
Trilobite palaeobiodiversity during the Devonian in the Ardennes Massif
11
Bull. Soc. géol. Fr., 2007, n o 6 Trilobite palaeobiodiversity during the Devonian in the Ardennes Massif CATHERINE CRÔNIER 1 and ALLART van VIERSEN 2 Key-words. – Trilobites, Devonian, Ardennes Massif, Biodiversity, Assemblages Abstract. – Trilobite occurrences and biodiversity from Devonian sections of the Ardennes Massif are presented. Fluc- tuations in diversity reflect environmental changes and event-stratigraphic turnovers throughout the Devonian. Peak di- versity is attained during the Eifelian (Middle Devonian) when the carbonate system is developed. Six trilobite associations have been delineated based on cluster analysis. Distribution of these associations is correlated with the de- position environments and age of the strata. Paléobiodiversité des trilobites en Ardenne occidentale au Dévonien Mots-clés. – Trilobites, Dévonien, Ardenne occidentale, Biodiversité, Assemblages Résumé. – Dans les environnements favorables du Dévonien de l’Ardenne occidentale, les trilobites sont des compo- sants majeurs du macrobenthos. Une analyse des changements de la paléobiodiversité et une modélisation de la réparti- tion des communautés à trilobites du massif des Ardennes au Dévonien sont proposées. Le pic de diversité est atteint à l’Eifélien (Dévonien moyen) quand le système carbonaté est développé. Six associations ont été reconnues à l’aide des analyses de classification hiérarchique. La distribution de ces associations est corrélée aux conditions écologiques et temporelles. INTRODUCTION The Ardennes Massif is part of a complex of Palaeozoic outcrops between the English Channel in the west and the Rhine River and beyond in the east. Originally part of Ava- lonia, a terrane located north of the Gondwana, it became an element of the southern margin of the Old Red Sandstone Continent in the Devonian and Carboniferous, when it suf- fered the effects of the Hercynian Orogeny. These global tectonic events, linked to climatic changes due to continen- tal drift, had profound consequences on the living orga- nisms of the Ardennes Massif. Recent works [Meilliez, 1989; Cibaj, 1992; Goffette, 1991; Lacquement, 2001] have reported knowledge of the tectonic and geodynamical context of the Ardennes area; they have allowed revision of the stratigraphy [Bultynck et al., 1991; Bultynck and Dejonghe, 2002] and of the geolo- gical map of Givet [Lacquement et al., 2003]. In addition, these works have shown the need to improve knowledge of the palaeoenvironments represented by numerous stratigra- phic units such as the formation cropping out at the ‘Mur des douaniers’ in the French Ardennes [Crônier and van Viersen, 2008]. This knowledge must be based on a revision of the sedimentological and faunal contents. Such fossilife- rous outcrops must be studied in the contexts not only of the taxa present where the biodiversity can be estimated but also of the paleoecology and paleoenvironments. Trilobites are important elements of the Devonian ma- crobenthos. During this period, one of the most severe res- trictions in trilobite diversity on a global scale occurred: at the end of the early Late Devonian, during the global Kell- wasser Crisis [Schindler, 1990]. At that time, most trilobites that were adapted to outer - shelf level - bottom habitats af- ter the mid-Givetian Taghanic Onlap, i.e. the major trans- gressive event of the Givetian Age [Johnson, 1970], were affected by repeated rises of sea-level leading to a world-wide ‘drowning’ of reef ecosystems and reductions of shallow water habitats [Schlager, 1981; Feist, 1991]. Long-lived orders such as the Corynexochida, Harpetida and Lichida disappeared, and the two orders surviving the crisis, the Phacopida and Proetida, were severely affected. In this context, we focussed our investigations on repre- sentative trilobite-bearing sections of the Devonian Ardennes Massif, in order to evaluate this biodiversity. This work, which started in the spring of 2003 by one of us (A.vanV.), is an account of numerous observations in the Belgian Arden- nes and a first estimate of Belgian Devonian trilobite paleo- biodiversity. These Belgian faunas appear to be highly diverse but are poorly documented, not being as well known as those from the German Eifel and French Ardennes. Bull. Soc. géol. Fr., 2007, t. 178, n o 6, pp. 473-483 1. Université de Lille 1, Sciences de la Terre, UMR 8014 du CNRS, Laboratoire de Paléontologie et Paléogéographie du Paléozoïque, F-59655 Villeneuve d’Ascq cedex. [email protected] 2. Graaf van Loonstraat 25, NL-6121 JS Born, The Netherlands. [email protected] Manuscrit déposé le 30 novembre 2006; accepté après révision le 21 mai 2007. Séance spécialisée Sédimentation et structuration du front nord varisque et évolution méso-cénozoïque : exemple de l’Ardenne Givet, 4-6 mai 2006
-
Upload
nhmmaastricht -
Category
Documents
-
view
1 -
download
0
Transcript of Trilobite palaeobiodiversity during the Devonian in the Ardennes Massif

Bull. Soc. géol. Fr., 2007, no 6
Trilobite palaeobiodiversity during the Devonian in the Ardennes MassifCATHERINE CRÔNIER1 and ALLART van VIERSEN2
Key-words. – Trilobites, Devonian, Ardennes Massif, Biodiversity, Assemblages
Abstract. – Trilobite occurrences and biodiversity from Devonian sections of the Ardennes Massif are presented. Fluc-tuations in diversity reflect environmental changes and event-stratigraphic turnovers throughout the Devonian. Peak di-versity is attained during the Eifelian (Middle Devonian) when the carbonate system is developed. Six trilobiteassociations have been delineated based on cluster analysis. Distribution of these associations is correlated with the de-position environments and age of the strata.
Paléobiodiversité des trilobites en Ardenne occidentale au Dévonien
Mots-clés. – Trilobites, Dévonien, Ardenne occidentale, Biodiversité, Assemblages
Résumé. – Dans les environnements favorables du Dévonien de l’Ardenne occidentale, les trilobites sont des compo-sants majeurs du macrobenthos. Une analyse des changements de la paléobiodiversité et une modélisation de la réparti-tion des communautés à trilobites du massif des Ardennes au Dévonien sont proposées. Le pic de diversité est atteint àl’Eifélien (Dévonien moyen) quand le système carbonaté est développé. Six associations ont été reconnues à l’aide desanalyses de classification hiérarchique. La distribution de ces associations est corrélée aux conditions écologiques ettemporelles.
INTRODUCTION
The Ardennes Massif is part of a complex of Palaeozoicoutcrops between the English Channel in the west and theRhine River and beyond in the east. Originally part of Ava-lonia, a terrane located north of the Gondwana, it became anelement of the southern margin of the Old Red SandstoneContinent in the Devonian and Carboniferous, when it suf-fered the effects of the Hercynian Orogeny. These globaltectonic events, linked to climatic changes due to continen-tal drift, had profound consequences on the living orga-nisms of the Ardennes Massif.
