
An ichthyofauna from the subsurface Devonian of northwestern Iowa and its biostratigraphic and...
13
Paleontological Society An Ichthyofauna from the Subsurface Devonian of Northwestern Iowa and Its Biostratigraphic and Paleoecologic Significance Author(s): Glenn W. Storrs Source: Journal of Paleontology, Vol. 61, No. 2 (Mar., 1987), pp. 363-374 Published by: Paleontological Society Stable URL: http://www.jstor.org/stable/1305328 . Accessed: 22/01/2014 11:00 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . Paleontological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal of Paleontology. http://www.jstor.org This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AM All use subject to JSTOR Terms and Conditions
-
Upload
cincymuseum -
Category
Documents
-
view
1 -
download
0
Transcript of An ichthyofauna from the subsurface Devonian of northwestern Iowa and its biostratigraphic and...

Paleontological Society
An Ichthyofauna from the Subsurface Devonian of Northwestern Iowa and ItsBiostratigraphic and Paleoecologic SignificanceAuthor(s): Glenn W. StorrsSource: Journal of Paleontology, Vol. 61, No. 2 (Mar., 1987), pp. 363-374Published by: Paleontological SocietyStable URL: http://www.jstor.org/stable/1305328 .
Accessed: 22/01/2014 11:00
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Paleontological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal ofPaleontology.
http://www.jstor.org
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

JOURNAL OF PALEONTOLOGY, V. 61, NO. 2, P. 363-374, 5 FIGS., MARCH 1987
AN ICHTHYOFAUNA FROM THE SUBSURFACE DEVONIAN OF NORTHWESTERN IOWA AND ITS BIOSTRATIGRAPHIC
AND PALEOECOLOGIC SIGNIFICANCE
GLENN W. STORRS Department of Geology and Geophysics and Peabody Museum of Natural History,
Yale University, New Haven, Connecticut 06511
ABSTRACT-A fauna of disarticulated Devonian fossil fish remains includes osteolepid, palaeo- niscoid, acanthodian, placoderm, and possible agnathan material. Lithologic and biostratigraphic evidence for a lacustrine or paludal environmental interpretation is examined. Palynomorphs from the deposit suggest a Middle (Givetian) to early Late (Frasnian) Devonian age which is compatible with the faunal evidence.
INTRODUCTION
DURING the course of recent hydrogeologic studies by the Iowa and U.S. Geological Sur- veys, a well-preserved assemblage of disar- ticulated fish remains was recovered from a well core taken near Hawarden, Sioux Coun- ty, Iowa (Figures 1, 2). This well (#D-7, NE?/4, NE/4, SE?4 sec. 5, T95N, R47W), part of the Dakota Aquifer Project of the I.G.S. and U.S.G.S., encountered at depths of 195.4- 197.0 m a thin, previously unknown deposit of silty shales above a thin sandstone over- lying Precambrian gneissic basement and overlain unconformably by the Nishnabotna Member of the Cretaceous Dakota Forma- tion (Ludvigson and Bunker, 1979). The unit presents a varve-like, laminated appearance and consists predominantly of light-brown to dark-grey, slightly silty, organic-rich shale, with some fine sand, disseminated pyrite, thin siltstone intercalations, and phosphatic lam- inae.
Portions of the 5.1 cm diameter rock core from the shale unit were repeatedly processed in household bleach to oxidize the organic shale binder, the residue screened and dried, and systematically picked. A moderately well- preserved palynomorph assemblage of low species diversity was obtained from the shale by standard processing techniques.
GEOLOGIC SETTING
The spores recovered from the Hawarden core are difficult to examine in detail because of the presence of amorphous kerogen in the samples but a Middle or Late Devonian age is suspected (Ravn, personal commun.). A
similar unit of brown, sandy, organic-rich shale was encountered approximately 20 km to the east, though not cored, in well D-44, Iowa Geological Survey, Boyden, Sioux County, Iowa (NE/4, SE/4, NE/4 sec. 9, T96N, R44W, Sioux Co.) (Figure 2). Here the shale overlies Cambrian dolomite, probably the Upper Cambrian Bonneterre Formation (Witzke, personal commun.). Although no fish debris was observed, spores from well cut- tings confirm both a Devonian age and a sim- iliarity to the Hawarden shale unit for these sediments. The lithologies and the spore as- semblages of both shales are so similar that the rocks probably represent the same strati- graphic unit.
Palynomorphs present in well D-44, all tri- lete spores, are relatively free of kerogen and include, most abundantly, Geminospora cf. G. svalbardiae (Vigran, 1964), but also Ver- rucosisporites premnus Richardson, 1965, Dictyotriletes sp., Samarisporites sp., and Convolutispora sp. (Figure 3.1-3.4). Verru- cosisporites premnus was originally reported from the Givetian (Old Red Sandstone) of Britain, while Geminospora svalbardiae is well known from the Givetian and the Frasnian. The entire floral assemblage thus seems to indicate a Middle to early Late Devonian, probably Givetian or Frasnian, age (Ravn, personal commun.). The lack of pollen in the sample supports this determination, as does the absence of the major components of the common Late Devonian (Famennian) Reti- spora lepidophyta-dominated assemblage. Ravn (personal commun.) also suggests that, while for paleoecologic reasons such data are far from conclusive, the present spore assem-
Copyright ? 1987, The Paleontological Society 363 0022-3360/87/0061-0363$03.00
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

364 GLENN W. STORRS
Devonian
Erosional Limits of Paleozoic (Pre-Devonian) Rocks
-r Strike and Dip of Paleozoic Rocks
7r-- Erosional Limits of Subsurface Devonian Rocks
Present Devonian Outcrop Pattern
Precambrian - Sioux Quartzite Ridge
SStudy Area
FIGURE I--Relationship of study area to Sioux Quartzite Ridge and the structural setting of mid- continental Devonian deposits (modified from Bunker, 198 1; and Ludvigson and Bunker, 1979).
blage differs markedly from those of the Em- sian and Eifelian of the Gulf Coast (see McGregor, 1979, for an overview of Devo- nian spore biostratigraphy).
