
Analysis of hybridoma mutants defective in synthesis of immunoglobulin M
11
Somatic Cell and Molecular Genetics, 1Iol. 12, No. 5, 1986, pp. 467-477 Analysis of Hybridoma Mutants Defective in Synthesis of Immunoglobulin M W.S. Trimble, 1'2 M.D. Baker, 2 G.L. Boulianne, 1'2 H. Murialdo, 2 N. Hozumi, 1 3 and M.J. Shulman 2'3 1Mount Sinai Hospital Research Institute, 600 University Ave., Toronto, Ontario, Canada M5G 1X5; and 2Depart- ment of Medical Genetics, 3Department of lmmunology, University of Toronto, Toronto, Ontario, Canada M5S 1.48 Received 7 April 1986 Final 21 May 1986 Abstract--Hybridoma mutants defective in the expression of IgM have been analyzed by molecular and somatic cell hybridization techniques. The frequency of K light-chain mutants in the hybridoma PC7 was much higher than for other cell lines. In contrast to the mutations which we observed previously, the K mutants examined here resulted from complete or partial deletion of the K gene. Mutants defective in # chain synthesis were of more diverse types including deletions, gross rearrangements, and more subtle changes. One mutant containing a cis-acting mutation resulting in reduced expression of the tz chain had an associated partial duplication of the # gene, while others making low or undetectable levels of# had no gross alterations in genetic structure. The usefulness of this approach to the study of gene structure and expression is discussed. INTRODUCTION It has become possible in recent years to apply the techniques of molecular and somatic cell genetics to study the molecular basis of immunoglobulin (Ig) synthesis and function. The advantages of this system are threefold: Ig-producing cell lines are especially amena- ble to genetic analysis because the Ig genes are haploid; molecular studies are facilitated because the [g genes are well characterized and expressed at high levels in a tissue-specific manner; and the availability of immortal myeloma and hybridoma cell lines provides large amounts of monoclonal antibody for studies of Ig production and function. Early studies using myeloma cell lines have indi- cated that the mutation rate of their Ig genes can be unusually high (up to 10 -4 per cell generation) (1-6), and it has been suggested that this high mutation rate is related to the diversity generated somatically by the normal immune system (7). The analysis of Ig gene mutations could provide valuable insights not only into the immune system but also in defining the relationship between the struc- ture and function of genes and their products. We have previously described a method for selecting [g-defective mutant cells from hapten-specific Immunoglobulin M (IgM) -producing hybridoma cell lines (6). From the selected population, we have obtained a vari- ety of rare structural mutants which produce abnormal IgM, defective in complement acti- vation, hapten binding, assembly, or glycosy- lation (8). However, the majority of the sur- viving cells produce no IgM, and in principle a portion of these mutants could be "regula- tory" in nature and might be useful for iden- tifying the molecular requirements of Ig 467 0740-7750/86/0900-046755.00/0 1986 Plenum Publishing Corporation
-
Upload
cardiacehealth -
Category
Documents
-
view
6 -
download
0
Transcript of Analysis of hybridoma mutants defective in synthesis of immunoglobulin M

Somatic Cell and Molecular Genetics, 1Iol. 12, No. 5, 1986, pp. 467-477
Analysis of Hybridoma Mutants Defective in Synthesis of Immunoglobulin M
W.S. Trimble, 1'2 M.D. Baker, 2 G.L. Boulianne, 1'2 H. Murialdo, 2 N. Hozumi, 1 3 and M.J. Shulman 2'3
1Mount Sinai Hospital Research Institute, 600 University Ave., Toronto, Ontario, Canada M5G 1X5; and 2Depart- ment of Medical Genetics, 3Department of lmmunology, University of Toronto, Toronto, Ontario, Canada M5S 1.48
Received 7 April 1986 Final 21 May 1986
Abstract--Hybridoma mutants defective in the expression of IgM have been analyzed by molecular and somatic cell hybridization techniques. The frequency of K light-chain mutants in the hybridoma PC7 was much higher than for other cell lines. In contrast to the mutations which we observed previously, the K mutants examined here resulted from complete or partial deletion of the K gene. Mutants defective in # chain synthesis were of more diverse types including deletions, gross rearrangements, and more subtle changes. One mutant containing a cis-acting mutation resulting in reduced expression of the tz chain had an associated partial duplication of the # gene, while others making low or undetectable levels o f# had no gross alterations in genetic structure. The usefulness of this approach to the study of gene structure and expression is discussed.
INTRODUCTION
It has become possible in recent years to apply the techniques of molecular and somatic cell genetics to study the molecular basis of immunoglobulin (Ig) synthesis and function. The advantages of this system are threefold: Ig-producing cell lines are especially amena- ble to genetic analysis because the Ig genes are haploid; molecular studies are facilitated because the [g genes are well characterized and expressed at high levels in a tissue-specific manner; and the availability of immortal myeloma and hybridoma cell lines provides large amounts of monoclonal antibody for studies of Ig production and function. Early studies using myeloma cell lines have indi- cated that the mutation rate of their Ig genes can be unusually high (up to 10 -4 per cell generation) (1-6), and it has been suggested
that this high mutation rate is related to the diversity generated somatically by the normal immune system (7). The analysis of Ig gene mutations could provide valuable insights not only into the immune system but also in defining the relationship between the struc- ture and function of genes and their products.
