
Análisis de las técnicas empleadas para el estudio de los restos óseos humanos
23
ANÁLISIS DE LAS TÉCNICAS EMPLEADAS PARA EL ESTUDIO DE RESTOS ÓSEOS HUMANOS Ángel Espino Herrera Trabajo para la asignatura: Prácticas en técnicas de Laboratorio Universidad Autónoma de Madrid RESUMEN Hoy en día, el campo de la investigación arqueológica recibe cada vez más la inestimable ayuda de la tecnología, pudiendo realizar tareas que hace años le serían impensables, como observar algo con miles de aumentos, saber de qué se alimentaban en el pasado o relacionar genéticamente poblaciones de diferente espacio o tiempo. Sin embargo, es necesario saber cómo hacer uso adecuadamente de esta tecnología, para interpretar el pasado de la manera más acertada posible. En este trabajo se desarrolla una síntesis de las principales técnicas en investigación arqueológica, centrando la atención en el estudio de los restos óseos e ilustrando cada técnica con ejemplos que puedan ayudar a su comprensión. Palabras clave: huesos humanos, colágeno, ADN, PCR, isótopos, paleodietas, ICP-MS, FRX, SEM ABSTRACT Nowadays, the field of archaeological research receives increasingly the priceless help of the technology, being able to solve tasks in a manner which would seem unthinkable years ago, as watching something thousands of times zoomed in, knowing what did the people from the past eat or relate genetically populations coming from different ages or locations. However, knowledge of the correct use of this technology is needed, in order to read the past in the most accurate way. On this paper, a synthesis of the main techniques for archaeological research is developed, focusing the attention on the study of skeletal remains and giving examples for the application of each technique to provide its understanding. Keywords: human bones, collagen, DNA, PCR, isotopes, paleodiets, ICP-MS, XRF, SEM
Transcript of Análisis de las técnicas empleadas para el estudio de los restos óseos humanos

ANÁLISIS DE LAS TÉCNICAS EMPLEADAS PARA EL ESTUDIO DE RESTOS ÓSEOS HUMANOS
Ángel Espino Herrera
Trabajo para la asignatura: Prácticas en técnicas de Laboratorio
Universidad Autónoma de Madrid
RESUMEN
Hoy en día, el campo de la investigación arqueológica recibe cada vez más la inestimable ayuda de la tecnología, pudiendo realizar tareas que hace años le serían impensables, como observar algo con miles de aumentos, saber de qué se alimentaban en el pasado o relacionar genéticamente poblaciones de diferente espacio o tiempo. Sin embargo, es necesario saber cómo hacer uso adecuadamente de esta tecnología, para interpretar el pasado de la manera más acertada posible. En este trabajo se desarrolla una síntesis de las principales técnicas en investigación arqueológica, centrando la atención en el estudio de los restos óseos e ilustrando cada técnica con ejemplos que puedan ayudar a su comprensión.
Palabras clave: huesos humanos, colágeno, ADN, PCR, isótopos, paleodietas, ICP-MS, FRX, SEM
ABSTRACT
Nowadays, the field of archaeological research receives increasingly the priceless help of the technology, being able to solve tasks in a manner which would seem unthinkable years ago, as watching something thousands of times zoomed in, knowing what did the people from the past eat or relate genetically populations coming from different ages or locations. However, knowledge of the correct use of this technology is needed, in order to read the past in the most accurate way. On this paper, a synthesis of the main techniques for archaeological research is developed, focusing the attention on the study of skeletal remains and giving examples for the application of each technique to provide its understanding.
Keywords: human bones, collagen, DNA, PCR, isotopes, paleodiets, ICP-MS, XRF, SEM

1. INTRODUCCIÓN
A menudo, uno de los objetivos más presentes en el campo de la investigación arqueológica es derribar la concepción de arqueólogo que trabaja por sí solo sin valerse de la ayuda de profesionales de otras disciplinas: el arqueólogo que excava, que conserva y restaura, que analiza, que investiga, que interpreta... Saber colaborar de manera adecuada en el estudio de una determinada pieza es fundamental para adentrarse de verdad en el conocimiento que esta pueda ofrecer.
Uno de los objetivos de este trabajo es, por tanto, ilustrar en qué medida la ayuda de los demás puede ser útil: un restaurador que mantenga la pieza en un estado idóneo, un químico que analice la pieza para facilitarnos los datos que necesitamos, un historiador que, apoyado en fuentes de todo tipo, pueda aportar una interpretación plausible de la información obtenida... En otras palabras, colaborar en conjunto por un objetivo común: el conocimiento del pasado a través de una determinada muestra.
De igual manera, en este trabajo se pretende hacer un acercamiento al estudio de las técnicas empleadas por físicos y químicos en el análisis de piezas arqueológicas, y cerrando aún más el círculo, de restos óseos humanos.
Conocer su información genética estableciendo haplogrupos que relacionen conjuntos poblacionales o correlaciones de linajes que conecten el pasado con nuestros días, determinar en qué basaban su alimentación gracias al estudio de los isótopos estables o examinar su composición elemental para evaluar el estado del hueso o su grado de fosilización, todo ello sin menospreciar la importancia de la propia observación y la información que puede facilitarnos.
Dado que para todos estos análisis es necesario el uso de una máquina con sus respectivas y complejas técnicas y resultados -a excepción de la observación, en las que a veces con mirarlo detenidamente basta para sacar conclusiones-, el propósito del presente trabajo será adentrarse en el conocimiento de cada una para determinar qué técnica debemos usar en función de lo que deseemos buscar.
2. ADN
2.1. Introducción al estudio del ADN antiguo:
La paleogenética, arqueogenética (Izagirre, Alonso y de la Rúa, 2005: 328) o recuperación de material genético a partir de restos biológicos antiguos ofrece un gran abanico de posibilidades: caracterización genética de nuestros antepasados próximos, análisis del material genético de especies extintas para esclarecer su relación con las actuales, o el seguimiento de migraciones históricas o prehistóricas.
Este tipo de análisis comienza en 1984, con el trabajo pionero de Higuchi y sus colaboradores, quienes obtuvieron por primera vez ADN de una especie extinta: el

cuagga, un équido sudafricano extinto hace unos 140 años (Fernández Domínguez, Pérez-Pérez, Turbón y Arroyo-Pardo, 2003: 27; Izagirre et al., 2005, 328) . Al año siguiente, Paabo recuperó ADN humano de una momia de 2.400 años de antigüedad (Izagirre 2005: 328). El procedimiento aplicado a humanos, sin embargo, enfrentó cierto escepticismo, pues había quien defendía que las secuencias de ADN antiguo serían en realidad moderno, introducido en la muestra por fallos de procedimiento, lo que se denomina “contaminación”, y cuyas posibilidades de solución veremos más adelante.
2.2. PCR
Generalmente, al lograr una muestra de ADN antiguo (en adelante ADNa) no se dispone de la suficiente para hacer las pruebas e interesa tener ADN de reserva. La denominada PCR o Reacción en Cadena de la Polimerasa permite aumentar esa cantidad, obteniendo así información genética con cantidades ínfimas y muy fragmentadas de ADN, y los secuenciadores automáticos permiten comparar muestras antiguas y modernas, estableciendo patrones filogéneticos (Fernández Domínguez et al. 2003: 28). Este proceso consiste en abrir la doble hebra de la que se compone cada ADN y hacer copias de cada una, mediante varios ciclos: desnaturalizado (donde se abre la doble hélice y se separan las hebras), polimerización (copiando cada una de las dos mitades) e hibridación (formando por complementariedad una nueva cadena de doble hebra, siendo cada una de ellas una copia del original). Esta copia del PCR se realiza por complementariedad de bases, es decir, que en cada ADN hay cuatro tipos de nucleótidos (A o Adenanina, T o Timina, G o Guanina y C o Citosina), complementarios por parejas entre sí (A-T y G-C).
2.3. Ventajas
El estudio de ADN en hueso en lugar de en tejidos conlleva una serie de ventajas, como una mejor conservación frente a la degradación gracias a la unión entre moléculas de ADN y cristales de hidroxiapatita (presentes en hueso y diente), mayor eficiencia en cuanto a cantidad disponible para amplificación de ADN, y que no es necesario destruir toda la pieza, pues el ADN extraído de la raíz del diente es suficiente para los análisis moleculares, dejando la corona intacta (Fernández Domínguez et al. 2003: 29).
2.4. Inconvenientes
Asimismo, existen una serie de inconvenientes para el uso de la PCR. Uno de los principales es la presencia de procesos bioquímicos degradativos tras la muerte del individuo, lo que hace que este se descomponga en conjunción con los químicos del suelo, produciéndose mutaciones o variaciones en la información genética del ADN. Sin embargo, ciertas condiciones pueden ralentizar o detener este proceso de