Recent works [Meilliez, 1989; Cibaj, 1992; Goffette,1991; Lacquement, 2001] have reported knowledge of thetectonic and geodynamical context of the Ardennes area;they have allowed revision of the stratigraphy [Bultynck etal., 1991; Bultynck and Dejonghe, 2002] and of the geolo-gical map of Givet [Lacquement et al., 2003]. In addition,these works have shown the need to improve knowledge ofthe palaeoenvironments represented by numerous stratigra-phic units such as the formation cropping out at the ‘Murdes douaniers’ in the French Ardennes [Crônier and vanViersen, 2008]. This knowledge must be based on a revisionof the sedimentological and faunal contents. Such fossilife-rous outcrops must be studied in the contexts not only of the
taxa present where the biodiversity can be estimated butalso of the paleoecology and paleoenvironments.
Trilobites are important elements of the Devonian ma-crobenthos. During this period, one of the most severe res-trictions in trilobite diversity on a global scale occurred: atthe end of the early Late Devonian, during the global Kell-wasser Crisis [Schindler, 1990]. At that time, most trilobitesthat were adapted to outer - shelf level - bottom habitats af-ter the mid-Givetian Taghanic Onlap, i.e. the major trans-gressive event of the Givetian Age [Johnson, 1970], wereaffected by repeated rises of sea-level leading to aworld-wide ‘drowning’ of reef ecosystems and reductionsof shallow water habitats [Schlager, 1981; Feist, 1991].Long-lived orders such as the Corynexochida, Harpetidaand Lichida disappeared, and the two orders surviving thecrisis, the Phacopida and Proetida, were severely affected.
In this context, we focussed our investigations on repre-sentative trilobite-bearing sections of the Devonian ArdennesMassif, in order to evaluate this biodiversity. This work,which started in the spring of 2003 by one of us (A.vanV.), isan account of numerous observations in the Belgian Arden-nes and a first estimate of Belgian Devonian trilobite paleo-biodiversity. These Belgian faunas appear to be highlydiverse but are poorly documented, not being as well knownas those from the German Eifel and French Ardennes.
Bull. Soc. géol. Fr., 2007, t. 178, no 6, pp. 473-483
1. Université de Lille 1, Sciences de la Terre, UMR 8014 du CNRS, Laboratoire de Paléontologie et Paléogéographie du Paléozoïque, F-59655 Villeneuved’Ascq cedex. [email protected]. Graaf van Loonstraat 25, NL-6121 JS Born, The Netherlands. [email protected] déposé le 30 novembre 2006; accepté après révision le 21 mai 2007.
Séance spécialiséeSédimentation et structuration
du front nord varisque et évolutionméso-cénozoïque : exemple de l’Ardenne
Givet, 4-6 mai 2006

GEOLOGICAL SETTING AND MATERIAL
Several sections from the Devonian Ardennes Massif, loca-ted in the ‘Allochthonous Ardennes’ [Meilliez and Mansy,1990; Mansy and Meilliez, 1993], along the southern andnorthern-eastern marging of the Dinant synclinorium (fig. 1)are known to yield trilobites. These sections include the fa-mous fossiliferous site known as the ‘Mur des douaniers’[van Viersen, 2006a; Crônier and van Viersen, 2008]. The tri-lobites from these sites have remained largely undocumen-ted. Moreover, most of these trilobite-bearing sections havenot yet been described with respect to their sedimentologi-cal/faunal content and bioevent stratigraphy. Here, ten sec-tions with trilobite records of varying quality – some of thesample sizes are small – are documented in order to obtain apreliminary assessment of paleobiodiversity in the westernArdennes (fig. 1). The main focus is on the evolution of trilo-bite associations from the Early to Late Devonian and pos-sible correlations with sea-level variations.
BIODIVERSITY ANALYSIS THROUGH DEVONIANTIME
The study of palaeobiodiversity, especially at regional andlocal scales, is meaningful even though estimate of Phane-rozoic global biodiversity is limited by biases that may bepresent in the fossil record. Important biases include un-der-representation of some areas, inaccuracies in taxonomic
data and uneven sampling intensity across groups of orga-nisms, environments, and time intervals [Badgley, 2003].Recognition of these methodological problems has resultedin notable improvements in the Phanerozoic diversity data-base [Adrain and Westrop, 2000; Alroy et al., 2001].
Fluctuations in biodiversity
In the Ardennes Massif, trilobites are mainly represented bynine families: Styginidae (Corynexochida), Lichidae andOdontopleuridae (Lichida), Homalonotidae, Phacopidaeand Acastidae (Phacopida), Proetidae, Tropidocoryphidaeand Aulacopleuridae (Proetida). Another family, Harpetidae(Harpetida), is rare. For comparison, trilobite paleobiodi-versity from Early to Late Devonian at the global and regio-nal scales (Ardennes Massif) is given in figure 2A-B.
Lower Devonian
Trilobites seem to be poorly diverse macrofaunal consti-tuents in the Lower Devonian of the Ardennes Massif, incontrast with their worldwide diversity (fig. 2A-B). At theglobal scale, trilobite diversity is relatively poor (fig. 2A,2C). Only a marginal decrease in diversity during this timeseems to exist. In fact, according to Chlupá� [1994], this lo-wered trilobite diversity persists from the Silurian into theLochkovian with no drastic changes, only minor extinctionsand more important migrations. The Basal Pragian Event ofregressive character, i.e. an eustatic sea level lowering[Chlupá� and Kukal, 1986] contributed to widening of shal-
474 CRÔNIER C. and VAN VIERSEN A.
Bull. Soc. géol. Fr., 2007, no 6
FIG. 1. – Geological map showing the location of several Devonian fossiliferous sections in the Ardennes Massif, southern Belgium [modified after De-jonghe, 2001].FIG. 1. – Localisations géographique et géologique des différents sites fossilifères du Dévonien en Ardennes occidentale, Wallonie, Belgique [modifiéd’après Dejonghe, 2001].

low marine realms with carbonate sedimentation favourablefor trilobites. This led to an increase of the generic diversity[Chlupá�, 1994] and, at least, to a preservation of the familydiversity which lasted with some changes until the early Ei-felian.
For the regional scale, at the beginning of the DevonianPeriod, a major marine transgression from the south-westreached the northern borders of the Ardennes and Eifel[Ziegler, 1982; Steemans, 1989]. Erosion of several ancientmassifs, such as the Rocroi Massif, led to important shaly
Bull. Soc. géol. Fr., 2007, no 6
TRILOBITE PALAEOBIODIVERSITY IN THE ARDENNES MASSIF 475
FIG. 2. – Total occurrences of trilobite taxa (A) at the global scale [modified from Feist, 1991; Chlupá1, 1994], (B) in the Ardennes Massif; total familyoccurrences for each trilobite order (C) at the global scale [modified from Feist, 1991; Chlupá1, 1994], (D) in the Ardennes Massif; total (E) subfamilyand (F) genus occurrences of each trilobite order in the Ardennes Massif, from Early to Late Devonian.FIG. 2. – Nombre total de trilobites (A) à l’échelle mondiale [modifié d’après Feist, 1991 ; Chlupá1, 1994], (B) dans le massif des Ardennes ; nombre totalde familles pour chaque ordre de trilobites (C) à l’échelle mondiale [modifié d’après Feist, 1991 ; Chlupá1, 1994], (D) dans le massif des Ardennes ;nombre total de (E) sous-familles et (F) genres pour chaque ordre de trilobites dans le massif des Ardennes, au Dévonien.

