
Leak Detection in Pipelines using the Damping of Fluid Transients

www.elsevier.com/locate/agee
Agriculture, Ecosystems and Environment 106 (2005) 189–198
Analysis of chlorophyll fluorescence transients in mandarin
leaves during a photo-oxidative cold shock and recovery
F. Pietrinia, D. Chaudhurib, A.P. Thapliyalc, A. Massaccia,*
aInstitute of Agro-Environment and Forest Biology of National Research Council,
Via Salaria Km 29,300, 00016 Monterotondo Scalo (Roma), ItalybRubber Research Institute of India, Regional Research Station, Rubber Board, Guwahati, AssamcRubber Research Institute of India, Regional Research Station, Rubber Board, Tura, Meghalaya
Accepted 30 June 2004
Abstract
In temperate climates, plants are more frequently exposed to sudden and strong decrease of temperature combined with high
light intensity that compromise the photosynthetic efficiency and, often, the development and their survival. To investigate
deeper the effects of such a photo-oxidative cold shock on photosynthesis of mandarin (Citrus reticulata Blanco) we used
transient fluorescence analysis and gas exchange techniques. Mandarin plants were acclimated at 13 8C or grown at 25 8C for 30
days, then single leaves were exposed for 6 h at 5 8C and 1200 mmol photons m�2 s�1 and again exposed for 72 h under growth
conditions. Acclimated leaves showed lower photosynthesis and higher total carotenoid contents than non-acclimated leaves.
The photo-oxidative cold shock inhibited photosystem II (PSII) efficiency, as suggested by the reduction of Fv/Fm ratios, in all
leaves. Carotenoid content was reduced in non-acclimated leaves. A time-dependent increase in the initial fluorescence level
(F0) and a faster decline in Fv/Fm ratio indicated a stronger PSII inhibition in non-acclimated leaves in comparison with
acclimated leaves. The chlorophyll a fluorescence kinetic showed in all leaves complex changes in the O–J, J–I and I–P phases.
The I and P steps of the fluorescence rise were particularly affected in non-acclimated leaves. Photosynthesis, fluorescence and
pigment contents, measured in leaves maintained for 2 h in the dark and then returned for 72 h to growth conditions, showed a
different pattern of recovery between leaves. Photosynthesis and fluorescence completely recovered in acclimated leaves after
72 h under growth conditions. On the contrary, non-acclimated leaves still showed very low values of photosynthesis and Fv/Fm,
suppression of the sigmoidal shape of fluorescent transients and further reduction of chlorophyll contents, indicating the
presence of irreversible damages to PSII and, probably, an anticipated senescence. It is suggested that carotenoids have a relevant
role in protecting leaves of mandarin from the effects of a photo-oxidative cold shock.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Chlorophyll fluorescence; Cold-acclimation; Photosystem II; Photo-oxidative cold shock; Recovery; Mandarin (Citrus reticulata
Blanco)
* Corresponding author. Tel.: +39 0690672537; fax: +39 069064492.
E-mail address: [email protected] (A. Massacci).
0167-8809/$ – see front matter # 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.agee.2004.10.007

F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198190
1. Introduction
The occurrence of unexpected frosts, numerous in
the last decades, causes serious damages to many
cultivated species (Zinoni et al., 2002). The higher
frequency of the phenomenon seems to be attributed to
the underway process of hearth warming (EPA
WBBsite, 2004). The higher temperatures, in fact,
anticipate the growth season so that the new leaves are
emitted when the risk of frost is elevated. Never-
theless, if frosts are associated with high light
damaging effects are even more severe (Long et al.,
1994). In these conditions, light energy arriving to
chloroplasts is not utilisable for photosynthesis and
this energy cannot be all convertible into heat. The
formation of singlet oxygen and other active oxygen
species becomes, thus, inevitable and, when the
capacity of the scavenging mechanisms is overloaded,
these species are damaging to pigments, thylakoid
membranes and various components of the photo-
chemical process (Asada and Takahashi, 1987). The
protective conversion of excess light energy into heat
is, then, an extremely important process. It is
considered as a safety valve for photosynthesis and
it is associated with a prompt regulatory reduction of
PSII activity in response to either a sudden or slow
build-up of high transthylakoid DpH (Niyogi, 2000).
When stress factors are removed, PSII function can,
however, recover with a half time depending on the
intensity of the perturbation: 7–10 min or, even,
several hours. In both cases, xanthophylls and
phosphatase inhibitors seem to be involved in a
mechanistically similar process that through confor-
mational changes can induce such rapid or sustained
energy dissipations (Gilmore and Ball, 2000; Xu et al.,
1999). Xanthophylls represent about 80% of total
carotenoids and are mostly associate with polypep-
tides of minor antenna such as CP24, CP26 and CP29
(Bassi et al., 1993) while b-carotene, the other major
component of total carotenoids, is associated with the
core reaction centres of PSII and PSI (Peter and
Thomber, 1991). Studies on an overwintering ever-
green (snow gum) suggested that the conversion
capacity of excess energy into heat increased when
leaves were acclimated to low temperatures (Ottander
et al., 1995). The lower request of energy absorption
and electron transport at PSII induced also in maize a
different organisation and pigment composition of the
photochemical apparatus (Haldimann et al., 1995).
Mandarin, a species growing in temperate climates,
seems to possess some chilling acclimation capacity
but it suffers particularly photo-oxidative cold shock
episodes in early spring. Newly emitted leaves are, in
fact, severely damaged under such a condition
(Yelenosky, 1982; Cakmak et al., 1995). Changes
induced by acclimation to winter temperatures could
involve all cell components particularly chloroplasts.
The capacity of energy dissipation and of other
protective mechanisms (e.g. antioxidants) in these
organelles, in fact, must be increased to prevent
irreversible damages to the photochemical process
that might trigger anticipated senescence. Chlorophyll
a fluorescence is considered as a proper technique to
study the extent of such damaging modifications in
acclimated or stressed leaves (Leipner et al., 1997).
Particularly, the Fv/Fm ratio obtained by the measured
basal F0 and maximum Fm fluorescence emission [Fv/
Fm = (Fm � F0)/Fm] is recognised as a good indicator
of photoinhibitory or photo-oxidative effects on PSII
(Maxwell and Johnson, 2000; Baker et al., 1994).
