
Late Devonian conodonts and the global Frasnian-Famennian extinction event, Thong Pha Phum, western...
14
Palaeoworld 15 (2006) 171–184 Research paper Late Devonian conodonts and the global Frasnian-Famennian extinction event, Thong Pha Phum, western Thailand Norman M. Savage a,∗ , Apsorn Sardsud b , Werner Buggisch c a Department of Geology, University of Oregon, Eugene, OR 97403, USA b Department of Mineral Resources, Geological Survey Division, Bangkok 10400, Thailand c Geologie u. Mineralogie, Universitat Erlangen, Erlangen D-91054, Germany Received 10 May 2005; received in revised form 7 July 2006; accepted 17 July 2006 Available online 1 September 2006 Abstract The study of Late Devonian Frasnian and Famennian conodonts in western Thailand confirms these species are mostly cos- mopolitan. The Thailand section includes the Upper Kellwasser Event recorded in Europe, North America, China, and elsewhere, and probably also includes the upper part of the Lower Kellwasser Event. Stable isotope geochemistry for this interval shows a positive d13C excursion in the Late rhenana conodont Zone followed by a gradual return to normal, followed by a sudden positive excursion near the conodont extinction event. The latter has been chosen elsewhere as the Frasnian-Famennian boundary. The extinction event seems a consequence of unsteady conditions that could include eutrophic, oxygen, carbon dioxide, eustatic, and temperature fluctuations. There was a regional disappearance or extinction of several conodont species at the Frasnian-Famennian boundary followed by recovery and the spread of new species during the early Famennian. © 2006 Nanjing Institute of Geology and Palaeontology, CAS. Published by Elsevier Ltd. All rights reserved. Keywords: Thailand; Late Devonian; Extinction; Conodonts; Isotopes 1. Introduction The Late Devonian conodonts described here were obtained during field work in western Thailand, in an area 250 km northwest of Bangkok (Fig. 1) where some preliminary mapping and collecting was done by Thai and German geologists during the years 1965–1971, although a map and palaeontological lists were published much later (Hahn and Siebenh¨ uner, 1982). The prelim- inary mapping was published at a scale of 1:250,000 and gives a general interpretation of the stratigraphy and structure. The work presented herein results from ∗ Corresponding author. E-mail address: [email protected] (N.M. Savage). several visits to the Thong Pha Phum area in the years 2003–2004. Initial spot samples from many roadside and creek exposures were processed and led to the identifica- tion of several conodont-yielding beds. The conodonts described here are from a new road section not avail- able to the earlier workers. Our detailed collecting and processing from this section shows that the examined Late Devonian sequence spans the Frasnian-Famennian boundary and includes the extinction crisis and isotope excursions characteristic of that boundary. 2. Occurrence and age of collections The section studied in this present research is exposed in a road cutting 11 km north of the town of Thong Pha Phum (Fig. 2). The road at this point is running approx- 1871-174X/$ – see front matter © 2006 Nanjing Institute of Geology and Palaeontology, CAS. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.palwor.2006.07.005
Transcript of Late Devonian conodonts and the global Frasnian-Famennian extinction event, Thong Pha Phum, western...

A
mapeetb©
K
1
oapaamiaa
1
Palaeoworld 15 (2006) 171–184
Research paper
Late Devonian conodonts and the global Frasnian-Famennianextinction event, Thong Pha Phum, western Thailand
Norman M. Savage a,∗, Apsorn Sardsud b, Werner Buggisch c
a Department of Geology, University of Oregon, Eugene, OR 97403, USAb Department of Mineral Resources, Geological Survey Division, Bangkok 10400, Thailand
c Geologie u. Mineralogie, Universitat Erlangen, Erlangen D-91054, Germany
Received 10 May 2005; received in revised form 7 July 2006; accepted 17 July 2006Available online 1 September 2006
bstract
The study of Late Devonian Frasnian and Famennian conodonts in western Thailand confirms these species are mostly cos-opolitan. The Thailand section includes the Upper Kellwasser Event recorded in Europe, North America, China, and elsewhere,
nd probably also includes the upper part of the Lower Kellwasser Event. Stable isotope geochemistry for this interval shows aositive d13C excursion in the Late rhenana conodont Zone followed by a gradual return to normal, followed by a sudden positivexcursion near the conodont extinction event. The latter has been chosen elsewhere as the Frasnian-Famennian boundary. The
xtinction event seems a consequence of unsteady conditions that could include eutrophic, oxygen, carbon dioxide, eustatic, andemperature fluctuations. There was a regional disappearance or extinction of several conodont species at the Frasnian-Famennianoundary followed by recovery and the spread of new species during the early Famennian.2006 Nanjing Institute of Geology and Palaeontology, CAS. Published by Elsevier Ltd. All rights reserved.
eywords: Thailand; Late Devonian; Extinction; Conodonts; Isotopes
. Introduction
The Late Devonian conodonts described here werebtained during field work in western Thailand, in anrea 250 km northwest of Bangkok (Fig. 1) where somereliminary mapping and collecting was done by Thaind German geologists during the years 1965–1971,lthough a map and palaeontological lists were publisheduch later (Hahn and Siebenhuner, 1982). The prelim-
nary mapping was published at a scale of 1:250,000nd gives a general interpretation of the stratigraphynd structure. The work presented herein results from
∗ Corresponding author.E-mail address: [email protected] (N.M. Savage).
