
An alternative ELISA for T4 determination based on idiotype anti-idiotype interaction and a latex...
13
JOURNAL OF IMMU~ICAL METHOIJS ELSEVIER Journal of Immunological Methods 192 (1996) 73-85 An alternative ELISA for T4 determination based on idiotype anti-idiotype interaction and a latex method for anti-idiotype monoclonal antibody selection Maria In& Becker a~cT*, Juan E. Aguayo a, Adolf0 Jamett ‘, Franklin Juica ‘, Arturo Yudelevich ‘, Arnaldo Foradori b, Alfred0 E. De Ioannes ’ ’ Laboratorio de Inmunologia. Bias Chile lngenieria GeriCricrr S.A.. Santiago, Chile h Lclboratorio de Medicina Nuclear. Centro de Diagndstico (CEDIUCI. Ponti’cia Unil,er.sidad Cat6lica de Chile. Santiago. Chile ’ Unidud de Inmunolo,@z. Facultad de Ciencias Biol6gicas. Pontijicia Unit,ersidad Catrilico de Chile. Santiago. Chile Received 18 August 1995: revised 1 I January 1996: accepted 25 January 1996 Abstract This paper is the first report on the use of an idiotype-anti-idiotype monoclonal antibody reaction to develop an enzyme immunoassay for thyroxine (T4). We have developed a monoclonal antibody against T4, named IF10 of IgGl subclass and K, 5.21 X IO8 M- ’ which was used to obtain anti-idiotypic monoclonal antibodies. Anti-idiotypic antibodies were selected by a novel method. a passive agglutination assay with the idiotype monoclonal lFl0 absorbed on latex particles and subsequently characterized by RIA. One of these anti-idiotype antibodies, named 5B3 - type /3 antibody - of IgGl subclass, was used to develop an enzyme-linked T4 idiotype-anti-idiotype immunosorbent assay. The T4 calibration curve. using the lFl0 idiotypic antibody adsorbed to solid phase and the 5B3 anti-idiotypic antibody conjugated to alkaline phosphatase (ALP), shows adequate performance in the range between 0.7-25 kg% of the analyte. The reliability of the proposed method is demonstrated by the correlation coefficient r = 0.74, found between T4 measured by RIA and our assay, with a panel of sera from euthyroid, hypothyroid and hyperthyroid individuals. The correlation coefficient was r = 0.93 within assays and r = 0.88 between assays. These results provide the basis for a new non isotopic assay for the study and diagnosis of T4-related human disease and provides a model to develop immunoassays for other haptens and small molecules of clinical interest. Kewords: T4: Idiotype-anti-idiotype monoclonal antibody; Latex: ELISA Abbreviations: ANS. 8-anilino-I-naphthalenesulfonic acid; ALP, alkaline phosphatase; PBS, phosphate-buffered saline; BSA, bovine serum albumin; CAP, American College of Pathology: CFC. complete Freund’s adjuvant: IFC, incomplete Freund’s adjuvant; D-MEM, Dulbecco’s medium; ELISA, enzyme-linked immunosorbent assay(s); id, idiotype; K,, affinity constant; KLH, keyhole limpet hemocyanin: OD. optical density; OVO, ovalbumin from chicken eggs; PEG. polyethylene glycol: p-NPP, p-nitrophenylphosphate; r. correlation coefficient: RIA. radioimmunoassay; T3. triiodothyronine: T4, thyroxine. ’ Corresponding author. At: Bios Chile I.G.S.A., Av. Marathon 1943. Santiago, Chile. Tel,: (56-2)238-1878: Fax: (56-2) 239-4250. Elsevier Science B.V. PfI SOO22- 1759(96)000’3-3
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of An alternative ELISA for T4 determination based on idiotype anti-idiotype interaction and a latex...

JOURNAL OF IMMU~ICAL METHOIJS
ELSEVIER Journal of Immunological Methods 192 (1996) 73-85
An alternative ELISA for T4 determination based on idiotype anti-idiotype interaction and a latex method for anti-idiotype
monoclonal antibody selection
Maria In& Becker a~cT*, Juan E. Aguayo a, Adolf0 Jamett ‘, Franklin Juica ‘, Arturo Yudelevich ‘, Arnaldo Foradori b, Alfred0 E. De Ioannes ’
’ Laboratorio de Inmunologia. Bias Chile lngenieria GeriCricrr S.A.. Santiago, Chile
h Lclboratorio de Medicina Nuclear. Centro de Diagndstico (CEDIUCI. Ponti’cia Unil,er.sidad Cat6lica de Chile. Santiago. Chile
’ Unidud de Inmunolo,@z. Facultad de Ciencias Biol6gicas. Pontijicia Unit,ersidad Catrilico de Chile. Santiago. Chile
Received 18 August 1995: revised 1 I January 1996: accepted 25 January 1996
Abstract
This paper is the first report on the use of an idiotype-anti-idiotype monoclonal antibody reaction to develop an enzyme immunoassay for thyroxine (T4). We have developed a monoclonal antibody against T4, named IF10 of IgGl subclass and
K, 5.21 X IO8 M- ’ which was used to obtain anti-idiotypic monoclonal antibodies. Anti-idiotypic antibodies were selected by a novel method. a passive agglutination assay with the idiotype monoclonal lFl0 absorbed on latex particles and subsequently characterized by RIA. One of these anti-idiotype antibodies, named 5B3 - type /3 antibody - of IgGl subclass,
was used to develop an enzyme-linked T4 idiotype-anti-idiotype immunosorbent assay. The T4 calibration curve. using the
lFl0 idiotypic antibody adsorbed to solid phase and the 5B3 anti-idiotypic antibody conjugated to alkaline phosphatase
(ALP), shows adequate performance in the range between 0.7-25 kg% of the analyte. The reliability of the proposed method is demonstrated by the correlation coefficient r = 0.74, found between T4 measured by RIA and our assay, with a
panel of sera from euthyroid, hypothyroid and hyperthyroid individuals. The correlation coefficient was r = 0.93 within assays and r = 0.88 between assays. These results provide the basis for a new non isotopic assay for the study and diagnosis of T4-related human disease and provides a model to develop immunoassays for other haptens and small molecules of clinical interest.
Kewords: T4: Idiotype-anti-idiotype monoclonal antibody; Latex: ELISA
Abbreviations: ANS. 8-anilino-I-naphthalenesulfonic acid; ALP, alkaline phosphatase; PBS, phosphate-buffered saline; BSA, bovine
serum albumin; CAP, American College of Pathology: CFC. complete Freund’s adjuvant: IFC, incomplete Freund’s adjuvant; D-MEM,
Dulbecco’s medium; ELISA, enzyme-linked immunosorbent assay(s); id, idiotype; K,, affinity constant; KLH, keyhole limpet hemocyanin:
OD. optical density; OVO, ovalbumin from chicken eggs; PEG. polyethylene glycol: p-NPP, p-nitrophenylphosphate; r. correlation coefficient: RIA. radioimmunoassay; T3. triiodothyronine: T4, thyroxine.
’ Corresponding author. At: Bios Chile I.G.S.A., Av. Marathon 1943. Santiago, Chile. Tel,: (56-2)238-1878: Fax: (56-2) 239-4250.
Elsevier Science B.V.
PfI SOO22- 1759(96)000’3-3

1. Introduction
Human thyroid hormones are a key factor in the maintenance of basal metabolism. Alterations in the
normal concentration of T3 and T4 generate a num-
ber of pathologies and affect the energetic metabolism in human beings (Wilson and Forter, 1985). The
availability of sensitive and accurate methods for the quantitation of these hormones in serum samples is therefore of clinical importance. Pioneering work
described methods based solely on chemical (Pind,
1957) or radiochemical approaches (Arons and Hy- dovitz, 1959). Later. some methods were developed on the basis of the physico-chemical behavior of
these molecules, such as equilibrium dialysis (Fang and Selenkow, 1970), ultrafiltration (Sophianopoulos
et al., 1980) and gas chromatography (Petersen et al., 1977). Although all these methods are sensitive, they
are technically complex. hence not easily applicable in the routine of clinical laboratories.