Further, other fluorescence parameters, derived by the
theory of fluxes, have been suggested to describe
changes of absorbed, dissipative, trapping, and
electron transport fluxes (Lazar, 1999; Strasser et
al., 2000; Force et al., 2003). The analysis of these
parameters, named JIP test, is performed on the typical
fluorescence induction from the basal emission F0 (0)
to a maximum emission level Fm (P) through two
intermediate steps called, according to a recent
terminology, J and I (Strasser and Govindjee, 1992;
Strasser et al., 1995). The fluorescence induction has a
typical sigmoidal shape evidencing three main phases:
O–J, J–I and I–P. Specifically, the first phase O–J is
determined by the charge separation reactions, the
photochemical event that leads to the reduction of the
primary quinone acceptor QA, and seems influenced
by the S-state of the oxygen evolving complex (OEC).
The following J–I phase is sensitive to temperature
and, as well as the I–P phase, reflects the accumulation
of reduced quinones Q�AQ�
B and of Q�AQ�2
B , respec-
tively (Neubauer and Schreiber, 1987; Schreiber and
Krieger, 1996). The heterogeneity of PSII could
account for the occurrence of two phases related to the
reduction of secondary quinones. The time needed to
reach the P step from time zero, when fluorescence
rises, is that needed to reduce the plastoquinone pool

F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198 191
(PQ) and is limited by the slow events in the electron
transport at PSII which include the S-states of OEC
(Stirbet et al., 1998). Although a satisfactory inter-
pretation relating all phases to specific photochemical
events is not yet available the analysis of fluorescence
induction could be, however, considered as an
interesting complement to the information obtainable
by the various classical and well investigated
fluorescence parameters (e.g. F0, Fv, Fv/Fm, qE, qI,
and DF/Fm) (Govindjee, 1995; Force et al.,
2003).
In this study we exposed newly developed leaves at
13 and 25 8C to a photo-oxidative shock at 5 8C and
1200 mmol photons m�2 s�1 and to a recovery period
of 72 h under their growth conditions. The effects of
this treatment were analysed by measuring gas
exchange, fluorescence transients and pigment con-
tents. Our results show that changes induced by low
temperature acclimation allowed mandarin leaves to
tolerate the photo-oxidative shock and to recover
completely photosynthesis and photochemical activ-
ities. The increase of carotenoid contents is suggested
to play an important role in this tolerance.
2. Materials and methods
2.1. Plant material
One-year-old leaves of mandarin (Citrus reticulata
Blanco) were placed in climatic chambers under a
photoperiod of 14 h (600 mmol photons m�2 s�1) at
25 � 2 8C (non-acclimated) for 45 days. At the end of
this period, some leaves were moved to a second
chamber under the same conditions except that the
temperature was set at 13 � 2 8C. Approximately 30
days later, new leaves emitted at this temperature were
fully expanded.
2.2. Photo-oxidative cold shock and recovery
The central part of a last fully expanded leaf was
enclosed in a cuvette in the dark for 20 min at 25 or
13 8C and then for 40 min at 5 8C. After the dark
adaptation at 5 8C, the leaf was exposed at
1200 mmol photons m�2 s�1 for 6 h. Recovery from
this treatment was induced for 2 h in the dark and for
72 h under growth conditions (25 or 13 8C and
600 mmol photons m�2 s�1 for 14 h). This experiment
was replicated four times.
2.3. Fluorescence measurements
Fluorescence transients (OJIP) were measured at
room temperature by a Plant Efficiency Analyser
(Hansatech, King’s Lynn, Norfolk, UK) according to
Strasser et al. (1995). The transients were induced by a
light provided by an array of six light-emitting diodes
(peak 650 nm), which focused on the sample surface
to give homogenous illumination over the exposed
area (4 mm in diameter). The chlorophyll a fluores-
cence signals were recorded for 5 s with data
acquisition rate of 10 ms for the first 2 and 1 ms for
the rest of the time. The fluorescence signal at 50 ms,
the earliest measurement free of any artefacts related
to the electronics of the instrument (Haldimann and
Strasser, 1999), was considered as F0. The maximal
measured fluorescence Fm and the variable fluores-
cence, Fv (obtained by the difference between the
initial fluorescence, F0, and the maximal fluorescence,
Fm of dark adapted leaves), were used to calculate the
Fv/Fm ratios. The duration of dark adaptation required
to obtain the relaxation of the rapid energy-dependent
non-photochemical quenching was experimentally
established to 20 min. At least four repetitions were
done for each measurement.
2.4. Measurements of CO2 assimilation
Measurements of CO2 gas exchange were made
using a LI-6400 (LI-COR, Inc., Lincoln, NE, USA)
portable IRGA analyser. A central part of the leaf was
clamped into a cuvette and illuminated with red light
of 600 mmol m�2 s�1. Measurements were taken
under steady CO2 uptake maintained for 5 min, at
the constant relative humidity of 50% and at the
temperature of 25 8C. Four replicates were done for
this measurement.
2.5. Pigment analysis
Two square centimeters of the last full-expanded
leaf used for photosynthesis and fluorescence mea-
surements were ground under dim light in a mortar
containing liquid N2. When the leaf was reduced to a
fine powder, 2 ml of acetone–water (80%, v/v) were
F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198192
added to extract the pigments. The samples were
centrifuged at 12 000 � g and 5 8C for 10 min, and the
supernatant was removed and used for pigment
determinations. Absorbance was measured at 470,
646.8 and 663.2 nm with a spectrophotometer (Perkin
Elmer, Norwalk, CT, USA). The extinction coeffi-
cients and the equations reported by Lichtenthaler
(1987) were used to calculate the chlorophyll a and b
and total carotenoid contents. Four replicates were
done for each measurement.
2.6. Statistical analysis
In any one experiment, all the measurements were
made on at least four replicates. To evaluate the
significance of photosynthesis activity, fluorescence
and contents of pigments data the Student–Newman–
Keuls test was performed.
