
Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine ...

www.elsevier.com/locate/vph
Vascular Pharmacology
An ethyl acetate fraction obtained from a Southern Brazilian red wine
relaxes rat mesenteric arterial bed through hyperpolarization and
NO-cGMP pathway
Elke Zuleika Schuldta,*, Angela Cristina Beta, Mariana Appel Horta, Carla Ianssenb,
Marcelo Maraschinb, Karina Cklessc, Rosa Maria Ribeiro-do-Vallea
aDepartamento de Farmacologia, CCB, UFSC, Florianopolis, BrazilbLaboratorio de Fitotecnia e Morfogenese Vegetal, CCA, UFSC, Florianopolis, Brazil
cDepartment of Pathology, University of Vermont, HRSF Building, Room 214, Burlington, VT 05405, USA
Received 29 December 2004; received in revised form 23 March 2005; accepted 22 April 2005
Abstract
A number of studies suggest that moderate consumption of red wine may be more effective than other alcoholic beverages in decreasing
the risk of coronary heart disease (CAD). In this study, we investigated the effect of a crude extract (CE), as well as an ethyl acetate fraction
(EAF) obtained from a Brazilian red wine in the mesenteric arterial bed (MAB) from rats.
Our results showed that after the tonus of MAB was increased with phenylephrine (PE), increasing concentrations of CE induced a
concentration-dependent relaxation; moreover, EAF was more potent in relaxing the MAB when compared with CE. In vessels depolarized
with KCl (80 mM) or treated with the Na+/K+-ATPase pump inhibitor, ouabain (OUA; 100 AM), or with the K+ channel blockers: barium
(BaCl2, 100 AM) and tetraethylammonium (TEA; 500 AM ), the effect of EAF was significantly reduced. However, this effect was not altered
by the ATP-dependent K+ (KATP) channel blocker, glibenclamide (GLI; 100 AM) as well as Charybdotoxin (ChTx 10 nM), a nonselective
inhibitor of KCa channels of large and intermediate conductance plus Apamin (Apamin 100 nM), a specific inhibitor of KCa channels of small
conductance. The residual vasodilator effect of EAF observed in vessels pretreated with L-NOARG (100 AM), 1H-[1,2,4,] oxadiazolo[4,3-
alfa]quinoxalin, ODQ (10 AM) or KCl (80 mM), given separately, was reduced by the administration of KCl (40 mM) plus L-NOARG (100
AM). The present study demonstrates that the vasodilator effect of EAF is partially dependent upon membrane hyperpolarization in
combination with nitric oxide (NO) release.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Wine; Mesenteric arterial bed; NO; Hyperpolarization
1. Introduction
Consumption of polyphenol-rich foods, such as fruits
and vegetables, and beverages derived from plants, such as
cocoa, red wine and tea, may represent a beneficial diet in
terms of cardiovascular protection (Yamakoshi et al., 1999;
1537-1891/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.vph.2005.04.002
* Corresponding author. Universidade Federal de Santa Catarina-UFSC,
Coordenadoria Especial de Farmacologia/BLOCO ‘‘D’’, Campus Universi-
tario-Trindade-Florianopolis-SC, CEP: 88049-900, Caixa Postal: 476,
Brazil. Tel.: +55 48 331 9491, +55 48 331 9764; fax: +55 48 337 5479.
E-mail address: [email protected] (E.Z. Schuldt).
Stoclet et al., 2004). Among the numerous plausible
mechanisms by which polyphenols may confer cardiovas-
cular protection, improvement of the endothelial function
and inhibition of angiogenesis and cell migration and
proliferation in blood vessels have been the focus of recent
studies. (Howard et al., 2002; Pal et al., 2002; Stoclet et al.,
2004).
Recently, it was reported that incubation of endothelial
cells with French red wines up-regulated endothelial-type
nitric oxide synthase (eNOS) expression and eNOS activity,
but in contrast, red wines from Germany showed little or no
effect on eNOS expression (Wallerath et al., 2003). Red
43 (2005) 62 – 68

E.Z. Schuldt et al. / Vascular Pharmacology 43 (2005) 62–68 63
wine contains a range of polyphenols derived from the skin
of the grape and these include flavonols (quercetin and
myricetin), gallic acid, condensed tannins (catechin and
epicatechin polymers) and polymeric anthocyanins (Frankel
et al., 1995). Variations in the concentration of these
constituents among red wines may be responsible for the
range of biological effects and this suggests that not all red
wines might have the same pharmacological properties
(Ritchey and Waterhouse, 1999; Leighton et al., 1999;
Howard et al., 2002; Soares de Moura et al., 2004).
Red wine is particularly rich in polyphenol substances
compared to white wine. Polyphenols are potent in
preventing lower density lipoprotein oxidation, the corner-
stone event in the oxidation theory of atherogenesis and may
play an important role in preventing cardiovascular disease
(Schuldt et al., 2000; Vinson et al., 2001).