With the advent of immunodiagnostic methods, RIA became the method of choice for the quantita-
tion of T4, initially with polyclonal antibodies
(Chopra et al.. 1971) and later with monoclonal antibodies (Wang et al., 1981). Since T4 is an hapten (M, 777.9) it is necessary to use T4-protein conju- gates for antibody preparation and specific markers
including radioactive T4 and enzyme T4 conjugate which enzymes like malic dehydrogenase (Ullman et
al., 1975), peroxidase (Schall et al., 1978). P-galac- tosidase (Monji et al., 1978). acetylcholinesterase (Schall et al., 1978) and alkaline phosphatase (Miyai et al., 1980). T4 RIA has been used in extensive
screening programs to detect congenital hypothy- roidism. The fact that this type of programs needs the processing of a huge number of samples, has led scientists to develop alternative technologies based on non-isotopic methods for thyroid hormone deter-
mination, e.g., ELISA (Ullman et al.. 1975). lumi-
nescence (Christofides and Sheehan, 1995) and latex immunoassays (Mareschal et al.. 1990).
We describe here the development of a mono- clonal antibody against T4. its type syngeneic anti- idiotype and its use for the development of a new enzyme-linked immunosorbent assay for T4 determi- nation. The use of idiotype-anti-idiotype antibodies interactions, first described by Jerne (1974). has been reported in different systems (Pang et al., 1995). In
particular. the concept of the internal image of the antigen has been successfully applied to characterize receptors (Sege and Peterson, 1978; Colucci et al.,
1987). possible vaccines against pathogens (Kennedy et al., 1986;; McClintock et al., 1986) or related cancer antigens (Herlyn et al., 1986; Schmitt et al.,
1994). Also. anti-idiotypic antibodies are useful tools in the prognosis of human autoimmune diseases
(Poskitt et al.. 1991) and in the detection of a parasite antigens (Potocnjak et al., 1982; Thanavala,
1989). In recent years. anti-idiotypes against haptens including insecticides. herbicides (Spinks et al., 1993) mycotoxins and phycotoxins (Chanh et al.. 1989;
Shestowsky et al.. 1992) have been successfully generated.
We also report here the use of a novel assay to screen anti-idiotype monoclonal antibodies based in passive agglutination reaction. This method can be
used in the selection of hybridomas secreting anti-id- iotypic antibodies for other haptens and small molecules of clinical interest. Our results show that
the anti-idiotype ELISA for T4 provides a new alter-
native method for the study and diagnosis of T4 related human diseases.
2. Materials and methods
2.1. DeL:elopment of anti-T4 morloclorlal arltibodies
2. I. I. Mice immuriiratiorl 3-month-old female BALB/c mice were immu-
nized as follows: on day 1, they received a subcuta- neous and intraperitoneal injection of 50 (ug of
porcine thyroglobulin (American Laboratories) in complete Freund’s coadjuvant (CFC): on days 15 and 30, the same immunization was carried out, but
with incomplete Freund’s coadjuvant (IFC). 10 days after each injection, the animals were bled by the tail to cbtain the serum and the humoral response against T4 was determined by a direct ELISA using T4
coupled to BSA (T4-BSA).
2.1.2. T4-BSA coupling The procedure described by Marchand et al.
(1987) was used. In brief, 8 mg of T4 (Sigma) were coupled to 10 mg of BSA using 30 mg carbodiimide (Aldrich). The mixture was stirred for 24 h at 4°C

M.I. Becker et ul./Journal qflmmunological Methods 192 (1996) 73-85 7.5
then centrifuged at 10 000 rpm for 15 min. The monoclonals, an ELISA was run in parallel using
supematant was dialyzed against 0.9% NaCl for 96 h T3-BSA, BSA treated with carbodiimide and BSA
and stored at - 20 C. without modifications as control.
2.1.3. T4-BSA ELISA The procedure described by Crowther and Abu-
Elzeein (1980) was used. In brief, 96-well
polystyrene plates (Nunc) were activated with 50 ~1 of a solution of 10 pg/ml TbBSA for 2 h at room temperature and blocked with a 2% PBS-ovalbumin
(PBS-OVO) solution for 90 min at room tempera- ture. Then, serial half dilutions of immune serum in
2% PBS-OVO were added to the plates. and incu- bated for 2 h at room temperature. The plates were washed three times for 2 min with 0.02% PBS-Tween
20, and then 50 ~1 of goat serum mouse anti-IgG conjugated to ALP (Sigma) diluted l/1000 in 0.2% PBS-OVO was added to the wells. After incubating
for 30 min at room temperature, the plates were washed as described above. To start the enzymatic
reaction, SO ~1 of p-NPP 1 mg/ml in a buffer solution containing 0.1 M Tris. 0.1 M NaCI, 5 mM MgCl,, pH 9.5 was added to the wells and the plates were incubated at 37°C for 30 min. The reaction was
stopped with 50 ~1 3 N NaOH and was read spec- trophotometrically at 405 nm.
2. I .5. Pur$cation of anti-T4 I FlO tnonoclonal anti-
body The anti-T4 antibody was recloned twice and
produced in ascites. It was then purified by affinity chromatography with Protein G-Sepharose CL-4B (Pharmacia) as described by Harlow and Lane (1988).
2.1.6. Molecular e.rciusion chromatography The general procedure described by Johnstone
and Thorpe ( 1988) was used. In brief. a IO X 0.5 cm
Sephadex G-25 (Pharmacia) column was prepared. The exclusion volume was determined with 500 ~1
of 1% Blue Dextran. Later, 1 ml containing 0.96 mg/ml of purified lFl0 monoclonal antibody was
applied after being previously incubated during 24 h with 360000 cpm of T4l’“l. For the elution of the
antibody 0.05 M PBS and 0.1% sodium azide pH 7.5 was used. Elution of T4 was done with 3 N NaOH.
Radioactivity present in each fraction was deter- mined in a Gamma Counter. As control. a chro- matography with only T4”‘I was run.
2.1.7. Detertnination of the qffinig constant (K, )
2.1.4. Hybridoma production The protocol described by Kiihler and Milstein
(1975) was used. An intravenous booster with 100
pg of porcine thyroglobulin was administered to the mouse exhibiting the higher titer against T4. After 3 days the animal was killed by cervical dislocation
and the spleen was removed. The splenic lympho-
cytes were fused to NS0/2 myeloid cells (provided by C. Milstein, from MRC Laboratory of Molecular
Biology, Cambridge, England). The fusion was car- ried out with 4000 PEG (Merck) and the cells were cultured in a HAT selection medium as described by Becker et al. ( 1994). The supematants of all wells
that presented hybridoma growth were assayed by ELISA using T4-BSA, as described above.
Clones secreting antibodies to T4 were expanded
to Wwell plates, in 1 ml of complete D-MEM medium previously conditioned with spleen cells from a non-immune mouse. The isotype of the monoclonals was determined with a commercial as- say (Serotec). To determine the specificity of the
The K, of the lFl0 antibody was determined
using a classical RIA (Johnstone and Thorpe, 1988).
The assay was performed in polypropylene tubes as follows: a volume of 0.1 ml of serum samples con-
taining known amounts of T4 were mixed with 0.1 ml of lFl0 antibody (3.7 Fg/ml) and 0.2 ml T4”‘I
(approximately 30000 cpm) in 0.075 M sodium barbital, 0.05% ANS, 2.0% mouse serum and 0.1%
sodium azide, pH 8.6. The mixture was stirred and incubated for I h at 37°C. Then. 0.2 ml of 0.5 mg/ml anti-mouse IgG developed in rabbit in PBS
and 0.1 M EDTA pH 7.8 were added, followed by 0.5 ml 6% PEG 8000 in PBS. The preparations were stirred and centrifuged at 2000 rpm for 15 min at
4°C. The supematants were discarded and the pellets were counted in a Gamma Counter. The results were analyzed by the Scatchard method (Paul. 1989).