and sandy deposits [Godefroid et al. 1994] in which trilobi-tes are rare. The paleobiodiversity is low during the Loch-kovian-Pragian, characterized exclusively by Phacopida(fig. 2D-F). Only two families occur, and those are rare:Acastidae and Homalonotidae. New discoveries in the eas-tern Ardennes (Treveropyge, Dunopyge and Acastava; un-pub. data from van Viersen) corroborate the occurrences oftwo subfamilies of the Acastidae: Acastavinae and Astero-pyginae, as mentioned by Maillieux [1933] and Asselberghs[1946]. Homalonotinae are represented by Digonus ornatusdisornatus RICHTER & RICHTER and Scabrella asselberghsi(MAILLIEUX), among others. Wenndorf [1990] resolvedmany of the synonyms within this subfamily in the Arden-nes and Eifel areas. Additional study and fieldwork are re-quired to revise and complete the list of Early Devoniantrilobites in the Ardennes.
Trilobite diversity is greater in the Emsian than in theLochkovian and Pragian but is not particularly high(fig. 2D-F). Increase in diversity is linked to the opening ofthe marine habitat during the Early Devonian. During theMiddle Emsian, a regressive episode occurred which resul-ted in characteristic red shales [Godefroid et al., 1994].Three additional families were recorded by Maillieux[1933]: Proetidae, Phacopidae and Odontopleuridae. So far,none of these records could be corroborated although itseems likely that these families actually occur in the Bel-gian Emsian. Treveropyge is among the most common Aste-ropyginae of the Ardennes-Eifel area during this time.Locally, another common trilobite genus is Digonus (Homa-lonotinae).
Middle Devonian
At the global scale (fig. 2A, 2C), the general decline in tri-lobites, marked by stepwise diversity falls, was strongly ac-celerated by the Middle Devonian Kacák and Taghanicevents in the late Eifelian and middle Givetian respectively[Chlupá�, 1994].
At the regional scale, a more major transgressive phase[Ziegler, 1982; Bultynck et al., 1991] leads to more diversi-fied trilobites (fig. 2D-F). Middle Eifelian limestones inparticular yield a rich trilobite fauna, including a number ofspecies known also from the German Eifel [Struve, 1982;Basse, 1996, 1997, 1998; Magrean and van Viersen, 2005;van Viersen, 2007]. The initiation of carbonate deposition,characterised by shallow, warm and clear seas, favoured theappearance of Corynexochida, Proetida and Lichida (Li-chidae being generally linked to neritic and peri-reefal envi-ronments [Feist, 1991]), and the diversification ofPhacopida among trilobites. Stromatoporids [Lecompte,1951-1952], corals [Tourneur, 1985], crinoids and brachio-pods [Godefroid, 1995] also occur. Peak diversity was attai-ned during the Eifelian when typically Rhenish faunasflourished. Their generic diversity is three times that duringthe Emsian (fig. 2D-F). All orders present at the globalscale are present also in the Ardennes Massif, with Phacopi-da and Proetida dominant. Harpetida are rare.
Eifelian-Givetian transitional sequences are comparati-vely well exposed in some places, especially in the Roche-fort area. Trilobites from the Givetian are mostly poorlydocumented (fig. 2B, D-F). Calycoscutellum, Dechenella,Gerastos and Nyterops have been identified, and notably
Dechenella, Gerastos and Nyterops in the Rochefort biotaduring the Early Givetian (see the next section, fig. 4).
Upper Devonian
The Belgian Upper Devonian is not particularly rich in tri-lobites although fragmentary material may be abundant lo-cally. The basal Frasnian is represented by a mainly shalyunit that corresponds to a major transgressive phase [Bul-tynck et al. 2000]. At some localities on the southern borderof the Dinant Synclinorium, reef cycles may be observed[Boulvain, 2006]; each period of reefal growth is interrup-ted by a phase of sea-level increase leading to terrigenousdeposits [Boulvain et al., 1999]. Trilobites are mostly res-tricted to Middle Frasnian units, especially for Proetida andPhacopida (fig. 2D-F). A third remaining order, Corynexo-chida seems to be present (figs. 2D-F). Indeed, Richter andRichter [1926] and Maillieux [1927] pointed out some re-cords of Scutellum. Nevertheless, no new samples were col-lected in this study.
The Frasnian-Famennian crisis led to the extinction ofalmost all reefs and their associated trilobites (Lichidae),and a significant decrease in other trilobite taxa.
The Famennian is associated with an important regres-sive phase with terrigenous deposits on a shallow platformin the process of filling [Thorez et al. 2006; Bultynck andDejonghe, 2002]. A record of a Famennian trilobite, Pha-cops (Omegops) accipitrinus maretiolensis RICHTER &RICHTER, 1933, seems to exist [Rome, 1936; Struve, 1976].Nevertheless, no new samples were collected in this study.