degradación del ADN, como una rápida deshidratación, impidiendo la actuación de microorganismos, la presencia de elevadas concentraciones de sales o las altas temperaturas (Fernández Dominguez et al., 2003: 28). También hay que tener en cuenta la existencia de una serie de inhibidores del PCR, reconocibles por dos factores (color marrón y fluorescencia): ácidos húmicos y fúlvicos, residuos de porfirinas, productos de degradación del ADN (Fernández Domínguez et al. 2003: 31-32)... Hay estrategias para eliminar o atenuar esa inhibición, como aumentar la pureza del ADN extraído o contrarrestar el efecto inhibidor durante la propia fase de amplificación, pero la mayoría de estas actuaciones incrementan el riesgo de contaminación con ADN moderno, pues aumentan la manipulación por parte del investigador. Sin embargo, hay otras actuaciones que no tienen ese problema, como almacenar la muestra en frío varios días antes de amplificarla (Fernández Domínguez et al. 2003: 31-32). Para evitar esta contaminación de ADN exógeno moderno, existen una serie de criterios básicos (Fernández Domínguez et al. 2003: 30):
Esterilización extensiva de las áreas de trabajo con HCl, seguido de exposición a luz UV. El HCl causa la hidrólisis del ADN, y la radiación UV, la formación de aductos covalentes no válidos para la reacción de PCR, o en otras palabras, funciona a modo de “antibloqueo” del PCR, para que esta técnica se pueda emplear correctamente.
Delimitación de áreas de trabajo: separar áreas de extracción del ADN de las áreas de amplificación, para evitar el carry-over, arrastre de moléculas de ADN provenientes de amplificaciones anteriores.
Uso de campanas de flujo laminar o de seguridad biológica, que impiden la contaminación de las muestras en el área de trabajo.
Además, existen protocolos particulares: Ward y Stringer consideran necesario repetir las amplificaciones, para confirmar los resultados (Fernández Domínguez et al. 2003: 30), y en los análisis de Oslo lavaron el polvo de huesos con hipoclorito de sodio para eliminar cualquier posible resto contaminante de ADN moderno (Krzewinska et al. , 2014: 2-3)
2.5. Ejemplo de uso de ADNmt: el caso de Priego de Córdoba
El motivo del análisis viene justificado por las oleadas provenientes del Norte de África a Priego en el s. VIII y la implantación del Imperio Almohade en el s. XII, lo que hace que se quiera averiguar hasta qué punto ha cambiado la población medieval, y si tiene relación genética con el Norte de África a causa de estas oleadas (Casas, 2005: 138).
El ADN empleado para este estudio fue ADNmt o mitocondrial, en lugar de ADN nuclear, dado que este no puede determinar con tanta fiabilidad el parentesco o relación genética de las muestras analizadas. La razón es que el ADN nuclear es más susceptible de mutaciones por mitosis en el núcleo, mientras que el ADNmt no recombina, por lo que se mantiene intacto a través de generaciones sucesivas

(Martínez-Marignac, Bravi, Lahitte y Bianchi, 1999: 282). Sin embargo, el ADN mitocondrial procede exclusivamente de la madre biológica, por lo que todas las personas que tengan el mismo linaje materno deberán tener el mismo ADNmt.
Fundamental antes de la extracción del ADN es asegurarse de que no va a haber contaminación de ADN actual, por lo que antes de extraer el ADNmt de la superficie ósea, quitaron 1 mm de superficie de la muestra, que posteriormente fue pulverizada, tratada con diferentes agentes químicos y almacenada en tubos estériles (Casas et al. 2006: 540).
Así pues, examinan el ADNmt extraído de los restos óseos y dentarios de 71 individuos procedentes de tres áreas de necrópolis musulmana de los ss. XII-XIII en Priego de Córdoba (El Palenque, La Cava y El Castillo), y se comparan con las muestras de saliva tomadas de 108 voluntarios de esa población actual (Casas, 2005: 137) y con el ADNmt de poblaciones actuales del Noroeste de África y Sur de la Península Ibérica (Casas, 2005: 139).
De estos análisis se pudieron extraer una importante cantidad de resultados de todo tipo (Casas, 2005: 139). En primer lugar, hubo 7 secuencias o haplotipos medievales y 7 actuales que no habían sido detectados nunca en ninguna población. Por otro lado, ambas poblaciones de Priego (medievales y actuales) comparten 12 haplotipos en común, pero no en exclusiva, puesto que todos estos haplotipos se han encontrado también en otras áreas geográficas. Sin embargo estos haplotipos, aunque no son exclusivos, sí son poco frecuentes, lo que deja ver una relativa continuidad genética de la población de Priego desde el Medievo. Además, se comprobó que los habitantes medievales de Priego tenían una mayor similitud genética con las poblaciones actuales del Noroeste de África que con las actuales de Priego de Córdoba. La población prieguense actual es más similar al resto del Sur de la Península que a la población medieval de Priego. Por último, también se descubrió que el porcentaje de haplogrupos de origen africano era superior en el Priego medieval que en todo el Sur de la Península Ibérica.
2.6. Ejemplo de uso de ADNmt: expansión noruega en época vikinga
Para ayudar a conocer las afinidades genéticas de los antiguos nórdicos, y su contribución a la diversidad genética de otros europeos, se analizaron los marcadores de ADN con restos esqueléticos de la Edad del Hierro Tardía en Noruega. El objetivo de este análisis era caracterizar los linajes maternos de noruegos a través de análisis de polimorfismos de ADNmt presentes en el ADN obtenido de restos humanos de la Edad del Hierro Tardía (550-1050), dando como resultado una prueba de los movimientos de los nórdicos por las islas durante la época vikinga.
El material empleado para este estudio fueron 80 muestras de hueso y dientes en distintos estados de conservación. La mayoría provienen de enterramientos en el norte