2.1.8. Anti-T4 IF10 antibody evaluation n+th a panel f3f serum samples
A similar RIA for the determination of K, was performed on a 60 panel serum samples from nor-

mal, hypothyroid and hyperthyroid patients. All sam- ples were previously quantified for T4 by RIA at the Laboratorio de Medicina Nuclear, Pontificia Univer-
sidad Cat6lica de Chile. using the Wiersinga and Chopra (1982) method. consisting in a classic RIA
with a rabbit polyclonal serum against T4 and pre- cipitation of the complex with goat serum against rabbit IgG. The standard used was L-thyroxine
(Sigma). The methodology was carried out with
internal quality guarantee (three reference sera, with low, medium and high T4 concentration) and exter-
nal quality guarantee (Program of the American Col-
lege of Pathologists No. 1 1042-03-01-01).
2.2. Der?elopment of anti-idyotypic monoclonal unti- bodies
2.2.1. Coupling of anti-T4 IF10 antibody to KLH The protocol described by Harlow and Lane
(1988) was used. In brief, 500 pg AMC 1 F10 plus 500 pg of hemocyanin (KLH from Sigma) were mixed in the presence of citrate buffer pH 6.5 and
glutaraldehyde at a final concentration of 0.1%. The mixture was left in the dark at room temperature for 1 h and then dialyzed overnight against PBS.
2.2.2. Prepurution of latex particles coated with IF10 antibody
We used the protocol described by Hudson and Hay (1980). 1 ml of 0.8 pm polystyrene latex beads (Sigma) were resuspended to 1% in 50 ml 0.054 M
glycine-saline buffer pH 8.2 and centrifuged at 12 500 X g for 15 min. The pellet was resuspended in the same buffer. Then, the 1FlO monoclonal
antibody (previously dialyzed against 0.054 M glycine saline buffer. pH 8.2) was added to a final concentration of 150 pg/ml of 1% latex. The mix-
ture was stirred at room temperature for 30 min and centrifuged at 12 500 X g for 15 min. The super- natant was discarded and the pellet was resuspended
in 50 ml of 0.2% BSA glycine-saline buffer pH 8.2.
2.2.3. Mice immunization BALB/c female mice were immunized with IF10
monoclonal antibody coupled to KLH, according to the protocol described by Jaureguiberry et al. (1988). In brief, on day 1, 25 pg 1 F10 antibody coupled to KLH in CFC was injected intraperitoneally; on days 15 and 30. 25 pg in IFC and boosted on day 45 with
an intraperitoneal injection of 25 pg 1FI 0 mono- clonal antibody in glucose-phosphate buffer. 10 days after each injection, bleeding trials were performed
and the antibody response was evaluated with an agglutination assay using latex beads coated with anti-T4 IF10 monoclonal antibody.
2.2.4. Lutex agglutination assay ,for screening of anti-idiotype untibodies
Mouse serum was serially diluted. An aliquot of
20 ~1 of each dilution or of hybridoma supernatant was placed on a dark slide and 20 ~1 of the latex
preparation was added. The reagents were mixed and spread on the slide, which was gently rocked for 2
min, then agglutination was scored. Aliquots of the appropriate dilutions of pre-immune serum or super-
natant from the parental hybridomas cell line were used as negative controls.
A reaction was considered as positive when a granular latex agglutination pattern was observed
within 2 min. Agglutination was evaluated on a + 1
to +3 scale. The maintenance of the smooth latex suspension at the end of the 2 min period was
considered as a negative reaction.
22.5. Hvbridomu production The same immunization schedule described to
obtain anti-T4 monoclonal antibodies was used. We selected the mouse with the best humoral immune
response to IF10 monoclonal antibody. The final endovenous booster was 100 pg of uncoupled 1 FIO
monoclonal antibody. The primary selection of hy- bridomas secreting anti-idiotype monoclonal antibod- ies against anti-T4 IF10 antibody was performed with the latex agglutination assay described above. The specificity of all positive supernatants was tested by using latex activated with a unrelated monoclonal
antibody belonging to the same subclass of anti-T4 IF10 antibody, i.e. an IgGl. For latex preparation the same conditions used to prepare the anti-T4
IFlO-latex were used. All supernatants which agglu- tinated this irrelevant IgG I -latex. were discarded and
those that did not agglutinate were selected to be further characterized by RIA.
22.6. Inhibition of T4 binding in RIA by anti-idio- type untibodies
This assay was performed by adding 0.1 ml IF 10 hybridoma supernatant (about of 5 pg/tube of the
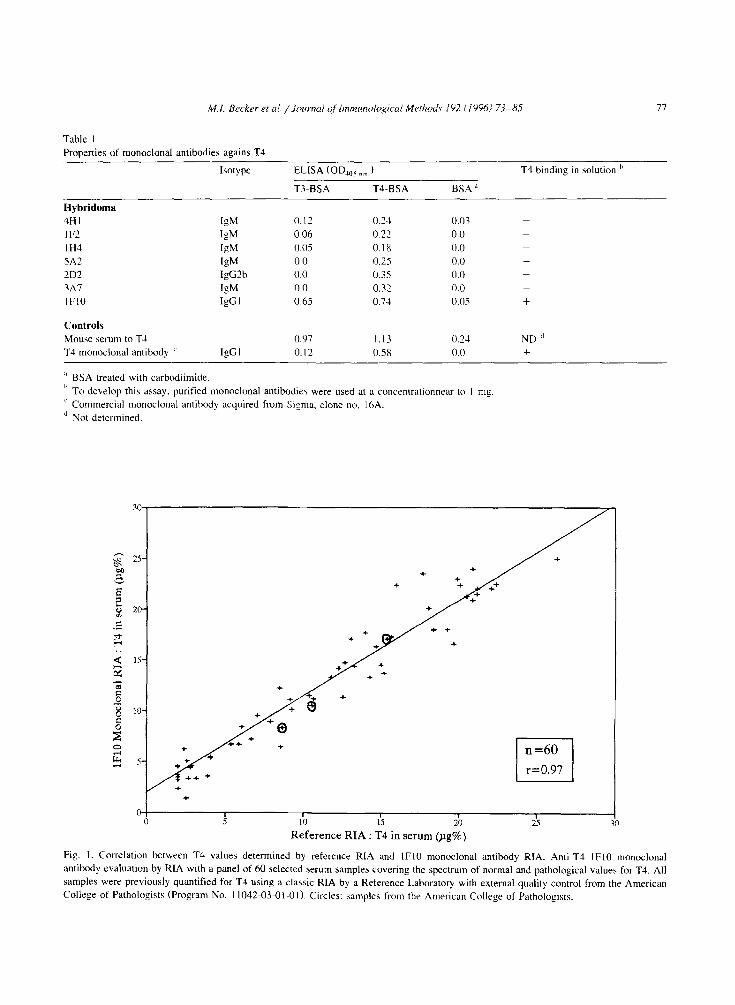
77
Table I Properties of monoclonal antibodies agains T4
Isotype
Hybridoma
4HI
IF?
IH4
5A2
‘D2
3A7
IF10
Controls
Mouse serum to T-!
T4 monoclonal antibody ’
IgM
‘gM
IgM
W IgG2b
IgM
IgG I
IgCl
HASA (OD,,,, or,, )
T?-BSA T4-BSA BSA *
T4 binding in solution h
0.17 02-i 0.03 _
0.06 0.27 0.0 _
0.05 0.18 0.0 00 0.25 0.0 _
0 0 0.35 0.0 _
0 0 0.3’ 0.0 _
0 65 0.74 0.05 +
0.97 1.13 0.21 ND “
0. I2 0.58 0.0 +
” BSA treated with carhodiimide.
h To develop this assay, purified monoclonal antihodk were used at a concentrationnear to I mg. ’ Commercial monoclonal antibody acquired from Sigma. clone no. 16A.
’ Not determined.