We have previously described a method for selecting [g-defective mutant cells from hapten-specific Immunoglobulin M (IgM) -producing hybridoma cell lines (6). From the selected population, we have obtained a vari- ety of rare structural mutants which produce abnormal IgM, defective in complement acti- vation, hapten binding, assembly, or glycosy- lation (8). However, the majority of the sur- viving cells produce no IgM, and in principle a portion of these mutants could be "regula- tory" in nature and might be useful for iden- tifying the molecular requirements of Ig
467
0740-7750/86/0900-046755.00/0 �9 1986 Plenum Publishing Corporation

468 Trimble et al.
expression. On the other hand, it might be expected that the karyotype of the parental hybridoma cell line would be so unstable (9, 10) that loss of a chromosome bearing an Ig gene would be the predominant cause of the Ig-negative phenotype. To test this expecta- tion we have analyzed several hybridoma mutants which are deficient in the production of either the K light chain or # heavy chain and found that in about half the cases the Ig genes have not been entirely deleted. Some of the mutants bear gross rearrangements of an Ig gene, whereas others bear more subtle (point?) mutations. We conclude from this analysis that, despite its chromosomal over- load, this hybridoma cell line can be used to obtain interesting regulatory mutations.
MATERIALS AND METHODS
Cell Lines. The origin of the wild-type hybridoma PC7 and the enrichment of mutants by suicide selection have been described (8). The mutants were selected from four independent sublines of PC7 called PC700, PC704, PC705, and PC700 Oul. 1. All cell lines were grown as described pre- viously (6). Somatic hybrids were generated as described (11), and hybrid cells were selected for growth in the presence of 1 mM ouabain and HAT (0.1 mM hypoxanthine, 0.8 pM aminopterin, and 16 #M thymidine).
Bacterial Strains and Plasmids. Es- cherichia coli strain K803 (r k , mk, SupE, SupF) was obtained from G. Matthysens and was used for plasmid preparation. Plasmids containing a cDNA clone of the # gene (12), the VKPC gene (13), and the V~PC gene were kindly provided by Drs. J Adams, M. Scharff, and P. Leder, respectively. Genomic ~ and # genes have been previously described (14, 15).
DATA Preparation. High-molecular- weight DNA was prepared from the various hybridoma cell lines, and from Balb/c kid- neys, by the SDS/proteinase K method of Gross-Bellard et al (16). Plasmid DNA was
prepared as described (17), with the final purification step involving isopycnic CsC1- EtBr gradient centrifugation. The DNA probes used in Southern and RNA blot hybridizations were prepared by digestion of plasmid DNA with appropriate restriction enzymes and isolation by electrophoresis through 2% agarose or 5% polyacrylamide followed by electroelution within dialysis membranes. A 2-kb BamHI/EcoRI fragment encoding JH3~, was isolated from the germ-line # gene. A #-specific probe of 949 bp was isolated by SmaI/PstI digestion of the # cDNA clone. The VHPC probe used in these studies was a 625-bp HinfI fragment contain- ing VnPC plus the JH1-2 coding regions. The VKPC probe used in these studies was a 680-bp XbaI/AvalI fragment and the CK probes used were either a 900-bp HaelII fragment or a 413-bp XmnI fragment.
Electrophoresis and Hybridiza- tions. Restriction enzymes were purchased from New England Biolabs, Boehringer- Mannheim, and Bethesda Research Laborato- ries. All enzymatic reactions were performed as recommended by the manufacturers. Digests involving more than one enzyme were performed either sequentially or in combina- tion. Following digestion, DNA was electro- phoresed through horizontal or vertical gels composed of 0.7 1.3% agarose using HindllI digested 2,CamI85757 DNA as a molecular- weight marker. The preparation of gels and subsequent blotting of DNA onto nitrocellu- lose was performed by the method of Southern (18). DNA blot hybridizations were per- formed at 65~ in 4x SSC (4x SSC = 0.6 M NaC1, 0.06 M Na Citrate, pH 7.0) or in 50% formamide and 4 x SSC at 42~ as described (17), using 3zp-labeled nick-translated DNA probes of specific activity between 5 and 10 x 10 7 cpm/#g. RNA blots and hybridizations were performed as described by Thomas (19). Final washes were performed at 65~ in 0.1 x SSC and 0.2% SDS.
Measurement of IgM. The parental hybridoma PC7 produces IgM(K) specific for

lgM Mutants 469
the hapten phosphorylcholine. Total IgM was assayed by an enzyme-linked immunosorbent assay (ELISA) or by hemagglutination (6); the # and K chains were assayed by SDS gel electrophoresis of immunoprecipitated bio- synthetically labeled culture material (8).
RESULTS
Mutant Isolation. Using the suicide selection system described previously (6, 8), we have enriched for mutant cells defective in the normal production of the phosphorylcho- line (PC) -specific IgM. The enrichment involved covalent attachment of a reactive derivative of the hapten PC to the surface of the hybridoma cells, followed by incubation in the presence of complement. Suicide selection occurred because the PC specific IgM binds preferentially to the surface of cells from which it was secreted. Thus, cells secreting normal amounts of normal cytolytic IgM are more readily lysed by complement than mutant cells. After several enrichment cycles, the enriched population, which contained less than 0.1% normal (plaque-forming) cells, was subcloned, and the sublines were tested for secretion of lgM. Cells secreting much less than (0-10%) normal amounts of IgM were
examined for intracellular t~ and K chains by immunoprecipitation of biosynthetically la- beled proteins (Table 1). The cells isolated could be classified according to their pheno- type into four groups: (1) undetectable ~z but normal ~ (u-K+); (2) normal t~ but undetect- able K (#+K); (3) undetectable ~ and K (#-K-); and (4) low levels of/~ (1-10%) but normal K (#LK+). We have examined representatives from each group to determine if gross rearran- gements in the # and K genes could account for the reductions in IgM production.