Both shales apparently represent an outlier approximately 40 km northwest of the main body of Devonian shales and carbonates found throughout most of the subsurface of western Iowa (Figures 1, 2). No Devonian rocks are exposed at the surface in western Iowa. However, the Devonian marine se- quence onlaps to the northwest in Iowa and is probably entirely Late Devonian in age near its northwestern margin. Middle Devonian rocks are found in central and southwestern Iowa. Additionally, both new sections are lo- cated near to the southern margin of the Sioux Quartzite Ridge. The quartzite body forming this paleotopographic high is composed of gently folded, highly resistant Proterozoic rocks which locally formed the topographi- cally highest portion of the Transcontinental Arch throughout the Paleozoic and Mesozoic (Bunker, 1981; Johnson, 1970). The presence of quartzite clasts in the basal sands (Devo-
:????????.~i~iii~i~iQuortzjiieiii "i~iiiiiiii.~ !!
S:iuxi i oux::f:::?::~i 1
....
. . ..:
. ....:.:
:!:':" '" : ::" "o
~Sequence
N 0 20 40 km
..........I . . . . . . . . ..:??: ??: . . . . . . . . ..~.: ?::: ??:: ? ?
Sioux
Devonian
Marine
Sequence
0 20 40km FIGURE 2--Northwest Iowa study area enlarged
from Figure 1 showing subcrop patterns of pre- Mississippian age rocks. These rocks are now largely overlain by widespread Cretaceous sed- iments; only the Sioux Quartzite Ridge is pres- ently exposed. -C, undifferentiated Cambrian rocks (subsurface); 0, undifferentiated Ordovi- cian rocks (subsurface); P-C, undifferentiated Precambrian rocks (subsurface); 1, Iowa Geo- logical Survey well #D-7, Hawarden; 2, Iowa Geological Survey well #D-44, Boyden.
nian) of the Hawarden core suggests the past existence of fluvial networks draining the ridge. Because of its high local relief, the ridge was probably an island during the maximum extent of the Late Devonian marine trans- gressions in the area. Erosional stripping has since removed all Devonian carbonates from around the Sioux Ridge (Witzke, personal commun.).
DESCRIPTION OF FAUNA
The faunal assemblage consists of a wide range of skeletal elements but is particularly rich in scales, teeth, and miscellaneous frag- ments of bone. All specimens are small (1.0- 4.0 mm) and their delicate nature makes ex- amination difficult. Nevertheless, remains of several fish taxa can be recognized, although some specimens, due to their fragmentary na- ture, are indeterminate. All specimens are in the paleontology repository of the Depart- ment of Geology, University of Iowa, lot number 51487.
Acanthodii. --Acanthodians are represent-
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

DE VONIAN ICHTHYOFA UNA FROM IOWA 365
ed in the fauna by two sections of fin spine and dozens of scales (Figure 3.5-3.8). Only slight proportional and size variation is pres- ent in the scales, which are the nonimbricat- ing studs peculiar to the Acanthodii. All are less than 1.0 mm in length. The flat, dia- mond-shaped crown of each scale is separat- ed from the bulbous base by a uniformly wide constriction or neck. The crown is areally larger than, and extends caudad to, the domed, ovate base and thus the latter cannot be viewed in dorsal aspect. The crown is smooth and unsculptured. Concentric growth rings can sometimes be seen under high mag- nification on the surface of the bony base. The scales are distinguishable from the gross- ly similar scales of thelodonts on the basis of the lack of a pulp cavity in the present spec- imens. Additionally, they are of the Acan- thodes histologic "type" (Gross, 1947; Orvig, 1951, 1967a, 1967b) in possessing a crown of true dentine and an acellular base, as op- posed to the Nostolepis condition (Gross, 1971; Moy-Thomas and Miles, 1971).
These scales are identical to, and can be assigned with certainty to, Acanthodes? dub- linensis Stauffer, 1938. This species was pro- posed on the basis of scales recovered from the Upper Devonian (Frasnian) Olentangy Formation of central Ohio (Denison, 1979; Stauffer, 1938). It seems unlikely, however, that this species actually belongs to the Car- boniferous genus Acanthodes Agassiz, 1833. According to Denison (1979), the scales of A.? dublinensis resemble those of Acanthodes but are "surely not Acanthodes." Acanthodes scales can perhaps be distinguished by their somewhat concave coronal surface and their smaller posterior overhang of the crown (Wells, 1944). However, Gross (1973), in ex- amining the scale histology ofA.? dublinensis, indicates that Wells (1944) inadvisedly re- ferred the Devonian scales to Acanthoides, a genus improperly erected by Brotzen (1934), and which is now considered a synonym for Gomphonchus Gross, 1971. Wells (1944) proposed two new scale form species ofAcan- thoides, both of which probably represent body region scale variants of Acanthodes? dublinensis.
Acanthodes? dublinensis is also known from Eifelian bone beds in the Boyle Formation of east-central Kentucky, the Jeffersonville For- mation of southeastern Indiana, and the Del-
aware and Columbus formations of central Ohio (Denison, 1979; Wells, 1944). Similar scales have been reported from the Givetian Marcellus Formation of central New York (Smith, 1910). According to Gross (1973), scales of this species are also present in the uppermost Devonian (Famennian) Maple Mill Formation of Iowa. In Canada, this species has been found in the Emsian Eids and Blue Fiord formations of Bathurst and Ellesmere islands, respectively (Vieth, 1980). Valyukyavichyus (1979) reports the presence of Acanthodes? dublinensis from the Eifelian Forkdalen Formation of Spitsbergen and the Lyaday and Kyarnava formations of the So- viet Baltic region.