I 1 I
10 1.5 20
Reference RIA : T4 in serum @g%)
Fig. I. Correlation between T4 values determined by reference RIA and IF10 monoclonal antibody RIA. Anti-T4 lFl0 monoclonal
antibody evaluation by RIA with a panel of 60 selected berum samples covering the spectrum of normal and pathological values for T4. All
samples were previously quantified for TJ using a classic RIA by a Reference Laboratory with external quality control from the American
College of Pathologists (Program No. 11042-03-01-01) Circles: samples from the American College of Pathologists.
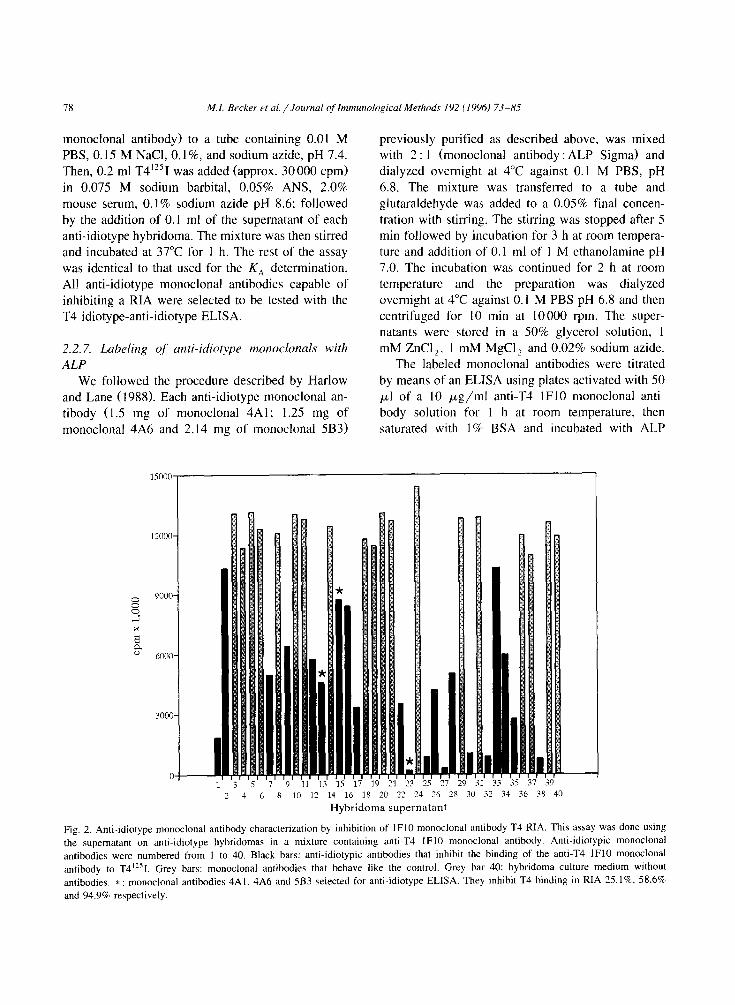
78 M. I. Becker et al. /Journal of Immunological Methods 192 C I9961 73-85
monoclonal antibody) to a tube containing 0.01 M PBS, 0.15 M NaCl, O.l%, and sodium azide, pH 7.4.
Then, 0.2 ml T4’151 was added (approx. 30000 cpm)
in 0.075 M sodium barbital, 0.05% ANS, 2.0% mouse serum, 0.1% sodium azide pH 8.6; followed
by the addition of 0.1 ml of the supernatant of each anti-idiotype hybridoma. The mixture was then stirred and incubated at 37°C for 1 h. The rest of the assay
was identical to that used for the K, determination.
All anti-idiotype monoclonal antibodies capable of inhibiting a RIA were selected to be tested with the T4 idiotype-anti-idiotype ELISA.
2.2.7. Labeling of unti-idiotype monoclonals with ALP
We followed the procedure described by Harlow
and Lane (1988). Each anti-idiotype monoclonal an- tibody (I .5 mg of monoclonal 4Al; 1.25 mg of
monoclonal 4A6 and 2.14 mg of monoclonal 5B3)
15000~
12000~
3 9000
5 -i k-c
B 9
6000
3000
previously purified as described above, was mixed with 2 : 1 (monoclonal antibody: ALP Sigma) and
dialyzed overnight at 4°C against 0.1 M PBS. pH
6.8. The mixture was transferred to a tube and glutaraldehyde was added to a 0.05% final concen- tration with stirring. The stirring was stopped after 5 min followed by incubation for 3 h at room tempera-
ture and addition of 0.1 ml of 1 M ethanolamine pH 7.0. The incubation was continued for 2 h at room temperature and the preparation was dialyzed
overnight at 4°C against 0.1 M PBS pH 6.8 and then centrifuged for 10 min at 10000 ‘pm. The super- natants were stored in a 50% glycerol solution, 1 mM ZnCl,. 1 mM MgCl, and 0.02% sodium azide.
The labeled monoclonal antibodies were titrated
by means of an ELISA using plates activated with 50 ~1 of a 10 pg/ml anti-T4 lFl0 monoclonal anti-
body solution for 1 h at room temperature, then saturated with 1% BSA and incubated with ALP
0
2 4 6 10 17 14 16 18 20 23 24 26 12 14 3b 38 4V
Hybridoma supernatant
Fig. 2. Anti-idiotype monoclonal antibody characterization by inhibition of IF10 monoclonal antibody T4 RIA. This assay was done using
the supematant on anti-idiotype hybridomas in a mixture containing anti-T4 IF10 monoclonal antibody. Anti-idiotypic monoclonal
antibodies were numbered from I to 40. Black bars: anti-idiotypic antibodies that inhibit the binding of the anti-T4 IF10 monoclonal
antibody to T4”sI. Grey bars: monoclonal antibodies that behave like the control. Grey bar 40: hybridoma culture medium without
antibodies. * : monoclonal antibodies 4A1. 4A6 and 5B3 selected for anti-idiotype ELISA. They inhibit T4 binding in RIA 25. lR, 58.6%
and 94.9% respectively.
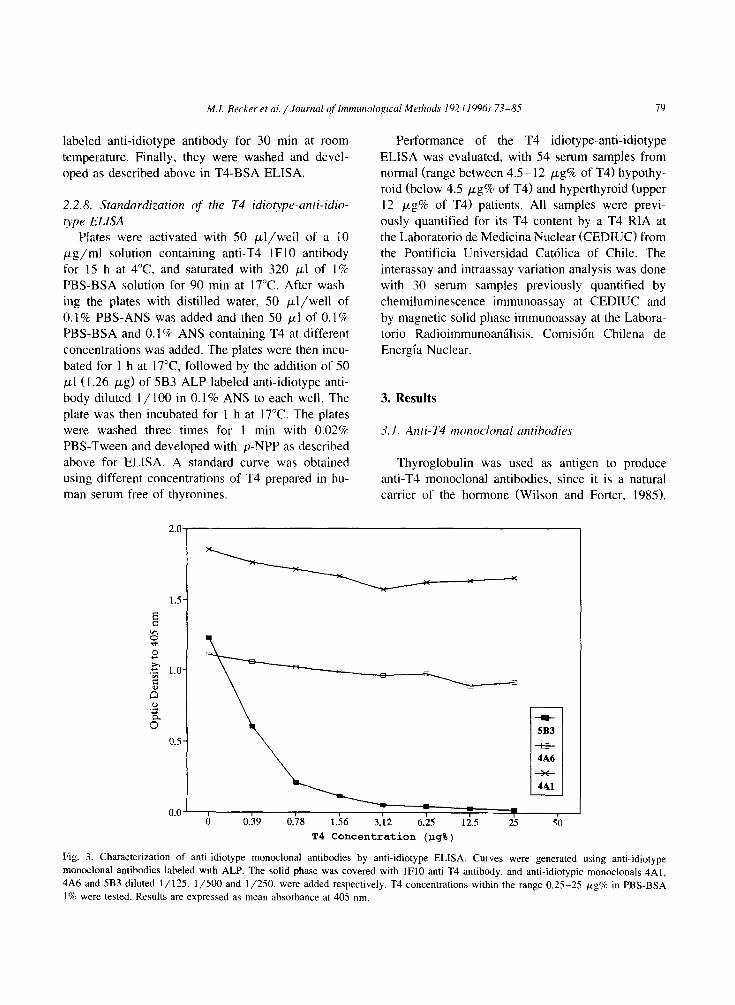
M.I. Becker et al. / Journal cf Immunnlo~ical Methods 192 (lY96J 73-85 79
labeled anti-idiotype antibody for 30 min at room
temperature. Finally, they were washed and devel-
oped as described above in T4-BSA ELISA.