Kappa Nonexpression. In order to screen the mutants for structural alterations of the K genes, mutant and wild-type cellular DNA was digested with various restriction enzymes, either alone or in combination, and analyzed by Southern blot techniques using a 32p-labeled probe for the K gene constant (Ca) region. The results of a BamHI/EcoRI DNA digestion are shown in Fig. 1A. In the germ line of the Balb/c mouse, the DNA encoding the K chain J and constant region (J~-CK) gene is contained within a 6.8-kb EcoRI/BamHI fragment (20) and is present in all of the cell lines examined. As a consequence of V,-JK joining, the size of the fragments bearing the germ-line C~ gene should be altered, so that digestion with these two enzymes can be used to identify fragments bearing rearranged K
Table 1. Origin and Phenotype o f l g M - M u t a n t s of PC7 a
Parental line u-K + U+K U-K- uLK +
PC700 1" ,10 ,37 PC700.0ul.1 350" ,400" ,529" 574* 343* PC704 259* 261" PC705 132",134,1102, 1106",1134,
1115",1122" 1178" 1123, 1133" 1145 1183, 1192 1147,1170,1172 1175,1179
"Mutants defective in IgM synthesis were isolated after limiting dilution as described previously. I g M - mutaniLs were examined for production of u and K chains by SDS-PAGE analysis of immunoprecipitated intracellular lysates. Cells producing undetectable (<1% relative to wild-type levels) u or K chains were classed u-K + and u+K -, respectively. Those which made normal K but between 1 and 10% normal u levels were classed as uLK +. Mutants denoted with an asterisk were further studied herein.
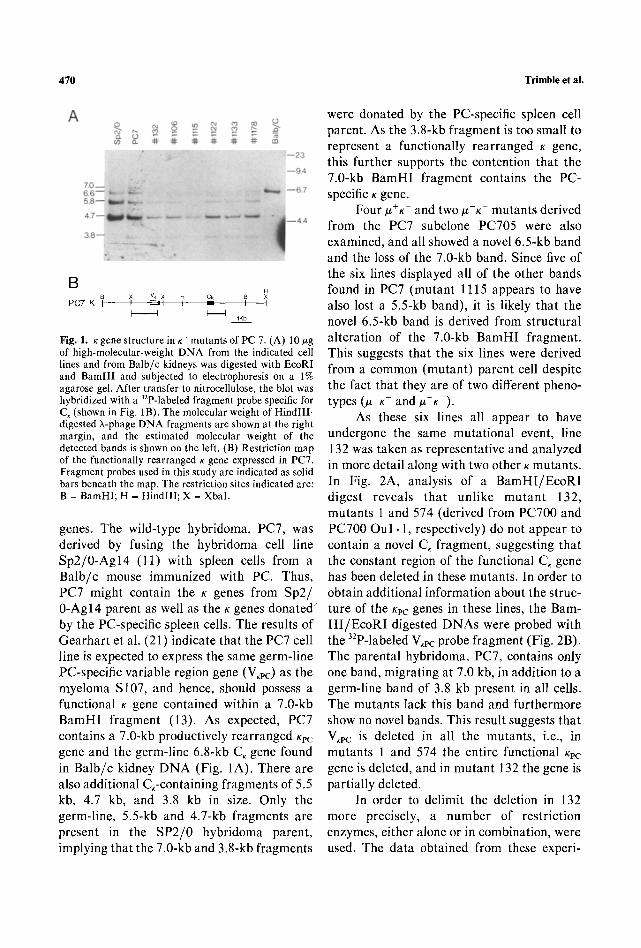
470 Trimble et al.
B B X Vk X H Ck H
P C 7 K i i " - " ', ', - " ~ I I I I I I K b .
Fig. 1. ~ gene structure in K- mutants of PC 7. (A) 10 #g of high-molecular-weight DNA from the indicated cell lines and from Balb/c kidneys was digested with EcoRl and BamHI and subjected to electrophoresis on a 1% agarose gel. After transfer to nitrocellulose, the blot was hybridized with a 32P-labeled fragment probe specific for C, (shown in Fig. 1B). The molecular weight of Hindlll- digested X-phage DNA fragments are shown at the right margin, and the estimated molecular weight of the detected bands is shown on the left. (B) Restriction map of the functionally rearranged K gene expressed in PC7. Fragment probes used in this study are indicated as solid bars beneath the map. The restriction sites indicated are: B = BamHI; H = Hindlll; X - Xbal.
genes. The wild-type hybridoma, PC7, was derived by fusing the hybridoma cell line Sp2 /0 -Agl4 (11) with spleen cells from a Balb/c mouse immunized with PC. Thus, PC7 might contain the K genes from Sp2/ 0-Agl4 parent as well as the K genes dona ted by the PC-specific spleen cells. The results of Gearhart et al. (21) indicate that the PC7 cell line is expected to express the same germ-line PC-specific variable region gene (V,pc) as the myeloma S107, and hence, should possess a functional K gene contained within a 7.0-kb BamHI fragment (13). As expected, PC7 contains a 7.0-kb productively rearranged Kpc gene and the germ-line 6.8-kb C~ gene found in Balb/c kidney D N A (Fig. 1A). There are also additional C,-containing fragments of 5.5 kb, 4.7 kb, and 3.8 kb in size. Only the germ-line, 5.5-kb and 4.7-kb fragments are present in the SP2/0 hybridoma parent, implying that the 7.0-kb and 3.8-kb fragments
were donated by the PC-specific spleen cell parent. As the 3.8-kb fragment is too small to represent a functionally rearranged K gene, this further supports the contention that the 7.0-kb BamHI fragment contains the PC- specific K gene.
Four #+K and two #-K- mutants derived from the PC7 subclone PC705 were also examined, and all showed a novel 6.5-kb band and the loss of the 7.0-kb band. Since five of the six lines displayed all of the other bands found in PC7 (mutant 1115 appears to have also lost a 5.5-kb band), it is likely that the novel 6.5-kb band is derived from structural alteration of the 7.0-kb BamHI fragment. This suggests that the six lines were derived from a common (mutant) parent cell despite the fact that they are of two different pheno- types 0t+K - and #-K ).