Associative and ecological structures in biodiversity
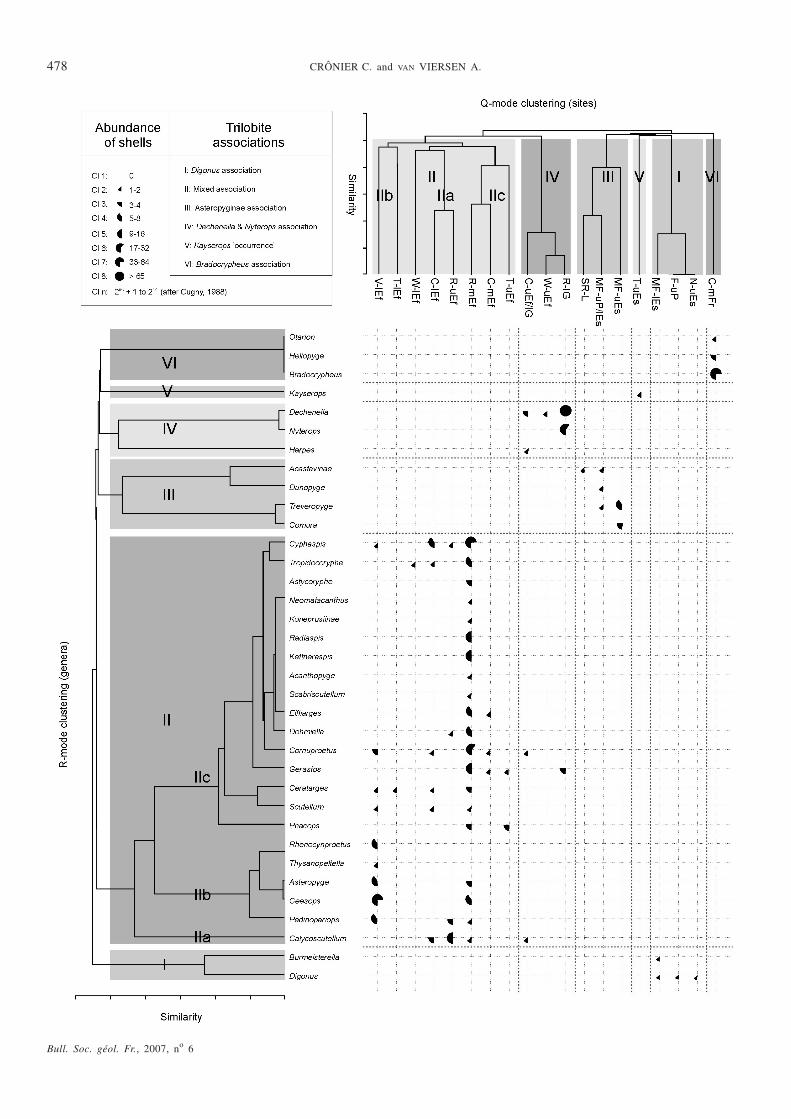
In order to understand the biodiversity better, a preliminaryanalysis of trilobite associations from the Devonian of theArdennes Massif has been undertaken. The term associationis used here for assemblages of trilobites with similar taxo-nomic composition [Brenchley and Harper, 1998; Thomasand Lane, 1999; Chlupá�, 1987]. Assemblages are generallycharacterised both by their species composition and the re-lative abundance of individuals of the different species pre-sent. This analysis has focused on the quantitative aspect,especially cluster analyses, which are commonly applied topalaeoecological problems [Cugny, 1988; Brenchley andHarper, 1998; Botquelen et al. 2006], notably on Cam-brian-Ordovician trilobites [Ludvigsen and Westrop, 1983],and helped in the identification of associations.
The biodiversity of sampled faunas is represented in ourstudy by 33 genera and two subfamilies that refer to asingle, undetermined genus (fig. 4), for 19 samples fromLochkovian to Frasnian strata (fig. 1). Data have been ana-lysed with hierarchical cluster analyses. The relative abun-dances of the 35 genera/subfamilies have been quantified ina separate way in eight classes (fig. 4). Hierarchical clusteranalysis based on the trilobite abundance classes was achie-ved using PAST 1.04 software [Hammer et al., 2001]. Clus-ter analysis is an agglomerating technique that groupstogether objects by level of similarity [Davis, 1986; Harper,1999]. Results of Q-mode clustering (grouping of samples)and R-mode clustering (groupings of taxa) are presentedas dendrograms, which were achieved using paired-groupmethod with the Pearson Similarity Coefficient. Groups of
Bull. Soc. géol. Fr., 2007, no 6
476 CRÔNIER C. and VAN VIERSEN A.

Bull. Soc. géol. Fr., 2007, no 6
TRILOBITE PALAEOBIODIVERSITY IN THE ARDENNES MASSIF 477
FIG. 3. – (A) Changes in trilobite faunal diversity,dominance and evenness through 19 sites from Earlyto Upper Devonian of the Ardennes Massif. (B) Re-lative abundance of trilobite taxa in the ‘Rochefort’site from the Middle Eifelian.FIG. 3. – (A) Changement dans la diversité, la do-minance et la régularité des faunes trilobitiques de19 sites provenant du Dévonien des Ardennes.(B) Abondance relative des taxons de trilobites dansle gisement de ’Rochefort’ de l’Eifélien moyen.
Bull. Soc. géol. Fr., 2007, no 6
478 CRÔNIER C. and VAN VIERSEN A.

genera with a high probability of mutual occurrence arethus grouped together.
In order to complete the palaeoecological informationof these assemblages, three ecological indices have beenused (fig. 3): diversity, dominance and evenness indices[Brenchley and Harper, 1998]. Diversity measures (Marga-lef Div = S-1/log N; S = number of species, N = number ofspecimens) are usually standardized against the sample sizewhereas dominance measures (Dom = S (ni/N)2; ni = num-ber of ith species), based on relative abundance, have highvalues for assemblages with a few common elements, andlow values where species are more or less evenly represen-ted; measures of evenness (Even = 1/S (ni/N)2), are thususually the inverse of dominance.
Symbols have been used to identify each site and age: C= Couvin, F = Ferrières, MF = Marche-en-Famenne, N =Nassogne, R = Rochefort, SR = Sougné-Remouchamps, T =Treignes, V = Vireux-Molhain, W = Wellin; L = Lochko-vian, P = Pragian, Es = Emsian, Ef = Eifelian, G = Givetian,Fr = Frasnian; l = lower, m = middle, u = upper.
Ecological structure of samples
Cluster analysis performed on palaeoecological parametershas permitted recognition of assemblage types on the basisof the relative abundance and diversity of the trilobite ma-crofauna. Thus, four specific assemblage types are definedas follows (fig. 3A):
– type A characterised by only 1 species, from the Lo-wer Devonian: SR-L, F-uP, N-uEs, T-uEs;
– type B characterised by an important dominance fromthe Lower Devonian, lower and upper parts of the MiddleDevonian and Upper Devonian: MF-lEs, MF-uEs, W-lEf,R-uEf, R-lG, C-mFr. For example, R-uEf biota is dominatedby Calycoscutellum, R-lG by Nyterops and C-mFr by Bra-docryphaeus;
– type C characterised by a moderate diversity andevenness: MF-uP/lEs, V-lEf, C-lEf, C-mEf, C-uEf/lG;
– type D characterised by an important diversity andevenness: Rochefort site from Middle Devonian (R-mEf).
The distinction between ecological assemblages andthose that arise from transport can be difficult. Assemblagesthat are either in-situ or nearly in-situ because they show nosigns of prolonged transport could represent a former com-munity or a palaeocommunity [Brenchley and Harper,1998]. Species within a community are associated partly be-cause they have similar tolerances of physical aspects of theenvironment and partly because they interact with one ano-ther through a food chain or through complementary nicherequirements [Brenchley and Harper, 1998]. Moreover, in‘pioneer’ communities, there occur one or two very abun-dant, opportunistic species. By contrast, in ‘equilibrium’communities, there is a relatively high diversity of taxa
more or less equally present [Brenchley and Harper, 1998].Some sites from the Eifelian (types C-D) could represent‘equilibrium’ communities: for example, V-lEF [Crônierand van Viersen, 2008] or R-mEf (see the next section).