2.2.8, Standardization of the T4 idiotype-anti-idio- type ELISA
Plates were activated with 50 @/well of a 10 pg/ml solution containing anti-T4 lFl0 antibody
for 15 h at 4°C and saturated with 320 ~1 of 1% PBS-BSA solution for 90 min at 17°C. After wash- ing the plates with distilled water, 50 pi/well of
0.1% PBS-ANS was added and then 50 ~1 of 0.1% PBS-BSA and 0.1% ANS containing T4 at different
concentrations was added. The plates were then incu-
bated for 1 h at 17°C followed by the addition of 50 ~1 ( 1.26 pg) of 5B3 ALP-labeled anti-idiotype anti- body diluted l/ 100 in 0.1% ANS to each well. The
plate was then incubated for 1 h at 17°C. The plates were washed three times for 1 min with 0.02% PBS-Tween and developed with p-NPP as described
above for ELISA. A standard curve was obtained using different concentrations of T4 prepared in hu- man serum free of thyronines.
Performance of the T4 idiotype-anti-idiotype
ELISA was evaluated, with 54 serum samples from normal (range between 4.5-12 pg% of T4) hypothy- roid (below 4.5 pg% of T4) and hyperthyroid (upper 12 pg% of T4) patients. All samples were previ-
ously quantified for its T4 content by a T4 RIA at the Laboratorio de Medicina Nuclear (CEDIUC) from the Pontificia Universidad Catolica of Chile. The
interassay and intraassay variation analysis was done with 30 serum samples previously quantified by
chemiluminescence immunoassay at CEDIUC and
by magnetic solid phase immunoassay at the Labora- torio Radioimmunoanalisis, Comision Chilena de Energia Nuclear.
3. Results
3. I. Anti-T4 monoclonal antibodies
Thyroglobulin was used as antigen to produce anti-T4 monoclonal antibodies, since it is a natural carrier of the hormone (Wilson and Forter, 1985).
K
T4 Concentration (Pg%)
-m- 5B3
-5- 4A6
++ 4Al
Fig. 3. Characterization of anti-idiotype monoclonal antibodies by anti-idiotype ELISA. Curves were generated using anti-idiotype
monoclonal antibodies labeled with ALP. The solid phase was covered with lFl0 anti-T4 antibody, and anti-idiotypic monoclonals 4A1,
4A6 and 5B3 diluted l/125. l/500 and l/250, were added respectively. T4 concentrations within the range 0.25-25 WgW in PBS-BSA
1% were tested. Results are expressed as mean absorbance at 405 nm.
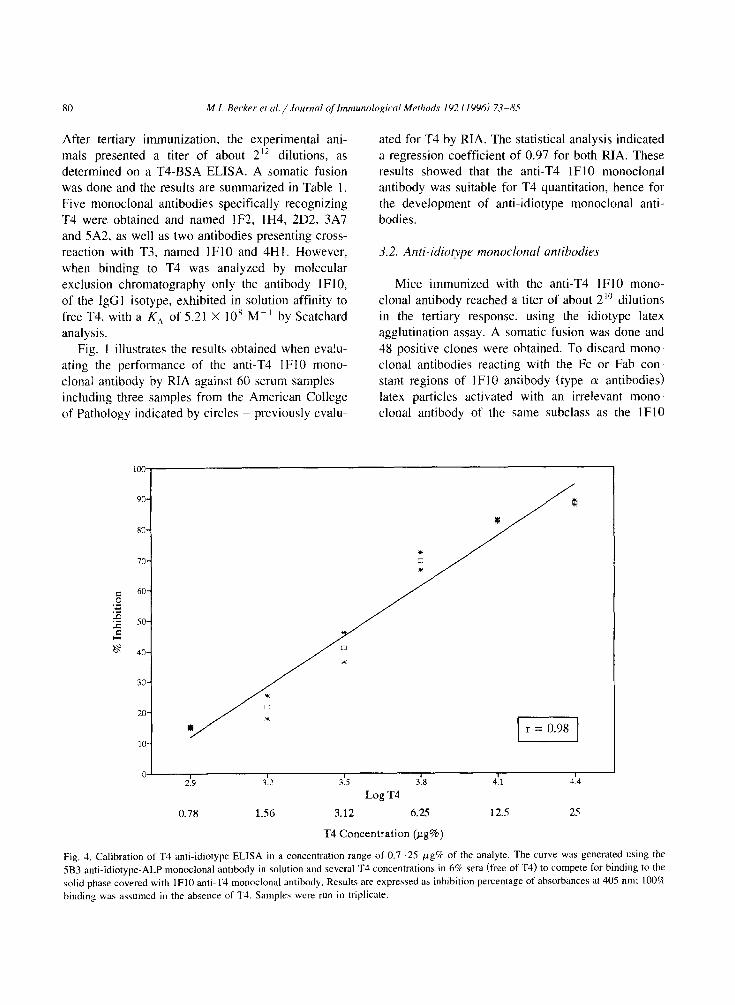
After tertiary immunization, the experimental ani- mals presented a titer of about 2” dilutions, as determined on a T4-BSA ELISA. A somatic fusion
was done and the results are summarized in Table 1. Five monoclonal antibodies specifically recognizing T4 were obtained and named 1 F2, lH4, 2D2, 3A7
and 5A2, as well as two antibodies presenting cross- reaction with T3. named IF10 and 4HI. However,
when binding to T4 was analyzed by molecular
exclusion chromatography only the antibody 1 FlO, of the IgGl isotype, exhibited in solution affinity to
free T4, with a K, of 5.21 X 10’ Mm’ by Scatchard
analysis.
Fig. 1 illustrates the results obtained when eva!u- ating the performance of the anti-T4 IF10 mono- clonal antibody by RIA against 60 serum samples -
including three samples from the American College of Pathology indicated by circles - previously evalu-
ated for T4 by RIA. The statistical analysis indicated
a regression coefficient of 0.97 for both RIA. These results showed that the anti-T4 lFl0 monoclonal antibody was suitable for T4 quantitation, hence for
the development of anti-idiotype monoclonal anti- bodies.
3.2. Anti-idiotype morwclonul antibodies
Mice immunized with the anti-T4 lFl0 mono-
clonal antibody reached a titer of about 2’” dilutions
in the tertiary response. using the idiotype latex agglutination assay. A somatic fusion was done and
48 positive clones were obtained. To discard mono- clonal antibodies reacting with the Fc or Fab con- stant regions of IF10 antibody (type (Y antibodies) latex particles activated with an irrelevant mono- clonal antibody of the same subclass as the 1 FlO
0’ I I 1 a , I
2.9 3.2 3.5 3.8 4.1 4.4
Log T4
0.78 1.56 3.12 6.25 12.s 25
T4 Concentration @g%)
Fig, 4. Calibration of T4 anti-idiotype ELISA in a concentration range of 0.7-25 pg% of the analyte. The curve was generated using the
5B3 anti-idiotype-ALP monoclonal antibody in solution and several T4 concentrations in 6% sera (free of T3) to compete for binding to the
solid phase covered with lFl0 anti-T4 monoclonal antibody. Results are expressed as inhibition percentage of absorbances at 405 nm: 100%
binding was assumed in the absence of T4. Samples were run in triplicate.