871-174X/$ – see front matter © 2006 Nanjing Institute of Geology and Paldoi:10.1016/j.palwor.2006.07.005
several visits to the Thong Pha Phum area in the years2003–2004. Initial spot samples from many roadside andcreek exposures were processed and led to the identifica-tion of several conodont-yielding beds. The conodontsdescribed here are from a new road section not avail-able to the earlier workers. Our detailed collecting andprocessing from this section shows that the examinedLate Devonian sequence spans the Frasnian-Famennianboundary and includes the extinction crisis and isotopeexcursions characteristic of that boundary.
2. Occurrence and age of collections
The section studied in this present research is exposedin a road cutting 11 km north of the town of Thong PhaPhum (Fig. 2). The road at this point is running approx-
aeontology, CAS. Published by Elsevier Ltd. All rights reserved.
172 N.M. Savage et al. / Palaeoworld 15 (2006) 171–184
Fig. 1. Location of the Thong Pha Phum field area in western Thailand.

N.M. Savage et al. / Palaeoworld 15 (2006) 171–184 173
f Thon
irwtta
sais
Fig. 2. Study area 11 km north o
mately north-south and the cutting extends along theoadside for about 100 m, with the beds dipping north-ard at up to 18◦. However, numerous minor faults cause
he section to be repeated several times so that the totalhickness appears to be no more than about 7 m, althought first sight it might seem much greater.
Samples were collected during successive visits to the
ite during 2003 and 2004. The early collecting in Febru-ry 2003 was from near the south end of the cutting andncluded Bed 4 (not shown in Fig. 3 but considered theame as Bed 30 in the north section—see below). Bed 4g Pha Phum, western Thailand.
yielded a good fauna of Late rhenana Zone P elements.On a subsequent visit, in late 2003, we decided to collectfrom a section farther north where the cutting seemed toextend stratigraphically higher. The stratigraphic thick-ness is 6.6 m at this northern part of the cutting, withour Bed 26 at the base and Bed 42 at the top (Fig. 3).Conodont samples weighing 2 kg were collected from
each of these beds and these samples were divided withone part kept for reference in Bangkok and the otherpart processed in Oregon for conodonts. A notable fea-ture of the conodont collections from all the beds is
174 N.M. Savage et al. / Palaeoworld 15 (2006) 171–184
amenn
Fig. 3. Late Devonian (Frasnian-Fthe almost complete absence of elements other than P1elements.
When the conodonts were examined they suggestedthat Beds 26–30 at the base of the cutting are of Laterhenana Zone age, Beds 32–37 are probably of lin-guiformis Zone age (although Palmatolepis linguiformisis not known from this section nor from another Frasnian-
Fammennian currently being studied 360 km farthernorth), Beds 38–39 are of Early triangularis Zone age,and Beds 40–42 are of possible Middle triangularis Zoneage. Thus, the section was shown to include the Frasnian-ian) Section at Thong Pha Phum.
Famennian boundary. It was because of this that wereturned in early 2004 to collect closely spaced isotopesamples from this measured section. The results of thiswork are discussed below.
All the sampled beds yielded some conodonts butyields were low in Beds 26–28 and Beds 34–37, butmuch higher in Beds 29–33 and Beds 38–40. Exami-
nation of the conodont faunas suggests that the high-yielding Bed 4 at the southern end of the cutting is thesame horizon as high-yielding Bed 30 farther north inthe measured section. This horizon includes the species
laeowor
AcYY1
Fat
N.M. Savage et al. / Pa
ncyrodella nodosa Ulrich et Bassler, 1926, Ancyrodella
urvata Branson and Mehl, 1934, Ancyrodella gigasounquist, 1947, Ancyrognathus calvini Miller andoungquist, 1947, Palmatolepis aff. prominens Muller,956 (a species later assigned by some workers to Pa.ig. 4. d13C isotope excursions. Note positive value at the base of the sectiot 3.25 m above the section base (Bed 38A) extending through to 42D. Afteriangularis Zone (within Bed 38) conodonts gradually reappear. This occurs
ld 15 (2006) 171–184 175
bogartensis or Pa. gigas, or Pa. hassi, but the holo-
types of these species are distict from the holotype ofPa. prominens), Palmatolepis hassi Muller and Muller,1957, Icriodus alternatus alternatus Branson and Mehl,1934, and Polygnathus decorosus Stauffer, 1938, and isn then a gradual return to normal. Then a sudden positive excursionr the sudden return to positive d13C levels at the base of the Earlyin the early Famennian.

laeowo
176 N.M. Savage et al. / Pajudged to be of Late rhenana Zone age. Bed 31, immedi-ately above, is also thought to be in this zone and includesPolygnathus aspelundi Savage and Funai, 1980, and Pal-
matolepis thanisi n. sp. in addition to most of those inBed 30. Yields drop off gradually in Beds 32–37 and thenpick up in Bed 38 with the appearance of specimens ofPalmatolepis triangularis Sannemann, 1955 and Palma-rld 15 (2006) 171–184
tolepis ultima Ziegler, 1958. Beds 39 and 40 have yieldednumerous Palmatolepis triangularis Sannemann, 1955,indicating the the boundary between the Frasnian and
Famennian stages, and in Bed 40 there are severaladditional species including Palmatolepis werneri Jiand Ziegler, 1993, Palmatolepis delicatula delicatulaBranson and Mehl, 1934, and Palmatolepis subperlobata
laeowor
B4uip
3
boTtfwmd
rtPdNisartiepiatcpRpc
and Somchai Charoensuk. Figs. 1–3 are prepared by
FO(PaBSMl1lPv
N.M. Savage et al. / Pa
ranson and Mehl, 1934. Yields appear to drop off in Bed2 although this may be a result of sampling from annusally thick bed where sedimentation rates may havencreased and diluted the number of conodonts recovereder kg. This bed marks the top of the exposure.