3. Results
3.1. Effects of cold-acclimation
3.1.1. Chlorophyll a fluorescence
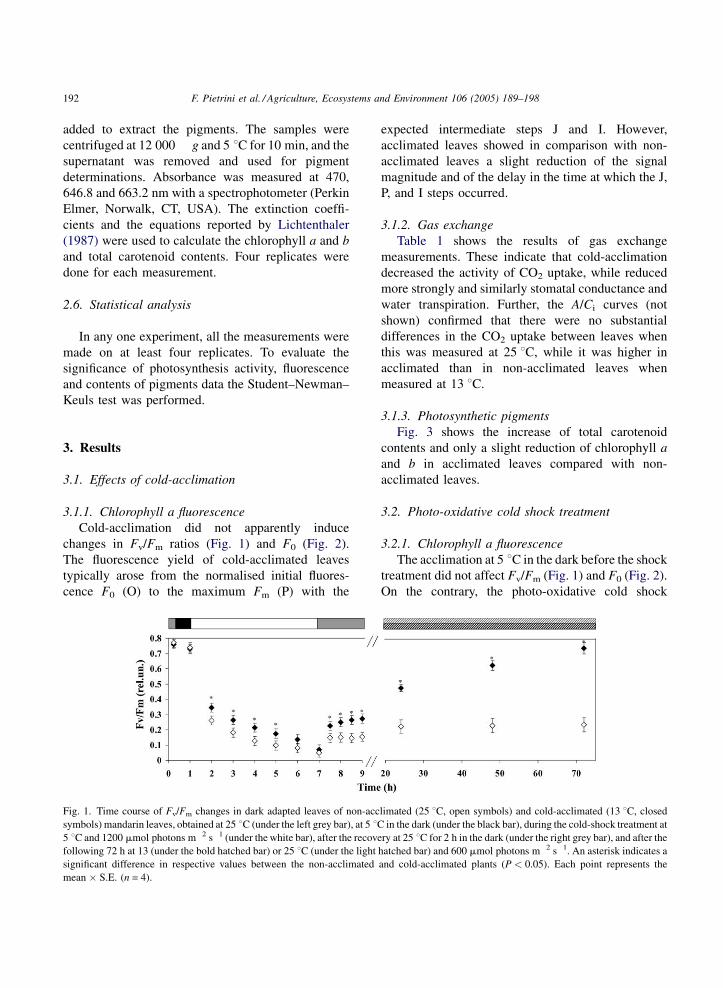
Cold-acclimation did not apparently induce
changes in Fv/Fm ratios (Fig. 1) and F0 (Fig. 2).
The fluorescence yield of cold-acclimated leaves
typically arose from the normalised initial fluores-
cence F0 (O) to the maximum Fm (P) with the
Fig. 1. Time course of Fv/Fm changes in dark adapted leaves of non-acc
symbols) mandarin leaves, obtained at 25 8C (under the left grey bar), at 5 8C5 8C and 1200 mmol photons m�2 s�1 (under the white bar), after the recove
following 72 h at 13 (under the bold hatched bar) or 25 8C (under the light
significant difference in respective values between the non-acclimated
mean � S.E. (n = 4).
expected intermediate steps J and I. However,
acclimated leaves showed in comparison with non-
acclimated leaves a slight reduction of the signal
magnitude and of the delay in the time at which the J,
P, and I steps occurred.
3.1.2. Gas exchange
Table 1 shows the results of gas exchange
measurements. These indicate that cold-acclimation
decreased the activity of CO2 uptake, while reduced
more strongly and similarly stomatal conductance and
water transpiration. Further, the A/Ci curves (not
shown) confirmed that there were no substantial
differences in the CO2 uptake between leaves when
this was measured at 25 8C, while it was higher in
acclimated than in non-acclimated leaves when
measured at 13 8C.
3.1.3. Photosynthetic pigments
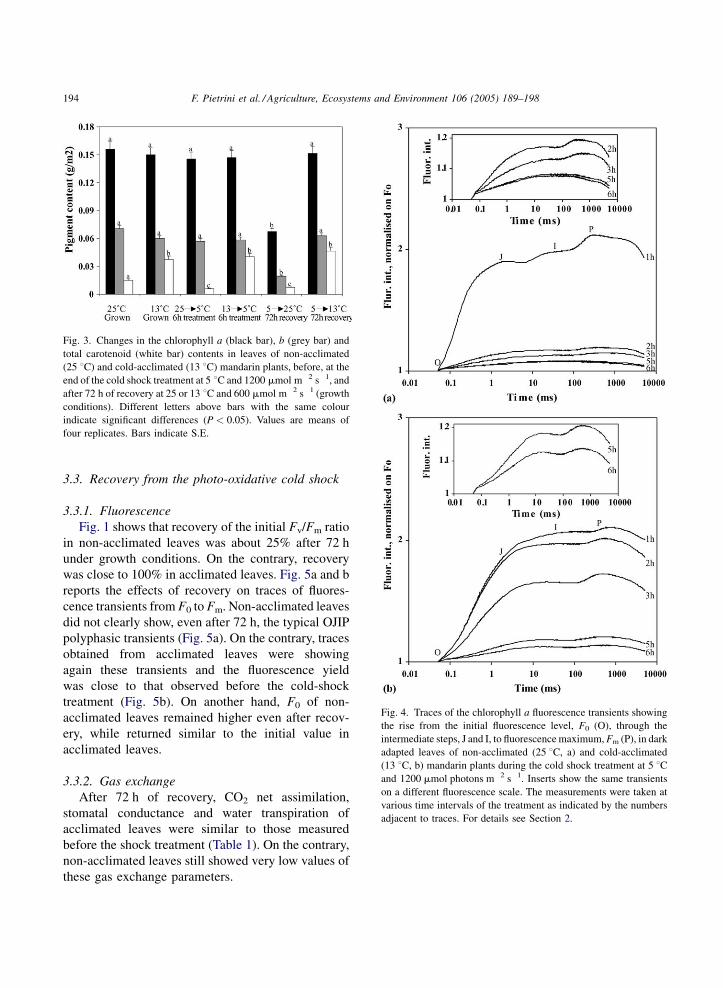
Fig. 3 shows the increase of total carotenoid
contents and only a slight reduction of chlorophyll a
and b in acclimated leaves compared with non-
acclimated leaves.
3.2. Photo-oxidative cold shock treatment
3.2.1. Chlorophyll a fluorescence
The acclimation at 5 8C in the dark before the shock
treatment did not affect Fv/Fm (Fig. 1) and F0 (Fig. 2).