The only well known Devonian trilobite fauna from theArdennes is that of the famous ‘Mur des douaniers’ near Vi-reux-Molhain (V-lEf), southern Ardennes, France [vanViersen, 2006a; Crônier and van Viersen, 2008]. Neverthe-less, of particular interest is the Middle Eifelian Rochefortsite yielding a rich, diverse trilobite fauna that shows a re-markable resemblance to the trilobite fauna of the GermanEifel [van Viersen, 2007]. This fauna is dominated by mem-bers of the Proetida, which is represented by six genera oftrilobites, representing half the total number of specimenscollected (51%, fig. 3B). The other groups with a moderaterelative abundance are Phacopida (27%) with five genera,Odontopleuridae (15%) and Lichidae (6%). Corynexochidais uncommon (abundance 3%).
Associative structure of samples
The hierarchical cluster analysis performed on all taxa al-lows us to delineate six discrete trilobite associations re-flecting their taxonomic affinities and partly related tospatial and temporal gradients (fig. 4).
I) The Digonus Association characterises the Lower De-vonian: Praguian-Emsian (Nassogne, Ferrières, Marche-en-Famenne). This genus is typically found in sandstonesthat were deposited near-shore. Because of this turbulentenvironment favoured by Digonus, articulated specimensare particularly rare. Mass occurrences are not exceptionalin homalonotines, as is also the case in other regions[Gandl, 1972; Wenndorf, 1990]. Nevertheless, Sandford[2005] described a complex relationship between homalo-notid trilobite faunal composition and taphonomy. Sandfordrecognized a recurrent relationship between homalono-tid-dominated low diversity assemblages and high diversityassemblages in relatively shallower-water facies.
II) The mixed association characterises the Middle De-vonian, and especially the Lower-Middle Eifelian. Threemain sub-associations can be observed:
(IIa) association dominated by Calycoscutellum repre-sented at the Couvin, Lower Eifelian (C-lEf) and Rochefort,Upper Eifelian (R-uEf) sites. The C-lEf biota, confined toargillaceous limestone beds, is characterised by mostly di-sarticulated exoskeletons with most specimens of Calycos-cutellum also being fragmented. Complete specimens arerare, but surprisingly they are of taxa that are usually notfound complete elsewhere, such as Tropidocoryphe, Cy-phaspis and Dohmiella [van Viersen, 2006b]. Accumulationof small individuals occurs locally in the bed, with othermacrofaunal elements. The R-uEf is dominated by Calycos-cutellum , which is known from numerous fragments and
Bull. Soc. géol. Fr., 2007, no 6
TRILOBITE PALAEOBIODIVERSITY IN THE ARDENNES MASSIF 479
FIG. 4. – R and Q mode cluster analysis of data set (35 genera and 19 samples) of Devonian trilobites from the Ardennes Massif. The R and Q mode den-drograms have been plotted against the original data, modified to show relative abundance classes of taxa, to indicate the generic and locality group be-hind the clusters. 6 clusters or associations (I to VI) are recognized.FIG. 4. – Classification hiérarchique ascendante réalisée sur 35 genres et 19 sites provenant du massif des Ardennes. La proportion des taxa a été quan-tifiée de manière disjonctive en 8 classes d’abondance relative. 6 clusters ou associations ont été identifés (I à VI).

disarticulated exoskeletons, rarely accumulated. No articu-lated trilobites are known from this locality.
Calycoscutellum is possibly among the most commontrilobites in the Upper Eifelian although usually representedby disarticulated sclerites. So far, complete specimens haveonly been reported from the Lower and Middle Eifelian;
(IIb) association with dominant Geesops, representedby the Vireux-Molhain biota [see Crônier and van Viersen,2008];
(IIc) association with dominant Cyphaspis (Aulaco-pleuridae), Radiaspis (Odontopleuridae), Kettneraspis(Odontopleuridae), Cornuproetus (Proetidae) and Geras-tos (Proetidae), represented by the Rochefort biota. Ecolo-gical indices and preservation state indicate that the
Rochefort biota is an equilibrium community probablyfound in situ. Trilobites are abundant and diverse, compri-sing both disarticulated exoskeletons and a significant por-tion of fully articulated ones. Accumulation, especially ofsmall odontopleurid and aulacopleurid parts (librigenae),seems to occur locally in the bed where trilobites are trans-ported together with other typical macrofaunal constituentssuch as bryozoans, crinoid ossicles and especially brachio-pods. For the Couvin Middle Eifelian biota, the faunalcontent is less rich than the Rochefort biota but has thesame state of preservation.
III) The Asteropyginae Association, dominated by Tre-veropyge, is representative of the Lower Devonian. Only di-sarticulated exoskeletons of asteropygines are usually found
Bull. Soc. géol. Fr., 2007, no 6
480 CRÔNIER C. and VAN VIERSEN A.
FIG. 5. – Localisation of samples through time and a bathymetric profile and comparison with diversity index curve, the global sea-level curve and the re-gressions/transgressions at the regional scale.FIG. 5. – Positionnement des affleurements au cours du temps et le long d’un profil bathymétrique et comparaison avec la courbe obtenue d’après l’indiceécologique de diversité, la courbe eustatique globale et les régressions/transgressions connues à l’échelle régionale.

in the Ardennes-Eifel; articulated specimens are representedonly by one complete enrolled Comura and a fully articula-ted Treveropyge thoracopygidium from Marche-en-Famenne(MF-uEs), confined to sandstone with calcareous cement. Inolder strata at the same locality (MF-uP/lEs), trilobites arefragmented and confined to sandy layers intercalated withdark blue slates. These represent post-mortem accumula-tions of trilobites and macrofaunal elements such as bra-chiopods, crinoids and corals.
IV) The Dechenella (Dechenellinae, Proetidae) and Ny-terops (Phacopinae, Phacopidae) Association characterisesthe Upper Eifelian-Lower Givetian. Dechenellinae appearto be slightly more common during the Upper Eifelian inthe Ardennes than they are in contemporaneous strata in theadjacent German Eifel [Basse, 2002].
At the Couvin locality (C-uEf/lG) all trilobites areknown from disarticulated exoskeletons and fragments frommainly crinoidal limestones, as is also the case at the Wellin(W-uEf) and Rochefort localities (R-lG).
V) The Kayserops ‘occurrence’ is restricted to theUpper Emsian of the Treignes area. Only one well-preser-ved cranidium has been sampled for this locality. Conse-quently, this single ‘endemic’ individual cannot constitutean association. In this study, the term of ‘occurrence’ hasbeen used until the discovery of new material.
VI) The Bradocryphaeus Association, dominated byBradocryphaeus supradevonicus (69%), is restricted here togreenish shales at the Middle Frasnian Couvin. The remai-ning species are Bradocryphaeus maillieuxi (17%), Otarioncf. ibergium (2%) and the highly endemic Heliopyge helios(5%). Bradocryphaeus supradevonicus occurs in both Bel-gium and Germany, whereas Heliopyge helios and Brado-cryphaeus maillieuxi are only found in Belgium. Otarion isextremely rare in the Ardennes, being known only from theFrasnian in the area between Couvin and Olloy-sur-Viroin[Richter and Richter, 1926; Maillieux, 1927]. In this area,specimens are usually fragmented, complete specimens arevery rare, and accumulations often occur.