Since it has been hypothesized that other mediators than
NO may be involved in vasodilator responses in small
vessels (such as coronary and mesenteric arteries), and
because the mechanisms of these mediators may involve K+
channels (Soares de Moura et al., 2004; Dabisch et al.,
2004; Ndiaye et al., 2004), the current study was undertaken
to investigate the role of NO, as well as K+ channels in
mediating the vasodilator responses to an ethyl acetate
fraction (EAF) obtained from a Southern Brazilian red wine,
in vitro.
2. Materials and methods
2.1. Alcohol-free extract and organosolvent fraction
Red wine (variety Bordeaux-Vitis labrusca L.), thought
to have originated from Southern Brazil (Vale do Rio do
Peixe, Tangara-Santa Catarina state), from the 2000 vintage.
The bottles were kept in the dark and at 20–22 -C prior to
analysis.
Aliquots of 300 mL of Bordeaux red wine were collected
and the alcohol-free extract (CE) was obtained under
reduced pressure (80 -C, ¨50 min), following measurement
of the ethanol content using an alcoholometer apparatus
(Incoterm Co.). The polyphenols in the alcohol-free extract
(ca. 100 mL) were isolated by phase partitioning with ethyl
acetate (EtOAc–300 mL, 24 h incubation at room temper-
ature/in the dark). The organic phase (residue) was collected
and the EtOAc was removed by evaporation using an N2
stream. During the preparation of the alcohol-free extract
and the organosolvent fraction (EAF), the samples were
constantly protected from light to avoid photochemical
isomerization of any polyphenol compound.
Since the pharmacological protocols could not be carried
out in the presence of solvents, it was necessary to
determine the level of dry matter beginning with the
obtained residue. To this end, the residue was transferred
to Petri dishes and maintained in an incubator (;60 -C) untilit had reached constant weight. Through the resuspension of
the dry matter in deionized water, samples of CE and EAF
were obtained, which were used in the subsequent pharma-
cological experiments.
2.2. Pharmacological assays—Mesenteric Arterial Bed
The animal care research protocols were in accordance
with the guidelines of our university and were approved by
our Institutional Ethics Committee.
Male Wistar rats were anesthetized with a mixture of
ketamine (Francotar\) and 2-(2,6-xilidino)-5,6-dihydro-4H-
1,3-tiazine chlorhydrate (Rompum\) and a stainless poly-
ethylene cannula (PE-50) was inserted into the superior
mesenteric artery, then flushed with 10 mL of PBS
containing heparin (10 IU/mL) to remove possible blood
clots. After ligating all branches except the 4 or 5 directed to
the terminal ileum, the whole mesentery was cut along the
ileal border and transferred to a humidifying chamber
(McGregor, 1965), where it was perfused with 5 mL of
warm (37 -C) Krebs solution (composition in mM): NaCl,
113; KCl, 4.7; CaCl2, 2.5; KH2PO4, 0.9; NaHCO3, 25;
MgSO4, 1.1; glucose, 11; pH 7.4, saturated with 5% CO2 in
O2. To investigate the possible influence of endogenous
eicosanoids, concentration–response curves (CRC) to CE
an EAF were conducted in the presence of indomethacin.
According to our experiments, indomethacin does not affect
the vasodilator effect of CE as well as EAF (data not
shown), and for this reason, all subsequent experiments
were conducted in the presence of indomethacin (10 AM),
added to Krebs solution.
The perfusion pressure (PP) was measured by a trans-
ducer connected to a preamplifier and chart recorder (Digi-
Med, model 190, USA). Since the flow rate was constant
throughout the whole experimental period (4.0 mL/min),
any pressure alteration reflected changes in vascular
resistance. Drugs were either dissolved in Krebs solution
and perfused at the desired concentration or were adminis-
tered as bolus injections directly into the perfusion stream
(volume=100 AL). All preparations were allowed to
equilibrate for 30 min before the start of the experiments.
After this period, isolated injections of CE or EAF (30–
3000 Ag; bolus) were administered until consistent
responses were obtained. The basal pressure after the
equilibration period was 19.85T0.30 mm Hg (n =20).
Concentrations of CE or EAF were injected after the PP
had been elevated (70–80 mm Hg) with phenylephrine (PE,
750–1500 Ag; added to the perfusion fluid and adjusted to
maintain the same increase in PP). Acetylcholine (ACh, 200
nmol, bolus) was also injected to test endothelium function-
ality before concentration–response curves to CE and EAF
were obtained. All vasodilator responses to ACh, CE and
EAF were obtained in vessels precontracted with PE. As CE
and EAF induced a long-lasting inhibition of the PE
constrictor effect (Register 1), only one dose–response
curve to CE or EAF was obtained in each arterial bed
preparation.