A large section of an acanthodian fin spine is characterized by a rough, tuberculated sur- face and a serrated edge (Figure 3.7). As no other acanthodian species are represented in the assemblage by scales, the spine might be assumed to belong to Acanthodes? dublinen- sis. If so, it represents the first find of a spine of this species. However, a portion of a sec- ond, possibly distinct, spine may also prove to be of acanthodian origin. This is the distal tip of a pronged spine whose external surface is smooth (Figure 3.8). Although only a frac- tion of the entire spine, it most closely re- sembles specimens that have been referred to the acanthodian organ genus Gyracanthus (Eastman, 1907; Wells, 1944). Denison (1979) notes that such spines range from the Lower Devonian through the Carboniferous and that several genera are probably represented by them.
Palaeoniscidae.--A major component of the assemblage is the large number of pa- laeoniscoid flank scales (Figures 4, 5.1, 5.2). These primarily rhomboidal scales are often fragmentary, probably as a result of the pro- cessing method. A size range of 1.0-3.0 mm is estimated. The scales bear the typically variable ribbed external surface and spiked posterior margin of palaeoniscoid actinop- terygians and possess both a thick isopedin base and a transparent ganoine crown, all of which are primitive features.
The ridges on the glossy enamel crown run horizontally from the posterior spines for- ward until they are deflected dorsally along the anterior margin of the scale. This is in contrast with, for example, the diamond- shaped scales of Cheirolepis in which each
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions
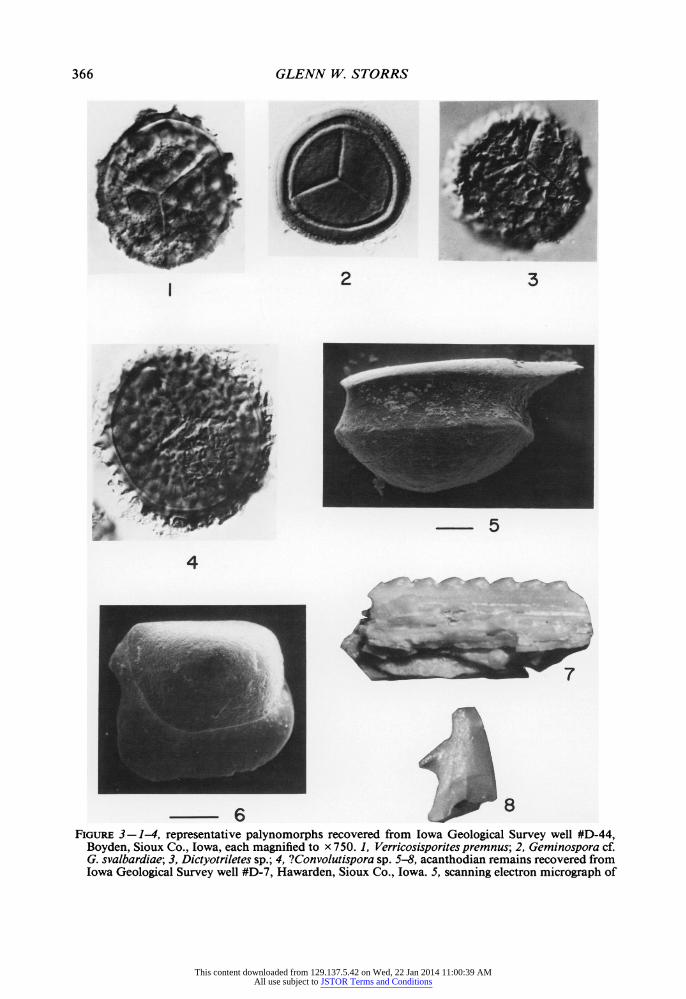
366 GLENN W. STORRS
2 3
Itr
5
'1'
4
% i
,1
N'
6 "
6 ,:•8 ? :
FIGURE 3-1-4, representative palynomorphs recovered from Iowa Geological Survey well #D-44, Boyden, Sioux Co., Iowa, each magnified to x 750. 1, Verricosisporites premnus; 2, Geminospora cf. G. svalbardiae; 3, Dictyotriletes sp.; 4, ?Convolutispora sp. 5-8, acanthodian remains recovered from Iowa Geological Survey well #D-7, Hawarden, Sioux Co., Iowa. 5, scanning electron micrograph of
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

DE VONIAN ICHTHYOFA UNA FROM IOWA 367
ridge intersects the anterior margin (Gross, 1947; Pearson and Westoll, 1979). On the present scales the anterior upturned portion of each ridge is itself ornamented with small, subhorizontal striae (Figures 4, 5.1). Each scale exhibits a dorsal articulatory peg, when not broken, and a low median ridge runs an- terodorsally-posteroventrally across the smooth inner surface. No prominent articu- latory facet is found along the anterior border of the scale.
The overall scale morphology of the Iowa material differs dramatically from those of most known Devonian palaeoniscoid genera, whose scales are either not rhomboidal in outline (e.g., Cheirolepis and Orvikuina), are not of similar proportions (e.g., Ligulalepis), or else display striking and unique ornamen- tation (e.g., Dialipina, Mimia, and Oso- rioichthys (Stereolepis)) (Casier, 1952, 1954; Gardiner, 1984; Giffin, 1980; Pearson and Westoll, 1979; Schultze, 1968, 1977). Con- versely, the Iowa scales are most similar to taxa such as Moythomasia (Aldingeria) or Stegotrachelus, although a wide range of scale ornamentation is known from the many species assigned to the first of these genera. Tegeolepis (Actinophorus) is known from the Famennian Ohio Shale (Gardiner, 1963; Dunkle and Schaeffer, 1973) and has rhom- boidal scales, but these are very poorly known and are perhaps too thin to be equated with the Iowa specimens.