As these six lines all appear to have undergone the same mutational event, line 132 was taken as representative and analyzed in more detail along with two other K mutants. In Fig. 2A, analysis of a B a m H I / E c o R I digest reveals that unlike mutant 132, mutants 1 and 574 (derived from PC700 and PC700 Oul �9 1, respectively) do not appear to contain a novel C, fragment, suggesting that the constant region of the functional C, gene has been deleted in these mutants. In order to obtain additional information about the struc- ture of the Kpc genes in these lines, the Bam- H I / E c o R I digested DNAs were probed with the 32p-labeled V,pc probe fragment (Fig. 2B). The parental hybridoma, PC7, contains only one band, migrating at 7.0 kb, in addition to a germ-line band of 3.8 kb present in all cells. The mutants lack this band and furthermore show no novel bands. This result suggests that V,pc is deleted in all the mutants, i.e., in mutants 1 and 574 the entire functional Kpc gene is deleted, and in mutant 132 the gene is partially deleted.
In order to delimit the deletion in 132 more precisely, a number of restriction enzymes, either alone or in combination, were used. The data obtained from these experi-
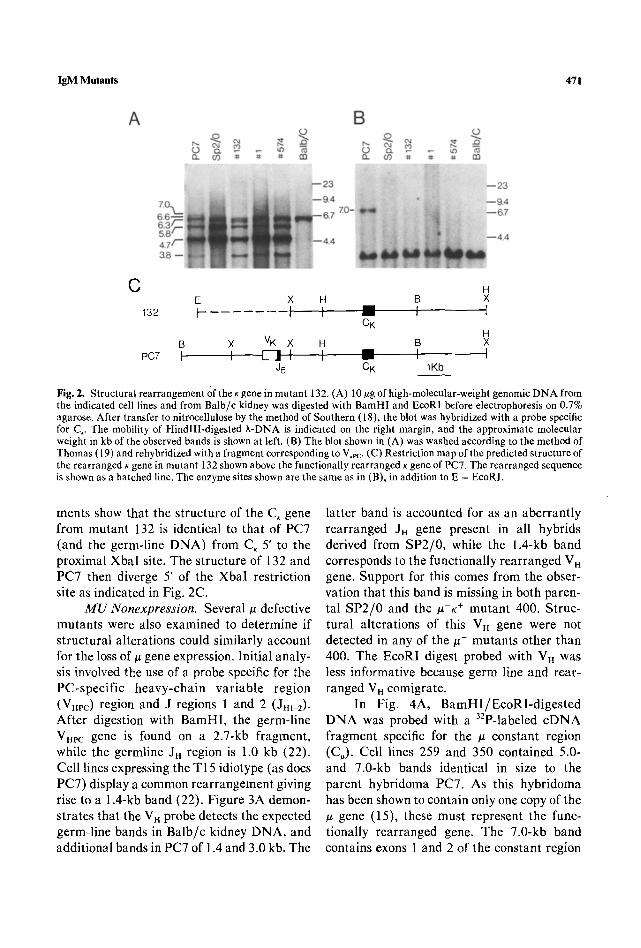
IgM Mutants 471
C H E X H B X
132 t t t --,,,-- t 1 CK
H B X VK X H B X
PC7 F I F 7 1 1 I ~ ,' I J5 C K 1Kb
Fig. 2. Structuralrearrangement0ftheKgeneinmutant 132, (A) 10~gofhigh-molecular-weight genomicDNA from the indicated cell lines and from Balb/c kidney was digested with BamHI and EcoRI before electrophoresis on 0.7% agarose. After transfer to nitrocellulose by the method of Southern (18), the blot was hybridized with a probe specific for C,. The mobility of HindlII-digested X-DNA is indicated on the right margin, and the approximate molecular weight in kb of the observed bands is shown at left. (B) The blot shown in (A) was washed according to the method of Thomas (19) and rehybridized with a fragment corresponding to V,pc. (C) Restriction map of the predicted structure of the rearranged K gene in mutant 132 shown above the functionally rearranged K gene of PC7. The rearranged sequence is shown as a hatched line. The enzyme sites shown are the same as in (B), in addition to E = EcoRI.
ments show that the structure of the C, gene from mutant 132 is identical to that of PC7 (and the germ-line DNA) from C~ 5' to the proximal XbaI site. The structure of 132 and PC7 then diverge 5' of the XbaI restriction site as indicated in Fig. 2C.
MU Nonexpression. Several # defective mutants were also examined to determine if structural alterations could similarly account for the loss of u gene expression. Initial analy- sis involved the use of a probe specific for the PC-specific heavy-chain variable region (VHpc) region and J regions 1 and 2 (JHl-2). After digestion with BamHI, the germ-line VHp c gene is found on a 2.7-kb fragment, while the germline Jn region is 1,0 kb (22). Cell lines expressing the T15 idiotype (as does PC7) display a common rearrangement giving rise to a 1.4-kb band (22). Figure 3A demon- strates that the VH probe detects the expected germ-line bands in Balb/c kidney DNA, and additional bands in PC7 of 1.4 and 3.0 kb. The
latter band is accounted for as an aberrantly rearranged J . gene present in all hybrids derived from SP2/0, while the 1.4-kb band corresponds to the functionally rearranged V H gene. Support for this comes from the obser- vation that this band is missing in both paren- tal SP2/0 and the ~-~+ mutant 400. Struc- tural alterations of this VH gene were not detected in any of the #- mutants other than 400. The EcoRI digest probed with VH was less informative because germ line and rear- ranged Vn comigrate.