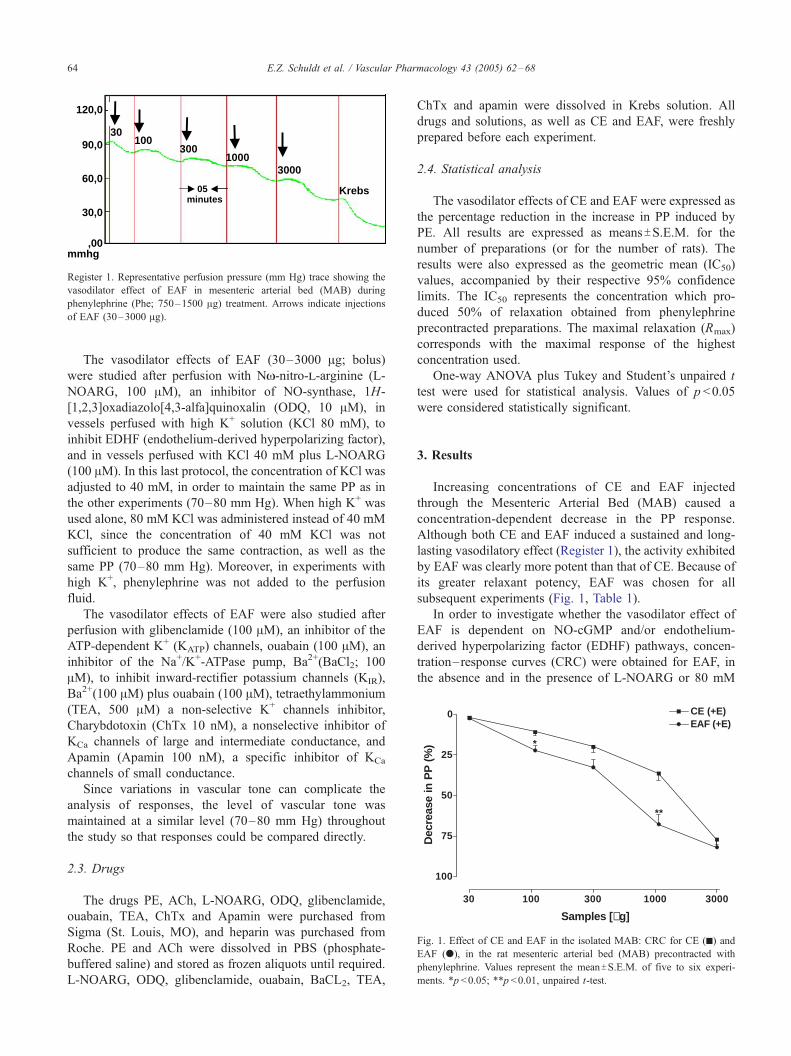
120,0
90,0
60,0
30,0
,00mmhg
30100
300
30001000
Krebs05minutes
Register 1. Representative perfusion pressure (mm Hg) trace showing the
vasodilator effect of EAF in mesenteric arterial bed (MAB) during
phenylephrine (Phe; 750–1500 Ag) treatment. Arrows indicate injections
of EAF (30–3000 Ag).
30 100 300 1000 3000
0
25
50
75
100
CE (+E)EAF (+E)
*
**
Samples [µg]
Dec
reas
e in
PP
(%
)
Fig. 1. Effect of CE and EAF in the isolated MAB: CRC for CE (h) andEAF (?), in the rat mesenteric arterial bed (MAB) precontracted with
phenylephrine. Values represent the meanTS.E.M. of five to six experi-
ments. *p <0.05; **p <0.01, unpaired t-test.
E.Z. Schuldt et al. / Vascular Pharmacology 43 (2005) 62–6864
The vasodilator effects of EAF (30–3000 Ag; bolus)
were studied after perfusion with NN-nitro-l-arginine (L-
NOARG, 100 AM), an inhibitor of NO-synthase, 1H-
[1,2,3]oxadiazolo[4,3-alfa]quinoxalin (ODQ, 10 AM), in
vessels perfused with high K+ solution (KCl 80 mM), to
inhibit EDHF (endothelium-derived hyperpolarizing factor),
and in vessels perfused with KCl 40 mM plus L-NOARG
(100 AM). In this last protocol, the concentration of KCl was
adjusted to 40 mM, in order to maintain the same PP as in
the other experiments (70–80 mm Hg). When high K+ was
used alone, 80 mM KCl was administered instead of 40 mM
KCl, since the concentration of 40 mM KCl was not
sufficient to produce the same contraction, as well as the
same PP (70–80 mm Hg). Moreover, in experiments with
high K+, phenylephrine was not added to the perfusion
fluid.
The vasodilator effects of EAF were also studied after
perfusion with glibenclamide (100 AM), an inhibitor of the
ATP-dependent K+ (KATP) channels, ouabain (100 AM), an
inhibitor of the Na+/K+-ATPase pump, Ba2+(BaCl2; 100
AM), to inhibit inward-rectifier potassium channels (KIR),
Ba2+(100 AM) plus ouabain (100 AM), tetraethylammonium
(TEA, 500 AM) a non-selective K+ channels inhibitor,
Charybdotoxin (ChTx 10 nM), a nonselective inhibitor of
KCa channels of large and intermediate conductance, and
Apamin (Apamin 100 nM), a specific inhibitor of KCa
channels of small conductance.