Stegotrachelus is known only from a single species, S. finlayi, from the Givetian of Scot- land (Woodward and White, 1926). While Gross (1942) suggested that this fish is pos- sibly congeneric with Moythomasia, Gardi- ner (1963) maintained, on the basis of skull morphology, that the two are quite distinct, although he placed them together in the fam- ily Stegotrachelidae, along with the Missis- sippian Kentuckia, and later with Mimia from the Frasnian of Australia (Gardiner, 1984; Gardiner and Bartram, 1977). The rhom- boidal flank scales of Stegotrachelus are sim- ilar proportionally to those of Moythomasia and to those of the present specimens, but
?~??~
r: ?..,. ?-- '?
?.
-?~.. Y~ '`?
~Y~Li~.
~c
FIGURE 4--Composite drawing of representative palaeoniscoid flank scale with undamaged ar- ticulatory peg, possibly ?Moythomasia sp., an- terior to left, approximately 2.5 mm long.
are each marked by a single, coarse, diagonal rib. A typical dentine vascular system exists internally (Aldinger, 1937).
Two poorly known species of what appears to be Moythomasia are known from North America from the Frasnian "Portage shales" of western New York (Clarke, 1885; East- man, 1907; Gardiner, 1963; Hussakof and Bryant, 1918; Williams, 1886). However, the scales of neither Mothythomasia devonica (Clarke) nor M. antiqua (Williams) (both originally described as Palaeoniscus, and lat- er as Rhadinichthys) exhibit external orna- mentation which is identical to that of the well core specimens. The scales of Moytho- masia devonica are characterized by numer- ous punctae that are arranged in single rows between the horizontal ribbing. These punc- tae are an external expression of the promi- nent vascular canals inhabiting the median dentine layer of the scale (Orvig, 1957). This is a pattern similar to that seen in M. nitida (Gross, 1953; Jessen, 1968) and M. perforata (Gross, 1942, 1953) of the Middle Devonian of central Europe, and in M. durgaringa of the Upper Devonian of Australia (Gardiner,
Acanthodes? dublinensis flank scale, lateral aspect, anterior to left, bar scale equals 100 microns; 6, Acanthodes? dublinensis flank scale, ventral aspect, anterior to northeast, bar scale equals 100 microns; 7, regular light photograph of acanthodian fin spine fragment, approximately 2.5 mm long; 8, fin spine tip, ?Gyracanthus sp., approximately 1 mm long.
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

368 GLENN W. STORRS
44
4jt4
5--
hA
5t 7 , . -
. .
711*10 12i
' " . ,,A•,
6' ,.
9
...•, p•
? • .. •. , . .. S .." 'h . -
i. 5 i :re& -pQ. tJ. ?
I r,
10-Ct.• ii I
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

DE VONIAN ICHTHYOFA UNA FROM IOWA 369
1984), but no punctae are found in the pres- ent specimens. The scales of M. antiqua are apparently dissimilar in that they are essen- tially smooth with little or no ribbing, much as in the German M. laevigata (Gross, 1953), while possessing relatively few punctae. Moy- thomasia antiqua is known only from a few scales and, if distinct, remains a problematic species. Moythomasia striata (Gross, 1933a, 1933b, 1953; Jessen, 1968), like M. laevigata from the lower Upper Devonian of Germany, has no punctae and displays scale ribbing similar to that of the Iowa material, but the horizontal ribs are not upturned at their an- terior ends. The scales of M. striata also pos- sess large anterior articulatory facets which are noticeably lacking from the Iowa speci- mens.
While a large degree of body scale vari- ability is known among palaeoniscoids, it is apparent that the Iowa scales cannot be equated with any known species of Moytho- masia. However, because of the general sim- ilarity of the present specimens to scales of this genus, they can be most prudently clas- sified as ?Moythomasia sp.
Devonian palaeoniscoids, as the earliest representatives of the group, are extremely important from an evolutionary and a paleo- biogeographical point of view. Eastman (1907) indicated that indeterminate Devo- nian palaeoniscoid scales have also been re- ported from Kentucky, and scales and teeth can be locally common in the acetic acid res- idues of many Devonian rocks (Schultze, per- sonal commun.). However, the Devonian forms are still poorly understood, especially those known only from isolated scales. Fur- ther interpretations must await better mate- rial and, with it, the opportunity for a more comprehensive study.
A delicate fragment of tooth-bearing bone (Figure 5.3) is present in the Iowa sample. This is presumably palaeoniscoid in origin, and probably pertains to the same taxon as the scales. Three teeth, probably part of the marginal dentition, are preserved, each with the characteristic acrodin enameloid cap of palaeoniscoids.
Osteolepidae. - Some of the largest and most common elements in the assemblage are the scales of rhipidistian crossopterygians (Figure 5.4-5.6). All such scales are noncy- cloid and belong to the family Osteolepidae. They all bear a full external covering of cos- mine as is typical of most members of the family and the minute pore openings are clearly visible (for details of cosmine mor- phology see Gross, 1956; Meinke, 1984; Or- vig, 1969a; Thomson, 1975). The scales are largely rhomboidal and range from approx- imately 1.0 to 3.0 mm in length.
The anterior border of each scale usually corresponds in shape to the margin of the finished cosmine surface. An interesting fea- ture of the cosmine margin is the ribbed pat- tern observed at high magnification (Figure 5.5). The smooth medial surface of the scale displays the typical diagonal ridge of Devo- nian osteolepids.
A single bone fragment from the well core appears to be part of an osteolepid scale in which cosmine resorption has taken place, leaving a single tubercle of cosmine on an otherwise naked bone surface. Other osteo- lepid scales in the sample possess portions of the lateral line system as a string of large pits recessed into the external surface of the scale (Figure 5.6).