y centro de Noruega, donde la conservación es más favorable. 15 de los enterramientos individuales se conocen por hallazgos casuales. Es importante tener en cuenta que estos individuos no tuvieron por qué vivir necesariamente en la misma época, y que entre unos y otros puede existir una diferencia cronológica de varios siglos, pero para el objeto de este estudio se los analiza como pertenecientes a una misma población (Krzewinska et al., 2014: 6).
Antes de analizar la muestra, era necesario prepararla adecuadamente, por lo que las piezas pequeñas de hueso o dientes eran separados con sumo cuidado del conjunto para evitar daños excesivos. Si era posible, las cuñas de huesos largos (hasta 3 g), eran cortadas con una sierra para metales. Esta preparación fue llevada a cabo por un técnico protegido por todo el cuerpo, para evitar contaminar el ADN antiguo.
Tras esto, la superficie del hueso fue limpiada mediante arenado o chorreado abrasivo con arena de alúmina, lo que suele conocerse con el vocablo inglés de sandblasting, tras lo que son expuestos a rayos ultravioleta durante 15-30 minutos por cada lado (Krzewinska, 2014: 2). Los huesos fueron pulverizados usando un molino congelador, refrigerado con nitrógeno líquido, y lavados con hipoclorito de sodio. Las secuencias de ADNa obtenidas se compararon con una base de datos de ADN actual.
Finalmente, se consiguió extraer ADNmt de 69 de las 80 muestras, aunque en algunas de estas el material genético era muy inconsistente, de manera que se redujo la cifra a 45 secuencias de ADN. Curiosamente, estas muestras definitivas provenían de huesos excavados de media unos 10 años más tarde que las muestras descartadas, indicando que el tiempo de almacenamiento también influye a la hora de recuperar ADN que pueda servir.
Analizando ese ADN antiguo, se descubrió que los noruegos de época vikinga tenían una frecuencia genética de los haplogrupos K, I, V y U más alta que los actuales habitantes del Norte de Europa, pero más baja en cuanto a los haplogrupos T y H. Al llevar estos datos al análisis de haplotipos en común, se comprobó que los antiguos noruegos compartían la mayor parte de sus haplotipos con las actuales Noruega, Shetland, Orcadas, Francia e Inglaterra. Sin embargo, estas pruebas de comparación de haplotipos demostraron que los antiguos noruegos e islandeses son más similares genéticamente hablando a los actuales islandeses y noruegos, de forma cruzada, que a los actuales habitantes de sus respectivos territorios (Krzewinska et al., 2014: 3).
Para evitar una representación excesiva de linajes pertenecientes a posibles mezclas entre diferentes familias, se retiraron las secuencias duplicadas, lo que dejó 43 secuencias individuales, en representación de 34 haplotipos de ADNmt (FIG. 1), de los que 24 se encontraron en la base de datos de ADNmt moderno, 9 se identificaron en bases de datos más amplias, con un espectro a nivel mundial. y sólo un haplotipo permanece sin relación alguna con individuos actuales (Krzewinska et al., 2014: 4-6).
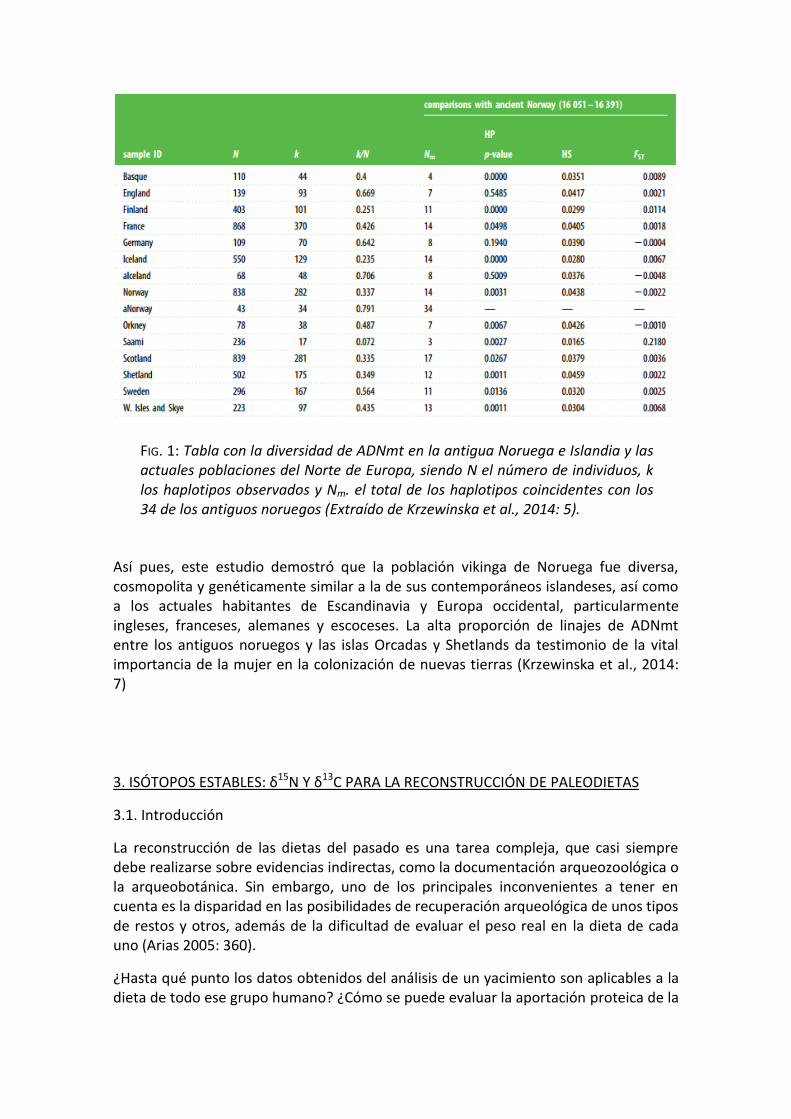
Así pues, este estudio demostró que la población vikinga de Noruega fue diversa, cosmopolita y genéticamente similar a la de sus contemporáneos islandeses, así como a los actuales habitantes de Escandinavia y Europa occidental, particularmente ingleses, franceses, alemanes y escoceses. La alta proporción de linajes de ADNmt entre los antiguos noruegos y las islas Orcadas y Shetlands da testimonio de la vital importancia de la mujer en la colonización de nuevas tierras (Krzewinska et al., 2014: 7)
3. ISÓTOPOS ESTABLES: δ15N Y δ13C PARA LA RECONSTRUCCIÓN DE PALEODIETAS
3.1. Introducción
La reconstrucción de las dietas del pasado es una tarea compleja, que casi siempre debe realizarse sobre evidencias indirectas, como la documentación arqueozoológica o la arqueobotánica. Sin embargo, uno de los principales inconvenientes a tener en cuenta es la disparidad en las posibilidades de recuperación arqueológica de unos tipos de restos y otros, además de la dificultad de evaluar el peso real en la dieta de cada uno (Arias 2005: 360).
¿Hasta qué punto los datos obtenidos del análisis de un yacimiento son aplicables a la dieta de todo ese grupo humano? ¿Cómo se puede evaluar la aportación proteica de la
FIG. 1: Tabla con la diversidad de ADNmt en la antigua Noruega e Islandia y las actuales poblaciones del Norte de Europa, siendo N el número de individuos, k los haplotipos observados y Nm. el total de los haplotipos coincidentes con los 34 de los antiguos noruegos (Extraído de Krzewinska et al., 2014: 5).

carne en relación con otros alimentos como verdura o pescado? Para dar respuesta a estas cuestiones, se han producido una serie de avances significativos a través de análisis químicos, destacando la medición de los isótopos estables de carbono y nitrógeno en el colágeno, método que comenzó a desarrollarse en la década de 1980 (Arias 2005: 360). Esta técnica permite valorar el peso relativo en la dieta (o más exactamente, en la ingesta de proteínas) de los alimentos de origen marino o terrestre y de ciertas plantas.
Pero el colágeno no es la única materia presente en los restos humanos con los que se pueden realizar análisis isotópicos. También pueden ser objetos de estudio los minerales biogénicos como el apatito, a partir del cual se analiza el 18O del fosfato, que permite saber la abundancia de esta especie en la precipitación a escala regional (Fernández Mosquera y Vila Taboada, 2000: 52). Para ambos análisis, es imprescindible que el material a analizar no haya sufrido alteraciones en su estructura isotópica. En el caso del 18O del apatito, esto se comprueba por DRX, observando que no existan recristalizaciones, mientras que para el colágeno se debe usar la relación atómica C/N como índice de conservación (Fernández Mosquera y Vila Taboada, 2000: 52).
Paralelamente a estos primeros estudios de δ13C se desarrollan los de δ15N, para indicar el nivel de un organismo en la cadena trófica, pudiendo emplearse para averiguar el origen terrestre o marino de las proteínas, ya que las cadenas tróficas de los ecosistemas marinos son más complejas que las de los terrestres, y en consecuencia tienen valores más elevados de δ15N, valores que pueden ser cotejados en referencias como la que menciona Arias (2005: 360).
3.2. Ventajas
Los análisis isotópicos son un gran aliado de la arqueología, al ser capaces de ofrecer información directa sobre los estudios de las paleodietas, datos que de otro modo sólo podrían ser obtenidos por evidencias indirectas, como el estudio de la flora y la fauna presente en el entorno circundante al individuo. Por otro lado, este tipo de análisis permite aislar fenómenos con determinados marcos espaciotemporales, y desde ahí dar prueba de diferentes situaciones (Salazar-García, 2012: 270).
3.3. Inconvenientes
Es un error, sin embargo, considerar que estos análisis isotópicos son válidos como evidencia por sí sola a la hora de justificar una determinada interpretación, pues habrá que combinarlos con métodos como la antropología física, estudiando el impacto que los procesos biológicos del entorno causan sobre un determinado individuo o población.
El carbono del colágeno óseo procede casi exclusivamente de proteínas, enmascarando el aporte de los hidratos de carbono y las grasas, de modo que es difícil valorar en estas dietas alimentos con estos componentes dominantes (como tubérculos). Una posible solución son los análisis de 13C en la bioapatita, dado que en

ella hay también lípidos y glúcidos. En este aspecto, el nitrógeno presenta menos problemas, dado que los animales sólo tienen este elemento en las proteínas (Arias 2005: 360).
Combinando ambos métodos (carbono y nitrógeno) se encuentra el instrumento más efectivo para el estudio de las paleodietas.
Generalmente, los organismos con una dieta de origen terrestre tienen unos valores de δ13C más negativos que aquellos que obtienen sus proteínas del mar. Arneborg et al. (1999: 158) menciona valores extremos de -21 ‰ para dietas totalmente terrestres y de -12,5 ‰ para dietas totalmente marinas, aunque la cifra exacta puede variar en función de cada autor (FIG. 2). Sin embargo, hay que tener en cuenta también que estos valores pueden verse alterados por el consumo de plantas C3 o C4. Las plantas C3 como el trigo o el arroz, dan unos valores de δ13C más negativos que las plantas C4, como el maíz.
Otros problemas son la necesidad de contar con un número representativo de muestras, lo que no siempre es posible, y de considerar los efectos metabólicos por la edad, la fisiología o la población a la que pertenece el individuo, así como tener una buena datación absoluta de la que partir (Fernández Mosquera y Vila Taboada, 2000: 52).
FIG. 2: Cuadro con los niveles de δ13C y δ15N, en función de la alimentación (extraído de Schulting, 1998: 2005). Este cuadro está elaborado usando como ejemplos animales cuya dieta es bien conocida, pero puede ser aplicable perfectamente al ser humano, en función de su dieta.