Since variations in vascular tone can complicate the
analysis of responses, the level of vascular tone was
maintained at a similar level (70–80 mm Hg) throughout
the study so that responses could be compared directly.
2.3. Drugs
The drugs PE, ACh, L-NOARG, ODQ, glibenclamide,
ouabain, TEA, ChTx and Apamin were purchased from
Sigma (St. Louis, MO), and heparin was purchased from
Roche. PE and ACh were dissolved in PBS (phosphate-
buffered saline) and stored as frozen aliquots until required.
L-NOARG, ODQ, glibenclamide, ouabain, BaCL2, TEA,
ChTx and apamin were dissolved in Krebs solution. All
drugs and solutions, as well as CE and EAF, were freshly
prepared before each experiment.
2.4. Statistical analysis
The vasodilator effects of CE and EAF were expressed as
the percentage reduction in the increase in PP induced by
PE. All results are expressed as meansTS.E.M. for the
number of preparations (or for the number of rats). The
results were also expressed as the geometric mean (IC50)
values, accompanied by their respective 95% confidence
limits. The IC50 represents the concentration which pro-
duced 50% of relaxation obtained from phenylephrine
precontracted preparations. The maximal relaxation (Rmax)
corresponds with the maximal response of the highest
concentration used.
One-way ANOVA plus Tukey and Student’s unpaired t
test were used for statistical analysis. Values of p <0.05
were considered statistically significant.
3. Results
Increasing concentrations of CE and EAF injected
through the Mesenteric Arterial Bed (MAB) caused a
concentration-dependent decrease in the PP response.
Although both CE and EAF induced a sustained and long-
lasting vasodilatory effect (Register 1), the activity exhibited
by EAF was clearly more potent than that of CE. Because of
its greater relaxant potency, EAF was chosen for all
subsequent experiments (Fig. 1, Table 1).
In order to investigate whether the vasodilator effect of
EAF is dependent on NO-cGMP and/or endothelium-
derived hyperpolarizing factor (EDHF) pathways, concen-
tration–response curves (CRC) were obtained for EAF, in
the absence and in the presence of L-NOARG or 80 mM

Table 1
Mean IC50 and maximal relaxation (Rmax) values obtained with CE and
EAF, with and without different solutions and drugs, in MAB of
normotensive Wistar rats
Group IC50 (a) (Ag) Rmax (b) (%)
CE 891.25 (538.10–1475.70)*** 78.40T3.67EAF (control) 390.84 (252.27–605.53) 81.88T4.04
+KCl 40+L-NOARG) 1059.25 (906.06–1238.34)*** 49.68T1.69***
+KCl 80 767.36 (457.35–1287.49)*** 61.11T3.64**
+L-NOARG 736.20 (330.17–1641.53)*** 70.57T4.38+GLI 252.34 (128.77–494.51) 82.01T6.05
+OUA 665.27 (474.14–933.45)*** 64.29T1.80*
+BaCl2 638.26 (275.43–1479.02)*** 86.53T1.96
+BaCl2+OUA 743.02 (312.70–1765.51)*** 71.16T5.64+TEA 851.13 (543.30–1333.80)*** 68.60T4.92
+ChTx+Apamin 421.70 (210.71–843.94) 98.01T6.45
+ODQ 881.04 (370.13–2097.20)*** 83.71T3.62
(a) Mean IC50 with the respective 95% confidence limits. ***p <0.001:
significantly different from the EAF (control) group. ANOVA/Tukey test.
(b) MeanTS.E.M. *p <0.05; **p <0.01; ***p <0.001: significantly different
from the EAF (control) group. ANOVA/Tukey test.
30 100 300 1000 3000
0
25
50
75
100
+ GLI 100 µM+ OUA 100 µM+ TEA 500 µM
Control
EAF [µg]
Dec
reas
e in
PP
(%
)
**
**
(A)
(B)
E.Z. Schuldt et al. / Vascular Pharmacology 43 (2005) 62–68 65
KCl. The pretreatment of the vessels with both inhibitors
(alone) significantly reduced, to the same extent, the
vasodilator effect of EAF. However, the combination of L-
NOARG with 40 mM KCl markedly shifted the CRC for
EAF to the right, when compared with the control (Fig. 2,
Table 1).
For this purpose, in order to investigate if the mecha-
nisms by which the EAF exerts its vasodilatory effect occur
mainly by hyperpolarization, CRC for EAF were obtained
in the MAB, after perfusion of GLI, OUA, TEA, barium,
ChTx and Apamin. The perfusion with GLI produced a
30 100 300 1000 3000
0
25
50
75
100
+ KCl 80
+ L-NOARG 100 µM
+ KCl 40 + L-NOARG 100 µM
Control
***
***** ***
******
*****
**
+ ODQ 10 µM
*
***
EAF [µg]
Dec
reas
e in
PP
(%
)
Fig. 2. Effect of EAF in the presence of L-NOARG, ODQ and high K+:
CRC for EAF in the rat mesenteric arterial bed (MAB), in the absence (?)