On the contrary, the photo-oxidative cold shock
limated (25 8C, open symbols) and cold-acclimated (13 8C, closed
in the dark (under the black bar), during the cold-shock treatment at
ry at 25 8C for 2 h in the dark (under the right grey bar), and after the
hatched bar) and 600 mmol photons m�2 s�1. An asterisk indicates a
and cold-acclimated plants (P < 0.05). Each point represents the
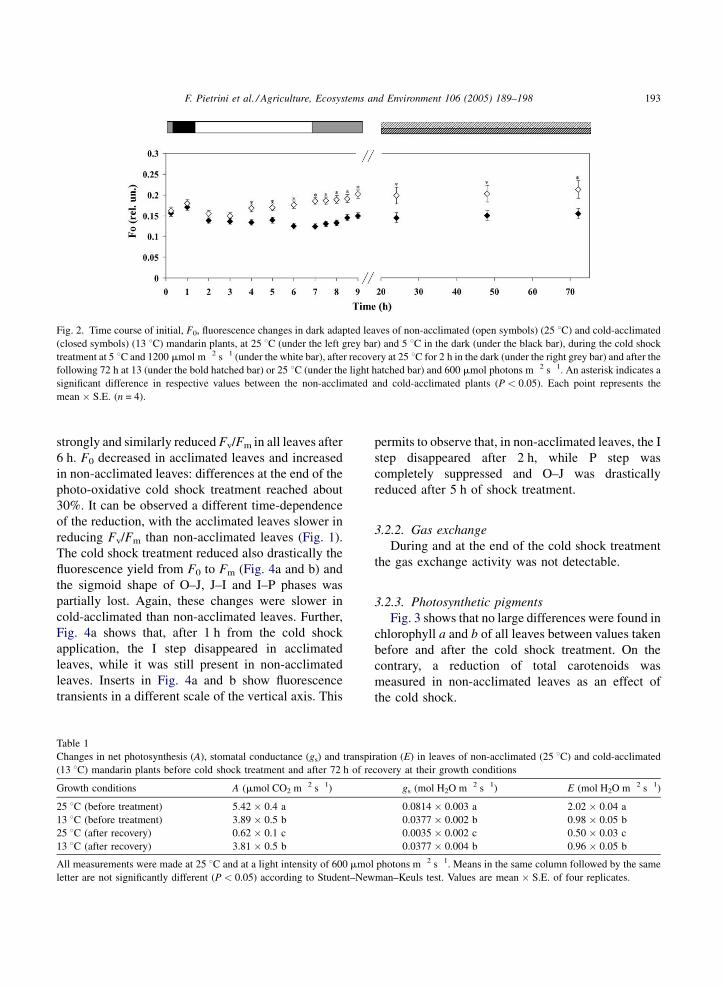
F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198 193
Fig. 2. Time course of initial, F0, fluorescence changes in dark adapted leaves of non-acclimated (open symbols) (25 8C) and cold-acclimated
(closed symbols) (13 8C) mandarin plants, at 25 8C (under the left grey bar) and 5 8C in the dark (under the black bar), during the cold shock
treatment at 5 8C and 1200 mmol m�2 s�1 (under the white bar), after recovery at 25 8C for 2 h in the dark (under the right grey bar) and after the
following 72 h at 13 (under the bold hatched bar) or 25 8C (under the light hatched bar) and 600 mmol photons m�2 s�1. An asterisk indicates a
significant difference in respective values between the non-acclimated and cold-acclimated plants (P < 0.05). Each point represents the
mean � S.E. (n = 4).
strongly and similarly reduced Fv/Fm in all leaves after
6 h. F0 decreased in acclimated leaves and increased
in non-acclimated leaves: differences at the end of the
photo-oxidative cold shock treatment reached about
30%. It can be observed a different time-dependence
of the reduction, with the acclimated leaves slower in
reducing Fv/Fm than non-acclimated leaves (Fig. 1).
The cold shock treatment reduced also drastically the
fluorescence yield from F0 to Fm (Fig. 4a and b) and
the sigmoid shape of O–J, J–I and I–P phases was
partially lost. Again, these changes were slower in
cold-acclimated than non-acclimated leaves. Further,
Fig. 4a shows that, after 1 h from the cold shock
application, the I step disappeared in acclimated
leaves, while it was still present in non-acclimated
leaves. Inserts in Fig. 4a and b show fluorescence
transients in a different scale of the vertical axis. This
Table 1
Changes in net photosynthesis (A), stomatal conductance (gs) and transpi
(13 8C) mandarin plants before cold shock treatment and after 72 h of re
Growth conditions A (mmol CO2 m�2 s�1)
25 8C (before treatment) 5.42 � 0.4 a
13 8C (before treatment) 3.89 � 0.5 b
25 8C (after recovery) 0.62 � 0.1 c
13 8C (after recovery) 3.81 � 0.5 b
All measurements were made at 25 8C and at a light intensity of 600 mmol
letter are not significantly different (P < 0.05) according to Student–New
permits to observe that, in non-acclimated leaves, the I
step disappeared after 2 h, while P step was
completely suppressed and O–J was drastically
reduced after 5 h of shock treatment.
3.2.2. Gas exchange
During and at the end of the cold shock treatment
the gas exchange activity was not detectable.
3.2.3. Photosynthetic pigments
Fig. 3 shows that no large differences were found in
chlorophyll a and b of all leaves between values taken
before and after the cold shock treatment. On the
contrary, a reduction of total carotenoids was
measured in non-acclimated leaves as an effect of
the cold shock.
ration (E) in leaves of non-acclimated (25 8C) and cold-acclimated
covery at their growth conditions
gs (mol H2O m�2 s�1) E (mol H2O m�2 s�1)
0.0814 � 0.003 a 2.02 � 0.04 a
0.0377 � 0.002 b 0.98 � 0.05 b
0.0035 � 0.002 c 0.50 � 0.03 c
0.0377 � 0.004 b 0.96 � 0.05 b
photons m�2 s�1. Means in the same column followed by the same
man–Keuls test. Values are mean � S.E. of four replicates.