. Isotope excursions
Taking samples for isotope analysis was difficultecause of the need to collect several samples from eachf the thick limestone beds within the measured section.he beds were marked in situ with the subdivisions and
hen removed as blocks to the laboratory in Bangkokor sawing into slabs for analysis. The 17 beds (26–42)ere subdivided into a total of 126 slabs and these wereailed to Werner Buggisch in Germany for d13C and
18O analysis.It was decided by Werner Buggisch that the d18O
esults were unreliable because of contamination buthat the d13C results were meaningful. The Thong Phahum section includes the global Upper Kellwasser13C isotope positive excursion known from Europe,orth America, China, and elsewhere, and probably also
ncludes the upper part of the Lower Kellwasser excur-ion. Stable isotope geochemistry for this interval showspositive d13C excursion in the lower part of the Late
henana conodont Zone followed by a gradual returno normal followed by a sudden positive excursion dur-ng the later part of that zone and the subsequent timequivalent of the Palmatolepis linguiformis Zone. Theositive excursion continues in Beds 38–41 through whats probably the Early triangularis Zone and then gradu-lly returns toward normal in Bed 42, of possible Middleriangularis Zone age. There are many theories on theause or causes of the Upper Kellwasser d13C isotope
ositive excursion and several of these are discussed byacki (2005). It is possibly linked to enhanced primaryroduction in high nutrient regimes related to sea levelhanges. In general the sedimentary, biotic, and rela-ig. 5. Conodonts from Bed 4 (southern continuation of Bed 30 in roadside mral and lateral views of P element UOPS 176, ×27. C–I. Palmatolepis hassi M
D–F) oral, lateral, and aboral views of P element UOPS 178, ×25; (G) oral velement UOPS 220, ×32. (J and K) Palmatolepis aff. eureka Ziegler and San
nd M) Icriodus alternatus alternatus Branson and Mehl, 1934. Oral and lateranson and Mehl, 1934. (N–P) Oral, lateral, and aboral views of P element U) Ancyrodella aff. And. nodosa Ulrich and Bassler, 1926. Oral and aboral viiller and Youngquist, 1947. Oral and lateral views of P element UOPS 18
ateral, and aboral views of P element UOPS 186, ×19. (Y and Z) Ancyrodel87, ×25. (AA–FF) Polygnathus decorosus Stauffer, 1938. (AA–CC) Oral, laateral, and aboral views of P element 189, ×30. (GG–II) Polygnathus sp. Oolygnathus. aff. webbi Stauffer, 1938. (JJ and KK) Oral and aboral views of siews of dextral P element UOPS 192, ×30. (OO–QQ) Polygnathus n. sp. Or
ld 15 (2006) 171–184 177
tively rapid isotope changes during this time suggestunsteady eutrophic, oxygen, carbon dioxide, eustatic,and temperature conditions (Yudina et al., 2002; Savage,2002).
4. Conclusions
Late Devonian Frasnian and Famennian conodontsin western Thailand confirm that recognized speciesare mostly cosmopolitan. The Thong Pha Phum sectionincludes the global Upper Kellwasser extinction eventrecorded in other parts of the world and probably alsopart of the Lower Kellwasser event. Stable isotope geo-chemistry for this interval shows a positive d13C excur-sion in the Late rhenana conodont Zone followed bya gradual return to normal during the Palmatolepis lin-guiformis conodont Zone preceding the extinction event,followed by a sudden positive excursion at the extinctionevent, which corresponds with the appearance of Pal-matolepis triangularis. In the Thong Pha Phum section,as elsewhere, there was a disappearance or extinctionof several conodont species at or just preceding theFrasnian-Famennian boundary followed by recovery andthe appearance of other species during the early Famen-nian. The extinction event corresponds with a positived13C excursion in the composition of seawater and theatmosphere (Fig. 4; Table 1).
Acknowledgements
For help in the field we wish to thank Depart-ment of Mineral Resources geologists Doungrutai Sae-saengseerung and Worakit Khaochan, geological tech-nician Mana Rakbumrung, and drivers Shan Subyuyen,
Worakit Khaochan. Most of the funding for field workwas provided by the Department of Mineral Resources,Bangkok. We thank John Talent and Thomas Becker forconstructive criticism of the manuscript.
easured section). (A and B) Palmatolepis aff. prominens Muller, 1956.uller and Muller, 1957. (C) Oral view of P element UOPS 177, ×24;
iew of P element UOPS 179, ×25; (H and I) oral and lateral views ofdberg, 1990. Oral and lateral views of P element UOPS 180, ×42. (Lral views of P element UOPS 181, ×47. (N–Q) Ancyrodella curvataOPS 182, ×35; (Q) oral view of P element UOPS 183, ×26. (R and
ews of P element UOPS 184, ×22. (T and U) Ancyrognathus calvini5, ×26. (V–X) Ancyrodella nodosa Ulrich and Bassler, 1926. Oral,la gigas Younquist, 1947. Oral and aboral views of P element UOPSteral, and aboral views of P element UOPS 188, ×28; (DD–FF) oral,ral, lateral, and aboral views of P element UOPS 190, ×26. (JJ–NN)inistral P element UOPS 191, ×25; (LL–NN) oral, lateral, and aboralal, lateral, and aboral views of P element UOPS 193, ×40.

