp), a single mutation was detected (DNA polymerase
error rate ¼ 1Æ5 · 10)4/bp). Comparisons between the ob-
tained sequences and those of the various germ line IgVHDJH
genes were performed with the Ig basic local alignment search
tool (IgBLAST) directory (http://www.ncbi.nlm.nih.gov/ig-
blast) utilizing the MacVector 7.1 sequence analysis software
(Accelrys; Symantec Co., Accelrys European Headquarters,
Cambridge, UK). According to this algorithm, a stretch of
seven or more nucleotides was required for D and JH
assignment. Only when the same rearrangement was identified
in at least 5–10 clones (three or four clones in 11 cases), a given
IgVH sequence was further analysed. As described (Gurrieri
et al, 2002), alignment of the IgVH sequences available for each
patient revealed, along with mutations shared by all the
transcripts analysed, a number of partially shared or unique
mutations. For this reason, all mutational analyses (see below)
M. Degan et al
30 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42

were carried out in each IgVH transcript separately, and the per
cent mutation assigned to a given B-CLL was the mean value of
the per cent mutations found in each individual transcript.
IgVH mutation analysis
A census of somatic point mutations was performed by solely
considering the IgVH sequence, i.e. from FR1 to FR3. As
evidence of antigen-driven selection (Chang & Casali, 1994;
Dorner et al, 1998a; Lossos et al, 2000; Gurrieri et al, 2002),
the probability that an excess or scarcity of R mutations
occurred by chance in IgVH CDR or FR, respectively, was
computed by applying the available binomial and multinomial
distribution models (Chang & Casali, 1994; Lossos et al, 2000).
Both these models calculate the expected frequency of
mutations by taking into account the base composition of a
given germ line IgVH sequence (Chang & Casali, 1994; Lossos
et al, 2000). In the absence of a positive selective pressure
operated by the antigen, nucleotide changes yielding R or
S mutation will be distributed throughout the coding sequence
with a pattern superimposible to that of randomly occurring
mutations (P > 0Æ05). Conversely, a skewing of R mutation
from FR to CDR and/or of S mutations from CDR to FR will
indicate a positive selection operated by the antigen; in this
case, the probability that these mutations are due by chance
will be <0Æ05 (Chang & Casali, 1994; Lossos et al, 2000). In the
present study, P-values were computed by applying the
multinomial distribution model, taking advantage of the
ready-to-use computer program at the http://www-stat.stan-
ford.edu/immunoglobulin website (Lossos et al, 2000). Due to
some flaws of the available JAVA applet, the multinomial
model was not appropriate when analysing specific IgVH (3–53
and 4–39, overall accounting for nine cases of 77 M B-CLL in
our series). In these cases, calculations of biased R/S mutations
were made according to the binomial distribution model
utilizing the reported formula (Chang & Casali, 1994). The
expected values of transitions or transversions, of mutations
targeting specific nucleotides as well as the RGYW (i.e. purine/
G/pyrimidine/A or T) or its inverse (WRCY) motifs were
computed as reported (Smith et al, 1996; Dorner et al, 1998b;
Neuberger et al, 1998).
Expression of translesion DNA polymerases g and f andAID
Expression of DNA pol g and f was evaluated by quantitative
real-time polymerase chain reaction (QRT-PCR), while AID
expression was investigated by conventional reverse transcrip-
tion-PCR (RT-PCR) on a selected cohort of B-CLL cases (52
cases). To avoid interference with normal B and T cells that
allegedly express these enzymes, only cases with more than 95%
of neoplastic cells in PB samples were analysed. Briefly, cDNAs
from B-CLL cells were amplified with primer pairs specific for
pol g, pol f and the housekeeping gene b2-microglobulin
(b2M), as described (Zan et al, 2001; Spessotto et al, 2002). The
incorporation of the SYBR� Green dye (Applied Biosystems)
into the PCR products was monitored in real-time with a
sequence detection system (ABI PRISM 7700), resulting in a
calculation of threshold cycles (Ct value), i.e. the PCR cycle
number corresponding to the beginning of the exponential
growth of PCR products. Ct values were converted into
attomoles by means of QRT-PCR experiments carried out
with serial dilution of known concentrations of pol g, pol f and
b2M-specific amplicons. Results were reported as the ratio of
the relative expression levels of pol g/b2M or pol f/b2M.
Activation-induced cytidine deaminase expression was
checked in the same cohort of cases essentially as described
(Albesiano et al, 2003; McCarthy et al, 2003; Oppezzo et al,
2003). Briefly, cDNAs were amplified with primers chosen to
anneal to sequences corresponding to the first (sense, position
45–64) and the fifth (antisense, position 675–653) exons of
AID-specific cDNA (Albesiano et al, 2003; McCarthy et al,
2003; Oppezzo et al, 2003). Amplified products (10 ll) were
run on 1Æ5% ethidium bromide-stained agarose gels and
analysed under ultra-violet light.
Statistical analysis
Complete survival data were available for 141 patients. The
mean follow-up was 67 months (range 8–220 months). Patient
survivals were analysed using the Kaplan–Meier survival curves
and log-rank test (Armitage & Berry, 1987). Correlation
between IgVH mutational status and CD38 expression was
performed by the chi-square test (Armitage & Berry, 1987).
Differences between expected and observed values of transi-
tions and transversions, mutations targeting specific nucleo-
tides, as well as RGYW/WRCY motifs in IgVH sequences were
assessed by the Student’s t-test for paired samples, by
separately evaluating each transcript (Armitage & Berry,
1987). Differences between the relative expression levels of
pol f/g ratios were assessed by the Student’s t-test for unpaired
samples. Statistical significance was always accepted when
P < 0Æ05.
Results
IgVH per cent mutations, biased IgVH usage, intraclonalIgVH diversification, CD38 expression and survivals
The mutational status of IgVH genes was investigated in 124 of
141 B-CLLs by sequencing at least 5–10 transcripts (three to
four in 11 cases) for each patient. Values of IgVH gene
mutations (Fig 1), represent the mean values of per cent
mutations due to a certain degree of intraclonal IgVH
diversification (see below). According to this analysis, 47
B-CLL cases (38%) had neoplastic cells bearing less than 2%
average mutations when compared with the nearest germ line
sequence (UM B-CLL), while 77 cases (62%) displayed ‡2%
average mutations and were classified as M B-CLLs (Damle
et al, 1999; Hamblin et al, 1999; Maloum et al, 2000; Krober
IgVH Gene Mutations and Prognosis in B-CLL
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 31

et al, 2002). Tables I and II summarize VHDJH assignments
and the means of per cent mutations for UM and M B-CLLs
respectively.
In agreement with previous reports (Keating et al, 2003;
Messmer et al, 2004), a higher number of B-CLLs expressing
the VH1 family was found in the UM, when compared with the
M, B-CLL subgroups (37Æ5% vs. 10Æ1%; Table III). Conversely,
M B-CLLs preferentially expressed VH genes belonging to the
VH3 family (58Æ2% vs. 31Æ3%; Table III). No differences were
observed between UM and M B-CLLs regarding the expression
of other IgVH families, including VH4 (about 20% in both
subgroups) and other rarely expressed IgVH genes, such as
VH2, VH5 and VH6 (Table III).
Table II also reports a detailed analysis of IgVH point
mutations for the M B-CLL group. According to Gurrieri et al
(2002), they were divided into ‘shared’ (i.e. mutations found in
all the IgVH transcripts analysed), ‘partially shared’ (i.e.
mutations present in a part of the analysed IgVH transcripts)
and ‘unique’ (i.e. mutations found only in distinct IgVH
transcripts) mutations (Tables II and IV). Among 79 IgVH
(449 IgVH transcripts), 63 (about 80%) exhibited either no
intraclonal IgVH diversification (i.e. shared mutations only, 13
IgVH, 16Æ5%) or a low number of unique and/or partially
shared mutations, always in the presence of a great excess of
shared mutations (Tables II and IV). Conversely, in 16 of 79
IgVH, the sum of unique and partially shared mutations
exceeded the number of shared mutations (Tables II and IV),
thus revealing a higher degree of intraclonal IgVH diversifica-
tion.