Scales of this general type are common among Devonian osteolepiform genera. Jar- vik (1948) has pointed out that few differ-
FIGURE 5-1-3, palaeoniscoid remains. 1, scanning electron micrograph of representative flank scale minus articulatory peg, possibly ?Moythomasia sp., anterior to left, bar scale equals 100 microns; 2, regular light photograph of similar scale, anterior to left, approximnately 3 mm long; 3, fragment of tooth-bearing bone, approximately 1.5 mm long. 4-8, osteolepid \remains. 4, representative flank scale of osteolepid, genus indeterminate, anterior to right, approximately 3 mm long; 5, scanning electron micrograph of anterior cosmine margin of similar scale, anterior to northwest, bar scale equals 10 microns; 6, scanning electron micrograph of osteolepid scale with portion of lateral line system, anterior to left; bar scale equals 100 microns; 7, indeterminate osteolepid tusk, approximately 1 mm long; 8, indeterminate cosmine-bearing bone, possibly part of an osteolepid lepidotrichium, approximately 2.5 mm long. 9-12, problematic bone fragments. 9-11, possible placoderm fragments, 9 and 10, approximately 1 mm long, 11, approximately 2.5 mm long; 12, possible agnathan fragment, approximately 1 mm long.
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

370 GLENN W. STORRS
ences exist among the scales of known osteo- lepid taxa, and that such isolates are almost useless in taxonomy. Glyptopomus from the Famennian of Scotland (Agassiz, 1844) and Panderichthys from the Frasnian of Latvia (Gross, 1941) can be ruled out in this instance because the scales of these fish are charac- terized by bony ornaments and no cosmine. Megistolepis from the Frasnian of Russia (Obruchev, 1955) is similarly unusual in that it is a very large form having scales averaging 2.0 cm in length.
The known Middle Devonian genera all bore small, nondescript, rhomboidal scales with cosmine. Gyroptychius, Osteolepis, and Thursius of the Eifelian and Givetian had such scales, are part of the circum-North At- lantic fauna discussed by Thomson (1976), and could theoretically have been present in Iowa. All were first reported from Scotland, but Gyroptychius is particularly well known from Greenland (Jarvik, 1950). Our knowl- edge of Devonian osteolepid distributions is limited by the presently small number of use- ful fossil localities. A similar North Atlantic taxon is the Frasnian Latvius which, in ad- dition to the Baltic region, has also been re- ported from New Brunswick (Greiner, 1977). Devonian osteolepid occurrences have pre- viously been reported in North America from Pennsylvania (Thomson, 1968, 1972), Ne- vada (Gregory et al., 1977), and Colorado (Denison, 1951). All of these are from the Upper Devonian, few have been generically identifiable, and in none are the scales re- markably different from typical osteolepids. The Iowa scales must also be considered ge- nus indeterminate.
Numerous isolated teeth in the collection (2.0 mm or less in length) can be attributed to crossopterygian fish. Most were probably contributed by osteolepids. Figure 5.7 shows a palatal tusk presumed to have come from an osteolepid. As is typical for Middle De- vonian osteolepids (Schultze, 1970), there is no labyrinthine infolding of the outer enam- eloid layer. One additional bone or scale in the sample bears an external cosmine layer. This is a relatively long, narrow element which may be part of an osteolepid lepidotrichium (Figure 5.8).
Placodermi. -A small number of problem- atic bone scraps in the collection seemingly
belong to placoderms, but because of their fragmentary nature these are of uncertain af- finities. One fragment of bony exoskeleton (Figure 5.9) displays several raised cylindri- cal tubercles on its surface. The bases of these tubercles are stellate, or supported by radial struts. This pattern is superficially similar to the peculiar dermal ornamentation of Tra- chosteus clarkii Newberry, 1889, a poorly known placoderm from the Famennian Cleveland Shale of Ohio. Two additional der- mal bone fragments (Figure 5.10, 5.11) dis- play rounded tubercles or nodes on their ex- ternal surfaces which are very like those found in antiarchs such as Pterichthyodes from the Middle Devonian, although certain arthro- dires, for example Coccosteus (also Middle Devonian), exhibit similar nodes.
?Agnatha. -What may be a small cephalic shield polygon is the only indication of pos- sible agnathans in the sample (Figure 5.12). This polygonal plate displays six raised tu- bercles on its outer surface which are smooth and hemispherical. While the plate appears similar to those figured by Stensi5 (1932) for various species of Cephalaspis, its isolated nature prevents identification. There is also some question as to whether the plates of Cephalaspis (well known from the Lower and Middle Devonian of the circum-North At- lantic region) constitute independent tesserae or rather only polygonal areas of the cephalic shield which have become separate through fracture (Wells, 1944). The cephalic shield exoskeleton is known to have been contin- uous in most cephalaspids.
Ohioaspis tumulosus Wells, 1944, of the Middle Devonian of the Midwest and Canada displays similar isolated plates, but these pos- sess quite different stellate tubercles. Ohioas- pis may not be a cephalaspid but is perhaps congeneric with the rhenanid placoderm As- terosteus (Gross, 1973; Orvig, 1969b). Rhen- anids do possess isolated tesserae and it is therefore possible that the Iowa plate is from a placoderm. Histological studies have not been made on placoderm tesserae (Denison, 1978).
DISCUSSION
As the Devonian stratigraphic sequence of northwestern Iowa is virtually unstudied, this assemblage presents potentially important
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

DE VONIAN ICHTHYOFA UNA FROM IOWA 371
information for analysis of mid-continental paleoenvironments and paleogeography. Un- fortunately, while there is ongoing progress in the field, Devonian vertebrate biostratig- raphy is currently a crucial but poorly refined science. Devonian stage determinations, for example, are always difficult to make, and accurate correlations between marine and nonmarine sections are still needed (Dineley, 1984). Similarly, the biostratigraphic reso- lution provided by the Iowa faunal assem- blage is, at present, insufficient to distinguish between a Middle and a Late Devonian age for the Hawarden shale unit. However, the recovered fish material is certainly compat- ible with the Givetian-Frasnian age suggest- ed by the preserved palynomorphs, and this interpretation should be accepted.