178 N.M. Savage et al. / Palaeoworld 15 (2006) 171–184
Table 1Numbers of P1 element conodonts from Beds 29 to 42 (Beds 26–28 yielded very few or no elements)
Bed number 29 30 (=4) 31 32 33 34 35 36 37 38 39 40 41 42
TaxaPalmatolepis prominens 2 6 1 0 0 0 0 0 0 0 0 0 0 0Palmatolepis hassi 20 138 14 13 0 5 0 0 0 0 0 0 0 0Palmatolepis aff. Pa. eureka 5 30 0 0 0 0 0 0 0 0 0 0 0 0Icriodus alternatus alternatus 0 2 0 0 0 0 0 0 0 0 6 7 3 0Ancyrodella curvata 28 43 9 0 0 0 0 0 0 0 0 0 0 0Ancyrodella aff. And. nodosa 0 1 0 0 0 0 0 0 0 0 0 0 0 0Ancyrognathus calvini 1 2 0 0 0 0 0 0 0 0 0 0 0 0Ancyrodella nodosa 4 66 8 12 0 0 0 0 0 0 0 0 0 0Ancyrodella gigas 4 1 0 1 0 0 0 0 0 0 0 0 0 0Polygnathus decorosus 42 249 16 7 0 5 0 0 2 9 34 27 5 6Polygnathus sp. 0 2 0 0 0 0 0 0 0 0 0 0 0 0Polygnathus aff. Po. webbi (sinist.) 0 10 0 0 0 0 0 0 0 0 0 0 0 0Polygnathus n. sp. 0 5 0 0 0 0 0 0 0 0 0 0 0 0Palmatolepis triangularis 0 0 0 0 0 0 0 0 0 9 9 33 2 1Palmatolepis thanisi 0 0 3 0 0 0 0 0 0 0 0 0 0 0Polygnathus aspelundi 1 2 2 1 0 0 0 0 0 0 0 0 0 0Polygnathus aff. Po. webbi (dextral) 0 10 2 0 0 0 0 0 0 0 0 0 0 0Polygnathus aff. Po. decorosus 0 1 0 0 0 0 0 0 0 0 0 0 0 0Palmatolepis werneri 0 0 0 0 0 0 0 0 0 0 6 17 0 3Palmatolepis delicatula delicatula 0 0 0 0 0 0 0 0 0 0 1 3 0 0Palmatolepis subperlobata 0 0 0 0 0 0 0 0 0 0 4 11 0 2Palmatolepis delicatula platys 0 0 0 0 0 0 0 0 0 0 1 4 0 0Palmatolepis ultima 0 0 0 0 0 0 0 0 0 2 0 0 0 0
Appendix A. Systematic Palaeontology
The conodont faunas recovered from this section arecomprised almost wholly of P1 elements and thus onlyP1 elements are discussed and illustrated herein. Thespecimen numbers are those of the of the University ofOregon Paleontological Collection (UOPS).
Genus Ancyrodella Ulrich and Bassler, 1926
Type species: Ancyrodella nodosa Ulrich and Bassler,1926.
Ancyrodella curvata Branson and Mehl, 1934
Fig. 5N–QDiscussion: Both illustrated specimens have the pos-
terior part of the outer lateral lobe broken but closelyresemble the holotype, which is itself broken with theposterior ends of both lateral lobes missing. The distinc-tive features of the species include the concave marginof the outer lateral lobe with a secondary carina and sec-
ondary keel, and the convex margin of the inner laterallobe.Material: Figured specimen UOPS 182–183 fromBed 4 (Late rhenana Zone).
Ancyrodella nodosa Ulrich and Bassler, 1926
Figs. 5V–X, 6G, H, Q, W–YDiscussion: This is the type species of the genus.
The lectotype of the species is broken and lacks theanterior blade but the distinctive features are the trian-gular platform outline with the lateral lobes constrictedposteriorly, particularly on the inner side. Of the fourThong Pha Phum specimens illustrated here, three (Figs.5V–X, 6G, H, Q) are very typical of the species. Thefourth (Fig. 6W–Y), which is quite large, has a markedlystraight margin to the outer lobe, probably connected toits occlusional relationship with the anterior blade of theopposing element.
Material: Figured specimens UOPS 186 from Bed 4;UOPS 196 from Bed 31; UOPS 202, 205 from Bed 30(all Late rhenana Zone).
Ancyrodella aff. nodosa Ulrich and Bassler, 1926
Fig. 5R and S
Discussion: This specimen differs from charac-teristic specimens of the species in having a finernodose surface, particularly along the platform marg-ins.