Data on CD38 expression was available for 140 of 141
patients, of which 51 cases (36Æ4%) were CD38high B-CLL, and
the remaining 89 cases (63Æ6%) were classified as CD38low
B-CLL (Damle et al, 1999; Del Poeta et al, 2001). In agreement
with some of the current literature (Damle et al, 1999), we
found a statistically significant correlation between CD38high
and low mutations of IgVH genes by applying a chi-square test
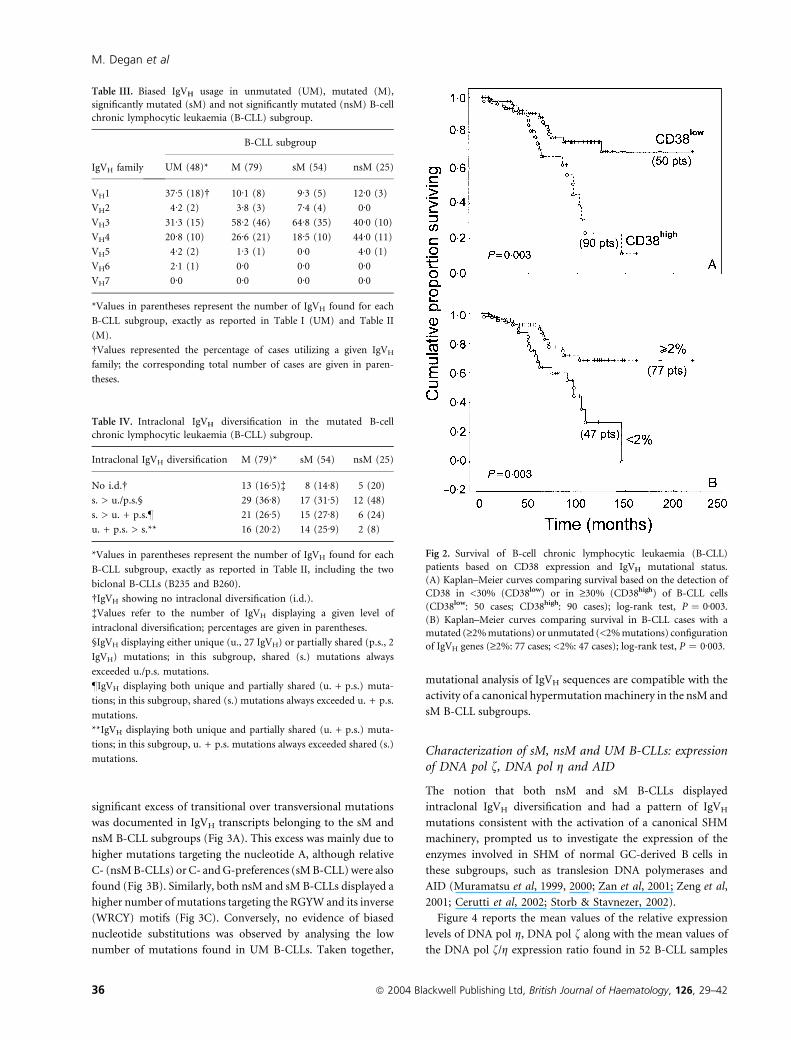
(P < 0Æ01; Fig 1).
A comparison of patient survivals with CD38 expression
(140 cases) or IgVH mutational status (124 cases) also
demonstrated the positive impact of CD38low and IgVH
mutations above the standard cut-off of 2% on prognosis
(Fig 2) (Damle et al, 1999; Del Poeta et al, 2001; Krober et al,
2002; Lin et al, 2002).
Altogether, IgVH per cent mutations and usage, intraclonal
IgVH diversification, CD38 expression and survivals, as found
on our cohort of B-CLL patients, were in keeping with the
above reports.
Evidence of antigen-driven selection identified three B-CLLsubsets
The hypothesis that M B-CLL cases may derive as a transfor-
mation of a competent B cell (Naylor & Capra, 1999), led us to
investigate whether the IgVH gene mutational status found in
Fig 1. CD38 expression and IgVH mutational status in B-cell chronic lymphocytic leukaemia (B-CLL). B-CLL cases were arranged according to the
IgVH mutational status, expressed as per cent mutation (upper panel); the lower panel indicates, for each B-CLL case, the corresponding expression
level for CD38, reported as percentage of positive B cells. Cases for which IgVH mutational status and CD38 expression data were not simultaneously
available are reported as open histograms. Dotted lines refer to the established cut-offs of 2% mutation (upper panel) and 30% CD38+ cells (lower
panel). Asterisks indicate three cases (B241, B235, B260) with biclonal disease, whose per cent mutations were separately reported. A chi-square
correlation between IgVH mutational status (cut-off: 2% mutations) and CD38 expression (cut-off: 30% of positive cells) is also reported (P < 0Æ01).
n.a., not available.
M. Degan et al
32 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42

B-CLL cells was consistent with that of B cells positively
selected by the antigen (Chang & Casali, 1994; Dorner et al,
1998a; Lossos et al, 2000). Within IgVH genes, sequences
encoding for the antigen-contacting CDRs are inherently more
prone to R mutations than those encoding for the respective
FRs (Chang & Casali, 1994), this susceptibility being further
stressed by antigen selection (Chang & Casali, 1994; Dorner
et al, 1998a; Lossos et al, 2000). In our study, among 77 M
B-CLLs, 449 IgVH transcripts were analysed by applying the
multinomial distribution model to assess the probability that
biased R/S mutations in IgVH gene CDR/FR arise randomly
(Lossos et al, 2000). When the multinomial model gave a
result that was not appropriate (52/449 IgVH transcripts,
11Æ5%), we took advantage of the binomial distribution model
according to Chang and Casali (1994). Overall, a given IgVH
transcript was defined as having evidence of antigen-driven
selection if the applied statistical model yielded a significant
result (P < 0Æ05) either in CDRs or FRs (Dorner et al, 1998a;
Sakai et al, 2000).
By using such an approach, we identified 52 of 77 M B-CLLs
for which evidence of antigen-driven selection was found in all
or in part of the IgVH transcripts analysed (Table II). These
B-CLL cases were classified as ‘significantly mutated’ (sM
B-CLL). Table II summarizes the characteristics of the sM B-CLL
cases, including VHDJH gene assignment, per cent mutations,
number of IgVH transcripts analysed and number of IgVH
transcripts showing evidence of antigen-driven selection in
CDR and/or FR. Among 289 analysed IgVH transcripts
(average 5Æ7, range 3–10), 247 showed a significant excess of
R mutations in CDR (25 IgVH transcripts) or a scarcity of
R mutations in FR (114 IgVH transcripts) or both (108 IgVH
transcripts), while the remaining 42 IgVH transcripts did not
display evidence of antigen-driven selection. Taken together,
36 of 52 sM B-CLLs (69Æ2%) showed evidence of antigen-
driven selection in all of the transcripts analysed, while in 16
cases this evidence was documented only in some IgVH
transcripts (Table II). The remaining B-CLL cases were
classified as ‘not significantly mutated’ (nsM, 25 cases) or
‘unmutated’ (UM, 47 cases) when the per cent mismatch was
above or below the 2% cut-off respectively. For all these cases,
the average number of IgVH transcripts analysed for each
B-CLL patients was 6Æ1 (range 5–10) and none showed
evidence of antigen-driven selection using the two available
statistical models.
Characterization of sM and nsM B-CLLs: biased IgVH
usage and intraclonal IgVH diversification
The sM subgroup of B-CLL cases displayed a pattern of IgVH
usage similar to that reported for M B-CLLs (Table III).
However, when compared with M B-CLL, a more evident
skewing towards the preferential usage of IgVH3 family was
documented in sM B-CLL. In fact, almost 2/3 of sM B-CLLs
rearranged IgVH genes of the VH3 family, whereas nsM B-CLLs
Table I. Mutation characteristics of IgVHDJH genes in the unmutated
B-cell chronic lymphocytic leukaemia (B-CLL) subgroup.
CLL case
VHDJH gene
% mutation*VH D JH
B 48 1-18 3-10 4 0Æ0B 201 3-15 3-3 4 0Æ0B 206 3-11 6-6 6 0Æ0B 265 3-73 3-22 3 0Æ0B 210 3-53 3-3 4 0Æ1B 16 3-13 3-3 4 0Æ2B 209 1-2 6-19 4 0Æ2B 232 4-39 6-13 5 0Æ2B 169 1-2 1-26 4 0Æ3B 205 3-21 5-24 6 0Æ3B 47 1-18 3-10 4 0Æ4B 149 4-59 3-22 6 0Æ4B 42 4-39 6-13 5 0Æ5B 255 5-51 3-22 4 0Æ5B 249 5-51 3-22 4 0Æ5B 202 1-69 3-16 3 0Æ6B 179 4-39 6-13 4 0Æ7B 191 3-11 3-9 6 0Æ7B 241 2-26 6-13 4 0Æ7
2-5 1-7 4 1Æ9B 116 1-69 2-2 2 0Æ8B 173 3-30 7-21 1 0Æ8B 220 1-69 3-22 6 0Æ8B 128 3-33 3-3 3 0Æ9B 159 6-1 2-2 4 0Æ9B 21 3-23 5-10 4 0Æ9B 196 4-34 6-13 6 1Æ0B 223 1-69 n.a. 3 1Æ0B 118 1-2 2-21 6 1Æ1B 18 1-2 1-26 6 1Æ1B 176 1-69 6-13 3 1Æ1B 83 3-23 2-8 6 1Æ1B 119 4-31 3-3 5 1Æ1B 166 4-39 4-17 5 1Æ1B 158 1-69 7-27 1 1Æ1B 244 3-30 n.a. 5 1Æ1B 161 4-59 3-22 4 1Æ2B 183 1-69 3-16 4 1Æ3B 245 3-33 7-27 4 1Æ3B 174 1-69 3-3 6 1Æ3B 17 1-46 3-10 6 1Æ4B 14 4-34 3-3 6 1Æ6B 110 1-18 6-19 4 1Æ6B 15 3-21 5-18 6 1Æ6B 184 3-48 2-8 4 1Æ7B 165 1-2 3-10 5 1Æ7B 39 1-2 6-13 4 1Æ7B 75 4-39 2-8 4 1Æ7
n.a., not assigned.
*Values represent the mean value of 5–10 transcripts analysed for each
B-CLL case.