In Fig. 4A, BamHI/EcoRI-d iges ted DNA was probed with a 3zp-labeled cDNA fragment specific for the # constant region (C,). Cell lines 259 and 350 contained 5.0- and 7.0-kb bands identical in size to the parent hybridoma PC7. As this hybridoma has been shown to contain only one copy of the
gene (15), these must represent the func- tionally rearranged gene. The 7.0-kb band contains exons 1 and 2 of the constant region
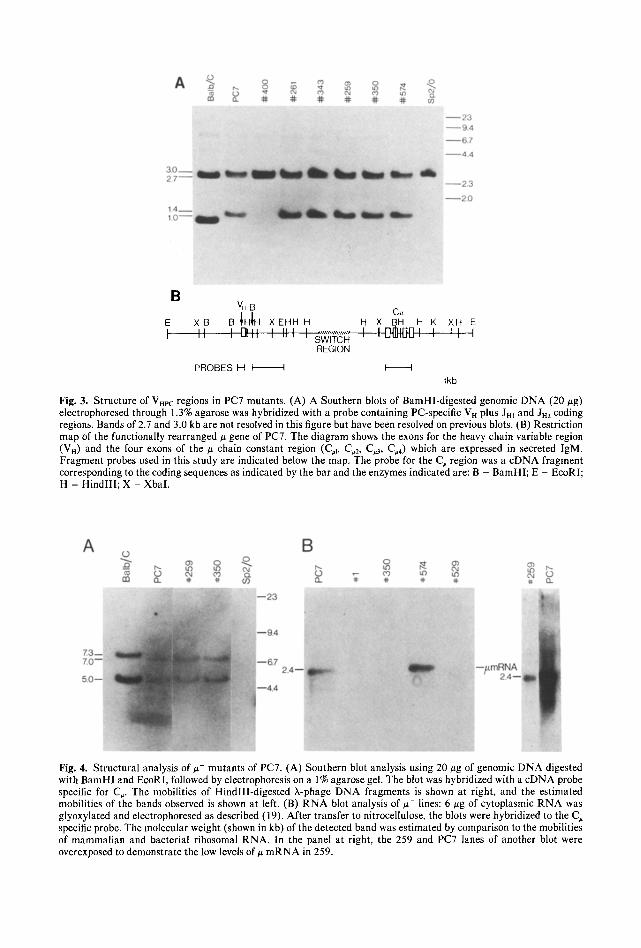
B V . B
C~ E X B B IHIH X EHH H H X BH H K XH E
I " I gl tt ', ',I I ', SWITCH , , ................. I I I ] - t : l }~J I I I I REGION
P R O B E S H I I I I l kb
Fig. 3. Structure of VHr.c regions in PC7 mutants. (A) A Southern blots of BamHI-digested genomic DNA (20 #g) electrophoresed through 1.3% agarose was hybridized with a probe containing PC-specific V H plus Jm and Jm coding regions. Bands of 2.7 and 3.0 kb are not resolved in this figure but have been resolved on previous blots. (B) Restriction map of the functionally rearranged # gene of PC7. The diagram shows the exons for the heavy chain variable region (VH) and the four exons of the # chain constant region (C,1, C,2, C,3, C~4) which are expressed in secreted IgM. Fragment probes used in this study are indicated below the map, The probe for the C, region was a cDNA fragment corresponding to the coding sequences as indicated by the bar and the enzymes indicated are: B = BamH1; E = EcoRI; H = HindlIl; X - XbaI.
Fig. 4. Structural analysis of # - mutants of PC7. (A) Southern blot analysis using 20 #g of genomic DNA digested with BamHl and EcoRI, followed by electrophoresis on a 1% agarose gel. The blot was hybridized with a cDNA probe specific for C~. The mobilities of HindlII-digested X-phage DNA fragments is shown at right, and the estimated mobilities of the bands observed is shown at left. (B) R N A blot analysis of ~- lines: 6 gg of cytoplasmic R N A was glyoxylated and electrophoresed as described (19). After transfer to nitrocellulose, the blots were hybridized to the C~ specific probe. The molecular weight (shown in kb) of the detected band was estimated by comparison to the mobilities of mammalian and bacterial ribosomal RNA. In the panel at right, the 259 and PC7 lanes of another blot were overexposed to demonstrate the low levels of # mRNA in 259.
IgM Mutants 473
(C#1_2) and is slightly smaller than the 7.3-kb band found in the germ-line configuration, possibly due to a deletion within the switch region (12). The C~ gene of mutant 529 was also found to be intact, while mutants 1 and 400 were previously shown to lack C~ DNA upon similar analysis (reference 15, and N. Pennell, personal communication) and appear to have deleted the V n gene.
The production of ~t-specific transcripts in the #- mutants was also examined (Fig. 4B). As expected, mutant 1, which lacks the tt gene, produces no detectable u-specific mRNA. Mutants 350 and 529, for which we detected no rearrangements in their/z DNA, also show no # specific mRNA. Mutant 259 makes about 10% the normal amount of # mRNA. However, this RNA is apparently defective, because it is not converted to detect- able protein (i.e., the culture supernatant of mutant 259 contains < 1% the normal amount of IgM, as measured by ELISA).