3.4. Análisis de isótopos estables sobre restos humanos de la región Cantábrica
Para verificar análisis realizados con anterioridad, se llevó a cabo un análisis sistemático con muestras prehistóricas de la región Cantábrica en los siguientes yacimientos: Los Canes, Los Azules, La Poza l'Egua, Colomba, Cuartamentero, J3, Cotero de la Mina y todos los niveles sepulcrales de La Garma A (Arias, 2005 361). Tanto en Los Azules como en Cuartamentero no se encontró suficiente colágeno como para obtener una datación, pudiendo hacerse análisis sólo de δ13C. Con el resto de yacimientos, tomando sus niveles de δ13C y δ15N se estableció una gráfica (Arias 2005: 366) donde se distinguen claramente dos grupos (FIG. 3):
El primero de ellos (Colomba, La Poza l'Egua y J3) se caracteriza por valores elevados de δ13C y δ15N. Arias (2005: 366) atribuye los valores de δ13C al consumo de proteínas de origen marino, y dado que los tres yacimientos tienen niveles similares, se sugiere que los tres individuos consumirían proporciones similares de proteínas procedentes del mar y la tierra. Por otro lado, los valores elevados de δ15N apuntan a la posibilidad de que las proteínas de origen marino procediesen en mayor medida del consumo de pescado que de la explotación de invertebrados.
En el otro grupo (Los Canes, Cotero de la Mina y Garma A), sin embargo, los valores de δ13C se sitúan entre los márgenes de variabilidad de las dietas que extraen las proteínas de alimentos terrestres, sugiriendo que en los últimos 5-10 años de su vida, estos individuos apenas consumieron alimentos de origen marino. En cuanto a los valores de δ15N, se acercan a los niveles de los depredadores, de modo que podría suponerse que estos individuos llevaban una dieta mayoritariamente basada en el consumo de carne y algún vegetal. Esta teoría estaría reforzada por las patologías bucodentales halladas en la mujer de la tumba I de Los Canes, dado que las caries suelen atribuirse a las dietas basadas en hidratos de carbono (Arias 2005: 366-367).
Aunque posiblemente, el ejemplo más llamativo lo haría al añadir a todos estos yacimientos otras mediciones de δ13C y dataciones de 14C de diversas épocas desde el Mesolítico hasta la Edad Moderna (Arias 2005: 368-369, FIG. 4). El resultado de esta comparación indicaría que en el Paleolítico y en el Aziliense tendrían una dieta plenamente terrestre, situación que cambia en el Mesolítico, donde aparece una clara indicación de ingesta de alimentos de origen marino, que constituye aproximadamente la mitad de proteínas consumidas por los grupos costeros cantábricos.
FIG. 3: Resultados de los análisis de isótopos
estables sobre las muestras de la región
Cantábrica (Extraído de Arias, 2005: 366).
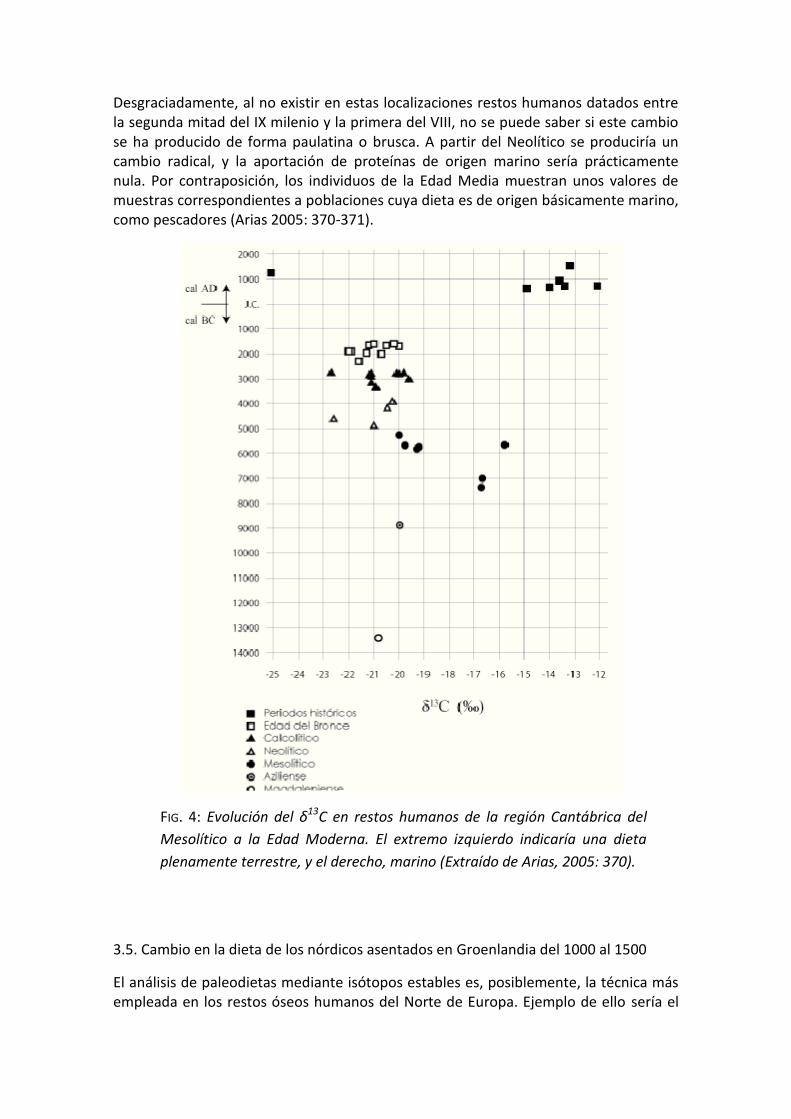
Desgraciadamente, al no existir en estas localizaciones restos humanos datados entre la segunda mitad del IX milenio y la primera del VIII, no se puede saber si este cambio se ha producido de forma paulatina o brusca. A partir del Neolítico se produciría un cambio radical, y la aportación de proteínas de origen marino sería prácticamente nula. Por contraposición, los individuos de la Edad Media muestran unos valores de muestras correspondientes a poblaciones cuya dieta es de origen básicamente marino, como pescadores (Arias 2005: 370-371).
3.5. Cambio en la dieta de los nórdicos asentados en Groenlandia del 1000 al 1500
El análisis de paleodietas mediante isótopos estables es, posiblemente, la técnica más empleada en los restos óseos humanos del Norte de Europa. Ejemplo de ello sería el
FIG. 4: Evolución del δ13C en restos humanos de la región Cantábrica del
Mesolítico a la Edad Moderna. El extremo izquierdo indicaría una dieta
plenamente terrestre, y el derecho, marino (Extraído de Arias, 2005: 370).
estudio realizado por Arneborg et al. (1999), donde se analiza el cambio de dieta de los vikingos establecidos en Groenlandia a partir de análisis de isótopos de carbono y datación por radiocarbono.
El principal objetivo consistía en, mediante el uso de referencias extremas de dieta 100% terrestre y 100% marina (Arneborg et al. 1999: 158), calcular el porcentaje de dieta marina en cada uno de los 27 individuos, y tras datarlos mediante 14C, extraer conclusiones acerca del cambio de dieta a lo largo de sus siglos de asentamiento.
Las excavaciones indican que la subsistencia de los nórdicos dependía básicamente de la ganadería y la caza de focas, aunque es difícil reforzar esta idea con el estudio de sus dietas, dado que los desperdicios de comida hallados en granjas y muladares pueden no ser suficientemente representativos (Arneborg et al., 1999: 159-160). Si el pescado formó o no parte sustancial de su dieta, no se puede demostrar por el momento con análisis isotópicos (Arneborg et al., 1999: 165).
El muestreo se realizó en tumbas de 6 cementerios al suroeste de Groenlandia, 5 del Asentamiento Oriental y uno del Occidental, cuyas fechas oscilan entre el s. X y el XV (Arneborg et al., 1999: 160).
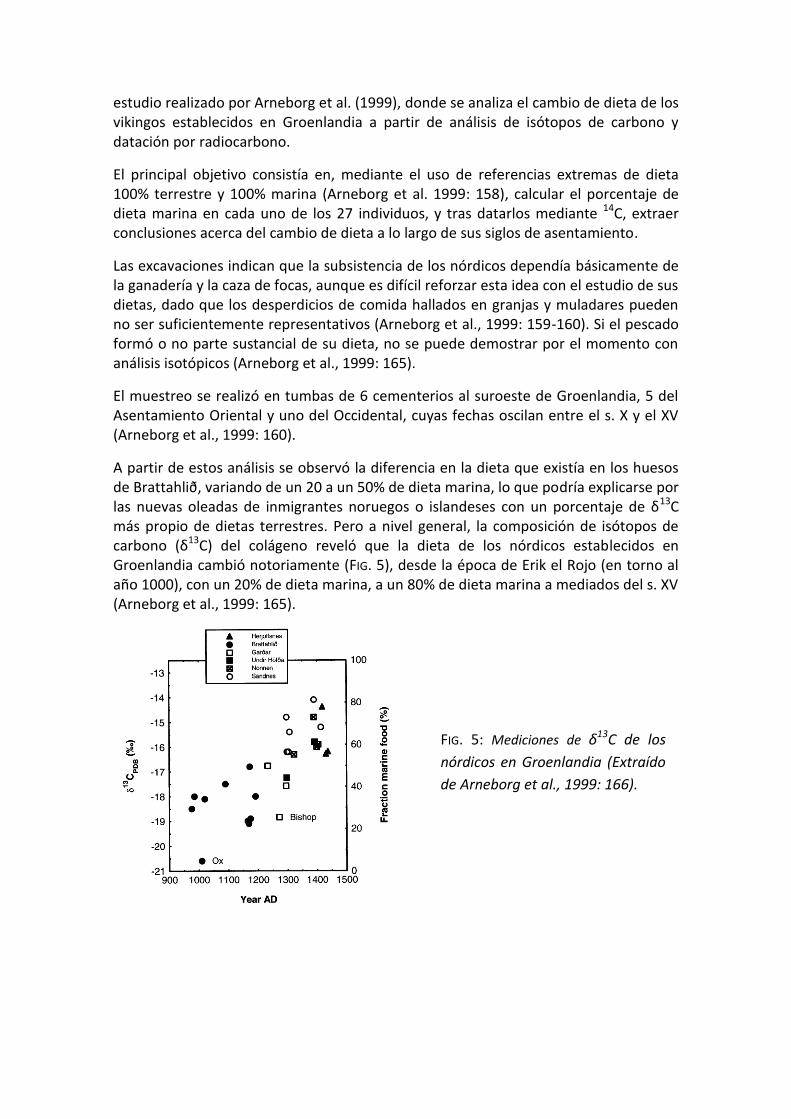
A partir de estos análisis se observó la diferencia en la dieta que existía en los huesos de Brattahlið, variando de un 20 a un 50% de dieta marina, lo que podría explicarse por las nuevas oleadas de inmigrantes noruegos o islandeses con un porcentaje de δ13C más propio de dietas terrestres. Pero a nivel general, la composición de isótopos de carbono (δ13C) del colágeno reveló que la dieta de los nórdicos establecidos en Groenlandia cambió notoriamente (FIG. 5), desde la época de Erik el Rojo (en torno al año 1000), con un 20% de dieta marina, a un 80% de dieta marina a mediados del s. XV (Arneborg et al., 1999: 165).
FIG. 5: Mediciones de δ13C de los
nórdicos en Groenlandia (Extraído
de Arneborg et al., 1999: 166).