and in the presence of L-NOARG (r), ODQ (>), L-NOARG plus 40 mM
KCl (h) and 80 mM KCl (g). Values represent the meanTS.E.M. of four to
six experiments. *p <0.05; **p <0.01; ***p <0.001 significant difference
when compared with the control. ANOVA/Tukey test.
small leftward shift in the CRC for EAF. However,
perfusion with OUA or TEA significantly reduced the
vasodilatory effects of EAF (Fig. 3A). Fig. 3B illustrates the
CRC for EAF, obtained with BaCl2, BaCl2+plus OUA and
ChTx plus apamin. Barium and OUA, perfused alone or
simultaneously, displayed inhibitory effects characterized as
parallel rightward shifts of the EAF control curve. However,
ChTx and apamin were not able to promote an inhibitory
effect in EAF-induced relaxation in the MAB (Fig. 3B).
30 100 300 1000 3000
0
20
40
60
80
100
+ BaCl2 100 µM
Control
+ BaCl2 100 µM + OUA 100 µM
*
**
+ ChTx 10 nM + Apamin 100 nM
#
#
EAF [µg]
Dec
reas
e in
PP
(%
)
Fig. 3. Effect of EAF in the presence of the Na+/K+-ATPase pump inhibitor
and K+ channel blockers: CRC for EAF in the rat mesenteric arterial bed
(MAB) precontracted with phenylephrine, in the absence (?) and in the
presence of (A): OUA (h), GLI (r) and TEA (g). (B): BaCl2 (h), BaCl2plus OUA (g) and ChTx plus Apamin (>). Values represent the
meanTS.E.M. of five to six experiments. *p <0.05; **p <0.01 indicate a
significant difference when compared with the control. #p <0.05 indicate
significant difference between BaCl2 and BaCl2+OUA; ANOVA/Tukey
test.

0
20
40
60
80
100
120 Control+ L-NOARG+ ODQ+ KCl40 + L-NOARG+ KCl 80+ BaCl2+ BaCl2 + OUA + ChTx + Apamin+ GLI+ OUA+ TEA
****
***
***
*
Acethylcholine [200 nmol]
Dec
reas
e in
Per
fusi
on
Pre
ssu
re (
%)
Fig. 4. Effect of ACh-mediated relaxation in MAB: effect of a single injection of ACh (200 nmol) in rat mesenteric arterial bed (MAB), in the absence (control)
and in the presence of different solutions and drugs. Values represent the meanTS.E.M. of five to six experiments. *p <0.05; **p <0.01; ***p <0.001 indicate
significant difference when compared with the control. ANOVA/Tukey test.
E.Z. Schuldt et al. / Vascular Pharmacology 43 (2005) 62–6866
In all experiments, after the consistent precontraction of
the preparations with PE or KCl, a single dose of ACh (200
nmol) was administered in the MAB as a positive control.
According to Fig. 4, a high concentration of K+ as well as
Na+/K+-ATPase pump inhibitor and K+ channel blockers
(except GLI and BaCl2) were more efficient in decreasing
the vasodilatory effect of ACh in the MAB, when compared
to control or L-NOARG groups. ODQ was not able to
decrease the vasodilatory effect of ACh in the MAB.
However, the protocol in which the NO-synthase inhibitor
(L-NOARG) was perfused simultaneously with KCl 40 mM
was the more potent in inhibiting the decrease in PP induced
by ACh.
Table 1 shows the IC50 values with their respective
confidence limits (95%) and maximal relaxation (Rmax).
According to Table 1, in all experiments where the EDHF
mechanism was investigated (in protocols with high K+,
Na+/K+-ATPase pump inhibitor or K+ channel blockers,
except glibenclamide), there was an increase in IC50 values,
when compared with the IC50 control group.
4. Discussion
This study was performed in order to investigate the
mechanisms by which an ethyl acetate fraction (EAF),
obtained from a Brazilian red wine, induces a decrease in
PP, in MAB from normotensive Wistar rats. According to
our results, the EAF, and also to some extent the CE, was
effective in inducing a significant decrease in PP, in isolated
vessels of experimental animals (Fig. 1).
Previous data from our laboratory revealed that by
contrast with CE, EAF possesses a high concentration of
phenolic compounds; these compounds include gallic acid,
m-coumaric acid, chlorogenic acid, quercetin and resvera-
trol. In fact, EAF, an enriched polyphenol fraction obtained
by phytochemical partition of CE with ethyl acetate, showed
interesting results towards the scavenging of reactive
oxygen species (ROS), probably as a result of its higher
concentration of phenolic compounds (data not shown).
Therefore, the concentration-dependent relaxant response
induced by EAF in the MAB may be partly due to a radical
scavenging property, mainly superoxide radical. Superoxide
radical can react with NO at a high rate constant, generating
the oxidant peroxynitrite (Malinski et al., 1993; Demiryurek
et al., 1998), that has been implicated in the pathophysiol-
ogy of a variety of diseases including inflammation,
atherosclerosis and arthritis (White et al., 1994). The
antioxidant property of phenolic compounds could increase
the half-life of NO (Fitzpatrick et al., 1995; Dell’Agli et al.,
2004) and in this way contribute to the relaxing effect
verified in our experiment.