F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198194
Fig. 3. Changes in the chlorophyll a (black bar), b (grey bar) and
total carotenoid (white bar) contents in leaves of non-acclimated
(25 8C) and cold-acclimated (13 8C) mandarin plants, before, at the
end of the cold shock treatment at 5 8C and 1200 mmol m�2 s�1, and
after 72 h of recovery at 25 or 13 8C and 600 mmol m�2 s�1 (growth
conditions). Different letters above bars with the same colour
indicate significant differences (P < 0.05). Values are means of
four replicates. Bars indicate S.E.
Fig. 4. Traces of the chlorophyll a fluorescence transients showing
the rise from the initial fluorescence level, F0 (O), through the
intermediate steps, J and I, to fluorescence maximum, Fm (P), in dark
adapted leaves of non-acclimated (25 8C, a) and cold-acclimated
(13 8C, b) mandarin plants during the cold shock treatment at 5 8Cand 1200 mmol photons m�2 s�1. Inserts show the same transients
on a different fluorescence scale. The measurements were taken at
various time intervals of the treatment as indicated by the numbers
adjacent to traces. For details see Section 2.
3.3. Recovery from the photo-oxidative cold shock
3.3.1. Fluorescence
Fig. 1 shows that recovery of the initial Fv/Fm ratio
in non-acclimated leaves was about 25% after 72 h
under growth conditions. On the contrary, recovery
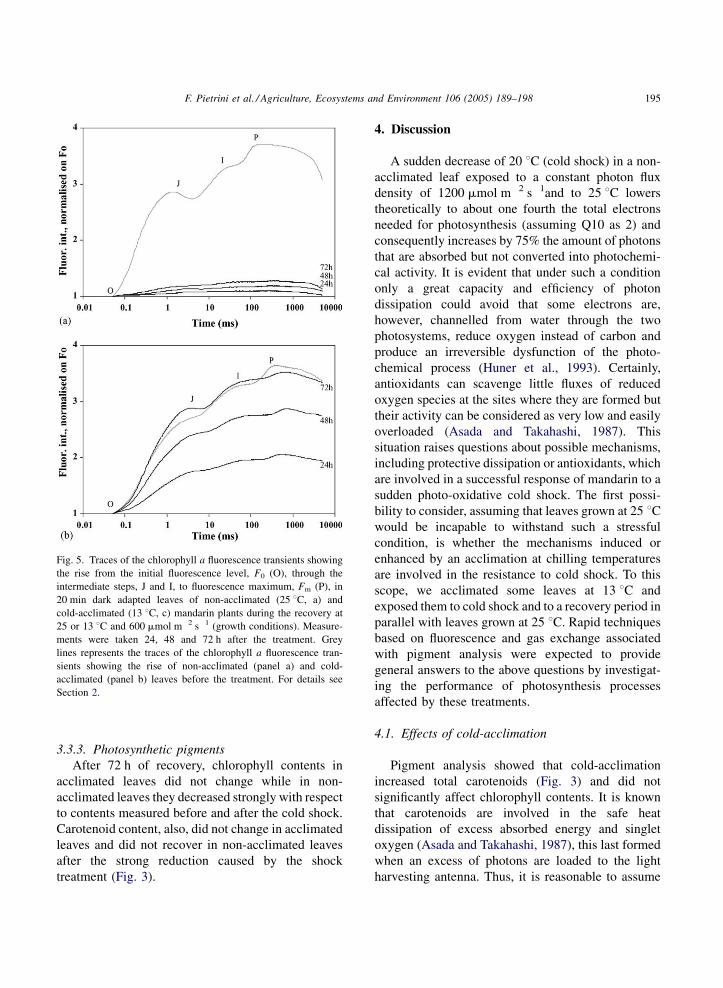
was close to 100% in acclimated leaves. Fig. 5a and b
reports the effects of recovery on traces of fluores-
cence transients from F0 to Fm. Non-acclimated leaves
did not clearly show, even after 72 h, the typical OJIP
polyphasic transients (Fig. 5a). On the contrary, traces
obtained from acclimated leaves were showing
again these transients and the fluorescence yield
was close to that observed before the cold-shock
treatment (Fig. 5b). On another hand, F0 of non-
acclimated leaves remained higher even after recov-
ery, while returned similar to the initial value in
acclimated leaves.
3.3.2. Gas exchange
After 72 h of recovery, CO2 net assimilation,
stomatal conductance and water transpiration of
acclimated leaves were similar to those measured
before the shock treatment (Table 1). On the contrary,
non-acclimated leaves still showed very low values of
these gas exchange parameters.
F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198 195
Fig. 5. Traces of the chlorophyll a fluorescence transients showing
the rise from the initial fluorescence level, F0 (O), through the
intermediate steps, J and I, to fluorescence maximum, Fm (P), in
20 min dark adapted leaves of non-acclimated (25 8C, a) and
cold-acclimated (13 8C, c) mandarin plants during the recovery at
25 or 13 8C and 600 mmol m�2 s�1 (growth conditions). Measure-
ments were taken 24, 48 and 72 h after the treatment. Grey
lines represents the traces of the chlorophyll a fluorescence tran-
sients showing the rise of non-acclimated (panel a) and cold-
acclimated (panel b) leaves before the treatment. For details see
Section 2.
3.3.3. Photosynthetic pigments
After 72 h of recovery, chlorophyll contents in
acclimated leaves did not change while in non-
acclimated leaves they decreased strongly with respect
to contents measured before and after the cold shock.
Carotenoid content, also, did not change in acclimated
leaves and did not recover in non-acclimated leaves
after the strong reduction caused by the shock
treatment (Fig. 3).