IgVH Gene Mutations and Prognosis in B-CLL
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 33

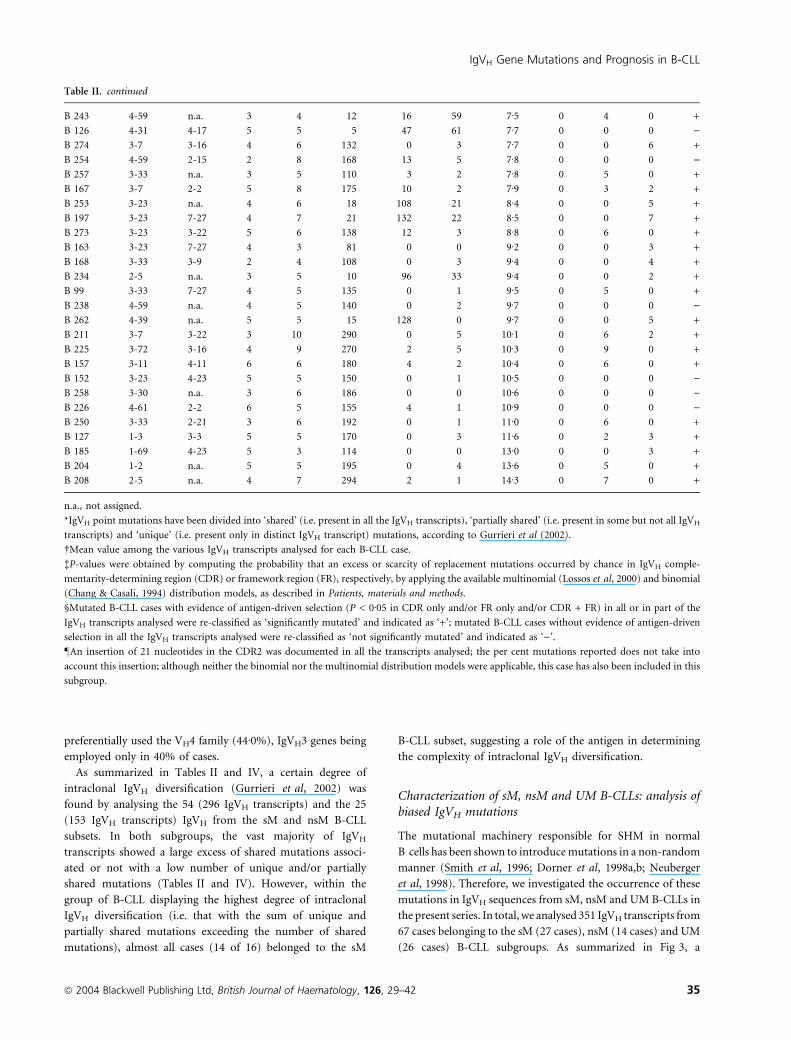
Table II. Mutation characteristics of IgVHDJH genes in the mutated B-cell chronic lymphocytic leukaemia (B-CLL) subgroup.
CLL case
VHDJH genesTranscripts
analysed
IgVH point mutations*
% mutation�
IgVH transcripts with P < 0Æ05�
Ag selection§VH D JH Shared Partially shared Unique CDR only FR only CDR + FR
B 186 1-69 3-16 4 5 30 0 0 2Æ0 0 0 0 )B 49 3-30 3-21 6 5 10 16 4 2Æ1 0 0 0 )B 180 3-23 2-15 5 7 42 0 0 2Æ2 n.a.– n.a. n.a. +
B 153 3-30 5-12 4 4 20 3 2 2Æ3 0 1 1 +
B 216 3-53 7-27 4 7 49 0 0 2Æ4 0 0 0 )B 217 4-39 3-22 4 7 49 0 1 2Æ4 7 0 0 +
B 224 3-33 3-9 3 5 35 2 1 2Æ6 0 0 0 )B 67 1-46 3-10 6 6 6 21 25 2Æ9 0 0 2 +
B 156 1-58 3-10 3 8 64 7 0 3Æ0 0 0 0 )B 182 3-53 4-11 4 5 45 4 3 3Æ4 5 0 0 +
B 227 3-48 6-19 5 4 40 0 0 3Æ4 0 4 0 +
B 218 3-64 2-2 6 10 90 11 2 3Æ5 0 0 0 )B 188 3-7 4-23 3 5 55 0 0 3Æ7 0 0 0 )B 194 4-39 6-25 4 5 55 0 2 3Æ7 0 0 0 )B 195 4-39 6-25 4 5 55 0 4 3Æ9 0 0 0 )B 219 3-48 n.a. 5 5 13 33 13 4Æ0 0 3 0 +
B 68 4-34 6-19 4 5 60 0 1 4Æ1 0 0 0 )B 189 3-21 n.a. 6 5 50 6 5 4Æ2 0 0 0 )B 114 3-48 6-19 5 6 66 4 2 4Æ2 0 3 2 +
B 98 4-28 3-22 3 7 91 0 0 4Æ4 0 0 7 +
B 125 4-34 6-19 4 6 78 0 1 4Æ6 0 0 0 )B 236 3-48 5-24 4 6 36 35 10 4Æ6 0 6 0 +
B 154 4-39 6-13 4 5 65 0 3 4Æ7 0 0 0 )B 261 3-21 6-13 4 5 60 4 5 4Æ7 0 0 5 +
B 229 3-23 5-12 5 4 56 0 0 4Æ8 0 0 4 +
B 231 3-23 5-12 5 6 84 0 1 4Æ8 0 0 6 +
B 198 3-48 3-22 4 6 84 2 0 4Æ9 0 0 6 +
B 221 3-7 3-3 4 6 84 0 6 5Æ1 0 0 0 )B 164 3-21 5-24 6 3 42 0 3 5Æ1 0 0 3 +
B 177 3-23 2-15 5 4 56 0 3 5Æ1 4 0 0 +
B 215 4-4 7-27 4 10 150 0 3 5Æ2 0 3 0 +
B 240 3-11 3-21 4 7 105 0 5 5Æ4 0 0 7 +
B 207 4-59 2-15 3 9 144 0 2 5Æ5 0 0 0 )B 242 2-5 n.a. 5 5 10 50 22 5Æ5 0 1 1 +
B 80 4-28 3-3 4 5 85 0 0 5Æ7 0 0 5 +
B 13 4-28 3-10 5 6 96 5 2 5Æ8 3 2 2 +
B 235 4-59 2-15 2 5 85 0 1 5Æ9 0 5 0 +
3-9 3-3 6 4 132 0 1 11Æ4 4 0 0 +
B 222 3-72 2-21 4 5 85 2 3 6Æ0 0 4 0 +
B 84 5-51 3-3 4 5 90 0 0 6Æ1 0 0 0 )B 181 4-31 7-27 4 5 35 53 0 6Æ1 0 5 0 +
B 264 3-33 n.a. 6 5 5 64 22 6Æ2 0 0 1 +
B 263 4-39 n.a. 4 6 12 81 19 6Æ3 1 0 3 +
B 259 4-39 2-8 3 7 125 0 2 6Æ4 0 0 0 )B 239 1-3 6-13 4 9 171 0 4 6Æ7 0 0 0 )B 230 2-5 n.a. 4 5 95 0 6 6Æ7 0 1 0 +
B 260 3-33 n.a. 3 6 24 48 46 6Æ7 0 2 3 +
1-69 n.a. 4 5 15 48 55 8Æ0 0 0 4 +
B 237 3-7 3-4 4 5 95 4 2 6Æ9 0 3 2 +
B 107 3-23 5-24 4 4 64 9 11 7Æ1 0 4 0 +
B 271 3-23 7-27 4 7 140 6 4 7Æ3 0 0 0 )B 170 3-15 3-3 3 3 66 0 0 7Æ3 0 3 0 +
B 155 3-11 2-21 6 5 100 4 1 7Æ3 1 0 2 +
M. Degan et al
34 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42
preferentially used the VH4 family (44Æ0%), IgVH3 genes being
employed only in 40% of cases.
As summarized in Tables II and IV, a certain degree of
intraclonal IgVH diversification (Gurrieri et al, 2002) was
found by analysing the 54 (296 IgVH transcripts) and the 25
(153 IgVH transcripts) IgVH from the sM and nsM B-CLL
subsets. In both subgroups, the vast majority of IgVH
transcripts showed a large excess of shared mutations associ-
ated or not with a low number of unique and/or partially
shared mutations (Tables II and IV). However, within the
group of B-CLL displaying the highest degree of intraclonal
IgVH diversification (i.e. that with the sum of unique and
partially shared mutations exceeding the number of shared
mutations), almost all cases (14 of 16) belonged to the sM
B-CLL subset, suggesting a role of the antigen in determining
the complexity of intraclonal IgVH diversification.