The present study suggests that the vasodilator effect of
the EAF partially depends on the release of NO from the
endothelial cells, since the perfusion of the MAB with L-
NOARG, an inhibitor of NO-synthase and ODQ, an
inhibitor of guanylate cyclase, significantly reduced the
effect of EAF (Fig. 2, Table 1). Similar results were found in
a recent study which showed that another inhibitor of NO-
synthase, NG-nitro-l-arginine methyl ester (L-NAME),
significantly reduced the vasodilator effect of a Brazilian
red wine, in the rat MAB (Soares de Moura et al., 2004).
The vasorelaxation induced by NO is dependent on a
reduction in intracellular calcium in smooth muscle cells,
due not only to the activation of soluble guanylate cyclase
(GC), but also to hyperpolarization induced by the opening
of K+ channels (Waldman and Murad, 1987; Tare et al.,
1990). For this purpose, several pieces of evidence suggest
that NO may be only one of several EDRFs mediating the
vasodilator response in small vessels. When NO and
prostaglandin synthesis are blocked, stimulation of the
vascular endothelium is still capable of eliciting vaso-
dilatation in various vascular preparations (Parkington et al.,
2002; Savage et al., 2003).

E.Z. Schuldt et al. / Vascular Pharmacology 43 (2005) 62–68 67
According to Fig. 2, the vasodilator effect of EAF may
involve hyperpolarization of the cells through K+ channels,
since 80 mM KCl also reduced the decrease in PP induced
by EAF in the MAB (Fig. 2, Table 1). In our experiment, the
inhibitory effect of L-NOARG and high K+ on the EAF-
induced vasodilatory activity was further enhanced when
those inhibitors were perfused simultaneously. According to
Fig. 2 and IC50 values described in Table 1, the decrease in
PP brought about by EAF might be due to a combination of
an activation of GC enzyme plus hyperpolarization by the
opening of K+ channels.
Some authors (Li et al., 1997; Naderali et al., 1997) have
reported that flavonoids such as quercetin and resveratrol
are able to open Ca2+-activated K+ channels (KCa).
Resveratrol, a polyphenolic phytoalexin that is generated
in response to stress in specific plants and also grape skins,
has a stimulatory effect on KCa (large-conductance potas-
sium channel) found in human vascular endothelial cells
(Chen and Pace-Asciak, 1996). It is postulated that
increased K+ efflux following the activation of KCa channels
by resveratrol may increase K+ concentration in the
myoendothelial space followed by hyperpolarization of
smooth muscle cells, leading to the dilation of blood vessels
(Bhat et al., 2001).
Another piece of evidence which indicates that EAF
could be involved in the EDHF pathway is illustrated in Fig.
3A and B, since the vasodilator effect of EAF is inhibited by
the perfusion of the MAB with the Na+/K+-ATPase pump
inhibitor, ouabain (OUA) and the different K+ channel
blockers, including tetraethylammonium (TEA) and barium.
The perfusion of the MAB with OUA or TEA markedly
inhibited the decrease in PP induced by EAF, and to the
same extent as in the presence of 80 mM KCl. According to
Soares de Moura et al. (2004), KATP channels are probably
not involved in wine polyphenols-induced vasodilatation in
the MAB; in our experiment, EAF-induced vasorelaxation
was also not inhibited by glibenclamide (Fig. 3A).
The decrease in PP induced by EAF was also partially
inhibited by the perfusion of ouabain plus barium. Even
though the perfusion of the MAB with barium (alone) did
not significantly shift the CRC for EAF to the right, when
barium and ouabain were perfused in the same solution, this
inhibitory effect was more evident (Fig. 3B).
Our results are in accordance with Edwards et al. (1998),
who have shown that the relaxations stimulated by K+ were
similarly inhibited by barium and ouabain in rat hepatic and
mesenteric arteries. These authors proposed that K+ entering
the subendothelial space after efflux through endothelial Kca
channels stimulates smooth muscle hyperpolarization by
activation of KIR and the Na+/K+-ATPase pump on the
smooth muscle cells. In our experiments, although barium
plus ouabain significantly shifted the EAF control curve to
the right, ChTx plus Apamin were not able to inhibit the
decrease in PP induced by EAF (Fig. 3B; Table 1).
Eventually, the pharmacological similarities of the hyper-
polarization activity and the vasodilator effect induced by
EAF (both inhibited by high K+ and some K+ channel
blockers) suggest that the EAF effect could be partially
mediated by activation of KIR and Na+/K+-ATPase pump.