Relative paleobathymetric position of samples
Each of the trilobite associations has a distinctive taxono-mic composition and abundance structure that reflects someecological conditions, ranged across shelves. Trilobite com-munities have already proved useful in recognizing marineenvironments of different depths [Fortey, 1975].
Palaeocological indices, state of preservation and natureof sediment allow us to locate fossiliferous sites along a ba-thymetric profile [modified from Fortey and Owens, 1997and Brenchley and Harper, 1998]. Additionally, Erben[1964] recognized two dominant magnafacies, i.e. nearshoreclastic and offshore carbonates, in the Devonian of Europe.
The shoreline environment (0-15 m), characterised byshallow, proximal and agitated deposits includes possibly aDigonus-dominated biota always known from fragmentedexoskeletons in sandstones. Diversity, though generally low,may be increased by introduction of allochthonous faunasby wave activity.
The upper offshore or inner shelf environment is cha-racterised by mudstones interbedded with sandstones. It in-cludes possibly the Bradocrypheus-dominated biota knownfrom disarticulated and fragmented exoskeletons and/or ascoquinas.
The median offshore or middle shelf environment cha-racterised by relatively deep deposits includes moderatelyto highly diverse assemblages. Their abundance is generallyhigh but decreases with depth. Mudstones may contain insitu trilobites with well-developed eyes, clumped or disper-sed sclerites that are commonly articulated, and various sta-ges. Quieter hydrodynamic conditions are favourable to thedevelopment of faunas and/or to conditions for their preser-vation. Mixed biotas are typically found in such environ-ments.
The lower offshore or outer shelf environment is loca-ted below effective storm wave-base (from 50-60 m). Trilo-bites are rare, often blind or with reduced-eyes and occur asarticulated exoskeletons. No sampled locality in the Arden-nes represents this environmental type.
Spatial and temporal interpretation of associations
The distribution of these associations recognised in theArdennes Massif and the diversity curve (fig. 5) are correla-ted with the transgressive/regressive trends in the ArdennesMassif [from Godefroid et al. 1994; Steemans, 1989; Gou-wy and Bultynck, 2000]. Three trends can be observed: 1)the Digonus-dominated association spreads out during aphase of low relative sea-level and is known from the shore-line domain; 2) the mixed associations flourished during aperiod of rising sea-level and they are known from the me-dian offshore domain; 3) peaks of diversity coincide withdeepening of the environment (rising sea-level).
The distribution of the trilobite associations is related topalaeobathymetric domains and sea-level variations. Anassociation is potentially a rather precise palaeoenviron-mental indicator. Trilobites are considered as vagrant epi-benthos, living mobile above the substratum whereconditions are little uniform. Trilobites are sensitive tosea-level variations, resulting in numerous association re-placements due to rapid alteration of the ecosystems. Thereis a continuous turnover in species, so the composition ofpalaeocommunities changes with time and their taxonomiccomposition and environmental ranges.
CONCLUSION
Fluctuation in biodiversity
At all sampled localities, trilobites increase in both numberand diversity in the Middle Devonian. The relative variationof trilobite abundance may be related to changes in ecologi-cal conditions influenced by substratum and palaeobathy-metry. The data show that the trilobites are most diversifiedand abundant in a peri-reefal context.
Taxonomic structuration
Five trilobite associations and one ‘occurrence’ delineatedamong the 35 genera/subfamilies were recognised from theEarly to Late Devonian, across environmental gradients, re-lated to water depth and substrate. Trilobites that mostly be-long to the vagrant epibenthos seem to be sensitive topalaeoenvironmental fluctuations. Sections of the same en-vironmental gradient are very similar to each other concer-ning the trilobite content, even if faunal replacement seemsto exist through time (for example, possible replacement ofthe Dechenella and Nyterops Association from the Early Gi-
Bull. Soc. géol. Fr., 2007, no 6
TRILOBITE PALAEOBIODIVERSITY IN THE ARDENNES MASSIF 481

vetian by the Bradocrypheus Association from the Frasnianfor the same environmental gradient).
Lower Devonian strata are characterised by abundantAsteropyginae or Homalonotinae. Subsequently, these sub-families are replaced by Phacopinae and Proetinae in theMiddle Devonian. The mixed middle Eifelian fauna at Cou-vin is much less diversified than that at Rochefort and re-presentatives of Cyphaspis, Radiaspis and Kettneraspishave not been found by us. This is due to sampling biassince a rich trilobite fauna was reported from the middle Ei-felian of this area by Maillieux [1904, 1907, 1912]. The‘Mur des douaniers’ Lagerstatte offers a glimpse of what isabsent from the Lower Eifelian.
At the Ardennes regional scale, changes in species rich-ness and ecological diversity of trilobites are in relation tophysical environmental gradients. Changes in climate andtopography limit most species ranges, resulting in conside-rable spatial turnover in species richness and ecologicalstructure.
Acknowledgments. – With this paper, the authors would like to pay homageto the eminent Professor J.L. Mansy (University of Lille) who was workingon the structural geology of the French Ardennes Massif; we miss him.This is a contribution of UMR 8014 of Centre National de la RechercheScientifique.
References
ADRAIN J. M. & WESTROP S. R. (2000). – An empirical assessment of taxicpaleobiology. – Science, 289, 110-112.
ALROY J., MARSHALL C. R., BAMBACH R. K., BEZUSKO K., FOOTE M.,FÜRSICH F. T., HANSEN T.A., HOLLAND S. M., IVANY L. C., JA-
BLONSKI D., JACOBS D. K., JONES D. C., KOSNIK M. A., LIDGARD
S., LOW S., MILLER A. I., NOVACK-GOTTSHALL P. M., OLSZEWSKI
T. D., PATZKOWSKY M. E., RAUP D. M., ROY K., SEPKOSKI JR. J.J., SOMMERS M. G., WAGNER P. J. & WEBBER A. (2001). –Effects of sampling standardization on estimates of Phanerozoicmarine diversification. – Proceedings of the National Academyof Sciences USA, 98, 6261–6266.
ASSELBERGHS E. (1946). – L’Eodévonien de l’Ardenne et des régions voisi-nes. – Mém. Inst. géol. Univ. Louvain, 14, 1-598.
BADGLEY C. (2003). – The multiple scales of biodiversity. – Paleobiology,29, 11–13.
BASSE M. (1996). – Trilobiten aus mittlerem Devon des Rhenohercyni-kums: I. Corynexochida und Proetida (1). – Palaeontographica(A), 239, 89-182.