4. Discussion
A sudden decrease of 20 8C (cold shock) in a non-
acclimated leaf exposed to a constant photon flux
density of 1200 mmol m�2 s�1and to 25 8C lowers
theoretically to about one fourth the total electrons
needed for photosynthesis (assuming Q10 as 2) and
consequently increases by 75% the amount of photons
that are absorbed but not converted into photochemi-
cal activity. It is evident that under such a condition
only a great capacity and efficiency of photon
dissipation could avoid that some electrons are,
however, channelled from water through the two
photosystems, reduce oxygen instead of carbon and
produce an irreversible dysfunction of the photo-
chemical process (Huner et al., 1993). Certainly,
antioxidants can scavenge little fluxes of reduced
oxygen species at the sites where they are formed but
their activity can be considered as very low and easily
overloaded (Asada and Takahashi, 1987). This
situation raises questions about possible mechanisms,
including protective dissipation or antioxidants, which
are involved in a successful response of mandarin to a
sudden photo-oxidative cold shock. The first possi-
bility to consider, assuming that leaves grown at 25 8Cwould be incapable to withstand such a stressful
condition, is whether the mechanisms induced or
enhanced by an acclimation at chilling temperatures
are involved in the resistance to cold shock. To this
scope, we acclimated some leaves at 13 8C and
exposed them to cold shock and to a recovery period in
parallel with leaves grown at 25 8C. Rapid techniques
based on fluorescence and gas exchange associated
with pigment analysis were expected to provide
general answers to the above questions by investigat-
ing the performance of photosynthesis processes
affected by these treatments.
4.1. Effects of cold-acclimation
Pigment analysis showed that cold-acclimation
increased total carotenoids (Fig. 3) and did not
significantly affect chlorophyll contents. It is known
that carotenoids are involved in the safe heat
dissipation of excess absorbed energy and singlet
oxygen (Asada and Takahashi, 1987), this last formed
when an excess of photons are loaded to the light
harvesting antenna. Thus, it is reasonable to assume
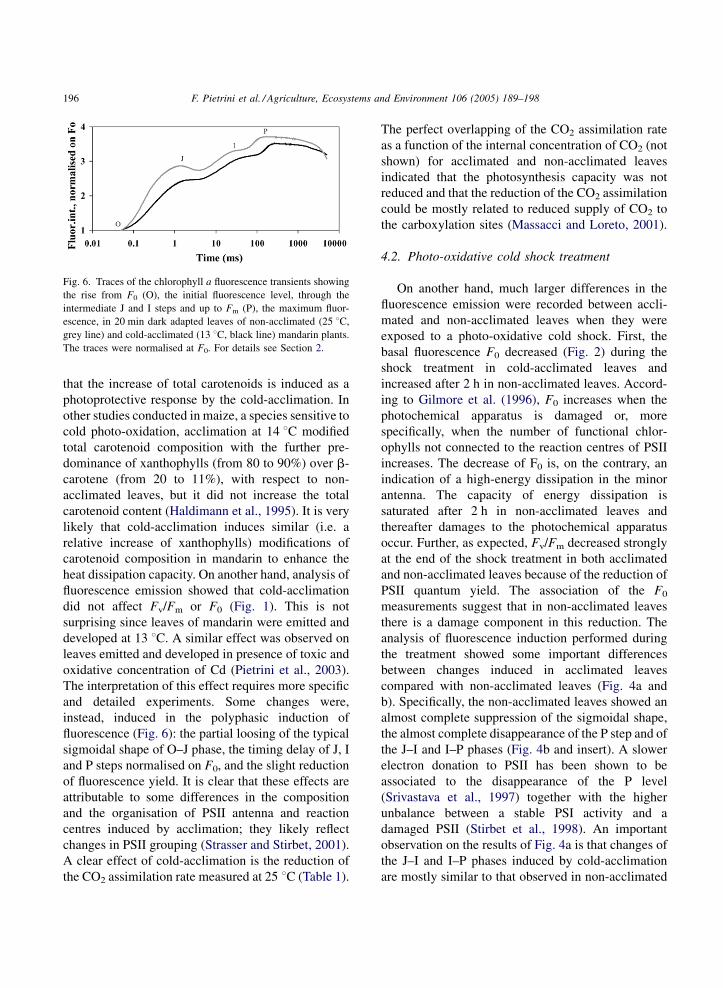
F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198196
Fig. 6. Traces of the chlorophyll a fluorescence transients showing
the rise from F0 (O), the initial fluorescence level, through the
intermediate J and I steps and up to Fm (P), the maximum fluor-
escence, in 20 min dark adapted leaves of non-acclimated (25 8C,
grey line) and cold-acclimated (13 8C, black line) mandarin plants.
The traces were normalised at F0. For details see Section 2.
that the increase of total carotenoids is induced as a
photoprotective response by the cold-acclimation. In
other studies conducted in maize, a species sensitive to
cold photo-oxidation, acclimation at 14 8C modified
total carotenoid composition with the further pre-
dominance of xanthophylls (from 80 to 90%) over b-
carotene (from 20 to 11%), with respect to non-
acclimated leaves, but it did not increase the total
carotenoid content (Haldimann et al., 1995). It is very
likely that cold-acclimation induces similar (i.e. a
relative increase of xanthophylls) modifications of
carotenoid composition in mandarin to enhance the
heat dissipation capacity. On another hand, analysis of
fluorescence emission showed that cold-acclimation
did not affect Fv/Fm or F0 (Fig. 1). This is not
surprising since leaves of mandarin were emitted and
developed at 13 8C. A similar effect was observed on
leaves emitted and developed in presence of toxic and
oxidative concentration of Cd (Pietrini et al., 2003).
The interpretation of this effect requires more specific
and detailed experiments. Some changes were,
instead, induced in the polyphasic induction of
fluorescence (Fig. 6): the partial loosing of the typical
sigmoidal shape of O–J phase, the timing delay of J, I
and P steps normalised on F0, and the slight reduction
of fluorescence yield. It is clear that these effects are
attributable to some differences in the composition
and the organisation of PSII antenna and reaction
centres induced by acclimation; they likely reflect
changes in PSII grouping (Strasser and Stirbet, 2001).
A clear effect of cold-acclimation is the reduction of
the CO2 assimilation rate measured at 25 8C (Table 1).
The perfect overlapping of the CO2 assimilation rate
as a function of the internal concentration of CO2 (not
shown) for acclimated and non-acclimated leaves
indicated that the photosynthesis capacity was not
reduced and that the reduction of the CO2 assimilation
could be mostly related to reduced supply of CO2 to
the carboxylation sites (Massacci and Loreto, 2001).