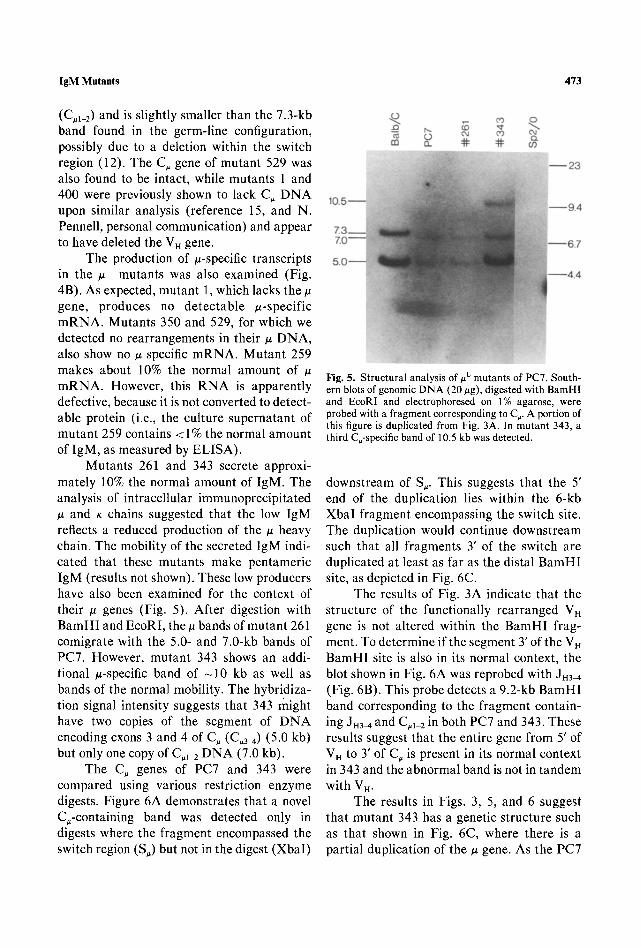
Mutants 261 and 343 secrete approxi- mately 10% the normal amount of IgM. The analysis of intracellular immunoprecipitated ~z and K chains suggested that the low IgM reflects a reduced production of the # heavy chain. The mobility of the secreted IgM indi- cated that these mutants make pentameric IgM (results not shown). These low producers have also been examined for the context of their # genes (Fig. 5). After digestion with BamHI and EcoRI, the/z bands of mutant 261 comigrate with the 5.0- and 7.0-kb bands of PC7. However, mutant 343 shows an addi- tional ~t-specific band of - 1 0 kb as well as bands of the normal mobility. The hybridiza- tion signal intensity suggests that 343 might have two copies of the segment of DNA encoding exons 3 and 4 of C, (C,3~) (5.0 kb) but only one copy of C,1 2 DNA (7.0 kb).
The C, genes of PC7 and 343 were compared using various restriction enzyme digests. Figure 6A demonstrates that a novel C,-containing band was detected only in digests where the fragment encompassed the switch region (S,) but not in the digest (XbaI)
Fig. 5. Structural analysis of #L mutants of PC7. South- ern blots of genomic DNA (20 #g), digested with BamHI and EcoRI and electrophoresed on 1% agarose, were probed with a fragment corresponding to C~. A portion of this figure is duplicated from Fig. 3A. In mutant 343, a third C~-specific band of 10.5 kb was detected.
downstream of S~. This suggests that the 5' end of the duplication lies within the 6-kb XbaI fragment encompassing the switch site. The duplication would continue downstream such that all fragments 3' of the switch are duplicated at least as far as the distal BamHI site, as depicted in Fig. 6C.
The results of Fig. 3A indicate that the structure of the functionally rearranged VH gene is not altered within the BamHI frag- ment. To determine if the segment 3' of the V . BamHI site is also in its normal context, the blot shown in Fig. 6A was reprobed with Jn3~ (Fig. 6B). This probe detects a 9.2-kb BamHI band corresponding to the fragment contain- ing JH3M and C#I 2 in both PC7 and 343. These results suggest that the entire gene from 5' of V . to 3' of C, is present in its normal context in 343 and the abnormal band is not in tandem with VH.
The results in Figs. 3, 5, and 6 suggest that mutant 343 has a genetic structure such as that shown in Fig. 6C, where there is a partial duplication of the ~ gene. As the PC7

474 Trimble et al.
C E XB B B XEX X K X E B I II I P,,', I ' 4 .................... 4 ~ 4 ] ' [ 3 I I I I - -
SWITCH REGION
:1:1:3431a x B K X E B
. . . . . . . . . . . . F I ~ I I I I='--
1Kb
Fig. 6. Rearranged t~ gene in mutant 343. (A) Southern blot analysis of 343 compared to PC7:20 ~tg of each genomic DN A was digested with the indicated restriction enzymes and electrophoresed on 1% agarose. The blot was hybridized with a C~-specific probe. (B) The blot shown in (A) was washed as described by Thomas (19) and rehybridized with a probe specific for JH (Fig. 3C) revealing the presence of one functionally rearranged J. segment in both PC7 and 343. (C) Restriction map of the predicted ~t gene content of 343.
wild-type parent has only one copy of the # gene, the additional #-specific DNA in mutant 343 must have been derived in some way from the functional PC-specific ~ gene. We do not, however, know how closely the duplicated region is linked to the functional PC-specific gene.
In order to test whether the low produc- tion of ~ by mutant 343 reflects a mutation in the # gene itself (rather than in a trans-acting regulatory factor), mutant 343 was fused to the hybridoma igm-12. This hybridoma pro- duces a normal amount of a TNP-specific # chain which lacks exon Cul (6) and can be readily distinguished by SDS gel electropho- resis from the full-length u chain produced by 343. As shown in Fig 7, the fusion product of igm-12 and PC7 produces comparable amounts of the two ~ chains. By contrast, the fusion of igm-12 to 343 yields cell lines which
make much less of the 343 u chain that igm-12 # chain, implying that the defect in 343 is cis-acting. The relationship between the pres- ence of abnormal ~ DNA and the low level of expression of the functional ~t gene remains to be elucidated.