Since it has been hypothesized that other mediators than
NO may be involved in the vasodilator responses to ACh
and because the mechanisms of these mediators may
involve K+ channels (Dabisch et al., 2004), Fig. 4 was
obtained as our positive control, in order to demonstrate the
ACh-mediated relaxation, in all different protocols. Since
the perfusion of L-NOARG and ODQ do not inhibit the
ACh-mediated relaxation in the MAB, this demonstrates
that the ACh mechanism possesses a significant component
that is independent of NO production. These observations
are consistent with those reported in the literature (Wood-
man et al., 2000; Savage et al., 2003), in which NO makes a
small but significant contribution to ACh-mediated vaso-
dilatation with a more substantial role for EDHF, in small
resistance arteries. However, comparing the results of Fig. 4
with the CRC for EAF obtained in the presence of L-
NOARG and ODQ (Fig. 2), EAF appears to be more NO-
dependent when compared with ACh, since the perfusion
with both drugs significantly shifted the CRC for EAF to the
right.
5. Conclusions
This study showed that NO appears to make an important
contribution to EAF-mediated vasodilatation, either via the
NO-cGMP pathway or through hyperpolarization induced
by the opening of K+ channels. The phenolic compounds
present in EAF (such as quercetin and resveratrol) may exert
their effects by activation of KIR and the Na+/K+ ATPase
pump. The present investigation also demonstrated that EAF
induces a vasodilator effect that does not involve the
opening of KATP channels and potassium conductance
(KCa) channels.
In concert with other systemic beneficial effects of red
wine, this could contribute to the cardiovascular protection
enjoyed by moderate drinkers of red wine.
Acknowledgments
The authors are grateful to CNPq (Brazil) for financial
support.
References
Bhat, K.P.L., Kosmeder II, J.W., Pezzuto, J.M., 2001. Biological effects of
resveratrol. Antiox. Red. Signal. 3 (6), 1041–1064.
Chen, C.K., Pace-Asciak, C.R., 1996. Vasorelaxing activity of resveratrol
and quercetin in isolated rat aorta. Gen. Pharmacol. 27, 363–366.
Dabisch, P.A., Liles, J.T., Taylor, J.T., Sears, B.W., Saenz, R., Kadowitz,
P.J., 2004. Role of potassium channels in the nitric oxide-independent
vasodilator response to acetylcholine. Pharmacol. Res. 49, 207–215.

E.Z. Schuldt et al. / Vascular Pharmacology 43 (2005) 62–6868
Dell’Agli, M., Busciala, A., Bosisio, E., 2004. Vascular effects of wine
polyphenols. Cardiovasc. Res. 63, 593–602.
Demiryurek, A.T., Cakici, I., Kanzik, I., 1998. Peroxynitrite: a putative
cytotoxin. Pharmacol. Toxicol. 82, 113–117.
Edwards, G., Dora, K.A., Gardener, M.J., Garland, C.J., Weston, A.H.,
1998. K+ is an endothelium-derived hyperpolarizing factor in rat
arteries. Nature 396, 269–272.
Fitzpatrick, D.F., Hirschfield, S.L., Ricci, T., Coffey, R.G., 1995.
Endothelium-dependent vasorelaxation caused by various plant
extracts. J. Cardiovasc. Pharmacol. 26, 90–95.
Frankel, E.N., Waterhouse, A.L., Teissedre, P.L., 1995. Principal
phenolic phytochemicals in selected California wines and their
antioxidant activity in inhibiting oxidation of human low-density
lipoproteins. J. Agric. Food Chem. 43, 890–894.
Howard, A., Chopra, M., Thurnham, D.I., Strain, J.J., Fuhrman, B.,
Aviram, M., 2002. Red wine consumption and inhibition of LDL
oxidation: what are the important components? Med. Hypotheses 59
(1), 101–104.
Leighton, F., Cuevas, A., Guasch, V., Perez, D.D., Strobel, P., San Martın,
A., Urzua, U., Dıez, M.S., Fonsea, R., Castillo, O., Mizon, C.,
Espinoza, M.A., Urquiaga, I., Rozowski, J., Maiz, A., Germain, A.,
1999. Plasma polyphenols and antioxidants, oxidative DNA damage
and endothelial function in a diet and wine intervention study in
humans. Drugs Exp. Clin. Res. XXV (2/3), 133–141.
Li, Y., Starrett, J.E., Meanwell, N.A., Johnson, G., Harte, W.E., Dworetzky,
S.I., Boissard, C.G., Gribkoff, V.K., 1997. The discovery of novel
openers of Ca2+-dependent large-conductance potassium channels:
Pharmacophore search and physiological evaluation of flavonoids.
Biorg. Med. Chem. Lett. 7 (7), 759–762.
Malinski, T., Bailey, F., Zhang, Z.G., Chopp, M., 1993. Nitric oxide
measured by a porphyrinic microsensor in rat brain after transient
middle cerebral artery occlusion. J. Cereb. Blood Flow Metab. 13,
355–358.
McGregor, D.D., 1965. The effect of sympathetic nerve stimulation on
vasoconstrictor responses in perfused mesenteric blood vessels of the
rat. J. Physiol. 177, 21–30.
Naderali, E.K., Buttell, N., Taggart, M.J., Bullock, A.J., Eisner, D.A., Wray,
S., 1997. The role of the sarcolemmal Ca(2+)-ATPase in the pH
transients associated with contraction in rat smooth muscle. J. Physiol.