Acanthodes? dublinensis is known com- monly, though not exclusively, from the Mid- dle Devonian, and Moythomasia is found in both Middle and Upper Devonian rocks. Os- teolepiform rhipidistians are especially com- mon in Middle Devonian faunas, although the work of various authors has begun to doc- ument a broad diversity of osteolepiforms in the Late Devonian as well (Denison, 1951; Lebedev, 1983; Obruchev, 1955; Thomson, 1968, 1972; Vorobyeva, 1977). Among os- teolepids possessing scales similar to the present specimens, Latvius, for example, is from the Frasnian but Gyroptychius, Osteo- lepis, and Thursius are all Middle Devonian in age. Thomson (1976) has suggested that the traditional paucity of cosmine-bearing osteolepids from the Upper Devonian rocks of Europe and Greenland (where the osteo- lepids are well represented in the Middle De- vonian) is perhaps related to environmental factors and may have bearing on the func- tional and evolutionary significance of cos- mine. The Cephalaspida is one of the few agnathan groups to have survived into the Givetian but it disappears in the Frasnian (Westoll, 1979). As previously stated, the present agnathan occurrence is questionable.
The paleoenvironment represented by the Hawarden shale unit cannot be determined through faunal evidence alone, as the habitat of few Paleozoic fish groups is known un- equivocally. Acanthodians were marine fish during the Silurian, but they had invaded freshwater habitats by the beginning of the
Devonian and became very successful there (Moy-Thomas and Miles, 1971). Denison (1979) noted that the Acanthodidae, to which Acanthodes? dublinensis may perhaps belong, was a predominantly freshwater family. However, A.? dublinensis has been found to date only in presumed nearshore marine de- posits. After the Lower Devonian, crossop- terygians are believed to have been over- whelmingly freshwater forms, although Thomson (1969) pointed out that they were not exclusively so and has suggested that crossopterygians may actually have been pri- marily associated with proximal marine en- vironments in the Devonian (Thomson, 1980). The presence of cosmine-bearing os- teolepids may, at times, indicate a close as- sociation with nearshore marine conditions (Thomson, 1976). Palaeoniscoids also are typically found in both freshwater (e.g., Ste- gotrachelus) and marine (e.g., Tegeolepis and Moythomasia) deposits.
Certain placoderm groups were, as far as is known, restricted to marine environments but others thrived in fresh water, particularly in the Middle Devonian. Many arthrodires lived in fresh water as did most antiarchs (Denison, 1978). While the Agnatha began as marine organisms (Denison, 1956; Rob- ertson, 1957; White, 1958), some became freshwater forms in the Silurian or Devonian, and osteostracans were predominantly so. In either case, freshwater or marine provenance, the preservation and delicacy of many of the Iowa specimens would seem to preclude the possibility of long-distance transport.
Lithologic and stratigraphic evidence is more useful and argues in favor of a restricted lacustrine or paludal facies for the well core shale. The "varved," low energy appearance of the unit, its dissimilarity to the Devonian shale/carbonate marine sequence to the south, and its position as an outlier on basement so near to the Sioux Ridge are relevant. So too is the richness of organic material and the abundance of disseminated pyrite, suggesting not only bottom sediment anoxia, but also a rich supply of terrestrially derived iron. The presence of terrestrial spores and the lack of typical marine invertebrates and conodonts also favor a fresh water interpretation. The possibility that the true environment of de- position was part of a similarly restricted
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

372 GLENN W. STORRS
nearshore marine or estuarine environment cannot, however, be ruled out at this time.
ACKNOWLEDGMENTS
I want to thank particularly K. S. Thom- son, who suggested the topic for this paper and provided invaluable advice and assis- tance during its preparation. B. J. Witzke kindly brought the existence of this deposit to our attention, provided the samples, and supplied extremely helpful information throughout the course of my research. Thanks are due also to R. L. Ravn, who generously furnished the palynomorph identifications and photographs, and to H.-P. Schultze, L. J. Hickey, B. Schaeffer, P. Forey, and M. Wil- liams, who contributed useful help and/or advice. A. S. Pooley, W. K. Sacco, R. Board- man, and R. Purdy provided technical assis- tance. The manuscript was kindly reviewed by K. S. Thomson, B. J. Witzke, D. L. Dine- ley, and H.-P. Schultze. This study was sup- ported by National Science Foundation grant no. DEB 79-21746 to K. S. Thomson, by a Yale University Louis Pirrson Fellowship, and by the Department of Geology and Geo- physics, Yale University.
REFERENCES
AGASSIZ, L. 1833-1844. Recherches sur les Pois- sons Fossiles. Neuchitel, 5 vols., 1,420 p.
ALDINGER, H. 1937. Permische Ganoidfische aus Ostgrinland. Meddelanden om Gronland, 102(3):1-392.
BROTZEN, F. 1934. Die Morphologie und His- tologie der Proostea-(Acanthodiden) Schuppen. Arkiv fdr Zoologi, 26A(23):1-27.
BUNKER, B. J. 1981. The tectonic history of the Transcontinental Arch and Nemaha Uplift and their relationship to the Cretaceous rocks of the central midcontinent region. Iowa Geological Survey Guidebook, 4, 24 p.
CASIER, E. 1952. Contributions & l'atude des poissons fossiles de la Belgique, X. Un palaeo- niscoide du Famennian inf'erieur de la Fagne: Stereolepis marginis n. gen., n. sp. Bulletin, In- stitut Royal de Sciences Naturelles de Belgique (Brussels), 28(47):1-17.
1954. Contributions & l'atude des poissons fossiles de la Belgique, XI. Note additionnelle relative a "Stereolepis" (=Osorioichthys nov. nom.) et & l'origine de l'interoperculaire. Bul- letin, Institut Royal de Sciences Belgique (Brus- sels), 30(2): 1-12.
CLARKE, J. M. 1885. On the Higher Devonian fauna of Ontario County, New York. U.S. Geo- logical Survey, Bulletin, 3(16):39-120.