BASSE M. (1997). – Trilobiten aus mittlerem Devon des Rhenohercyni-kums: II. Proetida (2), Ptychopariida, Phacopida (1). – Palaeon-tographica (A), 246, 53-142.
BASSE M. (1998). – Trilobiten aus mittlerem Devon des Rhenohercyni-kums: III. Proetida (3), Phacopida (2), Lichida (Lichoidea,Odontopleuroidea) und ergänzende Daten. – Palaeontographica(A), 249, 1-162.
BASSE M. (2002). – Eifel-Trilobiten 1. Proetida. – Goldschneck-Verlag,Korb, 152 pp.
BOTQUELEN A., LE MENN J., GOURVENNEC R. & LOI A. (2006). – Crinoidcolumnal associations and sequence stratigraphy architecture:the Le Faou Formation, Lower Devonian of the Massif Armori-cain (France). – Bull. Soc. géol. Fr., 177, 1, 19-26.
BOULVAIN F. (2006). – Atolls et récifs du Frasnian du Synclinorium de Di-nant (Belgique, France): sédimentologie et implications paléo-céanographiques. – Géol. France, 1-2, 53-57.
BOULVAIN F., BULTYNCK P., COEN M., COEN-AUBERT M., HELSEN S., LA-
CROIX D., LALOUX M., CASIER J.G., DEJONGHE L., DUMOULIN V.,GHYSEL P., GODEFROID J., MOURAVIEFF N., SARTENAER P., TOUR-
NEUR F. & VANGUESTAINE M. (1999). – Les formations du Fras-nien de la Belgique. – Mem. Geol. Survey of Belgium, 44, 1-125.
BRENCHLEY P. J. & HARPER D. A. T. (1998). – Palaeoecology. Ecosystems,environments and evolution. – Chapman & Hall, London.
BULTYNCK P., COEN-AUBERT M., DEJONGHE L., GODEFROID J., HANCE L.,LACROIX D., PRÉAT A., STAINIER P., STEEMANS P., STREEL M. &TOURNEUR F. (1991). – Les formations du Dévonien moyen de laBelgique. – Mém. Explic. cartes géol. min. Belg., 30, 106 p.
BULTYNCK P., COEN-AUBERT M. & GODEFROID J. (2000). – Summary of thestate of correlation in the Devonian of the Ardennes (Bel-gium-NE France) resulting from the decisions of the SDS. –Cour. Forsch.-Inst. Senckenberg, 225, 91-114.
BULTYNCK P. & DEJONGHE L. (2002). – Guide to a revised lithostratigraphicscale of Belgium. – Geol. Belgica, 4, 1-168.
CHLUPÁ0 I. (1987). – Ecostratigraphy of Silurian trilobite assemblages ofthe Barrandian area, Czechoslovakia. – Newsletters on Stratigra-phy, 17, 169-186.
CHLUPÁ0 I. (1994). – Devonian trilobites. Evolution and events. – Geobios,27, 487-505.
CHLUPÁ0 I. & KUKAL Z. (1986). – Reflection of possible global Devonianevents in the Barrandian area, CSSR. In: O. H. WALLISER, Ed.,Global bio-events. – Lecture Notes in Earth Sciences, 8,169-179.
CIBAJ I. (1992). – Structures sédimentaires dans les grès de Vireux (Dévo-nien inférieur). La coupe du Mont Vireux, Ardenne méridionale.– Ann. Soc. géol. Nord, 2ème série, 1, 195-203.
CRÔNIER C. & van VIERSEN A. (2008). – The ’Mur des douaniers’, an ex-ceptionally well-preserved Early Eifelian fossil site. – Bull. Soc.géol. France, 179, 1 (in press)
CUGNY P. (1988). – Analyse quantitative des faciès de diverses formationscrétacées des marges néotéthysiennes et atlantiques. Associa-tions paléontologiques et paléoenvironnementales. – Thèse Uni-versité Paul Sabatier de Toulouse. – Strata, série 1, 10, 331 p.
DAVIS J.C. (1986). – Statistics and data analysis in geology. – John Wiley& Sons, New York.
DEJONGHE l. (2001). – Guide de lecture des cartes géologiques de Wallonie.– Ministère de la région wallonne, 2ème éd., 51p.
ERBEN (1964). – Facies developments in the marine Devonian of the OldWorld. – Proceedings of the Ussher Society, 1, 92-118.
FEIST R. (1991). – The late Devonian trilobite crises. – Historical Biol., 5,197-214.
FORTEY R. A. (1975). – Early Ordovician trilobite communities. – Fossilsand Strata, 4, 331-352.
FORTEY R. A. & OWENS R. M. (1997). – Evolutionary history. In: R. L.KAESLER, Ed., Treatise on Invertebrate Paleontology. Part O.Arthropoda 1, Trilobita 1 (revised). – Geological Society ofAmerica and University of Kansas Press, Lawrence, 249-287.
GANDL J. (1972). – Die Acastavinae und Asteropyginae (Trilobita) Keltibe-riens (NE-Spanien). – Abh. Senck. Naturforsch. Gesellsch., 530,1-183.
GODEFROID J. (1995). – Les brachiopodes (Pentamerida, Atrypida et Spiri-ferida) de la fin de l’Eifelien début du Givétien à Pondrôme,bord sud du synclinorium de Dinant. – Inst. Roy. Sci. Nat. Belg.,Bull., 65, 69-116.
GODEFROID J., BLIECK A., BULTINCK K., DEJONGHE L., GERIENNE P., HANCE
L., MEILLEZ F., STAINIER P. & STEEMANS P. (1994). – Les forma-tions du Dévonien inférieur du massif de la Vesdre, de la fenêtrede Theux et du synclinorium de Dinant (Belgique, France). –Mém. Expl. Cartes géol. et minières de la Belg., 38, 1-78.
Bull. Soc. géol. Fr., 2007, no 6
482 CRÔNIER C. and VAN VIERSEN A.

GOFFETTE O. (1991). – Le magmatisme varisque en Ardenne méridionale:un marqueur de l’évolution géodynamique d’une paléomarge. –Thèse doct., Université Lille 1, Villeneuve d’Ascq, 309 p.
GOUWY S. & BULTYNCK P. (2000). – Graphic correlation of Frasnian sec-tions (Upper Devonian) in the Ardennes, Belgium. – Bull. Inst.Roy. Sci. Nat. Belg., Sciences de la Terre, 70, 25-52.
HAMMER Ø., HARPER D.A.T. & RYAN P.D. (2001). – Past: Paleontologicalstatistics software package for education and data analysis. –Palaeontologia Electronica, 4, 1-9.
HARPER D.A.T. (1999). – Numerical palaeobiology. – John Wiley & Sons,New York.
JOHNSON J. G. (1970). – Taghanic onlap and the end of North AmericanDevonian provinciality. – Geol. Soc. Amer. Bull., 81, 2077-2105.