4. ICP-MS
4.1. Introducción
La Espectrometría de Masas con fuente de Plasma de Acoplamiento Inductivo (ICP-MS) es una técnica empleada para determinar elementos. Fue presentada en 1983, siendo rápidamente adoptada por los laboratorios de geoquímica, dadas sus especiales capacidades de detección (Wolf, 2005).
Para analizar las muestras por ICP-MS, deben ser diluidas e introducidas en el plasma del ICP en forma de aerosol (Wolf, 2005), que es sometida a temperaturas de entre 8.000 y 10.000 Kelvin (7.700-9.700 ° C) para ionizarla. Los iones resultantes pasan a la cámara de vacío incluida en el sistema de ICP-MS, donde son acelerados, pasando por una serie de lentes, un analizador electrostático y un imán, que separa los iones en función de su proporción masa/carga, pasando finalmente por el detector (Lienhop, 2003).
Otra posibilidad es analizarlo por LA-ICP-MS (ICP-MS por ablación con láser), para lo cual, la muestra debe colocarse en un soporte donde tendrá lugar la ablación, que varía en función del tamaño de la muestra, pero que no suele tener un área mayor de 1x1 mm y 0,1 mm de profundidad (Lienhop, 2003). El resto del proceso es el mismo que con el método estándar.
4.2. Ventajas
El ICP-MS tiene una serie de ventajas sobre otras técnicas de análisis elemental, como la posibilidad de obtener información isotópica, un alto rendimiento o la ausencia casi completa de interferencias gracias a las altas temperaturas (Wolf, 2005).
LA-ICP-MS) permite análisis en puntos específicos muy pequeños. Además, justo antes de la ablación y recolección de datos realizada por el láser, este mismo limpia adecuadamente la muestra, reduciendo los riesgos de analizar una muestra contaminada.
4.3. Inconvenientes
Los iones formados por la descarga del ICP son generalmente positivos, por lo que elementos tendentes a formar iones negativos, como el cloro, el yodo o el flúor son muy difíciles de determinar por esta técnica (Wolf, 2005).
Si el total de sólidos disueltos es muy alto, es probable que los orificios queden bloqueados, aminorando la sensibilidad del detector (Wolf, 2005). Por este motivo, muestras como suelos o rocas deben ser necesariamente disueltas en ácido antes de ser analizadas por el método estándar (Wolf, 2005; Lienhop, 2003). Esto no sucede con
el LA-ICP-MS, donde sólo se necesita trabajar la muestra para que quepa en la cámara de ablación (Lienhop, 2003).
4.4. ¿Cómo puede aplicarse al estudio de los restos óseos?
El Bario (Ba) es un metal alcalino presente en el hueso a través del tracto intestinal. En zonas terrestres, el Bario y el Estroncio (Sr) tienen una abundancia similar, pero en zonas marinas forma un precipitado (sólido creado en una disolución por efecto de cristalización) indisoluble por los altos contenidos en sulfato del agua salada. De esta manera, la proporción Ba/Sr puede servir para indicar la dieta y posición trófica de un individuo: las dietas marinas provocarán una baja proporción Ba/Sr, al contrario que las dietas terrestres. Tanto los huesos como los dientes pueden ser analizados con esta técnica, si bien el esmalte de los dientes lo protege más que al hueso frente al efecto de la diagénesis, y por ello suele ser más fiable (Lienhop, 2003).
4.5. Ejemplo de aplicación de LA-ICP-MS en huesos humanos
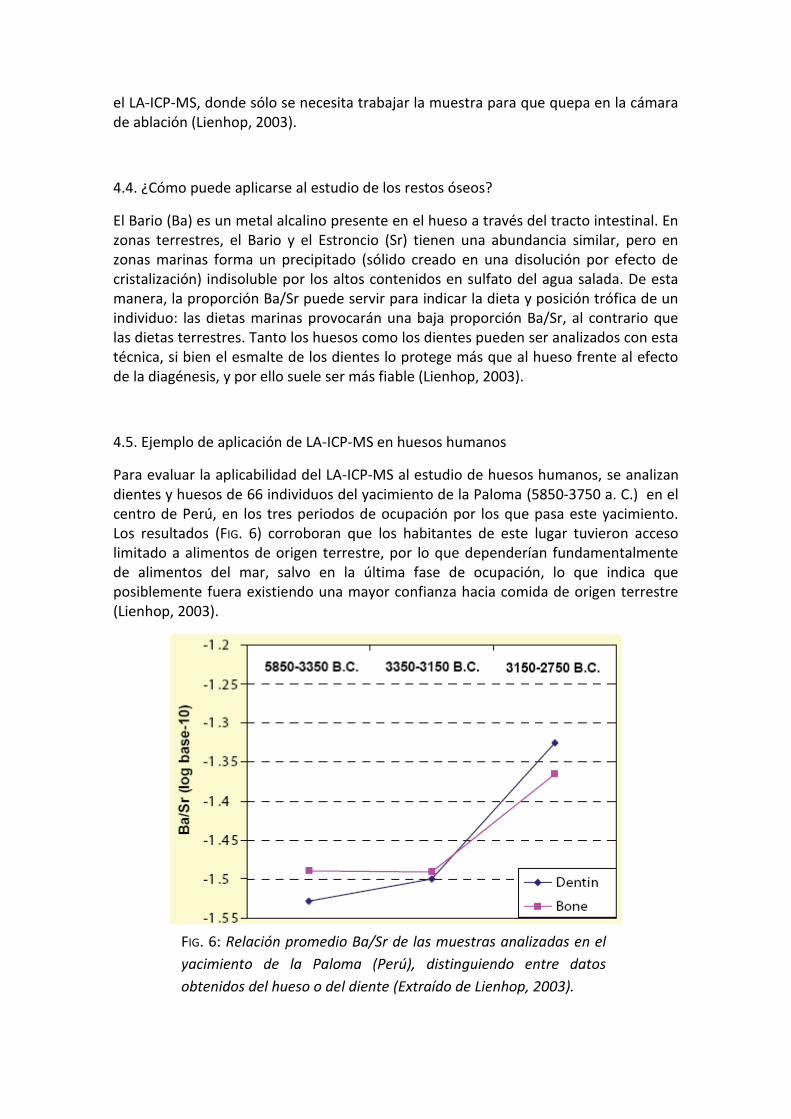
Para evaluar la aplicabilidad del LA-ICP-MS al estudio de huesos humanos, se analizan dientes y huesos de 66 individuos del yacimiento de la Paloma (5850-3750 a. C.) en el centro de Perú, en los tres periodos de ocupación por los que pasa este yacimiento. Los resultados (FIG. 6) corroboran que los habitantes de este lugar tuvieron acceso limitado a alimentos de origen terrestre, por lo que dependerían fundamentalmente de alimentos del mar, salvo en la última fase de ocupación, lo que indica que posiblemente fuera existiendo una mayor confianza hacia comida de origen terrestre (Lienhop, 2003).
FIG. 6: Relación promedio Ba/Sr de las muestras analizadas en el
yacimiento de la Paloma (Perú), distinguiendo entre datos
obtenidos del hueso o del diente (Extraído de Lienhop, 2003).