4.2. Photo-oxidative cold shock treatment
On another hand, much larger differences in the
fluorescence emission were recorded between accli-
mated and non-acclimated leaves when they were
exposed to a photo-oxidative cold shock. First, the
basal fluorescence F0 decreased (Fig. 2) during the
shock treatment in cold-acclimated leaves and
increased after 2 h in non-acclimated leaves. Accord-
ing to Gilmore et al. (1996), F0 increases when the
photochemical apparatus is damaged or, more
specifically, when the number of functional chlor-
ophylls not connected to the reaction centres of PSII
increases. The decrease of F0 is, on the contrary, an
indication of a high-energy dissipation in the minor
antenna. The capacity of energy dissipation is
saturated after 2 h in non-acclimated leaves and
thereafter damages to the photochemical apparatus
occur. Further, as expected, Fv/Fm decreased strongly
at the end of the shock treatment in both acclimated
and non-acclimated leaves because of the reduction of
PSII quantum yield. The association of the F0
measurements suggest that in non-acclimated leaves
there is a damage component in this reduction. The
analysis of fluorescence induction performed during
the treatment showed some important differences
between changes induced in acclimated leaves
compared with non-acclimated leaves (Fig. 4a and
b). Specifically, the non-acclimated leaves showed an
almost complete suppression of the sigmoidal shape,
the almost complete disappearance of the P step and of
the J–I and I–P phases (Fig. 4b and insert). A slower
electron donation to PSII has been shown to be
associated to the disappearance of the P level
(Srivastava et al., 1997) together with the higher
unbalance between a stable PSI activity and a
damaged PSII (Stirbet et al., 1998). An important
observation on the results of Fig. 4a is that changes of
the J–I and I–P phases induced by cold-acclimation
are mostly similar to that observed in non-acclimated

F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198 197
leaves but are time-dependent, they appears with a
delay of about 3 h. This suggests that acclimation
enhances the photoprotection but it cannot completely
remove some causes of the progressive damage to the
photochemical apparatus. An interesting aspect of this
cold shock treatment is the decrease of carotenoid
contents in non-acclimated leaves. Very likely the
energy load to the photochemical apparatus is so high
that singlet oxygen and other oxygen active species are
formed in large amount causing also destruction of b-
carotenes and of xantophylls. In maize, a species
sensitive to chilling, the exposure to 14 8C induced a
decrease of b-carotenes compensated by a similar
increase of xantophylls (Haldimann et al., 1995).
4.3. Recovery from the photo-oxidative cold shock
The return to growth conditions evidenced that all
effects of the cold treatment on the photosynthesis
process were reversible in cold-acclimated leaves and
irreversible in non-acclimated leaves (Table 1 and Fig.
1). Cold-acclimation seems to induce physiological
and biochemical changes that confer mandarin the
capacity to tolerate a sudden and intense photo-
oxidative event. The increase of total carotenoids has
certainly some positive role in this acquired tolerance
as indicated by many works on xantophylls, the major
components of total carotenoids (Leipner et al., 1997;
Niyogi et al., 1997). On another hand, the high
susceptibility of mandarin to cold shocks is clearly
evidenced in the measurements of non-acclimated
leaves. In fact, chlorophyll contents were further
reduced during 72 h recovery. Photosynthesis
appeared irreversibly inhibited with damages to the
photochemical apparatus as indicated by the low Fv/Fm
(Fig. 1). F0 continued decreasing (Fig. 2). Dysfunctions
at donor and acceptor sides of PSII were maintained and
reflected in the shapeless fluorescence transients (Fig.
5a). Some studies have already related reduction of
photosynthesis with degradation of rubisco and chlor-
ophylls by active oxygen species as symptoms of
senescence (Pietrini et al., 1999; Desimone et al., 1998).
5. Conclusions
These results seem to indicate that the mechanisms
enhanced by cold-acclimation conferred mandarin the
capacity to tolerate a sudden and intense photo-
oxidative shock to leaves. The increase of total
carotenoids after cold-acclimation and their decrease
in non-acclimated leaves during the cold shock
indicates that these molecules might have an
important role in this capacity.
References
Asada, K., Takahashi, M., 1987. Production and scavenging of
active oxygen in photosynthesis. In: Kyle, D.J., Osmond,
C.B., Arntzen, C.D. (Eds.), Photoinhibition. Topics in Photo-
synthesis, vol. 9. Elsevier, Amsterdam, pp. 227–287.
Baker, N.R., Farage, P.K., Stirling, C.M., Long, S.P., 1994. Photo-
inhibition of crop photosynthesis in the field at low temperatures.
In: Baker, N.R., Bowyer, J.R. (Eds.), Photoinhibition of Photo-
synthesis from Molecular Mechanisms to the Field. bios Scien-
tific Publisher, Oxford, pp. 349–364.
Bassi, R., Pineau, B., Danese, P., Marquardt, J., 1993. Carotenoid-
binding proteins of photosystem II. Eur. J. Biochem. 212, 297–
303.
Cakmak, I., Atli, M., Kaya, R., Evliya, H., Marschner, H., 1995.
Association of high light and zinc deficiency in cold-induced
leaf chlorosis in grapefruit and mandarin trees. J. Plant Physiol.
146, 355–360.
Desimone, M., Wagner, E., Joanningmeier, U., 1998. Degradation of
active-oxygen-modified ribulose-1,5-bisphosphate carboxylase/
oxygenase by chloroplastic proteases requires ATP-hydrolysis.
Planta 205, 459–466.
EPA WBBsite, 2004. http://www.epa.gov/globalwarming/.
Force, L., Critcley, C., van Rensen, J.J.S., 2003. New fluorescence
parameters for monitoring photosynthesis in leaves. Photosynth.
Res. 78, 17–33.
Gilmore, A.M., Ball, M.C., 2000. Protection and storage of chlor-
ophyll in over-wintering evergreens. Proc. Natl. Acad. Sci.
U.S.A. 97, 11098–11101.
Gilmore, A.M., Hazlett, T.L., Debrunner, P.G., Govindjee, 1996.
Comparative time-resolved photosystem II chlorophyll a fluor-
escence analyses reveal distinctive differences between photo-
inhibitory reaction center damage and xanthophyll cycle-
dependent energy dissipation. Photochem. Photobiol. 64,
552–563.
Govindjee, 1995. Sixty-three years since Kautsky: chlorophyll a
fluorescence. Aust. J. Plant Physiol. 22, 131–160.
Haldimann, P., Fracheboud, Y., Stamp, P., 1995. Carotenoid com-
position in Zea mays developed at sub-optimal temperature and
different light intensities. Physiol. Planta. 95, 409–414.