Characterization of sM, nsM and UM B-CLLs: analysis ofbiased IgVH mutations
The mutational machinery responsible for SHM in normal
B cells has been shown to introduce mutations in a non-random
manner (Smith et al, 1996; Dorner et al, 1998a,b; Neuberger
et al, 1998). Therefore, we investigated the occurrence of these
mutations in IgVH sequences from sM, nsM and UM B-CLLs in
the present series. In total, we analysed 351 IgVH transcripts from
67 cases belonging to the sM (27 cases), nsM (14 cases) and UM
(26 cases) B-CLL subgroups. As summarized in Fig 3, a
Table II. continued
B 243 4-59 n.a. 3 4 12 16 59 7Æ5 0 4 0 +
B 126 4-31 4-17 5 5 5 47 61 7Æ7 0 0 0 )B 274 3-7 3-16 4 6 132 0 3 7Æ7 0 0 6 +
B 254 4-59 2-15 2 8 168 13 5 7Æ8 0 0 0 )B 257 3-33 n.a. 3 5 110 3 2 7Æ8 0 5 0 +
B 167 3-7 2-2 5 8 175 10 2 7Æ9 0 3 2 +
B 253 3-23 n.a. 4 6 18 108 21 8Æ4 0 0 5 +
B 197 3-23 7-27 4 7 21 132 22 8Æ5 0 0 7 +
B 273 3-23 3-22 5 6 138 12 3 8Æ8 0 6 0 +
B 163 3-23 7-27 4 3 81 0 0 9Æ2 0 0 3 +
B 168 3-33 3-9 2 4 108 0 3 9Æ4 0 0 4 +
B 234 2-5 n.a. 3 5 10 96 33 9Æ4 0 0 2 +
B 99 3-33 7-27 4 5 135 0 1 9Æ5 0 5 0 +
B 238 4-59 n.a. 4 5 140 0 2 9Æ7 0 0 0 )B 262 4-39 n.a. 5 5 15 128 0 9Æ7 0 0 5 +
B 211 3-7 3-22 3 10 290 0 5 10Æ1 0 6 2 +
B 225 3-72 3-16 4 9 270 2 5 10Æ3 0 9 0 +
B 157 3-11 4-11 6 6 180 4 2 10Æ4 0 6 0 +
B 152 3-23 4-23 5 5 150 0 1 10Æ5 0 0 0 )B 258 3-30 n.a. 3 6 186 0 0 10Æ6 0 0 0 )B 226 4-61 2-2 6 5 155 4 1 10Æ9 0 0 0 )B 250 3-33 2-21 3 6 192 0 1 11Æ0 0 6 0 +
B 127 1-3 3-3 5 5 170 0 3 11Æ6 0 2 3 +
B 185 1-69 4-23 5 3 114 0 0 13Æ0 0 0 3 +
B 204 1-2 n.a. 5 5 195 0 4 13Æ6 0 5 0 +
B 208 2-5 n.a. 4 7 294 2 1 14Æ3 0 7 0 +
n.a., not assigned.
*IgVH point mutations have been divided into ‘shared’ (i.e. present in all the IgVH transcripts), ‘partially shared’ (i.e. present in some but not all IgVH
transcripts) and ‘unique’ (i.e. present only in distinct IgVH transcript) mutations, according to Gurrieri et al (2002).
�Mean value among the various IgVH transcripts analysed for each B-CLL case.
�P-values were obtained by computing the probability that an excess or scarcity of replacement mutations occurred by chance in IgVH comple-
mentarity-determining region (CDR) or framework region (FR), respectively, by applying the available multinomial (Lossos et al, 2000) and binomial
(Chang & Casali, 1994) distribution models, as described in Patients, materials and methods.
§Mutated B-CLL cases with evidence of antigen-driven selection (P < 0Æ05 in CDR only and/or FR only and/or CDR + FR) in all or in part of the
IgVH transcripts analysed were re-classified as ‘significantly mutated’ and indicated as ‘+’; mutated B-CLL cases without evidence of antigen-driven
selection in all the IgVH transcripts analysed were re-classified as ‘not significantly mutated’ and indicated as ‘)’.
–An insertion of 21 nucleotides in the CDR2 was documented in all the transcripts analysed; the per cent mutations reported does not take into
account this insertion; although neither the binomial nor the multinomial distribution models were applicable, this case has also been included in this
subgroup.
IgVH Gene Mutations and Prognosis in B-CLL
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 35
significant excess of transitional over transversional mutations
was documented in IgVH transcripts belonging to the sM and
nsM B-CLL subgroups (Fig 3A). This excess was mainly due to
higher mutations targeting the nucleotide A, although relative
C- (nsM B-CLLs) or C- and G-preferences (sM B-CLL) were also
found (Fig 3B). Similarly, both nsM and sM B-CLLs displayed a
higher number of mutations targeting the RGYW and its inverse
(WRCY) motifs (Fig 3C). Conversely, no evidence of biased
nucleotide substitutions was observed by analysing the low
number of mutations found in UM B-CLLs. Taken together,
mutational analysis of IgVH sequences are compatible with the
activity of a canonical hypermutation machinery in the nsM and
sM B-CLL subgroups.
Characterization of sM, nsM and UM B-CLLs: expressionof DNA pol f, DNA pol g and AID
The notion that both nsM and sM B-CLLs displayed
intraclonal IgVH diversification and had a pattern of IgVH
mutations consistent with the activation of a canonical SHM
machinery, prompted us to investigate the expression of the
enzymes involved in SHM of normal GC-derived B cells in
these subgroups, such as translesion DNA polymerases and
AID (Muramatsu et al, 1999, 2000; Zan et al, 2001; Zeng et al,
2001; Cerutti et al, 2002; Storb & Stavnezer, 2002).
Figure 4 reports the mean values of the relative expression
levels of DNA pol g, DNA pol f along with the mean values of
the DNA pol f/g expression ratio found in 52 B-CLL samples
Table III. Biased IgVH usage in unmutated (UM), mutated (M),
significantly mutated (sM) and not significantly mutated (nsM) B-cell
chronic lymphocytic leukaemia (B-CLL) subgroup.
IgVH family
B-CLL subgroup
UM (48)* M (79) sM (54) nsM (25)
VH1 37Æ5 (18)� 10Æ1 (8) 9Æ3 (5) 12Æ0 (3)
VH2 4Æ2 (2) 3Æ8 (3) 7Æ4 (4) 0Æ0VH3 31Æ3 (15) 58Æ2 (46) 64Æ8 (35) 40Æ0 (10)
VH4 20Æ8 (10) 26Æ6 (21) 18Æ5 (10) 44Æ0 (11)
VH5 4Æ2 (2) 1Æ3 (1) 0Æ0 4Æ0 (1)
VH6 2Æ1 (1) 0Æ0 0Æ0 0Æ0VH7 0Æ0 0Æ0 0Æ0 0Æ0
*Values in parentheses represent the number of IgVH found for each
B-CLL subgroup, exactly as reported in Table I (UM) and Table II
(M).
�Values represented the percentage of cases utilizing a given IgVH
family; the corresponding total number of cases are given in paren-
theses.
Table IV. Intraclonal IgVH diversification in the mutated B-cell
chronic lymphocytic leukaemia (B-CLL) subgroup.
Intraclonal IgVH diversification M (79)* sM (54) nsM (25)
No i.d.� 13 (16Æ5)� 8 (14Æ8) 5 (20)
s. > u./p.s.§ 29 (36Æ8) 17 (31Æ5) 12 (48)
s. > u. + p.s.– 21 (26Æ5) 15 (27Æ8) 6 (24)
u. + p.s. > s.** 16 (20Æ2) 14 (25Æ9) 2 (8)
*Values in parentheses represent the number of IgVH found for each
B-CLL subgroup, exactly as reported in Table II, including the two
biclonal B-CLLs (B235 and B260).
�IgVH showing no intraclonal diversification (i.d.).
�Values refer to the number of IgVH displaying a given level of
intraclonal diversification; percentages are given in parentheses.
§IgVH displaying either unique (u., 27 IgVH) or partially shared (p.s., 2
IgVH) mutations; in this subgroup, shared (s.) mutations always
exceeded u./p.s. mutations.
–IgVH displaying both unique and partially shared (u. + p.s.) muta-
tions; in this subgroup, shared (s.) mutations always exceeded u. + p.s.
mutations.
**IgVH displaying both unique and partially shared (u. + p.s.) muta-
tions; in this subgroup, u. + p.s. mutations always exceeded shared (s.)
mutations.
Fig 2. Survival of B-cell chronic lymphocytic leukaemia (B-CLL)
patients based on CD38 expression and IgVH mutational status.
(A) Kaplan–Meier curves comparing survival based on the detection of
CD38 in <30% (CD38low) or in ‡30% (CD38high) of B-CLL cells
(CD38low: 50 cases; CD38high: 90 cases); log-rank test, P ¼ 0Æ003.
(B) Kaplan–Meier curves comparing survival in B-CLL cases with a
mutated (‡2% mutations) or unmutated (<2% mutations) configuration
of IgVH genes (‡2%: 77 cases; <2%: 47 cases); log-rank test, P ¼ 0Æ003.
M. Degan et al
36 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42

(UM ¼ 21 cases; nsM ¼ 10 cases; sM ¼ 21 cases), all selected
as having a very low contamination by normal B and T cells.
Notwithstanding a certain heterogeneity also within the same
B-CLL subgroup, significantly higher expression levels of DNA
pol g could be observed both in the nsM and sM B-CLL
subgroups, when compared with UM B-CLLs (Fig 4). Con-
versely, the mean value of DNA pol f expression did not change
in the various subgroups (Fig 4), hence the DNA pol f/gexpression ratio, which was high in UM B-CLLs, was signifi-
cantly down-regulated both in sM and nsM B-CLLs (Fig 4).