5. FRX
5.1. Introducción
La Espectrometría de Fluorescencia de Rayos-X o simplemente Fluorescencia de Rayos-X (FRX) es una técnica aplicada para determinar la composición elemental de una muestra, lo que en nuestro ámbito puede trasladarse a la detección de concentraciones de elementos en los huesos y la evaluación de la diagénesis o los procesos de fosilización (Piga et al., 2014: 647). Se basa en las transiciones de electrones de los átomos producidos cuando una radiación electromagnética incide con la muestra en cuestión, produciendo una excitación del átomo, que pasa de estable a inestable, expulsando un electrón (Meléndez y Dávila, 2009: 2). Para estabilizar el átomo, los electrones adyacentes rellenan el espacio que dejó el electrón expulsado, emitiéndose una energía en forma de radiación de rayos X. Este proceso de emisión de rayos X es la fluorescencia (Meléndez y Dávila, 2009: 2-3).
Esa energía liberada por el electrón expulsado se puede cuantificar, convirtiendo la energía de los rayos X en longitud de onda y determinando así de qué elemento se trata y en qué cantidad se encuentra (Meléndez y Dávila, 2009: 2). Estos análisis pueden realizarse usando la energía o la longitud de onda. Si se emplea este último, la técnica se conoce por sus siglas en inglés WDXRF (FRX de dispersión de longitud de onda), mientras que si se usa la energía, se conoce como EDXRF o FRX de dispersión de energía, técnica empleada en equipos portátiles, y más barata que la WDXRF (Meléndez y Dávila, 2009: 3).
5.2. Ventajas
El análisis elemental se puede realizar bien por absorción elemental o por inducción de plasma acoplado. En ambos casos se trata de una técnica destructiva, lo que se traduce en la pérdida de la muestra. En consecuencia, cuando la muestra es muy pequeña o de suma importancia, conviene tener una técnica no destructiva, y el FRX es una de ellas (Meléndez y Dávila, 2009: 2).
La longitud de onda es única para cada elemento, lo que permite hacer una inequívoca identificación del elemento, y además permite hacer análisis cuantitativos, ya que la intensidad de los rayos X es proporcional a la concentración del elemento (Meléndez y Dávila, 2009: 3). Además, podrían citarse otras ventajas: determinación rápida, interpretación sencilla, posibilidad de determinaciones multielementales y en cualquier estado (sólido, líquido o gaseoso), preparación de la muestra mínima o inexistente, etc. (Meléndez y Dávila, 2009: 4)
5.3. Desventajas
Pocas desventajas se mencionan al empleo del FRX, destacando quizá la necesidad de disponer de patrones generalmente caros, la posibilidad de interferencias en el análisis

al realizar una penetración bastante baja o la limitada sensibilidad del método en la detección de elementos ligeros (Martínez et al., 2006: 793; Roquet, 2012: 8)
5.4. Ejemplo de uso de FRX en los huesos de Pedro III y Blanca de Anjou
En 2010 Pedro III, Jaime II y Blanca de Anjou fueron exhumados por un proyecto de restauración e investigación de sus tumbas, y sus huesos fueron analizados. Dado que el FRX es una técnica de análisis elemental no destructiva, se estudian con ella fragmentos de hueso de Pedro III y Blanca de Anjou para evaluar la composición elemental de sus huesos, eligiendo en ambos casos fragmentos de costilla y fémur, y también del metacarpo en el caso de Blanca de Anjou,, que fueron sometidos a análisis de FRX, así como de DRX y FTIR (Piga et al., 2014: 648-649).
Por un lado, el DRX y FTIR dan pistas acerca del estado de conservación de los huesos a través del estudio de sus fases mineralógicas y propiedades vibracionales (Piga et al., 2014: 649). Mediante el DRX se comprueba que la hidroxiapatita es el componente principal en los huesos de ambos individuos. El FTIR por su parte demuestra que ambos tenían una escasa proporción señal/intensidad, teniendo bandas de fosfato que colapsan con otras (entre los rangos 500-650 y 1000-1200), posiblemente debido a la presencia de colágeno u otra materia orgánica (Piga et al., 2014: 649). La diferencia en la gráfica es notable al compararla con un fragmento de hueso perteneciente a la necrópolis de S’Illot de Porros, que ha sufrido un proceso diagenético traducido en la pérdida del colágeno (Piga et al., 2014: 650).
A su vez, el rango 1300-1700 es similar entre los dos reyes y un fragmento de hueso actual, del que se sabe que no ha pasado ningún proceso diagenético. Esto sugiere que la parte orgánica de los huesos de ambos reyes fuera la causante de su buena preservación en el interior de las tumbas (Piga et al., 2014: 650).
En cuanto al uso de FRX, uno de los resultados más sorprendentes obtenidos fue que entre los componentes presentes en Pedro III (FIG. 7) había azufre y en una medida considerable, lo que se explica teniendo en cuenta que la mortaja que lo cubría llevaba yeso, material que incluye azufre entre sus elementos (Piga et al., 2014: 651). En cuanto a los niveles bajos, pero presentes, de hierro, zinc y plomo, puede explicarse por los hábitos de comida y el empleo de vajillas que contenían muchos de los metales de transición (Piga et al., 2014: 651).
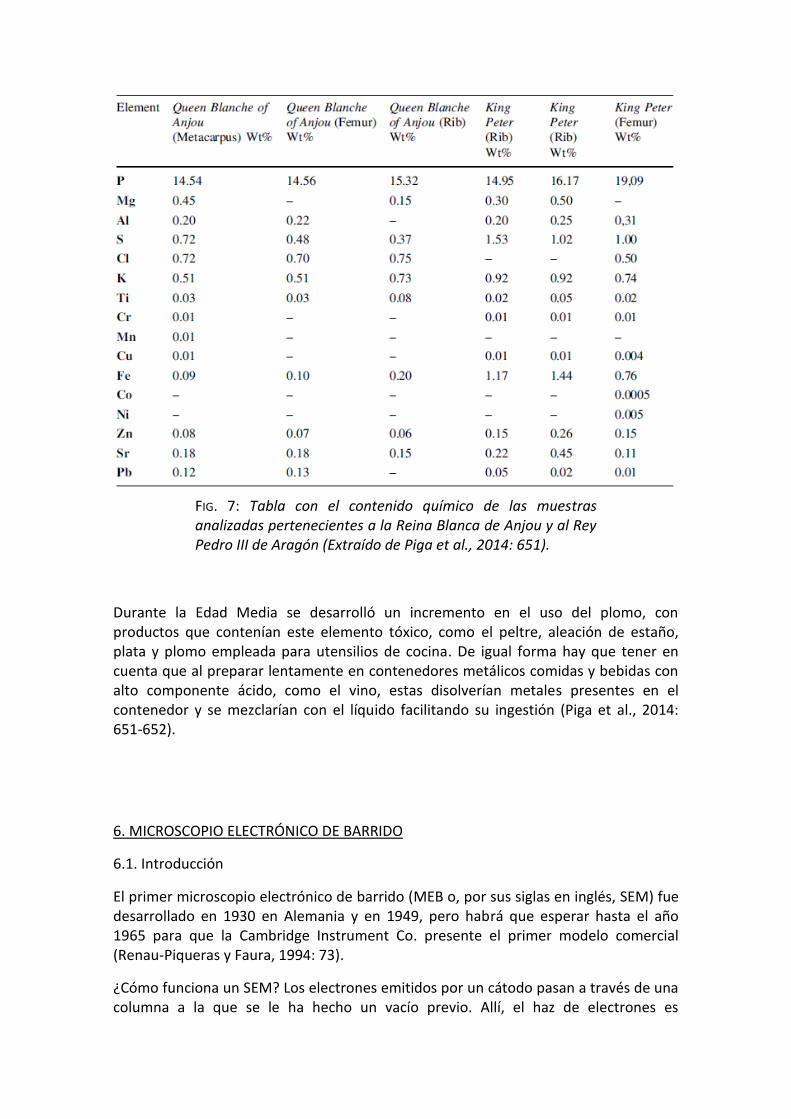
Durante la Edad Media se desarrolló un incremento en el uso del plomo, con productos que contenían este elemento tóxico, como el peltre, aleación de estaño, plata y plomo empleada para utensilios de cocina. De igual forma hay que tener en cuenta que al preparar lentamente en contenedores metálicos comidas y bebidas con alto componente ácido, como el vino, estas disolverían metales presentes en el contenedor y se mezclarían con el líquido facilitando su ingestión (Piga et al., 2014: 651-652).
6. MICROSCOPIO ELECTRÓNICO DE BARRIDO
6.1. Introducción
El primer microscopio electrónico de barrido (MEB o, por sus siglas en inglés, SEM) fue desarrollado en 1930 en Alemania y en 1949, pero habrá que esperar hasta el año 1965 para que la Cambridge Instrument Co. presente el primer modelo comercial (Renau-Piqueras y Faura, 1994: 73).
¿Cómo funciona un SEM? Los electrones emitidos por un cátodo pasan a través de una columna a la que se le ha hecho un vacío previo. Allí, el haz de electrones es
FIG. 7: Tabla con el contenido químico de las muestras analizadas pertenecientes a la Reina Blanca de Anjou y al Rey Pedro III de Aragón (Extraído de Piga et al., 2014: 651).