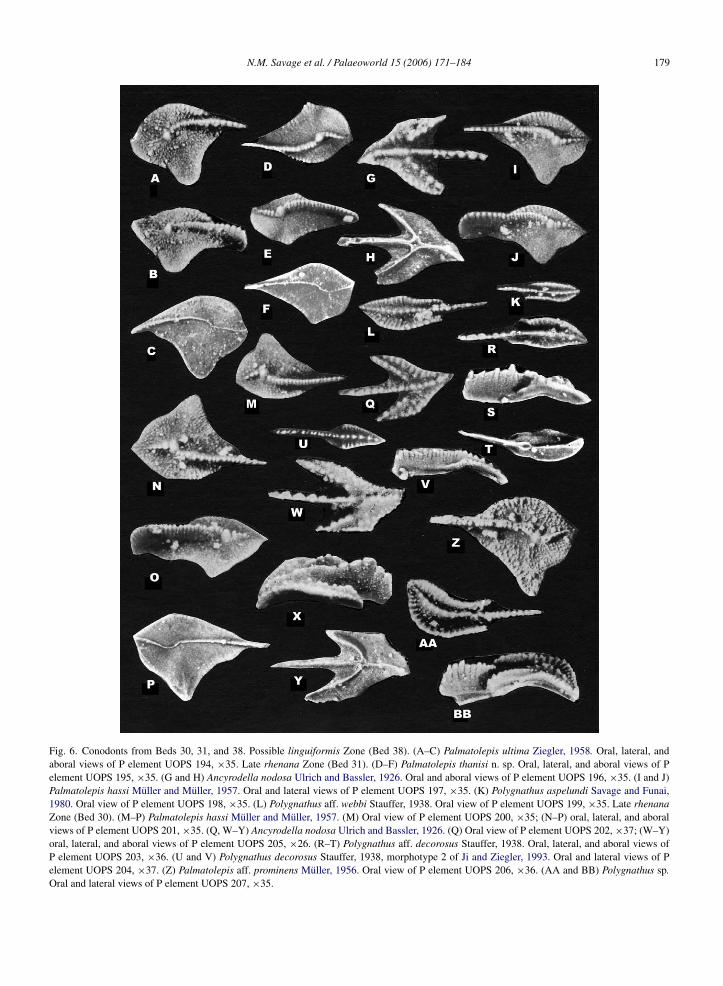
N.M. Savage et al. / Palaeoworld 15 (2006) 171–184 179
Fig. 6. Conodonts from Beds 30, 31, and 38. Possible linguiformis Zone (Bed 38). (A–C) Palmatolepis ultima Ziegler, 1958. Oral, lateral, andaboral views of P element UOPS 194, ×35. Late rhenana Zone (Bed 31). (D–F) Palmatolepis thanisi n. sp. Oral, lateral, and aboral views of Pelement UOPS 195, ×35. (G and H) Ancyrodella nodosa Ulrich and Bassler, 1926. Oral and aboral views of P element UOPS 196, ×35. (I and J)Palmatolepis hassi Muller and Muller, 1957. Oral and lateral views of P element UOPS 197, ×35. (K) Polygnathus aspelundi Savage and Funai,1980. Oral view of P element UOPS 198, ×35. (L) Polygnathus aff. webbi Stauffer, 1938. Oral view of P element UOPS 199, ×35. Late rhenanaZone (Bed 30). (M–P) Palmatolepis hassi Muller and Muller, 1957. (M) Oral view of P element UOPS 200, ×35; (N–P) oral, lateral, and aboralviews of P element UOPS 201, ×35. (Q, W–Y) Ancyrodella nodosa Ulrich and Bassler, 1926. (Q) Oral view of P element UOPS 202, ×37; (W–Y)oral, lateral, and aboral views of P element UOPS 205, ×26. (R–T) Polygnathus aff. decorosus Stauffer, 1938. Oral, lateral, and aboral views ofP element UOPS 203, ×36. (U and V) Polygnathus decorosus Stauffer, 1938, morphotype 2 of Ji and Ziegler, 1993. Oral and lateral views of Pelement UOPS 204, ×37. (Z) Palmatolepis aff. prominens Muller, 1956. Oral view of P element UOPS 206, ×36. (AA and BB) Polygnathus sp.Oral and lateral views of P element UOPS 207, ×35.

laeowo
180 N.M. Savage et al. / PaMaterial: Figured specimen UOPS 184 from Bed 4(Late rhenana Zone).
Ancyrodella gigas Younquist, 1947
Fig. 5Y and ZDiscussion: In its general outline and nodose oral
surface the Thong Pha Phum material is close to whatmost authors have named Ancyrodella gigas althoughconfident identification is made difficult becausethe holotype of the species is laterally compressed.Generally the species is diagnosed to have a platformthat is wider and less constricted posteriorly than thatof Ancyrodella nodosa, and to have a more randomlynodose oral surface.
Material: Figured specimen UOPS 187 from Bed 4(Late rhenana Zone).
Genus Ancyrognathus Branson and Mehl, 1934
Type species: Ancyrognathus symmetricus Branson andMehl, 1934.
Ancyrognathus calvini Miller and Youngquist, 1947
Fig. 5T and UDiscussion: The illustrated specimen differs from
Ancyrognathus tsiensi Mouravieff in being wider andhaving a less acute angle between the posterior cari-nae. It is assigned to Ancyrognathus calvini becauseof its short, high anterior blade, comparable platformoutline and the similar angle between the posteriorcarinae.
Material: Figured specimen UOPS 185 from Bed 4(Late rhenana Zone).
Genus Icriodus Branson and Mehl, 1938
Type species: Icriodus expansus Branson and Mehl,1938.
Icriodus alternatus alternatus Branson and Mehl, 1934
Figs. 5L and M, 7CC–GGDiscussion: A few specimens of Icriodus occur in
samples throughout the section. All have the alterna-tion of medial and lateral nodes characteristic of Icrio-dus alternatus and within that species none has beenobserved with the alignment of the posterior cusp with alateral row that Sandberg and Dreesen (1984) observedin distinguishing Icriodus alternatus helmsi.
Material: Figured specimens UOPS 181 from Bed 4
(Late rhenana Zone); UOPS 218-219 from Bed 40 (Mid-dle triangularis Zone).Genus Palmatolepis Ulrich and Bassler, 1926
rld 15 (2006) 171–184
Type species: Palmatolepis perlobata Ulrich and Bassler,1926.
Palmatolepis delicatula Branson and Mehl, 1934
Palmatolepis delicatula aff. delicatula Branson andMehl, 1934
Fig. 7Z-BBDiscussion: This subspecies is distinguished from
Palmatolepis delicatula platys Ziegler and Sandberg inhaving almost equal areas of the outer platform presentanterior and posterior of a line between the node andthe tip of the lobe. It may be close to juveniles of Pa.triangularis but these are generally more elongate.