Expression of AID was investigated by a conventional
RT-PCR strategy, utilizing primers located in exons 1 and 5 of
the AID gene. As shown in Fig 5 this strategy identified, in all
AID-positive cases, four spliced variants: (i) a 924-bp fragment
corresponding to the described high-molecular weight variant
and due to the retention of intron 4; (ii) a 631-bp fragment
corresponding to the wild-type form of AID; (iii) a fragment of
601 bp due to a deletion of 30 nucleotides of the initial portion
of exon 4; and (iv) a 515-bp fragment due to the omission of
exon 4 (Albesiano et al, 2003; McCarthy et al, 2003; Oppezzo
et al, 2003; Degan et al, 2004). AID expression was found in 21
(40%) of 52 B-CLL cases; when stratified by IgVH mutational
status, AID-positive cases accounted for 57% (12/21) of UM B-
CLLs and 29% (9/31) of the sM and nsM B-CLL cases together,
Fig 3. Mutational analysis of IgVH sequences in unmutated (UM), not
significantly mutated (nsM) and significantly mutated (sM) B-cell
chronic lymphocytic leukaemia (B-CLL). (A) Comparison between
expected and observed number of transitions (trs) or transversions
(trv). The expected number (open bars) of trs or trv was computed by
considering that randomly occurring point mutations should be 1/3
transitions and 2/3 transversions. Closed bars refer to observed
mutations found in the three B-CLL subgroups. (B) Comparison be-
tween expected and observed number of mutations targeting specific
nucleotides in transitional mutations. The expected number of muta-
tions targeting specific nucleotides (open bars) was computed as fol-
lows: frequency of occurrence of the nucleotide of interest in a given
germ line IgVH sequence multiplied by the total number of observed
mutations and the result divided by three, as, theoretically, each
nucleotide has the same probability to be mutated into one of the other
three nucleotides. Closed bars refer to observed mutations found in the
three B-CLL subgroups. (C) The expected frequency of mutations
targeting the RGYW (i.e. purine/G/pyrimidine/A or T) or its inverse
(WRCY) motifs (open bars) is the percentage of RGYW/WRCY motifs
in a given IgVH sequence. The actual frequency of mutations targeting
the motifs of interest (closed bars) was computed by taking into ac-
count the total number of mutations in a given IgVH sequence and the
number of mutations targeting the motifs. Differences between
expected and observed values of transitions and transversions, muta-
tions targeting specific nucleotides, as well as RGYW/WRCY motifs in
IgVH sequences were assessed by means of the Student’s t-test for
paired samples by separately evaluating each transcript. Asterisks
indicate conditions in which were statistically significant (P < 0Æ05).
Fig 4. Expression of translesion DNA pol g and f by QRT-PCR.
cDNAs from unmutated (UM), not significantly mutated (nsM) and
significantly mutated (sM) B-cell chronic lymphocytic leukaemia
(B-CLL) (52 cases) were amplified with primer pairs specific for pol g,
pol f and the housekeeping gene b2M. The incorporation of the
SYBR� Green dye into the PCR products was monitored in real-time
and the resulting threshold cycles (Ct; i.e. the PCR cycle number
corresponding to the beginning of the exponential growth of PCR
products) were computed. Ct values were converted into attomoles by
means of QRT-PCR experiments carried out with serial dilution of
known concentrations of pol g, pol f and b2M-specific amplicons.
Results for each B-CLL subgroup represent the mean values of the
relative expression levels of pol g/b2M (open bars), pol f/b2M (striped
bars) and pol f/g ratio (closed bars). Reported P-values refer to dif-
ferences between the relative expression levels of pol f/g ratios, as
assessed by the Student’s t-test for unpaired samples.
IgVH Gene Mutations and Prognosis in B-CLL
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 37
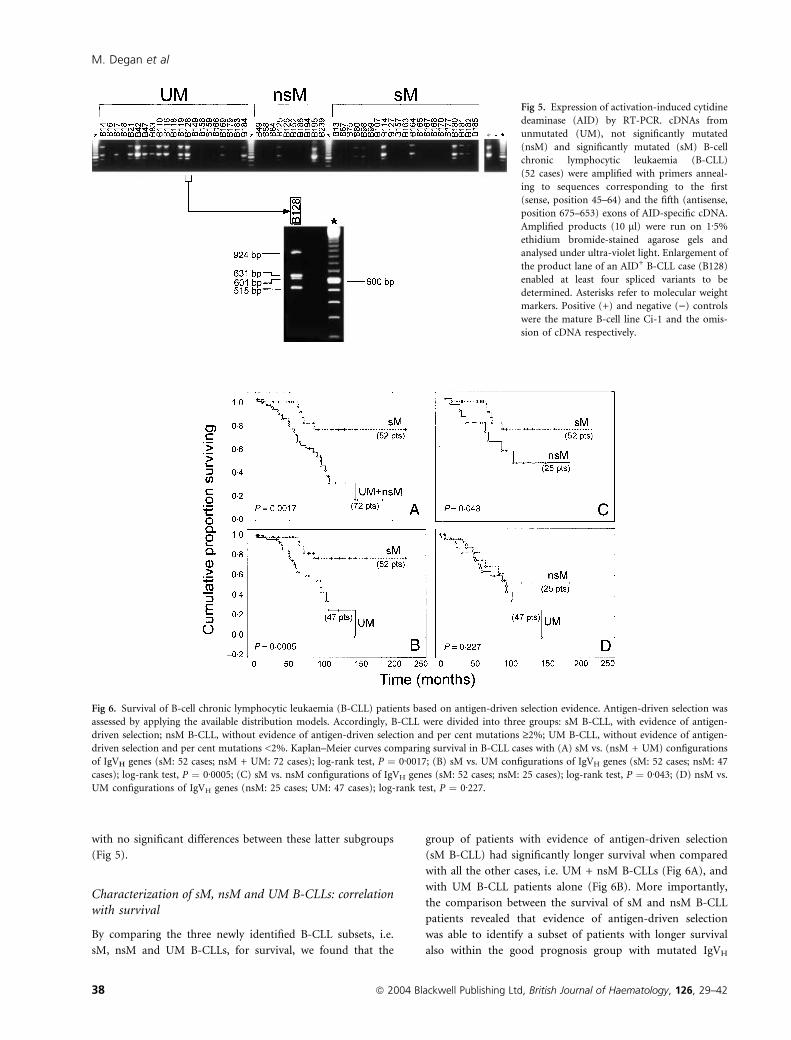
with no significant differences between these latter subgroups
(Fig 5).
Characterization of sM, nsM and UM B-CLLs: correlationwith survival
By comparing the three newly identified B-CLL subsets, i.e.
sM, nsM and UM B-CLLs, for survival, we found that the
group of patients with evidence of antigen-driven selection
(sM B-CLL) had significantly longer survival when compared
with all the other cases, i.e. UM + nsM B-CLLs (Fig 6A), and
with UM B-CLL patients alone (Fig 6B). More importantly,
the comparison between the survival of sM and nsM B-CLL
patients revealed that evidence of antigen-driven selection
was able to identify a subset of patients with longer survival
also within the good prognosis group with mutated IgVH
Fig 6. Survival of B-cell chronic lymphocytic leukaemia (B-CLL) patients based on antigen-driven selection evidence. Antigen-driven selection was
assessed by applying the available distribution models. Accordingly, B-CLL were divided into three groups: sM B-CLL, with evidence of antigen-
driven selection; nsM B-CLL, without evidence of antigen-driven selection and per cent mutations ‡2%; UM B-CLL, without evidence of antigen-
driven selection and per cent mutations <2%. Kaplan–Meier curves comparing survival in B-CLL cases with (A) sM vs. (nsM + UM) configurations
of IgVH genes (sM: 52 cases; nsM + UM: 72 cases); log-rank test, P ¼ 0Æ0017; (B) sM vs. UM configurations of IgVH genes (sM: 52 cases; nsM: 47
cases); log-rank test, P ¼ 0Æ0005; (C) sM vs. nsM configurations of IgVH genes (sM: 52 cases; nsM: 25 cases); log-rank test, P ¼ 0Æ043; (D) nsM vs.
UM configurations of IgVH genes (nsM: 25 cases; UM: 47 cases); log-rank test, P ¼ 0Æ227.
Fig 5. Expression of activation-induced cytidine
deaminase (AID) by RT-PCR. cDNAs from
unmutated (UM), not significantly mutated
(nsM) and significantly mutated (sM) B-cell
chronic lymphocytic leukaemia (B-CLL)
(52 cases) were amplified with primers anneal-
ing to sequences corresponding to the first
(sense, position 45–64) and the fifth (antisense,
position 675–653) exons of AID-specific cDNA.
Amplified products (10 ll) were run on 1Æ5%
ethidium bromide-stained agarose gels and
analysed under ultra-violet light. Enlargement of
the product lane of an AID+ B-CLL case (B128)
enabled at least four spliced variants to be
determined. Asterisks refer to molecular weight
markers. Positive (+) and negative ()) controls
were the mature B-cell line Ci-1 and the omis-
sion of cDNA respectively.