DENISON, R. H. 1951. Late Devonian freshwater fishes from the western United States. Fieldiana, Geology, 11(5):221-261.
. 1956. A review of the habitat of the earliest vertebrates. Fieldiana, Geology, 11(8):359-457.
. 1978. Placodermi. Handbook of Pa- leoichthyology, 2. 128 p.
. 1979. Acanthodii. Handbook of Pa- leoichthyology, 5. 62 p.
DINELEY, D. L. 1984. Devonian vertebrates in biostratigraphy. Proceedings of the Linnean So- ciety of New South Wales, 107(3):185-196 (1983).
DUNKLE, D. H. AND B. SCHAEFFER. 1973. Te- geolepis clarki (Newberry), a palaeonisciform from the Upper Devonian Ohio Shale. Palae- ontographica, ser. A, 143:151-158.
EASTMAN, C. R. 1907. Devonian fishes of Iowa. Iowa Geological Survey, Annual Report, 18:29- 386.
GARDINER, B. G. 1963. Certain palaeoniscoid fishes and the evolution of the snout in acti- nopterygians. Bulletin of the British Museum (Natural History), Geology, 8(6):255-325.
. 1984. The relationships of the paleoniscid fishes, a review based on new specimens of Mimia and Moythomasia from the Upper De- vonian of Western Australia. Bulletin of the British Museum (Natural History), Geology, 37(4): 173-428.
- AND A. W. H. BARTRAM. 1977. The ho- mologies of ventral cranial fissures in osteich- thyans, p. 227-245. In S. M. Andrews, R. S. Miles and A. Walker (eds.), Problems in Ver- tebrate Evolution. Academic Press, London.
GIFFIN, E. B. 1980. Devonian vertebrates from Australia. Postilla, 180:1-15.
GREGORY, J. T., T. G. MORGAN AND J. W. REED. 1977. Devonian fishes in central Nevada, p. 112-120. in M. A. Murphy, W. B. N. Berry and C. A. Sandberg (eds.), Western North America: Devonian. University of California, Riverside, Museum Contributions, 4.
GREINER, H. R. 1977. Crossopterygian fauna from the Albert Formation, New Brunswick, Canada, and its stratigraphical-paleoecological significance. Journal of Paleontology, 51:44-56.
GROSS, W. 1933a. Die Fische des baltischen Devons. Palaeontographica, ser. A, 79:1-72.
1933b. Die Wirbeltiere des rheinischen Devons. Abhandlungen Preussische Geolo- gische Landesanstalt, (N.F.), 154:5-84.
. 1941. Uber den Unterkiefer einiger de- vonischer Crossopterygier. Abhandlungen Preussische Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse, 7: 1-51.
. 1942. Die Fischfauna des baltischen De- vons und ihre biostratigraphische Bedeutung. Korrespondenzblatt der Naturforscher-Vereins zu Riga, 64:373-436.
. 1947. Die Agnathen und Acanthodier des
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

DE VONIAN ICHTHYOFA UNA FROM IOWA 373
obersilurischen Beyrichienkalks. Palaeonto- graphica, ser. A, 96:91-161.
. 1953. Devonische Palaeonisciden-Reste in Mittel- und Osteuropa. Paldiontologische Zei- tung, 27(1/2):85-112.
. 1956. Uber Crossopterygier und Dipnoer aus dem baltischen Oberdevon im Zusammen- hang einer vergleichenden Untersuchung des Porekanalsystems paliozoischer Agnathen und Fische. Kungliga Svenska Vetenskapsakade- miens Handlingar, ser. 4, 5:1-140.
. 1971. Downtonische und dittonische Acanthodier-reste des Ostseegebietes. Palaeon- tographica, ser. A, 136:1-82.
. 1973. Kleinschuppen, Flossenstacheln und Ziihne von Fischen aus europiiischen und nord- amerikanischen bonebeds des Devons. Palae- ontographica, ser. A, 142:51-155.
HUSSAKOF, L. AND W. L. BRYANT. 1918. Catalog of the fossil fishes in the Museum of the Buffalo Society of Natural Science. Bulletin of the Buf- falo Society of Natural Science, 12:1-346.
JARVIK, E. 1948. On the morphology and tax- onomy of the Middle Devonian osteolepid fish- es of Scotland. Kungliga Svenska Vetenskaps- akademiens Handlingar, ser. 3, 25(1):1-301.
. 1950. Middle Devonian vertebrates from Canning Land and Wegeners Halv6 (East Greenland). Part II. Crossopterygii. Meddelan- den om Gronland, 96(4):1-132.
JESSEN, H. 1968. Moythomasia nitida Gross und M. cf. striata Gross, devonische Palaeonisciden aus dem Oberen Plattenkalk der Bergisch-Glad- bach-Paffrather Mulde (Rheinisches Schiefer- gebirge). Palaeontographica, ser. A, 128:87-114.
JOHNSON, J. G. 1970. Taghanic onlap and the end of North American Devonian provinciality. Geological Society ofAmerica Bulletin, 8 1:2077- 2106.
LEBEDEV, O. A. 1983. A new crossopterygian fish from the central Devonian region. Paleontolo- gicheskiy Zhurnal, 4:68-75 (in Russian).
LUDVIGSON, G. A. AND B. J. BUNKER. 1979. Sta- tus of hydrogeologic studies in northwest Iowa. Iowa Geological Survey Open File Report, Sept. 1979, 37 p.
MCGREGOR, D. C. 1979. Spores in Devonian stratigraphical correlation. Palaeontological As- sociation Special Papers, 23:163-184.
MEINKE, D. K. 1984. A review of cosmine: its structure, development, and relationship to oth- er forms of the dermal skeleton in osteichthyans. Journal of Vertebrate Paleontology, 4:457-470.
MoY-THOMAS, J. A. AND R. S. MILES. 1971. Pa- laeozoic Fishes. W. B. Saunders Co., Philadel- phia, 259 p.