Material: Figured specimen UOPS 217 from Bed 40(Middle triangularis Zone).
Palmatolepis delicatula platys Ziegler and Sandberg,1990
Fig. 7V and WDiscussion: This subspecies is distinguished from
Palmatolepis delicatula delicatula by having the largerpart of the outer platform anterior of the node.
Material: Figured specimen UOPS 215 from Bed 40(Middle triangularis Zone).
Palmatolepis aff. eureka Ziegler and Sandberg, 1990
Fig. 5J and KDiscussion: This is a small, probably juvenile speci-
men.Material: Figured specimen UOPS 180 from Bed 4
(Late rhenana Zone).
Palmatolepis hassi Muller and Muller, 1957
Figs. 5C–I, 6I, J, M–PDiscussion: As commonly diagnosed this is a long
ranging species with considerable variation in outlineand other features. The more characteristic specimensof such as here in Fig. 6 I, J, N–P, have a large outer lobedemarcated by distinct sinuses anteriorly and posteriorlyand with a convex bulge farther to the posterior. Theposterior tip of the element is pointed, with a small sinuseither side. Some specimens have a less pointed posteriortip, as in here in Figs. 5C–I, 6M. The anterior carinais gently curved, as in the holotype, or almost straight.The surface is nodose, particularly toward the platformmargins.
Material: Figured specimens UOPS 177–179 fromBed 4; UOPS 200, 201 from Bed 30; UOPS 197 fromBed 31; UOPS 220 from Bed 4 (all Late rhenanaZone).

N.M. Savage et al. / Palaeoworld 15 (2006) 171–184 181
Fig. 7. Conodonts from Bed 40. (A–I, S–U) Palmatolepis triangularis Sannemann, 1955. (A–C) Oral, lateral, and aboral views of P element UOPS208, ×40; (D–F) oral, lateral, and aboral views of P element UOPS 209, ×37; (G–I) oral, lateral, and aboral views of P element UOPS 210, ×39;(S–U) oral, lateral, and aboral views of P element UOPS 214, ×39. (J–R) Palmatolepis werneri Ji and Ziegler, 1993. (J–L) Oral, lateral, and aboralviews of P element UOPS 211, ×39; (M–O) oral, lateral, and aboral views of possible juvenile P element UOPS 212, ×38; (P–R) oral, lateral, andaboral views of possible juvenile P element UOPS 213, ×38. (V and W) Palmatolepis delicatula platys Ziegler and Sandberg, 1990. Oral and lateralviews of P element UOPS 215, ×39. (X and Y) Palmatolepis subperlobata Branson and Mehl, 1934. Oral and aboral views of P element UOPS216, ×39. (Z-BB) Palmatolepis delicatula aff. delicatula Branson and Mehl, 1934. Oral, lateral, and aboral views of P element UOPS 217, ×40.(CC–GG) Icriodus alternatus alternatus Branson and Mehl, 1934. (CC–EE) Oral, lateral, and aboral views of P element UOPS 218, ×33; (FF andGG) oral and aboral views of P element UOPS 219, ×39.

laeowo
182 N.M. Savage et al. / PaPalmatolepis aff. prominens Muller, 1956
Figs. 5A and B, 6ZDiscussion: The illustrated specimen from Bed 31
(Fig. 6Z) is very like the holotype of Palmatolepis promi-nens and that from Bed 4 (Fig. 5A and B) is very similarbut with a straight anterior margin to the outer plat-form. Ziegler and Sandberg (1990) made the holotypeof P. prominens a synonym of P. gigas gigas Miller andYoungquist, 1947, but the latter taxon differs in hav-ing a marked sinus anterior of the outer lobe. Klapperand Foster (1993) used evidence from the fused clusterof Lange (1968) from the Upper Kellwasser Limestoneat Steinbruch Schmidt, and the P2 (Pb) element thatStauffer (1938) designated as the holotype of his speciesNothognathella bogartensis, to describe the multiele-ment species Palmatolepis bogartensis (Staufer, 1938).In this assessment they included the P1 holotype of P.prominens Muller, 1956, as a synonym of of P. boga-rtensis. However, the P1 elements of P. bogartensis theyillustrate as representaive of the species are differentfrom the holotype of P. prominens in that the lobe isgenerally directed slightly to the anterior and the plat-form extends anteriorly to the carina tip or almost to thecarina tip. Neither of these features, nor the presenceof uniform nodes on the platform, occur on the holo-type of P. prominens. Therefore, we do not agree thatP. prominens is a junior synonym of P. bogartensis. Ourspecimens are close to P. prominens, but as noted above,differ in having a relatively straight outer anterior plat-form margin. Possibly these Thailand specimens couldbe described as a new species but they are insufficient innumber and so close to P. prominens that the presentassignment seems warranted, at least for the present.The P1 elements of P. wildungensis Muller, 1956, illus-trated by that author, including the holotype, differfrom his illustrations of P. prominens, and our specimenfrom Thailand, in having a much less prominent outerlobe.
Material: Figured specimen UOPS 176 from Bed 4;UOPS 206 from Bed 30 (all Late rhenana Zone).
Palmatolepis werneri Ji and Ziegler, 1993
Fig. 7J–RDiscussion: There seems a transition from Pa. trian-
gularis to Pa. werneri in our collection from Bed 40. Thespecimens illustrated here in Fig. 7A–I appear to belongto Pa. triangularis and those in Fig. 7M–R to belong to
Pa. werneri, whereas that in Fig. 7S–U, is somewhat tran-sitional but assigned herein to triangularis whilst that inFig. 7J–L, is assigned to werneri.rld 15 (2006) 171–184
Material: Figured specimens UOPS 211–213 fromBed 40 (Middle triangularis Zone).