M. Degan et al
38 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42

gene configuration (Fig 6C). Consistently, no differences in
terms of overall survival were detected between those patients
affected by a disease characterized by more than 2% IgVH
gene mutations but lacking evidence of antigen-driven
selection (nsM B-CLL) and the bad prognosis B-CLL patient
subset displaying less than 2% mutations of IgVH genes
(Fig 6D).
Discussion
By combining the analysis of IgVH gene per cent mutations
with the application of algorithms evaluating skewing of R/S
mutations in FR/CDR as evidence of antigen-driven selection
(Chang & Casali, 1994; Smith et al, 1996; Dorner et al,
1998a,b; Neuberger et al, 1998; Lossos et al, 2000) in a large
series of B-CLL cases, we were able to identify at least three
B-CLL subsets with different prognoses and usage of the
canonical SHM machinery.
The strategy chosen for determining the IgVH mutational
status included the use of RNA as source material and the
sequencing of RT-PCR products upon their cloning into
appropriate plasmids. This approach, although still not widely
applied (Damle et al, 1999; Hamblin et al, 1999; Maloum et al,
2000; Jelinek et al, 2001; Krober et al, 2002; Lin et al, 2002;
McCarthy et al, 2003; Oppezzo et al, 2003), has the advantages
of reducing the possibility of amplifying aberrantly rearranged
VH genes (Hamblin et al, 1999) and of revealing those cases in
which an intraclonal IgVH diversification has occurred (Gurrieri
et al, 2002). In this regard, we found that a certain degree of
intraclonal IgVH diversification was a common event in M B-
CLL, occurring in about 80% of cases in our series. These results
are consistent with those reported by Gurrieri et al (2002),
although obtained by using a different approach. The possibility
that our finding might be biased by errors introduced by our
PCR and cloning strategy could be ruled out by the observation
that this approach yielded a DNA polymerase error rate of less
than one base out of 3000–6000 bp. In keeping with previous
studies (Damle et al, 1999; Hamblin et al, 1999; Maloum et al,
2000; Krober et al, 2002; Lin et al, 2002), our results included a
significant correlation between low mutations and CD38high
expression (Damle et al, 1999; Del Poeta et al, 2001; Lin et al,
2002). CD38, however, could not be considered a reliable
surrogate of IgVH mutational status (Hamblin et al, 2000;
Matrai et al, 2001; Thunberg et al, 2001) also in our study. In
addition, both CD38 and IgVH mutations had prognostic
relevance as, at the respective standard cut-off values of 30%
positive cells and 2% mutations, we were able to split patients
into subgroups with different survivals, as previously reported
by several authors (Damle et al, 1999; Hamblin et al, 1999; Del
Poeta et al, 2001; Jelinek et al, 2001; Matrai et al, 2001; Krober
et al, 2002; Lin et al, 2002).
The usage of a certain value of IgVH per cent mutations to
separate M from UM B-CLL cases (Damle et al, 1999; Hamblin
et al, 1999), surely of undiscussed clinical relevance, represents
an arbitrary method that only partially takes into account the
physiology of the SHM process, as it occurs during
T-dependent GC differentiation of normal B cells (Chang &
Casali, 1994; Smith et al, 1996; Dorner et al, 1998a,b; Neu-
berger et al, 1998; Lossos et al, 2000). Since IgVH mutations,
introduced as the result of a physiological SHM process, fulfil
precise criteria due to the antigenic stimulation acting in
concert with the hypermutation machinery (Chang & Casali,
1994; Smith et al, 1996; Dorner et al, 1998a,b; Neuberger et al,
1998; Lossos et al, 2000), we speculated that these factors
might be important in defining specific clinical and biological
features of B-CLL. Thus, we investigated whether our B-CLL
cases displayed evidence of positive selection operated by the
antigen by evaluating the excess and/or scarcity of R mutations
in CDR or FR of a given IgVH sequence respectively. This was
accomplished by utilizing a multinomial statistical algorithm
(Dorner et al, 1998a; Lossos et al, 2000; Sakai et al, 2000) or
the binomial distribution model described by of Chang and
Casali (1994). The use of this latter method was restricted to
those cases (about 11% of IgVH transcripts from the M B-CLL
subgroup) in which analyses were not amenable with the JAVA
applet available for the application of the multinomial
algorithm. According to this strategy, we identified three B-
CLL subsets: sM, with evidence of antigen-driven selection,
nsM and UM, without this evidence and IgVH gene per cent
mutations above or below the 2% cut-off.
These three newly identified B-CLL subsets were also
characterized for the biased usage of IgVH families, the
complexity of intraclonal IgVH diversification, the preference
of mutations to target-specific nucleotides or hotspots, as well
as for the expression of enzymes involved in SHM process of
normal B cells (Muramatsu et al, 1999, 2000; Zan et al, 2001;
Zeng et al, 2001; Cerutti et al, 2002; Storb & Stavnezer, 2002;
Albesiano et al, 2003; McCarthy et al, 2003; Oppezzo et al,
2003). Data accumulated in the present study favours the idea
of a mechanism similar to the canonical IgVH mutation
machinery of normal B cells operating in both sM and nsM
B-CLLs (Messmer et al, 2004). This conclusion is supported by
the demonstration of a similar degree of intraclonal IgVH
diversification, along with an excess of transitions over
transversions or of mutations clustering within RGWY/WRCY
motifs in both sM and nsM B-CLL subsets (Gurrieri et al,
2002; Messmer et al, 2004). Consistently, high levels of both
DNA pol f and g were found in nsM and sM B-CLL but not in
UM B-CLL. This data, if associated with the finding of a
relative A-preference in nsM and sM B-CLLs, is in agreement
with the notion that DNA pol g is an A-T mutator during the
SHM process (Zeng et al, 2001). The additional expression, in
all B-CLL subgroups, of high levels of the error-prone DNA
pol i, a known G-C mutator (Faili et al, 2002), could explain
the finding of a certain degree of G- and/or C-preference in
nsM and sM B-CLLs (R. Bomben, unpublished data).
Specific sets of IgVH genes were preferentially expressed by
sM, nsM and UM B-CLLs. In agreement with previous studies
(Albesiano et al, 2003; Keating et al, 2003; Messmer et al,
2004), UM and M B-CLLs preferentially utilized the IgVH1
IgVH Gene Mutations and Prognosis in B-CLL
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 39

and IgVH3 families respectively. However, when considering
the sM and nsM B-CLL subsets, a more striking skewing
towards the usage of IgVH3 family was documented in sM
B-CLLs, while nsM cases more frequently employed genes
from the IgVH4 family. A different behaviour between sM and
nsM B-CLLs was also observed when analysing the intraclonal
IgVH diversification. In this regard, although the vast majority
of IgVH transcripts displayed an excess of shared mutations
associated or not with few unique and/or partially shared
mutations, the small subset of IgVH transcripts characterized
by the highest degree of intraclonal IgVH diversification almost
exclusively belonged to the sM B-CLL subset. Intraclonal IgVH
diversification has been so far described by a single study
reporting only B-CLL cases lacking evidence of antigen-driven
selection (Gurrieri et al, 2002). In addition to confirming in a
wider cohort of cases that a certain degree of intraclonal IgVH
diversification is not an unfrequent event in B-CLL, we
provided evidence that a positive selection operated by the
antigen can play a role in determining the intrinsic complexity
of intraclonal IgVH diversification.
Strikingly, by examining the prognostic value of antigen-
driven selection, we were able to demonstrate a significantly
longer survival of sM B-CLL patients even within the good
prognosis subgroup with more than 2% mutations of IgVH
genes. Consistently, those B-CLL patients affected by a disease
with more than 2% mutations but lacking evidence of antigen-
driven selection had survivals comparable with bad prognosis
patients, i.e. displaying less than 2% mutations of IgVH genes.
The reasons for this different clinical behaviour remain to be
established. A putative hypothesis could be that the neoplastic
clone from sM B-CLLs continues to be under the positive
pressure exerted by the antigen also following the neoplastic
transformation, while, in nsM B-CLL, the leukaemic growth
has mostly become independent from any external factor,
including the antigen. The higher complexity level of intra-
clonal IgVH diversification found in sM B-CLLs, when
compared with nsM cases, is in keeping with such a view.
Alternatively, the notion that nsM and sM B-CLL cells
preferentially use different IgVH genes may support the
hypothesis that these B-CLL subsets are the result of the
neoplastic transformation of normal B-cell clones selected by
different antigens (Keating et al, 2003; Messmer et al, 2004).
Finally, it cannot be excluded that leukaemic cells from nsM B-
CLLs might represent the neoplastic counterpart of B cells
discarded during a T-dependent SHM process that, by
dysfunction of specific apoptotic pathways (Caligaris-Cappio
& Hamblin, 1999; Lin et al, 2002), are not eliminated by
apoptosis and are more resistant to therapy.