JOHNSON J. G., KLAPPER G. & SANDBERG C. A. (1985). – Devonian eustaticfluctuations in Euramerica. – Geol. Soc. Amer. Bull., 96,567-587.
LACQUEMENT F. (2001). – L’Ardenne varisque. Déformation progressived’un prisme sédimentaire préstructuré; de l’affleurement au mo-dèle de chaîne. – Soc. Géol. Nord, Publ., 29, 285p.
LACQUEMENT F., MANSY J.-L., MEILLIEZ F., VAN VLIET LANOË B., COEN M.,CORNEILLE J.-P., DUMOULIN V., HANOT F., LEMONNE E., OUDOIRE
T. & PENISSON J.-P. (2003). – Notice explicative de la carte géo-logique de la France (1/50 000), feuille no 40 Givet 2ème édi-tion. – Editions du BRGM, Orléans.
LECOMPTE M. (1951-1952). – Les stromatoporoïdes du Dévonien moyen etsupérieur du bassin de Dinant. – Mém. Inst. Roy. Hist. Nat. Belg.Première partie [1951], 117, 1-216. Deuxième partie [1952],117, 217-259.
LUDVIGSEN R. & WESTROP S. R. (1983). – Trilobite biofacies of the Cam-brian-Ordovician boundary interval in North America. – Alche-ringa, 7, 301-319.
MAGREAN B. & VAN VIERSEN A. P. (2005). – A revision of Devonian trilobi-tes from Belgium – Part 1. The genera Cornuproetus and Ra-diaspis. – Bull. Inst. Roy. Sci. Nat. Belg., Sciences de la Terre,75, 87-93.
MAILLIEUX E. (1904). – Quelques mots sur les trilobites du Couvinien desenvirons de Couvin. – Bull. Soc. belge Géol., Paléontol. et Hy-drol., 17, 579-582.
MAILLIEUX E. (1907). – Compte rendu de l’excursion dans les environs deCouvin les 14 et 15 août 1906. – Bull. Soc. belge Géol., Paléon-tol. et Hydrol., 21, 133-167.
MAILLIEUX E. (1912). – Texte explicatif du levé géologique de la plan-chette de Couvin. – Service géol. Belg., Bruxelles, 70 pp.
MAILLIEUX E. (1927). – Sur les trilobites du Frasnien de la Belgique. –Bull. Soc. belge Géol., Paléontol. et Hydrol., 37, 77-87.
MAILLIEUX E. (1933). – Terrains, roches et fossiles de la Belgique. –Musée royal d’Histoire naturelle de Belgique, Bruxelles, 217 p.
MANSY J.-L. & MEILLIEZ F. (1993). – Eléments d’analyse structurale à par-tir d’exemples pris en Ardenne-Avesnois. – Ann. Soc. géol.Nord, 2, 45-60.
MEILLIEZ F. (1989). – Importance de l’événement calédonien dans l’alloch-tone ardennais; essai sur une cinématique paléozoïque del’Ardenne dans la chaîne Varisque. – Thèse doct. Etat, Universi-té du Maine, Le Mans, 518 p.
MEILLIEZ F. & MANSY J.-L. (1990). – Déformation pelliculaire différenciéedans une série à lithologie hétérogène: le Dévono-Carbonifèrede l’Ardenne. – Bull. Soc. géol. France, 8, VI, 177-188.
RICHTER R. & RICHTER E. (1926). – Die Trilobiten des Oberdevons.Beiträge zur Kenntnis devonischer Trilobiten. IV. – Abh. Geol.Bund., Neue Folge, 99, 1-314.
ROME D. R. (1936). – Note sur la microstructure de l’appareil tégumentairede Phacops (Ph.) accipitrinus maretiolensis R. & E. RICHTER. –Bull. Mus. Roy. Hist. Nat. Belg., XII, 31, 1-7.
SANDFORD A.C. (2005). – Homalonotid tribolites from the Silurian and lo-wer Devonian of southeastern Australia and New Zealand(Arthropoda, Trilobita, Homalonotidae). – Mem. Museum Victo-ria, 68, 1-66.
SCHINDLER E. (1990). – The late Frasnian (Upper Devonian) KellwasserCrisis. – Lecture Notes Earth Sci., 30, 151-159.
SCHLAGER W. (1981). – The paradox of drowned reefs and carbonate plat-forms. – Geol. Soc. Amer. Bull., 92, 197-211.
STEEMANS P. (1989). – Paléogéographie de l’Eodévonien ardennais et desrégions limitrophes. – Ann. Soc. Géol. Belg., 112, 103-119.
STRUVE W. (1976). – Phacops (Omegops) n. sg. (Trilobita; Ober-Devon). –Senckenberg. lethaea, 56, 429-451.
STRUVE W. (1982). – Beiträge zur Kenntnis der Phacopina (Trilobita), 9:Phacops (Omegops) n. sg. (Trilobita; Ober-devon). – Sencken-berg. lethaea, 56, 429-451.
THOMAS A. T. & LANE P. D. (1999). – Trilobite assemblages of the NorthAtlantic region. In: A. J. BOUCOT and J. D. LAWSON, Eds., Paleo-communities: a case study from the Silurian and Lower Devo-nian. – Cambridge University Press, 444-457.
THOREZ J., STREEL M., BOUCKAERT J. & BLESS M. (2006). – Stratigraphie etpaléogéographie de la partie orientale du synclinorium de Di-nant (Belgique) au Famennien supérieur: un modèle de basin sé-dimentaire reconstitué par analyse pluridisciplinairesédimentologique et micropaléontologique. – Meded. Rijks.Geol. Dienst., 28, 17-28.
TOURNEUR F. (1985). – Contribution à l’étude des tabulés du Dévonienmoyen de la Belgique. – Thèse Doct. Sc. Université CatholiqueLouvain-la-Neuve, 568 p.
VIERSEN van A. P. (2006a). – New Middle Devonian trilobites from Vi-reux-Molhain (Ardennes, northern France). – Senckenberg. le-thaea, 86, 63-75.
VIERSEN van A. P. (2006b). – Dohmiella LÜTKE 1990 (Trilobita), a validproetid genus, with two new species from Belgium and Germa-ny. – Senckenberg. lethaea, 86, 229-241.
VIERSEN van A. P. (2007). – Kettneraspis, Radiaspis and Ceratarges (Trilo-bita) from the Middle Devonian of the Rochefort area (Arden-nes, Belgium). – Scripta Geologica, 134, 1-18.
WENNDORF K. (1990). – Homalonotinae (Trilobita) aus dem RheinischenUnter-Devon. – Palaeontogr. Abt. A, 211, 1–184.
ZIEGLER P. A. (1982). – Geological atlas of the western and central Europe.– Shell Internationale Petroleum Maatschappij B.V., 130 p.
Bull. Soc. géol. Fr., 2007, no 6
TRILOBITE PALAEOBIODIVERSITY IN THE ARDENNES MASSIF 483




