concentrado por lentes electromagnéticas, disminuyendo el diámetro del haz de 25.000-50.000 nm a tan sólo 10, casi puntual (Renau-Piqueras y Faura, 1994: 77). Este haz puntual se desplaza una y otra vez por la superficie de la muestra. Renau-Piqueras y Faura (1994: 78) realizan una analogía al respecto, comparándolo con "un pincel que iría barriendo la muestra con continuas idas y venidas". Al chocar el haz con la superficie de la muestra, se producen nuevos electrones que son captados por un detector, dando origen a varios fotones (Renau-Piqueras y Faura, 1994: 78), que son dirigidos hasta un fotomultiplicador donde cada uno crea electrones secundarios. En otras palabras, lo que se consigue con esta técnica es una amplificación de los electrones, que son los que nos dan la información de la muestra analizada (Renau-Piqueras y Faura, 1994: 78). A más electrones creados, más información sobre la muestra.
Los electrones secundarios pasarán finalmente por un videoamplificador, produciendo la imagen.
6.2. Ventajas
Gracias a una serie de fórmulas se ha podido comprobar que, mediante la utilización de un haz de electrones acelerados se pueden conseguir longitudes de onda 105 veces menores que cuando se usa luz visible (Renau-Piqueras y Faura, 1994: 76).
Por otro lado, Renau-Piqueras y Faura (1994: 73-74) mencionan una serie de ventajas en comparación con su antecesor el TEM o microscopio electrónico de transmisión, siendo quizá las más destacables la imagen tridimensional (el TEM ofrece una imagen en dos dimensiones) y la posibilidad de mover el haz electrónico (a diferencia del haz fijo del TEM) y de colocar muestras íntegras, mientras que en el TEM era necesario preparar la muestra para extraer secciones ultrafinas.
6.3. Inconvenientes
A pesar de las mejoras del SEM con respecto a su antecesor, también existen ciertos inconvenientes, como la pérdida de aumentos, siendo de 500.000 en el TEM y de 140.000 en el SEM (Renau-Piqueras y Faura, 1994: 74). Por otro lado, aunque la muestra pueda colocarse íntegra necesita de una preparación previa, donde es necesario realizar una serie de pasos particularmente críticos que hay que controlar o conocer para interpretar adecuadamente los resultados. Habrá que prestar atención a la temperatura de lavado y fijación o el secado de la muestra, en el que podrían producirse algunos problemas como la rotura o alteración de parte de la superficie (Renau-Piqueras y Faura, 1994: 88-89).
Tampoco hay que olvidar la presencia de astigmatismo en el SEM, que da como resultado diferentes intensidades en las dos direcciones que conforman un ángulo recto, de forma que mientras una está enfocada, la otra está desenfocada. Sin embargo, este problema puede corregirse con una lente adecuada, y en general la

mayoría de inconvenientes de esta técnica pueden solucionarse estableciendo las condiciones de trabajo óptimas (Renau-Piqueras y Faura, 1994: 89).
6.4. Ejemplo de uso del SEM en el análisis del pH del suelo
El objetivo de estos análisis era esclarecer si la escasa proporción de huesos humanos recuperados de dos yacimientos de Huelva y su estado fragmentario se debe a un expolio sistemático, limpieza periódica o enterramiento secundario, o si por el contrario es efecto de condiciones físico-químicas adversas (Nieto et al., 2002: 182).
Para ello, en primer lugar se mide el pH del suelo en las inmediaciones de los restos óseos (Nieto et al., 2002: 182): el Dolmen de la Paloma (5.95, lo que se puede clasificar como moderadamente ácido), y el Dolmen de la Venta (5.52, fuertemente ácido). Para establecer una comparativa con otros yacimientos coetáneos de otras áreas geográficas cuyos restos óseos se encontrasen bien conservados, miden también el pH de Cabezo Juré (8.25) y La Cueva (8.31, ambos moderadamente alcalinos). Y para evaluar el factor tiempo, se analiza también el pH del enterramiento de época moderna de la Venta, dando el resultado más bajo de todos (5.40), pero el esqueleto se conservaba, aunque con importantes muestras de corrosión apreciables con lupa binocular. Esto pone de manifiesto que en suelos ácidos los restos óseos sufren un proceso de disolución, cuyo grado está en función del pH del suelo y del tiempo que lleve enterrado (Nieto et al., 2002: 182-183).
Gracias al uso del MEB se han podido realizar microanálisis microelementales, y el estudio por DRX de huesos y suelos permitió conocer la composición mineralógica, y los cambios que sufren los restos óseos en función del pH del suelo (Nieto et al., 2002: 183).
En el Dolmen de la Venta pudo observarse una estructura ósea muy transformada, con grietas, fisuras, descamaciones superficiales…mientras que en el enterramiento moderno, los daños son mucho menores, pero hay descamaciones, grietas y aspecto terroso en un hueso que se encuentra descalcificado. Al observar este hueso con MEB, se ve que el tejido trabecular está siendo reemplazado por minerales de arcilla (Nieto et al., 2002: 183-184).
Como puede observarse en la tabla (FIG. 8), la vermiculita y la haloisita no sólo se encuentran en los huesos de La Venta, sino también en el propio suelo. De esta manera, se concluye que para el estudio adecuado de los restos óseos es fundamental el análisis del entorno en el que se encuentra (Nieto et al., 2002: 184).