Unmutated B-CLLs, frequently expressing CD38 and AID
(Albesiano et al, 2003; McCarthy et al, 2003; Oppezzo et al,
2003; Degan et al, 2004) also in our series, might be viewed as
the neoplastic counterpart of an ‘early-GC’ B cell. The demon-
stration that somatic point mutation of BCL6 gene, a marker of
GC passage (Migliazza et al, 1995), may also occur in UM B-
CLLs (Sahota et al, 2000), are in agreement with this hypothe-
sis. Moreover, the expression of low levels of translesion DNA
pol g are in keeping with a still incomplete activation of the
SHM machinery (Zeng et al, 2001) and may explain the low
mutation rate of this B-CLL subset (Klein et al, 1998). However,
the finding that UM B-CLLs preferentially expressed IgVH genes
from the IgVH1 family again underscore the role of a putative
selection by specific antigens also for this B-CLL subset. In this
regard, it cannot be excluded that UM B-CLLs may derive from
memory B cells selected by specific antigen via a T-independent
pathway, thus expressing none or few mutations not fulfilling
the features of a canonical T-dependent SHM process (Klein
et al, 2001; Rosenwald et al, 2001).
From a clinical standpoint, this study underscores the role of
the antigen in defining the clinical behaviour of B-CLLs and
suggests a certain caution in the application of the per cent
mutations as a marker of prognostic value in the absence of
evidence of antigen-driven selection. The availability of ready-
to-use computer programs aimed at evaluating the biased R/S
mutations within the various IgVH segments (Chang & Casali,
1994; Lossos et al, 2000) could be of help for the rapid
detection of good prognosis B-CLL subsets during routine
diagnostic/prognostic procedures.
Acknowledgments
The authors thank M. Maio (Cancer Bioimmunotherapy,
CRO, Aviano) and A. Cerutti (Weill Medical College of
Cornell University, New York) for helpful discussion. This
work was supported in part by the Associazione Italiana per la
Ricerca sul Cancro (AIRC), Milan, Italy; and the Ministero
della Salute (Ricerca Finalizzata IRCCS and ‘Alleanza Contro il
Cancro’), Rome, Italy.
References
Albesiano, E., Messmer, B.T., Damle, R.N., Allen, S.L., Rai, K.L. &
Choirazzi, N. (2003) Activation-induced cytidine deaminase in
chronic lymphocytic leukemia B cells: expression as multiple forms in
a dynamic, variably sized fraction of the clone. Blood, 102, 3333–3339.
Armitage, P. & Berry, G. (1987) Statistical Methods in Medical Research.
Blackwell Scientific, London, UK.
Caligaris-Cappio, F. & Hamblin, T.J. (1999) B-cell chronic lympho-
cytic leukemia: a bird with a different feather. Journal of Clinical
Oncology, 17, 399–408.
Cerutti, A., Zan, H., Kim, EC., Shah, S., Schattner, E.J., Schaffer, A. &
Casali, P. (2002) Ongoing in vivo immunoglobulin class switch
DNA recombination in chronic lymphocytic leukemia B cells.
Journal of Immunology, 169, 6594–6603.
Chang, B. & Casali, P. (1994) The CDR1 sequences of a major pro-
portion of human germline Ig VH genes are inherently susceptible to
amino acid replacement. Immunology Today, 15, 367–373.
Chiorazzi, N. & Ferrarini, M. (2003) Chronic lymphocytic leukemia:
lessons learned from studies of the B cell antigen receptor. Annual
Review of Immunology, 21, 841–894.
Damle, R.N., Wasil, T., Fais, F., Ghiotto, F., Valetto, A., Allen, S.L.,
Buchbinder, A., Budman, D., Dittmar, K., Kolitz, J., Lichtman, S.M.,
M. Degan et al
40 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42

Schulman, P., Vinciguerra, V.P., Rai, K.R., Ferrarini, M. & Chior-
azzi, N. (1999) IgV gene mutation status and CD38 expression as
novel prognostic indicators in chronic lymphocytic leukemia. Blood,
94, 1840–1847.
Damle, R.N., Ghiotto, F., Valetto, A., Albesiano, E., Fais, F., Yan, X.J.,
Sison, C.P, Allen, S.L., Kolitz, J., Schulman, P., Vinciguerra, V.P.,
Budde, P., Frey, J., Rai, K.R., Ferrarini, M. & Chiorazzi, N. (2002)
B-cell chronic lymphocytic leukemia cells express a surface mem-
brane phenotype of activated, antigen-experienced B lymphocytes.
Blood, 99, 4087–4093.
Degan, M., Mazzocco, F.T., DiFrancia, R., Rossi, F.M., Pinto, A. &
Gattei, V. (2000) Normalizing complementary DNA by quantitative
reverse transcriptase-polymerase chain reaction of beta2-micro-
globulin: molecular monitoring of minimal residual disease in acute
promyelocytic leukemia. Diagnostic Molecular Pathology, 9, 98–109.
Degan, M., Zucchetto, A., Bomben, R., Dal Bo, M., Stefanon, A.,
Rupolo, M., Canton, E., Nanni, P., Attadia, V., Del Poeta, G.,
Colombatti, A. & Gattei, V. (2004) Activation-induced cytidine
deaminase (AID) and CD38 expression in B-cell chronic lympho-
cytic leukemia (B-CLL). Clinical Lymphoma, in press.
Del Poeta, G., Maurillo, L., Venditti, A., Buccisano, F., Epiceno, A.M.,
Capelli, G., Tamburini, A., Suppo, G., Battaglia, A., Del Principe,
M.I., Del Moro, B., Masi, M. & Amadori, S. (2001) Clinical
significance of CD38 expression in chronic lymphocytic leukemia.
Blood, 98, 2633–2639.
Dorner, T., Foster, S.J., Brezinschek, H.P. & Lipsky, P.E. (1998a) Analysis
of the hypermutational machinery and the impact of subsequent se-
lection on the distribution of nucleotide changes in human VHDJH
rearrangements. Immunological Reviews, 162, 161–171.
Dorner, T., Foster, S.J., Farner, N.L. & Lipsky, P.E. (1998b) Somatic
hypermutation of immunoglobulin heavy chain genes: targeting of
RGYW motifs on both strands. European Journal of Immunology, 28,
3384–3396.
Faili, A., Aoufouchi, S., Flatter, E., Gueranger, Q., Reynaud, C.A. &
Weill, J.C. (2002) Induction of somatic hypermutation in
immunoglobulin genes is dependent on DNA polymerase iota.
Nature, 419, 944–947.
Fais, F., Ghiotto, F., Hashimoto, S., Sellars, B., Valetto, A., Allen, S.L.,
Schulman, P., Vinciguerra, V.P., Rai, K., Rassenti, L.Z., Kipps, T.J.,
Dighiero, G., Schroeder, Jr, H.W., Ferrarini, M. & Chiorazzi, N.
(1998) Chronic lymphocytic leukemia B cells express restricted sets
of mutated and unmutated antigen receptors. Journal of Clinical
Investigation, 102, 1515–1525.
Gattei, V., Degan, M., Gloghini, A., De Iuliis, A., Improta, S., Rossi, F.M.,
Aldinucci, D., Perin, V., Serraino, D., Babare, R., Zagonel, V., Gruss,
H.J., Carbone, A. & Pinto, A. (1997) CD30 ligand is frequently
expressed in human hematopoietic malignancies of myeloid and
lymphoid origin. Blood, 89, 2048–2059.
Gattei, V., Rupolo, M., Zucchetto, A., Berretta, M., Nanni, P., Tumolo, S.,
Doliana, R., Attadia, V., Colombatti, A. & Degan, M. (2003) Mutational
status of IgVH genes in B-cell chronic lymphocytic leukemia: evidence
of antigen-driven selection has prognostic impact instead of percent of
mutation. Journal of Clinical Oncology, 22, 564 (abstract no. 2267).
Gurrieri, C., McGuire, P., Zan, H., Yan, X.J., Cerutti, A., Albesiano, E.,
Allen, S.L., Vinciguerra, V., Rai, K.R., Ferrarini, M., Casali, P. &
Chiorazzi, N. (2002) Chronic lymphocytic leukemia B cells undergo
somatic hypermutation and intraclonal immunoglobulin VHDJH
gene diversification. Journal of Experimental Medicine, 196, 629–639.
Hamblin, T.J. (2002) Chronic lymphocytic leukaemia: one disease or
two? Annals of Hematology, 81, 299–303.
Hamblin, T.J., Davis, Z., Gardiner, A., Oscier, D.G. & Stevenson, F.K.
(1999) Unmutated Ig V(H) genes are associated with a more
aggressive form of chronic lymphocytic leukemia. Blood, 94, 1848–
1854.
Hamblin, T.J., Orchard, J.A., Gardiner, A., Oscier, D.G., Davis, Z.,
Stevenson, F.K. (2000) Immunoglobulin V genes and CD38
expression in CLL. Blood, 95, 2455–2457.
Jelinek, D.F., Tschumper, S., Geyer, S.M., Bone, N.D., Dewald, G.W.,
Hanson, C.A., Stenson, M.J., Witzig, T.E., Tefferi, I.A. & Kay, N.E.
(2001) Analysis of clonal B-cell CD38 and immunoglobulin variable
region sequence status in relation to clinical outcome for B-chronic
lymphocytic leukaemia. British Journal of Haematology, 115, 854–
861.
Keating, M.J., Chiorazzi, N., Messmer, B., Damle, R.N., Allen, S.L.,
Rai, K.R., Ferrarini, M. & Kipps, T.J. (2003) Biology and treatment
of chronic lymphocytic leukemia. In: Hematology (American Society
of Hematology. Education Program), pp. 153–175. American Society
of Hematology, Washington, DC, USA.