Palmatolepis ultima Ziegler, 1958
Fig. 6A–CMaterial: Figured specimens UOPS 194 from Bed 38
(Early triangularis Zone).
Palmatolepis triangularis Sannemann, 1955
Fig. 7A–I, S–UDiscussion: The species is distinguished by its
upflexed posterior platform, weak posterior carina, largecentral node, and strongly raised anterior inner margin.The outline of the species varies markedly, with somespecimens tending to be more elongate and to have a pos-terior carina that curves inward rather than being shortand straight as in the wider specimens. In our collectionfrom Bed 40 there is some transition to Pa. werneri, asdiscussed above.
Material: Figured specimens UOPS 208–210, 214from Bed 40 (Middle triangularis Zone).
Palmatolepis thanisi n. sp.
Fig. 6D–FDiagnosis: A palmatolepid with the P1 element hav-
ing a long, ovate platform with a small but distinct angu-lar projection on the outer lobe just anterior of the centralnode and with a surface that is smooth apart from smallnodes around the edges. The anterior platform marginsmeet the blade at very small angles and almost at theanterior of the blade. The anterior blade and carina iscomprised of equi-sized denticles. The posterior carinaextends to the posterior platform margin.
Discussion: The species has some resemblance to Pa.linguiformis Muller, 1956 and to Pa. ederi Ziegler andSandberg, 1990, but differs from both its less sinuousgeneral outline, its possession of a small angular outerlobe, and its straighter anterior carina and blade.
Material: Figured specimen UOPS 195 (holotype)from Bed 31 (Late rhenana Zone).
Palmatolepis subperlobata Branson and Mehl, 1934
Fig. 7X and YDiscussion: This species is distinguished from Pa. tri-
angularis by its smoother platform and more sigmoidalcarina and blade.
Material: Figured specimen UOPS 216 from Bed 40(Middle triangularis Zone).
Genus Polygnathus Hinde, 1879

laeowor
T
P
F
msw
(
P
F
satsFtppg
B
P
F
iapfllf
(
P
F
mpaI(1t
(
N.M. Savage et al. / Pa
ype species: Polygnathus dubius Hinde, 1879.
olygnathus aspelundi Savage and Funai, 1980
ig. 6KDiscussion: The illustrated specimen is a sinistral ele-
ent and is typical of the species first described fromoutheastern Alaska but since recognized widely else-here, including southern China.Material: Figured specimen UOPS 198 from Bed 31
Late rhenana Zone).
olygnathus decorosus Stauffer, 1938
igs. 5AA–FF, 6U, VDiscussion: These elements are characteristic of the
pecies in possessing a blade about equal in length tosmall platform, with the lower platform margin dis-
inctly higher than the lower blade margin. There isome variation in the illustrated specimens with that inig. 5DD–FF in having a relatively shorter blade whereas
hat in Fig. 6U and V has a shorter and more posteriorlyointed platform. None has clearly developed marginallatform nodes used by Ji and Ziegler (1993), to distin-uish their morphotype 2 of this species.
Material: Figured specimen UOPS 188–189 fromed 4; UOPS 204 from Bed 30 (all Late rhenana Zone).
olygnathus aff. decorosus Stauffer, 1938
ig. 6R–TDiscussion: The illustrated specimen differs from typ-
cal elements of Polygnathus decorosus in having a rel-tively long platform. It has some resemblance to Po.acificus Savage and Funai, 1980, but has the downwardexure of the platform at about one third the platform
ength rather than nearer the junction of the and plat-orm as in that species.
Material: Figured specimen UOPS 203 from Bed 30Late rhenana Zone).
olygnathus sp.
ig. 5GG–IIRemarks: This may be a new species but more speci-
ens are needed to decide if the features are consistentlyresent. It differs from Po. decorosus in its shorter bladend platform ornament of numerous fine marginal ridges.t differs from the material described by Ji and Ziegler1993), morphotype 1 of Po. evidens Klapper and Lane,985, in having a longer carina that extends to the pos-
erior of the platform.Material: Figured specimen UOPS 190 from Bed 4Late rhenana Zone).
ld 15 (2006) 171–184 183
Polygnathus n. sp.
Fig. 5OO–QQDiscussion: The illustrated specimen is a left curved
P1 element with a free blade about one quarter the ele-ment length and a platform that expands and is roundedposteriorly. The free blade has four or five stout, highdenticles. The platform has a weak, gently curved nodosecarina that barely reaches the posterior tip, and an orna-ment of faint transverse ridges that become nodosetoward the carina, particularly in the posterior third ofthe platform. The outer margin of the platform is gentlypinched for the anterior one third of its length. In aboralview, the basal pit is situated about one third the plat-form length and extends a short distance antereiorly as anarrow groove.
There are insufficient specimens to make a meaning-ful comparison, and it is not possible to distinguish theright-curved element of the species. It seems likely this isa new species as it is not close to any currently describedspecies of middle to upper Frasnian age.
Material: Figured specimen UOPS 193 from Bed 4(Late rhenana Zone).
Polygnathus aff. webbi Stauffer, 1938
Figs. 5JJ–NN, 6LDiscussion: Synonymy of Polygnathus webbi and Po.
normalis, representing dextral and sinistral P1 elementsof the same species, was first suggested by Klapper,1971. Both dextral and sinistral P1 elements have cari-nae that extend slightly posterior of the platform, freeblades about three-sevenths the element length that bearabout eight denticles, and platform margins that are ser-rated to reflect the short transverse ridges. The dextralelements have platforms with outer and inner marginsthat are gently and evenly curved. The sinistral elementshave platforms with outer margins that are slightly flat-tened. In lower view the basal pit is situated at a pointabout one-quarter the distance from the anterior end ofthe platform, with a wide basal groove extending fromthere to about midway along the free blade.