DISCUSSION
In this study we have isolated mutants from sublines of the PC-specific hybridoma PC7. The frequency of independent # and K expression mutants detected was approxi- mately equal and in the range of 1%. Failure to produce IgM was due either to loss of u or K production, or in some cases the loss of both. In the cases studied thus far, ~ expression in this cell line was terminated due to gross structural alterations of the gene, resulting
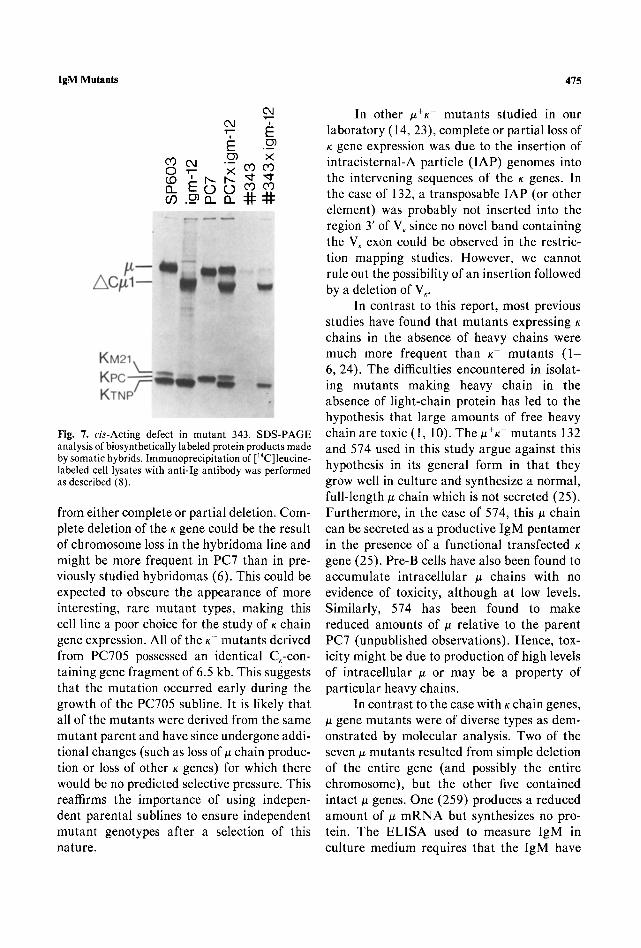
IgM Mutants 475
Cq Cq "7, E E .~
o~ .~ x o •
13_ COCO m . _ a . a_
Fig. 7. cis-Acting defect in mutant 343. SDS-PAGE analysis of biosynthetically labeled protein products made by somatic hybrids. Immunoprecipitation of [14C]leucine- labeled cell lysates with anti-Ig antibody was performed as described (8).
from either complete or partial deletion. Com- plete deletion of the K gene could be the result of chromosome loss in the hybridoma line and might be more frequent in PC7 than in pre- viously studied hybridomas (6). This could be expected to obscure the appearance of more interesting, rare mutant types, making this cell line a poor choice for the study of K chain gene expression. All of the K- mutants derived from PC705 possessed an identical C,-con- taining gene fragment of 6.5 kb. This suggests that the mutation occurred early during the growth of the PC705 subline. It is likely that all of the mutants were derived from the same mutant parent and have since undergone addi- tional changes (such as loss of ~ chain produc- tion or loss of other K genes) for which there would be no predicted selective pressure. This reaffirms the importance of using indepen- dent parental sublines to ensure independent mutant genotypes after a selection of this nature.
In other #+K- mutants studied in our laboratory (14, 23), complete or partial loss of K gene expression was due to the insertion of intracisternal-A particle (IAP) genomes into the intervening sequences of the K genes. In the case of 132, a transposable IAP (or other element) was probably not inserted into the region 3' of V, since no novel band containing the VK exon could be observed in the restric- tion mapping studies. However, we cannot rule out the possibility of an insertion followed by a deletion of V,.
In contrast to this report, most previous studies have found that mutants expressing K chains in the absence of heavy chains were much more frequent than K mutants (1- 6, 24). The difficulties encountered in isolat- ing mutants making heavy chain in the absence of light-chain protein has led to the hypothesis that large amounts of free heavy chain are toxic (1, 10). The~+K mutants 132 and 574 used in this study argue against this hypothesis in its general form in that they grow well in culture and synthesize a normal, full-length # chain which is not secreted (25). Furthermore, in the case of 574, this # chain can be secreted as a productive IgM pentamer in the presence of a functional transfected K gene (25). Pre-B cells have also been found to accumulate intracellular ~ chains with no evidence of toxicity, although at low levels. Similarly, 574 has been found to make reduced amounts of # relative to the parent PC7 (unpublished observations). Hence, tox- icity might be due to production of high levels of intracellular # or may be a property of particular heavy chains.
In contrast to the case with K chain genes, # gene mutants were of diverse types as dem- onstrated by molecular analysis. Two of the seven ~ mutants resulted from simple deletion of the entire gene (and possibly the entire chromosome), but the other five contained intact # genes. One (259) produces a reduced amount of ~ m R N A but synthesizes no pro- tein. The ELISA used to measure IgM in culture medium requires that the IgM have

476 Trimble et al.
PC binding activity and at least some anti- genic determinants of the # constant region. The phenotype of mutant 259 suggests that it might have a nonsense or frameshift muta- tion, perhaps near the amino terminus. This hypothesis is consistent with results reported elsewhere (26), in which various frameshift mutants were found to contain less than nor- mal amounts of g mRNA. In this case, non- sense or frameshift mutations could account for the phenotype and might result in decreased RNA stability. Mutants 350 and 529 differ from 259 in that they produce no detectable g mRNA. These might represent additional frameshift mutants with substan- tially reduced mRNA levels or might be defective in some step of RNA synthesis or processing. Characterization of mutants of this type should be useful in further elucidat- ing the molecular requirements for tran- scription or RNA processing.