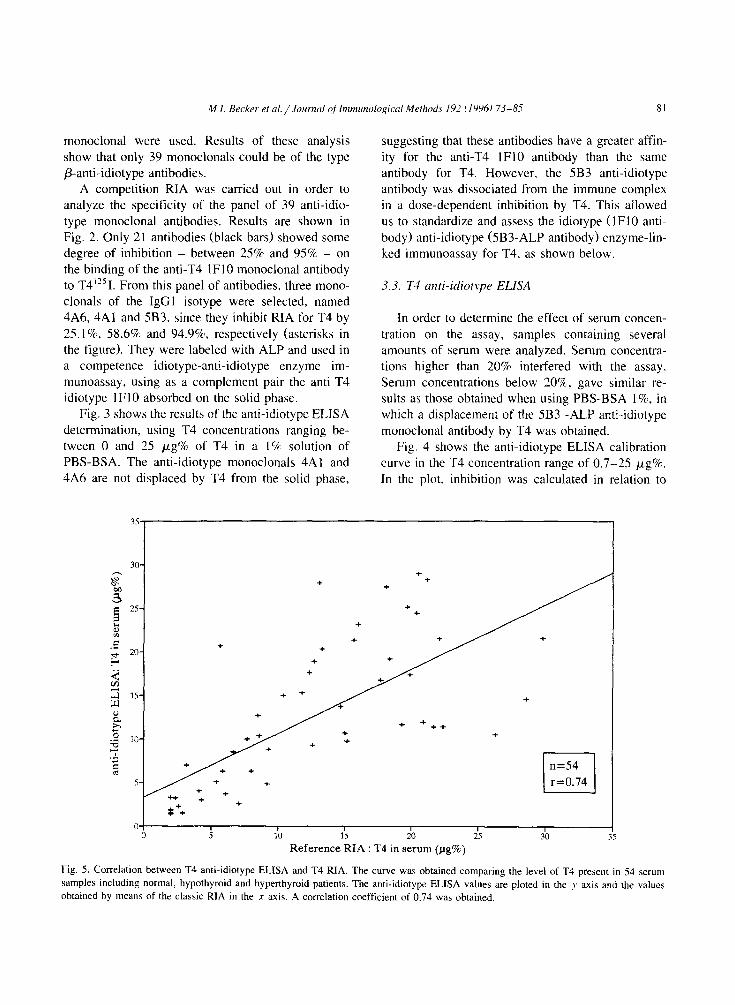
M.I. Becker et al. / Joumd of Inmuno/ogicol Methods 197 f IYY6) 73-85 81
monoclonal were used. Results of these analysis
show that only 39 monoclonals could be of the type /3-anti-idiotype antibodies.
A competition RIA was carried out in order to analyze the specificity of the panel of 39 anti-idio-
type monoclonal antibodies. Results are shown in
Fig. 2. Only 21 antibodies (black bars) showed some degree of inhibition - between 25% and 95% - on the binding of the anti-T4 IF10 monoclonal antibody to T4”‘I. From this panel of antibodies. three mono-
clonals of the IgGl isotype were selected, named 4A6, 4Al and 5B3. since they inhibit RIA for T4 by
25.1%, 58.6% and 94.9%. respectively (asterisks in the figure). They were labeled with ALP and used in a competence idiotype-anti-idiotype enzyme im-
munoassay, using as a complement pair the anti-T4 idiotype IF10 absorbed on the solid phase.
Fig. 3 shows the results of the anti-idiotype ELISA
determination. using T4 concentrations ranging be- tween 0 and 25 pg% of T4 in a 1% solution of PBS-BSA. The anti-idiotype monoclonals 4Al and
4A6 are not displaced by T4 from the solid phase,
suggesting that these antibodies have a greater affin-
ity for the anti-T4 lFl0 antibody than the same antibody for T4. However, the 5B3 anti-idiotype
antibody was dissociated from the immune complex in a dose-dependent inhibition by T4. This allowed us to standardize and assess the idiotype (lFl0 anti-
body) anti-idiotype (5B3-ALP antibody) enzyme-lin- ked immunoassay for T4, as shown below.
3.3. T3 anti-idiotyx ELISA
In order to determine the effect of serum concen-
tration on the assay, samples containing several amounts of serum were analyzed. Serum concentra- tions higher than 20% interfered with the assay. Serum concentrations below 20%, gave similar re-
sults as those obtained when using PBS-BSA I%, in which a displacement of the 5B3 -ALP anti--idiotype
monoclonal antibody by T4 was obtained. Fig. 4 shows the anti-idiotype ELISA calibration
curve in the T4 concentration range of 0.7--25 pg%.
In the plot, inhibition was calculated in relation to
3.5
30-
7 + b + +
s +
E - 2s + +
? +
s .E +
?O- +
g
$ 3 IS- + 0
R 5 _ < 10
*
‘: ‘3
5 S-
I I I I I
10 1s 20 25 30
Reference RIA : T4 in serum (pg%)
Fig. 5. Correlation between T4 anti-idiotype ELBA and T4 RIA. The curve was obtained comparing the level of T4 present in 54 serum
samples including normal, hypothyroid and hyperthyroid patients. The anti-idiotypr ELISA values are ploted in the y axis and the values
obtained by means of the classic RIA in the x axis. A correlation coefficient of 0.74 was obtained.
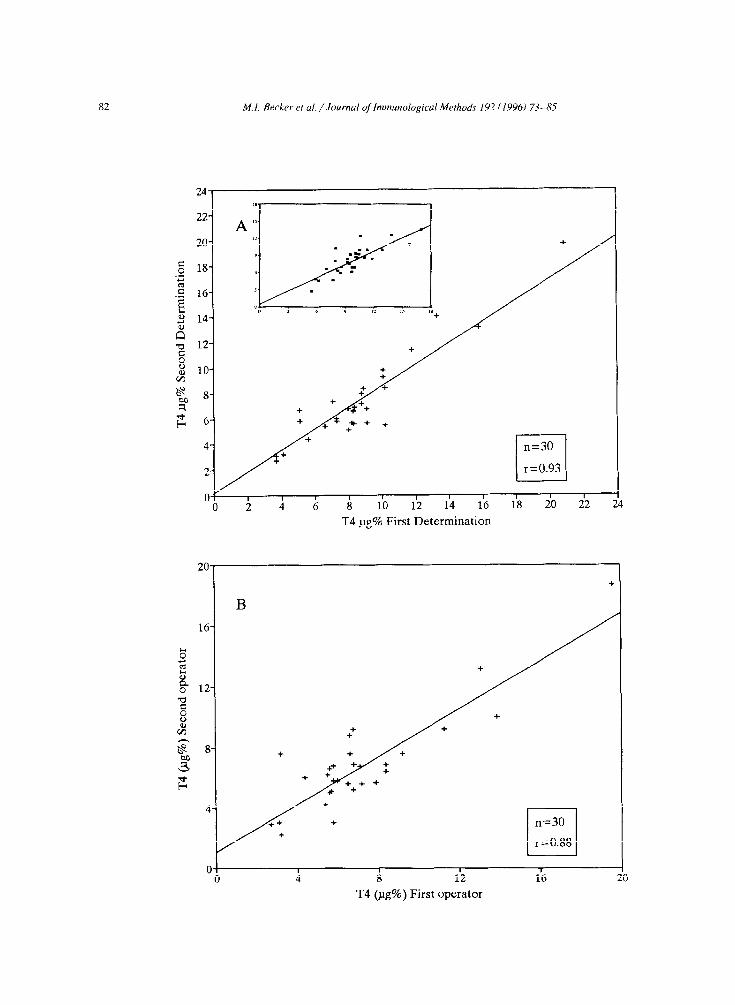
82 MI. Becker et ai. /Journal of Imnwzological Methods 192 (19961 73-85
T4 pg% First Determination
+
lG-
O- I 0 4
1 I 8 12
T4 (pg%) First operator
1 16

M.I. Becker et ~1. /Journal of bnmunological Methods 192 i 19961 73-85 83
the values obtained without T4, when the maximum
binding between lFl0 idiotype antibody and 5B3- ALP anti-idiotype antibody is obtained. The plot shows that the assay is close to linearity in the range
of I.5 12 pg% of T4. A correlation value of 0.74 (Fig. 5) was obtained with the panel of 54 samples used to characterize the anti-T4 IF10 idiotype anti-
body. Finally, to further determine the accuracy of the
anti-idiotype ELISA, a panel of 30 serum samples
was tested. Results are shown in Fig. 6. The intraas- say variation (Fig. 6A1, carried out in duplicate in
two different days by the same operator, was r =
0.93. The interassay variation (Fig. 6Bl performed in duplicate by two different operators was r = 0.88. It is worth noting that this panel of samples had been
previously evaluated by two reference centers. by means of a luminescence immunoassay and by a magnetic solid phase immunoassay; the correlation between both determinations was r = 0.82, as shown
in the insert of Fig. 6A.