Klein, U., Goossens, T., Fischer, M., Kanzler, H., Braeuninger, A.,
Rajewsky, K. & Kuppers, R. (1998) Somatic hypermutation in
normal and transformed human B cells. Immunological Reviews,
162, 261–280.
Klein, U., Stolovitzky, G.A., Mattioli, M., Cattoretti, G., Husson, H.,
Freedman, A., Inghirami, G., Cro, L., Baldini, L., Neri, A., Califano, A.
& Dalla-Favera, R. (2001) Gene expression profiling of B chronic
lymphocytic leukemia reveals a homogeneous phenotype related to
memory B cells. Journal of Experimental Medicine, 194, 1625–1638.
Krober, A., Seiler, T., Benner, A., Bullinger, L., Bruckle, E., Lichter, P.,
Dohner, H. & Stilgenbauer, S. (2002) VH mutational status, CD38
expression level, genomic aberrations, and survival in chronic lym-
phocytic leukemia. Blood, 100, 1410–1416.
Lin, K., Sherrington, P.D., Dennis, M., Matrai, Z., Cawley, J.C. &
Pettitt, A.R. (2002) Relationship between p53 dysfunction, CD38
expression, and IgVH mutation in chronic lymphocytic leukemia.
Blood, 100, 1404–1409.
Lossos, I.S., Tibshirani, R., Narasimhan, B. & Levy, R. (2000) The
inference of antigen selection on Ig genes. Journal of Immunology,
165, 5122–5126.
Maloum, K., Davi, F., Merle-Beral, H., Pritsch, O., Magnac, C., Vuil-
lier, F., Dighiero, G., Troussard, X., Mauro, F.F. & Benichou, J.
(2000) Expression of unmutated VH genes is a detrimental prog-
nostic factor in chronic lymphocytic leukemia. Blood, 96, 377–379.
Matrai, Z., Lin, K., Dennis, M., Sherrington, P., Zuzel, M., Pettitt, A.R.
& Cawley, J.C. (2001) CD38 expression and Ig VH gene mutation in
B-cell chronic lymphocytic leukemia. Blood, 97, 1902–1903.
Matutes, E., Owusu-Ankomah, K., Morilla, R., Garcia Marco, J.,
Houlihan, A., Que, T.H. & Catovsky, D. (1994) The immunological
profile of B cell disorders and proposal of a scoring system for the
diagnosis of CLL. Leukemia, 8, 1640–1645.
McCarthy, H., Wierda, W.G., Barron, L.L., Cromwell, C.C., Wang, J.,
Coombes, K.R., Rangel, R., Elenitoba-Johnson, K.S., Keating, M.J. &
Abruzzo, L.V. (2003) High expression of activation-induced cytidine
deaminase (AID) and splice variants is a distinctive feature of poor-
prognosis chronic lymphocytic leukemia. Blood, 101, 4903–4908.
Messmer, B.T., Albesiano, E., Messmer, D. & Chiorazzi, N. (2004) The
pattern and distribution of immunoglobulin VH gene mutations in
IgVH Gene Mutations and Prognosis in B-CLL
ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42 41

chronic lymphocytic leukemia B cells are consistent with the
canonical somatic hypermutation process. Blood, 103, 3490–3495.
Migliazza, A., Martinotti, S., Chen, W., Fusco, C., Ye, B.H., Knowles,
D.M., Offit, K., Chaganti, R.S. & Dalla-Favera, R. (1995) Frequent
somatic hypermutation of the 5¢ noncoding region of the BCL6 gene
in B-cell lymphoma. Proceedings of the National Academy of Sciences
of the United States of America, 92, 12520–12524.
Muramatsu, M., Sankaranand, V.S., Anant, S., Sugai, M., Kinoshita, K.,
Davidson, N.O. & Honjo, T. (1999) Specific expression of activa-
tion-induced cytidine deaminase (AID), a novel member of the
RNA-editing deaminase family in germinal center B cells. Journal of
Biological Chemistry, 274, 18470–18476.
Muramatsu, M., Kinoshita, K., Fagarasan, S., Yamada, S., Shinkai, Y. &
Honjo, T. (2000) Class switch recombination and hypermutation
require activation-induced cytidine deaminase (AID), a potential
RNA editing enzyme. Cell, 102, 553–563.
Naylor, M. & Capra, J.D. (1999) Mutational status of Ig VH genes
provides clinically valuable information in B-cell chronic lympho-
cytic leukemia. Blood, 94, 1837–1839.
Neuberger, M.S., Ehrenstein, M.R., Klix, N., Jolly, C.J., Yelamos, J.,
Rada, C. & Milstein, C. (1998) Monitoring and interpreting the
intrinsic features of somatic hypermutation. Immunological Reviews,
162, 107–116.
Oppezzo, P., Vuillier, F., Vasconcelos, Y., Dumas, G., Magnac, C.,
Payelle-Brogard, B., Pritsch, O. & Dighiero, G. (2003) Chronic
lymphocytic leukemia B cells expressing AID display a dissociation
between class switch recombination and somatic hypermutation.
Blood, 101, 4029–4032.
Oscier, D.G., Thompsett, A., Zhu, D. & Stevenson, F.K. (1997) Dif-
ferential rates of somatic hypermutation in V(H) genes among
subsets of chronic lymphocytic leukemia defined by chromosomal
abnormalities. Blood, 89, 4153–4160.
Pasqualucci, L., Migliazza, A., Fracchiolla, N., William, C., Neri, A.,
Baldini, L., Chaganti, R.S., Klein, U., Kuppers, R., Rajewsky, K. &
Dalla-Favera, R. (1998) BCL-6 mutations in normal germinal center
B cells: evidence of somatic hypermutation acting outside Ig loci.
Proceedings of the National Academy of Sciences of the United States of
America, 95, 11816–11821.
Rai, K.R., Sawitsky, A., Cronkite, E.P., Chanana, A.D., Levy, R.N. &
Pasternack, B.S. (1975) Clinical staging of chronic lymphocytic
leukemia. Blood, 46, 219–234.
Rosenwald, A., Alizadeh, A.A., Widhopf, G., Simon, R., Davis, R.E., Yu,
X., Yang, L., Pickeral, O.K., Rassenti, L.Z., Powell, J., Botstein, D.,
Byrd, J.C., Grever, M.R., Cheson, B.D., Chiorazzi, N., Wilson, W.H.,
Kipps, T.J., Brown, P.O. & Staudt, L.M. (2001) Regulation of gene
expression phenotype to immunoglobulin mutation genotype in
B cell chronic lymphocytic leukemia. Journal of Experimental
Medicine, 194, 1639–1647.
Sahota, S.S., Davis, Z., Hamblin, T.J. & Stevenson, F.K. (2000) Somatic
mutation of bcl-6 genes can occur in the absence of VH mutations
in chronic lymphocytic leukemia. Blood, 95, 3534–3540.
Sakai, A., Marti, G.E., Caporaso, N., Pittaluga, S., Touchman, J.W.,
Fend, F. & Raffeld, M. (2000) Analysis of expressed immunoglobulin
heavy chain genes in familial B-CLL. Blood, 95, 1413–1419.
Schroeder, Jr, H.W. & Dighiero, G. (1994) The pathogenesis of chronic
lymphocytic leukemia: analysis of the antibody repertoire.
Immunology Today, 15, 288–294.
Smith, D.S., Creadon, G., Jena, P.K., Portanova, J.P., Kotzin, B.L. &
Wysocki, L.J. (1996) Di- and trinucleotide target preferences of
somatic mutagenesis in normal and autoreactive B cells. Journal of
Immunology, 156, 2642–2652.
Spessotto, P., Rossi, F.M., Degan, M., Di Francia, R., Perris, R.,
Colombatti, A. & Gattei, V. (2002) Hyaluronan-CD44 interaction
hampers migration of osteoclast-like cells by down-regulating
MMP-9. Journal of Cell Biology, 158, 1133–1144.
Storb, U. & Stavnezer, J. (2002) Immunoglobulin genes: generating
diversity with AID and UNG. Current Biology, 12, R725–R727.
Thunberg, U., Johnson, A., Roos, G., Thorn, I., Tobin, G., Sallstrom, J.,
Sundstrom, C. & Rosenquist, R. (2001) CD38 expression is a poor
predictor for VH gene mutational status and prognosis in chronic
lymphocytic leukemia. Blood, 97, 1892–1894.
Zan, H., Komori, A., Li, Z., Cerutti, A., Schaffer, A., Flajnik, M.F., Diaz, M.
& Casali, P. (2001) The translesion DNA polymerase f plays a major
role in Ig and bcl-6 somatic hypermutation. Immunity, 14,
643–653.
Zeng, X., Winte, D.B., Kasmer, C., Kraemer, K.H., Lehmann, A.R. &
Gearhart, P.J. (2001) DNA polymerase g is an A-T mutator in
somatic hypermutation of immunoglobulin variable genes. Nature
Immunology, 2, 537–541.
M. Degan et al
42 ª 2004 Blackwell Publishing Ltd, British Journal of Haematology, 126, 29–42
Copyright © 2022 FDOKUMEN




