Material: Figured specimens UOPS 191–192 fromBed 4 (Late rhenana Zone).
Polygnathus sp.
Fig. 6AA–BBDiscussion: This form is characterized by the strongly
flexed platform carina and in this respect differs from
the typical dextral specimens of Po. webbi (=Po. nor-malis Miller and Youngquist, 1947), in which the carinais more gently curved.
laeowo
184 N.M. Savage et al. / PaMaterial: Figured specimen UOPS 207 from Bed 30(Late rhenana Zone).
References
Branson, E.B., Mehl, M.G., 1934. Conodonts from the Grassy CreekShale of Missouri. University of Missouri Studies 8, pp. 171–259.
Branson, E.B., Mehl, M.G., 1938. The conodont genus Icriodus andits stratigraphic distribution. J. Paleontol. 12, 156–166.
Hahn, L., Siebenhuner, M., 1982. Explanatory notes (Paleontology) onthe geological maps of northern and western Thailand 1:250,000.Bundesanstalt fur geowissenschaften und rohstoffe, Hanover, pp.3–76.
Hinde, G.J., 1879. On conodonts from the Chazy and Cincinnati Groupof the Cambro-Silurian, and from the Hamilton and Genesee-Shaleof the Devonian, in Canada and the United States. Q. J. Geol. Soc.Lond. 35, 351–369.
Ji, Q., Ziegler, W., 1993. The Lali section: an excellent reference sectionfor Upper Devonian in South China. Courier ForschungsinstitutSenckenberg 157, 1–183.
Klapper, G., 1971. Sequence within the conodont genus Polygnathus inthe New York lower Middle Devonian. Geol. Palaeontol. 5, 59–79.
Klapper, G., Lane, H.R., 1985. Upper Devonian (Frasnian) conodontsof the Polygnathus biofacies, N.W.T., Canada. J. Paleontol. 59,904–951.
Klapper, G., Foster Jr., C.T., 1993. Shape Analysis of Frasnian Speciesof the Late Devonian Conodont Genus Palmatolepis. Paleontolog-ical Society Memoir 32 (J. Paleontol. 67 (4) Supplement), 35 pp.
Lange, F.G., 1968. Conodonten-Gruppenfunde aus Kalken des tieferenOberdevon. Geol. Palaeontol. Sonderband 1, 97–114.
Miller, A.K., Youngquist, W.L., 1947. Conodonts from the type sectionof the Sweetland Creek Shale in Iowa. J. Paleontol. 21, 501–517.
Muller, K.J., 1956. Zur Kenntnis der Conodonten-Fauna des europais-chen Devons, 1; Die Gattung Palmatolepis. SenckenbergischeNaturforschende Gesellschaft Abhandlungen 494, 70 pp.
rld 15 (2006) 171–184
Muller, K.J., Muller, E.M., 1957. Early Upper Devonian (Indepen-dence) conodonts from Iowa, Part 1. J. Paleontol. 31, 1069–1108.
Racki, G., 2005. Toward understanding Late Devonian global events:few answers, many questions. In: Over, D.J., Morrow, J.R., Wig-nall, P.B. (Eds.), Understanding Late Devonian and Permian-Triassic Biotic and Climatic Events: Towards an IntegratedApproach. Elsevier, pp. 5–36.
Sandberg, C.A., Dreesen, R., 1984. Late Devonian Icriodontid Biofa-cies Models and Alternative Shallow-Water Conodont Zonation.Geological Society of America Special Paper 196, pp. 143–177.
Sannemann, D., 1955. Beitrag zur Untergliederung des Oberdevonsnach Conodonten. Neues Jahrbuch fur Geologie und Palaontologie,Abhandlungen 100, 324–331.
Savage, N.M., 2002. The Upper Devonian Frasnian-Famennian GlobalExtinction Event. In: Proceedings of the Symposium on the Geol-ogy of Thailand, Bangkok, Thailand, pp. 24–28.
Savage, N.M., Funai, C.A., 1980. Devonian conodonts of probableearly Frasnian age from the Coronados Islands of southeasternAlaska. J. Paleontol. 54, 806–813.
Stauffer, C.R., 1938. Conodonts of the Olentangy Shale. J. Paleontol.12, 411–443.
Ulrich, E.O., Bassler, R.S., 1926. A classification of the toothlikefossils, conodonts, with descriptions of American Devonian andMississippian species. Proc. U.S. National Museum 68 (12), 63.
Younquist, W.L., 1947. A new Upper Devonian conodont fauna fromIowa. J. Paleontol. 21, 95–112.
Yudina, A.B., Racki, G., Savage, N.M., Racki, M., Malkowski, K.,2002. The Frasnian-Famennian events in a deep-shelf succession,Subpolar Urals: biotic, depositional, and geochemical records.Acta Palaeontol. Polonica 47, 355–372.
Ziegler, W., 1958. Conodontenfeinstratigraphische Untersuchungen an
der Grenze Mitteldevon/Oberdevon und in der Adorfstufe. Hessis-che Landesamt Bodenforschung Notizblatt 87, pp. 7–77.Ziegler, W., Sandberg, C.A., 1990. The Late Devonian standardconodont zonation. Courier Forschungsinstitut Senckenberg 121,1–115.