4. Discussion
The determination of thyroid hormones is a tool of great diagnostic value in humans, hence the im-
portance of developing alternative methods to the traditional RIA, avoiding the use of dangerous ra- dioisotopes and reducing costs to the users. Since the
development of the first immunoenzymatic assays for T4, several fairly accurate methods have been made available.
In this work, a new ELISA format - type II, or ‘analyte observed’ (Porstmann and Kiessig, 1992) - was developed to quantitate T4. The rationale of the assay is the following: T4 present in serum competes
with a 5B3-ALP anti-idiotype monoclonal antibody for its binding to the lFl0 anti-T4 monoclonal anti-
body adhered to solid phase. In serum samples with- out T4, the maximum binding between both mono-
clonal antibodies occurs. In serum samples with T4,
the binding of both antibodies decreases in relation to hormone concentration. The method is based on two specific reactions which take place at, or close, to the antigen-antibody combining site, namely the idiotype anti-idiotype binding and the ability of the
analyte to interfere this reaction. The feasibility of the assay strongly depends on the relative affinities
of both reactions. The idiotype anti-idiotype reaction, does not need the two separate - or repeated -
epitopes required for an ELISA sandwich assay,
since T4 is a very small molecule (777.9 Dl. It is worth to note that idiotype anti-idiotype inhibition
ELISA has been used in the past for the quantitation
of a hepatitis B virus surface antigen (Kim et al., 1989) and recently, for the quantitation of the afla-
toxin hapten (Hsu and Chu, 1994). In both cases, results correlated well with those obtained by RIA.
To develop the T4 enzyme immunoassay reported in this study, was necessary to produce an anti-T4 monoclonal antibody with the proper affinity and specificity to determine T4 by RIA. The monoclonal
antibody lFl0 meet these requisites, as shown by
testing it against a panel of samples previously eval- uated by RIA (see Fig. I> and by using a non
classical RIA. in which this antibody was bound to a magnetic solid phase (data not shown). Although the
anti-T4 1 FlO monoclonal exhibits some cross-reac- tion with T3 (1.5 1%) by RIA, it is suitable for the
quantitation of T4 since the cross-reacting hormone is present in plasma at a concentration of 12 ng/ml,
so that the interference of T3 under the assay condi- tions is negligible. It is worth noting, although that all the monoclonal antibodies directed against thy- roid hormone described so far exhibit some cross-re-
action with both hormones (Marchand et al., 19871, they have been traditionally used for quantification
by RIA. The obtention of a monoclonal with full specificity to T3 or T4 is not easy to obtain. in view
of the great similarity and the small size of both hormones. On the other hand, rabbit or goat anti-T4
Fig. 6. Anti-idiotype ELISA intraassay and interassay comparison. To determine the intraassay variation, a panel of 30 serum samples was
evaluated by anti-idiotype ELISA in duplicate in two different days by the same operator, obtaining an r = 0.93 (A). The interassay
variation in duplicate was run by two different operators, obtaining an r = 0.88 (B). The inset in A, shows the analysis of the above 30
serum samples simultaneously analyzed by two reference centers, by luminescence immunoassay (X axis) at CEDIUC and magnetic solid
phase immunoassay (!: axis) at Laboratorio de Endocrinologia, Hospital J.J. Aguirre. A correlation coefficient of 0.82 was obtained,

polyclonal antibodies cross-reactive with T3. were extensively used in the past, before monoclonal anti- bodies were available.
To select anti-idiotype antibodies we developed a passive agglutination assay, based on the use of latex
particles coated with IF IO monoclonal antibody. This
procedure - simple, rapid and inexpensive - may be applied to the selection of anti-idiotype monoclonals for the quantitation of other haptens or small pep-
tides of clinical interest.
One attractive features of the T4 anti-idiotype ELISA is that the hapten does not have to be purified
or labeled, without concern about the stability of the
T4-enzyme conjugate in the assay kits. Also, since the assay uses monoclonal antibodies. it has a lower susceptibility to disturbing influences normally pre-
sent in polyclonal antibodies. In this respect, even purified hyperimmune serum to one antigen, may be contaminated by leaking of the non-related antigen
from the affinity column (Brown et al., 1989). In addition, it should be mentioned that monoclonals
are homogeneous, maintain their properties with time
and can be obtained in unlimited amounts; such properties make them the reagent of choice in differ-
ent areas of human clinical diagnosis (Waldmann,
1991; Becker et al., 1994; Castillo et al., 1995). Finally. our idiotype anti-idiotype assay for T4 is
comparable with a conventional RIA and magnetic bead RIA immunoassays, since anti-idiotype ELISA can measure with sensitivity, accuracy and repro-
ducibility the serum levels of T4 at all T4 concentra- tion ranges and therefore can be used to identify euthyroid, hypothyroid and hyperthyroid patients.
Acknowledgements
We are grateful to Dr. Patricia Contreras, Labora-
torio de Endocrinologia, Hospital J.J. Aguirre and Comision Chilena de Energia Nuclear. for evaluation of anti-T4 IF10 monoclonal antibody, Mr. J.J. Elizalde for collaboration in the tests to determine the accuracy of the assay, Dr. Pablo Valenzuela and Dr. Edith Hevia for revision of the manuscript. We also thank Miss Ignacia Aguirre for assistance in preparing the manuscript, Miss Marcela Cabezas for secretarial help and Mr. Alvaro Morales for animal care.
This work was partially supported by Grant no. 14008 from Corporation de Foment0 de la Produc- ci6n de Chile (CORFO).
References
Arons, W.L. and Hydovitz, J.D. (1959) The serum pattern of
thyroid hormones in euthyroidism and hyperthyroidism. J.
Clin. Endocrinol. 19, 548.
Becker, MI.. Juica, F.. Jamett. A., Tzichinovsky. S., Barros, S.,
Aguayo, J. and De Ioannes, A.E. (1994) Development of
anti-human B blood group monoclonal antibodies suitable as a
blood typing reagent. Hybridoma 13, 303.
Brown, D.W.. Kim. Y.T. and Siskind. G.W. (1989) The influence
of immune complexes. steric effects, and antigen-antigen in-
teractions on the sensitivity of enzyme-linked immunosorbent
assay. J. Immunol. Methods 116. 45.
Castillo, L.. Castillo, D.. Silva. W., Zapata. L., Reid. M., Ulloa,
M.T.. Seoane. M.. Maldonado, A., Valenzuela, M.T., Bustos,
R.. Tamplin, M.L.. Martinez, M.A., De Ioannes. A.E.. Jamett.
A.. Yudelevich. A. and Becker, MI. (1995) Development of
highly specific monoclonal antibodies for the diagnosis of
Vihrio cholrme 01. Hybridoma 14. 271.
Chanh, T.H.. Huot, R.I.. Schick. M.R.. Hewetson. F.J. (1989)
Anti-idiotypic antibodies against a monoclonal antibody spe-
cific for the trichothecene mycotoxin T-2, Toxicol. Appl.
Pharmacol. 100, 201.
Chopra, I.J.. Nelson, J.C.. Solomon. D.H. and Beall. G.N. (1971)
Production of antibodies specifically binding triiodothyronine
and thyroxine. J. Clin. Endocrinol. 32, 299.
Christofides. N.D. and Sheehan, C.P. (1995) Enhanced chemilu-
miniscence labeled-antibody immunoassay (Amerlite-MAB)
for free thyroxine: Desing, development, and technical valida-
tion. Clin. Chem. 31, 17.