505 (Pt 2), 329–336.
Ndiaye, M., Chataigneau, T., Chataigneau, M., Schini-Kerth, V.B., 2004.
Red wine polyphenols induce EDHF-mediated relaxations in porcine
coronary arteries through the redox-sensitive activation of the PI3-
kinase/Akt pathway. Br. J. Pharmacol. 142 (7), 1131–1136.
Pal, S., Ho, N., Santos, C., Dubois, P., Mamo, J., Croft, K., Allister, E.,
2002. Red wine polyphenolics increase LDL receptor expression and
activity and suppress the secretion of apoB100 from human HepG2
cells. Am. Soc. Nutr., 700–706.
Parkington, H.C., Chow, J.M., Evans, R.G., Coleman, H.A., Tare, M.,
2002. Role of endothelium-derived hyperpolarizing factor in vascular
tone in rat mesenteric and hindlimb circulations in vivo. J. Physiol. 542,
929–937.
Ritchey, J., Waterhouse, A.L., 1999. A standard red wine: monomeric
phenolic analysis of commercial Cabernet Sauvignon wines. Am. J.
Enol. Viticult. 50, 91–100.
Savage, D., Perkins, J., Hong Lim, C., Bund, S.J., 2003. Functional
evidence that K+ is the non-nitric oxide, non-prostanoid endothelium-
derived relaxing factor in rat femoral arteries. Vascul. Pharmacol. 40 (1),
23–28.
Schuldt, E.Z., Kcless, K., Simas, M.E., Farias, M.R., Ribeiro-do-Valle,
R.M., 2000. Butanolic fraction from Cuphea carthagenensis Jacq Mc
Bride relaxes rat thoracic aorta through endothelium-dependent and
endothelium-independent mechanisms. J. Cardiovasc. Pharmacol. 35
(2), 234–239.
Soares de Moura, R., Miranda, D.Z., Pinto, A.C.A., Sicca, R.F., Souza,
M.A.V., Rubenich, L.M.S., Carvalho, L.C.R.M., Rangel, B.M., Tano,
T., Madeira, S.V.F., Resende, A.C., 2004. Mechanism of the endothe-
lium-dependent vasodilation and the antihypertensive effect of Brazilian
red wine. J. Cardiovasc. Pharmacol. 44 (3), 302–309.
Stoclet, J-C., Chataigneau, T., Ndiaye, M., Ho Oak, M., El Bedoui, J.,
Chataigneau, M., Schini-Kerth, V.B., 2004. Vascular protection by
dietary polyphenols. Eur. J. Pharmacol. 500 (1–3), 299–313.
Tare, M., Parkington, H.C., Coleman, H.A., Neild, T.O., Dusting, G.J.,
1990. Hyperpolarization and relaxation of arterial smooth muscle
caused by nitric oxide derived from endothelium. Nature 346, 69–71.
Vinson, J.A., Teufel, K., Wu, N., 2001. Red wine, dealcoholized red wine,
and especially grape juice, inhibit atherosclerosis in a hamster model.
Atherosclerosis 156, 67–72.
Waldman, S.A., Murad, F., 1987. Cyclic GMP synthesis and function.
Pharmacol. Rev. 39, 163–196.
Wallerath, T., Polleo, D., Li, H., Forstermann, U., 2003. Red wine increases
the expression of human endothelial nitric oxide synthase. J. Am. Coll.
Cardiol. 41, 471–478.
White, C.R., Brock, T.A., Chang, L.Y., Crapo, J., Briscoe, P., Ku, D.,
Bradley, W.A., Gianturco, A.S., Gore, J., Freeman, B.A., Tarpey, M.M.,
1994. Superoxide and peroxynitrite in atherosclerosis. Prod. Natl. Acad.
Sci. 91, 1044–1048.
Woodman, O.L., Wongsawatkul, O., Sobey, C.G., 2000. Contribution of
nitric oxide, cyclic GMP and K+ channels to acetylcholine-induced
dilatation of the rat conduit resistance arteries. Clin. Exp. Pharmacol.
Physiol. 27, 34–40.
Yamakoshi, J., Kataoka, S., Koga, T., Ariga, T., 1999. Proanthocyani-
din-rich extract from grape seeds attenuates the development of
aortic atherosclerosis in cholesterol-fed rabbits. Atherosclerosis 142,
139–149.
Copyright © 2022 FDOKUMEN
















![Synthesis, Single Crystal X-Ray Structure, and Antimicrobial Activity of 6-(1,3-Benzodioxol-5-ylmethyl)-5-ethyl-2-{[2-(morpholin-4-yl)ethyl]sulfanyl}pyrimidin-4(3H)-one](https://static.fdokumen.com/doc/165x107/6324d4c2c9c7f5721c01c4b5/synthesis-single-crystal-x-ray-structure-and-antimicrobial-activity-of-6-13-benzodioxol-5-ylmethyl-5-ethyl-2-2-morpholin-4-ylethylsulfanylpyrimidin-43h-one.jpg)