Haldimann, P., Strasser, R.J., 1999. Effects of anaerobiosis as probed
by the polyphasic chlorophyll a fluorescence rise kinetic in pea
(Pisum sativum L.). Photosynth. Res. 62, 67–83.
Huner, N.P.A., Oquist, G., Hurry, V.M., Krol, M., Falk, S., Griffith,
M., 1993. Photosynthesis, photoinhibition and low temperature
acclimation in cold tolerant leaves. Photosynth. Res. 37,
19–39.

F. Pietrini et al. / Agriculture, Ecosystems and Environment 106 (2005) 189–198198
Lazar, D., 1999. Chlorophyll a fluorescence induction. Biochim.
Biophys. Acta 1412, 1–28.
Leipner, J., Fracheboud, Y., Stamp, P., 1997. Acclimation by sub-
optimal growth temperature diminishes photo-oxidative damage
in maize leaves. Plant Cell Environ. 20, 366–372.
Lichtenthaler, H.K., 1987. Chlorophylls and carotenoids: pigments
of photosynthetic biomembranes. Methods in Enzymology, vol.
148, Academic Press, London, pp. 350–382.
Long, S.P., Humphries, S., Falkowski, P.G., 1994. Photoinhibition of
photosynthesis in nature. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 45, 633–662.
Massacci, A., Loreto, F., 2001. Diffusive resistances to CO2 entry in
the leaves and their limitations to photosynthesis. In: Pessarakli,
M. (Ed.), Handbook of Plant and Crop Physiology. Marcel
Dekker, New York, pp. 327–336.
Maxwell, K., Johnson, G.N., 2000. Chlorophyll fluorescence—a
practical guide. J. Exp. Bot. 51, 659–668.
Neubauer, C., Schreiber, U., 1987. The polyphasic rise of chlor-
ophyll fluorescence upon onset of strong continuous illumina-
tion. I. Saturation characteristics and partial control by the
photosystem Il acceptor side. Z. Naturforsch. 42c, 1246–1254.
Niyogi, K.K., 2000. Safety valves for photosynthesis. Curr. Opin.
Plant Biol. 3, 455–460.
Niyogi, K.K., Bjorkman, O., Grossman, A.R., 1997. The roles of
specific xanthophylls in photoprotection. Proc. Natl. Acad. Sci.
U.S.A. 94, 14162–14167.
Ottander, C., Campbell, D., Oquist, G., 1995. Seasonal changes in
photosystem II organization and pigment composition in Pinus
sylvestris. Planta 197, 176–183.
Peter, G.F., Thomber, J.P., 1991. Biochemical composition and
organization pf higher plant photosystem light-harvesting pig-
ment-protein. J. Biol. Chem. 266, 16745–16754.
Pietrini, F., Iannelli, M.A., Battistelli, A., Moscatello, S., Loreto, F.,
Massacci, A., 1999. Effects on photosynthesis, carbohydrate
accumulation, and re-growth induced by temperature increase in
maize genotypes with different sensitivity to low temperature.
Aust. J. Plant Physiol. 26, 367–373.
Pietrini, F., Iannelli, A.M., Pasqualini, S., Massacci, A., 2003.
Interaction of cadmium with glutathione and photosynthesis
in developing leaves and chloroplasts of Phragmites australis
Trin. ex Steudel. Plant Physiol. 133 (2), 829–837.
Schreiber, U., Krieger, A., 1996. Two fundamentally different
variable chlorophyll fluorescence in vivo. FEBS Lett. 397,
131–135.
Srivastava, A., Guisse, B., Greppin, H., Strasser, R.J., 1997. Reg-
ulation of antenna structure and electron transport in photosys-
tem II of Pisum sativum under elevated temperature probed by
the fast polyphasic chlorophyll a fluorescence transient: OKJIP.
Biochim. Biophys. Acta 1320, 95–106.
Stirbet, A., Govindjee, Strasser, B.J., Strasser, R.J., 1998. Chlor-
ophyll a fluorescence induction in higher leaves: modelling and
numerical simulation. J. Theor. Biol. 193, 131–151.
Strasser, R.J., Govindjee, 1992. On the O–J–I–P fluorescent tran-
sient in leaves and D1 mutants of Chlamydomonas reinhardtii.
In: Murata, N. (Ed.), Research in Photosynthesis, vol. 4.
Kluwer Academic Publishers, Dordrecht, The Netherlands,
pp. 29–32.
Strasser, R.J., Srivastava, A., Govindjee, 1995. Polyphasic chlor-
ophyll a fluorescence transient in plant and cyanobacteria.
Photochem. Photobiol. 61, 32–42.
Strasser, R.J., Srivastava, A., Tsimilli-Michael, M., 2000. The
fluorescence transient as a tool to characterize and screen
photosynthetic samples. In: Yunus, M., Pathre, U., Mohanty,
P. (Eds.), Probing Photosynthesis: Mechanisms, Regulation
and Adaptation. Taylor and Francis, London, pp. 445–
483.
Strasser, R.J., Stirbet, A., 2001. Estimation of the energetic con-
nectivity of PSII centres in leaves using the fluorescence rise O–
J–I–P. Fitting of experimental data to three different PSII
models. Math. Comput. Simulat. 56, 451–461.
Xu, C.C., Jeon, Y.A., Hwang, H.J., Lee, C.H., 1999. Suppression of
zeaxanthin epoxidation by chloroplast phosphatase inhibitors in
rice leaves. Plant Sci. 146, 27–34.
Yelenosky, G., 1982. Chilling injury in leaves of citrus leaves at
1.7 8C. Hort. Sci. 17, 385–387.
Zinoni, F., Antolini, G., Campisi, T., Marletto, V., Rossi, F., 2002.
Characterization of Emilia Romagna in relation with late frost
risk. Chem. Phys. Earth 27, 1091–1102.
Copyright © 2022 FDOKUMEN










![Near-Membrane [Ca2+] Transients Resolved Using the Ca2+ Indicator FFP18](https://static.fdokumen.com/doc/165x107/631286873ed465f0570a4533/near-membrane-ca2-transients-resolved-using-the-ca2-indicator-ffp18.jpg)