In addition to mutants unable to produce any IgM, the enrichment procedure used in this study also selects for mutants making low amounts of pentameric IgM. Two mutants of this type (261 and 343) were studied and found to be different from the low producing K mutant IgK-I which resulted from the trans- position of an IAP (14). In one case (261) no obvious structural changes can be detected at the DNA level, suggesting that a subtle cis- or a trans-acting mutation has resulted in decreased expression of the u gene. However, in the case of mutant 343, a duplication of the # gene extending through the entire constant region downstream of the switch region appears to have occurred. As the structure of the functionally rearranged ~ gene does not appear to have been altered by this duplica- tion event, it is not clear at present what relationship exists between the low level of IgM production and presence of additional DNA. In this case, the mutation acts in cis with the functionally rearranged gene as dem- onstrated by the analysis of gene expression in hybrid cells. Although we cannot as yet rule
out the possibility that partial gene duplica- tion and low levels of ~ chain production might be unrelated, models can be proposed which could account for this phenotype. It has previously been shown by a number of groups (27-29) that the g gene enhancer is not required in cis to maintain the expression of endogenous ~ genes. This has led to specula- tion that other long-range cis-acting elements might exist elsewhere on chromosome 12 and be sufficient for expression of the endogenous genes. Duplication of the heavy chain locus could displace the functionally rearranged gene away from this active locus, and if so, this cell line could be valuable in demonstrat- ing and localizing such an element.
We have previously demonstrated the utility of the PC hapten-specific hybridoma cell line PC7 in the isolation of structural mutants expressing defective IgM antibodies. In the present study we have shown that, despite its chromosomal overload, a large per- centage of the nonexpressing mutants isolated still contain the Ig ~ genes in a form which is not grossly altered. Analysis of mutants of this type should permit us to identify novel ele- ments and mechanisms involved in gene expression.
ACKNOWLEDGMENTS
We thank Dr. G. D'Agostaro, N. Pen- nell, Dr. P. Leder, Dr. M. Scharff, Dr. S.-P. Kwan, and Dr. J. Adams for DNA clones used in these studies. This work was supported by grants from the Medical Research Council of Canada, the National Cancer Institute of Canada, the Arthritis Society of Canada, the Wellesley Hospital Research Institute and the Allstate Foundation. W.S.T. and G.L.B. were supported by studentships from the Medical Research Council and the National Cancer Institute, respectively. M.D.B. was supported by a fellowship from the Medical Research Council of Canada.

IgM Mutants 477
L I T E R A T U R E CITED
1. Coffino, P., and Scharff, M. (1971). Proc. Natl. Acad. Sci. U.S.A. 68:219-224.
2. Baumal, R., Birstein, B., Coffino, P., and Scharff, M. (1973). Science 182:164-166.
3. Laskov, R., and Scharff, M.D. (1974). J. Exp. Med. 140:1112-1116.
4. Baumal, R., and Scharff, M. (1976). J. Immunol. 116:65-74.
5. Mosmann, T., Baumal, R., and Williamson, A. (1979). Eur. J. Immunol. 9:511-516.
6. Kohler, G., and Shulman, M.J. (1980). Eur. J. Immunol. 10"467-476.
7. Morrison, S.L., and Scharff, M.D. (1981). CRC Crit. Rev. Immunol. 3:1-22.
8. Shulman, M.J., Heusser, C., Filkin, C., and Kohler, G. (1982). Mol. Cell. Biol. 2:1033-1043.
9. Hengartner, H., Meo, T., and Muller, E. (1978). Proe. Natl. Acad. Sci. U.S.A. 75:4494-4498.
10. Kohler, G. (1980). Proc. Natl. Acad. Sci U.S.A. 77:2197-2199.
11. Shulman, M., Wilde, C.D., and Kohler, G. (1978). Nature 276:269-270.
12. Gough, N.M., Kemp, D.J., Tyler, M.B., Adams, .I.M., and Cory, S. (1980). Proc. Natl. Acad. Sci. U.S.A. 77:554-558.
13. Kwan, S.-P., Rudikoff, S., Seidman, J.G., Leder, P., and Scharff, M.D. (1981). J. Exp. Med. 153:1366- 1370.
14. Hawley, R.G., Shulman, M.J., Murialdo, H., Gib- son, D.M., and Hozumi, N. (1982). Proc. Natl. Acad. Sci U.S.A. 79:7425-7429.
15. Baczynsky, W.O.T., Sugii, S., Murialdo, H., Pen- nell, N., Filkin, C., Hozumi, N., and Shulman, M.J. (1983). Nucleic Acids Res. 11:7471-7485.
16. Gross-Bellard, M., Dudet, P., and Chambon, P. (1973). Eur. J. Biochem. 36:32-38.
17. Maniatis, T., Fritsch, E.F., and Sambrook, J. (1982). Molecular Cloning--A Laboratory Manu- al, (Cold Spring Harbor Laboratory, New York).
18. Southern E.M. (1975). J. Mol. Biol. 98:503-517. 19. Thomas, P. (1980). Proc. Natl. Acad. Sci. U.S.A.
77:5201-5205. 20. Wilson, R., Miller, J., and Storb, U. (1979). Bio-
chemistry 18:5013-5021. 21. Gearhart, P., Johnson, N.D., Douglas, R., and
Hood, L. (1981). Nature 291:29-34. 22. Crews, S., Griffin, J., Huang, H., Kalame, K., and
Hood, L. (1981). Cell 25:59-66. 23. Kuff, E.L., Feenstra, A., Leuders, K., Smith, L.,
Hawley, R.G., Hozumi, N., and Shulman, M.J. (1983). Proc. Natl. Acad. Sci. U.S.A. 80:1992- 1996.
24. Wims, L., and Morrison, S. (1981). Mutat. Res. 81:215-228.
25. Pepe, V.H., Sonenshein, G.E., Yoshimura, M.I., and Shulman, M.J. (1986). J. Immunol. (in press).
26. Baumann, B., Potash, M.J., and Kohler, G. (1985). E M B O J. 4:351-359.
27. Wabl, M.R., and Burrows, P.D. (1984). Proc. Natl. Acad. Sci. U.S.A. 81:2452-2455.
28. Klein, S., Sablitsky, F., and Radbruch, A. (1984). E M B O J. 3:2473-2476.
29. Zaller, D.M., and Eckhardt, L.A. (1985). Proc. Natl. Acad. Sci. U.S.A. 82:5088-5092.




