7. DISCUSIÓN
Lo primero que salta a la vista tan sólo con la búsqueda bibliográfica es el largo camino que queda por delante, dado que, aunque algunas técnicas han recibido bastante más análisis que otras, en general apenas hay estudios suficientes como para centrar la atención en un periodo concreto. Muchas de los que publican estos estudios mencionan como motivación la falta de información al respecto, y prácticamente todas las conclusiones, en una forma u otra, dejan ver que todavía hay mucho por investigar.
De igual forma, otro de los aspectos a mejorar es la colaboración entre disciplinas que ya se mencionó en la introducción. Autores como Krzewinska (2014) limitan su estudio a interpretar los datos obtenidos de los análisis químicos, pero se echa en falta la colaboración de un profesional en el campo de las fuentes históricas que, empleando las interpretaciones nacidas de los análisis químicos, dé la correspondiente recreación histórica del pasado, o dicho de otro modo, que explique o trate de explicar la relación cruzada que vincula genéticamente a islandeses y noruegos, aunque también es necesario reconocer que en ocasiones, las fuentes históricas o literarias no pueden aportar más datos de los obtenidos en ese estudio, como sucede en el caso de Arneborg (1999).
En cuanto a las paleodietas, el punto a mejorar está bastante claro: tratar de descifrar en qué manera el consumo de plantas C3 o C4 afecta a los resultados obtenidos mediante el estudio de los isótopos estables.
Tampoco se puede descuidar, por superficial que parezca, la importancia del tema económico. Técnicas como el ICP-MS o las instalaciones de FRX suponen un considerable gasto económico –tanto para la realización de la técnica en sí como para la creación o mantenimiento de las máquinas encargadas-, que debe subsanarse para poder llevar a cabo dichas analíticas. En el caso del FRX, deberían asumirse sus gastos
FIG. 8: Valores de pH del suelo y análisis mineralógicos realizados por DRX
de las muestras de hueso y suelo, marcando con asteriscos la abundancia
del material en cuestión (Extraído de Nieto et al., 2002: 182).

inclusive por encima del de otras técnicas, dado su carácter no destructivo y la gran variedad de datos que puede ofrecer acerca de la composición elemental de una pieza.
Por último, cabe señalar la cantidad de técnicas cuyos inconvenientes podrían ser total o parcialmente solventados de establecerse las condiciones de trabajo óptimas para saber qué técnicas emplear en función del tipo de datos que esperemos obtener, cómo contrastar dichos datos, evitar contaminar en el proceso la pieza con ADN moderno o provocar grietas en su composición que dificulten su análisis o conservación, y prepararlas de manera adecuada para que no oculten su detección o identificación.
8. CONCLUSIÓN
De todos estos datos y analíticas, si hubiese que extraer una sola conclusión sería, posiblemente, la necesidad de establecer una serie de parámetros que guíen al mejor conocimiento y conservación de la muestra, parámetros que redundan finalmente en la adecuada estructuración del trabajo en todas sus vertientes: desde los profesionales de diversos ámbitos con los que colaborar, considerándose obsoleta la idea de “arqueólogo autónomo”, hasta los procedimientos de tratamiento de la muestra desde el mismo momento en que se encuentra, sin olvidar la necesidad de tener las ideas bien claras en lo que respecta a la obtención de información a partir de la pieza y a cómo obtener cada clase de información.
Tener en cuenta que de cada técnica puede obtenerse información absolutamente diferente, y que la gran mayoría de las veces no bastará con un solo ejemplo o una sola técnica, siendo necesario coordinar analíticas de todo tipo, es fundamental para que el arqueólogo pueda obtener los datos que busca, sin más deterioro de la pieza que el estrictamente necesario.
9. BIBLIOGRAFÍA
- Arias Cabal, P. (2005): Determinación de isótopos estables en restos humanos de la región Cantábrica. Aportación al estudio de la dieta de las poblaciones del Mesolítico y el Neolítico. Munibe, 57, 359-374
- Arneborg, J., Heinemeier, J., Lynnerup, N., Nielsen, H. L., Rud, N. y Sveinbjörnsdóttir, Á. E. (1999): Change of diet of the greenland vikings determined from stable carbon isotope analysis and 14C dating of their bones. Radiocarbon, 41 (2), 157-168
- Casas, M. J. (2005): El estudio de ADN humano de época almohade y actual revela la influencia migratoria norteafricana en Priego de Córdoba. Antiqvitas, 17, 137-141

- Casas, M. J., Hagelberg, E., Fregel, R., Larruga, J. M. y González, A. M. (2006): Human Mitochondrial DNA Diversity in an Archaeological Site in al-Andalus: Genetic Impact of Migrations from North Africa in Medieval Spain. American Journal of Physical Anthropology, 131, 539-551
- Fernández Domínguez, E., Pérez-Pérez, A., Turbón, D. y Arroyo-Pardo, E. (2003): ADN antiguo: química y aplicaciones. Anales de la Real Sociedad Española de Química, 3, 27-37
- Fernández Mosquera, D. y Vila Taboada, M. (2000): Consideraciones metodológicas acerca del estudio de isótopos estables en restos óseos fósiles. Cadernos Lab. Xeolóxico de Laxe, 25, 51-53
- Izagirre, N. Durán, L. M. y De la Rúa, C. (2005): Genética y Arqueología: Análisis molecular de ADN procedente de restos esqueléticos. Munibe, 50, 3-14
- Krzewinska, M., Bjørnstad, G., Skoglund, P., Olason, P. I., Bill, J., Götherström, A. y Hagelberg, E. (2014): Mitochondrial DNA variation in the Viking age population of Norway. Philosophical Transactions B, 370
- Lienhop, K. M., Speakman, R. J. y Glascock, M. D. (2003): Reconstruction of prehistoric human dietary patterns by laser ablation ICP-MS. Columbia.
- Martínez, D., Gil Novoa, O. D., y Barón González, A. J. (2006): Espectrometría de fluorescencia de Rayos X. Revista Colombiana de Física, 38 (2), 790-793.
- Martínez-Marignac, V. L., Bravi, C. M., Lahitte, H. B. y Bianchi, N. O. (1999): Estudio del ADN mitocondrial de una muestra de la Ciudad de la Plata. Revista Argentina de Antropología Biológica, 2 (1), 281-300
- Nieto, J. M., Nocete, F., Sáez, R., y Franco, F. (2002): Cambios mineralógicos en restos óseos en función de las condiciones de pH del suelo. Geogaceta, 31, 181-184.
- Piga, G., Brunetti, A., Lasio, B., Enzo, S. y Malgosa, A. (2014): XRF investigation on skeletal remains from King Peter III of Aragon (1239–1285 A.D.) and Queen Blanche of Anjou (1280–1310 A.D.). Applied Physics A., 114, 647-653.
-Renau-Piqueras, J. y Faura, M. (1994): Principios básicos del microscopio electrónico de barrido. En Méndez Felpeto, J., Rodríguez Torres, A. M., y Cerdán Villanueva, E. (eds.), II Reunión Científica en Biología Celular y Molecular: técnicas de microscopía, manipulación y análisis de imagen (73-92). A Coruña: Universidade da Coruña, Servicio de Publicacións.
-Schulting, R. M. (1998): Slightling the sea: stable isotopic evidence for the transition to farming in northwestern Europe. Documenta Praehistorica, 25, 203-218
- Salazar-García, D. C. (2012): Reconstrucción directa de la dieta mediante análisis de isótopos estables del carbono y del nitrógeno en individuos del Paleolítico Medio y Superior europeo. En Turbón Borrega, D., Fañanás Saura, L., Rissech Badalló, C., y Rosa de la Cruz, A. (eds.), Biodiversidad humana y Evolución (270-276). Barcelona: Sociedad Española de Antropología Física.

Recursos electrónicos
- Meléndez Pizarro, C. O. y Camacho Dávila, A. A. (2009): Espectrometría de Fluorescencia de Rayos X. Recuperado el 12 de enero de 2015, del Sitio web de la Universidad Autónoma de Chihuahua: http://www.uach.mx/extension_y_difusion/synthesis/2009/08/20/espectrometria_de_flourescencia_de_rayos_X.pdf
- Roquet, M. B. (2012): Métodos analíticos en geoquímica (DRX-FRX) y Pegmatitas. Recuperado el 12 de enero de 2015, del Sitio web de la Universidad de San Luis: http://www0.unsl.edu.ar/~geo/materias/geoquimica/documentos/teorias/tema-VI-FRX-Pegmatitas.pdf
- Wolf, R. E. (2005): What is ICP-MS?...and more importantly, what can it do?. Recuperado el 12 de enero de 2015, del Sitio web de Crustal Geophysics and Geochemistry Science Center: http://crustal.usgs.gov/laboratories/icpms/intro.html




