NEWBERRY, J. S. 1889. The Paleozoic fishes of North America. U.S. Geological Survey, Mono- graph 16, 340 p.
OBRUCHEV, O. P. 1955. Upper Devonian fishes of central Kazakhstan. Sovyetskaya Geologiya, 45:84-99 (in Russian).
ORVIG, T. 1951. Histologic studies of placo- derms and fossil elasmobranchs. 1. The endo- skeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv fdr Zoologi, 2(2):321-454.
1957. Palaeohistological notes. I. On the structure of the bone tissue in the scales of cer- tain Palaeonisciformes. Arkiv fdr Zoologi, 2(10): 481-490.
. 1967a. Some new acanthodian material from the Lower Devonian of Europe. Journal of the Linnean Society (Zoology), 47:131-153.
1967b. Phylogeny of tooth tissues: evo- lution of some calcified tissues in early verte- brates, p. 45-110. In A. E. W. Miles (ed.), Struc- tural and Chemical Organization of Teeth. Academic Press, New York.
. 1969a. Cosmine and cosmine growth. Le- thaia, 2:241-260.
. 1969b. Vertebrates from the Wood Bay Group and the position of the Emsian-Eifelian boundary in the Devonian of Vestspitsbergen. Lethaia, 2:273-328.
PEARSON, D. M. AND T. S. WESTOLL. 1979. The Devonian actinopterygian Cheirolepis Agassiz. Transactions, Royal Society of Edinburgh, 70(13-14):337-399.
RICHARDSON, J. B. 1965. Middle Old Red Sand- stone spore assemblages from the Orcadian Ba- sin, north-east Scotland. Palaeontology, 7:559- 605.
ROBERTSON, J. D. 1957. The habitat of the ear- liest vertebrates. Biology Review, 32:156-187.
SCHULTZE, H.-P. 1968. Palaeoniscoidea-Schup- pen aus dem Unterdevon Australiens und Ka- nadas und aus dem Mitteldevon Spitzbergens. Bulletin of the British Museum (Natural His- tory), Geology, 16(7):343-368.
. 1970. Folded teeth and the monophyletic origin of tetrapods. American Museum Novi- tates, 2408:1-10.
.1977. Ausgangsform und Entwicklung der rhombischen Schuppen der Osteichthyes (Pisces). Palaontologische Zeitung, 51(3/4): 152-168.
SMITH, B. 1910. Notes on some little-known fish- es from the New York Devonian. Proceedings, Academy of Natural Sciences of Philadelphia, 62:656-663.
STAUFFER, C. R. 1938. Conodonts of the Olen- tangy Shale. Journal of Paleontology, 12:411- 433.
STENSI6, E. A. 1932. The cephalaspids of Great Britain. British Museum (Natural History), London, 220 p.
THOMSON, K. S. 1968. A new Devonian fish (Crossopterygii, Rhipidistia) considered in re- lation to the origin of the Amphibia. Postilla, 124:1-13.
1969. The environment and distribution of Paleozoic sarcopterygian fishes. American Journal of Science, 269:457-464.
. 1972. New evidence on the evolution of
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions

374 GLENN W. STORRS
the paired fins of Rhipidistia and the origin of the tetrapod limb, with description of a new genus of Osteolepidae. Postilla, 157:1-7.
. 1975. On the biology of cosmine. Bulletin of the Peabody Museum of Natural History, 40: 1-57.
1976. The faunal relationships of rhipi- distian fishes (Crossopterygii) from the Catskill (Upper Devonian) of Pennsylvania. Journal of Paleontology, 50:1203-1208.
. 1980. The ecology of Devonian lobe-finned fishes, p. 187-222. In A. L. Panchen (ed.), The Terrestrial Environment and the Origin of Land Vertebrates. Academic Press, New York.
VALYUKYAVICHYUS, Y. Y. 1979. Acanthodian scales from the Eifelian of Spitsbergen. Paleon- tologicheskiy Zhurnal, 4:101-111 (in Russian).
VIETH, J. 1980. Thelodontier-, Acanthodier-, und Elasmobranchier-Schuppen aus dem Unter- Devon der kanadischen Arktis (Agnatha, Pisces). Gittinger Arbeiten zur Geologie und Paliion- tologie, 23:1-69.
VIGRAN, J. 0. 1964. Spores from Devonian de- posits, Mimerdalen, Spitsbergen. Norsk Polar- institut, Skrifter, 132:1-32.
VOROBYEVA, E. I. 1977. The phylogenetic con- nections of the osteolepiform Crossopterygii and
their systematic position, p. 71-88. In V. V. Menner (ed.), Ocherki po filogenii i sistematike iskopayemykh ryb i beschelyustnykh. Izda- tyelstva "Nauka," Moscow (in Russian).
WELLS, J. W. 1944. Fish remains from the Mid- dle Devonian bone beds of the Cincinnati Arch region. Palaeontographica Americana, 3(16): 1- 62.
WESTOLL, T. S. 1979. Devonian fish biostratig- raphy. Palaeontological Association Special Pa- pers, 23:341-353.
WHITE, E. I. 1958. Original environment of the craniates, p. 212-234. In T. S. Westoll (ed.), Studies on Fossil Vertebrates. The Athlone Press, London.
WILLIAMS, H. U. 1886. Notes on the fossil fishes of the Genesee and Portage Black Shales. Bul- letin of the Buffalo Society of Natural Science, 5:81-84.
WOODWARD, A. S. AND E. I. WHITE. 1926. The fossil fishes of the Old Red Sandstone of the Shetland Islands. Transactions, Royal Society of Edinburgh, 54:567-571.
MANUSCRIPT RECEIVED 21 AUGUST 1985 REVISED MANUSCRIPT RECEIVED 19 DECEMBER 1985
This content downloaded from 129.137.5.42 on Wed, 22 Jan 2014 11:00:39 AMAll use subject to JSTOR Terms and Conditions




