Colucci. G.. Beazer, Y. and Waksal, S. (1987) Interaction between
hepatitis B virus and polymeric human albumin. II. Develop-
ment of syngenic monoclonal anti-antiidiotypes which mimic
hepatitis B surface antigen in the induction of immune respon-
siveness. Eur. J. Immunol. 17. 371.
Crowther, J.R. and Abu-Elzeein, E.M. (1980) Detection of anti-
bodies against foot-and-mouth-desease virus using purified
Srnphyloc~occus A protein conjugated with alkaline phos-
phatase. J. Immunol. Methods 34. 261.
Fang, V.S. and Selenkow, H.A. (1970) Determination of free
thyroxine in serum by low temperature equilibrium dialysis.
Clin. Chem. 16. 185. Harlow, E. and Lane. D. (1988) Antibodies. A Laboratory Man-
ual. Cold Spring Harbor Laboratory, Cold Spring Harbor. NY.
Herlyn, D.. Ross. A.H. and Koprowski, H. (1986) Anti-idiotypic
antibodies bear the internal image of a human tumor antigen.
Science 232, 100.
Hudson, L. and Hay, F.C. f 1980) Practical Immunology, 2nd edn.
Blackwell Scientific Publications, Oxford.
Hsu, K.-H., Chu, F.S. (1994). Production and characterization of
anti-idiotype and anti-anti-idiotype antibodies from a mono-

M.I. Becker et (11. / Jortrnal of hnuncdo,~ical Methods 192 f lY96J 73-85 85
clonal antibody against aflatoxin. J. Agric. Food Chem. 42.
2353.
Jaureguiberry, B.. Liao, L., Kuchoo. V.. Dorf, M.E. and Diamond.
B. (1988) Monoclonal anti-idiotypic antibodies to an I-J inter-
acting molecule inhibit suppression in an H-2 restricted way.
J. Immunol. 140. 3286.
Jerne. N.K. (1974) Towards a network theory of the immune
system. Ann. Immunol. 125C, 373.
Johnstone. A. and Thorpe, R. (1988) Immunochemistry in Prac-
tice, 2nd edn. Blackwell Sctentific Publications, Oxford.
Kennedy, R.C.. Eichberg, J.W.. Lanford, R.E. and Dreesman.
G.R. (1986) Anti-idiotypic antibody vaccine for type B viral
hepatitis in chimpanzees. Science 232, 220.
Kim. W.B.. Jo, Y.W., Choi, E.C. and Kim. B.K. ( 1989) Inhibition
ELISA for hepatitis B surface antigen (HBaAgJ using mono-
clonal idiotype- anti-idiotype interaction. J. Immunol. Methods
125. 273.
Kohler, G. and Milstein, C. (1975) Continuous cultures of fused
cells secreting antibody of predefined specificity. Nature 256.
495,
Marchand, J.. Gorce, A.. Badouaille. G., Bras, J., Simon. D. and
Pau. B. (1987) Production and partial characterization of
monoclonal antibodies against 3,3’,5-triiodo-L-thyronine. Hy-
bridoma 6. 97.
Mareschal, J.C., Cambiaso, CL., Janssens J., Limet. J.N., Tasi-
aux. N. and Collet-Cassart, D. (1990) Homogeneous latex
immunoassay for thyroid hormone testing determination of
thyroxine and triidothyronine. J. Immunol. Methods 13 I, 137.
McClintock, P.R.. Prabhakar. B.S. and Notkins. A.L. (1986)
Anti-idiotypic antibodies to monoclonal antibodies that neu-
tralize Coxsackievirus B, do not recognize viral receptors.
Virology 150. 352.
Miyai. K.. Ishibashi. K. and Kawashima, M. (1980) Heterologous
enzyme immunoassay for measurement of thyroxine in dried
blood sample on filter paper. Endocrinol. Jpn. 27, 186.
Monji, N.. Nalkus. H. and Castro. A. (1978) Maleimide derivative
of hapten for coupling to enzyme. A new method in enzyme
immunoassay. Biochem. Biophys. Res. Commun. 85. 671.
Pang. Y.. Yuhaaz, S.C. and Amrel, L.M. (1995) Anti-idiotypic
antibodies: biological function and structural studies. FASEB
J. 9. 43.
Paul, W.E. ( 1989) Fundamental Immunology. Raven Press. New
York.
Petersen, B.A.. Giese. R.W.. Larsen, P.R. and Karger, B.L. (1977)
Measurements of free thyroid hormones in serum by dialysis
and gas chromatography. Clin. Chem. 23. 1389.
Pind, K. t 1957) Paper-chromatographic determination of thyroid
hormone (3.5.3’-triiodothyronine) in serum without radio-
iodine. Acta Endocrinol. 26. 263.
Porstmann, T. and Kiessig. S.T. (1992) Enzyme immunoassay
techniques. An overview. J. Immunol. Methods 150. 5.
Poskitt. D.C.. Jean-Francois, M.J.B., Turnbull, S.. MacDonald, L.
and Yasmeen, D. ( 1991) The nature of immunoglobulin idio-
types and idiotype-anti-idiotype interactions in immunological
networks. lmmunol. Cell Biol. 69. 61.
Potocnjak. P.. Zavala, F.. Nussenzweig. R.S. and Nussenzweig, V.
t 1987) Inhibition of idiotype-anti-idiotype interactions for de-
tection of a parasite antigen: A new immunoassay. Science
215. 1637.
Sege. K. and Peterson, P.A. (1978) Use of anti-idiotypic antibod-
ies as ceil-surface receptor probes. Proc. Natl. Acad. Sci. USA
75, 2443.
Schail. R.F.. Fraser. Jr.. R.. Hansen. H.W.. Kern. C.V. and
Tenoso. H.J. (1978) A sensitive manual enzyme immunoassay
for thyroxine. Clin. Chem. 24, 1801.
Schmitt. H.. Khazaeli. M.B.. Rodenburg, C.M.. Kaul. S. and
Bastert. G. (1994) Production of mouse monoclonal anti-idio-
typic antibodies specific to epitopes of tumor-associated mucin
TAG- 12. Hybridoma 13. 389.
Shestowsky. W.S.. Quilliam, M.A. and Sikorska. H.M. t 1992) An
idiotypic-anti-idiotypic competitive immunoassay for quantita-
tion of okadaic acid. Toxicon 30. 1441,
Sophianopoulos, J., Jerkunica. I.. Lee. C.N. and Sgoutas, D.
( 1980) An improved ultrafiltration method for free thyroxine
and triiodothyronine in serum. Clin. Chem. 26, 159.
Spinks, C.A.. Wang, B., Mills, E.N.C. and Morgan. M.R.A.
t 1993) Production and characterization of monoclonal anti-idi-
otype antibody mimics for the pyrethroid insecticides and
herbicide paraquat. Food Agric. Immunol. 5. 13.
Thanavala, Y. (1989) Anti-idiotypes vaccines. Trends Biotechnol.
7, 62.
Ullman. E.F., Blakermore, J.. Leute. R.K., Eimstad. W. and
Jaklitsch. A. (1975) Homogeneous enzyme immunoassay for
thyroxine. Clin. Chem. 2 I, IO1 I.
Waldmann. T.A. (1991) Monoclonal antibodies in diagnosis and
therapy. Science 252, 1657.
Wang. L.. Hexter. C.S. and Inbar. M. (1981) Monoclonal antibod-
ies to thyroid hormones anti-thyroxine and anti-tri-
iodothyronine. In: G.J. Hammerling, G.H. Hammerling and
J.F. Kearney (Eds.). Monoclonal Antibodies and T Cell Hy-
bridomas. Elsevier, Amsterdam, pp. 357.
Wiersinga. W.M. and Chopra, J. (1982) Radioimmunoassay of
thyroxine (T4) 3.5.3’-triiodothyronine, 3.3’,5-triiodothyronine
(reverse T3. rT3) and 3.3’-diiodothyronine. Methods Enzymol.
84, 273.
Wilson, J.D. and Forter. D.W. (1985) Textbook of Endocrinology.
7th edn. Williams. London. Ch. 5.




















